ΠΑΝΕΠ. ΕΤΟΣ Αριθµ. 1767
|
|
|
- Ἡρὼ Παχής
- 6 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΦΥΣΙΟΛΟΓΙΑΣ-ΦΑΡΜΑΚΟΛΟΓΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΠΕΙΡΑΜΑΤΙΚΗΣ ΦΥΣΙΟΛΟΓΙΑΣ ΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΡΙΑ ΟΛΥΜΠΙΑ ΓΚΙΜΠΑ-ΤΖΙΑΜΠΙΡΗ ΠΑΝΕΠ. ΕΤΟΣ Αριθµ Η ΕΠΙ ΡΑΣΗ ΤΗΣ ΡΙΒΑΣΤΙΓΜΙΝΗΣ ΣΕ ΑΝΩΤΕΡΕΣ ΦΛΟΙΙΚΕΣ ΛΕΙΤΟΥΡΓΙΕΣ (ΣΥΜΠΕΡΙΦΟΡΑ) ΕΠΙΜΥΩΝ, ΚΑΤΟΠΙΝ ΚΑΤΑΣΤΡΟΦΗΣ ΤΟΥ ΒΑΣΙΚΟΥ ΠΥΡΗΝΑ ΤΟΥ MEYNERT Ι ΑΚΤΟΡΙΚΗ ΙΑΤΡΙΒΗ ΓΕΩΡΓΙΟΥ Α. ΜΕΛΛΙΟΥ ΙΑΤΡΟΥ-ΦΑΡΜΑΚΟΠΟΙΟΥ ΥΠΟΒΛΗΘΗΚΕ ΣΤΗΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΥ ΑΡΙΣΤΟΤΕΛΕΙΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΘΕΣΣΑΛΟΝΙΚΗΣ ΘΕΣΣΑΛΟΝΙΚΗ 2006
2 1
3 2
4 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΦΥΣΙΟΛΟΓΙΑΣ-ΦΑΡΜΑΚΟΛΟΓΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΠΕΙΡΑΜΑΤΙΚΗΣ ΦΥΣΙΟΛΟΓΙΑΣ ΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΡΙΑ ΟΛΥΜΠΙΑ ΓΚΙΜΠΑ-ΤΖΙΑΜΠΙΡΗ ΠΑΝΕΠ. ΕΤΟΣ Αριθµ Η ΕΠΙ ΡΑΣΗ ΤΗΣ ΡΙΒΑΣΤΙΓΜΙΝΗΣ ΣΕ ΑΝΩΤΕΡΕΣ ΦΛΟΙΙΚΕΣ ΛΕΙΤΟΥΡΓΙΕΣ (ΣΥΜΠΕΡΙΦΟΡΑ) ΕΠΙΜΥΩΝ, ΚΑΤΟΠΙΝ ΚΑΤΑΣΤΡΟΦΗΣ ΤΟΥ ΒΑΣΙΚΟΥ ΠΥΡΗΝΑ ΤΟΥ MEYNERT Ι ΑΚΤΟΡΙΚΗ ΙΑΤΡΙΒΗ ΓΕΩΡΓΙΟΥ Α. ΜΕΛΛΙΟΥ ΙΑΤΡΟΥ-ΦΑΡΜΑΚΟΠΟΙΟΥ ΥΠΟΒΛΗΘΗΚΕ ΣΤΗΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΥ ΑΡΙΣΤΟΤΕΛΕΙΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΘΕΣΣΑΛΟΝΙΚΗΣ ΘΕΣΣΑΛΟΝΙΚΗ
5 Η ΤΡΙΜΕΛΗΣ ΕΠΙΤΡΟΠΗ ΟΛΥΜΠΙΑ ΓΚΙΜΠΑ-ΤΖΙΑΜΠΙΡΗ, ΚΑΘΗΓΗΤΡΙΑ ΚΩΝΣΤΑΝΤΙΝΟΣ ΚΑΛΛΑΡΑΣ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΗΜΗΤΡΙΟΣ ΚΟΥΤΣΟΝΙΚΟΛΑΣ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ Η ΕΠΤΑΜΕΛΗΣ ΕΠΙΤΡΟΠΗ ΟΛΥΜΠΙΑ ΓΚΙΜΠΑ-ΤΖΙΑΜΠΙΡΗ, ΚΑΘΗΓΗΤΡΙΑ ΚΩΝΣΤΑΝΤΙΝΟΣ ΚΑΛΛΑΡΑΣ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΗΜΗΤΡΙΟΣ ΚΟΥΤΣΟΝΙΚΟΛΑΣ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΒΑΣΙΛΙΚΗ ΜΗΡΤΣΟΥ-ΦΙ ΑΝΗ, ΚΑΘΗΓΗΤΡΙΑ ΒΑΣΙΛΕΙΟΣ ΚΟΚΚΑΣ, ΚΑΘΗΓΗΤΗΣ ΛΟΥΙΖΑ ΑΝ ΡΙΟΠΟΥΛΟΥ-ΟΙΚΟΝΟΜΟΥ, ΚΑΘΗΓΗΤΡΙΑ ΜΑΡΙΑ ΑΛΜΠΑΝΗ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΡΙΑ Η έγκριση της ιδακτορικής ιατριβής από την Ιατρική Σχολή του Αριστοτελείου Πανεπιστηµίου Θεσσαλονίκης, δεν υποδηλώνει αποδοχή των γνωµών του συγγραφέα. (Νόµος 5343/32, άρθρο και νόµος 1268/82, άρθρο 50 8) 4
6 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΡΟΕ ΡΟΣ ΤΜΗΜΑΤΟΣ ΙΩΑΝΝΗΣ ΜΠΟΝΤΗΣ 5
7 6
8 Στην οικογένειά µου Στους δασκάλους µου 7
9 8
10 ΠΕΡΙΕΧΟΜΕΝΑ Σελ. ΠΡΟΛΟΓΟΣ 13 ΓΕΝΙΚΟ ΜΕΡΟΣ Α. ΕΓΚΕΦΑΛΟΣ ΚΑΙ ΣΥΜΠΕΡΙΦΟΡΑ Νευρική βάση της µνήµης - Γνωστικές λειτουργίες και µνήµη Μνήµη και µάθηση Μετατετανική διευκόλυνση και παρατεινόµενη ενίσχυση Ο ρόλος της συναπτικής διευκόλυνσης των συνάψεων «Εθισµός» και «ευαισθητοποίηση» της συναπτικής διαβίβασης Ταξινόµηση της µνήµης Άµεση µνήµη Βραχυπρόθεσµη µνήµη Μοριακός µηχανισµός της βραχυπρόθεσµης µνήµης Μακροπρόθεσµη µνήµη οµικές µεταβολές των συνάψεων Μεταβολή του αριθµού των νευρώνων και των 31 συνάψεών τους 6.4. Εµπέδωση της µνήµης Κωδικοποίηση της µνήµης κατά την εµπέδωση 33 Β. ΝΕΥΡΟ ΙΑΒΙΒΑΣΤΕΣ Σύνθεση και αποθήκευση των νευροδιαβιβαστών Αποµάκρυνση των νευροδιαβιβαστών από τη συναπτική 37 σχισµή 3. Κατηγορίες νευροδιαβιβαστών Ακετυλοχολίνη 42 Γ. ΒΑΣΙΚΟΣ ΠΥΡΗΝΑΣ ΤΟΥ ΜEYNERT Χολινεργική νεύρωση του εγκεφαλικού φλοιού Στοιχεία φυσιολογίας της συµπεριφορικής εξειδίκευσης του 55 βασικού πυρήνα του Meynert 3. Αλλαγές στη φλοιϊκή χολινεργική νεύρωση κατά το γήρας και 59 τη νόσο Alzheimer 9
11 . Η ΧΟΛΙΝΕΡΓΙΚΗ ΥΠΟΘΕΣΗ ΤΗΣ ΜΝΗΜΗΣ ΚΑΙ ΤΗΣ ΑΝΟΙΑΣ ΣΤΗ ΝΟΣΟ ALZHEIMER 1. Νευροχηµικές και ιστοπαθολογικές αλλαγές σε χολινεργικούς και µη-χολινεργικούς νευρώνες στη νόσο Alzheimer Ε. ΦΑΡΜΑΚΟΛΟΓΙΚΗ ΘΕΡΑΠΕΙΑ ΤΗΣ ΝΟΣΟΥ ALZHEIMER Η λογική της χολινεργικής θεραπείας Αναστολείς χολινεστεράσης Χαρακτηριστικά αναστολέων της ακετυλοχολινεστεράσης Η Ριβαστιγµίνη (ΕΝΑ-713) Φαρµακοδυναµική Κλινική φαρµακοκινητική Απορρόφηση Κατανοµή Μεταβολισµός Αλληλεπιδράσεις µε άλλα φάρµακα Απέκκριση Κλινική σηµασία Μελλοντικές κατευθύνσεις της έρευνας 90 ΣΤ. ΟΚΙΜΑΣΙΕΣ ΣΥΜΠΕΡΙΦΟΡΑΣ Λαβύρινθος του Morris Λαβύρινθος ανοικτού πεδίου οκιµασία παθητικής αποφυγής 95 ΕΙ ΙΚΟ ΜΕΡΟΣ I. ΥΛΙΚΟ ΚΑΙ ΜΕΘΟ ΟΙ Πειραµατόζωα Πειραµατικό πρότυπο Οµάδες πειραµατοζώων Αισθητικοκινητικές δοκιµασίες Χορήγηση ριβαστιγµίνης Πειράµατα συµπεριφοράς Λαβύρινθος του Morris
12 6.2. Λαβύρινθος ανοικτού πεδίου οκιµασία παθητικής αποφυγής Ιστολογική µελέτη Στατιστική ανάλυση 119 II. ΑΠΟΤΕΛΕΣΜΑΤΑ Αισθητικοκινητικές δοκιµασίες Μονόζυγο Αντανακλαστικό έκτασης των πρόσθιων άκρων Αντίσταση των πρόσθιων άκρων σε ραχιαία ώθηση Αντανακλαστικό κάµψης Αντανακλαστικό σύλληψης Αντανακλαστικό επαναφοράς στην ορθή θέση Επαγρύπνηση Ακουστικό αντανακλαστικό Αντανακλαστικό του κερατοειδούς Εκκριτικά σηµεία Συνολική αξιολόγηση αισθητικοκινητικών δοκιµασιών Αξιολόγηση των χολινεργικών ανεπιθύµητων ενεργειών Πειράµατα συµπεριφοράς Βάρος σώµατος Λαβύρινθος του Morris Λαβύρινθος ανοικτού πεδίου οκιµασία παθητικής αποφυγής Συγκρίσεις µεταξύ των οµάδων Λαβύρινθος του Morris Λαβύρινθος ανοικτού πεδίου Μνήµη αναφοράς Λειτουργική µνήµη Συνολικός χρόνος οκιµασία παθητικής αποφυγής Ιστολογική µελέτη
13 III. ΣΥΖΗΤΗΣΗ Αισθητικοκινητικές δοκιµασίες Χολινεργικές ανεπιθύµητες ενέργειες Βασικός πυρήνας του Meynert και συµπεριφορά Σηµασία του βασικού πυρήνα του Meynert σε σχέση µε τα 179 υπόλοιπα ανατοµικά στοιχεία που εµπλέκονται στην παθογένεια της νόσου Alzheimer 4. οκιµασίες συµπεριφοράς Λαβύρινθος του Morris Λαβύρινθος ανοικτού πεδίου οκιµασία παθητικής αποφυγής Οι συµπεριφορικές µεταβολές ως αποτέλεσµα της 186 πειραµατικής καταστροφής του βασικού πυρήνα του Meynert 6. Οι συµπεριφορικές µεταβολές ως αποτέλεσµα της διαταραχής 187 της χολινεργικής οδού 7. Ειδικότητα των συµπεριφορικών δοκιµασιών για τις γνωστικές 188 διεργασίες 8. Αξιοποίηση του χρησιµοποιηθέντος πειραµατικού προτύπου 188 στη µελέτη της συµπεριφοράς IV. ΠΕΡΙΛΗΨΗ-ΣΥΜΠΕΡΑΣΜΑΤΑ V. SUMMARY-CONCLUSIONS ΒΙΒΛΙΟΓΡΑΦΙΑ
14 ΠΡΟΛΟΓΟΣ Η κατανόηση του µυστηρίου του ανθρώπινου εγκεφάλου και των µηχανισµών της µνήµης και της µάθησης αποτέλεσε διαχρονικό στόχο της φιλοσοφίας, των βιολογικών επιστηµών και της ψυχολογίας. Τις τελευταίες δεκαετίες, όµως, η πρόοδος των βιολογικών επιστηµών και η συνεργασία ανάµεσα στους διαφόρους κλάδους της Ιατρικής, οδήγησαν στην ενοποίηση της έρευνας σε ένα νέο πλαίσιο, αυτό των νευροεπιστηµών. Βασική αρχή των νευροεπιστηµών θεωρείται η συσχέτιση της συµπεριφοράς µε τις λειτουργίες του εγκεφάλου, καθώς και η κατανόηση και ερµηνεία των διαταραχών της συµπεριφοράς µε βάση αντίστοιχες διαταραχές των λειτουργιών του εγκεφάλου. Οι µηχανισµοί της µνήµης και της µάθησης, οι γνωστικές διαταραχές που συνδέονται µε το γήρας και η παθοφυσιολογία των διαφόρων µορφών άνοιας αποτέλεσαν βασικούς στόχους της έρευνας των νευροεπιστηµόνων. Έτσι, η «χολινεργική υπόθεση», που διατυπώθηκε πριν από 20 χρόνια περίπου 1, διευκρίνισε ότι η σηµαντική απώλεια της χολινεργικής λειτουργίας στο κεντρικό νευρικό σύστηµα (ΚΝΣ) συντελεί καθοριστικά στην εµφάνιση των γνωστικών διαταραχών, οι οποίες σχετίζονται µε την προχωρηµένη ηλικία και τη νόσο Alzheimer. Πριν από τη διατύπωσή της δεν υπήρχαν θεωρητικές ερµηνείες για τα κύρια συµπτώµατα της νόσου Alzheimer και για τα διαγνωστικά της κριτήρια. Η χολινεργική υπόθεση παρείχε το θεωρητικό υπόβαθρο για την ανάπτυξη θεραπευτικών στρατηγικών για τη νόσο Alzheimer. Αυτό το ερευνητικό πλαίσιο προήλθε από τον επιτυχή συνδυασµό δεδοµένων φυσιολογίας, βιοχηµείας, φαρµακολογίας και κλινικών µελετών. Και µάλιστα, σε αντίθεση µε άλλα σύγχρονα πεδία έρευνας, η χολινεργική υπόθεση στηρίχθηκε σε πειραµατικά µοντέλα και συµπεριφορικά πρωτόκολλα, τα οποία αναπτύχθηκαν και εξελίχθηκαν κατά τη διάρκεια της πλούσιας ιστορίας της βασικής έρευνας των νευροεπιστηµών. Η ιδέα της χρησιµοποίησης πειραµατοζώων στην προσπάθεια µελέτης της παθοφυσιολογίας µίας νόσου, που επηρεάζει τη µνήµη και της εύρεσης µίας κατάλληλης θεραπευτικής στρατηγικής, θεωρήθηκε στην καλύτερη περίπτωση ότι δηµιουργεί επιστηµονική αντιπαράθεση, ενώ δεν ήταν λίγοι αυτοί που τη χαρακτήρισαν αιρετική. 13
15 Επιστήµονες που προέρχονταν από παλαιότερες σχολές σκέψης, θεώρησαν ανθρωποµορφική τη χρήση του όρου «µνήµη» για την περιγραφή της συµπεριφοράς των ζώων. Έτσι, αρχικά ήταν δύσκολο να χρησιµοποιηθούν πειραµατόζωα, για να προσοµοιωθούν διεργασίες της ανθρώπινης µνήµης, καθώς και διαταραχές που σχετίζονται µε συγκεκριµένες παθήσεις του νευρικού συστήµατος του ανθρώπου. Οι µελέτες, όµως, που πραγµατοποιήθηκαν µε βάση τη χολινεργική υπόθεση, καθώς και τα πειραµατικά µοντέλα που χρησιµοποιήθηκαν, συνέβαλαν όχι µόνο στην καθιέρωση της θεωρίας και των πρώτων θεραπευτικών στρατηγικών που βασίστηκαν σε αυτή, αλλά συνετέλεσαν και στη γενικότερη πρόοδο των νευροεπιστηµών. Εξαιτίας µάλιστα αυτού του έντονου ερευνητικού ενδιαφέροντος, δηµιουργήθηκε ενθουσιασµός στην επιστηµονική κοινότητα για την ανάπτυξη φαρµακολογικών θεραπειών για τις νευροεκφυλιστικές παθήσεις, ενώ η χρήση εκλεκτικών πειραµατικών µοντέλων σε αυτό το πεδίο της έρευνας δε θεωρείται πια λανθασµένη ή αιρετική. Η ιδέα της χρήσης µίας φαρµακολογικής ουσίας για την αντιµετώπιση των συµπτωµάτων της νόσου Alzheimer ή ακόµη και την αναστολή της εξέλιξής της, δεν είναι καινούρια. Απέκτησε, όµως, µία συγκεκριµένη έκφραση στη δεκαετία του 1990 µε την εισαγωγή στη θεραπευτική πρακτική των αναστολέων της χολινεστεράσης, φαρµάκων βασισµένων στη θεωρία της χολινεργικής υπόθεσης, που όµως δεν αποτελούν οριστική λύση στην αντιµετώπιση της νόσου. Κατά την κρατούσα άποψη για τη θέση των φαρµάκων στη σύγχρονη αντιµετώπιση της νόσου Alzheimer θεωρείται απαραίτητη η χρήση τους για την άµβλυνση των συµπτωµάτων από τα πρώτα στάδια της νόσου. Παράλληλα, ενθαρρύνονται οι ερευνητικές προσπάθειες για την πλήρη αποσαφήνιση της παθοφυσιολογίας της νόσου. Οι µελλοντικές προσεγγίσεις περιλαµβάνουν τη γεφύρωση του χάσµατος ανάµεσα στην πρόοδο της νευροεπιστήµης και στο σχεδιασµό και την ανάπτυξη νέων φαρµάκων, κάτι που θα οδηγήσει στην ταχύτερη εισαγωγή στη θεραπευτική νέων φαρµακολογικών ουσιών. Η επικέντρωση της έρευνας σε κάποιο παθοφυσιολογικό µηχανισµό και συνεπακόλουθα ο σχεδιασµός, η ανάπτυξη και η εισαγωγή στη θεραπευτική ενός φαρµάκου θεωρούνται στο σύνολό τους αποτέλεσµα της σύζευξης της θεωρητικής νευροεπιστήµης µε την εφαρµοσµένη έρευνα, που χρησιµοποιεί σε µεγάλο βαθµό πειραµατικά µοντέλα προσοµοίωσης της νόσου για τη 14
16 δοκιµή της αποτελεσµατικότητας των φαρµακολογικά δραστικών ουσιών, πριν καταλήξει στις κλινικές µελέτες τους. Ο σκοπός της παρούσας µελέτης ήταν ο σχεδιασµός και η ανάπτυξη ενός πειραµατικού µοντέλου, το οποίο θα µπορούσε να χρησιµοποιηθεί για την προσοµοίωση γνωστικών διαταραχών, που σχετίζονται µε παθήσεις του κεντρικού νευρικού συστήµατος, καθώς και για την αξιολόγηση φαρµακολογικά δραστικών ουσιών, που επιδρούν στη µνήµη και τη µάθηση. Σηµαντικό ρόλο στο σχεδιασµό του πειραµατικού πρωτοκόλλου έπαιξε η υπάρχουσα εµπειρία από τη διεθνή βιβλιογραφία, ώστε να επιβεβαιωθεί ο σηµαίνων ρόλος του βασικού πυρήνα του Meynert στις γνωστικές λειτουργίες µε βάση τη χολινεργική υπόθεση της µνήµης, αλλά και την εµπλοκή αυτού του πυρήνα στην παθοφυσιολογία της νόσου Alzheimer. Είναι όµως περιορισµένος ο αριθµός των συµπεριφορικών µελετών, που χρησιµοποιούν αλληλουχία συµπεριφορικών δοκιµασιώνπειραµάτων για τη µελέτη της µνήµης και της µάθησης, καθώς και παράλληλο έλεγχο της απόκτησης και ανάκλησης της µνήµης πριν και µετά την καταστροφή του βασικού πυρήνα του Meynert 2. Βασική, εξάλλου, φαρµακολογική επιλογή για τη νόσο Alzheimer αποτελούν οι αναστολείς της χολινεστεράσης, η θεραπευτική χρήση των οποίων βασίζεται στη θεωρία της χολινεργικής υπόθεσης. Η ριβαστιγµίνη (ENA713), αποτελεί έναν από τους τελευταίους αναστολείς που χρησιµοποιούνται στη θεραπευτική, έχει εκλεκτική δράση και συµβάλλει στην άµβλυνση των συµπτωµάτων από τα πρώτα στάδια της νόσου. Στη διεθνή βιβλιογραφία υπάρχουν αναφορές για µελέτη της ριβαστιγµίνης σε συµπεριφορικά πρωτόκολλα, στα οποία προκαλείται ελάττωση της χολινεργικής δραστηριότητας µε τη χρήση σκοπολαµίνης 3,4. Επειδή, όµως, δεν υπήρχαν συµπεριφορικές µελέτες µε χορήγηση της ριβαστιγµίνης σε πειραµατόζωα µετά από καταστροφή του βασικού πυρήνα του Meynert, θεωρήθηκε σηµαντική η µελέτη της επίδρασης της ριβαστιγµίνης σε ανώτερες φλοιικές λειτουργίες (συµπεριφορά) επίµυων, µετά από καταστροφή του βασικού πυρήνα του Meynert µε τη χρήση ιµποτενικού οξέος (ibotenic acid), το οποίο αποτελεί νευροτοξίνη που καταστρέφει τους νευρώνες και όχι διερχόµενες νευρικές ίνες ή αγγεία της περιοχής του πυρήνα. Στο σηµείο αυτό θα ήθελα να αναφερθώ σε όλους όσους συνέβαλαν µε άµεσο ή έµµεσο τρόπο στο σχεδιασµό και την ολοκλήρωση της έρευνας, καθώς και στη συγγραφή της διδακτορικής διατριβής. 15
17 Στην Καθηγήτρια Φυσιολογίας της Ιατρικής Σχολής του Α.Π.Θ. κ. Ολυµπία Γκίµπα-Τζιαµπίρη, επιβλέπουσα καθηγήτρια της τριµελούς συµβουλευτικής επιτροπής, θα ήθελα να εκφράσω τις θερµές µου ευχαριστίες και την ειλικρινή µου ευγνωµοσύνη για την καθοριστική συµβολή της στην εκπόνηση της διατριβής. Οι πολύτιµες συµβουλές της και η στήριξή της διαχρονικά κατά τη διάρκεια των σπουδών µου, συνέβαλαν ουσιαστικά στην ολοκλήρωσή τους, καθώς και σε πολλές από τις επιλογές µου όσον αφορά την ενασχόλησή µου µε το γνωστικό πεδίο των επιστηµών υγείας. Τον αναπληρωτή Καθηγητή Φυσιολογίας της Ιατρικής Σχολής του Α.Π.Θ. κ. Κωνσταντίνο Καλλαρά, µέλος της τριµελούς συµβουλευτικής επιτροπής, ευχαριστώ, επίσης, για τη διαρκή καθοδήγησή του και την ουσιαστική συµβολή του στην ολοκλήρωση της διδακτορικής διατριβής. Τον αναπληρωτή Καθηγητή Φυσιολογίας της Ιατρικής Σχολής του Α.Π.Θ. κ. ηµήτριο Κουτσονικόλα, µέλος της τριµελούς συµβουλευτικής επιτροπής, ευχαριστώ για τη συµβολή του στην εκπόνηση της διδακτορικής διατριβής. Στη φίλη και συνάδελφο Νατάσα Παζαΐτη-Καραµανλή θα ήθελα να εκφράσω τις θερµές µου ευχαριστίες για τη συµβολή της στην ενασχόλησή µου µε θέµατα µνήµης, µάθησης και συµπεριφορικής έρευνας. Ευχαριστώ, επίσης, τη φίλη και συνάδελφο ωροθέα Καπουκρανίδου, Λέκτορα Φυσιολογίας της Ιατρικής Σχολής του Α.Π.Θ., για τη συµπαράστασή της κατά τη λήψη και επεξεργασία των ιστολογικών δειγµάτων. Κατά τη διάρκεια των ετών της εκπόνησης µίας διδακτορικής διατριβής καθοριστικό ρόλο παίζει η «παρέα του εργαστηρίου», που συµπαρίσταται στις ώρες των πειραµάτων, εµψυχώνει σε περιόδους απαισιοδοξίας και πιεστικών ωραρίων και συµµετέχει στη χαρά της ολοκλήρωσης της έρευνας. Έτσι, θα ήθελα να ευχαριστήσω τη ήµητρα Φαρδή, το Γιώργο Παπαζήση και τον Κώστα Λίλη, που αποτέλεσαν για εµένα τους φίλους, συναδέλφους και συνεργάτες κατά την περίοδο εκπόνησης της διδακτορικής διατριβής στο Εργαστήριο Φυσιολογίας της Ιατρικής Σχολής του Α.Π.Θ. Θα ήθελα, επίσης, να εκφράσω τις ευχαριστίες µου στα µέλη ΕΠ και το προσωπικό του Εργαστηρίου Φυσιολογίας της Ιατρικής Σχολής του Α.Π.Θ. για τη διακριτική συµπαράστασή τους. 16
18 Τέλος, θα ήθελα να ευχαριστήσω τους γονείς µου Αναστάσιο και Παρασκευή, την αδελφή µου Ελένη και τη θεία µου Μαρία για την αµέριστη συµπαράστασή τους κατά τη διάρκεια των πολυετών σπουδών µου. Συµπαράσταση ηθική και υλική, χωρίς όρια και ανταλλάγµατα, ακόµη και σε περιόδους όπου λόγω αυξηµένων υποχρεώσεων δεν είχα το χρόνο να ανταποδώσω το ενδιαφέρον και την αγάπη τους. 17
19 18
20 ΓΕΝΙΚΟ ΜΕΡΟΣ 19
21 20
22 Α. ΕΓΚΕΦΑΛΟΣ ΚΑΙ ΣΥΜΠΕΡΙΦΟΡΑ Στις µεγαλύτερες διαχρονικές προκλήσεις των επιστηµών ανήκει η κατανόηση της βιολογικής βάσης της συµπεριφοράς και των νοητικών διεργασιών µε τις οποίες αντιλαµβανόµαστε, ενεργούµε, µαθαίνουµε και θυµόµαστε. Η διατύπωση µίας γενικώς αποδεκτής θεωρίας για τις γνωστικές διεργασίες, καθώς και η τοπογραφική εντόπισή τους στον ανθρώπινο εγκέφαλο αποτέλεσαν το ερευνητικό κίνητρο για σηµαντικούς επιστήµονες της ιστορίας, ξεκινώντας από την κλασική ελληνική φιλοσοφία. Πριν από την εφεύρεση του σύνθετου µικροσκοπίου τον 18 ο αιώνα, ο νευρικός ιστός χαρακτηριζόταν αδενικός ως προς τη λειτουργία, άποψη που µπορεί να ανιχνευθεί στην αρχαιότητα και στην πεποίθηση του Γαληνού ότι τα νεύρα είναι αγωγοί µεταφοράς υγρού, το οποίο εκκρίνεται από τον εγκέφαλο και το νωτιαίο µυελό προς την περιφέρεια του σώµατος. Η κρατούσα αντίληψη για τα νευρικά κύτταρα, τον εγκέφαλο και τη συµπεριφορά προέκυψε το 19 ο αιώνα από πέντε κύριους πειραµατικούς κλάδους: την ανατοµία, την εµβρυολογία, τη φυσιολογία, τη φαρµακολογία και την ψυχολογία. Φηµισµένοι ερευνητές όπως ο Ιταλός ιατρός Golgi, ο Ισπανός ιστολόγος Cajal, ο Αµερικανός εµβρυολόγος Harrison, ο Ιταλός ιατρός και φυσικός Galvani, ο Γερµανός ιατρός και νευροανατόµος Gall, ο Bernard στη Γαλλία, ο Ehrlich στη Γερµανία και ο Langley στην Αγγλία, οι Γερµανοί φυσιολόγοι Dubois-Reymond, Muller και von Helmholtz, ο Descartes, ο Locke, ο Hume, ο Darwin, ο Γερµανός νευρολόγος Wernicke, ο Pavlov, o Penfield και ο Broca είναι ορισµένοι από τους επιστήµονες, που ασχολήθηκαν µε τον εγκέφαλο και τη συµπεριφορά. Τις τελευταίες δεκαετίες σηµειώθηκε µία αξιόλογη συνεργασία µεταξύ των διαφόρων βιολογικών επιστηµών, η οποία είχε ως αποτέλεσµα την ένωση της νευροεπιστήµης µε την κυτταρική και µοριακή βιολογία. Το επόµενο, και ακόµη πιο σηµαντικό, βήµα στην προοδευτική αυτή ενοποίηση ήταν η ένωση της µελέτης της συµπεριφοράς (της επιστήµης της νόησης και της ψυχολογίας) µε τη νευροεπιστήµη (την επιστήµη του εγκεφάλου). Η κεντρική αρχή αυτής της ενοποίησης είναι ότι αυτό που συνήθως ονοµάζουµε συµπεριφορά είναι µία σειρά λειτουργιών, οι οποίες εκτελούνται από τον εγκέφαλο. Οι ενέργειες του εγκεφάλου αποτελούν τη 21
23 βάση του συνόλου της συµπεριφοράς, όχι µόνο των σχετικά απλών κινητικών συµπεριφορών και ζωτικών λειτουργιών, όπως είναι η βάδιση και η λήψη τροφής, αλλά και όλων των σύνθετων γνωστικών διεργασιών, τις οποίες συνδέουµε κατεξοχήν µε την ανθρώπινη συµπεριφορά, όπως είναι η σκέψη, ο λόγος και η δηµιουργία έργων τέχνης. Με βάση τη συγκεκριµένη αντίληψη οι ανωµαλίες της συµπεριφοράς, οι οποίες χαρακτηρίζουν τις ψυχικές ασθένειες, είναι διαταραχές της λειτουργίας του εγκεφάλου. Σκοπός της νευροεπιστήµης είναι να ερµηνεύσει τη συµπεριφορά µε βάση τις λειτουργίες του εγκεφάλου, να εξηγήσει πώς εκατοµµύρια ξεχωριστά νευρικά κύτταρα στον εγκέφαλο λειτουργούν για την «παραγωγή» συµπεριφοράς και πώς αντιστρόφως τα κύτταρα αυτά επηρεάζονται από το περιβάλλον, στο οποίο περιλαµβάνεται και η συµπεριφορά των άλλων ανθρώπων 5,6. 1. Νευρική βάση της µνήµης - Γνωστικές λειτουργίες και µνήµη Κατά την ανάλυση των γνωστικών λειτουργιών και της µνήµης δεν είναι απολύτως γνωστά τα νευρωνικά κυκλώµατα που ενεργοποιούνται για να γίνει µία σκέψη, καθώς και οι διεργασίες απόκτησης και ανάκλησης της µνήµης. Είναι γνωστό ότι η καταστροφή µεγάλων περιοχών του φλοιού δεν εµποδίζει το άτοµο να σκέφτεται, µειώνει όµως την ικανότητά του να αντιλαµβάνεται το περιβάλλον. Για να γίνει µία σκέψη απαιτείται σχεδόν ταυτόχρονη ενεργοποίηση πολλών περιοχών του εγκεφαλικού φλοιού, του οπτικού θαλάµου, του µεταιχµιακού συστήµατος και του δικτυωτού σχηµατισµού του εγκεφαλικού στελέχους. Ορισµένες αδρές µορφές σκέψης-αντίληψης πιθανώς εξαρτώνται σχεδόν αποκλειστικά από κατώτερα κέντρα. Ένα τέτοιο παράδειγµα πιθανόν να αποτελεί η αντίληψη του πόνου, δεδοµένου ότι ηλεκτρικός ερεθισµός του φλοιού στον άνθρωπο σπανιότατα προκαλεί οτιδήποτε άλλο εκτός από ελαφρό πόνο, ενώ ο ερεθισµός ορισµένων περιοχών του υποθαλάµου και του µεσεγκεφάλου συνήθως προκαλεί ανυπόφορο πόνο. Αντίθετα, µία µορφή σκέψης-αντίληψης, που απαιτεί κυρίως το φλοιό, είναι η όραση δεδοµένου ότι η απώλεια του οπτικού φλοιού προκαλεί πλήρη αδυναµία διάκρισης του σχήµατος και των χρωµάτων των αντικειµένων. Οι γνωστικές λειτουργίες, µε βάση το νευρωνικό τους µηχανισµό, είναι αποτέλεσµα της συγχρονισµένης ενεργοποίησης πολλών τµηµάτων του 22
24 κεντρικού νευρικού συστήµατος. Η ενεργοποίηση αυτή γίνεται µε συγκεκριµένη αλληλουχία, που περιλαµβάνει τον εγκεφαλικό φλοιό, τον οπτικό θάλαµο, το µεταιχµιακό σύστηµα και το ανώτερο τµήµα του δικτυωτού σχηµατισµού του εγκεφαλικού στελέχους. Ο ορισµός αυτός αποτελεί την ειδική ολιστική θεωρία της εγκεφαλικής λειτουργίας. Η παλαιότερη γενικευµένη ολιστική θεωρία της φλοιικής ειδικά λειτουργίας έχει από καιρό εγκαταληφθεί. Η θεωρία αυτή υποστήριζε ότι όταν ένα οποιοδήποτε τµήµα του φλοιού καταστρεφόταν, µπορούσε αυτόµατα να αντικατασταθεί από κάποιο παρακείµενο τµήµα. Οι περιοχές του µεταιχµιακού συστήµατος, του οπτικού θαλάµου, και του δικτυωτού σχηµατισµού που ενεργοποιούνται, πιστεύεται ότι προσδιορίζουν τις γενικές ιδιότητες της σκέψης, όπως ευχαρίστηση ή δυσαρέσκεια, πόνο ή αίσθηµα άνεσης, καθώς και αδρά αισθητικά χαρακτηριστικά, όπως η εντόπιση στο χώρο. Επίσης, καθορίζουν τις συγκεκριµένες ιδιότητες της σκέψης, κυρίως αισθητικού τύπου, όπως η τοπογραφική εντόπιση αισθήµατος στο σώµα, ή αντικειµένου στο οπτικό πεδίο. Η ενεργοποίηση του φλοιού προσφέρει στη σκέψη άλλες µορφές αισθητικού προσδιορισµού του περιβάλλοντος, όπως είναι η αντίληψη του τετράγωνου σχήµατος ενός τοίχου, του υλικού και ύφανσης ενός χαλιού και άλλες συγκεκριµένες ιδιότητες και πληροφορίες για το περιβάλλον, οι οποίες συµµετέχουν στη σφαιρική αντίληψη του περιβάλλοντος σε κάθε δεδοµένη στιγµή 5. Σύµφωνα µε όλα τα παραπάνω, η µνήµη, ως επαναφορά µίας προηγούµενης σκέψης, πρέπει να θεωρείται ότι αντιστοιχεί σε µία συγχρονισµένη λειτουργία πολλών περιοχών του εγκεφάλου. Υπάρχουν βέβαια πειράµατα κατά τα οποία ο ηλεκτρικός ερεθισµός µίας µικρής περιοχής, π.χ. στον κροταφικό φλοιό, προκαλεί αναπόληση ολόκληρων περιστατικών ή σκέψεων. Αυτό όµως δε σηµαίνει ότι η προκείµενη µνήµη ήταν εναποθηκευµένη στη θέση αυτή, αλλά απλώς ότι η ηλεκτρική διέγερση στη συγκεκριµένη περιοχή ενεργοποίησε το κύκλωµα εγγραµµάτων της συγκεκριµένης µνήµης, το οποίο ήταν βέβαια κατανεµηµένο σε διάφορα άλλα σηµεία του εγκεφάλου 7. 23
25 2. Μνήµη και µάθηση Η ανάκληση στη συνειδητότητα µίας πληροφορίας ή ενός γεγονότος, που έγινε στο παρελθόν, ονοµάζεται µνήµη. Η εξέλιξη της λειτουργίας της µνήµης γίνεται σε τρία στάδια που αφορούν την πρόσληψη και επεξεργασία των πληροφοριών από το νευρικό σύστηµα, την αποθήκευσή τους και τελικά την ανάκλησή τους στη συνειδητότητα. Η µάθηση, αντιστοίχως, δεν αποτελεί µία ενιαία διεργασία και εµφανίζεται µε δύο µορφές. Τη δηλωτική µάθηση, στην οποία απαιτείται συνειδητή συµµετοχή και τη µη δηλωτική µάθηση, στην οποία δεν απαιτείται συνειδητή συµµετοχή. Η µάθηση συνοδεύεται από µεταβολές της αποτελεσµατικότητας των νευρωνικών συνδέσεων Μετατετανική διευκόλυνση και παρατεινόµενη ενίσχυση Η µετατετανική διευκόλυνση (facilitation) που εµφανίζεται κυρίως σε συνάψεις του ΚΝΣ, αποτελεί µία πρωτογενή µορφή µάθησης και οφείλεται σε αύξηση του επιπέδου των ιόντων ασβεστίου στις τελικές απολήξεις του προσυναπτικού νευρώνα, όταν αυτός εµφανίζει αλλεπάλληλες σειρές δυναµικών δράσεως (ώσεων). Η αύξηση αυτή οδηγεί σε αυξηµένη απελευθέρωση νευροδιαβιβαστικής ουσίας, δηλαδή σε µία έµµεση πτώση της βαλβίδας της σύναψης. Το φαινόµενο αυτό µπορεί να διαρκέσει επί µακρό (µερικές ώρες), µετά την αρχική του ενεργοποίηση. Ανάλογη παροδική αύξηση του ενδοκυττάριου ασβεστίου µπορεί να παρατηρηθεί και στο µετασυναπτικό νευρώνα. Στην περίπτωση αυτή αυξάνεται το ύψος του ερεθιστικού µετασυναπτικού δυναµικού στη σύναψη. Το φαινόµενο αυτό χαρακτηρίζεται παρατεινόµενη (µετασυναπτική) διευκόλυνση ή ενίσχυση (Long term potentiation, LTP), εµφανίζεται µετά την άφιξη στο µετασυναπτικό νευρώνα ειδικών συνδυασµών προσυναπτικών ώσεων και έχει συχνά µεγάλη διάρκεια, δηλαδή µπορεί να διατηρηθεί αρκετά 24ωρα µετά την αρχική του εκδήλωση. Λειτουργικά, η συναπτική ενίσχυση οποιουδήποτε τύπου διευκολύνει την εκτέλεση µίας κεντρικής νευρικής διαδικασίας, ως αποτέλεσµα εξάσκησης, δηλαδή µπορεί να θεωρηθεί ως µία µαθησιακή διαδικασία 7. 24
26 4. Ο ρόλος της συναπτικής διευκόλυνσης των συνάψεων Η µνήµη είναι αποτέλεσµα µεταβολής της συναπτικής µετάδοσης από ένα νευρικό κύτταρο στο επόµενο, η οποία είναι αποτέλεσµα προηγούµενης νευρωνικής δραστηριότητας. Οι µεταβολές αυτές, στη συνέχεια προκαλούν την ανάπτυξη νέων οδών, είτε συναπτική διευκόλυνση υφισταµένων οδών διαβίβασης των ώσεων από τα νευρωνικά κυκλώµατα του εγκεφάλου. Οι καινούριες οδοί διαβίβασης ή οι οδοί που υπέστησαν διευκόλυνση, ονοµάζονται µνηµονικά εγγράµµατα. Η απλούστερη εξήγηση της δηµιουργίας εγγραµµάτων είναι η νευροηλεκτρική, δηλαδή η δηµιουργία ενός συνεχώς λειτουργούντος ταλαντούµενου κυκλώµατος νευρώνων, το οποίο εφόσον εξακολουθεί να ευρίσκεται σε ενέργεια, διατηρεί και τη µνήµη. Τα µνηµονικά αυτά εγγράµµατα έχουν πολύ µεγάλη σηµασία, διότι από τη στιγµή που θα εγκατασταθούν, µπορούν να ενεργοποιούνται εκ νέου από το σκεπτόµενο εγκέφαλο και να αναπαράγουν τις µνήµες 6,7. Πειράµατα σε κατώτερα ζώα απέδειξαν ότι µνηµονικά εγγράµµατα είναι δυνατό να εγκαθίστανται σε όλα τα επίπεδα του νευρικού συστήµατος. Ακόµη και νωτιαία αντανακλαστικά µπορούν να τροποποιούνται, έστω και λίγο, µετά από επανειληµµένη ενεργοποίηση του νωτιαίου µυελού, πράγµα που αποτελεί µέρος διεργασίας µνήµης. Επίσης, ακόµη και ορισµένες µακροπρόθεσµες µνήµες είναι δυνατό να οφείλονται σε µεταβολή της συναπτικής διαβίβασης σε κατώτερα κέντρα του εγκεφάλου 5. Υπάρχουν επαρκείς ενδείξεις ότι ο µεγαλύτερος όγκος µνήµης, που συσχετίζεται µε τις νοητικές διεργασίες, είναι αποτέλεσµα µνηµονικών εγγραµµάτων κυρίως στον εγκεφαλικό φλοιό «Εθισµός» και «ευαισθητοποίηση» της συναπτικής διαβίβασης Αν και συνήθως αναφερόµαστε στη µνήµη, ως µία θετική ανάκληση προηγουµένων σκέψεων και εµπειριών, πιθανόν το µεγαλύτερο ποσοστό των µνηµονικών εγγραµµάτων είναι αρνητικές µνήµες και όχι θετικές. ηλαδή, ο εγκέφαλος κατακλύζεται µε αισθητικές πληροφορίες από όλες τις αισθήσεις. Αν ο εγκέφαλος δοκίµαζε να αποθηκεύσει όλες τις αισθητικές πληροφορίες, η ικανότητά του για αποµνηµόνευση θα εξαντλούνταν σε λίγα µόνο λεπτά. Ευτυχώς, όµως, ο εγκέφαλος έχει την ιδιαίτερη ικανότητα να 25
27 µαθαίνει να αγνοεί τις πληροφορίες, που είναι ασήµαντες. Το φαινόµενο αυτό προκαλείται µε αναστολή της συναπτικής διαβίβασης γι αυτόν τον τύπο πληροφοριών, και το αποτέλεσµα ονοµάζεται «εθισµός» (habituation). Το φαινόµενο αυτό κατά κάποιο τρόπο αποτελεί µορφή αρνητικής µνήµης. Αντίθετα, για εκείνα τα είδη των πληροφοριών που είναι σηµαντικές, όπως είναι ο πόνος ή ένα ευχάριστο αίσθηµα, ο εγκέφαλος έχει επίσης την ικανότητα να ενισχύει αυτόµατα και να αποθηκεύει τα µνηµονικά εγγράµµατα. Είναι εµφανές ότι το φαινόµενο αυτό αποτελεί µορφή θετικής µνήµης. Η µορφή αυτή της µνήµης είναι αποτέλεσµα διευκόλυνσης (facilitation) της συναπτικής διαβίβασης, και η διεργασία αυτή ονοµάζεται «ευαισθητοποίηση» (sensitization) της µνήµης. Έχει, επίσης, αποδειχθεί ότι συγκεκριµένες περιοχές, που ανήκουν στο µεταιχµιακό σύστηµα, προσδιορίζουν αν µία πληροφορία είναι σηµαντική ή ασήµαντη και αποφασίζουν υποσυνείδητα αν θα αποθηκευθεί µε τη µορφή ενισχυµένου µνηµονικού εγγράµµατος ή θα κατασταλεί 6. Κατά την τελευταία δεκαετία έχουν γίνει εκτενείς µελέτες των φαινοµένων αυτών της µάθησης και της µνήµης σε ασπόνδυλα µαλάκια (aplysia) και έχουν επισηµανθεί αρκετές λεπτοµέρειες σχετικές µε τους µηχανισµούς χάρη στους οποίους ορισµένες συνάψεις ή υφίστανται εθισµό (ξεχνούν) ή αντίθετα ευαισθητοποιούνται (θυµούνται) έναντι κάποιων εξωτερικών ερεθισµάτων και αυτό ανάλογα µε την ένταση του αρχικού ερεθίσµατος, αλλά και την ενεργοποιητική παρεµβολή διάµεσων νευρώνων. Τα ευρήµατα αυτά σχετίζονται µε διακυµάνσεις στην ενδοκυττάρια στάθµη των ιόντων ασβεστίου στον επηρεαζόµενο νευρώνα και εποµένως και στην ποσότητα της νευροδιαβιβαστικής ουσίας, η οποία εκκρίνεται όταν αυτός διεγερθεί 7. 26
28 6. Ταξινόµηση της µνήµης Είναι σε όλους γνωστό ότι µερικές πληροφορίες διατηρούνται στη µνήµη για µερικά δευτερόλεπτα και άλλες διατηρούνται για ώρες, µέρες, µήνες ή και για χρόνια. Η µνήµη διαθέτει διάφορα αλληλοεξαρτώµενα στάδια ή τύπους µνηµονικών διεργασιών. Σύµφωνα µε τις νεότερες απόψεις τα στάδια της µνήµης είναι τα ακόλουθα: (1) άµεση µνήµη (immediate memory), η οποία περιλαµβάνει τη διατήρηση πληροφοριών στη µνήµη για µερικά δευτερόλεπτα ή το πολύ λεπτά, εκτός εάν µετατραπούν σε βραχυπρόθεσµη µνήµη, (2) βραχυπρόθεσµη µνήµη (short-term memory), η οποία διαρκεί µερικές µέρες ή εβδοµάδες, αλλά τελικά χάνεται, και (3) µακροπρόθεσµη µνήµη (long-term memory), η οποία από τη στιγµή που θα αποθηκευθεί, µπορεί να ανακαλείται επί χρόνια ή και για ολόκληρη τη ζωή του ατόµου 6, Άµεση µνήµη Η άµεση µνήµη είναι βραχείας διάρκειας. Οι πληροφορίες, που αποθηκεύονται σε αυτή, διατηρούνται µόνο για µερικά δευτερόλεπτα µέχρι µερικά λεπτά, και κατόπιν χάνονται (λήθη), ενώ η χωρητικότητά της είναι σχετικά µικρή. Πολλοί φυσιολόγοι έχουν διατυπώσει την άποψη ότι η άµεση µνήµη οφείλεται σε συνεχή νευρωνική δραστηριότητα, που συνίσταται σε νευρικές ώσεις οι οποίες κινούνται κυκλικά σε ανατροφοδοτικά κυκλώµατα, τα οποία προσωρινά µετατρέπονται σε µνηµονικά εγγράµµατα. Μία άλλη πιθανή ερµηνεία της άµεσης µνήµης δίδεται µε το µηχανισµό της προσυναπτικής διευκόλυνσης ή αναστολής. Το φαινόµενο αυτό παρατηρείται σε συνάψεις που σχηµατίζονται όχι µε το επόµενο κύτταρο, αλλά µε τις προσυναπτικές απολήξεις. Οι νευροδιαβιβαστικές ουσίες, που απελευθερώνονται σε αυτές τις συνάψεις, προκαλούν συνήθως παρατεταµένη διευκόλυνση ή αναστολή των απολήξεων (η µορφή της επίδρασης εξαρτάται από το είδος της νευροδιαβιβαστικής ουσίας), η οποία διαρκεί µερικά δευτερόλεπτα ή και µερικά λεπτά. Είναι φανερό ότι η µορφή αυτή της νευρωνικής δραστηριότητας θα µπορούσε να εξηγήσει την άµεση µνήµη. Τέλος, µία άλλη πιθανή ερµηνεία της άµεσης µνήµης αποτελεί ο µηχανισµός της συναπτικής ενίσχυσης, µε τον οποίο ενισχύεται η αγωγή 27
29 στη σύναψη. Το φαινόµενο αυτό µπορεί να εµφανίζεται µετά από άθροιση µεγάλου αριθµού ιόντων ασβεστίου στις προσυναπτικές απολήξεις. Όταν δηλαδή µία αλληλουχία νευρικών ώσεων διέρχεται από το προσυναπτικό κοµβίο, το ποσό των ιόντων ασβεστίου της απόληξης αυξάνεται µε την κάθε ώση. Όταν το ποσό του ασβεστίου γίνει µεγαλύτερο από εκείνο που µπορούν να δεσµεύσουν τα µιτοχόνδρια και το ενδοπλασµατικό δίκτυο, το επιπλέον ασβέστιο παρατείνει την απελευθέρωση της νευροδιαβιβαστικής ουσίας στη σύναψη. Γι αυτό και ο συγκεκριµένος µηχανισµός θα µπορούσε να χρησιµοποιείται για την άµεση µνήµη Βραχυπρόθεσµη µνήµη Η βραχυπρόθεσµη µνήµη (short term memory), αφορά στις µνηµονικές διεργασίες µε τις οποίες οι πληροφορίες διατηρούνται για µικρό χρονικό διάστηµα (µνήµη µικρής διάρκειας ή λειτουργική µνήµη). Η διάρκειά της είναι από λίγα sec έως µερικές ώρες. Στη συνέχεια, οι πληροφορίες αυτές είτε χάνονται, είτε µεταπίπτουν στο στάδιο της µακροπρόθεσµης µνήµης 8. Πειράµατα σε κατώτερα ζώα έδειξαν ότι είναι δυνατό να εγκατασταθούν µνήµες αυτής της µορφής, µετά από πρόσκαιρες χηµικές ή φυσικές µεταβολές ή και από συνδυασµό χηµικών και φυσικών µεταβολών, που επιτελούνται είτε στις προσυναπτικές απολήξεις, είτε στη µετασυναπτική µεµβράνη και συνήθως διαρκούν από λίγα λεπτά ως µερικές εβδοµάδες Μοριακός µηχανισµός της βραχυπρόθεσµης µνήµης α) Μηχανισµός «εθισµού» Σε µοριακό επίπεδο, ο εθισµός της αισθητικής απόληξης είναι αποτέλεσµα του σταδιακού αποκλεισµού των διαύλων ασβεστίου της µεµβράνης της απόληξης. Μολονότι ο µηχανισµός µε τον οποίο γίνεται ο αποκλεισµός αυτός δεν είναι απόλυτα γνωστός, το ποσό των ιόντων ασβεστίου που εισέρχεται στην απόληξη µε κάθε δυναµικό δράσεως είναι πολύ µικρότερο από το φυσιολογικό και κατά συνέπεια το ποσό της νευροδιαβιβαστικής ουσίας που απελευθερώνεται, είναι και αυτό µικρότερο, δεδοµένου ότι η εισροή ασβεστίου αποτελεί το ερέθισµα για την απελευθέρωσή της. 28
30 β) Μηχανισµός διευκόλυνσης Στην περίπτωση της διευκόλυνσης, ο µοριακός µηχανισµός πιστεύεται ότι είναι ο ακόλουθος: 1. Η διέγερση του νευρώνα που υφίσταται διευκόλυνση της διαβίβασης ταυτόχρονα µε τη διέγερση της αισθητικής απόληξης προκαλεί απελευθέρωση σεροτονίνης στη σύναψη, η οποία σχηµατίζεται στην αισθητική προσυναπτική απόληξη. 2. Η σεροτονίνη συνδέεται µε τους υποδοχείς σεροτονίνης της µεµβράνης της αισθητικής απόληξης και ενεργοποιεί την αδενυλική κυκλάση στο εσωτερικό της µεµβράνης. Η ενεργοποίηση αυτή προκαλεί σχηµατισµό κυκλικής µονοφωσφορικής αδενοσίνης (camp) στο εσωτερικό της αισθητικής προσυναπτικής απόληξης. 3. Η camp ενεργοποιεί µία πρωτεϊνική κινάση, η οποία φωσφορυλιώνει µια πρωτεΐνη, που αποτελεί τµήµα του διαύλου καλίου της µεµβράνης της αισθητικής απόληξης. Αυτό αναστέλλει την αγωγιµότητα των διαύλων καλίου για χρονικό διάστηµα που κυµαίνεται από µερικά λεπτά ως µερικές εβδοµάδες. 4. Η απουσία αγωγιµότητας καλίου προκαλεί πολύ µεγάλη παράταση του δυναµικού δράσεως της προσυναπτικής απόληξης, αφού η εκροή ιόντων καλίου είναι απαραίτητη για την επαναπόλωση της µεµβράνης µετά την εκπόλωσή της από το δυναµικό δράσης. 5. Το παρατεταµένο δυναµικό δράσης προκαλεί παράταση της ενεργοποίησης των διαύλων ασβεστίου, από τους οποίους εισέρχονται πολύ µεγάλες ποσότητες ιόντων ασβεστίου στην αισθητική απόληξη. Στη συνέχεια, τα ιόντα ασβεστίου προκαλούν απελευθέρωση πολύ µεγάλου ποσού νευροδιαβιβαστικής ουσίας και µε αυτό τον τρόπο διευκολύνεται η συναπτική µετάδοση. Έτσι, µε έµµεσο τρόπο η συνδυασµένη επίδραση της διέγερσης του νευρώνα που υφίσταται διευκόλυνση της διαβίβασης, συγχρόνως µε τη διέγερση του αισθητικού νευρώνα, προκαλεί µία παρατεταµένη µεταβολή στην αισθητική απόληξη, που εγκαθιστά το µνηµονικό έγγραµµα 6. 29
31 6.3. Μακροπρόθεσµη µνήµη Στη µακροπρόθεσµη µνήµη (long term memory) οι ασταθείς αρχικά µνηµονικές πληροφορίες, που προέρχονται από τη βραχυπρόθεσµη µνήµη, σταθεροποιούνται µε την επανάληψη. Οι πληροφορίες που περιέχει η µακροπρόθεσµη µνήµη αναζητούνται και ανασυντίθενται είτε µε απλή ανάκληση είτε µε αναγνώριση. Αντιπροσωπεύεται σε διάφορες περιοχές του κεντρικού νευρικού συστήµατος και απαιτεί σύνθεση νέων πρωτεϊνών και δηµιουργία νέων συναπτικών συνδέσεων 8. ιαχωριστική γραµµή µεταξύ των παρατεταµένων τύπων βραχυπρόθεσµης και µακροπρόθεσµης µνήµης δεν υπάρχει. Γι αυτό και η διάκριση είναι σχηµατική. Ωστόσο, πιστεύεται ότι η µακροπρόθεσµη µνήµη γενικά οφείλεται σε µορφολογικές µεταβολές των συνάψεων, οι οποίες προκαλούν είτε αύξηση είτε µείωση της µετάδοσης του σήµατος από αυτές οµικές µεταβολές των συνάψεων Φωτογραφίες µε ηλεκτρονικό µικροσκόπιο σε ασπόνδυλα ζώα έδειξαν ότι κατά τη διάρκεια εγκατάστασης µακροπρόθεσµων µνηµονικών εγγραµµάτων, επέρχονται πολλαπλές φυσικές δοµικές µεταβολές στις συνάψεις. Οι δοµικές αυτές µεταβολές δεν παρατηρούνται εφόσον χορηγηθούν φαρµακολογικές ουσίες που εµποδίζουν το DNA να επάγει τον πολλαπλασιασµό πρωτεϊνικών µορίων στον προσυναπτικό νευρώνα. Στις περιπτώσεις αυτές µνηµονικό έγγραµµα δεν αναπτύσσεται. Συνεπώς, συµπεραίνεται ότι η ανάπτυξη γνήσιας µακροπρόθεσµης µνήµης εξαρτάται από τη φυσική αναδόµηση των ίδιων των συνάψεων, κατά τρόπον ώστε να αυξάνεται η ευαισθησία τους για τα διαβιβαζόµενα νευρικά σήµατα. Οι σηµαντικότερες φυσικές µεταβολές που προκαλούνται είναι: 1. Αύξηση του αριθµού των θέσεων απελευθέρωσης κυστιδίων για έκκριση νευροδιαβιβαστικής ουσίας. Κάθε προσυναπτική απόληξη διαθέτει στη µεµβράνη της ειδικές θέσεις απελευθέρωσης, από τις οποίες απελευθερώνεται νευροδιαβιβαστική ουσία προς τη συναπτική σχισµή όταν η απόληξη διεγείρεται. Σε λίγα λεπτά µετά την έναρξη συνεδρίας εξάσκησης µνήµης διαπιστώνεται µε το ηλεκτρονικό µικροσκόπιο έναρξη αύξησης του αριθµού αυτών των θέσεων απελευθέρωσης νευροδιαβιβαστικής ουσίας στην απόληξη. Οι καινούριες θέσεις δηµιουργούνται µε την ανάπτυξη κυστιδίων απελευθέρωσης πρωτεΐνης 30
32 στην έσω επιφάνεια της προσυναπτικής µεµβράνης. Στη συνέχεια, µετά την άφιξη του δυναµικού δράσης στην προσυναπτική απόληξη, οι επιπλέον αυτές θέσεις προκαλούν µεγάλη αύξηση της εξωκυττάρωσης των κυστιδίων µε προώθηση της νευροδιαβιβαστικής ουσίας προς τη συναπτική σχισµή. 2. Αύξηση του αριθµού των κυστιδίων µε νευροδιαβιβαστική ουσία. Παρατηρείται όχι µόνο αύξηση του αριθµού των θέσεων, αλλά και του αριθµού των κυστιδίων σε κάθε προσυναπτική απόληξη. 3. Αύξηση του αριθµού των προσυναπτικών απολήξεων. Με έντονη εξάσκηση αυξάνεται ακόµα και ο αριθµός των προσυναπτικών απολήξεων, σε µερικές περιπτώσεις µέχρι και στο διπλάσιο του φυσιολογικού. Παράλληλα µε την αύξηση των απολήξεων, οι δενδρίτες του επόµενου νευρώνα αυξάνονται σε µήκος για να χωρέσουν οι επιπρόσθετες συνάψεις. Με αυτόν τον τρόπο, η δοµική ικανότητα των συνάψεων να µεταδίδουν σήµατα αυξάνεται κατά την εγκατάσταση των εγγραµµάτων µακροπρόθεσµης µνήµης Μεταβολή του αριθµού των νευρώνων και των συνάψεών τους Από τις πρώτες εβδοµάδες και πιθανώς µέχρι το πρώτο έτος της ζωής σε διάφορες περιοχές του εγκεφάλου αναπτύσσονται νευρώνες, που διαθέτουν κλάδους από το νευράξονά τους για σύνδεσή τους µε άλλους νευρώνες. Εάν οι νέοι νευράξονες δεν µπορέσουν να συνδεθούν µε άλλους νευρώνες ή µυϊκά και αδενικά κύτταρα, τότε «διαλύονται» σε λίγες εβδοµάδες. Οι συνδέσεις καθορίζονται από ποικίλους ειδικούς αυξητικούς παράγοντες των νεύρων, που απελευθερώνονται από τους νευρώνες. Εάν ο τύπος του απελευθερούµενου αυξητικού παράγοντα δεν είναι ο κατάλληλος, ο αυξανόµενος νευράξονας δε συνδέεται και σύντοµα νεκρώνεται. Επιπρόσθετα, σε περιπτώσεις που οι συνδέσεις δεν είναι ικανοποιητικές, ολόκληρος ο νευρώνας από τον οποίο προέρχεται ο νευράξονας εξαφανίζεται. Με αυτό τον τρόπο στα πρώτα στάδια της ζωής ισχύει η αρχή «χρησιµοποίηση ή απώλεια», η οποία ισχύει για τον τελικό αριθµό των νευρώνων και των συνδέσεών τους στις διάφορες περιοχές του εγκεφάλου. Αυτό αποτελεί ένα είδος «µάθησης». Για παράδειγµα, εάν η όραση από τον έναν οφθαλµό νεογέννητου ζώου αποφραχθεί για µερικές εβδοµάδες µετά τη γέννηση του, πολλοί από τούς νευρώνες στις εναλλασσόµενες οπτικές ταινίες στον οπτικό φλοιό, δηλαδή οι νευρώνες που φυσιολογικά 31
33 συνδέονται µε τον καλυπτόµενο οφθαλµό, εκφυλίζονται και ο οφθαλµός εµφανίζει µερική είτε και ολική τύφλωση για το υπόλοιπο της ζωής του ζώου. Κατά συνέπεια, σε πολλές περιοχές του εγκεφαλικού φλοιού, το 50% ή και περισσότερο, από τους αρχικούς νευρώνες εξαφανίζονται. Αυτός ο µηχανισµός αποτελεί ένα είδος µακροπρόθεσµης µάθησης. Επιπρόσθετα, η πλαστικότητα του νευρικού συστήµατος κατά τα πρώτα στάδια της ζωής αποδεικνύει τη σηµασία της παροχής στο νεαρό άτοµο όσο το δυνατό µεγαλύτερης έκθεσης σε πολλαπλές εµπειρίες για µάθηση. Έτσι, στη συνέχεια αυτό το παιδί έχει το καλύτερο δυνατό θεµέλιο για µεταγενέστερη µόρφωση κατά το υπόλοιπο της ζωής του Εµπέδωση (consolidation) της µνήµης Για να µετατραπεί η άµεση µνήµη είτε σε βραχυπρόθεσµη είτε σε µακροπρόθεσµη, ώστε να µπορεί να ανακαλείται µετά από την παρέλευση ενός ικάνου χρονικού διαστήµατος, πρέπει να υποστεί µία ορισµένη διεργασία, την εµπέδωση (consolidation). Ειδικότερα, πρέπει να επέλθουν µε κάποιο τρόπο χηµικές, φυσικές και ανατοµικές µεταβολές στις συνάψεις, οι οποίες είναι υπεύθυνες για τη µακροπρόθεσµη µνήµη. Για τη διεργασία αυτή απαιτούνται 5 ως 10 λεπτά για τον ελάχιστο βαθµό εµπέδωσης και µία ώρα ή και περισσότερο για το µέγιστο. Αν επιδράσει στον εγκέφαλο ένα ισχυρό αισθητικό ερέθισµα και µετά από αυτό ακολουθήσει, σε ένα περίπου λεπτό, µία τεχνητή επιληπτική κρίση (µε ηλεκτρικό ερεθισµό), η αισθητική εµπειρία δεν ανακαλείται στη µνήµη. Αντιστοίχως, µία εγκεφαλική διάσειση, ή απότοµη εφαρµογή βαθιάς γενικής αναισθησίας ή οποιαδήποτε άλλη διεργασία, η οποία αναστέλλει πρόσκαιρα την εγκεφαλική λειτουργία εµποδίζει την εµπέδωση της µνήµης. Ωστόσο, αν η εφαρµογή του ισχυρού ηλεκτρικού ερεθίσµατος καθυστερήσει περισσότερο από 10 λεπτά, µέρος τουλάχιστο του µνηµονικού εγγράµµατος θα εγκατασταθεί, και αν το ηλεκτρικό ερέθισµα καθυστερήσει για πάνω από µία ώρα, η µνήµη θα έχει σχεδόν πλήρως εµπεδωθεί 6. Ψυχολογικές µελέτες έδειξαν ότι η επανάληψη της ίδιας πληροφορίας πολλές φορές (επανάληψη-probe test), επιταχύνει και ενισχύει το βαθµό µετατροπής της άµεσης σε βραχυπρόθεσµη και στη συνέχεια σε µακροπρόθεσµη µνήµη, και εποµένως επιταχύνει και ενισχύει την εµπέδωση. Ο εγκέφαλος έχει την τάση να επαναλαµβάνει από µόνος του τις 32
34 νέες πληροφορίες, και ιδιαίτερα εκείνες που του προκαλούν ενδιαφέρον. Εποµένως, µε την πάροδο του χρόνου τα σηµαντικά στοιχεία αισθητικής εµπειρίας καθηλώνονται όλο και περισσότερο σε αποθήκες µνήµης. Αυτό εξηγεί γιατί ένα άτοµο µπορεί να θυµάται καλύτερα µικρό όγκο πληροφοριών τις οποίες µελέτησε σε βάθος, παρά µεγάλο όγκο πληροφοριών που µελέτησε επιφανειακά. Επίσης, εξηγεί και το λόγο που ένα άτοµο που βρίσκεται σε εγρήγορση εµπεδώνει καλύτερα από κάποιο άλλο άτοµο που βρίσκεται σε κατάσταση πνευµατικού καµάτου Κωδικοποίηση της µνήµης κατά την εµπέδωση Ένα από τα σηµαντικότερα στάδια στη διεργασία της εµπέδωσης είναι το γεγονός ότι οι µνήµες κωδικοποιούνται σε διάφορες τάξεις πληροφοριών. Κατά τη διεργασία της κωδικοποίησης, η επεξεργασία των νέων πληροφοριών υποβοηθείται µε την ανάκληση από τη µνήµη παρόµοιων πληροφοριών. Οι νεότερες και οι παλαιότερες πληροφορίες συγκρίνονται για πιθανές οµοιότητες και διαφορές, και µέρος της διεργασίας εµπέδωσης συνίσταται στην αποθήκευση της πληροφορίας για τις οµοιότητες και τις διαφορές µάλλον παρά την αποθήκευση της πληροφορίας χωρίς επεξεργασία. Άρα, κατά τη διεργασία της εµπέδωσης, η αποθήκευση της µνήµης στον εγκέφαλο δεν είναι τυχαία, αλλά αντίθετα γίνεται πάντα σε συσχετισµό µε προηγούµενες εµπειρίες του ιδίου τύπου. Είναι φανερό ότι η παραπάνω διεργασία είναι απαραίτητη για να µπορεί κανείς να ανατρέχει στις αποθήκες της µνήµης του σε µεταγενέστερο χρόνο, και να βρίσκει τις πληροφορίες που χρειάζεται 6. 33
35 Β. ΝΕΥΡΟ ΙΑΒΙΒΑΣΤΕΣ Βάση για την απάντηση σηµαντικών ερωτηµάτων σχετικών µε τον εγκέφαλο και τη συµπεριφορά αποτελούν έρευνες, που σχετίζονται µε τη χηµεία της συναπτικής διαβίβασης στον εγκέφαλο (της επικοινωνίας µεταξύ των νευρώνων). Οι διεργασίες αποθήκευσης της µνήµης, η βιολογική βάση των ψυχικών νοσηµάτων και η παθοφυσιολογία των νευροεκφυλιστικών νοσηµάτων είναι ορισµένα από αυτά. Η ιεράρχηση των συνδέσεων των κυκλωµάτων των χηµικών συνάψεων αποτελεί µία ιδιαίτερα πολύπλοκη διαδικασία, αλλά σε αυτόν τον τοµέα έχει γίνει σηµαντική πρόοδος, όπως για παράδειγµα µε την ταυτοποίηση των κυκλωµάτων που εµπλέκονται στις νόσους Parkinson και Alzheimer. Η µελέτη των χηµικών διαβιβαστών και των νευρικών κυκλωµάτων στον εγκέφαλο, καθώς και των επιδράσεων φαρµακολογικώς δραστικών ουσιών σε αυτά, ξεκίνησε µόλις πριν λίγες δεκαετίες και έχει εξελιχθεί ως το σηµαντικότερο πεδίο έρευνας της νευροεπιστήµης. Οι πληροφορίες τις οποίες µεταφέρει ένας νευρώνας είναι κωδικοποιηµένες σε ηλεκτρικά σήµατα (νευρικές ώσεις), που διατρέχουν το νευράξονα και καταλήγουν στη νευρική απόληξη. Στη σύναψη, τα σήµατα αυτά µεταφέρονται στην άλλη πλευρά της συναπτικής σχισµής από έναν ή περισσότερους χηµικούς αγγελιοφόρους. Κανένας από τους χηµικούς διαβιβαστές δε µεταφέρει µοναδικές πληροφορίες, όπως συµβαίνει µε το RNA και το DNA. Πράγµατι, ορισµένοι νευροδιαβιβαστές έχουν και άλλες κυτταρικές λειτουργίες, ως µεταβολίτες άλλων βιοχηµικών οδών. Για παράδειγµα, τα αµινοξέα πολυµερίζονται και σχηµατίζουν πρωτεΐνες. Το γλουταµινικό οξύ και το GABA δρουν ως υποστρώµατα στον ενδιάµεσο µεταβολισµό και η ΑΤΡ είναι ο κύριος παράγοντας µεταφοράς µεταβολικής ενέργειας. Για να επιτελέσουν διαβιβαστική λειτουργία, τα µόρια αυτά πρέπει να απελευθερωθούν στη συναπτική σχισµή και να προσδεθούν σε µεµβρανικούς υποδοχείς στο µετασυναπτικό κύτταρο. Μετά την πρόσδεση των µορίων νευροδιαβιβαστή, ο υποδοχέας παράγει ηλεκτρικά ή µεταβολικά σήµατα στο µετασυναπτικό κύτταρο. Η συναπελευθέρωση διαφόρων νευροδραστικών ουσιών κοντά σε κατάλληλους µετασυναπτικούς υποδοχείς επιτρέπει µία εκπληκτική ποικιλία µεταφοράς πληροφοριών στην κάθε σύναψη. Εποµένως, η χηµική συναπτική διαβίβαση αποτελεί µία διεργασία που 34
36 πραγµατοποιείται σε δύο επίπεδα, ένα προσυναπτικό και ένα µετασυναπτικό. Συγκεκριµένα, σε προσυναπτικό επίπεδο πραγµατοποιείται η σύνθεση και η αποθήκευση του νευροδιαβιβαστή, καθώς και η απελευθέρωσή του στη συναπτική σχισµή. Σε µετασυναπτικό επίπεδο πραγµατοποιείται η αλληλεπίδραση του νευροδιαβιβαστή µε τους υποδοχείς του στη µετασυναπτική µεµβράνη. Το τελικό στάδιο αποτελεί η διάσπαση ή η επαναπρόσληψη ή η διάχυση του νευροδιαβιβαστή, µε αποτέλεσµα την αποµάκρυνσή του από τη συναπτική σχισµή. Η έννοια του χηµικού αγγελιοφόρου-διαβιβαστή και της χηµικής νευροδιαβίβασης χρησιµοποιήθηκε στα µέσα της δεκαετίας του 1920, όταν ο Otto Loewi απέδειξε ότι οι απολήξεις του πνευµονογαστρικού νεύρου απελευθερώνουν ακετυλοχολίνη (ACh) και αφού ο Henry Dale δηµοσίευσε τις εργασίες του σχετικά µε τη χολινεργική και την αδρενεργική διαβίβαση. Οι όροι «χολινεργικός» και «αδρενεργικός» εισήχθησαν για να δηλώσουν αν ένας νευρώνας χρησιµοποιεί ως νευροδιαβιβαστή την ακετυλοχολίνη ή τη νοραδρεναλίνη (ή την αδρεναλίνη). Επειδή, όµως, από την εποχή εκείνη ανακαλύφθηκαν πολλές νέες ουσίες, οι οποίες λειτουργούν ως νευροδιαβιβαστές, η έννοια υπέστη διαρκείς µεταβολές για να συµπεριλάβει νέες πληροφορίες σχετικά µε την κυτταρική βιολογία των νευρώνων και τη φαρµακολογία των υποδοχέων. Ως νευροδιαβιβαστής χαρακτηρίζεται µία ουσία, η οποία απελευθερώνεται από έναν νευρώνα σε µία σύναψη και η οποία επιδρά είτε σε έναν άλλο νευρώνα είτε σε ένα κύτταρο άλλου ιστού µε συγκεκριµένο τρόπο. Όπως συµβαίνει και µε πολλές άλλες λειτουργικές έννοιες στη βιολογία, η έννοια του νευροδιαβιβαστή είναι αρκετά σαφής στο κέντρο της και κάπως ασαφής στις παρυφές της. Γενικώς, ένας µικρός αριθµός ουσιών µικρού µοριακού βάρους θεωρούνται νευροδιαβιβαστές, αλλά πολλές άλλες ουσίες έχουν γίνει δεκτές ως πιθανοί νευροδιαβιβαστές, χωρίς να υπάρχει οµοφωνία. Συχνά, είναι δύσκολο να αποδειχθεί ότι µία ουσία λειτουργεί ως νευροδιαβιβαστής, ακόµη και για εκείνες τις ουσίες που θεωρούνται από πολλούς ότι είναι νευροδιαβιβαστές 5,9. 35
37 Για να χαρακτηριστεί µία χηµική ουσία ως νευροδιαβιβαστής πρέπει να πληρεί τα ακόλουθα κριτήρια: 1. Να συντίθεται στο νευρώνα 2. Να υπάρχει στην προσυναπτική απόληξη και να απελευθερώνεται σε επαρκείς ποσότητες, ώστε να ασκήσει ορισµένη δράση στο µετασυναπτικό νευρώνα ή σε ένα όργανο-τελεστή 3. Όταν χορηγηθεί εξωγενώς να µπορεί να µιµηθεί τη δράση του ενδογενώς απελευθερούµενου νευροδιαβιβαστή (δηλαδή να ενεργοποιήσει τους ίδιους διαύλους ιόντων ή την ίδια οδό δεύτερου αγγελιοφόρου στο µετασυναπτικό κύτταρο) 4. Να υπάρχει ειδικός µηχανισµός αποµάκρυνσης της ουσίας από τη συναπτική σχισµή είτε µε διάσπασή της είτε µε επαναπρόσληψη ή διάχυσή της Ο τελικός χαρακτηρισµός µίας ενδογενούς ουσίας ως νευροδιαβιβαστή αποτελεί µία εξαιρετικά πολύπλοκη και δύσκολη διαδικασία, µε αποτέλεσµα για πολλές ενδογενείς ουσίες να υπάρχουν θεωρητικές ενδείξεις, αλλά όχι πειραµατικές αποδείξεις για αυτήν την ιδιότητά τους. 1. Σύνθεση και αποθήκευση των νευροδιαβιβαστών Ο νευροδιαβιβαστής, που παράγεται σε κάθε νευρώνα, καθορίζεται από την παρουσία ενός ειδικού βιοσυνθετικού ενζύµου, το οποίο καταλύει τη βιοσυνθετική οδό του νευροδιαβιβαστή, αλλά δε χαρακτηρίζει και την εξειδίκευσή του. Οι νευροδιαβιβαστές είναι συσσωρευµένοι σε κυστίδια στο εσωτερικό του προσυναπτικού νευρώνα. Τα κυστίδια συµµετέχουν µε διαφορετικό τρόπο στις διεργασίες παραγωγής και αποθήκευσης των δύο µεγάλων κατηγοριών χηµικών αγγελιοφόρων, των µικροµοριακών νευροδιαβιβαστών και των νευροδραστικών πεπτιδίων. Οι µικροµοριακοί νευροδιαβιβαστές, αφού συντεθούν στο κυτταρόπλασµα, προσλαµβάνονται και συγκεντρώνονται µέσα σε συναπτικά κυστίδια, τα οποία τους προστατεύουν από τα ενδοκυτταρικά ένζυµα αποδόµησης, που συντελούν στη διατήρηση ενός σταθερού επιπέδου νευροδιαβιβαστή στο κυτταρόπλασµα. Επειδή οι µικροµοριακοί νευροδιαβιβαστές απελευθερώνονται σε σχετικώς µεγάλες ποσότητες µε ταχύ ρυθµό, το µεγαλύτερο µέρος των µικροµοριακών νευροδιαβιβαστών του νευρώνα συντίθεται στις απολήξεις. Εποµένως, οι 36
38 νευρικές απολήξεις περιέχουν αυξηµένη συγκέντρωση αυτών των συναπτικών κυστιδίων. Σε αντίθεση µε τη σύνθεση των µικροµοριακών νευροδιαβιβαστών, τα πρόδροµα πρωτεϊνικά µόρια των νευροδραστικών πεπτιδίων συντίθενται µόνο στο κυτταρικό σώµα. Εκεί, συσσωρεύονται σε εκκριτικά και συναπτικά κυστίδια, τα οποία µεταφέρονται από το κυτταρικό σώµα στις απολήξεις. Έτσι, αντίθετα µε τα κυστίδια που περιέχουν µικροµοριακούς νευροδιαβιβαστές, τα κυστίδια αυτά δεν αναγοµώνονται στη νευρική απόληξη. Η κατανόηση του µοριακού µηχανισµού της χηµικής συναπτικής διαβίβασης αρχίζει µε την ταυτοποίηση του περιεχοµένου των συναπτικών κυστιδίων. Στη συνέχεια θεωρείται απαραίτητο για µία χηµική ουσία να µπορεί να απελευθερωθεί από τα κυστίδια, ώστε να έχει και το δυναµικό να ενεργοποιήσει έναν υποδοχέα. Ωστόσο, τα µόρια που απελευθερώνονται από έναν νευρώνα δεν είναι όλα χηµικοί διαβιβαστές. Μόνον εκείνα που έχουν την ικανότητα να προσδεθούν στους κατάλληλους υποδοχείς µπορούν να χρησιµεύσουν ως νευροδιαβιβαστές. Έτσι, ενώ ορισµένες πλευρές της λειτουργίας των κυστιδίων ποικίλλουν αρκετά µεταξύ διαφόρων τύπων νευρώνων, µία πλευρά είναι κοινή στους περισσότερους από αυτούς. Με εξαίρεση τους νευρώνες που απελευθερώνουν νευροδιαβιβαστές µέσω διάχυσης ή µέσω µηχανισµών µεταφοράς, τα κυστίδια απελευθερώνουν το νευροδιαβιβαστή µε εξωκυττάρωση 5,9. 2. Αποµάκρυνση των νευροδιαβιβαστών από τη συναπτική σχισµή Η έγκαιρη αποµάκρυνση των νευροδιαβιβαστών από τη συναπτική σχισµή έχει ιδιαίτερη σηµασία για τη συναπτική διαβίβαση. Η συνεχής παρουσία του νευροδιαβιβαστή στη συναπτική σχισµή µετά την απελευθέρωσή του, θα εµπόδιζε τη µεταβίβαση νέων νευρικών ώσεων και η σύναψη θα γινόταν ανθεκτική, κυρίως λόγω της απευαισθητοποίησης των υποδοχέων από την παρατεταµένη επαφή τους µε το νευροδιαβιβαστή. Έχουν αναγνωριστεί τρεις µηχανισµοί, µε τους οποίους ο νευρικός ιστός απαλλάσσεται από διαλυτούς ή µη προσδεδεµένους νευροδιαβιβαστές. Αυτοί είναι η διάχυση, η ενζυµική αποδόµηση και η επαναπρόσληψη του χηµικού αγγελιοφόρου. 37
39 Η διάχυση αποµακρύνει αρκετούς από το σύνολο των νευροδιαβιβαστών και αποτελεί σηµαντικό τρόπο καθαρισµού της συναπτικής σχισµής από την περίσσεια των νευροδιαβιβαστών. Η ενζυµική αποδόµηση των νευροδιαβιβαστών εντοπίζεται κυρίως στο χολινεργικό σύστηµα. Το εξωκυτταρικό ένζυµο που παρεµβαίνει είναι η ακετυλοχολινεστεράση (AChE). Στη νευροµυϊκή σύναψη, οι ενεργές ζώνες της προσυναπτικής νευρικής απόληξης εντοπίζονται ακριβώς επάνω από τις συναπτικές πτυχές της µεµβράνης του µυϊκού κυττάρου, όπου βρίσκονται οι υποδοχείς της ACh. Οι υποδοχείς συγκεντρώνονται στην επιφάνεια του µυϊκού κυττάρου και δεν επεκτείνονται στο βάθος των πτυχών, ενώ η AChE είναι προσδεδεµένη στο βασικό υµένα µόνο στο εσωτερικό των πτυχών. Αυτή η ανατοµική διάταξη των µορίων εξυπηρετεί δύο λειτουργίες. εδοµένου ότι τα µόρια της ACh, µετά τον αποχωρισµό τους από τον υποδοχέα, έχουν µεγάλη πιθανότητα να διαχυθούν στο σχετικά µεγάλο όγκο των συναπτικών πτυχών και να υδρολυθούν προς χολίνη και οξικό οξύ, τα µόρια νευροδιαβιβαστή θα χρησιµεύσουν µόνο µία φορά. Έτσι, µία λειτουργία της AChE είναι η ταχεία στίξη του συναπτικού µηνύµατος. Η δεύτερη λειτουργία είναι η επαναπρόσληψη της χολίνης, η οποία µπορεί αλλιώς να χανόταν µε διάχυση από τη συναπτική σχισµή. Μετά την υδρόλυσή της από την AChE, η χολίνη παραµένει σε χαµηλή συγκέντρωση στη δεξαµενή, η οποία αποτελείται από τις συναπτικές πτυχές, και επαναπροσλαµβάνεται αργότερα από τις χολινεργικές νευρικές απολήξεις µέσω ενός ειδικού µηχανισµού υψηλής συγγένειας. ιάφορες άλλες ενζυµικές οδοί αποδοµούν τους νευροδιαβιβαστές στο νευρώνα και σε µη νευρικούς ιστούς, χωρίς να παρεµβαίνουν ειδικά στη διακοπή της συναπτικής διαβίβασης. Οι οδοί αυτές είναι δυνατό να έχουν σηµασία για τον έλεγχο της συγκέντρωσης του νευροδιαβιβαστή µέσα στο νευρώνα ή για την απενεργοποίηση νευροδιαβιβαστών, που έχουν διαχυθεί από τη συναπτική σχισµή. Πολλές έχουν κλινική σηµασία και παρέχουν σηµεία δράσης φαρµακευτικών ουσιών, καθώς και διαγνωστικές δυνατότητες. Για παράδειγµα, οι αναστολείς της µονοαµινοοξειδάσης, οι οποίοι εµποδίζουν την αποδόµηση των αµινικών νευροδιαβιβαστών µέσα στο κύτταρο, χρησιµεύουν σήµερα για τον έλεγχο της αρτηριακής υπέρτασης, καθώς και για την αντικαταθλιπτική αγωγή. Ένα άλλο παράδειγµα είναι το ενδοκυτταρικό ένζυµο κατεχολο-ο- µεθυλοτρανσφεράση, το οποίο είναι σηµαντικό για την αποδόµηση 38
40 βιογενών αµινών. Βρίσκεται στο κυτταρόπλασµα των περισσότερων κυττάρων, συµπεριλαµβανοµένων των νευρώνων, αλλά κυρίως στο ήπαρ και στο νεφρό. Οι συγκεντρώσεις των µεταβολιτών του ενζύµου αυτού στα υγρά του σώµατος παρέχουν µία διαγνωστική ένδειξη της αποτελεσµατικότητας των φαρµάκων, τα οποία επηρεάζουν τη σύνθεση ή την αποδόµηση των βιογενών αµινών στο νευρικό ιστό. Τέλος, πρωταρχικό ρόλο στην αντιµετώπιση της νόσου Alzheimer παίζουν οι αναστολείς της ακετυλοχολινεστεράσης, οι οποίοι αυξάνουν τα επίπεδα της ακετυλοχολίνης στα σηµεία δράσης της στο κεντρικό νευρικό σύστηµα. Τα νευροδραστικά πεπτίδια αποµακρύνονται από τη συναπτική σχισµή µε πιο βραδύ ρυθµό από τους µικροµοριακούς νευροδιαβιβαστές. Μοναδικοί µηχανισµοί αποµάκρυνσης των πεπτιδίων είναι πιθανώς η διάχυση και η πρωτεόλυση από εξωκυτταρικές πεπτιδάσες. Η βραδεία αποµάκρυνση των πεπτιδίων συµβάλλει στην αύξηση της διάρκειας της δράσης τους και κάνει το µεταβολισµό τους να µοιάζει µάλλον µε το µεταβολισµό των ορµονών. Η επαναπρόσληψη του νευροδιαβιβαστή είναι ο πιο κοινός µηχανισµός απενεργοποίησης. Υπάρχουν µηχανισµοί στενής συγγένειας για την επαναπρόσληψη νευροδιαβιβαστών, που έχουν απελευθερωθεί και οι οποίοι εξαρτώνται από µεταφορικά µόρια της µεµβράνης των νευρικών απολήξεων και των κυττάρων της νευρογλοίας µε σταθερές πρόσδεσης της τάξης των 25µΜ ή και λιγότερο. Οι µηχανισµοί επαναπρόσληψης στενής συγγένειας περιγράφηκαν αρχικά για τη νοραδρεναλίνη, την ντοπαµίνη και τη σεροτονίνη. Παρόµοιοι µηχανισµοί επαναπρόσληψης αµινοξέωννευροδιαβιβαστών, όπως είναι το γλουταµινικό οξύ, το GABA, η γλυκίνη και η χολίνη (αλλά όχι η ACh), ανακαλύφθηκαν αργότερα. Καθένας από τους ειδικούς νευρώνες έχει τους δικούς του χαρακτηριστικούς µηχανισµούς επαναπρόσληψης. Για παράδειγµα, οι µη χολινεργικοί νευρώνες δεν επαναπροσλαµβάνουν τη χολίνη µε υψηλή συγγένεια. Ορισµένα ισχυρά ψυχοτρόπα φάρµακα µπορούν να αναστείλουν τις διεργασίες επαναπρόσληψης (η κοκαΐνη αναστέλλει την επαναπρόσληψη της νοραδρεναλίνης, και τα τρικυκλικά αντικαταθλιπτικά την επαναπρόσληψη της σεροτονίνης). Η εφαρµογή των κατάλληλων φαρµάκων για την αναστολή της επαναπρόσληψης µπορεί να παρατείνει και να ενισχύσει τη δράση των βιογενών αµινών και του GABA 5,9. 39
41 3. Κατηγορίες νευροδιαβιβαστών Το νευρικό σύστηµα χρησιµοποιεί δύο κύριες κατηγορίες χηµικών ουσιών για τη µετάδοση σηµάτων: µικροµοριακούς νευροδιαβιβαστές και νευροδραστικά πεπτίδια, τα οποία είναι βραχείες αλυσίδες αµινοξέων. Στη συνέχεια αναφέρονται µόνο οι µικροµοριακοί νευροδιαβιβαστές και αναλύεται εκτενέστερα η ακετυλοχολίνη (ACh), λόγω της συµµετοχής της στα νευρωνικά κυκλώµατα και τις διεργασίες, που σχετίζονται µε τη µνήµη. Από τις µικροµοριακές ουσίες, που χαρακτηρίζονται ως νευροδιαβιβαστές, εννέα είναι αµίνες. Από αυτές οκτώ είναι αµινοξέα ή παράγωγά τους. Η ουσία που δεν είναι αµίνη είναι η ΑΤΡ και τα προϊόντα της αποδόµησής της. Οι αµινικοί χηµικοί διαβιβαστές έχουν πολλές βιοχηµικές οµοιότητες. Όλοι είναι φορτισµένα µικρά µόρια, που σχηµατίζονται σε σχετικώς βραχείες βιοσυνθετικές οδούς και όλοι συντίθενται από πρόδροµα µόρια, που παράγονται από τα µείζονα υδατανθρακικά υποστρώµατα του ενδιάµεσου µεταβολισµού. Όπως ισχύει και για πολλές οδούς του ενδιάµεσου µεταβολισµού, η σύνθεση αυτών των νευροδιαβιβαστών καταλύεται από ένζυµα, τα οποία σχεδόν χωρίς εξαίρεση είναι κυτοσολιακά. Στις βιοσυνθετικές οδούς των αµινικών διαβιβαστών, όπως και σε κάθε βιοσυνθετική οδό, η συνολική σύνθεση ρυθµίζεται από µία ενζυµική αντίδραση. Το ένζυµο, που ελέγχει τον ρυθµό σύνθεσης, χαρακτηρίζει συνήθως έναν τύπο νευρώνα και κατά κανόνα λείπει από άλλους τύπους νευρώνων (Πίνακας 1). 40
42 Πίνακας 1. Nευροδιαβιβαστές και βιοσυνθετικά ένζυµα 5 Νευροδιαβιβαστής Ένζυµο Ακετυλοχολίνη Βιογενείς αµίνες Ντοπαµίνη Νοραδρεναλίνη (νορεπινεφρίνη) Αδρεναλίνη (επινεφρίνη) Σεροτονίνη Ισταµίνη Αµινοξέα γ-αµινοβουτυρικό οξύ Γλυκίνη Γλουταµινικό οξύ Τριφωσφορική αδενοσίνη (ΑΤΡ) Χολινοακετυλοτρανσφεράση (ειδική) Υδροξυλάση της τυροσίνης (ειδική) Υδροξυλάση της τυροσίνης και υδροξυλάση β της ντοπαµίνης (ειδική) Υδροξυλάση της τυροσίνης και υδροξυλάση β της ντοπαµίνης (ειδική) Υδροξυλάση της τρυπτοφάνης (ειδική) Αποκαρβοξυλάση της ιστιδίνης (αβέβαιη εξειδίκευση) Αποκαρβοξυλάση του γλουταµινικού οξέος (πιθανώς ειδική) Γενικός µεταβολισµός (ακαθόριστη ειδική οδός) Γενικός µεταβολισµός (ακαθόριστη ειδική οδός) ΑΤΡ συνθετάσες 41
43 3.1. Ακετυλοχολίνη Η ακετυλοχολίνη είναι ο νευροδιαβιβαστής που έχει µελετηθεί περισσότερο από κάθε άλλο. Η µελέτη της ξεκίνησε το 1924 όταν ο Otto Loewi, σε ένα από τα κλασικά πειράµατα της νευροεπιστήµης, ανακάλυψε την ύπαρξή της και απάντησε στο ερώτηµα για το αν η συναπτική διαβίβαση από το πνευµονογαστρικό νεύρο στον καρδιακό µυ (και σε άλλες συνάψεις) ήταν ηλεκτρική ή χηµική. Το πείραµα του Loewi αποτελεί ένα πρότυπο απλότητας και συγκροτηµένου ερευνητικού σχεδιασµού. Το πνευµονογαστρικό νεύρο είναι ένα από τα κύρια νεύρα που ελέγχουν την καρδιά. Στο βάτραχο το πνευµονογαστρικό νεύρο και η καρδιά µπορούν να αφαιρεθούν και να διατηρηθούν ζωντανά µέσα σε διάλυµα Ringer (το διάλυµα Ringer περιέχει ανάλογα ιοντικά συστατικά µε το αίµα). Ηλεκτρική διέγερση του πνευµονογαστρικού νεύρου επιβράδυνε τον καρδιακό ρυθµό τόσο στο ζωντανό ζώο όσο και στο παρασκεύασµα πνευµονογαστρικού-καρδιάς. Ο Loewi διέγειρε το πνευµονογαστρικό νεύρο του αποµονωµένου παρασκευάσµατος πολλές φορές, µε αποτέλεσµα την επαναλαµβανόµενη επιβράδυνση του καρδιακού ρυθµού. Στη συνέχεια έλαβε ποσότητα διαλύµατος Ringer από το αποµονωµένο παρασκεύασµα που είχε διεγείρει. Αυτήν την ποσότητα διαλύµατος Ringer τη µετάγγισε σε ένα άλλο παρασκεύασµα πνευµονογαστρικού-καρδιάς, όπου επίσης διαπίστωσε επιβράδυνση του καρδιακού ρυθµού. Με αυτό το απλό πείραµα απέδειξε ότι η συναπτική διαβίβαση είναι χηµική. Ο Loewi ονόµασε την άγνωστη χηµική ουσία Vagusstoff, ουσία που σύντοµα αποδείχθηκε ότι ήταν η ακετυλοχολίνη (ACh). Η βιοσυνθετική οδός της ACh αποτελείται από µία και µόνη ενζυµική αντίδραση, η οποία καταλύεται από τη χολινοακετυλοτρανσφεράση (ChAT). Η τρανσφεράση αυτή είναι το ένζυµο, που χαρακτηρίζει τη βιοσύνθεση της ACh. Ακετυλοσυνένζυµο Α + χολίνη Ω Ακετυλοχολίνη Ο νευρικός ιστός δεν έχει την ικανότητα να συνθέτει χολίνη, την οποία προσλαµβάνει µέσω της τροφής και φθάνει στους νευρώνες µέσω της κυκλοφορίας του αίµατος. Το συνυπόστρωµα, το ακετυλοσυνένζυµο Α, συµµετέχει σε πολλές οδούς του γενικού µεταβολισµού και δεν υπάρχει 42
44 µόνο στους χολινεργικούς νευρώνες, αλλά εντοπίζεται σε µεγάλες ποσότητες στα µιτοχόνδρια όλων των κυττάρων. Η ακετυλοχολίνη είναι ο νευροδιαβιβαστής, τον οποίο χρησιµοποιούν οι νευρώνες του νωτιαίου µυελού και απελευθερώνεται σε όλες τις νευροµυϊκές συνάψεις των σπονδυλωτών. Στο αυτόνοµο νευρικό σύστηµα η ACh αποτελεί το νευροδιαβιβαστή όλων των προγαγγλιακών νευρώνων, καθώς και των µεταγαγγλιακών παρασυµπαθητικών νευρώνων και χρησιµεύει σε πολλές συνάψεις του εγκεφάλου. Ειδικότερα, πολλά κυτταρικά σώµατα που συνθέτουν ACh βρίσκονται στο βασικό πυρήνα του Meynert, ο οποίος έχει εκτεταµένες προβολές στο φλοιό των εγκεφαλικών ηµισφαιρίων 5,9. 43
45 Γ. ΒΑΣΙΚΟΣ ΠΥΡΗΝΑΣ ΤΟΥ MEYNERT Στα µέσα της δεκαετίας του 1970 η χολινεργική νεύρωση του εγκεφαλικού νεοφλοιού ήταν ελάχιστα κατανοητή και ακόµη υπό αµφισβήτηση. Τα τελευταία 30 χρόνια, όµως, έχουν προκύψει δραµατικές αλλαγές. Η χολινεργική νεύρωση του εγκεφαλικού φλοιού αποτελεί µία από τις πιο δυναµικές περιοχές έρευνας και προσφέρει σηµαντικές ευκαιρίες για τη διερεύνηση της χηµικής νευροανατοµίας της µάθησης, των σχετιζόµενων µε τη γήρανση διαταραχών της µνήµης και της νόσου Alzheimer. Η χολινεργική οδός από τη βάση του πρόσθιου εγκεφάλου περιέχει τη µεγαλύτερη πυκνότητα νευρικών ινών από όλες τις εξω-θαλαµικές προβολές του εγκεφαλικού φλοιού. Νέα στοιχεία, που βασίζονται σε έρευνες µε ηλεκτρονικό µικροσκόπιο, ανοσοϊστοχηµεία, αντισώµατα ανασυνδυασµένων υποτύπων υποδοχέων, in situ υβριδισµό και εκλεκτικές νευροτοξικές καταστροφές, έχουν προσφέρει µία πληθώρα νέων πληροφοριών για την οργάνωση και τη λειτουργία της χολινεργικής οδού σε διάφορα είδη, συµπεριλαµβανοµένου του ανθρώπου 10. Ο βασικός πρόσθιος εγκέφαλος περιέχει τέσσερις κύριες οµάδες κυττάρων, οι οποίες προβάλλουν σε άλλες δοµές του τελεγκεφάλου. Αυτές οι χολινεργικές οµάδες κυττάρων δε χαρακτηρίζονται από τα παραδοσιακά όρια των πυρήνων και τα δοµικά τους νευρικά κύτταρα αναµειγνύονται µε µη χολινεργικούς νευρώνες. Έτσι, χρησιµοποιείται η κατάταξη Ch1-Ch4 (Εικόνα 1) για να περιγραφούν οι χολινεργικοί νευρώνες (αυτοί που περιέχουν χολινο-ακετυλοτρανσφεράση ChAT) αυτών των τεσσάρων οµάδων νευρικών κυττάρων 11,12,13. Σύµφωνα µε αυτήν την κατάταξη η οµάδα Ch1 περιγράφει τα χολινεργικά κύτταρα που σχετίζονται κυρίως µε τον έσω πυρήνα του διαφράγµατος, η Ch2 µε τον κάθετο πυρήνα της διαγώνιας ταινίας, η Ch3 µε τη διαγώνια ζώνη της διαγώνιας ταινίας και η Ch4 µε το βασικό πυρήνα του Meynert. Πειράµατα που διεξήχθησαν σε διάφορα ζωικά είδη, έδειξαν ότι οι οµάδες Ch1 και Ch2 παρέχουν την κύρια χολινεργική νεύρωση στο σύµπλεγµα του ιππόκαµπου, η Ch3 στον οσφρητικό βολβό και η Ch4 στο υπόλοιπο του εγκεφαλικού φλοιού και την αµυγδαλή
46 Εικόνα 1. Κατάταξη των χολινεργικών νευρώνων της βάσης του πρόσθιου εγκεφάλου 12 (AMG = αµυγδαλή, HC = ιππόκαµπος, NC = νεοφλοιός, OB = οσφρητικός βολβός, TH = θάλαµος) Στον εγκέφαλο των ανώτερων θηλαστικών, η οµάδα Ch4 περιέχει ένα συµπαγές-βασικό στοιχείο στο βασικό πυρήνα του Meynert και διάµεσα στοιχεία ευρισκόµενα στην έσω κάψα, την ωχρά σφαίρα, την ansa peduncularis (πολύπλοκη δέσµη νευρικών ινών, που συνδέει τον αµυγδαλοειδή πυρήνα, την απιδοειδή περιοχή και τον πρόσθιο υποθάλαµο µε διάφορους θαλαµικούς πυρήνες) και τη φακοειδή αγκύλη (ansa lenticularis) 11,13,14,15. Ακόµη και στο βασικό πυρήνα του Meynert, οι χολινεργικοί νευρώνες Ch4 αναµειγνύονται µε έναν ετερογενή πληθυσµό µη-χολινεργικών νευρώνων 16. Εποµένως, οι όροι «Ch4» και «βασικός πυρήνας του Meynert» δεν είναι συνώνυµοι. Ο όρος «βασικός πυρήνας του Meynert» µπορεί να χρησιµοποιείται για την περιγραφή όλων των στοιχείων του πυρήνα, ενώ ο πιο περιοριστικός όρος «Ch4» χρησιµοποιείται για τους βασικούς και διάµεσους χολινεργικούς νευρώνες, όπως προσδιορίζονται µε ChAT ανοσοϊστοχηµεία. Ο βασικός πυρήνας του Meynert (Εικόνες 2α και 2β) στον άνθρωπο εκτείνεται από το επίπεδο του οσφρητικού φύµατος µέχρι την οπίσθια αµυγδαλή, εκτεινόµενος σε µία απόσταση 13-14mm στον προσθιοπίσθιο άξονα και έχοντας µεσοπλάγιο βάθος 18mm µέσα στη ανώνυµη ουσία. Οι Arendt και συν. (1985) 17 εκτίµησαν ότι ο βασικός πυρήνας του Meynert στον άνθρωπο περιέχει περίπου νευρώνες σε κάθε ηµισφαίριο. 45
 18 Τοπογραφικά ο βασικός πυρήνας του Meynert στον άνθρωπο (και η οµάδα Ch4 που")
47 Εικόνα 2α. Ιστολογικό παρασκεύασµα διατοµής εγκεφάλου σε µετωπιαίο επίπεδο, διερχόµενο από το βασικό πυρήνα του Meynert (βέλος) 18 Εικόνα 2β. Σχηµατική απεικόνιση διατοµής του εγκεφάλου σε µετωπιαίο επίπεδο Επισηµαίνεται ο βασικός πυρήνας του Meynert (B, βέλος) 18 Τοπογραφικά ο βασικός πυρήνας του Meynert στον άνθρωπο (και η οµάδα Ch4 που περιέχει) µπορεί να υποδιαιρεθεί σε τοµείς που περιλαµβάνουν την πρόσθια έσω (anteromedial nbch4am), την προσθιοπλάγια (anterolateral 46
48 nb-ch4al), την προσθιοδιάµεση (anterointermediate nb-ch4ai), τη διάµεση (intermediate nb-ch4i) και την οπίσθια (posterior nb-ch4p) οµάδα νευρικών κυττάρων 13. εν υπάρχουν αυστηρά όρια µεταξύ του βασικού πυρήνα του Meynert και των παρακείµενων οµάδων κυττάρων, όπως αυτά του οσφρητικού φύµατος, της προοπτικής περιοχής, του υποθαλαµικού πυρήνα, του πυρήνα της διαγώνιας ζώνης, του αµυγδαλοειδούς πυρήνα και της ωχράς σφαίρας. Επιπρόσθετα, σε αυτήν την ανοιχτή δοµή του πυρήνα, οι νευρώνες του βασικού πυρήνα του Meynert παρουσιάζουν φυσιολογική και µορφολογική ετερογένεια. Η πλειονότητα των Ch4 νευρώνων έχουν ισοδενδριτική µορφολογία µε επικαλυπτόµενα δενδριτικά πεδία, πολλά από τα οποία εκτείνονται µέσα σε οδούς νευρικών ινών διατρέχοντας τη βάση του πρόσθιου εγκεφάλου. Αυτά τα χαρακτηριστικά είναι επίσης παρόντα στον πυρήνα του δικτυωτού σχηµατισµού του εγκεφαλικού στελέχους και έχουν οδηγήσει στην υπόθεση ότι το σύµπλεγµα nb-ch4 µπορεί να θεωρηθεί ως η τελεγκεφαλική προέκταση του δικτυωτού σχηµατισµού του εγκεφαλικού στελέχους 19. Το περικάρυο (το κυτταρικό σώµα του νευρώνα) των Ch4 νευρώνων περιέχει µεγάλα ποσά κυτταροπλάσµατος µε περίσσεια οργανιδίων και καλά ανεπτυγµένο αδρό ενδοπλασµατικό δίκτυο, διπλασιαζόµενο πυρήνα και εµφανείς πυρηνίσκους 20,21,22. Η συναπτική διέγερση στο περικάρυο και στους γειτονικούς δενδρίτες είναι αραιή, αλλά αυξάνεται αποκεντρωτικά. Οι συναπτικές εξειδικεύσεις είναι κυρίως ασύµµετρες στους δενδρίτες και συµµετρικές στο σώµα. Στον ανθρώπινο εγκέφαλο, επιπροσθέτως της ChAT, οι Ch4 νευρώνες εκφράζουν την ακετυλοχολινεστεράση AChE, την calbindin-d28k, τον trka υψηλής συγγένειας υποδοχέα του παράγοντα ανάπτυξης των νεύρων (NGFr) και τον p75 χαµηλής συγγένειας υποδοχέα του παράγοντα ανάπτυξης των νεύρων (NGFr) 16,23. Μία µικρή µειονότητα Ch4 νευρώνων είναι NGFr-αρνητικοί και προβάλλουν επιλεκτικά στην αµυγδαλή 24,25,26. Ο βασικός πυρήνας του Meynert περιέχει, επίσης, ένα σύνθετο µωσαϊκό µηχολινεργικών νευρώνων, οι οποίοι είναι NADPHd θετικοί, GABAνεργικοί, πεπτιδεργικοί, και θετικοί στην υδροξυλάση της τυροσίνης (TH) 27,28,29,30,31,32. Οι GABAνεργικοί νευρώνες του βασικού πυρήνα του Meynert ίσως είναι όσοι και οι χολινεργικοί µε τους οποίους αναµειγνύονται. Ορισµένοι από αυτούς τους νευρώνες είναι διάµεσοι νευρώνες (interneurons), ενώ άλλοι προβάλλουν στον εγκεφαλικό φλοιό και 47
49 νευρώνουν φλοιϊκούς ανασταλτικούς διάµεσους νευρώνες 32,33. Οι θετικοί στην υδροξυλάση της τυροσίνης (TH) νευρώνες έχουν ιδιαίτερο ενδιαφέρον, αφού φαίνεται να είναι πολυάριθµοι σε αρκετά µη-ανθρώπινα είδη και µπορεί να προσφέρουν µία εναλλακτική πηγή µονοαµινεργικής νεύρωσης του βασικού πυρήνα του Meynert και του εγκεφαλικού φλοιού. Αυτοί οι νευρώνες δεν έχουν ακόµη ανιχνευθεί στον ανθρώπινο εγκέφαλο, αλλά από προκαταρκτικές µελέτες φαίνεται ότι οι θετικοί στην υδροξυλάση της τυροσίνης (TH) νευρώνες είναι αρκετά σπάνιοι στον ανθρώπινο βασικό πυρήνα του Meynert. Το κύκλωµα των νευροδιαβιβαστών του βασικού πυρήνα του Meynert είναι πολύπλοκο. Μελέτες µε ηλεκτρονικό µικροσκόπιο δείχνουν ότι οι GABAεργικές τελικές απολήξεις δηµιουργούν συνάψεις µε Ch4 χολινεργικούς νευρώνες και ότι οι χολινεργικές τελικές απολήξεις δηµιουργούν συνάψεις µε GABAεργικούς νευρώνες του βασικού πυρήνα του Meynert 34. Σε κυτταρικές καλλιέργειες νευρώνων του βασικού πυρήνα του Meynert βρέθηκε ότι οι νευρώνες αυτοί είναι ευαίσθητοι σε ακετυλοχολίνη, νευροτενσίνη, ουσία P και L-γλουταµινικό 35,36. Η µονοαµινεργική νεύρωση του βασικού πυρήνα του Meynert έχει επίσης προσελκύσει σηµαντικό ερευνητικό ενδιαφέρον. Ο βασικός πυρήνας του Meynert περιέχει θέσεις υποδοχέων για 5HT, ντοπαµίνη και νορεπινεφρίνη 37. Στον επίµυ έχουν περιγραφεί προβολές από ντοπαµινεργικούς κοιλιακούς νευρώνες της καλύπτρας, από σεροτονινεργικούς νευρώνες της ραφής και από νοραδρενεργικούς νευρώνες του υποµέλανα τόπου, αλλά η ταυτότητα των νευρώνων του βασικού πυρήνα του Meynert που συµµετέχουν σε αυτές, καθώς και οι λεπτοµέρειες της συναπτικής µορφολογίας παραµένουν αδιευκρίνιστες 38. Στον ανθρώπινο εγκέφαλο η ανοσοδραστικότητα της υδροξυλάσης της τυροσίνης (TH) και της βήτα-υδροξυλάσης της ντοπαµίνης (DBH) έχει µελετηθεί µε οπτικό µικροσκόπιο στη διαφραγµατική περιοχή, αλλά όχι στο βασικό πυρήνα του Meynert 39. Αξιόλογη ανοσοδραστικότητα της σεροτονίνης είναι δύσκολο να ανιχνευθεί στον ανθρώπινο εγκέφαλο. Στον εγκέφαλο του πιθήκου έχει ανιχνευθεί ένα πυκνό πλέγµα σεροτονινοανοσοδραστικών νευραξόνων µέσα στο βασικό πυρήνα του Meynert 40. Εποµένως στα ανώτερα θηλαστικά ο βασικός πυρήνας του Meynert αποτελεί µία θέση εκτεταµένων αλληλεπιδράσεων χολινεργικών- µονοαµινεργικών νευροδιαβιβαστών. 48
50 O βασικός πυρήνας του Meynert (NBM) λαµβάνει επίσης χολινεργική νεύρωση. Μελέτες ανοσοεπισηµασµένης ChAT µε ηλεκτρονικό µικροσκόπιο σε ιστούς του NBM αποκαλύπτουν την ύπαρξη χολινεργικών νευρικών απολήξεων. Η µεγάλη πλειονότητα αυτών των απολήξεων δηµιουργεί συνάψεις µε µη χολινεργικούς νευρώνες 41. Η ακριβής πηγή της χολινεργικής νεύρωσης του NBM είναι άγνωστη, αλλά πιθανόν να περιλαµβάνει πλευρικούς κλάδους από χολινεργικούς νευρώνες των οµάδων Ch1-Ch4 της βάσης του πρόσθιου εγκεφάλου ή ανιούσες προβολές από τις οµάδες Ch5-Ch6 του γεφυροµεσεγκεφαλικού χολινεργικού πυρήνα 38. Μελέτες σε νευρώνες του NBM και µε τεχνικές patch clamp αποδεικνύουν την παρουσία ρευµάτων που σχετίζονται µε νικοτινικούς και µε M2 µουσκαρινικούς υποδοχείς 42. Οι νευρώνες του βασικού πυρήνα στον επίµυ εκφράζουν mrna m2 (µοριακά οριζόµενων) υποδοχέων και περιέχουν m2 και m3 υποδοχείς. Ο m2 υποδοχέας φαίνεται να είναι ο επικρατών χολινεργικός υποδοχέας στο βασικού πυρήνα του Meynert 43,44,45. Αυτός ο υπότυπος του υποδοχέα εκφράζεται από σχεδόν το ένα τρίτο των Ch4 νευρώνων και από πολλούς µη-χολινεργικούς νευρώνες του βασικού πυρήνα του Meynert. Ανοσοκυτταροχηµεία µε αντισώµατα, έναντι ανασυνδυασµένης πρωτεΐνης m2 υποδοχέα, απoδεικνύει ότι ο NBM περιέχει αρκετές m2 ανοσοδραστικές νευραξονικές απολήξεις, οι οποίες συνάπτονται κυρίως µε µη επισηµασµένους δενδρίτες και κυτταρικά σώµατα, διαµέσου ασύµµετρων συνάψεων 46. Αυτές οι παρατηρήσεις δείχνουν ότι ο m2 υπότυπος υποδοχέα µπορεί να λειτουργεί ως προσυναπτικός, αλλά και ως µετασυναπτικός υποδοχέας για χολινεργικούς και µη-χολινεργικούς νευρώνες του NBM. Φαίνεται, επίσης, ότι η χολινεργική νευροδιαβίβαση στον NBM κυριαρχείται από τον m2 υπότυπο, ενώ στον εγκεφαλικό φλοιό από τον m1 υπότυπο. Η έρευνα στο βασικό πυρήνα του Meynert παραδοσιακά επικεντρώνεται στη χολινεργική εξάρτηση των νευρώνων. Οι µη-χολινεργικοί νευρώνες πιθανόν να παίζουν σηµαντικό ρόλο στη συναπτική οργάνωση του τοπικού κυκλώµατος και µπορεί να αποτελούν ένα δοµικό τµήµα των αποµακρυσµένων από το φλοιό προβολών, που πηγάζουν από αυτόν τον πυρήνα. Η σηµασία αυτού του µη-χολινεργικού τµήµατος έχει επισηµανθεί και από έρευνες µε εκλεκτικές νευροτοξικές καταστροφές χολινεργικών 49
51 νευρώνων στον NBM, οι οποίες προκαλούν λιγότερο σοβαρές διαταραχές από ό,τι οι καταστροφές που επιδρούν σε όλα τα είδη κυττάρων 47,48. Επισηµαίνεται ότι στα κατώτερα θηλαστικά χρησιµοποιείται και ο όρος βασικός µεγαλοκυτταρικός πυρήνας (nucleus basalis magnocellularis), ο οποίος είναι οµόλογος µε το βασικό πυρήνα του Meynert (nucleus basalis of Meynert) των ανώτερων θηλαστικών 49,50,51, Χολινεργική νεύρωση του εγκεφαλικού φλοιού Πειραµατικές νευροανατοµικές µέθοδοι σε εγκέφαλο πιθήκων απέδειξαν ότι συγκεκριµένες φλοιϊκές περιοχές δέχονται την κύρια χολινεργική τους νεύρωση από διαφορετικές περιοχές του nb-ch4 συµπλέγµατος. Έτσι, η Ch4am περιοχή παρέχει χολινεργική νεύρωση σε µέσες φλοιϊκές περιοχές συµπεριλαµβανοµένης της υπερµεσολόβιας έλικας (έλικας του προσαγωγίου), η Ch4al σε µετωποβρεγµατικές περιοχές της καλύπτρας της νήσου και του αµυγδαλοειδή πυρήνα, η Ch4i σε πλαγιοπίσθιες µετωποβρεγµατικές, ραβδωτές και µέσες κροταφικές περιοχές και η Ch4p σε ανώτερες κροταφικές περιοχές και τους κροταφικούς πόλους 11. Οι πειραµατικές µέθοδοι που εφαρµόζονται για αυτόν τον τοπογραφικό προσδιορισµό, δεν µπορούν να εφαρµοστούν στον ανθρώπινο εγκέφαλο. Ωστόσο, στοιχεία έµµεσης τεκµηρίωσης για την ύπαρξη αντίστοιχης τοπογραφικής κατανοµής στον ανθρώπινο εγκέφαλο, µπορούν να συλλεχθούν από ασθενείς της νόσου Alzheimer. Έχουν περιγραφεί ασθενείς στους οποίους διαπιστώθηκε ότι η εκτεταµένη απώλεια χολινεργικών ινών στους κροταφικούς πόλους, αλλά όχι στον πρόσθιο φλοιό της καλύπτρας της νήσου, συνδέεται µε απώλεια κυττάρων στον πρόσθιο (Ch4p), αλλά όχι στον οπίσθιο (Ch4am+Ch4al) τοµέα του Ch4 13. Αυτή η σχέση είναι συµβατή µε την τοπογραφία των προβολών στον εγκέφαλο του πιθήκου. Η ανοσοκυτταροχηµεία για χολινοακετυλοτρανσφεράση (ChAT) παρέχει ένα από τα πιο σηµαντικά εργαλεία για τη µορφολογική µελέτη των χολινεργικών οδών, αφού η ChAT είναι εκλεκτικός και συγκεκριµένος δείκτης για χολινεργικά περικάρυα και νευράξονες. Χολινεργικοί (ChATανοσοδραστικοί) νευράξονες είναι παρόντες σε όλες τις περιοχές του ανθρώπινου εγκεφαλικού φλοιού, αλλά εµφανίζουν επίσης µία οργανωµένη µεταβολή της πυκνότητάς τους, η οποία ακολουθεί τη συναπτική ιεραρχία των συστηµάτων επεξεργασίας των πληροφοριών 53,54. Η πυκνότητα των 50
52 χολινεργικών νευραξόνων είναι υψηλότερη στα επιφανειακά στρώµατα του εγκεφαλικού φλοιού, υποδηλώνοντας ότι οι νευράξονες, που εισέρχονται στο φλοιό από την υποκείµενη φαιά ουσία, διακλαδίζονται καθώς πορεύονται προς τη χοριοειδή επιφάνεια, µία ερµηνεία που υποστηρίζεται από στοιχεία φυσιολογίας 54,55. Στον επίµυ και στη γάτα, οι ChAT ανοσοδραστικές νευρικές ίνες είναι κυρίως αµύελες, παρουσιάζουν πολυάριθµες διογκώσεις και δηµιουργούν κυρίως συµµετρικές συνάψεις µε περικάρυα και δενδρίτες πυραµοειδών και µη-πυραµοειδών νευρώνων 56,57,58. Η ακετυλοχολίνη ACh, η οποία απελευθερώνεται από χολινεργικούς νευράξονες, ασκεί τη δράση της στον εγκεφαλικό φλοιό αλληλεπιδρώντας µε µουσκαρινικούς και νικοτινικούς υποδοχείς. Αντισώµατα κατά ετερογενών µουσκαρινικών και νικοτινικών υποδοχέων έχουν αποκαλύψει µεγάλο αριθµό ανοσοθετικών, προφανώς χολινορυθµιζόµενων, φλοιϊκών νευρώνων στον ανθρώπινο εγκέφαλο 59,60. Στον εγκέφαλο των θηλαστικών, οι φαρµακολογικά οριζόµενοι M1 (ή µοριακά οριζόµενοι m1) υπότυποι του µουσκαρινικού υποδοχέα αποτελούν το πιο πολυάριθµο είδος των χολινεργικών υποδοχέων στον εγκεφαλικό φλοιό. Με µεθόδους βασισµένες σε αυτοραδιογραφία (autoradiography) έχουν ανιχνευθεί περιοχικές και σε στιβάδες διαφοροποιήσεις στην κατανοµή των υποτύπων των µουσκαρινικών υποδοχέων στον εγκέφαλο του ανθρώπου και του πιθήκου 61,62,63. Οι µελέτες για τους χολινεργικούς υποδοχείς έχουν περάσει σε µία νέα και παραγωγική περίοδο µε το µοριακό προσδιορισµό και την κλωνοποίηση των πέντε υποτύπων (m1-m5) των µουσκαρινικών υποδοχέων 64,65,66,67. Και οι πέντε υπότυποι υποδοχέων είναι συνδεόµενοι µε πρωτεΐνη G: οι m1, m3 και m5 επιλεκτικά ενεργοποιούν τη φωσφολιπάση C, ενώ οι m2 και m4 εµποδίζουν τη δράση της αδενυλκυκλάσης 66. Η ύπαρξη ολιγονουκλεοτιδικών δεικτών και εκλεκτικών αντισωµάτων για ανασυνδυασµένους υποτύπους υποδοχέων έχει συντελέσει στη µελέτη αυτών των υποδοχέων µε µεθόδους in situ υβριδισµού και ανοσοκυτταροχηµείας 43,68,69. Με τις παραπάνω µεθόδους βρέθηκε ότι o m1 υπότυπος υπάρχει στην πλειονότητα των νευρώνων του φλοιού, κατανοµή που είναι συµβατή µε το ρόλο του, ως του κύριου µετασυναπτικού χολινεργικού υποδοχέα στον εγκεφαλικό φλοιό 43,68. Aνοσοδραστικότητα τύπου m1 στο περικάρυο παρατηρείται όχι µόνο σε σχέση µε συµµετρικές 51
53 συνάψεις, χαρακτηριστικές των φλοιϊκών χολινεργικών οδών, αλλά και σε σχέση µε ασύµµετρες συνάψεις, χαρακτηριστικές των οδών των διεγερτικών αµινοξέων. Αυτό αυξάνει την πιθανότητα η ακετυλοχολίνη ACh να παίζει ρυθµιστικό ρόλο στη νευροδιαβίβαση µε διεγερτικά αµινοξέα στον εγκεφαλικό φλοιό, µέσω θέσεων των m1 υποδοχέων 70. Στο φλοιό οι m2 υπότυποι εντοπίζονται κυρίως στο νευροπίληµα (neuropil: σύµπλεγµα συνυφαινόµενων δενδριτών, νευραξώνων και νευρογλοιακών κυττάρων στο κεντρικό νευρικό σύστηµα και σε ορισµένα τµήµατα του περιφερικού νευρικού συστήµατος), αλλά και σε κάποια περικάρυα, υποδηλώνοντας ότι αυτός ο υπότυπος υποδοχέα µπορεί να λειτουργεί ταυτόχρονα ως προσυναπτικός υποδοχέας (αυτο-υποδοχέας, autoreceptor) και ως µετασυναπτικός υποδοχέας 43. Ο m2 υπότυπος σχετίζεται επίσης µε µη-χολινεργικές νευρικές απολήξεις, γεγονός που ενισχύει την άποψη ότι µπορεί να δρα και ως προσυναπτικός ετεροϋποδοχέας (heteroceptor), µέσω του οποίου η ακετυλοχολίνη ACh µπορεί να ρυθµίζει την απελευθέρωση άλλων νευροδιαβιβαστών 70. Ο m3 υπότυπος ανιχνεύεται σε πολλές περιοχές του πρόσθιου εγκεφάλου, συµπεριλαµβανοµένων της πυραµοειδούς στιβάδας του ιπποκάµπου, του ρινεγκεφάλου και των ανώτερων στρωµάτων του εγκεφαλικού φλοιού 68. Σε ανοσοκυτταροχηµικά πειράµατα εντοπίζεται κυρίως στο νευροπίληµα και ελάχιστα σε περικάρυα, υποδηλώνοντας ότι ο m3 υπότυπος µπορεί να λειτουργεί ως προ- αλλά και ως µετα- συναπτικός υποδοχέας στον εγκεφαλικό φλοιό 45. Αυτές οι παρατηρήσεις δείχνουν ότι οι υπότυποι του µουσκαρινικού υποδοχέα µπορούν να λειτουργούν όχι µόνο ως µετασυναπτικοί υποδοχείς σε παραδοσιακά χολινορυθµιζόµενες θέσεις, αλλά και ως µετασυναπτικοί ρυθµιστές µη-χολινεργικής νευροδιαβίβασης και ως προσυναπτικοί υποδοχείς (αυτο-υποδοχείς, autoreceptors), που επηρεάζουν την απελευθέρωση της ακετυλοχολίνης ACh και άλλων νευροδιαβιβαστών. Οι εισερχόµενοι χολινεργικοί νευράξονες νευρώνουν µεγάλο αριθµό φλοιϊκών νευρώνων. Μία πλήρης µορφολογική και κυτταροχηµική κατάταξη των φλοιϊκών χολινορυθµιζόµενων νευρώνων δεν είναι ακόµη εφικτή. Ο µόνος δείκτης που φαίνεται να υπάρχει σε όλους τους χολινεργικούς νευρώνες είναι η ακετυλοχολινεστεράση AChE, το ένζυµο που διακόπτει τη χολινεργική νευροδιαβίβαση διαµέσου ταχείας υδρόλυσης της ακετυλοχολίνης ACh σε οξικό και χολίνη. Αν και όλοι οι χολινεργικοί νευρώνες πιθανόν εκφράζουν την ακετυλοχολινεστεράση AChE, µόνο ένα 52
54 υποσύνολό τους παράγει ποσότητα τέτοια, ώστε να είναι πλούσιοι ιστοχηµικώς στην ακετυλοχολινεστεράση AChE. Στον εγκεφαλικό φλοιό του ενήλικου επίµυος, τέτοια χαρακτηριστικά έχουν λίγοι πολυµορφικοί νευρώνες 71,72,73. Η κατάσταση είναι δραµατικά διαφορετική στον ανθρώπινο εγκέφαλο, ο οποίος περιέχει ένα πυκνό δίκτυο πλούσιων σε ακετυλοχολινεστεράση AChE φλοιϊκών νευρώνων, κυρίως στις στιβάδες III και V του προκινητικού και αισθητικού φλοιού 13,14,74,75,76. Η πυκνότητα και η ένταση της χρώσης αυτών των νευρώνων είναι υψηλότερη στον ανθρώπινο εγκέφαλο από κάθε άλλο είδος που έχει µελετηθεί, συµπεριλαµβανοµένων των πιθήκων macaque και baboon. Αυτοί οι νευρώνες παρουσιάζουν, επίσης, ένα ασυνήθιστο οντογενετικό προφίλ: η πλούσια σε ακετυλοχολινεστεράση AChE χρώση τους δεν είναι ανιχνεύσιµη µέχρι το δέκατο έτος της ηλικίας, ανιχνεύεται όµως πλήρως κατά την ενηλικίωση. Αυτοί οι πλούσιοι σε ακετυλοχολινεστεράση AChE νευρώνες είναι αρνητικοί σε ChAT και εποµένως µη-χολινεργικοί. Παραλιµβικές (paralimbic) και µεταιχµιακές- λιµβικές (limbic) περιοχές του ανθρώπινου εγκεφάλου δέχονται µία πολύ πυκνή χολινεργική νεύρωση, αλλά έχουν πολύ λίγους νευρώνες πλούσιους σε ακετυλοχολινεστεράση AChE. Επιπλέον, πλούσιοι σε ακετυλοχολινεστεράση AChE ενδοφλοιϊκοί νευρώνες είναι σπάνιοι κατά τη νηπιακή ηλικία, όταν ο εγκεφαλικός φλοιός περιέχει ένα πυκνό δίκτυο προσαγωγών χολινεργικών ινών και υποθετικά έναν αντιστοίχως µεγάλο αριθµό χολινορυθµιζόµενων νευρώνων. Εποµένως, είναι λογικό να υποθέσουµε ότι οι πλούσιοι σε ακετυλοχολινεστεράση AChE φλοιϊκοί νευρώνες αποτελούν ένα ειδικό υποσύνολο των χολινορυθµιζόµενων νευρώνων και ότι η υψηλή περιεκτικότητά τους σε ακετυλοχολινεστεράση AChE µπορεί να αντανακλά λειτουργίες, που ξεπερνούν τις απαιτήσεις της φυσιολογικής χολινεργικής νευροδιαβίβασης. Συµβατή µε αυτήν την πιθανότητα είναι και η άποψη ότι η έκφραση της ακετυλοχολινεστεράσης AChE µπορεί να εµπλέκεται σε έναν αριθµό µη-χολινεργικών φαινοµένων, όπως την προαγωγή της ανάπτυξης του νευράξονα, την αύξηση του µήκους του, τη συναπτογένεση, τη νευρωνική διαφοροποίηση και πλαστικότητα, τις νευροτροφικές επιδράσεις και την κυτταρική προσκόλληση 77,78,79. Οι µηχανισµοί αυτών των λειτουργιών παραµένουν αινιγµατικοί, αλλά φαίνεται ότι είναι µη-ενζυµικοί, αφού δεν καταργούνται από αναστολείς της ενζυµικής δραστηριότητας της ακετυλοχολινεστεράσης AChE. Η έντονη 53
55 δραστηριότητα της ακετυλοχολινεστεράσης AChE στους πυραµοειδείς ενδοφλοιϊκούς νευρώνες µπορεί να αποτελεί δείκτη ειδικών ενεργών δράσεων όπως η αναδιοργάνωση, η πλαστικότητα και η ωρίµανση, που παρατηρούνται σχετικά αργά στην ανάπτυξη, σε χρόνο που αναπτύσσονται οι ανώτερες συµπεριφορικές και γνωστικές δεξιότητες. Οι πλούσιοι σε ακετυλοχολινεστεράση AChE χολινεργικοί νευρώνες του εγκεφαλικού φλοιού, οι οποίοι αναγνωρίζονται µε µονοκλωνικά αντισώµατα για ανθρώπινη ακετυλοχολινεστεράση AChE, είναι πλούσιοι σε µηφωσφορυλιωµένες νευροϊνιδιακές πρωτεΐνες και η πυκνότητά τους παραµένει σταθερή στο προχωρηµένο γήρας, αλλά µόνο σε άτοµα µε άθικτη νοητική κατάσταση 13,75,80. Αυτοί οι νευρώνες βλάπτονται σοβαρά σε νευροεκφυλιστικές παθήσεις, που επηρεάζουν τη νοητική κατάσταση, όπως η νόσος Alzheimer 81. Φαίνεται ότι οι συγκεκριµένοι νευρώνες µπορεί να παίζουν σηµαντικό ρόλο σε ανώτερα στάδια φλοιϊκής και νοητικής ανάπτυξης και ότι η εκφύλισή τους µπορεί να αποτελεί το ανατοµικό υπόστρωµα της άνοιας. 54
56 2. Στοιχεία φυσιολογίας της συµπεριφορικής εξειδίκευσης του βασικού πυρήνα του Meynert Ο βασικός πυρήνας του Meynert είναι φυλογενετικά εξελιγµένος πυρήνας, ο οποίος παρουσιάζει τη µεγαλύτερη διαφοροποίησή του στον εγκέφαλο κητωδών ζώων και του ανθρώπου. Ακόµη και στον ανθρώπινο εγκέφαλο, όπου αποκτά το µεγαλύτερο µέγεθός του, δεν περιέχει περισσότερα από νευρικά κύτταρα σε κάθε ηµισφαίριο. Αν και λίγοι σε αριθµό, αυτοί οι νευρώνες παρέχουν σε κάθε κυτταροαρχιτεκτονικό τµήµα και σε κάθε φλοιϊκή στιβάδα του εγκεφαλικού φλοιού, µία πλούσια νεύρωση χολινεργικών νευραξόνων ανά mm. Η υψηλή πυκνότητα των χολινεργικών νευραξόνων, κυρίως σε παραλιµβικές (paralimbic) και µεταιχµιακές-λιµβικές (limbic) περιοχές είναι πραγµατικά εντυπωσιακή. Το µέγεθός της από µόνο του πιθανόν να αποτελεί ένδειξη ότι αυτή η οδός αποτελεί το µοναδικό σηµαντικό ρυθµιστικό προσαγωγό σύστηµα του εγκεφαλικού φλοιού. Με βάση απόψεις συγκριτικής νευροανατοµίας, ο βασικός πυρήνας του Meynert µπορεί να θεωρηθεί ως η τελεγκεφαλική προέκταση του ισοδενδριτικού δικτυωτού πυρήνα του εγκεφαλικού στελέχους και επίσης ως τµήµα του λιµβικού φλοιού 13,28. Σύµφωνα µε αυτές τις ανατοµικές απόψεις, οι δύο σηµαντικότερες συµπεριφορικές εξειδικεύσεις, που έχουν αποδοθεί στο βασικό πυρήνα του Meynert, είναι αυτές της προσοχής-εγρήγορσης και της µνήµης-µάθησης. Η κύρια επίδραση της ακετυλοχολίνης ACh στους φλοιϊκούς νευρώνες ρυθµίζεται διαµέσου των m1 µουσκαρινικών υποδοχέων και προκαλεί µία σχετικά παρατεταµένη µείωση στην αγωγιµότητα του καλίου, µε αποτέλεσµα να µειώνει τη διεγερσιµότητα των φλοιϊκών χολινορυθµιζόµενων νευρώνων από άλλες διεγερτικές επιδράσεις. Έχουν αναγνωριστεί, επίσης, σύνθετες ανασταλτικές επιδράσεις, τόσο άµεσες όσο και έµµεσες διαµέσου της ρύθµισης GABAεργικών διάµεσων νευρώνων 82. ιέγερση του βασικού πυρήνα του Meynert προκαλεί ηλεκτροεγκεφαλογραφική (EEG) ενεργοποίηση διαµέσου µουσκαρινικών υποδοχέων, εκπολώνει τους φλοιϊκούς νευρώνες και προκαλεί µία αλλαγή στις διακυµάνσεις του δυναµικού µεµβράνης από µεγάλου εύρους αργά κύµατα σε µικρού εύρους ταχέα κύµατα (20-40Hz) 83. Λαµβάνοντας υπόψη την καθολική κατανοµή της φλοιϊκής χολινεργικής οδού, δεν είναι παράδοξο ότι όλες οι λειτουργίες του φλοιού µπορούν να ρυθµιστούν 55
57 διαµέσου της χολινεργικής νευροδιαβίβασης. Στον πρωτεύοντα οπτικό φλοιό, για παράδειγµα, η χολινεργική διέγερση αυξάνει την πιθανότητα ο νευρώνας να ανταποκριθεί ως αντίδραση στο ειδικό ερέθισµα 84. Παρόµοια ρυθµιστική δράση έχει περιγραφεί στο σωµατοαισθητικό και ακουστικό φλοιό 85,86. Οι νευρώνες του βασικού πυρήνα του Meynert είναι ευαίσθητοι σε νέα και σχετιζόµενα µε κίνητρα αισθητηριακά γεγονότα 87,88, οδηγώντας στην υπόθεση ότι η φλοιϊκή χολινεργική νεύρωση µπορεί να ρυθµίζει την επίπτωση των αισθητηριακών γεγονότων στη φλοιϊκή λειτουργία, µε τρόπο που αντανακλά τη συµπεριφορική τους σηµαντικότητα. Σύµφωνα µε τα παραπάνω, τα προκύπτοντα δυναµικά P300 στον ανθρώπινο εγκεφαλικό φλοιό µπορούν να ανασταλούν µε τη χορήγηση χολινεργικών αναστολέων 89. Βλάβες στο βασικό πυρήνα του Meynert καταστέλλουν τη χαµηλού δυναµικού ταχεία EEG δραστηριότητα, µειώνουν τη φλοιϊκή χρήση γλυκόζης και επιδρούν σε δοκιµασίες προσοχής-εγρήγορσης 90,91,92. Αυτές οι παρατηρήσεις παρέχουν σηµαντική υποστήριξη στην άποψη ότι οι κατευθυνόµενες προς το φλοιό προβολές από το βασικό πυρήνα του Meynert συµπεριλαµβάνονται στο ανιόν δικτυωτό ενεργοποιητικό σύστηµα και ότι εµπλέκονται στη ρύθµιση των φλοιϊκών επιπέδων εγρήγορσης. Πολλές παρατηρήσεις σχετίζουν τις φλοιϊκές χολινεργικές οδούς µε τη νευρωνική οργάνωση της µνήµης και της µάθησης. Η συστηµατική χορήγηση του µουσκαρινικού αποκλειστή σκοπολαµίνη προκαλεί διαταραχές της µνήµης σε διάφορα ζωικά είδη και στον άνθρωπο 93. Κατά τη διάρκεια Pavlovian συνθηκών µάθησης, σχεδόν οι µισοί νευρώνες του βασικού πυρήνα του Meynert παρουσιάζουν σηµαντικά µεγαλύτερη αλλαγή της δραστηριότητάς τους ως απάντηση σε έναν ήχο που αντιστοιχεί στην παρουσία ενός επώδυνου ερεθίσµατος, από ό,τι σε έναν διαφορετικό ήχο 94. Φαρµακολογικά πειράµατα υποδηλώνουν ότι η χολινεργική νεύρωση της αµυγδαλής, παίζει σπουδαίο ρόλο στην εµπέδωση της µνήµης 95. Πειραµατικές βλάβες στο βασικό πυρήνα του Meynert, οι οποίες περιλαµβάνουν τόσο τα χολινεργικά όσο και τα µη-χολινεργικά στοιχεία του, προκαλούν διαταραχές στη µνήµη σε κάποιες δοκιµασίες 96, αλλά όχι σε κάποιες άλλες 92. Η πρόσφατη χρησιµοποίηση της IgG-saporin (µίας νευροτοξίνης που απενεργοποιεί τα ριβοσώµατα, συνδεδεµένης µε ένα αντίσωµα που αναγνωρίζει τον υποδοχέα NGFr) έχει κάνει εφικτή την εκλεκτική καταστροφή µόνο των χολινεργικών νευρώνων στο βασικό πυρήνα του Meynert. Τέτοια πειράµατα δείχνουν ότι η καταστροφή της 56
58 φλοιϊκής χολινεργικής νεύρωσης προκαλεί σοβαρή βλάβη στη χωρική µάθηση, αλλά όχι στις δοκιµασίες καθυστερηµένης αλλαγής (delayed alteration) και παθητικής αποφυγής 47,48. Η εξειδίκευση στη συµπεριφορά και στη φυσιολογία των µη-χολινεργικών νευρώνων του βασικού πυρήνα του Meynert παραµένει αδιευκρίνιστη. Οι µηχανισµοί, που συνδέουν τις χολινεργικές προβολές από τις οµάδες νευρικών κυττάρων Ch1-Ch4 µε τις λειτουργίες της µνήµης, δεν είναι πλήρως κατανοητοί και έχουν οδηγήσει σε διάφορες υποθέσεις. Ο ρόλος της ακετυλοχολίνης ACh στην παρατεταµένη ενίσχυση (LTP) στον ιππόκαµπο 97,98 µπορεί να παρέχει τον κυτταρικό µηχανισµό που εξηγεί τη σχέση των χολινεργικών οδών µε τη µνήµη. Πειράµατα σε εγκεφαλικές τοµές του απιοειδή φλοιού του επίµυος, έχουν δείξει ότι η ακετυλοχολίνη ACh µπορεί να αναστείλει εκλεκτικά την ενδογενή συναπτική νευροδιαβίβαση διαµέσου ενός προσυναπτικού µηχανισµού, ενώ αφήνουν ανεπηρέαστη την εξωγενή προσαγωγό νευροδιαβίβαση. Η συγκεκριµένη εκλεκτική αναστολή µπορεί να προλάβει την παρεµβολή από προηγούµενα αποθηκευµένες γνώσεις κατά τη διάρκεια απόκτησης νέων γνώσεων. Αυτός µπορεί να είναι ένας νέος µηχανισµός διαµέσου του οποίου η φλοιϊκή χολινεργική νεύρωση µπορεί να συµµετέχει στη µάθηση 99. Επίσης, έχει προταθεί ένα µοντέλο σύµφωνα µε το οποίο η χολινεργική νεύρωση, ιδίως του συµπλέγµατος του ιπποκάµπου, παίζει κύριο ρόλο στην αλλαγή µεταξύ της on-line επεξεργασίας, που χαρακτηρίζεται από θήτα ιπποκάµπιο ρυθµό, και της off-line εµπέδωσης, που χαρακτηρίζεται από δραστηριότητα οξέων κυµάτων 100. Ένας άλλος µηχανισµός που συνδέει τους χολινεργικούς νευράξονες µε τη µνήµη και τη µάθηση, ενδέχεται να σχετίζεται µε την τοπογραφική διαφοροποίηση της φλοιϊκής χολινεργικής νεύρωσης. Πειραµατικά στοιχεία οδηγούν στο συµπέρασµα ότι οι αισθητηριο-µεταιχµιακές (λιµβικές) οδοί παίζουν βασικό ρόλο σε ένα µεγάλος εύρος συµπεριφορών που σχετίζονται µε το συναίσθηµα, το κίνητρο και ειδικά τη µνήµη 101,102. Η επεξεργασία αρχίζει εντός των πρωτοταγών-ανώτερων περιοχών του εγκεφαλικού φλοιού, οι οποίες αποτελούν µία πύλη για την είσοδο των αισθητηριακών πληροφοριών στο φλοιϊκό κύκλωµα. Κεντρικές λιµβικές περιοχές, όπως η αµυγδαλή και ο ιππόκαµπος, περιέχουν την υψηλότερη πυκνότητα χολινεργικής νεύρωσης. Αυτή η διαφοροποίηση της πυκνότητας οδηγεί στην υπόθεση ότι οι αισθητηριακές πληροφορίες είναι πιθανόν να 57
59 προκύπτουν κάτω από προοδευτικά µεγαλύτερη χολινεργική επίδραση κατά την πορεία τους διαµέσου πολυσυναπτικών οδών προς το µεταιχµιακό (λιµβικό) σύστηµα. Ως συνέπεια αυτής της διάταξης, η φλοιϊκή χολινεργική νεύρωση µπορεί να βοηθήσει τη διοχέτευση των αισθητηριακών πληροφοριών προς και από το µεταιχµιακό (λιµβικό) σύστηµα µε τρόπο που είναι ευαίσθητος στη συµπεριφορική εµπειρία του σχετικού πειράµατος 103. Οι διαταραχές της µνήµης, που προκύπτουν µετά από την καταστροφή στις οµάδες νευρικών κυττάρων Ch1-Ch4 ή µετά από τη συστηµατική χορήγηση χολινεργικών ανταγωνιστών, µπορεί να αντανακλά µία διαταραχή των αισθητηριο-µεταιχµιακών (λιµβικών) αλληλεπιδράσεων, οι οποίες είναι απαραίτητες για αποτελεσµατική µνήµη και µάθηση. 58
60 3. Αλλαγές στη φλοιϊκή χολινεργική νεύρωση κατά το γήρας και τη νόσο Alzheimer Οι σχετιζόµενες µε τη γήρανση αλλαγές της χολινεργικής νεύρωσης στον ανθρώπινο εγκεφαλικό φλοιό είναι τοπικά εκλεκτικές και µάλλον περιορισµένες. Φαίνεται ότι οι αµυγδαλοειδείς πυρήνες και ο φλοιός του προσαγωγίου δεν παρουσιάζουν αλλαγές, ενώ ο κατώτερος κροταφικός και ο φλοιός του ρινικού εγκεφάλου εµφανίζουν σηµαντική αλλά σχετικά περιορισµένη απώλεια περίπου του 20%, όταν συγκρίνονται οι πυκνότητες των χολινεργικών νευραξόνων σε δείγµατα ανθρώπων ετών µε αντίστοιχα ανθρώπων πάνω από 68 ετών 104,105. Οι σχετιζόµενες µε τη γήρανση αλλαγές στο βασικό πυρήνα του Meynert είναι γενικά περιορισµένες και δεν είναι ανιχνεύσιµες µέχρι το προχωρηµένο γήρας 106. Παρατηρείται µία αρχική υπερτροφία του περικαρύου γύρω στην ηλικία των 60 ετών, η οποία ακολουθείται από συρρίκνωση και κυτταρική απώλεια. Αν και περιορισµένες, αυτές οι αλλαγές στη φλοιϊκή χολινεργική νεύρωση µπορεί να παρέχουν ένα σηµαντικό ανατοµικό υπόστρωµα για τις σχετιζόµενες µε τη γήρανση αλλαγές των λειτουργιών της µνήµης. Οι νευρώνες του βασικού πυρήνα του Meynert, καθώς και οι φλοιϊκοί χολινεργικοί νευράξονες πλήττονται σοβαρά στη νόσο Alzheimer 106,107,108,109,110. Η απώλεια της φλοιϊκής χολινεργικής νεύρωσης στη νόσο αυτή έχει αναφερθεί σε περισσότερες από 30 δηµοσιεύσεις 106. Αυτή η διαταραχή είναι σοβαρή (76-85%) στον κατώτερο κροταφικό, το µέσο κροταφικό και το φλοιό του ρινικού εγκεφάλου και σε τµήµατα της αµυγδαλής. Είναι περιορισµένη (40-67%) στον προµετωπιαίο, οπίσθιο βρεγµατικό, κογχοµετωπιαίο, νησιδιακό, οπίσθιου προσαγωγίου, πρωτοταγή ακουστικό και ιπποκάµπειο φλοιό. Είναι ήπια (4-28%) στον πρωτοταγή οπτικό, το σωµατοαισθητικό, τον πρωτοταγή κινητικό, τον προκινητικό και στο φλοιό του πρόσθιου προσαγωγίου 105,106. Η χολινεργική διαταραχή, γενικώς, τείνει να επικεντρώνεται στον κροταφικό λοβό, συµπεριλαµβανοµένων των µεταιχµιακών-λιµβικών και παραλιµβικών περιοχών. Ακόµη και σε πολύ προχωρηµένα στάδια της νόσου Alzheimer, αλλά και σε περιοχές µε σοβαρή διαταραχή της χολινεργικής νεύρωσης, ο εγκεφαλικός φλοιός περιέχει ορισµένους χολινεργικούς νευράξονες. Οι τοπικές πυκνότητες αυτών των χολινεργικών νευραξόνων στον κροταφικό φλοιό 59
61 φαίνεται να αντανακλούν διαφορές στα επίπεδα νεύρωσης πριν από την εγκατάσταση της νόσου. Έτσι, περιοχές µε σχετικά χαµηλή χολινεργική νεύρωση, όπως οι κατώτερες, µέσες και ανώτερες κροταφικές συνειρµικές περιοχές, φαίνεται πως απογυµνώνονται από χολινεργικές ίνες. Αντίθετα η περιοχή του ρινικού εγκεφάλου, η οποία χάνει αντίστοιχο ποσοστό της χολινεργικής νεύρωσης, αλλά έχει υψηλότερη προνοσηρή πυκνότητα χολινεργικής νεύρωσης, διατηρεί µία σηµαντικά υψηλότερη υπολειµµατική νεύρωση 106. Επιπροσθέτως της απώλειας των φλοιϊκών χολινεργικών νευραξόνων, στη νόσο Alzheimer παρατηρείται απώλεια µουσκαρινικών m2 υποδοχέων και νικοτινικών υποδοχέων, ενώ οι παραµένοντες χολινορυθµιζόµενοι νευρώνες µπορεί να αποτυγχάνουν να εκφράζουν νικοτινικούς υποδοχείς και AChE ενζυµική δραστηριότητα 106. Σε αρκετές ερευνητικές εργασίες στην προσιτή διεθνή βιβλιογραφία σχετικά µε τη νόσο Alzheimer, αναφέρεται µία µείωση κατά 30-90% του αριθµού των νευρώνων του βασικού πυρήνα του Meynert. Σε σχετική ανασκόπηση καταγράφεται ότι η απώλεια των χολινεργικών νευρώνων είναι µόλις κάτω του 50% στον πρόσθιο (nb-ch4a) και το διάµεσο (nb- CH4i) τοµέα του βασικού πυρήνα του Meynert, ενώ η απώλεια αυτή αυξάνεται πάνω από 60% στον οπίσθιο (nb-ch4p) τοµέα 106. Ο Ch4p τοµέας είναι η κύρια πηγή των χολινεργικών προβολών στον κροταφικό λοβό και τις άνω κροταφικές έλικες 11, δύο περιοχές που σταθερά παρουσιάζουν µία σοβαρή απώλεια των χολινεργικών νευραξόνων. Το σχετικά µεγαλύτερο ποσοστό κυτταρικής απώλειας σε αυτόν τον τοµέα είναι σύµφωνο µε τη συνδεσιµότητα του Ch4 και την τοπική κατανοµή της απώλειας των χολινεργικών νευραξόνων στη νόσο Alzheimer. Με βάση τη σχετικά περιορισµένη απώλεια της χολινεργικής νεύρωσης στον ιππόκαµπο και στην υποκείµενη στηρικτική δοµή, η κυτταρική απώλεια στους τοµείς Ch1 και Ch2 αναφέρεται συνήθως λιγότερο σηµαντική σε σύγκριση µε τον Ch4. Στη νόσο Alzheimer καθώς και κατά τη γήρανση, οι νευρώνες του βασικού πυρήνα του Meynert φαίνεται να παρουσιάζουν µία αρχική υπερτροφία του περικαρύου, η οποία ακολουθείται από συρρίκνωση και κυτταρική απώλεια. Πολλοί νευρώνες του πυρήνα σε ασθενείς µε τη νόσο αυτή εµφανίζουν νευροϊνιδιακό εκφυλισµό. Η νόσος Alzheimer οδηγεί σε αποδιοργάνωση των εξωθαλαµικών κατευθυνόµενων προς το φλοιό προβολών, που εκπορεύονται όχι µόνο από το βασικό πυρήνα του Meynert, αλλά και από τον ιππόκαµπο 60
62 (ισταµινεργικές), την κοιλιακή περιοχή της καλύπτρας (ντοπαµινεργικές), τον πυρήνα της ραφής (σεροτονινεργικές) και τον υποµέλανα τόπο (νοραδρενεργικές). Ωστόσο, η απώλεια της φλοιϊκής χολινεργικής νεύρωσης και ο σχετιζόµενος µε αυτήν εκφυλισµός του βασικού πυρήνα του Meynert, παρουσιάζεται νωρίτερα στην πορεία της νόσου και είναι µεγαλύτερης βαρύτητας 106. Ορισµένοι συγγραφείς υποστηρίζουν ότι ο βασικός πυρήνα του Meynert και η απώλεια της φλοιϊκής χολινεργικής νεύρωσης παίζουν παθογενετικό ρόλο στη νόσο Alzheimer. Έτσι, έχει αναφερθεί ότι πειραµατικές καταστροφές του βασικού πυρήνα του Meynert προκαλούν το σχηµατισµό αποθέσεων στον εγκεφαλικό φλοιό, αντίστοιχων µε τις γεροντικές πλάκες 111. Επίσης, ότι οι νευρώνες του βασικού πυρήνα του Meynert µπορεί να υπερεκφράζουν την πρόδροµη πρωτεΐνη του αµυλοειδούς, σε βαθµό που να προάγουν το σχηµατισµό γεροντικών πλακών στη νόσο Alzheimer 112. Άλλοι συγγραφείς δεν µπόρεσαν να επαναλάβουν τα αποτελέσµατα των Arendash και συν. (1987) 111, καθώς και τη σχέση µεταξύ της τοπικής πυκνότητας των πλακών και είτε της σοβαρότητας της απώλειας της χολινεργικής νεύρωσης είτε της προνοσηρής πυκνότητας της χολινεργικής νεύρωσης 106. Τέλος, έχει υποστηριχθεί ότι η µουσκαρινική διέγερση των φλοιϊκών νευρώνων προάγει τη µη-αµυλοειδική οδό επεξεργασίας της πρόδροµης πρωτεΐνης του αµυλοειδούς, οδηγώντας στην υπόθεση ότι η χολινεργική διαταραχή µπορεί να προάγει την αµυλοειδογένεση 113, γεγονός που διερευνάται σε ορισµένα ερευνητικά κέντρα. Η αιτία της εκλεκτικής τρωτότητας της χολινεργικής νεύρωσης στη νόσο Alzheimer δεν είναι ακόµη πλήρως κατανοητή. Η χολίνη είναι πρόδροµη ουσία τόσο της ακετυλοχολίνης ACh όσο και της φωσφατιδυλοχολίνης 114. Μία αρχική χολινεργική διαταραχή µπορεί να οδηγήσει σε αντισταθµιστική υπερδραστηριότητα των υπολοίπων χολινεργικών νευρώνων µε τρόπο που µπορεί να αποστερεί τη χολίνη από τη σύνθεση της µεµβράνης, για να τη διαθέσει στη σύνθεση της ακετυλοχολίνης ACh. Αυτό µπορεί να οδηγήσει σε αυτοκανιβαλισµό των χολινεργικών νευρώνων και µπορεί να αποτελεί το µηχανισµό της εκλεκτικής επιτάχυνσης του εκφυλισµού των οδών των χολινεργικών προβολών. Η µεγάλη πλειονότητα των χολινεργικών νευρώνων στο βασικό πυρήνα του Meynert εξαρτά την επιβίωσή της από την ανάστροφη µεταφορά του παραγόµενου στο φλοιό παράγοντα ανάπτυξης των νεύρων NGF. Στην 61
63 πράξη, πειραµατικά προκαλούµενες βλάβες στο φλοιό, που εµποδίζουν την ανάστροφη µεταφορά του NGF, προκαλούν παθολογικές αλλαγές στους Ch4 νευρώνες που σχετίζονται µε αυτές τις φλοιϊκές περιοχές 115. Αν και δεν υπάρχουν ενδείξεις ότι ο εγκεφαλικός φλοιός στη νόσο Alzheimer παρουσιάζει έλλειµµα NGF ή ότι οι Ch4 νευρώνες αποτυγχάνουν να εκφράσουν τους υποδοχείς NGFr, η παθολογία του φλοιού, συµπεριλαµβανοµένης της εξωκυτταρικής εναπόθεσης αµυλοειδούς, µπορεί να επιδρά στην επαρκή απελευθέρωση NGF, τη σύνδεσή του µε τους NGFr, καθώς και µε την ανάστροφη µεταφορά του. Οι προβολές από το βασικό πυρήνα του Meynert στον εγκεφαλικό φλοιό είναι σηµαντικά πυκνότερες συγκρινόµενες µε τις κατευθυνόµενες προς το φλοιό προβολές από την κοιλιακή περιοχή της καλύπτρας (ντοπαµινεργικές), τον πυρήνα της ραφής (σεροτονινεργικές) ή τον υποµέλανα τόπο (νοραδρενεργικές). Υπάρχουν ενδείξεις ότι ο βασικός πυρήνας του Meynert ίσως είναι µία από τις πιο ευαίσθητες περιοχές του εγκεφάλου στην υπερφωσφορυλίωση της µικροσωληναριακής πρωτεΐνης tau και στο σχηµατισµό νευροϊνιδιακών αλλοιώσεων στην πορεία της γήρανσης. Εποµένως, οι φλοιϊκές χολινεργικές προβολές είναι σε διπλό κίνδυνο: έχουν µεγαλύτερο αριθµό νευραξόνων εκτεθειµένων στη χολινεργική παθολογία των νευροεκφυλιστικών νόσων και τα κύτταρα από τα οποία προέρχονται στο βασικό πυρήνα του Meynert βρίσκονται σε µία περιοχή επιλεκτικά ευάλωτη στη σχετιζόµενη µε τη γήρανση νευρωνική παθολογία. Μία εκλεκτική απώλεια της ανοσοδραστικότητας της calbindind28k εµφανίζεται σε Ch4 νευρώνες ηλικιωµένων ατόµων χωρίς συµπτώµατα άνοιας, οδηγώντας στο συµπέρασµα ότι αλλαγές στην ενδοκυτταρική ρύθµιση του ασβεστίου µπορεί να παρέχουν ένα µηχανισµό για την εκλεκτική ευαισθησία αυτών των νευρώνων στις σχετιζόµενες µε τη γήρανση παθολογικές διεργασίες της νόσου Alzheimer 116. Η χολινεργική οδός, που εκπορεύεται από τη βάση του πρόσθιου εγκεφάλου, συνιστά µία από τις πιο σηµαντικές ρυθµιστικές προσαγωγές οδούς του εγκεφαλικού φλοιού των θηλαστικών. Η αρχική προσδοκία ότι η χολινεργική διαταραχή θα παρείχε µία ενοποιηµένη παθοφυσιολογική θεωρία για τη νόσο Alzheimer και ότι οι χολινεργικές θεραπείες θα θεράπευαν την άνοια, ήταν εµφανώς ιδιαίτερα αισιόδοξες. Ωστόσο, η φλοιϊκή χολινεργική απονεύρωση παραµένει µία από τις πρωταρχικές, ιδιαίτερα σοβαρές και δοµικές αλλαγές των νευροδιαβιβαστών στην πορεία 62
64 της νόσου. Η χολινεργική διαταραχή µπορεί να παρέχει το βασικό υπόστρωµα των νευροφυσιολογικών χαρακτηριστικών της νόσου Alzheimer και µπορεί τελικά να οδηγήσει σε σηµαντικά συµπεράσµατα για την παθογένεσή της. 63
65 . Η ΧΟΛΙΝΕΡΓΙΚΗ ΥΠΟΘΕΣΗ ΤΗΣ ΜΝΗΜΗΣ ΚΑΙ ΤΗΣ ΑΝΟΙΑΣ ΣΤΗ ΝΟΣΟ ALZHEIMER Φαρµακολογικές µελέτες κατά τη διάρκεια των τελευταίων δεκαετιών επιβεβαιώνουν τη θεωρητική και κλινική άποψη, ότι οι χολινεργικοί νευρώνες στον εγκέφαλο εµπλέκονται στη µνήµη και τη µάθηση. Από τις διαταραχές της συµπεριφοράς, που παρατηρούνται στους ηλικιωµένους, η µειωµένη γνωστική ικανότητα θεωρείται η πιο σοβαρή και µόνιµη διαταραχή. Πολυάριθµες ερευνητικές µελέτες αποδεικνύουν ότι η πλειονότητα των υγιών ηλικιωµένων ανθρώπων εµφανίζει σηµαντική µείωση της γνωστικής ικανότητας µε την πάροδο του χρόνου 117, διαταραχή κοινή και σε άλλα είδη θηλαστικών συµπεριλαµβανοµένων των µυώνmice 118,119, επίµυων-rats 120,121 και των πιθήκων 122,123,124,125. Στον άνθρωπο το πρόβληµα αυτό συχνά επιδεινώνεται µε την εµφάνιση γεροντικής άνοιας, η οποία εκτιµάται ότι επηρεάζει πάνω από δύο εκατοµµύρια ανθρώπων µόνο στις ΗΠΑ και αναµένεται να λάβει µορφή επιδηµίας µε την πάροδο των ετών 126. Σε ορισµένες περιπτώσεις γεροντικής άνοιας οι γνωστικές διαταραχές συχνά υποβαθµίζουν την ποιότητα ζωής του ασθενούς, απαιτούν ολοκληρωµένη και διαρκή ιδρυµατική φροντίδα του και προσθέτουν οικονοµικά και συναισθηµατικά βάρη στις οικογένειες και την κοινωνία. Παρά τους κοινωνικούς, πολιτισµικούς, οικονοµικούς και ψυχολογικούς παράγοντες, που πιθανόν συντελούν στη γνωστική αποδιοργάνωση, η ιατρική κοινότητα θεωρεί ότι οι διαταραχές, που σχετίζονται µε τη γήρανση στο κεντρικό νευρικό σύστηµα (ΚΝΣ), είναι αυτές που παίζουν τον κυρίαρχο ρόλο. Οι προσπάθειες για να διευκρινιστεί ποιες αλλαγές στο ΚΝΣ παίζουν κυρίαρχο ρόλο στη γνωστική απώλεια έχουν ενταθεί τα τελευταία χρόνια. Αν και η συσχέτιση µεταξύ συγκεκριµένων αλλαγών στο ΚΝΣ λόγω ηλικίας και των γνωστικών διαταραχών είναι πολύπλοκη, φαίνεται ότι ένας κυρίαρχος παράγοντας είναι η διαταραχή στο χολινεργικό σύστηµα νευροδιαβιβαστών. Η «χολινεργική υπόθεση» έχει καταλάβει ένα σηµαντικό τµήµα στη σχετική µε τη γηριατρική βιβλιογραφία και έχει οδηγήσει σε κλινικές µελέτες για τη φαρµακολογική αποκατάσταση αυτής της χολινεργικής διαταραχής. Η νόσος Alzheimer αφορά περίπου 15 εκατοµµύρια ανθρώπων διεθνώς και είναι η κύρια αιτία άνοιας στους ηλικιωµένους. Με το ποσοστό των ηλικιωµένων στο γενικό πληθυσµό να αυξάνεται σταθερά, η επίπτωση της 64
66 ασθένειας στους παρέχοντες τη φροντίδα στους ασθενείς, καθώς και στις εθνικές οικονοµίες, αναµένεται να αυξηθεί σηµαντικά τις επόµενες 2-3 δεκαετίες. Η νόσος Alzheimer είναι µία προοδευτική νευροεκφυλιστική διαταραχή µε µέση διάρκεια, µεταξύ της εµφάνισης των κλινικών συµπτωµάτων και του θανάτου, τα 8,5 έτη. Περιοχές του εγκεφάλου που σχετίζονται µε ανώτερες νοητικές λειτουργίες, κυρίως ο νεοφλοιός και ο ιππόκαµπος, είναι αυτές που επηρεάζονται περισσότερο από τη χαρακτηριστική παθολογία της νόσου. Αυτή περιλαµβάνει την εξωκυτταρική εναπόθεση β-αµυλοειδούς (που προέρχεται από την APP πρόδροµη πρωτεΐνη του αµυλοειδούς) στις γεροντικές πλάκες, τον ενδοκυτταρικό σχηµατισµό νευροϊνιδιακών αλλοιώσεων (που περιλαµβάνουν µία µη-φυσιολογικά φωσφορυλιωµένη µορφή της µικροσωληναριακής πρωτεΐνης tau) και την απώλεια νευρικών συνάψεων και πυραµοειδών νευρώνων. Οι αλλαγές αυτές προκαλούν την τυπική συµπτωµατολογία της νόσου Alzheimer, η οποία περιλαµβάνει βαριά και προοδευτική απώλεια της γνωστικής λειτουργίας, που συχνά συνοδεύεται από διαταραχές της συµπεριφοράς όπως επιθετικότητα, κατάθλιψη και άσκοπη περιπλάνηση. Οι διαταραχές της συµπεριφοράς δηµιουργούν τα µεγαλύτερα προβλήµατα στους παρέχοντες τη φροντίδα στους ασθενείς και αποτελούν την αιτία για την ιδρυµατοποίηση των ασθενών 127. Η συστηµατική βιοχηµική ανάλυση εγκεφάλων ασθενών µε νόσο Alzheimer ξεκίνησε στα τέλη της δεκαετίας του 1960 και στις αρχές της δεκαετίας του Η ελπίδα ήταν να βρεθεί µία εµφανής νευροχηµική διαταραχή, η οποία θα αποτελούσε τη βάση για µία θεραπευτική παρέµβαση ανάλογη µε τη levodopa στη νόσο Parkinson. Συνέχεια σε αυτήν την προοπτική δόθηκε στα µέσα της δεκαετίας του 1970, µε αναφορές για σηµαντικές διαταραχές στη χολινο-ακετυλοτρανσφεράση ChAT, του ενζύµου που είναι υπεύθυνο για τη σύνθεση της ακετυλοχολίνης ACh 108,109,128,129. Ακόλουθες ανακαλύψεις για µειωµένη πρόσληψη χολίνης 130, µειωµένη απελευθέρωση ακετυλοχολίνης ACh 131 και απώλεια χολινεργικών περικάρυων στο βασικό πυρήνα του Meynert 132, επιβεβαιώνουν τη σηµαντική προσυναπτική χολινεργική διαταραχή. Οι παραπάνω µελέτες µαζί µε τον εµφανή ρόλο της ακετυλοχολίνης ACh στη µνήµη και τη µάθηση 93, οδηγούν στη Χολινεργική Υπόθεση της νόσου Alzheimer. Έτσι, υποστηρίχθηκε ότι η αποδιοργάνωση των χολινεργικών 65
67 νευρώνων στη βάση του πρόσθιου εγκεφάλου και η σχετιζόµενη µε αυτήν απώλεια χολινεργικής νευροδιαβίβασης στον εγκεφαλικό φλοιό και σε άλλες περιοχές, συντελούν σηµαντικά στην αποδιοργάνωση της γνωστικής λειτουργίας που παρατηρείται στους ασθενείς της νόσου Alzheimer 1. Κατά τη διάρκεια των 30 περίπου ετών από τη διατύπωση της χολινεργικής υπόθεσης, έχουν προστεθεί στην αναζήτηση της ορθότητας αυτής της υπόθεσης τα στοιχεία πολλών µελετών για την ερµηνεία του συνδρόµου της άνοιας της νόσου Alzheimer. 1. Νευροχηµικές και ιστοπαθολογικές αλλαγές σε χολινεργικούς και µηχολινεργικούς νευρώνες στη νόσο Alzheimer Μεταθανάτιες έρευνες σε ασθενείς µε νόσο Alzheimer αναφέρουν απώλεια νευρώνων και σχηµατισµό νευροϊνιδιακών αλλοιώσεων σε εντοπισµένες περιοχές του νεοφλοιού και του ιππόκαµπου, που επηρεάζουν τα πυραµοειδή κύτταρα και τις συνάψεις τους 133,134. Υποφλοιώδεις πυρήνες ειδικοί για συγκεκριµένους νευροδιαβιβαστές, οι οποίοι προβάλλουν στο φλοιό, επηρεάζονται επίσης από νευροεκφυλιστικές διαταραχές και περιλαµβάνουν το χολινεργικό βασικό πυρήνα του Meynert, το σεροτονινεργικό πυρήνα της ραφής και το νοραδρενεργικό υποµέλανα τόπο. Βιοχηµικές έρευνες σε ιστούς από βιοψίες, που ελήφθησαν από ασθενείς µε νόσο Alzheimer µε µέσο όρο 3,5 έτη από την εµφάνιση των κλινικών συµπτωµάτων, έδειξαν ότι εµφανίζεται µία επιλεκτική παθολογία στους νευροδιαβιβαστές από τα πρώτα στάδια της πορείας της νόσου 135. Συγκεκριµένα εµφανίζονται οµοιόµορφα ελαττωµένοι οι προσυναπτικοί δείκτες του χολινεργικού συστήµατος (Εικόνα 3). Παρατηρείται, δηλαδή, µείωση της δραστηριότητας της ChAT, καθώς και µείωση της σύνθεσης της ACh, που συνδέονται στενά µε τις γνωστικές διαταραχές σε ασθενείς µε νόσο Alzheimer

