ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ. Τοµέας Γενετικής, Βιολογίας Κυττάρου και Ανάπτυξης
|
|
|
- Διώνη Κανακάρης-Ρούφος
- 7 χρόνια πριν
- Προβολές:
Transcript
1 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ Τοµέας Γενετικής, Βιολογίας Κυττάρου και Ανάπτυξης ΜΕΛΕΤΗ ΤΩΝ ΜΗΧΑΝΙΣΜΩΝ ΠΡΟΚΛΗΣΗΣ ΑΝΕΥΠΛΟΕΙ ΙΑΣ ΣΤΟΝ ΑΝΘΡΩΠΟ ΑΠΟ ΤΗ ΦΑΡΜΑΚΕΥΤΙΚΗ ΕΝΩΣΗ Υ ΡΟΧΛΩΡΟΘΕΙΑΖΙ ΙΟ (HCTZ) ΜΕ ΜΟΡΙΑΚΕΣ ΚΥΤΤΑΡΟΓΕΝΕΤΙΚΕΣ ΤΕΧΝΙΚΕΣ in vitro Ι ΑΚΤΟΡΙΚΗ ΙΑΤΡΙΒΗ ΤΟΥ ΚΩΝΣΤΑΝΤΙΝΟΥ ΑΝ ΡΙΑΝΟΠΟΥΛΟΥ ΒΙΟΛΟΓΟΥ, MSc ΠΑΤΡΑ 25

2 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ Τοµέας Γενετικής, Βιολογίας Κυττάρου και Ανάπτυξης ΜΕΛΕΤΗ ΤΩΝ ΜΗΧΑΝΙΣΜΩΝ ΠΡΟΚΛΗΣΗΣ ΑΝΕΥΠΛΟΕΙ ΙΑΣ ΣΤΟΝ ΑΝΘΡΩΠΟ ΑΠΟ ΤΗ ΦΑΡΜΑΚΕΥΤΙΚΗ ΕΝΩΣΗ Υ ΡΟΧΛΩΡΟΘΕΙΑΖΙ ΙΟ (HCTZ) ΜΕ ΜΟΡΙΑΚΕΣ ΚΥΤΤΑΡΟΓΕΝΕΤΙΚΕΣ ΤΕΧΝΙΚΕΣ in vitro Ι ΑΚΤΟΡΙΚΗ ΙΑΤΡΙΒΗ ΤΟΥ ΚΩΝΣΤΑΝΤΙΝΟΥ ΑΝ ΡΙΑΝΟΠΟΥΛΟΥ ΒΙΟΛΟΓΟΥ, MSc ΠΑΤΡΑ 25

3 ΣΥΜΒΟΥΛΕΥΤΙΚΗ ΕΠΙΤΡΟΠΗ Επιβλέπουσα Καθηγήτρια: Γ. Στεφάνου Αναπληρώτρια Καθηγήτρια Τµήµα Βιολογίας, Πανεπιστήµιο Πατρών Μέλη: Ν. ηµόπουλος Καθηγητής Τµήµα Βιολογίας, Πανεπιστήµιο Πατρών Σ. Αλαχιώτης Καθηγητής Τµήµα Βιολογίας, Πανεπιστήµιο Πατρών ΕΞΕΤΑΣΤΙΚΗ ΕΠΙΤΡΟΠΗ Σ. Αλαχιώτης Καθηγητής Τµήµα Βιολογίας, Πανεπιστήµιο Πατρών Γ. Γιαννόπουλος Καθηγητής Τµήµα Βιολογίας, Πανεπιστήµιο Πατρών Ν. ηµόπουλος Καθηγητής Τµήµα Βιολογίας, Πανεπιστήµιο Πατρών Π. Κορδοπάτης Καθηγητής Τµήµα Φαρµακευτικής, Πανεπιστήµιο Πατρών Ι. Ματσούκας Καθηγητής Τµήµα Χηµείας, Πανεπιστήµιο Πατρών Γ. Στεφάνου Αναπληρώτρια Καθηγήτρια Τµήµα Βιολογίας, Πανεπιστήµιο Πατρών. Τσαµπάος Καθηγητής Τµήµα Ιατρικής, Πανεπιστήµιο Πατρών Η έγκριση της διδακτορικής διατριβής από το Τµήµα Βιολογίας του Πανεπιστηµίου Πατρών δεν υποδηλώνει την αποδοχή των γνωµών του συγγραφέα. Ν. 5343/1932, άρθρο 22.

4 Στους γονείς µου, ελάχιστο δείγµα εκτίµησης και αγάπης Στον αδελφό µου, ιονύση

5 ΠΡΟΛΟΓΟΣ Η παρούσα διατριβή πραγµατοποιήθηκε στο Εργαστήριο Γενετικής του Τοµέα Γενετικής, Βιολογίας Κυττάρου και Ανάπτυξης του Τµήµατος Βιολογίας του Πανεπιστηµίου Πατρών. Η ανάθεση του θέµατος και η επίβλεψη της εργασίας έγινε από την Αναπληρώτρια Καθηγήτρια κ. Στεφάνου Γεωργία την οποία ευχαριστώ ιδιαίτερα για τη φιλική συνεργασία της, τις συνεχείς υποδείξεις της κατά τη διεξαγωγή των πειραµάτων και για τα σχόλια και προτάσεις της κατά τη συγγραφή της διατριβής. Στην Επιβλέπουσα Καθηγήτρια οφείλεται η υποστήριξη της εργασίας µε υποτροφία για το χρονικό διάστηµα από το πρόγραµµα "Καραθεοδωρή" της Επιτροπής Ερευνών του Πανεπιστηµίου Πατρών. Η εµπιστοσύνη που µου έδειξε µε τιµά, ενώ η υποµονή και η καλοσύνη της µε βοήθησαν να ξεπερνώ τα όποια προβλήµατα προέκυπταν. Τον Καθηγητή κ. Νικόλαο ηµόπουλο, µέλος της συµβουλευτικής επιτροπής, ευχαριστώ θερµά για την αµέριστη συµπαράσταση και αδιάκοπη καθοδήγησή του, παράλληλα µε την κ. Γ. Στεφάνου, σε όλη τη διάρκεια της διατριβής. Τον ευχαριστώ ιδιαίτερα για την ευκαιρία που µου έδωσε, ως επιστηµονικός υπεύθυνος, να συµµετάσχω σε δύο ερευνητικά προγράµµατα χρηµατοδοτούµενα από την Ευρωπαϊκή Ένωση. Η επιστηµονική του κατάρτιση, η απλότητα, η πραότητα και η ειλικρίνεια που τον διακρίνουν συνιστούν το πρότυπο επιστήµονα και ανθρώπου. Τον Καθηγητή Γενετικής κ. Σταµάτη Αλαχιώτη ευχαριστώ για την αποδοχή του να οριστεί µέλος της τριµελούς συµβουλευτικής επιτροπής και για τις συµβουλές του. Τον Καθηγητή του Τµήµατος Βιολογίας κ. Γεώργιο Γιαννόπουλο, τον Καθηγητή του Τµήµατος Φαρµακευτικής κ. Παύλο Κορδοπάτη, τον Καθηγητή του Τµήµατος Χηµείας κ. Ιωάννη Ματσούκα και τον Καθηγητή του Τµήµατος Ιατρικής κ. ιονύση Τσαµπάο ευχαριστώ θερµά για τη συµµετοχή τους στην Εξεταστική Επιτροπή και τις εύστοχες παρατηρήσεις τους. Ευχαριστίες επίσης οφείλω στους δότες του πειραµατικού υλικού της διατριβής και στον Αναπληρωτή Καθηγητή του Τµήµατος Βιολογίας κ. Π. Κατσώρη ο οποίος έκανε τις αιµοληψίες. Τον Επίκουρο Καθηγητή του Τµήµατος Μαθηµατικών κ. Φ. Αλεβίζο ευχαριστώ για τις χρήσιµες συµβουλές του σε θέµατα Στατιστικής. Την κ. ήµητρα Θωµαΐδου, Ερευνήτρια Γ του Ινστιτούτου Παστέρ ευχαριστώ για τη δυνατότητα χρήσης του Συνεστιακού Μικροσκοπίου. Την Καθηγήτρια του Τµήµατος Βιολογίας κ. C. Tanzarella του Πανεπιστηµίου "Roma Tre" της Ιταλίας και τους ερευνητές ρ. Α. Antoccia και ρ. A. Sgura ευχαριστώ για τη βοήθεια τους σε πειραµατικές διαδικασίες και για τη φιλοξενία τους στα πλαίσια του προγράµµατος INTERREG II (M.5-2). Επιπρόσθετα θα ήθελα να ευχαριστήσω τους φίλους και συνεργάτες του Εργαστηρίου M. Τυράκη, MSc, Α. Κουλουµέντα, MSc, Μ. Ευθυµίου, MSc, Σ. Σαλαµαστράκη και. Ουρανού για την άψογη και δηµιουργική συνεργασία που είχαµε. Τη Μ. Ευθυµίου ευχαριστώ θερµά για την ηθική συµπαράσταση και κατανόηση που έδειξε και η οποία συνέβαλε µε το δικό της ιδιαίτερο τρόπο στην ολοκλήρωση της παρούσας διατριβής. -I-

6 Ένα πολύ µεγάλο ευχαριστώ ανήκει στους γονείς µου και στον αδελφό µου ιονύση για τη συνεχή παρότρυνση και βοήθεια, ηθική και οικονοµική, στην ολοκλήρωση των σπουδών µου. Στους γονείς µου οφείλω τη µέχρι τώρα πορεία µου, αποτέλεσµα της αµέριστης συµπαράστασης και αγάπης τους. Με την ολοκλήρωση της διατριβής µου, νοιώθω την ανάγκη να εκφράσω την απόλυτη ευγνωµοσύνη προς το οικογενειακό µου περιβάλλον που µε βοήθησαν και που εξακολουθούν να µε στηρίζουν στις αποφάσεις µου. εκέµβριος, 25, Πάτρα Κωνσταντίνος Ανδριανόπουλος -II-

7 ΠΕΡΙΕΧΟΜΕΝΑ ΕΙΣΑΓΩΓΗ ΥΠΕΡΤΑΣΗ Γενικά Μηχανισµός ρύθµισης της αρτηριακής πίεσης Τύποι υπέρτασης Γενετική αιτιολογία της υπέρτασης Αντιϋπερτασικά φάρµακα ιουρητικά Αγγειοδιασταλτικά Παράγοντες οι οποίοι τροποποιούν τη λειτουργία του νευρικού συστήµατος Παράγοντες οι οποίοι επιδρούν άµεσα στο Κ.Ν.Σ Παράγοντες οι οποίοι επιδρούν στο περιφερικό νευρικό σύστηµα Παράγοντες που αναστέλλουν την παραγωγή ή τη δράση της αγγειοτασίνης ΙΙ ΙΟΥΡΗΤΙΚΑ ΦΑΡΜΑΚΑ-ΘΕΙΑΖΙ ΙΑ Γενικά Τύποι διουρητικών φαρµάκων Αναστολείς ανθρακικής ανυδράσης Ωσµωτικά διουρητικά Αναστολείς συµµεταφορέα Na + -Κ + -2Cl (διουρητικά αγκύλης ή ισχυρά διουρητικά) Αναστολείς συµµεταφορέα Na + -Cl - (θειαζίδια και θειαζιδικά ανάλογα) Αναστολείς νεφρικών επιθηλιακών διαύλων Na + (καλιοπροστατευτικά διουρητικά) Ανταγωνιστές αλατοκορτικοειδών υποδοχέων (ανταγωνιστές αλδοστερόνης-καλιοπροστατευτικά διουρητικά) Γενετικοί παράγοντες που επηρεάζουν τη δράση διουρητικών φαρµάκων Μεταβολισµός διουρητικών φαρµάκων Φάση Ι µεταβολισµού διουρητικών φαρµάκων Φάση ΙΙ µεταβολισµού διουρητικών φαρµάκων Αλληλεπιδράσεις διουρητικών φαρµάκων µε κυτταρικές πρωτεΐνες Σύστηµα ρενίνης-αγγειοτασίνης Πρωτεΐνη αντουκίνη Συνθετάση µονοξειδίου του αζώτου (NO) Υ ΡΟΧΛΩΡΟΘΕΙΑΖΙ ΙΟ (CAS No ) Φυσικές και χηµικές ιδιότητες Παραγωγή Ενδείξεις-Εµπορική Ονοµασία- οσολογία Μηχανισµός δράσης του υδροχλωροθειαζιδίου (HCTZ) Πρωτεΐνες-µεταφορείς Ανθρακική ανυδράση Περιφερικός τύπος υποδοχέα των βενζοδιαζεπινών Κατεχολαµινεργικές ορµόνες/β2-αδρενεργικοί υποδοχείς Προστακυκλίνη (PGI2) Καλλικρεΐνη III-
... 12 2.2.4 Αναστολείς συµµεταφορέα Na + -Cl - (θειαζίδια και θειαζιδικά ανάλογα).")
8 3.5 Φαρµακοκινητικές ιδιότητες Βιοδιαθεσιµότητα Κατανοµή Μεταβολισµός Απέκκριση Γενετική δράση του υδροχλωροθειαζιδίου In vitro µελέτες Προκαρυωτικοί οργανισµοί Ευκαρυωτικοί οργανισµοί Κύτταρα Θηλαστικών Άνθρωπος In vivo µελέτες Έντοµα Θηλαστικά Καρκινογένεση Τρωκτικά Άνθρωπος ΜΙΚΡΟΠΥΡΗΝΕΣ Ιστορική αναδροµή Μέθοδος αναστολής της κυτταροκίνησης-κυτταροχαλασίνη-β Προσδιορισµός της συχνότητας µικροπυρήνων- Κριτήρια αναγνώρισης Κυτταροτοξικότητα Αναγνώριση του µηχανισµού δηµιουργίας µικροπυρήνων και ανίχνευση γεγονότων λανθασµένου χρωµοσωµατικού αποχωρισµού Εφαρµογές Παράγοντες που επηρεάζουν τη συχνότητα µικροπυρήνων Πρόσθετοι βιολογικοί δείκτες Μειονεκτήµατα της µεθόδου αναστολής της κυτταροκίνησης ΧΡΩΜΟΣΩΜΑΤΙΚΗ ΑΣΤΑΘΕΙΑ - ΑΝΕΥΠΛΟΕΙ ΙΑ Αριθµητικές χρωµοσωµατικές ανωµαλίες Επιπτώσεις ανευπλοειδίας Μηχανισµοί ανευπλοειδίας Χρωµοσωµατικός µη αποχωρισµός Χρωµοσωµατική απώλεια Στόχοι δράσης ανευπλοειδογόνων ενώσεων Παράγοντες που διαταράσσουν το σωστό διαχωρισµό του γενετικού υλικού και σχετίζονται µε βασικά και ρυθµιστικά στοιχεία της µιτωτικής συσκευής Μικροσωληνίσκοι Κινητικές πρωτεΐνες Κεντρόσωµα Πυρηνική µεµβράνη Χρωµοσώµατα Κινητοχώρος Κεντροµέρος Τελοµέρη Πυρηνίσκος Κυτταροκίνηση Σηµεία ελέγχου της µίτωσης ΣΚΟΠΟΣ ΤΗΣ ΙΑΤΡΙΒΗΣ IV-

9 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟ ΟΙ ΚΥΤΤΑΡΙΚΕΣ ΚΑΛΛΙΕΡΓΕΙΕΣ Ανθρώπινα λεµφοκύτταρα ολικού αίµατος Λεµφοκύτταρα Συλλογή δειγµάτων Πηγή λεµφοκυττάρων Συνθήκες καλλιέργειας λεµφοκυττάρων Κυτταρικές σειρές H.Ep.2 (Human Epidermoid Carcinoma) H.F.F.F.2 (Human Caucasian F(o)etal Foreskin Fibroblast) C2C12 (Mouse, C3H, Muscle, Myoblast) ΜΕΛΕΤΗ ΚΥΤΤΑΡΟΤΟΞΙΚΟΤΗΤΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΒΛΑΒΗΣ ΜΕΘΟ ΟΣ ΑΝΑΣΤΟΛΗΣ ΤΗΣ ΚΥΤΤΑΡΟΚΙΝΗΣΗΣ (CYTOKINESIS-BLOCK METHOD/CBMN) Λεµφοκύτταρα Συνθήκες καλλιέργειας Αποµόνωση λεµφοκυττάρων Κυτταρική σειρά Hep Κυτταρική σειρά HFFF ΜΕΛΕΤΗ ΧΡΩΜΟΣΩΜΑΤΙΚΗΣ ΑΠΩΛΕΙΑΣ (CL) ΚΑΙ ΧΡΩΜΟΣΩΜΑΤΙΚΟΥ ΜΗ ΑΠΟΧΩΡΙΣΜΟΥ (ND) ΜΕΘΟ ΟΣ ΑΝΑΣΤΟΛΗΣ ΤΗΣ ΚΥΤΤΑΡΟΚΙΝΗΣΗΣ ΣΕ ΣΥΝ ΥΑΣΜΟ ΜΕ IN SITU ΥΒΡΙ ΟΠΟΙΗΣΗ ΜΕ ΦΘΟΡΟΧΡΩΜΑΤΑ (CBMN/FISH) In Situ υβριδοποίηση µε φθοροχρώµατα (FISH) - Χρήση πανκεντροµερικού ανιχνευτή για τον προσδιορισµό του µηχανισµού δηµιουργίας µικροπυρήνων Προετοιµασία παρασκευασµάτων (Slide preparation) Μονιµοποίηση-Ωρίµανση παρασκευασµάτων (Fixation-Aging) Επεξεργασία του κυτταρικού υλικού στις αντικειµενοφόρους (Pretreatment) Επίδραση µε HCl Επίδραση µε πεπσίνη Πειραµατική πορεία ιαλύµατα Αποδιάταξη DNA στόχου και DNA ανιχνευτή (Denaturation) Πειραµατική πορεία Υβριδοποίηση Θερµοκρασία ph Μονοσθενή κατιόντα Οργανικοί διαλύτες Πειραµατική πορεία ιαλύµατα Εκπλύσεις µετά την υβριδοποίηση (Posthybridization washes) Πειραµατική πορεία Ανοσοεντοπισµός του ανιχνευτή µε φθορίζουσες ουσίες (Ιmmunocytochemistry) Πειραµατική πορεία ιαλύµατα V-
 ΚΑΙ ΧΡΩΜΟΣΩΜΑΤΙΚΟΥ ΜΗ ΑΠΟΧΩΡΙΣΜΟΥ (ND) 96 8.")
10 8.2 ιπλή In Situ υβριδοποίηση µε φθοροχρώµατα (DUAL FISH) - Συµµετοχή των χρωµοσωµάτων Χ, Υ και 8 σε ανευπλοειδικά φαινόµενα Επεξεργασία του κυτταρικού υλικού στις αντικειµενοφόρους Επίδραση µε πεπσίνη Αποδιάταξη DNA ανιχνευτών Υβριδοποίηση Ανιχνευτές Ανοσοεντοπισµός των ανιχνευτών µε φθορίζουσες ουσίες Συνδυασµένη σήµανση του κινητοχώρου και του κεντροµέρους (CREST - FISH) Τεχνική CREST Μονιµοποίηση Πειραµατική πορεία ιαλύµατα Τεχνική FISH Μονιµοποίηση-Ωρίµανση παρασκευασµάτων Ανιχνευτής, Appligene Oncor CP595-DG Εντοπισµός του ανιχνευτή µε φθορίζουσες ουσίες ιαλύµατα ΜΕΛΕΤΗ ΤΗΣ ΑΚΕΡΑΙΟΤΗΤΑΣ ΤΗΣ ΜΙΤΩΤΙΚΗΣ ΣΥΣΚΕΥΗΣ Σε καλλιέργειες λεµφοκυττάρων Συνθήκες καλλιέργειας Αποµόνωση λεµφοκυττάρων Χρώση των παρασκευασµάτων Σε καλλιέργειες κυτταρικών σειρών µε τη µέθοδο διπλού ανοσοφθορισµού για την α- και γ- τουµπουλίνη (HEp2, HFFF2, C2C12) Κυτταρική σειρά HEp Κυτταρική σειρά HFFF Κυτταρική σειρά C2C Ανοσοχηµική ανίχνευση α- και γ- τουµπουλίνης ιαλύµατα ΜΙΚΡΟΣΚΟΠΙΚΗ ΠΑΡΑΤΗΡΗΣΗ ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ 12 ΑΠΟΤΕΛΕΣΜΑΤΑ ΕΠΑΓΩΓΗ ΜΙΚΡΟΠΥΡΗΝΩΝ ΩΣ ΕΙΚΤΗΣ ΓΕΝΕΤΙΚΗΣ ΒΛΑΒΗΣ - ΜΕΛΕΤΗ ΚΥΤΤΑΡΟΤΟΞΙΚΟΤΗΤΑΣ ιεγερµένα λεµφοκύτταρα Μη διεγερµένα λεµφοκύτταρα ΜΕΛΕΤΗ ΤΟΥ ΜΗΧΑΝΙΣΜΟΥ ΠΡΟΕΛΕΥΣΗΣ ΜΙΚΡΟΠΥΡΗΝΩΝ Λεµφοκύτταρα ανθρώπου ιεγερµένα λεµφοκύτταρα Μη διεγερµένα λεµφοκύτταρα Ανθρώπινες κυτταρικές σειρές HFFF HEp VI-

11 14. ΜΕΛΕΤΗ ΑΝΕΥΠΛΟΕΙ ΙΚΩΝ ΦΑΙΝΟΜΕΝΩΝ ΜΕ ΤΗ ΣΥΜΜΕΤΟΧΗ ΤΩΝ ΧΡΩΜΟΣΩΜΑΤΩΝ Χ, Υ ΚΑΙ ιεγερµένα λεµφοκύτταρα Καλλιέργειες λεµφοκυττάρων ατόµων νεαρής ηλικίας Χρωµόσωµα Χ Χρωµόσωµα Υ Χρωµόσωµα Καλλιέργειες λεµφοκυττάρων ατόµων ώριµης ηλικίας Χρωµόσωµα Χ Χρωµόσωµα Υ Χρωµόσωµα Μη διεγερµένα λεµφοκύτταρα Χρωµόσωµα Χ Χρωµόσωµα ΙΕΡΕΥΝΗΣΗ ΤΟΥ ΜΗΧΑΝΙΣΜΟΥ ΠΡΟΚΛΗΣΗΣ ΑΝΕΥΠΛΟΕΙ ΙΚΩΝ ΦΑΙΝΟΜΕΝΩΝ Κυτταρικές σειρές ΗFFF2, HEp2 και ανθρώπινα λεµφοκύτταρα Κυτταρική σειρά C2C ΣΥΖΗΤΗΣΗ..196 ΠΕΡΙΛΗΨΗ.218 SUMMARY...22 ΒΙΒΛΙΟΓΡΑΦΙΑ VII-


12
13 Εισαγωγή Υπέρταση 1. ΥΠΕΡΤΑΣΗ 1.1 Γενικά Η συνεχής εντός στενών ορίων ρύθµιση της αρτηριακής πίεσης πραγµατοποιείται, προκειµένου να παρέχει επαρκή αιµάτωση των ιστών, χωρίς να προκαλείται βλάβη στο αγγειακό σύστηµα, ιδιαίτερα στον εσωτερικό χιτώνα των αρτηριών. Η αρτηριακή πίεση είναι ευθέως ανάλογη του γινοµένου της καρδιακής παροχής και των περιφερικών αγγειακών αντιστάσεων. Ως υπέρταση ορίζεται η αρτηριακή πίεση που ξεπερνά τα ανώτερα φυσιολογικά όρια (14/9mmHg) και οφείλεται σε αυξηµένο τόνο των λείων µυών των περιφερικών αγγείων, που οδηγεί σε αυξηµένες αντιστάσεις των αρτηριδίων και µειωµένη χωρητικότητα του φλεβικού συστήµατος µε αποτέλεσµα µετατόπιση αίµατος από τις συστηµατικές φλέβες στην καρδιά. Η χρόνια υπέρταση µπορεί να οδηγήσει σε συµφορητική καρδιακή ανεπάρκεια, έµφραγµα του µυοκαρδίου, νεφρική βλάβη και αγγειακά εγκεφαλικά επεισόδια. Η εγκεφαλική συµφόρηση αποτελεί τη δεύτερη κύρια αιτία θανάτου σε ολόκληρο τον κόσµο, γεγονός που οφείλεται αποκλειστικά στην υψηλή αρτηριακή πίεση. Στις Η.Π.Α. η υψηλή αρτηριακή πίεση είναι µια πολύ κοινή διαταραχή, που παρουσιάζεται περίπου στο 25% του πληθυσµού, ενώ το αντίστοιχο ποσοστό στην Ευρώπη είναι 44%. Επιπλέον στις Η.Π.Α. η διάγνωση γίνεται έγκαιρα στο 7% του πληθυσµού και µολονότι το 6% λαµβάνει θεραπευτική αγωγή µόνο σε ένα ποσοστό 35% η υπέρταση θεραπεύεται κατάλληλα. Ερευνητές του Πανεπιστηµίου Loyola στο Μίσιγκαν των Η.Π.Α. σύγκριναν τα ήδη δηµοσιευµένα αποτελέσµατα εξέτασης της αρτηριακής πίεσης σε αντιπροσωπευτικά δείγµατα οκτώ διαφορετικών χωρών. Σε αυτές, εκτός των Η.Π.Α. και του Καναδά που έφερε και το µεγαλύτερο δείγµα άτοµα, συµπεριλαµβάνονται οι ευρωπαϊκές χώρες Ιταλία, Γερµανία, Ισπανία, Φιλανδία, Αγγλία και Σουηδία µε το µικρότερο δείγµα των 18 ατόµων. Η αναλογία των ατόµων που λαµβάνει αντιϋπερτασική φαρµακευτική αγωγή είναι σχεδόν διπλάσια στην οµάδα της Βορείου Αµερικής (44%) συγκρινόµενη µε αυτήν της Ευρώπης (27%) και παρ όλα αυτά µόνο το 8% των Ευρωπαίων καταφέρνουν να ελέγξουν και να επαναφέρουν την αρτηριακή πίεση στα φυσιολογικά επίπεδα. Αυτό συµβαίνει για το 23% των ατόµων της οµάδας της Βορείου Αµερικής. Ένα άλλο σηµαντικό στοιχείο που κατέδειξε η συγκριτική αυτή µελέτη είναι η στενή σχέση της υψηλής αρτηριακής πίεσης µε τα αγγειακά εγκεφαλικά επεισόδια και τους καρδιακούς θανάτους. Έτσι στην Ευρώπη η µέση αναλογία θνησιµότητας οφειλόµενη στην εγκεφαλική συµφόρηση-αποπληξία είναι 41.2 στα 1. άτοµα και στη Βόρειο Αµερική 27.6 στα 1. άτοµα, αναλογίες όµοιες µε τις συχνότητες εµφάνισης υπέρτασης σε αυτές τις χώρες. Γνωρίζοντας ότι περιβαλλοντικοί παράγοντες, όπως αγχώδης τρόπος ζωής, υψηλή διαιτητική πρόσληψη νατρίου, παχυσαρκία, έλλειψη φυσικής άσκησης και κάπνισµα οι οποίοι προδιαθέτουν στην εµφάνιση της υπέρτασης σ ένα άτοµο δεν διαφέρουν αρκετά µεταξύ των δύο οµάδων, η έγκαιρη διάγνωση, η κατάλληλη θεραπευτική αγωγή και η στενή παρακολούθηση των νοσούντων είναι οι κύριοι λόγοι της σηµαντικής διαφοράς νοσηρότητας και θνησιµότητας στην Ευρώπη και στη Βόρειο Αµερική (Η.Π.Α. και Καναδάς) (Wolf-Maier et al., 23)
 και οφείλεται σε αυξηµένο τόνο των λείων µυών των περιφερικών αγγείων, που οδηγεί σε αυξηµένες αντιστάσεις")
14 Εισαγωγή Υπέρταση 1.2 Μηχανισµός ρύθµισης της αρτηριακής πίεσης Υπάρχουν δύο κύριοι τύποι συστηµάτων ελέγχου της αρτηριακής πίεσης: 1. Το πρώτο σύστηµα αποτελεί την πρώτη γραµµή άµυνας εναντίον των παθολογικών πιέσεων και αποτελείται από νευρικούς µηχανισµούς παλίνδροµης ρύθµισης οι οποίοι λαµβάνουν χώρα µέσα σε δευτερόλεπτα. Πρόκειται για τον αντανακλαστικό µηχανισµό των αρτηριακών τασεοϋποδοχέων και για τον ισχαιµικό µηχανισµό του κεντρικού νευρικού συστήµατος. Στα τοιχώµατα όλων σχεδόν των µεγάλων αρτηριών βρίσκονται ελάχιστοι σε αριθµό υποδοχείς διάτασης (τασεοϋποδοχείς), ενώ σε ιδιαίτερα αυξηµένο αριθµό υπάρχουν στο τοίχωµα του αορτικού τόξου και στα τοιχώµατα των έσω καρωτίδων. Οι τασεοϋποδοχείς είναι τελοδενδριτικού τύπου νευρικές απολήξεις οι οποίες διεγείρονται όταν τα τοιχώµατα των αρτηριών διατείνονται. Τα ερεθίσµατα αυτά µεταβιβάζονται µέσω του γλωσσοφαρυγγικού (καρωτιδικοί βολβοί) και του πνευµονογαστρικού (αορτικό τόξο) νεύρου στην προµηκική περιοχή του εγκεφάλου. Οι τασεοϋποδοχείς διεγείρονται από πιέσεις >6 mmhg και ανταποκρίνονται προοδευτικά για να φθάσουν στην ανώτατη ανταπόκριση τους στα 18-2mmHg. Κατά συνέπεια, η διέγερση των τασεοϋποδοχέων από την υψηλή πίεση στις αρτηρίες αναστέλλει το αγγειοσυσταλτικό κέντρο του προµήκη και προκαλεί αγγειοδιαστολή όλου του περιφερικού κυκλοφορικού συστήµατος και ελάττωση της καρδιακής συχνότητας και έντασης της συστολής επαναφέροντας την αρτηριακή πίεση στα φυσιολογικά επίπεδα. Αντίθετα µια πτώση της πίεσης του αίµατος αποτελεί ερέθισµα για τους τασεοευαίσθητους νευρώνες που προκαλεί αντανακλαστικά αυξηµένες συµπαθητικές ώσεις προς την καρδιά και τα αγγεία, µε αποτέλεσµα την αγγειοσύσπαση και την αύξηση της αιµατικής ροής. Η σηµασία του συστήµατος των τασεοϋποδοχέων για την µακροπρόθεσµη ρύθµιση της αρτηριακής πίεσης είναι επουσιώδης, διότι σε µία έως τρεις ηµέρες οι τασεοϋποδοχείς προσαρµόζονται σε οποιοδήποτε επίπεδο πίεσης έχουν εκτεθεί χάνοντας ουσιαστικά την ικανότητα τους να ανταποκρίνονται. Η αύξηση της αρτηριακής πίεσης ως αντίδραση προς την ελάττωση της αιµατικής ροής προς τον εγκέφαλο είναι γνωστή ως ισχαιµική αντίδραση του Κ.Ν.Σ. Η αδυναµία του αίµατος, καθώς κινείται αργά, να αποµακρύνει το διοξείδιο του άνθρακα από το αγγειοκινητικό κέντρο του εγκεφάλου προκαλεί ελάττωση της θρέψης του (ισχαιµία) µε ταυτόχρονη διέγερση του συµπαθητικού νευρικού συστή- µατος. Η αντίδραση Cushing είναι ειδικού τύπου ισχαιµική αντίδραση του Κ.Ν.Σ., που οφείλεται σε αύξηση της ενδοκρανιακής πίεσης η οποία συµπιέζοντας τις εγκεφαλικές αρτηρίες διακόπτει την αιµατική ροή. Εκτός από τους ταχέως δρώντες νευρικούς µηχανισµούς ελέγχου της αρτηριακής πίεσης υπάρχουν δύο κύριοι ορµονικοί µηχανισµοί που ενεργοποιούνται σε διάστηµα από τριάντα λεπτά έως µερικές ώρες εξασφαλίζοντας ένα σχετικά ταχύ έλεγχο: (α) Ο αγγειοσυσταλτικός µηχανισµός νοραδρεναλίνης-αδρεναλίνης. Τα διάφορα αντανακλαστικά που ρυθµίζουν την αρτηριακή πίεση διεγείροντας το συµπαθητικό νευρικό σύστηµα, προκαλούν αύξηση της πίεσης µε δύο τρόπους: µε άµεση διέγερση των αιµοφόρων αγγείων και της καρδιάς και µε έµµεσο ερεθισµό απελευθερώνοντας νοραδρεναλίνη και αδρεναλίνη από τη µυελώδη µοίρα των επινεφριδίων στην κυκλοφορία. Οι δύο αυτές ορµόνες φθάνουν και σε τµήµατα της κυκλοφορίας που δεν έχουν συµπαθητική νεύρωση και ασκούν στο κυκλοφορικό σύστηµα την ίδια δράση µε την άµεση διέγερση του συµπαθητικού, δηλαδή διεγείρουν την καρδιά και προκαλούν συστολή των περισσοτέρων αγγείων
, ενώ σε ιδιαίτερα αυξηµένο αριθµό υπάρχουν στο τοίχωµα του αορτικού τόξου και στα")
15 Εισαγωγή Υπέρταση Η αδρεναλίνη δεσµεύεται πιο αποδοτικά µε τους β1-αδρενεργικούς υποδοχείς της καρδιάς αυξάνοντας την καρδιακή παροχή, και η νοραδρεναλίνη ενεργοποιεί τους α1-αδρενεργικούς υποδοχείς των λείων µυϊκών κυττάρων των αγγειακών τοιχω- µάτων αυξάνοντας τις περιφερικές αντιστάσεις, µε άµεσο επακόλουθο την ανύψωση της πίεσης του αίµατος. (β) Ο αγγειοσυσταλτικός µηχανισµός ελέγχου της αρτηριακής πίεσης µε το σύστηµα ρενίνης-αγγειοτασίνης. Όταν η αρτηριακή πίεση ελαττωθεί σηµαντικά µε αποτέλεσµα την πτώση της νεφρικής αιµατικής ροής, τα κύτταρα της παρασπειραµατικής συσκευής εκκρίνουν ρενίνη. Η ρενίνη είναι ένα ένζυµο που παραµένει στο αίµα για τριάντα λεπτά έως µια ώρα και διασπά την πρωτεΐνη του πλάσµατος, η οποία ονοµάζεται υπόστρωµα ρενίνης απελευθερώνοντας ένα δεκαπεπτίδιο, την αγγειοτασίνη Ι. Ένα άλλο ένζυµο, το ένζυµο µετατροπής (ΜΕΑ/ΑCE) καταλύει την απόσπαση δύο ακόµα αµινοξέων από την αγγειοτασίνη Ι σχηµατίζοντας ένα οκταπεπτίδιο την αγγειοτασίνη ΙΙ. Η αγγειοτασίνη ΙΙ παραµένει στο αίµα µόνο για διάστηµα περίπου ενός λεπτού, διότι αδρανοποιείται γρήγορα από διάφορα ένζυµα του αίµατος και των ιστών µε το γενικό όνοµα αγγειοτασινάση. Μία ταχεία επίδραση της αγγειοτασίνης ΙΙ είναι η αγγειοσύσπαση ιδιαίτερα των αρτηριδίων και λιγότερο, αλλά ταυτόχρονα, των φλεβών µε άµεση συνέπεια την αύξηση της αρτηριακής πίεσης σε φυσιολογικά επίπεδα. Εκτός από τους νευρικούς και ορµονικούς µηχανισµούς γρήγορου ελέγχου της αρτηριακής πίεσης, στον έλεγχο της συµβάλλει και ένας ενδογενής φυσικός µηχανισµός της κυκλοφορίας αυτός της µετακίνησης υγρών από τα τριχοειδή. Εάν για παράδειγµα η αρτηριακή πίεση αυξηθεί πολύ, παρόµοια αύξηση συνοδεύει και την τριχοειδική πίεση µε αποτέλεσµα την έξοδο υγρών διαµέσου της µεµβράνης των τριχοειδών από το αίµα στους διάµεσους χώρους. Το αποτέλεσµα είναι η ελάττωση του όγκου του αίµατος και η επάνοδος της αρτηριακής πίεσης στα φυσιολογικά επίπεδα. Αντίστροφα µείωση της αρτηριακής πίεσης προκαλεί απορρόφηση των διαµέσων υγρών προς το αίµα και η αύξηση του όγκου του παίζει σηµαντικό ρόλο στην άνοδο της αρτηριακής πίεσης σε φυσιολογικά επίπεδα. 2. Το δεύτερο σύστηµα µακροπρόθεσµου ελέγχου της πίεσης βασίζεται κυρίως στο µηχανισµό νεφρών - σωµατικών υγρών. Εµφανίζει µεγάλη µακροχρόνια ισχύ και η λειτουργία του υποβοηθείται από ένα µεγάλο αριθµό επικουρικών µηχανισµών, που περιλαµβάνει ειδικές επιδράσεις του συστήµατος ρενίνης-αγγειοτασίνης, του συστήµατος της αλδοστερόνης και του νευρικού συστήµατος. Οι κύριες λειτουργίες των νεφρών είναι δύο: η απέκκριση των περισσοτέρων από τα τελικά προϊόντα του µεταβολισµού και ο έλεγχος των συγκεντρώσεων των περισσοτέρων από τα συστατικά των υγρών του σώµατος. Οι νεφρώνες ως κύριες λειτουργικές µονάδες των νεφρών συνίστανται από το νεφρικό σωµάτιο, από το εγγύς εσπειραµένο σωληνάριο, από την αγκύλη Henle, από το άπω εσπειραµένο σωληνάριο και από το σύστηµα των αθροιστικών σωληναρίων. Η δοµή και η λειτουργία των νεφρών είναι άρρηκτα συνδεδεµένες. Η απαλλαγή ή κάθαρση του πλάσµατος του αίµατος από ανεπιθύµητες ουσίες, καθώς αυτό περνά από το νεφρό πραγµατοποιείται µε τη σπειραµατική διήθηση, την επαναρρόφηση των ουσιών (νερό και αρκετοί ηλεκτρολύτες) από το σωληναριακό υγρό στο πλάσµα των περισωληναριακών τριχοειδών (αίµα) και την έκκριση ουσιών από το αίµα απευθείας στο σωληναριακό υγρό. Αυτές οι τρεις διεργασίες καθορίζουν την τελική σύνθεση και τον όγκο των ούρων. Η νεφρική ροή αίµατος και ο ρυθµός σπειραµατικής διήθησης διατηρούνται σταθερά, παρά τις µεταβολές της αρτηριακής πίεσης µεταξύ 9 και - 4 -

16 Εισαγωγή Υπέρταση 18mmHg, εξαιτίας του µηχανισµού που αναφέρεται ως αυτορρύθµιση της ροής του αίµατος. Η άνοδος της αρτηριακής πίεσης προκαλεί µεγάλη αύξηση της ταχύτητας µε την οποία οι νεφροί απεκκρίνουν νερό και νάτριο (διούρηση και νατριούρηση από πίεση) µε αποτέλεσµα την απώλεια σηµαντικού όγκου εξωκυττάριου υγρού που έχει ως συνέπεια την ελάττωση του όγκου του αίµατος και την επαναφορά της στα απολύτως φυσιολογικά επίπεδα. Ένα χαρακτηριστικό του µηχανισµού ελέγχου της αρτηριακής πίεσης µε το σύστηµα νεφρού - σωµατικών υγρών, είναι σε πόσο µικρές µεταβολές της ποσότητας των υγρών του σώµατος προκαλούνται σηµαντικές µεταβολές της πίεσης. Η χρόνια αύξηση του εξωκυττάριου υγρού κατά λίγες εκατοντάδες ml, µπορεί να οδηγήσει σε υπέρταση, η οποία θεραπεύεται µε τη χορήγηση κάποιου νατριουρητικού φαρµάκου, το οποίο προκαλεί αύξηση της απέκκρισης νερού και νατρίου. Όταν αυξάνεται η αρτηριακή πίεση, εκτός από τη νατριούρηση και τη διούρηση από πίεση, παράγοντες που αυξάνουν την αποβολή νατρίου και νερού είναι η καταστολή της έκκρισης ρενίνης από τους νεφρούς, η ελάττωση της έκκρισης αλδοστερόνης από το φλοιό των επινεφριδίων λόγω απουσίας της αγγειοτασίνης ΙΙ και η µείωση των συµπαθητικών ώσεων προς τους νεφρούς. Με τις δράσεις αυτές ο ρυθµός σπειραµατικής διήθησης αυξάνεται, γεγονός που αυξάνει την διηθούµενη ποσότητα Na +, ενώ η επαναρρόφηση Na + στο εγγύς εσπειραµένο και στο αθροιστικό σωληνάριο µειώνεται. Όλες αυτές οι µεταβολές έχουν ως συνολικό αποτέλεσµα την αύξηση της απέκκρισης Na + και νερού από τους νεφρούς. Ένας τύπος υπέρτασης µε σηµαντικές διαφορές από την τύπου υπερφόρτισης του όγκου υγρών, είναι ή υπέρταση που προκαλούν η συνεχής έγχυση αγγειοτασίνης ΙΙ ή οι όγκοι της παρασπειραµατικής συσκευής που εκκρίνουν ρενίνη. Ένας άλλος τύπος αγγειοσυσπαστικής υπέρτασης προκαλείται από το φαιοχρωµοκύτωµα, έναν όγκο της µυελώδους µοίρας των επινεφριδίων που εκκρίνει µεγάλες ποσότητες αδρεναλίνης και νοραδρεναλίνης. Επιπρόσθετα, υπέρταση από πρωτοπαθή αλδοστερονισµό προκαλεί ένας µικρός όγκος που αναπτύσσεται στα επινεφρίδια εκκρίνοντας µεγάλες ποσότητες αλδοστερόνης. Η αλδοστερόνη αυξάνει το ρυθµό επαναπορρόφησης νατρίου και νερού στα άπω εσπειραµένα νεφρικά σωληνάρια, µειώνοντας σε µεγάλο βαθµό την ταχύτητα απέκκρισης της µε επακόλουθο την ανάπτυξη µικρού έως µέτριου βαθµού υπέρτασης. Αντίθετα η αναστολή της έκκρισης αλδοστερόνης (υποαλδοστερονισµός) προκαλεί µείωση της επαναρρόφησης νατρίου που έχει ως αποτέλεσµα την απώλεια νατρίου στα ούρα. Η αύξηση του συµπαθητικού τόνου, των επιπέδων ρενίνης, της αγγειοτασίνης ΙΙ και της αντιδιουρητικής ορµόνης (ADH), ως απάντηση στο µειωµένο όγκο του εξωκυττάριου υγρού, προκαλούν τη συστολή των αρτηριδίων και την αύξηση της πίεσης του αίµατος. Η νευρογενής υπέρταση χαρακτηρίζεται από πολλές νευρικές διαταραχές που µπορούν να προκαλέσουν παροδική, και µερικές φορές µόνιµη, υπέρταση. Για να προκληθεί µακροχρόνια υπέρταση τα νεφρικά αρτηρίδια θα πρέπει να συσπώνται συνεχώς από τη διέγερση του συµπαθητικού, για ηµέρες κάθε φορά, µε αποτέλεσµα ο νεφρικός µηχανισµός ελέγχου της πίεσης να διατηρεί την πίεση σε υψηλότερα επίπεδα. Το συνεχές άγχος ή πόνος συντηρεί αυτή την παθολογική διέγερση των νεφρών στους οποίους βαθµιαία προκαλείται διαταραχή στην αρχιτεκτονική του παρεγχύµατος τους. Χαρακτηριστικό είναι, ότι η υπέρταση παραµένει ακόµα και εάν απουσιάσει η διέγερση του συµπαθητικού (Berne και Levy, 1996, Guyton, 21)

17 Εισαγωγή Υπέρταση 1.3 Τύποι υπέρτασης Περίπου 95% από τα υπερτασικά άτοµα χαρακτηρίζονται ως ασθενείς µε ιδιοπαθή υπέρταση (essential hypertension), µε την έννοια της υπέρτασης άγνωστης αιτιολογίας που προσβάλει το µηχανισµό ρύθµισης της αρτηριακής πίεσης. Μέχρι πριν από λίγα χρόνια πολλοί από τους ασθενείς, που σήµερα είναι γνωστό ότι πάσχουν από κάποιο τύπο υπέρτασης που περιγράφηκε, θα αναφέρονταν ως ασθενείς µε ιδιοπαθή υπέρταση. Αντίθετα η δευτεροπαθής υπέρταση (secondary hypertension) εµφανίζεται σε ένα µικρό αριθµό ασθενών (περίπου 2%-5%), όταν υπάρχει βλάβη στη φυσιολογική λειτουργία ενός οργάνου (ενδοκρινικές διαταραχές, αγγειακά προβλήµατα, καρδιακή δυσλειτουργία) και θεραπεύεται µε την αποκατάσταση της λειτουργίας του οργάνου (Beevers et al., 21). 1.4 Γενετική αιτιολογία της υπέρτασης Η ανάπτυξη ιδιοπαθούς υπέρτασης φαίνεται να σχετίζεται µε γενετικούς παράγοντες. Πιθανόν η δράση πολλών γονιδίων (πολυγονίδια) τα οποία επηρεάζονται σε σχετικά µεγάλο βαθµό από το περιβάλλον (διατροφικές συνήθειες, γεωγραφική περιοχή, κοινωνικοοικονοµική κατάσταση, ηλικία) αυξάνουν την πιθανότητα σε ένα άτοµο να εµφανίσει υπερτασική νόσο. Επιδηµιολογικές µελέτες έδειξαν, ότι υπάρχει µία ποικιλότητα στις τιµές αρτηριακής πίεσης σε διάφορους πληθυσµούς. Επιπλέον δεν θα πρέπει να αγνοηθεί το οικογενειακό ιστορικό και ο σηµαντικός ρόλος της κληρονοµικότητας, αφού η πιθανότητα εµφάνισης υπέρτασης διπλασιάζεται σε οικογένειες µε ένα ή δύο υπερτασικούς γονείς. Η ιδιοπαθής υπέρταση εµφανίζεται τέσσερις φορές συχνότερα σε µαύρους απ ότι σε λευκούς και πιθανόν να συµβαίνει συχνότερα σε µεσήλικες άρρενες απ ότι σε µεσήλικες θήλεις. Αναλυτικότερα, η εµφάνιση υπέρτασης είναι σπάνια σε αγροτικές περιοχές ή σε φυλές της Αφρικής, αλλά είναι αρκετά κοινή στις σύγχρονες Αφρικανικές πόλεις και στους µαύρους πληθυσµούς της Μ. Βρετανίας και των Η.Π.Α.. Πειραµατικά µοντέλα που εξέτασαν τη γενετική βάση της υπέρτασης έδειξαν, ότι το νεφρό είναι υπεύθυνο για την κληρονοµούµενη τάση για υπέρταση. Η µεταµόσχευση ενός νεφρού από έναν υπερτασικό δότη προκαλεί άνοδο της αρτηριακής πίεσης και αυξάνει την ανάγκη χορήγησης αντιϋπερτασικών φαρµάκων σε µη υπερτασικούς δέκτες. Η εµφάνιση στο πλάσµα αυξηµένων επιπέδων αγγειοτασινογόνου (υπόστρωµα ρενίνης), το οποίο παράγεται στο ήπαρ, έχουν επίσης αναφερθεί τόσο σε υπερτασικούς ασθενείς, όσο και στους απογόνους υπερτασικών οικογενειών. Πρόκειται για µονογονιδιακή διαταραχή σχετίζοντας άµεσα το γονίδιο του αγγειοτασινογόνου µε την υπέρταση. Η γενετική αιτιολογία της υπέρτασης ενισχύεται και από τη δηµιουργία µεταλλάξεων σε ειδικούς γενετικούς τόπους, οι οποίες όµως συµβαίνουν σπάνια. Μεταλλάξεις του γονιδίου του µετατρεπτικού ενζύµου της αγγειοτασίνης Ι στα αγγειακά ενδοθηλιακά κύτταρα των πνευµόνων, όπου απαντάται, φαίνεται να προκαλούν υπερτροφία της αριστερής κοιλίας ή υπερτασική νεφροπάθεια. Μία γενετική διαταραχή η οποία σχετίζεται µε δέκα διαφορετικές µεταλλάξεις του γονιδίου CYP11B1, είναι υπεύθυνη για τη συγγενή υπερπλασία των επινεφριδίων οφειλόµενη σε ανεπάρκεια της 11-β-υδροξυλάσης και εντοπίζεται στο µεγάλο βραχίονα του χρωµοσώµατος 8 (8q21)

18 Εισαγωγή Υπέρταση Γενικότερα, οι γενετικές νόσοι της φλοιώδους µοίρας των επινεφριδίων είναι σε θέση να µεταβάλουν τα επίπεδα της αλδοστερόνης και να παρεµποδίσουν την ικανότητα των νεφρών να διατηρούν σε φυσιολογικά επίπεδα τον εξωκυττάριο όγκο. Το σύνδροµο Liddle s µε κύρια χαρακτηριστικά τα χαµηλά επίπεδα ρενίνης και αλδοστερόνης (υποαλδοστερονισµός), όπως και το σύνδροµο Gordon s που πιθανόν σχετίζεται µε το µεγάλο βραχίονα του χρωµοσώµατος 17 (ψευδουποαλδοστερονισµός) συνιστούν διαταραχές οι οποίες έχουν άµεση σχέση µε την υπέρταση (Beevers et al., 21). Μοριακές τροποποιήσεις της πρωτεΐνης του κυτταροσκελετού αντουκίνης σχετίζονται γενετικά µε την εµφάνιση υπέρτασης στον άνθρωπο, εξαιτίας της αυξανόµενης επαναρρόφησης ιόντων Na +. Οι αναδιατάξεις στον κυτταροσκελετό των νεφρικών επιθηλιακών κυττάρων που προκαλούνται παρουσία της µεταλλαγµένης µορφής αντουκίνης τροποποιούν κύρια την ενζυµική ενεργότητα (αύξηση) και κατανο- µή των αντλιών Na + -K + στο βασεοπλευρικό τµήµα της µεµβράνης. Επιπλέον, µειώνουν το χρόνο παραµονής της αντλίας Na +, K + -ATPάσης στην επιφάνεια της κυτταρικής µεµβράνης εξαιτίας ενός µηχανισµού ανακύκλωσης (internalization) µε επιταχυνόµενο ρυθµό. Η όλη διαδικασία πιθανόν να οδηγεί σε λανθασµένο σήµα ενίσχυσης για τη σύνθεση των α και β υποµονάδων του ενζύµου µε σκοπό να α- ποφευχθεί η έλλειψη του από το κύτταρο (Ferrandi et al., 1996, Ferrandi et al., 1999). Τα πειραµατικά αποτελέσµατα επιβεβαιώνουν µελέτες, οι οποίες συσχετίζουν την παρουσία του αλληλοµόρφου µεταλλαγµένου τύπου (ADD-W) σε οµάδες πληθυσµών που εµφανίζουν ήπια µορφή υπέρτασης (>14/95 mmhg). Άτοµα είτε οµοζυγωτικά (Τrp/Trp) είτε ετεροζυγωτικά (Gly/Trp) για το αλληλόµορφο ADD-W χαρακτηρίζονται από µειωµένη απέκκριση ιόντων λιθίου και ουρικού οξέος σε σύγκριση µε οµοζυγωτικά άτοµα (Gly/Gly) για το αλληλόµορφο αγρίου τύπου ADD- G (Ferrandi et al., 1999). Η µελέτη δεικτών σε υπερτασικές οµάδες που δεν λαµβάνουν αντιϋπερτασική φαρµακευτική αγωγή αποδεικνύει ότι η παρουσία της ισοµορφής της α-αντουκίνης, που φέρει το αµινοξύ τρυπτοφάνη στην πολυπεπτιδική αλυσίδα ευθύνεται για την αυξηµένη επαναρρόφηση ιόντων νατρίου από το εγγύς εσπειραµένο σωληνάριο µε αποτέλεσµα τη µείωση της απέκκρισης ιόντων νατρίου, λιθίου και νερού από τους νεφρούς, που οδηγεί σε αύξηση του όγκου του εξωκυττάριου υγρού και άνοδο της αρτηριακής πίεσης (Manunta et al., 1999). 1.5 Αντιϋπερτασικά φάρµακα Το πρωταρχικό βήµα στη θεραπεία της υπέρτασης είναι µη φαρµακολογικό και βασίζεται στη µείωση της διαιτητικής πρόσληψης νατρίου και του σωµατικού βάρους. Η ήπια υπέρταση (14-159/9-99mmHg) ελέγχεται µε ένα µόνο φάρµακο (διουρητικό, β-ανταγωνιστή, αναστολέα του Μ.Ε.Α. ή αναστολέα διαύλων Ca ++ ), ανάλογα µε το συγκεκριµένο ασθενή και λαµβάνοντας υπόψη συνυπάρχουσες νόσους. Εξαιτίας των πολλαπλών µηχανισµών ρύθµισης της αρτηριακής πίεσης και των ιδιαίτερων χαρακτηριστικών των ασθενών, η αντιϋπερτασική θεραπεία δεν δύναται να βασισθεί συχνά στη χορήγηση ενός µόνο φαρµάκου. Η µέτρια (16-179/1-19mmHg) έως βαρύτερη ( 18/11mmHg) υπέρταση απαιτεί τη θεραπεία µε διαφορετικές κατηγορίες φαρµάκων που επιλέγονται µε σκοπό να ελαχιστοποιηθούν οι ανεπιθύµητες ενέργειες της συνδυασµένης αγωγής
 συνιστούν διαταραχές οι οποίες έχουν άµεση σχέση µε την υπέρταση (Beevers et al., 21).")
19 Εισαγωγή Υπέρταση Όλοι οι αντιϋπερτασικοί παράγοντες αλληλεπιδρούν µε τα φυσιολογικά συστήµατα ελέγχου της αρτηριακής πίεσης και κατηγοριοποιούνται σύµφωνα µε τη βασική ρυθµιστική περιοχή ή µε τους µηχανισµούς στους οποίους ασκούν τη δράση τους. Παρακάτω, γίνεται µία συνοπτική αναφορά σε κατηγορίες φαρµάκων που χρησι- µοποιούνται στη θεραπεία της υπέρτασης (Benowitz, 1998) ιουρητικά Σε αυτήν την κατηγορία ανήκουν οι φαρµακευτικές ενώσεις: βουµετανίδη, φουροσεµίδη, υδροχλωροθειαζίδη, σπιρονολακτόνη και τριαµτερένη. Τα διουρητικά θειαζίδια προτείνονται ως φαρµακευτική αγωγή πρώτης γραµµής για την υπέρταση και είναι αποτελεσµατικότερη η χορήγηση τους σε ηλικιωµένα άτοµα, σε µαύρους ασθενείς και σε εκείνους µε χρόνια νεφρική νόσο. Η µείωση της αρτηριακής πίεσης βασίζεται στην αύξηση της απέκκρισης νατρίου και νερού. Αυτό προκαλεί µείωση του εξωκυττάριου όγκου που οδηγεί σε ελάττωση της καρδιακής παροχής και της νεφρικής αιµατικής ροής. Προκαλούν σηµαντικές διαταραχές στην ηλεκτρολυτική ισορροπία, µε αποτέλεσµα το 7% των ασθενών να παρουσιάζει υποκαλιαιµία, µειώνοντας τα επίπεδα στο αίµα τόσο του K +, όσο και του Mg ++, ενώ το Ca ++ κατακρατείται στον οργανισµό Αγγειοδιασταλτικά Σε αυτήν την κατηγορία ανήκουν κύρια οι φαρµακευτικές ενώσεις: υδραλαζίνη, µινοξιδίλη, νιτροπρωσσικό νάτριο, διαζοξίδη. Τα αµέσως δρώντα διασταλτικά των λείων µυών, όπως η υδραλαζίνη (hydralazine) και η µινοξιδίλη (minoxidil) δεν χρησιµοποιούνται ως πρώτης επιλογής φάρµακα στη θεραπεία της υπέρτασης. Προκαλούν χάλαση των αγγειακών λείων µυών µειώνοντας τις αρτηριακές αντιστάσεις και κατ επέκταση την αρτηριακή πίεση. Άµεσα δυσµενή συµπτώµατα είναι η ενεργοποίηση των τασεοϋποδοχέων µε επακόλουθη αύξηση της συσταλτικότητας του µυοκαρδίου και της καρδιακής συχνότητας. Οι δράσεις αυτές είναι δυνατόν να προκαλέσουν στηθάγχη, έµφραγµα του µυοκαρδίου ή καρδιακή ανεπάρκεια σε άτο- µα µε σχετική προδιάθεση. Τα αγγειοδιασταλτικά αυξάνουν επίσης τη συγκέντρωση ρενίνης στο πλάσµα, γεγονός που οδηγεί σε κατακράτηση νατρίου και νερού µε επακόλουθο την αύξηση του εξωκυττάριου όγκου. Σχεδόν πάντα τα φάρµακα αυτά χορηγούνται σε συνδυασµό µε ένα β-ανταγωνιστή και µε ένα διουρητικό για να ανασταλούν οι ανεπιθύµητες ενέργειες της θεραπείας. Οι αναστολείς διαύλων ασβεστίου δεσµεύονται στους ευαίσθητους στις διαφορές δυναµικού (voltage sensitive) τύπου-l διαύλους στην καρδιά και στους λείους µυς των στεφανιαίων και των περιφερικών αγγείων αναστέλλοντας την προς το εσωτερικό του κυττάρου κίνηση του ασβεστίου. Αυτό έχει ως αποτέλεσµα τη µείωση του τόνου των λείων µυών προκαλώντας αγγειοδιαστολή κυρίως των αρτηριδίων και του µυοκαρδίου. ιακρίνονται σε τρεις χηµικές κατηγορίες: (1) διφαινυλοαλκυλαµίνες (βεραπαµίλη) (2) βενζοθειαζεπίνες (διλτιαζέµη) και (3) διυδροπυριδίνες. Η τελευταία κατηγορία περιλαµβάνει την νιφεδιπίνη µε υψηλή συχνότητα ανεπιθύµητων ενεργειών (δυσκοιλιότητα, κεφαλαλγία, ίλιγγο, κόπωση) και πέντε νέες φαρµακευτικές ενώσεις: αµλοδιπίνη, ισραδιπίνη, φελοδιπίνη, νικαρδιπίνη και νισολδιπίνη. Όλες οι διυδροπυριδίνες έχουν πολύ µεγαλύτερη συγγένεια µε τους διαύλους ασβεστίου των αγγείων, απ ότι της καρδιάς γεγονός που τις καθιστά περισσότερο κατάλληλες για την θεραπεία της υπέρτασης. Γενικότερα, οι αναστολείς των διαύλων ασβεστίου παρουσιάζουν µια εγγενή νατριουρητική δράση, γι αυτό δεν απαιτείται η προσθήκη διουρητικού φαρµάκου

20 Εισαγωγή Υπέρταση Παράγοντες οι οποίοι τροποποιούν τη λειτουργία του νευρικού συστήµατος Σε αυτήν την κατηγορία ανήκουν κύρια οι φαρµακευτικές ενώσεις: α- µεθυλντόπα, κλονιδίνη, προπρανολόλη, µετοπρολόλη, λαβεταλόλη, ναδολόλη, ατενολόλη, πραζοσίνη, δοξαζοσίνη, τεραζοσίνη Η καταλληλότερη φαρµακευτική αγωγή για τη θεραπεία της µέτριας µέχρι σοβαρής µορφής υπέρτασης περιλαµβάνει αντιϋπερτασικούς παράγοντες, οι οποίοι αναστέλλουν τη λειτουργία του συµπαθητικού νευρικού συστήµατος. Η οµαδοποίηση τους γίνεται σύµφωνα µε τα σηµεία δράσης τους στο συµπαθητικό αντανακλαστικό τόξο Παράγοντες οι οποίοι επιδρούν άµεσα στο Κ.Ν.Σ. Η κλονιδίνη (α2-αγωνιστής) και η α-µεθυλντόπα µέσω του ενεργού µεταβολίτη της (α-µεθυλνορεπινεφρίνη) µειώνουν τις αδρενεργικές ώσεις από το Κ.Ν.Σ., µε αποτέλεσµα την ελάττωση των περιφερικών αγγειακών αντιστάσεων και τη µείωση της αρτηριακής πίεσης. Η βασική δράση τους αναφέρεται σε διαφορετικούς πληθυσµούς αδρενεργικών νευρώνων στα αγγειοκινητικά κέντρα του εγκεφαλικού στελέχους, οι οποίοι τροποποιούν τα αντανακλαστικά των τασεοϋποδοχέων. Οι ανεπιθύµητες ενέργειες είναι η ψυχοκινητική καταστολή, η ξηρότητα του ρινικού βλεννογόνου και οι διαταραχές του ύπνου Παράγοντες οι οποίοι επιδρούν στο περιφερικό νευρικό σύστηµα. Πρόκειται για φαρµακευτικές ενώσεις οι οποίες διακρίνονται σε: (1) εκλεκτικούς ανταγωνιστές των β-υποδοχέων της καρδιάς (β-blockers), µε κύριο αντιπρόσωπο την προπρανολόλη. Η ελάττωση της αρτηριακής πίεσης προέρχεται κατά κύριο λόγο από τη µείωση της καρδιακής παροχής. Οι β-ανταγωνιστές είναι περισσότερο αποτελεσµατικοί στη θεραπεία της υπέρτασης σε λευκούς παρά σε µαύρους ασθενείς, και σε νέους συγκριτικά µε ηλικιωµένους. Προκαλούν ανεπιθύ- µητες ενέργειες, όπως λήθαργο, αϋπνία, ψευδαισθήσεις. µειώνουν την libido και διαταράσσουν το µεταβολισµό των λιπιδίων αυξάνοντας τα τριγλυκερίδια του πλάσµατος. (2) α-αδρενεργικούς ανταγωνιστές (πραζοσίνη, δοξαζοσίνη, τεραζοσίνη), οι οποίοι προκαλούν συναγωνιστικό αποκλεισµό των α1-αδρενεργικών υποδοχέων των λείων µυών (α1-blockers). (3) παράγοντες οι οποίοι προκαλούν συναγωνιστικό αποκλεισµό των νικοτινιδικών χολινεργικών υποδοχέων στις παρασυµπαθητικές και συµπαθητικές µεταγαγγλιακές ίνες (ganglion-blocking agents). Η αντιϋπερτασική δράση βασίζεται στην αύξηση της χωρητικότητας των αγγείων. Η χρήση τους έχει διακοπεί εξαιτίας της υψηλής τοξικότητας που προκαλούν (τριµεθαφάνη). (4) παράγοντες οι οποίοι αναστέλλουν τη σύνθεση (ρεσερπίνη) ή την απελευθέρωση (γουανεθιδίνη) αδρενεργικών διαβιβαστών (νοραδρεναλίνη) από τις συµπαθητικές µεταγαγγλιακές απολήξεις. Η ρεσερπίνη (reserpine), εξαιτίας της ικανότητας της να διαπερνά τον αιµατοεγκεφαλικό φραγµό, προκαλεί κατάθλιψη και καταπράϋνση των πνευµατικών λειτουργιών, ενώ έντονα εµφανίζονται τα συµπτώµατα της ασθένειας Parkinson

