ΚΕΦΑΛΑΙΟ 9. Ο πυρήνας. Ευάγγελος Κωλέττας
|
|
|
- Αγάθη Σπανού
- 6 χρόνια πριν
- Προβολές:
Transcript
1 ΚΕΦΑΛΑΙΟ 9 Ο πυρήνας Ευάγγελος Κωλέττας Αναπληρωτής Καθηγητής Μοριακής Κυτταρικής Βιολογίας Εργαστήριο Γενικής Βιολογίας, Τμήμα Ιατρικής Σχολή Επιστημών Υγείας, Πανεπιστήμιο Ιωαννίνων και Συμβεβλημένο Μέλος ΔΕΠ - Ερευνητής Τμήμα Βιοϊατρικών Ερευνών Ινστιτούτο Μοριακής Βιολογίας & Βιοτεχνολογίας Ίδρυμα Τεχνολογίας & Έρευνας (ΒΕ-ΙΜΒΒ/ΙΤΕ), Ιωάννινα Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
2 Ο πυρηνικός φάκελος και η κυκλοφορία μορίων ανάμεσα στον πυρήνα και το κυτταρόπλασμα Η επιλεκτική διακίνηση πρωτεϊνών και RNA διαμέσου των πυρηνικών πόρων αφενός καθορίζει την εσωτερική σύσταση του πυρήνα και αφετέρου έχει καθοριστικό ρόλο στη ρύθμιση της γονιδιακής έκφρασης των ευκαρυωτών. Η δομή του πυρηνικού φακέλου Ο πυρήνας περιβάλλεται από ένα σύστημα δύο ομόκεντρων πυρηνικών μεμβρανών που σχηματίζουν τον πυρηνικό φάκελο, ο οποίος φέρει τα σύμπλοκα των πυρηνικών πόρων (NPC): Εξωτερική πυρηνική μεμβράνη - αποτελεί συνέχεια του ενδοπλασματικού δικτύου (ΕΔ) και φέρει ριβοσώματα, έτσι ο περιπυρηνικός χώρος ανάμεσα στην εσωτερική και την εξωτερική πυρηνική μεμβράνη να συνδέεται άμεσα με τον αυλό του ΕΔ. Εσωτερική πυρηνική μεμβράνη φέρει πρωτεΐνες που προσδένονται στην πυρηνικά λάμινα. Πυρηνική λάμινα - ένα ινώδες δίκτυο πρωτεϊνών που παρέχει στον πυρήνα δομική στήριξη και επενδύει εσωτερικά τον πυρήνα, αλληλοεπιδρώντας με την πυρηνοπλασματική επιφάνεια της εσωτερικής πυρηνικής μεμβράνης. 2
 Φωτογραφία ΗΜ που απεικονίζει: (Α) έναν")

 Σχηματική αναπαράσταση του")
3 ΕΙΚΟΝΑ 9.1: Ο ΠΥΡΗΝΙΚΌΣ ΦΆΚΕΛΟΣ (40-50 nm) Φωτογραφία ΗΜ που απεικονίζει: (Α) έναν πυρήνα. Η εσωτερική και η εξωτερική πυρηνική μεμβράνη ενώνονται στα σύμπλοκα των πυρηνικών πόρων (βέλη), και (Β) τη συνέχεια της εξωτερικής πυρηνικής μεμβράνης με το ενδοπλασματικό δίκτυο (Γ) Σχηματική αναπαράσταση του πυρηνικού φακέλου. Η εσωτερική πυρηνική μεμβράνη επενδύεται από την πυρηνική λάμινα, η οποία χρησιμεύει ως θέση προσάρτησης της χρωματίνης. 3