68 Εικόνα 3. Σχηµατική απεικόνιση της ελάττωσης των δεικτών του χολινεργικού συστήµατος 132 Αν και ορισµένοι από τους νοραδρενεργικούς και σεροτονινεργικούς δείκτες εµφανίζουν µεταβολές, το ίδιο δε συµβαίνει µε την ντοπαµίνη, το γ- αµινοβουτυρικό οξύ (GABA) και τη σωµατοστατίνη 135,136,137,138. Νεκροτοµικές µελέτες σε εγκεφάλους ασθενών, που βρίσκονταν σε προχωρηµένο στάδιο της νόσου, αποκαλύπτουν ότι πολλά περισσότερα συστήµατα νευροδιαβιβαστών επηρεάζονται σε µεγαλύτερη έκταση. Αυτά περιλαµβάνουν το GABA 139,140 και τη σωµατοστατίνη 141,142 και µπορεί να αποτελούν ένδειξη ότι οι νευρώνες του φλοιού επηρεάζονται σε πιο προχωρηµένο στάδιο της πορείας της νόσου. Ωστόσο, οι αλλαγές στη σεροτονινεργική νευροδιαβίβαση συσχετίζονται περισσότερο µε τις διαταραχές στη συµπεριφορά, όπως η κατάθλιψη, παρά µε τις γνωστικές διαταραχές 143,144. Με βάση τα παραπάνω, η χολινεργική νεύρωση του νεοφλοιού φαίνεται να επηρεάζεται σε αρχικό στάδιο της νόσου, συµπέρασµα που αποδεικνύεται από στοιχεία για παρόµοιες αλλαγές σε ασθενείς που έχουν εµφανίσει κλινικά σηµεία της νόσου λιγότερο από ένα χρόνο 145. Ωστόσο, παρά το ότι η διαταραχή της χολινεργικής λειτουργίας συνδέεται µε τις γνωστικές διαταραχές της νόσου Alzheimer, η συσχέτιση αυτή δε φαίνεται να έχει απόλυτη σχέση αιτίου-αιτιατού. Άλλες αιτίες, που συνδέονται µε τις γνωστικές διαταραχές της νόσου, είναι η απώλεια συνάψεων και περικαρύου πυραµοειδών κυττάρων 146. Επίσης, ένας µικρός αριθµός 67

