ΒΙΟΠΟΙΚΙΛΟΤΗΤΑ ΚΑΙ ΒΙΟΓΕΩΓΡΑΦΙΚΑ ΠΡΟΤΥΠΑ ΤΩΝ ΙΧΘΥΩΝ ΤΩΝ ΕΣΩΤΕΡΙΚΩΝ ΥΔΑΤΩΝ ΤΗΣ ΒΑΛΚΑΝΙΚΗΣ ΧΕΡΣΟΝΗΣΟΥ
|
|
|
- Ἄτλας Δυοβουνιώτης
- 7 χρόνια πριν
- Προβολές:
Transcript
1 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΒΙΟΛΟΓΙΚΩΝ ΕΦΑΡΜΟΓΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΩΝ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑ ΚΑΙ ΒΙΟΓΕΩΓΡΑΦΙΚΑ ΠΡΟΤΥΠΑ ΤΩΝ ΙΧΘΥΩΝ ΤΩΝ ΕΣΩΤΕΡΙΚΩΝ ΥΔΑΤΩΝ ΤΗΣ ΒΑΛΚΑΝΙΚΗΣ ΧΕΡΣΟΝΗΣΟΥ ΑΝΘΗ ΟΙΚΟΝΟΜΟΥ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΙΩΑΝΝΙΝΑ 2016
2
3 UNIVERSITY OF IOANNINA SCHOOL OF HEALTH SCIENCES DEPARTMENT OF BIOLOGICAL APPLICATIONS AND TECHNOLOGY BIODIVERSITY AND BIOGEOGRAPHICAL PATTERNS OF FRESHWATER FISHES OF THE BALKAN PENINSULA ANTHI OIKONOMOU PHD THESIS IOANNINA 2016
4
5 «Η έγκριση της διδακτορικής διατριβής από το Τμήμα Βιολογικών Εφαρμογών και Τεχνολογιών της Σχολής Επιστημών Υγείας του Πανεπιστημίου Ιωαννίνων δεν υποδηλώνει αποδοχή των γνωμών του συγγραφέα» (Ν. 5343/32, Παρ.2, Άρθρο 202)
6
7
8
9
10
11
12
13
14
15 ΕΡΓΑΣΙΕΣ ΠΟΥ ΠΡΟΕΚΥΨΑΝ ΑΠΟ ΤΗ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ Oikonomou A., Leprieur F. & Leonardos I., 2014b. Biogeography of freshwater fishes of the Balkan Peninsula. Hydrobiologia, 738: Oikonomou A., Anastasiadou Ch., Taskoudis Th. & Leonardos I., 2014a. Length- weight relations of seven native fish species (Actinopterygii) from the Louros River, Greece. Acta Ichthyologica et Piscatoria, 44 (2): ΑΛΛΕΣ ΕΡΓΑΣΙΕΣ Ntakis A., Liasko R., Oikonomou A & Leonardos I., Growth pattern of an endemic and endangered West Balkan Peninsula trout species (Salmo lourosensis, Delling, 2010). Journal of Applied Ichthyology, 31: Leprieur F. & Oikonomou A., The need for richness-independent measures of turnover when delineating biogeographical regions. Journal of Biogeography, 41: Gkenas C., Oikonomou A., Economou A., Kiosse F. & Leonardos I., Life history pattern and feeding habits of the invasive mosquitofish, Gambusia holbrooki, in Lake Pamvotis (NW Greece). Journal of Biological Research, 17: ΠΡΟΦΟΡΙΚΕΣ ΚΑΙ ΓΡΑΠΤΕΣ ΑΝΑΚΟΙΝΩΣΕΙΣ ΠΟΥ ΣΧΕΤΙΖΟΝΤΑΙ ΜΕ ΤΗ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ Oikonomou A., Leprieur F. & Leonardos I., Freshwater fishes in Greek lakes: Species richness and body size patterns. XV European Congress of Ichthyology, Oporto. Oikonomou A., Leprieur F. & Leonardos I., Delineating the biogeographical regions of the Balkan Peninsula. 7th HELECOS Congress, Mitilini. 15
16 Oikonomou A., Leprieur F. & Leonardos I., Exploring the species- area relationship (SAR) in the freshwater ecosystems of the Balkan Peninsula. 7th HELECOS Congress, Mitilini. Oikonomou A., Leprieur F. & Leonardos I., Conservation biogeography of the freshwater fishes of the Balkan Peninsula. 15th Panhellenic Congress of Ichthyologists, Thessaloniki. Oikonomou A., Leprieur F. & Leonardos I., Patterns of endemism and Conservation Status of the Balkan freshwater fish fauna. 35th Congress EEVE, Nafplio. Oikonomou A., Leprieur F. & Leonardos I., Freshwater fishes of the Balkan Peninsula: Delineating biogeographical regions. XIV European Congress of Ichthyology, Liège. Taskoudis T., Oikonomou A. & Leonardos I., Ichthyofauna of Louros River. 34th Congress EEVE, Trikala. Ntakis A., Oikonomou A., Liasko R. & Leonardos I., The use of non-lethal methods for the study of freshwater fishes. The case study of the endemic and endangered trout of Louros River.34th Congress EEVE, Trikala. Oikonomou A., Ntakis A., Anastasiadou C. & Leonardos I., Salmo lourosensis, Delling, 2010: An endangered species and its conservation status. 33rd Congress EEVE, Edessa. ΑΛΛΕΣ ΠΡΟΦΟΡΙΚΕΣ ΚΑΙ ΓΡΑΠΤΕΣ ΑΝΑΚΟΙΝΩΣΕΙΣ Sapounidis A., Liousia V., Tryfonopoulos G., Ntislidou Ch., Oikonomou A., Petriki O., Bobori D., Koutrakis E. & Leonardos I., Aphanius almiriensis: an established and breeding population in Moustos wetland. 13th ICZEGAR, Heraklion. Anastasiadou Ch., Christodoulou M. & Oikonomou A., Biodiversity and biogeographical patterns of the epigean freshwater shrimps (Atyidae, Palaemonidae) and amphipods (Gammaridae) in Greece. 13th ICZEGAR, Heraklion. 16
17 Papadopoulou P., Oikonomou A., Georgakli A., Koutrakis E. & Leonardos I., Age and growth of european eel (Anguilla anguilla, Linnaeus, 1758) in three lagoons of Western Greece. 37th Congress EEVE, Volos. Vardakas L., Kalogianni E., Kommatas D., Stoumboudi M.T., Mentzafou A., Oikonomou A. & Skoulikidis N. Th., Habitat use by three range-restricted endemic fish species in reference and impacted conditions (Evrotas River, Southern Greece) - Preliminary results. 11th Panhellenic Symposium of Oceanography & Fisheries, Mitilini. 17
18
19 Σε κάθε άνθρωπο που πιστεύει και αγωνίζεται για την ομορφιά των ονείρων του
20
21 Πίνακας Περιεχομένων Πίνακας Περιεχομένων Πρόλογος και ευχαριστίες Εκτεταμένη περίληψη Extended abstract Structure of the thesis- Outline of chapters Κεφάλαιο 1 Γενική εισαγωγή Περί βιογεωγραφίας και μακροοικολογίας Κατανομή, σύνθεση και δομή βιοκοινοτήτων στη μακρο- και μικρο- κλίμακα Πολλαπλές πτυχές της βιοποικιλότητας Ποικιλότητα οικοσυστημάτων Ποικιλότητα ειδών Ενδοειδική ποικιλότητα Γενετική ποικιλότητα Λειτουργική ποικιλότητα Τα οικοσυστήματα των εσωτερικών και υφάλμυρων υδάτων ως θερμά σημεία («hotspots») της βιοποικιλότητας Δράσεις διατήρησης στα εσωτερικά ύδατα Τα ψάρια των εσωτερικών υδάτων Τα ψάρια των εσωτερικών υδάτων ως δείκτες σε βιογεωγραφικές αναλύσεις ΜΕΡΟΣ I Περιφερειακό επίπεδο: Η Βαλκανική χερσόνησος Κεφάλαιο 2 Βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου Εισαγωγή Oριοθέτηση των βιογεωγραφικών περιοχών Η χρήση της β- ποικιλότητας για την οριοθέτηση των βιογεωγραφικών περιοχών Oριοθέτηση των βιογεωγραφικών περιοχών βάσει των κατανομών των ψαριών Είδη-ενδείκτες σε βιογεωγραφικές περιοχές Oριοθέτηση των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου βάσει των κατανομών των ψαριών των εσωτερικών υδάτων Παλαιογεωγραφική αναδρομή της Βαλκανικής χερσονήσου
22 2.2 Στόχος του κεφαλαίου Υλικά και Μέθοδοι Βάση δεδομένων κατανομής των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου Προτεινόμενοι δείκτες β- ποικιλότητας για την οριοθέτηση βιογεωγραφικών περιοχών Επιλογή του καταλληλότερου δείκτη β-ποικιλότητας για την οριοθέτηση των βιογεωγραφικών περιοχών στη Βαλκανική χερσόνησο Δημιουργώντας το δενδρόγραμμα για την οριοθέτηση των βιογεωγραφικών περιοχών Πόσες βιογεωγραφικές περιοχές ορίζονται από ένα δενδρόγραμμα; Είδη-ενδείκτες των Βαλκανικών βιογεωγραφικών περιοχών Στατιστική επεξεργασία και πακέτα Αποτελέσματα Βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου Είδη-ενδείκτες της ιχθυοπανίδας στις βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου Συζήτηση Oριοθέτηση των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου Βιογεωγραφική απόδοση από τα είδη-ενδείκτες των εσωτερικών υδάτων της Βαλκανικής χερσονήσου Κεφάλαιο 3 Πρότυπα βιοποικιλότητας των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου Εισαγωγή Θερμά σημεία της βιοποικιλότητας ή σημεία ενδιαφέροντος («Biodiversity Hotspots») Τα πρότυπα του αριθμού των ειδών και του ενδημισμού στα εσωτερικά ύδατα Σχέση ανάμεσα στον αριθμό των ειδών και την έκταση Στόχος του κεφαλαίου Υλικά και Μέθοδοι Πρότυπα αριθμού ειδών, ενδημισμού και των απειλούμενων ειδών (κατηγορίες IUCN) στις βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου Επιλογή του μοντέλου με την καλύτερη προσαρμογή για τη περιγραφή της σχέσης αριθμού ειδών-έκτασης στα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου Στατιστική επεξεργασία και πακέτα Αποτελέσματα Σύνθεση και πρότυπα αριθμού ειδών της Βαλκανικής ιχθυοπανίδας Πρότυπα ενδημισμού και απειλούμενων ειδών (IUCN Κατηγορίες) της Βαλκανικής ιχθυοπανίδας SARs στα οικοσυστήματα των εσωτερικών υδάτων στα Βαλκάνια Συζήτηση Σύνθεση και πρότυπα της Βαλκανικής ιχθυοπανίδας
23 3.5.2 Διαχειριστικές προτάσεις ΜΕΡΟΣ II Τοπικό επίπεδο: Ελλάδα Κεφάλαιο 4 Βιογεωγραφικά πρότυπα των ψαριών στα εσωτερικά ύδατα της Ελλάδας Εισαγωγή Τα εσωτερικά ύδατα της Ελλάδας Τα εσωτερικά ύδατα της Ελλάδας ως αποτέλεσμα της παλαιογεωγραφικής ιστορίας Οι Ελληνικές λίμνες Η Ελληνική ιχθυοπανίδα και οι βιογεωγραφικές ιδιαιτερότητές της Απειλές της Ελληνικής ιχθυοπανίδας Παράγοντες που καθορίζουν την ποικιλότητα των ειδών ψαριών σε λίμνες ιαβάθµιση του Μεγέθους του Σώματος Κατά Γεωγραφικό Πλάτος Στόχος του κεφαλαίου Υλικά και Μέθοδοι Σύνθεση, πρότυπα ποικιλότητας και απειλούμενων ειδών (κατηγορίες IUCN) της Ελληνικής ιχθυοπανίδας Βάση δεδομένων των Ελληνικών λιμνών Στατιστική επεξεργασία και πακέτα Αποτελέσματα Πρότυπα ποικιλότητας, ενδημισμού και των απειλούμενων ειδών (κατηγορίες IUCN) της Ελληνικής ιχθυοπανίδας Σχέση μεταξύ του αριθμού και του μεγέθους των ειδών ψαριών με τα περιβαλλοντικά χαρακτηριστικά των Ελληνικών λιμνών Συζήτηση Η Ελληνική ιχθυοπανίδα των εσωτερικών υδάτων Βιογεωγραφικοί παράγοντες που επηρεάζουν τον αριθμό των ειδών ψαριών και το μέγεθός τους σε λιμναία οικοσυστήματα στην Ελλάδα ΜΕΡΟΣ III Επίπεδο οικοσυστήματος: Ποταμός Λούρος Κεφάλαιο 5 Πρότυπα ποικιλότητας στον ποταμό Λούρο Εισαγωγή Βιοκοινότητες ψαριών σε ποτάμια οικοσυστήματα Λειτουργική ποικιλότητα σε ποτάμια οικοσυστήματα Λειτουργική ποικιλότητα των ψαριών των εσωτερικών υδάτων Ποια και πόσα λειτουργικά χαρακτηριστικά των ψαριών πρέπει να χρησιμοποιήσουμε; Στόχος του κεφαλαίου Υλικά και Μέθοδοι Κριτήρια επιλογής του οικοσυστήματος μελέτης
24 5.3.2 Περιοχή μελέτης: Ποταμός Λούρος Σταθμοί δειγματοληψίας Μεθοδολογία συλλογής ιχθυολογικού υλικού Περιβαλλοντικοί περιγραφείς Εργαστηριακή επεξεργασία ιχθυολογικού υλικού Ανάλυση δεδομένων λειτουργικής ποικιλότητας Στατιστική επεξεργασία και πακέτα Αποτελέσματα Λειτουργική ποικιλότητα του ποταμού Λούρου Συζήτηση Περιβαλλοντική διαβάθμιση στον ποταμό Λούρο Λειτουργικός χώρος των ψαριών του ποταμού Λούρου Σχέσεις μεταξύ των οικο-μορφολογικών και των περιβαλλοντικών χαρακτηριστικών Λειτουργική ποικιλότητα στον ποταμό Λούρο Λειτουργικά μοναδικά είδη της ιχθυοπανίδας του ποταμού Λούρου Περιβαλλοντικό φιλτράρισμα ως κανόνας συγκρότησης των ιχθυοκοινοτήτων Μειονεκτήματα και πλεονεκτήματα των επιλεγμένων αναλύσεων και διαχειριστικές προεκτάσεις Κεφάλαιο 6 Γενικά συμπεράσματα Βιβλιογραφικές αναφορές Παραρτήματα Παράρτημα 1 Βιβλιογραφικές πηγές βάσης δεδομένων κατανομής των ψαριών των εσωτερικών υδώτων της Βαλκανικής Χερσονήσου Παράρτημα 2 Λίστα ειδών ψαριών της Βαλκανικής χερσονήσου Παράρτημα 3 Συσχέτιση του αριθμού ειδών ψαριών με τα λιμνολογικά χαρακτηριστικά και τις φυσικοχημικές παραμέτρους του νερού σε 13 Ελληνικές λίμνες Παράρτημα 4 Αναλυτική περιγραφή σταθμών δειγματοληψίας στον ποταμό Λούρο Παράρτημα 5 Διακύμανση φυσικοχημικών παραμέτρων και συγκέντρωσης θρεπτικών αλάτων και χλωροφύλλης-α στο νερό του ποταμού Λούρου Αβιοτικά στοιχεία Θρεπτικά άλατα Χλωροφύλλη-α Παράρτημα 6 Ιχθυοπανίδα του ποταμού Λούρου
25 Πρόλογος και ευχαριστίες Το παρόν κείμενο αποτελεί το αποτέλεσμα ομαδικής προσπάθειας κατά την εκπόνηση της διδακτορικής διατριβής μου στο Εργαστήριο Ζωολογίας του Τμήματος Βιολογικών Εφαρμογών και Τεχνολογιών του Πανεπιστημίου Ιωαννίνων υπό την επίβλεψη του Καθηγητή του Πανεπιστημίου Ιωαννίνων, Ιωάννη Λεονάρδου. Σε αυτόν, λοιπόν, οφείλω ένα ειλικρινές και μεγάλο ευχαριστώ για την ευκαιρία που μου έδωσε να εκπονήσω το διδακτορικό μου, μια ευκαιρία που κάποιος δύσκολα θα έδινε στις αντίξοες συνθήκες των Ελληνικών Πανεπιστημίων, σε μία φαινομενικά άγνωστη φοιτήτρια που επικοινώνησε μαζί του. Τον ευχαριστώ βαθειά που πίστεψε ότι θα μπορούσα να ανταπεξέλθω στις ευθύνες αυτής της έρευνας, που με ανέχτηκε όταν τα πράγματα δεν ήταν εύκολα και τέλος, για την ευκαιρία που μου έδωσε να συμμετάσχω στα ερευνητικά προγράμματα του εργαστηρίου, που συνέβαλαν στη διεύρυνση του ερευνητικού μου αντικείμενου. Θα ήθελα να πω ένα «mille merci» στον Αναπληρωτή Καθηγητή του Πανεπιστημίου Montpellier 2, Fabien Leprieur, για την καθοριστική συνεισφορά του, για την άψογη συνεργασία μας κατά τη διάρκεια αυτών των 5 χρόνων και για τη μαθητεία δίπλα του κατά την παραμονή μου στο Montpellier. Οφείλω, επίσης, ένα μεγάλο ευχαριστώ στον Δρ. Άλκη Οικονόμου, Τακτικό Ερευνητή του Ινστιτούτου Θαλασσίων Βιολογικών Πόρων και Εσωτερικών Υδάτων του Ελληνικού Κέντρου Θαλασσίων Ερευνών (ΕΛ.ΚΕ.Θ.Ε.), για την εμπιστοσύνη που μου έδειξε όταν συμφώνησε να γίνει μέλος της Τριμελούς Συμβουλευτικής μου Επιτροπής, για τις πολύ εποικοδομητικές συζητήσεις που κάναμε και τις εύστοχες υποδείξεις και παρατηρήσεις του σε κείμενα μου. Πέραν όμως από τους τρεις ανθρώπους που συνέβαλαν καθοριστικά στην τελική μορφή αυτού του κειμένου, θα ήθελα να ευχαριστήσω και αυτούς που τυχαία ή μη, συνέβαλαν σε αυτό που είμαι σήμερα. Σε αυτό το σημείο, λοιπόν, θα ήθελα να ευχαριστήσω τα τέσσερα μέλη της Εξεταστικής μου Επιτροπής: τον Καθηγητή του Α.Π.Θ. και Διευθυντή του Ινστιτούτου Θαλασσίων Βιολογικών Πόρων και Εσωτερικών Υδάτων του Ελληνικού Κέντρου Θαλασσίων Ερευνών (ΕΛ.ΚΕ.Θ.Ε.), Κωνσταντίνο Στεργίου, γιατί ήταν ο άνθρωπος που κάποτε αναφέρθηκε στην οικολογική αξία που έχουν τα εσωτερικά ύδατα σε μία εκδρομή με το μάθημα της Ζωολογίας το 2006, ένα γεγονός που αποτέλεσε το ερέθισμα για να εμβαθύνω σε αυτό το αντικείμενο. Από το εργαστήριο Ιχθυολογίας 25
26 του τμήματος Βιολογίας του Α.Π.Θ., ευχαριστώ την Επίκουρη Καθηγήτρια Δήμητρα Μπόμπορη, για τις εύστοχες διορθώσεις και τόσο για τη διδασκαλία της στο Λιμνοποτάμιο κατά τα φοιτητικά μου χρόνια όσο και για τη συμπαράσταση τα χρόνια της γνωριμίας μας. Δε θα μπορούσα να παραλείψω να ευχαριστήσω τους Δρ. Μάνο Κουτράκη, Τακτικό Ερευνητή του Ινστιτούτου Αλιευτικής Έρευνας (ΙΝΑΛΕ), για τα θετικά του σχόλια και τις υποδείξεις του και τον Επίκουρο Καθηγητή του Πανεπιστημίου Πατρών, Γιώργο Κεχαγιά, για τις διορθώσεις επί του κειμένου και τις ενδιαφέρουσες προτάσεις του. Θα ήθελα, τέλος, να ευχαριστήσω τον Επίκουρο Καθηγητή του Α.Π.Θ., Θανάση Τσίκληρα, γιατί ήταν ο άνθρωπος που με προέτρεψε το 2009 να ξεκινήσω διδακτορικό στο Πανεπιστήμιο Ιωαννίνων και αυτή του η συμβουλή έπαιξε καθοριστικό ρόλο στη μετέπειτα ζωή μου. Ευχαριστώ βαθειά την «οικογένεια» του Εργαστηρίου Ζωολογίας που με αγκάλιασε, με υποστήριξε, με καθοδήγησε αλλά και ανέχτηκε τις φωνές και τη γκρίνια μου. Ευχαριστώ, λοιπόν, θερμά, τη σχεδόν αδερφική αλλά και πολλές φορές μητρική μου φίλη και συνεργάτιδα Δρ. Χρύσα Αναστασιάδου, το συνάδελφο Αλέξανδρο Ντάκη, την αγαπητή Δρ. Μιράντα Τσουμάνη και τον ιντερνετικό φίλο και συνεργάτη Δρ. Αργύρη Σαπουνίδη, για την καθοδήγηση, την άψογη συνεργασία στα ερευνητικά προγράμματα και τις ευχάριστες στιγμές στις δειγματοληψίες, αλλά και εκτός εργαστηρίου. Ευχαριστώ πολύ τα πρώην μέλη Δρ. Δότη Πανπαδημητρίου, Δρ. Χρήστο Γκένα και Δρ. Βέρα Λίουσια για το θερμό καλωσόρισμα και τις συμβουλές τους κατά τα πρώτα έτη στο εργαστήριο. Ευχαριστώ, επίσης, το Δρ. Ρομάν Λιάσκο για τα «εύκολα και τα δύσκολα» και το «παιδί του εργαστηρίου», Βούλα Παπαδοπούλου, για το χαμόγελο και τη θετική της ενέργεια. Ευχαριστώ το Φορέα Διαχείρισης Υγροτόπων Αμβρακικού και τον οδηγό του Πανεπιστημίου Ιωαννίνων, Παναγιώτη Βλάχο, για τη βοήθεια τους στις δειγματοληψίες μου στον ποταμό Λούρο. Τέλος, ευχαριστώ το φίλο Βασίλη Τάχο για την καθοδήγηση και τις εποικοδομητικές συζητήσεις κατά την εφαρμογή της Οδηγίας 92/43 στα Ελληνικά εσωτερικά ύδατα. Κατά την εκπαίδευση μου στο Πανεπιστήμιο του Montpellier, ευχαριστώ το Ίδρυμα Κρατικών υποτροφιών (Ι.Κ.Υ.) για τη χρηματοδότηση και τα μέλη του εργαστηρίου MARine Biodiversity, Exploitation and Conservation (Marbec) του Πανεπιστημίου του Montpellier, François Guilhaumon, Arthur Escalas, Eva Marie, Luisa Manna και Sébastien Villeger, που με βοήθησαν στις αναλύσεις αλλά και με υποστήριξαν συναισθηματικά. Οφείλω επίσης ένα μεγάλο ευχαριστώ στους Βαλκάνιους συναδέλφους μου, Milorad Mrakovčić, Marko Ćaleta, Davor Zanella, Zoran Marčić, Simo Georgiev, Predrag Simonović, Avdo Sofradžija, Avdul Adrović, Trajce Talevski, Tomislav Treer, Apostolos Apostolou, Pero Tutman, Luchezar Pehlivanov, Mirjana Lenhardt και Tichomir Stefanov, διότι η συμβολή τους ήταν καθοριστική στο 2 ο και 3 ο κεφάλαιο της διδακτορικής μου διατριβής. Τα περισσότερα και τα πιο μεγάλα ευχαριστώ ανήκουν όμως στους δικούς μου ανθρώπους. Είναι αυτοί που μου κρατούσαν το χέρι και πολλές φορές με έκαναν να 26
27 σφίξω τα δόντια με την τόση αγάπη τους, είναι εκείνοι που με άκουγαν πάντα και ας άλλαζα ψυχολογία κάθε εβδομάδα, ακόμη και όταν τους παραμελούσα. Είναι η πατρική μου οικογένεια και ιδιαίτερα η Έλλη, ο Κώστας, είναι η Κωνσταντίνα με τη νέα Χίπι, η Νίκη, η Ελένη και η Φανή, η Μαρία, η Γιώτα, η Jo και οι Τρικαλινοί μου Κωστής και Μπάμπης. Και ίσως από όλους εσάς, να χρωστάω το μεγαλύτερο ευχαριστώ στον Κώστα, στο φίλο μου, συνάδελφο και συνοδοιπόρο, που με βοήθησε τόσο πρακτικά αλλά και ψυχικά, που με καθοδήγησε, που έδειξε τόσο υπομονή και αποτελεί για μένα το πρότυπο ερευνητικής δεοντολογίας και επαγγελματικής αφοσίωσης. Σας ευχαριστώ. Κλείνοντας, μπορώ να πω πως κάνοντας ένα ξεκαθάρισμα συναισθημάτων και αναμνήσεων, κατέληξα πως αυτή η διαδικασία με άλλαξε ως άνθρωπο, με βοήθησε να βρω τα φυσικά και ψυχικά μου όρια, με ωρίμασε και με οδήγησε σε δρόμους αυτογνωσίας, προσωπικής αναθεώρησης και μελλοντικής εξέλιξης. Αυτό το «παιδί», λοιπόν, που γεννήθηκε ως μία ιδέα αλλά τελικά βρήκε έδαφος στο παρόν αρχείο, σίγουρα άξιζε ως μονοπάτι, θα το κουβαλάω μέσα μου για πάντα και αποτελεί για μένα σημείο αναφοράς. Σας ευχαριστώ λοιπόν για όλα xxx 2/2016 Για μένα, το λοιπόν, το πιο εκπληκτικό, πιο επιβλητικό, πιο μυστηριακό και πιο μεγάλο, είναι ένας άνθρωπος που τον μποδίζουν να βαδίζει, είναι ένας άνθρωπος που τον αλυσοδένουνε. Ναζίμ Χικμέτ 27
28
29 Εκτεταμένη περίληψη Η μελέτη των μηχανισμών που διαμορφώνουν τα πρότυπα κατανομής των οργανισμών και της διαβάθμισης της βιοποικιλότητας αποτελεί ένα από τα θεμελιώδη αντικείμενα της οικολογίας, της βιογεωγραφίας και της μακροικολογίας. Τα οικοσυστήματα των εσωτερικών υδάτων αποτελούν μία εξαιρετική περίπτωση για τη διερεύνηση των οικολογικών θεωριών διότι φιλοξενούν ένα ιδιαίτερα υψηλό ποσοστό της παγκόσμιας βιοποικιλότητας. Ενώ, καταλαμβάνουν μόνο το 0.8% της επιφάνειας της Γης, εκτιμάται ότι προσφέρουν καταφύγιο στο 12% της παγκόσμιας πανίδας. Ταυτόχρονα, τείνουν να παρουσιάζουν το υψηλότερο ποσοστό των ειδών που απειλούνται με εξαφάνιση, δεδομένου ότι ένα μεγάλο μέρος των ενδιαιτημάτων τους είτε έχει καταστραφεί είτε έχει υποβαθμιστεί από τις ανθρώπινες δραστηριότητες. Ανάμεσα στους οργανισμούς που διαβιούν στα εσωτερικά ύδατα, τα ψάρια αποτελούν έναν από τους χρησιμότερους οργανισμούς-μοντέλα για τη διερεύνηση της βιογεωγραφικής ιστορίας, των μακροοικολογικών προτύπων και των οικολογικών υποθέσεων, και αυτό διότι θεωρούνται πολύ καλοί δείκτες βιολογικής απομόνωσης μιας υδάτινης μάζας. Τα ψάρια των εσωτερικών υδάτων είναι καλοί ζωογεωγραφικοί δείκτες, διότι δε μπορούν να διασχίσουν το εμπόδιο του θαλασσινού νερού, έχουν γενικά μειωμένη ικανότητα διασποράς και η κατανομή τους περιορίζεται στο επίπεδο της λεκάνης απορροής, παρουσιάζοντας έτσι υψηλό ενδημισμό. Τα ψάρια έχουν χρησιμοποιηθεί ευρέως για την οριοθέτηση των βιογεωγραφικών περιοχών στα εσωτερικά ύδατα εξαιτίας της υψηλής ποικιλότητάς τους και της επαρκούς και τεκμηριωμένης γνώσης τόσο της συστηματικής τους όσο και της γεωγραφικής κατανομής των περισσότερων ειδών. Η οριοθέτηση των βιογεωγραφικών περιοχών διακρίνει τις διαφορετικές περιοχές με βάση την 29
30 ομοιότητα των πανίδων τους, και ως εκ τούτου, τα οικοσυστήματα εντός των βιογεωγραφικών περιοχών παρουσιάζουν όμοια πανιδική σύνθεση. Το γεγονός αυτό αποτελεί το πρώτο βήμα στη διερεύνηση των ζωογεωγραφικών προτύπων. Σε συνδυασμό με τη μελέτη της διακύμανσης του αριθμού των ειδών, του ενδημισμού και των απειλούμενων ειδών, επιτρέπει στους ερευνητές την αναγνώριση και τον προσδιορισμό των «θερμών σημείων» της βιοποικιλότητας και συνεπώς προσφέρει ένα πολύτιμο εργαλείο για τον προσδιορισμό των προστατευόμενων περιοχών. Οι τελευταίες βιογεωγραφικές και οικολογικές προσπάθειες σε ευρωπαϊκό ή παγκόσμιο επίπεδο, βασισμένες στις κατανομές ψαριών των εσωτερικών υδάτων, δεν έχουν συμπεριλάβει δεδομένα από τη Βαλκανική χερσόνησο, αν και η περιοχή αυτή φιλοξενεί σημαντικό μέρος της Ευρωπαϊκής βιοποικιλότητας. Παράλληλα, οι έως σήμερα, διαθέσιμες μελέτες προσδιορισμού των βιογεωγραφικών περιοχών της Βαλκανικής, έχουν καταλήξει σε αντικρουόμενα πρότυπα, καθώς είτε έχουν επικεντρωθεί μόνο στο νότιο τμήμα της, είτε έχουν βασιστεί σε εμπειρικές προσεγγίσεις. Βάσει των ανωτέρων διαπιστώσεων, οι πρώτοι βασικοί στόχοι της παρούσας διδακτορικής διατριβής αφορούν την πρώτη συστηματική συγκέντρωση και διαχείριση των διαθέσιμων βιβλιογραφικών δεδομένων για τις κατανομές των ψαριών των εσωτερικών υδάτων στο σύνολο της περιοχής, στη συνέχεια την οριοθέτηση των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου και τέλος τη μελέτη των προτύπων του αριθμού των ειδών σε σχέση με τον ενδημισμό, των απειλούμενων με εξαφάνιση ειδών και την έκταση των οικοσυστημάτων. Έπειτα από ενδελεχή ανασκόπηση 364 βιβλιογραφικών πηγών και αξιολόγηση της εγκυρότητάς τους, δημιουργήθηκε η πιο εκτεταμένη και πλήρης βάση δεδομένων κατανομής παρουσίας- απουσίας για 193 αυτόχθονα είδη ψαριών σε 78 λεκάνες απορροής ποταμών και 19 φυσικές λίμνες της Βαλκανικής χερσονήσου. Μετά τη δημιουργία της βάσης δεδομένων, το επόμενο βήμα κατά την οριοθέτηση των βιογεωγραφικών περιοχών είναι η επιλογή της κατάλληλης μετρικής για τη δημιουργία της μήτρας των αποστάσεων μεταξύ των οικοσυστημάτων. Ανάμεσα στις μετρικές που χρησιμοποιούνται κατά την οριοθέτηση των βιογεωγραφικών περιοχών, οι δείκτες β-ποικιλότητας έχουν ευρέως χρησιμοποιηθεί στη βιβλιογραφία καθώς εκφράζουν την αλλαγή στη σύνθεση των ειδών μεταξύ δύο περιοχών. 30
31 Ειδικότερα, επιλέχθηκε ο δείκτης βjtu, για να εντοπιστούν οι κύριες βιογεωγραφικές περιοχές των Βαλκανίων με βάση την αλλαγή της σύνθεσης των ειδών καθώς δεν επηρεάζεται από τις διαφορές του αριθμού των ειδών ανά οικοσύστημα. Με βάση τη μήτρα ανομοιότητας που προέκυψε από την εφαρμογή του δείκτη, στη συνέχεια εφαρμόστηκε η ιεραρχική ανάλυση ομαδοποίησης κατά συστάδες (HCA) που αποτελεί την πιο διαδεδομένη στατιστική ανάλυση για τον καθορισμό διακριτών ομάδων. Για τον εντοπισμό του αριθμού των διακριτών ομάδων (διακριτές βιογεωγραφικές περιοχές), στο δενδρόγραμμα που προέκυψε, χρησιμοποιήθηκε το κριτήριο Kelley - Gardner - Sutcliffe. Τέλος, προσδιορίστηκαν τα είδη- ενδείκτες τα οποία μπορούν να χρησιμοποιηθούν ως εκπρόσωποι των βιογεωγραφικών περιοχών. Χρησιμοποιώντας την κατανομή των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου ανά λεκάνη απορροής ποταμού και λίμνη, υπολογίστηκαν τα ποσοστά των ενδημικών ειδών και ο αριθμός των ειδών που κινδυνεύουν με εξαφάνιση σύμφωνα με την Κόκκινη Λίστα της Διεθνούς Ένωσης για τη Διατήρηση της Φύσης (IUCN) ανά βιογεωγραφική περιοχή. Τέλος, τα οχτώ βασικότερα μαθηματικά μοντέλα που έχουν προταθεί έως σήμερα για την περιγραφή της σχέσης του αριθμού ειδών και της έκτασης (SAR) εφαρμόστηκαν στα δεδομένα κατανομών των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου ανά λεκάνη απορροής ποταμού και λίμνη, ώστε να προσδιοριστεί το μοντέλο που περιγράφει αποτελεσματικότερα τη σχέση, σύμφωνα με το διορθωμένο Akaike Κριτήριο Πληροφορίας (AICc). Από τα αποτελέσματα των ανωτέρω αναλύσεων προέκυψε ότι η Βαλκανική χερσόνησος είναι η πιο άφθονη περιοχή της Ευρώπης, φιλοξενώντας 193 αυτόχθονα ψάρια των εσωτερικών υδάτων, η οποία χαρακτηρίζεται από υψηλά επίπεδα ενδημισμού (56%) και μεγάλα ποσοστά απειλούμενων ειδών (33%). Στην αυτόχθονη και ενδημική ιχθυοπανίδα της Βαλκανικής χερσονήσου, τα Κυπρινοειδή είναι η επικρατέστερη οικογένεια. Μετά την εφαρμογή της μετρικής β-ποικιλότητας (βjtu) προσδιορίστηκαν οκτώ κύριες βιογεωγραφικές περιοχές: 1. Ιόνιο Πέλαγος, 2. Νότια Αδριατική θάλασσα, 3. Βόρεια Αδριατική θάλασσα. 4. Λεκάνη απορροής του ποταμού Neretva, 5. 31
32 Αττικοβοιωτία, 6. Αιγαίο Πέλαγος, 7. Ποντιακή επαρχία και 8. Πρέσπες. Αυτές οι βιογεωγραφικές περιοχές μπορούν να ταξινομηθούν περαιτέρω στη δυτική ομάδα (Ιόνιο Πέλαγος, Νότια και Βόρεια Αδριατική θάλασσα και λεκάνη απορροής του ποταμού Neretva), την ανατολική ομάδα (Αττικοβοιωτία, Αιγαίο Πέλαγος και Ποντιακή επαρχία) και την ανεξάρτητη ομάδα (Πρέσπες). Η μεγάλη ανομοιότητα (0.81 ± 0.22 SD) της σύνθεσης των ψαριών των εσωτερικών υδάτων αυτών των οκτώ βιογεωγραφικών περιοχών είναι το αποτέλεσμα των ανεξάρτητων βιογεωγραφικών ιστοριών τους. Πιο συγκεκριμένα, οι νότιες και δυτικές βιογεωγραφικές περιοχές των Βαλκανίων δεν καλύφτηκαν με παγετώνες κατά το Πλειστόκαινο και χαρακτηρίζονται από μεγαλύτερο βαθμό απομόνωσης, εξαιτίας της αλπικής ορογένεσης που εμπόδισε τη διασπορά των ψαριών προς τα βόρεια και ανατολικά. Επίσης, η ανατολική ομάδα χαρακτηρίζεται από παραδουνάβιας καταγωγής είδη τα οποία διασπάρθηκαν μέσω της ποτάμιας σύνδεσης ανάμεσα στο Δούναβη και τον Αξιό και των παλαιογεωγραφικών συνδέσεων του Αιγαίου με τη Μαύρη Θάλασσα. Η ανάλυσης προσδιορισμού των ειδών- ενδεικτών των βιογεωγραφικών περιοχών έδειξε ότι 33 είδη (17%) παρουσίασαν στατιστικά σημαντική συσχέτιση με τις βιογεωγραφικές περιοχές. Από τα 33 είδη, τα 28 ανήκουν στην οικογένεια των Κυπρινοειδών. Επιπλέον, τρία είδη του γένους Cobitis, δύο ενδημικοί Αμμογωβιοί (Knipowitschia croatica και Economidichthys pygmaeus) και το ενδημικό είδος Valencia letourneuxi, που ανήκει στη μοναδική ενδημική οικογένεια της Ευρώπης Valenciidae αναδείχθηκαν ως είδη-ενδείκτες της Βαλκανικής χερσονήσου. Σύμφωνα με τα αποτελέσματα της διαδικασίας επιλογής του καταλληλότερου μοντέλου για την περιγραφή της σχέσης αριθμού ειδών και έκτασης, το δυναμικό μοντέλο (Power) παρουσίασε τις καλύτερες επιδόσεις και καταλαμβάνει την πρώτη θέση σε όλες τις περιπτώσεις τόσο για το σύνολο των δεδομένων όσο και για τις 75 λεκάνες απορροής ποταμών, ξεχωριστά. Ωστόσο, το συνολικό ποσοστό ερμηνείας από το δυναμικό μοντέλο για το σύνολο των δεδομένων (R 2 <0.36) και για τις λεκάνες απορροής ποταμών (R 2 <0.52) ήταν αρκετά χαμηλό. 32
33 Οι περιοχές με τα υψηλότερα επίπεδα ενδημισμού είναι αυτές που παρουσιάζουν το χαμηλότερο αριθμό ειδών, ένα γεγονός που πιθανόν σχετίζεται με το φαινόμενο ή την επίδραση της χερσονήσου (peninsula effect), το μέγεθος των οικοσυστημάτων και την ασθενέστερη συνδεσιμότητα των λεκανών απορροής των νότιων περιοχών που απέτρεψε τη διασπορά των ειδών σε μεγάλες αποστάσεις, αυξάνοντας έτσι τον τοπικό ενδημισμό. Οι περιοχές της Αττικοβοιωτίας, του Ιονίου και των λιμνών των Πρεσπών χαρακτηρίζονται ως σημεία υψηλού ενδιαφέροντος καθώς φιλοξενούν τα υψηλότερα ποσοστά ενδημικών και απειλούμενων ειδών. Στις βιογεωγραφικές περιοχές του Αιγαίου Πελάγους, της νότιας Αδριατικής θάλασσας, της λεκάνης απορροής του ποταμού Neretva και της βόρειας Αδριατικής Θάλασσας, η ανάγκη για κοινές διασυνοριακές δράσεις διατήρησης μεταξύ των χωρών είναι επιτακτική, καθότι οι διοικητικές διαιρέσεις δε συμπίπτουν με αυτές τις βιογεωγραφικές περιοχές. Επομένως, η βιογεωγραφική ιστορία που σχετίζεται με τη γεωλογική εξέλιξη, η συνδεσιμότητα και η έκταση των οικοσυστημάτων είναι από τους πιο σημαντικούς παράγοντες καθορισμού των προτύπων της σύνθεσης και του αριθμού των αυτόχθονων και ενδημικών ειδών των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου. Δεδομένου ότι η αναγνώριση των προτύπων σε διαφορετικές ιεραρχικές χωρικές κλίμακες είναι απαραίτητη για την κατανόηση της διαμόρφωσης της βιοποικιλότητας εν γένει, περνώντας σε τοπικό επίπεδο, η παρούσα διατριβή έθεσε ως ένα επόμενο στόχο τη μελέτη των προτύπων ποικιλότητας, ενδημισμού και απειλούμενων με εξαφάνιση ειδών ψαριών στα Ελληνικά εσωτερικά ύδατα. Για την αξιολόγηση των ειδών που κινδυνεύουν με εξαφάνιση, υπολογίστηκε ο αριθμός των ειδών που αντιστοιχεί σε κάθε κατηγορία του καθεστώτος διατήρησης των ειδών σύμφωνα με τη Διεθνή Ένωση για τη Διατήρηση της Φύσης (IUCN) και το Κόκκινο Βιβλίο Απειλούμενων Ζώων της Ελλάδας. Επιπλέον, διερευνήθηκαν τα πρότυπα της διαβάθμισης της ποικιλότητας στα Ελληνικά λιμναία οικοσυστήματα διότι αποτελούν μια μοναδική ευκαιρία διερεύνησης της επίδρασης των περιβαλλοντικών παραγόντων στη διαμόρφωση του αριθμού και της κατανομής του μεγέθους των ειδών των ψαριών. Δημιουργήθηκε μία βάση δεδομένων κατανομής των ψαριών των εσωτερικών υδάτων της Ελλάδας, η οποία περιλαμβάνει δεδομένα από 42 αυτόχθονα είδη ψαριών σε 13 φυσικές λίμνες, με βάση τη γεωγραφική 33
34 αντιπροσωπευτικότητα και την πληρότητα των διαθέσιμων δεδομένων. Για κάθε λιμναίο οικοσύστημα, καταγράφηκαν ο αριθμός των ειδών των ψαριών και το μέγιστο μήκος των μόνιμων ειδών από βιβλιογραφικές πηγές. Το μέγιστο μήκος των ειδών επιλέχτηκε ως βιολογικό αντικατάστατο του μεγέθους του σώματος. Επίσης, καταγράφηκαν το γεωγραφικό πλάτος και μήκος, τέσσερα λιμνολογικά χαρακτηριστικά και έξι φυσικοχημικές παράμετροι του νερού. Για να ελεγχθεί ποιοι περιβαλλοντικοί παράγοντες ερμηνεύουν τη διακύμανση του αριθμού και του μεγέθους των ειδών των ψαριών σε Ελληνικές λίμνες, εφαρμόστηκαν πολλαπλές γραμμικές παλινδρομήσεις και το πιο αντιπροσωπευτικό σύνολο των μεταβλητών προσδιορίστηκε με την εφαρμογή του Akaike Κριτηρίου Πληροφορίας (AIC), έπειτα από έλεγχο της επίδρασης της συγγραμικότητας, με την εφαρμογή του συντελεστή πληθωρισμού διακύμανσης (VIF- Variation Inflation Factor). Η Ελληνική ιχθυοπανίδα των εσωτερικών υδάτων είναι η πιο πλούσια της Ευρώπης και χαρακτηρίζεται από πολύ υψηλά επίπεδα ενδημισμού (42% των αυτόχθονων ψαριών), κάτι που καταφαίνεται και από τον υψηλό ενδημισμό που παρουσιάζουν τα εσωτερικά ύδατα της Ελλάδας, ο οποίος σχετίζεται άμεσα με την ιδιαίτερα κατακερματισμένη γεωγραφική τους κατανομή και την έντονη γεωλογική δραστηριότητα κατά το Τριτογενές και το Τεταρτογενές. Η Ελληνική ιχθυοπανίδα αντιπροσωπεύεται από ένα σχετικά μικρό αριθμό οικογενειών, με τα περισσότερα είδη να ανήκουν στην οικογένεια των Κυπρινοειδών (53%). Τρία περιβαλλοντικά (υψόμετρο, έκταση, γεωγραφικό πλάτος) και πέντε φυσικοχημικά χαρακτηριστικά (αγωγιμότητα, ολικός φώσφορος, ph, διαλυμένο ανόργανο άζωτο, συγκέντρωση χλωροφύλλης-α) χρησιμοποιήθηκαν ως ανεξάρτητες μεταβλητές στη σταδιακή πολλαπλή παλινδρόμηση, έπειτα από τον έλεγχο της συγγραμικότητας. Το καταλληλότερο μοντέλο εμπεριέχει τις μεταβλητές του υψομέτρου, της έκτασης και του ολικού φωσφόρου και εξηγεί το 94% της διακύμανσης του αριθμού των ειδών. Ο ολικός φώσφορος, ο οποίος σχετίζεται έμμεσα με την πρωτογενή παραγωγικότητα, έχει τη μικρότερη συνεισφορά, αφού έπειτα από τη συμπερίληψή του, η επεξηγηματικότητα του μοντέλου αυξήθηκε κατά 4%. Πιο συγκεκριμένα, ο αριθμός των ψαριών στα λιμναία οικοσυστήματα της 34
35 Ελλάδας μειώνεται με την αύξηση του υψομέτρου και αυξάνεται με την αύξηση της έκτασης του οικοσυστήματος. Τα αποτελέσματα σε σχέση με το μέσο μέγεθος των ειδών των ψαριών, έδειξαν ότι το καταλληλότερο μοντέλο αφορά μόνο τη μεταβλητή του γεωγραφικού πλάτους και εξηγεί το 40% της διακύμανσης του μεγέθους των ειδών. Ως εκ τούτου, το μέγεθος του σώματος των ψαριών των εσωτερικών υδάτων στις Ελληνικές λίμνες αυξάνεται με το γεωγραφικό πλάτος, εξαιτίας της διαφορετικής ικανότητας διασποράς των ειδών και της διαφορετικής γεωλογικής ιστορίας των υπό μελέτη συστημάτων. Η βορειοανατολική Ελλάδα φιλοξενεί ευρέως κατανεμημένα, μεγαλόσωμα είδη με μεγαλύτερη ικανότητα εποικισμού στη μεταπαγετώδη περίοδο, σε σύγκριση με τη νοτιοδυτική, όπου διαβιούν μικρόσωμα ενδημικά είδη ή είδη με περιορισμένη κατανομή. Η δυτική-ανατολική διάκριση των Ελληνικών εσωτερικών υδάτων και η διαφορετική σύνθεση τους αποδίδεται στη μακροχρόνια παρουσία του γεωγραφικού εμπόδιου της Πίνδου και την επακόλουθη απομόνωση που προκλήθηκε. Η βιογεωγραφία και η μακροοικολογία βασίστηκαν αρχικά, ως πεδία έρευνας, στην ανάλυση της ταξινομικής ποικιλότητας και παραδοσιακά επικεντρώνονται σε μετρικές όπως ο αριθμός των ειδών. Πέρα όμως από το περιφερειακό και τοπικό επίπεδο μελέτης των βιογεωγραφικών και μακροοικολογικών προτύπων, ειδικότερα από το τέλος του περασμένου αιώνα, αναδύθηκε η ανάγκη μελέτης της λειτουργικής ποικιλότητας των βιοκοινοτήτων, δηλαδή του αριθμού, του τύπου και της κατανομής των λειτουργιών που εκτελούνται από τους οργανισμούς μέσα σε ένα οικοσύστημα. Ως αποτέλεσμα της μακράς εξελικτικής τους ιστορίας, της υψηλής ποικιλότητας και των πολύπλοκων στρατηγικών ζωής στα διάφορα οντογενετικά τους στάδια, τα ψάρια των εσωτερικών υδάτων είναι ιδανικά για την εξέταση της σχέσης μεταξύ των χαρακτηριστικών τους με τη χρήση των ενδιαιτημάτων. Για τη διερεύνηση, επομένως, των προτύπων της λειτουργικής ποικιλότητας και των συσχετίσεων μεταξύ των περιβαλλοντικών χαρακτηριστικών και της λειτουργικής σύνθεσης των ιχθυοκοινοτήτων, ο ποταμός Λούρος επιλέχθηκε ως σύστημα μελέτης διότι αποτελεί ένα τυπικό πεδινό ομαλού πηγαίου τύπου ποτάμιο οικοσύστημα. Ανήκει στην Ιόνια βιογεωγραφική περιοχή, είναι ένα από τα λιγότερο 35
36 μελετημένα οικοσυστήματα στην Ελλάδα, με μακρά ιστορία ανθρώπινης παρουσίας και παρέμβασης. Χαρακτηρίζεται από αυξημένη ποικιλότητα μεσογειακών ποτάμιων ενδιαιτημάτων, τα οποία φιλοξενούν εξελικτικά απομονωμένη ιχθυοπανίδα, η οποία χαρακτηρίζεται από χαμηλό αριθμό ειδών, αλλά ταυτόχρονα από υψηλά επίπεδα ενδημισμού και μεγάλο αριθμό απειλούμενων ειδών. Τα δεδομένα για τη διερεύνηση των προτύπων λειτουργικής ποικιλότητας προέκυψαν έπειτα από τη συλλογή του ιχθυολογικού υλικού και τη μέτρηση των περιβαλλοντικών παραμέτρων σε 9 σταθμούς στο σύστημα μελέτης, σε διμηνιαία βάση για ένα έτος. Από κάθε άτομο ψαριού λήφθηκαν 18 μορφολογικές μετρήσεις που οδήγησαν στην εκτίμηση 13 οικο-μορφολογικών χαρακτηριστικών που σχετίζονται με τις λειτουργίες της σύλληψης τροφής και της μετακίνησης. Οι τελικές μετρήσεις των λειτουργικών χαρακτηριστικών κάθε είδους προέκυψαν από το μέσο όρο των τιμών των χαρακτηριστικών όλων των ενήλικων ατόμων. Για την ανάλυση των δεδομένων της λειτουργικής ποικιλότητας, κατασκευάστηκαν και χρησιμοποιήθηκαν τρεις πίνακες, ο πίνακας των περιβαλλοντικών παραμέτρων που χαρακτηρίζουν κάθε σταθμό, ο πίνακας της αφθονίας των ειδών σε κάθε σταθμό και ο πίνακας των λειτουργικών χαρακτηριστικών των ειδών. Στόχος της διδακτορικής διατριβής, ως προς τη λειτουργική ποικιλότητα στον ποταμό Λούρο, ήταν η διερεύνηση των συσχετίσεων μεταξύ των περιβαλλοντικών χαρακτηριστικών και της λειτουργικής σύνθεσης των ιχθυοκοινοτήτων. Η ανάλυση κύριων συνιστωσών (PCA) απεικόνισε τις περιβαλλοντικές διαβαθμίσεις στον ποταμό Λούρο και αποτύπωσε το λειτουργικό χώρο των ειδών στο δισδιάστατο επίπεδο. Στη συνέχεια, η πολυμεταβλητή ανάλυση διακύμανσης με μετάθεση (permanova) χρησιμοποιήθηκε προκειμένου να διαπιστωθεί ποιες είναι οι στατιστικά σημαντικότερες περιβαλλοντικές παράμετροι που διαμορφώνουν τη σύνθεση και την αφθονία των ειδών. Για να διερευνηθεί η υπόθεση ότι τα λειτουργικά χαρακτηριστικά ενός είδους σχετίζονται με τις συγκεκριμένες περιβαλλοντικές συνθήκες όπου διαβιεί, ακολουθήθηκε η μοναδική μεθοδολογική προσέγγιση που έχει προταθεί μέχρι σήμερα που συνιστά την εφαρμογή της πολυμεταβλητής ανάλυσης RLQ και της συμπληρωματικής μεθόδου fourth-corner. Η 36
37 στατιστική σημαντικότητα των συσχετίσεων που προέκυψαν από αυτές τις δύο αναλύσεις αξιολογήθηκε μέσω της τυχαιοποίησης με µεταθέσεις. Η λειτουργική ποικιλότητα κάθε ιχθυοκοινότητας υπολογίστηκε με τους δείκτες του λειτουργικού πλούτου (FRic), της λειτουργικής ισομέρειας (FEve), της λειτουργικής απόκλισης (FDiv), της λειτουργικής διασποράς (FDis), του δείκτη του Rao (Q) και απεικονίστηκε γραφικά με τη χρήση του δείκτη του πλούτου των λειτουργικών ομάδων (FGR). Για να εξεταστεί αν οι δείκτες της λειτουργικής ποικιλότητας επηρεάζονται από τον αριθμό των ειδών στις ιχθυοκοινότητες και κατά συνέπεια, αν η λειτουργική ποικιλότητα αυξάνεται με την προσθήκη περισσότερων ειδών, εφαρμόστηκαν γραμμικές παλινδρομήσεις μεταξύ των δεικτών και του αριθμού των ειδών. Σε συνέχεια των αναλύσεων της λειτουργικής ποικιλότητας, αναδείχθηκαν τα πιο οικο-μορφολογικά διακριτά είδη του ποταμού Λούρου που χρήζουν άμεσης προστασίας, μέσω του υπολογισμού των επτά δεικτών της λειτουργικής μοναδικότητας των ειδών που έχουν προταθεί έως σήμερα. Τέλος, ελέγχθηκε η επίδραση των δύο βασικών και αντίρροπων διεργασιών που αφορούν τη διαμόρφωση των ιχθυοκοινοτήτων, με βάση τα λειτουργικά γνωρίσματα των ειδών, το περιβαλλοντικό φιλτράρισμα και τις βιοτικές αλληλεπιδράσεις. Ακολουθώντας τη μεθοδολογική προσέγγιση που εφαρμόζεται σε φυλογενετικές αναλύσεις, πραγματοποιήθηκε σύγκριση μεταξύ της λειτουργικής ποικιλότητας των πραγματικών ιχθυοκοινοτήτων του ποταμού Λούρου και της λειτουργικής ποικιλότητας τυχαίων ιχθυοκοινοτήτων που προέκυψαν από το περιφερειακό αποθεματικό των ειδών. Τα αποτελέσματα των αναλύσεων έδειξαν ότι τα ενδιαιτήματα του ποταμού Λούρου χαρακτηρίζονται από αυξανόμενο μέσο βάθος, από χαμηλότερο υψόμετρο και από υπόστρωμα λεπτόκοκκου ιζήματος όσο προχωράμε από τις πηγές προς τις εκβολές του ποταμού. Από το σύνολο των περιβαλλοντικών παραμέτρων που εξετάστηκαν, το πλάτος, το μέσο βάθος και ο τύπος υποστρώματος (χονδρόκοκκο), ερμηνεύουν τη μεγαλύτερη μεταβλητότητα της αφθονίας των ιχθυοκοινοτήτων. Από τις αναλύσεις της λειτουργικής ποικιλότητας, προέκυψε ότι τα είδη των ψαριών του ποταμού Λούρου διαχωρίστηκαν κυρίως ως προς το σχήμα του ουραίου 37
38 πτερυγίου, τη θέση των θωρακικών πτερυγίων τους στον κάθετο άξονα του σώματός τους, το σχήμα του σώματος και τη σχετική θέση του στόματος ως προς τον κάθετο άξονα της κεφαλής. Επιπλέον, οι περιβαλλοντικοί παράγοντες περιορίζουν την κατανομή ειδών με συγκεκριμένα λειτουργικά χαρακτηριστικά οδηγώντας σε διαφορετικά επίπεδα λειτουργικής ποικιλότητας των ιχθυοκοινοτήτων στον ποταμό Λούρο. Οι σταθμοί ανάντη χαρακτηρίζονται από μικρό βάθος και χοντρόκοκκο υπόστρωμα και φιλοξενούν είδη με πιο υδροδυναμικό σχήμα σώματος, μεγαλύτερη κολυμβητική αντοχή και διχαλωτή ουρά. Αντίθετα, οι σταθμοί κατάντη χαρακτηρίζονται από αυξημένο λειτουργικό χώρο, καθώς φιλοξενούν είδη από ένα ευρύ φάσμα οικογενειών και με ακραίες τιμές των οικομορφολογικών χαρακτηριστικών τους. Για παράδειγμα, τα απειλούμενα είδη Valencia letourneuxi (Ζουρνάς) και Cobitis hellenica (Λουροβελονίτσα) χαρακτηρίζονται από την υψηλότερη λειτουργική μοναδικότητα μεταξύ των ειδών που διαβιούν στον ποταμό Λούρο και χρήζουν άμεσων διαχειριστικών ενεργειών, διότι κάθε παρέμβαση ή και καταστροφή του ενδιαιτήματός τους, μπορεί να έχει άμεσες και σημαντικές επιπτώσεις στους πληθυσμούς τους. Τέλος, η λειτουργική ποικιλότητα στον ποταμό Λούρο, κατά κανόνα αυξάνεται με την αύξηση του αριθμού των ειδών, καθώς η αύξηση του αριθμού των ειδών συνεπάγεται αύξηση της ποικιλότητας των λειτουργικών χαρακτηριστικών των ιχθυοκοινοτήτων. Εντούτοις, σημαντικό ρόλο παίζει και η σύνθεση των ειδών, όπως σε περιπτώσεις σταθμών με όμοια λειτουργικά είδη, όπου παρά την αυξημένη ταξινομική ποικιλότητα, η λειτουργική ποικιλότητα είναι χαμηλότερη. Οι ιχθυοκοινότητες ανάντη χαρακτηρίζονται από υψηλά επίπεδα πλεονασμού ως προς τη σύνθεση των ειδών και είναι λειτουργικά ομαδοποιημένες σε σύγκριση με τυχαίες ιχθυοκοινότητες που θα προέκυπταν από το περιφερειακό αποθεματικό των ειδών. Αυτό το πρότυπο αναδεικνύει το περιβαλλοντικό φιλτράρισμα ως το κυρίαρχο μηχανισμό συγκρότησης των ιχθυοκοινοτήτων στον ποταμό Λούρο. Εν κατακλείδι, η εκτίμηση της λειτουργικής ποικιλότητας στα ποτάμια οικοσυστήματα είναι απαραίτητη τόσο για την πληρέστερη μελέτη της βιοποικιλότητας όσο και τη βελτίωση της αποτελεσματικότητας των προσπαθειών διατήρησης των σπάνιων ειδών και των οικοσυστημάτων που χρήζουν άμεσης 38
39 προστασίας. Η μεθοδολογική προσέγγιση που ακολουθήθηκε καθώς και τα πρότυπα που περιγράφηκαν στην παρούσα διατριβή θα μπορούσαν να συμβάλουν στη σύνθεση των θεωριών της οικολογίας βιοκοινοτήτων και να αποτελέσουν τη βάση για τη δημιουργία διαχειριστικών εργαλείων που βρίσκουν εφαρμογή σε διαφορετικές ταξινομικές ομάδες και γεωγραφικές περιοχές. Η παρούσα διδακτορική διατριβή υπογραμμίζει επίσης την επείγουσα ανάγκη για τη δημιουργία βάσεων δεδομένων σχετικά με τα λειτουργικά χαρακτηριστικά των ιχθυοκοινοτήτων. Η παρούσα μελέτη εκτίμησης της βιοποικιλότητας σε ταξινομικό και λειτουργικό επίπεδο μπορεί να συνεισφέρει στη διασφάλιση της μακροπρόθεσμης διατήρησης της βιοποικιλότητας της Βαλκανικής χερσονήσου, και ως εκ τούτου, τα αποτελέσματα της έχουν δυνητικά, αντίκτυπο στην ευρωπαϊκή ερευνητική ατζέντα σχετικά με τα οικοσυστήματα των εσωτερικών υδάτων. Τόσο οι μεθοδολογικές προσεγγίσεις όσο και τα αποτελέσματα της παρούσας διδακτορικής διατριβής θα συνεισφέρουν σε μελλοντικές μελέτες σχετικά με τη βιοποικιλότητα και τη λειτουργία των οικοσυστημάτων των εσωτερικών υδάτων, καθώς και σε μελέτες που θα διερευνήσουν τη λειτουργική ποικιλότητα ειδών και βιοκοινοτήτων σε πολλαπλά χωρικά επίπεδα, με στόχο μια ολιστική κατανόηση της βιοποικιλότητας, κυρίως στη λιγότερο μελετημένη Βαλκανική Χερσόνησο. 39
40
41 Extended abstract One of the fundamental aims of ecology, biogeography and macroecology is the study of the mechanisms that establish the distributional patterns of organisms and species diversity. The last decades, the study of biodiversity at multiple spatial scales is continuously gaining space to international literature pointing towards to a more holistic understanding of biodiversity and thus, to a more effective conservation strategy. Freshwater ecosystems are an excellent model system for introducing and testing ecological theories since they host an extremely high percentage of global biodiversity. While occupying only 0.8% of Earth s surface, it is estimated that they harbor at least 12% of the global animal species richness. At the same time, freshwater ecosystems disproportionately host the highest percentage of imperiled faunas worldwide, as a large proportion of the habitats of these faunas has been lost or has been degraded by intense human activities. Amongst the taxa found in freshwater ecosystems, fish are the ideal organismmodel for testing and refining shared biogeographical histories, macroecological patterns and ecological hypotheses, since they are considered as very good biological indicators of water bodies isolation, in space and time. Additionally, freshwater fish are good zoogeographical indicators because they cannot cross the saltwater barrier, inherently they have reduced dispersal ability and their distribution is usually confined to drainage, leading to high local endemism. As a result of their high diversity, well-resolved systematics and welldocumented geographical distribution, fish have been widely used for the delineation of biogeographic regions of freshwaters. The delineation of biogeographical regions aims to identify different regions based on biotic similarity, and therefore, the ecosystems within each biogeographic region are expected to present higher 41
42 similarity in terms of their biotic composition, compared to ecosystems of other biogeographic regions. The delineation of biogeographic regions is a primary step towards investigating of biogeographic patterns. In combination with the spatial patterns of species diversity, endemism and the proportion of endangered species, it can provide a robust framework for the identification of biodiversity hotspots and thus, provide a valuable tool for delimitating areas in need of protection and necessary actions for their protection. Recent biogeographic and ecological efforts for large-scale analyses of freshwater fish at the European and the global scale have excluded the Balkan Peninsula, although it is considered as a European hotspot, mainly to the absence of adequate data. At the same time, previous ecoregional mapping studies in the Balkan Peninsula either focused only on the southern part of the peninsula or have been based on empirical approaches. Therefore, the first and major objective of the current thesis refers to the systematic collection and analysis of the available bibliographic data on freshwater fish occurrences, the delineation of biogeographic regions of the Balkan Peninsula and finally, the study of species richness patterns taking into account endemism, endangered species and area of ecosystems. Based upon the most extensive literature survey conducted to date, involving 364 bibliographical sources, and after the evaluation of their completeness, we established the most extensive and complete database of the freshwater fishes of the Balkan Peninsula, comprising of 193 native fish species and their presence in 78 river basins and 19 natural lakes. The next step for delimitating the biogeographic regions is the selection of the appropriate metric, through which beta diversity or turnover pairwise distances between ecosystems are estimated. A large number of such indices is available in the literature to estimate change in community composition. In the current study, we applied a compositional dissimilarity index that measures species replacement without any influence of differences in species richness, specifically the bjtu index, to identify the main biogeographic regions of the Balkans. Based on the compositional dissimilarity matrix, we applied the hierarchical clustering analysis (HCA). We employed the Kelley Gardner Sutcliffe penalty function (KGS) to identify distinct clusters of drainage basins (distinct biogeographical regions). Finally, we 42
43 performed the indicator species analysis, which consists of a list of species that are significantly associated with a given cluster and are often used as ecological representatives of biogeographic regions. Using the database of occurrences of 193 native fish species, we quantified the relative degree of endemism and the number of species assigned to each Red List category (IUCN) for each of the previously identified biogeographical regions. Finally, we applied the eight most-widely applied mathematical models that have been proposed to describe the species-area relationship (SAR), to the distributional data of freshwater fish of the Balkan Peninsula. Species-area models fit was compared using the corrected Akaike s information criterion (AICc). The results showed that the Balkan Peninsula is the most speciose region of Europe, hosting 193 native freshwater fish and is characterized by high levels of endemism (56%), while a high proportion of the fish fauna of the area is now threatened with extinction (33%). Cyprinids dominate the freshwater native and endemic fish fauna of the Balkan Peninsula. After the application of the richness-independent measure of species turnover, i.e., the bjtu index, eight main biogeographic regions were identified: 1. Ionian Sea, 2. southern Adriatic Sea, 3. northern Adriatic Sea, 4. Neretva watershed, 5. Attikobeotia, 6. Aegean Sea, 7. Pontic province and 8. Prespa Lakes. These eight biogeographical regions can be further classified into western (Ionian Sea, southern Adriatic Sea, Neretva watershed and northern Adriatic Sea), eastern (Attikobeotia, Aegean Sea and Pontic Province) and independent (Prespa Lakes) groupings. The high compositional dissimilarity (0.81 ± 0.22 SD) of these biogeographical regions results from their independent biogeographical histories. More specifically, the southern and western biogeographic regions of the Balkans were unglaciated during the Pleistocene and are characterized by a high degree of isolation mainly due to the Alpine orogenesis, which prevented most of freshwater species from moving east and northwards. The eastern group is represented by species of Danubian origin which dispersed, during the paleogeographical connection between the Black and Aegean Sea but also because Danube and Axios rivers were inter-connected. 43
44 The results of the indicator species analysis have shown that 33 species (17%) were associated significantly with the biogeographic regions; 28 out of the 33 species belong to the Cyprinidae family. In total, three Cobitis species, two endemic freshwater sand gobies (Knipowitschia croatica and Economidichthys pygmaeus) and the endemic species Valencia letourneuxi, belonging to the endemic Mediterranean family Valenciidae, were typical of the Balkan Peninsula. According to the results of the model selection, the power model is the most appropriate for describing the species-area relationship and ranks as first in all cases, i.e. the entire data and the 75 river basins separately, although the R 2 was quite low (R 2 =0.36, R 2 =0.52, respectively). In the Balkan Peninsula, the areas with the highest endemism are the speciespoorest areas, a pattern that can be assigned to the peninsula effect, the size of ecosystems and the weaker connectivity of drainage basins which prevented longdistance dispersal, thus increasing local endemism. The biogeographic regions of Attikobeotia, Ionian Sea and Prespa Lakes are biodiversity hotspots because of their high endemism and at the same time the high vulnerability of the species they host. Therefore, the biogeographic history related to the geological events, the connectivity and the area of ecosystems are among the most important factors determining the patterns of species composition, richness and endemism of the Balkan freshwater fish fauna. Since the elucidation of biogeographical patterns at different hierarchical spatial scales is critical for understanding species richness gradients, the next aim of this thesis was the study of spatial patterns of richness, endemism and vulnerability on a more local scale; i.e. Greece s freshwater fishes. We first identified patterns of endemism and the Greek fish species at a risk of extinction, by quantifying the number of species assigned to each Red List category according to the International Union for Conservation of Nature (IUCN) and the Red Data Book of Endangered Animals of Greece. Moreover, Greek lacustrine ecosystems were used for the study of certain biodiversity patterns, since they provide a unique opportunity to investigate the influence of environmental factors on shaping species richness along with the variation of freshwater fishes body size at the ecosystem level. 44
45 We created a database of 42 native freshwater fish species occurences distributed in 13 natural lakes, based on the criteria of geographical representation and completeness of available data. For each lake ecosystem, we assembled datasets of fish species richness, maximum length of resident species, latitude and longitude and four limnological and six water physicochemical parameters from an extensive literature search. Species maximum length was used as surrogate of body size. Multiple linear regressions were applied to determine the best set of explanatory variables for species richness and body size. We checked for multicollinearity among factors using a diagnostic measure of collinearity, the Variation Inflation Factor (VIF) and we chose the best model based on the Akaike Information Criterion (AIC). The Greek freshwater fish fauna is the most speciose in Europe and is characterized by high levels of endemism (42% of native fish), a pattern which is related to the highly fragmented hydrographic networks and the intense geological activity during the Tertiary and Quaternary. The Greek freshwater fish fauna is represented by a relatively small number of families, with most species belonging to Cyprinidae (53%). Three environmental (altitude, area, latitude) and five physicochemical (conductivity, total phosphorus, ph, dissolved inorganic nitrogen and chlorophyll-a) variables were selected according to AIC values, after the calculation of VIF. The bestfitted model contains the variables of altitude, area and total phosphorus and explains 94% of variation in species richness. Total phosphorus, which is indirectly related to primary productivity, has the weakest influence, increasing model s predictive power by just 4%. Thus, the two main variables selected were area and altitude; fish species richness in Greek lakes decreases with increasing altitude and increases in ecosystems with larger area. Our results identified latitude as the best explanatory variable of the variation in fish body size (R 2 =0.40). Therefore, body size of Greek lacustrine fish increases with latitude. The observed pattern can be assigned to species different dispersal ability and the influence of the major biogeographical events that took place in the studied systems. Northeastern Greece hosts widely distributed large-bodied fish species showing greater ability of postglacial colonization, compared to the southern 45
46 ecosystems which harbor small-bodied species which are either endemic or species exhibit restricted distributions. The west-east division of Greek freshwaters and consequently, their different species composition are the result of the longstanding geographical barriers to dispersal, such as the Pindos mountain range. Βiogeography and macroecology have been based initially on taxonomic diversity and have focused traditionally on measures such as species richness. However, it is increasingly recognized that the concept of biodiversity is multidimensional, including taxonomic, phylogenetic, and functional dimensions. Herein, apart from the study of biogeographical and macroecological patterns at the regional and local scale, we also evaluated the role of functional trait diversity, i.e. the number, type and distribution of functions performed by the organisms within an ecosystem. As a result of their long evolutionary history, high diversity and complex life strategies in their different ontogenetic stages, freshwater fish are ideal for examining trait environment relationships. To study functional diversity patterns and trait environment relationships of freshwater fish, Louros river was selected as the study system. It is a typical spring fed, lowland riverine ecosystem, belonging to the Ionian biogeographical region and is one of the Greek understudied ecosystems, with a long history of human presence and intervention. Louros river is characterized by high diversity of Mediterranean riverine habitats and a generally isolated fish fauna showing low species richness, but at the same consisting of a large number of endemic and endangered species. For the study of functional diversity patterns, we collected fish samples and recorded environmental data in nine riverine sampling stations, on a bimonthly basis for one year. A set of 18 measurements of morphological traits of individuals was taken from each specimen and was the basis for the quantification of 13 ecomorphological characteristics, associated with the functions of locomotion and food acquisition. The functional traits values were estimated as the average of the measurements on all adult individuals collected from each species. For functional diversity analysis, we constructed and used three major tables; the first matrix contained species abundances at the sampling sites, the second matrix displayed 46
47 environmental data of each station and the third matrix referred to the functional trait values of fish species. Regarding the functional diversity in Louros river, the main aim was the evaluation of the relationship between the functional composition of fish communities with the environmental factors. Principal component analysis (PCA) was applied to describe environmental gradients in Louros river and functional space of fish species, in the two-dimensional space. Then, permutational multivariate analysis of variance (permanova) was performed to determine which environmental parameters affect the composition and abundance of fish species. We tested the hypothesis that functional traits determine the spatial distribution of fish species by assessing the relationship between fish distributions, fish species traits and habitat environmental characteristics, following the only methodological approach available to date, which is the application of the multivariate RLQ analysis with the additional fourth-corner method. The statistical significance of the relationships between traits and environmental variables was tested by permutations. Trait diversity was calculated for each fish community using five functional diversity indices: functional richness (FRic), functional eveness (FEve), functional divergence (FDiv), functional dispersion (FDis) and Rao s quadratic entropy (Q) and was plotted using a posteriori functional group richness index (FGR). To test if functional diversity indices are influenced by species richness in fish communities and therefore, if functional diversity increases by the addition of species, linear regressions were applied. In addition to the functional diversity analysis, seven indices that have been developed to estimate the originality of species were calculated to highlight the most eco-morphologically distinct species of Louros river, in order to provide an hierarchical classification of species priority to protection measures. Finally, we tested the influence of the fundamental and opposing processes structuring fish communities in terms of functional traits; environmental filtering and biotic interactions. Following the methodological approach used in phylogenetic analyses, we measured trait dissimilarity among co-occurring species, and compared the observed trait diversity to a null model of a random community drawn by the available regional species pool. 47
48 Our results showed that Louros riverine habitats are characterized by increased mean depth, lower altitude and substrate of finer sediment as we move downstream. Among the environmental variables studied, width, mean depth and substrate type (rocky) explain most of fish abundances variation. Functional diversity analysis revealed that fish species in Louros river are separated mainly by traits such as the aspect ratio of the caudal fin, pectoral fin position, body transversal shape and oral gape position. Furthermore, environmental factors limit the distribution of species with specific functional traits resulting in different levels of functional diversity of fish communities in Louros river. The stations upstream that are related to shallow mean depth and coarse substrate are associated with fish species with a more streamlined body shape, swimming endurance and a forked caudal fin. On the contrary, the stations downstream that are characterized by increased functional space, host fish species from a wide range of families and with extreme values of their ecomorphological traits. The endangered species Valencia letourneuxi (Zournas) and Cobitis hellenica (Lourovelonitsa) show the highest functional uniqueness among Louros fish fauna and require immediate management actions, since any intervention or destruction of their habitat may have direct and significant impact on their populations size and thus survival ability. Ιn general, functional diversity increases with species richness, since each species added to a community results in an increase of trait diversity. Nevertheless, species composition plays a significant role in cases where functionally similar species coexist, for instance, in stations with high taxonomic diversity but low functional diversity. Upstream fish communities are characterized by high levels of trait redundancy and species traits are packed compared to random fish communities drawn by the regional species pool. This pattern shows that environmental filtering is the dominant mechanism structuring Louros riverine fish fauna upstream. In conclusion, functional diversity assessment in riverine ecosystems is essential both for the adequate study of biodiversity and the effectiveness of conservation efforts for both rare species and ecosystems in need of immediate protection. The methodological approach followed and the patterns described in this 48
49 thesis could contribute further to community ecology theory and its application to conservation actions taken upon different taxonomic groups and geographical regions. For example, our results give prominence to joint transboundary conservation efforts among stakeholders to increase the validity of the conservation actions in the Aegean Sea, southern Adriatic Sea, Neretva Watershed and northern Adriatic Sea regions, since administrative divisions do not coincide with these biogeographical regions. This thesis also highlights the need for a world-wide database on fish functional traits. Finally, the current study on the taxonomic and functional components of biodiversity can contribute to the development of a longterm research agenda for the conservation of the Balkan Peninsula s biodiversity; in the wider context of the biodiversity of the European freshwater ecosystems. 49
50
51 Structure of the thesis- Outline of chapters In the first chapter of the thesis (Chapter 1), the theoretical framework and terminology used thereafter are introduced. The structure and the aims of this thesis have been inspired and based on the facts that: 1) ecologists have begun to give more attention to biogeographic patterns over a hierarchy of spatial scales (Ricklefs, 2004) 2) biodiversity is a multifaceted concept and comparative analyses focusing on different components of biodiversity (taxonomic diversity, functional diversity, genetic and ecosystem diversity) are needed (Loreau et al. 2001) 3) the multifaceted approach may unravel the different drivers of community structure (e.g. competition or environmental filtering) (Webb et al. 2002) 4) although freshwater ecosystems present unique opportunities to test biogeographical patterns and hypotheses, freshwater fishes have been used in few biogeographic studies (Olden et al. 2010) 5) recent biogeographic and ecological efforts for large-scale analyses of freshwater fish at the European and the global scale have excluded the Balkan Peninsula, although it is considered as a European hotspot, mainly to the absence of adequate data At the regional level, the first and major objectives of the current thesis refer to the systematic collection and analysis of the available bibliographic data on freshwater fish occurrences and the delineation of biogeographic regions of the Balkan Peninsula (Chapter 2). Following up from the delineation of biogeographic regions, fish community composition, patterns of species richness, species-area relationships, levels of endemic and endangered species are described (Chapter 3). 51
. The next aim of this thesis was the study of spatial patterns of richness, endemism and vulnerability on a more local scale; i.e. Greece s freshwater fishes.")
52 It is increasingly apparent that knowledge of the roles of processes in shaping patterns at different spatial scales is at the very heart of an understanding of variation in biodiversity (Gaston, 2000). The next aim of this thesis was the study of spatial patterns of richness, endemism and vulnerability on a more local scale; i.e. Greece s freshwater fishes. Furthermore, Greek lacustrine ecosystems were used to investigate the influence of environmental factors on shaping species richness along with the variation of freshwater fishes body size at the ecosystem level (Chapter 4). The assessment of biodiversity patterns across local, environmental gradients is important to providing insights into ecological interactions and processes that regulate species diversity at the community level. At the same time, we acknowledge that conservation planning has traditionally focused on identifying priority areas that ensure taxonomic diversity, using metrics such as species richness (see Chapters 3 52
53 and 4). However, it is now well established that the functional role of species and their functional interactions with the environment and other species, are amongst the key ecological and evolutionary determinants of biodiversity, overall. Subsequently, the impact of the global loss of biodiversity is increasingly attributed to the loss of functional rather than taxonomic diversity. So far, the exploration of the functional diversity of the freshwater fishes of Greece has been limited. Therefore, to study functional diversity patterns and trait environment relationships of freshwater fish (Chapter 5), Louros river, a typical spring-fed lowland river, was selected as the casestudy system. We tested the hypothesis that functional traits determine the spatial distribution of fish species by assessing the relationship among fish distributions, fish species traits and the habitat environmental characteristics. We also tested the influence of the fundamental and opposing processes structuring fish communities functional diversity, i.e.; environmental filtering and biotic interactions. Finally, we also calculated the originality of each single species to highlight the most ecomorphologically distinct species of Louros river, in order to provide an hierarchical classification of species priority to conservation and protection measures. The main conclusions of the thesis are presented in Chapter 6. This work is innovative in assessing biodiversity through multiple metrics, e.g. species numbers and functional diversity, along with the multi-scale approach of biodiversity patterns. It is now well-established that single-scale and single-metric studies of biodiversity are not able to describe the multi-facet nature of biodiversity. Our understanding of species richness and functional diversity patterns at different spatial scales is of extreme importance if we want to answer the questions that are currently being asked by society, including the present and future effects of invasive species and natural habitats fragmentation on the maintenance of aquatic biodiversity. 53
54
55 Κεφάλαιο 1 Γενική εισαγωγή Most of these recent calls for increasing the spatiotemporal scale of ecological investigations could be encapsulated under the new research field of macroecology (Brown 1995), which urges ecologists to think big. (Hugueny, Oberdorff & Tedesco 2010)
56
57 1.1 Περί βιογεωγραφίας και μακροοικολογίας Η διερεύνηση και αποκάλυψη των βαθύτερων αιτιών που διαμορφώνουν τα διαφορετικά χωρικά πρότυπα διαβάθμισης της βιοποικιλότητας παραμένει ένας από τους πλέον θεμελιώδεις στόχους της Οικολογίας από τον 19 ο αιώνα (Darwin 1839, Wallace 1878) μέχρι σήμερα. Η βιογεωγραφία αποτελεί τον επιστημονικό κλάδο που διερευνά τα χωρικά πρότυπα και εντοπίζει τις διεργασίες (τους αιτιακούς μηχανισμούς παραγωγής των προτύπων) της βιοποικιλότητας 1, μελετώντας την κατανομή των οργανισμών, στο παρελθόν και στο παρόν. Σήμερα, ο κλάδος της βιογεωγραφίας χωρίζεται σε τρεις κύριους τομείς ανάλογα με το επιστημονικό πεδίο μελέτης και αναφοράς: την ιστορική βιογεωγραφία, την οικολογική βιογεωγραφία και τη βιογεωγραφία διατήρησης. I. Η ιστορική βιογεωγραφία (ή αλλιώς παλαιοβιογεωγραφία) μελετά τις παρελθοντικές κατανομές των ειδών, εξετάζοντας την εξελικτική ιστορία τους, ενώ συχνά χρησιμοποιεί και πληροφορίες από απολιθώματα ειδών καθώς και τις κλιματικές συνθήκες για να ερμηνεύσει τα σημερινά πρότυπα της ποικιλότητας (π.χ. Araújo et al. 2008, Hortal et al. 2011, Dias et al. 2014). Διερευνά την προέλευση, τη διασπορά όπως και την εξαφάνιση των τάξων. Με άλλα λόγια, η ιστορική βιογεωγραφία στηρίζεται κυρίως στην επίδραση της φυσικής απομόνωσης και της εξελικτικής διαφοροποίησης των ειδών (βλέπε Darwin 1859). II. Η οικολογική βιογεωγραφία εξετάζει τους οικολογικούς παράγοντες που καθορίζουν τη χωρική κατανομή των ειδών σήμερα (π.χ. Bonada et al. 2005). Αυτοί οι παράγοντες μπορούν να χωριστούν σε βιοτικούς (σχέσεις ανάμεσα στους οργανισμούς) και σε αβιοτικούς (επίδραση των περιβαλλοντικών παραγόντων στους οργανισμούς π.χ. θερμοκρασία, υγρασία, αλατότητα). 1 Ο όρος βιοποικιλότητα σύμφωνα με τη Σύμβαση για τη Βιοποικιλότητα, στην οποία κατέληξε η Συνδιάσκεψη των Ηνωμένων Εθνών για το Περιβάλλον και την Ανάπτυξη, στο Ρίο ντε Τζανέιρο το 1992, έχει αποδοθεί ως «η παραλλακτικότητα («variability») των ζωντανών οργανισμών κάθε προέλευσης, συμπεριλαμβανομένων, μεταξύ άλλων, των χερσαίων, θαλασσίων, και άλλων υδάτινων οικοσυστημάτων, και οικολογικών συμπλεγμάτων των οποίων αποτελούν τμήμα. Στον όρο βιοποικιλότητα συμπεριλαμβάνεται η ποικιλότητα εντός των ειδών, μεταξύ ειδών καθώς και η ποικιλότητα των οικοσυστημάτων» 57
58 III. Η βιογεωγραφία διατήρησης προτάθηκε ως κλάδος της βιογεωγραφίας μόλις το 2005 (Whittaker et al. 2005) και αποτελεί σήμερα ένα από τα πιο αναπτυσσόμενα πεδία της. Ορίζεται ως η εφαρμογή των βιογεωγραφικών αρχών, θεωριών, αναλύσεων και δεδομένων από άλλα ερευνητικά πεδία, όπως η ιστορική βιογεωγραφία, η βιολογία εισβολής («invasion biology»), η οικολογία τοπίου («landscape ecology»), η οικολογία κοινοτήτων («community ecology») και η εξελικτική βιολογία («evolutionary biology»), με απώτερο στόχο τη διατήρηση της βιοποικιλότητας (Whittaker et al. 2005). Παρά την παρουσία των προαναφερόμενων επιστημονικών τομέων έρευνας των προτύπων κατανομής και του πλήθους των θεωριών που αναπτύχθηκαν για κάθε έναν από αυτούς, η βιογεωγραφική μελέτη σε μικρές ή τοπικές κλίμακες, πολλές φορές δεν είναι σε θέση να ερμηνεύσει πλήρως τον αριθμό των ειδών 2 και τη γεωγραφική κατανομή τους, ενώ η προσέγγιση πολύπλοκων οικοσυστημάτων μπορεί να επιτευχτεί µόνο με αναγωγή σε υψηλότερη κλίμακα (π.χ. περιφερειακό επίπεδο). Οι πράξεις, σε ότι όπως ο Hutchinson (1965) αποκάλεσε «οικολογικό θέατρο», παίζονται σε διάφορες κλίμακες του χώρου και του χρόνου. Ως αποτέλεσμα, για να καταλάβουμε το δράμα πρέπει να το δούμε στην κατάλληλη κλίμακα (Wiens, 1989). Αυτή η ανάγκη, λοιπόν, οδήγησε στην εμφάνιση ενός συμπληρωματικού επιστημονικού κλάδου της βιογεωγραφίας: της μακροοικολογίας 3. Σήμερα, η μακροοικολογία διερευνά γενικές αρχές και γενικευμένα πρότυπα της βιοποικιλότητας, σε μεγάλες χωρικές και χρονικές κλίμακες (Brown 1995). Σύμφωνα με τους Beck et al. (2012) η μακροικολογία επιδιώκει (βλέπε επίσης Πίνακα 1.1): 2 Ο αγγλικός όρος «species richness» αποδίδεται ως «αριθμός ειδών». Ο όρος «αφθονία ειδών» αναφέρεται στον όρο «species abundance» 3 Σύμφωνα με τον Brown (1995): «Η μακροοικολογία είναι η μη πειραματική, στατιστική έρευνα της σχέσης μεταξύ των αλληλεπιδράσεων των πληθυσμών των ειδών που συνήθως μελετάται σε μικρές κλίμακες από τους οικολόγους με τις διαδικασίες της ειδογένεσης, εξαφάνισης και της αλλαγής των κατανομών των ειδών που έχουν διερευνηθεί σε πολύ μεγαλύτερες κλίμακες από τους βιογεωγράφους, τους παλαιοντολόγους και τους μακροεξελικτικούς. Χρησιμοποιεί ταυτόχρονα γεωγραφικά και ιστορικά δεδομένα, με σκοπό την καλύτερη κατανόηση της τοπικής αφθονίας, της κατανομής και της ποικιλότητας των ειδών και εφαρμόζει οικολογικές θεωρίες για την απόκτηση γνώσης σχετικά με την ιστορία και τη σύνθεση των ειδών» 58
59 I. Να ενσωματώσει παλαιοντολογικά, φυλογενετικά και δεδομένα της ιστορικής βιογεωγραφίας σε αναλύσεις χωρικής και χρονικής μακροκλίμακας, για την κατανόηση των σημερινών προτύπων της βιοποικιλότητας. II. III. Να εξετάσει τις τοπικές διεργασίες που συμβαίνουν σε μικρές χωρικές κλίμακες και οδηγούν στα παρατηρούμενα πρότυπα μεγαλύτερης χωρικής κλίμακας, επιτρέποντας έτσι την καλύτερη πρόβλεψη των μελλοντικών προτύπων (π.χ. κάτω από συνθήκες περιβαλλοντικών αλλαγών),διότι και οι τοπικές διεργασίες επηρεάζουν ως ένα βαθμό τα πρότυπα κατανομής των ειδών σε μεγάλη κλίμακα (Kissling et al. 2012). Να χρησιμοποιήσει λεπτομερή δεδομένα υψηλής ποιότητας από ένα ευρύ φάσμα τάξων και περιοχών. Εδώ, θα πρέπει να επισημάνουμε ότι, έως τώρα, η μακροοικολογία έχει επικεντρωθεί κυρίως στα χερσαία σπονδυλωτά, με τα θαλάσσια και τα οικοσυστήματα των εσωτερικών υδάτων καθώς και τους οργανισμούς που διαβιούν σε αυτά, να εκπροσωπούνται ελάχιστα στη βιβλιογραφία (Heino 2011). 59
60 Πίνακας 1.1. Κύρια ερωτήματα της μακροοικολογίας (τροποποιημένο από Gaston & Blackburn (2000)) που διερευνώνται στην παρούσα μελέτη Table 1.1. Principal concerns of macroecology (modified from Gaston & Blackburn (2000)) investigated in the present study Σχέση Αριθμού Ειδών και Έκτασης (Species-area relationship) Αριθμος Ειδών (Species richness) ιαβάθµιση του Αριθμού Ειδών με το Γεωγραφικό Πλάτος (Latitudinal gradient in species richness) ιαβάθµιση του Αριθμού Ειδών με το Γεωγραφικό Μήκος (Longitudinal gradient in species richness) ιαβάθµιση του Αριθμού Ειδών με το Υψόμετρο (Altitudinal gradient in species richness) Εύρος της γεωγραφικής κατανοµής (Range size) Εγκιβωτισμός της Παρουσίας των Ειδών (Nestedness of species occurrence) Χωρική Αντικατάσταση των Ειδών (Spatial turnover in species identities) Μέγεθος σώματος (Body size) Κατανομή Ειδών και Μεγέθους του Σώματος των Ειδών (Species-body size distribution) ιαβάθµιση του Μεγέθους του Σώματος Κατά Γεωγραφικό Πλάτος - Κανόνας του Bergmann (Latitudinal gradient in body size - Bergmann s rule) 60
61 1.2 Κατανομή, σύνθεση και δομή βιοκοινοτήτων στη μακρο- και μικρο- κλίμακα Όπως αναφέρθηκε ήδη, η διερεύνηση του βαθμού που οι σύγχρονοι και οι ιστορικοί παράγοντες διαμορφώνουν τα πρότυπα κατανομής των ειδών σε μικρή και μεγάλη κλίμακα, αποτελεί ένα από τα σπουδαιότερα ερωτήματα στη βιογεωγραφία και τη μακροικολογία (Olden et al. 2010). Επίσης, θέμα προβληματισμού για τους ερευνητές για πολλές δεκαετίες είναι αν οι βιοκοινότητες αποτελούν διακριτά-προβλέψιμα και όχι τυχαία σύνολα και κατά πόσο η σύνθεση των ειδών διαμορφώνεται από βιοτικούς ή αβιοτικούς παράγοντες (βλέπε τις πρώτες έρευνες των Hutchinson 1961, Connor & Simberloff 1979). Αναφορικά με τα πρότυπα που διέπουν την κατανομή και την αφθονία των ειδών καθώς και τη σύνθεση των βιοκοινοτήτων σε μεγάλες χωρικές αλλά και τις χρονικές κλίμακες, έχουν προταθεί μέχρι σήμερα μια σειρά υποθέσεων και θεωριών. Μια από αυτές τις θεωρίες, η οποία βασίζεται στην ανάλυση των οικολογικών θώκων («niche 4 assembly theory») και τη διαφοροποίηση των ειδών («species differentiation»), προτείνει πως οι φυσικές διεργασίες και οι βιολογικές αλληλεπιδράσεις είναι αυτές που καθορίζουν τελικά τη συνύπαρξη των ειδών (MacArthur & Wilson 1967). Αντίθετα, η δομή και σύνθεση μιας βιοκοινότητας καθορίζεται από την ικανότητα διασποράς των οικολογικά ισοδύναμων ειδών, σε συνδυασμό με την ειδογένεση και την ύπαρξη τυχαίων γεγονότων, όπως για παράδειγμα η γέννηση και ο θάνατος σύμφωνα με τη θεωρία της ουδερότητας της βιοποικιλότητας και βιογεωγραφίας («Unified neutral theory of biodiversity and biogeography») (Hubbell 2001). Έτσι, εάν οι βιοικοινότητες είναι πράγματι τυχαίες, τα πρότυπα που παρατηρούνται δεν παρέχουν καμία πληροφορία στις οικολογικές θεωρίες. Εντούτοις, η αξιολόγηση κατά πόσο οι βιοκοινότητες είναι τυχαίες ή δομημένες (εναλλακτικά, στοχαστικές ή ντετερμινιστικές) σχετίζεται άμεσα με το χρόνο και την έκταση της δειγματοληψίας (Jackson, Peres-Neto & Olden 2001), δηλαδή των διαθέσιμων δεδομένων μέσα στο χρόνο. Επιπλέον, το μέγεθος των χωρικών και χρονικών κλιμάκων που χρησιμοποιούνται στις αναλύσεις μας μπορεί 4 Σύμφωνα με τoν Hutchinson (1957) θεωρούμε ότι o οικολογικός θώκος περιγράφει το σύνολο των βιοτικών και αβιοτικών συνθηκών που επηρεάζουν τη βιωσιμότητα ενός πληθυσμού, ή αλλιώς είναι τα ιδεατά ή μέγιστα όρια επιβίωσης 61
62 να είναι σημαντικό, δεδομένου ότι ο παράγων μεταβλητότητα στις περιβαλλοντικές συνθήκες και στη σύνθεση των ειδών παρατηρείται σε διαφορετικές κλίμακες, επηρεάζοντας έτσι τα συμπεράσματά μας (Brown & Maurer 1989). Για παράδειγμα, σε παγκόσμια κλίμακα, η διαβάθµιση του αριθμού των ειδών με το γεωγραφικό πλάτος είναι ένα από τα πιο συχνά παρατηρούμενα πρότυπα (Whittaker, Willis & Field 2001), ενώ σε περιφερειακό επίπεδο ο αριθμός των ειδών καθορίζεται κυρίως από παράγοντες, όπως η απόσταση από τα θερμά σημεία («hotspots») βιοποικιλότητας (π.χ. Bellwood et al. 2005). Σε τοπικό επίπεδο, ο αριθμός των ειδών και η κατανομή τους ρυθμίζεται από τις αβιοτικές συνθήκες (π.χ. φυσικοχημικοι παράγοντες) και τις αλληλεπιδράσεις μεταξύ των ειδών (π.χ. ανταγωισμός, θήρευση) που ενεργούν ταυτόχρονα (Mouillot, Dumay & Tomasini 2007). Σύμφωνα με τη σχετική βιβλιογραφία, αναφορικά με τα γεωγραφικά ασυνεχή πρότυπα κατανομής των ειδών που παρατηρούνται σήμερα, έχουν προταθεί δύο θεωρίες που για δεκαετίες θεωρούνταν αλληλοσυγκρουόμενες: 1) η θεωρία της διασποράς και 2) ο βικαριανισμός 5. H υπόθεση της διασποράς υποστηρίζει ότι οι απογονικές μορφές είναι προϊόν τυχαίας διασποράς σε μεγάλη απόσταση πάνω από έναν προϋπάρχοντα φραγμό (π.χ. οροσειρά), ενώ ο βικαριανισμός θεωρεί ότι οι περιοχές της ευρείας κατανομής των ειδών κατακερματίστηκαν και χωρίστηκαν μεταξύ τους από την εμφάνιση ενός νέου φράγματος, με αποτέλεσμα σε πολλές περιπτώσεις να ακολουθήσει αλλοπάτρια ειδογένεση στους ξεχωριστούς πλέον πληθυσμούς (Myers & Giller 1988). Για παράδειγμα, ένα είδος που χαρακτηρίζεται από μεγάλη ικανότητα διασποράς («long-distance dispersal») κατανέμεται όπου υπάρχει διαθέσιμος κατάλληλος οικολογικός χώρος, αρκεί αυτός να μην έχει καταληφθεί από άλλους ανταγωνιστές (Lavin et al. 2004). Συνεπώς, εξελικτικά (και λειτουργικά) κοντινά είδη τείνουν να καταλαμβάνουν το ίδιο ενδιαίτημα, καθιστώντας δυνατή την πρόβλεψη της γεωγραφικής κατανομής τους, υπό γνωστές περιβαλλοντικές συνθήκες (Lomolino et al. 2010). 5 Κατά τη διασπορά, μια ταξινομική μονάδα αποκτά τη σημερινή της κατανομή μέσω ενεργητικής ή παθητικής διασποράς από ένα ή περισσότερα προγονικά κέντρα προέλευσης. Υπό βικαριανιστικές συνθήκες, η αλλοπάτρια ειδογένεση συντελείται χωρίς τη μετανάστευση ατόμων. Οι πληθυσμοί διαχωρίζονται όταν η συνεχής κατανομή της προγονικής μορφής κατακερματίζεται από απροσπέλαστους φραγμούς που διαχωρίζουν τους πληθυσμούς, όπως π.χ. ορογένεση. 62
63 Η σύνθεση και δομή των βιοκοινοτήτων έχει προταθεί ότι είναι αποτέλεσμα δύο διαφορετικών διαδικασιών: 1) του περιβαλλοντικού φιλτραρίσματος («environmental filtering»), σύμφωνα με το οποίο μία ομάδα ειδών που μπορεί δυνητικά να αποικίσει μια περιοχή, δηλαδή το δυνητικό αποθεματικό ειδών («species pool») επιλέγεται (ή αλλιώς φιλτράρεται) από μια σειρά περιβαλλοντικών παραμέτρων, με αποτέλεσμα τα είδη που μπορούν πραγματικά να επιβιώσουν σε αυτή την περιοχή να είναι περισσότερο λειτουργικά όμοια απ ό, τι αναμενόταν από τυχαίους παράγοντες (π.χ. Ruhí et al. 2014). Η επιλογή αυτή μπορεί να βασίζεται στις αβιοτικές συνθήκες που επικρατούν στο περιβάλλον, στα διαθέσιμα ενδιαιτήματα, στις κλιματικές συνθήκες και σε άλλους παράγοντες. 2) του περιορισμού ομοιότητας («limiting similarity»), σύμφωνα με τον οποίο τα είδη που απαντώνται, σε τοπική κλίμακα, διαφέρουν σημαντικά το ένα από το άλλο σε συγκεκριμένα λειτουργικά χαρακτηριστικά, που σχετίζονται με την απόκτηση ή/και τη χρήση των διαθέσιμων πόρων, ώστε να μειώνεται ο ανταγωνισμός (π.χ. Grime 2006). Η επίδραση των δύο παραπάνω αλλά και συνάμα αντίθετων διεργασιών οδηγεί σε δυο βασικά πρότυπα. Ωστόσο πολλές μελέτες μέχρι σήμερα έχουν δείξει πως η δομή των βιοκοινοτήτων μπορεί να είναι το αποτέλεσμα και των δύο αυτών μηχανισμών που δρουν ταυτόχρονα (Cornwell & Ackerly 2009). Πρότυπο 1 ο ) Αν υποθέσουμε ότι το περιβάλλον είναι ο κύριος παράγοντας που διαμορφώνει τις βιοκοινότητες, οργανισμοί με παρόμοια λειτουργικά χαρακτηριστικά θα έχουν παρόμοιες οικολογικές απαιτήσεις και επομένως θα κατανέμονται σε παρόμοια περιβάλλοντα. Τα είδη με ακατάλληλα χαρακτηριστικά για τις τοπικές περιβαλλοντικές συνθήκες δεν μπορούν να είναι εύρωστα. Συνεπώς, η μεταβλητότητα (ή αλλιώς ποικιλότητα) των χαρακτηριστικών των ειδών θα ποικίλει λιγότερο σε σύγκριση με μια τυχαία βιοκοινότητα, με τυχαία σύνθεση ειδών και τελικά τα χαρακτηριστικά των ειδών θα εμφανίζουν σύγκλιση («trait convergence») (π.χ. Lebrija-Trejos et al. 2010). 63
64 Πρότυπο 2 ο ) Αν υποθέσουμε ότι η μεταβλητότητα των χαρακτηριστικών μιας βιοκοινότητας είναι αυξημένη σε σχέση με μια τυχαία βιοκοινότητα, τότε ως μηχανισμός συγκρότησης βιοκοινοτήτων θα μπορούσαν να οριστούν οι βιοτικές αλληλεπιδράσεις, όπως ο αποκλειόμενος ανταγωνισμός («competitive exclusion») και η θήρευση («predation»). Σε αυτήν την περίπτωση, τα χαρακτηριστικά των ειδών θα εμφανίζουν απόκλιση («trait divergence»), αφού η θεωρία του ανταγωνισμού αναφέρει ότι υπάρχει ένα όριο για το πόσα όμοια είδη μπορούν να συνυπάρχουν (MacArthur & Levins 1967). Ως αποτέλεσμα, τα είδη που συνυπάρχουν είναι ανόμοια και επομένως συμπληρωματικά. 1.3 Πολλαπλές πτυχές της βιοποικιλότητας Τις τελευταίες δεκαετίες, η μελέτη των προτύπων του αριθμού ειδών στη μάκρο και μικρο-κλίμακα αμφισβητείται έντονα σχετικά με τη δυνατότητά της να αποτελέσει ισχυρή βάση αναφοράς για τον επιστημονικό κλάδο της διατήρησης της βιοποικιλότητας. Για παράδειγμα, ενώ υπάρχουν ενδείξεις ότι η απώλεια ποικιλότητας ειδών («diversity loss») επηρεάζει σημαντικά τις λειτουργίες των οικοσυστημάτων («ecosystem functioning») (Hooper et al. 2012), η δημοσιευμένη πληροφορία, ως προς την κατανόηση της σχέσης αυτής, είναι εξαιρετικά περιορισμένη (βλέπε McCann 2000 για την ανασκόπηση της δημοσιευμένης πληροφορίας). Μια πιθανή θετική συσχέτιση μεταξύ των λειτουργιών του οικοσυστήματος και της ποικιλότητας των ειδών (π.χ. Tilman 1994) αποτελεί το γεγονός ότι η σταθερότητα («stability») ενός οικοσυστήματος αυξάνεται με την αύξηση του αριθμού ειδών («species richness») (MacArthur 1955). Η θεωρία αυτή δεν υποστηρίζει ότι η ταξινομική ποικιλότητα είναι η κινητήρια δύναμη αυτής της σχέσης, δηλαδή ότι ο αυξανόμενος αριθμός ειδών οδηγεί από μόνος του στην σταθερότητα των οικοσυστημάτων. Αντ 'αυτού, η ποικιλότητα ειδών μπορεί να θεωρηθεί ως έμμεση απόδειξη σημαντικών οικολογικών μηχανισμών που δρουν στα οικοσυστήματα. Μία άλλη πιθανή σχέση μεγάλης σημασίας είναι η κατανόηση του τρόπου, πώς δηλαδή η απώλεια (ή προσθήκη) των ειδών επηρεάζει τη σταθερότητα και τη λειτουργία των οικοσυστημάτων (Tilman et al. 1997). Για παράδειγμα, η απώλεια της βιοποικιλότητας δε συνδέεται μόνο με τη μείωση της 64
65 ταξινομικής ποικιλότητας (δηλαδή τον αριθμό ειδών) αλλά και με την αντικατάσταση ειδών, με ιδιαίτερα χαρακτηριστικά, από είδη με διαφορετικά χαρακτηριστικά (Loreau et al. 2001). Αυτή η υπόθεση υποστηρίζει ότι η ταυτότητα των ειδών που εξαφανίζονται (ή η σειρά με την οποία έχουν χαθεί) μπορεί να είναι πιο σημαντική από τον αριθμό των ειδών που απομένει. Σε παγκόσμιο επίπεδο, η απώλεια της βιοποικιλότητας αποδίδεται κυρίως στην απώλεια των λειτουργικών και όχι των ταξινομικών ομάδων, δείχνοντας έτσι το δρόμο προς το πεδίο της λειτουργικής οικολογίας και ποικιλότητας (Bellwood et al. 2002), (βλέπε Ενότητα Λειτουργική ποικιλότητα). Ως αποτέλεσμα, οι βιολόγοι διατήρησης έχουν διευρύνει τους στόχους τους, περιλαμβάνοντας πολλαπλές πτυχές της ποικιλότητας, με σκοπό τη διερεύνηση των προτύπων του αριθμού, της κατανομής και της σύνθεσης των ειδών για τη διατήρηση των οικοσυστημικών λειτουργιών (Naeem, Duffy & Zavaleta 2012). Προς την κατεύθυνση της διατήρησης των οικοσυστημικών λειτουργιών, η νέο εισαχθείσα έννοια των πολλαπλών πτυχών της βιοποικιλότητας ενσωματώνει, παράλληλα, διαφορετικά ιεραρχικά επίπεδα της οργάνωσης, από το μοριακό έως το επίπεδο του οικοσυστήματος, και ως εκ τούτου οδηγεί σε ολιστικές προσεγγίσεις για τη διατήρηση. Ο ορισμός της βιοποικιλότητας, πέρα από το επίπεδο των ειδών («species diversity»), εμπερικλείει ένα ευρύτερο φάσμα εννοιών και πεδίων ανάλυσης (Matthews et al. 2001) που εκτείνονται από το επίπεδο ολόκληρων οικοσυστημάτων, στα ενδιαιτήματα και τους οικοτόνους («ecosystem diversity»), τη γενετική και τη λειτουργική ενδοειδική διαφοροποίηση («intraspecific diversity») καθώς και τη γενετική («genetic diversity») και λειτουργική ποικιλότητα («functional diversity») εντός των πληθυσμών και των βιοκοινοτήτων (Eικόνα 1.1). 65
66 Εικόνα 1.1. Ιεραρχική απεικόνιση των πολλαπλών πτυχών της βιοποικιλότητας (τροποποιημένο από Geist 2011) Figure 1.1. Hierarchical levels of multiple facets of biodiversity (modified from Geist 2011) Ποικιλότητα οικοσυστημάτων Η ποικιλότητα των οικοσυστημάτων ορίζεται ως το σύνολο των διαφορετικών τύπων των ενδιαιτημάτων και των οικολογικών διεργασιών που τα χαρακτηρίζουν και μπορεί να ιδωθεί ως συνώνυμο της ετερογένειας του τοπίου («landscape spatial heterogeneity») (Naeem 2002). Συγκεκριμένα είδη επικρατούν σε κάθε οικοσύστημα, ωστόσο, η οριοθέτηση του κάθε οικοσυστήματος και η κατανόησή των λειτουργιών του είναι τις περισσότερες φορές µια αρκετά δύσκολη υπόθεση, καθώς οι αλληλεπιδράσεις που λαμβάνουν χώρα ανάμεσα στους οργανισμούς και με το περιβάλλον τους δεν έχουν ξεκάθαρα όρια. 66
67 1.3.2 Ποικιλότητα ειδών Ιστορικά, από την πρωτοποριακή έρευνα του Alfred Russel Wallace στα τέλη του 19ου αιώνα (Wallace 1878), οι ερευνητές για να μελετήσουν τη βιοποικιλότητα και τις γεωγραφικές κατανομές των ειδών της χλωρίδας και πανίδας βασίστηκαν σε ταξινομικές μετρικές. Έτσι, από τη Διάσκεψη Κορυφής του Ρίο το 1992 για το Περιβάλλον και την Ανάπτυξη και μετέπειτα, η βιοποικιλότητα (ή «βιολογική ποικιλότητα») συνήθως ορίζεται ως ο αριθμός των ειδών σε ένα δεδομένο οικοσύστημα ή οικότοπο, με την έννοια του αριθμού των ειδών (ή αλλιώς πλούτος ειδών) να χρησιμοποιείται τελικά ως το πιο απλό μέτρο της βιοποικιλότητας (Krebs 2009) Ενδοειδική ποικιλότητα Η ενδοειδική ποικιλότητα έχει αναγνωριστεί ως μια από τις σημαντικές πτυχές της βιοποικιλότητας τις τελευταίες δεκαετίες (Fox, Ades & Bi 2001, Moritz 2002) και θεωρείται αναπόσπαστο μέρος της έννοιας αυτής. Περιλαμβάνει τη φαινοτυπική, γενετική, ηθολογική και λειτουργική ποικιλότητα των πληθυσμών, εντός των ειδών («within species diversity»). Η ποικιλότητα όσο αφορά τη γενετική σύσταση (π.χ. Gamfeldt et al. 2005) αλλά και η διαφοροποίηση των λειτουργικών χαρακτήρων (π.χ. Albert et al. 2010) εντός των πληθυσμών των ειδών, συνδέεται άμεσα με την κατανόηση της δομής και σύνθεσης των βιοκοινοτήτων, της επιλογής του ενδιαιτήματος και των οικοσυστημικών λειτουργιών. Εντούτοις, η ενδοειδική ποικιλότητα συχνά δεν συμπεριλαμβάνεται στις μελέτες γενετικής (Gugerli et al. 2008) και λειτουργικής ποικιλότητας (Mason et al. 2007, Cianciaruso et al. 2009, de Bello et al. 2011) Γενετική ποικιλότητα Η γενετική ποικιλότητα είναι απαραίτητη για τη προσαρμογή ενός είδους στις διαρκώς μεταβαλλόμενες περιβαλλοντικές συνθήκες, καθώς αντανακλά τη 67
68 δυνατότητα εξέλιξης του. Για παράδειγμα, είδη ή πληθυσμοί που έχουν υποστεί μείωση της γενετικής τους ποικιλότητας έχουν μειωμένη πιθανότητα προσαρμογής και επιβίωσης στο περιβάλλον και τις μεταβολές του (Frankham 2005) Λειτουργική ποικιλότητα Όπως ήδη αναφέρθηκε, η βιογεωγραφία και η μακροοικολογία βασίστηκαν αρχικά, ως πεδία έρευνας, στην ανάλυση της ταξινομικής ποικιλότητας. Από το τέλος, ωστόσο, του περασμένου αιώνα, οι οικολόγοι άρχισαν να δίνουν μεγαλύτερη έμφαση στα χαρακτηριστικά των ειδών μιας βιοκοινότητας παρά στον αριθμό των ταξινομικών ειδών (Tilman et al. 1997, Loreau 1998, Díaz & Cabido 2001, Petchey & Gaston 2002). Αυτή η επιστημονική τάση καταδεικνύεται και από τη συχνότητα της εμφάνισης των λέξεων-κλειδιά «λειτουργική ποικιλότητα» και «λειτουργική οικολογία» στις ερευνητικές δημοσιεύσεις, που όπως φαίνεται στην Εικόνα 1.2, ο όρος γίνεται όλο και περισσότερο αποδεκτός. Επιπλέον, γίνεται όλο και περισσότερο εμφανές πως ο αντίκτυπος της απώλειας της παγκόσμιας βιοποικιλότητας σχετίζεται όλο και περισσότερο με την απώλεια των λειτουργικών και όχι ταξινομικών ομάδων (Bellwood et al. 2002). Η λειτουργική ποικιλότητα (FD) σύμφωνα με τον Tilman (2001) «περιγράφει τα συστατικά της βιοποικιλότητας που επηρεάζουν το πώς ένα οικοσύστημα λειτουργεί» και έχει αναδειχθεί ως μια σημαντική πτυχή της βιοποικιλότητας για τη ποσοτική εκτίμηση της αξίας και το εύρος των λειτουργικών χαρακτηριστικών 6 και των οικοσυστημικών λειτουργιών (Díaz & Cabido 2001, Bellwood et al. 2002, Mims et al. 2010). Λειτουργικά, ένα είδος αποτελείται από μια συλλογή από άτομα με όμοια φαινοτυπικά, φυσιολογικά και ηθολογικά χαρακτηριστικά τα οποία και καθορίζουν πότε και πώς τα άτομα αυτά αλληλεπιδρούν με άλλα είδη (McGill et al. 2006). Αυτή η άποψη αναδεικνύει ότι, ενώ η λειτουργική ποικιλότητα αντιπροσωπεύει τη ποικιλότητα των χαρακτηριστικών, εμμέσως αντιπροσωπεύει μια μηχανιστική σχέση μεταξύ της ποικιλότητας των ειδών με τους οικολογικούς θώκους ή/και τις 6 Ως λειτουργικά χαρακτηριστικά αποδίδονται τα χαρακτηριστικά που επηρεάζουν σε μεγάλο βαθμό την απόδοση («performance») του οργανισμού (η την ικανότητά του να διατηρηθεί κατά τη διάρκεια πολλών γενεών) (Frimpong & Angermeier 2010) 68
69 οικοσυστημικές λειτουργίες (McGill et al. 2006, Petchey & Gaston 2006, Villéger, Mason & Mouillot 2008). Ένα σημαντικό πλεονέκτημα της μελέτης της λειτουργικής ποικιλότητας έγκειται στο γεγονός ότι μπορεί να ελεγχθεί κατά πόσο μία βιοκοινότητα που αποτελείται από μικρό αριθμό λειτουργικά ανόμοιων ειδών, με χαμηλές αφθονίες, παρουσιάζει υψηλότερη λειτουργική ποικιλότητα από μια άλλη βιοκοινότητα με μεγαλύτερο αριθμό ειδών, που είναι λειτουργικά όμοια και σε υψηλότερες αφθονίες (Petchey & Gaston 2006). Συνεπώς, η λειτουργική ποικιλότητα εκφράζει τον αριθμό των λειτουργικών ρόλων που απαντώνται σε ένα οικοσύστημα (και όχι την ταξινομική τους ταυτότητα και τον αριθμό των ειδών) και λαμβάνει υπόψη της την συμπληρωματικότητα και τον πλεονασμό των ειδών. Μπορεί να προσδιορίσει, συνεπώς, πώς οι βιοκοινότητες των ειδών ανταποκρίνονται στις ανθρωπογενείς διαταραχές, τόσο σε τοπικό όσο και σε περιφερειακό επίπεδο, και πώς αυτές επηρεάζουν τις λειτουργίες του οικοσυστήματος (Díaz & Cabido 2001, Petchey & Gaston 2002, Mouillot et al. 2013a) (Εικόνα 1.2). Με άλλα λόγια, όταν τα λειτουργικά χαρακτηριστικά κάποιων ειδών σχετίζονται με τις λειτουργίες των βιοκοινοτήτων, τότε η απώλειά τους θα επιφέρει σημαντική μείωση στο ρυθμό της οικοσυστημικής λειτουργίας. 69
, σε σύγκριση με το ετήσιο")
70 Εικόνα 1.2. Το ετήσιο ποσοστό των δημοσιεύσεων χρησιμοποιώντας το ISI Web of Science, χρησιμοποιώντας τις λέξεις-κλειδιά «λειτουργική ποικιλότητα» και «οικολογία» (σκούρες ράβδοι), σε σύγκριση με το ετήσιο ποσοστό των δημοσιεύσεων που βρέθηκαν με τη χρήση του όρου "οικολογίας κοινοτήτων" (διάφανες ράβδοι) σε όλες τις δημοσιεύσεις για το διάστημα (από Cadotte, Carscadden & Mirotchnick 2011) Figure 1.2. The yearly percentage of publications found using ISI Web of Science using the keywords functional diversity and ecology (darkbars), compared with the yearly percentage of publications found using community ecology (transparent light bars) for all papers published (from Cadotte, Carscadden & Mirotchnick 2011) 1.4 Τα οικοσυστήματα των εσωτερικών και υφάλμυρων υδάτων 7 ως θερμά σημεία («hotspots») της βιοποικιλότητας Αν και ορισμένες μελέτες έχουν συγκρίνει διαφορετικές πτυχές της ποικιλότητας των ειδών (π.χ. Wilsey et al. 2005, Petchey et al. 2007), μόνο προκαταρκτικές προσπάθειες έχουν γίνει για τη μελέτη της λειτουργικής ποικιλότητας των εσωτερικών υδάτων (π.χ. Heino 2005, Heino, Mykrä & Kotanen 2008, Villéger et al. 2010, Pool, Grenouillet & Villéger 2014). Γενικότερα, η πλειονότητα των μελετών των εσωτερικών υδάτων είναι προσανατολισμένη στη διερεύνηση της σχέσης 7 Οι ξενόγλωσσοι όροι «Inland waters» ή «Freshwaters», «Les Eaux Douces», «Innerliches Wasser», «Aqua dolce», «Aguas doulces» κτλ. αποδίδονται στο παρόν κείμενο στα ελληνικά ως «εσωτερικά ύδατα». Ο όρος «γλυκό νερό» περιγράφει τις μη θαλάσσιες υδατοσυλλογές, οι οποίες βρίσκονται σε χερσαίες περιοχές και εμφανίζουν πολύ χαμηλές συγκεντρώσεις διαλυμένων αλάτων 70
