Fiksacija dušika i biosinteza aminokiselina
|
|
|
- Ἀρτεμίδωρος Σπηλιωτόπουλος
- 6 χρόνια πριν
- Προβολές:
Transcript

1 Fiksacija dušika i biosinteza aminokiselina Boris Mildner Dušik u aminokiselinama, purinima, pirimidinima i drugim molekulama potječe od atmosferskog dušika, N 2. Biosintetski proces započinje redukcijom N 2 u NH 3, a taj proces nazivamo fiksacijom dušika. Bakterije koje su u čvorićima korijena djetelina pretvaraju atmosferski dušik u amonijak. Bakterije pretvaraju N 2 u amonijak koji se prvo ugrađuje u glutamat a zatim u ostale aminokiseline. 1
 pretvaraju plinoviti N 2 u amonijak kojeg biokemijski onda može koristiti većina organizama.")
2 Fiksacija dušika (pretvorba N 2 u NH 3 ) U atmosferi se nalazi 80% N 2, ali N 2 ne može iskoristiti većina organizama. Nekoliko organizama, kao npr. diazotrofne bakterije (bakterije koje fiksiraju dušik) pretvaraju plinoviti N 2 u amonijak kojeg biokemijski onda može koristiti većina organizama. Haber-Boschovim postupkom industrijski se dobiva amonijak. Munjama se dobiva amonijak. Kruženje dušika (ciklus dušika) Većina živih bića korisiti amonijak. U tlu postoje nitrificirajuće bakterije koje pretvaraju amonijak (oksidiraju) u nitrite i nitrate. Većina biljaka koristi nitrite i nitrate jer posjeduju nitrit- i nitrat-reduktaze. U tlu postoje i denitrificirajuće bakterije koje nitrite i nitrate pretvaraju u N 2. Samo određene vrste bakterija i arheje direktno fiksiraju N 2 i reduciraju ga u amonijak. U te vrste spadaju cijanobakterije, metanogene arheje i druge bakterije koje žive u tlu. Postoje i simbiotske bakterije, Rhizobium bakterije, koje fiksiraju dušik, a žive u nodulima (čvorićima) korijena leguminoza. 2
3 Redukcija dušika u amonijak N Ξ N veza je jako stabilna, energija veze je 930 kj. mol -1. Iako je redukcija dušika u amonijak egzergoni proces, N H 2 2 NH 3 DG o = - 33,5 kj. mol -1 reakcija se mora provesti pri o C i nekoliko desetaka tisuća kpa (300 atm) Haber Boschov postupak. U osnovi, za redukciju N 2 u NH 3 potrebno je 6 elektrona: N 2 + 6e H + 2 NH 3 Kako se u biološkim reakcijama uvijek stvara i 1 mol H 2 uz 2 NH 3 u postupku redukcije N Ξ N, to je jednadžba reakcije: N 2 + 8e H + 2 NH 3 + H 2 U većini mikroorganizama koji fiksiraju dušik, 8 elektrona koje donira feredoksin, potječu ili od fotosinteze ili od oksidacijskih procesa. Ujedno za svaki elektron hidrolizira se 2 ATP, pa je ukupna reakcija: N 2 + 8e ATP + 10 H + 2NH H ADP + 16 P i Kompleks nitrogenaze fiksira N 2 (reducira N 2 u amonijak) U diazotrofnim organizmima kompleks nitrogenaze fiksira N 2 u NH 3. Kompleks se sastoji od dva enzima: 1. Reduktaze koja donira energijom bogate elektrone nitrogenazi feredoksin je protein koji dovodi elektrone reduktazi. 2. Nitrogenaze koja koristi elektrone za redukciju N 2 u NH 3 prema jednadžbi: N 2 + 8e ATP + 10 H + 2NH H ADP + 16 P i U vodenim otopinama NH 3 veže H + te nastaje NH 4+. ATP je potreban kako bi se egzergona redukcija odvijala dovoljno brzo. 3



4 Fiksacija dušika u kompleksu nitrogenaze. N 2 + 8e ATP + 10 H + 2NH H ADP + 16 P i Hidroliza ATP se ne koristi za termodinamičke potrebe reakcije. Ona je potrebna za smanjenje energije aktivacije enzima. Tok elektrona je od feredoksina na reduktazu, Fe-S protein, te na nitrogenazu, Fe-Mo protein, kako bi se reducirao dušik u amonijak. Hidroliza ATP potpomaže potrebne konformacijske promjene reduktaze i nitrogenaze kako bi protok elektrona bio efikasan. Enzimi i kofaktori u kompleksu nitrogenaze. Podjedinice nitrogenaze prikazane su sivo i ljubičasto, a podjedinice reduktaze obojane su zeleno i plavo. Vezani ADP je označen crveno, 4Fe-4S kompleks (Fe atomi su narančasti, a S atomi su žuti), a u Fe-Mo kompleksu Mo je označen crno, S atomi su žuti, Fe atomi narančasti a homocitrat je označen sivo. 4


5 Kompleks nitrogenaze fiksira N 2 Molibden-željezo kofaktor u nitrogenazi veže i reducira atmosferski dušik. Reduktaza i nitrogenaza su željezo-sumpor proteini. Reduktaza, naziva se i željeznim proteinom, prenosi elektrone s feredoksina na nitrogenazu. Mjesto na reduktazi, zvano P-klaster, prihvaća elektrone s feredoksina. Iz P-klastera elektroni putuju do Mo-Fe središta gdje se N 2 reducira u NH 3. Mo-Fe središte sadrži i molibden i željezo i oba su potrebna za redukciju N 2. Prikaz podjedinica reduktaze i nitrogenaze. Kompleks nitrogenaze je heterodimer a 2 b 2 podjedinica Elektoni s reduktaze dolaze na P-klastere koji su između a i b podjedinica nitrogenaze. Uloga P-klastera je da čuva elektrone dok se oni ne mogu efikasno iskoristiti za redukciju dušika na FeMo kofaktoru. FeMo kofaktor je mjesto fiksacije dušika. Prikaz podjedinice reduktaze s 4Fe-4S. Reduktaza spada u skupinu NTP-aza s P-petljama koje vežu ATP P-klaster se sastoji od 8 atoma Fe i 7 sulfida povezanih na protein preko 6 cisteina. FeMo kofaktor sastoji se od Mo, 7 Fe, 9 SH, središnjeg atoma, homocitrata, a na protein je vezan preko jednog cisteina i jednog histidina. 5