69 ασθενών δεν εµφανίζει σηµαντική µείωση της δραστηριότητας της ChAT, αν και εµφανίζει µείωση στην αµυγδαλή 147. Έτσι, αν και η µειωµένη δραστηριότητα της ChAT συσχετίζεται µε τη νόσο Alzheimer, επιπρόσθετοι παράγοντες διαφορετικοί από τη µειωµένη χολινεργική δραστηριότητα, είναι πιθανόν να µετέχουν στη µείωση της γνωστικής λειτουργίας (Εικόνα 4). Εικόνα 4. Η χολινεργική υπόθεση 148 (VAChT= κυστίδια ακετυλοχολίνης, NGF= παράγοντας ανάπτυξης των νεύρων, M2AChR= M2 µουσκαρινικοί υποδοχείς, M1AChR= M1 µουσκαρινικοί υποδοχείς, ACh= ακετυλοχολίνη, TrKA= υποδοχέας τυροσινικής κινάσης, nachr= νικοτινικός υποδοχέας, ChAT= χολινοακετυλοτρανσφεράση, AChE= ακετυλοχολινεστεράση) Άλλες µελέτες έχουν αποδείξει µείωση στον αριθµό των νικοτινικών 149 και µουσκαρινικών (Μ2) υποδοχέων ACh σε εγκεφάλους ασθενών µε νόσο Alzheimer, οι περισσότεροι από τους οποίους εντοπίζονται σε προσυναπτικές χολινεργικές απολήξεις, αλλά έχουν επίσης αποδείξει µία σχετική διατήρηση των µετασυναπτικών υποδοχέων (Μ1, Μ3) 150. Υπάρχουν όµως υπάρχουν κάποια στοιχεία για διαταραχή στη σύνδεση µεταξύ των µουσκαρινικών Μ1 υποδοχέων, των G-πρωτεϊνών τους και 68
70 συστηµάτων δεύτερου αγγελιοφόρου 151. Επιπρόσθετα της χολινεργικής δυσλειτουργίας, υπάρχει συσχέτιση της άνοιας µε χηµικούς και ιστοπαθολογικούς δείκτες των διεγερτικών αµινοξέων EAA (excitatory amino acids), τα οποία απελευθερώνονται από πυραµοειδείς νευρώνες του φλοιού. Αυτοί οι νευρώνες φαίνεται να συµβάλλουν στη φυσιολογική γνωστική λειτουργία αυτόνοµα, ενώ έχουν σηµαντικό ρόλο στη χολινεργική λειτουργία αφού είναι χολινορυθµιζόµενοι 152,153,154,155. Αν και νευροχηµικές µελέτες των νευροδιαβιβαστών ΕΑΑ έχουν αποτύχει να δείξουν εµφανείς ή εκτεταµένες αλλαγές στη συγκέντρωσή τους στους νευρώνες 135, αυτό µπορεί να οφείλεται στη δυσκολία διαχωρισµού των νευροδιαβιβαστών ασπαρτικού και γλουταµινικού από αυτά που προέρχονται από το µεταβολισµό. Παρόλα αυτά, η συγκέντρωση του γλουταµινικού σε δείγµατα βιοψίας από τον κροταφικό λοβό ασθενών µε νόσο Alzheimer εµφανίζεται µειωµένη κατά 14%. Η πρόσληψη D-ασπαρτικού, ενός πιθανού δείκτη των ΕΑΑ νευρικών συνάψεων, είναι επίσης µειωµένη σε πολλές φλοιικές περιοχές εγκεφάλων ασθενών µε νόσο Alzheimer 156,157,158. In vivo απεικονιστικές µελέτες, που πραγµατοποιήθηκαν σε ασθενείς µε νόσο Alzheimer, υποστηρίζουν την εµπλοκή των πυραµοειδών νευρώνων στην πάθηση, αφού ο τοπικός υποµεταβολισµός συνδυάζεται µε την απώλεια και ατροφία νευρώνων, το σχηµατισµό νευροϊνιδιακών αλλοιώσεων και την απώλεια συνάψεων 133,159,160,161. Απώλεια πυραµοειδών κυττάρων του φλοιού 146,162,163, απώλεια συνάψεων 162 και µειωµένη συγκέντρωση γλουταµινικού 140 µαζί µε νευροϊνιδιακές αλλοιώσεις 164, συνδέονται µε τη σοβαρότητα της άνοιας. Αυτά τα στοιχεία οδηγούν στο συµπέρασµα ότι τα πυραµοειδή νευρικά κύτταρα και ο νευροδιαβιβαστής τους το γλουταµινικό (και/ή το ασπαρτικό), παίζουν σηµαντικό ρόλο στις γνωστικές διαταραχές της νόσου Alzheimer και για αυτό µπορεί να αποτελέσουν έναν ακόµη θεραπευτικό στόχο. Ωστόσο, αυτοί οι νευρώνες είναι χολινορυθµιζόµενοι και είναι πιθανόν µία από τις δράσεις των χολινοµιµητικών φαρµάκων, που χρησιµοποιούνται στη θεραπεία, να αποτελεί η αύξηση της δραστηριότητας των ΕΑΑ νευρώνων µέσω µουσκαρινικών και νικοτινικών υποδοχέων, που υπάρχουν σε αυτά τα κύτταρα 152. Αυτό ενισχύεται και από ηλεκτροφυσιολογικές µελέτες, οι οποίες έδειξαν τις διεγερτικές δράσεις των χολινοµιµητικών φαρµάκων στα πυραµοειδή κύτταρα του φλοιού ανθρώπων και επίµυων 153,154. Έτσι, ως αποτέλεσµα της απώλειας νευρώνων, η εµφανής µείωση της ΕΑΑ 69
71 νευροδιαβίβασης θα οδηγήσει σε πυραµοειδή υποδραστηριότητα, η οποία συνοδεύεται από διατήρηση των επιπέδων της ανασταλτικής δράσης των GABAεργικών νευρώνων. Συνεπώς, µπορεί να υποτεθεί ότι επιπροσθέτως της νευρωνικής απώλειας και του σχηµατισµού νευροϊνιδιακών αλλοιώσεων, παρατηρείται και µία αλλαγή στην ισορροπία της νευροδιαβίβασης στον εγκέφαλο ασθενών µε νόσο Alzheimer, µε αποτέλεσµα τη µειωµένη νευρωνική δραστηριότητα 135. Αυτό µπορεί να αντανακλάται στον υποµεταβολισµό που εντοπίζεται µε απεικονιστικές µεθόδους στον εγκέφαλο ασθενών, αν και ένα ποσοστό µπορεί να οφείλεται στην νευρωνική ατροφία 165. Παροµοίως, είναι ενδιαφέρον ότι η τοπική εγκεφαλική ροή αίµατος µπορεί να είναι αυξηµένη σε ασθενείς που λαµβάνουν αναστολείς της χολινεστεράσης, όπως η φυσοστιγµίνη 166,167. Η ανακάλυψη ότι σπάνιες µεταλλάξεις στο γονίδιο που κωδικοποιεί την APP (πρόδροµη πρωτεΐνη του αµυλοειδούς) προκαλούν την εµφάνιση της νόσου Alzheimer σε µέλη οικογενειών που φέρουν τη µετάλλαξη, οδήγησε στη διατύπωση της υπόθεσης του «αµυλοειδούς καταρράκτη» για την ερµηνεία της νόσου Alzheimer

.")

