ΚΑΘΗΛΩΣΗ ΠΕΠΤΙΔΙΩΝ ΒΙΟΛΟΓΙΚΟΥ ΕΝΔΙΑΦΕΡΟΝΤΟΣ ΣΕ ΝΑΝΟΣΩΛΗΝΕΣ ΑΝΘΡΑΚΑ ΚΑΙ ΜΕΛΕΤΗ ΤΗΣ ΔΡΑΣΤΙΚΟΤΗΤΑΣ ΤΟΥΣ
|
|
|
- reek Πολίτης
- 6 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΧΗΜΕΙΑΣ ΤΟΜΕΑΣ ΒΙΟΧΗΜΕΙΑΣ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΝΑΝΟΕΠΙΣΤΗΜΕΣ ΚΑΙ ΝΑΝΟΤΕΧΝΟΛΟΓΙΕΣ» ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΤΟΥ ΣΙΣΚΟΥ ΑΡΓΥΡΙΟΥ ΚΑΘΗΛΩΣΗ ΠΕΠΤΙΔΙΩΝ ΒΙΟΛΟΓΙΚΟΥ ΕΝΔΙΑΦΕΡΟΝΤΟΣ ΣΕ ΝΑΝΟΣΩΛΗΝΕΣ ΑΝΘΡΑΚΑ ΚΑΙ ΜΕΛΕΤΗ ΤΗΣ ΔΡΑΣΤΙΚΟΤΗΤΑΣ ΤΟΥΣ ΕΠΙΒΛΕΠΩΝ ΚΑΘΗΓΗΤΗΣ: ΚΥΡΙΑΚΙΔΗΣ ΔΗΜΗΤΡΙΟΣ ΘΕΣΣΑΛΟΝΙΚΗ 2008
2 ΠΕΡΙΕΧΟΜΕΝΑ ΠΡΟΛΟΓΟΣ.8 ΠΕΡΙΛΗΨΗ..9 SUMMARY ΣΥΝΤΜΗΣΕΙΣ 11 ΚΕΦΑΛΑΙΟ 1 ΘΕΩΡΗΤΙΚΟ ΜΕΡΟΣ Νανοτεχνολογία Σύντομη παρουσίαση νανοδομημένων υλικών Παρασκευή νανοδομημένων υλικών Μηχανικές ιδιότητες νανοδομημένων υλικών Ηλεκτρικές ιδιότητες νανοδομημένων υλικών Μαγνητικές ιδιότητες νανοδομημένων υλικών Οπτικές ιδιότητες νανοδομημένων υλικών Χημικές ιδιότητες νανοδομημένων υλικών Εφαρμογές νανοδομημένων υλικών Νανοσωματίδια Παρουσίαση μορφών άνθρακα Νανοΐνες άνθρακα Φουλερένια Τύποι νανοσωλήνων άνθρακα Τεχνικές σύνθεσης νανοσωλήνων άνθρακα Καταλυτική χημική απόθεση από ατμό Καθαρισμός από προσμίξεις Δομή νανοσωλήνων άνθρακα Ηλεκτρονικές ιδιότητες νανοσωλήνων άνθρακα Μηχανικές και άλλες ιδιότητες νανοσωλήνων άνθρακα. 31 2
3 Μηχανισμοί αποθήκευσης υδρογόνου σε νανοσωλήνες άνθρακα Φυσική προσρόφηση Χημειορρόφηση Αποθήκευση σε νανοσωλήνες άνθρακα προσμίξεων αλκαλίων Βασικές αρχές κατάλυσης ένζυμα Πρωτεολυτικά ένζυμα πρωτεϊνόλυση Καταβολισμός πρωτεϊνών πρωτεάσες Οι πρωτεάσες διευκολύνουν δύσκολες αντιδράσεις Η χυμοθρυψίνη έχει ένα υψηλά ενεργό κατάλοιπο σερίνης Η δράση της χυμοθρυψίνης προχωρά σε δύο βήματα που συνδέονται από ένα ομοιοπολικά προσδεμένο ενδιάμεσο Η σερίνη είναι μέρος μιας καταλυτικής τριάδας η οποία περιλαμβάνει επίσης ιστιδίνη και ασπαραγινικό οξύ Καταλυτικές τριάδες απαντούν σε άλλα υδρολυτικά ένζυμα Άλλες κύριες τάξεις ενζύμων που διασπούν πεπτίδια είναι οι ασπαρτυλοπρωτεάσες, οι κυστεϊνοπρωτεάσες και οι μεταλλοπρωτεάσες Οι αναστολείς των πρωτεασών είναι σημαντικά φάρμακα Μερικά πρωτεολυτικά ένζυμα έχουν ειδικούς αναστολείς Βιοτεχνολογικές εφαρμογές πρωτεασών Ένζυμα με βιοτεχνολογικό ενδιαφέρον - Θερμοάντοχα ένζυμα Σταθεροποίηση των ενζύμων Καθήλωση ενζύμων Καθήλωση με ομοιοπολικό δεσμό μέσω των -ΟΗ, -SH, -H 2 του ενζύμου (Έποξυ ενεργοποιημένη σεφαρόζη) Ομοιοπολική σύνδεση με το υπόστρωμα μέσω των SH, -H 2 του ενζύμου Πεπτιδική σύνθεση Προστατευτικές ομάδες αμινοξέων Προστασία α-αμινομάδας αμινοξέων Τριτοταγής βουτοξυ-καρβονυλομάδα (Boc) φλουορενυλ-μεθοξυ-καρβονυλομάδα (Fmoc) Τριφαινυλομάδα (Trt) Ακετυλομάδα (Ac) Προστασία α-καρβοξυλομάδας αμινοξέων 79 3
4 Βενζυλεστέρες (Bzl) Πλάγιες προστατευτικές ομάδες αμινοξέων Μέθοδοι σύζευξης Μέθοδος των καρβαμιδίων Μέθοδος των ασύμμετρων ή μικτών ανυδριτών Μέθοδος των ενεργών εστέρων Μέθοδος των φωσφονικών και ουρονικών παραγώγων Μεθοδολογίες πεπτιδικής σύνθεσης Boc/Bzl μέθοδος Fmoc/Bu t μέθοδος Τεχνικές πεπτιδικής σύνθεσης Σύνθεση σε υγρή φάση Σύνθεση σε στερεά φάση Στερεά υποστρώματα (ρητίνες) Συνδέτες (Linkers) Πρόσδεση πρώτου αμινοξέος στη ρητίνη Επιμήκυνση πεπτιδικής αλυσίδας Απόσπαση πεπτιδίου από το πολυμερές και απομόνωσή του Πεπτιδομιμητές Κατηγορίες πεπτιδομιμητών με τροποποίηση πεπτιδικού σκελετού Πεπτοειδή Αζαπεπτίδια Διάφορες δράσεις των αζαπεπτιδίων Υδραζινοπεπτίδια και Ν α -υδραζινο-πεπτοειδή Διάφορες δράσεις υδραζινοπεπτιδίων Εισαγωγή στη μικροσκοπία σαρώσεως ακίδας (SPM) Αρχή λειτουργίας μικροσκοπίας ατομικής δύναμης και οργανολογία Μέθοδοι - τεχνικές της μικροσκοπίας ατομικής δύναμης (AFM) Εφαρμογές της μικροσκοπίας ατομικής δύναμης (AFM)
5 ΚΕΦΑΛΑΙΟ 2 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 2.1 Υλικά Μέθοδοι Σύνθεση αζαπεπτιδίων Σύνθεση υδραζινοπεπτιδίων Γενική μέθοδος σύζευξης με DIC/HBt (στερεά φάση) Σύζευξη με τη μέθοδο του DIC/HBt Έλεγχος της αντίδρασης σύζευξης Διαδικασία απομάκρυνσης ομάδων Boc Έποξυ ενεργοποιημένη σεφαρόζη 6B (Ακινητοποίηση ενζύμου) Προσδιορισμός δραστικότητας πρωτεϊνάσης Χρωματομετρική μέθοδος κατά Bradford τροποποιημένη κατά Bearden Ενζυμικός προσδιορισμός τρυψίνης Προσδιορισμός αμινοξέων με τη μέθοδο της νινυδρίνης Φασματοσκοπία Φασματοσκοπία και ηλεκτρομαγνητικό φάσμα Φασματοσκοπία υπερύθρου οργανικών μορίων Ερμηνεία φασμάτων IR Φάσματα υπερύθρου των υδρογονανθράκων Αλκάνια Αλκένια Αλκύνια Φάσματα υπερύθρου άλλων λειτουργικών ομάδων Αλκοόλες Αμίνες Καρβονυλικές ενώσεις.130 5
6 ΚΕΦΑΛΑΙΟ 3 ΠΕΙΡΑΜΑΤΙΚΟ ΜΕΡΟΣ 3.1 Μελέτη νανοσωλήνων που μιμούνται τη δράση ενζύμων - πρωτεασών Προσδιορισμός δραστικότητας πρωτεϊνάσης K σε υπόστρωμα καζεΐνης Δραστικότητα πρωτεϊνάσης Κ συναρτήσει του χρόνου επώασης Δραστικότητα πρωτεϊνάσης Κ συναρτήσει της συγκέντρωσής της σε υπόστρωμα αλβουμίνης Προσδιορισμός δραστικότητας των τροποποιημένων υλικών Προσδιορισμός δραστικότητας των τροποποιημένων υλικών σε διάφορα ph Προσδιορισμός της βιολογικής δραστικότητας του τροποποιημένου νανοσωλήνα (K54) Επαναχρησιμοποίηση του τροποποιημένου νανοσωλήνα (ΝΚ54) Έλεγχος της δραστικότητας του μη τροποποιημένου νανοσωλήνα και επαναχρησιμοποίηση του τροποποιημένου νανοσωλήνα (K54) Έλεγχος δραστικότητας του τροποποιημένου νανοσωλήνα (K28) Απεικόνιση με μικροσκόπιο ατομικών δυνάμεων (AFM) των τροποποιημένων νανοσωλήνων και του μη τροποποιημένου νανοσωλήνα Mελέτη νανοσωλήνων άνθρακα (CT) ως φορείς βιολογικών μορίων Προσδιορισμός της βιολογικής δραστικότητας των προστατευμένων αναλόγων 31 και Προσδιορισμός της βιολογικής δραστικότητας του αναλόγου 31 και του καθηλωμένου αναλόγου 31 σε CT Δραστικότητα του καθηλωμένου αναλόγου 31 σε CT μετά από επαναλαμβανόμενη χρήση του Απεικόνιση με μικροσκόπιο ατομικών δυνάμεων (AFM) τoυ αναλόγου 31 και του καθηλωμένου αναλόγου 31 σε CT Προσδιορισμός της βιολογικής δραστικότητας των εξαπεπτιδίων 3.1, 3.2, 3.3, 4.1, 4.2 και Δέσμευση βιολογικών μορίων σε κλασικά υλικά Προσδιορισμός της βιολογικής δραστικότητας για το καθηλωμένο 6
7 και το ελεύθερο ένζυμο Προσδιορισμός της βιολογικής δραστικότητας του καθηλωμένου και του ελεύθερου ενζύμου σε συνάρτηση με την θερμοκρασία Προσδιορισμός της βιολογικής δραστικότητας του καθηλωμένου και του ελεύθερου ενζύμου συναρτήσει του ph Σταθερότητα του καθηλωμένου και του ελεύθερου ενζύμου (τρυψίνη) μετά από παρατεταμένη θέρμανσή τους ΣΥΜΠΕΡΑΣΜΑΤΑ..161 ΒΙΒΛΙΟΓΡΑΦΙΑ
8 ΠΡΟΛΟΓΟΣ Σκοπός της παρούσας διπλωματικής εργασίας είναι η μελέτη νανοσωλήνων άνθρακα (CT) ως φορείς βιολογικών μορίων και συγκεκριμένα μικρών πεπτιδίων και αμινοξέων με βιολογική δράση όπως επίσης και νανοσωλήνων που μιμούνται τη δράση ενζύμων-πρωτεασών. Η διπλωματική εργασία εκπονήθηκε στο εργαστήριο βιοχημείας, του τμήματος χημείας, του Αριστοτελείου Πανεπιστημίου Θεσσαλονίκης υπό την επίβλεψη του καθηγητή κ. Κυριακίδη Δημητρίου τον οποίο ευχαριστώ θερμά. Για να πραγματοποιηθεί η εργασία οι νανοσωλήνες συντέθηκαν στο ερευνητικό εργαστήριο του κ. Ν. Ταγματάρχη στο Ινστιτούτο Θεωρητικής και Φυσικής Χημείας του Εθνικού Ιδρύματος Ερευνών (ΕΙΕ) και τα πεπτίδια στο ερευνητικό εργαστήριο του κ. Γ. Σταυρόπουλου, τους οποίους ευχαριστώ θερμά. Επίσης θα ήθελα να ευχαριστήσω τον κ. Λογοθετίδη και την κα. Λιακοπούλου για τη συνεργασία. Θα ήθελα τέλος να ευχαριστήσω την μεταδιδακτορική ερευνήτρια του εργαστηρίου βιοχημείας Παπή Ρηγίνη για τη συνεχή καθοδήγηση μου σ όλη την διάρκεια των πειραμάτων και τους υπόλοιπους ερευνητές του εργαστηρίου βιοχημείας. 8
9 Περίληψη Στην παρούσα εργασία μελετήθηκε η χρήση των νανοσωλήνων άνθρακα για την καθήλωση βιολογικών μορίων όπως ολιγοπεπτιδίων και αμινοξέων. Στην πρώτη περίπτωση χρησιμοποιήθηκε το διπεπτίδιο {Βοc-D-Trp-H-(Bzl)-C-D-Trp}, το οποίο είχε μελετηθεί ως προς τη βιολογική του δράση και είχε διαπιστωθεί ότι δρα ως αναστολέας τρυψίνης. Στην παρούσα εργασία το διπεπτίδιο καθηλώθηκε σε νανοσωλήνα άνθρακα και διαπιστώθηκε ότι με την καθήλωση σταθεροποιείται η δράση του και είναι δυνατή η χρησιμοποίηση του για πολλούς κύκλους. Με σκοπό την καθήλωση και άλλων ολιγοπεπτιδίων σε νανοσωλήνες άνθρακα μελετήθηκαν το διπεπτίδιο: {Boc-H-(Bzl)-CH 2 -C-D-Trp-Leu-H} και τα εξαπεπτίδια {Glp 1 -Phe 2 -Gly 3 -[H-(Bzl)-CH 2 -C] 4 -D-Trp 5 -Leu 6 -H}, {Glp 1 -Ala 2 - Gly 3 -[H-(Bzl)-CH 2 -C] 4 -D-Trp 5 -Leu 6 -H}, {Glp 1 -Tic 2 -Gly 3 -[H-(Bzl)-CH 2 -C] 4 - D-Trp 5 -Leu 6 -H}, {Glp 1 -Phe 2 -Gly 3 -D-Trp 4 -[H-(Bzl)-C] 5 -D-Trp 6 -H}, {Glp 1 - Ala 2 -Gly 3 -D-Trp 4 -[H-(Bzl)-C] 5 -D-Trp 6 -H} και {Glp 1 -Tic 2 -Gly 3 -D-Trp 4 -[H- (Bzl)-C] 5 -D-Trp 6 -H}. Από την παραπάνω ομάδα επιλέχθηκαν τα πιο δραστικά ολιγοπεπτίδια τα οποία θα καθηλωθούν και θα μελετηθούν περαιτέρω. Επίσης πραγματοποιήθηκε δέσμευση τρυψίνης σε κλασικά υλικά όπως είναι η εποξυ-ενεργοποιημένη σεφαρόζη 6Β και μελετήθηκε η δραστικότητά του. Τέλος οι νανοσωλήνες άνθρακα χρησιμοποιήθηκαν για την καθήλωση κατάλληλων αμινοξέων με σκοπό την προσομοίωση του ενεργού κέντρου των πρωτεασών. Επιλέχθηκαν τα αμινοξέα τυροσίνη, ασπαραγινικό οξύ και γλουταμινικό οξύ και αφού έγινε η προσκόλλησή τους πάνω σε νανοσωλήνα άνθρακα ελέγχθηκε αν ο τροποποιημένος νανοσωλήνας εμφανίζει ενζυμική δραστικότητα. Μελετήθηκε η επίδραση του τροποποιημένου νανοσωλήνα σε υπόστρωμα αλβουμίνης καθώς και σε συνθετικό υπόστρωμα μεθυλεστέρα της π-τολουεν-σουλφονυλ-l αργινίνης (TAME) και προέκυψε ότι εμφανίζει δράση παρόμοια με αυτή των πρωτεασών. Ένα πρόσθετο συμπέρασμα είναι ότι ο τροποποιημένος νανοσωλήνας έχει δυνατότητα επαναχρησιμοποίησης χωρίς απώλεια της δραστικότητας. 9
10 SUMMARY In this work the immobilization of biological molecules, such as oligopeptides and amino acids, on carbon nanotubes (CTs) was studied. The dipeptide Βοc-D-Trp-H- (Bzl)-C-D-Trp} that present trypsin inhibitor activity was immobilized on a carbon nanotudes. This modification stabilizes the dipeptide s activity and finally the bound form can be used in many cycles without losing any activity. With the intention to immobilize other oligopeptides on CTS, a group of synthetic dipeptide and hexapeptides were studied for their protease inhibitor activity: {Boc-H-(Bzl)-CH 2 -C-D-Trp-Leu-H} and the six hexapeptides {Glp 1 -Phe 2 -Gly 3 - [H-(Bzl)-CH 2 -C] 4 -D-Trp 5 -Leu 6 -H}, {Glp 1 -Ala 2 -Gly 3 -[H-(Bzl)-CH 2 -C] 4 -D- Trp 5 -Leu 6 -H}, {Glp 1 -Tic 2 -Gly 3 -[H-(Bzl)-CH 2 -C] 4 -D-Trp 5 -Leu 6 -H}, {Glp 1 - Phe 2 -Gly 3 -D-Trp 4 -[H-(Bzl)-C] 5 -D-Trp 6 -H}, {Glp 1 -Ala 2 -Gly 3 -D-Trp 4 -[H- (Bzl)-C] 5 -D-Trp 6 -H} and {Glp 1 -Tic 2 -Gly 3 -D-Trp 4 -[H-(Bzl)-C] 5 -D-Trp 6 -H}. The immobilization of a conventional enzyme, such as trypsin, on a commercial material (epoxy-activated sepharose 6B) was studied as well. Finally, carbon nanotubes were used as a carrier of specific amino acids in an attempt to mimic the enzymic active site. This was achieved through the attachment of the amino acids Tyr, Asp and Glu, residues of the known protease active site, on the surface of CTs. The modified CT was tested with substrates albumin or the synthetic p- toluenesulphonyl-l-argine methyl ester (TAME). The modified CT with the three amino acids, mimics the proteases active site and exerts photolytic activity. Ιnterested enough the modified CTs can be used for many cycles with no loss of its activity. 10
11 ΣΥΝΤΜΗΣΕΙΣ AcH ΑFM Boc Βzl DCM DDT DIC DMF DMS [Ε] Εt 2 Fmoc ΗΟΒt i-prh ΜeH [S] TAME TCA ΤΕS ΤFΑ ΤFE Tris Οξικό οξύ Atomic Force Microscopy Τριτοταγής βουτοξυκαρβονυλομαδα Βενζυλομάδα Διχλωρομεθάνιο Διχλωροδιφαινυλοτριχλωροαιθάνιο, -διισοπροπυλοκαρβοδιιμίδιο Διμεθυλοφορμαμίδιο Διμεθυλοσουλφοξείδιο Ένζυμο Διαιθυλαιθέρας 9-φλουορενυλ-μεθοξυκαρβονυλομάδα 1-υδροξυ-βενζοτριαζόλιο Ισοπροπανόλη Μεθανόλη Υπόστρωμα p-toluenesulphonyl-l-argine methyl ester Τριχλωροξικό οξύ Τριαιθυλοσιλάνιο Τριφθοροξικό οξύ Τριφθοροαιθανόλη Υδροξυμεθυλοαμινομεθάνιο 11
12 ΚΕΦΑΛΑΙΟ 1 ΘΕΩΡΗΤΙΚΟ ΜΕΡΟΣ Νανοτεχνολογία Νανοτεχνολογία είναι ένας όρος ο οποίος χρησιμοποιείται για να περιγράψει τη δημιουργία και χρήση λειτουργικών δομών μεγέθους μεταξύ 1 και 100 νανομέτρων (nm), της τάξεως δηλαδή του 10-9 μέτρων. Οι διαστάσεις γίνονται ευκολότερα αντιληπτές αναφέροντας πως ένα νανόμετρο ισούται περίπου με το 1/80000 μιας ανθρώπινης τρίχας ή με το μήκος 10 ατόμων υδρογόνων σε σειρά. Κατά παρόμοιο τρόπο ορίζεται και ο όρος νανοεπιστήμη αναφερόμενος σε επιστήμες οι οποίες μελετούν φαινόμενα στην κλίμακα αυτή. Στη μέχρι τώρα ανάπτυξη της σημαντικό ρόλο έπαιξαν η σημαντική βελτίωση του ηλεκτρονικού μικροσκοπίου ενώ σταθμοί μπορούν να θεωρηθούν οι ανακαλύψεις δομών άνθρακα σε μορφή σφαίρας γνωστές ως φουλερένια (buckminister fullerenes ή fullerenes ή buckyballs) καθώς και σε μορφή σωλήνα γνωστές ως νανοσωλήνες άνθρακα (carbon nanotubes) με ιδιαίτερες ιδιότητες το καθένα. Ο όρος νανοτεχνολογία χαρακτηρίζεται από μεγάλη ευρύτητα όντας πολύ γενικός για να περιγράψει οτιδήποτε συμβαίνει στις διαστάσεις του νανομέτρου. Κατά συνέπεια, μπορεί να χωρισθεί σε πιο ειδικά θέματα όπως αυτό της νανοηλεκτρονικής, των νανοϋλικών καθώς και άλλων. Οι εφαρμογές της είναι αναρίθμητες ενώ οι επιπτώσεις γίνονται αντιληπτές σε πολλαπλά επίπεδα κατά κύριο λόγο στον οικονομικό τομέα επηρεάζοντας παγκόσμιες βιομηχανίες και οικονομίες, αλλά και στο κοινωνικό βελτιώνοντας το επίπεδο της ζωής μας. Οι παρακάτω εικόνες αν και περισσότερο αισθητικού περιεχομένου παρά επιστημονικού είναι χαρακτηριστικές των δυνατοτήτων της νανοτεχνολογίας. Αυτή μας δίνει τα μέσα να «χτίσουμε» δομές σε ατομικό επίπεδο ή να «δούμε» τις δομές αυτές σε ατομική κλίμακα. Οι επιφάνειες οι οποίες απεικονίζονται στις παρακάτω εικόνες μελετήθηκαν με τη χρήση STM μικροσκοπίου ενώ το ίδιο χρησιμοποιήθηκε και για την τοποθέτηση των ατόμων. Δε θα πρέπει ωστόσο να φανταστεί κάποιος πως η νανοτεχνολογία πρόκειται για επιστημονική επανάσταση. Τα περισσότερα θέματα που αυτή περικλείει προκύπτουν σαν λογική συνέπεια της εξέλιξης της ικανότητας της επιστήμης και της τεχνολογίας να ερευνά και να εργάζεται σε όλο και μικρότερη κλίμακα. Εξάλλου, η κατάλυση, ένα φαινόμενο που ανέκαθεν χαρακτηριζόταν από νανομετρικές διαστάσεις αποτελεί επιστημονικό κλάδο ο οποίος αναπτύσσεται πολλές δεκαετίες. Επιπλέον, ολόκληρα επιστημονικά πεδία όπως η χημεία, ή η 12


13 βιολογία ανέκαθεν δούλευαν σε τέτοιες διαστάσεις παρόλο που ο όρος νανοεπιστήμη εισήχθη μόλις πρόσφατα. Η νανοτεχνολογία στο επίπεδο των νανοδομημένων υλικών αποτελεί καθοριστικό παράγοντα στην ανάπτυξη της οικονομίας υδρογόνου. Υλικά για την αποθήκευση υδρογόνου, για ηλεκτρόδια κυψελών καυσίμου ή για την κατάλυση η οποία είναι καθοριστική σε όλα τα στάδια από αυτό της παραγωγής μέχρι της χρήσης βελτιώνουν την επίδοση τους ραγδαία όταν αυτά αποκτήσουν νανοκρυσταλλική δομή. Επομένως, η μελέτη των διεργασιών και η κατανόηση τους σε αυτό το επίπεδο μας ενδιαφέρει άμεσα. Αυτό δικαιολογεί τη μεγάλη πειραματική δραστηριότητα στον τομέα αλλά και τις προσπάθειες ανάπτυξης θεωρητικών μοντέλων με τη χρήση προσομοίωσης [2-5,18,19,25]. Εικόνα 1: (Αριστερά) STM εικόνα 48 ατόμων σιδήρου σε επιφάνεια Cu (111), (δεξιά πάνω) STM εικόνα 35 ατόμων Xe σε επιφάνεια i (110) και (δεξιά κάτω) STM εικόνα επιφάνειας Si [2-5] Σύντομη παρουσίαση νανοδομημένων υλικών Τα νανοϋλικά αποτελούν για τους λόγους που είπαμε ιδιαίτερο πεδίο έρευνας τα τελευταία χρόνια. Νανοϋλικά ονομάζονται τα υλικά των οποίων οι δομικοί λίθοι ανήκουν στην τάξη του νανομέτρου. Σε αυτά τα χαρακτηριστικά οφείλονται και οι ιδιαίτερες ιδιότητες των νανοϋλικών, ιδιότητες κατά πολύ ανώτερες αυτών των συμβατικών υλικών που χρησιμοποιούνται κατά κύριο λόγο αυτή τη στιγμή στη βιομηχανία. Στα πολυκρυσταλλικά νανοδομημένα υλικά ή αλλιώς νανοκρυσταλλικά υλικά (nanocrystalline materials) αναφερόμαστε στο μέγεθος ενός κρυσταλλίτη ή αλλιώς κόκκου (grain), ο οποίος αποτελείται από ένα μικρό αριθμό ατόμων. Συνέπεια αυτού είναι ένα μεγάλο ποσοστό (τα μισά ή περισσότερα) αυτών των ατόμων να βρίσκονται στην επιφάνεια του. Από άποψη 13
14 φυσικής, αυτό κάνει τα φαινόμενα επιφάνειας (φαινόμενα μεταφοράς μάζας, διαμόρφωση επιφανειακών ενεργειακών σταθμών κ.τ.λ.) να παίζουν πρωταρχικό ρόλο στη μελέτη των νανοδομημένων υλικών. Όπως είναι λογικό, οι εφαρμογές των νανοϋλικών είναι ανάλογες σε αριθμό των δυνατοτήτων τους. Οι βελτιωμένες ηλεκτρικές, οπτικές, φυσικές, χημικές, μαγνητικές και μηχανικές ιδιότητες τους είναι και το κίνητρο για την έρευνα που γίνεται πάνω σε αυτά ενώ έχουν αναπτυχθεί πολυάριθμες μέθοδοι παρασκευής τους, αν και όχι όλοι κατάλληλοι για την επιθυμητή από τη βιομηχανία, μαζική παραγωγή τους. Ένας δεύτερος λόγος που δικαιολογεί το ενδιαφέρον για τα νανοϋλικά είναι ότι η φυσική που τα χαρακτηρίζει διαφέρει ριζικά από αυτή που πάνω της στηρίζεται η σύγχρονη βιομηχανία (για παράδειγμα η μικροηλεκτρονική) και απαιτούνται νέες μέθοδοι προσέγγισης, που όμως μόλις πρόσφατα άρχισαν να αναπτύσσονται. Ενδεικτικές χρήσεις των νανοϋλικών είναι, καλύτερα υλικά για υπολογιστές νέας γενιάς, καλύτερα μονωτικά υλικά, καλύτερα κοπτικά εργαλεία, μπαταρίες υψηλής χωρητικότητας, εξαιρετικά ισχυροί μαγνήτες κτλ [2-5] Παρασκευή νανοδομημένων υλικών Καθώς ο όρος "νανοϋλικά" είναι ιδιαίτερα ευρύς καλύπτοντας μία πολύ μεγάλη κατηγορία υλικών δεν μπορούμε να μιλήσουμε για κάποια γενική μέθοδο παρασκευής τους. Υπάρχουν δύο γενικές κατηγορίες παρασκευής υλικών, η "από πάνω προς τα κάτω" και η "από κάτω προς τα πάνω". Χαρακτηριστικό παράδειγμα των πρώτων είναι η λιθογραφία στις διάφορες μορφές της. Στην πιο απλή της μορφή, ηλεκτρόνια ή φωτόνια κατάλληλων ενεργειών χρησιμοποιούνται για να μορφοποιήσουν μια επιφάνεια με τη βοήθεια μιας μάσκας. Την κατηγορία των "από κάτω προς τα πάνω" αντιπροσωπεύει επάξια ή τεχνικής της αυτοσυναρμολόγησης όπου μοριακά είδη αυτο-οργανώνονται σε περιοδικές δομές σε μια προσπάθεια ελαχιστοποίησης της ελεύθερης ενέργειας του συστήματος. Αρκετά δημοφιλής είναι η και λεγόμενη sol - gel synthesis η οποία ανήκει στην κατηγορία των χημικών μεθόδων και είναι ικανή να παράγει ικανοποιητικά μεγάλες ποσότητες, να συνθέσει μεγάλο αριθμό υλικών, να δώσει μέχρι και 99,99% καθαρό προϊόν ενώ προσφέρει ακριβή έλεγχο πάνω στη δομή του τελικού προϊόντος κτλ. Εξίσου χρησιμοποιείται η τεχνική του mechanical alloying (ή ball milling) κυρίως για να δημιουργήσει μεταλλικά κράματα νανοκρυσταλλικής δομής. Αρχικά τοποθετείται μείγμα σκόνης διαφορετικών στερεών ουσιών σε κατάλληλο κλειστό μηχανισμό ο οποίος εκτελεί ταχεία περιστροφική κίνηση γύρω από έναν ή περισσότερους άξονες. Τότε συνεχείς συγκρούσεις της 14

15 σκόνης με μεταλλικές μπάλες (λόγω φυγοκέντρου) μεγάλων σχετικά ενεργειών έχει ως αποτέλεσμα να προκύψουν σωματίδια μικρών διαστάσεων αναλογίας ουσιών ίδιας με του αρχικού μείγματος. Ο χώρος συνήθως βρίσκεται σε κενό ή σε αδρανή ατμόσφαιρα. Εικόνα 2: Mηχανισμός ball milling [2-5]. Στην εικόνα 2 βλέπουμε κάτοψη ενός μηχανισμού ball milling. Το μέγεθος των σωματιδίων εξαρτάται από τη χρονική διάρκεια της κατεργασίας. Για μεγάλους χρόνους παίρνουμε σωματίδια εξαιρετικά μικρών διαστάσεων μέχρι και λίγων νανομέτρων. Με αυτή τη τεχνική είναι δυνατόν να δημιουργηθούν κράματα μη αναμιγνύσιμων μετάλλων. Πλεονέκτημα της είναι η ευκολία της και η μικρή τιμή του εξοπλισμού που απαιτεί. Μειονέκτημα αποτελεί η δυσκολία συνένωσης των παραγόμενων σωματιδίων σε μεγαλύτερα στερεά με παράλληλη διατήρηση της νανοδομημένης φύσης τους [2-5,18,19,25] Μηχανικές ιδιότητες νανοδομημένων υλικών Αναφερόμενοι σε νανοκρυσταλλικά υλικά, καθώς ο όγκος του κόκκου μειώνεται, αυξάνεται σημαντικά ο αριθμός των ατόμων που βρίσκονται στην επιφάνεια του. Η απόδοση ενός κρυσταλλικού υλικού, κατά κανόνα, όσων αφορά κάποια ιδιότητα του είναι αντιστρόφως ανάλογη του μεγέθους των κόκκων, μέχρι ένα κατώτατο όριο φυσικά όπου μετά από αυτό δεν έχει νόημα πια η έννοια του πολυκρυσταλλικού υλικού αλλά μιλάμε πλέον για άμορφα υλικά. Αυτό γιατί τα περισσότερα ενεργά άτομα και αυτά στα οποία οφείλονται κυρίως οι ιδιότητες τους, είναι τα άτομα που βρίσκονται στην επιφάνεια του κρυσταλλίτη. Τα νανοκρυσταλλικά υλικά παρουσιάζουν πολύ υψηλή σκληρότητα. Για τα νανοκρυσταλλικά μέταλλα κόκκων μεγέθους περίπου 10 nm, για παράδειγμα, μπορεί να παρατηρηθεί από 2 έως και 7 φορές περισσότερη σκληρότητα από μέταλλα με μεγαλύτερους κόκκους μεγέθους (περισσότερο από 1μm). Εξαίρεση αποτελούν τα λεπτά υμένια (thin films) με κόκκους μικρότερους των 6 nm όπου η σκληρότητα μειώνεται καθώς μειώνεται το μέγεθος του κόκκου [2-5,18,19,25]. 15
16 Ηλεκτρικές ιδιότητες νανοδομημένων υλικών Η πιο χαρακτηριστική χρήση των νανοϋλικών με τέτοιο τρόπο ώστε να εκμεταλλεύονται οι ηλεκτρικές ιδιότητες τους είναι οι νανοσωλήνες άνθρακα, τα οποία έχουν ευρεία χρήση στη νανοηλεκτρονική και στην ηλεκτρονική μικροσκοπία. Οι ηλεκτρικές ικανότητες των νανοσωλήνων άνθρακα εξαρτώνται από τη διάμετρο τους και το προσανατολισμό των ανθράκων ως προς τον κεντρικό άξονα τους. Ανάλογα μπορούν και συμπεριφέρνονται ως μεταλλικά ή ως ημιαγώγιμα υλικά [2-5,18,19,25] Μαγνητικές ιδιότητες νανοδομημένων υλικών Από τις πιο σημαντικές ιδιότητες των νανοϋλικών είναι οι μαγνητικές. Η παραμένουσα μαγνήτιση των διφασικών σκληρών/μαλακών μαγνητικών προϊόντων νανοκρυσταλλικής δομής έχει βιομηχανικά πάρα πολλές χρήσεις. Μαλακή σιδηρομαγνητική συμπεριφορά εμφανίζουν για παράδειγμα διαλύματα μαγνητικών νανοσωματιδίων. Επιπλέον τα νανοκρυσταλλικά ελαφρά μαγνητικά προϊόντα παρουσιάζουν τις μικρότερες απώλειες ενέργειας από οποιοδήποτε άλλο υλικό. Στα νανοϋλικά επίσης μπορούμε να παρατηρήσουμε και το φαινόμενο της γιγαντιαίας μαγνητοαντίστασης (GMR) κατά το οποίο η ηλεκτρική αντίσταση ενός υλικού μειώνεται όταν το υλικό εκτεθεί σε μαγνητικό πεδίο και αναφέρθηκε πρώτη φορά για πολυστρωματικά λεπτά υμένια [2-5,18,19,25] Οπτικές ιδιότητες νανοδομημένων υλικών Οι οπτικές ιδιότητες ενός υλικού εξαρτώνται και από τις ηλεκτρονιακές του καταστάσεις. Η δομή των νανοϋλικών είναι τέτοια ώστε αυτές να παρουσιάζουν μεγάλες διαφορές σε σύγκριση με ένα κοινό υλικό. Μιλώντας για νανοκρυσταλλικά υλικά, ανάλογα με το προς χρήση υλικό και το μέγεθος των κόκκων του μπορεί να επιτευχθούν διαφορετικές τιμές ανακλαστικότητας ή οπτικής διαφάνειας για διαφορετικά μήκη κύματος. Συνέπεια αυτού του γεγονότος, οι πάρα πολλές τεχνολογικές εφαρμογές κυρίως σαν επιστρώσεις με τη μορφή λεπτών υμενίων [2-5,18,19] Χημικές ιδιότητες νανοδομημένων υλικών Τέλος πρέπει να αναφερθούμε στη χαρακτηριστική ικανότητα των νανοϋλικών να απορροφούν/αποθηκεύουν μεγάλες ποσότητες υδρογόνου καθώς αντιδρούν φυσικά ή χημικά με 16
17 αυτό. Αυτό τους δίνει συν τοις άλλοις τη δυνατότητα να χρησιμοποιηθούν ως πολύ καλοί καταλύτες σε αντιδράσεις που παίρνει μέρος υδρογόνο, κάτι αρκετά σημαντικό, αφού βρίσκει εφαρμογές σε πολλές ενεργειακές εφαρμογές όπως η λειτουργία των κυψελών καυσίμου. Γενικά τα νανοκρυσταλλικά υλικά, λόγω της μεγάλης επιφάνειας των δομικών λίθων τους (αναφερόμενοι είτε σε κρυσταλλίτες είτε στους μονοκρυστάλλους νανοσωματιδίων) μπορούν να δράσουν καταλυτικά και μάλιστα βέλτιστα σε πάρα πολλές περιπτώσεις [2-5,18,19,25] Εφαρμογές νανοδομημένων υλικών Όπως εύκολα μπορεί να καταλάβει κάποιος, υλικά με ιδιότητες όπως τις παραπάνω είναι επιθυμητά σε μία πληθώρα εφαρμογών. Αναφέρονται μερικές ενδεικτικά: Η ταχύτητα των υπολογιστικών συστημάτων είναι αντιστρόφως ανάλογη με το μέγεθος των μικροεπεξεργαστών τους. Ωστόσο με τη σημερινή τεχνολογία το κατώτατο όριο του μεγέθους αυτού δε θα αργήσει να σταματήσει κάθε περαιτέρω βελτίωση. Τα νανοϋλικά είναι αυτά που με τη χρήση τους μπορούμε να πετύχουμε το ελάχιστο δυνατό όριο. Βελτίωση επίσης της αποδοτικότητας και μείωσης της συχνότητας εμφάνισης λάθους μπορεί να γίνει με τη χρήση καθαρών υλικών όπως τα νανοϋλικά. Δεύτερον, τα νανοκρυσταλλικά υλικά μπορούν να παρασκευαστούν με τέτοιο τρόπο ώστε να καταστούν αρκετά πορώδη για να λειτουργήσουν ως πολύ καλοί μονωτές με τον αέρα να εγκλωβίζεται στα διάκενα του. Τέτοια υλικά δίνει η sol - gel τεχνική που αναφέρθηκε και παραπάνω. Επιπλέον, νανοκρυσταλλικά υλικά μπορούν να χρησιμοποιηθούν σε οπτικές συσκευές που απαιτείται μεγάλη ευκρίνεια. H υψηλής ευκρίνειας τηλεοράσεις με χρήση νανοϋλικών αντί για των συμβατικών υλικών που χρησιμοποιούνται σήμερα είναι ένα παράδειγμα αυτού. Δεύτερο παράδειγμα είναι οι ηλεκτροχρωμικές οπτικές συσκευές των οποίων η φωτεινότητα και χρωματική αντίθεση τους εξαρτάται αποκλειστικά από το μέγεθος του κόκκου του υλικού που χρησιμοποιούν. Με τη χρήση νανοϋλικών μπορούν να παρασκευαστούν, επίσης, ανθεκτικότερα και σκληρότερα κοπτικά εργαλεία. Σημαντική χρήση τέτοιων εργαλείων αρκετά μικρών διαστάσεων και στην κατασκευή μικροηλεκτρικών κυκλωμάτων. Ακόμα, νανοϋλικά, προϊόντα της sol - gel τεχνικής αποτελούν άριστα υλικά για την κατασκευή μπαταριών, μεγαλύτερης χωρητικότητας και στην περίπτωση των επαναφορτιζόμενων, μικρότερης συχνότητας επαναφόρτισης. 17
18 Οι βελτιωμένες μαγνητικές ιδιότητες αυτών των υλικών μπορούν να δώσουν αρκετά υψηλής ενέργειας μαγνήτες ώστε να βρουν χρήση σε γεννήτριες ρεύματος, κινητήρες πλοίων καθώς επίσης μπορούν να χρησιμοποιηθούν για την κατασκευή εξαιρετικά ευαίσθητων αναλυτικών οργάνων. Τα νανοϋλικά είναι εξαιρετικά ευαίσθητα στις αλλαγές του περιβάλλοντος τους, λόγω του μικρού μεγέθους του κόκκου τους. Χρήση τους μπορεί να γίνει και στη βελτίωση των υπαρχόντων αισθητήριων οργάνων, δεδομένου του ότι η αρχή λειτουργίας τους στηρίζεται στο ότι κατά την ανίχνευση ο ανιχνευτής αλληλεπιδρά με κάποιο τρόπο με τον περιβάλλον ώστε μια συνεχώς μετρούμενη ιδιότητα του να αλλάξει (για παράδειγμα η ηλεκτρονική αγωγιμότητα του ή η χημική του σύνθεση ή η χωρητικότητα του κτλ.). Στο ίδιο πλαίσιο, νανοαισθητήρες της παραπάνω μορφής μπορούν να αποτελέσουν μέρος ολοκληρωμένων συστημάτων ικανά να συλλέξουν, να επεξεργαστούν και να μεταδώσουν ογκώδη ποσά δεδομένων με ελάχιστο μέγεθος, βάρος και κατανάλωση ενέργειας. Συνεχίζοντας, εκμεταλλευόμενοι τη σκληρότητα των νανοϋλικών μπορούμε να έχουμε ουσιαστική χρήση τους σε αεροδιαστημικές εφαρμογές αφού τα υλικά που χρησιμοποιούνται σήμερα για αυτές τις χρήσεις μπορούν να αντικατασταθούν από άλλα ανθεκτικότερα και ελαφρύτερα. Χρήση τα νανοϋλικά μπορούν να βρουν και στην κατασκευή κεραμικών τα οποία χαρακτηρίζονται από ιδιαίτερη ευθραυστότητα αν και σκληρά δίνοντας κεραμικά εξίσου σκληρά αλλά με πλαστικές ικανότητες. Ακόμα μπορούν να επιτευχθούν υψηλότερες συχνότητες μετάδοσης πληροφορίας με πολλαπλάσια εύρος ζώνης καθώς και πιο αποτελεσματική χρήση του οπτικού φάσματος για αυτό το σκοπό, χάρη στις οπτικές ιδιότητες τους [2-5,18,19,25] Νανοσωματίδια Τα νανοσωματίδια είναι σωματίδια διαμέτρου μερικών νανομέτρων. Υβριδικά νανοσωματίδια με πυρήνα ανόργανης φάσης τροποποιημένης επιφάνειας με οργανικά μόρια είναι ένα παράδειγμα. Αμφίφυλα μπλοκ συμπολυμερή σε κατάλληλες συνθήκες ph σχηματίζουν μικύλλια και επομένως μπορούν επίσης να χαρακτηριστούν ως νανοσωματίδια. Μεταλλικά ή ημιαγώγιμα κρυσταλλικά νανοσωματίδια μπορούν να βρεθούν στη θέση υψηλά διεσπαρμένης ενεργού φάσης ενός στηριζόμενου καταλύτη. Σε τέτοιους καταλύτες όπως θα δούμε και παρακάτω το μέσο μέγεθος ενός μεταλλικού καταλυτικού σωματιδίου μπορεί να ανήκει στη τάξη του 18

19 νανομέτρου. Τα νανοσωματίδια αυτά συνήθως είναι μεμονωμένοι κρύσταλλοι παρόμοιους με αυτούς που ανήκουν στο εσωτερικό ενός πολυκρυσταλλικού υλικού. Το μέγεθος αυτών των κρυστάλλων καθορίζεται κυρίως από την συνεισφορά της επιφανειακής ενέργειας στην ολική. Καθώς η πιο σταθερή δομή είναι αυτή με τη μικρότερη ενέργεια ο κρύσταλλος τείνει να αποκτήσει μορφή που περιγράφεται από: Όσο το δυνατό μικρότερη επιφάνεια Πλευρές χαμηλής επιφανειακής ενέργειας Μελετώντας ένα υλικό μακροσκοπικά και συνεπώς σαν συνεχές μέσο θα λέγαμε ότι τις προϋποθέσεις αυτές πληρεί η σφαιρική μορφή. Ωστόσο, σε νανομετρικές διαστάσεις δε μπορούμε να αγνοήσουμε τη διακριτή τοποθέτηση των ατόμων στο χώρο. Εικόνα 3: Κυβο-οκταεδρική μορφή (αριστερά) και μορφή οκταέδρου (δεξιά) [2-5]. Η ενέργεια των επιφανειακών επιπέδων είναι υψηλότερη από αυτή του εσωτερικού καθώς χαρακτηρίζονται από θετική ελεύθερη ενέργεια σχηματισμού. Αυτό το καταλαβαίνουμε εύκολα φανταζόμενοι ένα μακροσκοπικό υλικό το οποίο τεμαχίζεται προκειμένου να δημιουργηθεί μία νέα ελεύθερη επιφάνεια. Θα πρέπει τότε να δοθεί ενέργεια στο σύστημα προκειμένου να σπάσουν οι δεσμοί που συγκρατούσαν το στερεό κατά το επίπεδο όπου εμείς το τεμαχίζουμε. Το πλεγματικό επίπεδο ενός f c c μετάλλου με την μικρότερη επιφανειακή ενέργεια είναι η (111). Κατασκευάζοντας ένα νανοσωματίδιο με (111) επίπεδα μόνο καταλήγουμε σε μία οκταεδρική μορφή. Ωστόσο αυτή η δομή δεν χαρακτηρίζεται από τη μέγιστη δυνατή επιφάνεια. Αν "κόψουμε" τις ακμές του οκταέδρου τότε θα πάρουμε μία δομή αποτελούμενη από 8 (111) πλεγματικά επίπεδα και 6 (100). Τα πλεγματικά αυτά επίπεδα αν και υψηλότερης ενέργειας συνεισφέρουν κατά τέτοιο τρόπο όπου ο νέος κρύσταλλος να χαρακτηρίζεται από μικρότερο λόγο επιφάνειας - όγκου και επομένως μειωμένη συνολική ενέργεια άρα και αυξημένη σταθερότητα. Η δομή αυτή συνήθως υιοθετείται ως μοντέλο για τα καταλυτικά σωματίδια τα οποία δεν αλληλεπιδρούν ισχυρά με το υπόστρωμα ώστε να επηρεαστεί η μορφολογία τους. Νανοσωματίδια μπορούν να παρασκευαστούν με πολλές τεχνικές όπως η sol - gel, η ball milling ή άλλες χημικές μεθόδους. Καθώς οι παράμετροι κάθε τεχνικής καθορίζουν τη τελική μορφή του νανοσωματιδίου μπορούμε να παρασκευάσουμε νανοσωματίδια βελτιστοποιημένα για κάποια συγκεκριμένη χρήση. Όσων αφορά την ενέργεια υδρογόνου, το ενδιαφέρον για τα 19
20 νανοσωματίδια έγγυται κυρίως στις καταλυτικές τους ιδιότητες για τη χρήση τους σε κυψέλες καυσίμου αλλά και γενικότερα [2-5,18,19,25] Παρουσίαση μορφών άνθρακα Το διαμάντι και ο γραφίτης είναι δύο πολύ γνωστές μορφές άνθρακα. Το διαμάντι σχηματίζεται από άτομα άνθρακα κάθε ένα από τα οποία χαρακτηρίζεται από 4 υβριδικά τροχιακά sp 3 και έτσι σχηματίζεται ένα τρισδιάστατο δίκτυο ατόμων. Στο γραφίτη τα άτομα άνθρακα ενώνονται μεταξύ τους μέσω τριών sp 2 υβριδικών τροχιακών σχηματίζοντας έτσι επίπεδα φύλλα από εξαμελείς δακτυλίους. Τα φύλλα αυτά είναι συγκρατημένα μεταξύ τους μέσω δυνάμεων Van der Waals. Εκτός όμως από τις δύο αυτές μορφές άνθρακα υπάρχουν και άλλες, όπως οι νανοΐνες άνθρακα, τα φουλερένια και οι νανοσωλήνες άνθρακα, τις οποίες θα εξετάσουμε παρακάτω. Όλες αυτές οι ενώσεις χαρακτηρίζονται από νανοκρυσταλλική δομή. Το ενδιαφέρον για αυτές όσων αφορά την οικονομία υδρογόνου έγγυται στο ότι είναι ελαφρά και μπορούν να χρησιμοποιηθούν (όπως και τα υδρίδια μετάλλων) ως μέσα αποθήκευσης υδρογόνου, με περισσότερη ασφάλεια από αυτή των μεθόδων αέριας ή υγρής αποθήκευσης του. Δεν μπορούμε όμως να μιλήσουμε συγκεκριμένα για το πώς γίνεται η αποθήκευση σε αυτά χωρίς πρώτα να αναφερθούμε στις σημαντικότερες ιδιότητες τους [2-5,18,19,25] Νανοΐνες άνθρακα Οι νανοΐνες άνθρακα (GFs) αποτελούνται από στρώματα γραφίτη τοποθετημένα με συγκεκριμένο προσανατολισμό ως προς τον άξονα της ίνας. Χαρακτηρίζονται από μεγάλης έκτασης επιφάνεια οι οποίες με τη σειρά τους αποτελούν θέσεις για χημική ή φυσική προσρόφηση αερίου. Η ελεύθερη επιφάνεια κυμαίνεται μεταξύ 300 έως και 700 m 2 ανά γραμμάριο υλικού. Οι νανοΐνες άνθρακα περιγράφονται γενικά από μήκη μερικών μm έως και μερικών mm και διαμέτρους από 0,4 έως 500 nm. Η απόσταση μεταξύ των φύλλων είναι περίπου 3,4 Å. Συνοπτικά, ο μηχανισμός δημιουργίας μιας νανοΐνας άνθρακα περιλαμβάνει τα εξής: Αέριος υδρογονάνθρακας ή κατάλληλο μείγμα ανθρακικών ενώσεων προσροφάται σε μεταλλική καταλυτική επιφάνεια (Α). Οι συνθήκες όπου επικρατούν εκεί ευνοούν τη διάσπαση των δεσμών μεταξύ των ανθράκων στα μόρια. Ο ατομικός άνθρακας διαχέεται στη συνέχεια διαμέσου του μετάλλου (Β). 20

 και τη θερμοκρασία.")
21 Τέλος, η νανοδομή του άνθρακα σχηματίζεται στην αντίθετη όψη του μετάλλου από αυτή όπου αρχικά εισήλθε ο άνθρακας (C). Η γεωμετρία του καταλύτη καθορίζει και τον προσανατολισμό των επιπέδων. Εικόνα 4: Herringbone νανοΐνα γραφίτη [2-5]. Το μοντέλο αυτό για τη δημιουργία των νανοϊνών χρησιμοποιείται και για την περιγραφή ενός από τους μηχανισμούς σύνθεσης των νανοσωλήνων, όπως θα δούμε παρακάτω με περισσότερη λεπτομέρεια. Η κρυσταλλική τελειότητα της εναποτιθέμενης ίνας καθορίζεται από τη φύση του καταλύτη, τη φύση του αντιδρώντος αερίου (ή με άλλα λόγια την αναλογία άνθρακα - υδρογόνου του αερίου) και τη θερμοκρασία. Στο τέλος της διαδικασίας τα σωματίδια του μεταλλικού καταλύτη απομακρύνονται με τη χρήση κατάλληλου όξινου διαλύματος. Ο προσανατολισμός των στρωμάτων ως προς τον άξονα της ίνας, τη χαρακτηρίζει ανάλογα. Στο σχήμα παρακάτω βλέπουμε κάθετο (ribbon) και παράλληλο (platelet) προσανατολισμό ενώ στο προηγούμενο το προσανατολισμό "herringbone" όπου η γωνία μεταξύ των στρωμάτων και του άξονα της ίνας είναι ίση με 45 μοίρες. Η σκούρα περιοχή παριστάνει τα μεταλλικά σωματίδια καταλύτη [2-5,18,19,25]. Εικόνα 5: Platelet νανοΐνα γραφίτη (αριστερά). Ribbon νανοΐνα γραφίτη (δεξιά). Φαίνεται μία μακροσκοπική εικόνα της κάθε ίνας και η μικροσκοπική της δομή με εικόνα ηλεκτρονικού μικροσκοπίου [2-5]. 21
.")
22 1.1.7 Φουλερένια Μια ακόμη αλλοτροπική μορφή άνθρακα είναι τα φουλερένια (fullerenes). Τα φουλερένια είναι ανθρακικές δομές με σφαιρικό σχήμα και ανακαλυφθήκαν το 1985 από τους Harold Kroto et al. (βραβείο Νόμπελ Χημείας 1996). Το πιο γνωστό φουλερένιο είναι αυτό το οποίο αποτελείται από 60 άτομα άνθρακα (C 60 ή buckyball) αλλά επίσης κοινά είναι αυτά με 70, 76 και 84 άτομα άνθρακα (C 70, C 76, C 84 fullerenes). Eικόνα 6: Φουλερένιο C 60 [2-5]. Τα φουλερένια είναι γενικά σταθερά μόρια και απαιτούν θερμοκρασίες τάξεως 1000 o C και άνω για να διασπαστούν οι δεσμοί μεταξύ των ανθράκων οι οποίοι το αποτελούν. Ανάλυση με ακτίνες Χ ή σκέδαση νετρονίων ενός μοριακού κρυστάλλου μπορεί να φανερώσει τη σφαιρική δομή του μορίου C 60 καθώς και τη διάμετρο του. Επιπλέον η φασματοσκοπία MR δίνει ένα φάσμα μίας μόνο κορυφής, υποδεικνύοντας πως όλα τα άτομα άνθρακα του μορίου είναι ισοδύναμα γεγονός το οποίο επίσης παραπέμπει σε σφαιρική δομή. Τρία ηλεκτρόνια από κάθε άνθρακα συμμετέχοντας σε sp 2 υβριδικά τροχιακά ενώνουν τους άνθρακες μεταξύ τους με σ δεσμούς. Υπάρχουν δύο είδη δεσμών στο μόριο, ένας μεταξύ ανθράκων στην κοινή ακμή μιας πενταγωνικής και μιας εξαγωνικής έδρας καθώς και ένας στην κοινή ακμή δύο εξαγωνικών εδρών. Η σκέδαση με νετρόνια μας δίνει μήκη δεσμών 0,1455 nm και 0,1391 nm αντίστοιχα. Τα άτομα άνθρακα σε ένα φύλλο γραφίτη χαρακτηρίζονται επίσης από ίδιου τύπου υβριδικά τροχιακά sp 2 όπως το φουλερένιο. Αυτά τα υβριδικά τροχιακά δίνουν τρεις ισχυρούς σ δεσμούς πάνω στο ίδιο επίπεδο και έναν ασθενή π σε κάθετη διεύθυνση. Καθώς οι σ δεσμοί είναι κορεσμένοι ο γραφίτης θεωρείται χημικά αδρανής. Στην περίπτωση του φουλερενίου ωστόσο, λόγω της κυρτότητας του μορίου, οι δεσμοί παύουν να βρίσκονται στο ίδιο επίπεδο ενώ επίσης το νέφος των π ηλεκτρονίων παραμορφώνεται κάνοντας το φουλερένιο περισσότερο ενεργό χημικά. 22




23 Η διάμετρος του βρίσκεται από διάφορες φασματοσκοπικές μεθόδους ίση με 0,710 nm. Προκειμένου να βρούμε την ενεργό διάμετρο του μορίου θα πρέπει να συνυπολογίσουμε το εύρος του ηλεκτρονικού νέφους των π-ηλεκτρονίων το οποίο είναι ίσο με 0,335 nm υπολογίζοντας τελικά τη διάμετρο του μορίου 1,380 nm. Αν και μεγάλα μόρια όσον αφόρα τις διαστάσεις, τα φουλερένια παραμένουν συγκριτικά με τα οργανικά μόρια αρκετά μικρά. Η σφαιρικότητα του φουλερενίου επιτυγχάνεται με την ύπαρξη πενταγωνικών εδρών καθώς είναι αυτά τα οποία εισάγουν κυρτότητα στο μόριο. Η ύπαρξη δύο γειτονικών πενταγώνων σε ένα φουλερένιο, επιβαρύνει τη σταθερότητα του μορίου καθώς έχει σαν αποτέλεσμα αυξημένη κυρτότητα και κατ' επέκταση μεγαλύτερη παραμόρφωση. Είναι επομένως ενεργειακά συμφέρον κάθε πεντάγωνο να βρίσκεται απομονωμένο και αυτό υποδεικνύει ο κανόνας του απομονωμένου πενταγώνου. Σταθεροποίηση μη σταθερών φουλερενίων μπορούμε να επιτύχουμε με άτομα της ΙΑ ομάδας του περιοδικού πίνακα αφού σχηματίζουν δεσμούς με τους άνθρακες του μορίου. Για μεγάλο αριθμό ανθράκων το φουλερένιο τείνει να αποκτήσει μηδενική καμπυλότητα κατά τη διαδρομή που ενώνει 2 οποιουσδήποτε πενταμελείς δακτυλίους. Η ηλεκτρονική μορφή ενός μορίου φουλερενίου χαρακτηρίζεται από HM με πέντε εκφυλισμένα ενεργειακά επίπεδα και LUM με τρία εκφυλισμένα ενεργειακά επίπεδα. Καθώς τα πέντε ενεργειακά επίπεδα της HM είναι κατειλημμένα από δέκα ηλεκτρόνια (και πιο συγκεκριμένα π ηλεκτρόνια) το ατομικό φουλερένιο θεωρείται μονωτής. Τα φουλερένια αποτελούν δομές με ιδιαίτερα μορφολογικά χαρακτηριστικά. Το φουλερένιο C 60 εμφανίζει 3 είδη αξόνων συμμετρίας, 2ης, 3ης και 5ης τάξεως, συνολικά σε αριθμό 21. Εμφανίζει επίσης 15 διαφορετικά επίπεδα συμμετρίας. Τέλος παραμένει αναλλοίωτο σε αντιστροφή των αξόνων του συστήματος αναφοράς. Συνδυάζοντας όλα τα παραπάνω βρίσκουμε πως είναι πιθανές 120 διαφορετικές διαδικασίες συμμετρίας σημείου. Αυτό καθιστά το μόριο του C 60 το περισσότερο συμμετρικό [2-5,18,19,25] Τύποι νανοσωλήνων άνθρακα Εικόνα 7: Από αριστερά προς τα δεξιά βλέπουμε έναν μονοφλοιϊκό νανοσωλήνα, έναν πολυφλοιϊκό νανοσωλήνα, μία συστοιχία νανοσωλήνων και μία εικόνα συστοιχίας νανοσωλήνων με SEM [2-5]. 23
24 Οι νανοσωλήνες άνθρακα είναι ομόκεντροι κύλινδροι γραφίτη, κλειστοί σε κάθε άκρο με πενταμελείς δακτυλίους και ανακαλυφθήκαν το 1991 από τον Sumio Iijima. Οι νανοσωλήνες μπορεί να είναι πολυφλοιϊκοί (MWT) με ένα κεντρικό σωλήνα να περιβάλλεται από ένα ή περισσότερα στρώματα γραφίτη ή μονοφλοιϊκοί (SWT) όπου υπάρχει μόνο ένας σωλήνας και καθόλου επιπλέον στρώματα γραφίτη. Όταν νανοσωλήνες ομαδοποιούνται έχουμε τις λεγόμενες συστοιχίες νανοσωλήνων [2-5,18,19,25] Τεχνικές σύνθεσης νανοσωλήνων άνθρακα Οι νανοσωλήνες άνθρακα μπορούν να παρασκευαστούν με τις παρακάτω τεχνικές: 1. Εξάχνωση ηλεκτροδίων άνθρακα με τη χρήση ηλεκτρικού τόξου εκκένωσης. 2. Φωτοδιάσπαση γραφίτη με τη χρήση laser. Ένα κομμάτι άνθρακα εξατμίζεται με ακτινοβολία από laser σε υψηλή θερμοκρασία και αδρανή ατμόσφαιρα. Οι παραγόμενοι σωλήνες έχουν μικρή διασπορά ως προς τη διάμετρο. 3. Καταλυτική χημική απόθεση από ατμό (CCVD). Αέριες ενώσεις του άνθρακα (συνήθως υδρογονανθράκων ή μονοξειδίου του άνθρακα) διασπώνται καταλυτικά με τη χρήση μεταλλικών καταλυτών (Fe, Co, i) υποστηριγμένων σε υποστρώματα οξειδίων μετάλλων ή αιωρούμενων στην αέρια φάση. Τα προϊόντα μπορεί να είναι πολυφλοιϊκοί νανοσωλήνες ή μονοφλοιϊκοί ανάλογα με τις παραμέτρους της μεθόδου. Γενικά, ενώ οι πολυφλοιϊκοί νανοσωλήνες μπορούν να συντεθούν και χωρίς τη χρήση καταλύτη, οι μονοφλοιϊκοί απαιτούν την παρουσία του. Το μέγεθος μάλιστα των καταλυτικών σωματιδίων καθορίζει και τη διάμετρο του νανοσωλήνα. Θα πρέπει να αναφερθεί ότι υπάρχουν και άλλες μέθοδοι παρασκευής, ωστόσο εδώ αναφέρθηκαν οι πιο σημαντικές [2-5,19] Καταλυτική χημική απόθεση από ατμό Στη συνέχεια, από τις μεθόδους σύνθεσης οι οποίες αναφέρθηκαν παραπάνω θα εξεταστεί αναλυτικότερα η καταλυτική χημική απόθεση από ατμό. Είναι αυτή η οποία αναφέρθηκε και παραπάνω για τη σύνθεση νανοϊνών άνθρακα. Η σημαντικότητα της έγγυται στα πλεονεκτήματα χρήσης της έναντι των άλλων, δηλαδή της εξάχνωσης ηλεκτροδίων άνθρακα μέσω εκκένωσης ηλεκτρικού τόξου και της φωτοδιάσπασης με χρήση laser. Τα πλεονεκτήματα αυτά είναι: 24