6 Karakteristika kompleksa nitrogenaze je da je nitrogenaza izuzetno osjetljiva na kisik, pa se reakcija provodi u striktno anaerobnim uvjetima. Simbioza bakterija u nodulima korijena leguminoza korisna je i za dobivanje energije a i za uklanjanje kisika. Bakterije u nodulima dobivaju energiju od biljke u obliku ugljikohidrata i međuprodukta citratnog ciklusa. Za uklanjanje kisika, bakterija je u otopini proteina koji veže kisik leghemoglobina, kojeg sintetizira biljka, a hem proizvodi bakterija. Leghemoglobin veže sav kisik pa se fiksacija dušika nesmetano provodi. Isto tako leghemoglobin dovodi kisik do bakterijskog sustava za prijenos elektrona. Elektronmikrogfija koja je artificijelno obojana. Bakterije su obojane crveno, okružene su peribakteriodnom membranom, plavo, a jezgra biljne stanice je žutozelena. Bakterije koje žive aerobno, a mogu fiksirati dušik, uklanjaju kisik tako da prekidaju tok elektrona koji se koristi za sintezu ATP. Na ovaj način povisuje se temperatura, te kisik izgara čim uđe u stanicu Ovim načinom uklanjanja kisika bakterija Azotobacter vinelandii. 6
7 Amonijev ion asimilira se u stanicu putem glutamata i glutamina Reakciju vezanja amonijaka na a-ketoglutarat katalizira glutamat dehidrogenaza. Reakcija se provodi u dva koraka, U prvom koraku nastaje Schiffova baza, koja se u drugom reakcijskom stupnju protonira. Schiffove baze između karbonilne skupine i amina ključne su reakcije u biosintezi aminokiselina. Reakcija protoniranja Schiffove baze ključna je za stereokemiju a-c atoma u glutamatu. Enzim, glutamat dehidrogenaza veže a-ketoglutarat tako da se hidridni ion s NADPH veže tako da akiralna Schiffova baza postane L-glutamat. Glutamat je prvi spoj u koji je ugrađen NH 3. NH a-ketoglutarat + NAD(P)H + H + glutamat + NAD(P) + H 2 O 7
8 Naredni amonijev ion ugrađuje se u glutamat, te nastaje glutamin. Reakciju katalizira glutamin sintetaza, a potrebna je i hidroliza ATP. Glutamin je univerzalni donor dušika. Regulacija glutamin sintetaze ključna je reakcija kojom se kontrolira metabolizam dušika. Većina prokariota i biljaka imaju i glutamat sintazu koja evolucijski nije srodna glutamin sintetazi Glutamat sintaza katalizira reduktivnu aminaciju a-ketoglutarata u glutamat: a-ketoglutarat + glutamin + NADPH + H + 2 glutamat + NADP + Glutamin sintetaza: glutamat + ATP + NH 4 + glutamin + ADP + P i Sumarna reakcija je: NH a-ketoglutarat + NADPH + ATP glutamat + NADP + + ADP + P i Kada nema dovoljno NH 4 + većina glutamata nastaje slijedom reakcija glutamin sintetaze i glutamat sintaze. Prokarioti koriste ovaj put jer je K M glutamat dehidrogenaze za NH 4 + velik (>> 1 mol dm -3 ) pa enzim nije zasićen kada je nestašica NH 4+. Nasuprot tome, glutamin sintetaza ima vrlo visoki afinitet za NH 4+, ali da se iskoristi NH 4 + mora se utrošiti ATP. 8


9 Glutamin sintetaza je prvi korak koji se kontrolira u metabolizmu dušika Glutamin sintetaza E. coli ima 12 podjedinica. Svih 12 podjedinica su identične. Zbog lakšeg raspoznavanja, podjedinice su obojane. Mr = Pogled sa strane (bokocrt) Pogled odozgora (nacrt) Enzim se regulira i alosterički i kovalentnom modifikacijom, kao i transkripcijom gena. Alosterička regulacija glutamin sintetaze Alanin, glicin i barem 6 produkata metabolizma glutamina alosterički su inhibitori glutamin sintetaze. Svaki spoj zasebno izaziva samo djelomičnu inhibiciju, ali inhibicije su aditivne, tako da 8 inhibitora zajedno u potpunosti inhibiraju aktivnost enzima. Alanin i glicin vjerojatno služe kao indikatori koncentracija aminokiselina u stanici. 9

10 Pretpostavljeni mehanizam glutamin amidotransferaza Više od deset reakcija u biosintezi aminokiselina koriste glutamin kao izvor amino skupina. Sve te reakcije provode glutamin amidotransferaze. U domeni koja veže glutamin je Cys koji je nukleofil, pa pomaže cijepanju amidne veze glutamina te radi kovalentni međuprodukt s enzimom. Oslobođeni NH 3 u ovoj reakciji se ne otpušta već odlazi u drugu domenu enzima gdje reagira sa supstratom. Biosinteza aminokiselina Aminokiseline se sintetiziraju iz međuprodukata glavnih metaboličkih putova 10
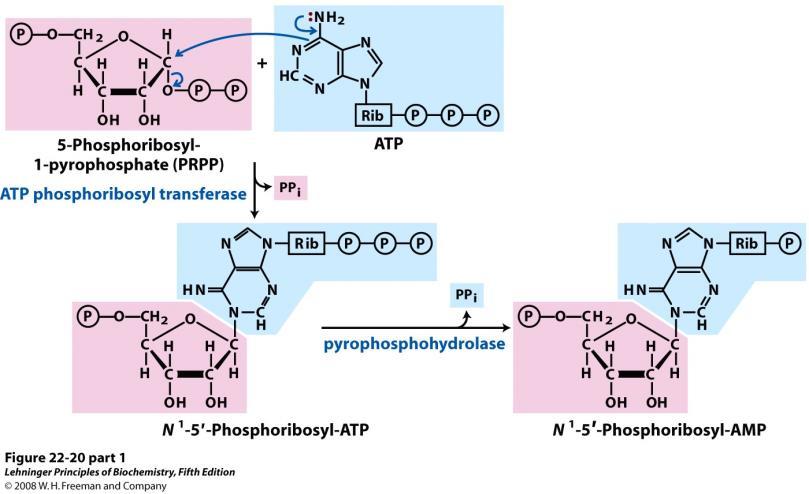
, citratnog ciklusa (svjetlo plavo) i puta pentoza fosfata (tamno plavo) 5-fosforibozil-1-pirofosfat")
11 Pregled metabolizma aminokiselina. Ugljikovi atomi aminokiselina potječu od puta glikolize (ružičasto), citratnog ciklusa (svjetlo plavo) i puta pentoza fosfata (tamno plavo) 5-fosforibozil-1-pirofosfat Riboza-5-fosfat + ATP 5-fosforibozil-1-pirofosfat Nastaje u pentoza fosfat metaboličkom putu. Ovaj spoj je međuprodukt putova sinteze histidina i nukleotida. 11