72 Εικόνα 5. Σχηµατισµός πλακών β-αµυλοειδούς 169 Έτσι, η βλάβη στο µεταβολισµό της APP, που οδηγεί σε αύξηση της παραγωγής β-αµυλοειδούς, θεωρήθηκε ως ο κρίσιµος παράγοντας για την οικογενή και σποραδική νόσο Alzheimer, µε τις υπόλοιπες αλλαγές-όπως την απώλεια νευρώνων και συνάψεων, τις νευροϊνιδιακές αλλοιώσεις και τη διαταραχή της νευροδιαβίβασης-να αποτελούν επακόλουθό της (Εικόνα 5). Η χολινεργική νευροδιαβίβαση ίσως είναι ο κύριος στόχος του β- αµυλοειδούς, αφού έχει αποδειχθεί ότι µειώνει τόσο την πρόσληψη χολίνης όσο και την απελευθέρωση ακετυλοχολίνης σε in vitro έρευνες 170,171. Είναι ενδιαφέρον ότι οι αλλαγές στον εγκέφαλο ασθενών µε νόσο Alzheimer επικεντρώνονται στα πυραµοειδή κύτταρα, που χάνονται κατά την πορεία της νόσου εξαιτίας νευροϊνιδιακών αλλοιώσεων, αποτελούν µία µεγάλη πηγή της APP (και γι αυτό είναι περιοχή της ανώµαλης λειτουργίας του µεταβολισµού µε αυξηµένη παραγωγή β-αµυλοειδούς) και ρυθµίζονται από την ακετυλοχολίνη, η οποία επηρεάζεται πολύ νωρίς στην πορεία της νόσου (Εικόνες 6 και 7). 71
73 Εικόνα 6. Σχηµατική απεικόνιση των αλλαγών στα πυραµοειδή κύτταρα 172 (ACh= ακετυλοχολίνη, AChE= ακετυλοχολινεστεράση, Glu= γλουταµινικό οξύ, machr= µουσκαρινικός υποδοχέας, nachr= νικοτινικός υποδοχέας, Aβ= β-αµυλοειδές, tau= πρωτεΐνη tau, APP= πρόδροµη πρωτεΐνη του αµυλοειδούς) Εποµένως, αυτοί οι νευρώνες φαίνεται να έχουν κεντρικό ρόλο στην εµφάνιση των κλινικών συµπτωµάτων, καθώς και στην παθοφυσιολογία της νόσου. Παρατηρήσεις σε κυτταρικές σειρές και καλλιέργειες πρόδροµων νευρικών κυττάρων έδειξαν ότι η ενεργοποίηση µουσκαρινικών, µεταβοτροπικών (metabotropic) γλουταµινικών και συνδεόµενων µε φωσφολιπάση-c υποδοχέων, ευνοεί τη µετατροπή της APP σε µηαµυλοειδή προϊόντα 173 και φαίνεται ότι οι φαρµακολογικές ουσίες που χρησιµοποιούνται για τη συµπτωµατική θεραπεία της νόσου πιθανόν να δρουν και µέσω της µείωσης της παραγωγής β-αµυλοειδών. Επιπλέον, η νευροτοξικότητα των β-αµυλοειδών µετριάζεται από µουσκαρινικούς αγωνιστές 174. Όµως, προς το παρόν δεν έχουν δηµοσιευτεί στοιχεία που αφορούν πιθανή ευεργετική δράση των χολινοµιµητικών φαρµάκων, που χορηγούνται σε ασθενείς της νόσου Alzheimer, µέσω αύξησης της APP ή µείωσης της παραγωγής β-αµυλοειδούς. Υπάρχουν, ωστόσο κάποια στοιχεία για µείωση της APP στο εγκεφαλονωτιαίο υγρό καταθλιπτικών ασθενών, που λαµβάνουν φάρµακα µε αντιχολινεργικές παρενέργειες
74 Εικόνα 7. Σχηµατική απεικόνιση της παραγωγής β-αµυλοειδούς 176 Άλλες µελέτες απέδειξαν ότι η φωσφορυλίωση της πρωτεΐνης tau, φαίνεται να αποτελεί ένα σηµαντικό βήµα στο σχηµατισµό νευροϊνιδιακών αλλοιώσεων (που παρουσιάζονται κυρίως στους ΕΑΑ πυραµοειδείς νευρώνες του φλοιού), οι οποίες µπορεί επίσης να επηρεάζονται από το σύστηµα δευτέρου αγγελιοφόρου της φωσφολιπάσης C 177. Έτσι, µετά από διέγερση των µουσκαρινικών χολινεργικών υποδοχέων, η ενεργοποίηση της πρωτεϊνικής κινάσης C, µπορεί να οδηγήσει σε in vitro απενεργοποίηση µίας πρωτεϊνικής κινάσης GSK-3, η οποία φωσφορυλιώνει την πρωτεΐνη tau, µε τρόπο παρόµοιο µε αυτό που παρατηρείται στη νόσο Alzheimer 177. Σε στήριξη αυτής της θεωρίας, η προσθήκη χολινεργικών αγωνιστών σε καλλιέργειες νευρικών κυττάρων µε Μ1 µουσκαρινικούς υποδοχείς προκάλεσε µείωση της φωσφορυλίωσης της πρωτεΐνης tau
75 Εικόνα 8. Σχηµατική απεικόνιση της υπερφωσφορυλίωσης της πρωτεΐνης tau 176 Ως αποτέλεσµα της µειωµένης χολινεργικής δραστηριότητας, η µειωµένη ενεργοποίηση της πρωτεϊνικής κινάσης C, µπορεί να οδηγήσει σε µεγαλύτερη ενεργοποίηση της GSK-3 και εποµένως σε υπερφωσφορυλίωση της πρωτεΐνης tau (Εικόνα 8). Συµπερασµατικά, αν αυτές οι αλληλεπιδράσεις µεταξύ νευροδιαβιβαστών-πρωτεϊνών συντελούνται στον εγκέφαλο ασθενών της νόσου Alzheimer, τότε δεν είναι απίθανο οι αλλαγές στην οµοιόσταση των νευροδιαβιβαστών στον εγκέφαλο στη νόσο Alzheimer, να συντελούν στην αύξηση της φωσφορυλίωσης της πρωτεΐνης tau, στην αύξηση παραγωγής β-αµυλοειδούς και εποµένως να προκαλούν νευροεκφύλιση σε συγκεκριµένες ευαίσθητες περιοχές. Επιπρόσθετα, είναι πιθανόν τα χολινοµιµητικά φάρµακα να µειώνουν τις ιστοπαθολογικές αλλοιώσεις της εξέλιξης της νόσου. Με βάση πρόσφατες έρευνες, προτάθηκε η «γλουταµινεργική υπόθεση», ως βοηθητική της «χολινεργικής υπόθεσης» για την ερµηνεία της νόσου 135,179. Με τον τρόπο αυτό, η χολινεργική υπόθεση µπορεί να αναθεωρηθεί ώστε να συµπεριλαµβάνει την ιδέα ότι ένας κύριος στόχος της χολινοµιµητικής δράσης είναι οι ΕΑΑ πυραµοειδείς νευρώνες και ότι η χολινεργική διαταραχή προκαλεί την απώλεια της δράσης των ΕΑΑ. Tα δύο συστήµατα µπορεί να είναι υπεύθυνα για τις νευροφυσιολογικές διαταραχές και την 74
76 εξελισσόµενη παθολογία της νόσου Alzheimer στους ασθενείς. Αυτή η αναθεωρηµένη χολινεργική υπόθεση παρέχει µία ισχυρή βάση για την ανάπτυξη χολινοµιµητικών φαρµάκων για τη συµπτωµατική θεραπεία της νόσου Alzheimer. 75
77 Ε. ΦΑΡΜΑΚΟΛΟΓΙΚΗ ΘΕΡΑΠΕΙΑ ΤΗΣ ΝΟΣΟΥ ALZHEIMER Το µεγαλύτερο µέρος της έρευνας για την ανάπτυξη νέων φαρµάκων και του σχεδιασµού της φαρµακολογικής θεραπείας της νόσου Alzheimer, έχει βασιστεί στη χολινεργική υπόθεση της νόσου. Οι στρατηγικές για τη βασική θεραπεία της άνοιας, περιλαµβάνουν την άµβλυνση των γνωστικών διαταραχών, την επιβράδυνση ή την αναστολή της πορείας της νόσου και τέλος την οριστική θεραπεία ή ακόµη και την πρόληψή της. Τα τελευταία χρόνια υπάρχουν αρκετά αποτελεσµατικά φάρµακα για την άµβλυνση των γνωστικών διαταραχών. Ο στόχος της επιβράδυνσης ή της αναστολής της πορείας της νόσου δεν έχει ακόµη επιτευχθεί, αλλά φαίνεται εφικτός µε τη χρήση υπαρχόντων φαρµάκων και άλλων που βρίσκονται υπό ανάπτυξη. Αν και µια θεραπεία για την εγκατεστηµένη, συµπτωµατική νόσο Alzheimer µπορεί να µην είναι εφικτή λόγω της µη-αναστρεψιµότητας των εγκεφαλικών βλαβών, η προληπτική θεραπεία ίσως είναι εφικτή. Υπάρχουν βάσιµες ελπίδες στηριζόµενες στην υπάρχουσα επιστηµονική γνώση, ότι οι τεράστιες επιπτώσεις τόσο σε προσωπικό επίπεδο όσο και σε επίπεδο δηµόσιας υγείας, µπορούν να περιοριστούν µε τις υπάρχουσες και τις αναπτυσσόµενες θεραπευτικές στρατηγικές. Ωστόσο, παρά την αισιοδοξία σχετικά µε την πιθανότητα ελέγχου της νόσου Alzheimer, υπάρχει σκεπτικισµός για την πλήρη αποδοχή των θεραπειών για την άµβλυνση των γνωστικών διαταραχών. Το αυξανόµενο ενδιαφέρον από την ακαδηµαϊκή κοινότητα και τη φαρµακοβιοµηχανία για την ανάπτυξη νέων φαρµάκων και τη διενέργεια κλινικών δοκιµών θεραπευτικών παραγόντων, θα επιτρέψει τη θέσπιση κοινά αποδεκτών θεραπευτικών στρατηγικών. 1. Η λογική της χολινεργικής θεραπείας Αν και τα ακριβή αίτια της νόσου Alzheimer δεν είναι πλήρως καθορισµένα, η χρήση των χολινεργικών παραγόντων στην αντιµετώπιση της νόσου στηρίζεται στο ότι η χολινεργική διαταραχή είναι η σηµαντικότερη διαταραχή των νευροδιαβιβαστών και στο ότι οι γνωστικές διαταραχές οφείλονται σε αυτή 109,180. Η ελάττωση στους χολινεργικούς δείκτες έχει συνδεθεί µε την πυκνότητα των γεροντικών πλακών στις 76
78 περιοχές του φλοιού και µε τις κλινικές κλίµακες της άνοιας. Πρόσφατα ερευνητικά στοιχεία φαίνεται να συνδέουν τη χολινεργική διαταραχή µε την παραγωγή µη-φυσιολογικού αµυλοειδούς και συσχετίζουν τη χολινεργική ενίσχυση µε την προαγωγή της ανάπτυξης του νευρίτη. Ωστόσο, είναι σηµαντικό να συνυπολογίσουµε το ότι η ακριβής σχέση µεταξύ της χολινεργικής διαταραχής και της παθοφυσιολογίας της νόσου παραµένει αδιευκρίνιστη και ότι ο βαθµός της χολινεργικής διαταραχής µπορεί να διαφέρει µεταξύ των ασθενών, αλλά και στον ίδιο ασθενή στα διάφορα στάδια της νόσου. Επίσης, πρέπει να αναφερθεί ότι υπάρχουν και άλλες νευροχηµικές και δοµικές διαταραχές που συντελούν στην παθογένεια της νόσου Alzheimer 181. Έτσι, η θεραπεία αποκατάστασης της χολινεργικής λειτουργίας δεν πρέπει να θεωρείται ότι θα αναστρέψει ή θα αµβλύνει όλα τα κλινικά συµπτώµατα της νόσου. 2. Αναστολείς χολινεστεράσης Η συσχέτιση µεταξύ της χολινεργικής διαταραχής και της νόσου Alzheimer είναι επαρκώς τεκµηριωµένη. Οι αναστολείς της χολινεστεράσης αποτελούν τη µόνη βασική θεραπευτική επιλογή, που έχει την έγκριση του FDA από το Με βάση θεωρητικές υποθέσεις και πειραµατικές αποδείξεις ότι ο εγκέφαλος ασθενών µε νόσο Alzheimer παρουσιάζει µειωµένη χολινεργική δραστηριότητα, είτε λόγω καταστροφής χολινεργικών νευρώνων είτε λόγω µείωσης της χολινοακετυλο-τρανσφεράσης (ChAΤ) και ελαττωµένης σύνθεσης της ακετυλοχολίνης (ACh), επιχειρήθηκε η χορήγηση αντιχολινεστερασικών ουσιών, οι οποίες διαπερνούν τον αιµατοεγκεφαλικό φραγµό, όπως η φυσοστιγµίνη 182. Αυτές οι φαρµακολογικά δραστικές ουσίες µπορούσαν να αυξήσουν την ελαττωµένη συγκέντρωση της ακετυλοχολίνης (ACh) στη συναπτική σχισµή, ώστε να µπορέσει να δράσει στους χολινεργικούς υποδοχείς (Εικόνα 9). 77

79 Εικόνα 9. Σχηµατική απεικόνιση του µηχανισµού δράσης των αναστολέων AChE 183 Προηγούµενες προσπάθειες για αύξηση της σύνθεσης της Ach µε τη χορήγηση µέσω της διατροφής προδρόµων ουσιών της, της χλωριούχου χολίνης και φωσφατιδυλοχολίνης (λεκιθίνης), δεν προσέφεραν σηµαντική κλινική βελτίωση Χαρακτηριστικά των αναστολέων της ακετυλοχολινεστεράσης Ο ιδανικός αναστολέας της ακετυλοχολινεστεράσης για τη θεραπεία της ΝΑ θα πρέπει να είναι καλά ανεκτός µέσα στο φάσµα της κλινικά δραστικής δόσης, µε ελάχιστες ανεπιθύµητες κεντρικές ή περιφερικές χολινεργικές επιδράσεις (ναυτία, έµετος, κεφαλαλγία, διάρροια, ίλιγγος, µειωµένη όρεξη, κοιλιακό άλγος), απουσία τοξικότητας των οργάνων (παθολογικό ΗΚΓ, αυξηµένα επίπεδα των ηπατικών ενζύµων) και θα πρέπει να είναι κατάλληλος για άτοµα µε συνυπάρχοντα νοσήµατα (Πίνακας 2). Λαµβάνοντας υπόψη την προχωρηµένη ηλικία των περισσότερων ατόµων που πάσχουν από ΝΑ, η χορήγηση οποιασδήποτε θεραπείας για µεγάλα χρονικά διαστήµατα δεν θα πρέπει να έχει ανεπιθύµητες καρδιαγγειακές επιδράσεις και αλληλεπιδράσεις µε συγχορηγούµενα φάρµακα. Το φάρµακο θα πρέπει να έχει όσο το δυνατόν 78
80 λιγότερες αλληλεπιδράσεις µε άλλα φάρµακα, καθώς τα περισσότερα άτοµα άνω των 65 ετών πάσχουν από ένα ή περισσότερα συνυπάρχοντα νοσήµατα για τα οποία ήδη λαµβάνουν µακροχρόνια φαρµακευτική αγωγή 185,186,187,188. Ένα ιδανικό φάρµακο δεν θα πρέπει να προκαλεί ή να επιδεινώνει τη δυσφορία, το διαταραγµένο συναίσθηµα και την εξασθένιση της µνήµης. Η φαρµακοκινητική και η διάρκεια της δράσης θα πρέπει να συµβαδίζει µε ένα απλό θεραπευτικό σχήµα, µε κατά προτίµηση από του στόµατος χορήγηση µία ή δύο φορές ηµερησίως. Πίνακας 2. Χαρακτηριστικά του ιδανικού αναστολέα AChE 185,186,187,188 Παρέχει σηµαντικό κλινικό όφελος Είναι καλά ανεκτός Είναι ιδιαίτερα εκλεκτικός Προκαλεί ελάχιστες κεντρικές ή περιφερικές χολινεργικές ανεπιθύµητες ενέργειες εν παρουσιάζει καµία τοξικότητα οργάνων εν προκαλεί καµία ανεπιθύµητη καρδιαγγειακή επίδραση Έχει ελάχιστες αλληλεπιδράσεις µε άλλα φάρµακα εν προκαλεί καµία ανεπιθύµητη καρδιαγγειακή επίδραση εν προκαλεί εξασθένιση της µνήµης Έχει απλό, ευέλικτο δοσολογικό σχήµα Γνωρίζουµε δύο είδη ενζύµων της χολινεστεράσης, η AChE που είναι το ένζυµο-στόχος στη θεραπεία της ΝΑ, βρίσκεται κυρίως στον εγκέφαλο, στους γραµµωτούς µυς και στα ερυθροκύτταρα, ενώ η βουτυλοχολινεστεράση (BChE) βρίσκεται κατά κύριο λόγο στον καρδιακό µυ, τους λείους µύες, στο δέρµα, σε κάποιους αδένες, στον ορό και τις πλάκες που σχετίζονται µε τη ΝΑ. Ένα ιδανικό σκεύασµα θα πρέπει να είναι εκλεκτικό ως προς το ένζυµο-στόχο, ενώ θα πρέπει να εµφανίζει ελάχιστες αλληλεπιδράσεις µε άλλα ένζυµα (Εικόνες 10α, 10β, 10γ). 79

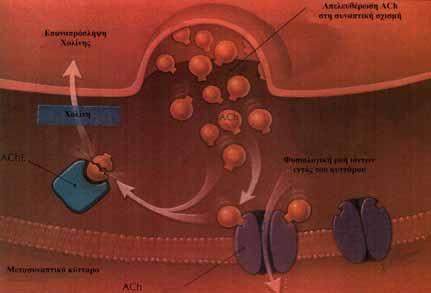
81 Εικόνα 10α Εικόνα 10β 80

82 Εικόνα 10γ Εικόνες 10α, 10β, 10γ. Χολινεργική διαβίβαση και µηχανισµός δράσης των αναστολέων AChE 189 Υπάρχουν αρκετοί δοµικοί σχηµατισµοί της σφαιρικής (G) µορφής της AChE. Η G4 είναι η µορφή που απαντάται σε µεγαλύτερη αφθονία στον εγκέφαλο, ενώ τη µορφή G1 τη συναντούµε σε µικρότερες ποσότητες. Ωστόσο, η ποσότητα της G4 στον εγκέφαλο µειώνεται µε την ηλικία και η µείωση αυτή είναι ακόµα ταχύτερη σε άτοµα που πάσχουν από τη ΝΑ. Αντίθετα, η ποσότητα της G1 παραµένει ίδια ή µειώνεται µε πολύ βραδύτερους ρυθµούς. Ένα ιδανικό σκεύασµα θα πρέπει να αναστέλλει επιλεκτικά τη µορφή G1 του ενζύµου, καθώς στη ΝΑ αυτή η µορφή διατηρείται, η αναλογία της ως προς τις άλλες µορφές AChE αυξάνεται και καθίσταται κατά αυτόν τον τρόπο περισσότερο σηµαντική για το γενικότερο έλεγχο της χολινεργικής δραστηριότητας του εγκεφάλου των ηλικιωµένων ατόµων. Τέλος, το σκεύασµα θα πρέπει να εµφανίζει ουσιαστική αποτελεσµατικότητα, σύµφωνα µε τα καθιερωµένα κριτήρια. Η κλινική αποτελεσµατικότητα του φαρµάκου δε θα πρέπει να έχει ευεργετική επίδραση µόνο στη γνωστική λειτουργία, αλλά και να συνεπάγεται ένα φανερό όφελος στη λειτουργική ικανότητα, κυρίως επηρεάζοντας και βελτιώνοντας τις καθηµερινές δραστηριότητες του ασθενή. 81
 να αποτελέσει υπόστρωµα για το ένζυµο, ενώ ταυτόχρονα αποκλείει την περαιτέρω ενζυµικώς καταλυόµενη υδρόλυση της ACh για ένα µεγάλο χρονικό διάστηµα (Εικόνα 11). Εικόνα 11.")
83 3. Η Ριβαστιγµίνη Η ριβαστιγµίνη (ΕΝΑ-713) είναι ένας ψευδο-µη-αναστρέψιµος αναστολέας της ακετυλοχολινεστεράσης (AChE) µε φαινυλοκαρβαµυλική δοµή, ο οποίος δεσµεύει το ένζυµο και δεν επιτρέπει στην ακετυλοχολίνη (ACh) να αποτελέσει υπόστρωµα για το ένζυµο, ενώ ταυτόχρονα αποκλείει την περαιτέρω ενζυµικώς καταλυόµενη υδρόλυση της ACh για ένα µεγάλο χρονικό διάστηµα (Εικόνα 11). Εικόνα 11. Σχηµατική απεικόνιση του µηχανισµού δράσης της ριβαστιγµίνης 185, Φαρµακοδυναµική Η ριβαστιγµίνη είναι ένα φάρµακο µακράς διάρκειας δράσης, κεντρικώς δρων και κεντρικώς εκλεκτικό. ιεισδύει σχεδόν αµέσως, µετά την παρεντερική ή από του στόµατος χορήγησή του, στο κεντρικό νευρικό σύστηµα (ΚΝΣ). Αναστέλλει εκλεκτικά την AChE, µε ελάχιστη περιφερική δράση 185. Η ριβαστιγµίνη µιµούµενη την ACh λειτουργεί ως υπόστρωµα για το ένζυµο AChE (Εικόνα 12). Συνήθως, η ACh σχηµατίζει ένα ακετυλιωµένο σύµπλοκο µε την AChE και στη συνέχεια υδρολύεται σε χολίνη και οξικό οξύ. Αντίθετα, η ριβαστιγµίνη δηµιουργεί ένα καρβαµυλιωµένο σύµπλοκο µε το ένζυµο, αναστέλλοντάς το µέχρι την αντικατάσταση της 82
 όσο και in vivo")
84 καρβαµυλικής οµάδας από την υδροξυλοµάδα στη θέση της εστεράσης 185. Εικόνα 12. Χηµική δοµή της ACh και της ριβαστιγµίνης-ena Από πειράµατα σε επίµυες βρέθηκε ότι η ριβαστιγµίνη είναι λιγότερο αποτελεσµατικός αναστολέας της AChE τόσο in vitro (100 φορές) όσο και in vivo (10 φορές) από τη φυσοστιγµίνη, αλλά παρουσιάζει εκλεκτικότητα για τον ιππόκαµπο και το φλοιό (περιοχές που επηρεάζονται περισσότερο στη νόσο Alzheimer) σε σύγκριση µε το ραβδωτό σώµα και τη γέφυρα. Έχει επίσης εκλεκτικότητα για τον εγκέφαλο και όχι για την καρδιά ή τους σκελετικούς µύες, µε αποτέλεσµα περισσότερη κεντρική (5 φορές περισσότερη από τη φυσοστιγµίνη) και λιγότερη περιφερική χολινεργική δράση 191,192,193,194. Αναστέλλει την AChE και BuChE του ΕΝΥ µε δοσοεξαρτώµενο τρόπο και αναστέλλει λιγότερο σταθερά τη BuChE (10 φορές µικρότερη συγγένεια από την G1AChE και µικρή συγγένεια για την AChE των ερυθροκυττάρων σε όλες τις περιοχές). Η ριβαστιγµίνη αυξάνει τα µειωµένα επίπεδα της ACh και της χολινοακετυλοτρανφεράσης ChAT. Επίσης, αυξάνει τον αριθµό των Μ1 υποδοχέων στο µετωπιαίο φλοιό ηλικιωµένων επίµυων και βελτιώνει την ελαττωµένη δεσµευτική ικανότητα 83
85 των φλοιωδών µουσκαρινικών υποδοχέων στην ACh, χωρίς όµως να επηρεάζει τη χηµική συγγένεια. Εξάλλου σε βλάβες της βάσης του πρόσθιου εγκεφάλου επιµύων, όπου παρατηρείται µείωση της µνήµης και της µαθήσης, η ριβαστιγµίνη βελτιώνει την απόδοση της δοκιµασίας λαβυρίνθου νερού και της δοκιµασίας παθητικής αποφυγής. Όµως, η ριβαστιγµίνη στερείται συγγενείας προς τους µουσκαρινικούς, α- ή β- αδρενεργικούς, ντοπαµινεργικούς ή οπιοειδείς υποδοχείς Κλινική φαρµακοκινητική Τα βασικά φαρµακοκινητικά χαρακτηριστικά της ριβαστιγµίνης είναι τα εξής: ταχεία και πλήρης απορρόφηση, αύξηση της βιοδιαθεσιµότητας µε την αύξηση της δόσης, ασθενής δέσµευση µε τις πρωτεΐνες του πλάσµατος, καλή διείσδυση στον εγκέφαλο, απενεργοποίησή της σε ένα µη δραστικό φαινολικό προϊόν από την AChE, ελάχιστη συµµετοχή των ισοενζύµων του P450 στο µεταβολισµό της, καµία αλληλεπίδραση µε άλλα φάρµακα σε σχέση µε το κυττόχρωµα P450, ταχύτατη απέκκριση µέσω των νεφρών του προϊόντος της διάσπασης (ΝΑΡ226-90), καµία συσσώρευση, εξατοµίκευση της ηµερήσιας δόσης, χορήγηση µαζί µε την τροφή προκαλεί αύξηση της περιοχής κάτω από την καµπύλη (AUC) κατά περίπου 30%. Η ριβαστιγµίνη απορροφάται ταχέως και σχεδόν πλήρως, δε δεσµεύεται πρακτικά από τις πρωτεΐνες του πλάσµατος και υδρολύεται σε ένα µη δραστικό προϊόν. Το φάρµακο χορηγείται εύκολα και είναι καλώς ανεκτό από τα ηλικιωµένα άτοµα, καθώς απεκκρίνεται πλήρως µέσω των νεφρών µε ελάχιστη συµµετοχή του συστήµατος ισοενζύµων του κυτοχρώµατος Ρ450 και δεν αλληλεπιδρά µε την πλειονότητα των πιο πιθανών συγχορηγούµενων φαρµάκων 186,187,188,
86 3.3. Απορρόφηση Η ριβαστιγµίνη απορροφάται ταχέως και πλήρως. Οι µέγιστες συγκεντρώσεις του πλάσµατος επιτυγχάνονται σε περίπου 1 ώρα. Η απόλυτη βιοδιαθεσιµότητα µίας δόσης 3mg είναι περίπου 36% ± 13%. Η χορήγησή της συγχρόνως µε την τροφή καθυστερεί τη µέγιστη απορρόφηση κατά 90 λεπτά και αυξάνει την AUC κατά περίπου 30%. Η απορρόφηση µίας εφάπαξ από του στόµατος δόσης της ριβαστιγµίνης σε υγιείς εθελοντές έχει αποδειχθεί ταχεία και σχεδόν ολική (Μελέτη Β151). Στη συγκεκριµένη µελέτη, το ποσοστό της απορρόφησης που επιτεύχθηκε ήταν πάνω από 96%, η ηµιπερίοδος απορρόφησής της (κάψουλες) ήταν 0,17-0,19 ώρες και ο tmax ήταν 0,8-1,1 ώρες. Η απορρόφηση γίνεται µε βραδύτερους ρυθµούς στα ηλικιωµένα άτοµα. Ο χρόνος µέχρι να σηµειωθούν οι µέγιστες συγκεντρώσεις στο πλάσµα ήταν 1-2 ώρες στις µελέτες Α103 και ENA/AD-VD/P-1A. Η µεταξύ των ατόµων διακύµανση των φαρµακοκινητικών παραµέτρων ήταν περίπου 20% για το µητρικό φάρµακο (συντελεστές µεταβλητότητας 22,7% για την AUC και 18,2% για τη Cmax). Οι σωµατικές διαφορές µεταξύ των ατόµων παίζουν σηµαντικό ρόλο στη µεταβλητότητα της AUC. Έτσι, ένα άτοµο µε µικρή σωµατική διάπλαση (40kg), που πάσχει από ΝΑ, φαίνεται ότι έχει τρεις φορές µεγαλύτερη AUC της ριβαστιγµίνης από ό,τι ένα άτοµο µεγαλύτερης σωµατικής διάπλασης (90kg) που λαµβάνει την ίδια δόση φαρµάκου. Η AUC για τη ριβαστιγµίνη αυξάνεται περισσότερο από αναλογικά µε την αύξηση της δόσης (µεταξύ των δόσεων 1,5-6,0mg δύο φορές ηµερησίως), γίνεται δηλαδή 1,5 φορά µεγαλύτερη από αυτή που αναµένεται για νέα υγιή άτοµα και για ασθενείς µε ΝΑ. Η βιοδιαθεσιµότητα αυξάνεται µε την αύξηση της δόσης, υποδηλώνοντας κεκορεσµένο µεταβολισµό πρώτης διόδου. Στη µελέτη Β 151 η βιοδιαθεσιµότητα ήταν 3% για τη δόση 1,0mg και 11% για τη δόση 2,5mg και η απόλυτη βιοδιαθεσιµότητα ήταν 35,5% για τη δόση 3mg (µελέτη W361) Η λήψη τροφής προκαλεί µέτρια αύξηση της απορρόφησης του φαρµάκου σε υγιείς εθελοντές. Παρατηρήθηκε αύξηση κατά 39% της AUC για µία δόση 1mg και αύξηση κατά 23% για µία δόση 2,5mg (µελέτη W101). Η ταχύτητα της απορρόφησης µειώνεται κατά 2,8-3,4 φορές µε τη λήψη τροφής. Για παράδειγµα οι τιµές της tmax που επιτεύχθηκαν µετά από 85
87 νηστεία και µετά από λήψη τροφής ήταν αντίστοιχα για µία δόση 1mg 0,78±0,30 και 2,16±0,75 ώρες. Συνιστάται λοιπόν η ριβαστιγµίνη να λαµβάνεται σε συνδυασµό µε τα γεύµατα 186,187,188, Κατανοµή Η ριβαστιγµίνη δεσµεύεται ασθενώς (περίπου 40%) µε τις πρωτεΐνες του πλάσµατος (µελέτη W104). ιαπερνά γρήγορα τον αιµατοεγκεφαλικό φραγµό. Ο θεωρητικός όγκος κατανοµής κυµαίνεται µεταξύ 1,8-2,7L/kg. Η αναλογία αίµατος/πλάσµατος είναι 0,8-0,9, µε ένα ποσοστό 40-50% του φαρµάκου να συσχετίζεται µε τα ερυθροκύτταρα, ανεξάρτητα από τη συγκέντρωση. Υπολογίζεται ότι ο όγκος κατανοµής της ριβαστιγµίνης µετά από ενδοφλέβια χορήγηση κυµαίνεται µεταξύ 1,8-2,7L/kg (δηλαδή µεγαλύτερος από τη συνολική ποσότητα του νερού στο σώµα) υποδηλώνοντας ότι η ριβαστιγµίνη κατανέµεται εξωαγγειακά. Το ίδιο ισχύει και για το αποκαρβαµυλιωµένο προϊόν της διάσπασης της ριβαστιγµίνης (ΝΑΡ226-90). Η διείσδυσή της στον εγκέφαλο είναι ιδιαίτερα µεγάλη. Στους ποντικούς, το 70% της ριβαστιγµίνης διεισδύει στον εγκεφαλικό ιστό σε αντίθεση µε το αντίστοιχο ποσοστό 19% του µεταβολίτη (ΝΑΡ226-90). Σε υγιείς εθελοντές, ήταν δυνατή η ανίχνευση µόνο του µεταβολίτη στο ΕΝΥ µετά από µία εφάπαξ από του στόµατος δόση 3mg, καθώς τα επίπεδα της ριβαστιγµίνης ήταν κάτω από τα χαµηλότερα ανιχνεύσιµα επίπεδα. Η κατανοµή στο ΕΝΥ αποδείχθηκε ιδιαίτερα µεγάλη στους ασθενείς µε υδροκέφαλο φυσιολογικής πίεσης (ΥΦΠ), ενώ στους ασθενείς µε ΝΑ ανιχνεύθηκε ένα ενδιάµεσο επίπεδο (µεταξύ αυτού των υγιών ατόµων και των ασθενών µε ΥΦΠ) του µητρικού φαρµάκου στο ΕΝΥ. Αυτές οι διαφοροποιήσεις που παρατηρούνται στους ασθενείς µε ΥΦΠ και στους ασθενείς µε ΝΑ, σε αντίθεση µε τα υγιή άτοµα, ενδέχεται να οφείλονται στην αυξηµένη διαβατότητα του αιµατοεγκεφαλικού φραγµού και στη µειωµένη δραστηριότητα χολινεστεράσης αντίστοιχα. Ωστόσο, είναι σαφές ότι στα άτοµα µε ΝΑ το φάρµακο φτάνει στον ιστό-στόχο και έχει κεντρική δράση 186,187,188,
88 3.5. Μεταβολισµός Ο µεταβολισµός της ριβαστιγµίνης είναι ταχύς και ολικός, κυρίως µέσω της υδρόλυσης από τη χολινεστεράση προς τον αποκαρβαµυλιωµένο µεταβολίτη (ΝΑΡ226-90). Ο µεταβολίτης αναστέλλει ελάχιστα την AChE. Τα κύρια ισοένζυµα P450 συµµετέχουν σε µικρό βαθµό στο µεταβολισµό της. εν έχουν παρατηρηθεί σε ανθρώπους αλληλεπιδράσεις φαρµάκων που να σχετίζονται µε το κυττόχρωµα P450. Η χορήγησή της σε άτοµα µε ΝΑ, τα οποία ήδη ελάµβαναν τις συνήθεις φαρµακευτικές αγωγές, δεν επέφερε καµία αλλαγή στην κινητική του συγκεκριµένου φαρµάκου, ούτε αύξησε τον κίνδυνο εµφάνισης κλινικά σχετιζόµενων ανεπιθύµητων ενεργειών. Η ριβαστιγµίνη αδρανοποιείται ταχύτατα, καθώς η ηµιπερίοδος ζωής της στο πλάσµα είναι περίπου 1-2 ώρες. Ο αποκαρβαµυλιωµένος µεταβολίτης (ΝΑΡ226-90) ενδέχεται να υποστεί Ν-αποµεθυλίωση ή/και θειική σύζευξη. Σηµαντική συµµετοχή σε αυτή τη διαδικασία έχουν οι χολινεστεράσες του ήπατος καθώς επίσης σε µικρότερο βαθµό και τα εντερικά ένζυµα. Μελέτες in vitro καθώς και µελέτες σε πειραµατόζωα αποδεικνύουν ότι γίνεται µικρής σηµασίας µεταβολισµός µέσω των κύριων ισοενζύµων του κυττοχρώµατος P ,187,188, Αλληλεπιδράσεις µε άλλα φάρµακα Σύµφωνα µε in vitro µελέτες, δεν αναµένεται να λαµβάνουν χώρα φαρµακοκινητικές αλληλεπιδράσεις µε φάρµακα που αδρανοποιούνται από τα ακόλουθα συστήµατα ισοενζύµων P450: CYP1A2, CYP2D6, CYP3A4/5, CYP2E1, CYP2C9, CYP2C8, ή CYP2C19. Επίσης, δεν παρατηρήθηκε φαρµακοκινητική αλληλεπίδραση µεταξύ της ριβαστιγµίνης και της διγοξίνης, γουαρφαρίνης, διαζεπάµης ή φλουοξετίνης σε µελέτες που πραγµατοποιήθηκαν σε υγιείς εθελοντές. Η χορήγηση ριβαστιγµίνης σε ασθενείς µε ΝΑ συγχρόνως µε τα συνήθη συνταγογραφούµενα φάρµακα, όπως είναι τα αντιόξινα, τα αντιεµετικά, τα αντιδιαβητικά, τα αντιυπερτασικά, οι β-αποκλειστές, οι αναστολείς των διαύλων του ασβεστίου, τα ινότροπα φάρµακα, τα αντιστηθαγχικά, τα µη στεροειδή αντιφλεγµονώδη φάρµακα (NSAID), τα οιστρογόνα, τα αναλγητικά, οι βενζοδιαζεπίνες και τα αντιισταµινικά, δε συσχετίστηκε µε µεταβολή της κινητικής της ριβαστιγµίνης ή µε αυξηµένο κίνδυνο εµφάνισης σχετικών ανεπιθύµητων ενεργειών 186,187,188,
89 3.7. Απέκκριση Ο αδρανής µεταβολίτης της ριβαστιγµίνης (ΝΑΡ226-90) απεκκρίνεται κυρίως µέσω των νεφρών. Η απέκκριση είναι ταχεία, µε ηµιπερίοδο ζωής στο πλάσµα λιγότερο από 1 ώρα µετά από µια δόση 3mg και πρωτίστως µέσω των νεφρών. Ένα ποσοστό µικρότερο από 1% της χορηγούµενης δόσης αποβάλλεται µε τα κόπρανα. Μετά από χορήγηση 14C-rivastigmine, η απέκκριση µέσω των νεφρών ήταν ταχεία µε ποσοστό απέκκρισης 90% µέσα σε διάστηµα 24 ωρών. Συσσώρευση της ριβαστιγµίνης σε άτοµα που πάσχουν από τη ΝΑ δεν παρατηρήθηκε σε χρόνια χορήγηση του φαρµάκου (2-12mg/ηµέρα, δύο φορές ηµερησίως) 186,187,188, Κλινική σηµασία Η απορρόφηση της ριβαστιγµίνης είναι ταχεία και πλήρης και η από του στόµατος χορήγηση δόσης προκαλεί παρατεταµένη και σηµαντική αναστολή της AChE στον εγκέφαλο µε µικρή αναστολή του περιφερικού ενζύµου. Το γεγονός ότι η φαρµακολογική αδρανοποίηση και η απέκκριση του φαρµάκου είναι ανεξάρτητες από το ηπατικό σύστηµα P450, σε συνδυασµό µε το γεγονός ότι εφαρµόζεται τιτλοποίηση της δόσης ανάλογα µε τις ανάγκες και απαιτήσεις κάθε ασθενή, συνεπάγεται ότι το φάρµακο µπορεί εύκολα να χρησιµοποιηθεί σε ασθενείς µε ήπια έως µέτρια ηπατική ανεπάρκεια (Εικόνα 13). Επιπλέον, σε κλινικές µελέτες που πραγµατοποιήθηκαν για το φάρµακο δεν παρατηρήθηκε ηπατοτοξικότητα. Σε σύγκριση µε το γενικό πληθυσµό τα ηλικιωµένα άτοµα χρειάζονται συχνότερα επείγουσα ιατρική παρέµβαση. Τα άτοµα που λαµβάνουν ριβαστιγµίνη είναι λιγότερο πιθανό να δηµιουργήσουν προβλήµατα στην παροχή επείγουσας περίθαλψης, και αυτό γιατί η φυσιολογική λειτουργία της AChE αποκαθίσταται µέσα σε 24ώρες από τη διακοπή λήψης του φαρµάκου. εν ισχύει το ίδιο και για τη χορήγηση µακράς δράσης αναστολέων της AChE, όπως είναι τα µη αναστρέψιµα οργανοφωσφορικά σκευάσµατα και τα αναστρέψιµα παράγωγα της πιπεριδίνης 186,187,188,

90 Εικόνα 13. Σχηµατική απεικόνιση του φαρµακοκινητικού µοντέλου για τη ριβαστιγµίνη 185,186,187,188 89
91 3.9. Μελλοντικές κατευθύνσεις της έρευνας Η αρχική κλινική επιτυχία των αναστολέων της χολινεστεράσης, που στηρίχθηκε στη θεωρία της χολινεργικής υπόθεσης, είναι αποτέλεσµα έρευνας δεκαετιών πολλών ερευνητών από ανεξάρτητες µεταξύ τους ερευνητικές οµάδες. Η χολινεργική υπόθεση βασιζόµενη σε ουσιαστική θεωρητική βάση, καθώς και σε πληθώρα δεδοµένων από ποικίλα πειραµατικά µοντέλα, συνέβαλε στην εξέλιξη και παγιοποίηση της µόνης υπάρχουσας θεραπευτικής αντιµετώπισης των γνωστικών διαταραχών της νόσου Alzheimer. Ωστόσο, η επιστηµονική κοινότητα συνεχίζει να εργάζεται σε θεωρητικό και πειραµατικό επίπεδο για τη βελτίωση της συγκεκριµένης κατηγορίας φαρµακολογικών ουσιών, καθώς και για την ανάπτυξη νέων. Για παράδειγµα, οι αγωνιστές των µουσκαρινικών υποδοχέων αποτελούν µία από τις κατηγορίες φαρµακολογικών ουσιών, η χρήση των οποίων στην αντιµετώπιση των γνωστικών διαταραχών της νόσου Alzheimer διερευνάται από ορισµένους ερευνητές 197. Αντιστοίχως, διερευνάται ο ρόλος των νικοτινικών υποδοχέων και η πιθανότητα χρήσης αγωνιστών των νικοτινικών υποδοχέων σε συνδυασµό µε άλλους χολινεργικούς παράγοντες. Αν και η πρόοδος που έχει επιτευχθεί µε τη χρήση των αναστολέων της χολινεστεράσης είναι εντυπωσιακή, καθώς αποτελούν τη µόνη αποτελεσµατική θεραπευτική αντιµετώπιση των συµπτωµάτων της νόσου Alzheimer, θεωρείται περισσότερο σηµαντική η διαπίστωση ότι παράλληλα µε τη βελτίωση των γνωστικών λειτουργιών παρατηρείται καθυστέρηση στο ρυθµό εξέλιξης της συγκεκριµένης νευροεκφυλιστικής νόσου. Θεωρείται ότι η αναστολή της ακετυλοχολινεστεράσης περιορίζει τη σύνθεση παθολογικού β-αµυλοειδούς
92 ΣΤ. ΟΚΙΜΑΣΙΕΣ ΣΥΜΠΕΡΙΦΟΡΑΣ Η συµπεριφορική νευροεπιστήµη είναι η µελέτη των µηχανισµών του εγκεφάλου και της συµπεριφοράς. Καλύπτει ένα τεράστιο επιστηµονικό πεδίο, όπου όµως υπάρχουν αρκετά θέµατα προς διερεύνηση. Μάλιστα, όταν προστεθούν και οι πολυπλοκότητες της νευροεπιστήµης, τότε το µέγεθος του προβλήµατος αυξάνει υπερβολικά. Ωστόσο, έχει επιχειρηθεί από διάφορους επιστηµονικούς κύκλους να οριστεί το ερευνητικό τους πεδίο πιο αυστηρά. Όροι όπως ψυχοφαρµακολογία, συµπεριφορική φαρµακολογία, ψυχονευρο-ενδοκρινολογία, νευροψυχοφαρµακολογία και νευροψυχολογία, έχουν χρησιµοποιηθεί για να ορίσουν ένα συγκεκριµένο πεδίο της συµπεριφορικής νευροεπιστήµης. Το ουσιώδες, όµως, είναι ότι όλοι οι συµπεριφορικοί νευροεπιστήµονες µοιράζονται ένα κοινό στρατηγικό σκοπό, την παρατήρηση και τη µέτρηση της συµπεριφοράς σε ζώα και στον άνθρωπο και το συσχετισµό αυτής µε συγκεκριµένες εγκεφαλικές λειτουργίες. Για να επιτευχθεί αυτό, είναι συνήθως απαραίτητη η επίδραση στο κεντρικό νευρικό σύστηµα µέσω φαρµάκων, ηλεκτρικής διέγερσης και πειραµατικών καταστροφών εγκεφαλικών δοµών. Οι επιδράσεις αυτές µετρώνται µε τη χρήση ειδικά σχεδιασµένων δοκιµασιών, των πειραµάτων-δοκιµασιών συµπεριφοράς. Για παράδειγµα, µπορεί να ενδιαφερόµαστε για τον καθορισµό των επιδράσεων ενός συγκεκριµένου φαρµάκου στη συµπεριφορά, ή για τη συµβολή µίας συγκεκριµένης οµάδας νευρικών κυττάρων στις γνωστικές διεργασίες, ή τέλος για το αν η πειραµατική καταστροφή µίας εγκεφαλικής δοµής επιδρά στη µνήµη. Τυπικά η αλληλουχία του πειραµατικού σχεδιασµού περιλαµβάνει: επιλογή του θέµατος προς διερεύνηση, σχεδιασµό της πιο κατάλληλης δοκιµασίας συµπεριφοράς, καθορισµό των πειραµατικών παραµέτρων (µέγεθος των οµάδων, εύρος της δόσης και θέση της πειραµατικής καταστροφής), επιλογή των στατιστικών µεθόδων που θα χρησιµοποιηθούν, διεξαγωγή του πειράµατος κάτω από αυστηρά ελεγχόµενες συνθήκες, ιστολογική ή νευροχηµική επιβεβαίωση, ερµηνεία των δεδοµένων. 91
93 1. Ο λαβύρινθος του Morris Ένα από τα πιο δηµοφιλή και ευρέως χρησιµοποιούµενα πειράµατα συµπεριφοράς για τον έλεγχο της µνήµης και της µάθησης είναι µία δοκιµασία προσανατολισµένης κολύµβησης, η οποία αναπτύχθηκε για τους επίµυες από τον Richard G.M. Morris 199. Εξαιτίας της τεχνικής απλότητας και της αξιοπιστίας της, η δοκιµασία έχει τύχει ευρύτατης εφαρµογής σε µελέτες νευροφαρµακολογίας και φυσιολογίας. Προσφάτως, έχει καθιερωθεί ως το κύριο πείραµα για τον έλεγχο αλλαγών στη µνήµη και τη µάθηση και για την ταυτοποίηση των νευρωνικών παραµέτρων των συµπεριφορικών αλλαγών. Ο λαβύρινθος του Morris (water maze) είναι µία απλή και δηµοφιλής διαδικασία για τον έλεγχο της γνωστικής λειτουργίας σε τρωκτικά 200,201. Χρησιµοποιείται ευρέως σε προκλινικές µελέτες, για τον έλεγχο του θεραπευτικού δυναµικού φαρµακολογικά δραστικών ενώσεων σε σχετιζόµενες µε την ηλικία γνωστικές διαταραχές 202,203,204,205,206,207, 208,209,210,211,212,213. Όπως σε κάθε δοκιµασία για τον έλεγχο της γνωστικής λειτουργίας, έτσι και στο λαβύρινθο του Morris η καταγραφή των παραµέτρων αποτελεί µία έµµεση µέτρηση µίας εγκεφαλικής-ψυχολογικής λειτουργίας. Πολυάριθµες µελέτες υποστηρίζουν την αξιοπιστία του λαβύρινθου του Morris στη µελέτη της γνωστικής λειτουργίας 211,214,215,216,217,218,219,220,221,222. Το πειραµατικό πρωτόκολλο περιλαµβάνει µία κυλινδρική δεξαµενή ύδατος (διαµέτρου περίπου 1,5m), η οποία είναι γεµάτη µε ύδωρ βάθους 30cm και θερµοκρασίας 22±1 C. Η δεξαµενή βρίσκεται σε ένα χώρο (Εικόνα 14) µε σταθερή θερµοκρασία 22 C και επαρκή σταθερό τεχνητό φωτισµό, στους τοίχους του οποίου βρίσκονται πίνακες-φωτογραφίες υψηλής χρωµατικής αντίθεσης 3,