4 Οι πυρηνικές μεμβράνες δρουν ως φραγμός που διαχωρίζει το περιεχόμενο του πυρήνα από το κυτταρόπλασμα. Η εσωτερική και εξωτερική πυρηνική μεμβράνη ενώνονται στα σύμπλοκα των πυρηνικών πόρων, που είναι οι μοναδικοί δίαυλοι διαμέσου των οποίων μπορούν να διαπεράσουν τον πυρηνικό φάκελο μικρά πολικά μόρια και μακρομόρια, δηλαδή ευθύνονται για την επιλεκτική μεταφορά πρωτεϊνών και RNA ανάμεσα στον πυρήνα και το κυτταρόπλασμα. Πυρηνικοί Πόροι ΕΙΚΟΝΑ 9.2: Φωτογραφία ΗΜ που δείχνει πυρηνικούς πόρους. Σε αυτό το παρασκεύασμα πυρηνικού φακέλου που έχει υποστεί τεμαχισμό υπό ψύξη διακρίνονται πολλοί πυρηνικοί πόροι (βέλη). 4


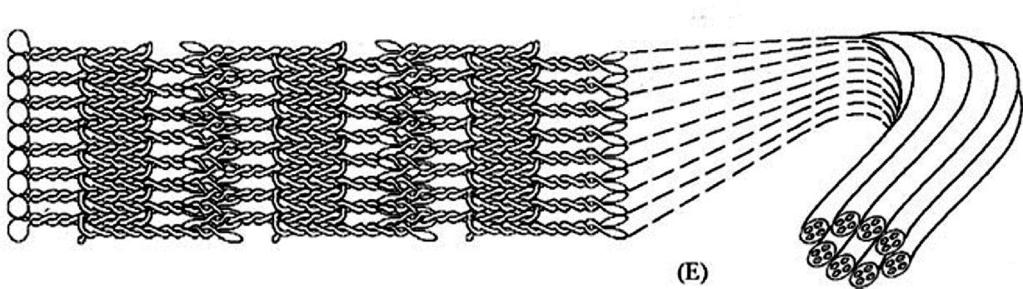
5 Η πυρηνική λάμινα, ένα ινώδες δίκτυο πρωτεϊνών, που παρέχει στον πυρήνα δομική στήριξη, επενδύει την πυρηνοπλασματική επιφάνεια της εσωτερικής πυρηνικής μεμβράνης και αποτελείται από ινώδεις πρωτεΐνες ΜΒ kda, τις λαμίνες, και από πρωτεΐνες που συνδέονται με αυτές. Οι λαμίνες είναι μια κατηγορία πρωτεϊνών των ενδιάμεσων ινιδίων πάχους nm. Οι άλλες κατηγορίες πρωτεϊνών των ενδιάμεσων ινιδίων συναντώνται στον κυτταροσκελετό. Τα κύτταρα των θηλαστικών διαθέτουν τρία γονίδια για τις λαμίνες, τα Α/C, Β1 και B2, που κωδικοποιούν τουλάχιστον επτά διακριτές πρωτεΐνες. ΕΙΚΟΝΑ 9.3: Φωτογραφία ΗΜ που δείχνει την πυρηνική λάμινα. Η λάμινα είναι ένα δίκτυο ινιδίων που βρίσκεται κάτω από την εσωτερική πυρηνική μεμβράνη και εκτείνεται στο εσωτερικό του πυρήνα. Ο κυτταροσκελετός αποτελείται από ένα σύστημα ινιδίων: τα λεπτά ινίδια ή ινίδια ακτίνης (actin filaments) πάχους 7-10 nm, τα παχύτερα ινίδια, τους μικροσωληνίσκους (microtubules) (25 nm) και τα ενδιάμεσα ινίδια (intermediate filaments), με ενδιάμεσο πάχος nm. 5
6 6
 και προκύπτουν με εναλλακτικό μάτισμα.")
 και B2 (LMNB2), ενώ η λαμίνη B3 εκφράζεται στους όρχεις και κωδικοποιείται από το γονίδιο LMNB2 με εναλλακτικό μάτισμα.")
7 Πυρηνικές λαμίνες, ινώδεις πρωτεΐνες ΜΒ kda, που σχηματίζουν ενδιάμεσα ινίδια τύπου V, πάχους nm. Δύο τύποι πυρηνικών λαμινών με όμοια δομή που κωδικοποιούνται από 3 γονίδια: Λαμίνες τύπου Α: A, C, C2 και ΑΔ10 που κωδικοποιούνται από το γονίδιο A/C (LMNA) και προκύπτουν με εναλλακτικό μάτισμα. Οι λαμίνες A και C είναι οι κύριοι τύποι, ενώ η C2 εκφράζεται στους όρχεις και η ΑΔ10 στα σωματικά κύτταρα με άγνωστο ρόλο. Λαμίνες τύπου Β: B1, B2 και B3. Οι Β1 και Β2 κωδικοποιούνται από τα γονίδια Β1 (LMNB1) και B2 (LMNB2), ενώ η λαμίνη B3 εκφράζεται στους όρχεις και κωδικοποιείται από το γονίδιο LMNB2 με εναλλακτικό μάτισμα. Οι λαμίνες φέρουν μια μικρή Ν-τελική περιοχή καταλοίπων, την κεφαλή, μια κεντρική α-ελικοειδή επικράτεια που φέρει 4 περιοχές 1Α, 1Β, 2Α και 2Β σπειρωμένου σπειράματος που διαχωρίζονται από τις συνδετικές περιοχές L1, L12 & L2, και μια μεγάλη C-τελική περιοχή (ουρά) που φέρει ένα NLS, μια Ig σφαιρική επικράτεια και ένα διατηρημένο πλαίσιο CAAX (όπου C, cys) που υπόκειται φαρνεσυλίωση. 7