12 Metaboličke preteče označene su plavo, aminokiselene koje su preteče drugim amininokiselinama označene su žuto, a esencijalne aminokiseline su označene masnim slovima Ljudi sintetiziraju neke aminokiseline, a druge aminokiseline moraju se unositi hranom Aminokiseline koje može sintetizirati ljudski organizam nazivaju se neesencijalnim aminokiselinama. Neesencijalne aminokiseline sintetiziraju se jednostavnim reakcijama. Aminokiseline koje dobivamo hranom nazivaju se esencijalnim aminokiselinama. Za sintezu ovih aminokiselina potrebni su kompleksni sintetski putovi i mi ih ne možemo provoditi. Nedostatak samo i jedne esencijalne aminokiseline dovodi do ozbiljnih pato-fizioloških posljedica. 12

13 Esencijalne i neesencijalne aminokiseline. Neke aminokiseline su neesencijalne za ljude jer ih se može sintetizirati u malom broju reakcijskih koraka. One aminokiseline za čiju biosintezu je potreban velik broj reakcijskih koraka su esencijalne i potrebno ih je uzimati hranom. Neke aminokiseline nastaju jednostavnim transaminacijskim reakcijama Transaminacijske reakcije kataliziraju transaminaze (aminotransferaze). Za aktivnost transaminaza neophodan je piridoksalfosfat koji potječe od piridoksamina (vitamina B 6 ). 13
14 Biosinteza prolina i arginina u bakterijama Biosinteza arginina - 1 Sinteza arginina

15 Biosinteza arginina - 2 Sinteza prolina odnosno ornitina kod sisavaca 15

16 Serin, cistein i glicin nastaju iz 3-fosfoglicerata Produkt glikolitičkog puta, 3- fosfoglicerat je preteča serina. Serin se metabolički pretvara u cistein i glicin. Za sintezu glicina neophodan je kofaktor tetrahidrofolat. Sinteza serina i glicina 16

17 Sinteza cisteina u sisavaca Iz piruvata i oksaloacetata sintetiziraju se tri nesencijalne i šest esencijalnih aminokiselina 17
18 Sinteza asparagina Biosinteza lizina, metionina, treonina, valina, izoleucina i leucina u bakterijama preteča za sintezu lizina, metionina i treonina Sinteza lizina Sinteza metionina 18
 Tijekom doniranja metilne skupine nastaje homocistein, preteča cisteina Ciklus")
19 Sinteza S-adenozilmetionina važnog donora metilnih skupina SAM nastaje iz metionina i ATP u neuobičajenoj reakciji u kojoj se ATP cijepa na PP i i P i. U sisavcima, sinteza cistena započinje razgradnjom S- adenozilmetionina (SAM) Tijekom doniranja metilne skupine nastaje homocistein, preteča cisteina Ciklus homocistein - metionin 19
20 Visoke koncentracije homocisteina koreliraju s vaskularnim bolestima Pacijenti koji imaju visoke koncentracije homocisteina imaju veći rizik za kardiovaskularne bolesti. Najčešći uzrok povišenoj koncentraciji homocisteina u krvi je nedostatak aktivnosti cistation sintaze, enzima koji pretvara homocistein u cistein. Sinteza aromatskih aminokiselina 20
21 Sinteza aromatskih aminokiselina - 1 Sinteza aromatskih aminokiselina - 2 Korizmat je ključni međuprodukt u biosintezi aromatskih aminokiselina. Glifozat inhibira adiciju PEP-a. 21
22 Sinteza fenilalanina i tirozina Sinteza triptofana 22

23 Sinteza histidina

24 Sinteza histidina - 2 Sinteza histidina
25 Sažetak sinteza neesencijalnih aminokiselina u sisavaca oksaloacetat + glutamat aspartat + a-ketoglutarat piruvat + glutamat alanin + a-ketoglutarat Sažetak -2 sinteza neesencijalnih aminokiselina u sisavaca arginin serin + tetrahidrofolat glicin + metilentetrahidrofolat 25
26 Regulacija biosinteze aminokiselina Brzina sinteze aminokiselina ovisi o količini biosintetskih enzima i o njihovoj aktivnosti. U biosintetskom putu prva ireverzibilna reakcija, koju nazivamo odlučujući korak, važno je regulacijsko mjesto. U inhibiciji povratnom spregom, krajnji produkt metaboličkog puta Z inhibira enzime koji kataliziraju odlučujući korak (A B). Ovakva kontrola je bitna za očuvanje metabolita koji su međuprodukti za sintezu biomolekula, kao i za očuvanje metaboličke energije. Odlučujući korak je uobičajeno regulacijsko mjesto za inhibiciju povratnom spregom 3-fosfoglicerat dehidrogenaza katalizira odlučujući korak u biosintezi serina iz 3-fosfoglicerata. Serin, krajnji produkt, inhibira 3-fosfoglicerat dehidrogenazu. 26