94 Εικόνα 14. Σχηµατική απεικόνιση του λαβύρινθου του Morris 223 Ο στόχος για το πειραµατόζωο είναι να βρει την εξέδρα διαφυγής (παραλληλόγραµµης επιφάνειας διαµέτρου 10 10cm, κατασκευασµένης από διαφανές plexiglas), η οποία είναι κρυµµένη 1,0cm κάτω από την επιφάνεια του νερού. Αρχικά το πειραµατόζωο κολυµπά κοντά στα τοιχώµατα της δεξαµενής. Αυτή η ενστικτώδης συµπεριφορά ονοµάζεται θιγµοταξία (thigmotaxis), από τις ελληνικές λέξεις «θίγµα» (επαφή) και «τάξις» (διάταξη). Μία άλλη αυθόρµητη αντίδραση είναι ότι το πειραµατόζωο επιπλέει ακίνητο, κάτι που δεν είναι συνηθισµένο στους επίµυες, που χρησιµοποιούνται στο εργαστήριο. Επειδή η εξέδρα διαφυγής είναι τοποθετηµένη σε απόσταση τουλάχιστον 30cm από το τοίχωµα της δεξαµενής, καµία από τις παραπάνω συµπεριφορές δεν επιτρέπει την ενεργητική απόδραση του πειραµατόζωου. 93
95 Μετά από 180sec κολύµβησης, ο παρατηρητής αποµακρύνει από το ύδωρ το πειραµατόζωο και το τοποθετεί στην εξέδρα διαφυγής για 20sec και στη συνέχεια το αποµακρύνει από τη δεξαµενή. Ύστερα από δύο δοκιµασίες τα περισσότερα πειραµατόζωα τείνουν να αναπτύξουν στρατηγικές κολύµβησης. Τελικώς, έρχονται τυχαία σε επαφή µε την εξέδρα και εάν ανεβούν επάνω της και παραµείνουν εκεί για µία µικρή χρονική περίοδο, αποµακρύνονται από τον παρατηρητή και αφού στεγνώσουν, ώστε να αποφευχθεί η υποθερµία, τοποθετούνται στο κλουβί τους. 2. Λαβύρινθος ανοικτού πεδίου Η χρήση λαβύρινθων στη µελέτη της µνήµης σε πειραµατόζωα έχει µεγάλη ιστορία. Οι µετρήσεις κατηγοριοποιούνται σε απόκτηση (acquisition) και ανάκληση (retention) της µνήµης, όπου το πειραµατόζωο πρέπει απλώς να µάθει και να θυµάται τη σωστή διαδροµή-διαδικασία στο λαβύρινθο. Το πρώτο µετρήσιµο δεδοµένο στους λαβυρίνθους ήταν ο συνολικός χρόνος εκτέλεσης της δοκιµασίας, ενώ δεν υπήρχε καµία συστηµατική κατηγοριοποίηση των λαθών. Μετέπειτα βελτιώσεις περιελάµβαναν αλλαγές στη διαδροµή επίτευξης του στόχου, µε αποτέλεσµα να απαιτείται από το πειραµατόζωο να εµφανίσει µία ευελιξία-προσαρµοστικότητα στη µάθηση κάθε νέου δεδοµένου στη διαδικασία του λαβύρινθου. Λαβύρινθοι δύο επιλογών, όπως ο T-maze ή τα αυτοµατοποιηµένα ανάλογά του, χρησιµοποιήθηκαν για πειράµατα διάκρισης-αναγνώρισης περιορισµένων επιλογών. Πιο πρόσφατες εξελίξεις στο σχεδιασµό των λαβυρίνθων εισήγαγαν σηµαντικές καινοτοµίες σε σύγκριση µε τα πρώιµα πειραµατικά πρωτόκολλα. Συγκεκριµένα, ο σχεδιασµός τους αποσκοπεί κυρίως στην αξιολόγηση της ακρίβειας των επιλογών κατά τη διάρκεια της ηµερήσιας δοκιµασίας, και δευτερευόντως στον έλεγχο της γνώσης που έχει συσσωρευθεί κατά τη διάρκεια των ηµερήσιων επαναλήψεων. Αυτοί οι δύο διαφορετικοί τύποι κατάταξης των δοκιµασιών καταγραφής της µνήµης, αναφέρονται ως παράµετροι λειτουργικής (working) µνήµης και µνήµης αναφοράς (reference). Ο λαβύρινθος ανοιχτού πεδίου (open field maze) χρησιµοποιείται για τον έλεγχο της λειτουργικής (working) µνήµης και της µνήµης αναφοράς 94
96 (reference) των πειραµατόζωων. Οι επίµυες πρέπει να βρουν την τροφή χρησιµοποιώντας σηµεία προσανατολισµού από το περιβάλλον (spatial memory). Κατά τη διάρκεια του πειράµατος η ηµερήσια χορηγούµενη ποσότητα τροφής µειώνεται κατά 20-25%, ώστε οι επίµυες να είναι νηστικοί και να επιδεικνύουν έντονο ενδιαφέρον για αναζήτηση τροφής. Μετά το τέλος της δοκιµασίας, τους χορηγείται ελεύθερα τροφή για δύο ώρες. Η δοκιµασία του λαβυρίνθου ανοικτού πεδίου πραγµατοποιείται για 15 συνεχόµενες ηµέρες κατά τη διάρκεια της φωτεινής περιόδου µεταξύ των ωρών και οκιµασία παθητικής αποφυγής Οι δοκιµασίες παθητικής αποφυγής (passive avoidance) χρησιµοποιούνται ευρέως για τη µέτρηση των γνωστικών µεταβολών, που ακολουθούν τη χορήγηση φαρµάκων και τις πειραµατικές καταστροφές εγκεφαλικών δοµών και παρά την κριτική από ορισµένους επιστήµονες, εξακολουθούν να αποτελούν ένα από τα πιο δηµοφιλή εργαλεία της συµπεριφορικής έρευνας. Αρκετοί συγγραφείς έχουν αναφερθεί στην παθητική αποφυγή 224,225, υποστηρίζοντας ότι τα πειραµατόζωα «θυµούνται» ότι µία συγκεκριµένη επιλογή-αντίδραση τερµατίζεται µε ένα «δυσάρεστο» συµβάν και γι αυτό θα διστάσουν να επαναλάβουν τη συγκεκριµένη επιλογή στο µέλλον. Εποµένως, η επακόλουθη αύξηση του χρόνου αντίδρασης των πειραµατοζώων, φαίνεται να αντανακλά την ένταση της µνηµονικής εγγραφής του «δυσάρεστου» συµβάντος. Οι δοκιµασίες παθητικής αποφυγής έχουν αρκετά πλεονεκτήµατα: είναι απλές στην εφαρµογή και απαιτούν λίγο και απλό εξοπλισµό, το σύνολο του πειράµατος µπορεί να ολοκληρωθεί στη διάρκεια µερικών ωρών (συνήθως σε 24ώρες), τα φάρµακα µπορούν να χορηγηθούν µετά τη δοκιµασία απόκτησης της µνήµης και τα πειραµατόζωα µπορούν να ελεγχθούν χωρίς να βρίσκονται υπό την επίδραση του φαρµάκου (για παράδειγµα 24 ώρες µετά). 95
97 Υπάρχουν, όµως, και ορισµένα µειονεκτήµατα επειδή: τα πειραµατόζωα διαφέρουν στην ικανότητά τους να ανέχονται τα δυσάρεστα ερεθίσµατα, οι χρόνοι αντίδρασης συχνά διαφέρουν και είναι απαραίτητη η χρήση µεγάλων οµάδων πειραµατόζωων, δεν είναι ξεκάθαρο τι µπορεί να προσδιορίσει αυτή η δοκιµασία, γιατί είναι σαφές ότι µετράται η µνήµη, εµπλέκονται όµως και άλλοι γνωστικοί και µη-γνωστικοί παράγοντες. 96
98 ΕΙ ΙΚΟ ΜΕΡΟΣ 97
99 98
100 I. ΥΛΙΚΟ ΚΑΙ ΜΕΘΟ ΟΙ 1. Πειραµατόζωα Στην παρούσα µελέτη χρησιµοποιήθηκαν ένηβοι άρρενες επίµυς φυλής Wistar, οι οποίοι προέρχονταν από την καλλιέργεια του Εργαστηρίου Πειραµατικής Φυσιολογίας της Ιατρικής Σχολής του Αριστοτέλειου Πανεπιστηµίου Θεσσαλονίκης. Οι επίµυες διαβιούσαν σε οµάδες των 4-5 πειραµατοζώων, µέσα σε ειδικά κλουβιά κατασκευασµένα από plexiglas. Οι συνθήκες στο χώρο φύλαξης των πειραµατοζώων (animal house) ήταν σταθερές, µε θερµοκρασία 22±2 C και φωτοπερίοδο 12 ωρών (12 ώρες φως /12 ώρες σκότος). Οι επίµυες είχαν ελεύθερη πρόσβαση (ad libitum) σε νερό και τροφή (βώλοι ειδικής πλήρους ζωοτροφής για επίµυες). Η έρευνα πραγµατοποιήθηκε σύµφωνα µε τις διατάξεις του Ν.2015/92 περί «Κυρώσεως της Ευρωπαϊκής Σύµβασης για την προστασία των σπονδυλωτών ζώων που χρησιµοποιούνται για πειραµατικούς ή άλλους επιστηµονικούς σκοπούς», το Προεδρικό ιάταγµα 160/91 «Περί προστασίας των ζώων που χρησιµοποιούνται για πειραµατικούς ή άλλους επιστηµονικούς σκοπούς», την οδηγία 86/609 της Ευρωπαϊκής Ένωσης και τη διακήρυξη του Ευρωπαϊκού Ιδρύµατος Επιστήµης ESF (European Science Foundation) για την ορθή ερευνητική πρακτική (Αριθµός πρωτοκόλλου άδειας: 13/12307, ιεύθυνση Κτηνιατρικής Νοµαρχίας Θεσσαλονίκης). 2. Πειραµατικό πρότυπο Το πειραµατικό πρότυπο βασίστηκε στην αµφοτερόπλευρη καταστροφή του βασικού πυρήνα του Meynert µε έγχυση ιµποτενικού οξέος (ibotenic acid, alpha-amino-3-hydroxy-5-isoxazoleacetic acid) διαµέσου µικροσύριγγας Hamilton 1,0µl, µε τη χρήση στερεοταξικής διάταξης. Χρησιµοποιήθηκε το πειραµατικό πρότυπο των Wenk και συν. (1984) 226 που περιγράφει τις βέλτιστες συντεταγµένες και τον ιδανικό όγκο εγχεόµενου διαλύµατος για την καταστροφή του βασικού πυρήνα του Meynert. Το συγκεκριµένο πρότυπο αναφέρεται ως η καλύτερη µέθοδος για την αµφοτερόπλευρη καταστροφή του βασικού πυρήνα του Meynert, µε τον υψηλότερο δείκτη 99
, σε δόση 1ml/100g βάρους σώµατος.")
101 επιβίωσης των πειραµατοζώων, στην ανασκόπηση των Pepeu και συν. (1986) 227, και του Smith (1988) 228. Το πειραµατόζωο αναισθητοποιούνταν µε ενδοπεριτοναϊκή έγχυση διαλύµατος ένυδρης χλωράλης (4,5%w/v σε φυσιολογικό ορό), σε δόση 1ml/100g βάρους σώµατος. Στη συνέχεια αποψιλωνόταν η περιοχή του κρανίου και ο επίµυς τοποθετούνταν στη στερεοταξική συσκευή (της εταιρείας Narishige, Japan), που ρυθµιζόταν 2mm κάτω από τη διωτιαία (interaural) γραµµή (Εικόνα 15). Εικόνα 15. Στερεοταξική συσκευή Narishige, Japan Με βάση τις συντεταγµένες της έγχυσης, που είναι 0,9mm οπίσθια του βρέγµατος και ±2,6mm πλάγια της µέσης γραµµής, γινόταν διάτρηση του κρανίου, ώστε να είναι δυνατή η είσοδος της βελόνας της έγχυσης. Οι συντεταγµένες της έγχυσης είναι 0,9mm οπίσθια του βρέγµατος, ±2,6mm πλάγια της µέσης γραµµής και 6,8mm κάτω από την επιφάνεια του εγκεφάλου. Το ιµποτενικό οξύ (ibotenic acid 25nmole) διαλυµένο σε ρυθµιστικό διάλυµα φωσφορικών όγκου 0,6µl, εγχέεται διαµέσου µικροσύριγγας 1,0µl Hamilton, µε ταχύτητα έγχυσης 0,1µl/min. Στην οµάδα 100
102 των επίµυων µε εικονική καταστροφή (sham-operated) του βασικού πυρήνα έγινε έγχυση ρυθµιστικού διαλύµατος φωσφορικών αντίστοιχου όγκου. Η βελόνα έγχυσης παραµένει στη θέση της για 5 min. Όλες οι διαδικασίες γίνονται αµφοτερόπλευρα. Οι επίµυες µετά από εικονική καταστροφή του βασικού πυρήνα είχαν καλή ανάρρωση, ενώ µετά από αµφοτερόπλευρη νευροτοξική καταστροφή του βασικού πυρήνα, το 5% (2/40) των πειραµατοζώων δεν επέζησε µετά τη χειρουργική διαδικασία. Έτσι, ο αριθµός των ζώων αυτών των οµάδων αντανακλά τη συγκεκριµένη διόρθωση 229. Μετά τη χειρουργική διαδικασία χορηγήθηκε υποδόρια αντιβιωτικό ενροφλοξασίνη (Baytril 5% enrofloxacin) σε δόση 5mg/kg βάρους σώµατος. Στη συνέχεια τα πειραµατόζωα τοποθετούνταν σε ειδικά κλουβιά το καθένα µόνο του, όπου είχαν ελεύθερη πρόσβαση (ad libitum) σε ύδωρ και τροφή. 101
103 3. Οµάδες πειραµατοζώων Χρησιµοποιήθηκαν 60 πειραµατόζωα, που χωρίστηκαν σε έξι (6) οµάδες (Πίνακας 3): Πίνακας 3. Οµάδες πειραµατοζώων Σύνολο πειραµατοζώων (n=60) ΟΜΑ Α Α Γενικοί µάρτυρες (controls) (n=10) ΟΜΑ Α Β Ειδικοί µάρτυρες µε εικονική εγχείρηση (sham operated) (n=10) ΟΜΑ Α Γ Πειραµατική καταστροφή του βασικού πυρήνα του Meynert. οκιµασίες διατήρησης µνήµης (retention tests) (n=10) ΟΜΑ Α Πειραµατική καταστροφή του βασικού πυρήνα του Meynert + χορήγηση ριβαστιγµίνης. οκιµασίες διατήρησης µνήµης (retention tests) (n=10) ΟΜΑ Α Ε Πειραµατική καταστροφή του βασικού πυρήνα του Meynert. οκιµασίες απόκτησης µνήµης (acquisition tests) (n=10) ΟΜΑ Α ΣΤ Πειραµατική καταστροφή του βασικού πυρήνα του Meynert + χορήγηση ριβαστιγµίνης. οκιµασίες απόκτησης µνήµης (acquisition tests) (n=10) Η πρώτη οµάδα (Α) περιελάµβανε δέκα (10) πειραµατόζωα, στα οποία δεν εφαρµόστηκε το πειραµατικό πρότυπο και χρησίµευσαν ως γενικοί µάρτυρες (controls). Στα πειραµατόζωα πραγµατοποιήθηκαν κατά τις ηµέρες 1 έως 20 δοκιµασίες συµπεριφοράς (acquisition tests), την 25η ηµέρα έγιναν αισθητικοκινητικές δοκιµασίες και τις ηµέρες 27, 28 και 29 έγινε έλεγχος διατήρησης της µνήµης των δοκιµασιών συµπεριφοράς (retention tests). Η δεύτερη οµάδα (Β) περιελάµβανε δέκα (10) πειραµατόζωα, στα οποία έγινε εικονική εγχείρηση χωρίς καταστροφή του βασικού πυρήνα (shamoperated) την 20η ηµέρα, για να ελεγχθούν οι τυχόν επιδράσεις της εγχειρητικής διαδικασίας και χρησίµευσαν ως ειδικοί µάρτυρες. Στα πειραµατόζωα πραγµατοποιήθηκαν οι ίδιες δοκιµασίες (όπως στα πειραµατόζωα της οµάδας Α) στα ίδια χρονικά διαστήµατα. Η τρίτη οµάδα (Γ) περιελάµβανε δέκα (10) πειραµατόζωα, στα οποία εφαρµόστηκε το πειραµατικό πρότυπο. Στα πειραµατόζωα αυτά, αφού πραγµατοποιήθηκαν κατά τις ηµέρες 1 έως 20 οι δοκιµασίες συµπεριφοράς, έγινε στερεοταξική καταστροφή του βασικού πυρήνα του Meynert την 20η ηµέρα. Την 25η ηµέρα έγιναν οι αισθητικοκινητικές δοκιµασίες και τις 102
104 ηµέρες 27, 28 και 29 έγινε έλεγχος διατήρησης της µνήµης των δοκιµασιών συµπεριφοράς µετά από υποδόρια έγχυση φυσιολογικού ορού. Η τέταρτη οµάδα ( ) περιελάµβανε δέκα (10) πειραµατόζωα, στα οποία εφαρµόστηκε το πειραµατικό πρότυπο. Στα πειραµατόζωα αυτά πραγµατοποιήθηκαν οι ίδιες, όπως στα πειραµατόζωα της οµάδας Γ, δοκιµασίες και στα ίδια χρονικά διαστήµατα, µε τη διαφορά ότι ο έλεγχος της διατήρησης της µνήµης των δοκιµασιών συµπεριφοράς γινόταν µετά από υποδόρια έγχυση διαλύµατος ριβαστιγµίνης. Η πέµπτη οµάδα (Ε) περιελάµβανε δέκα (10) πειραµατόζωα, στα οποία εφαρµόστηκε το πειραµατικό πρότυπο. Στα πειραµατόζωα αυτά έγινε στερεοταξική καταστροφή του βασικού πυρήνα του Meynert. Την πέµπτη (5η) ηµέρα µετά την καταστροφή, πραγµατοποιήθηκαν οι αισθητικοκινητικές δοκιµασίες. Μετά τη συµπλήρωση 7ήµερης ανάρρωσης, ακολούθησαν οι δοκιµασίες συµπεριφοράς κατά τις πειραµατικές ηµέρες 1 έως 20. Κατά τη διάρκεια των δοκιµασιών γινόταν υποδόρια έγχυση φυσιολογικού ορού µε βάση το πειραµατικό πρότυπο. Η έκτη οµάδα (ΣΤ) περιελάµβανε δέκα (10) πειραµατόζωα, στα οποία εφαρµόστηκε το πειραµατικό πρότυπο. Στα πειραµατόζωα αυτά έγινε στερεοταξική καταστροφή του βασικού πυρήνα του Meynert. Πραγµατοποιήθηκαν οι ίδιες, όπως στα πειραµατόζωα της οµάδας Ε, δοκιµασίες και στα ίδια χρονικά διαστήµατα, µε τη διαφορά ότι κατά τη διάρκεια της περιόδου των δοκιµασιών, γινόταν υποδόρια έγχυση διαλύµατος ριβαστιγµίνης. 103
105 Οµάδες Α, Β, Γ και Λαβύρινθος ανοικτού πεδίου πειραµατική ηµέρα 5 Λαβύρινθος ανοικτού πεδίου, δοκιµασία παθητικής αποφυγής (acquisition) πειραµατική ηµέρα 19 Αισθητικοκινητικές δοκιµασίες Οµάδες Α, Β, Γ, πειραµατική ηµέρα 25 Οµάδες Ε και ΣΤ πειραµατική ηµέρα 1 Λαβύρινθος του Morris πειραµατική ηµέρα 4 Λαβύρινθος του Morris πειραµατική ηµέρα 20 οκιµασία παθητικής αποφυγής (retention), καταστροφή ΝΒΜ Οµάδες Γ και, εικονική καταστροφή Οµάδα Β πειραµατικές ηµέρες 27, 28, 29 Λαβύρινθος του Morris, λαβύρινθος ανοικτού πεδίου, δοκιµασία παθητικής αποφυγής (retention) Καταστροφή ΝΒΜ Οµάδες Ε και ΣΤ Λαβύρινθος του Morris πειραµατική ηµέρα 1 Λαβύρινθος ανοικτού πεδίου πειραµατική ηµέρα 5 Λαβύρινθος ανοικτού πεδίου, δοκιµασία παθητικής αποφυγής (acquisition) πειραµατική ηµέρα 19 5η µετεγχειρητική ηµέρα Αισθητικοκινητικές δοκιµασίες Οµάδες Ε και ΣΤ πειραµατική ηµέρα 4 Λαβύρινθος του Morris πειραµατική ηµέρα 20 οκιµασία παθητικής αποφυγής (retention) Σχήµα 1. Σχηµατική απεικόνιση του πειραµατικού πρωτόκολλου (NBM: Nucleus Basalis of Meynert) 104
106 4. Αισθητικοκινητικές δοκιµασίες Μετά την ανάρρωση των πειραµατοζώων από τη χειρουργική διαδικασία και πριν από τη διενέργεια των πειραµάτων-δοκιµασιών συµπεριφοράς, οι επίµυες υποβλήθηκαν σε µία σειρά αισθητικοκινητικών δοκιµασιών για την εξακρίβωση της αισθητικοκινητικής τους συµπεριφοράς 229,230. Σκοπός αυτών των δοκιµασιών ήταν η αποφυγή χρησιµοποίησης στα πειράµατα συµπεριφοράς επιµύων µε αισθητικοκινητικές διαταραχές, κάτι που θα οδηγούσε σε λανθασµένα συµπεράσµατα. Τα πειράµατα διεξήχθησαν την πέµπτη (5η) ηµέρα µετά τη χειρουργική καταστροφή του βασικού πυρήνα του Meynert. Οι αισθητικοκινητικές δοκιµασίες που χρησιµοποιήθηκαν ήταν οι ακόλουθες: 4.1. Μονόζυγο Τα πειραµατόζωα τοποθετούνταν σε µία οριζόντια ράβδο διαµέτρου 0,6cm, στερεωµένη 45cm πάνω από µία µαλακή επιφάνεια. Οι επίµυες κρέµονταν από τη ράβδο µε τα πρόσθια άκρα τους (horizontal bar-grip strength), µε µέγιστο χρόνο παραµονής 60sec. Το test επαναλαµβανόταν δύο φορές και καταγραφόταν ο µέσος όρος του χρόνου παραµονής στη ράβδο Αντανακλαστικό έκτασης των πρόσθιων άκρων Το πειραµατόζωο κρεµόταν από την ουρά και σε κατακόρυφη θέση πάνω από την άκρη ενός τραπεζιού, όπου έπρεπε να εκτείνει τα πρόσθια άκρα προς την επιφάνεια του τραπεζιού (placing reflex). Καταγραφόταν η παρουσία ή η απουσία του συγκεκριµένου αντανακλαστικού Αντίσταση των πρόσθιων άκρων σε ραχιαία ώθηση Το πειραµατόζωο συγκρατούνταν από τη ραχιαία επιφάνεια, έχοντας επαφή µε την επιφάνεια ενός τραπεζιού και ωθούνταν προς την άκρη του τραπεζιού (placed on the table and pushed to the edge). Αξιολογήθηκε η αντίσταση στην πίεση και η ικανότητα συγκράτησης µε τα πρόσθια άκρα στην άκρη του τραπεζιού. 105
107 4.4. Αντανακλαστικό κάµψης Τα πειραµατόζωο συγκρατούνταν από τον αυχένα και ένα από τα πρόσθια άκρα πιεζόταν ελαφρώς µε µία λαβίδα (flexion reflex). Η αντίδραση ήταν η παρουσία ή η απουσία άµεσης απόσυρσης-κάµψης του άκρου Αντανακλαστικό σύλληψης Το πειραµατόζωο συγκρατούνταν από τον αυχένα και η παλαµιαία επιφάνεια των πρόσθιων άκρων αγγιζόταν µε ένα οριζόντιο σύρµα (grasping reflex). Αξιολογήθηκε η σύλληψη ή όχι του σύρµατος µε τα πρόσθια άκρα Αντανακλαστικό επαναφοράς στην ορθή θέση Το πειραµατόζωο τοποθετούνταν ανάστροφα µε τη ραχιαία επιφάνειά του σε ένα τραπέζι (righting reflex). Αξιολογήθηκε η επαναφορά ή όχι στην ορθή θέση Επαγρύπνηση Αξιολογήθηκε αν το πειραµατόζωο ήταν σε επαγρύπνηση (alertness) και ενδιαφερόταν για τα ερεθίσµατα του περιβάλλοντος (0: απουσία ενδιαφέροντος, 1: µέτριο ενδιαφέρον, 2: ενεργό ενδιαφέρον) Ακουστικό αντανακλαστικό Το πειραµατόζωο τοποθετούνταν σε ένα τραπέζι και προκαλούνταν ένα έντονο ηχητικό ερέθισµα (π.χ. χειροκρότηµα). Αξιολογήθηκε η αντίδραση ή η αδιαφορία για το ερέθισµα (auditory reflex) Αντανακλαστικό του κερατοειδούς Το πειραµατόζωο συγκρατούνταν από τον αυχένα και µε ένα τολύπιο βάµβακος αγγιζόταν ο κερατοειδής. Αξιολογήθηκε η παρουσία ή η απουσία του αντανακλαστικού του κερατοειδούς (corneal reflex) Εκκριτικά σηµεία Το πειραµατόζωο ελεγχόταν για εκκριτικά σηµεία (secretory signs) στο στόµα και τους οφθαλµούς. Αξιολογήθηκε η φυσιολογική εκκριτική λειτουργία ή η µη φυσιολογική σιελόρροια και δακρύρροια. 106
108 5. Χορήγηση ριβαστιγµίνης Οι αναστολείς της χολινεστεράσης είναι αποτελεσµατικοί σε ένα περιορισµένο εύρος δόσης, έχοντας σχέση δόσης-αποτελέσµατος ανεστραµµένου U 231,232. Οι αιτίες αυτής της σχέσης δεν είναι ξεκάθαρες, αλλά αντίστοιχη σχέση έχει παρατηρηθεί µε τη φυσοστιγµίνη 233 και την επτυλφυσοστιγµίνη 234 σε ασθενείς µε νόσο Alzheimer. Φαίνεται, επίσης, ότι προκαλείται µείωση της θεραπευτικής αποτελεσµατικότητας, όταν η αναστολή της χολινεστεράσης ξεπερνά το 40%. Η ριβαστιγµίνη εµφανίζει εκλεκτικότητα στην αναστολή της ακετυλοχολινεστεράσης στο φλοιό του εγκεφάλου και τον ιππόκαµπο σε δόσεις υποδόριας χορήγησης 0,5-1,5 mg/kg 3,192,235, ενώ έχει σηµαντικά µικρότερη δράση στους σκελετικούς µύες και το ραβδωτό σώµα (Πίνακας 4). Υψηλότερες δόσεις ριβαστιγµίνης (µεγαλύτερες από 2,5 mg/kg) προκαλούν τρόµο και διάρροια, καθώς και εµφάνιση συµπεριφοράς διέγερσης σε ακινησία 235. Πίνακας 4. Ποσοστά αναστολής χολινεστεράσης 3. Με έντονα στοιχεία φαίνεται η δόση της ριβαστιγµίνης που χρησιµοποιήθηκε, καθώς και τα ποσοστά (%) αναστολής που επιτυγχάνονται στον εγκέφαλο και τους µύες. Ποσοστό (%) αναστολής της χολινεστεράσης 30min από την έγχυση ριβαστιγµίνης οσολογία ριβαστιγµίνης Φλοιός Ιππόκαµπος Ραβδωτό σώµα Μύες 0,5mg/kg 30,1±5,5 21,0±6,7 11,6±4,9 15,4±4,9 0,75mg/kg 41,1±5,2 39,9±1,9 12,7±4,0 26,2±8,7 1,5mg/kg 49,2±1,8 46,0±3,7 36,2±4,6 36,7±3,3 2,5mg/kg 64,5±2,2 62,3±5,9 57,1±3,3 63,2±4,0 Εποµένως, έπρεπε να επιλεγεί µία δοσολογία υποδόριας χορήγησης της ριβαστιγµίνης, η οποία να εµφανίζει εκλεκτικότητα στην αναστολή της ακετυλοχολινεστεράσης στο φλοιό του εγκεφάλου και τον ιππόκαµπο και να µην προκαλεί τις χολινεργικές ανεπιθύµητες ενέργειες. Η δόση της υποδόριας χορήγησης της ριβαστιγµίνης, που χρησιµοποιήθηκε σε αυτό το πείραµα, ήταν 0,75mg/kg ανά 12ωρο, σε εγχεόµενο όγκο 1ml/kg. Η χορήγηση γινόταν κάθε ηµέρα σε χρονικές στιγµές, που δε σχετίζονταν µε τις δοκιµασίες-πειράµατα συµπεριφοράς. Η ακολουθούµενη 107
109 διαδικασία έχει το πλεονέκτηµα ότι οι επιδόσεις στις δοκιµασίες συµπεριφοράς δεν επηρεάζονταν από πιθανές επιδράσεις των φαρµάκων στην κινητική δραστηριότητα, τον ουδό του πόνου και την ικανότητα προσανατολισµού των πειραµατοζώων 3,236. Επιλέχθηκε να µη χορηγηθεί ριβαστιγµίνη στις οµάδες Α και Β (controls και sham-operated), γιατί µε βάση το σχεδιασµό της έρευνας και το µηχανισµό δράσης της ριβαστιγµίνης, η χρήση της στοχεύει στην αποκατάσταση της χολινεργικής διαταραχής, που δηµιουργείται µε την καταστροφή του βασικού πυρήνα του Meynert και δε χρησιµοποιήθηκε για να αυξήσει τη φυσιολογική γνωστική λειτουργία. Στις οµάδες Α, Β, Γ και Ε χορηγούνταν υποδόρια, σε αντιστοιχία µε τις οµάδες ριβαστιγµίνης και ΣΤ, 1ml/kg διαλύµατος φυσιολογικού ορού Αξιολόγηση των χολινεργικών ανεπιθύµητων ενεργειών Η απουσία ή η παρουσία σηµείων χολινεργικής υπερδραστηριότητας, όπως πτώση οπίσθιων άκρων και διάρροια, παρατηρήθηκε 15 min, 30 min και 45 min µετά την ένεση της ριβαστιγµίνης. Η ένταση του σωµατικού τρόµου βαθµολογήθηκε επίσης µε την εξής κλίµακα: 0=απουσία, 1=ήπιος, 2=µέτριος, 3=έντονος τρόµος
 είναι µία κυλινδρική δεξαµενή διαµέτρου 1,5m, η οποία είναι γεµάτη µε νερό βάθους 30cm και θερµοκρασίας 22±1 C (Εικόνα 16).")
110 6. Πειράµατα Συµπεριφοράς 6.1. Λαβύρινθος του Morris Ο λαβύρινθος του Morris (water maze) είναι µία κυλινδρική δεξαµενή διαµέτρου 1,5m, η οποία είναι γεµάτη µε νερό βάθους 30cm και θερµοκρασίας 22±1 C (Εικόνα 16). Τα πειραµατόζωα µπορούν να «δραπετεύσουν» από το νερό ανεβαίνοντας στην εξέδρα διαφυγής (παραλληλόγραµµη επιφάνεια, διαστάσεων 10 10cm, κατασκευασµένη από διαφανές plexiglas), η οποία είναι κρυµµένη 1,0cm κάτω από την επιφάνεια του νερού, ώστε να µην είναι ορατή από την επιφάνεια του νερού. Η θέση της εξέδρας κατά τη διάρκεια µίας δοκιµασίας ήταν σταθερή. Η δεξαµενή βρίσκεται σε ένα δωµάτιο µε σταθερή θερµοκρασία 22 C και επαρκή σταθερό τεχνητό φωτισµό. Στο δωµάτιο υπάρχουν ράφια, παράθυρα, κουρτίνες και πίνακες-φωτογραφίες υψηλής χρωµατικής αντίθεσης, που αποτελούν τα σηµεία προσανατολισµού των πειραµατοζώων. Η πειραµατική διαδικασία που εφαρµόστηκε στην παρούσα µελέτη στηρίχθηκε στο πρότυπο των Bejar και συν. (1999) 3, που αποτελεί παραλλαγή του πειραµατικού πρωτοκόλλου του Morris 199. Εικόνα 16. Σχηµατική απεικόνιση του λαβύρινθου του Morris Η δεξαµενή χωρίζεται νοητά σε τέσσερις περιοχές ίσου εµβαδού (ΒΑ, Β, ΝΑ, Ν ). Η εξέδρα διαφυγής τοποθετήθηκε 1,0cm κάτω από την επιφάνεια του νερού, στο µέσον ανάµεσα στο κέντρο και το τοίχωµα της δεξαµενής, στη ΒΑ περιοχή. Το πειραµατόζωο εισάγεται στη Ν περιοχή της 109
111 δεξαµενής και καταγράφεται ο χρόνος που χρειάζεται για να βρει την εξέδρα διαφυγής. Αν αποτύχει να ανακαλύψει την εξέδρα διαφυγής µέσα σε χρονικό διάστηµα 180 sec, τότε τοποθετείται από τον παρατηρητή στην εξέδρα για 20 sec και µετά αποµακρύνεται από τη δεξαµενή. Τα πειραµατόζωα υποβάλλονταν σε δύο ηµερήσιες δοκιµασίες για τέσσερις ηµέρες, µε µεσοδιάστηµα 15min µεταξύ των ηµερήσιων δοκιµασιών. Το σηµείο εισόδου του πειραµατόζωου στη δεξαµενή και η θέση της εξέδρας διαφυγής παρέµεναν ίδια µεταξύ των δύο ηµερήσιων δοκιµασιών, αλλά διαφοροποιούνταν κάθε µέρα. Η µείωση του χρόνου διαφυγής από ηµέρα σε ηµέρα στην πρώτη ηµερήσια δοκιµασία αντιπροσωπεύει τη µνήµη αναφοράς (reference memory) ή µνήµη µακράς διαρκείας, ενώ η αντίστοιχη µείωση µεταξύ των δύο ηµερήσιων δοκιµασιών σχετίζεται µε τη λειτουργική µνήµη (working memory) ή µνήµη µικρής διάρκειας
112 6.2. Λαβύρινθος ανοικτού πεδίου Ο λαβύρινθος ανοιχτού πεδίου (open field maze) χρησιµοποιήθηκε για τον έλεγχο της λειτουργικής (working) και αναφορικής µνήµης (reference memory) των πειραµατοζώων. Οι επίµυς έπρεπε να βρουν την τροφή χρησιµοποιώντας σηµεία προσανατολισµού από το περιβάλλον (spatial memory). Κατά τη διάρκεια του πειράµατος η ηµερήσια χορηγούµενη ποσότητα τροφής είχε µειωθεί κατά 20-25%, ώστε οι επίµυς να είναι νηστικοί και να επιδεικνύουν έντονο ενδιαφέρον αναζήτησης τροφής. Μετά το τέλος της δοκιµασίας, τους χορηγούνταν ελεύθερα τροφή για δύο ώρες. Η δοκιµασία του λαβυρίνθου ανοικτού πεδίου πραγµατοποιήθηκε για 15 συνεχόµενες ηµέρες κατά τη διάρκεια της φωτεινής περιόδου µεταξύ των ωρών και Η συσκευή του λαβυρίνθου, καθώς και η πειραµατική διαδικασία σχεδιάστηκαν από τον καθηγητή Ψυχολογίας του Πανεπιστηµίου Tufts της Βοστόνης Robert Cook το ,239. Η συσκευή του λαβυρίνθου ανοικτού πεδίου είναι κατασκευασµένη από λευκή µελαµίνη, έχει σχήµα κυβοειδές και έχει διαστάσεις 150cm 150cm 50cm. Οι διαστάσεις αυτές παρέχουν στα πειραµατόζωα τη δυνατότητα εξερεύνησης, επαρκούς οπτικού πεδίου και αποµόνωσης από τα εξωτερικά ερεθίσµατα κατά τη διάρκεια αναζήτησης της τροφής. Ο φωτισµός της συσκευής είναι 75 lux και προέρχεται από την οροφή του χώρου πειραµατισµού, ώστε να µη δηµιουργούνται φωτοσκιάσεις. Στο έδαφος της συσκευής είναι τοποθετηµένα σε κυκλική διάταξη έξι (6) τρυβλία σε ίσες αποστάσεις µεταξύ τους. Τα τρυβλία είναι γεµάτα µε άµµο και σε τέσσερα από αυτά υπάρχει τροφή (ένα κοµµάτι σοκολάτας γάλακτος). Στις τέσσερις γωνίες της συσκευής είναι τοποθετηµένα τέσσερα (4) αντικείµενα διαφορετικού ύψους και χρώµατος, τα οποία αποτελούν τα σηµεία προσανατολισµού των πειραµατοζώων. Τα αντικείµενα αυτά είναι: α) ένα µεταλλικό κυλινδρικό κουτί ροζ χρώµατος ύψους 20cm και διαµέτρου 12cm, β) ένα µεταλλικό κουτί καφέ χρώµατος cm, γ) ένας χάρτινος κύλινδρος πράσινου χρώµατος ύψους 30cm και διαµέτρου 8cm και δ) ένα σφαιρικό µπουκάλι χρώµατος µπλε και διαµέτρου 18cm. Η συσκευή είναι τοποθετηµένη στο κέντρο του δωµατίου πειραµατισµού, όπου υπάρχουν ελάχιστα έπιπλα µε σταθερή 111
.")
113 διάταξη στο χώρο, ικανοποιητική ηχοµόνωση και σταθερές συνθήκες θερµοκρασίας 22±2 C (Εικόνες 17α, 17β, 17γ). Εικόνα 17α Εικόνα 17β 112

114 Εικόνα 17γ Εικόνες 17α, 17β, 17γ. Λαβύρινθος ανοικτού πεδίου Πειραµατική διαδικασία λαβυρίνθου ανοικτού πεδίου Τα πειραµατόζωα πριν από την έναρξη της πειραµατικής διαδικασίας τοποθετούνταν στο λαβύρινθο για δύο συνεχόµενες ηµέρες και για 10min το καθένα, ώστε να εξοικειωθούν µε τη συσκευή. Κατά τη διάρκεια της περιόδου εξοικείωσης όλα τα τρυβλία περιείχαν τροφή (ένα κοµµάτι σοκολάτας γάλακτος). Κατά τη διάρκεια της 15νθήµερης δοκιµασίας του λαβυρίνθου ανοικτού πεδίου η τροφή τοποθετούνταν σε τέσσερα µόνο τρυβλία, πάντα στα ίδια. Κάθε πειραµατόζωο ασκούνταν για µέγιστο χρόνο 10 λεπτών ηµερησίως και η συσκευή καθαριζόταν επιµελώς µετά από κάθε δοκιµασία, ώστε να µην επηρεάζονται τα πειραµατόζωα από τις οσµές και τα περιττώµατα των προηγουµένων. Το πειραµατόζωο τοποθετούνταν στο κέντρο της συσκευής και αφηνόταν ελεύθερο να εξερευνήσει το χώρο για την ανεύρεση τροφής. Σκοπός της δοκιµασίας ήταν να εκπαιδευτεί το πειραµατόζωο για το ότι: i) Σε τέσσερα (4) συγκεκριµένα τρυβλία υπήρχε τροφή και µόνο σε αυτά (έλεγχος λειτουργικής µνήµης) ii) Αποκλειστικά και µόνο στην πρώτη επίσκεψη στο τρυβλίο υπήρχε ενίσχυση (τροφή) και για αυτό δεν έπρεπε να επαναλαµβάνουν τις επισκέψεις στο ίδιο τρυβλίο (έλεγχος αναφορικής µνήµης). 113
115 Όταν το πειραµατόζωο επισκεπτόταν τρυβλίο που περιείχε τροφή, σηµειωνόταν ως σωστή επιλογή. Αντίθετα, κάθε επίσκεψη σε τρυβλίο το οποίο το είχε ήδη επισκεφθεί ή δεν περιείχε τροφή, σηµειωνόταν ως λανθασµένη επιλογή. Η δοκιµασία ολοκληρωνόταν όταν το πειραµατόζωο έβρισκε όλα τα τρυβλία στα οποία υπείρχε τροφή ή όταν παρερχόταν ο µέγιστος χρόνος παραµονής στο λαβύρινθο (10min). Στη δοκιµασία αυτή καταγράφονταν οι σωστές και οι λανθασµένες επιλογές, ο χρόνος που µεσολαβούσε ανάµεσα στις επιλογές και ο συνολικός χρόνος παραµονής του πειραµατοζώου στη συσκευή. Μετά το τέλος της πειραµατικής διαδικασίας τα πειραµατόζωα επέστρεφαν στην ελεύθερη διατροφή (ad libitum). 114
116 6.3. οκιµασία παθητικής αποφυγής Η δοκιµασία παθητικής αποφυγής (passive avoidance test) χρησιµοποιείται για τον έλεγχο της µνήµης αποτελώντας µία από τις απλούστερες και πιο αξιόπιστες µεθόδους της συµπεριφορικής έρευνας. Χρησιµοποιείται ένα επώδυνο ερέθισµα για την απόκτηση της µνήµης (acquisition) και ελέγχεται τη διατήρηση της µνήµης (retention) σε 24ώρες. Η δοκιµασία πραγµατοποιήθηκε στην αυτόµατη συσκευή παθητικής αποφυγής (passive avoidance) της εταιρείας Ugo Basile. Η συσκευή αποτελείται από το κλουβί παθητικής αποφυγής και από τη µονάδα ρύθµισης, αυτόµατης λειτουργίας και καταγραφής (Εικόνες 18α, 18β). Το κλουβί παθητικής αποφυγής αποτελείται από το φωτεινό θάλαµο (θάλαµο εκκίνησης) και από το σκοτεινό θάλαµο (θάλαµο τιµωρίας). Ο θάλαµος εκκίνησης είναι κατασκευασµένος από λευκό plexiglas, έχει διαστάσεις 22x25cm, φωτίζεται από ένα λαµπτήρα 10W και καλύπτεται από διαφανές plexiglas, ώστε ο ερευνητής να έχει οπτική επαφή µε το πειραµατόζωο. Ο θάλαµος τιµωρίας είναι κατασκευασµένος από µαύρο plexiglas, έχει διαστάσεις 22x25cm, καλύπτεται από µαύρο plexiglas και είναι σκοτεινός. Οι δύο θάλαµοι επικοινωνούν µε αυτόµατη συρόµενη πόρτα δύο φύλλων, η οποία βρίσκεται στο κοινό τους τοίχωµα. Το δάπεδο των δύο θαλάµων αποτελείται από σαράντα µεταλλικές ράβδους διαµέτρου 0,3cm και µήκους 22cm, οι οποίες είναι τοποθετηµένες εν σειρά, απέχουν µεταξύ τους 1,2cm και σχηµατίζουν µία σχάρα. Το µεταλλικό δάπεδο είναι στηριγµένο σε άξονα που βρίσκεται στο ενδιάµεσο των δύο θαλάµων και στο άκρο ενός εκ των δύο θαλάµων. Έτσι, όταν το πειραµατόζωο κινείται από το θάλαµο εκκίνησης προς το θάλαµο τιµωρίας, το δάπεδο µετακινείται προς το θάλαµο τιµωρίας και ενεργοποιείται µία µαγνητική επαφή που διοχετεύει ηλεκτρικό ρεύµα χαµηλής έντασης στο δάπεδο, δηµιουργώντας ένα ήπιο επώδυνο ερέθισµα. Κάτω από το δάπεδο βρίσκεται ένα συρτάρι για τη συλλογή των περιττωµάτων των πειραµατοζώων. 115
 και")
.")
117 Εικόνες 18α, 18β. Συσκευή δοκιµασίας παθητικής αποφυγής. Φωτεινός και σκοτεινός θάλαµος, συσκευή ελέγχου. H µονάδα ρύθµισης, αυτόµατης λειτουργίας και καταγραφής περιλαµβάνει διακόπτες για τη ρύθµιση της έντασης (shock intensity) και της διάρκειας (shock duration) του ηλεκτρικού ρεύµατος, της καθυστέρησης του ανοίγµατος της πόρτας (door delay) και του µέγιστου χρόνου του πειράµατος (cut-off time). Περιλαµβάνει, επίσης, λυχνίες ενδείξεων για την πόρτα (door indicator) και τη διοχέτευση ηλεκτρικού ρεύµατος (shock 116
118 indicator), καθώς και µετρητή του χρόνου καθυστέρησης εισόδου (latency time) του πειραµατόζωου στο θάλαµο τιµωρίας. Τέλος, περιλαµβάνει έξοδο µεταφοράς των δεδοµένων σε ηλεκτρονικό υπολογιστή µε κατάλληλο λογισµικό επεξεργασίας των αποτελεσµάτων, καθώς και χειριστήριο ποδός, ώστε ο ερευνητής να µπορεί να συντονίζει ταχύτερα και µε µεγαλύτερη ακρίβεια τους χειρισµούς του. Το πρωτόκολλο της δοκιµασίας παθητικής αποφυγής (passive avoidance test) περιλαµβάνει δύο δοκιµασίες. Τη δοκιµασία απόκτησης µνήµης (acquisition test) και τη δοκιµασία ανάκλησης της µνήµης (retention test). Κατά τη δοκιµασία απόκτησης µνήµης το πειραµατόζωο τοποθετείται από τον ερευνητή στο φωτεινό θάλαµο (θάλαµο εκκίνησης), σε τέτοια θέση ώστε να µην έχει άµεση οπτική επαφή µε την πόρτα. Η πόρτα ανοίγει µετά από καθυστέρηση 3sec και το πειραµατόζωο έχει την επιλογή να περάσει στο σκοτεινό θάλαµο. Μόλις ο επίµυς εισέλθει στο σκοτεινό θάλαµο (θάλαµο τιµωρίας), η πόρτα κλείνει και ταυτόχρονα διοχετεύεται ηλεκτρικό ρεύµα έντασης 1,0mA και διάρκειας 3sec. Καταγράφεται ο χρόνος καθυστέρησης εισόδου του πειραµατόζωου στο θάλαµο τιµωρίας. Ο µέγιστος χρόνος παραµονής του επίµυος στο φωτεινό θάλαµο-θάλαµο εκκίνησης είναι 300sec. Μετά το τέλος του πειράµατος το πειραµατόζωο επανέρχεται στο κλουβί του, όπου έχει ελεύθερη πρόσβαση (ad libitum) σε τροφή και νερό. Κατά τη δοκιµασία ανάκλησης της µνήµης ακολουθείται η παραπάνω διαδικασία και καταγράφεται ο χρόνος καθυστέρησης εισόδου του πειραµατόζωου στο θάλαµο τιµωρίας. Η δοκιµασία ανάκλησης της µνήµης διενεργείται 24 ώρες µετά τη δοκιµασία απόκτησης µνήµης. Στις οµάδες Γ- εφαρµόσθηκε το πρωτόκολλο που περιγράφηκε µε τη µόνη διαφορά ότι η δοκιµασία ανάκλησης διενεργούνταν 7 ηµέρες µετά τη δοκιµασία απόκτησης της µνήµης. 117
119 7. Ιστολογική µελέτη Μετά το πέρας των πειραµάτων-δοκιµασιών συµπεριφοράς τα πειραµατόζωα θυσιάζονταν, ώστε να ληφθεί ο εγκέφαλός τους για ιστολογική µελέτη, για την επιβεβαίωση της βλάβης από το ιµποτενικό οξύ στο βασικό πυρήνα του Meynert. Συγκεκριµένα το πειραµατόζωο αναισθητοποιούνταν µε ενδοπεριτοναϊκή έγχυση διαλύµατος ένυδρης χλωράλης (4,5% w/v σε φυσιολογικό ορό), σε δόση 1ml/100g βάρους σώµατος και τοποθετούνταν σε ειδική χειρουργική τράπεζα. Με µία τοµή του δέρµατος από το ύψος της σφαγής του στέρνου µέχρι την υπερηβική χώρα, καθώς και µε διατοµή του θωρακικού τοιχώµατος και του διαφράγµατος, αποκαλύπτονταν η καρδιά και οι πνεύµονες. Ακολουθούσε έγχυση ηπαρινισµένου φυσιολογικού ορού στην αριστερή κοιλία, µέσω βελόνας προσαρµοσµένης σε συσκευή χορήγησης. Στη συνέχεια γινόταν έγχυση µονιµοποιητικού διαλύµατος φορµαλδεϋδης 10%. Κατά τη διάρκεια της έγχυσης των διαλυµάτων ήταν εφικτή η ελεύθερη ροή του αίµατος και των διαλυµάτων έξω από το κυκλοφορικό σύστηµα, µε µία τοµή στο δεξιό κόλπο. Μετά τη µονιµοποίηση, ο εγκέφαλος αφαιρούνταν από την κοιλότητα του κρανίου και διατηρούνταν για τουλάχιστον δύο ηµέρες σε φιαλίδια, τα οποία περιείχαν το µονιµοποιητικό υγρό. Οι προς εξέταση περιοχές του εγκεφάλου σκηνώνονταν και εγκλείονταν σε κύβους παραφίνης, ώστε να ληφθούν τοµές µε µικροτόµο παραφίνης, πάχους 4-10µm. Σε όλες τις τοµές πραγµατοποιήθηκε χρώση αιµατοξυλίνης-εωσίνης. 118
120 8. Στατιστική ανάλυση Οι καταγραφές των αποτελεσµάτων στις οµάδες των πειραµατοζώων περιγράφονται µε το µέσο όρο (ΜΟ), την τυπική απόκλιση (SD) και/ή το τυπικό σφάλµα (SEM). Για τον έλεγχο των βιοστατιστικών υποθέσεων και την ερµηνεία των αποτελεσµάτων µεταξύ των οµάδων, καθώς και για την εξαγωγή των συµπερασµάτων, χρησιµοποιήθηκε η ανάλυση διακύµανσης µε έναν παράγοντα (one way ANOVA) και η ανάλυση διακύµανσης επανειλληµένων µετρήσεων (repeated measures ANOVA RMANOVA), ακολουθούµενη από το Tukey-Kramer test πολλαπλών συγκρίσεων, όταν το F ήταν στατιστικά σηµαντικό. Ως επίπεδο σηµαντικότητας θεωρήθηκε το p=0,05. Για τη στατιστική ανάλυση χρησιµοποιήθηκε το πρόγραµµα NCSS
121 II. ΑΠΟΤΕΛΕΣΜΑΤΑ 1. Αισθητικοκινητικές δοκιµασίες Οι αισθητικοκινητικές δοκιµασίες διεξήχθησαν την πέµπτη (5η) ηµέρα µετά τη χειρουργική επέµβαση Μονόζυγο Κατά τη στατιστική επεξεργασία των καταγραφών στις διάφορες οµάδες µε ανάλυση της διακύµανσης ANOVA, δε διαπιστώθηκαν στατιστικά σηµαντικές διαφορές (σε επίπεδο P=0,05) µεταξύ των οµάδων για την παραµονή των πειραµατοζώων στο µονόζυγο-οριζόντια ράβδο. Στο γράφηµα 1 φαίνονται οι µέσοι όροι ±SEM των χρόνων παραµονής των πειραµατοζώων κάθε οµάδας στο µονόζυγο και στον πίνακα 5 οι χρόνοι παραµονής µε τους µέσους όρους ±SD Χρόνος παραµονής (sec) A B Γ E ΣΤ Οµάδες πειραµατοζώων Γράφηµα 1. Μέσοι όροι (±SEM) των χρόνων (sec) παραµονής των πειραµατοζώων στο µονόζυγο για κάθε πειραµατική οµάδα 120
122 Πίνακας 5. Χρόνος (sec) παραµονής στο µονόζυγο, ΜΟ και τυπική απόκλιση ( X ± SD) Οµάδα Α Χρόνος(sec) Οµάδα Β Χρόνος(sec) Οµάδα Γ Χρόνος(sec) Α1 60 Β1 60 Γ1 52,8 Α2 58,4 Β2 60 Γ2 60 Α3 60 Β3 60 Γ3 60 Α4 60 Β4 56,6 Γ4 54,4 Α5 60 Β5 60 Γ5 60 Α6 60 Β6 60 Γ6 60 Α7 60 Β7 60 Γ7 60 Α8 57,8 Β8 60 Γ8 55,7 Α9 60 Β9 58,4 Γ9 56,2 Α10 60 Β10 60 Γ10 60 Μ.Ο.=59,62± 0, Μ.Ο.=59,5±1, Μ.Ο.=57,91±2, Οµάδα Χρόνος(sec) Οµάδα Ε Χρόνος(sec) Οµάδα ΣΤ Χρόνος(sec) 1 60 Ε1 54,9 ΣΤ Ε2 60 ΣΤ ,4 Ε3 52,3 ΣΤ3 55,9 4 58,5 Ε4 60 ΣΤ Ε5 60 ΣΤ5 58, Ε6 57,6 ΣΤ ,2 Ε7 60 ΣΤ7 56, Ε8 60 ΣΤ8 56, Ε9 54,2 ΣΤ ,8 Ε10 57,4 ΣΤ10 60 Μ.Ο.=58,39±2, Μ.Ο.=57,64±2, Μ.Ο.=58,73±1,
123 1.2. Αντανακλαστικό έκτασης των πρόσθιων άκρων Στον πίνακα 6 φαίνεται η παρουσία ή η απουσία αντανακλαστικού έκτασης των πρόσθιων άκρων (placing reflex) σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 6. Αντανακλαστικό έκτασης των πρόσθιων άκρων Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Π Β1 Π Γ1 Π Α2 Π Β2 Π Γ2 Π Α3 Π Β3 Π Γ3 Π Α4 Π Β4 Π Γ4 Π Α5 Π Β5 Π Γ5 Π Α6 Π Β6 Π Γ6 Π Α7 Π Β7 Π Γ7 Π Α8 Π Β8 Π Γ8 Π Α9 Π Β9 Π Γ9 Π Α10 Π Β10 Π Γ10 Π Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Π Ε1 Π ΣΤ1 Π 2 Π Ε2 Π ΣΤ2 Π 3 Π Ε3 Π ΣΤ3 Π 4 Π Ε4 Π ΣΤ4 Π 5 Π Ε5 Π ΣΤ5 Π 6 Π Ε6 Π ΣΤ6 Π 7 Π Ε7 Π ΣΤ7 Π 8 Π Ε8 Π ΣΤ8 Π 9 Π Ε9 Π ΣΤ9 Π 10 Π Ε10 Π ΣΤ10 Π (Π: παρουσία αντανακλαστικού έκτασης των πρόσθιων άκρων) 122
124 1.3. Αντίσταση των πρόσθιων άκρων σε ραχιαία ώθηση Στον πίνακα 7 φαίνεται η συγκράτηση ή η αδυναµία συγκράτησης µε τα πρόσθια άκρα µετά από ραχιαία ώθηση (placed on the table and pushed to the edge) προς την άκρη του τραπεζιού σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 7. Αντίσταση των πρόσθιων άκρων σε ραχιαία ώθηση Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Φ Β1 Φ Γ1 Φ Α2 Φ Β2 Φ Γ2 Φ Α3 Φ Β3 Φ Γ3 Φ Α4 Φ Β4 Φ Γ4 Φ Α5 Φ Β5 Φ Γ5 Φ Α6 Φ Β6 Φ Γ6 Φ Α7 Φ Β7 Φ Γ7 Φ Α8 Φ Β8 Φ Γ8 Φ Α9 Φ Β9 Φ Γ9 Φ Α10 Φ Β10 Φ Γ10 Φ Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Φ Ε1 Φ ΣΤ1 Φ 2 Φ Ε2 Φ ΣΤ2 Φ 3 Φ Ε3 Φ ΣΤ3 Φ 4 Φ Ε4 Φ ΣΤ4 Φ 5 Φ Ε5 Φ ΣΤ5 Φ 6 Φ Ε6 Φ ΣΤ6 Φ 7 Φ Ε7 Φ ΣΤ7 Φ 8 Φ Ε8 Φ ΣΤ8 Φ 9 Φ Ε9 Φ ΣΤ9 Φ 10 Φ Ε10 Φ ΣΤ10 Φ (Φ: αντίσταση στην πίεση-συγκράτηση µε τα πρόσθια άκρα) 123
125 1.4. Αντανακλαστικό κάµψης Στον πίνακα 8 φαίνεται η παρουσία ή η απουσία άµεσης απόσυρσηςκάµψης (flexion reflex) του άκρου στο αντανακλαστικό κάµψης σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 8. Αντανακλαστικό κάµψης Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Π Β1 Π Γ1 Π Α2 Π Β2 Π Γ2 Π Α3 Π Β3 Π Γ3 Π Α4 Π Β4 Π Γ4 Π Α5 Π Β5 Π Γ5 Π Α6 Π Β6 Π Γ6 Π Α7 Π Β7 Π Γ7 Π Α8 Π Β8 Π Γ8 Π Α9 Π Β9 Π Γ9 Π Α10 Π Β10 Π Γ10 Π Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Π Ε1 Π ΣΤ1 Π 2 Π Ε2 Π ΣΤ2 Π 3 Π Ε3 Π ΣΤ3 Π 4 Π Ε4 Π ΣΤ4 Π 5 Π Ε5 Π ΣΤ5 Π 6 Π Ε6 Π ΣΤ6 Π 7 Π Ε7 Π ΣΤ7 Π 8 Π Ε8 Π ΣΤ8 Π 9 Π Ε9 Π ΣΤ9 Π 10 Π Ε10 Π ΣΤ10 Π (Π: παρουσία άµεσης απόσυρσης-κάµψης του άκρου) 124
126 1.5. Αντανακλαστικό σύλληψης Στον πίνακα 9 φαίνεται η ύπαρξη ή όχι αντανακλαστικού σύλληψης (grasping reflex) σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 9. Αντανακλαστικό σύλληψης Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Σ Β1 Σ Γ1 Σ Α2 Σ Β2 Σ Γ2 Σ Α3 Σ Β3 Σ Γ3 Σ Α4 Σ Β4 Σ Γ4 Σ Α5 Σ Β5 Σ Γ5 Σ Α6 Σ Β6 Σ Γ6 Σ Α7 Σ Β7 Σ Γ7 Σ Α8 Σ Β8 Σ Γ8 Σ Α9 Σ Β9 Σ Γ9 Σ Α10 Σ Β10 Σ Γ10 Σ Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Σ Ε1 Σ ΣΤ1 Σ 2 Σ Ε2 Σ ΣΤ2 Σ 3 Σ Ε3 Σ ΣΤ3 Σ 4 Σ Ε4 Σ ΣΤ4 Σ 5 Σ Ε5 Σ ΣΤ5 Σ 6 Σ Ε6 Σ ΣΤ6 Σ 7 Σ Ε7 Σ ΣΤ7 Σ 8 Σ Ε8 Σ ΣΤ8 Σ 9 Σ Ε9 Σ ΣΤ9 Σ 10 Σ Ε10 Σ ΣΤ10 Σ (Σ: σύλληψη) 125
127 1.6. Αντανακλαστικό επαναφοράς στην ορθή θέση Στον πίνακα 10 φαίνεται η ύπαρξη ή όχι φυσιολογικού αντανακλαστικού επαναφοράς (righting reflex) στην ορθή θέση σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 10. Αντανακλαστικό επαναφοράς στην ορθή θέση Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Ε Β1 Ε Γ1 Ε Α2 Ε Β2 Ε Γ2 Ε Α3 Ε Β3 Ε Γ3 Ε Α4 Ε Β4 Ε Γ4 Ε Α5 Ε Β5 Ε Γ5 Ε Α6 Ε Β6 Ε Γ6 Ε Α7 Ε Β7 Ε Γ7 Ε Α8 Ε Β8 Ε Γ8 Ε Α9 Ε Β9 Ε Γ9 Ε Α10 Ε Β10 Ε Γ10 Ε Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Ε Ε1 Ε ΣΤ1 Ε 2 Ε Ε2 Ε ΣΤ2 Ε 3 Ε Ε3 Ε ΣΤ3 Ε 4 Ε Ε4 Ε ΣΤ4 Ε 5 Ε Ε5 Ε ΣΤ5 Ε 6 Ε Ε6 Ε ΣΤ6 Ε 7 Ε Ε7 Ε ΣΤ7 Ε 8 Ε Ε8 Ε ΣΤ8 Ε 9 Ε Ε9 Ε ΣΤ9 Ε 10 Ε Ε10 Ε ΣΤ10 Ε (Ε: επαναφορά στην ορθή θέση) 126
128 1.7. Επαγρύπνηση Στον πίνακα 11 φαίνεται η ύπαρξη ή όχι επαγρύπνησης (alertness) και ενδιαφέροντος για τα ερεθίσµατα του περιβάλλοντος σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 11. Επαγρύπνηση Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 2 Β1 2 Γ1 2 Α2 2 Β2 2 Γ2 2 Α3 2 Β3 2 Γ3 2 Α4 2 Β4 2 Γ4 2 Α5 2 Β5 2 Γ5 2 Α6 2 Β6 2 Γ6 2 Α7 2 Β7 2 Γ7 2 Α8 2 Β8 2 Γ8 2 Α9 2 Β9 2 Γ9 2 Α10 2 Β10 2 Γ10 2 Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 2 Ε1 2 ΣΤ Ε2 2 ΣΤ Ε3 2 ΣΤ Ε4 2 ΣΤ Ε5 2 ΣΤ Ε6 2 ΣΤ Ε7 2 ΣΤ Ε8 2 ΣΤ Ε9 2 ΣΤ Ε10 2 ΣΤ10 2 (0:απουσία ενδιαφέροντος, 1:µέτριο ενδιαφέρον, 2:ενεργό ενδιαφέρον) 127
129 1.8. Ακουστικό αντανακλαστικό Στον πίνακα 12 φαίνεται το ακουστικό αντανακλαστικό (auditory reflex), δηλαδή η αντίδραση ή η αδιαφορία σε ακουστικό ερέθισµα κάθε πειραµατόζωου στις έξι οµάδες. Πίνακας 12. Ακουστικό αντανακλαστικό Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Π Β1 Π Γ1 Π Α2 Π Β2 Π Γ2 Π Α3 Π Β3 Π Γ3 Π Α4 Π Β4 Π Γ4 Π Α5 Π Β5 Π Γ5 Π Α6 Π Β6 Π Γ6 Π Α7 Π Β7 Π Γ7 Π Α8 Π Β8 Π Γ8 Π Α9 Π Β9 Π Γ9 Π Α10 Π Β10 Π Γ10 Π Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Π Ε1 Π ΣΤ1 Π 2 Π Ε2 Π ΣΤ2 Π 3 Π Ε3 Π ΣΤ3 Π 4 Π Ε4 Π ΣΤ4 Π 5 Π Ε5 Π ΣΤ5 Π 6 Π Ε6 Π ΣΤ6 Π 7 Π Ε7 Π ΣΤ7 Π 8 Π Ε8 Π ΣΤ8 Π 9 Π Ε9 Π ΣΤ9 Π 10 Π Ε10 Π ΣΤ10 Π (Π: παρουσία ακουστικού αντανακλαστικού) 128
130 1.9. Αντανακλαστικό του κερατοειδούς Στον πίνακα 13 φαίνεται η παρουσία ή η απουσία αντανακλαστικού του κερατοειδούς (corneal reflex) σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 13. Αντανακλαστικό του κερατοειδούς Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Π Β1 Π Γ1 Π Α2 Π Β2 Π Γ2 Π Α3 Π Β3 Π Γ3 Π Α4 Π Β4 Π Γ4 Π Α5 Π Β5 Π Γ5 Π Α6 Π Β6 Π Γ6 Π Α7 Π Β7 Π Γ7 Π Α8 Π Β8 Π Γ8 Π Α9 Π Β9 Π Γ9 Π Α10 Π Β10 Π Γ10 Π Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Π Ε1 Π ΣΤ1 Π 2 Π Ε2 Π ΣΤ2 Π 3 Π Ε3 Π ΣΤ3 Π 4 Π Ε4 Π ΣΤ4 Π 5 Π Ε5 Π ΣΤ5 Π 6 Π Ε6 Π ΣΤ6 Π 7 Π Ε7 Π ΣΤ7 Π 8 Π Ε8 Π ΣΤ8 Π 9 Π Ε9 Π ΣΤ9 Π 10 Π Ε10 Π ΣΤ10 Π (Π: παρουσία αντανακλαστικού του κερατοειδούς) 129
131 1.10. Εκκριτικά σηµεία Στον πίνακα 14 φαίνεται η ύπαρξη φυσιολογικής ή παθολογικής εκκριτικής λειτουργίας, δηλαδή σιελόρροιας και δακρύρροιας (secretory signs), σε κάθε πειραµατόζωο στις έξι οµάδες. Πίνακας 14. Εκκριτικά σηµεία Οµάδα Α Καταγραφή Οµάδα Β Καταγραφή Οµάδα Γ Καταγραφή Α1 Φ Β1 Φ Γ1 Φ Α2 Φ Β2 Φ Γ2 Φ Α3 Φ Β3 Φ Γ3 Φ Α4 Φ Β4 Φ Γ4 Φ Α5 Φ Β5 Φ Γ5 Φ Α6 Φ Β6 Φ Γ6 Φ Α7 Φ Β7 Φ Γ7 Φ Α8 Φ Β8 Φ Γ8 Φ Α9 Φ Β9 Φ Γ9 Φ Α10 Φ Β10 Φ Γ10 Φ Οµάδα Καταγραφή Οµάδα Ε Καταγραφή Οµάδα ΣΤ Καταγραφή 1 Φ Ε1 Φ ΣΤ1 Φ 2 Φ Ε2 Φ ΣΤ2 Φ 3 Φ Ε3 Φ ΣΤ3 Φ 4 Φ Ε4 Φ ΣΤ4 Φ 5 Φ Ε5 Φ ΣΤ5 Φ 6 Φ Ε6 Φ ΣΤ6 Φ 7 Φ Ε7 Φ ΣΤ7 Φ 8 Φ Ε8 Φ ΣΤ8 Φ 9 Φ Ε9 Φ ΣΤ9 Φ 10 Φ Ε10 Φ ΣΤ10 Φ (Φ: φυσιολογική εκκριτική λειτουργία) 130
132 1.11. Συνολική αξιολόγηση των αισθητικοκινητικών δοκιµασιών Τα αποτελέσµατα των αισθητικοκινητικών δοκιµασιών χρησιµοποιήθηκαν για την εκτίµηση της αισθητικοκινητικής συµπεριφοράς των επίµυων. Από την ανάγνωση των παραπάνω καταγραφών διαπιστώνεται ότι τηρείται ο σκοπός αυτών των δοκιµασιών, που ήταν η αποφυγή χρησιµοποίησης στα πειράµατα συµπεριφοράς επίµυων µε αισθητικοκινητικές διαταραχές, γεγονός που θα οδηγούσε σε λανθασµένα συµπεράσµατα 229, Αξιολόγηση των χολινεργικών ανεπιθύµητων ενεργειών Μετά τη χορήγηση ριβαστιγµίνης στα πειραµατόζωα αξιολογούνταν η απουσία ή η παρουσία χολινεργικής υπερδραστηριότητας στα 15 min, 30 min και 45 min µετά την ένεση. Όλες οι καταγραφές, κατά τη διάρκεια της περιόδου χορήγησης της ριβαστιγµίνης, ήταν αρνητικές για την παρουσία πτώσης οπίσθιων άκρων, διάρροιας και σωµατικού τρόµου. Έτσι, εξασφαλίστηκε ότι οι επιδόσεις των πειραµατοζώων στα πειράµατα συµπεριφοράς δεν οφείλονταν σε πιθανή επίδραση της ριβαστιγµίνης στην αισθητικοκινητική τους συµπεριφορά, επίδραση που θα εµπόδιζε την αξιολόγηση των επιδράσεων στις γνωστικές λειτουργίες
133 3. Πειράµατα Συµπεριφοράς 3.1. Βάρος σώµατος Στους πίνακες που ακολουθούν, αναφέρονται οι µέσοι όροι (Μ.Ο.) και οι τυπικές αποκλίσεις (SD) του βάρους (g) των πειραµατοζώων όλων των οµάδων την πρώτη και την τελευταία ηµέρα των δοκιµασιών. Πίνακας 15. Βάρος (g) πειραµατοζώων της οµάδας Α (γενικοί µάρτυρες) Ηµέρα 1 Ηµέρα 29 Μέσος όρος 262,84 293,07 Τυπική απόκλιση 7, , Πίνακας 16. Βάρος (g) πειραµατοζώων της οµάδας Β (ειδικοί µάρτυρες) Ηµέρα 1 Ηµέρα 29 Μέσος όρος 240,96 270,12 Τυπική απόκλιση 8, , Πίνακας 17. Βάρος (g) πειραµατοζώων της οµάδας Γ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης) Ηµέρα 1 Ηµέρα 29 Μέσος όρος 250,33 272,9 Τυπική απόκλιση 12, , Πίνακας 18. Βάρος (g) πειραµατοζώων της οµάδας (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης, µε χορήγηση ριβαστιγµίνης) Ηµέρα 1 Ηµέρα 29 Μέσος όρος 253,21 275,65 Τυπική απόκλιση 9, ,
134 Πίνακας 19. Βάρος (g) πειραµατοζώων της οµάδας Ε (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης) Ηµέρα καταστροφής Ηµέρα 20 του πυρήνα Μέσος όρος 269,66 292,11 Τυπική απόκλιση 12, , Πίνακας 20. Βάρος (g) πειραµατοζώων της οµάδας ΣΤ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης, µε χορήγηση ριβαστιγµίνης) Ηµέρα καταστροφής Ηµέρα 20 του πυρήνα Μέσος όρος 257,08 279,42 Τυπική απόκλιση 15, ,
135 3.2. Λαβύρινθος του Morris Στους πίνακες που ακολουθούν, αναφέρονται οι µέσοι όροι (Μ.Ο.) και οι τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων όλων των οµάδων στο λαβύρινθο του Morris. Πίνακας 21. Μέσοι όροι (Μ.Ο.) και τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Α (γενικοί µάρτυρες) στο λαβύρινθο του Morris Χρόνος (sec) Ηµέρα 1 Ηµέρα 2 Ηµέρα 3 Ηµέρα 4 Ηµέρα 27 οκιµασία 1 οκιµασία 2 Μ.Ο. SD Μ.Ο. SD 164,22 9, ,32 4, ,61 2, ,48 3, ,54 3, ,9 3, ,28 3, ,09 2, ,54 2, ,32 3, Πίνακας 22. Μέσοι όροι (Μ.Ο.) και τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Β (ειδικοί µάρτυρες) στο λαβύρινθο του Morris Χρόνος (sec) Ηµέρα 1 Ηµέρα 2 Ηµέρα 3 Ηµέρα 4 Ηµέρα 27 οκιµασία 1 οκιµασία 2 Μ.Ο. SD Μ.Ο. SD 170,86 7, ,2 3, ,57 2, ,92 2, ,66 2, ,77 2, ,54 2, ,31 2, ,75 2, ,06 3,08264 Πίνακας 23. Μέσοι όροι (Μ.Ο.) και τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Γ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης) στο λαβύρινθο του Morris Χρόνος (sec) Ηµέρα 1 Ηµέρα 2 Ηµέρα 3 Ηµέρα 4 Ηµέρα 27 οκιµασία 1 οκιµασία 2 Μ.Ο. SD Μ.Ο. SD 169,04 7, ,39 3, ,88 3, ,43 3, ,12 3, ,91 3, ,85 2, ,23 2, ,54 2, ,75 3,
136 Πίνακας 24. Μέσοι όροι (Μ.Ο.) και τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης, µε χορήγηση ριβαστιγµίνης) στο λαβύρινθο του Morris Χρόνος (sec) Ηµέρα 1 Ηµέρα 2 Ηµέρα 3 Ηµέρα 4 Ηµέρα 27 οκιµασία 1 οκιµασία 2 Μ.Ο. SD Μ.Ο. SD 167,43 7, ,72 4, ,68 4, ,1 2, ,95 2, ,15 3, ,75 2, ,81 2, ,34 5, ,95 3, Πίνακας 25. Μέσοι όροι (Μ.Ο.) και τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Ε (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης) στο λαβύρινθο του Morris Χρόνος (sec) Ηµέρα 1 Ηµέρα 2 Ηµέρα 3 Ηµέρα 4 οκιµασία 1 οκιµασία 2 Μ.Ο. SD Μ.Ο. SD 175,14 5, ,86 6, ,98 5, ,62 6, ,75 7, ,85 8, ,3 6, ,7 5, Πίνακας 26. Μέσοι όροι (Μ.Ο.) και τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας ΣΤ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης, µε χορήγηση ριβαστιγµίνης) στο λαβύρινθο του Morris Χρόνος (sec) Ηµέρα 1 Ηµέρα 2 Ηµέρα 3 Ηµέρα 4 οκιµασία 1 οκιµασία 2 Μ.Ο. SD Μ.Ο. SD 172,65 5, ,78 6, ,56 4, ,18 4, ,37 5, ,33 4, ,08 4, ,53 3,
137 3.3. Λαβύρινθος ανοικτού πεδίου Στους πίνακες αναφέρονται οι µέσοι όροι (Μ.Ο.) και οι τυπικές αποκλίσεις (SD) των καταγραφών των πειραµατοζώων όλων των οµάδων κατά τη διάρκεια των ηµερών πραγµατοποίησης της δοκιµασίας του λαβυρίνθου ανοικτού πεδίου, για την αναφορική µνήµη, τη λειτουργική µνήµη και το συνολικό χρόνο (total time). Υπενθυµίζεται ότι ο µέγιστος χρόνος παραµονής στο λαβύρινθο ήταν 600sec. Πίνακας 27. Μέσοι όροι και τυπικές αποκλίσεις των λαθών αναφορικής και λειτουργικής µνήµης και του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Α (γενικοί µάρτυρες) στο λαβύρινθο ανοικτού πεδίου Αναφορική µνήµη (Λάθη) Λειτουργική µνήµη (Λάθη) Συνολικός χρόνος (sec) Ηµέρα Μ.Ο. SD Μ.Ο. SD Μ.Ο. SD 5 2,2 0, ,3 0, ,7 44, ,5 0, ,1 0, ,7 29, ,4 0, ,8 1, ,1 20, ,2 0, ,4 0, ,4 22, ,7 0, ,8 0, ,1 14, ,3 0, ,3 0, ,2 13, ,8 0, ,9 0, ,3 15, ,5 0, , , ,4 0, ,9 0, ,2 6, ,5 0, ,8 0, ,4 6, ,3 0, ,9 0, ,2 11, ,4 0, , ,6 7, ,5 0, ,8 0, ,6 9, ,3 0, ,8 0, ,7 10, ,4 0, ,8 0, ,1 7, , ,1 0, ,7 10,
138 Πίνακας 28. Μέσοι όροι και τυπικές αποκλίσεις των λαθών αναφορικής και λειτουργικής µνήµης και του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Β (ειδικοί µάρτυρες) στο λαβύρινθο ανοικτού πεδίου Αναφορική µνήµη (Λάθη) Λειτουργική µνήµη (Λάθη) Συνολικός χρόνος (sec) Ηµέρα Μ.Ο. SD Μ.Ο. SD Μ.Ο. SD 5 2,1 1, ,4 0, ,2 35, ,4 0, , ,3 18, ,6 0, ,7 0, ,6 20, ,3 0, ,5 0, ,6 16, ,6 0, ,9 0, ,5 16, ,3 0, ,3 0, ,9 13, ,9 0, ,8 0, ,6 13, ,6 0, ,8 0, ,5 10, ,4 0, , ,3 8, ,5 0, ,9 0, , ,5 0, , ,8 8, ,3 0, ,9 0, , ,3 0, ,9 0, ,2 8, ,5 0, ,8 0, ,6 8, ,4 0, ,9 0, ,6 7, ,1 0, ,2 0, ,7 8,
139 Πίνακας 29. Μέσοι όροι και τυπικές αποκλίσεις των λαθών αναφορικής και λειτουργικής µνήµης και του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Γ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης) στο λαβύρινθο ανοικτού πεδίου Αναφορική µνήµη (Λάθη) Λειτουργική µνήµη (Λάθη) Συνολικός χρόνος (sec) Ηµέρα Μ.Ο. SD Μ.Ο. SD Μ.Ο. SD 5 2,3 0, ,3 0, ,6 44, ,5 0, ,1 0, ,7 30, ,4 0, ,8 0, ,2 28, ,2 0, ,6 0, ,9 22, ,7 0, ,7 0, ,3 20, ,2 0, ,2 0, ,8 19, ,8 0, ,9 0, ,3 17, ,6 0, ,9 0, ,2 12, ,3 0, , ,9 7, ,4 0, , ,2 6, ,3 0, ,9 0, ,6 9, ,4 0, ,9 0, , ,5 0, ,9 0, ,4 6, ,4 0, , ,1 6, ,3 0, ,8 0, ,9 9, ,8 0, ,4 1, ,2 21,
140 Πίνακας 30. Μέσοι όροι και τυπικές αποκλίσεις των λαθών αναφορικής και λειτουργικής µνήµης και του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης, µε χορήγηση ριβαστιγµίνης) στο λαβύρινθο ανοικτού πεδίου Αναφορική µνήµη (Λάθη) Λειτουργική µνήµη (Λάθη) Συνολικός χρόνος (sec) Ηµέρα Μ.Ο. SD Μ.Ο. SD Μ.Ο. SD 5 2,3 0, ,5 0, ,9 37, ,4 0, , ,8 27, ,4 0, ,9 0, ,5 20, ,3 0, ,6 0, ,1 20, ,6 0, ,8 0, , ,2 0, ,3 0, ,8 19, ,8 0, ,8 0, ,7 15, ,5 0, ,8 0, ,8 15, ,4 0, ,9 0, , ,4 0, , ,3 8, ,4 0, ,9 0, ,6 6, ,3 0, ,9 0, , ,3 0, ,8 0, ,2 7, ,3 0, , ,2 8, ,4 0, ,9 0, , ,3 0, ,8 0, ,7 11,
141 Πίνακας 31. Μέσοι όροι και τυπικές αποκλίσεις των λαθών αναφορικής και λειτουργικής µνήµης και του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Ε (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης) στο λαβύρινθο ανοικτού πεδίου Αναφορική µνήµη (Λάθη) Λειτουργική µνήµη (Λάθη) Συνολικός χρόνος (sec) Ηµέρα Μ.Ο. SD Μ.Ο. SD Μ.Ο. SD 5 2,1 0, ,7 0, ,5 25, ,7 1, ,8 0, ,3 17, ,7 0, ,6 0, ,2 19, ,9 0, ,3 1, ,2 16, ,2 0, ,1 0, ,3 11, ,9 0, ,2 0, ,5 14, ,8 0, ,1 0, ,8 16, ,7 0, ,1 0, ,2 22, ,7 0, ,3 0, ,5 17, ,6 0, ,2 0, ,3 8, ,7 0, ,4 0, ,4 9, ,6 0, ,4 0, , ,6 0, ,2 0, ,8 12, ,4 0, ,4 0, ,8 9, ,7 0, ,3 0, ,6 10,
142 Πίνακας 32. Μέσοι όροι και τυπικές αποκλίσεις των λαθών αναφορικής και λειτουργικής µνήµης και του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας ΣΤ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης, µε χορήγηση ριβαστιγµίνης) στο λαβύρινθο ανοικτού πεδίου Αναφορική µνήµη (Λάθη) Λειτουργική µνήµη (Λάθη) Συνολικός χρόνος (sec) Ηµέρα Μ.Ο. SD Μ.Ο. SD Μ.Ο. SD 5 2,2 0, ,5 0, ,1 22, ,6 0, ,4 0, ,6 20, ,7 0, ,1 0, , ,6 0, ,4 0, ,2 17, ,2 0, ,8 0, ,8 16, ,9 0, ,1 0, , ,6 0, ,7 0, ,4 23, ,5 0, ,4 0, ,8 21, ,2 0, ,4 0, ,6 10, ,8 0, ,2 0, ,8 14, ,6 0, , ,5 8, ,5 0, ,1 0, ,8 8, ,6 0, ,8 0, ,4 10, ,3 0, , ,4 12, ,5 0, ,9 0, ,6 11,
143 3.4. οκιµασία παθητικής αποφυγής Στους πίνακες που ακολουθούν, αναφέρονται οι µέσοι όροι (Μ.Ο.) και οι τυπικές αποκλίσεις (SD) του χρόνου παραµονής (sec) των πειραµατοζώων στο θάλαµο εκκίνησης (φωτεινό θάλαµο) της δοκιµασίας παθητικής αποφυγής. Υπενθυµίζεται ότι ο µέγιστος χρόνος παραµονής στο θάλαµο εκκίνησης (cut-off time) ήταν 300sec. Πίνακας 33. Μέσοι όροι (±SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Α (γενικοί µάρτυρες) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής Χρόνος παθητικής αποφυγής (sec) Ηµέρα 19 Ηµέρα 20 Ηµέρα 29 Μ.Ο. 10,42±4, Πίνακας 34. Μέσοι όροι (±SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Β (ειδικοί µάρτυρες) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής Χρόνος παθητικής αποφυγής (sec) Ηµέρα 19 Ηµέρα 20 Ηµέρα 29 Μ.Ο. 11,07±5, Πίνακας 35. Μέσοι όροι (±SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Γ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής Χρόνος παθητικής αποφυγής (sec) Ηµέρα 19 Ηµέρα 20 Ηµέρα 29 Μ.Ο. 10,54±4, ,19±8, Πίνακας 36. Μέσοι όροι (±SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης, µε χορήγηση ριβαστιγµίνης) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής Χρόνος παθητικής αποφυγής (sec) Ηµέρα 19 Ηµέρα 20 Ηµέρα 29 Μ.Ο. 10,68±5, ,04±109,
144 Πίνακας 37. Μέσοι όροι (±SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας Ε (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής Χρόνος παθητικής αποφυγής (sec) Ηµέρα 19 Ηµέρα 20 Μ.Ο. 11,86±5, ,5±8, Πίνακας 38. Μέσοι όροι (±SD) του χρόνου παραµονής (sec) των πειραµατοζώων της οµάδας ΣΤ (µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης, µε χορήγηση ριβαστιγµίνης) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής Χρόνος παθητικής Ηµέρα 19 Ηµέρα 20 αποφυγής (sec) Μ.Ο. 10,97±6, ,61±105,
145 4. Συγκρίσεις µεταξύ των οµάδων 4.1. Λαβύρινθος του Morris Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Β, Γ και έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. (στατιστικά σηµαντικά) µεταξύ τους (F(3,36)=1769,023, P<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(4,144)=4765,70, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(12,144)=100,03, p<0,0001), οµάδας και αριθµού ηµερήσιας δοκιµασίας (F(3,36)=2,14, p<0,001), ηµέρας και αριθµού ηµερήσιας δοκιµασίας (F(4,144)=1130,32, p<0,0001), καθώς και οµάδας, ηµέρας και αριθµού ηµερήσιας δοκιµασίας (F(12,144)=3,25, p<0,001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι οι χρόνοι παραµονής των πειραµατοζώων των οµάδων στο λαβύρινθο του Morris, δε διαφέρουν Σ.Σ. µεταξύ τους µέχρι την 4 η ηµέρα. Την 27 η ηµέρα που έγινε έλεγχος διατήρησης της µνήµης (retention) παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Γ και (Γ> >Α), ανάµεσα στις οµάδες Γ και (Γ> ), ενώ δεν παρατηρήθηκε Σ.Σ. διαφορά µεταξύ των οµάδων Α και Β. Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε εικονική καταστροφή του βασικού πυρήνα του Meynert (sham-operated) την 20 ηµέρα στην οµάδα Β, καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ την 20 ηµέρα στις οµάδες Γ και, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα. Συµπεραίνεται ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert στα πειραµατόζωα της οµάδας Β (ειδικοί µάρτυρες) δεν επέδρασε στις διεργασίες της µνήµης, αφού δεν παρατηρήθηκαν Σ.Σ. διαφορές µε τα πειραµατόζωα της οµάδας Α (γενικοί µάρτυρες). Οι Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Γ και αποτελούν απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τον αυξηµένο χρόνο παραµονής στο λαβύρινθο του Morris (Γ> >Α) κατά τις δοκιµασίες διατήρησης της µνήµης για τις οµάδες Γ και σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Και µάλιστα, παρατηρείται ότι η µνήµη αναφοράς στα πειραµατόζωα της οµάδας Γ διατηρείται σε µεγάλο βαθµό. Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Γ και 144
146 (Γ> ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως τη διατήρηση της µνήµης, τόσο της µνήµης αναφοράς όσο και της λειτουργικής µνήµης. Στα γραφήµατα 2 και 3 που ακολουθούν, φαίνονται οι µέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο λαβύρινθο του Morris για τις δύο ηµερήσιες δοκιµασίες (κατά τις ηµέρες 1, 2, 3, 4 και 27), στις οµάδες Α, Β, Γ και. Χρόνος παραµονής (sec) Λαβύρινθος του Morris οκιµασία 1 Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Γ Οµάδα Οµάδα Β Ηµέρες * * *=p<0,05 Γράφηµα 2. Μέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο λαβύρινθο του Morris για την ηµερήσια δοκιµασία 1, στις οµάδες Α, Β, Γ και 145
147 Χρόνος παραµονής (sec) Λαβύρινθος του Morris οκιµασία 2 Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Γ Οµάδα Οµάδα Β Ηµέρες * * *=p<0,05 Γράφηµα 3. Μέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο λαβύρινθο του Morris για την ηµερήσια δοκιµασία 2, στις οµάδες Α, Β, Γ και Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Ε και ΣΤ έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(2,27)=157791,9, p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(3,81)=1794,17, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(6,81)=418,35, p<0,0001), οµάδας και αριθµού ηµερήσιας δοκιµασίας (F(2,27)=134,55, p<0,0001), ηµέρας και αριθµού ηµερήσιας δοκιµασίας (F(3,81)=58,63, p<0,0001), καθώς και οµάδας, ηµέρας και αριθµού ηµερήσιας δοκιµασίας (F(6,81)=82,64, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι οι χρόνοι παραµονής των πειραµατοζώων των οµάδων στο λαβύρινθο του Morris διαφέρουν Σ.Σ. µεταξύ τους. Παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Ε και ΣΤ (Ε>ΣΤ>Α), καθώς και ανάµεσα στις οµάδες Ε και ΣΤ (Ε>ΣΤ). Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ στις οµάδες Ε και ΣΤ πριν από τις δοκιµασίες απόκτησης µνήµης, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα ΣΤ. Συµπεραίνεται ότι οι Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Ε και ΣΤ 146
148 αποτελούν απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ πριν από τις δοκιµασίες απόκτησης µνήµης, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τον αυξηµένο χρόνο παραµονής στο λαβύρινθο του Morris (Ε>ΣΤ>Α) κατά τις δοκιµασίες του λαβυρίνθου του Morris για τις οµάδες Ε και ΣΤ σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Ε και ΣΤ (Ε>ΣΤ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας ΣΤ συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως την απόκτηση και τη διατήρηση της µνήµης, τόσο της µνήµης αναφοράς όσο και της λειτουργικής µνήµης. Στα γραφήµατα 4 και 5 που ακολουθούν, φαίνονται οι µέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο λαβύρινθο του Morris για τις δύο ηµερήσιες δοκιµασίες (κατά τις ηµέρες 1, 2, 3 και 4), στις οµάδες Α, Ε και ΣΤ. Χρόνος παραµονής (sec)_ Λαβύρινθος του Morris οκιµασία 1 Οµάδες πειραµατόζωων * Οµάδα Α Οµάδα Ε Οµάδα ΣΤ Ηµέρες * *=p<0,05 Γράφηµα 4. Μέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο λαβύρινθο του Morris για την ηµερήσια δοκιµασία 1, στις οµάδες Α, Ε και ΣΤ 147
 του χρόνου παραµονής (sec) στο λαβύρινθο του Morris για την ηµερήσια δοκιµασία 2, στις οµάδες Α, Ε και ΣΤ Στο γράφηµα 6 αναφέρονται οι συνολικοί χρόνοι παραµονής (±SEM) στο")
149 Χρόνος παραµονής (sec) Λαβύρινθος του Morris οκιµασία 2 Οµάδες πειραµατόζωων * Οµάδα Α Οµάδα Ε Οµάδα ΣΤ Ηµέρες * *=p<0,05 Γράφηµα 5. Μέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο λαβύρινθο του Morris για την ηµερήσια δοκιµασία 2, στις οµάδες Α, Ε και ΣΤ Στο γράφηµα 6 αναφέρονται οι συνολικοί χρόνοι παραµονής (±SEM) στο λαβύρινθο του Morris (sec) για κάθε ηµερήσια δοκιµασία (κατά τις ηµέρες 1, 2, 3, 4 και 27), στις οµάδες Α, Β, Γ και. * * * * *=p<0,05 Γράφηµα 6. Συνολικός χρόνος παραµονής (±SEM) στο λαβύρινθο του Morris (sec) για κάθε ηµερήσια δοκιµασία στις οµάδες Α, Β, Γ και 148