25 Οι θερμοκρασίες που τη χαρακτηρίζουν είναι αρκετά χαμηλότερες (τάξεως ο Κ) από αυτές των άλλων (>3000 ο Κ). Χαρακτηρίζεται από καλύτερη ικανότητα ελέγχου της δομής του τελικού προϊόντος. Μπορεί να χρησιμοποιηθεί επομένως για να δώσει δομές νανοσωλήνων άνθρακα κατάλληλες για διάφορες χρήσεις σε αντίθεση με τις άλλες που δίνουν υψηλά εναγκαλισμένες δομές με αρκετές προσμίξεις. Ωστόσο θα πρέπει να σημειωθεί ότι σημαντική έρευνα γίνεται πάνω στην επίτευξη μεγαλύτερου ελέγχου. Η ουσία που αντιδρά μετατρέπεται σε αέρια μορφή με πυρόλυση και είναι συνήθως το ακετυλένιο (HC CH), το αιθυλένιο (H 2 C=CH 2 ), το μεθάνιο (CH 4 ) ή το μονοξείδιο του άνθρακα (C). Οι χαρακτηριστικοί χρόνοι της CVD μπορεί να είναι από λεπτά έως ώρες εν αντιθέσει με τις άλλες μεθόδους που είναι μικροί (μs ms). Κάτω από τις κατάλληλες συνθήκες (για παράδειγμα με χρήση μεταλλικών καταλυτών ενσωματωμένων στους μεσοπόρους διοξειδίου του πυριτίου) δίνει δέσμες νανοσωλήνων άνθρακα, προσανατολισμένους μεταξύ τους. Νανοσωλήνες τέτοιας δομής χρησιμοποιούνται σαν διατάξεις εκπομπής πεδίου σε ηλεκτρονικά κυκλώματα. Μέθοδοι για παραγωγή μεγάλων ωστόσο ποσοτήτων τέτοιας μορφής δεν έχουν επιτευχθεί ακόμα. Στην εικόνα 8 βλέπουμε CCVD συσκευές. Στην πρώτη περίπτωση (a) τα καταλυτικά σωματίδια βρίσκονται επί υποστρώματος μέσα σε οριζόντιο φούρνο και οι νανοσωλήνες (ή οι νανοΐνες) σχηματίζονται κάθετα στο υπόστρωμα. Οι κατακόρυφοι φούρνοι των εικόνων (b) και (c) είναι πιο κατάλληλοι για μαζική παραγωγή ενώ βλέπουμε πως στον φούρνο ( b ) ο καταλύτης δεν είναι στηριζόμενος. Εικόνα 8: (a) οριζόντιος και (b), (c) κατακόρυφοι φούρνοι CCVD. Στους (a) και (c) η καταλυτική φάση είναι στηριζόμενη ενώ στον (b) όχι [2-5]. 25
![Μέχρι σήμερα, δεν έχει κατανοηθεί πλήρως ο μηχανισμός ανάπτυξης νανοσωλήνων αλλά και νανοϊνών με τη CVD μέθοδο. Εικόνα 9: (a) ανάπτυξη κορυφής και (b) ανάπτυξη βάσης [2-5].](/docs-images/72/67903182/images/26-0.jpg "Έχουμε δύο μηχανισμούς ανάπτυξης νανοσωλήνων στην περίπτωση που ο καταλύτης είναι στηριζόμενος.")
26 Μέχρι σήμερα, δεν έχει κατανοηθεί πλήρως ο μηχανισμός ανάπτυξης νανοσωλήνων αλλά και νανοϊνών με τη CVD μέθοδο. Εικόνα 9: (a) ανάπτυξη κορυφής και (b) ανάπτυξη βάσης [2-5]. Έχουμε δύο μηχανισμούς ανάπτυξης νανοσωλήνων στην περίπτωση που ο καταλύτης είναι στηριζόμενος. Κατά τον πρώτο, το καταλυτικό σωματίδιο απομακρύνεται από το υπόστρωμα καθώς ο νανοσωλήνας σχηματίζεται σταδιακά μεταξύ αυτού και του υποστρώματος εικόνα 9(a). Κατά το δεύτερο, το καταλυτικό σωματίδιο παραμένει επί του υποστρώματος και ο νανοσωλήνας αναπτύσσεται με άνθρακες να προστίθενται συνεχώς στη βάση του εικόνα 9(b). Ο πρώτος μηχανισμός λέγεται ανάπτυξη κορυφής και ο δεύτερος ανάπτυξη βάσης. Ένας από τους πιο αποδεκτούς μηχανισμούς έχει προταθεί από τον Baker et al. Σύμφωνα με τον μηχανισμό αυτό, αέριο υδρογονάνθρακα διασπάται στην επιφάνεια του μεταλλικού καταλύτη. Παράγεται υδρογόνο και άνθρακας, και ο άνθρακας διαχέεται μέσα στο καταλυτικό σωματίδιο μέχρι την πίσω όψη του. Καθώς η διάσπαση είναι εξώθερμη δημιουργείται μια βαθμίδα θερμοκρασίας κατά μήκος του σωματιδίου. Καθώς ο συντελεστής διάχυσης του άνθρακα στο μέταλλο είναι ανάλογος της θερμοκρασίας, θα παρατηρηθεί συσσώρευση του στην πιο ψυχρή πλευρά, στην πίσω δηλαδή όψη του σωματιδίου. Ο άνθρακας θα συνεχίζει να εναποτίθεται στην πίσω μεριά του σωματιδίου και να χτίζει τη δομή όσο η μπροστινή πλευρά του καταλυτικού σωματιδίου είναι ενεργή. Στη περίπτωση που ο άνθρακας καλύψει ολοκληρωτικά αυτή τη πλευρά, ο καταλύτης δηλητηριάζεται και η ανάπτυξη της δομής του άνθρακα σταματά. Η παραπάνω διαδικασία δικαιολογεί το πώς το μέγεθος του καταλυτικού σωματιδίου καθορίζει και τη διάμετρο του νανοσωλήνα ή της ίνας. Για σωματίδια μέσης διαμέτρου μεταξύ 1 και 5 nm ευνοείται η δημιουργία μονοφλοιϊκών νανοσωλήνων ενώ για διαμέτρους σωματιδίων μεταξύ 5 και 25 nm ευνοούνται οι πολυφλοιϊκοί. Ο ανάπτυξη κορυφής μπορεί να εξηγηθεί από αυτό το μοντέλο. Το μοντέλο αυτό δεν είναι τέλειο. Καταρχάς, δεν είναι όλες οι αντιδράσεις διάσπασης εξώθερμες (όπως συμβαίνει με τη διάσπαση του μεθανίου), ωστόσο παρατηρείται δημιουργία καρβονικών δομών. Επιπλέον, η θερμοβαθμίδα την οποία δέχεται το μοντέλο δεν μπορεί να δικαιολογηθεί 26
27 καθώς τα μέταλλα παρουσιάζουν μεγάλη θερμική αγωγιμότητα και μία μικρή βάθμωση της θερμοκρασίας θα είχε ως αποτέλεσμα πολύ μεγάλη ροή θερμότητας μέσω του σωματιδίου, πράγμα το οποίο δεν ισχύει. Ένα επίσης κοινό μοντέλο, το οποίο προτάθηκε από τον berlin συνιστά ότι ο άνθρακας δε διαχέεται διαμέσου του μεταλλικού σωματιδίου αλλά στην επιφάνεια του. Τότε μία κυλινδρική δομή αρχίζει και σχηματίζεται ξεκινώντας από την περιφέρεια του καταλυτικού σωματιδίου. Αυτός ο μηχανισμός μπορεί να εφαρμοστεί εξίσου στην ανάπτυξη κορυφής όσο και στην ανάπτυξη βάσης. Οι δύο παραπάνω μηχανισμοί εφαρμόζονται και σε καταλυτική φάση με κολλοειδή μορφή. Ο άνθρακας καταρχάς καλύπτει το ένα ημισφαίριο του καταλύτη και ο νανοσωλήνας άνθρακα αναπτύσσεται καθώς άτομα άνθρακα αρχίζουν να προστίθενται στην περιφέρεια του καταλυτικού σωματιδίου. Καθώς γίνεται αυτό το σωματίδιο απομακρύνεται όλο και περισσότερο από την αρχικά σχηματισμένη ημισφαιρική επιφάνεια άνθρακα ενώ η ανάπτυξη του νανοσωλήνα τερματίζεται όταν το σωματίδιο καλυφθεί τελικά εντελώς από άνθρακες. Μοντέλα που ο νανοσωλήνας σχηματίζεται γύρω από φουλερένια τα οποία φέρουν δεσμούς με καταλυτικά άτομα (i ή Co) έχουν επίσης προταθεί καθώς υπάρχουν φουλερένια σε εικόνες ηλεκτρονικής μικροσκοπίας εγκλωβισμένα σε νανοσωλήνες [2-5,18,19,25] Καθαρισμός από προσμίξεις Τα προϊόντα όσο καθαρά και αν είναι, πάντα απαιτούν επιπλέον επεξεργασία προκειμένου να απομακρυνθούν οι προσμίξεις στο μεγαλύτερο βαθμό που γίνεται. Οι μηχανισμοί ανάπτυξης που είδαμε παραπάνω είναι δυνατόν να τερματιστούν με τέτοιο τρόπο ώστε το καταλυτικό σωματίδιο να μείνει εγκλωβισμένο μέσα στο νανοσωλήνα. Άλλες προσμίξεις εκτός από τα καταλυτικά μεταλλικά σωματίδια μπορεί να είναι φουλερένια, γραφίτης ή άμορφος άνθρακας. Τα μεταλλικά σωματίδια και τα φουλερένια απομακρύνονται σχετικά εύκολα ενώ ο γραφίτης και ο άμορφος άνθρακας απομακρύνονται δύσκολα. Ο καθαρισμός γίνεται με χημικές ή μηχανικές μεθόδους. Οι χημικές μέθοδοι έχουν το πλεονέκτημα του χαμηλού κόστους και της ευκολίας χρήσης. Οι μηχανικές προσφέρονται πιο πολύ για μεγάλης κλίμακας σύνθεσης νανοσωλήνων αλλά είναι απαιτητικές σε χρόνο. Καθαρισμός μπορεί να γίνει και με τον συνδυασμό τους. Για παράδειγμα, με φυγοκέντρηση απομακρύνονται τα μεγαλύτερα σωματίδια γραφίτη ενώ με κατάλληλα διαλύματα, μέσω διάλυσης απομακρύνονται τα φουλερένια και τα μεταλλικά σωματίδια. Τα μεταλλικά σωματίδια συνήθως απομακρύνονται απλά με τη χρήση υψηλών θερμοκρασιών. 27
28 Ωστόσο, δεν υπάρχει κάποια γενική αποτελεσματική και απλή μέθοδος για τον καθαρισμό των προϊόντων αλλά ανάλογα με τις απαιτήσεις επιλέγεται η κατάλληλη [2-5,18,19,25] Δομή νανοσωλήνων άνθρακα Στους πολυφλοιϊκούς νανοσωλήνες οι εσωτερικοί σωλήνες έχουν διάμετρο της τάξης μερικών nm ενώ οι εξωτερικοί μπορεί να έχουν διαμέτρους πολλές φορές πολλαπλάσιους. Στα άκρα των κυλίνδρων γραφίτη δεν υπάρχουν ελεύθεροι δεσμοί αλλά αυτά καλύπτονται από κατάλληλες ημισφαιρικές δομές παρόμοιες με αυτές των φουλερενίων. Θεωρούμε ένα στρώμα γραφίτη όπως στο σχήμα παρακάτω. Φανταζόμαστε έναν νανοσωλήνα σαν ένα στρώμα γραφίτη το οποίο έχει τυλιχτεί κατά τέτοιο τρόπο ώστε η αρχή και το τέλος του χειραλικού διανύσματος (chiral vector) C r h να συμπέσουν. Προκειμένου επομένως να περιγράψουμε τη δομή ενός νανοσωλήνα αρκεί να ορίσουμε το διάνυσμα C r h. Στην παραπάνω εικόνα φαίνεται ένα επίσης σημαντικό μέγεθος για τη περιγραφή της δομής, η χειραλική γωνία (chiral angle) θ. Η σχέση η οποία δίνει το C r h είναι η r r r Ch = na1 + ma 2 όπου a r 1 και a r 2 είναι τα μοναδιαία διανύσματα σε ένα δυσδιάστατο πλέγμα γραφίτη και n, m ακέραιοι. Η γωνία θ ορίζεται από τη σχέση: 28

29 Ισχύει 0 θ 30 όπου θ είναι δηλαδή η γωνία μεταξύ του διανύσματος C r h και του 1 a r 2 έτσι ώστε να ισχύει 30 0 θ. a r ή του Το μέτρο του C r h είναι ίσο με τη περίμετρο του νανοσωλήνα και μπορούμε να υπολογίσουμε από τη γεωμετρία του σχήματος τόσο τη διάμετρο d όσο και την ακριβή σχέση που δίνει το θ. Η διάμετρος d υπολογίζεται ως εξής: r r Έστω α 1 = α 2 = α. Αν α c-c η απόσταση μεταξύ δύο ανθράκων στο πλέγμα (α c-c = 1,4210 Å σε θερμοκρασία δωματίου) τότε: Για το μέτρο του π α = 2α c c 2α c c cos = 2α c c 2α c c = 3α c c α = 3α (1) c c 3 2 C r h έχουμε: Οπότε: Στη συνέχεια υπολογίζουμε τη σχέση η οποία μας δίνει τη γωνία θ. Από το νόμο των ημιτόνων και συνημίτονων έχουμε αντίστοιχα: και Διαιρώντας κατά μέλη παίρνουμε: Εικόνα 10: Παραδείγματα armchair, zigzag και chiral νανοσωλήνων a ) (5, 5)-, b ) (9, 0)- και c ) (10, 5)- νανοσωλήνας [2-5]. 29
30 Αν ισχύει n = m (κι επομένως θ=30 ο ) ο νανοσωλήνας είναι τύπου " armchair ". Αν πάλι ισχύει n = 0, m 0 ή n 0, m = 0 (και επομένως θ=0) ο νανοσωλήνας είναι τύπου "zigzag". Και τα δύο παραπάνω ανήκουν στην κατηγορία των achiral νανοσωλήνων. Οποιαδήποτε άλλη τιμής της γωνίας θ χαρακτηρίζει νανοσωλήνες του γενικού τύπου chiral, νανοσωλήνων τέτοιας δομής δηλαδή που δε μπορεί να ταυτιστεί με την κατοπτρική της με κανέναν τρόπο. Ανάλογα με τον τύπο του νανοσωλήνα διαφορετική τιμή παίρνει και η απόσταση μεταξύ των διαδοχικών στρωμάτων άνθρακα στους πολυφλοιϊκούς νανοσωλήνες. Για (n, n) και (n, 0) νανοσωλήνες έχουμε αντίστοιχα 3,38 Å και 3,41 Å. Εδώ επίσης θα πρέπει να αναφερθεί ότι τα στρώματα του γραφίτη στους πολυφλοιϊκούς νανοσωλήνες δεν εμφανίζουν την τέλεια διάταξη ΑΒΑΒ που παρουσιάζει ο κρυσταλλικός γραφίτης μεταξύ των στρωμάτων του. Όπως είδαμε οι νανοσωλήνες χαρακτηρίζονται τόσο από πενταμελείς όσο και εξαμελείς δακτυλίους άνθρακα. Ωστόσο, απόκλιση από τον εξαμελή αποτελούν και οι επταμελείς δακτύλιοι οι οποίοι σε αντίθεση με τους πενταμελείς δίνουν αρνητική καμπυλότητα στο νανοσωλήνα. Αν πενταμελείς δακτύλιοι βρίσκονται απέναντι από επταμελείς τότε ο νανοσωλήνας αποκτά κυρτότητα. Επίσης σωστός συνδυασμός πενταμελών και επταμελών δακτυλίων μπορούν να κάνουν εφικτή την ένωση ενός νανοσωλήνα με κάποιον άλλον, διαφορετικής δομής. Κατά αυτό τον τρόπο μπορούν να δημιουργηθούν ετεροεπαφές όπως μετάλλου - ημιαγωγού ή p - n εξ' ολοκλήρου από νανοσωλήνες. Για ημιαγώγιμους νανοσωλήνες τύπου p ή n θα μιλήσουμε παρακάτω. Τα άκρα στους πολυφλοιϊκούς σωλήνες μπορούν να απομακρυνθούν με οξείδωση από διοξείδιο του άνθρακα ή οξυγόνο σε υψηλές θερμοκρασίες καθώς και με διάλυμα H 3 υψηλής συγκέντρωσης. Έτσι γίνεται εφικτό το γέμισμα του νανοσωλήνα με άλλο υλικό. Αν για παράδειγμα μαζί με το διάλυμα H 3 είναι παρόν κάποιο μεταλλικό άλας όπως το i( 3 ) 2 τότε το δεύτερο μετατρέπεται κατά τη διαδικασία σε οξείδιο. Αναγωγή με υδρογόνο τελικά σε θερμοκρασία 400 ο C δίνει μέταλλο μέσα στο σωλήνα. Ομοίως σε μονοφλοιϊκούς χρησιμοποιούνται για τον ίδιο σκοπό ήπια όξινα διαλύματα [2-5,18,19,25] Ηλεκτρονικές ιδιότητες νανοσωλήνων άνθρακα Οι ηλεκτρονικές ιδιότητες των νανοσωλήνων παρουσιάζουν ιδιαίτερο ενδιαφέρον λόγω των πολλών τεχνολογικών εφαρμογών που έχουν. Όπως και στα φουλερένια, η καμπυλότητα των φύλλων γραφίτη επηρεάζει σημαντικά τις ηλεκτρονικές ιδιότητες των νανοσωλήνων. Ο κυριότερος παράγοντας ο οποίος καθορίζει τις ηλεκτρονικές ιδιότητες ενός νανοσωλήνα είναι ότι τα ηλεκτρόνια του είναι χωρικά περιορισμένα. Μπορούν να κινηθούν μόνο στο χώρο του γραφίτη και όχι κατά την κάθετη στον άξονα διεύθυνση στο φύλλο του γραφίτη. 30
31 Επιπλέον, καθώς το μήκος του είναι πολύ μεγαλύτερο από τη διάμετρο του οι επιτρεπόμενες ηλεκτρονικές καταστάσεις κατά την αξονική διεύθυνση θα είναι πολύ περισσότερες από αυτές κατά την περιφερειακή. Μπορούμε επομένως να θεωρήσουμε πως τα ηλεκτρόνια του νανοσωλήνα χαρακτηρίζονται από κυματανύσματα αξονικής διεύθυνσης. Η πεπερασμένη περιφέρεια και μήκος του νανοσωλήνα εισάγουν οριακές συνθήκες οι οποίες έχουν ως αποτέλεσμα διακριτές ενεργειακές στάθμες και οι νανοσωλήνες παρουσιάζουν τόσο μεταλλική όσο και ημιαγώγιμη συμπεριφορά. Για νανοσωλήνες μικρής διαμέτρου, η σχέση μεταξύ των n και m τα οποία περιγράφουν τη δομή του νανοσωλήνα, είναι ένα μέτρο για το αν ο νανοσωλήνας θα παρουσιάζει μεταλλική ή ημιαγώγιμη συμπεριφορά. Για έναν (n, m) νανοσωλήνα, αν ισχύει n m = 3q, όπου q ακέραιος, τότε ο νανοσωλήνας ανήκει στα μέταλλα. Αν όχι, ο νανοσωλήνας ανήκει στους ημιαγωγούς. Σύμφωνα επομένως με την παραπάνω σχέση, όλοι οι armchair νανοσωλήνες και το ένα τρίτο των zigzag είναι μέταλλα. Το ενεργειακό χάσμα σε έναν ημιαγώγιμο νανοσωλήνα είναι αντιστρόφως ανάλογο της διαμέτρου του. Επίσης, έχει δειχθεί ότι στους πολυφλοιϊκούς νανοσωλήνες το ρεύμα άγεται μόνο από το εξωτερικό στρώμα και όχι από όλο τον όγκο του νανοσωλήνα. Τέλος, είναι δυνατόν να εισάγουμε προσμίξεις στους νανοσωλήνες αντικαθιστώντας άτομα άνθρακα από άλλα άτομα, όπως το βόριο (B - C, συνήθως με αναλογία η οποία περιγράφεται από τον τύπο C 35 B) ή το άζωτο (C -, με αντίστοιχο τύπο C 33 ) προκειμένου να δοθούν στο νανοσωλήνα ημιαγώγιμες ιδιότητες τύπου p ή n αντίστοιχα. Συνδυασμοί των δύο παραπάνω είναι επίσης εφικτοί ( B - C - ) [2-5,18,19,25] Μηχανικές και άλλες ιδιότητες νανοσωλήνων άνθρακα Οι νανοσωλήνες είναι εξαιρετικά ανθεκτικοί κατά μήκος του άξονα τους. Η ιδιότητα τους αυτή βρίσκει χρήση στην ενίσχυση άλλων υλικών. Αυτός είναι ένας επιπλέον λόγος που επιζητούμε σύνθεση δεσμών νανοσωλήνων σε μεγάλες ποσότητες. Το μέτρο του Young τόσο για πολυφλοιϊκούς όσο και μονοφλοιϊκούς σωλήνες είναι άνω του 1 TPa (1TPa=10 12 m -2 ) και μειώνεται καθώς η διάμετρος τους αυξάνει. Πιέσεις μέχρι και 30 GPa δε καταστρέφουν τη βασική δομή του σωλήνα. Η πυκνότητα τους εξαρτάται από την ακριβή δομή τους. Ενδεικτικά, η πυκνότητα ενός zigzag (17,0) νανοσωλήνα υπολογίζεται 1,34 gr/cm 3. Όσον αφόρα την ειδική θερμοχωρητικότητα η οποία τους χαρακτηρίζει, για τους μεν πολυφλοιϊκούς βρίσκεται παρόμοια με αυτή του γραφίτη ενώ για τους μονοφλοιϊκούς μεγαλύτερη. Αυτό είναι αναμενόμενο καθώς τα φωτόνια του κρυστάλλου χαρακτηρίζονται από μεγαλύτερη μέση ελεύθερη διαδρομή (περίπου 100 nm) αφού δεν υπάρχουν επιπλέον στρώματα γραφίτη για να επηρεάσουν την κίνηση τους μέσω της μεταξύ τους αλληλεπίδρασης. 31
32 Η μετακίνηση των νανοσωλήνων στο χώρο πάνω σε κάποιο υπόστρωμα μπορεί να γίνει με τη χρήση της ακίδας ενός AFM μικροσκοπίου. Αυτή η τεχνική έχει χρησιμοποιηθεί και για άλλους σκοπούς, όπως η μέτρηση της φύσης της αλληλεπίδρασης μεταξύ ενός νανοσωλήνα και του υλικού του υποστρώματος. Τέλος, οι ίδιοι οι νανοσωλήνες έχουν πολύ καλή εφαρμογή σαν ακίδες AFM μικροσκοπίων λόγω των αναλογιών μήκους - διαμέτρου εισχωρώντας στις βαθιές ανωμαλίες των επιφανειών. Δεν σπάνε και όντας εξαιρετικά ελαστικοί αποφεύγονται φαινόμενα προσκόλλησης της ακίδας στο δείγμα. Ένα STM μικροσκόπιο μπορεί επίσης να χρησιμοποιηθεί για το χειρισμό νανοσωλήνων. Στέλνοντας έναν δυνατό παλμό δυναμικού μέσω της ακίδας στο νανοσωλήνα, αυτός μπορεί να τεμαχιστεί τοπικά. Τέλος η μεγάλη τους ηλεκτρική αγωγιμότητα τους κάνει κατάλληλους σαν ακίδες STM μικροσκοπίων [2-5,18,19,25] Μηχανισμοί αποθήκευσης υδρογόνου σε νανοσωλήνες άνθρακα Το υδρογόνο αποθηκεύεται στις νανοΐνες άνθρακα ανάμεσα στα επίπεδα του γραφίτη που την αποτελούν και συγκρατείται με δυνάμεις Van der Waals. Ανάλογα με τον προσανατολισμό των επιπέδων αλλάζει και η μέγιστη ποσότητα που μπορεί να αποθηκευτεί. Ωστόσο, αν και οι νανοΐνες άνθρακα παρουσιάζουν ιδιαίτερο ενδιαφέρον για την αποθήκευση υδρογόνου, η περισσότερη έρευνα, πειραματικά και θεωρητικά, σήμερα εστιάζεται στην αποθήκευση υδρογόνου σε νανοσωλήνες άνθρακα. Η αποθήκευση στους νανοσωλήνες γίνεται με δύο τρόπους, ή με φυσική προσρόφηση του υδρογόνου σε αυτούς ή με χημική προσρόφηση. Όπως γνωρίζουμε, στην πρώτη περίπτωση αυτό γίνεται με το υδρογόνο να διατηρεί τη μοριακή του δομή και να "δένεται" στην επιφάνεια με δυνάμεις Van der Waals. Στη δεύτερη γίνεται με το ατομικό υδρογόνο να δημιουργεί χημικούς δεσμούς με τους άνθρακες των νανοσωλήνων. Οι ακριβείς μηχανισμοί ωστόσο που κατευθύνουν το πώς γίνεται η προσρόφηση του υδρογόνου στους νανοσωλήνες δεν είναι εξακριβωμένοι ακόμα. Συχνά υπάρχει ασυμφωνία μεταξύ πειράματος και θεωρητικών προβλέψεων. Αυτό μπορεί να οφείλεται στην μη καθαρότητα των δειγμάτων αλλά επίσης σημαντικό ρόλο παίζει και η δυσκολία να πραγματοποιηθούν ακριβείς μετρήσεις σε ατομικό επίπεδο. Εξάλλου τα μοντέλα στα οποία στηρίζεται η θεωρία (κυρίως μέσω εξομοιώσεων) είναι ιδανικά και η παραμετροποίηση τους (όπου αυτή απαιτείται) είναι πολλές φορές μη ρεαλιστική. Επομένως είναι δύσκολο να καθορίσουμε πότε το υδρογόνο προσροφάται αποκλειστικά φυσικά ή λαμβάνουν χώρα και χημικές διεργασίες. Το υδρογόνο μπορεί να αποθηκευτεί στους νανοσωλήνες με τους παρακάτω τρόπους: 1. Στο εσωτερικό τους. Το υδρογόνο σε αυτή τη περίπτωση μπορεί να σχηματίζει μονόστρωμα κυλινδρικής μορφής ενώ για μεγαλύτερες ποσότητες σχηματίζονται 32
33 αλλεπάλληλα κυλινδρικά στρώματα που είναι ομοκεντρικά αν δεν αναφερόμαστε σε πολύ μεγάλες ποσότητες. 2. Στην επιφάνεια των μονοφλοιϊκών ή στην εξωτερική επιφάνεια των πολυφλοιϊκών. 3. Μεταξύ των εσωτερικών επιφανειών των πολυφλοιϊκών. Τότε, η ποσότητα του προσροφημένου υδρογόνου δεν επηρεάζεται από τη διάμετρο του νανοσωλήνα. 4. Μεταξύ των νανοσωλήνων όταν αυτοί βρίσκονται υπό τη μορφή συστοιχίας. Αυτό είναι αρκετά σημαντικό διότι η συγκεκριμένη ιδιότητα κυρίως δίνει στους νανοσωλήνες καλύτερες αποθηκευτικές ιδιότητες από ότι τα απλά στρώματα γραφίτη. Για το διαχωρισμό των νανοσωλήνων της συστοιχίας απαιτούνται υψηλές πιέσεις. Τότε, το υδρογόνο εισχωρεί μέσα σε αυτή και προσροφάται από όλες τις εκτεθειμένες επιφάνειες άνθρακα [2-5,18,19,25] Φυσική προσρόφηση Αναφερόμενοι στη φυσική προσρόφηση [14-20] υπολογίζεται ότι η θέση ισορροπίας για το προσροφημένο μόριο υδρογόνου βρίσκεται περίπου μία ατομική ακτίνα από την επιφάνεια του νανοσωλήνα είτε αυτό βρίσκεται εξωτερικά είτε εσωτερικά υπό τη μορφή μονοστρώματος. Είναι αναμενόμενο το υδρογόνο να προσροφάται πιο εύκολα σε έναν νανοσωλήνα άνθρακα παρά σε ένα στρώμα γραφίτη (χωρίς αυτό όμως να σημαίνει ότι και ποσοτικά οι νανοσωλήνες υπερτερούν πάντα του γραφίτη). Αυτό γιατί στην περίπτωση των νανοσωλήνων, λόγω της καμπυλότητας τους, τα πεδία από τα αντικρινά άτομα άνθρακα αλληλεπικαλύπτονται και έτσι η ελκτική δύναμη αυξάνεται συγκρινόμενη με αυτή μιας ελεύθερης επιφάνειας. Σε ένα zigzag νανοσωλήνα (13,0) το δυναμικό του νανοσωλήνα στο εσωτερικό του είναι 25% φορές μεγαλύτερο από αυτό την επιφάνεια ενός απλού φύλλου γραφίτη. Στο εξωτερικό ενός νανοσωλήνα, αρχικά το αέριο του υδρογόνου προσροφάται σε αυτόν. Όταν αρχίζει να σχηματίζεται σε κάποιο βαθμό ένα μονόστρωμα υδρογόνου τα επιπλέον μόρια θα προσροφόνται είτε στο στερεό νανοσωλήνα ή στο σχηματισθέν μονόστρωμα υγρού, μιλώντας πάντα για θερμοκρασίες κάτω από το σημείο εξατμίσεως. Σε υψηλότερες θερμοκρασίες σχηματίζεται μόνο ένα στρώμα προσροφημένου υδρογόνου. Αυτός είναι ο λόγος που το ποσοστό του προσροφημένου υδρογόνου βρίσκεται πολύ μεγάλο σε χαμηλές θερμοκρασίες αλλά ταυτόχρονα μη κατάλληλες για πρακτική εφαρμογή. Ο υπολογισμός της ποσότητας του υδρογόνου η οποία προσροφάται πάνω στο νανοσωλήνα γίνεται σύμφωνα με τους παρακάτω υπολογισμούς. Καταρχάς δεχόμαστε ότι το υδρογόνο στην επιφάνεια του νανοσωλήνα συμπεριφέρεται σαν υγρό. Δεδομένης της πυκνότητας του προσροφημένου υγρού P liq και της μοριακής μάζας του M ads, ο όγκος που καταλαμβάνει κάθε μόριο, είναι: 33
34 M ads V ad = P όπου A ο αριθμός Avogadro ( A = 6, mol -1 ). Θεωρώντας πως η μοριακή δομή του μονοστρώματος υδρογόνου περιγράφεται από πλέγμα f c c πυκνής κατάληψης, ο όγκος που καταλαμβάνεται από τα μόρια αν αυτά ήταν σφαιρικά υπολογίζεται σύμφωνα mε την εξίσωση: liq A Ο παράγοντας π ισούται με το ποσοστό κατάληψης μιας f c c κυψελίδας (74,05%). Από τον 3 2 όγκο μπορούμε να υπολογίσουμε την ακτίνα του μορίου: και τελικά την επιφάνεια του κάθε μορίου: Αντικαθιστώντας τις τιμές ειδικά για υδρογόνο προσροφημένο σε μονοφλοιϊκό νανοσωλήνα ειδικής επιφάνειας S spec = 1315 m 2 g -1 (η οποία είναι και η μέγιστη δυνατή) προκύπτει S M = m 2 mol -1. Η αναλογία επομένως μεταξύ υδρογόνου και άνθρακα είναι: από την οποία υπολογίζεται η συγκέντρωση C υδρογόνου στον άνθρακα: Χημειορρόφηση m C = m H C 1 = m 1+ m 2 = C H 2 3,3% Όπως αναφέρθηκε παραπάνω, νανοσωλήνες οι οποίοι χαρακτηρίζονται από χειραλική γωνία 0 ή 30 μοίρες είναι μεταλλικοί. Αυτού του τύπου νανοσωλήνες προσφέρονται για χρήση σαν ηλεκτρόδια σε ηλεκτροχημικά στοιχεία. Πρωτόνια μένουν προσκολλημένα στην επιφάνεια του νανοσωλήνα, ο οποίος αποτελεί το καθοδικό ηλεκτρόδιο, καθώς ηλεκτρόνια μεταφέρονται από αυτόν στο διάλυμα. Η αντίδραση η οποία περιγράφει το φαινόμενο είναι η: Αυτά τα άτομα μπορούν να ενωθούν μεταξύ τους και να δημιουργήσουν μόρια υδρογόνου. Τα μόρια αυτά είναι αρκετά ευκίνητα όσων αφορά την κίνηση τους πάνω στην 34

35 επιφάνεια του σωλήνα. Για να δημιουργηθεί ένα σταθερό στρώμα υδρογόνου θα πρέπει η κινητικότητα των μορίων να παρεμποδιστεί. Αυτό γίνεται καθώς μόρια νερού σχηματίζουν ένα δεύτερο προσροφημένο στρώμα το οποίο δημιουργεί ένα φράγμα δυναμικού στην κίνηση του υδρογόνου. Υπάρχουν ωστόσο και άλλοι πιθανοί μηχανισμοί για το πώς το υδρογόνο μπορεί να αποθηκευτεί σε νανοσωλήνες με αυτή τη μέθοδο. Μάλιστα αυτοί οι μηχανισμοί είναι δυνατόν να λαμβάνουν χώρα ταυτόχρονα με τη φυσική προσρόφηση. Αν και δεν αποτελούν επιβεβαιωμένα μοντέλα αλλά έχουν προκύψει από εξομοιώσεις αξίζει να αναφερθούν συνοπτικά: Το πρώτο μοντέλο θέλει το ατομικό υδρογόνο να καλύπτει την επιφάνεια ενός armchair νανοσωλήνα δημιουργώντας χημικούς δεσμούς με τους άνθρακες του. Το ατομικό υδρογόνο σπρώχνει το άτομο του άνθρακα προς το εσωτερικού του σωλήνα. Τότε αλληλεπιδρά με τα ηλεκτρονικά νέφη των δύο πιο κοντινών του ανθράκων. Τέλος πραγματοποιείται μία περιστροφή του συστήματος των ανθράκων και του υδρογόνου, το υδρογόνο μεταφέρεται στο εσωτερικό του σωλήνα και ο δεσμός μεταξύ των ανθράκων αποκαθίσταται. Αυτή η διαδικασία (flip - in process) επαναλαμβάνεται και για τα άλλα άτομα. Τελικά ενεργειακά ευνοείται η ένωση των ατομικών υδρογόνων στο εσωτερικό του σωλήνα και τη δημιουργία μοριακού, φυσικά προσροφημένου. Στην παρακάτω εικόνα 11 φαίνονται παραστατικά τα παραπάνω ( I και II ). Εικονα 11: Μοντέλα απορρόφησης υδρογόνου από νανοσωλήνες άνθρακα [2-5]. 35
36 Κατά ένα δεύτερο μοντέλο ενεργειακά ευνοείται η μεταφορά κάθε δεύτερου υδρογόνου στο εσωτερικού του σωλήνα. Κάθε νέο άτομο υδρογόνου το οποίο πλησιάζει το σωλήνα χημειορροφάται από κάποιο άτομο άνθρακα το οποίο είναι ενωμένο με άτομο υδρογόνου στο εσωτερικό του σωλήνα. Το άτομο αυτό προκειμένου να αλληλεπιδράσει ο άνθρακας με το εξωτερικό άτομο αποσπάται στο εσωτερικό του σωλήνα (kick - in process). Αν η διαδικασία αυτή συνδυαστεί με αυτή της προηγούμενης παραγράφου (μεταφοράς του υδρογόνου στο εσωτερικό) εναλλάξ θα έχουμε για άλλη μία φορά τη συσσώρευση μοριακού υδρογόνου στο εσωτερικό του σωλήνα (I, III και IV στην εικόνα 11). Η εκρόφηση του υδρογόνου γίνεται με παρόμοιο τρόπο ακολουθώντας τα παραπάνω βήματα κατά αντίθετη σειρά. Οι μηχανισμοί αυτοί μπορούν να εξηγήσουν εύκολα πως το υδρογόνο μπορεί να αποθηκευτεί εσωτερικά σε έναν νανοσωλήνα. Εξάλλου, με το να αφαιρέσουμε απλά τα κλειστά άκρα ενός νανοσωλήνα ώστε να εκτεθεί το εσωτερικό του στο υδρογόνο δεν σημαίνει ότι θα έχουμε ιδιαίτερη αύξηση στο προσροφημένο αέριο εξαιτίας του μεγάλου μήκους του νανοσωλήνα [2-5,18,19,25] Αποθήκευση σε νανοσωλήνες άνθρακα προσμίξεων αλκαλίων Οι νανοσωλήνες θεωρούνται ιδανικά υλικά για την αποθήκευση υδρογόνου. Ωστόσο, μέχρι στιγμής, τα ποσά υδρογόνου που αποθηκεύονται σε νανοσωλήνες (τα οποία έχουν επιβεβαιωθεί πειραματικά) απέχουν από τις θεωρητικές εκτιμήσεις τουλάχιστον σε θερμοκρασία περιβάλλοντος και σε χαμηλές πιέσεις. Ενθαρρυντικά όμως, είναι τα αποτελέσματα τα οποία δίνουν πειράματα σε τέτοιες συνθήκες με νανοσωλήνες άνθρακα και προσμίξεων αλκαλίων, όπως Li ή K. Το αλκαλικό άτομο βρίσκεται σε κάποια απόσταση πάνω από το κέντρο ενός εξαγωνικού δακτυλίου. Μία πιθανή εξήγηση για αυτό το φαινόμενο είναι ότι το μεταλλικό σωματίδιο πολώνεται θετικά καθώς λαμβάνει χώρα μεταφορά φορτίου από αυτό στο νανοσωλήνα. Το θετικά φορτισμένο αυτό σωματίδιο με τη σειρά του πολώνει το μοριακό υδρογόνο και τελικά δημιουργείται ένας ασθενής δεσμός μεταξύ των δύο [2-5,18,19,25]. 36
37 1.2.1 Bασικές αρχές κατάλυσης από ένζυμα Η ενζυμική κατάλυση αρχίζει με την πρόσδεση του υποστρώματος. Η ενέργεια της πρόσδεσης είναι η ελεύθερη ενέργεια που απελευθερώνεται κατά τον σχηματισμό ενός μεγάλου αριθμού ασθενών αλληλεπιδράσεων μεταξύ του ενζύμου και του υποστρώματος. Η ενέργεια αυτή της πρόσδεσης εξυπηρετεί δύο σκοπούς: εγκαθιστά την εξειδίκευση υποστρώματος και αυξάνει την καταλυτική ικανότητα. Μόνο το σωστό υπόστρωμα μπορεί να συμμετέχει στις περισσότερες ή όλες τις αλληλεπιδράσεις με το ένζυμο και έτσι μεγιστοποιεί την ενέργεια πρόσδεσης, εξηγώντας την εξαιρετική εξειδίκευση για το υπόστρωμα που εμφανίζουν πολλά ένζυμα. Επιπλέον, η πλήρης συμπληρωματικότητα τέτοιων αλληλεπιδράσεων σχηματίζεται μόνον όταν το υπόστρωμα είναι στη μεταβατική κατάσταση. Έτσι, αλληλεπιδράσεις μεταξύ του ενζύμου και του υποστρώματος όχι μόνον ευνοούν την πρόσδεση του υποστρώματος αλλά σταθεροποιούν τη μεταβατική κατάσταση, μειώνοντας με τον τρόπο αυτό την ενέργεια ενεργοποίησης. Η ενέργεια της πρόσδεσης μπορεί επίσης να προαγάγει δομικές αλλαγές και στο ένζυμο και στο υπόστρωμα που διευκολύνουν την κατάλυση, μια πορεία που αναφέρεται ως επαγόμενη προσαρμογή. Τα ένζυμα χρησιμοποιούν μία από τις ακόλουθες στρατηγικές για να καταλύσουν ειδικές αντιδράσεις: 1. Ομοιοπολική κατάλυση. Στην ομοιοπολική κατάλυση, το ενεργό κέντρο περιέχει μια ενεργό ομάδα, συνήθως ένα ισχυρό πυρηνόφιλο το οποίο στην πορεία της κατάλυσης προσωρινά τροποποιείται ομοιοπολικά. Το πρωτεολυτικό ένζυμο χυμοθρυψίνη αποτελεί ένα εξαιρετικό παράδειγμα του μηχανισμού αυτού. 2. Γενική οξεοβασική κατάλυση. Στη γενική οξεοβασική κατάλυση, ένα μόριο διαφορετικό από το νερό παίζει τον ρόλο ενός δότη ή δέκτη πρωτονίων. Η χυμοθρυψίνη χρησιμοποιεί ένα κατάλοιπο ιστιδίνης ως βασικό καταλύτη για να αυξήσει την πυρηνοφιλική ισχύ της σερίνης. 3. Κατάλυση ιόντος μετάλλου. Τα ιόντα μετάλλων μπορούν να λειτουργήσουν καταλυτικά με αρκετούς τρόπους. Για παράδειγμα, ένα ιόν μετάλλου μπορεί να χρησιμεύσει ως ηλεκτρονιόφιλος καταλύτης, σταθεροποιώντας ένα αρνητικό φορτίο σε ένα ενδιάμεσο της αντίδρασης. Εναλλακτικά, το ιόν μετάλλου μπορεί να παραγάγει ένα πυρηνόφιλο αυξάνοντας την οξύτητα ενός γειτονικού μορίου, όπως το νερό στην ενυδάτωση του C 2 από την ανθρακική ανυδράση. Τελικά, το ιόν μετάλλου μπορεί να προσδεθεί σε ένα υπόστρωμα, αυξάνοντας τον αριθμό των αλληλεπιδράσεων με το ένζυμο και έτσι την ενέργεια πρόσδεσης. Η στρατηγική αυτή χρησιμοποιείται από τις κινάσες. 4. Κατάλυση με προσέγγιση. Πολλές αντιδράσεις περιλαμβάνουν δύο ξεχωριστά υποστρώματα. Στις περιπτώσεις αυτές, η ταχύτητα της αντίδρασης μπορεί να αυξηθεί αξιοσημείωτα από την πρόσδεση και των δύο υποστρωμάτων μαζί σε μια μοναδική 37
38 επιφάνεια πρόσδεσης επάνω στο ένζυμο. Οι κινάσες ΝΜΡ φέρουν μαζί δύο νουκλεοτίδια για να διευκολύνουν τη μεταφορά μιας φωσφορικής ομάδας από το ένα νουκλεοτίδιο στο άλλο [11] Πρωτεολυτικά ένζυμα - πρωτεϊνόλυση Τα ένζυμα επιταχύνουν τις αντιδράσεις κατά ένα εκατομμύριο φορές ή περισσότερο. Πράγματι, στα βιολογικά συστήματα οι περισσότερες αντιδράσεις δεν γίνονται καν σε αντιληπτή ταχύτητα χωρίς την παρουσία ενζύμων. Ακόμη και μια απλή αντίδραση, όπως είναι η ενυδάτωση του διοξειδίου του άνθρακα, καταλύεται από ένα ένζυμο, την ανθρακική ανυδράση. Η μεταφορά του C 2, από τους ιστούς στο αίμα και στη συνέχεια στον κυψελιδικό αέρα χωρίς το ένζυμο αυτό δεν θα ήταν πλήρης. Στην πραγματικότητα, η ανθρακική ανυδράση είναι ένα από τα ταχύτερα ένζυμα που γνωρίζουμε. Κάθε μόριο ενζύμου μπορεί να ενυδατώνει 10 6 μόρια C 2 ανά δευτερόλεπτο. Η αντίδραση που καταλύεται είναι 10 7 φορές ταχύτερη από εκείνη που δεν καταλύεται. Τα ένζυμα έχουν υψηλό βαθμό εξειδίκευσης τόσο στην αντίδραση που καταλύουν όσο και στην επιλογή των αντιδρώντων, που ονομάζονται υποστρώματα. Ένα ένζυμο καταλύει συνήθως μια απλή χημική αντίδραση ή μια σειρά από πολύ συγγενικές αντιδράσεις. Στις ενζυμικές αντιδράσεις σπανίως συμβαίνουν παράπλευρες αντιδράσεις που οδηγούν στον άχρηστο σχηματισμό παραπροϊόντων, σε αντίθεση με τις αντιδράσεις που δεν καταλύονται. Ας πάρουμε ως παράδειγμα τα πρωτεολυτικά ένζυμα. Τα ένζυμα αυτά, in vivo, καταλύουν την πρωτεϊνόλυση, την υδρόλυση ενός πεπτιδικού δεσμού: Τα περισσότερα πρωτεολυτικά ένζυμα καταλύουν επίσης την υδρόλυση ενός εστερικού δεσμού, in vitro, που είναι μια διαφορετική αλλά σχετική αντίδραση. Τέτοιες αντιδράσεις παρακολουθούνται πιο εύκολα από ότι η πρωτεϊνόλυση και είναι χρήσιμες στην πειραματική έρευνα των ενζύμων αυτών. 38
39 Τα πρωτεολυτικά ένζυμα διαφέρουν σημαντικά στον βαθμό εξειδίκευσης που έχουν ως προς το υπόστρωμα. Η σουμπτιλυσίνη, η οποία βρέθηκε σε ορισμένα βακτήρια, δεν κάνει διακρίσεις: θα διασπάσει οποιονδήποτε δεσμό δίνοντας λίγη σημασία στη φύση των παρακείμενων πλευρικών αλυσίδων. Η θρυψίνη, ένα ένζυμο της πέψης, είναι απολύτως εξειδικευμένη στο να καταλύει μόνο τη διάσπαση των πεπτιδικών δεσμών στο καρβοξυ-τελικό άκρο των καταλοίπων της λυσίνης και της αργινίνης (Εικόνα 12A). Η θρομβίνη, ένα ένζυμο που συμμετέχει στην πήξη του αίματος, είναι περισσότερο εξειδικευμένη από τη θρυψίνη. Καταλύει την υδρόλυση των δεσμών Arg-Gly μόνο σε ειδικές αλληλουχίες πεπτιδίων (Εικόνα 12B). Εικόνα 12: Εξειδίκευση ενζύμου. (Α) Η θρυψίνη διασπά τους δεσμούς στο καρβοξυ-τελικό άκρο των καταλοίπων αργινίνης και λυσίνης, ενώ (Β) η θρομβίνη διασπά τους δεσμούς Arg-Gly ειδικά σε συγκεκριμένες αλληλουχίες [11]. Η DΝΑ πολυμεράση I, ένα ένζυμο που κατευθύνεται από ένα εκμαγείο, είναι ένας άλλος καταλύτης με υψηλή εξειδίκευση. Προσθέτει νουκλεοτίδια σε μια αλυσίδα DA που συντίθεται με αλληλουχία η οποία προσδιορίζεται από την αλληλουχία των νουκλεοτιδίων στην άλλη αλυσίδα DΝΑ που χρησιμεύει ως εκμαγείο. Η DA πολυμεράση I μεταφέρει τις πληροφορίες που δίνονται από το εκμαγείο με αξιοθαύμαστη ακρίβεια. Εισάγει το λάθος νουκλεοτίδιο στη νεοσχηματιζόμενη αλυσίδα DA με συχνότητα μικρότερη από ένα εκατομμύριο φορές. Η εξειδίκευση ενός ενζύμου οφείλεται στην ακριβή αλληλεπίδραση του υποστρώματος και του ενζύμου. Η ακρίβεια αυτή είναι αποτέλεσμα της πολύπλοκης τριδιάστατης δομής της εvζυμικής πρωτεΐνης [11]. 39
40 1.2.3 Καταβολισμός πρωτεϊνών - πρωτεάσες Οι πρωτεΐνες καταβολίζονται από υδρολυτικά ένζυμα που καλούνται πρωτεϊνάσες ή πρωτεάσες ή πεπτιδάσες. Σε αναλογία με τα αμυλολυτικά και νουκλεολυτικά ένζυμα, τα πρωτεολυτικά ένζυμα διακρίνονται σε ενδοπεπτιδάσες και εξωπεπτιδάσες. Οι ενδοπεπτιδάσες υδρολύουν πεπτιδικούς δεσμούς προς το εσωτερικό του μορίου με αποτέλεσμα τα προϊόντα υδρόλυσης να είναι κυρίως ολιγοπεπτίδια. Πολλά απ αυτά τα ένζυμα παρουσιάζουν και εξειδίκευση σε άλλοτε άλλο βαθμό ως προς τη φύση των αμινοξέων που συμμετέχουν στον πεπτιδικό δεσμό που υδρολύεται. Έτσι λ.χ. η τρυψίνη υδρολύει τους πεπτιδικούς δεσμούς εκείνους που η καρβονυλομάδα ανήκει στ αμινοξέα αργινίνη και λυσίνη. Η χυμοτρυψίνη προτιμά στην ίδια θέση φαινυλαλανίνη, τυροσίνη και τρυπτοφάνη κυρίως αλλά και δεσμούς που γειτονεύουν με λευκίνη, μεθειονίνη, ασπαραγίνη και ιστιδίνη. Η πεψίνη έχει παρόμοια εξειδίκευση όπως και η χυμοτρυψίνη ενώ η παπαΐνη υδρολύει όλους σχεδόν τους δεσμούς και αποφεύγει εκείνους μόνο που γειτονεύουν με όξινα αμινοξέα. Οι εξωπεπτιδάσες υδρολύουν πεπτιδικούς δεσμούς με την προϋπόθεση ότι η αμινομάδα ή η καρβοξυλομάδα ενός των δύο αμινοξέων που συμμετέχουν στον πεπτιδικό δεσμό θα είναι ελεύθερη. Και εάν απαιτούν την αμινομάδα ελεύθερη ονομάζονται αμινοπεπτιδάσες και υδρολύουν πεπτίδια αποσπώντας από ένα αμινοξύ από το Ν-τελικό άκρο, ενώ εάν απαιτούν ελεύθερη καρβοξυλομάδα ονομάζονται καρβοξυπεπτιδάσες και υδρολύουν πεπτίδια με σταδιακή απόσπαση ενός αμινοξέος από το C-τελικό άκρο. Εξειδίκευση ως προς τη φύση του αμινοξέος παρουσιάζουν και οι εξωπεπτιδάσες. Έτσι ενώ η καρβοξυπεπτιδάση Α υδρολύει όλα τα C-τελικά αμινοξέα πλην της αργινίνης, της λυσίνης και της προλίνης, η καρβοξυπεπτιδάση Β υδρολύει μόνο την αργινίνη και τη λυσίνη. Η λευκιναμινοπεπτιδάση πάλι προτιμά αμινοξέα με υδρόφοβα R. Γι αυτό και δρα καλύτερα, αν και όχι αποκλειστικά, σε πεπτίδια που έχουν λευκίνη σαν αμινοτελικό αμινοξύ. Υπάρχουν τέλος και ορισμένες διπεπτιδάσες, όπως εκείνη της γλυκυλογλυκίνης που υδρολύουν ειδικά ορισμένα διπεπτίδια. Οι ενδοπεπτιδάσες κατατάσσονται σε 4 κατηγορίες ανάλογα με το καταλυτικό αμινοξύ που βρίσκεται στο ενεργό κέντρο. Έτσι υπάρχουν πρωτεϊνάσες σερίνης, κυστεΐνης, ασπαραγινικού και μεταλλοπρωτεϊνάσες. Οι τελευταίες περιέχουν κάποιο μεταλλικό ιόν (συνήθως ψευδάργυρο) στο ενεργό κέντρο. Ο καταλυτικός μηχανισμός των πρωτεϊνασών σερίνης και κυστεΐνης περιλαμβάνει ομοιοπολική δέσμευση του υποστρώματος στις ομάδες R της σερίνης και κυστεΐνης των ενεργών κέντρων αντίστοιχα. Οι πρωτεϊνάσες ασπαραγινικού περιέχουν τις ομάδες R δύο ασπαραγινικών οξέων στο καταλυτικό τους κέντρο και λειτουργούν χωρίς τη δημιουργία ενδιάμεσου συμπλόκου με το υπόστρωμα. 40
41 Η ενδοκυττάρια αποικοδόμηση των πρωτεϊνών οφείλεται κυρίως σε όξινες πρωτεϊνάσες, που εντοπίζονται στα λυσοσώματα και ονομάζονται καθεψίνες. Άλλες ενδοκυττάριες πρωτεϊνάσες προκειμένου να δράσουν απαιτούν και την παρουσία ΑΤΡ το οποίο υδρολύεται ή όχι, αναλόγως. Οι καλπαΐνες είναι μια ενδιαφέρουσα οικογένεια ενδοκυττάριων πρωτεϊνασών που απαιτούν Ca 2+ προκειμένου να ενεργοποιηθούν και να δράσουν. Τέλος υπάρχει και ένα σύμπλοκο πρωτεολυτικών ενζύμων το οποίο αναγνωρίζει και υδρολύει μόνον εκείνες τις πρωτεΐνες που είναι ομοιοπολικά δεσμευμένες με μια ευρύτατα διαδεδομένη στη φύση μικρή πρωτεΐνη (μ.β. 8500) που λέγεται ουβικιτίνη. Η ουβικιτινυλίωση μιας πρωτεΐνης που πρόκειται να υδρολυθεί γίνεται με ταυτόχρονη υδρόλυση ΑΤΡ προς ΑΜΡ και ΡΡi. Υδρόλυση ΑΤΡ παρατηρείται και κατά τη συγκρότηση του συμπλόκου των πρωτεολυτικών ενζύμων το οποίο υδρολύει τις ουβικιτινυλιωμένες πρωτεΐνες [20] Οι πρωτεάσες διευκολύνουν δύσκολες αντιδράσεις Ο ρυθμός μετατροπής των πρωτεϊνών είναι μια σημαντική πορεία στους ζώντες οργανισμούς. Οι πρωτεΐνες οι οποίες εκπλήρωσαν την αποστολή τους πρέπει να αποδομηθούν έτσι ώστε τα αμινοξέα τους να ανακυκλωθούν για τη σύνθεση νέων πρωτεϊνών. Οι πρωτεΐνες που προσλαμβάνονται με τις τροφές πρέπει να διασπαστούν σε μικρά πεπτίδια και αμινοξέα για να απορροφηθούν στο έντερο. Επιπλέον οι πρωτεολυτικές αντιδράσεις είναι σημαντικές στο να ρυθμίζουν τη δραστικότητα μερικών ενζύμων και άλλων πρωτεϊνών. Οι πρωτεάσες διασπούν τις πρωτεΐνες με μια αντίδραση υδρόλυσης, δηλαδή προσθήκης ενός μορίου ύδατος σε έναν πεπτιδικό δεσμό: Αν και η υδρόλυση των πεπτιδικών δεσμών ευνοείται θερμοδυναμικά, οι αντιδράσεις αυτές είναι πάρα πολύ αργές. Απουσία ενός καταλύτη, η ημιζωή για την υδρόλυση ενός τυπικού πεπτιδίου σε ουδέτερο ph υπολογίζεται ότι είναι από 10 έως 1000 χρόνια. Εντούτοις, σε μερικές βιοχημικές πορείες οι πεπτιδικοί δεσμοί πρέπει να υδρολυθούν σε χιλιοστά του δευτερολέπτου. Οι χημικοί δεσμοί στους πεπτιδικούς δεσμούς είναι υπεύθυνοι για την κινητική τους σταθερότητα. Ειδικά, η δομή συντονισμού εξηγεί το γιατί τα άτομα ενός πεπτιδικού δεσμού βρίσκονται στο ίδιο επίπεδο και κάνει επίσης τους δεσμούς αυτούς ανθεκτικούς στην υδρόλυση. Αυτή η δομή συντονισμού προικίζει τον πεπτιδικό δεσμό με χαρακτήρα μερικού διπλού δεσμού: 41