71 βιοκοινοτήτων- περιβαλλοντικών συνθηκών στο επίπεδο της λεκάνης απορροής, παραλείποντας μία συνολικότερη προσέγγιση σε μακροοικολογικό επίπεδο. Εντούτοις, τα οικοσυστήματα των εσωτερικών υδάτων αποτελούν μία εξαιρετική περίπτωση για την κατανόηση προτύπων, τη διερεύνηση των οικολογικών θεωριών και την προώθηση της μακροοικολογικής έρευνας (Hugueny, Oberdorff & Tedesco 2010). Τα οικοσυστήματα των εσωτερικών υδάτων (δηλ. λίμνες, ποτάμια, ρέματα, πηγές και υγροβιότοποι) είναι ευρέως αναγνωρισμένα ως «θερμά σημεία» βιοποικιλότητας και ενδημισμού 8, αν και καταλαμβάνουν μόλις το 0.8% της επιφάνειας της Γης (Dudgeon et al. 2006). Μολονότι η εκτίμηση της ποικιλότητας των οργανισμών που διαβιούν σε αυτά δεν έχει ακόμα ολοκληρωθεί, υπoλογίζεται πως το 12% της παγκόσμιας πανίδας και το 1/3 ( είδη) των σπονδυλωτών φιλοξενούνται στο ετερογενές περιβάλλον των εσωτερικών υδάτων (Balian et al. 2008), που χαρακτηρίζεται από απομονωμένα ενδιαιτήματα («habitat isolation») και περιορισμένη διασπορά («dispersal limitation») των ειδών τους. Αυτός ο αριθμός είναι σίγουρα πολύ μεγαλύτερος, λαμβάνοντας υπόψη το γεγονός ότι σε ετήσια βάση πολλά νέα είδη περιγράφονται, ακόμα και στην περίπτωση των πιο πλήρως μελετημένων ομάδων (ψάρια και αμφίβια). Επιπρόσθετα, εκτός από τον υψηλό αριθμό ειδών, τα οικοσυστήματα των εσωτερικών υδάτων παρέχουν καταφύγιο σε ένα μεγάλο ποσοστό ενδημικών τάξων (Tisseuil et al. 2013). Πρόσφατες μελέτες έδειξαν πως τα υψηλότερα ποσοστά των απειλούμενων ειδών παγκοσμίως παρατηρούνται στα εσωτερικά ύδατα, καθώς ένα μεγάλο μέρος των ενδιαιτημάτων τους είτε έχει καταστραφεί (Millennium Ecosystem Assessment, 2005) είτε πιεστεί από ανθρώπινες παρεμβάσεις. Ο κίνδυνος, λοιπόν, εξαφάνισης ειδών στα εσωτερικά ύδατα είναι υψηλότερος από ό, τι στους άλλους τύπους οικοσυστημάτων (Collen et al. 2014). Πιο συγκεκριμένα, το 30,1% των ειδών των εσωτερικών υδάτων, που έχουν αξιολογηθεί από τη Διεθνή Ένωση Προστασίας της Φύσης (International Union for Conservation of Nature - IUCN), ταξινομούνται ως απειλούμενα (κατηγορίες Κρισίμως κινδυνεύοντα- CR, Κινδυνεύοντα- EN, Τρωτά- VUL), (IUCN 2013) ενώ το 25% των ψαριών των εσωτερικών υδάτων απειλούνται με εξαφάνιση (Vié et al. 2008). Έως το 2070, το 75% των ψαριών σε τοπικό επίπεδο 8 Ο όρος ενδημικό («endemic») αναφέρεται σε ένα είδος (ή τάξο) το οποίο απαντάται σε μία συγκεκριμένη γεωγραφική περιοχή και πουθενά αλλού (Morrone 1994) 71
72 προβλέπεται να εξαφανιστεί εξαιτίας των αλλαγών σε χαρακτηριστικά όπως, η ροή και η παροχή νερού, κυρίως λόγω της κλιματικής αλλαγής και της υπεράντλησης νερού (Xenopoulos et al. 2005). Μέχρι σήμερα, έχει καταγραφεί μια πληθώρα ανθρωπογενών πιέσεων που απειλούν αυτά τα οικοσυστήματα, με τις σημαντικότερες να αφορούν τη ρύπανση, την άντληση νερού, την τροποποίηση της ροής, την υπερεκμετάλλευση των ιχθυοαποθεμάτων, τον κατακερματισμό των ενδιαιτημάτων και τον εμπλουτισμό με ξενικά, χωροκατακτητικά είδη (Dudgeon 2010, Collen et al. 2014). Πιο συγκεκριμένα, η ρύπανση των υδάτων από οικιακές και βιομηχανικές πηγές έχει μειωθεί σημαντικά στις περισσότερες βιομηχανικές χώρες, αλλά εξακολουθεί να αποτελεί μεγάλη πρόκληση για στις μεσογειακές χώρες. Αν και η ποιότητα του νερού των ποταμών της Ευρώπης έχει βελτιωθεί σταθερά τις τελευταίες δεκαετίες (Aarts, Van den Brink & Nienhuis 2004), ο κατακερματισμός των ενδιαιτημάτων αποτελεί συχνό φαινόμενο, ως αποτέλεσμα της αποψίλωσης των παραποτάμιων δασών, της τροποποίησης της ροής και της κατασκευής φραγμάτων που αναγκάζουν τα υδρόβια είδη σε μειωμένες δυνατότητες μετανάστευσης κατά συνέπεια και σε μειωμένη ροή γενετικού υλικού. Η υπερεκμετάλλευση επηρεάζει κυρίως τα ψάρια με μεγάλη οικονομική αξία και, σε μικρότερο βαθμό, τα μαλάκια και τα καρκινοειδή. Τέλος, η εισαγωγή κοσμοπολίτικων, μη αυτόχθονων ειδών, σε συνδυασμό με την απώλεια των αυτόχθονων (συχνά ενδημικών) ειδών, προκαλεί την αύξηση της γενετικής, ταξινομικής και λειτουργικής ομοιότητας των βιοκοινοτήτων, μια διαδικασία που ονομάζεται βιοτική ομογενοποίηση («biotic homogenization») (Vitousek et al. 1997, McKinney & Lockwood 1999, Olden & Poff 2004). Για παράδειγμα, σε μια μελέτη, ο Rahel (2000) συνέκρινε την ομοιότητα της ιχθυοπανίδας των εσωτερικών υδάτων στις Ηνωμένες Πολιτείες πριν και μετά την ευρωπαϊκή εγκατάσταση και βρήκε πως οι περιοχές, ανά ζεύγος, φιλοξενούν 15 περισσότερα κοινά είδη απ ότι στο παρελθόν (αύξηση της ομοιότητας της ιχθυοπανίδας κατά 7,2%). Ως εκ τούτου, η μελέτη της βιοτικής ομογενοποίησης αποτελεί μία μοναδική πρόκληση στη βιογεωγραφία διατήρησης, συνδυάζοντας τις αρχές της βιολογίας εισβολής, την εξαφάνιση των ειδών και την περιβαλλοντική αλλοίωση (McKinney & Lockwood 1999). 72
73 1.5 Δράσεις διατήρησης στα εσωτερικά ύδατα Ο σχεδιασμός δράσεων μεγάλης κλίμακας για τη διατήρηση της βιοποικιλότητας στα εσωτερικά ύδατα, με βάση τις κατανομές των ειδών, τα πρότυπα ενδημισμού και τις ανθρωπογενείς πιέσεις, απουσιάζει από τη σχετική βιβλιογραφία (αλλά βλέπε Abell et al. 2008, Darwall et al. 2011, Clavero & Hermoso 2011, Tedesco et al. 2012, Collen et al. 2014). Επιπλέον, έχει δειχθεί ότι τα οικοσυστήματα των εσωτερικών υδάτων δεν περιλαμβάνονται επαρκώς στο παγκόσμιο δίκτυο των προστατευόμενων περιοχών (π.χ. Herbert et al. 2010). Σε ευρωπαϊκό επίπεδο, μόλις το 30% των ποταμών χαρακτηρίζεται «καλής οικολογικής κατάστασης», σύμφωνα με την Οδηγία Πλαίσιο για τα Ύδατα (Garcia-Moreno et al. 2014). Ένα ακόμα προβληματικό σημείο της διατήρησης των εσωτερικών υδάτων σχετίζεται με το γεγονός ότι βασίζεται, έως σήμερα, κυρίως σε στρατηγικές που εφαρμόζονται με επιτυχία σε χερσαία οικοσυστήματα (Olden et al. 2010) και οι οποίες έχουν στηριχθεί όλο και περισσότερο σε χαρισματικά τάξα από άποψη καταγραφής (π.χ. πουλιά, θηλαστικά) (π.χ. Rodrigues et al. 2004). Εντούτοις, οι στρατηγικές διατήρησης των χερσαίων οικοσυστημάτων είναι φτωχά υποκατάστατα των εσωτερικών υδάτων (Darwall et al. 2011). Επίσης, δεν γνωρίζουμε ακόμη κατά πόσο τα «θερμά σημεία» των χερσαίων οικοσυστημάτων και των εσωτερικών υδάτων επικαλύπτονται (Strayer & Dudgeon 2010), ώστε η διατήρηση τους να μπορεί να συμπεριληφθεί σε ένα κοινό διαχειριστικό σχέδιο. Για παράδειγμα, για τη διατήρηση των χερσαίων οικοσυστημάτων δίνεται έμφαση σε περιοχές με πλούσια ενδιαιτήματα και σημαντικούς βιοτόπους που μπορούν να ενοποιηθούν και να προστατευθούν. Αντιθέτως, οι στρατηγικές διατήρησης των εσωτερικών υδάτων απαιτούν μεγαλύτερη ακρίβεια σε μικρότερη κλίμακα για α) την προστασία των ανάντη τμημάτων των ποταμών και της παρόχθιας ζώνης, β) τη μελέτη των επιπτώσεων των φραγμάτων στην περίπτωση των ανάδρομων ειδών καθώς επίσης και γ) τη μελέτη του ανταγωνισμού των εισαχθέντων ειδών με τα αυτόχθονα. Ως αποτέλεσμα, υπάρχει μια επιτακτική ανάγκη για προσεκτικό σχεδιασμό των προστατευόμενων περιοχών για τη διατήρηση της βιοποικιλότητας των εσωτερικών υδάτων. 73
74 1.6 Τα ψάρια 9 των εσωτερικών υδάτων Ο ακριβής αριθμός των υπαρχόντων ειδών των ψαριών των εσωτερικών υδάτων δεν έχει ακόμη καθοριστεί. Πράγματι, το ποσοστό και ο ρυθμός της συστηματικής περιγραφής των ειδών καταδεικνύει σαφώς ότι πολλά είδη θα περιγραφούν στα επόμενα χρόνια (Pelayo-Villamil et al. 2014). Μέχρι τώρα η ομάδα αυτή περιλαμβάνει σχεδόν είδη (πρωτογενή, δευτερογενή και διάδρομα) που ανήκουν σε περίπου 207 οικογένειες, το μεγαλύτερο μέρος των οποίων κατατάσσονται στις τάξεις Characiformes, Cypriniformes, Siluriformes, Gymnotiformes, Perciformes και Cyprinodontiformes (Εικόνα 1.3, Lévêque et al. 2008, Mikšik & Schraml 2014). Σε ευρωπαϊκό επίπεδο, τα είδη ιχθύων ανήκουν κυρίως στις οικογένειες των Cyprinidae, Salmonidae, Coregonidae, Gobiidae και Cobitidae (Lévêque et al. 2008). Παρόλα αυτά, η ευρωπαϊκή ιχθυοπανίδα εσωτερικών υδάτων είναι φτωχή, σε σχέση με τις άλλες ηπείρους, πιθανώς ως αποτέλεσμα των πρόσφατων παγετώνων (Reyjol et al. 2007, Lévêque et al. 2008). Πιο συγκεκριμένα, τα βιογεωγραφικά στοιχεία δείχνουν ότι οι ευρύτερες περιοχές που καλύφτηκαν με πάγο κατά τις παγετώδεις περιόδους εποικίστηκαν μετέπειτα κυρίως από είδη προερχόμενα από την περιοχή του Πόντου και της Κασπίας, και ιδιαίτερα από τα μεσαία και κατώτερα τμήματα του Δούναβη, ο οποίος χρησίμευσε ως σημαντικό καταφύγιο ειδών (Bănărescu 1992, Griffiths 2006). Άλλα καταφύγια, κατά τη διάρκεια της τελευταίας εποχής των παγετώνων περιλαμβάνουν περιοχές στις νότιες χερσονήσους (Ιβηρική, Ιταλική και Βαλκανική) (Εικόνα 1.4). Ωστόσο, η διασπορά προς τη βόρεια και την κεντρική Ευρώπη των ειδών των νότιων χερσονήσων εμποδίστηκε από τα Πυρηναία, τις Άλπεις και τις Διναρίδες οδηγώντας έτσι την πανίδα των τριών αυτών χερσονήσων της Μεσογείου σε υψηλή γεωγραφική απομόνωση και ως εκ τούτου, σε υψηλό επίπεδο ενδημισμού (Bănărescu 1992, Reyjol et al. 2007, Oikonomou, Leprieur & Leonardos 2014). 9 Ως ψάρια ορίζουμε τα ποικιλόθερμα, υδρόβια χορδωτά με αμφίπλευρη συμμετρία, που φέρουν πτερύγια, τα οποία ως κύριο αναπνευστικό όργανο έχουν βράγχια και των οποίων το σώμα είναι συνήθως καλυμμένο με λέπια. Έχουν δίχωρη καρδιά και ένα ειδικό αισθητήριο όργανο που ονομάζεται πλευρική γραμμή 74
75 Εικόνα 1.3. Διάγραμμα ταξινόμησης για τα είδη ψαριών των εσωτερικών υδάτων (συμπεριλαμβανομένων των ειδών που δεν χρησιμοποιούν μόνο το γλυκό νερό) (τροποποιημένο από Nelson 2006) Figure 1.3. Classification diagram for the freshwater fish species (including species not using only freshwater) (modified from Nelson 2006)
 ποσοστό των απειλούμενων ειδών σε επίπεδο χώρας, Β) αριθμός των ειδών ανά χώρα και C) ποσοστό των εισαχθέντων ειδών σε")
76 Εικόνα 1.4. Χάρτης κατανομής των ψαριών των εσωτερικών υδάτων σε πανευρωπαϊκό επίπεδο Α) ποσοστό των απειλούμενων ειδών σε επίπεδο χώρας, Β) αριθμός των ειδών ανά χώρα και C) ποσοστό των εισαχθέντων ειδών σε κάθε χώρα (τροποποιημένο από European Environment Agency, Figure 1.4. Map showing distribution of freshwater fishes in the pan-european region A) percentage of threatened species of freshwater fishes at country level, B) shows number of species per country and C) percentage of introduced species in each country (modified by European Environment Agency, Τα ψάρια των εσωτερικών υδάτων ως δείκτες σε βιογεωγραφικές αναλύσεις Μεταξύ των τάξων που διαβιούν στα εσωτερικά ύδατα, τα ψάρια είναι το πιο ενδιαφέρον μοντέλο για τη διερεύνηση της βιογεωγραφικής ιστορίας, των μακροοικολογικών προτύπων και διάφορων οικολογικών υποθέσεων και αυτό διότι θεωρούνται πολύ καλοί δείκτες βιολογικής απομόνωσης μιας υδάτινης μάζας. Πιο συγκεκριμένα, τα ψάρια έχουν γενικά μειωμένη ικανότητα διασποράς και η κατανομή τους περιορίζεται στο επίπεδο της λεκάνης απορροής, παρουσιάζοντας έτσι υψηλό ενδημισμό. Εξίσου ενδιαφέρον είναι ότι, σε περιφερειακό αλλά και σε παγκόσμιο επίπεδο, τα περισσότερα ψάρια των εσωτερικών υδάτων καταλαμβάνουν μόνο ένα τμήμα από τα οικοσυστήματα όπου θα μπορούσαν εν 76
77 δυνάμει να φιλοξενηθούν (Darlington, 1948). Επιπλέον, τα εσωτερικά ύδατα μπορούν να χαρακτηριστούν ως «βιογεωγραφικά νησιά» (Hugueny 1989, Hugueny, Oberdorff & Tedesco 2010) κατά συνέπεια, η μελέτη της κατανομής των ψαριών που διαβιούν σε αυτά, να υπόκειται στις αρχές τις νησιωτικής βιογεωγραφίας. Επιπλέον, οι περισσότερες κλιματικές ή γεωμορφολογικές μεταβολές έχουν σημαντικό αντίκτυπο στα οικοσυστήματα των εσωτερικών υδάτων, υπό την έννοια ότι οι συνδέσεις και οι αποσυνδέσεις που συμβαίνουν σε αυτά (π.χ. συνδέσεις ποταμών) οδηγούν σε βικαριανιστικά γεγονότα (Burridge, Craw & Waters 2006). Τα πρωτογενή ψάρια, δηλαδή εκείνα που κατάγονται από τα εσωτερικά νερά, ζουν αποκλειστικά σε αυτά και δεν ανέχονται την αλατότητα. Θεωρούνται, δε, οι καλύτεροι δείκτες βιολογικής απομόνωσης των λεκανών απορροής, διότι δεν μπορούν να υπερβούν το φράγμα του αλμυρού νερού και να μετακινηθούν δια θαλάσσης, παρά μόνο με τη συνένωση ποταμών («σύλληψη ποταμών») στα ανάντη, είτε με την ένωση των εκβολών στα κατάντη όταν τα ποτάμια εκβάλλουν σε κοινή ακτή. Συνεπώς τα είδη περιορίζονται στις λεκάνες απορροής, αναδεικνύοντας έτσι τη συνδεσιμότητα του υδρολογικού δικτύου («hydrological connectivity»). Παρόμοιους οικολογικούς περιορισμούς αντιμετωπίζουν και τα δευτερογενή είδη, δηλαδή εκείνα που προσαρμόστηκαν με επιτυχία στα εσωτερικά ύδατα, αν και οι πρόγονοί τους ή τα συγγενικά τους είδη ζούσαν ή ζουν σήμερα στη θάλασσα, ενώ η κατανομή τους αντανακλά, εκτός των άλλων, τη δυνατότητα διασποράς και μέσω των παράκτιων υδάτων (Bianco & Nordlie 2008). Εν κατακλείδι, σε περίπτωση απουσίας της ανθρώπινης παρέμβασης (π.χ. μεταφορά και εμπλουτισμός ειδών), οι κατανομές των ψαριών αντανακλούν συνήθως το αποτέλεσμα γεωτεκτονικών και άλλων γεγονότων, όπως οι συλλήψεις ποταμών, οι πλημμύρες που δημιουργούν διαδρόμους διασποράς («dispersal corridors») και η ακούσια κίνηση από χερσαίους οργανισμούς (Bănărescu 1990). Η βιογεωγραφική μελέτη των ψαριών των εσωτερικών υδάτων εμφανίζει ομοιότητες με τη νησιωτική βιογεωγραφία, όπου οι πανίδες των νησιών αντανακλούν την απομόνωση και την ηλικία των οικοσυστημάτων. Για παράδειγμα, όπως και στη νησιωτική βιογεωγραφία, πολλές υποθέσεις έχουν προταθεί για να εξηγήσουν τον αριθμό των ειδών σε ευρεία χωρική κλίμακα (Oberdorff et al. 2011, 77
78 Griffiths, McGonigle & Quinn, 2014). Αυτές μπορούν να ομαδοποιηθούν σε τρεις βασικές οικολογικές υποθέσεις (Εικόνα 1.5): I. Η Σχέση Αριθμού Ειδών και Έκτασης αναφέρει ότι ο αριθμός των ειδών αυξάνει ως συνάρτηση του εμβαδού επιφανείας (έκτασης) σύμφωνα με το δυναμικό μοντέλο S = ca z (όπου S είναι ο αριθμός των ειδών, A είναι η έκταση (ή η επιφάνεια) και c και z παράμετροι) (MacArthur & Wilson 1967) κυρίως εξαιτίας δύο γεγονότων: 1) O αριθμός των ειδών αυξάνει με την έκταση per se (MacArthur & Wilson 1967) και 2) O αριθμός των ειδών αυξάνει με την έκταση λόγω της αύξησης της ετερογένειας του τοπίου (Triantis et al. 2003) II. Η υπόθεση της παραγωγικότητας προβλέπει ότι ο αριθμός των ειδών σε μία περιοχή παρουσιάζει θετική συσχέτιση με τη συνολική διαθέσιμη ενέργεια (Wright, 1983) III. Η «ιστορική» υπόθεση (Whittaker 1977, Ricklefs 2004) προτείνει ότι τα πρότυπα ποικιλότητας είναι αποτέλεσμα των εξελικτικών διεργασιών, όπως η ειδογένεση και εξαφάνιση, σε συνδυασμό με την ικανότητα διασποράς των ειδών ως αποτέλεσμα των παρελθοντικών και παρόντων περιβαλλοντικών συνθηκών 78
79 Εικόνα 1.5. Πιθανοί αιτιακοί μηχανισμοί μεταξύ περιβαλλοντικών παραγόντων και διεργασιών που επηρεάζουν θετικά ( + ) ή αρνητικά ( - ) τον αριθμό των ειδών στη λεκάνη απορροής ενός ποταμού (από Oberdorff et al. 2011) Figure 1.5. Potential causal mechanisms between environmental factors and processes affecting positively (+) or negatively ( ) the species richness of a river drainage (from Oberdorff et al. 2011) Ανάμεσα σε αυτές τις υποθέσεις, η έκταση per se εξηγεί το μεγαλύτερο ποσοστό του αριθμού των ειδών (αύξηση της έκτασης της λεκάνης απορροής συνεπάγεται αύξηση αριθμού ειδών) (Hugueny 1989). Εντούτοις, η παραγωγικότητα και η ιστορική υπόθεση έχει ερμηνεύσει αρκετές φορές τα πρότυπα ποικιλότητας σε αρχαίες λίμνες και ευρωπαϊκά ποτάμια οικοσυστήματα (Oberdorff, Guégan, & Hugueny 1995, Tedesco et al. 2005). Οι Oberdorff et al. (2011) έδειξαν, για πρώτη φορά, ότι τα πρότυπα του αριθμού των ψαριών των εσωτερικών υδάτων είναι το αποτέλεσμα των παρελθοντικών περιβαλλοντικών συνθηκών σε παγκόσμια κλίμακα, ενισχύοντας την ιστορική υπόθεση (βλέπε επίσης παρόμοια αποτελέσματα για τα πρότυπα της β ποικιλότητας από Leprieur et al. 2011). Τα ψάρια των εσωτερικών υδάτων έχουν ευρέως χρησιμοποιηθεί για τη διερεύνηση και άλλων υποθέσεων που έχουν προταθεί στα πεδία της ιστορικής βιογεωγραφίας και της μακροοικολογίας. Ενδεικτικά, οι Leprieur et al. (2009) χρησιμοποίησαν τις κατανομές των ψαριών στις μεγαλύτερες λεκάνες απορροής της Ευρώπης και περιέγραψαν το βιογεωγραφικό φαινόμενο της φθίνουσας απόστασης 79
80 («distance decay») 10, ότι δηλαδή η ομοιότητα στη σύνθεση των βιοκοινοτήτων μεταξύ τοποθεσιών (ή περιοχών) συχνά μειώνεται όσο η απόσταση μεταξύ των τοποθεσιών αυξάνει (Nekola & White 1999). Αυτό το πρότυπο οφείλεται στη μειωμένη ικανότητα διασποράς (π.χ. λόγω γεωγραφικών εμποδίων), αλλά και στις διαφορετικές περιβαλλοντικές συνθήκες που επικρατούν στα οικοσυστήματα αυτά. Επιπλέον, σε μικρότερες χωρικές κλίμακες (π.χ. μέσα στα όρια λεκανών απορροής ποταμών), οι κατανομές των ψαριών καθορίζονται από το συνδυασμό των βιοτικών και αβιοτικών παραγόντων που ποικίλουν ανά υδρογραφικό δίκτυο (Jackson, Peres- Neto & Olden 2001), χαρακτηρίζοντας έτσι τα είδη ιχθύων ως ιδανικούς δείκτες για την εξέταση του ρόλου των οικολογικών παραγόντων στη δομή και σύνθεση των βιοκοινοτήτων (Griffiths 2006). Ως εκ τούτου, μέσα στις λεκάνες απορροής των ποταμών η διερεύνηση των προτύπων ποικιλότητας των ψαριών παρέχει πολύτιμη πληροφορία αφενός σε θεωρητικό επίπεδο, αφετέρου σε επίπεδο διαχειριστικών ενεργειών διότι είναι απαραίτητη για το σχεδιασμό των προστατευόμενων περιοχών και της ορθής διαχείρισης αυτών των οικοσυστημάτων (Morlon et al. 2008, Leprieur et al. 2011, Dias et al. 2014, Pool, Grenouillet, & Villéger 2014) (Κεφάλαιο 5). 10 Το φαινόμενο αυτό διατυπώθηκε για πρώτη φορά από τον Tobler (1970) όπου τα πάντα σχετίζονται μεταξύ τους αλλά και τα πλησιέστερα σχετίζονται περισσότερο μεταξύ τους από τα απομακρυσμένα («Everything is related to everything else, but near things are more related than distant things») 80
81 ΜΕΡΟΣ I Περιφερειακό επίπεδο: Η Βαλκανική χερσόνησος
82
83 Κεφάλαιο 2 Βιογεωγραφικές της Βαλκανικής χερσονήσου περιοχές In considering the distribution of organic beings over the face of the globe, the first great fact which strikes us is, that neither the similarity nor the dissimilarity of the inhabitants of various regions can be accounted for by their climatal and other physical conditions. (Darwin 1859)
84 2.1 Εισαγωγή Oριοθέτηση των βιογεωγραφικών περιοχών Η οριοθέτηση των βιογεωγραφικών περιοχών ή περιφερειοποίηση 11 κατά Wallace (1894) είναι η προσπάθεια διάκρισης διαφορετικών περιοχών με βάση την (αν) ομοιότητα των πανίδων και χλωρίδων τους (Kreft & Jetz 2010) και ως εκ τούτου, τα οικοσυστήματα εντός των βιογεωγραφικών περιοχών παρουσιάζουν (αν) όμοια πανιδική και χλωριδική σύνθεση. Η οριοθέτηση των βιογεωγραφικών περιοχών θεωρείται ένα πολύτιμο εργαλείο στον τομέα της βιογεωγραφίας διατήρησης (Brooks 2010, Κεφάλαιο 1), παρέχοντας σημαντικές πληροφορίες για τον προσδιορισμό των προστατευόμενων περιοχών και των ευρύτερων προτεραιοτήτων της βιολογίας της διατήρησης (Holt et al. 2013). Επιτρέπει την τεκμηρίωση των χωρικών προτύπων της βιοποικιλότητας μεταξύ διαφορετικών ταξινομικών ομάδων (π.χ. Linder et al. 2012) και αποκαλύπτει τις σχέσεις μεταξύ των περιφερειών (Gonzàlez-Orozco et al. 2014). Επίσης, δίνει τη δυνατότητα εξερεύνησης του ρόλου των οικολογικών, εξελικτικών και ιστορικών παραγόντων στη σύνθεση των ειδών σε μεγάλες χωρικές κλίμακες (Ladle & Whittaker 2011). Τέλος, σε συνδυασμό με τη διερεύνηση των χωρικών προτύπων του αριθμού των ειδών, του ενδημισμού και των απειλούμενων ειδών επιτρέπει στους ερευνητές την αναγνώριση και τον προσδιορισμό των «θερμών σημείων» βιοποικιλότητας. Ο καθορισμός των βιογεωγραφικών περιοχών σε μεγάλη χωρική ή παγκόσμια κλίμακα αντιμετωπίζει μια σειρά από προκλήσεις: 1) Τη δυσκολία συγκέντρωσης αξιόπιστων δεδομένων κατανομής των οργανισμών και τη δυσκολία επεξεργασίας μεγάλου όγκου πληροφοριών σχετικά με την παρουσία και την κατανομή τους. Αυτό έχει ως αποτέλεσμα, ο καθορισμός των βιογεωγραφικών περιοχών να γίνεται εμπειρικά, χωρίς τη χρήση συγκεκριμένης 11 Σύμφωνα με τον Wallace (1894) οι περιφέρειες πρέπει να έχουν 3 χαρακτηριστικά: 1) Θα πρέπει να στηρίζονται, κατά προσέγγιση, στα μεγάλα πρωτογενή γεωγραφικά τμήματα της γης 2) Θα πρέπει να είναι πλούσιες σε πανιδικά είδη 3) Θα πρέπει να διαθέτουν μεγάλη «ατομικότητα», να χαρακτηρίζονται δηλαδή από πολλά ιδιόμορφα είδη, γένη ή οικογένειες ή από την απουσία των ειδών, γενών ή οικογενειών οι οποίες είναι άφθονες και ευρέως διαδεδομένες σε παρακείμενες περιοχές 84
85 στατιστικής μεθοδολογίας και κατά συνέπεια οι πολλαπλές οριοθετήσεις διαφέρουν ως προς τον αριθμό, το μέγεθος και τα όρια των βιογεωγραφικών περιοχών. 2) Ακόμη και όταν αξιόπιστα δεδομένα κατανομής είναι διαθέσιμα, δεν υπάρχει ενιαίος τρόπος για τον καθορισμό βιογεωγραφικών περιοχών βάσει της σύνθεσης των ειδών, δεδομένου ότι η οριοθέτηση μπορεί να εξαρτάται κάθε φορά από το ερώτημα που τίθεται. Για παράδειγμα, οι βιογεωγραφικές περιοχές μπορούν να χρησιμοποιηθούν σε μελέτες ιστορικής βιογεωγραφίας που επικεντρώνονται κυρίως στην επίδραση της φυσικής απομόνωσης και της εξελικτικής διαφοροποίησης των ειδών για την ερμηνεία των προτύπων, της οικολογικής βιογεωγραφίας που εξετάζουν τους οικολογικούς παράγοντες που καθορίζουν τη χωρική κατανομή των ειδών σήμερα αλλά και σε μελέτες της βιογεωγραφίας διατήρησης που στοχεύουν στη διατήρηση της βιοποικιλότητας. Επιπλέον, η μέθοδος που χρησιμοποιείται μπορεί να αλλάξει αν κάποιος επιλέξει για μονάδα μέτρησης τα ποσοστά ενδημισμού, τη σύνθεση των βιοκοινοτήτων, την ομοιότητα των βιοκοινοτήτων, τον αριθμό των ειδών- ενδεικτών («indicator species») ή τον αριθμό των ειδών ανά βιογεωγραφική περιοχή. 3) Οι ιδιαιτερότητες των διάφορων ταξινομικών ομάδων δυσκολεύουν τη συλλογή και επεξεργασία των δεδομένων καθώς και την ερμηνεία των προτύπων. Για παράδειγμα, τα πρότυπα της ποικιλότητας και του ενδημισμού των ταξινομικών ομάδων που διαβιούν στα εσωτερικά ύδατα δεν βρίσκονται σε αντιστοιχία. Ως εκ τούτου, οι στρατηγικές διατήρησης δεν είναι αποτελεσματικές για όλα τα τάξα (Collen et al. 2014). Στη σύγχρονη βιβλιογραφία παρόλα αυτά έχει αναγνωριστεί ένας αριθμός κοινών ιδιοτήτων που είναι επιθυμητές στις μεθόδους που χρησιμοποιούνται. Για παράδειγμα, οι ποσοτικές αναλύσεις περιφεριοποίησης δεν πρέπει να είναι υπερβολικά ευαίσθητες σε μικρές αλλαγές στη σύνθεση των ειδών ή και στον αριθμό των ειδών ανά οικοσύστημα (Wallace 1894). Οι ποσοτικές προσεγγίσεις πρέπει, επίσης, να παρέχουν μια βάση για την ανάπτυξη υποθέσεων που αφορούν την εξελικτική ιστορία και τις διαδικασίες που διέπουν τα πρότυπα της διαφοροποίησης και της κατανομής των ειδών και να μην βασίζονται σε υποκειμενικές προσεγγίσεις (π.χ. Abell et al. 2008). 85
86 2.1.2 Η χρήση της β- ποικιλότητας για την οριοθέτηση των βιογεωγραφικών περιοχών Ενώ ο συνολικός αριθμός των ειδών καθώς και ο αριθμός των ενδημικών ή απειλούμενων ειδών δίνει έμφαση στη σημασία μιας περιοχής (Myers et al. 2000, Qian 2009), το ποσοστό της αλλαγής της σύνθεσης των ειδών μεταξύ των περιοχών αναδεικνύει την ανάγκη σχεδιασμoύ δράσεων διατήρησης. Η β-ποικιλότητα ή αλλιώς «species turnover» έχει ευρέως χρησιμοποιηθεί στη βιβλιογραφία για την οριοθέτηση των βιογεωγραφικών περιοχών καθώς αντανακλά την αλλαγή στη σύνθεση των ειδών μεταξύ δύο περιοχών ή αλλιώς το συνολικό αριθμό ειδών που διαφοροποιούν δύο γειτονικές γεωγραφικά περιοχές που χαρακτηρίζονται από διαφορετικές συνθήκες ως αποτέλεσμα της επίδρασης των περιβαλλοντικών, ιστορικών και οικολογικών παραγόντων (Εικόνα 2.1) (Kreft & Jetz 2010). Η β- ποικιλότητα μαζί με την α- και γ- ποικιλότητα αποτελούν τις τρεις συνιστώσες της ποικιλότητας των ειδών (Whittaker 1960). Η α-ποικιλότητα περιγράφει τον αριθμό των ειδών που συναντώνται σε μία γεωγραφική περιοχή ή σε ένα οικοσύστημα ή μία βιοκοινότητα. Τέλος, η γ-ποικιλότητα αφορά το συνολικό αριθμό των ειδών σε περιφερειακό («regional») επίπεδο. 86
87 Εικόνα 2.1a. Απεικόνιση της β-ποικιλότητας. Το τμήμα a περιλαμβάνει το συνολικό αριθμό των κοινών ειδών μεταξύ των δύο περιοχών, το τμήμα b περιλαμβάνει το συνολικό αριθμό των ειδών που εμφανίζονται στη περιοχή b αλλά όχι στη c και τo τμήμα c περιλαμβάνει το συνολικό αριθμό των ειδών της περιοχής c, αλλά όχι της γειτονικής b (από Koleff, Gaston & Lennon 2003) b. Διάγραμμα με τον αριθμό των δημοσιευμένων άρθρων με τη λέξη κλειδί «β-ποικιλότητα» για τη χρονική περίοδο , με βάση το ISIs Web of Science (από Anderson et al. 2011) Figure 2.1a. Depiction of beta diversity. Component a comprises the total number of common species between the two regions, the component b comprises the total number of species occurring in the area b but not in c and the component c contains the total number of species in the region c, but not in b (from Koleff, Gaston & Lennon 2003) b. Plot showing the number of published articles with «βdiversity» as a keyword from 1974 to 2009, based on ISIs Web of Science database (from Anderson et al. 2011) Η β-ποικιλότητα σχετίζεται με το βαθμό διαφοροποίησης της βιοκοινότητας σε σχέση με μια περιβαλλοντική διαβάθμιση ή σε διαφορετικές περιβαλλοντικές συνθήκες και συνεπώς, με μια πληθώρα διεργασιών που αφορούν α) την αλληλεπίδραση των ειδών (π.χ. επικάλυψη οικολογικού θώκου) και β) τα χαρακτηριστικά των οικοσυστημάτων (π.χ. κλιματικές συνθήκες, τοπογραφική πολυπλοκότητα και απομόνωση). Ως αποτέλεσμα, η μελέτη της β-ποικιλότητας αντανακλά τις παρελθοντικές διαφοροποιήσεις σε περιβαλλοντικό, οικολογικό και βιογεωγραφικό επίπεδο (π.χ. εμπόδια διασποράς) (π.χ. Koleff, Gaston & Lennon 2003, Qian, Ricklefs & White 2005, Leprieur et al. 2011). Επιπλέον, η μέτρηση της είναι μεγάλης σημασίας για την κατανόηση των βασικών διεργασιών που διαμορφώνουν τη δομή και τη σύνθεση της βιοκοινότητας σε πολλαπλές χωρικές κλίμακες, θέτοντας τη μελέτη της ως ένα από τους πιο σημαντικούς στόχους της βιογεωγραφίας και της μακροοικολογίας (McKnight et al. 2007). Για παράδειγμα, οι McKnight et al. (2007) προσδιόρισαν τις περιοχές με τις 87
88 υψηλότερες τιμές β-ποικιλότητας παγκοσμίως για αμφίβια, πουλιά και θηλαστικά χαρακτηρίζοντάς τις ως σημεία υψηλής διαχειριστικής αξίας, δεδομένου ότι οι περιοχές αυτές είναι πιο ευάλωτες στην κλιματική αλλαγή Oριοθέτηση των βιογεωγραφικών περιοχών βάσει των κατανομών των ψαριών Oι κατανομές των ψαριών έχουν χρησιμοποιηθεί ευρέως στην οριοθέτηση των βιογεωγραφικών περιοχών τόσο στα εσωτερικά ύδατα (Reyjol et al. 2007, Abell et al. 2008, Matamoros, Kreiser & Schaefer 2012) όσο και στα θαλάσσια οικοσυστήματα (Mouillot et al. 2013b, Kulbicki et al. 2013). Αποτελούν μία από τις ενδεικτικότερες ομάδες- εργαλεία για την οριοθέτηση των βιογεωγραφικών περιοχών εξαιτίας της ποικιλότητάς τους, με επαρκή ταξινόμηση και τεκμηριωμένη γεωγραφική κατανομή. Επιπλέον, τα οικοσυστήματα των εσωτερικών υδάτων φέρουν ιδιότητες που υποστηρίζουν πολλές αρχές της βιογεωγραφίας διατήρησης και παρέχουν ένα πλαίσιο εντός του οποίου μπορεί να εφαρμοστεί η περιφερειοποίηση (Griffiths 2006, Leprieur et al. 2011, Tedesco et al. 2012). Για παράδειγμα, οι Reyjol et al. (2007) οριοθέτησαν μεγάλες βιογεωγραφικές περιοχές της Ευρώπης, και στη συνέχεια οι Abell et al. (2008) παρουσίασαν τον πρώτο χάρτη βιογεωγραφικών περιοχών των εσωτερικών υδάτων του πλανήτη βάσει της γνώσης των ειδικών για τα ψάρια των εσωτερικών υδάτων Είδη-ενδείκτες σε βιογεωγραφικές περιοχές Οι βιολόγοι της διατήρησης («conservation biologists)», με στόχο την εύρεση αποτελεσματικών στρατηγικών διατήρησης των ειδών, οδηγούνται, συχνά, στην ανάδειξη ειδών για να προάγουν το συνολικό στόχο της διατήρησης της βιοποικιλότητας. Οι Hall & Grinnell (1919) ήταν από τους πρώτους που χρησιμοποίησαν την έννοια των ειδών-ενδεικτών συνδέοντας φυτικά και ζωικά είδη σε συγκεκριμένες «ζώνες ζωής», δηλαδή μεγάλες γεωγραφικές περιοχές με παρόμοια 88
89 δομή και σύνθεση. Από τότε, τα είδη-ενδείκτες έχουν χρησιμοποιηθεί ευρέως σε έρευνες σχετιζόμενες με τη βιοποικιλότητα και τη διατήρηση της. Ανάλογα με το αντικείμενο της μελέτης, τα είδη-ενδείκτες είναι είδη που, λόγω εξειδικευμένων προτιμήσεων ή ιστορικών παραγόντων, μπορούν να χρησιμοποιηθούν ως οικολογικοί δείκτες βιοκοινοτήτων, ενδιαιτημάτων ή βιογεωγραφικών περιοχών. Η ανάλυση των ειδών-ενδεικτών επιτρέπει το χαρακτηρισμό των περιβαλλοντικών προτιμήσεων των ειδών-στόχων με πολλές εφαρμογές, συμπεριλαμβανομένης της διατήρησης, της χαρτογράφησης των τοπίων, ή το σχεδιασμό των προστατευόμενων περιοχών (Oikonomou, Leprieur & Leonardos 2014). Συνήθως, προσδιορίζονται χρησιμοποιώντας μια ανάλυση της σχέσης μεταξύ της παρατηρούμενης παρουσίαςαπουσίας ειδών σε οικοσυστήματα (ή θέσεων δειγματοληψίας) και την ταξινόμησή αυτών (Dufrêne & Legendre 1997) Oριοθέτηση των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου βάσει των κατανομών των ψαριών των εσωτερικών υδάτων Η Βαλκανική χερσόνησος προσφέρει μια μοναδική ευκαιρία για τη διερεύνηση των βιογεωγραφικών θεωριών, διότι χαρακτηρίζεται από σχετικά πρόσφατη γεωλογία με έντονα γεωγραφικά εμπόδια και εξαιρετικά κατακερματισμένα υδρογραφικά δίκτυα (Bănărescu 2004). Τα περιφερειακά πρότυπα της βιοποικιλότητας καταδεικνύουν ότι η Βαλκανική χερσόνησος αποτελεί θερμό σημείο («hotspot») της Ευρωπαϊκής βιοποικιλότητας (Kryštufek & Reed 2004) με ξεχωριστή πανίδα και ενδημικά είδη που συγκεντρώνονται σε εξαιρετικά μικρές περιοχές με μικρά πληθυσμιακά μεγέθη και πολλά απειλούμενα είδη (Bănărescu 2004). Για παράδειγμα, το 52% των μαλακίων και 28% των ψαριών των εσωτερικών υδάτων που βρίσκονται στη Βαλκανική χερσόνησο απειλούνται με εξαφάνιση (Freyhof 2012). Η βιογεωγραφική περιφερειοποίηση των Βαλκανίων παραμένει ασαφής, επειδή τα υποκείμενα βιογεωγραφικά πρότυπα διακόπτονται από τα έντονα χαρακτηριστικά της περιοχής (π.χ., πολυάριθμες λίμνες, η επίδραση της 89
90 χερσονήσου 12 ) (Skoulikidis et al. 2009). Επιπλέον, η γνώση σχετικά με την κατανομή των ψαριών των εσωτερικών υδάτων στην περιοχή είναι ελλιπής και χαρακτηρίζεται από το λεγόμενο έλλειμμα του Wallace («Wallacean shortfall»), το οποίο ορίζεται ως «η ανεπαρκής γνώση των κατανομών των ειδών» (Lomolino 2004). Κατά συνέπεια, οι τελευταίες βιογεωγραφικές και οικολογικές προσπάθειες μεγάλης κλίμακας (σε ευρωπαϊκό ή παγκόσμιο επίπεδο), βασισμένες σε κατανομές ψαριών των εσωτερικών υδάτων, δεν έχουν συμπεριλάβει τη Βαλκανική χερσόνησο (Reyjol et al. 2007, Logez, Pont & Ferreira 2010, Tedesco et al. 2012, Tisseuil et al. 2013). Οι Economidis & Bănărescu (1991) στην πρώτη προσπάθεια περιφεριοποίησης της Βαλκανικής χερσονήσου προσδιόρισαν τέσσερις κύριες διαιρέσεις (Εικόνα 2.2a). Αυτές είναι: 1) Πόντο-Αιγαίο, 2) Αττικό-Βοιωτία, 3) Νότια Αδριατική-Ιόνιο, και 4) Δαλματία και τις τρεις υποδιαιρέσεις: 1α) Ανατολική Βουλγαρία, 1β) Θράκη- Ανατολική Μακεδονία και 1γ) Μακεδονία-Θεσσαλία. Στη συνέχεια, ο Bănărescu (2004) μελετώντας την κατανομή της πανίδας της Βαλκανικής χερσονήσου διέκρινε επτά (7) βιογεωγραφικές περιοχές (Εικόνα 2.2b): 1) Λεκάνη απορροής του Δούναβη, 2) Ανατολική Βαλκανική 3) Θράκη 4) Μακεδονία- Θεσσαλία, 5) Δαλματία, 6) Νότια Αδριατική και Ιονίο και 7) Αττικο-Βοιωτία. Οι Abell et al. (2008) παρουσίασαν ένα παγκόσμιο χάρτη που οριοθετεί για πρώτη φορά τις βιογεωγραφικές περιοχές των οικοσυστημάτων των εσωτερικών υδάτων οι οποίες βασίζονται στην κατανομή και τη σύνθεση των ειδών ψαριών σύμφωνα με τους Kottelat & Freyhof (2007) λαμβάνοντας υπόψη τη γνώμη ειδικών. Οι βιογεωγραφικές περιοχές που αφορούν τη Βαλκανική χερσόνησο είναι (Εικόνα 2.2c): 1) Δνείστερος-Κάτω Δούναβης, 2) Δαλματία, 3) Νοτιοανατολική Αδριατική, 4) Ιόνιο, 5) Βαρδάρης, 6) Θράκη και 7) Αιγαίο. Τέλος, οι Zogaris et al. (2009) καθόρισαν τέσσερις βιογεωγραφικές περιοχές στη νότια Βαλκανική χερσόνησο χρησιμοποιώντας δεδομένα από τις 23 μεγαλύτερες λεκάνες απορροής ποταμών, κυρίως της ηπειρωτικής Ελλάδας και τμημάτων των γειτονικών βαλκανικών χωρών. Αυτές είναι (Εικόνα 2.2d): 1) Νοτιο-ανατολική Αδριατική, 2) Ιόνιο 3) Ανατολικό Αιγαίο (Αττικό-Βοιωτία) και 4) Βόρειο Αιγαίο. 12 Η επίδραση της χερσονήσου περιγράφει τη µείωση του αριθμού των ειδών κατά µήκος των χερσονήσων όσο αυξάνεται η απόσταση από την ηπειρωτική τους σύνδεση 90
91 Παρ όλα ταύτα, οι προηγούμενες μελέτες οριοθέτησης των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου έχουν βασιστεί σε παλαιότερες βάσεις δεδομένων με διαφορετική ονοματολογία ειδών (Economidis & Bănărescu 1991, Bănărescu 2004), έχουν επικεντρωθεί μόνο στo νότιο τμήμα της χερσονήσου (Zogaris, Economou & Dimopoulos 2009) ή βασίζονται σε υποκειμενικές προσεγγίσεις (Abell et al. 2008). Οι διαφορές στα αποτελέσματα αυτών των μελετών καθώς και η επιστημονική αναγκαιότητα και σημαντικότητα της εξέτασης των βιογεωγραφικών περιοχών αποτελούν επιστημονική πρόκληση, επειδή αποτελέσματα σαν και αυτά είναι σημαντικά για την κατανόηση των χωρικών προτύπων των ειδών των εσωτερικών υδάτων και την εφαρμογή στρατηγικών διατήρησης. 91
92 Εικόνα 2.2. Προηγούμενες μελέτες οριοθέτησης των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου a. Ιχθυογεωγραφικές περιοχές της Βαλκανικής χερσονήσου (από Economidis & Bănărescu 1991) b. Περιοχές των Βαλκανίων με βάση την ιχθυοπανίδα και τα δίκτυα απορροής ποταμών (από Bănărescu 2004) c. Βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου (από Abell et al. 2008) d. Προκαταρκτικές οικολογικές περιφέρειες της Ελλάδας και των γειτονικών Βαλκανικών χωρών (από Zogaris et al. 2009) Figure 2.2a. Ichthyogeographical divisions in the Balkan Peninsula (from Economidis & Bănărescu 1991) b. Balkan regions based on fish fauna and drainage networks (from Bănărescu 2004) c. Freshwater ecoregions of the Balkan Peninsula (from Abell et al. 2008) d. Preliminary ecoregional units of Greece and its neighbouring Balkan states (from Zogaris et al. 2009) 92
93 2.1.6 Παλαιογεωγραφική αναδρομή της Βαλκανικής χερσονήσου Η Βαλκανική χερσόνησος σχηματίζει ένα ακανόνιστο, ανεστραμμένο τρίγωνο γης που εκτείνεται από την Κεντρική Ευρώπη στα βόρεια, την ανατολική Μεσόγειο στο νότο, και οριοθετείται από την Αδριατική, το Ιόνιο, το Αιγαίο και τη Μαύρη Θάλασσα. Η δημιουργία της Βαλκανικής χερσονήσου είναι αποτέλεσμα του Αλπικού γεωτεκτονικού κύκλου (ή απλά της Αλπικής ορογένεσης), όπως άλλωστε ολόκληρη η Νότια Ευρώπη, που έλαβε χώρα στο Μεσοζωικό (περίπου 200 Μa πριν) και κορυφώθηκε στο Τριτογενές (περίπου 30 Μa πριν). Οι οροσειρές της Νότιας Ευρώπης ανήκουν στην αλυσίδα όρεων, που αρχίζει από τα Πυρηναία και καταλήγει στα Ιμαλάια περιλαμβάνοντας τις οροσειρές των Άλπεων, τα Καρπάθια, τις Διναρίδες, Ελληνίδες κ.λ.π. Ο μηχανισμός της Αλπικής ορογένεσης συνδέεται στενά με την σύγκρουση των δύο ηπειρωτικών λιθοσφαιρικών πλακών, της Ευρασίας και της Αφρικανικής Πλάκας (Γκοντβάνας), τα ιζήματα των οποίων αποτέθηκαν στον ωκεανό της Τηθύος. H Tηθύς, όπου γινόταν η αποστράγγιση του ηπειρωτικού χώρου, βρισκόταν μεταξύ της Ευρασίας, του ενιαίου ηπειρωτικού χώρου της Ευρώπης και της Ασίας στο βορρά, και της Γκοντβάνα, της ενιαίας ηπείρου που περιλάμβανε την Αφρική, την Ινδία και την Αυστραλία στο νότο. Σχηματίστηκε κατά τη διάρκεια την περίοδο του Πέρμιου- Τριασικού (Παλαιοζωικό, περίπου 240 Ma). Οι συμπιεστικές τάσεις που επικρατούσαν στην Τηθύ οφείλονταν στη σύγκρουση των μεγάλων λιθοσφαιρικών πλακών και της καταστροφής του ωκεανού της. Οι μεταβολές των χερσαίων συνδέσεων και του κλίματος καθώς και οι μετατοπίσεις του γεωγραφικού πλάτους των ηπειρωτικών μαζών επηρεάσαν την εξέλιξη της Βαλκανικής πανίδας και χλωρίδας. Πιο συγκεκριμένα, στο τέλος Κρητιδικού - αρχές Τριτογενούς αρχίζει το στάδιο σύγκρουσης μεταξύ των δύο τεκτονικών πλακών και η κορύφωση της ορογένεσης με την ανύψωση των Αλπικών οροσειρών καθώς και η τελική ορογενετική διεργασία των Ελληνίδων που οδηγεί στην οριστική τους ανάδυση. Την ίδια περίοδο είναι αποδεκτό ότι οι πρόγονοι των περισσοτέρων Ευρωπαϊκών ή Παλεαρκτικών γενών και ειδών που έχουν Αμερικάνικη ή Σιβηρική προέλευση ή προέρχονται από την Ανατολική Ασία 93
94 (Economidis & Bănărescu 1991, Bănărescu 2004) ήρθαν στην Ευρώπη κατά το Παλαιόκαινο ή το πρώιμο Ηώκαινο (βλέπε επίσης Smith, Rose & Gingerich 2006). 94 Με το σχηματισμό των Αλπικών πτυχώσεων και οροσειρών από τον ωκεανό της Τηθύος δηµιουργήθηκε ένα νέο ανάγλυφο στο νότιο τµήµα της Ευρώπης. Το ανάγλυφο αυτό ευνόησε το σχηµατισµό δύο εσωτερικών θαλασσών, υπολειµµάτων του παλαιού ωκεανού, γνωστών µε τους όρους Παρατηθύς 13 και Μεσογέα. 14. Η ανάδυση της Πίνδου έγινε από το Ηώκαινο έως το Κάτω Ολιγόκαινο όταν οριστικά σχηματίσθηκε και η Παρατηθύς, που λειτούργησε στον εσωτερικό χώρο του Ευρασιατικού Περιθωρίου (Olteanu & Jipa 2006) (Εικόνα 2.3a, 2.3b, 2.3c). Κατά τη διάρκεια του Νεογενούς, η γεωτεκτονική εξέλιξη στην Πανόνια λεκάνη (βόρεια Σερβία και Κροατία στη Βαλκανική χερσόνησο) διαδραμάτησε σημαντικό ρόλο στην περιφερειακή χλωριδική σύνθεση (Erdei et al. 2007, Neubauer et al. 2015) (φάση του Σερραβάλλιου για την Μεσογέα) (Εικόνα 2.3e, 2.3f, 2.3g, 2.3h). Χαρακτηριστικό παράδειγμα της τεκτονικής αστάθειας και των ραγδαίων εξελίξεων εκείνη την εποχή, αποτελεί η περίοδος του Ύστερου Σερραβάλλιου ( ,5 Ma) όπου η μειωμένη αλατότητα και η αύξηση της αλκαλικότητας οδήγησε τα στενόαλα ενδημικά είδη της Σαρματικής θάλασσας σε εξαφάνιση (Pisera 1996). Το σημερινό γεωτεκτονικό καθεστώς του Αιγαίου και του Ελληνικού τόξου με τη βύθιση της Αφρικανικής πλάκας κάτω από την Ευρασιατική λιθόσφαιρα διαμορφώθηκε στο Μέσο Μειόκαινο, δηλαδή πριν 10 Ma (Εικόνα 2.3e, 2.3f, 2.3g). Στο Ανώτερο Μειόκαινο η ηπειρωτική Ελλάδα ήταν ενωμένη με το χώρο του Αιγαίου, που 13 Η Παρατηθύς, που ήταν χαμηλής αλατότητας θάλασσα, αποτέλεσε το καθοριστικό μέσο για την εξάπλωση των ειδών των εσωτερικών υδάτων (Bianco 1990, Reyjol et al. 2007, Neubauer et al. 2015). Κάλυπτε την περιοχή από τη σηµερινή Ουγγαρία (λεκάνη Πανονίας), Ρουµανία, Μαύρη Θάλασσα, Κριµαία, Κασπία Θάλασσα µέχρι και τη λίµνη Αράλη. Το τελευταίο στάδιο εξέλιξης της Παρατηθύος, που άρχισε µεταξύ Μ. και Α. Μειοκαίνου και ολοκληρώθηκε στο Πλειόκαινο ονοµάζεται Νεοπαρατηθύς. Την εποχή εκείνη άρχισε η διάσπαση της Παρατηθύος σε µικρές λεκάνες, αφού είχε διακοπεί οριστικά πλέον η σύνδεση της Παρατηθύος µε τον Ινδο-Ειρηνικό Ωκεανό. Η θαλάσσια επίκλυση της κλειστής Παρατηθέως περίπου πριν 8,5 εκ. έτη δημιουργεί το Αιγαίο πέλαγος και η Ελληνική χερσόνησος αρχίζει να σχηματίζεται με την μορφή που την γνωρίζουμε σήμερα (Εικόνα 2.3f). Η σηµερινή Μαύρη Θάλασσα, η Κασπία Θάλασσα και η λίµνη Αράλη, θεωρούνται υπολείµµατα της Παρατηθύος. 14 Η Μεσογέα και η σηµερινή Μεσόγειος εγκλωβισμένες µεταξύ των αλπικών οροσειρών και της αφρικανικής ηπείρου, αποτέλεσαν ένα νέο χώρο αποστράγγισης της ηπειρωτικής Ευρώπης. Η σύνδεση της Μαύρης Θάλασσας µε τη Μεσόγειο µέσω της βύθισης Αιγηίδας και το σχηµατισµό του Αιγαίου Πελάγους είναι αποτέλεσµα των γεωδυναµικών εξελίξεων του Τεταρτογενούς και αποτέλεσε την αιτία της διασποράς πολλών ειδών με προέλευση Μαύρης Θάλασσας στην Μεσογειακή Λεκάνη (Dermitzakis & Papanikolaou 1981)
95 επίσης ήταν ξηρά, καθώς και με τη Μικρά Ασία (Εικόνα 2.3h). Το νοτιότερο τμήμα αυτής της περιοχής, ονομάστηκε από τον Phillipson (1898) «Αιγηίδα». Η σύνδεση αυτή επιβεβαιώνεται και με τα παλαιοντολογικά ευρήματα που πιστοποιούν τη μετακίνηση μεγάλων θηλαστικών ζώων (Dermitzakis & Papanikolaou 1981) καθώς και με την ομοιότητα της ιχθυοπανίδας μεταξύ της βιογεωγραφικής περιοχής του Αιγαίου με αυτής της Μικράς Ασίας (Zogaris, Economou & Dimopoulos 2009, Oikonomou, Leprieur & Leonardos 2014). Τα Ασιατικά είδη εισέρχονταν στα Βαλκάνια από την Ολιγόκαινο έως και τη Μειόκαινο περίοδο μέσω Σιβηρίας. Κατά το τέλος του Μειοκαίνου, και συγκεκριμένα, κατά την φάση της «Κρίσης της Αλατότητας» (5.5 Ma) (Hsü et al. 1977), τα στενά του Γιβραλτάρ έκλεισαν, η Μεσόγειος Θάλασσα αποξηράνθηκε και γέμισε με νερό από την Παρατηθύ. Η «Κρίση της Αλατότητας» είχε τεράστιο αντίκτυπο στην πανιδική σύνθεση των Βαλκανίων, και συνοδεύτηκε από σημαντική διάβρωση των ποταμών, κοιλάδων και σπηλαίων εκτός από την πρόκληση μιας πλήρους αλλαγής στις θαλάσσιες συνθήκες (Reed, Kryštufek & Eastwood 2004). Σε αυτή τη φάση αποδίδεται η διασπορά πολλών υδρόβιων οργανισμών στη Περι-Μεσογειακή περιοχή (Bianco 1990). Το σενάριο της διασποράς της πανίδας της Παρατηθύος στην Μεσόγειο, μέσω της φάσης «Lago Mare», προβλέπει μια ταχεία περίοδο διαφοροποίησης της πανίδας και μια στενή συσχέτιση μεταξύ της πανίδας των χερσονήσων της Νοτίου Ευρώπης (Levy, Doadrio & Almada 2009, Oikonomou, Leprieur & Leonardos 2014, Neubauer et al για τα γαστερόποδα των εσωτερικών υδάτων κατά το Νεογενές). Εκτός αυτών, όμως, δημιουργήθηκαν και εκτεταμένες γέφυρες ξηράς (διάδρομοι διασποράς) που επέτρεψαν τη μετακίνηση πολλών ειδών χλωρίδας και πανίδας από την Αφρική στην Ευρώπη (Sonnenfeld 1985, Agustí, Rook & Andrews 1999). Παράλληλα, οι γέφυρες ξηράς που συνέδεαν το κεντρικό Αιγαίο έσπασαν σχηματίζοντας νησιά που αποτέλεσαν τις σημερινές Κυκλάδες (Dermitzakis 1990). Η έναρξη του Πλειοκαίνου σηματοδοτείται από την αποκατάσταση των θαλάσσιων συνθηκών σε ολόκληρη τη Μεσόγειο (θαλάσσια επίκλυση από τον Ατλαντικό Ωκεανό), όταν και σχηματίστηκαν οι μεγάλες τάφροι των Βαλκανικών Ποταμών (Εικόνα 2.3i). Αποτέλεσμα των συμπιέσεων του Πλειστόκαινου θεωρήθηκε η απόσυρση της θάλασσας από το Αιγαίο και ο σχηματισμός μεγάλων ποταμών και 95
96 λιμνών με πανίδα προέλευσης από την Κασπία και τη Μαύρη θάλασσα (για περισσότερες λεπτομέρειες βλέπε Σημείωμα 1). Οι βασικές αιτίες αλλαγής της γεωγραφίας της περιοχής ήταν ο ευστατισμός και ο τεκτονισμός. Οι ευστατικές κινήσεις, που ήταν αποτέλεσμα των παγετώδων και μεσοπαγετώδων περιόδων, προκαλούσαν επέκταση ή μείωση των χερσαίων περιοχών και μεταβολή των μεταξύ τους χερσαίων συνδέσεων καθώς και αυξομειώσεις στην στάθμη της θάλασσας. Οκτώ τέτοιοι κύκλοι παρατηρήθηκαν καθόλη τη διάρκεια του Πλειστόκαινου σύμφωνα με τους Beard, Sangree & Smith (1982), η τελευταία εκ των οποίων, η Βούρμια παγετώδης περίοδος, ξεκίνησε πριν από και ολοκληρώθηκε πριν από χρόνια. Στην Ευρώπη, κατά τη διάρκεια της τελευταίας παγετώδους περιόδου ( έως χρόνια πριν- «Last Glacial Maximum»), το κάλυμμα πάγου χώριζε την ήπειρο σε δύο τμήματα, το βόρειο μέρος που καλύπτονταν από πάγο, ενώ το νότιο τμήμα (κυρίως περιοχές γύρω από τη Μεσόγειο Θάλασσα) παρέμεινε ακάλυπτο (Hewitt 2000, Reyjol et al. 2007). Ως συνέπεια, πολλά είδη βρήκαν καταφύγιο σε αυτές τις περιοχές (δηλαδή στα Βαλκάνια, στον Καύκασο, στην Ιβηρική και στην Ιταλική χερσόνησο) και τα οποία εποίκισαν αργότερα την Ευρώπη (Hewitt 2000). 96

97 97
 Figure 2.3. Paleogeographic maps of Paratethys (from Popov et al. 2004) a. Late Eocene (37-34 Ma) b. Early Oligocene (34-32 Ma) c. Late Oligocene (29-24 Ma) d. Early Miocene (20.")
98 Εικόνα 2.3. Παλαιογεωγραφικοί χάρτες της Παρατηθέως (από Popov et al. 2004) a. Άνω Ηώκαινο(3734 Ma) b. Κάτω Ολιγόκαινο (34-32 Ma) c. Άνω Ολιγόκαινο (29-24 Ma) d. Άνω Μειόκαινο ( Ma) Figure 2.3. Paleogeographic maps of Paratethys (from Popov et al. 2004) a. Late Eocene (37-34 Ma) b. Early Oligocene (34-32 Ma) c. Late Oligocene (29-24 Ma) d. Early Miocene ( Ma) 98
 Figure 2.3. e. Middle Miocene- Middle Serravaillan (14-13 Ma) f.")
99 Εικόνα 2.3. e. Μέσο Μειόκαινο- Μέσο Σερραβάλλιο (14-13 Ma) f. Μέσο Μειόκαινο- Κάτω Σερραβάλλιο - Σαρμάτιο (12-11 Ma) Figure 2.3. e. Middle Miocene- Middle Serravaillan (14-13 Ma) f. Middle Miocene- Late SerravaillanSarmatian (12-11 Ma) 99
100 Εικόνα 2.3. e. Μέσο Μειόκαινο- Μέσο Σερραβάλλιο (14-13 Ma) f. Μέσο Μειόκαινο- Κάτω Σερραβάλλιο- Σαρμάτιο (12-11 Ma) g. Μέσο- Κάτω Μειόκαινο- Κάτω Τορτόνιο- Άνω Μεσσήνιο (8.5-7 Ma) h. Κάτω Μειόκαινο- Κάτω Μεσσήνιο- Κάτω Παννόνιο ( Ma) Figure 2.3. g. Mid- Late Miocene- Late Tortonian- Early Messinian (8.5-7 Ma) h. Latest Miocene- Late Messinian- Late Pannonian ( Ma) 100
 Figure 2.3. i. Middle-Late Pliocene (3.4-1.")
101 Εικόνα 2.3. i. Μέσω-Κάτω Πλειόκαινο ( Ma) Figure 2.3. i. Middle-Late Pliocene ( Ma) 101
102 2.2 Στόχος του κεφαλαίου Η Βαλκανική χερσόνησος είναι ένα σημείο μεγάλου ενδιαφέροντος της ευρωπαϊκής βιοποικιλότητας και προσφέρει μια μοναδική ευκαιρία για τη διερεύνηση διάφορων μακροοικολογικών και βιογεωγραφικών θεωριών. Στο παρόν κεφάλαιο, οριοθετήθηκαν οι βιογεωγραφικές περιοχές της Βαλκανικής χρησιμοποιώντας την πληρέστερη βάση δεδομένων κατανομής ψαριών των εσωτερικών υδάτων που είναι διαθέσιμη μέχρι σήμερα. Παράλληλα, μέσω της ανάλυσης των ειδών-ενδεικτών, εξετάστηκαν ποια είδη χαρακτηρίζουν τις βιογεωγραφικές περιοχές. Οι προηγούμενες μελέτες οριοθέτησης των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου έχουν προτείνει διαφορετικά πρότυπα και είτε έχουν επικεντρωθεί μόνο στο νότιο τμήμα της Βαλκανικής είτε έχουν βασιστεί σε εμπειρικές προσεγγίσεις. Αυτό το γεγονός καθώς και η επιστημονική αναγκαιότητα και σημαντικότητα της εξέτασης των βιογεωγραφικών περιοχών αποτελούν επιστημονική πρόκληση για την περιοχή των Βαλκανίων. Η χρησιμοποίηση βάσεων δεδομένων με ευρεία γεωγραφική αναφορά είναι κομβικής σημασίας για να μελετηθούν τα χωρικά πρότυπα των οργανισμών των εσωτερικών υδάτων αλλά και για να αναπτυχθούν οι βέλτιστες στρατηγικές προστασίας και διαχείρισης τους (Heino 2011). Αυτό το κεφάλαιο αποτελεί ένα πρώτο βήμα για την ανάπτυξη πρακτικών σχετικά με την διατήρηση των εσωτερικών υδάτων στην ευρύτερη περιοχή της Βαλκανικής υπό ένα ενιαίο πλαίσιο διακριτών βιογεωγραφικών περιοχών. 2.3 Υλικά και Μέθοδοι Βάση δεδομένων κατανομής των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου Η βάση δεδομένων (ΒΔ) κατανομής των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου περιλαμβάνει πανιδικές λίστες από λεκάνες απορροής ποταμών και φυσικές λίμνες που κατανέμονται στη Βαλκανική χερσόνησο. Τα 102
103 δεδομένα συλλέχθηκαν από εκτεταμένη έρευνα βιβλιογραφίας συμπεριλαμβανομένων 364 βιβλιογραφικών πηγών και χρησιμοποιήθηκαν έπειτα από αξιολόγηση της εγκυρότητάς τους (Παράρτημα 1). Οι βιβλιογραφικές πηγές, εκτός των επιστημονικών άρθρων, περιλαμβάνουν δεδομένα από άλλες βάσεις δεδομένων (όπως και βιβλία και γκρίζα βιβλιογραφία (όπως σχέδια δράσης, τεχνικές εκθέσεις και τοπικές εφημερίδες). Τα δεδομένα συγκεντρώθηκαν με βάση τα ακόλουθα κριτήρια: 1) να συμπεριλαμβάνονται τα μεγαλύτερα οικοσυστήματα των εσωτερικών υδάτων των Βαλκανίων 2) να αντιπροσωπεύονται όλες οι γεωγραφικές περιοχές και 3) να υπάρχουν διαθέσιμα και αξιόπιστα επιστημονικά δεδομένα για κάθε οικοσύστημα. Στη ΒΔ καταχωρήθηκαν δεδομένα από παραποτάμους που καταλήγουν σε ποτάμια και λίμνες, εξαιτίας του γεγονότος ότι η σύνθεση ειδών του κάθε παραποτάμου επηρεάζει τη συνολική σύνθεση της ιχθυοπανίδας και τη μοναδικότητα της λεκάνης απορροής (Hoagstrom & Berry Jr 2006). Στη ΒΔ δεν καταχωρήθηκαν δεδομένα από νησιά (Αιγαίο και Ιόνιο Πέλαγος και Αδριατική Θάλασσα) ή την ανατολική Πελοπόννησο (νότια Ελλάδα), λόγω έλλειψης αξιόπιστων πληροφοριών. Μόνο αυτόχθονα και πρωτογενή είδη ή δευτερογενή που η κατανομή τους ομοιάζει με αυτή των πρωτογενών ειδών εξετάστηκαν για κάθε σύστημα ώστε κάθε λεκάνη απορροής να θεωρείται ως ένα κλειστό σύστημα (Hugueny, Oberdorff & Tedesco 2010). Συστήματα με ελλιπείς καταλόγους των ειδών (δηλαδή, απουσία μελέτης σε ένα τμήμα του οικοσυστήματος ή πληροφορία που βασίζεται μόνο σε ένα συγκεκριμένο γένος) παραλείφθηκαν από τη ΒΔ και από τις αναλύσεις. Τα υποείδη της πέστροφας, Salmo trutta fario L., S. trutta lacustris L. και S. trutta trutta L. και τα είδη του γένους Caspiomyzon παραλείφθηκαν από τις αναλύσεις, λόγω ταξινομικής αβεβαιότητας. Είδη με μεγάλη οικονομική σημασία, όπως τα Carassius carassius, Cyprinus carpio, Rhodeus amarus, Silurus glanis και Tinca tinca δεν προστέθηκαν στις πανιδικές λίστες εξαιτίας της μεταφοράς τους για εμπλουτισμό των οικοσυστημάτων και ως εκ τούτου, εξαιτίας της αμφίβολης φυσικής τους παρουσίας. Όσον αφορά την ονοματολογία των ειδών, ακολουθήθηκε η ταξινόμηση που προτείνεται από τους 103
104 Kottelat & Freyhof (2007). Τα είδη που χρησιμοποιήθηκαν στις αναλύσεις παρουσιάζονται στο Παράρτημα 2. Συνολικά, η ΒΔ περιέχει δεδομένα κατανομής για 193 είδη σε 78 λεκάνες απορροής ποταμών και 19 φυσικές λίμνες (Πίνακας 2.1). 104
105 Πίνακας 2.1. Ονόματα ποταμών και λιμνών που χρησιμοποιήθηκαν στις αναλύσεις. Το σύμβολο * υποδηλώνει τις λίμνες Table 2.1. River and Lake IDs and names used in the analyses. * indicates Lakes ID Basin name ID Basin name ID Basin name ID Basin name 1 Prespa* 31 Doiran* 61 Danube 91 Bregava 2 Sperchios 32 Vegoritis* 62 Tisa 92 Krupa 3 Assopos 33 Aliakmon 63 Zasavica 93 Buna 4 Kifissos 34 Loudias 64 Matura 94 Neretva 5 Trichonis* 35 Axios/ Vardar 65 Brzaja 95 Cetina 6 Lyssimachia* 36 Vistonis* 66 Great 96 Zrmanja morava 7 Amvrakia* 37 Byala 67 ^Cesma 97 Krka 8 Ozeros* 38 Krumovitsa 68 Bosut 9 Mornos 39 Arda 69 Lonja 10 Acheloos 40 Evros/ Maritsa 70 Drina 11 Evinos 41 Nestos/ Mesta 71 Ilova 12 Louros 42 Vacha 72 Drenica 13 Acheron 43 Volvi* 73 Sitnica 14 Alfios 44 Strymon/ Struma 74 Kolubara 15 Arachthos 45 Ropotamo 75 Ibar 16 Kalamas 46 Dyavolska 76 Lab 17 Aoos/ Vjose 47 Karaagch 77 West morava 18 Shkumbin 48 Rezovo 78 Spreca 19 Drin 49 Veleka 79 Rasina 20 White Drin 50 Burgas* 80 Sava 21 Skadar* 51 Srebarna* 81 Una 22 Buna/Bojana 52 Durankulak* 82 Sana 23 Sas* 53 Kamchyia 83 Pek 24 Zeta 54 Yantra 84 Vrbas 25 Moraca 55 Carska Bara Bog* 85 Drava 26 Ohrid* 56 Vit 86 Bosna 27 Black drin 57 South Morava 87 Trebisnjica 28 Crna 58 Lim/Ljuca/ Grncar/Vermosh 88 Trebizat/ Matica 29 Babuna 59 Ukrina 89 Bacina* 30 Pinios The 60 Lonjsko polje* 90 Deran* 105
106 2.3.2 Προτεινόμενοι δείκτες β- ποικιλότητας για την οριοθέτηση βιογεωγραφικών περιοχών Η οριοθέτηση των βιογεωγραφικών περιοχών συνιστά στην ομαδοποίηση περιοχών, σύμφωνα με την ανομοιότητα της πανιδικής και χλωριδικής σύνθεσης τους, και κατά συνέπεια, αφορά τη διάκριση των περιοχών με διακριτές βιογεωγραφικές ιστορίες (Kreft & Jetz 2010). Οι αναλύσεις ταξιθέτησης με την κατασκευή δενδρογραμμάτων («clustering») παράγουν ποσοτική, ιεραρχική κατάταξη της (αν) ομοιότητας μεταξύ του συνόλου των ειδών κάθε περιοχής, αλλά είναι ευαίσθητες ανάλογα με τη μετρική (αν) ομοιότητας και τον αλγόριθμο της ταξιθέτησης που επιλέγεται (Kulbicki et al. 2013). Ως εκ τούτου, η επιλογή της κατάλληλης μετρικής για τη δημιουργία της μήτρας των αποστάσεων μεταξύ των οικοσυστημάτων αποτελεί το πρώτο βήμα κατά την οριοθέτηση των βιογεωγραφικών περιοχών. Μέχρι σήμερα, έχει προταθεί μια πληθώρα μετρικών για τον ποσοτικό προσδιορισμό της β- ποικιλότητας (βλέπε Koleff, Gaston & Lennon, 2003, Anderson et al για την ανασκόπηση της βιβλιογραφίας), αν και υπάρχουν διαφωνίες μεταξύ των επιστημόνων στη χρήση αυτών των μετρικών κατά την οριοθέτηση βιογεωγραφικών περιοχών (Leprieur & Oikonomou 2014) Ο δείκτης βsim Πρόσφατα, οι Kreft & Jetz (2010) πρότειναν ένα ποσοτικοποιημένο πλαίσιο για την οριοθέτηση των βιογεωγραφικών περιοχών, με βάση την ταξινόμηση και ταξιθέτηση (Σημείωμα 2). Συγκεκριμένα, τόνισαν ότι οι μετρικές για την αντικατάσταση των ειδών που ασθενώς επηρεάζονται από τις διαφορές στον αριθμό των ειδών είναι πιο ενδεικτικές για την οριοθέτηση των βιογεωγραφικών περιοχών από τις κλασικές μετρικές, όπως οι δείκτες ανομοιότητας Jaccard και Sørensen (Σημείωμα 2). Ως εκ τούτου, οι Kreft & Jetz (2010) πρότειναν τη χρήση του δείκτη βsim, ο οποίος είναι γνωστό ότι επηρεάζεται ασθενώς από τις διαφορές στον αριθμό των ειδών (Koleff, Gaston & Lennon, 2003, Baselga, 2010, Mouillot et al. 2013b). Για παράδειγμα, οι 106
107 Mouillot et al. (2013b) έδειξαν ότι ο δείκτης βsim ελαχιστοποιεί το ενδεχόμενο παραπλανητικών αποτελεσμάτων λόγω του σχετικού μεγέθους των περιοχών δειγματοληψίας κατά τον προσδιορισμό των βιογεωγραφικών περιοχών, καθώς μία ανάλυση οριοθέτησης συχνά περιλαμβάνει ευρεία διακύμανση στις περιοχές δειγματοληψίας που μπορεί από μόνη της να προκαλέσει μεγάλες διαφορές στον αριθμό των ειδών. Ο δείκτης βsim δίνεται από τη σχέση: β sim = ( min( b, c) ) + min( b, c) ( α ) (1) όπου α είναι ο αριθμός των ειδών που είναι κοινά στις δύο τοποθεσίες, b είναι ο αριθμός των ειδών που εμφανίζονται στην πρώτη τοποθεσία, αλλά όχι στη δεύτερη, και c είναι ο αριθμός των ειδών που εμφανίζονται στην δεύτερη τοποθεσία, αλλά όχι στην πρώτη. Ο δείκτης βsim κυμαίνεται από 0 (χαμηλή ανομοιότητα, πανομοιότυπες λίστες ειδών) και 1 (υψηλή ανομοιότητα, χωρίς κοινά τάξα) Ο δείκτης β-3 Εντούτοις, ο δείκτης βsim πρόσφατα αξιολογήθηκε αρνητικά από τους Carvalho, Cardoso & Gomes (2012), οι οποίοι υποστήριξαν ότι υπερεκτιμά την αντικατάσταση των ειδών διότι μετρά την αντικατάσταση σε σχέση με τις φτωχότερες σε είδη περιοχές (βλέπε παρανομαστή εξίσωσης 1) και όχι ως προς το σύνολο των ειδών που θα μπορούσαν να αντικατασταθούν. Στην συνέχεια, οι Carvalho, Cardoso & Gomes (2012) πρότειναν τη χρήση του δείκτη β - 3, ο οποίος προτάθηκε αρχικά από τους Cardoso, Borges & Veech (2009) ως: β 3 = ( min( b, c) ) ( a+ b+ c) (2) Σύμφωνα με τους Cardoso, Borges & Veech (2009), ο δείκτης β - 3 κυμαίνεται μεταξύ 0 (ταυτόσημα τάξα) και 1 (χωρίς κοινά τάξα) και δεν είναι ευαίσθητος στις διαφορές στον αριθμό των ειδών μεταξύ περιοχών. Ομοίως με το δείκτη βsim, ο 107
108 δείκτης β- 3 ισούται επίσης με το μηδέν όταν τα σύνολα είναι εγκιβωτισμένα (π.χ. a = 10, b = 0 και c = 5). Ωστόσο, ο Baselga (2012) υποστήριξε ότι ο δείκτης β - 3 υποεκτιμά την αντικατάσταση των ειδών (βλέπε παρανομαστή εξίσωσης β-3), επειδή αντιπροσωπεύει το συνολικό αριθμό των ειδών στον παρονομαστή και όχι το συνολικό αριθμό των ειδών που δυνητικά μπορούν να αντικατασταθούν Ο δείκτης βjtu Ως ανταπάντηση, ο Baselga (2012) πρότεινε μια τροποποιημένη εκδοχή του δείκτη βsim, το δείκτη βjtu, ο οποίος δίνεται από τη σχέση: ( 2 min( b, c) ) + 2 min( b, c) β jtu = (3) ( α ) Ο δείκτης βjtu μετρά το ποσοστό των ειδών που θα αντικατασταθούν μεταξύ των βιοκοινοτήτων αν αυτές είχαν τον ίδιο αριθμό ειδών και, κατά συνέπεια, αντιπροσωπεύει την αντικατάσταση των ειδών, χωρίς την επίδραση των διαφορών στον αριθμό των ειδών. Ο δείκτης βjtu κυμαίνεται μεταξύ 0 (χαμηλή ανομοιότητα, πανομοιότυπα τάξα) και 1 (υψηλή ανομοιότητα, χωρίς κοινά τάξα). 108

109 109
110 2.3.3 Επιλογή του καταλληλότερου δείκτη β-ποικιλότητας για την οριοθέτηση των βιογεωγραφικών περιοχών στη Βαλκανική χερσόνησο Όπως ήδη αναφέρθηκε και προηγουμένως, η επιλογή του καταλληλότερου δείκτη β- ποικιλότητας αποτελεί καθοριστικό βήμα κατά την οριοθέτηση των βιογεωγραφικών περιοχών, μιας και υπάρχουν διαφωνίες μεταξύ των επιστημόνων στη χρήση αυτών των μετρικών (Leprieur & Oikonomou, 2014). Για παράδειγμα, σε προηγούμενες μελέτες έχουν ευρέως χρησιμοποιηθεί οι δείκτες του Sørensen/ Bray- Curtis, Jaccard και Kulczynski που όμως επηρεάζονται έντονα από τις διαφορές του αριθμού των ειδών μεταξύ των οικοσυστημάτων (Lennon et al. 2001). Με αφορμή αυτό το γεγονός, οι Leprieur & Oikonomou (2014) διερεύνησαν τα πιθανά πλεονεκτήματα και μειονεκτήματα των δεικτών για τον προσδιορισμό των βιογεωγραφικών περιοχών. Πιο συγκεκριμένα, χρησιμοποιώντας αριθμητικά παράδειγματα (Σημείωμα 3) και την κατανομή 136 αυτόχθονων ψαριών σε 26 μεγάλες ευρωπαϊκές λεκάνες απορροής ποταμών, οι Leprieur & Oikonomou (2014) συνέκριναν τα αποτελέσματα της ταξιθέτησης από τη χρήση των δεικτών βsim, βjtu και β- 3. Τα αποτελέσματά τους έδειξαν ότι ο δείκτης β-3 τείνει να υποεκτιμά το επίπεδο της χωρικής αντικατάστασης των ειδών εξαιτίας των διαφορών στον αριθμό των ειδών στον παρονομαστή (Baselga 2012), γεγονός που μπορεί να οδηγήσει σε παραπλανητική ταξιθέτηση των τοποθεσιών με βάση την σύνθεση των ειδών τους. Οι δείκτες βsim και βjtu είναι πράγματι ανεξάρτητοι από τον αριθμό των ειδών κατά τη χωρική αντικατάσταση των ειδών και θεωρούνται οι πιο κατάλληλοι για τον προσδιορισμό των βιογεωγραφικών περιοχών. Αυτή η διαπίστωση δεν ισχύει για το δείκτη β-3 που μειώνεται όσο αυξάνεται ο αριθμός των ειδών ακόμα και όταν οι δυο τοποθεσίες δεν μοιράζονται κανένα είδος (Σημείωμα 3). 110

111 Επιπλέον, ο δείκτης β-3 παραβιάζει την ιδιότητα της «συμπληρωματικότητας» 15, η οποία αποτελεί προϋπόθεση κατά την ανάλυση των προτύπων της β- ποικιλότητας (Legendre & De Cáceres 2013). Για παράδειγμα, ο 15 Η ιδιότητα της «συμπληρωματικότητας» περιγράφει την κατάσταση όπου περιοχές χωρίς κοινά είδη έχουν τη μεγαλύτερη ανομοιότητα και θα πρέπει να μην εμφανίζονται σε κοντινές θέσεις κατά την ταξιθέτηση 111
112 δείκτης β-3 έδειξε ότι τα ζευγάρια Α-D και A-I είναι ίσα ως προς τη ανομοιότητα (β- 3=0.4, Σημείωμα 3). Αυτό σημαίνει ότι οι τοποθεσίες D και I είναι εξίσου πιθανό να ομαδοποιηθούν με τη τοποθεσία A κατά την ιεραρχική ομαδοποίηση. Ωστόσο, στο ζευγάρι Α-D υπάρχουν 10 κοινά είδη σε σύγκριση με το ζευγάρι Α-Ι που δεν μοιράζεται κανένα είδος. Με βάση αυτά τα αποτελέσματα (Σημείωμα 4), οι Leprieur & Oikonomou (2014) κατέληξαν ότι ο δείκτης β-3 δεν πρέπει να χρησιμοποιείται στην οριοθέτηση των βιογεωγραφικών περιοχών. Αντίθετα, συνέστησαν τη χρήση των δεικτών βsim και βjtu επειδή φέρουν επιθυμητές ιδιότητες για την οριοθέτηση των βιογεωγραφικών περιοχών. Ως αποτέλεσμα, οι δείκτες βsim και βjtu που έχουν προταθεί για την οριοθέτηση των βιογεωγραφικών περιοχών, είναι οι καταληλότεροι και φέρουν επιθυμητές ιδιότητες για την οριοθέτηση των βιογεωγραφικών περιοχών. Θα πρέπει να σημειωθεί εδώ ότι σύμφωνα με τον Baselga (2012) και τους Leprieur & Oikonomou (2014), οι δείκτες βjtu και βsim είναι ταυτόσημοι και παρέχουν σχεδόν παρόμοια αποτελέσματα σε δεδομένα παρουσίας/απουσίας. Ως εκ τούτου, στο παρόν κεφάλαιο, χρησιμοποιήθηκε ο δείκτης βjtu (Baselga 2012) για να εντοπιστούν οι κύριες βιογεωγραφικές περιοχές των Βαλκανίων με βάση την αλλαγή της σύνθεσης των ειδών («species turnover»), χωρίς την επιρροή των διαφορών στον αριθμό των ειδών ανά οικοσύστημα (βλέπε επίσης Σημειώματα 2, 3, 4). 112
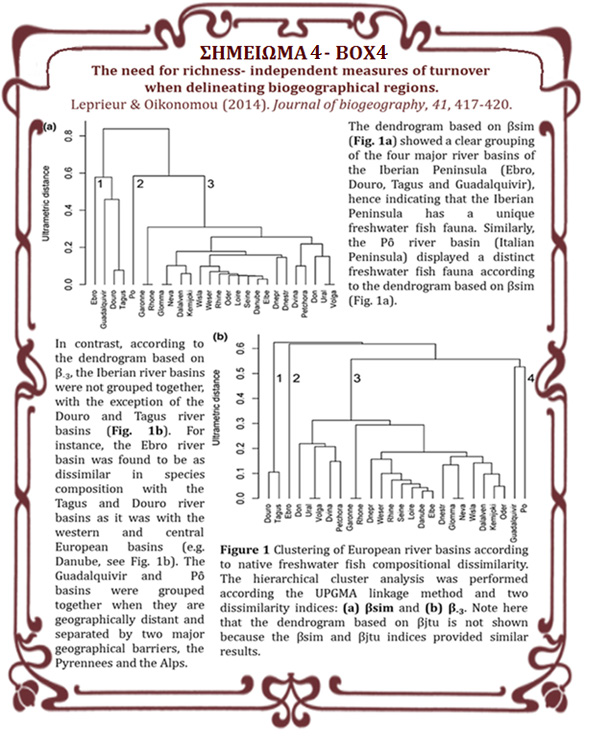
113 113
114 2.3.4 Δημιουργώντας το δενδρόγραμμα για την οριοθέτηση των βιογεωγραφικών περιοχών Η επιλογή του αλγόριθμου ομαδοποίησης είναι το δεύτερο σημαντικό βήμα κατά την οριοθέτηση των βιογεωγραφικών περιοχών και καθορίζεται από τους εκάστοτε στόχους της βιογεωγραφικής μελέτης. Για παράδειγμα, εάν ο στόχος της οριοθέτησης των βιογεωγραφικών περιοχών είναι η συσχέτιση με δεδομένα βλάστησης ή κλίματος (π.χ. Heikinheimo et al. 2007), προτείνεται η μη ιεραρχική ομαδοποίηση. Αντίθετα, αν ο στόχος είναι ο καθορισμός διακριτών ομάδων έπειτα από διαχωρισµό των οικοσυστημάτων µιας περιοχής σε υποσύνολα έτσι ώστε να υπάρχει οµοιογένεια µέσα στο κάθε υποσύνολο και ανοµοιογένεια µεταξύ των στοιχείων που ανήκουν σε διαφορετικά υποσύνολα, η ιεραρχική ανάλυση κατά συστάδες (hierarchical clustering) είναι η πιο διαδεδομένη στατιστική ανάλυση (Kreft & Jetz 2010). Εικόνα 2.4. Εννοιολογικό διάγραμμα των σταδίων ανάλυσης ροής δεδομένων σε πολυπαραγοντικές βιογεωγραφικές αναλύσεις (τροποποιημένο από Kreft & Jetz 2010) Figure 2.4. Conceptual diagram of analysis steps and data flow in multivariate biogeographical analyses (modified from Kreft & Jetz 2010) Στην παρούσα μελέτη, χρησιμοποιήθηκαν δεδομένα παρουσίας/απουσίας κατανομών των ψαριών στα εσωτερικά ύδατα της Βαλκανικής χερσονήσου και υπολογίστηκαν οι αποστάσεις κατά ζευγάρια (pairwise), με την εφαρμογή του δείκτη βjtu. Ο δείκτης βjtu δημιουργεί έναν πίνακα αποστάσεων ανομοιότητας μεταξύ των οικοσυστημάτων με βάση την αλλαγή στη σύνθεση των 114
115 ειδών. Με βάση τη μήτρα ανομοιότητας που προέκυψε από την εφαρμογή του δείκτη, στη συνέχεια εφαρμόστηκε η ιεραρχική ανάλυση ομαδοποίησης (HCA). Η διαδικασία της ιεραρχικής οµαδοποίησης συγχωνεύει τα πιο όμοια οικοσυστήματα για να δηµιουργήσει µια οµάδα. Τέλος, υπολογίστηκαν οι αποστάσεις ανάμεσα στην κάθε οµάδα και µεταξύ των άλλων οµάδων. Πρακτικά, η ιεραρχική ανάλυση ομαδοποίησης επιτρέπει την απεικόνιση της σχετικής απόστασης μεταξύ των οικοσυστημάτων με τα δεδομένα της σύνθεσης της πανίδας τους (δηλαδή οδηγεί στη δημιουργία ενός δενδρογράμματος). Στο τελευταίο στάδιο, χρησιμοποιήσαμε τη μέθοδο σύνδεσης «συσχέτιση µη σταθµισµένων οµάδων ανά ζεύγη µε αριθµητικούς µέσους όρους» (UPGMA- Unweighted Pair Group Method with Arithmetic Mean), όπως συνιστάται από τους Kreft & Jetz (2010) (Εικόνα 2.4). Πράγματι, με βάση τις προκαταρκτικές αναλύσεις των Mérigot, Durbec & Gaertner (2010) με το κριτήριο 2- norm, η μέθοδος σύνδεσης «συσχέτιση µη σταθµισµένων οµάδων ανά ζεύγη µε αριθµητικούς µέσους όρους» (UPGMA) παρείχε μια πιο πιστή αναπαράσταση του αρχικού πίνακα ανομοιότητας (Kreft & Jetz 2010) σε σχέση με άλλες μεθόδους σύνδεσης [συσχέτιση µη σταθµισµένων οµάδων ανά ζεύγη µε κεντροειδή (UPGMC- Unweighted Pair-Group Method using Centroids), συσχέτιση σταθµισµένων οµάδων ανά ζεύγη µε αριθµητικούς µέσους όρους (WPGMA- Weighted Pair Group Method with Arithmetic Mean), Ward, του κοντινότερου γείτονα (nearest neighbour ή single linkage) και του µακρινότερου γείτονα (furthest neighbour ή complete linkage)] Πόσες βιογεωγραφικές περιοχές ορίζονται από ένα δενδρόγραμμα; Ανάλογα µε την εκάστοτε µέθοδο που χρησιµοποιείται, ο αριθµός των οµάδων ενός δενδρογράμματος ποικίλλει. Στην πραγµατικότητα, οι ερευνητές αποφασίζουν τον αριθμό των βιογεωγραφικών περιοχών με κύριο άξονα την ερµηνεία που µπορούν να αποδώσουν σε αυτές. Με άλλα λόγια, ο καθορισμός του αριθμού των βιογεωγραφικών περιοχών γίνεται με υποκειμενικό και αυθαίρετο τρόπο (Reyjol et al. 2007, Abell et al. 2008, Zogaris, Economou & Dimopoulos 2009), ενώ η οριοθέτηση των βιογεωγραφικών περιοχών, έπειτα από έλεγχο με κριτήριο επιλογής του αριθµού των οµάδων, σπάνια εμφανίζεται στη βιβλιογραφία (αλλά βλέπε Carvalho et al. 2011, Leprieur & Oikonomou 2014, Oikonomou, Leprieur & Leonardos 2014). 115
116 Για τον εντοπισμό των διακριτών ομάδων (διακριτές βιογεωγραφικές περιοχές), στο δενδρόγραμμα που προέκυψε, χρησιμοποιήθηκε το κριτήριο Kelley - Gardner - Sutcliffe (KGS) (Kelley, Gardner & Sutcliffe 1996). Το κριτήριο KGS αποτελεί κριτήριο απεικόνισης των διαφορών μεταξύ των ομάδων και της συνεκτικότητας εντός αυτών με βάση τη μήτρα ανομοιότητας. Με άλλα λόγια, αποτελεί την πιο ενδεδειγμένη μέθοδο λήψης αποφάσεων για τον καθορισμό του αριθμού των διακριτών ομάδων, που συνθέτουν ένα ιεραρχικό δενδρόγραμμα. Το ελάχιστο του κριτηρίου KGS αντιστοιχεί στο βέλτιστο αριθμό των ομάδων Είδη-ενδείκτες των Βαλκανικών βιογεωγραφικών περιοχών Τα είδη-ενδείκτες είναι είδη που χρησιμοποιούνται συχνά ως εκπρόσωποι μιας βιογεωγραφικής περιοχής χρησιμοποιώντας δεδομένα κατανομής στις λεκάνες απορροής («basin specific») που βρίσκονται μόνο στην εν λόγω ενιαία βιογεωγραφική περιοχή δείχνοντας εξειδίκευση («specificity») και είναι παρόντα στα περισσότερα οικοσυστήματα της βιογεωγραφικής περιοχής διότι χαρακτηρίζονται από πιστότητα («fidelity»). Τα είδη-ενδείκτες προσδιορίζονται από την ανάλυση της σχέσης μεταξύ της παρουσίας των ειδών στα οικοσυστήματα μιας βιογεωγραφικής περιοχής και την ταξιθέτηση των οικοσυστημάτων αυτών σε ένα κλαδόγραμμα (Dufrêne & Legendre 1997). Τα αποτελέσματα της ανάλυσης προσδιορισμού των ειδών- ενδεικτών σε ένα κλάδο («cluster») αποτελούνται από έναν κατάλογο ειδών που είναι στατιστικά σημαντικά συσχετιζόμενα με αυτόν και παρουσιάζονται με σειρά φθίνουσας τιμής του δείκτη (IndVal) (De Cáceres et al. 2012). Στην ανάλυσή μας, η τιμή του δείκτη κάθε είδους αξιολογήθηκε μέσω μιας διαδικασίας τυχαιοποίησης με 999 µεταθέσεις («permutations»), λαμβάνοντας υπόψη το όριο της παρουσίας τουλάχιστον ενός είδους-ενδείκτη ανά βιογεωγραφική περιοχή (Εικόνα 2.5). 116
117 Εικόνα 2.5. Εννοιολογικό διάγραμμα των σταδίων ανάλυσης προσδιορισμού των ειδών-ενδεικτών κλαδόγραμμα (σύνθεση από Kreft & Jetz 2010 και Dufrêne & Legendre 1997) Figure 2.5. Conceptual diagram of steps in indicator value analysis κλαδόγραμμα (synthesis from Kreft & Jetz 2010 and Dufrêne & Legendre 1997) Στατιστική επεξεργασία και πακέτα Όλες οι αναλύσεις πραγματοποιήθηκαν στο «R statistical and programming environment» (R , R Development Core Team 2013) χρησιμοποιώντας τα πακέτα «betapart», «maptree» και «indicspecies». 117
118 2.4 Αποτελέσματα Βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου Σύμφωνα με την ιεραρχική ανάλυση ομαδοποίησης και τη χρήση του κριτηρίου Kelley - Gardner - Sutcliffe (KGS) (Kelley, Gardner & Sutcliffe 1996), με βάση το δείκτη βjtu προσδιορίστηκαν οκτώ ομάδες (Εικόνα 2.6). Εικόνα 2.6. Εφαρμογή του κριτηρίου Kelley - Gardner - Sutcliffe (KGS) (Kelley, Gardner & Sutcliffe 1996) για τον προσδιορισμό διακριτών βιογεωγραφικών περιοχών. Το ελάχιστο του κριτηρίου KGS αντιστοιχεί στο βέλτιστο αριθμό των ομάδων (ελάχιστο= 8) Figure 2.6. Kelley-Gardner-Sutcliffe penalty function (KGS) analysis to identify distinct biogeographical regions (Kelley, Gardner & Sutcliffe 1996). The minimum of KGS function corresponds to the optimal number of clusters (minimum= 8) H ανομοιότητα με βάση τη σύνθεση της ιχθυοπανίδας μεταξύ των λεκανών απορροής στη Βαλκανική χερσόνησο είναι υψηλή (μέση τιμή του δείκτη βjtu = 0,81 ± 0,22 SD) (Εικόνα 2.7). Οι οκτώ ομάδες που προσδιορίστηκαν αντιστοιχούν στις ακόλουθες διακριτές περιοχές της Βαλκανικής χερσονήσου (Εικόνα 2.7 και 2.8): 1) Πρέσπες, 2) Αττικοβοιωτία, 3) Ιόνιο,4) Νότια Αδριατική, 5) Αιγαίο, 6) Ποντιακή Επαρχεία, 7) Λεκάνη απορροής του ποταμού Neretva, 8) Βόρεια Αδριατική. 118
119 Εικόνα 2.7. Ιεραρχική ανάλυση ομαδοποίησης με βάση το δείκτη βjtu για τον καθορισμό των βιογεωγραφικών περιοχών στη Βαλκανική Χερσόνησο σε δεδομένα παρουσίας / απουσίας αυτόχθονων ειδών ψαριών. 1: Πρέσπες, 2-4: Αττικοβοιωτία, 5-16: Ιόνιο, 17-27: Νότια Αδριατική, 28-44: Αιγαίο, 45-86: Ποντιακή Επαρχεία, 87-94: Λεκάνη απορροής του π. Neretva, 95-97:Βόρεια Αδριατική. Το ποσοστό ανομοιότητας ισούται με 0.81 μεταξύ των λεκανών απορροής (± 0.22 SD) Figure 2.7. Hierarchical clustering analysis based on βjtu identifying the biogeographical re-gions of the Balkan Peninsula on native fish species presence/ absence data. 1: Prespa Lakes, 2-4: Attikobeotia Region, 5-16: Ionian Sea, 17-27: southern Adriatic Sea, 28-44: Aegean Sea, 45-86: Pontic Province, 87-94: Neretva Watershed, 95-97: northern Adriatic Sea. The level of dissimilarity between the drainage basins is at 0.81 (± 0.22 SD) 119
120 120
121 Εικόνα 2.8a. Ιεραρχική ανάλυση ομαδοποίησης για τον εντοπισμό των οκτώ βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου, σύμφωνα με τη ανομοιότητα στη σύνθεση των ψαριών των εσωτερικών υδάτων και b. ο αντίστοιχος χάρτης των βιογεωγραφικών περιοχών. Δύο υποομάδες της περιοχής του Αιγαίου μπορούν να αναγνωριστούν: η Θράκη (28-35) και η Μακεδονία-Θεσσαλία (36-44) (βλέπε κείμενο για περαιτέρω πληροφορίες). Η Αττικοβοιωτία, Ιόνιο Πέλαγος και οι Πρέσπες αποτελούν τα Βαλκανικά θερμά σημεία και χαρακτηρίζονται από τα υψηλά επίπεδα μοναδικότητας (ενδημισμού) και ευπάθειας (Κεφάλαιο 3, από Oikonomou, Leprieur & Leonardos 2014) Figure 2.8a. Hierarchical clustering analysis identifying eight biogeographical regions of the Balkan Peninsula according to freshwater fish compositional dissimilarity and b. the corresponding map showing these regions. Two subgroups of the Aegean Sea region can be recognized; i.e., Thrace (28-35) and Macedonia-Thessaly (36-44) (see text for further information). The Attikobeotia, Ionian Sea and Prespa Lakes regions constitute the Balkan hotspots being characterised by high levels of irreplaceability (endemism) and vulnerability (Chapter 3, Oikonomou, Leprieur & Leonardos 2014) Είδη-ενδείκτες της ιχθυοπανίδας στις βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου Από τα αποτελέσματα της ανάλυσης της ανάδειξης των ειδών- ενδεικτών στα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου προέκυψε ότι ανάμεσα στα 193 είδη ψαριών, 33 είδη (17%) παρουσίασαν στατιστικά σημαντική συσχέτιση με τις επτά βιογεωγραφικές περιοχές (Εικόνα 2.9, Πίνακας 2.2). Από τα 33 είδη, τα 28 ανήκουν στην οικογένεια των Cyprinidae, συμπεριλαμβανομένων των ειδών από τα ακόλουθα γένη: a) Alburnus, b) Aulopyge, c) Barbus, d) Blicca, e) Chondrostoma, f) Delminichthys, g) Luciobarbus, h) Pachychilon, i) Pelasgus, j) Rutilus, k) Scardinius, l) Squalius, m) Telestes και n) Vimba (Εικόνα 2.10). 121
122 Εικόνα 2.9. Κατανομή γενών- ενδεικτών σε όλες τις βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου. a-n. Cyprinidae: a. Alburnus b. Aulopyge c. Barbus d. Blicca e. Chondrostoma f. Delminichthys g. Luciobarbus h. Pachychilon i. Pelasgus j. Rutilus k. Scardinius l. Squalius m. Telestes n. Vimba, o. Cobitidae: Cobitis p. Valenciidae: Valencia q-r. Gobiidae q. Economidichthys r. Knipowitschia Figure 2.9. Distribution of Indicator genera across the biogeographical regions of the Balkan Peninsula. a-n. Cyprinidae: a. Alburnus b. Aulopyge c. Barbus d. Blicca e. Chondrostoma f. Delminichthys g. Luciobarbus h. Pachychilon i. Pelasgus j. Rutilus k. Scardinius l. Squalius m. Telestes n. Vimba, o. Cobitidae: Cobitis p. Valenciidae: Valencia q-r. Gobiidae q. Economidichthys r. Knipowitschia Πιο συγκεκριμένα, το είδος Pelasgus marathonicus (Vinciguerra 1921) κατανέμεται (IndVal=100%) σε όλα τα οικοσυστήματα της Αττικοβοιωτίας η οποία επίσης χαρακτηρίζεται από το Telestes beoticus (Stephanidis 1939) και το Luciobarbus graecus (Steindachner 1895). Οι ιχθυοκοινότητες της Αττικοβοιωτίας είναι παρόμοιες με αυτές του Ιονίου, οι οποίες χαρακτηρίζονται από είδη από τα ίδια 122
123 γένη: Luciobarbus albanicus Steindachner, 1870, Telestes pleurobipunctatus (Stephanidis 1939), Barbus peloponnesius Valenciennes 1842 και Pelasgus stymphalicus (Valenciennes 1844). Όσον αφορά την περιοχή της νότιας Αδριατικής Θάλασσας, το είδος Pachychilon pictum (Heckel & Kner 1858) απαντά σε όλα τα οικοσυστήματα (IndVal=100%), ενώ τέσσερα άλλα Κυπρινοειδή [Barbus rebeli Koller 1926, Telestes montenigrinus (Vukovic 1963), Rutilus ohridanus (Karaman 1924) και Chondrostoma nasus (Linnaeus 1758)], ταυτοποιήθηκαν ως είδη-ενδείκτες. Τα Chondrostoma vardarense Karaman 1928 και Vimba melanops (Heckel 1837) χαρακτηρίζουν τη περιοχή του Αιγαίου, ενώ τα Blicca bjoerkna (Linnaeus 1758) και Vimba vimba (Linnaeus 1758) σχετίζονται με την Ποντιακή επαρχία. Τέλος, τέσσερα Κυπρινοειδή είναι ενδεικτικά της βόρειας Αδριατικής Θάλασσας (Scardinius dergle Heckel & Kner 1858, Aulopyge huegelii Heckel 1843, Barbus plebejus Bonaparte 1839, Squalius illyricus Heckel & Kner 1858) και επτά Κυπρινοειδή [Alburnus neretvae Buj, Sanda & Perea 2010, Rutilus basak (Heckel 1843), Squalius svallize (Heckel & Kner 1858), Chondrostoma knerii Heckel 1843, Scardinius plotizza Heckel & Kner 1858, Delminichthys ghetaldii (Steindachner 1882), Delminichthys adspersus (Heckel 1843)] χαρακτηρίζουν τη λεκάνη απορροής του ποταμού Neretva. 123
124 Πίνακας 2.2. Τα αποτελέσματα της ανάλυσης ειδών-ενδεικτών που δείχνουν τις οικογένειες, τα γένη και τα είδη- ενδείκτες ανά βιογεωγραφική περιοχή, την τιμή του δείκτη (IndVal) και το επίπεδο στατιστικής σημαντικότητας (p). Στον πίνακα συμπεριλήφθηκαν μόνο τα είδη με τιμές του δείκτη IndVal πάνω από 48% (p<0,01) Table 2.2. Results of Indicator analysis showing family, genus and indicator species per biogeographical region, each Indicator Value (IndVal) and level of statistical significance (p). The table includes only spexies with IndVal above 48% (p<0,01) Family Genus Species Indicator Value p Biogeographical region (Indval) (%) Cyprinidae Chondrostoma Chondrostoma vardarense 74 0,0002 Aegean Sea Cobitidae Cobitis Cobitis strumicae 51 0,0030 Aegean Sea Cyprinidae Vimba Vimba melanops 71 0,0002 Aegean Sea Cyprinidae Luciobarbus Luciobarbus graecus 59 0,0036 Attikobeotia region Cyprinidae Pelasgus Pelasgus marathonicus 100 0,0002 Attikobeotia region Cyprinidae Telestes Telestes beoticus 67 0,0010 Attikobeotia region Cyprinidae Barbus Barbus peloponnesius 67 0,0008 Ionian Sea Gobiidae Economidichthys Economidichthys pygmaeus 92 0,0002 Ionian Sea Cyprinidae Luciobarbus Luciobarbus albanicus 83 0,0002 Ionian Sea Cyprinidae Pelasgus Pelasgus stymphalicus 67 0,0004 Ionian Sea Cyprinidae Telestes Telestes pleurobipunctatus 75 0,0006 Ionian Sea Valenciidae Valencia Valencia letourneuxi 67 0,0010 Ionian Sea Cyprinidae Alburnus Alburnus neretvae 88 0,0004 Neretva Watershed Cyprinidae Chondrostoma Chondrostoma knerii 63 0,0004 Neretva Watershed Cobitidae Cobitis Cobitis narentana 75 0,0002 Neretva Watershed Cyprinidae Delminichthys Delminichthys adspersus 50 0,0074 Neretva Watershed Cyprinidae Delminichthys Delminichthys ghetaldii 50 0,0072 Neretva Watershed Gobiidae Knipowitschia Knipowitschia croatica 50 0,0074 Neretva Watershed Cyprinidae Rutilus Rutilus basak 88 0,0002 Neretva Watershed Cyprinidae Scardinius Scardinius plotizza 63 0,0004 Neretva Watershed
125 Family Genus Species Indicator Value p Biogeographical region (Indval) (%) Cyprinidae Squalius Squalius svallize 88 0,0002 Neretva Watershed Cyprinidae Aulopyge Aulopyge huegelii 89 0,0002 northern Adriatic Sea Cyprinidae Barbus Barbus plebejus 67 0,0010 northern Adriatic Sea Cyprinidae Scardinius Scardinius dergle 100 0,0002 northern Adriatic Sea Cyprinidae Squalius Squalius illyricus 67 0,0014 northern Adriatic Sea Cyprinidae Blicca Blicca bjoerkna 48 0,0084 Pontic Province Cyprinidae Vimba Vimba vimba 48 0,0084 Pontic Province Cyprinidae Barbus Barbus rebeli 82 0,0002 southern Adriatic Sea Cyprinidae Chondrostoma Chondrostoma nasus 54 0,0002 southern Adriatic Sea Cobitidae Cobitis Cobitis ohridana 61 0,0006 southern Adriatic Sea Cyprinidae Pachychilon Pachychilon pictum 100 0,0002 southern Adriatic Sea Cyprinidae Rutilus Rutilus ohridanus 55 0,0024 southern Adriatic Sea Cyprinidae Telestes Telestes montenigrinus 64 0,0006 southern Adriatic Sea
126 Συνολικά, τρία είδη του γένους Cobitis αναδείχθηκαν ως είδη-ενδείκτες της Βαλκανικής χερσονήσου, και συγκεκριμένα της νότιας Αδριατικής θάλασσας (Cobitis ohridana Karaman 1928), του Αιγαίου (Cobitis strumicae Karaman 1955) και της λεκάνης απορροής του π. Neretva (Cobitis narentana Karaman 1928), ενώ δύο ενδημικοί αμμογωβιοί [Knipowitschia croatica Mrakovcic et al και Economidichthys pygmaeus (Holly 1929)] κυριαρχούν στις περιοχές της λεκάνης απορροής του π. Neretva και του Ιόνιου Πελάγους, αντίστοιχα. Τέλος, το ενδημικό είδος Valencia letourneuxi (Sauvage 1880), που ανήκει στη μοναδική ενδημική οικογένεια της Ευρώπης Valenciidae, χαρακτηρίζει το Ιόνιο Πέλαγος. Εικόνα Τα αποτελέσματα από την ανάλυση ειδών-ενδεικτών, για τις επτά βιογεωγραφικές περιοχές, που προέκυψαν μέσω της διαδικασίας τυχαιοποίησης με 999 μεταθέσεις. Τα είδη που αναφέρονται φέρουν τιμή του δείκτη πάνω από το όριο του 48% και p<0,01 (οι τιμές παρατίθενται στις παρενθέσεις). Η βιογεωγραφική περιοχή των Πρεσπών δεν εμφανίζεται, αφού περιλαμβάνει μόνο ένα οικοσύστημα Figure Results from the Indicator Species analysis for the seven biogeographical regions derived via a randomization procedure with 999 permutations. Species are only mentioned with indicator value above the threshold of 48% and p<0,01 (indicator value between parentheses). The biogeographical region of Prespa Lakes is not shown since it includes only one ecosystem 126
127 2.5 Συζήτηση Oριοθέτηση των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου Η ανάλυση οριοθέτησης βιογεωγραφικών περιοχών χρησιμοποιώντας τη μεγαλύτερη διαθέσιμη ΒΔ για τα υδάτινα οικοσυστήματα και τα ψάρια των εσωτερικών υδάτων των Βαλκανίων προσδιόρισε οκτώ κύριες βιογεωγραφικές περιοχές (Εικόνα 2.8a και 2.8b, Oikonomou, Leprieur, & Leonardos 2014). Αυτές οι οκτώ βιογεωγραφικές περιοχές μπορούν να θεωρηθούν αξιόπιστες δεδομένου ότι η παρούσα μελέτη βασίζεται σε στατιστική ανάλυση της ανομοιότητας της σύνθεσης των ειδών με τη χρήση της μετρικής β-ποικιλότητας (βjtu), που ενδείκνυται για περιοχές με υψηλά επίπεδα ενδημισμού, όπως η Βαλκανική χερσόνησος. Η Βαλκανική χερσόνησος δεν είναι χωρικά ομοιογενής ως προς την ιχθυοπανίδα της, καθώς χαρακτηρίζεται από υψηλό ποσοστό ανομοιότητας μεταξύ των βιογεωγραφικών περιοχών (Εικόνα 2.7). Αυτό το αποτέλεσμα επιβεβαιώνει τη διαπίστωση του Bănărescu (2004), ότι η Βαλκανική χερσόνησος χαρακτηρίζεται από κατακερματισμένο υδρογραφικό δίκτυο με ξεχωριστή πανίδα και ενδημικά είδη που συγκεντρώνονται σε εξαιρετικά μικρές περιοχές. Αυτά τα χαρακτηριστικά των εσωτερικών υδάτων της Βαλκανικής εμφανίζονται ως απόρροια της παρουσίας των μακροχρόνιων γεωγραφικών εμποδίων στη διασπορά των ψαριών και την επακόλουθη απομόνωση που προκλήθηκε από μεγάλα γεωλογικά γεγονότα, όπως η αλπική ορογένεση στα τέλη του Μειόκαινου, η οποία δημιούργησε τα Βαλκάνια και τις Διναρίδες-Ελληνίδες οροσειρές (Skoulikidis et al. 2009). Οι οκτώ βιογεωγραφικές περιοχές των εσωτερικών υδάτων της Βαλκανικής μπορούν να ταξινομηθούν στη δυτική ομάδα (Ιόνιο Πέλαγος, Νότια και Βόρεια Αδριατική θάλασσα και λεκάνη απορροής του ποταμού Neretva), την ανατολική ομάδα (Αττικοβοιωτία, Αιγαίο Πέλαγος και Ποντιακή επαρχία) και την ανεξάρτητη ομάδα (λίμνες Πρέσπες). Πράγματι, αυτή η δυτική-ανατολική διάκριση των Βαλκανίων έχει τεκμηριωθεί από πολλές φυλογενετικές μελέτες [Fritz et al. (2007) 127
128 για τα αμφίβια, Poulakakis et al. (2005) για τα ερπετά, Parmakelis et al. (2006) για τους σκορπιούς, Cooper, Ibrahim & Hewitt (1995) για τις ακρίδες, Albrecht et al. (2008) για τα υδρόβια σαλιγκάρια, Vanhove et al. (2012) για τα ψάρια των εσωτερικών υδάτων], παρέχοντας έτσι περαιτέρω υποστήριξη στα αποτελέσματα της βιογεωγραφικής μας ανάλυσης Δυτική ομάδα: Ιόνιο Πέλαγος, Νότια και Βόρεια Αδριατική θάλασσα και λεκάνη απορροής του ποταμού Neretva Ιόνιο Πέλαγος Η βιογεωγραφική περιοχή του Ιονίου περιλαμβάνει πολλά ακρωτήρια, καρστικά συστήματα, φαράγγια, μικρές λίμνες και μικρού μεγέθους ποτάμια από τη δυτική Ελλάδα μέχρι την Πελοπόννησο. Χαρακτηρίζεται από υψηλά επίπεδα ενδημισμού εξαιτίας της οροσειράς της Πίνδου, η οποία σχηματίστηκε στην αλπική ορογένεση και αποτελεί ένα διακριτό όριο για το βιοτικό διαχωρισμό και την απομόνωση της περιοχής αυτής από την ανατολική Ελλάδα (Αιγαίο πέλαγος). Τα έντονα γεωλογικά γεγονότα και η ύπαρξη πολλών παλαιο-λίμνών στην περιοχή έχουν επίσης συμβάλει στη μακροχρόνια απομόνωση και στον υψηλό ενδημισμό (Economou et al. 2007). Η οριοθέτηση της βιογεωγραφικής περιοχής του Ιονίου υποστηρίχτηκε πρώτα από τον Radoman (1985) και έχει αναφερθεί και σε πρόσφατες μελέτες σχετικά με τα ψάρια των εσωτερικών υδάτων, που είτε βασίζονται σε μια προσέγγιση από κάτω προς τα πάνω («bottom- up approach») (Abell et al. 2008) ή σε αυθαίρετο διαχωρισμό (Zogaris, Economou & Dimopoulos 2009). Εντούτοις, οι Economidis & Bănărescu (1991) και o Bănărescu (2004) έχουν αναγνωρίσει αυτή την περιοχή ως μια υποομάδα της περιοχής «νότια Αδριατική - Ιόνιο». Η διαφοροποίηση αυτή μπορεί να οφείλεται στην αλλαγή της ονοματολογίας των ειδών («split») και την αναθεώρηση της συστηματικής κατάταξης των ειδών. 128
129 Νότια Αδριατική Θάλασσα Η περιοχή της νότιας Αδριατικής θάλασσας εμπίπτει στην περιοχή της Αδριατικής ιχθυογεωγραφικής περιοχής των Δυτικών Βαλκανίων, όπως ορίστηκε από τους Economidis & Bănărescu (1991) και περιλαμβάνει τις λεκάνες απορροής ποταμών και τις λίμνες στην Αλβανία και το Μαυροβούνιο, από τον ποταμό Αώο-Vjose έως τον ποταμό Δρίνο-Drin. Χωρίστηκε από την Ιόνια περιοχή κατά το Τριτογενές και Τεταρτογενές (Perissoratis & Conispoliatis 2003), και ως εκ τούτου οι δύο περιοχές παρουσιάζουν διαφορετικά πρότυπα στον αριθμό των ειδών και στα ποσοστά ενδημισμού (Neubauer et al. 2015). Βόρεια Αδριατική Θάλασσα και λεκάνη απορροής του ποταμού Neretva Τα αποτελέσματα της παρούσας μελέτης προτείνουν για πρώτη φορά την οριοθέτηση της λεκάνης απορροής του π. Neretva, ως διακριτή βιογεωγραφική περιοχή και το διαχωρισμό της, από αυτή της βόρειας Αδριατικής Θάλασσας. Προηγούμενες μελέτες σχετικά με τα Βαλκάνια, από τους Economidis & Bănărescu (1991) και τον Bănărescu (2004) έχουν προτείνει την οριοθέτηση μιας ενιαίας περιοχής, της Δαλματίας. Πράγματι, οι δύο περιοχές (Βόρεια Αδριατική Θάλασσα και λεκάνη απορροής του π. Neretva) χαρακτηρίζονται από παρόμοια πολύπλοκα γεωλογικά και υδρολογικά φαινόμενα, συμπεριλαμβανομένων της παρουσίας υπόγειων υδάτων, καρστικών σχηματισμών, πηγών και λιμνών. Επιπλέον, η ενιαία περιοχή δεν ήταν καλυμμένη με πάγο κατά τη διάρκεια του Πλειστοκαίνου, παρέχοντας έτσι καταφύγια στα είδη των εσωτερικών υδάτων, οδηγώντας σε υψηλά επίπεδα ενδημισμού. Η ανύψωση των Διναρίδων μετά τις συμπιεστικές τεκτονικές τάσεις στα τέλη του Σερραβάλλιου (Pavelić & Belak 2008) επίσης συνέβαλε στην απομόνωση των βιοκοινοτήτων της περιοχής από τη δουνάβια πανίδα (Skoulikidis et al. 2009). Ωστόσο, τα αποτελέσματα της παρούσας μελέτης δείχνουν την υψηλή διαφοροποίηση της περιοχής της βόρειας Αδριατικής θάλασσας από την περιοχή της λεκάνης απορροής του π. Neretva, επιβεβαιώνοντας το «καταφύγια μέσα σε 129
130 καταφύγια» πρότυπο («refugia within refugia») που έχει αναφερθεί και στην Ιβηρική χερσόνησο (Gómez & Lunt 2007). Προηγούμενες φυλογενετικές μελέτες σε είδη των εσωτερικών υδάτων (Previšic et al. 2009, DeFaveri et al. 2012), θηλαστικών (Kryštufek et al. 2007) και φυτικών ειδών (Surina, Schönswetter & Schneeweiss 2011) έχουν δείξει, επίσης, αυτή τη διάκριση των πληθυσμών στην περιοχή του π. Neretva. Έτσι, γίνεται όλο και πιο σαφές ότι η λεκάνη απορροής του π. Neretva θα πρέπει να αντιμετωπίζεται ως ξεχωριστή βιογεωγραφική περιοχή Ανατολική ομάδα: Αττικοβοιωτία, Αιγαίο Πέλαγος και Ποντιακή επαρχία Αττικοβοιωτία Η οριοθέτηση της γεωγραφικά κατακερματισμένης περιοχής της Αττικοβοιωτίας υποστηρίζεται ισχυρά από τις προηγούμενες βιογεωγραφικές μελέτες (Economidis & Bănărescu 1991, Bănărescu 2004, Abell et al. 2008, Zogaris, Economou & Dimopoulos 2009). Αυτή η περιοχή παρουσιάζει φτωχή ιχθυοπανίδα, αλλά υψηλό βαθμό ενδημικών και απειλούμενων ειδών που επηρεάζονται σοβαρά από την υψηλή ανθρωπογενή πίεση που παρατηρείται στην περιοχή (βλέπε επίσης Κεφαλαιο 3). Αιγαίο Πέλαγος Η παρούσα μελέτη υποστηρίζει, επίσης, την ύπαρξη μίας ενιαίας βιογεωγραφικής περιοχής στο Αιγαίο (Radoman 1985, Bianco 1990), συμπεριλαμβανομένων των περιοχών της Θράκης, της Μακεδονίας και της Θεσσαλίας, από όπου διέρχονται τα μεγάλα ποτάμια που εκβάλλουν στο Αιγαίο (π.χ. Στρυμόνας-Struma, Αξιός-Vardar, Νέστος-Mesta και Έβρος-Maritsa). Ωστόσο, δύο υποομάδες μπορούν να αναγνωριστούν: η Θράκη και η Μακεδονία-Θεσσαλία (βλέπε τις υποομάδες του κλάδου στην Eικόνα 2.6a), σε συμφωνία με τα αποτελέσματα των Economidis & Bănărescu (1991), του Bănărescu (2004) και των Abell et al. (2008). Η διαίρεση αυτή μπορεί να εξηγηθεί επαρκώς από τις πρόσφατες γεωλογικές μελέτες στη ζώνη 130
131 μεταξύ του Βαρδάρη και των Σερβομακεδονικών όγκων και Ροδόπης (Himmerkus, Reischmann & Kostopoulos 2006). Σε γενικές γραμμές, η πανίδα της περιοχής του Αιγαίου παρουσιάζει εμφανείς δεσμούς με τη Δουνάβια πανίδα, δικαιολογώντας τον υψηλό αριθμό ειδών, καθώς οι ποταμοί Morava και Αξιός-Vardar ενώνονταν με τον ποταμό Δούναβη κατά τη διάρκεια της Πλειο - Πλειστόκαινης εποχής. Επιπλέον, οι λεκάνες απορροής των ποταμών στην περιοχή του Αιγαίου φιλοξενούν είδη με προέλευση τη Μαύρη Θάλασσα ως αποτέλεσμα της διείσδυσης των υδάτων της Μαύρης Θάλασσας (λιμναία φάση) στο όριο του Πλειόκαινου με το Πλειστόκαινο (Economou et al. 2007). Δείχνουν επίσης κοντινές σχέσεις με την παρακείμενη περιοχή της Μικράς Ασίας (βλέπε Durand et al. 2003, Zogaris, Economou & Dimopoulos 2009, Tsoumani, Apostolidis & Leonardos 2013 για το είδος Rutilus rutilus και Neubauer et al για τα υδρόβια σαλιγκάρια). Ποντιακή επαρχία Επιπλέον, τα αποτελέσματά μας επιβεβαιώνουν την παρουσία της Ποντιακής επαρχίας (λεκάνη απορροής Δούναβη), η οποία είναι η πιο πλούσια σε αριθμό ειδών και πιο φτωχή σε ενδημικά είδη μεταξύ των βαλκανικών περιοχών. Η δουνάβια πανίδα χαρακτηρίζεται από είδη με ευρεία κατανομή στην Ευρώπη και την Σιβηρία και φέρει δεσμούς με τη δυτική και κεντρική Ευρώπη (Economidis & Bănărescu 1991) Λίμνες Πρέσπες Η υψηλή ανομοιότητα των Πρεσπών, ως προς τη δυτική και ανατολική ομάδα των βιογεωγραφικών περιοχών, μας οδήγησαν στο συμπέρασμα ότι οι λίμνες Πρέσπες βρίσκονται στη διασταύρωση των δύο προαναφερθεισών σημαντικών ομάδωνβιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου και παρουσιάζουν το υψηλότερο επίπεδο διαφοροποίησης (Tsoumani, Apostolidis & Leonardos 2013, Berrebi et al. 2013). Προηγούμενες μελέτες (Bănărescu 2004, Zogaris, Economou & Dimopoulos 2009) πρότειναν ένα βιογεωγραφικό σενάριο σύμφωνα με το οποίο, οι 131
132 Πρέσπες και η γειτονική λίμνη Οχρίδα, ανήκουν στην ίδια ζωογεωγραφική διαίρεση (δυτική ομαδοποίηση). Πράγματι, ως μέλη της ομάδας των «Δασσαρητών», οι Πρέσπες και η Οχρίδα είναι από τις παλαιότερες λίμνες στην Ευρώπη (Wagner et al. 2010) καθώς είναι γεωγραφικά κοντά, συνδέονται υπόγεια με καρστικά κανάλια (Amataj et al. 2007) και έχουν κοινή προέλευση (Radoman, 1985). Αντιθέτως, οι Wilke et al. (2010) πρότειναν ένα εναλλακτικό σενάριο στο οποίο οι λίμνες των Πρεσπών σχετίζονται με τη λίμνη Βεγορίτιδα και τη λίμνη Δοϊράνη και ως εκ τούτου, θα πρέπει να συμπεριληφθούν στην περιοχή του Βαρδάρη (ανατολική ομαδοποίηση) (Albrecht & Wilke 2008). Αξίζει να σημειωθεί ότι, αυτή είναι η πρώτη μελέτη που υποδηλώνει την οριοθέτηση των Πρεσπών ως μία ταξινομικά διακριτή βιογεωγραφική περιοχή καθώς η ιχθυοπανίδα των Πρεσπών είναι διαφορετική σε σχέση με την ιχθυοπανίδα της Οχρίδας, της Δοϊράνης ή της Βεγορίτιδας. Για παράδειγμα, όσον αφορά τα Κυπρινοειδή, στις Πρέσπες συναντώνται τα ενδημικά είδη Alburnus belvica, Barbus prespensis, Rutilus prespensis και Squalius prespensis, ενώ αντίθετα στη λίμνη Οχρίδα, φιλοξενούνται τα είδη A. ohridanus, B. rebeli. R. ohridanus και S. squalus. Εντούτοις, τα είδη αυτά έχουν μεγάλη φυλογενετική συγγένεια με τα είδη των Πρεσπών (Geiger et al. 2014). Αντίθετα, στη Βεγορίτιδα και Δοϊράνη απαντώνται Κυπρινοειδή με ευρεία κατανομή όπως τα A. thessalicus και A. macedonicus αντίστοιχα, και τα B. balcanicus και R. rutilus ενώ δεν απαντούν είδη του γένους Pelasgus Η διαφορετική ιχθυοπανίδα των Πρεσπών μπορεί να συνδέεται με τον περιορισμό της διασποράς των ειδών (Schultheiß et al. 2008), τη γεωλογική ιστορία της περιοχής (Wagner et al. 2010, Wilke et al. 2010), τα υψηλά ποσοστά της εξαφάνισης ή/ και τις στρατηγικές ζωής των ειδών που διαβιούν στο οικοσύστημα, αλλά αυτές οι υποθέσεις δεν ελέγχθηκαν στην παρούσα μελέτη. 132
133 2.5.2 Βιογεωγραφική απόδοση από τα είδη-ενδείκτες των εσωτερικών υδάτων της Βαλκανικής χερσονήσου Τα Κυπρινοειδή είναι η επικρατέστερη οικογένεια των εσωτερικών υδάτων σε παγκόσμια κλίμακα και συνεπώς κυριαρχεί στην αυτόχθονη και ενδημική ιχθυοπανίδα της Βαλκανικής χερσονήσου. Πράγματι, ένα υψηλό ποσοστό (82%) των ειδών-ενδεικτών ανήκει στην οικογένεια Cyprinidae. Το ποσοστό αυτό είναι σύμφωνο με την υψηλή εκπροσώπηση των Κυπρινοειδών συνολικά στη Βαλκανική ιχθυοπανίδα (Κεφάλαιο 3). Μέχρι σήμερα έχουν προταθεί διάφορες υποθέσεις για την προέλευση, την ποικιλότητα και τους διαδρόμους διασποράς της οικογένειας. Πρόσφατα, οι Gaubert, Denys & Oberdorff (2009) επιβεβαίωσαν ότι αυτή η οικογένεια έχει ανατολίτικη καταγωγή προερχόμενη από την Ινδο-Μαλαισία και ότι τα Βαλκανικά Κυπρινοειδή διασπάρθηκαν μέσω διαδρόμων που συνέδεαν τις λεκάνες απορροής ποταμών της Κεντρικής Ευρώπης και της Σιβηρίας κατά τη διάρκεια των μεταναστεύσεων μεταξύ Ανατολής και Παλαιαρκτικής. Οι οικογένειες Cobitidae και Gobiidae έχουν πολλούς εκπροσώπους στη Βαλκανική ιχθυοπανίδα, όπως και στην υπόλοιπη Ευρώπη (Reyjol et al. 2007) και στην Ασία (Kang et al. 2013). Παρά το γεγονός ότι η γεωγραφική προέλευση της οικογένειας Cobitidae είναι άγνωστη, αυτή εκπροσωπείται στα Βαλκάνια από τρεις εξελικτικές γραμμές που χωρίστηκαν όταν η χερσαία σύνδεση μεταξύ της κεντρικής Ευρώπης και της Ανατολίας έσπασε (Bohlen et al. 2006). Τέλος, οι Βαλκανικοί αμμογωβιοί περιλαμβάνουν τα δύο γένη: Knipowitschia και Economidichthys (Vanhove et al. 2012), οι οποίοι εμφανίζουν ένα ευρύ φάσμα οικοτόπων και υψηλά επίπεδα ενδημισμού καθώς η απόσχιση του γένους Economidichthys από το γένος Knipowitschia και η διαφοροποίηση των ειδών τους συνέβη κατά το Πλειστόκαινο (Γκένας 2012). Μεταξύ των Βαλκανικών ειδών-ενδεικτών, ορισμένα είδη συντελούν ιδιαίτερα στην παρατήρηση των βιογεωγραφικών προτύπων και μας βοηθούν να κατανοήσουμε τις ομοιότητες της Βαλκανικής ιχθυοπανίδας. Για παράδειγμα, οι 133
134 λεκάνες απορροής της νότιας Αδριατικής χαρακτηρίζονται από το είδος Pachychilon pictum (Indval= 100%), αποδεικνύοντας ότι αυτό κατανέμεται σε όλα τα οικοσυστήματα (100% πιστότητα) αποκλειστικά αυτής της περιοχής (100% εξειδίκευση). Η διαφοροποίηση του είδους υπολογίζεται ότι συνέβη πριν από το Μεσσήνιο (Levy, Doadrio & Almada 2009), επιτρέποντας την ευρεία εξάπλωση του είδους στην περιορισμένη έκταση της απομονωμένης νότιας Αδριατικής περιοχής. Επιπλέον, το είδος Chondrostoma nasus, το οποίο είναι ευρέως κατανεμημένο στην περιοχή της νότιας Αδριατικής και στα Δυτικά Βαλκάνια, φέροντας προέλευση από την Παρατηθύ (Robalo et al. 2007), επιβεβαιώνει τη σύνδεση των λεκανών απορροής των ποταμών της νότιας Αδριατικής με το παραδουνάβιο καταφύγιο. Η παρουσία του ενδημικού είδους Alburnus neretvae και η απουσία των ενδημικών ειδών Aulopyge huegelii και Scardinius dergle στη λεκάνη απορροής του ποταμού Neretva τονίζουν το μέγεθος της επίδρασης των διεργασιών καρστικοποίησης οδηγώντας σε απομόνωση και υψηλή ειδογένεση των ειδών που διαβιούν σε αυτές τις περιοχές (βλέπε επίσης Perea et al. 2010), παρέχοντας ενδείξεις για την διάκριση της λεκάνης απορροής του Neretva από τη βόρεια Αδριατική. Επιπλέον, τα τρία είδη Κυπρινοειδών -ενδείκτες της Αττικοβοιωτίας (Pelasgus marathonicus, Telestes beoticus και Luciobarbus graecus) είναι συγγενικά με αυτά στην περιοχή του Ιονίου Πελάγους (Pelasgus stymphalicus, Telestes pleurobipunctatus και Luciobarbus albanicus) (Economou et al. 2007). Εξίσου ενδιαφέρουσα είναι η παρατήρηση, ότι η ευρεία κατανομή του γένους Vimba και του είδους Blicca bjoerkna υποδεικνύει τον ρόλο του Δούναβη στη μετανάστευση και στον εποικισμό (Hänfling et al. 2009), καθώς και το γενικότερο γνώρισμα της ευρείας κατανομής της παραδουνάβιας πανίδας. Τέλος, είναι πραγματικά ενδιαφέρον ότι το είδος Valencia letourneuxi, που απαντά στην περιοχή του Ιονίου, παρουσιάζει στενή συνάφεια με το Ιβηρικό είδος Valencia hispanica (Valenciennes 1846) (Zardoya & Doadrio 1999, Sanjur, Carmona & Doadrio 2003). Ομοίως, η παρουσία του είδους Chondrostoma knerii στη λεκάνη απορροής του ποταμού Neretva, το οποίο ανήκει στο ίδιο γένος με το ιταλικό είδος Chondrostoma soetta Bonaparte 1840 επιβεβαιώνει τις ανταλλαγές πολλών τάξων των εσωτερικών υδάτων διαμέσου της Αδριατικής μεταξύ των Ιταλικών και των Βαλκανικών ποταμών κατά τη διάρκεια της τελευταίας παγετώδους περιόδου του 134
135 Βούρμιου («Würmian στάδιο», βλέπε επίσης Šlechtová et al για τα είδη του γένους Cottus). Θα πρέπει να σημειωθεί εδώ ότι προηγούμενες έρευνες σε Βαλκανικούς πληθυσμούς της οικογένειας των Σαλμονοειδών (π.χ. Apostolidis et al. 1997, Delling 2010) ανέδειξαν ότι τα είδη παρουσιάζουν υψηλά επίπεδα γενετικής ποικιλότητας, υποδεικνύοντας την παρουσία ξεχωριστών γονιδιακών αποθεμάτων και τη μακροχρόνια απομόνωση των πληθυσμών τους. Ως αποτέλεσμα, η οικογένεια χαρακτηρίζεται από υψηλή εκπροσώπηση στη Βαλκανική ιχθυοπανίδα (Κεφάλαιο 3). Εντούτοις, η ανάλυση προσδιορισμού των ειδών-ενδεικτών παρήγαγε χαμηλές τιμές, αποκαλύπτοντας ότι η κατανομή κάθε είδους σολομονιδών είναι πολύ περιορισμένη σε κάθε βιογεωγραφική περιοχή, δηλαδή ότι τα είδη χαρακτηρίζονται από υψηλό τοπικό ενδημισμό. 135
136
137 Κεφάλαιο 3 Πρότυπα βιοποικιλότητας των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου The first objectives of historical biogeography are to identify and delimit the related biotas and to describe patterns they form in relation to present day geography. (Rosen 1975)
138
139 3.1 Εισαγωγή Θερμά σημεία της βιοποικιλότητας ή σημεία ενδιαφέροντος («Biodiversity Hotspots») Η μελέτη των προτύπων της ποικιλότητας και του ενδημισμού των ταξινομικών ομάδων σε παγκόσμιο επίπεδο είναι θεμελιώδης στην οικολογία (Πίνακας 1.1, Gaston & Blackburn 2000). Ο όρος «σημεία ενδιαφέροντος της βιοποικιλότητας» ή αλλιώς «θερμά σημεία της βιοποικιλότητας» αναφέρεται σε περιοχές υψηλής αξίας για τη διατήρηση της βιοποικιλότητας, έπειτα από διερεύνηση των προτύπων της (Εικόνα 3.1). Αυτές οι γεωγραφικές περιοχές χαρακτηρίζονται από μία ή περισσότερες από τις επόμενες παραμέτρους (Williams et al. 1996): α) Θερμά σημεία ή σημεία ενδιαφέροντος του αριθμού των ειδών («Richness hotspots») και αφορούν περιοχές, που παρουσιάζουν τον υψηλότερο αριθμό ειδών β) Θερμά σημεία ή σημεία ενδιαφέροντος σπανιότητας («Rarity hotspots»), τα οποία είναι τα πλουσιότερα σε είδη με περιορισμένα κατανομή γ) Θερμά σημεία ή σημεία ενδιαφέροντος απειλής («Threatspots»), τα οποία φιλοξενούν τον υψηλότερο αριθμό απειλούμενων ειδών δ) Θερμά σημεία ή σημεία ενδιαφέροντος ενδημισμού («Endemic hotspots»), όπου διαβιεί ο υψηλότερος αριθμός ενδημικών ειδών ε) Περιοχές συμπληρωματικού πλούτου («Complementary richness areas»), όπου συνδυαστικά εμφανίζεται ο μέγιστος αριθμός ειδών 139
 Ο σχεδιασμός για την προστασία κα τη διατήρηση της βιοποικιλότητας είναι άρρηκτα συνδεδεμένος με την ύπαρξη αξιόπιστων δεδομένων.")