150 Στο γράφηµα 7 αναφέρονται οι συνολικοί χρόνοι παραµονής (±SEM) στο λαβύρινθο του Morris (sec) για κάθε ηµερήσια δοκιµασία (κατά τις ηµέρες 1, 2, 3 και 4), στις οµάδες Α, Ε και ΣΤ. * * * * *=p<0,05 Γράφηµα 7. Συνολικός χρόνος παραµονής (±SEM) στο λαβύρινθο του Morris (sec) για κάθε ηµερήσια δοκιµασία στις οµάδες Α, Ε και ΣΤ 149
151 4.2. Λαβύρινθος ανοικτού πεδίου Μνήµη αναφοράς Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Β, Γ και έδειξε ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(1,36)=47,57, p<0,0001), ανεξάρτητα από την οµάδα, ενώ οι οµάδες δε διαφέρουν Σ.Σ. µεταξύ τους ανεξάρτητα από την ηµέρα πραγµατοποίησης της δοκιµασίας. Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι τα λάθη των πειραµατοζώων των οµάδων στη µνήµη αναφοράς (reference memory) στο λαβύρινθο ανοικτού πεδίου, δε διαφέρουν Σ.Σ. µεταξύ τους τη 19 η ηµέρα. Την 28 η ηµέρα που έγινε έλεγχος διατήρησης της µνήµης παρατηρήθηκε επίσης, µη Σ.Σ. διαφορά µεταξύ των οµάδων Α, Γ και, µεταξύ των οµάδων Γ και, όπως και µεταξύ των οµάδων Α και Β. Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε εικονική καταστροφή του βασικού πυρήνα του Meynert (sham-operated) την 20 ηµέρα στην οµάδα Β, καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ την 20 ηµέρα στις οµάδες Γ και, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα. Συµπεραίνεται ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert στα πειραµατόζωα της οµάδας Β (ειδικοί µάρτυρες) δεν επέδρασε στις διεργασίες της µνήµης, αφού δεν παρατηρήθηκαν Σ.Σ. διαφορές µε τα πειραµατόζωα της οµάδας Α (γενικοί µάρτυρες). Οι µη Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Γ και, αποδεικνύουν ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, δεν επιδρά Σ.Σ. στη µνήµη αναφοράς των πειραµατοζώων, όταν η απόκτηση της µνήµης (εκπαίδευση των πειραµατοζώων) γίνεται πριν την πειραµατική καταστροφή του πυρήνα. Η Σ.Σ. επίδραση της ηµέρας πραγµατοποίησης της δοκιµασίας θα µπορούσε να δικαιολογηθεί από το γεγονός ότι ο βέλτιστος χρόνος, που επιτυγχάνεται µετά από την 15ήµερη εκπαίδευση, δεν αναπαράγεται την 28η ηµέρα λόγω παρόδου 9ηµέρου από την προηγούµενη δοκιµασία. 150
152 Στο γράφηµα 8 φαίνονται οι µέσοι όροι (±SEM) των συνολικών λαθών της µνήµης αναφοράς (reference memory) των πειραµατοζώων των οµάδων Α, Β, Γ και στο λαβύρινθο ανοικτού πεδίου. Λάθη µνήµης αναφοράς Λαβύρινθος ανοικτού πεδίου Μνήµη αναφοράς Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Γ Οµάδα Οµάδα Β Ηµέρες Γράφηµα 8. Μέσοι όροι (±SEM) των συνολικών λαθών της µνήµης αναφοράς των πειραµατοζώων των οµάδων Α, Β, Γ και στο λαβύρινθο ανοικτού πεδίου 151
153 Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Ε και ΣΤ έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(2,27)=104,7356, p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(14,378)=23,53, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(28,378)=5,75, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι τα λάθη των πειραµατοζώων των οµάδων στη µνήµη αναφοράς στο λαβύρινθο ανοικτού πεδίου, διαφέρουν Σ.Σ. µεταξύ τους. Παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Ε και ΣΤ (Ε>ΣΤ>Α), καθώς και ανάµεσα στις οµάδες Ε και ΣΤ (Ε>ΣΤ). Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ στις οµάδες Ε και ΣΤ, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα ΣΤ. Συµπεραίνεται ότι οι Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Ε και ΣΤ αποτελούν απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τα αυξηµένα λάθη στη µνήµη αναφοράς κατά τις δοκιµασίες στο λαβύρινθο ανοικτού πεδίου (Ε>ΣΤ>Α) για τις οµάδες Ε και ΣΤ σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Ε και ΣΤ (Ε>ΣΤ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας ΣΤ συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως την απόκτηση και τη διατήρηση της µνήµης. Η βελτίωση των επιδόσεων της οµάδας ΣΤ, λόγω της χορήγησης ριβαστιγµίνης, φθάνει στο σηµείο σύγκλισης των επιδόσεων µε αυτές της οµάδας Α από την 14η ηµέρα και µετά. 152
154 Στο γράφηµα 9 φαίνονται οι µέσοι όροι (±SEM) των συνολικών λαθών της µνήµης αναφοράς των πειραµατοζώων των οµάδων Α, Ε και ΣΤ στο λαβύρινθο ανοικτού πεδίου. Λάθη µνήµης αναφοράς Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Ε Οµάδα ΣΤ * *=p<0, ΗΜΕΡΕΣ Γράφηµα 9. Μέσοι όροι (±SEM) των συνολικών λαθών της µνήµης αναφοράς των πειραµατοζώων των οµάδων Α, Ε και ΣΤ στο λαβύρινθο ανοικτού πεδίου 153
155 Λειτουργική µνήµη Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Β, Γ και έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(3,36)=11,57917, p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(1,36)=134,29, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(3,36)=16,10, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι τα λάθη των πειραµατοζώων των οµάδων στη λειτουργική µνήµη (working memory) στο λαβύρινθο ανοικτού πεδίου, δε διαφέρουν Σ.Σ. µεταξύ τους τη 19 η ηµέρα. Την 28 η ηµέρα που έγινε έλεγχος διατήρησης της µνήµης παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Γ και (Γ> >Α), ανάµεσα στις οµάδες Γ και (Γ> ), ενώ δεν παρατηρείται Σ.Σ. διαφορά µεταξύ των οµάδων Α και Β. Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε εικονική καταστροφή του βασικού πυρήνα του Meynert την 20 ηµέρα στην οµάδα Β, καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ την 20 ηµέρα στις οµάδες Γ και, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα. Συµπεραίνεται ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert στα πειραµατόζωα της οµάδας Β (ειδικοί µάρτυρες) δεν επέδρασε στις διεργασίες της µνήµης, αφού δεν παρατηρήθηκαν Σ.Σ. διαφορές µε τα πειραµατόζωα της οµάδας Α (γενικοί µάρτυρες). Οι Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Γ και αποτελούν απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τα αυξηµένα λάθη στη λειτουργική µνήµη στο λαβύρινθο ανοικτού πεδίου (Γ> >Α) κατά τις δοκιµασίες διατήρησης της µνήµης για τις οµάδες Γ και σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Γ και (Γ> ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως τη διατήρηση της µνήµης. 154
156 Στο γράφηµα 10 φαίνονται οι µέσοι όροι (±SEM) των συνολικών λαθών της λειτουργικής µνήµης των πειραµατοζώων των οµάδων Α, Β, Γ και στο λαβύρινθο ανοικτού πεδίου. Λάθη λειτουργικής µνήµης Λαβύρινθος ανοικτού πεδίου Λειτουργική µνήµη Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Γ * Οµάδα Οµάδα Β * *=p<0, Ηµέρες Γράφηµα 10. Μέσοι όροι (±SEM) των συνολικών λαθών της λειτουργικής µνήµης των πειραµατοζώων των οµάδων Α, Β, Γ και στο λαβύρινθο ανοικτού πεδίου 155
157 Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Ε και ΣΤ έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(2,27)=300,8422, p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(14,378)=23,91, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(28,378)=8,75, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι τα λάθη των πειραµατοζώων των οµάδων στη λειτουργική µνήµη στο λαβύρινθο ανοικτού πεδίου, διαφέρουν Σ.Σ. µεταξύ τους. Παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Ε και ΣΤ (Ε>ΣΤ>Α), καθώς και ανάµεσα στις οµάδες Ε και ΣΤ (Ε>ΣΤ). Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ στις οµάδες Ε και ΣΤ πριν από τις δοκιµασίες απόκτησης µνήµης, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα ΣΤ. Συµπεραίνεται ότι οι Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Ε και ΣΤ αποτελούν απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τα αυξηµένα λάθη στη λειτουργική µνήµη κατά τις δοκιµασίες στο λαβύρινθο ανοικτού πεδίου (Ε>ΣΤ>Α) για τις οµάδες Ε και ΣΤ σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Ε και ΣΤ (Ε>ΣΤ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας ΣΤ συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως την απόκτηση και τη διατήρηση της µνήµης. Η βελτίωση των επιδόσεων της οµάδας ΣΤ, λόγω της χορήγησης ριβαστιγµίνης, φθάνει στο σηµείο σύγκλισης των επιδόσεων µε αυτές της οµάδας Α από την 15η ηµέρα και µετά. 156
158 Στο γράφηµα 11 φαίνονται οι µέσοι όροι (±SEM) των συνολικών λαθών της λειτουργικής µνήµης των πειραµατοζώων των οµάδων Α, Ε και ΣΤ στο λαβύρινθο ανοικτού πεδίου. Λάθη λειτουργικής µνήµης Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Ε Οµάδα ΣΤ * *=p<0, ΗΜΕΡΕΣ Γράφηµα 11. Μέσοι όροι (±SEM) των συνολικών λαθών της λειτουργικής µνήµης των πειραµατοζώων των οµάδων Α, Ε και ΣΤ στο λαβύρινθο ανοικτού πεδίου 157
159 Συνολικός χρόνος παραµονής στο λαβύρινθο ανοικτού πεδίου Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Β, Γ και έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(3,36)=3331,879, p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(1,36)=257,49, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(3,36)=34,62, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι οι χρόνοι παραµονής των πειραµατοζώων των οµάδων στο λαβύρινθο ανοικτού πεδίου, δε διαφέρουν Σ.Σ. µεταξύ τους µέχρι τη 19 η ηµέρα. Την 28 η ηµέρα που έγινε έλεγχος διατήρησης της µνήµης παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α και Γ (Γ> Α), ανάµεσα στις οµάδες Γ και (Γ> ), ενώ δεν παρατηρείται Σ.Σ. διαφορά µεταξύ των οµάδων Α, Β και. Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε εικονική καταστροφή του βασικού πυρήνα του Meynert την 20 ηµέρα στην οµάδα Β, καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ την 20 ηµέρα στις οµάδες Γ και, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα. Συµπεραίνεται ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert στα πειραµατόζωα της οµάδας Β (ειδικοί µάρτυρες) δεν επέδρασε στις διεργασίες της µνήµης, αφού δεν παρατηρήθηκαν Σ.Σ. διαφορές µε τα πειραµατόζωα της οµάδας Α (γενικοί µάρτυρες). Η Σ.Σ. διαφορά, που παρατηρήθηκε µεταξύ των οµάδων Α και Γ, αποτελεί απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τον αυξηµένο χρόνο παραµονής στο λαβύρινθο ανοικτού πεδίου (Γ>Α) κατά τις δοκιµασίες διατήρησης της µνήµης για την οµάδα Γ σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Γ και (Γ> ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας συµβάλλει στην εντυπωσιακή βελτίωση των επιδόσεών τους, βελτιώνει εποµένως τη διατήρηση της µνήµης, σε σηµείο που να µην καταγράφονται Σ.Σ. διαφορές µεταξύ των οµάδων Α (γενικοί µάρτυρες) και. 158
160 Στο γράφηµα 12 φαίνονται οι µέσοι όροι (±SEM) του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων των οµάδων Α, Β, Γ και στο λαβύρινθο ανοικτού πεδίου. Συνολικός χρόνος (sec) Λαβύρινθος ανοικτού πεδίου Συνολικός χρόνος Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Γ Οµάδα Οµάδα Β * *=p<0, Ηµέρες Γράφηµα 12. Μέσοι όροι (±SEM) του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων των οµάδων Α, Β, Γ και στο λαβύρινθο ανοικτού πεδίου 159
161 Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Ε και ΣΤ έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(2,27)= , p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(14,378)=1579,69, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(28,378)=184,22, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι οι χρόνοι παραµονής των πειραµατοζώων των οµάδων στο λαβύρινθο ανοικτού πεδίου, διαφέρουν Σ.Σ. µεταξύ τους. Παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Ε και ΣΤ (Ε>ΣΤ>Α), καθώς και ανάµεσα στις οµάδες Ε και ΣΤ (Ε>ΣΤ). Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ στις οµάδες Ε και ΣΤ πριν από τις δοκιµασίες απόκτησης µνήµης, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα ΣΤ. Συµπεραίνεται ότι οι Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Ε και ΣΤ αποτελούν απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τον αυξηµένο χρόνο παραµονής (Ε>ΣΤ>Α) κατά τις δοκιµασίες στο λαβύρινθο ανοικτού πεδίου για τις οµάδες Ε και ΣΤ σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Ε και ΣΤ (Ε>ΣΤ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας ΣΤ συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως την απόκτηση και τη διατήρηση της µνήµης. 160
162 Στο γράφηµα 13 φαίνονται οι µέσοι όροι (±SEM) του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων των οµάδων Α, Ε και ΣΤ στο λαβύρινθο ανοικτού πεδίου. Συνολικός Χρόνος (sec) ΗΜΕΡΕΣ Οµάδες πειραµατόζωων Οµάδα Α Οµάδα Ε Οµάδα ΣΤ Γράφηµα 13. Μέσοι όροι (±SEM) του συνολικού χρόνου παραµονής (sec) των πειραµατοζώων των οµάδων Α, Ε και ΣΤ στο λαβύρινθο ανοικτού πεδίου * * *=p<0,05 161
163 4.3. οκιµασία παθητικής αποφυγής Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Β, Γ και έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(3,36)=88449,35, p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(1,36)=506,75, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(3,36)=57,36, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι οι χρόνοι παραµονής των πειραµατοζώων των οµάδων στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής, δε διαφέρουν Σ.Σ. µεταξύ τους τη 19 η ηµέρα. Την 29η ηµέρα, που έγινε έλεγχος διατήρησης της µνήµης, παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Γ και (Γ< <Α), ανάµεσα στις οµάδες Γ και (Γ< ), ενώ δεν παρατηρείται Σ.Σ. διαφορά µεταξύ των οµάδων Α και Β. Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε εικονική καταστροφή του βασικού πυρήνα του Meynert την 20η ηµέρα στην οµάδα Β, καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ την 20η ηµέρα στις οµάδες Γ και, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα. Συµπεραίνεται ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert στα πειραµατόζωα της οµάδας Β (ειδικοί µάρτυρες) δεν επέδρασε στις διεργασίες της µνήµης, αφού δεν παρατηρήθηκαν Σ.Σ. διαφορές µε τα πειραµατόζωα της οµάδας Α (γενικοί µάρτυρες). Οι Σ.Σ. διαφορές, που παρατηρήθηκαν µεταξύ των οµάδων Α, Γ και, αποτελεί απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα το µειωµένο χρόνο παραµονής στο θάλαµο εκκίνησης (Γ< <Α) κατά τις δοκιµασίες διατήρησης της µνήµης για την οµάδα Γ και σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Γ και (Γ< ), αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως τη διατήρηση της µνήµης. Η Σ.Σ. επίδραση της ηµέρας πραγµατοποίησης της δοκιµασίας θα µπορούσε να δικαιολογηθεί από το γεγονός ότι την ηµέρα απόκτησης της µνήµης παθητικής αποφυγής τα πειραµατόζωα αναµένεται να έχουν αντίστοιχους 162
164 χρόνους, αφού δεν έχουν δεχθεί το δυσάρεστο ερέθισµα και τείνουν να µετακινηθούν προς το σκοτεινό θάλαµο. Στο γράφηµα 14 φαίνονται οι µέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής των πειραµατοζώων των οµάδων Α, Β, Γ και. οκιµασία Παθητικής Αποφυγής Χρόνος παραµονής (sec) Οµάδες Πειραµατόζωων Οµάδα Α Οµάδα Γ Οµάδα Οµάδα Β *=p<0,05 * Ηµέρες Γράφηµα 14. Μέσοι όροι (±SEM) του χρόνου παραµονής (sec) των πειραµατοζώων των οµάδων Α, Β, Γ και στη δοκιµασία παθητικής αποφυγής 163
165 Η ανάλυση RMANOVA µεταξύ των οµάδων Α, Ε και ΣΤ έδειξε ότι οι οµάδες διαφέρουν Σ.Σ. µεταξύ τους (F(2,27)=106027,1, p<0,0001) ανεξάρτητα από τις ηµέρες πραγµατοποίησης της δοκιµασίας. Η RMANOVA έδειξε, επίσης, ότι η ηµέρα πραγµατοποίησης της δοκιµασίας επιδρά Σ.Σ. (F(1,27)=227,91, p<0,0001) ανεξάρτητα από την οµάδα. Έδειξε ακόµη ότι υπάρχει αλληλεπίδραση οµάδας και ηµέρας δοκιµασίας (F(2,27)=61,88, p<0,0001). Το Tukey-Kramer test (σε επίπεδο p=0,05) έδειξε ότι οι χρόνοι παραµονής των πειραµατοζώων των οµάδων στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής, διαφέρουν Σ.Σ. µεταξύ τους. Παρατηρήθηκε Σ.Σ. διαφορά ανάµεσα στις οµάδες Α, Ε και ΣΤ (Ε<ΣΤ<Α), καθώς και ανάµεσα στις οµάδες Ε και ΣΤ (Ε<ΣΤ). Για τη βιοστατιστική ερµηνεία των παραπάνω αποτελεσµάτων πρέπει να ληφθεί υπόψη ότι έγινε καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ στις οµάδες Ε και ΣΤ πριν από τις δοκιµασίες απόκτησης µνήµης, καθώς και χορήγηση ριβαστιγµίνης στην οµάδα ΣΤ. Συµπεραίνεται ότι οι Σ.Σ. διαφορές που παρατηρήθηκαν µεταξύ των οµάδων Α, Ε και ΣΤ αποτελούν απόδειξη ότι η καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, επιδρά στη συµπεριφορά των πειραµατοζώων και έχει ως αποτέλεσµα τον αυξηµένο χρόνο παραµονής (Ε<ΣΤ<Α) κατά τις δοκιµασίες στη συσκευή παθητικής αποφυγής για τις οµάδες Ε και ΣΤ σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες). Επίσης, η Σ.Σ. διαφορά που παρατηρείται µεταξύ των οµάδων Ε και ΣΤ (Ε<ΣΤ) αποδεικνύει ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα της οµάδας ΣΤ συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως τη διατήρηση της µνήµης. 164
166 Στο γράφηµα 15 αναφέρονται οι µέσοι όροι (±SEM) του χρόνου παραµονής (sec) στο θάλαµο εκκίνησης της δοκιµασίας παθητικής αποφυγής των πειραµατοζώων των οµάδων Α, Ε και ΣΤ. οκιµασία Παθητικής Αποφυγής Χρόνος παραµονής (sec) Οµάδες Πειραµατοζώων Οµάδα Α Οµάδα Ε Οµάδα ΣΤ *=p<0,05 * Ηµέρες Γράφηµα 15. Μέσοι όροι (±SEM) του χρόνου παραµονής (sec) των πειραµατοζώων των οµάδων Α, Ε και ΣΤ στη δοκιµασία παθητικής αποφυγής 165
167 Στο γράφηµα 16 αναφέρονται τα ποσοστά «συνολικής αποφυγής» των πειραµατοζώων όλων των οµάδων (incidence of avoidance: ποσοστό των πειραµατοζώων που παρουσιάζουν χρόνο παραµονής στο θάλαµο εκκίνησης µεγαλύτερο από 180sec) ΠΟΣΟΣΤΟ 60 ΣΥΝΟΛΙΚΗΣ 50 ΑΠΟΦΥΓΗΣ ΟΜΑ ΕΣ ΠΕΙΡΑΜΑΤΟΖΩΩΝ 70 ΟΜΑ Α Α ΟΜΑ Α Β ΟΜΑ Α Γ ΟΜΑ Α ΟΜΑ Α Ε ΟΜΑ Α ΣΤ Γράφηµα 16. Ποσοστά «συνολικής αποφυγής» των πειραµατοζώων στη δοκιµασία παθητικής αποφυγής 166

168 5. Ιστολογική µελέτη Οι προς εξέταση περιοχές του εγκεφάλου, αφού είχαν σκηνωθεί και εγκλειστεί σε κύβους παραφίνης, επεξεργάστηκαν σε µικροτόµο παραφίνης για τη λήψη τοµών πάχους 4-10µm. Σε όλες τις τοµές πραγµατοποιήθηκε χρώση αιµατοξυλίνης-εωσίνης. Κατά την εξέταση των τοµών διαπιστώθηκε απουσία αλλοιώσεων στα πειραµατόζωα της οµάδας Β. Εποµένως, συµπεραίνουµε ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert στα πειραµατόζωα της οµάδας Β (ειδικοί µάρτυρες) δεν προκάλεσε υπολειµµατικές βλάβες στην περιοχή του πυρήνα. Στα πειραµατόζωα των οµάδων Γ,, Ε και ΣΤ διαπιστώθηκαν αλλοιώσεις. Στη συνέχεια παρατίθενται αντιπροσωπευτικές εικόνες τοµών των εγκεφάλων των οµάδων των πειραµατοζώων. Εικόνα 19. Μετωπιαία διατοµή φυσιολογικού εγκεφάλου στο επίπεδο του βασικού πυρήνα του Meynert 167

 Εικόνα 21.")
169 Εικόνα 20. Μετωπιαία διατοµή φυσιολογικού εγκεφάλου (βέλος 3η κοιλία) στο επίπεδο του βασικού πυρήνα του Meynert (σε αντιστοιχία µε την εικόνα 19) Εικόνα 21. Μεγέθυνση ( 400) φυσιολογικών νευρώνων της εικόνας


170 Εικόνα 22. Μετωπιαία διατοµή εγκεφάλου που απεικονίζει την περιοχή της νευροτοξικής καταστροφής του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ Εικόνα 23. Μεγέθυνση ( 400) νεκρωµένων (βέλος) και φυσιολογικών νευρώνων (διπλό βέλος) της εικόνας


171 Εικόνα 24. Μετωπιαία διατοµή εγκεφάλου που απεικονίζει την περιοχή της νευροτοξικής καταστροφής του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ Εικόνα 25. Μεγέθυνση ( 400) νεκρωµένων (βέλος) και φυσιολογικών νευρώνων (διπλό βέλος) της εικόνας
 νεκρωµένων (βέλος) και φυσιολογικών νευρώνων")

