ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΒΙΟΣΥΣΤΗΜΑΤΙΚΗ ΜΕΛΕΤΗ ΤΟΥ ΓΕΝΟΥΣ ΗYMENONEMA CASS.
|
|
|
- Ελένη Βουρδουμπάς
- 6 χρόνια πριν
- Προβολές:
Transcript



1 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ - ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ - ΕΡΓΑΣΤΗΡΙΟ ΒΟΤΑΝΙΚΗΣ Μ.Π.Σ.«ΟΙΚΟΛΟΓΙΑ, ΔΙΑΧΕΙΡΙΣΗ ΚΑΙ ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΒΙΟΣΥΣΤΗΜΑΤΙΚΗ ΜΕΛΕΤΗ ΤΟΥ ΓΕΝΟΥΣ ΗYMENONEMA CASS. ΕΛΕΝΗ ΛΙΒΕΡΗ Α.Μ. 511 ΠΑΤΡΑ 2015
2 Τριμελής Εξεταστική Επιτροπή: Αργυρώ Λιβανίου-Τηνιακού, Επίκουρη Καθηγήτρια, Τμήμα Βιολογίας, Πανεπιστήμιο Πατρών (Επιβλέπουσα). Πανωραία Αρτελάρη, Αναπληρώτρια Καθηγήτρια, Τμήμα Βιολογίας, Πανεπιστήμιο Πατρών. Ελευθερία-Περδίκω Μπαρέκα, Λέκτορας, Τμήμα Επιστήμης Φυτικής Παραγωγής, Γεωπονικό Πανεπιστήμιο Αθηνών. I
3 ΠΡΟΛΟΓΟΣ Η παρούσα μεταπτυχιακή εργασία εκπονήθηκε στο Εργαστήριο Βοτανικής, του Τομέα Βιολογίας Φυτών του Πανεπιστημίου Πατρών, κατά τη χρονική περίοδο Μέρος της πειραματικής διαδικασίας εκπονήθηκε στο Institute of Plant Science του Regensburg University (Germany). Το ερευνητικό αυτό μέρος ενισχύθηκε οικονομικά από το πρόγραμμα «Erasmus + για Πρακτική Άσκηση». Με την ολοκλήρωση αυτής της μεταπτυχιακής εργασίας θα ήθελα να ευχαριστήσω τους ανθρώπους που με βοήθησαν να την φέρω εις πέρας και συνέβαλαν καθοριστικά σε αυτό το αποτέλεσμα. Αρχικά, θα ήθελα να ευχαριστήσω την Επίκ. Καθηγήτρια κ. Αργυρώ Λιβανίου- Τηνιακού, η οποία δέχτηκε να επιβλέψει την παρούσα εργασία. Την ευχαριστώ θερμά για την άψογη συνεργασία μας και τις λεπτομερείς διορθώσεις της καθώς και για την υποστήριξη, ενθάρρυνση και τις συμβουλές της όχι μόνο σε επίπεδο ερευνητικό αλλά και σε προσωπικό, οι οποίες αποτέλεσαν για μένα πολύ σημαντικά εφόδια. Ιδιαίτερες ευχαριστίες οφείλω και στα υπόλοιπα μέλη της Τριμελούς Επιτροπής: την Αναπλ. Καθηγήτρια κ. Ρέα Αρτελάρη, για την βοήθεια, την εμψύχωση και την υλική υποστήριξη που μου παρείχε και την Λέκτορα κ. Πέπη Μπαρέκα για την πολύ καλή συνεργασία μας, το ενδιαφέρον και την λεπτομερή διόρθωσή της. Ένα πολύ μεγάλο ευχαριστώ οφείλω στην Ομότιμη Καθηγήτρια κ. Γεωργία Καμάρη, η οποία εμπνεύστηκε το θέμα της εργασίας και με εμπιστεύτηκε για τη διεκπεραίωσή της. Την ευχαριστώ για την υπομονή της, την καθοδήγησή της, την ώθηση που μου δίνει να «κυνηγώ» τους ερευνητικούς μου στόχους και τη βοήθεια που μου παρέχει για να τους πετυχαίνω. Η συμβολή της στην ερευνητική μου πορεία από το προπτυχιακό επίπεδο έως τώρα είναι ανεκτίμητη. Ιδιαίτερη τιμή για μένα αποτελεί η συνεργασία με τον Ομότιμο Καθηγητή κ. Δημήτριο Φοίτο, και τον ευχαριστώ για τη διάθεση πολύτιμου χρόνου και τις συμβουλές του. Η αφοσίωση του στην επιστήμη της Βοτανικής αποτελεί παράδειγμα προς μίμηση για κάθε ερευνητή. Ευχαριστώ πολύ τον Prof. Dr. Christoph Oberprieler, που με δέχτηκε στην ερευνητική του ομάδα και με έκανε να αισθανθώ μέλος της από την πρώτη κιόλας μέρα. Η συνεργασία μας ήταν εξαιρετική, η μεθοδικότητα και η σχολαστικότητα του II
4 τρόπου εργασίας του είναι υποδειγματική ενώ η καθοδήγησή του στην πρώτη μου επαφή με την Μοριακή Φυλογενετική υπήρξε σπουδαία. Από το University of Regensburg θα ήθελα να ευχαριστήσω θερμά τον Διδάκτορα Salvatore Tomasello για την υπομονή του, την εξαιρετική συνεργασία μας και την πολύτιμη βοήθειά του για την προσαρμογή μου στη ζωή στην Γερμανία και για τον τρόπο εργασίας μου στο Εργαστήριο. Μαζί με τον Salvatore και τον Υποψ. Διδάκτορα David Gutiérrez-Larruscain μοιραστήκαμε τον ίδιο ερευνητικό χώρο και με κάνουν να θυμάμαι με τις καλύτερες αναμνήσεις τους τρεις μήνες που πέρασα στο Εργαστήριο. Ακόμα, θα ήθελα να ευχαριστήσω τον Λέκτορα κ. Γιώργο Μήτσαινα, ο οποίος με βοηθούσε κάθε φορά που αντιμετώπιζα πρόβλημα σε θέματα μικροσκοπίου, με συμβούλευσε σε θέματα αγγλικής ορολογίας και συνεργάστηκε μαζί μου για την απόδοση όλων των φωτογραφιών που περιλαμβάνονται στο υποκεφάλαιο των Μορφολογικών Παρατηρήσεων. Για πάνω από τρία χρόνια μοιραστήκαμε τον ίδιο ερευνητικό χώρο στο Εργαστήριο Βοτανικής με την συνάδελφο και φίλη, Υποψ. Διδάκτωρ κ. Σοφία Σαμαροπούλου, την οποία ευχαριστώ για τις όμορφες στιγμές που περάσαμε, για τις συζητήσεις μας επί ερευνητικών και προσωπικών ανησυχιών αλλά και για την συνεργασία μας σ αυτή τη μελέτη με την ανάλυση υλικού με κυτταρογενετικές μεθόδους. Ευχαριστώ τα υπόλοιπα μέλη του Εργαστηρίου Βοτανικής, την Γεωργία Ζαραφωνίτη και τη Μαρία Τσακίρη για τη συνεργασία μας καθ όλη τη διάρκεια των μεταπτυχιακών μου σπουδών και για τις ωραίες στιγμές στο Εργαστήριο. Ευχαριστώ τους Υποψ. Διδάκτορες κ. Χαράλαμπο Κυριακόπουλο και κ. Κώστα Κουγιουμουτζή για τη διάθεση υλικού και την βοήθεια που πρόθυμα μου παρείχαν. Σ αυτό το σημείο θα ήθελα να ευχαριστήσω τους ανθρώπους οι οποίοι χωρίς να σχετίζονται με τον ακαδημαϊκό χώρο μου έδωσαν σημαντικές πληροφορίες για τις συλλογές του υλικού μελέτης. Ευχαριστώ θερμά τον κ. Γιάννη Κοφινά που με συνόδευσε στις συλλογές και μου έστειλε υλικό και φωτογραφίες του Hymenonema laconicum, τον κ. Σαράντο Δολιανίτη για το υλικό από το Λεωνίδιο και τους κυρίους Παναγιώτη Κουμπέτσο και Ερωτόκριτο Καλογερόπουλο για τις οδηγίες τους για την εύρεση του Hymenonema graecum στην Τήνο. Τέλος, θα ήθελα να ευχαριστήσω τον συνοδοιπόρο μου σε όλες τις βοτανικές μου αναζητήσεις αλλά και στη ζωή μου, τον Βλάση, που με ανέχτηκε με υπομονή και III
5 αγάπη. Πάνω απ όλα, ένα μεγάλο ευχαριστώ στην οικογένεια μου, και ειδικά στους γονείς μου Γιάννη και Γεωργία, που με στήριξε και με στηρίζει για να φτάσω ως εδώ με απεριόριστη αγάπη! Ε. Λιβέρη Αύγουστος 2015 IV
6 ΠΕΡΙΛΗΨΗ Το γένος Hymenonema Cass. παρουσιάζει ιδιαίτερο ενδιαφέρον, καθώς είναι ένα από τα οκτώ ενδημικά γένη της Ελληνικής Χλωρίδας. Συγχρόνως είναι το μόνο από τα ενδημικά γένη που δεν είναι μονοτυπικό. Το γένος Hymenonema διακρίνεται σε δυο είδη, το Hymenonema laconicum Boiss. & Heldr. και H. graecum (L.) DC. Το H. laconicum Boiss. & Heldr. εξαπλώνεται στα χαμηλά υψόμετρα των ορέων της Πελοποννήσου (Ταΰγετος, Πάρνωνας και Μαίναλο) ενώ το H. graecum (L.) DC απαντά στα περισσότερα νησιά και βραχονησίδες των Κυκλάδων και στην ΒΔ. Κρήτη, αν και η εμφάνισή του στην τελευταία περιοχή θεωρείται αβέβαιη. Στα πλαίσια της παρούσας εργασίας το γένος Hymenonema Cass. μελετήθηκε από ταξινομική, μορφολογική, κυτταρολογική, κυτταρογενετική, μοριακή και φυλογενετική άποψη. Για την πραγματοποίηση αυτής της μελέτης, εξετάσθηκε υλικό, το οποίο καλύπτει μεγάλο τμήμα της περιοχής εξάπλωσης των δυο ειδών. Το υλικό που συλλέχθηκε, καλλιεργήθηκε στον πειραματικό κήπο του Εργαστηρίου Βοτανικής, του Πανεπιστημίου Πατρών. Η κατάταξη του γένους Hymenonema Cass. στα ταξινομικά επίπεδα της οικογένειας και του κλάδου (tribe) δεν παρουσίασε άξιες λόγου αλλαγές από την ανακάλυψη του γένους και μετά. Κατατάσσεται στην οικογένεια Compositae = Asteraceae και στoν κλάδο Cichorieae. Εντός του κλάδου διακρίνονται υποκλάδοι (subtribes), οι οποίοι διαφοροποιούνται ως προς τον αριθμό και τα γένη που περιλαμβάνουν ανάλογα με τους συγγραφείς. Σύμφωνα με τα πιο πρόσφατα δεδομένα, το γένος Hymenonema Cass. συνιστά μαζί με τα γένη Catanananche L., Scolymus L. και Gundelia L. την subtribe Scolyminae. Η μορφολογική μελέτη του γένους περιλαμβάνει παρατηρήσεις ως προς την ταξιανθία, τον καρπό και τους γυρεοκόκκους, καθώς και μετρήσεις δεκατριών μορφολογικών χαρακτήρων, που αφορούν το ύψος του φυτού, την μορφή και τις διαστάσεις της ταξιανθίας του (κεφαλίου), τον αριθμό των βρακτίων, τις διαστάσεις των φύλλων βάσης και βλαστού, τις διαστάσεις των σπερμάτων του (αχαινίων) και του πάππου. Από τις μορφολογικές παρατηρήσεις διαπιστώθηκε για πρώτη φορά ότι ορισμένοι πληθυσμοί (από τις νήσους Σύρο και Τήνο) του Hymenonema graecum (L.) DC εμφανίζουν ενδιάμεσους μορφολογικούς χαρακτήρες μεταξύ των δυο ειδών. Η ποικιλότητα των μορφολογικών χαρακτήρων μετρήθηκε μεταξύ των πληθυσμών του ιδίου είδους και μεταξύ των δυο ειδών. Η διαπληθυσμιακή ποικιλότητα δεν V
7 παρουσίασε έντονη διαφοροποίηση, αντίθετα με την ποικιλότητα σε επίπεδο ειδών, η οποία κατέδειξε στατιστικώς σημαντικές διαφορές σε επτά μορφολογικά γνωρίσματα, μερικά δε εξ αυτών φαίνεται να αποτελούν σημαντικά ταξινομικά γνωρίσματα μεταξύ των δυο ειδών, όπως είναι το μήκος των τριχών του πάππου, το μέγεθος των αχαινίων και το πλάτος του τελικού λοβού των φύλλων βάσης. Τα αποτελέσματα αυτά σε συνδυασμό με τις μορφολογικές παρατηρήσεις οδήγησαν στη δημιουργία νέων κλειδών προσδιορισμού των δύο ειδών και λεπτομερέστερης περιγραφής αυτών. Για το γένος Hymenonema Cass. οι κυτταρολογικές αναφορές ήσαν πολύ περιορισμένες, ενώ δεν είχε εξετασθεί μέχρι σήμερα με κυτταρογενετικές τεχνικές. Πραγματοποιήθηκε κυτταρολογική μελέτη των δυο ειδών με την κλασσική μέθοδο σύνθλιψης κυττάρων και την χρώση με το αντιδραστήριο Feulgen. Μελετήθηκαν και δόθηκαν, για πρώτη φορά, η ανάλυση της μορφολογίας του καρυοτύπου, των καρυογραμμάτων και των ιδιογραμμάτων, καθώς και η μέτρηση έξι χρωμοσωματικών δεικτών, των δύο ειδών του γένους Hymenonema Cass. Ο χρωμοσωματικός αριθμός 2n = 20, τον οποίο διαπιστώσαμε κατά την παρούσα μελέτη μας, συμφωνεί με ήδη προηγούμενες αναφορές. Ο καρυότυπος και για τα δυο είδη βρέθηκε διπλοειδής και συμμετρικός, αποτελούμενος από μετακεντρικά και υπομετακεντρικά χρωμοσώματα μεσαίου μεγέθους (1,92-4,48 μm), με το μικρότερο σε μέγεθος 10 ο ζεύγος να φέρει πάντοτε δορυφόρους (SAT-χρωμοσώματα). Σε όλους τους πληθυσμούς του H. laconicum Boiss. & Heldr. το δορυφορικό ζεύγος χρωμοσωμάτων είναι υπομετακεντρικό, ενώ στο H. graecum (L.) DC μπορεί να αποτελείται από δυο υπομετακεντρικά ή από ένα μετακεντρικό κι ένα υπομετακεντρικό χρωμόσωμα (δομική ετεροζυγωτία). Σημειώνεται ότι ο αριθμός των μετακεντρικών και υπομετακεντρικών χρωμοσωμάτων ποικίλλει μεταξύ των πληθυσμών. Η κυτταρογενετική τεχνική που χρησιμοποιήθηκε είναι η χρώση με φθορισμό με χρήση χρωμομυκίνης A 3 (CMA 3 ) για τον εντοπισμό των πλούσιων σε GC περιοχών. Τα αποτελέσματα έδειξαν φθορισμό σε ένα ζεύγος χρωμοσωμάτων, αλλά δεν ήταν δυνατό να διακριθεί συγκεκριμένη μορφή ζωνώσεων, ώστε να έχουμε αντλήσει περισσότερες πληροφορίες. Για την μοριακή και φυλογενετική μελέτη συμπεριλήφθηκαν αντιπρόσωποι από όλη την Subtribe Scolyminae, αλλά περισσότερα δείγματα από το γένος Hymenonema Cass. Η πειραματική προσέγγιση περιέλαβε τον πολλαπλασιασμό των αλληλουχιών με τη μέθοδο της αλυσιδωτής αντίδρασης της πολυμεράσης (PCR), τον προσδιορισμό VI
8 της αλληλουχίας τους και τη στατιστική και φυλογενετική ανάλυση των αποτελεσμάτων. Οι αλληλουχίες που χρησιμοποιήθηκαν αποτελούν ευρέως διαδεδομένους μοριακούς δείκτες του χλωροπλαστικού (trnl-f, rpl32-trnl), ριβοσωμικού πυρηνικού (ITS1, ITS2) και πυρηνικού (D10) DNA για μελέτες σε επίπεδο γενών και ειδών. Οι χλωροπλαστικοί μοριακοί δείκτες και ο D10 μελετώνται για πρώτη φορά για το γένος Hymenonema Cass. Οι αλληλουχίες επεξεργάστηκαν με δυο μεθόδους φυλογενετικής ανάλυσης, την Μέθοδο Μέγιστης Φειδωλότητας και τη Μπεϊσιανή Συμπερασματολογία. Τα αποτελέσματα που προέκυψαν από τη χρήση των μοριακών δεικτών και των δυο διαφορετικών φυλογενετικών μεθόδων συμφωνούν μεταξύ τους ως προς τις φυλογενετικές σχέσεις των γενών εντός της subtribe Scolyminae και ως προς την μονοφυλετικότητα του γένους Hymenonema Cass. Το πλέον συγγενές γένος του Hymenonema Cass. φαίνεται ότι είναι το γένος Scolymus L., καθώς σε όλα τα φυλογενετικά δένδρα τα δυο αυτά γένη συγκροτούν μια μονοφυλετικήν ομάδα. Επιπλέον, πραγματοποιήθηκε η εκτίμηση του χρόνου απόσχισης των γενεαλογικών γραμμών της subtribe Scolyminae με βάση την υπόθεση του μοριακού ρολογιού και λαμβάνοντας υπόψη τις αλληλουχίες όλων των μοριακών δεικτών. Η εμφάνιση των δυο ειδών του γένους Hymenonema Cass. υπολογίζεται ότι πραγματοποιήθηκε πριν από 2,3-0,4 εκατομμύρια χρόνια, δηλαδή κατά την περίοδο του Πλειστοκαίνου. VII
9 ABSTRACT The genus Hymenonema Cass. shows a special interest, because it is one of the eight endemic genera of Greece and the only one which consists of two species, i.e. Hymenonema laconicum Boiss & Heldr. and H. graecum (L.) DC. H. laconicum Boiss. & Heldr. grows at the lowlands of Peloponnisos mountains (Mts Taigetos, Parnonas and Mainalo), while H. graecum (L.) DC occurs on most Kiklades islands and islets and on NW Kriti, although its presence in the latter is considered doubtful. In the framework of this dissertation, the genus Hymenonema Cass. was studied from systematic, morphological, cytological, cytogenetic, molecular and phylogenetic points of view. For this study, material was examined, that covered most the two species distribution range. The collected material was cultivated in the experimental garden of the Botanical Institute, University of Patras. The systematic classification of the genus Hymenonema Cass. at the taxonomic levels of family and tribe hasn t changed much from the first discovery of the genus. It is classified in the family Compositae and in the tribe Cichorieae. The tribe contains several subtribes, which differ in the number and genera that they include, depending on the authors. According to the most recent data, the genus Hymenonema Cass. and the genera Catananache L., Scolymus L. and Gundelia L. constitute the subtribe Scolyminae. The morphological study of the genus Hymenonema Cass. includes morphological observations concerning the capitulum, the achene and the pollens, and measurements of 13 morphological features concerning the plant height, the capitulum size, the number of bracts, the size of the basal and cauline leaves, the size of the achenes and pappus. The morphological observations revealed for the first time H. graecum (L.) DC populations, which however displayed intermediate morphological features between the two species. The variation of the morphological features was calculated both on an intra- and inter-specific level. The intra-specific variation didn t show intense differentiation, in contrast to the inter-specific comparison, which showed statistically significant differences at seven morphological features, some of which being taxonomically important between the two species, such as the length of the pappus hairs, the size of the achenes and the width of the terminal lobe of the basal leaves. These results combined with the interesting morphological observations led to VIII
10 the development of new taxonomical or dichotomic keys and of more detailed species descriptions. For the genus Hymenonema Cass., previous karyological data were very restricted, whereas the genus hadn t been examined with any cytogenetic technique up to now. The classic cytological squash technique and Feulgen staining were performed. The analysis of the karyotype morphology, the preparation of karyograms, idiograms and the measurements of six chromosome markers were performed for the first time. The chromosome number 2n = 20, which was found agrees with previous references. Both species displayed a diploid and symmetrical karyotype, consisting of metacentric (m) and submetacentric (sm) chromosomes of medium size (1,92-4,48 μm) with the smallest in size pair (the 10 th ) always bearing a visible satellite pair (SATchromosomes). In all populations of H. laconicum Boiss. & Heldr. the SATchromosome pair consists of two submetacentric chromosomes, while in H. graecum one of the SAT-chromosomes is often submetacentric and the other metacentric (structural heterogeneity). It should be noted that the number of submetacentric chromosomes varies among the populations. The cytogenetic technique that was performed, consisted of fluorochrome staining, using chromomycin A 3 (CMA 3 ), in order to locate GC-rich areas. The results showed that one chromosome pair stained positively, but it was not possible to determine a specific banding pattern, that would be more informative. The molecular and the phylogenetic study included representatives from the whole subtribe Scolyminae, most sequences derived from the genus Hymenonema Cass. The experimental approach included sequence amplification, using polymerase chain reaction (PCR), determination of the individual sequences via automated sequencing and statistical and phylogenetic analysis of the results. The sequences that were used are widely used molecular chloroplast markers (trnl-f, rpl32-trnl), ribosomal nuclear (ITS1, ITS2) and nuclear (D10) DNA for studies at the taxonomic levels of genus and species. The chloroplast markers and D10 are used for the first time in the genus Hymenonema Cass. The sequences were analyzed with Maximum Parsimony and Bayesian Inference. The results from the five different molecular markers and the two phylogenetic methods, concerning the phylogenetic relationships among the genera in the subtribe Scolyminae and the monophyly of the genus Hymenonema Cass. are concordant. The genus Scolymus L. appears to be a sister taxon of the genus IX
11 Hymenonema Cass. and in all phylogenetic trees these two genera constitute a monophyletic group. In addition, an estimation of the divergence time of the lineages in the subtribe Scolyminae was performed, based on a molecular clock hypothesis and using the sequences from all the molecular markers. The appearance of the Hymenonema Cass. species is estimated at Mya, placing it within Pleistocene. X
12 ΠΕΡΙΕΧΟΜΕΝΑ ΠΕΡΙΛΗΨΗ... V ABSTRACT... VIII 1. ΕΙΣΑΓΩΓΗ Η ΕΛΛΗΝΙΚΗ ΧΛΩΡΙΔΑ ΙΣΤΟΡΙΚΗ ΠΟΡΕΙΑ ΤΗΣ ΕΛΛΗΝΙΚΗΣ ΧΛΩΡΙΔΑΣ ΕΝΔΗΜΙΣΜΟΣ Ο ΕΝΔΗΜΙΣΜΟΣ ΤΗΣ ΕΛΛΗΝΙΚΗΣ ΧΛΩΡΙΔΑΣ Hymenonema Cass. ΕΛΛΗΝΙΚΟ ΕΝΔΗΜΙΚΟ ΓΕΝΟΣ ΣΚΟΠΟΣ ΤΗΣ ΜΕΛΕΤΗΣ ΣΥΣΤΗΜΑΤΙΚΗ ΜΕΛΕΤΗ ΓΕΝΙΚΑ ΤΑΞΙΝΟΜΗΣΗ ΤΟΥ ΓΕΝΟΥΣ ΥΛΙΚΟ ΜΕΛΕΤΗΣ ΚΛΕΙΔΕΣ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΕΙΔΩΝ ΤΟΥ ΓΕΝΟΥΣ Hymenonema Cass ΠΕΡΙΓΡΑΦΕΣ ΕΙΔΩΝ ΜΟΡΦΟΛΟΓΙΚΗ ΜΕΛΕΤΗ ΜΟΡΦΟΛΟΓΙΑ ΥΛΙΚΑ & ΜΕΘΟΔΟΙ ΑΠΟΤΕΛΕΣΜΑΤΑ Μορφολογικές παρατηρήσεις Διαπληθυσμιακή ποικιλότητα εντός των ειδών Ποικιλότητα των μορφολογικών χαρακτήρων μεταξύ των δυο ειδών ΚΥΤΤΑΡΟΛΟΓΙΚΗ ΜΕΛΕΤΗ ΚΥΤΤΑΡΟΛΟΓΙΑ - ΚΥΤΤΑΡΟΓΕΝΕΤΙΚΗ ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Κυτταρολογικές μέθοδοι Κυτταρογενετικές μέθοδοι ΑΠΟΤΕΛΕΣΜΑΤΑ Κυτταρολογικά αποτελέσματα Κυτταρογενετικά αποτελέσματα ΜΟΡΙΑΚΗ ΜΕΛΕΤΗ ΜΟΡΙΑΚΗ ΦΥΛΟΓΕΝΕΤΙΚΗ ΥΛΙΚΑ & ΜΕΘΟΔΟΙ XI
13 Συλλογή υλικού & εκχύλιση DNA Επιλογή μοριακών δεικτών Μοριακές τεχνικές Επεξεργασία αλληλουχιών & ανάλυση δεδομένων ΑΠΟΤΕΛΕΣΜΑΤΑ Γονιδιακά δέντρα (gene trees) χλωροπλαστικό DNA (cpdna) Γονιδιακά δέντρα (gene trees) πυρηνικοί δείκτες (nrdna ITS & single-copy D10) Δέντρο ειδών (species tree) ΓΕΝΙΚΑ ΣΥΜΠΕΡΑΣΜΑΤΑ ΑΠΟΤΕΛΕΣΜΑΤΑ ΒΙΒΛΙΟΓΡΑΦΙΑ ΠΑΡΑΡΤΗΜΑ XII
14 1. ΕΙΣΑΓΩΓΗ 1.1. Η ΕΛΛΗΝΙΚΗ ΧΛΩΡΙΔΑ Ο ελλαδικός χώρος έχει αποτελέσει πόλον έλξης για τους Βοτανικούς ήδη πριν από την εποχή του Λινναίου (Carl Linné ). Η μεγάλη ποικιλία του σε φυτικά και ζωικά είδη, τύπους οικοτόπων και τοπία έχει εγείρει έντονο ερευνητικό ενδιαφέρον με αποτέλεσμα πληθώρα μελετών σε διάφορα επιστημονικά πεδία. Η εξερεύνηση και μελέτη της Ελληνικής Χλωρίδας άρχισε ήδη από τους κλασικούς χρόνους από τον Θεόφραστο τον Ερέσιο ( π.χ.) με τα έργα του «Περί φυτών ιστορίας» και «Περί φυτών αιτιών». Ακολούθησε ο Διοσκουρίδης (40-90 μ.χ.), ο οποίος κατέγραψε πολλά φυτά της Μεσογείου με έμφαση στην φαρμακευτική τους αξία. Έπειτα, η περίοδος ιδιαίτερου ενδιαφέροντος ξένων περιηγητών για την Ελλάδα ξεκίνησε τον 18 ο αιώνα με τους Tournefort ( ) και Sibthorp ( ), από τους πιο σημαντικούς Βοτανικούς που επισκέφθηκαν την χώρα μας. Ακολούθησαν κι άλλοι σπουδαίοι Βοτανικοί, όπως οι Boissier ( ), Grisebach ( ), Ορφανίδης ( ), Heldreich ( ), Halácsy ( ), Hayek ( ) και Rechinger ( ), οι οποίοι εξερεύνησαν και μελέτησαν την Ελληνική Χλωρίδα και από τις μελέτες τους εκδόθηκαν πολύ σημαντικά έργα. Κυρίως όμως τα τελευταία 150 έτη έχουν εκδοθεί πολλές εργασίες και μελέτες, που αφορούν γένη, περιοχές ορεινές, νησιωτικές, ή υγροτοπικές κ.ά., με αντικείμενο την μελέτη μέρους ή του συνόλου των φυτικών ειδών διαφόρων περιοχών της Ελλάδας (Πίνακας 1). Επίσης, έχουν δημοσιευθεί δύο έργα, τα οποία αφορούν τα απειλούμενα είδη της Ελληνικής Χλωρίδας (Phitos & al. 1995, Φοίτος & αλ. 2009). Προς διευκόλυνση της μελέτης της Ελληνικής Χλωρίδας έχει διακριθεί ο ελληνικός χώρος (Εικ. 1) σε 13 φυτογεωγραφικές περιοχές (Phitos & al και Strid & Tan 1997). Σύμφωνα με πρόσφατη μελέτη (Dimopoulos & al. 2013), η Ελληνική Χλωρίδα αριθμεί περί τα taxa αυτοχθόνων και εγκλιματισμένων Αγγειοφύτων (Πτεριδόφυτα, Γυμνόσπερμα και Αγγειόσπερμα).Τα taxa αυτά ανήκουν σε είδη και υποείδη ενώ χωρίζονται σε γένη και 185 οικογένειες. Η αυτοφυής χλωρίδα κυριαρχείται από ολαρκτικές οικογένειες, από τις οποίες οι πλουσιότερες σε είδη είναι οι: Asteraceae (749), Poaceae (439), Fabaceae (438), Caryophyllaceae (341), Brassicaceae (278), Lamiaceae (260), Apiaceae (230), 13
15 Πίνακας 1: Μερικά από τα έργα τα οποία αναφέρονται στην Ελληνική Χλωρίδα. ΤΙΤΛΟΣ ΧΡΟΝΟΛΟΓΙΑ ΣΥΓΓΡΑΦΕΑΣ/ΕΙΣ ConspectusFloraeGraecae (και 2 συμπληρώματα) , Halácsy 1908, 1912 Prodromus Florae Peninsulae Balcanicae (3 τόμοι) Hayek Flora Aegaea 1943 Rechinger Flora Europaea (5 τόμοι & επανέκδοση του τόμου 1) , 1993 Tutin, Heywood, Burges, Moore, Valentine, Walters, Webb Flora of Turkey and the East Aegean Islands (9 τόμοι& , Davis 2 συμπληρώματα) 1988, 2000 The Mountain Flora of Greece (2 τόμοι) 1986, 1991 Strid, Strid & Tan Red Data Book of Rare and Threatened Plants of Greece 1995 Phitos, Strid, Snogerup & Greuter Flora Hellenica (2 τόμοι) 1997, 2002 Strid & Tan Βιβλίο Ερυθρών Δεδομένων των Σπανίων και Απειλουμένων Φυτών της Ελλάδας (2 τόμοι) 2009 Φοίτος, Κωνσταντινίδης & Καμάρη Vascular plants of Greece: an annotated checklist 2013 Dimopoulos & al. Rosaceae (152), Boraginaceae (139), Rubiaceae (129). Οι πλουσιότερες οικογένειες σε αριθμό γενών είναι οι: Poaceae (125), Asteraceae (113), Apiaceae (75), Brassicaceae (65), Fabaceae (51), Lamiaceae (35), Caryophyllaceae (34), Boraginaceae (31), Rosaceae (26), Chenopodiaceae (25). Σε επίπεδο γένους, τα πλουσιότερα σε αριθμό ειδών γένη είναι τα: Centaurea (110), Silene (108), Allium (97), Trifolium (90), Limonium (87), Campanula (80), Taraxacum (77), Verbascum (76), Galium (72), Hieracium (66). Αξίζει να σημειωθεί ότι όταν η βιοποικιλότητα των γενών σε taxa (είδη, υποείδη ή χαμηλότερες ταξινομικές βαθμίδες) είναι μεγάλη, η παραπάνω σειρά αλλάζει λίγο ή πολύ, ειδικά στις περιπτώσεις απομικτικών γενών όπως π.χ. το γένος Hieracium το οποίο εμφανίζεται με 137 taxa. Οι οικογένειες και τα taxa τους δεν κατανέμονται ομοιόμορφα σε όλη την Ελλάδα. Η Βορειοανατολική χλωριδική περιοχή (ΝΕ) βρέθηκε να έχει τον μεγαλύτερο αριθμό ειδών, με είδη, ενώ τον μικρότερο παρουσιάζει η περιοχή των Κυκλάδων (Kik), με είδη. Πρέπει να τονισθεί ότι οι παραπάνω αριθμοί και γενικά οι αριθμοί που αναφέρονται στο πλήθος των taxa χλωριδικών περιοχών δεν πρέπει να θεωρούνται σταθεροί, αλλά κατά προσέγγιση, καθώς συχνά μεταβάλλονται λόγω νεότερων δεδομένων ή/και νεοτέρων ταξινομικών αναθεωρήσεων (Φοίτος & Καμάρη 2009). 14
, 7. Στερεά Ελλάς (StE), 8. Ιόνιοι νήσοι (ΙοΙ), 9. Νότια Πίνδος (SPi), 10. Βόρεια Πίνδος (NPi), 11. Ανατολική κεντρική περιοχή (EC), 12.")
16 Εικ.1: Οι 13 χλωριδικές περιοχές της Ελλάδας: 1. Κρήτη-Κάρπαθος (ΚΚ), 2. Κυκλάδες νήσοι (Kik), 3. Ανατολικές νήσοι του Αιγαίου (ΕΑe), 4. Δυτικές νήσοι του Αιγαίου (WAe), 5. Βόρειες νήσοι του Αιγαίου, 6. Πελοπόννησος (Pe), 7. Στερεά Ελλάς (StE), 8. Ιόνιοι νήσοι (ΙοΙ), 9. Νότια Πίνδος (SPi), 10. Βόρεια Πίνδος (NPi), 11. Ανατολική κεντρική περιοχή (EC), 12. Βόρεια κεντρική περιοχή (NC) και 13. Βορειανατολική περιοχή (NE) (από Phitos & al και Strid & Tan 1997) ΙΣΤΟΡΙΚΗ ΠΟΡΕΙΑ ΤΗΣ ΕΛΛΗΝΙΚΗΣ ΧΛΩΡΙΔΑΣ Η ιστορική πορεία της ελληνικής χλωρίδας είναι συνυφασμένη με την παλαιογεωγραφία και το παλαιοκλίμα του ελλαδικού χώρου. Η γνώση των παλαιοβιογεωγραφικών συνθηκών δια μέσου των γεωλογικών χρόνων αποτελεί πολύ σημαντικό εργαλείο για την ερμηνεία της σημερινής περιοχής εξάπλωσης των φυτικών taxa. Η σημερινή μορφή του ελλαδικού χώρου είναι το αποτέλεσμα μιας σειράς γεωτεκτονικών διεργασιών που έλαβαν χώρα στην ευρύτερη περιοχή. Ολόκληρος ο 15
, καλύφθηκαν και πάλι από την θάλασσα. Κατά το Ολιγόκαινο η περιοχή του Αιγαίου νοτίως της παλαιάς Κυκλαδικής μάζας ήταν ακόμα καλυμμένη από θάλασσα.")
, μιας πλατιάς, ορεινής, χερσαίας μάζας, η οποία καταλάμβανε ολόκληρο το Αιγαίο και εκτεινόταν ως τη Μικρά Ασία (Φοίτος & Καμάρη 2009).")
17 ελλαδικός χώρος αποτελούσε ως το τέλος της Ιουρασικής περιόδου τμήμα του βυθού της Τηθύος, οπότε άρχισαν να αναδύονται τα πρώτα τμήματα ξηράς, τα οποία όμως αργότερα, κατά το μέσον της Κρητιδικής περιόδου ( εκατ. έτη πριν), καλύφθηκαν και πάλι από την θάλασσα. Κατά το Ολιγόκαινο η περιοχή του Αιγαίου νοτίως της παλαιάς Κυκλαδικής μάζας ήταν ακόμα καλυμμένη από θάλασσα. Σ αυτή την περίοδο άρχισε η ορογένεση (αποτελούσε μέρος των μεγάλων αλπικών πτυχώσεων) η οποία περατώθηκε κατά τις αρχές του Μειοκαίνου (20 εκατ. έτη πριν). Το αποτέλεσμα αυτής ήταν η δημιουργία της Αιγαιΐδας (Εικ. 2Α), μιας πλατιάς, ορεινής, χερσαίας μάζας, η οποία καταλάμβανε ολόκληρο το Αιγαίο και εκτεινόταν ως τη Μικρά Ασία (Φοίτος & Καμάρη 2009). Α Β Εικ. 2: Η Αιγαιΐδα στα Α: 20 εκατ. χρόνια πριν και Β: 11 εκατ. χρόνια πριν (από internet). Στις γεωλογικές περιόδους που ακολούθησαν μετά το Μειόκαινο, η Αιγαιΐδα άρχισε να αλλάζει λόγω ευστατικών και τεκτονικών κινήσεων. Στο διάστημα μεταξύ Σερραβαλλίου και Τορτονίου, η μορφή του Ν. Αιγαίου άλλαξε εντελώς. Πριν από 11 εκατ. χρόνια όλη σχεδόν η Ν. Ελλάδα και μέρος της Κρήτης αποτελούσε μια ενιαία ξηρά με μερικές μικρές λίμνες (Εικ. 2Β). Η θάλασσα που διεισδύει χωρίζει τις Κυκλάδες από τα νησιά του Α. Αιγαίου και την Πελοπόννησο (Εικ. 3Α). Ωστόσο η περιοχή των Κυκλάδων παρέμεινε χέρσος ενωμένη με την Αττική και την Εύβοια, όπως και οι νήσοι του Α. Αιγαίου ενωμένες με τη Μικρά Ασία. Το κλίμα ήταν σαφώς θερμότερο του σημερινού με πλούσιες βροχοπτώσεις και η χλωρίδα συνίστατο κυρίως από υποτροπικά στοιχεία. Κατά το τέλος του Μειοκαίνου (5 εκατ. χρόνια πριν), η Μεσόγειος απομονώθηκε από τον Ατλαντικό και τον Ινδικό Ωκεανό με το κλείσιμο του Γιβραλτάρ. Αυτό είχε ως αποτέλεσμα την «κρίση αλατότητος» του Μεσσηνίου κατά τη διάρκεια της οποίας το κλίμα μετατράπηκε σε στεπικό με ανάλογες αλλαγές στη σύνθεση της χλωρίδας. Χαρακτηριστικά της κρίσης ήταν η 16
. Κατά τη διάρκεια του Μεσσηνίου (6-5,5 εκατ. χρόνια πριν), το μεγαλύτερο μέρος των Κυκλάδων ήταν ενωμένο με την Πελοπόννησο και τη Μ.")
, η κρίση αλατότητας έλαβε τέλος με τη διάνοιξη της οδού επικοινωνίας του Ατλαντικού και της Μεσογείου στο Γιβραλτάρ να επιτρέπει και πάλι την εισβολή της θάλασσας στο Αιγαίο και μάλιστα")
18 ανύψωση πολλών περιοχών και οι εκτεταμένες γέφυρες ξηράς που αποτέλεσαν διαδρόμους επικοινωνίας και μετακίνησης πολλών χλωριδικών και πανιδικών στοιχείων, καθώς και η δημιουργία μεγάλων, αλμυρών λιμνών και κλειστών θαλασσών στην περιοχή του Αιγαίου (Εικ. 3Β). Κατά τη διάρκεια του Μεσσηνίου (6-5,5 εκατ. χρόνια πριν), το μεγαλύτερο μέρος των Κυκλάδων ήταν ενωμένο με την Πελοπόννησο και τη Μ. Ασία (Φοίτος & Καμάρη 2009). Α Β Εικ. 3: Ο ελλαδικός χώρος τις περιόδους του Τορτονίου (Α) και του Μεσσηνίου (Β) (από internet). Κατά το Κατώτερο Πλειόκαινο (5,5-4,5 εκατ. χρόνια πριν), η κρίση αλατότητας έλαβε τέλος με τη διάνοιξη της οδού επικοινωνίας του Ατλαντικού και της Μεσογείου στο Γιβραλτάρ να επιτρέπει και πάλι την εισβολή της θάλασσας στο Αιγαίο και μάλιστα περισσότερο από κάθε άλλη φορά, αφού προχωρεί τώρα πολύ περισσότερο προς Βορρά. Νέες ανυψώσεις και καταβυθίσεις της ηπειρωτικής μάζας συνεχίστηκαν καθ όλο το Πλειόκαινο, δίνοντας στην Πελοπόννησο μια μορφή πολύκολπου νήσου και διαμελίζοντας το Ν. Αιγαίο σε πλήθος νήσων. Η Κρήτη αποχωρίζεται από την ηπειρωτική Ελλάδα, οι νότιες και κεντρικές Κυκλάδες απομονώνονται αποτελώντας ενιαία ξηρά, ενώ οι βόρειες ενώνονται μέσω της Εύβοιας με την ηπειρωτική Ελλάδα (Dermitzakis & Papanikolaou 1981, Poulakakis & Sfenthourakis 2008). Το κλίμα γινόταν βαθμιαία ψυχρότερο, ώστε κατά το τέλος του Πλειοκαίνου (Εικ. 4Α) πρέπει να ήταν όμοιο προς το σημερινό (Φοίτος & Καμάρη 2009). Μόλις κατά το τέλος του Πλειοκαίνου αρχές Τεταρτογενούς, μετά από αλλεπάλληλες ευστατικές κινήσεις έλαβε ο ελλαδικός χώρος για πρώτη φορά περίπου την σημερινή του μορφή. Το Πλειστόκαινο χαρακτηρίζεται από την ενναλαγή παγετωδών και μεσοπαγετωδών περιόδων, που έχουν ως αποτέλεσμα σημαντικές αλλαγές στη στάθμη της θάλασσας. Κατά το Κατώτερο Πλειστόκαινο, η Κρήτη 17