21 Εισαγωγή Υπέρταση Παράγοντες που αναστέλλουν την παραγωγή ή τη δράση της αγγειοτασίνης ΙΙ Σε αυτήν την κατηγορία ανήκουν οι φαρµακευτικές ενώσεις: καπτοπρίλη, εναλαπρίλη, µπεναζεπρίλη, κιναπρίλη, λοσαρτάνη, βαλσαρτάνη, σαραλασίνη. Οι αναστολείς του µετατρεπτικού ενζύµου της αγγειοτασίνης (Μ.Ε.Α.) συνιστώνται όταν τα αντιϋπερτασικά πρώτης γραµµής (διουρητικά ή β-ανταγωνιστές) αντενδείκνυνται ή είναι αναποτελεσµατικά. Η καπτοπρίλη (captopril) και άλλοι αναστολείς του Μ.Ε.Α. µιµούνται τη δοµή του υποστρώµατος (αγγειοτασίνη Ι) του συγκεκριµένου ενζύµου, µε αποτέλεσµα τη µείωση της αγγειοτασίνης ΙΙ. Η µείωση της αγγειοτασίνης ΙΙ στο πλάσµα έχει ως αποτέλεσµα: (α) τη µείωση της επαναρρόφησης NaCl και νερού, (β) την ελάττωση της έκκρισης αλδοστερόνης και (γ) επειδή η αγγειοτασίνη ΙΙ είναι ισχυρή αγγειοσυσταλτική ουσία σε συνδυασµό µε τα υψηλά επίπεδα βραδυκινίνης τα αρτηρίδια διαστέλλονται και η αρτηριακή πίεση µειώνεται. Η ρύθµιση της αρτηριακής πίεσης µε τη χορήγηση φαρµάκων αυτής της κατηγορίας οφείλεται στη µείωση των περιφερικών αγγειακών αντιστάσεων, χωρίς να επηρεάζεται η παροχή, ο ρυθµός ή η συσταλτικότητα της καρδιάς. Η φαρµακευτική αγωγή είναι αποτελεσµατική στη θεραπεία ασθενών µε χρόνια συµφορητική καρδιακή ανεπάρκεια και µετά από έµφραγµα του µυοκαρδίου. Συνήθεις ανεπιθύ- µητες ενέργειες είναι το δερµατικό εξάνθηµα, ο ξηρός βήχας, η υπερκαλιαιµία, η αλλοίωση της γεύσης, η πρωτεϊνουρία και η εµφάνιση αγγειονευρωτικού οιδήµατος. Οι αναστολείς του Μ.Ε.Α. είναι τοξικοί για το έµβρυο και δεν πρέπει να χορηγούνται σε έγκυες γυναίκες. Οι µη πεπτιδικές ενώσεις λοσαρτάνη (losartan) και βαλσαρτάνη (valsartan) είναι υψηλής εκλεκτικότητας ανταγωνιστές των υποδοχέων της αγγειοτασίνης ΙΙ. Ο µηχανισµός δράσης τους είναι παρόµοιος των αναστολέων του Μ.Ε.Α. στην πρόκληση αγγειοδιαστολής και στην αναστολή έκκρισης αλδοστερόνης
22 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια 2. ΙΟΥΡΗΤΙΚΑ ΦΑΡΜΑΚΑ-ΘΕΙΑΖΙ ΙΑ 2.1 Γενικά Τα διουρητικά φάρµακα αυξάνουν το ρυθµό της ροής των αποβαλλόµενων ούρων και είναι αναστολείς της µεταφοράς ιόντων οι οποίοι µειώνουν την επαναρρόφηση νατρίου σε διάφορες θέσεις του νεφρώνα. Το νάτριο (Νa + ) και τα συνοδά του ανιόντα (Cl - και HCO3 - ) είναι οι κύριοι καθοριστικοί παράγοντες της κατά βάρος ωσµωγραµµοµοριακότητας των υγρών του σώµατος. Η είσοδος ιόντων νατρίου, όπως και χλωρίου, στα ούρα σε ποσότητες µεγαλύτερες από τις φυσιολογικές σε συνδυασµό µε την παθητική µεταφορά ύδατος για να διατηρηθεί η ωσµωτική ισορροπία έχει ως αποτέλεσµα την αύξηση του όγκου των ούρων, τη µείωση του όγκου του εξωκυττάριου υγρού και συχνά τη µεταβολή του ph και της ιοντικής σύνθεσης των ούρων και του αίµατος (Berne και Levy, 1996). Οι κύριες κλινικές χρήσεις των διουρητικών αναφέρονται στην αντιµετώπιση διαταραχών που έχουν σχέση µε ανώµαλη κατακράτηση υγρών (οίδηµα) και στη θεραπεία της υπέρτασης. Η διουρητική τους δράση προκαλεί ελάττωση του όγκου του αίµατος που οδηγεί σε πτώση της αρτηριακής πίεσης κατά 1-15mmHg στους περισσότερους ασθενείς (Jackson, 1996). 2.2 Τύποι διουρητικών φαρµάκων Μια γενική ταξινόµηση των διουρητικών φαρµάκων γίνεται σύµφωνα µε την περιοχή δράσης τους κατά µήκος του νεφρώνα (διουρητικά της αγκύλης), την αποτελεσµατικότητα (άµεσα αγγειοδιασταλτικά διουρητικά), τη χηµική δοµή τους (διουρητικά θειαζίδια), τη δράση τους στην έκκριση του καλίου (καλιοπροστατευτικά διουρητικά) κ.α. (Jackson, 1996). Η εκτενής γνώση του µηχανισµού δράσης κάθε κατηγορίας διουρητικών οδήγησε στη λεπτοµερή οµαδοποίηση τους Αναστολείς ανθρακικής ανυδράσης Το ένζυµο ανθρακική ανυδράση εντοπίζεται ενδοκυτταρικά στο κορυφαίο τµήµα της µεµβράνης του επιθηλίου του εγγύς σωληναρίου και καταλύει την αντίδραση του CO2 και του H2O προς H + και HCO3 - (διττανθρακικό). Τα ιόντα υδρογόνου εκκρίνονται στο σωληναριακό υγρό µε µηχανισµό αντίθετης µεταφοράς (αντιµεταφορέας) Na + -H +, ενώ για τα διττανθρακικά, αν και η ηλεκτροχηµική κλίση επιτρέπει την παθητική τους έξοδο δια µέσου της βασεοπλευρικής µεµβράνης στο διάµεσο χώρο, η κίνησή τους είναι συζευγµένη µε την κίνηση άλλων ιόντων (συµµεταφορέας Na + -3HCO3 - και αντιµεταφορέας Cl - -HCO3 - ). Η παρουσία της ακεταζολαµίδης, εξαιτίας της µειωµένης ικανότητας ανταλλαγής Na + µε H +, έχει ως αποτέλεσµα µια ελαφρά διούρηση. Η κατακράτηση των διττανθρακικών στο σωληναριακό αυλό και η απώλεια τους προκαλεί µια σηµαντική άνοδο του ph των ούρων, υπερχλωραιµική µεταβολική οξέωση και µειωµένη διουρητική δράση ύστερα από ορισµένες ηµέρες θεραπείας Ωσµωτικά διουρητικά Απλές υδρόφιλες χηµικές ενώσεις οι οποίες διηθούνται από το αγγειώδες νεφρικό σπείραµα, όπως η µαννιτόλη, η γλυκερίνη και η ουρία επιφέρουν κάποιου βαθµού διούρηση. Μικρή ή καθόλου επαναρρόφηση της διηθούµενης ουσίας, σε συνδυασµό µε την παθητική µεταφορά ύδατος στο σωληναριακό υγρό προκαλεί αύξηση του παραγόµενου ποσού ούρων
23 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια Παρατηρούνται σηµαντικές ηλεκτρολυτικές διαταραχές, οι οποίες µεταβάλουν την κατά βάρος ωσµωγραµµοµοριακότητα των υγρών του σώµατος Αναστολείς συµµεταφορέα Na + -Κ + -2Cl (διουρητικά αγκύλης ή ισχυρά διουρητικά) Τα διουρητικά της αγκύλης αναστέλλουν την ταυτόχρονη µεταφορά 1Na + - 1Κ + -2Cl - από το σωληναριακό υγρό, δια µέσου της κορυφαίας µεµβράνης, στο εσωτερικό των επιθηλιακών κυττάρων, στο παχύ τµήµα του ανιόντος σκέλους της αγκύλης Henle. Η αγκύλη Henle (ανιόν σκέλος) επαναρροφά το 25-3% των διηθούµενων NaCl και Κ + και οι επόµενες θέσεις δεν είναι ικανές να αντισταθµίσουν το αυξηµένο φορτίο Na +. Γι αυτό το λόγο τα διουρητικά της αγκύλης θεωρούνται τα περισσότερο αποτελεσµατικά διουρητικά. Επίσης η αυξηµένη µεταφορά αλάτων από το παχύ ανιόν σκέλος της αγκύλης Henle αυξάνει τη θετική διαφορά δυναµικού στον αυλό η οποία αποτελεί σηµαντική κινητήρια δύναµη για την επαναρρόφηση αρκετών ειδών κατιόντων, όπως Na +, Κ +, Ca ++ και Mg ++ δια της παρακυτταρικής οδού. ιουρητικά φάρµακα αυτής της κατηγορίας, τα οποία χρησιµοποιούνται ευρέως στις Η.Π.Α., είναι η φουροσεµίδη (Lasix), η βουµετανίδη (Bumex), η τορσεµίδη (Demadex) και το αιθακρυνικό οξύ (Edecrin) (Kokko, 1984) Αναστολείς συµµεταφορέα Na + -Cl - (θειαζίδια και θειαζιδικά ανάλογα) Είναι σουλφοναµιδικά παράγωγα που παρουσιάζουν δοµικές αναλογίες µε τους αναστολείς της ανθρακικής ανυδράσης. Επειδή οι πρωταρχικοί αναστολείς του συµµεταφορέα Na + -Cl - ήταν ανάλογα της 1,2,4-βενζοθειαζιδίνης-1,1διοξείδιο, η κατηγορία αυτή ονοµάσθηκε διουρητικά θειαζίδια (Jackson, 1996). Η κύρια δράση τους εντοπίζεται στην κορυφαία µεµβράνη των επιθηλιακών κυττάρων του άπω εσπειραµένου σωληναρίου (Ellison et al., 1987). Τα διουρητικά της θειαζίδης αναστέλλουν τη συµµεταφορά των ιόντων Na + στο εσωτερικό των επιθηλιακών κυττάρων δρώντας συναγωνιστικά ή µη στη θέση πρόσδεσης των ιόντων Cl - µε αποτέλεσµα τη µείωση της επαναρρόφησης του NaCl (Beaumont et al., 1988). Η χλωροθειαζίδη είναι η πρώτη βενζοθειαδιαζίνη µε τη χαρακτηριστική αρωµατική σουλφοναµιδική δοµή και αποτελεί πρότυπο θειαζιδικό διουρητικό που φέρει την εµπορική ονοµασία Diuril (Oates, 1996) Αναστολείς νεφρικών επιθηλιακών διαύλων Na + (καλιοπροστατευτικά διουρητικά) Η τριαµτερένη (Dyrenium, Maxzide) και η αµιλορίδη (Damor) ανήκουν σε αυτήν την κατηγορία διουρητικών, τα οποία αναστέλλουν τη λειτουργία των διαύλων µεταφοράς Na + που εντοπίζονται στην κορυφαία µεµβράνη των κύριων επιθηλιακών κυττάρων του τελικού τµήµατος του άπω εσπειραµένου σωληναρίου και του αθροιστικού σωληναρίου. Ο κυτταρικός µηχανισµός έκκρισης Κ + από το αίµα στο σωληναριακό υγρό είναι µία διεργασία που συνεπάγεται: (α) την πρόσληψη Κ + από την Na + -Κ + -ATPάση της βασεοπλευρικής µεµβράνης των κυρίων κυττάρων και (β) τη διάχυση Κ + από το κύτταρο στο σωληναριακό υγρό. Η σωστή λειτουργία της Na + - Κ + -ATPάσης δηµιουργεί υψηλή ενδοκυττάρια [Κ + ], που αποτελεί την κινητήρια δύναµη (ηλεκτροχηµική κλίση) για την εκροή Κ + δια µέσου των διαύλων Κ + της κορυφαίας µεµβράνης (υψηλή διαπερατότητα). Η αµιλορίδη αναφέρεται ως διουρητικό που διασώζει τα Κ + διότι η αναστολή της επαναρρόφησης Na + µειώνει την αρνητική διαφορά διεπιθηλιακού δυναµικού στον αυλό, η οποία αποτελεί την
24 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια κινητήρια δύναµη για την παρακυτταρική επαναρρόφηση Cl -. Η µείωση της αρνητικής διαφοράς ηλεκτρικού δυναµικού του αυλού, προκαλεί αναστολή της έκκρισης Κ Ανταγωνιστές αλατοκορτικοειδών υποδοχέων (ανταγωνιστές αλδοστερόνηςκαλιοπροστατευτικά διουρητικά) Τα επιθηλιακά κύτταρα του τελικού τµήµατος του άπω εσπειραµένου σωληναρίου και του αθροιστικού σωληναρίου διαθέτουν κυτταροπλασµατικούς υποδοχείς των αλατοκορτικοειδών, οι οποίοι εµφανίζουν υψηλή συγγένεια µε την αλδοστερόνη. Οι υποδοχείς αυτοί είναι µέλη της υπεροικογένειας των υποδοχέων για τις στεροειδείς ορµόνες, τις θυρεοειδικές ορµόνες, τη βιταµίνη D και τα ρετινοειδή. Η σπιρονολακτόνη (Aldactone) είναι ένας συνθετικός ανταγωνιστής της αλδοστερόνης που την ανταγωνίζεται στους ενδοκυττάριους κυτταροπλασµατικούς υποδοχείς της. Το σύµπλοκο υποδοχέα-σπιρονολακτόνης είναι ανενεργό, δηλαδή εµποδίζει τη µετατόπιση και τη σύνδεση του συµπλόκου σε ειδικές ακολουθίες στο DNA του κυττάρου στόχου. Αυτό έχει ως αποτέλεσµα την αναστολή της σύνθεσης πρωτεϊνών επαγόµενων από την αλδοστερόνη. Σε φυσιολογικές συνθήκες, οι πρωτεΐνες-µεσολαβητές αυξάνουν τον αριθµό των µεµβρανικών διαύλων Na + στην επιφανειακή (σωληναριακή) πλευρά της µεµβράνης, ενεργοποιούν την Na + -Κ + - ATPάση στη βασική (τριχοειδική) επιφάνεια της κυτταρικής µεµβράνης, προκαλούν αλλαγές που επιφέρουν ανακατανοµή των διαύλων και των αντλιών Na + από το διαλυτό κυτταρόπλασµα στην κυτταρική µεµβράνη, µεταβάλλουν τη διαπερατότητα των παρακυτταρικών συνδέσεων και διεγείρουν τη δραστηριότητα ενζύµων των µιτοχονδρίων, τα οποία συµµετέχουν µέσα από τις αντιδράσεις του κύκλου Krebs, στην παραγωγή ATP. Η παρεµπόδιση της επαναρρόφησης Na + από τη σπιρονολακτόνη προκαλεί µείωση της αρνητικής διαφοράς διεπιθηλιακού δυναµικού στον αυλό η οποία αναστέλλει την απέκκριση Κ + και Η + στο σωληναριακό υγρό. Η ενέργεια της σπιρονολακτόνης οφείλεται σε µεγάλο βαθµό στη δράση του ενεργού µεταβολίτη της, κανρενόνης. 2.3 Γενετικοί παράγοντες που επηρεάζουν τη δράση διουρητικών φαρµάκων Οι ποικίλες αντιδράσεις στα φάρµακα µεταξύ ατόµων του ιδίου είδους ή και µεταξύ διαφορετικών ειδών µπορεί να οφείλονται τόσο σε πολυγονιδιακές διαφορές µεταξύ ειδών, φυλών, οµάδων, ατόµων, όσο και σε απλά γονίδια (επηρεασµός της δραστικότητας ενός ενζύµου). Γνωρίζοντας ότι οι γονιδιακές συχνότητες ποικίλουν µεταξύ ειδών και πληθυσµών, είναι πιθανό να διαφέρει, εξαιτίας των γενετικών πολυµορφισµών, και ο βαθµός αντίδρασης σε ένα φάρµακο. Οι περισσότερες ποικιλοµορφίες στην αλληλουχία του γονιδιώµατος παρατηρούνται σε µια συχνότητα στον πληθυσµό που είναι µεγαλύτερη από 1% τουλάχιστον. Αξιοσηµείωτο είναι, ότι οι περισσότεροι πολυµορφισµοί αφορούν αλλαγές στις αζωτούχες βάσεις του DNA, που συνήθως συµβαίνουν εκτός των κωδικών περιοχών των γονιδίων, οι οποίες δεν έχουν επιπτώσεις στη δοµή και στη λειτουργία της κωδικοποιούµενης πρωτεΐνης. Αλλαγές οι οποίες εµφανίζονται σε κωδικές περιοχές των γονιδίων (exons) έχουν ως αποτέλεσµα τη δηµιουργία λανθασµένων mrna και προκαλούν αλλαγές στην ενεργότητα των παραγόµενων πρωτεϊνών γεγονός που επιφέρει, είτε αύξηση, είτε ελάττωση, είτε έλλειψη της ενζυµικής έκφρασης
25 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια ύο είναι οι κύριοι µηχανισµοί µε τους οποίους γενετικοί παράγοντες επηρεάζουν το βαθµό απόκρισης ενός φαρµάκου: (α) τροποποίηση του µεταβολισµού (διάσπασης) του φαρµάκου και (β) µεταβολή της ευαισθησίας των ενζύµων ή των υποδοχέων διαµέσου των οποίων οι φαρµακολογικοί παράγοντες εµφανίζουν τη δράση τους (Stakos και Boudoulas, 22) Μεταβολισµός διουρητικών φαρµάκων Ο µεταβολισµός των φαρµάκων και άλλων εξωγενών ουσιών (ξενοβιοτικών) καταλύεται από οµάδες ενζύµων που σχετίζονται άµεσα µε το µεταβολισµό ενδογενών υποστρωµάτων όπως τα χολικά οξέα, οι στεροειδείς ορµόνες, τα λιπαρά οξέα και η χολερυθρίνη. Κυρίως συναντώνται σε περιοχές του οργανισµού που εκτίθενται σε υψηλές συγκεντρώσεις ξενοβιοτικών ουσιών όπως είναι το ήπαρ, οι πνεύµονες, το έντερο και οι νεφροί. Τα περισσότερα φάρµακα αποµακρύνονται από τον οργανισµό εφόσον έχουν υποστεί βιοχηµικές µετατροπές ή απεκκρίνονται µε διάφορες χηµικές µορφές, από τα νεφρά στα ούρα ή από το ήπαρ στη χολή. Το µέγεθος και η υψηλή συγκέντρωση καταβολικών ενζύµων καθιστά το ήπαρ το κυριότερο όργανο για το µεταβολισµό των φαρµάκων. Τα περισσότερα φάρµακα είναι λιπόφιλα µε αποτέλεσµα να διαπερνούν σχετικά εύκολα τις κυτταρικές µεµβράνες και να επαναρροφώνται στα άπω εσπειραµένα σωληνάρια όταν η συγκέντρωση του φαρµάκου στο διήθηµα γίνεται µεγαλύτερη από εκείνη στον περιαγγειακό χώρο, γεγονός που καθιστά δύσκολη την αποβολή τους από το νεφρό. Για να ελαχιστοποιηθεί αυτή η επαναρρόφηση, τα φάρµακα µετατρέπονται από τον οργανισµό σε πολικότερα µόρια µε δύο τύπους αντιδράσεων που ονοµάζονται Φάση Ι και Φάση ΙΙ του µεταβολισµού των φαρµάκων Φάση Ι µεταβολισµού διουρητικών φαρµάκων. Με τις αντιδράσεις οξείδωσης, αναγωγής ή και υδρόλυσης της Φάσης Ι σε ουσίες οι οποίες εισέρχονται στον οργανισµό προστίθενται ή αποκαλύπτονται µικρές πολικές λειτουργικές οµάδες (-OH, -NH2, -COOH). Τα ένζυµα του κυτοχρώµατος P45 (CYP) καταλύοντας αντιδράσεις οξείδωσης λαµβάνουν µέρος στο πρώτο στάδιο (Φάση Ι) του µεταβολισµού των φαρµάκων. Οι τρεις πρώτες οικογένειες (CYP1, CYP2, CYP3) του συστήµατος P45 διαδραµατίζουν ένα πολύ σηµαντικό ρόλο στην αποµάκρυνση φαρµακολογικών παραγόντων και τοξινών από τον οργανισµό (αποτοξίνωση). Στις υποοικογένειες CYP2C, CYP2 D, CYP3A, συναντώνται ισοµορφές και πολυµορφισµοί που εµφανίζονται σε αυτές, ικανές να µεταβολίσουν ένα σηµαντικό αριθµό αντιϋπερτασικών φαρµάκων. Συγκεκριµένα, η τορσεµίδη (Demadex), ένα ισχυρό διουρητικό, αποτελεί υπόστρωµα του ενζύµου CYP2C9, ενώ φαρµακευτικές ενώσεις που ανήκουν στην κατηγορία των σουλφοναµιδών εµφανίζουν ισχυρή ανασταλτική δράση. Πολυµορφισµοί στην κωδική περιοχή του γονιδίου CYP2C9, που εντοπίζεται στη χρωµοσω- µατική περιοχή 1q24.1, οφείλονται σε υποκαταστάσεις βάσεων και δηµιουργούν πολλαπλά αλληλόµορφα (CYP2C9*1, CYP2C9*2, CYP2C9*3), οι συχνότητες των οποίων ποικίλουν τόσο µεταξύ ατόµων του ιδίου πληθυσµού (Καυκάσιοι) όσο και µεταξύ διαφορετικών εθνοτήτων (Κινέζοι, Κορεάτες, Ευρωπαίοι, Νεγροαµερικάνοι). Η συντηρηµένη αντικατάσταση, έστω και ενός µόνο αµινοξέος (Ile σε Leu) σε θέσεις που αφορούν το ενεργό κέντρο του ενζύµου τροποποιεί δραµατικά την καταλυτική δραστικότητα και την ειδίκευση του υποστρώµατος των ενζυµικών πολυµορφισµών της ισοµορφής CYP2C
26 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια Άτοµα οµοζυγωτικά για το αλληλόµορφο Leu359 εµφανίζουν µειωµένη κάθαρση του φαρµάκου τορσεµίδη και χαρακτηρίζονται ως αργοί µεταβολίτες των υποστρω- µάτων του ενζύµου CYP2C9. Αντίθετα η δραστικότητα του ενζύµου δεν φαίνεται να επηρεάζεται από την αύξηση της ηλικίας και δεν υπάρχουν σαφείς ενδείξεις, τουλάχιστον για τις ανθρώπινες CYP ισοµορφές, για διαφορική έκφραση όσον αφορά το φύλο (Miners και Birkett, 1998) Φάση ΙΙ µεταβολισµού διουρητικών φαρµάκων. Με τις αντιδράσεις σύζευξης της Φάσης ΙΙ πραγµατοποιείται η πρόσδεση οξικού, ανόργανου θείου, σακχάρων ή αµινοξέων, µε αποτέλεσµα την πλήρη απενεργοποίηση και αποµάκρυνση της ουσίας από τον οργανισµό. Οι Ν-ακετυλοτρανσφεράσες (NAT) συµµετέχουν στη φάση ΙΙ του µεταβολισµού των φαρµάκων καταλύοντας τη µεταφορά µιας ακετυλοµάδας από το ακέτυλο-coas σε διάφορα υποστρώµατα όπως είναι οι αµίνες, οι υδραζίνες, η ισονιαζίδη, η προκαϊναµίδη, καθώς και σε ένα µεγάλο αριθµό σουλφοναµιδών που περιλαµβάνει τη σουλφαδιαζίνη, τη σουλφαµεθαζίνη, τη σουλφαπιριδίνη, τη σουλφαµεριδίνη και τη σουλφαδοξίνη (Meyer και Zanger, 1997). Στον κοντό βραχίονα του χρωµοσώµατος 8p εντοπίζονται στον άνθρωπο δύο γενετικοί τόποι οι οποίοι κωδικοποιούν λειτουργικές Ν-ακετυλοτρανσφεράσες (NAT1 και NAT2) (Blum et al., 199, Hickman et al., 1994). Τέσσερα διαφορετικά αλληλόµορφα του γονιδίου NAT1 προκύπτουν από αλλαγές οι οποίες συµβαίνουν στην τερµατική αλληλουχία µεταγραφής. Το NAT1*4 είναι το πιο συνηθισµένο και αποτελεί τον άγριο τύπο, ενώ το αλληλόµορφο NAT1*1, το οποίο προκύπτει µε δύο υποκαταστάσεις βάσεων από τον άγριο τύπο, σχετίζεται µε αυξηµένη ενζυµική δραστικότητα περίπου στο 3% των πληθυσµών µε ευρωπαϊκή καταγωγή. Ιδιαίτερα σηµαντικοί είναι οι πολυµορφισµοί του γονιδίου NAT2, διότι διαχωρίζουν τα άτοµα σε γρήγορους, ενδιάµεσους και αργούς αδρανοποιητές (acetylators). Με µεταλλάξεις του αρχικού γονιδίου NAT2 έχουν προκύψει περισσότερα από 25 αλληλόµορφα, από τα οποία το αλληλόµορφο αγρίου τύπου NAT2*4 είναι το πιο κοινό και ευθύνεται για το γρήγορο φαινότυπο. Οι αυξηµένοι ρυθµοί ακετυλίωσης εµφανίζονται σε οµόζυγους και ετερόζυγους φορείς του συγκεκριµένου αλληλο- µόρφου. Αντίθετα τα αλληλόµορφα NAT2*5A, B, C, NAT2*6A, NAT2 *7B και NAT 2*13 χαρακτηρίζουν το φαινότυπο αργή αδρανοποίηση και εµφανίζονται πάντα σε οµοζυγωτία στους διάφορους πληθυσµούς (Καυκάσιοι) (Taningher et al., 1999). Η µεταβολική ενεργοποίηση και η αδρανοποίηση µονοπατιών που σχετίζονται µε την ακετυλίωση αρωµατικών αµινών (π-αµινοβενζοϊκό οξύ και σουλφαµεθαζίνη) συνιστούν ένα εξαιρετικό µοντέλο πολυµορφισµών το οποίο ρυθµίζει τη διαδικασία καρκινογένεσης στην ουροδόχο κύστη (Badawai et al., 1995, Cascorbi, et al., 21). Ένα σηµαντικό µονοπάτι για την αδρανοποίηση ή την αποτοξίνωση του οργανισµού από ένα µεγάλο αριθµό φαρµάκων και ξενοβιοτικών ουσιών είναι οι αντιδράσεις µεθυλίωσης, οι οποίες πραγµατοποιούνται από µια ποικιλία µεθυλοτρανσφερασών (COMT, GAMT, HNMT, NNMT, PEMT, PNMT, TPMT). Η µεθυλοτρανσφεράση της θειοπουρίνης (TPMT) καταλύει την S-µεθυλίωση στη θέση C-6 του δακτυλίου της πουρίνης θειοπουρινικών αντιλευχαιµικών και ανοσοκατασταλτικών φαρµάκων όπως η 6-µερκαπτοπουρίνη, η αζαθειοπρίνη και η 6- θειογουανίνη µε αποτέλεσµα τη µεθυλίωση και την απενεργοποίηση τους. Η S- αδενόσυλ-l-µεθειονίνη (AdoMet) είναι ο κύριος δότης µεθυλοµάδας, ο οποίος εµπλέκεται στη µεθυλίωση σηµαντικών µορίων για το κύτταρο µε εξαιρετική
27 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια σηµασία στη διατήρηση του φυσιολογικού προτύπου µεθυλίωσης του DNA. Η µεθυλίωση των θειοπουρινών οδηγεί σε εξάντληση της AdoMet, ενώ οι µεθυλιω- µένοι θειοπουρινικοί µεταβολίτες αναστέλλουν τη de novo σύνθεση πουρινών και µειώνουν σηµαντικά το ενεργειακό µόριο ATP. Αυτές οι διαταραχές οδηγούν σε υποµεθυλίωση του DNA γεγονός που επηρεάζει τη γονιδιακή έκφραση και δηµιουργεί νέες θέσεις περιορισµού (Lambooy et al., 1998). Εχει βρεθεί ότι τα διουρητικά φάρµακα φουροσεµίδη, βενδροφλουµεθειαζίδη και τριχλωροµεθειαζίδη δρουν κύρια ως µη συναγωνιστικοί αναστολείς της µεθυλοτρανσφεράσης της θειοπουρίνης (TPMT) παρουσία του υποστρώµατος 6-µερκαπτοπουρίνης και του κύριου δότη µεθυλοµάδας AdoMet. Γνωρίζοντας ότι η S-µεθυλίωση είναι το κύριο µονοπάτι µεταβολισµού των θειοπουρινών υπάρχει άµεση αλληλεπίδραση µεταξύ διουρητικών και θειοπουρινικών φαρµάκων, η οποία γίνεται εµφανής σε ασθενείς µε ταυτόχρονη χορήγηση (Lysaa et al., 1996). Μία άλλη κατηγορία ενζύµων που συµµετέχουν στις αντιδράσεις του δευτέρου σταδίου (Φάση ΙΙ) του µεταβολισµού των φαρµάκων είναι οι S-τρανσφεράσες της γλουταθειόνης (GSTs), οι οποίες καταλύουν την προσθήκη του τριπεπτιδίου γλουταθειόνη (GSH) σε ηλεκτρονιόφιλα υποστρώµατα. Η σύζευξη της γλουταθειόνης σε µία µεγάλη ποικιλία εξωγενών ουσιών, η οποία περιλαµβάνει χηµικές ενώσεις που χρησιµοποιούνται στη χηµειοθεραπεία του καρκίνου (αλκυλιωτικοί παράγοντες), καρκινογόνες ουσίες που αποβάλλονται στο περιβάλλον, φυσικές ή χηµικές τοξίνες που σχετίζονται άµεσα µε ανθρώπινες δραστηριότητες και συνήθειες (δίαιτα, κάπνισµα, αλκοόλ) και ενεργά προϊόντα οξυγόνου, προκαλεί την απενεργοποίηση τους. Οι S-τρανσφεράσες της γλουταθειόνης (GSTs) είναι διµερείς πρωτεΐνες και συνιστούν µία εξαιρετικά πολυµορφική οικογένεια ενζύµων στην οποία διακρίνονται οι τάξεις: α (GSTA), µ (GSTM), π (GSTP) και θ (GSTT). Πληθυσµιακές µελέτες έδειξαν ότι ελλείµµατα των γονιδίων GSTM1 και GSTT1 σχετίζονται άµεσα µε αυξηµένη ευαισθησία στην εµφάνιση καρκινικών ασθενειών εξαιτίας της ελλιπής απενεργοποίησης τοξινών και άλλων ξενοβιοτικών από τον οργανισµό. Συγκεκριµένα, ο πολυµορφισµός του γονιδίου GSTM1 ο οποίος οφείλεται σε αντικατάσταση βάσης στην κωδική περιοχή 7 οδηγεί σε δύο αλληλό- µορφα, τα GSTM1*Α και GSTM1*Β. Άτοµα είτε οµοζυγωτικά για το ένα ή το άλλο αλληλόµορφο είτε ετεροζυγωτικά παράγουν τόσο οµοδιµερή (GSTM1a-1a ή GSTM1b-1b) όσο και ετεροδιµερή (GSTM1a-1b) ένζυµα µε παρόµοια καταλυτική ενεργότητα. Ο γενότυπος GSTM1(-/-) και GSTT1(-/-) (null genotype) εµφανίζεται πάντα σε οµοζυγωτία (Taningher et al., 1999). Η παντελής έλλειψη του ενζύµου αυξάνει την τοξικότητα ορισµένων αντικαρκινικών ενώσεων, ενώ υπερέκφραση των γονιδίων για τις ισοενζυµικές µορφές S-τρανσφερασών της γλουταθειόνης (GSTs) προκαλεί αυξηµένη απενεργοποίηση των αλκυλιωτικών παραγόντων µέσα στο κύτταρο, µε αποτέλεσµα την ανθεκτικότητα των καρκινικών κυττάρων και τη µειωµένη αποτελεσµατικότητα της χηµειοθεραπείας (Schultz et al., 1997). Σε µία προσπάθεια να παρακαµφθούν οι κυτταρικές αλλαγές που οδηγούν σε ανάπτυξη ανθεκτικότητας χρησιµοποιήθηκαν αναστολείς της καταλυτικής δράσης των κυριοτέρων τάξεων (alpha-, mu-, και pi-) τρανσφερασών της γλουταθειόνης, όπως το ισχυρό διουρητικό φάρµακο αιθακρυνικό οξύ (Edecrin). Η αναστολή όλων των ισοενζυµικών µορφών από το αιθακρυνικό οξύ (>8%) είναι συναγωνιστική παρουσία του υποστρώµατος 1-χλώρο-2,4-δινιτροβενζένιο και µη συναγωνιστική για τη γλουταθειόνη
28 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια Επιπρόσθετα η σύζευξη του αιθακρυνικού οξέος µε τη γλουταθειόνη προκαλεί ενζυµική αναστολή. Στη συγκεκριµένη µελέτη τα διουρητικά φουροσεµίδη και βουµετανίδη βρέθηκε να έχουν µικρότερη ανασταλτική δράση (Ahokas et al., 1985, Ploemen et al., 199). Πρόσφατες ερευνητικές µελέτες ταύτισαν την καταλυτική δράση S-τρανσφερασών της γλουταθειόνης µε αυτήν της σουλφοναµιδάσης, εξαιτίας της ικανότητας των ισοµορφών GSTA1, GSTM1, GSTP1 να διασπούν το σουλφονα- µιδικό δεσµό (-NH-SO2) µε διαφορετικούς ρυθµούς. Η ενζυµική διάσπαση των σουλφοναµιδών είναι ιδιαίτερα σηµαντική λαµβάνοντας υπόψη ότι φάρµακα µε τη χαρακτηριστική σουλφοναµιδική δοµή αποτελούν πιθανά υποστρώµατα των S- τρανσφερασών της γλουταθειόνης (Koeplinger et al., 1999, Zhao et al., 1999) Αλληλεπιδράσεις διουρητικών φαρµάκων µε κυτταρικές πρωτεΐνες Η κύρια δράση των αντιϋπερτασικών φαρµάκων εντοπίζεται στην αλληλεπίδραση τους µε πρωτεΐνες, όπως διαµεµβρανικοί υποδοχείς, ενδοκυτταρικά µηνυ- µατοφόρα µόρια και ένζυµα. Οι ενδοατοµικές διαφορές που παρουσιάζονται κατά τη θεραπεία µε διουρητικά φάρµακα οφείλονται κατά κύριο λόγο σε γενετικούς πολυµορφισµούς ενζύµων και υποδοχέων οι οποίοι τροποποιούν βιοχηµικά, ανατο- µικά και φυσιολογικά συστήµατα ελέγχου ρύθµισης της αρτηριακής πίεσης Σύστηµα ρενίνης-αγγειοτασίνης. Ιδιαίτερα σηµαντικοί είναι οι πολυµορφισµοί του αγγειοσυσταλτικού µηχανισµού ελέγχου της αρτηριακής πίεσης µε το σύστηµα ρενίνης-αγγειοτασίνης, οι οποίοι αφορούν τα γονίδια για τη ρενίνη, το υπόστρωµα ρενίνης, το ένζυµο µετατροπής και τους µεµβρανικούς υποδοχείς ΑΤ1 και ΑΤ2 της αγγειοτασίνης ΙΙ. Στο µακρύ βραχίονα του χρωµοσώµατος 17 (17q23) εντοπίζεται στον άνθρωπο ο γενετικός τόπος που κωδικοποιεί το ένζυµο µετατροπής της αγγειοτασίνης (ACE). Πολυµορφισµοί του γονιδίου ACE οι οποίοι χαρακτηρίζονται είτε από ένθεση (I) είτε από έλλειµµα (D) µίας νουκλεοτιδικής ακολουθίας 287 ζευγών βάσεων στη κωδική περιοχή 16 δηµιουργούν τα αλληλόµορφα I και D, η παρουσία των οποίων τροποποιεί τα επίπεδα του ενζύµου µετατροπής στο πλάσµα. Συγκεκριµένα τα επίπεδα του ενζύµου µετατροπής αυξάνουν ευθέως ανάλογα µε τον αριθµό των D αλληλοµόρφων και σε άτοµα µε το γενότυπο DD το ένζυµο εµφανίζει την υψηλότερη καταλυτική ενεργότητα µε αποτέλεσµα την ενίσχυση της παραγωγής της αγγειοσυσταλτικής ένωσης αγγειοτασίνη ΙΙ (Rigat et al., 199). Κατά τη θεραπεία της ήπιας και µέτριας µορφής υπέρτασης µε θειαζιδικά διουρητικά φάρµακα, όπως το υδροχλωροθειαζίδιο (HCTZ), υπάρχουν σαφείς ενδείξεις για διαφορική απόκριση των υπερτασικών ατόµων στο συγκεκριµένο φάρµακο όσον αφορά τη ρύθµιση τόσο της διαστολικής όσο και της συστολικής αρτηριακής πίεσης, η οποία σχετίζεται άµεσα µε την παρουσία του αλληλοµόρφου D και το φύλο (Schwartz et al., 22) Πρωτεΐνη αντουκίνη. Μία από τις γενεσιουργούς αιτίες της υπέρτασης και ταυτόχρονα της ευαισθησίας των ατόµων τόσο στη διαιτητική πρόσληψη NaCl όσο και στην απόκριση στα διουρητικά θειαζιδικά φάρµακα είναι η ποικιλοµορφία των µορίων της πρωτεΐνης του κυτταροσκελετού αντουκίνης, η οποία εντοπίζεται στις συνδέσεις σπεκτρίνης-ακτίνης. Η σπεκτρίνη είναι µία επιφανειακή ρυθµιστική πρωτεΐνη που προκαλεί την ένωση των ινιδίων της ακτίνης µε τις πρωτεΐνες της πλασµατικής
29 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια µεµβράνης. Μεταβολές στις υποµονάδες, που απαρτίζουν τις δεσµευµένες στην ακτίνη των ινιδίων πρωτεΐνες, οδηγούν σε αλλαγές στην οργάνωση του κυτταροσκελετού, οι οποίες σχετίζονται άµεσα µε αλλαγές του κυτταρικού σχήµατος, της κίνησης, της κυτταρικής προσκόλλησης και αλληλεπίδρασης, της κατανοµής και σταθερότητας διαµεµβρανικών πρωτεϊνών. Η αντουκίνη είναι ένα ετεροδιµερές πρωτεϊνικό µόριο, το οποίο απαρτίζεται από τις υποµονάδες α/β ή α/γ. Κάθε υποµονάδα φέρει τρεις διακριτές περιοχές: (α) µία αµινοτελική σφαιρική περιοχή, η οποία συνιστά την κεφαλή της αντουκίνης, (β) µία ενδιάµεση περιοχή και (γ) µία καρβοξυτελική ευαίσθητη σε πρωτεάσες περιοχή που καλείται ουρά. Το καρβοξυτελικό άκρο της αντουκίνης, ενώ είναι µοναδικό και ιδιαίτερα µεταβλητό σε κάθε µία από τις α και β υποµονάδες της, περιλαµβάνει µία πανοµοιότυπη ακολουθία 22 αµινοξέων, η οποία είναι η ίδια µε µία περιοχή των MARCKS (myristoylated alanine-rich C kinase substrate) πρωτεϊνών. Σε αυτή την περιοχή συναντώνται θέσεις φωσφορυλίωσης από τις κινάσες C των πρωτεϊνών (PKC) και από τις ειδικές κινάσες σερίνης/θρεονίνης (Rho-assosiated kinase), που ενεργοποιούνται από πρωτεΐνες της οικογένειας Rho (Rho, Rac, Cdc42), καθώς και θέσεις δέσµευσης για την καλµοντουλίνη, η οποία ενεργοποιείται µε την αύξηση των επιπέδων ασβεστίου. Σηµαντικό είναι να αναφέρουµε ότι οι πρωτεΐνες της οικογένειας Rho έχουν κεντρικό ρόλο στη ρύθµιση της οργάνωσης του κυτταροσκελετού της ακτίνης και ότι το σύµπλοκο ασβεστίου/καλµοντουλίνης ενεργοποιεί κινάσες που εξαρτώνται από το ασβέστιο και οι οποίες φωσφορυλιώνουν πλήθος πρωτεϊνών όπως ένζυµα µεταβολισµού, ιοντικούς διαύλους και µεταγραφικούς παράγοντες. Επιπλέον, οι ποικίλες περιοχές του καρβοξυτελικού άκρου της αντουκίνης εξαιτίας της υψηλής συγγένειας που εµφανίζουν µε το σύµπλεγµα σπεκτρίνης-ακτίνης συνενώνονται µε αποτέλεσµα περισσότερα µόρια σπεκτρίνης να κατευθύνονται µε τη βοήθεια της αντουκίνης στο τριµερές σύµπλοκο αντουκίνης-σπεκτρίνης-ακτίνης µε σκοπό την αύξηση της σταθερότητας του (Hughes και Bennett, 1995, Matsuoka et al., 2). Η ποικιλοµορφία της πρωτεΐνης αντουκίνης στους ανθρώπινους ιστούς οφείλεται στην ύπαρξη και διαφορική έκφραση των ADD1, ADD2 και ADD3 γονιδίων τα οποία χαρτογραφούνται σε διαφορετικά χρωµοσώµατα και κωδικοποιούν αντίστοιχα τις υποµονάδες α, β και γ. Η ανάλυση της οικογένειας γονιδίων της αντουκίνης έδειξε ότι πολυµορφισµοί που οφείλονται είτε σε απλές υποκαταστάσεις βάσεων είτε σε ενθέσεις/απαλοιφές νουκλεοτιδίων, σε συνδυασµό µε τους διαφορετικούς τρόπους επανένωσης των κωδικών αλληλουχιών του κάθε γονιδίου (εναλλακτικό µάτισµα-alternative splicing) παράγουν παραλλαγές των υποµονάδων µε παραπλήσια λειτουργικότητα. Σχετικά µε το γονίδιο ADD1, το οποίο στον άνθρωπο εντοπίζεται στην τελοµερική περιοχή του χρωµοσώµατος 4p16.3, βρέθηκε ότι εκφράζεται σε όλους τους ιστούς µε έντονη παρουσία στο νεφρικό και νευρικό ιστό σε αντίθεση µε το γονίδιο ADD2 (Joshi et al., 1991, Lin et al., 1995). Πληθυσµιακές µελέτες συσχετίζουν άµεσα έναν πολυµορφισµό του γονιδίου ADD1 µε την ανάπτυξη της πρωτοπαθούς υπέρτασης (primary hypertension) ως µονογονιδιακή διαταραχή και µε διακυµάνσεις της πτώσης της αρτηριακής πίεσης ως απόκριση στη φαρµακευτική αγωγή µε διουρητικά, όπως το υδροχλωροθειαζίδιο και η φουροσεµίδη. Συγκεκριµένα, η υποκατάσταση µιας νουκλεοτιδικής βάσης (G T) σε κωδική περιοχή του γονιδίου, προκαλεί την αντικατάσταση του αµινοξέος γλυκίνη από το αµινοξύ τρυπτοφάνη στην πολυπεπτιδική αλυσίδα της α-αντουκίνης. Το αποτέλεσµα της σηµειακής µετάλλαξης είναι η παρουσία στους πληθυσµούς δύο αλληλοµόρφων, αυτό του αγρίου τύπου (ADD-G) και του µεταλλαγµένου τύπου (ADD-W)
30 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια Ο σηµαντικός ρόλος της α-αντουκίνης στη ρύθµιση της αρτηριακής πίεσης βασίζεται στην ικανότητα της να χειρίζεται τη µετακίνηση ιόντων νατρίου διαµέσου της πλασµατικής µεµβράνης. Η λειτουργία αυτή επιτυγχάνεται µε τη ρύθµιση της συγκρότησης του συµπλέγµατος σπεκτρίνης-ακτίνης, το οποίο κατευθύνει τη δέσµευση των ινιδίων της ακτίνης σε διαµεµβρανικές πρωτεΐνες µεταφοράς ιόντων, όπως οι δίαυλοι Na +, o συµµεταφορέας Na + -Κ + -Cl - και η αντλία Na + -K + (Cantiello, 1995). Η επιµόλυνση φυσιολογικών επιθηλιακών νεφρικών κυττάρων αρουραίων (NRK-52E) µε τη µεταλλαγµένη µορφή της α-αντουκίνης επηρεάζει την οργάνωση του κυτταροσκελετού της ακτίνης και µεταβάλλει δραστικά το ρυθµό λειτουργίας και την έκφραση της αντλίας Na + -K + (Tripodi et al., 1996). Η µεταλλαγµένη µορφή της α-αντουκίνης σε υπερτασικά άτοµα σχετίζεται άµεσα, τόσο µε τη µη σωστή λειτουργία των µηχανισµών που εµπλέκονται στη διατήρηση του ισοζυγίου NaCl εξαιτίας της υψηλότερης ευαισθησίας που παρουσιάζουν στις διακυµάνσεις των προσλαµβανόµενων ποσοτήτων Na + (salt sensitivity), όσο και µε τη διαφορική απόκριση στο διουρητικό θειαζιδικό φάρµακο υδροχλωροθειαζίδιο (HCTZ). Η χρόνια χορήγηση υδροχλωροθειαζιδίου στην οµάδα των υπερτασικών ατόµων, οι οποίοι είναι φορείς του αλληλοµόρφου ADD-W έχει ως αποτέλεσµα µια απότοµη πτώση της αρτηριακής πίεσης έως και 15mmHg σε σύγκριση µε τους οµοζυγώτες φορείς του αλληλοµόρφου αγρίου τύπου ADD- G (Cusi et al., 1997). Η πλήρης κατανόηση του αποτελέσµατος που προκύπτει από τη δράση του αλληλοµόρφου ADD-W σε διάφορους πληθυσµούς κάνει επιτακτική την ανάγκη ανίχνευσης πολυµορφισµών άλλων γονιδίων που εµπλέκονται σε µηχανισµούς ρύθµισης τόσο της νεφρικής λειτουργίας όσο και της αρτηριακής πίεσης. Από την εξέταση των γενοτύπων µίας οµάδας ασθενών µε ήπια µορφή υπέρτασης για την ταυτόχρονη παρουσία πολυµορφισµών της α-αντουκίνης και του ενζύµου µετατροπής (ACE) του συστήµατος ρενίνης-αγγειοτασίνης δεν προέκυψαν διαφορές στις τιµές της αρτηριακής πίεσης που να σχετίζονται µε το συνδυασµό διαφορετικών αλληλοµόρφων των συγκεκριµένων γονιδίων. Οι ενδοατοµικές διαφορές που παρατηρούνται εντοπίζονται στην απότο- µη πτώση της αρτηριακής πίεσης ως απόκριση στη θεραπευτική αγωγή µε θειαζιδικά διουρητικά φάρµακα (HCTZ), η οποία εµφανίζεται εντονότερη σε ασθενείς που είναι ταυτόχρονα φορείς ενός τουλάχιστον αλληλοµόρφου ACE-I και ADD-W. Συγκεκριµένα το αποτέλεσµα του συνδυασµού των γενοτύπων DI ή II + Gly/Trp ή Trp/Trp στα υπό µελέτη άτοµα, όσον αφορά την πτώση της αρτηριακής πίεσης εξαιτίας της χορήγησης υδροχλωροθειαζιδίου είναι αθροιστικό και όχι επιστατικό. Αυτή η µορφή απόκρισης, ενώ σε οµοζυγωτικά άτοµα για το αλληλόµορφο ADD-W οφείλεται στην τροποποίηση του µηχανισµού διούρησης και νατριούρησης από πίεση, σε οµοζυγωτικά άτοµα για το αλληλόµορφο ACE-I οφείλεται στο µικρότερο βαθµό ενεργοποίησης του συστήµατος ρενίνης-αγγειοτασίνης (RAS) (Sciarrone et al., 23) Συνθετάση µονοξειδίου του αζώτου (NO). Το µονοξείδιο του αζώτου (NO) είναι ένα σηµαντικό ενδοκυτταρικό µήνυµα και ένας από τους σπουδαιότερους παράγοντες καθορισµού της αρτηριακής πίεσης. Από το αυτόνοµο νευρικό σύστηµα, που νευρώνει τα αιµοφόρα αγγεία, απελευθερώνεται ακετυλοχολίνη, η οποία δεσµεύεται στην επιφάνεια των ενδοθηλιακών κυττάρων που επενδύουν τα αιµοφόρα αγγεία. Η δέσµευση αυτή οδηγεί σε αύξηση της συγκέντρωσης των κυτταροπλασµατικών ιόντων ασβεστίου µε άµεσο επακόλουθο την ενεργοποίηση της συνθετάσης του NO. Το παραγόµενο µονοξείδιο του αζώτου διαχέεται εύκολα διαµέσου της πλασµατικής µεµβράνης από τα
31 Εισαγωγή ιουρητικά φάρµακα-θειαζίδια ενδοθηλιακά κύτταρα στα παρακείµενα λεία µυϊκά κύτταρα, όπου και επάγεται η σύνθεση του κυκλικού GMP µέσω της ενεργοποίησης του ενζύµου γουανυλική κυκλάση. Το cgmp προκαλεί χαλάρωση των µυϊκών κυττάρων και συνεπώς διαστολή των αιµοφόρων αγγείων. Η δυσλειτουργία της ενδοθηλιακής συνθετάσης του NO, η οποία προκύπτει µε αντικατάσταση του κανονικού αµινοξέος γλουταµίνη από την ασπαραγίνη, εντοπίζεται στην αυξηµένη πρωτεολυτική ευαισθησία του ενζύµου που οδηγεί σε µειωµένη παραγωγή µονοξειδίου του αζώτου. Πολυµορφισµοί που εξετάσθηκαν σε δύο διαφορετικές εθνικές οµάδες, οι οποίες απαρτίζονται από τους Νεγροαµερικάνους και τους Αµερικάνους, αφορούν τα γονίδια της α-αντουκίνης, της συνθετάσης του µονοξειδίου του αζώτου, των β1 και β2 αδρενεργικών υποδοχέων και της λιπάσης των λιποπρωτεϊνών. Η ανάλυση των γονιδιακών συχνοτήτων σε συνδυασµό µε τις µετρήσεις της αρτηριακής πίεσης έδειξε σηµαντικές διαφορές ανάµεσα σε αυτές τις εθνικές οµάδες. Στους Νεγροαµερικάνους η µέση τιµή της συστολικής αρτηριακής πίεσης είναι σηµαντικά υψηλότερη σε σύγκριση µε τους Αµερικάνους, ενώ η συχνότητα των αλληλοµόρφων που προκύπτουν από τους πολυµορφισµούς των γονιδίων που µελετήθηκαν είναι αντίστροφα υψηλότερη. Στην παρούσα µελέτη θεωρήθηκε ότι η παρουσία κάθε πολυµορφισµού στον πληθυσµό είναι σε θέση να χρησιµοποιηθεί ως πρόβλεψη στις αποκρίσεις της αρτηριακής πίεσης κατά την αντιϋπερτασική αγωγή. Η απότοµη πτώση της συστολικής αρτηριακής πίεσης ως απόκριση των ατόµων στη χορήγηση του υδροχλωροθειαζιδίου συνεχίζει να σχετίζεται θετικά µε κάθε έναν από τους πολυµορφισµούς των γονιδίων της α-αντουκίνης και της συνθετάσης του NO. Αντίθετα, εάν στη στατιστική ανάλυση η παρουσία των πολυµορφισµών συσχετισθεί µε δευτερεύουσες µεταβλητές, όπως η εθνικότητα, το φύλο και η ηλικία τότε, τόσο οι πολυµορφισµοί που αφορούν τα γονίδια της α-αντουκίνης, των β1 και β2 αδρενεργικών υποδοχέων και της λιπάσης των λιποπρωτεϊνών, όσο και η ταυτόχρονη παρουσία αλληλοµόρφων δύο ή περισσοτέρων πολυµορφικών γονιδίων που µελετήθηκαν δεν είναι σε θέση να επηρεάζουν τις ενδοατοµικές διαφορές που παρατηρούνται στις αποκρίσεις της αρτηριακής πίεσης εξαιτίας της χορήγησης του υδροχλωροθειαζιδίου. Παρ όλα αυτά η εθνικότητα, το φύλο και η ηλικία είναι υπεύθυνα σε ποσοστό 26% και 11% αντίστοιχα, για την ενδοατοµική ποικιλότητα στις αποκρίσεις της συστολικής και της διαστολικής αρτηριακής πίεσης (Turner et al., 23)
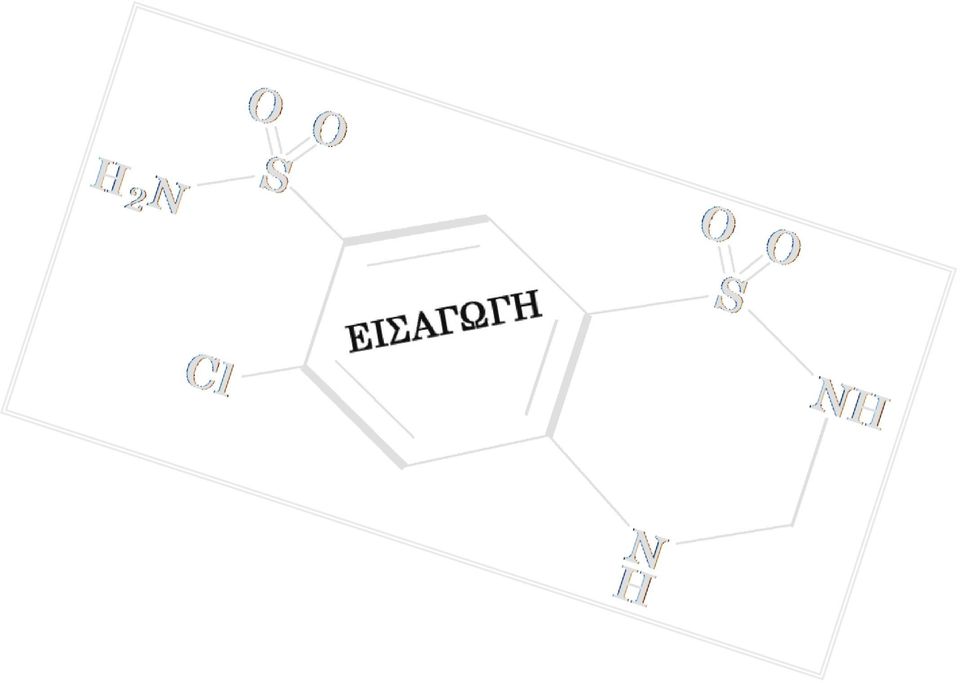
32 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ 3. Υ ΡΟΧΛΩΡΟΘΕΙΑΖΙ ΙΟ (CAS No ) 3.1 Φυσικές και χηµικές ιδιότητες Το υδροχλωροθειαζίδιο (HCTZ) είναι το 6-χλώρο-3,4-δίυδρο-2Η-1,2,4-βενζο θειαζίν-7-θειοαµίδ 1,1-διοξείδιο (6-Chloro-3,4-dihydro-2H-1,2,4-benzothiadiazine- 7-sulfonamide 1,1-dioxide) και η δοµή του παρουσιάζεται στην εικόνα 3-1. Άλλες συνώνυµες ονοµασίες που συναντώνται συχνά είναι οι ακόλουθες: α) 6-Chloro-3,4- dihydro-7-sulfamoyl-2h-1,2,4-benzothiadiazine1,1-dioxide, β) 6-Chloro-7-sulfamyl -3,4-dihydro-1,2,4-benzothiadiazine1,1-dioxide, γ) 3,4 dihydrochlorothiazide, δ) Chlorosulfonamidodihydrobenzothiadiazine dioxide και ε) Chlorosulthiadil. Ο µοριακός τύπος του υδροχλωροθειαζιδίου είναι C7H8CIN3O4S2 και έχει µοριακό βάρος 297,72. Το σηµείο τήξης είναι C. Είναι µια λευκή, άοσµη, µικροκρυσταλική σκόνη που έχει µια ελαφρά πικρή γεύση. Η χηµική ένωση είναι ελάχιστα διαλυτή στο νερό, ενώ είναι ευδιάλυτη σε αµµωνιακό διάλυµα, σε αραιό διάλυµα υδροξειδίου του νατρίου, σε αιθανόλη, σε µεθανόλη, σε ακετόνη και σε ακετονιτρίλιο, έχοντας µια σταθερά διάστασης pka ιαλυµένο σε υδατικό διάλυµα και σε ακραίες τιµές ph το υδροχλωροθειαζίδιο υδρολύεται σε φορµαλδεΰδη και 6-χλώρο-2,4-διθειοαµοϊλανιλίνη (IARC, 199). H 2 N O O S C H C C O O S NH Cl C C H C N H CH Παραγωγή Εικόνα 3-1. Χηµικός τύπος του υδροχλωροθειαζιδίου (HCTZ). Το υδροχλωροθειαζίδιο συντίθεται είτε µε την αντίδραση της παραφορµαλδεΰδης µε την 5-χλώρο-2,4-διθειοαµοϊλανιλίνη σε µη υδατικό µέσο είτε µε την αντίδραση της φορµαλδεΰδης µε το 6-χλώρο-7θειαµοϊλ-2Η-1,2,4-βενζοθειαζιδίνη-1, 1-διοξείδιο σε αλκαλικό υδατικό διάλυµα. εν είναι γνωστό εάν υπάρχει ως φυσικό προϊόν (IARC, 199). 3.3 Ενδείξεις-Εµπορική Ονοµασία- οσολογία Το υδροχλωροθειαζίδιο είναι ένα διουρητικό θειαζίδιο που χρησιµοποιείται ευρύτατα στη θεραπεία της υπέρτασης, από το Ο συνδυασµός υδροχλωροθειαζίδιο/τριαµτερένη (Dyazide) τη χρονική περίοδο βρίσκεται πρώτος στη λίστα συνταγογραφούµενων φαρµάκων στις Η.Π.Α., ενώ το υδροχλωροθειαζίδιο τη χρονική περίοδο είναι το έκτο ευρύτερα υποδεικνυόµενο φάρµακο. Το υδροχλωροθειαζίδιο χρησιµοποιείται ευρύτατα για τη µείωση οιδηµάτων που σχετίζονται µε την καρδιακή ανεπάρκεια, ως αντιϋπερτασικός παράγοντας, καθώς και σε ειδικές περιπτώσεις όπως η ασθένεια Ménière (van Deelen και Huizing, 1986), ο νεφρογενής άποιος διαβήτης (Kirchlechner et al., 1999) και η µείωση του σχηµατι
33 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ σµού λίθων οξαλικού ασβεστίου στην ουροφόρο οδό, σε άτοµα µε ιδιοπαθή υπερασβεστιουρία (Baggio et al., 1986). Οι συνήθεις εµπορικές ονοµασίες του φαρµάκου είναι: Aquarius, Apo- Hydro, Atenadon, Bremil, Cidrex, Dichlotride, Esidrix, Fluvin, Hydro-Diuril, Hypothiazide, Ivaugan, Oretic, Neoflumen, Ridaq, Thiuretic και πολλές άλλες, ανάλογα µε τη χώρα στην οποία κυκλοφορεί. Επιπρόσθετα, ευρέως διαδεδοµένη είναι η συνδυαστική χορήγηση του υδροχλωροθειαζιδίου, τόσο µε αναστολείς του µετατρεπτικού ενζύµου της αγγειοτασίνης (κιναπρίλη, λισινοπρίλη, εναλαπρίλη), όσο και µε ανταγωνιστές των υποδοχέων της αγγειοτασίνης ΙΙ (λοζαρτάνη, ιρµπεζαρτάνη), σε φαρµακευτικά σκευάσµατα που φέρουν την εµπορική ονοµασία Accuretic, Zestoretic, Renitec, Hyzaar, Karvezide και Avalide (Neutel et al., 1996). Σύµφωνα µε τις ιδιότητες του φαρµάκου (υδατοδιαλυτότητα, λιποδιαλυτότητα, ιονισµός) ακολουθείται η συνηθέστερη (εντερική) οδός χορήγησης από το στόµα, η οποία είναι η περισσότερο ασταθής και πολύπλοκη. Τα τελευταία σαράντα χρόνια υπάρχει µία εξέλιξη στην πορεία της προτεινόµενης δοσολογίας του υδροχλωροθειαζιδίου, ιδιαίτερα όσον αφορά τη θεραπεία της ιδιοπαθούς υπέρτασης. Συνεχώς ακολουθείται µία πτωτική τάση στην ηµερήσια δόση, η οποία από τα 2 mg στις αρχές της δεκαετίας του 196 µειώθηκε στα 6.25 mg το 1996, µε σκοπό να µειωθούν ανεπιθύµητα επεισόδια και διαταραχές σε µεταβολικά µονοπάτια, στα οποία εµπλέκεται άµεσα το υδροχλωροθειαζίδιο. Στη δεκαετία που διανύουµε, όσον αφορά τη θεραπευτική αγωγή που ακολουθείται για τη θεραπεία της ιδιοπαθούς υπέρτασης, ευρέως διαδεδοµένη είναι η συνδυαστική χορήγηση του υδροχλωροθειαζιδίου µε άλλους αντιϋπερτασικούς παράγοντες σε δισκία, στα οποία η ηµερήσια χορηγούµενη δόση κυµαίνεται µεταξύ mg. Η ίδια δοσολογία ακολουθείται, όταν χρησιµοποιείται µόνο του στην αντιϋπερτασική θεραπεία που αφορά τη χορήγηση αµιγούς διουρητικού φαρµάκου, εξαιτίας της δυνατότητας του σε αυτές τις συγκεντρώσεις να µειώνει ταυτόχρονα µε την αρτηριακή πίεση και τις αρνητικές επιπτώσεις που επιφέρει η εµπλοκή του στο µεταβολισµό των λιπιδίων. Πάντοτε, θα πρέπει να ακολουθείται εξατοµίκευση της θεραπευτικής δοσολογίας σύµφωνα µε την απόκριση του ασθενούς (Neutel, 1996, Reyes, 22). 3.4 Μηχανισµός δράσης του υδροχλωροθειαζιδίου (HCTZ) Ο ακριβής µοριακός µηχανισµός δράσης του υδροχλωροθειαζιδίου δεν έχει πλήρως αποσαφηνισθεί. Τα αποτελέσµατα της αντιϋπερτασικής αγωγής µε διουρητικά θειαζίδια (βενζοθειαδιαζίνες) επιβεβαιώνονται συνεχώς, αλλά ο ακριβής µηχανισµός διαφορετικής απόκρισης, που αναφέρεται σε οµάδες υπερτασικών ατόµων, κατά τη χρόνια χορήγηση τους, αποτελεί αναµφισβήτητα αντικείµενο ερευνητικών µελετών και επιστηµονικών συζητήσεων. Οι πρώτες µελέτες που αφορούσαν τη χρόνια χορήγηση υδροχλωροθειαζιδίου σε υπερτασικά άτοµα, σε ηµερήσιες δόσεις που κυµαίνονται από 5-1 mg, οδήγησαν την επιστηµονική κοινότητα στο συµπέρασµα ότι η ταυτόχρονη και συνεχιζόµενη, σε µικρότερο βαθµό, µείωση του όγκου του πλάσµατος και του εξωκυττάριου υγρού που επιτυγχάνονται µε τη νατριούρηση, παρά τα αυξηµένα επίπεδα ρενίνης που εντοπίζονται στο πλάσµα ως αποτέλεσµα της έλλειψης NaCl, καθορίζουν την ελάττωση της καρδιακής παροχής και της αρτηριακής πίεσης
34 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Πρωτεΐνες-µεταφορείς Η κύρια δράση του εντοπίζεται στην αναστολή της επαναρρόφησης του Na Cl στη φλοιώδη ουσία του νεφρού (άπω εσπειραµένο σωληνάριο), εξαιτίας της πιθανής ιδιότητας του να εµφανίζει υψηλή συγγένεια µε µία θέση δέσµευσης, η οποία σε φυσιολογικές συνθήκες καταλαµβάνεται από ανιόντα αλογόνων (Cl, Br, J) σε ένα εξαιρετικά ευαίσθητο στα θειαζίδια σύστηµα µεταφοράς ιόντων. Η υψηλής συγγένειας θέση δέσµευσης για τα θειαζίδια, η οποία συναντάται σε µία διαµεµβρανική πρωτεΐνη συµµεταφοράς ιόντων (Na +, Cl - ), πιθανώς αντιπροσωπεύει τον υποδοχέα των διουρητικών θειαζιδικού τύπου (Beaumont et al., 1988). Εικόνα 3-2. Eπαναρρόφηση NaCl στο άπω εσπειραµένο σωληνάριο και µηχανισµός δράσης των διουρητικών της θειαζίδης, υδροχλωροθειαζίδιο, τα οποία αναστέλλουν το συµµεταφορέα Na + -Cl -. Σε φυσιολογικές συνθήκες τα Na + εγκαταλείπουν το κύτταρο µε τη βοήθεια της αντλίας Na + -Κ + -ATPάση και τα Cl - µε διάχυση διαµέσου διαύλων (Berne και Levy, 1996, Jackson, 1996). Γενικότερα οι πρωτεΐνες-µεταφορείς, ανεξάρτητα από το µόριο που µεταφέρουν, την κατεύθυνση της µεταφοράς, το µηχανισµό µεταφοράς και τον οργανισµό συνιστούν µία πολυπληθή οικογένεια. Σε αυτή την οικογένεια διακρίνεται µία νέα κατηγορία µεταφορικών συστηµάτων σύµφωνα µε τo είδος του µεταφερόµενου κατιόντος (Na + -Cl - ή Na + -Κ + -2Cl - συµµεταφορείς), την ειδικότητα των αλληλεπιδράσεων µε αναστολείς διαφορετικού τύπου (συµµεταφορείς ευαίσθητοι στα θειαζίδια, TSC ή στη βουµετανίδη, BSC) και την καθορισµένη θέση τους στην κυτταροπλασµατική µεµβράνη των διαφόρων τύπων επιθηλιακών κυττάρων (κορυφαία ή βασεοπλευρική). Ο συµµεταφορέας Na + -Cl - στον άνθρωπο (htsc), στον αρουραίο (rtsc) και στα ψάρια (fltsh) εµφανίζει σηµαντική οµοιότητα τόσο στη νουκλεοτιδική ακολουθία (86% και 72%), όσο και στην αµινοξική αλληλουχία (92% και 78%). Αξιοσηµείωτο είναι ότι µεταξύ των διαφορετικών συµµεταφορέων Na + -Cl - και Na + - Κ + -2Cl - παρατηρείται εκτεταµένη οµολογία ανάµεσα στον άνθρωπο (htsc) και σε σχετικά αποµακρυσµένα είδη, όπως στις ζύµες, στα έντοµα, στα ψάρια και στον αρουραίο (rbsc), γεγονός που συνηγορεί στην άποψη ότι η διατήρηση αυτών των ακολουθιών αντανακλά στη διατήρηση της λειτουργίας σε διαφορετικά είδη και στην κοινή προέλευση των µεταφορικών αυτών πρωτεϊνών. Στον άνθρωπο, ο γενετικός τόπος που κωδικοποιεί το ευαίσθητο στα θειαζίδια σύστηµα µεταφοράς ιόντων (htsc) εντοπίζεται στο µακρύ βραχίονα του χρωµοσώµατος 16q13. Ένα εξαιρετικά ειδικό πρότυπο έκφρασης σε ανθρώπινους ιστούς διαπιστώνεται για τον htsc (τεχνική Northern), το οποίο αποκαλύπτει την ύπαρξη δύο µεταγραφηµάτων διαφορετικού µεγέθους (4.5kb, 6.5kb) και διαφορετικής έντασης στο νεφρό και την
35 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ παντελή απουσία αυτών στην καρδιά, στον εγκέφαλο, στους πνεύµονες, στο ήπαρ, στο πάγκρεας, στο θύµο αδένα, στα γεννητικά όργανα και στα λευκά αιµοσφαίρια του περιφερικού αίµατος. Η ανίχνευση µίας ασθενούς ζώνης που αφορά το µεταγράφηµα µεγαλύτερου µεγέθους (6.5kb) παρατηρείται στο λεπτό έντερο, στον πλακούντα, στον προστάτη, στο κόλον και στη σπλήνα και αντιπροσωπεύει χαµηλά επίπεδα έκφρασης για τον htsc σε αυτούς τους ιστούς (Chang et al., 1996). Τρεις διακριτές περιοχές χαρακτηρίζουν τη συγκεκριµένη πρωτεΐνη-µεταφορέα (121αα-112kDa) που βρίσκεται ενσωµατωµένη στις κυτταρικές µεµβράνες, από τις οποίες οι δύο είναι ενδοκυτταρικές και ισχυρά υδρόφιλες, εκπροσωπώντας το αµινοτελικό και καρβοξυτελικό άκρο της πρωτεΐνης και µία υδρόφοβη περιοχή που αποτελεί το µεσαίο τµήµα της πρωτεΐνης. Σε αυτό το τµήµα εντοπίζονται 12 περιοχές (διαµεµβρανικές), οι οποίες διαπερνούν το εσωτερικό του διµοριακού λιπιδικού στρώµατος. Μικρά τµήµατα, τα οποία προεξέχουν του στρώµατος προς τον εξωκυττάριο χώρο (extracellular loops) διαφέρουν ως προς την αµινοξική τους σύσταση µεταξύ των διαφορετικών συµµεταφορέων (TSC-BSC) και αποτελούν τις εξειδικευµένες θέσεις δέσµευσης διαφορετικής συγγένειας για τους διουρητικούς παράγοντες. Επίσης, στα δύο άκρα της πρωτεΐνης που βρίσκονται στην κυτταροπλασµατική πλευρά υπάρχουν πιθανές θέσεις φωσφορυλίωσης για διαφορετικές κινάσες πρωτεϊνών (PK). ιαφορές που παρατηρούνται στη λειτουργία των συµµεταφορέων διαφορετικού τύπου και εντοπίζονται σε αµινοξικές υποκαταστάσεις του αµινοτελικού άκρου τους, ιδιαίτερα στις θέσεις φωσφορυλίωσης, πιθανόν αντικατοπτρίζουν διαφορές στη ρύθµιση της ενεργοποίησης τους, καθώς εµπλέκονται διαφορετικοί ενδοκυτταρικοί µηχανισµοί (Mastroianni et al., 1996) Ανθρακική ανυδράση Οι αντιϋπερτασικές ιδιότητες του υδροχλωροθειαζιδίου σχετίζονται άµεσα µε την ικανότητα του να συνδέεται µε την πανταχού παρούσα ανθρακική ανυδράση την οποία αναστέλλει µη συναγωνιστικά. Η επιτάχυνση της αντίδρασης: CO2+H2O H2CO3 H + +HCO3 -, ανάλογα µε τη µερική πίεση του CO2 και τη συγκέντρωση των διττανθρακικών (HCO3 - ) οφείλεται στο ένζυµο ανθρακική ανυδράση. Μέρος της καταλυτικής αύξησης επιτυγχάνεται µε την παρουσία ενός ιόντος ψευδαργύρου που σχηµατίζει σύµπλοκο συναρµογής µε τα ιµιδαζόλια των τριών ιστιδινών του ενζύµου και βοηθά τόσο στη σωστή τοποθέτηση του CO2 όσο και σε µία τοπικά υψηλή συγκέντρωση ΟΗ - που προκύπτουν από τη διάσπαση του νερού. Σουλφοναµιδικά παράγωγα, όπως το υδροχλωροθειαζίδιο, δεσµεύονται πολύ κοντά στο ενεργό κέντρο του ενζύµου και διακόπτουν τις αλληλεπιδράσεις που αφορούν τη δέσµευση του νερού στο ιόν ψευδαργύρου και τη µετατροπή του σε ιόν υδροξυλίου αναστέλλοντας την ενεργότητα του ενζύµου ανθρακική ανυδράση. Στα θηλαστικά, η ανθρακική ανυδράση αποτελείται από µία πολυπεπτιδική αλυσίδα (µονοµερής) και ανήκει στην τάξη α της αρχαίας αυτής πολυµορφικής οικογένειας ενζύµων. Τουλάχιστον δέκα ελαφρά διαφορετικές ισοµορφές της α ανθρακικής ανυδράσης εντοπίζονται σε διάφορους ιστούς ή κυτταρικά διαµερίσµατα είτε σε διαλυτή µορφή είτε δεσµευµένες στις µεµβράνες ειδικών επιθηλιακών κυττάρων. Η ικανότητα των νεφρών, σε συνδυασµό µε διάφορα ρυθµιστικά συστήµατα (CO2/HCO3 - ) να ρυθµίζουν το ph των υγρών του σώµατος και να διατηρούν το οξεοβασικό ισοζύγιο σε σταθερή κατάσταση οφείλεται στην παρουσία της µεµβρανικής ισοµορφής IV, της α ανθρακικής ανυδράσης. Η αναστολή της συγκεκριµένης ισοµορ -φής στους νεφρούς από το υδροχλωροθειαζίδιο σχετίζεται έµµεσα µε την αναστολή του αντιµεταφορέα Na + -H + µε αποτέλεσµα τη µείωση της επαναρρόφησης των Na
36 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Η συνολική επαναρρόφηση των διαλελυµένων οργανικών ουσιών, των Cl -, των άλλων ιόντων (K +, Ca ++, Mg ++ ) και του ύδατος είναι στενά συνδεδεµένη µε την επαναρρόφηση των Na +. Κάθε µεταβολή στην επαναρρόφηση των Na + θα επηρεάσει την επαναρρόφηση του ύδατος και των διαλελυµένων σε αυτό ουσιών ιδιαίτερα από το εγγύς εσπειραµένο σωληνάριο. Η αναστολή της διακυτταρικής επαναρρόφησης των Na + (αντιµεταφορέας Na + -H + ) και Cl - (αντιµεταφορέας Cl - -Βάση - ) δια µέσου της κορυφαίας µεµβράνης των επιθηλιακών κυττάρων στο δεύτερο ήµισυ του εγγύς σωληναρίου µειώνουν σηµαντικά την ωσµωτική επαναρρόφηση του ύδατος µε αποτέλεσµα την πτώση της αρτηριακής πίεσης (Beaumont et al., 1988, Schaeffer et al., 199) Πρόσφατες ερευνητικές µελέτες έχουν αποδώσει στο υδροχλωροθειαζίδιο και γενικότερα στη χρόνια χορήγηση διουρητικών θειαζιδίων µία άµεση αγγειοδιασταλτική δράση, η οποία επιτυγχάνεται µε την αναστολή της ανθρακικής ανυδράσης που εντοπίζεται δεσµευµένη εξωκυτταρικά στη µεµβράνη των λείων µυϊκών κυττάρων των αιµοφόρων αγγείων. Είναι πιθανόν, αυτή η εξωκυτταρική ενζυµική ισοµορφή να τροποποιεί τη διαµεµβρανική µεταφορική πρωτεΐνη Cl/HCO3 - µε αποτέλεσµα µία εξασθένηση στην αποβολή των διττανθρακικών ιόντων που έχει άµεση επίδραση στο ενδοκυτταρικό ph. Συγκεκριµένα, η αύξηση του ph εξαιτίας της αναστολής της ανθρακικής ανυδράσης ενεργοποιεί τους διαύλους KCa, γεγονός που µεταβάλλει την εκπολωτική και επαναπολωτική φάση των δυναµικών ενεργείας των λείων µυϊκών κυττάρων. Η υπερπόλωση της κυτταρικής µεµβράνης σε συνδυασµό µε τη συνεχή πτώση της ενδοκυτταρικής [Ca ++ ] καθιστά τα λεία µυϊκά κύτταρα ανίκανα να πυροδοτήσουν δυναµικά ενεργείας και αδύναµα να συσπαστούν, µε αποτέλεσµα να επιτυγχάνεται µία συνεχή χαλάρωση που επιφέρει διαστολή των αιµοφόρων αγγείων, µείωση των περιφερικών αγγειακών αντιστάσεων και κατ επέκταση πτώση της αρτηριακής πίεσης (Calder et al., 1992, Pickkers et al., 1999) Περιφερικός τύπος υποδοχέα των βενζοδιαζεπινών Σηµαντική είναι η ικανότητα δέσµευσης του υδροχλωροθειαζιδίου µε τον περιφερικό τύπο υποδοχέα των βενζοδιαζεπινών (PBR), ο οποίος στα θηλαστικά κατανέµεται σε µία ποικιλία περιφερικών ιστών/οργάνων, όπως στους νεφρούς, στα επινεφρίδια, στους πνεύµονες, στο ήπαρ, στην καρδιά, στους όρχεις και στον εγκέφαλο. Συνήθως εντοπίζεται στην εξωτερική µεµβράνη των µιτοχονδρίων, ενώ εµφανίζεται συχνά τόσο στην επιφάνεια της κυτταροπλασµατικής µεµβράνης όσο και σε άλλες ενδοκυτταρικές τοποθεσίες σε ορισµένους κυτταρικούς τύπους και ιστούς (Casellas et al., 22). Πρόκειται για µία εξαιρετικά µικρή (18 kda) και εξελικτικά συντηρηµένη πρωτεΐνη, η οποία είναι στενά σχετιζόµενη µε το δίαυλο ανιόντων που ελέγχει τα δυναµικά ενεργείας των κυτταρικών µεµβρανών (VDAC) και τον ειδικό αντιµεταφορέα νουκλεοτιδίων αδενοσίνης (ANC), υπεύθυνο για την ανταλλαγή της ATP/ADP. Χαρακτηριστικές είναι οι υψηλής συγγένειας θέσεις δέσµευσης που εµφανίζει ο υποδοχέας (PBR), οι οποίες αφορούν δύο διακριτές κατηγορίες σύνθετων ενώσεων, τις βενζοδιαζεπίνες (BZs) και τις ισοκινολίνες (IQs). Πρόσφατες ερευνητικές µελέτες θεωρούν τον περιφερικό τύπο υποδοχέα των βενζοδιαζεπινών ως ένα πολυµερές σύµπλεγµα που δεν εµφανίζει στοιχεία µίας απλής πρωτεΐνης-υποδοχέα και στον οποίον η υποµονάδα 18 kda αφορά τη θέση δέσµευσης για τις ισοκινολίνες, ενώ η έκφραση της θέσης δέσµευσης για τις βενζοδιαζεπίνες αφορά την ταυτόχρονη παρουσία µίας δεύτερης υποµονάδας (34 kda) που είναι ο δίαυλος ανιόντων, VDAC
37 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Αντίθετα, ο ρόλος του αντιµεταφορέα νουκλεοτιδίων αδενοσίνης (ANC) δεν φαίνεται να είναι καθοριστικός στην απόκριση του υποδοχέα σε διαφορετικές φαρµακευτικές ενώσεις (Garnier et al., 1994). Στους νεφρούς, η συγκεκριµένη θέση δέσµευσης για τις βενζοδιαζεπίνες (PBBS) εντοπίζεται στο παχύ τµήµα του ανιόντος σκέλους της αγκύλης Henle και στο άπω εσπειραµένο σωληνάριο (Beaumont et al., 1984, Butlen, 1984) και η πυκνότητα του ρυθµίζεται επιλεκτικά από την παρουσία αναστολέων διαµεµβρανικών µεταφορικών πρωτεϊνών (διουρητικά θειαζίδια) που σχετίζονται µε την ανταλλαγή ιόντων (Basile et al., 1988). Αξιοσηµείωτο είναι ότι ο περιφερικός τύπος υποδοχέα των βενζοδιαζεπινών (PBR) αποτελεί κρίσιµο συστατικό ενός πολυπρωτεϊνικού συµπλέγµατος που είναι ένας εξαιρετικά µεγάλος δίαυλος (MPTP-mitochondrial permeability transition pore), ο οποίος εντοπίζεται σε θέσεις επαφής της εξωτερικής µε την εσωτερική µεµβράνη των µιτοχονδρίων. Ανάµεσα σε ένα µεγάλο αριθµό βιολογικών λειτουργιών στις οποίες εµπλέκεται άµεσα ή έµµεσα συµπεριλαµβάνονται η ρύθµιση της σύνθεσης στεροειδών ορµονών, του κυτταρικού πολλαπλασιασµού και της απόπτωσης, η µεταφορά των πορφυρινών και η βιοσύνθεση της αίµης, η τροποποίηση της ανοσοαπόκρισης και η µεταφορά ανιόντων. Η χρήση ειδικών µορίων που δεσµεύονται στον περιφερικό τύπο υποδοχέα των βενζοδιαζεπινών, τα οποία είναι σε θέση να τροποποιούν την ενεργοποίηση του και να µεταβάλλουν τις δραστηριότητες του πολυπρωτεϊνικού συµπλέγµατος MPTP, πιθανόν να εµφανίζουν θεραπευτικές εφαρµογές και να παρέχουν κλινικά οφέλη στη διαχείριση ενός µεγάλου φάσµατος διαφορετικών ασθενειών (καρκίνος, αυτοάνοσα και λοιµώδη νοσήµατα, νευροεκφυλιστικές νόσοι) (Galiegue et al., 23). Ο συγκεκριµένος υποδοχέας (PBR) εµφανίζει θέσεις υψηλής συγγένειας για την ταυτόχρονη δέσµευση φαρµακευτικών ενώσεων και µίας σηµαντικής κατηγορίας µεµβρανικών λιπιδίων, της χοληστερόλης, η οποία αποτελεί την πρόδροµη ένωση σε µία σειρά αντιδράσεων που αφορούν τη σύνθεση φλοιοκορτικοστεροειδών ορµονών. Υπάρχουν σαφείς ενδείξεις, οι οποίες συσχετίζουν την υπερέκφραση του υποδοχέα µε τη δυνατότητα πρόγνωσης επιθετικών µορφών καρκίνου του προστάτη, του παχέος εντέρου και του µαστού (Papadopoulo, 23). Επίσης, η αύξηση της πυκνότητας του υποδοχέα σε ειδικές περιοχές του εγκεφάλου (κροταφιαίο/µετωπιαίο φλοιό), σε άτοµα που πάσχουν από νευροεκφυλιστικές (Alzheimer, Huntington) και νευροψυχιατρικές (επιληψία) νόσους, δηλώνει το σηµαντικό ρόλο της µιτοχονδριακής λειτουργίας στην παθογένεση αρκετών νευρολογικών ασθενειών (Casellas et al., 22). Η in vivo χορήγηση υδροχλωροθειαζιδίου σε συγκεντρώσεις µικρότερες των 15 µμ επηρεάζει την έκφραση των θέσεων δέσµευσης για τις βενζοδιαζεπίνες (PB BS) στην εξωτερική µοίρα (φλοιώδη ουσία) του νεφρού. Αυτό έχει ως αποτέλεσµα, η πυκνότητα του περιφερικού τύπου υποδοχέα των βενζοδιαζεπινών (PBR) να παρουσιάζει µία σηµαντική αύξηση σε µία ευαίσθητη στα θειαζίδια περιοχή του νεφρώνα, η οποία αποτελεί χαρακτηριστική θέση επαναρρόφησης ιόντων που επηρεάζεται άµεσα από το υδροχλωροθειαζίδιο (Lukeman et al., 1987). Η ίδια ερευνητική οµάδα λαµβάνοντας υπόψη: (α) τις θέσεις δράσεις του υδροχλωροθειαζιδίου, (β) τις παρόµοιες συγκεντρώσεις (1-4 Μ) της φαρµακευτικής ένωσης µε τις οποίες εκδηλώνονται τόσο οι δυνατότητες δέσµευσης της σε συγκεκριµένη θέση (PBBS) του υποδοχέα, όσο και οι διουρητικές επιδράσεις της (νατριουρητική αποτελεσµατικότητα) σε ηλεκτρικά ουδέτερα µεµβρανικά συστήµατα µεταφοράς ιόντων και (γ) την άµεση συµµετοχή του υποδοχέα PBR στη ροή των ιόντων Na +, K +, Cl - ή Ca ++ διαµέσου της κυτταρικής µεµβράνης ή των επιθηλίων, υπέθεσε, ότι η
38 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ δοµή του περιφερικού τύπου υποδοχέα των βενζοδιαζεπινών στους νεφρούς πιθανόν είναι η ίδια, είτε σχετίζεται στενά µε τον υποδοχέα των διουρητικών θειαζιδικού τύπου (TSC) (Lukeman και Fanestil, 1987) Κατεχολαµινεργικές ορµόνες/β2-αδρενεργικοί υποδοχείς Ένας άλλος πιθανός µηχανισµός δράσης του υδροχλωροθειαζιδίου που σχετίζεται µε τη µείωση των περιφερικών αγγειακών αντιστάσεων σε οµάδες υπερτασικών ατόµων µε την καθορισµένη µορφή απόκρισης, στις οποίες ανιχνεύονται υψηλά επίπεδα κατεχολαµινεργικών ορµονών στο πλάσµα φαίνεται να είναι µία εξασθένηση στην ευαισθησία οργάνων-στόχων έναντι των ορµονών επινεφρίνης-νορεπινεφρίνης και πιθανόν της αγγειοτασίνης ΙΙ (Fernandez et al., 1987). Οι αντιϋπερτασικές ιδιότητες του υδροχλωροθειαζιδίου αποδίδονται στην εξίσου σηµαντική ικανότητα που παρουσιάζει να αλληλεπιδρά µε τους β2-αδρενεργικούς υποδοχείς των µονοπύρηνων κυττάρων του περιφερικού αίµατος και να τροποποιεί την έκφραση τους. Κατεχολαµινεργικές ορµόνες (επινεφρίνη, νορεπινεφρίνη) συνθέτονται από το αµινοξύ τυροσίνη µέσω µίας σειράς ενζυµικών αντιδράσεων σε ένα εξειδικευµένο συµπαθητικό γάγγλιο, το µυελό των επινεφριδίων και αποθηκεύονται σε εκκριτικά κοκκία στα χρωµιόφιλα κύτταρα της µυελώδους µοίρας. Εκκρίνονται κατευθείαν στην κυκλοφορία του αίµατος ως απόκριση στη δραστηριότητα των προγαγγλιακών χολινεργικών ινών του συµπαθητικού νευρικού συστήµατος και είναι σε θέση να ρυθµίζουν την αρτηριακή πίεση µε τη διαµεσολάβηση ειδικών διαµεµβρανικών υποδοχέων (β1, β2, β3, α1 και α2). Ιδιαίτερα η νορεπινεφρίνη, εµφανίζει µία επιπλέον δράση ως βιογενής αµίνη, είναι ο κύριος νευροδιαβιβαστής στους µεταγαγγλιακούς συµπαθητικούς νευρώνες. Η ιστοειδική έκφραση των αδρενεργικών υποδοχέων που αφορά ορισµένους τύπους κυττάρων (λεία µυϊκά κύτταρα) αντανακλά διαφορές στις οδούς µεταγωγής σήµατος ως απόκριση στη δέσµευση συγκεκριµένων διαβιβαστών (αγωνιστές). Οι υποδοχείς β1 και β2 είναι συζευγµένοι µε τη διεγερτική της αδενυλικής κυκλάσης υποµονάδα αs της ετεροτριµερούς πρωτεΐνης G και η σύνδεση της επινεφρίνης µε τον υποδοχέα ανυψώνει τα επίπεδα του κυκλικού AMP µε επακόλουθο έναν "καταρράκτη" φωσφορυλιώσεων πρωτεϊνών (ένζυµα ή ιοντικοί δίαυλοι) που καταλύεται από την πρωτεϊνική κινάση Α. Η χρόνια χορήγηση ηµερήσιας δόσης 5 mg υδροχλωροθειαζιδίου σε υπερτασικά άτοµα προκαλεί σηµαντική µείωση της πυκνότητας (4%) των β2-αδρενεργικών υποδοχέων χωρίς να επηρεάζει την υψηλή συγγένεια των θέσεων δέσµευσης του υποδοχέα για τις κατεχολαµινικές ορµόνες. Η αντιϋπερτασική δράση του υδροχλωροθειαζιδίου εστιάζεται στην παραδοχή ότι οι β2-αδρενεργικοί υποδοχείς που εντοπίζονται στην κυτταρική µεµβράνη των λεµφοκυττάρων πιθανόν εκπροσωπούν προσυναπτικούς β2-αδρενεργικούς υποδοχείς, η µειορρύθµιση (down regulation) των οποίων, παρουσία της φαρµακευτικής ένωσης, έχει ως αποτέλεσµα τη µείωση της απελευθέρωσης αδρενεργικού διαβιβαστή σε µετασυναπτικές θέσεις (αιµοφόρα αγγεία). Το γεγονός αυτό θα µπορούσε να εξηγήσει µία εξασθενηµένη συστολή των περισσοτέρων αγγείων ως απόκριση σε ορµόνες που ρυθµίζουν την αρτηριακή πίεση και ακολούθως την ελάττωση της. Έχει βρεθεί ότι κατά τη χρόνια χορήγηση διουρητικών φαρµάκων η µέτρηση τόσο της πυκνότητας, όσο και της συγγένειας των θέσεων δέσµευσης για τις κατεχολα- µινικές ορµόνες, των αδρενεργικών υποδοχέων µπορούν να χρησιµοποιηθούν ως δείκτες της αδρενεργικής ευαισθησίας µε σκοπό την επίτευξη ενός βέλτιστου θεραπευτικού αποτελέσµατος (Middeke et al., 1985). Παρά τις ελλιπείς γνώσεις, που αφορούν τη µειορρύθµιση των β2-αδρενεργικών υποδοχέων έχει διαπιστωθεί
39 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ µία θετική συσχέτιση της πυκνότητας τους, σε άθικτα µονοπύρηνα κύτταρα και σε λεµφοκύτταρα του περιφερικού αίµατος, µε τη µέση αρτηριακή πίεση. Συγκεκριµένα σε ασθενείς µε ιδιοπαθή υπέρταση διπλασιάζεται ο αριθµός των υποδοχέων σε σύγκριση µε τα φυσιολογικά άτοµα, µε αποτέλεσµα µία άνιση κατανοµή των εξωκυτταρικών και ενδοκυτταρικών θέσεων δέσµευσης που εντοπίζονται στην κυτταρική επιφάνεια µε αποτέλεσµα να τροποποιούνται φυσιολογικές λειτουργίες που σχετίζονται είτε µε τη συστολή είτε µε τη διαστολή των αιµοφόρων αγγείων (Middeke et al., 1983, Brodde et al, 1985). Πρόσφατα διατυπώθηκε η άποψη ότι µία ελλιπής αγγειοδιασταλτική λειτουργία, η οποία προκαλεί αύξηση των περιφερικών αντιστάσεων θεωρείται εξίσου σηµαντική τόσο στην παθογένεση και διατήρηση της υπερτασικής κατάστασης, όσο και στην προδιάθεση των ηλικιωµένων ατόµων να εµφανίσουν υπέρταση. Σε αυτήν την περίπτωση το δυναµικό πρότυπο ρύθµισης της απόκρισης των β-αδρενεργικών υποδοχέων δεν σχετίζεται µε την επαγόµενη από αγωνιστές απευαισθητοποίηση τους (Feldman et al., 1995). Η τροποποίηση της λειτουργίας των κινασών του β-αδρενεργικού υποδοχέα (βark), που αποτελούν µέλη µίας ευρύτερης οικογένειας κινασών σε υποδοχείς συζευγµένους µε ρυθµιστικές πρωτεΐνες δέσµευσης GTP (GRKs) υποκινεί τη φωσφορυλίωση καταλοίπων σερίνης και θρεονίνης που εντοπίζονται στο ενδοκυττάριο C-άκρο του, µε αποτέλεσµα την απευαισθητοποίηση του και τη µειωµένη απόκριση στον αγωνιστή. Η αυξηµένη δραστικότητα και έκφραση των GRKs που παρατηρείται σε λεµφοκύτταρα του περιφερικού αίµατος σε υπερτασικά άτοµα προκαλεί µείωση της ενεργοποίησης του ενζύµου αδενυλικής κυκλάσης που σχηµατίζει το κυκλικό AMP. Τo camp είναι ένας δεύτερος αγγελιοφόρος στις οδούς ενδοκυτταρικής µεταβίβασης σηµάτων, που η δράση του, στη πλειονότητα των περιπτώσεων, επιτελείται µεσω της κινάσης Α, η οποία φωσφορυλιώνει διάφορα ένζυµα του µεταβολισµού στο κυτταρόπλασµα και τον παράγοντα µεταγραφής CREB διαχεόµενη στον πυρήνα. Εποµένως το camp έχει σηµαντικό ρόλο στη ρύθµιση του µεταβολισµού, του κυτταρικού πολλαπλασιασµού, της διαφοροποίησης και της επιβίωσης µίας µεγάλης ποικιλίας κυτταρικών τύπων. Πράγµατι έχει παρατηρηθεί, µείωση της ικανότητας πολλαπλασιασµού των T-λεµφοκυττάρων, σε υπερτασικά άτοµα, παρουσία µιτογόνων ενώσεων (PHA, Con-A), σε σύγκριση µε φυσιολογικά άτοµα (Shasha et al., 1991). Επιπλέον η ρύθµιση της αδενυλικής κυκλάσης από οδούς στις οποίες διαµεσολαβούν πρωτεΐνες G, ως απόκριση στη δέσµευση είτε διεγερτικών είτε ανασταλτικών ουσιών σε β-αδρενεργικούς υποδοχείς στα λεµφοκύτταρα είναι παρόµοια µε ανάλογη ρύθµιση που αφορά την απόκριση των υποδοχέων στα αιµοφόρα αγγεία. Πιθανόν η αυξηµένη δράση των GRKs δηλώνει τη µειωµένη απόκριση των απευαισθητοποιηµένων β-αδρενεργικών υποδοχέων, γεγονός που αντανακλά στην εξασθένιση της φυσιολογικής λειτουργίας των αγγείων, χαρακτηριστικό της υπερτασικής κατάστασης (Gros et al., 1997) Προστακυκλίνη (PGI2) Σε αρκετές οµάδες ασθενών, η αρχική µείωση του όγκου επαναρρόφησης υγρών σε συνδυασµό µε τα χαµηλά επίπεδα ρενίνης, η οποία συµβαίνει στον πρώτο µήνα της θεραπείας, δε συνεχίζεται µε αποτέλεσµα να µην παρατηρείται οριστική ελάττωση της καρδιακής παροχής. Σύµφωνα µε την υπόθεση που διατυπώθηκε στις αρχές του 197 η δράση των θειαζιδίων θα πρέπει να αναζητηθεί και σε µηχανισµούς µείωσης των περιφερικών αγγειακών αντιστάσεων (Leth, 197, Tarazi et al., 197). Επόµενες µελέτες έδειξαν ότι πράγµατι η αρχική πτώση της αρτηριακής πίεσης σχετίζεται άµεσα µε τη µείωση του όγκου του εξωκυττάριου υγρού και την
40 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ ελάττωση της καρδιακής παροχής, ενώ ταυτόχρονα παρατηρείται µία αύξηση των περιφερικών αγγειακών αντιστάσεων. Με την πάροδο της αντιϋπερτασικής αγωγής, ενώ τα επίπεδα των εξωκυττάριων υγρών και της αρτηριακής πίεσης παραµένουν χαµηλά, οι περιφερικές αγγειακές αντιστάσεις φθίνουν και η καρδιακή παροχή αυξάνεται, προσεγγίζοντας τις φυσιολογικές τιµές. Τα στοιχεία αυτά συνηγορούν στην άποψη που απορρίπτει ένα άµεσο αγγειοδιασταλτικό αποτέλεσµα, το οποίο προκύπτει από τη δράση των διουρητικών θειαζιδίων (Shah et al., 1978). Αντίθετα η µειωµένη απόκριση (πτώση της αρτηριακής πίεσης<1%), µίας µεγάλης οµάδας υπερτασικών ατόµων (non responders), στη χρόνια χορήγηση υδροχλωροθειαζιδίου σχετίζεται µε µία παρατεταµένη και σε µεγαλύτερο βαθµό ελάττωση της καρδιακής παροχής και του όγκου των εξωκυττάριων υγρών που έχει ως αποτέλεσµα την αύξηση των περιφερικών αγγειακών αντιστάσεων, εξαιτίας της αντανακλαστικής ενεργοποίησης είτε του συµπαθητικού νευρικού συστήµατος είτε του αγγειοσυσταλτικού µηχανισµού ρενίνης-αγγειοτασίνης-αλδοστερόνης. Αδιαµφισβήτητα, η λειτουργία των αντισταθµιστικών µηχανισµών σε αυτήν την οµάδα (non responders), παρά τη σπουδαιότητα της, δεν είναι σε θέση να αιτιολογήσει την πτώση των περιφερικών αγγειακών αντιστάσεων που παρατηρείται στην οµάδα των υπερτασικών ατόµων (responders) µε την καθορισµένη απόκριση (πτώση της αρτηριακής πίεσης>1%). Επιπρόσθετα, οι διακυµάνσεις της καρδιακής παροχής στις δύο διακριτές αυτές οµάδες δεν θεωρούνται καθοριστικές στην τελική µείωση των περιφερικών αγγειακών αντιστάσεων που συµβαίνει στην οµάδα των ασθενών που εµφανίζουν την καθορισµένη απόκριση, κατά τη χρόνια χορήγηση διουρητικών θειαζιδίων (van Brummelen et al., 198). Η άποψη της επιστηµονικής κοινότητας που θεωρεί απαραίτητη για την επίτευξη της αντιϋπερτασικής δράσης των διουρητικών θειαζιδίων την ικανότητα των νεφρών να ρυθµίζουν την απέκκριση του NaCl και του ύδατος διαταράχθηκε µε τις συνεχόµενες αναφορές κλινικών περιπτώσεων ασθενών µε χρόνια νεφρική ανεπάρκεια στους οποίους η χορήγηση διουρητικών θειαζιδίων (χλωροθειαζίδιο) προκαλεί πτώση της αρτηριακής πίεσης. Σε αυτές τις περιπτώσεις, ο µηχανισµός της αντιϋπερτασικής δράσης τους δεν σχετίζεται µε την απέκκριση του NaCl και τις µεταβολές του όγκου του εξωκυττάριου υγρού, αλλά µε αλλαγές που προκύπτουν στις περιφερικές αγγειακές αντιστάσεις, γεγονός που ενισχύει την άποψη της έµµεσης αγγειοδιασταλτικής δράσης (Jones και Nanra, 1979). Πράγµατι η χορήγηση, ενός µη στεροειδικού αντιφλεγµονώδους φαρµάκου (NSAID), της ινδοµεθακίνης, σε υπερτασικά άτοµα που ακολουθούν χρόνια θεραπευτική αγωγή µε διουρητικά θειαζίδια εξασθενεί σηµαντικά την πτώση της αρτηριακής πίεσης, ενώ ταυτόχρονα παρατηρείται µείωση στα επίπεδα ρενίνης-αλδοστερόνης στο πλάσµα (Lopez-Ovejero et al., 1978, Wing et al., 1981). Η δράση της ινδοµεθακίνης εντοπίζεται στην αναστολή του πρώτου σταδίου της βιοσύνθεσης των προσταγλανδινών από το αραχιδονικό οξύ (C2:4 λιπαρό οξύ), εξαιτίας της ικανότητας της να απενεργοποιεί την ενζυµική υποµονάδα (κυκλοξυγονάση 1 και 2) ενός πολύπλοκου ενζυµικού συµπλόκου, της συνθετάσης των προσταγλανδινών. Παράλληλες ερευνητικές µελέτες, έδειξαν ότι στους µηχανισµούς πτώσης της αρτηριακής πίεσης κατά τη χρόνια αντιϋπερτασική αγωγή µε διουρητικά θειαζί δια (βενδροφλουµεθειαζίδη, υδροχλωροθειαζίδιο) εµπλέκονται οι προσταγλανδίνες. Πρόκειται για κυκλικά ακόρεστα λιπαρά οξέα, τα οποία θα µπορούσαµε να τα χαρακτηρίσουµε ως τοπικές ορµόνες που στις κύριες επιδράσεις τους, τόσο σε κύτταρα στα οποία συντίθενται, όσο και σε γειτονικά κύτταρα, περιλαµβάνονται η ισχυρή αναστολή των λιπολυτικών επιδράσεων (αδενυλική κυκλάση-camp) των
41 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ ορµονών επινεφρίνης, γλυκαγόνης, φλοιοτρόπου και θυρεοειδοτρόπου ορµόνης, η διέγερση της φλεγµονής, η ρύθµιση της ροής του αίµατος, ο έλεγχος της µεµβρανικής µεταφοράς ιόντων και η τροποποίηση της συναπτικής διαβίβασης. Η αντιϋπερτασική δράση του υδροχλωροθειαζιδίου επιτυγχάνεται µε την αύξηση της νεφρικής σύνθεσης προστακυκλίνης (PGI2), η οποία καθορίζει, ως ισχυρός αγγειοδιασταλτικός παράγοντας, τη χάλαση των λείων µυών των νεφρικών αρτηριδίων (αύξηση του camp) µε αποτέλεσµα τη µείωση των περιφερικών αγγειακών αντιστάσεων που οδηγεί σε πτώση της αρτηριακής πίεσης. Αξιοσηµείωτο είναι ότι οι κύριες δράσεις της ενδοαγγειακής χορήγησης διουρητικών της αγκύλης (φουροσεµίδη), οι οποίες, µεταξύ των άλλων, περιλαµβάνουν την παροδική νεφρική αγγειοδιαστολή και µία απότοµη αύξηση των επιπέδων ρενίνης στο πλάσµα, είναι ανεξάρτητες της διούρησης, ενώ σχετίζονται µε την αυξηµένη απελευθέρωση πρόδροµων ενώσεων των προσταγλανδινών από τα µεµβρανικά φωσφολιπίδια µε την ενεργοποίηση φωσφολιπασών και την έντονη βιοσύνθεση των προσταγλανδινών, συµπεριλαµβανο- µένης της προστακυκλίνης (PGI2). Η απότοµη αύξηση των επιπέδων ρενίνης, η οποία παρατηρείται µετά την ενδοαγγειακή χορήγηση της φουροσεµίδης και κατά τη χρόνια χορήγηση διουρητικών θειαζιδίων και η οποία αναστέλλεται παρουσία της ινδοµεθακίνης συσχετίζεται θετικά µε την αυξηµένη συγκέντρωση στο πλάσµα και απέκκριση στα ούρα ενός προϊόντος υδρόλυσης της PGI2, της 6-κέτο-PGF1α, γεγονός το οποίο δηλώνει τη διέγερση της σύνθεσης προστακυκλίνης στην εξωτερική µοίρα (φλοιώδη ουσία) του νεφρού. Αντίθετα, η ενδοαγγειακή χορήγηση διουρητικών θειαζιδίων (υδροχλωροθειαζίδιο, βενδροφλουµεθειαζίδη) δεν επάγει τη νεφρική αγγειοδιαστολή, η οποία θεωρείται αποτέλεσµα της χρόνιας χορήγησής τους σε υπερτασικά άτοµα µε την καθορισµένη απόκριση και σε ηµερήσιες δόσεις, ιδιαίτερα για το υδροχλωροθειαζίδιο, που δεν ξεπερνούν τα 5mg. Σε αυτούς τους ασθενείς πιστεύεται ότι η µείωση της επαναρρόφησης του NaCl µε τις σηµαντικές µεταβολές που επιφέρει στο ισοζύγιο Νa + ενεργοποιεί τη µεµβρανική απελευθέρωση µεγάλων ποσών αραχιδονικού οξέος. Σε ασθενείς µε ιδιοπαθή υπέρταση που εµφανίζουν µειωµένη απόκριση σε συγκεκριµένη δόση (5mg) υδροχλωροθειαζιδίου, η αύξηση της χορηγούµενης ηµερήσιας δοσολογίας σε 1 mg δεν προκαλεί περαιτέρω πτώση της αρτηριακής πίεσης, ενώ έντονες παρουσιάζονται οι γνωστές ανεπιθύµητες ενέργειες, όπως η υποκαλιαιµία και η υπερουριχαιµία. Πιθανόν οι αυξηµένες συγκεντρώσεις ρενίνης και αγγειοτασίνης ΙΙ που ανιχνεύονται στο πλάσµα, εξαιτίας µηχανισµών που δεν διαµεσολαβούν οι προσταγλανδίνες, σχετίζονται µε τη διατήρηση της αρτηριακής πίεσης σε υψηλά επίπεδα (Webster et al., 198, Wilson, 1986) Καλλικρεΐνη Για την πτώση της αρτηριακής πίεσης κατά τη χρόνια χορήγηση υδροχλωροθειαζιδίου σε µία οµάδα αρσενικών υπερτασικών ατόµων, που παρουσιάζει την καθορισµένη µορφή απόκρισης στην οποία δεν εµφανίζονται οι γνωστές µεταβολές του ενδοαγγειακού όγκου και µε το ρυθµό σπειραµατικής διήθησης ή νεφρικής ροής πλάσµατος να διατηρείται αµετάβλητος, υπεύθυνη θεωρείται η µειωµένη νεφρική αγγειακή αντίσταση, πιθανόν των προσαγωγών αρτηριδίων. Η αυξηµένη απέκκριση της καλλικρεΐνης, µίας αγγειοδραστικής ουσίας µε σηµαντικό ρόλο στη ρύθµιση και στην τροποποίηση της νεφρικής ροής αίµατος, σχετίζεται πλέον µε τα φυσιολογικά εύρη τιµών σε σύγκριση µε τα χαµηλά επίπεδα της υπερτασικής κατάστασης. Η καλλικρεΐνη είναι ένα πρωτεολυτικό ένζυµο που παράγεται στους νεφρούς και διασπά την κυκλοφορούσα πρωτεΐνη κινινιγόνο σε κινίνη (βραδυκινίνη), η οποία - 3 -
42 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ µπορεί να δράσει τοπικά ενισχύοντας την παραγωγή του µονοξειδίου του αζώτου και των προσταγλανδινών και να προκαλέσει διαστολή των νεφρικών αρτηριδίων αυξάνοντας την ροή του αίµατος ως απόκριση στη χορήγηση διουρητικών θειαζιδίων (O Connor et al., 1981). 3.5 Φαρµακοκινητικές ιδιότητες Ο όρος φαρµακοκινητική αποδίδεται στις ποσοτικές µεταβολές που επέρχονται µε την πάροδο του χρόνου στη συγκέντρωση και στη συνολική ποσότητα του φαρµάκου στο πλάσµα και στο σώµα αντίστοιχα, µετά τη χορήγηση του από διαφορετικές οδούς. Ο προσδιορισµός της φαρµακοκινητικής ενός φαρµάκου έχει ιδιαίτερη σηµασία και αποσκοπεί τόσο στη διαπίστωση παραγόντων που επηρεάζουν την παραµονή και τα επίπεδα του φαρµάκου στον οργανισµό όσο και στην ακριβή ρύθµιση του θεραπευτικού σχήµατος για φάρµακα που έχουν αυξηµένες πιθανότητες τοξικότητας. Αξίζει να σηµειωθεί ότι στις αλληλεπιδράσεις σηµαντικών διεργασιών που καθορίζουν άµεσα τις φαρµακοκινητικές ιδιότητες ενός φαρµάκου περιλαµβάνονται η απορρόφηση, η κατανοµή, ο µεταβολισµός και η απέκκριση (Holford και Benet, 1998) Βιοδιαθεσιµότητα Η µεταφορά ενός φαρµάκου από τη θέση χορήγησης του στην κυκλοφορία του αίµατος ονοµάζεται απορρόφηση. Πρόκειται για µία διεργασία της οποίας η ταχύτητα και η αποδοτικότητα εξαρτώνται από την οδό χορήγησης. Η βιοδιαθεσιµότητα (bioavailability) εκφράζεται ως το ποσοστό του χορηγούµενου φαρµάκου που εισέρχεται στη συστηµατική κυκλοφορία χωρίς να υποστεί καµία χηµική µετατροπή και προσδιορίζεται συγκρίνοντας τα επίπεδα του φαρµάκου στο πλάσµα µετά τη χορήγηση του από µία συγκεκριµένη οδό µε εκείνα που επιτυγχάνονται µετά από ενδοαγγειακή έγχυση. Η απορρόφηση του υδροχλωροθειαζιδίου συµβαίνει στο δωδεκαδάκτυλο, στο ανώτερο τµήµα της νήστιδας και σε µία µικρή επιφάνεια του στοµάχου και το 95% της χορηγούµενης δόσης µε ενδοαγγειακή έγχυση αποβάλλεται από τον οργανισµό σχεδόν αµετάβλητο µέσω των νεφρών µε αποτέλεσµα το συνολικό ποσό απορρόφησης να αντιστοιχεί στην ποσότητα του φαρµάκου που απεκκρίνεται στα ούρα (Beer mann et al., 1976, Meyer και Whyatt, 1976). Η αποδοτικότητα της απορρόφησης ηµερήσιας δόσης υδροχλωροθειαζιδίου (25-, 5-, 1- και 2mg) που χορηγείται µε τη µορφή δισκίων ή υγρού διαλύµατος, σε αντίθεση µε αυτή του χλωροθειαζιδίου είναι ανεξάρτητη από το µέγεθος της δόσης (Patel et al., 1984). Οι φυσικοχη- µικές ιδιότητες του υδροχλωροθειαζιδίου διαδραµατίζουν ένα σηµαντικό ρόλο στον καθορισµό µίας αναλογικής σχέσης ανάµεσα σε χορηγούµενες δόσεις και στη βιοδιαθεσιµότητα του φαρµάκου. Αυτή η διαφορά που παρατηρείται στη συµπεριφορά απορρόφησης αποδίδεται στην υψηλή διαλυτότητα που οφείλεται µερικώς σε υψηλές τιµές pka µε αποτέλεσµα το υδροχλωροθειαζίδιο στη µη φορτισµένη µορφή του να απορροφάται από το 9% της επιφάνειας του δωδεκαδακτύλου και της νήστιδας. Συγκεκριµένα, από το έντερο απορροφάται το 68% περίπου της χορηγούµενης στοµατικής δόσης που κυµαίνεται από mg (Dressman et al., 1984). Πιθανότατα, η µεταφορά του φαρµάκου από το γαστρεντερικό σωλήνα να πραγµατοποιείται µε το µηχανισµό της παθητικής διάχυσης, ενώ η δοσοεξαρτώµενη απορρόφηση του χλωροθειαζιδίου που συµβαίνει µε σταθερό ρυθµό να αντανακλά την ύπαρξη ενός συστήµατος ενεργητικής µεταφοράς που παρουσιάζει κινητική
43 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ κορεσµού και εξυπηρετεί την είσοδο του φαρµάκου στην κυκλοφορία του αίµατος (Patel et al., 1984, Redalieu et al., 1985). Έχει αποδειχθεί ότι σε περιπτώσεις που η χορηγούµενη δόση συνοδεύεται µε πρόσληψη τροφής, συγκρινόµενες µε καταστάσεις νηστείας, το ποσοστό του υ- δροχλωροθειαζιδίου που απορροφάται αυξάνεται µέχρι 75% πιθανότατα εξαιτίας της αυξηµένης παραµονής στο λεπτό έντερο (Beermann και Groschinsky-Grind, 1978a), ενώ αντίθετα υπάρχουν δεδοµένα σύµφωνα µε τα οποία τα επίπεδα του φαρµάκου στο πλάσµα µειώνονται σηµαντικά (Barbhaiya et al., 1982). Εάν προηγηθεί θεραπεία µε 6mg προπανθελίνης αυξάνεται το ποσό της χορηγούµενης δόσης υδροχλωροθειαζιδίου που απορροφάται (~9%), γεγονός που οφείλεται στο µειωµένο ρυθµό γαστρικής εκκένωσης (Beermann και Groschinsky-Grind, 1978b). Αξιοσηµείωτο είναι το γεγονός που αφορά την τροποποίηση της απορρόφησης του υδροχλωροθειαζιδίου σε καταστάσεις σοβαρών ασθενειών, η οποία µειώνεται σηµαντικά σε άτοµα µε συµφορητική καρδιακή ανεπάρκεια (Beermann και Groschinsky -Grind, 1979) και σε παχύσαρκα άτοµα που έχουν υποστεί εντερική παράκαµψη κατόπιν χειρουργικής επέµβασης (Backman et al., 1979). Η σύγκριση της βιοδιαθεσιµότητας του υδροχλωροθειαζιδίου σε περιπτώσεις που αφορούν τη χορήγηση του σε δισκία και σε υγρό διάλυµα στις δόσεις 25-, 5-, 1- και 2mg, έδειξε παρόµοια απορρόφηση του φαρµάκου που πραγµατοποιείται µε πολύ γρήγορο ρυθµό µε αποτέλεσµα η συγκέντρωση του στο πλάσµα να εµφανίζει τη µέγιστη τιµή περίπου δύο ώρες µετά τη χορήγηση (Patel et al., 1984). Με υγρή χρωµατογραφία υψηλής απόδοσης επιβεβαιώνεται ο ευεργετικός ρόλος διαφορετικών χηµικών ενώσεων (µαλτοδεξτράνη, πολυεθυλενογλυκόλη) στην αύξηση της βιοδιαθεσιµότητας του υδροχλωροθειαζιδίου, οι οποίες περιέχονται σε λυοφιλισµένα δισκία χορηγούµενης δόσης 25mg και ευθύνονται για την ταχύτατη διάσπαση και απορρόφηση του φαρµάκου σε σύγκριση µε τα συµβατικά εµπορικά σκευάσµατα (Corveleyn και Remon, 1998) Κατανοµή Πρόκειται για τη διαδικασία µε την οποία ένα φάρµακο εγκαταλείπει αντιστρεπτά την κυκλοφορία του αίµατος και εισέρχεται στο διάµεσο χώρο (εξωκυττάριο υγρό) ή/και στα κύτταρα των ιστών. Η µεταφορά του φαρµάκου από το πλάσµα στο διάµεσο χώρο εξαρτάται κυρίως από την αιµατική ροή, την τριχοειδική διαπερατότητα, το βαθµό σύνδεσης του µε τις πρωτεΐνες του πλάσµατος και από τη χηµική του δοµή (αφόρτιστη ή φορτισµένη µορφή-υδρόφοβη ή υδρόφιλη φύση). Το υδροχλωροθειαζίδιο δεν εµπλέκεται στον ηπατικό µεταβολισµό και δεν δεσµεύεται µε συστατικά του ηπατικού ιστού, γεγονός που επιτρέπει µετά την απορρόφηση του να εµφανίζει µία οµοιογενή κατανοµή στον οργανισµό, η οποία δεν συµβαίνει άµεσα διότι δεσµεύεται στα ερυθροκύτταρα (Abdelhameed et al., 1993). Το υδροχλωροθειαζίδιο συσσωρεύεται στα ερυθροκύτταρα σε αναλογία µε το πλάσµα 3.5:1 (Beermann et al., 1976) και το ποσοστό που αφορά τη δεσµευµένη µορφή του σε πρωτεΐνες του πλάσµατος αντιστοιχεί σε 4% (Beermann, 1984). Η δυνατότητα συσσώρευσης του υδροχλωροθειαζιδίου στα ερυθροκύτταρα οφείλεται αποκλειστικά σε φυσικοχηµικές και λειτουργικές αλληλεπιδράσεις µεταξύ της φαρµακευτικής ένωσης και του ενζύµου ανθρακική ανυδράση. Συγκεκριµένα, ο βαθµός συγγένειας που παρουσιάζει ο αρωµατικός υποκαταστάτης σουλφοναµιδικών παραγώγων καθορίζει την ικανότητα δέσµευσης του υδροχλωροθειαζιδίου µε διαφορετικές ισοµορφές του ένζυµου (B και C) που εντοπίζεται σε υψηλές συγκεντρώσεις στα ερυθροκύτταρα (Matsumoto et al., 1989)
44 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Πιθανότατα, µία άγνωστη πρωτεΐνη των ερυθρών αιµοσφαιρίων δεσµεύεται εξίσου ισχυρά µε τα σουλφοναµιδικά παράγωγα (ζονισαµίδη) µε χαµηλότερη συγγένεια συγκριτικά µε την ανθρακική ανυδράση και υψηλότερη σε σχέση µε την εξωκυτταρική αλβουµίνη (Matsumoto et al., 1989). Γενικότερα, οι φαρµακευτικές ενώσεις προσδεµένες σε πρωτεΐνες του πλάσµατος χαρακτηρίζονται φαρµακολογικά αδρανείς και µόνο η ελεύθερη µορφή του φαρµάκου είναι σε θέση να δράσει στις θέσεις-στόχους των ιστών και να προκαλέσει βιολογική απάντηση. Επιπρόσθετα, τα ερυθροκύτταρα σε συνδυασµό µε τη δεσµευµένη µορφή του υδροχλωροθειαζιδίου σε πρωτεΐνες του πλάσµατος (Gilman et al., 199) συνιστούν ένα φυσικό εµπόδιο στη µετακίνηση του ανάµεσα στα τρία λειτουργικά διακριτά διαµερίσµατα του οργανισµού µε αποτέλεσµα να παρατηρείται µία αρκετά αργή εισροή υδροχλωροθειαζιδίου από το πλάσµα στα ερυθροκύτταρα και µία εξαιρετικά αργή αντίστροφη εκροή (Chen et al., 1992). Μία µεγάλη απόκλιση έχει διαπιστωθεί για το υδροχλωροθειαζίδιο στο χρόνο που απαιτείται για να επιτευχθεί συγκέντρωση του φαρµάκου ίση µε το 5% της τελικής συγκέντρωσης σταθερής κατάστασης (Csteady-state), ο οποίος αντιστοιχεί στο χρόνο ηµίσειας ζωής (half time ή t1/2). Η χορήγηση του σε καθορισµένες δόσεις (12. 5-, 25-, 5- και 75mg) πραγµατοποιείται σε υγιείς δότες µε τη µορφή δισκίων στα πλαίσια του σχήµατος που περιλαµβάνει σταθερή δόση σε σταθερά χρονικά µεσοδια στήµατα και η κύρια αιτία εντοπίζεται στο γεγονός της διφασικής αποµάκρυνσης του υδροχλωροθειαζιδίου από το πλάσµα. Ο χρόνος ηµίσειας ζωής κυµαίνεται από ώρες και χαρακτηρίζεται από την εµφάνιση υψηλής ενδοατοµικής ποικιλότητας, ενώ η συγκέντρωση του υδροχλωροθειαζιδίου στο πλάσµα εµφανίζει τη µεγιστη τιµή (7-376ng/ml) περίπου ώρες µετά τη χορήγηση δόσης (Beermann και Groshinsky-Grind, 1977). Πρόσφατες ερευνητικές µελέτες αποδίδουν το χρόνο ηµίσειας ζωής του υδροχλωροθειαζιδίου σε 1-12 ώρες (Beermann, 1984, Chen και Chiou, 1992) και ο φαινοµενικός όγκος κατανοµής (Vd) του φαρµάκου σε περιπτώσεις στοµατικής χορήγησης αντιστοιχεί σε.83l/kg (Gilman et al., 199). Επιπρόσθετα, συγκεντρώσεις υδροχλωροθειαζιδίου ανιχνεύθηκαν στο πλάσµα εγκύων γυναικών και στο πλάσµα του οµφάλιου λώρου σε σχετικά παρό- µοια επίπεδα (Beermann et al., 198), οι οποίες είναι χαµηλότερες από αντίστοιχες που παρατηρήθηκαν στο αµνιακό υγρό (Mulley et al., 1978). Ιδιαίτερα σηµαντική είναι η παρουσία του φαρµάκου στο µητρικό γάλα, ως αποτέλεσµα θεραπευτικής αγωγής εγκύων γυναικών, ενώ δεν παρατηρούνται µετρήσιµες συγκεντρώσεις (όριο ανίχνευσης, 2ng/ml) σε νεογέννητα βρέφη κατά τη χρονική περίοδο του θηλασµού (Miller et al., 1982)
45 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Μεταβολισµός Ο µεταβολισµός των φαρµακευτικών ενώσεων από το ήπαρ, τους νεφρούς ή άλλους ιστούς αποσκοπεί στη µετατροπή τους σε πολικότερα µόρια µε αποτέλεσµα να διευκολύνεται η αποβολή τους από τον οργανισµό. Ιδιαίτερα σηµαντικές είναι οι αντιδράσεις της Φάσης Ι. του µεταβολισµού σύµφωνα µε τις οποίες η φαρµακολογική δραστικότητα ενός φαρµάκου είτε αυξάνεται είτε µειώνεται είτε παραµένει σχετικά αµετάβλητη. Το υδροχλωροθειαζίδιο µεταβολίζεται σε εξαιρετικά µικρό βαθµό στους αρουραίους και στους πιθήκους, ενώ δεν υφίσταται σχεδόν καµία βιοµετατροπή στον άνθρωπο προκειµένου να αποµακρυνθεί από τον οργανισµό µε αποτέλεσµα µετά από στοµατική χορήγηση να απεκκρίνεται αµετάβλητο µέσω των νεφρών (EMEA, 1999). Στη βιβλιογραφία εντοπίσθηκε η περίπτωση ενός θηλυκού ατόµου ώριµης ηλικίας µε νεφρική νόσο (νέφρωση), στο οποίο η συνολική ηµερήσια χορήγηση 1mg υδροχλωροθειαζιδίου (Esidrex, 25mg) σε δισκία είχε ως αποτέλεσµα να ανιχνευθεί µε υγρή χρωµατογραφία υψηλής απόδοσης η παρουσία της ετεροκυκλικής ένωσης ACBS (2-Amino-4-chloro-m-benzenedisulfonamide) στα ούρα, στο πλάσµα και στα ερυθροκύτταρα (Okuda et al., 1987). Η ένωση ACBS αναγνωρίσθηκε ως µεταβολίτης του υδροχλωροθειαζιδίου και προκύπτει in vivo ως προϊόν υδρόλυσης του φαρµάκου µε αποτέλεσµα εικοσιτέσσερις ώρες µετά τη χορήγηση του να ανιχνεύεται σε εξαιρετικά µικρή ποσότητα στα ούρα. Συγκεκριµένα, η ποσότητα της ACBS που αποβάλλεται αντιστοιχεί σε 4.3 % της ποσότητας υδροχλωροθειαζιδίου που απεκκρίνεται στα ούρα και εµφανίζει σαφώς χαµηλότερη συχνότητα απέκκρισης σε σύγκριση µε αυτή της υδρόλυσης του φαρµάκου στο µεταβολίτη του (Okuda et al., 1987). Πράγµατι, η συγκέντρωση του υδροχλωροθειαζιδίου στο πλάσµα είναι δέκα φορές µεγαλύτερη από αυτή του µεταβολίτη του (Okuda et al., 1987), γεγονός το οποίο επιβεβαιώνεται και σε επόµενες µελέτες (Yamazaki et al., 199). Αντίθετα, η παρουσία του υδροχλωροθειαζιδίου και του µεταβολίτη του σε ισοδύναµες συγκεντρώσεις στα ερυθροκύτταρα υποδηλώνει µία πολύ περισσότερο ισχυρή δεσµευτική ικανότητα της ACBS σε σύγκριση µε το φάρµακο (Okuda et al., 1987). Από πειράµατα σε ερυθροκύτταρα λαγού προσδιορίσθηκε η συγκέντρωση δέσµευσης της ACBS σε τρεις διαφορετικές περιοχές, που είναι είκοσι φορές υψηλότερη από αυτή που παρατηρείται στο πλάσµα. Ο µεταβολίτης εµφανίζει πέντε φορές µεγαλύτερη συγγένεια µε την ανθρακική ανυδράση της µεµβράνης των ερυθροκυττάρων συγκριτικά µε το υδροχλωροθειαζίδιο, ενώ όσον αφορά τη περιοχή δέσµευσης, αυτή αντιστοιχεί στο 1/5 της θέσης δέσµευσης του υδροχλωροθειαζιδίου (Yamazaki et al., 199)
46 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Απέκκριση Η πιο σηµαντική οδός για την αποµάκρυνση φαρµακευτικών ενώσεων από τον οργανισµό θεωρείται αυτή που ακολουθείται από τα περισσότερα αντιυπερτασικά µέσω της παραγωγής ούρων από τους νεφρούς. Άλλες οδοί περιλαµβάνουν την αποµάκρυνση από τη χολή, το έντερο, τους πνεύµονες και το µητρικό γάλα. Οι βασικές διεργασίες που καθορίζουν την απέκκριση του υδροχλωροθειαζιδίου διαµέσου της νεφρικής οδού περιλαµβάνουν τη σπειραµατική διήθηση και τη σωληναριακή έκκριση (Beerman et al., 1976b, Barbhaiya et al., 1982). Η απέκκριση αντιστοιχεί σε 5-7% χορηγούµενης στοµατικής δόσης (25-65mg) και πραγµατοποιείται µετά από 24 ώρες, ενώ συνήθως ολοκληρώνεται στις επόµενες ώρες (Meyer και Whyatt, 1976). Η νεφρική πλασµατοκάθαρση αντιπροσωπεύει τον όγκο του πλάσµατος, από τον οποίο η φαρµακευτική ένωση υδροχλωροθειαζίδιο αποµακρύνεται και αποβάλλεται στα ούρα στη µονάδα του χρόνου και είναι αρκετά υψηλή, περίπου 335ml/min, ύστερα από χορήγηση mg σε άτοµα µε φυσιολογική νεφρική λειτουργία, γεγονός το οποίο κύρια υποδηλώνει σωληναριακή έκκριση (Beermann και Groschinsky-Grind, 1977 ). Η σωληναριακή έκκριση των διουρητικών θειαζιδίων από τα περισωληναριακά τριχοειδή στον αυλό του εγγύς σωληναρίου επιτυγχάνεται µε το σύστηµα ενεργητικής µεταφοράς που περιλαµβάνει τους µεταφορείς οργανικών ανιόντων (hoats) και θεωρείται ιδιαίτερα κρίσιµη για την ανάπτυξη φαρµακολογικής δράσης (Hasannejad et al., 24). Επιπρόσθετα, µε ενδοφλέβια χορήγηση µικρής ποσότητας επισηµασµένου µε ραδιενεργό άνθρακα (C 14 ) φαρµάκου είναι δυνατή η ανίχνευση του σε περιττώµατα µε αποτέλεσµα να συµβαίνει έκκριση του υδροχλωροθειαζιδίου από το έντερο σε ελάχιστη ποσότητα, ενώ σε καµία περίπτωση δεν παρατηρείται απέκκριση από τη χοληδόχο κύστη (Beerman et al., 1976). Η τροποποίηση της απέκκρισης του υδροχλωροθειαζιδίου σχετίζεται άµεσα µε περιπτώσεις µη φυσιολογικής νεφρικής λειτουργίας, µε την αύξηση της ηλικίας και µε την ταυτόχρονη χορήγηση φαρµακευτικών ενώσεων διαφορετικής κατηγορίας σε περιπτώσεις συνδυαστικής αντιϋπερτασικής θεραπείας. Χαρακτηριστική είναι η αύξηση των συγκεντρώσεων υδροχλωροθειαζιδίου που παρατηρείται στο πλάσµα σε άτοµα µε νεφρική ανεπάρκεια, η οποία συνοδεύεται από αντίστοιχη µείωση της νεφρική κάθαρσης και απέκκρισης του φαρµάκου στα ούρα (Niemeyer et al., 1983). Επιπλέον µε την πάροδο της ηλικίας παρατηρείται ανάλογη µείωση του ρυθµού αποµάκρυνσης του φαρµάκου από τον οργανισµό που πιθανότατα οφείλεται σε µειωµένη νεφρική λειτουργία (Williams et al., 1986). Η κινητική του υδροχλωροθειαζιδίου επηρεάζεται σηµαντικά σε υγιείς δότες παρουσία άλλων φαρµάκων µε αποτέλεσµα η διουρητική συµπεριφορά και η δράση του να µεταβάλλεται µε διαφορετικό τρόπο από δύο µη στεροειδή αντιφλεγµονώδη φάρµακα (NSAIDs). Η αρτηριακή πίεση, το βάρος του σώµατος, η συγκέντρωση των ιόντων καλίου και η δραστικότητα της ρενίνης στο πλάσµα, η κρεατινίνη και η αλβουµίνη, ο αιµατοκρίτης, καθώς επίσης η εικοσιτετράωρη απέκκριση των ιόντων νατρίου και καλίου προσδιορίστηκαν, προκειµένου να διευκρινιστεί η επίδραση που ασκείται στη δράση του υδροχλωροθειαζιδίου. Παρουσία ινδοµεθακίνης αυξάνεται το βάρος του σώµατος, η συγκέντρωση των ιόντων καλίου και µειώνεται η δραστικότητα της ρενίνης στο πλάσµα, πιθανότατα εξαιτίας αναστολής της σύνθεσης νεφρικής προστακυκλίνης, χωρίς να επηρεάζεται η κινητική του υδροχλωροθειαζιδίου. Αντίθετα, παρουσία ενός άλλου αντιφλεγµονώδους φαρµάκου (sulindac) µειώνεται η νεφρική κάθαρση του υδροχλωροθειαζιδίου και αυξάνονται τα επίπεδα του στο πλάσµα (Koopmans et al., 1985)
47 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ 3.6 Γενετική δράση του υδροχλωροθειαζιδίου In vitro µελέτες Τα δεδοµένα που προκύπτουν από in vitro πειραµατικές µελέτες γονιδιοτοξι -κότητας του υδροχλωροθειαζιδίου είναι αµφιλεγόµενα Προκαρυωτικοί οργανισµοί. Η µεταλλαξιγόνος δράση φαρµακευτικών ενώσεων που χαρακτηρίζονται ως αµίνες ή αµίδια και των παραγώγων τους που προκύπτουν, ως προϊόντα αλληλεπίδρασης µε νιτρώδη οξέα ελέγχθηκαν σε τέσσερα µη παθογόνα στελέχη του βακτηρίου Salmonella typhimurium (ΤΑ1535, ΤΑ1, ΤΑ1538, ΤΑ98) παρουσία ή απουσία εξωγενούς (µικροσωµικό κλάσµα S9 αρουραίου) µεταβολικού συστήµατος ενεργοποίησης. Από τα αποτελέσµατα παρατηρείται σηµαντική αύξηση του αριθµού των επαναµεταλλαγµένων αποικιών σε ένα µόνο στέλεχος (ΤΑ98), µε και χωρίς µεταβολική ενεργοποίηση, από το παράγωγο Ν-νιτροζουδροχλωροθειαζίδιο (NNO- ΗCTZ), γεγονός το οποίο υποδηλώνει τη δυνατότητα της νιτροζοένωσης να επάγει σηµειακές µεταλλάξεις µετατόπισης αναγνωστικού πλαισίου (Andrews et al., 1984). Σε επόµενες µελέτες, που αφορούν τον έλεγχο της µεταλλαξιγόνου δράσης σαράντα δύο χηµικών ενώσεων, το υδροχλωροθειαζίδιο φαίνεται να προκαλεί απουσία εξωγενούς µεταβολικής ενεργοποίησης µικρή δοσοεξαρτώµενη αύξηση των επαναµεταλλαγµάτων στο στέλεχος ΤΑ98, µε αποτέλεσµα η δράση του να χαρακτηρίζεται αµφίβολη (Mortelmans et al., 1986) και σε επανεκτίµηση από άλλη ερευνητική οµάδα θεωρείται πλέον, µη µεταλλαξιγόνος παράγοντας (Zeiger, 199). Η χορήγηση υδροχλωροθειαζιδίου (.26mM) δεν επάγει επαναµεταλλάξεις σε στελέχη Hs3R argf(am) του βακτηρίου Escherichia coli, ενώ αντίθετα ακτινοβόληση µε υπεριώδες του ίδιου στελέχους παρουσία του φαρµάκου προκαλεί τη δη- µιουργία αναστροφών που ανιχνεύονται µε την αποµόνωση πολυάριθµων πρωτότροφων στελεχών (arg + ), γεγονός που οφείλεται στην έκφραση µεταλλαξιγόνου δράσης (Fujita, 1985) Ευκαρυωτικοί οργανισµοί. Η πρώτη αναφορά για τη γενετική δράση του υδροχλωροθειαζιδίου έγινε το 1974 και απεκάλυψε σε υψηλή συχνότητα (96%) το επαγόµενο από τη δράση του φαρµάκου (Esidrex) φαινόµενο του χρωµοσωµατικού µη αποχωρισµού σε διπλοειδή στελέχη του µύκητα Aspergillus nidulans (Bignami et al., 1974) Κύτταρα Θηλαστικών. Μία θετική απόκριση στην επαγωγή µεταλλάξεων (forward mutation) από το υδροχλωροθειαζίδιο παρατηρήθηκε σε κύτταρα L5178YΤΚ ± λεµφώµατος ποντικών (MOLY), στα οποία ο γενετικός τόπος κινάση της θυµιδίνης (ΤΚ) εκφράζεται σε ετεροζυγωτία. Σε συγκεντρώσεις µεγαλύτερες των 5µg/ml, η παρουσία της φαρµακευτικής ένωσης σε L5178YΤΚ ± κύτταρα λεµφώµατος, απουσία εξωγενούς µεταβολικού συστήµατος επάγει µεταλλάξεις ανθεκτικές στην τριφθοροθυµιδίνη (TFT + cells) και παρουσιάζει έντονη κυτταροτοξική δράση (Myhr et al., 199). Η κυτταρογενετική δράση διαφόρων χηµικών ενώσεων (42) µελετήθηκε σε CHO (Chinese hamster ovary) κύτταρα µε τη χρήση µεθόδων ανάλυσης χρωµοσω- µατικών ανωµαλιών (CA) και ανταλλαγών αδελφών χρωµατιδίων (SCE) (Anderson et al., 199)
48 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Ανάµεσα σε αυτές µε θετική δράση, το υδροχλωροθειαζίδιο επάγει αυξηµένες συχνότητες ανταλλαγών µεταξύ αδελφών χρωµατιδίων, παρουσία και απουσία εξωγενούς µεταβολικού συστήµατος, αλλά δεν φαίνεται να προκαλεί δοµικές χρωµοσωµατικές ανωµαλίες (Galloway et al., 1987, Anderson et al., 199). Σε CHL (Chinese hamster lung) κύτταρα ανιχνεύθηκαν φαινόµενα πολυπλοειδίας 48 ώρες µετά από τη χορήγηση υδροχλωροθειαζιδίου (5µg/ml), ενώ δεν παρατηρήθηκαν δοµικές χρωµοσωµατικές ανωµαλίες (Ishidate et al., 1988). Επιπρόσθετα, η χορήγηση υδροχλωροθειαζιδίου ( M) δεν επηρέασε τη σύνθεση του DNA και τον πολλαπλασιασµό καλλιεργούµενων λείων µυϊκών αορτικών κυττάρων (VSMC) αρουραίου (Sprague-Dawley rat) (Kang et al., 1997). Η προσθήκη της φαρµακευτικής ένωσης σε συγκεντρώσεις (1-1µM) σε καλλιέργειες κυττάρων οστεοβλαστών αρουραίου (UMR-16) εµφανίζει παρόµοια δράση (Hall και Schaueblin, 1994) Άνθρωπος. Η προσθήκη υδροχλωροθειαζιδίου σε συγκεντρώσεις που κυµαίνονται από 25-3µM δεν φαίνεται να επηρεάζει το ρυθµό πολλαπλασιασµού της ανθρώπινης κυτταρικής σειράς MG-63 που θεωρείται ανάλογη µε τους οστεοβλάστες, ενώ αναστέλλει την έκκριση οστεοκαλσίνης και σε µεγαλύτερες συγκεντρώσεις (5µM) προκαλεί σηµαντική µείωση στη σύνθεση του DNA (Aubin et al., 1996). Αξιοση- µείωτη είναι η ικανότητα του υδροχλωροθειαζιδίου να αναστέλλει τον πολλαπλασιασµό και τη διαφοροποίηση καλλιεργούµενων κυττάρων οστεοβλαστών και πολυδύναµων προγονικών κυττάρων σε συγκεντρώσεις µεγαλύτερες από 1µM, παρουσία η απουσία εµπλουτισµένου θρεπτικού µέσου µε 1% ορό εµβρύου βοός (Lau et al., 1996). Επιπρόσθετα, η κυτταροτοξική δράση του φαρµάκου εκδηλώνεται µε ακτινοβόληση (UVA-UVB) καλλιεργούµενων κυττάρων που προέρχονται από καρκίνωµα µητρικού τραχήλου (NHIK 325) σε συγκεντρώσεις µεγαλύτερες από.25mm (Selvaag, 1997) In vivo µελέτες Τα αποτελέσµατα που προκύπτουν από in vivo πειραµατικές µελέτες γονιδιοτοξικότητας του υδροχλωροθειαζιδίου θεωρούνται αρνητικά Έντοµα. Υδατικό διάλυµα υδροχλωροθειαζιδίου (1mg/ml) που χορηγήθηκε µε την τροφή ή ενέθηκε σε αρσενικά άτοµα της Drosophila melanogaster δεν επάγει φυλοσύνδετες υποτελείς θανατογόνες µεταλλάξεις (Valencia et al., 1985) Θηλαστικά. Τα αποτελέσµατα που προκύπτουν από την εκτίµηση της γονιδιοτοξικότητας του υδροχλωροθειαζιδίου µε την εφαρµογή της µεθόδου των µικροπυρήνων και των ανταλλαγών µεταξύ αδελφών χρωµατιδίων σε Chinese hamsters είναι αρνητικά. Επιπρόσθετα, µετά τη χορήγηση του φαρµάκου δεν ανιχνεύονται δοµικές θανατογόνες µεταλλάξεις σε πρώιµα σπερµατοκύτταρα ποντικών και δεν φαίνεται να επηρεάζονται τα γεννητικά κύτταρα (EMEA, 1999)
49 Εισαγωγή Υδροχλωροθειαζίδιο-HCTZ Καρκινογένεση Τρωκτικά. Μελέτες για την πιθανότητα τοξικότητας και καρκινογένεσης από το υδροχλωροθειαζίδιο πραγµατοποιήθηκαν σε δύο διαφορετικές οµάδες τρωκτικών, που περιλαµβάνουν F (Fisher) 344/N αρουραίους και B6C3F1 ποντίκια για 15 ηµέρες, 13 εβδοµάδες, 1 χρόνο και 2 χρόνια. Η χρόνια χορήγηση της φαρµακευτικής ένωσης διαµέσου της τροφής αφορά σταθερές δόσεις, που κυµαίνονται από 25-2ppm για τους αρουραίους και από 25-5ppm για τα ποντίκια (Bucher et al., 199). Στις µελέτες των 15 ηµερών σε δόσεις που έφθαναν έως 5.ppm όλοι οι ποντικοί και αρουραίοι επέζησαν, ενώ παρατηρήθηκε µία µείωση του βάρους τους σε σχέση µε τους αντίστοιχους µάρτυρες. Αντίθετα, ο θάνατος που επήλθε σε 7/1 θηλυκά ποντίκια και 1/1 αρσενικά ποντίκια στις µελέτες των 13 εβδοµάδων σε παρόµοιες χορηγούµενες δόσεις δεν είναι σε θέση να εκτιµηθεί, εξαιτίας της εκδήλωσης πνευµονικής ασθένειας και κανιβαλισµού. Το πρωταρχικό όργανο στο οποίο εµφανίζεται η τοξική δράση του υδροχλωροθειαζιδίου ως αποτέλεσµα της χρόνιας χορήγησης του σε ποντικούς και σε αρουραίους είναι το νεφρό. Σε αρσενικούς και θηλυκούς αρουραίους παρουσία του φαρµάκου παρατηρείται αύξηση στην επικινδυνότητα των βλαβών κατά την ανάπτυξη χρόνιας νεφρικής νόσου, η οποία χαρακτηρίζεται από εκφυλισµό και αναγέννηση του επιθηλίου των νεφρικών σωληναρίων, σωληναριακή διάταση και ατροφία, λέπτυνση της βασικής µεµβράνης των επιθηλιακών κυττάρων, σκλήρυνση του νεφρικού σωµατίου (σπείραµα), ενδιάµεση ίνωση και χρόνια φλεγµονή. Επιπρόσθετα, δευτερογενείς βλάβες που παρουσιάζονται περιλαµβάνουν υπερπλασία των παραθυρεοειδών αδένων, ινώδη οστεοδυστροφία και αδενώµατα του ινώδη ιστού στο µαστικό αδένα σε θηλυκούς αρουραίους, ενώ αντίθετα δεν παρατηρείται υπερπλασία και αδενώµατα στο επιθήλιο των νεφρικών σωληναρίων. Η παρουσία ηπατοκυτταρικών νεοπλασιών (καρκινώµατα/αδενώµατα) σε αρσενικά ποντίκια σε στατιστικώς σηµαντικά επίπεδα (2-42%) σε σχέση µε τους αντίστοιχους µάρτυρες (15%) της συγκεκριµένης πειραµατικής µελέτης θεωρήθηκαν ασαφείς ενδείξεις καρκινογένεσης. Πράγµατι, η τυχαία επαγωγή παρόµοιων περιστατικών σε αρσενικά ποντίκια µάρτυρες, όπως προκύπτει από ιστορικά δεδοµένα που αφορούν µελέτες δύο ετών αντιστοιχεί σε ποσοστό 3±7.6% µε ένα εύρος τιµών να κυµαίνεται από 16-6%. Η τελική εκτίµηση της µελέτης αποδίδει στο υδροχλωροθειαζίδιο στοιχεία µη καρκινογόνου δράσης στους αρουραίους F344/N και σε θηλυκά ποντίκια B6C3F1, ενώ τα αντίστοιχα αποτελέσµατα είναι διφορούµενα για τα αρσενικά ποντίκια B6C3F1 (Bucher, 1989, Bucher et al., 199). Η ηµερήσια χορήγηση υδροχλωροθειαζιδίου σε θηλυκά άτοµα CD R αρουραίων και CD R -1 ποντικών που παραµένουν σε περίοδο κυοφορίας και σε δόσεις που κυµαίνονται από 3-3mg/Kg, µεταξύ της 6 ης και 15 ης ηµέρας δεν επέφερε τερατογόνες επιδράσεις (δυσπλασίες) στους απογόνους (Bucher, 1989, NTP, 1984)
50 Εισαγωγή Υδροχλωροθειαζίδιο Άνθρωπος. Σε επιδηµιολογικές µελέτες στον άνθρωπο για την εξέταση πιθανής καρκινογένεσης από διάφορες φαρµακευτικές ενώσεις παρατηρήθηκε µία πιθανή συσχέτιση µεταξύ χορήγησης υδροχλωροθειαζιδίου και εµφάνισης καρκίνου του προστάτη σε χρονικό διάστηµα επτά ετών από την έναρξη της φαρµακευτικής αγωγής ( ) (Friedman και Ury, 198). Τα αποτελέσµατα της επιδηµιολογικής µελέτης που προκύπτουν κατόπιν συνεχούς παρακολούθησης της ίδιας οµάδας ατόµων για µεγαλύτερο χρονικό διάστηµα αποδίδουν στη χορήγηση διουρητικών θειαζιδίων σε συνδυασµό µε άλλους παράγοντες επικινδυνότητας την εµφάνιση καρκίνου σε ένα µεγάλο αριθµό ζωτικών οργάνων του οργανισµού (Selby et al., 1989). Γενικότερα, η ερευνητική οµάδα και ο ιεθνής Οργανισµός Ερευνών Καρκίνου (IARC) θεώρησε τη θετική συσχέτιση τυχαία που πιθανόν να οφείλεται σε άλλους παράγοντες, ενώ η πραγµατική εκτίµηση θα πρέπει να γίνεται ανεξάρτητα από συσχετισµούς και για χρονικό διάστηµα µεγαλύτερο των είκοσι ετών (IARC, 199). Σε µεταγενέστερη µελέτη που περιλαµβάνει περιορισµένο αριθµό αρσενικών και θηλυκών ατόµων παρατηρήθηκε θετική συσχέτιση µεταξύ της χρήσης υδροχλωροθειαζιδίου και της εµφάνισης καρκινωµάτων σε νεφρικά κύτταρα στα θηλυκά άτοµα (Hiatt et al., 1994)
51 Εισαγωγή Μικροπυρήνες 4. ΜΙΚΡΟΠΥΡΗΝΕΣ 4.1 Ιστορική αναδροµή Σύµφωνα µε τον ορισµό που αποδίδεται στο βιβλίο "Glossary of Genetics and Cytogenetics" (Rieger et al., 1968), ο µικροπυρήνας είναι ένας "πυρήνας διαχωρισµένος από και επιπρόσθετος στον κύριο πυρήνα του κυττάρου, ο οποίος έχει δηµιουργηθεί κατά τη διάρκεια της τελόφασης της µίτωσης ή της µείωσης από καθυστερούµενα χρωµοσώµατα ή χρωµοσωµατικά τµήµατα, που προέρχονται από τυχαίες ή πειραµατικά επαγόµενες δοµικές χρωµοσωµατικές αλλαγές". Οι µικροπυρήνες περιγράφηκαν για πρώτη φορά από κάποιους πρωτοπόρους κυτταρογενετιστές στις δεκαετίες 3 και 4, ως αποτέλεσµα της έκθεσης σε ακτίνες Χ µίας ποικιλίας βιολογικού υλικού, που µεταξύ άλλων περιλαµβάνει νευροβλάστες ακρίδας και µεριστωµατικά κύτταρα ακροριζίου κρεµυδιού. Ο σχηµατισµός τους, τη δεδοµένη χρονική περίοδο αποδόθηκε στην παρουσία γεφυρών και τµηµάτων χρωµατίνης στην ανάφαση µε άµεση συνέπεια τον αποκλεισµό αυτών των τµηµάτων στην τελόφαση. Επιπρόσθετα, η έννοια µικροπυρήνες, γνωστή µε την ονοµασία "σωµατίδια Howell-Jolly", περιγράφηκε από τους αιµατολόγους ως πυρηνικά σωµατίδια θετικά σε χρώση Feulgen που εµφανίζονται σπάνια σε δικτυοερυθροκύτταρα υγιών ατόµων και τα οποία προκύπτουν είτε από κατάτµηση είτε από ελλιπή αποβολή του πυρήνα κατά τη διαδικασία της ερυθροποίησης πιθανότατα από γενετικό υλικό µη ενσωµατωµένο στη µιτωτική άτρακτο (Evans, 1997). Η άποψη που θεωρεί την παρουσία των µικροπυρήνων ως σαφή ένδειξη ύ- παρξης χρωµοσωµατικών ανωµαλιών προτάθηκε για πρώτη φορά από έναν καινοτό- µο κυτταρογενετιστή, τον Η. John Evans, κατόπιν επίπονων µελετών σε κύτταρα ακροριζίου της Vicia faba στα οποία επέδρασε µε νετρόνια και γ-ακτινοβολία παρουσία και απουσία οξυγόνου και οι οποίες αφορούν τη χρήση της επαγόµενης συχνότητας µικροπυρήνων για την ακριβή εκτίµηση κυτταρογενετικών βλαβών (Evans et al., 1959). Στις αρχές της δεκαετίας του 7 πραγµατοποιήθηκαν ανεξάρτητες ερευνητικές µελέτες από τον Werner Schmid και τους συνεργάτες του (Matter και Schmid, 1971, von Ledebur και Schmid, 1973, Schmid, 1973) στην Ελβετία και από τον John A. Heddle (Heddle, 1973) στον Καναδά σχετικά µε την επαγωγή µικροπυρήνων in vivo σε αιµοποιητικά κύτταρα ποντικού ή σε πρόδροµα τους, οι οποίες συνετέλεσαν στην εφαρµογή µίας αξιόπιστης, απλής και γρήγορης µεθόδου για την ανίχνευση και τον ποσοτικό προσδιορισµό χρωµοσωµατικών βλαβών που προκαλούνται σε µικρά θηλαστικά µετά από έκθεση σε µεταλλαξιγόνους παράγοντες, in vivo (Evans, 1997). Η συγκεκριµένη µεθοδολογία (micronucleus test - m.t. in vivo) βασίζεται σε γεγονότα κυτταρικών διαιρέσεων πολυδύναµων αρχέγονων αιµοποιητικών κυττάρων που συµβαίνουν στο µυελό των οστών µε αποτέλεσµα κατά τη διαδικασία ωρίµανσης οι ερυθροβλάστες, ενώ αποβάλλουν τον πυρήνα τους µετά την ολοκλήρωση της τελευταίας µίτωσης, διατηρούν τους µικροπυρήνες και ονοµάζονται πολυχρωµατικά ερυθροκύτταρα (PCEs) (Schmid, 1975). Η µέθοδος διαδόθηκε ταχύτατα στην επιστηµονική κοινότητα και αποτέλεσε έναν εναλλακτικό τρόπο εκτίµησης κυτταρογενετικών βλαβών σε µελέτες γενετικής τοξικολογίας σε συνδυασµό µε την ανίχνευση και ανάλυση χρωµοσωµατικών ανω- µαλιών. Στην αναγκαιότητα τροποποίησης της µεθόδου που αφορά τη χρήση της σε in vitro µελέτες γονιδιοτοξικότητας για τη σαφή εκτίµηση της δράσης πιθανόν µεταλλαξιγόνων παραγόντων συνέβαλαν σηµαντικές αδυναµίες, οι οποίες µεταξύ άλλων σχετίζονταν είτε µε τη δυσκολία λήψης δειγµάτων µυελού των οστών είτε µε την αδυναµία ευρείας εφαρµογής της σε άλλους κυτταρικούς πληθυσµούς
52 Εισαγωγή Μικροπυρήνες Πράγµατι στην εφαρµογή της µεθόδου, in vivo είναι πιθανό παράγοντες των οποίων η δράση µελετάται να µη φθάσουν στα κύτταρα στόχους στην αρχική τους µορφή και στη δεδοµένη συγκέντρωση, είτε γιατί στην πορεία µεταβολίζονται είτε γιατί υφίστανται αραίωση, µε αποτέλεσµα να µην είναι δυνατόν να εξακριβωθεί η δράση τους (Heddle et al., 1983). Αναµφισβήτητα, η ανίχνευση και η ανάλυση µικροπυρήνων σε πολυχρωµατικά ερυθροκύτταρα του µυελού των οστών και του περιφερικού αίµατος ποντικού παρέχει µέχρι και σήµερα σηµαντικές πληροφορίες για την εκτί- µηση της δράσης αρκετών χηµικών ενώσεων, οι οποίες λαµβάνονται υπόψη από τους διεθνείς οργανισµούς (NTP, IARC) (Mäki-Paakkanen και Norppa, 1987, Suzuki et al., 1989, Tice et al., 199, Mavournin et al., 199, Heddle at al., 1991, Morales-Ramìrez et al., 1997, Jackson et al., 2, Morales-Ramìrez et al., 24). Την ίδια χρονική περίοδο, η περιγραφή της κινητικής που αφορά την επαγωγή µικροπυρήνων και χαρακτηριστικών του κυτταρικού κύκλου στα λεµφοκύτταρα (Obe et al., 1975, Obe και Beek, 1975) σε συνδυασµό µε το γενικό φαινόµενο του σχηµατισµού µικροπυρήνων σε διαιρούµενους κυτταρικούς πληθυσµούς ώθησε τους Countryman και Heddle (Countryman και Heddle, 1976) στην εφαρµογή της µεθόδου των µικροπυρήνων σε καλλιέργειες ανθρώπινων λεµφοκυττάρων από ελάχιστες ποσότητες περιφερικού αίµατος µετά από έκθεση σε µεταλλαξιγόνα (ακτίνες-χ και µιτοµυκίνη-c). Ένα από τα µειονεκτήµατα της εφαρµογής αφορά την καταµέτρηση των επαγόµενων µικροπυρήνων σε λεµφοκύτταρα, που κατά τη χρονική διάρκεια της επεξεργασίας τους µε το υποτονικό διάλυµα καταστρέφεται το κυτταρόπλασµα τους µε αποτέλεσµα να παρατηρείται συχνά εκτόπιση των µικροπυρήνων από τα κύτταρα που πραγµατικά προέρχονται. Η εκτίµηση της µεθόδου από επόµενους ερευνητές επέφερε την επιθυµητή λύση µε την ανάπτυξη µίας νέας επεξεργασίας, που περιελάµβανε κύρια την τροποποίηση του υποτονικού διαλύµατος µε άµεση συνέπεια τη διατήρηση του κυτταροπλάσµατος των λεµφοκυττάρων (Iskandar, 1979, Högstedt, 1984). Το επόµενο σηµαντικό πρόβληµα που έπρεπε να αντιµετωπισθεί αφορούσε τη διαφορετική απόκριση των λεµφοκυττάρων στη µιτογόνο διέγερση και την παρουσία στην καλλιέργεια κυττάρων που δεν είχαν υποστεί κυτταρική διαίρεση και δεν υπήρχε η δυνατότητα σχηµατισµού µικροπυρήνων µε αποτέλεσµα η καταµέτρηση τους να µειώνει την πραγµατική συχνότητα των µικροπυρήνων. Οι προτεινόµενες µεθοδολογίες που διατυπώθηκαν από την Pinku και τους συνεργάτες της (Pinku et al., 1984) και τους Fenech και Morley (Fenech και Morley, 1985) αποσκοπούσαν στην επίλυση του προβλήµατος µε τον περιορισµό της ανίχνευσης των µικροπυρήνων σε κύτταρα που έχουν συµπληρώσει µόνο έναν κύκλο διαίρεσης και συνέβαλαν σηµαντικά στην εξέλιξη της µεθόδου των µικροπυρήνων. Με την προσθήκη βρωµοδεόξυουριδίνης (BrdU) σε καλλιέργειες αν θρώπινων λεµφοκυττάρων περιφερικού αίµατος, η οποία ενσωµατώνεται στο DNA κατά την αντιγραφή είναι δυνατή η αναγνώριση των κυττάρων που έχουν διαιρεθεί από τη διαφορετική χρώση µε Giemsa (Pinku et al., 1984). Το βασικό µειονέκτηµα της µεθόδου που συνετέλεσε στη µη αποδοχή της θεωρείται η αδυναµία διάκρισης µεταξύ του διαιρούµενου κυτταρικού πληθυσµού, που οφείλεται στο γεγονός ότι κύτταρα της φάσης S ή G2 του πρώτου κύκλου διαίρεσης µετρώνται επιπλέον ως κύτταρα του δεύτερου κύκλου (Fenech και Morley, 1985a). Επιπρόσθετα, η BrdU σε µεγάλες συγκεντρώσεις είναι δυνατόν να επάγει ανταλλαγές µεταξύ αδελφών χρωµατιδίων (Mazrimas και Stetka, 1978) καθώς και χρωµοσωµατικά ρήγµατα, τα οποία πιθανότατα να προκαλέσουν αύξηση του αριθµού των µικροπυρήνων (Raj και Heddle, 198). Η ανίχνευση και καταµέτρηση µικροπυρήνων σε καλλιέργειες λεµφοκυττάρων που αναγνωρίζονται πραγµατικά ως κύτταρα που έχουν διαιρεθεί µόνο µία φό
53 Εισαγωγή Μικροπυρήνες ρα καθίσταται δυνατή µε την ανάπτυξη δύο µεθόδων που διατυπώθηκαν από τους ερευνητές Fenech και Morley. Με την αυτοραδιογραφική µέθοδο γίνεται επίδραση µε [Η 3 ] θυµιδίνη 48 ώρες µετά την έναρξη της καλλιέργειας και στη συνέχεια σε κύτταρα, που έχουν υποστεί µία µίτωση και εκπροσωπούν ένα συγχρονισµένο στη φάση-s υποπληθυσµό λεµφοκυττάρων ανιχνεύονται οι µικροπυρήνες µε αυτοραδιογραφία στο χρονικό διάστηµα µεταξύ ωρών. Η εφαρµογή της µεθόδου δεν ενδείκνυται για τον προσδιορισµό προϋπαρχόντων in vivo χρωµοσωµατικών βλαβών διότι η ενσωµάτωση της τριτιωµένης θυµιδίνης ενέχεται στην επαγωγή µικροπυρήνων πιθανόν εξαιτίας της εκποµπής β-σωµατιδίων. Η διαπίστωση αυτή δεν αναιρεί τη χρησιµότητα της µεθόδου για την ποσοτική ανίχνευση και καταµέτρηση µικροπυρήνων οι οποίοι επάγονται από την ακτινοβόληση λεµφοκυττάρων in vitro. Με τη µέθοδο αναστολής της κυτταροκίνησης (Cytokinesis-Block method), που επιτυγχάνεται µε τη χρήση της κυτταροχαλασίνης-β καθίσταται εξαιρετικά εύκολη η αναγνώριση των κυττάρων που έχουν συµπληρώσει έναν κύκλο διαίρεσης, από τη χαρακτηριστική διπύρηνη εµφάνιση τους µε αποτέλεσµα η καταµέτρηση των µικροπυρήνων σε τέτοια κύτταρα να θεωρείται απλή και αξιόπιστη (Fenech και Morley, 1985). 4.2 Μέθοδος αναστολής της κυτταροκίνησης-κυτταροχαλασίνη-β Η µέθοδος αναστολής της κυτταροκίνησης βασίζεται αποκλειστικά στη χρήση της κυτταροχαλασίνης-β (Cyt-B), η δράση της οποίας εµποδίζει τη διαίρεση του κυτταροπλάσµατος-κυτταροκίνηση, χωρίς να επηρεάζει την πυρηνική διαίρεση-µίτωση µε αποτέλεσµα τη δηµιουργία διπύρηνων κυττάρων (Fenech και Morley, 1985 a, 1986, Fenech, 1993) ή πολυπύρηνων, ανάλογα µε το χρόνο παραµονής και τη συγκέντρωση της ουσίας στην καλλιέργεια (Ridler και Smith, 1968, Lindholm et al., 1991, Suralles et al., 1992). Η βιολογική δράση της κυτταροχαλασίνης-β αποκαλύφθηκε αρχικά σε καλλιεργούµενα κύτταρα ινοβλαστών (Earle s "L") ποντικού, από τον Carter, ο οποίος παρατήρησε ότι η παραµονή της στην καλλιέργεια σε χαµηλές συγκεντρώσεις (1 µg/ml) για 24 ώρες αναστέλλει την κυτταροπλασµατική διαίρεση, ενώ η πυρηνική διαίρεση που αφορά την εµφάνιση διπύρηνων κυττάρων ολοκληρώνεται φυσιολογικά (Carter, 1967). Οι πρώτες παρατηρήσεις που αφορούν τη χρήση της σε καλλιέργειες ανθρώπινων λεµφοκυττάρων αποδίδουν παρόµοια δράση στη συγκέντρωση 6µg/ml (Ridler και Smith, 1968). Γενικότερα, οι κυτταροχαλασίνες (Cytos:cell-Chalasis:relaxation) είναι χη- µικά συγγενείς µεταβολίτες που έχουν αποµονωθεί από διαφορετικά είδη µυκήτων, από τον Turner, και απoτελούν µία νέα κατηγορία ουσιών µε ασυνήθιστη βιολογική δράση. Συγκεκριµένα, οι Α και Β κυτταροχαλασίνες αποµονώθηκαν από καλλιέργειες του µύκητα Helminthosporium dematiodeum, ενώ οι C και D από το µύκητα Metarrhizium anisopliae. Αξιοσηµείωτη, είναι η παρόµοια λειτουργία που εµφανίζουν, ενώ διαφέρουν σηµαντικά ως προς την δραστικότητα, µε τις C και D να παρουσιάζουν δέκα φορές πιο ισχυρή δράση από τις Α και Β. Η κυτταροχαλασίνη-β που χρησιµοποιείται στη µέθοδο αναστολής της κυτταροκίνησης αποτελείται από έ- να δακτύλιο λακτόνης που είναι συνδεδεµένος σε ένα δικυκλικό σύστηµα λακτάµης (Carter, 1967). Με τη δέσµευσή της αποκλειστικά στο αυξανόµενο άκρο των ινιδίων της ακτίνης (F-ακτίνη) αναστέλλει τον πολυµερισµό τους, ο οποίος απαιτείται για το σχηµατισµό του συσταλτού δακτυλίου (δέσµη µικροϊνιδίων ακτίνης-µυοσίνης ΙΙ), µε αποτέλεσµα να µην καθίσταται δυνατή η περίσφιγξη που θα επιφέρει τη διαίρεση του κυτταροπλάσµατος µεταξύ των θυγατρικών κυττάρων κατά την κυτταροκίνηση (MacLean-Fletcher και Pollard, 198)