27 3-fosfoglicerat dehidrogenaza je odlučujući korak u biosintezi serina iz 3-fosfoglicerata. E. coli enzim je tetramer izgrađen od četiri identične podjedinice. Svaka podjedinica ima katalitičku domenu i regulatornu domenu koja veže serin. Vezanje serina za regulatornu domenu smanjuje V max enzima. Enzim na koji su vezana četiri serina je neaktivan te se 3-fosfoglicerat može iskoristiti za druge metabličke pretvorbe. Razgranani putovi podliježu sofisticiranim regulacijama Razgranani putovi reguliraju se na nekoliko načina: 1. Inhibicijom povratne sprege i aktivacijom: ako dva puta imaju zajednički početni korak, jedan put se inhibira vlastitim produktom, a stimulira se produktom drugog puta. (Treonin deaminaza je primjer ovog tipa regulacije). 2. Multiplikacijom enzima: odlučujući korak kataliziraju dva ili više enzima a ovi enzimi imaju različita regulacijska svojstva. U E. coli, tri različite aspartat kinaze kontroliraju sintezu treonina, metionina i lizina. 3. Kumulativnom inhibicijom povratne sprege: zajednički korak za nekoliko metaboličkih putova djelomično se inhibira sa svakim krajnjim produktom ovih različitih putova. Ovaj tip regulacije je primjer glutamin sintetaze koju inhibiraju mnogi produkti. 27
28 Kontrola razgrananih metaboličkih puteva. Kontrola razgrananih puteva je kompliciranija jer se mora voditi računa o koncentracijama dva produkta. Dva metabolička puta sa zajedničkim prvim korakom (odlučujućim korakom) mogu biti ihnibirana s vlastitim produktima, a aktivirana s produktom drugog puta. U biosintezi valina, leucina i izoleucina zajednički prvi međuprodukt je hidroksietil-tpp koji vodi do sinteze sve tri aminokiseline. Za sintezu izoleucina, hidroksietil- TPP reagira s a- ketobutiratom. Isto tako hidroksietil-tpp reagira s piruvatom u sintezama valina i leucina. Prema tome koncentracije a-ketobutirata i piruvata određuju koliko će se sintetizirati izoleucina u odnosu na valin i leucin. Treonin deaminaza koja katalizira nastajanje a-ketobutirata alosterički je inhibirana izoleucinom dok valin alosterički aktivira ovaj enzim, pa dolazi do sinteze valina i leucina. Regulacija treonin deaminaze. Treonin se pretvara u a-ketobutirat u odlučujućem koraku te dolazi do sinteze izoleucina. Enzim koji katalizira ovaj korak, treonin deaminazu, inhibira izoleucin, a aktivira je valin, produkt paralelnog metaboličkog puta. Treonin deaminaza izoleucin valin i leucin 28