140 Εικόνα 3.1. Τα 25 θερμά σημεία βιοποικιλότητας στη γη (Myers et al. 2000) Figure 3.1. The 25 hotspots on earth (Myers et al. 2000) Ο σχεδιασμός για την προστασία κα τη διατήρηση της βιοποικιλότητας είναι άρρηκτα συνδεδεμένος με την ύπαρξη αξιόπιστων δεδομένων. Έτσι, η συγκέντρωση, αξιολόγηση και σύνθεση της διαθέσιμης πληροφορίας αποτελεί προϋπόθεση για τη μελέτη των προτύπων της βιοποικιλότητας και της βιογεωγραφίας. Παρόλα αυτά, η γνώση σχετικά με την κατανομή των ειδών είναι πολλές φορές ελλιπής και χαρακτηρίζεται από δυο βασικά ελλείμματα, το λεγόμενο έλλειμμα του Wallace («Wallacean shortfall») το οποίο ορίζεται ως «η ανεπαρκής γνώση των κατανομών των ειδών» και το έλλειμμα του Linnaeus («Linnean shortfall»), που ορίζει ότι «τα περισσότερα είδη που ζουν στη Γη δεν έχουν ακόμη περιγραφεί επισήμως» (Lomolino 2004, Bini et al. 2006) (Κεφάλαιο 2). Για παράδειγμα, ο σχεδιασμός για τη διατήρηση της βιοποικιλότητας παρεμποδίζεται διότι η κατανομή των ψαριών των εσωτερικών υδάτων στη Βαλκανική χερσόνησο για πολλά είδη είναι άγνωστη και η ιχθυοπανίδα ορισμένων υδάτινων συστημάτων δεν έχει ακόμα μελετηθεί επαρκώς. Επιπλέον, αν και σε παγκόσμια κλίμακα, τα δεδομένα κατανομής των ψαριών των εσωτερικών υδάτων θεωρούνται επαρκή, πολλά νέα είδη περιγράφονται κάθε χρόνο. Από τις αρχές του 140
141 2008 έως το τέλος του 2013, περιγράφηκαν 1484 νέα είδη (Mikšik & Schraml 2014). Με άλλα λόγια, το 10% των ήδη γνωστών ειδών περιγράφηκε στο μικρό χρονικό διάστημα των 6 ετών (Pelayo-Villamil et al. 2014) (Εικόνα 3.2). Εικόνα 3.2. Αριθμός ειδών που έχουν περιγραφεί (σε μπλε χρώμα) και νεπέριος λογάριθμος του αθροιστικού αριθμού των ειδών που περιγράφονται (με κόκκινο χρώμα) για τα ψάρια των εσωτερικών υδάτων. Η διακεκομμένη γραμμή αντιπροσωπεύει τα εκτιμώμενα είδη που έχουν περιγραφεί σύμφωνα με το μοντέλο Morgan-Mercer-Flodin (Pelayo-Villamil et al. 2014) Figure 3.2. Number of described species (in blue) and Naperian logarithm of the accumulated number of described species (in red) by year of description for freshwater fishes. The dashed line represents the described estimated species according to the Morgan-Mercer-Flodin model (Pelayo-Villamil et al. 2014) Τα πρότυπα του αριθμού των ειδών και του ενδημισμού στα εσωτερικά ύδατα Στα εσωτερικά ύδατα, ο αριθμός και η σύνθεση των ειδών των ψαριών είναι αποτέλεσμα συχνά ιστορικών παραγόντων (Dias et al. 2014). Eπίσης, τα πρότυπα του αριθμού των ειδών έχουν σχετιστεί με ένα περιορισμένο αριθμό κλιματικών παραμέτρων (επίγεια πρωτογενή παραγωγικότητα, ετήσια βροχόπτωση, ισοθερμικότητα και μέση ετήσια θερμοκρασία) και με την παραγωγικότητα των 141
142 οικοσυστημάτων των εσωτερικών υδάτων (Tedesco et al. 2005, Oberdorff et al. 2011, Griffiths, McGonigle & Quinn 2014, Pelayo-Villamil et al. 2014). Από την άλλη, η γνώση μας σχετικά με τους παράγοντες που διαμορφώνουν τα πρότυπα ενδημισμού των ψαριών είναι ακόμα σε μεγάλο βαθμό ανεπαρκής. Πράγματι, ο ενδημισμός δεν έχει μέχρι τώρα συσχετιστεί με περιβαλλοντικές παραμέτρους αλλά έχει δειχθεί πως σχετίζεται με την απομόνωση και άρα σε μεγάλο βαθμό με τη σύνδεση των διαφορετικών λεκανών απορροής. Αναλυτικότερα, η απομόνωση και η συνεπαγόμενη διαδικασία της διαφοροποίησης των ειδών, λόγω της μειωμένης γονιδιακής ροής, μπορούν να ερμηνεύσουν την ύπαρξη περιοχών με υψηλά επίπεδα ενδημισμού και συνήθως με μικρό μέγεθος των πληθυσμών των ειδών τους. Τα είδη των περιοχών αυτών που εμφανίζουν υψηλούς ενδημισμούς και κατά κανόνα μικρά πληθυσμιακά μεγέθη εμφανίζουν και υψηλό κίνδυνο εξαφάνισης τους, ως αποτέλεσμα της ανθρωπογενούς παρέμβασης 16 (Hugueny, Movellan & Belliard 2011). Ο περιορισμένος πλούτος ειδών των περιοχών αυτών οφείλεται στο γεγονός ότι στις απομονωμένες ή «ασύνδετες» λεκάνες απορροής η μετανάστευση των ειδών και ο εποικισμός 17 είναι δυσκολότερος (Tedesco et al. 2012, Dias et al. 2013). Οι Tedesco et al. (2012) διερευνώντας τα πρότυπα του ενδημισμού των ψαριών των ποταμών σε παγκόσμιο επίπεδο κατέληξαν πως τα είδη μπορούν να είναι 1) νέο-ενδημικά («neo-endemics») που εξελίχθηκαν λόγω κλαδογένεσης ή διχοτομικής ειδογένεσης, δηλαδή ένα είδος εξελίσσεται σε νέα είδη τα οποία δεν κατανέμονται εκτός της περιοχής προέλευσης 2) παλαιο-ενδημικά («palaeo-endemics») που είναι είτε είδη που εξελίχθηκαν επί τόπου με αναγένεση ή φυλετική εξέλιξη λόγω ιστορικής απομόνωσης, δηλαδή ένα είδος εξελίσσεται σε ένα νέο είδος, είτε ευρέως κατανεμημένα στο παρελθόν που έχουν υποστεί μείωση της κατανομής τους. 16 Το παράδοξο αυτό περιγράφηκε πρόσφατα ως «Τα ψάρια των εσωτερικών υδάτων συνιστούν πιθανά θύματα της δικής τους επιτυχίας» από τους Pelayo-Villamil et al. (2014) 17 Για τους ελληνικούς όρους, καταλήξαμε στη χρήση του αποικισμός για το «immigration», εποικισμός για το «colonization» και μετανάστευση για το «migration» σύμφωνα με τον Σφενδουράκη (2009) [Νησιωτική βιογεωγραφία: Οικολογία, εξέλιξη και διατήρηση- επιστημονική επιμέλεια έκδοσης] 142
143 Οι παλαιο-συνδεδεμένες λεκάνες, δηλαδή οι λεκάνες απορροής που ήταν συνδεδεμένες μεταξύ τους στον γεωλογικό χρόνο, εμφανίζουν υψηλότερο αριθμό ειδών, αλλά χαμηλότερα επίπεδα ενδημισμού και β- ποικιλότητας από ό, τι οι νέοσυνδεδεμένες λεκάνες, γεγονός που υποδηλώνει ότι οι συνδέσεις ευνόησαν την ανταλλαγή των ειδών ψαριών (Dias et al. 2014). Η απουσία ενδημικών ειδών σε μια λεκάνη απορροής αποδίδεται στην α) εξαφάνιση των ενδημικών ειδών, β) στη διασπορά-μετανάστευση των ειδών αυτών και σε άλλες περιοχές (π.χ. Δουνάβια πανίδα, Reyjol et al. 2007) και γ) στην απουσία κατάλληλων συνθηκών για in situ ειδογένεση (π.χ. μικρή συνολική έκταση) (Losos & Schluter 2000) Σχέση ανάμεσα στον αριθμό των ειδών και την έκταση Η επίδραση της έκτασης μιας περιοχής στον αριθμό των ειδών που έχει αποδοθεί στη βιβλιογραφία με τον όρο «σχέση αριθμού ειδών-έκτασης (species-area relationship, SAR)» αποτελεί ένα από τα πιο μελετημένα παγκόσμια βιογεωγραφικά πρότυπα (Simberloff, 1974). Το πρότυπο αρχικά αναδείχθηκε από μελέτες σε νησιά (MacArthur & Wilson 1967, Diamond 1975, Rosenzweig 1995), αλλά η θετική σχέση μεταξύ της έκτασης μιας περιοχής και του αριθμού των ειδών έχει επίσης επιβεβαιωθεί για ηπειρωτικές περιοχές, περιλαμβάνοντας όλο το εύρος της χωρικής κλίμακας, από μικρής έκτασης περιοχές ως και τις παγκόσμιες βιογεωγραφικές περιοχές αλλά και για το σύνολο των οργανισμών τόσο σπονδυλωτών όσο και ασπόνδυλων (Eadie et al. 1986, Ricklefs & Schlüter 1993, Brown 1995). Επιπλέον, η σχέση έχει επιβεβαιωθεί και για ενδιατηματικά νησιά («habitat islands») και όχι μόνο για γεωγραφικά νησιά, όπως οι κορυφές των βουνών ή ενδιαιτήματα σε μεγάλο υψόμετρο και πολλούς τύπους οικοσυστημάτων (λίμνες, ποτάμια) (Eadie et al. 1986). Η σχέση αριθμού ειδών-έκτασης περιγράφει ότι «ο αριθμός των ειδών αυξάνεται καθώς αυξάνεται η έκταση» και εκφράστηκε για πρώτη φορά ως μαθηματική σχέση του αριθμού των ειδών και της έκτασης από τον Arrhenius το 1921, με την εξίσωση: S z = ca (4) 143
144 όπου S είναι ο αριθμός των ειδών σε μια περιοχή, Α η έκταση της περιοχής και τα c, z οι σταθερές της σχέσης. Η τιμή z δεν είναι απλώς μια ένδειξη της απομόνωσης του υπό μελέτη οικοσυστήματος, αλλά στην ουσία είναι ενδεικτική του αριθμού των ειδών και των προτύπων σύνθεσης τους (Rosenzweig 1995). Η τιμή c αντικατοπτρίζει τη φέρουσα ικανότητα του οικοσυστήματος ανά μονάδα επιφάνειας (Brown & Lomolino 1998). Συνήθως, η σχέση αυτή χρησιμοποιείται στη λογαριθμική της μορφή ώστε να γίνεται γραμμική (Εικόνα 3.3): log ( S ) = log ( c) + z log ( A) (5) Εκτός από την εξίσωση του Arrhenius, το λογαριθμικό μοντέλο του Gleason (1922): S = c + zlog( A) (6) ήταν το δεύτερο που περιγράφηκε και έχει χρησιμοποιηθεί ευρέως στη βιβλιογραφία. Εικόνα 3.3. Σχέση έκτασης (Α) - αριθµού ειδών (S) σε αριθµητική και λογαριθµική κλίµακα (από Brown & Lomolino 1998) Figure 3.3. Species (S) - Area (A) Relationship in arithmetic and logarithmic scale (from Brown & Lomolino 1998) 144
145 3.2 Στόχος του κεφαλαίου Στο παρόν κεφάλαιο, μελετήθηκαν τα πρότυπα του αριθμού των ειδών σε σχέση με τον ενδημισμό, τον αριθμό των απειλούμενων με εξαφάνιση ειδών και την έκταση, στη Βαλκανική χερσόνησο. Οι περισσότερες από τις διαθέσιμες μελέτες σχετικά με τα ψάρια των εσωτερικών υδάτων στη Βαλκανική χερσόνησο έχουν επικεντρωθεί σε επιμέρους βιολογικά, οικολογικά και φυλογενετικά πρότυπα, όπως π.χ. στις στρατηγικές ζωής των ειδών ή σε ταξινομικές, μορφομετρικές ή φυλογενετικές διαφοροποιήσεις, αλλά λιγοστές προσπάθειες έχουν εστιαστεί στην παράλληλη μελέτη των διάφορων προτύπων της βιοποικιλότητας που σχετίζονται με τον αριθμό, τη σύνθεση καθώς και τον κίνδυνο εξαφάνισης των ειδών σε χωρικές κλίμακες που ξεπερνούν την τοπική κλίμακα ενός ποτάμιου συστήματος ή μιας λίμνης. Τα παραπάνω πρότυπα είναι αλληλοσυμπληρούμενα και διαμορφώνονται από τις ίδιες βασικές οικολογικές και εξελικτικές διεργασίες και συνεπώς η ταυτόχρονη μελέτη τους μπορεί να αναδείξει τη συμπεριφορά των διεργασιών αυτών σε σχέση με τα χαρακτηριστικά της Βαλκανικής χερσονήσου. 3.3 Υλικά και Μέθοδοι Πρότυπα αριθμού ειδών, ενδημισμού και των απειλούμενων ειδών (κατηγορίες IUCN) στις βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου Για να συγκρίνουμε το σχετικό βαθμό ενδημισμού μεταξύ των προσδιορισμένων βιογεωγραφικών περιοχών σύμφωνα με το Κεφάλαιο 2, χρησιμοποιήθηκε η κατανομή των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου ανά λεκάνη απορροής ποταμού και λίμνη (Κεφάλαιο 2). Τα ποσοστά των ενδημικών ειδών υπολογίστηκαν διαιρώντας τον αριθμό των ενδημικών ειδών με το συνολικό αριθμό των αυτόχθονων ειδών σε κάθε βιογεωγραφική περιοχή. 145
146 Παράλληλα, για τον εντοπισμό των ειδών που κινδυνεύουν με εξαφάνιση, υπολογίστηκε ο αριθμός των ειδών που αντιστοιχούν σε κάθε κατηγορία της Κόκκινης Λίστας (IUCN 2013) (Εικόνα 3.4) και εκτιμήθηκε το ποσοστό κάθε κατηγορίας σε κάθε βιογεωγραφική περιοχή. Ως Κρισίμως Κινδυνεύοντα - Critically Endangered (CR) ορίζονται τα είδη που αντιμετωπίζουν εξαιρετικά υψηλό κίνδυνο εξαφάνισης στο άμεσο μέλλον. Ως Κινδυνεύοντα - Endangered (EΝ) είναι είδη που αντιμετωπίζουν εξαιρετικά υψηλό κίνδυνο εξαφάνισης στο φυσικό τους περιβάλλον στο εγγύς μέλλον. Ως τρωτά - Vulnerable (VU) είναι είδη τα οποία θεωρείται ότι αντιμετωπίζουν άμεσο κίνδυνο εξαφάνισης μεσοπρόθεσμα. Τα είδη που αξιολογούνται ως Κρισίμως Κινδυνεύοντα (CR), Κινδυνεύοντα (ΕΝ) ή Τρωτά (VU) αναφέρονται ως «απειλούμενα» στην παρούσα μελέτη. Εικόνα 3.4. Κατηγορίες της Κόκκινης Λίστας της IUCN (Economidis 2009) Figure 3.4. IUCN Red List Categories (Economidis 2009) 146
147 3.3.2 Επιλογή του μοντέλου με την καλύτερη προσαρμογή για τη περιγραφή της σχέσης αριθμού ειδών-έκτασης στα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου Τα τελευταία χρόνια ο αριθμός των μοντέλων, που έχουν προταθεί για την περιγραφή της σχέσης αριθμού ειδών-έκτασης, έχει αυξηθεί σημαντικά και φτάνει τα 20 (Πίνακας 3.1, Triantis, Guilhaumon & Whittaker 2012). Το ερώτημα λοιπόν που προκύπτει είναι, εάν υπάρχει ένα μαθηματικό μοντέλο, που να περιγράφει αποτελεσματικότερα τη σχέση σε διαθέσιμα εμπειρικά δεδομένα. Για παράδειγμα, ο Dengler (2009) χρησιμοποιώντας δεδομένα ηπειρωτικού και νησιωτικού τύπου συνέκρινε 23 διαφορετικές συναρτήσεις και κατέληξε ότι το δυναμικό μοντέλο [το μοντέλο του Arrhenius (1921) (εξίσωση 4)] είναι κατάλληλο για την περιγραφή της SAR ανεξάρτητα από τον τύπο των δεδομένων και την κλίμακα της ανάλυσης. Αντίστοιχα, οι Triantis, Guilhaumon & Whittaker (2012) κατέληξαν στο συμπέρασμα πως το μοντέλο του Arrhenius (1921) (εξίσωση 4) εμφάνισε την καλύτερη προσαρμογή σε δεδομένα γεωγραφικών νησιών. 147
148 Πίνακας 3.1. Τα 20 μοντέλα που έχουν προταθεί για την περιγραφή της σχέσης αριθμού ειδώνέκτασης (SAR) (από Triantis, Guilhaumon & Whittaker 2012) Table 3.1. The 20 models that have been proposed to describe the species-area relationship (SAR) (from Triantis, Guilhaumon & Whittaker 2012) Name Formula Shape Asymptotic Linear- Γραμμική S=c+zA Linear - Γραμμική NO Power- Συνάρτηση S = ca z Convex- Κυρτή NO δύναμης Power - Συνάρτηση δύναμης Rosenzweig S = k + ca z Convex- Κυρτή NO Extended Power 1- Επαυξημένη συνάρτηση δύναμης 1 Extended Power 2- Επαυξημένη συνάρτηση δύναμης 2 S = ca za d S = ca z (d/a) Sigmoid- Σιγμοειδής Convex- Κυρτή Sigmoid- Σιγμοειδής Persistence Function 1 S = ca z exp( da) Convex- Κυρτή NO Persistence Function 2 S = ca z exp( d/a) Sigmoid- NO Σιγμοειδής Exponential- Εκθετική S = c + z loga Convex- Κυρτή NO Kobayashi Logarithmic S = c log(1 + A/z) Convex- Κυρτή NO Monod S = d/(1 + ca 1 ) Convex- Κυρτή YES (d) Morgan-Mercer-Flodin S = d/(1 + ca z ) Sigmoid- YES (d) Σιγμοειδής Logistic- Λογιστική S = c/(f + A z ) Sigmoid- YES (c/f) Σιγμοειδής Negative Exponential- S = d[1 exp( za)] Convex- Κυρτή YES (d) Αρνητική εκθετική Chapman-Richards S = d[1 exp( za)] c Sigmoid- YES (d) Σιγμοειδής Weibull-3 S = d[1 exp( ca z )] Sigmoid- YES (d) Σιγμοειδής Weibull-4 S = d[1 Sigmoid- YES (d) exp( ca z )] d Σιγμοειδής Asymptotic- S = d cz A Convex- Κυρτή YES (d) Ασυμπτωτική Rational S = (c + za)/(1 + da Convex- Κυρτή YES (z/d) ) Gompertz S = d exp[ exp( z(a Σιγμοειδής YES (d) c))] Beta-P S = d[1 (1 + (A/c) z ) f ] Σιγμοειδής YES (d) NO NO 148
149 Tα 8 βασικότερα μαθηματικά μοντέλα (Δυναμικό, Εκθετικό, Αρνητικό εκθετικό, Monod, Rational, Λογιστικό, Lomolino και cumulative Weibull) (Πίνακας 3.2) που έχουν προταθεί έως σήμερα για τη περιγραφή της SAR (Guilhaumon et al. 2008, Guilhaumon et al. 2010) εφαρμόστηκαν στα δεδομένα κατανομών των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου ανά λεκάνη απορροής ποταμού και λίμνη. Για κάθε οικοσύστημα καταγράφηκε η έκταση και ο αριθμός των ειδών. Tα προτεινόμενα μοντέλα εφαρμόστηκαν στις καμπύλες έκτασης- αριθμού ειδών ώστε να προσδιοριστεί το μοντέλο που αποδίδει καλύτερα σύμφωνα με το διορθωμένο Akaike Κριτήριο Πληροφορίας (AICc) επιλέγοντας εκείνο με την καλύτερη προσαρμογή για τη μαθηματική περιγραφή τους. Τα 8 προτεινόμενα μοντέλα εφαρμόστηκαν στο σύνολο των δεδομένων, στις 78 λεκάνες απορροής ποταμών και 19 λίμνες, ξεχωριστά. Πίνακας 3.2. Τα 8 βασικότερα μαθηματικά μοντέλα που εφαρμόστηκαν στις καμπύλες αριθμού ειδών στα δεδομένα κατανομών των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου όπου S= αριθμός ειδών, A= έκταση και c, z, d και f= σταθερές (από Guilhaumon et al. 2010) Table 3.2. Functional forms for the SAR implemented in distributional data of the freshwater fishes of the Balkan Peninsula where S and A represent, respectively, species richness and area,while c, z, d and f are fitted parameters (from Guilhaumon et al. 2010) Name Formula Shape Asymptotic Reference Power- Συνάρτηση δύναμης S = ca z Convex- Κυρτή NO (Arrhenius, 1921) Exponential- Εκθετική S = c + zlog(a) Convex- Κυρτή NO (Gleason, 1922) Negative exponential- S = c(1 - exp(-za)) Convex- Κυρτή YES (Holdridge et al. 1971) Αρνητική εκθετική Monod S = (ca) / (z + A) Convex- Κυρτή YES (Monod, 1950) Rational S = (c + za) / (1 + fa) Convex- Κυρτή YES (Ratkowsky, 1990) Logistic- Λογιστική S = c / (1 + exp(- za+f) Sigmoid- Σιγμοειδής YES (Ratkowsky, 1990) Lomolino S = c / 1 + (z log(f/a) ) Sigmoid- YES (Lomolino, Σιγμοειδής cumulative Weibull S = c(1 - exp(-za f )) Sigmoid- Σιγμοειδής YES 2000) (Weibull, 1951) 149
150 3.3.3 Στατιστική επεξεργασία και πακέτα Όλες οι αναλύσεις της επιλογής του μοντέλου με την καλύτερη προσαρμογή για τη περιγραφή της σχέσης αριθμού ειδών-έκτασης στα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου πραγματοποιήθηκαν στο «R statistical and programming environment» (R , R Development Core Team 2013) χρησιμοποιώντας το πακέτο «mmsar». 3.4 Αποτελέσματα Σύνθεση και πρότυπα αριθμού ειδών της Βαλκανικής ιχθυοπανίδας Η οικογένεια των Cyprinidae είναι κυρίαρχη της Βαλκανικής ιχθυοπανίδας, αντιπροσωπεύοντας το 59% του συνόλου των ειδών, ακολουθούμενη από τις οικογένειες Cobitidae (10%), Salmonidae (10%) και Gobiidae (6%). Όσον αφορά τα ενδημικά είδη, τα περισσότερα από τα μισά ανήκουν στην οικογένεια των Cyprinidae (64%), ακολουθούμενα από είδη των οικογενειών Cobitidae και Salmonidae (9%) και Gobiidae (8%). Τέλος, τέσσερα ενδημικά είδη κατατάσσονται στην οικογένεια Nemacheilidae και δύο είδη στην οικογένεια Clupeidae, ενώ οι οικογένειες Blennidae, Gasterosteidae, Percidae και Siluridae εκπροσωπούνται από ένα μόνο είδος. Η Βαλκανική χερσόνησος φιλοξενεί τουλάχιστον 193 αυτόχθονα είδη. Μεταξύ των βιογεωγραφικών περιοχών που οριοθετήθηκαν με βάση το δείκτη βjtu, ο αριθμός των ειδών, όπως φαίνεται στην Εικόνα 3.5a, είναι υψηλότερος για τη Ποντιακή επαρχία (n=62), ακολουθούμενος από τη νότια Αδριατική θάλασσα (n=57), το Αιγαίο (n=55), τη λεκάνη απορροής του π. Neretva (n=39), το Ιόνιο Πέλαγος (n=28) και τη βόρεια Αδριατική (n=27). Τα λιγότερα είδη βρέθηκαν στην Αττικοβοιωτία (n=11) και τις Πρέσπες (n=9). 150
151 3.4.2 Πρότυπα ενδημισμού και απειλούμενων ειδών (IUCN Κατηγορίες) της Βαλκανικής ιχθυοπανίδας Ο ενδημισμός της Βαλκανικής χερσονήσου είναι εξαιρετικά υψηλός (56%). Το υψηλότερο ποσοστό ενδημισμού παρατηρήθηκε στην περιοχή των Πρεσπών (89%, n =8) και το Ιόνιο Πέλαγος (75%, n=21), ενώ χαμηλότερα ποσοστά ενδημικών ειδών παρατηρήθηκαν στην περιοχή της Αττικοβοιωτίας (54%, n=6), του Αιγαίου (45%, n=25), της νότιας Αδριατικής θάλασσας (40%, n=23), της βόρειας Αδριατικής Θάλασσας (37%, n=10) και στη λεκάνη απορροής του π. Neretva (36%, n=14). Στην Ποντιακή επαρχία δεν καταγράφηκαν ενδημικά είδη. Συνολικά, το 33% των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου έχουν χαρακτηριστεί ως απειλούμενα (Εικόνα 3.5b). Το μεγαλύτερο ποσοστό των ειδών που ανήκουν στα απειλούμενα σύμφωνα με την Κόκκινη Λίστα (IUCN, 2013) παρατηρήθηκε στις Πρέσπες (66%), την Αττικοβοιωτία (45%), το Ιόνιο Πέλαγος (39%), τη λεκάνη απορροής του ποταμού Neretva (38%), τη βόρεια Αδριατική Θάλασσα (33%) και το Αιγαίο (22%). Η νότια Αδριατική και η Ποντιακή επαρχία εμφάνισαν τα χαμηλότερα ποσοστά των απειλούμενων ειδών (14% και 5%, αντίστοιχα). 151
152 Εικόνα 3.5a. Ο απόλυτος αριθμός των αυτόχθονων και ενδημικών ειδών στις οκτώ βιογεωγραφικές περιοχές της Βαλκανικής χερσονήσου. Τα ποσοστά στην κορυφή των στηλών δείχνουν το βαθμό ενδημισμού b. Ποσοστά των αυτόχθονων ειδών που ανήκουν σε κάθε βιογεωγραφική περιοχή, σύμφωνα με κριτήρια της IUCN (ΝΕ: Δεν έχει αξιολογηθεί, DD: Ανεπαρκώς γνωστό, NT: Σχεδόν Απειλούμενο, LC: Μειωμένου ενδιαφέροντος, VUL: Τρωτό, EN: Κινδυνεύον, CR: Κρισίμως Κινδυνεύον, EX: Εξαφανισμένο). Τα ποσοστά δείχνουν τα ποσοστά των απειλούμενων ειδών Figure 3.5a. Species richness and absolute number of endemic species in the eight biogeographical regions of the Balkan Peninsula. Percentages in the top of the bars indicate degree of endemism b. Percentages of native species belonging to each biogeographical region according to IUCN criteria (NE: Not evaluated, DD: Data Deficient, NT: Near Threatened, LC: Least Concern, VUL: Vulnerable, EN: Endangered, CR: Critically Endangered, EX: Extinct). Percentages show proportion of species listed as threatened SARs στα οικοσυστήματα των εσωτερικών υδάτων στα Βαλκάνια Σύμφωνα με τα αποτελέσματα της διαδικασίας επιλογής του καταλληλότερου μοντέλου για τη περιγραφή της SAR σε δεδομένα κατανομής ψαριών στα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου, το δυναμικό 152
153 μοντέλο (Power) παρουσίασε τις καλύτερες επιδόσεις και καταλαμβάνει την πρώτη θέση σε όλες τις περιπτώσεις τόσο για το σύνολο των δεδομένων όσο και για τις 75 λεκάνες απορροής ποταμών, ξεχωριστά (Πίνακας 3.3). Το συνολικό ποσοστό ερμηνείας από το δυναμικό μοντέλο για το σύνολο των δεδομένων (R 2 =0,36), τις λεκάνες απορροής ποταμών (R 2 =0,52) και τις λίμνες (R 2 =0,23) ήταν αρκετά χαμηλό (Εικόνες 3.6, 3.7 και 3.8). Μετά την αφαίρεση των δύο μεγαλύτερων ποτάμιων οικοσυστημάτων (Δούναβης-Danube, Σάβα-Sava) το ποσοστό ερμηνείας από το δυναμικό μοντέλο παρέμεινε χαμηλό για το σύνολο των δεδομένων (R 2 =0,23) και τις λεκάνες απορροής ποταμών (R 2 =0,37). Πίνακας 3.3. Αποτελέσματα της διαδικασίας επιλογής του καταλληλότερου μοντέλου για τη περιγραφή της SAR σε δεδομένα κατανομής ψαριών στα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου. D-AICc είναι οι διαφορές του διορθωμένου Akaike Κριτηρίου Πληροφορίας, AICc-W οι τιμές βαρύτητας ισοδύναμες με την πιθανότητα το μοντέλο να ερμηνεύει το σύνολο των δεδομένων και R 2 το ποσοστό µεταβλητότητας του αριθμού των ειδών που ερμηνεύεται από το µοντέλο (για λεπτομέρειες βλέπε Guilhaumon et al. 2008) Table 3.3. Results of the model-selection process for the description of SAR in fish distribution data on freshwaters of the Balkan Peninsula. D-AICc are differences in AICc, AICc-W values correspond to model weights that are equivalent to model probabilities in being the best to fit the data set and R 2 are the coefficients of determination (for details see Guilhaumon et al. 2008) D-AICc AICc-W R 2 1) All systems power 0 0,75 0,36 weibull 2,19 0,25 0,36 2) Only rivers power 0 0,56 0,52 expo 8,30 0,01 0,46 logist 5,37 0,04 0,50 ratio 1,99 0,21 0,52 weibull 2,25 0,18 0,52 3) Lakes power 0 0,84 0,23 weibull 3,28 0,16 0,23 153
154 Εικόνα 3.6. Σχέση αριθμού ειδών και έκτασης (SAR) για τα ψάρια στο σύνολο των λεκανών απορροής ποταμών και λιμνών στη Βαλκανική χερσόνησο με βάση το μοντέλο με την καλύτερη προσαρμογή [το μοντέλο του Arrhenius (1921), (εξίσωση 4)] Figure 3.6. Species- area relationship (SAR) of fish distribution data in all drainages of the Balkan Peninsula using the model with the best fit [model Arrhenius (1921), (Equation 4)] 154
155 Εικόνα 3.7. Σχέση αριθμού ειδών και έκτασης (SAR) για τα ψάρια στο σύνολο των ποταμών στη Βαλκανική χερσόνησο με βάση το μοντέλο με την καλύτερη προσαρμογή [το μοντέλο του Arrhenius (1921), (εξίσωση 4)] Figure 3.7. Species- area relationship (SAR) of fish distribution data in rivers of the Balkan Peninsula using the model with the best fit [model Arrhenius (1921), (Equation 4)] 155
156 Εικόνα 3.8. Σχέση αριθμού ειδών και έκτασης (SAR) για τα ψάρια στο σύνολο των λιμνών στη Βαλκανική χερσόνησο με βάση το μοντέλο με την καλύτερη προσαρμογή [το μοντέλο του Arrhenius (1921), (εξίσωση 4)] Figure 3.8. Species- area relationship (SAR) of fish distribution data in lakes of the Balkan Peninsula using the model with the best fit [model Arrhenius (1921), (Equation 4)] 3.5 Συζήτηση Σύνθεση και πρότυπα της Βαλκανικής ιχθυοπανίδας Η Βαλκανική χερσόνησος είναι η πιο άφθονη περιοχή (υψηλότερος αριθμός ειδών) της Ευρώπης, όσον αφορά τα ψάρια των εσωτερικών υδάτων. Φιλοξενεί πάνω από 193 αυτόχθονα είδη ψαριών, ενώ η βόρεια, κεντρική και δυτική Ευρώπη (Reyjol et al. 156
157 2007), η Ιταλική (Bianco 2013) και η Ιβηρική χερσόνησος (Maceda-Veiga 2013) χαρακτηρίζονται από χαμηλή αφθονία ειδών ( 50 είδη). Ο ενδημισμός των ψαριών των Βαλκανικών εσωτερικών υδάτων είναι υψηλός (56%) σε παρόμοια επίπεδα με άλλα ευρωπαϊκά θερμά σημεία, όπως η Ιβηρική (73%, Maceda-Veiga 2013) και η Ιταλική χερσόνησος (27%, Ruffo & Stoch 2006). Οι ιχθυοπανίδες των νότιων χερσονήσων (Ιβηρική, Ιταλική και Βαλκανική) απομονώθηκαν η μία από την άλλη και από αυτές της Βόρειας Ευρώπης, από το Ολιγόκαινο έως το Μειόκαινο (Levy, Doadrio & Almada 2009), οδηγώντας σε υψηλά επίπεδα ενδημισμού. Ο ενδημισμός στα Βαλκάνια αυξάνεται από το βορρά προς το νότο και από την ανατολή προς τη δύση λόγω της παρουσίας των Βαλκανικών και των Διναρίδων-Ελληνίδων οροσειρών, οι οποίες εμπόδισαν τα περισσότερα είδη της Μεσογείου να κινηθούν προς τα βόρεια. Αυτός ο διαχωρισμός οδήγησε σε ανεξάρτητες βιογεωγραφικές ιστορίες, πολλαπλά συμβάντα βικαριανισμού και υψηλούς ρυθμούς διαφοροποίησης στις νότιες και δυτικές βιογεωγραφικές περιοχές των Βαλκανίων (βλέπε επίσης Oberdorff, Lek & Guégan 1999). Ο υψηλός ενδημισμός ( 50%) των περι-μεσογειακών βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου (δηλαδή, το Ιόνιο Πέλαγος και η Αττικοβοιωτία) δείχνει επίσης ότι τα είδη που απαντούν σε αυτά τα σημεία ενδιαφέροντος απομονώθηκαν από τις ιχθυοπανίδες των άλλων βιογεωγραφικών περιοχών και από τους μη μεσογειακούς συγγενείς τους (Ποντιακή Επαρχία) πριν από το Πλειστόκαινο. Αν δε συνέβαινε αυτό, οι Μεσογειακές και μη περιοχές θα είχαν κοινά τα περισσότερα είδη (Durand, Persat & Bouvet 1999, Perdices & Doadrio 2001, Šlechtová et al. 2004). Επιπλέον, οι παγετώδεις περίοδοι κατά το Πλειστόκαινο, όπως αναφέρθηκε προηγουμένως, έπαιξαν σημαντικό ρόλο στη δευτερεύουσα εποίκιση από το παραδουνάβιο καταφύγιο και στον καθορισμό της παρούσας ευρωπαϊκής κατανομής των ψαριών των εσωτερικών υδάτων (Sommerwerk et al. 2009). Έτσι, η παραδουνάβια λεκάνη και οι συλλήψεις ποταμών της (νότια Αδριατική θάλασσα, Ανατολική Βαλκανική χερσόνησος) προώθησαν την επέκταση και τη γενετική ομογενοποίηση των ειδών των εσωτερικών υδάτων σε όλη την Ευρώπη (Perea et al. 2010), οδηγώντας αυτές τις περιοχές σε χαμηλό ενδημισμό. 157
158 Τα αποτελέσματα του παρόντος κεφαλαίου δείχνουν ότι τα πρότυπα του αριθμού των ειδών και τα πρότυπα του ενδημισμού είναι ανεστραμμένα στη Βαλκανική χερσόνησο, όπου οι περιοχές με τα υψηλότερα επίπεδα ενδημισμού είναι οι περιοχές με τον χαμηλότερο αριθμό ειδών, μια παρατήρηση που έρχεται σε συμφωνία με μελέτες της ευρωπαϊκής ιχθυοπανίδας των εσωτερικών υδάτων (Peter 2006). Έχει δειχθεί ότι η ιστορία είναι ένας από τους πιο σημαντικούς παράγοντες καθορισμού των προτύπων του αριθμού των αυτόχθονων και ενδημικών ειδών (Heino 2011, Griffiths, McGonigle & Quinn 2014). Οι φτωχές λεκάνες απορροής που εμφανίζουν υψηλά επίπεδα ενδημισμού, όπως αυτές στην περιοχή των Πρεσπών, Αττικοβοιωτίας και Ιονίου αφορούν ασύνδετες, μικρές λεκάνες απορροής σε μια στενή γεωγραφική περιοχή. Η ασθενέστερη συνδεσιμότητα των λεκανών απορροής μπορεί να αποτρέψει τη διασπορά των ειδών σε μεγάλες αποστάσεις, αυξάνοντας έτσι τον τοπικό ενδημισμό (Heino 2011) και οδηγεί σε μια συνολικά αρνητική σχέση μεταξύ του συνολικού αριθμού ειδών με το ποσοστό των ενδημικών ειδών. Για παράδειγμα, ένας σημαντικός αριθμός ειδών μπορεί να έχει εξαφανιστεί από τα νότια καταφύγια μετά την τελευταία εποχή των παγετώνων, όταν το μεσογειακό κλίμα μετατράπηκε σε ημι-άνυδρο (Salinas, Blanca & Romero 2000). Ο χαμηλός αριθμός των ειδών των νότιων περιοχών (δηλαδή, στο Ιόνιο και στην Αττικοβοιωτία) μπορεί να σχετίζεται και με την επίδραση της χερσονήσου, η οποία σύμφωνα με τον Kryštufek (2004) έχει επίσης παρατηρηθεί στην ομάδα των θηλαστικών στη Βαλκανική χερσόνησο. Αντίθετα, η αυξημένη συνδεσιμότητα και το μεγαλύτερο μέγεθος των λεκανών απορροής και οι περιβαλλοντικές συνθήκες των οικοσυστημάτων αυτών, μπορεί να οδήγησαν στους υψηλούς αριθμούς ειδών στις βορειότερες περιοχές (Griffiths, McGonigle & Quinn 2014). Παράλληλα, ένα μεγάλο ποσοστό της ιχθυοπανίδας της Βαλκανικής απειλείται με εξαφάνιση (33%), όπως στη περίπτωση της Ιβηρικής και Ιταλικής χερσονήσου, όπου τα μισά από τα είδη έχουν χαρακτηριστεί ως κρισίμως κινδυνεύοντα, κινδυνεύοντα ή τρωτά, σύμφωνα με τα κριτήρια της IUCN (Maceda-Veiga 2013, Bianco 2013). Το ποσοστό των απειλούμενων ειδών είναι υψηλότερο στην περιοχή των Πρεσπών, του Ιονίου και της Αττικοβοιωτίας από ότι σε άλλες βιογεωγραφικές περιοχές με μεγάλες συνδεδεμένες λεκάνες απορροής (δηλαδή στην Ποντιακή επαρχία, την περιοχή του Αιγαίου και της νότιας Αδριατικής θάλασσας), τονίζοντας 158
159 ότι ο αριθμός των απειλούμενων ειδών είναι το αποτέλεσμα της συνέργειας διαφόρων παραγόντων, όπως η συνδεσιμότητα, το μέγεθος της λεκάνης απορροής, η ικανότητα διασποράς των ειδών και η ανθρωπογενής διατάραξη (Smith & Darwall 2006). Το συνολικό ποσοστό ερμηνείας από το δυναμικό μοντέλο (στο σύνολο των δεδομένων και στις λεκάνες απορροής ποταμών ξεχωριστά) για τη σχέση αριθμού ειδών- έκτασης ήταν αρκετά χαμηλό (R 2 =0,52) καταδεικνύοντας την πολυπλοκότητα των διεργασιών διαμόρφωσης του αριθμού των ειδών των ψαριών των εσωτερικών υδάτων στη Βαλκανική χερσόνησο (βλέπε επίσης Reyjol et al. 2007). H τιμή του συντελεστή z (z=0.26) είναι μεγαλύτερη από το εύρος των τιμών (z=0,19-0,24) που καταγράφηκε στα ποτάμια οικοσυστήματα της κεντρικής και δυτικής Ευρώπης (Hugueny 1989, Reyjol et al. 2007) ή της Αφρικής όπου η τιμή του συντελεστή εκτιμήθηκε ως z = 0,32 (Hugueny 1989). Εντούτοις, η τιμή αυτή είναι πολύ μικρότερη εν συγκρίσει με την τιμή του δείκτη για τα 47 μεγαλύτερα ποτάμια οικοσυστήματα σε παγκόσμιο επίπεδο που εκτιμήθηκε ως z =0,4 (Welcomme 1985), καθώς αφορά τόσο εύκρατα όσο και τροπικά ποτάμια οικοσυστήματα. Αν και, η έκταση per se εξηγεί το μεγαλύτερο ποσοστό της διακύμανσης του αριθμού των ειδών σε ποτάμια οικοσυστήματα (αύξηση της έκτασης της λεκάνης απορροής συνεπάγεται αύξηση αριθμού ειδών) (Hugueny 1989), εντούτοις, είναι αναγκαία η συνεκτίμηση και άλλων παραμέτρων πέραν της έκτασης, όπως η περιβαλλοντική ετερογένεια, η διαθέσιμη ενέργεια καθώς και η επίδραση της ειδογένεσης και εξαφάνισης (Hugueny, Oberdorff & Tedesco 2010). Αναλυτικότερα, τα ποτάμια εκτείνονται σε ένα μεγάλο εύρος γεωλογικών σχηματισμών και βιογεωγραφικών περιοχών, χαρακτηρίζονται από μεγάλη μεταβλητότητα των περιβαλλοντικών παραμέτρων, όπως η ροή, το υπόστρωμα και η θερμοκρασία του νερού σε σύγκριση με τις λίμνες που έχουν παρόμοιο μέγεθος και βάθος (Horwitz 1978). Πέραν όμως των παραμέτρων αυτών, είναι απαραίτητη και η αναφορά στην ίδια την γεωλογική δυναμική (Ricklefs 2004) κάθε περιοχής, όπως έχει καταδειχτεί σε ευρωπαϊκό και παγκόσμιο επίπεδο για τα ψάρια των εσωτερικών υδάτων (Oberdorff, Guégan & Hugueny 1995, Tedesco et al. 2005), αλλά και στη Βαλκανική χερσόνησο (Oikonomou, Leprieur & Leonardos 2014). 159
160 Με βάση αυτά τα αποτελέσματα, για να κατανοηθούν οι διαφορές στα πρότυπα της χωρικής διαβάθμισης της βιολογικής ποικιλότητας ανάμεσα στις περιοχές, οι ερευνητές θα πρέπει να εξετάζουν την ιστορία των οικοσυστημάτων και των βιογεωγραφικών περιοχών, τις καταγωγές των ειδών (Reyjol et al. 2007) καθώς και τα οικολογικά χαρακτηριστικά των ειδών (Hof, Brändle, & Brandl 2008). Τα αποτελέσματα του παρόντος κεφαλαίου αναδεικνύουν τη μεγάλη επίδραση της ιστορικότητας που σχετίζεται τόσο με την γεωλογική εξέλιξη μιας περιοχής όσο και με τα γεγονότα εποίκισης και εξαφάνισης. Φυλογενετικές και φυλογεωγραφικές αναλύσεις τόσο στοχευμένων κλάδων ψαριών και του συνόλου των ειδών της Βαλκανικής είναι βέβαιο πως θα αναδείξουν τα βασικά γεγονότα εποίκισης, εξαφάνισης και διαφοροποίησης σε σχέση και με την παλαιογεωγραφική διαφοροποίηση της περιοχής που διαμόρφωσαν τα παρατηρούμενα πρότυπα βιοποικιλότητας Διαχειριστικές προτάσεις Η διατήρηση της βιοποικιλότητας των εσωτερικών υδάτων έχει σε μεγάλο βαθμό αγνοηθεί μέχρι σήμερα (Holland, Darwall & Smith 2012). Έχουμε ακόμη ελλιπή γνώση του κατά πόσον τα πρότυπα μέσα στις-ταξινομικές ομάδες ή/ και μεταξύ των ταξινομικών ομάδων, όσον αφορά τον αριθμό των ειδών και τον ενδημισμό, συμφωνούν (Pimm 1999, Lamoreux et al. 2006) και, ως εκ τούτου, κατά πόσο τα διαχειριστικά σχέδια βάσει του αριθμού των ειδών («Richness hotspots»), θα διατηρήσουν επαρκώς τις περιοχές υψηλού ενδημισμού («Endemic hotspots») (Ricketts 2001). Κατά συνέπεια, από την παρούσα μέλετη προτείνεται η ιεράρχηση των «θερμών περιοχών» που χαρακτηρίζονται από υψηλά επίπεδα μοναδικότητας (ενδημισμού) («Endemic hotspots») και απειλούμενων ειδών («Threatspots») (Oberdorff, Lek & Guégan 1999), καθώς παρουσιάζουν ξεχωριστή εξελικτική ιστορία ως κέντρα διαφοροποίησης των ειδών (Jetz, Rahbek & Colwell 2004). Οι περιοχές της Αττικοβοιωτίας, του Ιονίου και των λιμνών των Πρεσπών θα πρέπει να ληφθούν υπόψη ως σημεία υψηλού ενδιαφέροντος κατά την ανάπτυξη ενός πλαισίου αποτελεσματικής διατήρησης και στοχευμένης έρευνας για τη βιοποικιλότητα. 160
161 Επιπλέον, τα αποτελέσματά της παρούσας μελέτης αναδεικνύουν την ανάγκη για κοινές διασυνοριακές προσπάθειες διατήρησης μεταξύ των χωρών, όπως για παράδειγμα στην περίπτωση των βιογεωγραφικών περιοχών του Αιγαίου Πελάγους, της νότιας Αδριατικής θάλασσας, της λεκάνης απορροής του ποταμού Neretva και της βόρειας Αδριατικής Θάλασσας, μιας και οι διοικητικές διαιρέσεις δε συμπίπτουν με αυτές τις βιογεωγραφικές περιοχές. Ωστόσο, στο μέλλον θα πρέπει να εξετάζονται πρόσθετα κριτήρια, όπως η φυλογενετική (βλέπε Holt et al. 2013) και η λειτουργική μοναδικότητα των ειδών (Olden et al. 2010), με απώτερο στόχο την βελτιστοποίηση των στρατηγικών διατήρησης εντός των καθορισμένων βιογεωγραφικών περιοχών. Εν κατακλείδι, η παρούσα μελέτη εκτίμησης της βιοποικιλότητας της Βαλκανικής χερσονήσου μπορεί να συνεισφέρει στη διασφάλιση της μακροπρόθεσμης διατήρησης της βιοποικιλότητας των Βαλκανίων, και με βάση αυτό, τα αποτελέσματα μας δυνητικά θα πρέπει να ληφθούν υπόψη στην ευρωπαϊκή ερευνητική ατζέντα σχετικά με τα οικοσυστήματα των εσωτερικών υδάτων. 161
162
163 ΜΕΡΟΣ II Τοπικό επίπεδο: Ελλάδα
164
165 Κεφάλαιο 4 Βιογεωγραφικά πρότυπα των ψαριών στα εσωτερικά ύδατα της Ελλάδας
166
167 4.1 Εισαγωγή Τα εσωτερικά ύδατα της Ελλάδας H Ελλάδα χαρακτηρίζεται από ένα πλήθος μικρών λεκανών απορροής ποταμών και λιμνών και είναι χώρα πλούσια σε υγροτόπους 18. Η απογραφή που έγινε το 1993 από το Ελληνικό Κέντρο Βιοτόπων-Υγροτόπων έδειξε ότι υπάρχουν περί τους 400 υγροτόπους στην ηπειρωτική Ελλάδα 19, συνολικής έκτασης πάνω από km 2 (1.6% της έκτασης της), οι οποίοι αντιπροσωπεύουν το μεγαλύτερο μέρος των κατηγοριών των υγροτόπων που υπάρχουν παγκοσμίως σύμφωνα με τη συνθήκη Ramsar (Ζαλίδης & Μαντζαβέλας 1994). Συνολικά, 91 ποταμοί έχουν καταμετρηθεί στην Ελλάδα αν και σε μόνο δέκα από αυτούς η εκροή υπερβαίνει το 1 km 3 /a (Skoulikidis, Bertahas & Koussouris 1998). Η μεγαλύτερη συγκέντρωση εσωτερικών υδάτων παρατηρείται στη Βόρεια Ελλάδα (48% του συνολικού εμβαδού) με τους περισσότερους ποταμούς να βρίσκονται στην Ανατολική Μακεδονία και Θράκη (Ζαλίδης & Μαντζαβέλας 1994). Εκτός από τους ποταμούς στα βόρεια της χώρας που διαθέτουν συνήθως συνεχή ροή καθ όλη τη διάρκεια του έτους, τα υπόλοιπα ποτάμια οικοσυστήματα χαρακτηρίζονται από μεγάλες διακυμάνσεις ροής με τις υψηλότερες τιμές να παρατηρούνται κατά τη διάρκεια του χειμώνα, στις ελάχιστες ροές είτε την πλήρη ξήρανση, κατά την περίοδο του καλοκαιριού. Επιπλέον, χαρακτηρίζονται από αυξημένη ετερογένεια, η κλίση τους είναι ιδιαίτερα απότομη στα ανάντη, συχνά διέρχονται μέσα από φαράγγια ή χαράδρες, ενώ όσον αφορά το υπόστρωμά τους χαρακτηρίζεται γενικά από πέτρες και χαλίκια, παρέχοντας κατ αυτόν τον τρόπο κάλυψη στα ψάρια (Skoulikidis et al. 2009). Από τις 56 Ελληνικές λίμνες με συνολική επιφάνεια περίπου 560 km 2 (επιφάνεια μικρότερη από 0,5% της συνολικής έκτασης της χώρας) (Skoulikidis, 18 Οι ποταμοί και οι λίμνες εντάσσονται στους υγροτόπους σύμφωνα και με τον ευρύτερα αποδεκτό ορισμό της "Σύμβασης για τους Υγροτόπους Διεθνούς Σημασίας Ειδικά ως Ενδιαιτήματος για Υδρόβια Πουλιά" (1971), γνωστής απλώς και ως Σύμβασης Ραμσάρ. Στους υγροτόπους συμπεριλαμβάνονται επίσης τα δέλτα, έλη λιμνοθάλασσες, πηγές, εκβολές και τεχνητές λίμνες 19 Η Ελλάδα είναι εξίσου πλούσια και σε νησιωτικούς υγροτόπους, οι οποίοι ανέρχονται στους 803 και κατανέμονται σε 75 νησιά σύμφωνα με το WWF Ελλάς ( 167
168 Bertahas & Koussouris 1998), οι περισσότερες βρίσκονται στο γεωγραφικό διαμέρισμα της Ηπείρου (Ζαλίδης & Μαντζαβέλας 1994). Οι περισσότερες λίμνες βρίσκονται εντός ή έχουν μια ιστορική σύνδεση με τις λεκάνες απορροής ποταμών αποτελώντας κέντρα ενδημισμού (π.χ. Πρέσπες, Oikonomou, Leprieur & Leonardos 2014) Τα εσωτερικά ύδατα της Ελλάδας ως αποτέλεσμα της παλαιογεωγραφικής ιστορίας Γενικότερα, ο Ελληνικός χώρος αποτελεί ένα ζωντανό γεωλογικό εργαστήρι όπου διαδραματίζονται σημαντικά γεωλογικά γεγονότα. Η γεωδυναμική εξέλιξη της Ελλάδας καθορίζεται από την κίνηση των πλακών της Ανατολικής Μεσογείου και κυρίως από τη σύγκλιση της Ευρασιατικής και της Αφρικανικής πλάκας. Η παλαιογεωγραφική ιστορία του Ελληνικού χώρου εντάσσεται σε αυτή της δημιουργίας της Βαλκανικής χερσονήσου (Κεφάλαιο 2, Σημείωμα 1). Η Βόρεια Ελλάδα υπάγεται στην πλάκα της Ευρασίας ενώ η Νότια ηπειρωτική Ελλάδα και το Αιγαίο εντάσσονται στην πλάκα του Αιγαίου. Οι Εξωτερικές Ελληνίδες τοποθετούνται στην μικροπλάκα της Απουλίας, που αποσπάσθηκε από την Γκοντβάνα (Μουντράκης 2010). Κατά το Τριαδικό, ο Ελληνικός χώρος βρίσκονταν κάτω από τη στάθμη της θάλασσας της Τηθύος. Την περίοδο του Ανωτέρου Ιουρασικού ξεκινά η πρώιμη ορογενετική δράση. Στο Μέσο Κρητιδικό, η στάθμη της θάλασσας ανήλθε και κάλυψε το χώρο των Εσωτερικών Ελληνικών γεωτεκτονικών ζωνών 20 (ή αλλιώς Ελληνίδες ζώνες) με τη λεγόμενη «επίκλυση του Κενομανίου». Ανάμεσα στα σημαντικότερα γεγονότα στην παλαιογεωγραφική ιστορία των εσωτερικών υδάτων της Ελλάδας είναι η ανάδυση των Ελληνίδων-Διναριδών οροσειρών και μετέπειτα στο Κάτω Ολιγόκαινο, η ανάδυση της οροσειράς της Πίνδου που ως φυσιογραφικό φράγμα διαχωρίζει τη Δυτική από την Ανατολική Ελλάδα. Όπως φαίνεται στην Εικόνα 2.3, κατά το Νέο-Τορτόνιο (8 εκ. έτη), συνέβη η εισβολή 20 Οι Ελληνικές γεωτεκτονικές ζώνες διακρίνονται σε τρεις κατηγορίες (Μουντράκης, 1985). Οι μάζες Ροδόπης και Σερβομακεδονικής αποτελούν την Ελληνική Ενδοχώρα. Οι ζώνες Περιροδοπική, Παιονίας, Πάικου, Αλμωπίας, Πελαγονική, Αττικοκυκλαδική και Υποπελαγονική ονομάζονται Εσωτερικές Ελληνίδες. Οι ζώνες Παρνασσού-Γκιώνας, Ωλονού-Πίνδου, Γαβρόβου-Τρίπολης, Αδριατικοϊόνιος και Παξών ονομάζονται Εξωτερικές Ελληνίδες 168
169 των νερών της Μεσογείου στην Κασπία και τη Μαύρη θάλασσα, ενώ κατά τη βύθιση της Αιγήιδας, η Ελληνική χερσόνησος απομονώνεται από τη Μ. Ασία και αρχίζει να αποκτά τη σημερινής της μορφή. Αμέσως μετά, το Μεσσηνίο επισφραγίζεται από την Κρίση Αλατότητας, δηλαδή την εκτεταμένη αποξήρανση της Μεσογείου η οποία αντικαταστάθηκε από ένα δίκτυο ποταμών και λιμνών γλυκού νερού (Hsü et al. 1977). Κατά το ύστερο Πλειόκαινο, οι ποταμοί της Θράκης έρχονται σε επαφή με τη Μαύρη Θάλασσα ένα γεγονός που εξηγεί την ομοιότητα της ιχθυοπανίδας της Θράκης με αυτή του Δούναβη (Bănărescu 2004, Skoulikidis et al. 2009, Oikonomou, Leprieur & Leonardos 2014). Στο Τεταρτογενές, την εποχή του Πλειστόκαινου και κατά τη διάρκεια των παγετωνικών περιόδων, η στάθμη της θάλασσας ήταν έως και 120 μ. χαμηλότερα (Dermitzakis & Papanikolaou 1981, Skoulikidis et al. 2009). Πολλές λεκάνες απορροής, που στο παρελθόν ήταν απομονωμένες, συνδέθηκαν κατά την περίοδο αυτή (Perissoratis & Conispoliatis 2003) και μέσω αυτών των συνδέσεων πολλά είδη διασπάρθηκαν. Τέλος, στο Ολόκαινο, η στάθμη της θάλασσας ανέβηκε και η περιοχή της Ελλάδας αποκτά σταδιακά τη σημερινή της γεωγραφία (Σημείωμα 1). Σε αντίθεση με τις περισσότερες ευρωπαϊκές χώρες, η Ελλάδα χαρακτηρίζεται από παράλληλη απομόνωση, τόσο χωρικά στο επίπεδο απορροής ποταμού ή λίμνης, όσο και χρονικά, ως βιοτικό καταφύγιο όταν ένα μεγάλο μέρος της ηπείρου καλύφθηκε από τους παγετώνες κατά το Πλειστόκαινο (Perissoratis & Conispoliatis 2003, Economou et al. 2007). Αναλυτικότερα, η ανάδυση της οροσειράς της Πίνδου, οι ποικίλες κλιματικές ζώνες, οι έντονοι θαλάσσιοι κόλποι και οι βαθιές θάλασσες οδήγησαν στην αυξημένη απομόνωση των Ελληνικών εσωτερικών υδάτων (Economou et al. 2007) Οι Ελληνικές λίμνες Οι περισσότερες Ελληνικές λίμνες βρίσκονται στο βορειοδυτικό τμήμα της χώρας, όπου η βροχόπτωση υπερβαίνει τα mm ανά έτος (Zacharias et al. 2002). Η μορφολογική εξέλιξη των Ελληνικών λιμνών είναι αρκετά περίπλοκη και συνδέεται 169
170 με τις τεκτονικές κινήσεις της αλπικής και μετα-αλπικής ορογένεσης και της διεισδυτικής ικανότητας του νερού στα ανθρακικά πετρώματα (Skoulikidis, Bertahas & Koussouris 1998). Έτσι, οι Ελληνικές λίμνες μπορούν να χωριστούν σε δύο ομάδες: σε αυτές που, εξαιτίας της διάβρωσης, βρίσκονται μεταξύ 0 και 145 m πάνω από το επίπεδο της θάλασσας και σε αυτές που ξεπερνούν τα 450 m και δημιουργήθηκαν λόγω του τεκτονισμού (Zacharias et al. 2002). Η προέλευση των περισσότερων λιμνών είναι συνδεδεμένη με τον τεκτονισμό και τα καρστικά φαινόμενα. Ως παράδειγμα, αναφέρουμε τις αρχαίες λίμνες Πρέσπες (Π.Γ.Δ.Μ.-Ελλάδα-Αλβανία) και των Ιωαννίνων (Παμβώτις), οι οποίες θεωρούνται ότι έχουν σχηματιστεί κατά το Ύστερο Τριτογενές (2-3 Ma). Άλλα παραδείγματα της έντονης γεωλογικής δραστηριότητας αποτελούν η Λίμνη Δοϊράνη (Π.Γ.Δ.Μ.--Ελλάδα) και η λίμνη της Καστοριάς (Ορεστιάς). Οι Ελληνικές λίμνες φέρουν κοινές ιστορίες εξέλιξης, ανήκουν σε διαφορετικές βιογεωγραφικές περιοχές και είναι κατά κανόνα απομεινάρια παλαιότερων μεγάλων λιμνών του Πλειοκαίνου (Skoulikidis, Bertahas & Koussouris 1998). Ενδεικτικά αναφέρουμε τις λίμνες του νομού Αιτωλοακαρνανίας (Τριχωνίδα, Λυσιμαχεία, Οζερός και Αμβρακία), οι οποίες αναπτύσσονται μέσα σε ταφροειδές βύθισμα και αποτελούσαν μία ενιαία αρχαία λίμνη, η οποία κατά το τέλος του Πλειοκαίνου κάλυπτε ολόκληρη την Αιτωλοακαρνανική λεκάνη (Κουσουρής 2014). Με την προοδευτική πτώση της στάθμης λόγω των εκροών μέσω του φαραγγιού της Κλεισούρας, η λίμνη Αμβρακία διαχωρίστηκε από την Τριχωνίδα και έγινε ανεξάρτητη. Αργότερα έλαβε χώρα και ο διαχωρισμός των άλλων λιμνών (Δημητρίου 2005). Αντίστοιχα, η λίμνη Παμβώτιδα (νομός Ιωαννίνων) αποτελεί το υπόλειμμα ενός μεγαλύτερου οικοσυστήματος, πριν την αποξήρανση της Λαψίστας. Ανήκει στην ευρύτερη υδρολογική λεκάνη του λεκανοπεδίου Ιωαννίνων που δημιουργήθηκε από τεκτονικές κινήσεις και στη συνέχεια διαμορφώθηκε με καρστικές διεργασίες από τα τέλη του Πλειόκαινου και κατά την διάρκεια του Τεταρτογενούς (Frogley & Preece 2004). Στην Ήπειρο συναντάμε και τη λίμνη του Ζηρού (νομός Πρεβέζης), η οποία δημιουργήθηκε από την διάβρωση του ρήγματος Ζηρού- Στεφάνης- Ζαλόγγου και κατακρήμνιση της οροφής (Κουσουρής 2014). Η λίμνη της Καστοριάς, είναι καρστικής προέλευσης, σχηματίστηκε πριν από περίπου 10 εκατομμύρια χρόνια, 170
171 κατά το Μειόκαινο και είναι η υπολειμματική μορφή των μεγάλων λιμνών, που δέσποζαν στο χώρο της Δυτικής Μακεδονίας κατά το Τεταρτογενές και, οι οποίες σχηματιστήκαν κατά το στάδιο των αλπικών πτυχώσεων (Βαφειάδης 1983). Βορειότερα, η αρχαία λίμνη των Δασσαρητών, που κατά το Ιουρασικό ήταν συνδεδεμένη και με την Αδριατική θάλασσα που διασπάστηκε στις λίμνες Μικρή και Μεγάλη Πρέσπα, Οχρίδα και την αποξηραμένη Μαλίκη (Stanković 1960) κατά το Πλειόκαινο λόγω της επίδρασης των καρστικών διαβρώσεων (Hollis & Stevenson 1997). Ένα άλλο σύνολο λιμνών που συνδέονται μεταξύ τους υδρολογικά είναι η Χειμαδίτιδα, η Ζάζαρη, η Πετρών και η Βεγορίτιδα, οι οποίες σχηματίστηκαν από μεγάλες γεωλογικές καθιζήσεις και θεωρούνται τεκτονικής προέλευσης, αποτελώντας τα υπολείμματα της παλιάς Εορδαίας λεκάνης (Τσιούρης 1996). Η διασυνοριακή λίμνη Δοϊράνη, ανάμεσα στην Ελλάδα και την ΠΓΔΜ, είναι μια σχεδόν κυκλική λεκάνη και αποτελεί το υπόλειμμα της Πλειστόκαινης λίμνης Παιονίας, που δημιουργήθηκε κατά το Ανώτερο Τεταρτογενές από ένα συνδυασμό ηφαιστειακής και τεκτονικής δραστηριότητας (Stojanov & Micevski 1989). Ανατολικότερα, η δεύτερη μεγαλύτερη λίμνη της Ελλάδας, η Βόλβη, είναι τεκτονικής προέλευσης, ανήκει στη Σερβοµακεδονική Ζώνη και αποτελεί μαζί με τη γειτονική λίμνη Κορώνεια, υπόλειμμα της μεγάλης λίμνης της Μυγδονίας που σχηµατίστηκε κατά το κατώτερο Πλειστόκαινο. Με το άνοιγμα των στενών της Ρεντίνας, όπου είναι σήμερα ο ποταμός Ρήχιος, άδειασε η λεκάνη της Μυγδονίας στο Στρυμονικό κόλπο και προέκυψαν οι δύο λίμνες (Ψιλοβίκος 1977) Η Ελληνική ιχθυοπανίδα και οι βιογεωγραφικές ιδιαιτερότητές της Η βιοποικιλότητα των εσωτερικών υδάτων της χώρας είναι ιδιαιτέρως υψηλή, κάτι που καταφαίνεται και από το μεγάλο αριθμό ενδημικών ψαριών (Economou et al. 2007, Oikonomou, Leprieur & Leonardos 2012), ασπονδύλων (Legakis 2009) και του σημαντικότατου, για τα ευρωπαϊκά δεδομένα, αριθμού παρυδάτιων πουλιών που διαχειμάζουν ή φωλιάζουν στις Ελληνικές λίμνες, λιμνοθάλασσες, δέλτα και ποτάμια (Chandrinos 2009). Ο υψηλός ενδημισμός που παρουσιάζουν τα εσωτερικά ύδατα της Ελλάδας σχετίζεται άμεσα με την ιδιαίτερα κατακερματισμένη γεωγραφική τους 171
172 κατανομή και την έντονη γεωλογική δραστηριότητα κατά το Τριτογενές και ιδιαίτερα το Τεταρτογενές (Σημείωμα 1). Η Ελληνική ιχθυοπανίδα έχει δεχθεί επιρροές από πολλαπλές περιοχές αποτελώντας την βιογεωγραφική γέφυρα μεταξύ της Μεσογείου, του Δούναβη, της Μαύρης Θάλασσας και της Ανατολής (Bănărescu 2004). Όσον αφορά τους διαδρόμους διασποράς των ψαριών των εσωτερικών υδάτων της Ελλάδας, δύο υποθέσεις έχουν προταθεί. Η πρώτη περισσότερο αποδεκτή υπόθεση (Economidis & Bănărescu 1991) περιγράφει τον αποικισμό μέσω ποτάμιων συνδέσεων στη ξηρά που ένωνε την Ανατολία με το Ιράν και τα Βαλκάνια κατά το Ολιγόκαινο και το Μειόκαινο με διαδοχικά κύματα αποικισμού. Τα είδη που διασπάρθηκαν κατά την περίοδο αυτή έφεραν Σιβηρική καταγωγή, πιθανώς, από την νοτιοδυτική Ασία. Το δεύτερο κύμα αποίκισης, σύμφωνα με τους ίδιους συγγραφείς, αφορά την περίοδο εναλλαγής από το Πλειόκαινο στο Πλειστόκαινο, όπου είδη με παραδουνάβια καταγωγή διασπάρθηκαν μέσω της ποτάμιας σύνδεσης ανάμεσα στο Δούναβη και τον Αξιό και τη σύνδεση του Αιγαίου με τη Μαύρη Θάλασσα (λιμναία φάση). Ένα εναλλακτικό σενάριο (Bianco 1990) προτείνει ότι είδη ψαριών αποίκισαν την Ελλάδα κατά τη διάρκεια της φάσης «Lago Mare», όταν η Μεσόγειος θάλασσα αποξηράνθηκε και γέμισε νερό από την Παρατηθύ. Αυτή η θεωρία προβλέπει μια ταχεία περίοδο διαφοροποίησης της πανίδας και μια στενή συσχέτιση μεταξύ της πανίδας των χερσονήσων της Νοτίου Ευρώπης (Durand et al. 2003, Levy, Doadrio & Almada 2009, Oikonomou, Leprieur & Leonardos 2014). 172
173 Εικόνα 4.1. Χάρτης του σχετικού βαθμού ενδημισμού ανά χώρα της Βαλκανικής χερσονήσου (Oikonomou, Leprieur & Leonardos 2012) Figure 4.1. Map showing relative degree of endemism among countries in the Balkan Peninsula (Oikonomou, Leprieur & Leonardos 2012) Απειλές της Ελληνικής ιχθυοπανίδας Όπως αναφέρθηκε νωρίτερα, τα ψάρια των εσωτερικών υδάτων κατέχουν το υψηλότερο ποσοστό απειλούμενων ειδών ανάμεσα στους υδρόβιους οργανισμούς (Εικόνα 4.2) γιατί έχουν γεωγραφικά περιορισμένες κατανομές και στενές οικολογικές απαιτήσεις. Λόγω της αυξανόμενης έντασης των ανθρώπινων δραστηριοτήτων, τα ψάρια των εσωτερικών υδάτων στην Ελλάδα αντιμετωπίζουν όλο και μεγαλύτερες απειλές από μια σειρά δραστηριοτήτων, όπως η κατασκευή φραγμάτων, η άντληση νερού, η ρύπανση, η υπεραλίευση και η εισαγωγή χωροκρατικών ειδών τα οποία ανταγωνίζονται τα αυτόχθονα είδη Ο ανταγωνισμός με τα αυτόχθονα ψάρια μπορεί να αφορά α) τους πόρους (διαθεσιμότητα τροφής), λόγω της αλληλεπικάλυψης στις τροφικές προτιμήσεις μεταξύ των ειδών,όπως το Gambusia holbrooki με το Valencia letourneux), β) την αναπαραγωγή, όπως συμβαίνει με την πεταλούδα (Carassius gibelio) που χρησιμοποιεί το σπέρμα του γριβαδιού προκειμένου να αναπαραχθεί και γ) τη θήρευση των αυτόχθονων ειδών, όπως συμβαίνει με το ηλιόψαρο (Lepomis gibbosus), που θηρεύει τα αυγά και το γόνο άλλων ειδών 173
174 Με βάση τα διαθέσιμα πληθυσμιακά δεδομένα, που δεν είναι εκτεταμένα, για τα είδη που κατανέμονται στην Ελλάδα, προκύπτει η γενική εκτίμηση ότι οι πληθυσμοί είναι σχετικά αραιοί και μικροί (Leonardos et al. 2008), ιδίως αν τους συγκρίνουμε με αντίστοιχους της κεντρικής Ευρώπης. 23 ξενικά είδη (17% των αυτόχθονων) έχουν επιβεβαιωμένη παρουσία στα Ελληνικά εσωτερικά ύδατα και 28 έχουν αναφερθεί ως εισαχθέντα σε αυτά προκαλώντας από κατάρρευση ως και εξαφάνιση των αυτόχθονων πληθυσμών (Barbieri et al. 2015). 11 ξενικά είδη ( 40%) έχουν εγκλιματιστεί στα Ελληνικά εσωτερικά νερά (Ameiurus cf. nebulosus, Carassius auratus, C. gibelio, C. langsdorfii, Chelon haematocheilus, Gambusia holbrooki, Gymnocephalus cernua, Lepomis gibosus, Oreochromis niloticus, Pseudorasbora parva και Poecilia latipinna,), είτε για χρήση σε υδατοκαλλιέργειες (π.χ. P. parva και O. niloticus), ενυδρία (π.χ. C. auratus, P. latipinna και L. gibbosus) είτε για την αντιμετώπιση της ελονοσίας (G. holbrooki) (Perdikaris, Gouva & Paschos 2010). Επιπρόσθετα, ένα ακόμα πρόβλημα αποτελεί η αποσπασματική γνώση περί της κατανομής των εισαχθέντων ειδών στα Ελληνικά εσωτερικά ύδατα εξαιτίας της έλλειψης εθνικής κλίμακας στρατηγικής, που εφαρμόζεται με στοχευμένες δράσεις εμπλουτισμών από τους τοπικούς φορείς και καταπολέμηση των παράνομων εμπλουτισμών από ιδιώτες. Μεταξύ των ειδών της Ελληνικής ιχθυοπανίδας ορισμένα έχουν και σημαντική εμπορική αξία. Τα κυριότερα εμπορεύσιμα είδη των εσωτερικών υδάτων της χώρας είναι: ο κοινός Κυπρίνος (Cyprinus carpio), η Πεταλούδα (Carassius gibelio), η Λεστιά (Abramis brama), το Τσιρόνι (Rutilus rutilus και R. prespensis), η Δρομίτσα (Rutilus panosi), η Μπριάνα και τα Μουστακάτα (Barbus spp. και Luciobarbus spp.), το Γλήνι (Tinca tinca), η Τσερούκλα (Scardinius acarnanicus), η Κοκκινοφτέρα (Scardinius erythrophthalmus), η Τούρνα (Esox lucius), η Πέρκα (Perca fluviatilis), η Αθερίνα (Atherina boyeri), το Χέλι (Anguilla anguilla), ο Γουλιανός (Silurus glanis) και το Γλανίδι (Silurus aristotelis). Η αλιεία, ιδιαίτερα κατά τη διάρκεια της περιόδου ωοτοκίας, θέτει σε σημαντικό κίνδυνο τους ιχθυοπληθυσμούς λόγω της απομάκρυνσης των θηλυκών ατόμων που δεν έχουν ακόμη γεννήσει, μειώνοντας έτσι τις πιθανότητες επιτυχούς αναπαραγωγής και συμπεριλαμβάνονται στις ανθρωπογενείς δραστηριότητες που θέτουν σε κίνδυνο την ιχθυοπανίδα (Economidis 2009). 174

175 Εικόνα 4.2. Αριθμοί ειδών ανά κατηγορία IUCN και ποσοστό ειδών που εντάσσονται σε κατηγορία κινδύνου, με βάση το σύνολο των ειδών που απαντώνται στην Ελλάδα (τροποποιημένο από Economidis 2009) Figure 4.2. Number of species per IUCN category and percentage of endangered species based on all species found in Greece (modified from Economidis 2009) Παράγοντες που καθορίζουν την ποικιλότητα των ειδών ψαριών σε λίμνες Το μέγεθος των οικοσυστημάτων είναι από τους κυριότερους παράγοντες που καθορίζουν την ποικιλότητα στα λιμναία οικοσυστήματα. Για παράδειγμα, ο Griffiths (1997) χρησιμοποιώντας μία βάση δεδομένων των κατανομών λιμνόφιλων ειδών ψαριών σε 651 λίμνες της Αμερικής, έδειξε πως η έκταση ερμηνεύει το 37% της διακύμανσης του αριθμού των ειδών ενώ χρησιμοποιώντας το υψόμετρο και το μέγιστο βάθος, το ποσοστό ερμηνείας αυξήθηκε μόλις στο 41%. Οι Drakou et al. (2009) χρησιμοποιώντας δεδομένα κατανομής ψαριών στις Ελληνικές λίμνες έδειξαν επίσης ότι ο αριθμός των ειδών αυξάνεται με το μέγεθος του οικοσυστήματος και μειώνεται με το υψόμετρο. Τα πρότυπα του αριθμού των ειδών των ψαριών σε λίμνες έχουν επίσης συνδεθεί με την περιβαλλοντική ετερογένεια. Οι Eadie et al. (1986) έδειξαν πως ο αριθμός των ειδών ψαριών σε 82 λίμνες του Καναδά ερμηνεύεται από την έκταση και την πολυπλοκότητα των ενδιαιτημάτων των οικοσυστημάτων. 175
176 Πράγματι, τα τοπογραφικά και αβιοτικά χαρακτηριστικά του οικοσυστήματος (π.χ., βάθος, γεωγραφικό πλάτος, γεωγραφικό μήκος, υψόμετρο, παραγωγικότητα, ph) συχνά καθορίζουν τον αριθμό των ειδών της ιχθυοπανίδας (Allen et al. 1999, Brucet et al. 2013). Επιπλέον, η παραγωγικότητα, αποτελεί ακόμα ένα παράγοντα διαμόρφωσης των προτύπων της ποικιλότητας των ψαριών, καθώς όπως έδειξαν οι Helminen et al. (2000), o αριθμός των ειδών συσχετίζεται θετικά με τη συγκέντρωση του ολικού φώσφορου σε 22 Φινλανδικές λίμνες. Και στις Ελληνικές λίμνες, η παραγωγικότητα (συγκέντρωση χλωροφύλλης-α) έχει μεγάλη επίδραση στη σύνθεση της ιχθυοπανίδας τους (Mazaris et al. 2010) ιαβάθµιση του Μεγέθους του Σώματος Κατά Γεωγραφικό Πλάτος Ένα από τα παρατηρούμενα πρότυπα σε διαφορετικές ομάδες ζώων - συμπεριλαμβανομένων τόσο των ποικιλόθερμων και των ομοιόθερμων- είναι ότι το μέγεθος του σώματος αυξάνεται με την αύξηση του γεωγραφικού πλάτους (Gaston & Blackburn 2000), ένα πρότυπο που μπορεί να διαμορφώνεται από τις διαφορετικές θερμοκρασίες που αντιστοιχούν στα διάφορα γεωγραφικά πλάτη. Το μέγεθος του σώματος έχει δειχθεί ότι αυξάνεται με το γεωγραφικό πλάτος σε 65% έως 71% των θηλαστικών και σε 72% έως 76% των πτηνών σε παγκόσμιο επίπεδο (Ashton, Tracy & de Queiroz 2000, Meiri & Dayan 2003). Όσον αφορά τα ψάρια των εσωτερικών υδάτων, πρώτος ο Ray (1960) διατύπωσε την άποψη ότι το μέγεθος του σώματος αυξάνεται με την αύξηση του γεωγραφικού πλάτους. Αργότερα, ο Griffiths (2012) κατέληξε στο συμπέρασμα ότι η μεταπαγετώδης εποίκιση από ευρείας εξάπλωσης, μεταναστευτικά είδη ήταν ο κύριος καθοριστικός παράγοντας της αύξησης του σώματος στα βόρεια ευρωπαϊκα οικοσυστήματα. Αντίθετα, η εισαγωγή ξενικών ειδών προκαλεί μείωση του μεγέθους των αυτόχθονων ειδών και αναστροφή του προτύπου όπως πρόσφατα έδειξαν οι Blanchet et al. (2010). Με την πάροδο των ετών, άλλες οικολογικές και εξελικτικές υποθέσεις προτάθηκαν για την εξήγηση του προτύπου. Ως σήμερα, η συσχέτιση του μεγέθους με το γεωγραφικό πλάτος και για τα ψάρια των εσωτερικών υδάτων, έχει αποδοθεί σε μια σειρά υποθέσεων (Gaston & Blackburn 2000, Denys et al. 2014), όπως: 176
177 (i) το μέγεθος είναι μεγαλύτερο κατά την ωρίμανση, η οποία επιτυγχάνεται σε χαμηλότερη θερμοκρασία (Υπόθεση θερμοκρασίας) (ii) τα μεγάλα είδη ευνοούνται σε υψηλότερα γεωγραφικά πλάτη, επειδή μεταβολίζουν τα αποθέματα λίπους με χαμηλότερο ρυθμό (Υπόθεση του περιβάλλοντος ή της εποχικότητας) (iii) τα μικρόσωμα taxa είναι κυρίαρχα σε χαμηλά γεωγραφικά πλάτη λόγω της αδυναμίας τους να εξαπλωθούν σε υψηλά γεωγραφικά πλάτη, ιδίως μετά τους Πλειστοκαινικούς παγετώνες (Υπόθεση της μεταναστευτικής ικανότητας) (iv) η διαθεσιμότητα της ενέργειας επιδρά θετικά στο μέγεθος του σώματος (Υπόθεση της παραγωγικότητας) (v) η αύξηση του διαειδικού ανταγωνισμού μπορεί να ευνοήσει τη συνύπαρξη των μικρότερων ειδών (Υπόθεση του ανταγωνισμού) (vi) τα μεγαλύτερα ενδιαιτήματα ευνοούν τη διατήρηση πληθυσμών μεγαλύτερου μεγέθους (Υπόθεση διαθεσιμότητας ενδιαιτήματος) 4.2 Στόχος του κεφαλαίου Ο στόχος του παρόντος κεφαλαίου είναι διττός. Αρχικά, κεντρικός σκοπός αυτού του κεφαλαίου ήταν η μελέτη των προτύπων ποικιλότητας, ενδημισμού και απειλούμενων με εξαφάνιση ειδών ψαριών στα Ελληνικά εσωτερικά ύδατα. Επιπλέον, μελετήθηκαν οι κύριες περιβαλλοντικές παράμετροι που επηρεάζουν και καθορίζουν τον αριθμό και το μέγεθος των ειδών των ψαριών σε 13 φυσικές Ελληνικές λίμνες. 177
178 4.3 Υλικά και Μέθοδοι Σύνθεση, πρότυπα ποικιλότητας και απειλούμενων ειδών (κατηγορίες IUCN) της Ελληνικής ιχθυοπανίδας Η βάση δεδομένων (ΒΔ) κατανομής των ψαριών των εσωτερικών υδάτων στην Ελλάδα περιλαμβάνει πανιδικές λίστες από λεκάνες απορροής ποταμών και φυσικές λίμνες. Τα δεδομένα συλλέχθηκαν από εκτεταμένη έρευνα βιβλιογραφίας (Economidis 1991, Economou et al. 2007, Economidis 2009, Koutsikos et al. 2012, Barbieri et al. 2015) (βλέπε επίσης Παράρτημα 1). Για τα δεδομένα, χρησιμοποιήθηκε η ταξινόμηση και η ονοματολογία που προτάθηκε από τους Kottelat & Freyhof (2007), μετά τις τροποποιήσεις και προσθήκες που προτάθηκαν από τους Economou et al. (2007), Koutsikos et al. (2012), Geiger et al. (2014) και Barbieri et al. (2015). Ο κατάλογος των ειδών περιλαμβάνει επίσης και κάποια είδη που ανήκουν στις οικογένειες Acipenseridae, Anguillidae, Atherinidae, Clupeidae, Cyprinodontidae, Gobiidae, Mugilidae, Moronidae, Pleuronectidae, Syngnathidae και Valencidae και συχνά συναντώνται στα εσωτερικά ύδατα της χώρας, είτε ως απομονωμένοι πληθυσμοί (π.χ. είδη των οικογενειών Atherinidae, Clupeidae και Valencidae) είτε στα εκβολικά συστήματα των ποταμών και τις λιμνοθάλασσες (π.χ. Cyprinodontidae, Gobiidae, Mugilidae, Moronidae, Pleuronectidae, Syngnathidae) είτε είναι ανάδρομα (είδη της οικογένειας Acipenseridae, Alosa fallax) και κατάδρομα (Anguilla anguilla). Για την αξιολόγηση των ειδών που κινδυνεύουν με εξαφάνιση, υπολογίστηκε ο αριθμός των ειδών που αντιστοιχεί σε κάθε κατηγορία του καθεστώτος διατήρησης των ειδών σύμφωνα με τη Διεθνή Ένωση για τη Διατήρηση της Φύσης (IUCN) και το Κόκκινο Βιβλίο Απειλούμενων Ζώων της Ελλάδας (Economidis 2009) (Εικόνα 3.4). Τα είδη που αξιολογούνται ως Κρισίμως Κινδυνεύοντα (CR), Κινδυνεύοντα (ΕΝ) ή Τρωτά (VU) αναφέρονται ως «απειλούμενα» στην παρούσα μελέτη. 178
179 4.3.2 Βάση δεδομένων των Ελληνικών λιμνών Συνολικά, η βάση δεδομένων που χρησιμοποιήθηκε στις αναλύσεις περιλαμβάνει δεδομένα από 42 είδη και 13 φυσικές λίμνες. Για κάθε λιμναίο οικοσύστημα, καταγράφηκαν ο αριθμός των ειδών των ψαριών, οι κατανομές τους και το μέγιστο μήκος των μόνιμων ειδών («resident species») από βιβλιογραφικές πηγές (Barbieri et al. 2015) και βάσεις δεδομένων (όπως διδακτορικές διατριβές (Τσουμάνη 2013, Πετρίκη 2015) και προσωπική πληροφόρηση (Μπόμπορη Δήμητρα, Α.Π.Θ.). Το μέγιστο αναφερθέν ολικό μήκος (maximum TL) επιλέχτηκε ως βιολογικό αντικατάστατο του μεγέθους του σώματος (βλέπε Griffiths 2012, Griffiths 2013). Στη βάση δεδομένων συμπεριλήφθηκαν μόνο αυτόχθονα είδη για κάθε σύστημα ενώ τα είδη που χαρακτηρίζονται με αμφίβολη φυσική παρουσία παραλείφθηκαν από τις περαιτέρω αναλύσεις. Η επιλογή των λιμναίων οικοσυστημάτων έγινε με βάση την πληρότητα των διαθέσιμων δεδομένων, λαμβάνοντας υπόψιν τον αριθμό των δειγματοληψιών, και της γεωγραφικής αντιπροσωπευτικότητας. Σε κάθε υδάτινο σώμα, καταγράφηκαν το γεωγραφικό πλάτος και μήκος με τη χρήση του Google Earth Pro (7.01), τέσσερα λιμνολογικά χαρακτηριστικά και έξι φυσικοχημικές παράμετροι του νερού (Πίνακας 4.1). Η επιλογή των παραπάνω λιμνολογικών χαρακτηριστικών και των φυσικοχημικών παραμέτρων έγινε μετά από ενδελεχή επισκόπηση της βιβλιογραφίας. Οι βιβλιογραφικές πηγές για τα περιβαλλοντικά δεδομένα, εκτός των επιστημονικών άρθρων (π.χ. Skoulikidis, Bertahas & Koussouris 1998, Zacharias et al. 2002, Kagalou & Leonardos 2009, Stephanidis & Papastergiadou 2012) και διδακτορικών διατριβών (Δράκου 2009, Τσουμάνη 2013), περιλαμβάνουν δεδομένα από άλλες βάσεις δεδομένων (όπως Οι λίμνες Βιστωνίδα και Βουλκαρία παραλείφθηκαν από τη βάση δεδομένων λόγω της αυξημένης αγωγιμότητάς τους ως αποτέλεσμα της σύνδεσής τους με τη θάλασσα. Οι λίμνες που χρησιμοποιήθηκαν στις αναλύσεις παρουσιάζονται στην Εικόνα
180 Πίνακας 4.1. Στατιστική σύνοψη του αριθμού των ειδών των ψαριών, των λιμνολογικών χαρακτηριστικών και των φυσικοχημικών παραμέτρων που χρησιμοποιήθηκαν (n = 13). Οι μεταβλητές που σημειώνονται με * λογαριθμήθηκαν (log 10) για τις μετέπειτα αναλύσεις Table 4.1. Summary for statistics of fish richness, limnological characteristics, and water physicochemical parameters used in the study (n = 13). The variables marked with * were log 10- transformed for subsequent analyses Variables Min Max Mean SD Skewness Kurtosis Species* Number of fish ,24 0,25-1,82 species Location Mean latitude (o) 38,55 41,20 39,88 1-0,24-1,82 Mean longitude (o) 20,83 23,46 21,62 0,73 1,29 0,72 Limnological characteristics* Lake altitude (m) ,9 270,45 0,15-2,02 Lake area (km 2 ) 0,5 69,5 26,83 23,89 0,69-1,11 Mean depth (m) 2, ,36 26,74 0,72-1,40 Lake volume 3, ,7 0,89 0,05-1,21 (X 10 6 m 3 ) Water physicochemical characteristics Total Phosphorus 0,03 0,20 0,10 0,06 0,09-1,63 (mg/l) Chlorophyll-a ,4 11,25 1,37 0,94 (µg/l) Dissolved inorganic 0,07 1,04 0,41 0,30 1,03-0,08 nitrogen (mg/l) Conductivity ,6 282,63 0,46-1,47 (µs/cm) ph 6,9 8,5 7,87 0,41-0,76 0 Secchi depth (m) 0,3 6 1,99 2,28 0,77-1,44 180

181 Εικόνα 4.3. Χάρτης που απεικονίζει τα 13 Ελληνικά λιμναία οικοσυστήματα που μελετήθηκαν Figure 4.3. Map showing the 13 Greek lake ecosystems that were studied Στατιστική επεξεργασία και πακέτα Μια προβληματική πτυχή σε εμπειρικές αναλύσεις της επίδρασης των περιβαλλοντικών παραγόντων στον αριθμό ή το μέγεθος των ειδών είναι οι μεταξύ τους αλληλεπιδράσεις (συγγραμικότητα των μεταβλητών) (Mela & Kopalle 2002). Για παράδειγμα, οι μεγάλες λίμνες τείνουν να έχουν μικρότερες συγκεντρώσεις σε θρεπτικά συστατικά από τις μικρές λίμνες. Έτσι, σε κάποιες περιπτώσεις η έκταση μπορεί να επιλεχθεί ως σημαντικός παράγοντας για τον καθορισμό του αριθμού των ειδών, αλλά αντίθετα σε άλλες μπορεί να εξαλειφθεί λόγω της αρνητικής της σχέσης με τις αβιοτικές παραμέτρους (π.χ. Hobaek, Manca & Andersen 2002). Η διαδικασία 181
182 επιλογής των παραμέτρων που θα συμπεριληφθούν σε μοντέλα παλινδρομήσεων αποδεικνύεται ιδιαίτερα σημαντική. Γι αυτό το λόγο, η συγγραμικότητα μεταξύ των μεταβλητών ελέγχθηκε με τη χρήση ενός μέτρου διάγνωσης της συγγραμμικότητας, του συντελεστή πληθωρισμού διακύμανσης (VIF- Variation Inflation Factor). Όταν η τιμή του δείκτη για μία μεταβλητή είναι μεγαλύτερη του πέντε, αυτό αποτελεί ένδειξη υψηλής συγγραμμικότητας και η μεταβλητή απαλείφεται. Για να ελεγχθεί ποιοι περιβαλλοντικοί παράγοντες επηρεάζουν τη διακύμανση του αριθμού και του μεγέθους των ειδών των ψαριών σε Ελληνικές λίμνες, εφαρμόστηκαν πολλαπλές γραμμικές παλινδρομήσεις με όλους τους πιθανούς συνδυασμούς των μεταβλητών. Το καλύτερο σύνολο των μεταβλητών προσδιορίστηκε με την εφαρμογή του Akaike Κριτηρίου Πληροφορίας (AIC), χρησιμοποιώντας DAIC <2 ως κριτήριο επιλογής (Grueber et al. 2011). Όλες οι αναλύσεις πραγματοποιήθηκαν στο «R statistical and programming environment» (R , R Development Core Team 2013). Πιο συγκεκριμένα, χρησιμοποιήθηκαν τα πακέτα «MuMln» «graphics» και «car». 4.4 Αποτελέσματα Πρότυπα ποικιλότητας, ενδημισμού και των απειλούμενων ειδών (κατηγορίες IUCN) της Ελληνικής ιχθυοπανίδας Η Ελλάδα φιλοξενεί τουλάχιστον 137 αυτόχθονα είδη που ανήκουν σε 14 τάξεις και 21 οικογένειες (Εικόνα 4.4). Η οικογένεια των Cyprinidae είναι κυρίαρχη στην Ελληνική ιχθυοπανίδα, αντιπροσωπεύοντας το 53 % του συνόλου των ειδών, ακολουθούμενη από τις οικογένειες Cobitidae (8%), Salmonidae (3,6%), Nemacheilidae (2,9%) και Blennidae (1,5%). Οι οικογένειες Gobiidae (5,8%), Petromyzontidae (2,9%), Clupeidae και Gasterosteidae (2,2%) περιλαμβάνουν τόσο πρωτογενή και δευτερογενή είδη ενώ για τα ευρύαλα είδη, οι οικογένειες των 182
183 Mugilidae (4,3%), Acipenseridae (2,9%) και Cyprinodontidae (1,5%) κατέχουν τα μεγαλύτερα ποσοστά (Εικόνα 4.5). Στην Ελλάδα έχουν επίσης καταγραφεί 23 ξενικά είδη. 183
184 Εικόνα 4.4. Διάγραμμα ταξινόμησης για τα αυτόχθονα είδη των εσωτερικών υδάτων της Ελλάδας (συμπεριλαμβανομένων των δευτερογενών ειδών που χρησιμοποιούν το γλυκό νερό). Οι αριθμοί δηλώνουν τα είδη από κάθε τάξη στον Ελληνικό χώρο Figure 4.4. Classification diagram for the native freshwater species of Greece (including secondary species using freshwater). The numbers indicate species from each class in Greece
 Figure 4.5.")
185 Εικόνα 4.5. Διάγραμμα πίτας για τις Οικογένειες των αυτόχθονων ψαριών των εσωτερικών υδάτων της Ελλάδας (συμπεριλαμβανομένων των δευτερογενών ειδών που χρησιμοποιούν το γλυκό νερό) Figure 4.5. Pie diagram οf the Families of native freshwater fishes of Greece (including secondary species using freshwater) Ο ενδημισμός των ψαριών των εσωτερικών υδάτων είναι εξαιρετικά υψηλός και αγγίζει το 42%. Συνολικά, το 32% των ψαριών των εσωτερικών υδάτων της Ελλάδας έχουν χαρακτηριστεί ως απειλούμενα σύμφωνα με την Κόκκινη Λίστα (IUCN 2013) και 34% σύμφωνα με το Κόκκινο Βιβλίο των Απειλούμενων Σπονδυλόζωων της Ελλάδας (Economidis 2009) (Εικόνα 4.6). 185
. Τα ποσοστά υποδηλώνουν τα απειλούμενα είδη.")