42 Ο δεσμός άνθρακα-αζώτου ισχυροποιείται από τον χαρακτήρα του διπλού δεσμού και το καρβονυλικό άτομο άνθρακα είναι λιγότερο ηλεκτρονιόφιλο και λιγότερο επιδεκτικό στην πυρηνοφιλική προσβολή από ότι είναι τα καρβονυλικά άτομα άνθρακα σε ενώσεις όπως οι καρβοξυλικοί εστέρες. Επομένως, για να προαχθεί η διάσπαση του πεπτιδικού δεσμού, ένα ένζυμο πρέπει να διευκολύνει την πυρηνοφιλική προσβολή σε μια κανονικά μη ενεργό καρβονυλική ομάδα [11] Η χυμοθρυψίνη έχει ένα υψηλά ενεργό κατάλοιπο σερίνης Στο πεπτικό σύστημα των θηλαστικών και άλλων οργανισμών ένας αριθμός πρωτεολυτικών ενζύμων συμμετέχουν στη διάσπαση των πρωτεϊνών. Ένα τέτοιο ένζυμο, η χυμοθρυψίνη, διασπά επιλεκτικά πεπτιδικούς δεσμούς στο καρβοξυ-τελικό άκρο ενός μεγάλου υδρόφοβου αμινοξέος όπως είναι η θρυπτοφάνη, η τυροσίνη, η φαινυλαλανίνη και η μεθειονίνη (Εικόνα 13). Η χυμοθρυψίνη είναι ένα καλό παράδειγμα της χρησιμοποίησης της ομοιοπολικής μετατροπής ως μιας καταλυτικής στρατηγικής. Το ένζυμο χρησιμοποιεί ένα ισχυρό πυρηνόφιλο για να επιτεθεί σε μια μη ενεργό καρβονυλική ομάδα του υποστρώματος. Το πυρηνόφιλο αυτό προσκολλάται ομοιοπολικά στο υπόστρωμα για μικρό χρονικό διάστημα κατά τη διάρκεια της κατάλυσης. Εικόνα 13: Εξειδίκευση της χυμοθρυψίνης. Η χυμοθρυψίνη διασπά πρωτεΐνες στο καρβοξυ-τελικό άκρο αρωματικών ή μεγάλων υδρόφοβων αμινοξέων (σκιασμένα με πορτοκαλί). Οι δεσμοί που πιθανώς διασπώνται δείχνονται με κόκκινο [11]. 42
43 Ποιο είναι το πυρηνόφιλο που χρησιμοποιεί η χυμοθρυψίνη για να επιτεθεί στην καρβονυλική ομάδα του υποστρώματος; Ένα συμπέρασμα βγήκε από το γεγονός ότι η χυμοθρυψίνη περιέχει ένα εξαιρετικά ενεργό κατάλοιπο σερίνης. Η επεξεργασία με οργανικά φθοριοφωσφορικά, όπως το διισοπροπυλοφωσφοφθορίδιο (DIPF), βρέθηκε να απενεργοποιεί μη αντιστρεπτά το ένζυμο (Εικόνα 14). Παρά το γεγονός ότι το ένζυμο έχει 28 κατάλοιπα σερίνης, τροποποιήθηκε μόνον ένα, η σερίνη 195, με αποτέλεσμα τη συνολική απώλεια της ενζυμικής δραστικότητας. Αυτή η αντίδραση χημικής τροποποίησης υποδηλώνει ότι αυτό το ασυνήθιστα ενεργό κατάλοιπο σερίνης παίζει κεντρικό ρόλο στον καταλυτικό μηχανισμό της χυμοθρυψίνης [11]. Εικόνα 14: Μια ασυνήθιστα αντιδραστική σερίνη της χυμοθρυψίνης. Η χυμοθρυψίνη απενεργοποιείται με επεξεργασία με διισοπροπυλο-φωσφοφθορίδιο (DIPF), το οποίο αντιδρά με τη σερίνη 195 ανάμεσα στα 28 πιθανά κατάλοιπα σερίνης [11] Η δράση της χυμοθρυψίνης προχωρά σε δύο βήματα που συνδέονται από ένα ομοιοπολικά προσδεμένο ενδιάμεσο Πώς μπορούμε να διευκρινίσουμε τον ρόλο της σερίνης 195 στη δράση της χυμοθρυψίνης; Μια κινητική μελέτη του ενζύμου έδωσε ένα δεύτερο συμπέρασμα για τον καταλυτικό μηχανισμό της χυμοθρυψίνης και τον ρόλο της σερίνης 195. Η κινητική της ενζυμικής δράσης συχνά καταγράφεται εύκολα βάζοντας το ένζυμο να δράσει σε ένα ανάλογο του υποστρώματος το οποίο σχηματίζει ένα έγχρωμο προϊόν. Για τη χυμοθρυψίνη ένα τέτοιο χρωμογόνο υπόστρωμα είναι ο Ν-ακετυλο-L-φαινυλαλανυλο-π-νιτροφαινυλο-εστέρας. Αυτό το υπόστρωμα είναι ένας εστέρας παρά ένα αμίδιο, αλλά πολλές πρωτεάσες υδρολύουν και εστέρες. Ένα από τα προϊόντα που σχηματίζεται από τη διάσπαση του υποστρώματος αυτού από τη χυμοθρυψίνη είναι το π-νιτροφαινολικό, το οποίο έχει κίτρινο χρώμα (Εικόνα 15). Μετρήσεις της απορρόφησης του φωτός αποκάλυψαν την ποσότητα του π-νιτροφαινολικού που παρήχθη. 43
44 Εικόνα 15: Χρωμογόνο υπόστρωμα. Ο Ν-ακετυλο-L-φαινυλαλανυλο-π-νιτροφαινυλο-εστέρας παράγει ένα έγχρωμο προϊόν, το π-νιτροφαινολικό, όταν διασπάται από τη χυμοθρυψίνη. Το π-νιτροφαινολικό σχηματίζεται από την αποπρωτονίωση της π-νιτροφαινόλης σε ph 7 [11]. Εικόνα 16: Ομοιοπολική κατάλυση. Η υδρόλυση από τη χυμοθρυψίνη λαμβάνει χώρα σε δύο στάδια: (Α) ακυλίωση για να σχηματίσει τα ενδιάμεσα του ακυλοενζύμου ακολουθούμενη από (Β) αποακυλίωση για να αναπαραχθεί το ελεύθερο ένζυμο [11]. Η υδρόλυση προχωρά σε δύο βήματα. Τα δύο βήματα εξηγούνται από την αντίδραση της πυρηνόφιλης σερίνης με το υπόστρωμα για να σχηματιστεί ένα ομοιοπολικά συνδεδεμένο ενδιάμεσο ενζύμου-υποστρώματος (Εικόνα 16). Πρώτον, η υδροξυλική ομάδα της πάρα πολύ ενεργού σερίνης 195 προσβάλλει την καρβονυλική ομάδα του υποστρώματος για να σχηματίσει το ενδιάμεσο του ακυλοενζύμου, απελευθερώνοντας την αλκοόλη π-νιτροφαινόλη (ή μια αμίνη εάν το υπόστρωμα είναι ένα αμίδιο και όχι εστέρας). Δεύτερον, το ενδιάμεσο του ακυλοενζύμου υδρολύεται για να απελευθερώσει το συστατικό καρβοξυλικό οξύ του υποστρώματος και να αναπαραγάγει το ελεύθερο ένζυμο. Έτσι, με την προσθήκη του υποστρώματος το π- νιτροφαινολικό παράγεται γρήγορα καθώς σχηματίζεται το ενδιάμεσο του ακυλοενζύμου, αλλά το ένζυμο χρειάζεται περισσότερο χρόνο για να επανέλθει στην αρχική κατάσταση με την υδρόλυση του ενδιαμέσου του ακυλοενζύμου [11]. 44

45 Η σερίνη είναι μέρος μιας καταλυτικής τριάδας η οποία περιλαμβάνει επίσης ιστιδίνη και ασπαραγινικό οξύ Ο προσδιορισμός της τριδιάστατης δομής της χυμοθρυψίνης από τον David Blow το 1967 αύξησε τις γνώσεις μας για τον μηχανισμό δράσης της. Γενικά, η χυμοθρυψίvη είναι σχεδόν σφαιρική και απαρτίζεται από τρεις πολυπεπτιδικές αλυσίδες ενωμένες με δισουλφιδικούς δεσμούς. Συντίθεται ως ένα μοναδικό πολυπεπτίδιο, που ονομάζεται χυμοθρυψινογόνο, το οποίο ενεργοποιείται από την πρωτεολυτική διάσπαση του πεπτιδίου για να παραχθούν οι τρεις αλυσίδες. Το ενεργό κέντρο της χυμοθρυψίνης, που είναι σημειωμένο από τη σερίνη 195, βρίσκεται σε μια σχισμή στην επιφάνεια του ενζύμου (Εικόνα 17). Η δομική ανάλυση αποκάλυψε τη χημική βάση της ειδικής δραστικότητας της σερίνης 195 (Εικόνα 18). Η πλευρική αλυσίδα της σερίνης 195 είναι συνδεδεμένη με δεσμό υδρογόνου με τον ιμιδαζολικό δακτύλιο της ιστιδίνης 57. Η ομάδα -ΝΗ αυτού του ιμιδαζολικού δακτυλίου με τη σειρά της ενώνεται με δεσμό υδρογόνου με την καρβοξυλική ομάδα του ασπαραγινικού 102. Η ομάδα αυτή των καταλοίπων αναφέρεται ως η καταλυτική τριάδα. Πώς αυτή η διάταξη των καταλοίπων οδηγεί στην υψηλή δραστικότητα της σερίνης 195; Το κατάλοιπο της ιστιδίνης χρησιμεύει στην τοποθέτηση της πλευρικής αλυσίδας της σερίνης και στην πόλωση της υδροξυλικής ομάδας. Εικόνα 17: Τριδιάστατη δομή της χυμοθρυψίνης. Οι τρεις αλυσίδες δείχνονται σε μορφή κορδέλας σε καφέ, μπλε και πράσινο. Οι πλευρικές αλυσίδες των καταλοίπων της καταλυτικής τριάδας, συμπεριλαμβανομένης της σερίνης 195, παριστάνονται με σφαίρες και ράβδους, όπως είναι δύο δισουλφιδικοί δεσμοί της ενδοαλυσίδας και της μεσοαλυσίδας [11]. Έτσι, το κατάλοιπο δρα ως ένας γενικός βασικός καταλύτης, ένας δέκτης ιόντων υδρογόνου, διότι η πολωμένη υδροξυλική ομάδα του καταλοίπου της σερίνης βρίσκεται σε ετοιμότητα για αποπρωτονίωση. Η αφαίρεση ενός πρωτονίου από την υδροξυλική ομάδα παράγει ένα ιόν αλκοξειδίου, το οποίο είναι πολύ πιο ισχυρό πυρηνόφιλο από ότι είναι η αλκοόλη. Το κατάλοιπο 45
46 του ασπαραγινικού βοηθά στον προσανατολισμό του καταλοίπου της ιστιδίνης και στο να το κάνει έναν καλύτερο δέκτη πρωτονίων μέσω ηλεκτροστατικών επιδράσεων. Εικόνα 18: Η καταλυτική τριάδα. Η καταλυτική τριάδα, στα αριστερά, μετατρέπει τη σερίνη 195 σε ένα δυνητικό πυρηνόφιλο, όπως φαίνεται στα δεξιά [11]. Εικόνα 19: Υδρόλυση πεπτιδίου από τη χυμοθρυψίνη. Ο μηχανισμός υδρόλυσης πεπτιδίων δείχνει εμφανώς τις αρχές της ομοιοπολικής και οξεοβασικής κατάλυσης. Οι διακεκομμένες πράσινες γραμμές δείχνουν τις ευνοούμενες αλληλεπιδράσεις μεταξύ του αρνητικά φορτισμένου καταλοίπου ασπαραγινικού και του θετικά φορτισμένου καταλοίπου ιστιδίνης, οι οποίες κάνουν το κατάλοιπο ιστιδίνης μια πιο ισχυρή βάση [11]. Οι παρατηρήσεις αυτές υποδηλώνουν έναν μηχανισμό για την υδρόλυση του πεπτιδίου (Εικόνα 19). Μετά από την πρόσδεση του υποστρώματος (βήμα 1), η αντίδραση αρχίζει με την 46

47 πυρηνοφιλική προσβολή της υδροξυλικής ομάδας της σερίνης 195 στο καρβονυλικό άτομο άνθρακα του υποστρώματος (βήμα 2). Η πυρηνοφιλική προσβολή αλλάζει τη γεωμετρία από επίπεδη τριγωνική σε τετραεδρική γύρω από αυτό το άτομο άνθρακα. Το εκ φύσεως ασταθές τετραεδρικό ενδιάμεσο που σχηματίζεται έχει ένα τυπικό αρνητικό φορτίο στο άτομο οξυγόνου που προέρχεται από την καρβοξυλική ομάδα. Αυτό το φορτίο σταθεροποιείται με αλληλεπιδράσεις με ομάδες ΝΗ από την πρωτεΐνη σε μια θέση που καλείται οπή οξυανιόντος (Εικόνα 20): Εικόνα 20: Η οπή οξυανιόντος. Η δομή σταθεροποιεί το τετραεδρικό ενδιάμεσο της αντίδρασης της χυμοθρυψίνης. Δεσμοί υδρογόνου (δείχνονται με πράσινο) ενώνουν τις ομάδες ΝΗ του πεπτιδίου με το αρνητικά φορτισμένο οξυγόνο [11]. Επίσης, οι αλληλεπιδράσεις αυτές βοηθούν στη σταθεροποίηση της μεταβατικής κατάστασης που προηγείται του σχηματισμού του τετραεδρικού ενδιαμέσου. Στη συνέχεια, αυτό το τετραεδρικό ενδιάμεσο καταρρέει για να παραχθεί το ακυλοένζυμο (βήμα 3). Αυτό το βήμα διευκολύνεται από τη μεταφορά ενός πρωτονίου από το θετικά φορτισμένο κατάλοιπο της ιστιδίνης στην αμινική ομάδα που σχηματίζεται από τη διάσπαση του πεπτιδικού δεσμού. Το αμινικό συστατικό είναι τώρα ελεύθερο να αποχωριστεί από το ένζυμο (βήμα 4) και αντικαθίσταται από ένα μόριο ύδατος (βήμα 5). Στη συνέχεια υδρολύεται η εστερική ομάδα του ακυλοενζύμου από μια πορεία που είναι ουσιαστικά επανάληψη των βημάτων 2 έως 4. Το μόριο ύδατος προσβάλλει την καρβονυλική ομάδα ενώ ταυτόχρονα αφαιρείται ένα πρωτόνιο από το κατάλοιπο της ιστιδίνης, το οποίο τώρα δρα ως ένας γενικός όξινος καταλύτης, σχηματίζοντας ένα τετραεδρικό ενδιάμεσο (βήμα 6). Αυτή η δομή διασπάται για να σχηματίσει το προϊόν καρβοξυλικό οξύ (βήμα 7). Τελικά, η απελευθέρωση του προϊόντος ετοιμάζει το ένζυμο για έναν άλλο κύκλο κατάλυσης. Ο μηχανισμός αυτός εξηγεί όλα τα χαρακτηριστικά της δράσης της χυμοθρυψίνης, εκτός από την παρατηρούμενη προτίμηση για τη διάσπαση πεπτιδικών δεσμών αμέσως μετά από κατάλοιπα με μεγάλες υδρόφοβες αλυσίδες. Η εξέταση της τριδιάστατης δομής με ανάλογα υποστρώματος και αναστολείς ενζύμων αποκάλυψε την παρουσία μιας βαθιάς και σχετικά υδρόφοβης θήκης, η οποία ονομάζεται θήκη S 1, μέσα στην οποία μπορούν να ταιριάξουν οι 47
: Εικόνα 21: Η υδρόφοβη θήκη της χυμοθρυψίνης.")
48 μακριές αφόρτιστες πλευρικές αλυσίδες των καταλοίπων φαινυλαλανίνης και θρυπτοφάνης. Η πρόσδεση μιας κατάλληλης πλευρικής αλυσίδας μέσα στη θήκη αυτή τοποθετεί τον παρακείμενο πεπτιδικό δεσμό μέσα στο ενεργό κέντρο για διάσπαση (Εικόνα 21): Εικόνα 21: Η υδρόφοβη θήκη της χυμοθρυψίνης. Η υδρόφοβη θήκη της χυμοθρυψίνης είναι υπεύθυνη για την εξειδίκευση του υποστρώματος. Τα καθοριστικά αμινοξέα που απαρτίζουν τη θέση πρόσδεσης είναι σημασμένα, συμπεριλαμβανομένου του καταλοίπου της σερίνης του ενεργού κέντρου (πλαισιωμένο). Η θέση ενός αρωματικού δακτυλίου προσδεμένου στη θήκη δείχνεται με πράσινο [11]. Η εξειδίκευση της χυμοθρυψίνης εξαρτάται ολοκληρωτικά από το ποιο αμινοξύ βρίσκεται αμέσως μετά από το αμινο-τελικό άκρο του πεπτιδικού δεσμού που πρόκειται να διασπαστεί. Άλλες πρωτεάσες έχουν πιο περίπλοκο τύπο εξειδίκευσης, όπως εμφανώς δείχνεται στην Εικόνα 22. Τέτοια ένζυμα έχουν στις επιφάνειές τους επιπρόσθετες θήκες για την αναγνώριση και άλλων καταλοίπων του υποστρώματος. Κατάλοιπα στο αμινο-τελικό άκρο του πεπτιδικού δεσμού που πρόκειται να διασπαστεί σημαίνονται ως Ρ 1, Ρ 2, Ρ 3, και ούτω καθεξής, δείχνοντας τη θέση τους σε σχέση με τον δεσμό που πρόκειται να διασπαστεί. Παρομοίως, κατάλοιπα στο καρβοξυ-τελικό άκρο του δεσμού που πρόκειται να διασπαστεί σημαίνονται ως Ρ 1, Ρ 2, Ρ 3 και ούτω καθεξής. Οι περιοχές που αντιστοιχούν επάνω στο ένζυμο αναφέρονται ως S 1, S 2 ή S 1, S 2 και ούτω καθεξής. Εικόνα 22: Ονοματολογία εξειδίκευσης για αλληλεπιδράσεις πρωτεάσης υποστρώματος. Οι δυνητικές περιοχές της αλληλεπίδρασης του υποστρώματος με το ένζυμο ορίζονται ως Ρ (δείχνονται με κόκκινο) και αντίστοιχες περιοχές πρόσδεσης επάνω στο ένζυμο ορίζονται ως S. Ο δεσμός που διασπάται (επίσης δείχνεται με κόκκινο) είναι το σημείο αναφοράς [11]. 48

49 Καταλυτικές τριάδες απαντούν σε άλλα υδρολυτικά ένζυμα Έχει βρεθεί στη συνέχεια ότι πολλές άλλες πρωτεΐνες περιέχουν καταλυτικές τριάδες παρόμοιες με εκείνη που ανακαλύφθηκε στη χυμοθρυψίνη. Μερικές, όπως η θρυψίνη και η ελαστάση, είναι εμφανή ανάλογα της χυμοθρυψίνης. Οι αλληλουχίες των πρωτεϊνών αυτών είναι περίπου 40% πανομοιότυπες με εκείνη της χυμοθρυψίνης και η συνολική δομή τους είναι σχεδόν η ίδια (Εικόνα 23): Εικόνα 23: Δομική ομοιότητα της θρυψίνης και της χυμοθρυψίνης. Μια επικάλυψη της δομής της θρυψίνης (μπλε) από τη χυμοθρυψίνη (κόκκινο) δείχνει τον υψηλό βαθμό ομοιότητας. Δείχνονται μόνο οι θέσεις των ατόμων α-άνθρακα. Η μέση απόκλιση στη θέση μεταξύ των αντίστοιχων ατόμων α-άνθρακα είναι 1,7 Å [11]. Οι πρωτεΐνες αυτές λειτουργούν με πανομοιότυπους μηχανισμούς με εκείνον της χυμοθρυψίνης. Εντούτοις, έχουν πολύ διαφορετικές εξειδικεύσεις υποστρώματος. Η θρυψίνη διασπά τον πεπτιδικό δεσμό μετά από κατάλοιπα με μακριές θετικά φορτισμένες πλευρικές αλυσίδες, όπως η αργινίνη και η λυσίνη, ενώ η ελαστάση διασπά τον πεπτιδικό δεσμό μετά από αμινοξέα με μικρές πλευρικές αλυσίδες, όπως η αλανίνη και η σερίνη. Σύγκριση των θηκών S 1, των ενζύμων αυτών αποκαλύπτει τη βάση της εξειδίκευσης. Στη θρυψίνη ένα κατάλοιπο ασπαραγινικού (Asp 189) υπάρχει στον πυθμένα της θήκης S 1, στη θέση ενός καταλοίπου σερίνης στη χυμοθρυψίνη. Το κατάλοιπο ασπαραγινικού προσβάλλει και σταθεροποιεί ένα θετικά φορτισμένο κατάλοιπο αργινίνης ή λυσίνης του υποστρώματος. Στην ελαστάση, δύο κατάλοιπα στην κορυφή της θήκης στη χυμοθρυψίνη και στη θρυψίνη έχουν αντικατασταθεί από βαλίνη (Val 190 και 216). Τα κατάλοιπα αυτά κλείνουν το στόμιο της θήκης έτσι ώστε μόνο μικρές πλευρικές αλυσίδες να μπορούν να εισέλθουν (Εικόνα 24). 49

50 Εικόνα 24: Οι θήκες S 1 της χυμοθρυψίνης, της θρυψίνης και της ελαστάσης. Κάποια κατάλοιπα παίζουν καθοριστικούς ρόλους στον προσδιορισμό της εξειδίκευσης αυτών των ενζύμων. Οι πλευρικές αλυσίδες των καταλοίπων αυτών καθώς επίσης και εκείνων της σερίνης του ενεργού κέντρου, δείχνονται έγχρωμες [11]. Άλλα μέλη της οικογένειας της χυμοθρυψίνης περιλαμβάνουν μια συλλογή πρωτεϊνών που παίζουν ρόλο στην πήξη του αίματος. Επιπροσθέτως, βρέθηκε στα βακτήρια και στους ιούς μια μεγάλη ποικιλία πρωτεασών που επίσης ανήκουν στην ομάδα αυτή. Επιπλέον, άλλα ένζυμα που δεν είναι ομόλογα με τη χυμοθρυψίνη βρέθηκε ότι περιέχουν πολύ όμοια ενεργά κέντρα, η παρουσία πολύ όμοιων ενεργών κέντρων σε αυτές τις διαφορετικές οικογένειες πρωτεϊνών είναι ένα επακόλουθο της συγκλίνουσας εξέλιξης. Η σουμπτιλισίνη, μια πρωτεάση βακτηρίων όπως ο Bacillus amyloliquefaciens, είναι ένα ιδιαιτέρως καλά χαρακτηρισμένο παράδειγμα. Το ενεργό κέντρο του ενζύμου αυτού περιέχει και την καταλυτική τριάδα και την οπή οξυανιόντος. Εντούτοις, μία από τις ομάδες ΝΗ που σχηματίζουν την οπή οξυανιόντος προέρχεται από τις πλευρικές αλυσίδες ενός καταλοίπου ασπαραγίνης και όχι από τον κορμό του πεπτιδίου (Εικόνα 25). Η σουμπτιλισίνη είναι το ιδρυτικό μέλος μιας άλλης μεγάλης οικογένειας πρωτεασών που περιλαμβάνει αντιπροσώπους από τα αρχαία, τα βακτήρια και τα ευκάρυα. Εικόνα 25: Η καταλυτική τριάδα και η οπή οξυανιόντος της σουμπτιλισίνης. Ο πεπτιδικός δεσμός που προσβάλλεται από την πυρηνόφιλη σερίνη 221 της καταλυτικής τριάδας θα αναπτύξει ένα αρνητικό φορτίο, το οποίο σταθεροποιείται από τις ομάδες ΝΗ του ενζύμου (και τα δύο στον κορμό και στην πλευρική αλυσίδα της Asn 155) που είναι τοποθετημένες στην οπή οξυανιόντος [11]. 50
.")
51 Επιπλέον, ένα άλλο παράδειγμα της καταλυτικής τριάδας βρέθηκε στην καρβοξυπεπτιδάση IΙ από το σιτάρι. Η δομή του ενζύμου αυτού δεν είναι σημαντικά παρόμοια με εκείνη της χυμοθρυψίνης ή της σουμπτιλισίνης (Εικόνα 26). Η πρωτεΐνη αυτή είναι μέλος μιας ενδιαφέρουσας οικογένειας ομόλογων πρωτεϊνών που περιλαμβάνει εστεράσες, όπως η εστεράση της ακετυλοχολίνης (ακετυλοχολινεστεράση) και αρκετές λιπάσες. Όλα αυτά τα ένζυμα χρησιμοποιούν πυρηνόφιλα ενεργοποιημένα από την ιστιδίνη, αλλά τα πυρηνόφιλα κατάλοιπα μπορεί να είναι κυστεΐνης παρά σερίνης. Τελικά, ανακαλύφθηκαν και άλλες πρωτεάσες που περιέχουν στο ενεργό κέντρο ένα κατάλοιπο σερίνης ή θρεονίνης που δεν ενεργοποιούνται από ένα ζεύγος ιστιδίνης-ασπαραγινικού αλλά από μια βασική αμινική ομάδα από την πλευρική αλυσίδα της λυσίνης ή την αμινο-τελική ομάδα της πολυπεπτιδικής αλυσίδας. Έτσι, η καταλυτική τριάδα στις πρωτεάσες αναδύθηκε τουλάχιστον τρεις φορές κατά την πορεία της εξέλιξης. Μπορούμε να συμπεράνουμε ότι αυτή η καταλυτική στρατηγική πρέπει να είναι μια ιδιαίτερα αποτελεσματική προσέγγιση στην υδρόλυση των πεπτιδίων και σχετιζόμενων δεσμών [11]. Εικόνα 26: Καρβοξυπεπτιδάση II. Η δομή της καρβοξυπεπτιδάσης ΙΙ από το σιτάρι (δεξιά) εικονίζεται με τις δύο αλυσίδες της (μπλε και κόκκινο). Η καταλυτική τριάδα της καρβοξυπεπτιδάσης ΙΙ (αριστερά) αποτελείται από τα ίδια αμινοξέα με εκείνα της χυμοθρυψίνης, παρά το γεγονός ότι τα ένζυμα δεν εμφανίζουν δομική ομοιότητα. Τα κατάλοιπα που σχηματίζουν την οπή οξυανιόντος τονίζονται με κίτρινο [11] Άλλες κύριες τάξεις ενζύμων που διασπούν πεπτίδια είναι οι ασπαρτυλοπρωτεάσες, οι κυστεϊνοπρωτεάσες και οι μεταλλοπρωτεάσες Όλες οι πρωτεάσες δεν χρησιμοποιούν στρατηγικές βασισμένες στα ενεργοποιημένα κατάλοιπα σερίνης. Ανακαλύφθηκαν τρεις τάξεις πρωτεϊνών που χρησιμοποιούν τρεις εναλλακτικές προσεγγίσεις για την υδρόλυση των πεπτιδικών δεσμών (Εικόνα 27). Οι τάξεις αυτές είναι (1) οι κυστεϊνοπρωτεάσες, (2) οι ασπαρτυλοπρωτεάσες και (3) οι μεταλλοπρωτεάσες. 51

52 Σε κάθε περίπτωση, η στρατηγική παράγει ένα πυρηνόφιλο που επιτίθεται στην καρβονυλική ομάδα του πεπτιδίου (Εικόνα 28). Εικόνα 27: Τρεις τάξεις πρωτεασών και τα ενεργά κέντρα τους. Αυτά τα παραδείγματα μιας κυστεϊνοπρωτεάσης, μιας ασπαρτυλοπρωτεάσης και μιας μεταλλοπρωτεάσης χρησιμοποιούν αντίστοιχα ως πυρηνόφιλο ένα κατάλοιπο κυστεΐνης ενεργοποιημένο από ιστιδίνη, ένα ενεργοποιημένο μόριο ύδατος και ένα ενεργοποιημένο από μέταλλο μόριο ύδατος. Τα δύο μισά της ρενίνης είναι σε μπλε και σε κόκκινο για να τονιστεί η κατά προσέγγιση συμμετρία δευτέρας τάξεως των ασπαρτυλοπρωτεασών [11]. Εικόνα 28: Οι στρατηγικές ενεργοποίησης των τριών τάξεων των πρωτεασών. Η καρβονυλική ομάδα του πεπτιδίου προσβάλλεται από (Α) μια κυστεΐνη ενεργοποιημένη από ιστιδίνη, στις κυστεϊνοπρωτεάσες (Β) 52
53 ένα μόριο ύδατος ενεργοποιημένο από ασπαραγινικό, στις ασπαρτυλοπρωτεάσες και (Γ) ένα μόριο ύδατος ενεργοποιημένο από μέταλλο, στις μεταλλοπρωτεάσες. Για τις μεταλλοπρωτεάσες το γράμμα Β αντιπροσωπεύει μια βάση (συχνά ένα γλουταμινικό) η οποία βοηθά στην αποπρωτονίωση του δεσμευμένου στο μέταλλο μορίου ύδατος [11]. Η στρατηγική που χρησιμοποιείται από τις κυστεϊνοπρωτεάσες μοιάζει περισσότερο με εκείνη που χρησιμοποιείται από την οικογένεια της χυμοθρυψίνης. Στα ένζυμα αυτά, ένα κατάλοιπο κυστεΐνης που ενεργοποιείται από ένα κατάλοιπο ιστιδίνης παίζει τον ρόλο του πυρηνόφιλου που επιτίθεται στον πεπτιδικό δεσμό (Εικόνα 28), με έναν τρόπο εντελώς ανάλογο με εκείνον του καταλοίπου σερίνης στις σερινοπρωτεάσες. Ένα ιδεώδες παράδειγμα των πρωτεϊνών αυτών είναι η παπαΐνη, ένα ένζυμο που απομονώνεται από το φρούτο παπάγια. Έχουν ανακαλυφθεί πρωτεάσες θηλαστικών που είναι ομόλογες με την παπαΐνη, περισσότερο αξιοσημείωτες είναι οι καθεψίνες που έχουν κάποιο ρόλο στο ανοσοποιητικό και άλλα συστήματα. Το ενεργό κέντρο που είναι βασισμένο στην κυστεΐνη προέκυψε τουλάχιστον δύο φορές κατά τη διάρκεια της εξέλιξης, οι κασπάσες, ένζυμα που παίζουν κύριο ρόλο στην απόπτωση, έχουν ενεργά κέντρα παρόμοια με εκείνο της παπαΐνης, αλλά οι συνολικές δομές τους δεν σχετίζονται. Η δεύτερη τάξη περιλαμβάνει τις ασπαρτυλοπρωτεάσες. Το κύριο χαρακτηριστικό των ενεργών κέντρων είναι ένα ζεύγος καταλοίπων ασπαραγινικού τα οποία δρουν μαζί για να επιτρέπουν σε ένα μόριο ύδατος να επιτεθεί στον πεπτιδικό δεσμό. Ένα κατάλοιπο ασπαραγινικού (στην αποπρωτονιωμένη μορφή του) ενεργοποιεί το επιτιθέμενο μόριο ύδατος θέτοντάς το σε ετοιμότητα για αποπρωτονίωση, ενώ το άλλο κατάλοιπο ασπαραγινικού (στην πρωτονιωμένη μορφή του) πολώνει το καρβονύλιο του πεπτιδίου, αυξάνοντας την ευαισθησία του για επίθεση (Εικόνα 28). Στα μέλη της τάξης αυτής περιλαμβάνονται η ρενίνη, ένα ένζυμο που παίζει ρόλο στη ρύθμιση της πίεσης του αίματος, και το πεπτικό ένζυμο πεψίνη. Τα ένζυμα αυτά έχουν κατά προσέγγιση συμμετρία δευτέρας τάξεως, υποδηλώνοντας ότι τα δύο μισά σχετίζονται εξελικτικά. Ένα ελκυστικό σενάριο είναι ότι δύο αντίγραφα του αρχέγονου γονιδίου συγχωνεύθηκαν για να σχηματίσουν ένα μοναδικό γονίδιο που κωδίκευε ένα ένζυμο με μια αλυσίδα. Κάθε αντίγραφο του γονιδίου θα είχε συνεισφέρει στο ενεργό κέντρο ένα κατάλοιπο ασπαραγινικού. Ο ιός της ανθρώπινης ανοσοανεπάρκειας (HIV) και άλλοι ρετροϊοί περιέχουν μια μη συγχωνευμένη διμερή ασπαρτυλοπρωτεάση η οποία είναι παρόμοια με τη συγχωνευμένη πρωτεΐνη, αλλά οι αλυσίδες δεν είναι ενωμένες για να σχηματίσουν μια μοναδική αλυσίδα (Εικόνα 29). Η παρατήρηση αυτή είναι σύμφωνη με την ιδέα ότι το ένζυμο μπορεί να υπήρξε αρχικά ως ξεχωριστές υπομονάδες. 53

54 Εικόνα 29: Η δομή και η θήκη πρόσδεσης της πρωτεάσης του HIV. Η πρωτεάση είναι ένα διμερές πανομοιότυπων υπομονάδων (δείχνονται με μπλε και κίτρινο), που η κάθε μία απαρτίζεται από 99 αμινοξέα. Τα κατάλοιπα του ασπαραγινικού οξέος του ενεργού κέντρου, ένα από κάθε αλυσίδα, δείχνονται με σφαίρες και ράβδους. Τα πτερύγια θα κλείσουν επάνω στην θήκη πρόσδεσης μετά την πρόσδεση του υποστρώματος [11]. Οι μεταλλοπρωτεάσες αποτελούν την τελική κύρια τάξη των ενζύμων που διασπούν πεπτίδια. Το ενεργό κέντρο μιας τέτοιας πρωτεΐνης περιέχει ένα προσδεμένο ιόν μετάλλου, σχεδόν πάντοτε ψευδάργυρο, το οποίο ενεργοποιεί ένα μόριο ύδατος να δράσει ως πυρηνόφιλο για να επιτεθεί στην καρβονυλική ομάδα του πεπτιδίου. Το βακτηριακό ένζυμο θερμολυσίνη και το ένζυμο της πέψης καρβοξυπεπτιδάση Α είναι κλασικά παραδείγματα των πρωτεασών ψευδαργύρου. Η θερμολυσίνη, αλλά όχι η καρβοξυπεπτιδάση Α, είναι μέλος μιας μεγάλης και ποικίλης οικογένειας ομόλογων πρωτεασών ψευδαργύρου η οποία περιλαμβάνει τις μεταλλοπρωτεάσες εξωκυτταρικής μήτρας που καταλύουν τις αντιδράσεις ανακατασκευής και αποικοδόμησης ιστών. Σε κάθε μία από τις τρεις τάξεις των ενζύμων, τα ενεργά κέντρα περιλαμβάνουν χαρακτηριστικά που επιτρέπουν την ενεργοποίηση του νερού ή ενός άλλου πυρηνόφιλου, καθώς επίσης την πόλωση της καρβονυλικής ομάδας του πεπτιδίου και την ακόλουθη σταθεροποίηση του τετραεδρικού ενδιαμέσου (Εικόνα 28) Οι αναστολείς των πρωτεασών είναι σημαντικά φάρμακα Ενώσεις που παρεμποδίζουν ή τροποποιούν τις δραστικότητες των πρωτεασών μπορούν να έχουν εντυπωσιακές βιολογικές επιδράσεις. Οι περισσότεροι αναστολείς πρωτεασών είναι παρόμοιοι στη δομή με τα υποστρώματα των ενζύμων που αναστέλλουν. Αρκετά σημαντικά φάρμακα είναι αναστολείς πρωτεασών. Παραδείγματος χάριν, η καπτοπρίλη, ένας αναστολέας της μεταλλοπρωτεάσης που μετατρέπει την αγγειοτενσίνη, έχει χρησιμοποιηθεί για τη ρύθμιση της πίεσης του αίματος. Το crixivan, ένας αναστολέας της πρωτεάσης του HIV, χρησιμοποιείται για τη θεραπεία του AIDS. Οι πρωτεάσες αυτές διασπούν τις ιικές πρωτεΐνες στις ενεργές μορφές 54
55 τους, παρεμποδίζοντας εντελώς την πορεία αυτή, εμποδίζεται ο ιός από το να είναι μολυσματικός (Εικόνα 29). Για να εμποδιστούν οι μη επιθυμητές παρενέργειες, οι αναστολείς πρωτεασών που χρησιμοποιούνται ως φάρμακα πρέπει να είναι ειδικοί για ένα ένζυμο χωρίς να αναστέλλουν άλλες πρωτεΐνες μέσα στο σώμα. Ας εξετάσουμε με περισσότερη λεπτομέρεια την αλληλεπίδραση του crixivan με την πρωτεάση του HIV. Το crixivan είναι κατασκευασμένο γύρω από μια αλκοόλη που μιμείται το τετραεδρικό ενδιάμεσο άλλες ομάδες είναι παρούσες για να προσδεθούν στις περιοχές αναγνώρισης S 2, S 1, S 1 και S 2 επάνω στο ένζυμο (Εικόνα 30). Εικόνα 30: Crixivan, ένας αναστολέας πρωτεάσης του HIV. Η δομή του crixivan δείχνεται σε σύγκριση με εκείνη ενός υποστρώματος πεπτιδίου της πρωτεάσης του HIV [11]. Τα αποτελέσματα μελετών κρυσταλλογραφίας με ακτίνες Χ αποκάλυψαν τη δομή του συμπλόκου ενζύμου-crixiνan, δείχνοντας ότι το crixivan υιοθετεί μια στερεοδιάταξη που προσεγγίζει τη διπλή συμμετρία του ενζύμου (Εικόνα 31). Το ενεργό κέντρο της πρωτεάσης του HIV καλύπτεται από δύο εύκαμπτα πτερύγια τα οποία διπλώνονται στην κορυφή του προσδεμένου αναστολέα. Η υδροξυλική ομάδα της κεντρικής αλκοόλης αντιδρά με δύο κατάλοιπα ασπαραγινικού του ενεργού κέντρου, ένα σε κάθε υπομονάδα. Επιπροσθέτως, δύο καρβονυλικές ομάδες του αναστολέα ενώνονται με δεσμούς υδρογόνου με ένα μόριο ύδατος (δεν δείχνεται), το οποίο διαδοχικά ενώνεται με δεσμούς υδρογόνου σε μια ομάδα ΝΗ του πεπτιδίου σε κάθε ένα από τα πτερύγια. Αυτή η αλληλεπίδραση του αναστολέα με το νερό και το ένζυμο δεν είναι δυνατή με κυτταρικές ασπαρτυλοπρωτεάσες, όπως η ρενίνη, και έτσι ίσως συνεισφέρει στην εξειδίκευση του crixivan και άλλων αναστολέων για την πρωτεάση του HIV [11]. 55
 Το φάρμακο έχει περιστραφεί για να αποκαλύψει την κατά προσέγγιση στερεοδιάταξη με συμμετρία δευτέρας τάξεως [11]. 1.2.4.")
56 Εικόνα 31: Σύμπλοκο πρωτεάσης HIV-crixivan. (Αριστερά) Η πρωτεάση του HIV δείχνεται με τον αναστολέα Crixivan προσδεδεμένο στο ενεργό κέντρο. (Δεξιά) Το φάρμακο έχει περιστραφεί για να αποκαλύψει την κατά προσέγγιση στερεοδιάταξη με συμμετρία δευτέρας τάξεως [11] Μερικά πρωτεολυτικά ένζυμα έχουν ειδικούς αναστολείς Ένας ακριβής τρόπος για να ξεκινήσει η ενζυμική δραστικότητα είναι η μετατροπή του ζυμογόνου σε πρωτεάση με τη διάσπαση ενός μοναδικού πεπτιδικού δεσμού. Εντούτοις, το βήμα αυτό της ενεργοποίησης δεν είναι αντιστρεπτό και γι αυτό χρειάζεται ένας διαφορετικός μηχανισμός για να σταματήσει η πρωτεόλυση. Αυτό γίνεται από ειδικούς αναστολείς των πρωτεασών. Παραδείγματος χάριν, ο αναστολέας της παγκρεατικής θρυψίνης, που είναι μια πρωτεΐνη με μοριακό βάρος 6 kd, αναστέλλει τη θρυψίνη με το να προσδένεται πολύ ισχυρά στο ενεργό κέντρο της. Η σταθερά διάστασης του συμπλόκου είναι 0,1 pμ, η οποία αντιστοιχεί σε μια πρότυπη ελεύθερη ενέργεια πρόσδεσης -18 kcal mol -1 (-57 kj mol -1 ) περίπου. Αυτό το σύμπλοκο, σε αντίθεση με όλα σχεδόν τα γνωστά σύμπλοκα πρωτεϊνών, δεν διαχωρίζεται στα συστατικά του με αποδιατακτικούς παράγοντες όπως 8 Μ ουρίας ή 6 Μ υδροχλωρικής γουανιδίνης. Η αιτία για την εξαιρετική σταθερότητα του συμπλόκου είναι ότι ο αναστολέας της παγκρεατικής θρυψίνης είναι ένα πολύ αποτελεσματικό ανάλογο του υποστρώματος. Η ανάλυση με ακτίνες Χ έδειξε ότι ο αναστολέας βρίσκεται στο ενεργό κέντρο του ενζύμου, τοποθετημένος έτσι ώστε η πλευρική αλυσίδα της λυσίνης 15 του αναστολέα αυτού να αλληλεπιδρά με την πλευρική αλυσίδα του ασπαραγινικού στην κοιλότητα εξειδίκευσης του ενζύμου. Επιπλέον, υπάρχουν πολλοί δεσμοί υδρογόνου μεταξύ της κύριας αλυσίδας της θρυψίνης και της αλυσίδας του αναστολέα. Περαιτέρω, η καρβονυλική ομάδα της λυσίνης 15 και άτομα που την περιβάλλουν προσαρμόζονται καλά στο ενεργό κέντρο του ενζύμου. Σύγκριση της δομής του αναστολέα που είναι προσδεμένος στο ένζυμο με εκείνη του ελεύθερου αναστολέα αποκαλύπτει ότι η δομή ουσιαστικά δεν αλλάζει με την πρόσδεση στο ένζυμο (Εικόνα 32). Έτσι ο αναστολέας οργανώνεται εκ των προτέρων σε μια δομή η οποία είναι σε μεγάλο βαθμό συμπληρωματική στο ενεργό κέντρο του ενζύμου. Πράγματι, ο πεπτιδικός δεσμός μεταξύ της λυσίνης 15 και της 56

57 αλανίνης 16 στον αναστολέα της παγκρεατικής θρυψίνης διασπάται, αλλά με πολύ μικρότερη ταχύτητα. Αυτό το σύμπλοκο της θρυψίνης με τον αναστολέα έχει ημιζωή μερικών μηνών. Ουσιαστικά, ο αναστολέας είναι ένα υπόστρωμα, αλλά η δομή του είναι τόσο καλά συμπληρωματική στο ενεργό κέντρο του ενζύμου που προσδένεται πολύ ισχυρά και γυρίζει πίσω πολύ αργά [11]. Εικόνα 32: Αλληλεπίδραση της θρυψίνης με τον αναστολέα της. Δομή του συμπλόκου της θρυψίνης (κίτρινο) με τον αναστολέα της παγκρεατικής θρυψίνης (κόκκινο). Η λυσίνη 15 του αναστολέα εισδύει στο ενεργό κέντρο του ενζύμου και σχηματίζει μια γέφυρα άλατος με το ασπαραγινικό 189. Ο προσδεμένος και ο ελεύθερος αναστολέας έχουν σχεδόν πανομοιότυπη δομή [11] Βιοτεχνολογικές εφαρμογές πρωτεασών Οι πρωτεάσες σήμερα κατέχουν την πρώτη θέση στα ένζυμα που παρασκευάζονται για βιοτεχνολογικούς σκοπούς. Η ετήσια παραγωγή ξεπερνά τους 500 τόνους. Οι περισσότερες πρωτεάσες χρησιμοποιούνται στις βιομηχανίες απορρυπαντικών. Αλκαλικές πρωτεάσες βοηθούν την υδρολυτική διάσπαση πρωτεϊνών που προκαλούν λεκέδες και είναι δύσκολο να απομακρυνθούν. Όξινες πρωτεάσες χρησιμοποιούνται στη βιομηχανία τυριών και πουλιούνται ως μίγμα μαζί με τη ρενίνη. Οι πρωτεάσες επίσης χρησιμοποιούνται ευρέως στην υδρόλυση των πρωτεϊνών της σόγιας και στη βιομηχανία ζυμαρικών. Η ως επί το πλείστον χρησιμοποιούμενη είναι η αλκαλική πρωτεάση σερίνης του Β. picheniformis γνωστή με το εμπορικό όνομα subtilisin. Υδρολύει πολλούς πεπτιδικούς δεσμούς και συνεπώς εύκολα διαλυτοποιεί πρωτεΐνες. 57
58 Επίσης στις βιομηχανίες γαλακτοκομικών προϊόντων χρησιμοποιούνται σήμερα η μικροβιακή πυτία που είναι όξινη πρωτεάση ασπαραγινικού οξέος [22] Ένζυμα με βιοτεχνολογικό ενδιαφέρον - Θερμοάντοχα ένζυμα Έχουν ανακαλυφθεί μικροοργανισμοί που αναπτύσσονται σε θερμοκρασίες γύρω ή και ψηλότερα των 100 C. Οι υπερθερμόφιλοι αυτοί οργανισμοί ζουν σε ηφαιστιογενείς θαλάσσιες περιοχές και θεωρούνται ως οι πιο αρχέγονοι μικροοργανισμοί. Για την ανάπτυξή τους απαιτούν αναερόβιες συνθήκες και εξαρτώνται από την αναγωγή του S σε H 2 S. Οι μικροοργανισμοί αυτοί θεωρούνται ως πολύ καλές πηγές ενζύμων μια και τα ένζυμα που περιέχουν είναι θερμοάντοχα και μπορούν να καταλύουν αντιδράσεις σε ψηλές θερμοκρασίες. Σήμερα περισσότερα από 30 τέτοια ένζυμα έχουν απομονωθεί και χαρακτηριστεί. Όλα τα ένζυμα έχουν ψηλή βέλτιστη θερμοκρασία κατάλυσης και παρουσιάζουν χρόνο ημιζωής πολλές μέρες. Οι μικροοργανισμοί αυτοί περιέχουν ένζυμα πολλών κατηγοριών, όπως διάφορες πρωτεάσες, αμυλάσες, γλυκανάσες, κυτταρινάσες, ξυλανάσες, δεϋδρογονάσες και DA πολυμεράσες. Για παράδειγμα, υπάρχει μεγάλη ζήτηση για θερμοάντοχες DA πολυμεράσες, ένζυμα που χρησιμοποιούνται για την αλυσιδωτή αντίδραση πολυμεράσης του DA (PCR) ως εργαλεία ρουτίνας σε εργαστήρια βιοχημείας και μοριακής βιολογίας [21]. Μικροοργανισμοί - Ένζυμα Χρόνος ημιζωής (μικρότερος/μεγαλύτερος) σε Καταλυτική δράση λεπτά Pyrococcus furiosus Πρωτεάση 33/98 Υδρόλυση πεπτιδίων Αμυλάση 2/120 Υδρόλυση αμύλου α-γλυκοσιδάση 48/98 Υδρόλυση μαλτόζης Υδρογονάση 2/100 Παραγωγή H 2 DA πολυμεράση 20/95 Διπλασιασμός του DA Thermococcus litoralis Φερρεδοξίνη 24/95 Μεταφορά ηλεκτρονίων DA πολυμεράση 7/95 Διπλασιασμός του DA Pyrococcus woesei Αμυλάση 6/100 Υδρόλυση αμύλου Pyrodictium brockii Υδρογονάση 1/98 Οξείδωση του Η 2 58
59 Desulfurococcus mucosus Πρωτεάση 2/95 Υδρόλυση πεπτιδίων Thermotoga maritima 4-α-Γλυκονοτρανσφεράση 3/80 Υδρόλυση αμύλου Υδρογονάση 1/90 Παραγωγή H 2 Δεϋδρογονάση του γαλακτικού 2/90 Μετατροπή πυροσταφυλικού σε γαλακτικό και αντίστροφα Thermotoga sp. (FjSS3-B.1) Ξυλανάση 1.5/95 Υδρόλυση ξυλανίου Κελλοβιοϋδρολάση 1.1/108 Υδρόλυση κυτταρίνης Πίνακας 1: Ιδιότητες των ενζύμων που απομονώθηκαν από υπερθερμόφιλους οργανισμούς [21] Σταθεροποίηση των ενζύμων Υπάρχουν πολλοί τρόποι που μπορούν να εφαρμοσθούν ώστε να παρασκευασθούν ένζυμα με μεγαλύτερη σταθερότητα. Ποιος τρόπος θα ακολουθηθεί εξαρτάται από το κόστος και την αποτελεσματικότητα της μεθόδου. Πολλές φορές γίνεται συνδυασμός διαφόρων μεθόδων (εικόνα 33). Εικόνα 33: Τρόποι σταθεροποίησης της δομής των ενζύμων [22]. 59
60 Παράγοντες που επηρεάζουν την σταθεροποίηση της δομής των ενζύμων: 1) Επιλογή και γενετική μηχανική 2) Ενζυμομηχανική 3) Το περιβάλλον της αντίδρασης 4) Χημική τροποποίηση της δομής των ενζύμων 5) Διαμοριακή σύνδεση 6) Επαναδίπλωση του μορίου με θέρμανση. Δεδομένου ότι η σταθερότητα που παρουσιάζουν τα ένζυμα στη θερμοκρασία είναι αποτέλεσμα υδρόφοβων αλληλεπιδράσεων, προτάθηκε ότι τα ένζυμα μπορούν να σταθεροποιηθούν με το να αυξηθούν αυτές οι αλληλεπιδράσεις. Για να δημιουργηθούν περισσότερες υδρόφοβες αλληλεπιδράσεις το ένζυμο πρώτα ξεδιπλώνεται π.χ. με ουρία ή με δισουλφίδια και στη συνέχεια επαναδιπλώνεται σε υψηλότερη θερμοκρασία που είναι μια μη φυσική κατάσταση. Η τρυψίνη μετά το ξεδίπλωμα στους 50 C και επαναθέρμανση στους 80 C παρουσιάζει πέντε φορές υψηλότερη δραστικότητα απ αυτή που είχε στους 20 C. 7) Καθήλωση. Για τα βιομηχανικά ένζυμα το σπουδαιότερο χαρακτηριστικό είναι η σταθερότητα και η μεγαλύτερη επιβίωσή τους. Βρέθηκε ότι πολλά ένζυμα όταν καθηλώνονται σε αδιάλυτους φορείς καθίστανται περισσότερο σταθερά. Για παράδειγμα οι πρωτεάσες όταν καθηλωθούν χάνουν την ικανότητά τους να αυτοϋδρολύονται και έτσι σταθεροποιούνται. Η μεγαλύτερη ανενεργοποίηση που υφίστανται τα ένζυμα στους βιοαντιδραστήρες οφείλεται στην αύξηση της θερμοκρασίας που τις περισσότερες φορές είναι επιβεβλημένη. Σε υψηλότερες θερμοκρασίες αφενός μεν η απόδοση της αντίδρασης είναι μεγαλύτερη αφετέρου δε εμποδίζεται η ανάπτυξη μολυσματικών μικροβίων. Δεν υπάρχει αμφιβολία ότι η ανενεργοποίηση περιλαμβάνει αλλαγές στην δομή του ενζύμου που το τελικό αποτέλεσμα είναι η καταστροφή των ανωτέρων δομών του ενζύμου όπως φαίνεται στην εικόνα 33. Θα πρέπει να φανταστούμε ότι ένα ένζυμο που είναι καθηλωμένο, συνδέεται με το υπόστρωμα με διάφορους χημικούς δεσμούς. Η δομή ενός τέτοιου ενζύμου συνεπώς είναι πολύ πιο σταθερή απ ότι όταν βρίσκεται στη φυσική του ελεύθερη κατάσταση. Το απλό αυτό μοντέλο φαίνεται να είναι αρκετά ρεαλιστικό γιατί βρέθηκε ότι σε ένζυμα που εγκλωβίζονται π.χ. σε πολυακρυλαμίδιο όπου δημιουργούνται πολλοί δεσμοί, αυξάνεται δραματικά ή αντοχή τους στη θερμοκρασία. Το ίδιο συμβαίνει όταν το ξεδίπλωμα του μορίου γίνεται με αλλαγές ph, οργανικούς διαλύτες ή άλλα μετουσιωτικά αντιδραστήρια [22]. 60
61 Καθήλωση ενζύμων Τα ένζυμα είναι οι κατά κύριο λόγο πρωτεΐνες που καταλύουν τις διάφορες βιοχημικές αντιδράσεις (βιοκαταλύτες) οι οποίες γίνονται μέσα στα κύτταρα κάτω από πολύ ήπιες συνθήκες θερμοκρασίας, πιέσεως και pη. Οι μεταβολικές διαδικασίες των φυτικών και ζωικών κυττάρων είναι πολύπλοκες σειρές αντιδράσεων και κάθε στάδιο ελέγχεται από ένα ειδικό ένζυμο. Η σπουδαιότητα των ενζύμων ως καταλύτες και στις χημικές διεργασίες σε βιομηχανική ή και εργαστηριακή κλίμακα είναι εξίσου σημαντική. Τα ένζυμα εμφανίζουν μια σειρά χαρακτηριστικών τα οποία καθιστούν την χρήση τους πλεονεκτικότερη έναντι των συμβατικών χημικών καταλυτών: i. Εμφανίζουν εξαιρετική καταλυτική απόδοση, πολλές φορές πολλαπλάσια των χημικών καταλυτών. ii. Εμφανίζουν υψηλό βαθμό εξειδίκευσης όχι μόνο ως προς το είδος της αντίδρασης αλλά και ως προς το υπόστρωμα (εξειδίκευση υποστρώματος). Η εξειδίκευση τους είναι τόσο υψηλή ώστε μπορούν να διακρίνουν ακόμα και μεταξύ όμοιων περιοχών των μορίων (εξειδίκευση περιοχής) καθώς επίσης και μεταξύ οπτικών ισομερών (στερεοεξειδίκευση). Με τον τρόπο αυτό εξασφαλίζουν τον αποκλεισμό παράπλευρων αντιδράσεων, καταλύουν την παραγωγή ενός αποκλειστικού επιθυμητού προϊόντος, παρέχουν υψηλές αποδόσεις στις αντιδράσεις περιορίζοντας το κόστος των υλικών. iii. Λειτουργούν σε ήπιες συνθήκες θερμοκρασίας, πίεσης και pη με ρυθμούς αντίδρασης οι οποίοι στην περίπτωση χρήσης χημικών καταλυτών θα απαιτούσαν ακραίες συνθήκες. Με τον τρόπο αυτό περιορίζεται σε μεγάλο βαθμό η κατανάλωση ενέργειας και το λειτουργικό κόστος παραγωγής. iv. Τα ένζυμα δεδομένου ότι είναι κατά κύριο λόγο πρωτεΐνες ή πεπτίδια, είναι βιοαποικοδομήσιμα και μπορούν εύκολα να απομακρυνθούν από τα απόβλητα. Αυτός ο μοναδικός συνδυασμός χαρακτηριστικών των ενζύμων ως καταλύτες, γίνεται εκμεταλλεύσιμος από το 1960 και ένας μεγάλος αριθμός προϊόντων έχουν παραχθεί από αντίστοιχες βιοχημικές διεργασίες κατά τις οποίες χρησιμοποιούνται καταλύτες ένζυμα στα πλαίσια της βιοτεχνολογίας. Μεταξύ των βιοτεχνολογικών προϊόντων ανήκουν πολλά τρόφιμα, φαρμακευτικά σκευάσματα, αγροχημικά καθώς και οργανικά χημικά συνθετικά. Πέρα όμως από τα αναμφισβήτητα πλεονεκτήματα των ενζύμων υπάρχει και ένας αριθμός πρακτικών προβλημάτων που προκύπτουν κατά την χρήση των ενζύμων. Μεταξύ αυτών είναι: i. Το υψηλό κόστος απομόνωσης και καθαρισμού των ενζύμων. ii. Φαινόμενα αστάθειας της δομής τους δεδομένου ότι απομακρύνονται από το φυσικό τους περιβάλλον. 61
62 iii. Η ευαισθησία τους στις συνθήκες των διεργασιών οι οποίες είναι διαφορετικές από αυτές της βέλτιστης λειτουργία τους. iv. Η ευαισθησία τους ακόμα και σε ιχνοποσότητες ουσιών που μπορούν να αναστείλουν την λειτουργία τους. Τα δύο τελευταία προβλήματα καθιστούν τα ένζυμα σε ενεργούς καταλύτες μικρής διάρκειας. Επιπλέον, σε αντίθεση με τους ετερογενείς χημικούς καταλύτες, τα περισσότερα ένζυμα λειτουργούν διαλυμένα στο νερό σε συστήματα ομοιογενούς κατάλυσης μολύνοντας το προϊόν και καθιστώντας δύσκολη την ανάκτησή τους από το μίγμα της αντίδρασης σε δραστική μορφή. Διάφοροι μέθοδοι έχουν προταθεί προκειμένου να επιλυθούν πολλά από τα παραπάνω προβλήματα μεταξύ των οποίων είναι και η καθήλωση των ενζύμων σε αδιάλυτους φορείς (Tischer and Wedekjnd, 1999, Κυριακίδης, 2000) [3]. Ως καθήλωση ορίζουμε τον εγκλωβισμό ενός ενζύμου σε συγκεκριμένο υλικό ώστε να μπορεί να έρχεται σε επαφή με την κυρίως φάση που είναι το υπόστρωμα, ενεργοποιητές ή αναστολείς. Η κινητικότητα των εγκλωβισμένων μορίων του ενζύμου είναι συνήθως ένα μεγάλο μοριακού βάρους υδρόφιλο πολυμερές (π.χ. κυτταρίνη ή πολυακρυλαμίδιο). Η καθήλωση ενζύμων σε βιομηχανική κλίμακα άρχισε στα τέλη της δεκαετίας του 70 [11]. Η καθήλωση των ενζύμων πετυχαίνεται με δέσμευση του ενζύμου πάνω ή μέσα σε κάποιο στερεό υλικό υπόστρωμα προκειμένου να λαμβάνονται ετερογενείς καθηλωμένα ενζυμικά συστήματα. Με τον τρόπο αυτό μιμούμαστε τον φυσικό τρόπο σταθεροποίησης της δομής των ενζύμων στα ζωντανά κύτταρα όπου τα ένζυμα στις περισσότερες περιπτώσεις είναι προσκολλημένα σε κυτταρικές μεμβράνες. Σε σύγκριση με τα ελεύθερα ένζυμα, τα καθηλωμένα ενζυμικά συστήματα: i. Είναι πιο ανθεκτικά σε αλλαγές του περιβάλλοντος. ii. Επιτρέπουν την εύκολη ανάκτηση τόσο του ενζύμου όσο και του προϊόντος. iii. Επιτρέπουν την πολλαπλή χρήση του ενζύμου. iv. Εξασφαλίζουν την συνεχή λειτουργία των ενζυμικών διεργασιών. v. Διευκολύνουν τον γρήγορο τερματισμό των ενζυμικών αντιδράσεων. vi. Μπορούν να λειτουργήσουν σε μια μεγάλη ποικιλία βιοαντιδραστήρων [13]. Η συγκεκριμένη δομή και το ενεργό κέντρο του ενζύμου είναι δύο απαραίτητα στοιχεία για την καταλυτική του δράση. Το ενεργό κέντρο αποτελείται από δύο περιοχές με διαφορετικές λειτουργικότητες. Μία είναι αντιδρώσα ή καταλυτική θέση που συμμετέχει στην καταλυτική δράση, ενώ η άλλη η ειδική θέση δέσμευσης του υποστρώματος που ελέγχει την εξειδίκευση του ενζύμου ως προς το υπόστρωμα. Οι δυο αυτές περιοχές αποτελούνται από πολλά αμινοξέα που βρίσκονται σε συγκεκριμένες θέσεις στο χώρο. Συνεπώς για να διατηρήσει ένα ένζυμο τη δραστικότητά του όταν καθηλωθεί, είναι απαραίτητο να διατηρεί τη φυσική του δομή όσο το δυνατόν περισσότερο. Για να παρασκευαστούν καθηλωμένα ένζυμα που να συνεχίζουν να 62
63 διατηρούν τη δραστικότητά τους, θα πρέπει η καθήλωσή τους να γίνεται με ήπιες και ελεγχόμενες συνθήκες. Λειτουργικές ομάδες που συμμετέχουν συνήθως στην καθήλωση των ενζύμων είναι οι ελεύθερες αμινο- και καρβοξυ-ομάδες, η σουλφυδρυλομάδα της κυστεΐνης, το ιμιδαζόλιο της ιστιδίνης, οι φαινολικές ομάδες και οι υδροξυλομάδες της σερίνης και θρεονίνης. Όταν γίνει η καθήλωση ενός ενζύμου είναι απαραίτητο αυτές οι λειτουργικές ομάδες του ενεργού κέντρου που οδηγούν στη σύνδεση του ενζύμου με τον αδιάλυτο φορέα να μη συμμετέχουν στην αντίδραση. Επίσης αντιδράσεις σε υψηλές θερμοκρασίες ή με ισχυρά οξέα ή βάσεις πρέπει να αποφεύγονται γιατί οι ασθενείς δευτερεύοντες δεσμοί (υδρογόνου, υδρόφοβοι, ιονικοί) που διατηρούν την τεταρτοταγή δομή του ενζύμου καταστρέφονται, με αποτέλεσμα τη μείωση ή και εξαφάνιση της δραστικότητας του ενζύμου [22]. Οι μέθοδοι καθήλωσης των ενζύμων μπορούν να ταξινομηθούν στις παρακάτω κατηγορίες: 1) Σύνδεση ενζύμων σε αδιάλυτους φορείς με ιονικούς δεσμούς α) Μέθοδος φυσικής προσρόφησης β) Μέθοδος ιονικής σύνδεσης 2) Μέθοδος ομοιοπολικής σύνδεσης α) Διαζω-μέθοδος β) Πεπτιδο-μέθοδος γ) Μέθοδος αλκυλίωσης δ) Σύνδεση σε πορώδες γυαλί ε) Σύνδεση με ενεργοποιημένη κυτταρίνη στ) Σύνδεση με ενεργοποιημένο άμυλο 3) Διαμοριακή σύνδεση ενζύμων 4) Εγκλωβισμός ενζύμων σε δικτυωτά πολυμερή ή μικροκάψουλες 5) Εγκλωβισμός ενζύμων σε λιποσώματα ή άλλα υδρόφοβα πολυμερή [22] Η καθήλωση επιφέρει μια σειρά από μεταβολές στο ένζυμο προκαλώντας την μείωση της απόδοσής του (Chaplin and Bucke, 1990). Σε σύγκριση με τα περισσότερα ένζυμα, τα καθηλωμένα ένζυμα εμφανίζουν: i. χαμηλότερη δραστικότητα και ii. μεγαλύτερες σταθερές Michaelis-Menten. Αυτές οι μεταβολές στα χαρακτηριστικά των καθηλωμένων ενζύμων οφείλονται σε μεταβολές στην δομή τους κατά την διαδικασία καθήλωσης καθώς και στη δημιουργία ενός μικροπεριβάλλοντος στο οποίο λειτουργούν τα καθηλωμένα ένζυμα και το οποίο είναι διαφορετικό από αυτό της κύριας μάζας του διαλύματος. Το τελευταίο εξαρτάται ισχυρά από την αντίδραση που λαμβάνει χώρα και την φύση του υποστρώματος. Επιπλέον, τα συστήματα 63
64 καθηλωμένων ενζύμων ως συστήματα δύο φάσεων υποφέρουν από ανεπιθύμητους περιορισμούς σε φαινόμενα μεταφοράς μάζας, τα οποία προκαλούν ανεπιθύμητους περιορισμούς στην ολική απόδοση του συστήματος. Προκειμένου να χρησιμοποιηθεί μια βιοχημική διεργασία για την παραγωγή ενός συγκεκριμένου προϊόντος με ελεύθερο ή καθηλωμένο ένζυμο έναντι μιας συμβατικής χημικής διεργασίας με έναν κατάλληλο χημικό καταλύτη, θα πρέπει να ληφθούν υπόψη όλα τα παραπάνω καθώς και το κατά πόσο η διεργασία αυτή είναι οικονομικά βιώσιμη. Μέχρι σήμερα πολλές βιοχημικές διεργασίες με καθηλωμένα ένζυμα έχουν αποδειχτεί οικονομικά βιώσιμες και μια μεγάλη ποικιλία προϊόντων παράγεται με τον τρόπο αυτό. Παρακάτω δίνεται ένας ενδεικτικός πίνακας (πίνακας 2) με τέτοια προϊόντα. Άλλες εφαρμογές πέρα από την χρήση των καθηλωμένων ενζύμων σε βιομηχανική κλίμακα είναι και η χρήση τους για την οργανικές συνθέσεις σε εργαστηριακή κλίμακα καθώς και σε αναλυτικές και ιατρικές εφαρμογές. Τα ένζυμα μπορούν να απλοποιούν τις χημικές διεργασίες μειώνοντας τον αριθμό των σταδίων σύνθεσης, μπορούν να ενισχύουν την καθαρότητα των προϊόντων και το πιο σημαντικό μπορούν να καταλύουν αντιδράσεις στερεοεξειδίκευσης. Ένζυμο Υπόστρωμα Προϊόν Ισομεράση της γλυκόζης Γλυκόζη Φρουκτόζη β- Γαλακτοσιδάση Λακτόζη Γλυκόζη και γαλακτόζη Λιπάση Τριγλυκερίδια Υποκατάστατα βουτύρου κακάου Νίτριλο-υδρατάση Ακρυλονιτρίλιο 3-Κυανοπυριδίνη Αδιπονιτρίλιο Ακρυλαμίδιο Νικοταμίδιο 5-Κυανοβαλεραμίδιο Αμυλοακυλάση D,L-αμινοξέα L-αμινοξέα (μεθειονίνη, αλανίνη, φαινυλαλανίνη, τρυπτοφάνη, βαλίνη) Ραφινάση Ραφινόζη Γαλακτόζη και σουκρόζη Ιμβερτάση Σουκρόζη Μίγμα γλυκόζης/φρουκτόζης L-ασπαρτικό οξύ (χρησιμοποιείται για την Λυάση της αμμωνίας του Αμμωνία και φουμαρικό οξυ παραγωγή συνθετικών γλυκαντικών ουσιών ασπαραγινικού οξέος ασπαρτάμης) Θερμολυσίνη Πεπτίδια Ασπαρτάμη Γλυκοαμυλάση Άμυλο D-γλυκόζη Παπαΐνη Πρωτεΐνες Απομάκρυνση του chill haze στη μπύρα Υδαντοϊνάση D,L-αμινοξέα ιδαντοϊνών D,L-αμινοξέα Αμιδάση πενικιλίνης Πενικιλίνες V και G 6-αμινοπενικιλανικό οξύ (πρόδρομη ένωση για την παραγωγή ημισυνθετικών πενικιλινών π.χ. αμπικιλίνη) 64
65 β-τυροσινάση Πυροκατεχόλη (pyrocatechol) L-DPA Πίνακας 2: Μερικές από τις σημαντικότερες βιομηχανικές εφαρμογές των καθηλωμένων ενζύμων (Krajewska, 2004) [13]. Σε αναλυτικές εφαρμογές τα καθηλωμένα ένζυμα χρησιμοποιούνται κυρίως για την ανάπτυξη βιοαισθητήρων (Van de Velde et al., 2002). Η λειτουργία των βιοαισθητήρων στηρίζεται στην ολοκλήρωση βιολογικών συστημάτων με μετασχηματιστές. Ένα χημικό σήμα λόγω απόκρισης του βιολογικού συστήματος στο περιβάλλον, μετασχηματίζεται σε ένα αναλυτικό μετρούμενο σήμα. Έχουν χρησιμοποιηθεί διάφορα είδη μετασχηματιστών όπως ποτενσιομετρικοί, αμπερομετρικοί, οπτικοί και πιεζοηλεκτρικοί. Τα ένζυμα τις περισσότερες φορές είναι απευθείας καθηλωμένα στην επιφάνεια του μετασχηματιστή ή σε κάποιο κατάλληλο υλικό (πολυμερές) που περιβάλλει τον μετασχηματιστή. Δεδομένου ότι τα ένζυμα εμφανίζουν πολύ μεγάλη εξειδίκευση και ευαισθησία μπορούν να χρησιμοποιηθούν καθηλωμένα σε βιοαισθητήρες προκειμένου να ανιχνευτούν τόσο το υπόστρωμα όσο και οι αναστολείς του ενζύμου. Οι βιοαισθητήρες είναι πολύτιμα εργαλεία στην ιατρική για βιοιατρικές και φαρμακευτικές αναλύσεις καθώς επίσης και στην ανάλυση τροφίμων και περιβαλλοντικών δειγμάτων (απόβλητων κ.ά). Οι ιατρικές εφαρμογές των καθηλωμένων ενζύμων περιλαμβάνουν την χρήση τους τόσο στην διάγνωση ός και στην θεραπεία ασθενειών (Piskin, 1993, Bertucci et al., 2003). Χαρακτηριστικό παράδειγμα είναι η επικάλυψη τεχνικών οργάνων και άλλων υλικών που εισάγονται στον ανθρώπινο οργανισμό με κατάλληλα ένζυμα ώστε να εμφανίζουν καλύτερη βιοσυμβατότητα. Παρακάτω δίνεται ένας πίνακας (πίνακας 3) με παραδείγματα καθηλωμένων ενζύμων που έχουν χρησιμοποιηθεί για την ανάπτυξη βιοαισθητήρων [13]. Ένζυμο Υπόστρωμα Εφαρμογή Οξειδάση της γλυκόζης Γλυκόζη Διάγνωση και θεραπεία του διαβήτη, επιστήμη τροφίμων, βιοτεχνολογία Υπεροξειδάση H 2 2 Βιολογικές και βιομηχανικές εφαρμογές, προσδιορισμός μέσω αναστολής βαρέων μετάλλων και εντομοκτόνων Τυροσινάση Προσδιορισμός φαινολικών συστατικών στα Φαινόλες και τρόφιμα, προσδιορισμός carbamate εντομοκτόνου πολυφαινόλες μέσω αναστολής Οξειδάση του γλουταμινικού οξέος Γλουταμινικό Επιστήμη τροφίμων, βιοτεχνολογία Ουρεάση Ουρία Ιατρικές διαγνωστικές τεχνικές, τεχνητό νεφρό 65