8 Οι λαμίνες, όπως και άλλες πρωτεΐνες των ενδιάμεσων ινιδίων, συνδέονται μεταξύ τους σχηματίζοντας δομές με υψηλότερο βαθμό οργάνωσης. Το πρώτο στάδιο σύνδεσης των λαμινών είναι η αλληλεπίδραση δύο λαμινών που οδηγεί στον σχηματισμό ενός διμερούς. Στο διμερές λαμινών, οι περιοχές α-έλικας των 2 πολυπεπτιδικών αλυσίδων τυλίγονται η μία γύρω από την άλλη σχηματίζοντας μια δομή που ονομάζεται σπειρωμένο σπείραμα (coiled coil). Στη συνέχεια, τα διμερή λαμίνης συνδέονται μεταξύ τους σχηματίζοντας την πυρηνική λάμινα. ΕΙΚΟΝΑ 9.4: Μοντέλο συναρμολόγησης των λαμινών. Τα πολυπεπτίδια λαμίνης σχηματίζουν διμερή στα οποία οι κεντρικές περιοχές α-έλικας δύο πολυπεπτιδικών αλυσίδων τυλίγονται η μία γύρω από την άλλη. Στην περαιτέρω συναρμολόγηση μπορεί να συνεισφέρουν η κατά σειρά σύνδεση των διμερών, με την οποία σχηματίζονται γραμμικά πολυμερή, και η πλευρική σύνδεση των πολυμερών, με την οποία σχηματίζονται δομές υψηλότερης οργάνωσης. 8
9 Οι λαμίνες, όπως και άλλες πρωτεΐνες των ενδιάμεσων ινιδίων, συνδέονται μεταξύ τους σχηματίζοντας δομές με υψηλότερο βαθμό οργάνωσης. Το πρώτο στάδιο σύνδεσης των λαμινών είναι η αλληλεπίδραση δύο λαμινών που οδηγεί στον σχηματισμό ενός διμερούς. Στο διμερές λαμινών, οι περιοχές α-έλικας των 2 πολυπεπτιδικών αλυσίδων τυλίγονται η μία γύρω από την άλλη σχηματίζοντας μια δομή που ονομάζεται σπειρωμένο σπείραμα (coiled coil). X Στη συνέχεια, τα διμερή λαμίνης συνδέονται μεταξύ τους σχηματίζοντας την πυρηνική λάμινα. ΕΙΚΟΝΑ 9.4: Μοντέλο συναρμολόγησης των λαμινών. Τα πολυπεπτίδια λαμίνης σχηματίζουν διμερή στα οποία οι κεντρικές περιοχές α-έλικας δύο πολυπεπτιδικών αλυσίδων τυλίγονται η μία γύρω από την άλλη. Στην περαιτέρω συναρμολόγηση μπορεί να συνεισφέρουν η κατά σειρά σύνδεση των διμερών, με την οποία σχηματίζονται γραμμικά πολυμερή, και η πλευρική σύνδεση των πολυμερών, με την οποία σχηματίζονται δομές υψηλότερης οργάνωσης. 9
 αλληλοεπιδρούν και σχηματίζουν διμερή στα οποία οι κεντρικές περιοχές α-έλικας 2 πολυπεπτιδικών αλυσίδων διευθετούνται παράλληλα και τυλίσσονται η μία γύρω από την άλλη")
. Το τετραμερές είναι η ευδιάλυτη υπομονάδα των ενδιάμεσων ινιδίων.")