186 Εικόνα 4.6. Διαγράμματα πίτας για τις κατηγορίες των ειδών των ψαριών των εσωτερικών υδάτων της Ελλάδας σύμφωνα με την a. Κόκκινη Λίστα (IUCN 2013) και b. το Κόκκινο Βιβλίο των Απειλούμενων Σπονδυλόζωων της Ελλάδας (Economidis 2009). Τα ποσοστά υποδηλώνουν τα απειλούμενα είδη. (ΝΕ: Δεν έχει αξιολογηθεί, DD: Ανεπαρκώς γνωστό, NT: Σχεδόν Απειλούμενο, LC: Μειωμένου ενδιαφέροντος, VUL: Τρωτό, EN: Κινδυνεύον, CR: Κρισίμως Κινδυνεύον, EX: Εξαφανισμένο) Figure 4.6. Pie charts for categories of fish species in freshwaters of Greece according to a. the Red List (IUCN 2013) and b. the Red Book of Threatened Vertebrates of Greece (Economidis 2009). The percentages indicate the endangered species. (NE: Not evaluated, DD: Data Deficient, NT: Near Threatened, LC: Least Concern, VUL: Vulnerable, EN: Endangered, CR: Critically Endangered, EX: Extinct) 186
187 4.4.2 Σχέση μεταξύ του αριθμού και του μεγέθους των ειδών ψαριών με τα περιβαλλοντικά χαρακτηριστικά των Ελληνικών λιμνών Η επιλογή των μεταβλητών που χρησιμοποιήθηκαν στη σταδιακή πολλαπλή παλινδρόμηση για τον καθορισμό των παραγόντων που διαμορφώνουν τον αριθμό των ειδών των ψαριών στις Ελληνικές λίμνες και του μεγέθους του σώματος των ειδών ψαριών, βασίστηκε στην αφαίρεση της επίδρασης της συγγραμικότητας, έπειτα από εφαρμογή του συντελεστή πληθωρισμού διακύμανσης (VIF- Variation Inflation Factor) (μέγιστο VIFalt = 4,00 και VIFalt= 4,48 αντίστοιχα). Ως αποτέλεσμα, χρησιμοποιήθηκαν τρία περιβαλλοντικά χαρακτηριστικά (υψόμετρο, έκταση, γεωγραφικό πλάτος) και πέντε φυσικοχημικά χαρακτηριστικά (αγωγιμότητα, ολικός φώσφορος, ph, διαλυμένο ανόργανο άζωτο, συγκέντρωση χλωροφύλλης-α) ως ανεξάρτητες μεταβλητές στη σταδιακή πολλαπλή παλινδρόμηση. Τα αποτελέσματα σχετικά με τη μεταβλητότητα του αριθμού των ειδών έδειξαν ότι το καταλληλότερο μοντέλο εμπεριέχει τις μεταβλητές του υψομέτρου, της έκτασης και του ολικού φωσφόρου και εξηγεί το 94% της διακύμανσης του αριθμού των ειδών (R 2adj = 0,91) (Πίνακας 4.2). Ο ολικός φώσφορος έχει τη μικρότερη συνεισφορά (Πίνακας 4.3) αφού έπειτα από τη συμπερίληψή του η επεξηγηματικότητα του μοντέλου αυξήθηκε κατά 4%. 187
188 Πίνακας 4.2. Αποτελέσματα της επιλογής μεταβλητών της δημιουργίας μοντέλου πολλαπλής γραμμικής παλινδρόμησης μεταξύ του αριθμού των ειδών ως συνάρτηση οκτώ επεξηγηματικών μεταβλητών (υψόμετρο, έκταση, γεωγραφικό πλάτος, αγωγιμότητα, ολικός φώσφορος, ph, διαλυμένο ανόργανο άζωτο, συγκέντρωση χλωροφύλλης-α) Table 4.2. Results of the model selection analysis assessing species richness as a function of eight explanatory variables (altitude, area, latitude, conductivity, TP, Dissolved inorganic nitrogen, ph, Chlorophyll-a) Selected variables df loglik AICc delta weight R² Altitude + Area+ TP 5 14,92-11,26 0,000 0,32 0,94 Altitude + Area+ Latitude 5 14,78-10,99 0,270 0,048 0,93 Altitude + Area 4 11,95-10,90 0,37 0,029 0,90 Altitude + Area+ TP+Latitude 6 17,64-9,28 1,98 0,023 0,96 Πίνακας 4.3. Αποτελέσματα του μοντέλου πολλαπλής γραμμικής παλινδρόμησης μεταξύ του αριθμού των ειδών ως συνάρτηση των τριών επιλεγμένων περιβαλλοντικών μεταβλητών Table 4.3. Results of the multiple linear regression of species richness as a function of three selected environmental variables Selected variables Estimate Std, Error t value Pr(> t ) Intercept 1, , ,656 9,86e-07 *** Altitude -0, , ,784 2,75e-05 *** Area 0,3564 0, ,126 5,51e-05 *** TP -1, , ,282 0,0484 * *** p<0,001 * p<0,05 Τα αποτελέσματα σε σχέση με το μέσο όρο του μέγιστου μεγέθους των ειδών των ψαριών, έδειξαν ότι το καταλληλότερο μοντέλο αφορά μόνο τη μεταβλητή του γεωγραφικoύ πλάτους και εξηγεί το 40% της διακύμανσης του μεγέθους των ειδών (R 2adj = 0,35) (Πίνακας 4.4). Πράγματι, το μέγιστο μήκος διέφερε οριακά στατιστικά σημαντικά μεταξύ των βορειότερων και των νοτιότερων λιμναίων οικοσυστημάτων (Αποτελέσματα wilcox.test: P<0,05, Eικόνα 4.7). Πίνακας 4.4. Αποτελέσματα της επιλογής μεταβλητών της δημιουργίας μοντέλου πολλαπλής γραμμικής παλινδρόμησης μεταξύ του μεγέθους σώματος των ειδών ως συνάρτηση οκτώ επεξηγηματικών μεταβλητών (υψόμετρο, έκταση, γεωγραφικό πλάτος, αγωγιμότητα, ολικός φώσφορος, ph, διαλυμένο ανόργανο άζωτο, συγκέντρωση χλωροφύλλης-α) Table 4.4. Results of the model selection analysis assessing species body size as a function of eight explanatory variables (altitude, area, latitude, conductivity, TP, Dissolved inorganic nitrogen, ph, Chlorophyll-a) Selected variables df loglik AICc delta weight R² Latitude 3 5,05-1,44 0,000 0,055 0,40 Latitude+ ph 4 6,87-0,74 0,270 0,74 0,55 188
189 Εικόνα 4.7a. Σχέση μεταξύ μεγέθους των ειδών ψαριών με το γεωγραφικό πλάτος και b. θηκόγραμμα σύγκρισης του μέσου μεγέθους στις βορειότερες και νοτιότερες ομάδες λιμναίων οικοσυστημάτων Figure 0.7a. Relationship between size of fish species with latitude and b. boxplot comparing the body size in northern and southern groups of lake ecosystems 189
190 4.5 Συζήτηση Η Ελληνική ιχθυοπανίδα των εσωτερικών υδάτων Η Ελληνική ιχθυοπανίδα των εσωτερικών υδάτων είναι η πιο πλούσια της Ευρώπης. Συνολικά, έχουν καταγραφεί 160 είδη σύμφωνα με την παρούσα μελέτη και την τελευταία αναθεώρηση των Barbieri et al. (2015), εκ των οποίων 137 είναι αυτόχθονα (Economou et al. 2007, Koutsikos et al. 2012). Το 70% των καταγεγραμμένων ειδών συγκαταλέγονται στα πρωτογενή. Από τα υπόλοιπα, περίπου το 20% κατατάσσεται στα δευτερογενή και περίπου το 10% στα μεταναστευτικά γαμόδρομα και αγαμόδρομα (Economidis 2009). Η ιχθυοπανίδα στα εσωτερικά ύδατα της Ελλάδας αντιπροσωπεύεται από ένα σχετικά μικρό αριθμό οικογενειών, με τα περισσότερα είδη να ανήκουν στην οικογένεια των Cyprinidae. Αυτή η ομάδα αποίκησε την Ελλάδα κατά τις αρχές του Ολιγόκαινου μέσω των ποτάμιων συλλήψεων στη ξηρά, που συνέδεε την Ευρώπη με την Ασία (Perea et al. 2010). Στο Μειόκαινο, τα είδη της οικογένειας διαφοροποιήθηκαν και εξαπλώθηκαν στα εσωτερικά ύδατα της Μεσογειακής περιοχής. Κατά το Πλειόκαινο, οι πληθυσμοί των Κυπρινοειδών των εσωτερικών υδάτων απομονώθηκαν μέχρι και τις παγετώδεις περιόδους, δικαιολογώντας τα υψηλά επίπεδα ενδημισμού στα Πλειστοκαινικά καταφύγια. Στην Ελληνική ιχθυοπανίδα δεν υπάρχουν εκπρόσωποι με προέλευση την Τηθύ. Εντούτοις, κάποια είδη των οικογενειών Cobitidae, Cyprinodontidae, Gobiidae και Valenciidae φέρουν πολύ σημαντική βιογεωγραφική ιστορία. Για παράδειγμα, τα περισσότερα γένη των γωβιών του Αιγαίου και Ιονίου Πελάγους έχουν Ποντοκασπιακή προέλευση, δηλαδή από την Παρατηθύ (Γκένας 2012). Γενικότερα, τα είδη που προέρχονται από την Παρατηθύ είναι κυρίως ενδημικά σε αντίθεση με την ιχθυοπανίδα του Μεσσηνίου που είναι ευρέως εξαπλωμένη και αποτελεί ένα φτωχό και ταξινομικά περίπλοκο σύνολο (Por & Dimentman 1985). Πολύ ενδιαφέρουσα βιογεωγραφική ιστορία φέρoυν και τα είδη Salaria fluviatilis και Salaria economidisi (Blenniidae) που ενώ κατοικούν στα εσωτερικά ύδατα της χώρας μας, πολλοί ερευνητές διατύπωσαν την άποψη ότι εξελίχθηκαν από έναν θαλλασινό 190
191 πρόγονο, φυλογενετικά κοντά στο θαλασσινό είδος Salaria pavo, και έχουν προέλευση από το Μεσσήνιο (Zander 1972). Επίσης, ο πληθυσμός της αθερίνας (Atherina boyeri) στη λίμνη Τριχωνίδα και τα είδη της οικογένειας Clupeidae, όπως τα Alosa falax, Alosa macedonica και Alosa vistonica αποτελούν ενδιαφέροντα παραδείγματα για την Ελληνική ιχθυοπανίδα, που δημιούργησαν εσώκλειστους («landlocked») πληθυσμούς στις λίμνες Βόλβη και Βιστωνίδα. Άλλα παραδείγματα αποτελούν τα είδη της οικογένειας Mugilidae που δεν αναπαράγονται αλλά συναντωνται στα εσωτερικά ύδατα της χώρας, όπως επίσης το είδος Petromyzon marinus που εισέρχεται σε ποτάμια της Ελλάδας για αναπαραγωγή (Barbieri et al. 2015). Παράλληλα, πάνω από 42% των αυτόχθονων ψαριών των εσωτερικών υδάτων στην Ελλάδα είναι ενδημικά. Τα επίπεδα ενδημισμού της Ελληνικής ιχθυοπανίδας είναι παρόμοια με άλλα ευρωπαϊκά σημεία ενδιαφέροντος, όπως η Ιβηρική (73%, Maceda-Veiga 2013) χερσόνησος, η Κροατία (30%, Oikonomou, Leprieur & Leonardos 2014) και η Ιταλία (27%, Ruffo & Stoch 2006). Όπως ήδη αναφέρθηκε (Κεφάλαιο 3), οι ιχθυοπανίδες των νότιων χερσονήσων (Ιβηρική, Ιταλική και Βαλκανική) εμφανίζουν τα υψηλότερα ποσοστά ενδημισμού σε όλη την Ευρώπη, αφού οι παγετώνες κατά το Πλειστόκαινο έπαιξαν σημαντικό ρόλο στα υψηλά επίπεδα ενδημισμού. Ο υψηλός ενδημισμός της Ελλάδας οφείλεται κυρίως στη Δυτική Ελλάδα (Ιόνια βιογεωγραφική περιοχή), η οποία εμφανίζει σαφώς υψηλότερα επίπεδα ενδημισμού (75%, Oikonomou, Leprieur & Leonardos 2014) σε σύγκριση με την Ανατολική (βιογεωγραφική περιοχή του Αιγαίου) εξαιτίας των καταφυγίων που επικράτησαν στην περιοχή κατά τους παγετώνες. Μία ακόμα εξήγηση για αυτό το πρότυπο αποδίδεται στη συνδεσιμότητα των λεκανών απορροής και τη γεωγραφική απομόνωση (Oberdorff et al. 2011). Πράγματι, η ασθενέστερη συνδεσιμότητα των λεκανών απορροής στη Δυτική Ελλάδα μπορεί να απέτρεψε τη διασπορά σε μεγάλες αποστάσεις, αυξάνοντας έτσι τον τοπικό ενδημισμό (βλέπε επίσης Heino 2011). Οι περισσότερες απειλές για την Ελληνική ιχθυοπανίδα των εσωτερικών υδάτων προκύπτουν από ανθρωπογενείς αλλαγές των υδρολογικών και υδρομορφολογικών συνθηκών, την άντληση νερού, την απώλεια και υποβάθμιση 191
192 των ενδιαιτημάτων τους, την εισαγωγή ξενικών ειδών καθώς και την εγκατάσταση εμπόδιων στη μετανάστευση τους (π.χ. φράγματα). Αυτοί οι παράγοντες οδήγησαν πολλά είδη της Ελληνικής ιχθυοπανίδας των εσωτερικών υδάτων όπως τα Alburnus macedonicus, A. vistonicus, Alosa vistonica, Barbus euboicus, Cobitis stephanidisi, Caspiomyzon hellenicus, Ε. graecus, Pelasgus epiroticus, P. laconicus, Pungitius hellenicus και Valencia letourneuxi να χρήζουν άμεσης προστασίας αφού κατατάσσονται στα Κρισίμως κινδυνεύοντα (CR) (Economidis 2009). Εκτός αυτών, πολλά είδη χαρακτηρίζονται ως κινδυνεύοντα ή τρωτά και απειλούνται με εξαφάνιση αυξάνοντας το ποσοστό των απειλούμενων πάνω από το 30%, κάτι ανάλογο της Βαλκανικής, Ιβηρικής και Ιταλικής Χερσονήσου σύμφωνα με τα κριτήρια της IUCN (Maceda-Veiga 2013, Bianco 2013, Oikonomou, Leprieur & Leonardos 2014). Σε αυτό το σημείο, είναι σημαντικό να αναφερθεί η σπουδαία συμβολή των Α. Στεφανίδη 22 και Π. Οικονομίδη 23 στη γνώση μας για τις κατανομές των ψαριών των εσωτερικών υδάτων καθώς και τη συστηματική περιγραφή τους, η συνεισφορά των οποίων τους καθιστά από τους πιο σημαντικούς επιστήμονες στην Ελλάδα που έθεσαν τις βάσεις για τη βιογεωγραφική μελέτη στη χώρα μας. Επιπλέον, θα πρέπει να σημειωθεί πως, έως σήμερα, έχουν πραγματοποιηθεί 4 αναθεωρήσεις του κατάλογου της Ελληνικής ιχθυοπανίδας (Economidis 1991, Economou et al. 2007, Koutsikos et al. 2012, Barbieri et al. 2015), που αύξησαν τον αριθμό των ειδών από 105 σε 160 (δηλαδή πάνω από 60%) μέσα σε 20 χρόνια. Αυτή η διαφορά στο συνολικό αριθμό των ειδών μεταξύ των καταλόγων αφορά κυρίως την συστηματική κατάταξη των ειδών, την αξιοποίηση νέων και ποικίλων διαγνωστικών μεθόδων στην αναγνώριση και την αναθεώρηση έπειτα από φυλογενετικές αναλύσεις. Εντούτοις, ο αριθμός και η κατανομή των ειδών που φιλοξενούνται στα Ελληνικά εσωτερικά ύδατα εξακολουθεί να είναι προς διερεύνηση καθώς η γνώση μας χαρακτηρίζεται από δυο βασικά ελλείμματα, το έλλειμμα του Wallace («Wallacean shortfall») και το έλλειμμα του Linnaeus («Linnean shortfall») (Κεφάλαιο 3). 22 Στεφανίδης, A. (1939). Tα ψάρια του γλυκού νερού της Δ. Eλλάδος και της νήσου Kερκύρας. Διδακτορική διατριβή, Πανεπιστήμιο Aθηνών 23 Οικονομίδης, Π. Σ. (1974). Μορφολογική συστηματική και ζωογεωγραφική μελέτη των ιχθύων των γλύκεων υδάτων της Α. Μακεδονίας και Δ. Θράκης. Διδακτορική διατριβή, Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης 192
193 Αναλυτικότερα, σε εθνικό επίπεδο, ένας πλήρης κατάλογος της Ελληνικής ιχθυοπανίδας ανά λεκάνη απορροής ποταμού και λίμνη δεν έχει δημοσιευθεί ποτέ, και αυτό διότι η ιχθυοπανίδα ορισμένων υδάτινων σωμάτων δεν έχει μελετηθεί συστηματικά και επαρκώς. Είναι, επίσης, ενδεικτικό ότι η εφαρμογή του Άρθρου 8 που αφορά την υλοποίηση προγραμμάτων εποπτικής, λειτουργικής και διερευνητικής παρακολούθησης (monitoring) των ποσοτικών και ποιοτικών χαρακτηριστικών των υδάτινων σωμάτων σύμφωνα με τις απαιτήσεις της Ευρωπαϊκής Οδηγίας Πλαίσιο για τα Ύδατα 2000/60/ΕΕ ολοκληρώθηκε μόλις το Δεκέμβριο του Επίσης, σε σχέση με τον πλούτο της Ελληνικής ιχθυοπανίδας, η Οδηγία της ΕΕ για τους οικοτόπους (92/43/ΕΟΚ) δεν περιλαμβάνει σημαντικά είδη που χρήζουν άμεσης προστασίας και διαχείρισης. Πιο συγκεκριμένα, στην πρόσφατη εξαετή εθνική αναφορά ( ) για τα 71 είδη που περιλαμβάνονται στην Οδηγία για τους Οικοτόπους (92/43/ΕΟΚ) και τα 66 που αξιολογήθηκαν, η Ελλάδα δηλώνει άγνωστη Κατάσταση Διατήρησης (ΧΧ) για 6 είδη που ανήκουν στις οικογένειες Acipenseridae (Acipenser stellatus, A. sturio), Cyprinidae (Aspius aspius, Pelasgus thesproticus) και Petromyzontidae (Petromyzon marinus, Caspiomyzon graecus), ένα ποσοστό που ανέρχεται στο 10%. Από τα υπόλοιπα, 18 είδη (25%) αξιολογήθηκαν σε «ευνοϊκή» (FV), 23 είδη (32%) σε «ανεπαρκή» (U1) και 21 είδη (30%) σε «κακή» (U2) (Τάχος κ. ά. 2015). Εδώ θα πρέπει να σημειωθεί πως ο κατάλογος των προστατευόμενων ειδών ψαριών για την Ελλάδα που περιλαμβάνονται στα παραρτήματα της Οδηγίας της ΕΕ για τους οικοτόπους (92/43/ΕΟΚ) 24, περιλαμβάνει είδη με λανθασμένη ονοματολογία, εντούτοις δεν περιλαμβάνει ορισμένα είδη που απειλούνται με πολύ περιορισμένη κατανομή, δηλαδή είδη που έχουν προσδιοριστεί ως κινδυνεύοντα από το Κόκκινο Βιβλίο και τη Διεθνή Ένωση Προστασίας της Φύσης (IUCN) (Economou et al. 2007, Barbieri et al. 2015). Επιπλέον, η μεγάλη κλίμακας έρευνα στις κατανομές των ειδών σε επίπεδο χώρας έχει παραγκωνιστεί υπέρ ερευνών στο επίπεδο του είδους, γένους ή οικοσυστήματος. Οι ελλείψεις αυτές σε δεδομένα κατανομών καθώς και ο μεγάλος 24 Με στόχο τη διατήρηση και προστασία της βιοποικιλότητας συντάχθηκε η Οδηγία 92/43/ΕΟΚ «για τη διατήρηση των φυσικών οικοτόπων καθώς και της άγριας πανίδας και χλωρίδας», από το ευρωπαϊκό Συμβούλιο. Η Οδηγία αυτή κατοχυρώθηκε στην Ελλάδα βάσει της ΚΥΑ 33318/3028/1998. Η εποπτεία των τύπων οικοτόπων και των ειδών κοινοτικού ενδιαφέροντος στην Ελλάδα (περιοχές του Δικτύου NATURA 2000 και εθνικός χώρος) πραγματοποιείται σε ανταπόκριση της χώρας στις υποχρεώσεις που απορρέουν από τις Οδηγίες 92/43/ΕΟΚ και 2009/147/ΕΚ (η αλλιώς 79/409/ΕΟΚ) 193
194 αριθμός κρυπτικών, μη μελετημένων ή περιγραφέντων ειδών και σε συνδυασμό με το γεγονός ότι οι τροποποιήσεις στην ονοματολογία και στη συστηματική κατάταξη των ειδών είναι συνεχείς, έχουν αρνητικό αντίκτυπο στην επιστημονική έρευνα και διαχείριση των οικοσυστημάτων των εσωτερικών υδάτων στην Ελλάδα (Koutsikos et al. 2012) Βιογεωγραφικοί παράγοντες που επηρεάζουν τον αριθμό των ειδών ψαριών και το μέγεθός τους σε λιμναία οικοσυστήματα στην Ελλάδα Tα λιμναία οικοσυστήματα είναι ιδιαιτέρως σημαντικά για την εφαρμογή των αρχών της μακροοικολογίας αφού επιτρέπουν τη διερεύνηση όλων των προτύπων που έχουν προταθεί για τα νησιωτικά οικοσυστήματα (Hortal et al. 2014). Έως τώρα, έχει προταθεί πως ο αριθμός των ειδών στις λίμνες μειώνεται με το υψόμετρο- από τον ισημερινό προς τους πόλους- αλλά πολλές φορές επηρεάζεται και από ιστορικούς παράγοντες που αλληλεπιδρούν με τους κλιματικούς (Griffiths 2006). Ο αριθμός των ειδών στα λιμναία οικοσυστήματα της Ελλάδας μειώνεται με την αύξηση του υψομέτρου και αυξάνεται με την αύξηση της έκτασης του οικοσυστήματος (r 2 =0.90). Το ίδιο πρότυπο έχει παρατηρηθεί και σε άλλες μελέτες για τα ψάρια των εσωτερικών υδάτων στα Ελληνικά λιμναία οικοσυστήματα (Δράκου 2009), αν και οι Mazaris et al (2010) δε βρήκαν στατιστικά σημαντική γραμμική σχέση μεταξύ του αριθμού των ειδών και της έκτασης για διαφορετικές ομάδες οργανισμών (φυτοπλαγκτό, ζωοπλαγκτό και ψάρια). Έντούτοις, αυτό το γεγονός ίσως δεν αποδίδεται σε έλλειψη συσχέτισης της έκτασης με τον αριθμό των ειδών, αλλά μπορεί να οφείλεται στο μικρό αριθμό λιμναίων οικοσυστημάτων που χρησιμοποίηθηκαν στην ανάλυση τους (συνολικά επτά λιμναία οικοσυστήματα). Εκτός όμως από τα Ελληνικά οικοσυστήματα, το μέγεθος των οικοσυστημάτων είναι ο κυριότερος παράγοντας που καθορίζει την ποικιλότητα σε λιμναία οικοσυστήματα σε παγκόσμιο επίπεδο. Για παράδειγμα, το υψόμετρο και η έκταση έχουν αναδειχθεί ως οι σημαντικότεροι παράγοντες καθορισμού της ποικιλότητας των λιμναίων οικοσυστημάτων στην Ασία (Amarasinghe & Welcomme 194
195 2002, Zhao et al. 2006), ερμηνεύοντας περίπου το 75% της διακύμανσης του αριθμού των ειδών, της Ευρώπης (Brucet et al. 2013, Griffiths, McGonigle & Quinn 2014) και της Αμερικής (Jaramillo-Villa, Maldonado-Ocampo & Escobar 2010), καθώς και άλλων ομάδων οργασνισμών των εσωτερικών υδάτων, όπως του φυτοπλαγκτού, του ζωοπλαγκτού και των μακροφύτων (Dodson, Arnott & Cottingham 2000, Hobaek, Manca, & Andersen 2002). Η συγκέντρωση του ολικού φωσφόρου έχει στατιστικά σημαντική αλλά τη μικρότερη συνεισφορά στην ερμηνεία της διαβάθμισης του αριθμού των ειδών σε λίμνες. O φώσφορος είναι άρρηκτα συνδεδεμένος με την πρωτογενή παραγωγικότητα. Αυτή η παρατήρηση επιβεβαιώνεται από την ισχυρή αρνητική συσχέτιση του αριθμού των ειδών με τη συγκέντρωση της χλωροφύλλης (0.72), μία ακόμα ένδειξη της παραγωγικότητας (Παράρτημα 3). Ως εκ τούτου, τα αποτελέσματα της παρούσας μελέτης υποδηλώνουν ότι οι ανθρωπογενείς δραστηριότητες, που συχνά προκαλούν αυξημένες συγκεντρώσεις φωσφόρου, μπορεί να έχουν αρνητική επίδραση στα μακροοικολογικά πρότυπα της ποικιλότητας των ψαριών σε λίμνες. Καθώς η παραγωγικότητα στις Ελληνικές λίμνες έχει συνδεθεί άμεσα με την εισαγωγή των ξενικών ειδών (Kagalou, Papastergiadou & Leonardos 2008, Kagalou & Leonardos 2009), θα είχε πολύ μεγάλο επιστημονικό ενδιαφέρον, σε μία μελλοντική έρευνα, να συσχετιστεί η παραγωγικότητα με τον αριθμό των ξενικών ειδών και τον αριθμό των αυτόχθονων ειδών ώστε να ελεγχθεί αν και κατά πόσο επιφέρουν εξαφάνιση ειδών της αυτόχθονης ιχθυοπανίδας του συστήματος υποδοχής. Το μέγεθος του σώματος των ψαριών των εσωτερικών υδάτων στις Ελληνικές λίμνες αυξάνεται με το γεωγραφικό πλάτος. Αν και στην παρούσα μελέτη δεν ελέγχθηκε η συσχέτιση του μεγέθους με τη θερμοκρασία λόγω της έλλειψης αξιόπιστων δεδομένων μέτρησης θερμοκρασίας του νερού στα υπό εξέταση υδάτινα σώματα, η ανάδειξη του μακροοικολογικού προτύπου της αύξησης του σώματος με το γεωγραφικό πλάτος εντοπίζεται για πρώτη φορά στα Ελληνικά εσωτερικά ύδατα. Αυτή η ιδιαιτέρως ενδιαφέρουσα παρατήρηση έρχεται σε συμφωνία με πολλές έρευνες σε διαφορετικού τύπου οικοσυστήματα και περιοχές (Knouft 2004, Blanck & Lamouroux 2007, Griffiths 2012, Griffiths 2013). Παρόλα αυτά, με δεδομένο το μικρό 195
196 εύρος του γεωγραφικού πλάτους στον Ελληνικό χώρο, το συγκεκριμένο πρότυπο θα πρέπει να ελεχθεί και με τη χρήση άλλων οργανισμών και συστημάτων. Δεδομένων των διαφορετικών υποθέσεων που εμπλέκονται στην στήριξη αυτού του προτύπου, πιστεύουμε πως μέρος της ερμηνείας για τα ψάρια των εσωτερικών υδάτων στις Ελληνικές λίμνες συνδέεται με ιστορικούς παράγοντες και αποδίδεται στη διαφορετική ικανότητα διασποράς των ειδών και τη διαφορετική γεωλογική ιστορία των μελετούμενων συστημάτων. Πιο συγκεκριμένα, στην παρούσα μελέτη παρατηρήθηκε ότι το μέσο μέγεθος των ειδών στις νοτιότερες περιοχές διέφερε σημαντικά από αυτό στις βορειότερες. Αυτή η παρατήρηση συμπίπτει με αυτή του Griffiths (2010), ότι δηλαδή τα μικρόσωμα είδη είναι πιο πιθανό να εξακολουθούν να διαβιούν σε περιοχές που δεν επηρεάστηκαν από τους παγετώνες (π.χ. Ιόνια βιογεωγραφική περιοχή Oikonomou, Leprieur & Leonardos 2014). Επιπλέον, τα μικρά είδη τείνουν να καταλαμβάνουν γεωγραφικά περιορισμένες περιοχές, καθιστώντας τα πιο ευάλωτα για εξαφάνιση, αλλά επίσης, λόγω της μειωμένης γονιδιακής ροής, οδηγώντας τα σε μεγαλύτερη ειδογένεση (Griffiths 2013). Πράγματι, οι διαφορές του επιπέδου του ενδημισμού και απειλούμενων ειδών μεταξύ των νότιων και βόρειων περιοχών στον Ελληνικό χώρο είναι εμφανείς (Κεφάλαιο 3), ένα πρότυπο που αναδείχθηκε και κατά την οριοθέτηση των βιογεωγραφικών περιοχών της Βαλκανικής χερσονήσου (Κεφάλαιο 2 και Κεφάλαιο 3). Από τα αποτελέσματα της παρούσας μελέτης, μπορεί να συμπεράνει κάποιος ότι η συνδεσιμότητα και η έκταση των περιοχών παίζουν έναν καθοριστικό ρόλο και στο μέγεθος των ειδών. Για παράδειγμα, η βιογεωγραφική περιοχή του Αιγαίου και οι συλλήψεις ποταμών της ευνόησαν τον εποικισμό με Δουνάβια είδη ή με προέλευση τη Μαύρη Θάλασσα που είναι πιο μεγαλόσωμα (Economidis & Bănărescu 1991, Bănărescu 2004, Oikonomou, Leprieur, & Leonardos 2014). Επιπλέον, η Βορειοανατολική Ελλάδα φιλοξενεί περισσότερα λιμνόφιλα είδη, που κατά κανόνα είναι μεγαλύτερα από τα ρεόφιλα, σε σχέση με τη Δυτική καθώς στο παρελθόν οι μεγαλύτερες λίμνες της Ελλάδας δέσποζαν στη Μακεδονία και τη Θράκη. Ένας ακόμα παράγοντας, η έκταση των οικοσυστημάτων, αν και δε συσχετίστηκε σημαντικά με το μέγεθος των ειδών, θα μπορούσε να παίζει ένα σημαντικό ρόλο στην 196
197 περιβαλλοντική ετερογένεια και τη διαθέσιμη ενέργεια, ευνοώντας τα μεγαλύτερα είδη στα βορειότερα οικοσυστήματα. Εν κατακλείδι, η Βορειοανατολική Ελλάδα φιλοξενεί ευρέως κατανεμημένα, μεγαλόσωμα είδη με μεγαλύτερη ικανότητα μεταπαγετώδους εποικισμού σε σύγκριση με τη Νοτιοδυτική, όπου διαβιούν μικρόσωμα ενδημικά είδη ή είδη με περιορισμένη κατανομή. Είναι εξίσου ενδιαφέρον ότι η Βορειοανατολική Ελλάδα φιλοξενεί μεγαλύτερο αριθμό ψαριών σε σχέση με τη Δυτική, ένα γεγονός που μπορεί να οφείλεται στα διαφορετικά κύματα αποικισμού των δύο περιοχών μέσω των ποτάμιων συνδέσεων. Πιο συγκεκριμένα, η Δυτική Ελλάδα αποικίστηκε μία φορά από είδη που έφεραν Σιβηρική καταγωγή, πιθανώς, από την νοτιοδυτική Ασία, ενώ η Ανατολική Ελλάδα αποικίστηκε και μία δεύτερη φορά με είδη με παραδουνάβια καταγωγή που διασπάρθηκαν μέσω της ποτάμιας σύνδεσης ανάμεσα στο Δούναβη και τον Αξιό και τη σύνδεση του Αιγαίου με τη Μαύρη Θάλασσα (Economidis & Bănărescu 1991). Ως εκ τούτου, τα διαφορετικά πρότυπα ποικιλότητας, η δυτική-ανατολική διάκριση των Ελληνικών εσωτερικών υδάτων και η διαφορετική σύνθεση της Ελληνικής ιχθυοπανίδας αποδίδεται στην μακροχρόνια παρουσία του γεωγραφικού εμπόδιου της Πίνδου και την επακόλουθη απομόνωση που προκλήθηκε (Economou et al. 2007, Drakou et al. 2009) και έχει ήδη τεκμηριωθεί σε προηγούμενα κεφάλαια αυτής της διατριβής, παρέχοντας έτσι περαιτέρω υποστήριξη στα αποτελέσματα του παρόντος κεφαλαίου. 197
198
199 ΜΕΡΟΣ III Επίπεδο οικοσυστήματος: Ποταμός Λούρος
200
201 Κεφάλαιο 5 Πρότυπα ποικιλότητας στον ποταμό Λούρο The river fish are bulky in the middle because they move with their head and tail: the fishes of the torrents are traditionally well known by the possession of greatly flattened body on account of their habit of crawling with the chest and a relatively reduced anterior part of the body -Translation by Hora (1952) of a Sanskrit passage written between 600 B.C. and A.D. 500
202
203 5.1 Εισαγωγή Βιοκοινότητες ψαριών σε ποτάμια οικοσυστήματα Ένα βασικό ερώτημα στην οικολογία είναι τι καθορίζει τη σύνθεση και τη δομή των βιοκοινοτήτων (Nikolski 1933); Η απάντηση δεν είναι απλή επειδή τόσο η κατανομή όσο και η αφθονία των ειδών στα ποτάμια οικοσυστήματα διαμορφώνονται από πολλούς παράγοντες που δρουν σε διαφορετικές κλίμακες με αποτέλεσμα τα παρατηρούμενα πρότυπα να προκύπτουν από ένα συνδυασμό τοπικών και περιφερειακών διεργασιών, και κατά συνέπεια, να πρέπει να ερμηνεύονται μέσα στα εκάστοτε βιογεωγραφικά και ιστορικά πλαίσια (Ricklefs & Schlüter 1993, βλέπε Matthews 1998b για την επισκόπηση της βιβλιογραφίας). Ειδικότερα για τη σύνθεση των ιχθυοκοινοτήτων, σε μεγάλες χωρικές κλίμακες (περιφερειακό επίπεδο), οι κυριότεροι παράγοντες είναι το υψόμετρο, το μέγεθος του ποταμού και το κλίμα, μαζί με παράγοντες, όπως η σύνθεση του περιφερειακού αποθέματος των ειδών («regional species pool»), ο ρυθμός διαφοροποίησης και η ικανότητα διασποράς (Hugueny 1989) να συγκαταλέγονται στους κυριότερους (Σημείωμα 5). Εντός των ποτάμιων οικοσυστημάτων (τοπικό επίπεδο), οι αβιοτικές παράμετροι όπως ο τύπος υποστρώματος, η μορφολογία του ποταμού (π.χ. πλάτος και το βάθος) (Schlosser 1985) καθώς και οι φυσικο-χημικοί παράγοντες (π.χ. οξυγόνο, θερμοκρασία) σχετίζονται ισχυρά με τη σύνθεση των βιοκοινοτήτων (Hughes, Wang & Seelbach 2006). Επιπλέον, βιοτικές παράμετροι όπως η παρόχθια βλάστηση επιδρούν στις ιχθυοκοινότητες ενώ και οι ανθρωπογενείς αλλαγές επηρεάζουν τη σύνθεση (π.χ. φράγματα, εισαγωγή ξενικών ειδών) (Rowe, Pierce & Wilton 2009). Σε τοπικό επίπεδο, ο ανταγωνισμός (π.χ. Ross, Matthews & Echelle 1985) ή η θήρευση αλλά και η ποικιλότητα βιοτόπων (Grenouillet, Pont & Seip 2002) αλληλεπιδρούν και καθορίζουν τη σύνθεση των ιχθυοκοινοτήτων (Jackson, Peres- Neto & Olden 2001). Ένα από τα πιο ενδιαφέροντα παγκόσμια πρότυπα που παρατηρούνται και στα ποτάμια οικοσυστήματα, αναφέρεται στη διαμήκη μεταβολή των ιχθυοκοινοτήτων. Αυτό το πρότυπο έχει αποδοθεί σε δύο συμπληρωματικές διεργασίες: τη χωρική 203
204 αντικατάσταση (δηλαδή, σην αλλαγή στη σύνθεση των ειδών) και τη συνεχή προσθήκη ειδών (Rahel & Hubert 1991, Hoagstrom et al. 2007). Η χωρική αντικατάσταση των ειδών σχετίζεται με την μεταβολή των ενδιαιτήματων (Rahel & Hubert 1991) και συνδέεται με την «Αρχή της συνέχειας των ποταμών» («River Continuum Concept») (Vannote et al. 1980). Αυτή η θεωρία προτείνει αλλαγές στη δομή των βιοκοινοτήτων καθώς κινούμαστε κατάντη κυρίως λόγω των μεταβολών της ροής, της θερμοκρασίας, της διαθεσιμότητας τροφής και τις συνθήκες του υποστρώματος. Με άλλα λόγια, τα χαρακτηριστικά των ποταμών (π.χ. υψόμετρο, ταχύτητα ροής νερού, θερµοκρασία νερού, υπόστρωμα) παρουσιάζουν βαθμιαία μεταβολή που έχει ως συνέπεια την παρουσία διαφορετικών ιχθυοκοινοτήτων. Αντίθετα, η προσθήκη των ειδών αναφέρεται στην αύξηση της πολυπλοκότητας των ενδιαιτημάτων και των οικολογικών θώκων («habitat complexity and niche space») κατάντη (Sheldon 1968, Oberdorff, Guégan & Hugueny 1995, Smith, Quist & Hardy 2015), αν και έχει συνδεθεί επίσης με την αύξηση της έκτασης των ενδιαιτημάτων (ή της παροχής ή και του όγκου), καθώς και με την αύξηση της επίγειας παραγωγικότητας (Hugueny, Oberdorff & Tedesco 2010). 204

205 5.1.2 Λειτουργική ποικιλότητα σε ποτάμια οικοσυστήματα Όπως ήδη αναφέρθηκε, οι βιοκοινότητες ποικίλλουν μέσα στο χώρο και το χρόνο (Rosenzweig 1995) κυρίως εξαιτίας της μεταβλητότητας των διεργασιών που τις διαμορφώνουν. Η κατανόηση πώς αυτές οι εξελικτικές και οικολογικές διεργασίες επηρεάζουν τη βιοποικιλότητα στα διάφορα επίπεδα οργάνωσής της, αποτελεί ακόμα ένα από τα πιο σημαντικά ερωτήματα στην οικολογία διατήρησης. Οι οικολόγοι της διατήρησης παραδοσιακά διερευνούσαν τις σχέσεις μεταξύ των περιβαλλοντικών παραμέτρων και της πανιδικής σύνθεσης, χρησιμοποιώντας μετρικές, όπως ο αριθμός και η αφθονία των ειδών (δηλαδή στο επίπεδο της ποικιλότητας των ειδών) (Jackson, Peres-Neto & Olden 2001). Ωστόσο, το ερώτημα αυτό βρίσκει εφαρμογή στο πεδίο της λειτουργικής ποικιλότητας (Σημείωμα 6), δηλαδή στην πτυχή της βιοποικιλότητας που αντιπροσωπεύει τα λειτουργικά 205
206 χαρακτηριστικά των οργανισμών. Γενικότερα, η λειτουργική ποικιλότητα αντικατοπτρίζει τη χρήση των πόρων και την προτίμηση των ενδιαιτημάτων (Lavorel & Garnier 2002), διαμορφώνεται από τις συνθήκες του περιβάλλοντος και επηρεάζει και τις οικοσυστημικές λειτουργίες (Violle et al. 2011). Τις τελευταίες δεκαετίες έχει δειχθεί ότι τα λειτουργικά χαρακτηριστικά των ειδών και συνεπώς ο λειτουργικός τους ρόλος μέσα στο οικοσύστημα συνδέεται άμεσα με τη μορφολογία τους (Gatz 1979, Wainwright 1994). Σύμφωνα με αυτή τη συσχέτιση μεταξύ της μορφής και της λειτουργίας, η περιγραφή του λειτουργικούμορφολογικού χώρου των ειδών, δηλαδή η έκφραση των λειτουργικών χαρακτηριστικών σε πολυδιάστατο επίπεδο, αποτελεί ένα ισχυρό εργαλείο διερεύνησης της λειτουργικής δομής των βιοκοινοτήτων καθώς και τις απαντήσεις των ειδών στις περιβαλλοντικές μεταβολές. Συνεπώς, η μελέτη των λειτουργικών χαρακτηριστικών των ειδών και η κατανόηση του τρόπου με τον οποίο οι αβιοτικές παράμετροι και οι αλληλεπιδράσεις μεταξύ των ειδών (ανταγωνισμός, θήρευση) επηρεάζουν την κατανομή των χαρακτηριστικών των ειδών, κατά μήκος των περιβαλλοντικών διαβαθμίσεων, είναι σημαντική για τη μοντελοποίηση της κατανομής των ειδών (Thuiller et al. 2013) και τη διερεύνηση των οικολογικών διεργασιών που διαμορφώνουν τις βιοκοινότητες (Gatz 1979, Winemiller 1991). Οι δύο βασικές διεργασίες που έχουν παρατηρηθεί κατά τη μελέτη της συγκρότησης των ιχθυοκοινοτήτων αφορούν το περιβαλλοντικό φιλτράρισμα (αβιοτικοί παράγοντες που περιορίζουν είδη με μη-κατάλληλα χαρακτηριστικά να επιβιώσουν και να είναι εύρωστα σε μια συγκεκριμένη τοποθεσία) και τις βιοτικές αλληλεπιδράσεις (ανταγωνισμός, θήρευση) (Σημείωμα 5). Χρησιμοποιώντας δεδομένα που αφορούν τα λειτουργικά χαρακτηριστικά των ειδών, την κατανομή και την αφθονία τους και τις περιβαλλοντικές συνθήκες στις οποίες διαβιούν, δύναται να συγκρίνουμε τη σύνθεση των ειδών εξαιτίας της χρονικής και χωρικής διακύμανσης των αβιοτικών συνθηκών, του ανταγωνισμού και της θήρευσης (Winemiller 2005, McGill et al. 2006). Κατά τη διερεύνηση αυτών των δύο διεργασιών, η διασπορά των χαρακτηριστικών των ειδών μπορεί να παράσχει πληροφορίες σχετικά με τους μηχανισμούς συγκρότησης των βιοκοινοτήτων (Weiher & Keddy 1995). Πιο συγκεκριμένα, όταν τα χαρακτηριστικά των ειδών μιας βιοκοινότητας εμφανίζουν μικρότερη διασπορά (χαμηλή λειτουργική ποικιλότητα) σε σύγκριση με μια τυχαία 206
 σε σύγκριση με μία τυχαία, αποτελεί ένδειξη του αποκλειόμενου ανταγωνισμού και της θήρευσης")
207 βιοκοινότητα που προκύπτει από το δυνητικό αποθεματικό των ειδών, ως μηχανισμός συγκρότησης της βιοκοινότητας ορίζεται το περιβαλλοντικό φιλτράρισμα. Αντίθετα, η μεγαλύτερη διασπορά των χαρακτηριστικών μιας βιοκοινότητας (υψηλή λειτουργική ποικιλότητα) σε σύγκριση με μία τυχαία, αποτελεί ένδειξη του αποκλειόμενου ανταγωνισμού και της θήρευσης (Κεφάλαιο 1, Εικόνα 5.1). Μελέτες έχουν δείξει πως η σύνθεση των ιχθυοκοινοτήτων στα ποτάμια Εικόνα 5.1. Υποθετικές σχέσεις μεταξύ της λειτουργική ποικιλότητας και του αριθμού των ειδών. Η περιοχή με τη γκρι σκίαση είναι η αναμενόμενη κατανομή της λειτουργικής ποικιλότητας. Οι μικρές σφαίρες αντιπροσωπεύουν υποθετικά σύνολα ειδών. Βιοκοινότητες με υψηλή λειτουργική ποικιλότητα για ένα δεδομένο αριθμό ειδών (μπλε περιοχή) συγκροτούνται από είδη που παρουσιάζουν μεγαλύτερη διασπορά τιμών των λειτουργικών χαρακτηριστικών τους απ ότι αναμένεται από την τύχη. Βιοκοινότητες με χαμηλή λειτουργική ποικιλότητα για δεδομένο αριθμό ειδών (κίτρινη περιοχή) συγκροτούνται από είδη με παρόμοιες τιμές των λειτουργικών χαρακτηριστικών τους σε σύγκριση με τυχαίες βιοκοινότητες (τροποποιημένο από Petchey et al. 2007) Figure 5.1. Hypothetical relationships between functional diversity and species richness. The greyshaded region is the expected distribution of functional diversity. Small spheres represent hypothetical assemblages. Biocommunities with high functional diversity for a given level of species richness (blue region) contain species that are more dispersed in functional trait values than expected by chance. Assemblages with low functional diversity for their given level of species richness (yellow region) contain species that have more similar functional trait values than random biocommunities (modified from Petchey et al. 2007) 207
208 διαμορφώνεται κυρίως από το περιβαλλοντικό φιλτράρισμα παρά από τον ανταγωνισμό (Jackson, Peres-Neto & Olden 2001). Για παράδειγμα, ο Peres-Neto (2004) κατέληξε στο συμπέρασμα ότι η συνύπαρξη των ειδών των ψαριών σε Νεοτροπικά ποτάμια αποδίδεται στη χρήση παρόμοιων ενδιαιτημάτων. Εντούτοις, ο Winston (1995) έκανε ακριβώς την αντίθετη διαπίστωση, ότι δηλαδή ο αποκλειόμενος ανταγωνισμός διαμορφώνει τις ιχθυοκοινότητες. Αν και οι έρευνες σχετικά με το πώς οι αβιοτικοί και βιοτικοί παράγοντες διαμορφώνουν μια βιοκοινότητα αυξάνονται, η ικανότητα των επιστημόνων να ποσοτικοποιήσουν τους ρόλους του περιβαλλοντικού φιλτραρίσματος και των βιοτικών αλληλεπιδράσεων είναι περιορισμένη και αντανακλάται κυρίως στη χρήση θεωρητικών μοντέλων μηδενικής υπόθεσης που εφαρμόζονται στη φυλογένεση (Webb et al. 2002). Παρόλα αυτά, οι μελέτες διερεύνησης των μηχανισμών συγκρότησης των βιοκοινοτήτων με χρήση των λειτουργικών χαρακτηριστικών των ειδών σε ποτάμια οικοσυστήματα είναι σημαντικές για τον προσδιορισμό των ρόλων των βιοτικών και αβιοτικών παραγόντων στη διαδικασία εγκατάστασης των ειδών (Troia & Gido 2015). Συνοψίζοντας, οι μελέτες που βασίζονται στα χαρακτηριστικά των ειδών μπορούν να απαντήσουν μια σειρά ερωτημάτων όπως 1) πώς αποκρίνονται τα είδη και οι βιοικοινότητες στις αλλαγές των ενδιαιτημάτων 2) ποιες είναι οι πραγματικές διεργασίες που οδηγούν σε εξαφάνιση, εισβολή και βιοτική ομογενοποίηση 3) ποιοι είναι οι γενικότεροι κανόνες συγκρότησης των βιοκοινοτήτων (Frimpong & Angermeier 2010) (Σημείωμα 6). 208

209 209
210 5.1.3 Λειτουργική ποικιλότητα των ψαριών των εσωτερικών υδάτων Για τα ψάρια των εσωτερικών υδάτων, η λειτουργική προσέγγιση χρησιμοποιήθηκε για πρώτη φορά για τη διερεύνηση των οικολογικών διεργασιών που διαμορφώνουν τις ιχθυοκοινότητες σε πέντε γεωγραφικές περιοχές (Αλάσκα, Βόρεια Αμερική, Κεντρική Αμερική, Νότια Αμερική και Αφρική) χρησιμοποιώντας 30 μορφολογικά χαρακτηριστικά των ειδών ψαριών (Winemiller 1991). Σύμφωνα με τα αποτελέσματα αυτής της μελέτης, οι βιοτικές αλληλεπιδράσεις αποτελούν καθοριστικό παράγοντα διαμόρφωσης της δομής των ιχθυοκοινοτήτων και ο αριθμός των ειδών σχετίζεται θετικά με το ρυθμό διαφοροποίησης των οικολογικών θώκων («niche diversification») και την οικολογική εξειδίκευση («ecological specialization») των ειδών, οδηγώντας σε διαπεριφερειακή («interregional convergence») σύγκλιση των χαρακτηριστικών των ιχθυοκοινοτήτων τους. Αντίθετα, οι Irz et al. (2007) βρήκαν πρόσφατα ότι η αναπαραγωγική στρατηγική των ψαριών σε λίμνες της Γαλλίας και των Η.Π.Α. ανταποκρίνεται με ανάλογο τρόπο στις εκάστοτε περιβαλλοντικές συνθήκες. Ειδικότερα, οι θερμότερες λίμνες φιλοξενούν υψηλότερη αναλογία ειδών που αποθέτουν τα αβγά τους σε φυτά («phytophils») και ειδών που φυλάσσουν τα αβγά τους («guarders») από ότι οι ψυχρότερες. Η θερμοκρασία, επίσης, καταδείχτηκε ως ένας από τους πιο σημαντικούς παράγοντες που επηρεάζουν τη χωρική κατανομή των ειδών ψαριών και σχετίζεται ισχυρά με τις οικολογικές απαιτήσεις τους και στα ευρωπαϊκά ποτάμια οικοσυστήματα (Logez et al. 2013). Συν τοις άλλοις, τα αποτελέσματα από μελέτες λειτουργικής ποικιλότητας έχουν σημαντική συνεισφορά στην ανάπτυξη των βιολογικών δεικτών που συχνά βασίζονται σε λειτουργικά χαρακτηριστικά των ιχθυοκοινοτήτων (Karr & Chu 1999) και αποτελούν ένα εργαλείο πρόβλεψης για το πώς οι ιχθυοκοινότητες θα ανταποκριθούν σε μελλοντικές περιβαλλοντικές αλλαγές. Για παράδειγμα, οι Walser & Bart (1999) παρατήρησαν τη μείωση του αριθμού των ειδών των ψαριών που προτιμούν ενδιαιτήματα με χοντρόκοκκο υπόστρωμα, σε περιοχές με αυξημένη γεωργική δραστηριότητα, εξαιτίας της οποίας προκλήθηκε αύξηση της εναπόθεσης ιλύος στο υπόστρωμα και μείωση της πολυπλοκότητας στη σύνθεσή του. Η χρήση 210
211 γης σε λεκάνες απορροής της Βόρειας Αμερικής έχει καταδειχθεί μεταξύ των σημαντικότερων παραμέτρων που διαμορφώνουν τη σύνθεση των ειδών με βάση τα λειτουργικά χαρακτηριστικά τους (Pease et al. 2015, Parks, Quist & Pierce 2016). Σε μικρότερη κλίμακα, οι μελέτες της λειτουργικής ποικιλότητας που εκπονήθηκαν πρόσφατα, διερεύνησαν τις αποκρίσεις των ειδών με βάση τα περιβαλλοντικά χαρακτηριστικά μέσω της ποσοτικοποίησης της σχέσης μεταξύ των λειτουργικών χαρακτηριστικών των οργανισμών και των περιβαλλοντικών μεταβλητών (π.χ. Pease et al. 2012, Keck et al. 2014, Pease et al. 2015, Troia & Gido 2015). Για παράδειγμα, η παροχή του ποταμού έχει δειχθεί πως επηρεάζει τις αναπαραγωγικές στρατηγικές των ειδών (Mims & Olden 2012), ενώ το βάθος και η ταχύτητα της ροής σχετίζεται με την κολυμβητική ικανότητα, την ικανότητα ελιγμών και τη θέση στη στήλη του νερού (Sibbing & Nagelkerke 2001, Lamouroux, Poff & Angermeier 2002). Αφού παρόμοιες μελέτες λειτουργικής ποικιλότητας σε οικοσυστήματα εσωτερικών υδάτων στην Αμερική (Mims et al. 2010), την Ευρώπη (Lamouroux, Poff & Angermeier 2002), την Αυστραλία (Sternberg & Kennard 2013) και την Αφρική (Tedesco et al. 2008) υποστηρίζουν ότι τα πρότυπα είναι επαναλήψιμα μεταξύ των βιογεωγραφικών περιοχών, η κατανόηση της επίδρασης των περιβαλλοντικών και ανθρωπογενών παραγόντων στη διαμόρφωση της ποικιλότητας των ειδών, της λειτουργικής ποικιλότητας και της σύνθεσης των ιχθυοκοινοτήτων είναι απαραίτητη για την αποτελεσματική διαχείριση στα οικοσυστήματα των εσωτερικών υδάτων (Olden et al. 2010, Κεφάλαιο 1) Ποια και πόσα λειτουργικά χαρακτηριστικά των ψαριών πρέπει να χρησιμοποιήσουμε; Έως σήμερα, στις μελέτες λειτουργικής ποικιλότητας που αφορούν τα ψάρια έχουν χρησιμοποιηθεί διάφορα χαρακτηριστικά που συνδέονται με την προσαρμοστικότητα των ειδών («species performance») (Winemiller 2005, Πίνακας 5.1). Ενδεικτικά, οι Wainwright, Bellwood & Westneat (2002) και οι Bellwood et al. 211
212 (2002), οι οποίοι μελέτησαν υφάλους, χρησιμοποίησαν δύο ομάδες λειτουργικών χαρακτηριστικών: την ικανότητα κολύμβησης και την προτίμηση των ενδιαιτημάτων των ιχθυοκοινοτήτων. Το 2007, η Brind Amour συμπεριέλαβε χαρακτηριστικά που αφορούν τις διατροφικές συνήθειες, την αναπαραγωγική στρατηγική, τη θέση στη στήλη του νερού και την προτίμηση του υποστρώματος των ιχθυοκοινοτήτων της δυτικής ακτής της Γαλλίας. Οι Mouillot et al. (2005) και στη συνέχεια, οι Mason et al. (2008) μέτρησαν οκτώ και 13 συνεχή λειτουργικά χαρακτηριστικά αντίστοιχα, που περιγράφουν την ικανότητα κολύμβησης, τη θέση στη στήλη του νερού, το μέγεθος της λείας και τις διατροφικές συνήθειες. Με περισσότερη λεπτομέρεια, οι Villéger et al. (2010) χρησιμοποίησαν 17 συνεχή οικο-μορφολογικά χαρακτηριστικά που σχετίζονται με την τροφοληψία και τη μετακίνηση. Προκειμένου να ελέγξουν, αν οι κοινότητες των ψαριών της Μεσογείου και της Δυτικής Ευρώπης ανταποκρίνονται με παρόμοιο τρόπο σε περιβαλλοντικές διαβαθμίσεις, οι Logez, Pont & Ferreira (2010) χρησιμοποίησαν κατηγορικά δεδομένα σχετικά με τη διατροφή, τα ενδιαιτήματα διαβίωσης και αναπαραγωγής και τη μετακίνηση. Σε άλλη μία μελέτη, οι Mims et al. (2010) χρησιμοποίησαν επτά συνεχή γνωρίσματα των στρατηγικών ζωής για να μετρήσουν τη λειτουργική ποικιλότητα των αυτόχθονων ψαριών των εσωτερικών υδάτων στη Βόρεια Αμερική. Κατηγορικά λειτουργικά χαρακτηριστικά που σχετίζονται με τις στρατηγικές ζωής των ψαριών έχουν επίσης χρησιμοποιηθεί για τη μελέτη της απόκρισης των ειδών μετά από ανθρωπογενείς παρεμβάσεις (Keck et al. 2014, Parks, Quist & Pierce 2016). Πρόσφατα, οι Troia & Gido (2015) συμπεριέλαβαν εννιά λειτουργικά χαρακτηριστικά που σχετίζονται με την αναπαραγωγή, τη διατροφή και τη μετακίνηση των ψαριών κατά μήκος των περιβαλλοντικών διαβαθμίσεων και σε διαφορετικές χωρικές κλίμακες (βλέπε επίσης Pease et al. 2015). Τέλος, τα λειτουργικά χαρακτηριστικά που αφορούν το μεταβολισμό και τη συμπεριφορά των ειδών έχουν χρησιμοποιηθεί ελάχιστα στη βιβλιογραφία (Πίνακας 5.1) Από τα παραπάνω συμπεραίνουμε πως ο αριθμός των υποψηφίων βιολογικών γνωρισμάτων είναι δυνητικά μεγάλος και αφορά πολλές λειτουργίες όπως η μετακίνηση, η τροφοληψία, η αναπαραγωγή, η ανακύκλωση θρεπτικών στοιχείων, η ανοχή στο περιβαλλοντικό στρες και η συμπεριφορά (Πίνακας 5.1). Επίσης, συνεχώς προσθέτονται νέα ή τροποποιημένα χαρακτηριστικά στα ήδη υπάρχοντα (π.χ. 212
213 Sibbing & Nagelkerke 2001). Συνεπώς τίθεται το ερώτημα, ποια είναι τα κατάλληλα λειτουργικά χαρακτηριστικά που πρέπει να μετρήσουμε και να συμπεριλάβουμε στις αναλύσεις λειτουργικής ποικιλότητας των ψαριών των εσωτερικών υδάτων; Η απάντηση σε αυτή την ερώτηση έγκειται στην επιλογή των λειτουργικών χαρακτηριστικών που είναι αντιπροσωπευτικά για τις εκάστοτε υπό μελέτη λειτουργίες των ειδών. Η επιλογή ποσοτικών (και όχι κατηγορικών) λειτουργικών χαρακτηριστικών αποτελεί μία σημαντική προϋπόθεση επειδή μόνο οι ποσοτικές και συνεχείς μετρήσεις μπορούν να χρησιμοποιηθούν για τον υπολογισμό των πολυδιάστατων δεικτών λειτουργικής ποικιλότητας και ως εκ τούτου, των πολλαπλών διαστάσεων του οικολογικού θώκου. Η ευκολία απόκτησης των μετρήσεων στα άτομα των ιχθυοκοινοτήτων αποτελεί ένα ακόμα κριτήριο επιλογής των χαρακτηριστικών διότι η γνώση μας για τα λειτουργικά χαρακτηριστικά για πολλά είδη ψαριών είναι ελλιπής. Τέλος, τα χαρακτηριστικά που επιλέγονται θα πρέπει να είναι, σε μεγάλο βαθμό, ανεξάρτητα, δηλαδή να μην παρουσιάζουν υψηλή συσχέτιση μεταξύ τους, ώστε να μην οδηγούν σε προβληματικά πρότυπα και ερμηνείες. Συνοψίζοντας, προτείνουμε τη χρήση οικο-μορφολογικών χαρακτηριστικών που σχετίζονται με τις λειτουργίες της τροφοληψίας και της μετακίνησης καθώς: 1) η χρήση των ενδιαιτημάτων σχετίζεται ισχυρά με τις προσαρμογές της μορφής του σώματος και του στόματος (Keast & Webb 1966) 2) είναι τα πιο ευρέως χρησιμοποιούμενα σε μελέτες λειτουργικής ποικιλότητας διότι αφορούν τις πιο σημαντικές λειτουργίες των ψαριών των εσωτερικών υδάτων, που έχουν διερευνηθεί για αρκετές δεκαετίες από τις πρώτες προσπάθειες του Gatz (1979) και Winemiller (1991) και 3) υπολογίζονται εύκολα από συνεχείς μετρήσεις σε ένα μεγάλο αριθμό ατόμων των ειδών με τη χρήση ψηφιακών εικόνων και ψηφιακού λογισμικού επεξεργασίας. 213
214 Πίνακας 5.1. Χαρακτηριστικά που χρησιμοποιήθηκαν σε μελέτες λειτουργικής ποικιλότητας και περιγράφουν τις λειτουργίες της τροφοληψίας, της μετακίνησης, της ανακύκλωσης των θρεπτικών συστατικών, της αναπαραγωγής, της ανοχής στο στρες και της συμπεριφοράς των ψαριών Table 5.1. Traits used in functional diversity studies that describe food acquisition, locomotion, nutrient cycling, reproduction, stress tolerance and behavior for fishes Function Food acquisition Locomotion Functional traits Relevance Trait-based approach Eye size Visual acuity Mouillot et al. 2005, Mason et al. 2008, Villéger et al. 2010, Pease et al. 2015, Troia & Gido 2015 Barbel length Sediment Mason et al burrowing Oral gape Maximum prey Mouillot et al. 2005, Mason et al. 2008, surface size and filtering Villéger et al. 2010, Pease et al. 2015, ability Troia & Gido 2015 Oral gape Prey capture Mason et al. 2008, Villéger et al shape Oral gape position Gill raker Jaw protrusion Feeding habitat Feeding preference Gut length Eye position Body transversal shape Body elongation Body transversal surface Pectoral fin position Position of the prey in the water column Filtration or gill protection Mason et al. 2008, Brind Amour et al. 2011, Villéger et al. 2010, Pease et al. 2015, Troia & Gido, 2015 Mouillot et al. 2005, Mason et al. 2008, Villéger et al. 2010, Pease et al. 2015, Troia & Gido 2015 Suction Mouillot et al. 2005, Pease et al Position of the prey Prey capture and processing Relative to digestibility of food Position of fish and/ or its preys in the water column Position in the water column and hydrodynamism Mass repartition and hydrodynamism Mass repartition and hydrodynamism Logez, Pont & Ferreira 2010, Brind Amour et al Logez, Pont & Ferreira 2010, Brind Amour et al. 2011, Keck et al Mouillot et al. 2005, Mason et al. 2008, Villéger et al. 2010, Pease et al. 2015, Troia & Gido 2015 Mason et al. 2008, Villéger et al. 2010, Pease et al Brind Amour et al. 2011, Villéger et al Pease et al Mouillot et al. 2005, Mason et al. 2008, Villéger et al. 2010, Troia & Gido 2015 Manoeuvrability Villéger et al
215 Nutrient cycling Reproduction Pectoral fin aspect ratio Caudal peducle throtting Caudal fin aspect ratio Fins surface ratio Relative fins surface Affinity to flow velocity Nitrogent body concentration Phosphorous body concentration Nitrogent Phosphorous ratio Nitrogen excretion rate Phosphorous excretion rate N, P excretion ratio Maximum total length Mean length at maturation Maximum potential longevity Age at maturation Total fecundity per spawining season per female Relative to propopulsion and/ or manoeuvrability Endurance Villéger et al Endurance and manoeuvrability Type of swim Villéger et al Propulsion Villéger et al Endurance and manoeuvrability, type of swim Relative to digestibility of food and diet- Metabolism Life history strategies Wainwright, Bellwood & Westneat 2002, Bellwood et al. 2002, Mouillot et al. 2005, Villéger et al. 2010, Pease et al. 2015, Troia & Gido 2015 Mouillot et al. 2005, Mason et al. 2008, Villéger et al. 2010, Pease et al. 2015, Troia & Gido 2015 Logez, Pont & Ferreira 2010, Keck et al Vanni et al. 2002, McIntyre et al Mims et al. 2010, Keck et al. 2014, Pease et al. 2015, Troia & Gido 2015, Parks, Quist & Pierce 2016 Mims et al. 2010, Troia & Gido 2015, Parks, Quist & Pierce 2016 Mims et al. 2010, Keck et al. 2014, Pease et al. 2015, Troia & Gido 2015, Parks, Quist & Pierce 2016 Mims et al. 2010, Keck et al. 2014, Pease et al. 2015, Troia & Gido 2015, Parks, Quist & Pierce 2016 Mims et al. 2010, Keck et al. 2014, Parks, Quist & Pierce
216 Stress tolerance Behavior Mean diameter of egg fully yolked ovarian oocytes Degree of parental care based on placement of zygotes, parental protection and nutritional contribution Life history tactics (i.e. periodic, opportunistic, equilibrium strategists) Spawning substrate Mass of gonads Type of larval life Hypoxia tolerance Mims et al. 2010, Pease et al. 2015, Troia & Gido 2015, Parks, Quist & Pierce 2016 Mims et al. 2010, Parks, Quist & Pierce 2016 Miyazono et al. 2010, Parks, Quist & Pierce 2016 Logez, Pont & Ferreira 2010 Energy allocation Troia & Gido 2015 Parental care Keck et al Morphological adaptations Logez, Pont & Ferreira 2010, Brind Amour et al. 2011, Parks, Quist & Pierce 2016 Salinity tolerance Survival estimation Thermal tolerance Survival estimation Activity Temporal activity Brind Amour et al Migration Space occupancy Logez, Pont & Ferreira 2010, Brind Amour et al Social Relations with congeners and other species Leduc, da Silva & Rosenfeld
217 5.2 Στόχος του κεφαλαίου Στόχος του παρόντος κεφαλαίου είναι να εντοπίσει συσχετίσεις μεταξύ των περιβαλλοντικών χαρακτηριστικών και της λειτουργικής σύνθεσης των ιχθυοκοινοτήτων σε τοπικό επίπεδο και πιο συγκεκριμένα στο οικοσύστημα του ποταμού Λούρου. Ο ποταμός Λούρος αποτελεί ένα ενδιαφέρον οικοσύστημα για τη διερεύνηση της συσχέτισης των λειτουργικών χαρακτηριστικών των ειδών με περιβαλλοντικές παραμέτρους σε ένα ποτάμιο οικοσύστημα. Επιπλέον, στο παρόν κεφάλαιο μελετούνται οι διεργασίες μέσω των οποίων, τα λειτουργικά χαρακτηριστικά των ειδών καθορίζουν τις σχετικές θέσεις τους σε ένα οικοσύστημα. Εκτός από τον έλεγχο των συσχετίσεων μεταξύ των χαρακτηριστικών των ειδών με τα χαρακτηριστικά των ενδιαιτημάτων, χρησιμοποιήσαμε τη μοναδική μεθοδολογική προσέγγιση ελέγχου της στατιστικής σημασίας αυτών των συσχετίσεων που έχει προταθεί έως σήμερα. Εξετάσαμε τη σχέση μεταξύ της λειτουργικής ποικιλότητας και του αριθμού των ειδών και αναδείξαμε ποια από τα είδη του ποταμού Λούρου χρήζουν άμεσης προστασίας, έπειτα από εφαρμογή των προτεινόμενων δεικτών λειτουργικής μοναδικότητας. Τέλος, ελέγξαμε την επίδραση των δύο βασικών αντιτιθέμενων διεργασιών που αφορούν το περιβαλλοντικό φιλτράρισμα και τις βιοτικές αλληλεπιδράσεις στη διαμόρφωση των ιχθυοκοινοτήτων, με βάση τα λειτουργικά γνωρίσματα.των ειδών. Οι περισσότερες μελέτες στα ποτάμια οικοσυστήματα της Ελλάδας έχουν επικεντρωθεί σε συγκεκριμένα είδη ή στη σύνθεση των βιοκοινοτήτων σύμφωνα με την ταξινομική προσέγγιση. Στην παρούσα μελέτη προχωρήσαμε ένα βήμα παραπέρα εφαρμόζοντας ένα πολυεπίπεδο πλαίσιο μελέτης που συμπεριλαμβάνει και τα λειτουργικά χαρακτηριστικά. Η προσέγγιση που ακολουθείται στο παρόν κεφάλαιο μπορεί να αποτελέσει ένα ενιαίο πλαίσιο για τις στρατηγικές διατήρησης και προστασίας των ειδών, με βασική στόχευση την σύνδεση της προστασίας του αριθμού των ειδών, της λειτουργικής τους ποικιλότητας και των οικοσυστημικών διεργασιών εν γένει. 217
218 5.3 Υλικά και Μέθοδοι Κριτήρια επιλογής του οικοσυστήματος μελέτης Στην παρούσα μελέτη, επιλέχθηκε η διερεύνηση των προτύπων ποικιλότητας σε ένα ποτάμιο οικοσύστημα καθώς παρουσιάζει μια σειρά συγκριτικών πλεονεκτημάτων. Τα ποτάμια ανάμεσα στους διαφορετικούς τύπους οικοσυστημάτων παρουσιάζουν τις πιο αδρές περιβαλλοντικές αλλαγές. Επιπλέον, οι διεργασίες που διαμορφώνουν τη σύνθεση των βιοκοινοτήτων μπορούν να μελετηθούν τόσο σε χωρική και χρονική κλίμακα μέσα από την άμεση παρατήρηση και δειγματοληπτική προσπάθεια αφού είναι σαφέστατα πιο δύσκολο να εκτιμηθεί το εύρος των συνθηκών στις οποίες διαβιούν οι χερσαίοι και θαλάσσιοι ζωντανοί οργανισμοί. Επιπλέον, ορισμένα φυσιογραφικά χαρακτηριστικά των ποταμών (π.χ. πλάτος, βάθος, υπόστρωμα) είναι μετρήσιμα και συγκρίσιμα στις διάφορες χωρικές κλίμακες. Τα Μεσογειακά ποτάμια είναι από τα πιο ευάλωτα οικοσυστήματα σε παγκόσμια κλίμακα λόγω της μακράς ιστορίας των ανθρωπογενών επιδράσεων, όπως η υποβάθμιση των ενδιαιτημάτων και η ρύπανση των υδάτων τους παρά την αύξηση των εγκαταστάσεων επεξεργασίας λυμάτων κατά τις τελευταίες δεκαετίες (Sabater & Tockner 2010). Τα Μεσογειακά ποτάμια έχουν πολύ μεγάλη οικολογική αξία με σημαντικές μεταβολές του υδρολογικού τους καθεστώτος αντανακλώντας τα πρότυπα των κατακρημνίσεων και της θερμοκρασίας (Bonada & Resh 2013). Ως αποτέλεσμα, οι βιοκοινότητές τους υποβάλλονται σε εποχικές διαφοροποιήσεις εξαιτίας του κατακερματισμού και της συνδεσιμότητας των ενδιαιτημάτων, επηρεάζοντας επίσης τις λειτουργίες των οικοσυστημάτων (Hershkovitz & Gasith 2013). Όσον αφορά την ιχθυοπανίδα τους, τα ποτάμια της Μεσογείου αναγνωρίζονται ως «θερμά σημεία» της βιοποικιλότητας και του ενδημισμού (Myers et al. 2000). Παράλληλα, αποτελούν οικοσυστήματα με υψηλά ποσοστά, εκ προθέσεως και τυχαίας, εισαγωγής ξενικών ειδών. Χαρακτηρίζονται, λοιπόν, από τον υψηλότερο ρυθμό απώλειας ειδών σε παγκόσμιο επίπεδο (Moyle 1995, Marr et al. 2013), υψηλά ποσοστά ενδημισμού και μεγάλους αριθμούς ξενικών ειδών (Marr et al. 2010). 218
219 Στην παρούσα μελέτη, ο ποταμός Λούρος επιλέχθηκε ως σύστημα μελέτης επειδή είναι ένα πεδινό ομαλού πηγαίου τύπου ποτάμι. Αποτελεί ένα από τα λιγότερο μελετημένα οικοσυστήματα στην Ελλάδα, με μακρά ιστορία ανθρώπινης παρουσίας και παρέμβασης. Από οικολογική άποψη, έχει μεγάλη διαχειριστική αξία δεδομένου ότι ανήκει στην Ιόνια βιογεωγραφική περιοχή (Oikonomou, Leprieur & Leonardos 2014). Χαρακτηρίζεται από αυξημένη ποικιλότητα μεσογειακών ποτάμιων ενδιαιτημάτων, τα οποία φιλοξενούν εξελικτικά απομονωμένη ιχθυοπανίδα, η οποία χαρακτηρίζεται από χαμηλό αριθμό ειδών, αλλά ταυτόχρονα από υψηλά επίπεδα ενδημισμού και μεγάλο αριθμό απειλούμενων ειδών (Oikonomou et al. 2014) Περιοχή μελέτης: Ποταμός Λούρος Ο ποταμός Λούρος πηγάζει από το όρος Τόμαρος (ή Ολύτσικα, υψόμετρο 1976μ) του νομού Ιωαννίνων και εκβάλλει στον Αμβρακικό κόλπο στο νομό Πρέβεζας με κατεύθυνση ΒΒΔ-ΝΝΑ. Το συνολικό μήκος του ποταμού είναι 73 (km) ενώ η έκταση της λεκάνης απορροής του κυμαίνεται από 785 έως 961 (km 2 ) (Poulos et al. 2005, Υπουργείο Περιβάλλοντος Ενέργειας και Κλιματικής Αλλαγής 2013). Στη διαδρομή του ο ποταμός τροφοδοτείται από τις πηγές Καμπής και Χανόπουλου στην ανατολική πλευρά και τις πηγές Πριάλας και Σκάλας στη δυτική. Η μέση ροή είναι περίπου 19-28,6 m 3 /s (Konstantinou, Hela & Albanis 2006, Katsaounos et al. 2007, Πίνακας 5.2). Το 1954 κατασκευάστηκε από τη Δ.Ε.Η. στο χωριό Άγιος Γεώργιος, ταμιευτήρας χωρητικότητας 0,035 hm 3 και υδροηλεκτρικό φράγμα ενέργειας 10,3 MW, που τροφοδοτείται με νερό του ποταμού (Μαμάσης & Στεφανάκος 2010). Τα τελευταία χρόνια ο ταμιευτήρας έχει γίνει πολύ ρηχός εξαιτίας της ιζηματαπόθεσης. Το φράγμα αυτό είναι ουσιαστικά αδιαπέραστο από οποιοδήποτε ψάρι διότι στερείται ιχθυόσκαλων και ιχθυοδιαδρόμων. Μορφολογικά, το καρστικό σύστημα του Λούρου χαρακτηρίζεται από υψηλές επιμήκεις οροσειρές και στενές κοιλάδες, λόγω των γεωτεκτονικών και γεωλογικών συνθηκών της περιοχής και της λιθολογικής εναλλαγής μεταξύ των ασβεστόλιθων και της φλύσχης της Ιόνιας ζώνης (Fakiris et al. 2013). Ο Λούρος χαρακτηρίζεται από μικρές ποσότητες λεπτόκοκκου υλικού, κροκάλες και χαλίκια και η υδρογραφική του 219
220 λεκάνη αναπτύσσεται σε καρστικά πετρώματα από ασβεστόλιθους, δολομίτες και δευτερευόντως γύψων (Papadopoulou & Vriniotis 2007). Το βασικό γεωλογικό υπόστρωμα αποτελείται από αλλουβιακές αποθέσεις ασβεστιτικών και πυριτικών τριτογενών αποθέσεων καθώς και σκληρούς ασβεστόλιθους (Katsaounos et al. 2007). Αφετηρία των γεωµορφολογικών εξελίξεων της περιοχής, για τη σημερινή κατάληξη του ανάγλυφου, αποτέλεσε η έντονη γεωτεκτονική δραστηριότητα κατά το Τεταρτογενές και την εναλλαγή Πλειοκαίνου-Πλειστοκαίνου (βλέπε επίσης Σημείωμα 1). Στις αρχές του Πλειστοκαίνου σχηματίζεται ο παλαιοπόταμος των υδροδυναμικών συστημάτων των ποταμών Λούρου και Αράχθου στο βόρειο τμήμα του Αµβρακικού κόλπου, ο οποίος αναδύθηκε και απομονώθηκε από το Ιόνιο πριν από χρόνια, µε αποτέλεσμα τη δημιουργία των πεδινών εκτάσεων Άρτας- Πρέβεζας (Μερτζάνης 1992). Σύμφωνα με τον ίδιο συγγραφέα, η συμβολή αυτή των Λούρου και Αράχθου σε ενιαίο υδρογραφικό σύστημα δημιούργησε στη συνέχεια τα παλαιότερα εκτεταμένα δελταϊκά μέτωπα στις περιοχές της Κορωνησίας και της Σαλαώρας. Η ανύψωση των περιοχών του ρίσκου και του Ξηροβουνίου διαχώρισε τελείως τη ροή των δύο ποταμών (6000 χρόνια πριν) και δημιούργησε ξεχωριστά εκβολικά συστήματα ενώ οι εκβολές του Λούρου απωθήθηκαν όλο και περισσότερο προς τα δυτικά από τη μεγαλύτερη προσχωµατική ικανότητα του Αράχθου (Μερτζάνης 1992, Kapsimalis et al. 2005). Εντούτοις, στη σημερινή εποχή, το εκβολικό σύστημα του π. Λούρου ενώνεται με αυτό του π. Άραχθου και δημιουργούν ένα δέλτα λοβοειδούς μορφής με έκταση 350 km 2, το οποίο αποτελεί σύμπλεγμα πολλών υγροτόπων (Poulos et al. 2005). 220
221 Πίνακας 5.2. Φυσικά-γεωγραφικά χαρακτηριστικά του ποταμού Λούρου (από Poulos et al. 2005) Table 5.2. Physical-geographic characteristics of Louros river (after Poulos et al. 2005) River Louros Geomorphology Catchment area (km 2 ) 785 Maximum elevation (m) 2000 Deltaic plain area (km 2 ) 350 Type of river mouth Cuspate Maximum water depths in front of river mouth (m) 69 Lithology of catchment area Clastic (flysch, alluvial) sediment (%) 32,5 Carbonate rocks (%) 66,0 Metamorphic rocks (%) 1,5 Igneous rocks (%) 0 Climate Climatic variation Mean annual temperature ( C) 17,9 Mean annual precipitation (mm) 925 Water/sediment fluxes Mean annual water discharge (m 3 /s) 19 Mean annual suspended sediment flux (10 6 t) 0,8 Terrestrial Mediterranean to humid continental Ανθρωπογενείς δραστηριότητες και καθεστώς προστασίας στον ποταμό Λούρο Όπως έχει αναφερθεί και προηγουμένως, τα ποτάμια είναι ευάλωτα στις ανθρωπογενείς παρεμβάσεις. Πολλά από αυτά έχουν υποστεί αλλαγή των ενδιαιτημάτων τους από την εκτεταμένη κατασκευή φραγμάτων, καναλοποίηση, αποψίλωση των δασών, μεταφορά ιζήματος, ρύπανση και εισαγωγή ξενικών ειδών (Power et al. 1988, Κεφάλαιο 1). Από τα διαθέσιμα στοιχεία που συγκεντρώθηκαν για το Υδατικό Διαμέρισμα Ηπείρου, οι σημειακές πηγές ρύπανσης στον ποταμό Λούρο προέρχονται από δραστηριότητες του πρωτογενούς τομέα παραγωγής (πτηνο- κτηνοτροφία, ιχθυοκαλλιέργεια) και τη βιομηχανική δραστηριότητα, κυρίως 221
222 για την αξιοποίηση προϊόντων πρωτογενούς τομέα παραγωγής (Εικόνα 5.2). Πιο συγκεκριμένα, παρατηρείται υπερσυγκέντρωση των μονάδων ιχθυοκαλλιέργειας τύπου εσωτερικών υδάτων στον ποταμό Λούρο στην περιοχή Βουλιάστας- Μουσιωτίτσας. Σύμφωνα μάλιστα με τη Διεύθυνση Υδάτων Ηπείρου στην περιοχή αυτή έχουν γίνει ανεξέλεγκτες ανθρώπινες επεμβάσεις στις όχθες και στο ρου του ποταμού (διευθέτηση κοίτης, μικρά φράγματα) με αποτέλεσμα την εμφάνιση πλημμυρικών φαινομένων. Τα νερά του Λούρου χρησιμοποιούνται για τη λειτουργία και την υποστήριξη 23 μονάδων εντατικής εκτροφής πέστροφας οι οποίες καλύπτουν 34,120 τετραγωνικών μέτρων (m 2 ) υδάτινης επιφάνειας και παράγουν περίπου 547 τόνους (tn) της ιριδίζουσας πέστροφας (Oncorhynchus mykiss) ετησίως. Λόγω του μεγάλου αριθμού των μονάδων και της πυκνότητας των εκτρεφόμενων πληθυσμών επηρεάζεται η ποιότητα των νερών του ποταμού (ρυπαντές, διαφυγές εκτρεφόμενων οργανισμών, διασπορά παθογόνων οργανισμών). Σημαντική είναι και η πίεση από τους χώρους ανεξέλεγκτης διάθεσης απορριμμάτων που είναι διάσπαρτοι σε όλο το υδατικό διαμέρισμα και την απόρριψη σκουπιδιών κατά μήκος της λεκάνης απορροής του ποταμού Λούρου (Εικόνα 5.3) (Υπουργείο Περιβάλλοντος Ενέργειας και Κλιματικής Αλλαγής 2013). Για την αποτελεσματική προστασία του, ο ποταμός Λούρος και ο Αμβρακικός Κόλπος έχουν αναγνωρισθεί ως ευαίσθητες περιοχές με την ΚΥΑ 19661/1982/ (ΦΕΚ 1811/Β/ ). Επίσης, έχει θεσμοθετηθεί το Εθνικό Πάρκο Υγροτόπων Αμβρακικού (που περιλαμβάνει τις εκβολές Αράχθου και Λούρου) σύμφωνα με την ΚΥΑ 11989/08 (ΦΕΚ 123/Δ/ ) (GR ). 222
223 Εικόνα 5.2. Χάρτης των ανθρωπογενών πιέσεων στον ποταμό Λούρο (τροποποιημένο από Υπουργείο Περιβάλλοντος Ενέργειας και Κλιματικής Αλλαγής 2013) Figure 5.2. Map of anthropogenic pressures in Louros river (modified from Ministry of Environment, Energy and Climate Change 2013) 223
224 Εικόνα 5.3. Η ανεξέλεγκτη διάθεση απορριμμάτων είναι έντονη στη λεκάνη απορροής του ποταμού Λούρου Figure 5.3. The uncontrolled waste disposal is intense in Louros water basin Σταθμοί δειγματοληψίας Πριν ακόμα γίνει η επιλογή των σταθμών δειγματοληψίας, των θέσεων δηλαδή στις οποίες θα ακολουθούσε η συλλογή των ψαριών και η καταγραφή των διάφορων περιβαλλοντικών παραμέτρων, έγινε μια προκαταρκτική επισκόπηση της περιοχής, ώστε να εντοπιστούν τα πιθανά καταλληλότερα και αντιπροσωπευτικότερα σημεία δειγματοληψίας των ενδιαιτημάτων του ποταμού. Έτσι, η επιλογή των εννιά σταθμών δειγματοληψίας (Εικόνα 5.4, για την αναλυτική περιγραφή των σταθμών βλέπε Παράρτημα 4) έγινε με βάση τα παρακάτω κριτήρια: 1) Αντιπροσωπευτική ισοκατανομή σταθμών κατά μήκος των ποταμού και σε όλα τα τμήματά του (πηγή, άνω, μέσο, κάτω ρείθρο, άνω, μέσο και κάτω πόταμον και της εκβολής). Δηλαδή η μελέτη δεν εστιάστηκε μόνο σε ένα τμήμα ή δεν ήταν εντατικότερη στα βόρεια τμήματα από ότι στα νοτιότερα καθώς και πριν και μετά το φράγμα τoυ ποταμού. 224
 Καλή προσβασιμότητα σταθμών παρακολούθησης, ώστε να εξασφαλίζεται η ακριβής επαναληψιμότητα της διαδικασίας σε καθορισμένο τόπο και χρόνο, ανεξαρτήτως καιρικών συνθηκών.")
225 2) Κάλυψη όλου του φάσματος των ενδιαιτημάτων, δηλαδή ενδιαιτήματα περισσότερο ή λιγότερο προφυλαγμένα, με μικρότερη ή μεγαλύτερη ροή υδάτων, με πλούσια ή μέτρια υδρόβια βλάστηση, με χαρακτηριστικούς τύπους πυθμένων κτλ. 3) Καλή προσβασιμότητα σταθμών παρακολούθησης, ώστε να εξασφαλίζεται η ακριβής επαναληψιμότητα της διαδικασίας σε καθορισμένο τόπο και χρόνο, ανεξαρτήτως καιρικών συνθηκών. 4) Παρουσία σημειακών πηγών πιέσεων για την ιχθυοπανίδα της περιοχής, όπως φράγματα, μονάδες υδατοκαλλιέργειας, εκτάσεις εντατικής καλλιέργειας και άλλες ανθρωπογενείς δραστηριότητες. Εικόνα 5.4. Xάρτης των σταθμών δειγματοληψίας στον ποταμό Λούρο Figure 5.4. Map of sampling stations in Louros river 225
 σύμφωνα με τις οδηγίες που καθορίζονται από το CEN (2003).")
226 5.3.4 Μεθοδολογία συλλογής ιχθυολογικού υλικού Οι δειγματοληψίες πραγματοποιήθηκαν κατά τη χρονική περίοδο από το Δεκέμβριο του 2011 έως το Νοέμβριο του 2012 σε διμηνιαία βάση. Η συλλογή των ψαριών έγινε με τη χρήση ηλεκτραλιείας (Μοντέλο: HansGrassl. Τύπος IG 200/2) σύμφωνα με τις οδηγίες που καθορίζονται από το CEN (2003). Το πρωτόκολλο αυτό αποτελεί μία από τις πλέον διαδεδομένες, αποτελεσματικές και οικονομικές μεθόδους συλλογής ψαριών (CEN 2003). Με τον όρο ηλεκτραλιεία ορίζεται η αξιοποίηση ηλεκτρικού πεδίου για την σύλληψη ψαριών. Με την ηλεκτραλιεία (υπό κανονικές συνθήκες και φυσιολογική χρήση) δε θανατώνονται τα άτομα αλλά αναισθητοποιούνται. Αυτό έχει ως αποτέλεσμα να παρέχεται η δυνατότητα καταγραφής και μέτρησης των βιολογικών χαρακτηριστικών τους, ενώ στη συνέχεια και μετά την ανάνηψη τους, τα άτομα μπορούν να απελευθερωθούν. Η ηλεκτραλιεία βασίζεται στον σχηματισμό ενός ηλεκτρικού πεδίου στο νερό, που προκαλείται λόγω της διαφοράς δυναμικού Εικόνα 5.5. Απεικόνιση μεθόδου συλλογής ψαριών με ηλεκτραλιεία (Scottish Fisheries Co-ordination Centre, 2007) Figure 5.5. Demonstration of the method of electrofishing (Scottish Fisheries Co-ordination Centre, 2007) από τα ηλεκτρόδια, την άνοδο (+) και την κάθοδο (-). Η αποτελεσματικότητα της μεθόδου επηρεάζεται από διάφορους παράγοντες, όπως η αγωγιμότητα του νερού, πέραν της έντασης του παραγόμενου ρεύματος και της διαφοράς δυναμικού. Όταν η τιμή της αγωγιμότητας είναι χαμηλή (χαμηλότερη από αυτή σώματος του ψαριού π.χ. ύδατα ποταμών και λιμνών), τότε το ηλεκτρικό ρεύμα τείνει να διατρέχει το νερό, διαπερνά το σώμα του ατόμου, καθώς η διαφορά δυναμικού είναι υπεύθυνη για τη δημιουργία του ηλεκτρικού 226
227 φαινόμενου. Αντίθετα, όταν η αγωγιμότητα του νερού υψηλή (υψηλότερη από αυτή του σώματος π.χ. υφάλμυρα και αλμυρά νερά λιμνοθαλασσών και θάλασσας), τότε το ρεύμα τείνει να διατρέχει το νερό γύρω από το ψάρι (IMBRIW 2013). Το μήκος του κάθε σταθμού, στο οποίο εφαρμόστηκε το παραπάνω αλιευτικό εργαλείο ήταν περίπου 100m, ανάλογα με το πλάτος του ποταμού σε κάθε σταθμό. Η απόσταση αυτή θεωρείται αρκετά ικανοποιητική για τη συλλογή αντιπροσωπευτικού δείγματος ψαριών (Meador, Cuffney & Gurtz 1993). Η μεθοδολογία συλλογής με την ηλεκτραλιεία περιελάμβανε ένα πέρασμα αντίθετα στη ροή του ποταμού, σε πορεία μαιανδρική (Εικόνα 5.5) με σκοπό να καλυφθούν κατά το δυνατό τόσο οι όχθες όσο και το κέντρο του ποταμού και μόνο σε εκείνα τα σημεία που η χρήση της συσκευής ήταν ασφαλής για το χειριστή (<1.20 m βάθος). Πίσω από το χειριστή ακολουθούσε δεύτερο άτομο με απόχη, ώστε να συλλαμβάνει τα ψάρια που παρασύρονταν από τη ροή του ποταμού. Στην παρόχθια ζώνη χρησιμοποιήθηκε απόχη (με πλαίσιο διαστάσεων 30 cm x 35 cm και δίχτυ με άνοιγμα ματιού 2 mm) ώστε να συλλεχθούν μικρόσωμα είδη ή νεαρά άτομα ειδών που κρύβονται στην υδρόβια βλάστηση ή στον πυθμένα. Η καταμέτρηση και η αναγνώριση των ειδών έγινε κρατώντας τα ψάρια εντός του σάκου του δικτυού ώστε να διατηρούνται ζωντανά. Αμέσως μετά την καταμέτρηση κάποια από αυτά απελευθερώθηκαν και τα υπόλοιπα που ήταν αντιπροσωπευτικά τοποθετήθηκαν σε ειδικά ψυγεία, μεταφέρθηκαν στο εργαστήριο όπου και διατηρήθηκαν σε βαθιά ψύξη. Αξίζει να σημειωθεί ότι στους σταθμούς 7, 8, 9 η πρόσβαση αλλά και μέρος της διαδικασίας της δειγματοληψίας πραγματοποιήθηκε με χρήση βάρκας την οποία μας παραχώρησε ο Φορέας Διαχείρισης Υγροτόπων του Αμβρακικού. 227
228 5.3.5 Περιβαλλοντικοί περιγραφείς Μετρήσεις περιβαλλοντικών παραμέτρων in situ Κατά τη διάρκεια της δειγματοληψίας πραγματοποιήθηκαν φυσικoχημικές μετρήσεις στο νερό (Πίνακας 5.3). Σε κάθε δειγματοληψία λήφθηκαν οι τιμές της θερμοκρασίας του νερού (Temp) και του διαλυμένου στο νερό οξυγόνου (Ox) in situ, πάντοτε κοντά στη μέση του ποταμού λίγο κάτω από την επιφάνεια του νερού, στην έναρξη της δειγματοληψίας και πριν τη συλλογή των ψαριών, με στόχο να διατηρείται το ενδιαίτημα όσο το δυνατό αδιατάρακτο. Οι μετρήσεις πραγματοποιήθηκαν με φορητά όργανα μέτρησης (Μοντέλο YSI Τύποι 550 και 63) (Παράρτημα 5). Καταγράφηκαν επίσης χαρακτηριστικά του ενδιαιτήματος όπως η απόσταση από την πηγή (Dist), το υψόμετρο (Ele), το πλάτος (W), το μέσο βάθος (Depth) καθώς και το ποσοστό του κάθε τύπου του υποστρώματος σε κάθε σταθμό Δείγματα νερού Από κάθε θέση δειγματοληψίας συλλέχθηκαν δείγματα νερού από την επιφάνεια, σε ειδικές φιάλες του ενός λίτρου (1 lt), οι οποίες διατηρήθηκαν σε φορητό ψυγείο. Η θερμοκρασία των δειγμάτων διατηρήθηκε σε χαμηλά επίπεδα (5-10 C) με τη χρήση παγοκύστεων. Στη συνέχεια, τα δείγματα μεταφέρθηκαν στο εργαστήριο, όπου και πραγματοποιήθηκε ο ποσοτικός προσδιορισμός των θρεπτικών ανόργανων ενώσεων που περιέχονταν στα δείγματα του νερού: νιτρωδών (NO2 - ) 25 (Nitri), νιτρικών (NO3 - ) 26 (Nitra), αμμωνιακών (NH4 + ) 27 (Am) και ορθοφωσφορικών ιόντων (PO4 - ) 28 (Phos) 25 Ο προσδιορισμός της συγκέντρωσης των νιτρωδών ιόντων, πραγματοποιήθηκε με τη βοήθεια της μεθόδου διαζώτωσης. Σύμφωνα με αυτή τη μέθοδο, τα νιτρώδη ιόντα αντιδρούν με το σουλφανιλικό οξύ σχηματίζοντας ένα διαζωνιακό άλας. Στη συνέχεια λαμβάνει χώρα αντίδραση του διαζωνιακού άλατος με χρωμοτροπικό οξύ, σχηματίζοντας ένα έγχρωμο σύμπλοκο. Η ένταση του χρώματος μετράται στα 507 nm και είναι ανάλογη της συγκέντρωσης των νιτρωδών ιόντων στο δείγμα (APHA, AWWA & WEF 2012).Το εύρος των συγκεντρώσεων των νιτρωδών ιόντων που ανιχνεύει η συγκεκριμένη μέθοδος είναι 0,002-0,300 mg/ L 26 Ο προσδιορισμός της συγκέντρωσης των νιτρικών ιόντων πραγματοποιήθηκε βάσει της μεθόδου αναγωγής του καδμίου. Σύμφωνα με αυτή τη μέθοδο, το κάδμιο, χρησιμοποιείται για την αναγωγή των νιτρικών ιόντων σε νιτρώδη. Στη συνέχεια, τα νιτρώδη ιόντα αντιδρούν με το σουλφανιλικό οξύ 228
229 και της Χλωροφύλλης-α (Chl) σύμφωνα με τη μεθοδολογία που περιγράφεται στο εγχειρίδιο Standard Methods for the Examination of Water and Wastewater (APHA, AWWA & WEF 2012) (Πίνακας 5.3). Τα δείγματα διηθήθηκαν με αντλία κενού (συσκευή διήθησης νερού KNF LAB) και με τη χρήση ηθμών (microfiber) (Whatman GF/C/45μm). Από κάθε δείγμα λήφθηκαν υποδείγματα 100 ml, τα οποία χρησιμοποιήθηκαν για τον προσδιορισμό των θρεπτικών αλάτων. Για τη μέτρηση της χλωροφύλλης στα δείγματα έγινε εκχύλιση της από τον ηθμό σε διάλυμα ακετόνης συγκέντρωσης 90% για 24 ώρες. Μετά την πάροδο των 24 ωρών, έγινε φωτομέτρηση σε 4 μήκη κύματος (750 nm, 664 nm, 647 nm και 630 nm). Η τελική συγκέντρωση της χλωροφύλλης α (mg/l) προσδιορίστηκε σύμφωνα με την εξίσωση: Ca* Vex Chl a = (7) όπου Vsample [Vex= ο όγκος της κυψελίδας] [Vsample=ο όγκος δείγματος που διηθήθηκε] και με την εξίσωση των Jeffrey & Humphrey (1975): ( 11,85* 664) ( 1,54* 647) ( 0, 08* 630) Chl a = OD OD OD (8) όπου [OD664 = απορρόφηση 664nm- - απορρόφηση 750nm] [OD647 = απορρόφηση 667nm-απορρόφηση 750 nm] σχηματίζοντας ένα ενδιάμεσο διαζωνικό άλας. Το διαζωνικό άλας στη συνέχεια σχηματίζει ένα έγχρωμο σύμπλοκο με το χρωμοτροπικό οξύ. Η ένταση του χρώματος μετράται στα 430 nm και είναι ανάλογη με τη συγκέντρωση των νιτρικών ιόντων στο δείγμα (APHA, AWWA & WEF 2012). Το εύρος των συγκεντρώσεων των νιτρικών ιόντων που ανιχνεύει η συγκεκριμένη μέθοδος είναι 0,1-10,0 mg/ L 27 Για τον προσδιορισμό της συγκέντρωσης των αμμωνιακών ιόντων στα δείγματα του νερού χρησιμοποιήθηκε η μέθοδος Nessler. Σύμφωνα με αυτή τη μέθοδο, το αντιδραστήριο Nessler (K2HgI4) αντιδρά υπό ισχυρά αλκαλικές συνθήκες με την αμμωνία, σχηματίζοντας έγχρωμα προϊόντα. Η ένταση του χρώματος μετράται στα 425 nm και είναι ανάλογη με τη συγκέντρωση της αμμωνίας στο δείγμα (APHA, AWWA & WEF 2012). Το εύρος των συγκεντρώσεων των αμμωνιακών ιόντων που ανιχνεύει η συγκεκριμένη μέθοδος είναι 0,02-2,5 mg/l 28 Η μέθοδος προσδιορισμού των ορθοφωσφορικών ιόντων του νερού στηρίζεται στην συμπλοκοποίηση των φωσφορικών ιόντων με μολυβδαινικά ιόντα σε όξινες συνθήκες. Παρουσία ασκορβικού οξέος λαμβάνει χώρα αναγωγή του σχηματιζόμενου συμπλόκου, δίνοντας στο διάλυμα μπλε χρωματισμό. Η ένταση του χρώματος, η οποία είναι ανάλογη με τη συγκέντρωση του δείγματος σε φώσφορο, μετράται φασματοφωτομετρικά σε μήκος κύματος 890 nm (APHA, AWWA & WEF 2012). Το εύρος των συγκεντρώσεων των ορθοφωσφορικών ιόντων που ανιχνεύει η συγκεκριμένη μέθοδος είναι 0,02-2,5 mg/ L 229
230 [OD630 = απορρόφηση 630 nm - απορρόφηση 750 nm] Για τις μετρήσεις χρησιμοποιήθηκε το φασματοφωτόμετρο HACH DR/2400. Πίνακας 5.3. Περιβαλλοντικές μεταβλητές τοπικής κλίμακας που μετρήθηκαν στον ποταμό Λούρο Table 5.3. Local-scale environmental variables measured at Louros river Environmental variable Distance from source (km) Mean Depth (m) Elevation (m) Width (m) Rocks- Pebbles (>1.6 cm) (%) Substrate thick sand or gravel (2-16 mm) (%) Substrate fine sand ( mm) (%) Substrate mud or silt (<0.06 mm) (%) Dissolved Oxygen (mg/l) TH20 ( O C) Nitrate (mg/l) Nitrite (mg/l) Orthophosphate (mg/l) Ammonia (mg/l) Chlorophyll-a (mg/l) Code Dist Depth Ele W Rocks Thick_sand Fine_sand Mud Ox Temp Nitra Nitri Phos Am Chl 230
 (Παράρτημα 6).")