66 Δεϋδρογονάση της αιθυλικής αλκοόλης Αιθανόλη Επιστήμη τροφίμων, βιοτεχνολογία Ακετυλοχολινεστεράση Ακετυλοχολίνη, Προσδιορισμός οργανοφωσφόρου και εντομοκτόνων ακετυλοθειοχολίνη carbamate Οξειδάση της χολίνης Χολίνη Ένζυμα χρησιμοποιούμενα σε σύνδεση με ακετυλοχολινεστεράσες Δεϋδρογονάση του γαλακτικού οξέος Γαλακτικό οξύ Αθλητιατρική, επιστήμη τροφίμων, βιοτεχνολογία Οξειδάση της χοληστερόλης Χοληστερόλη Ιατρικές εφαρμογές Πενικιλινάση Πενικιλίνες Φαρμακευτικές εφαρμογές Αλλιϊνάση Σουλφοξείδια κυστεΐνης Βιομηχανία τροφίμων Πίνακας 3: Μερικά χαρακτηριστικά παραδείγματα καθήλωσης ενζύμων για την ανάπτυξη βιοαισθητήρων [13] Καθήλωση με ομοιοπολικό δεσμό μέσω των -ΟΗ, -SH, -H 2 του ενζύμου (Έποξυ ενεργοποιημένη σεφαρόζη) Πρόκειται για ένα προ-ενεργοποιημένο υλικό που χρησιμοποιείται ευρέως για την καθήλωση διαφόρων μορίων όπως σάκχαρα, άλλους υδρογονάνθρακες καθώς και πρωτεΐνες μέσω σταθερών αιθερικών δεσμών που αναπτύσσονται μεταξύ των εποξυ-ομάδων του υλικού και των υδροξυ- (ΟΗ-), άμινο- (-ΝΗ 2 ) και σουλφυδρυλομάδων (-SH) των προς καθήλωση μορίων. Αποτελείται από σφαιρίδια αγαρόζης τα οποία έχουν τροποποιηθεί κατάλληλα. Όπως είναι γνωστό, η αγαρόζη είναι ένα συμπολυμερές των μορίων D-γαλακτόζης και 3-6-άνυδρο L- γαλακτόζης τα οποία συνδέονται μεταξύ τους με γλυκοζιτικούς σχηματίζοντας τον ιστό του υλικού. Το υλικό αυτό στο εμπόριο είναι γνωστό με την ονομασία sepharose. Οι υδροξυ-ομάδες των σακχαριτών αντιδρούν εύκολα και σχηματίζουν ομοιοπολικούς δεσμούς με διάφορα μόρια συνδέσμων (ligands) δίνοντας ένα μεγάλο εύρος υλικών με διαφορετικές ενεργείς ομάδες [13,14]. Σχήμα 1: Η δομή της αγαρόζης [13]. 66
67 Η έποξυ ενεργοποιημένη σεφαρόζη σχηματίζεται συνήθως από την αντίδραση μεταξύ των σφαιριδίων αγαρόζης (σεφαρόζη) και του μορίου 1,4-δις-(2,3-εποξυ-πρόποξυ-) βουτάνιο δημιουργώντας έτσι ένα βραχίovα 12 ατόμων με ενεργείς εποξυ-ομάδες. Συγκεκριμένα αναπτύσσεται σταθερός αιθερικός δεσμός μεταξύ του υδρόφιλου συνδέσμου 1,4-δις-(2.3-εποξυπρόποξυ-)βουτάνιο και των υδροξυλίων του ιστού της αγαρόζης. Ανάλογα με το ποσοστό της αγαρόζης στην πηκτή από την οποία αποτελούνται τα σφαιρίδια του υλικού, υπάρχουν τρεις διαφορετικοί τύποι υλικών 2Β, 4Β και 6Β με περιεκτικότητα σε αγαρόζη 2%, 4% και 6% αντίστοιχα [13,14]. Sepharose CH 2 CH H CH 2 (CH 2 ) 4 CH 2 CH CH 2 Σχήμα 2: Δομή έποξυ-ενεργοποιημένης σεφαρόζης [13,14]. Υπάρχουν στο εμπόριο μια σειρά από τυποποιημένα υλικά αυτής της μορφής με διαφορετικό μέγεθος σφαιριδίων και πιθανώς με διαφορετικό μήκος βραχίονα. Τα βασικά χαρακτηριστικά της έποξυ-ενεργοποιημένης σεφαρόζης 6Β που χρησιμοποιήθηκε στα πειράματα είναι τα εξής: Ενεργή ομάδα: Έποξυ-ομάδα. Πυκνότητα ενεργούς ομάδας: μmole/ml ξηρού υλικού. Βραχίονας: 1,4-δις-(2.3-εποξυ-πρόποξυ-)βουτάνιο (12 άτομα). Δομή σφαιριδίων: 6% αγαρόζη. Μέγεθος σφαιριδίων: μm (μέσο μέγεθος 90 μm). Σταθερότητα σε τιμές ph: Χημική σταθερότητα: Σταθερό σε όλα τα κοινά υδατικά διαλύματα, σε μη ιοντικά επιφανειοδραστικά, μετουσιωτικές ουσίες και σε οργανικούς διαλύτες. Φυσική σταθερότητα: Αμελητέα αλλαγή όγκου λόγω μεταβολής της τιμής του ph ή της ιονικής ισχύος [13,14]. Επειδή το υλικό παρέχεται σε μορφή ξηρής σκόνης απαιτείται η ενυδάτωση του 1 gr υλικού μετά από διάλυση και ανάδευση με 100 ml νερού δίνει περίπου 3.5 ml όγκο υλικού. Η σύζευξη των διαφόρων μορίων στο υλικό εμφανίζεται βέλτιστη τις παρακάτω συνθήκες: Τιμή ph σύζευξης: 9-13, για σύζευξη μέσω των -ΟΗ του προς καθήλωση μορίου απαιτείται ισχυρά αλκαλικό περιβάλλον και τιμή ph μεγαλύτερη από 13. Διάλυμα σύζευξης: Η 2 Ο ή ρυθμιστικό διάλυμα βορικών ή φωσφορικών στα οποία η τιμή του ph ρυθμίζεται στην κατάλληλη τιμή με προσθήκη διαλύματος ah. Δεν 67
68 ενδείκνυται η χρήση ρυθμιστικού διαλύματος tris-hcl γιατί περιέχει αμινομάδες οι οποίες αντιδρούν με τις εποξυ-ομάδες του υλικού. Θερμοκρασία σύζευξης: C. Χρόνος επώασης κατά την σύζευξη: 16 hr. Συγκέντρωση προς καθήλωση μορίου: μmoles/ml υλικού (Affinity Chromatography, Pharmacia) [13,14] Ομοιοπολική σύνδεση με το υπόστρωμα μέσω των SH, -H 2 του ενζύμου Όπως έχει ήδη αναφερθεί τα υλικά καθήλωσης με ενεργοποιημένες εποξυ-ομάδες έχουν χρησιμοποιηθεί ευρέως για την καθήλωση πρωτεϊνών ακόμη και σε βιομηχανική κλίμακα δεδομένου ότι εμφανίζουν υψηλή σταθερότητα των ενεργών ομάδων, υψηλή σταθερότητα στην σύζευξη μεταξύ πρωτεΐνης-υποστρώματος, δυνατότητα απενεργοποίησης των ενεργών ομάδων που δεν έχουν δεσμεύσει πρωτεΐνες καθώς και δυνατότητα σταθεροποίησης των καθηλωμένων πρωτεϊνών μέσω πολλαπλής ομοιοπολικής προσέγγισης. Ωστόσο τα συμβατικά αυτά υλικά με τις ενεργές εποξυ-ομάδες έχουν υδρόφοβη φύση και η καθήλωση λαμβάνει χώρα σε υψηλές τιμές ιονικής ισχύος. Για τον λόγο αυτό, πολλές φορές απαιτείται η μερική τροποποίηση των εποξυ-ομάδων σε άλλου είδους ενεργές ομάδες οι οποίες μπορούν να προάγουν την φυσική και εξειδικευμένη προσρόφηση των πρωτεϊνών. Η καθήλωση τώρα δεν απαιτεί ούτε υψηλές τιμές ιονικής ισχύος ούτε υδρόφοβο υπόστρωμα. Ένα παράδειγμα τροποποιημένου εποξυ-υποστρώματος είναι και το Epoxy-Thiol reactive υπόστρωμα. Πρόκειται για ένα δι-λειτουργικό υλικό καθήλωσης το οποίο αποτελείται από: i. ένα μικρό ποσοστό ενεργών ομάδων οι οποίες αντιδρούν έντονα με τις σουλφυδρυλομάδες των πρωτεϊνών (-SH) και ii. από έναν υψηλό αριθμό ενεργών εποξυ-ομάδων που διατηρούνται στο αρχικό υλικό. Οι πρωτεΐνες αρχικά καθηλώνονται ομοιοπολικά μέσω σουλφυδρυλο (... SH) δισουλφιδικής (...S-S...) ανταλλαγής (thiol - disulfide exchange) μεταξύ των σουλφυδρυλομάδων που βρίσκονται στην επιφάνεια των πρωτεϊνών (surface thiol - groups) και των τροποποιημένων ενεργών ομάδων του υποστρώματος (thiol-reactive groups) και αντιστρόφως. Οι σουλφυδρυλομάδες είναι οι πιο πυρηνόφιλες ομάδες που απαντώνται στις πρωτεΐνες και ο ρυθμός αντίδρασης με τους ενεργούς δισουλφιδικούς δεσμούς του υλικού είναι υψηλός για ένα μεγάλο εύρος τιμών pη. Στη συνέχεια οι μη τροποποιημένες εποξυ-ομάδες του υλικού σχηματίζουν μη αντιστρεπτούς ομοιοπολικούς δεσμούς με τις αμινομάδες των πρωτεϊνών. Παράγοντας τροποποίησης του υλικού είναι η διθειοθρεϊτόλη (DΤΤ) σε συνδυασμό με το 2,2-διπυριδυλο δισουλφίδιο dipyridyl disulfide (2-PDS). Οι αντιδράσεις τροποποίησης και καθήλωσης έχουν ως εξής: αρχικά η διθειοθρεϊτόλη (DTT) αντιδρά με τις εποξυ-ομάδες του 68
69 υποστρώματος και το αποτέλεσμα αυτής της αντίδρασης είναι η ανάπτυξη δισουλφιδικού δεσμού μετά από αντίδραση με 2,2-διπυριδυλο- δισουλφίδιο (2-PDS) (Affinity Chromatography, Pharmacia) [13,14]. 1 ο στάδιο Sepharose CH 2 CH CH 2 (CH 2 ) 4 CH 2 CH CH 2 + HS CH 2 CH CH CH 2 SH H H H (DTT) Sepharose CH 2 CH CH 2 (CH 2 ) 4 CH 2 CH CH 2 S CH 2 CH CH CH 2 SH H H H H 2 ο στάδιο Sepharose SH + S S Sepharose S S + S 2-PDS H Σχήμα 3: Μηχανισμός τροποποίησης των εποξυ-ομάδων σε ενεργούς δισουλφιδικούς δεσμούς [13]. Ο δισουλφιδικός δεσμός S-S του τροποποιημένου υποστρώματος αντιδρά έντονα με τις σουλφυδρυλομάδες -SH των πρωτεϊνών. Στη συνέχεια αναπτύσσονται ομοιοπολικοί δεσμοί μεταξύ των εποξυ-ομάδων του υποστρώματος που δεν έχουν αντιδράσει με το DΤΤ και των αμινομάδων των πρωτεϊνών. Η ανάπτυξη των δεσμών αυτών απαιτεί αλκαλικό pη [13,14]. Το σχήμα απεικονίζει τους μηχανισμούς. 69
70 + H H SH SH DTT H S H H SH Enzyme S S thiopropionamide-enzyme H S H H S S H 2 Enzyme Epoxy/SH(DTT)-Sepabeads S S 2-PDS H S H H S S Enzyme thiol-enzyme SH H S H H S S H 2 Enzyme Epoxy/PyS 2 (DTT)-Sepabeads Σχήμα 4: Μερική τροποποίηση υλικού καθήλωσης με ενεργές εποξυ-ομάδες προς δι-λειτουργικό υλικό καθήλωσης με ενεργούς δισουλφιδικούς δεσμούς και μηχανισμός καθήλωσης πρωτεϊνών [13]. Η ανάπτυξη του δι-λειτουργικού αυτού υλικού καθήλωσης επιτρέπει: i. Μεγάλη αύξηση του εύρους των συνθηκών καθήλωσης (δεν απαιτείται υψηλή ιονική ισχύς ή η παρουσία οργανικών διαλυτών ή επιφανειοδραστικών σε τιμές ph μεγαλύτερες του 6). ii. Η εξειδικευμένη καθήλωση των πρωτεϊνών μέσω των σουλφυδρυλομάδων των κυστεϊνών φυσικών ή ανασυνδυασμένων πρωτεϊνών. iii. Η περαιτέρω καθήλωση των πρωτεϊνών μέσω ανάπτυξης μη αντιστρεπτών ομοιοπολικών δεσμών με τις εποξυ-ομάδες του υλικού. iv. Η ακαμψία των καθηλωμένων πρωτεϊνών λόγω της πολλαπλής ομοιοπολικής σύζευξης με το υλικό [13,14]. Προκειμένου να εξουδετερωθούν οι μη τροποποιημένες εποξυ-ομάδες του υποστρώματος γίνεται κατεργασία με 3 Μ γλυκίνης σε pη 8,5 ενώ προκειμένου να απομακρυνθούν από το υλικό πρωτεΐνες που είναι καθηλωμένες στο υπόστρωμα μόνο με δισουλφιδικούς δεσμούς, γίνεται κατεργασία με 400 mm DTT [13,14]. 70
71 H H H S S H S S CH CH 2 glycine H CH CH 2 glycine H + + Enzyme SH Enzyme SH H H H S S Enzyme H S S Enzyme H 2 CH CH 2 glycine H CH CH 2 glycine H DTT H H S S Enzyme 100% elution yield (monofunctional derivatives) CH CH 2 H H DTT low elution yield (difunctional derivatives) Grazu et al Mateo et al Σχήμα 5: Δι-λειτουργικό και μονο-λειτουργικό υλικό καθήλωσης [13]. 71
72 1.3 Πεπτιδική σύνθεση Οι βασικές αρχές της πεπτιδικής σύνθεσης, λαμβάνοντας σαν παράδειγμα τη σύνθεση ενός διπεπτιδίου (σχήμα 1), περιλαμβάνουν αρχικά την προστασία της αμινομάδας του Ν-τελικού αμινοξέος και της καρβοξυλομάδας του C-τελικού αμινοξέος, καθώς επίσης και των δραστικών ομάδων των πλευρικών αλυσίδων των αμινοξέων -εφόσον αυτές υπάρχουν- ώστε να μην εμπλέκονται στις αντιδράσεις σύζευξης. Στη συνέχεια απαιτείται η ενεργοποίηση της καρβοξυλομάδας του Ν-προστατευμένου αμινοξέος και ακολουθεί η αντίδραση σύζευξης αυτού με το C-προστατευμένο C-τελικό αμινοξύ, που οδηγεί στη δημιουργία του πεπτιδικού δεσμού. Τέλος απομακρύνονται οι προστατευτικές ομάδες από τα Ν- και C- τελικά αμινοξέα. H H 2 C CH R 1 A-X H H A H C CH H 2 C CH R 1 R 2 R'H BH A H H C R 1 CR' H + H 2 C CB A H CH C H CH R 2 R 1 R 2 CB H 2 CH C H CH CH Σχήμα 1: Σχηματική πορεία σύνθεσης διπεπτιδίου [12]. R 1 R Προστατευτικές ομάδες αμινοξέων Για κάθε συγκεκριμένη χαρακτηριστική ομάδα ενός αμινοξέος υπάρχουν περισσότερες από μια προστατευτικές ομάδες, παρέχοντας έτσι μεγάλη ευχέρεια επιλογής και επιτρέποντας χωρίς μεγάλη δυσκολία τη σχεδίαση της σύνθεσης ακόμη και εξαιρετικά πολύπλοκων μορίων. Για την εκλογή της πιο κατάλληλης προστατευτικής ομάδας λαμβάνεται υπόψη η φύση της υπό προστασία ομάδας, οι συνθήκες αντίδρασης, που θα ακολουθήσουν, ώστε να μπορέσει μεν να 72
73 παρουσιάσει το χαρακτήρα της η προστατευτική ομάδα, να είναι δε ανεκτές οι συνθήκες αυτές στο υπόλοιπο μόριο κατά την απομάκρυνση της ομάδας και φυσικά να μην προκαλείται ρακεμίωση αν υπάρχουν χειρόμορφα κέντρα [12] Προστασία α-αμινομάδας αμινοξέων Για να προστατευτεί μια αμινομάδα στα πλαίσια μιας αντίδρασης πεπτιδικής σύνθεσης, πρέπει να μειωθεί δραστικά ο πυρηνόφιλος χαρακτήρας της μετατοπίζοντας την ηλεκτρονιακή πυκνότητα του αζώτου σε έναν κατάλληλο αντικαταστάτη υδρογόνου ή αποκρύπτοντας την πίσω από κάποιον ογκώδη στερεοχημικό παρεμποδιστή. Οι ομάδες "ουρεθανικού τύπου" είναι μια μεγάλη κατηγορία προστατευτικών ομάδων, οι οποίες όταν προσδένονται στην αμινομάδα τη μετατρέπουν σε αλκοξυ-καρβονυλ-αμινοπαράγωγα, που ονομάζονται "ουρεθάνες" ή "καρβαμίδια". Τα παράγωγα αυτά είναι ταυτόχρονα και αμίδια και εστέρες. Σαν αμίδια έχουν μειωμένο πυρηνόφιλο χαρακτήρα στο άζωτο, ενώ σαν εστέρες μπορούν να μετατραπούν στα αντίστοιχα καρβαμιδικά οξέα. Αντιπροσωπευτικά δείγματα της κατηγορίας αυτής είναι η βενζυλοξυ-καρβονυλομάδα (Ζ), η τριτοταγής βουτοξυ-καρβονυλομάδα (Boc), η 2-(4-διφαινυλο-)ισοπροποξυ-καρβονυλομάδα (Bpoc) και η 9-φλουορενυλ-μεθοξυκαρβονυλομάδα (Fmoc). Οι αμινοπροστατευτικές αλκοξυλοκαρβονυλικές ομάδες προτιμήθηκαν στο πεδίο της προστασίας της αμινομάδας για πολλά χρόνια, λόγω της αποδοτικότητας τους. Ωστόσο υπάρχουν και άλλες ομάδες -μη ουρεθανικού τύπου- που είναι εξίσου χρήσιμες. Μερικές από αυτές είναι η τριφαιναιλομάδα (Trt), η 2-νιτρο-φαινυλο-σουλφαινυλομάδα (ps), η τοζυλομάδα (Tos), η φθαλιμιδομάδα κ.ά [12] Τριτοταγής βουτοξυ-καρβονυλομάδα (Boc) CH 3 H 3 C C C CH 3 Η Boc ομάδα εισήχθη στη βιβλιογραφία το 1957 από τον Carpino και άμεσα βρήκε εφαρμογή στην πεπτιδική σύνθεση. Λίγα χρόνια μετά ο Merrifield συνειδητοποίησε ότι ο διαφορετικός βαθμός αστάθειας, έναντι των οξέων, της Boc ομάδας σε σχέση με τις βενζυλ- τύπου ομάδες, θα μπορούσε να είναι συμβατός με ένα αποτελεσματικό σχήμα προστασίας που να παρέχει προσωρινή και μόνιμη προστασία αντίστοιχα, οπότε προέβη στη σύνθεση της βραδυκινίνης σε στερεά φάση χρησιμοποιώντας Boc- προστατευμένα αμινοξέα. 73
74 Η Boc ομάδα είναι απολύτως σταθερή σε συνθήκες καταλυτικής υδρογόνωσης (υδρογονόλυσης), και γενικότερα σε αναγωγικά μέσα, είναι όμως περισσότερο ασταθής από την Ζ ομάδα στα οξέα. Βασικά και πυρηνόφιλα αντιδραστήρια δεν ασκούν καμία επίδραση επάνω της, ακόμα και έπειτα από εκτεταμένη έκθεση της σε αυτά. Η εισαγωγή της παλαιότερα γίνονταν με επίδραση του t-βουτοξυ-καρβονυλο-αζιδίου πάνω σε άλατα αμινοξέων, σε υδατικό διάλυμα και ασθενώς αλκαλικό περιβάλλον. Η μέθοδος απαιτεί ιδιαίτερη προσοχή, λόγω του ότι το αζίδιο μπορεί να προκαλέσει έκρηξη, οπότε η χρήση του είναι περιορισμένη. Σήμερα προτιμάται η χρήση του ανυδρίτη Boc--Boc σε διάλυμα αμινοξέος, με μίγμα διοξάνης, νερού και υδροξειδίου του νατρίου ή τριαιθυλαμίνης (σχήμα 2). Παρόλο που πρόκειται για πιο ακριβή ουσία, είναι προφανώς ασφαλής, δίνει καλές αποδόσεις και είναι ένα στερεό που μπορεί να διατηρηθεί στο ψυγείο για μεγάλο χρονικό διάστημα. CH 3 CH 3 H C 2 H 3 C C C C C CH 3 + H 2 C CH CH 3 CH 3 R CH 3 H H CH 3 H 3 C C C C CH + H C CH 3 CH 3 R CH 3 Σχήμα 2: Εισαγωγή Boc ομάδας [12]. Η διάσπαση της Boc ομάδας επιτυγχάνεται με διάλυση του προστατευμένου αμινοξέος ή πεπτιδίου σε TFA (είτε καθαρό είτε διαλυμένο σε DCM) σε θερμοκρασία δωματίου για περίπου μισή ώρα και στη συνέχεια αραίωση με αιθέρα ή συμπύκνωση. Τα προϊόντα που απελευθερώνονται (ισοβουτένιο και διοξείδιο του άνθρακα) είναι πτητικά και απομακρύνονται από το μίγμα της αντίδρασης (σχήμα.3). Οι συνθήκες αυτές θεωρούνται ήπιες και αξιόπιστες. Στην περίπτωση που είναι παρόντα αμινοξέα με ευαίσθητες πλευρικές ομάδες (π.χ. Trp, Cys, Met) τότε προστίθενται δεσμευτές (scavengers), οι οποίοι αντιδρούν με τα αρχικά σχηματιζόμενα καρβοκατιόντα [12]. 74
75 CH 3 H H CH 3 H H H H 3 C C C C CH H + H 3 C C C C CH CH 3 R CH 3 R CH 3 H 3 C H 3 C C CH 2 H 3 C C CH 3 u H 3 C CH 3 C u CH 3 H H H H 3 C CH H C C CH R C 2 R Σχήμα 3: Απομάκρυνση Boc ομάδας [12] φλουορενυλ-μεθοξυ-καρβονυλομάδα (Fmoc) Λόγω της ευρείας διάδοσης της Fmoc/Bu t μεθοδολογίας στη στερεά φάση, η Fmoc ομάδα είναι από τις πιο σημαντικές προστατευτικές ομάδες της αμινομάδας που χρησιμοποιούνται σήμερα. Είναι σταθερή σε επίδραση οξέων και στην καταλυτική υδρογόνωση (υδρογονόλυση), ενώ διασπάται κάτω από ήπιες βασικές, μη υδρολυτικές συνθήκες. H C C H Η Fmoc ομάδα εισάγεται με αντίδραση της αμινομάδας με Fmοc-Cl σε μίγμα αλκαλικού διαλύματος διοξάνης (σχήμα 4), παρατηρείται όμως ταυτόχρονα ανεπιθύμητη παράπλευρη αντίδραση που οδηγεί στο σχηματισμό Fmoc-διπεπτιδίου, δύσκολα διαχωριζόμενου από το κύριο προϊόν. Έτσι χρησιμοποιούνται λιγότερο δραστικά αντιδραστήρια, εκ των οποίων το κυριότερο είναι ο ηλεκτριμιδικός εστέρας της 9-φλουορενυλ-μεθανόλης (Fmoc-Su) [12]. 75
76 H C C H Cl H + H 2 C Ca R a 2 C 3 HCl H C C H H H C CH R Σχήμα 4: Εισαγωγή Fmoc ομάδας [12]. Απομακρύνεται συνήθως με διάλυμα 20% πιπεριδίνης σε DMF (σχήμα 5), μπορούν όμως να χρησιμοποιηθούν και διαλύματα κυρίως δευτεροταγών αμινών (π.χ. διαιθυλαμίνη) σε οργανικούς διαλύτες. Η αμινομάδα που απελευθερώνεται μετά την απομάκρυνση της Fmoc ομάδας, είναι ελεύθερη και όχι πρωτονιωμένη, όπως συμβαίνει κατά την απόσπαση άλλων ομάδων με την επίδραση οξέων, ενώ το διβενζοφουλβένιο που σχηματίζεται αντιδρά με την πιπεριδίνη προς σχηματισμό μιας σταθερής τριτοταγούς βάσης, η οποία δεν παρεμποδίζει την αντίδραση και απομακρύνεται κατά τις πλύσεις [12]. H C C H CH 2 H H + + C H H H H H H H C + + C 2 + H 2 H H Σχήμα 5: Απομάκρυνση Fmoc ομάδας [12]. 76
77 Τριφαινυλομάδα (Trt) Η Trt ομάδα είναι σταθερή σε βασικά διαλύματα και ασταθής σε όξινα. Η τριτυλιωμένη αμινομάδα διατηρεί ασθενείς βασικές ιδιότητες, δεν έχει όμως πυρηνόφιλες, λόγω της στερεοχημικής παρεμπόδισης που της προσδίδει η Trt ομάδα. C Για την εισαγωγή της Trt ομάδας χρησιμοποιείται το τριτυλοχλωρίδιο (σχήμα 6). Η αντίδραση λαμβάνει χώρα συνήθως με απευθείας τριτυλίωση των αμινοξέων σε υδατικά διαλύματα παρουσία διαιθυλαμίνης. H H H C Cl + H 2 C CH C C CH + HCl R R Σχήμα 6: Εισαγωγή Trt ομάδας [12]. Η διάσπασή της είναι σχετικά εύκολη, λόγω της μεγάλης σταθερότητας του σχηματιζόμενου καρβονιόντος. Απομακρύνεται με επίδραση αραιού διαλύματος υδροχλωρικού οξέος ή άλλου ισχυρού οξέος με θέρμανση για 20 min σε διάλυμα 50% οξικού οξέος. Μπορεί επίσης να απομακρυνθεί με καταλυτική υδρογόνωση. Αξίζει να σημειωθεί ότι ανάλογα με τις συνθήκες αποπροστασίας, λαμβάνονται διάφορα παράγωγα, όπως φαίνεται στο σχήμα 7 [12]. 77
78 H 2 Ph 3 C H H Ph 3 C + H CH C R HCl/CH 3 H Ph 3 C CH 3 + H CH C Ph 3 C H CH C R R HCl/αιθ. Ph 3 C Cl + H CH C R H 2 /Pd Ph 3 C H + H CH C R Σχήμα 7: Απομάκρυνση Trt ομάδας [12] Ακετυλομάδα (Ac) Η ακετυλομάδα αναφέρεται ως προστατευτική ομάδα της αμινομάδας, καθώς όταν εισαχθεί καταστέλλει τον πυρηνόφιλα χαρακτήρα του αζώτου σε μεγάλου βαθμό και η απομάκρυνση της μπορεί να επιτευχθεί με υδρόλυση. Ωστόσο οι διάφορες συνθήκες υδρόλυσης οδηγούν ταυτόχρονα και στην αποικοδόμηση των περισσοτέρων πεπτιδικών αλυσίδων. CH 3 C Έτσι δεν μπορεί να χρησιμοποιηθεί για προσωρινή προστασία της αμινομάδας, παρά μόνο για μόνιμη. Συνήθως χρησιμοποιείται σε τελικό στάδιο σύνθεσης πεπτιδίων, καθώς προκύπτουν με τον τρόπο αυτό σταθερά παράγωγα. Η εισαγωγή της γίνεται συνήθως με οξικό ανυδρίτη (σχήμα.8) [12]. H 3 C C C CH 3 + H 2 CH CH R H 3 C C H CH CH + CH 3 CH R Σχήμα 8: Εισαγωγή Ac ομάδας [12]. 78
79 Προστασία α-καρβοξυλομάδας αμινοξέων Από την άποψη της ακυλίωσης, οι αμινομάδες των αμινοξέων και των πεπτιδίων είναι θεωρητικά πιο πυρηνόφιλες από τις καρβοξυλομάδες τους, οπότε αμινοξέα και πεπτίδια με αποπροστάτευτες καρβοξυλομάδες μπορούν να χρησιμοποιηθούν για σχηματισμό πεπτιδικού δεσμού, αν προστεθούν ξεχωριστά προενεργοποιημένα καρβοξυλικά συστατικά. Πολλές αντιδράσεις αυτού του τύπου έχουν λάβει χώρα με επιτυχία, όμως συχνά υιοθετείται η στρατηγική της μέγιστης δυνατής προστασίας, ακόμα και όταν δεν είναι απαραίτητο, διότι έτσι παρέχονται πλεονεκτήματα διαλυτότητας και διευκολύνεται η απομόνωση του προϊόντος. Ειδικά σε πειράματα όπου ενεργοποιείται το καρβοξυλικό συστατικό παρουσία του αμινοσυστατικού, τότε απαιτείται απόλυτα η εκλεκτική προστασία της α-καρβοξυλομάδας του τελευταίου. Σύνηθες μέσο καρβοξυ-προστασίας είναι η εστεροποίηση και συγκεκριμένα οι μεθυλ- (Me) και αιθυλεστέρες (ΟΕt), οι βενζυλ-(οβzl) και π-νιτροφαινυλεστέρες (ΟΝp), οι τριτοταγείς βουτυλεστέρες (Bu t ) και οι φαινυλεστέρες (Ph) [12] Βενζυλεστέρες (Bzl) CH 2 Οι βενζυλεστέρες παρασκευάζονται με εστεροποίηση άνυδρης βενζυλικής αλκοόλης και αμινοξέος παρουσία π-τολουολοσουλφονικού οξέος (που δρα σαν καταλύτης) σε διάλυμα βενζολίου (σχήμα 9), για την απομάκρυνση του παραγόμενου νερού αζεοτροπικά. H 2 H 2 CH R CH + HCH 2 + H 3 C S 3 H H 3 C S 3 H H 2 CH. C CH 2 R Σχήμα 9: Εισαγωγή Bzl ομάδας [12]. Η βενζυλομάδα απομακρύνεται με σαπωνοποίηση, με καταλυτική ή χημική υδρογονόλυση, με επίδραση διαλύματος HBr/AcH ή υγρό HF (σχήμα 10). Η καταλυτική υδρογονόλυση εφαρμόζεται όταν δεν υπάρχουν πλευρικές αλυσίδες που περιέχουν θείο (π.χ. Thr). Η 79
80 συμπεριφορά της Bzl ομάδας σε όξινο περιβάλλον μοιάζει με αυτή της Ζ ομάδας με μικρότερη ευαισθησία [12]. H - /H 2 H CH C + HCH 2 R H CH R C CH 2 H 2 /Pd ή a/h 3 HBr/AcH H H CH R CH CH CH + + HCH 2 HCH 2 R Σχήμα 10: Απομάκρυνση Bzl ομάδας [12] Πλάγιες προστατευτικές ομάδες αμινοξέων Όταν οι πλευρικές ομάδες δεν είναι δραστικές στις συνήθεις συνθήκες της πεπτιδικής σύνθεσης, τότε η προστασία τους δεν είναι αναγκαία. Έτσι πολλές φορές στην πεπτιδική σύνθεση χρησιμοποιούνται παράγωγα σερίνης ή ιστιδίνης με απροστάτευτη την πλευρική αλυσίδα. Άλλες όμως δραστικότερες ομάδες σαν την ε-αμινομάδα της λυσίνης ή το γ-καρβοξύλιο του γλουταμινικού οξέος πρέπει οπωσδήποτε να προστατεύονται. Στην περίπτωση των αμινομάδων ή των καρβοξυλομάδων που βρίσκονται στις πλευρικές αλυσίδες των αμινοξέων, χρησιμοποιούνται οι ίδιες μέθοδοι που εφαρμόζονται για την προστασία της α-αμινομάδας ή του α-καρβοξυλίου, αρκεί να γίνονται κατάλληλοι συνδυασμοί ώστε να απομακρύνεται η προστατευτική ομάδα, όταν είναι επιθυμητό. Για την προστασία της υδροξυλομάδας χρησιμοποιείται κυρίως η Bzl, η Trt και η Bu t ομάδα [12] Μέθοδοι σύζευξης Η καρβοξυλομάδα του καρβοξυλικού συστατικού είναι απαραίτητο να ενεργοποιηθεί προκειμένου να αντιδράσει με την αμινομάδα του αμινοσυστατικού. Αυτό γίνεται με την πρόσδεση στο καρβοξύλιο μιας ομάδας που έλκει ηλεκτρόνια, ώστε να αυξάνει την ηλεκτρονιακή ανεπάρκεια του άνθρακα με πόλωση του δεσμού C- και έτσι να ενισχύεται η δυνατότητα προσβολής του καρβονυλίου από την πυρηνόφιλη αμινομάδα του αμινοσυστατικού. Μετά την ενεργοποίηση η ομάδα αυτή αποσπάται εύκολα από το καρβοξύλιο στη διάρκεια της αντίδρασης. Παλαιότερα χρησιμοποιούνταν η "μέθοδος των χλωριδίων", η οποία εγκαταλείφθηκε διότι τα Ν- υποκαταστημένα χλωρίδια είναι πολύ δραστικά και οδηγούν σε παραπροϊόντα, αλλά και η "μέθοδος των αζιδίων", που επίσης δε χρησιμοποιείται, λόγω χαμηλών αποδόσεων. Έχουν 80
81 αναπτυχθεί διάφορες μέθοδοι σύζευξης με κυριότερες την μέθοδο των καρβαμιδίων, των μικτών ανυδριτών, των ενεργών εστέρων και των φωσφονικών και ουρονικών παραγώγων [12] Μέθοδος των καρβαμιδίων Τα καρβαμίδια είναι τα πιο διαδεδομένα αντιδραστήρια που χρησιμοποιούνται για ενεργοποίηση in situ στην πεπτιδική σύνθεση. Κύριος εκπρόσωπος της κατηγορίας είναι το Ν,Ν δικυκλοεξυλοκαρβοδιιμίδιο (DCC), μια κρυσταλλική ουσία, που λόγω των δυο συνεχόμενων διπλών δεσμών προσφέρεται σε αντιδράσεις πυρηνόφιλης προσθήκης. To DCC προστίθεται στο μίγμα του οξέος και της αμίνης που θα σχηματίσουν τον πεπτιδικό δεσμό και με απόσπαση ενός μορίου νερού παράγεται Ν,Ν-δικυκλοεξυλουρία (DCU) (σχήμα 11). Ενδιάμεσο της αντίδρασης σύζευξης είναι ένα είδος ενεργού εστέρα της ισοουρίας (αμινοακυλιωμένο παράγωγό της), το οποίο τελικά προκαλεί τη σύζευξη του αμινοσυστατικού. Από το ενδιάμεσο αυτό προϊόν είναι δυνατόν να σχηματιστεί ή πεπτιδικός δεσμός ή συμμετρικός ανυδρίτης, ο οποίος στη συνέχεια ακυλιώνει την αμινοένωση. Και στις δυο περιπτώσεις σχηματίζεται ως παραπροϊόν η δυσδιάλυτη Ν,Ν-δικυκλοεξυλουρία. Από την ισοουρία μπορεί να σχηματιστεί και ένα άλλο σταθερό παραπροϊόν, η Ν-ακυλουρία (σχήμα 12), γεγονός που αποτελεί μειονέκτημα της μεθόδου, οπότε προτιμάται η αντίδραση να γίνεται σε χαμηλή θερμοκρασία, προκειμένου να περιορίζεται ο σχηματισμός της. H C A H CH CH + C R 1 (DCC) A H CH C R 1 (-ακυλισοουρία) H 2 CH CB R 2 A H CH C H CH CB + H C H R 1 R 2 (DCU) Σχήμα 11: Μέθοδος σύζευξης με DCC [12]. 81
82 H C A H CH C R 1 (-ακυλισοουρία) A H CH R 1 C H C (Ν-ακυλουρία) Σχήμα 12: Σχηματισμός Ν-ακυλουρίας μέσω ενδομοριακής μετάθεσης Ο- ακυλισοουρίας [12]. Τo DCC είναι ένα πολύ ενεργό αντιδραστήριο που οδηγεί σε αντιδράσεις υψηλής απόδοσης. Χρησιμοποιείται δε και για την παρασκευή ενεργών εστέρων Ν-προστατευμένων αμινοξέων. Ωστόσο έχει αλλεργιογόνες ιδιότητες και δημιουργεί προβλήματα στον καθαρισμό των προϊόντων. Έτσι χρησιμοποιούνται σήμερα άλλα καρβοδιιμίδια που σχηματίζουν ευδιάλυτες ουρίες, όπως το Ν,Ν-διισοπροπυλοκαρβοδιιμίδιο (DΙC), το t-βουτυλομεθυλκαρβοδιιμίδιο, το t- βουτυλοαιθυλκαρβοδιιμίδιο κ.ά. Πιθανή ρακεμίωση περιορίζεται σημαντικά κατά το στάδιο της σύζευξης με τη χρήση πρόσθετων αντιδραστηρίων, όπως το 1-υδροξυ-βενζοτριαζόλιο (ΗΟΒt), που είναι και το ευρύτερα χρησιμοποιούμενο, το οποίο ως ασθενές οξύ, εμποδίζει την απόσπαση πρωτονίου από το ασύμμετρο άτομο άνθρακα του καρβοξυλικού συστατικού. Εκτός αυτού μπορεί να προστεθεί το Ν-υδροξυ-ηλεκτριμίδιο (HSu) ή η 3-υδροξυ-3,4-διυδρο-1,2,3-βενζοτριαζιν-4-όνη (HBt). Η συγκέντρωση του πρόσθετου, που δρα ως ένα δεύτερο πυρηνόφιλο, παραμένει σταθερή, αφού αναγεννάται συνεχώς κατά τη διάρκεια της σύζευξης. Επιπλέον επιταχύνει την όλη πορεία και μετατρέπει το πολύ δραστικό ενδιάμεσο της αντίδρασης του DCC στο λιγότερο δραστικό ενεργό εστέρα του 1-υδροξυ-βενζοτριαζολίου ή του Ν-υδροξυ-ηλεκτριμιδίου. Ο ενεργός αυτός εστέρας, σχηματιζόμενος in situ, συντελεί λιγότερο σε παράπλευρες αντιδράσεις και επιπλέον είναι αρκετά δραστικός, ώστε να εξασφαλίζει ικανοποιητικό ποσοστό ακυλίωσης (σχήμα 13) [12]. 82
83 H 2 R' RCHR ' RCH + C (DCC) H (HBt) CR H 2 R' RCHR ' + (HBt) H (HBt) H RCH (RC) 2 H 2 R' RCHR ' Σχήμα 13: Πορεία αντίδρασης του DCC παρουσία HBt [12] Μέθοδος των ασύμμετρων ή μικτών ανυδριτών Παλαιότερα παρασκευάζονταν οι συμμετρικοί ή απλοί ανυδρίτες από τα αντίστοιχα ακυλαμινοξέα με τη χρήση διαφόρων μέσων, συμπεριλαμβανομένου του DCC. Η αμινόλυση ενός συμμετρικού ανυδρίτη εμφανίζει το μειονέκτημα της αξιοποίησης μόνο της μισής ποσότητας του Ν-προστατευμένου αμινοξέος. αλλά και της συχνά δύσκολης απομάκρυνσης του οξέος που σχηματίζεται. Έτσι η μέθοδος τροποποιήθηκε με αποτέλεσμα να παρασκευάζονται οι ασύμμετροι ή μικτοί ανυδρίτες από το άλας του Ν-προστατευμένου αμινοξέος με τριτοταγή βάση (π.χ. TEA) και το χλωρίδιο του βοηθητικού οξέος. Μειονέκτημα της μεθόδου είναι το γεγονός ότι πρόκειται για ασταθείς ενώσεις, συνεπώς πρέπει να παρασκευάζονται σε χαμηλές θερμοκρασίες και να χρησιμοποιούνται αμέσως χωρίς να απομονωθούν. Απαραίτητη είναι και η απομάκρυνση του βοηθητικού οξέος, μετά τη σύζευξη, το οποίο πρέπει να δεσμεύεται από ισοδύναμη ποσότητα βάσης γιατί διαφορετικά θα σχηματιστεί άλας με την ελεύθερη αμινομάδα του αμινοσυστατικού και θα την αδρανοποιήσει. Περίσσεια βάσης όμως κατά τη σύζευξη προκαλεί ρακεμίωση και καλό είναι να αποφεύγεται. Για τους παραπάνω λόγους πολύ σημαντική προσφορά στην πεπτιδική σύνθεση είναι η εισαγωγή των μικτών ανυδριτών με παράγωγα του ανθρακικού οξέος, που παρασκευάζονται από τους χλωρομυρμηκικούς εστέρες (R'CCl) των αντίστοιχων αλκοολών. Η ομάδα (R'-) δρα ως δότης ηλεκτρονίων, όπου R' είναι συνήθως CH 3 CH 2 - ή CH 3 CH(CH 3 )CΗ 2 -. Σύμφωνα με τη μέθοδο, ελευθερώνεται ανθρακικός μονοεστέρας, που αμέσως διασπάται σε διοξείδιο του άνθρακα και αλκοόλη, οπότε δεν απαιτείται βάση, όπως παραπάνω (σχήμα 14). 83
84 A H CH C C R' + H 2 CH CB R 1 A H CH C H CH CB + R'H + C 2 R 1 R 2 R 2 Σχήμα 14: Μέθοδος σύζευξης με μικτούς ανυδρίτες-παράγωγα ανθρακικού οξέος [12]. Πλεονεκτήματα της μεθόδου είναι η ταχύτητα, η σχετική καθαρότητα των προϊόντων και η απόδοση, η οποία αυξάνει όταν χρησιμοποιείται χλωρομυρμηκικός ισοβουτυλεστέρας, οξικός αιθυλεστέρας ή τετραϋδροφουράνιο (ως διαλύτης) και θερμοκρασία -15 C. Στις συνθήκες αυτές ο σχηματισμός του ανυδρίτη γίνεται μέσα σε min, όταν η τριτοταγής βάση είναι μια μεθυλαμίνη. Μειονεκτήματα της μεθόδου είναι η πιθανότητα για στερεοχημικούς λόγους να επηρεαστεί η κατεύθυνση της αντίδρασης και να σχηματιστεί ως παραπροϊόν ένα παράγωγο ουρεθάνης (σχήμα 15), αλλά και η ρακεμίωση, η οποία εξαρτάται από τις ενώσεις που χρησιμοποιούνται. Συγκεκριμένα έχει παρατηρηθεί ότι η χρήση ενός ισοδύναμου Ν-μεθυλομορφολίνης, Ν- μεθυλοπιπεριδίνης ή Ν-μεθυλοπυρρολιδίνης περιορίζει σημαντικά τη ρακεμίωση [12]. A H CH C C R' + H 2 CH CB R 1 A H CH C H + R' C H CH R 1 R 2 R 2 CB Σχήμα 15: Παράπλευρη αντίδραση που δίνει παράγωγο ουρεθάνης [12] Μέθοδος των ενεργών εστέρων Η μέθοδος χρησιμοποιεί εστέρες, στους οποίους ο ηλεκτρονιόφιλος χαρακτήρας του καρβονυλικού άνθρακα ενισχύεται από ομάδες που έλκουν ηλεκτρόνια. Αντιπροσωπευτικό δείγμα αποτελούν οι εστέρες του 1-υδροξυ-1,2,3-βενζοτριαζολίου (Bt), του υδροξυ-ηλεκτριμιδίου (Su), οι 2,4,5-τριχλωροφαινυλεστέρες (Tcp), οι πενταφθοροφαινυλεστέρες (Pfp) και οι π- νιτροφαινυλεστέρες (ΟΝρ). Οι π-νιτροφαινυλεστέρες παρασκευάζονται από Ν-προστατευμένα αμινοξέα και π- νιτροφαινόλη. Πρόκειται για ενώσεις σταθερές, όταν φυλάσσονται με προσοχή και σε χαμηλή θερμοκρασία, κρυσταλλικές, που αμινολύονται με καλές αποδόσεις. Η παρουσία π-νιτροφαινόλης 84
85 ανιχνεύεται χάρη στο κίτρινο χρώμα του ανιόντος της σε αλκαλικό περιβάλλον. Η αντίδραση (σχήμα 16) λαμβάνει χώρα χωρίς ή με ελάχιστη ρακεμίωση, ενώ το ενεργοποιημένο παράγωγο του αμινοξέος απομονώνεται σε κρυσταλλική κατάσταση [12]. A-H CH C 2 R 1 + H 2 CH CB R 2 A-H CH C H CH CB + H 2 R 1 R 2 Σχήμα 16: Μέθοδος σύζευξης των ενεργών εστέρων [12] Μέθοδος των φωσφονικών και ουρονικών παραγώγων Διάφορα ακυλοφωσφονικά αντιδραστήρια, τα οποία μπορούν να δημιουργηθούν μέσω της προσβολής κατάλληλων φωσφονικών κατιόντων από καρβοξυλικά ανιόντα, αντιδρούν εύκολα με πυρηνόφιλα στον ακυλιωμένο άνθρακα του καρβονυλίου αμινοξέος ή πεπτιδίου και έχει αναπτυχθεί ένας αριθμός αλάτων κατιόντων προκειμένου να χρησιμοποιηθεί ως άμεσα αντιδραστήρια σύζευξης. Το εξαφθοροφωσφορικό άλας του βενζοτριαζολυλοξυ-τρις-διμεθυλαμινοφωσφονίου (ΒΟΡ) είναι ένα από τα πρώτα αντιδραστήρια σύζευξης που χρησιμοποιήθηκαν και ακολούθησαν άλλα, όπως το εξαφθοροφωσφορικό άλας του βενζοτριαζολυλοξυ-τρις-πυρρολιδινοφωσφονίου (PyBP), το εξαφθοροφωσφορικο άλας του 2-(1Η-βενζοτριαζολυλ)-1,1,3-τετραμεθυλοουρονίου (HBTU), το τετραφθοροβορικό άλας του 2-(1Η-βενζοτριαζολυλ)-1,1,3,3-τετραμεθυλοουρονίου (TBTU) και το εξαφθοροφωσφορικό άλας του βρωμο-τρις-πυρρολιδινοφωσφονίου PyBrP. Πρόκειται για φωσφονικά ή ουρονικά άλατα, τα οποία παρουσία μιας τριτοταγούς βάσης μετατρέπουν προστατευμένα αμινοξέα σε μια ποικιλία ενεργοποιημένων παραγώγων και βρίσκουν ευρεία εφαρμογή στην κλασσική πεπτιδική σύνθεση, τόσο σε στερεά όσο και σε υγρή φάση, ακόμα και σε δύσκολες περιπτώσεις. Για να πραγματοποιηθεί η σύζευξη απαιτείται η προσθήκη ισοδύναμης ποσότητας του μέσου σε ένα μίγμα 1:1 άμινο και καρβοξυσυστατικού, σε κατάλληλο αδρανή διαλύτη, μαζί με τριτοταγή βάση που εξασφαλίζει την παρουσία του καρβοξυλικού συστατικού σε ανιονική μορφή. Υπάρχουν διάφοροι τρόποι, ο κυριότερος όμως είναι μέσω σχηματισμού HBt εστέρων (σχήμα 17). 85
86 P [(CH 3 ) 2 ] 3 [(CH 3 ) 2 ] 3 P= RC RC + RC P[(CH 3 ) 2 ] 3 RC HR' (BP) R'H 2 Σχήμα 17: Μέθοδος σύζευξης με BP [12]. Τα αντιδραστήρια αυτά, που δρουν ως ενεργοποιητές in situ, είναι διαδεδομένα επειδή είναι εύχρηστα, δίνουν υψηλές αποδόσεις και ταχύτατες αντιδράσεις σύζευξης, ακόμα και σε περιπτώσεις που τα αμινοξέα είναι στερεοχημικά παρεμποδισμένα. Γενικά οδηγούν σε ελάχιστες παράπλευρες αντιδράσεις, ενώ τα παραπροϊόντα απομακρύνονται εύκολα. Όμως το BP τείνει να εγκαταλειφθεί λόγω του εξαμεθυλο-φωσφορικού τριαμιδίου [(Me 2 ) 3 P=)], που παράγεται σαν παραπροϊόν και το οποίο είναι καρκινογόνο, οπότε έχει αντικατασταθεί από το PyBP, που είναι εξίσου αποτελεσματικό. Το PyBrP χρησιμοποιείται ευρύτατα στα Ν-μεθυλ-αμινοξέα, καθώς επιτρέπει ταχύτατες και υψηλών αποδόσεων αντιδράσεις σύζευξης, χωρίς τον κίνδυνο της ρακεμίωσης. Αποτελεσματικά είναι και τα ουρονικά παράγωγα HBTU και TBTU, που είναι φθηνά και παρέχουν ταχύτατες συζεύξεις [12] Μεθοδολογίες πεπτιδικής σύνθεσης Το πρώτο βήμα, προκειμένου να επιτευχθεί μια πεπτιδική σύνθεση, είναι να αποφασιστεί η μεθοδολογία που θα ακολουθηθεί. Συγκεκριμένα πρέπει να επιλεχθεί ο σωστός συνδυασμός προστατευτικών ομάδων για την α-αμινομάδα και για τις πλευρικές ομάδες, ούτως ώστε να εξασφαλιστεί η ορθή σύνθεση της πεπτιδικής αλληλουχίας με το σχηματισμό των λιγότερο δυνατόν παραπροϊόντων. Οι κλασσικές μεθοδολογίες που έχουν αναπτυχθεί είναι η Boc/Βzl και η Fmoc/Bu t [12] Boc/Bzl μέθοδος Η μέθοδος χρησιμοποιήθηκε στην πεπτιδική σύνθεση σε υγρή φάση και σχεδόν αποκλειστικά τα πρώτα 15 χρόνια της SPPS και βασίζεται στη διαφορετικού βαθμού αστάθεια που εμφανίζουν οι συγκεκριμένες προστατευτικές ομάδες έναντι των οξέων. Σύμφωνα με αυτή 86
87 λοιπόν τη μέθοδο, οι ομάδες που απαιτούν προσωρινή προστασία (οι Ν α -αμινομάδες) προστατεύονται με την Boc ομάδα, ενώ για εκείνες που απαιτείται ημιμόνιμη προστασία (οι ομάδες των πλευρικών αλυσίδων) χρησιμοποιούνται οι Bzl τύπου ομάδες. Όσον αφορά την κατά στάδια σύνθεση σε στερεά φάση, χρησιμοποιεί TFA κατά τη διάρκεια της σύνθεσης για την απομάκρυνση της Boc ομάδας και HF τελικά για την απομάκρυνση της Bzl ομάδας και του πεπτιδίου από τη ρητίνη [12] Fmoc/Bu t μέθοδος Η εισαγωγή στην πεπτιδική σύνθεση της Fmoc ομάδας προσέφερε ευελιξία και οδήγησε στην ανάπτυξη της μεθοδολογίας Fmoc/Bu t, η οποία αποτελεί το πιο αντιπροσωπευτικό δείγμα "ορθογωνικής προστασίας". Ο όρος "ορθογωνική" εισήχθη το 1977 από τους Barany και Merrifield, για να περιγράψει κατηγορίες προστατευτικών ομάδων που απομακρύνονται με διαφορετικούς μηχανισμούς. Ως εκ τούτου μπορούν να απομακρυνθούν με οποιαδήποτε σειρά και παρουσία των υπόλοιπων ομάδων. Οι α-αμινομάδες λοιπόν προστατεύονται με την Fmoc ομάδα και οι ομάδες των πλευρικών αλυσίδων με Bzl τύπου ομάδες. Ο συνδυασμός Fmoc/Bu t είναι πράγματι ορθογωνικός σε αντίθεση με τον Boc/Bzl, τουλάχιστον όσον αφορά τις συνθήκες της SPPS. Στην Fmoc/Bu t μεθοδολογία χρησιμοποιείται TFA στο τελικό στάδιο για την απομάκρυνση του πεπτιδίου από τη ρητίνη, ενώ δε χρησιμοποιείται HF, γεγονός που καθιστά τη μέθοδο περισσότερο πρακτική [12] Τεχνικές πεπτιδικής σύνθεσης Η πεπτιδική σύνθεση μπορεί να εφαρμοστεί είτε σε διάλυμα (υγρή φάση) είτε σε στερεά φάση, ενώ πολλές φορές χρησιμοποιείται συνδυασμός και των δύο Σύνθεση σε υγρή φάση Περιλαμβάνει την πραγματοποίηση όλων των διεργασιών ανοικοδόμηση της αλυσίδας σε διάλυμα. Τα ενδιάμεσα προϊόντα απομονώνονται, ξηραίνονται και δύναται να καθαριστούν και να ταυτοποιηθούν μετά από κάθε στάδιο. Οι συνήθεις μέθοδοι καθαρισμού που χρησιμοποιούνται είναι εκχυλίσεις, καταβυθίσεις, ανακρυσταλλώσεις και χρωματογραφίες στήλης. Πλεονέκτημα της τεχνικής αποτελεί η υψηλή καθαρότητα του τελικού πεπτιδίου. Σημαντικά μειονεκτήματα όμως είναι η πιθανότητα να εμφανιστεί πρόβλημα διαλυτότητας 87
88 ενδιάμεσου προϊόντος, καθώς και ο μεγάλος χρόνος που απαιτείται για τη σύνθεση, οπότε η τεχνική καθίσταται δύσχρηστη για παρασκευή μεγάλων πεπτιδίων [12] Σύνθεση σε στερεά φάση Η τεχνική εισήχθη από το Merrifield και σύμφωνα με αυτή, η πεπτιδική αλυσίδα συνδέεται με το ένα άκρο της σε στερεό υπόστρωμα καθ όλη τη διάρκεια της σύνθεσης και αναπτύσσεται από το άλλο, ενώ διαχωρίζεται από τα διαλυτά αντιδραστήρια και διαλύτες με απλή διήθηση και πλύσεις. Μετά την επιμήκυνση της επιθυμητής αλυσίδας, το προϊόν αποδεσμεύεται από το στερεό υπόστρωμα, καθαρίζεται και ταυτοποιείται. Η σύνθεση σε στερεά φάση υπερτερεί έναντι εκείνης σε υγρή αναφορικά με τον χρόνο, που είναι σημαντικά μικρότερος, αλλά και από το γεγονός ότι μπορεί να είναι αυτοματοποιημένη διαδικασία. Εντούτοις δεν υπάρχει κατά τη συνήθη διαδικασία, η δυνατότητα καθαρισμού των ενδιάμεσων προϊόντων και είναι δύσκολη η ταυτοποίηση τους. Επίσης δεν είναι δυνατή η απομάκρυνση αδιάλυτων παραπροϊόντων, όπως πεπτιδίων μικρότερου μοριακού βάρους δεσμευμένων στο στερεό υπόστρωμα, λόγω μη ολοκληρωμένων συζεύξεων [12] Στερεά υποστρώματα (ρητίνες) Το στερεό υπόστρωμα είναι το υλικό που χαρακτηρίζει τη σύνθεση σε στερεά φάση και τη διαχωρίζει από την υγρή. Πρόκειται για αδιάλυτο πολυμερές με μορφή μικροπορώδους gel, το οποίο φέρει εντός και εκτός των πόρων του ενεργές ομάδες πάνω στις οποίες λαμβάνουν χώρα οι αντιδράσεις. Έτσι καθίσταται ιδιαίτερα σημαντική η καλή διόγκωση της ρητίνης, όπως και η δυνατότητα των αντιδραστηρίων να εισέρχονται στο εσωτερικό των πόρων, αλλά και να εξέρχονται από αυτό μέσω διάχυσης. Η ρητίνη θα. πρέπει να έχει δομική σταθερότητα στις συνθήκες της σύνθεσης, ταχεία και χωρίς στερεοχημικούς περιορισμούς επαφή της αναπτυσσόμενης αλυσίδας με τα αντιδραστήρια, καθώς και να διαχωρίζεται από την υγρή φάση σε κάθε στάδιο με απλή διήθηση [12] Συνδέτες Ο συνδέτης (linker) παρεμβάλλεται μεταξύ ρητίνης και πεπτιδίου και εξασφαλίζει μια αντιστρεπτή σύνδεση της αναπτυσσόμενης πεπτιδικής αλυσίδας με το στερεό υπόστρωμα, ενώ προστατεύει τη C-τελική α-καρβοξυλομάδα καθ όλη τη διαδικασία επιμήκυνσης της αλυσίδας. 88
89 Η επιλογή του linker καθορίζει τη C-τελική λειτουργική ομάδα στο τελικό προϊόν, καθώς μετά την απομάκρυνση του μπορεί να απελευθερώσει πεπτίδιο που να είναι παράγωγο οξέος ή αμίδιο [12] Πρόσδεση πρώτου αμινοξέος στη ρητίνη Το πρώτο βήμα κατά τη σύνθεση σε στερεά φάση είναι η σύνδεση της ρητίνης με το C- τελικό αμινοξύ. Η ικανοποιητική εκτέλεση αυτής της διαδικασίας είναι ιδιαίτερα σημαντική, επειδή, πρώτον, η έκταση της αντίδρασης αυτής θα καθορίσει την απόδοση του τελικού προϊόντος και, δεύτερον, θέσεις της ρητίνης που δεν έχουν αντιδράσει σε αυτή την αρχική διαδικασία δύναται να ακυλιωθούν σε ακόλουθους κύκλους αντιδράσεων, οδηγώντας στη δημιουργία σχετικών C-τελικών παραπροϊόντων [12] Επιμήκυνση πεπτιδικής αλυσίδας Για την ανοικοδόμηση της πεπτιδικής αλυσίδας απαιτείται η απομάκρυνση της Ν α - προστατευτικής ομάδας του C-τελικού αμινοξέος και στη συνέχεια ακολουθούν διαδοχικοί κύκλοι συζεύξεων-αποπροστασίας, μέχρις ότου επιτευχθεί ο επιθυμητός στόχος. Χρησιμοποιούνται μέθοδοι ενεργοποίησης και σύζευξης, οι οποίες βασίζονται κυρίως στο σχηματισμό ενεργών εστέρων είτε προσχηματισμένων είτε δημιουργημένων in situ [12] Απόσπαση πεπτιδίου από το πολυμερές και απομόνωσή του Όταν ολοκληρωθεί η σύνθεση του επιθυμητού πεπτιδίου ακολουθεί η απομάκρυνσή του από τη ρητίνη. Ανάλογα με τις συνθήκες που εφαρμόζονται, το πεπτίδιο μπορεί να απομακρυνθεί πλήρως αποπροστατευμένο, διατηρώντας όλες ή μέρος των προστατευτικών του ομάδων (της Ν α -αμινομάδας και των πλαγίων ομάδων). Προκειμένου να επιλεχθούν οι συνθήκες αυτές, λαμβάνεται υπόψη φυσικά το είδος του δεσμού μεταξύ πεπτιδίου και ρητίνης [12] Πεπτιδομιμητές Οι πεπτιδομιμητές είναι ενώσεις που έχουν προκύψει μετά από την τροποποίηση των αντίστοιχων πεπτιδίων,.με σκοπό την μεγαλύτερη σταθερότητα έναντι των πεπτιδασων. Υπάρχουν διάφορες κατηγορίες πεπτιδομιμητών, που περιγράφονται αναλυτικά παρακάτω, ανάλογα με την έκταση της τροποποίησης των πεπτιδίων από τα οποία προέρχονται. Για τη 89
90 σύνθεση τους απαιτείται ο συνδυασμός της κλασσικής πεπτιδικής χημείας και ενός ευρέος φάσματος αντιδράσεων συνθετικής οργανικής χημείας. Η σύνθεση μπορεί να λάβει χώρα σε υγρή, στερεά φάση ή και με συνδυασμό και των δύο φάσεων. Τα τελευταία χρόνια οι πεπτιδομιμητές έχουν αποκτήσει ιδιαίτερη σημασία στο βιολογικό, χημικό και φαρμακευτικό πεδίο, καθώς προσφέρουν μια σειρά πλεονεκτημάτων έναντι των φυσικά δραστικών πεπτιδίων, τα οποία κατέχουν σημαντικό ρόλο στον οργανισμό. Εκτός του ότι σε ορισμένες περιπτώσεις εμφανίζουν αυξημένη δραστικότητα συγκριτικά με τα πεπτίδια, έχουν επιπλέον καλύτερη βιοδιαθεσιμότητα σε από του στόματος χορήγηση και μεγαλύτερης διάρκειας δράση, διότι επιβραδύνεται η υδρόλυση τους από ένζυμα μέσα στον οργανισμό. Σήμερα παρέχεται η δυνατότητα συνδυάζοντας την υπολογιστική χημεία, τις εξελιγμένες τεχνικές του MR και την ανάλυση της δομής με ακτίνες-χ, να σχεδιαστούν και να συντεθούν φάρμακα, των οποίων η δομή να περιέχει ταυτόχρονα τροποποιημένα πεπτιδικά και μη-πεπτιδικά τμήματα. Η σύνθεση τέτοιων ενώσεων απαιτεί το συνδυασμό των τεχνικών της κλασσικής πεπτιδικής χημείας και ενός ευρέος φάσματος αντιδράσεων της συνθετικής οργανικής χημείας. Οι ενώσεις των πεπτιδομιμητών βασίζονται πρωταρχικά σε πληροφορίες για τη διαμόρφωση, τις τοποχημικές και ηλεκτρονιακές ιδιότητες των πεπτιδίων, αλλά και για τον υποδοχέα ή το ενεργό κέντρο του ενζύμου με το οποίο αλληλεπιδρούν. Ο σχεδιασμός των πεπτιδομιμητών, ως δυνητικά βιοδραστικών ουσιών, πρέπει να λαμβάνει λοιπόν υπόψη δύο παράγοντες: πρώτον, την ολική διαμόρφωση του αναλόγου σε σχέση με τη δομή του ενεργού κέντρου και δεύτερον, την τοποθέτηση συγκεκριμένων δομικών στοιχείων (π.χ. λειτουργικών ομάδων, πολικών και υδρόφοβων περιοχών) σε καθορισμένες θέσεις, έτσι ώστε να μπορούν vα λάβουν χώρα οι επιθυμητές αλληλεπιδράσεις (π.χ. δεσμοί υδρογόνου, ηλεκτροστατικές ή υδρόφοβες αλληλεπιδράσεις) [12] Κατηγορίες πεπτιδομιμητών με τροποποίηση πεπτιδικού σκελετού Υπάρχουν πολλές κατηγορίες πεπτιδομιμητών που έχουν προκύψει από την τροποποίηση του σκελετού των αντίστοιχων πεπτιδίων {-H-CH(R)-C-}. Οι πιο γνωστές από αυτές είναι τα πεπτοειδή (peptoids) {-(R)-CH 2 -C-}, τα αζαπεπτίδια {-ΝΗ-(R)-CΟ-}, τα αζα-πεπτοειδή {-(R)-H-C-}, τα υδραζινοπεπτίδια {-ΝΗ-H-CH(R)-C-}, τα α -υδραζινο-πεπτοειδή {-H- (R)-CH 2 -CΟ-}, τα retro-peptoids {-C-CH 2 -(R)-}, οι urea-πεπτιδομιμητές {-H-CH(R)-CH 2 - H-C-}, τα ureapeptoids {-(R)-CH 2 -CH 2 -H-C-}, τα sulfonamide peptoids {-(R)-CH 2 - CH 2 -S 2 -}, τα α-αμινοοξυ-πεπτίδια {-H--CH(R)-C-}, τα αμινοοξυ-peptoids {-(R)--CH 2 - C-}, πεπτίδια και πεπτοειδή με ενσωματωμένα σάκχαρα κ.ά [12]. 90
91 Πεπτοειδή Τα πεπτοειδή (peptoids) αποτελούν μια από τις πρώτες και πιο γνωστές κατηγορίες πεπτιδομιμητών που αναπτύχθηκαν και περιλαμβάνουν Ν-υποκατεστημένα παράγωγα γλυκίνης (Ν-αμινοξέα) (σχήμα 18). Παρακάτω διατηρείται ο όρος "peptoids" έναντι του μάλλον αδόκιμου όρου "πεπτοειδή". Συμβολίζονται όπως τα αμινοξέα τοποθετώντας αρχικά το γράμμα Ν (π.χ. το peptoid μονομερές της φαινυλαλαλίνης συμβολίζεται ως Phe). H H C C CH 2 C R α - αμινοξύ Ν - αμινοξύ R Σχήμα 18: Δομή αμινοξέος και Ν-υποκατεστημένου παραγώγου γλυκίνης [12]. Οι δομικές διαφορές ανάμεσα σε ένα πεπτίδιο και ένα peptoid φαίνονται στο σχήμα 19. Ουσιαστικά πρόκειται για την απουσία χειρομορφίας στα Ν-υποκατεστημένα παράγωγα γλυκίνης, με εξαίρεση την προλίνη. Κατά μία έννοια θα μπορούσε κανείς να θεωρήσει τα peptoids ως μιμητές πεπτιδίων, στους οποίους η πλευρική αλυσίδα έχει μετακινηθεί από το χειρόμορφο α- άτομο άνθρακα στο μη χειρόμορφο άτομο αζώτου. Αναμφίβολα αυτό επιφέρει συνέπειες ως προς τη βιολογική δραστικότητα των peptoids σε σχέση με τα αντίστοιχα πεπτίδια. Η σύνθεση ενός μεγάλου αριθμού peptoids και η σύγκριση της δράσης τους, με τα αντίστοιχα πάντα πεπτίδια, έχει αποδείξει oτι σε μεγάλο ποσοστό τα peptoids εμφανίζουν καλύτερη δράση από τα προγονικά πεπτίδια. Όμως κάτι τέτοιο δεν μπορεί να προβλεφθεί, αλλά θα πρέπει να εξετάζεται κάθε φορά, προκειμένου ενδεχομένως να επιβεβαιωθεί. Αντίθετα με τη βιολογική δράση, προβλέψεις με μεγαλύτερη ασφάλεια μπορούν να γίνουν για τις φυσικές και χημικές ιδιότητες των peptoids. Η απουσία ενός αμιδίου του τύπου C-H, που μπορεί να συμμετέχει σε δεσμό υδρογόνου, έχει ως συνέπεια τη μείωση της διαλυτότητας των peptoids σε διαλύτες ικανούς να σχηματίζουν δεσμούς υδρογόνου και αντίθετα αύξηση της διαλυτότητας τους σε περισσότερο άπολους διαλύτες. Επίσης η παρουσία τριτοταγών αμιδικών δεσμών έχει ως συνέπεια την εμφάνιση πολλών cis/trans ισομερών, λόγω της δυνατότητας περιστροφής γύρω από κάθε τριτοταγές αμίδιο στην αλληλουχία του peptoid, με αποτέλεσμα να δίνει περίπλοκα MR φάσματα. Ως εκ τούτου προτιμάται η χρήση φασματομετρίας μάζας για το χαρακτηρισμό των peptoids. Επιπλέον λόγω απουσίας του κλασσικού πεπτιδικού δεσμού, παρουσιάζουν μικρότερη ευαισθησία έναντι πρωτεολυτικής διάσπασης, όπως αποδεικνύεται από μελέτες που έχουν πραγματοποιηθεί. 91
92 ΠΕΠΤΙΔΙΟ R 2 R 4 H H H H H R 1 R 3 R 5 Ν-τελικό άκρο C-τελικό άκρο R 2 R 4 R 1 R 3 R 5 PEPTID Σχήμα 19: Δομή πεπτιδίου και peptoid [12]. Στα peptoid-peptide υβρίδια έχει αντικατασταθεί ένα ή περισσότερα αμινοξέα από Ν- υποκατεστημένο παράγωγο γλυκίνης. Η φιλοσοφία στην οποία βασίστηκε πρωταρχικά η σύνθεση τέτοιων αναλόγων ήταν η συστηματική αντικατάσταση ενός, δύο, τριών κ.λ.π. αμινοξέων της πεπτιδικής αλληλουχίας από τα αντίστοιχα Ν-υποκατεστημένα παράγωγα γλυκίνης, προκειμένου να εξεταστεί μέχρι ποιο βαθμό μπορεί να τροποποιηθεί μια πεπτιδική αλληλουχία, χωρίς να χάσει την ικανότητα πρόσδεσης στον κατάλληλο υποδοχέα [12] Αζαπεπτίδια Τα αζαπεπτίδια ανήκουν όπως και τα peptoids στην κατηγορία εκείνη των πεπτιδομιμητών που έχει υποστεί τροποποίηση ο πεπτιδικός σκελετός. Συγκεκριμένα έχει αντικατασταθεί το α- άτομο άνθρακα (α-ch) από ένα άτομο αζώτου (Ν), με αποτέλεσμα αυτό το άζωτο να φέρει την πλευρική αλυσίδα του αυθεντικού αμινοξέος (σχήμα 20). H H C C H C R α - αμινοξύ R αζα - αμινοξύ Σχήμα 20: Δομή αμινοξέος και αζα-αμινοξέος [12]. 92
93 Όταν η αντικατάσταση των α-αμινοξέων ενός πεπτιδίου από αζα-αμινοξέα είναι μερική, δηλαδή εξακολουθεί να υπάρχει κάποιο αμινοξύ στην αλληλουχία, τότε το παράγωγο που προκύπτει ονομάζεται "αζαπεπτίδιο", ενώ αν όλα τα αμινοξέα έχουν αντικατασταθεί, τότε ονομάζεται "αζατίδιο" (azatide). Οι δομικές διαφορές ανάμεσα σε ένα πεπτίδιο, ένα αζαπεπτίδιο και ένα αζατίδιο φαίνονται στο σχήμα 21. Τα αζα-αμινοξέα συμβολίζονται όπως και τα αμινοξέα τοποθετώντας αρχικά το πρόθεμα aza ή az (π.χ. η αζα-φαινυλαλαλίνη συμβολίζεται azaphe ή azphe). Ένα από τα πλεονεκτήματα της κατηγορίας αυτής είναι η άμεση και σε γενικές γραμμές μη προβληματική σύνθεση, γεγονός που επιτρέπει την αντικατάσταση σε μια πεπτιδική αλληλουχία οποιουδήποτε αμινοξέος ή οποιουδήποτε αριθμού αμινοξέων, από τα αντίστοιχα αζα-αμινοξέα, σε όποια θέση κι αν βρίσκονται. Το πρώτο αζαπεπτίδιο συντέθηκε από τον Stolle στις αρχές του 20 ου αιώνα, πέρασαν όμως αρκετές δεκαετίες μέχρι να ξεκινήσει η συστηματική σύνθεση των αζαπεπτιδίων. Το 1963 ο Hess αντικατέστησε ένα φυσικό αμινοξύ της αγγειοτασίνης II από το αντίστοιχο αζα-αμινοξύ, με αποτέλεσμα να προκύψει το τροποποιημένο παράγωγο [azaval 3 ]-αγγειοτασίνη II, το οποίο εμφανίζει παρατεταμένη δράση συγκριτικά με το αυθεντικό πεπτίδιο. Έτσι λοιπόν δόθηκε το έναυσμα σε πολλούς ερευνητές να ασχοληθούν με την κατηγορία αυτή των ενώσεων, ώστε τελικά να προκύψει ένας μεγάλος αριθμός αναλόγων με βιολογικό ενδιαφέρον, καθώς τα παράγωγα παρουσίαζαν υψηλή ανθεκτικότητα έναντι πρωτεασών. Ν-τελικό άκρο C-τελικό άκρο R 2 R 4 H H H H H ΠΕΠΤΙΔΙΟ R 1 R 3 R 5 R 2 R 4 H H H H H ΑΖΑΠΕΠΤΙΔΙΟ R 1 R 3 R 5 R 2 R 4 H H H H H ΑΖΑΤΙΔΙΟ R 1 R 3 R 5 Σχήμα 21: Δομή πεπτιδίου, αζαπεπτιδίου και αζατιδίου [12]. 93
94 Η βιολογική δράση των πεπτιδίων και των πρωτεϊνών συνδέεται άμεσα, όπως είναι γνωστό, με τη δομή τους. Η εισαγωγή του Ν α ατόμου οδηγεί σε δομική διαφοροποίηση του σκελετού των αζα-πεπτιδίων σε σχέση με τον τυπικό πεπτιδικό σκελετό. Η διαφοροποίηση αυτή συνίσταται στη συνύπαρξη ενός συστατικού υδραζίνης [-H- α (R)-C-] με ένα συστατικό ουρίας [-H- a (R)- C-]. Ωστόσο η αντικατάσταση της ομάδας a-ch από Ν, από καθαρά δομική άποψη, δεν επιφέρει μεγάλη αλλαγή όσον αφορά τη χωροδιάταξη των αζαπεπτιδίων σε σχέση με τα πεπτίδια, καθώς υπάρχει παρόμοια απόσταση των ατόμων και γωνίες δεσμών, ενώ η πλευρική αλυσίδα παραμένει η ίδια. Εντούτοις η παρουσία του α-ατόμου αζώτου, εκτός του ότι οδηγεί στην απουσία χειρομορφίας, έχει ως επιπλέον αποτέλεσμα να μπορούν να σχηματιστούν περισσότερες μεσομερείς δομές, ειδικά λόγω του ελεύθερου ζεύγους ηλεκτρονίων που διαθέτει. Μελέτες που έχουν χρησιμοποιήσει την τεχνική του κυκλικού διχρωισμού αποδεικνύουν πως εξαιτίας του περιορισμού της δυνατότητας περιστροφής, οι πλευρικές αλυσίδες λαμβάνουν θέση μεταξύ της D- και L- διαμόρφωσης των αντίστοιχων αμινοξέων. Επιπρόσθετα, σύμφωνα με μετρήσεις η οξύτητα της ΝΗ ομάδας που προσδένεται στο α-άτομο αζώτου είναι υψηλότερη από oτι στα πεπτίδια, γεγονός που διευκολύνει μεν το σχηματισμό περισσότερο ισχυρών δεσμών υδρογόνου, εμποδίζει δε την πρωτονίωσή της σε φυσιολογικό pη. Tα παραπάνω έχουν ως συνέπεια να επηρεάζεται η απορρόφηση, η μεταφορά, η κατανομή, η ικανότητα πρόσδεσης στο ενεργό κέντρο ενός ενζύμου και η μεταβολική σταθερότητα, ενός βιολογικά δραστικού αναλόγου της κατηγορίας αυτής στον οργανισμό. Υπό ορισμένες συνθήκες, όπως όταν ευνοείται ο σχηματισμός πολλαπλών δεσμών υδρογόνου, η παρουσία δεσμών Van der Waals ή οι σχέσεις μεταφοράς φορτίου, τότε η διαφοροποίηση ενός αζαπεπτιδίου γίνεται σε μεγαλύτερο βαθμό σε σχέση με το αντίστοιχο πεπτίδιο, και ειδικά στα μεγάλου μεγέθους αζαπεπτίδια [12] Διάφορες δράσεις των αζαπεπτιδίων Τα αζαπεπτίδια αναφέρονται στη βιβλιογραφία ως αναστολείς πρωτεασών. Ιδιαίτερα έχουν αναφερθεί περιπτώσεις που δρουν ως αναστολείς πρωτεασών σερίνης και κυστεΐνης. Τα τελευταία χρόνια έχει δημοσιευτεί η αντι-hiv (Human Immunodeficiency Virus) τύπου 1 (HIV-1) δραστικότητα του αζαπεπτιδίου με τον κωδικό BMS , το οποίο είναι αναστολέας πρωτεάσης του HIV-I. Παρατίθενται μερικά παραδείγματα μελέτης της βιολογικής δράσης αζα-αναλόγων, τα οποία εμφανίζουν από κοινού υψηλή ανθεκτικότητα έναντι πρωτεασών και επιπλέον σε ορισμένες περιπτώσεις έχουν και καλύτερη δράση σε σχέση με τα αντίστοιχα πεπτίδια. Συγκεκριμένα το αζαπεπτίδιο [azagly 9 ]-oxytocin είναι 1.5 φορά πιο δραστικό από την οκυτοκίνη, το αζαπεπτίδιο 94
95 [azaasn 5 ]-eledoisin εμφανίζει 3-4 φορές καλύτερη δράση από την ίδια τη φυσική ορμόνη, τα αζαπεπτίδια [azagly 6 ]-LHRH, [azagly 10 ]-LHRH και [azaala 6 ]-LHRH παρουσιάζουν 100 φορές υψηλότερη δράση από τη λουλιβερίνη (LHRH), ενώ χαρακτηριστικό είναι το παράδειγμα του [D- Ser(Bu t ) 6, azagly 10 ]-LHRH. το οποίο βρίσκεται εδώ και χρόνια στη φαρμακευτική αγορά με την εμπορική ονομασία "Zolatex" για τη θεραπεία του καρκίνου του προστάτη [12] Υδραζινοπεπτίδια και Ν α -υδραζινο-πεπτοειδή Τα υδραζινοπεπτίδια και τα Ν α -υδραζινο-πεπτοειδή είναι μέλη της κατηγορίας των πεπτιδομιμητών που έχουν υποστεί τροποποίηση στο σκελετό τους (όπως τα peptoids και τα αζαπεπτίδια) και αποτελούνται από υδραζινο-οξέα (hydrazinoacids) ή Ν α -υδραζινο-οξέα ( α - hydrazinoacids) αντίστοιχα (σχήμα 22). Τα τελευταία ονομάζονται και α -hydrazino-acetic acids. H H C C H β α H H C C β H α CH 2 C R α - αμινοξύ R υδραζινο-οξύ R Ν α -υδραζινο-οξύ Σχήμα 22: Δομή αμινοξέος, υδραζινο-οξέος και Ν α -υδραζινο-οξέος [12]. Ένα υδραζινο-οξύ διατηρεί τη χειρομορφία του αντίστοιχου αμινοξέος, αφού η πλευρική αλυσίδα (R) εξακολουθεί να βρίσκεται στο α-άτομο C και συμβολίζεται με το πρόθεμα h (π.χ. hphe). Η αντικατάσταση σε μια πεπτιδική αλυσίδα ενός τουλάχιστον αμινοξέος από το αντίστοιχο υδραζινο-οξύ οδηγεί στο σχηματισμό του υδραζινοπεπτιδίου. Ένα Ν α -υδραζινο-οξύ όμως στερείται χειρομορφίας, καθώς η πλευρική αλυσίδα μετατοπίζεται στο α-άτομο Ν και συμβολίζεται με το πρόθεμα α h (π.χ. α hphe). Με αντικατάσταση ενός τουλάχιστον αμινοξέος μιας πεπτιδικής αλληλουχίας από το αντίστοιχο Ν α -υδραζινο-οξύ προκύπτει Ν α -υδραζινο-peptoid (σχήμα 23). Η χημεία των ενώσεων των κατηγοριών αυτών δεν είναι ιδιαίτερα αναπτυγμένη, όπως είναι των αζαπεπτιδίων. Έχουν αναφερθεί μέθοδοι παρασκευής υδραζινο-οξέων και μάλιστα δίνεται ιδιαίτερη βαρύτητα στη σύνθεση οπτικά ενεργών ουσιών. Παρόλα αυτά φαίνεται πως είναι περιορισμένος ο αριθμός των πλευρικών αλυσίδων που είναι συμβατές με τις χημικές μεθόδους που χρησιμοποιούνται για το σκοπό αυτό. Επιπλέον, λόγω παρουσίας του επιπρόσθετου ατόμου αζώτου, εμφανίζονται πολλές φορές δυσκολίες τόσο κατά την παρασκευή των υδραζινο-οξέων, όσο και σε αντιδράσεις σύζευξης, καθώς δε δρουν πάντα ως τοποεκλεκτικά αντιδραστήρια. Αυτό 95
96 εξηγεί και το λόγο, για τον οποίο τα εμπορικά διαθέσιμα υδραζινο-οξέα είναι συνήθως Ν α - προστατευμένα [12]. R H H ΠΕΠΤΙΔΙΟ R H H H ΥΔΡΑΖΙΝΟΠΕΠΤΙΔΙΟ H H Ν α -ΥΔΡΑΖΙΝΟ-PEPTID R Σχήμα 23: Δομή πεπτιδίου, υδραζινοπεπτιδίου και Ν α -υδραζινο-peptoid [12] Διάφορες δράσεις υδραζινοπεπτιδίων Η ενσωμάτωση υδραζινο-οξέων σε πεπτιδικές αλυσίδες επιφέρει σταθερότητα στις αλληλουχίες σε σχέση με τα αντίστοιχα αυθεντικά πεπτίδια και αναφέρονται περιπτώσεις που οι ενώσεις της κατηγορίας αυτής χρησιμοποιούνται ως αναστολείς μεταβολικών ενζύμων διαφόρων αμινοξέων. Επιπλέον είναι γνωστή και η αντιβιοτική δράση ορισμένων αναλόγων της κατηγορίας, με πιο γνωστό εκπρόσωπο το αντιβιοτικό "negamycin". Σύμφωνα με άλλες μελέτες έχει αποδειχθεί η κυτταροστατική δράση in vitro διαφόρων συνθετικών υδραζινοπεπτιδίων έναντι της κυτταρικής καρκινικής σειράς L1210 (murine leukemia). Τέλος έχουν βρεθεί υδραζινο-αζα-διπεπτίδια. τα οποία παρουσία μικρής ποσότητας HCl σχηματίζουν θερμοαντιστρεπτά φυσικά gels σε αρωματικούς διαλύτες, {π.χ. τολουόλιο, ξυλόλιο, στυρένιο). Αυτά τα gels είναι σταθερά στους C και μπορούν να αποθηκευτούν για μήνες. Με ηλεκτρονικό μικροσκόπιο βρέθηκε πως, σε αρωματικούς διαλύτες, οι πεπτιδομιμητές αυτοί αυτοσυναρμολογούνται προς σχηματισμό ινών, που είναι επιμήκεις και πολύ λεπτές, οι οποίες 96
![στρέφονται με τέτοιο τρόπο που δημιουργούν τρισδιάστατα πλέγματα μέσα στα οποία εγκλωβίζονται μόρια διαλύτη [12]. 1.4.](/docs-images/72/67903182/images/97-0.jpg "1 Εισαγωγή στη μικροσκοπία σαρώσεως ακίδας (SPM) Κατά την δεκαετία του 80, επειδή οι μέχρι τότε τεχνικές παρόλο που χαρακτηρίζονταν από μεγάλη ευαισθησία δεν μπορούσαν να δώσουν άμεση εικόνα της")
97 στρέφονται με τέτοιο τρόπο που δημιουργούν τρισδιάστατα πλέγματα μέσα στα οποία εγκλωβίζονται μόρια διαλύτη [12] Εισαγωγή στη μικροσκοπία σαρώσεως ακίδας (SPM) Κατά την δεκαετία του 80, επειδή οι μέχρι τότε τεχνικές παρόλο που χαρακτηρίζονταν από μεγάλη ευαισθησία δεν μπορούσαν να δώσουν άμεση εικόνα της μελετώμενης επιφάνειας, εμφανίστηκαν και αναπτύχθηκαν με ραγδαίο ρυθμό οι SPM τεχνικές, οι οποίες έφεραν πραγματική επανάσταση στη μελέτη των επιφανειών των υλικών. Πρώτη παρουσιάσθηκε η Σαρωτική Μικροσκοπία Σήραγγας (Scanning Tunneling Microscopy - STM), το 1981 από τους Binning και Rohrer στη Ζυρίχη. Για την εφεύρεση τους αυτή οι δύο ερευνητές τιμήθηκαν με το βραβείο Νόμπελ Φυσικής το 1986, μαζί με τον Ruska, εφευρέτη του ηλεκτρονικού μικροσκοπίου. Το 1986 οι Binning, Quate και Gerber παρουσίασαν στο Stanford μία παραλλαγή της STM, την Σαρωτική Μικροσκοπία Ατομικών Δυνάμεων (Atomic Force Microscopy or Scanning Force Microscopy - AFM or SFM). Έκτοτε έχουν εμφανισθεί μερικές δεκάδες παραλλαγών των δύο αυτών τεχνικών, όπως Ballistic Electron Emission Microscopy, Scanning Ion-Conductance Microscopy, Friction (Lateral) Force Microscopy, Magnetic Force Microscopy, Electrostatic Force Microscopy, Scanning Thermal Microscopy, Scanning ear Field ptical Microscopy κ.λ.π. Αν και σύντομα επικράτησε να χαρακτηρίζονται όλες αυτές οι τεχνικές ως Scanning Probe Microscopies - SPM, (και τα όργανα Scanning Probe Microscopes) τελευταία χρησιμοποιούνται και οι όροι Scanning Point Probe Microscopies ή Scanning ear Field Probe Microscopies, οι οποίοι είναι προφανώς ορθότεροι. Πρόκειται, όπως διατυπώθηκε εύστοχα για μικροσκόπια χωρίς τη χρήση φακών. Σχήμα 24: Σχηματική παράσταση των τμημάτων σαρωτικού μικροσκοπίου ακίδας. Σε ένα τέτοιο μικροσκόπιο ο διερευνητής (probe) είναι μία πολύ λεπτή ακίδα, που σαρώνει την επιφάνεια του υλικού, με τη βοήθεια ευαίσθητων πιεζοηλεκτρικών κρυστάλλων, η 97
, με κατάλληλο ανιχνευτή, το φυσικό")
98 κίνηση των οποίων ελέγχεται μέσω ηλεκτρονικής μονάδας ελέγχου και ηλεκτρονικού υπολογιστή (σχήμα 24). Σε κάθε σημείο της επιφάνειας η ακίδα αλληλεπιδρά με το υλικό μέσω συγκεκριμένου φυσικού φαινομένου και εν συνεχεία καταγράφεται στον ηλεκτρονικό υπολογιστή (Η/Υ), με κατάλληλο ανιχνευτή, το φυσικό μέγεθος, που αποτελεί την απόκριση του δείγματος στην επίδραση του ανιχνευτή-διερευνητή. Ο Η/Υ, χρησιμοποιώντας κατάλληλο λογισμικό, αναπαράγει σε διδιάστατη ή τρισδιάστατη μορφή την εικόνα της επιφάνειας, με διακριτική ικανότητα, η οποία μπορεί υπό βέλτιστες συνθήκες να πλησιάζει το 0.1 nm. Για την επίτευξη αυτού του στόχου απαιτούνται τόσο υψηλής ποιότητας ηλεκτρονικά κυκλώματα, όσο και υψηλής ποιότητας πιεζοκρύσταλλοι. Απαιτείται επίσης εξαιρετική μηχανική σταθερότητα του όλου συστήματος. Σχήμα 25: Χρονολογικό διάγραμμα και μελλοντικές προσδοκίες της μικροσκοπίας ατομικής δύναμης (AFM) [17]. Σχήμα 26: Μετάβαση από το contact AFM του παρελθόντος στο non contact AFM του παρόντος και στο close contact (ή very close contact) AFM του μέλλοντος [17]. 98