54 Εισαγωγή Μικροπυρήνες Προσδιορισµός της συχνότητας µικροπυρήνων-κριτήρια αναγνώρισης Το βασικότερο πλεονέκτηµα της µεθόδου αναστολής της κυτταροκίνησης µε χρήση της κυτταροχαλασίνης-β σε διαιρούµενα ευκαρυωτικά κύτταρα, των οποίων η κινητική της κυτταρικής διαίρεσης είναι γνωστή, έγκειται στην αξιόπιστη αναγνώριση των κυττάρων που έχουν συµπληρώσει µία πυρηνική διαίρεση (Fenech, 2). Με την εφαρµογή της µεθόδου δεν υπάρχει αµφιβολία ότι η ανάλυση των µικροπυρήνων για την εκτίµηση των χρωµοσωµατικών βλαβών που προκαλούνται είτε in vivo είτε in vitro επαγόµενες από χηµικούς ή φυσικούς παράγοντες πραγµατοποιείται σε κύτταρα που έχουν ολοκληρώσει έναν κύκλο διαίρεσης και µπορούν να αναγνωριστούν από την διπύρηνη εµφάνιση τους. Οι µικροπυρήνες είναι δυνατόν να δηµιουργηθούν κατά την ανάφαση της πυρηνικής διαίρεσης, είτε από συσσώρευση άκεντρων χρωµοσωµατικών τµηµάτων, που προκύπτουν από χρωµοσωµατικά ρήγµατα, είτε από ολόκληρα χρωµοσώµατα, τα οποία αποτυγχάνουν να αποχωρισθούν κανονικά, καθυστερούν κατά την ανάφαση και αποκλείονται από τους θυγατρικούς πυρήνες (Fenech, 1997, 2). Αναστολή κυτταροκίνησης Εικόνα 4-1. Απεικόνιση της δηµιουργίας µικροπυρήνων µε τη µέθοδο αναστολής της κυτταροκίνησης µε χρήση της Cyt-B (Fenech, 2). Η συχνότητα των µικροπυρήνων στην αρχική µορφή της µεθόδου αναφέρεται σε κύτταρα ανεξάρτητα από το πρότυπο της πυρηνικής τους διαίρεσης µε αποτέλεσµα να µην αντανακλά τον πραγµατικό δείκτη χρωµοσωµατικής βλάβης. Οι τιµές που λαµβάνει σε αυτή την περίπτωση είναι σηµαντικά µικρότερες από την πραγµατική, διότι ακόµα και οι καλύτερες µέθοδοι συγχρονισµού των κυττάρων δεν κατάφεραν να επιτύχουν την ταυτόχρονη διαίρεση του κυτταρικού πληθυσµού. Με τη χρήση της κυτταροχαλασίνης-β, η πραγµατική εκτίµηση της χρωµοσω- µατικής βλάβης γίνεται πλέον εφικτή µε την καταµέτρηση των µικροπυρήνων αποκλειστικά σε διπύρηνα κύτταρα. Συγκεκριµένα, ο ρυθµός µε τον οποίο ο κυτταρικός πληθυσµός διαιρείται και η αναλογία των κυττάρων που έχουν συµπληρώσει µία πυρηνική διαίρεση ποικίλει µε την αύξηση της συγκέντρωσης χηµικών ενώσεων µε πιθανή τοξική δράση, ιδιαίτερα σε περιπτώσεις που µελετάται η δοσοεξαρτώµενη σχέση µεταξύ των δύο αυτών παραµέτρων. Επιπρόσθετα, η συχνότητα των µικροπυρήνων ανά κύτταρο σε διπύρηνα κύτταρα είναι διπλάσια από αυτή που θα παρατηρούσαµε στα θυγατρικά µονοπύρηνα κύτταρα, εάν επιτρέπαµε στο διπύρηνο κύτταρο να ολοκληρώσει την κυτταροκίνηση. Η αυξηµένη ευαισθησία της µεθόδου σε σύγκριση µε την αρχική µορφή εµφάνισης της αντικατοπτρίζεται από το διπλάσιο αριθµό µονοπύρηνων κυττάρων που θα πρέπει να αναλυθεί απουσία της κυτταροχαλασίνης-β για να παρατηρηθεί το ίδιο επίπεδο µικροπυρήνων που εµφανίζεται στα διπύρηνα κύτταρα (Fenech, 1997)
55 Εισαγωγή Μικροπυρήνες Η αρχική διατύπωση των κριτηρίων για τη διάκριση των µικροπυρήνων από τους Countryman και Heddle (1976) έπρεπε να αναθεωρηθεί µε βάση τα νέα δεδο- µένα που προέκυψαν από την εφαρµογή της µεθόδου της κυτταροχαλασίνης-β µε απoτέλεσµα ο Fenech (1993) να διαµορφώσει τις απαραίτητες προϋποθέσεις που θα πρέπει να πληρούν. Επιπρόσθετα, οι συνεργασίες ερευνητικών οµάδων στην Ευρώπη (Suralles και Natarajan, 1997) και σε διεθνές επίπεδο (Fenech et al., 1999a) µε τη συµµετοχή ενός µεγάλου αριθµού εργαστηρίων καθόρισαν µε σαφήνεια τα κριτήρια που πρέπει να ακολουθούνται αυστηρά για τη σωστή αναγνώριση και καταµέτρηση διπύρηνων κυττάρων και µικροπυρήνων ώστε να εξασφαλίζεται η ακριβής εκτίµηση της συχνότητας µικροπυρήνων σε κύτταρα που έχουν ολοκληρώσει έναν κυτταρικό κύκλο και να παρέχεται η δυνατότητα αναπαραγωγής και σύγκρισης των αποτελεσµάτων. Τα διπύρηνα κύτταρα τα οποία είναι αποτέλεσµα της δράσης της κυτταροχαλασίνης-β αναγνωρίζονται ως εξής: Οι δύο πυρήνες σε ένα διπύρηνο κύτταρο πρέπει να φέρουν άθικτες πυρηνικές µεµβράνες και να εντοπίζονται µέσα στα ίδια κυτταροπλασµατικά όρια. Οι δύο πυρήνες πρέπει να είναι περίπου παροµοίου µεγέθους, ιδίου χρω- µατικού προτύπου και χρωµατικής έντασης. Οι δύο πυρήνες δύναται να βρίσκονται σε επαφή και η µέτρηση κυττάρου που φέρει δύο επικαλυπτόµενους πυρήνες καθίσταται εφικτή µόνο εάν τα πυρηνικά όρια κάθε πυρήνα είναι διακριτά. Τα κυτταροπλασµατικά όρια ή η κυτταροπλασµατική µεµβράνη ενός διπύρηνου κυττάρου πρέπει να είναι άθικτα και ευδιάκριτα από τα κυτταροπλασµατικά όρια γειτονικών κυττάρων. Ένα διπύρηνο κύτταρο δεν θα πρέπει να περιέχει περισσότερους από έξι µικροπυρήνες (Fenech, 1993, 2). Οι µικροπυρήνες είναι µορφολογικά όµοιοι, αλλά µικρότεροι σε µέγεθος από τους κυρίως πυρήνες και θα πρέπει να ακολουθούν τα παρακάτω χαρακτηριστικά: Η ανάλυση των µικροπυρήνων γίνεται αποκλειστικά σε διπύρηνα κύτταρα. Το σχήµα του µικροπυρήνα καθορίζεται ως σφαιρικό ή ωοειδές. Η διάµετρός του στα ανθρώπινα λεµφοκύτταρα ποικίλει µεταξύ του 1/3 και 1/16 της διαµέτρου του κύριου πυρήνα, που αντιστοιχεί στο 1/256 και 1/9 της επιφάνειας ενός από τους κύριους πυρήνες σε ένα διπύρηνο κύτταρο. Οι µικροπυρήνες δεν πρέπει να εµφανίζουν διαθλαστικότητα µε συνέπεια να δύναται εύκολα να διαχωριστούν από άλλα στοιχεία, όπως σωµατίδια χρωστικής. Η σύνδεσή τους ή ένωση τους µε τον κυρίως πυρήνα θεωρείται ανασταλτικός παράγοντας για την καταµέτρησή τους. Ο µικροπυρήνας είναι σε θέση να ακουµπά ή να αλληλεπικαλύπτεται µε τον κύριο πυρήνα, µε την προϋπόθεση ότι τα όρια του µικροπυρήνα καθορίζονται σαφώς και είναι διακριτά από τα όρια του πυρήνα. Οι µικροπυρήνες συνήθως χαρακτηρίζονται από την ίδια ένταση χρώσης µε τον κύριο πυρήνα αλλά σε ορισµένες περιπτώσεις παρουσιάζεται πιο έντονα βαµµένος (Fenech, 2, Kirsch-Volders et al., 2, Bonassi et al., 21)
56 Εισαγωγή Μικροπυρήνες Κυτταροτοξικότητα Η µέτρηση των µονοπύρηνων, διπύρηνων και πολυπύρηνων κυττάρων (>2 πυρήνες) σε ένα καθορισµένο χρονικό σηµείο µετά την προσθήκη της Cyt-B αποτελεί ένα πρόσθετο πλεονέκτηµα της µεθόδου που επιτρέπει την εκτίµηση της έκτασης και εξέλιξης της πυρηνικής διαίρεσης σε ένα διαιρούµενο κυτταρικό πληθυσµό. Ο είκτης Πυρηνικής ιαίρεσης (Nuclear Division Index-NDI) αποδίδεται από τον τύπο: NDI=M1 +2M2+3M3+4M4/N, όπου M1-M4, ο αριθµός των κυττάρων που έχουν αντίστοιχα από έναν έως τέσσερις πυρήνες και Ν το σύνολο των κυττάρων που µελετήθηκαν. Η εκτίµηση του συγκεκριµένου δείκτη παρέχει σηµαντικές πληροφορίες για τις κυτταροστατικές επιδράσεις της υπό µελέτη χηµικής ένωσης, βοηθά στην αναγνώριση µορίων που διεγείρουν την κυτταρική διαίρεση και είναι σε θέση να εκτιµήσει την ανοσολογική ικανότητα του κυτταρικού πληθυσµού διαµέσου της απόκρισης σε µιτογόνους παράγοντες. Αξιοσηµείωτο είναι ότι όλες οι παραπάνω πληροφορίες µπορούν να εξαχθούν µε ταυτόχρονη ανάλυση των µικροπυρήνων στο ίδιο παρασκεύασµα, γεγονός το οποίο είναι ανέφικτο απουσία της Cyt-B (Eastmond και Tucker, 1989, Fenech, 1997). Επιπρόσθετα, προτεινόµενοι δείκτες µε σαφείς αδυναµίες όσον αφορά την εκτίµηση της κυτταροτοξικότητας που αντανακλά µερικά την πρόοδο του κυτταρικού κύκλου αναφέρονται είτε ως ποσοστό διπύρηνων κυττάρων (Fenech και Morley, 1985, Suralles et al., 1994) είτε ως ποσοστό πολυπύρηνων κυττάρων (Van Hummelen και Kirsch-Volders, 1992). Ο είκτης Πολλαπλασιασµού των Κυττάρων (Cytocinesis Block Proliferation Index - CBPI) αποδίδεται µε παρόµοιο τύπο σε σύγκριση µε το δείκτη πυρηνικής διαίρεσης µε τη διαφορά ότι χρησιµοποιείται το άθροισµα των τριπύρηνων και τετραπύρηνων κυττάρων 3(M3+M4), ως πολυπύρηνα κύτταρα που έχουν προέλθει από µία επιπλέον µιτωτική διαίρεση διπύρηνων κυττάρων µετά το σχηµατισµό µίας πολυπολικής (τρι- και τετραπολικής αντίστοιχα) ανάφασης (Schultz και Önfelt, 1994, Suralles et al., 1995a). Με τον προσδιορισµό των δύο δεικτών αξιολογείται µε εξαιρετική ακρίβεια ο ρυθµός πυρηνικών διαιρέσεων και γενικότερα η πρόοδος του κυτταρικού κύκλου και ενώ δεν έχουν βρεθεί ποσοτικές διαφορές σε ανάλογες µελέτες σύγκρισης, όσον αφορά την εκτίµηση της καθυστέρησης του κυτταρικού κύκλου, προτιµάται κύρια η χρήση του δείκτη πολλαπλασιασµού των κυττάρων (Suralles et al., 1995a). Μία πιθανή εξήγηση αποδίδεται στο γεγονός που θεωρεί το δείκτη πυρηνικής διαίρεσης ως έµµεση ένδειξη καθυστέρησης του κυτταρικού κύκλου διότι µε τη διάκριση τριπύρηνων και τετραπύρηνων κυττάρων αντιστοιχεί στη µέση τιµή των πυρήνων ανά κύτταρο και εκφράζει το ρυθµό πυρηνικών διαιρέσεων σε αντίθεση µε το δείκτη πολλαπλασιασµού των κυττάρων που αντιστοιχεί στο µέσο αριθµό των κυτταρικών κύκλων που έχει ολοκληρώσει ο κυτταρικός πληθυσµός και εκφράζει τον αριθµό των κυτταρικών διαιρέσεων ανά κύτταρο µε αποτέλεσµα µία σαφή εκτίµηση της έκτασης και εξέλιξης του κυτταρικού κύκλου (Suralles et al., 1995a, Suralles και Natarajan, 1997). Πρόσφατα διατυπώθηκε ένας νέος τύπος για τον υπολογισµό της κυτταροτοξικότητας (Cytotoxicity) που βασίζεται αποκλειστικά στις τιµές του δείκτη πολλαπλασιασµού των κυττάρων παρουσία και απουσία της επίδρασης µε χηµικές ενώσεις και αποδίδεται από τον τύπο : Cytotoxicity =1-1[CBPIT - 1/ CBPIC - 1] µε την παραδοχή ότι τιµή ίση µε τη µονάδα για το δείκτη CBPIT ισοδυναµεί µε 1% κυτταροτοξικότητα, γεγονός που δηλώνει την αδυναµία ολοκλήρωσης του κυτταρικού κύκλου και την παραµονή του κυτταρικού πληθυσµού σε µονοπύρηνο στάδιο (Kirsch- Volders et al., 23)
57 Εισαγωγή Μικροπυρήνες Αναγνώριση του µηχανισµού δηµιουργίας µικροπυρήνων και ανίχνευση γεγονότων λανθασµένου χρωµοσωµατικού αποχωρισµού Ο συνδυασµός της µεθόδου αναστολής της κυτταροκίνησης (CBMN) µε µοριακές κυτταρογενετικές τεχνικές προσδίδει σηµαντικές πληροφορίες για τη διαπίστωση του περιεχοµένου των µικροπυρήνων και συνεπώς για την εξακρίβωση του µηχανισµού που προηγήθηκε (χρωµοσωµατικά ρήγµατα - χρωµοσωµατική απώλεια) µε άµεσο αποτέλεσµα το σχηµατισµό τους (Fenech, 1997, 2). Η αναγνώριση ολόκληρων χρωµοσωµάτων στους µικροπυρήνες επιτυγχάνεται είτε µε αµιγείς ανοσοχηµικές µεθόδους (CREST) µε τη χρήση αντισωµάτων έναντι πρωτεϊνών του κινητοχώρου, οι οποίες αναµφίβολα συνιστούν αναπόσπαστο τµήµα της κεντροµερικής περιοχής ενεργών χρωµοσωµάτων (Thomson και Perry, 1988, Hennig et al., 1988, Eastmond και Tucker, 1989, Lynch και Parry, 1993, Stephanou et al., 1997), είτε µε in situ υβριδοποίηση µε φθοροχρώµατα (FISH) µε τη χρήση ανιχνευτών έναντι ειδικής αλληλουχίας που απαντάται στα κεντροµέρη (α-δορυφορικό DNA) όλων των ανθρώπινων χρωµοσωµάτων (Becker et al., 199, Migliore et al., 1993, Vlastos και Stephanou, 1998, Andrianopoulos et al., 2). Με την εφαρ- µογή των παραπάνω µεθόδων καθίσταται δυνατή η ακριβής διάκριση των µικροπυρήνων σε δύο τύπους, οι οποίοι µε την παρουσία ή απουσία σήµατος για την κεντρο- µερική περιοχή αποκαλύπτουν το µηχανισµό προέλευσής τους. Μικροπυρήνες που προέρχονται από τη συσσώρευση άκεντρων χρωµοσωµατικών τµηµάτων, εξαιτίας παραγόντων που προκαλούν ρήγµατα στο γενετικό υλικό (clastogens) χαρακτηρίζονται από απουσία σήµατος για την κεντροµερική περιοχή, ενώ µικροπυρήνες οι οποίοι περικλείουν άθικτα χρωµοσώµατα, ως αποτέλεσµα χρωµοσωµατικής καθυστέρησης κατά την ανάφαση επάγονται από παράγοντες που προκαλούν απώλεια ολόκληρων χρωµοσωµάτων (aneugens) και χαρακτηρίζονται από την παρουσία σήµατος για την κεντροµερική περιοχή (Eastmond και Rupa, 1995, Albertini et al., 2). Η διάκριση µικροπυρήνων µε ανοσοχηµικές µεθόδους βασίζεται στην εξειδίκευση των αντισωµάτων που εµφανίζονται στον ορό µίας οµάδας ασθενών µε χαρακτηριστικά που συνιστούν ένα αυτοάνοσο νόσηµα, το σύνδροµο CREST έναντι των πρωτεϊνών του πολυπρωτεϊνικού συµπλόκου του κινητοχώρου (CENPs A, B, C και D) σε µία ποικιλία κυτταρικών τύπων στα θηλαστικά (Moroi et al., 198, Brenner et al., 1981, Bernat et al., 1991) και σε διάφορα είδη ζωϊκών και φυτικών οργανισµών γεγονός που αποδεικνύει την οµολογία και συντηρητικότητα των συγκεκριµένων πρωτεϊνών (Sullivan et al., 1996). Σε σύγκριση µε την in situ υβριδοποίηση µε φθοροχρώµατα θεωρείται απλή και γρήγορη στην εφαρµογή της, ενώ η απουσία χρήσης οξικού οξέος κατά τη διαδικασία µονιµοποίησης, πρωτεϊνασών και ισχυρών απορρυπαντικών, εξαιτίας της ευαισθησίας των πρωτεϊνών του κινητοχώρου σε οξέα και γενικότερα σε συνθήκες οξείδωσης και υγρασίας, επιφέρει άριστη διατήρηση της κυτταρικής µορφολογίας µε αποτέλεσµα να επιτυγχάνεται µία υψηλή απόδοση στην ποιότητα του σήµατος που χαρακτηρίζει όλα τα χρωµοσώµατα. Αξιοσηµείωτες είναι οι περιπτώσεις στις οποίες µικροπυρήνες που περικλείουν ολόκληρα χρωµοσώµατα χαρακτηρίζονται από την απουσία σήµατος για την περιοχή του κινητοχώρου και θεωρούνται εσφαλµένα ως µικροπυρήνες που προέρχονται από τη συσσώρευση άκεντρων χρωµοσωµατικών τµηµάτων γεγονός που οφείλεται είτε σε σοβαρές βλάβες διαφορετικών επιτόπων (περιοχές αντιγόνου) είτε σε πλήρη αποκόλληση του πολυπρωτεϊνικού συµπλόκου του κινητοχώρου (Schuler et al., 1997). Η εφαρµογή της µεθόδου CREST δεν ενδείκνυται σε πληθυσµιακές µελέτες γονιδιοτοξικότητας ιδιαίτερα στην περίπτωση που περιλαµβάνονται ηλικιωµένα άτοµα διότι ένα µεγάλο ποσοστό µικροπυρήνων που σχηµατίζεται από τα φυλετικά
58 Εισαγωγή Μικροπυρήνες χρωµοσώµατα χαρακτηρίζεται από την απουσία σήµατος για την περιοχή του κινητοχώρου που εµφανίζει σοβαρές απώλειες στη λειτουργικότητα του (Hando et al., 1994, Nath et al., 1995). Επιπρόσθετα, η δράση φυσικών ή χηµικών παραγόντων που προκαλούν κυρίως ρήγµατα (ακτίνες-χ, µιτοµυκίνη) στο γενετικό υλικό είναι δυνατόν να αποσαφηνισθεί, εάν η παραπάνω εφαρµογή συνδυασθεί µε την in situ υβριδοποίηση µε φθοροχρώµατα γεγονός που αποδίδει στη µιτοµυκίνη µία ανευπλοειδογόνο ιδιότητα. Η ικανότητα δέσµευσης της στη θεµελιώδη ετεροχρωµατίνη προκαλεί αποκόλληση της τρίστοιβης δοµής του κινητοχώρου από τα κεντροµέρη των χρωµοσωµάτων µε αποτέλεσµα την αδυναµία µετακίνησης τους προς τους πόλους της ατράκτου κατά την ανάφαση και τη δηµιουργία µικροπυρήνων που χαρακτηρίζονται από την απουσία σήµατος για την περιοχή του κινητοχώρου και την παρουσία σήµατος για το κεντροµέρος (Brinkley et al., 1985, Miller et al., 1991, Sgura et al., 21). Με την εφαρµογή της µεθόδου FISH µικροπυρήνες οι οποίοι θα προέλθουν εξαιτίας ρήγµατος που περιλαµβάνει την κεντροµερική περιοχή µε αποτέλεσµα την απώλεια της λειτουργικότητας του θα χαρακτηρισθούν από την παρουσία σήµατος. Τα χρωµοσώµατα µε κεντροµέρος θα συνδεθούν φυσιολογικά µε τα ινίδια της µιτωτικής ατράκτου και θα ολισθήσουν προς τους πόλους, εκτός εάν έχει επέλθει απώλεια της λειτουργικότητας του κινητοχώρου. Η ανωτερότητα της µεθόδου FISH µε τη χρήση ανιχνευτών έναντι αλληλουχίας που απαντάται στα κεντροµέρη όλων των ανθρώπινων χρωµοσωµάτων βασίζεται στην άµεση ανίχνευση του κεντροµέρους χωρίς να επηρεάζεται από βλάβες που έχουν επιφέρει απώλεια της λειτουργικότητας είτε του κινητοχώρου είτε του κεντροµέρους. Οι µικροπυρήνες συνιστούν δύο κατηγορίες που περιλαµβάνουν σύµφωνα µε την προέλευση τους άκεντρα χρωµοσωµατικά τµήµατα ή ολόκληρα χρωµοσώµατα και η ανίχνευση ειδικών περιπτώσεων θεωρείται σχετικά σπάνιο γεγονός σε µη καρκινικά κύτταρα, που για να συµβεί απαιτείται είτε κάποιος επαγωγέας µε ειδικό τρόπο δράσης είτε η ταυτόχρονη εµφάνιση δύο ή περισσοτέρων ανεξάρτητων γεγονότων που η συνδυασµένη πιθανότητα συνύπαρξης τους θα έπρεπε να είναι εξαιρετικά χαµηλή (Schuler et al., 1997, Norppa και Falck, 23). Οι παραπάνω προσεγγίσεις δεν είναι σε θέση να προσδιορίσουν ανευπλοειδικά φαινόµενα που οφείλονται σε µη αποχωρισµό του χρωµοσωµατικού υλικού καθώς και τις συχνότητες χρωµοσωµατικής απώλειας συγκεκριµένων χρωµοσω- µάτων. Ένα µοναδικό χαρακτηριστικό της µεθόδου αναστολής της κυτταροκίνησης Ορθός αποχωρισµός Παρουσία Cyt-B Μη Αποχωρισµός 1. Μη Αποχωρισµός 2. Χρωµοσωµατική απώλεια Επίδραση χηµικού παράγοντα 3α. Χρωµοσωµατική απώλεια Απώλεια γενετικού υλικού 3β. Απώλεια άκεντρου χρωµοσω- µατικού τµήµατος Εικόνα 4-2. Η χρήση της µεθόδου αναστολής της κυτταροκίνησης (CBMN) για την ανίχνευση γεγονότων χρω- µοσωµατικής καθυστέρησης και µη αποχωρισµού κατά τη µίτωση. Οι πράσινες γραµµές ( ) αναπαριστούν ένα ζεύγος οµόλογων χρωµοσωµάτων, τα κόκκινα και πράσινα σφαιρίδια (, ) δύο εξειδικευµένους κεντροµερικούς ανιχνευτές διαφορετικών χρωµοσωµάτων και οι κίτρινοι αστερίσκοι ( ) αναπαριστούν πανκεντροµερικούς ανιχνευτές
59 Εισαγωγή Μικροπυρήνες σε συνδυασµό µε την εφαρµογή της in situ υβριδοποίησης µε φθοροχρώµατα επιτυγχάνεται µε τη χρήση ανιχνευτών εξειδικευµένων για τη σήµανση του κεντροµέρους κάθε ανθρώπινου χρωµοσώµατος και επιτρέπει την αναγνώριση ολόκληρου χρωµοσώµατος σε ένα µικροπυρήνα ενός διπύρηνου κυττάρου ταυτόχρονα µε την ανάλυση της κατανοµής τoυ ιδίου ή άλλων συγκεκριµένων χρωµοσωµάτων µεταξύ των θυγατρικών πυρήνων σε ένα διπύρηνο κύτταρo (Sgura et al., 1997, Kirsch- Volders et al., 1997, Bakou et al., 22). Επιπρόσθετα, η ανίχνευση χρωµοσωµατικού µη αποχωρισµού είναι εφικτή σε µεσοφασικά διπύρηνα κύτταρα που δεν φέρουν µικροπυρήνα και τα οποία προκύπτουν µε την επίδραση της κυτταροχαλασίνης-β. Αξιοσηµείωτο είναι ότι το διπύρηνο κύτταρο στο στάδιο της τελόφασης θεωρείται ιδανικό για την εκτίµηση γεγονότων λανθασµένου διαχωρισµού του χρωµοσωµατικού υλικού διότι µόνο σε αυτό το στάδιο είναι δυνατόν να εξακριβωθεί µε ακρίβεια εάν ο µη αποχωρισµός ή η χρωµοσωµατική απώλεια αποτελούν το µηχανισµό δράσης ενός φυσικού ή χηµικού παράγοντα (Fenech, 1997). Τελευταία επικρατεί η άποψη που θεωρεί την εκτίµηση της συχνότητας και την ανίχνευση του περιεχοµένου των µικροπυρήνων σε µονοπύρηνα κύτταρα σε σύγκριση µε τη χαµηλή επαγόµενη συχνότητα µικροπυρήνων σε διπύρηνα κύτταρα (και τρι-, τετραπύρηνα κύτταρα) ικανή να διακρίνει την ανευπλοειδογόνο δράση χηµικών ενώσεων (νοκοδαζόλη, κολχικίνη, βινκριστίνη). ιαταραχές στον πολυµερισµό - αποπολυµερισµό των µικροσωληνίσκων που συνιστούν βασικό δοµικό συστατικό της µιτωτικής ατράκτου έχει ως άµεση συνέπεια την αποτυχία διαχωρισµού και µετακίνησης των διπλασιασµένων χρωµατιδίων των χρωµοσωµάτων σε µονοπύρηνα κύτταρα που διέφυγαν της µίτωσης στην οποία δεν παρεµβάλλεται η πυρηνική διαίρεση και η κυτταροκίνηση µε µία διαδικασία που καλείται µιτωτική ολίσθηση (mitotic slippage). Με την εφαρµογή της in situ υβριδοποίησης µε φθοροχρώµατα καθίσταται δυνατή η ανίχνευση υψηλής συχνότητας τετραπλοειδών (4Ν) µονοπύρηνων κυττάρων τα οποία χαρακτηρίζονται από την παρουσία ή απουσία µικροπυρήνων. Επιπλέον, η αυξηµένη συχνότητα µικροπυρήνων σε διπλοειδή µονοπύρηνα κύτταρα (2Ν) πιθανότατα να οφείλεται στη δράση της χηµικής ένωσης, που επιτρέπει να ολοκληρωθεί η κυτταροκίνηση πριν την προσθήκη της κυτταροχαλασίνης-β, µε αποτέλεσµα τα κύτταρα αυτά παρουσία βλάβης να αποτυγχάνουν σε επόµενες διαιρέσεις. Μία άλλη εκδοχή είναι ότι κύτταρα που έχουν δεχθεί την επίδραση µετά την ολοκλήρωση µίας φυσιολογικής κυτταρικής διαίρεσης απευαισθητοποιούνται και παραµένουν ανεπηρέαστα στη δράση της κυτταροχαλασίνης-β µε αποτέλεσµα να προκύπτουν δύο διπλοειδή µονοπύρηνα κύτταρα έναντι ενός αναµενόµενου διπύρηνου κυττάρου. Η σηµασία αυτών των µηχανισµών που αντανακλούν την επαγόµενη δηµιουργία µικροπυρήνων σε µονοπύρηνα κύτταρα δεν έχει πλήρως αποσαφηνισθεί µέχρι σήµερα (Elhajoui et al., 1998, Rosefort et al., 24)
60 Εισαγωγή Μικροπυρήνες 4.3 Εφαρµογές Η ταχύτατη διάδοση και εφαρµογή της µεθόδου αναστολής της κυτταροκίνησης σε µία ποικιλία κυτταρικών τύπων στον άνθρωπο και σε πειραµατόζωα είτε in vitro είτε in vivo την καθιστά ένα εξαιρετικά χρήσιµο εργαλείο σε µελέτες Γενετικής Τοξικολογίας σχετικά µε την εκτίµηση της επίδρασης της ιονίζουσας και µη ιονίζουσας ακτινοβολίας σε ευκαρυωτικούς οργανισµούς, στην ανάλυση της ικανότητας χηµικών και φαρµακευτικών ενώσεων να προκαλούν χρωµοσωµατικά ρήγ- µατα ή απώλεια ολόκληρων χρωµοσωµάτων, καθώς και στην έρευνα των επιδράσεων της εργασιακής έκθεσης στον άνθρωπο. Η εκτίµηση της συχνότητας των µικροπυρήνων σε συνδυασµό µε την αποσαφήνιση του περιεχοµένου τους για την ανίχνευση βλαβών στο µόριο του DNA σε χρωµοσωµατικό επίπεδο, οι οποίες αναµφίβολα συνιστούν ένα καθοριστικό γεγονός στην πολύπλοκη εξέλιξη της καρκινογένεσης καθιστά δυνατό τον προσδιορισµό γενετικών αλλαγών που προκαλούνται από παράγοντες του περιβάλλοντος µε µακρόχρονη επίδραση στον ανθρώπινο πληθυσµό, όπως σε εργάτες βιοµηχανιών (Suralles et al., 1997, Srám et al., 1998, Pitarque et al., 22), ορυχείων (Kryscio et al., 21) ή πυρηνικών εργοστασίων (Thierens et al., 1999), προσωπικό νοσοκοµείων (Thierens et al., 2), κατοίκους περιοχών µε έντονη µόλυνση (Moore et al. 1996, Fenech et al., 1997, Chang et al., 1999). Πολυάριθµες ενδείξεις υποστηρίζουν ότι οι χρωµοσωµατικές ανωµαλίες αποτελούν την άµεση συνέπεια βλαβών στο µόριο του DNA µε κύριο χαρακτηριστικό την προέλευση χρωµοσωµατικών ρηγµάτων από µη επιδιορθωµένα δίκλωνα ρήγµατα στο DNA και χρωµοσωµατικών αναδιατάξεων από λάθη που συµβαίνουν κατά την επιδιόρθωση ρηγµάτων στο µόριο του DNA. Επιπρόσθετα, έχει αναγνωρισθεί ότι λανθασµένος διαχωρισµός των χρωµοσωµάτων που αποτελεί κυρίαρχο γεγονός στην ανάπτυξη του καρκίνου και τη γήρανση είναι δυνατόν να προκύψει από βλάβες στη µιτωτική άτρακτο, το κεντρόσωµα ή να αποτελεί συνέπεια της υποσυ- µπύκνωσης των χρωµοσωµάτων πριν το στάδιο της µετάφασης (Fenech, 2). Σε συνύπαρξη µε τις κλασσικές κυτταρογενετικές µεθόδους, που περιλαµβάνουν την άµεση µελέτη των χρωµοσωµάτων για την ανίχνευση και ανάλυση των χρωµοσωµατικών ανωµαλιών σε ένα µεγάλο αριθµό µεταφασικών κυττάρων από εξειδικευµένο επιστηµονικό προσωπικό, η ευρεία αποδοχή της µεθόδου αναστολής της κυτταροκίνησης σε συνδυασµό µε την in situ υβριδοποίηση µε φθοροχρώµατα (CBMN/FISH) αποτελεί έναν απλούστερο, ταχύτερο και κυρίως ασφαλέστερο τρόπο εκτίµησης της γενετικής βλάβης. Αναµφισβήτητα, η εκτίµηση της συχνότητας µικροπυρήνων θεωρείται πλέον ως κύριος βιολογικός δείκτης σε πληθυσµιακές µελέτες για την εκτίµηση της έκθεσης σε γονιδιοτοξικούς παράγοντες και τον καθορισµό αποδεκτών ορίων στη βιολογική δοσιµετρία και διαδραµατίζει πιθανότατα σηµαντικό ρόλο στη διερεύνηση προδιάθεσης για την εµφάνιση καρκίνου (Albertini et al., 2, Fenech, 22, Bonassi et al., 25). Σηµαντική είναι η συµβολή της µεθόδου αναστολής της κυτταροκίνησης που εφαρµόζεται σε καλλιέργειες ανθρώπινων λεµφοκυττάρων περιφερικού αίµατος σε κλινικές µελέτες. Σε συνδυασµό µε εξειδικευµένους ανιχνευτές για τα κεντροµέρη (13, 21) και τις περιοχές (21q22.2) συγκεκριµένων χρωµοσωµάτων είναι εφικτή η ανάλυση κυττάρων για την εκτίµηση ανευπλοειδικών φαινοµένων (τρισωµία 21) και δύναται επιτυχώς να χρησιµοποιηθεί για την αναγνώριση ατόµων που εµφανίζουν τη νευροεκφυλιστική ασθένεια Alzheimer (Migliore et al., 1999a). Επιτυχής είναι η χρήση της µεθόδου στο ίδιο βιολογικό σύστηµα για την αναγνώριση ατόµων που αναπτύσσουν καρκίνο του µαστού καθότι βρέθηκε ότι εµφανίζουν υψηλή ευαισθησία σε γενετικές βλάβες που προκαλεί η ιονίζουσα ακτινοβολία (Scott et al., 1998). Η ευαισθησία αυτή οφείλεται σε δίκλω
61 Εισαγωγή Μικροπυρήνες να ρήγµατα στο γενετικό υλικό που δεν επιδιορθώθηκαν και η οποία παρατηρήθηκε στις δέκα από τις έντεκα περιπτώσεις ατόµων που ήταν φορείς της µετάλλαξης BRCA1 (Rothfus et al., 2). Οι πληροφορίες αυτές συνηγορούν στην άποψη που θεωρεί τη µέθοδο αναστολής της κυτταροκίνησης ιδιαίτερα χρήσιµη τόσο ως ένα βιοδείκτη της γενετικής βλάβης όσο και ως µία ένδειξη εκτίµησης του φαινοτύπου που εκφράζεται µετά την ελλιπή δράση των επιδιορθωτικών µηχανισµών. Συνεπώς θεωρείται περισσότερο πρακτική η µελέτη του φαινοτύπου για την εξαγωγή άµεσων συµπερασµάτων σε πιθανούς φορείς της µετάλλαξης από επιβαρηµένες γενετικά οικογένειες, εάν ληφθεί υπόψη ο µεγάλος αριθµός διαφορετικών BRCA1 µεταλλάξεων που πρέπει να ανιχνευθεί σε συνδυασµό µε ένα εξίσου µεγάλο αριθµό γονιδίων (>12) που συµµετέχουν σε εξειδικευµένους µηχανισµούς επιδιόρθωσης του DNA (Rothfus et al., 2, Fenech, 22). Επιπρόσθετα, αυξηµένα επίπεδα µικροπυρήνων παρατηρήθηκαν σε λεµφοκύτταρα περιφερικού αίµατος σε άτοµα που εµφανίζουν αυτοάνοσα νοσήµατα (συστηµική σκλήρυνση-ss) (Migliore et al., 1999b), την ασθένεια Parkinson (Migliore et al., 22), καθώς και σε άτοµα µε ση- µαντικές βλάβες στην αναπνευστική αλυσίδα των µιτοχονδρίων µε αποτέλεσµα να εµφανίζουν εγκεφαλοµυοπάθειες (Naccarati et al., 2). Αναλυτικότερα, οι µικροπυρήνες έχουν χρησιµοποιηθεί ως in vivo σύστηµα σε πληθυσµιακές µελέτες για την εκτίµηση της δράσης χηµικών παραγόντων και ακτινοβολιών, εκτός από τα ανθρώπινα λεµφοκύτταρα περιφερικού αίµατος (παρουσία της κυτταροχαλασίνης-β) (Titenco-Holland et al., 1997, Pitarque et. al., 1999, Thierens et al., 1999, Leopardi et al., 23), σε απολεπισµένα ανθρώπινα επιθηλιακά κύτταρα που προέρχονται από το στοµατικό (buccal mucosa) (Suralles et al., 1997, Pastor et al., 23) και ρινικό βλεννογόνο (nasal mucosa) (Titenco-Holland, 1996), καθώς και σε ανάλογα κύτταρα της ουροδόχου κύστης (urothelial cells) (Moore et al., 1996, Fontana et al., 21). Η χρήση της συχνότητας µικροπυρήνων ως βιοδείκτη γενετικής βλάβης σε βραγχιακά κύτταρα του Mytilus galloprovincialis (Majone et al., 199) ενδείκνυται για την εκτίµηση της µόλυνσης θαλάσσιων οικοσυστηµάτων από βαρέα µέταλλα και πολυκυκλικούς αρωµατικούς υδρογονάνθρακες σε ακτές επιβαρηµένων περιβαλλοντικά περιοχών (Burgeot et al., 1996, Bolognesi et al., 24). Ειδικότερα, η µέθοδος αναστολής της κυτταροκίνησης (CBMN) εφαρ- µόσθηκε εκτεταµένα είτε in vitro είτε in vivo σε ποικίλους κυτταρικούς τύπους σε εργαστηριακά πειραµατόζωα, όπως σε σπληνοκύτταρα ποντικού (Dreosti et al., 199, Darroudi et al., 1992, Farooqi et al., 1993, Russo et al., 1996, Stephanou et al., 1997, Stephanou et al., 1998), σε κύτταρα ωοθηκών (Murray et al., 1991) και σε V79 κύτταρα πνευµόνων (Channarayappa et al., 1992) Chinese hamster, και στον άνθρωπο σε επιδερµικούς ινοβλάστες (Antoccia et al., 1993, Sgura et al., 21), σε καρκινικά κύτταρα ήπατος (Hep G2) (Darroudi et al., 1996), προστάτη (PC-3) και µαστού (MCF-7) (Kalantzi et al., 24), καθώς και σε γενετικά τροποποιηµένα λεµφοβλαστικά κύτταρα (AHH-1, MCL-5) (Parry et al., 1996, Parry et al., 22). Η συνεχώς αυξανόµενη γνώση, από πληθυσµιακές µελέτες της γενετικής δράσης µεταλλαξιγόνων χηµικών ενώσεων σε συγκεκριµένες οµάδες ατόµων σε συσχέτιση µε τη γενετική τους σύσταση προσδίδει µία νέα διάσταση στην εφαρµογή της µεθόδου αναστολής της κυτταροκίνησης και στη χρήση της συχνότητας µικροπυρήνων ως βιοδείκτη της γενετικής βλάβης. Η ανάλυση και η εκτίµηση της συχνότητας µικροπυρήνων σε συνδυασµό µε το γονιδιοτυπικό προσδιορισµό ατόµων για την αποκάλυψη πολυµορφισµών γονιδίων που σχετίζονται κυρίως µε ένζυµα που συµµετέχουν στη Φάση Ι (ένζυµα του κυτοχρώµατος P45-CYP) και φάση ΙΙ (Ν-ακετυλοτρανσφεράσες-ΝΑΤ, S-τρανσφεράσες της γλουταθειόνης-gsts) του µεταβο
62 Εισαγωγή Μικροπυρήνες λισµού (ενεργοποίηση/απενεργοποίηση) ενός µεγάλου αριθµού χηµικών ενώσεων (πολυκυκλικοί αρωµατικοί υδρογονάνθρακες-pahs, εποξείδια, στυρένιο, βενζένιο, βουταδιένιο-bd, διεποξυβουτάνιο-deb, ετεροκυκλικές αµίνες) αποκαλύπτει την ευαισθησία συγκεκριµένων οµάδων ατόµων στην πρόκληση γενετικών βλαβών από διάφορους περιβαλλοντικούς ή χηµικούς παράγοντες. Πρόσφατες µελέτες έχουν δείξει ότι η ευαισθησία ατόµων στο διεποξυβουτάνιο οφείλεται κατά κύριο λόγο σε ελλειµµατική µορφή του γονιδίου GSTT1 µε αποτέλεσµα να παρατηρείται απουσία ενεργότητας της τρανσφεράσης (null γενότυπος). Άτοµα στα οποία απουσιάζει η γλουταθειόνη της τρανσφεράσης από τα ερυθροκύτταρα τους είναι περισσότερο ευαίσθητα στη δηµιουργία χρωµοσωµατικών ανωµαλιών (Sorsa et al., 1996), ανταλλαγών µεταξύ αδελφών χρωµατιδίων (Kelsey et al., 1995) και στο σχηµατισµό µικροπυρήνων (Vlachodimitropoulos et al., 1997). Σηµαντική είναι η σύνδεση του πολυµορφισµού µε συγκεκριµένες µορφές καρκίνου, καθώς η απουσία του γονιδίου συνδέεται πιθανότατα µε την εµφάνιση µυελοδυσπλαστικών συνδρόµων (Chen et al., 1996) και έµµεσα µε αυξηµένο κίνδυνο για την εµφάνιση ορισµένων τύπων (αστεροειδές γλοίωµα, µηνιγγικός όγκος) καρκίνου του εγκεφάλου (Elexpuru- Camiruaga et al., 1995, Kelsey et al., 1997). Σε ανάλογες µελέτες που αφορούν την έκθεση εργατών σε χαµηλά επίπεδα στυρενίου έχει δειχθεί η σηµαντική συµµετοχή πολυµορφισµών σε ένζυµα µεταβολισµού, που σχετίζονται είτε µε το µετασχηµατισµό (CYP2E1) του στυρενίου σε οξείδιο του στυρενίου (SO) είτε µε την αποτοξικοποίηση (GSTM1, GSTT1, EPHX1) του SO στην ευαισθησία των ατόµων όσον αφορά την πρόκληση γενετικών βλαβών, η οποία εκτιµάται από τη συχνότητα µικροπυρήνων. Επιπρόσθετα, µία άλλη παράµετρος που µελετήθηκε στην ίδια οµάδα εργατών µε παρόµοιο τρόπο αφορά πολυµορφισµούς σε ένζυµα µηχανισµών επιδιόρθωσης του DNA που συµµετέχουν στην αποκατάσταση των γενετικών βλαβών (hogg1, XRCC1, XRCC3 ), οι οποίες προκαλούνται από το οξείδιο του στυρενίου (Godderis et al., 24). 4.4 Παράγοντες που επηρεάζουν τη συχνότητα µικροπυρήνων Πέρα από την εργασιακή και περιβαλλοντική έκθεση σε γονιδιοτοξικούς παράγοντες που σαφέστατα επηρεάζουν τη συχνότητα µικροπυρήνων αρκετοί άλλοι ενέχονται για τις ενδο- και δια-τοµικές διακυµάνσεις που παρατηρούνται κατά την εκτίµηση του συγκεκριµένου βιοδείκτη. ηµογραφικοί παράγοντες, όπως το φύλο και η ηλικία (Bolognesi et al., 1997, Peace και Succop, 1999), καθώς και ατοµικές συνήθειες, όπως το κάπνισµα (Tomanin et al., 1991, Ishikawa et al., 23), η κατάνάλωση αλκοόλ (Maffei et al., 2), η διατροφή (Schneider et al., 21, Botto et al., 23, Fenech et al., 25) επηρεάζουν σε µικρότερο ή µεγαλύτερο βαθµό τη συχνότητα µικροπυρήνων και θα πρέπει να εκτιµηθούν ανάλογα σε µελέτες, που αφορούν τη αποσαφήνιση του µηχανισµού γενετικής δράσης ενός φυσικού ή χηµικού παράγοντα. Η εκτεταµένη χρήση της µεθόδου αναστολής της κυτταροκίνησης σε ανθρώπινα λεµφοκύτταρα οδήγησε στη δηµιουργία ενός διεθνούς προγράµµατος µελέτης σχετικά µε τις παρατηρούµενες συχνότητες µικροπυρήνων σε ανθρώπινους πληθυσµούς (HUMN-Human MicroNucleus Project) που αποσκοπεί στη συγκρότηση µίας ενιαίας βάσης δεδοµένων. Οι αντικειµενικοί στόχοι αυτού του προγράµ- µατος είναι: (α) η συσχέτιση παραµέτρων, ανάµεσα στους οποίους περιλαµβάνονται οι δηµογραφικοί παράγοντες και αυτοί που προκύπτουν από την υιοθέτηση ενός συγκεκριµένου τρόπου ζωής µε το δείκτη της γενετικής βλάβης (β) η χρήση κατάλληλων βιοδεικτών στην αναγνώριση προδιάθεσης για την εµφάνιση καρκίνου,
63 Εισαγωγή Μικροπυρήνες επιλεγµένων γενετικών συνδρόµων και πιθανά άλλων ασθενειών (γ) οι πιθανές διακυµάνσεις της συχνότητας µικροπυρήνων σε οµάδες πληθυσµών που προέρχονται από διαφορετικές ηπείρους και (δ) ο καθορισµός σταθερών πρωτοκόλλων για την εφαρµογή της µεθόδου. Ο συγκεκριµένος φορέας περιλαµβάνει ένα δίκτυο 25 εργαστηρίων σε περισσότερες από 16 χώρες µε την υψηλότερη εκπροσώπηση στην Ευρώπη και τη σηµαντική συµµετοχή των υπολοίπων ηπείρων (Ασία, Αµερική και Αυστραλία) (Bonassi et al., 21). Τα πρώτα αποτελέσµατα έδειξαν θετική συσχέτιση της συχνότητας µικροπυρήνων µε την ηλικία, που ισχύει και για τα δύο φύλα. Η αύξηση αυτή παρουσιάζεται εντονότερη µετά την ηλικία των σαράντα ετών και συνεχίζει να ισχύει µε την αύξηση της ηλικίας. Το φύλο φαίνεται να επηρεάζει τη συχνότητα µικροπυρήνων σε µικρότερο βαθµό µε τα θηλυκά άτοµα σε ηλικία µεγαλύτερη των τριάντα ετών να εµφανίζουν υψηλότερες συχνότητες µικροπυρήνων σε σύγκριση µε τα αρσενικά (Bonassi et al., 21). Όσον αφορά τη συνήθεια του καπνίσµατος και την εκτίµηση της γενετικής βλάβης που προκαλεί µε τη χρήση του συγκεκριµένου βιοδείκτη σε ανθρώπινα λεµφοκύτταρα υπήρξε αδυναµία θετικής συσχέτισης, ενώ µία στατιστικά σηµαντική αύξηση στη συχνότητα µικροπυρήνων παρατηρείται σε βαρείς καπνιστές ( 3 τσιγάρα/ηµέρα). Ιδιαίτερη σηµασία θα πρέπει να αποδοθεί στη χρονική διάρκεια του καπνίσµατος και στην ηλικία έναρξης της βλαβερής για την ανθρώπινη υγεία συνήθειας, στοιχεία τα οποία δεν ήταν διαθέσιµα στη συγκεκριµένη αξιολόγηση (Bonassi et al., 23). Επιπλέον, οι διατροφικές συνήθειες και η επιρροή τους στις διακυµάνσεις της συχνότητας µικροπυρήνων έχουν αναλυτικά εκτιµηθεί σε εκτεταµένες πληθυσµιακές µελέτες που αφορούν συγκεκριµένες ηπείρους (Αυστραλία) (Fenech, 1999, Fenech et al., 25). Αναµφισβήτητα αρκετά ιχνοστοιχεία, όπως ο ψευδάργυρος, το µαγνήσιο και το ασβέστιο, αντιοξειδωτικές ενώσεις, όπως το β-καροτένιο, το ασκορβικό οξύ και η α-τοκοφερόλη καθώς και σηµαντικά στοιχεία διατροφής, όπως το φολικό οξύ και η βιταµίνη Β12 που διαδραµατίζουν σηµαντικό ρόλο στο µεταβολισµό του DNA ενέχονται σε διακυµάνσεις στις συχνότητες µικροπυρήνων που παρατηρούνται συχνά σε ανθρώπινους πληθυσµούς και θα πρέπει σε κάποιο βαθµό να λαµβάνονται υπόψη (Fenech, 22). Για τη συµµετοχή του γενοτύπου σε πιθανές διακυµάνσεις της συχνότητας µικροπυρήνων σε υγιείς πληθυσµιακές οµάδες ατόµων υπάρχουν περιορισµένα αποδεικτικά στοιχεία, ως αποτέλεσµα ερευνητικών µελετών που συσχετίζουν συγκεκριµένους ενζυµικούς πολυµορφισµούς (Norppa, 21, Bonassi et al., 25). Χαρακτηριστική είναι η µελέτη που αναφέρεται σε πολυµορφισµούς του γονιδίου CYP2E1 που παρατηρείται σε χαµηλή συχνότητα σε καυκάσιους πληθυσµούς. Το προϊόν του ενεργοποιεί προκαρκινογόνες ενώσεις, όπως νιτροζαµίνες, συστατικά του καπνού, χλωριούχους και µη χλωριούχους διαλύτες στους οποίους περιλαµβάνεται το βενζένιο, και δραστηριοποιείται στο µεταβολισµό της αλκοόλης και περισσότερων από 7 συνθετικών οργανικών ενώσεων τις οποίες µετατρέπει σε τοξικές. Ανάµεσα σε άλλους πολυµορφισµούς που µελετήθηκαν και περιελάµβαναν τα γονίδια CYP1A1, GSTM1 και GSTT1 η παρουσία του αλληλοµόρφου CYP2E1*3 σχετίζεται µε τη συχνότητα των µικροπυρήνων σε υγιείς οµάδες ατόµων της Ιαπωνίας (Ishikawa et al., 24). Ανάλογες ερευνητικές µελέτες στον ευρωπαϊκό χώρο δεν είναι σε θέση να καθορίσουν µε ακρίβεια το βαθµό στον οποίο οι πολυµορφισµοί των γονιδίων CYP1A1, GSTM1 και GSTT1 συµµετέχουν σε διακυµάνσεις του συγκεκριµένου βιοδείκτη σε υγιείς πληθυσµούς (Pavanello και Clonfero, 2, Norppa, 21). Επιπρόσθετα, η εκτίµηση διαφορετικών δεικτών (χρωµοσωµατικές ανωµαλίες και µικροπυρήνες) της γενετικής βλάβης που προκαλείται µετά από την έκθεση των ατόµων σε γονιδιοτοξικούς παράγοντες και η συσχέτιση τους µε
64 Εισαγωγή Μικροπυρήνες ενζυµικούς πολυµορφισµούς σε ανάλογες ερευνητικές µελέτες δεν βρίσκεται σε συµφωνία. Αναµφισβήτητα, συγκεκριµένοι γενότυποι είναι σε θέση να επηρεάσουν την εκτίµηση κυτταρογενετικών δεικτών γεγονός το οποίο θα πρέπει να αποτελέσει αντικείµενο έρευνας σε µελλοντικές µελέτες για να αποσαφηνισθούν πλήρως οι εµπλεκόµενοι µηχανισµοί (Bonassi et al., 25). 4.5 Πρόσθετοι βιολογικοί δείκτες Ένα επιπλέον χαρακτηριστικό της µεθόδου αναστολής της κυτταροκίνησης αφορά την ανίχνευση πυρηνοπλασµατικών γεφυρών (Nucleoplasmic Bridges- NPBs) που εµφανίζονται µεταξύ των δύο θυγατρικών πυρήνων ενός διπύρηνου κυττάρου µε τη µορφή πυρηνοπλασµατικής σύνδεσης, µετά από επίδραση παραγόντων που προκαλούν ρήγµατα στο γενετικό υλικό (ιονίζουσα ακτινοβολία, έλλειψη φολικού οξέος). Πιθανότατα η ανάλυση τους παρέχει ένα βιοδείκτη ασύµµετρων χρωµοσωµατικών αναδιατάξεων που θεωρείται ιδιαίτερα χρήσιµος στη βιολογική δοσιµετρία, είτε κατόπιν ατυχηµάτων έκλυσης ραδιενέργειας είτε χρόνιας έκθεσης στην κοσµική ακτινοβολία. Ο σχηµατισµός τους αποδίδεται στην πίεση προς τους αντίθετους πόλους του κυττάρου που δέχονται τα κεντροµέρη των δίκεντρων χρωµοσωµάτων ή χρωµατιδίων κατά την ανάφαση µε αποτέλεσµα η παρατήρηση πυρηνοπλασµατικών γεφυρών να καθίσταται αδύνατη σε µονοπύρηνα κύτταρα και να επιτυγχάνεται µόνο µε την αναστολή της κυτταροκίνησης µετά την ολοκλήρωση του σχηµατισµού της πυρηνικής µεµβράνης γύρω από τα χρωµοσώµατα (Fenech και Crott, 22, Thomas et al., 23). Σε πρόσφατες µελέτες, η ανάλυση πυρηνοπλασµατικών γεφυρών ως βιοδείκτης γενετικής βλάβης σε ανθρώπινα λεµφοβλαστικά κύτταρα WIL2-NS µετά από έκθεση σε µία ποικιλία ελευθέρων ριζών οξυγόνου (H2O2, Ο2-) και ενεργοποιηµένων ουδετερόφιλων κοκκιοκυττάρων (HOCl) έδειξε την ευαισθησία της µεθόδου. Ο υψηλός ρυθµός δηµιουργίας πυρηνοπλασµατικών γεφυρών συγκριτικά µε το σχηµατισµό µικροπυρήνων (NPBs/MN), κυρίως ως αποτέλεσµα της δράσης τοξικών παραγόντων (υποχλωριώδες οξύ) οφείλεται στην παρουσία δίκεντρων χρωµοσωµάτων και δακτυλίων (Umegaki και Fenech, 2). Σε επόµενες µελέτες, σε ανθρώπινα λεµφοκύτταρα περιφερικού αίµατος που έχουν εκτεθεί in vitro σε αυξανόµενες δόσεις γ-ακτινοβολίας στη Go φάση του κυτταρικού κύκλου γίνεται πλέον εµφανής η θετική συσχέτιση µεταξύ πυρηνοπλασµατικών γεφυρών-έκκεντρων δακτυλίων και δίκεντρων χρωµοσωµάτων, καθώς και µεταξύ µικροπυρήνων-άκεντρων χρωµοσωµατικών τµηµάτων και άκεντρων δακτυλίων. Για την αποσαφήνιση του µηχανισµού δηµιουργίας των NPBs έγιναν προκαταρκτικά πειράµατα µε συνδυασµένη χρήση της in situ υβριδοποίησης µε φθοροχρώµατα µε εξειδικευµένο ανιχνευτή για ένα επιλεγµένο χρωµόσωµα (chromosome painting), τα οποία έδειξαν ότι πιθανά προέρχονται από ασύµµετρες χρωµοσωµατικές αναδιατάξεις που συµβαίνουν στο ίδιο χρωµόσωµα ή µεταξύ οµολόγων χρωµοσω- µάτων (Thomas et al., 23). Με τη µέθοδο αναστολής της κυτταροκίνησης παρέχεται η δυνατότητα ανάλυσης αποπτωτικών και νεκρωτικών κυττάρων, που επάγονται από παράγοντες µε πιθανή τοξική δράση για το κύτταρο και είναι σε θέση να ενεργοποιήσουν µονοπάτια του αποπτωτικού µηχανισµού, χωρίς απαραίτητα να ευνοούν το σχηµατισµό µικροπυρήνων (Kirsch-Volders et al., 1997). Η απόπτωση είναι ένας γενετικά καθορισµένος, µη φλεγµονώδης, µηχανισµός καταστροφής του κυττάρου, στον οποίο συµµετέχουν αρκετά ογκογονίδια και ογκοκατασταλτικά γονίδια και η εκτίµηση ανάλογων παραµέτρων (νέκρωση) θεωρείται ιδιαίτερα σηµαντική για την περιγραφή του µηχανισµού δράσης και την ανάλυση της ευαισθησίας των κυττάρων σε
65 Εισαγωγή Μικροπυρήνες χηµικές ενώσεις ή στην έκθεση ακτινοβολίας. Σοβαρές βλάβες στο DNA ή σε άλλα κυτταρικά στοιχεία στη Go φάση δεν επιτρέπει στα κύτταρα να ολοκληρώσουν τον πρώτο κυτταρικό κύκλο, ώστε να σχηµατίσουν µικροπυρήνες µε συνέπεια να υφίστανται νέκρωση ή απόπτωση (Kirsch-Volders και Fenech, 21). Η νέκρωση αποτελεί ένα παθολογικό κυτταρικό θάνατο που συµβαίνει σε οµάδες κυττάρων και οφείλεται σε γενικευµένη έκπτωση των κυτταρικών ρυθµιστικών µηχανισµών µε αποτέλεσµα την απελευθέρωση υδρολυτικών ενζύµων, από τα λυοσώµατα του κυτταροπλάσµατος στον περιβάλλοντα χώρο και την πρόκληση φλεγµονής. Σηµαντικοί παράγοντες που κατευθύνουν τη διαδικασία της νέκρωσης σε ένα κύτταρο είναι η οξειδωτική του κατάσταση, τα επίπεδα της ATP και το µέγεθος της βλάβης της κυτταρικής µεµβράνης (Fenech, et al., 1999b). Η χρήση της κυτταροχαλασίνης-b καθιστά δυνατή την ανάλυση των αποπτωτικών κυττάρων διότι µε την αναστολή του σχηµατισµού των µικροϊνιδίων, τα οποία θεωρούνται απαραίτητα για τον κατακερµατισµό των κυττάρων παρεµποδίζεται η διάσπασή τους σε µικρότερα αποπτω τικά σωµάτια. Η παρατήρηση νεκρωτικών και αποπτωτικών κυττάρων αφορά κυτταρικές καλλιέργειες που δεν έχουν υποστεί επεξεργασία µε υποτονικό διάλυµα µε τα αποπτωτικά κύτταρα να παρουσιάζουν είτε διάσπαρτη συµπυκνωµένη χρωµατίνη µε άθικτα τα όρια κυτταροπλασµατικής και πυρηνικής µεµβράνης είτε µικρά πυρηνικά σωµάτια σε κύτταρα µε άθικτο κυτταρόπλασµα/κυτταροπλασµατική µεµβράνη. Αντίθετα τα νεκρωτικά κύτταρα, σε αρχικά στάδια παρουσιάζουν κενοτόπια, αποσύνθεση της κυτταροπλασµατικής µεµβράνης και απώλεια του κυτταροπλάσµατος, ενώ ο πυρήνας παραµένει αρχικά ανέπαφος (Fenech, 2). Η εφαρµογή της µεθόδου αναστολής της κυτταροκίνησης σε συνδυασµό µε τη χρήση της κυτοσίνης της αραβινοσίδης (ARA-C/CBMN) αποδίδει έναν επιπλέον βιοδείκτη για τη σαφή εκτίµηση της γενετικής βλάβης µε την ανίχνευση γεγονότων επιδιόρθωσης εκτοµής, ιδιαίτερα για µία ποικιλία γονιδιοτοξικών παραγόντων που επάγουν χαµηλή συχνότητα µικροπυρήνων σε σχέση µε την εκδήλωση ισχυρής κυτταροτοξικής δράσης (Fenech, 2). Παράγοντες, όπως η υπεριώδης ακτινοβολία και αρκετές χηµικές ενώσεις (N-µεθυλονιτροζοουρία, EMS) προκαλούν βλάβες σε βάσεις του DNA µε αποτέλεσµα τη δηµιουργία τροποποιηµένων νουκλεοτιδίων (DNA adducts). Οι µηχανισµοί που έχει αναπτύξει το κύτταρο για την επιδιόρθωση τροποποιηµένων νουκλεοτιδίων περιλαµβάνουν την επιδιόρθωση µε εκτοµή βάσεων ή νουκλεοτιδίων. Στην πρώτη περίπτωση αποµακρύνεται η βάση που φέρει τη βλάβη γεγονός που δηµιουργεί µία απουρινική ή απυριµιδινική θέση στον κλώνο του DNA, ενώ στη δεύτερη περίπτωση ενδονουκλεάσες αποµακρύνουν ένα ολιγονουκλεοτίδιο 24-3 βάσεων µε τη φερόµενη βλάβη. Χαρακτηριστικό είναι ότι και στις δύο περιπτώσεις δηµιουργούνται κενά (ρήγµατα) στο DNA, τα οποία συµπληρώνονται µε τη δράση πολυµερασών. Εάν για κάποιο λόγο αυτά τα ρήγµατα στο γενετικό υλικό δεν δύναται να επιδιορθωθούν πριν τη φάση S της αντιγραφής µετατρέπονται σε δίκλωνα ρήγµατα και εκφράζονται ως χρωµατιδικά ρήγµατα (chromatid breaks), τα οποία δηµιουργούν τους µικροπυρήνες µετά από έναν κύκλο σύνθεσης του DNA. Για να ελεγχθεί η ικανότητα φυσικών ή χηµικών παραγόντων να προκαλούν τέτοιου είδους βλάβες, που αναφέρονται, ως "excision repairable lesions" έγινε χρήση της κυτοσίνης της αραβινοσίδης (ARA-C), η οποία αναστέλλει το στάδιο πολυµερισµού (gap filling) µετά την εκτοµή µε αποτέλεσµα να µετατρέπει τη βλάβη στη βάση του DNA σε ένα µονόκλωνο ρήγµα που θα µετατραπεί σε ένα δίκλωνο µετά από έναν κύκλο αντιγραφής του DNA και θα οδηγήσει στη δηµιουργία ενός άκεντρου χρωµοσωµατικού τµήµατος που εκδηλώνεται ως µικροπυρήνας µέσα σε έναν κύκλο διαίρεσης (Fenech και Neville, 1992, Suralles et al., 1995b)
66 Εισαγωγή Μικροπυρήνες Αναµφισβήτητα, η ακριβής µέτρηση των βλαβών που επιδιορθώνονται µε εκτοµή, µε τη χρήση της κυτοσίνης της αραβινοσίδης, καθίσταται δυνατή µόνο µε την ταυτόχρονη εφαρµογή της µεθόδου αναστολής της κυτταροκίνησης διότι: (α) η µετατροπή των συγκεκριµένων βλαβών σε µικροπυρήνες είναι δυνατόν να συµβεί µόνο σε κύτταρα που έχουν ολοκληρώσει µία διαίρεση και (β) η προσθήκη της αραβινοσίδης είναι δυνατόν να οδηγήσει σε σηµαντική τροποποίηση της κινητικής της πυρηνικής διαίρεσης (Fenech, 1997). Με τη συνδυασµένη χρήση της µεθόδου (ARA-C/CBMN) σε ανθρώπινα λεµφοκύτταρα in vitro έγινε ακριβής εκτίµηση της δράσης µίας ποικιλίας χηµικών παραγόντων που ασκούν τη µέγιστη δράση τους στη S φάση του κυτταρικού κύκλου και περιλαµβάνουν ζιζανιοκτόνα και εντοµοκτόνα (Suralles et al., 1995b), καθώς και µία κατηγορία νεοσυντιθέµενων χηµειοθεραπευτικών ενώσεων που αποτελούν στεροειδικά παράγωγα του υπερίτη του αζώτου (nitrogen mustard) µε αλκυλιωτική δράση (Kouloumenta et al., 25). Επιπρόσθετα, η εφαρµογή της µεθόδου στο ίδιο βιολογικό σύστηµα σε ρυθµιστές κυκλοφορίας οχηµάτων για την εκτίµηση της εργασιακής έκθεσης σε αρωµατικές ρυπογόνες ενώσεις απέτυχε να αποδώσει θετική συσχέτιση της συχνότητας µικροπυρήνων που δηµιουργήθηκαν ως αποτέλεσµα βλαβών που επιδιορθώνονται µε εκτοµή µε την ατµοσφαιρική µόλυνση σε συνδυασµό µε το κάπνισµα, το φύλο και την ηλικία (Leopardi et al., 23). Το γονίδιο HPRT κωδικοποιεί το ένζυµο φωσφοριβοζυλοτρανσφεράση της υποξανθίνης-γουανίνης και θεωρείται υπεύθυνο για µία σειρά αντιδράσεων που κατευθύνει τη σύνθεση και παραγωγή πουρινών. Επιπρόσθετα µε τα φυσιολογικά υποστρώµατα, το ένζυµο καταλύει τη µετατροπή αναλόγων των πουρινών, όπως η 6- θειογουανίνη (6-TG) µε αποτέλεσµα η παρουσία τους να είναι κυτταροτοξική. Μεταλλάξεις του γονιδίου HPRT, ως αποτέλεσµα της έκθεσης σε µεταλλαξιγόνους παράγοντες είναι δυνατόν να αναγνωρισθούν σε Τ-λεµφοκύτταρα περιφερικού αίµατος, από την ανθεκτικότητα τους σε καλλιέργειες παρουσία αναλόγων των πουρινών. Η εκτίµηση της συχνότητας µεταλλάξεων του γονιδίου HPRT χρησι- µοποιείται συχνά ως βιοδείκτης γενετικής βλάβης σε πληθυσµιακές µελέτες για τον προσδιορισµό της in vivo γονιδιοτοξικότητας φυσικών ή χηµικών παραγόντων (Albertini et al., 2). Με την προσθήκη της κυτταροχαλασίνης-β σε ενεργοποιη- µένες µε φυτοαιµατογλουτινίνη καλλιέργειες περιφερικού αίµατος καθίσταται δυνατή η ανίχνευση Τ-λεµφοκυττάρων που φέρουν την υποτιθέµενη µετάλλαξη για το HPRT γονίδιο, από την ικανότητα τους να διαιρούνται τουλάχιστον µία φορά παρουσία 6-TG µε αποτέλεσµα να αναγνωρίζονται από τη διπύρηνη ή πολυπύρηνη εµφάνιση τους. Με την προσθήκη µίας επιπλέον καλλιέργειας απουσία 6-TG που επιτρέπει τη διαίρεση των φυσιολογικών για το HPRT γονίδιο λεµφοκυττάρων καθίσταται δυνατή η εκτίµηση της συχνότητας των µεταλλάξεων του γονιδίου που βασίζεται στην αναλογία της συχνότητας των διαιρούµενων (διπύρηνων και πολυπύρηνων) λεµφοκυττάρων παρουσία και απουσία 6-TG (Fenech, 1997). Μεταλλάξεις στο γονίδιο HPRT υποδηλώνουν µία µεγάλη ποικιλία γενετικών τροποποιήσεων στις οποίες περιλαµβάνονται υποκαταστάσεις νουκλεοτιδικών βάσεων, µεγάλα και µικρά ελλείµµατα, αναστροφές και µη οµόλογος χρωµοσωµατικός ανασυνδυασµός. Το γεγονός ότι ο συγκεκριµένος γονιδιακός τόπος εντοπίζεται στο χρωµόσωµα Χ αποδίδει έναν περιορισµό στην ανίχνευση µεταλλάξεων που συµβαίνουν στο φυλετικό χρωµόσωµα σχετικά µε τη συµµετοχή του σε οµόλογο χρωµοσωµατικό ανασυνδυασµό που οδηγεί σε απώλεια της ετεροζυγωτίας (Albertini at al., 2)

67 Εισαγωγή Μικροπυρήνες Εικόνα 4-3. Πιθανές βλάβες κυττάρων που έχουν εκτεθεί σε κυτταροτοξικούς και/ή γονιδιοτοξικούς παράγοντες µε τη µέθοδο αναστολής της κυτταροκίνησης (Fenech, 22). Πιθανότατα µελλοντικές προεκτάσεις της µεθόδου αναστολής της κυτταροκίνησης αφορούν την προσθήκη νέων βιολογικών δεικτών σε µελέτες γονιδιοτοξικότητας που σχετίζονται µε τη δυνατότητα διάκρισης γεγονότων είτε γονιδιακής ενίσχυσης είτε ανωµαλιών του κεντροσώµατος (Fenech, 2, 22). 4.6 Μειονεκτήµατα της µεθόδου αναστολής της κυτταροκίνησης Οι περιορισµοί που σχετίζονται µε την εφαρµογή της µεθόδου αναστολής της κυτταροκίνησης αφορούν κυρίως τον τρόπο δράσης της κυτταροχαλασίνης-β, η οποία αναστέλλει την κυτταροκίνηση µε την πρόσδεσή της σε σύµπλοκα µεγάλου µοριακού βάρους στην πλασµατική µεµβράνη που έχουν την ικανότητα να επάγουν τον πολυµερισµό της ακτίνης µε άµεση συνέπεια τη σωστή οργάνωση των µικροϊνιδίων (Fenech, 1997). Τα µικροϊνίδια θεωρούνται απαραίτητα για το σχηµατισµό της περίσφιξης (σχισµή αυλάκωσης) στον ισηµερινό του κυττάρου, γεγονός που επιφέρει την κυτταροπλασµατική διαίρεση και τον πλήρη διαχωρισµό των θυγατρικών κυττάρων. Η αποτελεσµατικότητα µε την οποία η κυτταροχαλασίνη-β αναστέλλει αυτή τη διαδικασία εξαρτάται από την συγκέντρωση της στην καλλιέργεια και η προσθήκη της σε ανάλογες ποσότητες θα πρέπει να εξασφαλίζει την αναστολή της κυτταροκίνησης στο µέγιστο βαθµό. Αξιοσηµείωτο είναι το γεγονός της απουσίας δοσοεξαρτώµενης σχέσης σχηµατισµού µικροπυρήνων σε διπύρηνα και σε πολυπύρηνα κύτταρα και της συγκέντρωσης της κυτταροχαλασίνης-β (Prosser et al., 1988, Lindholm et al., 1991, Suralles et al., 1992). Επιπρόσθετα, η χρήση της ένωσης καθιστά αδύνατη τη µελέτη χηµικών ενώσεων µε ανάλογη δράση, οι οποίες αναστέλλουν την κυτταροκίνηση ή τον πολυµερισµό των µικροϊνιδίων (Fenech, 1997). Η εκτίµηση της ανάλυσης µικροπυρήνων σε πολυπύρηνα κύτταρα δεν ενδείκνυται, ως ασφαλής τρόπος εκτίµησης της γονιδιοτοξικής δράσης χηµικών ενώσεων, πιθανότατα εξαιτίας της υψηλής συχνότητας γεγονότων χρωµοσωµατικής απώλειας που ακολουθούν µετά από επόµενες πυρηνικές διαιρέσεις (Lindholm et al., 1991, Schultz και Önfelt, 1994). Η µέθοδος αναστολής της κυτταροκίνησης εφαρµόζεται αποκλειστικά σε διαιρούµενους κυτταρικούς πληθυσµούς γεγονός το οποίο την αποκλείει ως µεθοδολογία για την εκτίµηση χρωµοσωµατικών βλαβών σε µη διαιρούµενα κύτταρα, όπως για παράδειγµα οι πρωτογενείς καλλιέργειες µυϊκού και νευρικού ιστού (εγκέφαλος) (Fenech, 1997)
68 Εισαγωγή Ανευπλοειδία 5. ΧΡΩΜΟΣΩΜΑΤΙΚΗ ΑΣΤΑΘΕΙΑ - ΑΝΕΥΠΛΟΕΙ ΙΑ 5.1 Αριθµητικές χρωµοσωµατικές ανωµαλίες Η σταθερότητα του καρυοτύπου ενός ευκαρυωτικού οργανισµού η οποία περιλαµβάνει τη διατήρηση του αριθµού και την ακεραιότητα της δοµής των χρω- µοσωµάτων εκφράζει τη σταθερότητα του γενετικού υλικού και εξαρτάται από τη συντονισµένη λειτουργία των κυτταρικών κύκλων διαίρεσης κατά τη διάρκεια της µίτωσης και της µείωσης. Στα σηµαντικά γεγονότα που σχετίζονται µε τη διατήρηση της σταθερότητας περιλαµβάνονται η πιστότητα του χρωµοσωµατικού διπλασιασµού (αντιγραφή του DNA) και η ακρίβεια του διαχωρισµού των χρωµοσωµάτων στα θυγατρικά κύτταρα. Οι διαδικασίες αυτές εξαρτώνται από τη λειτουργικότητα µίας ποικιλίας κυτταρικών οργανιδίων καθώς και από τη διακύµανση συγκεκρι- µένων µεταβολικών διεργασιών της κυτταρική διαίρεσης οι οποίες περιλαµβάνουν τη σύνθεση των πρωτεϊνών της κυτταρικής ατράκτου και την πρόσδεση και κίνηση των χρωµοσωµάτων στη µιτωτική συσκευή. Τροποποιήσεις στη συσκευή κυτταρικής διαίρεσης δύναται να οδηγήσουν σε ανώµαλο διαχωρισµό χρωµοσωµάτων µε αποτέλεσµα τη δηµιουργία θυγατρικών κυττάρων µε λανθασµένο αριθµό χρωµοσωµάτων είτε πολλαπλάσιο του φυσιολογικού (πολυπλοειδία) είτε διακυµάνσεις του κανονικού αριθµού (2N) στις οποίες διαπιστώνεται περίσσεια ή έλλειµµα συγκεκριµένων χρωµοσωµάτων σε χρωµοσωµατικές σειρές (ανευπλοειδία) (Parry et al., 1996). Αριθµητικές αλλαγές που συµβαίνουν κατά τη διάρκεια τη µείωσης δίνουν γένεση, εφόσον επακολουθήσει γονιµοποίηση σε έµβρυα που χαρακτηρίζονται ως ανευπλοειδή ή πολυπλοειδή. Ένα µεγάλο ποσοστό τέτοιων εµβρύων στον άνθρωπο δεν είναι σε θέση να επιβιώσει και έχει υπολογιστεί ότι το 5% των αποβολών στις δώδεκα εβδοµάδες της κύησης, σχετίζεται µε έµβρυα (96%) που φέρουν αριθµητικές χρωµοσωµατικές ανωµαλίες (Parry et al., 1996, Thompson et al., 21, Kirsch- Volders et al., 22). Γενικότερα οι γονιδιακές µεταλλάξεις, οι δοµικές χρωµοσω- µατικές αναδιατάξεις (ελλείµµατα, διπλασιασµοί, µετατοπίσεις, αναστροφές) και οι αριθµητικές αλλαγές (ανευπλοειδία, πολυπλοειδία) συµβάλλουν στη δηµιουργία γενετικών ανωµαλιών. Ένα επιβαρηµένο γενετικό φορτίο θεωρείται σαφώς υπεύθυνο για τις πρόωρες αποβολές εµβρύων, τα νεκρά νεογέννητα και την εµφάνιση απογόνων µε φυσικές, φυσιολογικές, διανοητικές και ψυχολογικές διαταραχές (Kirsch-Volders et al., 22) Επιπτώσεις ανευπλοειδίας Η απόκλιση από το φυσιολογικό χρωµοσωµατικό αριθµό (ανευπλοειδία) έχει σοβαρότατες επιπτώσεις στην ανθρώπινη υγεία. Σε αυτές περιλαµβάνονται γενετικές ασθένειες που σχετίζονται µε καθυστέρηση της σωµατικής και διανοητικής ανάπτυξης, µε τη στειρότητα και µε αυτόµατες αποβολές. Επιπρόσθετα, οι αριθµητικές χρωµοσωµατικές αλλαγές παρατηρούνται συχνά σε διάφορα στάδια της καρκινογένεσης στον άνθρωπο (Aardema et al., 1998). Ο Boveri (1914) ανέφερε τη συσχέτιση που υπάρχει µεταξύ αριθµητικών ανωµαλιών (ανευπλοειδία) και καρκίνου (Brinkley και Goepfert, 1998). Πρόκειται για µία θεωρία η οποία µελετάται από την επιστηµονική κοινότητα και σύµφωνα µε την οποία οποιεσδήποτε τροποποιήσεις επηρεάσουν την ακρίβεια του µηχανισµού διαχωρισµού των χρω- µοσωµάτων οδηγούν σε αστάθεια του καρυοτύπου. Η χρωµοσωµατική αστάθεια σε πρώιµα στάδια της καρκινογένεσης, ίσως αποτελεί την κινητήρια δύναµη που οδηγεί σε αλλαγή του αριθµού των αντιγράφων ενός ή περισσότερων γονιδίων που
69 Εισαγωγή Ανευπλοειδία ελέγχουν την κυτταρική αύξηση και πιθανότατα να επηρεάζει την έκφραση ογκογονιδίων και/ή ογκοκατασταλτικών γονιδίων (Duesberg et al., 1999, Parry et al., 22, Duesberg, 25). Κλινικές µελέτες σε ανθρώπινα γεννητικά κύτταρα υποδεικνύουν τη σηµασία των χρωµοσωµατικών ανωµαλιών οι οποίες ανιχνεύονται σε υψηλή συχνότητα σε γονιµοποιηµένα ωάρια. Ανάµεσα σε διάφορους τύπους που ανιχνεύθηκαν, η ανευπλοειδία (τρισωµία και µονοσωµία) παρατηρείται συχνότερα και θεωρείται, από κλινικής άποψης, αρκετά ενδιαφέρουσα. Σηµαντική είναι η αναφορά της συχνότητας ανευπλοειδικών φαινοµένων σε γαµέτες µε το υψηλότερο ποσοστό (2-25%) να εµφανίζεται σε ωοκύτταρα (Hassold et al., 1996). Επιπλέον, η ανευπλοειδία σε ποσοστό περίπου 7% αποτελεί τον κυριότερο παράγοντα τυχαίων αποβολών στον άνθρωπο (Thompson et al., 21, Kirsch-Volders et al., 22). Ο κυτταρογενετικός έλεγχος περίπου 6. νεογνών ανέδειξε την ανευπλοειδία ως την αριθµητική ανωµαλία που παρουσιάζεται στο.4% των γεννήσεων και τα άτοµα αυτά στην πλειονότητα τους παρουσιάζουν κάποια σηµαντική φυσική ή πνευµατική βλάβη. Η τρισωµία του 21 ου χρωµοσώµατος (Down s Syndrome) παρατηρείται συχνότερα (.1%) και αντιπροσωπεύει την κύρια αριθµητική χρωµοσωµατική ανωµαλία. (Hassold et al., 1996, Aardema et al., 1998, Thompson et al., 21). Συγκεκριµένα, στον άνθρωπο, οι πιο κοινές µορφές ανευπλοειδίας θεωρούνται οι τρισωµίες των χρωµοσωµάτων 13 (Σύνδροµο Patau), 18 (Σύνδροµο Edwards), 21, και η εµφάνιση ενός επιπλέον ή η απώλεια αντιγράφων των φυλετικών χρωµοσωµάτων (Parry et al., 1996). Αξιοσηµείωτη χαρακτηρίζεται η διαφοροποίηση που υπάρχει µεταξύ των διαφόρων τύπων τρισωµιών µε την τρισωµία 16 να είναι ο συνηθέστερος τύπος (3%) σε έµβρυα που αποβάλλονται, ενώ δεν παρατηρείται καθόλου στα νεογνά. Επιπρόσθετα σε τυχαίες αποβολές έχουν αναγνωρισθεί τρισωµίες για όλα τα χρωµοσώµατα εκτός από το χρωµόσωµα 1. Στην πραγµατικότητα αυτές οι διαφορές πιθανότατα αποδίδονται στη διαφορετική συχνότητα µη αποχωρισµού µεταξύ των διαφόρων χρωµοσωµάτων ή στη διαφορική επιλογή έναντι χαρακτηριστικών τρισωµιών. Σχετικά µε περιπτώσεις αυτοσωµατικής µονοσωµίας αυτές δεν παρατηρούνται σε τυχαίες αποβολές διότι η απουσία ενός αυτοσωµατικού χρωµοσώµατος από το ζυγωτό οδηγεί σε τόσο µεγάλη ανισορροπία του γενετικού υλικού, ώστε ακόµα και εάν δηµιουργηθούν τέτοια ζυγωτά, αποβάλλονται πολύ νωρίς. Πιθανόν στην ηµιζυγωτική κατάσταση της µονοσωµίας τα υποτελή θανατογόνα γονίδια εκφράζονται και οδηγούν στη διακοπή της ανάπτυξης του εµβρύου. Αντίθετα, η µονοσωµία του φυλετικού χρωµοσώµατος Χ, η οποία συνδέεται µε το σύνδροµο Turner (45,XΟ), εµφανίζεται στο 1.5% όλων των συλλήψεων και ανιχνεύεται σε υψηλή συχνότητα (1-18%) στο σύνολο των τυχαίων αποβολών που οφείλονται σε χρωµοσωµατικές ανωµαλίες. Σε σχέση µε άλλες ανευπλοειδίες των φυλετικών χρωµοσωµάτων (47,ΧΧΥ, 47,ΧΥΥ, 47,ΧΧΧ), το σύνδροµο Turner απαντάται λιγότερο συχνά σε νεογέννητα, ενώ εξαιτίας της απενεργοποίησης του χρωµοσώµατος Χ οι ανευπλοειδίες των φυλετικών χρωµοσω- µάτων είναι πιο συχνές σε σύγκριση µε τις ανευπλοειδίες των αυτοσωµάτων (Aardema et al., 1998, Thompson et al., 21). Πολυάριθµες πληροφορίες που προέρχονται από τη µελέτη όγκων σε διαφορετικούς ιστούς και κυτταρικούς τύπους υποστηρίζουν την άποψη που θεωρεί συγκεκριµένες αριθµητικές χρωµοσωµατικές αλλαγές (ανευπλοειδία) σε σωµατικά κύτταρα να διαδραµατίζουν σηµαντικό ρόλο στην παθολογία αρκετών µορφών καρκίνου. Συγκεκριµένα, στη χρόνια µυελογενή λευχαιµία (CML) έχει παρατηρηθεί τρισωµία 8, 17q, 19 και επιπλέον αντίγραφα της 9:22 µετατόπισης (χρωµόσωµα Ph 1 ), ενώ στη χρόνια λεµφοκυτταρική λευχαιµία (CLL) τρισωµία
70 Εισαγωγή Ανευπλοειδία Σε πρωτογενείς και δευτερογενείς µορφές της οξείας µη λεµφοκυτταρικής λευχαιµίας (ANL) συναντάται µονοσωµία των φυλετικών χρωµοσωµάτων Χ ή Υ και µονοσωµία 5, 7, ενώ στην οξεία λεµφοβλαστική λευχαιµία (ALL), η τρισωµία 21. Σε αρκετούς στερεούς όγκους η ανευπλοειδία είναι δυνατόν να συσχετισθεί µε εξειδικευµένους τύπους που περιλαµβάνουν µεγάλα καρκινώµατα του εντέρου (τρισωµία 7, 12), καρκινώµατα της ουροδόχου κύστης (τρισωµία 7, µονοσωµία 11p, 9, 6p, 14q, 17p) και των νεφρών (µονοσωµία 3p), καθώς και το µικροκυτταρικό (µονοσωµία 3p, 13q, 17p) και µη (τρισωµία 7) καρκίνωµα των πνευµόνων. Η µελέτη του ειδικού ρόλου της ανευπλοειδίας σε πρώιµα στάδια της καρκινογένεσης καθίσταται δυνατή από αναλύσεις γονιδίων που εµπλέκονται σε αυτή τη διαδικασία. Τα ογκοκατασταλτικά γονίδια χαρακτηρίζονται ως η κύρια κατηγορία γονιδίων που εµπλέκονται άµεσα στην καρκινογένεση και σε περιπτώσεις απώλειας και των δύο αλληλοµόρφων ογκοκατασταλτικών γονιδίων δεν έχουµε εκδήλωση της κατασταλτικής δράσης σε διαδικασίες που οδηγούν το κύτταρο σε αύξηση και διαίρεση. Αυτό που συµβαίνει είναι µία µετάλλαξη σε ένα αντίγραφο του φυσιολογικού γονιδίου που συνοδεύεται από απώλεια του άλλου (loss of heterozygosity) µε χρωµοσωµατικούς µηχανισµούς στους οποίους περιλαµβάνονται ο µη αποχωρισµός (χρωµοσωµατική απώλεια), ο µιτωτικός ανασυνδυασµός και το έλλειµµα. Πρόκειται για τους συνηθέστερους µηχανισµούς µετάλλαξης µέσω των οποίων πραγµατοποιείται η απώλεια του φυσιολογικού αλληλοµόρφου σε ετεροζυγώτες. Η ανάλυση πολυµορφισµών του DNA σε περιοχές που βρίσκονται κοντά στο γενετικό τόπο RB1, σε κληρονοµικούς καρκίνους ή µη, από ασθενείς µε γλοίωµα του αµφιβληστροειδούς (ρετινοβλάστωµα) έδειξε ότι ενώ άτοµα από τα οποία είχαν ληφθεί οι όγκοι ήταν ετερόζυγα σε πολλούς γενετικούς τόπους, οι όγκοι ήταν οµόζυγοι στους ίδιους γενετικούς τόπους και περιείχαν αλληλόµορφα του ενός µόνο από τα δύο οµόλογα του χρωµοσώµατος 13. Επιπρόσθετα, η απώλεια ετεροζυγωτίας αποτελεί χαρακτηριστικό του όγκου του Wilms και ενός µεγάλου αριθµού άλλων όγκων σε αρκετούς από τους οποίους εµπλέκονται τα ογκοκατασταλτικά γονίδια APC (adenomatous polyposis coli), DCC (deleted in colon carcinoma) και MCC (mutated in colorectal cancer). Τα ευρήµατα αυτά υποδεικνύουν την ανευπλοειδία ως ένα πρώιµο και αρχικό γεγονός σε διάφορες µορφές καρκίνου. Μία άλλη κατηγορία γονιδίων που συµπεριφέρονται ως υποτελή και ενέχονται στην καρκινογένεση πιθανότατα µέσω της ενεργοποίησης τους µε χρωµοσωµατική απώλεια θεωρούνται τα γονίδια µεταλλάκτες (mutator genes). Σε αντίθεση η ενεργοποίηση µίας άλλης σηµαντικής κλάσης γονιδίων που εµπλέκονται στην καρκινογένεση, τα πρωτοογκογονίδια, επιτυγχάνεται µε σηµειακή µετάλλαξη και µικρές ενθέσεις ή ελλείµµατα, ενώ θεωρείται απίθανο να συµβαίνει τουλάχιστον σε σηµαντική έκταση µε αριθµητικές χρωµοσωµατικές αλλαγές. Επιπρόσθετα, ενδείξεις του ρόλου της ανευπλοειδίας σε κατοπινά στάδια της καρκινογένεσης αποτελεί η περίπτωση της χρόνιας µυελογενούς λευχαιµίας που ως εναρκτήριο γεγονός θεωρείται η χρωµοσωµατική µετατόπιση µε αποτέλεσµα τη δηµιουργία του χρωµοσώµατος Φιλαδέλφεια, ενώ η εξέλιξη της ασθένειας σχετίζεται µε την επαγωγή ειδικών υπερπλοειδικών αλλαγών (Alberts et al., 1994, Parry et al., 1996, Aardema et al., 1998, Thompson et al., 21)
71 Εισαγωγή Ανευπλοειδία 5.2 Μηχανισµοί ανευπλοειδίας Η ανευπλοειδία αναφέρεται σε µία αλλαγή στον αριθµό των χρωµοσωµάτων σε ένα κύτταρο, η οποία προκύπτει ως αποτέλεσµα µηχανισµών που οδηγούν σε περίσσεια ή απώλεια ενός ή περισσοτέρων χρωµοσωµάτων κατά τη διάρκεια της κυτταρικής διαίρεσης. Αντίθετα, η πολυπλοειδία καθορίζεται από την αύξηση του αριθµού των χρωµοσωµάτων κατά ένα πολλαπλάσιο αριθµό του απλοειδούς χρω- µοσωµατικού αριθµού (3Ν, 4Ν,..). Υπάρχουν δύο κύριοι µηχανισµοί που οδηγούν σε ανευπλοειδία και είναι ο µη αποχωρισµός των χρωµοσωµάτων και η χρωµοσωµατική απώλεια. Επιπρόσθετα µε τους δύο κλασσικούς µηχανισµούς (χρωµοσωµατικός µη αποχωρισµός, χρωµοσω- µατική απώλεια), η ανευπλοειδία είναι δυνατόν να προκύψει και µέσω άλλων µηχανισµών στους οποίους περιλαµβάνονται: αδυναµία σύζευξης (non-conjunction) των χρωµοσωµάτων, σε περίπτωση αδυναµίας των οµολόγων χρωµοσωµάτων να τοποθετηθούν στο ισηµερινό επίπεδο ελαττωµατική διαίρεση των κεντροµερών που οδηγεί σε λανθασµένο αποχωρισµό στη µειωτική διαίρεση επιπρόσθετη αντιγραφή του χρωµοσώµατος που οφείλεται σε λάθος στην αντιγραφή του χρωµοσώµατος κατά τη µείωση, µε αποτέλεσµα την εµφάνιση ενός επιπλέον χρωµοσώµατος (Kirsch-Volders et al., 22) Χρωµοσωµατικός µη αποχωρισµός O χρωµοσωµατικός µη αποχωρισµός (chromosome non-disjunction) συµβαίνει κατά την ανάφαση της µίτωσης ή της µείωσης µε αποτέλεσµα το ένα θυγατρικό κύτταρο να είναι τρισωµικό και το άλλο µονοσωµικό. Στην περίπτωση που ο διαχωρισµός είναι συµµετρικός τότε προκύπτουν θυγατρικά κύτταρα από τα οποία το ένα κύτταρο περιέχει ένα επιπλέον χρωµόσωµα (τρισωµικό 2n+1 κατά τη µίτωση ή δισωµικό n+1 κατά τη µείωση) και στο άλλο παρατηρείται απώλεια ενός χρωµοσώµατος (µονοσωµικό 2n-1 στη µίτωση ή ασωµικό n-1 στη µείωση) (Parry et al., 1996). Μειωτικός µη αποχωρισµός. Η αποτυχία ενός ζεύγους οµολόγων χρωµοσωµάτων να διαχωριστούν φυσιολογικά δύναται να συµβεί κατά τη διάρκεια της µίας από τις δύο µειωτικές διαιρέσεις. Οι συνέπειες του µη αποχωρισµού είτε κατά τη µείωση Ι είτε κατά τη µείωση ΙΙ διαφέρουν. Εάν ο µη αποχωρισµός συµβεί στη µείωση Ι τα µισά θυγατρικά κύτταρα θα περιέχουν ταυτόχρονα τα πατρικά και τα µητρικά µέλη του ζεύγους (24 χρωµοσώµατα), ενώ τα υπόλοιπα µισά δεν θα περιέχουν κανένα (22 χρωµοσώµατα), έναντι του φυσιολογικού αριθµού (23 χρωµοσώµατα), µε αποτέλεσµα όλοι οι γαµέτες που θα προκύψουν να είναι ανώµαλοι. Το ζυγωτό που προέρχεται από τη σύντηξη ενός τέτοιου γαµέτη µε ένα φυσιολογικό θα φέρει αντίστοιχα 47 ή 45 χρωµοσώµατα και όλα τα κύτταρα του ατόµου που προκύπτουν από αυτό το ζυγωτό είναι ανευπλοειδή. Εάν ο µη αποχωρισµός συµβεί στη µείωση ΙΙ ο γαµέτης µε το επιπλέον χρωµόσωµα περιέχει δύο αντίγραφα είτε του πατρικού είτε του µητρικού χρωµοσώµατος µε κάποιες µικρές διαφορές µεταξύ των χρωµατιδίων εξαιτίας του ανασυνδυασµού που πραγµατοποιείται κατά την προγενέστερη µείωση Ι και σε αυτήν την περίπτωση οι µισοί γαµέτες είναι ανώµαλοι και οι µισοί φυσιολογικοί. Μιτωτικός µη αποχωρισµός: Μη αποχωρισµός δύναται να συµβεί σε µιτωτική διαίρεση µετά το σχηµατισµό ενός φυσιολογικού ζυγωτού. Τα αποτελέσµατα του µιτωτικού µη αποχωρισµού στον φαινότυπο εξαρτώνται από τα χρωµοσώµατα που - 6 -
72 Εισαγωγή Ανευπλοειδία λαµβάνουν µέρος και από το στάδιο ανάπτυξης στο οποίο συµβαίνει η ανευπλοειδία. Εάν τα θυγατρικά κύτταρα καταφέρουν να επιβιώσουν και συνεχίσουν να διαιρούνται δίνουν γένεση σε ανευπλοειδικά κύτταρα και το άτοµο που προκύπτει απαρτίζεται από κύτταρα µε διαφορετικό αριθµό χρωµοσωµάτων και ονοµάζεται µωσαϊκό. Σε περίπτωση που ο µιτωτικός µη αποχωρισµός συµβεί στην πρώτη αυλάκωση του ζυγωτού όλοι οι ιστοί του ατόµου αποτελούνται από ανευπλοειδικά κύτταρα (τρισωµικά και µονοσωµικά), ενώ εάν συµβεί στη δεύτερη αυλάκωση τα µισά από τα θυγατρικά κύτταρα είναι ανευπλοειδικά (Thompson et al., 21) Χρωµοσωµατική απώλεια Η καθυστέρηση της µετακίνησης ενός ή περισσοτέρων χρωµοσωµάτων στα ινίδια της ατράκτου και η αποτυχία ενσωµάτωσης τους στη µιτωτική συσκευή κατά την ανάφαση της πυρηνικής διαίρεσης έχει ως αποτέλεσµα την απώλεια και τον αποκλεισµό τους από τον πυρήνα του θυγατρικού κυττάρου. Με τη δηµιουργία πυρηνικής µεµβράνης κατά την τελόφαση είναι δυνατόν να σχηµατιστεί ένας µικροπυρήνας, ο οποίος παρατηρείται µε την εφαρµογή κυτταρογενετικών µεθόδων. Η χρωµοσωµατική απώλεια (chromosome loss) συµβαίνει κατά την κυτταρική διαίρεση και οδηγεί στη δηµιουργία ενός µονοσωµικού και ενός φυσιολογικού θυγατρικού κυττάρου. Εναλλακτικά, ο µικροπυρήνας µπορεί τυχαία να ενσωµατωθεί ξανά σε έναν από τους θυγατρικούς πυρήνες, έτσι ώστε το ένα θυγατρικό κύτταρο να είναι τρισωµικό και το άλλο µονοσωµικό ή και τα δύο θυγατρικά κύτταρα να περιέχουν το σωστό αριθµό χρωµοσωµάτων (διπλοειδή κύτταρα) (Parry et al., 22). Αξιοσηµείωτες είναι οι µελέτες που προσπαθούν να αποσαφηνίσουν την πορεία του µικροπυρήνα σύµφωνα µε τις οποίες είναι δυνατόν είτε να αποβληθεί από το κύτταρο είτε να ενσωµατωθεί σε έναν από τους πυρήνες κατά την µίτωση είτε να οδηγήσει σε κυτταρικό θάνατο. Γενικότερα, τα φαινόµενα χρωµοσωµατικής απώλειας (micronucleation) αντανακλούν την καθυστέρηση των χρωµοσωµάτων κατά τη φάση διαχωρισµού τους στην ανάφαση και δηµιουργούν µονοσωµίες (Parry et al., 22, Norppa και Falck, 23). 5.3 Στόχοι δράσης ανευπλοειδογόνων ενώσεων Η ανευπλοειδία είναι δυνατόν να προκληθεί είτε τυχαία είτε µέσω χηµικής ή φυσικής επαγωγής κατά τη διάρκεια της κυτταρικής διαίρεσης σε γεννητικά και σωµατικά κύτταρα. Οι χηµικές ενώσεις που προκαλούν ανευπλοειδία ή πολυπλοειδία ονοµάζονται ανευπλοειδογόνες (aneugens) και οι πιθανοί στόχοι τους είναι µόρια τα οποία: συµµετέχουν στην πρόσδεση και το διαχωρισµό των χρωµατιδίων (συ- µπύκνωση χρωµοσώµατος, διασκελισµός, κινητοχώροι, πρωτεΐνες συνάφειας χρωµατιδίων) αποτελούν µέρος βασικών δοµών του γενετικού υλικού (κεντροµέρη, τελοµέρη) συµµετέχουν στον έλεγχο του κυτταρικού κύκλου (κυκλίνες, Cdk s, p53, APC- Anaphase Promoting Complex) συνιστούν βασικά στοιχεία της µιτωτικής συσκευής (τουµπουλίνες, MAPs, κεντριόλια) συνδέονται έµµεσα µε τον κυτταρικό κύκλο (καλµοδουλίνη, κυτταρική ή πυρηνική µεµβράνη) (Kirsch-Volders et al., 22). Αναµφισβήτητα, η επαγόµενη από χηµικές ενώσεις ανευπλοειδία δεν δύναται να αποδοθεί σε έναν µόνο µοριακό µηχανισµό. Ο σωστός διαχωρισµός των
73 Εισαγωγή Ανευπλοειδία χρωµοσωµάτων εξαρτάται από αρκετά διαφορετικά οργανίδια και ελέγχεται από ένα αριθµό µεταβολικών οδών γεγονός που καθιστά την διαδικασία ευπρόσβλητη σε µία ποικιλία διαταραχών µε αποτέλεσµα η ανευπλοειδία να προκύπτει από διαφορετικούς µηχανισµούς κατά τη διάρκεια του κυτταρικού κύκλου (κυρίως κατά τη φάση Μ-Μίτωση). Σε αυτούς περιλαµβάνονται βλάβες σε βασικά στοιχεία της λειτουργίας του χρωµοσώµατος, µείωση της ικανότητας των χρωµοσωµάτων να ζευγαρώνουν, αλλαγές στη δοµή των χρωµοσωµάτων (χρωµοσωµατικές ενδοανταλλαγές), επιδράσεις στη συµπύκνωση των χρωµοσωµάτων, παραµονή των πυρηνίσκων στη µίτωση ή µείωση, αυξηµένη ικανότητα πρόσδεσης των χρωµοσωµάτων, βλάβες στα κεντριόλια ή στους κινητοχώρους, µεταβολές στη συγκέντρωση ιόντων (Ca ++ ) κατά τη µίτωση και βλάβη στην πυρηνική µεµβράνη. Τέτοιες τροποποιήσεις είναι σε θέση να προκαλούν ανωµαλίες στην τοποθέτηση των χρωµοσωµάτων στο ισηµερινό επίπεδο και να διαταράσσουν τους φυσιολογικούς µηχανισµούς διάσπασης της συνάφειας των αδελφών χρωµατιδίων επιφέροντας χρωµοσωµατική καθυστέρηση. Ιδιαίτερη σηµασία στην αδυναµία των χρωµοσωµάτων να διαχωριστούν µε ακρίβεια λαµβάνει η µιτωτική συσκευή µε τα κύρια λειτουργικά στοιχεία της να αποτελούν πιθανούς στόχους ανευπλοειδογόνων ενώσεων. Τροποποιήσεις στη σύνθεση και στη συγκρότηση των µικροσωληνίσκων που θεωρείται απαραίτητη διαδικασία για το σχηµατισµό µίας λειτουργικής κυτταρικής ατράκτου σε συνδυασµό µε διαταραχές στη σύνθεση, διαίρεση και λειτουργικότητα των κεντροσωµατίων, καθώς και στη συγκρότηση και λειτουργικότητα των πρωτεϊνών του κινητοχώρου και του κεντροµέρους των χρωµοσωµάτων επιφέρουν ανώµαλη κυτταρική διαίρεση µε αποτέλεσµα τη δηµιουργία κυττάρων µε λανθασµένο αριθµό χρωµοσωµάτων (ανευπλοειδία). Άλλα εξίσου σηµαντικά στοιχεία για την ορθή λειτουργία της κυτταρικής ατράκτου θεωρούνται οι µεµβράνες που περιβάλλουν προστατευτικά την περιφέρεια της ατράκτου, οι εξαρτηµένες από c-amp κινάσες και τα ιόντα ασβεστίου. Σχετικά µε τη µειωτική κυτταρική διαίρεση µία σειρά τροποποιήσεων σε πρόσθετους µηχανισµούς που περιλαµβάνουν τη σύναψη των οµολόγων χρωµοσωµάτων µε την εµφάνιση του συναπτονηµατικού συµπλέγµατος, το διασκελισµό και το διαφορετικής χρονικής διάρκειας µειωτικό κύκλο σε αρσενικά και θηλυκά άτοµα επιφέρουν το λανθασµένο αποχωρισµό των χρωµοσωµάτων (ανευπλοειδία) (Parry et al., 1996, Aardema et al., 1998). 5.4 Παράγοντες που διαταράσσουν το σωστό διαχωρισµό του γενετικού υλικού και σχετίζονται µε βασικά και ρυθµιστικά στοιχεία της µιτωτικής συσκευής Η σωστή οργάνωση και λειτουργία της µιτωτικής ατράκτου απαιτεί µία πολυσύνθετη ακολουθία συνδεδεµένων χρονικά και χωρικά γεγονότων και στηρίζεται σε µία αλυσίδα ρυθµιστικών µονοπατιών που ελέγχουν τα γεγονότα αυτά, τα οποία είναι κατάλληλα συγχρονισµένα κατά τη διάρκεια του κυτταρικού κύκλου. Τα γεγονότα που συµβάλλουν στην ορθή συγκρότηση της ατράκτου για την εκκίνηση της κυτταρικής διαίρεσης στα περισσότερα ανθρώπινα κύτταρα περιλαµβάνουν τη διάλυση του πυρηνικού φακέλου, τον αποπολυµερισµό των µεσοφασικών µικροσωληνίσκων, τη διαµεσολαβούµενη από κεντροσώµατα πυρήνωση νέων µικροσωληνίσκων, το διπλασιασµό και σωστό διαχωρισµό των κεντροσωµάτων και τη συµπύκνωση, ευθυγράµµιση, συνοχή και κίνηση των χρωµοσωµάτων (Pihan και Doxsey, 1999)
74 Εισαγωγή Ανευπλοειδία Μικροσωληνίσκοι Οι µικροσωληνίσκοι συνιστούν τα πιο βασικά στοιχεία για τη δόµηση και λειτουργία της µιτωτικής ατράκτου και φέρουν ως κύρια δοµική µονάδα το µόριο της τουµπουλίνης. Η πρόσδεση χηµικών ενώσεων σε ειδικές θέσεις στο µόριο της τουµπουλίνης διακόπτει αποτελεσµατικά τη διαδικασία της µιτωτικής διαίρεσης και κατά συνέπεια τον κυτταρικό πολλαπλασιασµό είτε µέσω αποπολυµερισµού των µικροσωληνίσκων είτε µέσω τροποποίησης της δυναµικής τους. Γενικότερα, έχουν αναγνωριστεί συγκεκριµένες θέσεις που αποτελούν θέσεις πρόσδεσης για δινουκλεοτίδια γουανοσίνης, για την κολχικίνη (δεµεκολσίνη) και αλκαλοειδή φυτών (βινκριστίνη, βινµπλαστίνη). Οι τελευταίες ονοµάζονται αντιµιτογόνες ενώσεις και ανήκουν στην κατηγορία των αλκαλοειδών που χρησιµοποιούνται ευρέως στη χηµειοθεραπεία του καρκίνου. Η πρόσδεση της κολχικίνης σε µία ειδική θέση δέσµευσης στη β υποµονάδα της τουµπουλίνης παρεµποδίζει την προσθήκη νέων υποµονάδων στο (+) άκρο του µικροσωληνίσκου µε αποτέλεσµα να διακόπτει τον πολυµερισµό τους και να αναστέλλει την ορθή συγκρότηση της µιτωτικής ατράκτου. Η βινµπλαστίνη και η βινκριστίνη δεσµεύονται σε µία θέση διαφορετική από αυτή της κολχικίνης και προκαλούν κρυσταλλοποίηση της τουµπουλίνης µε αποτέλεσµα τη συσσωµάτωση των µικροσωληνίσκων. Αντίθετα, η ταξόλη προκαλεί παύση του κυτταρικού κύκλου στη φάση της µίτωσης και πιθανόν να οδηγεί σε ανευπλοειδία µε µία χαρακτηριστική δράση που σχετίζεται µε την αύξηση του πολυµερισµού της τουµπουλίνης και τη σταθεροποίηση των µικροσωληνίσκων (Önfelt, 1986, Aardema et al., 1998, Pihan και Doxsey, 1999). Πολυάριθµες χηµικές ενώσεις που επηρεάζουν τη λειτουργία των µικροσωληνίσκων δύναται να επάγουν ανευπλοειδία, ενώ µικρού µεγέθους αλλαγές στις ιδιότητες των µικροσωληνίσκων είναι δυνατόν να προκαλέσουν µη αποχωρισµό των χρωµοσωµάτων. Επιπρόσθετα, κυτταρικά συστατικά που επηρεάζουν τον πολυµερισµό, τη δυναµική και τη σταθερότητα των µικροσωληνίσκων συµβάλλουν στη δυσλειτουργία της ατράκτου (Pihan και Doxsey, 1999). Το βενζιµιδαζολικό µυκητοκτόνο θειαβενδαζόλη (TBZ) (Pisano et al., 2), η νοκοδαζόλη (Musa et al., 23) και το νιτριλοτριοξικό οξύ (NTA), ένας χειλικός παράγοντας που συναντάται σε απορρυπαντικά οικιακής χρήσης, (Modesti et al., 1995) θεωρούνται ανευπλοειδογόνες ενώσεις που επηρεάζουν τον πολυµερισµό των τουµπουλινών και τη συγκρότηση των µικροσωληνίσκων. Η ένυδρη χλωράλη αναστέλλει το σχηµατισµό των πολικών µικροσωληνίσκων µε αποτέλεσµα να εµποδίζει την επιµήκυνση της ατράκτου και να προκαλεί ανευπλοειδία (Dellarco et al., 1986), ενώ οι διφαινόλες (BP-A) τροποποιούν τη διαµόρφωση του µορίου της τουµπουλίνης µε αποτέλεσµα η τροποποιηµένη τουµπουλίνη να σχηµατίζει πρωτοϊνίδια τα οποία δεν δύναται να συγκροτήσουν το φυσιολογικό µικροσωληνίσκο (Pfeiffer et al., 1997). Οι αλκυλιωτικοί παράγοντες (MNNG, 1,2-διχλωροαιθάνιο, κυκλοφωσµαµίδιο) ως ισχυρά ηλεκτρονιόφιλοι, εκτός από τη σηµαντική ικανότητα που εµφανίζουν να αντιδρούν µε πυρηνόφιλες θέσεις στο µόριο του DNA και να προκαλούν ένα χαρακτηρισµένο φάσµα µεταλλάξεων µε αποτέλεσµα τη δηµιουργία ρηγµάτων (χρωµοσωµατική θραύση) και χρωµοσωµατικών αναδιατάξεων είναι σε θέση να προσβάλλουν ανάλογες θέσεις σε πρωτεΐνες και επιπρόσθετα να προκαλούν ανευπλοειδία. Γενικότερα στα γεγονότα αυτά περιλαµβάνονται η αλκυλίωση πρωτεϊνών της κυτταρικής ατράκτου, του κινητοχώρου, MAP πρωτεϊνών και προδρόµων µορίων GTP (Dellarco et al., 1986). Μία νέα κατηγορία χηµειοθεραπευτικών παραγόντων (BCEU) σχεδιάστηκε µε χαρακτηριστική δράση που εντοπίζεται στην αλκυλίωση (µονοενεργός) που προκαλείται από το 2-χλωροαίθυλοαµινο τµήµα της χηµικής ένωσης
75 Εισαγωγή Ανευπλοειδία στη θέση Cys239 της β-τουµπουλίνης γεγονός που επιφέρει πλήρη αποπολυµερισµό των µικροσωληνίσκων και παύση του κυτταρικού κύκλου (G2/M) (Legault et al., 2). Σε ορισµένες χηµικές ενώσεις που πιθανότατα προκαλούν ανευπλοειδία εντοπίζεται η δράση τους στην τροποποίηση πρωτεϊνών που διαδραµατίζουν βασικό ρόλο κατά την πυρηνική διαίρεση. Η ένωση π-φθοριοφαινυλαλανίνη (p-fpa) στηρίζει τη δράση της ως ενός αναλόγου φαινυλαλανίνης που επηρεάζει την τεταρτοταγή δοµή της πρωτεΐνης και κατά συνέπεια τη λειτουργία της. Οργανικές ενώσεις που περιέχουν υδράργυρο (θυµεροζάλη) δεσµεύονται στις σουλφυδρυλικές (-SH) οµάδες των πρωτεϊνών και επηρεάζουν τη συγκρότηση των µικροσωληνίσκων (Aardema et al., 1998). Επιπρόσθετα, αλλαγές στην έκφραση των τουµπουλινών (υποµονάδων µικροσωληνίσκων) και µεταλλάξεις στα γονίδια τους προκαλούν ανω- µαλίες στον αποχωρισµό των χρωµοσωµάτων. Ένας µεγάλος αριθµός πρωτεϊνών που άµεσα ή έµµεσα αλληλεπιδρούν µε τους µικροσωληνίσκους, προκαλούν χρωµοσω- µατικό µη αποχωρισµό στην περίπτωση που είτε υπερεκφράζονται, είτε υφίστανται µεταλλάξεις και δυσλειτουργούν (Pihan και Doxsey, 1999). Αναµφισβήτητα, µεταβολές στη συγκρότηση της δοµής των µικροσωληνίσκων αντανακλούν ανωµαλίες στην οργάνωση της µιτωτικής ατράκτου µε συνέπεια την έκφραση ανευπλοειδικών φαινοµένων. Έχουν αναπτυχθεί µεθόδοι (ραδιοσήµανση, ανοσοκατακρήµνιση) παρακολούθησης της συγκρότησης της δοµής των µικροσωληνίσκων οι οποίες βασίζονται στην ικανότητα µιας χηµικής ένωσης να µεταβάλλει τη δοµή των µικροσωληνίσκων αναστέλλοντας της σύνθεση της β-τουµπουλίνης µε την παρουσία στο κυτταρόπλασµα µονοµερών τουµπουλίνης. Με τον τρόπο αυτό παρακολουθείται η αποσυναρµολόγηση των µικροσωληνίσκων παρουσία της υπό εξέταση χηµικής ένωσης. Συγκεκριµένα, παρουσία κολχικίνης και DES (diethylstilboestrol), µειώνεται ο ρυθµός σύνθεσης της β-τουµπουλίνης σε ποσοστό µικρότερο από το 3% συγκριτικά µε το µάρτυρα. Επιπρόσθετα, παρατηρήθηκε µειωµένη σύνθεση της β- τουµπουλίνης σε κύτταρα αρουραίου (Cl-1) παρουσία της ένωσης διαζεπάµης. Πρόκειται για ένα µη αναµενόµενο αποτέλεσµα αφού η κύρια δράση της ένωσης αφορά την αναστολή διαχωρισµού των κεντριολίων κατά το στάδιο της πρόφασης. Πιθανότατα, η µείωση που παρατηρείται να είναι αποτέλεσµα µίας έµµεσης επίδρασης στη συγκρότηση της δοµής των µικροσωληνίσκων και δεν αντανακλά την άµεση συνέπεια της κύριας δράσης της (Parry et al., 1996) Κινητικές πρωτεΐνες Οποιαδήποτε διάσπαση της φυσιολογικής δέσµευσης των κινητικών πρωτεϊνών (δυνεΐνες, κινεσίνες) των µικροσωληνίσκων που θα επιφέρει διαταραχές στην κίνηση των χρωµοσωµάτων και της κυτταρικής ατράκτου κατά τη µίτωση επάγει ανευπλοειδικά φαινόµενα. Ο κύριος ρόλος των κινητικών πρωτεϊνών είναι η παραγωγή κατευθυντήριας δύναµης σε κάθε άκρο του µικροσωληνίσκου και θεωρούνται υπεύθυνες για τη σύγκλιση και αποχωρισµό των χρωµοσωµάτων, την επιµήκυνση της ατράκτου κατά την ανάφαση-β, το διαχωρισµό των κεντροσωµάτων µετά το διπλασιασµό τους και τη θέση σχηµατισµού της ατράκτου. Ανωµαλίες σε κινητικές πρωτεΐνες που συµµετέχουν στη µετακίνηση των χρωµοσωµάτων είναι δυνατόν να επηρεάσουν την ευθυγράµµιση και τον αποχωρισµό των χρωµοσωµάτων. Μικροένεση αντισωµάτων έναντι της πρωτεΐνης (CENP-E) που εδράζεται στην εξωτερική επιφάνεια του κινητοχώρου και θεωρείται απαραίτητη για την κίνηση των χρωµοσωµάτων αναστέλλει την ευθυγράµµιση των χρωµοσωµάτων στο ισηµερινό επίπεδο. Παρόµοιο αποτέλεσµα επιφέρει η υπερέκφραση της δυναµιτίνης, που αποτελεί υποµονάδα του συµπλέγµατος της δυνακτίνης µε την αποµάκρυνση