231 5.3.6 Εργαστηριακή επεξεργασία ιχθυολογικού υλικού Η επεξεργασία του βιολογικού υλικού πραγματοποιήθηκε στο εργαστήριο μετά την απόψυξή του. Αφού διαχωρίστηκαν ανά σταθμό μελέτης, τα άτομα προσδιορίστηκαν σε επίπεδο είδους με τη χρήση ειδικών κλειδών (σύμφωνα με τους Kottelat & Freyhof 2007) (Παράρτημα 6). Κατόπιν, τοποθετήθηκαν με το ρύγχος κατευθυνόμενο προς το αριστερό χέρι του παρατηρητή και φωτογραφήθηκαν με ψηφιακή φωτογραφική μηχανή (NIKON S9300). Οι φωτογραφίες εισήχθησαν στο πρόγραμμα ανάλυσης εικόνας ImageJ (έκδοση 1.47) και σε κάθε άτομο λήφθηκαν 14 μορφολογικές μετρήσεις, όπως φαίνεται στην Εικόνα 5.6 ακολουθώντας το καινοτόμο πρωτόκολλο των Villéger et al. (2010). Οι μετρήσεις που σχετίζονται με τα πτερύγια λήφθηκαν στη μέγιστη έκτασή τους. Εικόνα 5.6. Τα μορφολογικά χαρακτηριστικά που μετρήθηκαν σε άτομα ψαριών και που χρησιμοποιήθηκαν στις αναλύσεις του λειτουργικού χαρακτηρισμού των ειδών (τροποιημένο σκίτσο από Barbieri et al. 2015) Figure 5.6. Mophological traits measured on fish individuals used in the analyses of functional characterization of fish species (the sketch is modified from Barbieri et al. 2015) 231
232 Επιπρόσθετα, καταγράφηκαν το πάχος του σώματος και το άνοιγμα του στόματος (κάθετο και οριζόντιο) (Εικόνα 5.6) με τη χρήση του ίδιου ψηφιακού παχυμέτρου (ακρίβειας ±1 mm), για να αποφευχθούν τυχόν σφάλματα. Τα άτομα ζυγίστηκαν με χρήση ζυγού ακριβείας (0,001 g). Συνολικά, όλες οι ανωτέρω 18 μετρήσεις οδήγησαν στην εκτίμηση 13 οικο-μορφολογικών χαρακτηριστικών για κάθε άτομο με σκοπό να εκτιμηθεί η λειτουργική ποικιλότητα μέσω της σύλληψης τροφής και της μετακίνησης (π.χ. Winemiller 1991, Πίνακας 5.4). Τα δείγματα, αφού μετρήθηκαν, τοποθετήθηκαν σε δοχεία με διάλυμα φορμόλης 4% για περαιτέρω συντήρηση. Πίνακας 5.4. Κατάλογος των 13 οικο-μορφολογικών χαρακτηριστικών, της κωδικής ονομασίας, των μεθόδων υπολογισμού και της οικολογικής τους ερμηνείας (τροποποιημένο από Villéger et al. 2010) Table 5.4. List of the 13 ecomorphological traits, their code, their calculation methods and the ecological meaning associated with them (modified from Villéger et al. 2010) Functional trait Code Formula Εcological meaning Mw x Md Relative to Oral gape surface Gsur maximum prey size Bw x Bd or ability to water filtering Md Relative to prey Oral gape shape Gsh shape and food Mw acquisition Mo Relative to position Oral gape position Gpos of preys in the Hd water column Eye size Es Ed Relative to prey Hd detection Eh Relative to position Eye position Epos in the water Hd column Bd Relative to position Body transversal shape Bsh in the water Bw column and hydrodynamism ln((π/4 x Bw x Bd)+1) Relative to mass Body transversal surface Bsur distribution along (ln Mass +1) the body and hydrodynamism PFi Relative to Pectoral fin position Pf maneuverability PFb and position in the water column 232
233 Aspect ratio of the pectoral fin Caudal peduncle throttling Aspect ratio of the caudal fin Fins surface ratio Fins surface to body size ratio Rpf Throt Rcf Fsur Frat PFl 2 Relative to PFs propulsion and/or maneuverability CFd Relative to CPd swimming endurance CFd Relative to endurance, CFs acceleration and/or maneuverability 2 x PFs Relative to the swimming type CFs (pectoral or caudal fin propulsion) (2 X PFs) + CFs Relative to endurance, π/4 x Bw x Bd acceleration and/or maneuverability Ανάλυση δεδομένων λειτουργικής ποικιλότητας Για την ανάλυση των δεδομένων της λειτουργικής ποικιλότητας, μόνο πρωτογενή είδη του ποταμού Λούρου εξετάστηκαν ώστε η λεκάνη απορροής να θεωρείται ως ένα κλειστό σύστημα (βλέπε Hugueny, Oberdorff & Tedesco 2010). Κατασκευάστηκαν και χρησιμοποιήθηκαν τρεις πίνακες, ένας πίνακας των περιβαλλοντικών παραμέτρων που χαρακτηρίζουν κάθε σταθμό (Πίνακας R), ένας πίνακας της αφθονίας των ειδών σε κάθε σταθμό (Πίνακας L) και ένας πίνακας των λειτουργικών χαρακτηριστικών των ειδών (Πίνακας Q) (Εικόνα 5.7). Οι μετρήσεις των λειτουργικών χαρακτηριστικών κάθε είδους προέκυψαν από το μέσο όρο των τιμών των χαρακτηριστικών όλων των ενήλικων ατόμων κάνοντας την παραδοχή ότι η ενδοειδική λειτουργική ποικιλότητα είναι μικρότερη από την διαειδική («intraspesific functional diversity < interspesific functional diversity»). Εντούτοις, εδώ θα πρέπει να τονίσουμε πως ο αριθμός των ατόμων των ειδών διέφερε κατά πολύ από το σύνηθες δείγμα αναλύσεων λειτουργικής ποικιλότητας (n 10), αφού για τον προσδιορισμό των λειτουργικών χαρακτηριστικών μετρήθηκαν συνολικά 785 άτομα που ανήκουν σε δέκα είδη (mean: 78,5, sd: 92, min: 6, max: 277) (Παράρτημα 6, Πίνακας 1) και ως αποτέλεσμα τα δεδομένα των 233
234 λειτουργικών χαρακτηριστικών θεωρούνται αντιπροσωπευτικά των πληθυσμών των ειδών. Επίσης, είναι σημαντικό να αναφερθεί πως ενώ οι περισσότερες μελέτες λειτουργικής ποικιλότητας των ιχθυοκοινοτήτων χρησιμοποιούν κατηγορικά λειτουργικά χαρακτηριστικά ή λειτουργικές ομάδες, στην παρούσα εργασία τα λειτουργικά χαρακτηριστικά είναι ποσοτικά και συνεχή ώστε να επιτρέπουν τον υπολογισμό πολυδιάστατων δεικτών λειτουργικής ποικιλότητας (π.χ. Villéger, Mason & Mouillot 2008, Laliberté & Legendre 2010). Οι τιμές των περιβαλλοντικών δεδομένων (Πίνακας R) και των λειτουργικών χαρακτηριστικών (Πίνακας Q) λογαριθμήθηκαν (log10) πριν από κάθε ανάλυση ώστε να επιτευχθεί η ομοιογένεια της διασποράς (ομοσκεδαστικότητα). Οι αφθονίες των ειδών (Πίνακας L) μετασχηματίστηκαν χρησιμοποιώντας την τετραγωνική ρίζα των τιμών ώστε να ενισχυθεί η συμβολή των σπάνιων ειδών και να μειωθεί η επιρροή των κυρίαρχων ειδών (Clarke & Warwick 1994) Ανάλυση κύριων συνιστωσών για την απεικόνιση της περιβαλλοντικής διαβάθμισης και του λειτουργικού χώρου Η πρώτη ανάλυση που εφαρμόσαμε ήταν η ανάλυση κύριων συνιστωσών (PCA) που ανήκει στην κατηγορία των πολυμεταβλητών στατιστικών αναλύσεων ταξινόμησης («Ordination»). Η ταξινόμηση περιλαμβάνει τεχνικές των οποίων η εφαρμογή σε οικολογικά δεδομένα έχει ως αποτέλεσμα τη δημιουργία μιας γραφικής παρουσίασης (συνήθως διαγραμμάτων διασποράς δύο ή το πολύ τριών διαστάσεων), πάνω στην οποία τοποθετούνται αντικείμενα με τέτοιο τρόπο ώστε η μεταξύ τους απόσταση να αντικατοπτρίζει την ομοιότητα ή ανομοιότητα τους. Μεταξύ των διαφορετικών τεχνικών ταξινόμησης, η ανάλυση κύριων συνιστωσών είναι η παλαιότερη, η απλούστερη και χρησιμοποιείται ευρέως εξαιτίας της επιτυχημένης απόδοσης των δεδομένων. Στόχος της ανάλυσης είναι η διαπλοκή των δεδομένων ώστε να παραχθούν άξονες οι οποίοι αποδίδουν τη μεταβλητότητα των δεδομένων και ελαχιστοποιούν 234
235 την απώλεια της πληροφορίας λόγω της μείωσης των αξόνων απεικόνισης των αντικειμένων. Η ανάλυση υπολογίζει ένα νέο σύστημα συντεταγμένων, στο οποίο ο πρώτος άξονας (η πρώτη κύρια συνιστώσα, PC1) μεγιστοποιεί τη διακύμανση των στοιχείων κατά μήκος του άξονα (δηλαδή αντιπροσωπεύει το μεγαλύτερο μέρος της μεταβλητότητας των δεδομένων). Η δεύτερη κύρια συνιστώσα (PC2) είναι ορθογώνια ως προς την πρώτη και έχει τη δεύτερη μέγιστη διακύμανση (το οποίο σημαίνει ότι οι πρώτες κύριες συνιστώσες είναι ασυσχέτιστες μεταξύ τους), και ούτω καθεξής. Ο αριθμός των κύριων συνιστωσών είναι μικρότερος ή το πολύ ίσος με τον αριθμό των αρχικών μεταβλητών και συνήθως επιλέγονται οι πρώτοι δύο άξονες που αποδίδουν ικανοποιητικά τη μεταβλητότητα των δεδομένων. Με βάση αυτές τις δύο πρώτες συνιστώσες, οι δειγματοληπτικές μονάδες τακτοποιούνται έτσι ώστε οι σχετικές τους θέσεις στους άξονες και μεταξύ τους να αντανακλούν τη μέγιστη πληροφορία για τις οικολογικές τους ομοιότητες. Με αυτό τον τρόπο δύναται να αποτυπωθούν εκείνοι οι περιβαλλοντικοί παράγοντες που χαρακτηρίζουν τους σταθμούς από τους οποίους λήφθηκαν τα δείγματα. Στη συγκεκριμένη περίπτωση και σύμφωνα με αυτή τη μέθοδο, δίνεται η δυνατότητα να απεικονιστούν οι περιβαλλοντικές διαβαθμίσεις στον ποταμό Λούρο χρησιμοποιώντας τους πρώτους δύο άξονες (PC1, PC2) της μήτρας συσχέτισης. Το ίδιο επιτυγχάνεται χρησιμοποιώντας τα λειτουργικά χαρακτηριστικά ώστε να αποτυπωθεί ο λειτουργικός χώρος («functional space») των ειδών στο δισδιάστατο επίπεδο Επιλογή των κυριότερων περιβαλλοντικών παραμέτρων και των οικο-μορφολογικών χαρακτηριστικών για τις αναλύσεις λειτουργικής ποικιλότητας Το πρώτο βήμα για την επιλογή των περιβαλλοντικών παραμέτρων και των οικομορφολογικών χαρακτηριστικών για τις αναλύσεις λειτουργικής ποικιλότητας ήταν ο έλεγχος της συγγραμικότητας των μεταβλητών σύμφωνα με το συντελεστή ιεράρχησης Spearman. Η συσχέτιση κατά Spearman είναι ένα μη-παραμετρικό μέτρο της στατιστικής εξάρτησης μεταξύ κάθε ζεύγους μεταβλητών. Τα αναλυτικά δεδομένα των λειτουργικών χαρακτηριστικών για το σύνολο των ατόμων κάθε 235
236 είδους (και όχι ο μέσος όρος τους ανά είδος) χρησιμοποιήθηκαν για τον υπολογισμό της μήτρας συσχετίσεων και ελέγχθηκε κατά πόσο τα λειτουργικά χαρακτηριστικά συσχετίζονται στατιστικά σημαντικά. Η ίδια μεθοδολογία ακολουθήθηκε και για τα περιβαλλοντικά χαρακτηριστικά. Πριν από την ανάλυση συσχέτισης κατά Spearman οι μεταβλητές λογαριθμήκαν (log10) για σταθεροποίηση της διασποράς των δεδομένων. Στη συνέχεια, προκειμένου να διαπιστωθεί ποιες είναι οι στατιστικά σημαντικότερες περιβαλλοντικές παράμετροι που διαμορφώνουν τη σύνθεση και την αφθονία των ειδών σε χωρική κλίμακα χρησιμοποιήθηκε η πολυμεταβλητή ανάλυση διακύμανσης με μετάθεση (permutational manova ή αλλιώς permanova) (Anderson 2001). Η ανάλυση αυτή αποτελεί προέκταση της απλής, μονομεταβλητής ανάλυσης διακύμανσης αφού περιλαμβάνει περισσότερες από μία εξαρτημένες γραμμικές μεταβλητές και χρησιμοποιεί πίνακα αποστάσεων μεταξύ των ζευγών των δειγμάτων των βιοκοινοτήτων ως ανεξάρτητη μεταβλητή. Η συγκεκριμένη μεθοδολογία είναι η πλέον χρησιμοποιούμενη σε αντίστοιχες μελέτες γιατί λαμβάνει υπόψη της την αλληλοεξάρτηση ανάμεσα στα πληθυσμιακά μεγέθη των ειδών που συγκροτούν τις ιχθυοκοινότητες. Για τη συγκεκριμένη ανάλυση, επιλέχθηκε ο δείκτης ανομοιότητας των Bray & Curtis (1957) μεταξύ των αφθονιών των ιχθυοκοινοτήτων των σταθμών δειγματοληψίας. Πριν τον υπολογισμό του δείκτη ανομοιότητας των Bray- Curtis, τα δεδομένα αφθονίας τροποποιήθηκαν κατάλληλα, ώστε να μειωθεί η επίδραση των άφθονων ειδών και να αυξηθεί η επίδραση των σπάνιων. Πιο συγκεκριμένα, χρησιμοποιήθηκε η τετραγωνική ρίζα της αφθονίας (Clarke & Warwick 1994). Για την πολυμεταβλητή ανάλυση διακύμανσης με μετάθεση ο αριθμός των επαναλήψεων των μεταθέσεων που ορίστηκε για τη δοκιμασία F ήταν Αναλύσεις RLQ και fourthcorner Για να διερευνηθεί η υπόθεση ότι τα λειτουργικά χαρακτηριστικά (ή κάποια από αυτά) επιτρέπουν σε ένα είδος να επιβιώσει κάτω από συγκεκριμένες 236
237 περιβαλλοντικές συνθήκες (Townsend & Hildrew 1994), η μοναδική στατιστική ανάλυση που έχει προταθεί μέχρι σήμερα είναι η πολυμεταβλητή ανάλυση RLQ. Η ανάλυση χρησιμοποιήθηκε για την απεικόνιση των σχέσεων μεταξύ των περιβαλλοντικών μεταβλητών και των λειτουργικών χαρακτηριστικών των ιχθυοκοινοτήτων, συνοψίζοντας γραφικά την κύρια δομή των τριών μητρών R, L και Q. Προηγούμενες μελέτες έχουν δείξει ότι η ανάλυση RLQ είναι ένα χρήσιμο εργαλείο για την αποκάλυψη των σχέσεων των λειτουργικών χαρακτηριστικών με τις περιβαλλοντικές συνθήκες των βιοκοινοτήτων (π.χ. Oldeland, Wesuls & Jürgens 2012, Spitz, Ridoux & Brind'Amour 2014, Pease et al. 2015). Η ανάλυση RLQ (Dolédec et al. 1996) είναι μία προέκταση των αναλύσεων συναδράνειας («co-inertia analyses») και εκτελεί ταυτόχρονα ταξινόμηση στους τρεις πίνακες αποκαλύπτοντας τη δομή των πινάκων (Εικόνα 5.7): 1) των περιβαλλοντικών μεταβλητών (R) μέσω της ταξινόμησης Hill και Smith («Hill and Smith ordination») (Hill & Smith 1976) για μεικτού τύπου δεδομένα (δηλαδή για ποσοτικές και κατηγορικές μεταβλητές) 2) των λειτουργικών χαρακτηριστικών των ειδών (Q) μέσω της ανάλυσης κύριων συνιστωσών 3) και των συσχετίσεων τους, μέσω της παραγοντικής ανάλυσης των αντιστοιχιών («Correspondence analysis») της αφθονίας των ειδών στους σταθμούς δειγματοληψίας (L) 237
238 Εικόνα 5.7. Εννοιολογικό διάγραμμα των σταδίων εφαρμογής των αναλύσεων RLQ και fourth-corner. Η ανάλυση RLQ χρησιμοποιήθηκε για την οικολογική ομαδοποίηση και την ερμηνεία των αποτελεσμάτων. Η μέθοδος fourth-corner χρησιμοποιήθηκε για να ελεγχθεί στατιστικά κάθε συνδυασμός των λειτουργικών χαρακτηριστικών με τις περιβαλλοντικές μεταβλητές (τροποποιημένο από Brind'Amour et al. 2011) Figure 5.7. Conceptual diagram of analysis steps of the RLQ and fourth- corner approaches. RLQ analysis was used to facilitate ecological grouping and interpretation of the results. The fourth-corner method was used to test statistically each combination of functional traits and environmental variables. (modified from Brind'Amour et al. 2011) Εδώ, αξίζει να αναφερθεί ότι η δοκιμασία rand («randtest») (Dray & Dufour 2007) εφαρμόστηκε για το στατιστικό έλεγχο, με βάση τη συνολική συναδράνεια της ανάλυσης RLQ, της σημαντικότητας των συσχετίσεων μεταξύ των R και Q πινάκων. Η στατιστική σημαντικότητα των συσχετίσεων αξιολογήθηκε μέσω δύο σταδίων τυχαιοποίησης με 999 µεταθέσεις («permutations»), όπου πρώτα ολόκληρες οι σειρές και, στη συνέχεια, ολόκληρες οι στήλες του πίνακα L τυχαιοποιούνται. Η μετάθεση των σειρών του πίνακα L ελέγχει την ανεξαρτησία των αφθονιών των βιοκοινοτήτων από τα περιβαλλοντικά χαρακτηριστικά του ενδιαιτήματος (Μοντέλο 2). Η μετάθεση των στηλών ελέγχει τη μηδενική υπόθεση ότι οι αφθονίες των ειδών είναι ανεξάρτητες από τα λειτουργικά χαρακτηριστικά τους (Μοντέλο 4) (Dray & Legendre 2008). 238
239 Στη συνέχεια, εφαρμόστηκε η μέθοδος fourth-corner (Legendre, Galzin & Harmelin- Vivien 1997) που χρησιμοποιείται συμπληρωματικά με την ανάλυση RQL. H fourth-corner χρησιμοποιεί τους τρεις πίνακες: της αφθονίας των ειδών (L), των περιβαλλοντικών μεταβλητών (R) και των λειτουργικών χαρακτηριστικών των ειδών (Q) και υπολογίζει τις συσχετίσεις των λειτουργικών χαρακτηριστικών με τις περιβαλλοντικές παραμέτρους σε μία τέταρτη μήτρα (D) (Εικόνα 5.7). Η στατιστική σημαντικότητα των συσχετίσεων αξιολογήθηκε, όπως και στην ανάλυση RLQ, μέσω δύο σταδίων τυχαιοποίησης με µεταθέσεις («permutations»), όπου πρώτα ολόκληρες οι σειρές και, στη συνέχεια, ολόκληρες οι στήλες του πίνακα L τυχαιοποιήθηκαν. Οι συνδυασμένες πιθανότητες των δύο μοντέλων χρησιμοποιούνται με σκοπό την αποφυγή Σφάλµατος Τύπου Ι (απόρριψη της αρχικής υπόθεσης ενώ είναι ορθή) και ελέγχουν την αρχική υπόθεση ότι τα χαρακτηριστικά των ειδών (Q) δε σχετίζονται με τα περιβαλλοντικά χαρακτηριστικά (R). Για τη διόρθωση των τιμών p των πολλαπλών συσχετίσεων της μήτρας D, χρησιμοποιήσαμε τη στατιστική μέθοδο false discovery rate (FDR) (Benjamini & Hochberg 1995) και έπειτα από τη διόρθωση, οι σημαντικές συσχετίσεις σε επίπεδο σημαντικότητας a= 0.05 χρησιμοποιήθηκαν για την οικολογική ερμηνεία. Ο αριθμός των μεταθέσεων ορίστηκε σε Δείκτες λειτουργικής ποικιλότητας των ειδών Από το 1999, έχουν προταθεί πολλοί δείκτες μέτρησης της λειτουργικής ποικιλότητας (για την ανασκόπηση της βιβλιογραφίας βλέπε Mouchet et al. 2010). Όταν χρησιμοποιούνται ποσοτικά δεδομένα των λειτουργικών χαρακτηριστικών, η λειτουργική ποικιλότητα μπορεί να εκτιμηθεί με συνεχείς μετρικές. Για παράδειγμα, ο Botta-Dukat (2005) ανέδειξε το δείκτη του Rao («Rao s quadratic entropy»- Q) ως ένα δείκτη μέτρησης της λειτουργικής ποικιλότητας φυτοκοινωνιών χρησιμοποιώντας δεδομένα αφθονίας και των χαρακτηριστικών των ειδών τους. Οι Mason et al. (2005) υπέδειξαν ότι η λειτουργική ποικιλότητα θα μπορούσε να διαιρεθεί σε τρία πρωτογενή στοιχεία - το λειτουργικό πλούτο («functional richness»), τη λειτουργική ισομέρεια («functional evenness») και τη λειτουργική απόκλιση («functional divergence»). Ο λειτουργικός πλούτος μετρά τον όγκο του 239
240 χώρου των λειτουργικών χαρακτηριστικών που καταλαμβάνει μια βιοκοινότητα. Η λειτουργική ισομέρεια περιγράφει την κατανομή της αφθονίας των ειδών μέσα σε αυτόν το χώρο και η λειτουργική απόκλιση αντικατοπτρίζει πόσο μακριά η αφθονία κατανέμεται από το κέντρο της βαρύτητας του λειτουργικού χώρου. Τα τρία αυτά στοιχεία είναι συμπληρωματικά και, από κοινού, δύναται να περιγράψουν την κατανομή και την αφθονία των ειδών στο λειτουργικό χώρο. Εντούτοις, μέχρι σήμερα, κανένας από τους δείκτες λειτουργικής ποικιλότητας δε μετρά τα τρία αυτά στοιχεία ταυτόχρονα και για το λόγο αυτό, οι Mouchet et al. (2010) πρότειναν τη χρήση ενός συνδυασμού των τριών δεικτών, ένα για κάθε στοιχείο. Έτσι, τα τρία αυτά στοιχεία μπορούν να υπολογιστούν χρησιμοποιώντας τους δείκτες που προτάθηκαν από τους Villéger, Mason & Mouillot (2008) και αφορούν το λειτουργικό πλούτο («functional richness»- FRic), τη λειτουργική ισομέρεια («functional evenness»- FEve) και τη λειτουργική απόκλιση («functional divergence» FDiv). Ο λειτουργικός πλούτος (FRic) αυξάνεται όσο αυξάνεται η μεταβλητότητα των λειτουργικών χαρακτηριστικών μιας βιοκοινότητας. Η λειτουργική ισομέρεια (FEve) κυμαίνεται από 0 έως 1, με τις βιοκοινότητες όπου κυριαρχούν λειτουργικά όμοια είδη να φέρουν χαμηλές τιμές του δείκτη. Τέλος, η λειτουργική απόκλιση (FDiv) κυμαίνεται από 0 έως 1, με υψηλές τιμές του δείκτη να χαρακτηρίζουν τις βιοκοινότητες, όπου τα πιο άφθονα είδη είναι και τα πιο ανόμοια λειτουργικά. Στη συνέχεια, οι Laliberté & Legendre (2010) πρότειναν έναν πολυδιάστατο δείκτη μέτρησης της λειτουργικής διασποράς («functional dispersion»- FDis) μιας κοινότητας S ειδών βάσει Τ ποσοτικών χαρακτηριστικών. Ο δείκτης αυτός σχετίζεται ισχυρά με το δείκτη του RaoQ και οδηγεί στην αξιόπιστη εκτίμηση της διασποράς στο T-διάστατο χώρο. Τα πλεονεκτήματα του έναντι του δείκτη Fric (Villéger, Mason & Mouillot 2008), που μετρά το εύρος των ειδών στο λειτουργικό χώρο, είναι ότι δεν είναι ευαίσθητος σε ακραίες τιμές και επιπλέον χρησιμοποιεί τις σχετικές αφθονίες των ειδών. Τέλος, ένας από τους πρώτους δείκτες που προτάθηκαν για την ποσοτικοποίηση της λειτουργικής ποικιλότητας βασίζεται στον διαχωρισμό των ειδών σε διάφορες λειτουργικές ομάδες σύμφωνα με μία a posteriori ταξιθέτηση. Ο δείκτης αυτός που ονομάστηκε δείκτης πλούτου λειτουργικών ομάδων («functional 240
241 group richness»- FGR) (Petchey & Gaston 2006), μετρά τον αριθμό των λειτουργικών ομάδων σε μια βιοκοινότητα και χρησιμεύει για περιγραφικούς σκοπούς ως προς τη λειτουργική μοναδικότητα των ειδών. Παρά την πληθώρα των δεικτών λειτουργικής ποικιλότητας, δεν υπάρχει ενιαίος τρόπος για να μετρήσουμε τις διαφορετικές πτυχές της λειτουργικής ποικιλότητας βάσει των χαρακτηριστικών των ειδών των βιοκοινοτήτων. Έτσι, η λειτουργική ποικιλότητα υπολογίστηκε για κάθε ιχθυοκοινότητα χρησιμοποιώντας τους δείκτες του λειτουργικού πλούτου (FRic), της λειτουργικής ισομέρειας (FEve) και της λειτουργικής απόκλισης (FDiv) ακολουθώντας τους Villéger, Mason & Mouillot (2008). Εκτιμήθηκε, επίσης, ο δείκτης λειτουργικής διασποράς (FDis), ο δείκτης του Rao (Q) και απεικονίστηκε γραφικά ο πλούτος των λειτουργικών ομάδων (FGR). Η μη παραμετρική ανάλυση Kruskal-Wallis χρησιμοποιήθηκε για τη σύγκριση των τιμών των δεικτών της λειτουργικής ποικιλότητας μεταξύ των σταθμών καθώς τα δεδομένα δεν υπόκεινται στις προϋποθέσεις εφαρμογής της Μονοδιάστατης Ανάλυσης Διακύμανσης (ANOVA). H συγκεκριμένη μέθοδος, ωστόσο, δε δείχνει ποιες ιχθυοκοινότητες διαφοροποιούνται. Για αυτό το λόγο, εφαρμόστηκε η μέθοδος Conover εκ των υστέρων (post hoc) πολλαπλών συγκρίσεων κατά ζεύγη (Conover & Iman 1979). Για τη διόρθωση των τιμών p των πολλαπλών συγκρίσεων χρησιμοποιήθηκε η στατιστική μέθοδος false discovery rate (FDR) (Benjamini & Hochberg 1995). Τέλος, για να εξετάσουμε αν οι δείκτες της λειτουργικής ποικιλότητας επηρεάζονται από τον αριθμό των ειδών στις ιχθυοκοινότητες και κατά συνέπεια, αν η λειτουργική ποικιλότητα αυξάνεται με την προσθήκη περισσότερων ειδών, εφαρμόστηκαν γραμμικές παλινδρομήσεις μεταξύ των δεικτών και του αριθμού των ειδών. 241
242 Δείκτες λειτουργικής μοναδικότητας των ειδών Αντιμέτωποι με την ταχύτατη εξαφάνιση των ειδών, οι οικολόγοι διατήρησης έχουν προτείνει πολλούς δείκτες οι οποίοι μετρούν τη μοναδικότητα ενός είδους (ή αλλιώς τη συμπληρωματικότητα) για την αξιολόγηση της απώλειας της βιοποικιλότητας από την τοπική έως και την παγκόσμια κλίμακα (Pavoine, Ollier & Dufour 2005). Η μοναδικότητα (δηλαδή ο αριθμός των μοναδικών χαρακτηριστικών που φέρει ένα είδος) συνδέεται στενά με την έννοια της σπανιότητας και της τρωτότητας (Faith 1992, Pavoine, Ollier & Dufour 2005). Γενικότερα, η τρωτότητα ενός είδους αξιολογείται από το μέγεθος των πληθυσμών του και από την ποιότητα των ενδιαιτημάτων στο οποίο ζει. Ωστόσο, πολυάριθμες πρόσφατες μελέτες έχουν δείξει ότι οι εξαφανίσεις των ειδών δεν είναι ανεξάρτητες από τις σχέσεις των ειδών και τα λειτουργικά χαρακτηριστικά τους (Witting & Loeschke 1995). Σε συνέχεια των αναλύσεων της λειτουργικής ποικιλότητας, διερευνήσαμε τη λειτουργική μοναδικότητα των ειδών και το πώς αυτή συνδέεται με την τρωτότητα αφού η απώλεια των λειτουργικά διαφορετικών ειδών θα επιφέρει σημαντική μείωση στο ρυθμό της οικοσυστημικής λειτουργίας. Χρησιμοποιήσαμε τους επτά δείκτες μέτρησης της μοναδικότητας των ειδών που έχουν προταθεί έως σήμερα: 1 = Vane-Wright, Humphries & Williams (1991) node-counting index 2 = May (1990) branch-counting index 3 = Nixon & Wheeler (1992) unweighted index 4 = Nixon & Wheeler (1992) weighted index 5 = Pavoine, Ollier & Dufour (2005) QE-based index 6 = Isaac et al. (2007) ED index 7 = Redding & Mooers (2006) Equal-split index Πιο συγκεκριμένα, ο δείκτης των Vane-Wright, Humphries & Williams (1991) (Node-counting index) είναι ανάλογος με το αντίστροφο του αριθμού των κόμβων μεταξύ ενός είδους και τη βάση του δενδρογράμματος που απεικονίζει τη λειτουργική ταξιθέτηση των ειδών. Με άλλα λόγια, τα είδη που βρίσκονται πιο κοντά 242
243 στη βάση του δενδρογράμματος («close to-root») είναι τα πιο διακριτά και εμφανίζουν τις υψηλότερες τιμές λειτουργικής μοναδικότητας. Ο May (1990) πρότεινε το δείκτη Branch-counting index που μετρά τον αριθμό των κλάδων αντί του αριθμού των κόμβων όπως συμβαίνει στην περίπτωση του Node-counting index. Αυτή η τροποποίηση οδηγεί στην παραδοχή ότι όσο περισσότερα είδη συνδέονται με ένα κόμβο τόσο λιγότερο λειτουργικά μοναδικά είναι. Για τους δύο δείκτες που προτάθηκαν από τους Nixon & Wheeler (1992) (Unweighted index, Weighted index), η λειτουργική μοναδικότητα ενός είδους είναι αντιστρόφως ανάλογη με τον αριθμό των ειδών που ταξιθετούνται στους κλάδους του δενδρογράμματος. Για παράδειγμα, τα είδη που ανήκουν στους πιο φτωχούς κλάδους (δηλαδή στους κλάδους με το μικρότερο αριθμό ειδών) έχουν την υψηλότερη λειτουργική πρωτοτυπία. Η διαφορά του σταθμισμένου δείκτη (Weighted index) από το μη- σταθμισμένο (Unweighted index) είναι ότι η πρωτοτυπία ενός είδους είναι αντιστρόφως ανάλογη με το άθροισμα του αριθμού των ειδών για κάθε υποκλάδο στον οποίο ανήκει το είδος αυτό. Αυτοί οι τέσσερις δείκτες που παρουσιάστηκαν ως τώρα αξιολογούν τη μοναδικότητα των ειδών χωρίς να λαμβάνουν υπόψη τα μήκη των κλάδων στο δενδρόγραμμα με βάση τα λειτουργικά χαρακτηριστικά των ειδών. Εντούτοις, το μήκος του κλάδου ενός είδους αποτελεί ένδειξη του αριθμού των μοναδικών χαρακτηριστικών που αυτό το είδος φέρει. Για το λόγο αυτό, κατασκευάστηκε ο δείκτης (QE-based index) από τους Pavoine, Ollier & Dufour (2005) που λαμβάνει υπόψη τα μήκη των κλάδων για τον υπολογισμό της λειτουργικής μοναδικότητας των ειδών. Στη συνέχεια, προτάθηκε ο δείκτης ED index (Isaac et al. 2007), ο υπολογισμός του οποίου επιτυγχάνεται με τη διαίρεση του μήκους κάθε κλάδου όπου υπάγεται ένα είδος με τον αριθμό των ειδών που ανήκουν στον κλάδο. Έτσι, η λειτουργική μοναδικότητα ενός είδους είναι το άθροισμα αυτών των τιμών για όλους τους κλάδους από τη βάση του δενδρογράμματος μέχρι την άκρη του κλάδου όπου το είδος βρίσκεται. Τέλος, προτάθηκε ο δείκτης Equal-split index (Redding & Mooers 2006) που λαμβάνει υπόψη εκτός από τα μήκη των κλάδων και τον αριθμό των ειδών σε κάθε κλάδο, τη θέση των ειδών στο δενδρόγραμμα, με τα πιο απομονωμένα είδη του δενδρογράμματος να χαρακτηρίζονται ως πιο λειτουργικά ανόμοια. 243
244 Οι επτά δείκτες μέτρησης της λειτουργικής μοναδικότητας των ειδών υπολογίστηκαν χρησιμοποιώντας τα λειτουργικά χαρακτηριστικά των ειδών των ψαριών του ποταμού Λούρου. Η μελέτη της λειτουργικής μοναδικότητας θα αναδείξει τα πιο λειτουργικά διαφορετικά είδη που χρήζουν άμεσης προστασίας με στόχο τη διασφάλιση της βιοποικιλότητας των ψαριών του ποταμού Λούρου, μιας και η μοναδικότητα των ειδών πρέπει να είναι ένα από τα κριτήρια που χρησιμοποιούνται για την λήψη των αποφάσεων για το ποια είδη πρέπει να προστατεύονται πρώτα Μοντέλα μηδενικής υπόθεσης («Null models») για τη μελέτη της συγκρότησης ιχθυοκοινοτήτων Η διερεύνηση των κανόνων συγκρότησης των βιοκοινοτήτων αποτελεί ένα από τα πιο ενδιαφέροντα ερωτήματα στην οικολογία των βιοκοινοτήτων (Weiher & Keddy 1999). Παρότι οι κανόνες συγκρότησης περιλαμβάνουν διάφορους μηχανισμούς, από τις διαειδικές αλληλεπιδράσεις μέχρι τις αλληλεπιδράσεις των ειδών με το περιβάλλον τους, οι Connor & Simberloff (1979) υποστήριξαν ότι η συνύπαρξη των ειδών στις βιοκοινότητες δεν είναι παρά αποτέλεσμα τυχαιότητας προτείνοντας τη χρήση μοντέλων μηδενικής υπόθεσης. Για τη διερεύνηση της επίδρασης των διεργασιών που διαμορφώνουν τις ιχθυοκοινότητες στον ποταμό Λούρο, αρχικά υπολογίστηκαν οι Ευκλείδειες αποστάσεις μεταξύ των ειδών βάσει των δεδομένων των λειτουργικών χαρακτηριστικών τους, αφού αυτά πρώτα λογαριθμήθηκαν (log10) και κανονικοποιήθηκαν (με μηδενική μέση τιμή και διακύμανση τη μονάδα). Οι αφθονίες σε κάθε ιχθυοκοινότητα μετασχηματίστηκαν χρησιμοποιώντας τη τετραγωνική ρίζα των τιμών, ώστε να ενισχυθεί η συμβολή των σπάνιων ειδών και να μειωθεί η επιρροή των κυρίαρχων ειδών (Clarke & Warwick 1994). Το επόμενο βήμα αφορούσε τη δημιουργία υποσυνόλων με τυχαία σύνθεση και τον ίδιο αριθμό ειδών όπως η πραγματική ιχθυοκοινότητα σε κάθε σταθμό. Τα είδη που συνιστούσαν αυτές τις τυχαίες ιχθυοκοινότητες επιλέχθηκαν από το περιφερειακό αποθεματικό των ειδών με τη χρήση του ανεξάρτητου αλγόριθμου 244
245 ανταλλαγής («independent swap algorithm») από την εφαρμογή του μοντέλου μηδενικής υπόθεσης μέσω τυχαιοποίησης (Gotelli 2000). Με τη χρήση της μήτρας των αποστάσεων και των αφθονιών, υπολογίστηκε η μέση απόσταση κατά ζεύγη («Mean pairwise distance» -MPD) για κάθε παρατηρούμενη (πραγματική) και τυχαία ιχθυοκοινότητα, ακολουθώντας τη μεθοδολογική προσέγγιση που εφαρμόζεται σε φυλογενετικές αναλύσεις (Webb et al. 2002). Η παράμετρος MPD υπολογίζει τη μέση απόσταση ανά ζεύγος μεταξύ όλων των ειδών σε κάθε βιοκοινότητα. Αυτή η διαδικασία επαναλήφθηκε 999 φορές. Στη συνέχεια, υπολογίστηκε το τυποποιημένο μέγεθος της επίδρασης («standardized effect size»- SES) για την παράμετρο MPD. Οι τιμές SES.MPD ισούται με τη διαφορά των τιμών της μετρικής MPD μεταξύ των παρατηρούμενων βιοκοινοτήτων από τη μέση τιμή των 999 μηδενικών βιοκοινοτήτων διαιρούμενη με την τυπική απόκλιση των τιμών της μετρικής MPD των 999 μηδενικών βιοκοινοτήτων. Με αυτό τον τρόπο, δύναται η σύγκριση της λειτουργικής ανομοιότητας μεταξύ των ειδών που συνυπάρχουν στις πραγματικές ιχθυοκοινότητες με τη λειτουργική ανομοιότητα των τυχαίων ιχθυοκοινοτήτων. Η μη παραμετρική ανάλυση Kruskal-Wallis χρησιμοποιήθηκε για τη σύγκριση των τιμών SES.MPD μεταξύ των σταθμών καθώς τα δεδομένα δεν υπόκεινται στις προϋποθέσεις εφαρμογής της Μονοδιάστατης Ανάλυσης Διακύμανσης (ANOVA). Eπιπλέον, εφαρμόστηκε η μέθοδος Conover εκ των υστέρων (post hoc) πολλαπλών συγκρίσεων κατά ζεύγη (Conover & Iman 1979) για τη διερεύνηση σε ποιες ιχθυοκοινότητες εντοπίζεται η διαφορά. Για τη διόρθωση των τιμών p των πολλαπλών συγκρίσεων χρησιμοποιήθηκε η στατιστική μέθοδος false discovery rate (FDR) (Benjamini & Hochberg 1995). Για να ελεγχθεί η υπόθεση ότι οι ιχθυοκοινότητες του ποταμού Λούρου αποτελούν τυχαία υποσύνολα του δυνητικού περιφερειακού αποθεματικού των ειδών, εφαρμόστηκε η μη- παραμετρική δοκιμασία Wilcoxon one-sample signed rank, αφού ο έλεγχος της κανονικότητας έδειξε ότι τα δεδομένα δεν υπακούουν στα κριτήρια της κανονικότητας. Η δοκιμασία Wilcoxon εφαρμόζεται για να ελέγξει την ισότητα του πληθυσμιακού μέσου μιας ποσοτικής μεταβλητής, με μία υποθετική 245
246 μέση τιμή, όταν δεν συντρέχουν οι προυποθέσεις εφαρμογής της δοκιμασίας- t. Στη συγκεκριμένη περίπτωση, ο μέσος όρος των τιμών SES.MPD των πραγματικών ιχθυοκοινοτήτων που δε διαφοροποιούνται συγκρίθηκε με το μηδέν, καθώς αυτή είναι η αναμενόμενη τιμή μιας τυχαίας ιχθυοκοινότητας (mpd.null.z= 0). Θετικές τιμές SES (mpd.obs.z> 0) δείχνουν υψηλή λειτουργική ισομέρεια και συμπληρωματικότητα των ειδών, η οποία οφείλεται στον αποκλειόμενο ανταγωνισμό ενώ αρνητικές τιμές SES (mpd.obs.z< 0) υποδηλώνουν υψηλή λειτουργική ομαδοποίηση και λειτουργικό πλεονασμό των ειδών λόγω του περιβαλλοντικού φιλτραρίσματος. Σε κάθε άλλη περίπτωση, οι παρατηρούμενες ιχθυοκοινότητες αποτελούν τυχαία υποσύνολα του δυνητικού περιφερειακού αποθεματικού των ειδών Στατιστική επεξεργασία και πακέτα Όλες οι αναλύσεις πραγματοποιήθηκαν στο «R statistical and programming environment» (R , R Development Core Team 2013). Πιο συγκεκριμένα, χρησιμοποιήθηκαν τα πακέτα «ggbiplot», «corrplot», «vegan», «FD», «Hmisc», «ade4» «plotrix», «PMCMR» και «picante». 246
247 5.4 Αποτελέσματα Λειτουργική ποικιλότητα του ποταμού Λούρου Ανάλυση κύριων συνιστωσών Σύμφωνα με την ανάλυση κύριων συνιστωσών, οι εννιά σταθμοί δειγματοληψίας στον ποταμό Λούρο χαρακτηρίζονται από διαφορετικούς περιβαλλοντικούς περιγραφείς (Εικόνα 5.8a). Οι πρώτες δύο κύριες συνιστώσες της ανάλυσης κύριων συνιστωσών αποδίδουν πάνω από το 50% της μεταβλητότητας των περιβαλλοντικών δεδομένων. Πιο συγκεκριμένα, η πρώτη κύρια συνιστώσα (PC1, 37%) σχετίστηκε ισχυρά με την απόσταση από την πηγή (Dist), το μέσο βάθος (Depth), το υψόμετρο (Ele) και την παρουσία ιλύος (Mud) και κροκάλας (Rocks) στο υπόστρωμα. Όπως φαίνεται χαρακτηριστικά και στον Πίνακα 5.5, καθώς κινούμαστε προς τις εκβολές (αυξανόμενη τιμή της παραμέτρου Dist), οι σταθμοί χαρακτηρίζονται από χαμηλότερο υψόμετρο, μεγαλύτερο βάθος και λεπτόκοκκα ιζήματα. Η δεύτερη κύρια συνιστώσα (PC2, 13,2%) σχετίστηκε αρνητικά ισχυρά με την παρουσία άμμου στο υπόστρωμα (Thick sand, Fine sand) και θετικά με την αύξηση της συγκέντρωσης του διαλυμένου οξυγόνου (Ox) και των νιτρωδών ιόντων (Nitri). Αναφορικά με τα λειτουργικά χαρακτηριστικά των ειδών των ψαριών, οι πρώτες δύο κύριες συνιστώσες, σύμφωνα με την ανάλυση, ερμηνεύουν πάνω από το 60% της διακύμανσης των οικο-μορφολογικών γνωρισμάτων των ειδών (Εικόνα 5.8b). Η πρώτη κύρια συνιστώσα (PC1, 37,5%) σχετίστηκε ισχυρά με το μέγεθος του ουραίου πτερυγίου (Rcf), το σχήμα του σώματος (Bsh), τη θέση του θωρακικού πτερυγίου (Pf), τη θέση (Gpos) και το άνοιγμα του στόματος (Gsur). Η δεύτερη κύρια συνιστώσα (PC2, 25,7%) διαχώρισε τα είδη με βάση την επιφάνεια του σώματος (Bsur), το μέγεθος και τη θέση του οφθαλμού (Es), την αναλογία της επιφάνειας των πτερυγίων με το μέγεθος του σώματος (Frat) και το σχήμα του στόματος (Gsh) (Εικόνα 5.8b). 247
248 Πίνακας 5.5. Συνιστώσες των μετασχηματισμένων μεταβλητών επί των δύο κύριων αξόνων της ανάλυσης κυρίων συνιστωσών για την απεικόνιση της περιβαλλοντικής διαβάθμισης και του λειτουργικού χώρου των ειδών του ποταμού Λούρου (για τις συντομογραφίες των μεταβλητών βλέπε Πίνακες 5.3 και 5.4) Table 5.5. Loadings of the standardized variables of the analysis of the two principal components showing the environmental gradients and functional space of the freshwater fish species of Louros river (for variables abbreviations see Tables 5.3 and 5.4) Environmental gradient Functional space Variable PC 1 PC 2 Dist 0,411-0,037 Depth 0,395 0,126 Mud 0,396-0,007 Rocks -0,377 0,237 Ele -0,384-0,166 W 0,291 0,175 Fine_sand 0,272-0,321 Thick_sand -0,134-0,455 Ox -0,024 0,447 Temp 0,179-0,109 Nitri 0,020 0,554 Phos -0,017 0,167 Am 0,029-0,089 Nitra -0,134 0,049 Chl -0,013 0,027 Rcf -0,420-0,022 Throt 0,284-0,066 Bsur -0,265, -0,355 Bsh 0,370 0,206 Rpf -0,096 0,117 Es -0,242-0,384 Pf -0,359-0,103 Frat 0,155-0,344 Fsur -0,222 0,214 Gsh -0,136 0,411 Gsur -0,313 0,174 Gpos -0,385 0,181 Epos 0,003 0,
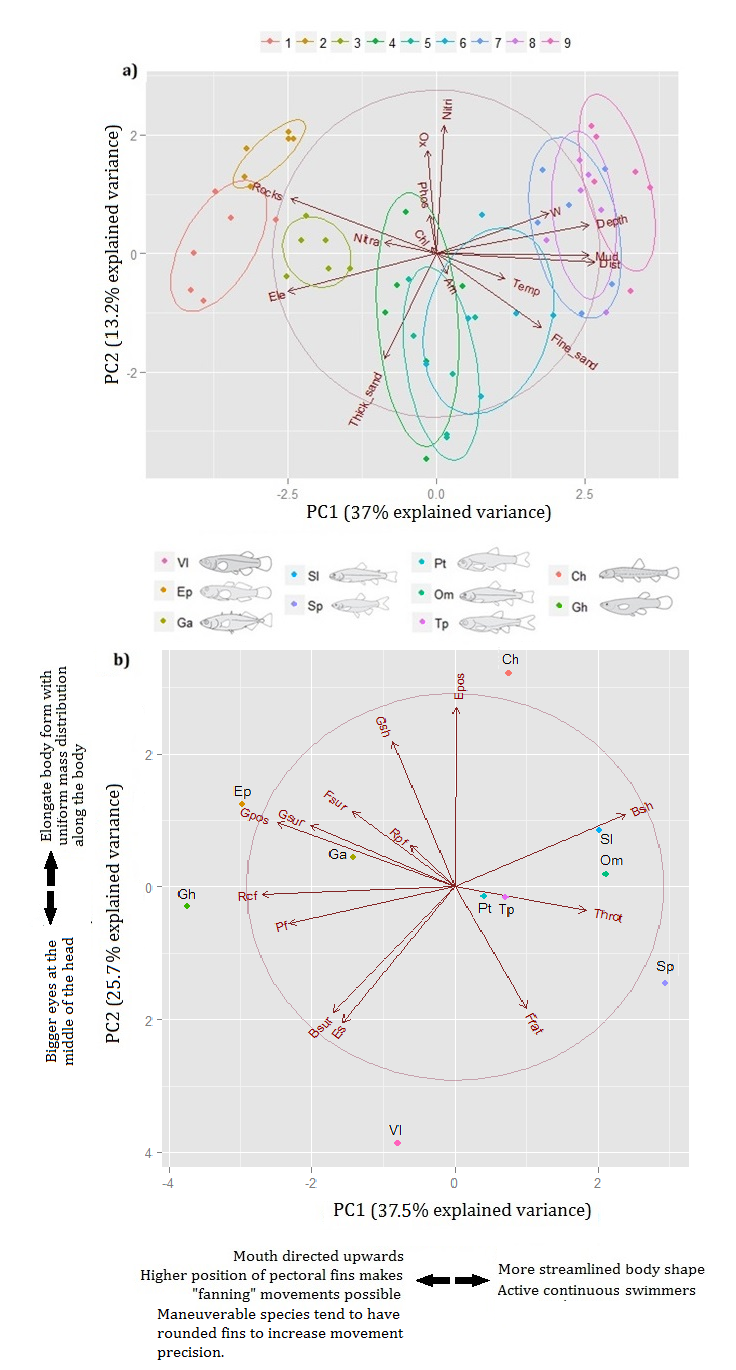
249 249
250 Εικόνα 5.8a. Διάγραμμα ταξινόμησης της ανάλυσης κύριων συνιστωσών του ποταμού Λούρου με βάση τις περιβαλλοντικές διαβαθμίσεις b. και τον λειτουργικό χώρο των ειδών. Οι συνιστώσες των μεταβλητών της περιβαλλοντικής διαβάθμισης και του λειτουργικού χώρου επί των δύο κύριων αξόνων (loadings) απεικονίζονται ως διανύσματα (βλέπε επίσης Πίνακα 5.5). Οι συντομογραφίες των μεταβλητών δίνονται στους Πίνακες 5.3 και 5. (τα σκίτσα των ειδών ανήκουν στη Ρ. Μπαρμπιέρι) Figure 5.8a. PCA ordination of Louros river based on environmental gradients b. and functional trait space of the freshwater fish species. Environmental and trait loadings on the two axes are depicted as vectors (see also Table 5.5). Abbreviations are shown in Tables 5.3 and 5.4. (species sketches belong to R. Barbieri) Επιλογή των κυριότερων περιβαλλοντικών παραμέτρων και των μορφολογικών χαρακτηριστικών για τις αναλύσεις λειτουργικής ποικιλότητας Από το σύνολο των 15 περιβαλλοντικών παραμέτρων, αφαιρέθηκαν αυτές που έδειξαν υψηλή συσχέτιση με βάση το συντελεστή συσχέτισης Spearman (r 0,80). Ένα παράδειγμα μιας παραμέτρου που είναι περιττή είναι η απόσταση από την πηγή (Εικόνα 5.9, Πίνακας 5.6). Αυτή η παράμετρος συσχετίστηκε ισχυρά με το μέσο βάθος (r=0,94), το υψόμετρο (r =- 0,983) το πλάτος του σταθμού (r=0,703), την παρουσία λάσπης (r =0,914) και κροκάλας (r =-0,821) στο υπόστρωμα και κατά συνέπεια, δε συμπεριλήφθηκε στις αναλύσεις. Μετά την αφαίρεση τριών μεταβλητών που έδειξαν στατιστικά υψηλή συσχέτιση μεταξύ τους [Απόσταση από την πηγή (Dist), Ιλυώδες υπόστρωμα (Mud) και Υψόμετρο (Ele)], 12 περιβαλλοντικές μεταβλητές χρησιμοποιήθηκαν στις μετέπειτα αναλύσεις [Μέσο βάθος (Depth), Πλάτος (W), Χονδρόκοκκο υπόστρωμα (Rocks), Συγκέντρωση διαλυμένου οξυγόνου (Ox), Αμμώδες υπόστρωμα (Fine sand), Θερμοκρασία (Temp), Χαλικώδες υπόστρωμα (Thick sand), συγκέντρωση νιτρωδών (Nitri), ορθοφωσφορικών (Phos), αμμωνιακών (Am), και νιτρικών ιόντων (Nitra) και συγκέντρωση χλωροφύλλης-α (Chl). 250

251 Εικόνα 5.9. Γραφική απεικόνιση του πίνακα συσχέτισης με βάση τα αποτελέσματα του συντελεστή συσχέτισης κατά Spearman για κάθε ζεύγος περιβαλλοντικών μεταβλητών Figure 5.9. Graphical display of the correlation matrix based on the results of Spearman's correlation coefficients for each pair of environmental variable 251
252 Πίνακας 5.6. Μήτρα συσχέτισης με βάση τα αποτελέσματα του συντελεστή συσχέτισης κατά Spearman για κάθε ζεύγος περιβαλλοντικών μεταβλητών Table 5.6. Correlation matrix based on the results of Spearman's correlation coefficients for each pair of environmental variable Dist Depth Mud Rocks Ele W Fine sand Thick sand Ox Temp Nitri Phos Am Nitra Chl Dist 1 Depth 0,940 1 Mud 0,914 0,850 1 Rocks -0,821-0,745-0, Ele -0,983-0,930-0,905 0,819 1 W 0,703 0,723 0,690 0,511-0,674 1 Fine 0,318 0,247 0,257 0,545-0,350 0,179 1 sand Thick -0,370-0,376-0,463 0,049 0,356-0,337 0,431 1 sand Ox -0,044-0,031 0,026 0,184 0,065 0,037-0,261-0,106 1 Temp 0,256 0,258 0,224-0,353-0,268-0,023 0,134-0,019-0,273 1 Nitri 0,113 0,168 0,173 0,189-0,096 0,189-0,237-0,367 0,350-0,112 1 Phos -0,004-0,010-0,020-0,004-0,011 0,020-0,183 0,042 0,065-0,025 0,080 1 Am 0,079 0,064 0,085-0,105-0,074 0,037-0,215-0,151-0,197-0,061-0,277 0,102 1 Nitra -0,295-0,286-0,268 0,291 0,283-0,051-0,018-0,003-0,040-0,275-0,077-0,235-0,101 1 Chl -0,018-0,024 0,027 0,057-0,015-0,045 0,042-0,084 0,098-0,268-0,143-0,025 0,131 0,237 1
253 Με βάση τα αποτελέσματα της πολυμεταβλητής ανάλυσης διακύμανσης με μετάθεση, από το σύνολο των περιβαλλοντικών παραμέτρων επιλέχθηκαν οι τρεις κυριότερες (βάθος, πλάτος και χονδρόκοκκος τύπος υποστρώματος) που ερμηνεύουν τη μεγαλύτερη διακύμανση της αφθονίας των ιχθυοκοινοτήτων (Πίνακας 5.7). Πίνακας 5.7. Αποτελέσματα της πολυμεταβλητής ανάλυσης διακύμανσης με μετάθεση (permanova) των Bray-Curtis αποστάσεων των αφθονιών των ψαριών σε σχέση με τις περιβαλλοντικές παραμέτρους, df = βαθμοί ελευθερίας, SS = άθροισμα των τετραγώνων, MS = μέσο άθροισμα των τετραγώνων, Μοντέλο F = τιμή F έπειτα από μετάθεση. Η έντονη γραφή υποδεικνύει στατιστική σημαντικότητα στο επίπεδο p<0,05 έπειτα από μεταθέσεις Table 5.7. Results of permanova analysis of Bray-Curtis dissimilarities in fish species abundances in relation to environmental parameters, Df = degrees of freedom; SS = sum of squares; MS = mean sum of squares; F. Model = F value by permutation. Boldface indicates statistical significance at p<0,05 based on permutations Df SS MS F, Model R 2 Pr(>F) Depth 1 2,437 2,437 9,215 0,133 0,000 W 1 0,640 0,640 2,418 0,035 0,039 Rocks 1 1,087 1,087 4,109 0,059 0,002 Ox 1 0,578 0,578 2,185 0,031 0,060 Fine_sand 1 0,580 0,580 2,193 0,032 0,056 Temp 1 0,541 0,541 2,044 0,029 0,071 Thick_sand 1 0,456 0,456 1,724 0,025 0,127 Nitri 1 0,269 0,269 1,019 0,015 0,408 Phos 1 0,154 0,154 0,582 0,008 0,718 Am 1 0,357 0,357 1,350 0,019 0,245 Nitra 1 0,143 0,143 0,539 0,008 0,768 Chl 1 0,284 0,284 1,072 0,015 0,376 Residuals 41 10,845 0,265 0,590 Total 53 18,371 1,000 Όσον αφορά τα λειτουργικά χαρακτηριστικά των ειδών, κανένα ζεύγος λειτουργικών χαρακτηριστικών δεν έδειξε ισχυρή συσχέτιση σύμφωνα με τη μη παραμετρική εκτίμηση του κατά Spearman συντελεστή συσχέτισης (Εικόνα 5.10, Πίνακας 5.8) και ως αποτέλεσμα, τα 13 υπό μελέτη λειτουργικά χαρακτηριστικά χρησιμοποιήθηκαν στις αναλύσεις λειτουργικής ποικιλότητας (Gsur, Gsh, Gpos, Es, Epos, Bsh, Bsur, Pf, Rpf, Throt, Rcf, Fsur και Frat). 253

254 Εικόνα Γραφική απεικόνιση του πίνακα συσχέτισης με βάση τα αποτελέσματα του συντελεστή συσχέτισης κατά Spearman για κάθε ζεύγος οικο- μορφολογικών χαρακτηριστικών Figure Graphical display of the correlation matrix based on the results of Spearman's correlation coefficients for each pair of eco-morphological traits 254
255 Πίνακας 5.8. Μήτρα συσχέτισης με βάση τα αποτελέσματα του συντελεστή συσχέτισης κατά Spearman για κάθε ζεύγος οικο- μορφολογικών χαρακτηριστικών Table 5.8. Correlation matrix based on the results of Spearman's correlation coefficients for each pair of eco-morphological traits Gsur Gsh Gpos Es Epos Bsh Bsur Pf Rpf Throt Rcf Fsur Frat Gsur 1 Gsh 0,252 1 Gpos 0,443-0,030 1 Es -0,166 0,038-0,136 1 Epos 0,280-0,054 0,483-0,465 1 Bsh -0,583-0,038-0,437 0,019-0,166 1 Bsur 0,215-0,202 0,409 0,116 0,150-0,389 1 Pf 0,323 0,010 0,504-0,054 0,480-0,418 0,345 1 Rpf 0,096 0,173-0,008 0,237-0,262-0,091-0,062-0,312 1 Throt -0,301-0,091-0,327-0,103-0,027 0,413-0,248-0,160-0,287 1 Rcf 0,385-0,028 0,467 0,180 0,126-0,547 0,461 0,493 0,127-0,221 1 Fsur 0,129-0,020 0,204-0,074 0,280-0,123 0,145 0,416-0,489-0,150 0,145 1 Frat -0,250-0,051-0,340-0,158-0,027 0,389-0,533-0,287-0,264 0,640-0,516-0,072 1
256 Με βάση τα παραπάνω στις αναλύσεις RLQ και fourth-corner χρησιμοποιήθηκαν τρεις περιβαλλοντικές παράμετροι και 13 λειτουργικά χαρακτηριστικά των ψαριών του ποταμού Λούρου όπως φαίνεται και στον Πίνακα 5.9. Πίνακας 5.9. Οι τρεις κυριότερες περιβαλλοντικές μεταβλητές και τα 13 λειτουργικά χαρακτηριστικά που χρησιμοποιήθηκαν στις αναλύσεις της λειτουργικής ποικιλότητας Table 5.9. The three main environmental variables and the 13 functional characteristics used in the analyses of functional diversity Environmental variable Mean Depth Substrate of Rocks-Pebbles (%) Width Functional trait Oral gape surface Oral gape shape Oral gape position Eye size Eye position Body transversal shape Body transversal surface Pectoral fin position Aspect ratio of the pectoral fin Caudal peduncle throttling Aspect ratio of the caudal fin Fins surface ratio Fins surface to body size ratio Code Depth Rocks W Code Gsur Gsh Gpos Es Epos Bsh Bsur Pf Rpf Throt Rcf Fsur Frat 256
257 Αναλύσεις RLQ και fourthcorner Τα αποτελέσματα από την εφαρμογή της ανάλυσης RLQ έδειξαν μια στατιστικά σημαντική συσχέτιση μεταξύ των περιβαλλοντικών παραμέτρων και των λειτουργικών χαρακτηριστικών των ψαριών (randtest: P<0,05, 999 μεταθέσεις) κυρίως για τον πρώτο άξονα της RLQ (0,60) ενώ η συσχέτιση ήταν ασθενέστερη για τον δεύτερο άξονα της RLQ (0.45) (Πίνακας 5.10). Οι πρώτοι δύο άξονες RLQ αποδίδουν πάνω από 98% της μεταβλητότητας των δεδομένων από τη μήτρα συσχέτισης των περιβαλλοντικών και των λειτουργικών χαρακτηριστικών των ιχθυοκοινοτήτων. Πίνακας Σύνοψη των αποτελεσμάτων της ανάλυσης RLQ Table Summarizing statistics for RLQ analysis RLQ Eigenvalue Covariance Inertia CorrL 1 3,76 1,94 96,47 0,60 2 0,10 0,32 2,51 0,45 The first two components of RLQ explain more than 95% of the variation and show high associations (corrl) with the sites-species table Πιο συγκεκριμένα, σύμφωνα με τον πρώτο άξονα (96,47%) και τη διάκριση του δεύτερου άξονα της RLQ (2,51%), ο τύπος του υποστρώματος και συγκεκριμένα το χονδρόκοκκο υπόστρωμα (Rocks) σχετίζεται θετικά με την αναλογία της επιφάνειας των πτερυγίων με το μέγεθος του σώματος (Frat), την κίνηση του ουραίου μίσχου (Throt) και το σχήμα του σώματος (Bsh), χαρακτηριστικά που αφορούν τη θέση στη στήλη του νερού, την υδροδυναμική, την κολυμβητική αντοχή, την επιτάχυνση και την ικανότητα ελιγμών. Αντίστοιχα, το πλάτος και το βάθος του σταθμού σχετίζονται θετικά με τα υπόλοιπα λειτουργικά χαρακτηριστικά, όπως η θέση του θωρακικού πτερυγίου (Pf), η αναλογία της επιφάνειας των πτερυγίων (Fsur), η επιφάνεια του σώματος (Bsur), το μέγεθος του θωρακικού (Rpf) και του ουραίου πτερυγίου (Rcf), η επιφάνεια (Gsur), το σχήμα (Gsh) και η θέση (Gpos) του στόματος και το μέγεθος (Es) και η θέση του ματιού (Epos) (Εικόνα 5.11). 257
258 Εικόνα 5.11a. Διάγραμμα κατάταξης σύμφωνα με την ανάλυση RLQ με βάση τα λειτουργικά χαρακτηριστικά των ειδών b. και τις περιβαλλοντικές μετρήσεις του ποταμού Λούρου. Οι συντομογραφίες των μεταβλητών δίνονται στον Πίνακα 5.9 Figure 5.11a. Ordination of RLQ scores of A functional traits of fish species and b. environmental variables of Louros river. Abbreviations are shown in Table 5.9
 συσχετίστηκε αρνητικά με το βάθος του σταθμού.")
259 Από την ανάλυση fourth-corner προέκυψε πως από τις σχέσεις που διαφαίνονται στο γράφημα της RLQ και περιγράφηκαν προηγουμένως, κάποιες εμφανίζουν στατιστικά σημαντική συσχέτιση. Πιο συγκεκριμένα, όπως φαίνεται και στην Εικόνα 5.12, η κίνηση του ουραίου μίσχου (Throt) συσχετίστηκε αρνητικά με το βάθος του σταθμού. Καθώς κινούμαστε προς τις πηγές όπου το υπόστρωμα γίνεται πιο χονδρόκοκκο, το μέγεθος του ουραίου πτερυγίου (Rcf) μικραίνει. Τέλος, το μέσο βάθος των σταθμών συσχετίστηκε αρνητικά με το υδρoδυναμικό σχήμα του σώματος (Bsh) (Πίνακας 5.11) Εικόνα Διάγραμμα απεικόνισης των σημαντικών συσχετίσεων που προσδιορίστηκαν μέσω της ανάλυσης fourth-corner. Οι στήλες αντιπροσωπεύουν τις περιβαλλοντικές μεταβλητές και οι σειρές τα λειτουργικά χαρακτηριστικά των ψαριών. Το μπλε χρώμα υποδεικνύει σημαντική αρνητική συσχέτιση (0.05 επίπεδο σημαντικότητας). Το γκρι υποδεικνύει μη στατιστικά σημαντικές συσχετίσεις Figure Plot of significant correlations determined through the fourth-corner analysis. The columns are environmental variables and rows are fish functional traits. Blue indicates significant negative correlation(at level of significance). Grey indicates non-significant correlations 259
260 Πίνακας Πίνακας συσχετίσεων βάση της ανάλυσης fourth-corner για κάθε ζεύγος περιβαλλοντικής μεταβλητής και λειτουργικού χαρακτηριστικού των ειδών των ψαριών του ποταμού Λούρου. Obs: παρατηρούμενες τιμές για κάθε δοκιμή, Std.obs: τυπική απόκλιση των παρατηρούμενών τιμών, Για τη διόρθωση των τιμών p των πολλαπλών συσχετίσεων χρησιμοποιήθηκε η στατιστική μέθοδος false discovery rate (FDR) (Benjamini & Hochberg 1995). Η έντονη γραφή υποδεικνύει στατιστική σημαντικότητα στο επίπεδο p<0,05 έπειτα από 9999 μεταθέσεις Figure Plot of significant correlations determined through the fourth-corner analysis for each pair of environmental variable and fish functional trait of Louros river. Obs: observed values for each test, Std.obs: standard deviation of observed values, p values of multiple correlations were corrected by false discovery rate (FDR) statistical method (Benjamini & Hochberg 1995). Boldface indicates statistical significance at p<0,05 based on permutations Test Obs Std,obs Adjusted P value Depth / Rcf 0,548 2,135 0,065 W / Rcf 0,410 2,328 0,065 Rocks / Rcf -0,535-2,325 0,033 Depth / Pf 0,398 1,553 0,259 W / Pf 0,308 1,781 0,171 Rocks / Pf -0,480-2,123 0,065 Depth / Throt -0,627-2,453 0,008 W / Throt -0,383-2,216 0,067 Rocks / Throt 0,427 1,872 0,140 Depth / Bsur 0,276 1,068 0,510 W / Bsur 0,161 0,883 0,523 Rocks / Bsur -0,259-1,115 0,508 Depth / Bsh -0,577-2,268 0,049 W / Bsh -0,274-1,572 0,250 Rocks / Bsh 0,413 1,837 0,142 Depth / Rpf 0,238 0,930 0,523 W / Rpf 0,169 0,959 0,523 Rocks / Rpf -0,046-0,207 0,889 Depth / Es -0,005-0,037 0,978 W / Es 0,151 0,848 0,523 Rocks / Es -0,088-0,372 0,790 Depth / Frat -0,330-1,325 0,387 W / Frat -0,368-2,070 0,073 Rocks / Frat 0,410 1,823 0,133 Depth / Fsur 0,160 0,628 0,668 W / Fsur 0,147 0,852 0,523 Rocks / Fsur -0,286-1,263 0,400 Depth / Gsh 0,216 0,859 0,535 W / Gsh 0,314 1,805 0,142 Rocks / Gsh -0,296-1,321 0,387 Depth / Gsur 0,455 1,757 0,142 W / Gsur 0,153 0,843 0,530 Rocks / Gsur -0,306-1,323 0,387 Depth / Gpos 0,529 2,054 0,
261 W / Gpos 0,335 1,922 0,140 Rocks / Gpos -0,462-2,023 0,077 Depth / Epos 0,243 0,968 0,523 W / Epos 0,084 0,491 0,716 Rocks / Epos -0,242-1,070 0, Δείκτες λειτουργικής ποικιλότητας των ειδών Ο υπολογισμός των δεικτών της λειτουργικής ποικιλότητας ανά σταθμό παρουσιάζεται στην Εικόνα Όσο κινούμαστε κατάντη, οι σταθμοί χαρακτηρίζονται από υψηλότερες τιμές του δείκτη RaoQ και συνεπώς, από μεγαλύτερο λειτουργικό χώρο σύμφωνα με τα αποτελέσματα της δοκιμασίας Kruskal-Wallis και της μεθόδου Conover εκ των υστέρων (post hoc) πολλαπλών συγκρίσεων κατά ζεύγη (Εικόνα 5.13, Πίνακας 5.12, Πίνακας 5.13). Παρόμοια αποτελέσματα παρατηρούμε και για το δείκτη FDis (r συσχέτισης των δύο δεικτών: 0,98) (Εικόνα 5.14). Εντούτοις, η μέση λειτουργική ποικιλότητα δε διαφέρει στατιστικά σημαντικά (P>0,05) ανά σταθμό μετά τον υπολογισμό του δείκτη FRic που δε λαμβάνει υπόψη τις αφθονίες των ειδών (mean= 1,21). Το ίδιο πρότυπο παρατηρήθηκε και για τους δείκτες FEve και FDiv, οι τιμές των οποίων κυμάνθηκαν σε παρόμοια επίπεδα (mean= 0,69, mean= 0,67 αντίστοιχα) καθώς δεν παρατηρήθηκαν στατιστικά σημαντικές διαφορές μεταξύ των σταθμών (P>0,05). Tέλος, θα πρέπει να αναφερθεί πως οι δείκτες λειτουργικού πλούτου (FRic), λειτουργικής ισομέρειας (FEve) και λειτουργικής απόκλισης (FDiv) δε δύναται να υπολογιστούν για τις ιχθυοκοινότητες με λίγοτερα από τρία λειτουργικά μοναδικά είδη (π.χ. Σταθμοί 2, 8 και 9). 261
262 Εικόνα Θηκογράμματα των αποτελεσμάτων του υπολογισμού των δεικτών λειτουργικής ποικιλότητας ανά βιοκοινότητα: λειτουργικός πλούτος («functional richness»- FRic), λειτουργική ισομέρεια («functional evenness»- FEve) και λειτουργική απόκλιση («functional divergence» FDiv), λειτουργική διασπορά («functional dispersion»- FDis) και δείκτης του Rao («Rao s quadratic entropy»- Q) ανά ιχθυοκοινότητα. Τα αποτελέσματα από τη μη παραμετρική δοκιμασία Kruskal- Wallis παρουσιάζονται πάνω δεξία Figure Boxplots of the results after the calculation of functional diversity indices per biocommunity: functional richness (FRic), functional evenness (FEve) and functional divergence (FDiv), functional dispersion (FDis) and Rao's quadratic entropy (Q) per fish community. The results οbtained from the nonparametric Kruskal- Wallis test are shown on the right top

263 Εικόνα Γραφική απεικόνιση του πίνακα συσχέτισης με βάση τα αποτελέσματα του συντελεστή συσχέτισης κατά Spearman για κάθε ζεύγος δεικτών λειτουργικής ποικιλότητας των ιχθυοκοινοτήτων του ποταμού Λούρου. FRic: λειτουργικός πλούτος («functional richness»), FEve: λειτουργική ισομέρεια («functional evenness») και FDiv: λειτουργική απόκλιση («functional divergence») (Villéger, Mason & Mouillot 2008), FDis: λειτουργική διασπορά («functional dispersion») (Laliberté & Legendre 2010) και RaoQ: δείκτης του Rao («Rao s quadratic entropy») (Botta-Dukát 2005) Figure Graphical display of the correlation matrix based on the results of Spearman's correlation coefficients for each pair of functional diversity indices of fish assemblages of Louros river. FRic: functional richness, FEve: functional evenness and FDiv: functional divergence (Villéger, Mason & Mouillot 2008), FDis: functional dispersion (Laliberté & Legendre 2010) and RaoQ: Rao s quadratic entropy (Botta-Dukát 2005) 263
264 Πίνακας Αποτελέσματα της μεθόδου Conover εκ των υστέρων (post hoc) πολλαπλών συγκρίσεων κατά ζεύγη σταθμών του ποταμού Λούρου για το δείκτη FDis. Η έντονη γραφή υποδεικνύει στατιστική σημαντικότητα στο επίπεδο p<0,05 Table Results of pairwise test for multiple comparisons of mean rank sums (Conover s-test) of stations of river Louros according to FDis index. Boldface indicates statistical significance at p<0, ,27 3 0,53 0,08 4 0,00 0,00 0,00 5 0,00 0,00 0,01 0,58 6 0,00 0,00 0,00 0,05 0,01 7 0,00 0,00 0,00 0,01 0,00 0,57 8 0,01 0,16 0,00 0,00 0,00 0,00 0,00 9 0,01 0,16 0,00 0,00 0,00 0,00 0,00 1,00 Πίνακας Αποτελέσματα της μεθόδου Conover εκ των υστέρων (post hoc) πολλαπλών συγκρίσεων κατά ζεύγη σταθμών του ποταμού Λούρου για το δείκτη RaoQ. Η έντονη γραφή υποδεικνύει στατιστική σημαντικότητα στο επίπεδο p<0,05 Table5.13. Results of pairwise test for multiple comparisons of mean rank sums (Conover s-test) of stations of river Louros according to RaoQ index. Boldface indicates statistical significance at p<0, ,32 3 0,33 0,05 4 0,00 0,00 0,01 5 0,00 0,00 0,02 0,68 6 0,00 0,00 0,00 0,06 0,02 7 0,00 0,00 0,00 0,02 0,01 0,55 8 0,02 0,18 0,00 0,00 0,00 0,00 0,00 9 0,02 0,18 0,00 0,00 0,00 0,00 0,00 1,00 Από τις γραμμικές παλινδρομήσεις μεταξύ των τιμών των δεικτών της λειτουργικής ποικιλότητας με τον αριθμό των ειδών προέκυψε πως μόνο οι τιμές των δεικτών FDis και RaoQ αυξάνονται στατιστικά σημαντικά με την αύξηση του αριθμού των ειδών (R 2 = 0,55, p<0,01 και R 2 = 0,42, p<0,01 αντίστοιχα) για τις ιχθυοκοινότητες του ποταμού Λούρου (Εικόνα 5.15), κάτι που δεν παρατηρήθηκε για το δείκτη FRic που υπολογίζει τη λειτουργική ποικιλότητα των βιοκοινοτήτων χωρίς την επιρροή της σχετικής αφθονίας των ειδών. Επίσης, η λειτουργική ισομέρεια (FEve) και η λειτουργική απόκλιση (FDiv) είναι ανεξάρτητες από τον αριθμό των 264
265 ειδών (p>0,05) και είναι επίσης ανεξάρτητες από το λειτουργικό πλούτο (Εικόνα 5.14). Εικόνα Σχέσεις μεταξύ του αριθμού των ειδών ψαριών με a. τη λειτουργική διασπορά («functional dispersion»- FDis) (Laliberté & Legendre 2010) και b. το δείκτη του RaoQ («Rao s quadratic entropy»- Q) (Botta-Dukát 2005) Figure Relationships between species richness and a. functional dispersion (FDis) (Laliberté & Legendre 2010) and b. Rao's quadratic entropy (Q) (Botta-Dukát 2005) Θέλοντας να αναπαραστήσουμε γραφικά μια λειτουργική ταξιθέτηση των ειδών του ποταμού Λούρου χρησιμοποιήσαμε το δείκτη πλούτου λειτουργικών ομάδων («functional group richness» -FGR) (Petchey & Gaston 2006) (Εικόνα 5.16) που πραγματοποιεί μία εκ των υστέρων ταξιθέτηση των ειδών. Με βάση αυτό το διάγραμμα βλέπουμε πως τα είδη διακρίνονται στους κλάδους με βάση την οικογένεια στην οποία ανήκουν. Όλα τα είδη διαφέρουν λειτουργικά, με τη μεγαλύτερη διαφοροποίηση να παρατηρείται για το είδος Valencia letourneuxi, εκτός από τα ζεύγη των ειδών Telestes pleurobipunctatus και Pelasgus thesproticus (Cyprinidae) και Salmo lourosensis και Oncorhynchus mykiss (Salmonidae) που ομαδοποιούνται λειτουργικά. 265
266 Εικόνα Αποτελέσματα της λειτουργικής ταξινόμησης των ειδών των ψαριών του ποταμού Λούρου έπειτα από υπολογισμό του δείκτη του πλούτου λειτουργικών ομάδων («functional group richness»- FGR) σύμφωνα με τους Petchey & Gaston (2006) [Valencia letourneuxi: (Vl), Gambusia holbrooki: (Gh), Economidichthys pygmaeus: (Ep), Gasterosteus aculeatus: (Ga), Cobitis hellenica: (Ch), Pelasgus thesproticus: (Pt), Telestes pleurobipunctatus: (Tp), Squalius pamvoticus: (Sp), Oncorhynchus mykiss: (Om), Salmo lourosensis: (Sl)] Figure Results of the functional clustering of the freshwater fish species of Louros river using the functional group richness index (FGR) following Petchey & Gaston (2006) [Valencia letourneuxi: (Vl), Gambusia holbrooki: (Gh), Economidichthys pygmaeus: (Ep), Gasterosteus aculeatus: (Ga), Cobitis hellenica: (Ch), Pelasgus thesproticus: (Pt), Telestes pleurobipunctatus: (Tp), Squalius pamvoticus: (Sp), Oncorhynchus mykiss: (Om), Salmo lourosensis: (Sl)] 266
267 Δείκτες λειτουργικής μοναδικότητας των ειδών Οι υψηλές τιμές λειτουργικής ποικιλότητας των σταθμών κατάντη μας οδήγησαν στη διερεύνηση του ερωτήματος ποια είδη είναι αυτά που προσδίδουν τόση μεγάλη μεταβλητότητα στα δεδομένα λειτουργικής ποικιλότητας. Για το σκοπό αυτό, χρησιμοποιήσαμε τους προτεινόμενους δείκτες μοναδικότητας των ειδών που αποτελούν εν δυνάμει κριτήρια δράσεων διατήρησης (Pavoine, Ollier & Dufour 2005). Από τον υπολογισμό των επτά δεικτών, παρατηρούμε ότι το είδος Valencia letourneuxi εμφανίζει τις υψηλότερες τιμές λειτουργικής μοναδικότητας και ακολουθείται από το είδος Cobitis hellenica, με εξαίρεση τα αποτελέσματα του δείκτη NWW όπου παρατηρείται το αντίστροφο πρότυπο (Εικόνα 5.17). Τα είδη Gasterosteus aculeatus, Squalius pamvoticus, Economidichthtys pygmaeus και Gambusia holbrooki καταλαμβάνουν τις επόμενες θέσεις. Τέλος, τα είδη Telestes pleurobipunctatus, Pelasgus thesproticus, Oncorhynchus mykiss και Salmo lourosensis παρουσίαζουν τις χαμηλότερες τιμές μοναδικότητας με βάση τα λειτουργικά χαρακτηριστικά τους (Πίνακας 5.14). 267
268 Πίνακας Αποτελέσματα της ανάλυσης μοναδικότητας για τα ψάρια του ποταμού Λούρου: 1 = Vane-Wright, Humphries & Williams (1991) node-counting index 2 = May (1990) branch-counting index 3 = Nixon & Wheeler (1992) unweighted index 4 = Nixon & Wheeler (1992) weighted index 5 = Pavoine, Ollier & Dufour (2005) QE-based index 6 = Isaac et al. (2007) ED index 7 = Redding & Mooers (2006) Equal-split index Table Results of the originality analysis of the freshwater fish species of Louros river: 1 = Vane-Wright, Humphries & Williams (1991) node-counting index 2 = May (1990) branch-counting index 3 = Nixon & Wheeler (1992) unweighted index 4 = Nixon & Wheeler (1992) weighted index 5 = Pavoine, Ollier & Dufour (2005) QE-based index 6 = Isaac et al. (2007) ED index 7 = Redding & Mooers (2006) Equal-split index Species Family IUCN Status VW 1 / M 2 NWU 3 NWW 4 QEbased 5 ED 6 Eqsplit 7 Valencia letourneuxi (Vl) Cobitis hellenica (Ch) Squalius pamvoticus (Sp) Pelasgus thesproticus (Pt) Telestes pleurobipunctatus (Tp) Oncorhynchus mykiss (Om) Salmo lourosensis (Sl) Gasterosteus aculeatus (Ga) Economidichthys pygmaeus (Ep) Gambusia holbrooki (Gh) Valenciidae CR 0,33 0,30 0,17 0,21 5,93 5,93 Cobitidae EN 0,16 0,15 0,21 0,19 5,77 5,84 Cyprinidae LC 0,08 0,08 0,09 0,09 4,01 4,57 Cyprinidae NT 0,05 0,05 0,07 0,05 2,97 2,80 Cyprinidae LC 0,05 0,05 0,07 0,05 2,97 2,80 Salmonidae - 0,05 0,05 0,07 0,06 3,33 3,16 Salmonidae 0,05 0,05 0,07 0,06 3,33 3,16 Gasterosteidae LC 0,08 0,10 0,09 0,11 4,54 4,80 Gobiidae LC 0,07 0,08 0,09 0,10 4,21 4,10 Poeciliidae - 0,07 0,08 0,09 0,10 4,21 4,10
269 Εικόνα Γραφική αναπαράσταση των αποτελεσμάτων της λειτουργικής μοναδικότητας για τα ψάρια του ποταμού Λούρου (VW = Vane-Wright, Humphries & Williams (1991) node-counting index, M = May (1990) branch-counting index, NWU = Nixon & Wheeler (1992) unweighted index, NWW = Nixon & Wheeler (1992) weighted index, QEbased= Pavoine, Ollier & Dufour (2005) QE-based index, ED = Isaac et al. (2007) ED index, eqsplit = Redding & Mooers (2006) Equal-split index) [Valencia letourneuxi: (Vl), Gambusia holbrooki: (Gh), Economidichthys pygmaeus: (Ep), Gasterosteus aculeatus: (Ga), Cobitis hellenica: (Ch), Pelasgus thesproticus: (Pt), Telestes pleurobipunctatus: (Tp), Squalius pamvoticus: (Sp), Oncorhynchus mykiss: (Om), Salmo lourosensis: (Sl)] Figure Graphical representation of the results of the functional uniqueness of the freshwater fishes of Louros river (VW = Vane-Wright, Humphries & Williams (1991) node-counting index, M = May (1990) branchcounting index, NWU = Nixon & Wheeler (1992) unweighted index, NWW = Nixon & Wheeler (1992) weighted index, QEbased= Pavoine, Ollier & Dufour (2005) QE-based index, ED = Isaac et al. (2007) ED index, eqsplit = Redding & Mooers (2006) Equal-split index) [Valencia letourneuxi: (Vl), Gambusia holbrooki: (Gh), Economidichthys pygmaeus: (Ep), Gasterosteus aculeatus: (Ga), Cobitis hellenica: (Ch), Pelasgus thesproticus: (Pt), Telestes pleurobipunctatus: (Tp), Squalius pamvoticus: (Sp), Oncorhynchus mykiss: (Om), Salmo lourosensis: (Sl)] 269
270 Κανόνες συγκρότησης των ιχθυοκοινοτήτων στον ποταμό Λούρο Για τη μελέτη των κανόνων συγκρότησης των ιχθυοκοινοτήτων στον ποταμό Λούρο εφαρμόστηκε το μοντέλο μηδενικής υπόθεσης με τη χρήση της παραμέτρου MPD που υπολογίζει τη μέση απόσταση ανά ζεύγος ειδών σε κάθε ιχθυοκοινότητα (Εικόνα 5.18). Σύμφωνα με τα αποτελέσματα αυτής της ανάλυσης, παρατηρήθηκε μία αυξητική τάση στις τιμές SES.MPD καθώς κινούμαστε προς τις εκβολές. Σύμφωνα με τα αποτελέσματα της μεθόδου Conover εκ των υστέρων πολλαπλών συγκρίσεων κατά ζεύγη, οι σταθμοί 1 έως 4 διαφοροποιούνται από τους σταθμούς 5 έως 7 όσον αφορά τις τιμές των τιμών SES.MPD (Πίνακας 5.15). Επιπλέον, τα αποτελέσματα της εφαρμογής της μη- παραμετρικής δοκιμασίας Wilcoxon one-sample signed αποκάλυψαν ότι ο μέσος όρος των SES.MPD των ιχθυοκοινοτήτων στους σταθμούς 1 έως 4 (μέσoς όρος:-0,76, p<0,05) είναι στατιστικά σημαντικά μικρότερος από το μηδέν, υποδεικνύοντας ότι αυτές οι ιχθυοκοινότητες τείνουν να είναι λειτουργικά ομαδοποιημένες σε σύγκριση με τις τυχαίες ιχθυοκοινότητες που θα προέκυπταν από το περιφερειακό αποθεματικό των ειδών. Αντίθετα, ο μέσος όρος των SES.MPD των ιχθυοκοινοτήτων στους σταθμούς 5 έως 7 (μέσoς όρος: -0,31, p> 0,05) δε διέφερε στατιστικά σημαντικά από το μηδέν. 270
271 Εικόνα Γραφική αναπαράσταση των τιμών SES.MPD των ιχθυοκοινοτήτων στους σταθμούς του ποταμού Λούρου. Τα αποτελέσματα της μη παραμετρικής δοκιμασίας Kruskal- Wallis μεταξύ των σταθμών εμφανίζονται πάνω δεξιά Figure Graphical representation of SES.MPD values of fish communities along the stations of Louros river. The results obtained from the nonparametric Kruskal- Wallis test among stations are shown on the right top panel Πίνακας Αποτελέσματα της μεθόδου Conover εκ των υστέρων (post hoc) πολλαπλών συγκρίσεων κατά ζεύγη σταθμών του ποταμού Λούρου βάσει των τιμών SES.MPD. Η έντονη γραφή υποδεικνύει στατιστική σημαντικότητα στο επίπεδο p<0,05 Table Results of pairwise test for multiple comparisons of mean rank sums (Conover s-test) of stations of river Louros according to SES.MPD values. Boldface indicates statistical significance at p<0, ,35 3 0,61 0,56 4 0,36 0,08 0,16 5 0,00 0,00 0,00 0,01 6 0,00 0,00 0,00 0,00 0,33 7 0,00 0,00 0,00 0,00 0,04 0,26 271
272 5.5 Συζήτηση Ως αποτέλεσμα της μακράς εξελικτικής τους ιστορίας, της υψηλής ποικιλότητας και των πολύπλοκων στρατηγικών ζωής στα διάφορα οντογενετικά τους στάδια, τα ψάρια αποτελούν ιδανικούς δείκτες για την εξέταση της σχέσης μεταξύ των χαρακτηριστικών των ειδών και της χρήσης των ενδιαιτημάτων (Wainwright 1994). Επιπλέον, τα ψάρια των ποταμών είναι κατάλληλα για τη διερεύνηση της υπόθεσης αν οι περιβαλλοντικές παράμετροι είναι παράγοντες διαμόρφωσης των προτύπων των ιχθυοκοινοτήτων (Lamouroux et al. 1999) καθώς μεταβλητές όπως η ταχύτητα του νερού, το βάθος και η γεωμορφολογία έχουν σχετιστεί με τα χαρακτηριστικά των ιχθυοκοινοτήτων (Gorman & Karr 1978, Angermeier & Winston 1999). Στην παρούσα μελέτη διερευνήσαμε τα πρότυπα των ιχθυοκοινοτήτων του ποταμού Λούρου χρησιμοποιώντας μία συνδυαστική μεθοδολογική προσέγγιση, της ταξινομικής και της λειτουργικής ποικιλότητας Περιβαλλοντική διαβάθμιση στον ποταμό Λούρο Με βάση την ανάλυση των κύριων συνιστωσών, οι σταθμοί στην περιοχή έρευνας διαχωρίστηκαν με βάση τέσσερα χαρακτηριστικά: α) την απόσταση από την πηγή, β) τα επίπεδα του υψομέτρου, γ) το μέσο βάθος και δ) τον τύπο του υποστρώματος, χαρακτηριστικά που σχετίζονται με τη γεωμορφολογία των σταθμών. Κατά μήκος αυτής της μελετούμενης περιβαλλοντικής διαβάθμισης, τα ενδιαιτήματα του ποταμού Λούρου χαρακτηρίζονται από αυξανόμενο μέσο βάθος, από χαμηλότερο υψόμετρο και από υπόστρωμα λεπτόκοκκου ιζήματος όσο προχωράμε από τις πηγές προς τις εκβολές του ποταμού. Με βάση τα αποτελέσματα της πολυμεταβλητής ανάλυσης διακύμανσης με μετάθεση, από το σύνολο των περιβαλλοντικών παραμέτρων που εξετάστηκαν, τρεις μεταβλητές, το πλάτος, το μέσο βάθος και ο τύπος υποστρώματος (χονδρόκοκκο), ερμηνεύουν τη μεγαλύτερη μεταβλητότητα της αφθονίας των ιχθυοκοινοτήτων. Εδώ αξίζει να αναφερθεί ότι οι περιβαλλοντικές παράμετροι συχνά εμφανίζουν υψηλή 272
273 συσχέτιση μεταξύ τους, γεγονός που αποδίδει μεγάλη δυσκολία στον προσδιορισμό των περιβαλλοντικών μεταβλητών που είναι υπεύθυνες για τη σύνθεση της βιοκοινότητας (Schlosser 1982). Για παράδειγμα, στην παρούσα μελέτη, το βάθος των σταθμών αντανακλά επίσης τη διαφοροποίηση του υψομέτρου των σταθμών καθώς οι δύο μεταβλητές συσχετίστηκαν ισχυρά. Για αυτό το λόγο, το υψόμετρο θα μπορούσε να αντικαταστήσει τη μεταβλητή του μέσου βάθους των σταθμών στην ερμηνεία της μεταβλητότητας της αφθονίας των ιχθυοκοινοτήτων. Τα αποτελέσματα της παρούσας μελέτης σχετίζονται με το μοντέλο της «συνέχειας των ποτάμιων οικοσυστημάτων» (Vannote et al. 1980) και με άλλες μελέτες που ακολούθησαν και περιγράφουν ότι το υπόστρωμα και τα μορφολογικά χαρακτηριστικά των ενδιαιτημάτων (π.χ. πλάτος, βάθος), που συνδέονται άμεσα με τα υδρολογικά χαρακτηριστικά (π.χ. ροή, θολερότητα) και τη διαθεσιμότητα της τροφής (μακροασπόνδυλα, οργανικό υλικό), σχετίζονται ισχυρά με την αφθονία των ψαριών και τη σύνθεση των ιχθυοκοινοτήτων σε ποτάμια οικοσυστήματα τόσο σε εύκρατες όσο και σε μεσογειακές λεκάνες απορροής (Oberdorff, Guilbert & Lucchetta 1993, Jackson, Peres-Neto & Olden 2001). Για παράδειγμα, οι Matthews & Robison (1988) και ο Paller (1994) βρήκαν σημαντική συσχέτιση μεταξύ του πλάτους, του βάθους και του υποστρώματος με τις αφθονίες των ψαριών σε λεκάνες απορροής στη Βόρεια Αμερική. Και στα μεσογειακά οικοσυστήματα, οι αλλαγές της θερμοκρασίας (μια μεταβλητή που επίσης αντανακλά τις υψομετρικές διαφοροποιήσεις), του βάθους και του πλάτους συσχετίστηκαν ισχυρά με τις αλλαγές των ιχθυοκοινοτήτων (Larsen et al. 2012). Παρομοίως στον Ελληνικό χώρο, οι βιοκοινότητες των ποτάμιων οικοσυστημάτων έχει δειχθεί πως διαφοροποιούνται, με το υψόμετρο και τον τύπο υποστρώματος να συγκαταλέγονται στους πιο σημαντικούς παράγοντες που διαμορφώνουν τις βιοκοινότητες μακροασπονδύλων, ακόμα και αν οι διαφοροποίησεις μεταξύ των σταθμών είναι μικρές (Skoulikidis, Karaouzas & Gritzalis 2009, Cheimonopoulou et al. 2011). 273
274 5.5.2 Λειτουργικός χώρος των ψαριών του ποταμού Λούρου Η ανάλυση κυρίων συνιστωσών αποκάλυψε τις κύριες οικο-μορφολογικές διαφορές μεταξύ των ειδών χρησιμοποιώντας δεδομένα που σχετίζονται με τη μετακίνηση και τη σύλληψη τροφής. Όσον αφορά τη μετακίνηση, η πρώτη κύρια συνιστώσα (PC1) διέκρινε τα είδη με βάση τις διαφορές σε δύο χαρακτηριστικά που σχετίζονται με τα πτερύγια. Πιο συγκεκριμένα, ως προς το σχήμα του ουραίου πτερυγίου, τα είδη που χαρακτηρίζονται από υψηλές τιμές της αναλογίας του μήκους του ουραίου πτερυγίου προς την επιφάνεια του (Rcf), δηλαδή είδη με αποστρογγυλεμένη ουρά, (π.χ. E. pygmaeus, G. holbrooki, V. letourneuxi, G. aculeatus) διαχωρίστηκαν από τα είδη με διχαλωτή ουρά (S. lourosensis, O. mykiss, P. thesproticus, T. pleurobipunctatus και S. pamvoticus). Τα είδη με αποστρογγυλεμένα πτερύγια έχουν μεγαλύτερη ικανότητα πραγματοποίησης ελιγμών ενώ τα είδη με διχαλωτή ουρά είναι ενεργοί και συνεχείς κολυμβητές καθώς το μικρότερο εμβαδό (χαμηλές τιμές του χαρακτηριστικού Rcf) οδηγεί σε μικρότερη οπισθέλκουσα αντίσταση (αντίρροπη δύναμη) κατά τη συνεχή και σταθερή κολύμβησή τους (Webb 1984, Balon et al. 1986). Ομοίως, τα είδη με υψηλότερη θέση των θωρακικών πτερυγίων τους στον κάθετο άξονα του σώματός τους (Pf) (E. pygmaeus, G. holbrooki, V. letourneuxi, G. aculeatus) χαρακτηρίζονται από αυξημένη ικανότητα πραγματοποίησης ελιγμών και δυνατότητα διατήρησής της σχετική τους θέσης στη στήλη του νερού κατά τη συνεχή κίνηση των θωρακικών πτερυγίων τους («fanning») (Harris 1937, Matthiews 1998b). Ένα ακόμα χαρακτηριστικό που διέκρινε τα είδη του ποταμού Λούρου είναι το σχήμα του σώματος (Bsh), όπου τα είδη διαχωρίστηκαν σε αυτά που έχουν πιο επίμηκες σχήμα σώματος (είδη της οικογένειας Salmonidae, Cyprinidae και Cobitidae) από αυτά με πιο επιπλατυσμένο (βλέπε την έντονη διαφοροποίηση των E. pygmaeus, G. holbrooki, V. letourneuxi, G. aculeatus) (Gatz 1979). Τέλος, η πρώτη κύρια συνιστώσα σχετίστηκε με χαρακτηριστικά που αφορούν τη σύλληψη της τροφής. Πιο συγκεκριμένα, τα είδη διαχωρίστηκαν με βάση τη σχετική θέση του στόματος ως προς τον κάθετο άξονα της κεφαλής (Gpos). 274
275 Ενδεικτικά, το ξενικό, επιθετικό και αρπακτικό κουνουπόψαρο (G. holbrooki) εκμεταλλεύεται τη θέση του στόματός του (Karpouzi & Stergiou 2003) και μετακινείται κοντά στην επιφάνεια του νερού ώστε να χρησιμοποιήσει την επιφανειακή αναπνοή για να ανταπεξέλθει στις συνθήκες υποξίας που συχνά παρατηρούμε στους σταθμούς που διαβιεί (προσωπική παρατήρηση) (βλέπε επίσης παρόμοια αποτελέσματα των Pease et al. 2015). Επιπλέον, η θέση του στόματός του είναι κατάλληλη για τη σύλληψη και κατάποση της λείας από την επιφάνεια ή τη στήλη του νερού, όντας ένα παμφάγο είδος που τρέφεται κυρίως με ζωοπλαγκτικούς οργανισμούς αλλά και έντομα που ζουν στην επιφάνεια (Gkenas et al. 2012). Αντίστοιχα με το G. holbrooki, η κεφαλή του V. letourneuxi είναι πιεσμένη νωτοκοιλιακά και το στόμα έχει κλίση προς τα πάνω, ένα χαρακτηριστικό που του επιτρέπει τη διατροφή από την επιφάνεια του νερού (Barbieri et al. 2000). Ομοίως, το είδος E. pygmaeus αν και βενθικό, φέρει στόμα με κλίση προς τα πάνω διότι δεν τρέφεται αποκλειστικά με βενθικές λείες αλλά επίσης με οργανισμούς από τη στήλη του νερού (Gkenas, Malavasi & Leonardos 2012). Όπως συνέβη και στην περίπτωση του σχήματος του σώματος (Bsh), η δέυτερη κύρια συσνιστώσα (PC2) διαχώρισε τα είδη με χαμηλές τιμές του χαρακτηριστικού της επιφάνειας του σώματος (Bsur) και της αναλογίας της επιφάνειας των πτερυγίων με το μέγεθος του σώματος (Frat), από αυτά που παρουσίασαν υψηλές τιμές των εν λόγω χαρακτηριστικών που σχετίζονται με τη λειτουργία της μετακίνησης. Πιο συγκεκριμένα, το πρώτο χαρακτηριστικό αντιπροσωπεύει την ομοιόμορφη κατανομή της βιομάζας κατά μήκος του σώματος και το δεύτερο την προώθηση στο μέσο του νερού. Συνεπώς, τα επιμήκη είδη εμφανίζουν αυξημένη υδροδυναμική (C. hellenica) σε αντίθεση με τα πιο συμπαγή (π.χ. V. letourneuxi, G. holbrooki) (Villéger 2008). Τέλος, το μέγεθος (Es) και η θέση (Epos) των οφθαλμών κατά μήκος της κεφαλής διέκρινε τα είδη σύμφωνα με τη θέση τους στη στήλη του νερού. Για παράδειγμα, το είδος C. hellenica ανήκει στα είδη με μικρούς οφθαλμούς τοποθετημένους στην κορυφή της κεφαλής που ζουν στο υπόστρωμα και τρέφονται με βενθικούς οργανισμούς (Robotham 1977). Αντίθετα, στο επιφανειακό («surfaceoriented») είδος V. letourneuxi, οι μεγάλοι του οφθαλμοί βρίσκονται περίπου στο 275
276 μέσο του κατακόρυφου άξονα της κεφαλής. Το είδος ζει και τρέφεται κοντά στην επιφάνεια (Barbieri et al. 2000) και χαρακτηρίζεται από υψηλή οπτική οξύτητα («visual acuity»), καθώς αυτή η ιδιότητα αυξάνεται με την αύξηση του μεγέθους των οφθαλμών (Northmore, Volkman & Yager 1978) Σχέσεις μεταξύ των οικο-μορφολογικών και των περιβαλλοντικών χαρακτηριστικών Η μελέτη της οικο-μορφολογίας των ειδών αποτελεί μια ολιστική προσέγγιση, που εμπερικλείει μεγάλο αριθμό οικο-μορφολογικών χαρακτηριστικών (συνήθως μέσω των πολυπαραγοντικών αναλύσεων) και συνδέει τα χαρακτηριστικά αυτά με τις λειτουργίες των ειδών (Douglas & Matthiews 1992). Σε ένα δεύτερο επίπεδο, η μελέτη των συσχετίσεων και κατ επέκταση, των αλληλεπιδράσεων μεταξύ των λειτουργικών οικο-μορφολογικών χαρακτηριστικών με τους περιβαλλοντικούς περιγραφείς των ενδιαιτημάτων όπου διαβιούν τα είδη, αποτελεί τη βάση σύνδεσης της μορφής των ειδών και του λειτουργικού τους ρόλου με τις διεργασίες συγκρότησης των βιοκοινοτήτων. Η βασική υπόθεση είναι πως αν οι σχέσεις μεταξύ των οικο-μορφολογικών χαρακτηριστικών των ειδών και των περιβαλλοντικών χαρακτηριστικών είναι ισχυρές, γίνεται σαφές ότι οι βιοκοινότητες είναι οικομορφολογικά δομημένες. Εάν αυτές οι σχέσεις είναι δυσδιάκριτες ή απουσιάζουν, οι βιοκοινότητες καθορίζονται από τις φυλογενετικές σχέσεις μεταξύ των ειδών ή αποτελούν τυχαία σύνολα (Mahon 1984, Motta, Norton & Luczkovich 1995). Τα αποτελέσματα από τις RLQ και fourth-corner αναλύσεις έδειξαν τον τρόπο με τον οποίο η σύνθεση των ειδών και συνεπώς η σύνθεση των οικο-μορφολογικών χαρακτηριστικών τους διαφέρουν με βάση τις περιβαλλοντικές συνθήκες στον ποταμό Λούρο. Πιο συγκεκριμένα, σε συμφωνία με προηγούμενες μελέτες σε άλλες περιοχές (π.χ. Lamouroux, Poff & Angermeier 2002), οι σταθμοί ανάντη που χαρακτηρίζονται από μικρότερο βάθος φιλοξενούν είδη με πιο υδροδυναμικό σχήμα σώματος (Body transversal shape) και μεγαλύτερη κολυμβητική αντοχή (Caudal peduncle throttling). Η αυξημένη υδροδυναμική και κολυμβητική αντοχή ενισχύουν την ικανότητα των ψαριών να καταλάβουν πιθανά ενδιαιτήματα για εύρεση 276
277 προστασίας (δηλαδή, καταφύγια) προς αποφυγή των θηρευτών, όσο και από τις απότομες αλλαγές στην ταχύτητα νερού (Townsend & Hildrew 1994). Επιπλέον, τα ατρακτόμορφα, υδροδυναμικά, είδη είναι πιο αποδοτικά στη σύλληψη τροφής με καταδίωξη σε λιγότερο πολύπλοκα ενδιαιτήματα με ανοιχτούς χώρους. Αντίθετα, τα είδη με πιο πεπλατυσμένη μορφή, που δίνει τη δυνατότητα για μεγαλύτερη ικανότητα ελιγμών, είναι πιο αποδοτικά σε πολύπλοκα μικροενδιαιτήματα με αυξημένο βάθος (Gatz 1979, βλέπε επίσης Ohlberger, Staaks & Hölker 2005). Τέλος, το σχήμα του ουραίου πτερυγίου (Ratio of caudal fin) συχετίστηκε με τον τύπο υποστρώματος. Πιο συγκεκριμένα, η παρουσία των ειδών με αποστρογγυλεμένη ουρά (π.χ. C. hellenica, E. pygmaeus, G. holbrooki, V. letourneuxi, G. aculeatus) συσχετίστηκε αρνητικά με το χοντρόκοκκο υπόστρωμα που παρατηρείται στους σταθμούς ανάντη. Μια πιθανή ερμηνεία της συσχέτισης αυτής μπορεί να αποδοθεί στο γεγονός ότι η μεγαλύτερη επιφάνεια του ουραίου πτερυγίου προσδίδει στα άτομα των ειδών αυτών το συγκριτικό πλεονέκτημα της καλύτερης πλευστότητας και πραγματοποίησης ελιγμών σε ενδιαιτήματα με ιλυώδεις πυθμένες και υδρόβια βλάστηση (προσωπική παρατήρηση). Επιπλέον, αυτή η μορφή της ουράς προσφέρει ένα πλεονέκτημα υπέρ της οπισθέλκουσας αντίστασης (αντίρροπη δύναμη) λόγω των ρευμάτων (Scarnecchia 1988) και είναι ιδιαιτέρως σημαντική για τα είδη που περνούν το χρόνο τους σε επαφή με τον πυθμένα (π.χ. C. hellenica, E. pygmaeus). Οι συσχετίσεις που προέκυψαν εμπίπτουν στη θεωρία των λειτουργικών ομάδων («functional guild concept») (Welcomme, Winemiller & Cowx 2006) που προτείνει πως η δομή της ιχθυοκοινότητας προσδιορίζεται από τη λειτουργική ποικιλότητα των ενδιαιτημάτων και τις περιβαλλοντικές συνθήκες που τα χαρακτηρίζουν, μία θεωρία ανάλογη με την «Αρχή της συνέχειας του πατάμιου οικοσυστήματος» (Vannote et al. 1980). Στο πλαίσιο αυτό, οποιεσδήποτε αλλαγές στα ποτάμια ενδιαιτήματα θα επιφέρουν αλλαγές στη λειτουργική ποικιλότητα και δομή των ιχθυοκοινοτήτων. 277
278 5.5.4 Λειτουργική ποικιλότητα στον ποταμό Λούρο Σύμφωνα με τα αποτελέσματα μετά τον υπολογισμό του δείκτη λειτουργικής ποικιλότητας του Rao («Rao s quadratic entropy»- Q) και του δείκτη της λειτουργικής διασποράς («functional dispersion»- FDis), οι σταθμοί καθώς κινούμαστε κατάντη στον ποταμό Λούρο χαρακτηρίζονται από αυξημένο λειτουργικό χώρο. Οι δείκτες αυτοί αυξάνουν σε σταθμούς που φιλοξενούν είδη με διαφορετικά χαρακτηριστικά. Με άλλα λόγια, στους σταθμούς κατάντη απαντώνται είδη από ένα ευρύ φάσμα οικογενειών και με ακραίες τιμές των οικο-μορφολογικών χαρακτηριστικών τους. Για παράδειγμα, οι ιχθυοκοινότητες κατάντη αποτελούνται από είδη με χαρακτηριστική μορφολογία ή στρατηγικές ζωής, όπως τα Valencia letourneuxi και Cobitis hellenica. Αντίθετα, οι ιχθυοκοινότητες ανάντη φιλοξενούν είδη που φέρουν παρόμοια χαρακτηριστικά που οδηγούν σε χαμηλές τιμές λειτουργικής ποικιλότητας. Το αποτέλεσμα αυτό μπορεί να εξηγηθεί από την επίδραση του περιβάλλοντος στη σύνθεση των ειδών, λειτουργώντας ως ένα φίλτρο (Zobel 1997). Κατά συνέπεια, σύμφωνα με τη θεωρία του περιβαλλοντικού φιλτραρίσματος («environmental filtering»), τα είδη που μπορούν να επιβιώσουν ανάντη είναι περισσότερο λειτουργικά όμοια. Η λειτουργική ποικιλότητα στον ποταμό Λούρο, κατά κανόνα αυξάνεται με τον αριθμό των ειδών, αλλά σημαντικό ρόλο παίζει και η σύνθεση των ειδών, μιας και σε περιπτώσεις πολλών όμοιων λειτουργικά ειδών, τότε παρά την αυξημένη ταξινομική ποικιλότητα, η λειτουργική ποικιλότητα είναι χαμηλότερη (πλεονασμός των λειτουργικών χαρακτηριστικών των ειδών). Για παράδειγμα, τα αποτελέσματα των γραμμικών παλινδρομήσεων του αριθμού των ειδών με τους δείκτες λειτουργικής ποικιλότητας ανά ιχθυοκοινότητα, υποδεικνύουν ότι μόνο οι δείκτες λειτουργικής ποικιλότητας του Rao («Rao s quadratic entropy»- Q) και του πολυδιάστατου δείκτη της λειτουργικής διασποράς («functional dispersion»- FDis) είναι ευαίσθητοι στις διακυμάνσεις του αριθμού των ειδών. Ως αποτέλεσμα, η λειτουργική ποικιλότητα εμφανίζει συσχέτιση με την ταξινομική ποικιλότητα καθώς η αύξηση του αριθμού των ειδών συνεπάγεται αύξηση της ποικιλότητας των λειτουργικών χαρακτηριστικών των βιοκοινοτήτων (Schmid, Joshi & Schläpfer 2002, Hooper et al. 278
279 2012). Παρόλα αυτά, όταν οι αφθονίες των ειδών δε ληφθούν υπόψη, κάποιες ιχθυοκοινότητες με ενδιάμεσα επίπεδα αριθμού ειδών χαρακτηρίζονται από μεγαλύτερο λειτουργικό χώρο απ ότι οι σταθμοί με μεγαλύτερο αριθμό ειδών (αποτελέσματα της γραμμικής παλινδρόμησης του δείκτη FRic με τον αριθμό των ειδών). Αυτή η παρατήρηση έρχεται σε συμφωνία με προηγούμενες μελέτες τόσο ψαριών (Pease et al. 2015) όσο και μακροασπόνδυλων (Gallardo et al. 2009), όπου η λειτουργική ποικιλότητα κορυφώθηκε όχι στους σταθμούς με τον μεγαλύτερο αριθμό ειδών, αλλά σε σταθμούς όπου η σύνθεση των ειδών [παρουσία τόσο γενικευμένων («generalists») όσο εξειδικευμένων («specialists») ειδών] ήταν ο καθοριστικότερος παράγοντας διαμόρφωσης της λειτουργικής ποικιλότητας των βιοκοινοτήτων. Ομοίως, σύμφωνα με τα αποτελέσματα των γραμμικών παλινδρομήσεων της λειτουργικής ισομέρειας (FEve) και της λειτουργικής απόκλισης (FDiv) με τον αριθμό των ειδών, δεν παρατηρήθηκαν στατιστικά σημαντικές συσχετίσεις, καθώς οι δείκτες αυτοί είναι ασθενώς ευαίσθητοι στον αριθμό των ειδών εξαιτίας των ιδιοτήτων τους κατά την κατασκευή τους (Villéger, Mason & Mouillot 2008) Λειτουργικά μοναδικά είδη της ιχθυοπανίδας του ποταμού Λούρου Kαθώς η λειτουργική ποικιλότητα αυξάνει με την αύξηση του λειτουργικού χώρου που καταλαμβάνεται από τα είδη (Mason et al. 2005), για τη διατήρηση της λειτουργικής ποικιλότητας πρέπει να προστατέψουμε τα πιο λειτουργικά διακριτά είδη, δηλαδή τα πιο ακραία είδη του λειτουργικού χώρου. Από τον υπολογισμό των επτά προτεινόμενων δεικτών λειτουργικής μοναδικότητας των ειδών, το V. letourneuxi (Ζουρνάς) χαρακτηρίζεται από την υψηλότερη λειτουργική μοναδικότητα μεταξύ των ειδών που διαβιούν στον ποταμό Λούρο. Το είδος V. letourneuxi μαζί με το είδος V. hispanica συνιστούν την οικογένεια των Valenciidae, που είναι και η μοναδική ενδημική οικογένεια της Ευρώπης. Αν και είναι ένα δευτερογενές είδος, αποτελεί είδος- ενδείκτη της βιογεωγραφικής περιοχής του Ιόνιου Πελάγους (Oikonomou, Leprieur & Leonardos 2014), καθώς η κατακερματισμένη γεωγραφική κατανομή του ομοιάζει με αυτή των πρωτογενών 279
280 ειδών («primary-like distribution») εξαιτίας της χαμηλής ανεκτικότητας του στην αλατότητα (Bianco & Nordlie 2008). Το V. letourneuxi κατατάσσεται στα Κρισίμως κινδυνεύοντα είδη (CR) του Κόκκινου Βιβλίου των απειλούμενων σπονδυλόζωων της Ελλάδας (Economidis 2009) και στην κόκκινη λίστα της IUCN (2013). Επίσης, περιλαμβάνεται στο παράρτημα ΙΙ της Οδηγίας των Οικοτόπων (Οδηγία 92/43/ΕΟΚ) και στο παράρτημα ΙΙ της Σύμβασης της Βέρνης, προστατεύεται από το Προεδρικό Διάταγμα 67/1981 του Ελληνικού Δημοσίου (Barbieri, Stoumboudi & Economou 2002) και έχειπρόσφατα συγκαταλεχθεί μεταξύ των «100 πιο απειλούμενων ειδών του κόσμου» (Baillie & Butcher 2012). Ζει σε ενδιαιτήματα που χαρακτηρίζονται από μεγάλο βάθος, χαμηλή ροή και πλούσια υδρόβια βλάστηση που παρέχει τροφή, προστασία από τα αρπακτικά είδη και κατάλληλο υπόστρωμα αναπαραγωγής (Barbieri et al. 2000). Εξαιτίας αυτής της υψηλής επιλεκτικότητας στη χρήση των ενδιαιτημάτων και των έντονων ανθρωπογενών δραστηριοτήτων στα ενδιαιτήματα όπου διαβιεί, όπως η υδροληψία, η ρύπανση και η καταστροφή της υδρόβιας βλάστησης, αντιμετωπίζει υψηλό κίνδυνο μείωσης των πληθυσμών του. Για παράδειγμα, η εντατική υδροληψία και τα υψηλά επίπεδα ρύπανσης προκάλεσαν την εξαφάνισή του από τα νησιά του Ιονίου (Κέρκυρα και Λευκάδα) (Economidis 1995). Επιπλέον, στις σημαντικές απειλές για το είδος συγκαταλέγεται και η εισαγωγή του είδους G. holbrooki (κουνουπόψαρο), το οποίο έχει εισβάλει σχεδόν στο σύνολο των ενδιαιτημάτων του V. letourneuxi, μειώνοντας τους πληθυσμούς του εξαιτίας του ανταγωνισμού και της θήρευσης (Barbieri et al. 2000, Kalogianni et al. 2010). Η παρατήρηση αυτή είναι σε συμφωνία με προηγούμενες μελέτες που δείχνουν ότι το κουνουπόψαρο, εξαιτίας των ιδιαίτερων λειτουργικών γνωρισμάτων του, αποτελεί έναν από τους πιο επιτυχημένους εισβολείς των εσωτερικών υδάτων (Pyke 2005). Πιο συγκεκριμένα, το είδος συγκαταλέγεται από την GISP (Global Invasive Species Program, database) στα 100 πιο επικίνδυνα χωροκρατικά είδη παγκοσμίως. Εμφανίζει αυξημένη πλαστικότητα και ικανότητα επιβίωσης σε ακραία περιβάλλοντα (Alcaraz & García-Berthou 2007). Επιπλέον, έχει δειχθεί ότι έχει προκαλέσει μείωση των διατροφικών ρυθμών και της αναπαραγωγικής επιτυχίας δύο ενδημικών ψαριών στην Ισπανία, του Valencia hispanica και Aphanius iberus 280
281 (Rincón et al. 2002) και συνεπώς, επιδρά αρνητικά στις λειτουργίες των οικοσυστημάτων που διαβιεί (Chapin et al. 2000). Στα Ελληνικά εσωτερικά ύδατα, σύμφωνα με τη συγκριτική μελέτη των διατροφικών συνηθειών των δύο ειδών που πραγματοποιήθηκε από τους Kalogianni et al. (2014), παρατηρήθηκε διαφορετική διατροφική στρατηγική μεταξύ τους. Αυτή η παρατήρηση αποδόθηκε από τους προαναφερθέντες ερευνητές ως μία μετατόπιση του οικολογικού θώκου του V. letourneuxi από την επιφάνεια- όπου τα δύο είδη συνυπάρχουν- προς αποφυγή του ανταγωνισμού. Ως αποτέλεσμα, παραδοσιακές πρακτικές διατήρησης, όπως η μετεγκατάσταση των πληθυσμών του Ζουρνά, δεν αποτελούν μια αποδοτική δράση λόγω των στενών οικολογικών απαιτήσεων (εξειδικευμένο είδος), καθώς και της μεγάλης γενετικής διαφοροποίησης μεταξύ των πληθυσμών του είδους (Vogiatzi et al. 2014). Έτσι, η διατήρηση και προστασία των υφιστάμενων ενδιαιτημάτων του προκύπτει ως ο πιο σημαντικός στόχος για την προστασία και τη διατήρηση του. Αντίστοιχα, το οικο-μορφολογικά διακριτό είδος C. hellenica (Λουροβελονίτσα) ανήκει στην οικογένεια Cobitidae που αποτελείται από μικρόσωμα είδη με μια χαρακτηριστική επιμήκη μορφή. Είναι κατά βάση νυχτόβιο είδος που την ημέρα παραμένει κρυμμένο μέσα στην ιλύ. Εξαιτίας της ιδιαίτερης μορφολογίας του και των μικρών κατακερματισμένων πληθυσμών του, η προστασία του είδους χρήζει άμεσων διαχειριστικών ενεργειών διότι κάθε παρέμβαση ή και καταστροφή του ενδιαιτήματός του, μπορεί να έχει άμεσες και σημαντικές επιπτώσεις στον πληθυσμό του ποταμού Λούρου. Επιπλέον, η εισαγωγή ξενικών ειδών (βλέπε το παράδειγμα ανταγωνισμού μεταξύ των Lepomis gibbosus και Cobitis jadovaensis, Mihinjač et al. 2015), η υπεράντληση νερού και οι αυξανόμενες περίοδοι ξηρασίας συνιστούν σημαντικές απειλές για το είδος. Τέλος, το Salmo lourosensis (Salmonidae), είναι ένα σολομοειδές των εσωτερικών υδάτων, ενδημικό είδος του ποταμού Λούρου που τις τελευταίες δύο δεκαετίες θεωρούνταν εξαφανισμένο. Έχει αναγνωριστεί ως υψηλής οικονομικής και διαχειριστικής αξίας. Εντούτοις, η ανθρωπογενής πίεση προς το είδος είναι αρκετά υψηλή, κυρίως λόγω της αστικοποίησης των παρακείμενων του ποταμού περιοχών, της ρύπανσης, του ανταγωνισμού και της επικάλυψης των διατροφικών συνηθειών με τη μη αυτόχθονη ιριδίζουσα πέστροφα (Oncorhynchus mykiss), η οποία διαφεύγει 281
282 από τις υδατοκαλλιέργειες της περιοχής (βλέπε επίσης τα αποτελέσματα του επικαλυπτόμενου λειτουργικού χώρου των δύο ειδών). Η κατασκευή του φράγματος στον ποταμό Λούρο που έχει περιορίσει την περιοχή κατανομής της πέστροφας σε ένα μικρό μέρος του ποτάμιου οικοσυστήματος (Oikonomou et al. 2011) και η χαμηλή πυκνότητα του πληθυσμού (1752 άτομα, Ntakis et al. 2015), σύμφωνα με τα κριτήρια που θέτει η κόκκινη λίστα της IUCN, οδηγούν στο συμπέρασμα πως το είδος πρέπει να προστατευτεί και να χαρακτηριστεί ως Κρισίμως κινδυνεύoν (Critically Endangered - CR). Παρόλα αυτά, το είδος δεν εμφανίζει υψηλή λειτουργική μοναδικότητα, αντιθέτως, εμφάνισε από τις χαμηλότερες τιμές. Η χαμηλή λειτουργική μοναδικότητα του μοναδικού ενδημικού είδους του ποταμού Λούρου, καταδεικνύει πως τα ενδημικά είδη που έχουν διαφοροποιηθεί λόγω της απομόνωσης του οικοσυστήματος, είναι πιθανόν να μην είναι μοναδικά από άποψη λειτουργικότητας και αυτό αποτελεί ένδειξη ότι η αξιολόγηση της σημαντικότητας των ειδών δεν μπορεί να γίνει μονοδιάστατα (π.χ. ενδημισμός), αλλά απαιτεί ολιστικές προσεγγίσεις (μελέτες ταξινομικής και λειτουργικής ποικιλότητας) Περιβαλλοντικό φιλτράρισμα ως κανόνας συγκρότησης των ιχθυοκοινοτήτων Η κατανόηση της οργάνωσης των βιοκοινοτήτων συνεισφέρει στη διερεύνηση της επίδρασης των ουδέτερων και των ντετερμινιστικών διαδικασιών (Hubbell 2001) στις πολλαπλές πτυχές της βιοποικιλότητας και είναι θεμελιώδης στην οικολογία και τη βιολογίας διατήρησης (Suding, Goldberg & Hartman 2003). Οι πρώτες προσπάθειες ερμηνείας των διεργασιών που διαμορφώνουν τις βιοκοινότητες βασίστηκαν στην ταξινομική ταυτότητα των οργανισμών (Tonn et al. 1990), χρησιμοποιώντας τον αριθμό των ειδών ως υποκατάστατο της βιοποικιλότητας (Magurran 1988). Εντούτοις, έχει δειχθεί πως οι ταξινομικές προσεγγίσεις αντιπροσωπεύουν μόνο περιορισμένα χαρακτηριστικά των βιοκοινοτήτων ενώ η μελέτη της λειτουργικής ποικιλότητας, δηλαδή η χρήση των βιολογικών δεδομένων- 282
283 χαρακτηριστικών (π.χ. διατροφικές συνήθειες, μορφολογία, στρατηγικές ζωής) των ειδών, συνεισφέρει αποτελεσματικότερα στην κατανόηση των σχέσεων μεταξύ της οικολογίας των ειδών με τις λειτουργίες των οικοσυστημάτων (Petchey & Gaston 2006, Webb et al. 2010). Συν τοις άλλοις, μιας και οι βιοκοινότητες συνιστούν ένα σύνολο αλληλεπιδράσεων μεταξύ των ειδών και των ειδών με το περιβάλλον (Palla et al. 2005), η διερεύνηση του τρόπου με τον οποίο τα λειτουργικά χαρακτηριστικά σχετίζονται με τις αποκρίσεις των ειδών σε ενδεχόμενες αλλαγές των ενδιαιτημάτων λόγω ανθρωπογενούς παρέμβασης, καθίσταται ιδιαιτέρως σημαντική (Erös et al. 2009). Όπως ήδη αναφέρθηκε, σύμφωνα με τα αποτελέσματα των δεικτών της λειτουργικής ποικιλότητας, οι περιβαλλοντικοί παράγοντες περιορίζουν την κατανομή ειδών με συγκεκριμένα λειτουργικά χαρακτηριστικά οδηγώντας σε διαφορετικά επίπεδα λειτουργικής ποικιλότητας των ιχθυοκοινοτήτων στον ποταμό Λούρο. Το συγκεκριμένο πρότυπο αναδύεται και από τα αποτελέσματα της εφαρμογής του μοντέλου μηδενικής υπόθεσης με τη χρήση της παραμέτρου MPD. Σύμφωνα με αυτή την ανάλυση, οι ιχθυοκοινότητες ανάντη χαρακτηρίζονται από υψηλά επίπεδα πλεονασμού ως προς τη σύνθεση των ειδών και είναι λειτουργικά ομαδοποιημένες σε σύγκριση με τυχαίες ιχθυοκοινότητες που θα προέκυπταν από το περιφερειακό αποθεματικό των ειδών. Αυτό το πρότυπο αναδεικνύει το περιβαλλοντικό φιλτράρισμα ως το κυρίαρχο μηχανισμό συγκρότησης των ιχθυοκοινοτήτων στον ποταμό Λούρο (Keddy 1992). Όταν επιδρά το περιβαλλοντικό φιλτράρισμα, κάποια είδη εξαιρούνται (φιλτράρονται) από τις βιοκοινότητες επειδή δεν μπορούν να προσαρμοστούν και να επιβιώσουν στις τοπικές περιβαλλοντικές συνθήκες (Grime 1973, Mayfield & Levine 2010). Ως αποτέλεσμα, τα είδη που μπορούν να επιβιώσουν είναι περισσότερο λειτουργικά όμοια απ ότι αναμενόταν από τυχαίους παράγοντες (Hoeinghaus, Winemiller & Birnbaum 2007). Πράγματι, όπως ήδη αναφέρθηκε, κάποιες αβιοτικές παράμετροι του ποταμού Λούρου (βάθος, πλάτος, υπόστρωμα) έχουν έντονη επίδραση στη σύνθεση των ιχθυοκοινοτήτων και σχετίζονται ισχυρά με τα λειτουργικά χαρακτηριστικά των ειδών. Για παράδειγμα, η πέστροφα κατανέμεται σε θέσεις του ποταμού με σχετικά μικρό βάθος, δυνατή ροή και αμμοχαλικώδη πυθμένα ενώ αντίθετα, τα βασικά ενδιαιτήματα του ζουρνά, της λουροβελονίτσας 283
284 και του λουρογωβιού, όπου τα είδη απαντώνται, είναι αμμώδεις ή ελώδεις εκτάσεις, με χαμηλή ροή, μεγάλο βάθος και πλούσια υδρόβια βλάστηση. Αυτό το πρότυπο μπορεί ασφαλώς να οφείλεται σε άλλους παράγοντες εκτός από τους αβιοτικούς που μελετήθηκαν στην παρούσα εργασία (π.χ. ταχύτητα ροής) και να συσχετίζεται με άλλα λειτουργικά χαρακτηριστικά των ειδών (π.χ. αναπαραγωγικές στρατηγικές, διατροφικές συνήθειες, ικανότητα ωσμορύθμισης), εντούτοις, υπάρχει έντονη ένδειξη φιλτραρίσματος στο οικοσύστημα μελέτης. Πράγματι, αυτή η ισχυρή συσχέτιση του ενδιαιτήματος με τα χαρακτηριστικά των ειδών των ψαριών του ποταμού Λούρου εμπίπτει στη θεωρία του ενδιαιτήματος («habitat templet theory») (Southwood 1977) που προτείνει πως η χωρική και χρονική διακύμανση των ενδιαιτημάτων οδηγεί στην επιλογή των χαρακτηριστικών. Η συγκεκριμένη διαπίστωση γίνεται για πρώτη φορά σε ιχθυοκοινότητες ποτάμιου οικοσυστήματος και έρχεται σε συμφωνία με προηγούμενες μελέτες όπου παρατηρήθηκε ισχυρή σύγκλιση των χαρακτηριστικών των ειδών υπό παρόμοιες περιβαλλοντικές συνθήκες μεταξύ των διαφόρων βιογεωγραφικών περιφερειών (Irz et al. 2007), όπως στην Αμερικανική ήπειρο (Jackson, Peres-Neto & Olden 2001, Peres-Neto 2004, Hoeinghaus, Winemiller & Birnbaum 2007) αλλά και την Ευρώπη (Erös et al. 2009). Σε μια πρόσφατη μελέτη, οι Mason et al. (2007), εφαρμόζοντας μοντέλα μηδενικής υπόθεσης, εξέτασαν τα πρότυπα ιχθυοκοινοτήτων σε λίμνες της Γαλλίας και κατέληξαν στο συμπέρασμα ότι το περιβαλλοντικό φιλτράρισμα ήταν ο κύριος ρυθμιστής της συγκρότησής τους ενώ οι ανταγωνιστικές αλληλεπιδράσεις και η ικανότητα διασποράς των ειδών είχαν ασθενέστερη επίδραση. Ωστόσο, στην παρούσα μελέτη, υπήρχαν κάποια αντιφατικά αποτελέσματα μεταξύ των διαφορετικών σταθμών, όπου δεν παρατηρήθηκε ισχυρή επίδραση του περιβαλλοντικού φιλτραρίσματος, ενώ αντίθετα, οι ιχθυοκοινότητες (σταθμοί 5-7) δε διέφεραν από τυχαίες ιχθυοκοινότητες. Αυτό το αποτέλεσμα μπορεί να οφείλεται στο γεγονός ότι κατά μήκος της περιβαλλοντικής διαβάθμισης (ανάντη- κατάντη), διαφορετικοί μηχανισμοί μπορούν να διαμορφώνουν τις ιχθυοκοινότητες (Mason et al. 2007). 284
285 5.5.7 Μειονεκτήματα και πλεονεκτήματα των επιλεγμένων αναλύσεων και διαχειριστικές προεκτάσεις H κατανόηση της δομής των ιχθυοκοινοτήτων σε σχέση με τα χαρακτηριστικά των ενδιαιτημάτων τους στα ποτάμια οικοσυστήματα, αποσκοπεί στη διερεύνηση του τρόπου με τον οποίο συγκροτούνται στις περιβαλλοντικές διαβαθμίσεις (Heino, Schmera & Erös 2013). Σε συνδυασμό με την παραδοσιακή ταξινομική (Angermeier & Winston 1999, Pyron et al. 2011) και τη φυλογενετική προσέγγιση (McGill et al. 2006), η διερεύνηση της λειτουργικής ποικιλότητας παρέχει μια πιο μηχανιστική κατανόηση, πώς οι περιβαλλοντικοί παράγοντες επηρεάζουν τις ιχθοκοινότητες, βελτιώνοντας την ικανότητά μας να προβλέψουμε τις οικολογικές αποκρίσεις τους μετά από την επίδραση των ανθρωπογενών διαταραχών ή περιβαλλοντικών αλλαγών (Dudgeon et al. 2006, Olden et al. 2010). Με άλλα λόγια, η εκτίμηση της λειτουργικής ποικιλότητας στα ποτάμια οικοσυστήματα είναι απαραίτητη τόσο για τη μελέτη της βιοποικιλότητας (Bady et al. 2005) όσο και την αξιολόγηση της επιτυχίας των προγραμμάτων προστασίας και αποκατάστασης (Cadotte, Carscadden & Mirotchnick 2011), βελτιώνοντας την απόδοση των προσπαθειών διατήρησης των σπάνιων ειδών και των οικοσυστημάτων που χρήζουν άμεσης προστασίας. Προς αυτή την κατεύθυνση, η ανάλυση RLQ σε συνδυασμό με τη fourth-corner συνδέει με επιτυχία τα περιβαλλοντικά δεδομένα και τα λειτουργικά χαρακτηριστικά των ειδών (Dray & Legendre 2008) αποτελώντας την πιο εξελιγμένη στατιστική μέθοδο για τη συσχέτισή τους που έχει προταθεί έως σήμερα. Ένα ακόμα πλεονέκτημα της παρούσας μελέτης συνιστά ότι η χρήση των οικο- μορφολογικών μετρήσεων από ένα μεγάλο αριθμό δειγμάτων (ατόμων) για κάθε είδος προσδίδει μεγαλύτερη στατιστική ισχύ στα αποτελέσματά μας αφού λαμβάνει υπόψη την ενδοειδική διακύμανση των λειτουργικών χαρακτηριστικών των ειδών, η οποία ενδέχεται να επηρεάζει την αξιολόγηση της λειτουργικής ποικιλότητας (π.χ. Zhao et al. 2014) και ειδικότερα τις σχέσεις μεταξύ των λειτουργικών και των περιβαλλοντικών χαρακτηριστικών (Brind'Amour et al. 2011). Επιπλέον, η χρήση μετρήσεων των λειτουργικών χαρακτηριστικών που αντιπροσωπεύουν πολλαπλές διαστάσεις του οικολογικού θώκου (ποσοτικές, συνεχείς μετρήσεις) αντί της 285
286 κατηγοριοποίησης των ειδών σε γενικές λειτουργικές ομάδες («functional guilds»), όπως παραδοσιακά συνηθίζεται στις αναλύσεις λειτουργικής ποικιλότητας των ψαριών των εσωτερικών υδάτων (π.χ. Schwartz, Simon & Klimetz 2011, Logez et al. 2013, Sternberg & Kennard 2014, Keck et al. 2014, Bergerot, Hugueny, & Belliard 2015) ενισχύει την ακρίβεια και την αξιοπιστία των αναλύσεων μας. Επιπλέον, τα αποτελέσματα της μέτρησης της λειτουργικής μοναδικότητας των ειδών της ιχθυοπανίδας του ποταμού Λούρου μπορούν να αποτελέσουν ένα από τα βασικά κριτήρια καθορισμού των προτεραιοτήτων διατήρησης και σημείο αναφοράς για τις μελλοντικές μελέτες με στόχο την προστασία του ποταμού Λούρου. Ενώ η μεθοδολογία και τα αποτελέσματά μας μπορούν να χρησιμοποιηθούν ως οδηγός για τη συσχέτιση της σύνθεσης των ειδών με βάση τα λειτουργικά χαρακτηριστικά τους σε διαφορετικές περιβαλλοντικές συνθήκες, είναι απαραίτητη περαιτέρω έρευνα σχετικά με το πώς αυτά τα χαρακτηριστικά σχετίζονται με την απόκριση των ειδών στην αλλοίωση των ενδιαιτημάτων, την τροποποίηση της ροής του ποταμού, τη ρύπανση και γενικότερα τις κλιματικές αλλαγές. Επιπλέον, αν και η στατιστική προσέγγιση που ακολουθήθηκε είναι πολλά υποσχόμενη για την ανάδυση των προτύπων οργάνωσης των ιχθυοκοινοτήτων, η αδυναμία εντοπισμού ισχυρών συσχετίσεων μεταξύ των οικο-μορφολογικών γνωρισμάτων και των χαρακτηριστικών των ενδιαιτημάτων υποδεικνύει κάποια μειονεκτήματα του σχεδιασμού της παρούσας μελέτης. Για παράδειγμα, οι περιβαλλοντικές μετρικές που εξετάστηκαν αντιπροσωπεύουν μόνο ένα μικρό ποσοστό μεταβλητότητας της ταξινομικής και λειτουργικής ποικιλότητας των ιχθυοκοινοτήτων και αυτό μπορεί να οφείλεται στο γεγονός ότι είτε ο αριθμός των ειδών στο σύστημα μελέτης είναι αρκετά μικρός (10 είδη) και η ιχθυοπανίδα αποτελείται από λίγα εξειδικευμένα είδη, είτε η περιβαλλοντική διαφοροποίηση των δειγματοληπτικών σταθμών είναι πολύ μικρή. Επομένως, θα ήταν ενδιαφέρον να εξεταστούν οι σχέσεις μεταξύ των λειτουργικών χαρακτηριστικών με τις περιβάλλοντικές συνθήκες σε οικοσυστήματα με μεγαλύτερη ποικιλότητα και πιο αδρές περιβαλλοντικές διαβαθμίσεις. Στην παρούσα μελέτη, η έλλειψη ενδείξεων του μηχανισμού του περιορισμού ομοιότητας (MacArthur & Levins 1967) ή του ανταγωνισμού (Grime 1973) μπορεί να οφείλεται στην επιλογή και χρήση οικο-μορφολογικών λειτουργικών χαρακτηριστικών και όχι πιο άμεσων γνωρισμάτων που υποδεικνύουν 286
287 αλληλεπίδραση, όπως για παράδειγμα η διατροφή. Παρ 'όλα αυτά, δύο είδη που εμφανίζουν να έχουν την ίδια δίαιτα, μπορεί να αλληλεπιδρούν ελάχιστα, εάν, για παράδειγμα, η σύλληψη της τροφής γίνεται σε διαφορετικά χρονικά διαστήματα (π.χ. ημέρα- νύχτα) καθιστώντας δύσκολη την επιλογή των καταλληλότερων υπό εξέταση χαρακτηριστικών. Επίσης, η οικολογική εξειδίκευση των ειδών είναι πιο δύσκολο να αναδειχθεί στα ποτάμια οικοσυστήματα, επειδή ο υδρολογικός κύκλος μεταβάλλει τη διαθεσιμότητα των πόρων (Winemiller 1989). Τέλος, η μεθοδολογική προσέγγιση που ακολουθήθηκε καθώς και τα πρότυπα που περιγράφηκαν στην παρούσα μελέτη θα μπορούσαν να συμβάλουν στη σύνθεση των θεωριών της οικολογίας βιοκοινοτήτων και να αποτελέσουν τη βάση για τη δημιουργία διαχειριστικών εργαλείων που βρίσκουν εφαρμογή στις διαφορετικές ταξινομικές ομάδες και γεωγραφικές περιοχές. Καθώς η παρούσα μελέτη αποτελεί την πρώτη προσέγγιση διερεύνησης των προτύπων λειτουργικής ποικιλότητας της ιχθυοπανίδας ενός ποταμού της Ελλάδας τόσο σε διαφορετικά χωρικά (διαφορετικοί σταθμοί κατά μήκος του ποταμού), όσο και χρονικά επίπεδα (διαφορετικές περίοδοι δειγματοληψίας), αντίστοιχες μελέτες και σε άλλα ποτάμια οικοσυστήματα της Ελλάδας, θα μας δώσουν την δυνατότητα αποτελεσματικότερης κατανόησης των μηχανισμών διαμόρφωσης της βιοποικιλότητας τόσο σε τοπικό όσο και περιφερειακό επίπεδο (Oikonomou, Leprieur & Leonardos 2014). 287
288
289 Κεφάλαιο 6 Γενικά συμπεράσματα
290