.")
99 1.4.2 Αρχή λειτουργίας μικροσκοπίας ατομικής δύναμης και οργανολογία Η λειτουργία ενός AFM μικροσκοπίου στηρίζεται εν γένει στις ίδιες αρχές με αυτές του STM μικροσκοπίου. Η διαφορά ωστόσο συνίσταται στο ότι αυτή τη φορά μετρούνται διαφορές στις δυνάμεις οι οποίες αναπτύσσονται μεταξύ ακίδας και επιφάνειας και όχι του ρεύματος το οποίο δημιουργείται λόγω του φαινομένου σήραγγας. Οι δυνάμεις αυτές εμφανίζονται ανεξαρτήτως από το αν η μελετώμενη επιφάνεια είναι αγώγιμη ή μη. Δηλαδή στην τεχνική αυτή η ακίδα προσεγγίζει την επιφάνεια μέχρι να την αγγίξει και να αισθανθεί έτσι την δύναμη από το σημείο επαφής (σχήμα 27). Η ακίδα είναι στερεωμένη σε ένα βραχίονα (cantilever), η επάνω επιφάνεια του οποίου έχει εξαιρετική ανακλαστικότητα. Ανάλογα με τη δύναμη που δέχεται η ακίδα, ο βραχίονας υφίσταται και αντίστοιχη κάμψη. Μία δέσμη laser, η οποία προσπίπτει στην ανακλαστική επιφάνεια του βραχίονα ανακλάται ανάλογα με την παραμόρφωση του βραχίονα και προσπίπτει σε διαφορετική κάθε φορά θέση ενός φωτοανιχνευτή. Σχήμα 27: Διάταξη AFM μικροσκοπίου. Το επαγόμενο κάθε φορά φωτόρευμα καταγράφεται στον Η/Υ. Επειδή το μέγεθος της δύναμης, αλλά και η φύση της (ελκτική ή απωστική) εξαρτώνται από τη φύση της ακίδας και του σημείου επαφής, η καταγραφή των ρευμάτων μας καταγράφει και πάλι την τοπογραφία της επιφάνειας. 99
. Η ακίδα βρίσκεται στο άκρο του ελάσματος.")

100 Το σύστημα εργάζεται είτε σε περιβάλλον ατμοσφαιρικού αέρα, είτε σε υγρό περιβάλλον ή υπό υψηλό κενό. Πλεονέκτημα της τεχνικής είναι το ότι μπορεί να δώσει πληροφορίες και για μη αγώγιμες επιφάνειες (πολυμερή, βιολογικά δείγματα). Η ακίδα βρίσκεται στο άκρο του ελάσματος. Όπως γνωρίζουμε από το νόμο του hooke η σχέση μεταξύ δύναμης F και παραμόρφωσης Δx ενός ελατηρίου δίνεται από τη σχέση: F = - K Δx Η κίνηση του ελάσματος παρακολουθείται καθώς μια δέσμη laser ανακλάται από την πίσω επιφάνεια του και ανιχνεύεται από έναν φωτοανιχνευτή. Για διαφορετικές θέσεις του ελάσματος το φως προσπίπτει και σε διαφορετικές περιοχές στο φωτοανιχνευτή. Ο φωτοανιχνευτής αυτός είναι σε θέση να ανιχνεύσει τις χωρικές μεταβολές της έντασης του προσπίπτοντας φωτός (position - sensitive detector, PSD). Ο ανιχνευτής αποτελείται από δύο μέρη, το πάνω (top (T)) και το κάτω (bottom (B)) (σχήμα 28, πάνω αριστερά). Καθώς το φως προσπίπτει σε αυτόν, η διαφορά τάσης μεταξύ των δύο αυτών περιοχών αποτελεί μέτρο της παραμόρφωσης του ελάσματος. Σχήμα 28: Τα δύο μέρη του ανιχνευτή, το πάνω (top (T)) και το κάτω (bottom (B)) [2]. 100

101 Όπως ένα STM μικροσκόπιο έτσι και ένα AFM μπορεί να λειτουργήσει με διάφορους τρόπους λειτουργίας. Καταρχάς, με τη χρήση του μηχανισμού ανάδρασης μπορούμε να διατηρήσουμε τη δύναμη σταθερή σε μία τιμή καθώς η παραμόρφωση του ελάσματος διατηρείται επίσης σταθερή (αφού όπως είδαμε η σχέση δύναμης - παραμόρφωσης είναι γραμμική). Ο μηχανισμός ανάδρασης εξυπηρετεί στο να διατηρεί την τάση που δίνεται από τον φωτοανιχνευτή σταθερή με το να μετακινεί το δείγμα ή το έλασμα κατά τον κατακόρυφο άξονα με τη βοήθεια πιεζοηλεκτρικού κρυστάλλου. Κατά αυτόν τον τρόπο, καθώς η ακίδα μετακινείται στο παράλληλο της επιφάνειας επίπεδο, αυτή ακολουθεί την τοπογραφία της επιφάνειας. Ο τρόπος λειτουργίας αυτός ονομάζεται λειτουργία σταθερής ανάκλασης (constant cantilever deflection mode) ή αλλιώς λειτουργία σταθερής δύναμης (constant force mode) (σχήμα 29 δεξιά). Δεύτερος τρόπος λειτουργίας είναι να διατηρείται σταθερή η απόσταση της ακίδας από την επιφάνεια ενώ μετρούνται οι αλλαγές στη δύναμη (ισοδύναμα, αλλαγές στην παραμόρφωση του ελάσματος) και ονομάζεται λειτουργία σταθερού ύψους (constant height mode) (σχήμα 9 αριστερά). Σχήμα 29: Οι δύο τρόποι λειτουργίας του contact mode. Αριστερά, σταθερού ύψους (Constant Height Mode) και δεξιά, σταθερής δύναμης (Constant Force Mode). Ένας τρίτος τρόπος είναι να τεθεί η ακίδα σε ταλάντωση καθώς αυτή σαρώνει την επιφάνεια. Κατά την κίνηση αυτή είναι πιθανό το πλάτος και η φάση της ταλάντωση αυτής να μεταβληθεί. Προκειμένου, για παράδειγμα, να παρατηρήσουμε τη χημική σύσταση σε ένα δείγμα, μπορούμε να διατηρήσουμε σταθερό το πλάτος με τη βοήθεια του μηχανισμού ανάδρασης, ενώ οι μεταβολές στη φάση θα μας δίνουν πληροφορίες για τη χημική σύσταση της επιφάνειας. Εξάλλου, η χημική σύσταση μπορεί να μελετηθεί και παρακολουθώντας την κίνηση της ακίδας καθώς αυτή βρίσκεται σε επαφή με την επιφάνεια (contact mode). Τότε το έλασμα παραμορφώνεται ανάλογα με τη δύναμη τριβής η οποία αναπτύσσεται και είναι διαφορετική για κάθε χημικό δεδομένο. Για αυτή τη λειτουργία, ωστόσο, απαιτείται η επιφάνεια να είναι επίπεδη. Όσων αφόρα την AFM μικροσκοπία, ιδιαίτερη προσοχή πρέπει να δοθεί στην κατασκευή του ελάσματος αφού θα πρέπει να επιτρέπει ικανοποιητική παραμόρφωση για δυνάμεις της τάξης του n και επομένως η σταθερά ελατηρίου θα πρέπει να είναι όσο μικρότερη γίνεται. Ταυτόχρονα 101
102 όμως θα πρέπει η συχνότητα συντονισμού ω ο να είναι αρκετά υψηλή προς αποφυγή κραδασμών. Συνδυάζοντας τις δύο αυτές απαιτήσεις και λαμβάνοντας υπόψη τη σχέση: βλέπουμε ότι αυτές ικανοποιούνται για πολύ μικρή μάζα κι επομένως πολύ μικρές γεωμετρικές διαστάσεις. Χαρακτηριστικές τιμές του Κ και των διαστάσεων είναι, 100 x 20 x 1 μm και με 1 /m αντίστοιχα. Προκειμένου δηλαδή το έλασμα να μετακινηθεί κατά 1 nm η δύναμη που πρέπει να του ασκηθεί είναι 1 n. Για την επιτυχή λειτουργία μιας διάταξης απαιτούνται μία σειρά από προϋποθέσεις: Ατομικά αιχμηρή ακίδα. Θα πρέπει η ακίδα να χαρακτηρίζεται από ατομική αιχμηρότητα. Ένας τρόπος για να επιτευχθεί αυτό είναι ο εξής. Σε πρώτη φάση, η ακίδα, η οποία συνήθως είναι πολυκρυσταλλικό βολφράμιο που λεπτύνεται με ηλεκτροχημικές μεθόδους διάβρωσης (etching) σε σημείο που η ακτίνα προσεγγίζει τα 100 nm. Στη συνέχεια η ακίδα αφήνεται να ακουμπήσει στην επιφάνεια και απομακρύνεται με τη βοήθεια του κατακόρυφου κρυστάλλου. Αυτή η κίνηση έχει ως αποτέλεσμα η ακίδα να αποκτήσει ατομικά αιχμηρή μορφή καθώς άτομα συγκρατούνται με δεσμούς από τα άτομα της επιφάνειας. Αν η ακίδα δεν είναι ατομικά αιχμηρή τότε μειώνεται η διακριτική ικανότητα του οργάνου όπως μπορούμε να καταλάβουμε εύκολα από το σχήμα που ακολουθεί: Σχήμα 30: Η αιχμηρότητα της ακίδας καθορίζει τη διακριτική ικανότητα του οργάνου. Πιεζοηλεκτρικοί κρύσταλλοι μεγάλης ευαισθησίας. Οι πιεζοηλεκτρικοί κρύσταλλοι είναι συνήθως κατασκευασμένοι από οξείδιο βαρίου τιτανίου. Προστασία του συστήματος από τυχόν κραδασμούς. Για αυτό το λόγο το σύστημα ακίδαςεπιφάνειας φτιάχνεται μικρό και συμπαγές ώστε η κανονική ρυθμοί ταλάντωσης του να βρίσκονται σε υψηλές συχνότητες (δεδομένου ότι οι μηχανικοί κραδασμοί χαρακτηρίζονται από χαμηλές συχνότητες). Επιπλέον, το όλο σύστημα βρίσκεται στερεωμένο σε κατάλληλα αντικραδασμικά υποστηρίγματα ενώ τέλος ο χώρος μέσα στο οποίο βρίσκεται το μικροσκόπιο φροντίζεται να είναι μονωμένος [2]. 102










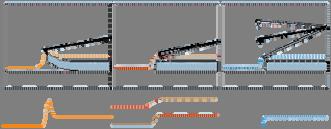
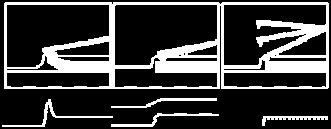
103 Σχήμα 31: Πραγματικές εικόνες από μικροσκόπιο ατομικής δύναμης - Atomic Force Microscope (AFM) Μέθοδοι - τεχνικές της μικροσκοπίας ατομικής δύναμης (AFM) Σχήμα 32: 1)Contact Mode (left), 2)on-contact Mode (middle) and 3)Tapping Mode (right) [2] 103
 στο δείγμα.")
 και του δείγματος.")
104 1) Contact Mode: Η συνηθέστερη μέθοδος που χρησιμοποιείται στο μικροσκόπιο ατομικής δύναμης είναι η μέθοδος επαφής στην οποία η ακίδα σαρώνει την επιφάνεια του δείγματος με επαφή. Η δύναμη στην ακίδα είναι απωστική με μέση τιμή 10-9 Ν. Τα προβλήματα στη μέθοδο αυτή προκαλούνται από τις υπερβολικές δυνάμεις που εφαρμόζονται από τον διερευνητή (probe) στο δείγμα. Μια μεγάλη κατηγορία δειγμάτων, συμπεριλαμβανομένων των ημιαγωγών και των μονωτών, μπορεί να παγιδεψει ηλεκτροστατικό φορτίο. Αυτό το φορτίο μπορεί να συμβάλει στις πρόσθετες ελκτικές δυνάμεις μεταξύ του διερευνητή (probe) και του δείγματος. Όλες αυτές οι δυνάμεις συνδυάζονται και καθορίζουν μια ελάχιστη δύναμη που μπορεί να εφαρμοστεί ελεγχόμενα από τον διερευνητή στο δείγμα. Αυτή η δύναμη δημιουργεί μια μεγάλη δύναμη τριβής καθώς ο διερευνητής (probe) σαρώνει το δείγμα. Στην πράξη, φαίνεται ότι αυτές οι δυνάμεις τριβής είναι πολύ πιο καταστρεπτικές από την κανονική δύναμη και μπορούν να βλάψουν το δείγμα και να διαστρεβλώσουν τα στοιχεία που προκύπτουν. Μια προσπάθεια να αποφευχθεί αυτό το πρόβλημα είναι η μέθοδος non-contact [2]. Σχήμα 33: Σχηματική απεικόνιση του a) contact mode του AFM και του b) non contact mode του AFM (C-AFM). Το a) δείχνει ότι το contact mode AFM είναι καταστρεπτικό και έχει μεγάλη περιοχή επαφής λόγω της μεγάλης απωστικής δύναμης ενώ το b) δείχνει ότι το C-AFM είναι μη καταστρεπτικό και μπορεί να παρατηρηθεί ακόμη και αν ένα ατομικό σημείο λείπει με μια ασθενή ελκτική δύναμη που μπορεί να ανιχνευθεί μέσω μεταβολής της συχνότητας που προκαλείται στην μηχανική ταλάντευση του cantilever [17]. 2) on-contact Mode. Μια νέα εποχή στην απεικόνιση άνοιξε όταν εισήχθη μικροσκοπικά ένα σύστημα για την μέθοδο non-contact το οποίο χρησιμοποιείται στις περιπτώσεις που η ακίδα επαφής μπορεί να μεταβάλλεται σε σχέση με το δείγμα. Σε αυτόν τον τρόπο η ακίδα αιωρείται Angstrom πάνω από την επιφάνεια των δειγμάτων. Οι ελκτικές δυνάμεις Van der Waals που ενεργούν μεταξύ της ακίδας και του δείγματος ανιχνεύονται και οι τοπογραφικές εικόνες δημιουργούνται με σάρωση της ακίδας πάνω από την επιφάνεια. Ένας μεγάλος αριθμός διαφορετικής φύσεως δυνάμεων μπορεί να εμφανιστεί όταν δύο σώματα έλθουν σε κοντινή απόσταση, όπως δυνάμεις Van Der Walls, απώθησης ιόντων, τριχοειδούς φύσεως, τριβής, δυνάμεις προερχόμενες από φυσική προσρόφηση, δυνάμεις ιοντικής ή ομοιοπολικής φύσεως κτλ. 104
105 Παρακάτω παρουσιάζονται συνοπτικά αυτές που παίζουν τον κυριότερο ρόλο στην AFM μικροσκοπία. Η αλληλεπίδραση Van Der Waals είναι ιδιαίτερα αισθητή σε αποστάσεις από λίγα έως μερικές εκατοντάδες Angstrom ενώ μπορεί να είναι ελκτικής ή απωστικής φύσεως. Αποτελεί συνισταμένη τριών επιμέρους δυνάμεων [2]. Της δύναμης αλληλεπίδρασης μεταξύ μόνιμα πολωμένων μορίων (δυνάμεις συντονισμού) Της δύναμης που αναπτύσσεται μεταξύ ενός δίπολου και ενός δίπολου το οποίο δημιουργείται από ένα κατά τα άλλα μη πολικό μόριο κατά την επαγωγή του από το πρώτο (δυνάμεις εξ επαγωγής) Της δύναμης που αναπτύσσεται καθώς μια στιγμιαία παραμόρφωση του ηλεκτρονικού νέφους ενός μορίου (ύστερα από σύγκρουση με άλλο μόριο) έχει ως αποτέλεσμα τη μετατόπιση του κέντρου βάρους του φορτίου και τη δημιουργία επομένως ηλεκτρικού πεδίου γύρω από αυτό 105
106 όπου με τη σειρά του θα πολώσει ένα δεύτερο μόριο (δυνάμεις διασποράς κατά London, London dispersion forces). Η τελευταία από αυτές είναι συνήθως η επικρατούσα. Σύμφωνα με τα παραπάνω επομένως το δυναμικό μεταξύ δύο ατόμων που μεταξύ τους αναπτύσσονται δυνάμεις Van Der Walls, μπορεί να γραφτεί σαν: με C vdw = C 1 + C 2 + C C i σταθερές οι οποίες εξαρτώνται από τις επιμέρους συνιστώσες και r η απόσταση μεταξύ των μορίων. Άλλες δυνάμεις είναι αυτές που εμφανίζονται λόγω επικάλυψης ηλεκτρονικών νεφών. Καθώς δύο άτομα πλησιάζουν μεταξύ τους σε αποστάσεις ίσες ή μικρότερες του 1 Angstrom τα ηλεκτρονικά νέφη τους επικαλύπτονται ισχυρά. Δεδομένης της απαγορευτικής αρχής του Pauli η θωράκιση του πυρηνικού φορτίου εξασθενεί και οι δύο πλέον πυρήνες αλληλεπιδρούν απωστικά μεταξύ τους. Οι δυνάμεις αυτές περιγράφονται από το νόμο: U r n με n ίσο ή μεγαλύτερο του 9 είτε γενικότερα από μία εκθετική σχέση. Οι απωστικής φύσεως δυνάμεις αυτές μπορούν να θεωρηθούν και σαν τέταρτη συνιστώσα της αλληλεπίδρασης Van Der Walls. Ένα ευρέως χρησιμοποιούμενο μοντέλο το οποίο περιγράφει ταυτόχρονα τις ελκτικές και απωστικές δυνάμεις είναι αυτό το οποίο στηρίζεται στο δυναμικό Lennard Jones: 3) Tapping Mode. Η μέθοδος ημιεπαφής (tapping mode) είναι μια βασική πρόοδος του AFM. Αυτή η αποτελεσματική τεχνική επιτρέπει υψηλή ανάλυση τοπογραφικής απεικόνισης επιφανειών δειγμάτων που φθείρονται εύκολα, που συγκρατούνται χαλαρά στο υπόστρωμά τους ή που δύσκολα απεικονίζονται από άλλες τεχνικές AFM. Η ημιεπαφή αυτή υπερνικά τα προβλήματα που συνδέονται με την τριβή, την προσκόλληση, τις ηλεκτροστατικές δυνάμεις και άλλες δυσκολίες μιας συμβατικής σάρωσης AFM. Με διαδοχικές τοποθετήσεις της ακίδας σε επαφή με την επιφάνεια, παρέχεται υψηλή ανάλυση αποφεύγοντας το σύρσιμο της επιφάνειας. Κατά τη διάρκεια της σάρωσης η κάθετα ταλαντευόμενη ακίδα έρχεται διαδοχικά σε επαφή με την επιφάνεια και ανασηκώνεται, γενικά με μια συχνότητα από έως κύκλων ανά δευτερόλεπτο. Κατά τη διάρκεια της λειτουργίας του tapping mode, το πλάτος ταλάντωσης του cantilever διατηρείτε σταθερό από ένα σύστημα ανατροφοδότησης πληροφοριών. Η επιλογή της βέλτιστης συχνότητας ταλάντωσης ρυθμίζεται με την βοήθεια κατάλληλου λογισμικού και η δύναμη στο δείγμα αυτόματα καθορίζεται και διατηρείται στο χαμηλότερο δυνατό επίπεδο. Όταν η ακίδα περνά από μια απότομη προεξοχή της επιφάνειας, το cantilever έχει λιγότερο χώρο για να 106
 μειώνει πολύ τις δυνάμεις προσκόλλησης ακίδας-δειγμάτων.")
![Η μέθοδος tapping mode επομένως αποτρέπει την ακίδα να κολλήσει στην επιφάνεια και την πρόκληση ζημίας κατά τη διάρκεια της σάρωσης [2]. 1.4.](/docs-images/72/67903182/images/107-1.jpg "4 Εφαρμογές της μικροσκοπίας ατομικής δύναμης (AFM) Η μικροσκοπία ατομικής δύναμης χρησιμοποιείται κυρίως στη μελέτη των επιφανειών των υλικών αφού όπως έχει είδη αναφερθεί γίνεται σάρωση της")
107 ταλαντευτεί και το πλάτος της ταλάντωσης μειώνεται. Αντιθέτως, όταν περνά η ακίδα από μια κοιλότητα-εσοχή, το cantilever έχει περισσότερο χώρο για να ταλαντευτεί και αυξάνεται το πλάτος ταλάντωσης. Όταν η άκρη έρχεται σε επαφή με την επιφάνεια η υψηλή συχνότητα ( khz) μειώνει πολύ τις δυνάμεις προσκόλλησης ακίδας-δειγμάτων. Η μέθοδος tapping mode επομένως αποτρέπει την ακίδα να κολλήσει στην επιφάνεια και την πρόκληση ζημίας κατά τη διάρκεια της σάρωσης [2] Εφαρμογές της μικροσκοπίας ατομικής δύναμης (AFM) Η μικροσκοπία ατομικής δύναμης χρησιμοποιείται κυρίως στη μελέτη των επιφανειών των υλικών αφού όπως έχει είδη αναφερθεί γίνεται σάρωση της επιφάνειας του υλικού και με την βοήθεια κατάλληλου λογισμικού ένας ηλεκτρονικός υπολογιστής αναπαράγει σε διδιάστατη ή τρισδιάστατη μορφή την εικόνα της επιφάνειας, με διακριτική ικανότητα, η οποία μπορεί υπό βέλτιστες συνθήκες να πλησιάζει το 0.1 nm. Το σύστημα εργάζεται είτε σε περιβάλλον ατμοσφαιρικού αέρα, είτε σε υγρό περιβάλλον ή υπό υψηλό κενό. Πλεονέκτημά της τεχνικής είναι το ότι μπορεί να δώσει πληροφορίες και για μη αγώγιμες επιφάνειες (πολυμερή, βιολογικά δείγματα). Η χημική σύσταση σε ένα δείγμα μπορεί να μελετηθεί αν διατηρηθεί σταθερό το πλάτος με τη βοήθεια του μηχανισμού ανάδρασης, ενώ οι μεταβολές στη φάση θα μας δίνουν πληροφορίες για τη χημική σύσταση της επιφάνειας. Η χημική σύσταση μπορεί να μελετηθεί και παρακολουθώντας την κίνηση της ακίδας καθώς αυτή βρίσκεται σε επαφή με την επιφάνεια (contact mode). Τότε το έλασμα παραμορφώνεται ανάλογα με τη δύναμη τριβής η οποία αναπτύσσεται και είναι διαφορετική για κάθε χημικό δεδομένο. Για αυτή τη λειτουργία, ωστόσο, απαιτείται η επιφάνεια να είναι επίπεδη. Ορισμένα παραδείγματα εμφανίζονται στα παρακάτω σχήματα: Σχήμα 34: Εικόνες AFM του βακτηριδίου Deinoccocus radiodurans Παρατηρούνται μεταβολές με το χρόνο της εξαγωνικής διάταξης των πρωτεϊνών [15]. Σχήμα 35: Εικόνες AFM της εξωκυτταρικής δομής του Ε. coli porin mpf που επηρεάζεται με την μεταβολή του ph: a) ph 7 b) ph 6 c) ph 2.5 και d) ph 4. Κάτω φαίνεται η σταδιακή διαφοροποίηση της δομής από αριστερά με ph 7 μέχρι δεξιά με ph 2.5 [15]. 107

 Περιγράμματα συνολικής πυκνότητας φορτίου ενός μορίου αδενίνης που προέκυψαν από υπολογισμούς των μοριακών τροχιακών [17]. Σχήμα 37: a) Εικόνα C-AFM του μορίου της θυμίνης σε επιφάνεια γραφίτη.")