76 Εισαγωγή Ανευπλοειδία της δυνεΐνης από τους κινητοχώρους. Οι πρωτεΐνες CENP-E και δυνεΐνη εµπλέκονται στο διαχωρισµό των χρωµοσωµάτων κατά την ανάφαση µε αποτέλεσµα βλάβη στις πρωτεΐνες αυτές να προκαλεί λανθασµένο διαχωρισµό των χρωµοσω- µάτων. Αντίθετα, βλάβες σε κινητικές πρωτεΐνες που δεν εδράζονται στους κινητοχώρους και είναι παρόµοιας δοµής µε τις κινεσίνες (Klps), όπως η πρωτεΐνη Νod, προκαλούν πρόωρη µετακίνηση των χρωµοσωµάτων προς τους πόλους. Στον άνθρωπο έχουν παρατηρηθεί σε κύτταρα ρετινοβλαστώµατος πρωτεΐνες παρόµοιας δοµής µε την κινεσίνη (χρωµοκινεσίνη) να εµφανίζονται απορυθµισµένες, ενώ δεν έχει διευκρινιστεί εάν γενικότερα οι κινητικές πρωτεΐνες διαδραµατίζουν σηµαντικό ρόλο στην καρκινογένεση. Σε µία άλλη διαδικασία κατά τη µίτωση που αφορά το διαχωρισµό των κεντροσωµάτων διαµεσολαβούν οι κινητικές πρωτεΐνες µε χαρακτηριστικές αλληλεπιδράσεις µε τους µικροσωληνίσκους που εδράζονται στο κεντρόσωµα. Η αδυναµία διαχωρισµού των κεντροσωµάτων πιθανότατα να προκαλεί τη δηµιουργία µονοπολικής ατράκτου µε αποτέλεσµα την αδυναµία ολοκλήρωσης της µίτωσης και την επαγωγή πολυπλοειδών και ανευπλοειδών κυττάρων, ενώ οποιαδήποτε διαταραχή στη διαδικασία θεωρείται υπεύθυνη για το σχηµατισµό ελαττωµατικής λειτουργικά διπολικής ατράκτου (Pihan και Doxsey, 1999). Αξιοσηµείωτη είναι η αναφορά για τη δράση συγκεκριµένου µυκητοκτόνου (griseofulvin) που διαταράσσει την κίνηση των χρωµοσωµάτων, εξαιτίας της ικανότητας του να αναστέλλει τη σύνδεση των µικροσωληνίσκων µε ειδικές πρωτεΐνες απαραίτητες για την ολίσθηση των χρωµοσωµάτων (Oshimura και Barrett, 1986) Κεντρόσωµα Το κεντρόσωµα θεωρείται το υπεύθυνο οργανίδιο για το σχηµατισµό και την κατανοµή των µικροσωληνίσκων και είναι γνωστό για τα κύτταρα των περισσοτέρων οργανισµών ως κέντρο οργάνωσης των µικροσωληνίσκων (MTOC). Ο ρόλος των κέντρων οργάνωσης των µικροσωληνίσκων είναι ο έλεγχος του αριθµού και της πολικότητας των µικροσωληνίσκων που σχηµατίζουν σε συνδυασµό µε τη χρονική διάρκεια και τη θέση του πολυµερισµού. Ανάµεσα σε µία πλειάδα πρωτεϊνών που εντοπίζονται στο κεντρόσωµα, η κοινή πρωτεΐνη που συµµετέχει ενεργά στην πυρήνωση και πολικότητα των µικροσωληνίσκων είναι η γ-τουµπουλίνη. Η σωστή λειτουργία και ακεραιότητα της πυρηνικής ατράκτου για την ακριβή κατανοµή των χρωµοσωµάτων στα θυγατρικά κύτταρα βασίζεται στο διπλασιασµό (φάση S) και στο σωστό διαχωρισµό και αποµάκρυνση των δύο κεντροσωµάτων (G2/M) προς τους πόλους του κυττάρου. ιπολική µιτωτική άτρακτος Κυτταροκίνηση Ωρίµανση και διαχωρισµός ιπλασιασµός Κεντρόσωµα Ζεύγος κεντριολίων Προσανατολισµός κεντριολίων Εικόνα 5-1. Σχηµατική απεικόνιση του κεντροσωµικού κύκλου, διπλασιασµός του κεντροσώ- µατος, σε σχέση µε τον κυτταρικό κύκλο (Fukasawa, 25)
77 Εισαγωγή Ανευπλοειδία Η παρουσία µη φυσιολογικού αριθµού κεντροσωµάτων (>2) διαταράσσει δραµατικά τη διαδικασία της µίτωσης και την κυτταροκίνηση διαµέσου του σχηµατισµού ατράκτου που φέρει περισσότερους από δύο πόλους µε αποτέλεσµα αυξηµένη συχνότητα λαθών κατά το διαχωρισµό των χρωµοσωµάτων γεγονός που επάγει χρωµοσωµατική αστάθεια (CIN) (Meraldi και Nigg, 22, Fukasawa, 25). Η ανευπλοειδογόνος δράση µίας ποικιλίας χηµικών ενώσεων οφείλεται σε ισχυρές αλληλεπιδράσεις που αναπτύσσουν µε τη γ-τουµπουλίνη η οποία εντοπίζεται στα κεντροσώµατα µε αποτέλεσµα την επαγωγή πολλαπλών κέντρων οργάνωσης των µικροσωληνίσκων (MTOCs) και το σχηµατισµό πολυπολικών µιτωτικών ατράκτων. Σε αυτές περιλαµβάνονται τα µεθυλιωµένα παράγωγα των ανόργανων αρσενικών (DMAA) (Ochi et al., 1999, Ochi, 2) και του υγραργύρου (MeHgCl) (Ochi, 22), τα οιστρογόνα (E2), οι διφαινόλες (Bp-A) (Ochi, 1999, Parry et al., 22) και τα στεροειδή οιστρογόνα (Ochi και Oda, 21). Επιπρόσθετα, η κολχικίνη (Sellito και Kuriyama, 1988) και οι ακτίνες-χ (Sato et al., 1983, Parry et al., 1985) προκαλούν ανωµαλίες στον αριθµό, στη δοµή και στην ενεργότητα πυρήνωσης των κέντρων οργάνωσης των µικροσωληνίσκων. Η διαζεπάµη (DZ), βασικό συστατικό αρκετών υπνωτικών και ηρεµιστικών φαρµάκων, αναστέλλει το διαχωρισµό των κεντριολίων µετά το διπλασιασµό τους κατά την προµετάφαση, ενώ έχουν συχνά αναφερθεί αναισθητικές ενώσεις οι οποίες είναι ικανές να επάγουν ανευπλοειδία µε την καταστροφή των κεντριολίων (Aardema et al., 1998). Ειδικότερα, η επίδραση της διαζεπάµης σε κύτταρα V79 επάγει κυρίως µονοπολικές µεταφάσεις, οι οποίες χαρακτηρίζονται είτε από ένα ή δύο διακριτά σήµατα γ-τουµπουλίνης σε πολύ µικρή απόσταση µεταξύ τους είτε από διασκορπισµένα/διάστικτα (punctate) σήµατα γ-τουµπουλίνης (Izzo et al., 1998). Πρόσφατες µελέτες υποστηρίζουν την υπόθεση που διατύπωσε στις αρχές του 2 ου αιώνα ο Theodor Boveri σύµφωνα µε την οποία ανωµαλίες στο κεντρόσωµα αποτελούν το θεµέλιο λίθο για την ανάπτυξη καρκίνου (Lingle και Salisbury, 2, Brinkley, 21). Με τη χρήση αντισωµάτων έναντι πρωτεϊνών (γ-τουµπουλίνη και περικεντρίνη) της θεµέλιας ουσίας του κεντροσώµατος έχει δειχθεί για τη συντριπτική πλειοψηφία κακοηθών όγκων να εµφανίζουν ανωµαλίες στο κεντρόσω- µα. Σε αυτούς περιλαµβάνονται καρκινώµατα στον προστάτη, στο στήθος, στους πνεύµονες και στο κόλον, καθώς και όγκοι στον εγκέφαλο. Οι ανωµαλίες του κεντροσώµατος περιλαµβάνουν πλεονάζοντα κεντροσώµατα, κεντροσώµατα απουσία κεντριολίων και κεντροσώµατα µε ανώµαλο µέγεθος και σχήµα και συνοδεύονται από δραµατικές αλλαγές στον αριθµό και στην κατανοµή των µικροσωληνίσκων που ξεκινούν από τις θέσεις πυρήνωσης. Οι ανωµαλίες στο κεντρόσωµα που παρατηρούνται σε όγκους και σε κυτταρικές σειρές, οι οποίες προέρχονται από όγκους, είναι δυνατόν να επάγουν δύο φαινόµενα που συµβάλλουν στην καρκινογένεση. Πρώτον, όλα τα κεντροσώµατα ανεξάρτητα από το µέγεθος, το σχήµα ή τον αριθµό είναι ικανά να συµµετέχουν στο σχηµατισµό δοµικά και λειτουργικά ανώµαλων µιτωτικών ατράκτων. εύτερον, ο µη αποχωρισµός παρατηρείται µε πολύ υψηλό ρυθµό σε κύτταρα µε ανώµαλα κεντροσώµατα µε αποτέλεσµα τη δηµιουργία ανευπλοειδικών κυττάρων µε εξαιρετικά διαφορετικούς αριθµούς χρωµοσωµάτων (χρωµοσωµατική αστάθεια). Ανάµεσα σε πιθανούς µηχανισµούς που συµµετέχουν στην εµφάνιση ενός ανώµαλου φαινοτύπου κεντροσωµάτων, η αύξηση των επιπέδων της περικεντρίνης και της γ-τουµπουλίνης που παρατηρείται σε καρκινικά κύτταρα θεωρείται να έχει ως αποτέλεσµα την έκτοπη συγκρότηση των πρωτεϊνών σε ανώµαλες και υπεράριθµες δοµές. Αξιοσηµείωτη είναι η παρατήρηση που αφορά την αδυναµία καρκινικών κυττάρων, τα οποία εµφανίζουν χαµηλές συγκεντρώσεις των πρωτεϊνών αυτών στην ικανότητα σχηµατισµού κεντροσωµάτων. Η ύπαρξη
78 Εισαγωγή Ανευπλοειδία πολλαπλών και ανώµαλων δοµικά κεντροσωµάτων στους πόλους της µιτωτικής ατράκτου συµβάλει στον ανώµαλο σχηµατισµό της και πιθανότατα οδηγεί σε καρκινογένεση. Ανώµαλα κεντροσώµατα στα καρκινικά κύτταρα προκύπτουν και ως αποτέλεσµα µη σωστής ρύθµισης του διπλασιασµού του κεντροσώµατος. Επιπρόσθετα, η αδυναµία πραγµατοποίησης της κυτταροκίνησης είναι δυνατόν να προκαλέσει αύξηση στον αριθµό (διπλάσιο) των δοµικά και λειτουργικά φυσιολογικών κεντροσωµάτων. Σε επόµενες διαιρέσεις η παρουσία περισσοτέρων από δύο κεντροσωµάτων σε ένα αρχικά σχηµατισµένο τετραπλοειδές κύτταρο (4N) διαταράσσει τη διαδικασία της µίτωσης και προκαλεί αποσταθεροποίηση των χρωµοσω- µάτων. Αρκετά ρυθµιστικά µόρια εµπλέκονται στη φυσιολογική λειτουργία του κεντροσώµατος και σε αυτά περιλαµβάνονται οι κινάσες Polo-Plk και Αurora A (ωρίµανση του κεντροσώµατος) στον άνθρωπο, Aurora 2 στη Drosophilla και η Nek 2. Η υπερέκφραση της aurora 2 προκαλεί µετασχηµατισµό των ινοβλαστών in vitro και τη δηµιουργία όγκου in vivo. Πρόσφατες µελέτες αποδίδουν τον έλεγχο του διπλασιασµού του κεντροσώµατος σε ένα πρωτεϊνικό σύµπλεγµα (cdk2-cyclin E) που συγκροτείται από την καταλυτική υποµονάδα κινάση 2 και τη ρυθµιστική υποµονάδα κυκλίνη Ε. Επιπρόσθετα, σε κύτταρα ποντικού (p53 - / - ) στα οποία έχει παύσει να συνθέτεται η πρωτεΐνη p53 (µεταγραφικός παράγοντας) παρατηρούνται πλεονάζοντα κεντροσώµατα γεγονός που υποδηλώνει τη σηµασία του ρυθµιστικού µορίου στη διαδικασία διπλασιασµού του κεντροσώµατος. Σε ανθρώπινα κύτταρα εξαιτίας της αυστηρότητας στον έλεγχο της έκφρασης της κυκλίνης Ε, η επαγωγή υπεράριθµων κεντροσωµάτων βασίζεται σε ένα συνδυαστικό αποτέλεσµα που προκαλεί ταυτόχρονα η απενεργοποίηση του ογκοκατασταλτικού γονιδίου p53 και η υψηλή έκφραση της κυκλίνης Ε (Pihan και Doxsey, 1999, Meraldi et al., 24, Fukasawa, 25) Πυρηνική µεµβράνη Η υπεύθυνη δοµή για τη διαµερισµατοποίηση του ευκαρυωτικού κυττάρου (κυτταρόπλασµα και πυρήνας) είναι η πυρηνική µεµβράνη. Αποτελεί σηµείο πρόσδεσης των χρωµοσωµάτων και παρέχει προστασία στο γενετικό υλικό µε αποτέλεσµα οποιαδήποτε διαταραχή της πυρηνικής µεµβράνης από χηµικές ενώσεις (αιθανόλη, δισόξινο χολικό νάτριο) να επάγει ανευπλοειδία (Aardema et al., 1998). Οι Oshimura και Barrett (1986) ανέφεραν ότι η πυρηνική µεµβράνη αποµακρύνει ιόντα ασβεστίου, τα οποία ρυθµίζουν τη µετάβαση από το στάδιο της µετάφασης σε αυτό της ανάφασης. Η δηµιουργία διαµερισµάτων στο κυτταρόπλασµα του κυττάρου εξακολουθεί να θεωρείται απαραίτητη κατά τη διάρκεια της κυτταρικής διαίρεσης και η πυρηνική µεµβράνη µε τη διάλυση της διαδραµατίζει ιδιαίτερο ρόλο στον έλεγχο και στη ρύθµιση της σωστής λειτουργίας της ατράκτου. Ειδικότερα, συµµετέχει σε ένα σύµπλοκο σύστηµα µεµβρανών που εντοπίζονται εσωτερικά και στην εξωτερική επιφάνεια της µιτωτικής συσκευής για την αποφυγή άνισης κατανοµής ρυθµιστικών ουσιών. Πράγµατι έχει διαπιστωθεί η ύπαρξη ενός επικαλύµµατος κυστιδίων που ανήκουν στο ενδοπλασµατικό δίκτυο και στην πυρηνική µεµβράνη και περιβάλλουν προστατευτικά την άτρακτο µε αποτέλεσµα µεγάλα οργανίδια (µιτοχόνδρια) τα οποία θα διατάραζαν τη µετακίνηση των χρωµοσωµάτων να παραµένουν στην εξωτερική επιφάνεια της µιτωτικής συσκευής. Επιπρόσθετα, ένας αστεροειδής σχηµατισµός κυστιδίων που δηµιουργείται γύρω από το κεντρόσωµα στους πόλους της ατράκτου σχετίζεται στενά µε τους µικροσωληνίσκους και διευκολύνει την επιµήκυνση της ατράκτου κατά την ανάφαση Β γεγονός που δηλώνει τις πιθανές αλληλεπιδράσεις των δύο δοµών κατά τη διάρκεια της κυτταρικής διαίρεσης
79 Εισαγωγή Ανευπλοειδία Συνεπώς οι µεµβράνες εµπλέκονται στη µετακίνηση των χρωµοσωµάτων και αποτελούν πιθανό στόχο χηµικών ενώσεων µε ανευπλοειδογόνο δράση. Συγκεκριµένα έχουν αναφερθεί µυκητοκτόνα (econazole, miconazole) που η δράση τους εντοπίζεται στην καταστροφή των µεµβρανών και προκαλούν ανευπλοειδία (Önfelt, 1986) Χρωµοσώµατα Η αστάθεια ενός χρωµοσώµατος που εισέρχεται στη µίτωση είναι σε θέση να επηρεάσει τον προσανατολισµό και τη σύνδεση του µε τη µιτωτική συσκευή µε αποτέλεσµα να επάγει ανευπλοειδικά φαινόµενα. Βασικά στοιχεία της δοµής των χρωµοσωµάτων που πιθανότατα επηρεάζουν το σωστό αποχωρισµό τους κατά την κυτταρική διαίρεση θεωρούνται η θέση και το µέγεθος του κεντροµέρους (θεµελιώδης ετεροχρωµατίνη) σε συνδυασµό µε τη σωστή λειτουργία αντιγραφής, η δοµή της χρωµατίνης, η συµπύκνωση των χρωµοσωµάτων, η ακέραια συγκρότηση των κινητοχώρων που φέρει κάθε χρωµατίδιο και η συνάφεια των αδελφών χρωµατιδίων (Pihan και Doxsey, 1999, Parry et al., 22). Κατά τη µείωση, παρεµβολές σε πρόσθετες διαδικασίες που δεν σχετίζονται µε τη λειτουργία της κυτταρικής ατράκτου, αλλά µε το ζευγάρωµα οµολόγων χρωµοσωµάτων (στάδιο σύναψης) και τον ανασυνδυασµό τους (χιασµατυπία) είναι δυνατόν να προκαλέσουν ανευπλοειδία. Σε ανευπλοειδικά φαινόµενα που συµβαίνουν κατά τη µειωτική διαίρεση συµµετέχουν η αδυναµία σύζευξης των οµολόγων χρωµοσωµάτων και ο µη αποχωρισµός τους, η πρώιµη διαίρεση των κεντροµερών στην πρώτη µειωτική διαίρεση, η αδυναµία των χρωµατιδίων να διαχωριστούν στη δεύτερη µειωτική διαίρεση, η επιπλέον αντιγραφή του χρωµοσώµατος και η χρωµοσωµατική απώλεια Το κυκλοφωσφαµίδιο είναι ένας αλκυλιωτικός παράγοντας που προκαλεί ρήγµατα στο γενετικό υλικό, επάγει ανευπλοειδία σε γεννητικά κύτταρα παρουσία µεταβολικού συστήµατος ενεργοποίησης και προκαλεί αδυναµία σύναψης µεταξύ οµολόγων χρωµοσωµάτων και τµηµατοποίηση του συναπτονηµατικού συµπλέγµατος (Aardema et al., 1998). Πρόσφατα δεδοµένα υποστηρίζουν την άποψη που θεωρεί βλάβες στη συµπύκνωση των χρωµοσωµάτων να επάγουν λανθασµένο αποχωρισµό των χρω- µοσωµάτων. Στη διαδικασία αυτή συµµετέχουν σηµαντικές πρωτεΐνες συµπεριλαµβανοµένης και της τοποϊσοµεράσης II, που συναντάται στον πυρήνα και η αναστολή της οποίας επάγει αριθµητικές και δοµικές ανωµαλίες (Pihan και Doxsey, 1999). Στις ζύµες, µεταλλαγµένες θερµοευαίσθητες µορφές του ενζύµου τοποϊσοµεράση ΙΙ σε περιοριστικές θερµοκρασίες διακόπτουν τον κυτταρικό κύκλο στο στάδιο της µίτωσης µε σηµαντικές ανωµαλίες στη συµπύκνωση των χρωµοσωµάτων. Εάν η λειτουργία του ενζύµου εµποδιστεί µετά από τη συµπύκνωση, τα κύτταρα παραµένουν στο στάδιο της µετάφασης και δεν πραγµατοποιείται διαχωρισµός των αδελφών χρωµατιδίων. Στην περίπτωση που ανασταλεί η δραστηριότητα της τοποϊσοµεράσης ΙΙ παρουσία αναστολέα του ενζύµου (ετοποσίδη) για συγκεκριµένο χρονικό διάστη- µα, σε κύτταρα αρουραίου (Cl-1) παρατηρείται λανθασµένος διαχωρισµός αδελφών χρωµατιδίων µε συνέπεια την επαγωγή ανευπλοειδίας (Parry et al., 1996). Μία άλλη κατηγορία µορίων που συµµετέχουν στη συµπύκνωση των χρωµοσωµάτων είναι τα µέλη της οικογένειας SMC πρωτεϊνών, οι οποίες σε συνδυασµό µε την κινάση cdk2-cyclin Β και την τοποϊσοµεράση Ι είναι δυνατόν να προκαλέσουν υπερελίκωση του DNA in vitro. Μεταλλάξεις σε κάποια από τα γονίδια που συµµετέχουν σε αυτή τη διαδικασία εµποδίζουν τη συµπύκνωση των χρωµοσωµάτων και προκαλούν τµηµατοποίηση και λανθασµένο αποχωρισµό των χρωµοσωµάτων κατά τη µίτωση (Pihan και Doxsey, 1999)
80 Εισαγωγή Ανευπλοειδία Η ύπαρξη συνάφειας µεταξύ των αδελφών χρωµατιδίων που έχουν προέλθει από την αντιγραφή του DNA επιτυγχάνεται µε αλληλεπιδράσεις µεταξύ πρωτεϊνών και οποιαδήποτε διαταραχή σε αυτή τη διαδικασία είναι δυνατόν να επάγει ανευπλοειδικά φαινόµενα. Ειδικότερα, σύµπλοκα κοχεσίνης (cohesin) δεσµεύονται στο διπλασιασµένο DNA και διατηρούν τα αδελφά χρωµατίδια σε άµεση συνάφεια µέχρι την έναρξη της ανάφασης, οπότε χάνεται η συνεκτικότητα τους και ξεκινά ο διαχωρισµός τους. Η απώλεια της συνεκτικότητας οφείλεται σε πρόσθετους πρωτεϊνικούς παράγοντες (APC) που προκαλούν τη σταδιακή διάσπαση των συµπλόκων της κοχεσίνης, η οποία φυσιολογικά ξεκινά από τα άκρα και καταλήγει στην περιοχή του κεντροµέρους. Μεταλλάξεις σε γονίδια που κωδικοποιούν αυτές τις πρωτεΐνες στις ζύµες εµποδίζουν την ύπαρξη συνάφειας µε αποτέλεσµα να προκαλείται πρόωρος διαχωρισµός των χρωµατιδίων και χρωµοσωµατικός µη αποχωρισµός, ενώ παρόµοιες βλάβες συµβάλλουν σε ανευπλοειδικά φαινόµενα που παρατηρούνται σε ανθρώπινους όγκους (Pihan και Doxsey, 1999). Επιπρόσθετα, η αυξηµένη ικανότητα πρόσδεσης των χρωµοσωµάτων, η παρουσία βλαβών στη δοµή τους (ακτινοµυκίνη D) και η ακατάλληλη τοποθέτηση τους στο ισηµερινό επίπεδο (νιτρώδες οξείδιο) που προκαλούνται από χηµικές ενώσεις ευθύνονται για την επαγωγή ανευπλοειδίας (Aardema et al., 1998). Αρκετές έρευνες υποστηρίζουν ότι χρωµοσώµατα που φέρουν επεκταµένες αλληλουχίες, υφίσταται σε µεγαλύτερη συχνότητα χρωµοσωµατικές αναδιατάξεις (Ottagio et al., 1988) και παρουσιάζουν αυξηµένες συχνότητες τυχαίας απώλειας (Miele et al., 1989). Οι επαναλαµβανόµενες αλληλουχίες DNA που εντοπίζονται στα χρωµοσώµατα συνιστούν ένα προτιµητέο στόχο από χηµικές ενώσεις για την επαγωγή ανευπλοειδίας. Πράγµατι, χρωµοσώ- µατα που φέρουν επεκταµένες περιοχές αποτελούν στόχο για τη δηµιουργία χρωµοσωµατικών ρηγµάτων και την επαγωγή χρωµοσωµατικής απώλειας από τις χηµικές ενώσεις BCNU και βινµπλαστίνη (Ottagio et al., 1993) Κινητοχώρος. Ο κινητοχώρος είναι ένας ειδικός πολυπρωτεϊνικός σχηµατισµός που συµµετέχει στην κίνηση των χρωµοσωµάτων κατά την κυτταρική διαίρεση και η αυτοσυγκρότηση του εξαρτάται αποκλειστικά από την αλληλουχία DNA του κεντροµέρους. Σε κάθε αδελφό χρωµατίδιο εντοπίζεται ένας κινητοχώρος µε αποτέλεσµα κάθε χρωµόσωµα να συνδέεται µε τους δύο πόλους της αναπτυσσόµενης ατράκτου διαµέσου της δέσµευσης των µικροσωληνίσκων µε τους δύο αδελφούς κινητοχώρους. Οι κινητοχώροι (αδελφοί κινητοχώροι) των αδελφών χρωµατιδίων είναι προσανατολισµένοι προς την αντίθετη κατεύθυνση (διαφορετικό προσανατολισµό) και συνεπώς δεσµεύουν µικροσωληνίσκους που προέρχονται από τους αντίθετους πόλους γεγονός το οποίο εξασφαλίζει τη σωστή κίνηση. Ειδικότερα, το σύµπλοκο κεντροµέρος-κινητοχώρος περικλείει την ετεροχρωµατίνη, τον κινητοχώρο και τις συνδεόµενες πρωτεΐνες και αποτελεί τη θέση των κινητήριων συστατικών (CENP- E, F, MCAK, δυνεΐνη) που συµβάλλουν στην ολίσθηση των χρωµοσωµάτων στους πόλους της ατράκτου εξασφαλίζοντας τον ισοµερή διαµοιρασµό των χρωµοσωµάτων κατά την κυτταρική διαίρεση (Wordeman και Mitchison, 1995, Sullivan et al., 1996, Preston, 1996). Μελέτες µε τη χρήση αντισωµάτων έναντι του κινητοχώρου (CREST) απεκάλυψαν την ύπαρξη τοποθετηµένων εστιών σε µεσοφασικούς πυρήνες που περιέχουν τουλάχιστον µερικά συστατικά του µιτωτικού κινητοχώρου (προκινητοχώροι) και εντοπίζονται συνδεδεµένοι είτε µε την επιφάνεια της πυρηνικής µεµβράνης είτε µε τους πυρηνίσκους. Οι κινητοχώροι κατά τη διάρκεια της G2 φάσης διπλασιάζονται και στην πρόφαση ανιχνεύονται πλέον ως ζεύγη, ενώ
81 Εισαγωγή Ανευπλοειδία αναγνωρίζονται µόλις τα χρωµοσώµατα αρχίσουν να συµπυκνώνονται (Moroi et al., 1981, Brenner et al., 1981). Ο σφαιροειδής σχηµατισµός του κινητοχώρου υφίσταται µία µικρή δοµική τροποποίηση κατά τη διάρκεια της πρόφασης µε αποτέλεσµα µετά τη ρήξη της πυρηνικής µεµβράνης να µεταπίπτει σε ένα δισκοειδή σχηµατισµό που θεωρείται απαραίτητη προϋπόθεση για την εµφάνιση µικροσωληνίσκων (Mitchison, 199). Οι δοµικές πρωτεΐνες (CENPs) της τρίστοιβης δοµής του συµπλόκου κεντροµέρους-κινητοχώρου περιλαµβάνουν τις CENP-A, B και C. H CENP-A εµφανίζει οµολογία ιστόνης (H3) και ευθύνεται για το διαφορετικό σχήµα της κεντροµερικής ετεροχρωµατίνης, ενώ η CENP-B αλληλεπιδρά άµεσα µε την κύρια τάξη του ανθρώπινου κεντροµερικού DNA, το α-δορυφορικό DNA και σχηµατίζει ένα διµερές συµβάλλοντας στην ανώτερη οργάνωση της χρωµατίνης. Η CENP -C περιέχει µία DNA συνδεόµενη αλληλουχία και εντοπίζεται στην εσωτερική επιφάνεια του µιτωτικού κινητοχώρου µε αποτέλεσµα να ανιχνεύεται στο συσπειρωµένο ενεργά κεντροµέρος και να θεωρείται απαραίτητη στη συγκρότηση του κινητοχώρου για τη δηµιουργία σταθερής σύνδεσης µε την κυτταρική άτρακτο (Sullivan et al., 1996). Εικόνα 5-2. Σχηµατική απεικόνιση της δοµής κινητοχώρου - κεντροµέρους σε ένα µεταφασικό χρωµόσωµα. Οι δισκοειδείς σχηµατισµοί αναπαριστούν τις πρωτεΐνες της εξωτερικής ( ) και εσωτερικής επιφάνειας ( ) του κινητοχώρου που εντοπίζεται στην κεντροµερική περιοχή ( ) του χρωµοσώ- µατος. Αξιοσηµείωτες είναι οι µελέτες που αφορούν τον κινητοχώρο και τον ενεργό ρόλο που διαδραµατίζει σε µηχανισµούς που σχετίζονται µε σηµεία ελέγχου της µιτωτικής διαδικασίας, ιδιαίτερα αυτόν που ελέγχει τη µετάβαση από το στάδιο της µετάφασης σε αυτό της ανάφασης. Η ακριβής συσσώρευση µικροσωληνίσκων στην περιοχή του κινητοχώρου και/ή η τάση που αυτοί παράγουν στον κινητοχώρο αποτελεί το σινιάλο για την απενεργοποίηση του σηµείου ελέγχου και τη µετάβαση στο επόµενο στάδιο της µίτωσης (Rieder and Salmon, 1998). Αναµφισβήτητα, οι περισσότερες πρωτεΐνες που συµµετέχουν στους µηχανισµούς ελέγχου συσσωρεύονται στον κινητοχώρο κατά τα αρχικά στάδια της µίτωσης (πρόφαση, προµετάφαση) και εξαφανίζονται από αυτόν όταν το κύτταρο εισέρχεται στην ανάφαση. Σε περίπτωση αδυναµίας είτε συγκρότησης της µιτωτικής ατράκτου είτε δέσµευσης ενός ή περισσοτέρων κινητοχώρων µε τους µικροσωληνίσκους, ενεργοποιείται ο - 7 -
82 Εισαγωγή Ανευπλοειδία µηχανισµός ελέγχου (Mad2, 3F3/2) και παρεµποδίζεται η είσοδος του κυττάρου στο στάδιο της ανάφασης µε αποτέλεσµα να αποτρέπεται ο ανώµαλος διαχωρισµός των χρωµοσωµάτων και ο σχηµατισµός ανευπλοειδών κυττάρων. Ωστόσο, είναι δυνατό, ο κινητοχώρος να µην προσανατολιστεί στη σωστή κατεύθυνση, αλλά το κύτταρο να εισέλθει στην ανάφαση. Η αναλυτική περιγραφή του φαινοµένου δείχνει ότι ενώ το χρωµατίδιο κάθε χρωµοσώµατος θα πρέπει να δέχεται τάση, µέσω του κινητοχώρου του, από τον ένα από τους δυο πόλους της µιτωτικής ατράκτου, στην περίπτωση µη σωστού προσανατολισµού, ο συγκεκριµένος κινητοχώρος αποκτά διπολικό προσανατολισµό (merotelic kinetochore orientation) µε αποτέλεσµα το χρωµατίδιο να δέχεται πίεση και από τους δυο πόλους και τελικά να αδυνατεί να µετακινηθεί προς τον ένα ή τον άλλο πόλο. Το φαινόµενο αυτό επεκτείνεται και στο άλλο αδελφό χρω- µατίδιο, µε αποτέλεσµα ολόκληρο το χρωµόσωµα να καθυστερεί στην ανάφαση και να µην ενσωµατώνεται στο θυγατρικό κύτταρο µε συνέπεια τη χρωµοσωµατική απώλεια (Cimini et al., 21, Cimini et al., 22). Σαφέστατα, αιτίες που προκαλούν ανευπλοειδία περιλαµβάνουν βλάβες στον κινητοχώρο (µιτοµυκίνη C) (Aardema et al., 1998), ενώ σε καρκινικά κύτταρα έχουν ανιχνευθεί κεντροµέρη στα οποία απουσιάζουν οι πρωτεΐνες του κινητοχώρου (Vig και Sternes, 1991), παρά το γεγονός ότι ελάχιστες είναι οι ενδείξεις που συνηγορούν στην άποψη που θεωρεί βλάβες στην περιοχή του κινητοχώρου να συµβάλλουν στην καρκινογένεση (Pihan και Doxsey, 1999) Κεντροµέρος. Συχνά συναντώνται θέσεις στο γονιδίωµα, γνωστές ως ''εύθραυστες θέσεις'' (Fragile Sites, FS), που αποτελούν θερµά σηµεία στη δηµιουργία ρηγµάτων, στις χρωµοσωµατικές αναδιατάξεις και στη δηµιουργία ελλειµµάτων (Cimprich, 23). Σε χρωµοσώµατα ποντικού έχουν αναφερθεί εύθραυστες θέσεις στην περιοχή του κεντροµέρους, ενώ από πειράµατα που έγιναν µε επίδραση MNU σε εµβρυϊκούς ινοβλάστες αρουραίου (Simi et al., 1998), έδειξαν να υπάρχει σαφής συσχέτιση της παρουσίας εύθραυστων θέσεων στην περιοχή του κεντροµέρους και της επαγωγής χρωµοσωµατικής απώλειας. Μια ιδιαίτερη προτίµηση για τη δηµιουργία ρηγµάτων στην ετεροχρωµατινική περιοχή του κεντροµέρους έχει αποδειχθεί στην περίπτωση της µιτοµυκίνης C, και σε αυτήν αποδίδεται η ανευπλοειδογόνος δράση της χηµικής ένωσης (aneuploidogenic activity), εκτός από την ιδιότητα της να προκαλεί ρήγµατα στο γενετικό υλικό (clastogenic activity) (Renzi et al.,1996). Επιπρόσθετα σε χρωµοσωµατική διαταραχή που σχετίζεται µε τον πρώιµο διαχωρισµό ετεροχρω- µατίνης του κεντροµέρους, η οποία συµβαίνει κατά τη µετάφαση οφείλεται το σύνδροµο Roberts (Tomkins, 1989, Thompson et al., 21). Ο πρώιµος διαχωρισµός του κεντροµέρους αποκλείει την επαφή του µε τους µικροσωληνίσκους της ατράκτου µε αποτέλεσµα την αποτυχία δηµιουργίας συµπλόκου που συµµετέχει στη µετακίνηση των χρωµοσωµάτων γεγονός που προκαλεί ανευπλοειδικά φαινόµενα (µη αποχωρισµό) (Vig, 1984, Vig και Paweletz, 1988) Τελοµέρη. Τα τελοµέρη είναι νουκλεοπρωτεϊνικά σύµπλοκα που καλύπτουν τα άκρα των ευκαρυωτικών χρωµοσωµάτων και δηµιουργούνται από µια αντίστροφη µεταγραφάση, την τελοµεράση, η οποία προσθέτει TTAGGG αλληλουχίες στο 3' άκρο (de Lange, 22). Εµφανίζουν ιδιαίτερη σηµασία για το χρωµοσωµατικό αποχωρισµό και τη διατήρηση της γονιδιωµατικής σταθερότητας, ενώ είναι γνωστό για τα φυσιολογικά σωµατικά κύτταρα ότι η διαδικασία γήρανσης σχετίζεται µε τη µείω
83 Εισαγωγή Ανευπλοειδία ση τελοµερικών επαναλαµβανόµενων αλληλουχιών (Pathak et al., 22), η οποία επιταχύνεται στην περίπτωση του απενεργοποιηµένου Χ χρωµοσώµατος (Surralles et al., 1999). Αναµφισβήτητα, καταστροφή ή/και ενίσχυση των επαναλαµβανόµενων περιοχών TTAGGG που εντοπίζονται στα άκρα των χρωµοσωµάτων προκαλεί αστάθεια του γενετικού υλικού. Άλλωστε τα τελοµέρη, εφόσον καθορίζουν την περιοχή κάθε χρωµοσώµατος ξεχωριστά µέσα στον πυρήνα και τα προστατεύουν από εσωτερικές και εξωτερικές επιδράσεις, η αποµάκρυνση τους θα έχει ως αποτέλεσµα την αποσταθεροποίηση του κυτταρικού καρυοτύπου (Pathak et al., 22). Ειδικότερα, σε τελοµερικά DNA µεταλλάγµατα της Tetrahymena thermophila, που δηµιουργήθηκαν µε την έκφραση ενός RNA µε τροποποιηµένη αλληλουχία, η διαίρεση του πυρήνα καθυστερεί σηµαντικά ή σταµατά στο στάδιο της ανάφασης, κατά τη µίτωση µε αποτέλεσµα τα µεταλλαγµένα χρωµατίδια να αδυνατούν να διαχωριστούν (Kirk et al., 1997). Παρόµοια, άλλες µελέτες υποδηλώνουν το σηµαντικό ρόλο που διαδραµατίζουν τα τελοµέρη και οι τελοµερικές πρωτεΐνες πρόσδεσης στον αποχωρισµό των χρωµοσωµάτων και στη µείωση (Hande et al., 1999). Επιπρόσθετα, η γενετική αστάθεια που προκύπτει ως αποτέλεσµα µεταβολών στην δυναµική των τελοµερικών περιοχών αποτελεί βασικό παράγοντα στην επαγωγή της καρκινογένεσης. Πιθανότατα, η δυναµική των περιοχών αυτών του γενετικού υλικού που σχετίζεται µε ενίσχυση των τελοµερικών περιοχών δεν συναντάται µόνο σε καρκινικά κύτταρα αλλά και σε λεµφοκύτταρα του περιφερικού αίµατος καρκινοπαθών σε περιπτώσεις µεταστάσεων. Μειωµένες ποσότητες τελοµερικού DNA εµφανίζονται σε κύτταρα που προέρχονται από αρκετά ανθρώπινα σύνδροµα (Down, Turner, Bloom, Werner, Fanconi), τα οποία είναι επιδεκτικά στην ανάπτυξη καρκίνου. Γενικότερα, η συσχέτιση της µείωσης των τελοµερικών περιοχών µε την ηλικία, που αποτελεί ένα παράγοντα κινδύνου για την ανάπτυξη καρκίνου στους ανθρώπινους πληθυσµούς, πιθανόν αντανακλά µεταβολές στα τελοµέρη, οι οποίες δίνουν το έναυσµα για την ανάπτυξη λευχαιµιών και καρκίνων σε ιστούς (Pathak et al., 22) Πυρηνίσκος. Ο πυρηνίσκος είναι µία ειδική περιοχή των µεσοφασικών χρωµοσωµάτων και θεωρείται υπεύθυνος για την παραγωγή και την ωρίµανση του rrna, καθώς και την αυτοσυγκρότηση των ριβοσωµατικών υποµονάδων. Είναι εµφανής µόνο κατά τη µεσόφαση και εξαφανίζεται προτού το κύτταρο εισέλθει στη φάση της µετάφασης. Έχει αναφερθεί συσχέτιση των πυρηνίσκων µε ανευπλοειδικά φαινόµενα σε περίπτωση παραµονής του πυρηνίσκου ή τµήµατος του, η οποία έχει ως αποτέλεσµα την αδυναµία των συναπτικών χρωµοσωµατικών ζευγών να διαχωριστούν µε άµεσο αντίκτυπο το µη αποχωρισµό των χρωµοσωµάτων (Aardema et al., 1998). Σε µία από τις υποθέσεις που έχουν διατυπωθεί για την επίδραση της ηλικίας της µητέρας στο µη αποχωρισµό του 21 ου χρωµοσώµατος αναφέρεται η επίµονη παρουσία των πυρηνίσκων στην πρόφαση Ι, η οποία παρατείνεται µε την αύξηση της ηλικίας (Gaulden, 1992) Κυτταροκίνηση Σε περίπτωση αποτυχίας της διαδικασίας της κυτταροκίνησης δηµιουργούνται κύτταρα που περιέχουν το διπλάσιο αριθµό χρωµοσωµάτων και κεντροσωµάτων µε αποτέλεσµα να σχηµατίζονται δυσλειτουργικές µιτωτικές άτρακτοι, οι οποίες σε επόµενες πυρηνικές διαιρέσεις οδηγούν στην παραγωγή πολυπλοειδικών κυττάρων (Pihan και Doxsey, 1999, Fukasawa, 25). Αξιοσηµείωτο
84 Εισαγωγή Ανευπλοειδία είναι ότι χηµικές ενώσεις που παρεµποδίζουν τη διαίρεση του κυτταροπλάσµατος (κυτταροχαλασίνη-β) συµβάλουν στο σχηµατισµό διπύρηνων κυττάρων. Η ανάκαµψη τέτοιων κυττάρων από τη χηµική επίδραση και η επακόλουθη διαίρεση τους έχει ως αποτέλεσµα το σχηµατισµό θυγατρικών κυττάρων που περιέχουν λανθασµένο αριθµό χρωµοσωµάτων (Schultz και Önfelt, 1994). Συνεπώς χηµικές ενώσεις ικανές να προκαλούν παροδική αναστολή της κυτταροκίνησης επάγουν ανευπλοειδικά φαινόµενα σε επόµενες διαιρέσεις. Eχουν αναφερθεί αρκετές χηµικές ενώσεις µε παρόµοια δράση σε V79 κύτταρα αρουραίου, οι οποίες χαρακτηρίζονται από ποικίλες φυσικοχηµικές και βιολογικές ιδιότητες και σε αυτές περιλαµβάνονται οι n-αλκοόλες, οι χλωριωµένες αλειφατικές ενώσεις, το διαµίδιο, η νεοµυκίνη, η σκοπαλαµίνη και η ατροπίνη. Επιπρόσθετα, για τον πολυκυκλικό αρωµατικό υδρογονάνθρακα βενζο(α)πυρένιο (B[α]P), που αποτελεί µία από τις πιο σηµαντικές τάξεις χηµικών ενώσεων η οποία ανιχνεύεται σε περιβαλλοντικά µολυσµένες ατµόσφαιρες, έχει αναφερθεί η δυνατότητα που εµφανίζει σε καλλιέργειες κυττάρων V79 (CYP1A1) να επάγει τροποποιήσεις στην κυτταρική διαίρεση και να προκαλεί ανευπλοειδία (Parry et al., 1996). Η αδυναµία πραγµατοποίησης της διαίρεσης του κυτταροπλάσµατος παρατηρείται συχνά σε καρκινικά κύτταρα πιθανότατα εξαιτίας απουσίας του λειτουργικού µονοπατιού p53/p21 µε αποτέλεσµα να αποφεύγεται η παραµονή στη φάση G2 του κυτταρικού κύκλου, τα κύτταρα να εισέρχονται στη µίτωση και να διαχωρίζουν τα χρωµοσώµατα τους αλλά να αδυνατούν να υποστούν κυτταροκίνηση και τα θυγατρικά κύτταρα να φέρουν διπλασιασµένο γενετικό υλικό. Πρόσφατα δεδοµένα ενοχοποιούν την υπερέκφραση της AIM-1, µίας πρωτεΐνης του ενδιάµεσου σώµατος µε παρόµοια δράση µε την πρωτεΐνη aurora Β, στην επαγωγή του σχηµατισµού πολλαπλών πυρήνων µε αποτέλεσµα να αυξάνεται η πολυπλοειδία, υποδεικνύοντας άµεση συσχέτιση µίας πρωτεΐνης που συµµετέχει στην κυτταροκίνηση και ενός καρκινικού φαινοτύπου (Pihan και Doxsey, 1999) Σηµεία ελέγχου της µίτωσης Οι µηχανισµοί ελέγχου του κυτταρικού κύκλου ρυθµίζουν την εξέλιξη των διακριτών βηµάτων από το ένα κυτταρικό στάδιο στο επόµενο (G1 S G2 M) ώστε να εξασφαλιστεί η πιστότητα στο διαχωρισµό των χρωµοσωµάτων (µιτωτικό σηµείο ελέγχου, spindle/mitotic checkpoint) και στην αντιγραφή του DNA (αντιγραφικό σηµείο ελέγχου, DNA replication checkpoint), καθώς και η διακοπή του κυτταρικού κύκλου και η επαγωγή της απόπτωσης σε περίπτωσης βλάβης του γενετικού υλικού (DNA damage checkpoint). Ο µηχανισµός απόκρισης στις βλάβες του DNA αποτελεί µια διαδικασία µεταγωγής σηµάτων, που συνίσταται από µόρια-αισθητήρες, µεταγωγείς και τελεστές, οι οποίοι απαρτίζουν ουσιαστικά ένα δίκτυο που περιλαµβάνει την αλληλεπίδραση διαφόρων µονοπατιών, µε τελικό αποτέλεσµα την οργανωµένη και ακριβή απόκριση του κυττάρου στη βλάβη του γενετικού του υλικού. Η διαταραχή οποιουδήποτε από τους παραπάνω µηχανισµούς ελέγχου που είναι πιθανόν να προκληθεί από χηµικά επαγόµενες µεταλλάξεις ή από χηµική αναστολή των εµπλεκόµενων πρωτεϊνικών παραγόντων δύναται να οδηγήσει σε λανθασµένο διαχωρισµό των χρωµοσωµάτων (ανευπλοειδία) και στη συσσώρευση µεταλλάξεων µε επακόλουθο τη γονιδιωµατική αστάθεια (Luch, 22). Η διαδικασία της µίτωσης παρακολουθείται στενά από τουλάχιστον δύο σηµεία ελέγχου, τα οποία ελέγχουν την είσοδο του κυττάρου στη µίτωση και τη µετάβαση από τη µετάφαση στην ανάφαση, ενώ θεωρείται πιθανή η ύπαρξη επιπρόσθετων σηµείων σε κύτταρα θηλαστικών που σχετίζονται µε την πρόοδο της
85 Εισαγωγή Ανευπλοειδία ανάφασης ή της κυτταροκίνησης (Pihan και Doxsey, 1999). Ειδικότερα, το πρωτεϊνικό σύµπλεγµα MPF (maturation- or mitosis- promoting factor) που συγκροτείται από την καταλυτική υποµονάδα CDK1 και τη ρυθµιστική υποµονάδα κυκλίνη B διαδραµατίζει ειδικό ρόλο στην είσοδο του κυττάρου από τη φάση G2 στη φάση Μ. Η αυξηµένη ενεργότητα του συµπλέγµατος στο τέλος της G2 φάσης επιφέρει την κατάλυση αντιδράσεων φωσφορυλίωσης σε διάφορες πρωτεΐνες στόχους (Η1, λαµίνες, ΜAPs) και ευθύνεται για τη συµπύκνωση των χρωµοσωµάτων, την αποδιοργάνωση της πυρηνικής µεµβράνης και τη συγκρότηση της µιτωτικής συσκευής (Luch, 22). Το επόµενο σηµείο ελέγχου που θεωρείται ιδιαίτερα σηµαντικό για τη µετάβαση από τη µετάφαση στην ανάφαση ενεργοποιείται από τους κινητοχώρους που δεν προσδένονται στην κυτταρική άτρακτο γεγονός που επιφέρει την αναστολή της ενεργοποίησης του συµπλόκου APC/C (anaphase promoting complex or cyclosome) µε αποτέλεσµα να παρεµποδίζεται ο πρόωρος διαχωρισµός των χρωµοσωµάτων και να εξασφαλίζεται µε ακρίβεια η συµµετοχή του γενετικού υλικού στα θυγατρικά κύτταρα. Συγκεκριµένα, πιθανές ανωµαλίες σχετικά µε την ορθή συγκρότηση της µιτωτικής συσκευής, τις οποίες αντιλαµβάνεται το σηµείο ελέγχου, σχετίζονται µε τις θέσεις δέσµευσης των µικροσωληνίσκων στον κινητοχώρο ή/και µε την τάση που αναπτύσσεται απέναντι από το ζεύγος των αδελφών χρωµατιδίων, η οποία οφείλεται στην ορθή πρόσδεση µικροσωληνίσκων του κινητοχώρου που φυσιολογικά προέρχονται από τους αντίθετους πόλους της ατράκτου. Ανάµεσα σε µόρια αισθητήρες του σηµείου ελέγχου που εντοπίζονται ως σύµπλεγµα στον κινητοχώρο στην προµετάφαση, στο ισηµερινό επίπεδο κατά την µετάφαση και στο ενδιάµεσο σώµα κατά την κυτταροκίνηση περιλαµβάνονται η Aurora B, οι εσωτερικές κεντροµερικές πρωτεΐνες (INCENP) και η Survinin. Η είσοδος στο στάδιο της ανάφασης απαιτεί τη διάσπαση της συνάφειας των αδελφών χρωµατιδίων, η οποία επιτυγχάνεται µε την πρωτεόλυση µίας υποµονάδας (Scc1) του πολυπρωτεϊνικού συµπλόκου κοχεσίνης από την πρωτεάση σεπαράση, που φυσιολογικά καθίσταται ανενεργή παρουσία αναστολέα (securin). Με την κατάλληλη πρόσδεση των µικροσωληνίσκων στους κινητοχώρους των αδελφών χρωµατιδίων επέρχεται η ικανοποίηση του σηµείου ελέγχου και η ενεργοποίηση του συµπλόκου APC/C, το οποίο µε δράση ουβικιτίνης προκαλεί την αποικοδόµηση της σεκουρίνης από το πρωτεόσωµα µε αποτέλεσµα την ενεργοποίηση της σεπαράσης και την έναρξη της ανάφασης. Ιδιαίτερο ρόλο στη µεταβίβαση της πληροφορίας και στην ενεργοποίηση του συστήµατος που προάγει την ανάφαση (APC/C) λαµβάνουν στοιχεία του σηµείου ελέγχου που εντοπίζονται σε πρωτεΐνες του συµπλέγµατος του κινητοχώρου (Mps1p, Mad και Bub πρωτεΐνες) και σε κυτταροπλασµατικά συστατικά του συστήµατος (Cdc2). Ειδικότερα, σε απόκριση βλάβης στη µιτωτική συσκευή, που σχετίζεται µε το σωστό προσανατολισµό των χρωµοσωµάτων και την ορθή δέσµευση των αδελφών χρωµατιδίων το σηµείο ελέγχου προάγει τη συγκρότηση πρωτεϊνικών συµπλεγµάτων (APC, Cdc2, Mad2, Bub3, BubR1) µε αποτέλεσµα την απενεργοποίηση του APC/C, γεγονός που επιφέρει τη διατήρηση της συνάφειας των αδελφών χρωµατιδίων και την καθυστέρηση του κυτταρικού κύκλου στη µετάφαση (Luch, 22, Bharadwaj και Yu, 24). Η χορήγηση αντισωµάτων έναντι πρωτεϊνών του συµπλέγµατος οδηγεί σε πρόωρη µετάβαση του κύκλου στο στάδιο της ανάφασης ακόµη και στην περίπτωση που οι κινητοχώροι δεν έχουν συνδεθεί µε την άτρακτο, ενώ έχει αναφερθεί σαφής συσχέτιση µεταξύ µη λειτουργικού σηµείου ελέγχου της µίτωσης (hbub1) και της ανάπτυξης χρωµοσωµατικής αστάθειας σε καρκίνο του ορθού (Pihan και Doxsey, 1999, Bharadwaj και Yu, 24). Συγκεκριµένα, σε κυτταρικές σειρές που εµφανί
86 Εισαγωγή Ανευπλοειδία ζουν χρωµοσωµατική αστάθεια δεν συµβαίνει παύση του κυτταρικού κύκλου στο στάδιο της µετάφασης, σε περίπτωση καταστροφής των µικροσωληνίσκων της µιτωτικής ατράκτου, σε αντίθεση µε διπλοειδείς κυτταρικές σειρές µε κανονικό αριθµό χρωµοσωµάτων (Cahill et al., 1998). Ογκογόνοι ιοί συχνά προκαλούν ανευπλοειδία και εκφράζουν πρωτεΐνες που αλληλεπιδρούν µε τον κυτταρικό κύκλο µε αποτέλεσµα στοιχεία ελέγχου της µίτωσης να αποτελούν πιθανούς στόχους ογκοπρωτεϊνών. Συγκεκριµένα, η HTLV1 TAX ογκοπρωτεΐνη στοχεύει τη Mad1 πρωτεΐνη του σηµείου ελέγχου της µίτωσης και επηρεάζει τη δράση της, γεγονός που προκαλεί πολυπυρήνωση και ανευπλοειδία, ενώ η υπερέκφραση ογκοπρωτεϊνών, όπως της πρωτεΐνης LMP-1 και του µεγάλου Τ αντιγόνου του SV4 επάγει το σχηµατισµό πολυπύρηνων κυττάρων. Συνεπώς η αποσταθεροποίηση του σηµείου ελέγχου της µίτωσης είναι δυνατόν να αποτελεί ένα κοινό µονοπάτι για την ογκογένεση από ιούς (Pihan και Doxsey, 1999). Αναµφισβήτητα, η πλήρης απενεργοποίηση συγκεκριµένων γονιδίων του σηµείου ελέγχου της µίτωσης (Mad2, Bub1) σε ανώτερους οργανισµούς σχετίζεται µε αυξηµένη εµβρυϊκή θνησιµότητα. Η απουσία Mad2 στον ποντικό έχει ως αποτέλεσµα τόσο την αποβολή νεκρών εµβρύων όσο και την εµφάνιση φαινοµένων χρωµοσωµατικού µη αποχωρισµού και απόπτωσης σε αρχικά στάδια της αναπτυξιακής τους πορείας. Μείωση των επιπέδων της πρωτεΐνης σε ετερόζυγα άτοµα για το γονίδιο αυτό οδηγεί σε απουσία λειτουργικού σηµείου ελέγχου της µίτωσης γεγονός που επιφέρει την εµφάνιση καρκίνου των πνευµόνων και επιβεβαιώνει την άποψη που θεωρεί µικρές τροποποιήσεις στα επίπεδα των πρωτεϊνών που ελέγχουν το στάδιο της µίτωσης να επάγουν την ανάπτυξη καρκίνου. Άλλωστε, εκτός από την πλήρη απενεργοποίηση τους, µερική καταστροφή αυτού του σηµείου ελέγχου θεωρείται περισσότερο πιθανό να παρατηρηθεί σε καρκίνους (Bharadwaj και Yu 24, Draviam et al., 24). Επιπρόσθετα, το p53 γονίδιο λαµβάνει καθοριστικό ρόλο σε γεγονότα του κυτταρικού κύκλου που υπόκεινται σε αυστηρή ρύθµιση και τα οποία εάν διαταραχθούν επάγουν ανευπλοειδικά φαινόµενα. Μεταλλάξεις στο γονίδιο αυτό προκαλούν αστάθεια του χρωµοσωµατικού υλικού in vitro και καρκινογένεση στον ανθρωπο. Πράγµατι ινοβλάστες Li-Fraumeni που προέρχονται από άτοµα που εµφανίζουν ετεροζυγωτία για το p53 γονίδιο είναι καρυοτυπικά ασταθείς in vitro. Η αλληλεπίδραση του γονιδίου p53 µε άλλα γονίδια τα οποία εάν εµφανίσουν µεταλλάξεις είναι δυνατόν να προκαλέσουν απώλεια της λειτουργίας του G1 ή G2 σηµείου ελέγχου του κυτταρικού κύκλου σχετίζεται µε πιθανή αστάθεια του γενετικού υλικού (Aardema et al., 1998). Παρά την πολυπλοκότητα της διαδικασίας της µίτωσης είναι δυνατόν σε φυσιολογικά κύτταρα να συµβούν µε κάποια συχνότητα ανώµαλες µιτώσεις µε αποτέλεσµα να προκύπτουν κύτταρα µε αριθµητικές χρωµοσωµατικές ανωµαλίες. Σε αυτήν την περίπτωση η πορεία που διαγράφεται περιλαµβάνει είτε το θάνατο εξαιτίας της απώλειας σηµαντικών για τη διατήρηση της ζωής γονιδίων είτε τη ''φυσιολογική'' ανάπτυξη είτε τη µετατροπή σε καρκινικά κύτταρα είτε τον προγραµµατισµένο κυτταρικό θάνατο. Ερευνητικές µελέτες συνηγορούν στην πιθανή παρουσία ενός προκαθορισµένου προαποπτωτικού µονοπατιού στη µίτωση το οποίο θα πρέπει ενεργά να ξεπερασθεί για την επιτυχή ολοκλήρωση της φυσιολογικής διαδικασίας. Στο µονοπάτι αυτό φαίνεται να συµµετέχει ένας αναστολέας της απόπτωσης (survinin) που εκφράζεται στη G2/M φάση και σχετίζεται µε τους µικροσωληνίσκους, ενώ η υπερέκφραση του αποτελεί ένα σηµαντικό µηχανισµό προόδου του κυτταρικού κύκλου αρκετών καρκινικών κυττάρων διαµέσου ανώµαλων µιτώσεων που αδυνατούν να υποστούν απόπτωση (Pihan και Doxsey, 1999)
87 Σκοπός της διατριβής ΣΚΟΠΟΣ ΤΗΣ ΙΑΤΡΙΒΗΣ Η ανισορροπία του γενετικού υλικού έχει σοβαρές συνέπειες στην ανθρώπινη υγεία µε τις δοµικές και αριθµητικές χρωµοσωµατικές ανωµαλίες-ανευπλοειδίες να χαρακτηρίζουν διάφορες παθολογικές καταστάσεις. Είναι σηµαντικό να γνωρίζουµε το ρόλο διαφόρων παραγόντων του περιβάλλοντος ως προς την ικανότητα πρόκλησης τέτοιων ανωµαλιών. Μία κατηγορία περιβαλλοντικών παραγόντων που επιδρούν στον άνθρωπο είναι οι φαρµακευτικές ενώσεις, οι οποίες θεωρούνται απαραίτητες για την αντιµετώπιση διαφόρων ασθενειών. Η υπέρταση θεωρείται ασθένεια του σύγχρονου τρόπου ζωής και ευθύνεται για την αυξηµένη εµφάνιση καρδιαγγειακών νοσηµάτων. Στη θεραπευτική αγωγή που ακολουθείται ευρέως διαδεδοµένη είναι η χορήγηση διουρητικών φαρµάκων. Το υδροχλωροθειαζίδιο ανήκει στα διουρητικά θειαζίδια και έχει την ικανότητα να αναπροσαρµόζει τον όγκο του αίµατος και να επαναφέρει την αρτηριακή πίεση σε φυσιολογικά επίπεδα. Η γενετική δράση του υδροχλωροθειαζιδίου δεν έχει πλήρως αποσαφηνισθεί. Έχει όµως αναφερθεί ότι προκαλεί µη αποχωρισµό σε διπλοειδή στελέχη του µύκητα Aspergillus nidulans, ενώ φαινόµενα πολυπλοειδίας ανιχνεύθηκαν µετά τη χορήγηση του σε CHL κύτταρα. Επιπλέον πειραµατικές ενδείξεις στο εργαστήριο µας υποδείκνυαν ότι η γενετική δράση του υδροχλωροθειαζιδίου σχετίζεται µε χρω- µοσωµατική καθυστέρηση σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro. Η παρούσα διατριβή έχει σκοπό τη µελέτη της γενετικής δράσης του υδροχλωροθειαζιδίου και τη διαλεύκανση των µηχανισµών µε τους οποίους εκδηλώνεται. Η µελέτη αφορά τον άνθρωπο και πραγµατοποιείται in vitro µε κύριο βιολογικό σύστηµα τα λεµφοκύτταρα του περιφερικού αίµατος. Επελέγησαν 32 υγιείς δότες οι οποίοι διαχωρίζονται σε οµάδες και αξιολογείται ο ρόλος της ηλικίας, του φύλου και του καπνίσµατος ως προς τη δράση της φαρµακευτικής ένωσης. Ως σύστηµα ελέγχου επιλέγεται η µέθοδος αναστολής της κυτταροκίνησης, CBMN, σε συνδυασµό µε in situ υβριδοποίηση µε φθοροχρώµατα, FISH. Η µέθοδος αυτή έχει ευρεία εφαρµογή στην εκτίµηση της γενετικής δράσης φαρµακευτικών ενώσεων τόσο in vitro όσο και in vivo. Αναλύονται γεγονότα χρωµοσωµατικής θραύσης αλλά και χρωµοσωµατικής καθυστέρησης µε επισήµανση της περιοχής του κεντροµέρους και του κινητοχώρου, FISH-CREST. Μελετώνται επίσης οι δύο κύριοι µηχανισµοί λανθασµένου χρωµοσωµατικού αποχωρισµού, µη αποχωρισµός και απώλεια, για τα επιλεγµένα χρωµοσώµατα Χ, Υ και 8, DUAL FISH. Επειδή ένας από τους στόχους των χηµικών ενώσεων που διαταράσσουν τον ισοµερή καταµερισµό των χρωµοσωµάτων είναι η µιτωτική συσκευή κρίνεται σκόπιµο να µελετηθεί η επίδραση του υδροχλωροθειαζιδίου στην ακεραιότητα και οργάνωση της. Η ανάλυση πραγµατοποιείται σε κυτταρικές σειρές ανθρώπου και ποντικού µε τη µέθοδο του διπλού ανοσοφθορισµού για την α- και γ- τουµπουλίνη, που αποτελούν βασικά συστατικά των µικροσωληνίσκων και των κεντροσωµάτων, αντίστοιχα της µιτωτικής ατράκτου
88
89 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες 6. ΚΥΤΤΑΡΙΚΕΣ ΚΑΛΛΙΕΡΓΕΙΕΣ Οι συνθήκες που απαιτούνται για την άριστη ανάπτυξη των κυτταρικών καλλιεργειών περιλαµβάνουν: Άσηπτο περιβάλλον εργασίας. Όλοι οι χειρισµοί και η προσθήκη των υλικών γίνονται σε εστία που διαθέτει λυχνία Bunsen και παροχή συνεχούς ροής αποστειρωµένου αέρα, η οποία επιτυγχάνεται µε τη διέλευσή του αέρα µέσα από ειδικά φίλτρα νιτροκυτταρίνης (d 2µm) (Laminar Downflow Work Station Type A, Microflow, MDH). Η ίδια εστία διαθέτει επίσης ειδική λάµπα ακτινοβολίας UV για την αποστείρωση του χώρου πριν τη χρήση. Αποστείρωση σε πολύ υψηλές θερµοκρασίες είτε σε ξηρό περιβάλλον (κλίβανος) µε τη παροχή θερµού αέρα, είτε σε υγρό περιβάλλον (αυτόκαυστο) µε τη παροχή ατµού µε πίεση, για την ενίσχυση των άσηπτων συνθηκών. Τα πλαστικά µικροαντικείµενα (ακροφύσια, φιαλίδια) καθώς και τα διαλύµατα που χρησιµοποιούνται αποστειρώνονται σε αυτόκαυστο κλίβανο (2 atm, C, 15-2 ), ενώ τα γυάλινα σκεύη και µεταλλικά αντικείµενα σε ξηρό κλίβανο (16 C, 6-9 ). οχεία καλλιέργειας ανάλογης επιφάνειας και χωρητικότητας για τη συγκεκριµένη κυτταρική καλλιέργεια. Κατάλληλα θρεπτικά µέσα και πρόσθετα. Επωαστικό θάλαµο για τη διατήρηση της βέλτιστης θερµοκρασίας ανάπτυξης, της απαιτούµενης υγρασίας και αέριας τάσης. Μετά την προσθήκη όλων των απαραίτητων συστατικών σε ειδικά αποστειρωµένα δοχεία, οι καλλιέργειες τοποθετούνται σε επωαστικό θάλαµο ανοιχτού τύπου (Water-Jacketed Incubator 325, USA Forma Scientific Inc.), µε παροχή 5% CO2/95% ατµοσφαιρικού αέρα, 9% σχετική υγρασία και θερµοκρασία 37 C. Για τον καθαρισµό του επωαστικού θαλάµου σε τακτά χρονικά διαστήµατα χρησιµοποιείται το διάλυµα Roccal II 1% (National Laboratories), ως χηµικό µέσο αποστείρωσης. Σταθερό pη καλλιέργειας ( ), ένα σηµαντικό παράγοντα ανάπτυξης ο οποίος πρέπει να ελέγχεται σε όλη τη διάρκεια της επώασης. 6.1 Ανθρώπινα λεµφοκύτταρα ολικού αίµατος Το βιολογικό σύστηµα που χρησιµοποιείται κύρια στην παρούσα εργασία για τη µελέτη του µηχανισµού πρόκλησης ανευπλοειδίας στον άνθρωπο από τη φαρµακευτική ένωση υδροχλωροθειαζίδιο είναι τα ανθρώπινα λεµφοκύτταρα του περιφερικού αίµατος. Πρόκειται για ένα ευρέως διαδεδοµένο και ταυτόχρονα ευαίσθητο σύστηµα ανίχνευσης αριθµητικών αλλαγών και δοµικών ανακατατάξεων χρωµοσωµάτων που εφαρµόζεται είτε σε ασθενείς για διαγνωστικές ιατρικές µελέτες είτε σε οµάδες ατόµων που εκτίθενται σε πιθανά επικίνδυνους παράγοντες είτε σε υγιείς δότες για τον έλεγχο της ενεργότητας τόσο περιβαλλοντικών παραγόντων όσο και χηµικών ενώσεων στην πρόκληση χρωµοσωµατικών ανωµαλιών. Η αξιοπιστία του συγκεκριµένου βιολογικού συστήµατος οφείλεται στο γεγονός ότι το αίµα είναι ένας από τους πιο ευπρόσιτους και ιδιαίτερους ιστούς του ανθρώπινου οργανισµού που τα κύτταρα του εµφανίζουν ένα σαφή, πλήρως χαρακτηρισµένο κυτταρικό κύκλο. Το αίµα είναι ένα εναιώρηµα διαφόρων τύπων κυττάρων σε ένα σύνθετο διάλυµα (πλάσµα), που περιέχει αέρια (Ο2, CO2, N2), άλατα, πρωτεΐνες (αλβουµίνη, ανοσοσφαιρίνες), υδρογονάνθρακες και λιπίδια
90 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες Τα κύτταρα του αίµατος των σπονδυλωτών διακρίνονται σε ερυθρά αιµοσφαίρια ή ερυθροκύτταρα, σε λευκά αιµοσφαίρια ή λευκοκύτταρα και σε αιµοπετάλια ή θροµβοκύτταρα, τα οποία προέρχονται από θραύσµατα του κυτταροπλάσµατος των µεγακαρυοκυττάρων (Berne και Levy, 1996). Τα λευκά αιµοσφαίρια, σε αντίθεση µε τα ερυθροκύτταρα είναι εµπύρηνα, γεγονός που τα καθιστά κατάλληλα σε κυτταρογενετικές µελέτες και ταξινοµούνται ανάλογα µε το µέγεθος τους (6-22µm σε διάµετρο), την παρουσία και τον τύπο κοκκίων, το σχήµα του πυρήνα και το χαρακτήρα του κυτταροπλάσµατος σε τρεις διακριτές κατηγορίες: (α) τα κοκκιοκύτταρα (granulocytes), τα οποία υποδιαιρούνται σε ουδετερόφιλα (πολυµορφοπύρηνα λευκοκύτταρα), βασεόφιλα και εωσινόφιλα, (β) τα µονοκύτταρα ή µονοπύρηνα (monocytes) και (γ) τα λεµφοκύτταρα (lymphocytes) (Alberts et al., 1994). Οι διαδοχικές διαιρέσεις πολυδύναµων αρχέγονων αιµοποιητικών κυττάρων που στο στάδιο της ενηλικίωσης στα θηλαστικά εντοπίζονται στο µυελό των οστών οδηγούν στη γένεση των διαφόρων κυτταρικών τύπων του περιφερικού αίµατος και αποτελούν µια διαδικασία γνωστή ως αιµοποίηση (hematopoiesis). Το πρώτο στάδιο ωρίµανσης των κυττάρων του αίµατος περιλαµβάνει τη διαίρεση των πρωταρχικών πολυδύναµων κυττάρων σε δύο κύριες κυτταρικές σειρές: τη µυελοειδή και τη λεµφοειδή. Η λεµφοειδής σειρά παράγει δύο κύριους κυτταρικούς τύπους που σχετίζονται άµεσα µε το ανοσοποιητικό σύστηµα, τα Β και τα Τ λεµφοκύτταρα (Alberts et al., 1994, Μαρµάρας και Λαµπροπούλου-Μαρµάρα, 2) Λεµφοκύτταρα Τα λεµφοκύτταρα συνιστούν µία κατηγορία σωµατικών κυττάρων τα οποία είναι υπεύθυνα για την επίκτητη ανοσία. Ένας µεγάλος αριθµός λεµφοκυττάρων παράγεται καθηµερινά (1 9 ) από τα πρωτογενή ή κεντρικά λεµφικά όργανα (µυελό των οστών και θύµο αδένα) τα οποία διαµέσου της κυκλοφορίας του αίµατος και της λέµφου µεταναστεύουν σε δευτερογενείς ή περιφερικούς λεµφικούς ιστούς και όργανα. Στο περιφερικό λεµφικό σύστηµα περιλαµβάνονται ο σπλήνας, οι λεµφαδένες, οι αµυγδαλές, η σκωληκοειδής απόφυση, οι πλάκες Peyer του λεπτού εντέρου και ο µυελός των οστών. Ο συνολικός αριθµός λεµφοκυττάρων στο ανθρώπινο σώµα είναι περίπου 2 Χ 1 12 κύτταρα και τα περισσότερα από αυτά είναι καθηλωµένα στα διάφορα όργανα µε αποτέλεσµα σε κάθε 1 κύτταρα του ανθρώπινου σώµατος, το ένα να είναι λεµφοκύτταρο. Επιπρόσθετα, σηµαντικό πλεονέκτηµα για τη διέγερση της ανοσοαπόκρισης είναι η δυνατότητα ανανέωσης που παρέχεται µε συνεχή κίνηση των λεµφοκυττάρων ανάµεσα στα διάφορα λεµφικά όργανα, µε αποτέλεσµα να αυξάνεται η πιθανότητα ένα λεµφοκύτταρο να έλθει σε επαφή µε οποιοδήποτε ξένο προς τον οργανισµό σώµα. Η µάζα που συγκροτούν στο σύνολο τους αποτελεί περίπου το 2% του βάρους του ανθρώπινου σώµατος. Στο περιφερικό αίµα ενός µέσου ανθρώπου η εκατοστιαία κατ όγκο αναλογία των λευκών αιµοσφαιρίων είναι 1% και από αυτά το 2-3% συνιστούν τα λεµφοκύτταρα. Συγκεκριµένα, ο συνολικός αριθµός λεµφοκυττάρων που εντοπίζεται στο αίµα φυσιολογικών ενήλικων ατόµων είναι Χ 1 9 κύτταρα/λίτρο αίµατος και από αυτά ένα υψηλό ποσοστό περίπου 7% είναι τύπου Τ (Roitt et al., 21, Gosden et al., 1992). Τα λεµφοκύτταρα ταξινοµούνται αρχικά σε δύο τύπους ανάλογα µε το µέγεθος (6-1µm σε διάµετρο) και τη µορφολογία. Οι βασικές διαφορές που παρατηρούνται εντοπίζονται στην αναλογία πυρήνα-κυτταροπλάσµατος (Ν:C ratio), στο σχήµα του πυρήνα και στην παρουσία ή απουσία κοκκίων. Η υποτυπώδης αυτή διάκριση περιλαµβάνει τα τυπικά µικρά λεµφοκύτταρα στα οποία ο πυρήνας είναι αρκετά µεγάλος και περιβάλλεται από µία µικρή ποσότητα κυτταροπλάσµατος που
91 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες δεν φέρει κοκκία, σε αντίθεση µε τα µεγάλα κοκκιώδη λεµφοκύτταρα (LGL). Η παρουσία συγκεκριµένων δεικτών στην πλασµατική µεµβράνη των λεµφοκυττάρων και η κύρια λειτουργία ανοσοαπόκρισης που επιτελούν τα διακρίνει σε δύο κύριους τύπους: τα Β-λεµφοκύτταρα τα οποία παρέχουν τη χυµική ανοσία και τα Τ- λεµφοκύτταρα υπεύθυνα για την κυτταρική ανοσία (Roitt et al., 21). Β-λεµφοκύτταρα Η µορφολογία των Β-λεµφοκυττάρων που βρίσκονται σε ηρεµία είναι χαρακτηριστική των τυπικών µικρών λεµφοκυττάρων και η θέση ωρίµανσης τους εντοπίζεται στο µυελό των οστών (bone marrow) στα ενήλικα θηλαστικά. Αποτελούν το 5-15% των λεµφοκυττάρων που βρίσκονται σε διαρκή κυκλοφορία διαµέσου του αίµατος και των λεµφαγγείων και αναγνωρίζονται από την παρουσία ανοσοσφαιρινών (κυρίως IgM και IgD) που παράγονται σε αυτά και εγκαθίστανται στην επιφάνεια της κυτταρικής τους µεµβράνης. Η αλληλεπίδραση ανοσοσφαιρινών µε βοηθητικά µόρια που αποτελούνται από τα ετεροδιµερή Ιgα (CD79a) και Ιgβ (CD79b) και εντοπίζονται αποκλειστικά στην επιφάνεια των Β-λεµφοκυττάρων συγκροτούν το σύµπλεγµα του αντιγονικού υποδοχέα των Β-κυττάρων (BCR). Η διέγερση των Β- λεµφοκυττάρων µε ένα αντιγόνο επάγει τη διαφοροποίηση τους αρχικά σε λεµφοβλάστες και στη συνέχεια µε συνεχείς διαιρέσεις σε κύτταρα µνήµης και σε πλασµατοκύτταρα τα οποία συνθέτουν και εκκρίνουν αντισώµατα σε µεγάλες ποσότητες που µεταφέρονται µε τη ροή του αίµατος στο σηµείο δράσης. Τ-λεµφοκύτταρα Μελέτες στο ηλεκτρονικό µικροσκόπιο έδειξαν το σωµάτιο Gall (Gall body) που αποτελείται από πρωτογενή λυοσώµατα και σταγονίδια λίπους, ως τη χαρακτηριστική λεπτή δοµή των Τ-λεµφοκυττάρων που βρίσκονται σε ηρεµία. Τα Τ-λεµφοκύτταρα προέρχονται από αρχέγονα αιµοποιητικά κύτταρα του µυελού των οστών, τα οποία διαφοροποιούνται στο θύµο αδένα (thymus gland). Ο κυριότερος δείκτης των Τ κυττάρων είναι ο αντιγονικός υποδοχέας TCR, ο οποίος αποτελείται από δύο πολυπεπτιδικές αλυσίδες παρόµοιου Μ.Β. τις α και β ή γ και δ που συνδέονται µεταξύ τους µε δισουλφιδικούς δεσµούς, εάν και ο πιο πρώιµος δείκτης των κυττάρων αυτών είναι ο Thy-1 που πρωτοεµφανίζεται κατά τη διάρκεια ωρίµανσης των Τ- κυττάρων στο θύµο αδένα. Οι αβ και γδ µορφές του υποδοχέα TCR σχετίζονται µε µία σειρά τεσσάρων πολυπεπτιδίων (συστατικό CD3) και συγκροτούν στην επιφάνεια των Τ-λεµφοκυττάρων το σύµπλεγµα TCR-CD3. Τα Τ-λεµφοκύτταρα ανάλογα µε τη λειτουργία την οποία επιτελούν στη διαδικασία της ανοσοαπόκρισης διακρίνονται κύρια σε Τ-κύτταρα αρωγοί (Τ helper cells-τη) και κυτταροτοξικά Τ- κύτταρα (T cytotoxic cells-tc). Στο περιφερικό αίµα τα ανθρώπινα Τ-λεµφοκύτταρα σε ποσοστό 9-95% εκφράζουν τους υποδοχείς α:β και περιλαµβάνουν δύο µη επικαλυπτόµενους υποπληθυσµούς (CD4, CD8) ανάλογα µε την παρουσία δεiκτών στην επιφάνεια τους. Τα Τ-κύτταρα αρωγοί (CD4) σε αναλογία 95% και τα κυτταροτοξικά Τ-κύτταρα (CD8) σε αναλογία 5% φέρουν την χαρακτηριστική µορφολογία των τυπικών µικρών λεµφοκυττάρων (Goldsby et al., 2, Roitt et al., 21). Τα ανθρώπινα λεµφοκύτταρα του περιφερικού αίµατος είναι ένας κυτταρικός πληθυσµός που στη µεγάλη του πλειονότητα βρίσκεται στη Go φάση του κυτταρικού κύκλου. Κατά τη διάρκεια αυτής της φάσης οι βιοχηµικές και φυσιολογικές δραστηριότητες των κυττάρων βρίσκονται σε καταστολή. Συγκεκριµένα γίνεται αντιγραφή DNA σε ένα ποσοστό λεµφοκυττάρων.2% ή και µικρότερο που πιθανόν εκπροσωπούν ενεργοποιηµένα Τ- ή Β- λεµφοκύτταρα (Natarajan και Obe, 1982). ιάφοροι παράγοντες όπως η ηλικία, το φύλο, το κάπνισµα, η λήψη φαρµάκων, η ακτινοβολία, η σωµατική καταπόνηση, οι ακραίες συναισθηµατικές - 8 -
92 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες καταστάσεις, η εγκυµοσύνη, αλλά και οι κλιµατικές συνθήκες επηρεάζουν σηµαντικά το ποσοστό των λευκών αιµοσφαιρίων (λεµφοκυττάρων) στο αίµα (Watt και Stephen, 1986). ιέγερση λεµφοκυττάρων Μερικές φυτικές ουσίες γνωστές ως λεκτίνες (γλυκοπρωτεΐνες) καθώς και κάποιοι βακτηριακοί πολυσακχαρίτες επάγουν βλαστογένεση και µίτωση σε λεµφοκύτταρα. Η δέσµευση τους σε ειδικούς υποδοχείς της κυτταρικής επιφάνειας είναι σε θέση να δηµιουργήσει τα κατάλληλα σήµατα που επιφέρουν σηµαντικές αλλαγές στο µεταβολισµό του κυττάρου. Το άµεσο επακόλουθο της ενεργοποίησης του πυρήνα είναι η αύξηση της σύνθεσης DNA και ο µετασχηµατισµός των λεµφοκυττάρων σε διαιρούµενους λεµφοβλάστες. Σηµαντικό βήµα στην ανάπτυξη της τεχνικής της καλλιέργειας των λεµφοκυττάρων in vitro υπήρξε η ανακάλυψη µιτογόνων ουσιών που έχουν τη δυνατότητα να διεγείρουν τον πολλαπλασιασµό και τη διαφοροποίηση των Τ λεµφοκυττάρων. Σε αυτή την κατηγορία ανήκει η φυτοαιµατογλουτινίνη (PHA), η οποία αποτελεί µία µιτογόνο ουσία που προέρχεται από τα κόκκινα φασόλια του είδους Phaseolus vulgaris. Η διέγερση των Τ-λεµφοκυττάρων από την PHA περιλαµβάνει την ενεργοποίηση πολύπλοκων µονοπατιών στα οποία εµπλέκεται ένας µεγάλος αριθµός ειδικών διαµεσολαβητών. Σε αυτούς περιλαµβάνονται το σύµπλεγµα TCR-CD3, MAP (Mitogen Activated Proteins) κινάσες και κυτταροκίνες. Η δέσµευση της PHA σε ειδικά σάκχαρα των επιφανειακών γλυκοπρωτεϊνών (CD3) του συµπλέγµατος TCR-CD3 προκαλεί την άµεση διέγερση των Τ-λεµφοκυττάρων και η διαδικασία ενεργοποίησης τους ολοκληρώνεται µε την επαγωγή της έκφρασης των γονιδίων της ιντερλευκίνης-2 (IL-2) και της υποµονάδας α του υποδοχέα της (IL-2 Rα). Οι πρώτες µιτωτικές διαιρέσεις παρατηρούνται 48 ώρες µετά την έναρξη της καλλιέργειας και επαναλαµβάνονται περιοδικά κάθε 24 ώρες (Gosden et al., 1992, O Donovan et al., 1995, Roitt et al., 21) Συλλογή δειγµάτων Στην παρούσα µελέτη συµπεριελήφθησαν συνολικά 32 άτοµα (πίνακας 6.1), τα οποία διαχωρίστηκαν σε οµάδες µε κριτήριο την ηλικία και το φύλο. Από αυτά τα 16 ήταν νεαρά αρσενικά και θηλυκά άτοµα ηλικίας έως 25 ετών. Τα υπόλοιπα 16 ήταν αρσενικά και θηλυκά άτοµα ώριµης ηλικίας άνω των 45 ετών. Από όλους τους δότες συµπληρώθηκε ερωτηµατολόγιο που αφορούσε την υγεία τους, τη λήψη θεραπευτικής αγωγής και τις προσωπικές τους συνήθειες όπως το κάπνισµα. Με αυτό τον τρόπο διασφαλίσαµε τη σωστή επιλογή ατόµων που κατά τη διάρκεια του τελευταίου τριµήνου πριν τη λήψη του αίµατος δεν είχαν υποστεί κάποια µορφή λοίµωξης, δεν τους χορηγήθηκαν φαρµακευτικά σκευάσµατα και δεν είχαν υποβληθεί σε ακτινολογική εξέταση
93 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες Πίνακας 6.1. ιαχωρισµός των ατόµων σε οµάδες µε κριτήριο το φύλο, την ηλικία και τη συνήθεια του καπνίσµατος. A.A. ΟΤΕΣ ΦΥΛΟ ΗΛΙΚΙΑ ΚΑΠΝΙΣΤΕΣ ΝΑΙ ΟΧΙ 1. Α.Χ. Α Α.Π. Α Α.Α. Α Κ.Γ. Α Σ.Η. Α Β.Α. Α Π.Ν. Α Τ.Μ. Α 23 + ΜΤ±ΤΣ 23.38± Κ.Μ. Θ Κ.Α. Θ Μ.Κ. Θ Ν.Κ. Θ Π.Ν. Θ Π.Π. Θ Π.Μ. Θ Σ.Κ. Θ 23 + ΜΤ±ΤΣ 23.63± Β.Α. Α Κ.Γ. Α Σ.Ν. Α Χ.Β. Α Γ.Χ. Α Γ.Θ. Α Ι.Γ. Α 52 (+) 24. Σ.Κ. Α 53 + ΜΤ±ΤΣ 52.38± Κ.Σ. Θ Λ.Μ. Θ Ν.Μ. Θ Π.Ε. Θ Σ.Σ. Θ Σ.Γ. Θ Χ.Ε. Θ 43 (+) 32. Χ.Α. Θ 51 + ΜΤ±ΤΣ 51.88± Πηγή λεµφοκυττάρων Ως πηγή λεµφοκυττάρων χρησιµοποιήθηκε φλεβικό αίµα που συλλέχθηκε από τους παραπάνω υγιείς δότες. Η αιµοληψία γίνεται µε αποστειρωµένη σύριγγα µίας χρήσης (της εταιρείας Becton Dickinson B-D Made in U.K.). Το δείγµα µεταφέρεται σε ηπαρινισµένο και αποστειρωµένο σωληνάριο χωρητικότητας 1 ml και διαστάσεων 16x1mm (BD Vacutainer, LH 143 I.U. Made in U.K.) αναδεύεται ήπια και παραµένει σε θερµοκρασία δωµατίου για µικρό χρονικό
94 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες διάστηµα (15-2 min), ενώ διατηρείται στους (+4) C, έως ότου χρησιµοποιηθεί. Στις καλλιέργειες µας το ανώτερο όριο παραµονής ήταν 1-2 ώρες µετά τη λήψη του αίµατος Συνθήκες καλλιέργειας λεµφοκυττάρων Απαραίτητα υλικά που χρησιµοποιήθηκαν για την καλλιέργεια ανθρώπινων λεµφοκυττάρων είναι τα εξής: Ειδικά αποστειρωµένα (sterile, DNAase, RNAase free) δοχεία καλλιέργειας µίας χρήσης (Cellstar R, Tissue Culture Flask-Greiner Bio One, GmbH) χωρητικότητας 5 ml και επιφανείας 25cm 2. Θρεπτικό µέσο (Nutrient mixture Ham s F1 w L-Glutamine/w/o L- Glutamine, Gibco BRL TM /Biochrom KG) που περιέχει ανόργανα άλατα, αµινοξέα (απαραίτητα και µη απαραίτητα), βιταµίνες σε χαµηλές συγκεντρώσεις και άλλα συστατικά (γλυκόζη, υποξανθίνη, ριβοφλαβίνη, θυµιδίνη, ερυθρό της φαινόλης ως δείκτη pη). Ορός εµβρύου βοός (Foetal Bovine Serum Heat/UV-light Inactivated, Gibco BRL TM /Biochrom KG) ως συµπλήρωµα του θρεπτικού µέσου, ο οποίος συλλέγεται µε καρδιακή παρακέντηση και ελέγχεται αυστηρά για ενδοτοξίνες, µυκόπλασµα, ιούς (BVD-MD, BHV-1, PI-3V), αντισώµατα έναντι αυτών των ιών και βακτήρια (L-Forms). Είναι ένα εξαιρετικά σύνθετο µείγµα το οποίο περιέχει πρωτεΐνες του πλάσµατος (α,β,γ σφαιρίνες, αιµοσφαιρίνη), παράγοντες ανάπτυξης, ορµόνες, µεγάλες ποσότητες ιόντων νατρίου, χλωρίου, διττανθρακικών ιόντων και µεταλλικά ιχνοστοιχεία. Φυτοαιµατογλουτινίνη (PHA lyophilised, Gibco BRL TM ), η οποία χρησιµοποιείται ως ανοσοδιεγέρτης της κυτταρικής διαίρεσης σε προτεινόµενες συγκεντρώσεις που δεν υπερβαίνουν το 1-2% του θρεπτικού µέσου και ενυδατώνεται µε αποστειρωµένο απεσταγµένο νερό. Παρ όλα αυτά η συγκεκριµένη συγκέντρωση µπορεί να ποικίλλει ανάλογα µε την καλλιέργεια κυττάρων για την οποία χρησιµοποιείται και την απόκριση του δότη. Στην ενυδατωµένη της µορφή τα αποθέµατα της φυτοαιµατογλουτινίνης φυλάγονται για διάστηµα 6 µηνών στους (-5) C - (-2) C και για δύο εβδοµάδες στους (+4) C. Γλουταµίνη (L-Glutamine, Serva), ως αµινοξύ το οποίο προστίθεται στο θρεπτικό µέσο, ενώ συντίθεται σε ικανοποιητικές ποσότητες (µη απαραίτητο) από τα κύτταρα των θηλαστικών, εξαιτίας της αστάθειας που παρουσιάζει σε διαλυτή µορφή στις συνήθεις συνθήκες αποθήκευσης. Η αστάθεια του οφείλεται στην ικανότητα που εµφανίζει να κυκλοποιείται και να σχηµατίζει τοξική αµµωνία. Παρ όλα αυτά, σε ένα πλήρες καλλιεργητικό µέσο που περιλαµβάνονται µη απαραίτητα αµινοξέα πιθανόν να ελαχιστοποιείται το µεταβολικό φορτίο των κυττάρων µε αποτέλεσµα να επάγεται ο πολλαπλασιασµός τους και κατ επέκταση ο ρυθµός ανάπτυξης της καλλιέργειας να είναι υψηλός. Πενικιλίνη και Στρεπτοµυκίνη (Penicillin-Streptomycin, Gibco BRL TM / Biochrom KG) είναι το µείγµα αντιβιοτικών έναντι κυρίως gram θετικών και gram αρνητικών βακτηρίων, αλλά µε ένα ευρύτερο φάσµα δράσης που χρησιµοποιείται στις καλλιέργειες λεµφοκυττάρων για την αποφυγή τυχόν επιµολύνσεων. Το πλήρες καλλιεργητικό υλικό που απαιτείται για τη σωστή ανάπτυξη των λεµφοκυττάρων σε καλλιέργεια περιλαµβάνει:
95 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες ΠΛΗΡΕΣ ΚΑΛΛΙΕΡΓΗΤΙΚΟ ΥΛΙΚΟ 1. Εµπλουτισµένο θρεπτικό µέσο Ham s F1 ΣΥΓΚΕΝΤΡΩΣΗ 8% (6.5ml) 2. Ορός εµβρύου βοός 2% (1.5ml) 3. Φυτοαιµατογλουτινίνη 2.5-3% (.26ml) 4. Φλεβικό αίµα.5ml Ο εµπλουτισµός του θρεπτικού µέσου πραγµατοποιείται µε την προσθήκη 2mM γλουταµίνης, 1 IU/ml πενικιλίνης και 1 µg/ml στρεπτοµυκίνης, ως µείγµα αντιβιοτικών σε 1 ml Ham s F1. Αξιοσηµείωτο είναι ότι η τελική συγκέντρωση των λεµφοκυττάρων σε καλλιέργειες ολικού αίµατος από µικρές ποσότητες (.3 -.5ml αίµατος) ποικίλει από X1 5 κύτταρα/ml, ανάλογα µε το δότη (Countryman και Heddle, 1976, Preston et al., 1987, Ellard και Parry, 1993). 6.2 Κυτταρικές σειρές Οι κυτταρικές σειρές (Hep 2, HFFF2, C2C12) που χρησιµοποιήθηκαν προέρχονται από κύτταρα πρωτογενών καλλιεργειών, τα οποία αποµονώθηκαν από διαφορετικούς ιστούς ή όργανα (φυσιολογικούς ή µη), ανακαλλιεργήθηκαν και επιλέχθηκαν για να αποτελέσουν οµοιογενείς πληθυσµούς. Αυτές είναι διαθέσιµες από συλλογές που δηµιουργήθηκαν στην Αµερική (ATCC), και στην Ευρώπη (ECACC, DSMZ). Για τις ανακαλλιέργειες κυττάρων διαφορετικού τύπου (καρκίνωµα του λάρυγγα, επιδερµικοί ινοβλάστες, µυοβλάστες) η επιλογή του κατάλληλου θρεπτικού µέσου σε συνδυασµό µε τον εµπλουτισµό του αποτελεί ένα σηµαντικό παράγοντα άριστης ανάπτυξης. Οι κύριες διαφορές εντοπίζονται στις συγκεντρώσεις των ανόργανων αλάτων, των βιταµινών, των απαραίτητων αµινοξέων, της γλυκόζης καθώς και στο ποσοστό των πρωτεϊνών του ορού. Μία άλλη παράµετρος αφορά τη διατήρηση των ζωικών κυττάρων για µεγάλο χρονικό διάστηµα που επιτυγχάνεται µε την αποθήκευση τους σε υγρό άζωτο σε θερµοκρασίες µεταξύ -175 C και -196 C. Η επιτυχής διατήρηση τους (cryopreservation) που συνδέεται άµεσα µε τη βαθµιαία διαδικασία ψύξης γίνεται για να αποφευχθεί η απώλεια της κυτταρικής σειράς από επιµολύνσεις, η γήρανση και ο µετασχηµατισµός σε σειρές µε πεπερασµένο αριθµό κυτταρικών διαιρέσεων (HFFF2, C2C12) και να ελαχιστοποιηθεί ο κίνδυνος να συµβούν γενετικές αλλαγές σε σειρές µε ακαθόριστο αριθµό κυτταρικών διαιρέσεων (Hep 2). Η διαδικασία που συνήθως ακολουθείται για τη βαθµιαία ψύξη (1 C/λεπτό) των κυττάρων, αφορά την προσθήκη 1% διµεθυλουσουλφοξειδίου (DMSO, sterile filtered, test for endotoxin, Sigma) σε πλήρες θρεπτικό µέσο (1ml) µε τη συγκέντρωση τους να ποικίλει από X1 6 κύτταρα/ml. Αρχικά το ειδικό αποστειρωµένο (sterile, DNAase, RNAase free) φιαλίδιο αποθήκευσης σε χαµηλές θερµοκρασίες (Cryogenic Storage Vial, Greiner Bio One) µε την επιθυµητή ποσότητα κυττάρων παραµένει στους (+4) C για 5, ενώ στη συνέχεια τοποθετείται σε µονωµένο δοχείο και µεταφέρεται στους (-7) C-(-9) C. Εκεί παραµένει για
96 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες ανάλογο χρονικό διάστηµα µέχρι να ολοκληρωθεί η µεταφορά και αποθήκευση του σε υγρό άζωτο. Με τη συγκεκριµένη διαδικασία αποφεύγεται ο σχηµατισµός πάγου στο εσωτερικό των κυττάρων, ελαχιστοποιείται η απώλεια νερού και η δηµιουργία υψηλών τοπικών συγκεντρώσεων DMSO, οι οποίες είναι τοξικές για τα κύτταρα. Αντίθετα, η διαδικασία απόψυξης των κυττάρων γίνεται ταχύτατα και περιλαµβάνει τη µεταφορά του φιαλιδίου µε το εναιώρηµα των κυττάρων από το δοχείο αποθήκευσης σε υδατόλουτρο (Memmert WB/OB 7-45) στους 37 C για 6-9, την τοποθέτηση των κυττάρων σε πλήρες θρεπτικό µέσο σε ειδικά δοχεία καλλιέργειας µίας χρήσης, την παραµονή των καλλιεργειών σε επωαστικό θάλαµο (θερµοκρασία 37 C και παροχή 5% CO2) και την αλλαγή του θρεπτικού µέσου µετά από ώρες επώασης. Τα κύτταρα παρουσιάζουν την ικανότητα προσκόλλησης στην επιφάνεια του δοχείου (γυάλινο ή πλαστικό), µία ιδιότητα που οφείλεται στην έκκριση κολλωδών και ινωδών πρωτεϊνών, όπως π.χ. λαµινίνης, φιµπρονεκτίνης και κολλαγόνου. Με την προσκόλληση τους αρχίζει η διαίρεση και κατά συνέπεια η επέκτασή τους (spreading) στην επιφάνεια του δοχείου καλλιέργειας. Συνήθως δηµιουργείται ένα στρώµα από κύτταρα πάχους ενός ή µερικών κυττάρων, ανάλογα µε τον κυτταρικό τύπο. Τα καρκινικά κύτταρα που έχουν συνήθως µικρή συγγένεια µε την επιφάνεια του δοχείου καλλιέργειας σχηµατίζουν ανώµαλες µάζες κυττάρων. Όταν η συνολική κάλυψη της επιφάνειας του δοχείου είναι ~7-9% (συνήθως µετά ώρες), ξεκινά η διαδικασία αποκόλλησης των κυττάρων, αποµόνωσης αυτών και επίστρωσής τους είτε σε τρυβλία petri (Cellstar R, Tissue Culture Dishes 35/1mm, Greiner Bio One, GmbH) µε τη χρησιµοποίηση καλυπτρίδων 22X22 mm για περαιτέρω επεξεργασία είτε σε νέα δοχεία καλλιέργειας που φέρουν ειδικό αεριζόµενο πώµα µε φίλτρο (25cm 2 Cell Culture Flask, 2µm Vent Cap-Corning), διαδικασία γνωστή ως ανακαλλιέργεια. Η παραπάνω διαδικασία πραγµατοποιείται ακολουθώντας τα εξής στάδια: Απόρριψη του υπερκειµένου. Έκπλυση των κυττάρων µε θρεπτικό µέσο ή µε Dulbecco s PBS (DPBS, Sigma). Επίδραση µε το διάλυµα αποκόλλησης. Αυτό περιέχει συνήθως τρυψίνη/ EDTA σε αναλογίες οι οποίες διαφέρουν ανάλογα µε τον τύπο της καλλιέργειας. Επίσης σηµαντικός είναι ο χρόνος παραµονής και η ποσότητα του διαλύµατος (1-3ml/25cm 2 ) που θα χρησιµοποιηθεί κατά τη διαδικασία αποκόλλησης για την αποφυγή κυτταρικής βλάβης. Η παρουσία EDTA κρίνεται απαραίτητη για τη δέσµευση του Ca 2+, από το οποίο εξαρτάται η πρόσφυση των κυττάρων. Σε κάποιες περιπτώσεις η αποκόλληση των κυττάρων επιτυγχάνεται µε λεπτούς χειρισµούς µίας ειδικής σπάτουλας (Scraper, Disposable Cell Scraper, Greiner Bio One, GmbH). Προσθήκη πλήρους θρεπτικού υλικού (η παρουσία ορού αναστέλλει τη δράση της τρυψίνης), µέτρηση του αριθµού των κυττάρων σε αιµοκυτταρόµετρο (πλάκα Neubauer) και διαχωρισµός (split) της καλλιέργειας H.Ep.2 (Human Epidermoid Carcinoma) Η πρωτογενής καλλιέργεια από την οποία προήλθε η συγκεκριµένη κυτταρική σειρά προέκυψε από µετασχηµατισµό εµβρύων ποντικού µε επιδερµικό καρκίνωµα του λάρυγγα που εµφανίσθηκε σε αρσενικό άτοµο ηλικίας 56 ετών(species: human, Caucasian male, Tissue: larynx, Tumor: carcinoma, epidermoid). Πρόκειται για µία κυτταρική σειρά (ECACC, Νο ) µε
97 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες απεριόριστο αριθµό κυτταρικών διαιρέσεων (continuous cell line). Εµφανίζουν µορφολογία παρόµοια µε αυτή των επιθηλιακών κυττάρων και σε άριστες συνθήκες καλλιέργειας (θερµοκρασία 37 C και παροχή 5% CO2) αναπτύσσονται ως µονόστιβο (monolayer) κυτταρικό στρώµα. Στη συγκεκριµένη κυτταρική σειρά, που εµφανίζει τα χαρακτηριστικά της ανθρώπινης σειράς HeLa, σηµαντική είναι η ικανότητα των κυττάρων να εκφράζουν ένζυµα µεταβολισµού ξενοβιοτικών ουσιών. Το πλήρες θρεπτικό µέσο που απαιτείται για την καλλιέργεια περιέχει τα εξής συστατικά: Eagle s Minimum Essential Medium w Earle s Salts, w/o L-Glutamine (EMEM, Gibco BRL TM ). Το ΜΕΜ είναι ένα απλό θρεπτικό µέσο που δηµιουργήθηκε από τον Eagle µε τροποποίηση του βασικού (BME) και θεωρείται κατάλληλο για την ανάπτυξη µίας µεγάλης ποικιλίας κυτταρικών τύπων, ιδιαίτερα στην περίπτωση που χρησιµοποιείται ορός ως συµπλήρωµα. Το ΜΕΜ µε την προσθήκη του ρυθµιστικού διαλύµατος Earle s σχεδιάστηκε για την ανάπτυξη καλλιεργειών σε επωαστικούς θαλάµους ανοιχτού τύπου µε παροχή 5%CO2/95% ατµοσφαιρικού αέρα. 5%-1% ορό εµβρύου βοός (FBS, Gibco BRL TM ). 3% διττανθρακικό νάτριο 7.5% (Sodium Bicarbonate 7.5%, Euroclone). Το διττανθρακικό νάτριο (NaHCO3) σε συνδυασµό µε τα κατάλληλα είδη ρυθµιστικών διαλυµάτων συµβάλλει στην αποφυγή ανεπιθύµητων διακυµάνσεων του ph, εξαιτίας του µεταβολισµού των κυττάρων, που πιθανόν να προκαλέσουν αναστολή της κυτταρικής ανάπτυξης ή να τροποποιήσουν τους βιολογικούς δείκτες που µελετώνται. 4mM γλουταµίνη (L-Glutamin 2mM, Biological Industries). 1 IU/ml πενικιλίνη και 1 µg/ml στρεπτοµυκίνη (Penicillin- Streptomycin 5IU/ML-5UG/ML, Gibco BRL TM ). ΠΛΗΡΕΣ ΘΡΕΠΤΙΚΟ ΜΕΣΟ 1. Εµπλουτισµένο (α,β,γ) θρεπτικό µέσο EMEM ΣΥΓΚΕΝΤΡΩΣΗ 95% 1α. ιττανθρακικό νάτριο 3% 1β. Γλουταµίνη 2% 1γ. Πενικιλίνη/Στρεπτοµυκίνη 2% 2. Ορός εµβρύου βοός 5% Η καλλιέργεια αναπτύσσεται σε δοχεία επιφανείας 25cm 2 που περιέχουν 5ml πλήρους θρεπτικού µέσου. Ο κατάλληλος αριθµός κυττάρων που προστίθεται για τη σωστή επέκταση τους σε τρυβλία petri και σε δοχεία καλλιέργειας είναι 1-3. κύτταρα/cm 2 επιφανείας. Για τις ανακαλλιέργειες χρησιµοποιήθηκε διάλυµα τρυψίνης/edta (.5%/.2%) σε PBS που δεν περιέχει Ca 2+ και Mg 2+ (Biological Industries). Η ποσότητα που προστίθεται είναι.5ml και ο χρόνος επώασης είναι 1-2 στους 37 C
98 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες Ο διαχωρισµός της καλλιέργειας πραγµατοποιείται µία φορά κάθε δύο ηµέρες σε αναλογία 1:3 έως 1:1. Οι δυνατότητες ανακαλλιέργειας της συγκεκριµένης κυτταρικής σειράς είναι απεριόριστες (~217) H.F.F.F.2 (Human Caucasian F(o)etal Foreskin Fibroblast) Πρόκειται για µία διπλοειδή κυτταρική σειρά (ECACC, Νο ) µε περιορισµένο αριθµό κυτταρικών διαιρέσεων (finite life span). Οι ινοβλάστες που εντοπίζονται στη δερµίδα αποτελούν τµήµα του συνδετικού ιστού του δέρµατος. Η αποµόνωση και καλλιέργεια ινοβλαστών από ανθρώπινο έµβρυο ηλικίας εβδοµάδων, έχει ως αποτέλεσµα κύτταρα της πρωτογενούς καλλιέργειας να ανακαλλιεργούνται διατηρώντας τις βασικές τους ιδιότητες. Το πλήρες θρεπτικό µέσο που απαιτείται για την καλλιέργεια περιέχει τα εξής συστατικά: Dulbecco s Modified Eagle s Medium (Low Glucose) w 1g/L D-Glucose, 1g/L Sodium Pyruvate, w/o L-Glutamine (DMEM, Gibco BRL TM ). 1% ορό εµβρύου βοός (FBS). 2mM γλουταµίνη (L-Glutamin 2mM). 1 IU/ml πενικιλίνη και 1 µg/ml στρεπτοµυκίνη (Penicillin- Streptomycin 5IU/ML-5UG/ML). ΠΛΗΡΕΣ ΘΡΕΠΤΙΚΟ ΜΕΣΟ 1. Εµπλουτισµένο (α,β) θρεπτικό µέσο DMEM ΣΥΓΚΕΝΤΡΩΣΗ 9% 1α. Γλουταµίνη 1% 1β. Πενικιλίνη/Στρεπτοµυκίνη 2% 2. Ορός εµβρύου βοός 1% Η καλλιέργεια αναπτύσσεται σε δοχεία επιφανείας 25cm 2 που περιέχουν 5ml πλήρους θρεπτικού µέσου. Ο κατάλληλος αριθµός κυττάρων που προστίθεται για τη σωστή επέκταση τους σε τρυβλία petri και σε δοχεία καλλιέργειας είναι 2-3. κύτταρα/cm 2 επιφανείας. Για τις ανακαλλιέργειες χρησιµοποιήθηκε διάλυµα τρυψίνης/edta (.5%/.2%) σε PBS που δεν περιέχει Ca 2+ και Mg 2+ (Biological Industries). Η ποσότητα που προστίθεται είναι.5ml και ο χρόνος επώασης είναι 3-5 στους 37 C. Ο διαχωρισµός της καλλιέργειας πραγµατοποιείται µία φορά την εβδοµάδα σε αναλογίες 1:2 έως 1:6. Οι δυνατότητες ανακαλλιέργειας της συγκεκριµένης κυτταρικής σειράς είναι περιορισµένες (~12) C2C12 (Mouse, C3H, Muscle, Myoblast) Η κυτταρική σειρά C2C12 (ATCC, Νο. CRL-1772) αποτελεί ένα διπλοειδή υποκλώνο (Blau et al., 1985) της πατρικής µυοβλαστικής σειράς C2, όπως αυτή καθιερώθηκε από τους Yaffe και Saxel (1977). Η αναγέννηση τραυµατισµένου µυϊκού ιστού που εντοπίζεται στο µηρό (γραµµωτοί
99 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες ή σκελετικοί µύες) φυσιολογικού ενήλικα ποντικού (στέλεχος C3H) είχε ως αποτέλεσµα τη διαίρεση και διαφοροποίηση πρόδροµων κυττάρων (satellite cells) σε µυοβλάστες. Η σειρά C2 προέκυψε από επιλεκτικές διαδοχικές ανακαλλιέργειες µυοβλαστών, οι οποίοι προέρχονται από το µηριαίο µυ, 7 ώρες µετά τον τραυµατισµό του. Στη βιβλιογραφία αναφέρεται η δυνατότητα διατήρησης µυοβλαστών στο στάδιο του πολλαπλασιασµού µε διαδοχικές ανακαλλιέργειες για χρονικό διάστηµα µεγαλύτερο από δύο χρόνια σε σταθερές συνθήκες καλλιέργειας (Yaffe, 1968). Σε αυτές τις συνθήκες, οι µυοβλάστες εµφανίζουν µορφολογία παρόµοια µε αυτή των ινοβλαστών του συνεκτικού ιστού. Για την αποφυγή της διαφοροποίησης τους σε συσταλτά µυϊκά ινίδια (µυϊκά κύτταρα) συνίσταται η χρήση ορού εµβρύου βοός (FBS), που προστίθεται σε υψηλές αναλογίες (~15-2%) στο πλήρες θρεπτικό µέσο της καλλιέργειας, αντί αλόγου (HS) και η παρουσία ενός καθοριστικού παράγοντα ανάπτυξης των ινοβλαστών (FGF). Επιπρόσθετα, σηµαντική παράµετρο αποτελεί η χρονική έναρξη των ανακαλλιεργειών, που ορίζεται από την µέτρια κάλυψη της επιφάνειας, στην οποία οι µυοβλάστες προσκολλούνται και πολλαπλασιάζονται. Tο πλήρες θρεπτικό µέσο που απαιτείται για την καλλιέργεια περιέχει τα εξής συστατικά: Dulbecco s Modified Eagle s Medium w 4.5g/L D-Glucose, 3.7g/L NaHCO3, w/o Sodium Pyruvate, L-Glutamine (DMEM, Biochrom KG). 2% ορό εµβρύου βοός (FBS, Biochrom KG). 4mM γλουταµίνη (L-Glutamin 2mM, Serva). 1 µg/ml γενταµικίνη (Gentamicin 1 mg/ml, GibcoBRL TM /BiochromKG).5 µg/ml αµιγώς µυκητοκτόνο αντιβιοτικό, Fungizone (Amphotericin B 25 µg/ml, GibcoBRL TM ). 1:2 εκχύλισµα εµβρύου όρνιθας, (CEE: Chick Embryo Extract 1 ml, Gibco BRL TM ), ως συµπλήρωµα παραγόντων ανάπτυξης στην καλλιέργεια για την αποφυγή διαφοροποίησης. ΠΛΗΡΕΣ ΘΡΕΠΤΙΚΟ ΜΕΣΟ 1. Εµπλουτισµένο (α,β,γ) θρεπτικό µέσο DMEM ΣΥΓΚΕΝΤΡΩΣΗ 79.5% 1α. Γλουταµίνη 2% 1β. Γενταµικίνη.1% 1γ. Fungizone.2% 2. Ορός εµβρύου βοός 2% 3. Εκχύλισµα εµβρύου όρνιθας.5% Η καλλιέργεια αναπτύσσεται σε δοχεία επιφανείας 25cm 2 που περιέχουν 3ml πλήρους θρεπτικού µέσου. Ο κατάλληλος αριθµός κυττάρων που προστίθεται για τη σωστή επέκταση τους σε τρυβλία petri και σε δοχεία καλλιέργειας είναι 2-2. κύτταρα/cm 2 επιφανείας
100 Υλικά και Μέθοδοι Κυτταρικές καλλιέργειες Για τις ανακαλλιέργειες χρησιµοποιήθηκε διάλυµα τρυψίνης/edta (.25%/1mM) σε ρυθµιστικό διάλυµα Hanks που δεν περιέχει Ca 2+ και Mg 2+ (Gibco BRL TM ). Η ποσότητα που προστίθεται είναι 1-2ml και ο χρόνος επώασης είναι 3-5 στους 37 C. Επιπρόσθετα, ενδείκνυται η χρήση διαλύµατος τρυψίνης/edta (.5%/.2%) σε PBS που δεν περιέχει Ca 2+ και Mg 2+ (Biochrom KG). Σε αυτή την περίπτωση για την αποκόλληση των κυττάρων, τα οποία αναπτύσσουν ισχυρές δυνάµεις πρόσφυσης χρησιµοποιήθηκε επιπρόσθετα µία ειδική σπάτουλα. Ο διαχωρισµός της καλλιέργειας πραγµατοποιείται µία φορά κάθε τέσσερις ηµέρες σε αναλογίες 1:3 έως 1:6. Η συνολική κάλυψη της επιφάνειας του δοχείου καλλιέργειας πριν το διαχωρισµό θα πρέπει να είναι ~6-7%. Οι δυνατότητες ανακαλλιέργειας της συγκεκριµένης κυτταρικής σειράς είναι περιορισµένες (~13)
101 Υλικά και Μέθοδοι Αναστολή της κυτταροκίνησης ΜΕΛΕΤΗ ΚΥΤΤΑΡΟΤΟΞΙΚΟΤΗΤΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΒΛΑΒΗΣ 7. ΜΕΘΟ ΟΣ ΑΝΑΣΤΟΛΗΣ ΤΗΣ ΚΥΤΤΑΡΟΚΙΝΗΣΗΣ (CYTOKINESIS-BLOCK METHOD/CBMN) Η εκτίµηση των χρωµοσωµατικών βλαβών σε διαιρούµενους κυτταρικούς πληθυσµούς µετά από έναν κύκλο διαίρεσης πραγµατοποιείται µε τη χρήση της µεθόδου αναστολής της κυτταροκίνησης (Cytocinesis-Block-CB), η οποία βασίζεται στη δράση της κυτταροχαλασίνης-β (Cytochalasin-B) (Fenech και Morley, 1985). Η ιδιότητα της κυτταροχαλασίνης-β (Cyt-B) να αναστέλλει την κυτταροκίνηση χωρίς να παρεµποδίζει την πυρηνική διαίρεση σε ευκαρυωτικούς οργανισµούς έχει ως αποτέλεσµα η καταµέτρηση των χρωµοσωµατικών βλαβών, που εκφράζονται ως µικροπυρήνες (Micronuclei-MN), να πραγµατοποιείται σε καλλιεργούµενα κύτταρα που έχουν διαιρεθεί µόνο µία φορά και αναγνωρίζονται από τη διπύρηνη τους εµφάνιση (Binucleated cell-bn). Οι µικροπυρήνες προκύπτουν από τη συσσώρευση χρωµοσωµατικού υλικού, το οποίο συνίσταται είτε από άκεντρα χρωµοσωµατικά τµήµατα είτε από άθικτα χρωµοσώµατα, που αδυνατούν να συνδεθούν µε τη µιτωτική συσκευή και να µετακινηθούν προς τους πόλους της µιτωτικής ατράκτου µε αποτέλεσµα να µην ενσωµατωθούν στους κύριους θυγατρικούς πυρήνες κατά τη διάρκεια της πυρηνικής διαίρεσης. Συγκεκριµένα, κατά την τελόφαση ένας πυρηνικός φάκελος σχηµατίζεται γύρω από το καθυστερούµενο χρωµοσωµατικό τµήµα ή το χρωµόσωµα, ο οποίος στη συνέχεια εκτυλίγεται και σταδιακά αποκτά τη µορφολογία ενός διακριτού µεσοφασικού πυρήνα που εντοπίζεται στο κυτταρόπλασµα και είναι µικρότερος σε µέγεθος από τους κυρίως πυρήνες. Η χρήση της κυτταροχαλασίνης-β επιτρέπει τη συσσώρευση ουσιαστικά του συνόλου των διαιρούµενων κυττάρων στο διπύρηνο στάδιο του διαιρούµενου κυτταρικού πληθυσµού, ανεξάρτητα από το βαθµό συγχρονισµού τους και την αναλογία των διαιρούµενων κυττάρων. Επιπρόσθετα, η καταµέτρηση των µικροπυρήνων µόνο σε άθικτα διπύρηνα κύτταρα επιτρέπει την αξιόπιστη σύγκριση της χρωµοσωµατικής βλάβης µεταξύ κυτταρικών πληθυσµών, οι οποίοι είναι δυνατόν να διαφέρουν ως προς την κινητική της κυτταρικής διαίρεσης (Fenech, 2). Ένα από τα σηµαντικότερα πλεονεκτήµατα της µεθόδου αποτελεί η γρήγορη και αξιόπιστη µέτρηση της κινητικής εξέλιξης του πολλαπλασιασµού των κυττάρων σε ένα διαιρούµενο κυτταρικό πληθυσµό, η οποία αποδίδεται από τον υπολογισµό του δείκτη πολλαπλασιασµού των κυττάρων (Cytocinesis Block Proliferation Index, CBPI=M1+2M2+3(M3+M4)/N). Αυτό επιτυγχάνεται µε τη µέτρηση των µονοπύρηνων (Μ1), διπύρηνων (Μ2) και πολυπύρηνων κυττάρων (> 2 πυρήνες) (Μ3+4) µετά από ένα καθορισµένο σηµείο, το οποίο επιλέγεται µετά την προσθήκη της Cyt-B, και Ν ο συνολικός αριθµός των κυττάρων που µετρήθηκαν. Ο δείκτης πολλαπλασιασµού των κυττάρων (CBPI) υπολογίζει το µέσο αριθµό των κυτταρικών κύκλων που έχει ολοκληρώσει ο κυτταρικός πληθυσµός και εκφράζει τον αριθµό των κυτταρικών διαιρέσεων που υφίσταται ένα κύτταρο µε δεδοµένο ότι τα τριπύρηνα και τετραπύρηνα κύτταρα έχουν ολοκληρώσει δύο κυτταρικές διαιρέσεις (Schultz και Önfelt, 1994, Suralles et al., 1995a)
102 Υλικά και Μέθοδοι Αναστολή της κυτταροκίνησης Επιπρόσθετα, κυτταροστατικά φαινόµενα αποδίδονται µε το δείκτη που αναφέρεται ως ποσοστό διπύρηνων κυττάρων (% of Binucleated cells-% BN) ο οποίος σε περιπτώσεις µελέτης που αφορά τον καθορισµό τοξικών δόσεων µίας χηµικής ένωσης δεν είναι σε θέση να διακρίνει τις διακυµάνσεις του ποσοστού των πολυπύρηνων κυττάρων σε ένα διαιρούµενο κυτταρικό πληθυσµό µε αποτέλεσµα, η εκτίµηση της έκτασης και εξέλιξης του κυτταρικού κύκλου να µην είναι ιδιαίτερα ακριβής και αξιόπιστη, όπως αυτή αποδίδεται επακριβώς µε το δείκτη πολλαπλασιασµού των κυττάρων (CBPI) (Suralles et al., 1994, Suralles et al., 1995a). 7.1 Λεµφοκύτταρα Συνθήκες καλλιέργειας Ο κυτταρικός κύκλος των ανθρώπινων λεµφοκυττάρων σε καλλιέργεια in vitro ξεκινά σε 24 ώρες µετά την έναρξη της καλλιέργειας παρουσία της φυτικής λεκτίνης φυτοαιµατογλουτινίνης (PHA) και χρειάζεται 24 ώρες περίπου για να ολοκληρωθεί. Η σύνθεση του DNA γίνεται στη διάρκεια της φάσης S, η οποία διαρκεί 6-8 ώρες, ενώ οι φάσεις G1 µε τη µεγαλύτερη χρονική διάρκεια και G2 διαρκούν αντίστοιχα 12 και 3-4 ώρες. Η χρονική διάρκεια της µίτωσης στα ανθρώπινα λεµφοκύτταρα είναι περίπου 1 ώρα (Watt και Stephen, 1986, Gosden et al., 1992). Η πειραµατική διαδικασία που ακολουθείται περιλαµβάνει δύο διαφορετικά πρωτόκολλα (Ι. και ΙΙ.) µε κύριο στόχο τον έλεγχο διαφοροποιηµένης γονιδιοτοξικής δράσης της χηµικής ένωσης υδροχλωροθειαζίδιο (HCTZ) σε σχέση µε την κατάσταση πολλαπλασιασµού των κυττάρων, γεγονός το οποίο αντανακλά επίδραση σε διαφορετικές φάσεις του κυτταρικού κύκλου των λεµφοκυττάρων. Σύµφωνα µε το πρωτόκολλο Ι. πραγµατοποιείται έκθεση των διεγερµένων µε PHA λεµφοκυττάρων (cycling/proliferating lymphocytes) στη φαρµακευτική ένωση υδροχλωροθειαζίδιο στη λογαριθµική φάση ανάπτυξης της καλλιέργειας, σε 41 ώρες από την έναρξή της, που αντιστοιχεί στη µετάπτωση από τη φάση S του κυτταρικού κύκλου στη φάση G2 (Watt και Stephen, 1986, Van Hummelen και Kirsch-Volders, 1992). Η παραµονή του υδροχλωροθειαζιδίου µέχρι τη λήξη της καλλιέργειας στις 72 ώρες κρίνεται απαραίτητη για να συµπεριληφθούν στη µελέτη κύτταρα που εκτέθηκαν σε όλες τις φάσεις του κυτταρικού κύκλου (G1/S/G2/M) και τα οποία µε τη δράση της κυτταροχαλασίνης-β αναγνωρίζονται από τη διπύρηνή τους εµφάνιση (Fenech, 2). Με το πρωτόκολλο Ι. εκτιµήθηκε η δράση της φαρµακευτικής ένωσης υδροχλωροθειαζίδιο σε δύο θηλυκά άτοµα νεαρής ηλικίας στις συγκεντρώσεις 5, 1, 2, 4 και 6 µg/ml και ακολούθως επιλέχθηκαν οι συγκεντρώσεις 5 και 4 µg/ml, που εξετάσθηκαν στο σύνολο των δοτών. Σε κάθε δότη και για κάθε συγκέντρωση του υδροχλωροθειαζιδίου πραγµατοποιήθηκαν παράλληλα δύο σειρές καλλιεργειών. Η απόλυτη αιθανόλη (Ethanol Absolute, Merck) χρησιµοποιείται ως διαλύτης, που περιλαµβάνει τις υπό εξέταση συγκεντρώσεις της φαρµακευτικής ένωσης, σε εξαιρε
103 Υλικά και Μέθοδοι Αναστολή της κυτταροκίνησης τικά χαµηλές περιεκτικότητες, οι οποίες αντιστοιχούν σε.2-.5% στον τελικό όγκο της καλλιέργειας λεµφοκυττάρων ολικού αίµατος (Preston et al., 1987). Επιπρόσθετα, για την αναστολή της κυτταροκίνησης η κυτταροχαλασίνη-β (Cyt-B, Sigma, St Louis, MO USA) προστίθεται στις 44 ώρες από την έναρξη της καλλιέργειας σε τελική συγκέντρωση 6 µg/ml. Η κυτταροχαλασίνη-β, ως πυκνό διάλυ- µα αποθήκευσης (2mg/ml) φυλάσσεται στους -8 C και βρίσκεται διαλυµένη στον οργανικό διαλύτη διµεθυλοσουλφοξείδιο (DMSO, Carlo Erba/Sigma), του οποίου η προσθήκη αντιστοιχεί σε συγκέντρωση.2% στον τελικό όγκο της καλλιέργειας. Η χρονική διάρκεια παραµονής της Cyt-Β (6µg/ml) στην καλλιέργεια, που ορίζεται στη διεθνή βιβλιογραφία στις 28 ώρες (44-72) και η οποία πραγµατοποιείται µε τη λήξη αυτής στις 72 ώρες θεωρείται ένας εξαιρετικά επιτυχηµένος συνδυασµός σε in vitro πληθυσµιακές µελέτες γονιδιοτοξικότητας, διότι προκύπτει ένας σχετικά υψηλός πληθυσµός διπύρηνων κυττάρων ( 6%) κατάλληλος για την καταµέτρηση των µικροπυρήνων (Van Hummelen και Kirsch-Volders, 199, Suralles et al., 1992, Fenech, 1993, Fenech, 2). Το χρονικό διάστηµα που µεσολαβεί µεταξύ της φαρµακευτικής ένωσης υδροχλωροθειαζίδιο (HCTZ) και της κυτταροχαλασίνης-β (Cyt-B), κατά την προγραµµατισµένη έκθεση των λεµφοκυττάρων στις συγκεκριµένες ενώσεις, κρίνεται απαραίτητο για την αποφυγή τυχόν αλληλεπιδράσεων, γνωρίζοντας ότι απαιτείται χρονικό διάστηµα περίπου 6 ωρών για να εκδηλωθεί η ανασταλτική δράση της κυτταροχαλασίνης-β (Fenech, 2). ιαγραµµατικά η εφαρµογή του Πρωτοκόλλου Ι. (διεγερµένα λεµφοκύτταρα) έχει ως εξής: t = Έναρξη καλλιέργειας παρουσία PHA t = 24 t = 41 Προσθήκη HCTZ t = 44 Προσθήκη Cyt-B t = 72 Ολοκλήρωση καλλιέργειας και αποµόνωση λεµφοκυττάρων Σύµφωνα µε το πρωτόκολλο ΙΙ. λαµβάνοντας υπόψη το πλεονέκτηµα ότι κατά τη συλλογή του φλεβικού αίµατος τα ανθρώπινα λεµφοκύτταρα αποτελούν στην πλειοψηφία τους ένα συγχρονισµένο πληθυσµό κυττάρων που βρίσκεται στην ίδια Gο φάση του κυτταρικού κύκλου πραγµατοποιήθηκε επίδραση µικρής σχετικά χρονικής διάρκειας (6 ώρες) σε καλλιέργειες λεµφοκυττάρων µε τη φαρµακευτική ένωση υδροχλωροθειαζίδιο, απουσία του ανοσοδιεγερτικού παράγοντα (PHA). Στο τέλος της χρονικής διάρκειας των έξι ωρών που αναλογεί σε µία µικρή περίοδο της προσυνθετικής Gο φάσης του κύκλου ζωής των λεµφοκυττάρων στις καλλιέργειες ολικού αίµατος (resting/unstimulated lymphocytes) ακολουθεί έκπλυση των κυττάρων για την αποµάκρυνση της φαρµακευτικής ένωσης (Fenech, 2). Η πορεία που ακολουθείται έχει ως εξής: 1. Μεταφορά του καλλιεργητικού υλικού σε αποστειρωµένους σωλήνες φυγοκέντρου (Cellstar R, Test Tubes 15ml, Greiner Bio One, GmbH). 2. Φυγοκέντρηση στις 15 στροφές για 1 λεπτά. 3. Προσεκτική αφαίρεση του υπερκειµένου χωρίς να διαταραχθεί η λεπτή επιφανειακή στοιβάδα των λεµφοκυττάρων µε πιπέτες µίας χρήσης (Stripette
104 Υλικά και Μέθοδοι Αναστολή της κυτταροκίνησης sterile, 1ml, Costar) και προσθήκη 3ml αποστειρωµένου ρυθµιστικού διαλύµατος PBS (37 C) µε ταυτόχρονη ελαφρά ανάδευση. Ακολουθεί νέα προσθήκη PBS µέχρι τελικού όγκου 8 ml. 4. Φυγοκέντρηση και επάναληψη της έκπλυσης των κυττάρων για την απόµάκρυνση υπολειµµάτων της φαρµακευτικής ένωσης µε θρεπτικό µέσο Ham s F1. 5. Αφαίρεση υπερκειµένου και προσθήκη 1 ml εµπλουτισµένου θρεπτικού µέσου Ham s F1 µε ταυτόχρονη αναδιάλυση του ιζήµατος των κυττάρων. 6. Μεταφορά του εναιωρήµατος των κυττάρων σε δοχεία καλλιέργειας, στα οποία έχει προηγούµενα τοποθετηθεί νέο καλλιεργητικό υλικό (6ml) που περιλαµβάνει 4.8ml εµπλουτισµένου θρεπτικού µέσου Ham s F1, 1.2ml FBS και τον ανοσοδιεγερτικό παράγοντα PHA σε αναλογία 2.5%. Με το πρωτόκολλο ΙΙ. εκτιµήθηκε η δράση της χηµικής ένωσης υδροχλωροθειαζίδιο σε έξι άτοµα νεαρής ηλικίας και διαφορετικού φύλου στις συγκεντρώσεις 5 και 4 µg/ml. Σε κάθε δότη και για κάθε συγκέντρωση του υδροχλωροθειαζιδίου πραγµατοποιήθηκαν παράλληλα δύο σειρές καλλιεργειών. Η προσθήκη της κυτταροχαλασίνης-β σε τελική συγκέντρωση 6 µg/ml για την επίτευξη αναστολής της κυτταροκίνησης πραγµατοποιείται στις 44 ώρες από την έναρξη της καλλιέργειας, η οποία σηµατοδοτείται µε την παρουσία της PHA και ολοκληρώνεται στις 72 ώρες. ιαγραµµατικά η εφαρµογή του Πρωτοκόλλου ΙΙ. (µη διεγερµένα λεµφοκύτταρα) έχει ως εξής: t = (- 6) t = Προσθήκη Αποµάκρυνση HCTZ HCTZ Έναρξη καλλιέργειας παρουσία PHA t = 24 t = 44 Προσθήκη Cyt-B t = 72 Ολοκλήρωση καλλιέργειας και αποµόνωση λεµφοκυττάρων Αποµόνωση λεµφοκυττάρων Οι µεθοδολογίες που περιγράφονται στη διεθνή βιβλιογραφία, όσον αφορά τη διαδικασία αποµόνωσης ανθρώπινων λεµφοκυττάρων και λήψης παρασκευασµάτων για την ακριβή ανίχνευση µικροπυρήνων στο κυτταρόπλασµα άθικτων διπύρηνων κυττάρων από καλλιέργειες µικρών ποσοτήτων φλεβικού αίµατος (.3-.5ml) (Human PBL/MN Assay) εστιάζουν την προσοχή τους στην άριστη διατήρηση του κυτταροπλάσµατος µε την ταυτόχρονη αποµάκρυνση των ερυθροκυττάρων. Ιδιαίτερη σηµασία δίνεται στη σύσταση και στο χρόνο επίδρασης του υποτονικού διαλύµατος, στην ταχύτητα φυγοκέντρησης και στον κατάλληλο χειρισµό του κυτταρικού ιζήµατος (Högstedt, 1984, Van Hummelen και Kirsch Volders, 199, Lee et al., 1994). Η διαδικασία αποµόνωσης λεµφοκυττάρων στο εργαστήριο µας πραγµατοποιείται µε κάποιες µικρές τροποποιήσεις που αποσκοπούν στην καλύτερη διατήρηση του κυτταροπλάσµατος µε αποτέλεσµα την αξιόπιστη διάκριση των µικροπυρήνων στα παρασκευάσµατά µας
105 Υλικά και Μέθοδοι Αναστολή της κυτταροκίνησης Κατά τη διαδικασία αποµόνωσης γίνεται χρήση των εξής διαλυµάτων: ΙΑΛΥΜΑΤΑ 1. Υποτονικό διάλυµα 2. Μονιµοποιητικό διάλυµα (Carnoy s) ΣΥΓΚΕΝΤΡΩΣΗ Ham s F1:dd H2O= 1:1 Το διάλυµα διατηρείται στους 37ºC Απαιτούνται περίπου 4ml ανά καλλιέργεια CH3OH (Merck) και CH3COOH (Merck) σε αναλογία 3:1 Το διάλυµα διατηρείται στους 4ºC Απαιτούνται περίπου 25 ml ανά καλλιέργεια Τα βήµατα που ακολουθούνται είναι τα παρακάτω: 1. Οι καλλιέργειες µεταφέρονται από τα δοχεία καλλιέργειας σε ειδικούς γυάλινους σωλήνες φυγοκέντρου (1ml) και ακολουθεί φυγοκέντρηση (Centrifuge 5416, Gerätebau Eppendorf, GmbH) στις 15 στροφές ανά λεπτό για 1 λεπτά. 2. Ακολουθεί αφαίρεση του υπερκειµένου, προσθήκη σε κάθε σωλήνα φυγοκέντρου 4ml υποτονικού διαλύµατος µε ταυτόχρονη ελαφρά ανάδευση (διαβάθµιση 2) σε συσκευή ανατάραξης (Vortex, Genie, SI) και παραµονή για 2 λεπτά σε θερµοκρασία δωµατίου. Η κατεργασία µε το υποτονικό διάλυµα προκαλεί την αιµόλυση των ερυθροκυττάρων και την ελαφρά διόγκωση των λεµφοκυττάρων. Ακολουθούν τρεις στιγµιαίες µονιµοποιήσεις 3. Προσθήκη 4ml µονιµοποιητικού διαλύµατος µε ταυτόχρονη πολύ καλή ανάδευση για την καλύτερη οµογενοποίηση του κυτταρικού εναιωρήµατος χωρίς να δηµιουργηθούν συσσωµατώµατα, µε σκοπό την καλύτερη δυνατή αποµάκρυνση των ερυθρών αιµοσφαιρίων σε συνδυασµό µε την άριστη διατήρηση της µορφολογίας των λεµφοκυττάρων (πρώτη µονιµοποίηση). 4. Φυγοκέντρηση στις 12 στροφές για 7 λεπτά. 5. Αποµάκρυνση του υπερκειµένου και κατεργασία του κυτταρικού ιζή- µατος µε 7ml νέου µονιµοποιητικού διαλύµατος µε ταυτόχρονη ανάδευση (δεύτερη µονιµοποίηση). 6. Φυγοκέντρηση του εναιώρηµατος των κυττάρων στις 12 στροφές για 7 λεπτά. 7. Μετά την αποµάκρυνση του υπερκειµένου ακολουθεί τρίτη µονιµοποίηση µε προσθήκη νέου µονιµοποιητικού διαλύµατος (7ml) και φυγοκέντρηση του εναιωρήµατος στις ίδιες συνθήκες. Τo εναιώρηµα των λεµφοκυττάρων διατηρείται σε µονιµοποιητικό διάλυµα (5ml) σε θερµοκρασία -2 C για αρκετά µεγάλο χρονικό διάστηµα. 7.2 Κυτταρική σειρά Hep2 Η πειραµατική διαδικασία που ακολουθείται για τις καλλιέργειες της κυτταρικής σειράς Hep2 περιλαµβάνει την επίστρωση των καλυπτρίδων µε X1 5 κύτταρα. Η φαρµακευτική ένωση υδροχλωροθειαζίδιο (HCTZ) εξετάσθηκε στις συγκεντρώσεις 5, 2, 4 και 6 µg/ml. Η επίδραση µε τις υπό µελέτη συγκεντρώσεις του υδροχλωροθειαζιδίου πραγµατοποιείται 41 ώρες µετά την έναρξη της καλλιέργειας, ενώ η κυτταροχαλασίνη-β προστίθεται στις 44 ώρες σε τελική συγκέντρωση 3µg/ml
106 Υλικά και Μέθοδοι Αναστολή της κυτταροκίνησης Με την ολοκλήρωση της καλλιέργειας στις 72 ώρες, σηµατοδοτείται η έναρξη κατάλληλων αντιδράσεων για τη συνδυασµένη σήµανση του κινητοχώρου (τεχνική CREST) και του κεντροµέρους (τεχνική FISH) όλων των ανθρώπινων χρωµοσωµάτων. ιαγραµµατικά το χρονοδιάγραµµα του πειράµατος έχει ως εξής: t = Έναρξη καλλιέργειας t = 41 Προσθήκη HCTZ t = 44 Προσθήκη Cyt-B t = 72 Τερµατισµός καλλιέργειας Μονιµοποίηση κυττάρων ιπλή σήµανση κινητοχώρουκεντροµέρους (CREST-FISH) 7.3 Κυτταρική σειρά HFFF2 Η πειραµατική διαδικασία που ακολουθείται για τις καλλιέργειες της κυτταρικής σειράς HFFF2 περιλαµβάνει την επίστρωση των καλυπτρίδων µε X1 5 κύτταρα. Η φαρµακευτική ένωση υδροχλωροθειαζίδιο (HCTZ) εξετάσθηκε στις συγκεντρώσεις 2, 4, 6 και 8 µg/ml και η χρονική διάρκεια παραµονής της στις καλλιέργειες εµβρυϊκών επιδερµικών ινοβλαστών αντιστοιχεί περίπου σε ένα κύκλο διαίρεσης. Επιπρόσθετα, για την αναστολή της κυτταροκίνησης η κυτταροχαλασίνη-β προστίθεται στις 26 ώρες από την έναρξη της καλλιέργειας σε τελική συγκέντρωση 3 µg/ml. Η χρονική διάρκεια παραµονής της Cyt-Β (3µg/ml) στην καλλιέργεια που ορίζεται στη διεθνή βιβλιογραφία στις 28 ώρες (26-54) και η οποία πραγµατοποιείται µε τη λήξη αυτής στις 54 ώρες θεωρείται ένας εξαιρετικά επιτυχηµένος συνδυασµός σε in vitro µελέτες γονιδιοτοξικότητας µε τη χρήση διαφόρων τύπων κυτταρικών σειρών, διότι προκύπτει ένας σχετικά υψηλός πληθυσµός διπύρηνων κυττάρων ( 5%) κατάλληλος για την καταµέτρηση των µικροπυρήνων (Fenech, 2, Sgura et al., 21, Parry et al., 22). Με την ολοκλήρωση της καλλιέργειας, σηµατοδοτείται η έναρξη κατάλληλων αντιδράσεων για την ταυτόχρονη σήµανση του κινητοχώρου (τεχνική CREST) και του κεντροµέρους (τεχνική FISH) όλων των ανθρώπινων χρωµοσωµάτων. ιαγραµµατικά το χρονοδιάγραµµα του πειράµατος έχει ως εξής: t = Έναρξη καλλιέργειας t = 23 Προσθήκη HCTZ t = 26 Προσθήκη Cyt-B t = 54 Τερµατισµός καλλιέργειας Μονιµοποίηση κυττάρων ιπλή σήµανση κινητοχώρουκεντροµέρους (CREST-FISH)
107 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH ΜΕΛΕΤΗ ΧΡΩΜΟΣΩΜΑΤΙΚΗΣ ΑΠΩΛΕΙΑΣ (CL) ΚΑΙ ΧΡΩΜΟΣΩΜΑΤΙΚΟΥ ΜΗ ΑΠΟΧΩΡΙΣΜΟΥ (ND) 8. ΜΕΘΟ ΟΣ ΑΝΑΣΤΟΛΗΣ ΤΗΣ ΚΥΤΤΑΡΟΚΙΝΗΣΗΣ ΣΕ ΣΥΝ ΥΑΣΜΟ ΜΕ IN SITU ΥΒΡΙ ΟΠΟΙΗΣΗ ΜΕ ΦΘΟΡΟΧΡΩΜΑΤΑ (CBMN/FISH) Η FISH (Fluorescence In Situ Hybridization), αποδίδεται ως in situ υβριδοποίηση µε φθοροχρώµατα, αποτελεί µία κυτταρογενετική-µοριακή τεχνική, η οποία χρησιµοποιεί ανιχνευτές DNA ή RNA επισηµασµένους µε µόρια-αναφοράς, τα οποία µε τη σειρά τους φθορίζουν και µπορούν µε τον τρόπο αυτό, να γίνονται ορατά µε παρατήρηση σε ένα µικροσκόπιο κατάλληλα εξοπλισµένο µε ειδικά φίλτρα φθορισµού. Η µέθοδος αυτή επιτρέπει τον προσδιορισµό του αριθµού και τον εντοπισµό της θέσης συγκεκριµένων ακολουθιών DNA στα ανθρώπινα χρωµοσώµατα. Η βασική αρχή της µεθόδου FISH στηρίζεται στο µοναδικό γεγονός της συµπληρω- µατικότητας ενός επισηµασµένου µονόκλωνου τµήµατος DNA (DNA ανιχνευτής) µε ένα µονόκλωνο-µετά από ειδική κατεργασία-dna (DNA στόχος) που εντοπίζεται σε ένα ή περισσότερα µορφολογικά διατηρηµένα χρωµοσώµατα µίας µετάφασης ή ενός µεσοφασικού πυρήνα.πρόκειται για µία δυναµική αντίδραση στην οποία απoδιαταγµένες αλληλουχίες-στόχοι και ανιχνευτές επανασυνδέονται για να σχη- µατίσουν σταθερά δίκλωνα υβριδικά µόρια. Υπάρχουν δύο τύποι µη ραδιενεργού υβριδοποίησης: η έµµεση (indirect) και η άµεση (direct) υβριδοποίηση. Στην έµµεση ο ανιχνευτής φέρει ένα µόριο αναφοράς, το οποίο χηµικά ή ενζυµικά δύναται να σηµανθεί µε άλλο φθορίζον µόριο (φθορόχρωµα) µέσω µίας εξειδικευµένης ανοσοχηµικής αντίδρασης, ενώ στην άµεση η φθορίζουσα χρωστική (το φθορόχρωµα) είναι ενωµένη απευθείας στον ανιχνευτή. Με την εφαρµογή της µεθόδου αναστολής της κυτταροκίνησης (CBMN) σε συνδυασµό µε in situ υβριδοποίηση µε φθοροχρώµατα (FISH) µε τη χρήση κατάλληλου DNA ανιχνευτή, είναι δυνατόν να προσδιορισθούν οι κύριοι µηχανισµοί δη- µιουργίας µικροπυρήνων σε µιτωτικά κύτταρα παρουσία της κυτταροχαλασίνης-β. Ο συγκεκριµένος ανιχνευτής συνίσταται από µία συνθετική ολιγονουκλεοτιδική αλληλουχία τριάντα βάσεων (SOαAllCen), για τον εντοπισµό µίας εξαιρετικά συντηρηµένης περιοχής (monomer unit) του υψηλά επαναλαµβανόµενου άλφα δορυφορικού DNA (α-satellite DNA) που εµφανίζεται στα κεντροµέρη όλων των ανθρώπινων χρωµοσωµάτων (Norppa et al., 1993b, Elhajouji et al., 1995). Μικροπυρήνες που έχουν σχηµατισθεί από τη συσσώρευση άκεντρων χρωµοσωµατικών τµηµάτων, ως αποτέλεσµα χρωµοσωµατικών ρηγµάτων διακρίνονται από εκείνους που περικλείουν ολόκληρα χρωµοσώµατα, ως αποτέλεσµα χρωµοσωµατικής καθυστέρησης κατά την ανάφαση, από την παρουσία ή απουσία των κεντροµερών των χρωµοσωµάτων που γίνεται ορατή µε την παρουσία ή απουσία σήµατος υβριδοποίησης. Επιπρόσθετα, η αναγνώριση ολόκληρων χρωµοσωµάτων στους µικροπυρήνες είναι εφικτή µε αµιγείς ανοσοχηµικές µεθόδους µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών του κινητοχώρου (CENPs), µέθοδος η οποία καλείται CREST, επειδή τα συγκεκριµένα αντισώµατα προέρχονται από τον ορό ασθενών µε σύνδροµο CREST (Calcinosis, Raynaud s phenomenon, Esophageal dysmotility, Sclerodactyly and Telangiectasia), ένα αυτοάνοσο νόσηµα που επηρεάζει πολλά οργανικά συστήµατα (Degrassi και Tanzarella, 1988, Eastmont και Tucker, 1989, Hennig, 199)