19 Α Β Εικ. 4: Ο ελλαδικός χώρος κατά τις εποχές Πλειόκαινο (Α) και Πλειστόκαινο (Β) (από internet). έλαβε το σημερινό της περίγραμμα ως αποτέλεσμα της ένωσης μιας ομάδας νήσων του Πλειοκαίνου και ακολούθησε ο αποχωρισμός νησιών του Αιγαίου από τις γειτονικές ηπειρωτικές μάζες. Κατά το μέσο Πλειστόκαινο, οι Κυκλάδες αποχωρίζονται από την Εύβοια και κατακερματίζονται. Τα νησιά του Α. Αιγαίου παραμένουν ενωμένα με τη Μικρά Ασία μέχρι και το Ολόκαινο, ενώ τα Κύθηρα είναι ακόμα συνδεδεμένα με την Πελοπόννησο. Περίπου χρ. πριν, τα νησιά των Κυκλάδων είχαν αποκοπεί προς δυσμάς από την ηπειρωτική Ελλάδα, αλλά παρέμειναν εν μέρει συνδεδεμένα με τη Μ. Ασία (Εικ. 4Β). Αργότερα ( χρ. πριν), οι Κυκλάδες έχουν ήδη απομονωθεί και αποτελούν κατά το μεγαλύτερο μέρος μια ενιαία ξηρά σχηματίζοντας τη λεγόμενη «Κυκλαδική ημι-χερσόνησο» (Van Andel & Shakleton 1982). Κατά το Ανώτερο Πλειστόκαινο αποκτούν τη σημερινή τους μορφή. Η τελευταία εποχή του Πλειστοκαίνου χαρακτηρίζεται από τις δημιουργηθείσες, λόγω των παγετώνων, ευστατικές μεταβολές στην στάθμη της θάλασσας, οπότε τεράστιες ποσότητες ύδατος συσσωρεύτηκαν στις Ηπείρους υπό μορφή πάγων, ενώ η στάθμη της θάλασσας κατήλθε αντίστοιχα (Φοίτος & Καμάρη 2009). Κατά το Ολόκαινο και ιδιαιτέρως κατά τα τελευταία έτη, διαπιστώνεται μια βραδεία ανύψωση της στάθμης της θάλασσας. Η ανύψωση της στάθμης της θάλασσας που ξεκίνησε περί τα χρ. πριν, με την έναρξη της μεταπαγετώδους περιόδου, δημιούργησε γύρω στα χρ. την ακτογραμμή του ελλαδικού χώρου περίπου όπως είναι διαμορφωμένη και σήμερα. Εν τω μεταξύ και εξ αιτίας της συνδυασμένης δράσης της μεταβολής του όγκου των υδάτων των ωκεανών και των ανοδικών ή καθοδικών κινήσεων διαφόρων τμημάτων του ελλαδικού χώρου, 18
20 ορισμένες περιοχές έχουν σήμερα αποκαλυφθεί ή αντιθέτως έχουν καλυφθεί από τη θάλασσα (Φοίτος & Καμάρη 2009) ΕΝΔΗΜΙΣΜΟΣ Ο ενδημισμός αποτελεί μια έννοια για την οποία οι επιστήμονες δεν έχουν καταλήξει σε έναν κοινά αποδεκτό ορισμό. Συνήθως ο ορισμός ποικίλλει ανά συγγραφέα, καθώς η έννοια του ενδημισμού είναι στενά συνδεδεμένη με την έκταση του χώρου που καταλαμβάνει έναν taxon. Κατά τον Good (1974), μια ταξινομική μονάδα (είδος ή γένος) μπορεί να χαρακτηριστεί ως ενδημική, εάν η γεωγραφική της εξάπλωση είναι μικρότερη της μέσης τιμής εξαπλώσεως των αντίστοιχων μονάδων. Το φαινόμενο του ενδημισμού δεν είναι στενά συνυφασμένο μόνο με την ερμηνεία της σημερινής εξαπλώσεως των ειδών, αλλά και με την ερμηνεία της δημιουργίας τους (ειδογένεση). Γενικά, ο ενδημισμός μπορεί να διακριθεί σε δυο κατηγορίες: τον παλαιοενδημισμό ή υπολειμματικό ενδημισμό και τον νεοενδημισμό ή προοδευτικό ενδημισμό. Στην πρώτη κατηγορία περιλαμβάνονται ενδημικά taxa τα οποία σε παλαιότερες γεωλογικές εποχές καταλάμβαναν περιοχές εξάπλωσης κατά πολύ ευρύτερες από τις σημερινές και τα οποία λόγω εξωτερικών ή βιολογικών αιτίων περιορίσθηκαν στις σημερινές περιοχές εξάπλωσής τους. Πρόκειται για γενετικώς παλαιά είδη, χωρίς άμεσους συγγενείς και ταξινομικώς απομονωμένα. Αντίθετα, τα νεοενδημικά taxa είναι συγκριτικά νεαρής ηλικίας και καταλαμβάνουν κατά κανόνα μικρές περιοχές, εφόσον δεν έχουν βρει τον χρόνο περαιτέρω εξάπλωσής τους. Αν ληφθεί υπόψη ο τρόπος σχηματισμού και οι σχέσεις των ενδημικών taxa με τα αμέσως συγγενή τους, μπορεί να κατηγοριοποιηθούν σε τέσσερις ομάδες (Favarger & Contandriopoulos 1961): α) Παλαιοενδημικά: μονοτυπικά γένη (δηλ. περιλαμβάνουν μόνο ένα είδος) ή είδη, τα οποία αποτελούν τον μοναδικό αντιπρόσωπο μιας συστηματικής μονάδας, μεγαλύτερης του γένους, όπως π.χ. η sectio. Τα παλαιοενδημικά δεν έχουν απαραιτήτως ως τόπο καταγωγής την σημερινή περιοχή εμφάνισής τους η οποία είναι υπολειμματική μιας άλλοτε πολύ ευρύτερης. Είναι συνήθως διπλοειδή, αλλά υπάρχουν και πολυπλοειδή. β) Σχιζοενδημικά: δυο ή περισσότερα είδη ή υποείδη, πολύ συγγενή μεταξύ τους, αλλά γεωγραφικά ή οικολογικά απομονωμένα. Έχουν προκύψει από το αυτό 19
21 πρωταρχικό (μητρικό) taxon με βραδεία και προοδευτική διαφοροποίηση των πληθυσμών του, σε διάφορες θέσεις της περιοχής εξαπλώσεώς του. Η διαφοροποίηση των αρχικών πληθυσμών μπορεί να προηγήθηκε του γεωγραφικού αποχωρισμού ή να προηγήθηκε η απομόνωση και έπειτα να επιταχύνθηκε η διαφοροποίηση. Τα σχιζοενδημικά taxa είναι κοινής καταγωγής κι εμφανίζουν τον ίδιο χρωμοσωματικό αριθμό. γ) Πατροενδημικά: taxa που έχουν δώσει γένεση σε συγγενή taxa γειτονικών περιοχών. Είναι διπλοειδή ή χαμηλού βαθμού πολυπλοειδίας. δ) Αποενδημικά: taxa που προέκυψαν με πολυπλοειδισμό από αρχικό, ευρέως εξαπλωμένο και διπλοειδές ή τουλάχιστον χαμηλότερης πολυπλοειδίας taxon. Αποτελούν την αντίστροφη περίπτωση από αυτή των πατροενδημικών. Ένα φαινόμενο που σχετίζεται άμεσα με τον ενδημισμό είναι αυτό της αντιστοιχίας. Αντίστοιχα χαρακτηρίζονται δύο ή περισσότερα taxa (συνήθως είδη του αυτού γένους ή υποείδη του αυτού είδους) στενώς συγγενή μεταξύ τους, όταν το ένα αποκλείει το άλλο στις περιοχές τις οποίες καταλαμβάνουν. Όταν τα αντίστοιχα είναι συγχρόνως ενδημικά, ονομάζονται ενδημο-αντίστοιχα. Τα γνήσια αντίστοιχα μάλιστα διαθέτουν πάντα τον ίδιο χρωμοσωματικό αριθμό (διπλοειδή ή πολυπλοειδή), διότι σχηματίζονται με βαθμιαία ειδογένεση από ένα πρωταρχικό είδος και σε διαφορετικά μέρη της εξαπλώσεώς του. Αυτή η ειδογένεση οφείλεται σε γονιδιακές μεταλλάξεις ή δομικές χρωμοσωματικές αλλαγές, που συνοδεύονται ενίοτε από δυσπλοειδία (π.χ. στην Crepis neglecta group, Kαμάρη 1976). Ο παραπάνω ορισμός των γνήσιων αντίστοιχων φαίνεται να συμπίπτει με αυτόν των σχιζοενδημικών. Πρέπει να τονισθεί ότι οι ρόλοι της γεωγραφίας (χώρος) και της οικολογίας (επιλογή) είναι πολύ σημαντικοί κατά τη διαδικασία της εξέλιξης. Η γεωγραφική απομόνωση διευκολύνει την αποκλίνουσα γενετική προσαρμογή και τον σχηματισμό των φυλών, που έχουν προέλθει υπό την επίδραση της επιλογής και της γενετικής εκτροπής, αποκλείοντας την ανάμιξη των αρχικών φυλών με διασταύρωση και ανασυνδυασμό, καθώς και τον άμεσο ανταγωνισμό τους. Έτσι, όταν στενώς συγγενείς αντίστοιχες φυλές σχηματίζονται σε γεωγραφικώς γειτονικές περιοχές, μπορεί να εξελιχθούν σε taxa ανώτερων ταξινομικών βαθμίδων και να καταλάβουν τελικά διαφορετικές περιοχές εξάπλωσης (αλλοπάτρια). Έτσι, η γεωγραφική απομόνωση λειτουργεί ως φραγμός διασταυρώσεων. Όμως, οι συγγενείς αυτές φυλές μπορεί να εξελιχθούν παραμένοντας στην ίδια περιοχή εξάπλωσης (συμπάτρια), 20
22 χρησιμοποιώντας μηχανισμούς αναπαραγωγικής απομόνωσης ως φραγμό διασταυρώσεων (Φοίτος & Καμάρη 2009). Συνοψίζοντας, η αλλοπάτρια ειδογένεση συμβαίνει ανάμεσα σε δυο (ή περισσότερους) πληθυσμούς, οι οποίοι είναι γεωγραφικά απομονωμένοι μεταξύ τους, ενώ η συμπάτρια αναφέρεται σε περιπτώσεις όπου η ειδογένεση συμβαίνει χωρίς γεωγραφική απομόνωση των πληθυσμών (Whittaker & Fernández-Palacios 2009) Ο ΕΝΔΗΜΙΣΜΟΣ ΤΗΣ ΕΛΛΗΝΙΚΗΣ ΧΛΩΡΙΔΑΣ Τα ενδημικά taxa της ελληνικής χλωρίδας φτάνουν τα από τα συνολικά 6.600, αγγίζοντας το ποσοστό του 22,2% της ελληνικής χλωρίδας. Αυτά διακρίνονται σε είδη (22,2% των ελληνικών ειδών) και 452 υποείδη (23,9% των ελληνικών υποειδών), ενώ δεν υπάρχουν ενδημικές οικογένειες στον ελλαδικό χώρο. Τα γένη με τον υψηλότερο αριθμό σε ενδημικά taxa είναι τα: Limonium (79), Centaurea (76), Hieracium (73), Campanula (60), Silene (53), Allium (52), Taraxacum (49), Dianthus (34), Asperula (31), Galium (29) (Dimopoulos & al. 2013). Οχτώ γένη χαρακτηρίζονται ως ενδημικά της Ελλάδας τα οποία αντιπροσωπεύονται από ένα μόνο είδος (μονοτυπικά), με εξαίρεση το γένος Hymenonema που διακρίνεται σε δυο είδη. Τα είδη που αντιπροσωπεύουν τα ελληνικά ενδημικά γένη είναι τα: Horstrissea dolinicola (Apiaceae), Hymenonema laconicum (Compositae-Asteraceae), Hymenonema graecum (Compositae-Asteraceae), Jankaea heldreichii (Gesneriaceae), Leptoplax emarginata (Brassicaceae), Lutzia cretica (Brassicaceae), Petromarula pinnata (Campanulaceae), Phitosia crocifolia (Compositae-Asteraceae), Thamnosciadium junceum (Apiaceae). Η κατανομή των ενδημικών taxa δεν είναι ομοιόμορφη στον ελλαδικό χώρο. Την πρώτη θέση σε αριθμό ενδημικών την καταλαμβάνει η Πελοπόννησος (Pe) με 468 taxa και ακολουθεί η Κρήτη (ΚΚ) με 395, ενώ την τελευταία θέση κατέχουν τα νησιά του Βόρειου Αιγαίου (NAe) με 57. Αν συγκριθεί όμως το ποσοστό των ενδημικών taxa της κάθε χλωριδικής περιοχής σε σχέση με τη συνολική χλωρίδα της τότε η Κρήτη ανέρχεται πρώτη με 17,6% και ακολουθεί η Πελοπόννησος (14,6%) (Dimopoulos & al. 2013). Σύμφωνα με τους Φοίτος & Καμάρη (2009), η σύγκριση του ποσοστού των ενδημικών ειδών των μεγάλων γεωγραφικών περιοχών της Ελλάδας δείχνει ότι αυτό αυξάνει προς Νότο με την Κρήτη να κατέχει την πρώτη θέση. Την ίδια αύξηση από Βορά προς Νότο ακολουθεί και η αναλογία στενότοπων ενδημικών ειδών, με την Κρήτη να έχει το 33,3% της ενδημικής χλωρίδας της με στενότοπα ενδημικά. 21
23 Όσον αφορά τα ενδιαιτήματα που προτιμούν τα ενδημικά του ελλαδικού χώρου, το 21,2% και το 20,7% των ενδημικών, εμφανίζονται ως στοιχείο της βλάστησης των υψηλών ορέων και σε απότομους βράχους και γκρεμνούς αντίστοιχα (Dimopoulos & al. 2013) Hymenonema Cass. ΕΛΛΗΝΙΚΟ ΕΝΔΗΜΙΚΟ ΓΕΝΟΣ Το γένος Hymenonema Cass., όπως αναφέρθηκε και ανωτέρω, φύεται αποκλειστικά στον ελληνικό χώρο και είναι το μοναδικό ελληνικό ενδημικό γένος που αντιπροσωπεύεται από δυο είδη. Τα είδη αυτά είναι τα Hymenonema laconicum Boiss. & Heldr.και Hymenonema graecum (L.) DC. Το H.laconicum εμφανίζεται στην κεντρική και νότια Πελοπόννησο και πιο συγκεκριμένα στα χαμηλά υψόμετρα των ορέων Ταΰγετος, Πάρνωνας και Μαίναλο. Το H.graecum φύεται στις Κυκλάδες και τη ΒΔ Κρήτη (?). Η πιο πρόσφατη καταγραφή για την Κρήτη προέρχεται από τον Zaffran την δεκαετία του 60 από το Ακρωτήριο Χανίων, αλλά παρότι αρκετοί ερευνητές έχουν αναζητήσει το είδος στην ευρύτερη περιοχή του Ακρωτηρίου Χανίων, αυτό δεν έχει επανευρεθεί! Οι αντιπρόσωποι του γένους Hymenonema δεν είναι γνωστοί μόνο στην επιστημονική κοινότητα, αλλά είναι διαδεδομένα φυτά και για το ευρύ κοινό. Τα δυο είδη, H. graecum και H. laconicum (Εικ. 5), συναντώνται στην ελληνική βιβλιογραφία και ως Υμενόνημα το ελληνικό και Υμενόνημα το λακωνικό, αντίστοιχα. Το H. graecum περιλαμβάνεται στο «Λεξικόν Φυτολογικόν» του Γεννάδιου (1914), ο οποίος το αναφέρει από τη νήσο Μύκονο με την κοινή ονομασία την «αδραλίδα» ενώ θεωρεί ότι είναι πιθανώς το «μικρόν Ιεράκιον» του Διοσκουρίδη. Ο Heldreich (1926) και ο Μαλακατές (1928α, 1928β) καταγράφουν στις εργασίες τους το H.graecum ως κοινά επονομαζόμενο «χοιροβοσκό». Από τον 19 ο αιώνα τουλάχιστον, ο χοιροβοσκός αποτελούσε βασικό μέρος της διατροφής των Κυκλαδιτών (Χρυσού-Καρατζά 2006). Στις μέρες μας, οι κάτοικοι της Σύρου και της Μυκόνου το αποκαλούν «αλεντρίδα» και το χρησιμοποιούν ακόμα ως συστατικό σε τοπικά εδέσματα. Η αλεντρίδα θεωρείται ένα ελληνικό εδώδιμο φυτό με υψηλή διατροφική αξία, ενώ παράλληλα παρουσιάζει ιδιαίτερο ενδιαφέρον η μελέτη της καλλιέργειας και παραγωγής του φυτού με σύγχρονες μεθόδους (Ακουμιανάκης κ.ά. 2009). 22


24 A B Εικ. 5: A. Hymenonema graecum (Bauer in: Flora Graeca) και B. Hymenonema laconicum (Johnsen in: Endemic Plants of Greece. The Peloponnese.) ΣΚΟΠΟΣ ΤΗΣ ΜΕΛΕΤΗΣ Ολοκληρωμένη μελέτη για το γένος Hymenonema μέχρι τώρα δεν έχει πραγματοποιηθεί. Υπάρχουν μόνο κάποιες αναφορές σε μελέτες ευρύτερου περιεχομένου. Σκοπός της εν λόγω εργασίας είναι να μελετηθεί το ενδημικό γένος Hymenonema από μορφολογική, κυτταρολογική και φυλογενετική σκοπιά. Η κατανομή του γένους στο χώρο μαρτυρά μια ενδιαφέρουσα προϊστορία, η οποία θα γίνει προσπάθεια να ερμηνευθεί μέσα από τον συνδυασμό δεδομένων διαφορετικών ερευνητικών πεδίων. Η συλλογή νέου υλικού και η μελέτη του παλαιότερου αποσκοπούν να δώσουν σημαντικές πληροφορίες για τη γεωγραφική εξάπλωση, τα ενδιαιτήματα, τα μορφολογικά χαρακτηριστικά, τη μορφολογία των χρωμοσωμάτων και το γονιδίωμα των αντιπροσώπων του γένους. 23
25 2. ΣΥΣΤΗΜΑΤΙΚΗ ΜΕΛΕΤΗ Η ταξινόμηση των φυτών έχει αλλάξει πολύ από την εποχή του Λινναίου έως σήμερα. Για την αποφυγή όμως σύγχυσης λόγω αλλαγών στην ταξινόμηση, κρίνεται σκόπιμο να αναφερθεί πως στην παρούσα εργασία χρησιμοποιούνται τα δεδομένα περί ονοματολογίας και συστηματικής κατάταξης από την Med-Checklist (Greuter & al , Greuter & Raab-Straube 2008), καθώς και από την ηλεκτρονική βάση της Euro+MedPlantBase the information resource for Euro-Mediterannean plant diversity ( ΓΕΝΙΚΑ Οικογένεια: Compositae Giseke in Prae. Ord. Nat. Pl = Asteraceae Bercht. & J. Presl. in Prir. Rostlin Η οικογένεια Compositae (Σύνθετα) ή Asteraceae (Αστεροειδή) αποτελεί τη μεγαλύτερη φυτική οικογένεια φτάνοντας σε αριθμό περίπου είδη, τα οποία έχουν περιγραφεί και επιβεβαιωθεί, ενώ αν συμπεριληφθούν κι αυτά που προβλέπεται να περιγραφούν, ο αριθμός αναμένεται να φτάσει τα είδη, σύμφωνα με τα στοιχεία των Funk & al. (2009). Καταλαμβάνει περίπου το 10% του φυτικού πλούτου της γης και περιλαμβάνει γένη, που κατανέμονται σε όλες τις ηπείρους εκτός της Ανταρκτικής. Κύριο χαρακτηριστικό της οικογένειας είναι ότι τα άνθη του φυτού (ανθίδια) οργανώνονται σε μια ταξιανθία που ονομάζεται κεφάλιο, το οποίο περιβάλλεται από βράκτια. Βασικά χαρακτηριστικά είναι ακόμα: ο δακτύλιος που σχηματίζουν οι ανθήρες και η παρουσία των σπερμάτων = αχαινίων, συνήθως φέροντα πάππο. Κατά τ άλλα, οι αντιπρόσωποι των Compositae παρουσιάζουν μεγάλη ποικιλομορφία σε όλους τους τομείς. Μπορεί να είναι μονοετή ή πολυετή, θάμνοι, πόες, αναρριχητικά, δέντρα, ακόμα και επίφυτα. Μπορούν να εποικήσουν πολλούς διαφορετικούς τύπους ενδιαιτημάτων, όπως, δάση, λιβάδια και υψηλού υψομέτρου εκτάσεις, αν και φαίνεται πως δεν επιλέγουν τα τροπικά, υγρά δάση. Μερικά από αυτά προτιμούν τα διαταραγμένα εδάφη, άλλα είναι επιβλαβή και άλλα χρήσιμα, εδώδιμα ή φαρμακευτικά. Μπορεί να είναι πολύ κοινά, να παρουσιάζουν μεγάλη εξάπλωση ή να είναι σπάνια και η εξάπλωσή τους να είναι αρκετά 24
26 περιορισμένη (Funk & al. 2009). Δεν είναι τυχαίο που τα 2 από τα 8 ενδημικά γένη της Ελληνικής Χλωρίδας ανήκουν στην οικογένεια Compositae. Το γένος Hymenonema είναι μέλος της μεγάλης οικογένειας των Compositae. Ιδρυτής της ταξινόμησης των Compositae θεωρείται ο Cassini (1816), καθώς σε όλη την καριέρα του ασχολήθηκε με αντιπροσώπους της οικογένειας, εξετάζοντας υλικό, περιγράφοντας αρκετά γένη, κατατάσσοντάς τα σε ομάδες, πολλές από τις οποίες εξακολουθούν να ισχύουν και σήμερα (Bremer 1994). Η συστηματική κατάταξη των Compositae έχει υποστεί πολλές αλλαγές από την εποχή του Cassini ως σήμερα. Ο Cassini το 1816 πρότεινε την ταξινόμηση της οικογένειας σε 19 tribes αλλά λόγω του τρόπου λειτουργίας των γαλλικών εκδόσεων η απήχηση της δουλειάς του τότε ήταν αυστηρά περιορισμένη. Το πρώτο βιβλίο που εκδόθηκε για τα Compositae ήταν από τον Lessing (1832) που διέκρινε 7 tribes. Το ίδιο μοτίβο ακολούθησε και ο De Candolle (1836). Ο Bentham (1873) ακολούθησε παρόμοιο τρόπο ταξινόμησης με τον Cassini, διακρίνοντας 13 tribes, όπως και ο Hoffmann (1890) που ουσιαστικά επανέλαβε την ταξινόμηση του πρώτου και υιοθετήθηκε από τους βοτανικούς για περίπου 100 χρόνια. Το 1976, ο Carlquist και ο Wagenitz (διαφωνώντας για τη θέση μιας εκ των tribes) διέκριναν ότι οι tribes χωρίζονται σε δυο κύριες ομάδες κι έτσι έγινε ομαδοποίηση της οικογένειας σε δυο υποοικογένειες (subfamilies), την Asteroideae και την Cichorioideae. Αυτό ήταν το πρώτο μεγάλο βήμα για την κατανόηση των σχέσεων μέσα στην οικογένεια. Το επόμενο σημαντικό βήμα πραγματοποιήθηκε με την ανάπτυξη των μοριακών τεχνικών και τη χρήση χλωροπλαστικού DNA (cpdna), γεγονός που οδήγησε τους Jansen & Palmer (1987) να παρατηρήσουν την έλλειψη μιας αναστροφής 22 kb σε 3 γένη της Cichorioideae, η οποία όμως είναι παρούσα σε όλους τους άλλους αντιπροσώπους. Έτσι, το 1992, οι Bremer & Jansen εγκαθίδρυσαν τρίτη υποοικογένεια, την Barnadesioideae. Ακολούθησε και τέταρτη υποοικογένεια, η Carduoideae (Bremer 1996) ενώ η tribe Mutisieae έμεινε στην ίδια ταξινομική βαθμίδα χρίζοντας περαιτέρω μελέτης λόγω της παραφυλετικότητας (Kim & Jansen 1995). Στη συνέχεια, ο αριθμός των υποοικογενειών αλλά και των tribes άρχισε να αυξομειώνεται ανάλογα με τη μελέτη: 11 υποοικογένειες και 35 tribes (Panero & Funk 2002) και σύμφωνα με πιο πρόσφατα στοιχεία 12 υποοικογένειες και 28 tribes (Panero & Funk 2008). Σύμφωνα όμως με την Euro+Med PlantBase, η οικογένεια Compositae περιλαμβάνει 15 tribes, όσον αφορά την εξάπλωσή της στη μεσογειακή περιοχή ενώ δεν αναφέρεται αριθμός υποοικογενειών ( 25
27 Tribe: Cichorieae Lam. & DC. (1806) = Lactuceae Cass. (1819), Catanancheae D. Don (1829), Crepideae Lindl. (1829), Hieracieae D. Don (1829), Hypochaerideae D. Don (1829), Scorzonereae D. Don (1829), Taraxaceae D. Don (1829), Gundelieae Lecoq & Juillet (1831), Hyoserideae Kostel. (1833), Scolymeae Kostel.(1833), Chondrilleae W.D.J. Koch (1837), Leontodonteae (Sch.Bip.) W.D.J. Koch (1834), Picrideae Sch.Bip. (1834), Tragopogoneae Sch.Bip.(1834), Urospermeae Sch.Bip. (1834) Σε επίπεδο tribe, το γένος Hymenonema κατατάσσεται στην Cichorieae. Είναι η πρώτη tribe που αναγνωρίστηκε μέσα στην οικογένεια και ίσως η πιο καλά μελετημένη. Τα μέλη της tribe διακρίνονται εύκολα από το γαλακτώδη χυμό που εκκρίνουν και το homogamous κεφάλιο τους με γλωσσοειδή = ταινιοειδή ανθίδια, που φέρουν 5 οδοντώσεις. Σύμφωνα με τους Kilian & al. (2009), η εύκολη αναγνώριση παρουσιάζει κι ένα σημαντικό μειονέκτημα, καθώς η tribe θεωρείται φτωχή σε μορφολογικά χαρακτηριστικά, πράγμα που δεν βοηθά στην διαλεύκανση της ταξινομικής κατάστασης των γενών που περιλαμβάνει. Επιπλέον, η εκτεταμένη παράλληλη εξέλιξη κάποιων χαρακτηριστικών, καθιστά ακόμα δυσκολότερη την ταξινόμηση των taxa εντός της tribe. Ο Tournefort (1694) ήταν ο πρώτος που διέκρινε και περιέγραψε αυτήν την ομάδα ως ταξινομική οντότητα δημιουργώντας την 13 η τάξη στο Βασίλειο των Φυτών. Στη συνέχεια, ο μαθητής του Vaillant (1719) ονόμασε αυτήν την τάξη Cichoracées και διέκρινε 5 sections βασισμένος στα χαρακτηριστικά: ενδιαίτημα, πάππος, δίσκος κεφαλίου (Vaillant 1723). Οι Lamarck & De Candolle(1806) επικύρωσαν το όνομα της tribe του Vaillant, αλλά δημιούργησαν 4 subtribes, βασισμένοι στη μελέτη του πάππου. Γενικά, οι παραπάνω βοτανικοί, αλλά και όσοι άλλοι [Cassini (1827, 1830), Don (1828), Lessing (1832), De Candolle (1838), Bentham (1873), Hoffmann ( )] ασχολήθηκαν στη διάρκεια του 19 ου αιώνα, βασίστηκαν αποκλειστικά σε μορφολογικά χαρακτηριστικά, δίνοντας πλούσιο και σημαντικό υλικό για την ανάλυση, την περιγραφή, την σύγκριση και την ταξινόμηση αυτής της ομάδας. Όμως, δε λήφθηκαν υπόψη εξελικτικά φαινόμενα, όπως η συγκλίνουσα εξέλιξη στον πάππο, με αποτέλεσμα τα συστήματα ταξινόμησης που προτάθηκαν μέχρι και τον Hoffmann 26
28 ( ), να μην περιλαμβάνουν σε μεγάλο βαθμό, πραγματικές ταξινομικές υποδιαιρέσεις φυτών. Έτσι, από την διαίρεση του Hoffmann σε 5 subtribes, οι Babcock & Stebbins (1938) αρχίζουν να επαναπροσδιορίζουν την ταξινόμηση της tribe συνδυάζοντας μορφολογικά, βιογεωγραφικά και κυτταρολογικά δεδομένα. Πολλοί ερευνητές ασχολήθηκαν με την κατάταξη σε tribes, ανάμεσά τους οι Poljakov (1967), Carlquist (1976), Wagenitz (1976), Cronquist (1977), Robinson (1983), Thorne (1983), με μεγαλύτερη απήχηση αυτή των Stebbins (1953) και Jeffrey (1966). Η ταξινόμηση του Stebbins (1953) αναγνώριζε 8 subtribes. Ο Jeffrey (1966), θέλοντας να βελτιώσει την εργασία του προηγούμενου, έλαβε υπόψη επιπρόσθετα κάποιους μικρομορφολογικούς χαρακτήρες και υποδιαίρεσε την tribe σε 5 ομάδες (groups), 8 υποομάδες (subgroups) και 18 σειρές (series). Η πρώτη κλαδιστική ταξινόμηση όμως πραγματοποιήθηκε από τον Bremer (1994) και είχε ως αποτέλεσμα 11 subtribes, καθώς και 2 γένη (Scolymus, Cichorium) απροσδιόριστα ως προς την subtribe. Την ταξινόμηση του Bremer (1994) ακολουθεί και ο Lack (2007), με μερικές τροποποιήσεις στις subtribes Sonchinae και Lactucinae. Η πιο πρόσφατη ταξινόμηση της tribe είναι των Kilian & al. (2009), σύμφωνα με την οποία διακρίνονται 11 subtribes και είναι αυτή που ακολουθείται στην παρούσα μελέτη ΤΑΞΙΝΟΜΗΣΗ ΤΟΥ ΓΕΝΟΥΣ Hymenonema Cass. in Bull. Sci. Soc. Philom. Paris 1817: Το γένος Hymenonema περιγράφηκε για πρώτη φορά από τον Γάλλο βοτανικό και δικαστικό Alexandre Henri Gabriel de Cassini ( ), στο Bulletin des Sciences / par la Société philomatique de Paris. Διάφοροι άλλοι ερευνητές που ασχολήθηκαν με αυτό, το κατέταξαν σε διάφορες ταξινομικές ομάδες (Πίνακας 2). Ο Stebbins (1953) το συμπεριέλαβε στην subtribe Cichoriinae, της οποίας κοινό χαρακτηριστικό των αντιπροσώπων της είναι ο πάππος, ο οποίος συνήθως φέρει λέπια ή είναι απών και περιλαμβάνει γένη του Παλαιού Κόσμου, όπως τα Tolpis, Arnoseris, Catananche, Hymenonema. Τα δύο πρώτα συνιστούν μία ομάδα γένους (generic group) και τα δυο τελευταία μία άλλη, επισημαίνεται όμως ότι μεταξύ τους οι δυο ομάδες δεν έχουν σχεδόν τίποτα κοινό. Ο Jeffrey (1966) χώρισε λίγο διαφορετικά την tribe, αλλά διατήρησε πολλά στοιχεία του προηγούμενου. Η υποομάδα του Catananche (Catananche subgroup) του Jeffrey (που ανήκει στην ευρύτερη ομάδα του Tolpis) ταυτίζεται με την ομάδα γένους Catananche-Hymenonema του Stebbins. Ο Bremer 27
29 (1993, 1994) κατατάσσει το γένος Hymenonema στην ίδια subtribe με τα Catananche και Rothmaleria και την ονομάζει Catananchinae. Τα τρία αυτά γένη μοιράζονται τους εξής κοινούς χαρακτήρες: βράκτια του κεφαλίου με μεμβρανώδη περιθώρια, σκληρότριχο-λεπιδωτό έως έχοντα παλαίες (paleate) δίσκο κεφαλίου και πάππο με λίγες μεγάλες φολίδες που προεκτείνονται καταλήγοντας σε σκληρές τρίχες (bristles). Θεωρούνται απομονωμένα μέσα στα Cichorieae, ενώ συνιστούν μονοφυλετική ομάδα σύμφωνα με τον Bremer. Η πιο πρόσφατη μελέτη (Kilian & al. 2009) παρουσιάζει λίγο διαφορετικά αποτελέσματα. Η subtribe Scolyminae αποτελεί έναν από τους τρεις κλάδους του φυλογενετικού δέντρου των Cichorieae και περιλαμβάνει τα γένη Catananche, Hymenonema, Scolymus, Gundelia. Το Catananche παρουσιάζεται ως αδελφή ομάδα των Hymenonema-Scolymus ενώ το γένος Gundelia ως αδελφή ομάδα της μονοφυλετικής ομάδας των Catananche, Hymenonema, Scolymus. Σχολιάζεται η σχέση του γένους Rothmaleria με τα παραπάνω γένη από παλυνολογικούς λόγους (Blackmore 1981), διατηρήθηκε εν μέρει και από άλλους ερευνητές, αλλά τα μοριακά δεδομένα το κατατάσσουν σε διαφορετικό κλάδο, μαζί με τα γένη Tolpis και Arnoseris. Πίνακας 2: Η ταξινόμηση του γένους Hymenonema στις διάφορες ταξινομικές κατηγορίες. Ταξινομικές κατηγορίες Βιβλιογραφία Subtribe Cichoriinae: Tolpis, Arnoseris, Catananche, Stebbins 1953 Hymenonema Catananche subgroup: Catananche, Hymenonema Jeffrey 1966 Subtribe Catananchinae: Catananche, Rothmaleria, Bremer 1993 Hymenonema Subtribe Scolyminae: Catananche, Hymenonema, Scolymus, Kilian & al. Gundelia ΥΛΙΚΟ ΜΕΛΕΤΗΣ Εξετάσθηκαν και συλλέχθηκαν 11 πληθυσμοί των Hymenonema laconicum και H. graecum από διάφορες περιοχές της εξάπλωσής τους (Εικ. 6 & 7). Στη συνέχεια καλλιεργήθηκαν φυτά στον Πειραματικό Κήπο του Πανεπιστημίου Πατρών, τα οποία 28
30 χρησιμοποιήθηκαν για κυτταρολογική, μορφολογική και μοριακή μελέτη. Το υλικό που συλλέχθηκε έχει κατατεθεί στο Βοτανικό Μουσείο του Πανεπιστημίου Πατρών (UPA) και μέρος του καλλιεργείται στον Πειραματικό Κήπο. Μελετήθηκαν επίσης συλλογές από το Βοτανικό Μουσείο του Πανεπιστημίου Πατρών (UPA), καθώς και συλλογές μέσω των ψηφιακών Herbaria, όπως τα: Herbarium of Lund University Botanical Museum (LD), Herbarium of the Royal Botanic Gardens Kew (RBGK), Herbarium of Botanic Garden and Botanical Museum Berlin-Dahlem (B), Herbarium of Karl Franzens University of Graz (GZU), Herbarium of the Muséum national d Histoire Naturelle Paris (MNHM), Herbarium of the Swedish Museum of Natural History (S), Herbarium of the Natural History Museum of Vienna (W) ΚΛΕΙΔΕΣ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΕΙΔΩΝ ΤΟΥ ΓΕΝΟΥΣ Hymenonema Cass. Από την προσεκτική εξέταση και μελέτη του υλικού που συλλέχθηκε για την παρούσα εργασία αλλά και από αποξηραμένο υλικό από τα Herbaria που αναφέρονται παραπάνω, προέκυψαν οι παρακάτω κλείδες προσδιορισμού ειδών του γένους Hymenonema. Πάππος έντονα ανισοϋψής με διαφορά μήκους μεταξύ των τριχών από 8-15 mm. Αχαίνια έντονα τριχωτά, μήκους 4,7-6 mm και πλάτους 1,4-2 mm. Σωλήνας ανθήρων σκούρος βυσσινί και έντονα τριχωτός. Γλωσσοειδή ανθίδια πορτοκαλοκίτρινα με σκούρα βυσσινί κηλίδα στη βάση. Φύλλα βάσης έντονα σχισμένα με τερματικό λοβό μήκους (9)10-35 mm. 1. Η.laconicum Πάππος σχεδόν ισοϋψής με διαφορά μήκους μεταξύ των τριχών από 0,5-4,8 mm. Αχαίνια αραιά τριχωτά, μήκους 3,5-5(6) mm. Σωλήνας ανθήρων κίτρινος και αραιά τριχωτός. Γλωσσοειδή ανθίδια κίτρινα χωρίς σκούρα βυσσινί κηλίδα στη βάση. Φύλλα βάσης εντονότατα σχισμένα με τερματικό λοβό μήκους 3-10(21) mm Η. graecum 29

31 Εικ. 6: Χάρτης εξάπλωσης του Hymenonema laconicum. Με κόκκινους κύκλους σημειώνονται οι περιοχές συλλογής του υλικού της μελέτης, με μαύρους οι περιοχές που αναφέρονται στην βιβλιογραφία και με μωβ κύκλους οι περιοχές που αναφέρονται σε αποξηραμένα δείγματα (Φωτ/φία από την εφαρμογή Google Earth). 30


32 Εικ. 7: Χάρτης εξάπλωσης του Hymenonema graecum. Με κόκκινους κύκλους σημειώνονται οι περιοχές συλλογής του υλικού της μελέτης, με μαύρους οι περιοχές που αναφέρονται στην βιβλιογραφία και με μωβ κύκλους οι περιοχές που αναφέρονται σε αποξηραμένα δείγματα (Φωτ/φία από την εφαρμογή Google Earth). 31
33 2.5. ΠΕΡΙΓΡΑΦΕΣ ΕΙΔΩΝ 1. Hymenonema laconicum Boiss. & Heldr. in Boiss., Fl. Or. 3: 715 (1875). Περιγραφή: Πολυετές. Βλαστοί όρθιοι, ύψους 30-75cm. Φύλλα βάσης μήκους cm και πλάτους 2,5-13,5 cm, έντονα σχισμένα με λοβωτά τμήματα, το τερματικό τμήμα πλάτους (9-)10-35 mm, μεγαλύτερο από τα πλευρικά τμήματα. Φύλλα βλαστού 1-6(-8), μήκους mm και πλάτους 3-36 mm, σαν της βάσης ή σαν βράκτια. Παρουσία τριχών σε βλαστό και σε φύλλα. Κεφάλια 1-4. Περίβλημα μήκους mm και πλάτους mm, βράκτια σε σειρές, επιμήκη-ωοειδή, λεία, με μεμβρανώδες περιθώριο 1-3 mm και οξύληκτο άκρο. Γλωσσοειδή ανθίδια πορτοκαλοκίτρινα με 5 οδοντώσεις στην άκρη, συνήθως με σκούρα βυσσινί κηλίδα στη βάση, νήματα ανθήρων 5 ενωμένα σε σωλήνα σκούρο βυσσινί και έντονα τριχωτό, στην άκρη του νήματος τριγωνική απόληξη ίδιου χρώματος, στίγμα κίτρινο, δισχιδές. Αχαίνια μήκους 4,7-6 mm και πλάτους 1,4-2 mm με 5 αυλακώσεις (ribs), έντονα τριχωτά με διακλαδισμένες τρίχες, στικτά, πάππος έντονα ανισοϋψής, με μια σειρά τριχών διακλαδισμένων, μήκους mm, περίπου το 1/3 της τρίχας του πάππου στο άκρο σκούρο βυσσινί. Περιγραφές για το Hymenonema laconicum έχουν δοθεί από τον Sell (1976) και από τους Tan & Iatrou (2001) (Εικ. 8). Στα περισσότερα χαρακτηριστικά υπάρχει συμφωνία με τους προηγούμενους ερευνητές. Διαφορές εντοπίστηκαν στο ύψος του βλαστού, όπου στην βιβλιογραφία δίνεται 20-70(-120), στον αριθμό των κεφαλίων όπου σημειώνονται 1-3, στα φύλλα βάσης, στα οποία το μήκος καταγράφεται 6-25 cm από τους Tan & Iatrou (2001), ενώ οι μετρήσεις μας μοιάζουν περισσότερο με αυτές της Flora Europaea ( mm). Όσον αφορά τις διαστάσεις του περιβλήματος και των αχαινίων, βρέθηκαν μικρές διαφορές σε σχέση με την υπάρχουσα βιβλιογραφία, που δίνει 15-24( ή 25) x mm για το περίβλημα και 4-5 mm για το μήκος αχαινίου. Οι μετρήσεις για το τερματικό τμήμα των φύλλων βάσης επίσης διαφέρουν συγκριτικά με αυτές που δίνονται από τη βιβλιογραφία, καθώς αναφέρεται το πλάτος του στα (10-)15-30 mm. Από τη μελέτη μας προέκυψαν νέες καταγραφές σχετικά με μετρήσεις και παρατηρήσεις για το είδος, σε σχέση με: το πλάτος των φύλλων βάσης, το μήκος και πλάτος φύλλων βλαστού, τον αριθμό βρακτίων περιβλήματος, το μήκος μεμβρανώδους περιθωρίου, το πλάτος αχαινίων, το μήκος τριχών πάππου και παρατηρήσεις σχετικά με το σωλήνα των ανθήρων, της 32