172 Εικόνα 26. Μεγέθυνση ( 400) νεκρωµένων (βέλος) και φυσιολογικών νευρώνων (διπλό βέλος) της εικόνας 24 Εικόνα 27. Μετωπιαία διατοµή εγκεφάλου που απεικονίζει την περιοχή της νευροτοξικής καταστροφής του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ 171
 νεκρωµένων (βέλος) και φυσιολογικών")
173 Εικόνα 28. Μεγέθυνση ( 400) νεκρωµένων (βέλος) και φυσιολογικών νευρώνων (διπλό βέλος) της εικόνας
174 III. ΣΥΖΗΤΗΣΗ Με τη «χολινεργική υπόθεση», που διατυπώθηκε πριν από 20 χρόνια περίπου, διευκρινίστηκε ότι η σηµαντική απώλεια της χολινεργικής λειτουργίας στο ΚΝΣ συντελεί καθοριστικά στην εµφάνιση των γνωστικών διαταραχών, που σχετίζονται µε την προχωρηµένη ηλικία και τη νόσο Alzheimer. Πριν από τη διατύπωσή της δεν υπήρχαν θεωρητικές ερµηνείες για τα κύρια συµπτώµατα της νόσου Alzheimer και για τα διαγνωστικά της κριτήρια 240,241,242. Ακόµη και η άποψη ότι συγκεκριµένες γνωστικές διαταραχές συνδέονται µε την προχωρηµένη ηλικία, ήταν σε περιορισµένο βαθµό αποδεκτή 122. Η αρχική µελέτη δηµοσίευση, στην οποία διατυπώθηκαν οι δοµικές αποδείξεις για τη χολινεργική υπόθεση 1, αποτέλεσε αφορµή για το σχεδιασµό και την πραγµατοποίηση πολυάριθµων νέων ερευνών. Η χολινεργική υπόθεση αναγνωρίζεται ως η µόνη θεωρία για τις γνωστικές διαταραχές, η οποία έχει οδηγήσει σε ουσιαστικό αποτέλεσµα, διαµέσου της ανάπτυξης των αναστολέων της χολινεστεράσης. Η αποτελεσµατικότητα των θεραπειών υποκατάστασης (µε τους αναστολείς χολινεστεράσης) και η συµβολή τους στην άµβλυνση των συµπτωµάτων και στη βελτίωση της ποιότητας ζωής των ασθενών, επιβάλλει τη συνέχιση της έρευνας πάνω σε αυτήν τη φαρµακολογική κατεύθυνση. Η χολινεργική υπόθεση, εποµένως, παρείχε το θεωρητικό υπόβαθρο για την ανάπτυξη θεραπευτικών προσεγγίσεων της νόσου Alzheimer, οι οποίες προήλθαν από τον επιτυχή συνδυασµό δεδοµένων της φυσιολογίας, της βιοχηµείας, της φαρµακολογίας και κλινικών µελετών. Και µάλιστα, σε αντίθεση µε άλλα σύγχρονα πεδία έρευνας, η χολινεργική υπόθεση στηρίχθηκε σε πειραµατικά µοντέλα και συµπεριφορικά πρωτόκολλα, τα οποία αναπτύχθηκαν-εξελίχθηκαν κατά τη διάρκεια της πλούσιας ιστορίας της βασικής έρευνας των νευροεπιστηµών. Η ιδέα της χρησιµοποίησης πειραµατοζώων στην προσπάθεια µελέτης των γνωστικών διαταραχών, που συνδέονται µε το γήρας και τη νόσο Alzheimer, και της εύρεσης µίας κατάλληλης θεραπευτικής αντιµετώπισης, θεωρήθηκε στην καλύτερη περίπτωση ότι δηµιουργεί επιστηµονική αντιπαράθεση, ενώ δεν ήταν λίγοι αυτοί που τη χαρακτήρισαν αιρετική. Και µάλιστα, οι επιστήµονες που προέρχονταν από παλαιότερες σχολές 173
175 σκέψης 243,244, θεώρησαν ανθρωποµορφική τη χρήση του όρου «µνήµη» για την περιγραφή της συµπεριφοράς των ζώων 245. Για να ερµηνευθούν οι συµπεριφορικές αλλαγές ως αποτέλεσµα µεταβολής µνηµονικών εγγραµµάτων είναι απαραίτητο να αποδεικνύεται ότι η συµπεριφορά οφείλεται σε προηγούµενη εµπειρία και δε συνδέεται µε ενεργούς τροποποιητικούς µηχανισµούς, που την επηρεάζουν 246. Παράδειγµα αποτελεί η επίδοση των πειραµατοζώων στη δοκιµασία παθητικής αποφυγής κατά τον έλεγχο διατήρησης της µνήµης µετά από χορήγηση ενός φαρµάκου, όπου είναι απαραίτητο να αποκλειστεί η επίδραση του φαρµάκου στη δραστηριότητα του πειραµατόζωου. Έτσι, η επίδοση ερµηνεύεται ως αποτέλεσµα της προηγούµενης δοκιµασίας απόκτησης µνήµης και της επίδρασης του φαρµάκου στις διεργασίες της µνήµης και µάθησης, και όχι ως συνέπεια της τροποποίησης της δραστηριότητας του πειραµατοζώου. Αντίστοιχο παράδειγµα αποτελεί η επίδοση του πειραµατοζώου στο λαβύρινθο ανοιχτού πεδίου, όπου η µη σωστή εφαρµογή των πρωτοκόλλων περιορισµού της τροφής πριν από τη δοκιµασία, µπορεί να αλλοιώσει τα αποτελέσµατα εξαιτίας της έλλειψης κινήτρου από το «χορτάτο» πειραµατόζωο. Η µνήµη που µελετάται στα πειράµατα συµπεριφοράς διακρίνεται σε δύο υποτύπους: τη µνήµη αναφοράς και τη λειτουργική µνήµη. Η µνήµη αναφοράς είναι ο υπότυπος, που σχετίζεται µε παραµέτρους της δοκιµασίας, οι οποίες παραµένουν σταθερές κατά τη διάρκεια των πειραµάτων συµπεριφοράς, όπως στο λαβύρινθο του Morris, στον οποίο το πειραµατόζωο πρέπει να βρει την εξέδρα διαφυγής, όταν αυτή βρίσκεται σε σταθερό σηµείο κάτω από την επιφάνεια του νερού. Αντιστοίχως, η λειτουργική µνήµη είναι ο υπότυπος, που σχετίζεται µε παραµέτρους της δοκιµασίας, οι οποίες αλλάζουν κατά τη διάρκεια των πειραµάτων συµπεριφοράς, όπως στο λαβύρινθο ανοιχτού πεδίου, όπου το πειραµατόζωο δεν πρέπει να επισκεφθεί για δεύτερη φορά τα τρυβλία µε την τροφή. Έτσι, αρχικά ήταν δύσκολο να χρησιµοποιηθούν πειραµατόζωα, για να προσοµοιωθούν πτυχές της ανθρώπινης µνήµης, καθώς και διαταραχές που σχετίζονται µε συγκεκριµένες παθήσεις του νευρικού συστήµατος του ανθρώπου. Οι µελέτες, όµως, που πραγµατοποιήθηκαν µε βάση τη χολινεργική υπόθεση, καθώς και τα πειραµατικά µοντέλα που χρησιµοποιήθηκαν, συνέβαλαν όχι µόνο στην καθιέρωση της θεωρίας και 174
176 των πρώτων θεραπευτικών στρατηγικών που βασίστηκαν σε αυτή, αλλά συνετέλεσαν και στη γενικότερη πρόοδο των νευροεπιστηµών. Εξαιτίας, µάλιστα, αυτού του έντονου ερευνητικού ενδιαφέροντος, δηµιουργήθηκε ενθουσιασµός στην επιστηµονική κοινότητα για την ανάπτυξη φαρµακολογικών θεραπειών για τις νευροεκφυλιστικές παθήσεις, ενώ η χρήση εκλεκτικών πειραµατικών µοντέλων σε αυτό το πεδίο της έρευνας δε θεωρείται πια λανθασµένη ή αιρετική. Στα εκλεκτικά πειραµατικά µοντέλα οι χηµικές καταστροφές µε νευροτοξίνες είναι πιο εκλεκτικές από τις αντίστοιχες ηλεκτρικές, επειδή καταστρέφουν µόνο τα νευρικά περικάρυα στο σηµείο της έγχυσης και αφήνουν ανέπαφες τις νευρικές ίνες που διέρχονται από την περιοχή 247. Για παράδειγµα, οι πειραµατικές καταστροφές µε ιµποτενικό οξύ (ibotenic acid), αλλά όχι και οι καταστροφές µε ηλεκτρικό ρεύµα, αφήνουν ανεπηρέαστη την ντοπαµινεργική και νοραδρενεργική νεύρωση του φλοιού, που διέρχεται διαµέσου της βάσης του πρόσθιου εγκεφάλου Αισθητικοκινητικές δοκιµασίες Οι νευροµυϊκές διαταραχές, που ενδεχοµένως παρατηρούνται τις πέντε πρώτες ηµέρες µετά την καταστροφή του βασικού πυρήνα του Meynert (NBM), περιλαµβάνουν τη µη φυσιολογική θέση των άκρων, την αδυναµία αντίστασης των άκρων στη ραχιαία πίεση, το επίπεδο επαγρύπνησης, την αντίδραση σε ακουστικά ερεθίσµατα, τα εκκριτικά σηµεία και τη διάρκεια παραµονής στο µονόζυγο. Οι Kassler και συν. (1986) 249 αναφέρουν ότι σε µία σειρά αισθητικοκινητικών δοκιµασιών, που πραγµατοποιήθηκαν στο χρονικό διάστηµα µέχρι τις επτά ηµέρες από την πειραµατική καταστροφή του βασικού πυρήνα του Meynert (NBM), µόνο ο χρόνος παραµονής στο µονόζυγο ήταν ελαφρώς ελαττωµένος, ενώ οι υπόλοιπες δοκιµασίες ήταν φυσιολογικές. Αναφέρεται επίσης 249,250,251,252,253, ότι παρά τις αλλαγές στις δοκιµασίες που µπορεί να παρατηρηθούν τις πρώτες τέσσερις ηµέρες µετά την πειραµατική καταστροφή του NBM, οι αισθητικοκινητικές λειτουργίες του πειραµατοζώου είναι αντίστοιχες µε εκείνες της οµάδας των γενικών µαρτύρων (controls) από την πέµπτη µέχρι την έβδοµη µετεγχειρητική ηµέρα, χρονική στιγµή έναρξης των δοκιµασιών συµπεριφοράς. 175
177 Στις αισθητικοκινητικές δοκιµασίες, που πραγµατοποιήθηκαν στην παρούσα µελέτη κατά την πέµπτη (5η) ηµέρα µετά τη στερεοταξική καταστροφή του βασικού πυρήνα του Meynert, δεν παρατηρήθηκαν διαφοροποιήσεις στις οµάδες σε σύγκριση µε εκείνη των γενικών, αλλά και των ειδικών µαρτύρων. Ο χρόνος παραµονής στο µονόζυγο στις οµάδες µε στερεοταξική καταστροφή του βασικού πυρήνα του Meynert (Γ,, Ε και ΣΤ) ήταν ελαφρώς ελαττωµένος, καταγραφή αντίστοιχη µε τα δεδοµένα της διεθνούς βιβλιογραφίας. Ωστόσο, οι διαφορές στους χρόνους παραµονής ήταν µη Σ.Σ. (p>0,05) σε σύγκριση µε την οµάδα των γενικών µαρτύρων. Οι υπόλοιπες καταγραφές των αισθητικοκινητικών δοκιµασιών για την ανίχνευση νευροµυϊκών διαταραχών ήταν, επίσης, αρνητικές. Έτσι, αποφεύχθηκε η χρησιµοποίηση, στα πειράµατα συµπεριφοράς, επιµύων µε αισθητικοκινητικές διαταραχές, γεγονός που θα οδηγούσε σε λανθασµένα συµπεράσµατα όπως υποστηρίζουν και άλλοι ερευνητές 229, Χολινεργικές ανεπιθύµητες ενέργειες Στα πειράµατα συµπεριφοράς, που χρησιµοποιούνται για τον έλεγχο των γνωστικών λειτουργιών, η απόδοση δεν εξαρτάται µόνο από τη χολινεργική νευροδιαβίβαση στον εγκεφαλικό φλοιό, αλλά και από τη φυσιολογική νευροµυϊκή λειτουργία, που επιτρέπει στο πειραµατόζωο να εκτελεί τις δοκιµασίες συµπεριφοράς µε ακρίβεια και ταχύτητα. Υπερβολική αναστολή της χολινεστεράσης στα βασικά γάγγλια και στους σκελετικούς µύες έχει ως αποτέλεσµα τη συσσώρευση ακετυλοχολίνης, που µπορεί να επηρεάσει την κινητική ικανότητα των πειραµατόζωων 3,192,235. Η ριβαστιγµίνη παρουσιάζει εκλεκτικότητα στην αναστολή της χολινεστεράσης στο φλοιό και τον ιππόκαµπο σε δόσεις 0,5-1,5mg/kg. Σε δόσεις 2,5-3,5mg/kg παρατηρείται αναστολή της χολινεστεράσης σε ποσοστό µεγαλύτερο του 50% στο ραβδωτό σώµα και τους σκελετικούς µύες, µε αποτέλεσµα χολινεργική υπερδραστηριότητα που εκδηλώνεται µε σηµεία κινητικών διαταραχών, όπως τρόµο, πτώση οπίσθιων άκρων και διάρροια, αλλά και εµφάνιση ακινησίας σε εγρήγορση (alert immobile behavior). Αυτές οι δράσεις της υπερβολικής συγκέντρωσης της ακετυλοχολίνης εκδηλώνονται διαµέσου µουσκαρινικών υποδοχέων στα βασικά γάγγλια και όχι διαµέσου νικοτινικών υποδοχέων στη νευροµυϊκή σύναψη
178 Στην παρούσα µελέτη µετά την υποδόρια ένεση ριβαστιγµίνης στα πειραµατόζωα, αξιολογούνταν η παρουσία ή η απουσία χολινεργικής υπερδραστηριότητας 15min, 30min και 45min µετά τη χορήγηση. Όλες οι παρατηρήσεις κατά την περίοδο διεξαγωγής των δοκιµασιών-πειραµάτων συµπεριφοράς, ήταν αρνητικές για την καταγραφή σηµείων χολινεργικής υπερδραστηριότητας. Έτσι, δεν καταγράφηκε σε κανένα από τα πειραµατόζωα πτώση οπίσθιων άκρων, διάρροια, ούτε και σωµατικός τρόµος. Τα αποτελέσµατα αυτά συµφωνούν µε ευρήµατα άλλων ερευνητών 3,4,192,235 για την εκλεκτικότητα της δράσης της ριβαστιγµίνης σε δόσεις 0,5-1,5mg/kg. Επίσης, τα αποτελέσµατα εξασφαλίζουν ότι οι επιδόσεις των πειραµατοζώων στις δοκιµασίες συµπεριφοράς δεν οφείλονται σε πιθανή επίδραση της ριβαστιγµίνης στην αισθητικοκινητική τους συµπεριφορά, µε αποτέλεσµα η αξιολόγηση τους και η εξαγωγή των συµπερασµάτων να είναι πιο αξιόπιστες Βασικός πυρήνας του Meynert και συµπεριφορά Σκοπός της παρούσας µελέτης ήταν ο σχεδιασµός και η ανάπτυξη ενός πειραµατικού µοντέλου, το οποίο θα µπορούσε να χρησιµοποιηθεί για την προσοµοίωση γνωστικών διαταραχών, που σχετίζονται µε παθήσεις του νευρικού συστήµατος, καθώς και για την αξιολόγηση φαρµακολογικά δραστικών ουσιών, που επιδρούν στη µνήµη και τη µάθηση. Επίσης, σηµαντικό ρόλο στο σχεδιασµό του πειραµατικού πρωτοκόλλου έπαιξε η υπάρχουσα εµπειρία από τη διεθνή βιβλιογραφία, ώστε να επιβεβαιωθεί ο σηµαίνων ρόλος του βασικού πυρήνα του Meynert (NBM) στις γνωστικές λειτουργίες µε βάση τη χολινεργική υπόθεση της µνήµης, αλλά και την εµπλοκή αυτού του πυρήνα στην παθοφυσιολογία της νόσου Alzheimer. Έτσι, κατά το σχεδιασµό του πειραµατικού προτύπου απαραίτητη προϋπόθεση θεωρήθηκε η συσχέτισή του µε γνωστικές διαταραχές, οι οποίες θα ανιχνεύονταν κατά την εκτέλεση των δοκιµασιών συµπεριφοράς, αλλά και θα µπορούσαν να αποδοθούν σε σχετιζόµενες µε το χολινεργικό σύστηµα διαταραχές, που θα επιχειρούνταν να αποκατασταθούν µε τη χορήγηση ενός αναστολέα της χολινεστεράσης, της ριβαστιγµίνης. Η καταστροφή του βασικού πυρήνα του Meynert επιτυγχάνεται είτε µε τοπική επίδραση ηλεκτρικού ρεύµατος 254 είτε µε τοπική έγχυση 177
179 νευροτοξίνης 255. Η ηλεκτρική καταστροφή παρά το ότι προκαλεί αλλαγές στις δοκιµασίες συµπεριφοράς, έχει το µειονέκτηµα ότι καταστρέφει µη εκλεκτικά τις ανατοµικές δοµές της περιοχής (νευρικές ίνες και αγγεία που διέρχονται διαµέσου αυτής) µε αποτέλεσµα την εµφάνιση αλλοιώσεων και σε αποµακρυσµένες περιοχές από αυτήν της καταστροφής. Η καταστροφή διαµέσου νευροτοξίνης είναι πιο εκλεκτική. Στην παρούσα µελέτη επιλέχθηκε το ιµποτενικό οξύ (ibotenic acid, alpha-amino-3-hydroxy-5- isoxazoleacetic acid), το οποίο προκαλεί νευρωνική νέκρωση διαµέσου υπερδιέγερσης των NMDA γλουταµινικών υποδοχέων, που οδηγεί σε υπερφόρτωση των κυττάρων µε ιόντα ασβεστίου 256. Η νευροτοξική δράση περιορίζεται στα σώµατα των νευρώνων και δεν επηρεάζει νευρικές ίνες και αγγεία που διέρχονται από την περιοχή της στερεοταξικής καταστροφής 256,257. Σε αντίθεση µάλιστα µε το καϊνικό οξύ (kainic acid), το οποίο συχνά προκαλεί επιληπτικές κρίσεις και αποµακρυσµένες νεκρώσεις, το ιµποτενικό οξύ δεν έχει επιληπτογόνο δράση, ενώ οι βλάβες περιορίζονται στην περιοχή της έγχυσης 258,259,260. Αυτά τα χαρακτηριστικά επιτρέπουν την υψηλής ακρίβειας πειραµατική καταστροφή του βασικού πυρήνα του Meynert. Επίσης, πρέπει να τονιστεί ότι η αµφοτερόπλευρη καταστροφή του βασικού πυρήνα του Meynert προσοµοιώνει καλύτερα τις γνωστικές διαταραχές, οι οποίες παρατηρούνται σε νευροεκφυλιστικές νόσους, ενώ η µονόπλευρη καταστροφή δε θεωρείται το ίδιο αξιόπιστη για τη µελέτη των διαταραχών της µνήµης και της µάθησης 2,227,
180 3.1. Σηµασία του βασικού πυρήνα του Meynert σε σχέση µε τα υπόλοιπα ανατοµικά στοιχεία που εµπλέκονται στην παθογένεια της νόσου Alzheimer Η χολινεργική οδός στη βάση του πρόσθιου εγκεφάλου περιέχει τη µεγαλύτερη πυκνότητα νευρικών ινών από όλες τις εξω-θαλαµικές προβολές του εγκεφαλικού φλοιού. Νέα στοιχεία, που βασίζονται σε έρευνες µε ηλεκτρονικό µικροσκόπιο, ανοσοϊστοχηµεία, αντισώµατα ανασυνδυασµένων υποτύπων υποδοχέων, in situ υβριδισµό και εκλεκτικές νευροτοξικές καταστροφές, έχουν προσφέρει πληθώρα νέων πληροφοριών για την οργάνωση και τη λειτουργία της χολινεργικής οδού σε διάφορα είδη, συµπεριλαµβανοµένου του ανθρώπου 10. Το χολινεργικό σύστηµα της βάσης του πρόσθιου εγκεφάλου είναι ένα σύστηµα µε διάφορα ανατοµικά στοιχεία (διαγώνια ζώνη, βασικός πυρήνας του Meynert, ραβδωτό σώµα), τα οποία προβάλλουν σε διάφορες περιοχές του εγκεφάλου (ιππόκαµπο, αµυγδαλή, νεοφλοιό). Στη νόσο Alzheimer φαίνεται ότι επηρεάζονται πολλές από τις παραπάνω περιοχές, µε αποτέλεσµα να θεωρείται ότι έχει δοθεί ιδιαίτερη έµφαση στη µελέτη µίας µόνο ανατοµικής δοµής, όπως είναι ο βασικός πυρήνας του Meynert. Οι νευρώνες του βασικού πυρήνα του Meynert, καθώς και οι φλοιϊκοί χολινεργικοί νευράξονες παραβλάπτονται στη νόσο Alzheimer 106,107,109, 110,145. Η απώλεια της φλοιϊκής χολινεργικής νεύρωσης στη νόσο Alzheimer αναφέρεται σε πληθώρα δηµοσιεύσεων 10,105,106. Ο ιδιαίτερα σηµαντικός ρόλος του βασικού πυρήνα του Meynert στην παθοφυσιολογία της νόσου Alzheimer και των σχετιζόµενων µε αυτήν γνωστικών διαταραχών επιβεβαιώνεται από τις υπάρχουσες σχετικές αναφορές στη διεθνή βιβλιογραφία 10,110,132,261,262. Από την παρούσα µελέτη διαπιστώνεται ότι η αµφοτερόπλευρη νευροτοξική καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ επιδρά στις γνωστικές διεργασίες των πειραµατοζώων. 179
181 4. οκιµασίες συµπεριφοράς Οι δοκιµασίες συµπεριφοράς πραγµατοποιήθηκαν σύµφωνα µε το πειραµατικό πρωτόκολλο σε όλες τις οµάδες των πειραµατοζώων. Εφαρµόστηκε µία αλληλουχία δοκιµασιών συµπεριφοράς, στηριζόµενη στη µεθοδολογία που περιγράφεται στη σύγχρονη βιβλιογραφία 3,199,230,239, ώστε να δοθεί η δυνατότητα να ελεγχθούν µε πολλαπλό τρόπο οι µηχανισµοί της µάθησης και οι διεργασίες της µνήµης, καθώς και η µνήµη αναφοράς και η λειτουργική µνήµη. Η πραγµατοποίηση των δοκιµασιών στα πειραµατόζωα των διαφόρων οµάδων οργανώθηκε µε βάση την καταστροφή του βασικού πυρήνα του Meynert, πριν ή µετά από τα πειράµατα συµπεριφοράς, σύµφωνα µε το πρωτόκολλο που έχει περιγραφεί στο τµήµα «υλικό και µέθοδοι» της παρούσας εργασίας. Η ριβαστιγµίνη χορηγήθηκε υποδόρια σε κατάλληλη δοσολογία, ώστε να εµφανίζει εκλεκτικότητα στην αναστολή της ακετυλοχολινεστεράσης στο φλοιό του εγκεφάλου και τον ιππόκαµπο, αλλά να µην προκαλεί χολινεργικές ανεπιθύµητες ενέργειες 3, Λαβύρινθος του Morris Η δοκιµασία του λαβυρίνθου του Morris χρησιµοποιεί τη φυσική ικανότητα των επιµύων στην κολύµβηση, καθώς και το ένστικτο της επιβίωσης και του φόβου του πνιγµού. Τα πειραµατόζωα «µαθαίνουν» να κολυµπούν στη δεξαµενή προς αναζήτηση της εξέδρας διαφυγής. Έχουν περιγραφεί διάφορα σχετικά πρωτόκολλα στη διεθνή βιβλιογραφία 3,199,200,201,263,264. Η πειραµατική διαδικασία, που εφαρµόστηκε στην παρούσα µελέτη, στηρίχθηκε στο πρότυπο των Bejar και συν. (1999) 3 και αποτελεί παραλλαγή του πειραµατικού πρωτοκόλλου του Morris (Morris 1984) 199. Όπως σε κάθε δοκιµασία για τον έλεγχο των γνωστικών λειτουργιών, έτσι και στο λαβύρινθο του Morris η καταγραφή των παραµέτρων αποτελεί µία έµµεση µέτρηση µίας εγκεφαλικής-ανώτερης φλοιικής λειτουργίας. Πολυάριθµες µελέτες υποστηρίζουν την αξιοπιστία του λαβύρινθου του Morris στη µελέτη των γνωστικών λειτουργιών 209,211,214,215,217,218, 219,220,221,222. Στην παρούσα µελέτη ελέγχθηκε τόσο η µνήµη αναφοράς όσο και η λειτουργική µνήµη των πειραµατόζωων. Από την ανάλυση των αποτελεσµάτων διαπιστώνεται ότι δε βρέθηκαν Σ.Σ. διαφορές µεταξύ των γενικών και των ειδικών µαρτύρων (οµάδες Α και Β, αντίστοιχα), γεγονός 180
182 που αποδεικνύει την απουσία επιδράσεων της πειραµατικής διαδικασίας της εικονικής καταστροφής του βασικού πυρήνα του Meynert στις γνωστικές λειτουργίες των πειραµατοζώων. Κατά τη σύγκριση των ληφθεισών καταγραφών από τις οµάδες των πειραµατοζώων µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο της διατήρησης ή απόκτησης της µνήµης (οµάδες Γ και Ε, αντίστοιχα) και από τους γενικούς µάρτυρες (οµάδα Α), διαπιστώθηκαν Σ.Σ. διαφορές (p<0,0001) τόσο για τη λειτουργική όσο και για τη µνήµη αναφοράς. Φαίνεται ότι πλήττονται τόσο οι διαδικασίες µάθησης όσο και οι διεργασίες της µνήµης στα πειραµατόζωα µε καταστροφή του πυρήνα. Εποµένως, η στερεοταξική καταστροφή του βασικού πυρήνα του Meynert, επιδρά στη συµπεριφορά-γνωστικές διεργασίες των πειραµατοζώων. Τα στοιχεία αυτά συµφωνούν µε τα ευρήµατα άλλων ερευνητών 3,4,251,261,265,266 και µάλιστα αποτελούν µία παράλληλη καταγραφή του ελέγχου τόσο της διατήρησης της µνήµης, που αποκτήθηκε πριν από την καταστροφή του βασικού πυρήνα του Meynert, όσο και της απόκτησης µνήµης µετά από την καταστροφή του NBM. Κατά τη σύγκριση των καταγραφών µεταξύ των οµάδων των πειραµατοζώων µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο της διατήρησης ή της απόκτησης µνήµης µε σύγχρονη χορήγηση ριβαστιγµίνης (οµάδες και ΣΤ αντίστοιχα) και των οµάδων των πειραµατοζώων µε καταστροφή του βασικού πυρήνα του Meynert (οµάδες Γ και Ε), διαπιστώθηκαν Σ.Σ. διαφορές (p<0,0001) τόσο στη λειτουργική όσο και στη µνήµη αναφοράς. Οι Σ.Σ. διαφορές που παρατηρούνται µεταξύ των οµάδων Ε και ΣΤ (Ε>ΣΤ), καθώς και µεταξύ των οµάδων Γ και (Γ> ) αποδεικνύουν ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα των οµάδων και ΣΤ συµβάλλει στη βελτίωση των επιδόσεών τους, βελτιώνει εποµένως την απόκτηση και τη διατήρηση της µνήµης. Με βάση την ανάλυση όλων των ευρηµάτων που προέκυψαν από τη δοκιµασία του λαβυρίνθου του Morris, διαπιστώθηκε ότι είναι πιο εύκολο να χρησιµοποιηθούν για την αξιολόγηση επιδράσεων φαρµακολογικά δραστικών ουσιών στις γνωστικές διεργασίες, τα πρωτόκολλα που περιλαµβάνουν δοκιµασίες συµπεριφοράς µε αξιολόγηση της απόκτησης της µνήµης από τα πειραµατόζωα. 181
183 4.2. Λαβύρινθος ανοικτού πεδίου Κατά τη διεξαγωγή των πειραµάτων στο λαβύρινθο ανοικτού πεδίου µελετήθηκαν η µνήµη αναφοράς και η λειτουργική µνήµη. Με βάση τις υπάρχουσες βιβλιογραφικές αναφορές η λειτουργική µνήµη επηρεάζεται σηµαντικά µετά από την καταστροφή του βασικού πυρήνα του Meynert 248,251,261,267,268, ενώ η µνήµη αναφοράς επηρεάζεται σε µικρότερο βαθµό 269,270. Σηµειώνεται η ύπαρξη αντικρουόµενων απόψεων 2,271,272,273 για το ποιά µορφή µνήµης επηρεάζεται (συνήθως επηρεάζονται και οι δύο, ενώ σε κάποιες µελέτες επηρεάζεται µόνο η µία από τις δύο), κάτι που επισηµαίνει ότι οι δύο αυτές µορφές µνήµης δεν είναι τόσο ανεξάρτητες, όσο υποδηλώνει η ονοµασία τους. Η µνήµη για συγκεκριµένες λεπτοµέρειες της πειραµατικής διαδικασίας εξαρτάται από το συνδυασµό πληροφοριών, που συλλέγονται σε κάθε δοκιµασία. Η µνήµη για τις µεταβαλλόµενες πληροφορίες-δεδοµένα είναι αξιοποιήσιµη µόνο µέσα στα πλαίσια της µνήµης για τα σταθερά δεδοµένα-πληροφορίες της δοκιµασίας. Αυτό σηµαίνει ότι µία επίδραση στη λειτουργική µνήµη θα επηρεάσει και τη µνήµη αναφοράς και αντιστρόφως. Πιθανόν και οι δύο διαταραχές να έχουν την ίδια αιτία. Γι αυτό, στην περίπτωση των πειραµατικών καταστροφών του βασικού πυρήνα του Meynert, είναι πιο δόκιµο να δεχθούµε ότι επηρεάζονται και οι δύο µορφές µνήµης, η λειτουργική µνήµη και η µνήµη αναφοράς. Η µνήµη των πειραµατοζώων µε πειραµατικές καταστροφές του βασικού πυρήνα του Meynert είναι ασταθής και ασύντακτη, δηλαδή δε γίνεται επαρκής επεξεργασία των πληροφοριών-ερεθισµάτων, ώστε να συνδυαστούν κατάλληλα µε άλλες πληροφορίες που υπάρχουν αποθηκευµένες στη µνήµη. Η ερµηνεία των διαφορετικών αποτελεσµάτων στις διάφορες µελέτες, σχετικά µε τις αλλαγές στη λειτουργική µνήµη, στη µνήµη αναφοράς ή και στις δύο ταυτόχρονα, βασίζεται στη δυσκολία της δοκιµασίας συµπεριφοράς 274 ή στη χρονική διάρκεια της εκπαίδευσης του πειραµατοζώου 228. Σύµφωνα µε την πρώτη ερµηνεία, δοκιµασίες µε απλούστερο πειραµατικό πρωτόκολλο χαρακτηρίζονται από λάθη στη λειτουργική µνήµη, ενώ πιο πολύπλοκο πρωτόκολλο οδηγεί σε αυξηµένα λάθη και στις δύο µορφές µνήµης. Σύµφωνα µε τη δεύτερη ερµηνεία, η εκπαίδευση πριν από την πειραµατική καταστροφή ελαττώνει τα λάθη της µνήµης αναφοράς, µε αποτέλεσµα τα λάθη της λειτουργικής µνήµης να 182
184 είναι αυξηµένα, αντιστοίχως η εκπαίδευση µετά την πειραµατική καταστροφή οδηγεί σε λάθη κυρίως της µνήµης αναφοράς 228,275. Ωστόσο, έχουν δηµοσιευθεί και περιπτώσεις στις οποίες λάθη της λειτουργικής µνήµης εµφανίζονται και µετά από εκπαίδευση του πειραµατοζώου µετά την καταστροφή του πυρήνα 274, καθώς και λάθη της µνήµης αναφοράς σε πειραµατόζωα που είχαν εκπαιδευτεί πριν από την καταστροφή του πυρήνα 270. Είναι απαραίτητο να διευκρινιστεί ότι οι αλλαγές στις επιδόσεις των δοκιµασιών συµπεριφοράς, που σχετίζονται µε τη λειτουργική µνήµη και τη µνήµη αναφοράς, εξαρτώνται από τη φύση και τη δυσκολία της δοκιµασίας. Εποµένως, οι συγκρίσεις-αναλύσεις των αποτελεσµάτων πρέπει να γίνονται µεταξύ των ίδιων δοκιµασιών συµπεριφοράς και όχι µε βάση τη µελετώµενη µορφή µνήµης. Στην παρούσα µελέτη διαπιστώθηκε ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert στα πειραµατόζωα της οµάδας Β (ειδικοί µάρτυρες) δεν επέδρασε στις διεργασίες της µνήµης, αφού δεν παρατηρήθηκαν Σ.Σ. διαφορές σε σχέση µε τα πειραµατόζωα της οµάδας Α (γενικοί µάρτυρες) στη µνήµη αναφοράς, στη λειτουργική µνήµη και στο χρόνο παραµονής στο λαβύρινθο ανοικτού πεδίου. Όσον αφορά στη µνήµη αναφοράς των πειραµατοζώων, διαπιστώθηκε ότι όταν η απόκτησή της (εκπαίδευση των πειραµατόζωων) γίνεται πριν από την πειραµατική καταστροφή του πυρήνα, τότε δεν παρατηρούνται Σ.Σ. διαφορές µεταξύ των οµάδων, ενώ όταν η καταστροφή του βασικού πυρήνα του Meynert γίνεται πριν από τη διεξαγωγή των δοκιµασιών συµπεριφοράς, τότε οι διαφορές µεταξύ των οµάδων στα λάθη της µνήµης αναφοράς είναι στατιστικά σηµαντικές. Οι καταγραφές αυτές είναι αντίστοιχες µε ανάλογες παρατηρήσεις άλλων ερευνητών 2,228,275. Η ανάλυση των αποτελεσµάτων από τις καταγραφές για τη λειτουργική µνήµη κατέδειξε ότι αυτή η µορφή της µνήµης επηρεάζεται ανεξάρτητα από τη χρονική εντόπιση της πειραµατικής καταστροφής του βασικού πυρήνα του Meynert (πριν ή µετά τη διεξαγωγή των δοκιµασιών συµπεριφοράς). Παρατηρήθηκαν Σ.Σ. διαφορές µεταξύ των οµάδων των πειραµατοζώων, καταγραφές παρόµοιες µε άλλων ερευνητών 2,248,251,261,267,268. Στις καταγραφές του συνολικού χρόνου παραµονής των πειραµατοζώων στο λαβύρινθο ανοικτού πεδίου παρουσιάστηκαν Σ.Σ. διαφορές µεταξύ των οµάδων. 183
185 Κατά την ανάλυση των αποτελεσµάτων διαπιστώθηκε ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα που υπέστησαν καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης ή απόκτησης µνήµης (οµάδες και ΣΤ αντίστοιχα) συνέβαλε στη βελτίωση των επιδόσεών τους. Η ριβαστιγµίνη, εποµένως, βελτίωσε τις διεργασίες απόκτησης της µνήµης και τους µηχανισµούς διατήρησης της µνήµης στα πειραµατόζωα µε καταστροφή του βασικού πυρήνα του Meynert οκιµασία παθητικής αποφυγής Με βάση την εµπειρία που έχει αποκτηθεί από τα διάφορα πειράµατα συµπεριφοράς, η δοκιµασία παθητικής αποφυγής (passive avoidance test) θεωρείται ότι εκπληρώνει τα κριτήρια ελέγχου της πρόσφατης µνήµης 262. Έτσι, παρά την απλότητα της συγκεκριµένης δοκιµασίας και τη φύση του µνηµονικού εγγράµµατος (αποφυγή επώδυνου ερεθίσµατος), ελέγχεται η πρόσφατη µνήµη των πειραµατοζώων και όταν καταγράφονται διαταραχές της, τα αποτελέσµατα µπορούν να οδηγήσουν στην εξαγωγή συµπερασµάτων, αλλά και να συνδυαστούν µε αυτά άλλων δοκιµασιών για τη µελέτη της µνήµης και της µάθησης. Η επίδραση της καταστροφής του βασικού πυρήνα του Meynert στην απόκτηση ή τη διατήρηση της µνηµονικής εγγραφής της δοκιµασίας παθητικής αποφυγής αποτέλεσε έναν από τους στόχους της παρούσας µελέτης. Αλλαγές στην απόκτηση, αλλά και στη διατήρηση της µνήµης της δοκιµασίας παθητικής αποφυγής, έχουν βρεθεί µετά από πειραµατικές καταστροφές του βασικού πυρήνα του Meynert από τις πρώτες απόπειρες µελέτης του συγκεκριµένου πυρήνα 250,276. Η επίδραση στη διατήρηση, µε καταγραφή ελαττωµένων χρόνων παραµονής των πειραµατoζώων στο θάλαµο εκκίνησης, αποτελεί το κύριο εύρηµα των περισσότερων ερευνητών 251,261,277. Στην παρούσα µελέτη επιχειρήθηκε, κατά το σχεδιασµό της πειραµατικής διαδικασίας, να ελεγχθούν όλες οι πτυχές της µνήµης που εµπλέκονται στη δοκιµασία παθητικής αποφυγής. Έτσι, στα πειραµατόζωα µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο διατήρησης µνήµης χωρίς ή µε χορήγηση ριβαστιγµίνης (οµάδες Γ και αντίστοιχα) ελέγχθηκε η διατήρηση της µνήµης, που είχε αποκτηθεί πριν από τη στερεοταξική καταστροφή του βασικού πυρήνα του Meynert, ενώ στα πειραµατόζωα µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης 184
186 µνήµης χωρίς ή µε χορήγηση ριβαστιγµίνης (οµάδες Ε και ΣΤ αντίστοιχα) ελέγχθηκε η διατήρηση της µνήµης, η απόκτηση της οποίας είχε γίνει µετά τη στερεοταξική καταστροφή. Κατά τη σύγκριση των γενικών και ειδικών µαρτύρων (οµάδων Α και Β) δε βρέθηκαν στατιστικά σηµαντικές διαφορές (p>0,05), στοιχείο που αποδεικνύει την απουσία επιδράσεων της χειρουργικής διαδικασίας στις γνωστικές λειτουργίες των πειραµατοζώων. Αντίθετα κατά τη σύγκριση των γενικών µαρτύρων και των οµάδων µε καταστροφή του πυρήνα (σύγκριση µεταξύ των οµάδων Α και Γ και των οµάδων Α και Ε), βρέθηκαν στατιστικά σηµαντικές διαφορές (p<0,0001) στους χρόνους παραµονής των πειραµατοζώων στο θάλαµο εκκίνησης. Έτσι, οι διαφορές µεταξύ των οµάδων Α και Γ αποδεικνύουν την επίδραση στη µνήµη, αφού είναι εµφανής η αδυναµία των πειραµατοζώων της οµάδας Γ να ανακαλέσουν τα µνηµονικά εγγράµµατα της δοκιµασίας απόκτησης της µνήµης. Αντιστοίχως, οι διαφορές µεταξύ των οµάδων Α και Ε αποδεικνύουν την επίδραση της καταστροφής του βασικού πυρήνα του Meynert στην πρόσφατη µνήµη, αφού είναι εµφανής η αδυναµία των πειραµατοζώων της οµάδας Ε να αποθηκεύσουν στη µνήµη και στη συνέχεια να ανακαλέσουν το δυσάρεστο ερέθισµα, που δέχθηκαν κατά τη διαδικασία µάθησης-απόκτησης της µνήµης. Εποµένως, η χολινεργική δυσλειτουργία µετά από τη νευροτοξική καταστροφή του βασικού πυρήνα του Meynert, επιδρά στους µηχανισµούς µνήµης και µάθησης, διαµέσου διαταραχών στην πρόσφατη µνήµη. Η χορήγηση ριβαστιγµίνης στις οµάδες και ΣΤ βελτίωσε τις επιδόσεις των πειραµατόζωων, αλλά δεν απεκατέστησε πλήρως τις γνωστικές λειτουργίες. Έτσι, µε βάση την ανάλυση του ποσοστού αποφυγής (incidence of avoidance) γίνεται εµφανές ότι η οµάδα ΣΤ παρουσιάζει ποσοστό αποφυγής 70%, ενώ η οµάδα 60%. Οι διαφορές που καταγράφηκαν τόσο µεταξύ των οµάδων Α και, όσο και µεταξύ Γ και ήταν στατιστικά σηµαντικές (p<0,05). Αντιστοίχως, καταγράφηκαν στατιστικά σηµαντικές διαφορές (p<0,05) τόσο µεταξύ των οµάδων Α και ΣΤ, όσο και µεταξύ Ε και ΣΤ. Η ανάλυση των αποτελεσµάτων των δοκιµασιών παθητικής αποφυγής καταδεικνύει ότι η διατήρηση της µνήµης επηρεάζεται σηµαντικά, ανεξάρτητα από τη χρονική εντόπιση της πειραµατικής καταστροφής του βασικού πυρήνα του Meynert (πριν ή µετά τη διεξαγωγή της δοκιµασίας 185
187 απόκτησης µνήµης της παθητικής αποφυγής). Με τις αισθητικοκινητικές δοκιµασίες έχει αποκλειστεί η επίδραση τυχόν νευροµυϊκών διαταραχών από την καταστροφή του πυρήνα στην αξιοπιστία των αποτελεσµάτων ενώ µε τις δοκιµασίες ελέγχου της χολινεργικής δραστηριότητας εξασφαλίστηκε η απουσία ανεπιθύµητων ενεργειών από τη χορήγηση ριβαστιγµίνης. Εποµένως, είναι δυνατή η συσχέτιση των αποτελεσµάτων µε τους µηχανισµούς της µνήµης και της µάθησης. 5. Οι συµπεριφορικές µεταβολές ως αποτέλεσµα της πειραµατικής καταστροφής του βασικού πυρήνα του Meynert Με βάση τον πειραµατικό σχεδιασµό και το πειραµατικό πρότυπο, το οποίο εφαρµόστηκε σε καθεµία από τις έξι οµάδες των πειραµατόζωων, είναι δυνατό να αποδοθούν οι αλλαγές, που παρατηρήθηκαν στα πειράµαταδοκιµασίες συµπεριφοράς, στην καταστροφή του βασικού πυρήνα του Meynert. ιαπιστώθηκε ότι στην οµάδα Β (ειδικοί µάρτυρες) δεν παρατηρήθηκαν Σ.Σ. διαφορές σε σύγκριση µε την οµάδα Α (γενικοί µάρτυρες) και συµπεραίνεται ότι η χειρουργική διαδικασία της εικονικής καταστροφής του πυρήνα δεν επέδρασε στη συµπεριφορά. Αντιστοίχως, από τη διεξαγωγή των αισθητικοκινητικών δοκιµασιών, διαπιστώθηκε ότι τα πειραµατόζωα των οµάδων Γ και Ε δεν παρουσίαζαν νευροµυϊκές διαταραχές. Εποµένως, οι Σ.Σ. διαφορές που παρατηρήθηκαν στα πειραµατόζωα αυτών των οµάδων, µπορούν να αποδοθούν στις πειραµατικές καταστροφές του βασικού πυρήνα του Meynert. 6. Οι συµπεριφορικές µεταβολές ως αποτέλεσµα της διαταραχής της χολινεργικής οδού Κατά την ανάλυση των αποτελεσµάτων των δοκιµασιών συµπεριφοράς, διαπιστώθηκε ότι η διαταραχή της χολινεργικής οδού, διαµέσου της καταστροφής του βασικού πυρήνα του Meynert, επέδρασε τόσο στους µηχανισµούς µάθησης-απόκτησης µνήµης, όσο και στις διεργασίες διατήρησης της µνήµης. ιαπιστώθηκαν, επίσης, διαταραχές τόσο της µνήµης αναφοράς, όσο και της λειτουργικής µνήµης στις οµάδες µε καταστροφή του βασικού πυρήνα του Meynert, στις οµάδες Γ και Ε, αντίστοιχα. 186
188 Η αναλογία των συµπεριφορικών αποτελεσµάτων µε εκείνα µελετών, στις οποίες το χολινεργικό σύστηµα ελέγχθηκε-τροποποιήθηκε φαρµακολογικά µε τη χρήση σκοπολαµίνης διαµέσου δέσµευσης µουσκαρινικών υποδοχέων 3,4,278,279,280, καταδεικνύει ότι οι αλλαγές που προκύπτουν µετά την νευροτοξική καταστροφή του βασικού πυρήνα του Meynert, σχετίζονται µε την ακετυλοχολίνη και το χολινεργικό σύστηµα. Επίσης, µελέτες που αφορούν ηλικιωµένους επίµυες, αναφέρουν διαταραχές στις γνωστικές λειτουργίες, που συνδυάζονται µε χολινεργική δυσλειτουργία 121,281,282. Η ισχυρότερη απόδειξη για την εµπλοκή του χολινεργικού συστήµατος στη µνήµη και τον έλεγχό του από τον βασικό πυρήνα του Meynert, είναι η σηµαντική βελτίωση των επιδόσεων των πειραµατόζωων στις δοκιµασίες συµπεριφοράς, µετά τη χορήγηση αναστολέων χολινεστεράσης. Αµφιβολίες έχουν διατυπωθεί κατά καιρούς για το πώς είναι δυνατόν να υπάρχει αποκατάσταση της χολινεργικής δυσλειτουργίας µε τη χορήγηση των αναστολέων, όταν έχουν καταστραφεί σε µεγάλο ποσοστό οι χολινεργικοί νευρώνες στο βασικό πυρήνα του Meynert. Ωστόσο, έχει βρεθεί 236 ότι οι ελάχιστοι παραµένοντες χολινεργικοί νευρώνες είναι δυνατό, παρουσία ενός αναστολέα της χολινεστεράσης, να αποκαταστήσουν τα λειτουργικά επίπεδα του νευροδιαβιβαστή ακετυλοχολίνη. Στην παρούσα µελέτη διαπιστώθηκε ότι η χορήγηση ριβαστιγµίνης στα πειραµατόζωα οµάδων, στα οποία είχε προκληθεί καταστροφή του βασικού πυρήνα του Meynert, στις οµάδες και ΣΤ αντίστοιχα, απεκατέστησε σε µεγάλο βαθµό τις επιδόσεις τους στις δοκιµασίες συµπεριφοράς. ιαπιστώθηκε βελτίωση τόσο της µνήµης αναφοράς, όσο και της λειτουργικής µνήµης. ηλαδή, η ριβαστιγµίνη απεκατέστησε τόσο τους µηχανισµούς µάθησης-απόκτησης µνήµης, όσο και τις διεργασίες διατήρησης της µνήµης. 187
189 7. Ειδικότητα των συµπεριφορικών δοκιµασιών για τις γνωστικές διεργασίες Με τις δοκιµασίες συµπεριφοράς αποδεικνύεται ότι η µνήµη επηρεάζεται στα πειραµατόζωα, στα οποία έγινε πειραµατική καταστροφή του βασικού πυρήνα του Meynert. Η συσχέτιση των µεταβολών των δοκιµασιών συµπεριφοράς µε τις διεργασίες της µνήµης και της µάθησης, δεν είναι απόλυτη. Ωστόσο, οι αλλαγές αυτές αποτελούν ενδείξεις για τις γνωστικές διαταραχές που προκύπτουν λόγω της δυσλειτουργίας του χολινεργικού συστήµατος. Στις διεργασίες της µνήµης και της µάθησης συµµετέχουν και άλλα συστήµατα, αλλά µε βάση την κρατούσα θεωρία της χολινεργικής υπόθεσης της νόσου Alzheimer, η χολινεργική διαταραχή είναι αυτή που οδηγεί στην έκπτωση των γνωστικών λειτουργιών του ασθενή. Γι αυτό και η αποκλειστική προς το παρόν θεραπευτική αντιµετώπιση των συµπτωµάτων της νόσου είναι η ενίσχυση του χολινεργικού συστήµατος. 8. Αξιοποίηση του χρησιµοποιηθέντος πειραµατικού προτύπου στη µελέτη της συµπεριφοράς Από την ανάλυση των αποτελεσµάτων σε όλες τις χρησιµοποιηθείσες δοκιµασίες συµπεριφοράς διαπιστώθηκε ότι το πειραµατικό πρότυπο προσοµοιάζει σε µεγάλο βαθµό αλλαγές στη συµπεριφορά των πειραµατοζώων, οι οποίες µπορούν να χαρακτηριστούν ως γνωστικές διαταραχές ή διαταραχές της µνήµης και της µάθησης. Επίσης, είναι σηµαντικό να τονισθεί ότι οι δοκιµασίες συµπεριφοράς µπορούν να αξιοποιηθούν στην αξιολόγηση των γνωστικών λειτουργιών και σε άλλα πειραµατικά µοντέλα, που αφορούν νευροεκφυλιστικές παθήσεις, αλλά και βλάβες του ΚΝΣ. Εποµένως, είναι δυνατή η αξιοποίηση του συγκεκριµένου πειραµατικού πρωτοκόλλου για την προκλινική αξιολόγηση φαρµακολογικά δραστικών ουσιών, που επιδρούν στους µηχανισµούς της µνήµης και της µάθησης. Για παράδειγµα, είναι δυνατή η αξιοποίηση του πρωτοκόλλου των δοκιµασιών συµπεριφοράς στην αξιολόγηση επιδράσεων των φαρµακολογικά δραστικών ουσιών που χρησιµοποιούνται στην αναισθησιολογία στις µνηµονικές διεργασίες. Τέλος, αξίζει να επισηµανθεί ότι οι αισθητικοκινητικές δοκιµασίες, που χρησιµοποιήθηκαν στην παρούσα µελέτη, είναι δυνατό να χρησιµοποιηθούν 188
190 στη αξιολόγηση πειραµατικών µοντέλων, που αφορούν νευροµυϊκές παθήσεις, αλλά και νευροµυϊκές διαταραχές, που οφείλονται σε φαρµακολογικά δραστικές ουσίες. Συµπερασµατικά, µπορεί να υποστηριχθεί ότι τα πειραµατικά µοντέλα δεν είναι δυνατό να αναπαραστήσουν πλήρως την αιτιολογία, τη νευροπαθολογία και τα συµπτώµατα των νόσων που προσοµοιάζουν. Αυτό, όµως, δεν πρέπει να οδηγήσει στην απόρριψη της χρήσης τους ως ερευνητικών εργαλείων. Κι αυτό, γιατί αποτελούν µεθόδους προσοµοίωσης βασικών χαρακτηριστικών των νόσων και συντελούν στην κατανόηση της παθοφυσιολογίας τους και στην ανάπτυξη νέων θεραπευτικών στρατηγικών. Ο συνδυασµός διαφόρων πειραµατικών µοντέλων µπορεί να προσεγγίσει συνεργιστικά την «ανθρώπινη» εικόνα µίας νόσου και να οδηγήσει τους ερευνητές σε αξιόπιστα συµπεράσµατα. 189
191 190
192 IV. ΠΕΡΙΛΗΨΗ-ΣΥΜΠΕΡΑΣΜΑΤΑ Η «χολινεργική υπόθεση», που διατυπώθηκε πριν από 20 χρόνια περίπου, υποστήριξε ότι η σηµαντική απώλεια της χολινεργικής λειτουργίας στο ΚΝΣ συντελεί καθοριστικά στην εµφάνιση των γνωστικών διαταραχών, οι οποίες σχετίζονται µε την προχωρηµένη ηλικία και τη νόσο Alzheimer. Οι µελέτες, που πραγµατοποιήθηκαν µε βάση τη χολινεργική υπόθεση, καθώς και τα πειραµατικά µοντέλα που χρησιµοποιήθηκαν, συνέβαλαν όχι µόνο στην καθιέρωση της θεωρίας, αλλά και των πρώτων θεραπευτικών στρατηγικών που βασίστηκαν σε αυτή. Η εισαγωγή στη θεραπευτική πρακτική των αναστολέων της χολινεστεράσης, φαρµάκων βασισµένων στη θεωρία της χολινεργικής υπόθεσης, συνέβαλε στην άµβλυνση των συµπτωµάτων και στη βελτίωση της ποιότητας ζωής των ασθενών. Ο σκοπός της παρούσας µελέτης ήταν ο σχεδιασµός και η ανάπτυξη ενός πειραµατικού µοντέλου, το οποίο θα µπορούσε να χρησιµοποιηθεί για την προσοµοίωση γνωστικών διαταραχών, που σχετίζονται µε παθήσεις του κεντρικού νευρικού συστήµατος (ΚΝΣ), καθώς και για την αξιολόγηση φαρµακολογικά δραστικών ουσιών, που επιδρούν στη µνήµη και τη µάθηση. Σηµαντικό ρόλο στο σχεδιασµό του πειραµατικού πρωτοκόλλου έπαιξε η υπάρχουσα σχετική βιβλιογραφία, που επιβεβαίωσε το σηµαίνοντα ρόλο του βασικού πυρήνα του Meynert τόσο στις γνωστικές λειτουργίες µε βάση τη χολινεργική υπόθεση της µνήµης, όσο και την εµπλοκή αυτού του πυρήνα στην παθοφυσιολογία της νόσου Alzheimer. Βέβαια είναι γεγονός ότι είναι περιορισµένος ο αριθµός των συµπεριφορικών µελετών, που χρησιµοποιούν αλληλουχία συµπεριφορικών δοκιµασιών-πειραµάτων για τη µελέτη της µνήµης και της µάθησης, καθώς και παράλληλο έλεγχο της απόκτησης και ανάκλησης της µνήµης πριν και µετά την καταστροφή του βασικού πυρήνα του Meynert. Βασική, εξάλλου, φαρµακολογική επιλογή για τη νόσο Alzheimer αποτελούν οι αναστολείς της χολινεστεράσης, η θεραπευτική χρήση των οποίων βασίζεται στη θεωρία της χολινεργικής υπόθεσης. Η ριβαστιγµίνη (ENA713), αποτελεί έναν από τους τελευταίους αναστολείς που χρησιµοποιούνται στη θεραπευτική, έχει εκλεκτική δράση και συµβάλλει στην άµβλυνση των συµπτωµάτων από τα πρώτα στάδια της νόσου. Επειδή στη διεθνή βιβλιογραφία δεν υπήρχαν συµπεριφορικές µελέτες µε χορήγηση της ριβαστιγµίνης σε πειραµατόζωα µετά από καταστροφή του βασικού πυρήνα του Meynert µε ιµποτενικό οξύ, 191
193 θεωρήθηκε σηµαντική η έρευνα της επίδρασης της ριβαστιγµίνης σε ανώτερες φλοιικές λειτουργίες (συµπεριφορά) επίµυων, µετά από καταστροφή του βασικού πυρήνα του Meynert µε τη χρήση ιµποτενικού οξέος (ibotenic acid), το οποίο αποτελεί νευροτοξίνη που καταστρέφει τους νευρώνες και όχι τις διερχόµενες νευρικές ίνες ή τα αγγεία της περιοχής του πυρήνα. Το πειραµατικό πρωτόκολλο της αµφοτερόπλευρης καταστροφής του βασικού πυρήνα του Meynert µε έγχυση ιµποτενικού οξέος (ibotenic acid) µε τη χρήση στερεοταξικής διάταξης, εφαρµόστηκε σε ένηβους άρρενες επίµυς φυλής Wistar. Τα πειραµατόζωα (n=60) κατανεµήθηκαν σε έξι (6) οµάδες. Η οµάδα Α περιελάµβανε δέκα (n=10) πειραµατόζωα, στα οποία δεν εφαρµόστηκε το πειραµατικό πρότυπο και χρησίµευσαν ως γενικοί µάρτυρες (controls). Η οµάδα Β περιελάµβανε δέκα (n=10) πειραµατόζωα, στα οποία έγινε εικονική εγχείρηση χωρίς καταστροφή του βασικού πυρήνα την 20 ηµέρα (sham-operated). Η οµάδα Γ περιελάµβανε δέκα (n=10) πειραµατόζωα, στα οποία αφού πραγµατοποιήθηκαν κατά τις ηµέρες 1 έως 20 οι δοκιµασίες συµπεριφοράς (acquisition tests), έγινε στερεοταξική καταστροφή του βασικού πυρήνα του Meynert την 20 ηµέρα. Την 25 ηµέρα έγιναν οι αισθητικοκινητικές δοκιµασίες και τις ηµέρες 27, 28 και 29 έγινε έλεγχος διατήρησης της µνήµης των δοκιµασιών συµπεριφοράς (retention tests) µετά από υποδόρια έγχυση φυσιολογικού ορού. Η οµάδα περιελάµβανε δέκα (n=10) πειραµατόζωα, στα οποία πραγµατοποιήθηκαν οι ίδιες, όπως στα πειραµατόζωα της οµάδας Γ, δοκιµασίες και στα ίδια χρονικά διαστήµατα, µε τη διαφορά ότι ο έλεγχος της διατήρησης της µνήµης των δοκιµασιών συµπεριφοράς (retention tests) γινόταν µετά από υποδόρια έγχυση διαλύµατος ριβαστιγµίνης. Η οµάδα Ε περιελάµβανε δέκα (n=10) πειραµατόζωα, στα οποία έγινε στερεοταξική καταστροφή του βασικού πυρήνα του Meynert. Την πέµπτη (5η) ηµέρα µετά την καταστροφή, πραγµατοποιήθηκαν οι αισθητικοκινητικές δοκιµασίες. Μετά τη συµπλήρωση 7ήµερης ανάρρωσης, ακολούθησαν οι δοκιµασίες συµπεριφοράς (acquisition tests) κατά τις πειραµατικές ηµέρες 1 έως 20. Κατά τη διάρκεια των δοκιµασιών γινόταν υποδόρια έγχυση φυσιολογικού ορού µε βάση το πειραµατικό πρότυπο. Η οµάδα ΣΤ περιελάµβανε δέκα (n=10) πειραµατόζωα, στα οποία έγινε στερεοταξική καταστροφή του βασικού πυρήνα του Meynert. Πραγµατοποιήθηκαν οι ίδιες, όπως στα πειραµατόζωα της οµάδας Ε, δοκιµασίες και στα ίδια χρονικά διαστήµατα, 192
194 µε τη διαφορά ότι κατά τη διάρκεια της περιόδου των δοκιµασιών, γινόταν υποδόρια έγχυση διαλύµατος ριβαστιγµίνης. Για τη µελέτη της µνήµης και της µάθησης χρησιµοποιήθηκε αλληλουχία συµπεριφορικών δοκιµασιών-πειραµάτων, που περιελάµβανε το λαβύρινθο του Morris, το λαβύρινθο ανοικτού πεδίου και τη δοκιµασία παθητικής αποφυγής. Στα πειραµατόζωα των οµάδων και ΣΤ (µε έλεγχο διατήρησης µνήµης και έλεγχο απόκτησης µνήµης αντίστοιχα) µε καταστροφή του βασικού πυρήνα του Meynert χορηγήθηκε ριβαστιγµίνη, ώστε να ελεγχθεί η επίδρασή της στους µηχανισµούς της µάθησης και στις διεργασίες της µνήµης. Για τη στατιστική επεξεργασία των καταγραφών των δοκιµασιών χρησιµοποιήθηκαν η ανάλυση διακύµανσης µε έναν παράγοντα (ANOVA) και η ανάλυση διακύµανσης επαναλαµβανοµένων µετρήσεων (RMANOVA), ακολουθούµενες από το Tukey-Kramer test, όταν το F ήταν στατιστικά σηµαντικό. Στα πειραµατόζωα πραγµατοποιήθηκαν αισθητικοκινητικές δοκιµασίες, οι οποίες απέβησαν αρνητικές για ύπαρξη νευροµυϊκών διαταραχών. Έτσι, εξασφαλίστηκε στα πειράµατα συµπεριφοράς η αποφυγή χρησιµοποίησης επιµύων µε αισθητικοκινητικές διαταραχές, γεγονός που θα οδηγούσε σε λανθασµένα συµπεράσµατα. Επίσης, η απουσία χολινεργικής υπερδραστηριότητας µετά τη χορήγηση ριβαστιγµίνης, εξασφαλίζει ότι οι επιδόσεις των πειραµατοζώων στις δοκιµασίες συµπεριφοράς δεν οφείλονται σε πιθανή επίδραση της ριβαστιγµίνης στην αισθητικοκινητική τους συµπεριφορά. Με βάση τα αποτελέσµατα των δοκιµασιών-πειραµάτων συµπεριφοράς της παρούσας µελέτης, τη στατιστική επεξεργασία τους και τη βιοστατιστική ερµηνεία τους συνάγονται τα παρακάτω συµπεράσµατα: 1. Μεταξύ των γενικών και των ειδικών µαρτύρων (οµάδες Α και Β αντίστοιχα) δεν παρατηρούνται στατιστικά σηµαντικές διαφορές, στοιχείο που αποδεικνύει ότι η πειραµατική διαδικασία της εικονικής καταστροφής του βασικού πυρήνα του Meynert δεν επιδρά στις γνωστικές λειτουργίες των πειραµατοζώων. 2. Η καταστροφή του βασικού πυρήνα του Meynert επέδρασε διαµέσου της διαταραχής της χολινεργικής οδού, τόσο στους 193
195 µηχανισµούς µάθησης-απόκτησης µνήµης, όσο και στις διεργασίες διατήρησης της µνήµης. 3. Οι οµάδες µε καταστροφή του βασικού πυρήνα του Meynert (οµάδες Γ,, Ε και ΣΤ) παρουσίασαν διαταραχές τόσο της µνήµης αναφοράς, όσο και της λειτουργικής µνήµης. 4. Σχετικά µε τη µνήµη αναφοράς, διαπιστώθηκε ότι όταν η απόκτησή της (εκπαίδευση των πειραµατόζωων) γίνεται πριν από την πειραµατική καταστροφή του βασικού πυρήνα του Meynert, τότε δεν παρατηρούνται στατιστικά σηµαντικές διαφορές στα λάθη της µνήµης αναφοράς µεταξύ των οµάδων, ενώ όταν η καταστροφή γίνεται πριν από τη διεξαγωγή των δοκιµασιών συµπεριφοράς, τότε οι διαφορές είναι στατιστικά σηµαντικές. 5. Η λειτουργική µνήµη επηρεάζεται ανεξάρτητα από τη χρονική εντόπιση της πειραµατικής καταστροφής του βασικού πυρήνα του Meynert (πριν ή µετά τη διεξαγωγή των δοκιµασιών συµπεριφοράς). 6. Η χορήγηση ριβαστιγµίνης στα πειραµατόζωα µε έλεγχο διατήρησης µνήµης και έλεγχο απόκτησης µνήµης, στα οποία είχε προκληθεί καταστροφή του βασικού πυρήνα του Meynert (οµάδες και ΣΤ, αντίστοιχα), αποκατέστησε σε µεγάλο βαθµό τις επιδόσεις τους στις δοκιµασίες συµπεριφοράς. ιαπιστώθηκε βελτίωση τόσο της µνήµης αναφοράς, όσο και της λειτουργικής µνήµης. ηλαδή, η ριβαστιγµίνη αποκατέστησε τόσο τους µηχανισµούς µάθησηςαπόκτησης µνήµης, όσο και τις διεργασίες διατήρησης της µνήµης. 7. Η βελτίωση των επιδόσεων στα πειραµατόζωα µε καταστροφή του βασικού πυρήνα του Meynert και έλεγχο απόκτησης µνήµης µε σύγχρονη χορήγηση ριβαστιγµίνης στο λαβύρινθο ανοικτού πεδίου (οµάδα ΣΤ), φθάνει σε σηµείο σύγκλισης των επιδόσεων µε αυτές των γενικών µαρτύρων (οµάδα Α), ως αποτέλεσµα της χορήγησης της ριβαστιγµίνης. 194
196 8. Με βάση την ανάλυση των αποτελεσµάτων στο λαβύρινθο του Morris, διαπιστώθηκε ότι τα πρωτόκολλα που περιλαµβάνουν δοκιµασίες συµπεριφοράς µε αξιολόγηση της απόκτησης της µνήµης από τα πειραµατόζωα είναι πιο εύκολο να χρησιµοποιηθούν για την αξιολόγηση επιδράσεων φαρµακολογικά δραστικών ουσιών στις γνωστικές διεργασίες. Συµπερασµατικά, τα αποτελέσµατα της παρούσας µελέτης επιβεβαιώνουν το σηµαίνοντα ρόλο του βασικού πυρήνα του Meynert στις γνωστικές λειτουργίες. Η αµφοτερόπλευρη καταστροφή του πυρήνα µε ιµποτενικό οξύ, προσοµοιώνει σε σηµαντικό βαθµό διαταραχές στους µηχανισµούς της µάθησης και στις διεργασίες της µνήµης και ως εκ τούτου µπορεί να χρησιµοποιηθεί ως πειραµατικό µοντέλο για την αξιολόγηση φαρµακολογικών ουσιών, οι οποίες επιδρούν στις γνωστικές διεργασίες. Επίσης, διαπιστώθηκε απουσία χολινεργικών ανεπιθύµητων ενεργειών της ριβαστιγµίνης στη δοσολογία που χρησιµοποιήθηκε, γεγονός που αποδεικνύει την εκλεκτικότητά της στην αναστολή της ακετυλοχολινεστεράσης στον εγκεφαλικό φλοιό και τον ιππόκαµπο. Τέλος, βρέθηκε ότι η χορήγηση ριβαστιγµίνης µετά από καταστροφή του βασικού πυρήνα του Meynert, επιφέρει βελτίωση των γνωστικών διεργασιών. 195
197 V. SUMMARY-CONCLUSIONS The cholinergic hypothesis, which was published almost 20 years ago, supported that the deterioration of the cholinergic function in the central nervous system was crucial for the development of cognitive dysfunction, related with aging and Alzheimer s disease. Studies based on this hypothesis and the experimental models that were used, contributed not only to the acceptance of this theory but also to the development of the first pharmacologic strategies for Alzheimer s disease. The use of cholinesterase inhibitors, drugs based on the theory of cholinergic hypothesis, has reduced the symptoms and improved the quality of life of the patients. The aim of this study was the development of an experimental model, which could be used for the simulation of cognitive disorders that are related with diseases of the central nervous system and could be also used for the evaluation of pharmacologic agents that affect learning and memory. Important role in the development of the experimental model played the already published studies which had confirmed the important role of the nucleus basalis of Meynert in cognitive functions according to the cholinergic hypothesis of geriatric memory dysfunction, as well as the relation of this nucleus with the pathophysiology of Alzheimer s disease. There are few behavioral studies that have used sequence of behavioral tests for the evaluation of learning and memory, as well as a parallel study of acquisition and retention of memory before and after the lesion of the nucleus basalis of Meynert. The main therapeutic option for Alzheimer s disease, based on the cholinergic hypothesis, is the use of cholinesterase inhibitors. Rivastigmine (ENA713) is one of the latest inhibitors which is used in therapeutics, has selective action and contributes to the amelioration of the symptoms from the first stages of the disease. In the published articles there were not behavioral studies in rats with nucleus basalis of Meynert ibotenic acid lesions and administration of rivastigmine, so it was considered important to study the effect of rivastigmine in the behavior of rats after nucleus basalis of Meynert lesions with ibotenic acid, which is a neurotoxin that destroys neurons and not the fibers and vessels that pass through the area of the nucleus. 196
198 The experimental protocol of the bilateral lesion of the nucleus basalis of Meynert with infusion of ibotenic acid with the use of a stereotaxic instrument was applied to mature male Wistar rats. The rats (n=10) were divided into six (6) groups. The first group (group A) included ten (n=10) rats in which the experimental protocol was not applied and served as controls animals. The second group (group B) included ten (n=10) rats in which was performed sham-operation without lesioning the nucleus basalis of Meynert on day 20. The third group (group C) included ten (n=10) rats which were trained on behavioral tests during days 1 to 20 (acquisition tests) and on day 20 nucleus basalis of Meynert was lesioned. On day 25 the sensomotory tests were performed and on days 27, 28 and 29 the behavioral tests for measuring the retention of memory (retention tests) were performed after SC infusion of normal saline. The fourth group (group D) included ten (n=10) rats in which was applied the same experimental protocol as in the animals of the group C with SC infusion of rivastigmine. The fifth group (group E) included ten (n=10) rats in which nucleus basalis of Meynert was lesioned. On day 5 the sensomotory tests were performed. After the 7 day recovery period the rats were trained on behavioral tests during experimental days 1 to 20 (acquisition tests). During the behavioral tests the rats were SC injected with normal saline according to the experimental protocol. The sixth group (group F) included ten (n=10) rats in which was applied the same experimental protocol as in the animals of the group E with SC infusion of rivastigmine. For studying learning and memory series of behavioral tests were used, including Morris water maze, open field maze and passive avoidance test. On the animals of groups D and F in which nucleus basalis of Meynert was lesioned, rivastignime was SC injected in order to study its effects on learning and memory processes. For the statistical analysis of the results, we used the one way ANOVA test and the repeated measures ANOVA test (RMANOVA) followed by Tukey- Kramer test, when F was significant. Rats were tested with sensomotory tests which were negative for neuromuscular disorders. The use of sensomotory tests secured that in behavioral tests were used animals without sensomotory defects and that the results would be reliable. 197
199 Also, the lack of cholinergic hyperactivity effects after the injection of rivastigmine, assured that the results of the behavioral tests are not due to possible effects of rivastigmine to the sensomotory behavior of the animals. Based on the results of the behavioral tests of this study, the statistical analysis and the biostatistical interpretation, the following conclusions were drawn: 1. There were not observed statistically significant differences between groups A and B, which proves that the experimental procedure of the sham-operation does not affect the cognitive function of the animals. 2. Nucleus basalis of Meynert lesions affect learning and memory through disturbance of the cholinergic system function. 3. In groups with nucleus basalis of Meynert lesions (groups C, D, E and F) disturbances in reference and also in working memory were observed. 4. When reference memory was acquired before the lesion of the nucleus basalis of Meynert, there were not observed statistically significant differences between the groups. When the lesion of the nucleus basalis of Meynert was performed before the behavioral tests, there were statistically significant differences on reference memory faults between the groups. 5. Working memory was affected independently the time of the lesion (before or after the behavioral tests training). 6. SC injection of rivastigmine in rats of the groups D and F restored their results on behavioral tests. Improvement in reference and working memory was observed. Rivastigmine restored learning and memory processes. 7. The improvement on behavioral tests results of the rats of group F tends to reach the results of the control animals of group A, as a result of SC injection of rivastigmine. 8. According the analysis of the results on the Morris water maze, it was concluded that the experimental protocols that include behavioral tests with evaluation of acquisition of the memory tasks, can be more accurately used for the evaluation of pharmacologically active substances that affect cognitive functions. 198
200 In conclusion, the results of this study indicate the importance of nucleus basalis of Meynert in cognitive functions.the bilateral lesion of the nucleus basalis of Meynert with ibotenic acid simulates disturbances in learning and memory processes and can be used as an experimental model for the evaluation of pharmacological substances that affect cognitive functions. Also, the lack of cholinergic adverse effects of rivastigmine in the dose that was used proves its selectivity in the inhibition of acetylcholinesterase in the cerebral cortex and the hippocampus. Finally, it was concluded that the administration of rivastigmine after lesions of the nucleus basalis of Meynert improves the cognitive functions of the rats. 199
201 200