108 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH Η δυνατότητα προσδιορισµού του µηχανισµού δηµιουργίας των µικροπυρήνων αποτέλεσε ένα σηµαντικό εργαλείο σε γενετικές µελέτες για την εκτίµηση παραγόντων που προκαλούν είτε ρήγµατα στο γενετικό υλικό (clastogen) είτε απώλεια ολόκληρων χρωµοσωµάτων (anuegen) που οφείλεται σε χρωµοσωµατική καθυστέρηση, φαινόµενα τα οποία οδηγούν αντίστοιχα σε δοµικές και αριθµητικές χρωµοσωµατικές µεταβολές (Kirsch Volders et al., 1997). Ένα σηµαντικό πλεονέκτηµα για τον καθορισµό του µηχανισµού πρόκλησης ανευπλοειδικών φαινοµένων από φυσικούς ή χηµικούς παράγοντες αποτέλεσε ο συνδυασµός της µεθόδου αναστολής της κυτταροκίνησης (CBMN) µε in situ υβριδοποίηση µε φθοροχρώµατα (FISH) που πραγµατοποιήθηκε µε τη χρήση DNA ανιχνευτών εξειδικευµένων για τα κεντροµέρη συγκεκριµένων χρωµοσωµάτων. Οι τελευταίοι αποτελούν ειδικές για το κεντροµέρος κάθε ανθρώπινου χρωµοσώµατος ολιγονουκλεοτιδικές αλληλουχίες, ώστε να υβριδοποιούνται µε την υψηλά επαναλαµβανόµενη άλφα δορυφορική περιοχή του DNA. Ανευπλοειδίες που οφείλονται σε µη αποχωρισµό είναι πλέον δυνατόν να ανιχνευθούν σε µεσοφασικά διπύρηνα κύτταρα, τα οποία προκύπτουν από την επίδραση της κυτταροχαλασίνης-β µε την ταυτόχρονη χρήση εξειδικευµένων ανιχνευτών για κάποιο συγκεκριµένο χρωµόσωµα. Αυτό το γεγονός αποτελεί ένα ιδιαίτερο χαρακτηριστικό της µεθόδου αναστολής της κυτταροκίνησης που δεν είναι δυνατό να ανιχνευθεί άµεσα µε άλλη µέθοδο και επιτρέπει τόσο την αναγνώριση ολόκληρου χρωµοσώµατος σε ένα µικροπυρήνα ενός διπύρηνου κυττάρου όσο και την ανάλυση της κατανοµής των χρωµοσωµάτων µεταξύ των θυγατρικών πυρήνων σε ένα διπύρηνο κύτταρο (Hando et al., 1994, Zijno et al., 1994). 8.1 In Situ υβριδοποίηση µε φθοροχρώµατα (FISH) - Χρήση πανκεντροµερικού ανιχνευτή για τον προσδιορισµό του µηχανισµού δηµιουργίας µικροπυρήνων Η τεχνική FISH αποτελεί αναµφισβήτητα µία σύνθετη διαδικασία. Τα στάδια της απλοποιηµένα είναι δυνατόν να συνοψισθούν ως εξής: 1. Παρασκευή και σήµανση των ανιχνευτών DNA. 2. Προετοιµασία των κυτταρολογικών δειγµάτων (µεταφασικών χρωµοσωµάτων ή κυτταρικών εναιωρηµάτων). 3. Αποδιάταξη των ανιχνευτών και των δειγµάτων (Denaturation). 4. Επιτόπια υβριδοποίηση (In situ hybridization). 5. Εκπλύσεις µετά την υβριδοποίηση (Posthybridization washes). 6. Ανοσοκυτταροχηµεία (Immunocytochemistry). 7. Μικροσκόπηση φθορισµού. Η µεθοδολογία η οποία αναπτύχθηκε αρχικά στα πειράµατα µας περιλαµβάνει τη χρήση κατάλληλου ανιχνευτή που εµφανίζει συµπληρωµατικότητα µε αλληλουχίες του υψηλά επαναλαµβανόµενου άλφα δορυφορικού DNA (α-satellite DNA) και εντοπίζεται στην κεντροµερική περιοχή όλων των ανθρώπινων χρωµοσωµάτων. Ο ανιχνευτής είναι επισηµασµένος µε ένα φυτικό παράγωγο, τη διγοξιγενίνη που ονοµάζεται απτένιο ή µόριο αναφοράς. Πρόκειται για ένα στεροειδές που αποµονώνεται από τα φυτά Digitalis purpurea και Digitalis linata. Η διγοξιγενίνη ενσωµατώνεται στον ανιχνευτή προσδεµένη σε τροποποιηµένα ανάλογα νουκλεοτιδίων (d-utp) µε ενδιάµεσα άτοµα άνθρακα. Η αντίδραση ανίχνευσης γίνεται έµµεσα µε φθοροχρώµατα µε αποτέλεσµα η διγοξιγενίνη να µεγεθύνεται µε επαναλαµβανόµενα βήµατα σήµανσης που περιλαµβάνουν
109 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH διαδοχικά στρώµατα αντισωµάτων, µε το τελευταίο να είναι συζευγµένο µε το φθορόχρωµα φθοροσκεΐνη Προετοιµασία παρασκευασµάτων (Slide preparation) Η προετοιµασία των αντικειµενοφόρων και των καλυπτρίδων περιλαµβάνει τον καθαρισµό που επιτυγχάνεται µε την παραµονή τους σε διάλυµα αιθανόλης: αιθέρα (1:1) ή 9% αιθανόλης και αποσκοπεί στην καλύτερη προσκόλληση του κυτταρικού υλικού και στην αποµάκρυνση τυχόν οργανικών υπολειµµάτων Μονιµοποίηση-Ωρίµανση παρασκευασµάτων (Fixation-Aging) Η µονιµοποίηση του κυτταρικού υλικού που θα χρησιµοποιηθεί για υβριδοποίηση κρίνεται απαραίτητη για να διατηρηθεί η µορφολογία του. Ειδικά για τη µεθοδολογία FISH δεν υπάρχει σαφής περιορισµός στον τύπο των µονιµοποιητικών τα οποία είναι δυνατόν να χρησιµοποιηθούν, διότι οι λειτουργικές οµάδες του DNA που συµµετέχουν στη σύζευξη των βάσεων προστατεύονται στη δίκλωνη έλικα του DNA. Ιδιαίτερη προσοχή πρέπει να δοθεί στην παρουσία πρωτεϊνών γύρω από τις ακολουθίες στόχους του DNA που είναι σε θέση να δηµιουργήσουν αρκετές αλληλεπιδράσεις, ώστε να επικαλύψουν το στόχο µε αποτέλεσµα να απαιτούνται συχνά διαδικασίες αύξησης της διαπερατότητας. Η διαδικασία ωρίµανσης αποσκοπεί: (α) στην καλύτερη στερέωση του κυτταρικού υλικού σε γυάλινες επιφάνειες και (β) στην σκληραγώγηση των χρωµοσωµάτων για την απόκτηση ανθεκτικής δοµής στην επακόλουθη και αρκετά επίπονη για τη µορφολογία τους διαδικασία αποδιάταξης του DNA. Σε παρασκευάσµατα στα οποία δεν έχει εφαρµοστεί η διαδικασία ωρίµανσης παρατηρείται είτε µερική απώλεια των πυρήνων/χρωµοσωµάτων κατά την αποδιάταξη του DNA είτε παραµόρφωση ("puffy" chromosomes) του σχήµατος των χρωµοσωµάτων. Επιπρόσθετα, ο παρατεταµένος χρόνος ωρίµανσης έχει ως αποτέλεσµα µία απότοµη πτώση στην απόδοση της υβριδοποίησης. Γενικότερα, η διαδικασία ωρίµανσης θα πρέπει να είναι ήπια και να µην επιτυγχάνεται µε την παραµονή των παρασκευασµάτων σε υψηλές θερµοκρασίες (dry heat aging). Η χρήση µονιµοποιητικών διαλυµάτων (chemical aging) και η παραµονή των παρασκευασµάτων για µικρό χρονικό διάστη- µα σε θερµοκρασία δωµατίου θεωρείται η καταλληλότερη διαδικασία ωρίµανσης. Αξιοσηµείωτο είναι ότι σε κυτταρικά εναιωρήµατα που παρέµειναν για χρονικό διάστηµα δέκα ετών σε συνήθη µονιµοποιητικά διαλύµατα (Carnoy s, αιθανόλη) εφαρµόσθηκε επιτυχώς η µεθοδολογία FISH. Το καλλιεργητικό υλικό (λεµφοκύτταρα) διατηρείται µε τη µορφή εναιωρή- µατος σε µονιµοποιητικό διάλυµα (µεθανόλης:οξικού, 3:1) στους -2 C για περίπου 3 µήνες από την ολοκλήρωση της καλλιέργειας. Η επίστρωση του υλικού σε καθαρές αντικειµενοφόρους πραγµατοποιείται µετά από φυγοκέντρηση για 1 στις 17 στροφές και επαναδιάλυση του ιζήµατος που προκύπτει σε.5ml µονιµοποιητικού διαλύµατος. Η επίστρωση µε ενστάλαξη γίνεται µε ιδιαίτερη προσοχή από ελάχιστη απόσταση και 2-3 σταγόνες από το εναιώρηµα κρίνονται ικανοποιητικές. Τα παρασκευάσµατα προφυλαγµένα από τη σκόνη αφήνονται να στεγνώσουν σε θερµοκρασία δωµατίου και ακολούθως τοποθετούνται στους (+4) C για τέσσερις ηµέρες για να ολοκληρωθεί η διαδικασία ωρίµανσης. Το µονιµοποιητικό διάλυµα αποµακρύνει τα λιπίδια της µεµβράνης του κυττάρου και τις πρωτεΐνες που είναι δεσµευµένες στο DNA. Με αυτήν την κατεργασία µειώνονται οι δυσκολίες που θα αντιµετωπίσει ο ανιχνευτής (probe) στην προσπάθεια του να προσεγγίσει την αλληλουχία στόχο και ταυτόχρονα αυξάνεται η διαπερατότητα του κυττάρου στα διάφορα αντιδραστήρια
110 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH Κατόπιν στο µικροσκόπιο, σε πεδίο αντίθεσης φάσεων, ελέγχεται η ποιότητα των παρασκευασµάτων που περιλαµβάνει την οµοιόµορφη κατανοµή ενός ικανοποιητικού αριθµού µεσοφασικών πυρήνων και τη µορφολογία του κυτταροπλάσµατος που επηρεάζει άµεσα το χρόνο παραµονής στο µονιµοποιητικό διάλυµα. Η ποιότητα των παρασκευασµάτων αποτελεί σηµαντική παράµετρο για µία σωστή και επιτυχηµένη υβριδοποίηση. Ακολούθως, σηµαδεύεται µε χάραξη (ειδικό στυλό Vicking) το οπτικό πεδίο στο οποίο θα πραγµατοποιηθεί υβριδοποίηση Επεξεργασία του κυτταρικού υλικού στις αντικειµενοφόρους (Pretreatment) Η συνήθης πρακτική που ακολουθείται περιλαµβάνει την ταυτόχρονη κατεργασία των παρασκευασµάτων µε πρωτεάσες παρουσία διαλύµατος HCl Επίδραση µε HCl. Πραγµατοποιείται επίδραση 1mM HCl για 1. Η ακριβής δράση του οξέος δεν είναι γνωστή, αλλά η αποµάκρυνση των πρωτεϊνών και η µερική υδρόλυση των ακολουθιών στόχων συµβάλλει σηµαντικά στη βελτίωση του σήµατος υβριδοποίησης Επίδραση µε πεπσίνη. Η συνήθης επεξεργασία περιλαµβάνει επώαση των παρασκευασµάτων για 1 στους 37 C σε διάλυµα 1mM HCl, που περιέχει 5-15µg/ml πεπσίνης. Η αύξηση της διαπερατότητας θεωρείται απαραίτητη για να διευκολυνθεί η δίοδος των αντιδραστηρίων υβριδοποίησης, συµπεριλαµβανοµένου και του ανιχνευτή, προς το DNA-στόχο. Η πεψίνη αποδίδει έξοχα αποτελέσµατα µε την πέψη και καταβολισµό των πρωτεϊνικών µορίων που περιβάλλουν τις αλληλουχίες στόχους Πειραµατική πορεία. 1. Προετοιµάζουµε το υδατόλουτρο στους 37 C και τοποθετούµε δοχείο µε 7ml 1mM HCl (ph 3). Η ρύθµιση του ph πραγµατοποιείται µε την προσθήκη.1n NaOH. 2. Προετοιµάζουµε τα ακόλουθα δοχεία: ένα µε αποστειρωµένο ddh2o, ένα µε PBS χωρίς Mg ++ & Ca ++, ένα µε 1% φορµαλδεΰδη/pbs (4 C). 3. Προετοιµάζουµε πέντε δοχεία µε αυξανόµενες συγκεντρώσεις αιθανόλης (3%, 5%, 7%, 9% & 1%). 4. Ελέγχουµε τη θερµοκρασία στο δοχείο που περιέχει 7 ml διαλύµατος 1mM HCl και προσθέτουµε 2-5µl πεπσίνης 2mg/ml. Ακολουθεί επώαση των παρασκευασµάτων για 1 στους 37 C. 5. Σύντοµες εκπλύσεις (2 ) των παρασκευασµάτων µε ταυτόχρονη ανάδευση µε ddh2o που ακολουθείται από PBS σε θερµοκρασία δωµατίου. Ως αρχή και τέλος του χρόνου θεωρούµε το πρώτο παρασκεύασµα. 6. Μονιµοποίηση των παρασκευασµάτων µε 1%φορµαλδεΰδη/PBS για 7-1 στους (+4) C. 7. Επανάληψη των εκπλύσεων (2 ), πρώτα µε PBS και κατόπιν µε ddη2ο. 8. Συνεχίζουµε µε σταδιακές αφυδατώσεις και τα παρασκευάσµατα τοποθετούνται σε διαδοχικές συγκεντρώσεις αιθανόλης σε θερµοκρασία δωµατίου χωρίς ανάδευση. Στις συγκεντρώσεις των 3, 5, 7% παραµένουν για 2 και στις 9 και 1% για Τα παρασκευάσµατα αφήνονται να στεγνώσουν προφυλαγµένα από τη σκόνη
111 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH ιαλύµατα. 1. 1mM (.1N) HCl (ph 3) 1ml.1Ν HCl:.83ml 37% w/w (12.7N) HCl (Carlo Erba) 99.17ml ddh2o. 7ml.1N HCl: 7ml.1Ν HCl 63ml ddh2o. 2..1N NaOH 1Ν NaOH: 4gr NaOH (Sigma) σε 1ml ddh2o. 7ml.1N NaOH: 7ml 1Ν NaOH 63ml ddh2o. 3. Πεψίνη Pepsin, Lyophilised powder, 25mg, (Sigma P712). Ενδιάµεσο πυκνό διάλυµα αποθήκευσης 2mg/ml: 8mg πεπσίνης σε.4ml ddh2o ιάλυµα εργασίας 15µg/ml: 52.5µl πεπσίνης 2mg/ml σε 69.95ml HCl 1mM. 4. PBS (Phosphate Buffered Saline) Σε 1L ddh2o προσθέτουµε: 8.gr χλωριούχο νάτριο, (.14Μ) NaCl (MERCK).2gr χλωριούχο κάλιο, (2.7mM) KCl (MERCK).2gr δισόξινο φωσφορικό κάλιο (1.5mM) KH2PO4 (MERCK) 1.15gr άνυδρο όξινο φωσφορικό νάτριο (8.1mM) Na2HPO4 (MERCK) Ρύθµιση σε ph % φορµαλδεΰδη/pbs 7ml διαλύµατος περιέχει: 1.9ml 37% HCHO (MERCK) 68.1ml PBS. 6. Απόλυτη αιθανόλη CH3CH2OH (MERCK) Παρασκευάζουµε πέντε διαλύµατα µε αυξανόµενες συγκεντρώσεις αιθανόλης σε τελικό όγκο 7 ml. Αιθανόλη: 3% 5% 7% 9% 1% Αιθανόλη απόλυτη (ml): ddh2o (ml): Αποδιάταξη DNA στόχου και DNA ανιχνευτή (Denaturation) Για να επιτευχθεί υβριδοποίηση θα πρέπει να πραγµατοποιηθεί µε επιτυχία η αποδιάταξη όλων των δίκλωνων µορίων (ανιχνευτών και στόχων). Οι συνήθεις τρόποι αποδιάταξης του DNA είναι δύο και περιλαµβάνουν τη χρήση αλκαλικών διαλυµάτων ή υψηλής θερµοκρασίας. Η θερµική αποδιάταξη σε 9-1 C (5 ) παρουσία.1-.2μ Νa + είναι απλή και αποδίδει πολύ καλή υβριδοποίηση αλλά πρακτικά επιφέρει σηµαντικές µορφολογικές απώλειες. Σηµαντική παράµετρο αποτελεί το γεγονός ότι ο ανιχνευτής και ο στόχος πρέπει να αποδιαταχθούν ταυτόχρονα. ιαφοροποιήσεις σχετικά µε το χρόνο και τη θερµοκρασία (8 C για 2 ) είναι δυνατόν να αποδώσουν καλύτερες συνθήκες αποδιάταξης
112 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH Οι άριστες συνθήκες αποδιάταξης επιτυγχάνονται µε συνδυασµό θερµοκρασίας και του οργανικού διαλύτη φορµαµιδίου (7% φορµαµίδιο, 7 C), που µειώνει σηµαντικά το σηµείο τήξης (Τm) του δίκλωνου DNA Πειραµατική πορεία. Αποδιάταξη DNA στόχου 1. Αποδιατάσσουµε το κυτταρικό DNA των παρασκευασµάτων (συνήθως τέσσερα) τοποθετώντας τα σε 7% φορµαµίδιο/2χssc ph 7 στους 72 C για Τοποθετούµε τα παρασκευάσµατα απ ευθείας σε 7% αιθανόλη (η οποία διατηρείται σε πάγο) για Αφυδατώνουµε ακόµη περισσότερο τα παρασκευάσµατα µας αφήνοντας τα από 2 σε 9 και 1% αιθανόλη σε θερµοκρασία δωµατίου, και κατόπιν αφήνονται να στεγνώσουν τελείως πριν την υβριδοποίηση. Αποδιάταξη DNA ανιχνευτή 4. Προθέρµανση του ανιχνευτή (-2 C) στους 37 C για Ακολουθεί ήπια ανάδευση σε συσκευή ανατάραξης (vortex) και φυγοκέντρηση (cytospin) για Παρασκευάζουµε το διάλυµα υβριδοποίησης που αποτελείται από 65% φορµαµίδιο/2χssc, ph 7 (Hybrisol VI). 7. Αναµιγνύουµε σε µικροφιαλίδιο.5µl (1µl:1ng) ανιχνευτή µε 1µl διαλύµατος υβριδοποίησης και αναδεύουµε ήπια για να επιτευχθεί οµογενοποίηση. 8. Αποδιατάσουµε τον ανιχνευτή µε την τοποθέτηση του µικροφιαλιδίου στους 72 C (±2 C) για Μετά το πέρας του χρόνου ακολουθεί σύντοµη φυγοκέντρηση (cytospin) για 2-3 και το µικροφιαλίδιο τοποθετείτε σε πάγο για τη διατήρηση της αποδιάταξης Υβριδοποίηση Η υβριδοποίηση εξαρτάται από την ικανότητα του αποδιαταγµένου DNA να επανασυνδέεται µε τις συµπληρωµατικές αλυσίδες σε ένα περιβάλλον µε θερµοκρασία λίγο χαµηλότερη από το σηµείο (Τm) τήξης του. Η σταθερότητα του DNA εξαρτάται άµεσα από την περιεκτικότητα σε GC βάσεις µε αποτέλεσµα όσο υψηλότερη είναι η αναλογία των ζευγών GC, τόσο υψηλότερη είναι η Τm. Οι κυριότερες παράµετροι που επηρεάζουν την Τm και την επαναδιάταξη του DNA είναι η θερµοκρασία, το ph, η συγκέντρωση µονοσθενών κατιόντων και η παρουσία οργανικών διαλυτών Θερµοκρασία. Η µεγαλύτερη ταχύτητα επαναδιάταξης-υβριδοποίησης (renaturationhybridization) επιτυγχάνεται σε θερµοκρασία 25 C χαµηλότερη από την Τm. Η βέλτιστη σχέση επαναδιάταξης και θερµοκρασίας κυµαίνεται µεταξύ C χαµηλότερα από την Τm ph. Σε τιµές ph που κυµαίνονται από 6-8 ο ρυθµός επαναδιάταξης είναι αρκετά ανεξάρτητος από το ph. Αντίθετα, σε υψηλότερο ή χαµηλότερο ph προκαλείται αλκαλική αποδιάταξη και υδρόλυση του DNA. Συνήθως καταλληλότερα
113 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH ρυθµιστικά διαλύµατα θεωρούνται αυτά που περιέχουν 2-5mM φωσφορικά µε ph Μονοσθενή κατιόντα. Μονοσθενή κατιόντα (ιόντα νατρίου) αλληλεπιδρούν ηλεκτροστατικά µε τα νουκλεϊκά οξέα (κυρίως µε τις φωσφορικές οµάδες) µε αποτέλεσµα η ηλεκτροστατική απώθηση µεταξύ των δύο αλυσίδων του δίκλωνου DNA να µειώνεται µε την αύξηση της συγκέντρωσης αλάτων. Αυτό πρακτικά σηµαίνει, ότι όσο υψηλότερη είναι η συγκέντρωση του άλατος, τόσο περισσότερο αυξάνεται η σταθερότητα του υβριδίου. Επιπρόσθετα, χαµηλές συγκεντρώσεις ιόντων νατρίου επηρεάζουν ση- µαντικά την Τm και το ρυθµό επαναδιάταξης, σε αντίθεση µε συγκεντρώσεις νατρίου υψηλότερες από.4μ. ισθενή κατιόντα σε ελεύθερη µορφή σταθεροποιούν δραµατικά το διπλό DNA και πρέπει να αποµακρύνονται µε τη χρήση κατάλληλων παραγόντων (EDTA, κιτρικό) Οργανικοί διαλύτες. Οι οργανικοί διαλύτες µειώνουν τη θερµοσταθερότητα των δίκλωνων πολυνουκλεοτιδίων, ώστε η υβριδοποίηση να επιτυγχάνεται σε αρκετά χαµηλότερες θερµοκρασίες µε αποτέλεσµα τη διατήρηση της µορφολογίας µε ταυτόχρονη αύξηση της σταθερότητας του ανιχνευτή. Το φορµαµίδιο έχει αποδειχθεί ένας εξαιρετικός οργανικός διαλύτης που µειώνει την Τm DNA:DNA και DNA:RNA γραµµικά κατά.72 C για κάθε 1% προστιθέµενου φορµαµιδίου. Η υβριδοποίηση είναι δυνατόν να πραγµατοποιηθεί σε 3-45 C µε την παρουσία 5% φορµαµιδίου. Επιπλέον, η χρήση υψηλής συγκέντρωσης φορµαµιδίου ( 95%) δεν ενδείκνυται διότι τα σχηµατιζόµενα υβρίδια είναι σχετικά ασταθή Πειραµατική πορεία. Η πειραµατική πορεία της επιτόπιας υβριδοποίησης που αποτελεί συνέχεια της αποδιάταξης του DNA (ανιχνευτή και στόχου) και πραγµατοποιείται την πρώτη ηµέρα εφαρµογής της τεχνικής FISH, περιγράφεται ως εξής: 1. Καλυπτρίδες διαστάσεων 22Χ22 mm που φυλάσσονται σε απόλυτη αιθανόλη (4 C) αφήνονται να στεγνώσουν σε θερµοκρασία δωµατίου. 11. Τοποθετούµε 1-15µl (2-3 µl/cm 2 ) διαλύµατος υβριδοποίησης που περιέχει 5-7ng αποδιαταγµένου ανιχνευτή σε κάθε παρασκεύασµα και κατόπιν καλύπτουµε προσεκτικά µε καλυπτρίδα µε τη βοήθεια λαβίδας, ώστε να αποφευχθεί η δηµιουργία µικροφυσαλίδων αέρα. 12. Οι καλυπτρίδες σφραγίζονται περιµετρικά µε ειδική κόλλα (Velo Gummi Iosung, Aspeg). 13. Αφήνουµε να επιτευχθεί υβριδοποίηση των παρασκευασµάτων µε τον ανιχνευτή κατά τη διάρκεια της νύχτας για ώρες (overnight) σε υγρή ατµόσφαιρα στους 37 C ιαλύµατα. 1. 2XSSC (Saline Sodium Citrate) Σε 1L ddh2o προσθέτουµε: gr χλωριούχο νάτριο, (3M) NaCl (MERCK) 88.23gr κιτρικό νάτριο, (.3M) HOC(COONa)(CH2COONa)2 (MERCK) Ρύθµιση σε ph
114 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH 2. 7% φορµαµίδιο/2χssc 7ml διαλύµατος περιέχει: 49ml φορµαµίδιο (MERCK) 7ml 2ΧSSC 14ml ddh2o Ρύθµιση σε ph Ανιχνευτής, Appligene Oncor CP595-DG.5 All Human Centromeres (α-satellite) digoxigenin labelled,.25µg (25µl). 4. ιάλυµα υβριδοποίησης (Hybrisol VI) 1ml διαλύµατος 65% φορµαµίδιο/2χssc (-2 C) περιέχει: 65µl φορµαµίδιο (MERCK) 1µl 2ΧSSC 25µl ddh2o Ρύθµιση σε ph Εκπλύσεις µετά την υβριδοποίηση (Posthybridization washes) Ο επισηµασµένος ανιχνευτής είναι δυνατόν να υβριδοποιηθεί µη ειδικά σε αλληλουχίες, οι οποίες δεν είναι πλήρως οµόλογες µε την αλληλουχία του ανιχνευτή. Τέτοια υβρίδια είναι περισσότερο ασταθή από τα πλήρως οµόλογα συζευγµένα και µπορούν να αποδιαταχθούν µε εκπλύσεις, οι οποίες κυµαίνονται ως προς την δυναµικότητα τους ανάλογα µε τη συγκέντρωση του φορµαµιδίου, των αλάτων και τη θερµοκρασία. Εκπλύσεις σε διάλυµα 2ΧSSC που περιέχει 5% φορµαµίδιο, ph 7 θεωρούνται αρκετά ικανοποιητικές. Η χρήση διαλυµάτων µε τιµές ph που κυµαίνονται υψηλότερα από 6-8 αντιστοιχεί σε αυστηρές συνθήκες έκπλυσης. Επιπρόσθετα, χαµηλές συγκεντρώσεις αλάτων σε συνδυασµό µε υψηλές θερµοκρασίες καθορίζουν άµεσα την αυστηρότητα (stringency) των συνθηκών έκπλυσης. Γενικότερα, µεγαλύτερη εξειδίκευση επιτυγχάνεται, όταν η υβριδοποίηση πραγµατοποιείται σε αυστηρές συνθήκες και οι εκπλύσεις, οι οποίες ακολουθούν είναι παρόµοιας ή χαµηλότερης δυναµικότητας Πειραµατική πορεία. 1. Προετοιµάζουµε το υδατόλουτρο στους 37 C και τοποθετούµε τα ακόλουθα δοχεία: δύο µε 5% φορµαµίδιο/2xssc, ph 7, δύο µε 2XSSC, ph 7 και τρία µε 4ΧSSC/.5% Tween Αφαιρούµε προσεκτικά την κόλλα από τα παρασκευάσµατα που εντοπίζεται περιµετρικά της καλυπτρίδας, µε τη βοήθεια λαβίδας. 3. Πραγµατοποιούµε δύο εκπλύσεις σε 5% φορµαµίδιο/2xssc, στους 37 C από 7, µε συνεχή ελαφρά ανακίνηση. Κατά τη διάρκεια της πρώτης έκπλυσης οι καλυπτρίδες αποµακρύνονται οµαλά. 4. Ακολουθούν δύο εκπλύσεις σε 2XSSC, στους 37 C από Ανοσοεντοπισµός του ανιχνευτή µε φθορίζουσες ουσίες (Ιmmunocytochemistry) Οι αντιδράσεις ανίχνευσης πραγµατοποιούνται έµµεσα µε τη βοήθεια µακροµοριακών ενώσεων (αντισώµατα) στις οποίες προσδένονται φθορίζουσες ουσίες (φθοροσκεΐνη) µε αποτέλεσµα τον εντοπισµό του µορίου αναφοράς (διγοξιγενίνη)
115 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH στον ανιχνευτή, γεγονός που επιτρέπει την αναγνώριση των θέσεων του σχηµατισµένου υβριδίου µετά την υβριδοποίηση. Επιπλέον, η χρήση αρκετών ειδικών αντισωµάτων που αντιδρούν µε αντιγόνα, κατά την έµµεση µέθοδο, δίνει τη δυνατότητα ενίσχυσης του σήµατος υβριδοποίησης. Σηµαντική παράµετρος πριν τη διαδικασία προσθήκης των αντισωµάτων αποτελεί η εφαρµογή της αντίδρασης φραγµού (blocking reaction) που µειώνει σηµαντικά την πιθανότητα µη ειδικής δέσµευσης των αντιδραστηρίων ανίχνευσης µε µη ειδικές θέσεις υβριδοποίησης. Με το ίδιο σκεπτικό χρησιµοποιούνται αραιώσεις των αντισωµάτων σε ανάλογα διαλύµατα (λευκωµατίνη ορού βοός, αποβουτυρωµένο γάλα). Επιπρόσθετα, η χρήση µη ιοντικών (nonionic) απορρυπαντικών (Tween 2) αποµακρύνει τα εναποµείναντα, από προηγούµενες κατεργασίες κυτταρικά υπολείµµατα και προκαλεί ήπια διάσπαση της διπλοστοιβάδας λιπιδίων της µεµβράνης, εξαιτίας της απελευθέρωσης και διαλυτοποίησης µεµβρανικών πρωτεϊνών, µε αποτέλεσµα τη διάνοιξη διόδων, ώστε οι θέσεις υβριδοποίησης να είναι περισσότερο ευπρόσιτες σε αντιδραστήρια ανίχνευσης Πειραµατική πορεία. Η πειραµατική πορεία εντοπισµού του ανιχνευτή µε κατάλληλα αντισώ- µατα που αποτελεί συνέχεια της αποµάκρυνσης των µη ειδικών υβριδοποιήσεων και πραγµατοποιείται τη δεύτερη ηµέρα εφαρµογής της τεχνικής FISH, περιγράφεται ως εξής: 5. Καλυπτρίδες διαστάσεων 24Χ24 mm που φυλάσσονται σε απόλυτη αιθανόλη (4 C) αφήνονται να στεγνώσουν σε θερµοκρασία δωµατίου. 6. Προσθήκη 3-5µl (5µl/cm 2 ) 5% αποβουτυρωµένου γάλακτος/4χssc και κάλυψη του παρασκευάσµατος µε καλυπτρίδα. Η επώαση διαρκεί 3 και πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C. 7. Έκπλυση των παρασκευασµάτων σε 4ΧSSC/.5% Tween-2 στους 37 C για Προσθήκη πρώτου αντισώµατος έναντι της διγοξιγενίνης και κάλυψη του παρασκευάσµατος µε καλυπτρίδα. Η επώαση πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C και διαρκεί Ακολουθούν τρεις εκπλύσεις µε 4ΧSSC/.5% Tween-2 στους 37 C από 2, µε ελαφρά ανακίνηση. Οι εκπλύσεις αυτές επαναλαµβάνονται µε νέο διάλυ- µα 4ΧSSC/.5% Tween-2 µετά την επώαση των παρασκευασµάτων µε το δεύτερο και τρίτο αντίσωµα. 1. Προσθήκη δεύτερου αντισώµατος και κάλυψη του παρασκευάσµατος µε καλυπτρίδα. Η επώαση πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C και διαρκεί Προσθήκη τρίτου αντισώµατος, που είναι συζευγµένο µε το φθορόχρωµα φθοροσκεΐνη και κάλυψη του παρασκευάσµατος µε καλυπτρίδα. Η επώαση πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C και διαρκεί Για την παρατήρηση των µεσοφασικών κυττάρων ακολουθεί χρώση των παρασκευασµάτων σε διάλυµα 2ΧSSC που περιέχει την κυανή φθορίζουσα χρωστική DAPI και το ερυθρό ιωδιούχο προπίδιο (PI). Η χρώση πραγµατοποιείται για 1 σε θερµοκρασία δωµατίου. 13. Έκπλυση των παρασκευασµάτων µε 4ΧSSC/.5% Tween-2 σε θερµοκρασία δωµατίου για Προσθήκη 1-15µl ειδικού διαλύµατος (Anti-fading reagent - Mounting Medium) για την ενίσχυση και διατήρηση του σήµατος και προσεκτική κάλυψη
116 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH των παρασκευασµάτων µε καλυπτρίδες. 15. Ακολουθεί σφράγιση των καλυπτρίδων περιµετρικά µε διαφανές υλικό (nail polish) και µικροσκοπική εξέταση των παρασκευασµάτων µετά την παραµονή τους τουλάχιστον για 3-6 στους 4 C ιαλύµατα. 1. 5% φορµαµίδιο/2χssc 7ml διαλύµατος περιέχει: 35ml φορµαµίδιο (MERCK) 7ml 2ΧSSC 28ml ddh2o Ρύθµιση σε ph XSSC, 4XSSC (Saline Sodium Citrate) Παρασκευάζουµε τα διαλύµατα 2XSSC και 4XSSC από πυκνό διάλυµα 2XSSC µε αραιώσεις 1:1 και 1:5, σε τελικό όγκο 5 ml. ιάλυµα: 2XSSC 4XSSC 2ΧSSC (ml): 5 1 ddh2o (ml): 45 4 Ρύθµιση σε ph % αποβουτυρωµένο γάλα/4xssc.5gr αποβουτυρωµένου γάλακτος (Regilait, UCA) σε1ml 4XSSC. 4..5% αποβουτυρωµένο γάλα/4xssc 5ml διαλύµατος περιέχει: 5ml 5% αποβουτυρωµένο γάλα/4xssc 45ml 4ΧSSC. 5. 4XSSC/.5% Tween-2 5ml διαλύµατος περιέχει: ml 4ΧSSC.25ml Tween-2 Polyoxyethylenesorbitan Monalaurate (Sigma P1379). 6. Αντισώµατα Πρώτο αντίσωµα α) Mouse Monoclonal Anti-Digoxigenin (Clone DI-22, IgG1). Sigma D8156. Χρησιµοποιείται σε αραίωση 1:25 (.4µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (.3µg). εύτερο αντίσωµα β) Anti-mouse IgG (whole molecule) F(ab')2 fragment-r-phycoerythrin conjugate from sheep. Sigma P8547. Χρησιµοποιείται σε αραίωση 1:2 (25µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (2µg). Τρίτο αντίσωµα γ) Anti-sheep IgG (whole molecule) FITC conjugate from donkey. Sigma F7634. Χρησιµοποιείται σε αραίωση 1:2 (5µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (4µg)
117 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH 7. DAPI DAPI, (4,6-Diamidino-2-phenylindole dihydrochloride), powder, 1mg (Aldrich) Ενδιάµεσα πυκνά διαλύµατα αποθήκευσης: DAPI 5mg/ml: 1mg διαµιδίνης σε 2ml ddh2o DAPI.1mg/ml: 2µl DAPI 5mg/ml σε 9.8ml PBS ιάλυµα εργασίας DAPI.2µg/ml: 14µl DAPI 1µg/ml σε 69.86ml 2ΧSSC. 8. PI PI, (propidium iodide), powder, 1mg, (Sigma) Ενδιάµεσο πυκνό διάλυµα αποθήκευσης: PI 5mg/ml: 1mg ιωδιούχου προπιδίου σε 2 ml ddh2o ιάλυµα εργασίας PI 1µg/ml: 14µl PI 5mg/ml σε 69.99ml 2ΧSSC. 9. Vectashield Mounting Medium (Vector H-1) 8.2 ιπλή In Situ υβριδοποίηση µε φθοροχρώµατα (DUAL FISH) - Συµµετοχή των χρωµοσωµάτων Χ, Υ και 8 σε ανευπλοειδικά φαινόµενα Η εφαρµογή της συγκεκριµένης µεθοδολογίας που επιτυγχάνεται µε τη χρησιµοποίηση εξειδικευµένων ανιχνευτών που εµφανίζουν συµπληρωµατικότητα µε αλληλουχίες του υψηλά επαναλαµβανόµενου άλφα δορυφορικού DNA (αsatellite DNA) και εντοπίζονται στην κεντροµερική περιοχή των χρωµοσωµάτων Χ, Υ και 8 βασίζεται στο γεγονός ότι η οργάνωση της α-δορυφορικής περιοχής είναι µοναδική για κάθε χρωµόσωµα. O ειδικός ανιχνευτής για την κεντροµερική περιοχή του χρωµοσώµατος Χ είναι επισηµασµένος µε το φυτικό παράγωγο διγοξιγενίνη και ο εντοπισµός του περιλαµβάνει επαναλαµβανόµενα βήµατα σήµανσης µε διαδοχικά στρώµατα αντισωµάτων, µε το τελευταίο να είναι συζευγµένο µε το φθορόχρω- µα φθοροσκεΐνη. Για την επισήµανση των ανιχνευτών που αφορούν την κεντροµερική περιοχή των χρωµοσωµάτων Υ και 8 χρησιµοποιείται µία φυσική βιταµίνη, η βιοτίνη και ο εντοπισµός τους πραγµατοποιείται µε τη χρήση αβιδίνης, η οποία είναι συζευγµένη µε το φθορόχρωµα ροδαµίνη. Το πειραµατικό πρωτόκολλο της διπλής in situ υβριδοποίησης µε φθοροχρώµατα περιλαµβάνει τα απλοποιηµένα στάδια της τεχνικής FISH (8.1) µε τις εξής τροποποιήσεις: Επεξεργασία του κυτταρικού υλικού στις αντικειµενοφόρους Επίδραση µε πεπσίνη. Η συνήθης επεξεργασία περιλαµβάνει επώαση των παρασκευασµάτων για 1 στους 37 C σε διάλυµα 1mM HCl, που περιέχει 3-5µg/ml πεπσίνης Αποδιάταξη DNA ανιχνευτών Αναµιγνύουµε σε µικροφιαλίδιο 1µl (1ng) µείγµατος ανιχνευτών (αsatellite probes) για την κεντροµερική περιοχή των χρωµοσωµάτων Χ και 8 ή Χ και Υ σε αναλογία 1:1 µε 9µl διαλύµατος υβριδοποίησης (Hybrisol VI), ώστε να επιτευχθεί τελική συγκέντρωση 1ng/µl
118 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH Υβριδοποίηση Σε κάθε παρασκεύασµα τοποθετούµε 1-12µl διαλύµατος υβριδοποίησης που περιέχει 5-6ng από τον κάθε αποδιαταγµένο ανιχνευτή Ανιχνευτές. α) Για το χρωµόσωµα Χ -Appligene Oncor CP56-DG.5 Chromosome X a-satellite (DXZ1) digoxigenin labeled,.5µg (5µl) β) Για το χρωµόσωµα Υ -Appligene Oncor CP565-B.5 Chromosome Y a-satellite (DYZ3) biotin labeled,.25µg (25µl) γ) Για το χρωµόσωµα 8 -Appligene Oncor CP513-B.5 Chromosome 8 a-satellite (D8Z2) biotin labeled,.25µg (25µl) Ανοσοεντοπισµός των ανιχνευτών µε φθορίζουσες ουσίες Ανίχνευση κεντροµερικής περιοχής χρωµοσώµατος Χ Πρώτο αντίσωµα α) Mouse Monoclonal Anti-Digoxigenin (Clone DI-22, IgG1). Sigma D8156. Χρησιµοποιείται σε αραίωση 1:25 (.4µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (.3µg). εύτερο αντίσωµα β) Anti-mouse Ig Digoxigenin conjugated secondary antibody [F(ab')2 fragments] from sheep. Chemicon AQ3D. Χρησιµοποιείται σε αραίωση 1:1 (2µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (1.6µg). Τρίτο αντίσωµα γ) Anti-digoxigenin-fluorescein, Fab fragments [FITC]. Roche Χρησιµοποιείται σε αραίωση 1:1 (2µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (1.6µg). Ανίχνευση κεντροµερικής περιοχής χρωµοσώµατος Υ, 8 Πρώτο αντίσωµα α) Rhodamine6 Avidin D. Vector A25. Χρησιµοποιείται σε αραίωση 1:1 (5µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (4µg). εύτερο αντίσωµα β) Mouse Monoclonal Anti-Avidin Biotin conjugate. Sigma B9655. Χρησιµοποιείται σε αραίωση 1:1 (22µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (1.8µg). Τρίτο αντίσωµα γ) Rhodamine6 Avidin D. Vector A25. Χρησιµοποιείται σε αραίωση 1:1 (5µg/ml) σε.5% αποβουτυρωµένο γάλα και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 8 µl (4µg). Για την παρατήρηση των µεσοφασικών κυττάρων ακολουθεί χρώση των παρασκευασµάτων σε διάλυµα 2ΧSSC που περιέχει την κυανή φθορίζουσα χρωστική DAPI (διάλυµα εργασίας.2µg/ml)
119 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH 8.3 Συνδυασµένη σήµανση του κινητοχώρου και του κεντροµέρους (CREST - FISH) Η µεθοδολογία CREST-FISH αποτελεί µία σύνθετη διαδικασία που αποσκοπεί στη συνδυασµένη σήµανση του κινητοχώρου και του κεντροµέρους όλων των ανθρώπινων χρωµοσωµάτων και η οποία περιλαµβάνει την ταυτόχρονη εφαρµογή δύο ανεξάρτητων τεχνικών µε την εξής σειρά: Τεχνική CREST Πρόκειται για τον ανοσολογικό προσδιορισµό των πρωτεϊνών του εξειδικευµένου πολυπρωτεϊνικού συµπλόκου του κινητοχώρου (CENPs A, B και C) µε τη χρήση ορού (αντισωµάτων CREST) που λαµβάνεται από µία οµάδα ασθενών κατά τη διάρκεια της περιόδου στην οποία αναπτύσσονται προοδευτικά τα συ- µπτώµατα του συνδρόµου CREST. Η ταυτοποίηση του κινητοχώρου συνίσταται στη χρήση ενός δεύτερου αντισώµατος σηµασµένου µε τη φθορίζουσα χρωστική φθοροσκεΐνη που συνδέεται µε τα εξειδικευµένα για τις πρωτεΐνες του κινητοχώρου, αντισώµατα CREST Μονιµοποίηση. Η ακινητοποίηση του κυτταρικού υλικού επιτυγχάνεται µε τη χρήση κατάλληλων µονιµοποιητικών διαλυµάτων κατακρήµνισης. Η χρήση απόλυτης µεθανόλης σε χαµηλές θερµοκρασίες (-1) C-(-2) C εξασφαλίζει τη διατήρηση της µορφολογίας ενδοκυτταρικών δοµών (πρωτεΐνες κινητοχώρου) και της λειτουργικότητας των επίτοπων ή αντιγονικών καθοριστών, γεγονός που καθιστά δυνατή την εφαρµογή αµιγών ανοσοχηµικών µεθόδων Πειραµατική πορεία. Τα βήµατα που ακολουθούνται στη µέθοδο ανίχνευσης πρωτεϊνών του κινητοχώρου σε διπύρηνα µεσοφασικά κύτταρα, τα οποία προκύπτουν µε την επίδραση της κυτταροχαλασίνης-β είναι τα εξής: 1. Απόρριψη καλλιεργητικού υλικού και στιγµιαία προσθήκη παγωµένου (4 C) ρυθµιστικού διαλύµατος φωσφορικών αλάτων (PBS) που πραγµατοποιείται µε ιδιαίτερα αργό ρυθµό στα άκρα του τρυβλίου petri για την αποφυγή ενόχλησης των κυττάρων. 2. Ακολουθεί µονιµοποίηση των κυττάρων µε παγωµένη απόλυτη µεθανόλη (-2 C) για 3 λεπτά. 3. Τρεις διαδοχικές εκπλύσεις µε PBS (4 C) από 3 λεπτά, µε ελαφρά ανάδευση. 4. Προσθήκη πρώτου αντισώµατος (CREST antibody). Η επώαση πραγµατοποιείται κατά τη διάρκεια της νύχτας για 12 ώρες σε υγρή ατµόσφαιρα στους 4 C. Για εξοικονόµηση χρόνου, η επώαση είναι δυνατόν να πραγµατοποιηθεί σε υγρή ατµόσφαιρα στους 37 C και διαρκεί 2 ώρες. 5. Ακολουθούν πέντε εκπλύσεις µε PBS (4 C) που περιέχει 1% BSA από 5 λεπτά, µε ελαφρά ανάδευση. 6. Προσθήκη δεύτερου αντισώµατος (Anti-Human antibody) που είναι σηµασµένο µε το φθορόχρωµα φθοροσκεΐνη. Η επώαση διαρκεί 45 λεπτά και πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C. 7. Πέντε διαδοχικές εκπλύσεις µε PBS (4 C) που περιέχει 1% BSA από 5 λεπτά, µε ελαφρά ανάδευση
120 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH 8. Ακολουθούν πέντε νέες εκπλύσεις µε PBS (4 C) από 5 λεπτά, µε ελαφρά ανάδευση. 9. Τα παρασκευάσµατα αφήνονται να στεγνώσουν σε θερµοκρασία δωµατίου σε περιβάλλον προστατευµένο από το φως και τη σκόνη ιαλύµατα. 1. PBS (Phosphate Buffer Solution) 2. Μεθανόλη απόλυτη CH3OH (Sigma, St.Louis, Mo., USA) 3. PBS/BSA 1% 1gr Bovine Serum Albumin-BSA (Sigma, A453) σε 1ml PBS 4. PBST (PBS/.1% Tween 2) 1µl Tween-2 σε 1ml PBS 5. Αντισώµατα Πρώτο αντίσωµα Antinuclear Antibody-Positive Control Serum (Centromere). Antibodies Inc., Davis, CA Χρησιµοποιείται σε αραίωση 1:1 σε PBS που περιέχει.1% Tween-2 (PBST) και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl. εύτερο αντίσωµα Anti-Human-Polyvalent Immunoglobulins (whole molecule)-fitc from rabbit. Sigma F4637. Χρησιµοποιείται σε αραίωση 1:8 σε PBS που περιέχει 1% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl Τεχνική FISH Πρόκειται για την ανίχνευση επαναλαµβανόµενων ειδικών αλληλουχιών κεντροµερικού DNA (α-δορυφορικού DNA) που πραγµατοποιείται µετά τον ανοσολογικό προσδιορισµό των πρωτεϊνών του εξειδικευµένου πολυπρωτεϊνικού συµπλόκου του κινητοχώρου. Στο πειραµατικό πρωτόκολλο που εφαρµόζεται δεν είναι εφικτή η επεξεργασία του κυτταρικού υλικού µε πρωτεάσες παρουσία διαλύµατος HCl, ενώ περιλαµβάνονται τα υπόλοιπα απλοποιηµένα στάδια της τεχνικής FISH (3.1) µε τις εξής τροποποιήσεις: Μονιµοποίηση-Ωρίµανση παρασκευασµάτων. Η νέα µονιµοποίηση του κυτταρικού υλικού πραγµατοποιείται µε την επώαση των παρασκευασµάτων σε διάλυµα µεθανόλης:οξικού (3:1) για µία ώρα, ενώ η διαδικασία ωρίµανσης επιτυγχάνεται µε την παραµονή τους για µία εβδοµάδα σε θερµοκρασία δωµατίου σε περιβάλλον προστατευµένο από το φως και τη σκόνη Ανιχνευτής, Appligene Oncor CP595-DG.5. All Human Centromeres (α-satellite) digoxigenin labelled,.5µg/15µl Εντοπισµός του ανιχνευτή µε φθορίζουσες ουσίες. Ο εντοπισµός του µορίου αναφοράς στον ανιχνευτή γίνεται έµµεσα µε φθο ροχρώµατα µε αποτέλεσµα η διγοξιγενίνη να µεγεθύνεται µε επαναλαµβανόµενα
121 Υλικά και Μέθοδοι Συνδυασµός CBMN/CREST-FISH βήµατα σήµανσης που περιλαµβάνουν διαδοχικά στρώµατα αντισωµάτων, µε το τελευταίο να είναι συζευγµένο µε το φθορόχρωµα ροδαµίνη. Για την αντίδραση φραγµού (blocking raction) χρησιµοποιείται διάλυµα 3% λευκωµατίνης ορού βοός (BSA)/4XSSC που περιέχει.1% Tween-2, ενώ για τις αραιώσεις των αντισωµάτων η προτεινόµενη αναλογία είναι 1% BSA/4XSSC (.1% Tween-2). Επιπρόσθετα, για τις εκπλύσεις που ακολουθούν κάθε φορά µετά την προσθήκη των αντισωµάτων χρησιµοποιείται διάλυµα 4XSSC/.1% Tween ιαλύµατα. 1. 3% BSA/4XSSC (.1% Tween 2) 5ml διαλύµατος περιέχει: 15mg BSA 1ml 2ΧSSC 3.95ml H2O 5µl Tween 2 1% 2. 1% BSA/4XSSC (.1% Tween 2) 5ml διαλύµατος περιέχει: 5mg BSA 1ml 2ΧSSC 3.95ml H2O 5µl Tween 2 1% 3. Tween 2 1%.5ml διαλύµατος περιέχει: 5µl Tween 2 45µl ddh2 4. 4XSSC/.1% Tween 2 5ml διαλύµατος περιέχει: 499.5ml 4ΧSSC.5ml Tween-2 5. Αντισώµατα Πρώτο αντίσωµα α) Mouse Monoclonal Anti-Digoxigenin (Clone , IgG1). Boehringer Mannheim, Χρησιµοποιείται σε αραίωση 1:2 (.5µg/ml) σε 1% BSA/ 4XSSC (.1% Tween-2) και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl (.3µg). εύτερο αντίσωµα β) Anti-mouse Ig-Digoxigenin, F(ab)2 fragment from sheep. Boehringer Mannheim, Χρησιµοποιείται σε αραίωση 1:1 (2µg/ml) σε 1% BSA/ 4XSSC (.1 % Tween-2) και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl (.1µg). Τρίτο αντίσωµα γ) Anti-digoxigenin-rhodamine, Fab fragments. Boehringer Mannheim, Χρησιµοποιείται σε αραίωση 1:8 (25µg/ml) σε 1% BSA/4XSSC (.1% Tween- 2) και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl (1.25µg). Για την παρατήρηση των µεσοφασικών κυττάρων ακολουθεί χρώση των παρασκευασµάτων σε διάλυµα 2ΧSSC που περιέχει την κυανή φθορίζουσα χρωστική DAPI (διάλυµα εργασίας.2µg/ml)
122 Υλικά και Μέθοδοι Μιτωτική συσκευή 9. ΜΕΛΕΤΗ ΤΗΣ ΑΚΕΡΑΙΟΤΗΤΑΣ ΤΗΣ ΜΙΤΩΤΙΚΗΣ ΣΥΣΚΕΥΗΣ 9.1 Σε καλλιέργειες λεµφοκυττάρων Για τη µελέτη των ανώµαλων µιτωτικών κυττάρων και την παρατήρηση του χρωµοσωµατικού υλικού σε διάφορες φάσεις της µίτωσης εφαρµόσθηκε η µέθοδος διαφορικής χρώσης µε εξειδικευµένες φθορίζουσες χρωστικές (διαµιδίνη και ιωδιούχο προπίδιο) σε ανθρώπινα λεµφοκύτταρα ολικού αίµατος. Ως πηγή λεµφοκυττάρων χρησιµοποιήθηκε φλεβικό αίµα που συλλέχθηκε από νεαρό αρσενικό άτοµο (Τ.Μ.) Συνθήκες καλλιέργειας Η πειραµατική διαδικασία περιλαµβάνει την προετοιµασία δύο σειρών καλλιεργειών στις οποίες η φαρµακευτική ένωση υδροχλωροθειαζίδιο (HCTZ) εξετάσθηκε στις συγκεντρώσεις 4, 6 και 8 µg/ml. Η επίδραση µε τις υπό µελέτη συγκεντρώσεις του υδροχλωροθειαζιδίου πραγµατοποιείται 24 ώρες µετά την έναρξη της καλλιέργειας, χρονική διάρκεια που αντιστοιχεί περίπου στη µέση της G1 φάσης του κυτταρικού κύκλου των λεµφοκυττάρων (Buckton και Evans, 1973). Μετά την προσθήκη όλων των απαραίτητων συστατικών, οι καλλιέργειες τοποθετούνται σε επωαστικό θάλαµο (θερµοκρασία 37 C και παροχή 5% CO2). Η καλλιέργεια λεµφοκυττάρων ολοκληρώνεται στις 72 ώρες και ακολουθεί η διαδικασία αποµόνωσης των λεµφοκυττάρων και λήψης παρασκευασµάτων για την παρατήρηση µιτωτικών φάσεων (Ford και Congedi, 1987). ιαγραµµατικά το χρονοδιάγραµµα του πειράµατος έχει ως εξής: t = Έναρξη καλλιέργειας t = 24 Προσθήκη των υπό µελέτη συγκεντρώσεων HCTZ t = 72 Ολοκλήρωση καλλιέργειας και αποµόνωση λεµφοκυττάρων Αποµόνωση λεµφοκυττάρων Η διαδικασία αποµόνωσης των λεµφοκυττάρων περιλαµβάνει τα ίδια στάδια, όπως περιγράφονται στο υποκεφάλαιο (7.1.2) στα Υλικά και Μέθοδοι µε µία µικρή τροποποίηση. Για την ταυτόχρονη ανάδευση του υλικού κατά την προσθήκη του υποτονικού και του µονιµοποιητικού διαλύµατος δεν χρησιµοποιήθηκε η συσκευή ανατάραξης (vortex), αλλά ακολουθήσαµε επιδέξιους λεπτούς χειρισµούς µε σκοπό την καλύτερη οµογενοποίηση του κυτταρικού ιζήµατος (Lee et al., 1994). Επιπρόσθετα, για την καλύτερη διατήρηση του κυτταροπλάσµατος ενδείκνυται η χρήση µεθανόλης:οξικoύ σε αναλογία 9:1 που εφαρµόζεται στο τελευταίο στάδιο µονιµοποίησης (Lindholm et al., 1991)
123 Υλικά και Μέθοδοι Μιτωτική συσκευή Χρώση των παρασκευασµάτων Η χρώση των παρασκευασµάτων γίνεται σε ρυθµιστικό διάλυµα PBS που περιέχει.5 µg/ml DAPI (4,6-διαµιδινο-2-φαινυλοινδόλιο-2-HCL) και 1 µg/ml PI (ιωδιούχο προπίδιο). Η χρονική διάρκεια παραµονής των παρασκευασµάτων στο διάλυµα είναι 1 σε θερµοκρασία δωµατίου και σε σκοτεινό περιβάλλον. Ακολουθεί έκπλυση µε PBS/.1%Tween 2 (PBST), προσθήκη 2-25 µl ειδικού διαλύ- µατος (Anti-fading reagent-mounting medium) και κάλυψη των παρασκευασµάτων µε καλυπτρίδες διαστάσεων 25Χ5mm. 9.2 Σε καλλιέργειες κυτταρικών σειρών µε τη µέθοδο διπλού ανοσοφθορισµού για την α- και γ- τουµπουλίνη (HEp2, HFFF2, C2C12) Για την µελέτη της ακεραιότητας της µιτωτικής συσκευής εφαρµόσθηκε η µέθοδος του διπλού ανοσοφθορισµού για την α- (ως κύριο συστατικό της µιτωτικής ατράκτου) και γ- τουµπουλίνη (ως βασικό συστατικό του κεντροσώµατος) και ως βιολογικό σύστηµα χρησιµοποιήθηκαν τρεις κυτταρικές σειρές (HEp2, HFFF2, και C2C12). Το κοινό χαρακτηριστικό τους είναι ότι εµφανίζουν ένα άριστα ανεπτυγµένο δίκτυο µικροσωληνίσκων, γεγονός που τις καθιστά κατάλληλο πειραµατικό µοντέλο για τη µελέτη µορίων που συµµετέχουν στη συγκρότηση της ατράκτου και της µιτωτικής συσκευής και κατ επέκταση στο συµµετρικό και ακριβή διαχωρισµό των χρωµοσωµάτων. Η επιφάνεια στην οποία θα αναπτυχθεί η καλλιέργεια της κάθε κυτταρικής σειράς σε συνδυασµό µε το ρυθµό πολλαπλασιασµού καθορίζει τον ακριβή αριθµό των κυττάρων που θα πρέπει να χρησιµοποιηθούν για την επίστρωση της, γεγονός που αντανακλά στην προσκόλληση, διαίρεση και επέκταση τους, ως µονοπύρηνα κύτταρα, ώστε να αναπτυχθεί άριστα το δίκτυο των µικροσωληνίσκων. Για την επίστρωση των κυττάρων χρησιµοποιούνται γυάλινες αποστειρωµένες καλυπτρίδες, διαστάσεων 22x22mm, τις οποίες τοποθετούµε σε τρυβλία petri, διαστάσεων 35/1mm. Το επόµενο στάδιο περιλαµβάνει την προσθήκη κατάλληλου πλήρους θρεπτικού µέσου για κάθε κυτταρική σειρά σε ποσότητα που δεν πρέπει να υπερβαίνει τα 2ml για τα συγκεκριµένα τρυβλία petri. Ακολουθεί επώαση των καλλιεργειών σε θάλαµο σταθερής θερµοκρασίας 37 C µε σταθερή παροχή 5% CO2. Επιπρόσθετα, επιβάλλεται η παρακολούθηση τους ανά τακτά χρονικά διαστήµατα σε ανάστροφο µικροσκόπιο, προκειµένου να πιστοποιηθεί η σωστή µορφολογία των κυττάρων και να ελεγχθεί η πυκνότητα και η κάλυψη της επιφανείας στις καλυπτρίδες. Η επίδραση µε την υπό µελέτη φαρµακευτική ένωση υδροχλωροθειαζίδιο (HCTZ) πραγµατοποιείται ~24 ώρες µετά την έναρξη της καλλιέργειας, που θεωρείται απαραίτητο χρονικό διάστηµα για την προσκόλληση των κυττάρων, ενώ διαφορές εντοπίζονται στις συγκεντρώσεις που εξετάσθηκαν και στη χρονική διάρκεια παραµονής της στις καλλιέργειες κάθε κυτταρικής σειράς. Επιπρόσθετα, το υδροχλωροθειαζίδιο (HCTZ) είναι ευδιάλυτο σε απόλυτη αιθανόλη (Ethanol Absolute, Merck) και χρησιµοποιείται σε εξαιρετικά χαµηλές περιεκτικότητες που αντιστοιχούν σε.1-.8% στον τελικό όγκο της καλλιέργειας, ως διαλύτης που περιλαµβάνει τις υπό εξέταση συγκεντρώσεις της χηµικής ένωσης
124 Υλικά και Μέθοδοι Μιτωτική συσκευή Κυτταρική σειρά HEp2 Η πειραµατική διαδικασία που ακολουθείται για τις καλλιέργειες της κυτταρικής σειράς Hep 2 περιλαµβάνει την επίστρωση των καλυπτρίδων µε X1 5 κύτταρα. Η φαρµακευτική ένωση υδροχλωροθειαζίδιο εξετάσθηκε στις συγκεντρώσεις 5, 2, 4, 6 και 8 µg/ml και η διάρκεια παραµονής της στις καλλιέργειες είναι 24 ώρες. Με τη διακοπή των κυτταροκαλλιεργειών, που πραγµατοποιείται στις 72 ώρες, σηµατοδοτείται η έναρξη των ανοσοχηµικών αντιδράσεων για την ταυτόχρονη σήµανση της α- και γ- τουµπουλίνης. ιαγραµµατικά το χρονοδιάγραµµα του πειράµατος έχει ως εξής: t = Έναρξη καλλιέργειας t = 24 Προσθήκη των υπό µελέτη συγκεντρώσεων HCTZ t = 48 Αποµάκρυνση της χηµικής ένωσης HCTZ t = 72 Τερµατισµός καλλιέργειας Μονιµοποίηση κυττάρων ιπλός ανοσοφθορισµός Κυτταρική σειρά HFFF2 Η πειραµατική διαδικασία που ακολουθείται για τις καλλιέργειες της κυτταρικής σειράς HFFF2 περιλαµβάνει την επίστρωση των καλυπτρίδων µε X1 5 κύτταρα. Η φαρµακευτική ένωση υδροχλωροθειαζίδιο εξετάσθηκε στις συγκεντρώσεις 2, 4, 6 και 8 µg/ml και η χρονική διάρκεια παραµονής της στις καλλιέργειες εµβρυϊκών επιδερµικών ινοβλαστών αντιστοιχεί περίπου σε ένα κύκλο διαίρεσης. Με τη διακοπή των κυτταροκαλλιεργειών, που πραγµατοποιείται στις 54 ώρες, σηµατοδοτείται η έναρξη των ανοσοχηµικών αντιδράσεων για την ταυτόχρονη σήµανση της α- και γ- τουµπουλίνης. ιαγραµµατικά το χρονοδιάγραµµα του πειράµατος έχει ως εξής: t = Έναρξη καλλιέργειας t = 23 Προσθήκη των υπό µελέτη συγκεντρώσεων HCTZ t = 54 Τερµατισµός καλλιέργειας Μονιµοποίηση κυττάρων ιπλός ανοσοφθορισµός Κυτταρική σειρά C2C12 Η πειραµατική διαδικασία που ακολουθείται για τις καλλιέργειες της κυτταρικής σειράς C2C12 περιλαµβάνει την επίστρωση των καλυπτρίδων µε X1 5 κύτταρα
125 Υλικά και Μέθοδοι Μιτωτική συσκευή Η φαρµακευτική ένωση υδροχλωροθειαζίδιο εξετάσθηκε στις συγκεντρώσεις 4, 6 και 8 µg/ml και η χρονική διάρκεια παραµονής της στις καλλιέργειες είναι 48 ώρες. Επιπρόσθετα, στις καλλιέργειες C2C12 χρησιµοποιήθηκε, ως θετικός µάρτυρας, η γνωστή για τις ανευπλοειδογόνες ιδιότητες χηµική ένωση δεµεκολσίνη, η οποία εξετάσθηκε στις συγκεντρώσεις.1 και.5 µg/ml. Με τη διακοπή των κυτταροκαλλιεργειών, που πραγµατοποιείται στις 72 ώρες, σηµατοδοτείται η έναρξη των ανοσοχηµικών αντιδράσεων για την ταυτόχρονη σήµανση της α- και γ- τουµπουλίνης. ιαγραµµατικά το χρονοδιάγραµµα του πειράµατος έχει ως εξής: t = Έναρξη καλλιέργειας t = 24 Προσθήκη των υπό µελέτη συγκεντρώσεων HCTZ t = 72 Τερµατισµός καλλιέργειας Μονιµοποίηση κυττάρων ιπλός ανοσοφθορισµός Ανοσοχηµική ανίχνευση α- και γ- τουµπουλίνης Με τη διακοπή των καλλιεργειών ξεκινούν οι ανοσοχηµικές αντιδράσεις για τη σήµανση των πρωτεϊνών α- και γ- τουµπουλίνης µε πρώτο βήµα τη σωστή µονιµοποίηση των κυττάρων. Η επιλογή κατάλληλων µονιµοποιητικών διαλυµάτων κατακρήµνισης (precipitating fixatives) είναι ιδιαίτερα σηµαντική όσον αφορά τη διατήρηση της ακεραιότητας ενδοκυτταρικών δοµών (κυτταροσκελετός) και την ακινητοποίηση τους για την εφαρµογή µεθόδων ανοσοφθορισµού. Η µεθανόλη, η οποία χαρακτηρίζεται ως συγκολλητικός παράγοντας (coagulant) είναι ένας οργανικός διαλύτης που προκαλεί µερική εκχύλιση των λιπιδίων της µεµβράνης µε αποτέλεσµα να διευκολύνεται η πρόσβαση στις υπό µελέτη ενδοκυτταρικές δοµές. Επιπρόσθετα, το γεγονός ότι οι χαµηλές θερµοκρασίες (4 C-12 C) είναι εξαιρετικά επιζήµιες για την ακεραιότητα της δοµής των µικροσωληνίσκων διότι προκαλείται αποπολυµερισµός των τουµπουλινών κάνει επιτακτική τη χρήση µονιµοποιητικών διαλυµάτων σε πολύ χαµηλότερες θερµοκρασίες (-1) C-(-2) C. Τα βήµατα που ακολουθούνται στη µέθοδο διπλού ανοσοφθορισµού είναι τα εξής: 1. Απόρριψη καλλιεργητικού υλικού και στιγµιαία προσθήκη ρυθµιστικού διαλύµατος φωσφορικών αλάτων (PBS) που πραγµατοποιείται µε ιδιαίτερα αργό ρυθµό στα άκρα του τρυβλίου petri για την αποφυγή ενόχλησης των κυττάρων. Η συγκεκριµένη διαδικασία σκοπό έχει την αποµάκρυνση διαλυτών κυτταρικών πρωτεϊνών (προϊόντα µεταβολισµού) και την αποφυγή µονιµοποίησης τους. Επιπρόσθετα, ενδείκνυται η χρήση θρεπτικού µέσου απουσία ορού ως αρχικό µέσο έκπλυσης. 2. Ακολουθεί µονιµοποίηση των κυττάρων µε παγωµένη απόλυτη µεθανόλη. Η επώαση πραγµατοποιείται στους -2 C για 3 λεπτά. Με ιδιαίτερη προσοχή εάν πρόκειται για πλαστικά υλικά είναι δυνατή η χρήση ενός δεύτερου οργανικού διαλύτη, της ακετόνης (-2 C) για Έκπλυση µε PBS (4 C) για 5 λεπτά, µε ελαφρά ανάδευση. 4. Προσθήκη πρώτου αντισώµατος για τη γ- τουµπουλίνη. Για
126 Υλικά και Μέθοδοι Μιτωτική συσκευή εξοικονόµηση χρόνου, η επώαση πραγµατοποιείται κατά τη διάρκεια της νύχτας για ώρες (overnight) σε υγρή ατµόσφαιρα στους 4 C. Η επώαση είναι δυνατόν να πραγµατοποιηθεί σε υγρή ατµόσφαιρα στους 37 C και διαρκεί 45-6 λεπτά. 5. Ακολουθούν τρεις εκπλύσεις µε PBS (4 C) από 5 λεπτά, µε ελαφρά ανάδευση. 6. Προσθήκη δεύτερου αντισώµατος για τη γ- τουµπουλίνη που είναι σηµασµένο µε το φθορόχρωµα φθοροσκεΐνη. Η επώαση διαρκεί 45 λεπτά και πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C. 7. Ακολουθούν τρεις εκπλύσεις µε PBS (4 C) από 5 λεπτά, µε ελαφρά ανάδευση. 8. Προσθήκη ορού ζωικού οργανισµού που επιλέγεται κατάλληλα, ώστε να λειτουργεί ως αντιδραστήριο φραγµός (blocking reagent) µε κύριο σκοπό να επιτευχθεί πλήρης κορεσµός των µη ειδικών θέσεων δέσµευσης και να µαταιωθούν αλληλεπιδράσεις µε τα αντισώµατα που ακολουθούν (διασταυρωτές αντιδράσεις). Η επώαση διαρκεί 5 λεπτά και πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C. 9. Ακολουθούν τρεις εκπλύσεις µε PBS (4 C) από 5 λεπτά, µε ελαφρά ανάδευση. 1. Προσθήκη πρώτου αντισώµατος για την α- τουµπουλίνη. Για εξοικονόµηση χρόνου, η επώαση πραγµατοποιείται κατά τη διάρκεια της νύχτας για ώρες (overnight) σε υγρή ατµόσφαιρα στους 4 C. Η επώαση είναι δυνατόν να πραγµατοποιηθεί σε υγρή ατµόσφαιρα στους 37 C και διαρκεί 45-6 λεπτά. 11. Ακολουθούν τρεις εκπλύσεις µε PBS (4 C) από 5 λεπτά, µε ελαφρά ανάδευση. 12. Προσθήκη δεύτερου αντισώµατος για την α- τουµπουλίνη, που είναι σηµασµένο µε φθορόχρωµα ερυθρού χρώµατος, όπως η ροδαµίνη ή το Texas Red. Η επώαση διαρκεί 45 λεπτά και πραγµατοποιείται σε υγρή ατµόσφαιρα στους 37 C. 13. Ακολουθούν τρεις εκπλύσεις µε PBS (4 C) από 5 λεπτά, µε ελαφρά ανάδευση. 14. Για την ανίχνευση και παρατήρηση του χρωµοσωµατικού υλικού πραγµατοποιείται χρώση των παρασκευασµάτων σε ρυθµιστικό διάλυµα PBS που περιέχει.2 µg/ml DAPI (από ενδιάµεσο πυκνό διάλυµα αποθήκευσης 1 µg/ml). Η χρονική διάρκεια παραµονής των παρασκευασµάτων στο διάλυµα είναι 1 σε θερµοκρασία δωµατίου και σε σκοτεινό περιβάλλον. 15. Με την απόρριψη του διαλύµατος DAPI ακολουθούν δύο στιγµιαίες εκπλύσεις µε PBS (4 C). 16. Προσθήκη 1-15µl ειδικού διαλύµατος (Anti-fading reagent - Mounting Medium) σε καθαρές και στεγνές αντικειµενοφόρους που εµποδίζει την εξασθένιση και αποχρωµατισµό των φθοροχρωµάτων κατά τη µικροσκοπική παρατήρηση και τη φύλαξη των παρασκευασµάτων. Επιπρόσθετα, η χρώση µπορεί να πραγµατοποιηθεί µε ένα ενιαίο διάλυµα που περιλαµβάνει το συνδυασµό της φθορίζουσας χρωστικής DAPI µε το αντιδραστήριο Antifade σε αναλογία 1:1 και σε τελική συγκέντρωση.2 µg/ml. 17. Αποµάκρυνση µε κατάλληλη χρήση διηθητικού χαρτιού της περίσσειας διαλύµατος που παραµένει στις καλυπτρίδες µετά τις εκπλύσεις και κάλυψη των αντικειµενοφόρων, που πραγµατοποιείται προσεκτικά για την αποφυγή φυσαλίδων µε την επιφάνεια της καλυπτρίδας στην οποία αναπτύσσονται τα κύτταρα. 18. Ακολουθεί σφράγιση των καλυπτρίδων περιµετρικά µε διαφανές υλικό (nail polish) και µικροσκοπική εξέταση των παρασκευασµάτων µετά την παραµονή τους τουλάχιστον για 4-6 λεπτά στους 4 C
127 Υλικά και Μέθοδοι Μιτωτική συσκευή ιαλύµατα. 1. PBS (Phosphate Buffer Solution) 2. Απόλυτη Μεθανόλη CH3OH (Sigma, St.Louis, Mo., USA) 3. Ακετόνη CH3COCH3 (Carlo Erba) 4. Αντισώµατα Σήµανση γ- τουµπουλίνης Πρώτο αντίσωµα α) Κυτταρική σειρά C2C12. Mouse Monoclonal Anti-γ-Tubulin (IgG1). Sigma T Χρησιµοποιείται σε αραίωση 1:2 σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl. Κυτταρικές σειρές ΗEp2 και HFFF2. TU-3 Mouse Monoclonal Anti-γ-Tubulin (IgG2b). Το αντίσωµα TU-3 (Novakova et al., 1996) προσεφέρθη από τον κ. Pavel Dráber, ιευθυντή του Ινστιτούτου Μοριακής Γενετικής της Ακαδηµίας Επιστηµών της Τσεχίας. Χρησιµοποιείται αδιάλυτο σε ποσότητα που δεν υπερβαίνει τα 3 µl. εύτερο αντίσωµα β) Anti-mouse IgG (whole molecule) F(ab')2 fragment-fitc from sheep. Sigma F2266. Χρησιµοποιείται σε αραίωση 1:2 σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl. Anti-mouse Ig Fluorescein-Linked whole Ab from sheep. Amersham N131. Χρησιµοποιείται σε αραίωση 1:15 σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 3 µl. Fluorescein Anti-mouse IgG (H+L) from horse. Vector FI-2. Χρησιµοποιείται σε αραίωση 1:15 σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 3 µl. Σήµανση α- τουµπουλίνης Πρώτο αντίσωµα γ) Κυτταρική σειρά C2C12.Mouse Monoclonal Anti-α-Tubulin (IgG1). Sigma T5168. Χρησιµοποιείται σε αραίωση 1:1 σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl. Κυτταρικές σειρές ΗEp2 και HFFF2. Mouse Monoclonal Anti-α-Tubulin. Amersham N35(9)6 Χρησιµοποιείται σε αραίωση 1:5 σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl. εύτερο αντίσωµα δ) Anti-mouse IgG (Fab specific) TRITC from goat. Sigma T7782. Χρησιµοποιείται σε αραίωση (1:1) - (1:15) σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 5 µl. Anti-mouse Ig Texas Red-Linked whole Ab from sheep. Amersham N231. Χρησιµοποιείται σε αραίωση (1:15) - (1:2) σε PBS που περιέχει 2% BSA και προστίθεται σε ποσότητα που δεν υπερβαίνει τα 4 µl
128 Υλικά και Μέθοδοι Μιτωτική συσκευή 5. Bovine Serum Albumin-BSA (Sigma A453) 6. Goat Serum (Sigma G923) 7. DAPI (Aldrich) 8. Vectashield Mounting Medium (Vector H-1)
129 Υλικά και Μέθοδοι Μικροσκοπική Παρατήρηση 1. ΜΙΚΡΟΣΚΟΠΙΚΗ ΠΑΡΑΤΗΡΗΣΗ Η µελέτη των παρασκευασµάτων πραγµατοποιείται σε κατάλληλο οπτικό µικροσκόπιο φθορισµού (Zeiss Axioskop) προσπίπτοντος φωτισµού, το οποίο διαθέτει λάµπα υδραργύρου (HBO 5W), ως πηγή ακτινοβολίας. Επιπρόσθετα, διαθέτει ένα µετακινούµενο φορέα στον οποίο είναι ενσωµατωµένα τα απαραίτητα φίλτρα µε τα οποία γίνονται ορατές οι φθορίζουσες χρωστικές. Αυτές χαρακτηρίζονται από ένα εξειδικευµένο φάσµα απορρόφησης ή διέγερσης (excitation) και εκποµπής (emission) της ακτινοβολίας, που συµβαίνει σε διαφορετικές περιοχές του ορατού φάσµατος (4-75nm), εξαιτίας της µετατόπισης κατά Stokes (Stokes shift). Οι φθορίζουσες χρωστικές που χρησιµοποιήθηκαν στα πειράµατα µας µε τις φασµατοσκοπικές ιδιότητες διαχωρισµού που τις χαρακτηρίζουν (πίνακας 1.1), αναγνωρίζονται παρουσία ειδικών φίλτρων είτε ως κυανό χρώµα (DAPI), είτε ως πράσινο χρώµα (FITC), είτε ως ερυθρά χρώµατα (PI, RHODOS, TRITC, XRITC, Texas Red). Επιπλέον, για την ταυτόχρονη ανίχνευση τριών διαφορετικών φθοροχρω- µάτων DAPI, FITC, TRITC ή XRITC χρησιµοποιείται φορέας που διαθέτει φίλτρο τριπλής διέλευσης. Πίνακας 1.1. Φασµατοσκοπικές ιδιότητες φθοροχρωµάτων (Sigma, Boehringer Mannheim, Vector) και ειδικά φίλτρα (Zeiss) που χρησιµοποιήθηκαν στη µέθοδο της in situ υβριδοποίησης (FISH) και του διπλού ανοσοφθορισµού. ΦΘΟΡΟΧΡΩΜΑΤΑ ΜΕΓΙΣΤΟ ΜΗΚΟΣ ΚΥΜΑΤΟΣ (nm) ΙΕΓΕΡΣΗ ΕΚΠΟΜΠΗ ΧΡΩΜΑ ΕΙ ΙΚΑ ΦΙΛΤΡΑ (ZEISS) DAPI /45 κυανό 488(2) FITC 494/ /525 πράσινο 488(9) PI ερυθρό 488(15) RHODOS-Rhodamine ερυθρό 488() TRITC ερυθρό 488(15) XRITC-Rhodamine ερυθρό 488(15) TEXAS RED 59/ /62 ερυθρό 488() DAPI/FITC/TRITC ή XRITC 36/485/56 46/52/6 κυ/πρα/ερ 121(4) Αξιοσηµείωτο είναι ότι η κυανή φθορίζουσα χρωστική διαµιδίνη (DAPI), εξαιτίας της ιδιότητας της να δεσµεύεται ισχυρά µε ορισµένες ακολουθίες που εντοπίζονται στη µικρή αύλακα του DNA και οι οποίες περιέχουν τρία έως τέσσερα γειτονικά ΑΤ ζεύγη βάσεων, είναι εξαιρετικά χρήσιµη για την παρατήρηση χρωµοσωµατικού υλικού (πυρήνες, µικροπυρήνες) και αποτελεί τη συνηθέστερη χρωστική, ως υπόστρωµα όλων των φθοροχρωµάτων. Το ερυθρό ιωδιούχο προπίδιο (PI), που έχει παρόµοια δοµή µε τη διαµιδίνη, συνδέεται µη ειδικά µε το δίκλωνο DNA και το RNA και συνεπώς η παρουσία του είναι έντονα ορατή στους πυρήνες και µικροπυρήνες, ενώ αµυδρά εµφανίζεται το κυτταρόπλασµα. Συνήθως, χρησι- µοποιείται ως υπόστρωµα για την φθοροσκεΐνη και η κοινή χρήση τους επιτρέπεται διότι και τα δύο µόρια διεγείρονται στο ίδιο µήκος κύµατος, ενώ εκπέµπουν σε διαφορετικά µήκη
130 Υλικά και Μέθοδοι Μικροσκοπική Παρατήρηση Τα φθορίζοντα σήµατα και οι φθορίζουσες χρωστικές υποστρώµατος καταγράφονται µε ενσωµατωµένη κάµερα µικροσκοπίου (MC 8 Microscope Camera, Zeiss) και για τη φωτογράφηση χρησιµοποιείται έγχρωµο φιλµ ASA 4 µε χρόνο έκθεσης απο Για τη λήψη ορισµένων εικόνων φθορισµού χρησι- µοποιήθηκε ειδική κάµερα, γνωστή ως CCD camera (Charge Coupled Device, Photometrics) και η ηλεκτρονική καταγραφή τους γίνεται µε τη χρήση του προγράµµατος Adobe Photoshop 5.. Ιδιαίτερα σηµαντική για την εξαγωγή συµπερασµάτων είναι η παρατήρηση παρασκευασµάτων τα οποία προέκυψαν µε τη µέθοδο του διπλού ανοσοφθορισµού για την α- και γ- τουµπουλίνη σε συνεστιακό µικροσκόπιο Leica TCS-SP CM, εξοπλισµένο µε δέσµη ιόντων αργού και ηλίου-νέου το οποίο καλύπτει ένα ευρύ φάσµα σε µήκη κύµατος από 45-65nm. Η τρισδιάστατη απεικόνιση και η επανατοποθέτηση των λεπτών οπτικών τοµών ενός µεσοφασικού πυρήνα ή µιτωτικού κυττάρου λαµβάνονται µε το πρόγραµµα ανάλυσης εικόνας Leica Confocal Software (LCS)
131 Υλικά και Μέθοδοι Στατιστική ανάλυση 11. ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ Η στατιστική ανάλυση των αποτελεσµάτων για την εκτίµηση της κυτταρογενετικής δράσης του υδροχλωροθειαζιδίου σε ανθρώπινα λεµφοκύτταρα περιφερικού αίµατος βασίζεται σε ελέγχους σύγκρισης στις µέσες τιµές που λαµβάνουν οι βιολογικοί δείκτες που προσδιορίσθηκαν. Σε αυτούς περιλαµβάνονται, για την εκτί- µηση της κυτταροτοξικότητας, ο δείκτης CBPI και της πρόκλησης γενετικής βλάβης, οι δείκτες ΜΝ, C + MN και C - MN. Για την εκτίµηση γεγονότων λανθασµένου χρωµοσωµατικού αποχωρισµού ακολουθείται η ίδια στατιστική ανάλυση στις µέσες τιµές που λαµβάνουν οι δείκτες που αναφέρονται στην απώλεια -ΜΝΧ +, ΜΝΥ +, ΜΝ8 + - και στο µη αποχωρισµό -NDX, NDY, ND8- των χρωµοσωµάτων Χ, Υ και 8. Εφαρµόζεται ένας παραµετρικός έλεγχος, t-test, που αφορά τη σύγκριση της µέσης τιµής δύο πληθυσµών σε ανεξάρτητα δείγµατα. Η ύπαρξη κανονικής κατανοµής στον πληθυσµό από τον οποίο προέρχονται τα ανεξάρτητα δείγµατα ελέγχεται µε τη δοκιµασία Shapiro-Wilk Normality test. Για τη σύγκριση των µέσων τιµών σε περισσότερα από δύο δείγµατα εφαρµόζεται η ανάλυση διασποράς µε έναν παράγοντα one-way ANOVA. Ελέγχεται η µηδενική υπόθεση ότι οι µέσες τιµές των οµάδων ενός πληθυσµού είναι ίσες και οι δειγµατικές διαφορές οφείλονται σε τυχαία µεταβλητότητα. Ο έλεγχος βασίζεται στη µεταβλητή F η οποία συγκρίνεται στη συνέχεια µε το κρίσιµο σηµείο α που αναφέρεται στο επίπεδο σηµαντικότητας.5. Ταυτόχρονα γίνεται έλεγχος της ισότητας των διασπορών, Levene s test, διότι η ανάλυση διασποράς στηρίζεται στην υπόθεση ότι οι µεταβλητές ακολουθούν κανονική κατανοµή και οι διασπορές των οµάδων είναι ίσες και ακολουθεί η σύγκριση των µέσων τιµών στις οµάδες ανά ζεύγη µε την εφαρµογή πολλαπλών συγκρίσεων Tukey. Με την πολυπαραγοντική ανάλυση παλινδρόµησης, Multiple Regression, εκτιµάται η ικανότητα των παραµέτρων ηλικία, φύλο, κάπνισµα και επίδραση HCTZ να επηρεάζουν τους προαναφερθέντες βιολογικούς δείκτες σε µεγαλύτερο ή µικρότερο βαθµό. Ο συντελεστής προσδιορισµού εκφράζει το ποσοστό της συνολικής µεταβλητότητας του βιολογικού δείκτη που υπολογίζεται από τις παραµέτρους και παίρνει τιµές στο διάστηµα [, 1]. Όσο πιο κοντά στη µονάδα βρίσκονται οι τιµές που λαµβάνει ο συντελεστής προσδιορισµού τόσο καλύτερο θεωρείται το µοντέλο που σχεδιάζεται. Στην περίπτωση της µελέτης του µηχανισµού της ανευπλοειδογόνου δράσης που εµφανίζει το υδροχλωροθειαζίδιο χρησιµοποιείται το G-test στις τιµές που λαµβάνουν οι βιολογικοί δείκτες που εκτιµήθηκαν στην ανάλυση των µιτωτικών κυττάρων. Αποτελεί µία τροποποιηµένη µορφή της δοκιµασίας x 2 για πίνακες συνάφειας (contingency tables) µε σύνολα γραµµών ή στηλών καθορισµένα που ανιχνεύει την οµοιογένεια του πληθυσµού. Εάν η τιµή της πιθανότητας είναι µικρότερη από το επίπεδο σηµαντικότητας.5 τότε υπάρχει στατιστικά σηµαντική διαφορά και τα αποτελέσµατα κρίνονται ανοµοιογενή. Επιπρόσθετα, εφαρµόζεται η ανάλυση παλινδρόµησης, Regression analysis, για να ελεγχθεί εάν υπάρχει γραµµική σχέση µεταξύ των συγκεντρώσεων της φαρµακευτικής ένωσης και των ανώµαλων µιτωτικών φάσεων. Η ίδια στατιστική ανάλυση χρησιµοποιείται και στην περίπτωση της µελέτης της ικανότητας του υδροχλωροθειαζιδίου να προκαλεί χρωµοσωµατική καθυστέρηση σε κυτταρικές σειρές. Τα στατιστικά προγράµµατα που χρησιµοποιήθηκαν είναι το Origin 7. (ttest, one-way ANOVA, Regression analysis) και το StatView 5.2 (G-test)
132
133 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα 12. ΕΠΑΓΩΓΗ ΜΙΚΡΟΠΥΡΗΝΩΝ ΩΣ ΕΙΚΤΗΣ ΓΕΝΕΤΙΚΗΣ ΒΛΑΒΗΣ - ΜΕΛΕΤΗ ΚΥΤΤΑΡΟΤΟΞΙΚΟΤΗΤΑΣ 12.1 ιεγερµένα λεµφοκύτταρα Η εκτίµηση της συχνότητας µικροπυρήνων σε ανθρώπινα λεµφοκύτταρα περιφερικού αίµατος θεωρείται ως κύριος δείκτης γενετικής βλάβης και χρησι- µοποιείται ευρύτατα σε κυτταρογενετικές µελέτες που αποσκοπούν στη διερεύνηση της γενετικής δράσης χηµικών και φαρµακευτικών ενώσεων. Στην παρούσα µελέτη επιλέχθηκαν οι συγκεντρώσεις 5 και 4 µg/ml της φαρµακευτικής ένωσης υδροχλωροθειαζίδιο (HCTZ) που εξετάσθηκαν στο σύνολο των δοτών, 32 άτοµα, αφού προηγούµενα εκτιµήθηκε η δράση της σε δύο θηλυκά άτοµα νεαρής ηλικίας στις συγκεντρώσεις 5, 1, 2, 4 και 6 µg/ml. Τα άτοµα που συµπεριελήφθησαν στη µελέτη διαχωρίστηκαν σε οµάδες µε κριτήριο την ηλικία και το φύλο. Αναλυτικότερα, τα στοιχεία των δοτών αναφέρονται στον πίνακα 6.1 στο Κεφάλαιο Κυτταρικές καλλιέργειες. Στους πίνακες 12.1, 12.2 και 12.3 αναφέρονται τα 16 νεαρά αρσενικά και θηλυκά άτοµα ηλικίας έως 25 ετών, ενώ στους πίνακες 12.4, 12.5 και 12.6 τα 16 αρσενικά και θηλυκά άτοµα ώριµης ηλικίας, δηλαδή άνω των 45 ετών. Συγκεκριµένα µελετήθηκε η τυχαία επαγωγή µικροπυρήνων σε καλλιέργειες λεµφοκυττάρων ολικού αίµατος σε δύο οµάδες ατόµων διαφορετικής ηλικίας (πίνακες 12.1, 12.4), οι οποίες θεωρούνται ως καλλιέργειες-µάρτυρες. Επίσης εξετάσθηκαν οι διαφορές στην επαγωγή µικροπυρήνων στις ίδιες οµάδες µετά από επίδραση HCTZ στις συγκεντρώσεις 5 µg/ml (πίνακες 12.2, 12.5) και 4 µg/ml (πίνακες 12.3, 12.6). Η συχνότητα µικροπυρήνων αφορά τον αριθµό των µικροπυρήνων (ΜΝ) σε 1 διπύρηνα κύτταρα, ενώ η συχνότητα διπύρηνων κυττάρων που εµφανίζουν µικροπυρήνα ( ΠΜΝ) αφορά τον αριθµό των διπύρηνων κυττάρων που περικλείουν µικροπυρήνα σε 1 διπύρηνα κύτταρα. Στους πίνακες που ακολουθούν στην πρώτη στήλη εµφανίζονται οι δότες που εξετάσθηκαν, στη δεύτερη ο αριθµός των διπύρηνων κυττάρων που αναλύθηκαν, στην τρίτη ο αριθµός των διπύρηνων κυττάρων που περιέχουν µικροπυρήνα και στην τέταρτη ο αριθµός των µικροπυρήνων. Στην τρίτη και τέταρτη στήλη έχουν υπολογισθεί οι συχνότητες των διπύρηνων κυττάρων µε µικροπυρήνα και των µικροπυρήνων σε διπύρηνα κύτταρα. Στην οµάδα ατόµων νεαρής ηλικίας η αύξηση που παρατηρείται στις συχνότητες των ΠΜΝ (8.69±.51) και ΜΝ (9.8±.56) στην ελάχιστη συγκέντρωση (5 µg/ml) HCTZ (πίνακας 12.2) είναι στατιστικώς σηµαντική σε σχέση µε τις καλλιέργειες-µάρτυρες (7.37±.27 και 7.67±.28, αντίστοιχα), όπως αποκαλύπτεται από τη σύγκριση των µέσων τιµών των δύο πληθυσµών. Στην ίδια ηλικιακή οµάδα, τη µεγαλύτερη αύξηση στις συχνότητες ΠΜΝ και ΜΝ σε σχέση µε τις καλλιέργειες- µάρτυρες την προκαλεί η συγκέντρωση 4 µg/ml HCTZ (13.29±.53 και 14.24±.56, αντίστοιχα), όπως προκύπτει από τη µέση τιµή των νεαρών δοτών που αναλύθηκαν. Επιπρόσθετα, η στατιστική ανάλυση των αποτελεσµάτων προσδιόρισε τη σηµαντική επίδραση της συγκέντρωσης 4 µg/ml HCTZ (πίνακας 12.3) στους βιολογικούς δείκτες που µελετήθηκαν σε σχέση τόσο µε τις καλλιέργειες-µάρτυρες όσο και µε τις καλλιέργειες HCTZ 5 µg/ml
134 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Πίνακας Συχνότητες διπύρηνων κυττάρων µε µικροπυρήνες ( ΠΜΝ), συχνότητες µικροπυρήνων (ΜΝ) και δείκτης πολλαπλασιασµού των κυττάρων (CBPI) σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας ( 25 ετών) και διαφορετικού φύλου, in vitro. ΜΑΡΤΥΡΕΣ ότες ιπύρηνα κύτταρα που αναλύθηκαν Α.Χ. ( ) 3641 Α.Π. ( ) 63 Α.Α. ( ) 358 Κ.Γ. ( ) 2882 Σ.Η. ( ) 3848 Β.Α. ( ) 2879 Π.Ν. ( ) 2245 Τ.Μ. ( ) 3324 ιπύρηνα κύτταρα µε µικροπυρήνες ΠΜΝ ( ) 29 (7.96) 43 (7.16) 26 (7.41) 26 (9.2) 32 (8.32) 14 (4.86) 15 (6.68) 21 (6.32) Μικροπυρήνες ΜΝ ( ) 29 (7.96) 47 (7.83) 27 (7.7) 28 (9.71) 34 (8.84) 15 (5.21) 16 (7.13) 21 (6.32) CBPI Μ.Τ.±Τ.Σ ± ± ±.4 Κ.Μ. ( ) 1857 Κ.Α. ( ) 25 Μ.Κ. ( ) 345 Ν.Κ. ( ) 245 Π.Ν. ( ) 242 Π.Π. ( ) 5943 Π.Μ. ( ) 34 Σ.Κ. ( ) (8.62) 19 (7.6) 26 (7.54) 19 (7.75) 18 (7.49) 35 (5.89) 2 (6.66) 5 (8.69) 16 (8.62) 19 (7.6) 26 (7.54) 19 (7.75) 19 (7.91) 37 (6.23) 22 (7.32) 52 (9.4) Μ.Τ.±Τ.Σ ± ± ±.5 Σ Μ.Τ.±Τ.Σ ± ± ±
135 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Πίνακας Συχνότητες διπύρηνων κυττάρων µε µικροπυρήνες ( ΠΜΝ), συχνότητες µικροπυρήνων (ΜΝ) και µεταβολή του δείκτη πολλαπλασιασµού των κυττάρων (CBPI) που επάγονται από τη δράση του υδροχλωροθειαζιδίου (HCTZ) σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας ( 25 ετών) και διαφορετικού φύλου, in vitro. ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν Α.Χ. ( ) 3699 Α.Π. ( ) 5346 Α.Α. ( ) 3151 Κ.Γ. ( ) 545 Σ.Η. ( ) 3637 Β.Α. ( ) 2749 Π.Ν. ( ) 2612 Τ.Μ. ( ) 3234 ιπύρηνα κύτταρα µε µικροπυρήνες ΠΜΝ ( ) 34 (9.19) 4 (7.48) 31 (9.84) 53 (9.81) 34 (9.35) 13 (4.73) 14 (5.36) 21 (6.49) Μικροπυρήνες ΜΝ ( ) 36 (9.73) 44 (8.23) 32 (1.15) 57 (1.55) 36 (9.9) 13 (4.73) 14 (5.36) 21 (6.49) CBPI Μ.Τ.±Τ.Σ ± ± ±.4 Κ.Μ. ( ) 174 Κ.Α. ( ) 39 Μ.Κ. ( ) 1272 Ν.Κ. ( ) 397 Π.Ν. ( ) 248 Π.Π. ( ) 4864 Π.Μ. ( ) 132 Σ.Κ. ( ) (8.8) 37 (12.3) 1 (7.86) 34 (1.98) 24 (9.97) 42 (8.63) 1 (7.58) 63 (1.68) 15 (8.8) 4 (13.29) 11 (8.65) 34 (1.98) 25 (1.38) 45 (9.25) 1 (7.58) 66 (11.19) Μ.Τ.±Τ.Σ ± ± ±.4 Σ Μ.Τ.±Τ.Σ * ± * ± ±.3 * p<.5 σε σύγκριση µε τις καλλιέργειες µάρτυρες (Student's t-test)
136 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Πίνακας Συχνότητες διπύρηνων κυττάρων µε µικροπυρήνες ( ΠΜΝ), συχνότητες µικροπυρήνων (ΜΝ) και µεταβολή του δείκτη πολλαπλασιασµού των κυττάρων (CBPI) που επάγονται από τη δράση του υδροχλωροθειαζιδίου (HCTZ) σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας ( 25 ετών) και διαφορετικού φύλου, in vitro. ότες ιπύρηνα κύτταρα που αναλύθηκαν Α.Χ. ( ) 3837 Α.Π. ( ) 311 Α.Α. ( ) 3481 Κ.Γ. ( ) 392 Σ.Η. ( ) 42 Β.Α. ( ) 2884 Π.Ν. ( ) 273 Τ.Μ. ( ) 3338 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml ιπύρηνα κύτταρα µε µικροπυρήνες ΠΜΝ ( ) 57 (14.85) 37 (11.93) 54 (15.51) 48 (15.52) 57 (14.24) 29 (1.5) 27 (9.89) 37 (11.8) Μικροπυρήνες ΜΝ ( ) 59 (15.38) 43 (13.87) 6 (17.24) 54 (17.46) 59 (14.74) 32 (11.1) 32 (11.72) 38 (11.38) CBPI Μ.Τ.±Τ.Σ ± ± ±.4 Κ.Μ. ( ) 1394 Κ.Α. ( ) 2 Μ.Κ. ( ) 1378 Ν.Κ. ( ) 2 Π.Ν. ( ) 2583 Π.Π. ( ) 4171 Π.Μ. ( ) 1458 Σ.Κ. ( ) (11.48) 31 (15.5) 17 (12.34) 29 (14.5) 39 (15.1) 48 (11.51) 19 (13.3) 91 (16.16) 16 (11.48) 31 (15.5) 18 (13.6) 29 (14.5) 42 (16.26) 51 (12.23) 21 (14.4) 99 (17.58) Μ.Τ.±Τ.Σ ± ± ±.4 Σ Μ.Τ.±Τ.Σ , ± , ± , ±.3 p<.5 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test) p<.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (Student's t-test)
137 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Στην οµάδα ατόµων ώριµης ηλικίας η αύξηση που παρατηρείται στις συχνότητες των ΠΜΝ (19.35±1.44) και ΜΝ (21.8±1.77) στην ελάχιστη συγκέντρωση (5 µg/ml) HCTZ (πίνακας 12.5) δεν διαφέρει στατιστικά από τις καλλιέργειες-µάρτυρες (17.8±1.36 και 2.44±1.65, αντίστοιχα). Στην ίδια ηλικιακή οµάδα, η αύξηση που προκαλείται στις συχνότητες ΠΜΝ (28.52±2.41) και ΜΝ (33.14±2.77) από την επίδραση της συγκέντρωσης 4 µg/ml HCTZ (πίνακας 12.6) είναι στατιστικώς σηµαντική σε σχέση τόσο µε τις καλλιέργειες-µάρτυρες όσο και µε τις καλλιέργειες HCTZ 5 µg/ml, όπως προκύπτει µετά την εφαρµογή των δύο στατιστικών µεθόδων ελέγχου. Πράγµατι, η επίδραση του HCTZ προκαλεί αύξηση στις συχνότητες των διπύρηνων κυττάρων µε µικροπυρήνα και των µικροπυρήνων η οποία ισχύει για όλες τις οµάδες των ατόµων που εξετάσθηκαν, διαφορετικής ηλικίας και διαφορετικού φύλου, ενώ η αύξηση αυτή είναι εντονότερη στη συγκέντρωση 4 µg/ml. Από τη σύγκριση των δύο ηλικιακών οµάδων φαίνεται ότι οι συχνότητες των ΠΜΝ και ΜΝ είναι υψηλότερες στα άτοµα ώριµης ηλικίας γεγονός που ισχύει τόσο για τις καλλιέργειες-µάρτυρες όσο και για κάθε πειραµατικό σηµείο που ελέγχθηκε. Προκειµένου να διερευνηθεί εάν η επίδραση της φαρµακευτικής ένωσης επηρεάζει τη διάρκεια του κυτταρικού κύκλου υπολογίσθηκε, σε σύνολο 2 κυττάρων, ο δείκτης πολλαπλασιασµού των κυττάρων (CBPI). Σε όλους τους προαναφερθέντες πίνακες ( ) η τιµή του δείκτη πολλαπλασιασµού των κυττάρων αναγράφεται στην τελευταία στήλη. Στις καλλιέργειες-µάρτυρες ο δείκτης εµφανίζει τιµή 1.9±.3 για τα νεαρά άτοµα (πίνακας 12.1) και 1.76±.2 για τα άτοµα ώριµης ηλικίας (πίνακας 12.4), η οποία µειώνεται µε την επίδραση 4 µg/ml HCTZ (πίνακες 12.3, 12.6) στην τιµή 1.79±.3 και 1.67 ±.2, αντίστοιχα. Από τη στατιστική ανάλυση προκύπτει ότι η µείωση αυτή είναι σηµαντική σε σχέση µε τις καλλιέργειες-µάρτυρες τόσο στα νεαρά όσο και στα ηλικιωµένα άτοµα. Επιπρόσθετα, αποκαλύπτεται η σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml (πίνακες 12.2, 12.5). Από τη µείωση του δείκτη πολλαπλασιασµού των κυττάρων σε σχέση µε το µάρτυρα προκύπτει η ικανότητα του HCTZ να προκαλεί καθυστέρηση του κυτταρικού κύκλου, ιδιαίτερα στη συγκέντρωση 4 µg/ml. Οι διακυµάνσεις στις τιµές του δείκτη πολλαπλασιασµού των κυττάρων, παρουσία και απουσία της επίδρασης HCTZ, αντανακλούν ένα ποσοστό κυτταροτοξικότητας, όπως αποδίδεται από τον τύπο: Cytotoxicity = 1-1[CBPIT-1/CBPIC-1]. Συγκεκριµένα, το ποσοστό κυτταροτοξικότητας που προκύπτει µετά την επίδραση HCTZ 4 µg/ml είναι 12.22% για τα νεαρά και 11.84% για τα ηλικιωµένα άτοµα, ενώ µετά την επίδραση HCTZ 5 µg/ml οι τιµές είναι 3.33% και 2.63% αντίστοιχα. Πράγµατι, η επίδραση του HCTZ προκαλεί µικρή κυτταροτοξικότητα στις καλλιέργειες από νεαρά και ηλικιωµένα άτοµα. Η µεγαλύτερη παρατηρείται στη συγκέντρωση 4 µg/ml και δεν φαίνεται να υπάρχει διαφορά ανάµεσα στις δύο ηλικιακές οµάδες που µελετήθηκαν
138 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Πίνακας Συχνότητες διπύρηνων κυττάρων µε µικροπυρήνες ( ΠΜΝ), συχνότητες µικροπυρήνων (ΜΝ) και δείκτης πολλαπλασιασµού των κυττάρων (CBPI) σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων ώριµης ηλικίας ( 45 ετών) και διαφορετικού φύλου, in vitro. ΜΑΡΤΥΡΕΣ ότες ιπύρηνα κύτταρα που αναλύθηκαν Β.Α. ( ) 3976 Κ.Γ. ( ) 2267 Σ.Ν. ( ) 2526 Χ.Β. ( ) 3492 Γ.Χ. ( ) 534 Γ.Θ. ( ) 1338 Ι.Γ. ( ) 1533 Σ.Κ. ( ) 1252 ιπύρηνα κύτταρα µε µικροπυρήνες ΠΜΝ ( ) 79 (19.87) 44 (19.41) 52 (2.59) 44 (12.6) 96 (19.7) 13 (9.72) 16 (1.44) 12 (9.58) Μικροπυρήνες ΜΝ ( ) 92 (23.14) 51 (22.5) 63 (24.94) 49 (14.3) 19 (21.65) 15 (11.21) 18 (11.74) 12 (9.58) CBPI Μ.Τ.±Τ.Σ ± ± ±.2 Κ.Σ. ( ) 2647 Λ.Μ. ( ) 2497 Ν.Μ. ( ) 128 Π.Ε. ( ) 481 Σ.Σ. ( ) 1774 Σ.Γ. ( ) 735 Χ.Ε. ( ) 163 Χ.Α. ( ) (27.96) 44 (17.62) 18 (14.9) 87 (21.32) 21.33± (19.73) 181 (25.73) 27.15± (16.84) 67 (17.79) 17.94± (31.73) 52 (2.82) 19 (15.73) 11 (24.75) 24.81± (21.98) 213 (3.27) 31.8± (19.96) 78 (2.71) 21.43± Μ.Τ.±Τ.Σ ± ±2. 1.8±.3 Σ Μ.Τ.±Τ.Σ ± ± ±
139 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Πίνακας Συχνότητες διπύρηνων κυττάρων µε µικροπυρήνες ( ΠΜΝ), συχνότητες µικροπυρήνων (ΜΝ) και µεταβολή του δείκτη πολλαπλασιασµού των κυττάρων (CBPI) που επάγονται από τη δράση του υδροχλωροθειαζιδίου (HCTZ) σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων ώριµης ηλικίας ( 45 ετών) και διαφορετικού φύλου, in vitro. ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν Β.Α. ( ) 363 Κ.Γ. ( ) 286 Σ.Ν. ( ) 2632 Χ.Β. ( ) 3356 Γ.Χ. ( ) 4378 Γ.Θ. ( ) 1894 Ι.Γ. ( ) 168 Σ.Κ. ( ) 1472 ιπύρηνα κύτταρα µε µικροπυρήνες ΠΜΝ ( ) 87 (23.96) 5 (23.97) 63 (23.94) 46 (13.71) 87 (19.87) 21 (11.9) 17 (1.57) 16 (1.87) Μικροπυρήνες ΜΝ ( ) 94 (25.89) 56 (26.85) 72 (27.36) 49 (14.6) 12 (23.3) 23 (12.14) 19 (11.82) 18 (12.23) CBPI Μ.Τ.±Τ.Σ ± ± ±.2 Κ.Σ. ( ) 276 Λ.Μ. ( ) 2531 Ν.Μ. ( ) 187 Π.Ε. ( ) 285 Σ.Σ. ( ) 3732 Σ.Γ. ( ) 2126 Χ.Ε. ( ) 156 Χ.Α. ( ) (29.71) 48 (18.96) 16 (14.72) 45 (21.58) 73 (19.56) 55 (25.87) 28 (18.59) 48 (22.72) 11 (36.59) 56 (22.13) 17 (15.64) 49 (23.5) 81 (21.7) 63 (29.63) 3 (19.92) 54 (25.56) Μ.Τ.±Τ.Σ ± ± ±.3 Σ Μ.Τ.±Τ.Σ ± ± ±
140 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Πίνακας Συχνότητες διπύρηνων κυττάρων µε µικροπυρήνες ( ΠΜΝ), συχνότητες µικροπυρήνων (ΜΝ) και µεταβολή του δείκτη πολλαπλασιασµού των κυττάρων (CBPI) που επάγονται από τη δράση του υδροχλωροθειαζιδίου (HCTZ) σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων ώριµης ηλικίας ( 45 ετών) και διαφορετικού φύλου, in vitro. ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν Β.Α. ( ) 34 Κ.Γ. ( ) 2197 Σ.Ν. ( ) 2357 Χ.Β. ( ) 4146 Γ.Χ. ( ) 3693 Γ.Θ. ( ) 24 Ι.Γ. ( ) 293 Σ.Κ. ( ) 1486 ιπύρηνα κύτταρα µε µικροπυρήνες ΠΜΝ ( ) 118 (34.71) 74 (33.68) 85 (36.6) 75 (18.9) 125 (33.85) 27 (13.47) 33 (15.77) 22 (14.8) Μικροπυρήνες ΜΝ ( ) 138 (4.59) 87 (39.6) 97 (41.15) 9 (21.71) 139 (37.64) 33 (16.47) 41 (19.59) 26 (17.5) CBPI Μ.Τ.±Τ.Σ ± ± ±.2 Κ.Σ. ( ) 259 Λ.Μ. ( ) 2626 Ν.Μ. ( ) 1198 Π.Ε. ( ) 2389 Σ.Σ. ( ) 1431 Σ.Γ. ( ) 2314 Χ.Ε. ( ) 1562 Χ.Α. ( ) (44.79) 77 (29.32) 25 (2.87) 82 (34.32) 42 (29.35) 94 (4.62) 36 (23.5) 76 (33.52) 137 (52.9) 96 (36.56) 28 (23.37) 96 (4.18) 46 (32.14) 16 (45.81) 39 (24.97) 91 (4.14) Μ.Τ.±Τ.Σ ± ± ±.2 Σ Μ.Τ.±Τ.Σ , ± , ± , ±.2 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
141 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα 12.2 Μη διεγερµένα λεµφοκύτταρα Προκειµένου να διερευνηθεί εάν η γενετική δράση του υδροχλωροθειαζιδίου είναι διαφορετική σε περίπτωση επίδρασης της φαρµακευτικής ένωσης σε µη διαιρούµενα λεµφοκύτταρα σχεδιάσθηκαν πειράµατα κατά τα οποία το HCTZ προστίθεται σε µη διεγερµένα λεµφοκύτταρα και αποµακρύνεται πριν αυτά διεγερθούν προς διαίρεση. Αναλυτικότερα, η επίδραση του υδροχλωροθειαζιδίου στις συγκεντρώσεις 5 και 4 µg/ml πραγµατοποιήθηκε στην προσυνθετική Gο φάση του κυτταρικού κύκλου απουσία του ανοσοδιεγερτικού παράγοντα PHA µε αποτέλεσµα οι καλλιέργειες να θεωρούνται ως µη διεγερµένες. Στην παρούσα µελέτη συµπεριελήφθησαν 3 αρσενικά και 3 θηλυκά άτοµα που προέρχονται από την οµάδα ατόµων νεαρής ηλικίας εξαιτίας της ευαισθησίας που παρουσιάζουν τα νεαρά άτοµα σε σύγκριση µε τα ηλικιωµένα στη δράση του υδροχλωροθειαζιδίου στις καλλιέργειες διεγερµένων λεµφοκυττάρων. Στον πίνακα που ακολουθεί (πίνακας 12.7) αναφέρονται αναλυτικά οι τιµές που λαµβάνουν οι βιολογικοί δείκτες, οι οποίοι µελετήθηκαν σε καλλιέργειες µη διεγερµένων λεµφοκυττάρων που προέρχονται από 6 νεαρά άτοµα. Η αύξηση που παρατηρείται στις συχνότητες των ΠΜΝ (1.24±.29) και ΜΝ (1.65±.31) στην ελάχιστη συγκέντρωση (5 µg/ml) HCTZ είναι στατιστικώς σηµαντική σε σχέση µε τις καλλιέργειες-µάρτυρες (8.63±.35 και 8.85±.41, αντίστοιχα). Τη µεγαλύτερη αύξηση στις συχνότητες ΠΜΝ και ΜΝ σε σχέση µε τις καλλιέργειες-µάρτυρες την προκαλεί η συγκέντρωση 4 µg/ml HCTZ (14.1±.86 και 14.96±.87, αντίστοιχα). Η στατιστική ανάλυση των αποτελεσµάτων επιβεβαίωσε τη σηµαντική επίπτωση της συγκέντρωσης 4 µg/ml HCTZ στους βιολογικούς δείκτες που µελετήθηκαν σε σχέση τόσο µε τις καλλιέργειες-µάρτυρες όσο και µε τις καλλιέργειες HCTZ 5 µg/ml. Πράγµατι, η επίδραση του HCTZ σε µη διεγερµένες καλλιέργειες προκαλεί αύξηση στις συχνότητες των διπύρηνων κυττάρων µε µικροπυρήνα και των µικροπυρήνων, η οποία ισχύει για κάθε πειραµατικό σηµείο που ελέγχθηκε και για τα δύο φύλα, ενώ η αύξηση αυτή είναι εντονότερη στη συγκέντρωση 4 µg/ml. Επιπρόσθετα, ελέγχθηκε η ικανότητα του HCTZ να προκαλεί καθυστέρηση του κυτταρικού κύκλου σε µη διεγερµένες καλλιέργειες από τη µείωση των τιµών του δείκτη πολλαπλασιασµού των κυττάρων (CBPI) σε σχέση µε τις καλλιέργειες- µάρτυρες. Συγκεκριµένα, στις καλλιέργειες-µάρτυρες ο δείκτης εµφανίζει τιµή 1.97±.2, η οποία µειώνεται µε την επίδραση 5 µg/ml HCTZ στην τιµή 1.94±.1, ενώ εµφανίζει την µεγαλύτερη πτώση (1.89±.1) µε την επίδραση 4 µg/ml HCTZ. Η στατιστική ανάλυση αποκαλύπτει τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση τόσο µε τις καλλιέργειες-µάρτυρες όσο και µε τις καλλιέργειες HCTZ 5 µg/ml. Τα αποτελέσµατα αυτά επιβεβαιώνονται από τον υπολογισµό του ποσοστού κυτταροτοξικότητας που εµφανίζεται ελαφρά υψηλότερο στις καλλιέργειες HCTZ 4 µg/ml (8.25%) σε σύγκριση µε τις καλλιέργειες HCTZ 5 µg/ml (3.9%), γεγονός που αποκαλύπτει τη µικρή κυτταροτοξικότητα που προκαλεί το υδροχλωροθειαζίδιο και η οποία φαίνεται να ισχύει και για τους δύο τύπους καλλιεργειών που µελετήθηκαν. Αξιοσηµείωτο είναι ότι οι παρατηρήσεις που αφορούν την επαγωγή µικροπυρήνων και την κυτταροτοξικότητα συµφωνούν µε τα αποτελέσµατα που προέκυψαν από τα πειράµατα µε καλλιέργειες διεγερµένων λεµφοκυττάρων, τα οποία περιγράφηκαν στο προηγούµενο υποκεφάλαιο
142 Αποτελέσµατα Μικροπυρήνες-κυτταροτοξικότητα Πίνακας Συχνότητες διπύρηνων κυττάρων µε µικροπυρήνες ( ΠΜΝ), συχνότητες µικροπυρήνων (ΜΝ) και µεταβολή του δείκτη πολλαπλασιασµού των κυττάρων (CBPI) σε µη διεγερµένες µε PHA κατά την επίδραση µε υδροχλωροθειαζίδιο (HCTZ) καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας ( 25 ετών) και διαφορετικού φύλου, in vitro. ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπύρηνα κύτταρα µε µικροπυρήνες ΠΜΝ ( ) Μικροπυρήνες ΜΝ ( ) CBPI ΜΑΡΤΥΡΕΣ Α.Π. ( ) (8.21) 19 (8.21) 1.99 Κ.Γ. ( ) (1.11) 54 (1.7) 1.95 Σ.Η. ( ) (8.44) 19 (8.44) 1.98 Μ.Τ.±Τ.Σ ± ± ±.1 Κ.Α. ( ) 2 17 (8.5) 17 (8.5) 2.1 Ν.Κ. ( ) (7.55) 2 (7.95) 1.98 Σ.Κ. ( ) (8.99) 3 (9.3) 1.9 Μ.Τ.±Τ.Σ ± ± ±.3 Σ Μ.Τ.±Τ.Σ ± ± ±.2 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml Α.Π. ( ) (9.67) 26 (1.48) 1.97 Κ.Γ. ( ) (1.) 25 (1.42) 1.95 Σ.Η. ( ) (9.91) 26 (9.91) 1.94 Μ.Τ.±Τ.Σ ± ± ±.1 Κ.Α. ( ) (11.66) 27 (11.66) 1.96 Ν.Κ. ( ) (9.95) 21 (9.95) 1.95 Σ.Κ. ( ) (1.23) 28 (11.46) 1.89 Μ.Τ.±Τ.Σ ± ± ±.2 Σ Μ.Τ.±Τ.Σ * ± * ± ±.1 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml Α.Π. ( ) (11.7) 28 (12.4) 1.91 Κ.Γ. ( ) (17.29) 46 (18.5) 1.87 Σ.Η. ( ) (13.25) 29 (13.72) 1.9 Μ.Τ.±Τ.Σ ± ± ±.1 Κ.Α. ( ) (14.37) 35 (15.24) 1.92 Ν.Κ. ( ) (12.95) 28 (13.94) 1.89 Σ.Κ. ( ) (15.14) 37 (16.) 1.83 Μ.Τ.±Τ.Σ ± ± ±.3 Σ Μ.Τ.±Τ.Σ , ± , ± , ±.1 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student s t-test) p<.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student s t-test) * p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (Student s t-test)
143 Αποτελέσµατα Μηχανισµός µικροπυρήνων 13. ΜΕΛΕΤΗ ΤΟΥ ΜΗΧΑΝΙΣΜΟΥ ΠΡΟΕΛΕΥΣΗΣ ΤΩΝ ΜΙΚΡΟΠΥΡΗΝΩΝ 13.1 Λεµφοκύτταρα ανθρώπου Η επαγωγή του σχηµατισµού µικροπυρήνων αποτέλεσε το έναυσµα για την περαιτέρω µελέτη και αποσαφήνιση του µηχανισµού κυτταρογενετικής δράσης της φαρµακευτικής ένωσης υδροχλωροθειαζίδιο. Συγκεκριµένα, για να προσδιορισθεί το περιεχόµενο των µικροπυρήνων που επάγονται στις δύο ηλικιακές οµάδες µετά την επίδραση του υδροχλωροθειαζιδίου, εάν δηλαδή περιέχουν άκεντρα χρωµοσω- µατικά τµήµατα ή ολόκληρα χρωµοσώµατα, ώστε να εξακριβωθεί ο µηχανισµός προέλευσης τους, χρωµοσωµατικά ρήγµατα-χρωµοσωµατική απώλεια, εφαρµόσθηκε η µέθοδος αναστολής της κυτταροκίνησης σε συνδυασµό µε in situ υβριδοποίηση µε φθοροχρώµατα (FISH). Η µεθοδολογία πραγµατοποιήθηκε µε τη χρήση ανιχνευτή συµπληρωµατικού του α-δορυφορικού DNA που συναντάται στα κεντροµέρη όλων των ανθρώπινων χρωµοσωµάτων (εικόνα 13-1). Για την παρουσία ή απουσία κεντροµερικού σήµατος σε κάθε πειραµατικό σηµείο αναλύθηκαν τουλάχιστον 5 µικροπυρήνες (56-11 στην οµάδα νεαρών ατόµων και στην οµάδα ατόµων ώριµης ηλικίας). Η παρουσία σήµατος υβριδοποίησης για την κεντροµερική περιοχή στον µικροπυρήνα (C + MN) υποδηλώνει την παρουσία ολόκληρου χρωµοσώµατος (χρωµοσωµατική απώλεια), ενώ η απουσία σήµατος υβριδοποίησης (C - MN) σηµαίνει την παρουσία άκεντρων χρωµοσωµατικών τµηµάτων (χρωµοσωµατικά ρήγµατα). Η ανάλυση σήµατος γίνεται αποκλειστικά σε διπύρηνα κύτταρα των οποίων οι πυρήνες παρουσιάζουν κεντροµερικά σήµατα, ενώ για την ταυτοποίηση του µικροπυρήνα ως C + MN απαιτείται η ίδια ένταση σήµατος µε αυτήν που παρατηρείται στους κυρίως πυρήνες του διπύρηνου κυττάρου. Περισσότερα του ενός σήµατα για να θεωρηθούν διαφορετικά θα πρέπει να είναι σαφώς διαχωρισµένα, ενώ ως ένα σήµα λαµβάνονται περιπτώσεις που αφορούν την ανίχνευση δύο σηµάτων που βρίσκονται σε επαφή και είτε αλληλεπικαλύπτονται είτε συνδέονται µεταξύ τους µε µία λεπτή γέφυρα (Eastmont et al., 1995). Η αποδοτικότητα της υβριδοποίησης για την παρουσία κεντροµερικού σήµατος σε όλα τα χρωµοσώµατα ελέγχθηκε µε την εφαρ- µογή της µεθόδου FISH σε µεταφασικά παρασκευάσµατα ιεγερµένα λεµφοκύτταρα Στην παρούσα µελέτη συµπεριελήφθησαν 2 άτοµα από το σύνολο των δοτών, τα οποία διαχωρίστηκαν σε οµάδες µε κριτήριο την ηλικία και το φύλο. Από αυτά τα 1 ήταν νεαρής ηλικίας, 5 αρσενικά και 5 θηλυκά και 1 ήταν ώριµης ηλικίας, 5 αρσενικά και 5 θηλυκά. Στους πίνακες που ακολουθούν παρουσιάζονται τα αποτελέσµατα που αφορούν τις συχνότητες µικροπυρήνων παρουσία ή απουσία σήµατος κεντροµέρους, καθώς και την κατανοµή των µικροπυρήνων που περιέχουν κεντροµερικά σήµατα στην οµάδα ατόµων νεαρής (πίνακες 13.1) και ώριµης ηλικίας (πίνακες 13.2). Στην πρώτη στήλη αναφέρονται οι δότες που εξετάσθηκαν, στη δεύτερη ο αριθµός των διπύρηνων κυττάρων, στην τρίτη ο αριθµός των µικροπυρήνων που αναλύθηκαν, στην τέταρτη ο αριθµός των µικροπυρήνων που παρουσιάζουν σήµα κεντροµέρους (C + MN), στην πέµπτη στήλη ο αριθµός των µικροπυρήνων που δεν παρουσιάζουν σήµα κεντροµέρους (C - MN), ενώ στις δύο επόµενες στήλες γίνεται διαχωρισµός µεταξύ των µικροπυρήνων που παρουσιάζουν ένα κεντροµερικό σήµα (C 1+ MN) και δύο ή περισσότερα σήµατα (C 2+ MN). Οι συχνότητες µικροπυρήνων που αναγράφονται στις τέσσερις τελευταίες στήλες έχουν υπολογισθεί σε 1 διπύρηνα κύτταρα
144 Αποτελέσµατα Μηχανισµός µικροπυρήνων Πίνακας Συχνότητες µικροπυρήνων που περιέχουν σήµα κεντροµέρους (C + MN) ή όχι (C - MN) και κατανοµή µικροπυρήνων που περιέχουν ένα και δύο ή περισσότερα σήµατα κεντροµέρους που επάγονται από τη δράση του υδροχλωροθειαζιδίου (HCTZ), σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας ( 25 ετών) και διαφορετικού φύλου, in vitro. ότες ιπύρηνα κύτταρα που αναλύθηκαν ΜΝ C + ΜΝ ( ) C - ΜN ( ) C 1+ ΜΝ ( ) C 2+ ΜN ( ) ΜΑΡΤΥΡΕΣ Α.Χ. ( ) (2.67) 8 (5.33) 4 (2.67) () Α.Π. ( ) (2.12) 7 (4.95) 3 (2.12) () Α.Α. ( ) (3.) 6 (6.) 3 (3.) () Κ.Γ. ( ) (3.91) 7 (5.48) 4 (3.13) 1 (.78) Σ.Η. ( ) (3.45) 8 (5.52) 5 (3.45) () Μ.Τ.±Τ.Σ ± ± ±.23.16±.16 Κ.Α. ( ) (2.67) 7 (4.67) 4 (2.67) () Ν.Κ. ( ) (2.76) 7 (4.83) 4 (2.76) () Π.Ν. ( ) (4.) 5 (5.) 4 (4.) () Π.Π. ( ) (3.2) 9 (3.6) 7 (2.8) 1 (.4) Σ.Κ. ( ) (4.54) 14 (5.79) 9 (3.72) 2 (.83) Μ.Τ.±Τ.Σ ± ± ±.28.25±.16 Σ Μ.Τ.±Τ.Σ ± ± ±.18.2±.11 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml Α.Χ. ( ) (3.33) 1 (6.67) 5 (3.33) () Α.Π. ( ) (4.67) 8 (5.33) 7 (4.67) () Α.Α. ( ) (4.) 7 (7.) 4 (4.) () Κ.Γ. ( ) (4.31) 22 (5.92) 13 (3.5) 3 (.81) Σ.Η. ( ) (4.) 6 (6.) 4 (4.) () Μ.Τ.±Τ.Σ ± ± ±.23.16±.16 Κ.Α. ( ) (7.) 13 (6.5) 11 (5.5) 3 (1.5) Ν.Κ. ( ) (5.) 12 (6.) 9 (4.5) 1 (.5) Π.Ν. ( ) (5.) 6 (6.) 5 (5.) () Π.Π. ( ) (4.67) 8 (5.33) 6 (4.) 1 (.67) Σ.Κ. ( ) (6.7) 18 (6.43) 14 (5.) 3 (1.7) Μ.Τ.±Τ.Σ ± ± ±.25.75±.25 Σ Μ.Τ.±Τ.Σ ± * ± ±.22.45±.17 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml Α.Χ. ( ) (8.) 12 (8.) 9 (6.) 3 (2.) Α.Π. ( ) (8.) 7 (7.) 7 (7.) 1 (1.) Α.Α. ( ) (11.) 1 (1.) 9 (9.) 2 (2.) Κ.Γ. ( ) (1.83) 11 (7.) 13 (8.28) 4 (2.55) Σ.Η. ( ) (8.67) 1 (6.67) 1 (6.67) 3 (2.) Μ.Τ.±Τ.Σ ± ± ± ±.25 Κ.Α. ( ) (1.) 6 (6.) 7 (7.) 3 (3.) Ν.Κ. ( ) (8.) 5 (5.) 7 (7.) 1 (1.) Π.Ν. ( ) (1.) 7 (7.) 9 (9.) 1 (1.) Π.Π. ( ) (1.) 5 (5.) 8 (8.) 2 (2.) Σ.Κ. ( ) (11.91) 21 (8.94) 24 (1.21) 4 (1.7) Μ.Τ.±Τ.Σ ± ± ± ±.37 Σ Μ.Τ.±Τ.Σ , ± ± , ± , ±.21 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student s t-test) p<.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student s t-test) * p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (Student s t-test)
145 Αποτελέσµατα Μηχανισµός µικροπυρήνων Στην οµάδα ατόµων νεαρής ηλικίας (πίνακας 13.1), η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων, που δεν εµφανίζουν σήµα υβριδοποίησης στην περιοχή του κεντροµέρους (C - MN), στην ελάχιστη συγκέντρωση 5 µg/ml HCTZ (6.12±.17) είναι στατιστικώς σηµαντική σε σχέση µε τις καλλιέργειες- µάρτυρες (5.12±.22). Αντίθετα στα ηλικιωµένα άτοµα (πίνακας 13.2), η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων C - MN στην ελάχιστη συγκέντρωση HCTZ (1.28±.84) δεν διαφέρει στατιστικά από τις καλλιέργειες-µάρτυρες (9.73±.65). Σχετικά µε τη δράση του HCTZ στη µέγιστη συγκέντρωση (4 µg/ml) παρατηρείται αύξηση τόσο στα νεαρά (7.6±.5) όσο και στα ηλικιωµένα άτοµα (13.68±1.8) και η στατιστική ανάλυση έδειξε τη σηµαντική διαφορά που υπάρχει σε σχέση µε τις αντίστοιχες καλλιέργειες-µάρτυρες (5.12±.22 και 9.73±.65) και στις δύο ηλικιακές οµάδες. Επιπρόσθετα, στα ηλικιωµένα άτοµα µε την εφαρµογή των δύο στατιστικών µεθόδων ελέγχου, όσον αφορά τις συχνότητες µικροπυρήνων που δεν εµφανίζουν σήµα κεντροµέρους, επιβεβαιώνεται η σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml (13.68±1.8) σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml (1.28±.84), γεγονός το οποίο δεν ισχύει για τα νεαρά άτοµα. Στην οµάδα ατόµων νεαρής ηλικίας (πίνακας 13.1), η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων που περιέχουν κεντροµερικό σήµα (C + MN) στις συγκεντρώσεις 5 µg/ml (4.8±.34) και 4 µg/ml HCTZ (9.64±.44) είναι στατιστικώς σηµαντική σε σχέση µε τις καλλιέργειες-µάρτυρες (3.23±.23), όπως αποκαλύπτεται από την εφαρµογή των δύο στατιστικών µεθόδων ελέγχου µε τη σύγκριση των µέσων τιµών. Αντίθετα, στα ηλικιωµένα άτοµα (πίνακας 13.2) µόνο η επίδραση της µέγιστης συγκέντρωσης (4 µg/ml) HCTZ προκαλεί στατιστικά σηµαντική αύξηση (25.96± 2.2) σε σχέση µε τις καλλιέργειες-µάρτυρες (13.51±1.51). Επιπρόσθετα, η στατιστική ανάλυση των αποτελεσµάτων αποκαλύπτει τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml και στις δύο ηλικιακές οµάδες. Σύµφωνα µε τα ανωτέρω, είναι φανερό πως η επίδραση του HCTZ προκαλεί µεγαλύτερη αύξηση στις συχνότητες των µικροπυρήνων που περιέχουν κεντροµερικό σήµα (C + MN), ενώ ταυτόχρονα επάγει σε µικρότερο βαθµό το σχηµατισµό µικροπυρήνων απουσία κεντροµερικού σήµατος (C - MN). Η δράση αυτή ισχύει για όλες τις οµάδες ατόµων που εξετάσθηκαν, διαφορετικής ηλικίας και διαφορετικού φύλου και παρουσιάζεται εντονότερη στη συγκέντρωση 4 µg/ml HCTZ. Επιπρόσθετα, από τη σύγκριση των δύο ηλικιακών οµάδων προκύπτει ότι τα άτοµα ώριµης ηλικίας, ιδιαίτερα τα θηλυκά άτοµα, παρουσιάζουν µεγαλύτερη συχνότητα µικροπυρήνων µε κεντροµερικό σήµα για κάθε πειραµατικό σηµείο που ελέγχθηκε. Από τα παραπάνω διαφαίνεται ό,τι το HCTZ εκδηλώνει µία σχετικά ισχυρή ανευπλοειδογόνο δράση, ενώ ταυτόχρονα παρατηρείται µία λιγότερο έντονη δράση της φαρµακευτικής ένωσης ως προς την ικανότητα δηµιουργίας ρηγµάτων
146 Αποτελέσµατα Μηχανισµός µικροπυρήνων Στους ίδιους πίνακες, από την κατανοµή των µικροπυρήνων ανάλογα µε τα σήµατα υβριδοποίησης (1 ή 2) που παρουσιάζουν, παρατηρούµε, τόσο στις καλλιέργειες-µάρτυρες όσο και στις καλλιέργειες που δέχθηκαν την επίδραση των δύο συγκεντρώσεων υδροχλωροθειαζιδίου, οι συχνότητες των C 1+ ΜΝ να είναι υψηλότερες από τις αντίστοιχες των C 2+ ΜΝ. Φαίνεται πως την κυρίαρχη κατηγορία και στις δύο ηλικιακές οµάδες για κάθε πειραµατικό σηµείο που ελέγχθηκε απαρτίζουν οι µικροπυρήνες που εµφανίζουν ένα κεντροµερικό σήµα (C 1+ ΜΝ) και εποµένως περιέχουν ένα χρωµόσωµα. Συγκεκριµένα, στην οµάδα ατόµων νεαρής ηλικίας η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων C 1+ ΜΝ στις συγκεντρώσεις 5 µg/ml (4.35±.22) και 4 µg/ml HCTZ (7.82±.41) είναι στατιστικώς σηµαντική σε σχέση µε τις καλλιέργειες-µάρτυρες (3.3±.18). Αντίθετα, στα ηλικιωµένα άτοµα µόνο η επίδραση της µέγιστης συγκέντρωσης HCTZ προκαλεί στατιστικά σηµαντική αύξηση (18.17±1.1) σε σχέση µε τις καλλιέργειες-µάρτυρες (9.68±.66). Επιπρόσθετα, η στατιστική ανάλυση των αποτελεσµάτων αποκαλύπτει τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml και στις δύο ηλικιακές οµάδες. Αξιοσηµείωτη είναι η παρατήρηση που αφορά την αύξηση στη συχνότητα µικροπυρήνων που φέρουν δύο ή περισσότερα σήµατα υβριδοποίησης (C 2+ ΜΝ) στη συγκέντρωση 4 µg/ml HCTZ να διαφέρει στατιστικά τόσο µε τις καλλιέργειες- µάρτυρες όσο και µε τις καλλιέργειες HCTZ 5 µg/ml και στις δύο ηλικιακές οµάδες. Αναλυτικότερα, ενώ στις καλλιέργειες-µάρτυρες η µέση τιµή της συχνότητας των C 2+ ΜΝ στα νεαρά και ηλικιωµένα άτοµα είναι.2±.11 και 3.84±.9, µετά την επίδραση 5 µg/ml και 4 µg/ml HCTZ αυξάνεται και γίνεται αντίστοιχα.45±.17, 3.74±.84 και 1.82±.21, 7.79±1.3. Η παρατήρηση αυτή ενισχύει ακόµη περισσότερο την ανευπλοειδογόνο δραστικότητα του HCTZ, ιδιαίτερα στη µέγιστη συγκέντρωση
147 Αποτελέσµατα Μηχανισµός µικροπυρήνων Πίνακας Συχνότητες µικροπυρήνων που περιέχουν σήµα κεντροµέρους (C + MN) ή όχι (C - MN) και κατανοµή µικροπυρήνων που περιέχουν ένα και δύο ή περισσότερα σήµατα κεντροµέρους που επάγονται από τη δράση του υδροχλωροθειαζιδίου (HCTZ), σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων ώριµης ηλικίας ( 45 ετών) και διαφορετικού φύλου, in vitro. ότες ιπύρηνα κύτταρα που αναλύθηκαν ΜΝ C + ΜΝ ( ) C - ΜN ( ) C 1+ ΜΝ ( ) C 2+ ΜN ( ) ΜΑΡΤΥΡΕΣ Β.Α. ( ) (9.48) 14 (13.27) 8 (7.58) 2 (1.9) Κ.Γ. ( ) (12.78) 11 (1.5) 1 (9.13) 4 (3.65) Σ.Ν. ( ) (11.74) 19 (13.12) 13 (8.98) 4 (2.76) Χ.Β. ( ) (6.46) 9 (8.31) 7 (6.46) () Γ.Χ. ( ) (11.43) 15 (1.71) 13 (9.29) 3 (2.14) Μ.Τ.±Τ.Σ ± ± ± ±.6 Χ.Α. ( ) (12.6) 19 (7.39) 23 (8.95) 8 (3.11) Π.Ε. ( ) (17.44) 17 (8.72) 2 (1.26) 14 (7.18) Σ.Γ. ( ) (21.14) 45 (8.6) 72 (12.9) 46 (8.24) Λ.Μ. ( ) (11.79) 1 (8.42) 12 (1.11) 2 (1.68) Κ.Σ. ( ) (2.8) 12 (9.24) 17 (13.1) 1 (7.7) Μ.Τ.±Τ.Σ ± ± ± ±1.33 Σ Μ.Τ.±Τ.Σ ± ± ± ±.9 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml Β.Α. ( ) (13.) 15 (15.) 11 (11.) 2 (2.) Κ.Γ. ( ) (16.) 11 (11.) 12 (12.) 4 (4.) Σ.Ν. ( ) (13.33) 19 (12.67) 15 (1.) 5 (3.33) Χ.Β. ( ) (7.43) 9 (8.36) 8 (7.43) () Γ.Χ. ( ) (1.42) 13 (9.68) 12 (8.93) 2 (1.49) Μ.Τ.±Τ.Σ ± ± ± ±.7 Χ.Α. ( ) (17.) 1 (1.) 13 (13.) 4 (4.) Π.Ε. ( ) (17.) 7 (7.) 11 (11.) 6 (6.) Σ.Γ. ( ) (2.) 8 (8.) 14 (14.) 6 (6.) Λ.Μ. ( ) (13.) 9 (7.8) 13 (11.27) 2 (1.73) Κ.Σ. ( ) (23.7) 18 (13.33) 2 (14.81) 12 (8.89) Μ.Τ.±Τ.Σ ± ± ± ±1.19 Σ Μ.Τ.±Τ.Σ ± ± ± ±.84 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml Β.Α. ( ) (22.33) 22 (2.47) 19 (17.67) 5 (4.65) Κ.Γ. ( ) (23.59) 15 (13.61) 18 (16.33) 8 (7.26) Σ.Ν. ( ) (24.73) 19 (17.39) 21 (19.23) 6 (5.5) Χ.Β. ( ) (12.77) 14 (13.75) 11 (1.81) 2 (1.96) Γ.Χ. ( ) (22.16) 18 (15.34) 2 (17.5) 6 (5.11) Μ.Τ.±Τ.Σ ± ± ± ±.86 Χ.Α. ( ) (29.) 1 (1.) 2 (2.) 9 (9.) Π.Ε. ( ) (31.3) 9 (9.1) 19 (19.2) 12 (12.1) Σ.Γ. ( ) (34.) 11 (11.) 21 (21.) 13 (13.) Λ.Μ. ( ) (23.14) 15 (12.85) 21 (17.99) 6 (5.14) Κ.Σ. ( ) (36.88) 16 (13.41) 27 (22.63) 17 (14.25) Μ.Τ.±Τ.Σ ± ± ± ±1.63 Σ Μ.Τ.±Τ.Σ , ± , ± , ± , ±1.3 p<.5 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student s t-test) p<.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student s t-test)
148 Αποτελέσµατα Μηχανισµός µικροπυρήνων Προκειµένου να διερευνηθεί πληρέστερα η κυτταρογενετική δράση της φαρµακευτικής ένωσης, η οποία εκτιµάται από την ανάλυση των βιολογικών δεικτών που προσδιορίσθηκαν, κρίθηκε σκόπιµο να συνυπολογισθούν παράµετροι όπως ηλικία, φύλο, συνήθεια καπνίσµατος που δύναται να τους επηρεάσουν σε µικρότερο ή µεγαλύτερο βαθµό. Με το σχεδιασµό ενός γενικού γραµµικού µοντέλου (πίνακας 13.3) στο οποίο οι συγκεκριµένοι παράγοντες λαµβάνονται ως ανεξάρτητες µεταβλητές δύναται να εξαχθούν σηµαντικά συµπεράσµατα. Πίνακας 13.3 Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών CBPI, MN, C + MN, C - MN που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο, το κάπνισµα και την επίδραση του HCTZ ως ανεξάρτητες µεταβλητές. CBPI MN C + MN C - MN Ηλικία <.1 <.1 <.1 <.1 Φύλο Κάπνισµα Επίδραση HCTZ <.1 <.1 <.1 <.1 R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.45 P<.1 R 2 =.675 P<.1 R 2 =.786 P<.1 R 2 =.738 P<.1 Ο σχεδιασµός του γραµµικού µοντέλου µε το δείκτη πολλαπλασιασµού των κυττάρων να λαµβάνεται ως εξαρτηµένη µεταβλητή έδειξε τη σηµαντική εξάρτηση του CBPI από τους παράγοντες ηλικία και επίδραση υδροχλωροθειαζιδίου. Η τιµή του συντελεστή προσδιορισµού R 2 ισούται µε.45, δηλαδή το 4.5% της συνολικής µεταβλητότητας του CBPI ερµηνεύεται από τις µεταβλητές ηλικία και επίδραση HCTZ, σύµφωνα µε το προτεινόµενο µοντέλο. Επιπρόσθετα, ο έλεγχος ανάλυσης διασποράς (ANOVA) έδειξε F = µε σηµαντικότητα P<.1. Αντίθετα, η συχνότητα ΜΝ φαίνεται να επηρεάζεται από όλες τις παραµέτρους που εκτιµήθηκαν στις οποίες περιλαµβάνεται και το κάπνισµα. Το φύλο φαίνεται να επηρεάζει τη συχνότητα µικροπυρήνων σε µικρότερο βαθµό (P =.8) σε σύγκριση µε την ηλικία (P<.1). Πρόκειται για ένα φαινόµενο που παρατηρήθηκε και στους προαναφερθέντες αναλυτικούς πίνακες µε τα αρσενικά και θηλυκά άτοµα ώριµης ηλικίας να εµφανίζουν υψηλότερες συχνότητες µικροπυρήνων σε σύγκριση µε τα νεαρά. Σε αυτό το µοντέλο η τιµή του συντελεστή προσδιορισµού (R 2 =.675) είναι αρκετά κοντά στη µονάδα γεγονός που αποδίδει µία σχετικά καλή γραµµική συσχέτιση. Η ηλικία και η επίδραση HCTZ θεωρούνται οι κύριοι παράγοντες που επηρεάζουν τόσο τις συχνότητες µικροπυρήνων που περιέχουν κεντροµερικό σήµα (C + MN) όσο και το σχηµατισµό µικροπυρήνων απουσία κεντροµερικού σήµατος (C - MN), ενώ το φύλο φαίνεται να επηρεάζει σε µεγαλύτερη έκταση τους C + MN από ότι τους C - MN. Όσον αφορά τη συνήθεια του καπνίσµατος, ενώ επηρεάζει το σχηµατισµό µικροπυρήνων που περιέχουν άκεντρα χρωµοσωµατικά τµήµατα (P =.14), δεν επάγει το σχηµατισµό µικροπυρήνων που θεωρούνται προϊόν χρωµοσωµατικής απώλειας (P =.267). Επιπρόσθετα, η τιµή που λαµβάνει ο συντελεστής προσδιορισµού R 2 για τις εξαρτηµένες µεταβλητές C + MN και C - MN είναι αρκετά κοντά στη µονάδα (.786,.738) γεγονός που αποδίδει µία ισχυρή γραµµική συσχέτιση ως προς τις παραµέτρους που εξετάσθηκαν
149 Αποτελέσµατα Μηχανισµός µικροπυρήνων Το γενικό γραµµικό µοντέλο που περιγράφηκε στην προηγούµενη παράγρα φο (πίνακας 13.3) βασίζεται σε µία συνολική εκτίµηση των βιολογικών δεικτών από τις παραµέτρους που δύναται να τους επηρεάσουν και σε αυτές περιλαµβάνεται η ε- πίδραση του υδροχλωροθειαζιδίου ως ανεξάρτητη µεταβλητή. Με το σχεδιασµό ενός δεύτερου γραµµικού µοντέλου (πίνακας 13.4) µελετήθηκε η επίδραση των παραγόντων ηλικία, φύλο και συνήθεια του καπνίσµατος που λαµβάνονται ως ανεξάρτητες µεταβλητές στην ικανότητα του HCTZ να προκαλεί αυξηµένες συχνότητες µικροπυρήνων (ΜΝ, C + MN, C - MN) και διπύρηνων κυττάρων µε µικροπυρήνες ( Π ΜΝ). Για την πραγµατοποίηση του εκτιµήθηκε η επίδραση της ηλικίας, του φύλου και της συνήθειας του καπνίσµατος στην τυχαία επαγωγή µικροπυρήνων στις καλλιέργειες-µάρτυρες (πίνακας 13.4α). Στη συνέχεια υπολογίσθηκαν οι διαφορές στην επαγωγή µικροπυρήνων µετά από επίδραση µε HCTZ. Οι νέες συχνότητες προκύπτουν αφαιρώντας, για κάθε δότη, τη συχνότητα των µικροπυρήνων που παρατηρήθηκε στο µάρτυρα από την αντίστοιχη συχνότητα µικροπυρήνων που παρατηρήθηκε στη µέγιστη συγκέντρωση HCTZ. Η πολυπαραγοντική ανάλυση παλινδρόµησης εφαρµόσθηκε στις νέες συχνότητες µικροπυρήνων και εξετάσθηκε ο βαθµός στον οποίον οι παράγοντες που προαναφέρθηκαν τις επηρεάζουν (πίνακας 13.4β). Πίνακας 13.4α. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών ΠΜΝ, MN, C + MN, C - MN που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο και το κάπνισµα ως ανεξάρτητες µεταβλητές σε καλλιέργειες-µάρτυρες. ΠMN MN C + MN C - MN Ηλικία <.1 <.1 <.1 <.1 Φύλο Κάπνισµα R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.688 P<.1 R 2 =.687 P<.1 R 2 =.751 P<.1 R 2 =.799 P<.1 Σε καλλιέργειες-µάρτυρες η ηλικία (P<.1, P<.1) αποτελεί τον κυριότερο παράγοντα που επηρεάζει τις συχνότητες ΠΜΝ και ΜΝ, ενώ το φύλο φαίνεται να επηρεάζει οριακά (P =.48) τη συχνότητα ΠΜΝ. Η ηλικία σε µεγαλύτερη έκταση (P<.1, P<.1) και το φύλο σε µικρότερη έκταση (P =.28, P =.11) θωρούνται παράγοντες οι οποίοι επηρεάζουν τόσο τις συχνότητες µικροπυρήνων που περιέχουν κεντροµερικό σήµα (C + MN) όσο και το σχηµατισµό µικροπυρήνων απουσία κεντροµερικού σήµατος (C - MN). Oι συχνότητες µικροπυρήνων δεν φαίνεται να επηρεάζονται από τη συνήθεια του καπνίσµατος. Πίνακας 13.4β. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών ΠΜΝ, MN, C + MN, C - MN που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο και το κάπνισµα ως ανεξάρτητες µεταβλητές σε καλλιέργειες µετά από επίδραση µε HCTZ. ΠMN MN C + MN C - MN Ηλικία.3 <.1 <.1.5 Φύλο Κάπνισµα R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.366 P=.1 R 2 =.421 P=.3 R 2 =.721 P<.1 R 2 =.396 P=