108 Σχήμα 36: a) Εικόνα C-AFM του μορίου της αδενίνης σε επιφάνεια γραφίτη. Η περιοχή σάρωσης είναι 70Å Χ 70Å. Το άσπρο ορθογώνιο δείχνει την κρυσταλλική μονάδα του κυττάρου. Το ένθετο είναι μία μεγεθυσμένη εικόνα από ένα διμερές αδενίνης. b) Δομικό μοντέλο από ένα διμερές αδενίνης c) Υψηλής μεγέθυνσης εικόνα μορίου αδενίνης που είναι τοποθετημένο από πάνω ένα δομικό μοντέλο αδενίνης. d) Περιγράμματα συνολικής πυκνότητας φορτίου ενός μορίου αδενίνης που προέκυψαν από υπολογισμούς των μοριακών τροχιακών [17]. Σχήμα 37: a) Εικόνα C-AFM του μορίου της θυμίνης σε επιφάνεια γραφίτη. Η περιοχή σάρωσης είναι 120Å Χ 120Å. Το άσπρο ορθογώνιο δείχνει την κρυσταλλική μονάδα του κυττάρου. Το ένθετο είναι μία μεγεθυσμένη εικόνα από ένα διμερές θυμίνης που φαίνεται ότι οι δύο περιοχές είναι περιστραμένες κατά 120 ο η μία σε σχέση με την άλλη. b) Δομικό μοντέλο από ένα διμερές θυμίνης c) Υψηλής μεγέθυνσης εικόνα μορίου θυμίνης που είναι τοποθετημένο από πάνω ένα δομικό μοντέλο θυμίνης. d) Περιγράμματα συνολικής πυκνότητας φορτίου ενός μορίου θυμίνης που προέκυψαν από υπολογισμούς των μοριακών τροχιακών [17]. Σχήμα 38: a) Εικόνα C-AFM ενός φουλερενίου C 60. Η περιοχή σάρωσης είναι 20nm Χ 20nm. b) Σχηματικό διάγραμμα που επεξηγεί τον τρόπο που προέκυψε η εικόνα (a) του φουλερενίου C 60 [17]. 108


 Εικόνα C-AFM 20nm X 20nm b) Εικόνα C-AFM 5nm X 5nm c) Αντιπροσωπευτικές μετρήσεις (Height")
![[nm] Distance [nm]) και η γραφική παράστασή τους κατά μήκος της γραμμής Α-Β του (b) [17].](/docs-images/72/67903182/images/109-4.jpg "Σχήμα 41: Εικόνες C-AFM μονοστρωματικής μεμβράνης οκτανοθειόλης (αλκανοθειόλη: CH 3 (CH 2 ) n-1")
 Εικόνα C-AFM 60nm X 60nm, b) Εικόνα C-AFM 15nm X 15nm, c) Αντιπροσωπευτικές μετρήσεις (Height")
109 Σχήμα 39: Εικόνα AFM τοξίνης χολέρας δεσμευμένη σε μεμβράνη. Η πενταμερή δομή της υπομονάδας εμφανίζεται σε πολλά μόρια. Ο διερευνητής (probe) δηλαδή η πολύ λεπτή ακίδα σαρώνει με δύναμη 0,4 n και ταχύτητα 8,68 Hz [16]. Σχήμα 40: Εικόνες C-AFM μονοστρωματικής μεμβράνης δεκανοθειόλης (αλκανοθειόλη : CH 3 (CH 2 ) n-1 SH με n=10). a) Εικόνα C-AFM 20nm X 20nm b) Εικόνα C-AFM 5nm X 5nm c) Αντιπροσωπευτικές μετρήσεις (Height [nm] Distance [nm]) και η γραφική παράστασή τους κατά μήκος της γραμμής Α-Β του (b) [17]. Σχήμα 41: Εικόνες C-AFM μονοστρωματικής μεμβράνης οκτανοθειόλης (αλκανοθειόλη: CH 3 (CH 2 ) n-1 SH με n=8). a) Εικόνα C-AFM 60nm X 60nm, b) Εικόνα C-AFM 15nm X 15nm, c) Αντιπροσωπευτικές μετρήσεις (Height [nm] Distance [nm]) και η γραφική παράστασή τους κατά μήκος της γραμμής Α-Β του (b), d) Εικόνα C-AFM 6nm X 6nm και e) Αντιπροσωπευτικές μετρήσεις (Height [nm] Distance [nm]) και η γραφική παράστασή τους κατά μήκος της γραμμής C-D του (d) [17]. 109
 πολυστρωματική μορφή του διμέθυλο-πένταθειοφαινίου (DM5T) σε υπόστρωμα λευκοχρύσου πυριτίου και e) C-AFM τοπογραφική εικόνα του διμέθυλο-πέντα-θειοφαινίου (DM5T) σε")
110 Σχήμα 42: a) Μοριακή δομή του διμέθυλο-πέντα-θειοφαινίου (DM5T) b) σχηματική απεικόνιση της μοριακής διευθέτησης του διμεθυλο-πεντα-θειοφαινίου (DM5T) στο παρόν πείραμα c) σχηματική απεικόνιση κατά την σάρωση της ακίδας του AFM d) πολυστρωματική μορφή του διμέθυλο-πένταθειοφαινίου (DM5T) σε υπόστρωμα λευκοχρύσου πυριτίου και e) C-AFM τοπογραφική εικόνα του διμέθυλο-πέντα-θειοφαινίου (DM5T) σε υπόστρωμα λευκοχρύσου πυριτίου [17]. 110
111 ΚΕΦΑΛΑΙΟ 2 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 2.1 Υλικά Όλα τα χημικά αντιδραστήρια που χρησιμοποιήθηκαν ήταν αναλυτικής καθαρότητας των οίκων Merck (Dramstadt, Germany), Sigma-Aldrich (St. Louis, USA), Riedel de Haan (Seelzer, Germany), J.T. Baker (Deventer, Holland), Gibco BRL (Karlsruhe, Germany), BDH (Poole, England), Qiagen GmbH (Hilden, Germany), Amersham Pharmacia Biotech AB (Uppsala, Sweden) και Panreac Quimica SA (Barcelona, Spain). Χρησιμοποιήθηκαν επίσης τροποποιημένοι νανοσωλήνες και φουλερένια τα οποία συντέθηκαν στο ερευνητικό εργαστήριο του κ. Ν. Ταγματάρχη (Ινστιτούτο Θεωρητικής και Φυσικής Χημείας του Εθνικού Ιδρύματος Ερευνών) και ήταν τα ακόλουθα: CH H R Boc-Tyr-H Boc-Asp-H Boc-Glu-H K54 SWTs CF 3 C H 3 GP5 H 3 Cl K28 SWTs n 111
112 H 2 H SWTs H H 2 EDCI / HBt CH Ανάλογο 31 H H SWTs K133 H H Σχήμα 1: Τροποποιημένοι νανοσωλήνες και φουλερένια που χρησιμοποιήθηκαν στην παρούσα εργασία. 2.2 Μέθοδοι Σύνθεση αζαπεπτιδίων Η σύνθεση αζα-αμινοξέων και η ενσωμάτωση τους σε μια πεπτιδική αλυσίδα αποτελεί συνδυασμό της χημείας της υδραζίνης και της πεπτιδικής χημείας. Για το σκοπό αυτό ένα παράγωγο υδραζίνης αντιδρά με έναν ισχυρό "δότη καρβονυλομάδας" ή η αλυσίδα επιμηκύνεται χρησιμοποιώντας τις μεθόδους σύζευξης της κλασσικής πεπτιδικής σύνθεσης. Έχουν αναπτυχθεί διάφορες γενικές μέθοδοι σύνθεσης αζαπεπτιδίων, όπως η μέθοδος των ισοκυανικών εστέρων, των ενεργών εστέρων, των αζολιδίων, των χλωριδίων των οξέων και των 1,3,4-οξαζολ-2(3Η)-ονών. Η σύνθεση του αζαπεπτιδίου 31 προαπαιτεί την παρασκευή της αλληλουχίας Fmoc-D-Trp- H-(Bzl)-C-D-Trp--ρητίνη, που προέρχεται από την Fmoc-D-Trp-H-(Bzl)H και την 112
113 Ο 2 Ν-C 6 H 4 -C-D-Trp--ρητίνη. Η συνθετική πορεία του αναλόγου 31 παρατίθεται στο σχήμα 2. Μετά το σχηματισμό της αλληλουχίας Fmoc-D-Trp-H-(Bzl)-C-D-Trp--ρητίνη ακολούθησε διάσπαση της Fmoc ομάδας, απομάκρυνση του αζαπεπτιδίου από τη ρητίνη και καθαρισμός με ημι-παρασκευαστικό HPLC [12]. 1. Fmoc-D-Trp-H DCM / DIEA 2. pip / DMF, 2:8 Fmoc-D-Trp-H H-D-Trp- -10 o C H 2-H-Bzl / διοξάνη DMF / PyBP / DIEA 2 C 2 DMF Fmoc-D-Trp-H-H-Bzl + 2 C D Trp DMF DIEA Fmoc-D-Trp-H-(Bzl)-C-D-Trp- 1. pip / DMF, 2:8 2. DCM / TFE / AcH, 7:2:1 H-D-Trp-H-(Bzl)-C-D-Trp-H (31) Σχήμα 2: Συνθετική πορεία αζαπεπτιδίου 31 [12] Σύνθεση υδραζινοπεπτιδίων Στη βιβλιογραφία αναφέρονται τέσσερις κύριες μέθοδοι παρασκευής σε υγρή φάση χειρόμορφων α-υδραζινο-οξέων: η καταλυτική υδρογόνωση υδραζονών, η αναδιάταξη Schestakov, η ηλεκτρονιόφιλη Ν-αμίνωση α-αμινοξέων και η πυρηνόφιλη υποκατάσταση των nosyloxy εστέρων. Τα τελευταία χρόνια δημοσιεύθηκε και η πρώτη μέθοδος σύνθεσης σε στερεά 113
114 φάση, η οποία εμπεριέχει Ν-αλκυλοξυκαρβονυλαμινοφθαλιμίδια προσδεδεμένα σε πολυμερές που συμμετέχουν ως οξέα κατά την αντίδραση Mitsunobu και α-υδροξυεστέρες. Αντίστοιχα η συνένωση Fmoc-H-H-Bzl και Br-CH 2 -C-D-Trp-Leu--ρητίvη οδήγησε στην παρασκευή του αναλόγου 37 (σχήμα 3), στο οποίο ακολούθως διασπάσθηκε η Fmoc ομάδα, αποκόπηκε από τη ρητίνη και καθαρίσθηκε με ημι-παρασκευαστικό HPLC [12]. 1. Fmoc-Leu-H DCM / DIEA 2. pip / DMF, 2:8 H-Leu- 1. Fmoc-D-Trp-H DMF / DIC / HBt 2. pip / DMF, 2:8 H-D-Trp-Leu- Fmoc-Su -10 o C H 2-H-Bzl / διοξάνη 10% ahc 3 Fmoc-H-H-Bzl + Br-CH 2 -CH DMF / DIC Br-CH 2 -C-D-Trp-Leu- DMF Fmoc-H-(Bzl)-CH 2 -C-D-Trp-Leu- 1. pip / DMF, 2:8 2. DCM / TFE / AcH, 7:2:1 H 2 -(Bzl)-CH 2 -C-D-Trp-Leu-H (37) Σχήμα 3: Συνθετική πορεία Ν α -υδραζινο-πεπτοειδή 37 [12] Γενική μέθοδος σύζευξης με DIC/HBt (στερεά φάση) Ποσότητα Ν α -Fmoc-αμινοξέος (k mmol), όπου κ=3χ (υποκατάσταση ρητίνης) χ (μάζα ρητίνης), διαλύεται στην ελάχιστη ποσότητα DMF. Προστίθεται HBt (1.5 x k mmol, MW= 114
115 153.1) και ακολούθως DIC (1.1 x k mmol, MW= 120.2, d=0.81 g/ml). Το μίγμα προστίθεται αμέσως στη ρητίνη που έχει προηγουμένως διογκωθεί με DCM. Η αντίδραση σύζευξης λαμβάνει χώρα σε τουλάχιστον 3h σε θερμοκρασία δωματίου. Ακολουθεί έλεγχος για να διαπιστωθεί η ολοκλήρωση της αντίδρασης [12] Σύζευξη με τη μέθοδο του DIC/HBt Αρχικά ενεργοποιείται το αμινοξύ που θα ενσωματωθεί στην πεπτιδική αλυσίδα 3x ισοδύναμα Boc-AA 2 -H (τριπλάσια ποσότητα σε σχέση με την αρχική υποκατάσταση του πεπτιδίου ρητίνη) και 4,5x ισοδύναμα HBt, διαλύονται σε 5-10 ml διαλύματος DMF σε θερμοκρασία δωματίου Μετά από ένα λεπτό προστίθενται 3,3x ισοδύναμα DIC Το διάλυμα προστίθεται στη σφαιρική που περιέχει τη ρητίνη και το μίγμα αναδεύεται 2-4 ώρες εώς και 24 ώρες σε θερμοκρασία δωματίου, ανάλογα με το αμινοξύ που προστίθεται Μετά το πέρας της αντίδρασης η ρητίνη διηθείται υπό κενό και πλένεται με (~10 ml/gr ρητίνης): 3x DMF, 2x i-prh, 2x DMF, 3x i-prh και 1x Et 2 Το στερεό υπόλειμμα ξηραίνεται πάνω από P 2 5 Ο έλεγχος της ολοκλήρωσης της αντίδρασης γίνεται με το test Kaiser και T.L.C. o Η ποσότητα HBt που χρησιμοποιείται είναι 1,5x πεπτίδιο σε mol o Η ποσότητα DIC που χρησιμοποιείται είναι 1,1x πεπτίδιο σε mol Έλεγχος της αντίδρασης σύζευξης Είναι απαραίτητο να εκτελείται ο έλεγχος αυτός μετά από κάθε αντίδραση σύζευξης και απομάκρυνσης της Ν α -Fmoc-ομάδας, ώστε να αποφευχθούν οι ημιτελείς αντιδράσεις, οι οποίες οδηγούν στο σχηματισμό παραπροϊόντων και τελικά σε μειωμένες αποδόσεις. Ο έλεγχος μπορεί να γίνει είτε με test Kaiser είτε με χρωματογραφία λεπτής στοιβάδας (TLC). Η μέθοδος Kaiser ανιχνεύει ελεύθερες πρωτοταγείς αμινομάδες μέσω της έγχρωμης ένωσης που σχηματίζεται με αντίδραση της αμινομάδας με τη νινυδρίνη. Σύμφωνα με αυτή τη μέθοδο, πολύ μικρή ποσότητα πεπτιδίου-ρητίνης τοποθετείται σε δοκιμαστικό σωλήνα και προστίθενται δυο σταγόνες από τα παρακάτω διαλύματα: ΔΙΑΛΥΜΑ 1: 500 mg νινυδρίνης σε 10 ml 1-βουτανόλης ΔΙΑΛΥΜΑ 2: 80 g φαινόλης σε 20 ml 1-βουτανόλης ΔΙΑΛΥΜΑ 3: 2 ml διαλύματος KC Μ σε 98 ml πυριδίνης 115
116 Το μίγμα θερμαίνεται στους 100 C για 5 min και παρατηρείται το χρώμα του. Αν είναι μπλε, το test θεωρείται θετικό, δηλαδή υπάρχει ελεύθερη αμινομάδα του αμινοξέος ή του πεπτιδίου, η οποία έχει αντιδράσει με τη νινυδρίνη, για να δώσει το έγχρωμο παράγωγο. Στην περίπτωση αυτή η αντίδραση σύζευξης κρίνεται ημιτελής και απαιτείται επανασύζευξη με τις μισές περίπου ποσότητες αντιδραστηρίων σύζευξης. Αν όμως παραμείνει κίτρινο, το test θεωρείται αρνητικό, οπότε η αντίδραση σύζευξης έχει ολοκληρωθεί. Σύμφωνα με τη χρωματογραφία λεπτής στοιβάδας (TLC), πολύ μικρή ποσότητα ρητίνης τοποθετείται σε δοκιμαστικό σωλήνα και προστίθενται μερικές σταγόνες διαλύματος διάσπασης του πεπτιδίου από τη ρητίνη [DCM/TFΕ/AcH (7:2:1, ν/ν)], οπότε ύστερα από μερικά λεπτά απελευθερώνεται ένα ποσοστό πεπτιδίου στο διάλυμα. Τοποθετείται κηλίδα από αυτό το διάλυμα σε πλάκα χρωματογραφίας και αναπτύσσεται σε κατάλληλο σύστημα διαλυτών [συνήθως στο Tol/MeH/AcH, (7:1.5:1.5, ν/ν)]. Στη συνέχεια και αφού εξατμιστεί ο διαλύτης από την επιφάνεια της πλάκας, παρατηρείται το χρωματογράφημα σε θάλαμο UV και ψεκάζεται με διάλυμα νινυδρίνης ή/και αμύλου [12] Διαδικασία απομάκρυνσης ομάδων Boc Το διάλυμα που χρησιμοποιήθηκε για την απομάκρυνση των ομάδων Βοc ήταν: TFA: 50% DCM: 40% TES: 3% H 2 : 3% Phenol: 3% DDT: 1% Μέσα σε eppendorf στο οποίο υπάρχουν 2 mg πεπτιδίου ρίχνονται 250 μl από το διάλυμα αποπροστασίας. Το eppendorf ήταν συνέχεια σκεπασμένο με αλουμινόχαρτο και υπήρχαν συνέχεια συνθήκες χαμηλού φωτισμού επειδή η πορεία είναι φωτοευαίσθητη. Αμέσως μετά την προσθήκη των 250 μl διαλύματος αποπροστασίας διοχετεύθηκε στο eppendorf αέριο άζωτο ώστε να απομακρυνθεί η ποσότητα του οξυγόνου επειδή υπάρχει μεγάλος κίνδυνος οξείδωσης. Ακολούθησε ανάδευση για 15 min και εξατμίστηκε ο διαλύτης μέχρι σταγόνας με αέριο άζωτο. Στη συνέχεια προστέθηκε κρύο Εt 2 Ο και το eppendorf μεταφέρθηκε στην κατάψυξη για 24 ώρες. (Αν κατά την προσθήκη του Εt 2 Ο το διάλυμα δεν θολώσει δηλαδή δεν σχηματιστεί το πεπτίδιο, εξατμίζεται και πάλι ο διαλύτης με άζωτο μέχρι σταγόνας ώστε να απομακρυνθεί ο Εt 2 Ο και το TFA και ξαναπροστίθεται Εt 2 Ο). Μετά τις 24 ώρες ακολούθησε φυγοκέντρηση, απομάκρυνση του 116
117 Εt 2 Ο, προσθήκη νέου Εt 2 Ο, φυγοκέντρηση και απομάκρυνση του Εt 2 Ο. Έγινε έλεγχος με χρωματογραφία λεπτής στιβάδας σε διάλυμα ανάπτυξης AcC : H 2 με αναλογία 5:1 (το αποτέλεσμα φαίνεται με 1) UV light ή/και 2) με ψεκασμό με νινυδρίνη και παραμονή στους 100 ο C για 5 10 min. Επίσης έλεγχος έγινε και με υγρή χρωματογραφία υψηλής πίεσης (HPLC) Έποξυ ενεργοποιημένη σεφαρόζη 6B (Ακινητοποίηση ενζύμου) Διαλύματα: 1 M ethanolamine ph 8.0 (Το πυκνό έχει d=1.02 g/ml, F.W.=61.08 και 16,7 Μ) Buffer ph 4: 0.1 M CH 3 CH-CH 3 Ca ph 4 και 0.5 Μ ΝaCl Buffer ph 8: 0.1 M Tris-HCl ph 8 και 0.5 Μ ΝaCl Διαδικασία: Ζύγιση 0.3 gr epoxy-activated sepharose και διαλυτοποίηση σε 10 ml δις απιονισμένο Η 2 Ο. Αφήνεται για 15 min και αποχύνεται σε διηθητικό χαρτί. Ακολουθεί πλύση του υλικού στο διηθητικό χαρτί με 100 ml Η 2 Ο. Σε 20 ml δις απιονισμένο Η 2 Ο (ph με ah) προστίθεται 1 gr τρυψίνης και αναδεύεται μέχρι να διαλυθεί. Ανάμειξη των δύο παραπάνω καλά στο falcon. Ακολουθεί ανάδευση στον επωαστήρα για 16 ώρες. Μετά τις 16 ώρες που το διάλυμα πρέπει να είναι άσπρο και όχι κόκκινο, το falcon φυγοκεντρείται στις 3000 rpm ώστε να διαχωριστεί το υπερκείμενο από το ίζημα. Το ίζημα πλένεται με 20 ml δις απιονισμένου Η 2 Ο ph 9-10 με ah. Στη συνέχεια ακολουθεί πάλι ανάδευση στον επωαστήρα για 10 min και φυγοκέντρηση στις 3000 rpm για 10 min. Κατόπιν μέσα στο falcon που ήταν το ίζημα της τελευταίας φυγοκέντρησης ρίχνεται διάλυμα 1Μ αιθανολαμίνης ph 8.0 με ΗCl. Το falcon μεταφέρεται στον επωαστήρα στους 45 o C και rpm. Αφήνεται over night (~16h) και φυγοκεντρείται στις 3000 rpm Τέλος το ίζημα πλένεται διαδοχικά με buffer ph 4 και 8 στον επωαστήρα για 30 min στους 43.5 o C. Συνολικά από τρεις φορές το κάθε ph [14]. 117
118 2.2.7 Προσδιορισμός δραστικότητας πρωτεϊνάσης Τα αντιδραστήρια που χρησιμοποιούνται είναι: 0,2 M Tris-HCl ph 7 με 5 mm CaCl 2 (Buffer) 50 % T.C.A. Η πειραματική διαδικασία είναι: Το ένζυμο διαλυτοποιείται σε Tris-HCl, CaCl 2 buffer σε συγκέντρωση μg/ml. Σε δοκιμαστικούς σωλήνες που περιέχουν 50 mg casein προσθέτουμε 4,9 ml buffer και το επωάζουμε 5-10 min στους 37 ο C. Σε χρονική στιγμή 0 προσθέτουμε 0,1 ml διαφόρων αραιώσεων ενζύμου και επωάζουμε 20 min στους 37 ο C με ανάδευση ανά τακτά διαστήματα. Υπάρχει τυφλό με [E] + buffer και τυφλό με [S] + buffer. Μετά από 20 min προσθέτουμε 0,5 ml 50% Τ.C.A., φυγοκεντρούμε ή διηθούμε και μετράμε την απορρόφηση στα 280 nm [13]. [A Units / mg = sample - (A280 [E] blank A280 [S] blank)] x 1000 min x mg ενζύμου Χρωματομετρική μέθοδος κατά Bradford τροποποιημένη κατά Bearden Η μέθοδος βασίζεται στην ιδιότητα της χρωστικής Coomassie Brilliant Blue G 250 να συμπλέκεται με τις πρωτεΐνες σε όξινο περιβάλλον. Ο σχηματισμός του συμπλόκου έχει ως αποτέλεσμα τη μετατόπιση του μέγιστου της απορρόφησης της χρωστικής από τα 465 nm στα 595 nm. To αντιδραστήριο Bradford παρασκευάζεται με διάλυση της χρωστικής σε πυκνό φωσφορικό οξύ (85% v/v), σε συγκέντρωση 1 mg/ml. Το αντιδραστήριο αναδεύεται 12 εώς 16 ώρες και στη συνέχεια αραιώνεται με H 2 σε πενταπλάσιο όγκο και διηθείται. Χρησιμοποιείται σε αναλογία 1:1 με το δείγμα και μετράται η απορρόφηση στα 595 nm. Ο προσδιορισμός της συγκέντρωσης γίνεται βάσει πρότυπης καμπύλης αναφοράς με γνωστές ποσότητες αλβουμίνης ορού βοδιού (BSA). Πλεονεκτεί η μέθοδος ως προς την ακρίβεια και την ευαισθησία της, παρεμποδίζεται όμως από την παρουσία απορρυπαντικών και σουλφυδρυλο-ομάδων. Πειραματικά για να βελτιωθεί η ευαισθησία του προσδιορισμού αντικαταστάθηκε η μέτρηση της απορρόφησης στα 280 nm με τον προσδιορισμό των πρωτεϊνών και των ολιγοπεπτιδίων με τη μέθοδο Bradford. Κατασκευάστηκε πρότυπη καμπύλη ανάφορας χρησιμοποιώντας αλβουμίνη ως πρότυπη ουσία Οπότε σε δοκιμαστικούς σωλήνες που περιέχουν 118
119 2 ml διαφόρων συγκεντρώσεων αλβουμίνης προστέθηκαν 2 ml Βradford. Μετά από 5-10 min μετρήθηκαν οι απορροφήσεις των διαλυμάτων των δοκιμαστικών σωλήνων στα 595 nm. Στη συνέχεια πραγματοποιήθηκε η αντίδραση πρωτεόλυσης της πρωτεϊνάσης Κ και των τροποποιημένων υλικών με υπόστρωμα αλβουμίνης και σε διάφορες χρονικές στιγμές προσδιοριζόταν το ποσό των πρωτεϊνών με τη μέθοδο Bradford. Οι διαφοροποιήσεις που παρατηρήθηκαν δεν ήταν ιδιαίτερα ικανοποιητικές καθώς δεν μπορούσε να γίνει ο διαχωρισμός των πρωτεϊνών από τα ολιγοπεπτίδια που σχηματίζονται. Επομένως επιλέχθηκε να χρησιμοποιηθεί ως μέθοδος προσδιορισμού της δραστικότητας των τροποποιημένων υλικών η ενζυμική μέθοδος προσδιορισμού τρυψίνης (όπως αναφέρεται στην παράγραφο 2.2.9). Η μέθοδος αυτή είναι πιο ευαίσθητη και εξειδικευμένη αφού πραγματοποιείται με την βοήθεια ενός συνθετικού πεπτιδίου ως υπόστρωμα το ΤΑΜΕ (p-toluenesulphonyl-l-argine methyl ester) Ενζυμικός προσδιορισμός τρυψίνης Η τρυψίνη είναι μια πρωτεάση σερίνης του παγκρέατος. Το υπόστρωμά της εμφανίζει συγγένεια με θετικά φορτισμένες πλευρικές αλυσίδες λυσίνης και αργινίνης. Τα χαρακτηριστικά του ενζύμου που προέρχεται από πάγκρεας βοός είναι: Μοριακό βάρος: Τρυψινογόνου 24,000 και Τρυψίνης 23,800 Σύσταση: Το τρυψινογόνο έχει βρεθεί ότι έχει ενδογενή ενεργότητα. Ωστόσο, συνήθως θεωρείται ως ανενεργή πρόδρομη ουσία τρυψίνης, η οποία μπορεί να ενεργοποιηθεί με την απομάκρυνση ενός τελικού εξαπεπτιδίου, οπότε προκύπτει μονή αλυσίδα β-τρυψίνης. Με επόμενη περιορισμένη αυτόλυση παράγονται άλλες ενεργές μορφές, οι οποίες έχουν μια ή δύο αλυσίδες συνδεδεμένες με δισουλφιδικούς δεσμούς. Οι κύριες μορφές είναι η α- τρυψίνη που έχει δύο πεπτιδικές αλυσίδες και η β-τρυψίνη που έχει μια αλυσίδα. Οι δύο αυτές μορφές εμφανίζουν διαφορετική ενεργότητα και θερμική σταθερότητα. Βέλτιστο ph: Περίπου 8 1% Συντελεστής απόσβεσης: Ε 280 =14.3 Ισοηλεκτρικό σημείο: Τρυψινογόνου ph 9.3 και Τρυψίνης ph 10.5 Αναστολείς: Η δράση της τρυψίνη αναστέλλεται από οργανοφωσφορικές ενώσεις όπως διισοπροπυλο-φθοροφωσφορικές και φυσικούς αναστολείς παγκρέατος. Ενεργοποιητές: Ο ρυθμός μετατροπής τρυψινογόνου ενισχύεται με τη χρήση λανθανιδίου στη θέση ιόντων ασβεστίου Εξειδίκευση: Η ενεργότητα πρωτεάσης της τρυψίνης εμφανίζει υψηλή εξειδίκευση προς θετικά φορτισμένες πλευρικές αλυσίδες λυσίνης και αργινίνης. 119
120 Σταθεροποιητές: Ιόντα ασβεστίου επιβραδύνουν την αυτόλυση της τρυψίνης και προάγουν την ενεργοποίηση του τρυψινογόνου. Σταθερότητα: H τρυψίνη είναι σταθερή ως σκόνη στους 5 ο C για χρόνια. Προσδιορίστηκε η δραστικότητα τρυψίνης παρουσία αναστολέα τρυψίνης ή των υπό μελέτη ενώσεων για να διαπιστωθεί αν αυτές έχουν πιθανή δράση αναστολής πρωτεασών [24]. Τα διαλύματα που χρησιμοποιήθηκαν ήταν: Διάλυμα τρυψίνης: 0,2 mg/ml σε 0,001 M HCl Διάλυμα αναστολέα τρυψίνης (soya bean): 0,2 mg/ml σε buffer φωσφορικών 0,01 Μ ph 6,5 με 0,15 Μ acl Yπόστρωμα διάλυμα ΤΑΜΕ (p-toluenesulphonyl-l-argine methyl ester): 0,01 Μ σε H 2 Ρυθμιστικό διάλυμα : 0,046 M Tris HCl ph 8,1 με 0,0115 Μ CaCl 2 Το μίγμα της αντίδρασης επωάζεται στους 37 ο C για 10 min και είναι: 1,3 ml buffer (ρυθμιστικό διάλυμα) 0,150 ml TAME 50 μl enzyme Η μέτρηση της απορρόφηση του δείγματος γίνεται στα 247nm (A 247 ). Όταν προστίθεται ο αναστολέας χρησιμοποιείται σε ποσότητα 50 μl. Ο προσδιορισμός της ενζυμικής δράσης γίνεται βάσει του τύπου [24]: ΔΑ / min x 1000 U / mg = 540 x mg trypsin Προσδιορισμός αμινοξέων με τη μέθοδο της νινυδρίνης Τα αντιδραστήρια που χρησιμοποιούνται είναι [6]: 3 gr νινυδρίνης σε 100 ml 2-methoxyethanol Sodium acetate 2,65 M με ph 5,3-5,4 ρυθμισμένο με glacial CH 3 CH Acetate-cyanide buffer: προσθήκη διαλύματος KC 10mM σε buffer sodium acetate CH 3 CH 2,65 M με ph 5,3-5,4 σε αναλογία 1:49 v/v Diluent solution: 2 - προπανόλη σε H 2 (1:1) 120
121 Η πειραματική διαδικασία είναι [6]: 200 μl δείγματος Προστίθενται 100 μl cyanide acetate buffer Προστίθενται 100 μl νινυδρίνης Επώαση στους 100 ο C για 15 min Προσθήκη 1 ml diluent solution Μέτρηση απορρόφησης στα 570 nm (A 570 ) Παρατηρήσεις: Το διάλυμα CH 3 Ca CH 3 CH 2,65 M ph 5,4 φτιάχνεται ως εξής: Φτιάχνονται 2 διαλύματα CH 3 Ca 2,65 M και CH 3 CH 2,65 M. Στη συνέχεια ρυθμίζεται το ph του CH 3 Ca με προσθήκη του CH 3 CH 2,65 M. Το διάλυμα της νινυδρίνης είναι κίτρινο και φυλλάσεται στο ψυγείο σε σκοτεινό μπουκάλι Το μίγμα της αντίδρασης αρχικά είναι άχρωμο (ανοιχτό κίτρινο) και μετά την επώαση γίνεται ανοιχτό κόκκινο μπλε. Με την προσθήκη του diluent solution γίνεται πιο κυανό. Όσο περισσότερη ποσότητα αμινοξέος υπάρχει τόσο πιο κυανό είναι. Γίνεται πρότυπη καμπύλη αναφοράς με γνωστές συγκεντρώσεις μεθειονίνης και κατόπιν με την εξίσωση της καμπύλης αναφοράς γίνεται η επεξεργασία των δεδομένων: y(a 570 ) = a + bx(mm) Φασματοσκοπία Ο προσδιορισμός της δομής μιας οργανικής ένωσης αποτελούσε δύσκολο και χρονοβόρο εγχείρημα κατά τον 19 ο και στις αρχές του 20 ού αιώνα. Τις τελευταίες δεκαετίες, όμως, σημειώθηκαν καταπληκτικές πρόοδοι στον τομέα αυτό. Σήμερα υπάρχουν διαθέσιμες εκλεπτυσμένες τεχνικές μέθοδοι, που απλοποιούν σημαντικά το πρόβλημα του προσδιορισμού κάποιας δομής όπως είναι: η φασματομετρία μαζών (MS), η φασματοσκοπία υπερύθρου (IR), η φασματοσκοπία πυρηνικού μαγνητικού συντονισμού (MR) και η φασματοσκοπία υπεριώδους (UV). Οι πληροφορίες που παρέχει η κάθε μέθοδος είναι [23]: Φασματομετρία μαζών: Ποιο είναι το μέγεθος και ο μοριακός τύπος; Φασματοσκοπία υπερύθρου: Ποιες είναι οι λειτουργικές ομάδες; Φασματοσκοπία υπεριώδους: Αν υπάρχει κάποιο συζυγιακό ή ηλεκτρονικό σύστημα; Φασματοσκοπία πυρηνικού μαγνητικού συντονισμού: Με ποιον τρόπο συνδέονται οι άνθρακες με τα υδρογόνα; 121

122 Φασματοσκοπία και ηλεκτρομαγνητικό φάσμα Οι φασματοσκοπίες υπερύθρου, υπεριώδους και πυρηνικού μαγνητικού συντονισμού αποτελούν τεχνικές που διαφέρουν από τη φασματομετρία μαζών κατά το ότι καταγράφουν την αλληλεπίδραση μορίων με ηλεκτρομαγνητική ενέργεια και όχι με μια υψηλής ενέργειας ηλεκτρονική δέσμη. Το ορατό φως, οι ακτίνες Χ, τα μικροκύματα, τα ραδιοκύματα κ.τ.λ. αποτελούν διαφορετικά είδη της ηλεκτρομαγνητικής ακτινοβολίας. Όλα μαζί συγκροτούν το ηλεκτρομαγνητικό φάσμα που αναπαρίσταται στο σχήμα 4. Όπως φαίνεται, το ηλεκτρομαγνητικό φάσμα υποδιαιρείται αυθαίρετα σε διάφορες περιοχές, από τις οποίες η περιοχή του ορατού καταλαμβάνει μικρό μόνο μέρος του συνολικού φάσματος, σε μήκος κύματος από 3,8 x 10-5 cm έως 7,8 x 10-5 cm. Η περιοχή του ορατού περιβάλλεται από τις περιοχές του υπερύθρου και του υπεριώδους. Σχήμα 4: Το ηλεκτρομαγνητικό φάσμα [23]. Η ηλεκτρομαγνητική ακτινοβολία εμφανίζει διττή συμπεριφορά. Σε ορισμένες περιπτώσεις, παρουσιάζει ιδιότητες σωματιδίου (που αποκαλείται φωτόνιο), ενώ σε άλλες συμπεριφέρεται ως ενεργειακό κύμα που ταξιδεύει με την ταχύτητα του φωτός. Όπως όλα τα κύματα, η ηλεκτρομαγνητική ακτινοβολία χαρακτηρίζεται από κάποιο μήκος κύματος, κάποια συχνότητα και κάποιο πλάτος (σχήμα 5). Το μήκος κύματος, λ, είναι απλώς η απόσταση ανάμεσα στα μέγιστα δύο διαδοχικών κυμάτων. Η συχνότητα, V, είναι ο αριθμός που εκφράζει το πόσα μέγιστα κυμάτων διέρχονται από ορισμένο σημείο ανά μονάδα χρόνου και συνήθως δίνεται σε αντίστροφα δευτερόλεπτα (s -1 ) ή hertz, Hz (1 Ηz = 1 s -1 ). Πλάτος είναι το ύψος του κύματος, μετρούμενο από το μέσο σημείο έως το μέγιστο. Η ένταση της ενέργειας ακτινοβολίας, είτε πρόκειται για ασθενή δέσμη είτε για εκτυφλωτική, είναι ανάλογη προς το τετράγωνο του πλάτους του κύματος. 122
123 Σχήμα 5: (α) Το μήκος κύματος (λ) είναι η απόσταση μεταξύ δύο διαδοχικών μέγιστων κυμάτων. Πλάτος είναι το ύψος του κύματος, μετρούμενο από το κέντρο. (β)-(γ) Ότι αντιλαμβανόμαστε ως διαφορετικά είδη ηλεκτρομαγνητικής ακτινοβολίας είναι απλώς κύματα με διαφορετικά μήκη κύματος (ή συχνότητες) [23]. Πολλαπλασιάζοντας το μήκος ενός κύματος, σε εκατοστόμετρα (cm), με τη συχνότητά του, σε αντίστροφα δευτερόλεπτα (s -1 ), υπολογίζουμε την ταχύτητά του, σε εκατοστόμετρα ανά δευτερόλεπτο (cm/s). Η ταχύτητα κάθε ηλεκτρομαγνητικής ακτινοβολίας στο κενό είναι σταθερή και αποκαλείται «ταχύτητα του φωτός», συντομογραφικά c. Πρόκειται για μια από τις φυσικές σταθερές που γνωρίζουμε με μεγάλη ακρίβεια. Έχει αριθμητική τιμή 2, x cm/s, που συνήθως στρογγυλοποιείται στα 3,00 Χ cm/s. Μήκος κύματος x Συχνότητα = Ταχύτητα λ (cm) x ν(s -1 ) = c (cm/sec) που μπορεί να αναγραφεί και ως: λ = c/ν ή ν =c/λ Η ηλεκτρομαγνητική ενέργεια μεταδίδεται μόνο σε διακριτές ενεργειακές δέσμες, τα κβάντα. Η ποσότητα ενέργειας, ε, που αντιστοιχεί σε 1 κβάντο ενέργειας (ή 1 φωτόνιο) δεδομένης συχνότητας ν εκφράζεται με την εξίσωση: ε = hv = (hc)/λ όπου ε = Ενέργεια 1 φωτονίου (1 κβάντο) h = Σταθερά του Planck (6,62 x J s = 1,58 x cal s) ν = Συχνότητα (s -1 ) λ = Μήκος κύματος (cm) c = Ταχύτητα φωτός (3,00 x cm/s) Αυτή η εξίσωση μας λέει ότι η ενέργεια ενός φωτονίου μεταβάλλεται ευθέως ανάλογα προς τη συχνότητα ν, αλλά αντιστρόφως ανάλογα προς το μήκος κύματος λ. Υψηλές συχνότητες και μικρά 123

124 μήκη κύματος αντιστοιχούν σε ακτινοβολία υψηλής ενέργειας, όπως οι ακτίνες γ. Χαμηλές συχνότητες και μεγάλα μήκη κύματος αντιστοιχούν σε ακτινοβολία χαμηλής ενέργειας, όπως τα ραδιοκύματα. Αν πολλαπλασιάσουμε το ε με τον αριθμό Ανοgadrο Ν Α, καταλήγουμε στην ίδια εξίσωση, εκφρασμένη όμως σε μονάδες οικείες στους οργανικούς τύπους: -2-3 Ahc 1,20 x 10 kj/mol 2,86 x 10 kcal/mol E = = = λ λ (cm) λ (cm) όπου το Ε αντιπροσωπεύει την ενέργεια του αριθμού Ανοgadrο (ενός «mole») φωτονίων με μήκος κύματος λ. Όταν κάποια οργανική ένωση προσβληθεί από μια δέσμη ηλεκτρομαγνητικής ακτινοβολίας, απορροφά ενέργεια σε συγκεκριμένα μήκη κύματος, αλλά αφήνει να διέλθει ενέργεια σε διαφορετικά μήκη κύματος. Αν ακτινοβολήσουμε ένα δείγμα με ενέργεια πολλών διαφορετικών μηκών κύματος και εντοπίσουμε ποια απορροφώνται και ποια διέρχονται, μπορούμε να προσδιορίσουμε το φάσμα απορρόφησης της ένωσης. Τα αποτελέσματα απεικονίζονται σε ένα γράφημα που καταγράφει το μήκος κύματος σε σχέση με την διερχόμενη ακτινοβολία. Ένα παράδειγμα φάσματος απορρόφησης, και συγκεκριμένα της αιθυλικής αλκοόλης όταν εκτίθεται σε υπέρυθρη ακτινοβολία, φαίνεται στο σχήμα 6. Ο οριζόντιος άξονας καταγράφει το μήκος κύματος και ο κατακόρυφος την ένταση των διαφόρων απορροφήσεων ενέργειας ως εκατοστιαία διαπερατότητα (transmittance). Η γραμμή βάσης, που αντιστοιχεί σε απορρόφηση 0% (ή διαπερατότητα 100%), βρίσκεται στην άνω πλευρά του γραφήματος, έτσι η δημιουργία μιας κορυφής προς τα κάτω σημαίνει ότι σ αυτό το μήκος κύματος έλαβε χώρα απορρόφηση ενέργειας. Σχήμα 6: Το φάσμα απορρόφησης υπερύθρου της αιθυλικής αλκοόλης, CH 3 CH 2 H. Διαπερατότητα 100% σημαίνει ότι όλη η ενέργεια διέρχεται μέσα από το δείγμα ενώ χαμηλότερη διαπερατότητα σημαίνει ότι κάποιο ποσοστό ενέργειας απορροφάται. Έτσι, κάθε κορυφή προς τα κάτω αντιστοιχεί σε απορρόφηση ενέργειας [23]. 124

125 Η πρόσθετη ενέργεια που αποκτά ένα μόριο, όταν απορροφά ακτινοβολία, πρέπει να κατανεμηθεί με κάποιο τρόπο σε ολόκληρο το μόριο. Για παράδειγμα η απορρόφηση ακτινοβολίας μπορεί να αυξήσει την κινητική ενέργεια του μορίου, αναγκάζοντας τους δεσμούς να αποκτούν μεγαλύτερο μήκος ή να κάμπτονται περισσότερο. Εναλλακτικά, η απορρόφηση ακτινοβολίας μπορεί να αναγκάσει κάποιο ηλεκτρόνιο να μεταπηδήσει από ένα τροχιακό χαμηλής ενέργειας σε ένα τροχιακό υψηλότερης ενέργειας. Διαφορετικές συχνότητες ακτινοβολίας επιδρούν στα μόρια με διαφορετικούς τρόπους, αλλά όλες παρέχουν πληροφορίες για τη δομή του μορίου, εφόσον τα αποτελέσματα της επίδρασης τους ερμηνευθούν σωστά [23] Φασματοσκοπία υπερύθρου οργανικών μορίων Η περιοχή υπερύθρου (IR) του ηλεκτρομαγνητικού φάσματος καλύπτει την περιοχή αμέσως μετά το ορατό (7,8 x 10-5 cm), μέχρι τα 10-2 cm περίπου, αλλά μόνο η ενδιάμεση περιοχή, από τα 2,5 x 10-4 cm ως τα 2,5 x 10-3 cm, χρησιμοποιείται από τους οργανικούς χημικούς (σχήμα 7). Σχήμα 7: Η υπέρυθρη περιοχή του ηλεκτρομαγνητικού φάσματος [23]. Τα μήκη κύματος εντός της περιοχής IR δίνονται συνήθως σε μικρόμετρα (1 μm = 10-4 cm), ενώ οι συχνότητες εκφράζονται σε κυματάριθμους ( ~ ν ) μάλλον παρά σε hertz. Ο κυματάριθμος, που εκφράζεται σε μονάδες αντιστρόφων εκατοστόμετρων (cm -1 ), είναι απλώς το αντίστροφο του μήκους κύματος: ~ Κυματάριθμος, ν (cm -1 ) = 1/λ (cm) Έτσι, η χρήσιμη περιοχή του IR είναι από τα 4000 cm -1 ως τα 400 cm -1. Χρησιμοποιώντας την εξίσωση Ε = (1,20 x 10-2 kj/mοl)/λ, μπορούμε να υπολογίσουμε τα επίπεδα ενέργειας της ακτινοβολίας IR, που κυμαίνονται μεταξύ 48,0 kj/mol και 4,80 kj/mol (11,5-1,15 kcal/mol). 125
126 Γιατί ένα οργανικό μόριο απορροφά κάποια μήκη κύματος της ακτινοβολίας IR και άλλα όχι; Όλα τα μόρια διαθέτουν κάποια συγκεκριμένη ποσότητα ενέργειας κατανεμημένη σε όλη τη δομή τους, που προκαλεί στους δεσμούς δονήσεις (επιμηκύνσεις) και κάμψεις. Ταυτόχρονα, εξαιτίας της, τα άτομα πάλλονται και περιστρέφονται ενώ παρατηρούνται και διάφορες άλλες μοριακές δονήσεις. Μερικές επιτρεπτές μορφές δονήσεων και κάμψεων είναι οι παρακάτω: Η ποσότητα ενέργειας που περιλαμβάνει ένα μόριο δεν μεταβάλλεται κατά συνεχή τρόπο, αλλά είναι κβαντισμένη. Δηλαδή, ένα μόριο μπορεί να επιμηκύνεται, να κάμπτεται ή να πάλλεται, σε συγκεκριμένες συχνότητες που αντιστοιχούν σε συγκεκριμένα ενεργειακά επίπεδα. Ας πάρουμε για παράδειγμα την επιμήκυνση του δεσμού. Αν και συνήθως αναφερόμαστε στα μήκη δεσμών σαν να ήταν δεδομένα, οι αριθμοί που αναφέρονται αποτελούν μέσους όρους. Στην πραγματικότητα, οι δεσμοί συνεχώς δονούνται και κάμπτονται, με συνέπεια το μήκος τους να αυξάνεται και να μειώνεται. Έτσι, ένας τυπικός δεσμός C-H, με μέσο μήκος 1,10 Å, στην πραγματικότητα πάλλεται σε ορισμένη συχνότητα και διαδοχικά εκτείνεται και συμπιέζεται, σαν να συνέδεε τα δύο άτομα ένα ελατήριο. Όταν το μόριο δέχεται ηλεκτρομαγνητική ακτινοβολία, απορροφάται ενέργεια όταν η ενέργεια της ακτινοβολίας είναι ίδια με την ενεργειακή διαφορά μεταξύ δυο δονητικών συχνοτήτων. Όταν ένα μόριο απορροφά ακτινοβολία IR, η μοριακή δόνηση που έχει συχνότητα ίση με εκείνη της ακτινοβολίας αυξάνει το πλάτος της. Δηλαδή, το «ελατήριο» που συνδέει τα δυο άτομα διαδοχικά εκτείνεται και συμπιέζεται λίγο περισσότερο. Εφόσον κάθε συχνότητα που απορροφάται από ένα μόριο αντιστοιχεί σε μια προκαθορισμένη μοριακή κίνηση, μπορούμε να διαπιστώσουμε τις κινήσεις του μορίου, μελετώντας το φάσμα IR. Από την ερμηνεία αυτών των κινήσεων μπορούμε να συμπεράνουμε τι είδους δεσμοί (λειτουργικές ομάδες) υπάρχουν στο μόριο [12]. 126

127 Ερμηνεία φασμάτων IR Η πλήρης ερμηνεία ενός φάσματος ΙR είναι δύσκολη, επειδή τα περισσότερα οργανικά μόρια είναι τόσο μεγάλα, ώστε εμφανίζουν δεκάδες μορφές επιμήκυνσης και κάμψης. Έτσι, ένα φάσμα IR περιλαμβάνει δεκάδες περιοχές απορρόφησης. Αυτή η περιπλοκότητα είναι εξαιρετικά χρήσιμη, επειδή ένα φάσμα IR μπορεί να λειτουργήσει σαν ένα είδος δακτυλικού αποτυπώματος για μια συγκεκριμένη ένωση. Για το λόγο αυτό, η περιοχή του φάσματος IR μεταξύ 1500 cm -1 και 400 cm -1 περίπου αποκαλείται περιοχή δακτυλικού αποτυπώματος. Εάν δύο ενώσεις έχουν πανομοιότυπα φάσματα IR, τότε είμαστε σχεδόν βέβαιοι ότι είναι όμοιες. Πίνακας 1: Χαρακτηριστικές απορροφήσεις IR μερικών λειτουργικών ομάδων [23]. 127
128 Ευτυχώς, δεν είναι απαραίτητο να ερμηνεύσουμε πλήρως ένα φάσμα IR, προκειμένου να αποκτήσουμε χρήσιμες πληροφορίες για τη δομή του μορίου. Οι περισσότερες λειτουργικές ομάδες διαθέτουν συγκεκριμένες περιοχές απορρόφησης, που δεν διαφέρουν από ένωση σε ένωση. Η απορρόφηση C= μιας κετόνης εντοπίζεται σχεδόν πάντοτε στην περιοχή cm -1. Η απορρόφηση Ο-Η μιας αλκοόλης βρίσκεται σχεδόν πάντοτε στην περιοχή cm -1 και η απορρόφηση C=C ενός αλκενίου σχεδόν πάντα μεταξύ cm -1. Μαθαίνοντας πού εμφανίζονται οι χαρακτηριστικές απορροφήσεις των διαφόρων λειτουργικών ομάδων, είμαστε σε θέση να αποκτούμε δομικές πληροφορίες από τα φάσματα IR. Στον πίνακα 1 καταγράφονται οι χαρακτηριστικές απορροφήσεις IR μερικών συνηθισμένων λειτουργικών ομάδων. Για να θυμόμαστε τη θέση συγκεκριμένων απορροφήσεων IR, μπορούμε να χωρίσουμε την υπέρυθρη περιοχή, από τις 4000 cm -1 ως τα 400 cm -1, σε τέσσερα τμήματα, όπως απεικονίζονται στο σχήμα 8: 1. Η περιοχή από 4000 ως 2500 cm -1 αντιστοιχεί σε απορροφήσεις που προκαλούνται από δονήσεις επιμήκυνσης (τάσης) των απλών δεσμών Ν-Η, C-H και Ο-Η. Οι δεσμοί Ν-Η και Ο-Η απορροφούν στην περιοχή cm -1, ενώ ο δεσμός C-H απορροφά γύρω στα 3000 cm Στην περιοχή cm -1 λαμβάνει χώρα η δόνηση επιμήκυνσης (τάσης) του τριπλού δεσμού. Εδώ απορροφούν τα νιτρίλια (RC ) και τα αλκύνια. 3. Στην περιοχή από 2000 ως 1500 cm -1 απορροφούν όλοι οι διπλοί δεσμοί (C=, C= και C=C). Οι καρβονυλικές ομάδες απορροφούν γενικά στην περιοχή μεταξύ 1680 και 1750 cm -1, ενώ η επιμήκυνση του δεσμού των αλκενίων συμβαίνει συνήθως σε μια περιορισμένη περιοχή μεταξύ 1640 και 1680 cm Η περιοχή κάτω από τα 1500 cm -1 είναι η περιοχή του δακτυλικού αποτυπώματος. Εδώ εμφανίζονται πολλές απορροφήσεις, που οφείλονται σε μια ποικιλία απλών δεσμών C-C, C-Ο, C- και C-X. Σχήμα 8: Περιοχές του φάσματος υπερύθρου. Το φάσμα IR υποδιαιρείται σε τέσσερις περιοχές: μια περιοχή απλών δεσμών με υδρογόνο, μια περιοχή τριπλών δεσμών, μια περιοχή διπλών δεσμών και την περιοχή του δακτυλικού αποτυπώματος [23]. 128
129 Γιατί, όμως, απορροφούν στις παρατηρούμενες περιοχές οι διάφορες λειτουργικές ομάδες; Η καλύτερη αντιστοιχία που μπορούμε να χρησιμοποιήσουμε είναι δύο βάρη (άτομα) που συνδέονται με ένα ελατήριο (δεσμός). Βραχείς, ισχυροί δεσμοί δονούνται σε υψηλότερη συχνότητα και με μικρότερο μήκος κύματος από τους επιμήκεις, ασθενείς δεσμούς, όπως ακριβώς ένα βραχύ ισχυρό ελατήριο δονείται ταχύτερα από κάποιο επίμηκες και περισσότερο χαλαρό ελατήριο. Έτσι, οι τριπλοί δεσμοί απορροφούν σε υψηλότερη συχνότητα από τους διπλούς, οι οποίοι με τη σειρά τους απορροφούν υψηλότερα από τους απλούς δεσμούς. Επιπλέον, ελατήρια που συνδέουν μικρά βάρη δονούνται ταχύτερα από ελατήρια που συνδέουν μεγάλα βάρη [23] Φάσματα υπερύθρου των υδρογονανθράκων Αλκάνια Το υπέρυθρο φάσμα ενός αλκανίου δεν παρέχει ιδιαίτερες πληροφορίες, επειδή δεν υπάρχουν λειτουργικές ομάδες και όλες οι απορροφήσεις οφείλονται σε δονήσεις τάσης και κάμψης των δεσμών C-H και C-C. Οι δεσμοί C-H των αλκανίων παρουσιάζουν πάντοτε μια ισχυρή απορρόφηση στα cm -1, ενώ οι κορεσμένοι δεσμοί C-C παρουσιάζουν λιγότερο έντονες κορυφές, ταινίες στην περιοχή cm -1. Εφόσον όλες οι οργανικές ενώσεις περιέχουν τμήματα κορεσμένων ανθρακικών αλυσίδων, οι περισσότερες εμφανίζουν και αυτές τις χαρακτηριστικές απορροφήσεις IR [23] Αλκένια Τα αλκένια παρουσιάζουν αρκετές χαρακτηριστικές απορροφήσεις τάσης. Οι βινυλικοί δεσμοί =C-H απορροφούν μεταξύ 3020 και 3100 cm -1,ενώ οι δεσμοί C=C των αλκενίων συνήθως απορροφούν γύρω στα 1650 cm -1, αν και σε ορισμένες περιπτώσεις οι κορυφές μπορεί να είναι μάλλον χαμηλές και δυσδιάκριτες. Τα μονο- και διυποκατεστημένα αλκένια εμφανίζουν χαρακτηριστικές απορροφήσεις κάμψης εκτός επιπέδου των δεσμών =C-Η στην περιοχή cm -1, οι οποίες επιτρέπουν να προσδιορίσουμε το είδος της υποκατάστασης στον διπλό δεσμό. Τα μονοϋποκατεστημένα αλκένια, όπως το 1-εξένιο, παρουσιάζουν ισχυρές χαρακτηριστικές απορροφήσεις στα 910 και στα 990 cm -1 ενώ τα 2,2-διυποκατεστημένα αλκένια (R 2 C=CH 2 ) εμφανίζουν μια ισχυρή απορρόφηση στα 890 cm -1 [23]. 129
130 Αλκύνια Τα αλκύνια παρουσιάζουν μια απορρόφηση τάσης του δεσμού C C στα cm -1, η οποία είναι πολύ πιο έντονη στα ακραία αλκύνια παρά στα εσωτερικά. Στην πραγματικότητα, οι συμμετρικά υποκατεστημένοι τριπλοί δεσμοί δεν παρουσιάζουν καθόλου απορρόφηση. Τα ακραία αλκύνια εμφανίζουν επίσης μια χαρακτηριστική δόνηση τάσης του δεσμού C-H στα 3300 cm -1. Αυτή η απορρόφηση έχει διαγνωστικό χαρακτήρα για τα ακραία αλκύνια, επειδή είναι αρκετά έντονη και οξεία [23] Φάσματα υπερύθρου άλλων λειτουργικών ομάδων Αλκοόλες Η λειτουργική ομάδα -H των αλκοολών είναι εύκολο να εντοπισθεί στο IR. Οι αλκοόλες παρουσιάζουν μια χαρακτηριστική απορρόφηση στην περιοχή cm -1, συνήθως αρκετά ευρεία και ισχυρή. Εάν υπάρχει, είναι δύσκολο να την αγνοήσει κανείς ή να την συγχέει με κάτι άλλο [23] Αμίνες Η λειτουργική ομάδα Ν-Η των αμινών είναι επίσης εύκολο να εντοπιστεί στο IR, από τη χαρακτηριστική απορρόφησή της στην περιοχή cm -1. Παρόλο που στην ίδια περιοχή απορροφούν και οι αλκοόλες, μια απορρόφηση Ν-Η είναι πολύ οξύτερη και λιγότερο έντονη από την απορρόφηση της ομάδας Ο-Η [23] Καρβονυλικές ενώσεις Οι καρβονυλικές ομάδες εντοπίζονται ευκολότερα από όλες τις άλλες απορροφήσεις IR, λόγω της οξείας και ισχυρής κορυφής τους στα cm -1. Ακόμη σημαντικότερο είναι το γεγονός ότι συχνά η ακριβής θέση της απορρόφησης στη συγκεκριμένη περιοχή μπορεί να χρησιμοποιηθεί για τον προσδιορισμό του είδους του καρβονυλίου: αλδεΰδης, κετόνης, εστέρα, κ.λ.π. Αλδεΰδες. Οι κορεσμένες αλδεΰδες απορροφούν στα 1730 cm -1, ενώ οι αλδεΰδες δίπλα είτε σε διπλό δεσμό είτε σε αρωματικό δακτύλιο απορροφούν στα 1705 cm
131 Κετόνες. Οι κετόνες ανοικτής αλυσίδας και οι εξαμελείς κυκλικές κετόνες απορροφούν στα 1715 cm -1, οι πενταμελείς κυκλικές κετόνες στα 1750 cm -1 και οι κετόνες δίπλα σε διπλό δεσμό ή αρωματικό δακτύλιο στα 1690 cm -1 Εστέρες. Οι κορεσμένοι εστέρες απορροφούν στα 1735 cm -1 ενώ οι εστέρες με το καρβονύλιο δίπλα σε διπλό δεσμό ή αρωματικό δακτύλιο στα 1715 cm -1 Μια πρόσθετη σημαντική παρατήρηση για τη φασματοσκοπία IR: είναι δυνατό να αποκτήσουμε πληροφορίες για τη δομή ενός μορίου από ένα φάσμα IR παρατηρώντας και ποιες απορροφήσεις δεν υπάρχουν. Εάν το φάσμα μιας ένωσης δεν παρουσιάζει απορροφήσεις στα 3300 και 2150 cm -1, η ένωση δεν είναι ακραίο αλκύνιο. Εάν το φάσμα δεν εμφανίζει απορρόφηση κοντά στα 3400 cm -1, δεν πρόκειται για αλκοόλη κ.ο.κ [23]. 131
132 ΚΕΦΑΛΑΙΟ 3 ΠΕΙΡΑΜΑΤΙΚΟ ΜΕΡΟΣ 3.1 Μελέτη νανοσωλήνων που μιμούνται τη δράση ενζύμων - πρωτεασών Η παρασκευή των τροποποιημένων νανοσωλήνων άνθρακα έγινε στο ερευνητικό εργαστήριο του κ. Ταγματάρχη στο Ινστιτούτο Θεωρητικής και Φυσικής Χημείας του Εθνικού Ιδρύματος Ερευνών (ΕΙΕ). Ακολούθησε προσδιορισμός των βέλτιστων συνθηκών για τον υπολογισμό της ενζυμικής δραστικότητας της πρωτεϊνάσης Κ και κατόπιν προσδιορίστηκε η πιθανή δράση των τροποποιημένων νανοσωλήνων Προσδιορισμός δραστικότητας πρωτεϊνάσης K σε υπόστρωμα καζεΐνης Σύμφωνα με την παράγραφο σε δοκιμαστικούς σωλήνες που περιέχουν 50 mg υπόστρωμα καζεΐνης (10 mg/ml) προστέθηκαν 4,9 ml buffer και 0,1 ml διαφόρων συγκεντρώσεων ενζύμου πρωτεϊνάσης Κ. Επωάστηκαν για 20 min στους 37 ο C με ανάδευση. Μετά το τέλος της επώασης η αντίδραση σταμάτησε με 0,5 ml 50 % TCA, ακολούθησε 10 min φυγοκέντρηση και μετρήθηκε η απορρόφηση των διαλυμάτων στα 280 nm. Προέκυψε το παρακάτω διάγραμμα: 0,8 Απορρόφηση (280 nm) 0,4 0 y = 0,0037x Συγκέντρωση (μg/ml) Διάγραμμα 1: Δραστικότητα πρωτεϊνάσης Κ συναρτήσει της συγκέντρωσης. 132
133 3.1.2 Δραστικότητα πρωτεϊνάσης Κ συναρτήσει του χρόνου επώασης Η αντίδραση επαναλήφθηκε χρησιμοποιώντας αλβουμίνη ως υπόστρωμα του ενζύμου. Οπότε σε δοκιμαστικούς σωλήνες που περιέχουν 2 και 5 mg/ml υπόστρωμα αλβουμίνης προστέθηκαν 4,9 ml buffer και 0,1 ml από διάλυμα αρχικής συγκέντρωσης 1 mg/ml ενζύμου πρωτεϊνάσης Κ. Επωάστηκαν για 20, 30, 45, 60, 90 και 120 min στους 37 ο C με ανάδευση. Μετά το τέλος της επώασης η αντίδραση σταματούσε με την προσθήκη 0,1 ml 50 % TCA ανά ml διαλύματος, ακολουθούσε 10 min φυγοκέντρηση και μετριόταν η απορρόφηση των διαλυμάτων στα 280 nm. Προέκυψε το παρακάτω διάγραμμα: Απορρόφηση (280 nm) 0,2 0, Χρόνος επώασης (min) Διάγραμμα 2: Ενζυμική δραστικότητα πρωτεϊνάσης Κ συναρτήσει του χρόνου επώασης. Υπόστρωμα: (- -) 2 mg/ml αλβουμίνη και (- -) 5 mg/ml αλβουμίνη. Από το διάγραμμα προκύπτει ότι ο καλύτερος χρόνος επώασης είναι αυτός των 60 min, γι αυτό στη συνέχεια η επώαση θα γίνει για 60 min Δραστικότητα πρωτεϊνάσης Κ συναρτήσει της συγκέντρωσής της σε υπόστρωμα αλβουμίνης Σύμφωνα με την παράγραφο σε δοκιμαστικούς σωλήνες που περιέχουν 4 mg/ml υπόστρωμα αλβουμίνης προστέθηκαν 4,9 ml buffer και 0,1 ml διαφόρων 133
134 συγκεντρώσεων ενζύμου πρωτεϊνάσης Κ. Επωάστηκαν για 60 min (λόγω της παραγράφου 3.1.2) στους 37 ο C. Μετά το τέλος της επώασης η αντίδραση σταμάτησε με την προσθήκη 0,5 ml 50 % TCA, ακολούθησε 10 min φυγοκέντρηση και μετρήθηκε η απορρόφηση των διαλυμάτων στα 280 nm. Προέκυψε το παρακάτω διάγραμμα: Απορρόφηση (280 nm) 0,4 0,3 0,2 0,1 0 y = 0,0004x Συγκέντρωση (μg/ml) Διάγραμμα 3: Ενζυμική δραστικότητα πρωτεϊνάσης Κ σε συνάρτηση με την συγκέντρωση της (μg/ml) σε υπόστρωμα αλβουμίνης Προσδιορισμός δραστικότητας των τροποποιημένων υλικών Σύμφωνα με την παράγραφο σε δοκιμαστικούς σωλήνες που περιέχουν 4 mg/ml υπόστρωμα αλβουμίνης προστέθηκαν 4,9 ml buffer και 0,1 ml (10 και 20 μg/ml) διαλύματος ΝΚ54 και GP5. Επωάστηκαν για 1, 3, 5, 8 και 20 h στους 37 ο C με ανάδευση. Μετά το τέλος της επώασης η αντίδραση σταμάτησε με την προσθήκη 0,1 ml 50 % TCA ανά ml διαλύματος, ακολουθούσε 10 min φυγοκέντρηση και μετριόταν η απορρόφηση των διαλυμάτων στα 280 nm. Προέκυψε το παρακάτω διάγραμμα: 134
135 Απορρόφηση (280 nm) 0,2 0,12 0, Χρόνος επώασης (h) Διάγραμμα 4: Δραστικότητα των τροποποιημένων υλικών GP5 και ΝΚ54. (- -) ΝΚ54 (10 μg/ml), (- -) ΝΚ54 (20 μg/ml), (- -) GP5 (10 μg/ml), (- -) GP5 (20 μg/ml), (- -) control χωρίς αλβουμίνη με (10 μg/ml) GP5 και (- -) control με αλβουμίνη χωρίς τροποποιημένο υλικό. Στην περίπτωση αυτή ο χρόνος επώασης ήταν μεγαλύτερος από 60 min. Αφού από το διάγραμμα προκύπτει ότι ο καλύτερος χρόνος επώασης είναι αυτός των 8 h, γι αυτό στη συνέχεια η επώαση θα γίνει για 8 h Προσδιορισμός δραστικότητας των τροποποιημένων υλικών σε διάφορα ph Σύμφωνα με την παράγραφο σε δοκιμαστικούς σωλήνες που περιέχουν 4 mg/ml υπόστρωμα αλβουμίνης προστέθηκαν 4,9 ml buffer διαφορετικού ph (5.2, 7, 8.9 και 11) και 0,1 ml (20 μg/ml) από το ΝΚ54 και το GP5. Επωάστηκαν για 8 h (λόγω της παραγράφου 3.1.4) στους 37 ο C με ανάδευση. Μετά το τέλος της επώασης η αντίδραση σταμάτησε με την προσθήκη 0,1 ml 50 % TCA ανά ml διαλύματος, ακολουθούσε 10 min φυγοκέντρηση και μετριόταν η απορρόφηση των διαλυμάτων στα 280 nm. Προέκυψε το παρακάτω διάγραμμα: 135
136 Μεταβολή απορρόφησης (280nm) 0,08 0,07 0,06 0, ph Διάγραμμα 5: Καμπύλη ph των τροποποιημένων υλικών (- -) ΝΚ54 (20 μg/ml) και (- -) GP5 (20 μg/ml). Από το διάγραμμα προκύπτει ότι το βέλτιστο ph και για το ΝΚ54 και για το GP5 είναι μεταξύ 8 και Προσδιορισμός της βιολογικής δραστικότητας του τροποποιημένου νανοσωλήνα (K54) Πραγματοποιήθηκε προσδιορισμός της βιολογικής δραστικότητας για το K54 με την μέθοδο της τρυψίνης (λόγω της παραγράφου 2.2.8). Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν τρία διαλύματα: Δείγμα 1 Δείγμα 2 Δείγμα 3 Buffer 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) μl K54 0 0,01 mg 0 Η 2 Ο 100 μl 0 50 μl 136
137 Οι συγκεντρώσεις ήταν σύμφωνα με την παράγραφο Τα τρία διαλύματα επωάστηκαν σε διάφορα χρονικά διαστήματα στους 37 ο C. Πριν από κάθε φωτομέτρηση το δείγμα φυγοκεντρείται 1 min ώστε να απομακρυνθεί το τροποποιημένο υλικό ΝΚ54 και το υπερκείμενο να μετρηθει στα 247 nm. Προέκυψε το παρακάτω διάγραμμα: Απορρόφηση (247 nm) 0,8 0,6 0,4 0, Χρόνος επώασης (h) Διάγραμμα 7: Δραστικότητα του τροποποιημένου υλικού (ΝΚ54). (- -) ΝΚ54 και (- -) τρυψίνη. Το τροποποιημένο υλικό (K54) αρχικά τις πρώτες πέντε ώρες έχει μικρότερη δράση σε σχέση με την τρυψίνη αλλά στη συνέχεια ενώ η δραστικοτητα της τρυψίνης μειώνεται και τελικά μηδενίζεται, η δράση του ΝΚ54 αυξάνεται συνεχώς Επαναχρησιμοποίηση του τροποποιημένου νανοσωλήνα (ΝΚ54) Η ποσότητα του ΝΚ54 που είχε χρησιμοποιηθεί σε προηγούμενο προσδιορισμό αφού καθαρίστηκε (τρεις πλύσεις με 1 ml Η 2 Ο) και απομακρύνθηκαν οι προσμίξεις που είχε, δοκιμάστηκε αν μπορεί να ξαναχρησιμοποιηθεί δηλαδή αν συνεχίζει να εμφανίζει δραστικότητα μετά την πρώτη χρήση του. Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν τρία διαλύματα: 137
138 Δείγμα 1 Δείγμα 2 Δείγμα 3 Buffer 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) μl K54 0 0,01 mg 0 Η 2 Ο 100 μl 0 50 μl Οι συγκεντρώσεις ήταν σύμφωνα με την παράγραφο Τα τρία διαλύματα επωάστηκαν με συνεχή ανάδευση σε διάφορα χρονικά διαστήματα και μετά από 1 min φυγοκέντρηση ακολουθούσε μέτρηση της απορρόφησης στα 247 nm. Προέκυψε το παρακάτω διάγραμμα: Απορρόφηση (247 nm) 0,7 0, Χρόνος επώασης (h) Διάγραμμα 7: Δυνατότητα επαναχρησιμοποίησης του τροποποιημένου υλικού (ΝΚ54). (- -) ΝΚ54 και (- -) τρυψίνη. Το τροποποιημένο υλικό (ΝΚ54) αν καθαριστεί μπορεί να επαναχρησιμοποιηθεί με παραπλήσια δραστικότητα οπώς και στην αρχική του χρήση Έλεγχος της δραστικότητας του μη τροποποιημένου νανοσωλήνα και επαναχρησιμοποίηση του τροποποιημένου νανοσωλήνα (K54) Πραγματοποιήθηκε έλεγχος της βιολογικής δραστικότητας για το μη τροποποιημένο υλικό (Ρ0279) με την μέθοδο της τρυψίνης. Επίσης για το τροποποιημένο υλικό (ΝΚ54) που είχε χρησιμοποιηθεί σε δύο προηγούμενους προσδιορισμούς αφού 138
139 καθαρίστηκε (τρεις πλύσεις με 1 ml Η 2 Ο) και απομακρύνθηκαν οι προσμίξεις του, δοκιμάστηκε αν μπορεί να ξαναχρησιμοποιηθεί και τρίτη φορά δηλαδή αν συνεχίζει να εμφανίζει δραστικότητα. Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν τέσσερα διαλύματα: Δείγμα 1 Δείγμα 2 Δείγμα 3 Δείγμα 4 Buffer 1.3 ml 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) μl P mg 0 K mg 0 0 Η 2 Ο 50 μl Οι συγκεντρώσεις ήταν σύμφωνα με την παράγραφο Τα τέσσερα διαλύματα επωάστηκαν με συνεχή ανάδευση σε διάφορα χρονικά διαστήματα και μετά από 1 min φυγοκέντρηση ακολουθούσε μέτρηση της απορρόφησης στα 247 nm. Προέκυψε το παρακάτω διάγραμμα: 0,8 Απορρόφηση (247 nm) 0,6 0,4 0,2 0-0,2-0, Χρόνος επώασης (h) Διάγραμμα 8: Έλεγχος της δραστικότητας του μη τροποποιημένου υλικού (Ρ0279) και επαναχρησιμοποίηση του τροποποιημένου (ΝΚ54). (- -) ΝΚ54, (- -) P0279 και (- -) τρυψίνη. Από το διάγραμμα 8 φαίνεται ότι ενώ το μη τροποποιημένο υλικό (P0279) δεν εμφανίζει δραστικότητα, το τροποποιημένο (ΝΚ54) συνεχίζει να είναι δραστικό και μετά από την τρίτη χρήση του. 139
140 3.1.9 Έλεγχος δραστικότητας του τροποποιημένου νανοσωλήνα (K28) Πραγματοποιήθηκε έλεγχος δραστικότητας για το τροποποιημένο υλικό (ΝΚ28) με την μέθοδο της τρυψίνης. Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν τρία διαλύματα: Δείγμα 1 Δείγμα 2 Δείγμα 3 Buffer 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) μl K54 0 0,06 mg 0 Η 2 Ο 60 μl 0 10 μl Οι συγκεντρώσεις ήταν σύμφωνα με την παράγραφο Τα τρία διαλύματα επωάστηκαν με συνεχή ανάδευση σε διάφορα χρονικά διαστήματα και μετά από 1 min φυγοκέντρηση ακολουθούσε μέτρηση της απορρόφησης στα 247 nm. Προέκυψε το παρακάτω διάγραμμα: Απορρόφηση (247 nm) 0,45 0,25 0,05-0, Χρόνος επώασης (h) Διάγραμμα 9: Έλεγχος της δραστικότητας του τροποποιημένου υλικού (ΝΚ28). (- -) ΝΚ28 και (- -) τρυψίνη. Το τροποποιημένο υλικό (ΝΚ28) όπως φαίνεται στο διάγραμμα 9 δεν εμφανίζει δραστικότητα. 140
 του P0279 που είναι ο μη τροποποιημένος")
 κολλημένα στην επιφάνεια")
. Η περιοχή σάρωσης είναι 1x1 μm.")
 που δείχνει την παράλληλη στον άξονα x")
141 Απεικόνιση με μικροσκόπιο ατομικών δυνάμεων (AFM) των τροποποιημένων νανοσωλήνων και του μη τροποποιημένου νανοσωλήνα Πραγματοποιήθηκε απεικόνιση με μικροσκόπιο ατομικών δυνάμεων (AFM) του P0279 που είναι ο μη τροποποιημένος νανοσωλήνας δηλαδή το control, του ΝΚ28 που είναι ο νανοσωλήνας με ένα αμμωνιακό άλλας και του ΝΚ54 που είναι ο νανοσωλήνας με τρία διαφορετικά αμινοξέα (L-Τυροσίνη, L-Ασπαραγινικό οξύ και L-Γλουταμινικό οξύ) κολλημένα στην επιφάνεια του. Οι εικόνες που προέκυψαν παρουσιάζονται παρακάτω: Εικόνα 1: Εικόνα AFM του P0279 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite). Η περιοχή σάρωσης είναι 1x1 μm. Από αριστερά προς τα δεξιά φαίνεται η τοπογραφία της επιφάνειας (height), η φάση (phase) και η 3D απεικόνηση. Εικόνα 2: Εικόνα AFM του P0279 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει την παράλληλη στον άξονα x x τομή. Η περιοχή σάρωσης είναι 800x800 nm. Από την τομή φαίνεται ότι το ύψος του νανοσωλήνα είναι 4,65 nm. 141
 που δείχνει ότι το πλάτος του είναι 24,1 nm.")