291 Η παρούσα διατριβή αποτελεί την πρώτη προσέγγιση παράλληλης διερεύνησης των προτύπων της ποικιλότητας της ιχθυοπανίδας και των μηχανισμών διαμόρφωσής της, τόσο σε περιφερειακό όσο και τοπικό επίπεδο. Ειδικότερα, η μελέτη εστιάστηκε σε τρία βασικά χωρικά επίπεδα: στο περιφερειακό επίπεδο, το οποίο αναφέρεται στη Βαλκανική χερσόνησο, στο τοπικό επίπεδο, το οποίο αναφέρεται στην Ελλάδα και στο οικοσυστημικό επίπεδο, το οποίο αφορά το σύστημα του ποταμού Λούρου. Ακολούθως, παρουσιάζονται τα ειδικά αλλά και γενικά συμπεράσματα που εξάγονται από τα αποτελέσματα της μελέτης και για τα τρία επίπεδα 29. ΠΕΡΙΦΕΡΕΙΑΚΟ ΕΠΙΠΕΔΟ Σε περιφερειακό επίπεδο, δημιουργήθηκε και χρησιμοποιήθηκε η πληρέστερη μέχρι σήμερα, βάση δεδομένων κατανομής 193 ειδών ψαριών στα 97 μεγαλύτερα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου με στόχο την οριοθέτηση και μελέτη των βιογεωγραφικών περιοχών των Βαλκανίων, η οποία αποτέλεσε τον πρώτο στόχο της διδακτορικής διατριβής (Κεφάλαιο 2). Η ανάγκη μιας ολοκληρωμένης εξέτασης των βιογεωγραφικών περιοχών της Βαλκανικής δημιουργήθηκε διότι η επιστημονική αναγκαιότητα και σημαντικότητα της εξέτασης των βιογεωγραφικών περιοχών αποτελούν επιστημονική πρόκληση για την περιοχή και οι προηγούμενες μελέτες οριοθέτησης έχουν προτείνει διαφορετικά πρότυπα, καθώς είτε έχουν επικεντρωθεί μόνο στο νότιο τμήμα της, είτε έχουν βασιστεί σε εμπειρικές προσεγγίσεις. Επιπλέον, προσδιορίστηκαν τα είδη- ενδείκτες που, λόγω εξειδικευμένων προτιμήσεων, μπορούν να χρησιμοποιηθούν ως εκπρόσωποι μιας βιογεωγραφικής περιοχής. Δεύτερος στόχος της διδακτορικής διατριβής (Κεφάλαιο 3) ήταν η μελέτη των προτύπων του αριθμού των ειδών σε σχέση με τον ενδημισμό, των απειλούμενων με εξαφάνιση ειδών και την έκταση των οικοσυστημάτων, στη Βαλκανική χερσόνησο. Από το σύνολο των αποτελεσμάτων που αφορούν το περιφερειακό επίπεδο εξάγονται τα ακόλουθα: 1. Έπειτα από ενδελεχή ανασκόπηση 364 βιβλιογραφικών πηγών και αξιολόγηση της εγκυρότητάς τους, δημιουργήθηκε η πιο εκτεταμένη και πλήρης 29 Για λόγους συνοχής και λειτουργικότητας της συμπερασματικής σύνοψης, κρίθηκε σκόπιμο τα ειδικά συμπεράσματα στα επιμέρους κεφάλαια να παρουσιαστούν με συνεχή αρίθμηση 291
292 μέχρι σήμερα βάση δεδομένων κατανομής για 193 είδη ψαριών σε 78 λεκάνες απορροής ποταμών και 19 φυσικές λίμνες της Βαλκανικής χερσονήσου. 2. Η Βαλκανική χερσόνησος είναι η πιο άφθονη περιοχή της Ευρώπης, φιλοξενώντας 193 αυτόχθονα ψάρια των εσωτερικών υδάτων, χαρακτηρίζεται από υψηλά επίπεδα ενδημισμού (56%) και μεγάλα ποσοστά απειλούμενων ειδών (33%). Τα Κυπρινοειδή είναι η επικρατέστερη οικογένεια στην αυτόχθονη και ενδημική ιχθυοπανίδα της Βαλκανικής χερσονήσου. 3. Μετά την εφαρμογή της μετρικής β-ποικιλότητας (βjtu), χρησιμοποιώντας την ανομοιότητα της σύνθεσης των ψαριών στα μεγαλύτερα υδάτινα οικοσυστήματα της Βαλκανικής χερσονήσου, προσδιορίστηκαν οκτώ κύριες βιογεωγραφικές περιοχές: 1. Ιόνιο Πέλαγος, 2. Νότια Αδριατική θάλασσα, 3. Βόρεια Αδριατική θάλασσα. 4. Λεκάνη απορροής του ποταμού Neretva, 5. Αττικοβοιωτία, 6. Αιγαίο Πέλαγος, 7. Ποντιακή επαρχία και 8. Πρέσπες. Αυτές οι βιογεωγραφικές περιοχές μπορούν να ταξινομηθούν στη δυτική ομάδα (Ιόνιο Πέλαγος, Νότια και Βόρεια Αδριατική θάλασσα και λεκάνη απορροής του ποταμού Neretva), την ανατολική ομάδα (Αττικοβοιωτία, Αιγαίο Πέλαγος και Ποντιακή επαρχία) και την ανεξάρτητη ομάδα (Πρέσπες). 4. Η μεγάλη ανομοιότητα της σύνθεσης των ψαριών των εσωτερικών υδάτων των οκτώ βιογεωγραφικών περιοχών είναι το αποτέλεσμα των ανεξάρτητων βιογεωγραφικών ιστοριών τους. Οι νότιες και δυτικές βιογεωγραφικές περιοχές των Βαλκανίων δεν καλύφτηκαν με παγετώνες κατά το Πλειστόκαινο και χαρακτηρίζονται από μεγαλύτερο βαθμό απομόνωσης, εξαιτίας της αλπικής ορογένεσης που εμπόδισε τη διασπορά των ψαριών προς τα βόρεια και ανατολικά. 5. Μέσω της ανάλυσης προσδιορισμού των ειδών- ενδεικτών των βιογεωγραφικών περιοχών, 33 είδη (17%) παρουσίασαν στατιστικά σημαντική συσχέτιση με τις βιογεωγραφικές περιοχές. Από τα 33 είδη, τα 28 ανήκουν στην οικογένεια των Κυπρινοειδών. Επιπλέον, τρία είδη του γένους Cobitis, δύο ενδημικοί αμμογωβιοί (Knipowitschia croatica και Economidichthys pygmaeus) και το ενδημικό είδος Valencia letourneuxi, που ανήκει στη μοναδική ενδημική οικογένεια της Ευρώπης Valenciidae, αναδείχθηκαν ως είδη-ενδείκτες της Βαλκανικής χερσονήσου. 6. Οι περιοχές με τα υψηλότερα επίπεδα ενδημισμού είναι αυτές που παρουσιάζουν το χαμηλότερο αριθμό ειδών, ένα γεγονός που πιθανόν σχετίζεται με 292
293 την επίδραση της χερσονήσου, το μέγεθος των οικοσυστημάτων και την ασθενέστερη συνδεσιμότητα των λεκανών απορροής των νότιων περιοχών που απέτρεψε τη διασπορά των ειδών σε μεγάλες αποστάσεις, αυξάνοντας έτσι τον τοπικό ενδημισμό. 7. Οι περιοχές της Αττικοβοιωτίας, του Ιονίου και των λιμνών των Πρεσπών χαρακτηρίζονται ως σημεία υψηλού ενδιαφέροντος κατά την ανάπτυξη ενός πλαισίου αποτελεσματικής διατήρησης και στοχευμένης έρευνας για τη βιοποικιλότητα, καθώς φιλοξενούν τα υψηλότερα ποσοστά ενδημικών και απειλούμενων ειδών. 8. Στις βιογεωγραφικές περιοχές του Αιγαίου Πελάγους, της νότιας Αδριατικής θάλασσας, της λεκάνης απορροής του ποταμού Neretva και της βόρειας Αδριατικής Θάλασσας, η ανάγκη για κοινές διασυνοριακές προσπάθειες διατήρησης μεταξύ των χωρών είναι επιτακτική, καθότι οι διοικητικές διαιρέσεις δε συμπίπτουν με αυτές των βιογεωγραφικών περιοχών. 9. Σύμφωνα με τα αποτελέσματα της διαδικασίας επιλογής του καταλληλότερου μοντέλου για τη περιγραφή της SAR σε δεδομένα κατανομής ψαριών στα οικοσυστήματα των εσωτερικών υδάτων της Βαλκανικής χερσονήσου, το δυναμικό μοντέλο (Power) παρουσίασε τις καλύτερες επιδόσεις αν και το συνολικό ποσοστό συσχέτισης ανάμεσα στον αριθμό των ειδών και την έκταση της λεκάνης απορροής ήταν αρκετά χαμηλό (R 2 =0,52). Συμπεραίνοντας γενικότερα θα λέγαμε ότι «Η βιογεωγραφική ιστορία που σχετίζεται με τη γεωλογική εξέλιξη, η συνδεσιμότητα και η έκταση των οικοσυστημάτων είναι από τους πιο σημαντικούς παράγοντες καθορισμού των προτύπων της σύνθεσης και του αριθμού των αυτόχθονων και ενδημικών ειδών των ψαριών των εσωτερικών υδάτων της Βαλκανικής χερσονήσου». ΤΟΠΙΚΟ ΕΠΙΠΕΔΟ Η αναγνώριση των προτύπων σε διαφορετικές ιεραρχικές χωρικές κλίμακες είναι απαραίτητη για την κατανόηση της βιοποικιλότητας. Ως εκ τούτου, προκειμένου να διερευνήσουμε τη διαβάθμιση της ποικιλότητας και σε χαμηλότερη 293
294 κλίμακα, επικεντρωθήκαμε σε ένα πιο τοπικό επίπεδο, την Ελλάδα (Κεφάλαιο 4). Τα Ελληνικά λιμναία οικοσυστήματα αποτέλεσαν μια μοναδική ευκαιρία διερεύνησης της επίδρασης των περιβαλλοντικών παραγόντων στη διαμόρφωση του αριθμού και της κατανομής του μεγέθους των ειδών των ψαριών. 10. Η Ελληνική ιχθυοπανίδα των εσωτερικών υδάτων είναι η πιο πλούσια της Ευρώπης, χαρακτηρίζεται από πολύ υψηλά επίπεδα ενδημισμού (42% των αυτόχθονων ψαριών των εσωτερικών υδάτων) και αντιπροσωπεύεται από ένα σχετικά μικρό αριθμό οικογενειών, με τα περισσότερα είδη να ανήκουν στην οικογένεια των Κυπρινοειδών. Συνολικά, το 32% των ψαριών των εσωτερικών υδάτων της Ελλάδας έχουν χαρακτηριστεί ως απειλούμενα σύμφωνα με την Κόκκινη Λίστα και 34% σύμφωνα με το Κόκκινο Βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας. 11. Ο αριθμός των ψαριών στα λιμναία οικοσυστήματα της Ελλάδας μειώνεται με την αύξηση του υψομέτρου και αυξάνεται με την αύξηση της έκτασης του οικοσυστήματος. Η συγκέντρωση του ολικού φωσφόρου που σχετίζεται έμμεσα με την πρωτογενή παραγωγικότητα έχει τη μικρότερη συνεισφορά στην ερμηνεία της διαβάθμισης του αριθμού των ειδών σε λίμνες. 12. Το μέγεθος του σώματος των ψαριών των εσωτερικών υδάτων στις Ελληνικές λίμνες αυξάνεται με το γεωγραφικό, εξαιτίας της διαφορετικής ικανότητας διασποράς των ειδών και της διαφορετικής γεωλογικής ιστορίας των υπό μελέτη συστημάτων. Η βορειοανατολική Ελλάδα φιλοξενεί ευρέως κατανεμημένα, μεγαλόσωμα είδη με μεγαλύτερη ικανότητα εποικισμού στη μεταπαγετώδη περίοδο, σε σύγκριση με τη νοτιοδυτική, όπου διαβιούν μικρόσωμα ενδημικά είδη ή είδη με περιορισμένη κατανομή. Επομένως, «Η δυτική-ανατολική διάκριση των Ελληνικών εσωτερικών υδάτων και η διαφορετική σύνθεση τους αποδίδεται στη μακροχρόνια παρουσία του γεωγραφικού εμπόδιου της Πίνδου και την επακόλουθη απομόνωση που προκλήθηκε». 294
295 ΕΠΙΠΕΔΟ ΟΙΚΟΣΥΣΤΗΜΑΤΟΣ Η βιολογία της διατήρησης παραδοσιακά επικεντρώθηκε σε μετρικές βασισμένες στην ταξινομική ποικιλότητα, με τον αριθμό των ειδών να αποτελεί δείκτη-υποκατάστατο της βιοποικιλότητας (Κεφάλαια 2, 3 και 4). Ωστόσο, ιδιαίτερα σε τοπικό επίπεδο, είναι πλέον αποδεκτό ότι η λειτουργική ποικιλότητα των βιοκοινοτήτων, δηλαδή ο αριθμός, ο τύπος και η κατανομή των λειτουργιών που εκτελούνται από τους οργανισμούς μέσα σε ένα οικοσύστημα, μέσα από τη χρήση των οικο-μορφολογικών χαρακτηριστικών των ειδών, συνεισφέρει αποτελεσματικότερα στην κατανόηση των σχέσεων μεταξύ της οικολογίας των ειδών με τις λειτουργίες των οικοσυστημάτων. Για τη διερεύνηση των προτύπων της λειτουργικής ποικιλότητας (Κεφάλαιο 5), ως σύστημα μελέτης επιλέχθηκε ο ποταμός Λούρος διότι αποτελεί ένα τυπικό πεδινό ομαλού πηγαίου τύπου ποτάμιο οικοσύστημα. Επιπλέον, ο ποταμός Λούρος ανήκει στην Ιόνια βιογεωγραφική περιοχή, είναι ένα από τα λιγότερο μελετημένα οικοσυστήματα στην Ελλάδα, με μακρά ιστορία ανθρώπινης παρουσίας και παρέμβασης. Χαρακτηρίζεται από αυξημένη ποικιλότητα μεσογειακών ποτάμιων ενδιαιτημάτων, τα οποία φιλοξενούν εξελικτικά απομονωμένη ιχθυοπανίδα, η οποία χαρακτηρίζεται από χαμηλό αριθμό ειδών, αλλά ταυτόχρονα από υψηλά επίπεδα ενδημισμού και μεγάλο αριθμό απειλούμενων ειδών. Τα δεδομένα για τη διερεύνηση των ερωτημάτων του πέμπτου κεφαλαίου προέκυψαν έπειτα από συλλογή ιχθυολογικού υλικού και μέτρηση των περιβαλλοντικών παραμέτρων σε 9 σταθμούς στο σύστημα μελέτης, σε διμηνιαία βάση για ένα χρόνο. Οι στόχοι του τελευταίου κεφαλαίου της διδακτορικής διατριβής ήταν η διερεύνηση των συσχετίσεων μεταξύ των περιβαλλοντικών χαρακτηριστικών και της λειτουργικής σύνθεσης των ιχθυοκοινοτήτων. Η μελέτη των συσχετίσεων και κατ επέκταση, των αλληλεπιδράσεων μεταξύ των λειτουργικών οικο-μορφολογικών χαρακτηριστικών με τις περιβαλλοντικές συνθήκες των ενδιαιτημάτων όπου διαβιούν τα είδη, αποτελεί τη βάση σύνδεσης της μορφής των ειδών και του λειτουργικού τους ρόλου με τις διεργασίες διαμόρφωσης των βιοκοινοτήτων. Με αυτή τη στοχοθέτηση, μας δόθηκε η ευκαιρία να ελέγξουμε επίσης, την επίδραση των δύο βασικών αντιτιθέμενων διεργασιών που αφορούν το περιβαλλοντικό φιλτράρισμα και τις βιοτικές αλληλεπιδράσεις, στη διαμόρφωση των ιχθυοκοινοτήτων, με βάση τα λειτουργικά γνωρίσματα των ειδών. Επιπλέον, έπειτα 295
296 από εφαρμογή των προτεινόμενων δεικτών λειτουργικής μοναδικότητας, αναδείχθηκαν τα πιο οικο-μορφολογικά διακριτά είδη του ποταμού Λούρου που χρήζουν άμεσης προστασίας. 13. Τα ενδιαιτήματα του ποταμού Λούρου χαρακτηρίζονται από αυξανόμενο μέσο βάθος, από χαμηλότερο υψόμετρο και από υπόστρωμα λεπτόκοκκου ιζήματος όσο προχωράμε από τις πηγές προς τις εκβολές του ποταμού. 14. Από το σύνολο των περιβαλλοντικών παραμέτρων που εξετάστηκαν, το πλάτος, το μέσο βάθος και ο τύπος υποστρώματος (χονδρόκοκκο), ερμηνεύουν τη μεγαλύτερη μεταβλητότητα της αφθονίας των ιχθυοκοινοτήτων. 15. Τα είδη των ψαριών του ποταμού Λούρου διαχωρίστηκαν κυρίως ως προς το σχήμα του ουραίου πτερυγίου, τη θέση των θωρακικών πτερυγίων τους στον κάθετο άξονα του σώματός τους, το σχήμα του σώματος και τη σχετική θέση του στόματος ως προς τον κάθετο άξονα της κεφαλής. 16. Οι περιβαλλοντικοί παράγοντες περιορίζουν την κατανομή ειδών που εμφανίζουν συγκεκριμένα λειτουργικά χαρακτηριστικά οδηγώντας έτσι τις ιχθυοκοινότητες του ποταμού Λούρου να οργανώνονται σε διαφορετικά επίπεδα λειτουργικής ποικιλότητας. Οι σταθμοί ανάντη χαρακτηρίζονται από μικρό βάθος και χοντρόκοκκο υπόστρωμα και φιλοξενούν είδη με πιο υδροδυναμικό σχήμα σώματος, μεγαλύτερη κολυμβητική αντοχή και διχαλωτή ουρά. Αντίθετα, οι σταθμοί κατάντη χαρακτηρίζονται από αυξημένο λειτουργικό χώρο, καθώς φιλοξενούν είδη από ένα ευρύ φάσμα οικογενειών και με ακραίες τιμές των οικο-μορφολογικών χαρακτηριστικών τους. 17. Η λειτουργική ποικιλότητα στον ποταμό Λούρο, κατά κανόνα αυξάνεται με την αύξηση του αριθμού των ειδών, καθώς η αύξηση του αριθμού των ειδών συνεπάγεται και αύξηση της ποικιλότητας των λειτουργικών χαρακτηριστικών των ιχθυοκοινοτήτων. Εντούτοις, σημαντικό ρόλο παίζει και η σύνθεση των ειδών, όπως σε περιπτώσεις σταθμών με όμοια λειτουργικά είδη, όπου παρά την αυξημένη ταξινομική ποικιλότητα, η λειτουργική ποικιλότητα είναι χαμηλότερη. 18. Οι ιχθυοκοινότητες ανάντη χαρακτηρίζονται από υψηλά επίπεδα πλεονασμού ως προς τη σύνθεση των ειδών και είναι λειτουργικά ομαδοποιημένες σε σύγκριση με τυχαίες ιχθυοκοινότητες που θα προέκυπταν από το περιφερειακό αποθεματικό των 296
297 ειδών. Αυτό το πρότυπο αναδεικνύει το περιβαλλοντικό φιλτράρισμα ως το κυρίαρχο μηχανισμό συγκρότησης των ιχθυοκοινοτήτων στον ποταμό Λούρο. 19. Τα απειλούμενα είδη Valencia letourneuxi (Ζουρνάς) και Cobitis hellenica (Λουροβελονίτσα) χαρακτηρίζονται από την υψηλότερη λειτουργική μοναδικότητα μεταξύ των ειδών που διαβιούν στον ποταμό Λούρο και χρήζουν άμεσων διαχειριστικών ενεργειών, διότι κάθε παρέμβαση ή και καταστροφή του ενδιαιτήματός τους, μπορεί να έχει άμεσες και σημαντικές επιπτώσεις στους πληθυσμούς τους. Επομένως, «Η εκτίμηση της λειτουργικής ποικιλότητας στα ποτάμια οικοσυστήματα είναι απαραίτητη τόσο για τη πληρέστερη μελέτη της βιοποικιλότητας όσο και τη βελτίωση της αποτελεσματικότητας των προσπαθειών διατήρησης των σπάνιων ειδών και των οικοσυστημάτων που χρήζουν άμεσης προστασίας». Η μεθοδολογική προσέγγιση που ακολουθήθηκε καθώς και τα πρότυπα που περιγράφηκαν στην παρούσα μελέτη θα μπορούσαν να συμβάλουν στη σύνθεση των θεωριών της οικολογίας βιοκοινοτήτων και να αποτελέσουν τη βάση για τη δημιουργία διαχειριστικών εργαλείων που βρίσκουν εφαρμογή σε διαφορετικές ταξινομικές ομάδες και γεωγραφικές περιοχές. Η παρούσα μελέτη εκτίμησης της βιοποικιλότητας σε ταξινομικό και λειτουργικό επίπεδο μπορεί να συνεισφέρει στη διασφάλιση της μακροπρόθεσμης διατήρησης της βιοποικιλότητας της Βαλκανικής χερσονήσου, και ως εκ τούτου, τα αποτελέσματα της έχουν δυνητικά, αντίκτυπο στην ευρωπαϊκή ερευνητική ατζέντα σχετικά με τα οικοσυστήματα των εσωτερικών υδάτων. Πιστεύουμε πως τόσο οι μεθοδολογικές προσεγγίσεις όσο και τα αποτελέσματα της παρούσας διδακτορικής διατριβής θα συνεισφέρουν σε μελλοντικές μελέτες σχετικά με τη βιοποικιλότητα και τη λειτουργία των οικοσυστημάτων των εσωτερικών υδάτων, καθώς και σε μελέτες που θα διερευνήσουν τη λειτουργική ποικιλότητα ειδών και βιοκοινοτήτων σε πολλαπλά χωρικά επίπεδα, ώστε να αποκτήσουμε μια ολιστική κατανόηση της βιοποικιλότητας, κυρίως στη λιγότερο μελετημένη Βαλκανική Χερσόνησο. 297
298
299 Βιβλιογραφικές αναφορές
300
301 Aarts, B. G., Van den Brink, F. W., & Nienhuis, P. H. (2004). Habitat loss as the main cause of the slow recovery of fish faunas of regulated large rivers in Europe: the transversal floodplain gradient. River Research and Applications, 20, Abell, R., Thieme, M. L., Revenga, C., Bryer, M., Kottelat, M., Bogutskaya, N., Coad, B., Mandrak, N., Balderas, S. C., Bussing, W., Stiassny, M. L. J., Skelton, P., Allen, G. R., Unmack, P., Naseka, A., Sindorf, N., Robertson, J., Armijo, E., Higgins, J. V., Heibel, T. J., Wikramanayake, E., Olson, D., López, H. L., Reis, R. E., Lundberg, J. G., Pérez, M. H. S, & Petry, P. (2008). Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience, 58, Agustí, J., Rook, L., & Andrews, P. (1999). Evolution of Neogene Terrestrial Ecosystems in Europe. Cambridge: Cambridge University Press. Albert, C., Thuiller, W., Yoccoz, N., Soudant, A., Boucher, F., Saccone, P., & Lavorel, S. (2010). Intraspecific functional variability: extent, structure and sources of variation. Journal of Ecology, 98, Albrecht, C., & Wilke, T. (2008). Lake Ohrid: biodiversity and evolution. Hydrobiologia, 615, Albrecht, C., Wolff, C., Glöer, P., & Wilke, T. (2008). Concurrent evolution of ancient sister lakes and sister species: The freshwater gastropod genus Radix in lakes Ohrid and Prespa. Hydrobiologia, 615, Alcaraz, C., & García-Berthou, E. (2007). Life history variation of invasive mosquitofish (Gambusia holbrooki) along a salinity gradient. Biological Conservation, 139, Allen, A., Whittier, T., Kaufmann, P., Larsen, D. P., O Connor, R. J., Hughes, R. M., Stemberger, R. S., Dixit, S. S., Brinkhurst, R. O., Herlihy, A. T., & Paulsen, S. G. (1999). Concordance of taxonomic richness patterns across multiple assemblages in lakes of the northeastern United States. Canadian Journal of Fisheries and Aquatic Sciences, 56, Amarasinghe, U., & Welcomme, R. (2002). An analysis of fish species richness in natural lakes. Environmental Biology of Fishes, 65, Amataj, S., Anovski, T., Benischke, R., Eftimi, R., Gourcy, L. L., Kola, L., Leontiadis, I., Micevsky, E., Stamos, A., & Zoto, J. (2007). Tracer methods used to verify the hypothesis of Cvijić about the underground connection between Prespa and Ohrid Lake. Environmental Geology, 51, Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecology, 26,
302 Anderson, M. J., Crist, T. O., Chase, J. M., Vellend, M., Inouye, B. D., Freestone, A. L., Sanders, N. J., Cornell, H. V., Comita, L. S., Davies, K. F., Harrison, S. P., Kraft, N. J. B., Stegen, J. C., & Swenson, N. G. (2011). Navigating the multiple meanings of diversity: a roadmap for the practicing ecologist. Ecology Letters, 14, Angermeier, P., & Winston, M. (1999). Characterizing fish community diversity across Virginia landscapes: prerequisite for conservation. Ecological Applications, 9, APHA, AWWA, & WEF. (2012). Standard Methods for the Examination of Water and Wastewater, 22nd Edition. Washington DC: American Public Health Association, American Water Works Association, Water Environment Federation. Apostolidis, A. P., Triantaphyllidis, C., Kouvatsi, A., & Economidis, P. S. (1997). Mitochondrial DNA sequence variation and phylogeography among Salmo trutta L. (Greek brown trout) populations. Molecular Ecology, 6, Araújo, M. B., Nogués Bravo, D., Diniz Filho, J. A., Haywood, A. M., Valdes, P. J., & Rahbek, C. (2008). Quaternary climate changes explain diversity among reptiles and amphibians. Ecography, 31, Arrhenius, O. (1921). Species and area. Journal of Ecology, 9, Ashton, K., Tracy, M., & de Queiroz, A. (2000). Is Bergmann s rule valid for mammals? American Naturalist, 156, Bady, P., Dolédec, S., Fesl, C., Gayraud, S., Bacchi, M., & Schöll, F. (2005). Use of invertebrate traits for the biomonitoring of European large rivers: the effects of sampling effort on genus richness and functional diversity. Freshwater Biology, 50, Baillie, J., & Butcher, E. (2012). Priceless or Worthless? London: Zoological Society of London. Balian, E. V., Lévêque, C., Segers, H., & Martens, K. (2008). Freshwater Animal Diversity Assessment. Dordrecht: Springer. Balon, E. K. (1986). Types of feeding in the ontogeny of fishes and the life-history model. Environmental Biology of Fishes, 16, Bănărescu, P. M. (1990). Zoogeography of fresh waters. Volume 1. General distribution and dispersal of freshwater animals. Wiesbaden: Aula Verlag. 302
303 Bănărescu, P. M. (1992). Zoogeography of fresh waters. Volume 2. Distribution and dispersal of freshwater animals in North America and Eurasia. Wiesbaden: Aula- Verlag. Bănărescu, P. M. (2004). Distribution pattern of the aquatic fauna of the Balkan Peninsula. In H. J. Griffiths, B. Kryštufek, & J. M. Reed, Balkan Biodiversity, Pattern and Process in the European Hotspot (pp ). Dordrecht, Boston, London: Kluwer Academic Publishers. Barbieri, R., Daoulas, C., Psarras, T., Stoumboudi, M., & Economou, A. N. (2000). The biology and ecology of Valencia letourneuxi Sauvage 1880 (Valenciidae). Mediterranean Marine Science, 1, Barbieri, R., Stoumboudi, M., & Economou, A. N. (2002). Threatened fishes of the world: Valencia letourneuxi Sauvage, 1880 (Valenciidae). Environmental Biology of Fishes, 65, 46. Barbieri, R., Zogaris, S., Kalogianni, E., Stoumboudi, M., Chatzinikolaou, Y., Giakoumi, S., Kapakos, Y., Kommatas, D., Koutsikos, N., Tachos, V., Vardakas, L., & Economou, A. N. (2015). Freshwater fishes and lampreys of Greece: An annotated checklist. Monographs on Marine Sciences No. 8. Athens: Hellenic Centre for Marine Research. Baselga, A. (2010). Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography, 19, Baselga, A. (2012). The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Global Ecology and Biogeography, 21, Beard, J., Sangree, J., & Smith, L. (1982). Quaternary Chronology, Paleoclimate, Depositional Sequences and Eustatic Cycles. American Association of Petroleum Geologists, 66, Beck, J., Ballesteros-Mejia, L., Buchmann, C. M., Dengler, J., Fritz, S. A., Gruber, B., Hof, C., Jansen, F., Knapp, S., Kreft, H., Schneider, A.-K., Winter, M., & Dormann, C. F. (2012). What's on the horizon for macroecology? Ecography, 35, Bellwood, D. R., Hughes, T. P., Connolly, S. R., & Tanner, J. (2005). Environmental and geometric constraints on Indo-Pacific coral reef biodiversity. Ecology Letters, 8, Bellwood, D. R., Wainwright, P. C., Fulton, C., & Hoey, A. (2002). Assembly rules and functional groups at global biogeographical scales. Functional Ecology, 16,
304 Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society. Series B, 28, Bergerot, B., Hugueny, B., & Belliard, J. (2015). Relating life-history traits, environmental constraints and local extinctions in river fish. Freshwater Biology, 60, Berrebi, P., Tougard, C., Dubois, S., Shao, Z., Koutseri, I., Petkovski, S., & Crivelli, A. J. (2013). Genetic Diversity and Conservation of the Prespa Trout in the Balkans. International Journal of Molecular Sciences, 14, Bianco, P. G. (1990). Potential role of the paleohistory of the Mediterranean and Paratethys basins on the early dispersal of Euro-Mediterranean freshwater fishes. Ichthyological Exploration of Freshwaters, 1, Bianco, P. G. (2013). An update on the status of native and exotic freshwater fishes of Italy. Journal of Applied Ichthyology, 30, Bianco, P. G., & Nordlie, F. (2008). The salinity tolerance of Pseudophoxinus stymphalicus (Cyprinidae) and Valencia letourneuxi (Valenciidae) from western Greece suggests a revision of the ecological categories of freshwater fishes. Italian Journal of Zoology, 75, Bini, L. M., Diniz-Filho, J. A., Rangel, T. F., Bastos, R. P., & Pinto, M. P. (2006). Challenging Wallacean and Linnean shortfalls: knowledge gradients and conservation planning in a biodiversity hotspot. Diversity and Distributions, 12, Blanchet, S., Grenouillet, G., Beauchard, O., Tedesco, P. A., Leprieur, F., Dürr, H. H., Busson, F., Oberdorff, T., & Brosse, S. (2010). Non-native species disrupt the worldwide patterns of freshwater fish body size: implications for Bergmann's rule. Ecology Letters, 13, Blanck, A., & Lamouroux, N. (2007). Large-scale intraspecific variation in life-history traits of European freshwater fish. Journal of Biogeography, 34, Bohlen, J., Perdices, A., Doadrio, I., & Economidis, P. S. (2006). Vicariance, colonisation, and fast local speciation in Asia Minor and the Balkans as revealed from the phylogeny of spined loaches (Osteichthyes; Cobitidae). Molecular Phylogenetics and Evolution, 39, Bonada, N., & Resh, V. H. (2013). Mediterranean-climate streams and rivers: geographically separated but ecological comparable freshwater systems. Hydrobiologia, 719,
305 Bonada, N., Zamora-Muñoz, C., Rieradevall, M., & Prat, N. (2005). Ecological and historical filters constraining spatial caddisfly distribution in Mediterranean rivers. Freshwater Biology, 50, Botta-Dukát, Z. (2005). Rao s quadratic entropy as a measure of functional diversity based on multiple. Journal of Vegetation Science, 16, Bray, R. J., & Curtis, J. T. (1957). An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 27, Brind'Aamour, A. (2007). Functional diversity: a study on the Bay of Biscay nursery habitats. ICES Annual Science Conference. International Council for the Exploration of the Sea, Helsinki. Brind Amour, A., Boisclair, D., Dray, S., & Legendre, P. (2011). Relationships between species feeding traits and environmental conditions in fish communities: a three-matrix approach. Ecological Applications, 21, Brooks, T. (2010). Conservation planning and priorities. In N. S. Sodhi, & P. R. Ehrlich, Conservation Biology for All (pp ). Oxford: Oxford University Press. Brown, J. H., & Maurer, A. B. (1989). Macroecology: The division of food and space among species on continents. Science, 243, Brown, J. H. (1995). Macroecology. Chicago: The University of Chicago Press. Brown, J. H., & Lomolino, M. V. (1998). Biogeography. Sunderland (Massachusetts): Sinauer Associates. Brucet, S., Pédron, S., Mehner, T., Lauridsen, T. L., Argillier, C., Winfield, I. J., Volta, P., Emmrich, M., Hesthagen, T., Holmgren, K., Benejam, L., Kelly, F., Krause, T., Palm, A., Rask, M., & Jeppesen, E. (2013). Fish diversity in European lakes: geographical factors dominate over anthropogenic pressures. Freshwater Biology, 58, Burridge, C. P., Craw, D., & Waters, J. M. (2006). River capture, range expansion, and cladogenesis: the genetic signature of freshwater vicariance. Evolution, 60, Cadotte, M. W., Carscadden, K., & Mirotchnick, N. (2011). Beyond species: functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology, 48, Cardoso, P., Borges, P., & Veech, J. (2009). Testing the performance of beta diversity measures based on incidence data: the robustness to undersampling. Diversity and Distributions, 15,
306 Carvalho, J. C., Cardoso, P., & Gomes, P. (2012). Determining the relative roles of species turnover and species richness differences patterns. Global Ecology and Biogeography, 21, Carvalho, J. C., Cardoso, P., Crespo, L. C., Henriques, S., Carvalho, R., & Gomes, P. (2011). Biogeographic patterns of spiders in coastal dunes along a gradient of mediterraneity. Biodiversity and Conservation, 20, CEN (2003). Water quality sampling of fish with electricity. European Standard EN 14011: 2003 E. Brussels: Comité Européen de Normalisation. Chandrinos, G. (2009). Birds. In A. Legakis, & P. Maragou, The Red Data Book of Threatened Animals of Greece (pp ). Athens: Hellenic Zoological Society. Chapin, F. S., Zavaleta, E. S., Eviner, V. T., Naylor, R. L., Vitousek, P. M., Reynolds, H. L., Hooper, D. U., Lavorel, S., Sala, O. E., Hobbie, S. E., Mack, M. C. & Diaz, S. Z (2000). Consequences of changing biodiversity. Nature, 405, Cheimonopoulou, M. T., Bobori, D. C., Theocharopoulos, I., & Lazaridou. M. (2011). Assessing ecological water quality with macroinvertebrates and fish: a case study from a small Mediterranean river. Environmental Management, 47, Cianciaruso, M., Batalha, M., Gaston, K. J., & Petchey, O. (2009). Including intraspecific variability in functional diversity. Ecology, 90, Clarke, K. R., & Warwick, R. M. (1994). Similarity-based testing for community pattern: the two-way layout with no replication. Marine Biology, 118, Clavero, M., & Hermoso, V. (2011). Threatening processes and conservation management of endemic freshwater fish in the Mediterranean Basin: a review. Marine and Freshwater Research, 62, Collen, B., Whitton, F., Dyer, E. E., Baillie, J. E. M., Cumberlidge, N., Darwall, W. R. T., Pollock, C., Richman, N. I., Soulsby, A.-M., & Böhm, M. (2014). Global patterns of freshwater species diversity, threat and endemism. Global Ecology and Biogeography, 23, Connor, E. F., & Simberloff, D. (1979). The Assembly of Species Communities: Chance or competition. Ecology, 60, Conover, W. J., & Iman, R. L. (1979). On multiple-comparison procedures. New Mexico: Los Alamos Scientific Laboratory. 306
307 Cooper, S. J., Ibrahim, K. M., & Hewitt, G. M. (1995). Postglacial expansion and genome subdivision in the European grasshopper Chorthippus parallelus. Molecular Ecology, 4, Cornwell, W., & Ackerly, D. (2009). Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California. Ecological Monographs, 79, Darlington, P. J. (1948). The geographical distribution of cold blooded vertebrates. Quarterly Review of Biology, 23, Darwall, W. R. T., Holland, R. A., Smith, K. G., Allen, D., Brooks, E. G. E., Katarya, V., Pollock, C. M., Shi, Y., Clausnitzer, V., Cumberlidge, N., Cuttelod, A., Dijkstra, K.- D. B., Diop, M. D., García, N., Seddon, M. B., Skelton, P. H., Snoeks, J., Tweddle, D., & Vié, J.-C. (2011). Implications of bias in conservation research and investment for freshwater species. Conservation Letters, 4, Darwin, C. (1839). Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. London: Colburn. Darwin, C. (1859). On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: Murray. de Bello, F., Lavorel, S., Albert, C., Thuiller, W., Grigulis, K., Dolezal, J., Janeček, Š., & Lepš, J. (2011). Quantifying the relevance of intraspecific trait variability for functional diversity. Methods in Ecology and Evolution, 2, De Cáceres, M., Legendre, P., Valencia, R., Cao, M., Chang, L.-W., Chuyong, G., Condit, R., Hao, Z., Hsieh, C.-F., Hubbell, S., Kenfack, D., Ma, K., Mi, X., Supardi Noor, Md. N., Kassim, A. R., Ren, H., Su, S.-H., Sun, I.-F., Thomas, D., Ye, W., & He, F. (2012). The variation of tree beta diversity across a global network of forest plots. Global Ecology and Biogeography, 21, DeFaveri, J., Zanella, L. N., Zanella, D., Mrakovčić, M., & Merilä, J. (2012). Phylogeography of isolated freshwater threespine stickleback Gasterosteus aculeatus populations in the Adriatic Sea basin. Journal of Fish Biology, 80, Delling, B. (2010). Diversity of western and southern Balkan trouts, with the description of a new species from the Louros River, Greece (Teleostei: Salmonidae). Ichthyological Exploration of Freshwaters, 21, Dengler, J. (2009). Which function describes the species-area relationship best? - A review and empirical evaluation. Journal of Biogeography, 36,
308 Denys, G., Tedesco, P., Oberdorff, T., & Gaubert, P. (2014). Environmental correlates of body size distribution in Cyprinidae (Actinopterygians) depend on phylogenetic scale. Ecology of Freshwater Fish,25, Dermitzakis, M. (1990). Paleogeography, geodynamic processes and event stratigraphy during the Late Cenozoic of the Aegean area. Accademia Nazionale dei Lincei, 85, Dermitzakis, M., & Papanikolaou, D. (1981). Palaeogeography and geodynamics of the Aegean region during the Neogene. Annales Geologique des Pays Hellenique, 30, Diamond, J. M. (1975). The island dilemma: lessons of modern biogeographic studies for the design of natural reserves. Biological Conservation, 7, Dias, M. S., Cornu, J. F., Oberdorff, T., Lasso, C. A., & Tedesco, P. A. (2013). Natural fragmentation in river networks as a driver of speciation for freshwater fishes. Ecography, Dias, M. S., Oberdorff, T., Hugueny, B., Leprieur, F., Jézéquel, C., Cornu, J. F., Brosse, S., Grenouillet, G., & Tedesco, P. A. (2014). Global imprint of historical connectivity on freshwater fish biodiversity. Ecology Letters, 17, Díaz, S., & Cabido, M. (2001). Vive la differénce: plant functional diversity matters to ecosystem functioning. Trends in Ecology and Evolution, 16, Dodson, S., Arnott, S., & Cottingham, K. (2000). The relationship in lake communities between primary productivity and species richness. Ecology, 81, Dolédec, S., Chessel, D., Ter Braak, C., & Champely, S. (1996). Matching species traits to environmental variables: a new three-table ordination method. Environmental and Ecological Statistics, 3, Drakou, E., Bobori, D. C., Kallimanis, A., Mazaris, A., Sgardelis, S., & Pantis, J. (2009). Freshwater fish community structured more by dispersal limitation than by environmental heterogeneity. Ecology of Freshwater Fish, 18, Dray, S., & Dufour, A. B. (2007). The ade4 package: implementing the duality diagram for ecologists. Journal of Statistical Software, 22, Legendre, P. (2008). Testing the species traits-environment relationships: The fourthcorner problem revisited. Ecology, 89, Dudgeon, D. (2010). Prospects for sustaining freshwater biodiversity in the 21st century: linking ecosystem structure and function. Current Opinion in Environmental Sustainability, 2,
309 Dudgeon, D., Arthington, A., Gessner, M., Kawabata, Z.-I., Knowler, D. J., Lévêque, C., Naiman, R. J., Prieur-Richard, A.-H., Soto, D., Stiassny, M. L. J., & Sullivan, C. A. (2006). Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews, 81, Dufrêne, M., & Legendre, P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs, 67, Douglas, M. E., & W. J. Matthews (1992). Does morphology predict ecology? Hypotheses testing within a freshwater stream fish community. Oikos, 65, Durand, J. D., Bianco, P. G., Laroche, J., & Gilles, A. (2003). Insight into the origin of endemic Mediterranean ichthyofauna: phylogeography of Chondrostoma genus (Teleostei, Cyprinidae). Journal of Heredity, 94, Durand, J. D., Persat, H., & Bouvet, Y. (1999). Phylogeography and postglacial dispersion of the chub (Leuciscus cephalus) in Europe. Molecular Ecology, 8, Eadie, J., Hurly, T., Montgomerie, R., & Teather, K. (1986). Lakes and rivers as islands: species-area relationships in the fish faunas of Ontario. Environmental Biology of Fishes, 15, Economidis, P. S. (1991). Checklist of the freshwater fish of Greece (recent status of threat and protection). Athens: Bulletin of the Hellenic Society for the Protection of Nature. Economidis, P. S. (1995). Endangered freshwater fishes of Greece. Biological Conservation, 72, Economidis, P. S. (2009). Fishes of inland waters. In A. Legakis, & P. Maragou, The Red Data Book of Threatened Animals of Greece (pp ). Athens: Hellenic Zoological Society. Economidis, P. S., & Bănărescu, P. M. (1991). The distribution and origins of freshwater fishes in the Balkan Peninsula, especially in Greece. Internationale Revue Der Gesamten Hydrobiologie, 76, Economou, A. N., Giakoumi, S., Vardakas, L., Barbieri, R., Stoumboudi, M., & Zogaris, S. (2007). The freshwater ichthyofauna of Greece: an update based on a hydrographic basin survey. Mediterranean Marine Science, 8, Erdei, B., Hably, L., Kazmer, M., Utescher, T., & Bruch, A. (2007). Neogene flora and vegetation development of the Pannonian domain in relation to palaeoclimate 309
310 and palaeogeography. Palaeogeography, Palaeoclimatology, Palaeoecology, 253, Erös, T., Heino, J., Schmera, D. & Rask, M. (2009), Characterising functional trait diversity and trait environment relationships in fish assemblages of boreal lakes. Freshwater Biology, 54, European Environment Agency Faith, D. (1992). Conservation evaluation and phylogenetic diversity. Biological Conservation, 61, Fakiris, Ε., Zoura, D., Katsanou, K., Kriempardi, P., & Lambrakis, N. (2013). Using spectral analysis for missing values treatment in long-term, daily sampled rainfall time series. In N. Lambrakis, G. Stournaras, & K. Katsanou, Advances in the Research of Aquatic Environment (pp ). Berlin: Springer. FishBase, World Wide Web electronic publication. Version Fox, J., Ades, P., & Bi, H. (2001). Stochastic structure and individual-tree growth models. Forest Ecology and Management, 154, Frankham, R. (2005). Genetics and extinction. Biological Conservation, 126, Freyhof, J. (2012). Threatened freshwater fishes and mollusks of the Balkan: Potential Impact of hydropower projects. Berlin: ECA Watch Austria & Euronatur. Frimpong, E., & Angermeier, P. (2010). Traits-based approaches in stream fish community analysis. In D. Jackson, & K. Gido, Community ecology of stream fishes: concepts, approaches, and techniques (pp ). Bethesda, Maryland: American Fisheries Society, Symposium 73. Fritz, U., Guicking, D., Kami, H., Arakelyan, M., Auer, M., Ayaz, D., Ayres Fernández, D., Bakiev, A. G., Celani, A., Džukić, G., Fahd, S., Havaš, P., Joger, U., Khabibullin, V. F., Mazanaeva, L. F., Široký, P., Tripepi, S., Valdeón Vélez, A., Velo Antón, G., & Wink, M. (2007). Mitochondrial phylogeography of European pond turtles (Emys orbicularis, Emys trinacris) - an update. Amphibia-Reptilia, 28, Frogley, M., & Preece, R. (2004). A Faunistic Review of the Modern and Fossil Molluscan Fauna from Lake Pamvotis, Ioannina, an Ancient Lake in Nw Greece: Implications for Endemism in the Balkans. In H. Griffiths, B. Kryštufek, & J. Reed, Balkan Biodiversity, Pattern and Process in the European Hotspot (pp ). Dordrecht, Boston, London: Kluwer Academic Publishers. 310
311 Gallardo, B., Gascon, S., Cabezas, A., Gonzalez, M., Garcia, M., & Comin, F. (2009). Relationship between invertebrate traits and lateral environmental gradients in a Mediterranean river-floodplain. Fundamendall and Applied Limnology, 173, Gamfeldt, L., Wallen, J., Jonsson, P., Berntsson, K., & Havenhand, J. (2005). Increasing intraspecific diversity enhances settling success in a marine invertebrate. Ecology, 86, Garcia-Moreno, J., Harrison, I. J., Dudgeon, D., Clausnitzer, V., Darwall, W., Farrell, T., Savy, C., K. Tockner, K., & Tubbs, N. (2014). Sustaining Freshwater Biodiversity in the Anthropocene. In A. Bhaduri, J. Bogardi, J. Leentvaar, & S. Marx, The Global Water System in the Anthropocene (pp ). Cham Heidelberg New York Dordrecht London: Springer. Gaston, K. J., & Blackburn, T. M. (2000). Pattern and process in macroecology. Oxford: Blackwell Science. Gatz, A. (1979). Ecological morphology of freshwater stream fishes. Tulane Studies in Zoology and Botany, 21, Gaubert, P., Denys, G., & Oberdorff, T. (2009). Genus-level supertree of Cyprinidae (Actinopterygii: Cypriniformes), partitioned qualitative clade support and test of macro-evolutionary scenarios. Biological reviews of the Cambridge Philosophical Society, 84, Geiger, M. F., Herder, F., Monaghan, M. T., Almada, V., Barbieri, R., Bariche, M., Berrebi, P., Bohlen, J., Casal-Lopez, M., Delmastro, G. B., Denys, G. P. J., Dettai, A., Doadrio, I., Kalogianni, E., Kärst, H., Kottelat, M., Kovačić, M., Laporte, M., Lorenzoni, M., Marčić, Z., Özuluğ, M., Perdices, A., Perea, S., Persat, H., Porcelotti, S., Puzzi, C., Robalo, J., Šanda, R., Schneider, M., Šlechtová, V., Stoumboudi, M., Walter, S., & Freyhof, J. (2014). Spatial heterogeneity in the Mediterranean Biodiversity Hotspot affects barcoding accuracy of its freshwater fishes. Molecular Ecology Resources, 14, Geist, J. (2011). Integrative freshwater ecology and biodiversity conservation. Ecological Indicators, 11, Global Invasive Species Program (2015) Gkenas, Ch., & Leonardos, I. D. (2012). Journal of Applied Ichthyology. Life cycle of a small endemic goby Economidichthys pygmaeus (Family: Gobiidae) in Lake Pamvotis, northwestern Greece, 28, Gkenas, Ch., Malavasi, S., Georgalas, V., Leonardos, I. D., & Torricelli, P. (2010). The reproductive behavior of Economidichthys pygmaeus: secondary loss of sound 311
312 production within the sand goby group? Environmental Biology of Fishes, 87, Gkenas, Ch., Malavasi, S., & Leonardos, I. D. (2012). Diet and feeding habits of Economidichthys pygmaeus (Perciformes: Gobiidae) in Lake Pamvotis, NW Greece. Journal of Applied Ichthyology, 28, Gkenas, Ch., Oikonomou, A., Economou, A., Kiosse, F., & Leonardos, I. (2012). Life history pattern and feeding habits of the invasive mosquitofish, Gambusia holbrooki, in Lake Pamvotis (NW Greece). Journal of Biological Research- Thessaloniki, 17, Gleason, H. A. (1922). On the relationship between species and area. Ecology, 3, Gómez, A., & Lunt, D. H. (2007). Refugia within refugia: patterns of phylogeographic concordance in the Iberian Peninsula. In S. Weiss, & N. Ferrand, Phylogeography of Southern European Refugia (pp ). Dordrecht: Springer. Gonzàlez-Orozco, C. E., Thornhill, A. H., Knerr, N., Laffan, S., & Miller, J. T. (2014). Biogeographical regions and phytogeography of the eucalypts. Diversity and Distributions, 20, Google Earth (2015). Google. Version Googleplex, Gorman, O. T., & Karr, J. R. (1978). Habitat structure and stream fish communities. Ecology, 59, Gotelli, N. J. (2000). Null model analysis of species co-occurrence patterns. Ecology, 81, Grenouillet, G., Pont, D., & Seip, K. (2002). Abundance and species richness as a function of food resources and vegetation structure: juvenile fish assemblages in rivers. Ecography, 25, Griffiths, D. (1997). Local and regional species richness in North American lacustrine fish. Journal of Animal Ecology, 66, Griffiths, D. (2006). Pattern and process in the ecological biogeography of European freshwater fish. Journal of Animal Ecology, 75, Griffiths, D. (2012). Body size distributions in North American freshwater fish: largescale factors. Global Ecology and Biogeography, 21,
313 Griffiths, D. (2013). Body size distributions in North American fishes: small-scale factors and synthesis. Ecology of Freshwater Fish, 22, Griffiths, D., McGonigle, C., & Quinn, R. (2014). Climate and species richness patterns of freshwater fish in North America and Europe. Journal of Biogeography, 41, Grime, J. P. (1973). Control of species density in herbaceous vegetation. Journal of Environmental Management, 1, Grime, J. P. (2006). Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences. Journal of Vegetation Science, 17, Grueber, C., Nakagawa, S., Laws, R., & Jamieson, I. (2011). Multimodel inference in ecology and evolution: challenges and solutions. Journal of Evolutionary Biology, 24, Gugerli, F., Englisch, T., Niklfeld, H., Tribsch, A., Mirek, Z., Ronikier, M., Zimmermann, N. E., Holderegger, R., & Taberlet, P. (2008). Relationships among levels of biodiversity and the relevance of intraspecific diversity in conservation - a project synopsis. Perspectives in Plant Ecology, Evolution and Systematics, 10, Guilhaumon, F., Gimenez, O., Gaston, K. J., & Mouillot, D. (2008). Taxonomic and regional uncertainty in species-area relationships and the identification of richness hotspots. Proceedings of the National Academy of Sciences USA, 105, Guilhaumon, F., Mouillot, D., & Gimenez, O. (2010). mmsar: an R-package for multimodel species-area relationship inference. Ecography, 33, Hall, H. M., & Grinnell, J. (1919). Life-zone indicators in California. Proceedings of the California Academy of Sciences, 9, Hänfling, B., Dumpelmann, C., Bogutskaya, N. G., Brandl, R., & Brändle, M. (2009). Shallow phylogeographic structuring of Vimba vimba across Europe suggests two distinct refugia during the last glaciation. Journal of Fish Biology, 75, Harris, J. E.(1937). The mechanical significance of the position and movements of the paired fins in the Teleostei. Papers from the Tortugas Laboratory Carnegie Institution of Washington, 16,
314 Heikinheimo, H., Fortelius, M., Eronen, J. T. & Mannila, H. (2007). European land mammal species cluster into environmentally distinct and spatially coherent entities. Journal of Biogeography, 34, Heino, J. (2005). Functional biodiversity of macroinvertebrate assemblages along major ecological gradients of boreal headwater streams. Freshwater Biology, 50, Heino, J. (2011). A macroecological perspective of diversity patterns in the freshwater realm. Freshwater Biology, 56, Heino, J., Mykrä, H., & Kotanen, J. (2008). Weak relationships between landscape characteristics and multiple facets of stream macroinvertebrate biodiversity in a boreal drainage basin. Landscape Ecology, 23, Heino, J., Schmera, D., & Erös, T. (2013). A macroecological perspective of trait patterns in stream communities. Freshwater Biology, 58, Helminen, H., Karjalainen, J., Kurkilahti, M., Rask, M., & Sarvala, J. (2000). Eutrophication and fish biodiversity in Finnish lakes. Verhandlungen Internationale Vereinigung Limnologie, 27, Herbert, M. E., Mcintyre, P. B., Doran, P. J., Allan, J. D., & Abell, R. (2010). Terrestrial Reserve Networks Do Not Adequately Represent Aquatic Ecosystems. Conservation Biology, 24, Hershkovitz, Y., & Gasith, A. (2013). Resistance, resilience, and community dynamics in mediterranean-climate streams. Hydrobiologia, 719, Hewitt, G. M. (2000). The genetic legacy of the ice ages. Nature, 405, Hill, M. O., & Smith, A. J. E. (1976). Principal component analysis of taxonomic data with multi-state discrete characters. Taxon, 25, Himmerkus, F., Reischmann, T., & Kostopoulos, D. K. (2006). Late Proterozoic and Silurian basement units within the Serbo-Macedonian Massif, northern Greece: the significance of terrane accretion in the Hellenides. In A. H. Robertson, & D. Mountrakis, Tectonic Development of the Eastern Mediterranean Region (pp ). Bath: Geological Society Publishing House. Hoagstrom, C. W., & Berry Jr, C. R. (2006). Island biogeography of native fish faunas among Great Plains drainage basins: basin scale features influence composition. American Fisheries Society Symposium, 48, Hoagstrom, C. W., Gosch, N., DeWitte, A., Berry, Jr, C. R., & Duehr, J. (2007). Biodiversity, biogeography, and longitudinal fish faunal structure among 314
315 perennial, warmwater streams of the Cheyenne River drainage. Prairie Naturalist, 39, Hobaek, A., Manca, M., & Andersen, T. (2002). Factors influencing species richness in lacustrine zooplankton. Acta Oecologica, 23, Hoeinghaus, D. J., Winemiller, K. O., & Birnbaum, J. S. (2007). Local and regional determinants of stream fish assemblage structure: inferences based on taxonomic vs. functional groups. Journal of Biogeography, 34, Hof, C., Brändle, M., & Brandl, R. (2008). Latitudinal variation of diversity in European freshwater animals is not concordant across habitat types. Global Ecology and Biogeography, 17, Holdridge, L., Grenke, W., Hatheway, W., Liang, T., & Tosi, J. (1971). Forest environments in tropical life zone. Oxford: Pergamon Press. Holland, R. A., Darwall, W. R. T., & Smith, K. G. (2012). Conservation priorities for freshwater biodiversity: The Key Biodiversity Area approach refined and tested for continental Africa. Biological Conservation, 148, Hollis, G., & Stevenson, A. (1997). The physical basis of Lake Mikri Prespa systems: geology, climate and water quality. Hydrobiologia, 351, Holt, B. G., Lessard, J. P., Borregaard, M. K., Fritz, S. A., Araújo, M. B., Dimitrov, D., F., Fabre, P.-H., Graham, C. H., Graves, G. R., Jønsson, K. A., Nogues-Bravo, D., Wang, Z., Whittaker, R. J., Fjeldså, J., & Rahbek, C. (2013). An update of Wallace's zoogeographic regions of the World. Science, 339, Hooper, D. U., Adair, E. C., Cardinale, B. J., Byrnes, J. E., Hungate, B. A., Matulich, K. L., Gonzalez, A., Duffy, J. E., Gamfeldt, L., & O'Connor, M. I. (2012). A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature, 486, Hora, S. L. (1952). The Himalayan fishes. Himalaya, 1, Hortal, J., Diniz Filho, J., Bini, L., Rodríguez, M. A., Baselga, A., Nogués Bravo, D., Rangel, T. F., Hawkins, B. A., & Lobo, J. M. (2011). Ice age climate, evolutionary constraints and diversity patterns of European dung beetles. Ecology Letters, 14, Hortal, J., Nabout, J., Calatayud, J. C., Carneiro, F. M., Padial, A., Santos, A. M. C., Siqueira, T., Bokma, F., Bini, L. M., & Ventura, M. (2014). Perspectives on the use of lakes and ponds as model systems for macroecological research. Journal of limnology, 73,
316 Horwitz, R. J. (1978) Temporal variability patterns and the distributional patterns of stream fishes. Ecological Monographs, 48, Hsü, K. J., Montadert, L., Bernoulli, D., Cita, M. B., Erickson, A., Erickson, A., Garrison, R. E., Kidd, R. B., Mèlierés, F., Müller, C., & Wright, R. (1977). History of the Mediterranean salinity crisis. Nature, 267, Hubbell, S. (2001). The Unified Neutral Theory of Biodiversity and Biogeography. Princenton, NJ: Princeton University Press. Hughes, R., Wang, L., & Seelbach, P. (2006). Landscape influences on stream habitats and biological assemblages. Bethesda (Maryland): American Fisheries Society, Symposium 48. Hugueny, B. (1989). West African rivers as biogeographic islands: species richness of fish communities. Oecologia, 79, Hugueny, B., Movellan, A., & Belliard, J. (2011). Habitat fragmentation and extinction rates within freshwater fish communities: a faunal relaxation approach. Global Ecology and Biogeography, 20, Hugueny, B., Oberdorff, T., & Tedesco, P. (2010). Community ecology of river fishes: a large scale perspective. InIn K. B. Gido & D. A. Jackson, Community ecology of stream fishes: concepts, approaches, and techniques (pp ). Bethesda (Massachusetts): American Fisheries Society Symposium. Hutchinson, G. E. (1957). Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology, 22, Hutchinson, G. E. (1961). The paradox of the plankton. American Naturalist, 95, Hutchinson, G. E. (1965). The Ecological Theater and the Evolutionary Play. New Haven, Connecticut: Yale University Press. IMBRIW. (2013). Ιnland waters fish monitoring operations manual, electrofishinh health and safety / HCMR rapid fish sampling protocol. Anavyssos Attiki: Hellenic Centre for Marine Research. Irz, P., Michonneau, F., Oberdorff, T., Whittier, T., Lamouroux, N., Mouillot, D., & Argillier, C. (2007). Fish community comparisons along environmental gradients in lakes of France and north-east USA. Global Ecology and Biogeography, 16,
317 Isaac, N., Turvey, S., Collen, B., Waterman, C., & Baillie, J. (2007). Mammals on the EDGE: conservation priorities based on threat and phylogeny. PloS ONE, 2, e IUCN (2013). IUCN Red List of Threatened Species. Version Jackson, D. A., Peres-Neto, P. R., & Olden, J. D. (2001). What controls who is where in freshwater fish communities - the roles of biotic, abiotic, and spatial factors. Canadian Journal of Fisheries and Aquatic Sciences, 58, Jaramillo-Villa, U., Maldonado-Ocampo, J. A., & Escobar, F. (2010). Altitudinal variation in fish assemblage diversity in streams of the central Andes of Colombia. Journal of Fish Biology, 76, Jeffrey, S., & Humphrey, G. (1975). New spectrophotometric equations for determining chlorophyll a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochemie und Physiologie der Pflanzen, 167, Jetz, W., Rahbek, C., & Colwell, R. K. (2004). The coincidence of rarity and richness and the potential signature of history in centres of endemism. Ecology Letters, 7, Kagalou, I., & Leonardos, I. (2009). Typology, classification and management issues of Greek lakes: implication of the Water Framework Directive (2000/60/EC). Environmental Monitoring and Assessment, 150, Kagalou, I., Papastergiadou, E., & Leonardos, I. (2008). Long term changes on eutrophication process in a shallow Mediterranean lake ecosystem of W. Greece: response after the reduction of external load. Journal of Environmental Management, 87, Kalogianni, E., Giakoumi, S., Andriopoulou, A., & Chatzinikolaou, Y. (2014). Prey utilisation and trophic overlap between the non native mosquitofish and a native fish in two Mediterranean rivers. Mediterranean Marine Science, 15, Kalogianni, E., Giakoumi, S., Zogaris, S., Chatzinikolaou, Y., Zimmerman, B., & Economou, A. N. (2010). Current distribution and ecology of the critically endangered Valencia letourneuxi in Greece. Biologia, 65, Kang, B., Deng, J., Wu, Y., Chen, L., Zhang, J., Qiu, H., Lu, Y., & He, D. (2014 (2013). Mapping China s freshwater fishes: diversity and biogeography. Fish and Fisheries, 15,
318 Kapsimalis, V., Pavlakis, P., Poulos, S., Alexandri, S., Tziavos, C., Sioulas, A., Filippas, D., & Lykousis, V. (2005). Internal structure and evolution of the Late Qauternary sequence in a shallow embayment: The Amvrakikos Gulf, NW Greece. Marine Geology, , Karpouzi, V., & Stergiou, K. (2003). The relationships between mouth size and shape and body length for 18 species of marine fishes and their trophic implications. Journal of Fish Biology, 62, Karr, J. R.,& Chu, E. W. (1999). Restoring Life in Running Waters: Better Biological Monitoring. Washington: Island Press. Katsaounos, C., Giokas, D., Leonardos, I. D., & Karayannis, M. (2007). Speciation of phosphorus fractionation in river sediments by explanatory data analysis. Water Research, 41, Keast, A., & Webb, D. (1966). Mouth and body form relative to feeding ecology in the fish fauna of a small lake, Lake Opinicon, Ontario. Journal of the Fisheries Research Board of Canada, 23, Keck, B., Marion, Z., Martin, D., Kaufman, J., Harden, C., Schwartz, J., & Strange, R. (2014). Fish functional traits correlated with environmental variables in a temperate biodiversity hotspot. PLoS One, 9, e Keddy, P. A. (1992), Assembly and response rules: two goals for predictive community ecology. Journal of Vegetation Science, 3, Kelley, L. A., Gardner, S. P., & Sutcliffe, M. J. (1996). An automated approach for clustering an ensemble of NMR-derived protein structures into conformationally related subfamilies. Protein Engineering, 9, Kissling, W. D., Dormann, C. F., Groeneveld, J., Hickler, T., Kühn, I., McInerny, G. J., Montoya, J. M., Römermann, C., Schiffers, K., Schurr, F. M., Singer, A., Svenning, J.-C., Zimmermann, N. E., & O Hara, R. B. (2012). Towards novel approaches to modelling biotic interactions in multispecies assemblages at large spatial extents. Journal of Biogeography, 39, Knouft, J. (2004). Latitudinal variation in the shape of the species body size distribution: an analysis using freshwater fishes. Oecologia, 139, Koleff, P., Gaston, K., & Lennon, J. (2003). Measuring beta diversity for presenceabsence data. Journal of Animal Ecology, 72, Konstantinou, I., Hela, D., & Albanis, T. (2006). The status of pesticide pollution in surface waters (rivers and lakes) of Greece. Part I. Review on occurrence and levels. Environmental Pollution, 141,
319 Kottelat, M., & Freyhof, J. (2007). Handbook of European freshwater fishes. Publications Kottelat: Cornol. Koutsikos, N., Zogaris, S., Vardakas, L., Tachos, V., Kalogianni, E., Šanda, R., Chatzinikolaou, Y., Giakoumi, S., Economidis, P. S., & Economou, A. N. (2012). Recent contributions to the distribution of the freshwater ichthyofauna in Greece. Mediterranean Marine Science, 13, Krebs, C. (2009). Ecology The Experimental Analysis of Distribution and Abundance. San Francisco: Pearson Benjamin Cummings. Kreft, H., & Jetz, W. (2010). A framework for delineating biogeographical biogeographical regions based on species distributions. Journal of Biogeography, 37, Kryštufek, B. (2004). A quantitative assessment of Balkan mammal diversity. In H. J. Griffiths, B. Kryštufek, & J. M. Reed, Balkan Biodiversity, Pattern and Process in the European Hotspot (pp ). Dordrecht, Boston, London: Kluwer Academic Publisher. Kryštufek, B., & Reed, J. M. (2004). Pattern and process in Balkan biodiversity- an overview. In H. J. Griffiths, B. Kryštufek, & J. M. & Reed, Balkan Biodiversity, Pattern and Process in the European Hotspot (pp. 1-8). Dordrecht, Boston, London: Kluwer Academic Publishers. Kryštufek, B., Bužan, E., Hutchinson, W. F., & Hӓnfling, B. (2007). Phylogeography of the rare Balkan endemic Martino's vole, Dinaromys bogdanovi, reveals strong differentiation within the western Balkan Peninsula. Molecular Ecology, 16, Kulbicki, M., Parravicini, V., Arias-Gonzàlez, E., Chabanet, P., Floeter, S. R., Friedlander, A., McPherson, J., Myers, R. E., Vigliola, L., & Mouillot, D. (2013). Global Biogeography of Reef Fishes: A Hierarchical Quantitative Delineation of Regions. PLoS ONE, 8: e Ladle, R. J., & Whittaker, R. J. (2011). Conservation biogeography. Oxford: Wiley- Blackwell. Laliberté, E., & Legendre, P. (2010). A distance-based framework for measuring functional diversity from multiple traits. Ecology, 91, Lamoreux, J. F., Morrison, J. C., Ricketts, T. H., Olson, D. M., Dinerstein, E., McKnight, M. W., & Shugart, H. H. (2006). Global tests of biodiversity concordance and the importance of endemism. Nature, 440,
320 Lamouroux, N., Olivier, J.-M., Persat, H., PouilLy, M., Souchon, Y., & Statzner, B. (1999). Predicting community characteristics from habitat conditions: fluvial fish and hydraulics. Freshwater Biology, 42, Lamouroux, N., Poff, N., & Angermeier, P. (2002). Intercontinental convergence of stream fish community traits along geomorphic and hydraulic gradients. Ecology, 83, Larsen, S., Mancini, L., Pace, G., Scalici, M., & Tancioni, L. (2012). Weak concordance between fish and macroinvertebrates in Mediterranean streams. PLoS ONE, 7, e Lavin, M., Schrire, B. P., Lewis, G., Pennington, R. T., Delgado-Salinas, A., Thulin, M., Hughes, C. E., Matos, A. B., & Wojciechowski, M. F. (2004). Metacommunity process rather than continental tectonic history better explains geographically structured phylogenies in legumes. Philosophical Transactions of the Royal Society B, 359, Lavorel, S., & Garnier, E. (2002). Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology, 16, Lebrija-Trejos, E., Perez-Garcia, E., Meave, J., Bongers, F., & Poorter, L. (2010). Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology, 91, Leduc, A. O. H. C., Silva, E. M., & Rosenfeld, J. S. (2015). Effects of species vs. functional diversity: Understanding the roles of complementarity and competition on ecosystem function in a tropical stream fish assemblage. Ecological Indicators, 48, Legakis, A. (2009). Invertebrates. In A. Legakis, & P. Maragou, The Red Data Book of Threatened Animals of Greece (pp ). Athens: Hellenic Zoological Scociety. Legendre, P., & De Cáceres, M. (2013). Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecology Letters, 16, Legendre, P., Galzin, R., & Harmelin-Vivien, M. (1997). Relating behavior to habitat: Solutions to the fourth-corner problem. Ecology, 78, Lennon, J. J., Koleff, P., Greenwood, J. J. D., & Gaston, K. J. (2001). The geographical structure of British bird distributions: diversity, spatial turnover and scale. Journal of Animal Ecology, 70,
321 Leonardos, I., Kagalou, I., Tsoumani, M., & Economidis, P. (2008). Fish fauna in a Protected Greek lake: biodiversity, introduced fish species over a 80-year period and their impacts on the ecosystem. Ecology of Freshwater Fish, 17, Leprieur, F., & Oikonomou, A. (2014). The need for richness-independent measures of turnover when delineating biogeographical regions. Journal of Biogeography, 41, Leprieur, F., Olden, J. D., Lek, S., & Brosse, S. (2009). Contrasting patterns and mechanisms of spatial turnover for native and exotic freshwater fish in Europe. Journal of Biogeography, 36, Leprieur, F., Tedesco, P., Hugueny, B., Beauchard, O., Dür, H., Brosse, S., & Oberdorff, T. (2011). Partitioning global patterns of freshwater fish beta diversity reveals contrasting signatures of past climate changes. Ecology Letters, 14, Lévêque, C., Oberdorff, T., Paugy, D., Stiassny, M. L., & Tedesco, P. A. (2008). Global diversity of fish (Pisces) in freshwater. Hydrobiologia, 595, Levy, A., Doadrio, I., & Almada, V. C. (2009). Historical biogeography of European leuciscins (Cyprinidae): evaluating the Lago Mare dispersal hypothesis. Journal of Biogeography, 36, Linder, H. P., de Klerk, H. M., Born, J., Burgess, N. D., Fjeldså, J., & Rahbek, C. (2012). The partitioning of Africa: statistically defined biogeographical regions in sub- Saharan Africa. Journal of Biogeography, 39, Logez, M., Bady, P., Melcher, A., & Pont, D. (2013). A continental-scale analysis of fish assemblage functional structure in European rivers. Ecography, 36, Logez, M., Pont, D., & Ferreira, M. (2010). Do Iberian and European fish faunas exhibit convergent functional structure along environmental gradients. Journal of the North American Benthological Society, 29, Lomolino, M. V. (2000). Ecology s most general, yet protean pattern: the species-area relationship. Journal of biogeography, 27, Lomolino, M. V. (2004). Conservation biogeography. In: M. V. Lomolino, & L. R. Heaney, Frontiers of Biogeography: new directions in the geography of nature (pp ). Sunderland (Massachusetts): Sinauer Associates. Lomolino, M. V., Riddle, B. R., Whittaker, R. J., Brown, J. H. (2010). Biogeography. Sunderland (Massachusetts): Sinauer Associates. 321
322 Loreau, M. (1998). Biodiversity and ecosystem functioning: a mechanistic model. Proceedings of the National Academy of Sciences of the United States of America, 95, Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., Hooper, D. U., Huston, M. A., Raffaelli, D., Schmid, B., Tilman, D., Wardle, D. A. (2001). Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science, 294, Losos, J. B., & Schluter, D. (2000). Analysis of an evolutionary species-area relationship. Nature, 408, MacArthur, R. (1955). Fluctuations of animal populations and a measure of community stability. Ecology, 36, MacArthur, R. H., & Levins, R. (1967). The limiting similarity convergence and divergence of coexisting species. American Naturalist, 101, MacArthur, R., & Wilson, E. (1967). The Theory of Island Biogeography. Princeton, NJ: Princeton University Press. Maceda-Veiga, A. (2013). Towards the conservation of freshwater fish: Iberian Rivers as an example of threats and management practices. Reviews in Fish Biology and Fisheries, 23, Magurran, A. E. (1988). Ecological Diversity and its Measurement. Princeton: Princeton University Press. Mahon, R. (1984). Divergent structure in fish taxocenes of north temperate streams. Canadian Journal of Fisheries and Aquatic Sciences, 41, Marr, S. M., Marchetti, M. P., Olden, J. D., Morgan, D. L., García-Berthou, E., Arismendi, I., Day, J. A., Griffiths, C. L., & Skelton, P. H. (2010). Freshwater fish introductions in mediterranean-climate regions: are there commonalities in the conservation problem? Diversity and Distributions, 16, Marr, S. M., Olden, J. D., Leprieur, F., Arismendi, I., Ćaleta, M., Morgan, D. L., Nocita, A., Šanda, R., Tarkan, A. S., & García-Berthou, E. (2013). A global assessment of freshwater fish introductions in Mediterranean-climate regions. Hydrobiologia, 719, Mason, N., Irz, P., Lanoiselee, C., Mouillot, D., & Argillier, C. (2008). Evidence that niche specialization explains species-energy relationships in lake fish communities. Journal of Animal Ecology, 77,
323 Mason, N., Lanoiselee, C., Mouillot, D., Irz, P., & Argillier, C. (2007). Functional characters combined with null models reveal inconsistency in mechanisms of species turnover in lacustrine fish communities. Oecologia, 153, Mason, N., Mouillot, D., Lee, W., & Wilson, J. (2005). Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 111, Matamoros, W. A., Kreiser, B. R., & Schaefer, J. F. (2012). A delineation of Nuclear Middle America biogeographical provinces based on river basin faunistic similarities. Reviews in Fish Biology and Fisheries, 22, Matthews, J. A., Bridges, E. M., Caseldine, C. J., Luckman, A. J., Owen, G., Perry, A. H., Shakesby, R. A. Walsh, R. P. D., Whittaker, R. J. & Willis, K. J. (2001). The encyclopaedic dictionary of environmental change. London: Arnold. Matthews, W. J. (1998a). Influence of Global to Regional Zoogeography on Local Fish Assemblages. InIn W. Matthews, Patterns in Freshwater Fish Ecology (pp ). Dodrect: Springer US. Matthews, W. J. (1998b). Patterns in Freshwater Fish Ecology. Dodrect: Springer US Matthews, W. J., & Robison, H. W. (1998). Influence of drainage connectivity, drainage area and regional species richness on fishes of the interior highlands in Arkansas. American Midland Naturalist, 139, May, R. (1990). Taxonomy as destiny. Nature, 347, Mayfield, M. M., & Levine, J. M. (2010). Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecology Letters, 13, Mazaris, A., Moustaka-Gouni, M., Michaloudi, E., & Bobori, D. C. (2010). Biogeographical patterns of freshwater micro- and macroorganisms: a comparison between phytoplankton, zooplankton and fish in the eastern Mediterranean. Journal of Biogeography, 37, McCann, K. (2000). The diversity-stability debate. Nature, 405, McGill, B. J., Enquist, B. J., Weiher, E., & Westoby, M. (2006). Rebuilding community ecology from functional traits. Trends in Ecology and Evolution, 21, McIntyre, P., Flecker, A. S., Vanni, M. J., Hood, J. M., Taylor, B. W., & Thomas, S. A. (2008). Fish distributions and nutrient cycling in streams: can fish create biogeochemical hotspots? Ecology, 89,
324 McKinney, M. L., & Lockwood, J. L. (1999). Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends in Ecology and Evolution, 14, McKnight, M., White, P., McDonald, R., Lamoreux, J., Sechrest, W., Ridgely, R., & Stuart, S. (2007). Putting beta-diversity on the map: broad-scale congruence and coincidence in the extremes. PLoS Biology, 5, Meador, M., Cuffney, T., & Gurtz, M. (1993). Methods for sampling fish communities as part of the National Water-Quality Assessment Program. Reston (Virginia): Department of the Interior, U.S. Geological Survey. Meiri, S., & Dayan, T. (2003). On the validity of Bergmann s rule. Journal of Biogeography, 30, Mela, C., & Kopalle, P. (2002). The impact of collinearity on regression analysis: the asymmetric effect of negative and positive correlations. Applied Economics, 34, Mérigot, B., Durbec, J., & Gaertner, J. (2010). On goodness-of-fit measure for dendrogram-based analyses. Ecology, 91, Mihinjač, T., Marčić, Z., Mrakovčić, M., Mustafić, P., Zanella, D., & Buj, I. (2015). Threatened fishes of the world: Cobitis jadovaensis Mustafić & Mrakovčić, 2008 (Cobitidae). Croatian Journal of Fisheries, 73, Mikšik, M., & Schraml, E. (2014). World of fishes. Millennium Ecosystem Assessment (2005). Ecosystems and Human Well-being: Synthesis. Washington, DC: Island Press. Mims, M., & Olden, J. (2012). Life history theory predicts fish assemblage response to hydrologic regimes. Ecology, 93, Mims, M., Olden, J., Shattuck, Z., & Poff, N. (2010). Life history trait diversity of native freshwater fishes in North America. Ecology of Freshwater Fish, 19, Miyazono, S., Aycock, J. N., Miranda, L. E., & Tietjen, T. E. (2010). Assemblage patterns of fish functional groups relative to habitat connectivity and conditions in floodplain lakes. Ecology of Freshwater Fish, 19, Monod, J. (1950). La technique de culture continue, théorie et applications. Annales de l Institut Pasteur, 79, Moritz, C. (2002). Strategies to protect biological diversity and the evolutionary processes that sustain it. Systematic Biology, 51,
325 Morlon, H., Chuyong, G., Condit, R., Hubbell, S. P., Kenfack, D., & Thomas, D. (2008). A general framework for the distance-decay of similarity in ecological communities. Ecology Letters, 11, Morrone, J. J. (1994) On the identification of areas of endemism. Systematic Biology, 43, Motta, J. P., Norton, S. F., & Luczkovich, J. J. (1995). Perspective on the ecomorphology of bony fishes. Environmental Biology of Fishes, 44, Mouchet, M., Villéger, S., Mason, N., & Mouillot, D. (2010). Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 24, Mouillot, D., Dumay, O., & Tomasini, J. A. (2007). Limiting similarity, niche filtering and functional diversity in coastal lagoon fish communities. Estuarine, Coastal and Shelf Science, 71, Mouillot, D., Graham, N., Villéger, S., Mason, N., & Bellwood, D. (2013a). A functional approach reveals community responses to disturbances. Trends in Ecology and Evolution, 28, Mouillot, D., De Bortoli, J., Leprieur, F., Parravicini, V., Kulbicki, M., & Bellwood, D. R. (2013b). The challenge of delineating biogeographical regions: nestedness matters for Indo-Pacific coral reef fishes. Journal of Biogeography, 40, Mouillot, D., Stubbs, W., Faure, M., Dumay, O., Tomasini, J. A., Wilson, J.B., & Chi, T.D. (2005). Niche overlap estimates based on quantitative functional traits: a new family of non-parametric indices. Oecologia, 145, Moyle, P. B. (1995). Conservation of native freshwater fishes in the Mediterraneantype climate of California, USA: A review. Biological Conservation. 72, Myers, A. A., & Giller, P. S. (1988). Analytical biogeography. London: Chapman & Hall. Myers, N., Mittermeier, R., Mittermeier, C., da Fonseca, G., & Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature, 403, Naeem, S. (2002). Ecosystem consequences of biodiversity loss: the evolution of a paradigm. Ecology, 83, Naeem, S., Duffy, J., & Zavaleta, E. (2012). The functions of biological diversity in an age of extinction. Science, 336, Nekola, J. C., & White, P. S. (1999). The distance decay of similarity in biogeography and ecology. Journal of Biogeography, 26,
326 Nelson, J. S. (2006). Fishes of the World. Hoboken (New Jersey): John Wiley & Sons. Neubauer, T., Harzhauser, M., Kroh, A., Georgopoulou, E., & Mandic, O. (2015). A gastropod-based biogeographic scheme for the European Neogene freshwater systems. Earth-Science Reviews, 143, Nikolski, G. (1933). On the influence of the rate of flow on the fish fauna of the rivers of Central Asia. Journal of Animal Ecology, 2, Nixon, K., & Wheeler, Q. (1992). Measures of phylogenetic diversity. In M. Novacek, & Q. Wheeler, Extinction and Phylogeny (pp ). New York: Columbia University Press. Northmore, D., Volkman, F. C., & Yager, D. (1978). Vision in fishes: colour and pattern. In D. T. Mostofsky, Behaviour in Fish and other Aquatic Animals (pp ). New York: Academic Press. Ntakis, A., Liasko, R., Oikonomou, A., & Leonardos, I. (2015). Growth pattern of an endemic and endangered West Balkan Peninsula troutspecies (Salmo lourosensis, Delling, 2010). Journal of Applied Ichthyology, 31, Oberdorff, T. T., Hugueny, B., Leprieur, F., Beauchard, O., Brosse, S., & Dürr, H. H. (2011). Global and Regional Patterns in Riverine Fish Species Richness: A Review. International Journal of Ecology, 12, Article ID Oberdorff, T., Guégan, J. F., & Hugueny, B. (1995). Global scale patterns of fish species richness in rivers. Ecography, 18, Oberdorff, T., Guilbert, E., & Lucchetta, J. C. (1993). Patterns of fish species richness in the Seine River basin, France. Hydrobiologia, 259, Oberdorff, T., Lek, S., & Guégan, J.-F. (1999). Patterns of endemism in riverine fish of the Northern Hemisphere. Ecology Letters, 2, Ohlberger, J., Staaks, G., & Hölker, F. (2005). Swimming efficiency and the influence of morphology on swimming costs in fishes. Journal of Comparative Physiology B, 176, Oikonomou, A., Anastasiadou, C., Taskoudis, T., & Leonardos, I. (2014). Length- weight relations of seven native fish species (Actinopterygii) from the Louros River, Greece. Acta Ichthyologica et Piscatoria, 44, Oikonomou, A., Leprieur, F., & Leonardos, I. (2012). Freshwater fishes of the Balkan Peninsula: Delineating biogeographical regions. XIV European Congress of Ichthyology, (p. 162). Liege. 326
327 Oikonomou, A., Leprieur, F., & Leonardos, I. D. (2014). Biogeography of freshwater fishes of the Balkan Peninsula. Hydrobiologia, 738, Oikonomou, A., Ntakis, A., Anastasiadou, C., & Leonardos, I. (2011). Salmo lourosensis, Delling, 2010: An endangered species and its conservation status. 33rd Congress EEVE, Edessa. Oldeland, J., Wesuls, D., & Jürgens, N. (2012). RLQ and fourth-corner analysis of plant species traits and spectral indices derived from HyMap and CHRIS-PROBA imagery. International Journal of Remote Sensing, 33, Olden, J. D., & Poff, N. L. (2004). Ecological processes driving biotic homogenization: Testing a mechanistic model using fish faunas. Ecology, 85, Olden, J. D., Kennard, M. J., Leprieur, F., Tedesco, P. A., Winemiller, K. O., & García - Berthou, E. (2010). Conservation biogeography of freshwater fishes: recent progress and future challenges. Diversity and Distributions, 16, Olteanu, R., & Jipa, D. (2006). Dacian Basin environmental evolution during Upper Neogene within the Paratethys domain. Geo-Eco-Marina, 12, Palla, G., Derényi, I., Farkas, I., & Vicsek, T. (2005). Uncovering the overlapping community structure of complex networks in nature and society. Nature, 435, Paller, M. H. (1994). Relationships between fish assemblage structure and stream order in South Carolina coastal plain streams. Transactions of the American Fisheries Society, 12, Papadopoulou, K., & Vriniotis, D. (2007). Quality of soil and water in deltaic deposits of Louros and Arachthos rivers related to karstic rocks of the wider area. Proceedings of the 11th International Congress, (pp ). Athens: Geological Society of Greece. Parks, T. P., Quist, M. C., & Pierce, C. L. (2016). Anthropogenic Disturbance and Environmental Associations with Fish Assemblage Structure in Two Nonwadeable Rivers. River Research and Applications, 32, Parmakelis, A., Stathi, I., Chatzaki, M., Simaiakis, S., Spanos, L., Louis, C., & Mylonas, M. (2006). Evolution of Mesobuthus gibbosus (Brullé, 1832) (Scorpiones, Buthidae) in the north eastern Mediterranean region. Molecular Ecology, 15, Pavelić, D., & Belak, M. (2008). Dinarids in north Croatia and Bosnia. In T. McCann, The Geology of Central Europe, Volume 2: Mesosoic and Cenozoic (pp ). London: Geological Society. 327
328 Pavoine, S., Ollier, S., & Dufour, A. B. (2005). Is the originality of a species measurable? Ecology Letters, 8, Pease, A., Gonzalez-Diaz, A., Rodiles-Hernandez, R., & Winemiller, K. Ο. (2012). Functional diversity and trait-environment relationships of stream fish assemblages in a large tropical catchment. Freshwater Biology, 57, Pease, A., Taylor, J., Winemiller, K. Ο., & King, R. (2015). Ecoregional, catchment, and reach-scale environmental factors shape functional-trait structure of stream fish assemblages. Hydrobiologia, 753, doi: /s z Pelayo-Villamil, P., Guisande, C., Vari, R. P., Manjarrés-Hernández, A., García-Roselló, E., González-Dacosta, J., Heine, J., González Vilas, L., Patti, B., Quinci, E. M., Jiménez, L. F., Granado-Lorencio, C., Tedesco, P. A., & Lobo, J. M. (2015) (2014). Global diversity patterns of freshwater fishes - potential victims of their own success. Diversity and Distributions, 21, Perdices, A., & Doadrio, I. (2001). The molecular systematics and biogeography of the European cobitids based on mitochondrial DNA sequences. Molecular phylogenetics and evolution, 19, Perdikaris, C., Gouva, E., & Paschos, I. (2010). Alien fish and crayfish species in Hellenic freshwaters and aquaculture. Reviews in Aquaculture, 2, Perea, S., Böhme, M., Zupančič, P., Freyhof, J., Šanda, R., Özuluǧ, M., Abdoli, A., & Doadrio, I. (2010). Phylogenetic relationships and biogeographical patterns in Circum-Mediterranean subfamily Leuciscinae (Teleostei, Cyprinidae) inferred from both mitochondrial and nuclear data. BMC Evolutionary Biology, 10, 265. Peres-Neto, P. R. (2004). Patterns in the co-occurrence of fish species in streams: the role of site suitability, morphology and phylogeny versus species interactions. Oecologia, 140, Perissoratis, C., & Conispoliatis, N. (2003). The impacts of sea-level changes during latest Pleistocene and Holocene times on the morphology of the Ionian and Aegean seas (SE Alpine Europe). Marine Geology, 196, Petchey, O., & Gaston, K. J. (2002). Functional diversity (FD), species richness and community composition. Ecology Letters, 5, Petchey, O., & Gaston, K. J. (2006). Functional diversity: back to basics and looking forward. Ecology Letters, 9,
329 Petchey, O., Evans, K., Fishburn, I., & Gaston, K. J. (2007). Low functional diversity and no redundancy in British avian assemblages. Journal of Animal Ecology, 76, Peter, F. (2006). Biodiversity of European Freshwater Fish - Threats and Conservation Priorities at the Catchment Scale. Basel: Department of Geography. Phillipson, A. (1898). Bosporus und Hellespont. Geographische Zeitschrift, 4, Pimm, S. L. (1999). Seeing both the woods and the trees. Nature, 402, Pisera, A. (1996). Miocene reefs of the Paratethys; a review. In E. Franseen, M. Esteban, & W. R. Ward, Models for Carbonate Stratigraphy from Miocene Reef Complexes of Mediterranean Regions (pp ). Tulsa (Oklahoma): SEPM Concepts in Sedimentology and Paleontology. Pool, T. K., Grenouillet, G., & Villéger, S. (2014). Species contribute differently to the taxonomic, functional, and phylogenetic alpha and beta diversity of freshwater fish communities. Diversity and Distributions, 20, Popov, S., Rögl, F., Rozanov, A. Y., Steininger, F. F., Shcherba; I. G., & Kovac, M. (2004). Lithological-Paleogeographic maps of Paratethys. Frankfurt am Main: Courier Forschungsinstitut Senckenberg. Por, F., & Dimentman, C. (1985). Continuity of Messinian biota in the Mediterranean basin. In D. Stanley, & F. Wezel, Geological evolution of the Mediterranean basin (pp ). Berlin: Springer. Poulakakis, N., Lymberakis, P., Valakos, E., Zouros, E., & Mylonas, M. (2005). Phylogenetic relationships and biogeography of Podarcis species from the Balkan Peninsula, by Bayesian and maximum likelihood analyses of mitochondrial DNA sequences. Molecular Phylogenetics and Evolution, 37, Poulos, S., Kapsimalis, V., Tziavos, C., Pavlakis, P., Livaditis, G., & Collins, M. (2005). Sea-level stands and Holocene geomorphological evolution of the northern deltaic margin of the Amvrakikos Gulf, western Greece. Zeitchrift für Geomorphologie, 135, Power, M. E., Stout, R. J., Cushing, C. E., Harper, P. P., Hauer, F. R., Matthews, W. J., Moyle, P. B., Statzner, B., & Debadgen, I. R. W. (1988). Biotic and abiotic controls in river and stream communities. Journal of the North American Benthological Society, 7, Previšic, A., Walton, C., Kučinić, M., Mitrikeski, P. T., & Kerovec, M. (2009). Pleistocene divergence of Dinaric Drusus endemics (Trichoptera, Limnephilidae) in 329
330 multiple microrefugia within the Balkan Peninsula. Molecular Ecology, 18, Pyke, G. H. (2005). A review of the biology of Gambusia affinis and G. holbrooki. Reviews in Fish Biology and Fisheries, 15, Pyron, M., Williams, L., Beugly, J., & Acquemin, S. (2011). The role of trait-based approaches in understanding stream fish assemblages. Freshwater Biology, 56, Qian, H. (2009). Global comparisons of beta diversity among mammals, birds, reptiles, and amphibians across spatial scales and taxonomic ranks. Journal of Systematics and Evolution, 47, Qian, H., Ricklefs, R., & White, P. (2005). Beta diversity of angiosperms in temperate floras of eastern Asia and eastern North America. Ecology Letters, 8, R Development Core Team (2013). R: a language and environment for statistical computing. Version R Foundationfor Statistical Computing, Vienna. Radoman, P. (1985). Hydrobioidea a superfamily Prosobranchia (Gastropoda), II. Origin, Zoogeography, Evolution in the Balkans and Asia Minor. Belgrade: Monographs Institute of Zoology. Rahel, F. J. (2000). Homogenization of fish faunas across the United States. Science, 288, Rahel, F. J., & Hubert, W. (1991). Fish assemblages and habitat gradients in a Rocky Mountain-Great Plains stream: biotic zonation and additive patterns of community change. Transactions of the American Fisheries Society, 120, Ratkowsky, D. A. (1990). Handbook of nonlinear regression models. New York: Marcel Dekker. Ray, C. (1960). The application of Bergmann s and Allen s rules to poikilotherms. Journal of Morphology, 106, Redding, D., & Mooers, A. (2006). Incorporating evolutionary measures into conservation prioritization. Conservation Biology, 20, Reed, J., Kryštufek, B., & Eastwood, W. (2004). The physical geography of the Balkans and nomenclature of place names. In H. J. Griffiths, B. Kryštufek, & J. M. Reed, Balkan Biodiversity, Pattern and Process in the European Hotspot (pp. 9-22). Dordrecht, Boston, London: Kluwer Academic Publishers. 330
331 Reyjol, Y., Hugueny, B., Pont, D., Bianco, P. G., Beier, U., Caiola, N., Casals, F., Cowx, I., Economou, A. N., Ferreira, T., Haidvogl, G., Noble, R., De Sostoa, A., Vigneron, T., & Virbickas, T. (2007). Patterns in species richness and endemism of European freshwater fish. Global Ecology and Biogeography, 16, Ricketts, T. H. (2001). Aligning conservation goals: are patterns of species richness and endemism concordant at regional scales? Animal Biodiversity and Conservation, 24, Ricklefs, R. E. (2004). A comprehensive framework for global patterns in biodiversity. Ecology Letters, 7, Ricklefs, R. E., & Schlüter, D. (1993). Species diversity in ecological communities, historical and geographical perspectives. Chicago: The University of Chicago Press. Rincón, P., Correas, A., Morcillo, F., Risueño, P., & Lobón-Cerviá, J. (2002). Interaction between the introduced eastern mosquitofish and two autochthonous Spanish toothcarps. Journal of Fish Biology, 61, Robalo, J., Almada, V. C., Levy, A., & Doadrio, I. (2007). Re-examination and phylogeny of the genus Chondrostoma based on mitochondrial and nuclear data and the definition of 5 new genera. Molecular Phylogenetics and Evolution, 42, Robotham, P. W. J. (1977). Feeding habits and diet in two populations of spined loach, Cobitis taenia. Freshwater Biology, 7, Rodrigues, A. S. L., Akcakaya, H. R., Andelman, S. J., Bakarr, M. I., Boitani, L., Brooks, T. M., Chanson, J. S., Fishpool, L. D. C., Da Fonseca, G. A. B., Gaston, K. J., Hoffmann, M., Marquet, P. A., Pilgrim, J. D., Pressey, R. L., Schipper, J., Sechrest, W., Stuart, S. N., Underhill, L. G., Waller, R. W., Watts, M. E. J., & Yan, X. (2004). Global Gap Analysis: Priority Regions for Expanding the Global Protected-Area Network. Bioscience, 54, Rosen, D. E. (1975). A vicariance model of Caribbean biogeography. Systematic Zoology, 24, Rosenzweig, M. L. (1995). Species Diversity in Space and Time. Cambridge: Cambridge University Press. Ross, S. T., Matthews, W. J., & Echelle, A. A. (1985). Persistence of stream fish assemblages: effects of environmental change. American naturalist, 126, Rowe, D., Pierce, C., & Wilton, T. (2009). Physical habitat and fish assemblage relationships with landscape variables at multiple spatial scales in wadeable 331
332 Iowa streams. North American Journal of Fisheries Management, 29, Ruffo, S., & Stoch, F. (2006). Checklist and Distribution of the Italian Fauna. Memorie del Museo Civico di Storia Naturale di Verona. Ruhí, A., Chappuis, E., Escoriza, D., Jover, M., Sala, J., Boix, D., Gascón, S., & Gacia, E. (2014). Environmental filtering determines community patterns in temporary wetlands: a multi-taxon approach. Hydrobiologia, 723, Sabater, S., & Tockner, K. (2010). Effects of Hydrologic Alterations onthe Ecological Quality of River Ecosystems. In: S. Sabater, & D.Barceló, Water Scarcity in the Mediterranean: Prospectives Under Global Change (pp15 39). Berlin Heidelberg: Springer Verlag. Salinas, M., Blanca, G., & Romero, A. (2000). Riparian vegetation and water chemistry in a basin under semiarid Mediterranean climate, Andarax River, Spain. Environmental Management, 26, Sanjur, O. I., Carmona, J. A., & Doadrio, I. (2003). Evolutionary and biogeographical patterns within Iberian populations of the genus Squalius inferred from molecular data. Molecular Phylogenetics and Evolution, 29, Scarnecchia, D. (1988). The importance of streamlining in influencing fish community structure in channelized and unchannelized reaches of a prairie stream. Regulated Rivers: Research & Management, 2, Schlosser, I. J. (1982). Fish community structure and function along two habitat gradients in a headwater stream. Ecological Monographs, 52, Schlosser, I. J. (1985). Flow regime. juvenile abundance, and the assemblage structure of stream fishes. Ecology, 66, Schmid, B., Joshi, J., & Schläpfer, F. (2002). Empirical evidence for biodiversityecosystem functioning relationships. In A. Kinzig, D. Tilman, & P. Pacala, Consequences of Biodiversity: Experimental Progress and Theoretical Extensions (pp ). Princeton (New Jersey): University Press. Schultheiß, R., Albrecht, C., Bößneck, U., & Wilke, T. (2008). The neglected side of speciation in anacient lakes: phylogeography of an inconspicuous mollusc taxon in lakes Ohrid and Presp. Hydrobiologia, 615, Schwartz, J., Simon, A., & Klimetz, L. (2011). Use of fish functional traits to associate in-stream suspended sediment transport metrics with biological impairment. Environmental Monitoring and Assessment, 179,
333 Scottish Fisheries Co-ordination Centre (2007). Fisheries management SVQ Level 2: Catch Fish using Electrofishing Techniques. Inverness: Barony College. Sheldon, A. (1968). Species diversity and longitudinal succession in stream fishes. Ecology, 49, Sibbing, F., & Nagelkerke, L. (2001). Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics. Reviews in Fish Biology and Fisheries, 10, Simberloff, D. S. (1974). Equilibrium theory of island biogeography and ecology. Annual Review of Ecology, Evolution, and Systematics, 5, Skoulikidis, N., Economou, A. N., Gritzalis, K., & Zogaris, S. (2009). Rivers of the Balkans. In K. Tockner, U. Uehlinger, & C. C. Robinson, Rivers of Europe (pp ). Amsterdam: Academic Press, Elsevier. Skoulikidis, Ν., Bertahas, I., & Koussouris, T. (1998). The environmental state of freshwater resources in Greece (rivers and lakes). Environmental Geology, 36, Skoulikidis, N., Karaouzas, I., & Gritzalis, K. (2009). Identifying key environmental variables structuring benthic fauna for establishing a biotic typology for Greek running waters. Limnologica, 39, Šlechtová, V., Bohlen, J., Freyhof, J., Persat, H., & Delmastro, G. B. (2004). The Alps as a barrier to dispersal incold-adapted fishes? Phylogeographic history and taxonomic status of the bullhead in the Adriatic freshwater drainage. Molecular Phylogenetics and Evolution, 33, Smith, C. D., Quist, M. C., & Hardy, R. S. (2015). Fish assemblage structure and habitat associations in a large western river system. River Research and Applications. doi: /rra.2877 Smith, K. G., & Darwall, W. R. T. (2006). The Status and Distribution of Freshwater Fish Endemic to the Mediterranean Basin. Gland, Cambridge: IUCN. Smith, T., Rose, K., & Gingerich, P. (2006). Rapid Asia-Europe-North America geographic dispersal of earliest Eocene primate Teilhardina during the Paleocene-Eocene Thermal Maximum. Proceedings of the National Academy of Sciences of the United States of America, 103, Sommerwerk, N., Hein, T., Schneider-Jakoby, M., Baumgartner, C., Ostojić, A., Siber, R., Bloesch, J., Paunović, M., & Tockner, K. (2009). The Danube River Basin. In K. Tockner, U. Uehlinger, & C. C. Robinson, Rivers of Europe (pp ). Amsterdam: Academic Press, Elsevier. 333
334 Sonnenfeld, P. (1985). Models of late Miocene evaporite genesis in the Mediterranean region. In D. Stanley, & F. Wezel, Geological Evolution of the Mediterranean Basin (pp ). Berlin: Springer-Verlag. Southwood, T. (1977). Habitat, the templet for ecological strategies? Journal of Animal Ecology, 46, Spitz, J., Ridoux, V., & Brind'Amour, A. (2014). Let's go beyond taxonomy in diet description: testing a trait-based approach to prey-predator relationships. Journal of Animal Ecology, 83, Stanković, S. (1960). The Balkan Lake Ohrid and its living world. Monographiae Biologicae, Vol. IX. Den Haag: Dr.W. Junk. Stephanidis, K., & Papastergiadou, E. (2012). Relationships between lake morphometry, water quality, and aquatic macrophytes, in Greek lakes. Fresenius Environmental Bulletin, 21, Sternberg, D., & Kennard, M. (2013). Environmental, spatial and phylogenetic determinants of fish life-history traits and functional composition of Australian rivers. Freshwater Biology, 58, Sternberg, D., & Kennard, M. (2014). Phylogenetic effects on functional traits and life history strategies of Australian freshwater fish. Ecography, 37, Stojanov, R., & Micevski, E. (1989). Geology of Lake Doiran and its surrounding. Skopje: Macedonian Academy of Science and Art. Strayer, D. L., & Dudgeon, D. (2010). Freshwater biodiversity conservation: recent progress and future challenges. Journal of the North American Benthological Society, 29, Suding, K. N., Goldberg, D. E., & Hartman, K. M. (2003). Relationships among species traits: separating levels of response and identifying linkages to abundance. Ecology, 84, Surina, B., Schönswetter, P., & Schneeweiss, G. M. (2011). Quaternary range dynamics of ecologically contrasting species (Edraianthus serpyllifolius and E. tenuifolius, Campanulaceae) within the Balkan refugium. Journal of Biogeography, 38, Tedesco, P. A., Oberdorff, T., Lasso, C. A., Zapata, M., & Hugueny, B. (2005). Evidence of history in explaining diversity patterns in tropical riverine fish. Journal of Biogeography, 32,
335 Tedesco, P., Hugueny, B., Oberdorff, T., Durr, H., Merigoux, S., & de Mérona, B. (2008). River hydrological seasonality influences life history strategies of tropical riverine fishes. Oecologia, 156, Tedesco, P., Leprieur, F., Hugueny, B., Brosse, S., Dürr, H. H., Beauchard, O., Busson, F., & Oberdorff, T. (2012). Patterns and processes of global freshwater fish endemism. Global Ecology and Biogeography, 21, Thuiller, W., Munkemuller, T., Lavergne, S., Mouillot, D., Mouquet, N., Schiffers, K., & Gravel, D. (2013). A road map for integrating eco-evolutionary processes into biodiversity models. Ecology Letters, 16, Tilman, D. (1994). Competition and biodiversity in spatially structured habitats. Ecology, 75, Tilman, D. (2001). Functional diversity. In S. Levin, Encyclopedia of biodiversity (pp ). Waltham (Massachusetts): Academic Press. Tilman, D., Knops, J., Wedin, D., Reich, P., Ritchie, M., & Sieman, E. (1997). The influence of functional diversity and composition on ecosystem processes. Science, 277, Tisseuil, C., Cornu, J.-F., Beauchard, O., Brosse, S., Darwall, W. R. T., Holland, R., Hugueny, B., Tedesco, P. A., & Oberdorff, T. (2013). Global diversity patterns and cross-taxa convergence in freshwater systems. Journal of Animal Ecology, 82, Tobler, W. (1970). A computer model simulating urban growth in the Detroit region. Economic Geography, 46, Tonn, W., Magnuson, J., Rask, M., & Toivonen, J. (1990). Intercontinental comparison of small-lake fish assemblages: the balance between local and regional processes. American Naturalist, 136, Townsend, C., & Hildrew, A. (1994). Species traits in relation to a habitat templet for river systems. Freshwater Biology, 31, Triantis, K. A., Guilhaumon, F., & Whittaker, R. J. (2012). The island species-area relationship: biology and statistics. Journal of Biogeography, 39, Triantis, K. A., Mylonas, M., Lika, K., & Vardinoyannis, K. (2003). A model for the species-area-habitat relationship. Journal of Biogeography, 30, Troia, M., & Gido, K. (2015). Functional strategies drive community assembly of stream fishes along environmental gradients and across spatial scales. Oecologia, 177,
336 Tsoumani, M., Apostolidis, A. P., & Leonardos, I. D. (2013). Biogeography of Rutilus species of the southern Balkan Peninsula as inferred by multivariate analysis of morphological data. Journal of Zoology, 289, Vane-Wright, R., Humphries, C., & Williams, P. (1991). What to protect? Systematics and the agony of choice. Biological Conservation, 55, Vanhove, M. P., Economou, A. N., Zogaris, S., Larmuseau, M. H., Giakoumi, S., Kalogianni, E., Volckaert, F. A. M., & Huyse, T. (2012). Phylogenetics and biogeography of the Balkan sand gobies (Teleostei: Gobiidae): vulnerable species in need of taxonomic revision. Biological Journal of the Linnean Society, 105, Vanni, M. J. (2002). Nutrient cycling by animals in freshwater ecosystems. Annual Review of Ecology, Evolution, and Systematics, 33, Vannote, R., Minshall, G., Cummins, K., Sedell, J., & Cushing, C. (1980). The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 37, Vié, J., Hilton-Taylor, C., & Stuart, S. N. (2008). Wildlife in a changing world: an analysis of the 2008 IUCN Red List of Threatened Species. Gland: IUCN. Villéger, S. (2008). Dynamique de la diversitι fonctionnelle des communautιs de poissons (Lagune de Terminos, Mexique). Montpellier: SupAgro. Villéger, S., Mason, N., & Mouillot, D. (2008). New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 89, Villéger, S., Ramos Miranda, J., Flores, H. D., & Mouillot, D. (2010). Contrasting changes in taxonomic vs functional diversity of tropical fish assemblages after habitat degradation. Ecological Applications, 20, Violle, C., Bonis, A., Plantegenest, M., Cudennec, C., Damgaard, C., Marion, B., Le Cœur, D., & Bouzillé, J.-B. (2011). Plant functional traits capture species richness variations along a flooding gradient. Oikos, 120, Violle, C., Reich, P., Pacala, S., Enquist, B., & Kattge, J. (2014). The emergence and promise of functional biogeography. Proceedings of the National Academy of Sciences USA,, 111, Vitousek, P. M., Mooney, H. A., Lubchenco, J., & Melillo, J. M. (1997). Human domination of Earth s ecosystems. Science, 277,
337 Vogiatzi, E., Kalogianni, E., Zimmerman, B., Giakoumi, S., Barbieri, R., Paschou, P., Magoulas, A., Tsaparis, D., Poulakakis, N., & Tsigenopoulos, C. S. (2014). Reduced genetic variation and strong genetic population structure in the freshwater killifish. Biological Journal of the Linnean Society, 111, Wainwright, P. C. (1994). Functional morphology as a tool in ecological research. In P. C. Wainwright, & S. M. Reilly, Ecological Morphology: Integrative Organismal Biology (pp 42-59). Chicago: University Chicago Press. Wainwright, P. C., Bellwood, D. R., & Westneat, M. W. (2002). Ecomorphology of locomotion in labrid fishes. Environmental Biology of Fishes, 65, Walser, C. A., & Bart, H. L. (2009) Influence of agriculture on in-stream habitat and fish community structure in Piedmont watersheds of the Chattahoochee River system. Ecology of Freshwater Fish, 8, Wagner, B., Vogel, H., Zanchetta, G., & Sulpizio, R. (2010). Environmental change within the Balkan region during the past ca. 50 ka recorded in the sediments from lakes Prespa and Ohrid. Biogeosciences, 7, Wallace, A. R. (1878). Tropical nature, and other essays. London: Macmillan and Co. Wallace, A. R. (1894). What are zoological regions? Nature, 49, Webb, C., Ackerly, D., McPeek, M., & Donoghue, M. (2002). Phylogenies and community ecology. Annual Review of Ecology and Systematics, 33, Webb, C., Hoeting, J., Ames, G., Pyne, M., & LeRoy Poff, N. (2010). A structured and dynamic framework to advance traits-based theory and prediction in ecology. Ecology Letters, 13, Webb, P. (1984). Form and function in fish swimming. Scientific American, 251, Weibull, W. (1951). A statistical distribution function of wide applicability. Journal of Applied Mathematics, 18, Weiher, E., & Keddy, P. (1995). Assembly rules, null models, and trait dispersion: new questions from old patterns. Oikos,74, Welcomme, R. L. (1985) River fisheries. Rome: FAO Fisheries. Welcomme, R. L., Winemiller, K. O., & Cowx, I. G. (2006). Fish environmental guilds as a tool for assessment of ecological condition of rivers. Rivers Research and Applications, 22, Whittaker, R. H. (1960). Vegetation of the Siskiyou mountains, Oregon and California. Ecological Monographs, 30,