150 Αποτελέσµατα Μηχανισµός µικροπυρήνων Η ανάλυση των βιολογικών δεικτών που επαναπροσδιορίσθηκαν σε καλλιέργειες στις οποίες πραγµατοποιήθηκε επίδραση µε τη µέγιστη συγκέντρωση HCTZ έδειξε ότι η ηλικία (P =.3, P<.1) αποτελεί τον κύριο παράγοντα που επηρεάζει τις συχνότητες ΠΜΝ και ΜΝ σε αντίθεση µε το φύλο (P =.434, P =.624) και τη συνήθεια του καπνίσµατος (P =.72, P =.62). Η ηλικία σε µεγαλύτερη έκταση (P<.1, P =.5) και το φύλο σε µικρότερη έκταση (P =.42, P =.4) εξακολουθούν να θωρούνται παράγοντες οι οποίοι επηρεάζουν τόσο την επαγωγή µικροπυρήνων που θεωρούνται προϊόν χρωµοσωµατικής απώλειας όσο και το σχηµατισµό µικροπυρήνων που περιέχουν άκεντρα χρωµοσωµατικά τµήµατα. Όσον αφορά τη συνήθεια του καπνίσµατος δεν φαίνεται να επάγει κανέναν από τους δύο τύπους µικροπυρήνων. Από τη σύγκριση των αποτελεσµάτων που προκύπτουν µετά το σχεδιασµό του δεύτερου γραµµικού µοντέλου (πίνακας 13.4α, 13.4β) διαφαίνεται ότι η ηλικία και το φύλο επηρεάζουν καθοριστικά την ικανότητα γενετικής δράσης της φαρµακευτικής ένωσης Μη διεγερµένα λεµφοκύτταρα Ο προσδιορισµός του µηχανισµού προέλευσης των µικροπυρήνων διερευνήθηκε σε καλλιέργειες µη διεγερµένων λεµφοκυττάρων (πίνακας 13.5) για τους λόγους που αναφέρονται στο υποκεφάλαιο Η παρούσα µελέτη αφορά τους ίδιους δότες στους οποίους µελετήθηκε η επαγωγή µικροπυρήνων από τη δράση της φαρµακευτικής ένωσης σε καλλιέργειες µη διεγερµένων λεµφοκυττάρων (πίνακας 12.7). Πρόκειται για άτοµα νεαρής ηλικίας, τα οποία έχουν συµπεριληφθεί στα πειράµατα προσδιορισµού του περιεχοµένου των µικροπυρήνων σε καλλιέργειες διεγερµένων λεµφοκυττάρων (πίνακας 13.1). Τα αποτελέσµατα που προκύπτουν από την εφαρµογή της µεθοδολογίας FISH µε τη χρήση πανκεντροµερικού ανιχνευτή έδειξαν τη σηµαντική επίδραση της συγκέντρωσης 4 µg/ml HCTZ στην επαγωγή του σχηµατισµού µικροπυρήνων που χαρακτηρίζονται, είτε από την παρουσία (8.25±.76) είτε από την απουσία (7.85 ±.73) κεντροµερικού σήµατος σε σχέση µε τις καλλιέργειες-µάρτυρες (3.54±.29 και 5.22±.41, αντίστοιχα). Σε µικρότερο βαθµό παρατηρήθηκε αύξηση στο σχηµατισµό των δύο τύπων µικροπυρήνων, C + MN και C - MN (4.44±.4 και 6.7±.17, αντίστοιχα) στις καλλιέργειες HCTZ 5 µg/ml, η οποία δεν διέφερε στατιστικά από τις καλλιέργειες-µάρτυρες. Επιπρόσθετα, η στατιστική ανάλυση των αποτελεσµάτων αποκαλύπτει τη σηµαντική διαφορά που υπάρχει στους δύο τύπους µικροπυρήνων στις καλλιέργειες HCTZ 4 µg/ml σε σχέση τόσο µε τις καλλιέργειες-µάρτυρες όσο και µε τις καλλιέργειες HCTZ 5 µg/ml. Αξιοσηµείωτη είναι η παρατήρηση που προέρχεται από την κατανοµή των µικροπυρήνων ανάλογα µε τα σήµατα υβριδοποίησης σύµφωνα µε την οποία για κάθε πειραµατικό σηµείο που ελέγχθηκε την κυρίαρχη κατηγορία απαρτίζουν οι µικροπυρήνες που εµφανίζουν ένα κεντροµερικό σήµα, ενώ σηµαντική είναι η αύξηση των µικροπυρήνων που φέρουν δύο ή περισσότερα σήµατα υβριδοποίησης για την κεντροµερική περιοχή στη συγκέντρωση 4 µg/ml HCTZ. Αναλυτικότερα, ενώ στις καλλιέργειες-µάρτυρες η µέση τιµή της συχνότητας των C 2+ ΜΝ στα νεαρά άτοµα είναι.29±.13, µετά την επίδραση 5 µg/ml και 4 µg/ml HCTZ αυξάνεται και γίνεται αντίστοιχα.41±.18 και 1.54±.28. Τα παραπάνω αποτελέσµατα συµφωνούν µε αυτά που προκύπτουν από τις καλλιέργειες διεγερµένων λεµφοκυττάρων ατόµων νεαρής ηλικίας, γεγονός που φαίνεται να αποκλείει µία εξαρτώµενη δράση της φαρµακευτικής ένωσης από την κατάσταση διαίρεσης των κυττάρων
151 Αποτελέσµατα Μηχανισµός µικροπυρήνων Πίνακας 13.5 Συχνότητες µικροπυρήνων που περιέχουν σήµα κεντροµέρους (C + MN) ή όχι (C - MN) και κατανοµή µικροπυρήνων που περιέχουν ένα και δύο ή περισσότερα σήµατα κεντροµέρους σε µη διεγερµένες µε PHA κατά την επίδραση µε υδροχλωροθειαζίδιο (HCTZ) καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας ( 25 ετών) και διαφορετικού φύλου, in vitro. ότες ιπύρηνα κύτταρα που αναλύθηκαν ΜΝ C + ΜΝ ( ) C - ΜN ( ) C 1+ ΜΝ ( ) C 2+ ΜN ( ) ΜΑΡΤΥΡΕΣ Α.Π. ( ) (2.38) 7 (5.56) 3 (2.38) () Κ.Γ. ( ) (4.2) 24 (6.9) 12 (3.45) 2 (.57) Σ.Η. ( ) (3.23) 6 (4.84) 4 (3.23) () Μ.Τ.±Τ.Σ ± ±.6 3.2±.33.19±.19 Κ.Α. ( ) (4.) 4 (4.) 4 (4.) () Ν.Κ. ( ) (3.3) 7 (4.62) 4 (2.64) 1 (.66) Σ.Κ. ( ) (4.32) 1 (5.41) 7 (3.78) 1 (.54) Μ.Τ.±Τ.Σ ± ± ±.42.4±.2 Σ Μ.Τ.±Τ.Σ ± ± ±.26.29±.13 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml Α.Π. ( ) (3.52) 9 (6.34) 5 (3.52) () Κ.Γ. ( ) (4.53) 8 (6.4) 5 (3.77) 1 (.76) Σ.Η. ( ) (3.93) 9 (5.9) 6 (3.93) () Μ.Τ.±Τ.Σ ± ± ±.12.25±.25 Κ.Α. ( ) (4.96) 8 (6.62) 5 (4.13) 1 (.83) Ν.Κ. ( ) (3.61) 6 (5.42) 4 (3.61) () Σ.Κ. ( ) (6.9) 7 (6.9) 6 (5.22) 1 (.87) Μ.Τ.±Τ.Σ ± ± ±.47.57±.28 Σ Μ.Τ.±Τ.Σ ±.4 6.7± ±.25.41±.18 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml Α.Π. ( ) (5.8) 8 (6.63) 6 (4.97) 1 (.83) Κ.Γ. ( ) (1.) 11 (11.) 8 (8.) 2 (2.) Σ.Η. ( ) (7.) 7 (7.) 5 (5.) 2 (2.) Μ.Τ.±Τ.Σ ± ± ± ±.39 Κ.Α. ( ) (9.73) 8 (6.49) 9 (7.3) 3 (2.43) Ν.Κ. ( ) (7.) 7 (7.) 6 (6.) 1 (1.) Σ.Κ. ( ) (1.) 9 (9.) 9 (9.) 1 (1.) Μ.Τ.±Τ.Σ ± ± ± ±.48 Σ Μ.Τ.±Τ.Σ , ± , ± , ± , ±.28 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test) p<.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (Student's t-test)
152 Αποτελέσµατα Μηχανισµός µικροπυρήνων Εικόνα Εφαρµογή της µεθόδου FISH µε χρήση πανκεντροµερικού ανιχνευτή σε ανθρώπινα λεµφοκύτταρα. Α1-Α2: Μετάφαση στο φίλτρο του υπεριώδους (κυανό-α1) και στο φίλτρο της φθοροσκεΐνης (πράσινο-α2). Παρατηρείται το κεντροµερικό σήµα για κάθε χρωµόσωµα, γεγονός που επιβεβαιώνει την απόδοση της υβριδοποίησης. Β1-Β2: Πολυπολική ανατελόφαση. Παρατηρείται η καθυστέρηση στη µετακίνηση δύο χρωµοσωµάτων στα φίλτρα του υπεριώδους (Β1) και της φθοροσκεΐνης (Β2) µε αποτέλεσµα την απώλεια και τον αποκλεισµό τους από τους κυρίως πυρήνες του κυττάρου. Γ1-Γ2: ιπύρηνο κύτταρο µε δύο µικροπυρήνες στο φίλτρο του υπεριώδους (Γ1). Στους δύο µικροπυρήνες ανιχνεύεται κεντροµερικό σήµα στο φίλτρο της φθοροσκεΐνης (Γ2)
153 Αποτελέσµατα Μηχανισµός µικροπυρήνων 13.2 Ανθρώπινες κυτταρικές σειρές Η επαγωγή µικροπυρήνων που θεωρούνται προϊόν χρωµοσωµατικής απώλειας ως αποτέλεσµα της γενετικής δράσης της φαρµακευτικής ένωσης σε ανθρώπινα λεµφοκύτταρα περιφερικού αίµατος αποτέλεσε το έναυσµα για την περαιτέρω µελέτη της ικανότητας του υδροχλωροθειαζιδίου να προκαλεί χρωµοσωµατική καθυστέρηση. Για το λόγο αυτό µελετήθηκε η ακεραιότητα της κεντροµερικής περιοχής, βασικής δοµής του γενετικού υλικού. Συγκεκριµένα µε την εφαρµογή της µεθόδου αναστολής της κυτταροκίνησης σε ανθρώπινες κυτταρικές σειρές µελετήθηκε η επαγωγή µικροπυρήνων, ενώ µε τη συνδυασµένη σήµανση του κινητοχώρου (CREST) και του κεντροµέρους (FISH) όλων των ανθρώπινων χρωµοσω- µάτων διερευνήθηκε ο µηχανισµός προέλευσης τους και εξετάσθηκε η δυνατότητα δράσης του υδροχλωροθειαζιδίου σε πιθανούς στόχους που ενέχονται στην πρόκληση χρωµοσωµατικής απώλειας. Η µεθοδολογία CREST-FISH εφαρµόσθηκε σε δύο κυτταρικές σειρές από τις οποίες η µία είναι φυσιολογική, HFFF2 (πίνακας 13.6), και η άλλη είναι καρκινική, HEp2 (πίνακας 13.7). Η επιλογή της δεύτερης κυτταρικής σειράς έγινε διότι σε καρκινικά κύτταρα έχουν ανιχνευθεί κεντροµέρη στα οποία απουσιάζουν οι πρωτεΐνες του κινητοχώρου (Vig και Sternes, 1991). Με την εφαρµογή της µεθοδολογίας είναι δυνατή η ταυτόχρονη ανίχνευση του κεντροµερικού DNA και των πρωτεϊνών του κινητοχώρου στον ίδιο µικροπυρήνα µε αποτέλεσµα οι µικροπυρήνες που προκύπτουν να κατατάσσονται σε τέσσερις διακριτές κατηγορίες (Schuler et al., 1997). Αναλυτικότερα: (α) Μικροπυρήνες οι Εικόνα ιαφορετικοί µηχανισµοί που οδηγούν στο σχηµατισµό µικροπυρήνων προσδιορίζονται µε τη συνδυασµένη σήµανση του κινητοχώρου και του κεντροµέρους, CREST - FISH. (1) χρωµοσωµατική καθυστέρηση (2) ρήγµα εκτός κεντροµερικής περιοχής του χρωµοσώ- µατος (3) ρήγµα στην κεντροµερική περιοχή (4) αποκόλληση του κινητοχώρου από το κεντροµέρος (Schuler et al., 1997). οποίοι χαρακτηρίζονται από την ταυτόχρονη παρουσία σήµατος για τον κινητοχώρο και το κεντροµέρος, αναφέρονται ως K + C + MN. (β) Μικροπυρήνες οι οποίοι χαρακτηρίζονται από την απουσία σήµατος για τον κινητοχώρο, ενώ ταυτόχρονα περιέχουν σήµα κεντροµέρους αναφέρονται ως K - C + MN. Οι δύο αυτές κατηγορίες θεωρούνται προϊόν χρωµοσωµατικής απώλειας εξαιτίας της παρουσίας σήµατος υβριδοποίησης για την κεντροµερική περιοχή και κατατάσσονται περαιτέρω ως C + MN. (γ) Μικροπυρήνες οι οποίοι χαρακτηρίζονται από την παρουσία σήµατος για τον κινητοχώρο, ενώ ταυτόχρονα δεν περιέχουν σήµα κεντροµέρους αναφέρονται ως K + C - MN και σε συνδυασµό µε τους K + C + MN αποτελούν µία ευρύτερη κατηγορία µικροπυρήνων που περιέχουν σήµα κινητοχώρου και κατατάσσονται ως K + MN. (δ) Μικροπυρήνες οι οποίοι χαρακτηρίζονται από την ταυτόχρονη απουσία σήµατος για τον κινητοχώρο και το κεντροµέρος αναφέρονται ως K - C - MN. Οι µικροπυρήνες
154 Αποτελέσµατα Μηχανισµός µικροπυρήνων αυτοί θεωρούνται προϊόν δηµιουργίας χρωµοσωµατικών ρηγµάτων (εικόνες 13-2, 13-3). Στους πίνακες που περιγράφονται στο συγκεκριµένο υποκεφάλαιο στην πρώτη στήλη εµφανίζεται η συγκέντρωση της εξεταζόµενης χηµικής ένωσης, ενώ στη δεύτερη στήλη αναφέρεται ο αριθµός των διπύρηνων κυττάρων. Στην τρίτη στήλη αναφέρεται ο αριθµός των µικροπυρήνων που αναλύθηκαν, ενώ στις επόµενες στήλες γίνεται διαχωρισµός των µικροπυρήνων στις κατηγορίες που περιγράφηκαν. Οι συχνότητες των µικροπυρήνων έχουν υπολογισθεί σε 1 διπύρηνα κύτταρα. Η αποδοτικότητα της συνδυασµένης σήµανσης κινητοχώρουκεντροµέρους ελέγχθηκε σε καλλιέργειες-µάρτυρες της φυσιολογικής κυτταρικής σειράς HFFF2 αποδίδοντας ποσοστά K + MN, 45.45%, και C + MN, 36.36%. Τα αποτελέσµατα αυτά είναι σύµφωνα µε τα διεθνή βιβλιογραφικά δεδοµένα (Vlastos και Stephanou, 1998, Andrianopoulos et al., 2) HFFF2 H δράση του υδροχλωροθειαζιδίου µελετήθηκε στις συγκεντρώσεις 2, 4, 6 και 8 µg/ml σε καλλιέργειες της κυτταρικής σειράς HFFF2 (πίνακας 13.6). Παρατηρήθηκε αύξηση της συχνότητας µικροπυρήνων στις καλλιέργειες που πραγµατοποιήθηκε επίδραση HCTZ, η οποία εµφανίζεται εντονότερη από τη συγκέντρωση 4 µg/ml (1.5) σε σχέση µε την καλλιέργεια-µάρτυρα (5.5). Η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων στις καλλιέργειες HCTZ 6 µg/ml (16.4) και 8 µg/ml (17.) είναι στατιστικώς σηµαντική σε σχέση µε την καλλιέργεια µάρτυρα, γεγονός το οποίο φαίνεται να ισχύει οριακά (P =.5) για τη συγκέντρωση 4 µg/ml HCTZ, ενώ δεν ισχύει για τη συγκέντρωση 2 µg/ml HCTZ (6.). Η ανάλυση του περιεχοµένου των µικροπυρήνων έδειξε µία µικρή αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων που δεν περιέχουν σήµα κινητοχώρου και κεντροµέρους (K - C - MN). Η αύξηση αυτή στη συγκέντρωση HCTZ 8 µg/ml (7.) στατιστικά φαίνεται να διαφέρει οριακά (P =.5) από την καλλιέργεια-µάρτυρα (3.). Αντίθετα, σηµαντική είναι η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων που περιέχουν σήµα κινητοχώρου και κεντροµέρους (K + C + MN) στις συγκεντρώσεις 6 µg/ml (6.8) και 8 µg/ml HCTZ (6.) σε σχέση µε την καλλιέργεια-µάρτυρα (2.). Επιπρόσθετα, η δράση του HCTZ δεν φαίνεται να επηρεάζει σε σηµαντικό βαθµό τις δύο επόµενες κατηγορίες µικροπυρήνων K + C - MN και K - C + MN. Ιδιαίτερα οι συχνότητες των K - C + MN είναι µηδενικές τόσο στην καλλιέργεια-µάρτυρα όσο και στις καλλιέργειες στις οποίες πραγµατοποιήθηκε επίδραση µε τις συγκεντρώσεις HCTZ 2 και 4 µg/ml, ενώ παρατηρείται µικρή αύξηση µετά την επίδραση 6 µg/ml (.6) και 8 µg/ml HCTZ (1.). Στον ίδιο πίνακα, µε τον προσδιορισµό των συχνοτήτων µικροπυρήνων που περιέχουν αποκλειστικά είτε σήµα κεντροµέρους (C + MN) είτε σήµα κινητοχώρου (K + MN) επιβεβαιώθηκε από τη στατιστική ανάλυση η σηµαντική δράση των συγκεντρώσεων 6 µg/ml (7.4 και 9.1) και 8 µg/ml HCTZ (7. και 9.) προκαλώντας αύξηση στους βιολογικούς δείκτες που µελετήθηκαν σε σχέση µε την καλλιέργεια- µάρτυρα (2. και 2.5). Από τα παραπάνω αποτελέσµατα επιβεβαιώνεται η ικανότητα του HCTZ να προκαλεί χρωµοσωµατική απώλεια
155 Αποτελέσµατα Μηχανισµός µικροπυρήνων ΠΙΝΑΚΑΣ Συχνότητες µικροπυρήνων, κατανοµή µικροπυρήνων σε τέσσερις διακριτές κατηγορίες που προκύπτουν από την ταυτόχρονη εφαρµογή της µεθόδου σήµανσης του κινητοχώρου (CREST) και του κεντροµέρους (FISH), δείκτης πολλαπλασιασµού των κυττάρων (CBPI) και ποσοστό διπύρηνων κυττάρων σε καλλιέργειες κυττάρων HFFF2 στο µάρτυρα και µετά από επίδραση µε υδροχλωροθειαζίδιο (HCTZ). Συγκέντρωση HCTZ (µg/ml) ιπύρηνα κύτταρα που αναλύθηκαν Μικροπυρήνες ( ) K - C - ΜΝ ( ) K + C + MΝ ( ) K + C - ΜΝ ( ) K - C + ΜN ( ) K + ΜΝ ( ) C + ΜN ( ) CBPI % Ποσοστό διπύρηνων κυττάρων 4 22 (5.5) 12 (3.) 8 (2.) 2 (.5) () 1 (2.5) 8 (2.) (6.) 12 (3.) 8 (2.) 4 (1.) () 12 (3.) 8 (2.) (1.5)* 1 (5.) 7 (3.5) 4 (2.) () 11 (5.5) 7 (3.5) * (16.4)* 12 (6.8) 12 (6.8)* 4 (2.3) 1 (.6) 16 (9.1)* 13 (7.4)* * (17.)* 14 (7.)* 12 (6.)* 6 (3.)* 2 (1.) 18 (9.)* 14 (7.)* * *:.1 p<.5 σε σύγκριση µε το µάρτυρα (G-test)
156 Αποτελέσµατα Μηχανισµός µικροπυρήνων Προκειµένου να διερευνηθεί η ικανότητα του HCTZ να προκαλεί καθυστέρηση του κυτταρικού κύκλου σε ανθρώπινες κυτταρικές σειρές υπολογίσθηκαν, σε σύνολο 1 κυττάρων, ο δείκτης πολλαπλασιασµού των κυττάρων (CBPI) και ο δείκτης που αναφέρεται ως ποσοστό διπύρηνων κυττάρων. Το ποσοστό των διπύρηνων κυττάρων εκφράζει την εκατοστιαία αναλογία των κυττάρων που έχουν υποστεί µία πυρηνική διαίρεση (διπύρηνων) προς το σύνολο των διπύρηνων και µονοπύρηνων κυττάρων. Οι τιµές που λαµβάνουν οι δύο δείκτες αναγράφονται στις δύο τελευταίες στήλες του πίνακα Από τη µείωση των δεικτών σε σχέση µε την καλλιέργεια-µάρτυρα αποδίδεται στο υδροχλωροθειαζίδιο µία κυτταροστατική δράση, ενώ όπως φαίνεται από τον πίνακα δεν υπάρχουν σηµαντικές διαφορές στο CBPI και στο ποσοστό των διπύρηνων κυττάρων µεταξύ των συγκεντρώσεων. Συγκεκριµένα, στην καλλιέργεια-µάρτυρα οι δύο δείκτες εµφανίζουν αντίστοιχα τιµή 1.62 και 53.2 η οποία µειώνεται µε την επίδραση 2 µg/ml HCTZ στην τιµή 1.58 και 49.1, ενώ εµφανίζουν τη µεγαλύτερη πτώση (1.37 και 36.3) µε την επίδραση 8 µg/ml HCTZ. Από τη στατιστική ανάλυση προκύπτει ότι η µείωση αυτή είναι σηµαντική σε σχέση µε την καλλιέργεια-µάρτυρα στη συγκέντρωση 4 µg/ml HCTZ (1.5 και 42.2), ενώ εξίσου σηµαντική είναι η µείωση που παρατηρείται στις συγκεντρώσεις 6 µg/ml και 8 µg/ml HCTZ
157 Αποτελέσµατα Μηχανισµός µικροπυρήνων HEp2 H δράση του υδροχλωροθειαζιδίου µελετήθηκε στις συγκεντρώσεις 5, 2, 4 και 6 µg/ml σε καλλιέργειες της κυτταρικής σειράς HEp2 (πίνακας 13.7). Παρατηρήθηκε αύξηση της συχνότητας µικροπυρήνων στις καλλιέργειες που πραγµατοποιήθηκε επίδραση HCTZ. Η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων τόσο στην ελάχιστη συγκέντρωση (5 µg/ml) HCTZ (17.7) όσο και στη συγκέντρωση 2 µg/ml HCTZ (2.9) δεν διαφέρει στατιστικά από την καλλιέργεια-µάρτυρα (17.3). Επιπρόσθετα, η στατιστική ανάλυση έδειξε πως η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων στις καλλιέργειες HCTZ 4 µg/ml (31.4) και 6 µg/ml (41.6) είναι στατιστικώς σηµαντική σε σχέση µε την καλλιέργεια-µάρτυρα. Η ανάλυση του περιεχοµένου των µικροπυρήνων έδειξε µία µικρή αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων που δεν περιέχουν σήµα κινητοχώρου και κεντροµέρους (K - C - MN). Η αύξηση αυτή στη συγκέντρωση HCTZ 6 µg/ml (16.7) στατιστικά φαίνεται να διαφέρει οριακά (P =.5) από την καλλιέργεια-µάρτυρα (7.3). Αντίθετα, στην ίδια συγκέντρωση (6 µg/ml) HCTZ (18.5) η αύξηση που παρατηρείται στις συχνότητες µικροπυρήνων που περιέχουν σήµα κινητοχώρου και κεντροµέρους (K + C + MN) είναι στατιστικά σηµαντική σε σχέση µε την καλλιέργεια-µάρτυρα (8.). Η δράση του HCTZ δεν φαίνεται να επηρεάζει σε σηµαντικό βαθµό τις δύο επόµενες κατηγορίες µικροπυρήνων K + C - MN και K - C + MN, παρά τις αυξηµένες συχνότητες που παρατηρούνται. Ιδιαίτερα οι συχνότητες των K - C + MN κυµαίνονται από.7 στην καλλιέργεια-µάρτυρα έως 1.8 στην καλλιέργεια που πραγµατοποιήθηκε επίδραση µε τη συγκέντρωση 6 µg/ml HCTZ και η στατιστική ανάλυση έδειξε πως δεν υπάρχει σηµαντική διαφορά. Στον ίδιο πίνακα, µε τον προσδιορισµό των συχνοτήτων µικροπυρήνων που περιέχουν αποκλειστικά είτε σήµα κεντροµέρους (C + MN) είτε σήµα κινητοχώρου (K + MN) επιβεβαιώθηκε από τη στατιστική ανάλυση η σηµαντική δράση της συγκέντρωσης 6 µg/ml HCTZ (2.4 και 23.2) προκαλώντας αύξηση στους βιολογικούς δείκτες που µελετήθηκαν σε σχέση µε την καλλιέργεια-µάρτυρα (8.6 και 9.3). Από τα παραπάνω αποτελέσµατα επιβεβαιώνεται η ανευπλοειδογόνος δράση της φαρµακευτικής ένωσης. Η ικανότητα του HCTZ να επηρεάζει τη διάρκεια του κυτταρικού κύκλου στο διαιρούµενο κυτταρικό πληθυσµό της καρκινικής σειράς HEp2 ελέγχθηκε από την µείωση των τιµών του δείκτη που αναφέρεται ως ποσοστό διπύρηνων κυττάρων. Σε αυτές τις καλλιέργειες δεν εκτιµήθηκε ο δείκτης πολλαπλασιασµού των κυττάρων διότι ήταν αδύνατη η διάκριση των πολυπύρηνων κυττάρων σε τριπύρηνα και τετραπύρηνα. Το ποσοστό των διπύρηνων κυττάρων υπολογίσθηκε σε σύνολο 5 κυττάρων και η τιµή που λαµβάνει αναγράφεται στην τελευταία στήλη του πίνακα Στην καλλιέργεια-µάρτυρα ο δείκτης εµφανίζει τιµή 45.5, η οποία µειώνεται µε την επίδραση 5 µg/ml (44.) και 2 µg/ml (41.7) HCTZ, ενώ εµφανίζει µεγάλη πτώση στις συγκεντρώσεις 4 µg/ml (37.) και 6 µg/ml (3.4) HCTZ. Από τη στατιστική ανάλυση προκύπτει ότι η µείωση που παρατηρείται στις συγκεντρώσεις 4 και 6 µg/ml HCTZ είναι σηµαντική σε σχέση µε την καλλιέργεια-µάρτυρα. Σύµφωνα µε τα αποτελέσµατα που προκύπτουν από την ανάλυση των δεικτών για την εκτίµηση της έκτασης και εξέλιξης του κυτταρικού κύκλου στις ανθρώπινες κυτταρικές σειρές HFFF2 και HEp2 αποκαλύπτεται η ικανότητα της φαρµακευτικής ένωσης να προκαλεί σηµαντική καθυστέρηση του κυτταρικού κύκλου στις συγκεντρώσεις 4, 6 και 8 µg/ml HCTZ
158 Αποτελέσµατα Μηχανισµός µικροπυρήνων ΠΙΝΑΚΑΣ 13.7 Συχνότητες µικροπυρήνων, κατανοµή µικροπυρήνων σε τέσσερις διακριτές κατηγορίες που προκύπτουν από την ταυτόχρονη εφαρµογή της µεθόδου σήµανσης του κινητοχώρου (CREST) και του κεντροµέρους (FISH) και ποσοστό διπύρηνων κυττάρων σε καλλιέργειες κυττάρων HEp2 στο µάρτυρα και µετά από επίδραση µε υδροχλωροθειαζίδιο (HCTZ). Συγκέντρωση HCTZ (µg/ml) ιπύρηνα κύτταρα που αναλύθηκαν Μικροπυρήνες ( ) K - C - ΜΝ ( ) K + C + MΝ ( ) K + C - ΜΝ ( ) K - C + ΜN ( ) K + ΜΝ ( ) C + ΜN ( ) % Ποσοστό διπύρηνων κυττάρων (17.3) 11 (7.3) 12 (8.) 2 (1.3) 1 (.7) 14 (9.3) 13 (8.6) (17.7) 13 (7.7) 14 (8.2) 2 (1.2) 1 (.6) 16 (9.4) 15 (8.8) (2.9) 11 (9.6) 1 (8.7) 2 (1.7) 1 (.9) 12 (1.4) 11 (9.6) (31.4)* 15 (14.3) 13 (12.4) 4 (3.8) 1 (1.) 17 (16.2) 14 (13.3) 37.* (41.6)* 18 (16.7)* 2 (18.5)* 5 (4.6) 2 (1.8) 25 (23.2)* 22 (2.4)* 3.4* *:.5 p<.5 σε σύγκριση µε το µάρτυρα (G-test)
159 Αποτελέσµατα Μηχανισµός µικροπυρήνων Εικόνα Συνδυασµένη σήµανση του κινητοχώρου, CREST, και του κεντροµέρους, FISH, όλων των χρωµοσωµάτων σε ανθρώπινες κυτταρικές σειρές. Η παρατήρηση του πυρηνικού υλικού, πυρήνες-µικροπυρήνες, (κυανό) γίνεται στο φίλτρο του υπεριώδους (1), του κινητοχώρου (πράσινο) στο φίλτρο της φθοροσκεΐνης (2) και του κεντροµέρους (ερυθρό) στο φίλτρο της ροδαµίνης (3). A1-A3: ιπύρηνο κύτταρο που δεν φέρει ΜΝ, Β1-Β3: ιπύρηνο κύτταρο µε K - C - MN, Γ1-Γ3 και 1-3: ύο διπύρηνα κύτταρα που το καθένα φέρει από έναν K + C + MN
160 Αποτελέσµατα Ανευπλοειδικά φαινόµενα 14. ΜΕΛΕΤΗ ΑΝΕΥΠΛΟΕΙ ΙΚΩΝ ΦΑΙΝΟΜΕΝΩΝ ΜΕ ΤΗ ΣΥΜΜΕΤΟΧΗ ΤΩΝ ΧΡΩΜΟΣΩΜΑΤΩΝ Χ, Υ ΚΑΙ ιεγερµένα λεµφοκύτταρα Η αυξηµένη συχνότητα µικροπυρήνων που θεωρούνται προϊόν χρωµοσω- µατικής απώλειας απέδωσε στο υδροχλωροθειαζίδιο µία ανευπλοειδογόνο δράση. Η περαιτέρω µελέτη αφορά τη διαπίστωση και αποσαφήνιση της ανευπλοειδογόνου δράσης του, η οποία επιτυγχάνεται µε την ανάλυση των δύο κύριων µηχανισµών που οδηγούν σε ανευπλοειδία, του µη αποχωρισµού των χρωµοσωµάτων και της χρωµοσωµατικής απώλειας. Εφαρµόσθηκε η µέθοδος αναστολής της κυτταροκίνησης σε συνδυασµό µε τη διπλή in situ υβριδοποίηση µε φθοροχρώµατα, Dual FISH, µε τη χρήση εξειδικευµένων ανιχνευτών για ορισµένες ειδικές περιοχές (κεντροµέρος) συγκεκριµένων χρωµοσωµάτων. Αναλυτικότερα, η κατανοµή των σηµάτων υβριδοποίησης στους δύο πυρήνες ενός διπύρηνου µεσοφασικού κυττάρου ή/και σε υπάρχοντα µικροπυρήνα δίνει σηµαντικές πληροφορίες για τα φαινόµενα του τυχαίου και επαγόµενου από τη δράση του υδροχλωροθειαζιδίου χρωµοσω- µατικού µη αποχωρισµού ή/και της χρωµοσωµατικής απώλειας (εικόνα 14-1). Συγκεκριµένα, ελέγχθηκαν τα χρωµοσώµατα Χ, Υ και 8 για τη συµµετοχή τους στα φαινόµενα του µη αποχωρισµού και της χρωµοσωµατικής απώλειας σε άτοµα νεαρής (πίνακες ) και ώριµης ηλικίας (πίνακες ). Τα κριτήρια που σχετίζονται µε την επιλογή των χρωµοσωµάτων αφορούν πρώτιστα τη χρωµοσωµατική δοµή (µέγεθος, θέση κεντροµέρους, δοµή χρωµατίνης), η οποία πιθανότατα ενέχεται σε ανευπλοειδικά φαινόµενα. Πρόσθετα στοιχεία που λαµβάνονται υπόψη είναι η συχνότητα εµφάνισης τους σε ανευπλοειδίες που παρατηρούνται σε νεογέννητα και σε αποβολές, καθώς και η παρουσία γονιδίων που συµµετέχουν στην ανάπτυξη καρκίνου στον άνθρωπο (Parry et al., 1994, Parry et al., 22). Το φυλετικό χρωµόσωµα Χ και το αυτοσωµατικό χρωµόσωµα 8 ανήκουν στην ίδια οµάδα C που περιλαµβάνει µεγάλα έως µεσαία υποµετακεντρικά χρω- µοσώµατα, ενώ το φυλετικό χρωµόσωµα Υ είναι το µικρότερο σε µέγεθος και ανήκει στην οµάδα G που απαρτίζεται από τα µικρά ακροκεντρικά χρωµοσώµατα (21 και 22). Οι ανωµαλίες των φυλετικών χρωµοσωµάτων µε τις περισσότερο κοινές και χαρακτηρισµένες ανευπλοειδίες εµφανίζονται σε σχετικά µεγάλη συχνότητα σε αρσενικά και θηλυκά νεογέννητα. Στο χρωµόσωµα 8 εντοπίζονται δύο σηµαντικά ογκογονίδια, τα myc και mos, που η υπερέκφραση τους ενέχεται σε καρκινικές καταστάσεις (Yunis, 1983). Από τη µελέτη των φυλετικών χρωµοσωµάτων ιδιαίτερο ενδιαφέρον παρουσιάζει το χρωµόσωµα Χ εξαιτίας τόσο της παρουσίας αρκετών ση- µαντικών ασθενειών που σχετίζονται µε αυτό όσο και της ύπαρξης του ανενεργού Χ χρωµοσώµατος στα θηλυκά άτοµα. Στην παρούσα µελέτη συµπεριελήφθησαν τα 32 άτοµα που αποτελούν το σύνολο των δοτών και στα οποία πραγµατοποιήθηκαν τα πειράµατα προσδιορισµού της συχνότητας των µικροπυρήνων. Στους πίνακες που ακολουθούν παρουσιάζεται η τυχαία επαγωγή ανευπλοειδικών φαινοµένων σε καλλιέργειες ανθρώπινων λεµφοκυττάρων ολικού αίµατος σε δύο διαφορετικές ηλικιακές οµάδες αρσενικών και θηλυκών ατόµων, οι οποίες θεωρούνται ως καλλιέργειες-µάρτυρες. Επίσης, ελέγχθηκε η ικανότητα του υδροχλωροθειαζιδίου, στις συγκεντρώσεις 5 και 4 µg/ml, να επηρεάζει τη συχνότητα χρωµοσωµατικής απώλειας και µη αποχωρισµού των χρωµοσωµάτων Χ, Υ και
161 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Κατά την εφαρµογή της διπλής in situ υβριδοποίησης, τα συνήθη ζεύγη ειδικών ανιχνευτών που χρησιµοποιήθηκαν περιλαµβάνουν την κεντροµερική περιοχή των χρωµοσωµάτων, είτε Χ και 8 είτε Χ και Υ. Η αποδοτικότητα της υβριδοποίησης ελέγχθηκε σε µεταφασικά παρασκευάσµατα. Ειδικότερα, έγινε ανάλυση µεταφάσεων και ελέγχθηκαν τα σήµατα που παρουσιάζονται ως προς την ειδικότητά τους για τον ανάλογο συνδυασµό χρωµοσωµάτων που µελετήθηκε. Επιπρόσθετα, οι κανονισµοί που ακολουθούνται για την ανάλυση σήµατος υβριδοποίησης σε διπύρηνα κύτταρα και σε µικροπυρήνες έχουν ήδη αναφερθεί στο υποκεφάλαιο Τουλάχιστον, 1 διπύρηνα κύτταρα µε το σωστό αριθµό σηµάτων αναλύθηκαν σε κάθε πειραµατικό σηµείο για τον υπολογισµό του µη αποχωρισµού και της χρω- µοσωµατικής απώλειας. ιπύρηνα κύτταρα µε το σωστό αριθµό σηµάτων χαρακτηρίζονται ως φυσιολογικά διπλοειδή κύτταρα, εφόσον εµφανίζουν κανονική κατανοµή (normal) στους δύο θυγατρικούς πυρήνες. Η περίπτωση της χρωµοσω- µατικής καθυστέρησης (chromosome delay) ή απώλειας (chromosome loss) αναφέρεται στην παρουσία µικροπυρήνα σε διπύρηνο κύτταρο, στον οποίο εντοπίζονται ένα ή περισσότερα σήµατα υβριδοποίησης, ενώ η περίπτωση του µη αποχωρισµού (non-disjunction) αναφέρεται στην παρουσία µη ισορροπηµένης κατανοµής ση- µάτων µεταξύ των πυρήνων ενός διπύρηνου κυττάρου (Zijno et al., 1996). Στις περισσότερες περιπτώσεις, τα ανευπλοειδικά φαινόµενα που µελετήθηκαν αναλύθηκαν ανεξάρτητα για τα χρωµοσώµατα Χ, Υ και 8, ενώ καταγράφηκαν παράλληλα σε κάθε κύτταρο. Στους πίνακες που περιγράφονται στο συγκεκριµένο υποκεφάλαιο στην πρώτη στήλη εµφανίζονται οι δότες που εξετάσθηκαν, ενώ στη δεύτερη αναφέρεται ο αριθµός των διπύρηνων κυττάρων που αναλύθηκαν. Στην τρίτη στήλη εµφανίζεται ο αριθµός των διπύρηνων κυττάρων που χαρακτηρίζονται ως διπλοειδή κύτταρα και είναι, είτε κύτταρα µε τη σωστή κατανοµή σηµάτων είτε κύτταρα που παρουσιάζουν φαινόµενα λανθασµένου αποχωρισµού, ενώ στην τέταρτη η συχνότητα του µη αποχωρισµού (% ND). Στην επόµενη (πέµπτη) στήλη αναφέρεται ο αριθµός των διπύρηνων κυττάρων µε µικροπυρήνα, ο οποίος παρουσιάζει σήµα υβριδοποίησης για την κεντροµερική περιοχή του χρωµοσώµατος που µελετήθηκε και στην τελευταία στήλη ο αριθµός των µικροπυρήνων µε σήµα κεντροµέρους για το συγκεκριµένο χρωµόσωµα. Στην πέµπτη και έκτη στήλη έχουν υπολογισθεί οι συχνότητες των διπύρηνων κυττάρων µε µικροπυρήνα και των µικροπυρήνων σε διπύρηνα κύτταρα, οι οποίοι χαρακτηρίζονται από την παρουσία κεντροµερικού σήµατος
162 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Καλλιέργειες λεµφοκυττάρων ατόµων νεαρής ηλικίας Χρωµόσωµα Χ. Η µελέτη ανευπλοειδικών φαινοµένων που αφορούν το χρωµόσωµα Χ στην οµάδα ατόµων νεαρής ηλικίας περιγράφεται αναλυτικά στους πίνακες Η ανάλυση γίνεται αποκλειστικά σε διπύρηνα κύτταρα µε το σωστό αριθµό σηµάτων που για το χρωµόσωµα Χ είναι 2 στα αρσενικά άτοµα και 4 στα θηλυκά άτοµα, ανεξάρτητα από τον τύπο κατανοµής. ιπύρηνα κύτταρα µε την κανονική κατανοµή σήµατος αναφέρονται ως 1:1 για τα αρσενικά άτοµα και 2:2 για τα θηλυκά άτοµα, µε απλό µη αποχωρισµό ως 2: για τα αρσενικά και 3:1 για τα θηλυκά άτοµα και µε διπλό µη αποχωρισµό ως 4: για τα θηλυκά άτοµα. Παρουσία της ελάχιστης (5 µg/ml) (πίνακας 14.2) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ (πίνακας 14.3) παρατηρείται αύξηση στις συχνότητες του χρωµοσωµατικού µη αποχωρισµού από.57±.5 στις καλλιέργειες-µάρτυρες (πίνακας 14.1) σε 2.2±.21 και 3.67±.23 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα προκαλεί η µέγιστη συγκέντρωση HCTZ γεγονός που ισχύει τόσο για τα φαινόµενα απλού όσο και για το φαινόµενο διπλού µη αποχωρισµού. Θα πρέπει να αναφερθεί ότι ο απλός µη αποχωρισµός αποτελεί το συνηθέστερο τύπο µη αποχωρισµού που παρατηρήθηκε. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν µη αποχωρισµό του χρωµοσώ- µατος Χ, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Στους ίδιους πίνακες προσδιορίζεται η απώλεια του χρωµοσώµατος Χ, η οποία ανιχνεύεται από την παρουσία σήµατος υβριδοποίησης στο µικροπυρήνα. Η επίδραση της ελάχιστης συγκέντρωσης HCTZ προκαλεί σχετικά µικρή αύξηση στην παρουσία του χρωµοσώµατος Χ σε διπύρηνα κύτταρα µε µικροπυρήνες ( ΠΜΝΧ + ) και στους µικροπυρήνες (ΜΝΧ + ) µε αποτέλεσµα να µην παρουσιάζεται στατιστικά σηµαντική διαφορά στις αντίστοιχες συχνότητες (1.68±.37) σε σχέση µε τις καλλιέργειες-µάρτυρες (1.52±.38). Αντίθετα, η αύξηση που παρατηρείται στη συγκέντρωση 4 µg/ml HCTZ (3.57±.62 ) είναι σαφώς µεγαλύτερη σε σχέση µε τις καλλιέργειες-µάρτυρες. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα της µέγιστης συγκέντρωσης HCTZ να επάγει απώλεια του χρωµοσώµατος Χ, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Οι παρατηρήσεις οι οποίες αφορούν το µη αποχωρισµό και την απώλεια του χρωµοσώµατος Χ στο σύνολο των 16 νεαρών δοτών που µελετήθηκαν ισχύουν τόσο για τα θηλυκά όσο και για τα αρσενικά άτοµα. Σχετικά µε το φαινόµενο της παρουσίας του χρωµοσώµατος Χ σε µικροπυρήνες φαίνεται να συµβαίνει µε µεγαλύτερη ένταση στα θηλυκά άτοµα σε σχέση µε τα αρσενικά, τόσο για τις καλλιέργειες-µάρτυρες (2.81±.38,.24±.12) όσο και για τις καλλιέργειες λεµφοκυττάρων στις οποίες πραγµατοποιήθηκε επίδραση µε τις δύο συγκεντρώσεις του HCTZ, 5 µg/ml (2.93±.33,.42±.16) και 4 µg/ml (5.31±.77, 1.83±.42). Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος Χ, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού στις οµάδες που απαρτίζονται από τα δύο φύλα των νεαρών δοτών είναι υψηλότερες από τις αντίστοιχες της χρωµοσωµατικής απώλειας για κάθε πειραµατικό σηµείο που ελέγχθηκε
163 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες τυχαίου µη αποχωρισµού (ΝDΧ) για το χρωµόσωµα Χ και απώλειας του ίδιου χρωµοσώµατος (MNΧ + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν αντίστοιχα δύο και τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. ΜΑΡΤΥΡΕΣ ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 2: ΝDΧ (%) Α.Χ. ( ) Α.Π. ( ) Α.Α. ( ) Κ.Γ. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) ΠΜΝΧ + ( ) () 1 (.64) 1 (.84) () 1 (.42) () () () ΜΝΧ + ( ) () 1 (.64) 1 (.84) () 1 (.42) () () () Μ.Τ.±Τ.Σ ±.8.24±.12.24±.12 2:2 3:1 4: Κ.Μ. ( ) Κ.Α. ( ) Μ.Κ. ( ) Ν.Κ. ( ) Π.Ν. ( ) Π.Π. ( ) Π.Μ. ( ) Σ.Κ. ( ) (3.24) 4 (4.) 1 (2.9) 4 (4.) 4 (3.34) 2 (1.32) 7 (2.33) 2 (1.31) 4 (3.24) 4 (4.) 1 (2.9) 4 (4.) 4 (3.34) 2 (1.32) 7 (2.33) 2 (1.31) Μ.Τ.±Τ.Σ ± ± ±.38 Σ Μ.Τ.±Τ.Σ ± ± ±
164 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝDΧ) για το χρωµόσωµα Χ και απώλειας του ίδιου χρωµοσώµατος (MNΧ + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν αντίστοιχα δύο και τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 2: ΝDΧ (%) Α.Χ. ( ) Α.Π. ( ) Α.Α. ( ) Κ.Γ. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) ΠΜΝΧ + ( ) () 1 (.94) 1 (.97) 1 (.68) 2 (.79) () () () ΜΝΧ + ( ) () 1 (.94) 1 (.97) 1 (.68) 2 (.79) () () () Μ.Τ.±Τ.Σ ±.32.42±.16.42±.16 2:2 3:1 4: Κ.Μ. ( ) Κ.Α. ( ) Μ.Κ. ( ) Ν.Κ. ( ) Π.Ν. ( ) Π.Π. ( ) Π.Μ. ( ) Σ.Κ. ( ) (1.91) 4 (4.) 3 (2.44) 4 (4.) 3 (2.47) 5 (3.36) 2 (1.64) 5 (3.62) 2 (1.91) 4 (4.) 3 (2.44) 4 (4.) 3 (2.47) 5 (3.36) 2 (1.64) 5 (3.62) Μ.Τ.±Τ.Σ ± ± ±.33 Σ Μ.Τ.±Τ.Σ ± ± ±.37 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test)
165 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝDΧ) για το χρωµόσωµα Χ και απώλειας του ίδιου χρωµοσώµατος (MNΧ + ) σε διπύρηνα κύτταρα αρσενικών και θηλυ-κών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν αντίστοιχα δύο και τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 2: ΝDΧ (%) Α.Χ. ( ) Α.Π. ( ) Α.Α. ( ) Κ.Γ. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) ΠΜΝΧ + ( ) () 1 (1.) 2 (1.86) 4 (2.78) 3 (1.25) 2 (1.94) 4 (3.95) 2 (1.89) ΜΝΧ + ( ) () 1 (1.) 2 (1.86) 4 (2.78) 3 (1.25) 2 (1.94) 4 (3.95) 2 (1.89) Μ.Τ.±Τ.Σ ± ± ±.42 2:2 3:1 4: Κ.Μ. ( ) Κ.Α. ( ) Μ.Κ. ( ) Ν.Κ. ( ) Π.Ν. ( ) Π.Π. ( ) Π.Μ. ( ) Σ.Κ. ( ) (1.98) 9 (9.) 6 (4.87) 7 (7.) 6 (5.61) 5 (3.75) 4 (3.97) 7 (6.29) 2 (1.98) 9 (9.) 6 (4.87) 7 (7.) 6 (5.61) 5 (3.75) 4 (3.97) 7 (6.29) Μ.Τ.±Τ.Σ ± ± ±.77 Σ Μ.Τ.±Τ.Σ , ± ± , ±.62 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
166 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Χρωµόσωµα Υ. Η µελέτη ανευπλοειδικών φαινοµένων που αφορούν το χρωµόσωµα Υ στην οµάδα αρσενικών ατόµων νεαρής ηλικίας περιγράφεται αναλυτικά στον πίνακα Η ανάλυση γίνεται αποκλειστικά σε διπύρηνα κύτταρα µε το σωστό αριθµό σηµάτων που για το χρωµόσωµα Υ είναι 2 στα αρσενικά άτοµα, ανεξάρτητα από τον τύπο κατανοµής. ιπύρηνα κύτταρα µε την κανονική κατανοµή σήµατος αναφέρονται ως 1:1 και µε µη αποχωρισµό ως 2:. Παρουσία της ελάχιστης (5 µg/ml) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ παρατηρείται αύξηση στις συχνότητες του χρωµοσωµατικού µη αποχωρισµού από.8±.4 στις καλλιέργειες-µάρτυρες σε.35±.5 και.86±.13 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα προκαλεί η µέγιστη συγκέντρωση HCTZ. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν µη αποχωρισµό του χρωµοσώµατος Υ, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Στον ίδιο πίνακα από τον προσδιορισµό της απώλειας του χρωµοσώµατος Υ, ενώ φαίνεται η επίδραση της συγκέντρωσης 4 µg/ml HCTZ να προκαλεί µία σχετικά µεγάλη αύξηση (.6±.24) στην παρουσία του χρωµοσώµατος σε µικροπυρήνες σε σχέση µε τις καλλιέργειες-µάρτυρες (.34±.21) η στατιστική ανάλυση που εφαρµόσθηκε έδειξε πως δεν υπάρχει σηµαντική διαφορά. Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος Y, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού είναι υψηλότερες από τις αντίστοιχες της χρωµοσωµατικής απώλειας, γεγονός που ισχύει τόσο για τις καλλιέργειες-µάρτυρες όσο και για τις καλλιέργειες λεµφοκυττάρων στις οποίες πραγµατοποιήθηκε επίδραση µε τις δύο συγκεντρώσεις του HCTZ
167 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες τυχαίου και επαγόµενου µη αποχωρισµού (ΝDΥ) για το χρω- µόσωµα Υ και απώλειας του ίδιου χρωµοσώµατος (ΜΝΥ + ) σε διπύρηνα κύτταρα αρσενικών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν δύο σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 2: ΝDY (%) ΠΜΝY + ( ) ΜΝY + ( ) ΜΑΡΤΥΡΕΣ Α.Π. ( ) Σ.Η. ( ) 1 1 Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) (.7) () () 1 (.99) () 2 (.7) () () 1 (.99) () Μ.Τ.±Τ.Σ ±.4.34±.21.34±.21 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml Α.Π. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) () () () () () () () () () () Μ.Τ.±Τ.Σ * ±.5 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml Α.Π. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) () 1 (.91) () 1 (1.) 2 (1.7) () 1 (.91) () 1 (1.) 2 (1.7) Μ.Τ.±Τ.Σ , ±.13.6±.24.6±.24 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test) * p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (Student's t-test)
168 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Χρωµόσωµα 8. Η µελέτη ανευπλοειδικών φαινοµένων που αφορούν το χρωµόσωµα 8 στην οµάδα ατόµων νεαρής ηλικίας περιγράφεται αναλυτικά στους πίνακες Η ανάλυση γίνεται αποκλειστικά σε διπύρηνα κύτταρα µε το σωστό αριθµό σηµάτων που για το χρωµόσωµα 8 είναι 4 σε αρσενικά και θηλυκά άτοµα, ανεξάρτητα από τον τύπο κατανοµής. ιπύρηνα κύτταρα µε την κανονική κατανοµή σήµατος αναφέρονται ως 2:2, µε απλό µη αποχωρισµό ως 3:1 και µε διπλό µη αποχωρισµό ως 4:. Παρουσία της ελάχιστης (5 µg/ml) (πίνακας 14.6) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ (πίνακας 14.7) παρατηρείται αύξηση στις συχνότητες του χρωµοσωµατικού µη αποχωρισµού από.24±.4 στις καλλιέργειες-µάρτυρες (πίνακας 14.5) σε.94±.7 και 1.96±.14 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα (.26±.6,.22±.6) προκαλεί η µέγιστη συγκέντρωση HCTZ (1.76±.2, 2.16±.17) τόσο στα αρσενικά όσο και στα θηλυκά άτοµα αντίστοιχα. Ο συνηθέστερος τύπος λανθασµένου αποχωρισµού για το χρωµόσωµα 8 είναι ο απλός µη αποχωρισµός ανεξάρτητα από το φύλο ή από την επίδραση HCTZ, γεγονός που ισχύει και για τα φυλετικά χρωµοσώµατα. Η εµφάνιση του διπλού µη αποχωρισµού για το χρωµοσώµα 8 σε αρσενικά και θηλυκά άτοµα παρατηρείται σε πολύ µικρότερες συχνότητες σε σχέση µε το χρωµόσωµα Χ στα θηλυκά άτοµα. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν µη αποχωρισµό του χρωµοσώµατος 8, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Στους ίδιους πίνακες προσδιορίζεται η απώλεια του χρωµοσώµατος 8, η οποία ανιχνεύεται από την παρουσία σήµατος υβριδοποίησης στο µικροπυρήνα. Στα νεαρά άτοµα ανεξάρτητα από το φύλο δεν παρατηρείται στις καλλιέργειες-µάρτυρες παρουσία του χρωµοσώµατος 8 τόσο στα διπύρηνα κύτταρα µε µικροπυρήνες ( Π ΜΝ8 + ) όσο και στους µικροπυρήνες (ΜΝ8 + ) µε αντίστοιχες µηδενικές συχνότητες. Η επίδραση της ελάχιστης συγκέντρωσης HCTZ προκαλεί µικρή αύξηση στην παρουσία του χρωµοσώµατος 8 σε διπύρηνα κύτταρα µε µικροπυρήνες ( ΠΜΝ8 + ) και στους µικροπυρήνες (ΜΝ8 + ) µε αποτέλεσµα να µην παρουσιάζεται σηµαντική διαφορά στις αντίστοιχες συχνότητες (.15±.8) σε σχέση µε τις καλλιέργειες-µάρτυρες. Αντίθετα, η αύξηση των ΠΜΝ8 + και ΜΝ8 + που παρατηρείται στη συγκέντρωση 4 µg/ml HCTZ (1.8±.32) είναι σαφώς µεγαλύτερη σε σχέση µε τις αντίστοιχες συχνότητες στις καλλιέργειες µάρτυρες. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα της µέγιστης συγκέντρωσης HCTZ να επάγει απώλεια του χρωµοσώµατος 8, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Οι παρατηρήσεις οι οποίες αφορούν το µη αποχωρισµό και την απώλεια του χρωµοσώµατος 8 στο σύνολο των 16 νεαρών δοτών που µελετήθηκαν ισχύουν και για τα δύο φύλα. Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος 8, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού στις οµάδες που απαρτίζονται από τα νεαρά αρσενικά και θηλυκά άτοµα είναι υψηλότερες από τις αντίστοιχες της χρωµοσωµατικής απώλειας για κάθε πειραµατικό σηµείο που ελέγχθηκε
169 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες τυχαίου µη αποχωρισµού (ΝD8) για το χρωµόσωµα 8 και απώλειας του ίδιου χρωµοσώµατος (ΜΝ8 + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. ΜΑΡΤΥΡΕΣ ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 3:1 4: ΝD8 (%) Α.Χ. ( ) Α.Π. ( ) Α.Α. ( ) Κ.Γ. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) ΠΜΝ8 + ( ) () () () () () () () () ΜΝ8 + ( ) () () () () () () () () Μ.Τ.±Τ.Σ ±.6 Κ.Μ. ( ) Κ.Α. ( ) Μ.Κ. ( ) 1 1 Ν.Κ. ( ) Π.Ν. ( ) Π.Π. ( ) Π.Μ. ( ) Σ.Κ. ( ) () () () () () () () () () () () () () () () () Μ.Τ.±Τ.Σ ±.6 Σ Μ.Τ.±Τ.Σ ±
170 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝD8) για το χρωµόσωµα 8 και απώλειας του ίδιου χρωµοσώµατος (ΜΝ8 + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 2:2 3:1 4: ΝD8 (%) Α.Χ. ( ) Α.Π. ( ) Α.Α. ( ) Κ.Γ. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) ΠΜΝ8 + ( ) () () () 1 (.77) () () () () ΜΝ8 + ( ) () () () 1 (.77) () () () () Μ.Τ.±Τ.Σ ±.11.1±.1.1±.1 Κ.Μ. ( ) Κ.Α. ( ) Μ.Κ. ( ) Ν.Κ. ( ) Π.Ν. ( ) Π.Π. ( ) Π.Μ. ( ) Σ.Κ. ( ) () () 1 (.97) () () 1 (.67) () () () () 1 (.97) () () 1 (.67) () () Μ.Τ.±Τ.Σ ±.7.21±.14.21±.14 Σ Μ.Τ.±Τ.Σ ±.7.15±.8.15±.8 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test)
171 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝD8) για το χρωµόσωµα 8 και απώλειας του ίδιου χρωµοσώµατος (ΜΝ8 + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ) ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 2:2 3:1 4: ΝD8 (%) Α.Χ. ( ) Α.Π. ( ) Α.Α. ( ) Κ.Γ. ( ) Σ.Η. ( ) Β.Α. ( ) Π.Ν. ( ) Τ.Μ. ( ) ΠΜΝ8 + ( ) 1 (.94) () 1 (.99) 1 (.7) () 1 (.83) 1 (1.) () ΜΝ8 + ( ) 1 (.94) () 1 (.99) 1 (.7) () 1 (.83) 1 (1.) () Μ.Τ.±Τ.Σ ±.2.56±.17.56±.17 Κ.Μ. ( ) Κ.Α. ( ) Μ.Κ. ( ) Ν.Κ. ( ) Π.Ν. ( ) Π.Π. ( ) Π.Μ. ( ) Σ.Κ. ( ) () 4 (4.) 1 (.99) 4 (4.) () 1 (.73) 1 (.99) 3 (2.16) () 4 (4.) 1 (.99) 4 (4.) () 1 (.73) 1 (.99) 3 (2.16) Μ.Τ.±Τ.Σ ± ± ±.57 Σ Μ.Τ.±Τ.Σ , ± ± , ±.32 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
172 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Καλλιέργειες λεµφοκυττάρων ατόµων ώριµης ηλικίας Χρωµόσωµα Χ. Η µελέτη ανευπλοειδικών φαινοµένων που αφορούν το χρωµόσωµα Χ στην οµάδα ατόµων ώριµης ηλικίας περιγράφεται αναλυτικά στους πίνακες Παρουσία της ελάχιστης (5 µg/ml) (πίνακας 14.9) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ (πίνακας 14.1) παρατηρείται αύξηση στις συχνότητες του χρωµοσωµατικού µη αποχωρισµού από 1.15±.7 στις καλλιέργειες-µάρτυρες (πίνακας 14.8) σε 2.62±.21 και 4.1±.28 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα προκαλεί η µέγιστη συγκέντρωση HCTZ γεγονός που ισχύει τόσο για τα φαινόµενα απλού όσο και για το φαινόµενο διπλού µη αποχωρισµού. Θα πρέπει να αναφερθεί ότι ο απλός µη αποχωρισµός αποτελεί το συνηθέστερο τύπο µη αποχωρισµού που παρατηρήθηκε. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν µη αποχωρισµό του χρωµοσώ- µατος Χ, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Στους ίδιους πίνακες προσδιορίζεται η απώλεια του χρωµοσώµατος Χ, η οποία ανιχνεύεται από την παρουσία σήµατος υβριδοποίησης στο µικροπυρήνα. Η επίδραση της ελάχιστης συγκέντρωσης HCTZ προκαλεί σχετικά µικρή αύξηση στην παρουσία του χρωµοσώµατος Χ σε διπύρηνα κύτταρα µε µικροπυρήνες ( ΠΜΝΧ + ) και στους µικροπυρήνες (ΜΝΧ + ) µε αποτέλεσµα να µην παρουσιάζεται στατιστικά σηµαντική διαφορά στις αντίστοιχες συχνότητες (8.4±1.13) σε σχέση µε τις καλλιέργειες-µάρτυρες (8.7±1.3). Αντίθετα, η αύξηση που παρατηρείται στη συγκέντρωση 4 µg/ml HCTZ (13.6±1.56 ) είναι σαφώς µεγαλύτερη σε σχέση µε τις καλλιέργειες-µάρτυρες. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα της µέγιστης συγκέντρωσης HCTZ να επάγει απώλεια του χρωµοσώµατος Χ, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Οι παρατηρήσεις οι οποίες αφορούν το µη αποχωρισµό και την απώλεια του χρωµοσώµατος Χ στο σύνολο των 16 ηλικιωµένων δοτών που µελετήθηκαν ισχύουν τόσο για τα θηλυκά όσο και για τα αρσενικά άτοµα. Σχετικά µε το φαινόµενο της παρουσίας του χρωµοσώµατος Χ σε µικροπυρήνες φαίνεται να συµβαίνει µε µεγαλύτερη ένταση στα θηλυκά άτοµα σε σχέση µε τα αρσενικά, τόσο για τις καλλιέργειες-µάρτυρες (12.31±1.3, 3.83±.62) όσο και για τις καλλιέργειες λεµφοκυττάρων στις οποίες πραγµατοποιήθηκε επίδραση µε τις δύο συγκεντρώσεις HCTZ, 5 µg/ml (12.1±1.17, 4.79±.62) και 4 µg/ml (17.68±1.59, 8.43±1.35). Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος Χ, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού στις οµάδες που απαρτίζονται από τα δύο φύλα των ηλικιωµένων δοτών είναι υψηλότερες από τις αντίστοιχες της χρωµοσωµατικής απώλειας για κάθε πειραµατικό σηµείο που ελέγχθηκε. Επιπρόσθετα, παρατηρούνται υψηλότερες συχνότητες ανευπλοειδικών φαινοµένων, µη αποχωρισµού και απώλειας, για το χρωµόσωµα Χ στα ηλικιωµένα άτοµα σε σύγκριση µε τα νεαρά
173 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες τυχαίου µη αποχωρισµού (ΝDΧ) για το χρωµόσωµα Χ και απώλειας του ίδιου χρωµοσώµατος (MNΧ + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων ώριµης ηλικίας ( 45 ετών), που φέρουν αντίστοιχα δύο και τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. ΜΑΡΤΥΡΕΣ ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 2: ΝDΧ (%) Β.Α. ( ) Κ.Γ. ( ) Σ.Ν. ( ) Χ.Β. ( ) Γ.Χ. ( ) Γ.Θ. ( ) Ι.Γ. ( ) Σ.Κ. ( ) ΠΜΝΧ + ( ) 6 (4.74) 6 (5.28) 6 (5.77) 2 (1.93) 4 (3.82) 3 (2.54) 4 (4.) 2 (1.62) ΜΝΧ + ( ) 6 (4.74) 6 (5.28) 7 (6.73) 2 (1.93) 4 (3.82) 3 (2.54) 4 (4.) 2 (1.62) Μ.Τ.±Τ.Σ ± ± ±.62 2:2 3:1 4: Κ.Σ. ( ) Λ.Μ. ( ) Ν.Μ. ( ) Π.Ε. ( ) Σ.Σ. ( ) Σ.Γ. ( ) Χ.Ε. ( ) Χ.Α. ( ) (15.29) 9 (8.72) 12 (1.34) 21 (9.86) 16 (11.56) 26 (17.93) 1 (7.11) 1 (9.81) 2 (15.29) 1 (9.69) 12 (1.34) 24 (11.27) 16 (11.56) 29 (2.) 12 (8.53) 12 (11.78) Μ.Τ.±Τ.Σ ± ± ±1.3 Σ Μ.Τ.±Τ.Σ ± ± ±
174 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝDΧ) για το χρωµόσωµα Χ και απώλειας του ίδιου χρωµοσώµατος (MNΧ + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων ώριµης ηλικίας ( 45 ετών), που φέρουν αντίστοιχα δύο και τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 2: ΝDΧ (%) Β.Α. ( ) Κ.Γ. ( ) Σ.Ν. ( ) Χ.Β. ( ) Γ.Χ. ( ) Γ.Θ. ( ) Ι.Γ. ( ) Σ.Κ. ( ) ΠΜΝΧ + ( ) 7 (7.) 7 (6.58) 7 (6.38) 3 (2.9) 3 (3.) 4 (2.31) 5 (5.) 6 (4.15) ΜΝΧ + ( ) 7 (7.) 7 (6.58) 7 (6.38) 3 (2.9) 4 (4.) 4 (2.31) 5 (5.) 6 (4.15) Μ.Τ.±Τ.Σ ± ± ±.62 2:2 3:1 4: Κ.Σ. ( ) Λ.Μ. ( ) Ν.Μ. ( ) Π.Ε. ( ) Σ.Σ. ( ) Σ.Γ. ( ) Χ.Ε. ( ) Χ.Α. ( ) (13.47) 8 (7.65) 7 (6.65) 14 (13.23) 19 (12.74) 15 (14.8) 11 (8.44) 12 (11.25) 19 (15.5) 9 (8.6) 7 (6.65) 15 (14.18) 2 (13.41) 17 (15.96) 13 (9.98) 13 (12.18) Μ.Τ.±Τ.Σ ± ± ±1.17 Σ Μ.Τ.±Τ.Σ ± ±1. 8.4±1.13 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test)
175 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝDΧ) για το χρωµόσωµα Χ και απώλειας του ίδιου χρωµοσώµατος (MNΧ + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων ώριµης ηλικίας ( 45 ετών), που φέρουν αντίστοιχα δύο και τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 2: ΝDΧ (%) Β.Α. ( ) Κ.Γ. ( ) Σ.Ν. ( ) Χ.Β. ( ) Γ.Χ. ( ) Γ.Θ. ( ) Ι.Γ. ( ) Σ.Κ. ( ) ΠΜΝΧ + ( ) 11 (1.14) 16 (15.56) 1 (8.29) 7 (4.74) 9 (9.) 9 (4.95) 13 (8.19) 7 (4.92) ΜΝΧ + ( ) 11 (1.14) 17 (16.54) 1 (8.29) 7 (4.74) 9 (9.) 9 (4.95) 13 (8.19) 8 (5.62) Μ.Τ.±Τ.Σ ± ± ±1.35 2:2 3:1 4: Κ.Σ. ( ) Λ.Μ. ( ) Ν.Μ. ( ) Π.Ε. ( ) Σ.Σ. ( ) Σ.Γ. ( ) Χ.Ε. ( ) Χ.Α. ( ) (22.14) 15 (13.71) 13 (12.4) 2 (15.61) 2 (15.17) 26 (21.98) 15 (11.39) 2 (17.79) 29 (24.7) 16 (14.62) 14 (12.96) 23 (17.95) 22 (16.69) 27 (22.82) 16 (12.15) 22 (19.57) Μ.Τ.±Τ.Σ ± ± ±1.59 Σ Μ.Τ.±Τ.Σ , ± ± , ±1.56 p<.5 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
176 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Χρωµόσωµα Υ. Η µελέτη ανευπλοειδικών φαινοµένων που αφορούν το χρωµόσωµα Υ στην οµάδα αρσενικών ατόµων ώριµης ηλικίας περιγράφεται αναλυτικά στον πίνακα Παρουσία της ελάχιστης (5 µg/ml) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ παρατηρείται αύξηση στις συχνότητες του χρωµοσωµατικού µη αποχωρισµού από.59±.9 στις καλλιέργειες-µάρτυρες σε.82±.8 και 1.43±.17 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα προκαλεί η µέγιστη συγκέντρωση HCTZ. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν µη αποχωρισµό του χρωµοσώµατος Υ, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Στον ίδιο πίνακα από τον προσδιορισµό της απώλειας του χρωµοσώµατος Υ η επίδραση της ελάχιστης συγκέντρωσης HCTZ προκαλεί σχετικά µικρή αύξηση στην παρουσία του χρωµοσώµατος σε µικροπυρήνες µε αποτέλεσµα να µην παρουσιάζεται στατιστικά σηµαντική διαφορά στις αντίστοιχες συχνότητες (2.34±.29) σε σχέση µε τις καλλιέργειες-µάρτυρες (1.81±.37). Αντίθετα, η αύξηση που παρατηρείται στη συγκέντρωση 4 µg/ml HCTZ (4.69±1.3) είναι σαφώς µεγαλύτερη σε σχέση µε τις καλλιέργειες-µάρτυρες και η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα της µέγιστης συγκέντρωσης να επάγει απώλεια του χρωµοσώµατος Υ. Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος Y, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού είναι υψηλότερες από τις αντίστοιχες της χρωµοσωµατικής απώλειας, γεγονός που ισχύει τόσο για τις καλλιέργειες-µάρτυρες όσο και για τις καλλιέργειες λεµφοκυττάρων στις οποίες πραγµατοποιήθηκε επίδραση µε τις δύο συγκεντρώσεις του HCTZ. Επιπρόσθετα, παρατηρούνται υψηλότερες συχνότητες ανευπλοειδικών φαινοµένων, µη αποχωρισµού και απώλειας, για το χρωµόσωµα Υ στα ηλικιωµένα αρσενικά άτοµα σε σύγκριση µε τα νεαρά
177 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες τυχαίου και επαγόµενου µη αποχωρισµού (ΝDΥ) για το χρω- µόσωµα Υ και απώλειας του ίδιου χρωµοσώµατος (ΜΝΥ + ) σε διπύρηνα κύτταρα αρσενικών ατόµων ώριµης ηλικίας ( 45 ετών), που φέρουν δύο σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ότες ιπύρηνα κύτταρα που ιπλοειδή κύτταρα αναλύθηκαν 1:1 2: ΜΑΡΤΥΡΕΣ ΝDY (%) Β.Α. ( ) Χ.Β. ( ) Γ.Χ. ( ) Γ.Θ. ( ) Ι.Γ. ( ) Σ.Κ. ( ) ΠΜΝY + ( ) 2 (1.88) 1 (.97) 7 (3.45) 1 (1.) 2 (1.6) 2 (1.96) ΜΝY + ( ) 2 (1.88) 1 (.97) 7 (3.45) 1 (1.) 2 (1.6) 2 (1.96) Μ.Τ.±Τ.Σ ± ± ±.37 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml Β.Α. ( ) Χ.Β. ( ) Γ.Χ. ( ) Γ.Θ. ( ) Ι.Γ. ( ) Σ.Κ. ( ) (3.) 2 (1.98) 4 (3.12) 2 (1.39) 1 (.91) 2 (1.8) 3 (3.) 2 (1.98) 4 (3.12) 2 (1.39) 2 (1.83) 3 (2.7) Μ.Τ.±Τ.Σ ±.8 2.3± ±.29 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml Β.Α. ( ) Χ.Β. ( ) Γ.Χ. ( ) Γ.Θ. ( ) Ι.Γ. ( ) Σ.Κ. ( ) (4.98) 5 (3.62) 11 (9.33) 3 (2.33) 6 (3.71) 3 (2.92) 5 (4.98) 5 (3.62) 11 (9.33) 3 (2.33) 8 (4.94) 3 (2.92) Μ.Τ.±Τ.Σ , ± ± ±1.3 p<.5 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
178 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Χρωµόσωµα 8. Η µελέτη ανευπλοειδικών φαινοµένων που αφορούν το χρωµόσωµα 8 στην οµάδα ατόµων ώριµης ηλικίας περιγράφεται αναλυτικά στους πίνακες Παρουσία της ελάχιστης (5 µg/ml) (πίνακας 14.13) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ (πίνακας 14.14) παρατηρείται αύξηση στις συχνότητες του χρωµοσωµατικού µη αποχωρισµού από.52±.5 στις καλλιέργειες-µάρτυρες (πίνακας 14.12) σε 1.14±.9 και 2.21±.17 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα (.51±.9,.53±.7) προκαλεί η µέγιστη συγκέντρωση HCTZ (2.2±.17, 2.22±.27) τόσο στα αρσενικά όσο και στα θηλυκά άτοµα αντίστοιχα. Ο συνηθέστερος τύπος λανθασµένου αποχωρισµού για το χρωµόσωµα 8 είναι ο απλός µη αποχωρισµός ανεξάρτητα από το φύλο ή από την επίδραση HCTZ, γεγονός που ισχύει και για τα φυλετικά χρωµοσώµατα. Η εµφάνιση του διπλού µη αποχωρισµού για το χρωµοσώµα 8 σε αρσενικά και θηλυκά άτοµα παρατηρείται σε πολύ µικρότερες συχνότητες σε σχέση µε το χρωµόσωµα Χ στα θηλυκά άτοµα. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν µη αποχωρισµό του χρωµοσώµατος 8, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Στους ίδιους πίνακες προσδιορίζεται η απώλεια του χρωµοσώµατος 8, η οποία ανιχνεύεται από την παρουσία σήµατος υβριδοποίησης στο µικροπυρήνα. Στα ηλικιωµένα άτοµα, σε αντίθεση µε τα νεαρά αρσενικά και θηλυκά άτοµα, παρατηρείται στις καλλιέργειες-µάρτυρες η παρουσία του χρωµοσώµατος 8 σε µικροπυρήνες. Η επίδραση της ελάχιστης συγκέντρωσης HCTZ (.51±.23) φαίνεται να µην επηρεάζει την απώλεια του χρωµοσώµατος 8, γεγονός το οποίο ισχύει και για τα νεαρά άτοµα. Αντίθετα, ενώ η αύξηση των ΠΜΝ8 + και ΜΝ8 + που παρατηρείται στη συγκέντρωση 4 µg/ml HCTZ (1.7±.32) είναι σχετικά µεγαλύτερη σε σχέση µε τις αντίστοιχες συχνότητες στις καλλιέργειες µάρτυρες (.72±.22) η στατιστική ανάλυση που εφαρµόσθηκε έδειξε πως δεν υπάρχει σηµαντική διαφορά, γεγονός το οποίο δεν ισχύει για τα νεαρά άτοµα. Οι παρατηρήσεις οι οποίες αφορούν το µη αποχωρισµό και την απώλεια του χρωµοσώµατος 8 στο σύνολο των 13 ηλικιωµένων δοτών που µελετήθηκαν ισχύουν και για τα δύο φύλα. Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος 8, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού στις οµάδες που απαρτίζονται από τα ηλικιωµένα αρσενικά και θηλυκά άτοµα είναι υψηλότερες από τις αντίστοιχες της χρωµοσωµατικής απώλειας για κάθε πειραµατικό σηµείο που ελέγχθηκε. Επιπρόσθετα, παρατηρούνται υψηλότερες συχνότητες µη αποχωρισµού για το χρωµόσωµα 8 στα ηλικιωµένα άτοµα σε σύγκριση µε τα νεαρά, γεγονός το οποίο φαίνεται να ισχύει και για τη χρωµοσωµατική απώλεια µε εξαίρεση τις καλλιέργειες λεµφοκυττάρων θηλυκών ατόµων στις οποίες πραγµατοποιήθηκε επίδραση µε τη µέγιστη συγκέντρωση HCTZ
179 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες τυχαίου µη αποχωρισµού (ΝD8) για το χρωµόσωµα 8 και απώλειας του ίδιου χρωµοσώµατος (ΜΝ8 + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων ώριµης ηλικίας ( 45 ετών), που φέρουν τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. ΜΑΡΤΥΡΕΣ ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 3:1 4: ΝD8 (%) Β.Α. ( ) Κ.Γ. ( ) Σ.Ν. ( ) Χ.Β. ( ) Γ.Χ. ( ) ΠΜΝ8 + ( ) () 1 (.88) 3 (2.84) () 1 (.94) ΜΝ8 + ( ) () 1 (.88) 3 (2.84) () 1 (.94) Μ.Τ.±Τ.Σ ±.9.93±.52.93±.52 Κ.Σ. ( ) Λ.Μ. ( ) Ν.Μ. ( ) Π.Ε. ( ) Σ.Σ. ( ) Σ.Γ. ( ) Χ.Ε. ( ) Χ.Α. ( ) (.94) () 1 (.9) 3 (1.41) () 1 (.69) 1 (.8) () 1 (.94) () 1 (.9) 3 (1.41) () 1 (.69) 1 (.8) () Μ.Τ.±Τ.Σ ±.7.59±.18.59±.18 Σ Μ.Τ.±Τ.Σ ±.5.72±.22.72±
180 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝD8) για το χρωµόσωµα 8 και απώλειας του ίδιου χρωµοσώµατος (ΜΝ8 + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων ώριµης ηλικίας ( 45 ετών), που φέρουν τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 3:1 4: ΝD8 (%) Β.Α. ( ) Κ.Γ. ( ) Σ.Ν. ( ) Χ.Β. ( ) Γ.Χ. ( ) ΠΜΝ8 + ( ) 1 (.98) () 3 (2.88) () () ΜΝ8 + ( ) 1 (.98) () 3 (2.88) () () Μ.Τ.±Τ.Σ ±.13.77±.56.77±.56 Κ.Σ. ( ) Λ.Μ. ( ) Ν.Μ. ( ) Π.Ε. ( ) Σ.Σ. ( ) Σ.Γ. ( ) Χ.Ε. ( ) Χ.Α. ( ) () () 1 (1.) () 1 (.88) () () 1 (.95) () () 1 (1.) () 1 (.88) () () 1 (.95) Μ.Τ.±Τ.Σ ±.13.35±.17.35±.17 Σ Μ.Τ.±Τ.Σ ±.9.51±.23.51±.23 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test)
181 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝD8) για το χρωµόσωµα 8 και απώλειας του ίδιου χρωµοσώµατος (ΜΝ8 + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων ώριµης ηλικίας ( 45 ετών), που φέρουν τέσσερα σήµατα υβριδοποίησης σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro, µετά από επίδραση υδροχλωροθειαζιδίου (HCTZ). ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 3:1 4: ΝD8 (%) Β.Α. ( ) Κ.Γ. ( ) Σ.Ν. ( ) Χ.Β. ( ) Γ.Χ. ( ) ΠΜΝ8 + ( ) 1 (.97) () 3 (2.55) () 1 (.96) ΜΝ8 + ( ) 1 (.97) () 3 (2.55) () 1 (.96) Μ.Τ.±Τ.Σ ±.17.9±.46.9±.46 Κ.Σ. ( ) Λ.Μ. ( ) Ν.Μ. ( ) Π.Ε. ( ) Σ.Σ. ( ) Σ.Γ. ( ) Χ.Ε. ( ) Χ.Α. ( ) (1.84) () 1 (.99) 2 (1.94) 1 (.94) 1 (.82) () 2 (1.97) 2 (1.84) () 1 (.99) 2 (1.94) 1 (.94) 1 (.82) () 3 (2.96) Μ.Τ.±Τ.Σ ± ± ±.35 Σ Μ.Τ.±Τ.Σ , ±.17 1.± ±.28 p<.1 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.1 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
182 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Προκειµένου να διερευνηθεί πληρέστερα η ανευπλοειδογόνος δράση του υδροχλωροθειαζιδίου, η οποία εκτιµάται από την ανάλυση των βιολογικών δεικτών που προσδιορίσθηκαν, κρίθηκε σκόπιµο να συνυπολογισθούν παράµετροι όπως ηλικία, φύλο, συνήθεια καπνίσµατος που δύναται να τους επηρεάσουν σε µικρότερο ή µεγαλύτερο βαθµό. Με το σχεδιασµό ενός γενικού γραµµικού µοντέλου (πίνακας 14.15) επιτυγχάνεται µία συνολική εκτίµηση των ανευπλοειδικών φαινοµένων, χρωµοσωµατικής απώλειας (πίνακας 14.15α) και µη αποχωρισµού (πίνακας 14.15β) των χρωµοσωµάτων Χ, Υ και 8. Στην πολυπαραγοντική ανάλυση παλινδρόµησης που ακολουθεί οι βιολογικοί δείκτες που αναφέρονται στην απώλεια -Χ + ΜΝ, Υ + ΜΝ, 8 + ΜΝ- και στο µη αποχωρισµό -NDX, NDY, ND 8- των χρωµοσωµάτων Χ, Υ και 8 λαµβάνονται ως εξαρτηµένες µεταβλητές και οι συγκεκριµένοι παράγοντες ως ανεξάρτητες. Ο παράγοντας κάπνισµα δεν αξιολογήθηκε στην εκτίµηση ανευπλοειδικών φαινοµένων για το χρωµόσωµα Υ διότι ο αριθµός των καπνιστών που συµπεριελήφθησαν στη µελέτη είναι εξαιρετικά µικρός. Συγκεκριµένα σε σύνολο 11 ατόµων, 5 νεαρών και 6 ηλικιωµένων, 9 είναι µη καπνιστές και 2 είναι καπνιστές. Πίνακας 14.15α. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών X + MN, Y + MN και 8 + MN που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο, το κάπνισµα και την επίδραση του HCTZ ως ανεξάρτητες µεταβλητές. ΧΡΩΜΟΣΩΜΑΤΙΚΗ ΑΠΩΛΕΙΑ X + MN Y + MN 8 + MN Ηλικία <.1 <.1.67 Φύλο < Κάπνισµα Επίδραση HCTZ < R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.737 P<.1 R 2 =.546 P<.1 R 2 =.163 P=.8 Σχετικά µε το φαινόµενο της χρωµοσωµατικής απώλειας, από τα αποτελέσµατα της ανάλυσης όσον αφορά την απώλεια του χρωµοσώµατος Χ φαίνεται η σηµαντική εξάρτηση (P<.1) του δείκτη Χ + ΜΝ από τους παράγοντες ηλικία, φύλο και επίδραση HCTZ. Η τιµή του συντελεστή προσδιορισµού R 2 ισούται µε.737, δηλαδή το 73.7% της συνολικής µεταβλητότητας του Χ + ΜΝ ερµηνεύεται από τις µεταβλητές ηλικία, φύλο και επίδραση HCTZ, σύµφωνα µε το προτεινόµενο µοντέλο. Σε αυτό το µοντέλο η τιµή R 2 είναι αρκετά κοντά στη µονάδα γεγονός που αποδίδει µία ισχυρή γραµµική συσχέτιση µεταξύ της εξαρτηµένης µεταβλητής Χ + ΜΝ και των παραµέτρων που εξετάσθηκαν. Επιπρόσθετα, ο έλεγχος ανάλυσης διασποράς (ANOVA) έδειξε F = 67.4 µε σηµαντικότητα P<.1. Στην εκτίµηση της απώλειας του χρωµοσώµατος Υ σε αρσενικά άτοµα η ηλικία (P<.1) και η επίδραση HCTZ (P =.6) θεωρούνται οι κύριοι παράγοντες που επηρεάζουν τη συχνότητα Υ + ΜΝ. Η απώλεια του χρωµοσώµατος 8 επάγεται από την επίδραση HCTZ (P =.7), ενώ δεν φαίνεται να επηρεάζεται από την ηλικία (P =.67) και το φύλο (P =.374). Η συνήθεια του καπνίσµατος δεν επάγει απώλεια των χρω- µοσωµάτων Χ (P =.246) και 8 (P =.55)
183 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας 14.15β. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών NDX, NDY και ND8 που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο, το κάπνισµα και την επίδραση του HCTZ ως ανεξάρτητες µεταβλητές. ΜΗ ΑΠΟΧΩΡΙΣΜΟΣ NDX NDY ND8 Ηλικία.1 <.1.8 Φύλο Κάπνισµα Επίδραση HCTZ <.1 <.1 <.1 R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.746 P<.1 R 2 =.738 P<.1 R 2 =.767 P<.1 Σχετικά µε το φαινόµενο του χρωµοσωµατικού µη αποχωρισµού, από τα αποτελέσµατα της ανάλυσης όσον αφορά το µη αποχωρισµό του χρωµοσώµατος Χ φαίνεται η σηµαντική εξάρτηση του δείκτη NDΧ από τους παράγοντες ηλικία, κάπνισµα και επίδραση HCTZ. H ηλικία (P =.1) και η επίδραση HCTZ (P<.1) επηρεάζουν τη συχνότητα µη αποχωρισµού σε µεγαλύτερο βαθµό σε σύγκριση µε τη συνήθεια του καπνίσµατος (P =.12). Αντίθετα, το φύλο (P =.244) δεν θεωρείται παράγοντας που δύναται να επηρεάσει τη συχνότητα µη αποχωρισµού για το χρωµόσωµα Χ. Στην εκτίµηση του µη αποχωρισµού για το χρωµόσωµα Υ σε αρσενικά άτοµα η ηλικία (P<.1) και η επίδραση HCTZ (P<.1) θεωρούνται οι κύριοι παράγοντες που επηρεάζουν τη συχνότητα NDY. Όσον αφορά το µη αποχωρισµό του χρωµοσώµατος 8 φαίνεται η σηµαντική εξάρτηση του δείκτη ND8 από τους παράγοντες ηλικία (P =.8) και επίδραση HCTZ (P<.1). Αντίθετα, το φύλο (P =.199) και το κάπνισµα (P =.87) δεν επηρεάζουν τη συχνότητα µη αποχωρισµού για το χρωµόσωµα 8. Επιπρόσθετα, η τιµή που λαµβάνει ο συντελεστής προσδιορισµού R 2 για τις εξαρτηµένες µεταβλητές NDX, NDY και ND8 είναι αρκετά κοντά στη µονάδα (.746,.738,.767) γεγονός που αποδίδει µία ισχυρή γραµµική συσχέτιση ως προς τις παραµέτρους που εξετάσθηκαν. Το γενικό γραµµικό µοντέλο που περιγράφηκε στην προηγούµενη παράγραφο (πίνακας 14.15) βασίζεται σε µία συνολική εκτίµηση των βιολογικών δεικτών από τις παραµέτρους που δύναται να τους επηρεάσουν και σε αυτές περιλαµβάνεται η επίδραση του υδροχλωροθειαζιδίου ως ανεξάρτητη µεταβλητή. Με το σχεδιασµό ενός δεύτερου γραµµικού µοντέλου (πίνακας 14.16) µελετήθηκε η επίδραση των παραγόντων ηλικία, φύλο και συνήθεια του καπνίσµατος που λαµβάνονται ως ανεξάρτητες µεταβλητές στην ικανότητα του HCTZ να επάγει απώλεια των χρωµοσω- µάτων Χ, Υ και 8. Για την πραγµατοποίηση του εκτιµήθηκε η επίδραση της ηλικίας, του φύλου και της συνήθειας του καπνίσµατος στην τυχαία επαγωγή χρωµοσωµατικής απώλειας στις καλλιέργειες-µάρτυρες (πίνακας 14.16α). Στη συνέχεια υπολογίσθηκαν οι διαφορές στην επαγωγή των Χ + ΜΝ, Υ + ΜΝ και 8 + ΜΝ µετά από επίδραση µε HCTZ. Σε κάθε χρωµόσωµα οι νέες συχνότητες προκύπτουν αφαιρώντας, για κάθε δότη, τη συχνότητα των µικροπυρήνων που παρατηρήθηκε στο µάρτυρα από την αντίστοιχη συχνότητα µικροπυρήνων που παρατηρήθηκε στη µέγιστη συγκέντρωση HCTZ. Η πολυπαραγοντική ανάλυση παλινδρόµησης εφαρµόσθηκε στις νέες συχνότητες απώλειας των χρωµοσωµάτων Χ, Υ και 8 και εξετάσθηκε ο βαθµός στον οποίον οι παράγοντες που προαναφέρθηκαν τις επηρεάζουν (πίνακας 14.16β)
184 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας 14.16α. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών X + MN, Y + MN και 8 + MN που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο και το κάπνισµα ως ανεξάρτητες µεταβλητές σε καλλιέργειες-µάρτυρες. ΧΡΩΜΟΣΩΜΑΤΙΚΗ ΑΠΩΛΕΙΑ X + MN Y + MN 8 + MN Ηλικία < Φύλο < Κάπνισµα R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.73 P<.1 R 2 =.49 P=.9 R 2 =.281 P=.1 Σε καλλιέργειες-µάρτυρες η ηλικία (P<.1) και το φύλο (P<.1) θεωρούνται οι κύριοι παράγοντες που επάγουν την απώλεια του χρωµοσώµατος Χ. Η ηλικία (P =.9, P =.1) παραµένει σηµαντικός παράγοντας που επηρεάζει σε µικρότερη έκταση την απώλεια των χρωµοσωµάτων Υ και 8. Όσον αφορά τη συνήθεια του καπνίσµατος (P =.428, P =.419) δεν επάγει το σχηµατισµό µικροπυρήνων που θεωρούνται προϊόν απώλειας των χρωµοσωµάτων Χ και 8. Το φύλο (P =.49) δεν επηρεάζει την απώλεια του χρωµοσώµατος 8. Πίνακας 14.16β. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών X + MN, Y + MN και 8 + MN που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο και το κάπνισµα ως ανεξάρτητες µεταβλητές σε καλλιέργειες µετά από επίδραση µε HCTZ. ΧΡΩΜΟΣΩΜΑΤΙΚΗ ΑΠΩΛΕΙΑ X + MN Y + MN 8 + MN Ηλικία Φύλο Κάπνισµα R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.281 P=.6 R 2 =.462 P=.13 R 2 =.148 P=.72 Η ανάλυση των βιολογικών δεικτών που επαναπροσδιορίσθηκαν σε καλλιέργειες στις οποίες πραγµατοποιήθηκε επίδραση µε τη µέγιστη συγκέντρωση HCTZ έδειξε ότι η ηλικία (P =.9, P=.13) αποτελεί τον κύριο παράγοντα που επηρεάζει την απώλεια των χρωµοσωµάτων Χ και Υ. Το φύλο (P=.321) και το κάπνισµα (P=.522) δεν επάγουν απώλεια του χρωµοσώµατος Χ. Όσον αφορά την απώλεια του χρωµοσώµατος 8 δεν φαίνεται να επηρεάζεται από καµία από τις παραµέτρους που εκτιµήθηκαν. Από τη σύγκριση των αποτελεσµάτων που προκύπτουν µετά το σχεδιασµό του δεύτερου γραµµικού µοντέλου (πίνακας 14.16α, 14.16β) διαφαίνεται ότι η ηλικία επηρεάζει καθοριστικά την ικανότητα της φαρµακευτικής ένωσης να επάγει απώλεια των χρωµοσωµάτων Χ και Υ. Με το σχεδιασµό ενός τρίτου γραµµικού µοντέλου (πίνακας 14.17) µελετήθηκε η επίδραση των παραγόντων ηλικία, φύλο και συνήθεια του καπνίσµατος που λαµβάνονται ως ανεξάρτητες µεταβλητές στην ικανότητα του HCTZ να επάγει µη αποχωρισµό των χρωµοσωµάτων Χ, Υ και 8. Για την πραγµατοποίηση του εκτιµήθηκε η επίδραση της ηλικίας, του φύλου και της συνήθειας του καπνίσµατος στην τυχαία επαγωγή χρωµοσωµατικού µη αποχωρισµού στις καλλιέργειες
185 Αποτελέσµατα Ανευπλοειδικά φαινόµενα µάρτυρες (πίνακας 14.17α).Στη συνέχεια υπολογίσθηκαν οι διαφορές στην επαγωγή των συχνοτήτων NDX, NDY και ND8 µετά από επίδραση µε HCTZ. Σε κάθε χρω- µόσωµα και σε κάθε συγκέντρωση υδροχλωροθειαζιδίου, ελάχιστη και µέγιστη, οι νέες συχνότητες προκύπτουν αφαιρώντας, για κάθε δότη, τη συχνότητα µη αποχωρισµού που παρατηρήθηκε στο µάρτυρα από την αντίστοιχη συχνότητα µη αποχωρισµού που παρατηρήθηκε παρουσία HCTZ. Η πολυπαραγοντική ανάλυση παλινδρόµησης εφαρµόσθηκε στις νέες συχνότητες µη αποχωρισµού των χρωµοσωµάτων Χ, Υ και 8 και εξετάσθηκε ο βαθµός στον οποίον οι παράγοντες που προαναφέρθηκαν τις επηρεάζουν (πίνακας 14.17β). Πίνακας 14.17α. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών NDX, NDY και ND8 που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο και το κάπνισµα ως ανεξάρτητες µεταβλητές σε καλλιέργειες-µάρτυρες. ΜΗ ΑΠΟΧΩΡΙΣΜΟΣ NDX NDY ND8 Ηλικία < Φύλο Κάπνισµα R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.574 P<.1 R 2 =.71 P=.8 R 2 =.39 P=.1 Σε καλλιέργειες-µάρτυρες η ηλικία (P<.1, P =.8, P =.2) αποτελεί τον κύριο παράγοντα που επηρεάζει το µη αποχωρισµό των χρωµοσωµάτων Χ, Υ και 8. Το φύλο και η συνήθεια του καπνίσµατος δεν επάγουν χρωµοσωµατικό µη αποχωρισµό. Πίνακας 14.17β. Πολυπαραγοντική ανάλυση παλινδρόµησης των βιολογικών δεικτών NDX, NDY και ND8 που µελετήθηκαν σε σχέση µε την ηλικία, το φύλο και το κάπνισµα ως ανεξάρτητες µεταβλητές σε καλλιέργειες µετά από επίδραση µε HCTZ. ΜΗ ΑΠΟΧΩΡΙΣΜΟΣ NDX NDY ND8 Ηλικία Φύλο Κάπνισµα R 2 (Συντελεστής προσδιορισµού) Σηµαντικότητα R 2 =.12 P=.388 R 2 =-.49 P=.924 R 2 =-.29 P=.79 Η ανάλυση των βιολογικών δεικτών που επαναπροσδιορίσθηκαν σε καλλιέργειες στις οποίες πραγµατοποιήθηκε επίδραση HCTZ έδειξε ότι καµία από τις παραµέτρους που εκτιµήθηκαν δεν δύναται να τροποποιήσει την ικανότητα της φαρµακευτικής ένωσης να επάγει χρωµοσωµατικό µη αποχωρισµό. Από τη σύγκριση των αποτελεσµάτων που προκύπτουν µετά το σχεδιασµό του τρίτου γραµµικού µοντέλου (πίνακας 14.17α, 14.17β) διαφαίνεται ότι ενώ η ηλικία επηρεάζει καθοριστικά την τυχαία επαγωγή χρωµοσωµατικού µη αποχωρισµού δεν επηρεάζει την ικανότητα της φαρµακευτικής ένωσης να επάγει µη αποχωρισµό των χρωµοσω- µάτων Χ, Υ και
186 Αποτελέσµατα Ανευπλοειδικά φαινόµενα 14.2 Μη διεγερµένα λεµφοκύτταρα Ο προσδιορισµός ανευπλοειδικών φαινοµένων, µη αποχωρισµού των χρω- µοσωµάτων και χρωµοσωµατικής απώλειας, διερευνήθηκε σε καλλιέργειες µη διεγερµένων λεµφοκυττάρων µε τη συµµετοχή των χρωµοσωµάτων Χ (πίνακας 14.18) και 8 (πίνακας 14.19) για τους λόγους που αναφέρονται στο υποκεφάλαιο Η παρούσα µελέτη αφορά τους ίδιους δότες στους οποίους προσδιορίσθηκε ο µηχανισµός προέλευσης των µικροπυρήνων σε καλλιέργειες µη διεγερµένων λεµφοκυττάρων (πίνακας 13.5). Πρόκειται για άτοµα νεαρής ηλικίας, τα οποία έχουν συµπεριληφθεί στα πειράµατα αποσαφήνισης της ανευπλοειδογόνου δράσης του υδροχλωροθειαζιδίου σε καλλιέργειες διεγερµένων λεµφοκυττάρων µε τη συµµετοχή των χρωµοσωµάτων Χ (πίνακες ) και 8 (πίνακες ) Χρωµόσωµα Χ Στις συχνότητες µη αποχωρισµού του χρωµοσώµατος Χ παρατηρείται αύξηση παρουσία της ελάχιστης (5 µg/ml) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ, από.45±.6 στις καλλιέργειες-µάρτυρες σε 2.27±.26 και 3.62±.24 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα προκαλεί η µέγιστη συγκέντρωση HCTZ γεγονός που ισχύει τόσο για τα φαινόµενα απλού όσο και για το φαινόµενο διπλού µη αποχωρισµού. Θα πρέπει να αναφερθεί ότι ο απλός µη αποχωρισµός συνεχίζει να αποτελεί το συνηθέστερο τύπο µη αποχωρισµού που παρατηρήθηκε. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν χρωµοσωµατικό µη αποχωρισµό, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Οι παρατηρήσεις που αφορούν το µη αποχωρισµό του χρωµοσώµατος Χ στο σύνολο των 6 νεαρών δοτών που µελετήθηκαν ισχύουν τόσο για τα θηλυκά όσο και για τα αρσενικά άτοµα. Στον ίδιο πίνακα προσδιορίζεται η απώλεια του χρωµοσώµατος Χ και παρατηρείται η επίδραση της ελάχιστης συγκέντρωσης HCTZ να προκαλεί σχετικά µικρή αύξηση στην παρουσία του χρωµοσώµατος Χ σε διπύρηνα κύτταρα µε µικροπυρήνες ( ΠΜΝΧ + ) και στους µικροπυρήνες (ΜΝΧ + ) µε αποτέλεσµα να µην παρουσιάζεται στατιστικά σηµαντική διαφορά στις αντίστοιχες συχνότητες (1.55±.44) σε σχέση µε τις καλλιέργειες-µάρτυρες (1.31±.48). Αντίθετα, η αύξηση που παρατηρείται στη συγκέντρωση 4 µg/ml HCTZ (3.42±.77) είναι σαφώς µεγαλύτερη σε σχέση µε τις καλλιέργειες-µάρτυρες και η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα της µέγιστης συγκέντρωσης HCTZ να επάγει απώλεια του χρωµοσώµατος Χ. Σχετικά µε το φαινόµενο της παρουσίας του χρωµοσώµατος Χ σε µικροπυρήνες φαίνεται να συµβαίνει µε µεγαλύτερη ένταση στα θηλυκά άτοµα σε σχέση µε τα αρσενικά, τόσο για τις καλλιέργειες-µάρτυρες (2.18±.43,.43±.43) όσο και για τις καλλιέργειες λεµφοκυττάρων στις οποίες πραγµατοποιήθηκε επίδραση µε τις δύο συγκεντρώσεις του HCTZ, 5 µg/ml (2.44±.29,.66±.33) και 4 µg/ml (4.96±.56, 1.89±.53). Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος Χ, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού στις οµάδες που απαρτίζονται από τα νεαρά αρσενικά και θηλυκά άτοµα είναι υψηλότερες από τις αντίστοιχες της χρω- µοσωµατικής απώλειας για κάθε πειραµατικό σηµείο που ελέγχθηκε. Τα παραπάνω αποτελέσµατα συµφωνούν µε αυτά που προκύπτουν από τις καλλιέργειες διεγερµένων λεµφοκυττάρων ατόµων νεαρής ηλικίας
187 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝDΧ) για το χρωµόσωµα Χ και απώλειας του ίδιου χρωµοσώµατος (MNΧ + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν αντίστοιχα δύο και τέσσερα σήµατα υβριδοποίησης σε µη διεγερµένες µε PHA κατά την επίδραση µε υδροχλωροθειαζίδιο (HCTZ) καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro. ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα ΜΑΡΤΥΡΕΣ 1:1 2: ΝDΧ (%) ΠΜΝΧ + ( ) ΜΝΧ + ( ) Α.Π. ( ) () () Κ.Γ. ( ) (1.29) 2 (1.29) Σ.Η. ( ) () () Μ.Τ.±Τ.Σ ±.11.43±.43.43±.43 2:2 3:1 4: Κ.Α. ( ) (3.) 3 (3.) Ν.Κ. ( ) (2.) 2 (2.) Σ.Κ. ( ) (1.55) 2 (1.55) Μ.Τ.±Τ.Σ ± ± ±.43 Σ Μ.Τ.±Τ.Σ ± ± ±.48 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml 1:1 2: Α.Π. ( ) (.99) 1 (.99) Κ.Γ. ( ) (.99) 1 (.99) Σ.Η. ( ) () () Μ.Τ.±Τ.Σ ±.55.66±.33.66±.33 2:2 3:1 4: Κ.Α. ( ) (2.98) 3 (2.98) Ν.Κ. ( ) (1.99) 2 (1.99) Σ.Κ. ( ) (2.35) 3 (2.35) Μ.Τ.±Τ.Σ ± ± ±.29 Σ Μ.Τ.±Τ.Σ ± ± ±.44 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml 1:1 2: Α.Π. ( ) (1.) 1 (1.) Κ.Γ. ( ) (2.82) 4 (2.82) Σ.Η. ( ) (1.85) 2 (1.85) Μ.Τ.±Τ.Σ ± ± ±.53 2:2 3:1 4: Κ.Α. ( ) (5.93) 6 (5.93) Ν.Κ. ( ) (4.96) 5 (4.96) Σ.Κ. ( ) (3.98) 5 (3.98) Μ.Τ.±Τ.Σ ± ± ±.56 Σ Μ.Τ.±Τ.Σ , ± ± ±.77 p<.5 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p<.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
188 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Χρωµόσωµα 8 Στις συχνότητες µη αποχωρισµού του χρωµοσώµατος 8 παρατηρείται αύξηση παρουσία της ελάχιστης (5 µg/ml) και µέγιστης συγκέντρωσης (4 µg/ml) HCTZ, από.21±.4 στις καλλιέργειες-µάρτυρες σε.8±.9 και 1.51±.12 αντίστοιχα. Τη µεγαλύτερη αύξηση σε σχέση µε το µάρτυρα (.24±.8,.18±.4) προκαλεί η µέγιστη συγκέντρωση HCTZ (1.64±.23, 1.38±.7) τόσο στα αρσενικά όσο και στα θηλυκά άτοµα αντίστοιχα. Ο συνηθέστερος τύπος λανθασµένου αποχωρισµού για το χρωµόσωµα 8 είναι ο απλός µη αποχωρισµός ανεξάρτητα από το φύλο ή από την επίδραση HCTZ, γεγονός που ισχύει και για το φυλετικό χρωµόσωµα Χ. Η εµφάνιση του διπλού µη αποχωρισµού για το χρωµοσώµα 8 σε αρσενικά και θηλυκά άτοµα παρατηρείται σε πολύ µικρότερες συχνότητες σε σχέση µε το χρωµόσωµα Χ στα θηλυκά άτοµα. Η στατιστική ανάλυση έδειξε τη σηµαντική ικανότητα των δύο συγκεντρώσεων HCTZ να επάγουν µη αποχωρισµό του χρωµοσώµατος 8, ενώ ταυτόχρονα απεκάλυψε τη σηµαντική διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Οι παρατηρήσεις που αφορούν το µη αποχωρισµό του χρωµοσώµατος 8 στο σύνολο των 6 νεαρών δοτών που µελετήθηκαν ισχύουν τόσο για τα αρσενικά όσο και για τα θηλυκά άτοµα. Στον ίδιο πίνακα προσδιορίζεται η απώλεια του χρωµοσώµατος 8 και στα νεαρά άτοµα ανεξάρτητα από το φύλο δεν παρατηρείται στις καλλιέργειες-µάρτυρες παρουσία του χρωµοσώµατος 8 τόσο στα διπύρηνα κύτταρα µε µικροπυρήνες ( ΠΜΝ8 + ) όσο και στους µικροπυρήνες (ΜΝ8 + ) µε αντίστοιχες µηδενικές συχνότητες. Η επίδραση της ελάχιστης συγκέντρωσης HCTZ φαίνεται να µην επηρεάζει την απώλεια του χρωµοσώµατος 8. Αντίθετα, η αύξηση των ΠΜΝ8 + και ΜΝ8 + που παρατηρείται στη συγκέντρωση 4 µg/ml HCTZ (.44±.2) είναι µεγαλύτερη σε σχέση µε τις αντίστοιχες συχνότητες στις καλλιέργειες µάρτυρες. Η στατιστική ανάλυση έδειξε την οριακή ικανότητα της µέγιστης συγκέντρωσης HCTZ να επάγει απώλεια του χρωµοσώµατος 8, ενώ ταυτόχρονα απεκάλυψε την οριακή διαφορά που υπάρχει στις καλλιέργειες HCTZ 4 µg/ml σε σχέση µε τις καλλιέργειες HCTZ 5 µg/ml. Αξιοσηµείωτη είναι η παρατήρηση, που προκύπτει από τη σύγκριση των συχνοτήτων µη αποχωρισµού και απώλειας του χρωµοσώµατος 8, σύµφωνα µε την οποία οι συχνότητες µη αποχωρισµού στις οµάδες που απαρτίζονται από τα νεαρά αρσενικά και θηλυκά άτοµα είναι υψηλότερες από τις αντίστοιχες της χρωµοσω- µατικής απώλειας για κάθε πειραµατικό σηµείο που ελέγχθηκε. Τα παραπάνω αποτελέσµατα φαίνεται να συµφωνούν µε αυτά που προκύπτουν από τις καλλιέργειες διεγερµένων λεµφοκυττάρων ατόµων νεαρής ηλικίας
189 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Πίνακας Συχνότητες επαγόµενου µη αποχωρισµού (ΝD8) για το χρωµόσωµα 8 και απώλειας του ίδιου χρωµοσώµατος (MN8 + ) σε διπύρηνα κύτταρα αρσενικών και θηλυκών ατόµων νεαρής ηλικίας ( 25 ετών), που φέρουν τέσσερα σήµατα υβριδοποίησης σε µη διεγερµένες µε PHA κατά την επίδραση µε υδροχλωροθειαζίδιο (HCTZ) καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro. ότες ιπύρηνα κύτταρα που αναλύθηκαν ιπλοειδή κύτταρα 1:1 3:1 4: ΝD8 (%) ΠΜΝ8 + ( ) ΜΝ8 + ( ) ΜΑΡΤΥΡΕΣ Α.Π. ( ) () () Κ.Γ. ( ) () () Σ.Η. ( ) () () Μ.Τ.±Τ.Σ ±.8 Κ.Α. ( ) () () Ν.Κ. ( ) () () Σ.Κ. ( ) () () Μ.Τ.±Τ.Σ ±.4 Σ Μ.Τ.±Τ.Σ ±.4 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 5µg/ml Α.Π. ( ) () () Κ.Γ. ( ) () () Σ.Η. ( ) () () Μ.Τ.±Τ.Σ ±.18 Κ.Α. ( ) () () Ν.Κ. ( ) () () Σ.Κ. ( ) () () Μ.Τ.±Τ.Σ ±.6 Σ Μ.Τ.±Τ.Σ ±.9 ΣΥΓΚΕΝΤΡΩΣΗ HCTZ 4µg/ml Α.Π. ( ) () () Κ.Γ. ( ) (.71) 1 (.71) Σ.Η. ( ) () () Μ.Τ.±Τ.Σ ±.23.24±.24.24±.24 Κ.Α. ( ) (.96) 1 (.96) Ν.Κ. ( ) (.99) 1 (.99) Σ.Κ. ( ) () () Μ.Τ.±Τ.Σ ±.7.65±.33.65±.33 Σ Μ.Τ.±Τ.Σ , ±.12.44±.2.44, ±.2 p.5 σε σύγκριση µε τις καλλιέργειες µάρτυρες (One-Way ANOVA, Student's t-test) p.5 σε σύγκριση µε τις καλλιέργειες HCTZ 5µg/ml (One-Way ANOVA, Student's t-test)
190 Αποτελέσµατα Ανευπλοειδικά φαινόµενα Εικόνα ιπλή in situ υβριδοποίηση µε φθοροχρώµατα, DUAL FISH, µε χρήση ανιχνευτών για ειδικές περιοχές, κεντροµέρη, των χρωµοσωµάτων Χ, Υ και 8 σε καλλιέργειες ανθρώπινων λεµφοκυττάρων µάρτυρα (Α, Β, Γ, Α ) και παρουσία HCTZ (, Ε, B ), in vitro. Για την ταυτόχρονη παρατήρηση των φθοροχρωµάτων DAPI (κυανό), FITC (πράσινο) και TRITC (ερυθρό) χρησιµοποιείται φορέας που διαθέτει φίλτρο τριπλής διέλευσης. DUAL FISH Χ-8: Α. Μετάφαση, Β. Κανονική κατανοµή για τα χρωµοσώµατα Χ (2:2) και 8 (2:2), Γ. Κανονική κατανοµή για τα χρωµοσώµατα Χ (1:1:) και 8 (2:2:) σε διπύρηνο κύτταρο µε ΜΝ,. Μη αποχωρισµός για το χρωµόσωµα Χ (3:1) και κανονική κατανοµή για το χρωµόσωµα 8, Ε. Μη αποχωρισµός για το χρωµόσωµα Χ και απώλεια των χρωµοσω- µάτων Χ (3::1) και 8 (2:1:1) σε διπύρηνο κύτταρο µε τρεις ΜΝ. DUAL FISH Χ-Υ: Α. Μετάφαση, Β. Κανονική κατανοµή για το χρωµόσωµα Χ (1:1) και απώλεια του χρωµοσώµατος Υ (1::1) σε διπύρηνο κύτταρο µε ΜΝ
191 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας 15. ΙΕΡΕΥΝΗΣΗ ΤΟΥ ΜΗΧΑΝΙΣΜΟΥ ΠΡΟΚΛΗΣΗΣ ΑΝΕΥΠΛΟΕΙ ΙΚΩΝ ΦΑΙΝΟΜΕΝΩΝ Ο προσδιορισµός φαινοµένων λανθασµένου αποχωρισµού, χρωµοσωµατική απώλεια και µη αποχωρισµός των οµολόγων χρωµοσωµάτων, σε ανθρώπινα λεµφοκύτταρα µε in situ υβριδοποίηση µε τη χρήση κατάλληλων ανιχνευτών απέδωσε στο υδροχλωροθειαζίδιο µία ισχυρή ανευπλοειδογόνο δράση. Η µιτωτική συσκευή αποτελεί µία µοριακή µηχανή που διαδραµατίζει ουσιώδη ρόλο για τον ορθό προσανατολισµό και την ολίσθηση των χρωµοσωµάτων στους πόλους της ατράκτου κατά τη µιτωτική διαίρεση. Βλάβες σε στοιχεία της µιτωτικής συσκευής επηρεάζουν αρνητικά τον ισοµερή καταµερισµό των χρωµοσωµάτων µε αποτέλεσµα την επαγωγή ανευπλοειδικών φαινοµένων. Ο µηχανισµός πρόκλησης φαινοµένων λανθασµένου αποχωρισµού διερευνήθηκε µε τη µελέτη δύο πρωτεϊνικών µορίων που συµµετέχουν στη συγκρότηση της ατράκτου και της µιτωτικής συσκευής, και κατ επέκταση στο συµµετρικό και ισοµερή διαχωρισµό των χρωµοσωµάτων κατά τη µίτωση. Πρόκειται για την α- και γ- τουµπουλίνη, βασικά συστατικά των µικροσωληνίσκων και του κεντροσώµατος, αντίστοιχα. Σε αυτή τη µελέτη εφαρµόσθηκε η µέθοδος του διπλού ανοσοφθορισµού µε χρήση κατάλληλων µονοκλωνικών αντισω- µάτων έναντι της α- και γ- τουµπουλίνης σε διαιρούµενα κύτταρα που έχουν την ικανότητα προσκόλλησης σε υπόστρωµα ώστε να αναπτύσσεται ένα εκτεταµένο δίκτυο µικροσωληνίσκων. Κατάλληλο βιολογικό σύστηµα για την παραπάνω µελέτη αποτέλεσαν οι ανθρώπινες κυτταρικές σειρές HFFF2, HEp2 και η κυτταρική µυοβλαστική σειρά ποντικού C2C12. Τα ανθρώπινα λεµφοκύτταρα ολικού αίµατος αναπτύσσονται σε καλλιέργεια ως εναιώρηµα κυττάρων και δεν εκπληρώνουν τις απαιτούµενες προϋποθέσεις µελέτης σχετικά µε τη µορφή ενός δικτύου µικροσωληνίσκων. Σε ανάλογη µελέτη σε ανθρώπινα λεµφοκύτταρα για την παρατήρηση του χρωµοσωµατικού υλικού σε διάφορες φάσεις της µίτωσης εφαρµόσθηκε η µέθοδος διαφορικής χρώσης µε εξειδικευµένες φθορίζουσες χρωστικές (διαµιδίνη και ιωδιούχο προπίδιο). Η παρούσα µελέτη περιελάµβανε την ανάλυση µιτωτικών και µεσοφασικών κυττάρων. Οι βιολογικοί δείκτες που εκτιµήθηκαν στην ανάλυση των µιτωτικών κυττάρων είναι: α) Ο µιτωτικός δείκτης, ο οποίος εκφράζει την εκατοστιαία αναλογία των µιτωτικών κυττάρων προς το σύνολο των κυττάρων που αναλύθηκαν και αποδίδεται από τον τύπο: Μ = 1 [Μιτωτικά / Μιτωτικά + Μεσοφασικά κύτταρα]. Με τον τρόπο αυτό µελετήθηκε ο ρυθµός διαίρεσης των κυττάρων. Ο µιτωτικός δείκτης υπολογίζεται σε σύνολο 1 κυττάρων. β) Το ποσοστό των µιτωτικών φάσεων στο σύνολο των µιτωτικών κυττάρων. Οι µιτωτικές φάσεις διακρίθηκαν σε πρόφαση, µετάφαση, ανάφαση και τελόφαση από την οργάνωση του γενετικού υλικού και την ιδιαίτερη µορφολογία της µιτωτικής συσκευής. Το ποσοστό εµφάνισης κάθε µιτωτικής φάσης αποτελεί ένδειξη για την πρόοδο του κυτταρικού κύκλου. γ) Το ποσοστό των ανώµαλων µιτωτικών κυττάρων στο σύνολο των µιτωτικών κυττάρων. Συγκεκριµένα, σε κάθε φάση ελέγχθηκε η ακέραια διαµόρφωση της µιτωτικής συσκευής και η σωστή οργάνωση του γενετικού υλικού και έγινε διάκριση µεταξύ των φυσιολογικών και µη. δ) Το ποσοστό των ανώµαλων µεταφάσεων στο σύνολο των µεταφασικών κυττάρων. Τα µεταφασικά κύτταρα διακρίθηκαν σε φυσιολογικά και ανώµαλα µε κριτήριο τον αριθµό των κεντροσωµάτων (µελέτη γ- τουµπουλίνης), την οργάνωση των ινιδίων της µιτωτικής ατράκτου (µελέτη α- τουµπουλίνης) και του χρωµοσωµατικού υλικού (διαµιδίνη). Σε µεσοφασικά κύτταρα µελετήθηκε η ακεραιότητα του κυτταροσκελετού, ο αριθµός των σηµάτων για τη γ-τουµπουλίνη και η παρουσία µη ενσωµατωµένου χρωµοσωµατικού υλικού, µικροπυρήνες
192 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας 15.1 Κυτταρικές σειρές ΗFFF2, HEp2 και ανθρώπινα λεµφοκύτταρα Η επίδραση του υδροχλωροθειαζιδίου στη µιτωτική συσκευή διερευνήθηκε σε κυτταρικές σειρές HFFF2, HEp2 και σε ανθρώπινα λεµφοκύτταρα. Από την προηγούµενη µελέτη σε ανθρώπινα λεµφοκύτταρα αποδόθηκε στο υδροχλωροθειαζίδιο διπλή ανευπλοειδογόνο δράση στη συγκέντρωση 4 µg/ml. Σε ανάλογη µελέτη σχετικά µε τη δράση της φαρµακευτικής ένωσης σε ανθρώπινες κυτταρικές σειρές φάνηκε η ικανότητα του υδροχλωροθειαζιδίου να προκαλεί χρωµοσωµατική απώλεια στις συγκεντρώσεις 6 και 8 µg/ml. Σε προκαταρκτική µελέτη, σύµφωνα µε αυτές τις πληροφορίες, διερευνήθηκε ο µηχανισµός πρόκλησης ανευπλοειδικών φαινοµένων στις συγκεντρώσεις 4, 6 και 8 µg/ml HCTZ. Στο διάγραµµα 15.1 παρουσιάζεται το ποσοστό των ανώµαλων κυττάρων που βρίσκονται στο στάδιο της µετάφασης στο σύνολο των µεταφάσεων. Σε όλες τις συγκεντρώσεις HCTZ, 4, 6 και 8 µg/ml, παρατηρούνται ανώµαλες µορφές µεταφάσεων. Σε ανθρώπινες κυτταρικές σειρές HFFF2, HEp2 επικρατούν οι µονοπολικές και πολυπολικές µεταφάσεις (εικόνα 15-1). Σε ανθρώπινα λεµφοκύτταρα διακρίνονται, εκτός από µονοπολικές, ανώµαλες διπολικές µεταφάσεις µε µη ενσωµατω- µένο χρωµοσωµατικό υλικό που υποδηλώνει καθυστερούµενα χρωµοσώµατα. 2 HL Linear Fit HFFF2 Linear Fit HEp2 Linear Fit 18 % Ανώµαλες Μεταφάσεις Μάρτυρας HCTZ (µg/ml) ιάγραµµα Μεταβολή του ποσοστού των ανώµαλων µεταφάσεων σε ανθρώπινα λεµφοκύτταρα, HL, και σε κυτταρικές σειρές, HFFF2, HEp2, στο µάρτυρα και µετά από επίδραση µε HCTZ. Από το διάγραµµα διασποράς και την ανάλυση παλινδρόµησης σε προκαταρκτικά αποτελέσµατα από τα ανθρώπινα λεµφοκύτταρα και την κυτταρική σειρά HFFF2 βρέθηκε ότι υπάρχει θετική γραµµική σχέση µεταξύ της συγκέντρωσης HCTZ και του ποσοστού των ανώµαλων µεταφάσεων. Όπως φαίνεται από το σχήµα, η προσαρ
193 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας µογή της ευθείας γραµµικής παλινδρόµησης στα δεδοµένα είναι αρκετά καλή και επιβεβαιώνεται από την τιµή του συντελεστή προσδιορισµού R 2 τόσο για τα ανθρώπινα λεµφοκύτταρα (R 2 =89.68%, P=.35, treg=5.2) όσο και για την κυτταρική σειρά HFFF2 (R 2 =94.3%, P=.2, treg=6.95). Όσον αφορά την κυτταρική σειρά HEp2 δεν φαίνεται να υπάρχει δοσοεξαρτώµενη αύξηση των ανώµαλων µεταφάσεων (R 2 =1.8%, P=.416, treg=1.2). Στο διάγραµµα 15.2 παρουσιάζεται η κατανοµή των µιτωτικών φάσεων, µεταφάσεων και ανατελοφάσεων, ως εκατοστιαία αναλογία στο σύνολο των κυττάρων που αναλύθηκαν. Σε ανθρώπινα λεµφοκύτταρα και σε κυτταρικές σειρές η παρουσία HCTZ στη συγκέντρωση 8 µg/ml προκαλεί αύξηση στο ποσοστό των κυττάρων που βρίσκονται στο στάδιο της µετάφασης, η οποία ακολουθείται από µείωση του ποσοστού των ανατελοφάσεων. Από τα παραπάνω φαίνεται ότι το HCTZ εµποδίζει την πρόοδο του κυτταρικού κύκλου συσσωρεύοντας τα κύτταρα στο στάδιο της µετάφασης, δράση που χαρακτηρίζει τις ανευπλοειδογόνες ενώσεις. Το φαινόµενο αυτό δεν παρατηρήθηκε στις συγκεντρώσεις 4 και 6 µg/ml HCTZ πιθανότατα εξαιτίας του µικρού αριθµού µεσοφασικών και µιτωτικών κυττάρων που αναλύθηκαν. % Μεταφάσεις % Ανα τελοφάσεις Μάρτυρας Μάρτυρας 4 6 HCTZ (µg/ml) 4 6 HCTZ (µg/ml) 8 8 HL HFFF2 Hep2 HL HFFF2 Hep2 ιάγραµµα Μεταβολή του ποσοστού των µεταφάσεων και ανατελοφάσεων σε ανθρώπινα λεµφοκύτταρα, HL, και σε κυτταρικές σειρές, HFFF2, HEp2, στο µάρτυρα και µετά από επίδραση µε HCTZ. Η εξέταση µεσοφασικών κυττάρων µε τη σήµανση α- τουµπουλίνης έδειξε την ικανότητα του HCTZ να προκαλεί αποδιοργάνωση του δικτύου των µικροσωληνίσκων σε κυτταρικές σειρές HFFF2 και HEp2 (εικόνα 15-2). Τα αποτελέσµατα της προκαταρκτικής µελέτης αξιολογήθηκαν και οι πληροφορίες που προέκυψαν αποτέλεσαν το έναυσµα για περαιτέρω διερεύνηση της επίδρασης του HCTZ στη µιτωτική συσκευή και στον κυτταροσκελετό σε ένα διαφορετικό βιολογικό σύστηµα
194 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Εικόνα Μιτωτικές φάσεις κυττάρων HEp2 (Α, Β, Γ, ) και HFFF2 (A, B ) στο µάρτυρα και παρουσία HCTZ
195 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Εικόνα ίκτυο µικροσωληνίσκων (α- τουµπουλίνη) κυττάρων HFFF2 (Α) και HEp2 (B) στο µάρτυρα και µετά από την επίδραση HCTZ. Οι φωτογραφίες προέρχονται από µικροσκόπιο φθορισµού Zeiss Axioskop
196 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας 15.2 Κυτταρική σειρά C2C12 Στην παρούσα µελέτη διερευνήθηκε εκτενέστερα η επίδραση του υδροχλωροθειαζιδίου στη µιτωτική συσκευή σε κυτταρική µυοβλαστική σειρά ποντικού C2C12. H δράση του HCTZ µελετήθηκε στις συγκεντρώσεις 4, 6 και 8 µg/ml. Για το λόγο αυτό πραγµατοποιήθηκαν τρία ανεξάρτητα πειράµατα. Στον πίνακα 15.1 παρουσιάζονται τα αποτελέσµατα της µελέτης των µιτωτικών κυττάρων στο σύνολο των κυττάρων που αναλύθηκαν µετά την επίδραση µε υδροχλωροθειαζίδιο. Στην πρώτη στήλη εµφανίζεται η συγκέντρωση της εξεταζόµενης χηµικής ένωσης, ενώ στη δεύτερη στήλη ο συνολικός αριθµός των κυττάρων που αναλύθηκαν. Τα κύτταρα αυτά διακρίνονται σε µεσοφασικά και µιτωτικά. Στην τρίτη στήλη αναφέρεται το σύνολο των µεσοφασικών κυττάρων και ακολουθεί στις επόµενες στήλες, τέταρτη, πέµπτη, έκτη και έβδοµη, η κατανοµή των µιτωτικών κυττάρων στις µιτωτικές φάσεις πρόφαση, µετάφαση, ανάφαση και τελόφαση. Ο συνολικός αριθµός αναφάσεων και τελοφάσεων αναφέρεται στην επόµενη στήλη, όγδοη, ως ανατελοφάσεις. Στις δύο τελευταίες στήλες αναφέρεται το σύνολο των µιτωτικών κυττάρων και ο µιτωτικός δείκτης. Στις καλλιέργειες που πραγµατοποιήθηκε επίδραση HCTZ παρατηρείται πτώση του µιτωτικού δείκτη που αντανακλά στη µείωση του ρυθµού πολλαπλασιασµού των κυττάρων. Συγκεκριµένα, στην καλλιέργεια-µάρτυρα ο δείκτης εµφανίζει τιµή 7.4, η οποία µειώνεται µε την επίδραση 4 µg/ml (5.35) και 6 µg/ml HCTZ (6.9), ενώ εµφανίζει τη µεγαλύτερη πτώση (2.92) µε την επίδραση 8 µg/ml HCTZ. Από τη στατιστική ανάλυση προκύπτει ότι η µείωση αυτή είναι σηµαντική σε σχέση µε την καλλιέργεια-µάρτυρα στη συγκέντρωση 8 µg/ml HCTZ (x 2 =62.96, p=.1, G=67.76). Στον πίνακα 15.2 παρουσιάζεται η κατανοµή των µιτωτικών φάσεων στο σύνολο των µιτωτικών κυττάρων που αναλύθηκαν. Στην πρώτη στήλη εµφανίζεται η συγκέντρωση της εξεταζόµενης χηµικής ένωσης, στη δεύτερη στήλη ο συνολικός αριθµός των µιτωτικών κυττάρων που αναλύθηκαν και στις επόµενες στήλες, τρίτη, τέταρτη, πέµπτη και έκτη, ακολουθεί η κατανοµή των κυττάρων στις µιτωτικές φάσεις πρόφαση, µετάφαση, ανάφαση και τελόφαση. Ο συνολικός αριθµός αναφάσεων και τελοφάσεων αναφέρεται στην έβδοµη στήλη ως ανατελοφάσεις. Στην όγδοη και ένατη στήλη αναφέρονται τα ανώµαλα µιτωτικά κύτταρα και τα ανώµαλα µεταφασικά κύτταρα στο σύνολο των µιτωτικών κυττάρων. Σε καλλιέργειες που πραγµατοποιήθηκε επίδραση HCTZ παρατηρείται αύξηση του ποσοστού των µεταφάσεων, η οποία συνοδεύεται από µία αντίστοιχη µείωση του ποσοστού των ανατελοφάσεων. Το ποσοστό των µεταφάσεων από 3.34% στην καλλιέργεια-µάρτυρα αυξάνεται στο 3.62%, 4.17% και 44.59% µετά την επίδραση 4, 6 και 8 µg/ml HCTZ, αντίστοιχα. Από τη στατιστική ανάλυση προκύπτει ότι η αύξηση αυτή είναι σηµαντική σε σχέση µε την καλλιέργεια- µάρτυρα στις συγκεντρώσεις 6 µg/ml (x 2 =18.68, p=.1, G=18.91) και 8 µg/ml HCTZ (x 2 =19.99, p=.1, G=2.17). Το ποσοστό των ανατελοφάσεων από 64.57% στην καλλιέργεια-µάρτυρα µειώνεται στο 61.33%, 53.75% και 47.9% µετά την επίδραση 4, 6 και 8 µg/ml HCTZ, αντίστοιχα. Από τη στατιστική ανάλυση προκύπτει ότι η µείωση αυτή είναι σηµαντική σε σχέση µε την καλλιέργεια- µάρτυρα στις συγκεντρώσεις, 6 µg/ml (x 2 =22.72, p=.1, G=23.2) και 8 µg/ml HCTZ (x 2 =32.11, p=.1, G=32.37). Συνεπώς, η αύξηση της συγκέντρωσης HCTZ οδηγεί στη συγκράτηση των κυττάρων στο στάδιο της µετάφασης και προκαλεί διακοπή της µίτωσης (mitotic arrest). Το γεγονός αυτό οφείλεται σε ενεργοποίηση του σηµείου ελέγχου της µίτωσης και µόνο όσα κύτταρα πληρούν τις προϋποθέσεις ξεπερνούν το σηµείο αυτό και συνεχίζουν τον κυτταρικό κύκλο µε αποτέλεσµα να
197 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας ΠΙΝΑΚΑΣ Κατανοµή των µιτωτικών κυττάρων και µιτωτικός δείκτης σε καλλιέργειες κυττάρων C2C12 στο µάρτυρα και µετά από επίδραση µε υδροχλωροθειαζίδιο (HCTZ). Συγκέντρωση HCTZ (µg/ml) Σύνολο κυττάρων Μεσοφασικά κύτταρα Πρόφαση (%) Μετάφαση (%) Ανάφαση Τελόφαση Ανα- Τελοφάσεις (%) Σύνολο µιτωτικών κυττάρων Μιτωτικός δείκτης % (5.57) 5.52± (24.92) 24.91± (69.51) 69.58± ± (9.76) 9.29± (3.89) 31.33± (59.35) 59.38± ± (6.85) 6.59± (38.36) 38.54± (54.79) 54.88± ± (4.8) 3.85± (46.94) 49.14± (48.98) 47.1± *±.4 *: p<.5 σε σύγκριση µε το µάρτυρα (G-test)
198 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας ΠΙΝΑΚΑΣ Κατανοµή των µιτωτικών φάσεων στο σύνολο των µιτωτικών κυττάρων σε καλλιέργειες κυττάρων C2C12 στο µάρτυρα και µετά από επίδραση µε υδροχλωροθειαζίδιο (HCTZ). Συγκέντρωση HCTZ (µg/ml) Σύνολο µιτωτικών κυττάρων Πρόφαση (%) Μετάφαση (%) Ανάφαση Τελόφαση Ανα- Τελοφάσεις (%) Σύνολο ανώµαλων µιτωτικών κυττάρων (%) Σύνολο ανώµαλων µεταφάσεων (%) (5.34) 5.9± (29.4) 3.34± (65.26) 64.57± (5.6) 4.75± (4.5) 4.25± (8.8) 8.6± (3.55) 3.62± (61.37) 61.33± (14.38) 14.4*±.38 7 (9.59) 9.59*± (6.6) 6.8± (38.24) 4.17*± (55.16) 53.75*± (19.24) 19.9*± (14.25) 13.91*± (8.37) 7.51± (39.) 44.59*± (52.63) 47.9*± (26.32) 26.17*± (16.87) 19.3*±3.17 *: p<.5 σε σύγκριση µε το µάρτυρα (G-test)
199 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας τα παρατηρούµε στις ανατελοφάσεις. Αξιοσηµείωτη είναι η παρατήρηση που αφορά τα ανώµαλα µιτωτικά κύτταρα και τις ανώµαλες µεταφάσεις. Παρατηρείται σηµαντική αύξηση στο σύνολο των ανώµαλων µιτωτικών κυττάρων (x 2 =54.28, p=.1, G=52.13) από τη µικρότερη εξεταζόµενη συγκέντρωση, 4 µg/ml HCTZ, η οποία παρατηρείται και στην περίπτωση των ανώµαλων µεταφάσεων (x 2 =2.53, p=.1, G=2.24). Η ανάλυση παλινδρόµησης έδειξε ότι υπάρχει θετική γραµµική σχέση µεταξύ της συγκέντρωσης HCTZ και του ποσοστού των ανώµαλων µιτωτικών κυττάρων (R 2 = 98.74%, P=.4, treg=15.34), καθώς και µεταξύ της συγκέντρωσης HCTZ και του ποσοστού των ανώµαλων µεταφάσεων (R 2 =95.98%, P=.14, treg=8.52). Η µελέτη περιελάµβανε την ανάλυση των ανώµαλων κυττάρων που βρίσκονται στο στάδιο της µετάφασης µε βασικό κριτήριο τον αριθµό των πόλων της µιτωτικής ατράκτου που καθορίζεται από τα κέντρα οργάνωσης των µικροσωληνίσκων (MTOC). Σε αυτά εντοπίζεται η γ- τουµπουλίνη, η οποία συµµετέχει ενεργά στην πυρήνωση και πολικότητα των µικροσωληνίσκων, α- και β- τουµπουλίνη, και αποτελεί βασικό συστατικό του κεντροσώµατος. Η ανάλυση των σηµάτων γ- τουµπουλίνης αποδίδει σηµαντικές πληροφορίες για την οργάνωση και τη δοµή της µιτωτικής ατράκτου. Έτσι οι ανώµαλες µεταφάσεις διακρίνονται σε: (α) ιπολικές µη οµαδοποιηµένες οι οποίες φέρουν δύο πόλους, κεντροσώµατα, αλλά το χρωµοσωµατικό υλικό δεν είναι τοποθετηµένο σωστά στο ισηµερινό επίπεδο. (β) Μονοπολικές οι οποίες φέρουν ένα κεντρόσωµα ή δύο σε άµεση επαφή µεταξύ τους και το χρωµοσω- µατικό υλικό διατάσσεται κυκλικά γύρω από τον πόλο της µιτωτικής ατράκτου. Χαρακτηριστική είναι η ακτινωτή διάταξη των µικροσωληνίσκων γύρω από τον πόλο. (γ) Πολυπολικές οι οποίες φέρουν τρία ή περισσότερα κεντροσώµατα και σε κάθε πόλο διατάσσεται το χρωµοσωµατικό υλικό. Θα πρέπει να αναφερθεί ότι στη φυσιολογική διπολική µετάφαση σχηµατίζεται από δύο πόλους, κεντροσώµατα, µία συµµετρική µιτωτική άτρακτος και το χρωµοσωµατικό υλικό είναι σωστά τοποθετηµένο στο ισηµερινό επίπεδο (εικόνα 15-3). Στον πίνακα 15.3 που ακολουθεί στην πρώτη στήλη εµφανίζεται η συγκέντρωση της εξεταζόµενης χηµικής ένωσης, στη δεύτερη το σύνολο των µεταφάσεων που αναλύθηκαν και στην τρίτη οι φυσιολογικές διπολικές µεταφάσεις. Σε επόµενες στήλες, τέταρτη, πέµπτη και έκτη γίνεται διάκριση των ανώµαλων µεταφάσεων σε διπολικές µη οµαδοποιηµένες, µονοπολικές και πολυπολικές και στην έβδοµη στήλη αναφέρεται το σύνολο των ανώµαλων µεταφάσεων. Σε καλλιέργειες που πραγµατοποιήθηκε επίδραση HCTZ παρατηρείται µείωση του ποσοστού των φυσιολογικών µεταφάσεων και ταυτόχρονα αύξηση του ποσοστού όλων των ανώµαλων µορφών. Στη συγκέντρωση 4 µg/ml HCTZ παρατηρείται σηµαντική αύξηση στις διπολικές µη οµαδοποιηµένες (x 2 =4.94, p=.26, G=5.42) και στις µονοπολικές µεταφάσεις (x 2 =8.69, p=.3, G=9.9). Η δράση του HCTZ σε µεγαλύτερες συγκεντρώσεις, 6 και 8 µg/ml, επηρεάζει σε σηµαντικό βαθµό το σχηµατισµό τόσο των διπολικών µη οµαδοποιηµένων όσο και των µονοπολικών µεταφάσεων. Όσον αφορά τις πολυπολικές µεταφάσεις ενώ στην καλλιέργεια- µάρτυρα αποτελούν το 2.74% του συνόλου των µεταφάσεων, το ποσοστό αυτό ανέρχεται στο 6.3% µετά την επίδραση 4 µg/ml HCTZ και φθάνει στο 8.51% και 1.13% µετά την επίδραση 6 και 8 µg/ml HCTZ. Η αύξηση που παρατηρείται στις συγκεντρώσεις 6 µg/ml (x 2 =8.58, p=.3, G=9.61) και 8 µg/ml HCTZ (x 2 = 17.11, p=.1, G=18.29) είναι σηµαντική σε σχέση µε την καλλιέργεια-µάρτυρα. Ανάµεσα στους τύπους των ανώµαλων µεταφάσεων που προκαλούνται από τη δράση του HCTZ οι µονοπολικές εµφανίζονται σε µεγαλύτερη συχνότητα και φαίνεται να αποτελούν την επικρατέστερη µορφή
200 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας ΠΙΝΑΚΑΣ Κατανοµή των ανώµαλων µεταφάσεων στο σύνολο των µεταφασικών κυττάρων που αναλύθηκαν σε καλλιέργειες κυττάρων C2C12 στο µάρτυρα και µετά από επίδραση µε υδροχλωροθειαζίδιο (HCTZ). Συγκέντρωση HCTZ (µg/ml) Σύνολο µεταφάσεωνπροµεταφάσεων ιπολικές (2 σήµατα) ιπολικές (2 σήµατα) Άρθρο I. Μη οµαδοποιηµένες Μονοπολικές (1 ή 2 σήµατα) πολικές Σύνολο ανώµαλων µεταφάσεων (84.69) 85.29± (5.26) 5.±1.6 3 (7.18) 6.97± (2.87) 2.74±.5 64 (15.31) 14.71± (68.61) 68.58± (1.31) 1.31*± (14.8) 14.82*± (6.28) 6.3±1.4 7 (31.39) 31.43*± (62.73) 64.95± (11.82) 1.78*± (17.58) 15.75*± (7.88) 8.51*± (37.27) 35.5*± (56.75) 56.63± (11.7) 11.47*± (21.45) 21.78*± (1.73) 1.13*± (43.25) 43.37*±3.38 *:.1 p<.5 σε σύγκριση µε το µάρτυρα (G-test)
201 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Αξιοσηµείωτη είναι η παρατήρηση που αφορά το σύνολο των ανώµαλων µεταφάσεων. Το ποσοστό των ανώµαλων µεταφάσεων αυξάνεται σηµαντικά µε την αύξηση της συγκέντρωσης HCTZ και η ανάλυση παλινδρόµησης έδειξε τη θετική γραµµική σχέση που συνδέει αυτές τις µεταβλητές (R 2 =97.79%, P=.7, treg=11.55 ). Επιπρόσθετα, η ανάλυση παλινδρόµησης έδειξε ότι υπάρχει θετική γραµµική σχέση µεταξύ της συγκέντρωσης HCTZ και των µονοπολικών µεταφάσεων (R 2 = 94.14%, P=.2, treg= 7.1), καθώς και µεταξύ της συγκέντρωσης HCTZ και των πολυπολικών µεταφάσεων (R 2 =99.69%, P=.1, treg=31.16). Τα παραπάνω αποτελέσµατα συµφωνούν µε αυτά που προκύπτουν από τα ανθρώπινα λεµφοκύτταρα και την ανθρώπινη κυτταρική σειρά HFFF2. Ακολουθούν φωτογραφίες από µικροσκόπιο φθορισµού, από παρασκευάσµατα διπλού ανοσοφθορισµού για την α- και γ- τουµπουλίνη. Στις φωτογραφίες απεικονίζονται φυσιολογικές και ανώµαλες µορφές µεταφάσεων (εικόνα 15-3), αναφάσεων και τελοφάσεων (εικόνα 15-4)
202 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Εικόνα Μεταφάσεις κυττάρων C2C12 στο µάρτυρα (Α) και παρουσία HCTZ (Β, Γ,, Ε)
203 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Εικόνα Αναφάσεις και τελοφάσεις κυττάρων C2C12 στο µάρτυρα (Α) και παρουσία HCTZ (Β, Γ, ). Παρατηρείται µη ενσωµατωµένο χρωµοσωµατικό υλικό σε ανώµαλες µορφές µιτωτικών φάσεων παρουσία HCTZ
204 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Η µελέτη περιελάµβανε την παρατήρηση µεσοφασικών κυττάρων για την οργάνωση του κυτταροσκελετού, τον αριθµό σηµάτων γ- τουµπουλίνης και την παρουσία καθυστερούµενου χρωµοσωµατικού υλικού. Η επίδραση HCTZ προκαλεί σηµαντικές τροποποιήσεις στο δίκτυο των µικροσωληνίσκων, ενώ δεν φαίνεται να επηρεάζεται η γ-τουµπουλίνη διότι οι µεσοφασικοί πυρήνες φέρουν το σωστό αριθµό κεντροσωµάτων (εικόνες 15.5, 15.6). Στον πίνακα 15.4 παρουσιάζεται η κατανοµή των µεσοφασικών κυττάρων µε κριτήριο την παρουσία µη ενσωµατωµένου χρωµοσωµατικού υλικού, µικροπυρήνες. Στην πρώτη στήλη εµφανίζεται η συγκέντρωση της εξεταζόµενης χηµικής ένωσης, στη δεύτερη ο αριθµός των µεσοφασικών πυρήνων που αναλύθηκαν, στην τρίτη ο αριθµός των φυσιολογικών και στην τέταρτη ο αριθµός των πυρήνων που φέρουν µικροπυρήνα. Η επίδραση υδροχλωροθειαζιδίου προκαλεί αύξηση στο ποσοστό των µεσοφασικών πυρήνων µε µικροπυρήνα από τη µικρότερη εξεταζόµενη συγκέντρωση (.9%). Η αύξηση αυτή εµφανίζεται εντονότερη στη συγκέντρωση 8 µg/ml HCTZ (1.42%). Η στατιστική ανάλυση (G-test) έδειξε τη σηµαντική ικανότητα του HCTZ να επάγει ανώµαλους µεσοφασικούς πυρήνες σε όλες τις εξεταζόµενες συγκεντρώσεις, 4, 6 και 8 µg/ml, σε σχέση µε την καλλιέργεια-µάρτυρα (.27%). ΠΙΝΑΚΑΣ Κατανοµή µεσοφασικών πυρήνων σε καλλιέργειες κυττάρων C2C12 στο µάρτυρα και µετά από επίδραση µε υδροχλωροθειαζίδιο (HCTZ). Συγκέντρωση HCTZ (µg/ml) Μεσοφασικοί πυρήνες Φυσιολογικοί (%) Με ΜΝ (%) Άρθρο II (99.73) 99.73± (99.11) 99.1± (99.37) 99.39± (98.55) 98.58± (.27).27± (.89).9*± (.63).61*± (1.45) 1.42*±.64 *:.1 p <.5 σε σύγκριση µε το µάρτυρα (G-test). Από τη σύγκριση των αποτελεσµάτων µε αυτά που προκύπτουν από τα ανθρώπινα λεµφοκύτταρα και τις ανθρώπινες κυτταρικές σειρές και την εκτίµηση τους αποδίδεται στο υδροχλωροθειαζίδιο µία ισχυρή ανευπλοειδογόνος δράση
205 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Εικόνα ίκτυο µικροσωληνίσκων (α- τουµπουλίνη) κυττάρων C2C12 στο µάρτυρα (Α) και µετά από την επίδραση HCTZ (Β) και δεµεκολσίνης (Γ). Οι φωτογραφίες προέρχονται από µικροσκόπιο φθορισµού Zeiss Axioskop
206 Αποτελέσµατα Μηχανισµοί ανευπλοειδίας Εικόνα Μεσοφασικοί πυρήνες µε σήµανση α- (δίκτυο µικροσωληνίσκων) και γ- τουµπουλίνης (κεντρόσωµα) κυττάρων C2C12 στο µάρτυρα (Α) και µετά από την επίδραση HCTZ (Β). Οι φωτογραφίες προέρχονται από συνεστιακό µικροσκόπιο Leika TCS-SP εξοπλισµένο µε το σύστηµα ανάλυσης εικόνας Leica Confocal Software