, η")

142 Εικόνα 3: Εικόνα AFM του P0279 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει ότι το πλάτος του είναι 24,1 nm. Η περιοχή σάρωσης είναι 1x1 μm. Εικόνα 4: Εικόνα AFM του ΝΚ28 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite). Η περιοχή σάρωσης είναι 500x500 nm. Από αριστερά προς τα δεξιά φαίνεται η τοπογραφία της επιφάνειας (height), η φάση (phase) και η 3D απεικόνηση. Εικόνα 5: Εικόνα AFM του ΝΚ28 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει την παράλληλη στον άξονα x x τομή. Η περιοχή σάρωσης είναι 500x500 nm. Από την τομή φαίνεται ότι το ύψος του νανοσωλήνα είναι 3,4 nm. 142
 που δείχνει ότι το πλάτος του είναι 9,9 nm.")

, η")

143 Εικόνα 6: Εικόνα AFM του ΝΚ28 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει ότι το πλάτος του είναι 9,9 nm. Η περιοχή σάρωσης είναι 1x1 μm. Εικόνα 7: Εικόνα AFM του ΝΚ54 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite). Η περιοχή σάρωσης είναι 600x600 nm. Από αριστερά προς τα δεξιά φαίνεται η τοπογραφία της επιφάνειας (height), η φάση (phase) και η 3D απεικόνηση. Εικόνα 8: Εικόνα AFM του ΝΚ54 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει την παράλληλη στον άξονα x x τομή. Η περιοχή σάρωσης είναι 600x600 nm. Από την τομή φαίνεται ότι το ύψος του νανοσωλήνα είναι 13,2 nm. 143
 ως φορείς βιολογικών μορίων Στο ερευνητικό εργαστήριο του κ. Γ.")
144 Εικόνα 9: Εικόνα AFM του ΝΚ54 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει ότι το πλάτος του είναι 17,5 nm. Η περιοχή σάρωσης είναι 600x600 nm. Από τις εικόνες AFM είναι εμφανές ότι το Ρ0279, το ΝΚ28 και το ΝΚ54 είναι νανοσωλήνες. 3.2 Mελέτη νανοσωλήνων άνθρακα (CT) ως φορείς βιολογικών μορίων Στο ερευνητικό εργαστήριο του κ. Γ. Σταυρόπουλου έχουν μελετηθεί μερικά βιολογικά μόρια και συγκεκριμένα μικρά πεπτίδια και αμινοξέα με βιολογική δράση όπως είναι το ανάλογο 31 {Βοc-D-Trp-H-(Bzl)-C-D-Trp} και 37 {Boc-H-(Bzl)-CH 2 - C-D-Trp-Leu-H} [12] αφού λειτουργούν ως αναστολείς πρωτεασών. Αυτά τα ανάλογα καθηλώθηκαν σε νανοσωλήνες άνθρακα στο ερευνητικό εργαστήριο του κ. Ταγματάρχη στο Ινστιτούτο Θεωρητικής και Φυσικής Χημείας του Εθνικού Ιδρύματος Ερευνών (ΕΙΕ) και μελετήθηκαν περαιτέρω Προσδιορισμός της βιολογικής δραστικότητας των προστατευμένων αναλόγων 31 και 37 Πραγματοποιήθηκε προσδιορισμός βιολογικής δραστικότητας για το ανάλογο 31 {Βοc-D-Trp-H-(Bzl)-C-D-Trp} και 37 {Boc-H-(Bzl)-CH 2 -C-D-Trp-Leu-H}. Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν εννέα διαλύματα: 144
145 Δείγμα 1 Δείγμα 2 Δείγμα 3 Δείγμα 4 Δείγμα 5 Δείγμα 6 Δείγμα 7 Δείγμα 8 Δείγμα 9 Buffer 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) 0 50 μl 50 μl 50 μl 50 μl 50 μl 50 μl 50 μl 50 μl Trypsin inhibitor μl (0.2 mg/ml) Ανάλογο mg 0.07 mg 0.1 mg Ανάλογο mg 0.07 mg 0.1 mg Η 2 Ο 50 μl Τα εννέα διαλύματα επωάστηκαν σε διάφορα χρονικά διαστήματα και ακολούθησε μέτρηση της απορρόφησης στα 247 nm. Από τα αποτελέσματα προέκυψε ότι τα προστατευμένα ανάλογα δεν λειτουργούν ως αναστολείς πρωτεασών και συγκεκριμένα της τρυψίνης οπότε αποπροστατεύτηκαν από τις ομάδες Boc (σύμφωνα με την παράγραφο 2.2.5) και επαναλήφθηκε η διαδικασία προσδιορισμού της βιολογικής δραστικότητας Προσδιορισμός της βιολογικής δραστικότητας του αναλόγου 31 και του καθηλωμένου αναλόγου 31 σε CT Πραγματοποιήθηκε προσδιορισμός βιολογικής δραστικότητας για το ανάλογο 31 και το καθηλωμένο ανάλογο 31 σε CT (K133) αφού προηγουμένως είχαν απομακρυνθεί οι ομάδες Boc (σύμφωνα με την παράγραφο 2.2.5). Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν τα παρακάτω διαλύματα: Δείγμα 1 Δείγμα 2 Δείγμα 3 Δείγμα 4 Δείγμα 5 Δείγμα 6 Buffer 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) 0 50 μl 50 μl 50 μl 50 μl 50 μl Trypsin inhibitor (0.2 mg/ml) μl Ανάλογο mg 0.05 mg 0.1 mg Η 2 Ο 50 μl
146 Επαναλήφθηκε ο προσδιορισμός της δραστικότητας για το ΝΚ133 και για το ανάλογο 31. Η ποσότητα των 0,0348 mg αναλόγου 31 αντιστοιχεί στα 0,3 mg του υλικού K133, βάσει του αριθμού των moles που έχουν καθηλωθεί. πότε παρασκευάστηκαν τα ακόλουθα διαλύματα: Δείγμα 7 Δείγμα 8 Δείγμα 9 Δείγμα 10 Δείγμα 11 Buffer 1.3 ml 1.3 ml 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) 0 50 μl 50 μl 50 μl 50 μl Trypsin inhibitor (0.2 mg/ml) μl 0 0 Ανάλογο mg 0 K mg Η 2 Ο 50 μl Οι συγκεντρώσεις ήταν σύμφωνα με την παράγραφο Τα έντεκα διαλύματα επωάστηκαν με συνεχή ανάδευση σε διάφορα χρονικά διαστήματα και ακολούθησε μέτρηση της απορρόφησης στα 247 nm. Προέκυψαν τα παρακάτω διαγράμματα: Παραμένουσα δραστικότητα (%) Χρόνος επώασης (min) Διάγραμμα 10: Παραμένουσα δραστικότητα (%) της τρυψίνης παρουσία (- -) 0,01 mg αναστολέα τρυψίνης, (- -) 0,0348 mg ανάλογο 31, (- -) 0,05 mg ανάλογο 31, (- -) 0,1 mg ανάλογο 31 και (- -) control. 146
147 Παραμένουσα δραστικότητα (%) Χρόνος επώασης (min) Διάγραμμα 11: Παραμένουσα δραστικότητα (%) της τρυψίνης παρουσία (- -) 0,01 mg αναστολέα τρυψίνης, (- -) 0,0348 mg ανάλογο 31, (- -) 0,3 mg K133 και (- -) control Δραστικότητα του καθηλωμένου αναλόγου 31 σε CT μετά από επαναλαμβανόμενη χρήση του H ποσότητα του καθηλωμένου αναλόγου 31 σε CT (ΝΚ133) που είχε χρησιμοποιηθεί σε προηγούμενο προσδιορισμό, αφού καθαρίστηκε (τρεις πλύσεις με 1 ml Η 2 Ο) και απομακρύνθηκαν οι προσμίξεις του, δοκιμάστηκε αν μπορεί να ξαναχρησιμοποιηθεί δηλαδή αν συνεχίζει να εμφανίζει δραστικότητα. Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν τέσσερα διαλύματα: Δείγμα 1 Δείγμα 2 Δείγμα 3 Δείγμα 4 Buffer 1.3 ml 1.3 ml 1.3 ml 1.3 ml TAME 0.15 ml 0.15 ml 0.15 ml 0.15 ml Trypsin (0.2 mg/ml) 0 50 μl 50 μl 50 μl Trypsin inhibitor (0.2 mg/ml) μl 0 K mg Η 2 Ο 50 μl Τα τέσσερα διαλύματα επωάστηκαν με συνεχή ανάδευση σε διάφορα χρονικά διαστήματα και μετά από 1 min φυγοκέντρηση ακολούθησε μέτρηση της απορρόφησης 147
148 στα 247 nm. Όμοια επαναλήφθηκε το πείραμα άλλες τέσσερις φορές. Οπότε προέκυψαν τα παρακάτω διαγράμματα: Παραμένουσα δραστικότητα (%) Χρόνος επώασης (min) Διάγραμμα 12: Παραμένουσα δραστικότητα (%) της τρυψίνης παρουσία (- -) 0,01 mg αναστολέα τρυψίνης, (- -) 0,3 mg ΝΚ133 (πρώτη επαναχρησιμοποίηση), (- -) 0,3 mg ΝΚ133 (δεύτερη επαναχρησιμοποίηση), (- -) 0,3 mg ΝΚ133 (τρίτη επαναχρησιμοποίηση), (- -) 0,3 mg ΝΚ133 (τέταρτη επαναχρησιμοποίηση), (- -) 0,3 mg ΝΚ133 (πέμπτη επαναχρησιμοποίηση), (- -) 0,0348 mg ανάλογο 31 και (- -) control. Επομένως παρατηρείται ότι η ποσότητα των 0,3 mg του καθηλωμένου αναλόγου 31 σε CT (ΝΚ133) εμφανίζει πολύ καλύτερη δράση σε σχέση με την αντίστοιχη ποσότητα των 0,0348 mg του αναλόγου 31 (η αντιστοιχία υπολογίστηκε βάσει του αριθμού των moles που έχουν καθηλωθεί). Επίσης ποσότητα του καθηλωμένου αναλόγου 31 σε CT (ΝΚ133) μπορεί να επαναχρησιμοποιηθεί αφού παρατηρήθηκε ότι μετά από χρησιμοποίηση της 6 φορές εμφανίζει παραπλήσια δραστικότητα. 148
 του αναλόγου 31 που είναι ένα")

, η φάση (phase) και η 3D")
 που δείχνει")
149 3.2.4 Απεικόνιση με μικροσκόπιο ατομικών δυνάμεων (AFM) τoυ αναλόγου 31 και του καθηλωμένου αναλόγου 31 σε CT Πραγματοποιήθηκε απεικόνηση με μικροσκόπιο ατομικών δυνάμεων (AFM) του αναλόγου 31 που είναι ένα διαζωνιακό διπεπτίδιο {Βοc-D-Trp-H-(Bzl)-C-D-Trp} και του καθηλωμένου αναλόγου 31 σε CT (K133). Οι εικόνες που προέκυψαν παρουσιάζονται παρακάτω: Εικόνα 10: Εικόνα AFM του αναλόγου 31 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite). Η περιοχή σάρωσης είναι 600x600 nm. Από αριστερά προς τα δεξιά φαίνεται η τοπογραφία της επιφάνειας (height), η φάση (phase) και η 3D απεικόνηση. Εικόνα 11: Εικόνα AFM του αναλόγου 31 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει την παράλληλη στον άξονα x x τομή. Η περιοχή σάρωσης είναι 600x600 nm. Από την τομή φαίνεται ότι το ύψος του αναλόγου 31 είναι 28 nm. 149
 που δείχνει ότι το πλάτος του είναι 58,3 nm.")

 και δεξιά η φάση")

150 Εικόνα 12: Εικόνα AFM του αναλόγου 31 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει ότι το πλάτος του είναι 58,3 nm. Η περιοχή σάρωσης είναι 600x600 nm. Εικόνα 13: Εικόνα AFM του ΝΚ133 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite). Η περιοχή σάρωσης είναι 2x2 μm. Αριστερά φαίνεται η τοπογραφία της επιφάνειας (height) και δεξιά η φάση (phase). Εικόνα 14: Εικόνα AFM του ΝΚ133 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει την κάθετη στον άξονα x x τομή. Η περιοχή σάρωσης είναι 2x2 μm. Από την τομή φαίνεται ότι το ύψος του νανοσωλήνα είναι 95,2 nm. 150