34 A Γ Β Δ Εικ. 8: Hymenonema laconicum: Α, βιότοπος του φυτού στο Γεράκι Λακωνίας, Β, άτομο από το Ξηροκάμπι, Γ, ταξιανθία από τον Ταΰγετο και Δ, άτομα από το Γεράκι Λακωνίας (Φωτ. Α, Β, Δ από Ι. Κοφινάς και Γ από Χ. Κυριακόπουλος). 33
35 σκούρας βυσσινί κηλίδας στη βάση των ανθιδίων, το στικτό αχαίνιο και το σκούρο βυσσινί τερματικό τμήμα της τρίχας του πάππου. Χρωμοσωματικός αριθμός: 2n = 2x = 20 Άνθιση: Μάιος - Ιούλιος. Βιότοπος: Ξηρές και εγκαταλειμμένες εκτάσεις, ανάμεσα σε θάμνους Quercus, Pistacia, Cistus, περιθώρια δρόμων, ελαιώνες, σε ασβεστολιθικά πετρώματα και σε υψόμετρο m. Εξάπλωση: Κεντρική και Νότια Πελοπόννησος. Καθεστώς Προστασίας: Δεν αναφέρεται. Σύμφωνα με την τελική αναφορά/κείμενο διαβούλευσης για την «Οικολογική αποτύπωση του όρους Ταΰγετος για τη διατήρηση της βιοποικιλότητας» (υπό την επιστημονική αξιολόγηση του ΕΚΒΥ), το H. laconicum προτείνεται μεταξύ άλλων 47 ειδών, να χρησιμοποιηθεί ως είδος-δείκτης ή διαχειριστικός στόχος για τη λήψη των κατάλληλων μέτρων προστασίας του ενδιαιτήματός του και τη διαρκή αξιολόγηση της αποτελεσματικότητάς του. Εν δυνάμει θα μπορούσε να χρησιμεύσει στην ενημέρωση και ευαισθητοποίηση του κοινού, καθώς και στην προσέλκυση συγκεκριμένων ομάδων επισκεπτών, στο πλαίσιο ενός οργανωμένου οικοτουριστικού σχεδιασμού (Γεωργιακάκης & al. 2011). Στη βάση δεδομένων του NATURA 2000, το ενδημικό φυτό αναφέρεται σε 5 περιοχές NATURA τις: GR Όρος Μαίναλο, GR Όρος Πάρνωνας και περιοχή Μαλεβής (χαρακτηρίζεται ως πολύ σπάνιο), GR Μονή Ελώνας και χαράδρα Λεωνιδίου, GR Όρος Ταΰγετος (το μέγεθος πληθυσμού υπολογίζεται άτομα), GR Όρος Ταΰγετος Λαγκάδα Τρύπης. Εξετασθέντα δείγματα: GREECE: PELOPONNISOS: ARKADIA: c. 3.1 km from the small town of Leonidio towards Tsitalia village. Stony slopes with abandoned olive groves, together with Ceratonia siliqua, Pistacia lentiscus, Calicotome villosa, Cistus creticus etc. Large population., alt. c m, N, E, , Kalpoutzakis 1506 (UPA); ibid. alt. 113 m, N, E, , Kofinas & Dolianitis 129 (UPA); c. 6.8 km from Leonidion towards Poulithra, close to Poulithra. Roadsides and margins of abandoned fields. Florets orange., alt m, N, E, , Constantinidis & Kalpoutzakis (UPA). 34
36 LAKONIA: Geraki, road from the village to Alepohori, alt m, N, E, , Liveri & Ketsilis-Rinis 124 (UPA); ibid , Kofinas 126; ibid , Kofinas 128; Lagkada gorge, climbing region, alt. 800 m, 37 4'59.09"N, 22 18'39.30"E, , Charalampopoulos 1524 (UPA); ibid , Liveri & Kofinas 130; entrance of Kaiadas, , Kofinas 132 (UPA); Anavriti, alt. 440 m, N, E, , Charalampopoulos & Kofinas s.n. (UPA); prope pagum Krokeae, in argilosis, N, E, 22.VI.1980, Iatrou 588I (UPA); ibid.9.v.1965, Phitos 1459 (UPA); ibid , Liveri & Kofinas 131 (UPA); 1-2 km from Siderokastron, along road to Skofianitika and Skamnitsa. Quercus/Pistacia scrub. Latex white. Ligules orange-yellow with purplish-black base. Stigmas bright lemon-yellow., alt. 530 m, N, E, , Tan & Vold 9754 (UPA); ibid , Tan (B); inter oppidum Gythion et pagum Krokeae, ad viam in argilosis, N, E, , Tzanoudakis & Iatrou 5724 (UPA); Mt. Taigetos. NW of Arna, c. 25 km S of Sparti, 950 m, 22º 24' E, 36º 53' N, , Runemark & Snogerup ( LD); Inter Sparta et Skouras, Iter Graecum XI 1964, Rechinger (B); In regione [olearum] ad radices Taygeti in herbidis prope [Androuvista], 06/1844,Heldreich s.n. (RBGK); Taygetus, Heldreich s.n. (RBGK). MESSINIA: Laconia boreo-occidentalis: in regione litorali ad radices m. Selitza prope Kalamata, Zahn 1355 (RBGK); Ep. Kalamon: Pr. Selitza ad radices m. Taygeti, 22º 7' E, 37º 3' N, , Heldreich (37837 LD); ibid , Heldreich 1355 (37838 LD); 8 km SE of Kalamata. Locally common on roadsides along olive gr. (This spec. was taken above hotel Lida), , Emanuelsson 1886 ( S). 2. Hymenonema graecum (L.) DC., Prodr. 7: 116 (1838). = Catananche graeca L.= Scorzonera elongata Willd. Περιγραφή: Πολυετές. Βλαστοί τριχωτοί, όρθιοι, ύψους cm. Φύλλα βάσης μήκους 3,2-35,4 cm και πλάτους 0,7-7 cm, πολύ έντονα σχισμένα με οδοντωτά τμήματα, το τερματικό τμήμα πλάτους 3-15(-21) mm, μεγαλύτερο από τα πλευρικά τμήματα. Φύλλα βλαστού 1-6(-8), μήκους mm και πλάτους 3-38 mm, σαν της βάσης ή σαν βράκτια. Παρουσία τριχών σε βλαστό και σε φύλλα. Κεφάλια 1-4. Περίβλημα μήκους mm και πλάτους mm, βράκτια σε σειρές, 35
37 επιμήκη-ωοειδή, λεία, με μεμβρανώδες περιθώριο 0,8-2,5 mm με οξύληκτο άκρο. Γλωσσοειδή ανθίδια κίτρινα με 5 οδοντώσεις στην άκρη, νήματα ανθήρων 5 ενωμένα σε σωλήνα κίτρινο και αραιά τριχωτό, στην άκρη του νήματος τριγωνική απόληξη ίδιου χρώματος, στίγμα κίτρινο, δισχιδές. Αχαίνια μήκους 3,5-6 mm και πλάτους 0,8-1,8 mm με 5 αυλακώσεις (ribs), αραιά τριχωτά με διακλαδισμένες τρίχες, στικτά, πάππος σχεδόν ισοϋψής, από μια σειρά τριχών διακλαδισμένων μήκους mm. Αναφορές στην περιγραφή του είδους γίνονται στις ίδιες βιβλιογραφικές πηγές στις οποίες γίνονται αναφορές και στην περιγραφή του H. laconicum. Σ αυτές τις πηγές σημειώνεται ότι το H. graecum (Εικ. 9) παρουσιάζει τα ίδια χαρακτηριστικά με το H. laconicum, εκτός από 4 διαφορές: 1) ο τερματικός λοβός των φύλλων βάσης δεν ξεπερνά τα 10 mm, 2) τα ανθίδια μερικές φορές έχουν μια σκούρα βυσσινί κηλίδα στη βάση, 3) οι λάκκοι του δίσκου του κεφαλίου είναι λείοι ή με λίγες κοντές βλεφαρίδες, 4) τα αχαίνια έχουν σχεδόν ομοιόμορφο πάππο. Όμως στην παρούσα μελέτη, το 28% των τερματικών λοβών που μετρήθηκε για το H. graecum έχει πλάτος μεγαλύτερο των 10 mm, ενώ σκούρα βυσσινί κηλίδα δεν παρατηρήθηκε ποτέ σε κανέναν αντιπρόσωπο του είδους. Αντίθετα, η σκούρα βυσσσινί κηλίδα στη βάση του ανθιδίου αποτελεί χαρακτηριστικό του H. laconicum, στην περιγραφή του οποίου δεν αναφέρεται. Πρέπει να σημειωθεί, ότι σε μερικά άτομα του είδους των πληθυσμών από τη Σύρο και την Τήνο, παρατηρήθηκε διαφοροποίηση στο χρώμα του σωλήνα των ανθήρων και στη μορφολογία των αχαινίων, η οποία αναλύεται εκτενώς στο κεφάλαιο της μορφολογικής μελέτης. Χρωμοσωματικός αριθμός: 2n = 2x = 20 Άνθιση: Μάιος -Ιούλιος. Βιότοπος: περιθώρια παραλιακών αλατούχων εδαφών, παρυφές των δρόμων, πετρώδεις θέσεις, περιοχές με φρύγανα και garigue, κατοικημένες περιοχές, ασβεστολιθικά, σχιστολιθικά και γρανιτικά πετρώματα, σε υψόμετρο m. Εξάπλωση: Κυκλάδες και ΒΔ Κρήτη (?). Από την περιοχή των Κυκλάδων έχει βρεθεί σε πολλά νησιά και νησίδες. Παρ όλα αυτά, οι καταγραφές του H. graecum για τους πληθυσμούς από Αγκαθωπές (Σύρος), Μέγας Γυαλό (Σύρος), Κολυμπήθρα (Τήνος) και Μοναστήρι, Αρνάδος, Δυο 36




38 Β Α Γ Δ Εικ. 9: Hymenonema graecum: Α, άτομο από Μέριχα Κύθνου, Β, ταξιανθία από Κιόνια Τήνου, Γ, ταξιανθία από καλλιεργημένο υλικό από Μέριχα Κύθνου και Δ, βιότοπος στις Αγκαθωπές Σύρου (Φωτ. Α από Γ. Καμάρη). 37
 (Εικ. 10).")
39 Χωριά (Τήνος) είναι νέες. Η εμφάνισή της στην Κρήτη έχει επιβεβαιωθεί με δείγμα του Zaffran από την δεκαετία του 60 και έκτοτε δεν έχει επανευρεθεί. Σύμφωνα με αυτή την καταγραφή, το H. graecum βρέθηκε ανατολικά των Χανίων, στο δυτικότερο σημείο του κόλπου της Σούδας (Turland & al. 1993, προσ. επικ. Nick Turland) (Εικ. 10). Το δείγμα του Zaffran αρχικά βρισκόταν στο Herbarium της Ορθόδοξης Ακαδημίας στο Κολυμπάρι κι έπειτα στο ΜΑΙΧ. Στην Euro+Med PlantBase, η εμφάνιση του φυτού στην Κρήτη χαρακτηρίζεται ως αβέβαιη. Σύμφωνα με την ίδια πηγή, αβέβαιη θεωρείται η εμφάνιση του φυτού και στην Τουρκία. Ο ενδοιασμός αυτός προέκυψε, διότι το ένα δείγμα του Herb.-Boissier που βρίσκεται στην Τουρκία (Κωνσταντινούπολη), φαίνεται ότι ανήκει σε καλλιεργημένο υλικό (Γ. Καμάρη, προσ. επικ.). Εικ. 10: Η περιοχή εμφάνισης του Hymenonema graecum στην Κρήτη κατά Turland & al Καθεστώς Προστασίας: Π.Δ. 67/1981, WCMC. Στο Π.Δ. 67/1981 το H. graecum χαρακτηρίζεται ως αμφιβόλως ενδημικό. Εντοπίζεται σε 5 περιοχές NATURA τις: GR Νήσος Πολύαιγος - Κίμωλος, GR Κεντρική και νότια Νάξος: Ζας και Βίγλα έως Μαυροβούνι και θαλάσσια ζώνη (όρμος Καράδες όρμος Μουτσούνας), GR Μικρές Κυκλάδες: Ηρακλειά, Σχοινούσσα, Κουφονήσια, Κέρος, Αντικεριά και θαλάσσια ζώνη, GR Ανάφη: χερσόνησος Κάλαμος - Ρουκούνας, GR Σαντορίνη: Νέα και Παλιά Καμένη Προφήτης Ηλίας. Εξετασθέντα δείγματα: GREECE: INSULAEAEGEAE: KIKLADES: Ins. Anafi: Kalamos, 1 km E of the monastery, cliffs, garrigue, m, 36º21'N, 25º51'E, , Runemark & 38
40 Snogerup 8132 ( LD); ibid ( LD);alt. 322 m, substrate: calcareous rocks, N, E, 26.ΙV.2011, Kougioumoutzis 1172 (UPA); ibid. alt. 244 m, substrate: gabbrodiorites, N, E, 1431 (UPA); ibid. alt. 209 m, substrate: alluvial, N, E, 1499 (UPA); ibid. alt. 277 m, substrate: granite, N, E, 28.V.2011, 1842 (UPA); 3.VI.1898, Leonis s.n. (MNHN). Ins. Andros: Batsi, along the roadsides NE of the village, phrygana, alt m, N, E, , Liveri & Ketsilis-Rinis 114 (UPA); ibid , Snogerup & Snogerup 6666 ( LD); Batsi, residential area, Agios Filippos church, alt. 45 m, N, E, , Liveri & Ketsilis-Rinis 115 (UPA); oros Rakhi, m, , Snogerup & Bothmer ( LD); near the sea 2 km SW of Zaganiari, m, , Snogerup ( LD). Ins. Andikeros: cliffs, 36º51'N, 25º41'E, , Runemark & Snogerup ( LD). Ins. Anidros: the SW-part of the island, limestone, garigue, 36º38'N, 25º41'E, , Runemark & Snogerup 8261 ( LD); insula Anhydros (Amorgopulos), in saxosis, substr. calc., 36º37'N, 25º41'E, Rechinger 5260 ( LD). Ins. Folegandros: Adelfia Petra, the E-island, litoral limestone rocks, 36º37'N, 24º59'E, , Runemark & Nordenstam ( LD). Ins. Iraklia: S of the harbor, 36º50'N, 25º29'E, , Runemark 4146 ( LD); ormos Pegadi, the small valley W of the bay, on sandy soil, 36º50'N, 25º29'E, , Runemark & Snogerup 5297 ( LD); cycladum insula Heraklia: in saxosis, 36º51'N, 25º28'E, , Rechinger 4892 ( LD). Ins. Kardiotissa: garigue, 36º38'N, 25º1'E, , Runemark & Nordenstam ( LD). Ins. Kithnos: in collibus ad meridiem pagi Merichas spectantibus, alt. c. 50 m, in nanofruticetosis, solo schistoso, 20.V.2007, Phitos & Kamari (UPA); ibid , Liveri & Ketsilis-Rinis 100 (UPA); ibid m, , Strid ( LD); in collibus supra pagi Merichas ad viam versus pagum Dryopida, alt. c. 100 m, solo schistoso, 20.V.2007, Phitos & Kamari (UPA). Ins. Koufonisia: Kopria, hard limestone cliffs, 36º59'N, 25º38'E, , Runemark & Snogerup 10279a ( LD). Ins. Makares: Prasonisi, garigue, 0-80 m, 37º5'N, 25º42'E, , Runemark & Snogerup 9662 ( LD); ibid. ( LD).Ins. Mikonos: the valley S of Mt. Ag. Elias, m, 37º28'N, 25º20'E, , Runemark & Engstrand ( LD); S of Ag.Stefano, 37º28'N, 25º20'E, , Runemark & Engstrand (
41 LD); 2 km N of Ag. Stefano, 37º30'N, 25º19'E, , Runemark & Engstrand ( LD); the island of Stapodia, 37º25'N, 25º34'E, , Runemark & Engstrand ( LD); the island of Ag.Georgios, 37º27'N, 25º18'E, , Runemark & Engstrand ( LD). Ins. Milos: W and SW Chivadolimni, wet area,36 44'N, 24 26'E, , Runemark ( LD); S of Akr. Roma, in a valley, 36 42'N, 24 32'E, , Runemark & Bentzer ( LD); Urville s.n. (MNHM). Ins. Naxos: Prokopios, garrigue, 30 m, , Jäth s.n. (B); E of Mytria, along a small stream, limestone cliffs, garigue, m, 37º8'N, 25º27' E, , Runemark 2637 ( LD); ibid ( LD); ibid , 3635 ( LD); Metri N of Moni, m, 37º5'N, 25º30'E, , Runemark 3174 ( LD); Faneromeni, along a small stream, m, 37º9'N, 25º29'E, , Runemark 3299 ( LD); ENE of Skado, valley with a rill, 500 m, 37º8'N, 25º33'E, , Runemark 3736 ( LD); SSE of Axapsis, along a small stream, 37º7'N, 25º26'E, , Runemark 3909 ( LD); Kalando, 36º56'N, 25º28'E, , Runemark cult ( LD); ibid. ( LD); in the valley 0-3 km N of Ormos Agiasou, garigue, 5-60 m, 36º59'N, 25º26'E, , Runemark & Snogerup ( LD); Psiliamos Ormos, 37º1'N, 25º34'E, , Runemark & Snogerup ( LD); ibid ( LD); 2 km NNW of Ormos Liona, garigue near the sea, 37º9'N, 25º35'E, , Runemark & Snogerup 9052 ( LD); NW outskirts of the town of Naxos, calcareous soils, 25 m, 37º7'N, 25º24'E, , Franzén & Andersson 1009 ( LD); in monte Phanariotissa ad pagum Apiranthos, in saxosis calc., m, 37º8'N, 25º31'E, , Rechinger 2237 ( LD); in Cycladum insula Naxos, 37º5'N, 25º26'E, Leonis 4171 ( LD); Ostküste, Moutsouna, Landzunge S des Hafens, 10 m, , Karl s.n. ( GZU). Ins. Paros: Gaidouronisi, 30 m, 37º9'N, 25º16 E, , Runemark & Bentzer ( LD). Ins. Poliegos: the NW-part, garigue-field, 36º47'N, 24º37'E, , Runemark & Bentzer ( LD). Ins. Schinousa: between the principal village and the harbor, 36º52'N, 25º31'E, , Runemark 4355 ( LD); ibid. cult. 124 ( LD); ibid. ( LD). Ins. Serifopoula: 37º 15' N, 24º 36' E, , Runemark & Bentzer ( LD). Ins. Serifos: from the Taxiarchis closter downwards the Plati Gialos Bay, micaceous schist., N, E, , Tiniakou 478 (UPA); ibid. 494; Profitis Ilias, Tiniakou 3454 (UPA); Livadi, granitic rocks of 40
42 the port, N, E, , Tiniakou 545 (UPA); 1-2 km SW of Livadion, m, 37º 8' N, 24º 30' E, , Runemark & Bentzer ( LD); ibid ( LD); ibid m, ( LD); above the Kato Ampeli bay, alt m, granitic substrate, N, E, , Tiniakou, Artelari, Georgiou & Christodoulakis 2305 (UPA); at the Petrias Mt., 1-2 km after Pano Chora, alt m, granitic substrate, N, E, , Tiniakou, Artelari, Georgiou & Christodoulakis 2256 (UPA); at the strand and on the rocks of the Vagia-bay, N, E, , Tiniakou 285 (UPA); between the villages of Panagia and Kentarchos, alt. ca 300 m, micaceous schist., N, E, , Tiniakou 470 (UPA); Aspros Kavos; marbles, N, E, , Tiniakou 534 (UPA); Koutalas Tiniakou 3654 (UPA); ibid. 3585; ibid. 400 m, , Malicky s.n. (W); ibid , Runemark & Bentzer ( LD); vertical rocks facing NW, just N of the village of Serifos, m, 37º 9' N, 24º 30' E, , Strid ( LD); S of Panagia, 400 m, 37 11'N, 24 25' E, , Runemark ( LD). Ins. Sifnos: S of Kamares, 36º59'N, 24º40'E, , Runemark & Bentzer ( LD); limestone hill S of Ormos Kondos, garigue, m, 36º54'N, 24º42'E, , Runemark & Snogerup 8402 ( LD). Ins. Sikinos: N-exposed cliffs E of Kastro, m, 36º42'N, 25º8'E, ,Runemark & Bentzer 24645( LD); the E- promontory, S-exposed cliffs facing the sea, 36º43'N, 25º11'E, , Runemark & Bentzer ( LD); the S-slope of the highest mt., m, 36º40'N, 25º06'E, , Runemark & Bentzer ( LD). Ins. Siros: Agathopes, road towards to the beach, on the rocks very close to the sea, alt m, N, E, , Liveri & Ketsilis-Rinis 110 (UPA); Kini, along the roadsides from the beach to the village for a distance up to 1 km approximately, N, E, , Liveri & Ketsilis-Rinis 107 (UPA); ibid. 200 m, , Snogerup ( LD); Megas Gialos, alt. 24 m, N, E, , Ketsilis-Rinis 113 (UPA); the islet of Strongylo E of Didymi, 37º26'N, 24º59'E, , Snogerup & Bothmer ( LD); the island of Aspronisos, hard limestone, 50 m, 37º24'N, 25º0'E, , Snogerup & Bothmer ( LD); the islet of Ampelos E of Vari, 5 m, 37º23'N, 24º57'E, , Snogerup & Bothmer ( LD); the islet of Kommeno NE of Ermoupolis, 20 m, 37º28'N, 24º57'E, , 41
43 Snogerup & Bothmer ( LD); the islet of Varvarousa, limestone, 25 m, 37º28'N, 24º54'E, , Snogerup & Bothmer ( LD); Cycladum insulae Syros, Tenos, Naxos et Melos, legimus in ins. Syro, in collibus siccis et ad vias, 37º27'N, 24º55'E, , Heldreich 1055 ( LD); ibid. (MNHM). Ins. Tinos: Kionia bay, on the rocks, phrygana, alt m, N, E, , Liveri & Ketsilis-Rinis 121 (UPA); ibid , Landström 7346 ( LD); Kolimpithra bay, on the rocks, phrygana, alt m, N, E, , Liveri & Ketsilis-Rinis 123 (UPA); Monastiri, Arnados, Dio Xoria, along the roadsides connecting these three villages, alt m, N to N, E to E, , Liveri & Ketsilis-Rinis 117 (UPA); Platia Ammos, 37º33'N, 25º8'E, , Runemark & Engstrand ( LD); ad litore ins. Teni, 37º33'N, 25º6 E, , Sartori s.n. ( LD); ibid. (MNHM). 42
44 3. ΜΟΡΦΟΛΟΓΙΚΗ ΜΕΛΕΤΗ 3.1. ΜΟΡΦΟΛΟΓΙΑ Τα μορφολογικά χαρακτηριστικά χρησιμοποιούνται από την αρχαιότητα για να ερμηνεύσουν τις σχέσεις μεταξύ των οργανισμών. Πιο συγκεκριμένα, οι πρώτες προσπάθειες κατάταξης των φυτών σε ταξινομικές μονάδες βασίστηκαν σε παρατηρήσεις επί της εξωτερικής μορφολογίας των φυτών και προοδευτικώς σε πιο λεπτομερή μορφολογικά χαρακτηριστικά. Από τον πατέρα της Βοτανικής, Θεόφραστο ( π.χ.), ως τον θεμελιωτή της σύγχρονης ταξινομικής, Κάρολο Λινναίο (Carl Linné ), η μορφολογία των φυτών έπαιξε πρωταγωνιστικό ρόλο. Στη σημερινή εποχή, παρ ότι έχουν αναπτυχθεί σύγχρονες μέθοδοι για την ταξινόμηση και ανάλυση των φυτών, οι μορφολογικοί χαρακτήρες αποτελούν αναπόσπαστο κομμάτι μιας ολοκληρωμένης εργασίας για ένα φυτικό taxon και μας παρέχουν αναμφίβολα σημαντικές πληροφορίες. Η χρήση του όρου «μορφολογία», στην εν λόγω μελέτη, γίνεται με την ευρεία έννοια του όρου δηλ. περιλαμβάνονται αναπαραγωγικοί, βλαστητικοί, μορφολογικοί και ανατομικοί χαρακτήρες. Τα παρακάτω δεδομένα προέρχονται από την υπάρχουσα βιβλιογραφία για το γένος Hymenonema καθώς και για συγγενικά του είδη. Φύλλωμα: Η θέση των φύλλων είναι ένα από τα πιο χρήσιμα μορφολογικά χαρακτηριστικά σε επίπεδο γένους και tribe. Επίσης, ο βαθμός εγκόλπωσης και νεύρωσης των φύλλων, καθώς και η παρουσία ή όχι ακάνθων αποτελούν χρήσιμες μορφολογικές παρατηρήσεις. Τα φύλλα βάσης παρουσιάζουν έντονη εγκόλπωση, ενώ το πλάτος το τερματικού λοβού των φύλλων βάσης αποτελεί διαχωριστικό ταξινομικό κριτήριο για τα δυο είδη. Σύμφωνα με τον Sell (1976), τα 10 mm αποτελούν το μέγιστο πλάτος του τερματικού λοβού για το Hymenonema graecum, ενώ για το H. laconicum κυμαίνεται από 10 έως και 30 mm. Σε όλα τα φύλλα παρατηρούνται τρίχες (Tan & Iatrou 2001). Τρίχωση: Σε ολόκληρη την οικογένεια Compositae υπάρχουν αρκετοί τύποι τριχών όπως μονο- ή πολυ-κύτταρες, διακλαδισμένες ή όχι, σχήματος Τ ή Υ, αστεροειδείς, καθώς και αδενώδεις (Ramayya 1962). Για την tribe Lactuceae έχουν αναγνωριστεί συγκεκριμένα 8 τύποι τριχών, ενώ από τα δεδομένα αυτά έχει προταθεί και η ταξινόμησή τους σε επίπεδο γένους. Σύμφωνα μ αυτή τη μελέτη, οι τρίχες που παρατηρήθηκαν στα φύλλα των Hymenonema graecum και H. laconicum, 43
.")
. Α Εικ. 11: Α, Κοντή κωνική τρίχα του Stephanomeria panniculata.")
45 χαρακτηρίζονται ως κοντές, κωνικές, μη αδενώδεις, σε μια σειρά, όπως και στο είδος Stephanomeria panniculata (Εικ. 11Α). Στο συγγενικό με το γένος Hymenonema, Catanananche caerulea, οι τρίχες είναι του ίδιου τύπου, αλλά μακριές (Εικ. 11Β). Απαρτίζονται από 5 συνήθως κύτταρα με παχιά κυτταρικά τοιχώματα ενώ σε μερικές περιπτώσεις η βάση αποτελείται από 2 κύτταρα. Η ύπαρξη εφυμενιδικών μιρκοθηλών και παχιών κυτταρικών τοιχωμάτων είναι χαρακτηριστικά γι αυτό τον τύπο τρίχας (Krak & Mráz 2008). Α Εικ. 11: Α, Κοντή κωνική τρίχα του Stephanomeria panniculata. Κλίμακα = 20 μm. Β, Μακριά κωνική τρίχα του Catananche caerulea Κλίμακα = 200 μm (Krak & Mráz 2008). Ταξιανθία: Το κεφάλιο είναι ο τύπος ταξιανθίας που χαρακτηρίζει όλους τους αντιπροσώπους της οικογένειας Compositae. Το κεφάλιο αποτελείται από ανθίδια τα οποία μπορεί να είναι είτε σωληνοειδή είτε γλωσσοειδή είτε συνδυασμός και των δυο (Εικ. 12). Η στεφάνη είναι συμπέταλη. Το ανθίδιο μπορεί να φέρει 3 ή 5 οδοντώσεις στην άκρη. Οι στήμονες είναι 5, έχουν ελεύθερα νήματα και στερεώνονται στη στεφάνη (επιπέταλοι). Οι ανθήρες έχουν ακιδωτή απόληξη ενώ ενώνονται με τέτοιο τρόπο ώστε να σχηματίζουν ένα σωλήνα στον οποίο απελευθερώνεται η γύρη. Ο σωλήνας αυτός περιβάλλει το στύλο. Ο στύλος είναι μονήρης με δισχιδές στίγμα. Η ωοθήκη είναι υποφυής (Moore & al. 1976). Στο γένος Hymenonema παρατηρούνται μόνο γλωσσοειδή ανθίδια με 5 οδοντώσεις, τα οποία μάλιστα διαφέρουν στο χρωματισμό ανάμεσα στα δυο είδη. Καρπός: Ο καρπός είναι σημαντικό κριτήριο στα Compositae. Οι περισσότεροι εκπρόσωποι της οικογένειας, όπως και το γένος Hymenonema, χαρακτηρίζονται από σπέρματα μορφής αχαινίου (Εικ. 13). Πρόκειται για καρπό μικρό, ξηρό, μη 44
 και ένα σπέρμα / ωάριο (ovule), με το σπέρμα να συνδέεται με το τοίχωμα της ωοθήκης σε ένα μόνο σημείο (Harris & Woolf Harris 1994).")
, όπου χαρακτηρίζεται από ανισομεγέθεις τρίχες. A B Α Β Γ Εικ. 13: Αχαίνιο, πάππος και λεπτομέρειες του πάππου (Β), = 200 mm (Γ) (Kilian et al. 2009).")
. Με βάση την εξωτερική μορφολογία οι γυρεόκοκκοι μπορεί να χαρακτηριστούν ως echinate, echinolophate ή psilolophate, δηλαδή φέρουν μικρές τριγωνικές ακάνθους.")
46 Εικ. 12: Σχηματική απεικόνιση A, ταξιανθίας (Compositae) αποτελούμενη από τους δυο τύπους ανθιδίων ( και B, γλωσσοειδούς ανθιδίου (en.wikipedia.org/wiki/asteraceae). διανοιγόμενο, με μια κοιλότητα (locule) και ένα σπέρμα / ωάριο (ovule), με το σπέρμα να συνδέεται με το τοίχωμα της ωοθήκης σε ένα μόνο σημείο (Harris & Woolf Harris 1994). Η ομοιομορφία ή όχι του πάππου αποτελεί το δεύτερο γνώρισμα διάκρισης ανάμεσα στα δυο είδη Hymenonema, με το H. graecum να παρουσιάζει ομοιόμορφο πάππο με ισομεγέθεις τρίχες, σε σχέση με το H. laconicum (Sell 1976, Tan & Iatrou 2001), όπου χαρακτηρίζεται από ανισομεγέθεις τρίχες. A B Α Β Γ Εικ. 13: Αχαίνιο, πάππος και λεπτομέρειες του πάππου (Β), = 200 mm (Γ) (Kilian et al. 2009). Κλίμακες = 3 mm (Α), = 100 mm Γυρεόκοκκοι: Η μορφολογία των γυρεοκόκκων σε αντιπροσώπους των Compositae έχει μελετηθεί από πολλούς ερευνητές, όπως ο Blackmore (1982), που ασχολήθηκε μάλιστα με την tribe Lactuceae (Εικ. 14). Με βάση την εξωτερική μορφολογία οι γυρεόκοκκοι μπορεί να χαρακτηριστούν ως echinate, echinolophate ή psilolophate, δηλαδή φέρουν μικρές τριγωνικές ακάνθους. Το τοίχωμα του γυρεόκοκκου, επίσης, 45
 και Scolymus hispanicus (B). Κλίμακα = 10 μm. (Blackmore 1982)")
.")
47 μπορεί να δώσει σημαντικές πληροφορίες. Διακρίνεται σε δυο τύπους: caveate και ecaveate, με τη δεύτερη κατηγορία να είναι πιο συνηθισμένη στα Cichorioideae (Bremer 1994). Α Β Εικ. 14: Γυρεόκοκκοι από Catananche caerulea (A) και Scolymus hispanicus (B). Κλίμακα = 10 μm. (Blackmore 1982) ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Συνολικά έγιναν μετρήσεις σε 30 δείγματα από το Hymenonema graecum και 20 από το H. laconicum καλύπτοντας 7 και 4 πληθυσμούς για το κάθε είδος αντίστοιχα (Πίνακας 3). Οι μετρήσεις έγιναν σε αποξηραμένα δείγματα συλλογών από το Herbarium του Βοτανικού Μουσείου του Πανεπιστημίου Πατρών (UPA). Για τις μακροσκοπικές παρατηρήσεις και μετρήσεις χρησιμοποιήθηκε στερεοσκόπιο Stemi SV-6 (Zeiss), καθώς και το στερεοσκόπιο Stemi 2000-C (Zeiss), το οποίο συνδέθηκε με φωτογραφική μηχανή για τη λήψη των φωτογραφιών. Για τις μικροσκοπικές παρατηρήσεις, χρησιμοποιήθηκε το μικροσκόπιο AXIOPHOT (Zeiss). Οι μορφολογικοί χαρακτήρες που εξετάστηκαν είναι οι: ύψος φυτού, μήκος και πλάτος κεφαλίου, αριθμός βρακτίων, μήκος και πλάτος φύλλων βάσης, τερματικός λοβός φύλλων βάσης, μήκος και πλάτος φύλλων βλαστού, μήκος και πλάτος αχαινίου, μήκος πάππου (μετρήθηκε η μεγαλύτερη τρίχα του πάππου), εύρος του μήκους των τριχών του πάππου. Η χρώση των γυρεοκόκκων έγινε με cotton-blue. Μετρήθηκε το ποσοστό γονιμότητας σε 30 παρασκευάσματα γυρεοκόκκων. Οι γυρεόκοκκοι οι οποίοι βάφτηκαν έντονα θεωρούνται γόνιμοι. Ο υπολογισμός των στατιστικών δεδομένων έγινε με τη χρήση Microsoft Office Excel Για τη μελέτη της ποικιλότητας των μορφολογικών χαρακτήρων, έγιναν 46
48 διαγράμματα μέσης τιμής. Υπολογίζεται το τυπικό σφάλμα με διάστημα εμπιστοσύνης 95%. Πίνακας 3: Πληθυσμοί του γένους Hymenonema οι οποίοι μελετήθηκαν μορφολογικά. Taxon Πληθυσμός Pop. no H. laconicum LAKONIA Krokees, alt. 350 m, N, E, , Liveri & Kofinas 131 (UPA); ibid. 22.VI.1980, Iatrou 588I (UPA); ibid. 9.V.1965, Phitos 1459 (UPA); ibid (UPA), Tzanoudakis & Iatrou 5724 (UPA)» ARKADIA: SE of Leonidio, road from Leonidio to 2 Tsitalia village, alt. 113 m, N, E, , Kofinas & Dolianitis 129 (UPA); ibid , Constantinidis & Kalpoutzakis (UPA); ibid , Kalpoutzakis 1506 (UPA).» LAKONIA: Geraki, road from the village to 3 Alepohori, alt m, N, E, , Liveri & Ketsilis- Rinis 124 (UPA); ibid , Kofinas 128 (UPA); ibid , Kofinas 126 (UPA).» LAKONIA: Lagkada gorge, climbing region, alt. 800 m, 37 4'59.09"N, 22 18'39.30"E, , Charalampopoulos 1524 (UPA); ibid , Liveri & Kofinas 130 (UPA). 4 Hymenonema Ins. ANAFI: alt. 322 m, substrate: 5 graecum calcareous rocks, N, E, 26.ΙV.2011, Kougioumoutzis 1172 (UPA); ibid (UPA); ibid (UPA); ibid. 28.V (UPA).» Ins. ANDROS: Batsi, along the roadsides NE of the 6 village, phrygana, alt m, N, E, , Liveri & Ketsilis- Rinis 114 (UPA); ibid. alt. 45 m, 115.» Ins. KITHNOS: in collibus ad meridiem pagi 7 Merichas spectantibus, alt. c. 50 m, in nanofruticetosis, solo schistoso, 20.V.2007, Phitos & Kamari (UPA); ibid (UPA); ibid , Liveri & Ketsilis-Rinis 100 (UPA) 47
.» Ins. TINOS: Monastiri, Arnados, Dio Xoria, along the roadsides connecting these three villages, alt.")
49 » Ins. SERIFOS: Koutalas, Tiniakou 3645 (UPA); ibid » Ins. SERIFOS: above the Kalo Ampeli bay, alt m, granitic substrate, N, E, , Tiniakou, Artelari, Georgiou & Christodoulakis 2305 (UPA).» Ins. SIROS: Kini, along the roadsides from the beach to the village for a distance up to 1 km approximately, N, E, , Liveri & Ketsilis-Rinis 107 (UPA).» Ins. TINOS: Monastiri, Arnados, Dio Xoria, along the roadsides connecting these three villages, alt m, N to N, E to E, , Liveri & Ketsilis-Rinis 117 (UPA) ΑΠΟΤΕΛΕΣΜΑΤΑ Μορφολογικές παρατηρήσεις Ταξιανθία Τα ανθίδια στο H. graecum έχουν χρώμα κίτρινο, ενώ στο H. laconicum πορτοκαλοκίτρινο, με σκούρα μωβ κηλίδα λιγότερο ή περισσότερο έντονη στη βάση του ανθιδίου (Εικ ). Τα ανθίδια έχουν 5 λοβούς που αντιστοιχούν στα 5 πέταλα της συμπέταλης στεφάνης (Εικ. 15) και ο στύλος είναι δισχιδής και για τα δυο είδη. Ο σωλήνας των ανθήρων αλλά και η ακιδωτή απόληξη είναι σκούρο βυσσινί στο H. laconicum (Εικ. 18). Τρίχες περιβάλλουν το σωλήνα, και φαίνονται περισσότερες στο H. laconicum απ ότι στο H. graecum. Ανάμεσα στα άτομα του H. graecum που μελετήθηκαν, παρατηρήθηκαν 2 μορφές σωλήνα ανθήρων: 1) κίτρινος σωλήνας και κίτρινη ακιδωτή απόληξη και 2) σκούρος βυσσινί σωλήνας και κίτρινη ακιδωτή απόληξη (Εικ ). Εικ. 15: Τα 5 ενωμένα πέταλα της στεφάνης του Hymenonema graecum, από την Κύθνο. Κλίμακα = 1 mm. 48






50 Α Β Εικ. 16: Κεφάλιο του Hymenonema laconicum: Α, από το Γεράκι Λακωνίας (Φωτ. Ι. Κοφινάς) και Β, από τις Κροκεές. Α Β Εικ. 17: Κεφάλιο του Hymenonema graecum με κίτρινο σωλήνα ανθήρων: Α, καλλιεργημένο υλικό από την Κύθνο και Β, στο φυσικό του ενδιαίτημα από την Τήνο. Α Β Εικ. 18: Κεφάλιο του Hymenonema graecum με βυσσινί σκούρο σωλήνα ανθήρων από την Τήνο. 49

.")