29 Izoenzimi u regulaciji metaboličkih puteva. Odlučujući korak mogu katalizirati dva ili više enzima s različitim regulacijskim svojstvima. Fosforilacija aspartata je obvezni korak u biosintezi treonina, metionina i lizina. U E. coli tri različite aspartokinaze kataliziraju ovu reakciju. Katalitičke domene imaju 30% identičnost primarne strukture. Iako su im mehanizmi katalize gotovo identični, njihove aktivnosti su različito regulirane. Jedan enzim nije reguliran povratnom spregom, drugog inhibira treonin, a trećeg inhibira lizin. Složen regulacijski mehanizam može se razviti i duplikacijom gena koji kodiraju biosintetske enzime. Biosinteza lizina, metionina, treonina, valina, izoleucina i leucina u bakterijama. Biosintezu reguliraju različite aspartat kinaze - primjer multiplikacije enzima. Više enzima katalizira identičnu ili sličnu reakciju, ali svaki od njih ima drugačije alosteričke karakteristike i zbog toga mogu katalizirati odlučujući korak u određenom metaboličkom putu. preteča za sintezu lizina, metionina i treonina Sinteza lizina Sinteza metionina 29
Aminokiseline. Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina 22.12.2014
 Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Put pentoza fosfata. B. Mildner. Put pentoza fosfata
 Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
CIKLUS LIMUNSKE KISELINE (CLK)
") SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
Osnove biokemije Enzimska kinetika. Boris Mildner. Kinetika proučava brzine reakcija
 Osnove biokemije Enzimska kinetika Boris Mildner Kinetika proučava brzine reakcija Za reakciju: A P Brzina reakcije v je: v = - d[a]/dt = d[p]/dt (1) pri čemu d označava smanjenje koncentracije supstrata,
Osnove biokemije Enzimska kinetika Boris Mildner Kinetika proučava brzine reakcija Za reakciju: A P Brzina reakcije v je: v = - d[a]/dt = d[p]/dt (1) pri čemu d označava smanjenje koncentracije supstrata,
Glukoneogeneza i regulacija glukoneogeneze
 Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
Ciklus limunske kiseline-2
 Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
3.1 Granična vrednost funkcije u tački
 3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE
 Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
2 tg x ctg x 1 = =, cos 2x Zbog četvrtog kvadranta rješenje je: 2 ctg x
 Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
18. listopada listopada / 13
 18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović
 DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
Linearna algebra 2 prvi kolokvij,
 Linearna algebra 2 prvi kolokvij, 27.. 20.. Za koji cijeli broj t je funkcija f : R 4 R 4 R definirana s f(x, y) = x y (t + )x 2 y 2 + x y (t 2 + t)x 4 y 4, x = (x, x 2, x, x 4 ), y = (y, y 2, y, y 4 )
Linearna algebra 2 prvi kolokvij, 27.. 20.. Za koji cijeli broj t je funkcija f : R 4 R 4 R definirana s f(x, y) = x y (t + )x 2 y 2 + x y (t 2 + t)x 4 y 4, x = (x, x 2, x, x 4 ), y = (y, y 2, y, y 4 )
RIJEŠENI ZADACI I TEORIJA IZ
 RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
M086 LA 1 M106 GRP. Tema: Baza vektorskog prostora. Koordinatni sustav. Norma. CSB nejednakost
 M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
41. Jednačine koje se svode na kvadratne
 . Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
. Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
(P.I.) PRETPOSTAVKA INDUKCIJE - pretpostavimo da tvrdnja vrijedi za n = k.
 PRETPOSTAVKA INDUKCIJE - pretpostavimo da tvrdnja vrijedi za n = k.") 1 3 Skupovi brojeva 3.1 Skup prirodnih brojeva - N N = {1, 2, 3,...} Aksiom matematičke indukcije Neka je N skup prirodnih brojeva i M podskup od N. Ako za M vrijede svojstva: 1) 1 M 2) n M (n + 1) M,
1 3 Skupovi brojeva 3.1 Skup prirodnih brojeva - N N = {1, 2, 3,...} Aksiom matematičke indukcije Neka je N skup prirodnih brojeva i M podskup od N. Ako za M vrijede svojstva: 1) 1 M 2) n M (n + 1) M,
ELEKTROTEHNIČKI ODJEL
 MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
Trigonometrija 2. Adicijske formule. Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto
 Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
7 Algebarske jednadžbe
 7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
Riješeni zadaci: Nizovi realnih brojeva
 Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama.
 Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
PRAVA. Prava je u prostoru određena jednom svojom tačkom i vektorom paralelnim sa tom pravom ( vektor paralelnosti).
.") PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
PARCIJALNI IZVODI I DIFERENCIJALI. Sama definicija parcijalnog izvoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je,
 PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
1 Promjena baze vektora
 Promjena baze vektora Neka su dane dvije različite uredene baze u R n, označimo ih s A = (a, a,, a n i B = (b, b,, b n Svaki vektor v R n ima medusobno različite koordinatne zapise u bazama A i B Zapis
Promjena baze vektora Neka su dane dvije različite uredene baze u R n, označimo ih s A = (a, a,, a n i B = (b, b,, b n Svaki vektor v R n ima medusobno različite koordinatne zapise u bazama A i B Zapis
numeričkih deskriptivnih mera.
 DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
ENZIMI KOJI UČESTVUJU U "PLANT GROWTH PROMOTING" POTENCIJALU BAKTERIJA
 ENZIMI KOJI UČESTVUJU U "PLANT GROWTH PROMOTING" POTENCIJALU BAKTERIJA Klimatske promene Porast broja ljudi na planeti ZAŠTO JE OVO VAŽNO Veći zahtevi za poizvodnjom hrane, hrane za životinje, goriva posebno
ENZIMI KOJI UČESTVUJU U "PLANT GROWTH PROMOTING" POTENCIJALU BAKTERIJA Klimatske promene Porast broja ljudi na planeti ZAŠTO JE OVO VAŽNO Veći zahtevi za poizvodnjom hrane, hrane za životinje, goriva posebno
NOMENKLATURA ORGANSKIH SPOJEVA. Imenovanje aromatskih ugljikovodika
 NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
C kao nukleofil (Organometalni spojevi)