151 Εικόνα 15: Εικόνα AFM του ΝΚ133 σε επιφάνεια HPG (Highly riented Pyrolytic Graphite) που δείχνει ότι το πλάτος του είναι 17,6 nm. Η περιοχή σάρωσης είναι 1x1 μm. Από τις εικόνες AFM είναι εμφανές ότι το ΝΚ133 είναι νανοσωλήνας και διαφοροποιείται από το ανάλογο Προσδιορισμός της βιολογικής δραστικότητας των εξαπεπτιδίων 3.1, 3.2, 3.3, 4.1, 4.2 και 4.3 Στα πλαίσια της παρούσας διπλωματικής ελέχθηκαν και δύο νέες σειρές εξαπεπτιδίων με σκοπό την μελλοντική τους καθήλωση σε νανοφορείς. Οπότε προσδιορίστηκε η βιολογική δραστικότητα για τα εξαπεπτίδια 3.1 {Glp 1 -Phe 2 -Gly 3 - [H-(Bzl)-CH 2 -C] 4 -D-Trp 5 -Leu 6 -H}, 3.2 {Glp 1 -Ala 2 -Gly 3 -[H-(Bzl)-CH 2 -C] 4 - D-Trp 5 -Leu 6 -H}, 3.3 {Glp 1 -Tic 2 -Gly 3 -[H-(Bzl)-CH 2 -C] 4 -D-Trp 5 -Leu 6 -H}, 4.1 {Glp 1 -Phe 2 -Gly 3 -D-Trp 4 -[H-(Bzl)-C] 5 -D-Trp 6 -H}, 4.2 {Glp 1 -Ala 2 -Gly 3 -D- Trp 4 -[H-(Bzl)-C] 5 -D-Trp 6 -H} και 4.3 {Glp 1 -Tic 2 -Gly 3 -D-Trp 4 -[H-(Bzl)-C] 5 - D-Trp 6 -H}. Τα εξαπεπτίδια αυτά παρασκευάστηκαν στο ερευνητικό εργαστήριο του κ. Γ. Σταυρόπουλου. Οπότε σύμφωνα με την παράγραφο παρασκευάστηκαν εννέα διαλύματα: 151
Οι περισσότεροι μονοτοιχωματικοί νανοσωλήνες έχουν διάμετρο περί του 1 νανομέτρου (υπενθυμίζεται ότι 1nm = 10 Å).
.") 1 2 Οι περισσότεροι μονοτοιχωματικοί νανοσωλήνες έχουν διάμετρο περί του 1 νανομέτρου (υπενθυμίζεται ότι 1nm = 10 Å). Οι πολυτοιχωματικοί νανοσωλήνες άνθρακα αποτελούνται από δύο ή περισσότερους ομοαξονικούς
1 2 Οι περισσότεροι μονοτοιχωματικοί νανοσωλήνες έχουν διάμετρο περί του 1 νανομέτρου (υπενθυμίζεται ότι 1nm = 10 Å). Οι πολυτοιχωματικοί νανοσωλήνες άνθρακα αποτελούνται από δύο ή περισσότερους ομοαξονικούς
Nανοσωλήνες άνθρακα. Ηλεκτρονική δομή ηλεκτρικές ιδιότητες. Εφαρμογές στα ηλεκτρονικά
 Nανοσωλήνες άνθρακα Ηλεκτρονική δομή ηλεκτρικές ιδιότητες Εφαρμογές στα ηλεκτρονικά Νανοσωλήνες άνθρακα ιστορική αναδρομή Από το γραφίτη στους Νανοσωλήνες άνθρακα Στο γραφίτη τα άτομα C συνδέονται ισχυρά
Nανοσωλήνες άνθρακα Ηλεκτρονική δομή ηλεκτρικές ιδιότητες Εφαρμογές στα ηλεκτρονικά Νανοσωλήνες άνθρακα ιστορική αναδρομή Από το γραφίτη στους Νανοσωλήνες άνθρακα Στο γραφίτη τα άτομα C συνδέονται ισχυρά
ΣΥΝΘΕΣΗ ΝΑΝΟΣΩΛΗΝΩΝ ΑΝΘΡΑΚΑ ΜΕΣΩ ΘΕΡΜΟΛΥΣΗΣ ΟΡΓΑΜΟΜΕΤΑΛΛΙΚΗΣ ΕΝΩΣΗΣ ΣΕ ΣΤΕΡΕΑ ΚΑΤΑΣΤΑΣΗ
 ΣΥΝΘΕΣΗ ΝΑΝΟΣΩΛΗΝΩΝ ΑΝΘΡΑΚΑ ΜΕΣΩ ΘΕΡΜΟΛΥΣΗΣ ΟΡΓΑΜΟΜΕΤΑΛΛΙΚΗΣ ΕΝΩΣΗΣ ΣΕ ΣΤΕΡΕΑ ΚΑΤΑΣΤΑΣΗ Α.Μ. Νέτσου 1, Ε. Χουντουλέση 1, Μ.Περράκη 2, Α.Ντζιούνη 1, Κ. Κορδάτος 1 1 Σχολή Χημικών Μηχανικών, ΕΜΠ 2 Σχολή
ΣΥΝΘΕΣΗ ΝΑΝΟΣΩΛΗΝΩΝ ΑΝΘΡΑΚΑ ΜΕΣΩ ΘΕΡΜΟΛΥΣΗΣ ΟΡΓΑΜΟΜΕΤΑΛΛΙΚΗΣ ΕΝΩΣΗΣ ΣΕ ΣΤΕΡΕΑ ΚΑΤΑΣΤΑΣΗ Α.Μ. Νέτσου 1, Ε. Χουντουλέση 1, Μ.Περράκη 2, Α.Ντζιούνη 1, Κ. Κορδάτος 1 1 Σχολή Χημικών Μηχανικών, ΕΜΠ 2 Σχολή
Κεφάλαιο 2 Χημικοί Δεσμοί
 Κεφάλαιο 2 Χημικοί Δεσμοί Σύνοψη Παρουσιάζονται οι χημικοί δεσμοί, ιοντικός, μοριακός, ατομικός, μεταλλικός. Οι ιδιότητες των υλικών τόσο οι φυσικές όσο και οι χημικές εξαρτώνται από το είδος ή τα είδη
Κεφάλαιο 2 Χημικοί Δεσμοί Σύνοψη Παρουσιάζονται οι χημικοί δεσμοί, ιοντικός, μοριακός, ατομικός, μεταλλικός. Οι ιδιότητες των υλικών τόσο οι φυσικές όσο και οι χημικές εξαρτώνται από το είδος ή τα είδη
ΚΑΤΑΛΥΤΙΚΆ ΥΛΙΚΆ. 1. Η Δομή των Στερεών Καταλυτών. 2. Παρασκευή μη Στηριγμένων Καταλυτών
 ΚΑΤΑΛΥΤΙΚΆ ΥΛΙΚΆ 1. Η Δομή των Στερεών Καταλυτών 2. Παρασκευή μη Στηριγμένων Καταλυτών Οργάνωση της στερεάς ύλης Άτομα-Ιόντα Μόρια (Διαστάσεις στην περιοχή των Å) Συγκροτήματα ατόμων-ιόντων-μορίων / κρυσταλλικά
ΚΑΤΑΛΥΤΙΚΆ ΥΛΙΚΆ 1. Η Δομή των Στερεών Καταλυτών 2. Παρασκευή μη Στηριγμένων Καταλυτών Οργάνωση της στερεάς ύλης Άτομα-Ιόντα Μόρια (Διαστάσεις στην περιοχή των Å) Συγκροτήματα ατόμων-ιόντων-μορίων / κρυσταλλικά
ΧΗΜΕΙΑ ΑΜΕΤΑΛΛΩΝ «ΑΕΡΕΣ», «ΑΝΘΡΑΚΑΣ
 ΧΗΜΕΙΑ ΑΜΕΤΑΛΛΩΝ «ΑΕΡΕΣ», «ΑΝΘΡΑΚΑΣ Ο ΘΗΣΑΥΡΟΣ!!!!!!!!!!!!!!!!!!!!!!!!!!!!» Η ομάδα του άνθρακα Ξυλάνθρακας, αναγωγή μεταλλευμάτων Αιθάλη, παραγωγή μελάνης Αύξηση μεταλλικού χαρακτήρα από πάνω προς τα
ΧΗΜΕΙΑ ΑΜΕΤΑΛΛΩΝ «ΑΕΡΕΣ», «ΑΝΘΡΑΚΑΣ Ο ΘΗΣΑΥΡΟΣ!!!!!!!!!!!!!!!!!!!!!!!!!!!!» Η ομάδα του άνθρακα Ξυλάνθρακας, αναγωγή μεταλλευμάτων Αιθάλη, παραγωγή μελάνης Αύξηση μεταλλικού χαρακτήρα από πάνω προς τα
Τίτλος: ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ: Κατασκευάζοντας ένα Μοντέλο Φουλερενίου Θέματα: ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ: Κατασκευάζοντας ένα Μοντέλο Φουλερενίου
 Τίτλος: ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ: Κατασκευάζοντας ένα Μοντέλο Φουλερενίου Θέματα: ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ: Κατασκευάζοντας ένα Μοντέλο Φουλερενίου Χρόνος: 90 λεπτά (2 μαθήματα) Ηλικία: Α' Λυκείου 15 16 χρονών Διαφοροποίηση:
Τίτλος: ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ: Κατασκευάζοντας ένα Μοντέλο Φουλερενίου Θέματα: ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ: Κατασκευάζοντας ένα Μοντέλο Φουλερενίου Χρόνος: 90 λεπτά (2 μαθήματα) Ηλικία: Α' Λυκείου 15 16 χρονών Διαφοροποίηση:
ΚΕΦΑΛΑΙΟ ΕΚΤΟ ΤΕΧΝΟΛΟΓΙΚΕΣ ΔΙΕΡΓΑΣΙΕΣ ΣΤΕΡΕΑΣ ΚΑΤΑΣΤΑΣΗΣ. Περιληπτική θεωρητική εισαγωγή
 ΚΕΦΑΛΑΙΟ ΕΚΤΟ ΤΕΧΝΟΛΟΓΙΚΕΣ ΔΙΕΡΓΑΣΙΕΣ ΣΤΕΡΕΑΣ ΚΑΤΑΣΤΑΣΗΣ Περιληπτική θεωρητική εισαγωγή α) Τεχνική zchralski Η πιο συχνά χρησιμοποιούμενη τεχνική ανάπτυξης μονοκρυστάλλων πυριτίου (i), αρίστης ποιότητας,
ΚΕΦΑΛΑΙΟ ΕΚΤΟ ΤΕΧΝΟΛΟΓΙΚΕΣ ΔΙΕΡΓΑΣΙΕΣ ΣΤΕΡΕΑΣ ΚΑΤΑΣΤΑΣΗΣ Περιληπτική θεωρητική εισαγωγή α) Τεχνική zchralski Η πιο συχνά χρησιμοποιούμενη τεχνική ανάπτυξης μονοκρυστάλλων πυριτίου (i), αρίστης ποιότητας,
Νανο-τεχνολογία. Νανο-Επιστήμη. Προσέγγιση από κάτω προς τα πάνω
 Νανο-τεχνολογία Ο σχεδιασμός, ο χαρακτηρισμός, η παραγωγή και η εφαρμογή των δομών, συσκευών και συστημάτων, ελέγχοντας τη μορφή και το μέγεθος σε κλίμακα νανόμετρου Νανο-Επιστήμη Η μελέτη των φαινομένων
Νανο-τεχνολογία Ο σχεδιασμός, ο χαρακτηρισμός, η παραγωγή και η εφαρμογή των δομών, συσκευών και συστημάτων, ελέγχοντας τη μορφή και το μέγεθος σε κλίμακα νανόμετρου Νανο-Επιστήμη Η μελέτη των φαινομένων
Τμήμα Τεχνολογίας Τροφίμων. Ανόργανη Χημεία. Ενότητα 8 η : Υγρά, Στερεά & Αλλαγή Φάσεων. Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής.
 Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 8 η : Υγρά, Στερεά & Αλλαγή Φάσεων Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Πολικοί Ομοιοπολικοί Δεσμοί & Διπολικές Ροπές 2 Όπως έχει
Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 8 η : Υγρά, Στερεά & Αλλαγή Φάσεων Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Πολικοί Ομοιοπολικοί Δεσμοί & Διπολικές Ροπές 2 Όπως έχει
Χημικές αντιδράσεις καταλυμένες από στερεούς καταλύτες
 Χημικές αντιδράσεις καταλυμένες από στερεούς καταλύτες Σε πολλές χημικές αντιδράσεις, οι ταχύτητές τους επηρεάζονται από κάποια συστατικά τα οποία δεν είναι ούτε αντιδρώντα ούτε προϊόντα. Αυτά τα υλικά
Χημικές αντιδράσεις καταλυμένες από στερεούς καταλύτες Σε πολλές χημικές αντιδράσεις, οι ταχύτητές τους επηρεάζονται από κάποια συστατικά τα οποία δεν είναι ούτε αντιδρώντα ούτε προϊόντα. Αυτά τα υλικά
Χαρακτηρισμός και μοντέλα τρανζίστορ λεπτών υμενίων βιομηχανικής παραγωγής: Τεχνολογία μικροκρυσταλλικού πυριτίου χαμηλής θερμοκρασίας
 Χαρακτηρισμός και μοντέλα τρανζίστορ λεπτών υμενίων βιομηχανικής παραγωγής: Τεχνολογία μικροκρυσταλλικού πυριτίου χαμηλής θερμοκρασίας Υποψήφιος Διδάκτορας: Α. Χατζόπουλος Περίληψη Οι τελευταίες εξελίξεις
Χαρακτηρισμός και μοντέλα τρανζίστορ λεπτών υμενίων βιομηχανικής παραγωγής: Τεχνολογία μικροκρυσταλλικού πυριτίου χαμηλής θερμοκρασίας Υποψήφιος Διδάκτορας: Α. Χατζόπουλος Περίληψη Οι τελευταίες εξελίξεις
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ
 ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
Αγωγιμότητα στα μέταλλα
 Η κίνηση των ατόμων σε κρυσταλλικό στερεό Θερμοκρασία 0 Θερμοκρασία 0 Δ. Γ. Παπαγεωργίου Τμήμα Μηχανικών Επιστήμης Υλικών Πανεπιστήμιο Ιωαννίνων dpapageo@cc.uoi.gr http://pc164.materials.uoi.gr/dpapageo
Η κίνηση των ατόμων σε κρυσταλλικό στερεό Θερμοκρασία 0 Θερμοκρασία 0 Δ. Γ. Παπαγεωργίου Τμήμα Μηχανικών Επιστήμης Υλικών Πανεπιστήμιο Ιωαννίνων dpapageo@cc.uoi.gr http://pc164.materials.uoi.gr/dpapageo
ΤΕΧΝΙΚΑ ΠΡΟΒΛΗΜΑΤΑ ΚΑΤΑ ΤΗΝ ΑΞΙΟΠΟΙΗΣΗ ΤΗΣ ΓΕΩΘΕΡΜΙΑΣ
 ΤΕΧΝΙΚΑ ΠΡΟΒΛΗΜΑΤΑ ΚΑΤΑ ΤΗΝ ΑΞΙΟΠΟΙΗΣΗ ΤΗΣ ΓΕΩΘΕΡΜΙΑΣ Η αξιοποίηση της γεωθερμικής ενέργειας συναντά ορισμένα τεχνικά προβλήματα, Τα προβλήματα αυτά είναι: (α) ο σχηματισμός επικαθίσεων (ή καθαλατώσεις
ΤΕΧΝΙΚΑ ΠΡΟΒΛΗΜΑΤΑ ΚΑΤΑ ΤΗΝ ΑΞΙΟΠΟΙΗΣΗ ΤΗΣ ΓΕΩΘΕΡΜΙΑΣ Η αξιοποίηση της γεωθερμικής ενέργειας συναντά ορισμένα τεχνικά προβλήματα, Τα προβλήματα αυτά είναι: (α) ο σχηματισμός επικαθίσεων (ή καθαλατώσεις
Από πού προέρχεται η θερμότητα που μεταφέρεται από τον αντιστάτη στο περιβάλλον;
 3. ΗΛΕΚΤΡΙΚΗ ΕΝΕΡΓΕΙΑ Ένα ανοικτό ηλεκτρικό κύκλωμα μετατρέπεται σε κλειστό, οπότε διέρχεται από αυτό ηλεκτρικό ρεύμα που μεταφέρει ενέργεια. Τα σπουδαιότερα χαρακτηριστικά της ηλεκτρικής ενέργειας είναι
3. ΗΛΕΚΤΡΙΚΗ ΕΝΕΡΓΕΙΑ Ένα ανοικτό ηλεκτρικό κύκλωμα μετατρέπεται σε κλειστό, οπότε διέρχεται από αυτό ηλεκτρικό ρεύμα που μεταφέρει ενέργεια. Τα σπουδαιότερα χαρακτηριστικά της ηλεκτρικής ενέργειας είναι
Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά
 Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά Είναι απλή πολυπεπτιδική αλυσίδα 307 αμινοξέων Είναι συμπαγής και έχει σχήμα ελλειψοειδές διαστάσεων 50 x 42 x 38 A Περιέχει περιοχές α-έλικος 38%
Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά Είναι απλή πολυπεπτιδική αλυσίδα 307 αμινοξέων Είναι συμπαγής και έχει σχήμα ελλειψοειδές διαστάσεων 50 x 42 x 38 A Περιέχει περιοχές α-έλικος 38%
Νανοτεχνολογία ΧΑΡΙΣ ΑΣΗΜΑΚΟΠΟΥΛΟΣ ΕΦΑΡΜΟΓΕΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 2013 14
 Νανοτεχνολογία 1.Ορισμός της Νανοτεχνολογίας 2.Πότε έκανε την εμφάνιση της; 3.Τα επιτευγματά της 4.Σε τι ωφελει την ανθρωπότητα 5.Που τη συναντάμε στη φύση 6.Κριτικές 7.Τι περιμένουμε απο την εξέλιξη της
Νανοτεχνολογία 1.Ορισμός της Νανοτεχνολογίας 2.Πότε έκανε την εμφάνιση της; 3.Τα επιτευγματά της 4.Σε τι ωφελει την ανθρωπότητα 5.Που τη συναντάμε στη φύση 6.Κριτικές 7.Τι περιμένουμε απο την εξέλιξη της
Αγωγιμότητα στα μέταλλα
 Η κίνηση των ατόμων σε κρυσταλλικό στερεό Θερμοκρασία 0 Θερμοκρασία 0 Δ. Γ. Παπαγεωργίου Τμήμα Μηχανικών Επιστήμης Υλικών Πανεπιστήμιο Ιωαννίνων dpapageo@cc.uoi.gr http://pc164.materials.uoi.gr/dpapageo
Η κίνηση των ατόμων σε κρυσταλλικό στερεό Θερμοκρασία 0 Θερμοκρασία 0 Δ. Γ. Παπαγεωργίου Τμήμα Μηχανικών Επιστήμης Υλικών Πανεπιστήμιο Ιωαννίνων dpapageo@cc.uoi.gr http://pc164.materials.uoi.gr/dpapageo
Τμήμα Τεχνολογίας Τροφίμων. Ανόργανη Χημεία. Ενότητα 10 η : Χημική κινητική. Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής.
 Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 10 η : Χημική κινητική Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ταχύτητες Αντίδρασης 2 Ως ταχύτητα αντίδρασης ορίζεται είτε η αύξηση
Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 10 η : Χημική κινητική Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ταχύτητες Αντίδρασης 2 Ως ταχύτητα αντίδρασης ορίζεται είτε η αύξηση
ΘΕΩΡΗΤΙΚΟ ΜΕΡΟΣ. Εργαστήριο Φυσικής IΙ. Μελέτη της απόδοσης φωτοβολταϊκού στοιχείου με χρήση υπολογιστή. 1. Σκοπός. 2. Σύντομο θεωρητικό μέρος
 ΘΕΩΡΗΤΙΚΟ ΜΕΡΟΣ 1. Σκοπός Το φωτοβολταϊκό στοιχείο είναι μία διάταξη ημιαγωγών η οποία μετατρέπει την φωτεινή ενέργεια που προσπίπτει σε αυτήν σε ηλεκτρική.. Όταν αυτή φωτιστεί με φωτόνια κατάλληλης συχνότητας
ΘΕΩΡΗΤΙΚΟ ΜΕΡΟΣ 1. Σκοπός Το φωτοβολταϊκό στοιχείο είναι μία διάταξη ημιαγωγών η οποία μετατρέπει την φωτεινή ενέργεια που προσπίπτει σε αυτήν σε ηλεκτρική.. Όταν αυτή φωτιστεί με φωτόνια κατάλληλης συχνότητας
 Πανεπιστήμιο Πατρών Πολυτεχνική σχολή Τμήμα Χημικών Μηχανικών Ακαδημαϊκό Έτος 2007-20082008 Μάθημα: Οικονομία Περιβάλλοντος για Οικονομολόγους Διδάσκων:Σκούρας Δημήτριος ΚΑΤΑΛΥΤΙΚΗ ΑΝΤΙΔΡΑΣΗ ΠΑΡΑΓΩΓΗΣ
Πανεπιστήμιο Πατρών Πολυτεχνική σχολή Τμήμα Χημικών Μηχανικών Ακαδημαϊκό Έτος 2007-20082008 Μάθημα: Οικονομία Περιβάλλοντος για Οικονομολόγους Διδάσκων:Σκούρας Δημήτριος ΚΑΤΑΛΥΤΙΚΗ ΑΝΤΙΔΡΑΣΗ ΠΑΡΑΓΩΓΗΣ
ΠΡΩΤΕΪΝΕΣ. Φατούρος Ιωάννης Αναπληρωτής Καθηγητής
 ΠΡΩΤΕΪΝΕΣ Φατούρος Ιωάννης Αναπληρωτής Καθηγητής Θέματα Διάλεξης Δομή, αριθμός και διαχωρισμός των αμινοξέων Ένωση αμινοξέων με τον πεπτιδικό δεσμό για τη δημιουργία πρωτεΐνης Λειτουργίες των πρωτεϊνών
ΠΡΩΤΕΪΝΕΣ Φατούρος Ιωάννης Αναπληρωτής Καθηγητής Θέματα Διάλεξης Δομή, αριθμός και διαχωρισμός των αμινοξέων Ένωση αμινοξέων με τον πεπτιδικό δεσμό για τη δημιουργία πρωτεΐνης Λειτουργίες των πρωτεϊνών
ΘΕΜΑΤΑ ΤΕΛΙΚΩΝ ΕΞΕΤΑΣΕΩΝ ΣΤΗ ΦΥΣΙΚΟΧΗΜΕΙΑ (Α. Χημική Θερμοδυναμική) 1 η Άσκηση
 1 η Άσκηση") ΘΕΜΑΤΑ ΤΕΛΙΚΩΝ ΕΞΕΤΑΣΕΩΝ ΣΤΗ ΦΥΣΙΚΟΧΗΜΕΙΑ 004-05 (Α. Χημική Θερμοδυναμική) η Άσκηση Στερεό CO, βάρους 6 g, εισάγεται μέσα σε κενό δοχείο όγκου 00 cm 3 που βρίσκεται συνεχώς σε θερμοκρασία δωματίου (300
ΘΕΜΑΤΑ ΤΕΛΙΚΩΝ ΕΞΕΤΑΣΕΩΝ ΣΤΗ ΦΥΣΙΚΟΧΗΜΕΙΑ 004-05 (Α. Χημική Θερμοδυναμική) η Άσκηση Στερεό CO, βάρους 6 g, εισάγεται μέσα σε κενό δοχείο όγκου 00 cm 3 που βρίσκεται συνεχώς σε θερμοκρασία δωματίου (300
Ένωση Ελλήνων Φυσικών ΠΑΝΕΛΛΗΝΙΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΦΥΣΙΚΗΣ B Λυκείου
 Θεωρητικό Μέρος B Λυκείου 21 Απριλίου 2007 Θέμα 1 ο 1. Στο παρακάτω σχήμα φαίνονται οι δυναμικές γραμμές του ηλεκτρικού πεδίου το οποίο δημιουργείται μεταξύ δύο αντίθετων ηλεκτρικών φορτίων. Ένα ηλεκτρόνιο
Θεωρητικό Μέρος B Λυκείου 21 Απριλίου 2007 Θέμα 1 ο 1. Στο παρακάτω σχήμα φαίνονται οι δυναμικές γραμμές του ηλεκτρικού πεδίου το οποίο δημιουργείται μεταξύ δύο αντίθετων ηλεκτρικών φορτίων. Ένα ηλεκτρόνιο
ΔΟΜΗ ΚΑΙ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΚΕΡΑΜΙΚΩΝ. Χ. Κορδούλης
 ΔΟΜΗ ΚΑΙ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΚΕΡΑΜΙΚΩΝ Χ. Κορδούλης ΚΕΡΑΜΙΚΑ ΥΛΙΚΑ Τα κεραμικά υλικά είναι ανόργανα µη μεταλλικά υλικά (ενώσεις μεταλλικών και μη μεταλλικών στοιχείων), τα οποία έχουν υποστεί θερμική κατεργασία
ΔΟΜΗ ΚΑΙ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΚΕΡΑΜΙΚΩΝ Χ. Κορδούλης ΚΕΡΑΜΙΚΑ ΥΛΙΚΑ Τα κεραμικά υλικά είναι ανόργανα µη μεταλλικά υλικά (ενώσεις μεταλλικών και μη μεταλλικών στοιχείων), τα οποία έχουν υποστεί θερμική κατεργασία
Ε. Μαλαμίδου-Ξενικάκη
 Ε. Μαλαμίδου-Ξενικάκη Θεσσαλονίκη 2015 ΑΛΚΥΝΙΑ: C ν H 2ν-2 Ο τριπλός δεσμός άνθρακα άνθρακα Τριπλός δεσμός αλκυνίου ΑΛΚΥΝΙΑ Μόρια πρότυπα για «μοριακούς διακόπτες» Μικροσκοπία σάρωσης σήραγγας (Scanning
Ε. Μαλαμίδου-Ξενικάκη Θεσσαλονίκη 2015 ΑΛΚΥΝΙΑ: C ν H 2ν-2 Ο τριπλός δεσμός άνθρακα άνθρακα Τριπλός δεσμός αλκυνίου ΑΛΚΥΝΙΑ Μόρια πρότυπα για «μοριακούς διακόπτες» Μικροσκοπία σάρωσης σήραγγας (Scanning
ΚΕΦΑΛΑΙΟ 2 Ο H XHΜΕΙΑ ΤΗΣ ΖΩΗΣ. Χημεία της ζωής 1
 ΚΕΦΑΛΑΙΟ 2 Ο H XHΜΕΙΑ ΤΗΣ ΖΩΗΣ Χημεία της ζωής 1 2.1 ΒΑΣΙΚΕΣ ΧΗΜΙΚΕΣ ΕΝΝΟΙΕΣ Η Βιολογία μπορεί να μελετηθεί μέσα από πολλά και διαφορετικά επίπεδα. Οι βιοχημικοί, για παράδειγμα, ενδιαφέρονται περισσότερο
ΚΕΦΑΛΑΙΟ 2 Ο H XHΜΕΙΑ ΤΗΣ ΖΩΗΣ Χημεία της ζωής 1 2.1 ΒΑΣΙΚΕΣ ΧΗΜΙΚΕΣ ΕΝΝΟΙΕΣ Η Βιολογία μπορεί να μελετηθεί μέσα από πολλά και διαφορετικά επίπεδα. Οι βιοχημικοί, για παράδειγμα, ενδιαφέρονται περισσότερο
Μερικά χαρακτηριστικά του ενεργού κέντρου των ενζύμων
 Μερικά χαρακτηριστικά του ενεργού κέντρου των ενζύμων Το ενεργό κέντρο καταλαμβάνει σχετικά μικρό τμήμα του ολικού όγκου του ενζύμου Το ενεργό κέντρο είναι μια τρισδιάστατη ολότητα Η ειδικότητα δέσμευσης
Μερικά χαρακτηριστικά του ενεργού κέντρου των ενζύμων Το ενεργό κέντρο καταλαμβάνει σχετικά μικρό τμήμα του ολικού όγκου του ενζύμου Το ενεργό κέντρο είναι μια τρισδιάστατη ολότητα Η ειδικότητα δέσμευσης
Οργανική Χημεία. Κεφάλαια 12 &13: Φασματοσκοπία μαζών και υπερύθρου
 Οργανική Χημεία Κεφάλαια 12 &13: Φασματοσκοπία μαζών και υπερύθρου 1. Γενικά Δυνατότητα προσδιορισμού δομών με σαφήνεια χρησιμοποιώντας τεχνικές φασματοσκοπίας Φασματοσκοπία μαζών Μέγεθος, μοριακός τύπος
Οργανική Χημεία Κεφάλαια 12 &13: Φασματοσκοπία μαζών και υπερύθρου 1. Γενικά Δυνατότητα προσδιορισμού δομών με σαφήνεια χρησιμοποιώντας τεχνικές φασματοσκοπίας Φασματοσκοπία μαζών Μέγεθος, μοριακός τύπος
Α Ε Τ. ΤΕΙ Αθήνας. Στ. Μπογιατζής, επίκουρος καθηγητής ΤΕΙ Αθήνας. ΤΕΙ Αθήνας / ΣΑΕΤ / Στ. Μπογιατζής
 Στ. Μπογιατζής, επίκουρος καθηγητής Ομοιοπολικές χημικές ενώσεις Ενώσεις του άνθρακα Χαρακτηριστικές ομάδες Τετραεδρική μοριακή δομή Επίπεδη τριγωνική μοριακή δομή Ευθύγραμμη μοριακή δομή Τα οργανικά μόρια
Στ. Μπογιατζής, επίκουρος καθηγητής Ομοιοπολικές χημικές ενώσεις Ενώσεις του άνθρακα Χαρακτηριστικές ομάδες Τετραεδρική μοριακή δομή Επίπεδη τριγωνική μοριακή δομή Ευθύγραμμη μοριακή δομή Τα οργανικά μόρια
ΦΥΣΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ. Οι φυσικές καταστάσεις της ύλης είναι η στερεή, η υγρή και η αέρια.
 ΦΥΣΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ Οι φυσικές καταστάσεις της ύλης είναι η στερεή, η υγρή και η αέρια. Οι μεταξύ τους μεταβολές εξαρτώνται από τη θερμοκρασία και την πίεση και είναι οι παρακάτω: ΣΗΜΕΙΟ ΤΗΞΗΣ ΚΑΙ ΣΗΜΕΙΟ
ΦΥΣΙΚΕΣ ΚΑΤΑΣΤΑΣΕΙΣ Οι φυσικές καταστάσεις της ύλης είναι η στερεή, η υγρή και η αέρια. Οι μεταξύ τους μεταβολές εξαρτώνται από τη θερμοκρασία και την πίεση και είναι οι παρακάτω: ΣΗΜΕΙΟ ΤΗΞΗΣ ΚΑΙ ΣΗΜΕΙΟ
ΕΝΖΥΜΙΚΕΣ ΑΝΤΙΔΡΑΣΕΙΣ ΣΕ ΕΤΕΡΟΓΕΝΗ ΣΥΣΤΗΜΑΤΑ
 ΕΝΖΥΜΙΚΕΣ ΑΝΤΙΔΡΑΣΕΙΣ ΣΕ ΕΤΕΡΟΓΕΝΗ ΣΥΣΤΗΜΑΤΑ ΚΙΝΗΤΙΚΗ ΕΝΖΥΜΩΝ ΣΕ ΔΙΑΛΥΜΑ ΕΠΕΝΕΡΓΟΥΝΤΩΝ ΣΕ ΑΔΙΑΛΥΤΑ ΥΠΟΣΤΡΩΜΑΤΑ το υπόστρωμα σε στερεά (αδιάλυτη) μορφή κλασσική περίπτωση: η υδρόλυση αδιάλυτων πολυμερών
ΕΝΖΥΜΙΚΕΣ ΑΝΤΙΔΡΑΣΕΙΣ ΣΕ ΕΤΕΡΟΓΕΝΗ ΣΥΣΤΗΜΑΤΑ ΚΙΝΗΤΙΚΗ ΕΝΖΥΜΩΝ ΣΕ ΔΙΑΛΥΜΑ ΕΠΕΝΕΡΓΟΥΝΤΩΝ ΣΕ ΑΔΙΑΛΥΤΑ ΥΠΟΣΤΡΩΜΑΤΑ το υπόστρωμα σε στερεά (αδιάλυτη) μορφή κλασσική περίπτωση: η υδρόλυση αδιάλυτων πολυμερών
+ ^ + + Ε + Υ-ΕΥ-Π + Π + Ε. 8. Σημειώστε σωστό ή λάθος σε καθεμιά από τις απαντήσεις που ακολουθούν.
 Υποενότητα 3.2 1. Τι είναι ενέργεια ενεργοποίησης και ποια η σχέση ενός ενζύμου με αυτήν. Η απάντηση βρίσκεται στην παράγραφο του σχολικού βιβλίου «Μηχανισμός δράσης των ενζύμων». 2.0 τρόπος δράσης των
Υποενότητα 3.2 1. Τι είναι ενέργεια ενεργοποίησης και ποια η σχέση ενός ενζύμου με αυτήν. Η απάντηση βρίσκεται στην παράγραφο του σχολικού βιβλίου «Μηχανισμός δράσης των ενζύμων». 2.0 τρόπος δράσης των
Υλικά Ηλεκτρονικής & Διατάξεις
 Τμήμα Ηλεκτρονικών Μηχανικών Υλικά Ηλεκτρονικής & Διατάξεις 3 η σειρά διαφανειών Δημήτριος Λαμπάκης Τύποι Στερεών Βασική Ερώτηση: Πως τα άτομα διατάσσονται στο χώρο ώστε να σχηματίσουν στερεά? Τύποι Στερεών
Τμήμα Ηλεκτρονικών Μηχανικών Υλικά Ηλεκτρονικής & Διατάξεις 3 η σειρά διαφανειών Δημήτριος Λαμπάκης Τύποι Στερεών Βασική Ερώτηση: Πως τα άτομα διατάσσονται στο χώρο ώστε να σχηματίσουν στερεά? Τύποι Στερεών
ΠΕΤΡΕΛΑΙΟ ΥΔΡΟΓΟΝΑΝΘΡΑΚΕΣ ΚΑΥΣΗ και ΚΑΥΣΙΜΑ
 ΠΕΤΡΕΛΑΙΟ ΥΔΡΟΓΟΝΑΝΘΡΑΚΕΣ ΚΑΥΣΗ και ΚΑΥΣΙΜΑ Καύση ονομάζεται η αντίδραση μιας οργανικής ή ανόργανης ουσίας με το Ο 2, κατά την οποία εκλύεται θερμότητα στο περιβάλλον και παράγεται φως. Είδη καύσης Α.
ΠΕΤΡΕΛΑΙΟ ΥΔΡΟΓΟΝΑΝΘΡΑΚΕΣ ΚΑΥΣΗ και ΚΑΥΣΙΜΑ Καύση ονομάζεται η αντίδραση μιας οργανικής ή ανόργανης ουσίας με το Ο 2, κατά την οποία εκλύεται θερμότητα στο περιβάλλον και παράγεται φως. Είδη καύσης Α.
Θεωρία Μοριακών Τροχιακών (ΜΟ)
") Θεωρία Μοριακών Τροχιακών (ΜΟ) Ετεροπυρηνικά διατομικά μόρια ή ιόντα (πολικοί δεσμοί) Το πιο ηλεκτραρνητικό στοιχείο (με ατομικά τροχιακά χαμηλότερης ενεργειακής στάθμης) συνεισφέρει περισσότερο στο δεσμικό
Θεωρία Μοριακών Τροχιακών (ΜΟ) Ετεροπυρηνικά διατομικά μόρια ή ιόντα (πολικοί δεσμοί) Το πιο ηλεκτραρνητικό στοιχείο (με ατομικά τροχιακά χαμηλότερης ενεργειακής στάθμης) συνεισφέρει περισσότερο στο δεσμικό
Επιφανειακή οξεοβασική κατάλυση
 Επιφανειακή οξεοβασική κατάλυση Μια μεγάλη κατηγορία στερεών καταλυτών εκδηλώνουν επιφανειακή οξεοβασική συμπεριφορά γ-αl 2 O 3, SiO 2, TiO 2, MgO, SiO 2 -Al 2 O 3, ζεόλιθοι Στην επιφάνεια τέτοιων οξειδίων
Επιφανειακή οξεοβασική κατάλυση Μια μεγάλη κατηγορία στερεών καταλυτών εκδηλώνουν επιφανειακή οξεοβασική συμπεριφορά γ-αl 2 O 3, SiO 2, TiO 2, MgO, SiO 2 -Al 2 O 3, ζεόλιθοι Στην επιφάνεια τέτοιων οξειδίων
ΝΑΝΟΥΛΙΚΑ ΚΑΙ ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ ΣΤΕΛΛΑ ΚΕΝΝΟΥ ΚΑΘΗΓΗΤΡΙΑ
 ΣΤΕΛΛΑ ΚΕΝΝΟΥ ΚΑΘΗΓΗΤΡΙΑ 1 Ιδιότητες εξαρτώμενες από το μέγεθος Στην νανοκλίμακα, οι ιδιότητες εξαρτώνται δραματικά από το μέγεθος Για παράδειγμα, ΙΔΙΟΤΗΤΕΣ ΝΑΝΟΥΛΙΚΩΝ (1) Θερμικές ιδιότητες θερμοκρασία
ΣΤΕΛΛΑ ΚΕΝΝΟΥ ΚΑΘΗΓΗΤΡΙΑ 1 Ιδιότητες εξαρτώμενες από το μέγεθος Στην νανοκλίμακα, οι ιδιότητες εξαρτώνται δραματικά από το μέγεθος Για παράδειγμα, ΙΔΙΟΤΗΤΕΣ ΝΑΝΟΥΛΙΚΩΝ (1) Θερμικές ιδιότητες θερμοκρασία
ΕΙΣΑΓΩΓΗ ΣΤΗ ΜΕΤΑΔΟΣΗ ΘΕΡΜΟΤΗΤΑΣ
 ΕΙΣΑΓΩΓΗ ΣΤΗ ΜΕΤΑΔΟΣΗ ΘΕΡΜΟΤΗΤΑΣ Η Επιστήμη της Θερμοδυναμικής ασχολείται με την ποσότητα της θερμότητας που μεταφέρεται σε ένα κλειστό και απομονωμένο σύστημα από μια κατάσταση ισορροπίας σε μια άλλη
ΕΙΣΑΓΩΓΗ ΣΤΗ ΜΕΤΑΔΟΣΗ ΘΕΡΜΟΤΗΤΑΣ Η Επιστήμη της Θερμοδυναμικής ασχολείται με την ποσότητα της θερμότητας που μεταφέρεται σε ένα κλειστό και απομονωμένο σύστημα από μια κατάσταση ισορροπίας σε μια άλλη
Καταστάσεις της ύλης. Αέρια: Παντελής απουσία τάξεως. Τα µόρια βρίσκονται σε συνεχή τυχαία κίνηση σε σχεδόν κενό χώρο.
 Καταστάσεις της ύλης Αέρια: Παντελής απουσία τάξεως. Τα µόρια βρίσκονται σε συνεχή τυχαία κίνηση σε σχεδόν κενό χώρο. Υγρά: Τάξη πολύ µικρού βαθµού και κλίµακας-ελκτικές δυνάµεις-ολίσθηση. Τα µόρια βρίσκονται
Καταστάσεις της ύλης Αέρια: Παντελής απουσία τάξεως. Τα µόρια βρίσκονται σε συνεχή τυχαία κίνηση σε σχεδόν κενό χώρο. Υγρά: Τάξη πολύ µικρού βαθµού και κλίµακας-ελκτικές δυνάµεις-ολίσθηση. Τα µόρια βρίσκονται
ΜΙΑ ΔΙΔΑΚΤΙΚΗ ΠΕΡΙΟΔΟΣ ΤΗ ΒΔΟΜΑΔΑ ΔΙΔΑΚΤΕΑ ΥΛΗ ΣΤΟΧΟΙ ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΠΡΩΤΗ ΕΝΟΤΗΤΑ
 ΑΝΑΛΥΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΧΗΜΕΙΑΣ Γ ΤΑΞΗ ΓΥΜΝΑΣΙΟΥ ΥΠΟΥΡΓΕΙΟ ΠΑΙΔΕΙΑΣ ΚΑΙ ΠΟΛΙΤΙΣΜΟΥ ΓΡΑΦΕΙΑ ΕΠΙΘΕΩΡΗΤΩΝ ΜΕΣΗΣ ΕΚΠΑΙΔΕΥΣΗΣ ΛΕΥΚΩΣΙΑ ΣΧΟΛΙΚΗ ΧΡΟΝΙΑ 2007-2008 ΜΙΑ ΔΙΔΑΚΤΙΚΗ ΠΕΡΙΟΔΟΣ ΤΗ ΒΔΟΜΑΔΑ ΔΙΔΑΚΤΕΑ ΥΛΗ ΣΤΟΧΟΙ
ΑΝΑΛΥΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΧΗΜΕΙΑΣ Γ ΤΑΞΗ ΓΥΜΝΑΣΙΟΥ ΥΠΟΥΡΓΕΙΟ ΠΑΙΔΕΙΑΣ ΚΑΙ ΠΟΛΙΤΙΣΜΟΥ ΓΡΑΦΕΙΑ ΕΠΙΘΕΩΡΗΤΩΝ ΜΕΣΗΣ ΕΚΠΑΙΔΕΥΣΗΣ ΛΕΥΚΩΣΙΑ ΣΧΟΛΙΚΗ ΧΡΟΝΙΑ 2007-2008 ΜΙΑ ΔΙΔΑΚΤΙΚΗ ΠΕΡΙΟΔΟΣ ΤΗ ΒΔΟΜΑΔΑ ΔΙΔΑΚΤΕΑ ΥΛΗ ΣΤΟΧΟΙ
Ασκήσεις ακαδ. έτους
 Τμήμα Επιστήμης και Τεχνολογίας Υλικών Πανεπιστήμιο Κρήτης Επιστήμη Επιφανειών - Νανοϋλικών (ETY/METY 346) Μεταπτυχιακό: Νανοτεχνολογία για Ενεργειακές Εφαρμογές ¹ Nanomaterials for Energy (Νανοϋλικά για
Τμήμα Επιστήμης και Τεχνολογίας Υλικών Πανεπιστήμιο Κρήτης Επιστήμη Επιφανειών - Νανοϋλικών (ETY/METY 346) Μεταπτυχιακό: Νανοτεχνολογία για Ενεργειακές Εφαρμογές ¹ Nanomaterials for Energy (Νανοϋλικά για
ηλεκτρικό ρεύμα ampere
 Ηλεκτρικό ρεύμα Το ηλεκτρικό ρεύμα είναι ο ρυθμός με τον οποίο διέρχεται ηλεκτρικό φορτίο από μια περιοχή του χώρου. Η μονάδα μέτρησης του ηλεκτρικού ρεύματος στο σύστημα SI είναι το ampere (A). 1 A =
Ηλεκτρικό ρεύμα Το ηλεκτρικό ρεύμα είναι ο ρυθμός με τον οποίο διέρχεται ηλεκτρικό φορτίο από μια περιοχή του χώρου. Η μονάδα μέτρησης του ηλεκτρικού ρεύματος στο σύστημα SI είναι το ampere (A). 1 A =
ΧΗΜΕΙΑ Β ΓΥΜΝΑΣΙΟΥ ΕΝΟΤΗΤΑ: 1.2
 ΕΝΟΤΗΤΑ: 1.2 Η ύλη συναντάται σε τρεις φυσικές καταστάσεις: Στερεή: έχει καθορισμένη μάζα, σχήμα και όγκο. Υγρή: έχει καθορισμένη μάζα και όγκο, ενώ σχήμα κάθε φορά παίρνει το σχήμα του δοχείου που το
ΕΝΟΤΗΤΑ: 1.2 Η ύλη συναντάται σε τρεις φυσικές καταστάσεις: Στερεή: έχει καθορισμένη μάζα, σχήμα και όγκο. Υγρή: έχει καθορισμένη μάζα και όγκο, ενώ σχήμα κάθε φορά παίρνει το σχήμα του δοχείου που το
Θεωρία δεσµού σθένους - Υβριδισµός. Αντιδράσεις προσθήκης Αντιδράσεις απόσπασης. Αντιδράσεις υποκατάστασης Πολυµερισµός
 11 ο Μάθηµα: Θεωρία δεσµού σθένους - Υβριδισµός 12 ο Μάθηµα: Αντιδράσεις προσθήκης Αντιδράσεις απόσπασης 13 ο Μάθηµα: Αντιδράσεις υποκατάστασης Πολυµερισµός 14 ο Μάθηµα: Αντιδράσεις οξείδωσης - αναγωγής
11 ο Μάθηµα: Θεωρία δεσµού σθένους - Υβριδισµός 12 ο Μάθηµα: Αντιδράσεις προσθήκης Αντιδράσεις απόσπασης 13 ο Μάθηµα: Αντιδράσεις υποκατάστασης Πολυµερισµός 14 ο Μάθηµα: Αντιδράσεις οξείδωσης - αναγωγής
ΣΤΟΙΧΕΙΑ ΤΕΧΝΟΛΟΓΙΑΣ ΠΑΡΑΓΩΓΗΣ
 ΔΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΕΙΟ ΘΡΑΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΜΗΧΑΝΙΚΩΝ ΠΑΡΑΓΩΓΗΣ κ ΔΙΟΙΚΗΣΗΣ ΤΟΜΕΑΣ ΥΛΙΚΩΝ, ΔΙΕΡΓΑΣΙΩΝ κ ΜΗΧΑΝΟΛΟΓΙΑΣ ΣΤΟΙΧΕΙΑ ΤΕΧΝΟΛΟΓΙΑΣ ΠΑΡΑΓΩΓΗΣ Παραουσίαση μαθήματος με διαφάνειες στο
ΔΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΕΙΟ ΘΡΑΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΜΗΧΑΝΙΚΩΝ ΠΑΡΑΓΩΓΗΣ κ ΔΙΟΙΚΗΣΗΣ ΤΟΜΕΑΣ ΥΛΙΚΩΝ, ΔΙΕΡΓΑΣΙΩΝ κ ΜΗΧΑΝΟΛΟΓΙΑΣ ΣΤΟΙΧΕΙΑ ΤΕΧΝΟΛΟΓΙΑΣ ΠΑΡΑΓΩΓΗΣ Παραουσίαση μαθήματος με διαφάνειες στο
panagiotisathanasopoulos.gr
 Χημική Ισορροπία 61 Παναγιώτης Αθανασόπουλος Χημικός, Διδάκτωρ Πανεπιστημίου Πατρών Χημικός Διδάκτωρ Παν. Πατρών 62 Τι ονομάζεται κλειστό χημικό σύστημα; Παναγιώτης Αθανασόπουλος Κλειστό ονομάζεται το
Χημική Ισορροπία 61 Παναγιώτης Αθανασόπουλος Χημικός, Διδάκτωρ Πανεπιστημίου Πατρών Χημικός Διδάκτωρ Παν. Πατρών 62 Τι ονομάζεται κλειστό χημικό σύστημα; Παναγιώτης Αθανασόπουλος Κλειστό ονομάζεται το
και χρειάζεται μέσα στο ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. ενζύμων κύτταρο τρόπους
 Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
Επιστήμη των Υλικών. Πανεπιστήμιο Ιωαννίνων. Τμήμα Φυσικής
 Επιστήμη των Υλικών Πανεπιστήμιο Ιωαννίνων Τμήμα Φυσικής 2017 Α. Δούβαλης Άνθρακας και υβριδικά υλικά με βάση τον άνθρακα Graphite Diamond Fullerene Nanotube Ηλεκτρονική διαμόρφωση του C, υβριδισμός και
Επιστήμη των Υλικών Πανεπιστήμιο Ιωαννίνων Τμήμα Φυσικής 2017 Α. Δούβαλης Άνθρακας και υβριδικά υλικά με βάση τον άνθρακα Graphite Diamond Fullerene Nanotube Ηλεκτρονική διαμόρφωση του C, υβριδισμός και
κυματικής συνάρτησης (Ψ) κυματική συνάρτηση
 κυματική συνάρτηση") Στην κβαντομηχανική ο χώρος μέσα στον οποίο κινείται το ηλεκτρόνιο γύρω από τον πυρήνα παύει να περιγράφεται από μια απλή τροχιά, χαρακτηριστικό του μοντέλου του Bohr, αλλά περιγράφεται ο χώρος μέσα στον
Στην κβαντομηχανική ο χώρος μέσα στον οποίο κινείται το ηλεκτρόνιο γύρω από τον πυρήνα παύει να περιγράφεται από μια απλή τροχιά, χαρακτηριστικό του μοντέλου του Bohr, αλλά περιγράφεται ο χώρος μέσα στον
ΚΡΥΣΤΑΛΛΙΚΑ ΠΟΛΥΜΕΡΗ ΠΟΡΩΔΗ ΥΛΙΚΑ (MOFs) ΓΙΑ ΑΠΟΘΗΚΕΥΣΗ Η 2
 ΓΙΑ ΑΠΟΘΗΚΕΥΣΗ Η 2") ΚΡΥΣΤΑΛΛΙΚΑ ΠΟΛΥΜΕΡΗ ΠΟΡΩΔΗ ΥΛΙΚΑ (MOFs) ΓΙΑ ΑΠΟΘΗΚΕΥΣΗ Η 2 Επιστημονική Υπεύθυνη: Δρ.Αικατερίνη Ραπτοπούλου, Ερευνήτρια Β, ΙΕΥ Το υδρογόνο ως φορέας ενέργειας παρουσιάζει συγκεκριμένα Πλεονεκτήματα:α)
ΚΡΥΣΤΑΛΛΙΚΑ ΠΟΛΥΜΕΡΗ ΠΟΡΩΔΗ ΥΛΙΚΑ (MOFs) ΓΙΑ ΑΠΟΘΗΚΕΥΣΗ Η 2 Επιστημονική Υπεύθυνη: Δρ.Αικατερίνη Ραπτοπούλου, Ερευνήτρια Β, ΙΕΥ Το υδρογόνο ως φορέας ενέργειας παρουσιάζει συγκεκριμένα Πλεονεκτήματα:α)
Το Καφενείο της Επιστήμης (5 ος κύκλος) Ίδρυμα Ευγενίδου, Γαλλικό Ινστιτούτο, Βρετανικό Συμβούλιο
 Ίδρυμα Ευγενίδου, Γαλλικό Ινστιτούτο, Βρετανικό Συμβούλιο") Το Καφενείο της Επιστήμης (5 ος κύκλος) «Η Φωτοχημεία στην υπηρεσία της προστασίας του περιβάλλοντος: Νέα Φωτοβολταϊκά Συστήματα με χρήση καινοτόμων νανο-υλικών» Οικονομόπουλος Σόλων Ινστιτούτο Θεωρητικής
Το Καφενείο της Επιστήμης (5 ος κύκλος) «Η Φωτοχημεία στην υπηρεσία της προστασίας του περιβάλλοντος: Νέα Φωτοβολταϊκά Συστήματα με χρήση καινοτόμων νανο-υλικών» Οικονομόπουλος Σόλων Ινστιτούτο Θεωρητικής
Μάθημα 23 ο. Μεταλλικός Δεσμός Θεωρία Ζωνών- Ημιαγωγοί Διαμοριακές Δυνάμεις
 Μάθημα 23 ο Μεταλλικός Δεσμός Θεωρία Ζωνών- Ημιαγωγοί Διαμοριακές Δυνάμεις Μεταλλικός Δεσμός Μοντέλο θάλασσας ηλεκτρονίων Πυρήνες σε θάλασσα e -. Μεταλλική λάμψη. Ολκιμότητα. Εφαρμογή δύναμης Γενική και
Μάθημα 23 ο Μεταλλικός Δεσμός Θεωρία Ζωνών- Ημιαγωγοί Διαμοριακές Δυνάμεις Μεταλλικός Δεσμός Μοντέλο θάλασσας ηλεκτρονίων Πυρήνες σε θάλασσα e -. Μεταλλική λάμψη. Ολκιμότητα. Εφαρμογή δύναμης Γενική και
Τίτλος Μαθήματος: Βασικές Έννοιες Φυσικής. Ενότητα: Στερεά. Διδάσκων: Καθηγητής Κ. Κώτσης. Τμήμα: Παιδαγωγικό, Δημοτικής Εκπαίδευσης
 Τίτλος Μαθήματος: Βασικές Έννοιες Φυσικής Ενότητα: Στερεά Διδάσκων: Καθηγητής Κ. Κώτσης Τμήμα: Παιδαγωγικό, Δημοτικής Εκπαίδευσης 7. Στερεά Η επιβεβαίωση ότι τα στερεά σώματα αποτελούνται από μια ιδιαίτερη
Τίτλος Μαθήματος: Βασικές Έννοιες Φυσικής Ενότητα: Στερεά Διδάσκων: Καθηγητής Κ. Κώτσης Τμήμα: Παιδαγωγικό, Δημοτικής Εκπαίδευσης 7. Στερεά Η επιβεβαίωση ότι τα στερεά σώματα αποτελούνται από μια ιδιαίτερη
6. ΘΕΡΜΙΚΕΣ Ι ΙΟΤΗΤΕΣ ΤΩΝ ΠΟΛΥΜΕΡΩΝ
 6-1 6. ΘΕΡΜΙΚΕΣ Ι ΙΟΤΗΤΕΣ ΤΩΝ ΠΟΛΥΜΕΡΩΝ 6.1. ΙΑ ΟΣΗ ΤΗΣ ΘΕΡΜΟΤΗΤΑΣ Πολλές βιοµηχανικές εφαρµογές των πολυµερών αφορούν τη διάδοση της θερµότητας µέσα από αυτά ή γύρω από αυτά. Πολλά πολυµερή χρησιµοποιούνται
6-1 6. ΘΕΡΜΙΚΕΣ Ι ΙΟΤΗΤΕΣ ΤΩΝ ΠΟΛΥΜΕΡΩΝ 6.1. ΙΑ ΟΣΗ ΤΗΣ ΘΕΡΜΟΤΗΤΑΣ Πολλές βιοµηχανικές εφαρµογές των πολυµερών αφορούν τη διάδοση της θερµότητας µέσα από αυτά ή γύρω από αυτά. Πολλά πολυµερή χρησιµοποιούνται
ΠΕΡΙΘΛΑΣΗ ΗΛΕΚΤΡΟΝΙΩΝ
 ΠΕΡΙΘΛΑΣΗ ΗΛΕΚΤΡΟΝΙΩΝ Αποδείξαμε πειραματικά, με τη βοήθεια του φαινομένου της περίθλασης, ότι τα ηλεκτρόνια έχουν εκτός από τη σωματιδιακή και κυματική φύση. Υπολογίσαμε τις σταθερές πλέγματος του γραφίτη
ΠΕΡΙΘΛΑΣΗ ΗΛΕΚΤΡΟΝΙΩΝ Αποδείξαμε πειραματικά, με τη βοήθεια του φαινομένου της περίθλασης, ότι τα ηλεκτρόνια έχουν εκτός από τη σωματιδιακή και κυματική φύση. Υπολογίσαμε τις σταθερές πλέγματος του γραφίτη
Ανόργανη Χημεία. Τμήμα Τεχνολογίας Τροφίμων. Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή. Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής
 Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ο Ομοιοπολικός Δεσμός 2 Ο δεσμός Η Η στο μόριο Η
Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ο Ομοιοπολικός Δεσμός 2 Ο δεσμός Η Η στο μόριο Η
Εργαστήριο Τεχνολογίας Υλικών
 Εργαστήριο Τεχνολογίας Υλικών Εργαστηριακή Άσκηση 01 Κατηγοριοποιήση υλικών-επίδειξη δοκιμίων Διδάσκοντες: Δρ Γεώργιος Ι. Γιαννόπουλος Δρ Θεώνη Ασημακοπούλου Δρ ΘεόδωροςΛούτας Τμήμα Μηχανολογίας ΑΤΕΙ Πατρών
Εργαστήριο Τεχνολογίας Υλικών Εργαστηριακή Άσκηση 01 Κατηγοριοποιήση υλικών-επίδειξη δοκιμίων Διδάσκοντες: Δρ Γεώργιος Ι. Γιαννόπουλος Δρ Θεώνη Ασημακοπούλου Δρ ΘεόδωροςΛούτας Τμήμα Μηχανολογίας ΑΤΕΙ Πατρών
Βρέντζου Τίνα Φυσικός Μεταπτυχιακός τίτλος: «Σπουδές στην εκπαίδευση» ΜEd Email : stvrentzou@gmail.com
 1 2.4 Παράγοντες από τους οποίους εξαρτάται η αντίσταση ενός αγωγού Λέξεις κλειδιά: ειδική αντίσταση, μικροσκοπική ερμηνεία, μεταβλητός αντισ ροοστάτης, ποτενσιόμετρο 2.4 Παράγοντες που επηρεάζουν την
1 2.4 Παράγοντες από τους οποίους εξαρτάται η αντίσταση ενός αγωγού Λέξεις κλειδιά: ειδική αντίσταση, μικροσκοπική ερμηνεία, μεταβλητός αντισ ροοστάτης, ποτενσιόμετρο 2.4 Παράγοντες που επηρεάζουν την
Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ
 ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ") Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ Βιοενεργητική είναι ο κλάδος της Βιολογίας που μελετά τον τρόπο με τον οποίο οι οργανισμοί χρησιμοποιούν ενέργεια για να επιβιώσουν και να υλοποιήσουν τις
Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ Βιοενεργητική είναι ο κλάδος της Βιολογίας που μελετά τον τρόπο με τον οποίο οι οργανισμοί χρησιμοποιούν ενέργεια για να επιβιώσουν και να υλοποιήσουν τις
 NTSE - Nano Technology Science Education Project No: 511787-LLP-1-2010-1-TR-KA3-KA3MP ΜΕΡΟΣ I: ΜΕΛΕΤΗ ΠΡΙΝ ΤΟ ΠΕΙΡΑΜΑ ΟΔΗΓΙΕΣ ΓΙΑ ΜΑΘΗΤΕΣ ΝΑΝΟΣΩΛΗΝΕΣ ΑΝΘΡΑΚΑ Στους περισσότερους από εμάς δεν αρέσουν τα
NTSE - Nano Technology Science Education Project No: 511787-LLP-1-2010-1-TR-KA3-KA3MP ΜΕΡΟΣ I: ΜΕΛΕΤΗ ΠΡΙΝ ΤΟ ΠΕΙΡΑΜΑ ΟΔΗΓΙΕΣ ΓΙΑ ΜΑΘΗΤΕΣ ΝΑΝΟΣΩΛΗΝΕΣ ΑΝΘΡΑΚΑ Στους περισσότερους από εμάς δεν αρέσουν τα
Επιτροπάκη Ειρήνη. Xianghui Xu,Hui Yuan,Jing Chang,Bin He and Zhongwei Gu. Angew.Chem.Int.Ed. 2012,51,1-5
 Επιτροπάκη Ειρήνη ΑΜ:429 Xianghui Xu,Hui Yuan,Jing Chang,Bin He and Zhongwei Gu Angew.Chem.Int.Ed. 2012,51,1-5 Κατασκευή υπερμοριακών δομών με μεταφορικές λειτουργίες για χρήση σε ιατρικές εφαρμογές σε
Επιτροπάκη Ειρήνη ΑΜ:429 Xianghui Xu,Hui Yuan,Jing Chang,Bin He and Zhongwei Gu Angew.Chem.Int.Ed. 2012,51,1-5 Κατασκευή υπερμοριακών δομών με μεταφορικές λειτουργίες για χρήση σε ιατρικές εφαρμογές σε
ΠΕΡΙΟΔΙΚΟΣ ΠΙΝΑΚΑΣ ΠΕΡΙΟΔΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΑΤΟΜΙΚΗ ΑΚΤΙΝΑ ΕΝΕΡΓΕΙΑ ΙΟΝΤΙΣΜΟΥ
 ΠΕΡΙΟΔΙΚΟΣ ΠΙΝΑΚΑΣ ΠΕΡΙΟΔΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΑΤΟΜΙΚΗ ΑΚΤΙΝΑ ΕΝΕΡΓΕΙΑ ΙΟΝΤΙΣΜΟΥ Ν. ΜΠΕΚΙΑΡΗΣ ΕΙΣΑΓΩΓΗ Η ηλεκτρονιακή δομή και κυρίως τα ηλεκτρόνια σθένους (τελευταία ηλεκτρόνια) προσδίδουν στο άτομο τη χημική
ΠΕΡΙΟΔΙΚΟΣ ΠΙΝΑΚΑΣ ΠΕΡΙΟΔΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΑΤΟΜΙΚΗ ΑΚΤΙΝΑ ΕΝΕΡΓΕΙΑ ΙΟΝΤΙΣΜΟΥ Ν. ΜΠΕΚΙΑΡΗΣ ΕΙΣΑΓΩΓΗ Η ηλεκτρονιακή δομή και κυρίως τα ηλεκτρόνια σθένους (τελευταία ηλεκτρόνια) προσδίδουν στο άτομο τη χημική
1.1 Ηλεκτρονικές ιδιότητες των στερεών. Μονωτές και αγωγοί
 1. Εισαγωγή 1.1 Ηλεκτρονικές ιδιότητες των στερεών. Μονωτές και αγωγοί Από την Ατομική Φυσική είναι γνωστό ότι οι επιτρεπόμενες ενεργειακές τιμές των ηλεκτρονίων είναι κβαντισμένες, όπως στο σχήμα 1. Σε
1. Εισαγωγή 1.1 Ηλεκτρονικές ιδιότητες των στερεών. Μονωτές και αγωγοί Από την Ατομική Φυσική είναι γνωστό ότι οι επιτρεπόμενες ενεργειακές τιμές των ηλεκτρονίων είναι κβαντισμένες, όπως στο σχήμα 1. Σε
Χημεία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ. Τ μαθητ : Σχολικό Έτος:
 Χημεία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ Τ μαθητ : Σχολικό Έτος: 1 1.2 Καταστάσεις των υλικών 1. Συμπληρώστε το παρακάτω σχεδιάγραμμα 2 2. Πώς ονομάζονται οι παρακάτω μετατροπές της φυσικής κατάστασης; 3 1.3
Χημεία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ Τ μαθητ : Σχολικό Έτος: 1 1.2 Καταστάσεις των υλικών 1. Συμπληρώστε το παρακάτω σχεδιάγραμμα 2 2. Πώς ονομάζονται οι παρακάτω μετατροπές της φυσικής κατάστασης; 3 1.3
Μέθοδοι έρευνας ορυκτών και πετρωμάτων
 Μέθοδοι έρευνας ορυκτών και πετρωμάτων Μάθημα 9 ο Φασματοσκοπία Raman Διδάσκων Δρ. Αδαμαντία Χατζηαποστόλου Τμήμα Γεωλογίας Πανεπιστημίου Πατρών Ακαδημαϊκό Έτος 2017-2018 Ύλη 9 ου μαθήματος Αρχές λειτουργίας
Μέθοδοι έρευνας ορυκτών και πετρωμάτων Μάθημα 9 ο Φασματοσκοπία Raman Διδάσκων Δρ. Αδαμαντία Χατζηαποστόλου Τμήμα Γεωλογίας Πανεπιστημίου Πατρών Ακαδημαϊκό Έτος 2017-2018 Ύλη 9 ου μαθήματος Αρχές λειτουργίας
ΝΑΝΟΚΛΙΜΑΚΑ ΚΑΙ ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ
 NTSE - Nan Technlgy Science Educatin Prject N: 511787-LLP-1-2010-1-TR-KA3-KA3MP ΟΔΗΓΙΕΣ ΓΙΑ ΜΑΘΗΤΕΣ ΝΑΝΟΚΛΙΜΑΚΑ ΚΑΙ ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ Εικονικό εργαστήριο: http://vlab.ntse-nantech.eu/nanvirtuallab/ 1 ΜΕΛΕΤΗ
NTSE - Nan Technlgy Science Educatin Prject N: 511787-LLP-1-2010-1-TR-KA3-KA3MP ΟΔΗΓΙΕΣ ΓΙΑ ΜΑΘΗΤΕΣ ΝΑΝΟΚΛΙΜΑΚΑ ΚΑΙ ΝΑΝΟΤΕΧΝΟΛΟΓΙΑ Εικονικό εργαστήριο: http://vlab.ntse-nantech.eu/nanvirtuallab/ 1 ΜΕΛΕΤΗ
ΤΕΛΟΣ 1ΗΣ ΑΠΟ 7 ΣΕΛΙΔΕΣ
 ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΣΑΒΒΑΤΟ 8 ΣΕΠΤΕΜΒΡΙΟΥ 018 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΣΥΝΟΛΟ
ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΣΑΒΒΑΤΟ 8 ΣΕΠΤΕΜΒΡΙΟΥ 018 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΣΥΝΟΛΟ
Χημική Κινητική. Κωδ. Μαθήματος 718 Τομέας Φυσικοχημείας, Τμήμα Χημείας, ΕΚΠΑ. Μάθημα 12. Βίκη Νουσίου
 Χημική Κινητική Κωδ. Μαθήματος 718 Τομέας Φυσικοχημείας, Τμήμα Χημείας, ΕΚΠΑ Μάθημα 12 Βίκη Νουσίου Εαρινό εξάμηνο 2019 Ένζυμα 2 3 4 Ένζυμα Ένζυμα: Βιολογικοί καταλύτες Μακρομόρια ΜΒ 10 4-10 6 Πρωτεΐνες
Χημική Κινητική Κωδ. Μαθήματος 718 Τομέας Φυσικοχημείας, Τμήμα Χημείας, ΕΚΠΑ Μάθημα 12 Βίκη Νουσίου Εαρινό εξάμηνο 2019 Ένζυμα 2 3 4 Ένζυμα Ένζυμα: Βιολογικοί καταλύτες Μακρομόρια ΜΒ 10 4-10 6 Πρωτεΐνες
ΤΕΛΟΣ 1ΗΣ ΑΠΟ 7 ΣΕΛΙΔΕΣ
 ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 6 ΣΕΠΤΕΜΒΡΙΟΥ 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΣΥΝΟΛΟ
ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 6 ΣΕΠΤΕΜΒΡΙΟΥ 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΣΥΝΟΛΟ
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΦΥΣΙΚΗ ΙΙ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΦΥΣΙΚΗ ΙΙ 1. Οι δυναμικές γραμμές ηλεκτροστατικού πεδίου α Είναι κλειστές β Είναι δυνατόν να τέμνονται γ Είναι πυκνότερες σε περιοχές όπου η ένταση του πεδίου είναι μεγαλύτερη δ Ξεκινούν
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΦΥΣΙΚΗ ΙΙ 1. Οι δυναμικές γραμμές ηλεκτροστατικού πεδίου α Είναι κλειστές β Είναι δυνατόν να τέμνονται γ Είναι πυκνότερες σε περιοχές όπου η ένταση του πεδίου είναι μεγαλύτερη δ Ξεκινούν
ΦΥΣΙΚΗ ΚΑΤΕΥΘΥΝΣΗΣ. ΘΕΜΑ 1 ο
 ΦΥΣΙΚΗ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1 ο 1.1. Φορτισμένο σωματίδιο αφήνεται ελεύθερο μέσα σε ομογενές ηλεκτρικό πεδίο χωρίς την επίδραση της βαρύτητας. Το σωματίδιο: α. παραμένει ακίνητο. β. εκτελεί ομαλή κυκλική κίνηση.
ΦΥΣΙΚΗ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1 ο 1.1. Φορτισμένο σωματίδιο αφήνεται ελεύθερο μέσα σε ομογενές ηλεκτρικό πεδίο χωρίς την επίδραση της βαρύτητας. Το σωματίδιο: α. παραμένει ακίνητο. β. εκτελεί ομαλή κυκλική κίνηση.
Θέμα 1 ο (30 μονάδες)
") ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ-ΤΟΜΕΑΣ ΤΕΧΝΟΛΟΓΙΩΝ ΕΡΓΑΣΤΗΡΙΟ ΤΕΧΝΟΛΟΓΙΑΣ ΥΛΙΚΩΝ Θέμα 1 ο (30 μονάδες) (Καθ. Β.Ζασπάλης) Θεωρείστε ένα δοκίμιο καθαρού Νικελίου
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ-ΤΟΜΕΑΣ ΤΕΧΝΟΛΟΓΙΩΝ ΕΡΓΑΣΤΗΡΙΟ ΤΕΧΝΟΛΟΓΙΑΣ ΥΛΙΚΩΝ Θέμα 1 ο (30 μονάδες) (Καθ. Β.Ζασπάλης) Θεωρείστε ένα δοκίμιο καθαρού Νικελίου
Φασματοσκοπία Υπερύθρου (IR, FTIR)
") Φασματοσκοπία Υπερύθρου (IR, FTIR) Εργαστήριο Ανάλυσης ΤΕΙ Αθήνας 2016-2017 Διδάσκοντες Βασιλεία Σινάνογλου Παναγιώτης Ζουμπουλάκης Σωτήρης Μπρατάκος Γενικά Στην φασματοσκοπία υπερύθρου μελετάμε την απορρόφηση
Φασματοσκοπία Υπερύθρου (IR, FTIR) Εργαστήριο Ανάλυσης ΤΕΙ Αθήνας 2016-2017 Διδάσκοντες Βασιλεία Σινάνογλου Παναγιώτης Ζουμπουλάκης Σωτήρης Μπρατάκος Γενικά Στην φασματοσκοπία υπερύθρου μελετάμε την απορρόφηση
Γκύζη 14-Αθήνα Τηλ : 210.64.52.777
 ΘΕΜΑ Α Α1. α Α2. γ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 24 ΜΑΪΟΥ 2013 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΒΙΟΧΗΜΕΙΑ ΤΕΧΝΟΛΟΓΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ (ΚΥΚΛΟΣ ΤΕΧΝΟΛΟΓΙΑΣ ΚΑΙ ΠΑΡΑΓΩΓΗΣ) ΑΠΑΝΤΗΣΕΙΣ
ΘΕΜΑ Α Α1. α Α2. γ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 24 ΜΑΪΟΥ 2013 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΒΙΟΧΗΜΕΙΑ ΤΕΧΝΟΛΟΓΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ (ΚΥΚΛΟΣ ΤΕΧΝΟΛΟΓΙΑΣ ΚΑΙ ΠΑΡΑΓΩΓΗΣ) ΑΠΑΝΤΗΣΕΙΣ
ΓΓ/Μ ΣΥΣΤΗΜΑ ΠΑΙΔΕΙΑΣ ΟΡΟΣΗΜΟ. Τεύχος 2ο: Υδρογονάνθρακες Πετρέλαιο Προϊόντα από υδρογονάνθρακες Αιθανόλη - Ζυμώσεις
 ΓΓ/Μ2 05-06 ΣΥΣΤΗΜΑ ΠΑΙΔΕΙΑΣ ΟΡΟΣΗΜΟ Τεύχος 2ο: Υδρογονάνθρακες Πετρέλαιο Προϊόντα από υδρογονάνθρακες Αιθανόλη - Ζυμώσεις 140 ΧΗΜΕΙΑ: Υδρογονάνθρακες- Πετρέλαιο - Προιόντα από υδρογονάνθρακες - Αιθανόλη
ΓΓ/Μ2 05-06 ΣΥΣΤΗΜΑ ΠΑΙΔΕΙΑΣ ΟΡΟΣΗΜΟ Τεύχος 2ο: Υδρογονάνθρακες Πετρέλαιο Προϊόντα από υδρογονάνθρακες Αιθανόλη - Ζυμώσεις 140 ΧΗΜΕΙΑ: Υδρογονάνθρακες- Πετρέλαιο - Προιόντα από υδρογονάνθρακες - Αιθανόλη
1 η ΕΝΟΤΗΤΑ ΔΟΜΙΚΑ ΥΛΙΚΑ (ΕΙΣΑΓΩΓΗ)
") ΣΧΟΛΗ ΠΟΛΙΤΙΚΩΝ ΜΗΧΑΝΙΚΩΝ ΕΜΠ ΤΕΧΝΙΚΑ ΥΛΙΚΑ 1 η ΕΝΟΤΗΤΑ ΔΟΜΙΚΑ ΥΛΙΚΑ (ΕΙΣΑΓΩΓΗ) Ε. Βιντζηλαίου (Συντονιστής), Ε. Βουγιούκας, Ε. Μπαδογιάννης Άδεια Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
ΣΧΟΛΗ ΠΟΛΙΤΙΚΩΝ ΜΗΧΑΝΙΚΩΝ ΕΜΠ ΤΕΧΝΙΚΑ ΥΛΙΚΑ 1 η ΕΝΟΤΗΤΑ ΔΟΜΙΚΑ ΥΛΙΚΑ (ΕΙΣΑΓΩΓΗ) Ε. Βιντζηλαίου (Συντονιστής), Ε. Βουγιούκας, Ε. Μπαδογιάννης Άδεια Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
Θέµατα ιάλεξης ΠΡΩΤΕΪΝΕΣ - ΕΝΖΥΜΑ ΠΡΩΤΕΪΝΕΣ. ιαχωρισµός Αµινοξέων
 MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΠΡΩΤΕΪΝΕΣ - ΕΝΖΥΜΑ Θέµατα ιάλεξης οµή, αριθµός και διαχωρισµός των αµινοξέων Ένωση αµινοξέων µε τον πεπτιδικό δεσµό
MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΠΡΩΤΕΪΝΕΣ - ΕΝΖΥΜΑ Θέµατα ιάλεξης οµή, αριθµός και διαχωρισµός των αµινοξέων Ένωση αµινοξέων µε τον πεπτιδικό δεσµό
Τ, Κ Η 2 Ο(g) CΟ(g) CO 2 (g) Λύση Για τη συγκεκριμένη αντίδραση στους 1300 Κ έχουμε:
 CΟ(g) CO 2 (g) Λύση Για τη συγκεκριμένη αντίδραση στους 1300 Κ έχουμε:") ΘΕΜΑΤΑ ΤΕΛΙΚΩΝ ΕΞΕΤΑΣΕΩΝ ΣΤΗ ΦΥΣΙΚΟΧΗΜΕΙΑ - ΑΣΚΗΣΕΙΣ 5-6 (Α. Χημική Θερμοδυναμική) η Άσκηση Η αντίδραση CO(g) + H O(g) CO (g) + H (g) γίνεται σε θερμοκρασία 3 Κ. Να υπολογιστεί το κλάσμα των ατμών του
ΘΕΜΑΤΑ ΤΕΛΙΚΩΝ ΕΞΕΤΑΣΕΩΝ ΣΤΗ ΦΥΣΙΚΟΧΗΜΕΙΑ - ΑΣΚΗΣΕΙΣ 5-6 (Α. Χημική Θερμοδυναμική) η Άσκηση Η αντίδραση CO(g) + H O(g) CO (g) + H (g) γίνεται σε θερμοκρασία 3 Κ. Να υπολογιστεί το κλάσμα των ατμών του
Ανόργανη Χημεία. Τμήμα Τεχνολογίας Τροφίμων. Ενότητα 4 η : Ιοντικοί Δεσμοί Χημεία Κύριων Ομάδων. Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής
 Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 4 η : Ιοντικοί Δεσμοί Χημεία Κύριων Ομάδων Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Δόμηση Ηλεκτρονίων στα Ιόντα 2 Για τα στοιχεία
Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 4 η : Ιοντικοί Δεσμοί Χημεία Κύριων Ομάδων Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Δόμηση Ηλεκτρονίων στα Ιόντα 2 Για τα στοιχεία
2η Εργαστηριακή Άσκηση Εξάρτηση της ηλεκτρικής αντίστασης από τη θερμοκρασία Θεωρητικό μέρος
 2η Εργαστηριακή Άσκηση Εξάρτηση της ηλεκτρικής αντίστασης από τη θερμοκρασία Θεωρητικό μέρος Όπως είναι γνωστό από την καθημερινή εμπειρία τα περισσότερα σώματα που χρησιμοποιούνται στις ηλεκτρικές ηλεκτρονικές
2η Εργαστηριακή Άσκηση Εξάρτηση της ηλεκτρικής αντίστασης από τη θερμοκρασία Θεωρητικό μέρος Όπως είναι γνωστό από την καθημερινή εμπειρία τα περισσότερα σώματα που χρησιμοποιούνται στις ηλεκτρικές ηλεκτρονικές
Υλικά Ηλεκτρονικής & Διατάξεις
 Τμήμα Ηλεκτρονικών Μηχανικών Υλικά Ηλεκτρονικής & Διατάξεις 2 η σειρά διαφανειών Δημήτριος Λαμπάκης ΜΟΡΙΑΚΗ ΔΟΜΗ Μεμονωμένα άτομα: Μόνο τα ευγενή αέρια Μόρια: Τα υπόλοιπα άτομα σχηματίζουν μόρια, γιατί
Τμήμα Ηλεκτρονικών Μηχανικών Υλικά Ηλεκτρονικής & Διατάξεις 2 η σειρά διαφανειών Δημήτριος Λαμπάκης ΜΟΡΙΑΚΗ ΔΟΜΗ Μεμονωμένα άτομα: Μόνο τα ευγενή αέρια Μόρια: Τα υπόλοιπα άτομα σχηματίζουν μόρια, γιατί
Ο πυρήνας του ατόμου
 Ο πυρήνας του ατόμου Αρχές 19 ου αιώνα: Η ανακάλυψη της ραδιενέργειας, (αυθόρμητης εκπομπής σωματιδίων και / ή ακτινοβολίας από στοιχεία), βοήθησε τα μέγιστα στην έρευνα της δομής του ατόμου. Ποια είδη
Ο πυρήνας του ατόμου Αρχές 19 ου αιώνα: Η ανακάλυψη της ραδιενέργειας, (αυθόρμητης εκπομπής σωματιδίων και / ή ακτινοβολίας από στοιχεία), βοήθησε τα μέγιστα στην έρευνα της δομής του ατόμου. Ποια είδη
Εργασία για το μάθημα της Βιολογίας. Περίληψη πάνω στο κεφάλαιο 3 του σχολικού βιβλίου
 Εργασία για το μάθημα της Βιολογίας Περίληψη πάνω στο κεφάλαιο 3 του σχολικού βιβλίου Στο 3 ο κεφάλαιο του βιβλίου η συγγραφική ομάδα πραγματεύεται την ενέργεια και την σχέση που έχει αυτή με τους οργανισμούς
Εργασία για το μάθημα της Βιολογίας Περίληψη πάνω στο κεφάλαιο 3 του σχολικού βιβλίου Στο 3 ο κεφάλαιο του βιβλίου η συγγραφική ομάδα πραγματεύεται την ενέργεια και την σχέση που έχει αυτή με τους οργανισμούς
7 ο Κεφάλαιο Οργανική Χημεία. Δ. Παπαδόπουλος, χημικός
 7 ο Κεφάλαιο Οργανική Χημεία Δ. Παπαδόπουλος, χημικός Βύρωνας, 2015 Θεωρίες ερμηνείας του ομοιοπολικού δεσμού με βάση την κβαντική θεωρία. Θεωρία δεσμού σθένους. Θεωρία των μοριακών τροχιακών. Κάθε θεωρία
7 ο Κεφάλαιο Οργανική Χημεία Δ. Παπαδόπουλος, χημικός Βύρωνας, 2015 Θεωρίες ερμηνείας του ομοιοπολικού δεσμού με βάση την κβαντική θεωρία. Θεωρία δεσμού σθένους. Θεωρία των μοριακών τροχιακών. Κάθε θεωρία
ΧΗΜΕΙΑ Γ' ΓΥΜΝΑΣΙΟΥ. + SO 4 Βάσεις είναι οι ενώσεις που όταν διαλύονται σε νερό δίνουν ανιόντα υδροξειδίου (ΟΗ - ). NaOH Na
. NaOH Na") ΧΗΜΕΙΑ Γ' ΓΥΜΝΑΣΙΟΥ ΙΔΙΟΤΗΤΕΣ ΟΞΕΩΝ Αλλάζουν το χρώμα των δεικτών. Αντιδρούν με μέταλλα και παράγουν αέριο υδρογόνο (δες απλή αντικατάσταση) Αντιδρούν με ανθρακικά άλατα και παράγουν αέριο CO2. Έχουν όξινη
ΧΗΜΕΙΑ Γ' ΓΥΜΝΑΣΙΟΥ ΙΔΙΟΤΗΤΕΣ ΟΞΕΩΝ Αλλάζουν το χρώμα των δεικτών. Αντιδρούν με μέταλλα και παράγουν αέριο υδρογόνο (δες απλή αντικατάσταση) Αντιδρούν με ανθρακικά άλατα και παράγουν αέριο CO2. Έχουν όξινη
Ασκήσεις ακαδ. έτους
 Τμήμα Επιστήμης και Τεχνολογίας Υλικών Πανεπιστήμιο Κρήτης Επιστήμη Επιφανειών - Νανοϋλικών (ETY/METY 346) Μεταπτυχιακό: Νανοτεχνολογία για Ενεργειακές Εφαρμογές ¹ Nanomaterials for Energy (Νανοϋλικά για
Τμήμα Επιστήμης και Τεχνολογίας Υλικών Πανεπιστήμιο Κρήτης Επιστήμη Επιφανειών - Νανοϋλικών (ETY/METY 346) Μεταπτυχιακό: Νανοτεχνολογία για Ενεργειακές Εφαρμογές ¹ Nanomaterials for Energy (Νανοϋλικά για
ΠΥΡΗΝΙΚΟΣ ΜΑΓΝΗΤΙΚΟΣ ΣΥΝΤΟΝΙΣΜΟΣ ΚΑΙ ΔΟΜΗ ΤΟΥ ΑΤΟΜΟΥ. Του Αλέκου Χαραλαμπόπουλου ΕΙΣΑΓΩΓΗ
 ΠΥΡΗΝΙΚΟΣ ΜΑΓΝΗΤΙΚΟΣ ΣΥΝΤΟΝΙΣΜΟΣ ΚΑΙ ΔΟΜΗ ΤΟΥ ΑΤΟΜΟΥ Του Αλέκου Χαραλαμπόπουλου ΕΙΣΑΓΩΓΗ Ένα επαναλαμβανόμενο περιοδικά φαινόμενο, έχει μία συχνότητα επανάληψης μέσα στο χρόνο και μία περίοδο. Επειδή κάθε
ΠΥΡΗΝΙΚΟΣ ΜΑΓΝΗΤΙΚΟΣ ΣΥΝΤΟΝΙΣΜΟΣ ΚΑΙ ΔΟΜΗ ΤΟΥ ΑΤΟΜΟΥ Του Αλέκου Χαραλαμπόπουλου ΕΙΣΑΓΩΓΗ Ένα επαναλαμβανόμενο περιοδικά φαινόμενο, έχει μία συχνότητα επανάληψης μέσα στο χρόνο και μία περίοδο. Επειδή κάθε
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ-ΤΟΜΕΑΣ ΤΕΧΝΟΛΟΓΙΩΝ ΕΡΓΑΣΤΗΡΙΟ ΤΕΧΝΟΛΟΓΙΑΣ ΥΛΙΚΩΝ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ-ΤΟΜΕΑΣ ΤΕΧΝΟΛΟΓΙΩΝ ΕΡΓΑΣΤΗΡΙΟ ΤΕΧΝΟΛΟΓΙΑΣ ΥΛΙΚΩΝ Γραπτό τεστ (συν-)αξιολόγησης στο μάθημα: «ΔΙΑΓΝΩΣΤΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗ ΜΕΛΕΤΗ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ-ΤΟΜΕΑΣ ΤΕΧΝΟΛΟΓΙΩΝ ΕΡΓΑΣΤΗΡΙΟ ΤΕΧΝΟΛΟΓΙΑΣ ΥΛΙΚΩΝ Γραπτό τεστ (συν-)αξιολόγησης στο μάθημα: «ΔΙΑΓΝΩΣΤΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗ ΜΕΛΕΤΗ
Τελική γραπτή εξέταση «Επιστήμη και Τεχνολογία Υλικών ΙΙ»-Ιούνιος 2016
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ-ΤΟΜΕΑΣ ΤΕΧΝΟΛΟΓΙΩΝ ΕΡΓΑΣΤΗΡΙΟ ΤΕΧΝΟΛΟΓΙΑΣ ΥΛΙΚΩΝ ΘΕΜΑ 1 ο (25 Μονάδες) (Καθ. Β.Ζασπάλης) Δοκίμιο από PMMA (Poly Methyl MethAcrylate)
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ-ΤΟΜΕΑΣ ΤΕΧΝΟΛΟΓΙΩΝ ΕΡΓΑΣΤΗΡΙΟ ΤΕΧΝΟΛΟΓΙΑΣ ΥΛΙΚΩΝ ΘΕΜΑ 1 ο (25 Μονάδες) (Καθ. Β.Ζασπάλης) Δοκίμιο από PMMA (Poly Methyl MethAcrylate)
ΜΟΡΦΟΠΟΙΗΣΗ ΜΕ ΤΕΧΝΙΚΕΣ ΚΟΝΙΟΜΕΤΑΛΛΟΥΡΓΙΑΣ
 ΜΟΡΦΟΠΟΙΗΣΗ ΜΕ ΤΕΧΝΙΚΕΣ ΚΟΝΙΟΜΕΤΑΛΛΟΥΡΓΙΑΣ Η πρώτη ύλη με τη μορφή σωματιδίων (κόνεως) μορφοποιείται μέσα σε καλούπια, με μηχανισμό που οδηγεί σε δομική διασύνδεση των σωματιδίων με πρόσδοση θερμότητας.
ΜΟΡΦΟΠΟΙΗΣΗ ΜΕ ΤΕΧΝΙΚΕΣ ΚΟΝΙΟΜΕΤΑΛΛΟΥΡΓΙΑΣ Η πρώτη ύλη με τη μορφή σωματιδίων (κόνεως) μορφοποιείται μέσα σε καλούπια, με μηχανισμό που οδηγεί σε δομική διασύνδεση των σωματιδίων με πρόσδοση θερμότητας.
Δομές (Διαμορφώσεις) Πρωτεινικών μορίων
 Πρωτεινικών μορίων") Δομές (Διαμορφώσεις) Πρωτεινικών μορίων Πρωτοταγής δομή (αλληλουχία αμινοξέων) Δευτεροταγής δομή Η διάταξη της πεπτιδικής αλυσίδας στον χωρο αυτής καθ αυτής (χωρίς να ληφθούν υπ όψη οι ομάδες R) Τριτοταγής
Δομές (Διαμορφώσεις) Πρωτεινικών μορίων Πρωτοταγής δομή (αλληλουχία αμινοξέων) Δευτεροταγής δομή Η διάταξη της πεπτιδικής αλυσίδας στον χωρο αυτής καθ αυτής (χωρίς να ληφθούν υπ όψη οι ομάδες R) Τριτοταγής
Μεταλλικός δεσμός - Κρυσταλλικές δομές Ασκήσεις
 Μεταλλικός δεσμός - Κρυσταλλικές δομές Ασκήσεις Ποια από τις ακόλουθες προτάσεις ισχύει για τους μεταλλικούς δεσμούς; α) Οι μεταλλικοί δεσμοί σχηματίζονται αποκλειστικά μεταξύ ατόμων του ίδιου είδους μετάλλου.
Μεταλλικός δεσμός - Κρυσταλλικές δομές Ασκήσεις Ποια από τις ακόλουθες προτάσεις ισχύει για τους μεταλλικούς δεσμούς; α) Οι μεταλλικοί δεσμοί σχηματίζονται αποκλειστικά μεταξύ ατόμων του ίδιου είδους μετάλλου.
ΑΣΚΗΣΗ ΦΑΣΜΑΤΟΜΕΤΡΙΑΣ ΜΑΖΑΣ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΛΥΤΙΚΗΣ ΧΗΜΕΙΑΣ ΙΙ
 ΑΣΚΗΣΗ ΦΑΣΜΑΤΟΜΕΤΡΙΑΣ ΜΑΖΑΣ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΛΥΤΙΚΗΣ ΧΗΜΕΙΑΣ ΙΙ Τίτλος Εργαστηριακής Άσκησης: Προσδιορισμός Σχετικής Μοριακής Μάζας (Μ r ) Πρωτεΐνης με την Xρησιμοποίηση Φασματομετρίας Μάζας Ηλεκτροψεκασμού
ΑΣΚΗΣΗ ΦΑΣΜΑΤΟΜΕΤΡΙΑΣ ΜΑΖΑΣ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΛΥΤΙΚΗΣ ΧΗΜΕΙΑΣ ΙΙ Τίτλος Εργαστηριακής Άσκησης: Προσδιορισμός Σχετικής Μοριακής Μάζας (Μ r ) Πρωτεΐνης με την Xρησιμοποίηση Φασματομετρίας Μάζας Ηλεκτροψεκασμού
v = 1 ρ. (2) website:
 website:") Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης Τμήμα Φυσικής Μηχανική Ρευστών Βασικές έννοιες στη μηχανική των ρευστών Μαάιτα Τζαμάλ-Οδυσσέας 17 Φεβρουαρίου 2019 1 Ιδιότητες των ρευστών 1.1 Πυκνότητα Πυκνότητα
Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης Τμήμα Φυσικής Μηχανική Ρευστών Βασικές έννοιες στη μηχανική των ρευστών Μαάιτα Τζαμάλ-Οδυσσέας 17 Φεβρουαρίου 2019 1 Ιδιότητες των ρευστών 1.1 Πυκνότητα Πυκνότητα
ΑΝΑΠΤΥΞΗ ΝΑΝΟΔΟΜΗΜΕΝΩΝ ΥΛΙΚΩΝ ΜΕ ΝΑΝΟΣΩΛΗΝΕΣ ΑΝΘΡΑΚΑ ΓΙΑ ΧΡΗΣΗ ΣΕ ΕΦΑΡΜΟΓΕΣ ΥΨΗΛΗΣ ΑΝΤΟΧΗΣ
 ΑΝΑΠΤΥΞΗ ΝΑΝΟΔΟΜΗΜΕΝΩΝ ΥΛΙΚΩΝ ΜΕ ΝΑΝΟΣΩΛΗΝΕΣ ΑΝΘΡΑΚΑ ΓΙΑ ΧΡΗΣΗ ΣΕ ΕΦΑΡΜΟΓΕΣ ΥΨΗΛΗΣ ΑΝΤΟΧΗΣ Πετούσης Μάρκος, Δρ. Μηχανολόγος Μηχανικός Τμήμα Μηχανολόγων Μηχανικών Τ.Ε. ΤΕΙ Κρήτης Σύνθετα υλικά Σύνθετα υλικά
ΑΝΑΠΤΥΞΗ ΝΑΝΟΔΟΜΗΜΕΝΩΝ ΥΛΙΚΩΝ ΜΕ ΝΑΝΟΣΩΛΗΝΕΣ ΑΝΘΡΑΚΑ ΓΙΑ ΧΡΗΣΗ ΣΕ ΕΦΑΡΜΟΓΕΣ ΥΨΗΛΗΣ ΑΝΤΟΧΗΣ Πετούσης Μάρκος, Δρ. Μηχανολόγος Μηχανικός Τμήμα Μηχανολόγων Μηχανικών Τ.Ε. ΤΕΙ Κρήτης Σύνθετα υλικά Σύνθετα υλικά
ΤΕΣΤ 30 ΕΡΩΤΗΣΕΩΝ ΓΝΩΣΤΙΚΟΥ ΧΗΜΕΙΑΣ
 ΤΕΣΤ 30 ΕΡΩΤΗΣΕΩΝ ΓΝΩΣΤΙΚΟΥ ΧΗΜΕΙΑΣ ο αριθμός Avogadro, N A, L = 6,022 10 23 mol -1 η σταθερά Faraday, F = 96 487 C mol -1 σταθερά αερίων R = 8,314 510 (70) J K -1 mol -1 = 0,082 L atm mol -1 K -1 μοριακός
ΤΕΣΤ 30 ΕΡΩΤΗΣΕΩΝ ΓΝΩΣΤΙΚΟΥ ΧΗΜΕΙΑΣ ο αριθμός Avogadro, N A, L = 6,022 10 23 mol -1 η σταθερά Faraday, F = 96 487 C mol -1 σταθερά αερίων R = 8,314 510 (70) J K -1 mol -1 = 0,082 L atm mol -1 K -1 μοριακός
ρ ε υ ν α Οι ανάγκες για ενέργεια παγκοσμίως αυξάνονται συνεχώς και εκτιμάται ότι θα διπλασιασθούν
 Οργανικά Φωτοβολταϊκά Τμήμα Ηλεκτρολογίας & Κέντρο Τεχνολογίας Υλικών και Λέιζερ, ΤΕΙ Κρήτης των Δρ. Εμμανουήλ Κουδουμά, Δρ. Εμμανουηλ Κυμάκη Οι ανάγκες για ενέργεια παγκοσμίως αυξάνονται συνεχώς και εκτιμάται
Οργανικά Φωτοβολταϊκά Τμήμα Ηλεκτρολογίας & Κέντρο Τεχνολογίας Υλικών και Λέιζερ, ΤΕΙ Κρήτης των Δρ. Εμμανουήλ Κουδουμά, Δρ. Εμμανουηλ Κυμάκη Οι ανάγκες για ενέργεια παγκοσμίως αυξάνονται συνεχώς και εκτιμάται
ΣΤΟΙΧΕΙΑ ΚΡΥΣΤΑΛΛΟΓΡΑΦΙΑΣ
 ΣΤΟΙΧΕΙΑ ΚΡΥΣΤΑΛΛΟΓΡΑΦΙΑΣ 1. ΓΕΝΙΚΑ Από τις καταστάσεις της ύλης τα αέρια και τα υγρά δεν παρουσιάζουν κάποια τυπική διάταξη ατόμων, ενώ από τα στερεά ορισμένα παρουσιάζουν συγκεκριμένη διάταξη ατόμων
ΣΤΟΙΧΕΙΑ ΚΡΥΣΤΑΛΛΟΓΡΑΦΙΑΣ 1. ΓΕΝΙΚΑ Από τις καταστάσεις της ύλης τα αέρια και τα υγρά δεν παρουσιάζουν κάποια τυπική διάταξη ατόμων, ενώ από τα στερεά ορισμένα παρουσιάζουν συγκεκριμένη διάταξη ατόμων