51 Εικ. 19: Σωλήνας ανθήρων του Hymenonema laconicum από το Γεράκι Λακωνίας (από καλλιεργημένο υλικό). Κλίμακες = 1mm. Εικ. 20: Σωλήνας ανθήρων κίτρινος του Hymenonema graecum από την Κύθνο (καλλιεργημένο υλικό). Κλίμακες = 1mm. Εικ. 21: Σωλήνας ανθήρων μαύρος του Hymenonema graecum από τη Σύρο (καλλιεργημένο υλικό). Κλίμακες = 1mm. 50


52 Καρπός Ο καρπός, όπως έχει αναφερθεί, είναι αχαίνιο που φέρει τον χαρακτηριστικό πάππο (Εικ ). Το σπέρμα φέρει 5 αυλακώσεις, είναι στικτό και φέρει διακλαδισμένες τρίχες. Τα σπέρματα του Hymenonema laconicum φέρουν πυκνή τρίχωση με αποτέλεσμα να διακρίνονται δυσκολότερα οι αυλακώσεις και τα στίγματα. Για το H. graecum εξετάστηκαν σπέρματα και από τις δυο ομάδες που διαμορφώθηκαν σύμφωνα με το χρώμα του σωλήνα των ανθήρων (Ομάδα 1: κίτρινος σωλήνας Ομάδα 2: σκούρος βυσσινί σωλήνας). Τα σπέρματα της ομάδας 1 έχουν αραιή τρίχωση σε σχέση με το H. laconicum ενώ τα σπέρματα της ομάδας 2 παρουσιάζουν μια ενδιάμεση κατάσταση. Οι τρίχες του πάππου διαμορφώνονται σε μια σειρά και είναι διακλαδισμένες. Η άκρη της κάθε τρίχας του πάππου στο H. laconicum, φέρει εναπόθεση χρωματικών κοκκίων που δίνουν σε αυτό το τμήμα της τρίχας σκούρο μωβ χρώμα. Στο H. graecum δεν υπάρχει αυτός ο χρωματισμός στην τρίχα, απλά καταλήγει σε οξύληκτο άκρο του ίδιου χρώματος. Στον πάππο της ομάδας 2 παρατηρήθηκε ότι το άκρο αυτό είναι πιο έντονα τριχωτό (Εικ ). Α Β Εικ. 22: Πάπποι του Hymenonema laconicum από το Γεράκι (Β. φωτ. από Ι. Κοφινάς). 51


. Β Εικ.")
.")
53 Α Εικ. 23: Πάπποι του Hymenonema graecum από: Α, την Κύθνο και Β, την Άνδρο (ομάδα 1). Β Εικ. 24: Πάπποι του Hymenonema graecum από τη Σύρο (ομάδα 2). 52



.")
54 Εικ. 25: Αχαίνιο και λεπτομέρειες των τριχών του πάππου του Hymenonema laconicum (από καλλιεργημένο υλικό από Γεράκι). Κλίμακες = 1mm. 53


. Κλίμακες = 1mm. Εικ.")
.")

55 Εικ. 26: Αχαίνιο και λεπτομέρειες των τριχών του πάππου του Hymenonema graecum, ομάδα 1 (καλλιεργημένο υλικό από Κύθνο). Κλίμακες = 1mm. Εικ. 27: Αχαίνιο και λεπτομέρειες των τριχών του πάππου του Hymenonema graecum, ομάδα 2 (καλλιεργημένο υλικό από Σύρο). Κλίμακες = 1mm. 54
.Το ποσοστό των γόνιμων γυρεοκόκκων είναι πολύ υψηλό και πάντοτε μεγαλύτερο του 80%.")
56 Γυρεόκοκκοι Η μορφή των γυρεοκόκκων του γένους Hymenonema είναι η κλασσική μορφή των περισσοτέρων γενών των Compositae. Είναι αγκαθωτοί, τριαξονικοί και μοιάζουν με αυτούς του γένους Crepis (Kαμάρη 1976).Το ποσοστό των γόνιμων γυρεοκόκκων είναι πολύ υψηλό και πάντοτε μεγαλύτερο του 80%. Ειδικότερα για το Hymenonema laconicum μετρήθηκαν από 95,83-97,05%, για το H. graecum της ομάδας 1 από 90,7-100% και για το H. graecum της ομάδας 2, από 82,86-95,73%. Το μέγεθός τους φαίνεται ίδιο και στις 3 περιπτώσεις, με διάμετρο περίπου στα 0,06 mm (Εικ ). Εικ. 28: Γυρεόκοκκοι του Hymenonema laconicum (καλλιεργημένο υλικό από τη Λαγκάδα Λακωνίας). Με κόκκινα βέλη σημειώνονται οι άγονοι γυρεόκοκκοι. Κλίμακα = 0,1 mm. 55

.")
57 Εικ. 29: Γυρεόκοκκοι του Hymenonema graecum, ομάδα 1 (καλλιεργημένο υλικό από την Κύθνο). Με κόκκινα βέλη σημειώνονται οι άγονοι γυρεόκοκκοι. Με πράσινο βέλος σημειώνεται γυρεόκοκκος που βλαστάνει. Κλίμακα = 0,1 mm. Εικ. 30: Γυρεόκοκκοι του Hymenonema graecum, ομάδα 2 (καλλιεργημένο υλικό από τη Σύρο). Με κόκκινα βέλη σημειώνονται οι άγονοι γυρεόκοκκοι. Κλίμακα = 0,1 mm. 56
58 Διαπληθυσμιακή ποικιλότητα εντός των ειδών H. laconicum Boiss.& Heldr. Ύψος φυτού Ως προς το ύψος των φυτών, δεν παρατηρείται σημαντική διαπληθυσμιακή ποικιλότητα. Ο πληθυσμός no1 εμφανίζει τη μεγαλύτερη τιμή, με τους πληθυσμούς no 2 & 3 να ακολουθούν με σχεδόν ίσες τιμές (Εικ. 31Α). Μήκος κεφαλίου Το μήκος του κεφαλίου εμφανίζει παρόμοια διαπληθυσμιακή ποικιλότητα, με τον πληθυσμό no 2 να έχει λίγο μεγαλύτερη τιμή (Εικ. 32Α). Πλάτος κεφαλίου Οι τιμές για το πλάτος του κεφαλίου δεν παρουσιάζουν σημαντική ποικιλότητα ανάμεσα στους πληθυσμούς. Ο πληθυσμός no 3 έχει τη μικρότερη τιμή, με τους άλλους δυο να προηγούνται με παρόμοιες τιμές (Εικ. 32Β). Αριθμός βρακτίων Ο αριθμός των βρακτίων για τους πληθυσμούς no1 & 2 δεν αποκλίνει πολύ μεταξύ τους, ενώ ο πληθυσμός no 3 έχει σημαντικά τις μικρότερες τιμές (Εικ. 31Β). Μήκος φύλλων βάσης Ο χαρακτήρας αυτός εμφανίζει παρόμοια διαπληθυσμιακή ποικιλότητα, με τον πληθυσμό no 3 να παρουσιάζει τη μεγαλύτερη μέση τιμή (Εικ. 33Α). Πλάτος φύλλων βάσης Οι διαφορές μεταξύ των πληθυσμών για το πλάτος φύλλων βάσης δεν είναι σημαντικές. Η μεγαλύτερη τιμή παρουσιάζεται στον πληθυσμό no 2 (Εικ. 33Β). Πλάτος τερματικού λοβού φύλλων βάσης Ως προς το πλάτος του τερματικού λοβού των φύλλων βάσης δεν βρέθηκε σημαντική διαπληθυσμιακή ποικιλότητα (Εικ. 35Α). 57
59 cm Ύψος φυτού no Αριθμός βρακτίων Α Pop. no 25 Β Pop. no Εικ. 31: Ποικιλότητα του ύψους φυτών και του αριθμού βρακτίων σε διάφορους πληθυσμούς του Hymenonema laconicum. cm 2,6 2,4 2,2 2 1,8 1,6 1,4 1,2 1 Α Μήκος κεφαλίου Pop. no cm 2,4 2,2 2 1,8 1,6 1,4 1,2 1 Β Πλάτος κεφαλίου Pop. no Εικ. 32: Ποικιλότητα του μήκους και του πλάτους κεφαλίου σε διάφορους πληθυσμούς του Hymenonema laconicum. cm Α Μήκος φύλλων βάσης Pop. no cm Β Πλάτος φύλλων βάσης Pop. no Εικ. 33: Ποικιλότητα του μήκους και του πλάτους φύλλων βάσης σε διάφορους πληθυσμούς του Hymenonema laconicum. 58
60 Μήκος φύλλων βλαστού Το μήκος των φύλλων βλαστού παρουσιάζει παρόμοια διαπληθυσμιακή ποικιλότητα. Μεγαλύτερη μέση τιμή σημειώνεται στον πληθυσμό no 1 και μικρότερη στον no 3 (Εικ. 34Α). Πλάτος φύλλων βλαστού Το χαρακτηριστικό αυτό δεν εμφανίζει σημαντική ποικιλότητα ανάμεσα στους πληθυσμούς. Την πιο μεγάλη μέση τιμή έχει ο πληθυσμός no 2, ενώ τη μικρότερη ο no 3 (Εικ. 34Β). Μήκος αχαινίου Το μήκος του αχαινίου δεν παρουσιάζει σημαντικές διαφορές μεταξύ των πληθυσμών. Η μικρότερη μέση τιμή εμφανίζεται στον πληθυσμό no 2 και η μεγαλύτερη στον no 4 (Εικ. 36Α). Πλάτος αχαινίου Στο πλάτος αχαινίου διαφέρει σημαντικά ο πληθυσμός no 2, στον οποίο εμφανίζεται και η μικρότερη τιμή σε σχέση με τους υπόλοιπους. Οι πληθυσμοί no 3 & 4 δεν διαφέρουν σημαντικά μεταξύ τους. Τη μεγαλύτερη μέση τιμή έχει ο πληθυσμός no 4 (Εικ. 36Β). Μήκος πάππου Για τον χαρακτήρα αυτόν οι πληθυσμοί no 2 & 4 διαφέρουν σημαντικά μεταξύ τους καταλαμβάνοντας τη μεγαλύτερη και τη μικρότερη μέση τιμή αντίστοιχα (Εικ. 35Β). Εύρος μήκους τριχών πάππου (ομοιομορφία πάππου) Το εύρος του μήκους των τριχών του πάππου είναι παρόμοιο και στους τρεις πληθυσμούς. Οι πιο ομοιόμορφοι πάπποι παρουσιάζονται στον πληθυσμό no 3 (Εικ. 37). 59
61 Α Β Εικ. 34: Ποικιλότητα του μήκους και του πλάτους φύλλων βλαστού σε διάφορους πληθυσμούς του Hymenonema laconicum. Α Β Εικ. 35: Ποικιλότητα του πλάτους του τερματικού λοβού φύλλων βάσης και του μήκους πάππου σε διάφορους πληθυσμούς του Hymenonema laconicum. Α Εικ. 36: Ποικιλότητα του μήκους και του πλάτους αχαινίου σε διάφορους πληθυσμούς του Hymenonema laconicum. Β 60
62 Εικ. 37: Ποικιλότητα του εύρους του μήκους τριχών πάππου σε διάφορους πληθυσμούς του H. laconicum. H. graecum (L.)DC. Ύψος φυτού Στο ύψος των φυτών παρατηρείται μικρή διαπληθυσμιακή ποικιλότητα. Ο πληθυσμός no 5 διαφέρει σημαντικά από όλους τους άλλους πληθυσμούς εκτός από τους no 6 & 7. Οι πληθυσμοί no 6-11 δεν εμφανίζουν σημαντικές διαφορές μεταξύ τους. Ο πληθυσμός no 5 έχει τη μικρότερη μέση τιμή (Εικ. 38Α). Μήκος κεφαλίου Το μήκος του κεφαλίου εμφανίζει παρόμοια διαπληθυσμιακή ποικιλότητα, με τον πληθυσμό no 5 να έχει λίγο μικρότερη τιμή (Εικ. 39Α). Πλάτος κεφαλίου Το πλάτος κεφαλίου, όπως και το μήκος, εμφανίζει παρόμοια διαπληθυσμιακή ποικιλότητα (Εικ. 39Β). Αριθμός βρακτίων Ο αριθμός των βρακτίων δεν παρουσιάζει σημαντική ποικιλότητα μεταξύ των πληθυσμών. Μικρότερη μέση τιμή ως προς των αριθμών βρακτίων εντοπίζεται στον πληθυσμό no 5 (Εικ. 38Β). 61
63 cm A Ύψος φυτού Pop. no no Β Αριθμός βρακτίων Pop. no Εικ. 38: Ποικιλότητα του ύψους φυτών και του αριθμού βρακτίων σε διάφορους πληθυσμούς του Hymenonema graecum. cm 2,4 2,2 2 1,8 1,6 1,4 1,2 1 A Μήκος κεφαλίου Pop. no cm 2,4 2,2 2 1,8 1,6 1,4 1,2 1 Β Πλάτος κεφαλίου Pop. no Εικ. 39: Ποικιλότητα του μήκους και πλάτους κεφαλίου σε διάφορους πληθυσμούς του Hymenonema graecum. cm A Μήκος φύλλων βάσης Pop. no cm 6 Β Πλάτος φύλλων βάσης Pop. no Εικ. 40: Ποικιλότητα του μήκους και πλάτους φύλλων βάσης σε διάφορους πληθυσμούς του Hymenonema graecum. 62
64 Μήκος φύλλων βάσης Το μήκος φύλλων βάσης στον πληθυσμό no 8 διαφέρει σημαντικά από τους υπόλοιπους πληθυσμούς και έχει τις μεγαλύτερες τιμές. Επιπλέον, ο πληθυσμός no 11, ο οποίος έχει τη μικρότερη μέση τιμή, εμφανίζει σημαντική διαφορά από τους πληθυσμούς no 7 & 9 (Εικ. 40Α). Πλάτος φύλλων βάσης Για τον χαρακτήρα αυτό, οι πληθυσμοί no 9 & 11 παρουσιάζουν σημαντική διαφορά. Τη μικρότερη μέση τιμή έχει ο πληθυσμός no 11 και τη μεγαλύτερη ο no 8 (Εικ. 40Β). Πλάτος τερματικού λοβού φύλλων βάσης Το πλάτος τερματικού λοβού των φύλλων βάσης εμφανίζει αρκετές σημαντικές διαφορές. Τις πιο μεγάλες τιμές για τον τερματικό λοβό φαίνεται ότι έχουν οι πληθυσμοί no 8 & 9, που ανήκουν και οι δυο στο νησί της Σερίφου. Ο πληθυσμός no 8 διαφέρει σημαντικά από όλους τους πληθυσμούς, εκτός από τον no 9, ενώ ο πληθυσμός no 9 διαφέρει από τους no 5 & 7. Τις πιο μικρές μέσες τιμές παρουσιάζουν οι πληθυσμοί no 5 & 7 (Εικ. 42Α). Μήκος φύλλων βλαστού Η διαπληθυσμιακή ποικιλότητα για το μήκος των φύλλων βλαστού είναι παρόμοια σχεδόν για όλους τους πληθυσμούς. Σημαντική διαφορά εντοπίζεται ανάμεσα στους πληθυσμούς no 5 & 8 (Εικ. 41Α). Πλάτος φύλλων βλαστού Οι πληθυσμοί που εξετάστηκαν ως προς το πλάτος των φύλλων βλαστού δεν παρουσιάζουν σημαντική διαπληθυσμιακή ποικιλότητα (Εικ. 41Β). Μήκος αχαινίου Το μήκος του αχαινίου δεν παρουσιάζει σημαντικές διαφορές μεταξύ των πληθυσμών (Εικ. 43Α). 63
65 Μήκος φύλλων βλαστού cm A Pop. no Πλάτος φύλλων βλαστού cm 3 2,5 2 1,5 1 0, Β Pop. no Εικ. 41: Ποικιλότητα του μήκους και πλάτους φύλλων βλαστού σε διάφορους πληθυσμούς του Hymenonema graecum. cm 2 Πλάτος τερματικού λοβού φύλλων βάσης cm 1,5 1,4 Μήκος πάππου 1,5 1,3 1 1,2 0,5 1,1 A Pop. no Β Pop. no Εικ. 42: Ποικιλότητα του πλάτους του τερματικού λοβού φύλλων βάσης και του μήκους πάππου σε διάφορους πληθυσμούς του Hymenonema graecum. cm 0,6 0,55 0,5 0,45 0,4 0,35 0,3 A Μήκος αχαινίου Pop. no mm 1,7 1,5 1,3 1,1 0,9 0,7 0,5 Β Πλάτος αχαινίου Pop. no Εικ. 43: Ποικιλότητα του μήκους και πλάτους αχαινίου σε διάφορους πληθυσμούς του Hymenonema graecum. 64
66 Πλάτος αχαινίου Για το χαρακτηριστικό αυτό, ο πληθυσμός no 7 διαφέρει σημαντικά από τον no 10. Οι πληθυσμοί αυτοί εμφανίζουν την ανώτερη και κατώτερη μέση τιμή αντίστοιχα (Εικ. 43Β). Μήκος πάππου Το μήκος πάππου εμφανίζει σημαντική ποικιλότητα ανάμεσα στον πληθυσμό no 7 και τους πληθυσμούς no 10 & 11. Τη μικρότερη μέση τιμή έχει ο πληθυσμός no 7 ενώ τη μεγαλύτερη ο no 11 (Εικ. 42Β). Εύρος μήκους τριχών πάππου (ομοιομορφία πάππου) Το εύρος του μήκους των τριχών του πάππου δε διαφέρει σημαντικά μεταξύ των πληθυσμών (Εικ. 44). Εικ. 44: Ποικιλότητα του εύρους του μήκους τριχών πάππου σε διάφορους πληθυσμούς του Hymenonema graecum Ποικιλότητα των μορφολογικών χαρακτήρων μεταξύ των δυο ειδών Ύψος φυτών Το ύψος των φυτών παρουσιάζει στατιστικώς σημαντική διαφορά ανάμεσα στα δυο είδη. Το ύψος των φυτών που ανήκουν στο είδος Hymenonema laconicum είναι 65
67 μεγαλύτερο απ ότι στο H. graecum. Για το H. laconicum οι τιμές κυμάνθηκαν από 32,4-76,8 cm ενώ για το H. graecum από 14-66,7 cm (Εικ. 48Α). Μήκος κεφαλίου Ο χαρακτήρας αυτός εμφανίζει παρόμοιες τιμές στα δυο είδη. Στο H. laconicum παρατηρείται μεγαλύτερη μέση τιμή από το άλλο είδος, χωρίς όμως σημαντική διαφορά (Εικ. 46Α). Πλάτος κεφαλίου Ομοίως και για το πλάτος κεφαλίου (Εικ. 46Β). Αριθμός βρακτίων Ο αριθμός βρακτίων εμφανίζει παρόμοια ποικιλότητα και για τα δυο είδη. Μεγαλύτερη μέση τιμή εντοπίζεται στο H. laconicum (Εικ. 48Β). Μήκος φύλλων βάσης Η ποικιλότητα του μήκους φύλλων βάσης μεταξύ των δυο ειδών είναι παρόμοια, χωρίς να εντοπίζεται σημαντική διαφορά. Η μεγαλύτερη μέση τιμή παρατηρείται στο H. laconicum (Εικ. 47Α). Πλάτος φύλλων βάσης Το πλάτος φύλλων βάσης διαφέρει σημαντικά ανάμεσα στα δυο είδη, με τις μεγαλύτερες τιμές να παρατηρούνται στο H. laconicum, οι οποίες αρχίζουν από 2,4-13,5 cm. Οι αντίστοιχες τιμές για το H. graecum είναι 0,7-5,2 cm (Εικ. 47Β). Πλάτος τερματικού λοβού φύλλων βάσης Το πλάτος τερματικού λοβού φύλλων βάσης παρουσιάζει στατιστικώς σημαντική διαφορά ανάμεσα στα δυο είδη. Το αποτέλεσμα αυτό ενισχύει τη σημαντικότητα αυτού του χαρακτήρα ως προς τη διάκριση των δυο ειδών. Ο τερματικός λοβός φαίνεται πλατύτερος στο H. laconicum, με μέγιστη και ελάχιστη τιμή 0,9 και 3,5 cm αντίστοιχα. Για το H. graecum οι αντίστοιχες τιμές κυμαίνονται από 0,3-2,1 cm. Πρέπει να σημειωθεί, ότι σχεδόν όλες οι τιμές που ξεπερνούν το 1 cm για το H. graecum εντοπίζονται στα φυτά της Σερίφου (Εικ. 49Α). 66
68 cm 70 Ύψος φυτού no 45 Αριθμός βρακτίων H. graecum H. laconicum 20 H. graecum H. laconicum Α Taxa Β Taxa Εικ. 45: Ποικιλότητα του ύψους φυτών και του αριθμού βρακτίων ανάμεσα στα δυο είδη. cm Α 2,5 2 1,5 1 Μήκος κεφαλίου H. graecum H. laconicum Taxa cm Β 2,2 2 1,8 1,6 1,4 1,2 1 Πλάτος κεφαλίου H. graecum H. laconicum Taxa Εικ. 46: Ποικιλότητα του μήκους και πλάτους κεφαλίου ανάμεσα στα δυο είδη. cm Α Μήκος φύλλων βάσης H. graecum H. laconicum Taxa cm Πλάτος φύλλων βάσης H. graecum H. laconicum Εικ.47: Ποικιλότητα του μήκους και πλάτους φύλλων βάσης ανάμεσα στα δυο είδη. Β Taxa 67
69 Μήκος φύλλων βλαστού Η ποικιλότητα του μήκους φύλλων βλαστού είναι παρόμοια για τα δυο είδη, με μεγαλύτερη μέση τιμή στο H. laconicum (Εικ. 48Α). Πλάτος φύλλων βλαστού Ομοίως και για το πλάτος φύλλων βλαστού (Εικ. 48Β). Μήκος αχαινίου Το μήκος αχαινίου εμφανίζει σημαντική ποικιλότητα ανάμεσα στα δυο είδη. Το H. laconicum έχει μεγαλύτερα αχαίνια σε μήκος, με τιμές από 0,45-0,6 cm. Το H. graecum έχει ελάχιστη και μέγιστη τιμή για το χαρακτήρα αυτό 0,35 και 0,6 cm αντίστοιχα (Εικ. 50Α). Πλάτος αχαινίου Το πλάτος αχαινίου διαφέρει σημαντικά μεταξύ των δυο ειδών. Τις μεγαλύτερες τιμές εμφανίζει το H. laconicum με 1,4-2 mm, ενώ οι τιμές για το άλλο είδος κυμαίνονται μεταξύ 0,8-1,8 mm (Εικ. 50Β). Μήκος πάππου Το μήκος του πάππου στο H. laconicum διαφέρει σημαντικά σε σχέση με το H. graecum. Τις μεγαλύτερες τιμές για το χαρακτηριστικό αυτό καταλαμβάνει το H. laconicum. Πιο συγκεκριμένα, το μήκος της μεγαλύτερης τρίχας του πάππου στο πρώτο είδος κυμαίνεται μεταξύ 1,5-1,86 cm, ενώ στο δεύτερο μεταξύ 1-1,46 cm (Εικ. 49Β). Εύρος μήκους τριχών πάππου (ομοιομορφία πάππου) Ο πάππος είναι σημαντικά πιο ομοιόμορφος στο H. laconicum απ ότι στο H. graecum. Το εύρος του μήκους των τριχών του πάππου έχει πολύ μικρότερες τιμές στα φυτά του είδους H. graecum. Για το είδος αυτό, το εύρος του μήκους των τριχών αρχίζει από 0,05-0,48 cm, ενώ για το άλλο είδος οι τιμές αυτές ξεκινούν από 0,79-1,52 cm. Ο χαρακτήρας αυτός έρχεται σε συμφωνία με τη βιβλιογραφία, που το αναφέρει ως διακριτικό γνώρισμα για τα δυο είδη του γένους (Εικ. 51). 68
70 cm Μήκος φύλλων βλαστού H. graecum H. laconicum Taxa cm 2 Πλάτος φύλλων βλαστού 1,5 1 0,5 0 H. graecum H. laconicum Taxa Εικ. 48: Ποικιλότητα του μήκους και πλάτους φύλλων βλαστού ανάμεσα στα δυο είδη. Εικ. 49: Ποικιλότητα του πλάτους του τερματικού λοβού φύλλων βάσης και του μήκους πάππου ανάμεσα στα δυο είδη. cm 0,56 0,54 0,52 0,5 0,48 0,46 0,44 0,42 0,4 Μήκος αχαινίου H. graecum H. laconicum Taxa mm 1,9 1,7 1,5 1,3 1,1 0,9 0,7 0,5 Πλάτος αχαινίου H. graecum H. laconicum Taxa Εικ. 50: Ποικιλότητα του μήκους και πλάτους αχαινίου ανάμεσα στα δυο είδη. 69
71 Εικόνα 51: Ποικιλότητα του εύρους του μήκους τριχών πάππου ανάμεσα στα δυο είδη. 70
72 4. ΚΥΤΤΑΡΟΛΟΓΙΚΗ ΜΕΛΕΤΗ 4.1. ΚΥΤΤΑΡΟΛΟΓΙΑ - ΚΥΤΤΑΡΟΓΕΝΕΤΙΚΗ Η Κυτταρολογία αποτελεί έναν κλάδο της Βιολογίας με τον οποίο μελετάται η μορφή, η σύσταση και η λειτουργία του γενετικού υλικού. Η κυτταρολογική μελέτη μπορεί να παίξει σημαντικό ρόλο στην ακριβέστερη διάκριση πολλών μορφολογικά δυσδιάκριτων taxa. Η βοήθεια που προσφέρει στην ερμηνεία εξελικτικών, φυλογενετικών και βιογεωγραφικών σχέσεων των διάφορων φυτικών taxa αναγνωρίζεται έως τώρα. Η Κυτταρογενετική είναι ένας κλάδος της Βιολογίας που συνδυάζει την κυτταρολογία (μελέτη χρωμοσωμάτων και άλλων κυτταρικών συστατικών) και τη γενετική (μελέτη της κληρονομικότητας). Τα πεδία μελέτης της Κυτταρογενετικής περιλαμβάνουν διάφορες τεχνικές χρώσης, την λειτουργία και την κίνηση των χρωμοσωμάτων (κυτταρική διαίρεση: μίτωση, μείωση), τον αριθμό και τη μορφή των χρωμοσωμάτων (ανάλυση καρυοτύπου) και τις πολλές αλλαγές της δομής και συμπεριφοράς των χρωμοσωμάτων, καθώς αυτές σχετίζονται με τον ανασυνδυασμό, την μετατόπιση (μεταβίβαση) και την έκφραση γονιδίων. Οι κυτταρογενετικές μελέτες συμβάλλουν σημαντικά στην χαρτογράφηση των γονιδιωμάτων πολλών οργανισμών και χαρακτηρίζονται ως σημαντικά εργαλεία στην ερμηνεία φαινομένων κληρονομικότητας, ταξινόμησης και φυλογένεσης (Gill & Friebe 1998, Μπαρέκα 2007). Η μορφολογία και ο αριθμός των χρωμοσωμάτων είναι συνήθως σταθερά χαρακτηριστικά για κάθε είδος και αποτελούν τη βιολογική του ταυτότητα. Το σύνολο των χρωμοσωμάτων, με τη συγκεκριμένη μορφολογία τους, αποτελεί τον καρυότυπο του οργανισμού και αντιστοιχεί στη συνολική εικόνα του (φωτογραφία), όπως αυτή φαίνεται στο μικροσκόπιο. Ο αριθμός των χρωμοσωμάτων σε έναν καρυότυπο ποικίλλει από είδος σε είδος, σπανιότερα και μέσα στο ίδιο το είδος. Ο καρυότυπος διακρίνεται σε συμμετρικό και ασύμμετρο. Ο συμμετρικός καρυότυπος αποτελείται από χρωμοσώματα παραπλήσιου ή κανονικά διαβαθμισμένου μεγέθους και έχουν περίπου την ίδια μορφή ή έχουν τα περισσότερα το κεντρομέρος περίπου στη μέση (+ ισοβραχιόνια δηλ. m ή sm). Ασύμμετρος χαρακτηρίζεται ο καρυότυπος που αποτελείται από χρωμοσώματα μερικά εκ των οποίων είναι πολύ μεγάλα ή πολύ μικρά, χωρίς κάποιο από τα υπόλοιπα να έχει ενδιάμεσο μέγεθος και τα περισσότερα 71
73 χρωμοσώματά του είναι ανισοβραχίονα (δηλ. st ή t). Ο καρυότυπος των περισσοτέρων οργανισμών είναι διπλοειδής (2n) και αποτελείται από δυο όμοιες σειρές (n) χρωμοσωμάτων (μια σειρά από κάθε γονέα, η οποία καλείται απλοειδής). Απλοειδής είναι συνήθως ο αριθμός των χρωμοσωμάτων στα αναπαραγωγικά κύτταρα των οργανισμών που πολλαπλασιάζονται φυλετικά (γαμέτες). Τα ανά δυο όμοια χρωμοσώματα ενός οργανισμού, που προέρχονται από τους δυο γαμέτες (μητρικό και πατρικό), καλούνται ομόλογα. Το σύνολο των χρωμοσωμάτων της απλοειδούς σειράς καλείται απλοειδής χρωμοσωματικός αριθμός (n), ενώ βασικός αριθμός (x) ο παλαιότερος φυλογενετικά απλοειδής χρωμοσωματικός αριθμός, ο οποίος είναι συνήθως μικρότερος όταν έχουμε περισσότερους από ένα χρωμοσωματικούς αριθμούς σε ένα είδος ή γένος. Πολυπλοειδία καλείται οποιαδήποτε μεταβολή του χρωμοσωματικού αριθμού ενός taxon από τον κανονικό διπλοειδή χρωμοσωματικό αριθμό(ψαρράς κ.ά. 2002α, Ψαρράς κ.ά. 2002β). Η ταξινόμηση όλων των χρωμοσωμάτων 2n ενός καρυοτύπου σε σειρά μεγέθους, καλείται καρυόγραμμα. Η διαγραμματική απεικόνιση της απλοειδούς σειράς (n) ενός οργανισμού, που γίνεται από τη μέση τιμή πέντε τουλάχιστον μετρηθέντων καρυοτύπων (υπολογισμός r-index και R-length), αποτελεί το ιδιόγραμμα ενός taxon (Ψαρράς κ.ά. 2002α, Ψαρράς κ.ά. 2002β). Τα χρωμοσώματα εκτός από την πρωτογενή περίσφιγξη (κεντρομέρος) έχουν μερικές φορές και δευτερογενείς περισφίγξεις, μια ή περισσότερες. Όταν οι δευτερογενείς περισφίγξεις αποχωρίζουν ένα μικρό σφαιρικό τμήμα χρωμοσώματος αυτό καλείται δορυφόρος και τα αντίστοιχα χρωμοσώματα δορυφορικά χρωμοσώματα ή SAT-χρωμοσώματα. Οι δευτερογενείς περισφίγξεις των SATχρωμοσωμάτων αποτελούν και τις θέσεις σχηματισμού των οργανωτών των πυρηνίσκων (Ψαρράς κ.ά. 2002α, Ψαρράς κ.ά. 2002β) ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Κυτταρολογικές μέθοδοι Μελετήθηκαν κυτταρολογικά 10 πληθυσμοί (37 άτομα συνολικά) του γένους Hymenonema (Πίνακας 4). Από κάθε πληθυσμό καλλιεργήθηκαν 2-8 άτομα στον Πειραματικό Κήπο του Εργαστηρίου Βοτανικής, του Πανεπιστημίου Πατρών, τα οποία μοιράστηκαν σε γλάστρες, έτσι ώστε να βρίσκονται 1-2 άτομα σε κάθε μια. 72
74 Πίνακας 4: Πληθυσμοί του γένους Hymenonema που μελετήθηκαν κυτταρολογικά. Taxon Πληθυσμός Pop. no Hymenonema laconicum ARKADIA: SE of Leonidio, road from Leonidio to Tsitalia village, alt. 113 m, N, E, , Kofinas & Dolianitis 129 (UPA). 2» LAKONIA: Geraki, road from the village to Alepohori, alt m, N, E, , Liveri & Ketsilis-Rinis 124 (UPA).» LAKONIA: Lagkada gorge, climbing region, alt. 800 m, 37 4'59.09"N, 22 18'39.30"E, , Charalampopoulos 1524 (UPA) ; ibid , Liveri & Kofinas 130 (UPA). 3 4 Hymenonema graecum Ins. ANDROS: Batsi, residential area, Agios Filippos church, alt. 45 m, N, E, , Liveri & Ketsilis-Rinis 115 (UPA). 6» Ins. KITHNOS: Merichas Bay (SW side), phrygana, alt m, N, E, , Liveri & Ketsilis-Rinis 100 (UPA).» Ins. SIROS: Kini, along the roadsides from the beach to the village for a distance up to 1 km approximately, N, E, , Liveri & Ketsilis- Rinis 107 (UPA).» Ins. SIROS: Agathopes, road towards to the beach, on the rocks very close to the sea, alt m., N, E, , Liveri & Ketsilis-Rinis 110 (UPA).» Ins. TINOS: Monastiri, Arnados, DioXoria, along the roadsides connecting these three villages, alt m, N to N, E to E, , Liveri & Ketsilis-Rinis 117 (UPA).» Ins. TINOS: Kionia bay, on the rocks, phrygana, alt m, N, E, , Liveri & Ketsilis-Rinis 121 (UPA)
75 » Ins. TINOS: Kolimpithra bay, on the rocks, phrygana, alt m, N, E, , Liveri & Ketsilis-Rinis 123 (UPA). 14 Η μελέτη των καρυοτύπων έγινε σε ημιμόνιμα παρασκευάσματα ακρορριζίων, τα οποία παρασκευάστηκαν σύμφωνα με την τεχνική της σύνθλιψης (squash technique) των Östergren & Heneen (1992). Έγιναν σχετικές τροποποιήσεις στην τεχνική αυτή, ώστε να έχουμε όσον το δυνατό καλύτερα αποτελέσματα (Καμάρη 1976, Constandinidis & al. 2002). Η πειραματική διαδικασία που ακολουθήθηκε είναι η εξής: Προκατεργασία του υλικού: 1. Κατά τη διάρκεια της έντονης βλαστητικής ανάπτυξης των φυτών (Οκτώβριος-Μάρτιος), οπότε οι μιτωτικές διαιρέσεις των ακρορριζίων είναι περισσότερες, συλλέχθηκαν ακραία τμήματα ριζών μήκους 2-3 cm από τα καλλιεργούμενα άτομα. Η συλλογή τους έγινε νωρίς το πρωί ( π.μ.), ώστε να έχουμε ακόμα μεγαλύτερο αριθμό μιτωτικών διαιρέσεων στα ακρορρίζια. 2. Προκειμένου να παρεμποδιστεί ο σχηματισμός της μιτωτικής ατράκτου κατά το στάδιο της πυρηνικής διαίρεσης, έγινε προκατεργασία των ακρορριζίων σε μίγμα 2:1 v/v 2Mm 8-υδροξυκινολίνης 0,002% w/v:κολχικίνης 0,3% w/v ή σε μίγμα 1:1 v/v 2Mm 8-υδροξυκινολίνης 0,002% w/v:κολχικίνης 0,2% w/v. 3. Τα ακρορρίζια τοποθετήθηκαν σε θερμοκρασία 4 C για περίπου 6 ώρες. 4. Στη συνέχεια έγινε στερέωση των φάσεων της μίτωσης σε διάλυμα Carnoy (μίγμα 3:1 απόλυτης αλκοόλης και παγόμορφου οξικού οξέος) για περίπου 24h, σε θερμοκρασία 4 C. 5. Τα ακρορρίζια μεταφέρθηκαν σε διάλυμα αλκοόλης 70% και διατηρήθηκαν σε χαμηλή θερμοκρασία (-20 C) μέχρι το επόμενο στάδιο επεξεργασίας τους. Καρυομορφομετρική ανάλυση χρωμοσωμάτων: 1. Πριν τη χρώση τους τα ακρορρίζια υδρολύθηκαν στους 60 C σε διάλυμα 1 Ν ΗCl για 12 min. 2. Στη συνέχεια πραγματοποιήθηκε χρώση των χρωμοσωμάτων με αντιδραστήριο Feulgen (Darlington & La Cour 1969), όπου παρέμειναν σ αυτό για 3 74
76 h σε σκοτεινό μέρος. Το αντιδραστήριο αυτό βάφει επιλεκτικά το DNA κατά τη διάρκεια μιας αντίδρασης στον πυρήνα με τη δεσοξυριβόζη. Το κυτταρόπλασμα είναι αρνητικό στην χρώση Feulgen, ενώ η περιοχή της πρωτογενούς περίσφιξης χρωματίζεται αχνά. 3. Το ακραίο μέρος των ακρορριζίων που αντιστοιχεί στην περιοχή της μεριστωματικής ζώνης (όπου έχουμε πολλές μιτωτικές διαιρέσεις) και που βάφτηκε έντονα, αποκόπηκε και τοποθετήθηκε σε αντικειμενοφόρο πλάκα με μια σταγόνα οξικού οξέος 45%. Εκεί κατατεμαχίστηκε και καλύφθηκε με υάλινη καλυπτρίδα. Με διηθητικό χαρτί απομακρύνθηκε η περίσσεια του οξικού οξέος. Το ριζίδιο συνθλίφθηκε με μηχανικό τρόπο κάτω από στεγνό διηθητικό χαρτί που κάλυψε την καλυπτρίδα. Ασκώντας ισχυρή μηχανική πίεση με τον αντίχειρα σε κατακόρυφη θέση, το υλικό προς παρατήρηση τοποθετήθηκε σε ένα οπτικό επίπεδο. Χρωμοσωματικές μετρήσεις: Οι δυο παράμετροι που καθορίζουν τη μορφολογία των χρωμοσωμάτων είναι: i) η αναλογία των βραχιόνων (r-index), που ισούται με τον λόγο του μεγαλύτερου βραχίονα (l) προς τον μικρότερο (s) και καθορίζεται από τη θέση του κεντρομέρους (πρωτογενής περίσφιξη). Αναλόγως προκύπτει η μορφολογία των χρωμοσωμάτων. ii) το σχετικό μήκος (Relative length) των χρωμοσωμάτων, που ισούται με το επί τις εκατό (%) ποσοστό της αναλογίας του μήκους κάθε ενός χρωμοσώματος (l+s) προς το συνολικό μήκος των χρωμοσωμάτων της απλοειδούς σειράς Σn(l+s)/2. Εκτός των ανωτέρω δεικτών, μελετήθηκαν επίσης για κάθε taxon: α) το μήκος του μικρότερου χρωμοσώματος (min-cl: minimum Chromosome Length), β) το μήκος του μεγαλύτερου χρωμοσώματος (max-cl: maximum Chromosome Length), γ) ο κεντρομερικός δείκτης (centromeric index = l/l+s), δ) η αναλογία διαφοράς βραχιόνων (arm difference ratio = l-s/l+s), ε) το συνολικό μήκος όλων των χρωμοσωμάτων (TCL: Total Chromosome Length) και 75
77 στ) ο μέσος όρος του μήκους των χρωμοσωμάτων (ACL: Average Chromosome Length) (Ayodele 1999, Jee & Vijayavalli 1999, Martin & al. 2011, Funamoto & Kondo 2012, Sharma & al. 2012, Kocygit & Bona 2013). Κατά την κυτταρολογική μελέτη πραγματοποιήθηκε μέτρηση των χρωμοσωμάτων για την εξακρίβωση του χρωμοσωματικού αριθμού κάθε taxon και κατάταξη των χρωμοσωμάτων στις κατηγορίες που αναγράφονται στον Πίνακα 5, με βάση τη θέση της πρωτογενούς περίσφιξης (κεντρομέρος) και την αναλογία των βραχιόνων. Σε ότι αφορά την ορολογία της μορφολογίας των χρωμοσωμάτων λήφθηκαν υπόψη οι Levan & al. (1965), Stebbins (1971) και Καμάρη (1976). Για τη μελέτη και την ανάλυση της μορφολογίας των χρωμοσωμάτων παρατηρήθηκαν όσο το δυνατόν περισσότεροι πυρήνες στο στάδιο της μετάφασης, με τον ίδιο περίπου βαθμό συσπείρωσης και την ίδια πάντοτε κλίμακα. Πρέπει να σημειωθεί ότι στη μελέτη της αναλογίας των δυο βραχιόνων και του σχετικού μήκους κάθε χρωμοσώματος δεν λαμβάνεται υπόψη το μέγεθος του δορυφόρου, ούτε και η διαφορετική κάθε φορά απόστασή του από το βραχίονα. Τα καρυογράμματα και τα ιδιογράμματα των μελετηθέντων φυτών έγιναν κατά τη φθίνουσα σειρά του μεγέθους των χρωμοσωμάτων. Πίνακας 5: Πίνακας μορφολογίας χρωμοσωμάτων. Συμβολισμός Ορολογία Θέση κεντρομέρους Αναλογία βραχιόνων M, m Μετακεντρικό Μέση περιοχή 1,0-1,7 sm Υπομετακεντρικό Υπομέση περιοχή 1,7-3,0 st Ακροκεντρικό Υποτελική περιοχή 3,0-7,0 t Υποτελοκεντρικό Τελική περιοχή 7,0- T Τελοκεντρικό Τελικό σημείο Τα παρασκευάσματα παρατηρήθηκαν και φωτογραφήθηκαν σε AXIOPHOT (Zeiss) μικροσκόπιο, με καταδυτικό planapo φακό Χ40, Χ63, Χ100. Η επεξεργασία των καρυογραμμάτων έγινε με το πρόγραμμα Adobe Photoshop CS3 Extended. Ο 76
78 προσδιορισμός των δεικτών και η δημιουργία των ιδιογραμμάτων με τη χρήση του Microsoft Office Excel Κυτταρογενετικές μέθοδοι Η κυτταρογενετική μελέτη πραγματοποιήθηκε σε συνεργασία με τη συνάδελφο κ. Σοφία Σαμαροπούλου στο Εργαστήριο Laboratoire d Evolution et Systématique του Πανεπιστημίου του Παρισιού (Université Paris Sud XI), υπό την εποπτεία της Prof. S. Siljiak-Yakovlev. Μελετήθηκαν 2 πληθυσμοί του γένους Hymenonema, ένας πληθυσμός για κάθε είδος. Η τεχνική που χρησιμοποιήθηκε είναι η σήμανση των πλούσιων σε GC περιοχών με χρωμομυκίνη Α3. Η πειραματική διαδικασία που ακολουθήθηκε είναι η εξής: Προετοιμασία πρωτοπλαστών σύμφωνα με την τεχνική των Geber & Schweizer (1988) με ελάχιστες τροποιήσεις (Siljak-Yakovlev & al. 2014): 1. Πλύση των ακρορριζίων για τουλάχιστον 10 λεπτά σε 0,01 M citrate buffer ph = 4,6. 2. Μεταφορά των ακρορριζίων για λεπτά (ανάλογα με το είδος) στους 37 C σε μείγμα ενζύμων αποτελούμενο από 4% cellulace R10, 1 % pectolyase Y23 και 4% hemicellulace (Sigma Chemical Co) σε 0,05 Μ citrate buffer. 3.Τοποθέτηση του μεριστώματος σε αντικειμενοφόρο με μια σταγόνα φρέσκου οξικού οξέος45%, σύνθλιψη και κάλυψη με υάλινη καλυπτρίδα. 4.Παραμονή των αντικειμενοφόρων για τουλάχιστον 24 ώρες στους -80 C. 5.Αφαίρεση των καλυπτρίδων σύμφωνα με τους Conger & Fairchild (1953) και παραμονή των αντικειμενοφόρων για να στεγνώσουν σε θερμοκρασία δωματίου για τουλάχιστον 24 ώρες. Σήμανση των πλούσιων σε GC περιοχών με χρωμομυκίνη Α 3 σύμφωνα με τις τροποποιημένες τεχνικές των Schweizer (1976), Kondo & Hizume (1982) και των Siljak-Yakovlev & al. (2002): 1.Εμβάπτιση του υλικού σε Mac Ilvaine buffer + MgSO 4 ph = 7. 2.Στέγνωμα των αντικειμενοφόρων. Ακολούθησε τοποθέτηση μιας σταγόνας χρωμομυκίνης Α 3 και μεταφορά των αντικειμενοφόρων στο σκοτάδι για μια ώρα υπό συνθήκες υγρασίας. 3.Πλύση των αντικειμενοφόρων με Mac Ilvaine buffer ph = 7. 77
79 4.Επιχρωματισμός αντίθεσης των χρωμοσωμάτων με τη χρωστική πράσινο του μεθυλίου 0,1 % για 5-7 λεπτά στο σκοτάδι. 5.Πλύση των αντικειμενοφόρων με Mac Ilvaine buffer ph = 5,5. 6.Έπειτα από στέγνωμα των αντικειμενοφόρων, τοποθετήθηκε μια σταγόνα Citifluor και το υλικό καλύφθηκε με υάλινη καλυπτρίδα ΑΠΟΤΕΛΕΣΜΑΤΑ Κυτταρολογικά αποτελέσματα Στα πλαίσια αυτής της ερευνητικής εργασίας πραγματοποιήθηκε κυτταρολογική και καρυομορφομετρική μελέτη των δυο ειδών του γένους Hymenonema. Για κάθε πληθυσμό δίνονται ο καρυότυπος και ο χρωμοσωματικός αριθμός καθώς και καρυόγραμμα, όπου είναι εφικτό. Για κάθε taxon δίνονται οι χρωμοσωματικοί δείκτες: αναλογία βραχιόνων (r-index), σχετικό μήκος (R-length), ελάχιστο (min-cl) και μέγιστο (max-cl) μήκος χρωμοσωμάτων, κεντρομερικός δείκτης, αναλογία διαφοράς βραχιόνων, συνολικό μήκος (TCL) και το μέσο μέγεθος (ACL) χρωμοσωμάτων. Από τη μελέτη των καρυοτύπων ενός πληθυσμού από κάθε είδος έχει κατασκευασθεί ιδιόγραμμα. Για το H. graecum έχουν γίνει δυο ιδιογράμματα, ένα από πληθυσμό που εμφανίζει την τυπική μορφολογία του είδους (κίτρινος σωλήνας ανθήρων) και ένα επιπλέον από πληθυσμό που εμφανίζει σκούρο βυσσινί σωλήνα ανθήρων. Ένα μικρό μέρος από την κυτταρολογική εργασία έχει ήδη δημοσιευτεί (Liveri & al. 2014). 1. Hymenonema laconicum Boiss. &Heldr., 2n = 2x = 20. Ο χρωμοσωματικός αριθμός 2n= 20 αναφέρεται από τους Ιατρού (1986), Tan & Iatrou (2001) και Liveri & al. (2014) για τα όρη Ταΰγετος και Πάρνων. Ο ίδιος αριθμός βρέθηκε και στην παρούσα μελέτη, δίνοντας όμως για πρώτη φορά καρυοτύπους και καρυογράμματα για 2 πληθυσμούς από τον Ταΰγετο (nos 3 & 4) και 1 από τον Πάρνωνα (no 2) καθώς και ιδιόγραμμα για το είδος (Εικ. 54). Ο καρυότυπος του Hymenonema laconicum είναι διπλοειδής, συμμετρικός και αποτελείται κυρίως από μετακεντρικά (m) αλλά και υπομετακεντρικά (sm) χρωμοσώματα, μικρού έως μεσαίου μεγέθους. Το μικρότερο σε μέγεθος ζεύγος χρωμοσωμάτων είναι πάντα υπομετακεντρικό και δορυφορικό (sm-sat). Το μέγεθος 78
80 των δορυφόρων είναι περίπου όσο ο μικρός βραχίονας του χρωμοσώματος. Ο αριθμός των υπομετακεντρικών χρωμοσωμάτων (εκτός του δορυφορικού) ποικίλλει από 0-4 χρωμοσώματα. Επιπλέον, εντοπίζονται ζεύγη χρωμοσωμάτων τα οποία είναι δύσκολο να χαρακτηριστούν ως μετακεντρικά ή υπομετακεντρικά διότι η τιμή της αναλογίας βραχιόνων είναι οριακή μεταξύ των δυο κατηγοριών. Για τον πληθυσμό από το Λεωνίδιο του Πάρνωνα (no 2) δίνεται καρυότυπος, ο οποίος χαρακτηρίζεται ως διπλοειδής και συμμετρικός ενώ διακρίνεται καθαρά το ζεύγος των δορυφορικών χρωμοσωμάτων (Εικ. 52Α). Το μέγιστο μήκος χρωμοσωμάτων για τον πληθυσμό είναι 3,61 μm ενώ το ελάχιστο είναι 2,05 μm. Το συνολικό μήκος και το μέσο μέγεθος χρωμοσωμάτων ισοδυναμεί με 54,9 μm και 2,75 μm, αντιστοίχως. Από την περιοχή του Ταϋγέτου μελετήθηκαν δυο πληθυσμοί, ο no 3 από το Γεράκι και ο no 4 από το φαράγγι της Λαγκάδας. Ο καρυότυπος του πληθυσμού no 3 είναι διπλοειδής, συμμετρικός και αποτελείται από 2n = 14m + 2m/sm + 2sm + 2sm-SAT = 20 χρωμοσώματα (Εικ. 52Γ). Το 4 ο ζεύγος είναι υπομετακεντρικό ενώ το 7 ο παρουσιάζει δομική ετεροζυγωτία, δηλ. αποτελείται από ένα μετακεντρικό κι ένα υπομετακεντρικό χρωμόσωμα. Το μέγεθος των χρωμοσωμάτων ποικίλλει από 2,12-3,39 μm. Το TCL ισούται με 59,09 μm και το ACL με 2,95 μm. Ο πληθυσμός της Λαγκάδας παρουσιάζει, επίσης, συμμετρικό καρυότυπο με 2n = 16m + 2m/sm + 2sm-SAT= 20 χρωμοσώματα (Εικ. 52Β, 52Δ) με μέγεθος που ποικίλλει μεταξύ 2,2 μm και 4,13 μm. To TCL ισούται με 65,48 μm και το ACL με 1,43. Στο καρυόγραμμα (Εικ. 53Β)το 5 ο ζεύγος χρωμοσωμάτων χαρακτηρίζεται ως m/sm λόγω οριακής τιμής αναλογίας βραχιόνων. Γι αυτόν τον πληθυσμό δίνεται και ιδιόγραμμα (Εικ. 54, Πίνακας 7) το οποίο δείχνει πως η γενική μορφολογία καρυοτύπου για τον πληθυσμό της Λαγκάδας είναι 2n = 16m + 2sm + 2sm-SAT= 20 χρωμοσώματα με το 5 ο και 10 ο ζεύγος να είναι υπομετακεντρικά. 79
81 Πίνακας 6: Χρωμοσωματικός αριθμός, μέγεθος χρωμοσωμάτων και χρωμοσωματικοί δείκτες (TCL, ACL) για τους πληθυσμούς του Hymenonema laconicum. Είδος Pop. 2n Μέγεθος χρωμοσωμάτων (min-max) μm H. laconicum TCL ACL ,05-3,61 54,9 2,75» ,12-3,39 59,09 2,95» ,2-4,13 65,48 1,43 Πίνακας 7: Μορφομετρικά δεδομένα των χρωμοσωμάτων του Hymenonema laconicum (πληθυσμός no 4), 2n = 2x = 20. Ζεύγη χρωμ/των l (μm) s (μm) l+s l/s Κεντρομερικός δείκτης l/l+s Αναλογία διαφοράς βραχιόνων Σχετικό μήκος l+s/sn(l+s) l-s/l+s 1 2,1414 1,8233 3,9647 1,1886 0,5394 0,0788 0, ,0788 1,6937 3,7725 1,1875 0,54 0,0799 0, ,8763 1,6605 3,5367 1,1468 0,5314 0,0628 0, ,9996 1,4867 3,4863 1,3583 0,5719 0,1438 0, ,1004 1,2562 3,3567 1,7226 0,6297 0,2594 0, ,8549 1,4545 3,3094 1,2536 0,5551 0,1103 0, ,7754 1,4279 3,2033 1,256 0,555 0,11 0, ,6195 1,3775 2,997 1,1917 0,5411 0,0823 0, ,525 1,2951 2,8201 1,1638 0,5351 0,0702 0, ,4689 0,8491 2,2952 1,7967 0,6361 0,2722 0,