202 201
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ Ορίζουμε ως διαβιβαστή μια ουσία που απελευθερώνεται από έναν νευρώνα σε μια σύναψη και που επηρεάζει ένα άλλο κύτταρο, είτε έναν νευρώνα είτε ένα κύτταρο
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ Ορίζουμε ως διαβιβαστή μια ουσία που απελευθερώνεται από έναν νευρώνα σε μια σύναψη και που επηρεάζει ένα άλλο κύτταρο, είτε έναν νευρώνα είτε ένα κύτταρο
Γνωστική-Πειραµατική Ψυχολογία
 Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Μηχανισµοί της ΣΔ à Βάση διεργασιών όπως η αντίληψη, µάθηση, εκούσια κίνηση. 10.000 à Μέσος νευρώνας à 1000 (1011 1014). 2 θεµελιώδεις µηχανισµοί ΣΔς:
Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Μηχανισµοί της ΣΔ à Βάση διεργασιών όπως η αντίληψη, µάθηση, εκούσια κίνηση. 10.000 à Μέσος νευρώνας à 1000 (1011 1014). 2 θεµελιώδεις µηχανισµοί ΣΔς:
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
Σύναψη µεταξύ της απόληξης του νευράξονα ενός νευρώνα και του δενδρίτη ενός άλλου νευρώνα.
 ΟΙ ΝΕΥΡΩΝΕΣ ΕΠΙΚΟΙΝΩΝΟΥΝ ΜΕΣΩ ΤΗΣ ΣΥΝΑΨΗΣ Άντα Μητσάκου Αναπληρώτρια Καθηγήτρια, Ιατρική Σχολή, Πανεπιστήµιο Πατρών Γνωρίζουµε ότι είµαστε ικανοί να εκτελούµε σύνθετες νοητικές διεργασίες εξαιτίας της
ΟΙ ΝΕΥΡΩΝΕΣ ΕΠΙΚΟΙΝΩΝΟΥΝ ΜΕΣΩ ΤΗΣ ΣΥΝΑΨΗΣ Άντα Μητσάκου Αναπληρώτρια Καθηγήτρια, Ιατρική Σχολή, Πανεπιστήµιο Πατρών Γνωρίζουµε ότι είµαστε ικανοί να εκτελούµε σύνθετες νοητικές διεργασίες εξαιτίας της
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ
 Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α. Ο ηλεκτρονικός υπολογιστής του οργανισμού μας
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
Βιολογία Α Λυκείου Κεφ. 9. Νευρικό Σύστημα. Δομή και λειτουργία των νευρικών κυττάρων
 Βιολογία Α Λυκείου Κεφ. 9 Νευρικό Σύστημα Δομή και λειτουργία των νευρικών κυττάρων Νευρικό Σύστημα Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων φροντίζουν να διατηρείται σταθερό το εσωτερικό
Βιολογία Α Λυκείου Κεφ. 9 Νευρικό Σύστημα Δομή και λειτουργία των νευρικών κυττάρων Νευρικό Σύστημα Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων φροντίζουν να διατηρείται σταθερό το εσωτερικό
9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΝΕΥΡΙΚΩΝ. Νευρώνες
 9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος (ομοιόσταση), ελέγχοντας και συντονίζοντας τις λειτουργίες των
9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος (ομοιόσταση), ελέγχοντας και συντονίζοντας τις λειτουργίες των
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ. ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΣΥΣΠΑΣΗΣ
 ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
Συστήματα αισθήσεων. Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
 Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας 2018 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας 2018 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I)
") ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν
Συστήματα αισθήσεων. Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
 Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Θέµατα διάλεξης ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ. Τρόποι µετάδοσης των νευρικών σηµάτων. υναµικό Ηρεµίας. Νευρώνας
 Θέµατα διάλεξης MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ Τρόποι µετάδοσης νευρικών σηµάτων Ρόλος και λειτουργία των νευροδιαβιβαστών
Θέµατα διάλεξης MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ Τρόποι µετάδοσης νευρικών σηµάτων Ρόλος και λειτουργία των νευροδιαβιβαστών
Σκοπός του μαθήματος είναι ο συνδυασμός των θεωρητικών και ποσοτικών τεχνικών με τις αντίστοιχες περιγραφικές. Κεφάλαιο 1: περιγράφονται οι βασικές
 Εισαγωγή Ασχολείται με τη μελέτη των ηλεκτρικών, η λ ε κ τ ρ ο μ α γ ν η τ ι κ ώ ν κ α ι μ α γ ν η τ ι κ ώ ν φαινομένων που εμφανίζονται στους βιολογικούς ιστούς. Το αντικείμενο του εμβιοηλεκτρομαγνητισμού
Εισαγωγή Ασχολείται με τη μελέτη των ηλεκτρικών, η λ ε κ τ ρ ο μ α γ ν η τ ι κ ώ ν κ α ι μ α γ ν η τ ι κ ώ ν φαινομένων που εμφανίζονται στους βιολογικούς ιστούς. Το αντικείμενο του εμβιοηλεκτρομαγνητισμού
χρόνιου πόνου κι των συναισθημάτων. Μάλιστα, μεγάλο μέρος αυτού
 Το μαιτεχμιακό σύστημα συνδέεται με τμήματα του μετωπιαίου κι κροταφικού λοβού ( τμήματα των εγκεφαλικών ημισφαιρίων,ονομασμένα σύμφωνα με το κρανιακό οστό που τα καλύπτει). Το ίδιο σχετίζεται με τον έλεγχο
Το μαιτεχμιακό σύστημα συνδέεται με τμήματα του μετωπιαίου κι κροταφικού λοβού ( τμήματα των εγκεφαλικών ημισφαιρίων,ονομασμένα σύμφωνα με το κρανιακό οστό που τα καλύπτει). Το ίδιο σχετίζεται με τον έλεγχο
ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ
 ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Εγκέφαλος Μεγάλη αιµάτωση, πολύ σηµαντική για την λειτουργία του Επικοινωνία µε το περιβάλλον Χρησιµοποιεί το 20% του Ο 2 και ως πηγή ενέργειας γλυκόζη Στις χειρουργικές επεµβάσεις
ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Εγκέφαλος Μεγάλη αιµάτωση, πολύ σηµαντική για την λειτουργία του Επικοινωνία µε το περιβάλλον Χρησιµοποιεί το 20% του Ο 2 και ως πηγή ενέργειας γλυκόζη Στις χειρουργικές επεµβάσεις
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ
 ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
Η βιολογία της μάθησης και της μνήμης: Μακρόχρονη ενδυνάμωση/αποδυνάμωση
 Η βιολογία της μάθησης και της μνήμης: Μακρόχρονη ενδυνάμωση/αποδυνάμωση 1 Τι λέμε μνήμη? Η ικανότητα να καταγράφουμε, να αποθηκεύουμε και να ανακαλούμε πληροφορίες Είναι ένας πολύπλοκος συνδυασμός υποσυστημάτων
Η βιολογία της μάθησης και της μνήμης: Μακρόχρονη ενδυνάμωση/αποδυνάμωση 1 Τι λέμε μνήμη? Η ικανότητα να καταγράφουμε, να αποθηκεύουμε και να ανακαλούμε πληροφορίες Είναι ένας πολύπλοκος συνδυασμός υποσυστημάτων
MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΟΡΜΟΝΕΣ ΚΑΙ ΑΣΚΗΣΗ. ΘΑΝΑΣΗΣ ΤΖΙΑΜΟΥΡΤΑΣ, Ph.D., C.S.C.
 MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΟΡΜΟΝΕΣ ΚΑΙ ΑΣΚΗΣΗ ΘΑΝΑΣΗΣ ΤΖΙΑΜΟΥΡΤΑΣ, Ph.D., C.S.C.S Αδένες Έκκρισης Ορμονών Υπόφυση Θυρεοειδής Αδένας Παραθυροειδείς
MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΟΡΜΟΝΕΣ ΚΑΙ ΑΣΚΗΣΗ ΘΑΝΑΣΗΣ ΤΖΙΑΜΟΥΡΤΑΣ, Ph.D., C.S.C.S Αδένες Έκκρισης Ορμονών Υπόφυση Θυρεοειδής Αδένας Παραθυροειδείς
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ
 ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΑΠΕΛΕΥΘΕΡΩΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ ΥΠΟΔΟΧΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ 1 Ηλεκτρικές και Χημικές Συνάψεις 2 Ηλεκτρικές
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΑΠΕΛΕΥΘΕΡΩΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ ΥΠΟΔΟΧΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ 1 Ηλεκτρικές και Χημικές Συνάψεις 2 Ηλεκτρικές
3. Να συμπληρώσετε κατάλληλα τα μέρη από τα οποία αποτελείται ένας νευρώνας.
 ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 9 ο ΚΕΦΑΛΑΙΟ «ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ» ΜΕΡΟΣ Α: ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Να συμπληρώσετε το παρακάτω διάγραμμα. 2. Ποιος είναι ο ρόλος του
ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 9 ο ΚΕΦΑΛΑΙΟ «ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ» ΜΕΡΟΣ Α: ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Να συμπληρώσετε το παρακάτω διάγραμμα. 2. Ποιος είναι ο ρόλος του
Συστήματα επικοινωνίας Ανθρωπίνου σώματος. ενδοκρινολογικό νευρικό σύστημα
 Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Γνωστική-Πειραµατική Ψυχολογία
 Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Γνωστική λειτουργία & φλοιός. Γνωστική λειτουργία & φλοιός. Γνωστικές λειτουργίες à επεξεργασία πληροφοριών από διαφορετικές περιοχές (µεγαλύτερη ( αποκλειστική)
Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Γνωστική λειτουργία & φλοιός. Γνωστική λειτουργία & φλοιός. Γνωστικές λειτουργίες à επεξεργασία πληροφοριών από διαφορετικές περιοχές (µεγαλύτερη ( αποκλειστική)
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΨΥΧΟΛΟΓΙΑ με έμφαση στις γνωστικές λειτουργίες. Θεματική Ενότητα 2: Συμβολή της φυσιολογίας στην ψυχολογία
 ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΨΥΧΟΛΟΓΙΑ με έμφαση στις γνωστικές λειτουργίες Θεματική Ενότητα 2: Συμβολή της φυσιολογίας στην ψυχολογία Θεματική Ενότητα 2: Στόχοι: Η απόκτηση ενημερότητας, εκ μέρους των φοιτητών, για
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΨΥΧΟΛΟΓΙΑ με έμφαση στις γνωστικές λειτουργίες Θεματική Ενότητα 2: Συμβολή της φυσιολογίας στην ψυχολογία Θεματική Ενότητα 2: Στόχοι: Η απόκτηση ενημερότητας, εκ μέρους των φοιτητών, για
ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ. Βιολογία A λυκείου. Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος:
 ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ Βιολογία A λυκείου Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος: 2013-2014 Ένα αισθητικό σύστημα στα σπονδυλωτά αποτελείται από τρία βασικά μέρη: 1. Τους αισθητικούς υποδοχείς,
ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ Βιολογία A λυκείου Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος: 2013-2014 Ένα αισθητικό σύστημα στα σπονδυλωτά αποτελείται από τρία βασικά μέρη: 1. Τους αισθητικούς υποδοχείς,
Μεταιχμιακό Σύστημα του Εγκεφάλου
 Μεταιχμιακό Σύστημα του Εγκεφάλου Άρθρο του ΧΑΡΑΛΑΜΠΟY ΤΙΓΓΙΝΑΓΚΑ, MT, CST, MNT Το μεταιχμιακό σύστημα ελέγχει το κύκλωμα του χρόνιου πόνου και των συναισθημάτων, ενώ συνδέεται με τα βαθύτερα τμήματα του
Μεταιχμιακό Σύστημα του Εγκεφάλου Άρθρο του ΧΑΡΑΛΑΜΠΟY ΤΙΓΓΙΝΑΓΚΑ, MT, CST, MNT Το μεταιχμιακό σύστημα ελέγχει το κύκλωμα του χρόνιου πόνου και των συναισθημάτων, ενώ συνδέεται με τα βαθύτερα τμήματα του
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ. Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.
 2310-999185 (γραμματεία) anogian@auth.") ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Σύνοψη των όσων εξετάσαμε για τους ιοντικούς διαύλους: 1. Διαπερνούν
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Σύνοψη των όσων εξετάσαμε για τους ιοντικούς διαύλους: 1. Διαπερνούν
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 Page1 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Μαθητές: Ρουμπάνης Γιάννης και Οικονομίδης Αριστείδης Τάξη: Γ γυμνασίου Κερατέας Τμήμα: Γ 4 Οκτώβριος 2013 Page2 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί
Page1 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Μαθητές: Ρουμπάνης Γιάννης και Οικονομίδης Αριστείδης Τάξη: Γ γυμνασίου Κερατέας Τμήμα: Γ 4 Οκτώβριος 2013 Page2 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΨΥΧΟΛΟΓΙΑ με έμφαση στις γνωστικές λειτουργίες. Θεματική Ενότητα 10: Μελέτη του Εγκεφάλου
 ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΨΥΧΟΛΟΓΙΑ με έμφαση στις γνωστικές λειτουργίες Θεματική Ενότητα 10: Μελέτη του Εγκεφάλου Θεματική Ενότητα 10: Στόχοι: Η εισαγωγή των φοιτητών στις μεθόδους μελέτης του εγκεφάλου. Λέξεις κλειδιά:
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΨΥΧΟΛΟΓΙΑ με έμφαση στις γνωστικές λειτουργίες Θεματική Ενότητα 10: Μελέτη του Εγκεφάλου Θεματική Ενότητα 10: Στόχοι: Η εισαγωγή των φοιτητών στις μεθόδους μελέτης του εγκεφάλου. Λέξεις κλειδιά:
ΜΑΘΗΜΑ 10ο ΜΕΡΟΣ Γ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ
 ΜΑΘΗΜΑ 10ο ΜΕΡΟΣ Γ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ Πρόσφατες εργασίες έχουν αποδείξει ότι στη μνήμη παρεμβαίνουν πολλές περιοχές του εγκεφάλου Παρ όλα αυτά, υπάρχουν διαφορετικοί τύποι μνήμης και ορισμένες
ΜΑΘΗΜΑ 10ο ΜΕΡΟΣ Γ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ Πρόσφατες εργασίες έχουν αποδείξει ότι στη μνήμη παρεμβαίνουν πολλές περιοχές του εγκεφάλου Παρ όλα αυτά, υπάρχουν διαφορετικοί τύποι μνήμης και ορισμένες
ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ
 ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ Δρ. Κυριακή Σιδηροπούλου Λέκτορας Νευροφυσιολογίας Γραφείο: Γ316δ ΤΗΛ: 28103940871 (γραφείο) E- MAIL: sidirop@imbb.forth.gr Εισαγωγή Σιδηροπούλου - Νευροβιολογία 1 Δομή μαθήματος
ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ Δρ. Κυριακή Σιδηροπούλου Λέκτορας Νευροφυσιολογίας Γραφείο: Γ316δ ΤΗΛ: 28103940871 (γραφείο) E- MAIL: sidirop@imbb.forth.gr Εισαγωγή Σιδηροπούλου - Νευροβιολογία 1 Δομή μαθήματος
ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ
 ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 5-6/29-02-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ 1. ΕΙΣΑΓΩΓΗ 2. ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 3. ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 4. ΚΕΝΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΡΥΘΜΙΣΗ 1. ΕΙΣΑΓΩΓΗ διαβίβαση στο νευρικό
ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 5-6/29-02-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ 1. ΕΙΣΑΓΩΓΗ 2. ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 3. ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 4. ΚΕΝΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΡΥΘΜΙΣΗ 1. ΕΙΣΑΓΩΓΗ διαβίβαση στο νευρικό
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ. Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας (προσωπικό) (γραμματεία)
 (γραμματεία)") ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Αρχές της ηλεκτρικής διακυτταρικής επικοινωνίας Ή πως το νευρικό
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Αρχές της ηλεκτρικής διακυτταρικής επικοινωνίας Ή πως το νευρικό
Πρώτα μηνύματα: ορμόνες, νευροδιαβιβαστές, παρακρινείς/αυτοκρινείς παράγοντες που φθάνουν στηνκμαπότονεξωκυττάριοχώροκαιδεσμεύονται με ειδικούς
 Πρώτα μηνύματα: ορμόνες, νευροδιαβιβαστές, παρακρινείς/αυτοκρινείς παράγοντες που φθάνουν στηνκμαπότονεξωκυττάριοχώροκαιδεσμεύονται με ειδικούς κυτταρικούς υποδοχείς Δεύτερα μηνύματα: μη-πρωτεϊνικές ουσίες
Πρώτα μηνύματα: ορμόνες, νευροδιαβιβαστές, παρακρινείς/αυτοκρινείς παράγοντες που φθάνουν στηνκμαπότονεξωκυττάριοχώροκαιδεσμεύονται με ειδικούς κυτταρικούς υποδοχείς Δεύτερα μηνύματα: μη-πρωτεϊνικές ουσίες
Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας. Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ
 2016 Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ Εισαγωγή Η σύσπαση των σκελετικών μυών ελέγχεται από
2016 Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ Εισαγωγή Η σύσπαση των σκελετικών μυών ελέγχεται από
Βιοδυναμικά: Ασθενή ηλεκτρικά ρεύματα τα οποία παράγονται στους ιστούς των ζωντανών οργανισμών κατά τις βιολογικές λειτουργίες.
 Bιοηλεκτρισμός To νευρικό σύστημα Το νευρικό κύτταρο Ηλεκτρικά δυναμικά στον άξονα Δυναμικά δράσης Ο άξονας ως ηλεκτρικό καλώδιο Διάδοση των δυναμικών δράσης Δυναμικά δράσεις στους μύες Δυναμικά επιφανείας
Bιοηλεκτρισμός To νευρικό σύστημα Το νευρικό κύτταρο Ηλεκτρικά δυναμικά στον άξονα Δυναμικά δράσης Ο άξονας ως ηλεκτρικό καλώδιο Διάδοση των δυναμικών δράσης Δυναμικά δράσεις στους μύες Δυναμικά επιφανείας
+ - - εκπολώνεται. ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ
 ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ Στόχοι Κατανόησης: -Να σας είναι ξεκάθαρες οι έννοιες πόλωση, εκπόλωση, υπερπόλωση, διεγερτικό ερέθισμα, ανασταλτικό ερέθισμα, κατώφλιο δυναμικό, υποκατώφλιες εκπολώσεις, υπερκατώφλιες
ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ Στόχοι Κατανόησης: -Να σας είναι ξεκάθαρες οι έννοιες πόλωση, εκπόλωση, υπερπόλωση, διεγερτικό ερέθισμα, ανασταλτικό ερέθισμα, κατώφλιο δυναμικό, υποκατώφλιες εκπολώσεις, υπερκατώφλιες
ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ
 ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Νευρικό Σύστημα 2 Ανατομία ΝΣ ΝΣ ΚΝΣ (εγκέφαλος + ΝΜ) ΠΝΣ Προσαγωγό (αισθητικό) Απαγωγό Κινητικοί νευρώνες ΑΝΣ Συμπαθητικό Παρασυμπαθητικό 3 Οι διαταραχές της λειτουργίας
ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Νευρικό Σύστημα 2 Ανατομία ΝΣ ΝΣ ΚΝΣ (εγκέφαλος + ΝΜ) ΠΝΣ Προσαγωγό (αισθητικό) Απαγωγό Κινητικοί νευρώνες ΑΝΣ Συμπαθητικό Παρασυμπαθητικό 3 Οι διαταραχές της λειτουργίας
Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών
 Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών Τίτλος Ερευνητικού Έργου «Δομικές και Λειτουργικές Μελέτες Νικοτινικών
Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών Τίτλος Ερευνητικού Έργου «Δομικές και Λειτουργικές Μελέτες Νικοτινικών
Μνήμη και μάθηση. Συμπεριφορικό επίπεδο Κυτταρικό επίπεδο. Παρασκευή, 5 Απριλίου 13
 Μνήμη και μάθηση Συμπεριφορικό επίπεδο Κυτταρικό επίπεδο Κατηγορίες μνήμης Βραχύχρονη/Μακρόχρονη μνήνη Έκδηλη μνήμη (ιππόκαμπος/φλοιός) Άδηλη μνήμη (αμυγδαλή, παρεγκεφαλίδα) Συνειρμική Μη συνειρμική Διαδικαστική
Μνήμη και μάθηση Συμπεριφορικό επίπεδο Κυτταρικό επίπεδο Κατηγορίες μνήμης Βραχύχρονη/Μακρόχρονη μνήνη Έκδηλη μνήμη (ιππόκαμπος/φλοιός) Άδηλη μνήμη (αμυγδαλή, παρεγκεφαλίδα) Συνειρμική Μη συνειρμική Διαδικαστική
ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 7-8/ Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ
 ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 7-8/02-03-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ Νευροδιαβιβαστές α) παραγωγή, είδη β) αναγνώριση γ) σηματοδότηση Σε πρώτη προσέγγιση, ο ορισμός του διαβιβαστή είναι: Διαβιβαστής είναι μια ουσία που
ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 7-8/02-03-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ Νευροδιαβιβαστές α) παραγωγή, είδη β) αναγνώριση γ) σηματοδότηση Σε πρώτη προσέγγιση, ο ορισμός του διαβιβαστή είναι: Διαβιβαστής είναι μια ουσία που
Νευρικό σύστημα - εισαγωγή. Μιχάλης Ζωγραφάκης - Σφακιανάκης Νοσηλευτής ΠΕ, M.Sc. Καθηγητής Εφαρμ. Νοσηλευτικής ΤΕΙ ΚΡΗΤΗΣ
 Νευρικό σύστημα - εισαγωγή Μιχάλης Ζωγραφάκης - Σφακιανάκης Νοσηλευτής ΠΕ, M.Sc. Καθηγητής Εφαρμ. Νοσηλευτικής ΤΕΙ ΚΡΗΤΗΣ Δενδρίτες Κυτταρικό σώμα Προσυναπτικό κύτταρο Πυρήνας Άξονας Κόμβοι Ranvier Μυελώδες
Νευρικό σύστημα - εισαγωγή Μιχάλης Ζωγραφάκης - Σφακιανάκης Νοσηλευτής ΠΕ, M.Sc. Καθηγητής Εφαρμ. Νοσηλευτικής ΤΕΙ ΚΡΗΤΗΣ Δενδρίτες Κυτταρικό σώμα Προσυναπτικό κύτταρο Πυρήνας Άξονας Κόμβοι Ranvier Μυελώδες
Συστήµατα Αισθήσεων Σωµατικές Αισθήσεις
 Συστήµατα Αισθήσεων Σωµατικές Αισθήσεις ΟΡΓΑΝΩΣΗ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Φλοιός (Ανώτερος Εγκέφαλος) Κατώτερος Εγκέφαλος Ειδικές Αισθήσεις Εν τω Βάθει Αισθητικότητα Επί πολλής Αισθητικότητα Χυµικά Ερεθίσµατα
Συστήµατα Αισθήσεων Σωµατικές Αισθήσεις ΟΡΓΑΝΩΣΗ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Φλοιός (Ανώτερος Εγκέφαλος) Κατώτερος Εγκέφαλος Ειδικές Αισθήσεις Εν τω Βάθει Αισθητικότητα Επί πολλής Αισθητικότητα Χυµικά Ερεθίσµατα
Εγκέφαλος-Αισθητήρια Όργανα και Ορμόνες. Μαγδαληνή Γκέιτς Α Τάξη Γυμνάσιο Αμυγδαλεώνα
 Εγκέφαλος-Αισθητήρια Όργανα και Ορμόνες O εγκέφαλος Ο εγκέφαλος είναι το κέντρο ελέγχου του σώματος μας και ελέγχει όλες τις ακούσιες και εκούσιες δραστηριότητες που γίνονται μέσα σε αυτό. Αποτελεί το
Εγκέφαλος-Αισθητήρια Όργανα και Ορμόνες O εγκέφαλος Ο εγκέφαλος είναι το κέντρο ελέγχου του σώματος μας και ελέγχει όλες τις ακούσιες και εκούσιες δραστηριότητες που γίνονται μέσα σε αυτό. Αποτελεί το
Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών
 Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
ΕΡΩΤΗΣΕΙΣ Επιλέξτε τη σωστή απάντηση στις παρακάτω προτάσεις: 1) Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά
 Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά") ΕΡΩΤΗΣΕΙΣ Επιλέξτε τη σωστή απάντηση στις παρακάτω προτάσεις: 1) Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά συστατικά και να απομακρύνουν τις άχρηστες ουσίες. β. Να
ΕΡΩΤΗΣΕΙΣ Επιλέξτε τη σωστή απάντηση στις παρακάτω προτάσεις: 1) Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά συστατικά και να απομακρύνουν τις άχρηστες ουσίες. β. Να
Φαρμακοκινητική. Χρυσάνθη Σαρδέλη
 Φαρμακοκινητική Χρυσάνθη Σαρδέλη Αναπληρώτρια Καθηγήτρια Κλινικής Φαρμακολογίας Εργαστήριο Κλινικής Φαρμακολογίας Τμήμα Ιατρικής, Σχολή Επιστημών Υγείας, ΑΠΘ Φαρμακοκινητική Η Φαρμακοκινητική είναι η επιστήμη
Φαρμακοκινητική Χρυσάνθη Σαρδέλη Αναπληρώτρια Καθηγήτρια Κλινικής Φαρμακολογίας Εργαστήριο Κλινικής Φαρμακολογίας Τμήμα Ιατρικής, Σχολή Επιστημών Υγείας, ΑΠΘ Φαρμακοκινητική Η Φαρμακοκινητική είναι η επιστήμη
ΒΙΟΧΗΜΕΙΑ ΤΟΥ ΜΕΤΑΒΟΛΙΣΜΟΥ
 ΒΙΟΧΗΜΕΙΑ ΤΟΥ ΜΕΤΑΒΟΛΙΣΜΟΥ ΠΑΡΟΥΣΙΑΣΗ 6 Η ΚΑΤΑΝΑΛΩΣΗ ΥΔΑΤΑΝΘΡΑΚΩΝ ΚΑΙ ΡΥΘΜΙΣΗ ΤΗΣ ΓΛΥΚΑΙΜΙΑΣ 1 Έλεγχος της ενέργειας Τα πραγματικά «Βιοκαύσιμα» 2 Υδατανθρακούχα τρόφιμα 3 Σημασία της ρύθμισης κατανάλωσης
ΒΙΟΧΗΜΕΙΑ ΤΟΥ ΜΕΤΑΒΟΛΙΣΜΟΥ ΠΑΡΟΥΣΙΑΣΗ 6 Η ΚΑΤΑΝΑΛΩΣΗ ΥΔΑΤΑΝΘΡΑΚΩΝ ΚΑΙ ΡΥΘΜΙΣΗ ΤΗΣ ΓΛΥΚΑΙΜΙΑΣ 1 Έλεγχος της ενέργειας Τα πραγματικά «Βιοκαύσιμα» 2 Υδατανθρακούχα τρόφιμα 3 Σημασία της ρύθμισης κατανάλωσης
Μάθηση και Μνήμη. Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2018
 Μάθηση και Μνήμη Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2018 Μάθηση: η διεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας
Μάθηση και Μνήμη Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2018 Μάθηση: η διεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας
Β. Να επιλέξετε την ορθή απάντηση αναγράφοντας στον πίνακα της ακόλουθης
 Ονοματεπώνυμο:.. Βαθμός: Ωριαία γραπτή εξέταση Α Τετραμήνου στη Βιολογία [Κεφ. 9 ο, σελ. 153-158] Α. Να χαρακτηρίσετε τις ακόλουθες προτάσεις με το γράμμα Ο, εφόσον είναι ορθές, ή με το γράμμα Λ, αν είναι
Ονοματεπώνυμο:.. Βαθμός: Ωριαία γραπτή εξέταση Α Τετραμήνου στη Βιολογία [Κεφ. 9 ο, σελ. 153-158] Α. Να χαρακτηρίσετε τις ακόλουθες προτάσεις με το γράμμα Ο, εφόσον είναι ορθές, ή με το γράμμα Λ, αν είναι
Συνάψεις Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2018
 Συνάψεις Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2018 Σύναψη Σύναψη είναι μια ανατομικά εξειδικευμένη σύνδεση μεταξύ δύο νευρώνων. Σύναψη Στη σύναψη η ηλεκτρική δραστηριότητα
Συνάψεις Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2018 Σύναψη Σύναψη είναι μια ανατομικά εξειδικευμένη σύνδεση μεταξύ δύο νευρώνων. Σύναψη Στη σύναψη η ηλεκτρική δραστηριότητα
Ρύθµιση κυτταρικής λειτουργίας. Μεταγωγή σήµατος
 Ρύθµιση κυτταρικής λειτουργίας Μεταγωγή σήµατος 1 Εισαγωγή Η διαδικασία εξέλιξης των πολυκύτταρων οργανισµών (πρίν 2.5 δις χρόνια) άρχισε πολύ πιο αργά από την ύπαρξη των µονοκύτταρων οργανισµών (πρίν
Ρύθµιση κυτταρικής λειτουργίας Μεταγωγή σήµατος 1 Εισαγωγή Η διαδικασία εξέλιξης των πολυκύτταρων οργανισµών (πρίν 2.5 δις χρόνια) άρχισε πολύ πιο αργά από την ύπαρξη των µονοκύτταρων οργανισµών (πρίν
Αρχές Ηλεκτροθεραπείας Φυσική του Ηλεκτρισμού Ηλεκτροφυσιολογία Γαλβανικά ρεύματα Παλμικά-εναλλασσόμενα ρεύματα Μαγνητικά πεδία Υπέρηχοι Ακτινοβολιες
 Περιεχόμενα μαθήματος Αρχές Ηλεκτροθεραπείας Φυσική του Ηλεκτρισμού Ηλεκτροφυσιολογία Γαλβανικά ρεύματα Παλμικά-εναλλασσόμενα ρεύματα Μαγνητικά πεδία Υπέρηχοι Ακτινοβολιες - Laser Θερμοθεραπεία Υδροθεραπεία
Περιεχόμενα μαθήματος Αρχές Ηλεκτροθεραπείας Φυσική του Ηλεκτρισμού Ηλεκτροφυσιολογία Γαλβανικά ρεύματα Παλμικά-εναλλασσόμενα ρεύματα Μαγνητικά πεδία Υπέρηχοι Ακτινοβολιες - Laser Θερμοθεραπεία Υδροθεραπεία
Physiology of synapses
 ΜΟΡΦΟΛΟΓΙΑ ΚΑΙ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Physiology of synapses Consoulas Christos Medical School, Physiology NKUA An understanding of synaptic transmission is necessary to understand the operations
ΜΟΡΦΟΛΟΓΙΑ ΚΑΙ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Physiology of synapses Consoulas Christos Medical School, Physiology NKUA An understanding of synaptic transmission is necessary to understand the operations
Γνωστική Ψυχολογία Ι (ΨΧ32)
") Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 3 Η φυσιολογία των γνωστικών διεργασιών Πέτρος Ρούσσος Η νευροψυχολογική βάση των γνωστικών διεργασιών Γνωστική νευροεπιστήμη: μελετάει τους τρόπους με τους οποίους
Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 3 Η φυσιολογία των γνωστικών διεργασιών Πέτρος Ρούσσος Η νευροψυχολογική βάση των γνωστικών διεργασιών Γνωστική νευροεπιστήμη: μελετάει τους τρόπους με τους οποίους
11. ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ
 11. ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Στον ανθρώπινο οργανισμό υπάρχουν δύο είδη αδένων, οι εξωκρινείς και οι ενδοκρινείς. Οι εξωκρινείς (ιδρωτοποιοί αδένες, σμηγματογόνοι αδένες κ.ά.) εκκρίνουν το προϊόν τους στην επιφάνεια
11. ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Στον ανθρώπινο οργανισμό υπάρχουν δύο είδη αδένων, οι εξωκρινείς και οι ενδοκρινείς. Οι εξωκρινείς (ιδρωτοποιοί αδένες, σμηγματογόνοι αδένες κ.ά.) εκκρίνουν το προϊόν τους στην επιφάνεια
Αλκοόλ, Εθεβεία & Εγκέθαλορ. Γιώργος Παναγής Πανεπιστήμιο Κρήτης Τμήμα Ψυχολογίας Εργαστήριο Νευροεπιστημών & Συμπεριφοράς
 Αλκοόλ, Εθεβεία & Εγκέθαλορ Γιώργος Παναγής Πανεπιστήμιο Κρήτης Τμήμα Ψυχολογίας Εργαστήριο Νευροεπιστημών & Συμπεριφοράς Κατανάλωση οινοπνευματωδών στους Έλληνες μαθητές (2011) Στην Ελλάδα, τα αγόρια
Αλκοόλ, Εθεβεία & Εγκέθαλορ Γιώργος Παναγής Πανεπιστήμιο Κρήτης Τμήμα Ψυχολογίας Εργαστήριο Νευροεπιστημών & Συμπεριφοράς Κατανάλωση οινοπνευματωδών στους Έλληνες μαθητές (2011) Στην Ελλάδα, τα αγόρια
Συναπτική ολοκλήρωση. Η διαδικασία της άθροισης όλων των εισερχόμενων διεγερτικών και ανασταλτικών σημάτων σε ένα νευρώνα. Τετάρτη, 20 Μαρτίου 13
 Συναπτική ολοκλήρωση Η διαδικασία της άθροισης όλων των εισερχόμενων διεγερτικών και ανασταλτικών σημάτων σε ένα νευρώνα http://www.mpg.de/13795/learning_memory_perception?print=yes 2 Τοποθεσία συνάψεων
Συναπτική ολοκλήρωση Η διαδικασία της άθροισης όλων των εισερχόμενων διεγερτικών και ανασταλτικών σημάτων σε ένα νευρώνα http://www.mpg.de/13795/learning_memory_perception?print=yes 2 Τοποθεσία συνάψεων
Φαρμακολογία Τμήμα Ιατρικής Α.Π.Θ.
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΧΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Α.Π.Θ. Ενότητα 3: Φαρμακοδυναμική Μαρία Μυρωνίδου-Τζουβελέκη Α.Π.Θ. Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΧΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Α.Π.Θ. Ενότητα 3: Φαρμακοδυναμική Μαρία Μυρωνίδου-Τζουβελέκη Α.Π.Θ. Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης
ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΓΙΑ ΤΙΣ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ HELLENIC SOCIETY FOR NEUROSCIENCE
 Τ.Θ. 5, Πανε ιστήµιο Πατρών, 650 Πάτρα, P.O. Box 5, University of Patras, 6 50 Patra ΠΑΓΚΟΣΜΙΑ ΕΒ ΟΜΑ Α ΕΝΗΜΕΡΩΣΗΣ ΓΙΑ ΤΟΝ ΕΓΚΕΦΑΛΟ ΕΓΚΕΦΑΛΟΣ ΚΑΙ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ: ΤΙ ΓΝΩΡΙΖΟΥΝ ΟΙ ΤΕΛΕΙΟΦΟΙΤΟΙ ΤΗΣ ΥΠΟΧΡΕΩΤΙΚΗΣ
Τ.Θ. 5, Πανε ιστήµιο Πατρών, 650 Πάτρα, P.O. Box 5, University of Patras, 6 50 Patra ΠΑΓΚΟΣΜΙΑ ΕΒ ΟΜΑ Α ΕΝΗΜΕΡΩΣΗΣ ΓΙΑ ΤΟΝ ΕΓΚΕΦΑΛΟ ΕΓΚΕΦΑΛΟΣ ΚΑΙ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ: ΤΙ ΓΝΩΡΙΖΟΥΝ ΟΙ ΤΕΛΕΙΟΦΟΙΤΟΙ ΤΗΣ ΥΠΟΧΡΕΩΤΙΚΗΣ
ΠΡΩΤΟΚΟΛΛΟ ΕΡΕΥΝΑΣ ΜΥΟΚΑΡ ΙΑΚΗ ΑΝΑΓΕΝΝΗΣΗ ΜΕΤΑ ΤΟ ΕΜΦΡΑΓΜΑ ΕΡΕΥΝΗΤΙΚΗ ΟΜΑ Α
 ΠΡΩΤΟΚΟΛΛΟ ΕΡΕΥΝΑΣ ΜΥΟΚΑΡ ΙΑΚΗ ΑΝΑΓΕΝΝΗΣΗ ΜΕΤΑ ΤΟ ΕΜΦΡΑΓΜΑ ΕΡΕΥΝΗΤΙΚΗ ΟΜΑ Α ΘΕΟΦΙΛΟΣ ΚΩΛΕΤΤΗΣ ΑΠΟΣΤΟΛΟΣ ΠΑΠΑΛΟΗΣ ΓΙΑΝΝΗΣ ΜΠΑΛΤΟΓΙΑΝΝΗΣ ΗΜΗΤΡΙΟΣ ΟΙΚΟΝΟΜΙ ΗΣ ΜΑΡΙΑ ΑΓΓΕΛΑΚΗ ΑΝΤΩΝΙΟΣ ΒΛΑΧΟΣ ΑΓΑΘΟΚΛΕΙΑ ΜΗΤΣΗ
ΠΡΩΤΟΚΟΛΛΟ ΕΡΕΥΝΑΣ ΜΥΟΚΑΡ ΙΑΚΗ ΑΝΑΓΕΝΝΗΣΗ ΜΕΤΑ ΤΟ ΕΜΦΡΑΓΜΑ ΕΡΕΥΝΗΤΙΚΗ ΟΜΑ Α ΘΕΟΦΙΛΟΣ ΚΩΛΕΤΤΗΣ ΑΠΟΣΤΟΛΟΣ ΠΑΠΑΛΟΗΣ ΓΙΑΝΝΗΣ ΜΠΑΛΤΟΓΙΑΝΝΗΣ ΗΜΗΤΡΙΟΣ ΟΙΚΟΝΟΜΙ ΗΣ ΜΑΡΙΑ ΑΓΓΕΛΑΚΗ ΑΝΤΩΝΙΟΣ ΒΛΑΧΟΣ ΑΓΑΘΟΚΛΕΙΑ ΜΗΤΣΗ
Τι θα προτιμούσατε; Γνωστική Ψυχολογία Ι (ΨΧ32) 25/4/2012. Διάλεξη 5 Όραση και οπτική αντίληψη. Πέτρος Ρούσσος. Να περιγράψετε τι βλέπετε στην εικόνα;
 25/4/2012. Διάλεξη 5 Όραση και οπτική αντίληψη. Πέτρος Ρούσσος. Να περιγράψετε τι βλέπετε στην εικόνα;") Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 5 Όραση και οπτική αντίληψη Πέτρος Ρούσσος Να περιγράψετε τι βλέπετε στην εικόνα; Τι θα προτιμούσατε; Ή να αντιμετωπίσετε τον Γκάρι Κασπάροβ σε μια παρτίδα σκάκι; 1
Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 5 Όραση και οπτική αντίληψη Πέτρος Ρούσσος Να περιγράψετε τι βλέπετε στην εικόνα; Τι θα προτιμούσατε; Ή να αντιμετωπίσετε τον Γκάρι Κασπάροβ σε μια παρτίδα σκάκι; 1
«ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ» Λειτουργία Νευρικού Συστήματος
 ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ «ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ» Λειτουργία Νευρικού Συστήματος Κώστας Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2013 Ομοιόσταση Ορισμός: Το σύνολο των φυσιολογικών
ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ «ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ» Λειτουργία Νευρικού Συστήματος Κώστας Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2013 Ομοιόσταση Ορισμός: Το σύνολο των φυσιολογικών
Η νόσος του Parkinson δεν είναι µόνο κινητική διαταραχή. Έχει υπολογισθεί ότι µέχρι και 50% των ασθενών µε νόσο Πάρκινσον, µπορεί να βιώσουν κάποια
 ρ ZΩΗ ΚΑΤΣΑΡΟΥ Νευρολόγος ιευθύντρια ΕΣΥ Η νόσος του Parkinson δεν είναι µόνο κινητική διαταραχή. Έχει υπολογισθεί ότι µέχρι και 50% των ασθενών µε νόσο Πάρκινσον, µπορεί να βιώσουν κάποια µορφή κατάθλιψης,
ρ ZΩΗ ΚΑΤΣΑΡΟΥ Νευρολόγος ιευθύντρια ΕΣΥ Η νόσος του Parkinson δεν είναι µόνο κινητική διαταραχή. Έχει υπολογισθεί ότι µέχρι και 50% των ασθενών µε νόσο Πάρκινσον, µπορεί να βιώσουν κάποια µορφή κατάθλιψης,
ΥΠΟΔΟΧΕΙΣ ΣΩΜΑΤΙΚΕΣ ΑΙΣΘΗΣΕΙΣ
 ΥΠΟΔΟΧΕΙΣ ΣΩΜΑΤΙΚΕΣ ΑΙΣΘΗΣΕΙΣ ΑΙΣΘΗΤΙΚΟΙ ΥΠΟΔΟΧΕΙΣ (συγκεντρωμένοι ή διάσπαρτοι) ΝΕΥΡΙΚΕΣ ΟΔΟΙ ΕΓΚΕΦΑΛΙΚΟΣ ΦΛΟΙΟΣ Ειδικά κύτταρα - υποδοχείς, ευαίσθητα στις αλλαγές αυτές, είναι τα κύρια μέσα συλλογής
ΥΠΟΔΟΧΕΙΣ ΣΩΜΑΤΙΚΕΣ ΑΙΣΘΗΣΕΙΣ ΑΙΣΘΗΤΙΚΟΙ ΥΠΟΔΟΧΕΙΣ (συγκεντρωμένοι ή διάσπαρτοι) ΝΕΥΡΙΚΕΣ ΟΔΟΙ ΕΓΚΕΦΑΛΙΚΟΣ ΦΛΟΙΟΣ Ειδικά κύτταρα - υποδοχείς, ευαίσθητα στις αλλαγές αυτές, είναι τα κύρια μέσα συλλογής
M.Sc. Bioinformatics and Neuroinformatics
 M.Sc. Bioinformatics and Neuroinformatics Recording and Processing Brain Signals Μαρία Σαγιαδινού Ο ανθρώπινος εγκέφαλος Πιο πολύπλοκο δημιούργημα της φύσης Προιόν βιολογικής εξέλιξης εκατομμυρίων ετών
M.Sc. Bioinformatics and Neuroinformatics Recording and Processing Brain Signals Μαρία Σαγιαδινού Ο ανθρώπινος εγκέφαλος Πιο πολύπλοκο δημιούργημα της φύσης Προιόν βιολογικής εξέλιξης εκατομμυρίων ετών
ΦΑΡΜΑΚΟ ΥΝΑΜΙΚΗ ΙΙ. Γενικές έννοιες (Θεωρία υποδοχέων - Αγωνιστής ανταγωνιστής) Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων
 Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων") ΦΑΡΜΑΚΟ ΥΝΑΜΙΚΗ ΙΙ Γενικές έννοιες (Θεωρία υποδοχέων - Αγωνιστής ανταγωνιστής) Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων Υποδοχείς (φαρµάκων) και ενδοκυττάριες σηµατοδοτικές πορείες - Συστήµατα Μεταγωγής
ΦΑΡΜΑΚΟ ΥΝΑΜΙΚΗ ΙΙ Γενικές έννοιες (Θεωρία υποδοχέων - Αγωνιστής ανταγωνιστής) Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων Υποδοχείς (φαρµάκων) και ενδοκυττάριες σηµατοδοτικές πορείες - Συστήµατα Μεταγωγής
ηδιεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς
 Μάθηση και Μνήµη Μάθηση Μνήµη Μάθηση: Μνήµη: ηδιεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς ηαποθήκευσητωνπληροφοριώναυτών η ικανότητα να
Μάθηση και Μνήµη Μάθηση Μνήµη Μάθηση: Μνήµη: ηδιεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς ηαποθήκευσητωνπληροφοριώναυτών η ικανότητα να
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ. Κάντε κλικ για να επεξεργαστείτε τον υπότιτλο του υποδείγματος
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Κάντε κλικ για να επεξεργαστείτε τον υπότιτλο του υποδείγματος ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα θέτει σε επικοινωνία τον οργανισμό μας με τον έξω κόσμο. Μοιάζει με τηλεφωνικό δίκτυο,
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Κάντε κλικ για να επεξεργαστείτε τον υπότιτλο του υποδείγματος ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα θέτει σε επικοινωνία τον οργανισμό μας με τον έξω κόσμο. Μοιάζει με τηλεφωνικό δίκτυο,
Συνιστώνται για... Οι δονήσεις είναι αποτελεσματικές...
 ΠΕΔΙΑ ΕΦΑΡΜΟΓΗΣ Εκφυλιστικές αλλοιώσεις Αγγειακές παθήσεις Παθολογίες των πνευμόνων Ουρο-γυναικολογικές διαταραχές Καρδιακές παθήσεις Παθολογίες σπονδυλικής στήλης Παθολογίες αρθρώσεων Παθολογίες συνδέσμων
ΠΕΔΙΑ ΕΦΑΡΜΟΓΗΣ Εκφυλιστικές αλλοιώσεις Αγγειακές παθήσεις Παθολογίες των πνευμόνων Ουρο-γυναικολογικές διαταραχές Καρδιακές παθήσεις Παθολογίες σπονδυλικής στήλης Παθολογίες αρθρώσεων Παθολογίες συνδέσμων
ΜΕΤΑΒΟΛΙΣΜΟΣ ΝΗΣΤΙΚΟΥ ΚΑΙ ΤΡΑΦΕΝΤΟΣ ΟΡΓΑΝΙΣΜΟΥ Tον ανθρώπινο µεταβολισµό το χαρακτηρίζουν δύο στάδια. Tοπρώτοείναιηκατάστασητουοργανισµούµετά
 ΜΕΤΑΒΟΛΙΣΜΟΣ ΝΗΣΤΙΚΟΥ ΚΑΙ ΤΡΑΦΕΝΤΟΣ ΟΡΓΑΝΙΣΜΟΥ Tον ανθρώπινο µεταβολισµό το χαρακτηρίζουν δύο στάδια. Tοπρώτοείναιηκατάστασητουοργανισµούµετά απόκάποιογεύµα, οπότετοαίµαείναιπλούσιοσε θρεπτικές ύλες από
ΜΕΤΑΒΟΛΙΣΜΟΣ ΝΗΣΤΙΚΟΥ ΚΑΙ ΤΡΑΦΕΝΤΟΣ ΟΡΓΑΝΙΣΜΟΥ Tον ανθρώπινο µεταβολισµό το χαρακτηρίζουν δύο στάδια. Tοπρώτοείναιηκατάστασητουοργανισµούµετά απόκάποιογεύµα, οπότετοαίµαείναιπλούσιοσε θρεπτικές ύλες από
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ ΕΡΩΤΗΣΕΙΣ ΤΗΣ ΤΡΑΠΕΖΑΣ ΘΕΜΑΤΩΝ ΣΤΟ 11 Ο ΚΕΦΑΛΑΙΟ ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ ΘΕΜΑ Β
 ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ ΕΡΩΤΗΣΕΙΣ ΤΗΣ ΤΡΑΠΕΖΑΣ ΘΕΜΑΤΩΝ ΣΤΟ 11 Ο ΚΕΦΑΛΑΙΟ ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ ΘΕΜΑ Β 1. Το σύστημα των ενδοκρινών αδένων είναι το ένα από τα δύο συστήματα του οργανισμού μας που συντονίζουν και
ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ ΕΡΩΤΗΣΕΙΣ ΤΗΣ ΤΡΑΠΕΖΑΣ ΘΕΜΑΤΩΝ ΣΤΟ 11 Ο ΚΕΦΑΛΑΙΟ ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ ΘΕΜΑ Β 1. Το σύστημα των ενδοκρινών αδένων είναι το ένα από τα δύο συστήματα του οργανισμού μας που συντονίζουν και
5o Μάθηµα. Αισθητικά Συστήµατα
 5o Μάθηµα Αισθητικά Συστήµατα Γραφή Μπράιγ Σωµαταισθητικό Σύστηµα Αισθήσεις:Αφής,Ιδιοδεκτικότητας Πόνου,Θερµοκρασίας Οργάνωση:Ιεραρχική-Παράλληλη Τρείς Νευρώνες:Πρωτοταγής ευτεροταγής Τριτοταγής Ραχιαίο
5o Μάθηµα Αισθητικά Συστήµατα Γραφή Μπράιγ Σωµαταισθητικό Σύστηµα Αισθήσεις:Αφής,Ιδιοδεκτικότητας Πόνου,Θερµοκρασίας Οργάνωση:Ιεραρχική-Παράλληλη Τρείς Νευρώνες:Πρωτοταγής ευτεροταγής Τριτοταγής Ραχιαίο
Κεφάλαιο 3 ΜΕΤΑΒΟΛΙΣΜΟΣ
 Κεφάλαιο 3 ΜΕΤΑΒΟΛΙΣΜΟΣ 3.1 Ενέργεια και οργανισμοί Όλοι οι οργανισμοί, εκτός από αυτούς από αυτούς που έχουν την ικανότητα να φωτοσυνθέτουν, εξασφαλίζουν ενέργεια διασπώντας τις θρεπτικές ουσιές που περιέχονται
Κεφάλαιο 3 ΜΕΤΑΒΟΛΙΣΜΟΣ 3.1 Ενέργεια και οργανισμοί Όλοι οι οργανισμοί, εκτός από αυτούς από αυτούς που έχουν την ικανότητα να φωτοσυνθέτουν, εξασφαλίζουν ενέργεια διασπώντας τις θρεπτικές ουσιές που περιέχονται
Βιολογία. Θετικής κατεύθυνσης. Β λυκείου. ΑΡΓΥΡΗΣ ΓΙΑΝΝΗΣ Βιολόγος 3 ο λύκ. ηλιούπολης
 Βιολογία Β λυκείου Θετικής κατεύθυνσης ΑΡΓΥΡΗΣ ΓΙΑΝΝΗΣ Βιολόγος 3 ο λύκ. ηλιούπολης 1. Εισαγωγή Το κύτταρο αποτελεί τη βασική δομική και λειτουργική μονάδα των οργανισμών. 1.1 Το κύτταρο. 3ο λύκ. ηλιούπολης
Βιολογία Β λυκείου Θετικής κατεύθυνσης ΑΡΓΥΡΗΣ ΓΙΑΝΝΗΣ Βιολόγος 3 ο λύκ. ηλιούπολης 1. Εισαγωγή Το κύτταρο αποτελεί τη βασική δομική και λειτουργική μονάδα των οργανισμών. 1.1 Το κύτταρο. 3ο λύκ. ηλιούπολης
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ. Δομή και λειτουργία των νευρικών κυττάρων. Ηλιάνα Καρβουντζή Βιολόγος
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Δομή και λειτουργία των νευρικών κυττάρων Ρόλος του νευρικού συστήματος Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Δομή και λειτουργία των νευρικών κυττάρων Ρόλος του νευρικού συστήματος Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος
Εισαγωγή στην Ψυχολογία με έμφαση στις γνωστικές λειτουργίες
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Εισαγωγή στην Ψυχολογία με έμφαση στις γνωστικές λειτουργίες Συμβολή της φυσιολογίας στην ψυχολογία Διδάσκουσα: Επίκ. Καθ. Γεωργία Α. Παπαντωνίου Άδειες
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Εισαγωγή στην Ψυχολογία με έμφαση στις γνωστικές λειτουργίες Συμβολή της φυσιολογίας στην ψυχολογία Διδάσκουσα: Επίκ. Καθ. Γεωργία Α. Παπαντωνίου Άδειες
Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός. Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας
 Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Θάλαμος Ο θάλαμος: «πύλη προς τον εγκέφαλο» Είναι μια μεγάλη συλλογή νευρώνων στο διεγκέφαλο Παίρνει
Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Θάλαμος Ο θάλαμος: «πύλη προς τον εγκέφαλο» Είναι μια μεγάλη συλλογή νευρώνων στο διεγκέφαλο Παίρνει
Φυσιολογία ΙΙ Ενότητα 1:
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 1: Το Νευρικό Σύστημα Ανωγειανάκις Γεώργιος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 1: Το Νευρικό Σύστημα Ανωγειανάκις Γεώργιος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
ΦΥΣΙΟΛΟΓΙΑ ΑΝΑΠΝΕΥΣΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΦΥΣΙΟΛΟΓΙΑ ΑΝΑΠΝΕΥΣΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΙΩΑΝΝΑ ΔΗΜΟΠΟΥΛΟΥ ΚΑΘΗΓΗΤΡΙΑ ΕΡΓΑΣΤΗΡΙΟ ΠΕΙΡΑΜΑΤΙΚΗΣ ΦΥΣΙΟΛΟΓΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΚΠΑ ΤΟ ΑΝΑΠΝΕΥΣΤΙΚΟ ΣΕ ΚΑΤΑΣΤΑΣΕΙΣ STRESS ΦΥΣΙΟΛΟΓΙΑ ΜΕΓΑΛΟΥ ΥΨΟΜΕΤΡΟΥ Η ατμοσφαιρική
ΦΥΣΙΟΛΟΓΙΑ ΑΝΑΠΝΕΥΣΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΙΩΑΝΝΑ ΔΗΜΟΠΟΥΛΟΥ ΚΑΘΗΓΗΤΡΙΑ ΕΡΓΑΣΤΗΡΙΟ ΠΕΙΡΑΜΑΤΙΚΗΣ ΦΥΣΙΟΛΟΓΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΚΠΑ ΤΟ ΑΝΑΠΝΕΥΣΤΙΚΟ ΣΕ ΚΑΤΑΣΤΑΣΕΙΣ STRESS ΦΥΣΙΟΛΟΓΙΑ ΜΕΓΑΛΟΥ ΥΨΟΜΕΤΡΟΥ Η ατμοσφαιρική
2) την αριθ. 2925/ απόφαση της Συνεδρίασης της Συγκλήτου
 την αριθ. 2925/ απόφαση της Συνεδρίασης της Συγκλήτου") ΠΡΟΣΚΛΗΣΗ ΕΚ ΗΛΩΣΗΣ ΕΝ ΙΑΦΕΡΟΝΤΟΣ ΓΙΑ ΤΗΝ ΠΡΟΚΗΡΥΞΗ ΤΕΣΣΑΡΩΝ (4) ΚΕΝΩΝ ΘΕΣΕΩΝ ΓΙΑ ΤΗΝ ΠΡΟΣΛΗΨΗ ΚΑΘΗΓΗΤΩΝ ΣΤΗ ΒΑΘΜΙ Α ΤΟΥ ΑΝΑΠΛΗΡΩΤΗ ή ΕΠΙΚΟΥΡΟΥ ΚΑΘΗΓΗΤΗ ΣΤΟ ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ ΤΗΣ ΣΧΟΛΗΣ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
ΠΡΟΣΚΛΗΣΗ ΕΚ ΗΛΩΣΗΣ ΕΝ ΙΑΦΕΡΟΝΤΟΣ ΓΙΑ ΤΗΝ ΠΡΟΚΗΡΥΞΗ ΤΕΣΣΑΡΩΝ (4) ΚΕΝΩΝ ΘΕΣΕΩΝ ΓΙΑ ΤΗΝ ΠΡΟΣΛΗΨΗ ΚΑΘΗΓΗΤΩΝ ΣΤΗ ΒΑΘΜΙ Α ΤΟΥ ΑΝΑΠΛΗΡΩΤΗ ή ΕΠΙΚΟΥΡΟΥ ΚΑΘΗΓΗΤΗ ΣΤΟ ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ ΤΗΣ ΣΧΟΛΗΣ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
ΚΕΦΑΛΑΙΟ 11 ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ
 ΚΕΦΑΛΑΙΟ 11 ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ 1o ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11207, 96ο ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11303 Ι. Το σύστημα των ενδοκρινών αδένων είναι το ένα από τα δύο συστήματα του οργανισμού μας που συντονίζουν
ΚΕΦΑΛΑΙΟ 11 ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ 1o ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11207, 96ο ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11303 Ι. Το σύστημα των ενδοκρινών αδένων είναι το ένα από τα δύο συστήματα του οργανισμού μας που συντονίζουν
ΕΚ ΗΛΩΣΗ ΕΝΗΜΕΡΩΣΗΣ ΚΑΙ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗΣ ΤΟΥ ΚΟΙΝΟΥ ΣΤΙΣ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ. Πόλη Ηµεροµηνία Ώρα Αίθουσα ΠΡΟΓΡΑΜΜΑ
 ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΓΙΑ ΤΙΣ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ ΕΚ ΗΛΩΣΗ ΕΝΗΜΕΡΩΣΗΣ ΚΑΙ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗΣ ΤΟΥ ΚΟΙΝΟΥ ΣΤΙΣ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ Πόλη Ηµεροµηνία Ώρα Αίθουσα Σαλαµίνα 17 Μαρτίου 2012 09:30 πµ Κινηµατοθέατρο Ναυστάθµου
ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΓΙΑ ΤΙΣ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ ΕΚ ΗΛΩΣΗ ΕΝΗΜΕΡΩΣΗΣ ΚΑΙ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗΣ ΤΟΥ ΚΟΙΝΟΥ ΣΤΙΣ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ Πόλη Ηµεροµηνία Ώρα Αίθουσα Σαλαµίνα 17 Μαρτίου 2012 09:30 πµ Κινηµατοθέατρο Ναυστάθµου
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Μνήμη και Μάθηση Νόσος του Alzheimer
 ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Μνήμη και Μάθηση Νόσος του Alzheimer Περιγραφή της περίπτωσης (Ι) Γυναίκα 43 ετών διακομίσθηκε στο τμήμα επειγόντων περιστατικών από την οικογένειά της λόγω διαταραχών μνήμης. Η ασθενής
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Μνήμη και Μάθηση Νόσος του Alzheimer Περιγραφή της περίπτωσης (Ι) Γυναίκα 43 ετών διακομίσθηκε στο τμήμα επειγόντων περιστατικών από την οικογένειά της λόγω διαταραχών μνήμης. Η ασθενής
Διαταραχές Μνήμης. Πιλοτική μετάφραση και απόδοση του εγχειριδίου: Treating Memory Impairments
 Διαταραχές Μνήμης. Πιλοτική μετάφραση και απόδοση του εγχειριδίου: Treating Memory Impairments ΜΑΝΤΖΑΚΙΔΟΥ ΣΟΦΙΑ Α.Μ. 11523 ΕΠΙΒΛΕΠΟΥΣΑ ΚΑΘΗΓΗΤΡΙΑ: ΖΑΚΟΠΟΥΛΟΥ ΒΙΚΤΩΡΙΑ ΕΙΣΑΓΩΓΗ Η δυνατότητα που έχουμε
Διαταραχές Μνήμης. Πιλοτική μετάφραση και απόδοση του εγχειριδίου: Treating Memory Impairments ΜΑΝΤΖΑΚΙΔΟΥ ΣΟΦΙΑ Α.Μ. 11523 ΕΠΙΒΛΕΠΟΥΣΑ ΚΑΘΗΓΗΤΡΙΑ: ΖΑΚΟΠΟΥΛΟΥ ΒΙΚΤΩΡΙΑ ΕΙΣΑΓΩΓΗ Η δυνατότητα που έχουμε
Ν Ε Υ Ρ Ο Ψ Υ Χ Ο Λ Ο Γ Ι Κ Ο Ι Π Α Ρ Α Γ Ο Ν Τ Ε Σ Π Ο Υ Ε Π Η Ρ Ε Α Ζ Ο Υ Ν Τ Η Μ Α Θ Η Σ Η
 Ν Ε Υ Ρ Ο Ψ Υ Χ Ο Λ Ο Γ Ι Κ Ο Ι Π Α Ρ Α Γ Ο Ν Τ Ε Σ Π Ο Υ Ε Π Η Ρ Ε Α Ζ Ο Υ Ν Τ Η Μ Α Θ Η Σ Η 2 ΤΙ ΕΙΝΑΙ ΣΥΝΑΙΣΘΗΜΑ; Μηχανισμός επικοινωνίας που διατηρούν την κοινωνική τάξη / δομή Μαθημένη συμπεριφορά
Ν Ε Υ Ρ Ο Ψ Υ Χ Ο Λ Ο Γ Ι Κ Ο Ι Π Α Ρ Α Γ Ο Ν Τ Ε Σ Π Ο Υ Ε Π Η Ρ Ε Α Ζ Ο Υ Ν Τ Η Μ Α Θ Η Σ Η 2 ΤΙ ΕΙΝΑΙ ΣΥΝΑΙΣΘΗΜΑ; Μηχανισμός επικοινωνίας που διατηρούν την κοινωνική τάξη / δομή Μαθημένη συμπεριφορά
Η ΣΥΜΒΟΛΗ ΤΗΣ ΝΕΥΡΟΨΥΧΟΛΟΓΙΑΣ ΣΤΗΝ ΨΥΧΙΑΤΡΙΚΗ. Ευανθία Σούμπαση. Απαρτιωμένη Διδασκαλία
 Η ΣΥΜΒΟΛΗ ΤΗΣ ΝΕΥΡΟΨΥΧΟΛΟΓΙΑΣ ΣΤΗΝ ΨΥΧΙΑΤΡΙΚΗ Ευανθία Σούμπαση Απαρτιωμένη Διδασκαλία ΠΕΔΙΟ ΝΕΥΡΟΨΥΧΟΛΟΓΙΑΣ ΟΡΙΣΜΟΣ Η επιστήμη που ασχολείται με τον προσδιορισμό της λειτουργικής κατάστασης του εγκεφάλου
Η ΣΥΜΒΟΛΗ ΤΗΣ ΝΕΥΡΟΨΥΧΟΛΟΓΙΑΣ ΣΤΗΝ ΨΥΧΙΑΤΡΙΚΗ Ευανθία Σούμπαση Απαρτιωμένη Διδασκαλία ΠΕΔΙΟ ΝΕΥΡΟΨΥΧΟΛΟΓΙΑΣ ΟΡΙΣΜΟΣ Η επιστήμη που ασχολείται με τον προσδιορισμό της λειτουργικής κατάστασης του εγκεφάλου
Πως ο Νους Χειρίζεται το Φόβο
 Πως ο Νους Χειρίζεται το Φόβο Σύμφωνα με δύο σχετικά πρόσφατες έρευνες, οι μνήμες φόβου και τρόμου διαφέρουν σημαντικά από τις συνηθισμένες μνήμες. Οι διαφορές αυτές δεν συνίστανται μόνο στις εμφανείς
Πως ο Νους Χειρίζεται το Φόβο Σύμφωνα με δύο σχετικά πρόσφατες έρευνες, οι μνήμες φόβου και τρόμου διαφέρουν σημαντικά από τις συνηθισμένες μνήμες. Οι διαφορές αυτές δεν συνίστανται μόνο στις εμφανείς
3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ
 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Όλοι οι οργανισμοί προκειμένου να επιβιώσουν και να επιτελέσουν τις λειτουργίες τους χρειάζονται ενέργεια. Οι φυτικοί οργανισμοί μετατρέπουν την ηλιακή ενέργεια με τη διαδικασία
3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Όλοι οι οργανισμοί προκειμένου να επιβιώσουν και να επιτελέσουν τις λειτουργίες τους χρειάζονται ενέργεια. Οι φυτικοί οργανισμοί μετατρέπουν την ηλιακή ενέργεια με τη διαδικασία
Μεταβολικές ανάγκες ανοσοκυττάρων
 Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
ΑΠΕΙΚΟΝΙΣΗΣ ΤΩΝ ΟΓΚΩΝ
 2. ΜΕΤΑΒΟΛΙΚΟ ΜΟΝΤΕΛΟ ΑΠΕΙΚΟΝΙΣΗΣ ΤΩΝ ΟΓΚΩΝ Οι όγκοι χαρακτηρίζονται από πολλαπλές αλλαγές του μεταβολισμού. Η χαρακτηριστική μεταβολική λειτουργία μπορεί να μετρηθεί in vivo με τη βοήθεια ενός ραδιοσημασμένου
2. ΜΕΤΑΒΟΛΙΚΟ ΜΟΝΤΕΛΟ ΑΠΕΙΚΟΝΙΣΗΣ ΤΩΝ ΟΓΚΩΝ Οι όγκοι χαρακτηρίζονται από πολλαπλές αλλαγές του μεταβολισμού. Η χαρακτηριστική μεταβολική λειτουργία μπορεί να μετρηθεί in vivo με τη βοήθεια ενός ραδιοσημασμένου
Βετεράνοι αθλητές. Απόδοση & Ηλικία. Βασικά στοιχεία. Αθλητισμός Επιδόσεων στη 2η και 3η Ηλικία. Γενικευμένη θεωρία για τη
 Αθλητισμός Επιδόσεων στη 2η και 3η Ηλικία. Γενικευμένη θεωρία για τη Διατήρηση η της αθλητικής απόδοσης 710: 8 η Διάλεξη Μιχαλοπούλου Μαρία Ph.D. Περιεχόμενο της διάλεξης αυτής αποτελούν: Αγωνιστικός αθλητισμός
Αθλητισμός Επιδόσεων στη 2η και 3η Ηλικία. Γενικευμένη θεωρία για τη Διατήρηση η της αθλητικής απόδοσης 710: 8 η Διάλεξη Μιχαλοπούλου Μαρία Ph.D. Περιεχόμενο της διάλεξης αυτής αποτελούν: Αγωνιστικός αθλητισμός
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ
 ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Περιγραφή της περίπτωσης Άνδρας 43 ετών εισάγεται σε κλινική λόγω επιγαστραλγίας. Μετά από έλεγχο ετέθη η διάγνωση του πεπτικού
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Περιγραφή της περίπτωσης Άνδρας 43 ετών εισάγεται σε κλινική λόγω επιγαστραλγίας. Μετά από έλεγχο ετέθη η διάγνωση του πεπτικού
Μεταπτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΜΕΛΕΤΗ ΤΗΣ ΕΚΛΕΚΤΙΚΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΟΞΕΙΔΩΣΗΣ ΤΟΥ ΜΕΘΑΝΙΟΥ ΠΡΟΣ ΔΙΟΞΕΙΔΙΟ ΤΟΥ ΑΝΘΡΑΚΑ Βασιλική
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΜΕΛΕΤΗ ΤΗΣ ΕΚΛΕΚΤΙΚΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΟΞΕΙΔΩΣΗΣ ΤΟΥ ΜΕΘΑΝΙΟΥ ΠΡΟΣ ΔΙΟΞΕΙΔΙΟ ΤΟΥ ΑΝΘΡΑΚΑ Βασιλική
Μάθηση και Μνήµη 12/4/16. Ε. Παρασκευά, Εργ. Φυσιολογίας, Τμήμα Ιατρικής Π.Θ.
 Μάθηση και Μνήµη 1 Μάθηση: η διεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς Μνήµη: η αποθήκευση των πληροφοριών αυτών η ικανότητα να θυµόµαστε
Μάθηση και Μνήµη 1 Μάθηση: η διεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς Μνήµη: η αποθήκευση των πληροφοριών αυτών η ικανότητα να θυµόµαστε
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων. Μεταιχµιακό Σύστηµα
 Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Μεταιχµιακό Σύστηµα Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά Γάγγλια µεταιχµιακό (στεφανιαίο) σύστηµα διάµεσος εγκέφαλος
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Μεταιχµιακό Σύστηµα Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά Γάγγλια µεταιχµιακό (στεφανιαίο) σύστηµα διάµεσος εγκέφαλος
ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ. Οι ρυθμιστές του οργανισμού
 ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Οι ρυθμιστές του οργανισμού Είδη αδένων στον άνθρωπο o Εξωκρινείς αδένες: εκκρίνουν το προϊόν τους μέσω εκφορητικού πόρου είτε στην επιφάνεια του σώματος (π.χ. ιδρωτοποιοί και σμηγματογόνοι
ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Οι ρυθμιστές του οργανισμού Είδη αδένων στον άνθρωπο o Εξωκρινείς αδένες: εκκρίνουν το προϊόν τους μέσω εκφορητικού πόρου είτε στην επιφάνεια του σώματος (π.χ. ιδρωτοποιοί και σμηγματογόνοι