207
208 Συζήτηση ΣΥΖΗΤΗΣΗ Οι χρωµοσωµατικές ανωµαλίες τόσο οι αριθµητικές όσο και οι δοµικές συµβάλλουν στην εµφάνιση γενετικών ασθενειών στον άνθρωπο. Η ανευπλοειδία είναι η συνηθέστερα προσδιορισµένη ανωµαλία χρωµοσωµάτων στον άνθρωπο και αποτελεί έναν ακούσιο παράγοντα διακοπής της κύησης. Εκτιµάται ότι ένα ποσοστό 1-3% των γονιµοποιηµένων ανθρώπινων ωαρίων φέρουν λανθασµένο αριθµό χρωµοσωµάτων και αυτό έχει σηµαντικές κλινικές συνέπειες. Συγκεκριµένα, ανευπλοειδικά φαινόµενα χαρακτηρίζουν το ένα τρίτο όλων των τυχαίων αποβολών, ενώ αποτελούν τη γενετική αιτία των αναπτυξιακών ανικανοτήτων και της διανοητικής καθυστέρησης (Hassold και Hunt, 21, Kirsch-Volders et al., 22). Εκτεταµένες µελέτες έχουν καθορίσει το σηµαντικό ρόλο που διαδραµατίζουν οι γενετικές αλλαγές στην καρκινογένεση. Η ανευπλοειδία σε σωµατικά κύτταρα σχετίζεται µε διάφορες µορφές καρκίνου παρατήρηση γνωστή περισσότερο από έναν αιώνα πριν, όταν οι D. Hansemann (189) και T. Βoveri (1914) ανέφεραν τη σχέση που υπάρχει µεταξύ αριθµητικών ανωµαλιών, ασύµµετρων µιτώσεων και καρκίνου (Duesberg et al, 2, Brinkley, 21, Draviam et al., 24). Σύµφωνα µε αυτή τη σχέση τροποποιήσεις που διαταράσσουν την ακρίβεια του µηχανισµού διαχωρισµού των χρωµοσωµάτων οδηγούν σε χρωµοσωµατική αστάθεια. Η χρωµοσωµατική αστάθεια (CIN) σε πρώιµα στάδια της καρκινογένεσης πιθανότατα αποτελεί την κινητήρια δύναµη που οδηγεί σε απώλεια της ετεροζυγωτίας ενός ογκοκατασταλτικού γονιδίου ή/και ενισχύει αποτελεσµατικά ένα ογκογονίδιο µε το διπλασιασµό του χρωµοσώµατος στο οποίο βρίσκεται και προκαλεί αλλαγή στον αριθµό των αντιγράφων ενός ή περισσοτέρων γονιδίων που ελέγχουν την κυτταρική αύξηση (Parry at al., 22). Ενδείξεις οι οποίες συνηγορούν στην εµπλοκή ανευπλοειδικών φαινοµένων στην καρκινογένεση είναι το µη τυχαίο πρότυπο εµφάνισης ανευπλοειδίας σε ζωικούς και ανθρώπινους καρκίνους και η προδιάθεση για εµφάνιση καρκίνου σε άτοµα µε συγγενείς ανευπλοειδίες. Επιπλέον ανευπλοειδογόνες ιδιότητες έχουν προσδιορισθεί για διάφορους καρκινογόνους παράγοντες, οι οποίοι έχουν αναφερθεί ως µη µεταλλαξιγόνοι σε συστήµατα ελέγχου µεταλλάξεων (Kirsch-Volders et al., 22). Εποµένως είναι σηµαντικό να προσδιορίσουµε τη δράση περιβαλλοντικών παραγόντων ως προς την ικανότητα πρόκλησης φαινο- µένων που σχετίζονται µε ανισορροπία του γενετικού υλικού. Οι φαρµακευτικές ενώσεις αποτελούν µία κατηγορία παραγόντων του περιβάλλοντος που επιδρούν άµεσα στον άνθρωπο και σε αρκετές από αυτές έχουν αποδοθεί ανευπλοειδογόνες ιδιότητες σε γονιδιοτοξικές µελέτες. Αντικαρκινικά φάρµακα (Elhajouji et al., 1997, Decordier et al., 22, Bakou et al., 22, Kouloumenta et al., 25), βασικά συστατικά αντιβηχικών (Schuler et al., 1999, 23), αντιµυκητιακών (Pisano et al., 2, Pacchierotti et al., 22), ηρεµιστικών ή/και υπνωτικών φαρµάκων (Izzo et al., 1998, Vitale et al., 25), καθώς και αντιϋπερτασικά φάρµακα (Telez et al., 2, Telez et al., 21) έχουν χαρακτηρισθεί ως ανευπλοειδογόνα. Η παρούσα διατριβή αναφέρεται στη µελέτη της κυτταρογενετικής δράσης της φαρµακευτικής ένωσης υδροχλωροθειαζίδιο. Επιλέγονται οι συγκεντρώσεις 5 και 4 µg/ml HCTZ που εξετάζονται σε ανθρώπινα λεµφοκύτταρα περιφερικού αίµατος in vitro σε 32 υγιείς δότες. Τα άτοµα που συµπεριελήφθησαν στη µελέτη διαχωρίστηκαν σε οµάδες µε κριτήριο την ηλικία, το φύλο και το κάπνισµα
209 Συζήτηση Μελετήθηκε η επίδραση του υδροχλωροθειαζιδίου στον κυτταρικό κύκλο µε την εκτίµηση του δείκτη πολλαπλασιασµού των κυττάρων, CBPI, σε ανθρώπινα λεµφοκύτταρα. Ο δείκτης CBPI θεωρείται περισσότερο ακριβής και αξιόπιστος σε σχέση µε το δείκτη NDI και το ποσοστό διπύρηνων κυττάρων για την εκτίµηση της προόδου του κυτταρικού κύκλου (Suralles et al., 1994, Suralles et al., 1995a). H επίδραση του HCTZ προκαλεί µείωση του δείκτη πολλαπλασιασµού των κυττάρων σε νεαρά και ηλικιωµένα άτοµα (διάγραµµα 1). Η ικανότητα του HCTZ να προκαλεί καθυστέρηση του κυτταρικού κύκλου προκύπτει από τη µείωση των τιµών του είκτης πολλαπλασιασµού των κυττάρων Νεαρά Ηλικιωµένα :p<.5 σε σχέση µε το µάρτυρα :p<.5 σε σχέση µε 5 µg/ml HCTZ (One-way ANOVA, Student's t-test) :p<.5 σε σχέση µε 5 µg/ml HCTZ (Student's t-test),, HCTZ (µg/ml) ιάγραµµα 1. Μεταβολή του δείκτη πολλαπλασιασµού των κυττάρων (CBPI) στο µάρτυρα και µετά από επίδραση HCTZ σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. δείκτη CBPI σε νεαρά και ηλικιωµένα άτοµα. Περισσότερο κυτταροτοξική εµφανίζεται η συγκέντρωση 4 µg/ml HCTZ, η οποία προκαλεί τη µεγαλύτερη πτώση του δείκτη CBPI συγκριτικά µε το µάρτυρα και τη συγκέντρωση 5 µg/ml HCTZ. Επιπλέον ο δείκτης CBPI εµφανίζει χαµηλότερες τιµές σε ηλικιωµένα άτοµα συγκριτικά µε τα νεαρά. Πρόκειται για ένα φαινόµενο που έχει παρατηρηθεί σε ανθρώπινα λεµφοκύτταρα σε πληθυσµιακές µελέτες και αποδίδεται σε φυσιολογικές αιτίες (Pastor et al., 21). Συγκεκριµένα τα λεµφοκύτταρα ηλικιωµένων ατόµων εµφανίζουν είτε µειωµένη ικανότητα απόκρισης σε µιτογόνους παράγοντες είτε αποτυγχάνουν σε επόµενες διαιρέσεις µετά τη διέγερση τους (Lucivero et al., 1988). Από τον υπολογισµό του ποσοστού κυτταροτοξικότητας (Kirsch-Volders et al., 23) προκύπτει ότι η επίδραση 5 µg/ml HCTZ προκαλεί αδυναµία ολοκλήρωσης του κυτταρικού κύκλου σε ποσοστό 2.41%. Μετά την επίδραση 4 µg/ml HCTZ το ποσοστό αυτό ανέρχεται σε 12.5%. Η επίδραση του HCTZ προκαλεί µικρή κυτταροτοξικότητα στις καλλιέργειες από νεαρά και ηλικιωµένα άτοµα και δεν φαίνεται να υπάρχει διαφορά ανάµεσα στις δύο ηλικιακές οµάδες που µελετήθηκαν. Έχει αναφερθεί η ικανότητα του υδροχλωροθειαζιδίου να αναστέλλει τον πολλαπλασιασµό και τη διαφοροποίηση οστεοβλαστών και πολυδύναµων προγονικών κυττάρων σε συγκεντρώσεις µεγαλύτερες από 1 µm (3 µg/ml) (Lau et
210 Συζήτηση al., 1996). Η κυτταροτοξική δράση του HCTZ είναι δυνατόν να αποδοθεί στην ικανότητα δέσµευσης του µε τον περιφερικό τύπο υποδοχέα των βενζοδιαζεπινών (Lukeman και Fanestil, 1987), ο οποίος εµπλέκεται στη ρύθµιση του κυτταρικού πολλαπλασιασµού και της απόπτωσης (Casellas et al., 22). Η συγκεκριµένη θέση δέσµευσης για τις βενζοδιαζεπίνες έχει ανιχνευθεί στην πλασµατική µεµβράνη των ερυθρών αιµοσφαιρίων στον άνθρωπο (Olson et al., 1988) και σε άθικτα λεµφοκύτταρα του περιφερικού αίµατος (Garau et al., 1989). Η ανάλυση µικροπυρήνων αποτελεί πολύτιµο εργαλείο στη διερεύνηση της γενετικής δράσης χηµικών ενώσεων. Ο σχηµατισµός µικροπυρήνων οφείλεται σε βλάβες/τροποποιήσεις στο DNA, στα χρωµοσώµατα και σε πρωτεΐνες που άµεσα ή έµµεσα εµπλέκονται στον ορθό διαχωρισµό των χρωµοσωµάτων (Kirsch-Volders et al., 23). Οι µικροπυρήνες αποτελούνται από άκεντρα χρωµοσωµατικά τµήµατα ή ολόκληρα χρωµοσώµατα που δεν ενσωµατώθηκαν στη µιτωτική συσκευή και δεν µετακινήθηκαν προς τους πόλους του κυττάρου στο στάδιο της ανάφασης. Έτσι στο στάδιο της µεσόφασης ο µικροπυρήνας εντοπίζεται κοντά στον κυρίως πυρήνα υποδηλώνοντας την παρουσία γενετικής βλάβης. Η συχνότητα µικροπυρήνων σε καλλιέργειες-µάρτυρες και σε καλλιέργειες µετά από επίδραση HCTZ αποτελεί ένα δείκτη χρωµοσωµατικών ρηγµάτων και χρωµοσωµατικής καθυστέρησης (διάγραµµα 2). Τη µεγαλύτερη αύξηση στις συχνότητες ΜΝ σε σχέση µε τις καλλιέργειες-µαρ- Μικροπυρήνες *, Νεαρά Ηλικιωµένα :p<.5 σε σχέση µε το µάρτυρα :p<.5 σε σχέση µε 5 µg/ml HCTZ (One-way ANOVA, Student's t-test) * :p<.5 σε σχέση µε το µάρτυρα (Student's t-test), Μικροπυρήνες * HCTZ (µg/ml) HCTZ (µg/ml), :p<.5 σε σχέση µε το µάρτυρα :p<.5 σε σχέση µε 5 µg/ml HCTZ (One-way ANOVA, Student's t-test) * :p<.5 σε σχέση µε το µάρτυρα (Student's t-test),, Νεαρά Ηλικιωµένα C + MN C - MN ιάγραµµα 2. Μεταβολή στη συνολική συχνότητα των µικροπυρήνων (ΜΝ) και στη συχνότητα των µικροπυρήνων µε (C + MN) και χωρίς (C - MN) κεντροµερικό σήµα στο µάρτυρα και µετά από επίδραση HCTZ σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. -τυρες την προκαλεί η συγκέντρωση 4 µg/ml HCTZ σε νεαρά και ηλικιωµένα άτο
211 Συζήτηση µα. Στην οµάδα ατόµων νεαρής ηλικίας η αύξηση που παρατηρείται στις συχνότητες ΜΝ στην ελάχιστη συγκέντρωση 5 µg/ml HCTZ είναι στατιστικώς σηµαντική σε σχέση µε τις καλλιέργειες-µάρτυρες. Πιθανότατα οι διαφορές στη δράση της ελάχιστης συγκέντρωσης HCTZ στις δύο ηλικιακές οµάδες οφείλονται σε διαφορετικές κινητικές πολλαπλασιασµού των λεµφοκυττάρων που επηρεάζουν το σχηµατισµό µικροπυρήνων. Η δράση της µιτοµυκίνης-c, MMC, και των ακτίνων Χ εξαρτάται από την ηλικία και το ρυθµό πολλαπλασιασµού των κυττάρων. Εφόσον για το σχηµατισµό µικροπυρήνων θωρείται απαραίτητη η κυτταρική, πυρηνική, διαίρεση ο ρυθµός πολλαπλασιασµού των κυττάρων επηρεάζει σηµαντικά τις συχνότητες µικροπυρήνων (Fenech και Morley, 1985b). Προκειµένου να εξακριβωθεί ο µηχανισµός δηµιουργίας των µικροπυρήνων, χρωµοσωµατικά ρήγµατα-χρωµοσωµατική απώλεια, που επάγονται στις δύο ηλικιακές οµάδες µετά την επίδραση του υδροχλωροθειαζιδίου, εφαρµόσθηκε η µέθοδος αναστολής της κυτταροκίνησης, CBMN, σε συνδυασµό µε in situ υβριδοποίηση µε φθοροχρώµατα, FISH µε χρήση πανκεντροµερικού ανιχνευτή. Η γενετική δράση του HCTZ εκδηλώνεται µέσω δύο διαφορετικών µηχανισµών. Η επίδραση της συγκέντρωσης, 4 µg/ml HCTZ προκαλεί το φαινόµενο της χρωµοσωµατικής απώλειας, όπως αποδεικνύεται από την πλειονότητα των επαγόµενων µικροπυρήνων που περιέχουν κεντροµερικό σήµα (C + MN), που υποδηλώνει την παρουσία ολόκληρων χρωµοσωµάτων. Την κυρίαρχη κατηγορία και στις δύο ηλικιακές οµάδες αποτελούν οι µικροπυρήνες που εµφανίζουν ένα κεντροµερικό σήµα (C 1+ ΜΝ) και εποµένως περιέχουν ένα χρωµόσωµα. Παρουσία της µέγιστης συγκέντρωσης HCTZ η αύξηση που παρατηρείται στις συχνότητες C + MN σε σύγκριση µε τις καλλιέργειες-µάρτυρες είναι τριπλάσια σε νεαρά άτοµα και διπλάσια σε ηλικιωµένα άτοµα. Η ικανότητα του HCTZ να προκαλεί χρωµοσωµατική θραύση προκύπτει από την αύξηση που παρατηρείται σε µικροπυρήνες οι οποίοι χαρακτηρίζονται από την απουσία κεντροµερικού σήµατος (C - MN) και εποµένως δηµιουργούνται από τη συσσώρευση άκεντρων χρωµοσω- µατικών τµηµάτων. Η αύξηση αυτή συµβαίνει σε µικρότερο βαθµό, κατά 1.4 φορές σε σύγκριση µε τις καλλιέργειες-µάρτυρες, στις δύο ηλικιακές οµάδες παρουσία της µέγιστης συγκέντρωσης HCTZ. Το HCTZ εκδηλώνει µία σχετικά ισχυρή ανευπλοειδογόνο δράση, η οποία ισχύει για όλες τις οµάδες ατόµων που εξετάσθηκαν, διαφορετικής ηλικίας και διαφορετικού φύλου. Σε αρκετές φαρµακευτικές ενώσεις που χρησιµοποιούνται ευρέως στη θεραπεία της υπέρτασης έχουν αποδοθεί γονιδιοτοξικές και µεταλλαξιγόνες ιδιότητες. Αγγειοδιασταλτικά, όπως η υδραλαζίνη και η διυδραλαζίνη έχουν εκτεταµένα µελετηθεί, in vitro και in vivo, για την αποσαφήνιση του µηχανισµού της γενετικής δράσης τους (Kodama et al., 198, Williams et al., 198, De Flora et al., 1982, Kitamura et al., 1985). Με την εφαρµογή της µεθόδου των µικροπυρήνων σε L929 κύτταρα βρέθηκε ότι οι δύο φαρµακευτικές ενώσεις επάγουν το σχηµατισµό µικροπυρήνων απουσία µεταβολικής ενεργοποίησης (Chlopkiewicz, 21). Η µεταλλαξιγόνος δράση ελέγχθηκε σε στελέχη του βακτηρίου Salmonella typhimirium παρουσία ή απουσία εξωγενούς µεταβολικού συστήµατος ενεργοποίησης. Παρατηρείται σηµαντική αύξηση του αριθµού των επαναµεταλλαγµένων αποικιών στα στελέχη ΤΑ1 και ΤΑ1537 από την υδραλαζίνη γεγονός που υποδηλώνει την ικανότητα της ένωσης να επάγει σηµειακές µεταλλάξεις αντικατάστασης βάσεων και µετατόπισης αναγνωστικού πλαισίου Η διυδραλαζίνη προκαλεί µεταλλάξεις τύπου µετατόπισης αναγνωστικού πλαισίου. (Williams et al., 198). Επιπλέον η χλωριω- µένη µορφή της υδραλαζίνης, hydralazine hydrochloride, προκαλεί χρωµοσω- µατικές ανωµαλίες σε κύτταρα θηλαστικών (Kodama et al., 198). Ανταγωνιστές των β-αδρενεργικών υποδοχέων της καρδιάς (β-blockers) µε ευρεία χρήση στη - 2 -
212 Συζήτηση θεραπεία της υπέρτασης έχουν χαρακτηρισθεί σε in vivo µελέτες για την ανευπλοειδογόνο δράση τους. Η ατενολόλη ανήκει σε αυτή την κατηγορία και έχει βρεθεί ότι σε λεµφοκύτταρα περιφερικού αίµατος προκαλεί σηµαντική αύξηση στις συχνότητες µικροπυρήνων που περιέχουν ολόκληρα χρωµοσώµατα, ενώ δεν επάγει αυξηµένες συχνότητες ανταλλαγών µεταξύ αδελφών χρωµατιδίων (Telez et al., 2). Η γενετική δράση της προπρανολόλης µελετήθηκε σε σωµατικά και γεννητικά κύτταρα ποντικού. Η προπρανολόλη επάγει µικροπυρήνες in vivo σε αιµοποιητικά κύτταρα ποντικού και δεν προκαλεί χρωµοσωµατικές ανωµαλίες σε γεννητικά κύτταρα (Aruna και Krishnamurthy, 1986). Αναστολείς διαύλων ασβεστίου, όπως η βεραπαµίλη και η νιφεδιπίνη, έχουν εκτεταµένα µελετηθεί σε διάφορα βιολογικά συστήµατα (Friedman et al., 199, Nesterova et al., 1999, Telez et al., 21). Η νιφεδιπίνη εκδηλώνει µία ισχυρή ανευπλοειδογόνο δράση σε λεµφοκύτταρα περιφερικού αίµατος in vitro, όπως διαπιστώνεται από την επαγωγή µικροπυρήνων µε κεντροµερικό σήµα. Η δράση αυτή δεν παρατηρείται in vivo σε λεµφοκύτταρα ασθενών µετά τη χορήγηση της φαρµακευτικής ένωσης πιθανότατα είτε εξαιτίας της µετατροπής της σε ένα λιγότερο δραστικό µεταβολίτη είτε στην ικανότητα προσαρµογής των κυττάρων σε χαµηλές δόσεις (adaptive response) (Telez et al., 21). Η διευθέτηση των κεντροµερών των χρωµοσωµάτων, ο σχηµατισµός συµπλεγµάτων και η συσχέτιση τους µε τον πυρηνίσκο και την πυρηνική µεµβράνη είναι χαρακτηριστικά στοιχεία σε µεσοφασικούς πυρήνες κυττάρων στον άνθρωπο και εξαρτώνται από τις φάσεις του κυτταρικού κύκλου. Αυτή η εξειδικευµένη χωροταξική διάταξη των χρωµοσωµάτων σχετίζεται µε την έκφραση γονιδίων, την αντιγραφή του DNA, τη µεταγραφή και ωρίµανση του RNA και την κυτταρική διαίρεση (Weimer at al., 1992, Solovei et al., 24). Εποµένως, η οργάνωση των κεντροµερικών περιοχών σε µεσοφασικούς πυρήνες πιθανότατα παρουσιάζει σηµαντικό ρόλο στην κυτταρική απόκριση σε χηµικές ενώσεις, βινµπλαστίνη (Norppa et al., 1993a), µιτοµυκίνη-c (Renzi et al., 1996). Παράλληλα µε την εκτίµηση της γενετικής δράσης του υδροχλωροθειαζιδίου σε διεγερµένα λεµφοκύτταρα εξετάσθηκε εάν η φαρµακευτική ένωση εκδηλώνει διαφορετική δράση σε περίπτωση επίδρασης σε µη διαιρούµενα λεµφοκύτταρα. Το HCTZ προστίθεται σε µη διεγερµένα λεµφοκύτταρα και αποµακρύνεται πριν αυτά διεγερθούν προς διαίρεση. Αναλυτικότερα, η επίδραση του υδροχλωροθειαζιδίου στις συγκεντρώσεις 5 και 4 µg/ml πραγµατοποιήθηκε στην προσυνθετική Gο φάση του κυτταρικού κύκλου απουσία του ανοσοδιεγερτικού παράγοντα PHA µε αποτέλεσµα οι καλλιέργειες να θεωρούνται ως µη διεγερµένες. Η µελέτη περιλαµβάνει νεαρά άτοµα εξαιτίας της ευαισθησίας που παρουσιάζουν σε σύγκριση µε τα ηλικιωµένα στη δράση του υδροχλωροθειαζιδίου σε καλλιέργειες διεγερµένων λεµφοκυττάρων. Οι παρατηρήσεις που αφορούν την επαγωγή µικροπυρήνων και την κυτταροτοξικότητα συµφωνούν µε τα αποτελέσµατα σε καλλιέργειες διεγερµένων λεµφοκυττάρων. Σηµαντική είναι η επίδραση της συγκέντρωσης 4 µg/ml HCTZ στην επαγωγή του σχηµατισµού µικροπυρήνων που χαρακτηρίζονται, είτε από την παρουσία είτε από την απουσία κεντροµερικού σήµατος. Το γεγονός αυτό αποδίδει στο HCTZ την ικανότητα χρωµοσωµατικής καθυστέρησης και χρωµοσωµατικής θραύσης. Η αύξηση που προκαλείται στους C + MN και στους C - MN είναι 2.4 και 1.5, αντίστοιχα σε σχέση µε τις καλλιέργειες- µάρτυρες γεγονός που υποδηλώνει µία ισχυρή ανευπλοειδογόνο δράση σε συµφωνία µε τα αποτελέσµατα σε καλλιέργειες διεγερµένων λεµφοκυττάρων (διάγραµµα 3)
213 Συζήτηση C + MN MN HCTZ (µg/ml) 5 ιεγερµένα Μη διεγερµένα ιάγραµµα 3. Μεταβολή στη συνολική συχνότητα των µικροπυρήνων (ΜΝ) και στη συχνότητα των µικροπυρήνων µε κεντροµερικό σήµα (C + MN) σε διεγερµένες και µη διεγερµένες µε PHA κατά την επίδραση µε HCTZ καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας, in vitro. 2 Σύµφωνα µε τα παραπάνω αποτελέσµατα φαίνεται να αποκλείεται µία εξαρτώµενη δράση της φαρµακευτικής ένωσης από την κατάσταση διαίρεσης των κυττάρων και πιθανότατα από τη διαφορετική θέση των κεντροµερών των χρωµοσωµάτων σε διεγερµένα και µη διεγερµένα λεµφοκύτταρα. Η MMC γνωστή για την ικανότητα της να προκαλεί ρήγµατα στο γενετικό υλικό εµφανίζει ανευπλοειδογόνο δράση που οφείλεται σε ρήξη ή/και αποκόλληση του κινητοχώρου από το κεντροµέρος σε διεγερµένα σπληνοκύτταρα ποντικού. Παρόµοια δράση εµφανίζει σε µη διεγερµένα σπληνοκύτταρα η οποία δεν δύναται να αποδοθεί στην ίδια αιτία εξαιτίας της ακεραιότητας του συµπλόκου κινητοχώρου-κεντροµέρους, αλλά σε διαφορετική ευαισθησία των κυττάρων κατά τη χηµική επίδραση (Renzi et al., 1996). Η παρουσία βινµπλαστίνης σε καλλιέργειες λεµφοκυττάρων 24 ώρες πριν τη διέγερση τους σχετίζεται µε την εκδήλωση ανευπλοειδογόνου δράσης. Σε διεγερµένες καλλιέργειες η ίδια γενετική δράση παρατηρείται σε χαµηλότερες συγκεντρώσεις (Norppa et al., 1993). Αναµφισβήτητα, το υδροχλωροθειαζίδιο στη συγκέντρωση 4 µg/ml εµφανίζει µία ισχυρή ανευπλοειδογόνο δράση και ταυτόχρονα παρατηρείται µία λιγότερο έντονη δράση της φαρµακευτικής ένωσης ως προς την ικανότητα δηµιουργίας ρηγµάτων. Αρκετές χηµικές ενώσεις έχουν αναφερθεί για τη θραυσµατογόνο, clastogen, και ανευπλοειδογόνο δράση τους. Ελάχιστες είναι οι χηµικές ενώσεις που επάγουν αποκλειστικά C - MN ή C + MN και στις οποίες δύναται να αποδοθεί µία ιδιότητα (Schuler et al., 1997). Χηµικές ενώσεις που επιδρούν στη µιτωτική συσκευή, δεµεκολσίνη, βινκριστίνη, και χαρακτηρίζονται ως ανευπλοειδογόνες, είναι δυνατόν να προκαλούν µικρή αύξηση σε µικροπυρήνες που δεν φέρουν κεντροµερικό σήµα, C - MN (Bakou et al., 22). Η κολχικίνη και η δεµεκολσίνη βρέθηκε να επάγουν χρωµοσωµατικά ρήγµατα και άλλες δοµικές χρωµοσωµατικές ανωµαλίες σε CHO κύτταρα (Matsuoka et al., 1992, Arni και Hertner, 1997). Το βουταδιένιο και οι µεταβολίτες του, εποξυβουταδιένιο και διεποξυβουταδιένιο, εξετάσθηκε σε διαφορετικά βιολογικά συστήµατα στον ποντικό και στον άνθρωπο και βρέθηκε ότι
214 Συζήτηση εκτός από την ικανότητα πρόκλησης ρηγµάτων παρουσιάζουν επιπλέον ασθενή ανευπλοειδογόνο δράση (Xiao et al., 1996, Stephanou et al., 1997, 1998, Vlacho dimitropoulos et al., 1997). Σε αλκυλιωτικούς παράγοντες που ανήκουν στην κατηγορία των υπεριτών του αζώτου έχουν αποδοθεί ανευπλοειδογόνες ιδιότητες (Andrianopoulos et al., 2, Kouloumenta et al., 25). Αυτή η ιδιότητα πιθανά οφείλεται στην ικανότητα τους να αλκυλιώνουν πρωτεΐνες που συµµετέχουν στο διαµοιρασµό των χρωµοσωµάτων (Dellarco et al., 1986) µε αποτέλεσµα τη δηµιουργία οµοιοπολικών δεσµών µεταξύ νουκλεϊνικών οξέων και πρωτεϊνών. Επιπρόσθετα, η ανευπλοειδογόνος ιδιότητα χηµικών ενώσεων που προκαλούν κυρίως ρήγµατα στο γενετικό υλικό αποδίδεται σε ασύµµετρη χρωµοσωµατική ανταλλαγή η οποία οδηγεί σε χρωµοσωµατικές γέφυρες ή σε αλληλοσυνδεόµενα χρωµατίδια. Έτσι από τη συσσώρευση χρωµοσωµάτων στα οποία έχουν συµβεί τέτοιου είδους ανταλλαγές προκύπτουν µικροπυρήνες µε κεντροµερικό σήµα, C + MN (Abramsson Zetterberg, 1997). Το υδροχλωροθειαζίδιο εµφανίζει αυξηµένες συχνότητες ανταλλαγών µεταξύ αδελφών χρωµατιδίων σε CHO κύτταρα (Anderson et al., 199). Οι θεωρίες που έχουν προταθεί για την εξήγηση του µηχανισµού που προκαλεί ανταλλαγές αδελφών χρωµατιδίων έχουν ένα κοινό σηµείο αναφοράς. Σύµφωνα µε αυτό συµβαίνουν ανταλλαγές σε προϊόντα αντιγραφής του DNA σε οµόλογες θέσεις οι οποίες προϋποθέτουν τη δηµιουργία ρήγµατος και επανασύνδεσης. Τα γεγονότα αυτά συµβαίνουν κοντά στη διχάλα αντιγραφής του DNA και επηρεάζονται από τη δυνατότητα των κυττάρων να επιδιορθώνουν τις βλάβες πριν το DNA προχωρήσει στην αντιγραφή (Tucker et al., 1993). Η βεραπαµίλη ενισχύει τη θραυσµατογόνο δράση του κυκλοφωσφαµιδίου, της ακρυλαµίδης και της διοξιδίνης σε ποντικούς, γεγονός που οφείλεται σε αναστολή της π-γλυκοπρωτεΐνης (Nesterova et al., 1999). Η βεραπαµίλη και η µεθυλντόπα βρέθηκε ότι επάγουν δοµικές ανωµαλίες χρωµατιδικού τύπου σε ανθρώπινα λεµφοκύτταρα in vitro, ενώ παρόµοια δράση εµφανίζει η βεραπαµίλη σε µικρότερο βαθµό, in vivo (Friedman et al., 199, Grisolia και Takahashi, 1991). Οι επιδράσεις της ηλικίας στον άνθρωπο φαίνεται να είναι συνδυασµός επιρροών από γενετικά προγραµµατισµένα φαινόµενα και εξωγενείς περιβαλλοντικούς παράγοντες, οι οποίοι λαµβάνουν χώρα σε κυτταρικό επίπεδο, γήρανση, παρά σε επίπεδο οργανισµού. Η διαδικασία της κυτταρικής-αντιγραφικής γήρανσης προκαλείται από βλάβες του πυρηνικού και µιτοχονδριακού DNA ή/και από ελαττωµένη επιδιόρθωση του DNA (Campisi, 2, Hasty, 25). Η εκτίµηση της συχνότητας χρωµοσωµατικών ανωµαλιών και η ανάλυση µικροπυρήνων σε επιδη- µιολογικές µελέτες ενδείκνυνται ως βιολογικοί δείκτες της κυτταρικής γήρανσης. Η απώλεια του DNA, η οποία εκφράζεται µε το σχηµατισµό µικροπυρήνων σχετίζεται µε τη γήρανση, τον καρκίνο και την περιβαλλοντική έκθεση (Leach και Jackson-Cook, 21, Wojda και Witt, 23). Στην παρούσα εργασία παρατηρείται αυξηµένη συχνότητα µικροπυρήνων σε αρσενικά και θηλυκά ηλικιωµένα άτοµα. Το φύλο επηρεάζει τη συχνότητα µικροπυρήνων σε µικρότερο βαθµό σε σύγκριση µε την ηλικία µε τα θηλυκά άτοµα να εµφανίζουν υψηλότερες συχνότητες µικροπυρήνων σε σύγκριση µε τα αρσενικά. Τα αποτελέσµατα αυτά και οι στατιστικές αναλύσεις επιβεβαιώνουν την επίδραση της ηλικίας και του φύλου στην επαγωγή µικροπυρήνων σε ανθρώπινα λεµφοκύτταρα (Fenech και Morley, 1985b, Migliore et al., 1991, Fenech et al., 1994). Από τα ευρήµατα ενός διεθνούς προγράµµατος, HUMN, προκύπτει πως η αύξηση στις συχνότητες µικροπυρήνων παρουσιάζεται εντονότερη µετά την ηλικία των σαράντα ετών και συνεχίζει να ισχύει µε την αύξηση της ηλικίας (Fenech et al., 1999, Bonassi et al., 21). Ένα άλλο στοιχείο που προκύπτει από τη σύγκριση των δύο ηλικιακών οµάδων είναι ότι τα θηλυκά
215 Συζήτηση ηλικιωµένα άτοµα παρουσιάζουν µεγαλύτερη συχνότητα µικροπυρήνων µε κεντροµερικό σήµα σε σύγκριση µε τα αρσενικά, γεγονός το οποίο αποδίδεται κυρίως σε αυξηµένη απώλεια του χρωµοσώµατος Χ. Σε αρσενικά ηλικιωµένα άτοµα παρατηρείται αυξηµένη συχνότητα µικροπυρήνων µε κεντροµερικό σήµα που αποδίδεται στην απώλεια των φυλετικών χρωµοσωµάτων, Χ και Υ (Hando et al., 1994, Zijno et al., 1996c, Catalan et al., 1998, Bakou et al., 22). Σηµαντική είναι η επίδραση της ηλικίας στο σχηµατισµό µικροπυρήνων που περιέχουν άκεντρα χρωµοσωµατικά τµήµατα, γεγονός το οποίο οφείλεται σε συνάθροιση βλαβών στο DNA αδρανών λεµφοκυττάρων. Οι βλάβες αυτές µετατρέπονται σε χρωµοσω- µατικά ρήγµατα και εκφράζονται ως µικροπυρήνες οι οποίοι χαρακτηρίζονται από την απουσία κεντροµερικού σήµατος (Falck et al., 22, Bakou et al., 22, Norppa και Falck, 23). Απουσία υδροχλωροθειαζιδίου οι συχνότητες µικροπυρήνων δεν φαίνεται να επηρεάζονται από τη συνήθεια του καπνίσµατος. Tα µεταλλαξιγόνα συστατικά του καπνού, νιτροζαµίνες, αρωµατικοί και ετεροκυκλικοί υδρογονάνθρακες, αµίνες ευθύνονται για τις αυξηµένες συχνότητες χρωµοσω- µατικών βλαβών που παρατηρούνται σε καπνιστές σε σχέση µε τους µη καπνιστές. Σε αρκετές µελέτες τo κάπνισµα έχει σχετιστεί θετικά µε τη συχνότητα εµφάνισης µικροπυρήνων σε ανθρώπινους ιστούς. Αντίθετα, σε άλλες επιδηµιολογικές µελέτες οι οποίες διερευνούν την επίδραση της εργασιακής έκθεσης στον άνθρωπο, όπου το κάπνισµα δεν έχει συνυπολογισθεί ως επιπλέον παράµετρος δεν έχει βρεθεί παρόµοια σχέση. Πιθανόν οι χηµικές ενώσεις που εξετάζονται ενεργοποιούν είτε την έκφραση γονιδίων που σχετίζονται µε τα συστήµατα επιδιόρθωσης βλαβών του DNA είτε µηχανισµούς που προκαλούν την αποτοξικοποίηση τους. Οι κυτταρικές αυτές διαδικασίες προκαλούν εξασθένηση της γενετικής δράσης αρκετών µεταλλαξιγόνων χηµικών συστατικών που ανιχνεύονται στον καπνό του τσιγάρου. Από την ανάλυση ενός µεγάλου αριθµού δειγµάτων ανθρώπινων λεµφοκυττάρων, HUMN, παρατηρείται στατιστικά σηµαντική αύξηση στη συχνότητα µικροπυρήνων σε βαρείς καπνιστές που δεν έχουν εκτεθεί σε γονιδιοτοξικούς παράγοντες (Bonassi et al., 23). Επιπρόσθετα, οι βαρείς καπνιστές εµφανίζουν µειωµένη ικανότητα απορρόφησης φολικού οξέος και βιταµίνης Β12, και η έλλειψη τους συµβάλλει καθοριστικά στη δηµιουργία µικροπυρήνων. Οι βιταµίνες Α, C και Ε παρέχουν προστατευτικό ρόλο προκαλώντας σηµαντική µείωση στις συχνότητες των µικροπυρήνων σε καπνιστές, εάν και αµφισβητείται ο ρόλος της βιταµίνης C στην προστασία του κυττάρου από το οξειδωτικό στρες, ROS (Alaoui-Jamali et al., 1991, Odagiri και Uchida, 1998, Schneider et al., 21, Fenech et al., 25). Εάν η επίδραση του υδροχλωροθειαζιδίου και η συνήθεια του καπνίσµατος συνυπολογισθούν ως παράµετροι που δύναται να επηρεάσουν τις συχνότητες µικροπυρήνων σε ανθρώπινα λεµφοκύτταρα προκύπτει ότι το κάπνισµα επηρεάζει το σχηµατισµό µικροπυρήνων που περιέχουν άκεντρα χρωµοσωµατικά τµήµατα, ενώ δεν επάγει το σχηµατισµό µικροπυρήνων που θεωρούνται προϊόν χρωµοσωµατικής απώλειας. Έτσι αποδίδεται στο κάπνισµα µία θραυσµατογόνος δράση, η οποία επιβεβαιώνει την άποψη άλλων ερευνητών για τις βλάβες που είναι δυνατόν να ανιχνευθούν σε ανθρώπινα λεµφοκύτταρα (Norppa και Falck, 23). Ένα από τα πιο σηµαντικά συστατικά του αέρα των εσωτερικών χώρων είναι ο καπνός του τσιγάρου (ETS) που ευθύνεται για το παθητικό κάπνισµα. Σε µία ανασκόπηση της Husgafvel- Pursiainen (24) σχετικά µε την έκθεση στον περιβαλλοντικό καπνό του τσιγάρου (ETS) παρουσιάζονται τα αποτελέσµατα in vitro και in vivo µελετών. Οι in vitro µελέτες παρουσιάζουν επαγωγή ρηγµάτων στο DNA, σχηµατισµό τροποποιηµένων νουκλεοτιδίων, µεταλλαξιγένεση σε βακτηριακούς ελέγχους και κυτταρογενετικές βλάβες
216 Συζήτηση Τα αποτελέσµατα in vivo πειραµάτων σε τρωκτικά συσχετίζουν την ολόσωµη έκθεση σε ETS µε την πρόκληση µονόκλωνων ρηγµάτων σε αλυσίδες του DNA, οξειδωτικής βλάβης από το σχηµατισµό αρωµατικών παραπροϊόντων, χρωµοσω- µατικών ανωµαλιών και δηµιουργίας µικροπυρήνων. Σε παιδιά ηλικίας 3-15 ετών που βρίσκονται σε περιβάλλον παθητικού καπνίσµατος, η συχνότητα των µικροπυρήνων σε περιφερικά λεµφοκύτταρα παρατηρείται αυξηµένη σε σχέση µε συνοµήλικα παιδιά που δεν εκτίθενται σε παθητικό κάπνισµα (Baier et al., 22). Πρόσφατα, σε µία ευρείας κλίµακα µοριακή επιδηµιολογική µελέτη, AULIS, ερευνάται η σχέση µεταξύ της έκθεσης πληθυσµού σε ατµοσφαιρικούς ρυπαντές, PAHs και αιωρούµενο σωµατιδιακό υλικό µε διάφορους βιολογικούς και γενετικούς δείκτες. Η έκθεση σε ETS προκαλεί αύξηση στο σχηµατισµό τροποποιηµένων νουκλεοτιδίων και στις συχνότητες δοµικών χρωµοσωµατικών ανωµαλιών, που ανιχνεύονται σε λεµφοκύτταρα περιφερικού αίµατος νεαρών ατόµων. Επειδή αρκετοί από τους πολυκυκλικούς αρωµατικούς υδρογονάνθρακες είναι έµµεσα µεταλλαξιγόνα και για την εκδήλωση της δράσης τους απαιτείται µεταβολική ενεργοποίηση µελετώνται πολυµορφισµοί ενζύµων που συµµετέχουν στο πρώτο και δεύτερο στάδιο του µεταβολισµού ξενοβιοτικών ενώσεων. Πολυµορφισµοί των γονιδίων CYP1A1, GSTM1, GSTP1, EPHX σχετίζονται µε αυξηµένο σχηµατισµό τροποποιη- µένων νουκλεοτιδίων σε άτοµα που εκτίθενται σε βεβαρηµένο αέρα εσωτερικών χώρων από τον καπνό των τσιγάρων (Georgiadis et al., 24, 25). Πρόκειται για οµοιοπολικά σύµπλοκα που σχηµατίζονται µεταξύ γονιδιοτοξικών καρκινογόνων ενώσεων και βάσεων του DNA και αποτελούν ένα πρώιµο στάδιο στο µονοπάτι της χηµικής καρκινογένεσης (Luch, 22, Baird et al., 25). Οι µηχανισµοί που έχει αναπτύξει το κύτταρο για την επιδιόρθωση τροποποιηµένων νουκλεοτιδίων περιλαµβάνουν την επιδιόρθωση µε εκτοµή βάσεων, BER ή νουκλεοτιδίων, NER. Η αδυναµία επιδιόρθωσης τους πριν τη φάση S της αντιγραφής έχει ως συνέπεια τη δηµιουργία ρηγµάτων στο γενετικό υλικό. Αρκετά συστατικά του καπνού που εκλύονται είτε ως αέρια είτε ως συµπυκνωµένα σωµατίδια πίσσας ευθύνονται για παθολογικές αλλαγές που συµβαίνουν στο αναπνευστικό επιθήλιο και σχετίζονται µε την εµφάνιση καρκίνου των πνευµόνων. Οι πολυκυκλικοί αρωµατικοί υδρογονάνθρακες, PAHs ενέχονται στην πρόκληση µεταλλάξεων ενός σηµαντικού αριθµού ογκοκατασταλτικών γονιδίων, TP53, και ογκογονιδίων, KRAS. Αυτές θεωρούνται δείκτες αυξηµένου κινδύνου εµφάνισης καρκίνου των πνευµόνων σε άτοµα που εκτίθενται σε ETS (Vineis και Husgafvel-Pursiainen, 25). Η επαγωγή µικροπυρήνων και η δυνατότητα δράσης του υδροχλωροθειαζιδίου σε πιθανούς στόχους που ενέχονται στην πρόκληση χρωµοσωµατικής καθυστέρησης µελετήθηκε σε ανθρώπινες κυτταρικές σειρές. Αυξηµένες συχνότητες µικροπυρήνων παρατηρούνται σε καλλιέργειες κυττάρων HFFF2 και HEp2 µετά από την επίδραση µε HCTZ. Από τον προσδιορισµό του µηχανισµού δηµιουργίας των µικροπυρήνων µε τη συνδυασµένη σήµανση του κινητοχώρου και του κεντροµέρους προκύπτει ότι το HCTZ εµφανίζει ισχυρή ανευπλοειδογόνο δράση. Η επίδραση του HCTZ προκαλεί αύξηση σε µικροπυρήνες που χαρακτηρίζονται από την ταυτόχρονη παρουσία του κινητοχώρου και του κεντροµέρους, K + C + MN, ενώ παρόµοια αύξηση δεν ισχύει για τους Κ - C + MN. Το γεγονός αυτό υποδηλώνει την απώλεια ολόκληρων χρωµοσωµάτων που χαρακτηρίζονται από την ακεραιότητα της κεντροµερικής περιοχής και αποκλείει την πιθανότητα δράσης του HCTZ διαµέσου της δηµιουργίας ρηγµάτων στην κεντροµερική περιοχή ή της αποκόλλησης των κινητοχώρων. Οι µικροπυρήνες που περιέχουν άκεντρα χρωµοσωµατικά τµήµατα, K - C - ΜΝ, παρουσιάζουν µικρότερη αύξηση σε σχέση µε τους C + MN και διαφέρουν
217 Συζήτηση στατιστικά από τις καλλιέργειες-µάρτυρες γεγονός που αποδίδει στο HCTZ ασθενή ικανότητα πρόκλησης ρηγµάτων στο γενετικό υλικό. Οι Vlastos και Stephanou (1998) απέδωσαν στη διυδροχλωρική σετιριζίνη, αντισταµινική φαρµακευτική ένωση, διπλή γενετική δράση σε ανθρώπινα λεµφοκύτταρα. Η ανευπλοειδογόνος δράση της σετιριζίνης οφείλεται σε απώλεια ολόκληρων χρωµοσωµάτων και δεν σχετίζεται µε την αποκόλληση των κινητοχώρων από τα κεντροµέρη. H ακτινοβόληση µε ακτίνες Χ πρωτογενών ανθρώπινων κυττάρων in vitro στη G1 και G2 φάση προκαλεί ανευπλοειδικά φαινόµενα, που οφείλονται τόσο στη χρωµοσωµατική απώλεια, όσο και σε µη αποχωρισµό. Υψηλότερες συχνότητες χρωµοσωµατικής απώλειας παρατηρούνται όταν η ακτινοβόληση γίνεται στη G1 φάση, υποδηλώνοντας ότι η ανευπλοειδία κατά τη διάρκεια της G1 φάσης µπορεί να επιτυγχάνεται µέσω αλληλεπίδρασης µε στόχους στο µόριο του DNA (Kirsch-Volders et al., 1996). Η ανάλυση της δοµής του κινητοχώρου και του κεντροµέρους που χρησιµοποιήθηκε στη µελέτη χαµηλών δόσεων ακτινοβολίας σε ανθρώπινους ινοβλάστες αποσαφήνισε τη γενετική δράση των ακτίνων Χ. Οι ακτίνες Χ, εκτός από ισχυρή θραυσµατογόνο δράση εµφανίζουν την ικανότητα να προκαλούν απώλεια ολόκληρων χρωµοσωµάτων που χαρακτηρίζονται από την ακεραιότητα της κεντροµερικής περιοχής. Έτσι φαίνεται ότι αλληλεπιδρούν µε άλλους κυτταρικούς στόχους οι οποίοι εµπλέκονται στην πρόκληση φαινοµένων λανθασµένου αποχωρισµού. Αντίθετα η επαγόµενη, από 28.5 KeV πρωτόνια, χρωµοσωµατική απώλεια φαίνεται να είναι συνέπεια της δηµιουργίας ρηγµάτων στην κεντροµερική περιοχή ή της αποκόλλησης των κινητοχώρων που συνεπάγεται την απώλεια ολόκληρων χρωµοσωµάτων (Sgura et al., 21). Η κυτταροτοξική δράση του HCTZ σε ανθρώπινα λεµφοκύτταρα επιβεβαιώνεται σε ανθρώπινες κυτταρικές σειρές από την πτώση των τιµών του δείκτη πολλαπλασιασµού των κυττάρων, CBPI και του ποσοστού διπύρηνων κυττάρων. Η αυξηµένη συχνότητα µικροπυρήνων που θεωρούνται προϊόν χρω- µοσωµατικής απώλειας απέδωσε στο υδροχλωροθειαζίδιο ανευπλοειδογόνο δράση. Η χρωµοσωµατική απώλεια και ο µη αποχωρισµός είναι τα κύρια αποτελέσµατα της δράσης των ανευπλοειδογόνων ενώσεων είτε µε την άµεση είτε µε την έµµεση πρόσδεση τους στη µιτωτική άτρακτο, στις τουµπουλίνες, στις πυρηνικές πρωτεΐνες, στο κεντροµέρος ή σε περιοχές του κινητοχώρου (Elhajouji et al., 1997). Η διαπίστωση και αποσαφήνιση της ανευπλοειδογόνου δράσης του HCTZ επιτυγχάνεται µε την ανάλυση των δύο κύριων µηχανισµών που οδηγούν σε ανευπλοειδία, του µη αποχωρισµού των χρωµοσωµάτων και της χρωµοσωµατικής απώλειας. Απαραίτητη προϋπόθεση για τη µελέτη της δράσης χηµικών ενώσεων είναι η επιλογή των κατάλληλων χρωµοσωµάτων. Στις παραµέτρους που θα πρέπει να λαµβάνονται υπόψη είναι το µέγεθος του χρωµοσώµατος, η συχνότητας εµφάνισης του µε σχετιζόµενες µε την ανευπλοειδία ασθένειες ή αποβολές και η παρουσία σηµαντικών γονιδίων που συµµετέχουν στην καρκινογένεση στον άνθρωπο (Parry et al., 1994, Parry et al., 22). Η µελέτη µας πραγµατοποιήθηκε µε τη διπλή in situ υβριδοποίηση, DUAL FISH, µε τη συµµετοχή κεντροµερικών ανιχνευτών για τα χρωµοσώµατα Χ, Υ και 8. Το HCTZ προκαλεί µη αποχωρισµό των χρωµοσωµάτων Χ, Υ και 8 στις εξεταζόµενες συγκεντρώσεις 5 και 4 µg/ml και στις δύο ηλικιακές οµάδες. Η ηλικία, το φύλο και η συνήθεια του καπνίσµατος δεν τροποποιεί την ικανότητα του HCTZ να επάγει χρωµοσωµατικό µη αποχωρισµό. Ωστόσο η ηλικία αποτελεί σηµαντικό παράγοντα που επηρεάζει τον τυχαίο µη αποχωρισµό των φυλετικών χρωµοσωµάτων και του χρωµοσώµατος 8 σε αντίθεση µε το φύλο και τη συνήθεια του καπνίσµατος
218 Συζήτηση Σε αρκετές µελέτες έχει διαπιστωθεί ότι το φαινόµενο του χρωµοσωµατικού µη αποχωρισµού επηρεάζεται από την ηλικία τόσο σε αρσενικά όσο και σε θηλυκά άτοµα. Το χρωµόσωµα Χ εµφανίζει µεγαλύτερη ευαισθησία στο φαινόµενο αυτό συγκριτικά µε το χρωµόσωµα Υ και το χρωµόσωµα 8 ή άλλα αυτοσωµατικά χρωµοσώµατα (Carere et al., 1999, Catalan et al., 2b, Bakou et al., 22, Leopardi et al., 22). Με δεδοµένο τον αριθµό των δοτών που συµπεριελήφθησαν σε µία από τις µελέτες, 7 καπνιστές, 12 µη καπνιστές, το κάπνισµα δεν φαίνεται να ενέχεται σε φαινόµενα λανθασµένου αποχωρισµού (Carere et al., 1999). Τα αποτελέσµατα µας έδειξαν ότι συγκριτικά µε το χρωµόσωµα Υ και 8, το χρωµόσωµα Χ είναι περισσότερο ευαίσθητο σε ανευπλοειδικά φαινόµενα που προκαλούνται από το HCTZ τόσο στις νεαρές όσο και στις µεγαλύτερες ηλικιακές οµάδες (διάγραµµα 4). Η ευαισθησία του χρωµοσώµατος Χ συγκριτικά µε το χρωµόσωµα 8 ή άλλα αυτοσω- ΜΗ ΑΠΟΧΩΡΙΣΜΟΣ (%) Xρωµόσωµα Χ Χρωµόσωµα Υ Χρωµόσωµα 8 :p<.5 σε σχέση µε το µάρτυρα :p<.5 σε σχέση µε 5 µg/ml HCTZ (One-way ANOVA, Student's t-test) *:p<.5 σε σχέση µε το µάρτυρα (Student's t-test) * Νεαρά Ηλικιωµένα ΜΑΡΤΥΡΑΣ Νεαρά Ηλικιωµένα HCTZ 5 (µg/ml) Νεαρά Ηλικιωµένα HCTZ 4 (µg/ml). ιάγραµµα 4. Μεταβολή του µη αποχωρισµού των χρωµοσωµάτων Χ, Υ και 8 στο µάρτυρα και µετά από επίδραση HCTZ σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. µατικά χρωµοσώµατα (1, 2, 11, 17) µετά από επίδραση µε ανευπλοειδογόνους παράγοντες σε ανθρώπινα λεµφοκύτταρα έχει διαπιστωθεί σε αρκετές µελέτες. Η επίδραση µε µπενοµύλη και καρβενδαζίµη (Bentley et al., 2), µε γκρισεοφουλβίνη και βανάδιο (Migliore et al., 1999), µε δεµεκολσίνη και βινκριστίνη (Bakou et al., 22), µε κολχικίνη και βινµπλαστίνη (Zigno et al., 1996b, Leopardi et al., 22) έδειξε ότι το χρωµόσωµα Χ ήταν πιο ευαίσθητο στο φαινόµενο αυτό τόσο σε νεαρά όσο και σε ηλικιωµένα άτοµα ανεξάρτητα από το φύλο. Πρόκειται για ένα φαινό- µενο που εµφανίζεται εντονότερο µε την αύξηση της ηλικίας (Zijno et al, 1996c, Bakou et al, 22). Έχει αναφερθεί ότι ο χρωµοσωµατικός µη αποχωρισµός αποτελεί τον κυριότερο µηχανισµό ανευπλοειδίας όταν επάγεται από ενώσεις αναστολείς της µιτωτικής ατράκτου και συµβαίνει σε χαµηλότερες συγκεντρώσεις από ότι η χρωµοσωµατική απώλεια. Αναµφισβήτητα για την επαγωγή χρωµοσωµατικού µη αποχωρισµού από ανευπλοειδογόνες ενώσεις µικρότερος αριθµός στόχων τροποποιείται συγκριτικά µε την επαγωγή χρωµοσωµατικής απώλειας (Elhajouji et al.,
219 Συζήτηση 1997). Οι ενώσεις κολχικίνη, καρβενδαζίµη, µεβενδαζόλη και νοκοδαζόλη, αναστέλλουν τον πολυµερισµό των τουµπουλινών, εµποδίζοντας τη συγκρότηση των µικροσωληνίσκων µε τη δέσµευση τους στη β-τουµπουλίνη σε συγκεκριµένες συγκεντρώσεις. Έτσι βλάβες στα ινίδια της ατράκτου από τους αναστολείς αυτούς αυξάνονται µε την αύξηση της συγκέντρωσης, αλλά η επίδραση των χηµικών αυτών ενώσεων στη µετακίνηση των χρωµοσωµάτων είναι αποτελεσµατική µόνο σε εκείνες τις συγκεντρώσεις που προκαλούν βλάβες στους µικροσωληνίσκους σε πολύ µεγάλο βαθµό (Elhajouji et al., 1995). Πρόσφατες µελέτες υποστηρίζουν µία σχέση µεταξύ ανευπλοειδίας και ηλικίας στην οποία εµπλέκονται κυρίως τα φυλετικά χρωµοσώ- µατα. Πρόκειται για ένα φαινόµενο που γίνεται εντονότερο µε την πρόοδο της ηλικίας. Στα θηλυκά άτοµα το φαινόµενο αυτό αφορά κυρίως την απώλεια του χρω- µοσώµατος Χ, το οποίο παρουσιάζεται σε πολύ υψηλό βαθµό στους µικροπυρήνες θηλυκών ατόµων ο οποίος αυξάνεται ακόµη περισσότερο µε την πάροδο της ηλικίας. Στην παρούσα εργασία εκτός από το φαινόµενο του χρωµοσωµατικού µη αποχωρισµού µελετήθηκε και η χρωµοσωµατική απώλεια που αποτελεί έναν από τους κύριους µηχανισµούς που οδηγούν στην ανευπλοειδία. Το HCTZ προκαλεί απώλεια των χρωµοσωµάτων Χ, Υ και 8 στις εξεταζόµενες συγκεντρώσεις 5 και 4 µg/ml. Η χρωµοσωµατική απώλεια εµφανίζεται εντονότερη στη συγκέντρωση 4 µg/ml HCTZ και στις δύο ηλικιακές οµάδες. Η ηλικία είναι ο κύριος παράγοντας που επηρεάζει την ικανότητα απόκρισης των λεµφοκυττάρων στην επίδραση του HCTZ και επάγει απώλεια των φυλετικών χρωµοσωµάτων. Το φύλο δεν τροποποιεί την ικανότητα του HCTZ να επάγει απώλεια του χρωµοσώµατος Χ (διάγραµµα 5). Σε καλλιέργειες- ΧΡΩΜΟΣΩΜΑΤΙΚΗ ΑΠΩΛΕΙΑ ( ) Χρωµόσωµα Χ ( ) Χρωµόσωµα Χ ( ) Χρωµόσωµα Υ ( ) Νεαρά Ηλικιωµένα ΜΑΡΤΥΡΑΣ :p<.5 σε σχέση µε το µάρτυρα :p<.5 σε σχέση µε 5 µg/ml HCTZ (One-way ANOVA, Student's t-test) Νεαρά Ηλικιωµένα HCTZ 5 (µg/ml) Νεαρά Ηλικιωµένα HCTZ 4 (µg/ml) ιάγραµµα 5. Μεταβολή στη συχνότητα της απώλειας των φυλετικών χρωµοσωµάτων στο µάρτυρα και µετά από επίδραση HCTZ σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro. µάρτυρες η ηλικία και το φύλο αποτελεί σηµαντικό παράγοντα που επάγει το σχηµατισµό µικροπυρήνων που θεωρούνται προϊόν απώλειας του χρωµοσώµατος Χ. Τα θηλυκά ηλικιωµένα άτοµα εµφανίζουν τις υψηλότερες συχνότητες απώλειας του χρωµοσώµατος Χ. Σε αρσενικά άτοµα η απώλεια του χρωµοσώµατος Χ συµβαίνει
220 Συζήτηση µε χαµηλότερους ρυθµούς συγκριτικά µε τα θηλυκά άτοµα. Η ηλικία παραµένει σηµαντικός παράγοντας που επάγει την απώλεια του χρωµοσώµατος Υ. Η µελέτη του χρωµοσώµατος Χ παρουσιάζει σηµαντικό ενδιαφέρον και αποτελεί αντικείµενο ερευνητικής µελέτης εξαιτίας αρκετών σηµαντικών ασθενειών που σχετίζονται µε το συγκεκριµένο χρωµόσωµα και της ύπαρξης στα θηλυκά άτοµα του ανενεργού Χ χρωµοσώµατος. Τα αποτελέσµατα µας επιβεβαιώνουν αυτά άλλων ερευνητικών µελετών και επεκτείνουν τις γνώσεις σχετικά µε την απώλεια των φυλετικών χρω- µοσωµάτων στη διεθνή βιβλιογραφία. Σύµφωνα µε τον Richard και τους συνεργάτες του (1994) οι συχνότητες µικροπυρήνων που φέρουν το χρωµόσωµα Χ αυξάνονται µε την ηλικία σε θηλυκά άτοµα. Οι µικροπυρήνες που φέρουν κεντροµερικό σήµα είναι αυξηµένοι σε θηλυκά ηλικιωµένα άτοµα συγκριτικά µε τα νεαρά άτοµα. Σε ηλικιωµένα άτοµα το µεγαλύτερο ποσοστό µικροπυρήνων είναι προϊόν απώλειας του χρωµοσώµατος Χ (Catalan et al., 1995). Σηµαντική είναι η επίδραση της ηλικίας στην επαγωγή φαινοµένων λανθασµένου αποχωρισµού για το χρωµόσωµα Χ σε αρσενικά και θηλυκά άτοµα (Zijno et al., 1996a, c). Απώλεια του χρωµοσώµατος Χ συµβαίνει σε αρσενικά άτοµα αλλά σε χαµηλότερους ρυθµούς συγκριτικά µε τα θηλυκά άτοµα. Απώλεια του χρωµοσώµατος Υ συµβαίνει σε ηλικιωµένα αρσενικά άτοµα. Η συχνότητα Υ + ΜΝ εµφανίζεται σε ηλικιωµένα άτοµα κατά 8-11 φορές υψηλότερη συγκριτικά µε τα νεαρά άτοµα (Catalan et al., 1998). Αυξηµένες είναι οι συχνότητες µικροπυρήνων που φέρουν το χρωµόσωµα Χ (2.33 ) συγκριτικά µε τα αυτοσωµατικά χρωµοσώµατα 7, 11 και 18 (.2,.16 και.8, αντίστοιχα) σε νεαρά και ηλικιωµένα αρσενικά άτοµα. Η αύξηση της απώλειας του χρωµοσώµατος Χ σε ηλικιωµένα άτοµα φαίνεται να διαφέρει οριακά από τα νεαρά άτοµα (Carere et al., 1999). Η ηλικία και το φύλο δεν φαίνεται να επηρεάζουν καθοριστικά την ικανότητα του HCTZ να επάγει απώλεια του χρωµοσώµατος 8. Σε καλλιέργειες-µάρτυρες η ηλικία παραµένει σηµαντικός παράγοντας που επηρεάζει την απώλεια του χρωµοσώµατος 8 (διάγραµµα 6). Το φύλο δεν επάγει το σχηµατισµό µικροπυρήνων ΧΡΩΜΟΣΩΜΑΤΙΚΗ ΑΠΩΛΕΙΑ ( ) Χρωµόσωµα 8 Χρωµόσωµα Χ Χρωµόσωµα Υ :p<.5 σε σχέση µε το µάρτυρα :p<.5 σε σχέση µε 5 µg/ml HCTZ (One-way ANOVA, Student's t-test) Νεαρά Ηλικιωµένα ΜΑΡΤΥΡΑΣ Νεαρά Ηλικιωµένα HCTZ 5 (µg/ml) Νεαρά Ηλικιωµένα HCTZ 4 (µg/ml) ιάγραµµα 6. Μεταβολή στη συχνότητα της απώλειας των χρωµοσωµάτων Χ, Υ και 8 στο µάρτυρα και µετά από επίδραση HCTZ σε καλλιέργειες ανθρώπινων λεµφοκυττάρων, in vitro
221 Συζήτηση που θεωρούνται προϊόν απώλειας του χρωµοσώµατος 8. Το κάπνισµα δεν επάγει την απώλεια των φυλετικών χρωµοσωµάτων και του χρωµοσώµατος 8 τόσο σε καλλιέργειες-µάρτυρες όσο και σε καλλιέργειες µετά από επίδραση µε HCTZ. H απενεργοποίηση του ενός χρωµοσώµατος Χ προκάλεσε το ενδιαφέρον αρκετών ερευνητών προκείµενου να διευκρινισθεί εάν το χρωµόσωµα Χ το οποίο χάνεται ως αποτέλεσµα χρωµοσωµατικής απώλειας είναι το ενεργό ή το ανενεργό. ιάφορες µέθοδοι έχουν αναπτυχθεί για τη µελέτη της συµπεριφοράς και της συµµετοχής του ανενεργού Χ χρωµοσώµατος σε ανευπλοειδικά φαινόµενα. Αρχικά προτάθηκε ότι η ευαισθησία που παρουσιάζει το χρωµόσωµα Χ µε τη συµµετοχή του σε ανευπλοειδικά φαινόµενα οφείλεται σε πρώιµη διαίρεση του κεντροµέρους και γενικότερα σε εξασθενηµένη λειτουργία του κεντροµέρους που παρουσιάζεται µε την αύξηση της ηλικίας (Fitzgerald et al., 1983, Nakagome et al., 1984). Σύµφωνα µε τον Abruzzo και τους συνεργάτες του (1985) η απώλεια ή περίσσεια ενός Χ χρωµοσώµατος σε θηλυκά άτοµα 45,Χ και 47,ΧΧΧ οφείλεται στο απενεργοποιηµένο Χ χρωµόσωµα. Η ανάλυση του περιεχοµένου των µικροπυρήνων σε θηλυκά άτοµα µε µετατόπιση είτε για το ενεργό είτε για το ανενεργό Χ χρωµόσωµα και το χρωµόσωµα 9 έδειξε ότι η πλειονότητα των µικροπυρήνων σχηµατίζονται από το απενεργοποιηµένο Χ χρωµόσωµα (Tucker et al., 1996). Η παρουσία του χρωµοσώµατος Χ σε µικροπυρήνες εµφανίζεται αυξηµένη κατά 1 φορές σε θηλυκά άτοµα 46,ΧΧ συγκρινόµενη µε αρσενικά άτοµα 46,ΧΥ και θηλυκά άτοµα 45,Χ, υποδηλώνοντας την απώλεια του απενεργοποιηµένου Χ χρωµοσώµατος. Η ίδια ερευνητική οµάδα υποστηρίζει ότι η απώλεια του χρωµοσώµατος Χ είναι ανεκτή από το κύτταρο και δεν επηρεάζει τη βιωσιµότητα του εάν πρόκειται για το απενεργοποιηµένο Χ αφού πολύ λίγα γονίδια που βρίσκονται σε αυτό εκφράζονται (Hando et al., 1997). Άλλες ερευνητικές οµάδες υποστηρίζουν ότι δεν υπάρχει διαφορά όσον αφορά τη µικροπυρήνωση των δύο οµολόγων χρωµοσωµάτων. Ο Surrales και οι συνεργάτες του (1996) βασισµένοι στο γεγονός ότι στο ανενεργό χρωµόσωµα Χ η ιστόνη Η4 δεν είναι ακετυλιωµένη αποδεικνύουν µε ανοσοιστοχηµικές µεθόδους ότι όχι µόνο το ανενεργό αλλά και το ενεργό χρωµόσωµα Χ έχει την ικανότητα να χάνεται µε µία διαδικασία που βρίσκεται σε άµεση σχέση µε την πρόοδο της ηλικίας. Σε λεµφοκύτταρα θηλυκών ατόµων φαινόµενα χρωµοσωµατικής καθυστέρησης παρουσιάζουν και τα δύο Χ χρωµοσώ- µατα (Catalan et al, 2a). Εκτός από την παρουσία του ανενεργού χρωµοσώµατος Χ στα θηλυκά άτοµα, η µελέτη του συγκεκριµένου χρωµοσώµατος εµφανίζει ιδιαίτερο ενδιαφέρον διότι ανευπλοειδικά φαινόµενα που αφορούν το χρωµόσωµα Χ παρουσιάζονται σε µεγαλύτερη συχνότητα από ότι στα αυτοσωµατικά χρωµοσώµατα (Zijno et al., 1996c, Catalan et al., 2a). Συγκεκριµένα, τα χρωµοσώµατα Χ αποτελούν το ¼ των χρωµοσωµάτων που καθυστερούν να µετακινηθούν στο στάδιο της ανάφασης, γεγονός που αντανακλά στο ότι έχουν την τάση να υφίστανται απώλεια µε µεγαλύτερο ρυθµό σε σύγκριση µε τα αυτοσωµικά χρωµοσώµατα. Μία πιθανή εξήγηση αφορά την απόσταση των χρωµοσωµάτων από τους πόλους της µιτωτικής ατράκτου. Τα καθυστερηµένα χρωµοσώµατα Χ βρίσκονται σε µεγαλύτερη απόσταση (.18) από τους πόλους συγκριτικά µε τα αυτοσωµικά (.13). Σε λεµφοκύτταρα ενός θηλυκού ατόµου ώριµης ηλικίας οι µικροπυρήνες που περιέχουν άκεντρα χρωµοσωµατικά τµήµατα, αυτοσωµατικά χρωµοσώµατα και Χ χρωµοσώµατα εµφανίζουν ποσοστά 34.9, 22.3 και 42.9%, αντίστοιχα (Catalan et al., 2a, Falck et al., 22). Μια εναλλακτική εξήγηση υποστηρίζει ότι τα αυτοσω- µατικά χρωµοσώµατα περιέχουν σηµαντικά γονίδια για την επιβίωση του κυττάρου µε απoτέλεσµα να επεµβαίνει η φυσική επιλογή και τα κύτταρα αυτά να πεθαίνουν είτε πριν από το σχηµατισµό µικροπυρήνων είτε πριν τη µετάβαση στο στάδιο της ανάφασης. Συνεπώς δεν είναι δυνατόν να παρατηρήσουµε τα χρωµοσώµατα αυτά
222 Συζήτηση στους µικροπυρήνες (Catalan et al, 2a). Όµως η δράση της φυσικής επιλογής θα πρέπει να είναι παρόµοια και σε κύτταρα που έχουν χάσει το ενεργό Χ χρωµόσω- µα µε αποτέλεσµα να ευνοείται η επιλεκτική απώλεια του απενεργοποιηµένου Χ χρωµοσώµατος (Hando et al., 1994). Εντούτοις απώλεια του χρωµοσώµατος Χ συµβαίνει σε υψηλό ρυθµό σε αρσενικά άτοµα υποδηλώνοντας µία τάση µικροπυρήνωσης του χρωµοσώµατος Χ ανεξάρτητα από το απενεργοποιηµένο Χ χρωµόσωµα. Πιθανόν η απουσία πρωτεϊνών του κινητοχώρου η οποία έχει διαπιστωθεί στο απενεργοποιηµένο Χ χρωµόσωµα ή βλάβες στον κινητοχώρο που αντανακλούν στη µη σωστή δέσµευση του µε τους µικροσωληνίσκους να εµπλέκονται σε µηχανισµούς χρωµοσωµατικής απώλειας (Hando et al., 1994, Cimini et al., 21). Αναµφισβήτητα η παρουσία του χρωµοσώµατος Χ και των αυτοσωµατικών χρωµοσωµάτων στους µικροπυρήνες εξαρτάται άµεσα από την ηλικία. Αύξηση της ηλικίας οδηγεί στην αύξηση της συχνότητας εµφάνισης των χρωµοσωµάτων σε µικροπυρήνες (Catalan et al., 1995, Bakou et al., 22). Τα φυλετικά χρωµοσώµατα έχουν την τάση να περικλείονται σε µικροπυρήνες σε µεγάλης ηλικίας άτοµα. Το χρωµόσωµα Χ έχει την τάση να χάνεται µε µεγαλύτερη συχνότητα τόσο σε µικρής όσο και σε µεγάλης ηλικίας γυναίκες παρά σε άντρες (Catalan et al., 1998). Η απώλεια των φυλετικών χρωµοσωµάτων εµφανίζεται αυξηµένη σε ηλικιωµένα άτοµα. Σε αρσενικά άτοµα η απώλεια του χρωµοσώµατος Υ είναι εντονότερη συγκριτικά µε τα αυτοσωµατικά χρωµοσώµατα. Ανευπλοειδία του χρωµοσώµατος Υ δεν οδηγεί το κύτταρο σε θάνατο πιθανόν εξαιτίας του µικρού του µεγέθους και της απουσίας σηµαντικών γονιδίων για την επιβίωση του κυττάρου (Guttenbach et al., 1995). Εντούτοις έχει βρεθεί ότι η ανευπλοειδία του χρωµοσώµατος Υ σχετίζεται µε διάφορες µορφές καρκίνου, όπως µελάνωµα και καρκινώµατα της ουροδόχου κύστης (Ozisik, 1994, Sauter et al., 1995). Πρώιµη διαίρεση του κεντροµέρους του Υ χρωµοσώµατος αυξάνεται µε την αύξηση της ηλικίας, µία ένδειξη που υποστηρίζει ότι µη λειτουργικά κεντροµέρη σχετίζονται µε την ανευπλοειδία του χρωµοσώµατος Υ που εµφανίζεται σε ηλικιωµένα άτοµα. Άλλες πιθανές αιτίες που οδηγούν στην µικροπυρήνωση ενός καθυστερηµένου Υ χρωµοσώµατος αφορούν τη µη λειτουργική ή τη µικρή έκφραση πρωτεϊνών του κινητοχώρου, CENP-B και τη µείωση των τελοµερών µε την αύξηση της ηλικίας µε δραµατικές συνέπειες για ένα χρωµόσωµα µικρού µεγέθους (Cherry et al., 1987, Nath et al., 1995). Ελάχιστες είναι οι µελέτες σχετικά µε την απώλεια των αυτοσωµατικών χρωµοσωµάτων και ιδιαίτερα αυτές που αφορούν το χρωµόσωµα 8. Η απώλεια των ακροκεντρικών χρωµοσωµάτων που ανήκουν στις οµάδες Ε (13, 14 και 15) και G (21 και 22) ακολουθεί τις αναµενόµενες τιµές και δεν φαίνεται να επηρεάζεται από την ηλικία και το φύλο (Scarpato et al., 1996). Ο Shi και οι συνεργάτες του (2) συµφωνούν ότι η απώλεια του χρωµοσώµατος 21 δεν επηρεάζεται από το φύλο και την ηλικία. Αντίθετα ο µη αποχωρισµός του χρωµοσώ- µατος 21 εµφανίζεται αυξηµένος σε ηλικιωµένα άτοµα. ύο ανεξάρτητες µελέτες υποστηρίζουν ότι η απώλεια του χρωµοσώµατος 8 επηρεάζεται από την ηλικία και συµβαίνει σε αρκετά χαµηλότερους ρυθµούς συγκριτικά µε την απώλεια του Χ χρω- µοσώµατος (Bakou et al., 22, Leopardi et al., 22). Σύγκριση των διαγραµµάτων 4 και 6 δείχνει ότι το φαινόµενο του µη αποχωρισµού επάγεται από τo HCTZ σε µεγαλύτερη ένταση από τη χρωµοσωµατική απώλεια. Επιπλέον το φαινόµενο του χρωµοσωµατικού µη αποχωρισµού επάγεται από την ελάχιστη συγκέντρωση HCTZ. Η δράση αυτή είναι χαρακτηριστική των ανευπλοειδογόνων ενώσεων και γενικότερα φυσικών ή χηµικών παραγόντων που αλληλεπιδρούν µε στοιχεία της µιτωτικής συσκευής (Kirsch-Volders et al., 1996, Elhajouji et al., 1997, Sgura et al., 1997, Touil et al., 2, Bakou et al., 22). Η ικανότητα του HCTZ να επάγει ανευπλοειδικά φαινόµενα δεν φαίνεται να επηρεάζεται από την κατάσταση διαίρεσης
223 Συζήτηση των κυττάρων (διάγραµµα 7). Το HCTZ επάγει το µη αποχωρισµό των χρωµοσω- µάτων Χ και 8 στις συγκεντρώσεις 5 και 4 µg/ml HCTZ σε µη διεγερµένα λεµφοκύτταρα αρσενικών και θηλυκών νεαρών ατόµων. Η επίδραση της µέγιστης συγκέντρωσης HCTZ προκαλεί απώλεια του χρωµοσώµατος Χ, ενώ οριακή είναι η ικανότητα του HCTZ να επάγει απώλεια του χρωµοσώµατος 8. Πιθανότατα η δράση Χρωµόσωµα Χ Χρωµόσωµα 8 4 HCTZ (µg/ml) ιεγερµένα Μη διεγερµένα ιάγραµµα 7. Μεταβολή του µη αποχωρισµού των χρωµοσωµάτων Χ και 8 σε διεγερµένες και µη διεγερµένες µε PHA κατά την επίδραση µε HCTZ καλλιέργειες ανθρώπινων λεµφοκυττάρων ατόµων νεαρής ηλικίας, in vitro. Μη Αποχωρισµός % του HCTZ στη µη διεγερµένη κατάσταση των κυττάρων είναι αρκετά ισχυρή και φαίνεται να ισχύει µέχρι το στάδιο της µίτωσης. Η ανευπλοειδογόνος δράση του υδροχλωροθειαζιδίου ελέγχεται µε τη µελέτη πρωτεϊνικών µορίων-δεικτών που διαδραµατίζουν ουσιώδη ρόλο στη µιτωτική διαίρεση και συµβάλλουν στην ακεραιότητα της µιτωτικής συσκευής. Πρόκειται για την α- και γ- τουµπουλίνη συστατικά των ινιδίων της µιτωτικής ατράκτου και του κεντροσώµατος. Η µελέτη πραγµατοποιήθηκε µε διπλή ανοσοσήµανση έναντι των µορίων αυτών µε χρήση κατάλληλων αντισωµάτων σε κύτταρα που βρίσκονται σε διαδικασία διαίρεσης και έχουν την ικανότητα προσκόλλησης σε κατάλληλο υπόστρωµα ώστε να αναπτύσσεται το δίκτυο των µικροσωληνίσκων. Κατάλληλο βιολογικό σύστηµα αποτελούν οι κυτταρικές σειρές HFFF2, HEp2 και C2C12. Η µελέτη σε ανθρώπινα λεµφοκύτταρα απέδωσε στο HCTZ διπλή ανευπλοειδογόνο δράση στη συγκέντρωση 4 µg/ml. Σε ανθρώπινες κυτταρικές σειρές φάνηκε η ικανότητα του HCTZ να προκαλεί χρωµοσωµατική καθυστέρηση στις συγκεντρώσεις 6 και 8 µg/ml. Για τη διερεύνηση του µηχανισµού πρόκλησης φαινοµένων λανθασµένου αποχωρισµού επιλέγονται οι συγκεντρώσεις 4, 6 και 8 µg/ml HCTZ. Μελετάται η επίδραση του HCTZ στην ακεραιότητα του κυτταροσκελετού, στο σχηµατισµό των κέντρων οργάνωσης των µικροσωληνίσκων και στην ακεραιότητα της µιτωτικής συσκευής. Σε ανθρώπινα λεµφοκύτταρα και σε κυτταρικές σειρές το HCTZ βρέθηκε να εµποδίζει την πρόοδο του κυτταρικού κύκλου συσσωρεύοντας τα κύτταρα στο σταδιο της µετάφασης. Η επίδραση HCTZ σε κύτταρα C2C12 προκαλεί πτώση του µιτωτικού δείκτη που αντανακλά στη µείωση του ρυθµού διαίρεσης των κυττάρων. Η αύξηση στον αριθµό των µεταφάσεων συνοδεύεται από µείωση στον αριθµό των ανα
224 Συζήτηση τελοφάσεων. Φαίνεται πως το HCTZ διακόπτει τη µιτωτική διαδικασία και οδηγεί σε συγκράτηση των κυττάρων στο στάδιο της µετάφασης, δράση που χαρακτηρίζει τις ανευπλοειδογόνες ενώσεις. Το φαινόµενο αυτό οφείλεται στο σύστηµα ελέγχου του κυτταρικού κύκλου το οποίο ρυθµίζει τη σωστή λειτουργία κάθε συγκεκρι- µένου σταδίου. Βλάβες στο γενετικό υλικό ή/και αναστολή της λειτουργίας της µιτωτικής ατράκτου εξαιτίας της παρουσίας αναστολέων επιφέρουν την ενεργοποίηση του σηµείου ελέγχου της µίτωσης και την παραµονή των κυττάρων στη µεταβατική φάση µεταξύ µετάφασης και ανάφασης. Ειδικότερα αδυναµία του σωστού προσανατολισµού των χρωµοσωµάτων και της δέσµευσης των µικροσωληνίσκων του κινητοχώρου στα αδελφά χρωµατίδια προάγει τη συγκρότηση πρωτεϊνικών συµπλεγµάτων µε αποτέλεσµα την απενεργοποίηση του APC γεγονός που επιφέρει τη διατήρηση της συνάφειας των αδελφών χρωµατιδίων και την καθυστέρηση του κυτταρικού κύκλου στη µετάφαση. Η διαταραχή των µηχανισµών ελέγχου από χηµικά επαγόµενες µεταλλάξεις ή από χηµική αναστολή των εµπλεκόµενων πρωτεϊνικών παραγόντων είναι δυνατόν να οδηγήσει σε λανθασµένο διαχωρισµό των χρωµοσω- µάτων (Luch, 22, Bharadwaj και Yu, 24). Μελετώντας δοµικές αλλαγές στο σχηµατισµό της µιτωτικής ατράκτου και µορφολογικές τροποποιήσεις στο δίκτυο των κυττάρων C2C12, HFFF2 και HEp2, υπό την επίδραση του υδροχλωροθειαζιδίου, καταλήγουµε στο συµπέρασµα ότι επηρεάζεται η συγκρότηση της δοµής του δικτύου των µικροσωληνίσκων. Τροποποιήσεις στο σχηµατισµό του δικτύου των µικροσωληνίσκων συµβαίνει τόσο σε µιτωτικά κύτταρα όσο και σε µεσοφασικά κύτταρα. Συγκεκριµένα, χάνεται η συνέχεια του δικτύου των µικροσωληνίσκων και σε αρκετές περιπτώσεις παρατηρείται συρρίκνωση των µικροσωληνίσκων γύρω από τον πυρήνα του κυττάρου σε αντίθεση µε τις καλλιέργειες του µάρτυρα. Επιπρόσθετα, η παρουσία διπολικών µεταφάσεων µε µη ενσωµατωµένο χρωµοσωµατικό υλικό, διπολικές µη οµαδοποιηµένες µεταφάσεις, αποτελεί µία ένδειξη της επίδρασης του υδροχλωροθειαζιδίου στη δοµή των µικροσωληνίσκων. Στη συγκρότηση της δοµής του δικτύου των µικροσωληνίσκων συµβάλλουν τόσο η α- όσο και η β- τουµπουλίνη. Πιθανόν να επηρεάζεται η α- τουµπουλίνη η οποία έχει σηµανθεί, ενώ δεν αποκλείεται το HCTZ να εµφανίζει έµµεση επίδραση στο δίκτυο µέσω της β-τουµπουλίνης. Αναµφισβήτητα µε την επίδραση του HCTZ τροποποιείται το δίκτυο των µικροσωληνίσκων και συνεπώς ο σωστός σχηµατισµός της µιτωτικής ατράκτου. Η θέση, η µορφολογία και ο αριθµός των κεντροσωµάτων συµβάλλουν ση- µαντικά στη σωστή λειτουργία και δοµή της µιτωτικής ατράκτου. Η γ- τουµπουλίνη αποτελεί βασικό συστατικό του κεντροσώµατος και χρησιµοποιείται ως δείκτης για την παρατήρηση κατά τη διάρκεια του κυτταρικού κύκλου γεγονότων που σχετίζονται µε το κεντρόσωµα. Το υδροχλωροθειαζίδιο επηρεάζει την οργάνωση της µιτωτικής συσκευής αφού προκαλεί αύξηση των ανώµαλων µορφών µετάφασης. Ανάµεσα στους τύπους των ανώµαλων µεταφάσεων οι µονοπολικές εµφανίζονται σε µεγαλύτερη συχνότητα και φαίνεται να αποτελούν την επικρατέστερη µορφή (διάγραµµα 8). Μονοπολικές άτρακτοι προκύπτουν είτε στην περίπτωση που συµβούν µεταλλάξεις στο πρώτο στάδιο του κεντροσωµικού κύκλου, στο στάδιο του διπλασιασµού, είτε µετά από επίδραση µε χηµικές ενώσεις. Κατά συνέπεια δεν πραγµατοποιείται ο διπλασιασµός µε αποτέλεσµα το σχηµατισµό µόνο ενός λειτουργικού κεντροσώµατος. Τα κύτταρα αυτά συνεχίζουν τη σύνθεση του DNA και καταλήγουν στην µίτωση, σχηµατίζοντας µία µονοπολική µιτωτική άτρακτο. Εάν είναι λειτουργικό το σηµείο ελέγχου της µίτωσης τότε ο κυτταρικός κύκλος διακόπτεται. Αντίθετα, µεταλλάξεις σε γονίδια που σχετίζονται µε το σηµείο αυτό, επιτρέπουν την ολοκλήρωση του κυτταρικού κύκλου. Συγκεκριµένα, έχει
225 Συζήτηση αναφερθεί ότι συµβαίνει µία παροδική διακοπή του κυτταρικού κύκλου (transient mitotic arrest). Αυτή οφείλεται είτε σε µη λειτουργικό σηµείο ελέγχου, είτε στο γεγονός ότι η µονοπολική άτρακτος πληρεί τα κριτήρια για τη συνέχιση και ολοκλήρωση του εξαιτίας της δέσµευσης των κινητοχώρων στα ινίδια της µιτωτικής ατράκτου (Glover et al., 1995, Winey, 1996). Μονοπολικές µεταφάσεις προκύπτουν και ως αποτέλεσµα βλαβών στις κινητικές πρωτεΐνες και στη δοµή των µικροσωληνίσκων, µε αποτέλεσµα τα κεντροσώµατα να µην δύνανται να µετακινηθούν προς τους αντίθετους πόλους και να παραµένουν σε άµεση επαφή (Pihan και Doxsey, 1999). Έτσι κατά το σχηµατισµό της µονοπολικής µετάφασης είναι ορατά στο κέντρο δύο κεντροσώµατα πολύ κοντά το ένα στο άλλο και γύρω από αυτά σχη- µατίζονται τα ινίδια των µικροσωληνίσκων. Το φαινόµενο αυτό έχει παρατηρηθεί µετά από επίδραση µε τη χηµική ένωση νοκοδαζόλη (NOZ) η οποία καταστρέφει τους µικροσωληνίσκους και επηρεάζει µε τον τρόπο αυτό τη µετακίνηση των κεντροσωµάτων κατά την πρόφαση σε κύτταρα V79. Είναι προφανές ότι η µετακίνηση αυτή καθορίζεται από ένα εξαρτώµενο µηχανισµό από τους µικροσωληνίσκους (Ochi, 2). Η επίδραση διαζεπάµης (DZ), βασικό συστατικό υπνωτικών φαρµάκων, προκαλεί χρωµοσωµατική απώλεια και εµποδίζει τη σωστή οργάνωση του κεντροσώµατος σε Cl-1 κύτταρα µε αποτέλεσµα τη δηµιουργία πολυπολικών αναφάσεων. Συγκεκριµένα προκαλεί αναστολή του διαχωρισµού των κεντριολίων µετά το διπλασιασµό τους µε αποτέλεσµα να σχηµατίζονται µονοπολικές άτρακτοι (Anderson et al., 1981). Επιπρόσθετα σε ένα µεγάλο ποσοστό διακρίνονται µονοπολικές άτρακτοι µε διάστικτα σήµατα γ- τουµπουλίνης (punctate) που οφείλονται στην επίδραση του φαρµάκου στο στάδιο ωρίµανσης του κεντροσωµικού κύκλου στη φάση G2/M (Izzo et al., 1998). Φυσιολογικές Ανώµαλες ΠΜΟ ΜΠ ΠΠ *:.1<p<.5 σε σχέση µε το µάρτυρα (G-test) ΜΕΤΑΦΑΣΕΙΣ (%) * * * * * * * * * * * 2 1 ΜΑΡΤΥΡΑΣ HCTZ 4 (µg/ml) HCTZ 6 (µg/ml) HCTZ 8 (µg/ml) ιάγραµµα 8. Μεταβολή φυσιολογικών και ανώµαλων µεταφασικών κυττάρων και κατανοµή των ανώµαλων µεταφάσεων στο σύνολο των µεταφασικών κυττάρων σε καλλιέργειες κυττάρων C2C12 στο µάρτυρα και µετά από επίδραση µε HCTZ. ΠΜΟ: ιπολικές µη οµαδοποιηµένες, ΜΠ: Μονοπολικές, ΠΠ: Πολυπολικές µεταφάσεις
226 Συζήτηση Εκτός από την επαγωγή µονοπολικών µεταφάσεων το HCTZ προκαλεί το σχη- µατισµό πολυπολικών µεταφάσεων ως αποτέλεσµα της ύπαρξης µεγάλου αριθµού κεντροσωµάτων (διάγραµµα 8). Σύµφωνα µε πρόσφατες µελέτες έχουν προταθεί δύο µηχανισµοί για την εµφάνιση κυττάρων µε πολλαπλά κεντροσώµατα. Ο πρώτος αναφέρεται στην ανικανότητα του κυττάρου για κυτταροκίνηση. Εάν τα κύτταρα ολοκληρώσουν φυσιολογικά τον κυτταρικό κύκλο και δεν λάβει χώρα η διαδικασία της κυτταροκίνησης, τότε τα κύτταρα θα περιέχουν διπλάσια ποσότητα γενετικού υλικού καθώς επίσης και διπλάσιο αριθµό κεντροσωµάτων. Ο δεύτερος µηχανισµός αναφέρεται στην απορύθµιση του κεντροσωµικού κύκλου. Τα κύτταρα στα οποία συµβαίνουν πολλοί κεντροσωµικοί κύκλοι σε ένα κυτταρικό κύκλο, χαρακτηρίζονται από υπεράριθµα κεντροσώµατα, ενώ περιέχουν τη φυσιολογική ποσότητα γενετικού υλικού (Tarapore and Fukasawa, 22). Το γεγονός αυτό οδηγεί στο σχηµατισµό πολλαπλών µιτωτικών πόλων και στον ανώµαλο διαχωρισµό των χρω- µοσωµάτων κατά τη διαίρεση του κυττάρου (Duensing and Münger, 21). Συγκεκριµένα ένα κύτταρο µε τριπολική άτρακτο υφίσταται κυτταροκίνηση µε αποτέλεσµα το σχηµατισµό τριών θυγατρικών κυττάρων. Αρκετά από αυτά είναι µη βιώσιµα εξαιτίας βλαβερών τροποποιήσεων του καρυοτύπου, ενώ κάποια που είναι βιώσιµα χαρακτηρίζονται ανευπλοειδή. Αντίθετα ένα κύτταρο που η µιτωτική άτρακτος σχηµατίζεται από περισσότερα από τρία κεντροσώµατα αποτυγχάνει να διαιρεθεί και σχηµατίζει ένα διπύρηνο ή ένα µεγάλο µονοπύρηνο κύτταρο. Ωστόσο σε ορισµένες περιπτώσεις µε τη συνάθροιση των κεντροσωµάτων ή την επιλεκτική απενεργοποίηση κάποιων δύναται να σχηµατισθούν διπολικές µιτωτικές άτρακτοι που θα οδηγήσουν στο σχηµατισµό ανευπλοειδικών κυττάρων (Brinkley, 21, Fukasawa, 25). Σηµαντικό ρόλο διαδραµατίζει η πρωτεΐνη p53 που εντοπίζεται στα κεντροσώµατα και ελέγχει το διπλασιασµό του κεντροσώµατος µόνο µία φορά σε κάθε κυτταρικό κύκλο. Αυτή η ρύθµιση δεν απαιτεί την πρόσδεση της στο DNA, γεγονός που αντανακλά στο ότι η πρωτεΐνη p53 ρυθµίζει το διπλασιασµό του κεντροσώµατος µε ένα τρόπο ανεξάρτητα από το ρόλο της ως µεταγραφικός παράγοντας (Tarapore et al, 21). Έτσι απώλεια ή µετάλλαξη του p53 προκαλεί ανώµαλο διπλασιασµό του κεντροσώµατος εξαιτίας απορύθµισης του κεντροσωµικού κύκλου και αδυναµία διαίρεσης του κυτταροπλάσµατος (Fisk et al., 22). Το p53 ελέγχει το διπλασιασµό του κεντροσώµατος µέσω δύο µηχανισµών. Ο πρώτος αναφέρεται στη συµµετοχή του γονιδίου στο G1/S σηµείο ελέγχου του κυτταρικού κύκλου, όπου το p53 ελέγχει ταυτόχρονα την έναρξη του κεντροσωµικού κύκλου αλλά και την καταστολή επανεκκίνησης του και ο δεύτερος µέσω φυσικής δέσµευσης της p53 στο κεντρόσωµα. Παρατεταµένοι κύκλοι πολλαπλασιασµού του κεντροσώµατος µε επακόλουθο ανισοµερή διαχωρισµό των χρωµοσωµάτων στα θυγατρικά κύτταρα έχουν διαπιστωθεί σε εµβρυϊκούς ινοβλάστες ποντικού που χαρακτηρίζονται από απουσία λειτουργικότητας για την p53, p53 -/- (Fukasawa et al., 1996, Tarapore και Fukasawa, 22). Επιπρόσθετα είναι γνωστό ότι ο πολλαπλασιασµός του κεντροσώ- µατος στα σωµατικά κύτταρα συνοδεύεται και από άλλα γεγονότα στον κυτταρικό κύκλο, όπως η αντιγραφή του DNA. Μειωµένη ικανότητα είτε αντιγραφής του DNA είτε επιδιόρθωσης σηµαντικών βλαβών που προκαλείται από χηµικές ενώσεις επιφέρει παρατεταµένη παραµονή των κυττάρων στη φάση S και στο όριο µεταξύ G2/Μ φάσης του κυτταρικού κύκλου. Το γεγονός αυτό προκαλεί διάσπαση του κεντροσώµατος σε τµήµατα που περιέχουν µόνο ένα κεντριόλιο µε αποτέλεσµα το σχηµατισµό πολυπολικών ατράκτων (Hut et al., 23). Σε άλλες ερευνητικές µελέτες υποστηρίζεται ότι παρουσία κολχικίνης το περικεντροσωµικό υλικό (PCMs) που περιβάλλει το κεντρόσωµα διαχωρίζεται από τα κεντριόλια µε αποτέλεσµα την επαγωγή νέων αντιγράφων που αποτελούν θέσεις πυρήνωσης των µικροσωληνίσκων
227 Συζήτηση (Kerryer et al., 1984, Sellito και Kuriyama, 1988). Κατανοώντας τους παραπάνω µηχανισµούς, που έχουν ως αποτέλεσµα το σχηµατισµό µονοπολικών και πολυπολικών µεταφάσεων, οδηγούµαστε στην υπόθεση ότι η φαρµακευτική ένωση υδροχλωροθειαζίδιο προκαλεί αλλαγές στον κεντροσωµικό κύκλο. Πιθανόν να επιδρά είτε στη φάση G1/S του κυτταρικού κύκλου όπου ταυτόχρονα µε την έναρξη της αντιγραφής του γενετικού υλικού, ξεκινάει ο διπλασιασµός του κεντροσώµατος είτε στη µεταβατική φάση G2/M κατά τη διάρκεια του σταδίου διαχωρισµού των κεντροσωµάτων. Η απουσία πολλαπλών σηµάτων γ- τουµπουλίνης σε µεσοφασικούς πυρήνες ενισχύει την πιθανή δράση του HCTZ στη G2/M φάση. Επιπλέον είναι δυνατόν να προκαλεί βλάβες σε κινητικές πρωτεΐνες που συµµετέχουν στην κίνηση των κεντροσωµάτων αλλά και σε πρωτεΐνες που συµβάλλουν στη σωστή ευθυγράµ- µιση των χρωµοσωµάτων στο ισηµερινό επίπεδο κατά τη διάρκεια της µετάφασης, όπως οι πρωτεΐνες του κινητοχώρου των χρωµοσωµάτων ή πρωτεΐνες συνδεδεµένες µε τα ινίδια της ατράκτου, µε αποτέλεσµα την αδυναµία ενσωµάτωσης ολόκληρου του γενετικού υλικού στα θυγατρικά κύτταρα. Πιθανόν το HCTZ σε χαµηλές συγκεντρώσεις επιδρά στο πρώτο και τρίτο στάδιο του κεντροσωµικού κύκλου ενώ σε µεγαλύτερες επιδρά εκτός από τα στάδια αυτά και σε πρωτεΐνες που συµµετέχουν όχι µόνο στον κεντροσωµικό κύκλο αλλά και σε στάδια της µιτωτικής διαδικασίας. Έτσι παράλληλα µε την αύξηση των µονοπολικών µεταφάσεων παρατηρούνται σε µεγαλύτερες συγκεντρώσεις αυξηµένα ποσοστά πολυπολικών φάσεων. Η επίδραση ξενοβιοτικών ενώσεων σε συγκεκριµένες συγκεντρώσεις εµφανίζει δοσοεξαρτώµενο ανευπλοειδικό αποτέλεσµα που είναι µικρότερο για το χρωµοσωµατικό µη αποχωρισµό από ότι για τη χρωµοσωµατική απώλεια. Αναλυτικότερα η δράση της νοκοδαζόλης οφείλεται στην πρόσδεση της σε µόρια τουµπουλίνης επάγοντας τον αποπολυµερισµό τους. Σε µικρές συγκεντρώσεις η φαρµακευτική ένωση δεν προσδένεται σε αρκετά µόρια ώστε να εµποδίσει τον πολυµερισµό των µικροσωληνίσκων και δεν προκαλεί ουσιώδη προβλήµατα στη λειτουργία της µιτωτικής συσκευής. Αυξανοµένης της συγκέντρωσης, αυξάνονται και τα µόρια στόχοι που έ- χουν επηρεαστεί µε αποτέλεσµα να προκαλείται χρωµοσωµατικός µη αποχωρισµός και χρωµοσωµατική απώλεια. Ωστόσο η απουσία µερικών πρωτοϊνιδίων τουµπουλίνης από τον κινητοχώρο είναι αρκετή για να προκληθεί ο µη αποχωρισµός. Αντίθετα για την επαγωγή χρωµοσωµατικής απώλειας θα πρέπει οι κινητοχώροι ενός χρωµοσώµατος να αποσπασθούν από τη µιτωτική άτρακτο. Σε αυτή την περίπτωση απαιτείται η καταστροφή περισσοτέρων ινιδίων τουµπουλίνης. Η νοκοδαζόλη σε σχέση µε άλλες ανευπλοειδογόνες ενώσεις, κολχικίνη, καρβενδαζίµη, µεβενδαζόλη που δρουν µε τον ίδιο τρόπο πρέπει να επηρεάσει µεγαλύτερο αριθµό µορίων στοχων για την επαγωγή µη αποχωρισµού (Elhajouji et al., 1997). Η δράση αυτή δεν χαρακτηρίζει µόνο τις ανευπλοειδογόνες ενώσεις αλλά και τα ανευπλοειδικά φαινό- µενα που προκαλούνται από την ιονίζουσα ακτινοβολία. Σε µικρές δόσεις ακτινοβολίας δεν παρατηρείται δοσοεξαρτώµενο αποτέλεσµα, πιθανόν λόγω της επίδρασης της ακτινοβολίας στις τουµπουλίνες ή σε άλλους στόχους εκτός από το DNA. Όταν η ανευπλοειδία επάγεται ως αποτέλεσµα απώλειας ή µη αποχωρισµού, δοµές που σχετίζονται µε το διαχωρισµό των χρωµοσωµάτων αποτελούν τους κύριους στόχους (Touil et al., 2). Έχει µελετηθεί µία ποικιλία χηµικών ενώσεων που επιδρούν στη µιτωτική συσκευή και προκαλούν σηµαντικές βλάβες στο µηχανισµό της πυρηνικής και κυτταρικής διαίρεσης. Το βενζιµιδαζολικό αντιµυκητιακό φάρµακο θειαβενδαζόλη (TB) βρέθηκε ότι προκαλεί λανθασµένο χρωµοσωµατικό αποχωρισµό, ο οποίος προάγεται µέσω του σχηµατισµού ανώµαλης µιτωτικής ατράκτου (Pisano et al., 2). Η αναστολή της διαίρεσης του κυτταροπλάσµατος είναι δυνατόν να οδηγήσει στο σχηµατισµό πολυπύρηνων κυττάρων που παρουσιάζουν µικροπυρήνες
228 Συζήτηση διαφορετικών µεγεθών. Στην περίπτωση όµως που η κυτταροκίνηση πραγµατοποιηθεί τότε δηµιουργούνται θυγατρικά κύτταρα που εµφανίζουν φαινόµενα ανευπλοειδίας. Μέρος των κυττάρων αυτών δεν επιβιώνουν εξαιτίας της απώλειας χρω- µοσωµάτων που είναι αναγκαία για την επιβίωση, ενώ κάποια άλλα θα συνεχίσουν να διαιρούνται, δίνοντας µια νέα κυτταρική σειρά. Ο Ochi (1999) διαπίστωσε ότι η δράση αυτή ισχύει για τα οιστρογόνα διεθυλοστιλβεστρόλη (DES), οιστραδιόλη-17β (E2) και διφαινόλη Α (BP-A). Οι ενώσεις αυτές γνωστές για την ανευπλοειδογόνο δράση τους προκαλούν το σχηµατισµό πολυπολικών µιτωτικών ατράκτων, µε αποτέλεσµα το σχηµατισµό πολυπολικών µεταφάσεων. Ειδικότερα για τη διαφαινόλη Α βρέθηκε ότι τροποποιεί τη διαµόρφωση του µορίου της τουµπουλίνης µε αποτέλεσµα η τροποποιηµένη τουµπουλίνη να σχηµατίζει πρωτοϊνίδια τα οποία δεν δύνανται να συγκροτήσουν το φυσιολογικό µικροσωληνίσκο (Pfeiffer et al., 1997). Επιπρόσθετα η κολχικίνη και οι ακτίνες Χ προκαλούν ανωµαλίες στον αριθµό, στη δοµή και στην ενεργότητα πυρήνωσης των κέντρων οργάνωσης των µικροσωληνίσκων (Sato et al., 1983, Parry et al., 1985, Sellito και Kuriyama, 1988). Συµπερασµατικά µπορούµε να πούµε ότι το HCTZ προκαλεί ανευπλοειδία διεγείροντας και τους δύο µηχανισµούς, το µη αποχωρισµό και την απώλεια των χρωµοσωµάτων. Ο µη αποχωρισµός είναι ο κύριος µηχανισµός δηµιουργίας κυττάρων µε λανθασµένο αριθµό χρωµοσωµάτων. Από τις παραµέτρους, ηλικία, φύλο και κάπνισµα, που εκτιµήθηκαν στην πορεία διερεύνησης της γενετικής δράσης της φαρµακευτικής ένωσης, η ηλικία επηρεάζει την απόκριση των λεµφοκυττάρων έναντι του HCTZ. Η έκφραση ανευπλοειδικών φαινοµένων φαίνεται ότι οφείλεται σε δοµικές αλλαγές στην οργάνωση της µιτωτικής ατράκτου εξαιτίας αλληλεπιδράσεων του HCTZ µε τις τουµπουλίνες των µικροσωληνίσκων και του κεντροσώµατος. Τα αποτελέσµατα µας συµφωνούν µε ευρήµατα άλλων ερευνητικών οµάδων όπου η φαρµακευτική ένωση επάγει µη αποχωρισµό σε διπλοειδή στελέχη του µύκητα Aspergillus nidulans (Bignami et al., 1974) και µεταλλάξεις σε κύτταρα L518YTK ± λεµφώµατος ποντικού (Myhr et al., 199). Οι µεταλλάξεις αυτές είναι δυνατόν να προκαλούνται από µηχανισµούς µεταξύ των οποίων περιλαµβάνονται ο µιτωτικός ανασυνδυασµός, η χρωµοσωµατική απώλεια και ο λανθασµένος χρωµοσωµατικός αποχωρισµός (Henry et al., 1998)
229
230 Περίληψη ΠΕΡΙΛΗΨΗ Η φαρµακευτική ένωση υδροχλωροθειαζίδιο, HCTZ, ανήκει στην κατηγορία των διουρητικών θειαζιδίων και χρησιµοποιείται στη θεραπεία της υπέρτασης. Η πτώση της αρτηριακής πίεσης επιτυγχάνεται µε την αναστολή της επαναρρόφησης του χλωριούχου νατρίου στο άπω εσπειραµένο σωληνάριο του νεφρού. Η γενετική δράση του υδροχλωροθειαζιδίου δεν έχει αποσαφηνισθεί. Είναι γνωστό ότι προκαλεί µη αποχωρισµό σε διπλοειδή στελέχη του µύκητα Aspergillus nidulans και πολυπλοειδία σε CHL κύτταρα. Στόχος της παρούσας διατριβής είναι η µελέτη της γενετικής δράσης του HCTZ και οι µηχανισµοί µε τους οποίους εκδηλώνεται. Η µελέτη πραγµατοποιήθηκε σε καλλιέργειες ανθρώπινων λεµφοκυττάρων in vitro από 32 υγιείς δότες οι οποίοι διαχωρίστηκαν σε οµάδες µε κριτήριο την ηλικία, το φύλο και τη συνήθεια του καπνίσµατος και σε κυτταρικές σειρές. ιερευνήθηκε η ικανότητα του HCTZ να επάγει αυξηµένες συχνότητες µικροπυρήνων και µελετήθηκε ο µηχανισµός δηµιουργίας τους. Ως σύστηµα ελέγχου χρησιµοποιήθηκε η µέθοδος αναστολής της κυτταροκίνησης,cbmn, σε συνδυασµό µε in situ υβριδοποίηση µε φθοροχρώµατα, FISH-DUAL FISH, για την επισήµανση της κεντροµερικής περιοχής των χρωµοσωµάτων και µε ανοσοφθορισµό, CREST, για την επισήµανση των πρωτεϊνών του κινητοχώρου. Τα ευρήµατα που προέκυψαν από τη µελέτη της γενετικής δράσης της φαρµακευτικής ένωσης συνοψίζονται στα εξής: Αποκαλύφθηκε ότι το HCTZ είναι ισχυρός παράγοντας επαγωγής µικροπυρήνων. Οι αυξηµένες συχνότητες µικροπυρήνων είναι αποτέλεσµα δύο µηχανισµών, της χρωµοσωµατικής καθυστέρησης και σε χαµηλότερο βαθµό της χρωµοσωµατικής θραύσης. Η περαιτέρω διερεύνηση της ανευπλοειδογόνου ικανότητας του HCTZ έδειξε ότι ο µη αποχωρισµός είναι ο κύριος µηχανισµός διατάραξης του χρωµοσωµατικού διαχωρισµού, ενώ η χρωµοσωµατική καθυστέρηση συµβαίνει σε µικρότερη έκταση. Από τα µελετηθέντα χρωµοσώµατα Χ, Υ και 8, το Χ υπόκειται σε υψηλότερους ρυθµούς ανευπλοειδίας. Η ανευπλοειδογόνος δράση του HCTZ δεν φαίνεται να επηρεάζεται από την κατάσταση διαίρεσης των κυττάρων και συνεπώς από την διαφορετική τοπολογία της κεντροµερικής περιοχής σε διαιρούµενα λεµφοκύτταρα και σε λεµφοκύτταρα ευρισκόµενα σε φάση ηρεµίας (G). Η πολυδιάστατη στατιστική ανάλυση των αποτελεσµάτων έδειξε ότι από τις τρείς µεταβλητές που µελετήθηκαν, ηλικία, φύλο και συνήθεια καπνίσµατος, µόνο η ηλικία επηρεάζει την απόκριση των λεµφοκυττάρων στο HCTZ κατά θετικό τρόπο. Πειράµατα διπλού ανοσοφθορισµού σε κυτταρικές σειρές ανθρώπου και ποντικού για την α- και γ- τουµπουλίνη έδειξαν ότι το HCTZ εκφράζει την ανευπλοειδογόνο δράση του µέσω αποδιοργάνωσης του δικτύου των µικροσωληνίσκων και διατάραξης της ακεραιότητας ή/και του διπλασιασµού του κεντροσώµατος
231 Summary SUMMARY The diuretic drug hydrochlorothiazide, HCTZ, belongs to the class of thiazides and is widely used in the treatment of hypertension. The reduction of high blood-pressure is achieved by inhibiting the reabsorption of NaCl in the distal tubule of the kidney. Data on the in vitro genotoxicity of hydrochlorothiazide are equivocal. HCTZ (Esidrex) is capable to induce high frequency of nondisjunction in diploid strains of Aspergillus nidulans and polyploidy in Chinese hamster lung cells. In the present thesis we investigated the genotoxicity of HCTZ and an effort was made to elucidate the mechanisms leading to HCTZ-induced aneugenic phenomena. For this purpose 32 healthy adults were recruited in our study. Donors were divided into groups according to their age, gender and smoking habits. The Cytokinesis Blocked Micronucleus Assay (CBMN) was applied in human lymphocyte cultures, in vitro as well as in human cell lines. The method was combined with In Situ Fluorescence Hybridization (FISH/DUAL-FISH) and immunofluorescence with antikinetochore antibodies (CREST) to distinguish between aneugenic and clastogenic phenomena. In conclusion the results of our study can be summarized as follows: It was revealed that HCTZ is a strong inducer of micronuclei. FISH and CREST analysis indicated that HCTZ induces micronuclei principally through chromosome delay, with a lesser frequency of micronuclei induced through chromosome breakage. DUAL FISH analysis showed that HCTZ expresses its aneugenic activity mainly due to chromosome non-disjunction, with chromosome loss to take place in a lesser extend. Among the three studied chromosomes, X, Y and 8, the X chromosome was shown to be the more sensitive one. The genotoxic activity of HCTZ was not affected by the schedule of treatment, before or after PHA stimulation, and probably not by the different centromere topology in cycling and resting lymphocytes. Multifactorial regression analysis of our results indicated that under the experimental conditions used only age clearly affected the response of lymphocytes to HCTZ in a positive way. Further analysis of HCTZ aneugenicity, with combined application of double immunofluorescence method for α- and γ- tubulin in human and mouse cell lines, revealed that HCTZ induces structural alterations in microtubule network and affects the integrity and/or the centrosomes duplication