82 A B Γ Δ Εικ. 52: Καρυότυποι του Hymenonema laconicum από τους πληθυσμούς: Α, no 2. Β, no 4. Γ, no 3 και Δ, no 4. Κλίμακες = 10 μm. A B m m m sm m m m/sm m m sm-sat m m m m m/sm m m m m sm-sat Εικ. 53: Καρυογράμματα του Hymenonema laconicum από τους πληθυσμούς: Α, no 2. Β, no 4. Κλίμακες = 10 μm. 81
83 m m m m sm m m m m sm-sat Εικ. 54: Ιδιόγραμμα του Hymenonema laconicum (πληθυσμός no 4). 2. Hymenonema graecum (L.) DC., 2n = 2x = 20. Ο χρωμοσωματικός αριθμός 2n = 20 έχει βρεθεί να αναγράφεται σε δείγματα του Hymenonema graecum, κατατεθειμένα στο Herbarium του Πανεπιστημίου της Lund (Σουηδία). Τα δείγματα αυτά έχουν συλλεχθεί από τον Runemark (2 δείγματα με no 1212) και από τους Runermark & Snogerup (no 10649) και προέρχονται από τη Νάξο. Ο Ιατρού (1986) υποστηρίζει πως τα δυο είδη του γένους Hymenonema είναι σχιζοενδημικά και παραθέτει τον ίδιο σωματικό αριθμό και για τα δυο, χωρίς όμως να αναφέρει την περιοχή συλλογής του H. graecum. Ο χρωμοσωματικός αριθμός που βρέθηκε στην παρούσα μελέτη είναι ο ίδιος με αυτόν που αναφέρεται από τους προηγούμενους ερευνητές. Πρώτη φορά, όμως, δίνονται καρυότυποι και καρυογράμματα για διάφορους πληθυσμούς από τα νησιά των Κυκλάδων, αλλά και ιδιογράμματα. Όλοι οι πληθυσμοί που μελετήθηκαν έχουν διπλοειδή, συμμετρικό καρυότυπο, ο οποίος αποτελείται κυρίως από μετακεντρικά (m), αλλά μερικά υπομετακεντρικά (sm) χρωμοσώματα μικρού έως μεσαίου μεγέθους. Παρατηρούνται αρκετές διαφοροποιήσεις ανάμεσα στους πληθυσμούς. Η παρουσία ενός ζεύγους δορυφορικών χρωμοσωμάτων όμως είναι σταθερή σε κάθε πληθυσμό. Πρέπει να 82
84 σημειωθεί ότι το ζεύγος αυτό αποτελεί και το μικρότερο σε μέγεθος ζεύγος του καρυοτύπου και αποτελείται είτε από ένα μετακεντρικό και ένα υπομετακεντρικό χρωμόσωμα (δομική ετεροζυγωτία) είτε από 2 υπομετακεντρικά. Ο χαρακτηρισμός ως m/sm για ένα ζεύγος χρωμοσωμάτων μπορεί να υποδηλώνει φαινόμενο δομικής ετεροζυγωτίας ή να σημαίνει πως οι μετρήσεις για τον χαρακτηρισμό αυτών των χρωμοσωμάτων βρίσκονται στο όριο μεταξύ των δυο κατηγοριών. Ο πληθυσμός no 6 από το Μπατσί της Άνδρου παρουσιάζει καρυότυπο διπλοειδή, συμμετρικό και αποτελούμενο από 2n = 12m + 4m/sm + 2sm + 2SAT-m/sm = 20 χρωμοσώματα (Εικ. 55Α, 57Α). Το μέγεθος των χρωμοσωμάτων κυμαίνεται από 2,06-4,38 μm ενώ το μέσο μέγεθος ισούται με 3,39 μm. Το συνολικό μήκος χρωμοσωμάτων γι αυτόν τον πληθυσμό αντιστοιχεί σε 67,72 μm. Ο καρυότυπος από τον Μέριχα της Κύθνου (no 7) παρουσιάζει λίγο διαφορετική μορφολογία από τον προηγούμενο (Εικ. 55Β, 55Γ, 57Β) και είναι η ακόλουθη: 2n = 14m + 2m/sm + 2 sm + 2SAT-m/sm = 20. Το μέγεθος των χρωμοσωμάτων ποικίλλει από 2,3-4,48 μm. Το συνολικό μήκος των χρωμοσωμάτων του καρυοτύπου ισούται με 71,31 μm ενώ το μέσο μέγεθος με 1,79 μm. Από το ιδιόγραμμα που κατασκευάστηκε (Εικ. 58A, Πίνακας 8) συμπεραίνεται πως 7 ζεύγη χρωμοσωμάτων είναι μετακεντρικά, 2 ζεύγη χαρακτηρίζονται ως μετακεντρικά/υπομετακεντρικά και 1 δορυφορικό μετακεντρικό ζεύγος. Τα 2 m/sm ζεύγη εντοπίζονται στην 5 η και 8 η θέση του ιδιογράμματος. Πρέπει να σημειωθεί ότι ο πληθυσμός αυτός αντιπροσωπεύει το τυπικό Hymenonema graecum με τον κίτρινο σωλήνα ανθήρων. Από το νησί της Σύρου μελετήθηκαν κυτταρολογικά 2 πληθυσμοί, ο no 10 από το Κίνι και ο no 12 από τις Αγκαθωπές. Για τον πληθυσμό no 10 δίνεται καρυότυπος και καρυόγραμμα σύμφωνα με τα οποία η μορφολογία του είναι ως εξής: 2n = 10m + 8m/sm +2SAT-sm = 20 (Εικ. 55Δ, 56Α, 57Γ). Στο 6 ο ζευγάρι εκτός από δομική ετεροζυγωτία παρατηρείται και μια έντονη δευτερογενής περίσφιγξη. Το μέγεθος των χρωμοσωμάτων κυμαίνεται από 2,31-4,11 μm καθώς οι τιμές των δεικτών TCL και ACL ανέρχονται στα 65,89 μm και 1,16 μm αντίστοιχα. Στο ιδιόγραμμα (Εικ. 58B, Πίνακας 9) φαίνεται πως όλα τα ζεύγη χρωμοσωμάτων είναι μετακεντρικά εκτός από 2 ζεύγη, το 5 ο και το 10 ο ζευγάρι τα οποία χαρακτηρίζονται ως m/sm. Ο πληθυσμός no 10 αντιπροσωπεύει την ομάδα του Hymenonema graecum που εμφανίζει σκούρο σωλήνα ανθήρων. Ο καρυότυπος του πληθυσμού no 12 (Εικ. 56Β) παρέχει πληροφορίες για τον αριθμό των χρωμοσωμάτων (2n=20), την ύπαρξη των χαρακτηριστικών SAT-χρωμοσωμάτων και για το μέγεθος των χρωμοσωμάτων. Το 83
85 μέγεθος των χρωμοσωμάτων αυτού του πληθυσμού κυμαίνεται από 1,92-3,04 μm ενώ οι χρωμοσωματικοί δείκτες TCL και ACL ισοδυναμούν με 49,25 μm και 2,46 μm, αντίστοιχα. Οι πληθυσμοί (no 11, 13 & 14) από την Τήνο είναι συμμετρικοί και διπλοειδείς, παρουσιάζοντας βέβαια μικρές διαφορές μεταξύ τους. Ο πληθυσμός no 11, από την περιοχή που καλύπτουν τα χωριά Μοναστήρι, Αρνάδος, Δυο Χωριά, παρουσιάζει καρυότυπο αποτελούμενο μόνο από μετακεντρικά χρωμοσώματα, εκτός από το δορυφορικό υπομετακεντρικό ζευγάρι, μεγέθους 2,07-3,88 μm (Εικ. 56Ε, 57Δ). Το συνολικό μέγεθος χρωμοσωμάτων ισούται με 61,68 μm και το μέσο μέγεθος χρωμοσωμάτων με 3,08 μm. Για τους πληθυσμούς no 13 & 14 δίνεται καρυότυπος καθώς επίσης μετρήθηκαν το μέγεθος των χρωμοσωμάτων, το συνολικό μήκος και το μέσο μέγεθος των χρωμοσωμάτων. Για τον πληθυσμό από τα Κιόνια της Τήνου (no 13) το μέγεθος των χρωμοσωμάτων ποικίλλει μεταξύ 2,2-4,41 μm, το TCL ισούται με 64,87 μm και το ACL με 3,24 μm. Για τον πληθυσμό από την Κολυμπήθρα της Τήνου (no 14) το μέγεθος των χρωμοσωμάτων μετρήθηκε από 2,04-4,08 μm και οι τιμές των TCL και ACL ισοδυναμούν με 65,51 μm και 3,28 μm, αντίστοιχα. Και στους δυο καρυότυπους είναι ευκρινές το δορυφορικό ζεύγος χρωμοσωμάτων. Πίνακας 8 :Χρωμοσωματικός αριθμός, μέγεθος χρωμοσωμάτων και χρωμοσωματικοί δείκτες (TCL, ACL) για τους πληθυσμούς του Hymenonema graecum. Είδος Pop. 2n Μέγεθος χρωμοσωμάτων (min-max) μm TCL ACL H. graecum ,06-4,38 67,72 3,39» ,3-4,48 71,31 1,79» ,31-4,11 65,89 1,16» ,07-3,88 61,68 3,08» ,92-3,04 49,25 2,46 84
86 » ,2-4,41 64,87 3,24» ,04-4,08 65,51 3,28 Πίνακας 9: Μορφομετρικά δεδομένα του Hymenonema graecum (πληθυσμός no 7), 2n = 2x = 20. Ζεύγη χρωμ/των l (μm) s (μm) l+s l/s Κεντρομερικός δείκτης l/l+s Αναλογία διαφοράς βραχιόνων l-s/l+s Σχετικό μήκος l+s/sn(l+s) 1 2,4284 1,9542 4,3826 1,2562 0,5534 0,1068 0, ,2656 1,9452 4,2108 1,1744 0,5391 0,0782 0, ,1545 1,7824 3,937 1,2155 0,5471 0,0785 0, ,1725 1,6482 3,8207 1,3252 0,5689 0,1377 0, ,2371 1,4299 3,6669 1,6117 0,6123 0,2246 0, ,8522 1,6804 3,5326 1,1105 0,5252 0,0503 0, ,829 1,5371 3,3661 1,221 0,546 0,0919 0, ,972 1,2313 3,2033 1,68 0,6138 0,2276 0, ,6697 1,3063 3,0083 1,3111 0, ,1236 0, ,4854 1,0398 2,5251 1,5533 0,5948 0,1897 0,03988 Πίνακας 10: Μορφομετρικά δεδομένα του Hymenonema graecum (πληθυσμός no 10), 2n = 2x = 20. Ζεύγη χρωμ/των l (μm) s (μm) l+s l/s Κεντρομερικός δείκτης l/l+s Αναλογία διαφοράς βραχιόνων Σχετικό μήκος l+s/sn(l+s) l-s/l+s 1 2,3185 1,6956 4,0141 1,3813 0,5778 0,1556 0, ,0991 1,7406 3,8398 1,2375 0,548 0,096 0, ,9876 1,5568 3,5445 1,2689 0,5581 0,0968 0, ,807 1,6731 3,4801 1,1 0,522 0,044 0, ,1524 1,3245 3,4769 1,6198 0,6154 0,2308 0, ,8134 1,4488 3,2622 1,2789 0,5556 0,1111 0, ,8223 1,3718 3,1941 1,329 0,5702 0,1404 0, ,6313 1,4155 3,0467 1,1557 0,5332 0,0664 0, ,557 1,1808 2,7378 1,3361 0,5694 0,1388 0, ,4304 0,9009 2,3314 1,65 0,6177 0,2353 0,



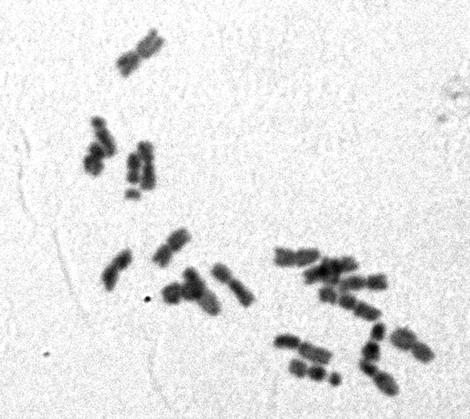
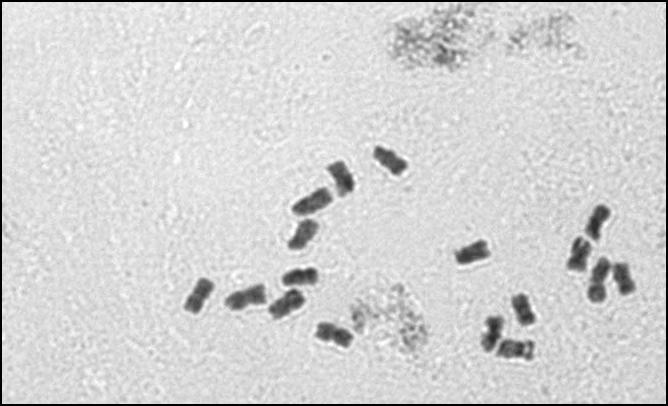
87 A Β Γ Δ Εικ. 55: Καρυότυποι του Hymenonema graecum από τους πληθυσμούς: Α, no 6. Β, no 7. Γ, no 7. Δ. no 10. Κλίμακες = 10 μm. 86






88 A Β Γ Δ Ε ΣΤ Εικ. 56: Καρυότυποι του Hymenonema graecum από τους πληθυσμούς: Α, no 10. Β, no 12. Γ, no 13. Δ, no 13. Ε, no 11. ΣΤ, no 14. Κλίμακες = 10 μm. 87


89 Α m m m m m m/sm m m/sm sm m/sm-sat Β m m m m sm m m m m/sm m/sm-sat Γ m/sm m m m m/sm m/sm m m m/sm sm-sat Δ m m m m m m m m m sm-sat Εικ. 57: Καρυογράμματα του Hymenonema graecum από τους πληθυσμούς: Α. no 6, Β. no 7, Γ. no 10, Δ. no 11. Κλίμακες = 10 μm. A m m m m m/sm m m m/sm m m-sat 88
90 B m m m m m/sm m m m m m/sm-sat Εικ. 58: Ιδιόγραμμα του Hymenonema graecum Α. πληθυσμός no 7, B. πληθυσμός no Κυτταρογενετικά αποτελέσματα Εκτός από την κλασσική κυτταρολογική μέθοδο χρώσης με Feulgen, πραγματοποιήθηκε και χρώση ζωνώσεων με φθορισμό με χρήση χρωμομυκίνης Α 3 (CMA 3 ). Παρόμοιες μελέτες έχουν γίνει για μερικούς αντιπροσώπους της tribe Cichorieae όπως για τα γένη Cichorium (Bernades 2010, Bernades & al. 2013), Crepis (Fregonezi & al. 2003), Hypochaeris (Cerbah & al. 1998, Cerbah & al. 1999, Weiss-Schneeweiss & al. 2003, Fiorin & al. 2013), Lactuca (Matoba & al. 2007) και Scorzonera (D Amato 2000). Για το γένος Hymenonema δεν υπάρχει καμία μελέτη που να χρησιμοποιεί σύγχρονες μεθόδους κυτταρογενετικής. Η CMA 3 χρώση κατέδειξε τις πλούσιες σε GC περιοχές, δίνοντας σήματα και στα δυο είδη. Στα περισσότερα δικότυλα φυτά οι θετικές σε χρωμομυκίνη περιοχές (CMA 3 +) περιορίζονται σε ετεροχρωματινικές περιοχές γύρω από το κεντρομέρος και ειδικά σε rdna ενώ έχουν παρατηρηθεί και σε δευτερογενείς περισφίγξεις (Weiss- Schneeweiss & al. 2003). Οι CMA 3 + εντοπίζονται σε 1 ζεύγος χρωμοσωμάτων, χωρίς ωστόσο να μπορούμε να διακρίνουμε συγκεκριμένο μοτίβο ζωνώσεων (Εικ 59, 60). Παρατηρούμε πως οι φθορίζουσες περιοχές εντοπίζονται στο μικρότερο ζεύγος χρωμοσωμάτων του καρυοτύπου και συγκεκριμένα στο μικρό βραχίονα του κάθε χρωμοσώματος. Από τα κυτταρολογικά δεδομένα, είναι γνωστό πως το μικρότερο 89
 και πιθανόν χρήση περισσότερων κυτταρογενετικών τεχνικών (π.")
91 ζεύγος χρωμοσωμάτων είναι δορυφορικό, επομένως οι φθορίζουσες περιοχές φαίνεται να συμπίπτουν με την δορυφορική περιοχή των χρωμοσωμάτων. Για να εξαχθούν συγκεκριμένα συμπεράσματα κρίνεται απαραίτητη η μελέτη περισσότερων πληθυσμών από κάθε είδος, χρώση με φθορίζουσα χρωστική για τον εντοπισμό των πλούσιων σε AT περιοχών (π.χ. DAPI) και πιθανόν χρήση περισσότερων κυτταρογενετικών τεχνικών (π.χ. in situ υβριδοποίηση φθορισμού σε ριβοσωμικά γονίδια). Με μια πιο ολοκληρωμένη κυτταρογενετική έρευνα για το γένος θα επιτραπεί η μελέτη της οργάνωσης του γονιδιώματος του γένους και θα αποκαλυφθούν πιθανές διαφοροποιήσεις μεταξύ των ειδών ή/και των πληθυσμών τους. Εικ. 59: Καρυότυπος με χρώση CMA 3 του Hymenonema laconicum από τον πληθυσμό no 4. Με κόκκινα βέλη σημειώνονται οι φθορίζουσες περιοχές. 90

92 Εικ. 60: Καρυότυπος με χρώση CMA 3 του Hymenonema graecum από τον πληθυσμό no 7. Με κόκκινα βέλη σημειώνονται οι φθορίζουσες περιοχές. 91
93 5. ΜΟΡΙΑΚΗ ΜΕΛΕΤΗ 5.1. ΜΟΡΙΑΚΗ ΦΥΛΟΓΕΝΕΤΙΚΗ Με τον όρο φυλογένεση εννοούμε την μελέτη των εξελικτικών σχέσεων των οργανισμών, όπως αυτές διαμορφώνονται με το πέρασμα του χρόνου. Η εξελικτική διαδικασία προχωρά όχι πάντοτε σε ευθεία γραμμή, συνήθως σχηματίζει διακλαδώσεις, καθώς οι πληθυσμοί μεταβάλλονται στο χρόνο και μπορούν να διαφοροποιούνται σε ξεχωριστούς κλάδους (ειδογένεση), να διασταυρώνονται ή να εξαφανίζονται. Από την εποχή του Δαρβίνου ήδη στόχος των ταξινόμων είναι η ταξινόμηση να αντανακλά όσο πιο πιστά γίνεται τις φυλογενετικές σχέσεις των οργανισμών. Έτσι, ο προσδιορισμός των φυλογενετικών σχέσεων συνδέεται πολύ στενά με την ταξινόμηση των οργανισμών. Παλαιότερα η περιγραφή των σχέσεων μεταξύ των οργανισμών βασιζόταν σε φαινοτυπικά χαρακτηριστικά. Οι αναλύσεις που στηρίζονταν στη μορφολογία των ατόμων παρουσίαζαν αρκετά προβλήματα κι έτσι οι ερευνητές στράφηκαν σε άλλες μεθόδους, όπως την κυτταροταξινόμηση, την χημειοταξινόμηση, κ.λπ. Τα τελευταία τουλάχιστον 30 χρόνια άρχισαν να χρησιμοποιούνται τα μοριακά δεδομένα και κυρίως οι αλληλουχίες του DNA στις φυλογενετικές αναλύσεις, κι έτσι είχαμε την ανάπτυξη των μοριακών τεχνικών. Πρέπει να σημειωθεί ότι όλες οι μέθοδοι που βασίζονται είτε σε μορφολογικά είτε σε κυτταρολογικά ή σε μοριακά δεδομένα παρουσιάζουν πλεονεκτήματα και μειονεκτήματα, οπότε η χρήση περισσότερων μεθόδων ταξινόμησης σίγουρα οδηγεί στην καλύτερη ερμηνεία των φυλογενετικών σχέσεων των οργανισμών. Η μοριακή προσέγγιση της ταξινόμησης των οργανισμών βασίζεται στη γενική αρχή ότι η αλληλουχία του DNA των έμβιων όντων κρύβει την εξελικτική τους ιστορία, καθώς ο βαθμός συγγένειας δυο οργανισμών είναι ανάλογος του βαθμού ομοιότητας των γονιδιωμάτων τους ή, καλύτερα, των πληροφοριακών γενετικών στοιχείων τους. Αυτή η αρχή διέπει και τον επιστημονικό κλάδο της Μοριακής Φυλογενετικής. Πιο συγκεκριμένα, αν τα γονιδιώματα εξελίσσονται με τη σταδιακή συσσώρευση μεταλλάξεων, τότε οι διαφορές στη νουκλεοτιδική αλληλουχία ενός ζεύγους γονιδιωμάτων θα υποδεικνύει το πόσο πρόσφατα αυτά τα γονιδιώματα μοιράζονται έναν κοινό πρόγονο. Δυο γονιδιώματα που διαχωρίστηκαν στο πρόσφατο παρελθόν, θα αναμένεται να έχουν λιγότερες διαφορές από δυο 92
94 γονιδιώματα που ο κοινός τους πρόγονος είναι παλαιότερος. Αυτό σημαίνει ότι, συγκρίνοντας τρία ή περισσότερα γονιδιώματα, είναι εφικτό να αποσαφηνιστούν οι εξελικτικές σχέσεις μεταξύ τους. Αυτό είναι το αντικείμενο με το οποίο ασχολείται η Μοριακή Φυλογενετική (Molecular Phylogenetics) (Κράιτσεκ 2006, Αλαχιώτης 2007). Η Μοριακή Φυλογενετική προέρχεται κατά κάποιο τρόπο από την παραδοσιακή μέθοδο ταξινόμησης των πληθυσμών, σύμφωνα με τις ομοιότητες και τις διαφορές, όπως έκανε ο Λινναίος τον 18 ο αιώνα. Ο Λινναίος ήταν συστηματικός και όχι εξελικτικός, αλλά χωρίς να το γνωρίζει έθετε τα θεμέλια για διάφορα θέματα που θα απασχολούσαν μετέπειτα τους εξελικτικούς, με το να διαχωρίζει ιεραρχικά τους διάφορους οργανισμούς σε ταξινομικές κατηγορίες. Οι φυσικοί του 18 ου και του 19 ου αιώνα προτίμησαν να αναφέρονται σε αυτή την ιεραρχική κατηγοριοποίηση ως «δένδρο της ζωής» (Tree of life), ένας χαρακτηρισμός που είχε υιοθετηθεί από το Darwin (1859) στο βιβλίο του THE ORIGIN OF THE SPECIES, προσπαθώντας να περιγράψει τις αλληλοσυνδεόμενες εξελικτικές ιστορίες των ζώντων οργανισμών. Η κατηγοριοποίηση που είχε γίνει από τον Λινναίο ερμηνευόταν αργότερα ως φυλογένεια, υποδεικνύοντας όχι μόνο τις ομοιότητες μεταξύ των ειδών αλλά και τις εξελικτικές τους σχέσεις (Brown 2002, Κράιτσεκ 2006). Η φυλογένεση (phylogeny) ορίζεται ως η εξελικτική αλληλουχία των οργανισμών, η ιστορία μιας ομάδας, ενός taxon ή διαφόρων taxa, που περιγράφεται σε μορφή εξελικτικού δέντρου, με κοινό πρόγονο. Μια ομάδα που περιλαμβάνει μέλη είδη τα οποία είναι απόγονοι ενός κοινού προγόνου λέγεται μονοφυλετική (monophyletic) και είναι συνώνυμη του κλάδου. Πολυφυλετική (polyphyletic) χαρακτηρίζεται μια ομάδα, όταν τα μέλη είδη της έχουν προέλθει κλαδογενετικά, φυλογενετικά από μερικές διαφορετικές προγονικές γραμμές, από τις οποίες έχουν προέλθει και είδη που ανήκουν σε άλλες ομάδες. Παραφυλετική (paraphyletic), είναι μια ομάδα στην οποία δεν κατατάσσονται όλα τα είδη που προήλθαν από τον πιο πρόσφατο κοινό πρόγονο, καθώς μερικά είδη βρίσκονται εκτός της ομάδας αυτής (Αλαχιώτης 2007). Ο στόχος των περισσότερων φυλογενετικών μελετών είναι η κατασκευή ενός φυλογενετικού δέντρου. Ένα φυλογενετικό δέντρο αποτελεί μια γραφική αναπαράσταση της φυλογένειας μιας ομάδας από taxa ή γονίδια. Οι αντικειμενικοί στόχοι της κατασκευής τους είναι: α) η αναπαράσταση των πραγματικών γενεαλογικών σχέσεων των οργανισμών και β) η χρονολόγηση της διάσπασης των ειδών από τον τελευταίο τους πρόγονο. Ανάλογα με το αν αντανακλά φυλογενετικές 93
95 σχέσεις ομάδας πληθυσμών ή ειδών, λέγεται φυλογενετικό δέντρο πληθυσμών ή ειδών. Όταν το φυλογενετικό δέντρο κατασκευάζεται με βάση τις νουκλεοτιδικές αλλαγές ενός ή πολύ λίγων γονιδίων (από κάθε είδος), τότε λέγεται γονιδιακό δέντρο (gene tree). Τα γονίδια ενός οργανισμού μπορεί να έχουν διαφορετικές εξελικτικές ιστορίες, οι οποίες μπορεί να συμφωνούν κατά ένα μέρος ή καθόλου με τη φυλογένεση του είδους. Έτσι συμβαίνει το γονιδιακό δέντρο να μην συμφωνεί πάνοτε με το δέντρο των ειδών (species tree). Τα φυλογενετικά δέντρα συνήθως είναι με ρίζα (rooted), η οποία δείχνει τον κοινό πρόγονο, όπως και την εξελικτική κατεύθυνση (Αλαχιώτης 2007). Οι μέθοδοι κατασκευής εξελικτικών δέντρων κατατάσσονται σε δυο μεγάλες κατηγορίες: 1) την κατηγορία των μητρών απόστασης (distance matrix methods), που περιλαμβάνει τις μεθόδους UPGMA, των μετασχηματισμένων αποστάσεων, των Fitsch-Margoliash και των γειτονικών ζευγαριών (neighbor joining) και 2) την κατηγορία που βασίζεται στην παρουσία ή απουσία πληροφοριακών χαρακτήρων και περιλαμβάνει τις μεθόδους της μέγιστης φειδωλότητας (maximum parsimony), της μέγιστης πιθανοφάνειας (maximum likelihood) και της μπεϊσιανής ανάλυσης (Bayes analysis). Στην πρώτη κατηγορία, οι εξελικτικές αποστάσεις (συνήθως νουκλεοτιδικές ή αμινοξικές διαφορές) υπολογίζονται για όλα τα ζευγάρια και χρησιμοποιείται ένας αλγόριθμος για την κατασκευή του δέντρου. Στην δεύτερη κατηγορία, και ειδικότερα στη μέγιστη φειδωλότητα, επιλέγονται οι μικρότερες (κάθε φορά) εξελικτικές αποστάσεις (νουκλεοτιδικές ή αμινοξικές υποκαταστάσεις) για την κατασκευή του φυλογενετικού δέντρου, το οποίο επιλέγεται με βάση αυτή τη συλλογιστική σε σχέση με όλα τα πιθανά δέντρα και θεωρείται το πιο πιθανό να είναι το σωστό δέντρο. Είναι σημαντικό να σημειωθεί ότι, οι διάφορες μέθοδοι κατασκευής εξελικτικών δέντρων χωρίς την εξαντλητική διερεύνηση των δυνατοτήτων τους, ενέχει τον κίνδυνο υπερεκτίμησης της κατάστασης. Για παράδειγμα, η μοριακή φυλογένεση στηρίζεται στους αριθμούς των νουκλεοτιδικών και αμινοξικών υποκαταστάσεων, οι οποίες διέπονται από πολύ περίπλοκες διεργασίες, που καθοδηγούνται από τη μετάλλαξη, τη φυσική επιλογή ή την τυχαία γενετική παρέκκλιση, οι οποίες αν δεν έχουν αποσαφηνιστεί και συμπεριληφθεί στη μελέτη, ενδέχεται να οδηγήσουν σε λάθη. Εξάλλου, η κατασκευή πολλών γονιδιακών δέντρων, τα οποία εκλαμβάνονται ως δέντρα ειδών, ενέχει τον κίνδυνο παρερμηνείας, αφού οι δυνατότητες των δυο τύπων δέντρων είναι διαφορετικές. Έτσι, μπορούν να 94
96 γενικευτούν εύκολα καταστάσεις και εξελικτικές ιστορίες, οι οποίες εμπεριέχουν σοβαρά λάθη (Αλαχιώτης 2007) ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Η μοριακή και φυλογενετική μελέτη πραγματοποιήθηκε στο Institute of Plant Science του University of Regensburg, υπό την εποπτεία του Prof. Dr. Christoph Oberprieler. Η πειραματική διαδικασία και η επεξεργασία των δεδομένων ολοκληρώθηκε στο Εργαστήριο της ερευνητικής ομάδας Plant Systematics and Evolution Group Συλλογή υλικού & εκχύλιση DNA Συλλογή υλικού Κατά τη διάρκεια των ετών 2013 και 2014 πραγματοποιήθηκαν συλλογές του γένους Hymenonema για τις ανάγκες της μοριακής μελέτης. Συλλέχθηκε υλικό, επίσης, και από τα υπόλοιπα γένη της subtribe Scolyminae, Catananche και Scolymus, τα οποία εμφανίζονται στον ελληνικό χώρο. Το υλικό αυτό συλλέχθηκε στη φύση ή/και καλλιεργήθηκε στον Πειραματικό Κήπο του Πανεπιστημίου Πατρών και μέρος αυτού είναι κατατεθειμένο στο Herbarium του Πανεπιστημίου Πατρών (UPA). Συλλέχτηκαν φρέσκα φύλλα, τα οποία τοποθετήθηκαν αμέσως σε silica gel για αποξήρανση. Προκειμένου να υπάρχουν αντιπρόσωποι από όλα τα είδη της subtribe, χρησιμοποιήθηκε και αποξηραμένο υλικό από το Herbarium of the Botanic Garden and Botanical Museum Berlin-Dahlem (B) για μερικά είδη καθώς και υλικό προηγούμενης μελέτης (Hammerschmid 2014) το οποίο βρίσκεται αποθηκευμένο στο Institute of Botany and Cell Biology of Regensburg University. Συνολικά, στην παρούσα μελέτη περιλαμβάνονται 31 δείγματα (Πίνακας 11, Εικ. 61). Για κάθε είδος χρησιμοποιήθηκαν 2-3 δείγματα, εκτός από τα Catananche montana, Gundelia aragatsii και Gundelia tournefortii για τα οποία χρησιμοποιήθηκε 1 δείγμα για το καθένα. Το γένος Gundelia χρησιμοποιήθηκε ως μέτρο φυλογενετικής συγγένειας δηλ. ως εξωτερική ομάδα ή ομάδα αναφοράς (outgroup). 95
97 Πίνακας 11: Πληθυσμοί των taxa που χρησιμοποιήθηκαν στη μοριακή μελέτη. Taxon Sample no Πληθυσμός Catananche arenaria Α146 MAROKKO: Anti-Atlas, road P 30 between Tiznit and Goulimine, Tizi Mighert, limestone, 1050 m, N, 9 48 W, , Vogt & Oberprieler 6333 (B ).» Α862 MAROKKO: Prov. Er-Rachidia, road P 32 between Er- Rachidia and Boudnib, c. 10 km E junction to Erfoud, stony plains, 1110m, N-4 16 W, , Vogt & Oberprieler 4848 (B ).» Α894 TUNISIA: Gouvernorat de Gafsa, Tunisie du Sud, road C 201 between Moulares and Redeyef, sandy plains of Oued Selja c. 7 km S Moulares, 400 m, N E, , Vogt & Oberprieler 7044 (B ). Catananche caerulea Α868 MAROKKO: Prov. Er-Rachidia, Hoher Atlas, P21 (Midelt - Er-Rachidia), Tunnel du Legionaire, steinige Hänge und Felsen, N-4 21 W, , Vogt 5397, Bayón & Oberprieler (B ).» Α863 SPANIEN: Prov. Huesca, Arro, 700 m, ERN 4272 & AL., cult. In Hort. Bot. Berlin Dahlem, , Oberprieler & Vogt, 2n = 18 (B )» Α871 TUNESIEN: Gouvernorat de Kasserine, Darsale Tunisienne, summit of Djebel Chambi W Kasserine, limestone, 1530 m, N E, , Vogt & Oberprieler (B ). Catananche caespitosa Α149 MAROKKO: Prov. Er-Rachidia, Mittlerer Atlas, Khenifra - Midelt (P 33), 4 km E der Passhöhe Tonout-ou-Fillali, Felder, 1910 m, N-5 24 W, , Vogt 5859, Baytón & Oberprieler (B ).» Α864 MAROKKO: Meknes, Middle Atlas, Timahdite, Col du Zad, c. 44 km from Azrou on road to Midelt, 2100 m, 33 2 N- 5 4 W, , Valdes, Vogt 9446, Oberprieler 3884 & al. [Iter Med. V: n ] (B ). Catananche lutea Α910 GREECE: Lakonia, port of Gerakas, olive groves, alt. 15 m, N, E, , Liveri & Ketsilis- Rinis s.n. (UPA).» Α147 TUNESIEN: Gouvernorat de El Kef, Dorsale Tunisienne, road between Kalaat Es Senan and Kalaat Khasba, 16 km NW Kalaat Khasba, road embankment, fields and oued, 880 m, N E, , Vogt & Oberprieler 6821 (B ).» Α865 Zypern, Pahos, Peyia, steinige Hänge an der Straße nach Polis, 96
98 Catananche montana Gundelia aragatsii Α311 Α m, Kalk, , Vogt 8716 (B ) ALGERIEN: Wilaya de Bouira, Parc National du Djurdjura, pentes sud de l Akouker, à environ 5 km au NE de Tikjda, 1600 m, , Dubuis & Maurel (B ) ARMENIA: Aragatsotn province, SE slopes of Mt. Aragats, track from water reservoir below Avtona to Kakavadzor, 1890 m, N E, , Vitek (B ). Gundelia tournefortii Hymenonema graecum Α872 ZYPERN: Giolou, SSE of village, old track to Stroumbi, c. 250 m from watertank towards St., grassy banks in vineyards, 360 m, N E, , Hand 5796 & Christodoulou (B ). Α901 GREECE: Kykladen, Naxos, Prokopios, garigue, 30 m, , Jäth s.n. (B ).» Α902 GREECE: Kiklades, Ins. Tinos, Monastiri, Arnados, Dio Xoria, along the roadsides connecting these three villages, alt m, N to N, E to E, , Liveri & Ketsilis-Rinis 117 (UPA).» Α903 GREECE: Kiklades, Ins. Andros, Batsi, residential area, Agios Filippos church, alt. 45 m, N, E, , Liveri & Ketsilis-Rinis 115 (UPA).» Α904 GREECE: Kiklades, Ins. Kithnos, Merichas Bay (SW side), phrygana, alt m, N, E, , Liveri & Ketsilis-Rinis 100 (UPA).» Α905 GREECE: Kiklades, Ins. Siros, Kini, along the roadsides from the beach to the village for a distance up to 1 km approximately, N, E, , Liveri & Ketsilis-Rinis 107 (UPA).» Α256 GRIECHENLAND: Kikladen, Serifos, am Bach oberhalb Koutalas, 400 m, , Malicky s.n. (W). Hymenonema laconicum Α906 GREECE: Lakonia, Lagkada gorge, climbing region, alt. 800 m, 37 4'59.09"N, 22 18'39.30"E, , Charalampopoulos 1524 (UPA)» Α907 GREECE: Lakonia, Geraki, road from the village to Alepohori, alt m, N, E, , Liveri & Ketsilis-Rinis 124 (UPA).» Α908 GREECE: Arkadia, SE of Leonidio, road from Leonidio to Tsitalia village, alt. 113 m, N, E, , Kofinas & Dolianitis 129 (UPA).» Α909 GREECE: Lakonia, Krokees, alt. 350 m, N, 97
99 Scolymus grandiflorus Α E, , Liveri & Kofinas 131 (UPA). MAROKKO: Peninsula of Melilla, road 8106 between Segangane and Aazanèn, c. 3 km W Segangane, 190 m, N, 3 4 W, , Vogt & Oberprieler 5355 (B ).» Α896 MOROCCO: Prov. Oujda, Monts de Beni-Snassen, track 5308 between Ain-Almou and road 5319 (Ahfir - Ain-Asa), surroundings of Tizi-n-Tirchete, field margins, 910 m, N W, , Vogt & Oberprieler 9600 (B ). Scolymus hispanicus Α898 GREECE: Lakonia, Velies, alt m, N, E, , Liveri & Ketsilis-Rinis s.n. (UPA).» Α897 MOROCCO: Prov. Marrakech, Marrakech - Beni-Mellal (P 24), 26 km nach El-Kelaa-Srarhna, Felder, 410 m, 32 9 N- 7 6 W, , VOGT 5770, Bayón & Oberprieler (B ).» Α150 TUNESIEN: Gouvernorat de Siliana, Dorsale Tunisienne, road C77 between El Hajeb and Makthar, c. 8 km S Makthar, fields and road margins, 910 m, N, E, , Vogt & Oberprieler 7772 (B ). Scolymus maculatus Α899 MOROCCO: Moyen Sebou Nord, Sefiane, Col de Rmel, c. 21 km from OOuezzane on road to Souk-el-Arba-du-Rharb (P 23), river banks of an Oued and N-facing slopes, 140 m, N-5 45 W, , Vogt 9617 & Oberprieler 4053 [= Iter. Med. V: n ] (B ).» Α151 SPANIEN: Prov. Cadiz, Tarifa, Dünen und Strand (bzw. Kiefernwald) ca. 10 km NW Tarifa, ca. 1-5 m, , Vogt 9276 & Prem-Vogt (B ) Εκχύλιση DNA (DNA extraction) Αρχικά, μέρος του φύλλου (10-20 mg) κάθε δείγματος ομογενοποιήθηκε με ειδική συσκευή (mortal mill - Retsch, Germany). Το DNA των δειγμάτων εκχυλίστηκε χρησιμοποιώντας τροποποιημένο πρωτόκολλο βασισμένο στην CTAB μέθοδο από τους Doyle & Doyle (1987). Η ποιότητα του εκχυλισθέντος DNA ελέγχθηκε με ηλεκτροφόρηση σε 1.5% ΤΒΕ gel αγαρόζης. Το εκχυλισθέν DNA φυλάσσεται στους -20 C. 98