338 Whittaker, R. H. (1977). Evolution of species diversity in land communities. Evolutionary Biology, 10, Whittaker, R. J., Araújo, M., Jepson, P., Ladle, R., Watson, J., & Willis, K. (2005). Conservation biogeography: assessment and prospect. Diversity and Distributions, 11, Whittaker, R. J., Willis, K. J., & Field, R. (2001). Scale and species richness: towards a general, hierarchical theory of species diversity. Journal of Biogeography, 28: Wiens, J. A. (1989). Spatial Scaling in Ecology. Functional Ecology, 3, Wilke, T., Schultheiß, R., Albrecht, C., Bornmann, N., Trajanovski, S., & Kevrekidis, T. (2010). Native Dreissena freshwater mussels in the Balkans: in and out of ancient lakes. Biogeosciences, 7, Williams, P. H., Gibbons, D. W., Margules, C. R., Rebelo, A. G., Humphries, C. J., & Pressey, R. L. (1996). A comparison of richness hotspots, rarity hotspots and complementary areas for conserving diversity using British birds. Conservation Biology,10, Wilsey, B. J., Chalcraft, D. R., Bowles, C. M., & Willig, M. R. (2005). Relationships among indices suggest that richness is an incomplete surrogate for grassland biodiversity. Ecology, 86, Winemiller, K.O. (1989). Patterns of variation in life history among South American fishes in seasonal environments. Oecologia, 81, Winemiller, K. Ο. (1991). Ecomorphological diversification in lowland freshwater fish assemblages from five biotic regions. Ecological Monographs, 61, Winemiller, K. Ο. (2005). Life history strategies, population regulation, and implications for fisheries management. Canadian Journal of Fisheries and Aquatic Sciences, 62, Winston, M. (1995). Co-occurrence of morphologically similar species of stream fishes. The American Naturalist, 145, Witting, L., & Loeschke, V. (1995). The optimization of biodiversity conservation. Biological Conservation, 71, Wright, D. H. (1983). Species-energy theory: an extension of species-area theory. Oikos, 41,
339 Xenopoulos, M., Lodge, D., Alcamo, J., Marker, M., Schulze, K., & Van Vuurens, D. (2005). Scenarios of freshwater fish extinctions from climate change and water withdrawal. Global Change Biology, 11, Zacharias, I., Bertahas, I., Skoulikidis, N., & Koussouris, T. (2002). Greek lakes: Limnological overview. Lakes & Reservoirs: Research and Management, 7, Zander, C. (1972). Evolution of Blennioidei in the Mediterranean Sea. Revue des Travaux de l'institut des Pêches Maritimes, 37, Zardoya, R., & Doadrio, I. (1999). Molecular evidence on the evolutionary and biogeographical patterns of European Cyprinids. Journal of Molecular Evolution, 49, Zhao, S., Fang, J., Peng, C., Tang, Z., & Piao, S. (2006). Patterns of fish species richness in China's lakes. Global Ecology and Biogeography, 15, Zhao, T., Villéger, S., Lek, S., & Cucherousset, J. (2014). High intraspecific variability in the functional niche of a predator is associated with ontogenetic shift and individual specialization. Ecology and Evolution, 4, Zobel, M. (1997). The relative role of species pools in determining plant species richness. An alternative explanation of species coexistence? Trends in Ecology and Evolution, 12, Zogaris, S., Economou, A. N., & Dimopoulos, P. (2009). Ecoregions in the southern Balkans: Should their boundaries be revised? Environmental Management, 43,
340 Ανοικτά γεωχωρικά δεδομένα και υπηρεσίες (2015). Βαφειάδης, Π. (1983). Υδρογεωλογική Μελέτη της Λεκάνης της Καστοριάς. Θεσσαλονίκη: Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης. Γκένας, Χ. (2012). Μελέτη της οικολογίας ειδών της οικογένειας Gobiidae στα εσωτερικά ύδατα της Δυτικής Ελλάδας με έμφαση στο είδος Economidichthys pygmaeus (Holly, 1929) της Λίμνης Παμβώτιδας. Ιωάννινα: Πανεπιστήμιο Ιωαννίνων. Δημητρίου, Η. (2005). Συμβολή στην βιώσιμη διαχείριση των υδατικών πόρων της υδρολογικής λεκάνης της λίμνης Τριχωνίδας. Αθήνα: Εθνικό και Καποδιστριακό Πανεπιστήμιο Αθηνών (ΕΚΠΑ). Δράκου, Ε. (2009). Χωρική ετερογένεια και ποικιλότητα ειδών στις προστατευόμενες περιοχές της Ελλάδας. Θεσσαλονίκη: Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης. Ζαλίδης, Χ., & Μαντζαβέλας, Α. (1994). Απογραφή των Ελληνικών υγροτόπων ως φυσικών πόρων (Πρώτη προσέγγιση). Αθήνα: Ελληνικό Κέντρο Βιοτόπων - Υγροτόπων (ΕΚΒΥ). Κουσουρής, Θ. (2014). Οι λίμνες στην Ελλάδα. Αθήνα: Εθνικό Κέντρο Κοινωνικών Ερευνών. Μαμάσης, Ν., & Στεφανάκος, I. (2010). Εισαγωγή στην Ενεργειακή Τεχνολογία. Αθήνα: Εθνικό Μετσόβιο Πολυτεχνείο. Μερτζάνης, Α. (1992). Γεωμορφολογική εξέλιξη του Αμβρακικού κόλπου. Αθήνα: Πανεπιστήμιο Αθήνας. Μουντράκης, Δ. (1985). Γεωλογία της Ελλάδας. Θεσσαλονίκη: University Studio Press. Μουντράκης, Δ. (2010). Γεωλογία και γεωτεκτονική εξέλιξη της Ελλάδας. Θεσσαλονίκη: University Studio Press. Πετρίκη, Ο. (2015). Διερεύνηση της δυνατότητας δημιουργίας ιχθυολογικού πολυμετρικού δείκτη για την εκτίμηση της οικολογικής ποιότητας λιμνών. Θεσσαλονίκη: Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης. Ραµσάρ, Σύµβαση (1971). Για την προστασία των διεθνούς ενδιαφέροντος υγροτόπων ιδίως ως υγροβιοτόπων. Ν.. 191/ 74, ΦΕΚ 350 Α, Σφενδουράκης, Σ. (2009). Νησιωτική βιογεωγραφία: Οικολογία, εξέλιξη και διατήρηση. Ηράκλειο: ΙΤΕ / Πανεπιστημιακές Εκδόσεις Κρήτης. 340
341 Τάχος, Β., Ζόγκαρης, Σ., Κουτράκης, Ε., Λεονάρδος, Ι., Μπόμπορη, Δ., Χατζηνικολάου, Γ., Αναγνόπουλος, Ν., Λουκάτος, Α., Μπουρδανιώτης, Ν., & Οικονόμου, Α. (2015). Αξιολόγηση της κατάστασης διατήρησης για κάθε είδος ιχθυοπανίδας κοιν. ενδιαφέροντος και σύνταξη των αντίστοιχων εντύπων αναφοράς του Άρθρου 17 της Οδηγίας 92/43/ΕΟΚ» στα πλαίσια της Μελέτης 6 «Εποπτεία και Αξιολόγηση της Κατάστασης Διατήρησης ειδών κοινο. Αθήνα: ΥΠΑΠΕΝ, Κοινοπραξία Αναγνόπουλος Νικ'ολαος -ΕΠΕΜ Α.Ε.-Κωνσταντινίδης Ηλίας. Τσιούρης, Σ. (1996). Ειδικό Διαχειριστικό Σχέδιο για την περιοχή Λίμνες Χειμαδίτιδα- Ζάζαρη (GR ). Θεσσαλονίκη: Ελληνικό Κέντρο Βιοτόπων-Υγροτόπων (ΕΚΒΥ). Τσουμάνη, Μ. (2013). Μελέτη της βιολογίας και των φυλογενετικών σχέσεων των ειδών Rutilus της οικογένειας Cyprinidae των εσωτερικών υδάτων της Ελλάδας. Ιωάννινα: Πανεπιστήμιο Ιωαννίνων. Υγροτόπιο νήσων (2015). Υπουργείο Περιβάλλοντος Ενέργειας και Κλιματικής Αλλαγής, Ε. Γ. (2013). Κ/ΞΙΑ Διαχείρισης Υδάτων Θεσσαλίας, Ηπείρου και Δυτικής Στερεάς Ελλάδας. Αθήνα. Ψιλοβίκος, Α. (1977). Παλαιογεωγραφική εξέλιξη της λεκάνης και της λίµνης της Μυγδονίας. Θεσσαλονίκη: Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης. 341
342
343 Παραρτήματα
344
345 Παράρτημα 1 Βιβλιογραφικές πηγές βάσης δεδομένων κατανομής των ψαριών των εσωτερικών υδώτων της Βαλκανικής Χερσονήσου 1. Adrović, A. & I. Skenderović, Allochthonous Ichthyofauna of Certain Water Bodies in North-Eastern Bosnia. Acta agriculturae Serbica 23: Adrović, A., I. Skednderović & S. Ibrišimović, Ekološke karakteristike rijeka Gostelje, Spreče i Brke. Zbornik radova PMF - a Tuzla 1: Adrović, A., R. Škrijelj & I. Skenderović, Ecologial Characteristices of the Roach (Rutilus rutilus L.) and Bream (Abramis brama L.) in the Reservoir of the Lake Modrac. Acta Agriculturae Serbica 27: Ahnelt, H., P. Tutman, J. Dulčić & B. Glamuzina, A new species of freshwater goby of the genus Knipowitschia (Teleostei: Gobiidae) from Hutovo Blato wetland (Neretva River basin), Bosnia and Herzegovina. 13th European congress of ichthyology, Book of Abstracts, Klaipeda, Lithuania, 6-11 September Apostolidis, A. P., D. Loukovitis & C. S. Tsigenopoulos, Genetic characterization of brown trout (Salmo trutta) populations from the Southern Balkans using mtdna sequencing and RFLP analysis. Hydrobiologia 600: Apostolidis A. P., M. T. Stoumboudi, E. Kalogianni, G. Cote & L. Bernatchez, Genetic divergence among native trout Salmo trutta populations from southern Balkans based on mitochondrial DNA and microsatellite variation. Journal of Fish Biology 79: Apostolidis, A. P., C. Triantafyllidis, A. Kouvatsi & P. S. Economidis, Mitochondrial DNA sequence variation and phylogeography among Salmo trutta L. (Greek brown trout) populations. Molecular Ecology 6: Apostolou, A., The ichthyofauna from the Bulgarian sector of the Mesta River. Acta zoologica bulgarica 57: Apostolou, A., M. Koutrakis, L. Pehlivanov, M. Vassilev, T. Stefanov & B. Velkov, Notes on the Fish Fauna Composition of Mesta (Nestos) River in regard to Management and Conservation. Acta zoologica bulgarica 62: Apostolou, A., T. Stefanov, M. Vassilev, A. Šedivá & L. Pehlivanov, Distribution of the stone loaches (Pisces: Balitoridae) in Bulgaria - past and present status. Acta zoologica bulgarica 2: Bănărescu, P. M., Zoogeography of Fresh Waters. Vol. 1. General Distribution and Dispersal of Freshwater Animals. AULA-Verlag, Wiesbaden. 345
346 12. Bănărescu, P. M., Distribution pattern of the aquatic fauna of the Balkan Peninsula. In Griffiths, H. J., B. Kryštufek & J. M. Reed (eds), Balkan Biodiversity, Pattern and Process in the European Hotspot. Kluwer Academic Publishers, Dordrecht, Boston, London: Barbieri, R., Cyprinids of Peloponnese. Distribution, ecology and threats. Pan-Hellenic Ichthyological Congress, Book of Abstracts, Messolonghi, Greece, January Barbieri, R., A. N. Economou, M. T. Stoumboudi & P. S. Economidis, Freshwater fishes of Peloponnese (Greece): Distribution, ecology and threats. In Collares- Pereira, M. J., M. M. Coelho & I. G. Cowx (eds), Freshwater Fish Conservation: options for the Future. Fishing News Book, Blackwell Science, Oxford: Barbieri, R., M. T. H. Stoumboudi & A. N. Economou, Threatened fishes of the world: Valencia letourneuxi Sauvage, 1880 (Valenciidae). Environmental Biology of Fishes 65: Beqiraj, S. & Dh. Dhora, Buna Bojana. APAWA, Kalimera, REC: Tirane. 17. Berrebi, P., C. Tougard, S. Dubois, Z. Shao, I. Koutseri, S. Petkovski & A. J. Crivelli, Genetic Diversity and Conservation of the Prespa Trout in the Balkans. International Journal of Molecular Sciences 14: Bianco, P. G., Potential role of the palaeohistory of the Mediterranean and Paratethys basins of the early dispersal of Euro-Mediterranean freshwater fishes. Ichthyological Exploration of Freshwaters: Bianco, P. G., The status of the twaite shad, Alosa agone, in Italy and the Western Balkans. Marine Ecology 23: Bianco P. G. & M. Kottelat Scardinius knezevici, a new species of rudd from Lake Skadar, Montenegro (Teleostei: Cyprinidae). Ichthyological Exploration of Freshwaters 16: Bianco, P. G. & R. R. Miller, First record of Valencia letourneuxi (Sauvage, 1880) in Peloponnese (Greece) and remarks on the mediterranean family Valenciidae (Cyprinodontiformes). Cybium 13: Bianco, P. G. & F. Nordlie, The salinity tolerance of Pseudophoxinus stymphalicus (Cyprinidae) and Valencia letourneuxi (Valenciidae) from western Greece suggests a revision of the ecological categories of freshwater fishes. Italian Journal of Zoology 75:
347 23. Bless, R. & R. Riehl, Biology and egg morphology of the Dalmatian barbell gudgeon Aulopyge huegeli, an endangered endemic species in Croatia. Environmental Biology of Fishes 63: Bobori, D. C., Bioaccumulation of heavy metals in the ecosystem of Lake Koronia, (Macedonia, Greece). Doctorate Thesis, Aristotle University, Thessaloniki. 25. Bobori, D. C. & P. S. Economidis, Freshwater fishes of Greece: their biodiversity, fisheries and habitats. Aquatic Ecosystem Health and Management 9: Bobori, D. C., P. S. Economidis & E. G. Maurakis, 2001a. Freshwater fish habitat science and management in Greece. Aquatic Ecosystem Health & Management 4: Bobori, D. C., E. T. Koutrakis & P. S. Economidis, 2001b. Shad species in Greek waters-an historical overview and present status. Bulletin Francais de la Peche et de la Protection des Milieux Aquatiques 362/363: Bobori, D. C., D. K. Moutopoulos, M. Bekri, I. Salvarina & A. I. P. Muñoz, Length-weight relationships of freshwater fish species caught in three Greek lakes. Journal of Biological Research- Thessaloniki 14: Bobori, D. C., I. Salvarina & E. Michaloudi, Fish dietary patterns in the eutrophic Lake Volvi (East Mediterranean). Journal of Biological Research- Thessaloniki 19: Bogutskaya, N. G., Petroleuciscus, a new genus for the Leuciscus borysthenicus species group (Teleostei: Cyprinidae). Zoosystematica Rossica, 11: Bogutskaya, N. G. & P. Zupančič, A re-description of Leuciscus zrmanjae (Karaman, 1928) and new data on the taxonomy of Leuciscus illyricus, L. svallize and L. cephalus (Pisces: Cyprinidae) in the West Balkans. Annalen des Naturhistorischen Museums in Wien. Serie B für Botanik und Zoologie 101: Bogutskaya, N. G. & P. Zupančič, Phoxinellus pseudalepidotus, a new species from the Neretva basin (Teleostei: Cyprinidae) with an overview of the morphology of Phoxinellus species of Croatia and Bosnia and Herzegovina. Ichthyological Exploration of Freshwaters 14: Bogutskaya, N. G. & P. Zupančič, Squalius janae, a new species of fish from the Adriatic Sea basin in Slovenia (Actinopterygii: Cyprinidae). Zootaxa 2536:
348 34. Bogutskaya, N. G., P. Zupančič & A. M. Naseka, Two new species of freshwater fishes of the genus Alburnoides, A. fangfangae and A. devolli (Actinopterygii: Cyprinidae), from the Adriatic Sea basin in Albania. Proceedings of the Zoological Institute Russian Academy of Sciences 314: Bohlen, J., A. Perdices, I. Doadrio & P. S. Economidis, Vicariance, colonisation, and fast local speciation in Asia Minor and the Balkans as revealed from the phylogeny of spined loaches (Osteichthyes; Cobitidae). Molecular Phylogenetics and Evolution 39: Bohlen, J., V. Šlechtová, R. Šanda, L. Kalous, J. Freyhof, J. Vukić & D. Mrdak, Cobitis ohridana and Barbatula zetensis in the River Morača basin, Montenegro: distribution, habitat, population structure and conservation needs. Folia Biologica 51: Buj, I., M. Podnar, M. Mrakovčić, L. Choleva, V. Šlechtová, N. Tvrtković, M. Ćaleta, P. Mustafić, Z. Marčić, D. Zanella & A. Brigić, Genetic diversity and phylogenetic relationships of spined loaches (genus Cobitis) in Croatia based on mtdna and allozyme analyses. Folia Zoologica 57: Buj, I., J. Vukić, R. Šanda, S. Perea, M. Ćaleta, Z. Marčić, I. Bogut, M. Povž & M. Mrakovčić, Morphological comparison of bleaks (Alburnus, Cyprinidae) from the Adriatic Basin with the description of a new species. Folia Zoologica 59: Cake, A., Data on the ichtiofauna of Shkumbini River and the influence of some abiotic factors in its distribution and density. Doctorate Thesis, University of Tirana, Tirana. 40. Cake, A. & A. Miho, Fishes from Shkumbini River (Central Albania): an ecological view. Journal of Environmental Protection and Ecology 2: Cakić, P., Growth of the bleaks (Alburnus alburnus L.) in the Velika Morava River. Ichthyologia, 15: Cakić, P., M. Lenhardt, J. Kolarevic, B. Mickovic & A. Hegedis, Distribution of the Asiatic cyprinid Pseudorasbora parva in Serbia and Montenegro. Journal of Fish Biology 65: Cakić, P., M. Lenhardt & Z. Petrovic, The first record of Gasterosteus aculeatus L (Pisces: Gasterosteidae) in the Yugoslav section of the Danube. Ichthyologia 32: Ćaleta, M., D. Jelić, I. Buj, D. Zanella, Z. Marčić, P. Mustafić & M. Mrakovčić, First record of the alien invasive species rotan (Perccottus glenii Dybowski, 1877) in Croatia. Journal of Applied Ichthyology 27:
349 45. Ćaleta, M., M. Mrakovčić, I. Buj, P. Mustafić, D. Zanella, & Z. Marčić, Threatened fishes of the world: Chondrostoma phoxinus Heckel, 1843 (Cyprinidae). Environmental Biology of Fishes 84: Catsadorakis, G., M. Malakou & A. J. Crivelli, 1996a. The Prespa barbel, Barbus prespensis, Karaman 1924, in the Prespa lakes basin, north-western Greece. Tour du Valat, Arles. 47. Catsadorakis, G., M. Malakou & A. J. Crivelli, 1996b. The effects of the drought on the colonial waterbirds nesting at Lake Mikri Prespa with special attention on pelicans. Colonial Waterbirds 19: Choleva L., A. Apostolou, P. Ráb & K. Janko, Making it on their own: sperm-dependent hybrid fishes (Cobitis) switch the sexual hosts and expand beyond the ranges of their original sperm donors. Philosophical Transactions of the Royal Society B: Biological Sciences 363: Crivelli, A. J., Fisheries decline in the freshwater lakes of northern Greece with special attention for Lake Mikri Prespa. In van Densen, W. L. T., B. Steinmetz & R. H. Hughes (eds), Management of Freshwater fisheries, Wageningen: Crivelli, A. J., Are fish introductions a threat to endemic fishes in the Northern Mediterranean region? Biological Conservation 72: Crivelli, A. J., The role of protected areas in freshwater fish conservation. In Collares- Pereira, M. J., M. M. Coelho & I. G. Cowx (eds), Freshwater Fish Conservation: options for the Future. Fishing News Book, Blackwell Science, Oxford: Crivelli A. J., G. Catsadorakis, M. Malakou & E. Rosecchi, Fish and fisheries of the Prespa lakes. Hydrobiologia 351: Crivelli, A. J., I. Koutseri & S. Petkovski, The Prespa trout, Salmo peristericus (Karaman, 1938), an endangered species in need of action. Society for the Protection of Prespa, Agios Germanos. 54. Crivelli, A. J. & T. W. Lee, Observation on the age, growth and fecundity of Cobitis meridionalis, an endemic loach of Prespa Lake (Greece). Folia Zoologica 49: Crivelli, A. J., M. Malakou, G. Catsadorakis & E. Rosecchi, The Prespa barbel, Barbus prespensis, a fish species endemic to the Prespa Lakes (North-western Greece). Folia Zoologica 45:
350 56. Crivelli, A. J., M. C. Ximenes, P. Grillas, J. M. Deslous-Paoli, Study on fishery improvement. European Commission PHARE programme: Karavasta lagoon wetland management project, Tour du Valat, Arles. 57. Çullaj, A, A. Hasko, A. Miho, F. Schanz, H. Brandl & R Bachofen, The quality of Albanian natural waters and the human impact. Environment International 31: Çullaj, A., A. Miho, B. Baraj, A. Hasko, R. Bachofen, H. Brandl & F. Schanz, Preliminary water quality report for some important Albanian Rivers. Journal of Environmental Protection and Ecology Special Issue: Daoulas, C. & P. S. Economidis, The feeding of Rutilus rubilio (Pisces, Cyprinidae) in Lake Trichonis Greece. Cybium 8: Delić, A., Ihtiofauna rijeke Ilove u području gornjeg Poilovlja (središnja Hrvatska). Ribarstvo 44: Delić, A., M. Kučinić, D. Marić & M. Bučar, New data about the distribution of Phoxinellus alepidotus (Heckel, 1843) and Aulopyge huegelii (Heckel, 1841). Natura Croatica 114: Delling, B., Species diversity and phylogeny of Salmo with emphasis on Southers Trouts (Teleostei, Salmonidae). Doctorate Thesis, University of Stockholm, Stockholm. 63. Delling, B., Diversity of western and southern Balkan trouts, with the description of a new species from the Louros River, Greece (Teleostei: Salmonidae). Ichthyological Exploration of Freshwaters, 21: Dhora, Dh., Liqeni i Shkodres. Botimet Camaj-Pipa: Shkodër. 65. Dhora, Dh., Regjistër i Specieve të Faunës së Shqipërisë Botimet Camaj-Pipa: Shkodër. 66. Dhora, Dh., S. Beqiraj & D. Dhora, Report on Biodiversity of River Buna. APAWA, Kalimera, REC: Tirane. 67. Djikanović V., M. Paunović, V. Nikolić, P. Simonović & P. Cakić, Parasitofauna of freshwater fishes in the Serbian open waters: a checklist of parasites of freshwater fishes in Serbian open waters. Reviews in Fish Biology and Fisheries 22: Dikov, T. & M. Zivkov, Abundance and Biomass of Fishes in the Veleka River, Bulgaria. Folia Zoologica 53:
351 69. Dimovski A. & R. Grupče, Nekoi problemi od sistematikata na plašicite (Alburnus) bo Makedonija. Acta Musei Macedonici Scientarium Naturalium 12: Dimovski A. & R. Grupče, Varijabilnost vrste Alburnus alburnus (L.) u različitim basenima Makedonije. Ichthyologia 7: Dulčić, J. & B. Glamuzina, Length-weight relationships for selected fish species from three eastern Adriatic estuarine systems (Croatia). Journal of Applied Ichthyology 22: Dulčić, J., B. Glamuzina & P. Tutman, First record of ruffe, Gymnocephalus cernuus (Percidae), in the Hutovo Blato wetland, Adriatic drainage system of Bosnia and Herzegovina. Cybium 29: Dulčić, J. P. Tutman, I. Prusina, S. Tomšić, B. Dragičević, E. Hasković & B. Glamuzina, Length-weight relationships for six endemic freshwater fishes from Hutovo Blato wetland (Bosnia and Herzegovina). Journal of Applied Ichthyology 25: Durand, J. D., P. G. Bianco, J. Laroche & A. Gilles, Insight into the origin of endemic Mediterranean ichthyofauna: phylogeography of Chondrostoma genus (Teleostei, Cyprinidae). Journal of Heredity 94: Durand, J. D., H. Persat & Y. Bouvet, Phylogeography and postglacial dispersion of the chub (Leuciscus cephalus) in Europe. Molecular Ecology 8: Economidis, P. S., Chalcalburnus belvica (Karaman, 1924) (Pisces, Cyprinidae), nouvelle combinaison taxonomique pour la population du lac Mikri Prespa (Macédoine, Grèce). Cybium 10: Economidis, P. S., Distribution pattern of the genus Barbus (Pisces, Cyprinidae) in the freshwaters of Greece. Travaux du Muséum National d Histoire Naturelle Grigore Antipa 30: Economidis, P. S., Check List of Freshwater Fishes of Greece, Recent Status of Threats and Protection. Hellenic Society for the Protection of Nature, Athens. 79. Economidis, P. S., Endangered freshwater fishes of Greece. Biological Conservation 72: Economidis, P. S., Leuciscus keadicus Stephanidis, 1971 (Pisces, Cyprinidae), a valid species from river Evrotas (Greece). Cybium 20: Economidis, P. S., Biology of rare and endangered non- migratory fish species: problems and constraints. In Collares- Pereira, M. J., M. M. Coelho & I. G. Cowx 351
352 (eds), Freshwater Fish Conservation: options for the Future. Fishing News Book, Blackwell Science, Oxford: Economidis, P. S., Barbatula pindus, a new species of stone loach from Greece (Teleostei: Balitoridae). Ichthyological Exploration of Freshwaters 16: Economidis, P. S., Fishes of inland waters of Greece- Check list. In Legakis, A. & P. Maragou (eds), The Red Data Book of Threatened Animals of Greece. Hellenic Zoological Society, Athens: Economidis, P. S. & P. M. Bănărescu, The distribution and origins of freshwater fishes in the Balkan Peninsula, especially in Greece. Internationale Revue Der Gesamten Hydrobiologie 76: Economidis, P. S., E. Dimitriou, R. Pagoni, E. Michaloudi & L. Natsis, 2000a. Introduced and translocated fish species in the inland waters of Greece. Fisheries Management and Ecology 7: Economidis, P. S., M. Koutrakis, A. Apostolou, M. Vassilev & L. Pehlivanov, Atlas of River Nestos Fish Fauna. Prefectural authority of Drama-Kavala-Xanthi, NAGREF-Fisheries Research Institute & Bulgarian Academy of Sciences, Kavala. 87. Economidis, P. S., M. T. Koutrakis & D. C. Bobori, 2000b. Distribution and conservation of Acipenser sturio and related species in the Greek waters. Boletin del Instituto Espanol de Oceanografia 16: Economidis, P. S. & P. J. Miller, Systematics of freshwater gobies from Greece (Teleostei: Gobiidae). Journal of Zoology 211: Economidis, P. S. & T. T. Nalbant, A study of the loaches of the genera Cobitis and Sabanejewia (Pisces, Cobitidae) of Greece, with description of six new taxa. Travaux du Muséum National d Histoire Naturelle Grigore Antipa 36: Economou, A. N., C. H. Daoulas, Th. Psarras & R. Barberi-Tseliki, Freshwater larval fish from Lake Trichonis (Greece). Journal of Fish Biology 45: Economou, A. N., S. Giakoumi, L. Vardakas, R. Barbieri, M. Stoumboudi, & S. Zogaris, The freshwater ichthyofauna of Greece: an update based on a hydrographic basin survey. Mediterranean Marine Science 8: Fet, V. & A. Popov, Biogeography and Ecology of Bulgaria. Springer, Dordrecht. 93. FishBase, World Wide Web electronic publication. Version Available at: 352
353 94. Fotis, G., A. Conides, T. Koussouris, A. Diapoulis & K. Gritzalis, Fishery potential of lakes in Macedonia, north Greece. Fresenius Environmental Bulletin 1: Freyhof, J., Threatened freshwater fishes and mollusks of the Balkan. Potential Impact of hydropower projects. Unpublished report, ECA Watch Austria & Euronatur, Berlin. 96. Freyhof, J., D. Lieckfeldt, N. G. Bogutskaya, C. Pitra & A. Ludwig, Phylogenetic position of the Dalmatian genus Phoxinellus and description of the newly proposed genus Delminichthys (Teleostei: Cyprinidae). Molecular Phylogenetics and Evolution 38: Gavrilović, Lj. & D. Đukić, Reke Srbije. Zavod za udžbenike i nastavna sredstva, Beograd. 98. Georgiev, D. G., The fish species participation in otters (Lutra lutra) diet in Maritza River, west of Plovdiv town (southern Bulgaria). Animalia 42: Georgiev, S. B., Ulov ribe na području općine Stuga OD DO Ribarstvo 57: Georgiev, S. B., 2000a. Stanje ihtiofaune rijeke Vardara u razdoblju od 1996 do Ribarstvo 58: Georgiev, S. B., 2000b. Taxonomical characteristics of chub Leuciscus cephalus (Linnaeus, 1758) from the River Babuna (Macedonia). Ribarstvo 58: Georgiev, S. B., 2003a. Length-weight relationship of six cyprinid fish in the River Vardar (Republic of Macedonia). Ribarstvo 61: Georgiev, S. B., 2003b. On the origin of the Balkan Peninsula salmonids. Ribarstvo 61: Georgiev, S. B., 2003c. Succession and structure of the fish in the Mavrovo Reservoir (The Republic of Macedonia). Ribarstvo 61: Georgiev, S. B., Sadašnje poznavanje endemizma ihtiofaune Republike Makedonije. Ribarstvo 62: Georgiev, S. B., Study of Salmo genus (Pisces, Salmonidae) on the Balkan Peninsula. Ribarstvo 69: Gilles, A., C. Costedoat, B. Barascud, A. Voisin, P. Banarescu, P. G. Bianco, P. S. Economidis, D. Marić & R. Chappaz, Speciation pattern of Telestes souffia complex (Teleostei, Cyprinidae) in Europe using morphological and molecular markers. Zoologica Scripta 39:
354 108. Gkenas, Ch., S. Malavasi & I. Leonardos, Diet and feeding habits of Economidichthys pygmaeus (Perciformes: Gobiidae) in Lake Pamvotis, NW Greece. Journal of Applied Ichthyology 28: Glamuzina, B. & V. Bartulović, Some characteristics of the endemic dentex trout, Salmo dentex (Heckel, 1851) from the Neretva River, Bosnia- Herzegovina. Ribarstvo 64: Glamuzina, B., V. Bartulović, J. Dulčić, A. J. Conides, P. Tutman, S. Matić-Skoko, A. Gavrilović, J. Jug-Dujaković, E. Hasković, A. Ivanc & N. Zovko, Some biological characteristics of the endemic Neretvan nase, Chondrostoma knerii Heckel, 1843, in the Hutovo Blato wetlands (Bosnia and Herzegovina). Journal of Applied Ichthyology 23: Glamuzina, B., J. Dulčić, P. Tutman, S. Matić-Skoko, S. Tomšić & V. Bartulović, Record size of Salmo dentex (Heckel, 1851) (Salmonidae) from Neretva River (Bosnia and Herzegovina). Cybium 33: Grapci-Kotor, L., F. Zhushi-Etemi, H. Sahiti, A. Gashi, R. Škrijelj & H. Ibrahimi, 2010a. The ichthyofauna of Drini i Dardhë river (Kosovo). Ribarstvo 68: Grapci-Kotor, L., F. Zhushi-Etemi, H. Sahiti, A. Gashi & H. Ibrahimi, 2010b. Assessing the Ecological Status of Lumebardhi i Pejes River (Drini i Bardhe River basin, Kosovo) Using Fish Assemblages. BALWOIS 2010 Book of Abstracts, Ohrid, Republic of Macedonia, May Grupcheva, G. I. & I. L. Nedeva, Ichthyofauna of the Zrebchevo reservoir (Bulgaria). Acta zoologica bulgarica 51: Guzina, N., Morphologically-taxonomical characteristics of the species Aulopyge hügeli Heckel, Doctorate Thesis, University of Sarajevo, Sarajevo Habeković, D., R. Safner, I. Aničić & T. Treer, Ihtiofauna djela rijeke Save. Ribarstvo 55: Habeković, D., M. Mrakovčić & M. Bogdan, Ihtiofauna djela rijeke Drave nakon izgradnje sustave HE Čakovec. Ribarstvo 41: Has-Schön, E., I. Bogut, G. Kralik, S. Bogut, J. Horvatić & I. Čačić, Heavy metal concentration in fish tissues inhabiting waters of Busko blato reservoir (Bosnia and Herzegovina). Environmental Monitoring and Assessment 144: Holčík, J., Changes in the fish fauna and fisheries in the Slovak section of the Danube River: a review. Annales de Limnologie: International Journal of Limnology 39:
355 120. Holčík, J. & A. Delić, New discovery of Ukrainian brook lamprey in Croatia. Journal of Fish Biology 56: Iliadou, K., Feeding of Scardinius acarnanicus Stephanidis, 1939 (Pisces: Cyprinidae) from lakes Lysimachia and Trichonis, Greece. Journal of Fish Biology 38: IUCN, IUCN Red List of Threatened Species. Version Available at: Ivanković, P., M. Piria, T. Treer & Z. Knezović, Meristic and morphometric characteristic of endemic Neretva chub, Squalius svallize from Neretva River area, Bosnia and Herzegovina. Journal of Applied Ichthyology 27: Ivanković, P., T. Treer, M. Piria & Z. Knezović, Diet and growth of the endemic Neretva chub, Squalius svallize from the Neretva river area, Bosnia and Herzegovina. Folia Zoologica 59: Janković, D., Serbia - Part of the south zone of the range of distribution of the European grayling (Thymallus thymallus L.). Archives of Biological Sciences 62: Jelić, D., P. Žutinić & M. Jelić, Characteristics and new data of river Ilova ichthyofauna (central Croatia). Ribarstvo 67: Kalogianni, E., S. Giakoumi, S. Zogaris, Y. Chatzinikolaou, B. Zimmerman & A. Economou, Current distribution and ecology of the critically endangered Valencia letourneuxi in Greece. Biologia 65: Karapetkova, M. & M. Živkov, Ribite v. Bulgaria. Geya Libris Publishers, Sofia Karapetkova, M., M. Živkov & K. Alexandrova-Kolemanova, The freshwater fishes of Bulgaria. In Meine, C. (ed.) Bulgaria s biological diversity: Conservation status and needs assessment. Pensoft Publishers, Sofia: Kerovec, M., M. Mrakovčić, D. Schneider & N. Tomašković, Occurrence of Chondrostoma phoxinus in Croatia. Folia Zoologica 47: Ketmaier, V., P. G. Bianco & J. D. Durand, Molecular systematics, phylogeny and biogeography of roaches (Rutilus, Teleostei, Cyprinidae). Molecular Phylogenetics and Evolution, 49: Ketmaier, V., P. G. Bianco, M. Cobolli, M. Krivokapic, R. Caniglia & E. De Matthaeis, Molecular phylogeny of two lineages of Leuciscinae cyprinids 355
356 (Telestes and Scardinius) from the peri-mediterranean area based on cytochrome b data. Molecular Phylogenetics and Evolution 32: Kleanthidis, P. K. & A. I. Sinis, Feeding habits of the Macedonian shad, Alosa macedonica (Vinciguerra, 1921) in Lake Volvi (Greece): seasonal and ontogenetic changes. Israel Journal of Zoology 47: Knežević, B., D. Kažić, D. Nedic, M. Kavarić & B. Ivanović, Unique characteristics of ichthyofauna and ichthyoparasites of Skadar Lake. Verhandlungen des Internationalen Verein Limnologie 20: Kohout, J., A. Šedivá, A. Apostolou, T. Stefanov, S. Marić, M. Gaffaroğlu & V. Šlechta, Genetic diversity and phylogenetic origin of brown trout Salmo trutta populations in eastern Balkans. Biologia 68: Kolaneci, V., Ç. Shegani, M. Nikolla & B. Kuka, Some Biological Properties of Two Cyprinid Fishes Living in Shkodra Lake. BALWOIS 2010 Book of Abstracts, Ohrid, Republic of Macedonia, May Kosovo environmental protection agency, Report on the state of nature Ministry of Environment and Spatial Planning, Kosovo Environmental Protection Agency, Kosovo Institute for Nature Protection, Prishtina Kostić, D. & S. Maletin, Contribution to the knowledge of ichthyofauna of some stagnant waters in Vojvodina. Ichthyologia 24: Kostov, V., 2008a. Results of ichthyofauna investigation in Macedonian part of Lake Dojran. 1st Symposium for Protection of Natural Lakes in Republic of Macedonia Book of Abstracts, Ohrid, Republic of Macedonia, 31 May-3 June Kostov, V. 2008b. First record of species Acipenser ruthenus Linnaeus 1758 into the waters of R. Macedonia. 1st Symposium for Protection of Natural Lakes in Republic of Macedonia Book of Abstracts, Ohrid, Republic of Macedonia, 31 May-3 June Kostov, V., M. Naumovski & R. Nastova-Gjorgjioska, Antropogenic influence on the ichthyofauna of river Vardar. International Eco-Conference Environmental protection of urban and suburban settlements Book of Abstracts, Novi Sad, Serbia, September Kostov, V., K. Rebok, V. Slavevska-Stamenković & M. Ristovska, Fish fauna of Bregalnica River (R. Macedonia). BALWOIS 2010 Book of Abstracts, Ohrid, Republic of Macedonia, May Kostov, V., M. Ristovska, D. Prelić & V. Slavevska-Stamenković, 2011a. Assessement of the ecological status of the Crna River based on the fish fauna - 356
357 contribution to the establishment of the monitoring system of rivers in R. Macedonia. Macedonian Journal of Animal Science 1: Kostov, V., M. Ristovska, V. Slavevska-Stamenković, B. Miljanović & M. Paunović, 2011b. Setting up a system for ecological status assessment based on Fish Fauna - the Pcinja River - case study. Macedonian Journal of Animal Science 1: Kostov, V. & M. Van der Knaap, The collapse of Fisheries of Lake Dojran - Reasons, Actual situation and Perspectives. IV International Conference Fishery Book of Abstracts, Zemun-Belgrade, Serbia, May Kotlík, P., C. S. Tsigenopoulos, P. Ráb & P. Berrebi, Two new Barbus species from the Danube River basin, with redescription of B. petenyi (Teleostei: Cyprinidae). Folia Zoologica 51: Kottelat, M. & J. Freyhof, Handbook of European freshwater fishes. Publications Kottelat, Cornol Kovačić, M., A new species of Knipowitschia (Gobiidae) from Dalmatia, Croatia. Cybium 29: Kovačić, M., The key for identification of Gobiidae (Pisces: Perciformes) in the Adriatic Sea. Acta Adriatica 49: Kovačić, M. & R. Šanda, A new species of Knipowitschia (Perciformes: Gobiidae) from southern Montenegro. Journal of the National Museum (Prague), Natural History Series 176: Krivokapic, M., Growth of Telestes montenegrinus from River Moraca, Montenegro (Cyprinidae), Italian Journal of Zoology 65: Krpo-Ćetković, J., A. Hegediš & M. Lenhardt, Diet and growth of asp, Aspius aspius (Linnaeus, 1758), in the Danube River near the confluence wirh the Sava River (Serbia). Journal of Applied Ichthyology 26: Larmuseau, M. H. D., J. Freyhof, F. A. M. Volckaert & J. K. J. Van Houdt, Matrilinear phylogeography and demographical patterns of Rutilus rutilus: implications for taxonomy and conservation. Journal of Fish Biology 75: Lenhardt, M., P. Cakic & J. Kolarevic, Influence of the HEPS Djerdap I and Djerdap II dam construction on catch of economically important fish species in the Danube River. Ecohydrology and Hydrobiology 4:
358 155. Lenhardt, M., G. Marković, A. Hegediš, S. Maletin, M. Ćirković & Z. Marković, Non-native and translocated fish species in Serbia and their impact on the native ichthyofauna. Reviews in Fish Biology and Fisheries Lenhardt, M., I. Jaric, A. Kalauzi & G. Cvijanovic, Assessment of extinction risk and reasons for decline in sturgeon. Biodiversity and Conservation 15: Leonardos, I. D., Life history traits of Scardinius acarnanicus (Stephanidis, 1939) (Pisces: Cyprinidae) in two Greek Lakes (Lysimachia and Trichonis). Journal of Applied Ichthyology 20: Leonardos, I. D., I. Paschos & M. Prassa, Threatened fishes of the world: Phoxinellus epiroticus (Steindachner, 1895) (Cyprinidae). Environmental Biology of Fishes 72: Leonardos, I. D., I. Kagalou & A. Triantafyllidis, Threatened fishes of the world: Silurus aristotelis (Agassiz, 1856) (Siluridae). Environmental Biology of Fishes 78: Leonardos, I. D., I. Kagalou, M. Tsoumani & P. S. Economidis, Fish fauna in a Greek lake: biodiversity, introduced fish species over a 80-year period and their impacts on the ecosystem. Ecology of Freshwater Fish 17: Leonardos, I. D., A. C. Tsikliras, I. Batzakas, A. Ntakis & V. Liousia, Lifehistory characteristics of the endangered Aristotle s catfish (Silurus aristotelis Garman, 1890), Lake Pamvotis, north-western Greece. Journal of Applied Ichthyology 25: Liasko, R., Ch. Anastasiadou, A. Ntakis, Ch. Gkenas & I. D. Leonardos, Morphological differentiation among native trout populations in North-Western Greece. Journal of Biological Research-Thessaloniki 17: Liousia, V., S. Batziakas, E. Koutrakis & I. D. Leonardos, Length- Weight relationships of 22 fish species from the littoral zone of the Eastern Ionian Sea, Greece. Acta Ichthyologica et Piscatoria 42: Lujić, J., D. Kostić, E. Popović, M. Ćirković, D. Ljubojević, N. Novakov & Z. Marinović. Presence of the most economically important fish species in the river Danube and its flood zones in Serbia. Ribarstvo 70: Marčić, Z., I. Buj, A. Duplić, M. Ćaleta, P. Mustafić, D. Zanella, P. Zupančič & M. Mrakovčić, A new endemic cyprinid species from the Danube drainage. Journal of Fish Biology 79:
359 166. Marić, D. S., Endemic Fish Species in Montenegro. Biological conservation 72: Marić, D. S., Polno sazrijevanje i polni ciklus gonada kod introdukovanog srebrnog karaša (Carassius auratus gibelio Bloch) u Skadarskom jezeru. Natura Montenegrina 1: Marić, D. S., Rutilus albus sp. n. (Teleostei: Cyprinidae) from Lake Skadar. Periodicum Biologorum 2: Marić, D. S. & D. Milošević, Katalog slatkovodnih riba (Osteichthyes) Crne Gore. Crnogorska akademija nauka i umjetnosti, Podgorica Marić, D. S. & V. Pavlović, First records and description of Cobitis elongata Keckel & Kner, 1858 (Cobitidae) in Montenegro. Natura Montenegrina 5: Marić, D. S. & D. Milošević, First records and description of the Goldside loach Sabanejewia balcanica (Cobitidae) in Montenegro. Periodicum Biologorum 2: Marić, D. S. & B. Radujković, Rutilus ohridanus (Karaman, 1924) from Ohrid, Prespa and Skadar Lakes. Natura Montenegrina 8: Marić, D. S. & V. N. Šorić, Nase (Chondrostoma) (Cyprinidae, Pisces) from Ohrid-Drim-Skadar system. Natura Montenegrina 8: Marić, S., A. Hegediš, V. Nikolić & P. Simonović, Conservation status of two Eastern Balkan endemic fish species in Serbia and proposal for their protection. Acta zoologica bulgarica 56: Marić, S., V. Nikolić, L. Tomović & P. Simonović, 2011a. Morphological differentiation of trout (subf. Salmoninae) based on characteristics of head skeleton. Italian Journal of Zoology 78: Marić, S., V. Nikolić, A. Tošić and P. Simonović, Record of the brown trout Salmo trutta L., 1758 in the main riverbed of the Serbian part of the Danube River. Journal of Applied Ichthyology 28: Marić, S., A. Razpet, V. Nikolić & P. Simonović, 2011b. Genetic differentiation of European grayling (Thymallus thymallus) populations in Serbia, based on mitochondrial and nuclear DNA analyses. Genetics Selection Evolution 43: Marić, S., A. Snoj, V. Nikolić & P. Simonović, 2006a. Genetic differentiation of trout (Salmo spp.) populations in Serbia ascertained using RFLP technique on PCR amplified control region of mitochondrial DNA. Acta Veterinaria 56:
360 179. Marić, S., S. Sušnik, P. Simonović & A. Snoj, 2006b. Phylogeographic study of brown trout from Serbia, based on mitochondrial DNA control region analysis. Genetics Selection Evolution 38: Marić S. D., J. V. Rakočević & D. S. Marić, Morphological characteristics of barbles (Barbus, Cyprinidae) from the water of Montenegro. Natura Montenegrina 11: Marinov, B., Taxonomy, binomial and faunistics of some species of the family Cyprinidae and Cotidae (Pisces) from Bulgaria. Doctorate Thesis, St. Kliment Ohridski Sofia University, Sofia Markova, S., R. Šanda, A. Crivelli, S. Shumka, I. F. Wilson, J. Vukić, P. Berrebi & P. Kotlík, Nuclear and mitochondrial DNA sequence data reveal the evolutionary history of Barbus (Cyprinidae) in the ancient lake systems of the Balkans. Molecular Phylogenetics and Evolution 55: Marković, G. S., V. M Simić, A. M. Ostojić & S. B. Simić, The diet of Leuciscus cephalus L. in the reservoir Medjuvrsje (Western Morava, Serbia). Acta agriculturae Serbica 16: Matić-Skoko, S., P. Tutman, J. Dulčić, I. Prusina, Ž. Đođo, J. Pavličević & B. Glamuzina, Growth pattern of the endemic Neretvan roach, Rutilus basak (Heckel, 1843) in the Hutovo Blato wetland. Journal of Applied Ichthyology 27: Maxhuni, Q., R. Skrijelj & L. Grapci - Kotori, The Influence of Ecological Factors on Fish Population Dynamics in River Drenica. BALWOIS 2010 Book of Abstracts, Ohrid, Republic of Macedonia, May, Memić, K. & A. Adrović, Populacije ciprinidnih riba rijeke Gostelje. Zbornik radova Filozofskog fakulteta u Tuzli 17: Mérő, T. O., Periodical appearance of pike (Esox lucius Linneaus, 1758) in the Danube River in northwest Vojvodina (N Serbia). Natura Croatica 20: Mikavica, D. & N. Savić, Ribe rijeke Drine. Poljoprivredni Fakultet u Banjoj Luci, Banja Luka Miller, P. J. & R. Šanda, A west Balkanian sand- goby (Teleostei: Gobiidae). Journal of Fish Biology 72: Milošević, D., V. Pešić, D. Petrović, A. Pavićević & D. Marić, Length-weight relationship and condition factor of two sympatric Rutilus (Rrafinesque, 1820) species from Lake Skadar (Montenegro). Archives of Biological Sciences 64:
361 191. Milošević, D., K. A. Winkler, D. Marić & S. Weiss, Genotypic and phenotypic evaluation of Rutilus spp. from Skadar, Ohrid and Prespa Lakes supports revision of endemic as well as taxonomic status of several taxa. Journal of Fish Biology 79: Mrakovčić, M., Ichthyological study of accumulation Butoniga, Istra. Archives of Fisheries Centre, Zagreb Mrakovčić, M., Status of Freshwater Fish in Croatian Adriatic River Systems. Biological Conservation 72: Mrakovčić, M., A. Brigić, I. Buj, M. Ćaleta, P. Mustafić & D. Zanella, Crvena knjiga slatkovodnih riba Hrvatske. Ministarstvo kulture, Državni zavod za zaštitu prirode, Zagreb Mrakovčić, M., A. Brigić, S. Mišetić, Z. Marčić & M. Ćaleta, Endemska ihtiofauna sliva rijeke Neretve i njenih pritoka i stanje ugroženosti prema IUCN. In Skaramuca, B. & J. Dulčić (eds), Ugrožene i endemske vrste riba u slivovima rijeka Neretve, Trebišnjice i Morače: zbornik radova znanstveno-stručnog skupa. Sveučilište u Dubrovniku, EastWest Institute, Dubrovnik: Mrakovčić, M., A. Duplić, P. Mustafić & Z. Marčić, Conservation status of the genus Cobitis and related genera in Croatia. Folia Zoologica, 57: Mrakovčić, M. & S. Mišetić, 1990a. Status, distribution and conservation of the salmonid, Salmotymus obtusirostris (Heckel) and the cyprinid, Aulopyge hugeli (Heckel) in Yugoslavia. Journal of Fish Biology 37: Mrakovčić, M. & S. Mišetić, 1990b. Značaj i karakteristike ihtiofaune rijeke Krke. In Kerovec, M. (ed), Nacionalni park Krka vol 2. Hrvatsko ekološko društvo: Mrakovčić, M., S. Mišetić & M. Povz, Status of freshwater fish in Croatian Adriatic river systems. Biological Conservation 72: Mrakovčić, M., D. Schneider & M. Kerovec, Freshwater gobies of Croatia. Periodicum Biologorum 96: Mrakovčić, M., D. Schneider, P. Mustafić & M. Kerovec, Status of the genus Cobitis and related species in Croatia. Folia Zoologica, 49: Mrdak, D., Environmental Risk Assessment of the Morača Dams: Fish Fauna of Morača River Canyon and Skadar Lake. Report committed by WWF MedPO and Green Home under the framework of Sharing Water Project - Skadar Lake component, Podgorica. 361
362 203. Mrdak, D., V. Nikolić, A. Tošić & P. D. Simonović, Molecular and ecological features of the soft-muzzled trout Salmo obtusirostris (Heckel, 1852) in the Zeta River, Montenegro. Biologia 67: Munari, C., U. Tessari, R. Rossi & M. Mistri, The ecological status of Karavasta Lagoon (Albania): closing the stable door before the horse has bolted? Marine Environmental Research 69: Mustafić, P., Z. Marčić, A. Duplić, M. Mrakovčić, M. Ćaleta, D. Zanella, I. Buj, M. Podnar & Z. Dolenec, A new loach species of the genus Cobitis in Croatia. Folia Zoologica 57: Mustafić, P., M. Mrakovcić, M. Ćaleta, I. Radić, D. Zanella, Z. Mihaljević & I. Ternjej, Loaches in long term study of the Drava River in Croatia. Folia Biologica 51: Naumovski, M., Ribite vo Makedonija. Zaki, Skopje Nikcevic, M., B. Mickovic, A. Hegedis & R. K. Andjus, Feeding habits of huchen Hucho hucho (Salmonidae) fry in the River Tresnjica, Yugoslavia. Italian Journal of Zoology 65: Nikolova, M., E. Uzunova, S. Studenkov, M. Georgieva, L. Pehlivanov & B. Velkov, Fееding patterns and seasonal variation in the diet of non -indigenous fish species Lepomis gibbosus L. from shallow eutrophic lakes along River Vit, Bulgaria. Natura Montenegrina 7: Oikonomou, A., Ch. Anastasiadou, Th. Taskoudis & I. D. Leonardos, Length-weight relations of seven native fish species (Actinopterygii) from the Louros River, Greece. Acta Ichthyologica et Piscatoria (in press) Pehlivanov, L. Z., State of the ichthyofauna in Ropotamo Reserve Complex: ecological, conservation and economic aspects. Water Science and Technology 39: Pehlivanov, L. Z., Ichthyofauna of the East Rhodopes (South Bulgaria): composition and distribution. Acta zoologica bulgarica 52: Pehlivanov, L. Z. & G. Atanasov Silurus aristotelis (Pisces: Siluridae): a New Species in the Bulgarian Ichthyofauna. Acta zoologica bulgarica 59: Pehlivanov L., E. Uzunova & M. Pavlova, Ichthyofauna of the Vit River (Danube Basin): composition, distribution and conservation significance. Biotechnology and Biotechnological Equipment 23:
363 215. Pehlivanov, L., M. Pavlova, M. Vassilev, A. Apostolou & B. Velkov, Ecological Quality Assessment of Salmonid Rivers in Bulgaria Using Ichthyologic Parameters. Acta zoologica bulgarica 4: Peja, N., A. Vaso, A. Miho, N. Rakaj & A. J. Crivelli, Characteristics of Albanian lagoons and their fisheries. Fishery Research 27: Perdices, A. & I. Doadrio, The molecular systematics and biogeography of the European cobitids based on mitochondrial DNA sequences. Molecular Phylogenetics and Evolution 19: Perdikaris, C., E. Gouva & I. Paschos, 2010a. Status and development prospects for indigenous freshwater finfish and crayfish farming in Greece. Reviews in Aquaculture 2: Perdikaris, C., E. Gouva & I. Paschos, 2010b. Alien fish and crayfish species in Hellenic freshwaters and aquaculture. Reviews in Aquaculture 2: Perdikaris, C., K. Nathanailides, E. Gouva, C. Karipoglou & I. Paschos, Collapse of Epirus minnow (P. epiroticus) population in Lake Pamvotis, Greece (Teleostei: Cyprinidae). Ichthyological Exploration of Freshwaters 16: Perea, S., M. Böhme, P. Zupančič, J. Freyhof, R. Šanda, M. Özuluǧ, A. Abdoli & I. Doadrio, Phylogenetic relationships and biogeographical patterns in Circum- Mediterranean subfamily Leuciscinae (Teleostei, Cyprinidae) inferred from both mitochondrial and nuclear data. BMC Evolutionary Biology 10: Petriki, O., E. Gousia & D. C. Bobori, Weight-length relationships of 36 fish species from the River Strymon system (northern Greece). Journal of Applied Ichthyology 27: Piria, M., D. Matulić, T. Treer, I. Aničić, R. Safner, N. Šprem, N & T. Tomljanović, Condition, length-weight relationship and morphological differences between Cobitis elongata and Cobitis elongatoides from the Sava River. 3rd International Conference Loaches of the Genus Cobitis and Related Genera, Book of Abstracts, Šibenik, Croatia, September Piria, M., N. Šprem, T. Tomljanović, M. Slišković, G. Jelić Mrčelić & T. Treer, Length weight relationships of the european eel Anguilla anguilla (Linnaeus, 1758) from six karst catchments of the Adriatic basin, Croatia. Ribarstvo 72: Piria, M., T. Treer, I. Aničić, R. Safner & T. Odak, The Natural Diet of Five Cyprinid Fish Species. Agriculturae Conspectus Scientificus 70:
364 226. Polačik, M., T. Trichkova, M. Janáč, M. Vassilev & P. Jurajda, The ichthyofauna of the shoreline zone in the longitudinal profile of the Danube River, Bulgaria. Acta zoologica bulgarica 60: Povz, M., Threatened fishes of the world: Umbra krameri Walbaum, 1792 (Umbridae). Environmental Biology of Fishes 43: Povz, M., S. Leiner, M. Mrakovčič & J. Popović, Rare and endangered fishes from Yugoslavian Adriatic rivers. Journal of Fish Biology 37: Povz, M., M. Mrakovčič & M. Kerovec, The first find of Balon s ruffe (Gymnocephalus baloni) in the River Drava in Slovenia and in Croatia. Folia Zoologica 46: Prpa, Z., T. Treer, M. Piria & N. Šprem, The condition of fish from some freshwaters of Croatia. Ribarstvo 65: Radović, D., V. Stevanović, D. Marković, S. Jovanović, G. Džukić & I. Radović, Implementation of GIS technologies in assessment and protection of natural values of Tara national park. Archives of Biological Science 57: Radulović, S., P. J. Boon, D. Laketić, P. Simonović, S. Puzović, M. Živković, T. Jurca, M. Ovuka, S. Malaguti & I. Teodorović, Preliminary check-lists for applying SERCON (System for Evaluating Rivers for Conservation) to rivers in Serbia. Archives of Biological Science 64: Raikova-Petrova, G., The ichthyofauna in the glacial water of the Rila mountains. In Golemanski, V. & V. Naĭdenov (eds), Biodiversity and evolution of glacial water ecosystems in the Rila mountains. Academic Publishing House, Sofia: Raikova-Petrova, G., N. I. Hamwi & I. K. Petrov, Changes in the ichthyofauna of the middle stream of Iskar River (Bulgaria). Forest Science 3: Raikova-Petrova, G. N., I. K. Petrov, N. I. Hamwi & D. M. Marinova, Growth rate and condition of riffle minnow (Alburnoides bipunctatus Bloch) from the middle stream of Iskar river (Bulgaria). Acta Zoologica Bulgarica 63: Rakaj, N., Iktiofauna E Shqipërisë. Shtëpia Botuese Libri Universitar, Tiranë Rakaj N. & A. Flloko, Conservation status of freshwater fish of Albania. Biological Conservation 72: Razpet, A., S. Sušnik, T. Jug & A. Snoj, Genetic variation among trout in the River Neretva basin, Bosnia and Herzegovina. Journal of Fish Biology 70:
365 239. Regional Environmental Center for Central and Eastern Europe (REC), Albania, Biodiversity database of Skadar/Shkodra Lake (Checklist of species with their conservation status). tydb.pdf 240. Rexhepi, A., K. Bërxholi, P. Scheinert, A. Hamidi & K. Sherifi, Study of viral diseases in some freshwater fish in the Republic of Kosovo. Veterinarski arhiv 81: Rosecchi, E., Crivelli, A. J. & G. Catsadorakis, The establishment and impact of Pseudorasbora parva, an exotic fish species introduced into Lake Mikri Prespa (northwestern Greece). Aquatic Conservation: Marine and Freshwater Ecosystems 3: Šanda, R., I. Bogut, I. Doadrio, J. Kohout, A. Perdices, S. Perea, A. Šedivá & J. Vukić, Distribution and relationships of spined loaches (Cobitidae) in the Neretva River basin in Bosnia and Herzegovina. Folia Zoologica 57: Šanda, R. & M. Kovačić, Freshwater gobies in the Adriatic drainage basin of the Western Balkans. Annales Series Historia Naturalis, 19: Sapounidis A., E. Koutrakis & I. D. Leonardos, Length-Weight relationships of 13 species from a flow regulated Balkan River. Journal of Applied Ichthyology 27: Šanda, R., V. Lusková & J. Vukić, Notes on the distribution and taxonomic status of Gobio gobio from the Morača River basin (Montenegro). Folia Zoologica 54: Schneider, D., M. Mrakovčić, P. Mustafić & M. Kerovec, Morphological differences in some Cobitis populations from Croatia. Folia Zoologica 49: Schneider, D., P. Mustafić, M. Mrakovčić & Z. Mihaljević, Some aspects of the biology of the Neretvan spined loach. Folia Zoologica 49: Šanda, R., J. Vukić, L. Choleva, J. Křížek, A. Šedivá, S. Shumka & I. F. Wilson, Distribution of loach fishes (Cobitidae, Nemacheilidae) in Albania, with genetic analysis of populations of Cobitis ohridana. Folia Zoologica 57: Schneider-Jacoby M., U. Schwarz, P. Sackl, D. Dhora, D. Saveljic & B. Stumberger, Rapid assessment of the Ecological Value of the Bojana-Buna Delta (Albania / Montenegro). Euronatur, Radolfzell Šedivá, A., A. Apostolou, J. Kohout, V. Kostov & R. Šanda, Genetic structure and distribution of Oxynoemacheilus bureschi (Balitoridae, Teleostei), and 365
366 its phylogenetic relationships with other European stone loaches. Folia Zoologica 57: Šedivá, A., A. Apostolou, J. Kohout & J. Bohlen, Molecular phylogenetic analyses of the loach Oxynoemacheilus bureschi reveal postglacial range extensions across major river systems of the Balkans. Journal of Fish Biology 76: Sekulić, N., Lj. Budakov & D. Branković, 1998a. Comparison of feeding spectra of some exotic and native fishes in Vojvodina (Serbia). Italian Journal of Zoology 65: Sekulić, N., Lj. Budakov & D. Branković, 1998b. Distribution of the European mudminnow Umbra krameri (Umbridae) in Serbia. Italian Journal of Zoology 65: Shumka, S., A. Cake & E. Nikleka, The fish composition and importance in the Drinos River ecosystem. Natura Montenegrina 9: Shumka, S., S. Grazhdani, A. Cake & S. Mali, 2008a. Fishery of some tributaries and reservoirs and zooplankton at the Drini Catchment: the needs of better conception of the monitoring practices at transboundary water bodies. Natura Montenegrina 8: Shumka, S., A. Paparisto & S. Grazhdani, 2008b. Identification of non-native fishes in Albania and assessment of their potential threats to the national biological freshwater diversity. BALWOIS 2008 Book of Abstracts, Ohrid, Republic of Macedonia, May, Shumka, S., L. Shuka & S. Mali, Rivers Water Life and the Responses of Possible Hydropower s to be Constructed in the Water Courses of Vjosa, Semani and Drini in Albania. BALWOIS 2008 Book of Abstracts, Ohrid, Republic of Macedonia, May, Simić, V., S. Simić, M. Paunović, P. Simonović, N. Radojković & A. Petrović, Scardinius knezevici Bianco & Kottelat, 2005 and Alburnus scoranza Bonaparte, 1845: new species of ichthyofauna of Serbia and the Danube basin. Archives of Biological Science 64: Simonović, P., Ribe Srbije. NNK International, Zavod za zaštitu prirode Srbije & Biološki fakultet, Beograd Simonović, P., Ihtiofauna. In Stevanović, M. (ed.), Drina. Zavod za udžbenike i nastavna sredstva, Beograd: Simonović, P., Ribe Morave. In Jovanović, N. (ed.), Morava. Zavod za udžbenike i nastavna sredstva, Beograd:
367 262. Simonović, P., T. Karan-Žnidaršič & V. Nikolić, Ichtyofauna of the upper course of Kolubara River and its tributaries. 36th International Conference of International Association for Danube Research (IAD), Book of Abstracts, Vienna, Austria, September Simonović, P. & S. Marić, Comments of atricle of Simić and Šorić about new data on ichthyofauna of Serbia. Archives of Biological Science 58: Simonović, P., S. Marić & V. Nikolić, Growth characteristics of huchen Hucho hucho (L.) from Rivers Drina, Una and Sana. Ekologija 35: Simonović, P. D. & V. P. Nikolić, Freshwater fish of Serbia: an annotated check list with some faunistic and zoogeographic considerations. Bios Thessaloniki 4: Simonović, P. D. & V. P. Nikolić, Density-dependence of growth characteristics and maturation in stream-dwelling resident brown trout, Salmo trutta, in Serbia. Fisheries Management and Ecology 14: Simonović, P. D., V. P. Nikolić, A. D. Tošić & S. P. Marić, Length-weight relationship in adult huchen Hucho hucho (L., 1758) from Drina River, Serbia. Biologia 66: Sinis, A. & D. Petridis, Age structure and reproductive pattern of Chalcalburnus belvica (Karaman, 1924) in Lake Micro Prespa (Northwestern Greece). Israel Journal of Zoology 41: Škrijelj, R., Populacija riba Neretvanskih Jezera. Ihtiološka monografija. Prirodno - Matematički Fakultet, Universitet u Sarajevu Škrijelj, R. & M. Masović, Populacija smudja (Stizostedion lucioperca L.) u Neretvanskim hidroakumulacijama. Ribarstvo, 59: Škrijelj, R., M. Mitrašinović, A. Ivanc, J. Krnić, S. ðug, S. Šljuka & A. Mušović, Ecological, morphological-taxonomic and chematological characterisation of pike perch (Sander lucioperca, Linnaeus 1758) from reservoirs on the River Neretva. Ribarstvo 69: Šlechtová, V., J. Bohlen, J. Freyhof, H. Persat & G. B. Delmastro, The Alps as a barrier to dispersal incold-adapted fishes? Phylogeographic history and taxonomic status of the bullhead in the Adriatic freshwater drainage. Molecular Phylogenetics and Evolution 33: Smederevac-Lalić, M., R. Pešić, S. Cvejić & P. D. Simonović, Socioeconomic features of commercial fishery in the bordering upper Danube River area of Serbia. Environmental Monitoring and Assessment 184:
368 274. Snoj, A., B. Glamuzina, A. Razpet, J. Zablocki, I. Bogut, E. Lerceteau-Kohler, N. Pojskić & S. Sušnik, Resolving taxonomic uncertainties using molecular systematics: Salmo dentex and the Balkan trout community. Hydrobiologia 651: Snoj, A., S. Marić, P. Berrebi, A. Crivelli, S. Shumka & S. Sušnik, Genetic architecture of trout from Albania as revealed by mtdna control region variation. Genetics Selection Evolution 41: Snoj, A., E. Melkič, S. Sušnik, S. Muhamedagić & P. Dovč, DNA phylogeny supports revised classification of Salmothymus obtusirostris. Biological Journal of the Linnaean Society 77: Sofradžija, A., Slatkovodne ribe Bosne i Hercegovine. Vijeće Kongresa bošnjačkih intelektualaca, Sarajevo Šorić, V., Rutilus rubilio (Cyprinidae) u Ohrid-Drim-Skadar sistem. Biosistematika 9: Šorić, V., Ichthyofauna of the Ohrid-Drim-Skadar system. Ichthyologia 22: Šorić, V., Osteological characteristics in six species of the genus Phoxinellus Heck., Ichthyologia 22: Šorić, V., Morphological characteristics of the minnow Phoxinus phoxinus in some waters of the Black Sea and the Adriatic Sea basin. Ichthyologia 26: Šorić, V., Ihtiofauna reke Gruže, pritoke Zapadne Morave (Dunavski sliv) I. Reproduktivni potencijal vrsta Leuciscus cephalus, Alburnus alburnus i Rutilus rutilus. Ichthyologia 28: Šorić, V., 1998a. Pollution of the Morava River (Danube basin Serbia) and changes of the Ichtyofauna under its influence. Ichthyologia 30: Šorić, V., 1998b. New data the taxonomy and distribution of lampreys (Petromyzontidae) in southern Europe. Ichthyologia 30: Šorić, V., 1999a. Growth of the Barbel (Barbus barbus) in the Middle Course of the Danube. Ichthyologia 31: Šorić, V., 1999b. First finding of Orthrias brandti (Cobitidae) in the drainage area of Vardar River (FYROM) and description of the subspecies Orthrias brandti macedonicus ssp. Nov. Ichthyologia 31:
369 287. Šorić, V., Intraspecific variations of stone loach Orthrias barbatulus (Cobitidae) in southeastern Europe and description of Orthrias barbatulus zetensis spp. nov. Ichthyologia 32: Šorić, V., New data on the taxonomy and distribution of lampreys (Petromyzontidae) in southern Europe: revision of data on the lampreys in the waters of Serbia and Montenegro. Ichthyologia 34: Šorić, V., Redescription and some ecological characteristics of Alburnus arborella (Bonapartae, 1844). Kragujevac Journal of Science 28: Šorić V. M. & K. R. Ilić, Systematical and ecological characteristics of Alburnoides bipunctatus (Bloch) in some waters of Yugoslavia. Ichthyologia 17: Šprem, N., D. Matulić, T. Treer & I. Aničić, A new maximum length and weight for Scardinius erythrophthalmus. Journal of Applied Ichthyology 26: Šprem, N., M. Piria & T. Treer, Morfološke osobine i dužinsko-maseni odnosi triju populacija bodorki (Rutilus rutilus) u sjeverozapadnoj Hrvatskoj. Ribarstvo 59: Stefanov, T., Fauna and distribution of fishes in Bulgaria. In Fet, V. & A. Popov (eds), Biogeography and Ecology of Bulgaria. Springer, Dordrecht: Stefanov, T. & J. Holčík, The lampreys of Bulgaria. Folia Zoologica 56: Stefanov, T. & T. Trichkova, Fish species diversity in the Eastern Rhodopes (Bulgaria). In: Beron P. & A. Popov (eds.). Biodiversity of Bulgaria. Biodiversity of Eastern Rhodopes (Bulgaria and Greece) Volume II. Pensoft Publishers & National Museum of Natural History, Sofia: Stefanov, T. & T. Trichkova, The ichthyofauna of the Western Rhodopes (Bulgaria). In Beron, P. (ed.), Biodiversity of Bulgaria, Biodiversity of Western Rhodopes (Bulgaria and Greece) Volume I. Pensoft Publishers & National Museum of Natural History, Sofia: Stefanova, E., E. Uzunova, T. Hubenova, P. Vasileva, D. Terziyski & I. Iliev, Age and growth of the chub, Leuciscus cephalus L. from the Maritza River (South Bulgaria). Bulgarian Journal of Agricultural Science 14: Stephanidis, A., Freshwater fish of western Greece and Corfu Island. Doctorate Thesis, University of Athens, Athens. 369
370 299. Štrbac, S., A. Šajnović, L. Budakov, N. Vasić, M. Kašanin-Grubin, P. D. Simonović & B. Jovančićević, Metals in the sediment and liver of four fish species from different trophic levels in Tisza River, Serbia. Chemistry and Ecology 30: Sušnik, S., A. Snoj, I. F. Wilson, D. Mrdak & S. Weiss, Historical demography of brown trout (Salmo trutta) in the Adriatic drainage including the putative S. letnica endemic to Lake Ohrid. Molecular Phylogenetics and Evolution 44: Tachos, A. V., The fish fauna of Aoos river. Postgraduate Doctorate Thesis, Aristotle University, Thessaloniki Talevska, M., D. Petrovic, D. Milosevic, T. Talevski, D. Maric & A. Talevska, Biodiversity of macrophyte vegetation from Lake Prespa, Lake Ohrid and Lake Skadar. Biotechnology and Biotechnological Equipment 23: Talevski, T., D. Milosevic, D. Maric, D. Petrovic, M. Talevska & A. Talevska, 2009a. Biodiversity of ichthyofauna from Lake Prespa, Lake Ohrid and Lake Skadar. Biotechnology and Biotechnological Equipment 23: Talevski, T., D. Milosevic, D. Maric, D. Petrovic, M. Talevska & A. Talevska, 2009b. Anthrophogenic Influence on Biodiversity of Ichthyofauna and Macrophyte Vegetation from Lake Ohrid and Lake Skadar. Journal of International Environmental Application & Science 4: Talevski, T., D. Milosevic & A. Talevska, 2010a. Anthropogenic influence and conservation status of authochthonous fish fauna from Lake Ohrid. BALWOIS 2010 Book of Abstracts, Ohrid, Republic of Macedonia, May, Talevski, T., D. Milosevic & A. Talevska, 2010b. Anthropogenic Influence and Conservation Status of Autochthonous Fish Fauna from Lake Prespa. BALWOIS 2010 Book of Abstracts, Ohrid, Republic of Macedonia, May, Talevski, T., M. Talevska, D. Milosevic & A. Talevska, 2010c. Anthropogenic Influence on Ichthyofauna and Macrophyte Diversity in the Crn Drim Ecosystem. BALWOIS 2010 Book of Abstracts, Ohrid, Republic of Macedonia, May, Teskeredžić, E., Z. Teskeredžić, E. McLean, M. Tomec & R. Čoz-Rakovac, Occurrence, Distribution, and Potential Future of Yugoslavian Salmonids. In Cloud, J. G. & G. H. Thorgaard (eds), Genetic Conservation of Salmonid Fishes. Plenum Press, New York: Treer, T., I. Aničić, R. Safner, T. Odak & M. Piria, 2008a. Growth and condition of endemic trout Salmothymus obtusirostris in Jadro, a Dalmatian river. In Nielsen J. L., J. J. Dodson, K. Friedland, T. R. Hamon, J. Musick & E. Verspoor (eds), Reconciling 370
371 fisheries with conservation: proceedings of the Fourth World Fisheries Congress. American Fisheries Society, Symposium 49, Bethesda, Maryland: Treer, T., I. Aničić, R. Safner, M. Piria & T. Odak, Post-spawning condition of endemic soft-muzzled trout Salmothymus obtusirostris in the Žrnovnica River. Ribarstvo 63: Treer, T., D. Habeković, I. Aničić, R. Safner & A. Kolak, Standard growth curve for chub (Leuciscus cephalus L. 1758) in Croatia. Ribarstvo 55: Treer, T., D. Habeković, I. Aničić, R. Safner & M. Piria, Growth of five spirlin (Alburnoides bipunctatus) populations from the Croatian rivers. Agriculturae Conspectus Scientificus 65: Treer, T., D. Matulić, G. Bogdanović, I. Aničić, R. Safner, M. Piria, N. Šprem & T. Tomljanović, The condition of allochtonous fishes in the Mediterranean Vransko Lake. Journal of Applied Ichthyology 27: Treer, T., A. Opačak, I. Aničić, R. Safner, M. Piria & T. Odak, 2003a. Growth of bream, Abramis brama, in the Croatian section of the Danube. Czech Journal of Animal Science 48: Treer, T., M. Piria, I. Aničić, R. Safner & T. Tomljanović, Diet and growth of spirlin, Alburnoides bipunctatus in the barbel zone of the Sava River. Folia Zoologica 55: Treer, T., R. Safner, I. Aničić, A. Kolak & M. Dražić, Morphological variation among four strains of common carp Cyprinus carpio in Croatia. Folia Zoologica 49: Treer, T., N. Šprem, H. Torcu-Koc, Y. Sun & M. Piria, 2008b. Length- weight relationships of freshwater fishes of Croatia. Journal of Applied Ichthyology 24: Treer, T., B. Varga, R. Safner, I. Aničić, M. Piria & T. Odak, 2003b. Growth of the common carp (Cyprinus carpio) introduced into the Mediterranean Vransko Lake. Journal of Applied Ichthyology 19: Triantafyllidis, A., D. Bobori, C. Koliamitra, E. Gbandi, M. Mpanti, O. Petriki & N. Karaiskou, DNA barcoding analysis of fish species diversity in four north Greek lakes. Mitochondrial DNA 22: Trichkova, A., M. Živkov & A. Apostolou, Some biological characteristics of gibel carp (Carassius gibelio) from three localities in the Bulgarian stretch of the Danube River. 17th International Symposium Ecology & Safety 2008, Sunny Beach, Bulgaria, 9-13 June
372 321. Trožić-Borovac, S. & R. Škrijelj, The Nutrition of a Pikeperch, Stizostedion lucioperca Linnaeus, 1758 in the River Neretva. Ribarstvo 65: Tsigenopoulos, C. S. & P. Berrebi, Molecular phylogeny of North Mediterranean freshwater Barbs (Genus Barbus: Cyprinidae) inferred from Cytochrome b sequences: biogeographic and systematic implications. Molecular Phylogenetics and Evolution 14: Tsigenopoulos, C. S., P. Kotlik & P. Berrebi, Biogeography and pattern of gene flow among Barbus species (Teleostei: Cyprinidae) inhabiting the Italian Peninsula and neighbouring Adriatic drainages as revealed by allozyme and mitochondrial sequence data. Biological Journal of the Linnean Society 75: Tsoumani, M., A. P. Apostolidis & I. D. Leonardos, 2013a. Length-weight relationships of Rutilus species from fifteen Greek lakes. Journal of Applied Ichthyology 29: Tsoumani, M., A. P. Apostolidis & I. D. Leonardos, 2013b. Biogeography of Rutilus species of the southern Balkan Peninsula as inferred by multivariate analysis of morphological data. Journal of Zoology 289: Turk, M., Hrvatsko slatkovodno ribartsvo u godini Ribartsvo 55: Tutman, P., B. Glamuzina & J. Dulčić, 2008a. Threatened fishes of the world: Chondrostoma knerii Heckel, 1843 (Cyprinidae). Environmental Biology of Fishes 83: Tutman, P., B. Glamuzina & J. Dulčić, 2008b. Threatened fishes of the world: Salmo dentex (Heckel 1852) (Salmonidae). Environmental Biology of Fishes 82: Tutman, P., B. Glamuzina, J., Dulčić, V. Bartulović & E. Hasković, Endemic fish fauna of the Hutovo Blato wetland (Neretva river basin, Bosnia and Herzegovina) and their conservation status. 13th European congress of ichthyology, Book of Abstracts, Klaipeda, Lithuania, 6-11 September Tutman, P., B. Glamuzina, J. Dulčić & N. Zovko, Ihtiofauna močvare Hutovo blato (donji tok rijeke Neretve, Bosna i Hercegovina); stanje i ugroženost. Ribarstvo 70: United States Agency for International Development, Biodiversity assessment of Kosovo, ARD/BIOFOR, Vermont. 372
373 332. Uzunov, Y., B. B. Georgiev, E. Varadinoiva, N. Ivanova, L. Pehlivanov & V. Vasilev, Ecosystems of the Biosphere Reserve Srebarna Lake. Professor Marin Drinov Academic Publishing House, Sofia Uzunova, E Survival, growth and maturation of triploid brook trout (Salvelinus fontinalis) induced by heat shock. Biotechnology and Biotechnological equipment 18: Uzunova, E., Role of the introduced freshwater fishes in Bulgaria: economical benefits and ecological consequences. Bulgarian Journal of Agriculture Science 12: Uzunova, E. & S. Zlatanova, A review of fish introductions in Bulgarian freshwaters. Acta Ichthyologica et Piscatoria 37: Valoukas, V. A. & P. S. Economidis, Growth, population composition and reproduction of Bream, Abramis brama (L.), in Lake Volvi, Macedonia, Greece. Ecology of Freshwater Fish 5: Vanhove, M. P. M, A. N. Economou, S. Zogaris, M. H. D. Larmuseau, S. Giakoumi, E. Kalogianni, F. A. M. Volckaert & T. Huyse, Phylogenetics and biogeography of the Balkan sand gobies (Teleostei: Gobiidae): vulnerable species in need of taxonomic revision. Biological Journal of the Linnean Society 105: Vassilev, M., Alteration of the ichthyofauna in the Shabla and Ezerets Lakes. In Golemansky, V. & W. Naidenow (eds), Biodiversity of Shabla Lake System. Prof. Marin Drinov Academic Publishing House, Sofia: Vassilev, M., Changes of ichthyofauna in the Durankulak Lake. Acta zoologica bulgarica 51: Vassilev, M., A. Apostolou, B. Velkov, D. Dobrev & V. Zarev, Atlas of the Bulgarian Gobies (Gobiidae). Pensoft publishing, Sofia Vassilev, M., A. Apostolou, B. Velkov, P. Ivanova, M. Panayotova, D. Dobrev & L. Pehlivanov, Status of Gobiid ichthyofauna (Gobiidae) in Bulgaria: taxonomical, conservative, ecological and social aspects. Natura montenegrina 10: Vassilev, M. & L. Z. Pehlivanov, Ichthyofauna of the Bulgarian section of the Struma River. Historia naturalis bulgarica 14: Vassilev, M. & L. Z. Pehlivanov, Checklist of Bulgarian freshwater fishes. Acta Zoologica Bulgarica 57: Vassilev, M., A. Trichkova, D. Ureche, I. Stoica, K. Battes & M. T. Zivkov, Distribution of Gobiid species (Gobiidae, Pisces) in the Yantra River (Danube basin, 373
374 Bulgaria). Anniversary scientific conference of ecology Book of Abstracts, Plovdiv, Bulgaria, 1 November Vasil eva, E. D. & V. P. Vasil ev, Cobitis pontica sp. nova a new spined loach species (Cobitidae) from the Bulgarian waters. Journal of Ichthyology 46: Velkov B., L. Z. Pehlivanov & M. Vassilev, Umbra krameri (Pisces: Umbridae): A Reinstated Species for the Bulgarian Ichthyofauna. Acta Zoologica Bulgarica 56: Velkova-Jordanoska, L., P. Ivanova, I. Dobrovolov, S. Stojanovski, G. Kostoski & G. Atanasov, Population-genetic structure of Barbus cyclolepis (Pisces, Cyprinidae). Biotechnology and Biotechnological Equipment 23: Vilizzi, L The common carp, Cyprinus carpio, in the Mediterranean region: origin, distribution, economic benefits, impacts and management. Fisheries Management and Ecology 19: Vuković, D., A. Tursi, R. Carlucci & R. Dekić, Ichthyofauna of the wetland ecosystem in the Bardača area (Bosnia and Herzegovina). Ribarstvo 66: Vuković, T., Ribe Bosne i Hercegovine. Svjetlost, Sarajevo Vuković, T. & B. Ivanović, Slatkovodne ribe Jugoslavije. Zemaljski muzej Bosne i Hercegovine, Sarajevo Zanella, D., Z. Mihaljević, M. Mrakovčić & M. Ćaleta, Ecology and diet of the endemic Telestes ukliva (Cyprinidae) in the Cetina River system, Croatia. Cybium 33: Zardoya, R. & I. Doadrio, Molecular evidence on the evolutionary and biogeographical patterns of European Cyprinids. Journal of Molecular Evolution 49: Zardoya R., P. S. Economidis & I. Doadrio, Phylogenetic relationships of Greek Cyprinidae: molecular evidence for at least two origins of the Greek cyprinid fauna. Molecular Phylogenetics and Evolution 13: Zaplata R. & Z. Taler, 1933: Ribe Sarajeva i okoline. Državna Štamparija, Sarajevo Zenetos, A., M. A. Pancucci-Papadopoulou, S. Zogaris, E. Papastergiadou, L. Vardakas, K. Aligizaki & A. N. Economou, Aquatic alien species in Greece (2009): tracking sources, patterns and effects on the ecosystem. Journal of Biological Research- Thessaloniki 12:
375 357. Zeqir, V., M. Behxhet, H. Avni & K. Zenel, Biodiversity conservation status in the Republic of Kosovo with focus on biodiversity centres. Journal of Environmental Biology 33: Živkov, M., Ichtyofauna and fish utilization of the Dospat reservoir. Hydrobiology 30: Živkov, M. & G. Raikova-Petrova, Comparative analysis of age composition, growth rate and condition of roach, Rutilus rutilus (L.), in three Bulgarian reservoirs. Acta zoologica bulgarica 53: Zogaris, S., L. Vardakas, A. N. Economou & P. S Economidis A preliminary list of fishes of the Dadia - Lefkimi - Soufli Forest National Park. In Catsadorakis G. & H. Kallander (eds), The Dadia- Lefkimi-Soufli Forest National Park, Greece: Biodiversity, Management, and Conservation. WWF-Greece, Athens: Zupančič, P., Rijetke i ugrožene ribe jadranskog slijeva Hrvatske, Slovenije i Bosne i Hercegovine. Atlas u boji. AZV d.o.o., Dolsko, Slovenia Zupančič, P. & N. G. Bogutskaya, Description of a new species, Phoxinellus dalmaticus (Cyprinidae: Leuciscinae), from the Cikola river in the Krka river system, Adriatic basin (Croatia). Natura Croatica 9: Zupančič, P. & N. G. Bogutskaya, Description of two new species, Phoxinellus krbavensis and P. jadovensis, re-description of P. fontinalis Karaman, 1972, and a discussion of the distribution of Phoxinellus species (Teleostei: Cyprinidae) in Croatia and Bosnia- Herzegovina. Natura Croatica 11: Zupančič, P., M. Mrakovčić, M. Marčić, A. M. Naseka & N. G. Bogutskaya, Identity of Squalius (Actinopterygii, Cyprinidae) from Istra Peninsula in Croatia (Adriatic Sea basin). ZooKeys 53:
376
377 Παράρτημα 2 Λίστα ειδών ψαριών της Βαλκανικής χερσονήσου Species Species 1. Abramis brama 101. Pelasgus minutus 2. Alburnoides bipunctatus 102. Pelasgus prespensis 3. Alburnoides ohridanus 103. Pelasgus stymphalicus 4. Alburnoides prespensis 104. Pelasgus thesproticus 5. Alburnus albidus 105. Pelecus cultratus 6. Alburnus alburnus 106. Petroleuciscus borysthenicus 7. Alburnus arborella 107. Phoxinellus alepidotus 8. Alburnus belvica 108. Phoxinellus pseudalepidotus 9. Alburnus chalcoides 109. Phoxinus lumaireul 10. Alburnus macedonicus 110. Phoxinus phoxinus 11. Alburnus maximus 111. Phoxinus strymonicus 12. Alburnus neretvae 112. Pomatoschistus canestrinii 13. Alburnus scoranza 113. Pomatoschistus montenegrensis 14. Alburnus sp. Volvi 114. Ponticola kessleri 15. Alburnus thessalicus 115. Proterorhinus semilunaris 16. Alburnus vistonicus 116. Pungitius hellenicus 17. Alburnus volviticus 117. Pungitius platygaster 18. Alosa agone 118. Rhodeus meridionalis 19. Alosa macedonica 119. Romanogobio albipinnatus 20. Alosa vistonica 120. Romanogobio elimeius 21. Aspius aspius 121. Romanogobio kesslerii 22. Aulopyge huegelii 122. Romanogobio uranoscopus 23. Ballerus ballerus 123. Romanogobio vladykovi 24. Ballerus sapa 124. Rutilus albus 25. Barbatula barbatula 125. Rutilus basak 26. Barbatula sturanyi 126. Rutilus frisii 27. Barbatula zetensis 127. Rutilus ohridanus 28. Barbus balcanicus 128. Rutilus panosi 29. Barbus barbus 129. Rutilus prespensis 30. Barbus cyclolepis 130. Rutilus rutilus 31. Barbus macedonicus 131. Rutilus sperchios 32. Barbus meridionalis 132. Rutilus virgo 33. Barbus peloponnesius 133. Rutilus ylikiensis 34. Barbus petenyi 134. Sabanejewia balcanica 35. Barbus plebejus 135. Sabanejewia bulgarica 36. Barbus prespensis 136. Salaria economidisi 37. Barbus rebeli 137. Salaria fluviatilis 38. Barbus sperchiensis 138. Salmo balcanicus 39. Barbus strumicae 139. Salmo farioides 40. Barbus tauricus 140. Salmo labrax 41. Blicca bjoerkna 141. Salmo letnica 42. Chondrostoma knerii 142. Salmo lourosensis 43. Chondrostoma nasus 143. Salmo lumi 377
378 44. Chondrostoma prespense 144. Salmo macedonicus 45. Chondrostoma scodrense 145. Salmo macrostigma 46. Chondrostoma vardarense 146. Salmo marmoratus 47. Cobitis arachthosensis 147. Salmo montenigrinus 48. Cobitis bilineata 148. Salmo obtusirostris 49. Cobitis dalmatina 149. Salmo ohridanus 50. Cobitis elongata 150. Salmo pelagonicus 51. Cobitis elongatoides 151. Salmo peristericus 52. Cobitis hellenica 152. Salmo taleri 53. Cobitis meridionalis 153. Salmo trutta 54. Cobitis narentana 154. Salmo zrmanjaensis 55. Cobitis ohridana 155. Sander lucioperca 56. Cobitis puncticulata 156. Sander volgensis 57. Cobitis punctilineata 157. Scardinius acarnanicus 58. Cobitis rhodopensis 158. Scardinius dergle 59. Cobitis stephanidisi 159. Scardinius erythrophthalmus 60. Cobitis strumicae 160. Scardinius graecus 61. Cobitis taenia 161. Scardinius knezevici 62. Cobitis trichonica 162. Scardinius plotizza 63. Cobitis vardarensis 163. Silurus aristotelis 64. Cottus gobio 164. Squalius cephalus 65. Cottus poecilopus 165. Squalius illyricus 66. Delminichthys adspersus 166. Squalius microlepis 67. Delminichthys ghetaldii 167. Squalius orpheus 68. Economidichthys pygmaeus 168. Squalius peloponnensis 69. Economidichthys trichonis 169. Squalius prespensis 70. Gasterosteus aculeatus 170. Squalius sp. Aoos 71. Gasterosteus gymnurus 171. Squalius sp. Evinos 72. Gobio bulgaricus 172. Squalius squalus 73. Gobio feraeensis 173. Squalius svallize 74. Gobio gobio 174. Squalius tenellus 75. Gobio ohridanus 175. Squalius vardarensis 76. Gobio skadarensis 176. Squalius zrmanjae 77. Gymnocephalus baloni 177. Telestes beoticus 78. Gymnocephalus cernua 178. Telestes croaticus 79. Gymnocephalus schraetser 179. Telestes metohiensis 80. Hucho hucho 180. Telestes montenigrinus 81. Knipowitschia croatica 181. Telestes pleurobipunctatus 82. Knipowitschia milleri 182. Telestes souffia 83. Knipowitschia montenegrina 183. Telestes turskyi 84. Knipowitschia mrakovcici 184. Telestes ukliva 85. Knipowitschia radovici 185. Thymallus thymallus 86. Knipowitschia thessala 186. Tropidophoxinellus hellenicus 87. Leucaspius delineatus 187. Umbra krameri 88. Leuciscus idus 188. Valencia letourneuxi 89. Leuciscus leuciscus 189. Vimba melanops 90. Lota lota 190. Vimba vimba 91. Luciobarbus albanicus 191. Zingel balcanicus 378
379 92. Luciobarbus graecus 192. Zingel streber 93. Misgurnus fossilis 193. Zingel zingel 94. Oxynoemacheilus bureschi 95. Oxynoemacheilus pindus 96. Pachychilon macedonicum 97. Pachychilon pictum 98. Pelasgus epiroticus 99. Pelasgus laconicus 100. Pelasgus marathonicus 379
380
 (Chla = χλωροφύλλη-α, DIN =")
381 Παράρτημα 3 Συσχέτιση του αριθμού ειδών ψαριών με τα λιμνολογικά χαρακτηριστικά και τις φυσικοχημικές παραμέτρους του νερού σε 13 Ελληνικές λίμνες Εικόνα 1. Διάγραμμα συσχέτισης του αριθμού ειδών ψαριών με τα λιμνολογικά χαρακτηριστικά και τις φυσικοχημικές παραμέτρους του νερού που χρησιμοποιήθηκαν στη μελέτη (n = 13) (Chla = χλωροφύλλη-α, DIN = Διαλυμένο ανόργανο άζωτο, Secchi = βάθος κατά Secchi) Figure 1. Correlation graph of fish richness, limnological characteristics and water physicochemical parameters used in the study (n = 13) (CHLa= Chlorophyll-a, DIN= Dissolved inorganic nitrogen, Secchi=Secchi depth) 381
382
383 Παράρτημα 4 Αναλυτική περιγραφή σταθμών δειγματοληψίας στον ποταμό Λούρο Σταθμός 1- Χάνι Τερόβου [ , ] Ο πρώτος σταθμός (Εικόνα 1) βρίσκεται μόλις 5 χιλιόμετρα νοτιότερα από τις αναβλυστικές πηγές του ποταμού Λούρου, σε υψόμετρο περίπου 246 μέτρων, και συγκεκριμένα στο Χάνι Τερόβου. Το πλάτος της κοίτης του ποταμού στο σημείο κυμαίνεται από 2 ως 5m και το πλάτος των πλημμυρικών πεδίων εκτείνεται μόλις στο 1 m, ενώ υπάρχουν σημεία με πολύ μικρό βάθος. Η έντονη ροή, ο πυθμένας ασβεστολιθικών πετρωμάτων και η παρόχθια βλάστηση με βυθισμένες ρίζες δέντρων δημιουργούν χαρακτηριστικά ενδιαιτήματα προσφέροντας καταφύγιο στην ενδημική πέστροφα του ποταμού Λούρου (Salmo lourosensis). Πλησίον του σταθμού δεν υπάρχουν λιβάδια και καλλιεργούμενες εκτάσεις, υπάρχουν όμως μονάδες εκτροφής της ιριδίζουσας πέστροφας (Oncorhynchus mykiss). Εικόνα 1. Ο πρώτος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 1. The first sampling station of Louros river 383
384 Σταθμός 2- Πλατανάκια [ , ] Ο δεύτερος σταθμός (Εικόνα 2) βρίσκεται 6 km νοτιότερα από τον πρώτο, στην περιοχή Πλατανάκια, σε υψόμετρο περίπου 206 μέτρων και αποτελεί θέση στην περιοχή του μέσου ρείθρου. Το πλάτος της κοίτης του ποταμού κυμαίνεται από 2 έως 6 μέτρα, ενώ αυτό των πλημυρρικών πεδίων εκτείνεται στα 3 m. Ο πυθμένας καλύπτεται με χοντρό χαλίκι και κροκάλες, ενώ η βλάστηση αποτελείται από λίγα υδρόβια μακρόφυτα. Η έντονη ροή, οι πολλοί μαίανδροι και οι αναβαθμίδες καθώς και το περιορισμένο εύρος των πλημμυρικών πεδίων έχουν ως αποτέλεσμα σε περιόδους έντονης βροχόπτωσης να παρατηρείται άνοδος της στάθμης των υδάτων και ανάπτυξη του όγκου του νερού μέσα στη στενή λεκάνη απορροής του ποταμού. Έτσι παρατηρούμε το φαινόμενο η στάθμη να ανέρχεται έως το επίπεδο του παραποτάμιου δρόμου. Χαρακτηριστική είναι η εκτεταμμένη απόρριψη σκουπιδιών κατά μήκος του ποταμού. Εικόνα 2. Ο δεύτερος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 2. The second sampling station of Louros river 384
385 Σταθμός 3- Κουκλέσι ( , ) Ο τρίτος σταθμός (Εικόνα 3) 5 km νοτιότερα από τον δεύτερο, λίγο πριν από το χωριό Κουκλέσι, σε υψόμετρο 177 μέτρων, χαρακτηρίζεται από ήπια ροή, χαμηλό βάθος και πυθμένα που καλύπτεται από χοντρό χαλίκι. Το πλάτος του ποταμού στο σημείο αυτό είναι 5-6 μέτρα και τα πλημμυρικά πεδία περίπου μισό μέτρο. Χαρακτηριστικό του σταθμού είναι η αυξομείωση της στάθμης των υδάτων ανάλογα με τις βροχοπτώσεις και η συχνή ανατάραξη του ιζήματος του πυθμένα. Πολλά βυθισμένα υδρόβια μακρόφυτα και παράκτια εποχικά φυτά χαρακτηρίζουν τα ενδιαιτήματα. Κοντά στο σταθμό υπάρχουν μόνιμες καλλιέργειες και κτηνοτροφικές μονάδες. Εικόνα 3. Ο τρίτος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 3. The third sampling station of Louros river 385
386 Σταθμός 4 Κερασώνας ( , ) Ο τέταρτος σταθμός (Εικόνα 4) 7 km νοτιότερα, βρίσκεται στην περιοχή του χωριού Κερασώνα σε υψόμετρο 141 μέτρων. Πρόκειται για ρηχό σταθμό με έντονη ροή υδάτων και ανατάραξη ιζήματος. Ιδιαίτερα, μετά από έντονη βροχόπτωση, κατά τους χειμερινούς μήνες, η θολερότητα των υδάτων είναι πολύ υψηλή. Το πλάτος του ποταμού στο σταθμό αυτό κυμαίνεται από 5-7 m ενώ το πλάτος των πλημμυρικών πεδίων φτάνει μόλις το μισό μέτρο. Ο σταθμός χαρακτηρίζεται από σημεία με βλάστηση, σκληρό πυθμένα με χαλίκι και από βαθύτερα σημεία με ιλυώδη πυθμένα. Ο αριθμός των μαιάνδρων είναι μικρότερος από τους προηγούμενους σταθμούς, ενώ στην περιοχή γίνονται προσχώσεις στην κοίτη του ποταμού από καλλιεργητές για να συγκρατήσουν τα νερά από τις καλλιεργειές τους. Η περιοχή γύρω από το ποτάμι καλύπτεται από γεωργικές εκτάσεις και κτηνοτροφικές μονάδες. Εικόνα 4. Ο τέταρτος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 4. The fourth sampling station of Louros river 386
387 Σταθμός 5- Άγιος Γεώργιος [ , ] Ο πέμπτος σταθμός (Εικόνα 5) 9 km νοτιότερα, στο χωριό Άγιος Γεώργιος βρίσκεται σε υψόμετρο 102 μέτρων. Πρόκειται για τον τελευταίο σταθμό ανάντη του υδροηλεκτρικού φράγματος. Νέες εισροές υδάτων από τις πηγές του Αγ. Γεωργίου σε συνδυασμό με την πλούσια υδρόβια βλάστηση και τον χοντρόκοκκο πυθμένα, συνεισφέρουν στο σχηματισμό ποικίλων ενδιαιτημάτων και προσφέρουν καταφύγιο σε πολλά είδη ψαριών. Η κοίτη του ποταμού σχηματίζει πολυάριθμους μαιάνδρους και νερόλακκους βάθους από 0,3 ως 1m. Στην περιοχή δεν υπάρχουν λιβάδια και γεωργικές εκτάσεις, υπάρχουν όμως μικρής έκτασης πεστροφοκαλλιέργειες και μία νεροτριβή. Εικόνα 5. Ο πέμπτος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 5. The fifth sampling station of Louros river 387
388 Σταθμός 6- Σκάλα [ , ] Ο έκτος σταθμός (Εικόνα 6) με την ονομασία Σκάλα βρίσκεται σε απόσταση 26 km από τον πέμπτο, μετά το υδροηλεκτρικό φράγμα, κοντά στο χωριό Λούρος, σε υψόμετρο 10 μέτρων. Στο σημείο αυτό ο Λούρος δέχεται τα νερά παραποτάμων στους πρόποδες του όρους Βαλαωρίτη. Έπειτα από ανθρωπογενή παρέμβαση, νερά του ποταμού συγκεντρώνονται σε μια υδατοσυλλογή και η επικοινωνία με την κύρια κοίτη του ποταμού γίνεται με σωλήνες. Καλάμια και υδρόβια φυτά που συναντούμε σε έλη κυριαρχούν ενώ ο πυθμένας καλύπτεται από λεπτόκοκκο ίζημα και ιλύ. Η περιοχή καλύπτεται από γεωργικές εκτάσεις ενώ κοντά στο σταθμό λειτουργεί και μία νεροτριβή. Εικόνα 6. Ο έκτος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 6. The sixth sampling station of Louros river 388
389 Σταθμός 7- «Jumbo» [ , ] Ο σταθμός επτά (Εικόνα 7) με την ονομασία Jumbo, βρίσκεται στο επίπεδο της θάλασσας και 5 km νοτιότερα από τον προηγούμενο σταθμό. Το πλάτος του ποταμού στο σημείο είναι μεγάλο και το βάθος του φτάνει τα 3 μέτρα. Η ήπια ροή, το μεγάλο βάθος και η πλούσια βλάστηση που αποτελείται κυρίως από υδρόβια φυτά δημιουργεί ενδιαιτήματα για μικρά και μεγαλόσωμα ψάρια. Κοντά στο σταθμό καλλιεργούνται οπωροφόρα δέντρα. Εικόνα 7. Ο έβδομος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 7. The seventh sampling station of Louros river 389
390 Σταθμός 8- Φλάμπουρα [ , ] Ο όγδοος σταθμός (Εικόνα 8) με την ονομασία Φλάμπουρα, βρίσκεται κοντά στις εκβολές, νοτιότερα 9 km από τον προηγούμενο. Το πλάτος του ποταμού στο σημείο φτάνει τα 7 μέτρα και το βάθος του φτάνει τα 3 μέτρα. Χαρακτηρίζεται από ήπια ροή, ιλυώδες υπόστρωμα και πλούσια βλάστηση. Οι μοναδικές επεμβάσεις στο σταθμό είναι η κατασκευή ξύλινων εξεδρών για την ευκολότερη προσβασιμότητα των ψαράδων στην κοίτη του ποταμού. Εικόνα 8. Ο όγδοος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 8. The eighth sampling station of Louros river 390
391 Σταθμός 9- Εκβολές [ , ] Ο σταθμός 9 (Εικόνα 9) βρίσκεται στις εκβολές του ποταμού Λούρου στον Αμβρακικό κόλπο. Το πλάτος του ποταμού στη θέση αυτή ξεπερνά τα 10 μέτρα. Η ροή είναι ήπια ενώ ο πυθμένας καλύπτεται από άμμο και ιλύ. Η βλάστηση αντιπροσωπεύεται από καλαμώνες και πολλά αλόφυτα είδη, που καλύπτουν μεγάλο ποσοστό της έκτασης. Σε απόσταση μεγαλύτερη των 5 km υπάρχουν παραδοσιακά διβάρια.. Εικόνα 9. Ο ένατος σταθμός δειγματοληψίας στον ποταμό Λούρο Figure 9. The ninth sampling station of Louros river 391
392
393 Παράρτημα 5 Διακύμανση φυσικοχημικών παραμέτρων και συγκέντρωσης θρεπτικών αλάτων και χλωροφύλλης-α στο νερό του ποταμού Λούρου Αβιοτικά στοιχεία Θερμοκρασία Η θερμοκρασία αναγνωρίζεται ως ο σημαντικότερος περιβαλλοντικός παράγοντας στα ποτάμια οικοσυστήματα, επηρεάζοντας τα πρότυπα κατανομής και τους μεταβολικούς ρυθμούς των οργανισμών που διαβιούν σε αυτά (Matthews, 1998b). Η μέγιστη θερμοκρασία του νερού καταγράφηκε το μήνα Ιούνιο (25 ο C) και η ελάχιστη το μήνα Δεκέμβριο (10,3 ο C) στον εκβολικό σταθμό 9 (μέσος όρος 15 ±0.8). Η θερμοκρασία παρουσίασε τις ανάλογες μεταβολές καθώς κινούμαστε από τον χειμώνα προς στο καλοκαίρι (Εικόνα 1). Πράγματι, η διακύµανση των τιµών της θερμοκρασίας δε διέφερε στατιστικά σηµαντικά ανάµεσα στους σταθµούς (P>0,05, F=1,23) αν και διέφερε στατιστικά σημαντικά ανάμεσα στους μήνες (P<0,05, F=8,88). 393
394 Εικόνα 1. Διμηνιαία διακύμανση της θερμοκρασίας στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου Figure 1. Bimonthly variation of temperature in nine sampling stations of Louros river ph Η μέτρηση του ph στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου έδειξε ότι κυμάνθηκε από 6,42 έως 8,35 με μέσο όρο 7,41 (±0.14) (Εικόνα 2). Η διακύµανση των τιµών του ph διέφερε στατιστικά σηµαντικά ανάµεσα στους σταθµούς (P<0,05, F=2,34) αν και το ίδιο δεν παρατηρήθηκε ανάμεσα στους μήνες (P>0,05, F=1,94). Η ενεργός οξύτητα αποτελεί έναν από τους σημαντικούς αβιοτικούς παράγοντες που επηρεάζουν την ποιότητα του υδάτινου οικοσυστήματος με άμεση συνέπεια τη βιωσιμότητα των ψαριών. Η αύξηση του ph ιδιαιτέρως στις εκβολές, οφείλεται στην αυξημένη φωτοσυνθετική δραστηριότητα με συνεπαγόμενη την υψηλή κατανάλωση του CO2 και την επακόλουθη αύξηση του ph (Wetzel 2001). 394
395 Εικόνα 2. Διμηνιαία διακύμανση της ενεργού οξύτητας στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου Figure 2. Bimonthly variation of ph in nine sampling stations of Louros river 395
396 Θρεπτικά άλατα Νιτρικά άλατα Όσον αφορά τη διακύμανση της συγκέντρωσης των νιτρικών ιόντων (N-NO3) του νερού στον ποταμό Λούρο, οι υψηλότερες τιμές των νιτρικών ιόντων καταγράφηκαν κατά τη διάρκεια των χειμερινών μηνών (4,35) στο μέσο του ποταμού (Σταθμός 5), ενώ μηδενική τιμή καταγράφηκε στο βορειότερο σταθμό κατά τη διάρκεια των χειμερινών μηνών (Εικόνα 3). Ο μέσος όρος των νιτρικών ιόντων ανήρθε στα 0,98 (±0,24). Η διακύµανση των τιµών των νιτρικών δε διέφερε στατιστικά σηµαντικά ανάµεσα στους σταθµούς (P>0,05, F=1,03) αν και διέφερε ανάμεσα στους μήνες (P<0,05, F=3,30). Εικόνα 3. Διμηνιαία διακύμανση των νιτρικών ιόντων στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου Figure 3. Bimonthly variation of nitrate in nine sampling stations of Louros river 396
397 Νιτρώδη άλατα Όσον αφορά τη διακύμανση της συγκέντρωσης των νιτρωδών ιόντων (N-NO2) του νερού στον ποταμό Λούρο, οι υψηλότερες τιμές καταγράφηκαν κατά τη διάρκεια των χειμερινών μηνών (0,05) στο άνω ρείθρο (σταθμός 5), ενώ η μικρότερη τιμή (0,001) καταγράφηκε στο μέσο του ποταμού (σταθμός 4) κατά τη διάρκεια των θερινών μηνών (Εικόνα 4). Ο μέσος όρος των νιτρικών ιόντων ανήρθε στα 0,015 (±0,003). Η διακύµανση των τιµών των νιτρωδών δε διέφερε στατιστικά σηµαντικά ανάµεσα στους σταθµούς (P>0,05, F=1,70) και στους μήνες (P>0,05, F=1,77) (Εικόνα 4). Εικόνα 4. Διμηνιαία διακύμανση των νιτρωδών ιόντων στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου Figure 4. Bimonthly variation of nitrite in nine sampling stations of Louros river 397
398 Ορθοφωσφορικές ρίζες Η διακύμανση της συγκέντρωσης των ορθοφωσφορικών ριζών (PO4-3 ) βρέθηκε μεταξύ 0,075 και 2,5 με μέσο όρο 0,89 (±0,21) (Εικόνα 5). Οι υψηλότερες τιμές καταγράφηκαν κατά τους θερινούς και φθινοπωρινούς μήνες. Εντούτοις, η διακύµανση των τιµών των ορθοφωσφορικών δε διέφερε στατιστικά σηµαντικά ανάµεσα στους σταθµούς (P>0,05, F=0,45) και στους μήνες (P>0,05, F=1,92). Εικόνα 5. Διμηνιαία διακύμανση των ορθοφωσφορικών ριζών στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου Figure 5. Bimonthly variation of orthophosphates in nine sampling stations of Louros river 398
399 Αμμωνία Η συγκέντρωση των αμμωνιακών ιόντων κυμάνθηκε από μηδενικές τιμές έως 1,98 κατά τους εαρινούς μήνες στους εκβολικούς σταθμούς με μέσο όρο 0,25 (±0,11) (Εικόνα 6). Η διακύµανση των τιµών της αμμωνίας δε διέφερε στατιστικά σηµαντικά ανάµεσα στους σταθµούς (P>0,05, F=0,60) και στους μήνες (P>0,05, F=1,71). Εικόνα 6. Διμηνιαία διακύμανση της αμμωνίας στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου Figure 6. Bimonthly variation of ammonia in nine sampling stations of Louros river 399
400 Χλωροφύλλη-α Σε ό,τι αφορά τη χλωροφύλλη-α, οι τιμές κυμάνθηκαν σε χαμηλά επίπεδα από μηδέν έως 0,027 με μέσο όρο 0,003 (±0,002). Η χλωροφύλλη ακολούθησε ένα εποχικό πρότυπο με αύξηση κατά τους εαρινούς μήνες και καλοκαιρινούς μήνες ενώ οι μικρότερες τιμές καταγράφηκαν τους χειμερινούς μήνες (Εικόνα 7). Η διακύµανση των τιµών της αμμωνίας δε διέφερε στατιστικά σηµαντικά ανάµεσα στους σταθµούς (P>0,05, F=0,50) αν και διέφερε στατιστικά σημαντικά ανάμεσα στους μήνες (P<0,05, F=2,57). Το πρότυπο αποδίδεται στην φωτοπερίοδο και στις συγκεντρώσεις των θρεπτικών στο νερό που καθορίζουν την παραγωγικότητα των εσωτερικών υδάτων (Wetzel 2001). Εικόνα 7. Διμηνιαία διακύμανση της χλωροφύλλης-α στους εννιά σταθμούς δειγματοληψίας του ποταμού Λούρου Figure 7. Bimonthly variation of chlorophyll-a in nine sampling stations of Louros river Βιβλιογραφικές αναφορές Matthews, W. J. (1998b). Patterns in Freshwater Fish Ecology. Dodrect: Springer US Wetzel, R. (2001). Limnology: Lake and River Ecosystems. San Diego: Academic Press. 400
401 Παράρτημα 6 Ιχθυοπανίδα του ποταμού Λούρου Συνολικά, 965 άτομα μετρήθηκαν που ανήκουν σε 13 οικογένειες και 18 είδη του ποταμού Λούρου. Station name Species 1. Chani Terovou Oncorhychus mykiss Salmo lourosensis Telestes pleurobipunctatus 2. Platanakia Oncorhychus mykiss Pelasgus thesproticus Telestes pleurobipunctatus 3. Kouklesi Gasterosteus aculeatus Luciobarbus albanicus Pelasgus thesproticus Salmo lourosensis Squalius pamvoticus Telestes pleurobipunctatus 4. Kerasonas Economidichthys pygmaeus Gasterosteus aculeatus Pelasgus thesproticus Salmo lourosensis Squalius pamvoticus Telestes pleurobipunctatus 5. Agios Georgios Economidichthys pygmaeus Gasterosteus aculeatus Oncorhychus mykiss Pelasgus thesproticus Salmo lourosensis Squalius pamvoticus Telestes pleurobipunctatus 6. Skala Economidichthys pygmaeus Gambusia holbrooki 401
402 Telestes pleurobipunctatus Valencia letourneuxi 7. Jumbo Anguilla anguilla Atherina boyeri Cobitis hellenica Economidichthys pygmaeus Gasterosteus aculeatus Gambusia holbrooki Pelasgus thesproticus 8. Flamboura Atherina boyeri Economidichthys pygmaeus Gasterosteus aculeatus Gambusia holbrooki Knipowitschia caucasica Pomatoschistus marmoratus Pelasgus thesproticus 9. Estuaries Anguilla anguilla Atherina boyeri Dicentrarchus labrax Gambusia holbrooki Knipowitschia caucasica Chelon ramada Pomatoschistus marmoratus Salaria fluviatilis Syngnathus abaster 402
403 Για τις αναλύσεις λειτουργικής ποικιλότητας μετρήθηκαν 785 άτομα που ανήκουν σε 10 είδη του ποταμού Λούρου όπως φαίνεται στον Πίνακα 1. Πίνακας 1. Σύνθεση των 10 ειδών που χρησιμοποιήθηκαν στις αναλύσεις λειτουργικής ποικιλότητας σε κάθε σταθμό του ποταμού Λούρου. Οι σχετικές αφθονίες των ειδών δίνονται με τα ποσοστά εντός των παρενθέσεων Table 1. Species Composition referring to the 10 species used in functional diversity analyses at Louros river stations. Species relative abundances are given as percentages in parentheses Station name Species 1. Chani Terovou Oncorhychus mykiss (17%) Salmo lourosensis (81%) Telestes pleurobipunctatus (2%) 2. Platanakia Oncorhychus mykiss (4%) Pelasgus thesproticus (15%) Telestes pleurobipunctatus (81%) 3. Kouklesi Gasterosteus aculeatus (1%) Pelasgus thesproticus (7%) Salmo lourosensis (1%) Squalius pamvoticus (2%) Telestes pleurobipunctatus (89%) 4. Kerasonas Economidichthys pygmaeus (5%) Gasterosteus aculeatus (35%) Pelasgus thesproticus (41%) Salmo lourosensis (1%) Telestes pleurobipunctatus (18%) 5. Agios Georgios Economidichthys pygmaeus (38%) Gasterosteus aculeatus (53%) Oncorhychus mykiss (1%) Pelasgus thesproticus (2%) Salmo lourosensis (1%) Squalius pamvoticus (1%) Telestes pleurobipunctatus (4%) 6. Skala Economidichthys pygmaeus (54%) Gambusia holbrooki (10%) 403
404 Telestes pleurobipunctatus (21%) Valencia letourneuxi (15%) 7. Jumbo Cobitis hellenica (16%) Economidichthys pygmaeus (72%) Gasterosteus aculeatus (1%) Gambusia holbrooki (10%) Pelasgus thesproticus (1%) 8. Flamboura Economidichthys pygmaeus (37%) Gasterosteus aculeatus (3%) Gambusia holbrooki (32%) Pelasgus thesproticus (29%) 9. Estuaries Economidichthys pygmaeus (70%) Gambusia holbrooki (30%) 404
.")