") C kao nukleofil (Organometalni spojevi) 1 Nastajanje nukleofilnih C atoma i njihova adicija na karbonilnu grupu Ukupan proces je jedan od najkorisnijih sintetskih postupaka za stvaranje C-C veze 2 Priroda
C kao nukleofil (Organometalni spojevi) 1 Nastajanje nukleofilnih C atoma i njihova adicija na karbonilnu grupu Ukupan proces je jedan od najkorisnijih sintetskih postupaka za stvaranje C-C veze 2 Priroda
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
INTEGRALNI RAČUN. Teorije, metodike i povijest infinitezimalnih računa. Lucija Mijić 17. veljače 2011.
 INTEGRALNI RAČUN Teorije, metodike i povijest infinitezimalnih računa Lucija Mijić lucija@ktf-split.hr 17. veljače 2011. Pogledajmo Predstavimo gornju sumu sa Dodamo još jedan Dobivamo pravokutnik sa Odnosno
INTEGRALNI RAČUN Teorije, metodike i povijest infinitezimalnih računa Lucija Mijić lucija@ktf-split.hr 17. veljače 2011. Pogledajmo Predstavimo gornju sumu sa Dodamo još jedan Dobivamo pravokutnik sa Odnosno
Zavrxni ispit iz Matematiqke analize 1
 Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Iskazna logika 3. Matematička logika u računarstvu. novembar 2012
 Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Matematička analiza 1 dodatni zadaci
 Matematička analiza 1 dodatni zadaci 1. Ispitajte je li funkcija f() := 4 4 5 injekcija na intervalu I, te ako jest odredite joj sliku i inverz, ako je (a) I = [, 3), (b) I = [1, ], (c) I = ( 1, 0].. Neka
Matematička analiza 1 dodatni zadaci 1. Ispitajte je li funkcija f() := 4 4 5 injekcija na intervalu I, te ako jest odredite joj sliku i inverz, ako je (a) I = [, 3), (b) I = [1, ], (c) I = ( 1, 0].. Neka
Teorijske osnove informatike 1
 Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Seminar 13.b. Glikogen GLIKOGEN. B. Mildner
 Seminar 13.b Glikogen B. Mildner GLIKOGEN 1 Glikogen Nereducirani kraj Glikogen je jako dostupni skladišni oblik glukoze; kao i jako velik, razgranat polimer; Glukozne jedinice su povezane α-1,4-glikozidnim
Seminar 13.b Glikogen B. Mildner GLIKOGEN 1 Glikogen Nereducirani kraj Glikogen je jako dostupni skladišni oblik glukoze; kao i jako velik, razgranat polimer; Glukozne jedinice su povezane α-1,4-glikozidnim
Linearna algebra 2 prvi kolokvij,
 1 2 3 4 5 Σ jmbag smjer studija Linearna algebra 2 prvi kolokvij, 7. 11. 2012. 1. (10 bodova) Neka je dano preslikavanje s : R 2 R 2 R, s (x, y) = (Ax y), pri čemu je A: R 2 R 2 linearan operator oblika
1 2 3 4 5 Σ jmbag smjer studija Linearna algebra 2 prvi kolokvij, 7. 11. 2012. 1. (10 bodova) Neka je dano preslikavanje s : R 2 R 2 R, s (x, y) = (Ax y), pri čemu je A: R 2 R 2 linearan operator oblika
Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner
 Boris Mildner") Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner Čestice glikogena u hepatocitima. Glikogen, čestice za pohranjivanje glukoze, u hepatocitima se nalazi u citoplazmi blizu tubula endoplazmatske
Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner Čestice glikogena u hepatocitima. Glikogen, čestice za pohranjivanje glukoze, u hepatocitima se nalazi u citoplazmi blizu tubula endoplazmatske
1.4 Tangenta i normala
 28 1 DERIVACIJA 1.4 Tangenta i normala Ako funkcija f ima derivaciju u točki x 0, onda jednadžbe tangente i normale na graf funkcije f u točki (x 0 y 0 ) = (x 0 f(x 0 )) glase: t......... y y 0 = f (x
28 1 DERIVACIJA 1.4 Tangenta i normala Ako funkcija f ima derivaciju u točki x 0, onda jednadžbe tangente i normale na graf funkcije f u točki (x 0 y 0 ) = (x 0 f(x 0 )) glase: t......... y y 0 = f (x
Operacije s matricama
 Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Neka je a 3 x 3 + a 2 x 2 + a 1 x + a 0 = 0 algebarska jednadžba trećeg stupnja. Rješavanje ove jednadžbe sastoji se od nekoliko koraka.
 Neka je a 3 x 3 + a x + a 1 x + a 0 = 0 algebarska jednadžba trećeg stupnja. Rješavanje ove jednadžbe sastoji se od nekoliko koraka. 1 Normiranje jednadžbe. Jednadžbu podijelimo s a 3 i dobivamo x 3 +
Neka je a 3 x 3 + a x + a 1 x + a 0 = 0 algebarska jednadžba trećeg stupnja. Rješavanje ove jednadžbe sastoji se od nekoliko koraka. 1 Normiranje jednadžbe. Jednadžbu podijelimo s a 3 i dobivamo x 3 +
HEMIJSKA VEZA TEORIJA VALENTNE VEZE
 TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
Pismeni ispit iz matematike Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: ( ) + 1.
 + 1.") Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
a M a A. Može se pokazati da je supremum (ako postoji) jedinstven pa uvodimo oznaku sup A.
 jedinstven pa uvodimo oznaku sup A.") 3 Infimum i supremum Definicija. Neka je A R. Kažemo da je M R supremum skupa A ako je (i) M gornja meda skupa A, tj. a M a A. (ii) M najmanja gornja meda skupa A, tj. ( ε > 0)( a A) takav da je a > M
3 Infimum i supremum Definicija. Neka je A R. Kažemo da je M R supremum skupa A ako je (i) M gornja meda skupa A, tj. a M a A. (ii) M najmanja gornja meda skupa A, tj. ( ε > 0)( a A) takav da je a > M
OM2 V3 Ime i prezime: Index br: I SAVIJANJE SILAMA TANKOZIDNIH ŠTAPOVA
 OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A
, grupa A") Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
XI dvoqas veжbi dr Vladimir Balti. 4. Stabla
 XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
OKSIDACIJSKA FOSFORILACIJA
 OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
Funkcije dviju varjabli (zadaci za vježbu)
") Funkcije dviju varjabli (zadaci za vježbu) Vidosava Šimić 22. prosinca 2009. Domena funkcije dvije varijable Ako je zadano pridruživanje (x, y) z = f(x, y), onda se skup D = {(x, y) ; f(x, y) R} R 2 naziva
Funkcije dviju varjabli (zadaci za vježbu) Vidosava Šimić 22. prosinca 2009. Domena funkcije dvije varijable Ako je zadano pridruživanje (x, y) z = f(x, y), onda se skup D = {(x, y) ; f(x, y) R} R 2 naziva
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu
 Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika. Monotonost i ekstremi. Katica Jurasić. Rijeka, 2011.
 Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
METABOLIZAM PROTEINA
 METABOLIZAM PROTEINA PREGLED METABOLIZMA AMINO KISELINA Hranom unijeti proteini se razgrađuju do amino kiselina, koje se apsorbuju, prenose cirkulacijom i preuzimaju u ćelije različitih tkiva. Amino kiseline
METABOLIZAM PROTEINA PREGLED METABOLIZMA AMINO KISELINA Hranom unijeti proteini se razgrađuju do amino kiselina, koje se apsorbuju, prenose cirkulacijom i preuzimaju u ćelije različitih tkiva. Amino kiseline
Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. 4/17/2013
 Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
3. razred gimnazije- opšti i prirodno-matematički smer ALKENI. Aciklični nezasićeni ugljovodonici koji imaju jednu dvostruku vezu.
 ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
Sume kvadrata. mn = (ax + by) 2 + (ay bx) 2.
 2 + (ay bx) 2.") Sume kvadrata Koji se prirodni brojevi mogu prikazati kao zbroj kvadrata dva cijela broja? Propozicija 1. Ako su brojevi m i n sume dva kvadrata, onda je i njihov produkt m n takoder suma dva kvadrata.
Sume kvadrata Koji se prirodni brojevi mogu prikazati kao zbroj kvadrata dva cijela broja? Propozicija 1. Ako su brojevi m i n sume dva kvadrata, onda je i njihov produkt m n takoder suma dva kvadrata.
Dijagonalizacija operatora
 Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
Elementi spektralne teorije matrica
 Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Ispitivanje toka i skiciranje grafika funkcija
 Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE)
") (Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
(Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
Strukture podataka i algoritmi 1. kolokvij 16. studenog Zadatak 1
 Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Matematika 1 - vježbe. 11. prosinca 2015.
 Matematika - vježbe. prosinca 5. Stupnjevi i radijani Ako je kut φ jednak i rad, tada je veza između i 6 = Zadatak.. Izrazite u stupnjevima: a) 5 b) 7 9 c). d) 7. a) 5 9 b) 7 6 6 = = 5 c). 6 8.5 d) 7.
Matematika - vježbe. prosinca 5. Stupnjevi i radijani Ako je kut φ jednak i rad, tada je veza između i 6 = Zadatak.. Izrazite u stupnjevima: a) 5 b) 7 9 c). d) 7. a) 5 9 b) 7 6 6 = = 5 c). 6 8.5 d) 7.
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
Pismeni ispit iz matematike GRUPA A 1. Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj, zatim naći 4 z.
 Pismeni ispit iz matematike 06 007 Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj z = + i, zatim naći z Ispitati funkciju i nacrtati grafik : = ( ) y e + 6 Izračunati integral:
Pismeni ispit iz matematike 06 007 Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj z = + i, zatim naći z Ispitati funkciju i nacrtati grafik : = ( ) y e + 6 Izračunati integral:
Kiselo bazni indikatori
 Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
Više dokaza jedne poznate trigonometrijske nejednakosti u trokutu
 Osječki matematički list 000), 5 9 5 Više dokaza jedne poznate trigonometrijske nejednakosti u trokutu Šefket Arslanagić Alija Muminagić Sažetak. U radu se navodi nekoliko različitih dokaza jedne poznate
Osječki matematički list 000), 5 9 5 Više dokaza jedne poznate trigonometrijske nejednakosti u trokutu Šefket Arslanagić Alija Muminagić Sažetak. U radu se navodi nekoliko različitih dokaza jedne poznate
Pošto pretvaramo iz veće u manju mjernu jedinicu broj 2.5 množimo s 1000,
 PRERAČUNAVANJE MJERNIH JEDINICA PRIMJERI, OSNOVNE PRETVORBE, POTENCIJE I ZNANSTVENI ZAPIS, PREFIKSKI, ZADACI S RJEŠENJIMA Primjeri: 1. 2.5 m = mm Pretvaramo iz veće u manju mjernu jedinicu. 1 m ima dm,
PRERAČUNAVANJE MJERNIH JEDINICA PRIMJERI, OSNOVNE PRETVORBE, POTENCIJE I ZNANSTVENI ZAPIS, PREFIKSKI, ZADACI S RJEŠENJIMA Primjeri: 1. 2.5 m = mm Pretvaramo iz veće u manju mjernu jedinicu. 1 m ima dm,
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
radni nerecenzirani materijal za predavanja
 Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Kažemo da je funkcija f : a, b R u točki x 0 a, b postiže lokalni minimum ako postoji okolina O(x 0 ) broja x 0 takva da je
Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Kažemo da je funkcija f : a, b R u točki x 0 a, b postiže lokalni minimum ako postoji okolina O(x 0 ) broja x 0 takva da je
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija
 Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
Mehanizmi djelovanja enzima i regulacija aktivnosti enzima
 Mehanizmi djelovanja enzima i regulacija aktivnosti enzima Boris Mildner Aktivnost enzima mijenja se promjenama temperature, ph, inhibitorima te vezanjem malih molekula ili iona Temperatura povećava brzinu
Mehanizmi djelovanja enzima i regulacija aktivnosti enzima Boris Mildner Aktivnost enzima mijenja se promjenama temperature, ph, inhibitorima te vezanjem malih molekula ili iona Temperatura povećava brzinu
Grafičko prikazivanje atributivnih i geografskih nizova
 Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
-NH 3. Degradación de aminoácidos. 1) Eliminación del NH 3. 2) Degradación de esqueletos carbonados. Ac. grasos c. cetónicos glucosa.
 Eliminación del NH 3. 2) Degradación de esqueletos carbonados. Ac. grasos c. cetónicos glucosa.") Degradación de aminoácidos 1) Eliminación del NH 3 interfiere polarización/despolariación nerviosa compite transportadores metales alcalinos -NH 3 + 2) Degradación de esqueletos carbonados Ac. grasos c.
Degradación de aminoácidos 1) Eliminación del NH 3 interfiere polarización/despolariación nerviosa compite transportadores metales alcalinos -NH 3 + 2) Degradación de esqueletos carbonados Ac. grasos c.
radni nerecenzirani materijal za predavanja R(f) = {f(x) x D}
 = {f(x) x D}") Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
PRIMJER 3. MATLAB filtdemo
 PRIMJER 3. MATLAB filtdemo Prijenosna funkcija (IIR) Hz () =, 6 +, 3 z +, 78 z +, 3 z +, 53 z +, 3 z +, 78 z +, 3 z +, 6 z, 95 z +, 74 z +, z +, 9 z +, 4 z +, 5 z +, 3 z +, 4 z 3 4 5 6 7 8 3 4 5 6 7 8