100 Εικ. 61: A. Gundelia aragatsii, B. Gundelia tournefortii, Γ. Scolymus maculatus, Δ. Scolymus grandiflorus, Ε. Catananche lutea, ΣΤ. Catananche caerulea, Ζ. Catananche arenaria, H. Catananche caespitosa (Φωτ. Α, Γ, Δ, ΣΤ, Ζ, Η από και Β, από 99
101 5.2.2 Επιλογή μοριακών δεικτών Χλωροπλαστικοί δείκτες Οι χλωροπλάστες και τα μιτοχόνδρια έχουν γονιδιώματα που κληρονομούνται με μη μεντελικό τρόπο. Τυπικά εμφανίζουν μητρική κληρονομικότητα. Στα φυτά, τα γονιδιώματα των ίδιων οργανιδίων υφίστανται σωματικό διαχωρισμό, που σημαίνει ότι οι γονικοί χαρακτήρες διαχωρίζονται στα σωματικά κύτταρα, εμφανίζοντας ετερογένεια στα κύτταρα του οργανισμού. (Lewin 2004). Το χλωροπλαστικό DNA (cpdna) περιλαμβάνει δυο μονάδες, τη μεγάλη (large single copy, LSC) και τη μικρή (small single copy, SSC), καθώς και δυο ανεστραμμένες επαναλήψεις (inverted repeats, IRA, IRB) και κωδικοποιεί τα δικά του γονίδια, καθώς και μερικά γονίδια του φωτοσυνθετικού μηχανισμού (Wakasugi et al. 2001). Περιοχές του cpdna έχουν χρησιμοποιηθεί επιτυχώς σε φυλογεωγραφικές μελέτες (Petit et al. 1997, Caron et al. 2000, Dutech at al. 2000, Raspé et al. 2000, Falchi et al. 2009), με πιο συχνά χρησιμοποιούμενες τις μη κωδικοποιημένες. Αυτό οφείλεται στο ότι στα ανώτερα φυτά οι κωδικοποιημένες περιοχές είναι πολύ συντηρημένες περιοχές, σε αντίθεση με τις μη κωδικοποιούσες που εμφανίζουν χαμηλό ρυθμό εξέλιξης και γι αυτό προτιμώνται σε μελέτες φυτών περί φυλογένειας, γενετικής δομής των πληθυσμών και φυλογεωγραφίας (Sang 2002). Βέβαια, αν κι έχουν χρησιμοποιηθεί από επίπεδο υποοικογένειας έως είδους (Mes et al. 2000), φαίνεται να είναι πιο σημαντικοί σε χαμηλά ταξινομικά επίπεδα (Taberlet et al. 1991, Shaw et al. 2014). Στην εν λόγω μελέτη, επιλέχθηκαν δυο μοριακοί δείκτες, που αποτελούν μη κωδικοποιημένες περιοχές του cpdna, οι trnl-f και rpl32-trnl (Εικ. 62). Ο trnl-f είναι intergenic spacer (IGS) και βρίσκεται ανάμεσα στα γονίδια trnl (κωδικοποιεί το trna-λευκίνη) και trnl-f (κωδικοποιεί το trna-φαινυλαλανίνη) (Wakasugi et al. 2001). Ο rpl32-trnl ανακαλύφθηκε σχετικά πρόσφατα για την πιθανή φυλογενετική του χρησιμότητα (Shaw et al. 2007, Timme et al. 2007) και είναι πρώτος στην κατάταξη για τους καλύτερους χλωροπλαστικούς δείκτες της ομάδας των euasterids II, όπου ανήκει και το μελετώμενο γένος (Shaw et al. 2014). Πυρηνικοί δείκτες Λόγω του γεγονότος ότι οι χλωροπλάστες περνούν στην επόμενη γενιά αποκλειστικά από το ωάριο (Corriveau & Coleman 1988), δίνουν φως στην φυλογενετική ιστορία 100
 γονίδια.")
102 από την μητρική πλευρά. Γι αυτό διερευνώνται μοριακοί δείκτες και από το πυρηνικό γονιδίωμα. Μια ομάδα πυρηνικών δεικτών που χρησιμοποιούνται ευρέως για την διαλεύκανση φυλογενετικών σχέσεων είναι τα ριβοσωμικά (rdna) γονίδια. Τα ριβοσώματα είναι σύμπλοκα σωματίδια, που αποτελούνται από 3 διαφορετικά μόρια RNA και πρωτεΐνες. Στους ευκαρυωτικούς οργανισμούς αποτελούνται από 2 Εικ. 62: cpdna του Arabidopsis thaliana. Με κόκκινα βέλη σημειώνονται οι χλωροπλαστικοί δείκτες που χρησιμοποιούνται στη μελέτη ( υπομονάδες. Η κάθε υπομονάδα αποτελείται από τουλάχιστον 1 μόριο RNA και πρωτεΐνες. Η μεγαλύτερη υπομονάδα περιλαμβάνει τα 28s και 5,8s μόρια rrna ενώ η μικρότερη το 18s. Όσον αφορά αυτά τα γονίδια (18s, 5.8s και 28s) υπάρχουν δυο εσωτερικά μεταγραφόμενα διαστήματα (ITS1, ITS2) που τα χωρίζουν και μια εξωτερική μεταγραφόμενη περιοχή (ETS) που βρίσκεται πριν το 18s (Εικ. 63). Οι περιοχές αυτές περιέχουν σήματα που οδηγούν το rrna μεταγραφής. Τα γειτονικά αντίγραφα των rdna μονάδων χωρίζονται από ένα μη μεταγραφόμενο διάστημα (NTS). Αυτή η περιοχή περιέχει στοιχεία που δρουν ως ενισχυτές της μεταγραφής 101
103 (Hillis & al. 1991). Οι περιοχές κωδικοποίησης και τα ενδιάμεσα διαστήματα, επαναλαμβάνονται πολλές φορές σε μορφή συνεχών επαναλήψεων (Αλαχιώτης 2007). Για την μελέτη αυτή, έχουν επιλεχθεί οι ITS1 και ITS2 ως πυρηνικοί δείκτες. Οι δείκτες αυτοί χρησιμοποιούνται ευρέως διότι θεωρείται ότι τα πυρηνικά rdna διαστήματα εξελίσσονται πιο γρήγορα από τις πυρηνικές rrna κωδικές περιοχές. Αυτό παρατηρείται γιατί οι υποκαταστάσεις που μπορεί να συμβούν στα διαστήματα (ITS, ETS) δεν έχουν θανατηφόρες συνέπειες στον οργανισμό και γι αυτό στις περιοχές αυτές παρατηρείται πιο γρήγορος εξελικτικός ρυθμός. Η σχετική υψηλή ποικιλότητα των συγκεκριμένων δεικτών που έχει αναφερθεί στα Compositae, φαίνεται να τους καθιστά κατάλληλους δείκτες ειδικά σε ενδοειδικό επίπεδο (Baldwin 1992). Επιπλέον, χρησιμοποιείται ως πυρηνικός μοριακός δείκτης το γονίδιο D10 που έχει ένα μόνο αντίγραφο (single copy gene), πρόσφατα χαρακτηρισμένο από τους Chapman et al. (2007). Εικ. 63: Οργάνωση μιας rdna υπομονάδας. Οι υπομονάδες είναι οργανωμένες σε γειτονικές επαναλήψεις. Κάθε μια αποτελείται από τα γονίδια 18s, 5.8s & 28s.(Muir & Schlötterer 1999). Εικ. 64: Θέση πρόσδεσης και κατεύθυνση των εκκινητών που χρησιμοποιήθηκαν για τους μοριακούς δείκτες ITS1 & ITS2 (από internet στο Hammerschmid 2014). 102
104 Μοριακές τεχνικές Ενίσχυση (amplification) PCR: Η Αλυσιδωτή αντίδραση της Πολυμεράσης (PCR) χρησιμοποιήθηκε για την ενίσχυση των δεικτών με χρήση του Taq DNA Polymerase Master-mix Red (Ampliqon, Odense, Denmark) και τελικό όγκο 12.5 μl, σύμφωνα με το προτεινόμενο πρωτόκολλο από την εταιρεία. Συνοπτικά τα στάδια της PCR έχουν ως εξής (Εικ. 65): 1) αποδιάταξη (denaturation) της μήτρας του δίκλωνου DNA σε θερμοκρασία 95 C, 2) ψύξη σε θερμοκρασία μεταξύ C ώστε οι εκκινητές να προσδεθούν στο DNA (annealing) και 3) επιμήκυνση (extension) στους 72 C. Στο στάδιο της επιμήκυνσης συμμετέχει η DNA-πολυμεράση και ο κύκλος αυτός επαναλαμβάνεται φορές. Για την διαδικασία της ενίσχυσης χρησιμοποιήθηκαν διαφορετικοί εκκινητές (primers) κατά περίπτωση. Η διαδικασία επιλογής της κατάλληλης θερμοκρασίας υβριδοποίησης (annealing temperature) έγινε με ειδικές συσκευές PCR (gradient PCR machines). Εικ. 65: Βήματα της PCR: 1. Αποδιάταξη, 2. Πρόσδεση εκκινητών και 3. Επιμήκυνση (en.wikipedia.org). Εκκινητές: Όσον αφορά την nrdna ITS περιοχή, οι υποπεριοχές ITS1 και ITS2 ενισχύθηκαν ξεχωριστά, χρησιμοποιώντας τους εκκινητές P1 και P2 (White 1990) για την ITS1, καθώς και τους P3 και P4 (White 1990) για την ITS2 (Εικ. 64). Μόνο για το Catananche caespitosa (Α864) χρησιμοποιήθηκαν διαφορετικοί εκκινητές, οι 18SF (Rydin 2004) και P2B (White et al. 1990) για την ITS1, και οι P3 (White et al. 103
105 1990) και SR (Blattner et al. 2001) για την ITS2. Για την περιοχή trnl-f χρησιμοποιήθηκαν οι εκκινητές trnl2(e) και trnf(f) (Taberlet et al. 1991) ενώ για την rpl32-trnl οι trnl (UAG) και rpl32-f (Shaw et al. 2007). Για τον πυρηνικό δείκτη D10 χρησιμοποιήθηκαν οι εκκινητές D10f και D10r (Chapman et al. 2007). Καθαρισμός: Ο καθαρισμός των προϊόντων της PCR είναι απαραίτητος, για την περίπτωση που ένα γονίδιο πολλαπλασιάζεται με σκοπό να αναλυθεί η αλληλουχία του. Αυτό οφείλεται στο ότι οι αναλυτές αλληλουχίας είναι πολύ ευαίσθητοι στις προσμίξεις που μειώνουν τη συγκέντρωση του DNA. Τα προϊόντα της PCR καθαρίστηκαν με τη μέθοδο Agencourt AMPure magnetic beads (Agencourt Bioscience Corporation, Beverly, Massachusetts, USA). Ηλεκτροφόρηση: Η ηλεκτροφόρηση βασίζεται στην κίνηση φορτισμένων μορίων μέσα σε ηλεκτρικό πεδίο. Πρέπει να σημειωθεί ότι, η τεχνική αυτή χρησιμοποιήθηκε για τον έλεγχο της ποιότητας και της ποσότητας του DNA που προέκυπτε από τον πολλαπλασιασμό των τμημάτων DNA με PCR, αλλά και μετά από κάθε καθαρισμό των προϊόντων. Η ηλεκτροφόρηση έγινε με τη χρήση 1,5% ΤΒΕ gel αγαρόζης, ενώ για την οπτικοποίηση το gel τοποθετούνταν σε πλάκα υπεριώδους φωτός (αφού είχε προστεθεί στο gel ειδική χρωστική). Η ύπαρξη μιας μόνο ευκρινούς ζώνης, δήλωνε την απουσία τυχόν παραπροϊόντων που θα μπορούσαν να προκύψουν. Ο έλεγχος της ποσότητας των ζωνών γινόταν με αλληλουχία γνωστού μεγέθους (ladder). Κλωνοποίηση (cloning) Σε 6 περιπτώσεις, η περιοχή D10 χρειάστηκε να κλωνοποιηθεί. Στην προκειμένη περίπτωση παρατηρήθηκαν περισσότερα του ενός αντίγραφα. Πιθανόν αυτό οφείλεται σε περισσότερα αλληλόμορφα για τον ίδιο γενετικό τόπο ή παράλογες (paralogous) αλληλουχίες. Με την κλωνοποίηση, κάθε ένα από αυτά τα μόρια λαμβάνεται από ένα βακτήριο (Escherichia coli), το οποίο με τις κατάλληλες συνθήκες θα δημιουργήσει μια αποικία. Παίρνοντας μια αποικία λοιπόν, είναι σίγουρο ότι σ αυτήν έχει πολλαπλασιαστεί ένα μόνο μόριο. Για κάθε είδος πάρθηκαν περίπου 12 αποικίες και ο έλεγχος των προϊόντων έγινε πάλι με ηλεκτροφόρηση. Βάσει αυτού επιλέχθηκαν ποιες αλληλουχίες θα σταλούν για αλληλούχιση και μετά από προσεκτική επεξεργασία επιλέχθηκαν ποιες θα χρησιμοποιηθούν στην μελέτη. Τα είδη για τα οποία χρειάστηκε κλωνοποίηση του D10 είναι τα: Gundelia aragatsii (Α900), Gundelia tournefortii (Α872), Hymenonema laconicum (A906), Scolymus 104
, σύμφωνα με τις οδηγίες της κατασκευαστικής εταιρείας.")
106 hispanicus (Α897, Α150), Scolymus grandiflorus (A152). Η κλωνοποίηση έγινε με τη χρήση του CloneJET PCR cloning kit (Fisher Scientific, Waltham, Massachusetts, USA), σύμφωνα με τις οδηγίες της κατασκευαστικής εταιρείας. Σε αυτό το kit περιλαμβάνεται ο φορέας κλωνοποίησης, ο οποίος φέρει γονίδιο για το ένζυμο περιορισμού (eco47ir), καθώς και γονίδιο ανθεκτικότητας στην αμπικιλίνη (ApR) και γονίδιο αντιγραφής (rep(pmb1)) (Εικ. 66). Εικ. 66: Ο φορέας κλωνοποίησης, τα γονίδια bla (ApR) και rep(mb1) και η περιοχή που εισέρχεται το τμήμα DNA (MCS) που πρόκειται να κλωνοποιηθεί ( Αλληλούχιση (sequencing) Η αλληλούχιση (cycle sequencing) έγινε με το DTCS Sequencing Kit (Beckman Coulter, Fullerton, California, USA), ακολουθώντας το πρωτόκολλο από τον κατασκευαστή. Οι αλληλουχίες αναλύθηκαν από το CEQ 8000 sequencer (Beckman Coulter, Fullerton, California, USA). Πρέπει να σημειωθεί ότι η αλληλούχιση για την επιλογή των μοριακών δεικτών έγινε στο προαναφερόμενο sequencer αλλά η αλληλούχιση των υπόλοιπων δειγμάτων κατά τη διάρκεια της μελέτης έγινε από την Macrogen (Amsterdam, Netherlands). Για τα δείγματα που αλληλουχήθηκαν στο sequencer του εργαστηρίου, μετά την αλληλούχιση πραγματοποιήθηκε καθίζηση του DNA (DNA precipitation). Πριν σταλούν τα δείγματα στην Macrogen, 105
107 πραγματοποιήθηκε μέτρηση της συγκέντρωσης DNA με τη χρήση του NanoDrop 2000 (Thermo Scientific). Τα αποτελέσματα της αλληλούχισης απεστάλησαν μέσω ηλεκτρονικού ταχυδρομείου σε μορφή χρωματογραφημάτων Επεξεργασία αλληλουχιών & ανάλυση δεδομένων Ποιοτικός έλεγχος χρωματογραφημάτων Η ανάλυση των χρωματογραφημάτων καθώς και ο έλεγχος για ασάφειες έγινε με το πρόγραμμα Chromas Lite 2.10 (Technelysium Pty Ltd., Tewantin, Australia; Συνήθως η ποιότητα τους μετά την αλληλούχιση δεν είναι καλή στις πρώτες βάσεις, ακολουθεί ένας αριθμός βάσεων με καλή ποιότητα, ο οποίος ποικίλλει ανάλογα με το μήκος του γονιδίου, ενώ μετά από αυτές τις βάσεις το σήμα φθίνει. Τα χρωματογραφήματα που πρόκειται να χρησιμοποιηθούν πρέπει να έχουν το αναμενόμενο μήκος βάσεων και μοναδική ευδιάκριτη κορυφή για κάθε σημείο της αλληλουχίας. Αυτός ο τύπος ελέγχου είναι εμπειρικός. Στοίχιση αλληλουχιών Η στοίχιση αποσκοπεί στον προσδιορισμό των ομόλογων θέσεων των αλληλουχιών, αφού με βάση τις διαφορές σ αυτές τις θέσεις υπολογίζονται οι εξελικτικές σχέσεις των υπό μελέτη αλληλουχιών. Οι κατάλληλες μετατροπές των αρχείων (fasta format, txt), αλλά και η επεξεργασία των αλληλουχιών με το χέρι, όπου κρίθηκε απαραίτητο, έγιναν με τη χρήση του Bioedit version ( Για τη στοίχιση των αλληλουχιών χρησιμοποιήθηκε το πρόγραμμα Clustal W (Thompson et al. 1994) και πραγματοποιήθηκε στο διαδικτυακό τόπο ενώ στη συνέχεια ελέγχθηκαν και διορθώθηκαν με το χέρι. Οι μέθοδοι Clustal χρησιμοποιούν προοδευτικούς αλγορίθμους όπου οι δυο συνολικά πιο ομόλογες στοιχίζονται πρώτες και ακολουθεί μια σειρά από νέες στοιχίσεις, προσθέτοντας σε κάθε βήμα την αμέσως συγγενέστερη αλληλουχία προς της ήδη στοιχισμένες. Τα κενά στις αλληλουχίες κωδικοποιήθηκαν ως δυαδικοί χαρακτήρες, χρησιμοποιώντας την απλή μέθοδο κωδικοποίησης κενών των Simmons & Ochoterena (2000), όπως εφαρμόζεται στο πρόγραμμα κωδικοποίησης GapCoder (Young & Healy 2003). Κατασκευή φυλογενετικών δέντρων 106
108 Χρησιμοποιήθηκαν δυο μέθοδοι φυλογενετικής ανάλυσης: η μέθοδος της Μέγιστης Φειδωλότητας (Maximum Parsimony) και η Μπεϊσιανή Συμπερασματολογία (Bayesian Inference). Οι χλωροπλαστικοί δείκτες trnl-f και rpl32-trnl αντιμετωπίστηκαν ως ενιαία αλληλουχία (cpdna), όπως και οι ITS1 & ITS2 (nrdna) ενώ ο D10 ξεχωριστά. Μέγιστη Φειδωλότητα (Maximum Parsimony): Η μέθοδος Μέγιστης Φειδωλότητας (ΜΦ) βασίζεται στην άποψη ότι όσο λιγότερα χρειάζεται να γνωρίζει κανείς για την εξελικτική διαδικασία, τόσο πιο μεγάλη εμπιστοσύνη μπορεί να έχει στα συμπεράσματά του (Sober 1988). Σε αυτή τη μέθοδο εξετάζονται όλες οι πιθανές τοπολογίες που μπορούν να συνδέουν τις αλληλουχίες και για κάθε τοπολογία υπολογίζεται ο αριθμός των εξελικτικών αλλαγών που πρέπει να έχουν γίνει στις αλληλουχίες για να προκύψει η συγκεκριμένη αλληλουχία. Η τοπολογία που θα έχει τον μικρότερο (φειδωλότερο) αριθμό βημάτων, αποτελεί και το δέντρο μέγιστης φειδωλότητας. Με άλλα λόγια, το δέντρο που απαιτεί τις λιγότερες αλλαγές είναι καλύτερο από αυτό που απαιτεί πολλές. Είναι πιθανό να μη προκύψει ένα μόνο φειδωλό δέντρο, αλλά πολλά περισσότερα, τα οποία θεωρούνται εξίσου φειδωλά (equally parsimony trees). Σε αυτή την ανάλυση δε χρησιμοποιούνται όλες οι νουκλεοτιδικές βάσεις, αλλά μόνο αυτές που διαφοροποιούνται μεταξύ των αλληλουχιών, ενώ οι σταθερές νουκλεοτιδικές βάσεις εξαιρούνται. Για να είναι μια θέση πληροφοριακή για την κατασκευή δέντρου ΜΦ, πρέπει να φέρει τουλάχιστον δυο διαφορετικά νουκλεοτίδια και το καθένα να εκπροσωπείται σε δυο τουλάχιστον αλληλουχίες. Αυτές οι θέσεις χαρακτηρίζονται πληροφοριακές θέσεις φειδωλότητας (parsimony informative sites). Όσο περισσότερες είναι αυτές οι θέσεις, τόσο πιο αξιόπιστο αναμένεται να είναι το δέντρο που θα προκύψει. Οι αναλύσεις της ΜΦ έγιναν με το πρόγραμμα PAUP* 4.0 version beta 10 (Swofford 2002) με τη χρήση της Branch and bound αναζήτησης. Μπεϊσιανή Συμπερασματολογία (Bayesian Inference): Η Μπεϊσιανή Συμπερασματολογία (ΜΣ) χρησιμοποιεί μια σημαντική αρχή που ονομάζεται επανασταθμισμένη πιθανότητα (posterior propability). Βασίζεται στο θεώρημα του Bayes: το οποίο χρησιμοποιείται για να συνδυάσει την πιθανότητα μιας υπόθεσης να είναι σωστή, προτού κάνουμε κάποια παρατήρηση (prior propability) της φυλογένεσης, 107
109 δηλαδή το Pr[Tree], με την πιθανοφάνεια (likelihood) των δεδομένων Pr[Data/Tree], ώστε να παραχθεί η επανασταθμισμένη ή εκ των υστέρων πιθανότητα (posterior propability) της αρχικής υπόθεσης, υπό το πρίσμα των δεδομένων που παρατηρήθηκαν. Η ΜΣ έχει δηλ. ως στόχο να γίνει αναφορά σε κάτι που δεν έχει παρατηρηθεί. Η ΜΣ χρησιμοποιεί τη μέθοδο Metropolis-coupled Markov Chain Monte Carlo (MCMCMC), που μπορεί να θεωρηθεί ως μια ομάδα ανεξάρτητων αναζητήσεων που περιστασιακά ανταλάσσουν πληροφορίες. Έχει σχεδιαστεί ως μέθοδος αναρρίχησης λόφων (hill-climbing), επιτρέποντας κατά την αναζήτηση τη διάσχιση από μια κοιλάδα, χωρίς να παγιδευτεί σε ένα μέγιστο λόφο. Το αποτέλεσμα είναι μια ομάδα δέντρων, η οποία έχει αξιολογηθεί επανηλλειμένα και η οποία συγκροτεί την κορυφή του λόφου (Xiong 2006). Η ΜΣ επανασταθμίζει τις πιθανοφάνειες σε πραγματικές πιθανότητες, έτσι ώστε το άθροισμα των πιθανοτήτων σε όλα τα δέντρα να είναι 1, και με αυτό τον τρόπο μας επιτρέπει να χρησιμοποιήσουμε την κλασσική θεωρία των πιθανοτήτων για να αναλύσουμε τα δεδομένα. Σε αυτή τη μέθοδο χρησιμοποιούνται όλες οι νουκλεοτιδικές θέσεις, και όχι μόνο οι πληροφοριακές, ενώ λαμβάνεται υπόψη η δυναμική της νουκλεοτιδικής υποκατάστασης. Γι αυτό το λόγο χρησιμοποιούνται εξελικτικά μοντέλα αλληλουχιών DNA, που βασίζονται σε υποθέσεις σχετικά με τη διαδικασία υποκατάστασης του DNA κι έτσι επιλέγεται ένα μοντέλο που ταιριάζει καλύτερα στην πληροφορία που δίνει η αλληλουχία για κάθε μοριακό δείκτη. Όλα τα μοντέλα χρησιμοποιούν 3 βασικούς τύπους παραμέτρων: 1) παράμετροι συχνότητας βάσεων, 2) παράμετροι υποκατάστασης βάσεων και 3) παράμετροι του ρυθμού ετερογένειας (συνήθως σχηματική παράμετρος α που καθορίζει τη μορφή της γ-κατανομής). Η επιλογή του μοντέλου βασίζεται στο Akaike Information Criterion (AIC) των MODELTEST v 3.7 (Posada & Crandall 1998) και jmodeltest (Darriba et al. 2012). Τα ακόλουθα μοντέλα βρέθηκαν να είναι τα καλύτερα για κάθε περίπτωση: 1) για το cpdna: TVM+G με παραμέτρους συχνοτήτων των βάσεων freqa = 0,3625, freqc = 0,1378, freqg = 0, 1479, freqt = 0, 3517, παράμετρο του ρυθμού ετερογένειας α = 1,0671 και παραμέτρους υποκατάστασης των βάσεων R[A-C] = 0,9832, R[A-G] = 1,0422, R[C-T] = 1,0422, R[A-T] = 0,1152, R[C-G] = 1,2344, R[G-T] = 1,0000, 2) για το ITS: GTR+G με freqa = 0,2197, freqc = 0,2729, freqg = 0,2724, freqt = 0,2350, α = 0,6551, R[A-C] = 0,7160, R[A-G] = 1,1938, R[C-T] = 3,5707, R[A-T] = 0,7629, R[C-G] = 0,1716, R[G-T] = 1,0000 και 3) για τον D10: 108
110 HKY+I+G με freqa = 0,3196, freqc = 0,1961, freqg = 0,1915, freqt = 0,2928, α = 0,6640 και παράμετρο μεταστροφής/μετάθεσης Ti/Tv ratio = 1,2076. Η ανάλυση ΜΣ πραγματοποιήθηκε με τη χρήση του προγράμματος MrBayes, version (Ronquist & Huelsenbeck 2003). Το πρόγραμμα διενέργησε 2 ανεξάρτητες αναλύσεις για αριθμό γενεών 5 x 10 6, γινόταν κατακράτηση ενός δέντρου κάθε γενεές και απορρίφθηκε το 25% των πρώτων δέντρων (burnin). Τελικά, στόχος είναι να παραχθεί το δέντρο που έχει τη μεγαλύτερη πιθανότητα, έχοντας ως βάση τα δεδομένα και το εξελικτικό μοντέλο. Έλεγχος αξιοπιστίας των φυλογενετικών δέντρων: Ο έλεγχος της αξιοπιστίας στην τοπολογία των κλάδων ενός δέντρου πραγματοποιήθηκε με τη μέθοδο επανασυμπίπτουσας ιχνηλάτησης (σε ελεύθερη απόδοση από τον Αλαχιώτη, 2007) ή bootstrap ανάλυση (Felsenstein 1985, Brown 2002). Αυτή η μέθοδος μιμείται τις επαναλαμβανόμενες δειγματοληψίες, μόνο που αντί να παίρνει δείγματα από τους πληθυσμούς κάνει δειγματοληψία πάλι από τα δείγματά μας (Εικ. 67). Κάθε νέα δειγματοληψία αποτελεί ένα ψευδοδείγμα. Από αυτό το ψευδοδείγμα υπολογίζεται εκ νέου η παράμετρος που μας ενδιαφέρει, δηλ. οι συνδέσεις των αλληλουχιών πάνω στο δέντρο (Page & Holmes 1998). Πρακτικά, 100 νέες αναδιατάξεις αλληλουχιών οδηγούν στην κατασκευή 100 δέντρων. Η τιμή (bootstrap value) που παίρνουμε για κάθε κόμβο, υποδεικνύει πόσες φορές προέκυψε η συγκεκριμένη τοπολογία στα δέντρα από τα ψευδοδείγματα. Αν η τιμή bootstrap είναι μεγαλύτερη από 75%, τότε μπορούμε να υποστηρίξουμε πως η συγκεκριμένη τοπολογία στο συγκεκριμένο κόμβο είναι αρκετά αξιόπιστη. Ο αριθμός των bootstrap επαναλήψεων που χρησιμοποιήθηκε είναι 100 επαναλήψεις. Η bootstrap ανάλυση πραγματοποιείται για τα φυλογενετικά δέντρα της ΜΦ. Για την στατιστική υποστήριξη της ΜΣ λαμβάνεται η τιμή της εκ των υστέρων πιθανότητας (Huelsenbeck & Ronquist 2001). Αν η τιμή αυτή είναι μεγαλύτερη από 0,95, τότε ο συγκεκριμένος κλάδος του δέντρου θεωρείται αξιόπιστος. Γενικά, η αξιοπιστία των φυλογενετικών δέντρων βελτιώνεται με τη χρήση μιας λειτουργικής ταξινομικής μονάδας που λειτουργεί ως μέτρο φυλογενετικής συγγένειας. Πρόκειται για την εξωτερική ομάδα, η οποία επιλέγεται ώστε να είναι γνωστή σε γενικές γραμμές η φυλογενετική της σχέση με τα εξεταζόμενα taxa. Για την παρούσα μελέτη, το γένος Gundelia αποτελεί εξωτερική ομάδα. 109
 & μοριακό ρολόι Δέντρο ειδών: Προκειμένου να συναχθεί ένα φυλογενετικό δέντρο που να περιλαμβάνει συνολικά πληροφορίες από όλους τους μοριακούς δείκτες και όλα")
111 Εικ. 67: Η αρχή της λειτουργίας της bootstrap analysis (Κράιτσεκ 2006). Κατασκευή δέντρου ειδών (species tree) & μοριακό ρολόι Δέντρο ειδών: Προκειμένου να συναχθεί ένα φυλογενετικό δέντρο που να περιλαμβάνει συνολικά πληροφορίες από όλους τους μοριακούς δείκτες και όλα τα είδη, υποβλήθηκε το πλήρες σύνολο δεδομένων για την κατασκευή του δέντρου και τη διαδικασία εκτίμησης του χρόνου απόκλισης (divergence time estimation). Η διαδικασία αυτή έγινε με τη χρήση του προγράμματος *BEAST (Heled & Drummod 2010). Ο φάκελος εισαγωγής δεδομένων (BEAST.xml input file) παράχθηκε από το BEAUti, version (Drummond et al. 2012) και απαρτίστηκε από 10 διαφορετικά τμήματα (partitions) δηλ. 5 αλληλουχίες για κάθε μοριακό δείκτη συν τις αντίστοιχες κωδικοποιημένες για τα κενά αλληλουχίες. Τα μοντέλα υποκατάστασης νουκλεοτιδίων επιλέχθηκαν βάσει του jmodeltest (Darriba et al. 2012) για κάθε μοριακό δείκτη ξεχωριστά. Σύμφωνα με το jmodeltest τα καταλληλότερα μοντέλα είναι τα εξής: 1) για τον trnl-f: TPM3uf+G με freqa = 0,3042, freqc = 0,1755, freqg = 0,1661, freqt = 0,3542, α = 0,1480, R[A-C] = 2,3453, R[A-G] = 4,1161, R[C-T] = 4,1161, R[A-T] = 1,0000, R[C-G] = 2,3453, R[G-T] = 1,0000, 2) για τον rpl32-trnl: TVM με freqa = 0,3803, freqc = 0,1258, freqg = 0,1430, freqt = 0,3509, R[A-C] = 0,9823, R[A-G] = 0,8280, R[C-T] = 0,8280, R[A-T] = 0,8796, R[C-G] = 1,0802, R[G-T] = 1,0000, 3) για τον ITS1: SYM+G με ίσες συχνότητες βάσεων, α = 0,6650, R[A-C] = 0,8235, R[A-G] = 1,0652, R[C-T] = 5,4059, R[A-T] = 1,0000, R[C-G] = 0,3077, R[G-T] = 1,0000, για τον ITS2: GTR+G με freqa = 110
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΤΟΓΕΩΓΡΑΦΙΑ
 ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΤΟΓΕΩΓΡΑΦΙΑ ΑΝΤΙΚΕΙΜΕΝΑ ΤΗΣ ΦΥΤΟΓΕΩΓΡΑΦΙΑΣ Χωρολογία των φυτών Εξελικτική ή ιστορική φυτογεωγραφία Φυτοκοινωνιολογία Οικολογική φυτογεωγραφία Η ΠΕΡΙΟΧΗ ΕΞΑΠΛΩΣΗΣ 1) Η ερμηνεία της περιοχής
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΤΟΓΕΩΓΡΑΦΙΑ ΑΝΤΙΚΕΙΜΕΝΑ ΤΗΣ ΦΥΤΟΓΕΩΓΡΑΦΙΑΣ Χωρολογία των φυτών Εξελικτική ή ιστορική φυτογεωγραφία Φυτοκοινωνιολογία Οικολογική φυτογεωγραφία Η ΠΕΡΙΟΧΗ ΕΞΑΠΛΩΣΗΣ 1) Η ερμηνεία της περιοχής
Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) Εισαγωγή. Δρ. Γεωργία Τοουλάκου
 Εισαγωγή. Δρ. Γεωργία Τοουλάκου") Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) 2018-2019 Εισαγωγή Δρ. Γεωργία Τοουλάκου Η Συστηματική Βοτανική είναι κλάδος της Βοτανικής. δηλαδή της επιστήμης η οποία ασχολείται
Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) 2018-2019 Εισαγωγή Δρ. Γεωργία Τοουλάκου Η Συστηματική Βοτανική είναι κλάδος της Βοτανικής. δηλαδή της επιστήμης η οποία ασχολείται
Νεοφυτικός αιώνας (περίοδος των Αγγειοσπέρμων)
") Νεοφυτικός αιώνας (περίοδος των Αγγειοσπέρμων) Η κυριαρχία των Αγγειοσπέρμων αρχίζει από το Mέσο Kρητιδικό (πριν 100 εκ. χρόνια) και συνεχίζεται μέχρι σήμερα. Υπάρχουν περίπου 250.000 είδη Αγγειοσπέρμων.
Νεοφυτικός αιώνας (περίοδος των Αγγειοσπέρμων) Η κυριαρχία των Αγγειοσπέρμων αρχίζει από το Mέσο Kρητιδικό (πριν 100 εκ. χρόνια) και συνεχίζεται μέχρι σήμερα. Υπάρχουν περίπου 250.000 είδη Αγγειοσπέρμων.
Εξελικτική πορεία της ελληνικής χλωρίδας παράδειγμα τα νησιά του Αιγαίου
 Εξελικτική πορεία της ελληνικής χλωρίδας παράδειγμα τα νησιά του Αιγαίου Καθηγητής Δημήτριος Χριστοδουλάκης Τμήμα Βιολογίας Τομέας Βιολογίας Φυτών E-mail: dkchrist@upatras.gr Τηλ.: 2610 997277 20 Αιγηίδα:
Εξελικτική πορεία της ελληνικής χλωρίδας παράδειγμα τα νησιά του Αιγαίου Καθηγητής Δημήτριος Χριστοδουλάκης Τμήμα Βιολογίας Τομέας Βιολογίας Φυτών E-mail: dkchrist@upatras.gr Τηλ.: 2610 997277 20 Αιγηίδα:
ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Κεντούλλα Πέτρου Αριθμός Φοιτητικής Ταυτότητας 2008761539 Κύπρος
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Κεντούλλα Πέτρου Αριθμός Φοιτητικής Ταυτότητας 2008761539 Κύπρος
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΟΡΙΑΚΗ ΚΑΙ ΦΑΙΝΟΤΥΠΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΚΥΠΡΙΑΚΟΥ ΠΛΗΘΥΣΜΟΥ ΣΚΛΗΡΟΥ ΣΙΤΑΡΙΟΥ ΠΟΥ ΑΠΟΚΤΗΘΗΚΕ ΑΠΟ ΤΡΑΠΕΖΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΟΡΙΑΚΗ ΚΑΙ ΦΑΙΝΟΤΥΠΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΚΥΠΡΙΑΚΟΥ ΠΛΗΘΥΣΜΟΥ ΣΚΛΗΡΟΥ ΣΙΤΑΡΙΟΥ ΠΟΥ ΑΠΟΚΤΗΘΗΚΕ ΑΠΟ ΤΡΑΠΕΖΑ
ΒΙΟΓΕΩΓΡΑΦΙΑ ΜΑΚΡΟΟΙΚΟΛΟΓΙΚΑ ΠΡΟΤΥΠΑ. Κώστας Κουγιουμουτζής, M.Sc., Ph.D.
 ΒΙΟΓΕΩΓΡΑΦΙΑ ΜΑΚΡΟΟΙΚΟΛΟΓΙΚΑ ΠΡΟΤΥΠΑ Κώστας Κουγιουμουτζής, M.Sc., Ph.D. Ενδημισμός Ενδημισμός Ποιος ο ορισμός του ενδημισμού; Ενδημισμός Ποιος ο ορισμός του ενδημισμού; Μια συστηματική μονάδα μπορεί να
ΒΙΟΓΕΩΓΡΑΦΙΑ ΜΑΚΡΟΟΙΚΟΛΟΓΙΚΑ ΠΡΟΤΥΠΑ Κώστας Κουγιουμουτζής, M.Sc., Ph.D. Ενδημισμός Ενδημισμός Ποιος ο ορισμός του ενδημισμού; Ενδημισμός Ποιος ο ορισμός του ενδημισμού; Μια συστηματική μονάδα μπορεί να
Department of Biological Sciences, Texas Tech University, MS 43131, Lubbock,Texas 79409-3131. 3
 1 Τμήμα Βιολογίας, Πανεπιστήμιο Κρήτης, Ηράκλειο Κρήτης. 2 Department of Biological Sciences, Texas Tech University, MS 43131, Lubbock,Texas 79409-3131. 3 Department of Biology, Faculty of Science, Dokuz
1 Τμήμα Βιολογίας, Πανεπιστήμιο Κρήτης, Ηράκλειο Κρήτης. 2 Department of Biological Sciences, Texas Tech University, MS 43131, Lubbock,Texas 79409-3131. 3 Department of Biology, Faculty of Science, Dokuz
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων Καθηγητής: Δρ. Νίκος Μίτλεττον Η ΣΧΕΣΗ ΤΟΥ ΜΗΤΡΙΚΟΥ ΘΗΛΑΣΜΟΥ ΜΕ ΤΗΝ ΕΜΦΑΝΙΣΗ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 2 ΣΤΗΝ ΠΑΙΔΙΚΗ ΗΛΙΚΙΑ Ονοματεπώνυμο: Ιωσηφίνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων Καθηγητής: Δρ. Νίκος Μίτλεττον Η ΣΧΕΣΗ ΤΟΥ ΜΗΤΡΙΚΟΥ ΘΗΛΑΣΜΟΥ ΜΕ ΤΗΝ ΕΜΦΑΝΙΣΗ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 2 ΣΤΗΝ ΠΑΙΔΙΚΗ ΗΛΙΚΙΑ Ονοματεπώνυμο: Ιωσηφίνα
Η Αφρική είναι η τρίτη σε μέγεθος ήπειρος του πλανήτη μας, μετά την Ασία και την Αμερική. Η έκτασή της είναι, χωρίς τα νησιά, 29,2 εκατομμύρια τετρ. χ
 Β. Π. Γ. Π. Η Αφρική είναι η τρίτη σε μέγεθος ήπειρος του πλανήτη μας, μετά την Ασία και την Αμερική. Η έκτασή της είναι, χωρίς τα νησιά, 29,2 εκατομμύρια τετρ. χιλιόμετρα, ενώ με τα νησιά φτάνει τα 30,2
Β. Π. Γ. Π. Η Αφρική είναι η τρίτη σε μέγεθος ήπειρος του πλανήτη μας, μετά την Ασία και την Αμερική. Η έκτασή της είναι, χωρίς τα νησιά, 29,2 εκατομμύρια τετρ. χιλιόμετρα, ενώ με τα νησιά φτάνει τα 30,2
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
Πολλαπλασιασμός των ειδών - Ειδογένεση. Αλλοπάτρια, παραπάτρια και συμπάτρια ειδογένεση. Μηχανισμοί αναπαραγωγικής απομόνωσης.
 Πολλαπλασιασμός των ειδών - Ειδογένεση. Αλλοπάτρια, παραπάτρια και συμπάτρια ειδογένεση. Μηχανισμοί αναπαραγωγικής απομόνωσης. Εισηγητής: Ν. Πουλακάκης Ειδογένεση H ειδογένεση αναφέρεται στο σχηματισμό
Πολλαπλασιασμός των ειδών - Ειδογένεση. Αλλοπάτρια, παραπάτρια και συμπάτρια ειδογένεση. Μηχανισμοί αναπαραγωγικής απομόνωσης. Εισηγητής: Ν. Πουλακάκης Ειδογένεση H ειδογένεση αναφέρεται στο σχηματισμό
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΨΥΧΟΚΟΙΝΩΝΙΚΗ ΥΓΕΙΑ ΤΟΥ ΠΑΙΔΙΟΥ ΣΕ ΣΧΕΣΗ ΜΕ ΤΗΝ ΚΟΙΝΩΝΙΚΟΟΙΚΟΝΟΜΙΚΗ ΚΑΤΑΣΤΑΣΗ ΤΗΣ ΟΙΚΟΓΕΝΕΙΑΣ Μαρία Χρίστου Λεμεσός 2012 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΨΥΧΟΚΟΙΝΩΝΙΚΗ ΥΓΕΙΑ ΤΟΥ ΠΑΙΔΙΟΥ ΣΕ ΣΧΕΣΗ ΜΕ ΤΗΝ ΚΟΙΝΩΝΙΚΟΟΙΚΟΝΟΜΙΚΗ ΚΑΤΑΣΤΑΣΗ ΤΗΣ ΟΙΚΟΓΕΝΕΙΑΣ Μαρία Χρίστου Λεμεσός 2012 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΕΛΕΓΧΟΣ ΓΟΝΙΔΙΑΚΩΝ ΣΥΧΝΟΤΗΤΩΝ ACAA2 ΣΤΟ ΕΞΩΝΙΟ 10 ΣΤΑ ΠΡΟΒΑΤΑ ΧΙΟΥ Μύρια Χατζηκώστα Λεμεσός, 2013
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΕΛΕΓΧΟΣ ΓΟΝΙΔΙΑΚΩΝ ΣΥΧΝΟΤΗΤΩΝ ACAA2 ΣΤΟ ΕΞΩΝΙΟ 10 ΣΤΑ ΠΡΟΒΑΤΑ ΧΙΟΥ Μύρια Χατζηκώστα Λεμεσός, 2013
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία ΑΓΧΟΣ ΚΑΙ ΚΑΤΑΘΛΙΨΗ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΑΓΧΟΣ ΚΑΙ ΚΑΤΑΘΛΙΨΗ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ ΧΡΥΣΟΒΑΛΑΝΤΗΣ ΒΑΣΙΛΕΙΟΥ ΛΕΜΕΣΟΣ 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΑΓΧΟΣ ΚΑΙ ΚΑΤΑΘΛΙΨΗ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ ΧΡΥΣΟΒΑΛΑΝΤΗΣ ΒΑΣΙΛΕΙΟΥ ΛΕΜΕΣΟΣ 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ. Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων
 ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων Φυλογένεση Η φυλογένεσης αφορά την ανεύρεση των συνδετικών εκείνων κρίκων που συνδέουν τα διάφορα είδη µεταξύ τους εξελικτικά, σε µονοφυλετικές
ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων Φυλογένεση Η φυλογένεσης αφορά την ανεύρεση των συνδετικών εκείνων κρίκων που συνδέουν τα διάφορα είδη µεταξύ τους εξελικτικά, σε µονοφυλετικές
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
 πανεπιστημιούπολη Καισαριανή Βύρωνας ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΟΥ ΚΗΠΟΥ Τοποθεσία Έτος ίδρυσης Έκταση Γεωγρ. Μήκος Γεωγρ. Πλάτος Υψόμετρο Προσανατολισμός Πέτρωμα Κατώτερη θερμοκρασία Ανώτερη θερμοκρασία Βροχόπτωση
πανεπιστημιούπολη Καισαριανή Βύρωνας ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΟΥ ΚΗΠΟΥ Τοποθεσία Έτος ίδρυσης Έκταση Γεωγρ. Μήκος Γεωγρ. Πλάτος Υψόμετρο Προσανατολισμός Πέτρωμα Κατώτερη θερμοκρασία Ανώτερη θερμοκρασία Βροχόπτωση
ΕΛΛΗΝΙΚΟ ΤΟΞΟ. Γεωλογική εξέλιξη της Ελλάδας Το Ελληνικό τόξο
 ΕΛΛΗΝΙΚΟ ΤΟΞΟ Γεωλογική εξέλιξη της Ελλάδας Το Ελληνικό τόξο ρ. Ε. Λυκούδη Αθήνα 2005 Γεωλογική εξέλιξη της Ελλάδας Ο Ελλαδικός χώρος µε την ευρεία γεωγραφική έννοια του όρου, έχει µια σύνθετη γεωλογικοτεκτονική
ΕΛΛΗΝΙΚΟ ΤΟΞΟ Γεωλογική εξέλιξη της Ελλάδας Το Ελληνικό τόξο ρ. Ε. Λυκούδη Αθήνα 2005 Γεωλογική εξέλιξη της Ελλάδας Ο Ελλαδικός χώρος µε την ευρεία γεωγραφική έννοια του όρου, έχει µια σύνθετη γεωλογικοτεκτονική
Χλωριδικές μονάδες βλάστησης και συστήματα
 Χλωριδικές μονάδες βλάστησης και συστήματα Α. Ποια τα βήματα της χλωριδικής έρευνας; Β. Δειγματοληψία με quadrat 1. Επιλογή quadrat 2. Σχήμα και μέγεθος quadrat 3. Αριθμός και διάταξη quadrats 4. Πρότυπο
Χλωριδικές μονάδες βλάστησης και συστήματα Α. Ποια τα βήματα της χλωριδικής έρευνας; Β. Δειγματοληψία με quadrat 1. Επιλογή quadrat 2. Σχήμα και μέγεθος quadrat 3. Αριθμός και διάταξη quadrats 4. Πρότυπο
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 19/5/2007
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Ανθεκτικότητα του κοινού τετράνυχου Tetranychus urticae Koch (Acari : Tetranychidae) στα εντοµοκτόνα/ακαρεοκτόνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Ανθεκτικότητα του κοινού τετράνυχου Tetranychus urticae Koch (Acari : Tetranychidae) στα εντοµοκτόνα/ακαρεοκτόνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Ο ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ Η ΣΧΕΣΗ ΤΟΥ ΜΕ ΤΟ ΚΑΡΚΙΝΟ ΤΟΥ ΜΑΣΤΟΥΣ ΣΤΙΣ ΓΥΝΑΙΚΕΣ ΠΟΥ ΕΙΝΑΙ ΦΟΡΕΙΣ ΤΟΥ ΟΓΚΟΓΟΝΙΔΙΟΥ BRCA1 ΚΑΙ BRCA2. Βασούλλα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Ο ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ Η ΣΧΕΣΗ ΤΟΥ ΜΕ ΤΟ ΚΑΡΚΙΝΟ ΤΟΥ ΜΑΣΤΟΥΣ ΣΤΙΣ ΓΥΝΑΙΚΕΣ ΠΟΥ ΕΙΝΑΙ ΦΟΡΕΙΣ ΤΟΥ ΟΓΚΟΓΟΝΙΔΙΟΥ BRCA1 ΚΑΙ BRCA2. Βασούλλα
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
Συστηματική και φυλογεωγραφία του συμπλέγματος ειδών Ablepharus kitaibelii στη περιοχή της ανατολικής Μεσογείου
 Συστηματική και φυλογεωγραφία του συμπλέγματος ειδών Ablepharus kitaibelii στη περιοχή της ανατολικής Μεσογείου Νίκος Πουλακάκης, Cetin Ilgaz, Yusuf Kumlutaş, Aziz Avcı, Ahmadzadeh Faraham, Jelca Crnobrnja
Συστηματική και φυλογεωγραφία του συμπλέγματος ειδών Ablepharus kitaibelii στη περιοχή της ανατολικής Μεσογείου Νίκος Πουλακάκης, Cetin Ilgaz, Yusuf Kumlutaş, Aziz Avcı, Ahmadzadeh Faraham, Jelca Crnobrnja
Μικροβιολογική ποιότητα έτοιμων προς κατανάλωση σαλατών
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία Μικροβιολογική ποιότητα έτοιμων προς κατανάλωση σαλατών Σοφία Στεργίου Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία Μικροβιολογική ποιότητα έτοιμων προς κατανάλωση σαλατών Σοφία Στεργίου Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ E - Z
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ E - Z ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ Ε - Z Επιτροπή Έκδοσης: Δημήτριος
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ E - Z ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ Ε - Z Επιτροπή Έκδοσης: Δημήτριος
Η ταυτότητα και η ετερότητά της
 Deutsch-griechisches Netzwerk: Regionen-Städte-Bürger Ελληνογερμανικό δίκτυο: Περιφέρειες Δήμοι - Πολίτες Περιφέρεια Ανατολικής Μακεδονίας & Θράκης Η ταυτότητα και η ετερότητά της ΕΥΑΝΘΙΑ ΤΖΗΜΟΥ ΕΥΑΝΘΙΑ
Deutsch-griechisches Netzwerk: Regionen-Städte-Bürger Ελληνογερμανικό δίκτυο: Περιφέρειες Δήμοι - Πολίτες Περιφέρεια Ανατολικής Μακεδονίας & Θράκης Η ταυτότητα και η ετερότητά της ΕΥΑΝΘΙΑ ΤΖΗΜΟΥ ΕΥΑΝΘΙΑ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ E - Z
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ E - Z ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ Ε - Z Επιτροπή Έκδοσης: Δημήτριος
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ E - Z ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΔΕΥΤΕΡΟΣ Ε - Z Επιτροπή Έκδοσης: Δημήτριος
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή διατριβή. Ονοματεπώνυμο: Αργυρώ Ιωάννου. Επιβλέπων καθηγητής: Δρ. Αντρέας Χαραλάμπους
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Διερεύνηση της αποτελεσματικότητας εναλλακτικών και συμπληρωματικών τεχνικών στη βελτίωση της ποιότητας της ζωής σε άτομα με καρκίνο
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Διερεύνηση της αποτελεσματικότητας εναλλακτικών και συμπληρωματικών τεχνικών στη βελτίωση της ποιότητας της ζωής σε άτομα με καρκίνο
ΕΚΔΟΣΗ ΤΟΥ ΒΙΒΛΙΟΥ: Vascular Plants of Greece: An Annotated Checklist
 ΕΚΔΟΣΗ ΤΟΥ ΒΙΒΛΙΟΥ: Vascular Plants of Greece: An Annotated Checklist Τα Φυτά της Ελλάδας Πραγματοποιήθηκε την Πέμπτη 3 Οκτωβρίου στη Θεσσαλονίκη στο πλαίσιο του 13ου Συνεδρίου της Ελληνικής Βοτανικής
ΕΚΔΟΣΗ ΤΟΥ ΒΙΒΛΙΟΥ: Vascular Plants of Greece: An Annotated Checklist Τα Φυτά της Ελλάδας Πραγματοποιήθηκε την Πέμπτη 3 Οκτωβρίου στη Θεσσαλονίκη στο πλαίσιο του 13ου Συνεδρίου της Ελληνικής Βοτανικής
(Μερος 2 ο ) Εισηγητής: Ν. Πουλακάκης
 Εισηγητής: Ν. Πουλακάκης") Ταξινομικοί χαρακτήρες και Φυλογενετική ανασύσταση. Σχολές ταξινόμησης. Θεωρίες για την Ταξινομική. Φυλογενετική ανάλυση: Μοριακή συστηματική. Οι κύριες διαιρέσεις της Ζωής. (Μερος 2 ο ) Εισηγητής: Ν.
Ταξινομικοί χαρακτήρες και Φυλογενετική ανασύσταση. Σχολές ταξινόμησης. Θεωρίες για την Ταξινομική. Φυλογενετική ανάλυση: Μοριακή συστηματική. Οι κύριες διαιρέσεις της Ζωής. (Μερος 2 ο ) Εισηγητής: Ν.
Περιεχόμενα. 1 Η ιστορία της εξελικτικής βιολογίας: Εξέλιξη και Γενετική 2 Η Προέλευση της Μοριακής Βιολογίας 3 Αποδείξεις για την εξέλιξη 89
 Περιεχόμενα Οι Συγγραφείς Πρόλογος της Ελληνικής Έκδοσης Πρόλογος της Αμερικανικής Έκδοσης Σκοπός και Αντικείμενο του Βιβλίου ΜΕΡΟΣ Ι ΜΙΑ ΕΠΙΣΚΟΠΗΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ 1 Η ιστορία της εξελικτικής
Περιεχόμενα Οι Συγγραφείς Πρόλογος της Ελληνικής Έκδοσης Πρόλογος της Αμερικανικής Έκδοσης Σκοπός και Αντικείμενο του Βιβλίου ΜΕΡΟΣ Ι ΜΙΑ ΕΠΙΣΚΟΠΗΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ 1 Η ιστορία της εξελικτικής
Διερεύνηση χαρτογράφησης Ποσειδωνίας με χρήση επιβλεπόμενης ταξινόμησης οπτικών δορυφορικών εικόνων
 Σχολή Μηχανικής και Τεχνολογίας Μεταπτυχιακή διατριβή Διερεύνηση χαρτογράφησης Ποσειδωνίας με χρήση επιβλεπόμενης ταξινόμησης οπτικών δορυφορικών εικόνων Αναστασία Υφαντίδου Λεμεσός, Νοέμβριος 2017 ΤΕΧΝΟΛΟΓΙΚΟ
Σχολή Μηχανικής και Τεχνολογίας Μεταπτυχιακή διατριβή Διερεύνηση χαρτογράφησης Ποσειδωνίας με χρήση επιβλεπόμενης ταξινόμησης οπτικών δορυφορικών εικόνων Αναστασία Υφαντίδου Λεμεσός, Νοέμβριος 2017 ΤΕΧΝΟΛΟΓΙΚΟ
Κωνσταντίνος Στεφανίδης
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης Οικολογική
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης Οικολογική
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή ΣΥΓΚΡΙΣΗ ΜΙΚΡΟΒΙΟΛΟΓΙΚΗΣ ΠΟΙΟΤΗΤΑΣ ΒΙΟΛΟΓΙΚΟΥ ΚΑΙ ΣΥΜΒΑΤΙΚΟΥ ΝΩΠΟΥ ΓΑΛΑΚΤΟΣ Λούκας Νεοφύτου Λεμεσός
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή ΣΥΓΚΡΙΣΗ ΜΙΚΡΟΒΙΟΛΟΓΙΚΗΣ ΠΟΙΟΤΗΤΑΣ ΒΙΟΛΟΓΙΚΟΥ ΚΑΙ ΣΥΜΒΑΤΙΚΟΥ ΝΩΠΟΥ ΓΑΛΑΚΤΟΣ Λούκας Νεοφύτου Λεμεσός
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΞΟΥΘΕΝΩΣΗ ΠΟΥ ΒΙΩΝΕΙ ΤΟ ΝΟΣΗΛΕΥΤΙΚΟ ΠΡΟΣΩΠΙΚΟ ΣΤΙΣ ΜΟΝΑΔΕΣ ΕΝΑΤΙΚΗΣ ΘΕΡΑΠΕΙΑΣ Άντρη Αγαθαγγέλου Λεμεσός 2012 i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΞΟΥΘΕΝΩΣΗ ΠΟΥ ΒΙΩΝΕΙ ΤΟ ΝΟΣΗΛΕΥΤΙΚΟ ΠΡΟΣΩΠΙΚΟ ΣΤΙΣ ΜΟΝΑΔΕΣ ΕΝΑΤΙΚΗΣ ΘΕΡΑΠΕΙΑΣ Άντρη Αγαθαγγέλου Λεμεσός 2012 i ΤΕΧΝΟΛΟΓΙΚΟ
Η ελληνική βιοποικιλότητα Ενας κρυμμένος θησαυρός. Μανώλης Μιτάκης Φαρμακοποιός Αντιπρόεδρος Ελληνικής Εταιρείας Εθνοφαρμακολογίας
 Η ελληνική βιοποικιλότητα Ενας κρυμμένος θησαυρός Μανώλης Μιτάκης Φαρμακοποιός Αντιπρόεδρος Ελληνικής Εταιρείας Εθνοφαρμακολογίας Ο μεγάλος αριθμός και η ποικιλομορφία των σύγχρονων μορφών ζωής στη Γη
Η ελληνική βιοποικιλότητα Ενας κρυμμένος θησαυρός Μανώλης Μιτάκης Φαρμακοποιός Αντιπρόεδρος Ελληνικής Εταιρείας Εθνοφαρμακολογίας Ο μεγάλος αριθμός και η ποικιλομορφία των σύγχρονων μορφών ζωής στη Γη
2. ΓΕΩΓΡΑΦΙΑ ΤΗΣ Υ ΡΟΣΦΑΙΡΑΣ
 2. ΓΕΩΓΡΑΦΙΑ ΤΗΣ Υ ΡΟΣΦΑΙΡΑΣ 2.1 Ωκεανοί και Θάλασσες. Σύµφωνα µε τη ιεθνή Υδρογραφική Υπηρεσία (International Hydrographic Bureau, 1953) ως το 1999 θεωρούντο µόνο τρεις ωκεανοί: Ο Ατλαντικός, ο Ειρηνικός
2. ΓΕΩΓΡΑΦΙΑ ΤΗΣ Υ ΡΟΣΦΑΙΡΑΣ 2.1 Ωκεανοί και Θάλασσες. Σύµφωνα µε τη ιεθνή Υδρογραφική Υπηρεσία (International Hydrographic Bureau, 1953) ως το 1999 θεωρούντο µόνο τρεις ωκεανοί: Ο Ατλαντικός, ο Ειρηνικός
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ. Πτυχιακή εργασία ΕΤΟΙΜΑΣΙΑ ΔΕΛΤΙΟΥ ΠΟΣΟΤΗΤΩΝ ΜΕ ΤΗ ΧΡΗΣΗ ΛΟΓΙΣΜΙΚΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ Πτυχιακή εργασία ΕΤΟΙΜΑΣΙΑ ΔΕΛΤΙΟΥ ΠΟΣΟΤΗΤΩΝ ΜΕ ΤΗ ΧΡΗΣΗ ΛΟΓΙΣΜΙΚΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ Χρύσω Κωνσταντίνου Λεμεσός 2016 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ Πτυχιακή εργασία ΕΤΟΙΜΑΣΙΑ ΔΕΛΤΙΟΥ ΠΟΣΟΤΗΤΩΝ ΜΕ ΤΗ ΧΡΗΣΗ ΛΟΓΙΣΜΙΚΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ Χρύσω Κωνσταντίνου Λεμεσός 2016 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
«Αξιολόγηση ατόμων με αφασία για Επαυξητική και Εναλλακτική Επικοινωνία, σύμφωνα με το μοντέλο συμμετοχής»
 Σχολή Επιστημών Υγείας Τμήμα Αποκατάστασης ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Αξιολόγηση ατόμων με αφασία για Επαυξητική και Εναλλακτική Επικοινωνία, σύμφωνα με το μοντέλο συμμετοχής» Χρυσάνθη Μοδέστου Λεμεσός, Μάιος,
Σχολή Επιστημών Υγείας Τμήμα Αποκατάστασης ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Αξιολόγηση ατόμων με αφασία για Επαυξητική και Εναλλακτική Επικοινωνία, σύμφωνα με το μοντέλο συμμετοχής» Χρυσάνθη Μοδέστου Λεμεσός, Μάιος,
Ο νοσηλευτικός ρόλος στην πρόληψη του μελανώματος
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Ο νοσηλευτικός ρόλος στην πρόληψη του μελανώματος Ονοματεπώνυμο: Αρτέμης Παναγιώτου Επιβλέπων καθηγητής: Δρ. Αντρέας Χαραλάμπους
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Ο νοσηλευτικός ρόλος στην πρόληψη του μελανώματος Ονοματεπώνυμο: Αρτέμης Παναγιώτου Επιβλέπων καθηγητής: Δρ. Αντρέας Χαραλάμπους
Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) Plantae. Το Βασίλειο των φυτών. Δρ.
 Plantae. Το Βασίλειο των φυτών. Δρ.") Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) 2018-2019 Plantae Το Βασίλειο των φυτών Δρ. Γεωργία Τοουλάκου Στη Συστηματική Βοτανική χρησιμοποιείται συχνά ο όρος κατάταξη ή συστηματική
Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) 2018-2019 Plantae Το Βασίλειο των φυτών Δρ. Γεωργία Τοουλάκου Στη Συστηματική Βοτανική χρησιμοποιείται συχνά ο όρος κατάταξη ή συστηματική
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΣΧΕΤΙΖΟΜΕΝΗ ΜΕ ΤΗΝ ΥΓΕΙΑ ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΑΤΟΜΩΝ ΜΕ ΝΕΑΝΙΚΗ ΙΔΙΟΠΑΘΗ ΑΡΘΡΙΤΙΔΑ Όνομα Φοιτήτριας: Μαρία Θωμά Αριθμός φοιτητικής ταυτότητας:2010221455
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΣΧΕΤΙΖΟΜΕΝΗ ΜΕ ΤΗΝ ΥΓΕΙΑ ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΑΤΟΜΩΝ ΜΕ ΝΕΑΝΙΚΗ ΙΔΙΟΠΑΘΗ ΑΡΘΡΙΤΙΔΑ Όνομα Φοιτήτριας: Μαρία Θωμά Αριθμός φοιτητικής ταυτότητας:2010221455
Κυτταρικός κύκλος: Μιτωτική και μειωτική διαίρεση του πυρήνα
 Κυτταρικός κύκλος: Μιτωτική και μειωτική διαίρεση του πυρήνα Ο κυτταρικός κύκλος περιλαμβάνει 5 φάσεις: Τη Μεσόφαση και τις 4 Φάσεις της Μιτωτικής διαίρεσης (Πρόφαση, μετάφαση, ανάφαση, τελόφαση) G1= grow
Κυτταρικός κύκλος: Μιτωτική και μειωτική διαίρεση του πυρήνα Ο κυτταρικός κύκλος περιλαμβάνει 5 φάσεις: Τη Μεσόφαση και τις 4 Φάσεις της Μιτωτικής διαίρεσης (Πρόφαση, μετάφαση, ανάφαση, τελόφαση) G1= grow
ΤΡΙΤΟ ΚΕΦΑΛΑΙΟ. Χλωρίδα και Πανίδα
 ΤΡΙΤΟ ΚΕΦΑΛΑΙΟ Χλωρίδα και Πανίδα ΕΡΩΤΗΣΕΙΣ ΚΛΕΙΣΤΟΥ ΤΥΠΟΥ Ερωτήσεις της µορφής σωστό-λάθος Σηµειώστε αν είναι σωστή ή λάθος καθεµιά από τις παρακάτω προτάσεις περιβάλλοντας µε ένα κύκλο το αντίστοιχο
ΤΡΙΤΟ ΚΕΦΑΛΑΙΟ Χλωρίδα και Πανίδα ΕΡΩΤΗΣΕΙΣ ΚΛΕΙΣΤΟΥ ΤΥΠΟΥ Ερωτήσεις της µορφής σωστό-λάθος Σηµειώστε αν είναι σωστή ή λάθος καθεµιά από τις παρακάτω προτάσεις περιβάλλοντας µε ένα κύκλο το αντίστοιχο
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
Η θέση ύπνου του βρέφους και η σχέση της με το Σύνδρομο του αιφνίδιου βρεφικού θανάτου. ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η θέση ύπνου του βρέφους και η σχέση της με το Σύνδρομο του αιφνίδιου βρεφικού θανάτου. Χρυσάνθη Στυλιανού Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η θέση ύπνου του βρέφους και η σχέση της με το Σύνδρομο του αιφνίδιου βρεφικού θανάτου. Χρυσάνθη Στυλιανού Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία Η ΚΑΤΑΘΛΙΨΗ ΣΕ ΕΦΗΒΟΥΣ ΜΕ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 1
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΚΑΤΑΘΛΙΨΗ ΣΕ ΕΦΗΒΟΥΣ ΜΕ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 1 ΑΝΔΡΕΑΣ ΑΝΔΡΕΟΥ Φ.Τ:2008670839 Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΚΑΤΑΘΛΙΨΗ ΣΕ ΕΦΗΒΟΥΣ ΜΕ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 1 ΑΝΔΡΕΑΣ ΑΝΔΡΕΟΥ Φ.Τ:2008670839 Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΓΕΩΓΡΑΦΙΑ ΘΕΜΑΤΑ. ήταν ο κάθε ένας από αυτούς και σε ποιον από αυτούς σχηματίστηκε η Ελλάδα;
 ΓΕΩΓΡΑΦΙΑ ΘΕΜΑΤΑ ΘΕΜΑ 1 ο (Μονάδες 3,3) 1. Ποια είναι η διοικητική ιεραρχία των πόλεων στην Ελλάδα; Πως λέγεται ο διοικητής του κάθε διοικητικού τομέα; 2. Ποιους γεωλογικούς αιώνες περιλαμβάνει η γεωλογική
ΓΕΩΓΡΑΦΙΑ ΘΕΜΑΤΑ ΘΕΜΑ 1 ο (Μονάδες 3,3) 1. Ποια είναι η διοικητική ιεραρχία των πόλεων στην Ελλάδα; Πως λέγεται ο διοικητής του κάθε διοικητικού τομέα; 2. Ποιους γεωλογικούς αιώνες περιλαμβάνει η γεωλογική
ΤΜΗΜΑ ΟΙΚΟΛΟΓΙΑΣ & ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΧΕΙΜΕΡΙΝΟ ΕΞΑΜΗΝΟ
 ΕΝΟΤΗΤΕΣ ΥΛΗΣ Δρ. Κώστας Ποϊραζίδης, Δασολόγος ΤΜΗΜΑ ΟΙΚΟΛΟΓΙΑΣ & ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΧΕΙΜΕΡΙΝΟ ΕΞΑΜΗΝΟ 2009-2010 ΕΝΟΤΗΤΕΣ ΥΛΗΣ ΕΝΟΤΗΤΑ 2.2: Ορισμός είδους, Συστηματική, Οικολογία και Εξέλιξη, Φυσική επιλογή
ΕΝΟΤΗΤΕΣ ΥΛΗΣ Δρ. Κώστας Ποϊραζίδης, Δασολόγος ΤΜΗΜΑ ΟΙΚΟΛΟΓΙΑΣ & ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΧΕΙΜΕΡΙΝΟ ΕΞΑΜΗΝΟ 2009-2010 ΕΝΟΤΗΤΕΣ ΥΛΗΣ ΕΝΟΤΗΤΑ 2.2: Ορισμός είδους, Συστηματική, Οικολογία και Εξέλιξη, Φυσική επιλογή
ΠΟΛΥΤΕΧΝΕΙΟ ΚΡΗΤΗΣ ΣΧΟΛΗ ΜΗΧΑΝΙΚΩΝ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΕΡΓΑΣΤΗΡΙΟ ΔΙΚΑΙΟΥ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΚΑΙ ΠΕΡΙΒΑΛΛΟΝΤΙΚΗΣ ΔΙΑΚΥΒΕΡΝΗΣΗΣ ΧΑΝΙΑ - ΙΟΥΛΙΟΣ 2014
 ΠΟΛΥΤΕΧΝΕΙΟ ΚΡΗΤΗΣ ΣΧΟΛΗ ΜΗΧΑΝΙΚΩΝ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΕΡΓΑΣΤΗΡΙΟ ΔΙΚΑΙΟΥ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΚΑΙ ΠΕΡΙΒΑΛΛΟΝΤΙΚΗΣ ΔΙΑΚΥΒΕΡΝΗΣΗΣ ΛΙΜΝΙΟΥ ΓΕΩΡΓΙΑ ΠΑΝΑΓΙΩΤΑ ΧΑΝΙΑ - ΙΟΥΛΙΟΣ 2014 ΤΟ ΡΥΘΜΙΣΤΙΚΟ ΠΛΑΙΣΙΟ ΛΕΙΤΟΥΡΓΙΑΣ
ΠΟΛΥΤΕΧΝΕΙΟ ΚΡΗΤΗΣ ΣΧΟΛΗ ΜΗΧΑΝΙΚΩΝ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΕΡΓΑΣΤΗΡΙΟ ΔΙΚΑΙΟΥ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΚΑΙ ΠΕΡΙΒΑΛΛΟΝΤΙΚΗΣ ΔΙΑΚΥΒΕΡΝΗΣΗΣ ΛΙΜΝΙΟΥ ΓΕΩΡΓΙΑ ΠΑΝΑΓΙΩΤΑ ΧΑΝΙΑ - ΙΟΥΛΙΟΣ 2014 ΤΟ ΡΥΘΜΙΣΤΙΚΟ ΠΛΑΙΣΙΟ ΛΕΙΤΟΥΡΓΙΑΣ
TreeTOPS. ένα εισαγωγικό παιχνίδι για τα φυλογενετικά δέντρα. Teacher s Guide. ELLS European Learning Laboratory for the Life Sciences