405 Αυτόχθονα είδη Valencia letourneuxi (Sauvage, 1880) (Ζουρνάς) Ανήκει στην οικογένεια Valenciidae. Είναι ενδημικό είδος της Ιόνιας και της Νότιας Αδριατικής βιογεωγραφικής περιοχής. Το είδος έχει μια κατακερματισμένη κατανομή. Το μέγιστο μήκος που έχει καταγραφεί είναι 7 cm (SL). Κατοικεί σε πηγές με πυκνή υδρόβια βλάστηση στην οποία αναζητά την τροφή του ή κρύβεται, σε ρυάκια με χαμηλή ροή και απαντάται σε ελώδεις εκτάσεις, που τροφοδοτούνται από πηγές και έχουν καθαρό στάσιμο ή με ελαφρά ροή νερό και χαμηλό βάθος (Economidis 2009). Τρέφεται με ασπόνδυλα και η αναπαραγωγή του γίνεται την άνοιξη και το καλοκαίρι. Απειλείται από υποβάθμιση των οικοτόπων και του ανταγωνισμού με το ξενικό είδος Gambusia holbrooki (Kalogianni et al. 2014). Η κόκκινη λίστα της IUCN (2013) αλλά και το Κόκκινο Βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας χαρακτηρίζουν το είδος ως Κρισίμως Κινδυνεύον (CR) (Economidis 2009). Cobitis hellenica Economidis & Nalbant, 1996 (Λουροβελονίτσα) 405
406 Ανήκει στην οικογένεια των Cobitidae. Είναι ενδημικό της Ιόνιας βιογεωγραφικής περιοχής και συγκεκριμένα των ποταμών Λούρου και Καλαμά ενώ εισήχθη και στην Παμβώτιδα (Leonardos et al. 2008). Το μέγιστο καταγεγραμμένο μήκος του είναι 11 cm (TL) (Barbieri et al. 2015). Είναι ένα βραχύβιο είδος του οποίου τα άτομα προτιμούν αμμώδες και ιλυώδες υπόστρωμα με βλάστηση και χαμηλή έως καθόλου ροή. Χαρακτηρίζεται από την κόκκινη λίστα της IUCN (2013) ως Κινδυνεύoν (ΕΝ) και από το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας ως Τρωτό (VU) (Economidis 2009). Economidichthys pygmaeus (Holly, 1929) (Λουρογωβιός) Ανήκει στην οικογένεια Gobiidae. Είναι ενδημικό είδος της Ιόνιας βιογεωγραφικής περιοχής και πρόσφατα εισήχθη στο Βοιωτικό Κηφισό, στην Υλίκη και στη Τάκα (Barbieri et al. 2015). Το μέγιστο καταγεγραμμένο μήκος του είναι 6cm (TL). Το είδος είναι βραχύβιο και ενδιαιτεί σε ρηχά ρέοντα και στάσιμα ύδατα με άφθονη βλάστηση, ζει κρυμμένο στον πυθμένα και σε σκιώδη σημεία. Τρέφεται με ασπόνδυλα, κυρίως με αμφίποδα και κωπήποδα. Χαρακτηρίζεται από την κόκκινη λίστα της IUCN (2013) και το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας ως Μειωμένου Ενδιαφέροντος (LC) (Economidis 2009). 406
 αλλά και στον κάτω ρου πολλών ποταμών της Δυτικής Ελλάδας, ενώ παρά το όνομά του")
407 Luciobarbus albanicus (Steindachner, 1870) (Στρωσίδι) Ανήκει στην οικογένεια των Cyprinidae. Είναι ένα ενδημικό είδος της Ιόνιας βιογεωγραφικής περιοχής και ενδιαιτεί σε λίμνες (Τριχωνίδα, Αμβρακία, Παμβώτιδα) αλλά και στον κάτω ρου πολλών ποταμών της Δυτικής Ελλάδας, ενώ παρά το όνομά του δεν απαντάται στην Αλβανία. Το μέγιστο καταγεγραμμένο μήκος του είναι 60cm (ΤL). Tα άτομα του είδους προτιμούν αμμώδες και ιλυώδες υπόστρωμα. Είναι παμφάγο είδος το οποίο τρέφεται με φύκη, φυτικό υλικό, προνύμφες εντόμων και θρύμματα. Οι πυκνότητες των πληθυσμών του είδους είναι σχετικά μεγάλες, έτσι δεν θεωρείται απειλούμενο και χαρακτηρίζεται από την κόκκινη λίστα της IUCN (2013) και το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας ως Μειωμένου Ενδιαφέροντος (LC) (Economidis 2009). Pelasgus thesproticus (Stephanidis, 1939) (Θεσπρωτικός Πελασγός) Ανήκει στην οικογένεια των Cyprinidae. Είναι αυτόχθονο είδος της Ιόνιας βιογεωγραφικής περιοχής (από Καλαμά έως Άραχθο, συμπεριλαμβανομένης της Κέρκυρας και της Λευκάδας). Κατανέμεται επίσης και στη νότια Αλβανία. Tο μέγιστο 407
408 μήκος του φτάνει τα 6 cm (TL). Ενδιαιτεί σε πηγές, υδάτινα ρέματα και λιμνούλες με χαμηλή στάθμη και πυκνή βλάστηση (Barbieri et al. 2015). Χαρακτηρίζεται από την κόκκινη λίστα της IUCN (2013) και το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας ως Σχεδόν Απειλούμενο (ΝΤ) (Economidis 2009). Squalius pamvoticus (Stephanidis, 1939) (Ποταμοκέφαλος Παμβώτιδας) Ανήκει στην οικογένεια των Cyprinidae. Είναι αυτόχθονο είδος της Ιόνιας βιογεωγραφικής περιοχής και πιθανότατα της Νότιας Αδριατικής βιογεωγραφικής περιοχής. Το μέγιστο καταγεγραμμένο μήκος του είναι 30 cm (TL). Είναι ποταμόδρομο και ενδιαιτεί σε υδάτινα ρέματα με ήπια ή πιο έντονης ροής διαυγή ύδατα. Τρέφεται κυρίως με ασπόνδυλα (σκουλήκια, έντομα κλπ), καθώς και προνύμφες εντόμων. Χαρακτηρίζεται από την κόκκινη λίστα της IUCN (2013) και το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας ως Μειωμένου Ενδιαφέροντος (LC) (Economidis 2009). Telestes pleurobipunctatus (Stephanidis, 1939) (Λιάρα) Ανήκει στην οικογένεια των Cyprinidae. Είναι αυτόχθονο είδος της Ιόνιας 408
409 βιογεωγραφικής περιοχής (από Καλαμά έως νοτιοδυτική Πελοπόννησο, συμπεριλαμβανομένης της Κέρκυρας). Το μέγιστο καταγεγραμμένο μήκος του είναι 22cm (TL). Ενδιαιτεί σε υδάτινα ρέματα με ήπια ή πιο έντονη ροή. Τρέφεται κυρίως με προνύμφες εντόμων. Στα τέλη του χειμώνα, μεταναστεύει ανάντη σε δροσερά ρυάκια για αναπαραγωγή (Barbieri et al. 2015). Χαρακτηρίζεται από την κόκκινη λίστα της IUCN (2013) και το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας ως Μειωμένου Ενδιαφέροντος (LC) (Economidis 2009). Gasterosteus aculeatus Linnaeus, 1758 (Αγκαθερό) Ανήκει στην οικογένεια Gasterosteidae. Κατανέμεται στις βιογεωγραφικές περιοχές του Ιονίου και του Αιγαίου. Το μέγιστο μήκος που έχει καταγραφεί είναι 11 cm (TL). Απαντάται σε λίμνες και ποτάμια με ήπια ροή και διαυγή ύδατα και σε περιοχές με πλούσια βλάστηση και αμμώδες υπόστρωμα. Τρέφεται κυρίως με μικρά ασπόνδυλα. Χαρακτηρίζεται από την κόκκινη λίστα της IUCN (2013) και το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας ως Μειωμένου Ενδιαφέροντος (LC) (Economidis 2009). 409

410 Salmo lourosensis Delling, 2011 (Πέστροφα Λούρου) Το είδος ανήκει στην οικογένεια των Salmonidae και είναι ενδημικό του ποταμού Λούρου. Λόγω του υδροηλεκτρικού φράγματος, οι μετακινήσεις της πέστροφας είναι πολύ περιορισμένες και έτσι ο πληθυσμός βρίσκεται στον άνω ρου του ποταμού αμέσως μετά τις πηγές, όπου το νερό είναι καθαρό και με δυνατή ροή ( 24 km, Oikonomou et al. 2011). Το μέγιστο μήκος που έχει καταγραφεί είναι 32 cm (SL) (Oikonomou et al. 2014). Είναι αρπακτικό είδος και τρέφεται με άλλα ψάρια ή αμφίβια και ερπετά και μεγάλα ασπόνδυλα (καραβίδες) (Economidis 2009). Η περίοδος αναπαραγωγής του ξεκινά το Νοέμβριο μέχρι τον Ιανουάριο. Η ανθρωπογενής παρέμβαση στο ενδιαίτημα του είδους καθώς και ο ανταγωνισμός ο οποίος προκύπτει από τα διαφεύγοντα άτομα καλλιεργούμενης ιριδίζουσας πέστροφας, καθιστούν αυτό το είδος απειλούμενο καθώς το Κόκκινο Βιβλίο των Απειλούμενων Σπονδυλοζώων της Ελλάδας το χαρακτηρίζει ως Κινδυνεύον (ΕΝ) (Economidis 2009). Salaria fluviatilis (Asso, 1801) (Ποταμοσαλιάρα) Το είδος Salaria fluviatilis ανήκει στην οικογένεια των Blenniidae. Απαντάται στους ποταμούς Λούρο, Καλαμά, Αχέροντα, Αχελώο και Αλιάκμονα, σε ποταμούς της 410
411 Κρήτης και της Πελοποννήσου και στις λίμνες Δοϊράνη, Βόλβη και Βιστωνίδα. Πρόκειται για κρυπτοβενθικό είδος κυρίως των εσωτερικών υδάτων, αν και απαντάνται σε υφάλμυρα ενδιαιτήματα όταν η αλατότητα τους είναι σε χαμηλά επίπεδα. Η διατροφή της ποταμοσαλιάρας αποτελείται από μικρού μεγέθους βενθικούς οργανισμούς και έντομα. Το μέγιστο καταγεγραμμένο μήκος της φτάνει τα 17 cm (SL) (Barbieri et al. 2015). Η αναπαραγωγική της περίοδος διαρκεί από τα μέσα της άνοιξης έως το τέλος του καλοκαιριού ή τις αρχές φθινοπώρου. Σύμφωνα με την κόκκινη λίστα της IUCN (2013) και το κόκκινο βιβλίο των απειλούμενων σπονδυλόζωων της Ελλάδας χαρακτηρίζεται ως Μειωμένου Ενδιαφέροντος (LC) (Economidis 2009). Στον ποταμό Λούρο έχει επίσης αναφερθεί το Caspiomyzon graecus (Renaud & Economidis, 2010) (Γκαβόχελο Ηπείρου) Το είδος ανήκει στην οικογένεια των Petromyzontidae και είναι ενδημικό του ποταμού Λούρου. Η κατανομή του είναι πολύ περιορισμένη και εκτείνεται κοντά στο χωριό Καμπή (Φιλιππιάδα). Βασική απειλή για το είδος αποτελεί η αλλοίωση των ενδιαιτημάτων, όπως η διευθέτηση του πυθμένα (Economidis 2009). Αλλόχθονα είδη Oncorhychus mykiss (Walbaum, 1792) (Ιριδίζουσα πέστροφα) Μέλος της οικογένειας των Salmonidae, υποτροπικό είδος, απαντάται σε κρύα νερά ποταμών και λιμνών. Η ιριδίζουσα πέστροφα εισήχθη στην Ελλάδα για εμπορικούς σκοπούς (ιχθυοκαλλιέργεια). Το μέγιστο μήκος που έχει καταγραφεί είναι 120 cm (TL). Τα άτομα της πέστροφας που διαφεύγουν από τις παραποτάμιες υδατοκαλλιέργειες ανταγωνίζονται ως προς την τροφή τους τοπικούς πληθυσμούς πέστροφας. Υπάρχουν κάποιες ενδείξεις για τοπική αναπαραγωγή στην άγρια φύση, αλλά αυτό δεν έχει ακόμα αποδειχθεί (Barbieri et al. 2015). Το είδος τρέφεται με ζωοπλαγκτό, υδρόβια και χερσαία έντομα, μαλάκια, καρκινοειδή, αβγά ψαριών και μικρά ψάρια. 411
412 Gambusia holbrooki Girard, 1859 (Κουνουπόψαρο) Ανήκει στην οικογένεια Poecilidae. Πρόκειται για βενθοπελαγικό είδος των εσωτερικών υδάτων το οποίο απαντάται και σε υφάλμυρα, εκβολικά συστήματα. Το μέγιστο μήκος που έχει καταγραφεί είναι 8 cm (TL). Ενδιαιτεί σε ύδατα με χαμηλή ροή ή στάσιμα και τρέφεται με θρύμματα, ζωοπλαγκτό, λάρβες και μικρά έντομα. Το είδος παρουσιάζει κοσμοπολιτική κατανομή καθώς έχει εισαχθεί σε πολλές χώρες από τη Βόρεια Αμερική για την καταπολέμηση των πληθυσμών των κουνουπιών. Στην Ελλάδα είναι το πιο διαδεδομένο ξενικό είδος στα εσωτερικά ύδατα. Έχει επίσης παρατηρηθεί πως ανταγωνίζεται ως προς την τροφή τους τοπικούς πληθυσμούς του Ζουρνά (αυξημένος διαειδικός ανταγωνισμός) (Kalogianni et al. 2014). Ευρύαλα είδη Στον ποταμό Λούρο απαντώνται επίσης τα ευρύαλα είδη Anguilla anguilla (Anguillidae) (Χέλι), Chelon ramada (Mugilidae) (Μαυράκι), Atherina boyeri (Atherinidae) (Aθερίνα), Syngnathus abaster (Syngnathidae) (Ταινιοσακοράφα) και Dicentrarchus labrax (Moronidae) (Λαβράκι). Βιβλιογραφικές αναφορές Barbieri, R., Zogaris, S., Kalogianni, E., Stoumboudi, M., Chatzinikolaou, Y., Giakoumi, S., Kapakos, Y., Kommatas, D., Koutsikos, N., Tachos, V., Vardakas, L., & Economou, A. N. (2015). Freshwater fishes and lampreys of Greece: An annotated checklist. Monographs on Marine Sciences No. 8. Athens: Hellenic Centre for Marine Research. Economidis, P. S. (2009). Fishes of inland waters. In A. Legakis, & P. Maragou, The Red Data Book of Threatened Animals of Greece (pp ). Athens: Hellenic Zoological Society. Kalogianni, E., Giakoumi, S., Andriopoulou, A., & Chatzinikolaou, Y. (2014). Prey utilisation and trophic overlap between the non native mosquitofish and a native fish in two Mediterranean rivers. Mediterranean Marine Science, 15, Leonardos, I., Kagalou, I., Tsoumani, M., & Economidis, P. (2008). Fish fauna in a Protected Greek lake: biodiversity, introduced fish species over a 80-year 412
413 period and their impacts on the ecosystem. Ecology of Freshwater Fish, 17, Oikonomou, A., Anastasiadou, C., Taskoudis, T., & Leonardos, I. (2014). Length- weight relations of seven native fish species (Actinopterygii) from the Louros River, Greece. Acta Ichthyologica et Piscatoria, 44, Oikonomou, A., Ntakis, A., Anastasiadou, C., & Leonardos, I. (2011). Salmo lourosensis, Delling, 2010: An endangered species and its conservation status. 33rd Congress EEVE, Edessa. 413
414 Εκτίμηση της ταξινομικής κατάστασης των ατόμων των ειδών της οικογένειας Gobiidae του ποταμού Λούρου 30 * Εισαγωγή Η βιογεωγραφική περιοχή του Ιονίου, ως θερμό σημείο της ευρωπαϊκής βιοποικιλότητας ψαριών (Oikonomou, Leprieur & Leonardos 2014) φιλοξενεί πολλά είδη της ομάδας «sand gobies», συμπεριλαμβανομένων των γενών Pomatoschistus, Knipowitschia και Economidichthys (Kottelat & Freyhof 2007). Παρά τις πολυάριθμες έρευνες της Ελληνικής ιχθυοπανίδας των εσωτερικών υδάτων της Ελλάδας, οι πληροφορίες σχετικά με την κατανομή των γωβιών στην Ιόνια βιογεωγραφική περιοχή είναι πολύ σπάνιες και βασίζονται μόνο στις στρατηγικές ζωής και σε βιολογικά δεδομένα του Economidichthys pygmaeus (Economou et al. 1999, Gkenas et al. 2010, Rampin et al. 2011, Gkenas & Leonardos 2012). Επίσης, για τη συγκεκριμένη ομάδα υπάρχουν πολλές ταξινομικές αντιπαραθέσεις, με βάση τα εξωτερικά μορφολογικά χαρακτηριστικά και την οστεολογία (δηλαδή χαρακτήρες που μπορεί να εμφανίζουν αυξημένη ενδοειδική φαινοτυπική πλαστικότητα), ένα γεγονός που οδηγεί σε μια δύσκολη ταυτοποίηση των ειδών. Για τη εκτίμηση της ταξινομικής κατάστασης των ατόμων των ειδών της οικογένειας Gobiidae του ποταμού Λούρου πραγματοποιήθηκε ανάλυση του μιτοχονδριακού DNA σε δείγμα δεκαοχτώ (18) ατόμων που δε μπορούσαν να ταυτοποιηθούν. Τα άτομα συλλέχθηκαν ακολουθώντας τη μεθοδολογία συλλογής ιχθυολογικού υλικού που περιγράφηκε στο Κεφάλαιο Υλικά και μέθοδοι Απομόνωση γενετικού υλικού 30 *Για την εξαγωγή αυτών των αποτελεσμάτων συνέβαλε καθοριστικά η Λιούσια Βαρβάρα 414

415 Η απομόνωση του γενετικού υλικού πραγματοποιήθηκε με τροποποίηση του πρωτοκόλλου της μεθόδου φαινόλης-χλωροφορμίου (Sambrook & Russel 2001). Το απομονωμένο γενετικό υλικό κατακρημνίστηκε από δ/μα καθαρής αλκοόλης και επαναιωρήθηκε σε 50 μl διαλύματος ΤΕ. Για τον έλεγχο της ποιότητας του DNA και κατ επέκταση, της διαδικασίας απομόνωσης του DNA πραγματοποιήθηκε ηλεκτροφόρηση 5 μl διαλύματος DNA σε πηκτή αγαρόζης 1% w/v. Ενίσχυση γενετικού υλικού με τη μέθοδο της αλυσιδωτής αντίδρασης πολυμεράσης και αλληλούχιση Η ταξινομική κατάσταση των ατόμων των ειδών της οικογένειας Gobiidae του ποταμού Λούρου διερευνήθηκε ενισχύοντας 655 ζεύγη βάσεων του γονιδιακού τμήματος της υπομονάδας Ι της οξειδάσης του κυτοχρώματος C (COI). Για την ενίσχυση του γενετικού τόπου χρησιμοποιήθηκε το ζεύγος των οικουμενικών (Universal) εκκινητών Fish F1 (5 - TCAACCAACCACAAAGACATTGGCAC-3 ) και FISH R1 (5 - TAGACTTCTGGGTGGCCAAAGAATCA-3 ) (Ward et al. 2005). Οι αντιδράσεις της PCR πραγματοποιήθηκαν σε τελικό όγκο 25 μl και οι βέλτιστες συνθήκες για την ενίσχυση του μιτοχονδριακού τμήματος των ατόμων που μελετήθηκαν παρουσιάζονται στον Πίνακα 2. Οι αντιδράσεις ενίσχυσης του γονιδιακού τμήματος πραγματοποιήθηκαν με τη χρήση του προγραμματιζόμενου θερμικού κυκλοποιητή (PTC - 200) της εταιρείας MJ Research. Πίνακας 2. Βέλτιστες συνθήκες για την ενίσχυση της υπομονάδας Ι της οξειδάσης το κυτοχρώματος C (COI), της παρούσας μελέτης Table 2. Optimal amplification conditions for the Cytochrome c oxidase subunit I at the present study Οι PCRs περιελάμβαναν τα εξής στάσια: 1. Αρχική αποδιάταξη των αλυσίδων του DNA στους 95 C για 2 min. 415
ΒΙΟΠΟΙΚΙΛΟΤΗΤΑ ΚΑΙ ΒΙΟΓΕΩΓΡΑΦΙΚΑ ΠΡΟΤΥΠΑ ΤΩΝ ΙΧΘΥΩΝ ΤΩΝ ΕΣΩΤΕΡΙΚΩΝ ΥΔΑΤΩΝ ΤΗΣ ΒΑΛΚΑΝΙΚΗΣ ΧΕΡΣΟΝΗΣΟΥ ΟΙΚΟΝΟΜΟΥ ΑΝΘΗ
 ΒΙΟΠΟΙΚΙΛΟΤΗΤΑ ΚΑΙ ΒΙΟΓΕΩΓΡΑΦΙΚΑ ΠΡΟΤΥΠΑ ΤΩΝ ΙΧΘΥΩΝ ΤΩΝ ΕΣΩΤΕΡΙΚΩΝ ΥΔΑΤΩΝ ΤΗΣ ΒΑΛΚΑΝΙΚΗΣ ΧΕΡΣΟΝΗΣΟΥ ΟΙΚΟΝΟΜΟΥ ΑΝΘΗ Institute of Marine Biological Resources and Inland Waters Hellenic Centre for Marine
ΒΙΟΠΟΙΚΙΛΟΤΗΤΑ ΚΑΙ ΒΙΟΓΕΩΓΡΑΦΙΚΑ ΠΡΟΤΥΠΑ ΤΩΝ ΙΧΘΥΩΝ ΤΩΝ ΕΣΩΤΕΡΙΚΩΝ ΥΔΑΤΩΝ ΤΗΣ ΒΑΛΚΑΝΙΚΗΣ ΧΕΡΣΟΝΗΣΟΥ ΟΙΚΟΝΟΜΟΥ ΑΝΘΗ Institute of Marine Biological Resources and Inland Waters Hellenic Centre for Marine
ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Κεντούλλα Πέτρου Αριθμός Φοιτητικής Ταυτότητας 2008761539 Κύπρος
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Κεντούλλα Πέτρου Αριθμός Φοιτητικής Ταυτότητας 2008761539 Κύπρος
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων Καθηγητής: Δρ. Νίκος Μίτλεττον Η ΣΧΕΣΗ ΤΟΥ ΜΗΤΡΙΚΟΥ ΘΗΛΑΣΜΟΥ ΜΕ ΤΗΝ ΕΜΦΑΝΙΣΗ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 2 ΣΤΗΝ ΠΑΙΔΙΚΗ ΗΛΙΚΙΑ Ονοματεπώνυμο: Ιωσηφίνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων Καθηγητής: Δρ. Νίκος Μίτλεττον Η ΣΧΕΣΗ ΤΟΥ ΜΗΤΡΙΚΟΥ ΘΗΛΑΣΜΟΥ ΜΕ ΤΗΝ ΕΜΦΑΝΙΣΗ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 2 ΣΤΗΝ ΠΑΙΔΙΚΗ ΗΛΙΚΙΑ Ονοματεπώνυμο: Ιωσηφίνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΟΥ ΚΛΙΜΑΤΟΣ ΑΣΦΑΛΕΙΑΣ ΤΩΝ ΑΣΘΕΝΩΝ ΣΤΟ ΝΟΣΟΚΟΜΕΙΟ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΟΥ ΚΛΙΜΑΤΟΣ ΑΣΦΑΛΕΙΑΣ ΤΩΝ ΑΣΘΕΝΩΝ ΣΤΟ ΝΟΣΟΚΟΜΕΙΟ ΑΝΔΡΕΑΣ ΛΕΩΝΙΔΟΥ Λεμεσός, 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΟΥ ΚΛΙΜΑΤΟΣ ΑΣΦΑΛΕΙΑΣ ΤΩΝ ΑΣΘΕΝΩΝ ΣΤΟ ΝΟΣΟΚΟΜΕΙΟ ΑΝΔΡΕΑΣ ΛΕΩΝΙΔΟΥ Λεμεσός, 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ
«Αξιολόγηση ατόμων με αφασία για Επαυξητική και Εναλλακτική Επικοινωνία, σύμφωνα με το μοντέλο συμμετοχής»
 Σχολή Επιστημών Υγείας Τμήμα Αποκατάστασης ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Αξιολόγηση ατόμων με αφασία για Επαυξητική και Εναλλακτική Επικοινωνία, σύμφωνα με το μοντέλο συμμετοχής» Χρυσάνθη Μοδέστου Λεμεσός, Μάιος,
Σχολή Επιστημών Υγείας Τμήμα Αποκατάστασης ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Αξιολόγηση ατόμων με αφασία για Επαυξητική και Εναλλακτική Επικοινωνία, σύμφωνα με το μοντέλο συμμετοχής» Χρυσάνθη Μοδέστου Λεμεσός, Μάιος,
Η θέση ύπνου του βρέφους και η σχέση της με το Σύνδρομο του αιφνίδιου βρεφικού θανάτου. ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η θέση ύπνου του βρέφους και η σχέση της με το Σύνδρομο του αιφνίδιου βρεφικού θανάτου. Χρυσάνθη Στυλιανού Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η θέση ύπνου του βρέφους και η σχέση της με το Σύνδρομο του αιφνίδιου βρεφικού θανάτου. Χρυσάνθη Στυλιανού Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ
Πτυχιακή διατριβή. Η επίδραση της τασιενεργής ουσίας Ακεταλδεΰδης στη δημιουργία πυρήνων συμπύκνωσης νεφών (CCN) στην ατμόσφαιρα
 στην ατμόσφαιρα") ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η επίδραση της τασιενεργής ουσίας Ακεταλδεΰδης στη δημιουργία πυρήνων συμπύκνωσης νεφών (CCN)
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η επίδραση της τασιενεργής ουσίας Ακεταλδεΰδης στη δημιουργία πυρήνων συμπύκνωσης νεφών (CCN)
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΠΡΟΣΔΙΟΡΙΣΜΟΣ ΔΕΙΚΤΩΝ ΚΑΤΑΝΑΛΩΣΗΣ ΕΝΕΡΓΕΙΑΣ ΣΤΑ ΑΝΤΛΙΟΣΤΑΣΙΑ ΤΟΥ ΤΜΗΜΑΤΟΣ ΑΝΑΠΤΥΞΕΩΣ ΥΔΑΤΩΝ Γεωργίου
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΠΡΟΣΔΙΟΡΙΣΜΟΣ ΔΕΙΚΤΩΝ ΚΑΤΑΝΑΛΩΣΗΣ ΕΝΕΡΓΕΙΑΣ ΣΤΑ ΑΝΤΛΙΟΣΤΑΣΙΑ ΤΟΥ ΤΜΗΜΑΤΟΣ ΑΝΑΠΤΥΞΕΩΣ ΥΔΑΤΩΝ Γεωργίου
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Ο ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ Η ΣΧΕΣΗ ΤΟΥ ΜΕ ΤΟ ΚΑΡΚΙΝΟ ΤΟΥ ΜΑΣΤΟΥΣ ΣΤΙΣ ΓΥΝΑΙΚΕΣ ΠΟΥ ΕΙΝΑΙ ΦΟΡΕΙΣ ΤΟΥ ΟΓΚΟΓΟΝΙΔΙΟΥ BRCA1 ΚΑΙ BRCA2. Βασούλλα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Ο ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ Η ΣΧΕΣΗ ΤΟΥ ΜΕ ΤΟ ΚΑΡΚΙΝΟ ΤΟΥ ΜΑΣΤΟΥΣ ΣΤΙΣ ΓΥΝΑΙΚΕΣ ΠΟΥ ΕΙΝΑΙ ΦΟΡΕΙΣ ΤΟΥ ΟΓΚΟΓΟΝΙΔΙΟΥ BRCA1 ΚΑΙ BRCA2. Βασούλλα
ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΠΑΙΔΙΚΗ ΠΑΧΥΣΑΡΚΙΑ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΠΑΙΔΙΚΗ ΠΑΧΥΣΑΡΚΙΑ Ονοματεπώνυμο: Μιχαέλλα Σάββα Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΠΑΙΔΙΚΗ ΠΑΧΥΣΑΡΚΙΑ Ονοματεπώνυμο: Μιχαέλλα Σάββα Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η ΣΥΓΚΕΝΤΡΩΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΣΤΟ ΕΔΑΦΟΣ ΚΑΙ ΜΕΘΟΔΟΙ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΤΟΥΣ Μιχαήλ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η ΣΥΓΚΕΝΤΡΩΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΣΤΟ ΕΔΑΦΟΣ ΚΑΙ ΜΕΘΟΔΟΙ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΤΟΥΣ Μιχαήλ
Μεταπτυχιακή Διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΟΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή Διατριβή Η ΕΠΙΔΡΑΣΗ ΤΟΥ ΣΤΑΘΜΟΥ ΑΦΑΛΑΤΩΣΗΣ ΣΤΟ ΒΑΣΙΛΙΚΟ ΣΤΟ ΘΑΛΑΣΣΙΟ ΟΙΚΟΣΥΣΤΗΜΑ Κατερίνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΟΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή Διατριβή Η ΕΠΙΔΡΑΣΗ ΤΟΥ ΣΤΑΘΜΟΥ ΑΦΑΛΑΤΩΣΗΣ ΣΤΟ ΒΑΣΙΛΙΚΟ ΣΤΟ ΘΑΛΑΣΣΙΟ ΟΙΚΟΣΥΣΤΗΜΑ Κατερίνα
Κωνσταντίνος Στεφανίδης
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης Οικολογική
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης Οικολογική
AΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΠΟΛΙΤΙΚΩΝ ΜΗΧΑΝΙΚΩΝ
 AΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΠΟΛΙΤΙΚΩΝ ΜΗΧΑΝΙΚΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΕΙΔΙΚΕΥΣΗΣ ΠΡΟΣΤΑΣΙΑ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΚΑΙ ΒΙΩΣΙΜΗ ΑΝΑΠΤΥΞΗ ΔΙΕΡΕΥΝΗΣΗ ΤΩΝ ΠΙΕΣΕΩΝ ΣΤΟ ΠΕΡΙΒΑΛΛΟΝ
AΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΠΟΛΙΤΙΚΩΝ ΜΗΧΑΝΙΚΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΕΙΔΙΚΕΥΣΗΣ ΠΡΟΣΤΑΣΙΑ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΚΑΙ ΒΙΩΣΙΜΗ ΑΝΑΠΤΥΞΗ ΔΙΕΡΕΥΝΗΣΗ ΤΩΝ ΠΙΕΣΕΩΝ ΣΤΟ ΠΕΡΙΒΑΛΛΟΝ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΤΜΗΜΑ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΤΜΗΜΑ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή Διατριβή Η ΣΥΝΕΙΣΦΟΡΑ ΤΟΥ CΗ 4 ΣΤΟ ΦΑΙΝΟΜΕΝΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΤΜΗΜΑ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή Διατριβή Η ΣΥΝΕΙΣΦΟΡΑ ΤΟΥ CΗ 4 ΣΤΟ ΦΑΙΝΟΜΕΝΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΚΑΤΑΣΚΕΥΗ ΦΩΤΟΑΝΤΙΔΡΑΣΤΗΡΑ (UV) ΓΙΑ ΠΕΡΙΒΑΛΛΟΝΤΙΚΕΣ ΕΦΑΡΜΟΓΕΣ Δημήτρης Δημητρίου Λεμεσός 2015
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΚΑΤΑΣΚΕΥΗ ΦΩΤΟΑΝΤΙΔΡΑΣΤΗΡΑ (UV) ΓΙΑ ΠΕΡΙΒΑΛΛΟΝΤΙΚΕΣ ΕΦΑΡΜΟΓΕΣ Δημήτρης Δημητρίου Λεμεσός 2015
Πτυχιακή Εργασία ΓΝΩΣΕΙΣ KAI ΣΤΑΣΕΙΣ ΤΩΝ ΕΠΑΓΓΕΛΜΑΤΙΩΝ ΥΓΕΙΑΣ ΓΙΑ ΤΗΝ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΚΘΕΣΗ ΣΤΟΝ HIV. Στυλιανού Στυλιανή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία ΓΝΩΣΕΙΣ KAI ΣΤΑΣΕΙΣ ΤΩΝ ΕΠΑΓΓΕΛΜΑΤΙΩΝ ΥΓΕΙΑΣ ΓΙΑ ΤΗΝ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΚΘΕΣΗ ΣΤΟΝ HIV Στυλιανού Στυλιανή Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία ΓΝΩΣΕΙΣ KAI ΣΤΑΣΕΙΣ ΤΩΝ ΕΠΑΓΓΕΛΜΑΤΙΩΝ ΥΓΕΙΑΣ ΓΙΑ ΤΗΝ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΚΘΕΣΗ ΣΤΟΝ HIV Στυλιανού Στυλιανή Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ. Πτυχιακή εργασία ΟΛΙΣΘΗΡΟΤΗΤΑ ΚΑΙ ΜΑΚΡΟΥΦΗ ΤΩΝ ΟΔΟΔΤΡΩΜΑΤΩΝ ΚΥΚΛΟΦΟΡΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ Πτυχιακή εργασία ΟΛΙΣΘΗΡΟΤΗΤΑ ΚΑΙ ΜΑΚΡΟΥΦΗ ΤΩΝ ΟΔΟΔΤΡΩΜΑΤΩΝ ΚΥΚΛΟΦΟΡΙΑΣ Χριστοδούλου Αντρέας Λεμεσός 2014 2 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ Πτυχιακή εργασία ΟΛΙΣΘΗΡΟΤΗΤΑ ΚΑΙ ΜΑΚΡΟΥΦΗ ΤΩΝ ΟΔΟΔΤΡΩΜΑΤΩΝ ΚΥΚΛΟΦΟΡΙΑΣ Χριστοδούλου Αντρέας Λεμεσός 2014 2 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
Σχολή Διοίκησης και Οικονομίας. Μεταπτυχιακή διατριβή
 Σχολή Διοίκησης και Οικονομίας Μεταπτυχιακή διατριβή Samsung και Apple: Αναλύοντας τη χρηματοοικονομική πληροφόρηση των ηγετών της τεχνολογίας και η επίδραση των εξωτερικών και ενδοεπιχειρησιακών παραγόντων
Σχολή Διοίκησης και Οικονομίας Μεταπτυχιακή διατριβή Samsung και Apple: Αναλύοντας τη χρηματοοικονομική πληροφόρηση των ηγετών της τεχνολογίας και η επίδραση των εξωτερικών και ενδοεπιχειρησιακών παραγόντων
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Χαμηλά επίπεδα βιταμίνης D σχετιζόμενα με το βρογχικό άσθμα στα παιδιά και στους έφηβους Κουρομπίνα Αλεξάνδρα Λεμεσός [2014] i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Χαμηλά επίπεδα βιταμίνης D σχετιζόμενα με το βρογχικό άσθμα στα παιδιά και στους έφηβους Κουρομπίνα Αλεξάνδρα Λεμεσός [2014] i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΞΟΥΘΕΝΩΣΗ ΠΟΥ ΒΙΩΝΕΙ ΤΟ ΝΟΣΗΛΕΥΤΙΚΟ ΠΡΟΣΩΠΙΚΟ ΣΤΙΣ ΜΟΝΑΔΕΣ ΕΝΑΤΙΚΗΣ ΘΕΡΑΠΕΙΑΣ Άντρη Αγαθαγγέλου Λεμεσός 2012 i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΞΟΥΘΕΝΩΣΗ ΠΟΥ ΒΙΩΝΕΙ ΤΟ ΝΟΣΗΛΕΥΤΙΚΟ ΠΡΟΣΩΠΙΚΟ ΣΤΙΣ ΜΟΝΑΔΕΣ ΕΝΑΤΙΚΗΣ ΘΕΡΑΠΕΙΑΣ Άντρη Αγαθαγγέλου Λεμεσός 2012 i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΨΥΧΟΚΟΙΝΩΝΙΚΗ ΥΓΕΙΑ ΤΟΥ ΠΑΙΔΙΟΥ ΣΕ ΣΧΕΣΗ ΜΕ ΤΗΝ ΚΟΙΝΩΝΙΚΟΟΙΚΟΝΟΜΙΚΗ ΚΑΤΑΣΤΑΣΗ ΤΗΣ ΟΙΚΟΓΕΝΕΙΑΣ Μαρία Χρίστου Λεμεσός 2012 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΨΥΧΟΚΟΙΝΩΝΙΚΗ ΥΓΕΙΑ ΤΟΥ ΠΑΙΔΙΟΥ ΣΕ ΣΧΕΣΗ ΜΕ ΤΗΝ ΚΟΙΝΩΝΙΚΟΟΙΚΟΝΟΜΙΚΗ ΚΑΤΑΣΤΑΣΗ ΤΗΣ ΟΙΚΟΓΕΝΕΙΑΣ Μαρία Χρίστου Λεμεσός 2012 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΚΑΠΝΙΣΤΙΚΕΣ ΣΥΝΗΘΕΙΕΣ ΓΟΝΕΩΝ ΚΑΙ ΕΠΙΡΡΟΗ ΤΟΥΣ ΣΤΗΝ ΕΝΑΡΞΗ ΤΟΥ ΚΑΠΝΙΣΜΑΤΟΣ ΣΤΟΥΣ ΕΦΗΒΟΥΣ Ονοματεπώνυμο Φοιτήτριας: Χριστοφόρου Έλενα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΚΑΠΝΙΣΤΙΚΕΣ ΣΥΝΗΘΕΙΕΣ ΓΟΝΕΩΝ ΚΑΙ ΕΠΙΡΡΟΗ ΤΟΥΣ ΣΤΗΝ ΕΝΑΡΞΗ ΤΟΥ ΚΑΠΝΙΣΜΑΤΟΣ ΣΤΟΥΣ ΕΦΗΒΟΥΣ Ονοματεπώνυμο Φοιτήτριας: Χριστοφόρου Έλενα
Περιεχόμενο Μαθημάτων
 Περιεχόμενο Μαθημάτων Χερσαία και Θαλάσσια Οικοσυστήματα Συντονιστές I. Καρακάσης M. Μυλωνάς Σύνολο ωρών 100 Διδάσκοντες Χ. Αρβανιτίδης, Ι. Καρακάσης, Κ. Κοτζαμπάσης, Ν. Λαμπαδαρίου, Μ. Μυλωνάς, Π. Πήττα
Περιεχόμενο Μαθημάτων Χερσαία και Θαλάσσια Οικοσυστήματα Συντονιστές I. Καρακάσης M. Μυλωνάς Σύνολο ωρών 100 Διδάσκοντες Χ. Αρβανιτίδης, Ι. Καρακάσης, Κ. Κοτζαμπάσης, Ν. Λαμπαδαρίου, Μ. Μυλωνάς, Π. Πήττα
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ. Πτυχιακή Εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή Εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΠΟΤΕΛΕΣΜΑΤΙΚΟΤΗΤΑΣ ΤΩΝ ΜΕΤΡΩΝ ΠΡΟΛΗΨΗΣ ΤΗΣ ΝΟΣΟΚΟΜΕΙΑΚΗΣ ΜΕΤΑΔΟΣΗΣ ΤΗΣ ΦΥΜΑΤΙΩΣΗΣ Βασιλική Αλκιβιαδου
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή Εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΠΟΤΕΛΕΣΜΑΤΙΚΟΤΗΤΑΣ ΤΩΝ ΜΕΤΡΩΝ ΠΡΟΛΗΨΗΣ ΤΗΣ ΝΟΣΟΚΟΜΕΙΑΚΗΣ ΜΕΤΑΔΟΣΗΣ ΤΗΣ ΦΥΜΑΤΙΩΣΗΣ Βασιλική Αλκιβιαδου
ΟΙΚΟΝΟΜΟΤΕΧΝΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΕΝΕΡΓΕΙΑΚΑ ΑΥΤΟΝΟΜΟΥ ΝΗΣΙΟΥ ΜΕ Α.Π.Ε
 Τμήμα Ηλεκτρονικών Μηχανικών Τ.Ε. ΟΙΚΟΝΟΜΟΤΕΧΝΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΕΝΕΡΓΕΙΑΚΑ ΑΥΤΟΝΟΜΟΥ ΝΗΣΙΟΥ ΜΕ Α.Π.Ε Πτυχιακή Εργασία Φοιτητής: Γεμενής Κωνσταντίνος ΑΜ: 30931 Επιβλέπων Καθηγητής Κοκκόσης Απόστολος Λέκτορας
Τμήμα Ηλεκτρονικών Μηχανικών Τ.Ε. ΟΙΚΟΝΟΜΟΤΕΧΝΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΕΝΕΡΓΕΙΑΚΑ ΑΥΤΟΝΟΜΟΥ ΝΗΣΙΟΥ ΜΕ Α.Π.Ε Πτυχιακή Εργασία Φοιτητής: Γεμενής Κωνσταντίνος ΑΜ: 30931 Επιβλέπων Καθηγητής Κοκκόσης Απόστολος Λέκτορας
Πτυχιακή Εργασία Η ΑΝΤΙΛΑΜΒΑΝΟΜΕΝΗ ΚΟΙΝΩΝΙΚΗ ΣΤΗΡΙΞΗ ΣΤΙΣ ΘΗΛΑΖΟΥΣΕΣ ΜΗΤΕΡΕΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΑΝΤΙΛΑΜΒΑΝΟΜΕΝΗ ΚΟΙΝΩΝΙΚΗ ΣΤΗΡΙΞΗ ΣΤΙΣ ΘΗΛΑΖΟΥΣΕΣ ΜΗΤΕΡΕΣ Ονοματεπώνυμο: Στέλλα Κόντζιαλη Αριθμός Φοιτητικής Ταυτότητας: 2010414838
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΑΝΤΙΛΑΜΒΑΝΟΜΕΝΗ ΚΟΙΝΩΝΙΚΗ ΣΤΗΡΙΞΗ ΣΤΙΣ ΘΗΛΑΖΟΥΣΕΣ ΜΗΤΕΡΕΣ Ονοματεπώνυμο: Στέλλα Κόντζιαλη Αριθμός Φοιτητικής Ταυτότητας: 2010414838
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή διατριβή. Ονοματεπώνυμο: Αργυρώ Ιωάννου. Επιβλέπων καθηγητής: Δρ. Αντρέας Χαραλάμπους
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Διερεύνηση της αποτελεσματικότητας εναλλακτικών και συμπληρωματικών τεχνικών στη βελτίωση της ποιότητας της ζωής σε άτομα με καρκίνο
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Διερεύνηση της αποτελεσματικότητας εναλλακτικών και συμπληρωματικών τεχνικών στη βελτίωση της ποιότητας της ζωής σε άτομα με καρκίνο
Ο νοσηλευτικός ρόλος στην πρόληψη του μελανώματος
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Ο νοσηλευτικός ρόλος στην πρόληψη του μελανώματος Ονοματεπώνυμο: Αρτέμης Παναγιώτου Επιβλέπων καθηγητής: Δρ. Αντρέας Χαραλάμπους
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Ο νοσηλευτικός ρόλος στην πρόληψη του μελανώματος Ονοματεπώνυμο: Αρτέμης Παναγιώτου Επιβλέπων καθηγητής: Δρ. Αντρέας Χαραλάμπους
Προσδιορισμός Σημαντικών Χαρακτηριστικών της Αυθόρμητης Δραστηριότητας Απομονωμένου Εγκεφαλικού Φλοιού in vitro
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΠΛΗΡΟΦΟΡΙΚΗΣ ΚΑΙ ΤΗΛΕΠΙΚΟΙΝΩΝΙΩΝ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ "ΤΕΧΝΟΛΟΓΙΕΣ ΠΛΗΡΟΦΟΡΙΚΗΣ ΣΤΗΝ ΙΑΤΡΙΚΗ ΚΑΙ ΤΗ ΒΙΟΛΟΓΙΑ"
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΠΛΗΡΟΦΟΡΙΚΗΣ ΚΑΙ ΤΗΛΕΠΙΚΟΙΝΩΝΙΩΝ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ "ΤΕΧΝΟΛΟΓΙΕΣ ΠΛΗΡΟΦΟΡΙΚΗΣ ΣΤΗΝ ΙΑΤΡΙΚΗ ΚΑΙ ΤΗ ΒΙΟΛΟΓΙΑ"
Μεταπτυχιακή διατριβή. Ανδρέας Παπαευσταθίου
 Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Μεταπτυχιακή διατριβή Κτίρια σχεδόν μηδενικής ενεργειακής κατανάλωσης :Αξιολόγηση συστημάτων θέρμανσης -ψύξης και ΑΠΕ σε οικιστικά κτίρια στην
Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Μεταπτυχιακή διατριβή Κτίρια σχεδόν μηδενικής ενεργειακής κατανάλωσης :Αξιολόγηση συστημάτων θέρμανσης -ψύξης και ΑΠΕ σε οικιστικά κτίρια στην
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ Ο ΠΡΟΩΡΟΣ ΤΟΚΕΤΟΣ ΚΑΙ Η ΣΥΣΧΕΤΙΣΗ ΤΟΥ ΜΕ ΤΟ ΣΥΝΔΡΟΜΟ ΑΝΑΠΝΕΥΣΤΙΚΗΣ ΔΥΣΧΕΡΕΙΑΣ Όνομα Φοιτήτριας: Χρυσοστομή Αγαθοκλέους Αριθμός φοιτητικής
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ Ο ΠΡΟΩΡΟΣ ΤΟΚΕΤΟΣ ΚΑΙ Η ΣΥΣΧΕΤΙΣΗ ΤΟΥ ΜΕ ΤΟ ΣΥΝΔΡΟΜΟ ΑΝΑΠΝΕΥΣΤΙΚΗΣ ΔΥΣΧΕΡΕΙΑΣ Όνομα Φοιτήτριας: Χρυσοστομή Αγαθοκλέους Αριθμός φοιτητικής
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία ΑΓΧΟΣ ΚΑΙ ΚΑΤΑΘΛΙΨΗ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΑΓΧΟΣ ΚΑΙ ΚΑΤΑΘΛΙΨΗ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ ΧΡΥΣΟΒΑΛΑΝΤΗΣ ΒΑΣΙΛΕΙΟΥ ΛΕΜΕΣΟΣ 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΑΓΧΟΣ ΚΑΙ ΚΑΤΑΘΛΙΨΗ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ ΧΡΥΣΟΒΑΛΑΝΤΗΣ ΒΑΣΙΛΕΙΟΥ ΛΕΜΕΣΟΣ 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΑ ΤΜΗΜΑ ΝΑΥΤΙΛΙΑΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΝΑΥΤΙΛΙΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΑ ΤΜΗΜΑ ΝΑΥΤΙΛΙΑΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΝΑΥΤΙΛΙΑ ΝΟΜΙΚΟ ΚΑΙ ΘΕΣΜΙΚΟ ΦΟΡΟΛΟΓΙΚΟ ΠΛΑΙΣΙΟ ΚΤΗΣΗΣ ΚΑΙ ΕΚΜΕΤΑΛΛΕΥΣΗΣ ΠΛΟΙΟΥ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ που υποβλήθηκε στο
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΑ ΤΜΗΜΑ ΝΑΥΤΙΛΙΑΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΝΑΥΤΙΛΙΑ ΝΟΜΙΚΟ ΚΑΙ ΘΕΣΜΙΚΟ ΦΟΡΟΛΟΓΙΚΟ ΠΛΑΙΣΙΟ ΚΤΗΣΗΣ ΚΑΙ ΕΚΜΕΤΑΛΛΕΥΣΗΣ ΠΛΟΙΟΥ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ που υποβλήθηκε στο
ΕΤΑΙΡΙΚΗ ΚΟΙΝΩΝΙΚΗ ΕΥΘΥΝΗ ΣΤΗΝ ΝΑΥΤΙΛΙΑΚΗ ΒΙΟΜΗΧΑΜΙΑ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΔΙΟΙΚΗΣΗΣ ΚΑΙ ΟΙΚΟΝΟΜΙΑΣ Μεταπτυχιακή διατριβή ΕΤΑΙΡΙΚΗ ΚΟΙΝΩΝΙΚΗ ΕΥΘΥΝΗ ΣΤΗΝ ΝΑΥΤΙΛΙΑΚΗ ΒΙΟΜΗΧΑΜΙΑ Ανδρούλα Γιαπάνη Λεμεσός, Φεβρουάριος 2018 0 i ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΔΙΟΙΚΗΣΗΣ ΚΑΙ ΟΙΚΟΝΟΜΙΑΣ Μεταπτυχιακή διατριβή ΕΤΑΙΡΙΚΗ ΚΟΙΝΩΝΙΚΗ ΕΥΘΥΝΗ ΣΤΗΝ ΝΑΥΤΙΛΙΑΚΗ ΒΙΟΜΗΧΑΜΙΑ Ανδρούλα Γιαπάνη Λεμεσός, Φεβρουάριος 2018 0 i ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΟΡΙΑΚΗ ΚΑΙ ΦΑΙΝΟΤΥΠΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΚΥΠΡΙΑΚΟΥ ΠΛΗΘΥΣΜΟΥ ΣΚΛΗΡΟΥ ΣΙΤΑΡΙΟΥ ΠΟΥ ΑΠΟΚΤΗΘΗΚΕ ΑΠΟ ΤΡΑΠΕΖΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΟΡΙΑΚΗ ΚΑΙ ΦΑΙΝΟΤΥΠΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΚΥΠΡΙΑΚΟΥ ΠΛΗΘΥΣΜΟΥ ΣΚΛΗΡΟΥ ΣΙΤΑΡΙΟΥ ΠΟΥ ΑΠΟΚΤΗΘΗΚΕ ΑΠΟ ΤΡΑΠΕΖΑ
ΠΑΝΔΠΗΣΖΜΗΟ ΠΑΣΡΩΝ ΓΗΑΣΜΖΜΑΣΗΚΟ ΠΡΟΓΡΑΜΜΑ ΜΔΣΑΠΣΤΥΗΑΚΩΝ ΠΟΤΓΩΝ «ΤΣΖΜΑΣΑ ΔΠΔΞΔΡΓΑΗΑ ΖΜΑΣΩΝ ΚΑΗ ΔΠΗΚΟΗΝΩΝΗΩΝ» ΣΜΖΜΑ ΜΖΥΑΝΗΚΩΝ Ζ/Τ ΚΑΗ ΠΛΖΡΟΦΟΡΗΚΖ
 ΠΑΝΔΠΗΣΖΜΗΟ ΠΑΣΡΩΝ ΓΗΑΣΜΖΜΑΣΗΚΟ ΠΡΟΓΡΑΜΜΑ ΜΔΣΑΠΣΤΥΗΑΚΩΝ ΠΟΤΓΩΝ «ΤΣΖΜΑΣΑ ΔΠΔΞΔΡΓΑΗΑ ΖΜΑΣΩΝ ΚΑΗ ΔΠΗΚΟΗΝΩΝΗΩΝ» ΣΜΖΜΑ ΜΖΥΑΝΗΚΩΝ Ζ/Τ ΚΑΗ ΠΛΖΡΟΦΟΡΗΚΖ ΣΜΖΜΑ ΖΛΔΚΣΡΟΛΟΓΩΝ ΜΖΥΑΝΗΚΩΝ ΚΑΗ ΣΔΥΝΟΛΟΓΗΑ ΤΠΟΛΟΓΗΣΩΝ ΣΜΖΜΑ
ΠΑΝΔΠΗΣΖΜΗΟ ΠΑΣΡΩΝ ΓΗΑΣΜΖΜΑΣΗΚΟ ΠΡΟΓΡΑΜΜΑ ΜΔΣΑΠΣΤΥΗΑΚΩΝ ΠΟΤΓΩΝ «ΤΣΖΜΑΣΑ ΔΠΔΞΔΡΓΑΗΑ ΖΜΑΣΩΝ ΚΑΗ ΔΠΗΚΟΗΝΩΝΗΩΝ» ΣΜΖΜΑ ΜΖΥΑΝΗΚΩΝ Ζ/Τ ΚΑΗ ΠΛΖΡΟΦΟΡΗΚΖ ΣΜΖΜΑ ΖΛΔΚΣΡΟΛΟΓΩΝ ΜΖΥΑΝΗΚΩΝ ΚΑΗ ΣΔΥΝΟΛΟΓΗΑ ΤΠΟΛΟΓΗΣΩΝ ΣΜΖΜΑ
Μελέτη των μεταβολών των χρήσεων γης στο Ζαγόρι Ιωαννίνων 0
 Μελέτη των μεταβολών των χρήσεων γης στο Ζαγόρι Ιωαννίνων 0 ΕΘΝΙΚΟ ΜΕΤΣΟΒΙΟ ΠΟΛΥΤΕΧΝΕΙΟ ΔΙΕΠΙΣΤΗΜΟΝΙΚΟ - ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ (Δ.Π.Μ.Σ.) "ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ" 2 η ΚΑΤΕΥΘΥΝΣΗ
Μελέτη των μεταβολών των χρήσεων γης στο Ζαγόρι Ιωαννίνων 0 ΕΘΝΙΚΟ ΜΕΤΣΟΒΙΟ ΠΟΛΥΤΕΧΝΕΙΟ ΔΙΕΠΙΣΤΗΜΟΝΙΚΟ - ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ (Δ.Π.Μ.Σ.) "ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ" 2 η ΚΑΤΕΥΘΥΝΣΗ
ΔΗΜΙΟΥΡΓΙΑ ΔΥΝΑΜΙΚΟΥ ΜΟΝΤΕΛΟΥ ΑΝΟΔΙΚΗΣ ΠΡΟΣΕΓΓΙΣΗΣ ΓΙΑ ΤΗΝ ΕΚΤΙΜΗΣΗ ΤΗΣ ΖΗΤΗΣΗΣ ΕΝΕΡΓΕΙΑΣ ΣΤΟΝ ΟΙΚΙΣΤΙΚΟ ΤΟΜΕΑ ΤΗΣ ΚΥΠΡΟΥ
 Σχολή Γεωτεχνικών Επιστημών & Διαχείρισης Περιβάλλοντος Μεταπτυχιακή διατριβή ΔΗΜΙΟΥΡΓΙΑ ΔΥΝΑΜΙΚΟΥ ΜΟΝΤΕΛΟΥ ΑΝΟΔΙΚΗΣ ΠΡΟΣΕΓΓΙΣΗΣ ΓΙΑ ΤΗΝ ΕΚΤΙΜΗΣΗ ΤΗΣ ΖΗΤΗΣΗΣ ΕΝΕΡΓΕΙΑΣ ΣΤΟΝ ΟΙΚΙΣΤΙΚΟ ΤΟΜΕΑ ΤΗΣ ΚΥΠΡΟΥ Γαλάτεια
Σχολή Γεωτεχνικών Επιστημών & Διαχείρισης Περιβάλλοντος Μεταπτυχιακή διατριβή ΔΗΜΙΟΥΡΓΙΑ ΔΥΝΑΜΙΚΟΥ ΜΟΝΤΕΛΟΥ ΑΝΟΔΙΚΗΣ ΠΡΟΣΕΓΓΙΣΗΣ ΓΙΑ ΤΗΝ ΕΚΤΙΜΗΣΗ ΤΗΣ ΖΗΤΗΣΗΣ ΕΝΕΡΓΕΙΑΣ ΣΤΟΝ ΟΙΚΙΣΤΙΚΟ ΤΟΜΕΑ ΤΗΣ ΚΥΠΡΟΥ Γαλάτεια
Πτυχιακή Εργασία. Παραδοσιακά Προϊόντα Διατροφική Αξία και η Πιστοποίηση τους
 ΑΛΕΞΑΝΔΡΕΙΟ ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΣΧΟΛΗ ΤΕΧΝΟΛΟΓΙΑΣ ΤΡΟΦΙΜΩΝ ΚΑΙ ΔΙΑΤΡΟΦΗΣ ΤΜΗΜΑ ΔΙΑΤΡΟΦΗΣ ΚΑΙ ΔΙΑΙΤΟΛΟΓΙΑΣ Πτυχιακή Εργασία Παραδοσιακά Προϊόντα Διατροφική Αξία και η Πιστοποίηση τους Εκπόνηση:
ΑΛΕΞΑΝΔΡΕΙΟ ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΣΧΟΛΗ ΤΕΧΝΟΛΟΓΙΑΣ ΤΡΟΦΙΜΩΝ ΚΑΙ ΔΙΑΤΡΟΦΗΣ ΤΜΗΜΑ ΔΙΑΤΡΟΦΗΣ ΚΑΙ ΔΙΑΙΤΟΛΟΓΙΑΣ Πτυχιακή Εργασία Παραδοσιακά Προϊόντα Διατροφική Αξία και η Πιστοποίηση τους Εκπόνηση:
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΤΟ ΚΑΠΜΝΙΣΜΑ ΩΣ ΠΑΡΑΓΟΝΤΑΣ ΥΨΗΛΟΥ ΚΙΝΔΥΝΟΥ ΓΙΑ ΠΡΟΚΛΗΣΗ ΥΠΟΓΟΝΙΜΟΤΗΤΑΣ ΣΤΟΥΣ ΑΝΔΡΕΣ Κατσαρής Γιάγκος Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΤΟ ΚΑΠΜΝΙΣΜΑ ΩΣ ΠΑΡΑΓΟΝΤΑΣ ΥΨΗΛΟΥ ΚΙΝΔΥΝΟΥ ΓΙΑ ΠΡΟΚΛΗΣΗ ΥΠΟΓΟΝΙΜΟΤΗΤΑΣ ΣΤΟΥΣ ΑΝΔΡΕΣ Κατσαρής Γιάγκος Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
Μικροβιολογική ποιότητα έτοιμων προς κατανάλωση σαλατών
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία Μικροβιολογική ποιότητα έτοιμων προς κατανάλωση σαλατών Σοφία Στεργίου Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία Μικροβιολογική ποιότητα έτοιμων προς κατανάλωση σαλατών Σοφία Στεργίου Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ
ΔΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΑΓΩΓΗΣ
 ΔΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΑΓΩΓΗΣ ΤΜΗΜΑ ΕΠΙΣΤΗΜΩΝ ΕΚΠΑΙΔΕΥΣΗΣ ΣΤΗΝ ΠΡΟΣΧΟΛΙΚΗ ΗΛΙΚΙΑ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Διαπολιτισμική Εκπαίδευση και Θρησκευτική Ετερότητα: εθνικές και θρησκευτικές
ΔΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΑΓΩΓΗΣ ΤΜΗΜΑ ΕΠΙΣΤΗΜΩΝ ΕΚΠΑΙΔΕΥΣΗΣ ΣΤΗΝ ΠΡΟΣΧΟΛΙΚΗ ΗΛΙΚΙΑ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Διαπολιτισμική Εκπαίδευση και Θρησκευτική Ετερότητα: εθνικές και θρησκευτικές
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία. Κόπωση και ποιότητα ζωής ασθενών με καρκίνο.
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Κόπωση και ποιότητα ζωής ασθενών με καρκίνο Μαργαρίτα Μάου Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Κόπωση και ποιότητα ζωής ασθενών με καρκίνο Μαργαρίτα Μάου Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ
þÿ ½ Á Å, ˆ»µ½± Neapolis University þÿ Á̳Á±¼¼± ¼Ìù±Â ¹ º à Â, Ç» Ÿ¹º ½ ¼¹ºÎ½ À¹ÃÄ ¼Î½ º±¹ ¹ º à  þÿ ±½µÀ¹ÃÄ ¼¹ µ À»¹Â Æ Å
 Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2016-08 þÿ µà±³³µ»¼±ä¹º ½ ÀÄž ÄÉ þÿµºà±¹ µåä¹ºî½ - ¹µÁµÍ½ à Äɽ þÿ³½îãµé½
Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2016-08 þÿ µà±³³µ»¼±ä¹º ½ ÀÄž ÄÉ þÿµºà±¹ µåä¹ºî½ - ¹µÁµÍ½ à Äɽ þÿ³½îãµé½
Επιβλέπων καθηγητής: Δρ Βασίλειος Ραφτόπουλος ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΣΕ ΜΕΤΕΜΜΗΝΟΠΑΥΣΙΑΚΕΣ ΓΥΝΑΙΚΕΣ ΜΕΤΑ ΑΠΟ ΟΣΤΕΟΠΟΡΩΤΙΚΑ ΚΑΤΑΓΜΑΤΑ ΣΠΟΝΔΥΛΙΚΗΣ ΣΤΗΛΗΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων καθηγητής: Δρ Βασίλειος Ραφτόπουλος ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΣΕ ΜΕΤΕΜΜΗΝΟΠΑΥΣΙΑΚΕΣ ΓΥΝΑΙΚΕΣ ΜΕΤΑ ΑΠΟ ΟΣΤΕΟΠΟΡΩΤΙΚΑ ΚΑΤΑΓΜΑΤΑ ΣΠΟΝΔΥΛΙΚΗΣ ΣΤΗΛΗΣ Από τη
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων καθηγητής: Δρ Βασίλειος Ραφτόπουλος ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΣΕ ΜΕΤΕΜΜΗΝΟΠΑΥΣΙΑΚΕΣ ΓΥΝΑΙΚΕΣ ΜΕΤΑ ΑΠΟ ΟΣΤΕΟΠΟΡΩΤΙΚΑ ΚΑΤΑΓΜΑΤΑ ΣΠΟΝΔΥΛΙΚΗΣ ΣΤΗΛΗΣ Από τη
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Γνώση και στάση νοσηλευτών στη διαχείριση του πόνου καρκινοπαθών που νοσηλεύονται Παναγιώτης Χαραλάμπους Λεμεσός, 2014 i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Γνώση και στάση νοσηλευτών στη διαχείριση του πόνου καρκινοπαθών που νοσηλεύονται Παναγιώτης Χαραλάμπους Λεμεσός, 2014 i ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΗΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία ΑΝΑΠΤΥΞΗ ΔΕΙΚΤΩΝ ΠΟΙΟΤΗΤΑΣ ΕΔΑΦΟΥΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΗΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΑΝΑΠΤΥΞΗ ΔΕΙΚΤΩΝ ΠΟΙΟΤΗΤΑΣ ΕΔΑΦΟΥΣ [Μαρία Μαρκουλλή] Λεμεσός 2015 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΗΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΑΝΑΠΤΥΞΗ ΔΕΙΚΤΩΝ ΠΟΙΟΤΗΤΑΣ ΕΔΑΦΟΥΣ [Μαρία Μαρκουλλή] Λεμεσός 2015 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή Εργασία ΕΠΙΛΟΧΕΙΑ ΚΑΤΑΘΛΙΨΗ: Ο ΡΟΛΟΣ ΤΗΣ ΚΑΤ ΟΙΚΟΝ ΝΟΣΗΛΕΙΑΣ. Φοινίκη Αλεξάνδρου
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία ΕΠΙΛΟΧΕΙΑ ΚΑΤΑΘΛΙΨΗ: Ο ΡΟΛΟΣ ΤΗΣ ΚΑΤ ΟΙΚΟΝ ΝΟΣΗΛΕΙΑΣ Φοινίκη Αλεξάνδρου Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΕΠΙΣΤΗΜΩΝ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία ΕΠΙΛΟΧΕΙΑ ΚΑΤΑΘΛΙΨΗ: Ο ΡΟΛΟΣ ΤΗΣ ΚΑΤ ΟΙΚΟΝ ΝΟΣΗΛΕΙΑΣ Φοινίκη Αλεξάνδρου Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΕΠΙΣΤΗΜΩΝ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Ανθεκτικότητα του κοινού τετράνυχου Tetranychus urticae Koch (Acari : Tetranychidae) στα εντοµοκτόνα/ακαρεοκτόνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Ανθεκτικότητα του κοινού τετράνυχου Tetranychus urticae Koch (Acari : Tetranychidae) στα εντοµοκτόνα/ακαρεοκτόνα
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΑΝΘΡΩΠΙΣΤΙΚΩΝ ΚΑΙ ΚΟΙΝΩΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΦΙΛΟΛΟΓΙΑΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΑΝΘΡΩΠΙΣΤΙΚΩΝ ΚΑΙ ΚΟΙΝΩΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΦΙΛΟΛΟΓΙΑΣ Π.Μ.Σ: «Σύγχρονες Προσεγγίσεις στη γλώσσα και στα κείμενα» ΚΑΤΕΥΘΥΝΣΗ ΓΛΩΣΣΟΛΟΓΙΑΣ ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ Το φωνηεντικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΑΝΘΡΩΠΙΣΤΙΚΩΝ ΚΑΙ ΚΟΙΝΩΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΦΙΛΟΛΟΓΙΑΣ Π.Μ.Σ: «Σύγχρονες Προσεγγίσεις στη γλώσσα και στα κείμενα» ΚΑΤΕΥΘΥΝΣΗ ΓΛΩΣΣΟΛΟΓΙΑΣ ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ Το φωνηεντικό
þÿ ³¹µ¹½ º±¹ ±ÃÆ»µ¹± ÃÄ ÇÎÁ
 Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2014 þÿ ³¹µ¹½ º±¹ ±ÃÆ»µ¹± ÃÄ ÇÎÁ þÿµá³±ã ±Â Äɽ ½ à º ¼µ ɽ : Georgiou,
Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2014 þÿ ³¹µ¹½ º±¹ ±ÃÆ»µ¹± ÃÄ ÇÎÁ þÿµá³±ã ±Â Äɽ ½ à º ¼µ ɽ : Georgiou,
Θέμα: Παχυσαρκία και κύηση:
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜ Α ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Θέμα: Παχυσαρκία και κύηση: επιπτώσεις στην έκβαση της κύησης και στο έμβρυο Ονοματεπώνυμο: Στέλλα Ριαλά Αριθμός
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜ Α ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Θέμα: Παχυσαρκία και κύηση: επιπτώσεις στην έκβαση της κύησης και στο έμβρυο Ονοματεπώνυμο: Στέλλα Ριαλά Αριθμός
Σχολή Γεωπονικών Επιστημών και Διαχείρισης Περιβάλλοντος. Πτυχιακή εργασία
 Σχολή Γεωπονικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία ΕΦΑΡΜΟΓΗ ΤΗΣ ΜΕΘΟΔΟΥ ΠΑΡΕΜΠΟΔΙΣΗΣ ΤΗΣ ΣΥΖΕΥΞΗΣ ΓΙΑ ΤΟΝ ΕΛΕΓΧΟ ΤΗΣ ΕΥΔΕΜΙΔΑΣ ΚΑΙ ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΠΑΡΟΥΣΙΑΣ ΩΦΕΛΙΜΩΝ ΕΝΤΟΜΩΝ ΩΣ ΥΠΟΣΤΗΡΙΚΤΙΚΗ
Σχολή Γεωπονικών Επιστημών και Διαχείρισης Περιβάλλοντος Πτυχιακή εργασία ΕΦΑΡΜΟΓΗ ΤΗΣ ΜΕΘΟΔΟΥ ΠΑΡΕΜΠΟΔΙΣΗΣ ΤΗΣ ΣΥΖΕΥΞΗΣ ΓΙΑ ΤΟΝ ΕΛΕΓΧΟ ΤΗΣ ΕΥΔΕΜΙΔΑΣ ΚΑΙ ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΠΑΡΟΥΣΙΑΣ ΩΦΕΛΙΜΩΝ ΕΝΤΟΜΩΝ ΩΣ ΥΠΟΣΤΗΡΙΚΤΙΚΗ
ΚΑΠΝΙΣΜΑ ΚΑΙ ΣΥΝΔΡΟΜΟ ΑΙΦΝΙΔΙΟΥ ΒΡΕΦΙΚΟΥ ΘΑΝΑΤΟΥ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΚΑΠΝΙΣΜΑ ΚΑΙ ΣΥΝΔΡΟΜΟ ΑΙΦΝΙΔΙΟΥ ΒΡΕΦΙΚΟΥ ΘΑΝΑΤΟΥ Ονοματεπώνυμο: Λοϊζιά Ελένη Λεμεσός 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΚΑΠΝΙΣΜΑ ΚΑΙ ΣΥΝΔΡΟΜΟ ΑΙΦΝΙΔΙΟΥ ΒΡΕΦΙΚΟΥ ΘΑΝΑΤΟΥ Ονοματεπώνυμο: Λοϊζιά Ελένη Λεμεσός 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ
ΝΑΥΤΙΛΙΑΚΟΙ ΚΥΚΛΟΙ ΚΑΙ ΧΡΗΜΑΤΟΔΟΤΗΣΗ ΝΑΥΤΙΛΙΑΚΩΝ ΕΠΕΝΔΥΣΕΩΝ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ Σχολή Διοίκησης και Οικονομίας Μεταπτυχιακή διατριβή ΝΑΥΤΙΛΙΑΚΟΙ ΚΥΚΛΟΙ ΚΑΙ ΧΡΗΜΑΤΟΔΟΤΗΣΗ ΝΑΥΤΙΛΙΑΚΩΝ ΕΠΕΝΔΥΣΕΩΝ ΔΗΜΗΤΡΗΣ ΤΡΥΦΩΝΟΣ Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ Σχολή Διοίκησης και Οικονομίας Μεταπτυχιακή διατριβή ΝΑΥΤΙΛΙΑΚΟΙ ΚΥΚΛΟΙ ΚΑΙ ΧΡΗΜΑΤΟΔΟΤΗΣΗ ΝΑΥΤΙΛΙΑΚΩΝ ΕΠΕΝΔΥΣΕΩΝ ΔΗΜΗΤΡΗΣ ΤΡΥΦΩΝΟΣ Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΒΕΛΤΙΩΣΗ ΔΙΕΡΓΑΣΙΩΝ ΕΡΓΑΣΤΗΡΙΟΥ ΕΛΕΓΧΟΥ ΠΟΙΟΤΗΤΑΣ ΚΑΙ ΕΦΑΡΜΟΓΗ ΕΡΓΑΛΕΙΩΝ ΔΙΑΣΦΑΛΙΣΗΣ ΠΟΙΟΤΗΤΑΣ ΣΕ ΜΕΤΑΛΛΟΒΙΟΜΗΧΑΝΙΑ
 Σχολή Mηχανικής και Τεχνολογίας Πτυχιακή εργασία ΒΕΛΤΙΩΣΗ ΔΙΕΡΓΑΣΙΩΝ ΕΡΓΑΣΤΗΡΙΟΥ ΕΛΕΓΧΟΥ ΠΟΙΟΤΗΤΑΣ ΚΑΙ ΕΦΑΡΜΟΓΗ ΕΡΓΑΛΕΙΩΝ ΔΙΑΣΦΑΛΙΣΗΣ ΠΟΙΟΤΗΤΑΣ ΣΕ ΜΕΤΑΛΛΟΒΙΟΜΗΧΑΝΙΑ Στέλιος Καράσαββας Λεμεσός, Μάιος 2017
Σχολή Mηχανικής και Τεχνολογίας Πτυχιακή εργασία ΒΕΛΤΙΩΣΗ ΔΙΕΡΓΑΣΙΩΝ ΕΡΓΑΣΤΗΡΙΟΥ ΕΛΕΓΧΟΥ ΠΟΙΟΤΗΤΑΣ ΚΑΙ ΕΦΑΡΜΟΓΗ ΕΡΓΑΛΕΙΩΝ ΔΙΑΣΦΑΛΙΣΗΣ ΠΟΙΟΤΗΤΑΣ ΣΕ ΜΕΤΑΛΛΟΒΙΟΜΗΧΑΝΙΑ Στέλιος Καράσαββας Λεμεσός, Μάιος 2017
ΑΞΙΟΛΟΓΗΣΗ ΑΦΗΓΗΜΑΤΙΚΩΝ ΙΚΑΝΟΤΗΤΩΝ ΜΕΣΩ ΧΟΡΗΓΗΣΗΣ ΤΟΥ ΕΡΓΑΛΕΙΟΥ ΜΑΙΝ ΣΕ ΤΥΠΙΚΩΣ ΑΝΑΠΤΥΣΣΟΜΕΝΑ ΠΑΙΔΙΑ ΣΤΗΝ ΚΥΠΡΟ
 Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΑΞΙΟΛΟΓΗΣΗ ΑΦΗΓΗΜΑΤΙΚΩΝ ΙΚΑΝΟΤΗΤΩΝ ΜΕΣΩ ΧΟΡΗΓΗΣΗΣ ΤΟΥ ΕΡΓΑΛΕΙΟΥ ΜΑΙΝ ΣΕ ΤΥΠΙΚΩΣ ΑΝΑΠΤΥΣΣΟΜΕΝΑ ΠΑΙΔΙΑ ΣΤΗΝ ΚΥΠΡΟ Γεωργίου Μύρια Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΑΞΙΟΛΟΓΗΣΗ ΑΦΗΓΗΜΑΤΙΚΩΝ ΙΚΑΝΟΤΗΤΩΝ ΜΕΣΩ ΧΟΡΗΓΗΣΗΣ ΤΟΥ ΕΡΓΑΛΕΙΟΥ ΜΑΙΝ ΣΕ ΤΥΠΙΚΩΣ ΑΝΑΠΤΥΣΣΟΜΕΝΑ ΠΑΙΔΙΑ ΣΤΗΝ ΚΥΠΡΟ Γεωργίου Μύρια Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή διατριβή Η ΚΑΤΑΘΛΙΨΗ ΩΣ ΠΑΡΑΓΟΝΤΑΣ ΚΙΝΔΥΝΟΥ ΓΙΑ ΑΠΟΠΕΙΡΑ ΑΥΤΟΚΤΟΝΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Η ΚΑΤΑΘΛΙΨΗ ΩΣ ΠΑΡΑΓΟΝΤΑΣ ΚΙΝΔΥΝΟΥ ΓΙΑ ΑΠΟΠΕΙΡΑ ΑΥΤΟΚΤΟΝΙΑΣ Παναγιώτου Νεοφύτα 2008969752 Επιβλέπων καθηγητής Δρ. Νίκος Μίτλεττον,
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή διατριβή Η ΚΑΤΑΘΛΙΨΗ ΩΣ ΠΑΡΑΓΟΝΤΑΣ ΚΙΝΔΥΝΟΥ ΓΙΑ ΑΠΟΠΕΙΡΑ ΑΥΤΟΚΤΟΝΙΑΣ Παναγιώτου Νεοφύτα 2008969752 Επιβλέπων καθηγητής Δρ. Νίκος Μίτλεττον,
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή διατριβή ΟΙ ΠΑΡΑΓΟΝΤΕΣ ΠΟΥ ΕΠΗΡΕΑΖΟΥΝ ΤΗΝ ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΣΧΕΤΙΖΟΜΕΝΗ ΜΕ ΤΗΝ ΥΓΕΙΑ ΣΕ ΗΛΙΚΙΩΜΕΝΟΥΣ ΜΕ ΚΑΡΔΙΑΚΗ ΑΝΕΠΑΡΚΕΙΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή διατριβή ΟΙ ΠΑΡΑΓΟΝΤΕΣ ΠΟΥ ΕΠΗΡΕΑΖΟΥΝ ΤΗΝ ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΣΧΕΤΙΖΟΜΕΝΗ ΜΕ ΤΗΝ ΥΓΕΙΑ ΣΕ ΗΛΙΚΙΩΜΕΝΟΥΣ ΜΕ ΚΑΡΔΙΑΚΗ ΑΝΕΠΑΡΚΕΙΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία Η ΚΑΤΑΘΛΙΨΗ ΣΕ ΕΦΗΒΟΥΣ ΜΕ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 1
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΚΑΤΑΘΛΙΨΗ ΣΕ ΕΦΗΒΟΥΣ ΜΕ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 1 ΑΝΔΡΕΑΣ ΑΝΔΡΕΟΥ Φ.Τ:2008670839 Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία Η ΚΑΤΑΘΛΙΨΗ ΣΕ ΕΦΗΒΟΥΣ ΜΕ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 1 ΑΝΔΡΕΑΣ ΑΝΔΡΕΟΥ Φ.Τ:2008670839 Λεμεσός 2014 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΔΗΜΙΟΥΡΓΙΑ ΣΥΣΤΗΜΑΤΟΣ ΔΕΙΚΤΩΝ ΑΣΤΙΚΗΣ ΒΙΩΣΙΜΟΤΗΤΑΣ ΓΙΑ ΤΗΝ ΕΠΑΡΧΙΑ ΛΕΜΕΣΟΥ
 Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Μεταπτυχιακή διατριβή ΔΗΜΙΟΥΡΓΙΑ ΣΥΣΤΗΜΑΤΟΣ ΔΕΙΚΤΩΝ ΑΣΤΙΚΗΣ ΒΙΩΣΙΜΟΤΗΤΑΣ ΓΙΑ ΤΗΝ ΕΠΑΡΧΙΑ ΛΕΜΕΣΟΥ Παρασκευή Νταϊλιάνη Λεμεσός, Μάιος, 2017 TΕΧΝΟΛΟΓΙΚΟ
Σχολή Γεωτεχνικών Επιστημών και Διαχείρισης Περιβάλλοντος Μεταπτυχιακή διατριβή ΔΗΜΙΟΥΡΓΙΑ ΣΥΣΤΗΜΑΤΟΣ ΔΕΙΚΤΩΝ ΑΣΤΙΚΗΣ ΒΙΩΣΙΜΟΤΗΤΑΣ ΓΙΑ ΤΗΝ ΕΠΑΡΧΙΑ ΛΕΜΕΣΟΥ Παρασκευή Νταϊλιάνη Λεμεσός, Μάιος, 2017 TΕΧΝΟΛΟΓΙΚΟ
þÿ ½ ÁÉÀ ºµ½ÄÁ¹º ÀÁ à ³³¹Ã Ä þÿ Á³±½Éù±º  ±»»±³  ¼ ÃÉ þÿà» Á Æ Á¹±º Í ÃÅÃÄ ¼±Ä Â.
 Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2016-02 þÿ ½ ÁÉÀ ºµ½ÄÁ¹º ÀÁ à ³³¹Ã Ä þÿ Á³±½Éù±º  ±»»±³  ¼ ÃÉ þÿà»
Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2016-02 þÿ ½ ÁÉÀ ºµ½ÄÁ¹º ÀÁ à ³³¹Ã Ä þÿ Á³±½Éù±º  ±»»±³  ¼ ÃÉ þÿà»
ΠΑΝΔΠΗΣΖΜΗΟ ΠΑΣΡΩΝ ΣΜΖΜΑ ΖΛΔΚΣΡΟΛΟΓΩΝ ΜΖΥΑΝΗΚΩΝ ΚΑΗ ΣΔΥΝΟΛΟΓΗΑ ΤΠΟΛΟΓΗΣΩΝ ΣΟΜΔΑ ΤΣΖΜΑΣΩΝ ΖΛΔΚΣΡΗΚΖ ΔΝΔΡΓΔΗΑ
 ΠΑΝΔΠΗΣΖΜΗΟ ΠΑΣΡΩΝ ΣΜΖΜΑ ΖΛΔΚΣΡΟΛΟΓΩΝ ΜΖΥΑΝΗΚΩΝ ΚΑΗ ΣΔΥΝΟΛΟΓΗΑ ΤΠΟΛΟΓΗΣΩΝ ΣΟΜΔΑ ΤΣΖΜΑΣΩΝ ΖΛΔΚΣΡΗΚΖ ΔΝΔΡΓΔΗΑ Γηπισκαηηθή Δξγαζία ηνπ Φνηηεηή ηνπ ηκήκαηνο Ζιεθηξνιόγσλ Μεραληθώλ θαη Σερλνινγίαο Ζιεθηξνληθώλ
ΠΑΝΔΠΗΣΖΜΗΟ ΠΑΣΡΩΝ ΣΜΖΜΑ ΖΛΔΚΣΡΟΛΟΓΩΝ ΜΖΥΑΝΗΚΩΝ ΚΑΗ ΣΔΥΝΟΛΟΓΗΑ ΤΠΟΛΟΓΗΣΩΝ ΣΟΜΔΑ ΤΣΖΜΑΣΩΝ ΖΛΔΚΣΡΗΚΖ ΔΝΔΡΓΔΗΑ Γηπισκαηηθή Δξγαζία ηνπ Φνηηεηή ηνπ ηκήκαηνο Ζιεθηξνιόγσλ Μεραληθώλ θαη Σερλνινγίαο Ζιεθηξνληθώλ
ΓΕΩΠΟΝΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΤΜΗΜΑ ΑΓΡΟΤΙΚΗΣ ΟΙΚΟΝΟΜΙΑΣ & ΑΝΑΠΤΥΞΗΣ
 ΓΕΩΠΟΝΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΤΜΗΜΑ ΑΓΡΟΤΙΚΗΣ ΟΙΚΟΝΟΜΙΑΣ & ΑΝΑΠΤΥΞΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΟΛΟΚΛΗΡΩΜΕΝΗ ΑΝΑΠΤΥΞΗ & ΔΙΑΧΕΙΡΙΣΗ ΤΟΥ ΑΓΡΟΤΙΚΟΥ ΧΩΡΟΥ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Οικονομετρική διερεύνηση
ΓΕΩΠΟΝΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΤΜΗΜΑ ΑΓΡΟΤΙΚΗΣ ΟΙΚΟΝΟΜΙΑΣ & ΑΝΑΠΤΥΞΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΟΛΟΚΛΗΡΩΜΕΝΗ ΑΝΑΠΤΥΞΗ & ΔΙΑΧΕΙΡΙΣΗ ΤΟΥ ΑΓΡΟΤΙΚΟΥ ΧΩΡΟΥ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Οικονομετρική διερεύνηση
Μεταπτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΣΥΣΧΕΤΙΣΜΟΙ ΠΡΑΓΜΑΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΥΦΙΣΤΑΜΕΝΩΝ ΦΩΤΟΒΟΛΤΑΪΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΑΝΑΛΟΓΑ ΜΕ ΤΗ ΤΟΠΟΘΕΣΙΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΣΥΣΧΕΤΙΣΜΟΙ ΠΡΑΓΜΑΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΥΦΙΣΤΑΜΕΝΩΝ ΦΩΤΟΒΟΛΤΑΪΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΑΝΑΛΟΓΑ ΜΕ ΤΗ ΤΟΠΟΘΕΣΙΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ. Πτυχιακή εργασία ΕΤΟΙΜΑΣΙΑ ΔΕΛΤΙΟΥ ΠΟΣΟΤΗΤΩΝ ΜΕ ΤΗ ΧΡΗΣΗ ΛΟΓΙΣΜΙΚΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ Πτυχιακή εργασία ΕΤΟΙΜΑΣΙΑ ΔΕΛΤΙΟΥ ΠΟΣΟΤΗΤΩΝ ΜΕ ΤΗ ΧΡΗΣΗ ΛΟΓΙΣΜΙΚΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ Χρύσω Κωνσταντίνου Λεμεσός 2016 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΜΗΧΑΝΙΚΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ Πτυχιακή εργασία ΕΤΟΙΜΑΣΙΑ ΔΕΛΤΙΟΥ ΠΟΣΟΤΗΤΩΝ ΜΕ ΤΗ ΧΡΗΣΗ ΛΟΓΙΣΜΙΚΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ Χρύσω Κωνσταντίνου Λεμεσός 2016 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ
ΠΕΡΙΕΧΟΜΕΝΑ. Μάρκετινγκ Αθλητικών Τουριστικών Προορισμών 1
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΤΗΣ ΔΙΟΙΚΗΣΗΣ ΤΜΗΜΑ ΔΙΟΙΚΗΣΗΣ ΕΠΙΧΕΙΡΗΣΕΩΝ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «Σχεδιασμός, Διοίκηση και Πολιτική του Τουρισμού» ΜΑΡΚΕΤΙΝΓΚ ΑΘΛΗΤΙΚΩΝ ΤΟΥΡΙΣΤΙΚΩΝ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΤΗΣ ΔΙΟΙΚΗΣΗΣ ΤΜΗΜΑ ΔΙΟΙΚΗΣΗΣ ΕΠΙΧΕΙΡΗΣΕΩΝ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «Σχεδιασμός, Διοίκηση και Πολιτική του Τουρισμού» ΜΑΡΚΕΤΙΝΓΚ ΑΘΛΗΤΙΚΩΝ ΤΟΥΡΙΣΤΙΚΩΝ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΠΟΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΠΟΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία Η ΕΦΑΡΜΟΓΗ ΤΟΥ ΣΥΣΤΗΜΑΤΟΣ HACCP ΣΕ ΜΙΚΡΕΣ ΒΙΟΤΕΧΝΙΕΣ ΓΑΛΑΚΤΟΣ ΣΤΗΝ ΕΠΑΡΧΙΑ ΛΕΜΕΣΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΠΟΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία Η ΕΦΑΡΜΟΓΗ ΤΟΥ ΣΥΣΤΗΜΑΤΟΣ HACCP ΣΕ ΜΙΚΡΕΣ ΒΙΟΤΕΧΝΙΕΣ ΓΑΛΑΚΤΟΣ ΣΤΗΝ ΕΠΑΡΧΙΑ ΛΕΜΕΣΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙ ΕΥΤΙΚΟ Ι ΡΥΜΑ ΚΡΗΤΗΣ ΣΧΟΛΗ ΙΟΙΚΗΣΗΣ ΚΑΙ ΟΙΚΟΝΟΜΙΑΣ ΤΜΗΜΑ ΙΟΙΚΗΣΗΣ ΕΠΙΧΕΙΡΗΣΕΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ
 ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙ ΕΥΤΙΚΟ Ι ΡΥΜΑ ΚΡΗΤΗΣ ΣΧΟΛΗ ΙΟΙΚΗΣΗΣ ΚΑΙ ΟΙΚΟΝΟΜΙΑΣ ΤΜΗΜΑ ΙΟΙΚΗΣΗΣ ΕΠΙΧΕΙΡΗΣΕΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Το franchising ( δικαιόχρηση ) ως µέθοδος ανάπτυξης των επιχειρήσεων λιανικού εµπορίου
ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙ ΕΥΤΙΚΟ Ι ΡΥΜΑ ΚΡΗΤΗΣ ΣΧΟΛΗ ΙΟΙΚΗΣΗΣ ΚΑΙ ΟΙΚΟΝΟΜΙΑΣ ΤΜΗΜΑ ΙΟΙΚΗΣΗΣ ΕΠΙΧΕΙΡΗΣΕΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Το franchising ( δικαιόχρηση ) ως µέθοδος ανάπτυξης των επιχειρήσεων λιανικού εµπορίου
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή Διατριβή Επιβλέπουσα καθηγήτρια: Κα Παναγιώτα Ταμανά ΣΤΑΣΕΙΣ ΚΑΙ ΠΕΠΟΙΘΗΣΕΙΣ ΑΤΟΜΩΝ ΑΠΕΝΑΝΤΙ ΣΤΟ ΕΜΒΟΛΙΟ ΚΑΤΑ ΤΟΥ ΚΑΡΚΙΝΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή Διατριβή Επιβλέπουσα καθηγήτρια: Κα Παναγιώτα Ταμανά ΣΤΑΣΕΙΣ ΚΑΙ ΠΕΠΟΙΘΗΣΕΙΣ ΑΤΟΜΩΝ ΑΠΕΝΑΝΤΙ ΣΤΟ ΕΜΒΟΛΙΟ ΚΑΤΑ ΤΟΥ ΚΑΡΚΙΝΟΥ
ΠΕΡΙΕΧΟΜΕΝΑ. Κεφάλαιο 1: Κεφάλαιο 2: Κεφάλαιο 3:
 4 Πρόλογος Η παρούσα διπλωµατική εργασία µε τίτλο «ιερεύνηση χωρικής κατανοµής µετεωρολογικών µεταβλητών. Εφαρµογή στον ελληνικό χώρο», ανατέθηκε από το ιεπιστηµονικό ιατµηµατικό Πρόγραµµα Μεταπτυχιακών
4 Πρόλογος Η παρούσα διπλωµατική εργασία µε τίτλο «ιερεύνηση χωρικής κατανοµής µετεωρολογικών µεταβλητών. Εφαρµογή στον ελληνικό χώρο», ανατέθηκε από το ιεπιστηµονικό ιατµηµατικό Πρόγραµµα Μεταπτυχιακών
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΣΧΕΤΙΖΟΜΕΝΗ ΜΕ ΤΗΝ ΥΓΕΙΑ ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΑΤΟΜΩΝ ΜΕ ΝΕΑΝΙΚΗ ΙΔΙΟΠΑΘΗ ΑΡΘΡΙΤΙΔΑ Όνομα Φοιτήτριας: Μαρία Θωμά Αριθμός φοιτητικής ταυτότητας:2010221455
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή Εργασία Η ΣΧΕΤΙΖΟΜΕΝΗ ΜΕ ΤΗΝ ΥΓΕΙΑ ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΑΤΟΜΩΝ ΜΕ ΝΕΑΝΙΚΗ ΙΔΙΟΠΑΘΗ ΑΡΘΡΙΤΙΔΑ Όνομα Φοιτήτριας: Μαρία Θωμά Αριθμός φοιτητικής ταυτότητας:2010221455
Εκτεταμένη έκθεση σε υπεριώδη ακτινοβολία στην παιδική και εφηβική ηλικία και εμφάνιση μελανώματος.
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Εκτεταμένη έκθεση σε υπεριώδη ακτινοβολία στην παιδική και εφηβική ηλικία και εμφάνιση μελανώματος. Ονοματεπώνυμο φοιτήτριας : Ελίνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Εκτεταμένη έκθεση σε υπεριώδη ακτινοβολία στην παιδική και εφηβική ηλικία και εμφάνιση μελανώματος. Ονοματεπώνυμο φοιτήτριας : Ελίνα
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΗΣ ΕΠΙΣΤΗΜΗΣ ΑΜΕΣΕΣ ΞΕΝΕΣ ΕΠΕΝΔΥΣΕΙΣ ΣΕ ΕΥΡΩΠΑΙΚΕΣ ΧΩΡΕΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΗΣ ΕΠΙΣΤΗΜΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΟΙΚΟΝΟΜΙΚΗ ΚΑΙ ΕΠΙΧΕΙΡΗΣΙΑΚΗ ΣΤΡΑΤΗΓΙΚΗ ΑΜΕΣΕΣ ΞΕΝΕΣ ΕΠΕΝΔΥΣΕΙΣ ΣΕ ΕΥΡΩΠΑΙΚΕΣ ΧΩΡΕΣ Αθανάσιος Νταραβάνογλου Διπλωματική
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΩΣ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΗΣ ΕΠΙΣΤΗΜΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΟΙΚΟΝΟΜΙΚΗ ΚΑΙ ΕΠΙΧΕΙΡΗΣΙΑΚΗ ΣΤΡΑΤΗΓΙΚΗ ΑΜΕΣΕΣ ΞΕΝΕΣ ΕΠΕΝΔΥΣΕΙΣ ΣΕ ΕΥΡΩΠΑΙΚΕΣ ΧΩΡΕΣ Αθανάσιος Νταραβάνογλου Διπλωματική
TEΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ
 TEΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΞΟΥΘΕΝΩΣΗ ΣΤΟΥΣ ΝΟΣΗΛΕΥΤΕΣ ΕΝΤΑΤΙΚΗΣ ΜΟΝΑΔΑΣ ΚΑΙ ΟΙ ΠΕΡΙΒΑΛΟΝΤΙΚΟΙ ΠΑΡΑΓΟΝΤΕΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: MANΩΛΗ
TEΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΞΟΥΘΕΝΩΣΗ ΣΤΟΥΣ ΝΟΣΗΛΕΥΤΕΣ ΕΝΤΑΤΙΚΗΣ ΜΟΝΑΔΑΣ ΚΑΙ ΟΙ ΠΕΡΙΒΑΛΟΝΤΙΚΟΙ ΠΑΡΑΓΟΝΤΕΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: MANΩΛΗ
Η Επίδραση των Events στην Απόδοση των Μετοχών
 Χρηματοοικονομικά και Διοίκηση Μεταπτυχιακή διατριβή Η Επίδραση των Events στην Απόδοση των Μετοχών Άντρεα Φωτίου Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΩΝ ΚΑΙ ΔΙΟΙΚΗΣΗΣ
Χρηματοοικονομικά και Διοίκηση Μεταπτυχιακή διατριβή Η Επίδραση των Events στην Απόδοση των Μετοχών Άντρεα Φωτίου Λεμεσός, Μάιος 2018 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΧΡΗΜΑΤΟΟΙΚΟΝΟΜΙΚΩΝ ΚΑΙ ΔΙΟΙΚΗΣΗΣ
Ανάλυση εισοδήματος των μισθωτών και παράγοντες που το επηρεάζουν
 Π Α Ν Ε Π Ι Σ Τ Η Μ Ι Ο Π Ε Ι Ρ Α Ι Ω Σ ΤΜΗΜΑ Σ Τ Α Τ Ι Σ Τ Ι Κ Η Σ ΚΑΙ ΑΣΦΑΛΙΣΤΙΚΗΣ Ε Π Ι Σ Τ Η Μ Η Σ Μ Ε Τ Α Π Τ Υ Χ Ι Α Κ Ο ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΣΤΗΝ Ε Φ Α Ρ Μ Ο Σ Μ Ε Ν Η Σ Τ Α Τ Ι Σ Τ Ι Κ Η Ανάλυση εισοδήματος
Π Α Ν Ε Π Ι Σ Τ Η Μ Ι Ο Π Ε Ι Ρ Α Ι Ω Σ ΤΜΗΜΑ Σ Τ Α Τ Ι Σ Τ Ι Κ Η Σ ΚΑΙ ΑΣΦΑΛΙΣΤΙΚΗΣ Ε Π Ι Σ Τ Η Μ Η Σ Μ Ε Τ Α Π Τ Υ Χ Ι Α Κ Ο ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΣΤΗΝ Ε Φ Α Ρ Μ Ο Σ Μ Ε Ν Η Σ Τ Α Τ Ι Σ Τ Ι Κ Η Ανάλυση εισοδήματος
Μεταπτυχιακή Διατριβή
 Μεταπτυχιακή Διατριβή ΣΧΗΜΑΤΙΣΜΟΣ ΚΑΙ ΕΝΤΟΠΙΣΜΟΣ ΒΙΟΦΙΛΜ ΣΤΙΣ ΜΕΜΒΡΑΝΕΣ ΑΝΤΙΣΤΡΟΦΗΣ ΩΣΜΩΣΗΣ ΣΤΗΝ ΑΦΑΛΑΤΩΣΗ ΛΕΜΕΣΟΥ ΚΥΠΡΟΣ ΜΙΧΑΗΛ Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ
Μεταπτυχιακή Διατριβή ΣΧΗΜΑΤΙΣΜΟΣ ΚΑΙ ΕΝΤΟΠΙΣΜΟΣ ΒΙΟΦΙΛΜ ΣΤΙΣ ΜΕΜΒΡΑΝΕΣ ΑΝΤΙΣΤΡΟΦΗΣ ΩΣΜΩΣΗΣ ΣΤΗΝ ΑΦΑΛΑΤΩΣΗ ΛΕΜΕΣΟΥ ΚΥΠΡΟΣ ΜΙΧΑΗΛ Λεμεσός, Μάιος 2017 1 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ
«ΑΓΡΟΤΟΥΡΙΣΜΟΣ ΚΑΙ ΤΟΠΙΚΗ ΑΝΑΠΤΥΞΗ: Ο ΡΟΛΟΣ ΤΩΝ ΝΕΩΝ ΤΕΧΝΟΛΟΓΙΩΝ ΣΤΗΝ ΠΡΟΩΘΗΣΗ ΤΩΝ ΓΥΝΑΙΚΕΙΩΝ ΣΥΝΕΤΑΙΡΙΣΜΩΝ»
 I ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΝΟΜΙΚΩΝ ΟΙΚΟΝΟΜΙΚΩΝ ΚΑΙ ΠΟΛΙΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ «ΔΙΟΙΚΗΣΗ ΚΑΙ ΟΙΚΟΝΟΜΙΑ» ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΝΟΜΙΚΗ
I ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΝΟΜΙΚΩΝ ΟΙΚΟΝΟΜΙΚΩΝ ΚΑΙ ΠΟΛΙΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ «ΔΙΟΙΚΗΣΗ ΚΑΙ ΟΙΚΟΝΟΜΙΑ» ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΝΟΜΙΚΗ
Διερεύνηση χαρτογράφησης Ποσειδωνίας με χρήση επιβλεπόμενης ταξινόμησης οπτικών δορυφορικών εικόνων
 Σχολή Μηχανικής και Τεχνολογίας Μεταπτυχιακή διατριβή Διερεύνηση χαρτογράφησης Ποσειδωνίας με χρήση επιβλεπόμενης ταξινόμησης οπτικών δορυφορικών εικόνων Αναστασία Υφαντίδου Λεμεσός, Νοέμβριος 2017 ΤΕΧΝΟΛΟΓΙΚΟ
Σχολή Μηχανικής και Τεχνολογίας Μεταπτυχιακή διατριβή Διερεύνηση χαρτογράφησης Ποσειδωνίας με χρήση επιβλεπόμενης ταξινόμησης οπτικών δορυφορικών εικόνων Αναστασία Υφαντίδου Λεμεσός, Νοέμβριος 2017 ΤΕΧΝΟΛΟΓΙΚΟ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΑΞΙΟΛΟΓΗΣΗ ΤΟΥ ΚΙΝΔΥΝΟΥ ΥΠΟΒΑΘΜΙΣΗΣ ΤΟΥ ΥΔΡΟΒΙΟΤΟΠΟΥ ΤΗΣ ΑΛΥΚΗΣ ΛΑΡΝΑΚΑΣ ΑΠΟ ΤΗΝ ΕΠΙΦΑΝΕΙΑΚΗ ΑΠΟΡΡΟΗ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΑΞΙΟΛΟΓΗΣΗ ΤΟΥ ΚΙΝΔΥΝΟΥ ΥΠΟΒΑΘΜΙΣΗΣ ΤΟΥ ΥΔΡΟΒΙΟΤΟΠΟΥ ΤΗΣ ΑΛΥΚΗΣ ΛΑΡΝΑΚΑΣ ΑΠΟ ΤΗΝ ΕΠΙΦΑΝΕΙΑΚΗ ΑΠΟΡΡΟΗ
ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ "ΠΟΛΥΚΡΙΤΗΡΙΑ ΣΥΣΤΗΜΑΤΑ ΛΗΨΗΣ ΑΠΟΦΑΣΕΩΝ. Η ΠΕΡΙΠΤΩΣΗ ΤΗΣ ΕΠΙΛΟΓΗΣ ΑΣΦΑΛΙΣΤΗΡΙΟΥ ΣΥΜΒΟΛΑΙΟΥ ΥΓΕΙΑΣ "
 ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΚΑΛΑΜΑΤΑΣ ΣΧΟΛΗ ΔΙΟΙΚΗΣΗΣ ΟΙΚΟΝΟΜΙΑΣ ΤΜΗΜΑ ΜΟΝΑΔΩΝ ΥΓΕΙΑΣ ΚΑΙ ΠΡΟΝΟΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ "ΠΟΛΥΚΡΙΤΗΡΙΑ ΣΥΣΤΗΜΑΤΑ ΛΗΨΗΣ ΑΠΟΦΑΣΕΩΝ. Η ΠΕΡΙΠΤΩΣΗ ΤΗΣ ΕΠΙΛΟΓΗΣ ΑΣΦΑΛΙΣΤΗΡΙΟΥ ΣΥΜΒΟΛΑΙΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΑΙΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΚΑΛΑΜΑΤΑΣ ΣΧΟΛΗ ΔΙΟΙΚΗΣΗΣ ΟΙΚΟΝΟΜΙΑΣ ΤΜΗΜΑ ΜΟΝΑΔΩΝ ΥΓΕΙΑΣ ΚΑΙ ΠΡΟΝΟΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ "ΠΟΛΥΚΡΙΤΗΡΙΑ ΣΥΣΤΗΜΑΤΑ ΛΗΨΗΣ ΑΠΟΦΑΣΕΩΝ. Η ΠΕΡΙΠΤΩΣΗ ΤΗΣ ΕΠΙΛΟΓΗΣ ΑΣΦΑΛΙΣΤΗΡΙΟΥ ΣΥΜΒΟΛΑΙΟΥ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΠΑΡΑΓΟΝΤΕΣ ΠΟΥ ΕΠΗΡΕΑΖΟΥΝ ΤΗΝ ΘΝΗΣΙΜΟΤΗΤΑ ΕΡΙΦΙΩΝ ΤΗΣ ΦΥΛΗΣ ΔΑΜΑΣΚΟΥ ΜΕΧΡΙ ΤΟΝ ΑΠΟΓΑΛΑΚΤΙΣΜΟ ΣΟΦΟΚΛΕΟΥΣ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΠΑΡΑΓΟΝΤΕΣ ΠΟΥ ΕΠΗΡΕΑΖΟΥΝ ΤΗΝ ΘΝΗΣΙΜΟΤΗΤΑ ΕΡΙΦΙΩΝ ΤΗΣ ΦΥΛΗΣ ΔΑΜΑΣΚΟΥ ΜΕΧΡΙ ΤΟΝ ΑΠΟΓΑΛΑΚΤΙΣΜΟ ΣΟΦΟΚΛΕΟΥΣ
þÿ ɺÁ Ä ÅÂ, ±»Î¼ Neapolis University þÿ Á̳Á±¼¼± ¼Ìù±Â ¹ º à Â, Ç» Ÿ¹º ½ ¼¹ºÎ½ À¹ÃÄ ¼Î½ º±¹ ¹ º à  þÿ ±½µÀ¹ÃÄ ¼¹ µ À»¹Â Æ Å
 Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2016 þÿ ͽ Á ¼ µà±³³µ»¼±ä¹º  þÿµ¾ Å ½Éà  ³º» ³¹ºÎ½ ½ à þÿ ɺÁ Ä ÅÂ,
Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2016 þÿ ͽ Á ¼ µà±³³µ»¼±ä¹º  þÿµ¾ Å ½Éà  ³º» ³¹ºÎ½ ½ à þÿ ɺÁ Ä ÅÂ,
þÿ¼ ½ ±Â : ÁÌ» Â Ä Å ÃÄ ²µ þÿä Å ÃÇ»¹º Í Á³ Å
 Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2015 þÿ ½»Åà Äɽ µ½½ ¹Î½ Ä Â þÿ±¾¹»ì³ à  º±¹ Ä Â þÿ±à ĵ»µÃ¼±Ä¹ºÌÄ Ä±Â
Neapolis University HEPHAESTUS Repository School of Economic Sciences and Business http://hephaestus.nup.ac.cy Master Degree Thesis 2015 þÿ ½»Åà Äɽ µ½½ ¹Î½ Ä Â þÿ±¾¹»ì³ à  º±¹ Ä Â þÿ±à ĵ»µÃ¼±Ä¹ºÌÄ Ä±Â
ΑΘΗΝΑ 2013 ΑΝΑΛΥΣΗ ΕΠΕΞΕΡΓΑΣΜΕΝΟΥ ΥΔΑΤΟΣ (ΛYΜΑΤΩΝ) FRAMME - LIFE08 NAT/GR/000533 ΡΟΔΟΣ
 FRAMME - LIFE08 NAT/GR/000533 ΡΟΔΟΣ") FRAMME LIFE 08 NAT//GR//000533 ΑΘΗΝΑ 2013 ΑΝΑΛΥΣΗ ΕΠΕΞΕΡΓΑΣΜΕΝΟΥ ΥΔΑΤΟΣ (ΛYΜΑΤΩΝ) FRAMME - LIFE08 NAT/GR/000533 ΡΟΔΟΣ Το FRAMME, "Μεθοδολογία Αποκατάστασης Πυρόπληκτων Μεσογειακών Δασών - Ασφάλεια & Αποδοτικότητα
FRAMME LIFE 08 NAT//GR//000533 ΑΘΗΝΑ 2013 ΑΝΑΛΥΣΗ ΕΠΕΞΕΡΓΑΣΜΕΝΟΥ ΥΔΑΤΟΣ (ΛYΜΑΤΩΝ) FRAMME - LIFE08 NAT/GR/000533 ΡΟΔΟΣ Το FRAMME, "Μεθοδολογία Αποκατάστασης Πυρόπληκτων Μεσογειακών Δασών - Ασφάλεια & Αποδοτικότητα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή εργασία ΠΑΡΑΓΟΝΤΕΣ ΠΟΥ ΕΠΗΡΕΑΖΟΥΝ ΤΗΝ ΑΝΑΠΤΥΞΗ ΤΗΣ ΠΑΙΔΙΚΗΣ ΠΑΧΥΣΑΡΚΙΑΣ Ραφαέλα Χριστοδούλου Α.Φ.Τ.: 2010335637 Λεμεσός,
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Πτυχιακή εργασία ΠΑΡΑΓΟΝΤΕΣ ΠΟΥ ΕΠΗΡΕΑΖΟΥΝ ΤΗΝ ΑΝΑΠΤΥΞΗ ΤΗΣ ΠΑΙΔΙΚΗΣ ΠΑΧΥΣΑΡΚΙΑΣ Ραφαέλα Χριστοδούλου Α.Φ.Τ.: 2010335637 Λεμεσός,
ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ
 Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ Άντρια Πολυκάρπου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ Άντρια Πολυκάρπου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
ΖΩΝΟΠΟΙΗΣΗ ΤΗΣ ΚΑΤΟΛΙΣΘΗΤΙΚΗΣ ΕΠΙΚΙΝΔΥΝΟΤΗΤΑΣ ΣΤΟ ΟΡΟΣ ΠΗΛΙΟ ΜΕ ΤΗ ΣΥΜΒΟΛΗ ΔΕΔΟΜΕΝΩΝ ΣΥΜΒΟΛΟΜΕΤΡΙΑΣ ΜΟΝΙΜΩΝ ΣΚΕΔΑΣΤΩΝ
 EΘΝΙΚΟ ΜΕΤΣΟΒΙΟ ΠΟΛΥΤΕΧΕΙΟ Τμήμα Μηχανικών Μεταλλείων-Μεταλλουργών ΖΩΝΟΠΟΙΗΣΗ ΤΗΣ ΚΑΤΟΛΙΣΘΗΤΙΚΗΣ ΕΠΙΚΙΝΔΥΝΟΤΗΤΑΣ ΜΕ ΤΗ ΣΥΜΒΟΛΗ ΔΕΔΟΜΕΝΩΝ ΣΥΜΒΟΛΟΜΕΤΡΙΑΣ ΜΟΝΙΜΩΝ ΣΚΕΔΑΣΤΩΝ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Κιτσάκη Μαρίνα
EΘΝΙΚΟ ΜΕΤΣΟΒΙΟ ΠΟΛΥΤΕΧΕΙΟ Τμήμα Μηχανικών Μεταλλείων-Μεταλλουργών ΖΩΝΟΠΟΙΗΣΗ ΤΗΣ ΚΑΤΟΛΙΣΘΗΤΙΚΗΣ ΕΠΙΚΙΝΔΥΝΟΤΗΤΑΣ ΜΕ ΤΗ ΣΥΜΒΟΛΗ ΔΕΔΟΜΕΝΩΝ ΣΥΜΒΟΛΟΜΕΤΡΙΑΣ ΜΟΝΙΜΩΝ ΣΚΕΔΑΣΤΩΝ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Κιτσάκη Μαρίνα
Η ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΤΩΝ ΗΛΙΚΙΩΜΕΝΩΝ ΑΣΘΕΝΩΝ ΜΕΤΑ ΤΗΝ ΕΞΟΔΟ ΤΟΥΣ ΑΠΟ ΤΗΝ ΜΟΝΑΔΑ ΕΝΤΑΤΙΚΗΣ ΘΕΡΑΠΕΙΑΣ. Στυλιανός Σολωμή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπουσα Καθηγήτρια: Δρ. Λαμπρινού Αικατερίνη Η ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΤΩΝ ΗΛΙΚΙΩΜΕΝΩΝ ΑΣΘΕΝΩΝ ΜΕΤΑ ΤΗΝ ΕΞΟΔΟ ΤΟΥΣ ΑΠΟ ΤΗΝ ΜΟΝΑΔΑ ΕΝΤΑΤΙΚΗΣ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπουσα Καθηγήτρια: Δρ. Λαμπρινού Αικατερίνη Η ΠΟΙΟΤΗΤΑ ΖΩΗΣ ΤΩΝ ΗΛΙΚΙΩΜΕΝΩΝ ΑΣΘΕΝΩΝ ΜΕΤΑ ΤΗΝ ΕΞΟΔΟ ΤΟΥΣ ΑΠΟ ΤΗΝ ΜΟΝΑΔΑ ΕΝΤΑΤΙΚΗΣ
ΑΝΑΠΤΥΞΗ ΤΟΥ ΠΤΥΧΙΑΚΗ. Λεμεσός
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΤΟ ΚΑΠΝΙΣΜΑ ΚΑΤΑ ΤΗΝ ΕΓΚΥΜΟΣΥΝΗ ΚΑΙ Η ΒΛΑΠΤΙΚΗ ΕΠΙΔΡΑ ΑΣΗ ΤΟΥ ΣΤΗΝ ΑΝΑΠΤΥΞΗ ΤΟΥ ΝΕΟΓΝΟΥ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Αγγελική Παπαπαύλου Αριθμός Φοιτητικής
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΤΟ ΚΑΠΝΙΣΜΑ ΚΑΤΑ ΤΗΝ ΕΓΚΥΜΟΣΥΝΗ ΚΑΙ Η ΒΛΑΠΤΙΚΗ ΕΠΙΔΡΑ ΑΣΗ ΤΟΥ ΣΤΗΝ ΑΝΑΠΤΥΞΗ ΤΟΥ ΝΕΟΓΝΟΥ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Αγγελική Παπαπαύλου Αριθμός Φοιτητικής
 Πτυχιακή Εργασία ηµιουργία Εκπαιδευτικού Παιχνιδιού σε Tablets Καλλιγάς ηµήτρης Παναγιώτης Α.Μ.: 1195 Επιβλέπων καθηγητής: ρ. Συρµακέσης Σπύρος ΑΝΤΙΡΡΙΟ 2015 Ευχαριστίες Σ αυτό το σηµείο θα ήθελα να
Πτυχιακή Εργασία ηµιουργία Εκπαιδευτικού Παιχνιδιού σε Tablets Καλλιγάς ηµήτρης Παναγιώτης Α.Μ.: 1195 Επιβλέπων καθηγητής: ρ. Συρµακέσης Σπύρος ΑΝΤΙΡΡΙΟ 2015 Ευχαριστίες Σ αυτό το σηµείο θα ήθελα να
ΕΘΝΙΚΗ ΣΧΟΛΗ ΔΗΜΟΣΙΑΣ ΔΙΟΙΚΗΣΗΣ ΙΓ' ΕΚΠΑΙΔΕΥΤΙΚΗ ΣΕΙΡΑ
 ΕΘΝΙΚΗ ΣΧΟΛΗ ΔΗΜΟΣΙΑΣ ΔΙΟΙΚΗΣΗΣ ΙΓ' ΕΚΠΑΙΔΕΥΤΙΚΗ ΣΕΙΡΑ ΤΜΗΜΑ ΤΟΠΙΚΗΣ ΑΥΤΟΔΙΟΙΚΗΣΗΣ ΚΑΙ ΠΕΡΙΦΕΡΕΙΑΚΗΣ ΑΝΑΠΤΥΞΗΣ ΤΕΛΙΚΗ ΕΡΓΑΣΙΑ: ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ: ΠΡΟΣΕΓΓΙΣΗ ΜΕΣΩ ΔΕΙΚΤΩΝ Επιβλέπων: Αθ.Δελαπάσχος
ΕΘΝΙΚΗ ΣΧΟΛΗ ΔΗΜΟΣΙΑΣ ΔΙΟΙΚΗΣΗΣ ΙΓ' ΕΚΠΑΙΔΕΥΤΙΚΗ ΣΕΙΡΑ ΤΜΗΜΑ ΤΟΠΙΚΗΣ ΑΥΤΟΔΙΟΙΚΗΣΗΣ ΚΑΙ ΠΕΡΙΦΕΡΕΙΑΚΗΣ ΑΝΑΠΤΥΞΗΣ ΤΕΛΙΚΗ ΕΡΓΑΣΙΑ: ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ: ΠΡΟΣΕΓΓΙΣΗ ΜΕΣΩ ΔΕΙΚΤΩΝ Επιβλέπων: Αθ.Δελαπάσχος
Χρηματοοικονομική Ανάπτυξη, Θεσμοί και
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΝΟΜΙΚΩΝ, ΟΙΚΟΝΟΜΙΚΩΝ ΚΑΙ ΠΟΛΙΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Τομέας Ανάπτυξης και Προγραμματισμού Χρηματοοικονομική Ανάπτυξη, Θεσμοί και Οικονομική
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΝΟΜΙΚΩΝ, ΟΙΚΟΝΟΜΙΚΩΝ ΚΑΙ ΠΟΛΙΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΟΙΚΟΝΟΜΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Τομέας Ανάπτυξης και Προγραμματισμού Χρηματοοικονομική Ανάπτυξη, Θεσμοί και Οικονομική
ΑΝΑΠΤΥΞΗ ΠΡΟΓΡΑΜΜΑΤΩΝ ΕΚΠΑΙΔΕΥΣΗΣ ΜΕ ΣΤΟΧΟ ΤΗΝ ΠΕΡΙΒΑΛΛΟΝΤΙΚΗ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗ ΑΤΟΜΩΝ ΜΕ ΕΙΔΙΚΕΣ ΑΝΑΓΚΕΣ ΚΑΙ ΤΗΝ ΚΟΙΝΩΝΙΚΗ ΤΟΥΣ ΕΝΣΩΜΑΤΩΣΗ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΠΑΙΔΑΓΩΓΙΚΟ ΤΜΗΜΑ ΕΙΔΙΚΗΣ ΑΓΩΓΗΣ ΧΡΙΣΤΙΝΑ Σ. ΛΑΠΠΑ ΑΝΑΠΤΥΞΗ ΠΡΟΓΡΑΜΜΑΤΩΝ ΕΚΠΑΙΔΕΥΣΗΣ ΜΕ ΣΤΟΧΟ ΤΗΝ ΠΕΡΙΒΑΛΛΟΝΤΙΚΗ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗ ΑΤΟΜΩΝ ΜΕ ΕΙΔΙΚΕΣ ΑΝΑΓΚΕΣ ΚΑΙ ΤΗΝ ΚΟΙΝΩΝΙΚΗ ΤΟΥΣ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΠΑΙΔΑΓΩΓΙΚΟ ΤΜΗΜΑ ΕΙΔΙΚΗΣ ΑΓΩΓΗΣ ΧΡΙΣΤΙΝΑ Σ. ΛΑΠΠΑ ΑΝΑΠΤΥΞΗ ΠΡΟΓΡΑΜΜΑΤΩΝ ΕΚΠΑΙΔΕΥΣΗΣ ΜΕ ΣΤΟΧΟ ΤΗΝ ΠΕΡΙΒΑΛΛΟΝΤΙΚΗ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗ ΑΤΟΜΩΝ ΜΕ ΕΙΔΙΚΕΣ ΑΝΑΓΚΕΣ ΚΑΙ ΤΗΝ ΚΟΙΝΩΝΙΚΗ ΤΟΥΣ
Η πρόληψη των κατακλίσεων σε βαριά πάσχοντες και η χρήση ειδικών στρωμάτων για την πρόληψη και αντιμετώπιση των κατακλίσεων
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η πρόληψη των κατακλίσεων σε βαριά πάσχοντες και η χρήση ειδικών στρωμάτων για την πρόληψη και αντιμετώπιση των κατακλίσεων Ονοματεπώνυμο
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η πρόληψη των κατακλίσεων σε βαριά πάσχοντες και η χρήση ειδικών στρωμάτων για την πρόληψη και αντιμετώπιση των κατακλίσεων Ονοματεπώνυμο
ΠΕΡΙΛΗΨΗ. Εισαγωγή. Σκοπός
 ΠΕΡΙΛΗΨΗ Εισαγωγή Η παιδική παχυσαρκία έχει φτάσει σε επίπεδα επιδημίας στις μέρες μας. Μαστίζει παιδιά από μικρές ηλικίες μέχρι και σε εφήβους. Συντείνουν αρκετοί παράγοντες που ένα παιδί γίνεται παχύσαρκο
ΠΕΡΙΛΗΨΗ Εισαγωγή Η παιδική παχυσαρκία έχει φτάσει σε επίπεδα επιδημίας στις μέρες μας. Μαστίζει παιδιά από μικρές ηλικίες μέχρι και σε εφήβους. Συντείνουν αρκετοί παράγοντες που ένα παιδί γίνεται παχύσαρκο
Σχολή Μηχανικής και Τεχνολογίας. Πτυχιακή διατριβή
 Σχολή Μηχανικής και Τεχνολογίας Πτυχιακή διατριβή ΠΕΙΡΑΜΑΤΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΑΝΤΙΚΑΤΑΣΤΑΣΗΣ ΜΕΡΟΥΣ ΤΟΥ ΚΑΥΣΙΜΟΥ ΠΟΥ ΚΑΤΑΝΑΛΩΝΕΙ ΒΕΝΖΙΝΟΚΙΝΗΤΗΡΑΣ ΜΕ ΥΔΡΟΓΟΝΟ ΤΟ ΟΠΟΙΟ ΘΑ ΠΑΡΑΓΕΤΑΙ ΜΕ ΑΝΑΚΤΗΣΗ ΕΝΕΡΓΕΙΑΣ ΚΑΤΑ
Σχολή Μηχανικής και Τεχνολογίας Πτυχιακή διατριβή ΠΕΙΡΑΜΑΤΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΑΝΤΙΚΑΤΑΣΤΑΣΗΣ ΜΕΡΟΥΣ ΤΟΥ ΚΑΥΣΙΜΟΥ ΠΟΥ ΚΑΤΑΝΑΛΩΝΕΙ ΒΕΝΖΙΝΟΚΙΝΗΤΗΡΑΣ ΜΕ ΥΔΡΟΓΟΝΟ ΤΟ ΟΠΟΙΟ ΘΑ ΠΑΡΑΓΕΤΑΙ ΜΕ ΑΝΑΚΤΗΣΗ ΕΝΕΡΓΕΙΑΣ ΚΑΤΑ