PRIMJER 3. MATLAB filtdemo Prijenosna funkcija (IIR) Hz () =, 6 +, 3 z +, 78 z +, 3 z +, 53 z +, 3 z +, 78 z +, 3 z +, 6 z, 95 z +, 74 z +, z +, 9 z +, 4 z +, 5 z +, 3 z +, 4 z 3 4 5 6 7 8 3 4 5 6 7 8
METABOLIZAM PROTEINA
 METABOLIZAM PROTEINA PREGLED METABOLIZMA AMINO KISELINA Hranom unijeti proteini se razgrađuju do amino kiselina, koje se apsorbuju, prenose cirkulacijom i preuzimaju u ćelije različitih tkiva. Amino kiseline
METABOLIZAM PROTEINA PREGLED METABOLIZMA AMINO KISELINA Hranom unijeti proteini se razgrađuju do amino kiselina, koje se apsorbuju, prenose cirkulacijom i preuzimaju u ćelije različitih tkiva. Amino kiseline
ZBIRKA POTPUNO RIJEŠENIH ZADATAKA
 **** IVANA SRAGA **** 1992.-2011. ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE POTPUNO RIJEŠENI ZADACI PO ŽUTOJ ZBIRCI INTERNA SKRIPTA CENTRA ZA PODUKU α M.I.M.-Sraga - 1992.-2011.
**** IVANA SRAGA **** 1992.-2011. ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE POTPUNO RIJEŠENI ZADACI PO ŽUTOJ ZBIRCI INTERNA SKRIPTA CENTRA ZA PODUKU α M.I.M.-Sraga - 1992.-2011.
REAKCIJE ELIMINACIJE
 REAKIJE ELIMINAIJE 1 . DEIDROALOGENAIJA (-X) i DEIDRATAIJA (- 2 O) su najčešći tipovi eliminacionih reakcija X Y + X Y 2 Dehidrohalogenacija (-X) X strong base + " X " X = l, Br, I 3 E 2 Mehanizam Ova
REAKIJE ELIMINAIJE 1 . DEIDROALOGENAIJA (-X) i DEIDRATAIJA (- 2 O) su najčešći tipovi eliminacionih reakcija X Y + X Y 2 Dehidrohalogenacija (-X) X strong base + " X " X = l, Br, I 3 E 2 Mehanizam Ova
STVARANJE VEZE C-C POMO]U ORGANOBORANA
![STVARANJE VEZE C-C POMO]U ORGANOBORANA](/thumbs/55/37829094.jpg "STVARANJE VEZE C-C POMO]U ORGANOBORANA") STVAAJE VEZE C-C PM]U GAAA 2 6 rojne i raznovrsne reakcije * idroborovanje alkena i reakcije alkil-borana 3, Et 2 (ili TF ili diglim) Ar δ δ 2 2 3 * cis-adicija "suprotno" Markovnikov-ljevom pravilu *
STVAAJE VEZE C-C PM]U GAAA 2 6 rojne i raznovrsne reakcije * idroborovanje alkena i reakcije alkil-borana 3, Et 2 (ili TF ili diglim) Ar δ δ 2 2 3 * cis-adicija "suprotno" Markovnikov-ljevom pravilu *
- pravac n je zadan s točkom T(2,0) i koeficijentom smjera k=2. (30 bodova)
 i koeficijentom smjera k=2. (30 bodova)") MEHANIKA 1 1. KOLOKVIJ 04/2008. grupa I 1. Zadane su dvije sile F i. Sila F = 4i + 6j [ N]. Sila je zadana s veličinom = i leži na pravcu koji s koordinatnom osi x zatvara kut od 30 (sve komponente sile
MEHANIKA 1 1. KOLOKVIJ 04/2008. grupa I 1. Zadane su dvije sile F i. Sila F = 4i + 6j [ N]. Sila je zadana s veličinom = i leži na pravcu koji s koordinatnom osi x zatvara kut od 30 (sve komponente sile
Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici
 Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri
 Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri 1 1 Zadatak 1b Čisto savijanje - vezano dimenzionisanje Odrediti potrebnu površinu armature za presek poznatih dimenzija, pravougaonog
Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri 1 1 Zadatak 1b Čisto savijanje - vezano dimenzionisanje Odrediti potrebnu površinu armature za presek poznatih dimenzija, pravougaonog
POVRŠINA TANGENCIJALNO-TETIVNOG ČETVEROKUTA
 POVRŠIN TNGENIJLNO-TETIVNOG ČETVEROKUT MLEN HLP, JELOVR U mnoštvu mnogokuta zanimljiva je formula za površinu četverokuta kojemu se istoobno može upisati i opisati kružnica: gje su a, b, c, uljine stranica
POVRŠIN TNGENIJLNO-TETIVNOG ČETVEROKUT MLEN HLP, JELOVR U mnoštvu mnogokuta zanimljiva je formula za površinu četverokuta kojemu se istoobno može upisati i opisati kružnica: gje su a, b, c, uljine stranica
Dvanaesti praktikum iz Analize 1
 Dvaaesti praktikum iz Aalize Zlatko Lazovi 20. decembar 206.. Dokazati da fukcija f = 5 l tg + 5 ima bar jedu realu ulu. Ree e. Oblast defiisaosti fukcije je D f = k Z da postoji ula fukcije a 0, π 2.
Dvaaesti praktikum iz Aalize Zlatko Lazovi 20. decembar 206.. Dokazati da fukcija f = 5 l tg + 5 ima bar jedu realu ulu. Ree e. Oblast defiisaosti fukcije je D f = k Z da postoji ula fukcije a 0, π 2.
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE GODINE 8. RAZRED TOČNI ODGOVORI
 21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
Heterogene ravnoteže taloženje i otapanje. u vodi u prisustvu zajedničkog iona u prisustvu kompleksirajućegreagensa pri različitim ph vrijednostima
 Heterogene ravnoteže taloženje i otapanje u vodi u prisustvu zajedničkog iona u prisustvu kompleksirajućegreagensa pri različitim ph vrijednostima Ako je BA teško topljiva sol (npr. AgCl) dodatkom
Heterogene ravnoteže taloženje i otapanje u vodi u prisustvu zajedničkog iona u prisustvu kompleksirajućegreagensa pri različitim ph vrijednostima Ako je BA teško topljiva sol (npr. AgCl) dodatkom
VJEŽBE 3 BIPOLARNI TRANZISTORI. Slika 1. Postoje npn i pnp bipolarni tranziostori i njihovi simboli su dati na slici 2 i to npn lijevo i pnp desno.
 JŽ 3 POLAN TANZSTO ipolarni tranzistor se sastoji od dva pn spoja kod kojih je jedna oblast zajednička za oba i naziva se baza, slika 1 Slika 1 ipolarni tranzistor ima 3 izvoda: emitor (), kolektor (K)
JŽ 3 POLAN TANZSTO ipolarni tranzistor se sastoji od dva pn spoja kod kojih je jedna oblast zajednička za oba i naziva se baza, slika 1 Slika 1 ipolarni tranzistor ima 3 izvoda: emitor (), kolektor (K)
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f
 D f") IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
EMISIJA ŠTETNIH SASTOJAKA U ATMOSFERU IZ PROCESA IZGARANJA IZGARANJE - IZVOR EMISIJE
 Prof. dr. sc. Z. Prelec INŽENJERSTO ZAŠTITE OKOLIŠA Poglavlje: (Emisija u atmosferu) List: 1 EMISIJA ŠTETNIH SASTOJAKA U ATMOSFERU IZ PROCESA IZGARANJA IZGARANJE - IZOR EMISIJE Izgaranje - najveći uzrok
Prof. dr. sc. Z. Prelec INŽENJERSTO ZAŠTITE OKOLIŠA Poglavlje: (Emisija u atmosferu) List: 1 EMISIJA ŠTETNIH SASTOJAKA U ATMOSFERU IZ PROCESA IZGARANJA IZGARANJE - IZOR EMISIJE Izgaranje - najveći uzrok