 TreeTOPS ένα εισαγωγικό παιχνίδι για τα φυλογενετικά δέντρα Teacher s Guide ELLS European Learning Laboratory for the Life Sciences 1 Γενικός σκοπός Το συγκεκριμένο παιχνίδι έχει ως στόχο να εισάγει τους
TreeTOPS ένα εισαγωγικό παιχνίδι για τα φυλογενετικά δέντρα Teacher s Guide ELLS European Learning Laboratory for the Life Sciences 1 Γενικός σκοπός Το συγκεκριμένο παιχνίδι έχει ως στόχο να εισάγει τους
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
Θέμα: Παχυσαρκία και κύηση:
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜ Α ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Θέμα: Παχυσαρκία και κύηση: επιπτώσεις στην έκβαση της κύησης και στο έμβρυο Ονοματεπώνυμο: Στέλλα Ριαλά Αριθμός
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜ Α ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Θέμα: Παχυσαρκία και κύηση: επιπτώσεις στην έκβαση της κύησης και στο έμβρυο Ονοματεπώνυμο: Στέλλα Ριαλά Αριθμός
ΑΞΙΟΛΟΓΗΣΗ ΑΦΗΓΗΜΑΤΙΚΩΝ ΙΚΑΝΟΤΗΤΩΝ ΜΕΣΩ ΧΟΡΗΓΗΣΗΣ ΤΟΥ ΕΡΓΑΛΕΙΟΥ ΜΑΙΝ ΣΕ ΤΥΠΙΚΩΣ ΑΝΑΠΤΥΣΣΟΜΕΝΑ ΠΑΙΔΙΑ ΣΤΗΝ ΚΥΠΡΟ
 Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΑΞΙΟΛΟΓΗΣΗ ΑΦΗΓΗΜΑΤΙΚΩΝ ΙΚΑΝΟΤΗΤΩΝ ΜΕΣΩ ΧΟΡΗΓΗΣΗΣ ΤΟΥ ΕΡΓΑΛΕΙΟΥ ΜΑΙΝ ΣΕ ΤΥΠΙΚΩΣ ΑΝΑΠΤΥΣΣΟΜΕΝΑ ΠΑΙΔΙΑ ΣΤΗΝ ΚΥΠΡΟ Γεωργίου Μύρια Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΑΞΙΟΛΟΓΗΣΗ ΑΦΗΓΗΜΑΤΙΚΩΝ ΙΚΑΝΟΤΗΤΩΝ ΜΕΣΩ ΧΟΡΗΓΗΣΗΣ ΤΟΥ ΕΡΓΑΛΕΙΟΥ ΜΑΙΝ ΣΕ ΤΥΠΙΚΩΣ ΑΝΑΠΤΥΣΣΟΜΕΝΑ ΠΑΙΔΙΑ ΣΤΗΝ ΚΥΠΡΟ Γεωργίου Μύρια Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Χαμηλά επίπεδα βιταμίνης D σχετιζόμενα με το βρογχικό άσθμα στα παιδιά και στους έφηβους Κουρομπίνα Αλεξάνδρα Λεμεσός [2014] i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Χαμηλά επίπεδα βιταμίνης D σχετιζόμενα με το βρογχικό άσθμα στα παιδιά και στους έφηβους Κουρομπίνα Αλεξάνδρα Λεμεσός [2014] i ΤΕΧΝΟΛΟΓΙΚΟ
ΕΙΔΙΚΑ ΜΑΘΗΜΑΤΑ ΒΟΤΑΝΙΚΗΣ. Κώστας Κουγιουμουτζής, Λέκτορας
 ΕΙΔΙΚΑ ΜΑΘΗΜΑΤΑ ΒΟΤΑΝΙΚΗΣ Κώστας Κουγιουμουτζής, Λέκτορας Ειδικά Μαθήματα Βοτανικής Πληροφορίες σχετικά με το μάθημα Ειδικά Μαθήματα Βοτανικής Πληροφορίες σχετικά με το μάθημα Σχεδιάγραμμα διδασκαλίας
ΕΙΔΙΚΑ ΜΑΘΗΜΑΤΑ ΒΟΤΑΝΙΚΗΣ Κώστας Κουγιουμουτζής, Λέκτορας Ειδικά Μαθήματα Βοτανικής Πληροφορίες σχετικά με το μάθημα Ειδικά Μαθήματα Βοτανικής Πληροφορίες σχετικά με το μάθημα Σχεδιάγραμμα διδασκαλίας
Ενδημισμός και ενδημικά φυτά
 Ενδημισμός και ενδημικά φυτά Το Αιγαίο πέλαγος θεωρείται εργαστήρι βιοποικιλότητας λόγω της πολύπλοκης παλαιογεωγραφίας, της γεωμορφολογίας και της θέσης του στο σταυροδρόμι τριών ηπείρων αλλά και λόγω
Ενδημισμός και ενδημικά φυτά Το Αιγαίο πέλαγος θεωρείται εργαστήρι βιοποικιλότητας λόγω της πολύπλοκης παλαιογεωγραφίας, της γεωμορφολογίας και της θέσης του στο σταυροδρόμι τριών ηπείρων αλλά και λόγω
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΣΧΕΣΗ ΤΗΣ ΠΑΧΥΣΑΡΚΙΑΣ ΜΕ ΤΟ ΣΑΚΧΑΡΩΔΗ ΔΙΒΗΤΗ ΚΥΗΣΗΣ Χρυστάλλα, Γεωργίου Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΣΧΕΣΗ ΤΗΣ ΠΑΧΥΣΑΡΚΙΑΣ ΜΕ ΤΟ ΣΑΚΧΑΡΩΔΗ ΔΙΒΗΤΗ ΚΥΗΣΗΣ Χρυστάλλα, Γεωργίου Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ
Η ΧΛΩΡΙΔΑ ΚΑΙ Η ΠΑΝΙΔΑ ΣΤΗΝ ΧΩΡΑ ΜΑΣ. ΟΜΑΔΑ 1 Κορμπάκη Δέσποινα Κολακλίδη Ναταλία Ζαχαροπούλου Φιλιππούλα Θανοπούλου Ιωαννά
 Η ΧΛΩΡΙΔΑ ΚΑΙ Η ΠΑΝΙΔΑ ΣΤΗΝ ΧΩΡΑ ΜΑΣ ΟΜΑΔΑ 1 Κορμπάκη Δέσποινα Κολακλίδη Ναταλία Ζαχαροπούλου Φιλιππούλα Θανοπούλου Ιωαννά ΓΙΑΤΙ ΤΟ ΕΠΙΛΕΞΑΜΕ: Η χλωρίδα και η πανίδα στην χώρα μας είναι ένα πολύ σημαντικό
Η ΧΛΩΡΙΔΑ ΚΑΙ Η ΠΑΝΙΔΑ ΣΤΗΝ ΧΩΡΑ ΜΑΣ ΟΜΑΔΑ 1 Κορμπάκη Δέσποινα Κολακλίδη Ναταλία Ζαχαροπούλου Φιλιππούλα Θανοπούλου Ιωαννά ΓΙΑΤΙ ΤΟ ΕΠΙΛΕΞΑΜΕ: Η χλωρίδα και η πανίδα στην χώρα μας είναι ένα πολύ σημαντικό
4. γεωγραφικό/γεωλογικό πλαίσιο
 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο /Ελληνικός χώρος Τα ελληνικά βουνά (και γενικότερα οι ορεινοί όγκοι της
4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο /Ελληνικός χώρος Τα ελληνικά βουνά (και γενικότερα οι ορεινοί όγκοι της
ΚΛΙΜΑΤΙΚΗ ΤΑΞΙΝΟΝΗΣΗ ΕΛΛΑΔΑΣ
 ΚΛΙΜΑΤΙΚΗ ΤΑΞΙΝΟΝΗΣΗ ΕΛΛΑΔΑΣ ΚΡΙΤΗΡΙΑ ΤΑΞΙΝΟΜΗΣΗΣ Το κλίμα μιας γεωγραφικής περιοχής διαμορφώνεται κατά κύριο λόγο από τους 3 παρακάτω παράγοντες: 1)το γεωγραφικό πλάτος 2)την αναλογία ξηράς/θάλασσας 3)το
ΚΛΙΜΑΤΙΚΗ ΤΑΞΙΝΟΝΗΣΗ ΕΛΛΑΔΑΣ ΚΡΙΤΗΡΙΑ ΤΑΞΙΝΟΜΗΣΗΣ Το κλίμα μιας γεωγραφικής περιοχής διαμορφώνεται κατά κύριο λόγο από τους 3 παρακάτω παράγοντες: 1)το γεωγραφικό πλάτος 2)την αναλογία ξηράς/θάλασσας 3)το
ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ
 Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ Άντρια Πολυκάρπου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ Άντρια Πολυκάρπου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
Γεωλογία - Γεωγραφία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ. Τ μαθητ : Σχολικό Έτος:
 Γεωλογία - Γεωγραφία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ Τ μαθητ : Σχολικό Έτος: 1 ΜΑΘΗΜΑ 1, Οι έννοιες «γεωγραφική» και «σχετική» θέση 1. Με τη βοήθεια του χάρτη στη σελ.12, σημειώστε τις παρακάτω πόλεις στην
Γεωλογία - Γεωγραφία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ Τ μαθητ : Σχολικό Έτος: 1 ΜΑΘΗΜΑ 1, Οι έννοιες «γεωγραφική» και «σχετική» θέση 1. Με τη βοήθεια του χάρτη στη σελ.12, σημειώστε τις παρακάτω πόλεις στην
Πτυχιακή Εργασία ΓΝΩΣΕΙΣ KAI ΣΤΑΣΕΙΣ ΤΩΝ ΕΠΑΓΓΕΛΜΑΤΙΩΝ ΥΓΕΙΑΣ ΓΙΑ ΤΗΝ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΚΘΕΣΗ ΣΤΟΝ HIV. Στυλιανού Στυλιανή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία ΓΝΩΣΕΙΣ KAI ΣΤΑΣΕΙΣ ΤΩΝ ΕΠΑΓΓΕΛΜΑΤΙΩΝ ΥΓΕΙΑΣ ΓΙΑ ΤΗΝ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΚΘΕΣΗ ΣΤΟΝ HIV Στυλιανού Στυλιανή Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία ΓΝΩΣΕΙΣ KAI ΣΤΑΣΕΙΣ ΤΩΝ ΕΠΑΓΓΕΛΜΑΤΙΩΝ ΥΓΕΙΑΣ ΓΙΑ ΤΗΝ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΚΘΕΣΗ ΣΤΟΝ HIV Στυλιανού Στυλιανή Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΚΕΦΑΛΑΙΟ 1. Η ΠΡΟΕΛΕΥΣΗ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΣΚΕΨΗΣ
 ΚΕΦΑΛΑΙΟ 1. Η ΠΡΟΕΛΕΥΣΗ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΣΚΕΨΗΣ Οι αρχές της εξελικτικής σκέψης Η προέλευση των ειδών Ορθές και λανθασµένες αντιλήψεις σχετικά µε τη θεωρία της εξέλιξης Η θεωρία της εξέλιξης
ΚΕΦΑΛΑΙΟ 1. Η ΠΡΟΕΛΕΥΣΗ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΣΚΕΨΗΣ Οι αρχές της εξελικτικής σκέψης Η προέλευση των ειδών Ορθές και λανθασµένες αντιλήψεις σχετικά µε τη θεωρία της εξέλιξης Η θεωρία της εξέλιξης
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ Ο ΠΡΟΩΡΟΣ ΤΟΚΕΤΟΣ ΚΑΙ Η ΣΥΣΧΕΤΙΣΗ ΤΟΥ ΜΕ ΤΟ ΣΥΝΔΡΟΜΟ ΑΝΑΠΝΕΥΣΤΙΚΗΣ ΔΥΣΧΕΡΕΙΑΣ Όνομα Φοιτήτριας: Χρυσοστομή Αγαθοκλέους Αριθμός φοιτητικής
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ Ο ΠΡΟΩΡΟΣ ΤΟΚΕΤΟΣ ΚΑΙ Η ΣΥΣΧΕΤΙΣΗ ΤΟΥ ΜΕ ΤΟ ΣΥΝΔΡΟΜΟ ΑΝΑΠΝΕΥΣΤΙΚΗΣ ΔΥΣΧΕΡΕΙΑΣ Όνομα Φοιτήτριας: Χρυσοστομή Αγαθοκλέους Αριθμός φοιτητικής
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης
Εξελικτική Οικολογία. Σίνος Γκιώκας Πανεπιστήμιο Πάτρας Τμήμα Βιολογίας 2014
 Εξελικτική Οικολογία Σίνος Γκιώκας Πανεπιστήμιο Πάτρας Τμήμα Βιολογίας 2014 Εξελικτική Οικολογία - Σίνος Γκιώκας - Παν. Πατρών - Τμ. Βιολογίας - 2014 1 Πού συναντιούνται τα δύο πεδία; Οι κιχλίδες των Μεγάλων
Εξελικτική Οικολογία Σίνος Γκιώκας Πανεπιστήμιο Πάτρας Τμήμα Βιολογίας 2014 Εξελικτική Οικολογία - Σίνος Γκιώκας - Παν. Πατρών - Τμ. Βιολογίας - 2014 1 Πού συναντιούνται τα δύο πεδία; Οι κιχλίδες των Μεγάλων
ΦΥΣΙΚΕΣ ΠΡΟΣΤΑΤΕΥΟΜΕΝΕΣ ΠΕΡΙΟΧΕΣ. Μαρία Κιτριλάκη ΠΕ04.04
 ΦΥΣΙΚΕΣ ΠΡΟΣΤΑΤΕΥΟΜΕΝΕΣ ΠΕΡΙΟΧΕΣ Μαρία Κιτριλάκη Διαχείριση φυσικών περιοχών Η σύγχρονη αντίληψη για τη διαχείριση των φυσικών περιοχών δεν κυριαρχείται από την παλαιότερη τακτική της εξάντλησης αλλά από
ΦΥΣΙΚΕΣ ΠΡΟΣΤΑΤΕΥΟΜΕΝΕΣ ΠΕΡΙΟΧΕΣ Μαρία Κιτριλάκη Διαχείριση φυσικών περιοχών Η σύγχρονη αντίληψη για τη διαχείριση των φυσικών περιοχών δεν κυριαρχείται από την παλαιότερη τακτική της εξάντλησης αλλά από
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η ΣΥΓΚΕΝΤΡΩΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΣΤΟ ΕΔΑΦΟΣ ΚΑΙ ΜΕΘΟΔΟΙ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΤΟΥΣ Μιχαήλ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η ΣΥΓΚΕΝΤΡΩΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΣΤΟ ΕΔΑΦΟΣ ΚΑΙ ΜΕΘΟΔΟΙ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΤΟΥΣ Μιχαήλ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Γνώση και στάση νοσηλευτών στη διαχείριση του πόνου καρκινοπαθών που νοσηλεύονται Παναγιώτης Χαραλάμπους Λεμεσός, 2014 i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Γνώση και στάση νοσηλευτών στη διαχείριση του πόνου καρκινοπαθών που νοσηλεύονται Παναγιώτης Χαραλάμπους Λεμεσός, 2014 i ΤΕΧΝΟΛΟΓΙΚΟ
Η ιστορική πατρότητα του όρου «Μεσόγειος θάλασσα» ανήκει στους Λατίνους και μάλιστα περί τα μέσα του 3ου αιώνα που πρώτος ο Σολίνος τη ονομάζει
 Η ιστορική πατρότητα του όρου «Μεσόγειος θάλασσα» ανήκει στους Λατίνους και μάλιστα περί τα μέσα του 3ου αιώνα που πρώτος ο Σολίνος τη ονομάζει χαρακτηριστικά «Mare Mediterraneum» ως μεταξύ δύο ηπείρων
Η ιστορική πατρότητα του όρου «Μεσόγειος θάλασσα» ανήκει στους Λατίνους και μάλιστα περί τα μέσα του 3ου αιώνα που πρώτος ο Σολίνος τη ονομάζει χαρακτηριστικά «Mare Mediterraneum» ως μεταξύ δύο ηπείρων
Η ΚΑΤΑΝΟΜΗ ΤΟΥ EΠIΠEΔOΥ ΕΚΠΑΙΔΕΥΣΗΣ ΣΤΟΥΣ ΝΟΜΟΥΣ ΤΗΣ ΕΛΛΑΔΑΣ ΣΥΜΦΩΝΑ ΜΕ ΤΗΝ ΑΠΟΓΡΑΦΗ ΤΟΥ 2001
 Ελληνικό Στατιστικό Ινστιτούτο Πρακτικά 18 ου Πανελληνίου Συνεδρίου Στατιστικής (2005) σελ.351-356 Η ΚΑΤΑΝΟΜΗ ΤΟΥ EΠIΠEΔOΥ ΕΚΠΑΙΔΕΥΣΗΣ ΣΤΟΥΣ ΝΟΜΟΥΣ ΤΗΣ ΕΛΛΑΔΑΣ ΣΥΜΦΩΝΑ ΜΕ ΤΗΝ ΑΠΟΓΡΑΦΗ ΤΟΥ 2001 Στέφος Ευστάθιος
Ελληνικό Στατιστικό Ινστιτούτο Πρακτικά 18 ου Πανελληνίου Συνεδρίου Στατιστικής (2005) σελ.351-356 Η ΚΑΤΑΝΟΜΗ ΤΟΥ EΠIΠEΔOΥ ΕΚΠΑΙΔΕΥΣΗΣ ΣΤΟΥΣ ΝΟΜΟΥΣ ΤΗΣ ΕΛΛΑΔΑΣ ΣΥΜΦΩΝΑ ΜΕ ΤΗΝ ΑΠΟΓΡΑΦΗ ΤΟΥ 2001 Στέφος Ευστάθιος
ΝΗΣΙΩΤΙΚΗ ΒΙΟΓΕΩΓΡΑΦΙΑ ΤΟ ΠΛΑΙΣΙΟ ΤΩΝ ΘΕΡΜΩΝ ΣΗΜΕΙΩΝ ΤΗΣ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ
 ΝΗΣΙΩΤΙΚΗ ΒΙΟΓΕΩΓΡΑΦΙΑ ΤΟ ΠΛΑΙΣΙΟ ΤΩΝ ΘΕΡΜΩΝ ΣΗΜΕΙΩΝ ΤΗΣ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ Τύποι νησιών Αν θεωρηθεί (κάπως αυθαίρετα) ότι η Νέα Γουινέα είναι το μεγαλύτερο νησί, τότε τα θαλάσσια νησιά αποτελούν το 3% της
ΝΗΣΙΩΤΙΚΗ ΒΙΟΓΕΩΓΡΑΦΙΑ ΤΟ ΠΛΑΙΣΙΟ ΤΩΝ ΘΕΡΜΩΝ ΣΗΜΕΙΩΝ ΤΗΣ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ Τύποι νησιών Αν θεωρηθεί (κάπως αυθαίρετα) ότι η Νέα Γουινέα είναι το μεγαλύτερο νησί, τότε τα θαλάσσια νησιά αποτελούν το 3% της
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή ΣΥΓΚΡΙΣΗ ΥΔΡΟΠΟΝΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΣΕ ΚΑΛΛΙΕΡΓΕΙΑ ΜΑΡΟΥΛΙΟΥ Νικόλας Χαραλάμπους Λεμεσός 2015 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή ΣΥΓΚΡΙΣΗ ΥΔΡΟΠΟΝΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΣΕ ΚΑΛΛΙΕΡΓΕΙΑ ΜΑΡΟΥΛΙΟΥ Νικόλας Χαραλάμπους Λεμεσός 2015 ΤΕΧΝΟΛΟΓΙΚΟ
Ταξιδεύοντας στην ηπειρωτική Ελλάδα. Τάξη Φύλλο Εργασίας 1 Μάθημα Ε Δημοτικού Διαιρώντας την Ελλάδα σε διαμερίσματα και περιφέρειες Γεωγραφία
 Ταξιδεύοντας στην ηπειρωτική Ελλάδα Τάξη Φύλλο Εργασίας 1 Μάθημα Ε Δημοτικού Διαιρώντας την Ελλάδα σε διαμερίσματα και περιφέρειες Γεωγραφία Ταξιδεύουμε στην ηπειρωτική Ελλάδα, χρησιμοποιώντας διαφορετικά
Ταξιδεύοντας στην ηπειρωτική Ελλάδα Τάξη Φύλλο Εργασίας 1 Μάθημα Ε Δημοτικού Διαιρώντας την Ελλάδα σε διαμερίσματα και περιφέρειες Γεωγραφία Ταξιδεύουμε στην ηπειρωτική Ελλάδα, χρησιμοποιώντας διαφορετικά
Μεταπτυχιακή Διατριβή
 Μεταπτυχιακή Διατριβή ΣΧΗΜΑΤΙΣΜΟΣ ΚΑΙ ΕΝΤΟΠΙΣΜΟΣ ΒΙΟΦΙΛΜ ΣΤΙΣ ΜΕΜΒΡΑΝΕΣ ΑΝΤΙΣΤΡΟΦΗΣ ΩΣΜΩΣΗΣ ΣΤΗΝ ΑΦΑΛΑΤΩΣΗ ΛΕΜΕΣΟΥ ΚΥΠΡΟΣ ΜΙΧΑΗΛ Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ
Μεταπτυχιακή Διατριβή ΣΧΗΜΑΤΙΣΜΟΣ ΚΑΙ ΕΝΤΟΠΙΣΜΟΣ ΒΙΟΦΙΛΜ ΣΤΙΣ ΜΕΜΒΡΑΝΕΣ ΑΝΤΙΣΤΡΟΦΗΣ ΩΣΜΩΣΗΣ ΣΤΗΝ ΑΦΑΛΑΤΩΣΗ ΛΕΜΕΣΟΥ ΚΥΠΡΟΣ ΜΙΧΑΗΛ Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ
ENOTHTA 1: ΧΑΡΤΕΣ ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ
 1 ENOTHTA 1: ΧΑΡΤΕΣ ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ Μάθημα 1: Οι έννοιες και θέση 1. Τι ονομάζεται σχετική θέση ενός τόπου; Να δοθεί ένα παράδειγμα. Πότε ο προσδιορισμός της σχετικής θέσης
1 ENOTHTA 1: ΧΑΡΤΕΣ ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ Μάθημα 1: Οι έννοιες και θέση 1. Τι ονομάζεται σχετική θέση ενός τόπου; Να δοθεί ένα παράδειγμα. Πότε ο προσδιορισμός της σχετικής θέσης
Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία
 ΣΥΓΧΡΟΝΕΣ ΜΕΘΟΔΟΛΟΓΙΕΣ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ - ΓΕΝΕΤΙΚΗΣ Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία ΠΑΠΑΝΙΚΟΛΑΟΥ ΕΛΕΝΗ, Ph.D. Λέκτορας Εργαστήριο Βιολογίας, Ιατρική Σχολή Αθηνών
ΣΥΓΧΡΟΝΕΣ ΜΕΘΟΔΟΛΟΓΙΕΣ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ - ΓΕΝΕΤΙΚΗΣ Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία ΠΑΠΑΝΙΚΟΛΑΟΥ ΕΛΕΝΗ, Ph.D. Λέκτορας Εργαστήριο Βιολογίας, Ιατρική Σχολή Αθηνών
ΑΠΟΜΑΚΡΥΝΣΗ ΜΕΓΑΛΩΝ ΣΥΓΚΕΝΤΡΩΣΕΩΝ ΦΩΣΦΟΡΟΥ ΑΠΟ ΑΠΟΒΛΗΤΑ ΧΡΗΣΙΜΟΠΟΙΩΝΤΑΣ ΔΙΑΠΥΡΩΜΕΝΑ ΤΣΟΦΛΙΑ ΑΥΓΩΝ ΚΑΙ Η ΣΥΝΘΕΣΗ BRUSHITE ΩΣ ΤΕΛΙΚΟ ΠΡΟΪΟΝ
 Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία ΑΠΟΜΑΚΡΥΝΣΗ ΜΕΓΑΛΩΝ ΣΥΓΚΕΝΤΡΩΣΕΩΝ ΦΩΣΦΟΡΟΥ ΑΠΟ ΑΠΟΒΛΗΤΑ ΧΡΗΣΙΜΟΠΟΙΩΝΤΑΣ ΔΙΑΠΥΡΩΜΕΝΑ ΤΣΟΦΛΙΑ ΑΥΓΩΝ ΚΑΙ Η ΣΥΝΘΕΣΗ BRUSHITE ΩΣ ΤΕΛΙΚΟ
Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία ΑΠΟΜΑΚΡΥΝΣΗ ΜΕΓΑΛΩΝ ΣΥΓΚΕΝΤΡΩΣΕΩΝ ΦΩΣΦΟΡΟΥ ΑΠΟ ΑΠΟΒΛΗΤΑ ΧΡΗΣΙΜΟΠΟΙΩΝΤΑΣ ΔΙΑΠΥΡΩΜΕΝΑ ΤΣΟΦΛΙΑ ΑΥΓΩΝ ΚΑΙ Η ΣΥΝΘΕΣΗ BRUSHITE ΩΣ ΤΕΛΙΚΟ
Πτυχιακή διατριβή. Η επίδραση της τασιενεργής ουσίας Ακεταλδεΰδης στη δημιουργία πυρήνων συμπύκνωσης νεφών (CCN) στην ατμόσφαιρα
 στην ατμόσφαιρα") ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η επίδραση της τασιενεργής ουσίας Ακεταλδεΰδης στη δημιουργία πυρήνων συμπύκνωσης νεφών (CCN)
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η επίδραση της τασιενεργής ουσίας Ακεταλδεΰδης στη δημιουργία πυρήνων συμπύκνωσης νεφών (CCN)
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ
 ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
ΒΙΒΛΙΟ ΕΡΥΘΡΩΝ ΔΕΔΟΜΕΝΩΝ ΤΩΝ ΣΠΑΝΙΩΝ & ΑΠΕΙΛΟΥΜΕΝΩΝ ΦΥΤΩΝ ΤΗΣ ΕΛΛΑΔΑΣ ΤΟΜΟΣ ΠΡΩΤΟΣ A - D Επιτροπή Έκδοσης: Δημήτριος Φοίτος, Θεοφάνης Κωνσταντινίδης & Γεωργία Καμάρη Επιμέλεια κειμένων: Πέπη Μπαρέκα ΕΛΛΗΝΙΚΗ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΤΟ ΚΑΠΜΝΙΣΜΑ ΩΣ ΠΑΡΑΓΟΝΤΑΣ ΥΨΗΛΟΥ ΚΙΝΔΥΝΟΥ ΓΙΑ ΠΡΟΚΛΗΣΗ ΥΠΟΓΟΝΙΜΟΤΗΤΑΣ ΣΤΟΥΣ ΑΝΔΡΕΣ Κατσαρής Γιάγκος Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΤΟ ΚΑΠΜΝΙΣΜΑ ΩΣ ΠΑΡΑΓΟΝΤΑΣ ΥΨΗΛΟΥ ΚΙΝΔΥΝΟΥ ΓΙΑ ΠΡΟΚΛΗΣΗ ΥΠΟΓΟΝΙΜΟΤΗΤΑΣ ΣΤΟΥΣ ΑΝΔΡΕΣ Κατσαρής Γιάγκος Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
Εύη Καραγιαννίδου Χημικός Α.Π.Θ. ΟΙ ΕΠΟΞΕΙΔΙΚΕΣ ΚΟΛΛΕΣ ΣΤΗΝ ΑΠΟΚΑΤΑΣΤΑΣΗ ΕΡΓΩΝ ΤΕΧΝΗΣ ΑΠΟ ΓΥΑΛΙ ή ΚΕΡΑΜΙΚΟ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΧΗΜΕΙΑΣ Εύη Καραγιαννίδου Χημικός Α.Π.Θ. ΟΙ ΕΠΟΞΕΙΔΙΚΕΣ ΚΟΛΛΕΣ ΣΤΗΝ ΑΠΟΚΑΤΑΣΤΑΣΗ ΕΡΓΩΝ ΤΕΧΝΗΣ ΑΠΟ ΓΥΑΛΙ ή ΚΕΡΑΜΙΚΟ ΜΕΛΕΤΗ ΤΗΣ ΚΙΝΗΤΙΚΗΣ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΧΗΜΕΙΑΣ Εύη Καραγιαννίδου Χημικός Α.Π.Θ. ΟΙ ΕΠΟΞΕΙΔΙΚΕΣ ΚΟΛΛΕΣ ΣΤΗΝ ΑΠΟΚΑΤΑΣΤΑΣΗ ΕΡΓΩΝ ΤΕΧΝΗΣ ΑΠΟ ΓΥΑΛΙ ή ΚΕΡΑΜΙΚΟ ΜΕΛΕΤΗ ΤΗΣ ΚΙΝΗΤΙΚΗΣ
ΠΡΩΤΟΚΟΛΛΟ ΑΞΙΟΛΟΓΗΣΗΣ ΠΑΙΔΙΩΝ ΜΕ ΑΝΑΠΤΥΞΙΑΚΗ ΛΕΚΤΙΚΗ ΑΠΡΑΞΙΑ
 Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΠΡΩΤΟΚΟΛΛΟ ΑΞΙΟΛΟΓΗΣΗΣ ΠΑΙΔΙΩΝ ΜΕ ΑΝΑΠΤΥΞΙΑΚΗ ΛΕΚΤΙΚΗ ΑΠΡΑΞΙΑ Χαρίκλεια Χαρωνίτη Λεμεσός, Μάϊος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ
Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΠΡΩΤΟΚΟΛΛΟ ΑΞΙΟΛΟΓΗΣΗΣ ΠΑΙΔΙΩΝ ΜΕ ΑΝΑΠΤΥΞΙΑΚΗ ΛΕΚΤΙΚΗ ΑΠΡΑΞΙΑ Χαρίκλεια Χαρωνίτη Λεμεσός, Μάϊος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ
ΠΙΛΟΤΙΚΗ ΕΦΑΡΜΟΓΗ ΑΥΤΟΝΟΜΩΝ ΣΥΣΤΗΜΑΤΩΝ ΠΛΟΗΓΗΣΗΣ ΓΙΑ ΤΗΝ ΠΑΡΑΓΩΓΗ ΥΨΗΛΗΣ ΑΝΑΛΥΣΗΣ ΟΡΘΟΦΩΤΟΓΡΑΦΙΩΝ ΓΕΩΡΓΙΚΩΝ ΕΚΤΑΣΕΩΝ
 Σχολή Μηχανικής & Τεχνολογίας Τμήμα Πολιτικών & Μηχανικών Γεωπληροφορικής Μεταπτυχιακή διατριβή ΠΙΛΟΤΙΚΗ ΕΦΑΡΜΟΓΗ ΑΥΤΟΝΟΜΩΝ ΣΥΣΤΗΜΑΤΩΝ ΠΛΟΗΓΗΣΗΣ ΓΙΑ ΤΗΝ ΠΑΡΑΓΩΓΗ ΥΨΗΛΗΣ ΑΝΑΛΥΣΗΣ ΟΡΘΟΦΩΤΟΓΡΑΦΙΩΝ ΓΕΩΡΓΙΚΩΝ
Σχολή Μηχανικής & Τεχνολογίας Τμήμα Πολιτικών & Μηχανικών Γεωπληροφορικής Μεταπτυχιακή διατριβή ΠΙΛΟΤΙΚΗ ΕΦΑΡΜΟΓΗ ΑΥΤΟΝΟΜΩΝ ΣΥΣΤΗΜΑΤΩΝ ΠΛΟΗΓΗΣΗΣ ΓΙΑ ΤΗΝ ΠΑΡΑΓΩΓΗ ΥΨΗΛΗΣ ΑΝΑΛΥΣΗΣ ΟΡΘΟΦΩΤΟΓΡΑΦΙΩΝ ΓΕΩΡΓΙΚΩΝ
ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΔΙΟΙΚΗΣΗ της ΥΓΕΙΑΣ» ΑΞΙΟΛΟΓΗΣΗ ΑΠΟΔΟΣΗΣ ΠΡΟΣΩΠΙΚΟΥ: ΜΕΛΕΤΗ ΠΕΡΙΠΤΩΣΗΣ ΙΔΙΩΤΙΚΟΥ ΝΟΣΟΚΟΜΕΙΟΥ ΠΑΡΑΓΙΟΥΔΑΚΗ ΜΑΓΔΑΛΗΝΗ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΗΣ ΕΠΙΣΤΗΜΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΔΙΟΙΚΗΣΗ της ΥΓΕΙΑΣ» ΑΞΙΟΛΟΓΗΣΗ ΑΠΟΔΟΣΗΣ ΠΡΟΣΩΠΙΚΟΥ: ΜΕΛΕΤΗ ΠΕΡΙΠΤΩΣΗΣ ΙΔΙΩΤΙΚΟΥ ΝΟΣΟΚΟΜΕΙΟΥ ΠΑΡΑΓΙΟΥΔΑΚΗ ΜΑΓΔΑΛΗΝΗ Διπλωματική
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΗΣ ΕΠΙΣΤΗΜΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΔΙΟΙΚΗΣΗ της ΥΓΕΙΑΣ» ΑΞΙΟΛΟΓΗΣΗ ΑΠΟΔΟΣΗΣ ΠΡΟΣΩΠΙΚΟΥ: ΜΕΛΕΤΗ ΠΕΡΙΠΤΩΣΗΣ ΙΔΙΩΤΙΚΟΥ ΝΟΣΟΚΟΜΕΙΟΥ ΠΑΡΑΓΙΟΥΔΑΚΗ ΜΑΓΔΑΛΗΝΗ Διπλωματική
Πτυχιακή εργασία. Ελένη Κυριάκου
 Σχολή Γεωπονικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία Βιοδιάσπαση Ιοντικών Υγρών σε Διαφορετικές Θερμοκρασίες από τον Sphingomonas sp. VITPTHJ Ελένη Κυριάκου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ
Σχολή Γεωπονικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία Βιοδιάσπαση Ιοντικών Υγρών σε Διαφορετικές Θερμοκρασίες από τον Sphingomonas sp. VITPTHJ Ελένη Κυριάκου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ
Κ ι λ µα µ τι τ κές έ Α λλ λ α λ γές Επι π πτ π ώ τ σει ε ς στη τ β ιοπο π ικιλό λ τη τ τα τ κ αι τ η τ ν ν ά γρια ζ ωή
 Επιπτώσεις στη βιοποικιλότητα και την άγρια ζωή Η παγκόσµια κλιµατική αλλαγή θεωρείται ως η σηµαντικότερη τρέχουσα απειλή για τη βιοποικιλότητα του πλανήτη. Παραδείγµατα από την Κυπριακή Φύση Μερικές από
Επιπτώσεις στη βιοποικιλότητα και την άγρια ζωή Η παγκόσµια κλιµατική αλλαγή θεωρείται ως η σηµαντικότερη τρέχουσα απειλή για τη βιοποικιλότητα του πλανήτη. Παραδείγµατα από την Κυπριακή Φύση Μερικές από
ΣΧΕΔΙΑΣΜΟΣ ΚΑΙ ΕΝΙΣΧΥΣΗ ΤΩΝ ΚΟΜΒΩΝ ΟΠΛΙΣΜΕΝΟΥ ΣΚΥΡΟΔΕΜΑΤΟΣ ΜΕ ΒΑΣΗ ΤΟΥΣ ΕΥΡΩΚΩΔΙΚΕΣ
 Σχολή Μηχανικής και Τεχνολογίας Πτυχιακή εργασία ΣΧΕΔΙΑΣΜΟΣ ΚΑΙ ΕΝΙΣΧΥΣΗ ΤΩΝ ΚΟΜΒΩΝ ΟΠΛΙΣΜΕΝΟΥ ΣΚΥΡΟΔΕΜΑΤΟΣ ΜΕ ΒΑΣΗ ΤΟΥΣ ΕΥΡΩΚΩΔΙΚΕΣ Σωτήρης Παύλου Λεμεσός, Μάιος 2018 i ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
Σχολή Μηχανικής και Τεχνολογίας Πτυχιακή εργασία ΣΧΕΔΙΑΣΜΟΣ ΚΑΙ ΕΝΙΣΧΥΣΗ ΤΩΝ ΚΟΜΒΩΝ ΟΠΛΙΣΜΕΝΟΥ ΣΚΥΡΟΔΕΜΑΤΟΣ ΜΕ ΒΑΣΗ ΤΟΥΣ ΕΥΡΩΚΩΔΙΚΕΣ Σωτήρης Παύλου Λεμεσός, Μάιος 2018 i ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΠΙΛΟΤΙΚΗ ΕΡΕΥΝΑ ΣΕ ΣΧΕΣΗ ΜΕ ΤΟ ΓΛΩΣΣΙΚΟ ΠΡΟΦΙΛ ΤΩΝ ΕΝΗΛΙΚΩΝ ΑΝΩ ΤΩΝ 65 ΕΤΩΝ ΜΕ ΑΝΟΙΑ
 Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΠΙΛΟΤΙΚΗ ΕΡΕΥΝΑ ΣΕ ΣΧΕΣΗ ΜΕ ΤΟ ΓΛΩΣΣΙΚΟ ΠΡΟΦΙΛ ΤΩΝ ΕΝΗΛΙΚΩΝ ΑΝΩ ΤΩΝ 65 ΕΤΩΝ ΜΕ ΑΝΟΙΑ Παναγιώτα Παπαϊωάννου Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ
Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΠΙΛΟΤΙΚΗ ΕΡΕΥΝΑ ΣΕ ΣΧΕΣΗ ΜΕ ΤΟ ΓΛΩΣΣΙΚΟ ΠΡΟΦΙΛ ΤΩΝ ΕΝΗΛΙΚΩΝ ΑΝΩ ΤΩΝ 65 ΕΤΩΝ ΜΕ ΑΝΟΙΑ Παναγιώτα Παπαϊωάννου Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ
Η άγρια βρώσιμη χλωρίδα της Κρήτης
 Σεμινάριο Βάμος 25-11-2008 Γεωργία- Διατροφή- Ποιότητα ζωής ΤΙΤΛΟΣ ΑΝΑΚΟΙΝΩΣΗΣ Η άγρια βρώσιμη χλωρίδα της Κρήτης Κλεόνικος Σταυριδάκης Υπεύθ. Α/θμιας Περιβαλλοντικής Εκπαίδευσης Ν. Ρεθύμνου Εισαγωγή Η
Σεμινάριο Βάμος 25-11-2008 Γεωργία- Διατροφή- Ποιότητα ζωής ΤΙΤΛΟΣ ΑΝΑΚΟΙΝΩΣΗΣ Η άγρια βρώσιμη χλωρίδα της Κρήτης Κλεόνικος Σταυριδάκης Υπεύθ. Α/θμιας Περιβαλλοντικής Εκπαίδευσης Ν. Ρεθύμνου Εισαγωγή Η
ΠΕΡΙΓΡΑΜΜΑ ΜΑΘΗΜΑΤΟΣ
 ΠΕΡΙΓΡΑΜΜΑ ΜΑΘΗΜΑΤΟΣ 1. ΓΕΝΙΚΑ ΣΧΟΛΗ ΑΓΡΟΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΥΠΟΔΟΜΩΝ ΚΑΙ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΤΜΗΜΑ ΕΠΙΣΤΗΜΗΣ ΦΥΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΕΠΙΠΕΔΟ ΣΠΟΥΔΩΝ Προπτυχιακό ΚΩΔΙΚΟΣ ΜΑΘΗΜΑΤΟΣ 3465 ΕΞΑΜΗΝΟ ΣΠΟΥΔΩΝ 1 Ο ΤΙΤΛΟΣ ΜΑΘΗΜΑΤΟΣ
ΠΕΡΙΓΡΑΜΜΑ ΜΑΘΗΜΑΤΟΣ 1. ΓΕΝΙΚΑ ΣΧΟΛΗ ΑΓΡΟΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΥΠΟΔΟΜΩΝ ΚΑΙ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΤΜΗΜΑ ΕΠΙΣΤΗΜΗΣ ΦΥΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΕΠΙΠΕΔΟ ΣΠΟΥΔΩΝ Προπτυχιακό ΚΩΔΙΚΟΣ ΜΑΘΗΜΑΤΟΣ 3465 ΕΞΑΜΗΝΟ ΣΠΟΥΔΩΝ 1 Ο ΤΙΤΛΟΣ ΜΑΘΗΜΑΤΟΣ
Hλίας Αθανασιάδης * Συγκριτική θέση της Ηπείρου ως προς τις υπόλοιπες περιοχές της Ελλάδας με κριτήριο τους δείκτες ευημερίας
 Hλίας Αθανασιάδης * Συγκριτική θέση της Ηπείρου ως προς τις υπόλοιπες περιοχές της Ελλάδας με κριτήριο τους δείκτες ευημερίας Στη σημερινή εποχή της ευρωπαϊκής ενοποίησης, τα οικονομικά κριτήρια σύγκρισης
Hλίας Αθανασιάδης * Συγκριτική θέση της Ηπείρου ως προς τις υπόλοιπες περιοχές της Ελλάδας με κριτήριο τους δείκτες ευημερίας Στη σημερινή εποχή της ευρωπαϊκής ενοποίησης, τα οικονομικά κριτήρια σύγκρισης
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΑΘΗΜΑΤΙΚΟΙ ΑΛΓΟΡΙΘΜΟΙ ΓΙΑ ΑΝΑΛΥΣΗ ΠΙΣΤΟΠΟΙΗΤΙΚΩΝ ΕΝΕΡΓΕΙΑΚΗΣ ΑΠΟΔΟΣΗΣ ΚΤΙΡΙΩΝ Εβελίνα Θεμιστοκλέους
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΑΘΗΜΑΤΙΚΟΙ ΑΛΓΟΡΙΘΜΟΙ ΓΙΑ ΑΝΑΛΥΣΗ ΠΙΣΤΟΠΟΙΗΤΙΚΩΝ ΕΝΕΡΓΕΙΑΚΗΣ ΑΠΟΔΟΣΗΣ ΚΤΙΡΙΩΝ Εβελίνα Θεμιστοκλέους
ΣΥΜΠΛΟΚΑ CU KAI FE ΜΕ ΤΗΝ ΕΞΩΚΥΤΤΑΡΙΑ ΠΟΛΥΜΕΡΗ ΟΥΣΙΑ (EPS) ΤΟΥ PHAEOBACTER GALLAECIENSIS
 ΤΟΥ PHAEOBACTER GALLAECIENSIS") Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία ΣΥΜΠΛΟΚΑ CU KAI FE ΜΕ ΤΗΝ ΕΞΩΚΥΤΤΑΡΙΑ ΠΟΛΥΜΕΡΗ ΟΥΣΙΑ (EPS) ΤΟΥ PHAEOBACTER GALLAECIENSIS Κυριάκος Δημητρίου Λεμεσός, Μάιος 2017
Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία ΣΥΜΠΛΟΚΑ CU KAI FE ΜΕ ΤΗΝ ΕΞΩΚΥΤΤΑΡΙΑ ΠΟΛΥΜΕΡΗ ΟΥΣΙΑ (EPS) ΤΟΥ PHAEOBACTER GALLAECIENSIS Κυριάκος Δημητρίου Λεμεσός, Μάιος 2017
Μεταπτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΜΕΛΕΤΗ ΤΗΣ ΕΚΛΕΚΤΙΚΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΟΞΕΙΔΩΣΗΣ ΤΟΥ ΜΕΘΑΝΙΟΥ ΠΡΟΣ ΔΙΟΞΕΙΔΙΟ ΤΟΥ ΑΝΘΡΑΚΑ Βασιλική
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΜΕΛΕΤΗ ΤΗΣ ΕΚΛΕΚΤΙΚΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΟΞΕΙΔΩΣΗΣ ΤΟΥ ΜΕΘΑΝΙΟΥ ΠΡΟΣ ΔΙΟΞΕΙΔΙΟ ΤΟΥ ΑΝΘΡΑΚΑ Βασιλική
Πτυχιακή Εργασία Η ΑΝΤΙΛΑΜΒΑΝΟΜΕΝΗ ΚΟΙΝΩΝΙΚΗ ΣΤΗΡΙΞΗ ΣΤΙΣ ΘΗΛΑΖΟΥΣΕΣ ΜΗΤΕΡΕΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΑΝΤΙΛΑΜΒΑΝΟΜΕΝΗ ΚΟΙΝΩΝΙΚΗ ΣΤΗΡΙΞΗ ΣΤΙΣ ΘΗΛΑΖΟΥΣΕΣ ΜΗΤΕΡΕΣ Ονοματεπώνυμο: Στέλλα Κόντζιαλη Αριθμός Φοιτητικής Ταυτότητας: 2010414838
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΑΝΤΙΛΑΜΒΑΝΟΜΕΝΗ ΚΟΙΝΩΝΙΚΗ ΣΤΗΡΙΞΗ ΣΤΙΣ ΘΗΛΑΖΟΥΣΕΣ ΜΗΤΕΡΕΣ Ονοματεπώνυμο: Στέλλα Κόντζιαλη Αριθμός Φοιτητικής Ταυτότητας: 2010414838