232
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕ ΡΙΑΣΗ Ν. Πέµπτη 28 Ιανουαρίου 2010
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕ ΡΙΑΣΗ Ν Πέµπτη 28 Ιανουαρίου 2010 ΘΕΜΑΤΑ Α. ΕΙ ΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 2917,2977 2. Αδεια απουσίας του Βουλευτή κ. Κ. Μητσοτάκη, σελ. 2961 3. Ανακοινώνεται ότι
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕ ΡΙΑΣΗ Ν Πέµπτη 28 Ιανουαρίου 2010 ΘΕΜΑΤΑ Α. ΕΙ ΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 2917,2977 2. Αδεια απουσίας του Βουλευτή κ. Κ. Μητσοτάκη, σελ. 2961 3. Ανακοινώνεται ότι
ΕΡΓΟ: «ΕΦΑΡΜΟΓΕΣ ΕΙΚΟΝΙΚΗΣ ΠΡΑΓΜΑΤΙΚΟΤΗΤΑΣ ΣΤΗ ΒΟΙΩΤΙΑ: ΜΑΝΤΕΙΟ ΤΡΟΦΩΝΙΟΥ ΚΑΙ ΜΥΚΗΝΑΪΚΗ ΘΗΒΑ»
 ΕΡΓΟ: «ΕΦΑΡΜΟΓΕΣ ΕΙΚΟΝΙΚΗΣ ΠΡΑΓΜΑΤΙΚΟΤΗΤΑΣ ΣΤΗ ΒΟΙΩΤΙΑ:» ΠΡΟΚΗΡΥΞΗ ΔΙΑΓΩΝΙΣΜΟΥ ΤΟΥ ΙΔΡΥΜΑΤΟΣ ΜΕΙΖΟΝΟΣ ΕΛΛΗΝΙΣΜΟΥ ΓΙΑ ΤΗΝ «ΑΝΑΠΤΥΞΗ ΕΦΑΡΜΟΓΩΝ ΕΙΚΟΝΙΚΗΣ ΑΝΑΠΑΡΑΣΤΑΣΗΣ ΜΝΗΜΕΙΩΝ ΒΟΙΩΤΙΑΣ, ΕΦΑΡΜΟΓΩΝ ΙΣΤΟΡΙΚΗΣ,
ΕΡΓΟ: «ΕΦΑΡΜΟΓΕΣ ΕΙΚΟΝΙΚΗΣ ΠΡΑΓΜΑΤΙΚΟΤΗΤΑΣ ΣΤΗ ΒΟΙΩΤΙΑ:» ΠΡΟΚΗΡΥΞΗ ΔΙΑΓΩΝΙΣΜΟΥ ΤΟΥ ΙΔΡΥΜΑΤΟΣ ΜΕΙΖΟΝΟΣ ΕΛΛΗΝΙΣΜΟΥ ΓΙΑ ΤΗΝ «ΑΝΑΠΤΥΞΗ ΕΦΑΡΜΟΓΩΝ ΕΙΚΟΝΙΚΗΣ ΑΝΑΠΑΡΑΣΤΑΣΗΣ ΜΝΗΜΕΙΩΝ ΒΟΙΩΤΙΑΣ, ΕΦΑΡΜΟΓΩΝ ΙΣΤΟΡΙΚΗΣ,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ο. Τετάρτη 8 Ιουλίου 2015
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ο Τετάρτη 8 Ιουλίου 2015 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 5ο και το 15ο Γυµνάσιο Περιστερίου, σελ. 4174 2. Η Ειδική
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ο Τετάρτη 8 Ιουλίου 2015 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 5ο και το 15ο Γυµνάσιο Περιστερίου, σελ. 4174 2. Η Ειδική
Π Ρ Ο Γ Ρ Α Μ Μ Α Τ Ι Κ Η Σ Υ Μ Β Α Σ Η ΠΡΩΙΝΟ ΧΑΜΟΓΕΛΟ
 Π Ρ Ο Γ Ρ Α Μ Μ Α Τ Ι Κ Η Σ Υ Μ Β Α Σ Η «ΠΡΩΙΝΟ ΧΑΜΟΓΕΛΟ» Στην Κέρκυρα σήμερα την... 2013 και στο κατάστημα της Περιφερειακής Ενότητας Κέρκυρας (Περιφέρεια Ιονίων Νήσων) που βρίσκεται στην οδό Σαμάρα 13,
Π Ρ Ο Γ Ρ Α Μ Μ Α Τ Ι Κ Η Σ Υ Μ Β Α Σ Η «ΠΡΩΙΝΟ ΧΑΜΟΓΕΛΟ» Στην Κέρκυρα σήμερα την... 2013 και στο κατάστημα της Περιφερειακής Ενότητας Κέρκυρας (Περιφέρεια Ιονίων Νήσων) που βρίσκεται στην οδό Σαμάρα 13,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕ ΡΙΑΣΗ ΡΙΣΤ. Παρασκευή 7 Μαΐου 2010
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕ ΡΙΑΣΗ ΡΙΣΤ Παρασκευή 7 Μαΐου 2010 ΘΕΜΑΤΑ Α. ΕΙ ΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 6859, 6893 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 2ο ηµοτικό Σχολείο
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕ ΡΙΑΣΗ ΡΙΣΤ Παρασκευή 7 Μαΐου 2010 ΘΕΜΑΤΑ Α. ΕΙ ΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 6859, 6893 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 2ο ηµοτικό Σχολείο
Αξιολόγηση των Επιδράσεων του Σχεδίου Τοποθέτησης Άνεργων Νέων Αποφοίτων Γυμνασίων, Λυκείων, Τεχνικών Σχολών και Μεταλυκειακής Εκπαίδευσης μέχρι και
 Αξιολόγηση των Επιδράσεων του Σχεδίου Τοποθέτησης Άνεργων Νέων Αποφοίτων Γυμνασίων, Λυκείων, Τεχνικών Σχολών και Μεταλυκειακής Εκπαίδευσης μέχρι και ιετούς ιάρκειας για Απόκτηση Εργασιακής Πείρας σε Επιχειρήσεις/Οργανισμούς
Αξιολόγηση των Επιδράσεων του Σχεδίου Τοποθέτησης Άνεργων Νέων Αποφοίτων Γυμνασίων, Λυκείων, Τεχνικών Σχολών και Μεταλυκειακής Εκπαίδευσης μέχρι και ιετούς ιάρκειας για Απόκτηση Εργασιακής Πείρας σε Επιχειρήσεις/Οργανισμούς
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΣ 2014 ΣΥΝΕΔΡΙΑΣΗ ΛΒ Πέµπτη 4 Σεπτεµβρίου 2014
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΣ 2014 ΣΥΝΕΔΡΙΑΣΗ ΛΒ Πέµπτη 4 Σεπτεµβρίου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 2493, 2569 2. Επί διαδικαστικού θέµατος,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΣ 2014 ΣΥΝΕΔΡΙΑΣΗ ΛΒ Πέµπτη 4 Σεπτεµβρίου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 2493, 2569 2. Επί διαδικαστικού θέµατος,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΚΣΤ. Τετάρτη 4 Μαΐου 2011
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΚΣΤ Τετάρτη 4 Μαΐου 2011 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 9434 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν μαθητές από το 9ο Δημοτικό Σχολείο Αλίμου,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΚΣΤ Τετάρτη 4 Μαΐου 2011 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 9434 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν μαθητές από το 9ο Δημοτικό Σχολείο Αλίμου,
ΑΠΟΣΠΑΣΜΑ. Από το υπ' αριθμ. 17/01-08-2014 Πρακτικό της Οικονομικής Επιτροπής Ιονίων Νήσων
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΕΡΙΦΕΡΕΙΑ ΙΟΝΙΩΝ ΝΗΣΩΝ ΟΙΚΟΝΟΜΙΚΗ ΕΠΙΤΡΟΠΗ ΑΠΟΣΠΑΣΜΑ Από το υπ' αριθμ. 17/01-08-2014 Πρακτικό της Οικονομικής Επιτροπής Ιονίων Νήσων Αριθμ. απόφασης 496-17/01-08-2014 ΠΕΡΙΛΗΨΗ: Έγκριση
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΕΡΙΦΕΡΕΙΑ ΙΟΝΙΩΝ ΝΗΣΩΝ ΟΙΚΟΝΟΜΙΚΗ ΕΠΙΤΡΟΠΗ ΑΠΟΣΠΑΣΜΑ Από το υπ' αριθμ. 17/01-08-2014 Πρακτικό της Οικονομικής Επιτροπής Ιονίων Νήσων Αριθμ. απόφασης 496-17/01-08-2014 ΠΕΡΙΛΗΨΗ: Έγκριση
Α Π Ο Σ Π Α Σ Μ Α Από το Πρακτικό της 03ης Τακτικής Συνεδρίασης του ηµοτικού Συµβουλίου Σκοπέλου
 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΜΑΓΝΗΣΙΑΣ ΗΜΟΣ ΣΚΟΠΕΛΟΥ Πληροφορίες: Σπυριδούλα Καρβέλη Τηλέφωνο: 2424350103 E-mail: dstech@otenet.gr ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΙΑ ΙΚΤΥΟ ΑΡΙΘΜ. ΑΠΟΦ: 31 /2013 Α.Π. 1181 Α Π Ο Σ Π Α Σ Μ Α Από
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΜΑΓΝΗΣΙΑΣ ΗΜΟΣ ΣΚΟΠΕΛΟΥ Πληροφορίες: Σπυριδούλα Καρβέλη Τηλέφωνο: 2424350103 E-mail: dstech@otenet.gr ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΙΑ ΙΚΤΥΟ ΑΡΙΘΜ. ΑΠΟΦ: 31 /2013 Α.Π. 1181 Α Π Ο Σ Π Α Σ Μ Α Από
ΕΘΝΙΚΗ ΣΥΝΟΜΟΣΠΟΝΔΙΑ ΕΛΛΗΝΙΚΟΥ ΕΜΠΟΡΙΟΥ ΜΗΤΡΟΠΟΛΕΩΣ 42, 105 63 ΑΘΗΝΑ
 ΕΘΝΙΚΗ ΣΥΝΟΜΟΣΠΟΝΔΙΑ ΕΛΛΗΝΙΚΟΥ ΕΜΠΟΡΙΟΥ ΜΗΤΡΟΠΟΛΕΩΣ 42, 105 63 ΑΘΗΝΑ ΙΝΣΤΙΤΟΥΤΟ ΕΜΠΟΡΙΟΥ & ΥΠΗΡΕΣΙΩΝ ΠΕΤΡΑΚΗ 16 Τ.Κ. 105 63 ΑΘΗΝΑ ΤΗΛ: 210. 32.59.197 FAX 32.59.229 8 Σεπτεμβρίου 2011 ΑΠΟΤΕΛΕΣΜΑΤΑ ΝΕΑΣ
ΕΘΝΙΚΗ ΣΥΝΟΜΟΣΠΟΝΔΙΑ ΕΛΛΗΝΙΚΟΥ ΕΜΠΟΡΙΟΥ ΜΗΤΡΟΠΟΛΕΩΣ 42, 105 63 ΑΘΗΝΑ ΙΝΣΤΙΤΟΥΤΟ ΕΜΠΟΡΙΟΥ & ΥΠΗΡΕΣΙΩΝ ΠΕΤΡΑΚΗ 16 Τ.Κ. 105 63 ΑΘΗΝΑ ΤΗΛ: 210. 32.59.197 FAX 32.59.229 8 Σεπτεμβρίου 2011 ΑΠΟΤΕΛΕΣΜΑΤΑ ΝΕΑΣ
Η ΚΑΤΑΝΑΛΩΣΗ ΤΩΝ ΕΜΠΟΡΕΥΜΑΤΩΝ
 Κ Ε Φ Α Λ Α Ι Ο Τ Ρ Ι Τ Ο Η ΚΑΤΑΝΑΛΩΣΗ ΤΩΝ ΕΜΠΟΡΕΥΜΑΤΩΝ Π Ε Ρ Ι Ε Χ Ό Μ Ε Ν Α 3.1 Η χρηστική αξία των εμπορευμάτων 32 3.2 Ανάγκες και Επιθυμίες 32 3.3 Είδη και Ταξινόμηση των Αναγκών 35 3.4 Τα Μέσα Ικανοποίησης
Κ Ε Φ Α Λ Α Ι Ο Τ Ρ Ι Τ Ο Η ΚΑΤΑΝΑΛΩΣΗ ΤΩΝ ΕΜΠΟΡΕΥΜΑΤΩΝ Π Ε Ρ Ι Ε Χ Ό Μ Ε Ν Α 3.1 Η χρηστική αξία των εμπορευμάτων 32 3.2 Ανάγκες και Επιθυμίες 32 3.3 Είδη και Ταξινόμηση των Αναγκών 35 3.4 Τα Μέσα Ικανοποίησης
ΕΝΩΠΙΟΝ ΠΑΝΤΟΣ ΑΡΜΟΔΙΟΥ ΔΙΚΑΣΤΗΡΙΟΥ Η ΑΡΧΗΣ ΕΞΩΔΙΚΗ ΔΙΑΜΑΡΤΥΡΙΑ - ΠΡΟΣΚΛΗΣΗ
 ΕΝΩΠΙΟΝ ΠΑΝΤΟΣ ΑΡΜΟΔΙΟΥ ΔΙΚΑΣΤΗΡΙΟΥ Η ΑΡΧΗΣ ΕΞΩΔΙΚΗ ΔΙΑΜΑΡΤΥΡΙΑ - ΠΡΟΣΚΛΗΣΗ ΦΑΡΜΑΣΩΝΗ Κωνσταντίνου του Νικολάου, Σκηνoθέτη, νoμίμου εκπροσώπου της Θεατρικής Εταιρείας «ΣΚΑΡΑΒΑΙΟΙ» με έδρα την οδό Φρύνης,
ΕΝΩΠΙΟΝ ΠΑΝΤΟΣ ΑΡΜΟΔΙΟΥ ΔΙΚΑΣΤΗΡΙΟΥ Η ΑΡΧΗΣ ΕΞΩΔΙΚΗ ΔΙΑΜΑΡΤΥΡΙΑ - ΠΡΟΣΚΛΗΣΗ ΦΑΡΜΑΣΩΝΗ Κωνσταντίνου του Νικολάου, Σκηνoθέτη, νoμίμου εκπροσώπου της Θεατρικής Εταιρείας «ΣΚΑΡΑΒΑΙΟΙ» με έδρα την οδό Φρύνης,
VESTA40 [ΟΔΗΓΙΕΣ ΓΙΑ ΤΗΝ ΤΟΠΟΘΕΤΗΣΗ, ΤΗ ΧΡΗΣΗ ΚΑΙ ΤΗ ΣΥΝΤΗΡΗΣΗ] Το εγχειρίδιο οδηγιών χρήσης αποτελεί αναπόσπαστο μέρος του προϊόντος
![VESTA40 [ΟΔΗΓΙΕΣ ΓΙΑ ΤΗΝ ΤΟΠΟΘΕΤΗΣΗ, ΤΗ ΧΡΗΣΗ ΚΑΙ ΤΗ ΣΥΝΤΗΡΗΣΗ] Το εγχειρίδιο οδηγιών χρήσης αποτελεί αναπόσπαστο μέρος του προϊόντος](/thumbs/33/16629899.jpg "VESTA40 [ΟΔΗΓΙΕΣ ΓΙΑ ΤΗΝ ΤΟΠΟΘΕΤΗΣΗ, ΤΗ ΧΡΗΣΗ ΚΑΙ ΤΗ ΣΥΝΤΗΡΗΣΗ] Το εγχειρίδιο οδηγιών χρήσης αποτελεί αναπόσπαστο μέρος του προϊόντος") VESTA40 [ΟΔΗΓΙΕΣ ΓΙΑ ΤΗΝ ΤΟΠΟΘΕΤΗΣΗ, ΤΗ ΧΡΗΣΗ ΚΑΙ ΤΗ ΣΥΝΤΗΡΗΣΗ] Το εγχειρίδιο οδηγιών χρήσης αποτελεί αναπόσπαστο μέρος του προϊόντος Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α ΚΕΦΑΛΑΙΟ ΤΙΤΛΟΣ ΣΕΛΙΔΑ Εισαγωγή 4 Σκοπός του
VESTA40 [ΟΔΗΓΙΕΣ ΓΙΑ ΤΗΝ ΤΟΠΟΘΕΤΗΣΗ, ΤΗ ΧΡΗΣΗ ΚΑΙ ΤΗ ΣΥΝΤΗΡΗΣΗ] Το εγχειρίδιο οδηγιών χρήσης αποτελεί αναπόσπαστο μέρος του προϊόντος Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α ΚΕΦΑΛΑΙΟ ΤΙΤΛΟΣ ΣΕΛΙΔΑ Εισαγωγή 4 Σκοπός του
ΕΞ.ΕΠΕΙΓΟΝ -FAX. Αθήνα, 15 Οκτωβρίου 2014
 ΕΞ.ΕΠΕΙΓΟΝ -FAX ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΚΩΝ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΗΜΟΣΙΩΝ ΕΣΟΔΩΝ ΓΕΝΙΚΗ Δ/ΝΣΗ ΦΟΡΟΛΟΓΙΚΗΣ ΔΙΟΙΚΗΣΗΣ ΔΙΕΥΘΥΝΣΗ ΕΛΕΓΧΩΝ ΤMHMA Α Αθήνα, 15 Οκτωβρίου 2014 ΠΟΛ.:1228 ΠΡΟΣ: Ως Π.Δ.
ΕΞ.ΕΠΕΙΓΟΝ -FAX ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΚΩΝ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΗΜΟΣΙΩΝ ΕΣΟΔΩΝ ΓΕΝΙΚΗ Δ/ΝΣΗ ΦΟΡΟΛΟΓΙΚΗΣ ΔΙΟΙΚΗΣΗΣ ΔΙΕΥΘΥΝΣΗ ΕΛΕΓΧΩΝ ΤMHMA Α Αθήνα, 15 Οκτωβρίου 2014 ΠΟΛ.:1228 ΠΡΟΣ: Ως Π.Δ.
Κύκλος Κοινωνικής Προστασίας ΠΟΡΙΣΜΑ
 Κύκλος Κοινωνικής Προστασίας ΠΟΡΙΣΜΑ (ΝΟΜΟΣ 2477/1997 Συνήγορος του Πολίτη και Σώµα Ελεγκτών-Επιθεωρητών ηµόσιας ιοίκησης Άρθρο 4, Παράγραφος 6) [ΑΥΤΕΠΑΓΓΕΛΤΗ ΕΡΕΥΝΑ 6487/18-5-2001] Θέµα: ΑΠΟΦΑΣΕΙΣ ΚΑΙ
Κύκλος Κοινωνικής Προστασίας ΠΟΡΙΣΜΑ (ΝΟΜΟΣ 2477/1997 Συνήγορος του Πολίτη και Σώµα Ελεγκτών-Επιθεωρητών ηµόσιας ιοίκησης Άρθρο 4, Παράγραφος 6) [ΑΥΤΕΠΑΓΓΕΛΤΗ ΕΡΕΥΝΑ 6487/18-5-2001] Θέµα: ΑΠΟΦΑΣΕΙΣ ΚΑΙ
Σέρρες 11.5.2015 Αριθ. Πρωτ.: 1387
 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ TEΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙ ΕΥΤΙΚΟ Ι ΡΥΜΑ ΚΕΝΤΡΙΚΗΣ ΜΑΚΕ ΟΝΙΑΣ ΤΕΡΜΑ ΜΑΓΝΗΣΙΑΣ - 614 ΣΕΡΡΕΣ ΠΡΟΚΗΡΥΞΗ Σέρρες 11.5.015 Αριθ. Πρωτ.: 1387 ΠΡΟΣΛΗΨΗΣ ΕΠΙΣΤΗΜΟΝΙΚΩΝ ΣΥΝΕΡΓΑΤΩΝ, ΕΡΓΑΣΤΗΡΙΑΚΩΝ ΣΥΝΕΡΓΑΤΩΝ,
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ TEΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙ ΕΥΤΙΚΟ Ι ΡΥΜΑ ΚΕΝΤΡΙΚΗΣ ΜΑΚΕ ΟΝΙΑΣ ΤΕΡΜΑ ΜΑΓΝΗΣΙΑΣ - 614 ΣΕΡΡΕΣ ΠΡΟΚΗΡΥΞΗ Σέρρες 11.5.015 Αριθ. Πρωτ.: 1387 ΠΡΟΣΛΗΨΗΣ ΕΠΙΣΤΗΜΟΝΙΚΩΝ ΣΥΝΕΡΓΑΤΩΝ, ΕΡΓΑΣΤΗΡΙΑΚΩΝ ΣΥΝΕΡΓΑΤΩΝ,
στο σχέδιο νόµου «Κύρωση του Πρωτοκόλλου µεταξύ το οποίο τροποποιεί τη Σύµβαση µεταξύ της Ελληνικής Δηµοκρατίας και της Ελβετικής Συνοµοσπονδίας
 ΑΙΤΙΟΛΟΓΙΚΗ ΕΚΘΕΣΗ στο σχέδιο νόµου «Κύρωση του Πρωτοκόλλου µεταξύ της Ελληνικής Δηµοκρατίας και της Ελβετικής Συνοµοσπονδίας το οποίο τροποποιεί τη Σύµβαση µεταξύ της Ελληνικής Δηµοκρατίας και της Ελβετικής
ΑΙΤΙΟΛΟΓΙΚΗ ΕΚΘΕΣΗ στο σχέδιο νόµου «Κύρωση του Πρωτοκόλλου µεταξύ της Ελληνικής Δηµοκρατίας και της Ελβετικής Συνοµοσπονδίας το οποίο τροποποιεί τη Σύµβαση µεταξύ της Ελληνικής Δηµοκρατίας και της Ελβετικής
Ε Φ Υ Μ Ν Ι O N. Παρατίθενται γνώµες και απόψεις. σχετικές µε το έργο του Πρωτοψάλτη, οράρχη και κδότη βιβλίων. κκλησιαστικής ουσικής
 1 Ε Φ Υ Μ Ν Ι O N Παρατίθενται γνώµες και απόψεις σχετικές µε το έργο του Πρωτοψάλτη, οράρχη και κδότη βιβλίων κκλησιαστικής ουσικής Φ ώ τ η Θ ε ο δ ω ρ α κ ό π ο υ λ ο υ Εκδοτικός Οίκος «Μυρίπνοον» 2
1 Ε Φ Υ Μ Ν Ι O N Παρατίθενται γνώµες και απόψεις σχετικές µε το έργο του Πρωτοψάλτη, οράρχη και κδότη βιβλίων κκλησιαστικής ουσικής Φ ώ τ η Θ ε ο δ ω ρ α κ ό π ο υ λ ο υ Εκδοτικός Οίκος «Μυρίπνοον» 2
ΑΘΗΝΑ, 12 Ιουλίου 2012. Αριθ. Πρωτ. : 775. ΠΡΟΣ : Περιφέρεια Αττικής. τον Περιφερειάρχη Αττικής κ. Γ. Σγουρό ΚΟΙΝ : όπως ο συνηµµένος πίνακας
 ΑΘΗΝΑ, 12 Ιουλίου 2012 Αριθ. Πρωτ. : 775 ΠΡΟΣ : Περιφέρεια Αττικής τον Περιφερειάρχη Αττικής κ. Γ. Σγουρό ΚΟΙΝ : όπως ο συνηµµένος πίνακας ΘΕΜΑ: ΚΑΤΑΓΓΕΛΙΑ για την καταστροφή του πρασίνου των οδικών αξόνων
ΑΘΗΝΑ, 12 Ιουλίου 2012 Αριθ. Πρωτ. : 775 ΠΡΟΣ : Περιφέρεια Αττικής τον Περιφερειάρχη Αττικής κ. Γ. Σγουρό ΚΟΙΝ : όπως ο συνηµµένος πίνακας ΘΕΜΑ: ΚΑΤΑΓΓΕΛΙΑ για την καταστροφή του πρασίνου των οδικών αξόνων
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΥΣ 2014 ΣΥΝΕΔΡΙΑΣΗ ΛZ Τρίτη 16 Σεπτεµβριου 2014
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΥΣ 2014 ΣΥΝΕΔΡΙΑΣΗ ΛZ Τρίτη 16 Σεπτεµβριου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 2923, 2974 2. Επί διαδικαστικού θέµατος,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΥΣ 2014 ΣΥΝΕΔΡΙΑΣΗ ΛZ Τρίτη 16 Σεπτεµβριου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 2923, 2974 2. Επί διαδικαστικού θέµατος,
ΥΠΗΡΕΣΙΕΣ ΕΡΓΑΣΙΩΝ ΙΑΧΕΙΡΙΣΗΣ ΑΣΤΙΚΩΝ ΣΤΕΡΕΩΝ ΑΠΟΒΛΗΤΩΝ Τ Ε Υ Χ Ο Σ Τ Ε Χ Ν Ι Κ Ω Ν Π Ρ Ο Ι Α Γ Ρ Α Φ Ω Ν
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΕΡΙΦΕΡΕΙΑ ΙΟΝΙΩΝ ΝΗΣΩΝ ΠΕΡΙΦΕΡΕΙΑΚΗ ΕΝΟΤΗΤΑ ΚΕΦΑΛΛΗΝΙΑΣ & ΙΘΑΚΗΣ ΕΤΑΙΡΕΙΑ ΔΙΑΧΕΙΡΙΣΗΣ ΑΠΟΒΛΗΤΩΝ ΚΕΦΑΛΟΝΙΑΣ & ΙΘΑΚΗΣ ΑΕ ΟΤΑ ΥΠΗΡΕΣΙΕΣ ΕΡΓΑΣΙΩΝ ΙΑΧΕΙΡΙΣΗΣ ΑΣΤΙΚΩΝ ΣΤΕΡΕΩΝ ΑΠΟΒΛΗΤΩΝ ΤΕΥΧΗ
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΕΡΙΦΕΡΕΙΑ ΙΟΝΙΩΝ ΝΗΣΩΝ ΠΕΡΙΦΕΡΕΙΑΚΗ ΕΝΟΤΗΤΑ ΚΕΦΑΛΛΗΝΙΑΣ & ΙΘΑΚΗΣ ΕΤΑΙΡΕΙΑ ΔΙΑΧΕΙΡΙΣΗΣ ΑΠΟΒΛΗΤΩΝ ΚΕΦΑΛΟΝΙΑΣ & ΙΘΑΚΗΣ ΑΕ ΟΤΑ ΥΠΗΡΕΣΙΕΣ ΕΡΓΑΣΙΩΝ ΙΑΧΕΙΡΙΣΗΣ ΑΣΤΙΚΩΝ ΣΤΕΡΕΩΝ ΑΠΟΒΛΗΤΩΝ ΤΕΥΧΗ
«12 Η ΓΙΟΡΤΗ ΤΩΝ ΣΠΟΡΩΝ»
 ΔΕΛΤΙΟ ΤΥΠΟΥ «12 Η ΓΙΟΡΤΗ ΤΩΝ ΣΠΟΡΩΝ» 10-12 Απριλίου 2009, ΚΥΡΙΑΚΗ ΤΩΝ ΒΑΪΩΝ ΑΙΘΡΙΟ ΓΥΜΝΑΣΙΟΥ & Λ. Τ. ΛΕΧΑΙΟΥ Μια γιορτή αφιερωμένη στη Φ Υ Σ Ι Κ Η Κ Α Λ Λ Ι Ε Ρ Γ Ε Ι Α και τις Π Ο Ι Κ Ι Λ Ι Ε Σ Α Ν Ο
ΔΕΛΤΙΟ ΤΥΠΟΥ «12 Η ΓΙΟΡΤΗ ΤΩΝ ΣΠΟΡΩΝ» 10-12 Απριλίου 2009, ΚΥΡΙΑΚΗ ΤΩΝ ΒΑΪΩΝ ΑΙΘΡΙΟ ΓΥΜΝΑΣΙΟΥ & Λ. Τ. ΛΕΧΑΙΟΥ Μια γιορτή αφιερωμένη στη Φ Υ Σ Ι Κ Η Κ Α Λ Λ Ι Ε Ρ Γ Ε Ι Α και τις Π Ο Ι Κ Ι Λ Ι Ε Σ Α Ν Ο
Ηµεροµηνία επίδοσης της 033149 πρόσκλησης στα µέλη της Οικονοµικής Επιτροπής η 9/7/2015. Ηµεροµηνία συνεδρίασης : 13 Ιουλίου 2015
 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΙΑ ΙΚΤΥΟ ΝΟΜΟΣ ΑΤΤΙΚΗΣ ΑΡ. ΠΡΩΤ.: ΗΜΟΣ ΑΜΑΡΟΥΣΙΟΥ ΜΟΝΑ Α Γ' ΥΠΟΣΤΗΡΙΞΗ ΟΙΚΟΝΟΜΙΚΗΣ ΕΠΙΤΡΟΠΗΣ Α Π Ο Σ Π Α Σ Μ Α Από το βιβλίο πρακτικών της Οικονοµικής Επιτροπής της 36ης
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΙΑ ΙΚΤΥΟ ΝΟΜΟΣ ΑΤΤΙΚΗΣ ΑΡ. ΠΡΩΤ.: ΗΜΟΣ ΑΜΑΡΟΥΣΙΟΥ ΜΟΝΑ Α Γ' ΥΠΟΣΤΗΡΙΞΗ ΟΙΚΟΝΟΜΙΚΗΣ ΕΠΙΤΡΟΠΗΣ Α Π Ο Σ Π Α Σ Μ Α Από το βιβλίο πρακτικών της Οικονοµικής Επιτροπής της 36ης
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ε. Παρασκευή 10 Οκτωβρίου 2014
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ε Παρασκευή 10 Οκτωβρίου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 273 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το Γενικό Λύκειο Βαθέος
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ε Παρασκευή 10 Οκτωβρίου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 273 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το Γενικό Λύκειο Βαθέος
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ]Β. Πέµπτη 20 Φεβρουαρίου 2014
![ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ]Β. Πέµπτη 20 Φεβρουαρίου 2014](/thumbs/39/18963772.jpg "ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ]Β. Πέµπτη 20 Φεβρουαρίου 2014") ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ]Β Πέµπτη 20 Φεβρουαρίου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 7631, 7671 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 3ο Δηµοτικό
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ]Β Πέµπτη 20 Φεβρουαρίου 2014 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 7631, 7671 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 3ο Δηµοτικό
A1. Να γράψετε στο τετράδιό σας την περίληψη του κειμένου που σας δόθηκε (100-120 λέξεις). Μονάδες 25
. Μονάδες 25") ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 28 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΝΕΟΕΛΛΗΝΙΚΗ ΓΛΩΣΣΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΙΜΕΝΟ Η «ανθρωπιά» είναι
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 28 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΝΕΟΕΛΛΗΝΙΚΗ ΓΛΩΣΣΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΙΜΕΝΟ Η «ανθρωπιά» είναι
ΕΔΡΑΣΗ Χ. ΨΑΛΛΙΔΑΣ ΑΝΩΝΥΜΟΣ ΤΕΧΝΙΚΗ, ΤΟΥΡΙΣΤΙΚΗ, ΕΜΠΟΡΙΚΗ και ΒΙΟΜΗΧΑΝΙΚΗ ΕΤΑΙΡΕΙΑ Συνοπτικές Ενδιάμεσες Οικονομικές Καταστάσεις για την περίοδο από
 1 Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Έκθεση Επισκόπησης Ενδιάμεσης Χρηματοοικονομικής Πληροφόρησης... 3 Ενοποιημένη και Εταιρική Κατάσταση Συνολικών Εσόδων... 6 Ενοποιημένη και Εταιρική Κατάσταση Οικονομικής Θέσης...
1 Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Έκθεση Επισκόπησης Ενδιάμεσης Χρηματοοικονομικής Πληροφόρησης... 3 Ενοποιημένη και Εταιρική Κατάσταση Συνολικών Εσόδων... 6 Ενοποιημένη και Εταιρική Κατάσταση Οικονομικής Θέσης...
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΙΣΤ. Πέµπτη 31 Ιανουαρίου 2013
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΙΣΤ Πέµπτη 31 Ιανουαρίου 2013 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 7055, 7129 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 1ο Γυµνάσιο
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΙΣΤ Πέµπτη 31 Ιανουαρίου 2013 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 7055, 7129 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 1ο Γυµνάσιο
Γ49/59 ΕΞ. ΕΠΕΙΓΟΝ Π Ρ Ο Σ :
 Αθήνα, 30-5-2012 Δ Ι Ο Ι Κ Η Σ Η ΓΕΝΙΚΗ Δ/ΝΣΗ ΔΙΟΙΚ/ΚΩΝ ΥΠΗΡΕΣΙΩΝ Δ/ΝΣΗ ΔΙΟΙΚΗΤΙΚΟΥ ΠΡΟΣΩΠΙΚΟΥ ΤΜΗΜΑ : ΕΡΓΑΣΙΑΚΩΝ ΣΧΕΣΕΩΝ Ταχ. Δ/νση : Αγ. Κωνσταντίνου 8 Ταχ. Κώδικας: 102 41 ΑΘΗΝΑ Τηλέφωνο : 210-215289,290,291,292
Αθήνα, 30-5-2012 Δ Ι Ο Ι Κ Η Σ Η ΓΕΝΙΚΗ Δ/ΝΣΗ ΔΙΟΙΚ/ΚΩΝ ΥΠΗΡΕΣΙΩΝ Δ/ΝΣΗ ΔΙΟΙΚΗΤΙΚΟΥ ΠΡΟΣΩΠΙΚΟΥ ΤΜΗΜΑ : ΕΡΓΑΣΙΑΚΩΝ ΣΧΕΣΕΩΝ Ταχ. Δ/νση : Αγ. Κωνσταντίνου 8 Ταχ. Κώδικας: 102 41 ΑΘΗΝΑ Τηλέφωνο : 210-215289,290,291,292
15PROC003313118 2015-11-16
 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΛΕΣΒΟΥ ΗΜΟΣ ΛΗΜΝΟΥ ΤΜΗΜΑ ΟΙΚ. ΥΠΗΡΕΣΙΩΝ ΓΡΑΦΕΙΟ ΠΡΟΜΗΘΕΙΩΝ Μύρινα, 13 Νοεµβρίου 2015 Αρ. Πρωτ. : 17043 ΙΑΚΗΡΥΞΗ Ο ΗΜΑΡΧΟΣ ΛΗΜΝΟΥ Έχοντας υπ όψη: 1. Τις διατάξεις: α. Των άρθρων
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΛΕΣΒΟΥ ΗΜΟΣ ΛΗΜΝΟΥ ΤΜΗΜΑ ΟΙΚ. ΥΠΗΡΕΣΙΩΝ ΓΡΑΦΕΙΟ ΠΡΟΜΗΘΕΙΩΝ Μύρινα, 13 Νοεµβρίου 2015 Αρ. Πρωτ. : 17043 ΙΑΚΗΡΥΞΗ Ο ΗΜΑΡΧΟΣ ΛΗΜΝΟΥ Έχοντας υπ όψη: 1. Τις διατάξεις: α. Των άρθρων
ΕΝΗΜΕΡΩΤΙΚΟ ΔΕΛΤΙΟ ΤΟΥ ΙΑΤΡΙΚΟΥ ΣΥΛΛΟΓΟΥ ΑΘΗΝΩΝ
 ΕΝΗΜΕΡΩΣΗ τ ω ν γ ι α τ ρ ω ν ΕΝΗΜΕΡΩΤΙΚΟ ΔΕΛΤΙΟ ΤΟΥ ΙΑΤΡΙΚΟΥ ΣΥΛΛΟΓΟΥ ΑΘΗΝΩΝ Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Δ ι μ η ν ι α ί α Έ κ δ ο σ η Τ Ε Υ Χ Ο Σ 2 1 8 (Τυπώνεται σε 25.500 αντίτυπα) ianoyaριοσ-φεβρουαριοσ
ΕΝΗΜΕΡΩΣΗ τ ω ν γ ι α τ ρ ω ν ΕΝΗΜΕΡΩΤΙΚΟ ΔΕΛΤΙΟ ΤΟΥ ΙΑΤΡΙΚΟΥ ΣΥΛΛΟΓΟΥ ΑΘΗΝΩΝ Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Δ ι μ η ν ι α ί α Έ κ δ ο σ η Τ Ε Υ Χ Ο Σ 2 1 8 (Τυπώνεται σε 25.500 αντίτυπα) ianoyaριοσ-φεβρουαριοσ
Ορθη επαναληψη ΔΙΑΚΗΡΥΞΗ ΠΡΟΧΕΙΡΟY ΜΕΙΟΔΟΤΙΚΟY ΔΙΑΓΩΝΙΣΜΟY ΜΕ ΣΦΡΑΓΙΣΜΕΝΕΣ ΠΡΟΣΦΟΡΕΣ ΓΙΑ ΤΗΝ ΠΡΟΜΗΘΕΙΑ ΔΙΑΦΟΡΩΝ ΕΙΔΩΝ ΔΙΑΤΡΟΦΗΣ ΓΙΑ ΤΟ ΕΤΟΣ 2011.
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΜΑΓΝΗΣΙΑΣ ΔΗΜΟΣ ΒΟΛΟΥ Βόλος : 5-8-2011 ΤΜΗΜΑ ΠΡΟΜΗΘΕΙΩΝ Αριθ. Πρωτ.: 846971/ΓΠ22817 Πληροφορίες : Φώτου Κωνσταντία Τηλέφωνο: : 2421350103 FAX : : 2421097610 Ορθη επαναληψη ΔΙΑΚΗΡΥΞΗ
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΜΑΓΝΗΣΙΑΣ ΔΗΜΟΣ ΒΟΛΟΥ Βόλος : 5-8-2011 ΤΜΗΜΑ ΠΡΟΜΗΘΕΙΩΝ Αριθ. Πρωτ.: 846971/ΓΠ22817 Πληροφορίες : Φώτου Κωνσταντία Τηλέφωνο: : 2421350103 FAX : : 2421097610 Ορθη επαναληψη ΔΙΑΚΗΡΥΞΗ
Ε.Ε. Π α ρ.ι(i), Α ρ.3638, 27/9/2002
, Α ρ.3638, 27/9/2002") Ο περί Εμπορικής Ναυτιλίας (Ελάχιστες Προδιαγραφές Ιατρικής Περίθαλψης στα Πλοία) Νόμος του 2002 εκδίδεται με δημοσίευση στην Επίσημη Εφημερίδα της Κυπριακής Δημοκρατίας σύμφωνα με το Άρθρο 52 του Συντάγματος.
Ο περί Εμπορικής Ναυτιλίας (Ελάχιστες Προδιαγραφές Ιατρικής Περίθαλψης στα Πλοία) Νόμος του 2002 εκδίδεται με δημοσίευση στην Επίσημη Εφημερίδα της Κυπριακής Δημοκρατίας σύμφωνα με το Άρθρο 52 του Συντάγματος.
ΠΕΡΙΛΗΨΗ Λήψη απόφασης σχετικά με το υπ' αριθμ. 5016/13-02-13 αίτημα της Ευαγγελίας Σκουντζούρη κλπ που αφορά στο ΟΤ 823.
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΑΠΟΣΠΑΣΜΑ ΝΟΜΟΣ ΑΤΤΙΚΗΣ Από το Πρακτικό της με αριθμό 6/2013 ΔΗΜΟΣ ΑΓΙΟΥ ΔΗΜΗΤΡΙΟΥ Τακτικής Συνεδρίασης του Δημοτικού Συμβουλίου ΑΡΙΘΜΟΣ ΑΠΟΦΑΣΗΣ: 45/2013 Γραφείο: Δημοτικού Συμβουλίου
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΑΠΟΣΠΑΣΜΑ ΝΟΜΟΣ ΑΤΤΙΚΗΣ Από το Πρακτικό της με αριθμό 6/2013 ΔΗΜΟΣ ΑΓΙΟΥ ΔΗΜΗΤΡΙΟΥ Τακτικής Συνεδρίασης του Δημοτικού Συμβουλίου ΑΡΙΘΜΟΣ ΑΠΟΦΑΣΗΣ: 45/2013 Γραφείο: Δημοτικού Συμβουλίου
Π Ρ Α Κ Τ Ι Κ Α Β Ο Υ Λ Η Σ
 Π Ρ Α Κ Τ Ι Κ Α Β Ο Υ Λ Η Σ Θ' ΠΕΡΙΟ ΟΣ (ΠΡΟΕ ΡΕΥΟΜΕΝΗΣ ΗΜΟΚΡΑΤΙΑΣ) ΣΥΝΟ ΟΣ Γ' ΣΥΝΕ ΡΙΑΣΗ Ξ ' Παρασκευή 22 Ιανουαρίου 1999 Αθήνα, σήµερα στις 22 Ιανουαρίου 1999, ηµέρα Παρασκευή και ώρα 10.35' συνήλθε
Π Ρ Α Κ Τ Ι Κ Α Β Ο Υ Λ Η Σ Θ' ΠΕΡΙΟ ΟΣ (ΠΡΟΕ ΡΕΥΟΜΕΝΗΣ ΗΜΟΚΡΑΤΙΑΣ) ΣΥΝΟ ΟΣ Γ' ΣΥΝΕ ΡΙΑΣΗ Ξ ' Παρασκευή 22 Ιανουαρίου 1999 Αθήνα, σήµερα στις 22 Ιανουαρίου 1999, ηµέρα Παρασκευή και ώρα 10.35' συνήλθε
ΠΑΝΕΛΛΗΝΙΑ ΟΜΟΣΠΟΝΔΙΑ ΣΩΜΑΤΕΙΩΝ ΕΡΓΑΖΟΜΕΝΩΝ Υ.ΠΕ.ΧΩ.Δ.Ε.
 ΠΑΝΕΛΛΗΝΙΑ ΟΜΟΣΠΟΝΔΙΑ ΣΩΜΑΤΕΙΩΝ ΕΡΓΑΖΟΜΕΝΩΝ Υ.ΠΕ.ΧΩ.Δ.Ε. (Π.Ο.Σ.Ε. Υ.ΠΕ.ΧΩ.Δ.Ε.) Ιπποκράτους 196 198 114 71 ΑΘΗΝΑ Τηλ. : 210 6440873 FAX : 210 6454223 Site: www.poseypexode.gr Email:poseypexode@otenet.gr
ΠΑΝΕΛΛΗΝΙΑ ΟΜΟΣΠΟΝΔΙΑ ΣΩΜΑΤΕΙΩΝ ΕΡΓΑΖΟΜΕΝΩΝ Υ.ΠΕ.ΧΩ.Δ.Ε. (Π.Ο.Σ.Ε. Υ.ΠΕ.ΧΩ.Δ.Ε.) Ιπποκράτους 196 198 114 71 ΑΘΗΝΑ Τηλ. : 210 6440873 FAX : 210 6454223 Site: www.poseypexode.gr Email:poseypexode@otenet.gr
Ημερομηνία δημοσίευσης στον Ελληνικό Τύπο Α Π Ο Φ Α Σ Η
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ ΕΘΝΙΚΟ ΚΕΝΤΡΟ ΑΜΕΣΗΣ ΒΟΗΘΕΙΑΣ ΠΑΡΑΡΤΗΜΑ ΚΑΒΑΛΑΣ ΤΜΗΜΑ ΠΡΟΣΩΠΙΚΟΥ & ΟΙΚΟΝΟΜΙΚΗΣ ΔΙΑΧΕΙΡΙΣΗΣ ΓΡΑΦΕΙΟ ΠΡΟΜΗΘΕΙΩΝ Ταχ. Δ/νση : Τέρμα Αργυροκάστρου Παιδόπολη Καβάλας Ταχ.
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ ΕΘΝΙΚΟ ΚΕΝΤΡΟ ΑΜΕΣΗΣ ΒΟΗΘΕΙΑΣ ΠΑΡΑΡΤΗΜΑ ΚΑΒΑΛΑΣ ΤΜΗΜΑ ΠΡΟΣΩΠΙΚΟΥ & ΟΙΚΟΝΟΜΙΚΗΣ ΔΙΑΧΕΙΡΙΣΗΣ ΓΡΑΦΕΙΟ ΠΡΟΜΗΘΕΙΩΝ Ταχ. Δ/νση : Τέρμα Αργυροκάστρου Παιδόπολη Καβάλας Ταχ.
...105 ...109...111...112 - ...117 9....118...119 ...130 1. ...133...136 10. ...138...146
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ...i ΚΕΦΑΛΑΙΟ 1. ΕΙΣΑΓΩΓΗ...4 ΚΕΦΑΛΑΙΟ 2. ΙΣΤΟΡΙΚΗ ΑΝΑ ΡΟΜΗ...7 1. Επιχειρήσεων των ΟΤΑ µέχρι τον Κώδικα ήµων και Κοινοτήτων του 1954...7 Α. Νοµική µορφή των επιχειρήσεων...7
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ...i ΚΕΦΑΛΑΙΟ 1. ΕΙΣΑΓΩΓΗ...4 ΚΕΦΑΛΑΙΟ 2. ΙΣΤΟΡΙΚΗ ΑΝΑ ΡΟΜΗ...7 1. Επιχειρήσεων των ΟΤΑ µέχρι τον Κώδικα ήµων και Κοινοτήτων του 1954...7 Α. Νοµική µορφή των επιχειρήσεων...7
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΜΔ. Κυριακή 4 Δεκεµβρίου 2011
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΜΔ Κυριακή 4 Δεκεµβρίου 2011 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 3ο Δηµοτικό Σχολείο Γιαννιτσών, σελ. 2269 2. Επί διαδικαστικού
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΜΔ Κυριακή 4 Δεκεµβρίου 2011 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 3ο Δηµοτικό Σχολείο Γιαννιτσών, σελ. 2269 2. Επί διαδικαστικού
Α Π Ο Φ Α Σ Η 4/459/27.12.2007. του ιοικητικού Συµβουλίου
 Α Π Ο Φ Α Σ Η 4/459/27.12.2007 του ιοικητικού Συµβουλίου ΘΕΜΑ: «Υπολογισµός κεφαλαιακών απαιτήσεων των Επιχειρήσεων Παροχής Επενδυτικών Υπηρεσιών για τον κίνδυνο αγοράς» ΤΟ ΙΟΙΚΗΤΙΚΟ ΣΥΜΒΟΥΛΙΟ ΤΗΣ ΕΠΙΤΡΟΠΗΣ
Α Π Ο Φ Α Σ Η 4/459/27.12.2007 του ιοικητικού Συµβουλίου ΘΕΜΑ: «Υπολογισµός κεφαλαιακών απαιτήσεων των Επιχειρήσεων Παροχής Επενδυτικών Υπηρεσιών για τον κίνδυνο αγοράς» ΤΟ ΙΟΙΚΗΤΙΚΟ ΣΥΜΒΟΥΛΙΟ ΤΗΣ ΕΠΙΤΡΟΠΗΣ
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΕΥΒΟΙΑΣ ΣΧΟΛΙΚΗ ΕΠΙΤΡΟΠΗ ΜΟΝΑΔΩΝ Α ΒΑΘΜΙΑΣ ΕΚΠ/ΣΗΣ ΕΡΓΑΣΙΑ:
 ΕΡΓΑΣΙΑ: Αναγόμωση συντήρηση Αναγόμωση συντήρηση Μονάδες Α Βάθμιας εκπ/σης ΠΕΡΙΕΧΟΜΕΝΑ 1. Τεχνική περιγραφή 2. Ενδεικτικός Προϋπολογισμός 3. Συγγραφή υποχρεώσεων 1 ΕΡΓΑΣΙΑ: Αναγόμωση συντήρηση Τεχνική
ΕΡΓΑΣΙΑ: Αναγόμωση συντήρηση Αναγόμωση συντήρηση Μονάδες Α Βάθμιας εκπ/σης ΠΕΡΙΕΧΟΜΕΝΑ 1. Τεχνική περιγραφή 2. Ενδεικτικός Προϋπολογισμός 3. Συγγραφή υποχρεώσεων 1 ΕΡΓΑΣΙΑ: Αναγόμωση συντήρηση Τεχνική
ΑΝΩΣΗ Α.E. ΕΤΗΣΙΑ ΟΙΚΟΝΟΜΙΚΗ ΕΚΘΕΣΗ
 ΑΝΩΣΗ Α.E. ΑΝΩΝΥΜΗ ΕΤΑΙΡΕΙΑ ΜΟΥΣΙΚΩΝ ΕΚΔΗΛΩΣΕΩΝ & ΠΑΡΑΓΩΓΩΝ ΕΤΗΣΙΑ ΟΙΚΟΝΟΜΙΚΗ ΕΚΘΕΣΗ 31 ης Δεκεμβρίου, 2009 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΑΝΑΦΟΡΑΣ (Δ.Π.Χ.Α.) Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α ΕΚΘΕΣΗ
ΑΝΩΣΗ Α.E. ΑΝΩΝΥΜΗ ΕΤΑΙΡΕΙΑ ΜΟΥΣΙΚΩΝ ΕΚΔΗΛΩΣΕΩΝ & ΠΑΡΑΓΩΓΩΝ ΕΤΗΣΙΑ ΟΙΚΟΝΟΜΙΚΗ ΕΚΘΕΣΗ 31 ης Δεκεμβρίου, 2009 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΑΝΑΦΟΡΑΣ (Δ.Π.Χ.Α.) Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α ΕΚΘΕΣΗ
Διοίκηση & Γραφεία 2. Επιστολή του Προέδρου 4. Μήνυμα από την Αν. Γενική Διευθύντρια 5. Σύσταση και αρμοδιότητες 6. Αναπτυξιακό Πρόγραμμα 7
 Ετήσια Έκθεση 2013 Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Διοίκηση & Γραφεία 2 Επιστολή του Προέδρου 4 Μήνυμα από την Αν. Γενική Διευθύντρια 5 Σύσταση και αρμοδιότητες 6 Αναπτυξιακό Πρόγραμμα 7 Σχέδια που έχουν συμπληρωθεί
Ετήσια Έκθεση 2013 Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Διοίκηση & Γραφεία 2 Επιστολή του Προέδρου 4 Μήνυμα από την Αν. Γενική Διευθύντρια 5 Σύσταση και αρμοδιότητες 6 Αναπτυξιακό Πρόγραμμα 7 Σχέδια που έχουν συμπληρωθεί
ΑΠΟΦΑΣΗ. Αθήνα, 03.11.2015 Αριθ. Πρωτ.: 112277
 Αθήνα, 03.11.2015 Αριθ. Πρωτ.: 112277 ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΑΣ, ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΤΟΥΡΙΣΜΟΥ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΕΜΠΟΡΙΟΥ ΚΑΙ ΠΡΟΣΤΑΣΙΑΣ ΚΑΤΑΝΑΛΩΤΗ ΓΕΝΙΚΗ ΔΙΕΥΘΥΝΣΗ ΠΡΟΣΤΑΣΙΑΣ ΚΑΤΑΝΑΛΩΤΗ ΚΑΙ ΕΠΟΠΤΕΙΑΣ ΑΓΟΡΑΣ Γραφείο
Αθήνα, 03.11.2015 Αριθ. Πρωτ.: 112277 ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΑΣ, ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΤΟΥΡΙΣΜΟΥ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΕΜΠΟΡΙΟΥ ΚΑΙ ΠΡΟΣΤΑΣΙΑΣ ΚΑΤΑΝΑΛΩΤΗ ΓΕΝΙΚΗ ΔΙΕΥΘΥΝΣΗ ΠΡΟΣΤΑΣΙΑΣ ΚΑΤΑΝΑΛΩΤΗ ΚΑΙ ΕΠΟΠΤΕΙΑΣ ΑΓΟΡΑΣ Γραφείο
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ρ. Τετάρτη 7 Μαρτίου 2012
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ρ Τετάρτη 7 Μαρτίου 2012 ΘΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 6733 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 1ο Γυµνάσιο Πειραιά,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ Ρ Τετάρτη 7 Μαρτίου 2012 ΘΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 6733 2. Ανακοινώνεται ότι τη συνεδρίαση παρακολουθούν µαθητές από το 1ο Γυµνάσιο Πειραιά,
Τηλ. : 210 4110919 ΣΕ ΕΥΡΩ - ΕΛΕΥΘΕΡΟ FAX : 210 4117347 ΓΙΑ ΤΟΝ ΦΟΡΕΑ: ΥΠΟΥΡΓΕΙΟ ΕΜΠΟΡΙΚΗΣ ΝΑΥΤΙΛΙΑΣ
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ Πειραιάς 08-3-2006 ΥΠΟΥΡΓΕΙΟ ΕΜΠΟΡΙΚΗΣ ΝΑΥΤΙΛΙΑΣ Αριθ. Διακήρυξης: 03/2006 ΓΕΝΙΚΗ Δ/ΝΣΗ ΔΙΟΙΚΗΤΙΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ Αριθ. Πρωτ.: 5321.1/03/2006 ΔΙΕΥΘΥΝΣΗ ΠΡΟΜΗΘΕΙΩΝ & ΚΤΙΡΙΑΚΩΝ ΕΓΚΑΤΑΣΤ.
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ Πειραιάς 08-3-2006 ΥΠΟΥΡΓΕΙΟ ΕΜΠΟΡΙΚΗΣ ΝΑΥΤΙΛΙΑΣ Αριθ. Διακήρυξης: 03/2006 ΓΕΝΙΚΗ Δ/ΝΣΗ ΔΙΟΙΚΗΤΙΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ Αριθ. Πρωτ.: 5321.1/03/2006 ΔΙΕΥΘΥΝΣΗ ΠΡΟΜΗΘΕΙΩΝ & ΚΤΙΡΙΑΚΩΝ ΕΓΚΑΤΑΣΤ.
ΑΠΟΣΠΑΣΜΑ ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ. ΗΜΟΣ ΙΟΝΥΣΟΥ Οικονοµική Επιτροπή Ταχ. /νση: Λ. Μαραθώνος 29 & Αθ. ιάκου 01 Άγιος Στέφανος..Αριθ. Απόφασης:..240/2015..
 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΑΤΤΙΚΗΣ ΣΥΝΕ ΡΙΑΣΗ:..13η/2015.. ΗΜΟΣ ΙΟΝΥΣΟΥ της 31ης-7-2015 Οικονοµική Επιτροπή Ταχ. /νση: Λ. Μαραθώνος 29 & Αθ. ιάκου 01 Άγιος Στέφανος..Αριθ. Απόφασης:..240/2015.. ΑΠΟΣΠΑΣΜΑ
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΑΤΤΙΚΗΣ ΣΥΝΕ ΡΙΑΣΗ:..13η/2015.. ΗΜΟΣ ΙΟΝΥΣΟΥ της 31ης-7-2015 Οικονοµική Επιτροπή Ταχ. /νση: Λ. Μαραθώνος 29 & Αθ. ιάκου 01 Άγιος Στέφανος..Αριθ. Απόφασης:..240/2015.. ΑΠΟΣΠΑΣΜΑ
Ο Τάσσος Σταθόπουλος, 44 ετών, είναι δηµοσιογράφος αθλητικός συντάκτης στην εφηµερίδα της Πάτρας «Πελοπόννησος». Είναι µέλος
 Πατραϊκή Πυγμαχία Ο Τάσσος Σταθόπουλος, 44 ετών, είναι δηµοσιογράφος αθλητικός συντάκτης στην εφηµερίδα της Πάτρας «Πελοπόννησος». Είναι µέλος του ΠΣΑΤ και της ΕΣΗΕΠΗΝ έχει εκδώσει στο παρελθόν τα βιβλία
Πατραϊκή Πυγμαχία Ο Τάσσος Σταθόπουλος, 44 ετών, είναι δηµοσιογράφος αθλητικός συντάκτης στην εφηµερίδα της Πάτρας «Πελοπόννησος». Είναι µέλος του ΠΣΑΤ και της ΕΣΗΕΠΗΝ έχει εκδώσει στο παρελθόν τα βιβλία
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΟΥ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ / Γ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ: 29/11/2015
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΟΥ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ / Γ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ: 29/11/2015 ΘΕΜΑ 1 Ο 1α. 2γ. 3γ. 4δ. 5β. ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 2 Ο Α. Οι αλληλουχίες 1,3 και 4 βρίσκονται μπροστά από τα δομικά γονίδια
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΟΥ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ / Γ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ: 29/11/2015 ΘΕΜΑ 1 Ο 1α. 2γ. 3γ. 4δ. 5β. ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 2 Ο Α. Οι αλληλουχίες 1,3 και 4 βρίσκονται μπροστά από τα δομικά γονίδια
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΔΗΜΟΣ ΠΟΛΥΓΥΡΟΥ ΑΡΙΘΜ. ΑΠΟΦ:
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΔΗΜΟΣ ΠΟΛΥΓΥΡΟΥ ΑΡΙΘΜ. ΑΠΟΦ: 1/2013 Απόσπασμα από το πρακτικό της 1/2013 συνεδρίασης του Δημοτικού Συμβουλίου ΟΡΘΗ ΕΠΑΝΑΛΗΨΗ Θέμα: Εκλογή Προεδρείου Δημοτικού Συμβουλίου και των μελών
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΔΗΜΟΣ ΠΟΛΥΓΥΡΟΥ ΑΡΙΘΜ. ΑΠΟΦ: 1/2013 Απόσπασμα από το πρακτικό της 1/2013 συνεδρίασης του Δημοτικού Συμβουλίου ΟΡΘΗ ΕΠΑΝΑΛΗΨΗ Θέμα: Εκλογή Προεδρείου Δημοτικού Συμβουλίου και των μελών
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΜ. Πέµπτη 7 Μαρτίου 2013
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΜ Πέµπτη 7 Μαρτίου 2013 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 8674 2. Άδεια απουσίας των Βουλευτών κ. κ. Γ. Ψαριανού και Γ. Παπανδρέου, σελ. 8647, 8753 3.
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΡΜ Πέµπτη 7 Μαρτίου 2013 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 8674 2. Άδεια απουσίας των Βουλευτών κ. κ. Γ. Ψαριανού και Γ. Παπανδρέου, σελ. 8647, 8753 3.
Αθήνα, 14 Ιουνίου 2006. Αρ. Πρωτ. 1891/05.2.3 Π Ο Ρ Ι Σ Μ Α
 Αθήνα, 14 Ιουνίου 2006 Αρ. Πρωτ. 1891/05.2.3 Π Ο Ρ Ι Σ Μ Α Ο Συνήγορος του Πολίτη στο πλαίσιο των αρµοδιοτήτων του, κατά το άρθρο 103 παρ. 9 του Συντάγµατος και το ν. 3094/2003, εξέτασε την αναφορά του
Αθήνα, 14 Ιουνίου 2006 Αρ. Πρωτ. 1891/05.2.3 Π Ο Ρ Ι Σ Μ Α Ο Συνήγορος του Πολίτη στο πλαίσιο των αρµοδιοτήτων του, κατά το άρθρο 103 παρ. 9 του Συντάγµατος και το ν. 3094/2003, εξέτασε την αναφορά του
ΔΕΛΤΙΟ ΕΛΕΓΧΟΥ ΟΧΙ ΥΠΟΚ/ΜΑ ΙΚΑ-ΕΤΑΜ ΗΜΕΡΟΜΗΝΙΑ:.ΩΡΑ.. ΙΔΙΟΤΗΤΑ ΗΜΕΡΟΜΗΝΙΑ... ΑΠΟΓΕΓΡΑΜΜΕΝΟΣ ΑΝΑΠΟΓΡΑΦΟΣ
 ΑΡΙΘ. ΠΡΩΤ. :... ΔΕΛΤΙΟ ΕΛΕΓΧΟΥ ΤΗΡΗΣΗ ΕΝΤΥΠΟΥ (Ε4) : ΝΑΙ ΟΧΙ ΥΠΟΚ/ΜΑ ΙΚΑ-ΕΤΑΜ ΑΠΟΓΕΓΡΑΜΜΕΝΟΣ ΑΝΑΠΟΓΡΑΦΟΣ ΗΜΕΡΟΜΗΝΙΑ:.ΩΡΑ.. ΕΠΩΝΥΜΙΑ ΕΠΙΧΕΙΡΗΣΗΣ :... ΚΩΔ.ΥΠΟΚ/ΤΟΣ / Α.Μ.Ε. :... /... ΕΙΔΟΣ ΕΠΙΧΕΙΡΗΣΗΣ :...
ΑΡΙΘ. ΠΡΩΤ. :... ΔΕΛΤΙΟ ΕΛΕΓΧΟΥ ΤΗΡΗΣΗ ΕΝΤΥΠΟΥ (Ε4) : ΝΑΙ ΟΧΙ ΥΠΟΚ/ΜΑ ΙΚΑ-ΕΤΑΜ ΑΠΟΓΕΓΡΑΜΜΕΝΟΣ ΑΝΑΠΟΓΡΑΦΟΣ ΗΜΕΡΟΜΗΝΙΑ:.ΩΡΑ.. ΕΠΩΝΥΜΙΑ ΕΠΙΧΕΙΡΗΣΗΣ :... ΚΩΔ.ΥΠΟΚ/ΤΟΣ / Α.Μ.Ε. :... /... ΕΙΔΟΣ ΕΠΙΧΕΙΡΗΣΗΣ :...
Θεσσαλονίκη 1.3.2007 Αρ.Πρωτ.216. Προς Τα μέλη του Συμβολαιογραφικού Συλλόγου Εφετείου Θεσσαλονίκης. Θέμα: Αναπροσαρμογή τιμών αντικειμενικού
 Θεσσαλονίκη 1.3.2007 Αρ.Πρωτ.216 Προς Τα μέλη του Συμβολαιογραφικού Συλλόγου Εφετείου Θεσσαλονίκης Θέμα: Αναπροσαρμογή τιμών αντικειμενικού Σας διαβιβάζουμε τις με αριθμ.πρωτ. 1020562/486/ΟΟΤΥ/Δ / ΠΟΛ.1033/27
Θεσσαλονίκη 1.3.2007 Αρ.Πρωτ.216 Προς Τα μέλη του Συμβολαιογραφικού Συλλόγου Εφετείου Θεσσαλονίκης Θέμα: Αναπροσαρμογή τιμών αντικειμενικού Σας διαβιβάζουμε τις με αριθμ.πρωτ. 1020562/486/ΟΟΤΥ/Δ / ΠΟΛ.1033/27
1932, πτώχευση. Οι πολίτες κλήθηκαν από πατριωτικό καθήκον να δώσουν τα κοσμήματά για να ενισχυθούν τα αποθέματα της χώρας σε χρυσό
 1932, πτώχευση Οι πολίτες κλήθηκαν από πατριωτικό καθήκον να δώσουν τα κοσμήματά για να ενισχυθούν τα αποθέματα της χώρας σε χρυσό τους Τέτοιες μέρες, τον Απρίλιο του 1932, η κυβέρνηση των Φιλελευθέρων
1932, πτώχευση Οι πολίτες κλήθηκαν από πατριωτικό καθήκον να δώσουν τα κοσμήματά για να ενισχυθούν τα αποθέματα της χώρας σε χρυσό τους Τέτοιες μέρες, τον Απρίλιο του 1932, η κυβέρνηση των Φιλελευθέρων
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΥΣ 2013 ΣΥΝΕΔΡΙΑΣΗ Ι Τρίτη 27 Αυγούστου 2013
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΥΣ 2013 ΣΥΝΕΔΡΙΑΣΗ Ι Τρίτη 27 Αυγούστου 2013 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 813 2. Άδεια απουσίας του Βουλευτή κ. Σ. Αναστασιάδη,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΤΜΗΜΑ ΔΙΑΚΟΠΗΣ ΕΡΓΑΣΙΩΝ ΤΗΣ ΒΟΥΛΗΣ ΘΕΡΟΥΣ 2013 ΣΥΝΕΔΡΙΑΣΗ Ι Τρίτη 27 Αυγούστου 2013 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Επικύρωση Πρακτικών, σελ. 813 2. Άδεια απουσίας του Βουλευτή κ. Σ. Αναστασιάδη,
ΑΔΑ: ΔΗΜΟΣΙΕΥΘΗΚΕ ΣΤΟ ΦΕΚ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΚΩΝ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΗΜΟΣΙΩΝ ΕΣΟΔΩΝ Ι.ΓΕΝΙΚΗ ΔΙΕΥΘΥΝΣΗ ΦΟΡΟΛΟΓΙΚΩΝ ΕΛΕΓΧΩΝ ΚΑΙ ΕΙΣΠΡΑΞΗΣ ΔΗΜΟΣΙΩΝ ΕΣΟΔΩΝ -Δ/ΝΣΗ ΠΑΡΑΚΟΛΟΥΘΗΣΗΣ ΝΟΜΙΚΩΝ ΥΠΟΘΕΣΕΩΝ ΕΛΕΓΧΟΥ ΚΑΙ ΑΝΑΓΚΑΣΤΙΚΗΣ
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΚΩΝ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΗΜΟΣΙΩΝ ΕΣΟΔΩΝ Ι.ΓΕΝΙΚΗ ΔΙΕΥΘΥΝΣΗ ΦΟΡΟΛΟΓΙΚΩΝ ΕΛΕΓΧΩΝ ΚΑΙ ΕΙΣΠΡΑΞΗΣ ΔΗΜΟΣΙΩΝ ΕΣΟΔΩΝ -Δ/ΝΣΗ ΠΑΡΑΚΟΛΟΥΘΗΣΗΣ ΝΟΜΙΚΩΝ ΥΠΟΘΕΣΕΩΝ ΕΛΕΓΧΟΥ ΚΑΙ ΑΝΑΓΚΑΣΤΙΚΗΣ
Οι στρατηγικές πολιτικές (διπλωµατικές) αρετές του Αγησιλάου (3 διδακτικές ώρες)
 αρετές του Αγησιλάου (3 διδακτικές ώρες)") Κεφάλαιο 1. 17-22 Οι στρατηγικές πολιτικές (διπλωµατικές) αρετές του Αγησιλάου (3 διδακτικές ώρες) Ενδεικτικοί διδακτικοί στόχοι 1. Να επισηµάνουν οι µαθητές τις στρατηγικές και πολιτικές ικανότητες του
Κεφάλαιο 1. 17-22 Οι στρατηγικές πολιτικές (διπλωµατικές) αρετές του Αγησιλάου (3 διδακτικές ώρες) Ενδεικτικοί διδακτικοί στόχοι 1. Να επισηµάνουν οι µαθητές τις στρατηγικές και πολιτικές ικανότητες του
(ΜΕ ΤΑ ΔΥΟ ΜΕΙΟΝΕΚΤΗΜΑΤΑ)
") 1 ΤΟ ΒΙΒΛΙΟ ΤΩΝ ΠΟΝΩΝ ΤΟΥ ΣΩΜΑΤΟΣ (ΜΕ ΤΑ ΔΥΟ ΜΕΙΟΝΕΚΤΗΜΑΤΑ) Η πραγματικότητα ξεπερνά και την πιο τολμηρή φαντασία. Επίκτητος Σοφός δεν είναι όποιος ξέρει πολλά, αλλά όποιος ξέρει χρήσιμα. Ηράκλειτος Οι
1 ΤΟ ΒΙΒΛΙΟ ΤΩΝ ΠΟΝΩΝ ΤΟΥ ΣΩΜΑΤΟΣ (ΜΕ ΤΑ ΔΥΟ ΜΕΙΟΝΕΚΤΗΜΑΤΑ) Η πραγματικότητα ξεπερνά και την πιο τολμηρή φαντασία. Επίκτητος Σοφός δεν είναι όποιος ξέρει πολλά, αλλά όποιος ξέρει χρήσιμα. Ηράκλειτος Οι
ΣΧΕ ΙΑΣΜΟΣ ΖΩΝΩΝ ΧΡΗΣΕΩΝ ΓΗΣ ΣΕ ΠΕΡΙΟΧΕΣ Γ.Π.Σ.
 ΤΕΧΝΙΚΟ ΕΠΙΜΕΛΗΤΗΡΙΟ ΕΛΛΑ ΟΣ ΤΜΗΜΑ ΚΕΝΤΡΙΚΗΣ ΜΑΚΕ ΟΝΙΑΣ ΣΧΕ ΙΑΣΜΟΣ ΖΩΝΩΝ ΧΡΗΣΕΩΝ ΓΗΣ ΣΕ ΠΕΡΙΟΧΕΣ Γ.Π.Σ. (Οµάδα Εργασίας σύµφωνα µε την 136/ 2005 απόφαση) Πρώτη Έκθεση Οµάδα Εργασίας: ΑΓΓΕΛΙ ΗΣ Ι. ΑΜ, πολεοδόµος
ΤΕΧΝΙΚΟ ΕΠΙΜΕΛΗΤΗΡΙΟ ΕΛΛΑ ΟΣ ΤΜΗΜΑ ΚΕΝΤΡΙΚΗΣ ΜΑΚΕ ΟΝΙΑΣ ΣΧΕ ΙΑΣΜΟΣ ΖΩΝΩΝ ΧΡΗΣΕΩΝ ΓΗΣ ΣΕ ΠΕΡΙΟΧΕΣ Γ.Π.Σ. (Οµάδα Εργασίας σύµφωνα µε την 136/ 2005 απόφαση) Πρώτη Έκθεση Οµάδα Εργασίας: ΑΓΓΕΛΙ ΗΣ Ι. ΑΜ, πολεοδόµος
Γ49/ 35 ΕΞ. ΕΠΕΙΓΟΝ Π Ρ Ο Σ :
 Αθήνα, 19 / 5 / 2010 Δ Ι Ο Ι Κ Η Σ Η ΓΕΝΙΚΗ Δ/ΝΣΗ ΔΙΟΙΚ/ΚΩΝ ΥΠΗΡΕΣΙΩΝ Δ/ΝΣΗ ΔΙΟΙΚΗΤΙΚΟΥ ΠΡΟΣΩΠΙΚΟΥ ΤΜΗΜΑ : ΕΡΓΑΣΙΑΚΩΝ ΣΧΕΣΕΩΝ Ταχ. Δ/νση : Αγ. Κωνσταντίνου 8 Ταχ. Κώδικας: 102 41 ΑΘΗΝΑ Τηλέφωνο : 210-215292,289,290,294
Αθήνα, 19 / 5 / 2010 Δ Ι Ο Ι Κ Η Σ Η ΓΕΝΙΚΗ Δ/ΝΣΗ ΔΙΟΙΚ/ΚΩΝ ΥΠΗΡΕΣΙΩΝ Δ/ΝΣΗ ΔΙΟΙΚΗΤΙΚΟΥ ΠΡΟΣΩΠΙΚΟΥ ΤΜΗΜΑ : ΕΡΓΑΣΙΑΚΩΝ ΣΧΕΣΕΩΝ Ταχ. Δ/νση : Αγ. Κωνσταντίνου 8 Ταχ. Κώδικας: 102 41 ΑΘΗΝΑ Τηλέφωνο : 210-215292,289,290,294
Ημερομηνία δημοσίευσης στον Ελληνικό Τύπο Α Π Ο Φ Α Σ Η
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ ΕΘΝΙΚΟ ΚΕΝΤΡΟ ΑΜΕΣΗΣ ΒΟΗΘΕΙΑΣ ΠΑΡΑΡΤΗΜΑ ΙΩΑΝΝΙΝΩΝ ΤΜΗΜΑ ΠΡΟΣΩΠΙΚΟΥ & ΟΙΚΟΝΟΜΙΚΗΣ ΔΙΑΧΕΙΡΙΣΗΣ ΓΡΑΦΕΙΟ ΠΡΟΜΗΘΕΙΩΝ Ταχ. Δ/νση : Γ.ΧΑΤΖΗΚΩΣΤΑ 1 Ταχ. Κώδικας : 45444 Πληροφορίες
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ ΕΘΝΙΚΟ ΚΕΝΤΡΟ ΑΜΕΣΗΣ ΒΟΗΘΕΙΑΣ ΠΑΡΑΡΤΗΜΑ ΙΩΑΝΝΙΝΩΝ ΤΜΗΜΑ ΠΡΟΣΩΠΙΚΟΥ & ΟΙΚΟΝΟΜΙΚΗΣ ΔΙΑΧΕΙΡΙΣΗΣ ΓΡΑΦΕΙΟ ΠΡΟΜΗΘΕΙΩΝ Ταχ. Δ/νση : Γ.ΧΑΤΖΗΚΩΣΤΑ 1 Ταχ. Κώδικας : 45444 Πληροφορίες
ηµοσιεύθηκε στο ΦΕΚ 1296/Β /30.6.2015
 ηµοσιεύθηκε στο ΦΕΚ 1296/Β /30.6.2015 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ Αθήνα, 30 Ιουνίου 2015 ΥΠΟΥΡΓΕΙΟ ΕΣΩΤΕΡΙΚΩΝ Αριθ. Πρωτ.: 22138 ΚΑΙ ΙΟΙΚΗΤΙΚΗΣ ΑΝΑΣΥΓΚΡΟΤΗΣΗΣ ΓΕΝ. ΙΕΥ/ΝΣΗ ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ ΚΑΙ ΙΟΙΚ/ΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ
ηµοσιεύθηκε στο ΦΕΚ 1296/Β /30.6.2015 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ Αθήνα, 30 Ιουνίου 2015 ΥΠΟΥΡΓΕΙΟ ΕΣΩΤΕΡΙΚΩΝ Αριθ. Πρωτ.: 22138 ΚΑΙ ΙΟΙΚΗΤΙΚΗΣ ΑΝΑΣΥΓΚΡΟΤΗΣΗΣ ΓΕΝ. ΙΕΥ/ΝΣΗ ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ ΚΑΙ ΙΟΙΚ/ΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΤΜΗΜΑ ΦΥΣΙΚΗΣ ΕΤΗΣΙΑ ΑΠΟΓΡΑΦΙΚΗ ΕΚΘΕΣΗ ΑΚΑΔ. ΕΤΟΥΣ 2011-2012
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΤΜΗΜΑ ΦΥΣΙΚΗΣ ΕΤΗΣΙΑ ΑΠΟΓΡΑΦΙΚΗ ΕΚΘΕΣΗ ΑΚΑΔ. ΕΤΟΥΣ 2011-2012 Το κάθε τµήµα µπορεί προαιρετικά να προσθέσει επιπλέον στήλες στους πίνακες όπου αυτό κρίνεται απαραίτητο. Πίνακας 1.
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΤΜΗΜΑ ΦΥΣΙΚΗΣ ΕΤΗΣΙΑ ΑΠΟΓΡΑΦΙΚΗ ΕΚΘΕΣΗ ΑΚΑΔ. ΕΤΟΥΣ 2011-2012 Το κάθε τµήµα µπορεί προαιρετικά να προσθέσει επιπλέον στήλες στους πίνακες όπου αυτό κρίνεται απαραίτητο. Πίνακας 1.
ΑΠΟΦΑΣΗ. Βαθμός Ασφαλείας... Μαρούσι 24-9-08 Αριθ. Πρωτ. 122459 /Γ2 Βαθ. Προτερ... ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ
 Βαθμός Ασφαλείας... ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ Μαρούσι 24-9-08 Αριθ. Πρωτ. 122459 /Γ2 Βαθ. Προτερ.... ΕΝΙΑΙΟΣ ΙΟΙΚΗΤΙΚΟΣ ΤΟΜΕΑΣ Π/ΘΜΙΑΣ & /ΘΜΙΑΣ ΕΚΠ/ΣΗΣ ΙΕΥΘΥΝΣΗ ΣΠΟΥ ΩΝ / ΘΜΙΑΣ ΕΚΠΑΙ ΕΥΣΗΣ ΤΜΗΜΑ Α' Αν. Παπανδρέου
Βαθμός Ασφαλείας... ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ Μαρούσι 24-9-08 Αριθ. Πρωτ. 122459 /Γ2 Βαθ. Προτερ.... ΕΝΙΑΙΟΣ ΙΟΙΚΗΤΙΚΟΣ ΤΟΜΕΑΣ Π/ΘΜΙΑΣ & /ΘΜΙΑΣ ΕΚΠ/ΣΗΣ ΙΕΥΘΥΝΣΗ ΣΠΟΥ ΩΝ / ΘΜΙΑΣ ΕΚΠΑΙ ΕΥΣΗΣ ΤΜΗΜΑ Α' Αν. Παπανδρέου
ΤΟ ΚΛΕΙΔΙ ΤΩΝ ΜΑΘΗΜΑΤΙΚΩΝ
 ΓΙΩΡΓΟΣ ΑΛΒΑΝΟΣ ΤΟ ΚΛΕΙΔΙ ΤΩΝ ΜΑΘΗΜΑΤΙΚΩΝ Α ΔΗΜΟΤΙΚΟΥ Άριστο βοήθημα για τους μαθητές Περιέχει πλήρη θεωρία για κάθε μάθημα του σχολικού βιβλίου και πολλές εμπεδωτικές ασκήσεις και προβλήματα. Εισαγωγικό
ΓΙΩΡΓΟΣ ΑΛΒΑΝΟΣ ΤΟ ΚΛΕΙΔΙ ΤΩΝ ΜΑΘΗΜΑΤΙΚΩΝ Α ΔΗΜΟΤΙΚΟΥ Άριστο βοήθημα για τους μαθητές Περιέχει πλήρη θεωρία για κάθε μάθημα του σχολικού βιβλίου και πολλές εμπεδωτικές ασκήσεις και προβλήματα. Εισαγωγικό
ΠΗΓΑΣΟΣ ΕΚΔΟΤΙΚΗ Α.Ε. ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ 31 ΜΑΡΤΙΟΥ, 2008 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΠΛΗΡΟΦΟΡΗΣΗΣ (Δ.Π.Χ.Π.
 ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ 31 ΜΑΡΤΙΟΥ, 2008 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΠΛΗΡΟΦΟΡΗΣΗΣ (Δ.Π.Χ.Π.) Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Σελίδα ΚΑΤΑΣΤΑΣΗ ΛΟΓΑΡΙΑΣΜΟΥ ΑΠΟΤΕΛΕΣΜΑΤΩΝ ΠΕΡΙΟΔΟΥ 1 ΙΣΟΛΟΓΙΣΜΟΣ
ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ 31 ΜΑΡΤΙΟΥ, 2008 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΠΛΗΡΟΦΟΡΗΣΗΣ (Δ.Π.Χ.Π.) Π Ε Ρ Ι Ε Χ Ο Μ Ε Ν Α Σελίδα ΚΑΤΑΣΤΑΣΗ ΛΟΓΑΡΙΑΣΜΟΥ ΑΠΟΤΕΛΕΣΜΑΤΩΝ ΠΕΡΙΟΔΟΥ 1 ΙΣΟΛΟΓΙΣΜΟΣ
ΑΡΤΟΒΙΟΜΗΧΑΝΙΑ ΚΑΡΑΜΟΛΕΓΚΟΣ Α.Ε. ΕΔΡΑ: ΘΕΣΗ ΤΖΗΜΑ ΚΟΡΩΠΙ ΕΤΗΣΙΕΣ ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ. Για την περίοδο από 1 Ιανουαρίου έως 31 Δεκεμβρίου 2006
 . ΕΔΡΑ: ΘΕΣΗ ΤΖΗΜΑ ΚΟΡΩΠΙ ΕΤΗΣΙΕΣ ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ Για την περίοδο από 1 Ιανουαρίου έως 31 Δεκεμβρίου 2006 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΠΛΗΡΟΦΟΡΗΣΗΣ (Δ.Π.Χ.Π.) 1/35 Βεβαιώνεται
. ΕΔΡΑ: ΘΕΣΗ ΤΖΗΜΑ ΚΟΡΩΠΙ ΕΤΗΣΙΕΣ ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ Για την περίοδο από 1 Ιανουαρίου έως 31 Δεκεμβρίου 2006 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΠΛΗΡΟΦΟΡΗΣΗΣ (Δ.Π.Χ.Π.) 1/35 Βεβαιώνεται
ΕΤΑΙΡΕΙΑ ΑΝΑΠΤΥΞΗΣ ΕΠΑΡΧΙΑΣ ΑΠΟΚΟΡΩΝΑ ΑΝΩΝΥΜΗ ΕΤΑΙΡΕΙΑ ΑΡ.Μ.Α.Ε. 11964/73/Β/86/23. ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ 31 ης Δεκεμβρίου 2006
 ΕΤΑΙΡΕΙΑ ΑΝΑΠΤΥΞΗΣ ΕΠΑΡΧΙΑΣ ΑΠΟΚΟΡΩΝΑ ΑΝΩΝΥΜΗ ΕΤΑΙΡΕΙΑ ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ 31 ης Δεκεμβρίου 2006 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΠΛΗΡΟΦΟΡΗΣΗΣ (Δ.Π.Χ.Π.) ΠΟΥ ΕΧΟΥΝ ΕΓΚΡΙΘΕΙ ΑΠΟ ΤΗΝ ΕΥΡΩΠΑΪΚΗ
ΕΤΑΙΡΕΙΑ ΑΝΑΠΤΥΞΗΣ ΕΠΑΡΧΙΑΣ ΑΠΟΚΟΡΩΝΑ ΑΝΩΝΥΜΗ ΕΤΑΙΡΕΙΑ ΟΙΚΟΝΟΜΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ 31 ης Δεκεμβρίου 2006 ΣΥΜΦΩΝΑ ΜΕ ΤΑ ΔΙΕΘΝΗ ΠΡΟΤΥΠΑ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΗΣ ΠΛΗΡΟΦΟΡΗΣΗΣ (Δ.Π.Χ.Π.) ΠΟΥ ΕΧΟΥΝ ΕΓΚΡΙΘΕΙ ΑΠΟ ΤΗΝ ΕΥΡΩΠΑΪΚΗ
ΚΑΝΟΝΙΣΜΟΣ ΛΕΙΤΟΥΡΓΙΑΣ ΔΗΜΟΤΙΚΩΝ ΚΟΙΜΗΤΗΡΙΩΝ ΔΗΜΟΥ ΘΕΡΜΗΣ
 ΚΑΝΟΝΙΣΜΟΣ ΛΕΙΤΟΥΡΓΙΑΣ ΔΗΜΟΤΙΚΩΝ ΚΟΙΜΗΤΗΡΙΩΝ ΔΗΜΟΥ ΘΕΡΜΗΣ Αριθμός Απόφασης Δημοτικού Συμβουλίου Θέρμης 428/2011 1 Πίνακας περιεχομένων Άρθρο 1 - Γενικά 3 Άρθρο 2 Πεδίο εφαρμογής 3 Άρθρο 3 Γενικοί κανόνες
ΚΑΝΟΝΙΣΜΟΣ ΛΕΙΤΟΥΡΓΙΑΣ ΔΗΜΟΤΙΚΩΝ ΚΟΙΜΗΤΗΡΙΩΝ ΔΗΜΟΥ ΘΕΡΜΗΣ Αριθμός Απόφασης Δημοτικού Συμβουλίου Θέρμης 428/2011 1 Πίνακας περιεχομένων Άρθρο 1 - Γενικά 3 Άρθρο 2 Πεδίο εφαρμογής 3 Άρθρο 3 Γενικοί κανόνες
I.Επί της Αρχής του σχεδίου Νόµου: ΙΙ. Επί των άρθρων του σχεδίου Νόµου: ΕΙΣΗΓΗΤΙΚΗ ΕΚΘΕΣΗ
 ΕΙΣΗΓΗΤΙΚΗ ΕΚΘΕΣΗ στο σχέδιο νόµου «Πρωτοβάθµιο Εθνικό Δίκτυο Υγείας (Π.Ε.Δ.Υ.), αλλαγή σκοπού Ε.Ο.Π.Υ.Υ. και λοιπές διατάξεις» Προς τη Βουλή των Ελλήνων I.Επί της Αρχής του σχεδίου Νόµου: Με τις διατάξεις
ΕΙΣΗΓΗΤΙΚΗ ΕΚΘΕΣΗ στο σχέδιο νόµου «Πρωτοβάθµιο Εθνικό Δίκτυο Υγείας (Π.Ε.Δ.Υ.), αλλαγή σκοπού Ε.Ο.Π.Υ.Υ. και λοιπές διατάξεις» Προς τη Βουλή των Ελλήνων I.Επί της Αρχής του σχεδίου Νόµου: Με τις διατάξεις
Θ Ε Μ Α «Έγκριση σύναψης τροποποιηµένης προγραµµατικής σύµβασης (Γεωτεχνική έρευνα Ευστάθειας Βραχωδών Πρανών στο.. Καβάλας) µε το Ι.Γ.Μ.Ε.
 µε το Ι.Γ.Μ.Ε.") ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΚΑΒΑΛΑΣ ΗΜΟΣ ΚΑΒΑΛΑΣ ΗΜΟΤΙΚΟ ΣΥΜΒΟΥΛΙΟ ΑΠΟΣΠΑΣΜΑ Από το Πρακτικό της µε αριθ. 31 ης /30 Οκτωβρίου 2008 Συνεδρίασης του ηµοτικού Συµβουλίου Καβάλας Αριθ. Αποφάσεως 628/2008 Θ Ε
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΚΑΒΑΛΑΣ ΗΜΟΣ ΚΑΒΑΛΑΣ ΗΜΟΤΙΚΟ ΣΥΜΒΟΥΛΙΟ ΑΠΟΣΠΑΣΜΑ Από το Πρακτικό της µε αριθ. 31 ης /30 Οκτωβρίου 2008 Συνεδρίασης του ηµοτικού Συµβουλίου Καβάλας Αριθ. Αποφάσεως 628/2008 Θ Ε
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΚΘ. Τρίτη 18 Σεπτεµβρίου 2012
 ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΚΘ Τρίτη 18 Σεπτεµβρίου 2012 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Άδεια απουσίας του Βουλευτή κ. Β. Κατριβάνου, σελ. 1053 2. Επί διαδικαστικού θέµατος, σελ. 1054, 1069, 1070, 1071,
ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΣΥΝΕΔΡΙΑΣΗ ΚΘ Τρίτη 18 Σεπτεµβρίου 2012 ΘΕΜΑΤΑ Α. ΕΙΔΙΚΑ ΘΕΜΑΤΑ 1. Άδεια απουσίας του Βουλευτή κ. Β. Κατριβάνου, σελ. 1053 2. Επί διαδικαστικού θέµατος, σελ. 1054, 1069, 1070, 1071,
Π Ι Ν Α Κ Α Σ Α Μ Ο Ι Β Ω Ν Ε Π Ι Δ Ο Σ Ε Ω Ν
 Π Ι Ν Α Κ Α Σ Α Μ Ο Ι Β Ω Ν Ε Π Ι Δ Ο Σ Ε Ω Ν ΔΙΚΑΣΤΙΚΩΝ ΕΠΙΜΕΛΗΤΩΝ ΕΦΕΤΕΙΩΝ ΑΘΗΝΩΝ & ΠΕΙΡΑΙΩΣ ΔΙΟΡΙΣΜΕΝΩΝ ΣΤΑ ΠΡΩΤΟΔΙΚΕΙΑ ΑΘΗΝΩΝ & ΠΕΙΡΑΙΩΣ ΜΕ ΕΔΡΑ ΤΗΝ ΑΘΗΝΑ Η χιλιομετρική απόσταση υπολογίσθηκε με σημείο
Π Ι Ν Α Κ Α Σ Α Μ Ο Ι Β Ω Ν Ε Π Ι Δ Ο Σ Ε Ω Ν ΔΙΚΑΣΤΙΚΩΝ ΕΠΙΜΕΛΗΤΩΝ ΕΦΕΤΕΙΩΝ ΑΘΗΝΩΝ & ΠΕΙΡΑΙΩΣ ΔΙΟΡΙΣΜΕΝΩΝ ΣΤΑ ΠΡΩΤΟΔΙΚΕΙΑ ΑΘΗΝΩΝ & ΠΕΙΡΑΙΩΣ ΜΕ ΕΔΡΑ ΤΗΝ ΑΘΗΝΑ Η χιλιομετρική απόσταση υπολογίσθηκε με σημείο
Η ΚΟΙΝΩΝΙΚΗ ΔΙΑΣΤΡΩΜΑΤΩΣΗ ΤΟΥ ΜΟΥΣΟΥΛΜΑΝΙΚΟΥ ΠΛΗΘΥΣΜΟΥ ΤΟΥ ΡΕΘΥΜΝΟΥ ΚΑΙ Η ΔΙΕΚΔΙΚΗΣΗ ΤΗΣ ΑΝΤΑΛΛΑΞΙΜΗΣ ΠΕΡΙΟΥΣΙΑΣ, 1924-1927
 ΣΠΥΡΟΣ ΔΗΜΑΝΟΠΟΥΛΟΣ Η ΚΟΙΝΩΝΙΚΗ ΔΙΑΣΤΡΩΜΑΤΩΣΗ ΤΟΥ ΜΟΥΣΟΥΛΜΑΝΙΚΟΥ ΠΛΗΘΥΣΜΟΥ ΤΟΥ ΡΕΘΥΜΝΟΥ ΚΑΙ Η ΔΙΕΚΔΙΚΗΣΗ ΤΗΣ ΑΝΤΑΛΛΑΞΙΜΗΣ ΠΕΡΙΟΥΣΙΑΣ, 924-927 Στη χώρα μας η μελέτη του αστικού φαινομένου εντάχθηκε στα
ΣΠΥΡΟΣ ΔΗΜΑΝΟΠΟΥΛΟΣ Η ΚΟΙΝΩΝΙΚΗ ΔΙΑΣΤΡΩΜΑΤΩΣΗ ΤΟΥ ΜΟΥΣΟΥΛΜΑΝΙΚΟΥ ΠΛΗΘΥΣΜΟΥ ΤΟΥ ΡΕΘΥΜΝΟΥ ΚΑΙ Η ΔΙΕΚΔΙΚΗΣΗ ΤΗΣ ΑΝΤΑΛΛΑΞΙΜΗΣ ΠΕΡΙΟΥΣΙΑΣ, 924-927 Στη χώρα μας η μελέτη του αστικού φαινομένου εντάχθηκε στα
Η κυπαρική θεωρία στη σύγχρονη εκδοχή της υποστηρίζει
 Μια από τις επιδιώξεις των Φυσικών Επιστημών είναι να περιγράψουν και να εξηγήσουν τη δομή και τις ι- διότητες της ύλης, ξεκινώντας από τα μικρότερα δομικά συστατικά της. Η ατομική θεωρία αποτελεί την
Μια από τις επιδιώξεις των Φυσικών Επιστημών είναι να περιγράψουν και να εξηγήσουν τη δομή και τις ι- διότητες της ύλης, ξεκινώντας από τα μικρότερα δομικά συστατικά της. Η ατομική θεωρία αποτελεί την
Φ Α Ρ Μ Α Κ Ε Υ Τ Ι Κ Ο Σ Σ ΥΛ Λ Ο Γ Ο Σ ΑΤ Τ Ι Κ Η Σ
 Σεπτέμβριος 2012 Editorial: Οι τελευταίες εξελίξεις Έκφραση αλληλεγγύης στους φαρμακοποιούς από την Ο.Σ.Φ.Ε. Απάντηση σε συκοφαντικούς ισχυρισμούς του Μega Απάντηση σε δηλώσεις του καθηγητή κ. Τσαρμουγκέλη
Σεπτέμβριος 2012 Editorial: Οι τελευταίες εξελίξεις Έκφραση αλληλεγγύης στους φαρμακοποιούς από την Ο.Σ.Φ.Ε. Απάντηση σε συκοφαντικούς ισχυρισμούς του Μega Απάντηση σε δηλώσεις του καθηγητή κ. Τσαρμουγκέλη
 08 π ε ρ ι ε χ ό μ ε ν α 12 Το γράμμα της σύνταξης 16 Πώς θα υποβάλλονται οι αποδείξεις λιανικής καυσίμων ως 300 ευρώ 20 Αλλαγές στην αγορά καυσίμων με το πολυνομοσχέδιο 30 Ελεγκτές πρατηρίων καυσίμων
08 π ε ρ ι ε χ ό μ ε ν α 12 Το γράμμα της σύνταξης 16 Πώς θα υποβάλλονται οι αποδείξεις λιανικής καυσίμων ως 300 ευρώ 20 Αλλαγές στην αγορά καυσίμων με το πολυνομοσχέδιο 30 Ελεγκτές πρατηρίων καυσίμων
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΣΕΡΡΩΝ ΔΗΜΟΣ ΣΕΡΡΩΝ ΔΙΕΥΘΥΝΣΗ ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ ΤΜΗΜΑ ΠΡΟΜΗΘΕΙΩΝ ΜΕΛΕΤΗ 9/2015
 ΔΙΕΥΘΥΝΣΗ ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ ΜΕΛΕΤΗ 9/2015 ΠΡΟΜΗΘΕΙΑ ΦΡΕΣΚΟΥ ΓΑΛΑΚΤΟΣ ΠΡΟΫΠΟΛΟΓΙΣΜΟΣ: 9.602,17 με ΦΠΑ & 8.497,5 χωρίς ΦΠΑ ΠΡΟΕΛΕΥΣΗ : ΕΣΟΔΑ ΔΗΜΟΥ ΣΥΝΤΑΞΑΣΑ : ΜΑΖΑΡΑΚΗ ΑΝΑΣΤΑΣΙΑ 1 Αρ. Μελέτης: 9/2015
ΔΙΕΥΘΥΝΣΗ ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ ΜΕΛΕΤΗ 9/2015 ΠΡΟΜΗΘΕΙΑ ΦΡΕΣΚΟΥ ΓΑΛΑΚΤΟΣ ΠΡΟΫΠΟΛΟΓΙΣΜΟΣ: 9.602,17 με ΦΠΑ & 8.497,5 χωρίς ΦΠΑ ΠΡΟΕΛΕΥΣΗ : ΕΣΟΔΑ ΔΗΜΟΥ ΣΥΝΤΑΞΑΣΑ : ΜΑΖΑΡΑΚΗ ΑΝΑΣΤΑΣΙΑ 1 Αρ. Μελέτης: 9/2015
ΗοΙdings. για τη περίοδο από 1η Ιανουαρίου 2008 έως 30η Σεπτεμβρίου 2008 σύμφωνα με τα Διεθνή Πρότυπα Χρηματοοικονομικής Πληροφόρησης
 (Τα ποσά είναι εκπεφρασμένα σε χιλιάδες Ευρώ, εκτός εάν αναφέρεται διαφορετικά) ΗοΙdings για τη περίοδο από 1η Ιανουαρίου 2008 έως 30η Σεπτεμβρίου 2008 σύμφωνα με τα Διεθνή Πρότυπα Χρηματοοικονομικής Πληροφόρησης
(Τα ποσά είναι εκπεφρασμένα σε χιλιάδες Ευρώ, εκτός εάν αναφέρεται διαφορετικά) ΗοΙdings για τη περίοδο από 1η Ιανουαρίου 2008 έως 30η Σεπτεμβρίου 2008 σύμφωνα με τα Διεθνή Πρότυπα Χρηματοοικονομικής Πληροφόρησης
ΚΟΙΝΟΠΟΙΗΣΗ : Ως συν/νος πίνακας ΘΕΜΑ : «Καταβολή Δωροσήμου Χριστουγέννων 2015 σε εργατοτεχνίτες οικοδόμους»
 Αθήνα 7/12/2015 Δ Ι Ο Ι Κ Η Σ Η ΓΕΝ.Δ/ΝΣΕΙΣ : ΑΣΦΑΛΙΣΤΙΚΩΝ ΥΠΗΡΕΣΙΩΝ : ΟΙΚΟΝΟΜΟΤΕΧΝΙΚΩΝ ΥΠΗΡΕΣΙΩΝ : ΠΛΗΡΟΦΟΡΙΚΗΣ Δ/ΝΣΕΙΣ : ΑΣΦΑΛΙΣΗΣ ΕΣΟΔΩΝ : ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ : ΕΦΑΡΜΟΓΩΝ Ταχ. Δ/νση : Αγ. Κων/νου
Αθήνα 7/12/2015 Δ Ι Ο Ι Κ Η Σ Η ΓΕΝ.Δ/ΝΣΕΙΣ : ΑΣΦΑΛΙΣΤΙΚΩΝ ΥΠΗΡΕΣΙΩΝ : ΟΙΚΟΝΟΜΟΤΕΧΝΙΚΩΝ ΥΠΗΡΕΣΙΩΝ : ΠΛΗΡΟΦΟΡΙΚΗΣ Δ/ΝΣΕΙΣ : ΑΣΦΑΛΙΣΗΣ ΕΣΟΔΩΝ : ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ : ΕΦΑΡΜΟΓΩΝ Ταχ. Δ/νση : Αγ. Κων/νου
Η ΕΠΑΓΓΕΛΜΑΤΙΚΗ & ΕΠΙΧΕΙΡΗΜΑΤΙΚΗ ΔΡΑΣΤΗΡΙΟΠΟΙΗΣΗ ΤΩΝ ΕΛΛΗΝΩΝ ΤΗΣ ΔΙΑΣΠΟΡΑΣ ΜΕΤΑ ΤΟΝ Β ΠΑΓΚΟΣΜΙΟ ΠΟΛΕΜΟ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΗΣ ΣΤΟ ΕΘΝΙΚΟ ΚΕΝΤΡΟ
 ΤΕΧΝΟΛΟΓΙΚΌ ΕΚΠΑΙΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΚΑΒΑΛΑΣ ΣΧΟΛΗ ΔΙΟΙΚΗΣΗΣ ΚΑΙ ΟΙΚΟΝΟΜΙΑΣ ΤΜΗΜΑ ΔΙΟΙΚΗΣΗ ΕΠΙΧΕΙΡΗΣΕΩΝ Η ΕΠΑΓΓΕΛΜΑΤΙΚΗ & ΕΠΙΧΕΙΡΗΜΑΤΙΚΗ ΔΡΑΣΤΗΡΙΟΠΟΙΗΣΗ ΤΩΝ ΕΛΛΗΝΩΝ ΤΗΣ ΔΙΑΣΠΟΡΑΣ ΜΕΤΑ ΤΟΝ Β ΠΑΓΚΟΣΜΙΟ ΠΟΛΕΜΟ
ΤΕΧΝΟΛΟΓΙΚΌ ΕΚΠΑΙΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΚΑΒΑΛΑΣ ΣΧΟΛΗ ΔΙΟΙΚΗΣΗΣ ΚΑΙ ΟΙΚΟΝΟΜΙΑΣ ΤΜΗΜΑ ΔΙΟΙΚΗΣΗ ΕΠΙΧΕΙΡΗΣΕΩΝ Η ΕΠΑΓΓΕΛΜΑΤΙΚΗ & ΕΠΙΧΕΙΡΗΜΑΤΙΚΗ ΔΡΑΣΤΗΡΙΟΠΟΙΗΣΗ ΤΩΝ ΕΛΛΗΝΩΝ ΤΗΣ ΔΙΑΣΠΟΡΑΣ ΜΕΤΑ ΤΟΝ Β ΠΑΓΚΟΣΜΙΟ ΠΟΛΕΜΟ
ΘΕΜΑ: «Ορισμός αριθμού εισακτέων κατά τις εισιτήριες εξετάσεις για τη Σχολή Αρχιπυροσβεστών της Πυροσβεστικής Ακαδημίας». Ο Υπουργός Εσωτερικών
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΕΣΩΤΕΡΙΚΩΝ ΑΡΧΗΓΕΙΟ ΠΥΡΟΣΒΕΣΤΙΚΟΥ ΣΩΜΑΤΟΣ Β ΚΛΑΔΟΣ ΔΙΟΙΚΗΤΙΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ ΔΙΕΥΘΥΝΣΗ ΠΡΟΣΩΠΙΚΟΥ ΤΜΗΜΑ 2ο ΥΠΑΞ/ΚΩΝ & ΠΥΡ/ΣΤΩΝ Ταχ.Δ/νση : Μουρούζη 4 Τ.Κ. 101 72 ΑΘΗΝΑ Τηλέφωνο
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΕΣΩΤΕΡΙΚΩΝ ΑΡΧΗΓΕΙΟ ΠΥΡΟΣΒΕΣΤΙΚΟΥ ΣΩΜΑΤΟΣ Β ΚΛΑΔΟΣ ΔΙΟΙΚΗΤΙΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ ΔΙΕΥΘΥΝΣΗ ΠΡΟΣΩΠΙΚΟΥ ΤΜΗΜΑ 2ο ΥΠΑΞ/ΚΩΝ & ΠΥΡ/ΣΤΩΝ Ταχ.Δ/νση : Μουρούζη 4 Τ.Κ. 101 72 ΑΘΗΝΑ Τηλέφωνο
ΑΡΙΘΜΟΣ 0501/2012 2013 ΣΥΜΒΑΣΗ ΕΠΙΧΟΡΗΓΗΣΗΣ Ι.ΝΕ.ΔΙ.ΒΙ.Μ. - ΣΧΟΛΙΚΗ ΕΠΙΤΡΟΠΗ ΠΡΩΤΟΒΑΘΜΙΑΣ ΕΚΠΑΙΔΕΥΣΗΣ ΔΗΜΟΥ ΣΚΟΠΕΛΟΥ
 ΑΡΙΘΜΟΣ 0501/2012 2013 ΣΥΜΒΑΣΗ ΕΠΙΧΟΡΗΓΗΣΗΣ Ι.ΝΕ.ΔΙ.ΒΙ.Μ. - ΣΧΟΛΙΚΗ ΕΠΙΤΡΟΠΗ ΠΡΩΤΟΒΑΘΜΙΑΣ ΕΚΠΑΙΔΕΥΣΗΣ ΔΗΜΟΥ ΣΚΟΠΕΛΟΥ Στην Αθήνα, σήμερα, 10/12/2012, οι υπογράφοντες τη παρούσα: Αφενός το Ν.Π.Ι.Δ. με την
ΑΡΙΘΜΟΣ 0501/2012 2013 ΣΥΜΒΑΣΗ ΕΠΙΧΟΡΗΓΗΣΗΣ Ι.ΝΕ.ΔΙ.ΒΙ.Μ. - ΣΧΟΛΙΚΗ ΕΠΙΤΡΟΠΗ ΠΡΩΤΟΒΑΘΜΙΑΣ ΕΚΠΑΙΔΕΥΣΗΣ ΔΗΜΟΥ ΣΚΟΠΕΛΟΥ Στην Αθήνα, σήμερα, 10/12/2012, οι υπογράφοντες τη παρούσα: Αφενός το Ν.Π.Ι.Δ. με την
Α Π Ο Σ Π Α Σ Μ Α. Από το αριθµ. 16/2014 Πρακτικό Συνεδρίασης Οικονοµικής Επιτροπής του ήµου Βοΐου. Αριθµός Απόφασης: 253/2014
 Α Π Ο Σ Π Α Σ Μ Α Από το αριθµ. 16/2014 Πρακτικό Συνεδρίασης Οικονοµικής Επιτροπής του ήµου Βοΐου Αριθµός Απόφασης: 253/2014 ΠΕΡΙΛΗΨΗ Καθορισµός όρων δηµοπρασίας Στη Σιάτιστα και στο ηµοτικό Κατάστηµα
Α Π Ο Σ Π Α Σ Μ Α Από το αριθµ. 16/2014 Πρακτικό Συνεδρίασης Οικονοµικής Επιτροπής του ήµου Βοΐου Αριθµός Απόφασης: 253/2014 ΠΕΡΙΛΗΨΗ Καθορισµός όρων δηµοπρασίας Στη Σιάτιστα και στο ηµοτικό Κατάστηµα
Π Ρ Ο Κ Η Ρ Υ Ξ Η Ο ΑΡΧΗΓΟΣ ΤΗΣ ΕΛΛΗΝΙΚΗΣ ΑΣΤΥΝΟΜΙΑΣ
 ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ ΑΔΑ : ΩΟΔΙ465ΦΘΕ-ΦΒΑ ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΕΣΩΤΕΡΙΚΩΝ ΚΑΙ ΔΙΟΙΚΗΤΙΚΗΣ ΑΝΑΣΥΓΚΡΟΤΗΣΗΣ ΑΡΧΗΓΕΙΟ ΕΛΛΗΝΙΚΗΣ ΑΣΤΥΝΟΜΙΑΣ ΚΛΑΔΟΣ ΔΙΟΙΚΗΤΙΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ ΚΑΙ ΑΝΘΡΩΠΙΝΟΥ ΔΥΝΑΜΙΚΟΥ
ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ ΑΔΑ : ΩΟΔΙ465ΦΘΕ-ΦΒΑ ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΕΣΩΤΕΡΙΚΩΝ ΚΑΙ ΔΙΟΙΚΗΤΙΚΗΣ ΑΝΑΣΥΓΚΡΟΤΗΣΗΣ ΑΡΧΗΓΕΙΟ ΕΛΛΗΝΙΚΗΣ ΑΣΤΥΝΟΜΙΑΣ ΚΛΑΔΟΣ ΔΙΟΙΚΗΤΙΚΗΣ ΥΠΟΣΤΗΡΙΞΗΣ ΚΑΙ ΑΝΘΡΩΠΙΝΟΥ ΔΥΝΑΜΙΚΟΥ
Ι Α Κ Η Ρ Υ Ξ Η Ο ΗΜΑΡΧΟΣ ΚΟΜΟΤΗΝΗΣ Σε εκτέλεση της αριθ. 267 / 2013 Απόφασης της Οικονοµικής Επιτροπής. 11 η Φεβρουαρίου 2014, ηµέρα της εβδοµάδας
 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΗΜΟΣ ΚΟΜΟΤΗΝΗΣ /ΝΣΗ ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ ΤΜΗΜΑ ΕΣΟ ΩΝ-ΠΕΡΙΟΥΣΙΑΣ ΑΡΙΘΜ. ΠΡΩΤ.: 4910 Ι Α Κ Η Ρ Υ Ξ Η Ο ΗΜΑΡΧΟΣ ΚΟΜΟΤΗΝΗΣ Σε εκτέλεση της αριθ. 267 / 2013 Απόφασης της Οικονοµικής Επιτροπής
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΗΜΟΣ ΚΟΜΟΤΗΝΗΣ /ΝΣΗ ΟΙΚΟΝΟΜΙΚΩΝ ΥΠΗΡΕΣΙΩΝ ΤΜΗΜΑ ΕΣΟ ΩΝ-ΠΕΡΙΟΥΣΙΑΣ ΑΡΙΘΜ. ΠΡΩΤ.: 4910 Ι Α Κ Η Ρ Υ Ξ Η Ο ΗΜΑΡΧΟΣ ΚΟΜΟΤΗΝΗΣ Σε εκτέλεση της αριθ. 267 / 2013 Απόφασης της Οικονοµικής Επιτροπής
Α. ΙΣΤΟΡΙΚΑ ΓΕΓΟΝΟΝΤΑ
 Μηχαήλ Κ. Νομικός Ο Μιχαήλ Νομικός με καταγωγή από την Αμοργό, αρχικά υπήρξε ιδιοκτήτης τυπογραφείου και εκδότης της εφημερίδας στην Αίγυπτο σε διάσημη εφημερίδα. Αφού επέστρεψε στην Ελλάδα το 1882 αγόρασε
Μηχαήλ Κ. Νομικός Ο Μιχαήλ Νομικός με καταγωγή από την Αμοργό, αρχικά υπήρξε ιδιοκτήτης τυπογραφείου και εκδότης της εφημερίδας στην Αίγυπτο σε διάσημη εφημερίδα. Αφού επέστρεψε στην Ελλάδα το 1882 αγόρασε
ΕΛΛΗΝΙΚΕΣ ΑΛΥΚΕΣ Α.Ε.
 ΕΛΛΗΝΙΚΕΣ ΑΛΥΚΕΣ Α.Ε. Δ Ι Α Κ Η Ρ Υ Ξ Η ΑΝΟΙΚΤΟΥ ΜΕΙΟΔΟΤΙΚΟΥ ΔΙΑΓΩΝΙΣΜΟΥ ΓΙΑ ΤΗΝ ΑΝΑΘΕΣΗ ΤΩΝ ΧΕΡΣAIΩΝ ΜΕΤΑΦΟΡΩΝ ΑΛΑΤΟΣ Η εταιρία «ΕΛΛΗΝΙΚΕΣ ΑΛΥΚΕΣ Α.Ε.» προκηρύσσει δημόσιο ανοιχτό μειοδοτικό διαγωνισμό,
ΕΛΛΗΝΙΚΕΣ ΑΛΥΚΕΣ Α.Ε. Δ Ι Α Κ Η Ρ Υ Ξ Η ΑΝΟΙΚΤΟΥ ΜΕΙΟΔΟΤΙΚΟΥ ΔΙΑΓΩΝΙΣΜΟΥ ΓΙΑ ΤΗΝ ΑΝΑΘΕΣΗ ΤΩΝ ΧΕΡΣAIΩΝ ΜΕΤΑΦΟΡΩΝ ΑΛΑΤΟΣ Η εταιρία «ΕΛΛΗΝΙΚΕΣ ΑΛΥΚΕΣ Α.Ε.» προκηρύσσει δημόσιο ανοιχτό μειοδοτικό διαγωνισμό,
ΔΙΑΚΗΡΥΞΗ Αριθμ. 19/2015 ΑΝΟΙΚΤΟΥ ΔΗΜΟΣΙΟΥ ΔΙΑΓΩΝΙΣΜΟΥ
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ & ΚΟΙΝΩΝΙΚΗΣ ΑΛΛΗΛΕΓΓΥΗΣ 4 η ΥΓΕΙΟΝΟΜΙΚΗ ΠΕΡΙΦΕΡΕΙΑ ΜΑΚΕΔΟΝΙΑΣ & ΘΡΑΚΗΣ ΓΕΝΙΚΟ ΝΟΣΟΚΟΜΕΙΟ ΔΡΑΜΑΣ Οικονομικό Τμήμα/ Γραφείο Προμηθειών Πληροφορίες: Αλεξιάδου Μαρία ΤΗΛ.:
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ & ΚΟΙΝΩΝΙΚΗΣ ΑΛΛΗΛΕΓΓΥΗΣ 4 η ΥΓΕΙΟΝΟΜΙΚΗ ΠΕΡΙΦΕΡΕΙΑ ΜΑΚΕΔΟΝΙΑΣ & ΘΡΑΚΗΣ ΓΕΝΙΚΟ ΝΟΣΟΚΟΜΕΙΟ ΔΡΑΜΑΣ Οικονομικό Τμήμα/ Γραφείο Προμηθειών Πληροφορίες: Αλεξιάδου Μαρία ΤΗΛ.:
ΤΑΚΤΙΚΗΣ ΣΥΝΕΔΡΙΑΣΗΣ ΤΟΥ ΔΙΟΙΚΗΤΙΚΟΥ ΣΥΜΒΟΥΛΙΟΥ ΤΟΥ ΔΗΜΟΤΙΚΟΥ ΛΙΜΕΝΙΚΟΥ ΤΑΜΕΙΟΥ ΧΑΝΙΩΝ
 ΟΡΘΗ ΕΠΑΝΑΛΗΨΗ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ ΑΡΙΘΜ. ΠΡΩΤ. : 1882 /19-11-2013 ΠΡΑΚΤΙΚΟ 17 ο /2013 ΤΑΚΤΙΚΗΣ ΣΥΝΕΔΡΙΑΣΗΣ ΤΟΥ ΔΙΟΙΚΗΤΙΚΟΥ ΣΥΜΒΟΥΛΙΟΥ ΤΟΥ ΔΗΜΟΤΙΚΟΥ ΛΙΜΕΝΙΚΟΥ ΤΑΜΕΙΟΥ ΧΑΝΙΩΝ Το Διοικητικό Συμβούλιο
ΟΡΘΗ ΕΠΑΝΑΛΗΨΗ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ ΑΡΙΘΜ. ΠΡΩΤ. : 1882 /19-11-2013 ΠΡΑΚΤΙΚΟ 17 ο /2013 ΤΑΚΤΙΚΗΣ ΣΥΝΕΔΡΙΑΣΗΣ ΤΟΥ ΔΙΟΙΚΗΤΙΚΟΥ ΣΥΜΒΟΥΛΙΟΥ ΤΟΥ ΔΗΜΟΤΙΚΟΥ ΛΙΜΕΝΙΚΟΥ ΤΑΜΕΙΟΥ ΧΑΝΙΩΝ Το Διοικητικό Συμβούλιο
Α Π Ο Φ Α Σ Η Ο ΑΝΑΠΛΗΡΩΤΗΣ ΥΠΟΥΡΓΟΣ ΟΙΚΟΝΟΜΙΚΩΝ
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΚΩΝ Π. ΕΠΕΙΓΟΝ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ Α.Δ.Α.: ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΗΜΟΣΙΟΝΟΜΙΚΗΣ ΠΟΛΙΤΙΚΗΣ ΓΕΝΙΚΟ ΛΟΓΙΣΤΗΡΙΟ ΤΟΥ ΚΡΑΤΟΥΣ ΓΕΝΙΚΗ ΔΙΕΥΘΥΝΣΗ ΔΗΜΟΣΙΟΝΟΜΙΚΩΝ ΕΛΕΓΧΩΝ Δ/ΝΣΗ
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΚΩΝ Π. ΕΠΕΙΓΟΝ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ Α.Δ.Α.: ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΗΜΟΣΙΟΝΟΜΙΚΗΣ ΠΟΛΙΤΙΚΗΣ ΓΕΝΙΚΟ ΛΟΓΙΣΤΗΡΙΟ ΤΟΥ ΚΡΑΤΟΥΣ ΓΕΝΙΚΗ ΔΙΕΥΘΥΝΣΗ ΔΗΜΟΣΙΟΝΟΜΙΚΩΝ ΕΛΕΓΧΩΝ Δ/ΝΣΗ
ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΙΑ ΙΚΤΥΟ ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΑΤΤΙΚΗΣ ΗΜΟΣ ΠΑΛΑΙΟΥ ΦΑΛΗΡΟΥ
 ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΑΤΤΙΚΗΣ ΗΜΟΣ ΠΑΛΑΙΟΥ ΦΑΛΗΡΟΥ Αρ. Aπόφασης 133 ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΙΑ ΙΚΤΥΟ Απόσπασµα Από το πρακτικό της 6 ης συνεδρίασης του ηµοτικού Συµβουλίου του ήµου Παλαιού Φαλήρου έτους 2012
ΕΛΛΗΝΙΚΗ ΗΜΟΚΡΑΤΙΑ ΝΟΜΟΣ ΑΤΤΙΚΗΣ ΗΜΟΣ ΠΑΛΑΙΟΥ ΦΑΛΗΡΟΥ Αρ. Aπόφασης 133 ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΙΑ ΙΚΤΥΟ Απόσπασµα Από το πρακτικό της 6 ης συνεδρίασης του ηµοτικού Συµβουλίου του ήµου Παλαιού Φαλήρου έτους 2012
Ε.Π. ΕΚΠΑΙΔΕΥΣΗ ΚΑΙ ΑΡΧΙΚΗ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΚΑΤΑΡΤΙΣΗ, (ΕΠΕΑΕΚ ΙΙ) 2000-2006 ΜΕΤΡΟ 2.5, ΕΝΕΡΓΕΙΑ 2.5.1, ΚΑΤΗΓΟΡΙΑ ΠΡΑΞΕΩΝ 2.5.1.α
 2000-2006 ΜΕΤΡΟ 2.5, ΕΝΕΡΓΕΙΑ 2.5.1, ΚΑΤΗΓΟΡΙΑ ΠΡΑΞΕΩΝ 2.5.1.α") Ε.Π. ΕΚΠΑΙΔΕΥΣΗ ΚΑΙ ΑΡΧΙΚΗ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΚΑΤΑΡΤΙΣΗ, (ΕΠΕΑΕΚ ΙΙ) 2000-2006 ΜΕΤΡΟ 2.5, ΕΝΕΡΓΕΙΑ 2.5.1, ΚΑΤΗΓΟΡΙΑ ΠΡΑΞΕΩΝ 2.5.1.α ΕΡΓΟ: «ΕΚΠΑΙΔΕΥΤΙΚΗ ΕΝΔΥΝΑΜΩΣΗ ΤΩΝ ΑΤΟΜΩΝ ΜΕ ΑΝΑΠΗΡΙΑ ΚΑΙ ΤΩΝ ΣΤΕΛΕΧΩΝ ΤΩΝ
Ε.Π. ΕΚΠΑΙΔΕΥΣΗ ΚΑΙ ΑΡΧΙΚΗ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΚΑΤΑΡΤΙΣΗ, (ΕΠΕΑΕΚ ΙΙ) 2000-2006 ΜΕΤΡΟ 2.5, ΕΝΕΡΓΕΙΑ 2.5.1, ΚΑΤΗΓΟΡΙΑ ΠΡΑΞΕΩΝ 2.5.1.α ΕΡΓΟ: «ΕΚΠΑΙΔΕΥΤΙΚΗ ΕΝΔΥΝΑΜΩΣΗ ΤΩΝ ΑΤΟΜΩΝ ΜΕ ΑΝΑΠΗΡΙΑ ΚΑΙ ΤΩΝ ΣΤΕΛΕΧΩΝ ΤΩΝ
Ε Λ Λ Η Ν Ι Κ Η ΔΗΜΟΚΡΑΤΙΑ
 Ε Λ Λ Η Ν Ι Κ Η ΔΗΜΟΚΡΑΤΙΑ Αναρτητέα στο διαδίκτυο: Α.Δ.Α.: Ε Λ Λ Η Ν Ι Κ Η ΑΣΤΥΝΟΜΙΑ ΓΕΝΙΚΗ ΑΣΤΥΝ.Δ/ΝΣΗ ΠΕΡΙΦΕΡΕΙΑΣ ΠΕΛΟΠΟΝΝΗΣΟΥ ΝΑΥΠΛΙΟ 13 Νοεμβρίου 2013 ΑΣΤΥΝΟΜΙΚΗ ΔΙΕΥΘΥΝΣΗ ΑΡΓΟΛΙΔΑΣ ΓΡΑΦΕΙΟ ΜΕΡΙΚΗΣ
Ε Λ Λ Η Ν Ι Κ Η ΔΗΜΟΚΡΑΤΙΑ Αναρτητέα στο διαδίκτυο: Α.Δ.Α.: Ε Λ Λ Η Ν Ι Κ Η ΑΣΤΥΝΟΜΙΑ ΓΕΝΙΚΗ ΑΣΤΥΝ.Δ/ΝΣΗ ΠΕΡΙΦΕΡΕΙΑΣ ΠΕΛΟΠΟΝΝΗΣΟΥ ΝΑΥΠΛΙΟ 13 Νοεμβρίου 2013 ΑΣΤΥΝΟΜΙΚΗ ΔΙΕΥΘΥΝΣΗ ΑΡΓΟΛΙΔΑΣ ΓΡΑΦΕΙΟ ΜΕΡΙΚΗΣ
ARVEN S.A. ARVEN ΑΝΩΝΥΜΟΣ ΤΕΧΝΙΚΗ ΕΜΠΟΡΙΚΗ ΒΙΟΜΗΧΑΝΙΚΗ ΕΤΑΙΡΕΙΑ ΑΝΑΠΤΥΞΗΣ & ΕΚΜΕΤΑΛΛΕΥΣΗΣ ΑΚΙΝΗΤΩΝ
 ARVEN ΑΝΩΝΥΜΟΣ ΤΕΧΝΙΚΗ ΕΜΠΟΡΙΚΗ ΒΙΟΜΗΧΑΝΙΚΗ ΕΤΑΙΡΕΙΑ ΑΝΑΠΤΥΞΗΣ & ΕΚΜΕΤΑΛΛΕΥΣΗΣ ΑΚΙΝΗΤΩΝ Ετήσιες Οικονοµικές Καταστάσεις της χρήσης α ό 1 Ιανουαρίου 2011 µέχρι 31 εκεµβρίου 2012 σύµφωνα µε τα ιεθνή Πρότυ
ARVEN ΑΝΩΝΥΜΟΣ ΤΕΧΝΙΚΗ ΕΜΠΟΡΙΚΗ ΒΙΟΜΗΧΑΝΙΚΗ ΕΤΑΙΡΕΙΑ ΑΝΑΠΤΥΞΗΣ & ΕΚΜΕΤΑΛΛΕΥΣΗΣ ΑΚΙΝΗΤΩΝ Ετήσιες Οικονοµικές Καταστάσεις της χρήσης α ό 1 Ιανουαρίου 2011 µέχρι 31 εκεµβρίου 2012 σύµφωνα µε τα ιεθνή Πρότυ
ΠΡΩΤΟΔΙΚΕΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ Εκλογικών
 ΠΡΩΤΟΔΙΚΕΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ Εκλογικών Χρήσιμο Β Ο Η Θ Η Μ Α Ο Δ Η Γ Ο Σ του Αντιπροσώπου της Δικαστικής Αρχής (Περιέχονται σχέδια και έντυπα για διευκόλυνση του έργου των Αντιπροσώπων της Δικαστικής Αρχής
ΠΡΩΤΟΔΙΚΕΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ Εκλογικών Χρήσιμο Β Ο Η Θ Η Μ Α Ο Δ Η Γ Ο Σ του Αντιπροσώπου της Δικαστικής Αρχής (Περιέχονται σχέδια και έντυπα για διευκόλυνση του έργου των Αντιπροσώπων της Δικαστικής Αρχής
Στις 20 Ιουνίου ψηφίζουμε ΔΑΚΕ, για το 16ο συνέδριο τής ΟΛΜΕ
 Στις 20 Ιουνίου ψηφίζουμε ΔΑΚΕ, για το 16ο συνέδριο τής ΟΛΜΕ Αγαπητοί Συνάδελφοι, 17 Ιουνίου 2013 Την Πέμπτη 20 Ιουνίου 2013, από τις 8 το πρωί έως τις 8 το βράδυ στο 1ο Λύκειο Παλλήνης, καλούμαστε να
Στις 20 Ιουνίου ψηφίζουμε ΔΑΚΕ, για το 16ο συνέδριο τής ΟΛΜΕ Αγαπητοί Συνάδελφοι, 17 Ιουνίου 2013 Την Πέμπτη 20 Ιουνίου 2013, από τις 8 το πρωί έως τις 8 το βράδυ στο 1ο Λύκειο Παλλήνης, καλούμαστε να
ΑΠΟΣΠΑΣΜΑ Από το υπ' αριθμ. 30/12-11-2012 Πρακτικό της Οικονομικής Επιτροπής Ιονίων Νήσων
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΕΡΙΦΕΡΕΙΑ ΙΟΝΙΩΝ ΝΗΣΩΝ ΟΙΚΟΝΟΜΙΚΗ ΕΠΙΤΡΟΠΗ ΑΠΟΣΠΑΣΜΑ Από το υπ' αριθμ. 30/12-11-2012 Πρακτικό της Οικονομικής Επιτροπής Ιονίων Νήσων Αριθμ. απόφασης 732-30/12-11-2012 ΠΕΡΙΛΗΨΗ: Έγκριση
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΕΡΙΦΕΡΕΙΑ ΙΟΝΙΩΝ ΝΗΣΩΝ ΟΙΚΟΝΟΜΙΚΗ ΕΠΙΤΡΟΠΗ ΑΠΟΣΠΑΣΜΑ Από το υπ' αριθμ. 30/12-11-2012 Πρακτικό της Οικονομικής Επιτροπής Ιονίων Νήσων Αριθμ. απόφασης 732-30/12-11-2012 ΠΕΡΙΛΗΨΗ: Έγκριση