10 Οι λαμίνες, όπως και άλλες πρωτεΐνες των ενδιάμεσων ινιδίων, συνδέονται μεταξύ τους σχηματίζοντας δομές με υψηλότερο βαθμό οργάνωσης. ΕΙΚΟΝΑ 9.4: Μοντέλο συναρμολόγησης των λαμινών. Τα πολυπεπτίδια λαμίνης (Α) αλληλοεπιδρούν και σχηματίζουν διμερή στα οποία οι κεντρικές περιοχές α-έλικας 2 πολυπεπτιδικών αλυσίδων διευθετούνται παράλληλα και τυλίσσονται η μία γύρω από την άλλη σχηματίζοντας μια δομή που ονομάζεται σπειρωμένο σπείραμα (coiled-coil) (Β). Στη συνέχεια 2 διμερή ολισθαίνουν και αλληλοεπιδρούν σχηματίζοντας ένα αντιπαράλληλο τετραμερές 4 πολυπεπτιδικών αλυσίδων λαμίνης (C). Το τετραμερές είναι η ευδιάλυτη υπομονάδα των ενδιάμεσων ινιδίων. Σε κάθε τετραμερές, τα διμερή αντισταθμίζονται μεταξύ τους έτσι ώστε να επιτρέπεται η αλληλοεπίδραση τους με το επόμενο τετραμερές (Πρωτοινίδιο) (D). Στο τελικό ινίδιο λάμίνης 10 nm, τα τετραμερή (πρωτο-ινίδια) πακετάρονται σχηματίζοντας μια ελικοειδή δομή υψηλής οργάνωσης, που έχει 16 διμερή (32 σπειρωμένα σπειράματα) (E). Μισά απ αυτά τα διμερή δείχνουν προς μια κατεύθυνση, ενώ τα άλλα μισά 10 προς την αντίθετη κατεύθυνση
11 11
 τψν λαμινών A/C (κόκκινο) και λαμίνης B1 (πράσινο) στους πυρήνες της ανθρώπινης κυτταρικής σειράς οστεοσαρκώματος U2OS και")
12 Πυρηνικές Λαμίνες (Nuclear lamins): Εντοπισμός λαμινών στην περιφέρεια του πυρήνα και στο νουκλεόπλασμα. Ανοσοφθορισμός (Immunofluorescence staining) τψν λαμινών A/C (κόκκινο) και λαμίνης B1 (πράσινο) στους πυρήνες της ανθρώπινης κυτταρικής σειράς οστεοσαρκώματος U2OS και εμβρυϊκών ινοβλαστών ποντικού, MEF (Mouse Embryonic Fibroblasts), αντίστοιχα. 12
 J Struct Biol 122:42-66].")
13 Πυρηνική Λάμινα και Λαμίνες Οι πυρηνικές λαμίνες σχηματίζουν ένα δίκτυο ινιδίων εντός της πυρηνικής λάμινα. (Α) Παρασκεύασμα πυρηνικού φακέλου από ωοκύτταρα Xenopus έχει υποστεί τεμαχισμό υπό ψύξη. Απεικονίζεται το δίκτυο της πυρηνικής λάμινας ανάμεσα στο οποίο διακρίνονται πολλοί πυρηνικοί πόροι [Stuurman et al (1988) J Struct Biol 122:42-66]. (B) Δομική απεικονιστική μικροσκοπία (Structured illumination microscopy; SIM) σε συνάρτηση με ανοσοφθορισμό αποκαλύπτει ένα ακανόνιστο δίκτυο της πυρηνικής λαμίνης Β (πράσινο). Το παρασκεύασμα έχει επίσης επωαστεί με αντισώματα για τους πυρηνικούς πόρους (κόκκινο) και με DAPI για το DNA/χρωματίνη (μπλέ) [Schermelleh L et al (2008) Science 320: ]. (C) Συνεστιακή μικροσκοπία με λέηζερ σε συνάρτηση με ανοσοφθορισμό (Confocal laser scanning immunofluorescence microscopy) για τον εντοπισμό των λαμινών A/C (πράσινο) και της λαμίνης B1 (κόκκινο) σε κύτταρα HeLa. Οι λαμίνες A.C και Β1 σε έναν πυρήνα κυττάρων HeLa όπως απεικονείζονται σε μια τομή (αριστερά) και στην επιφάνεια του πυρήνα (δεξιά). Οι περιοχές που απεικονίζονται σε λευκά πλαίσια μεγεθύνθηκαν κατά 5 φορές στο κάτω μέρος της εικόνας. Αυτές οι εικόνες καταδεικνύουν ότι οι λαμίνες βασικά σχηματίζουν διαφορετικά δίκτυα με ορισμένες αλληλοεπικαλυπτόμενες περιοχές [Shimi T et al (2008) Genes Dev 22: ]. Dechat T et al (2010) Nuclear lamins. Cold Spring Harbor Perspect Biol 2(11):a


14 Η σύνδεση των λαμινών με την ΕΣΠΜ διευκολύνεται από τη μετα-μεταφραστική προσθήκη λιπιδίων - πρενυλίωση των καρβοξυτελικών καταλοίπων κυστεΐνης. Οι λαμίνες προσδένονται σε πρωτεΐνες της ΕΣΠΜ, όπως η εμερίνη και ο LBR, που διαμεσολαβούν στην πρόσδεση τους στον πυρηνικό φάκελο. Η πυρηνική λάμινα προσδένεται στη χρωματίνη διαμέσου των ιστονών Η2Α & Η2Β, και άμεσα στο DNA. Οι λαμίνες σχηματίζουν ένα χαλαρό δίχτυ που εκτείνεται σε ολόκληρο το εσωτερικό του πυρήνα. Στις λαμίνες προσδένονται πολλές πυρηνικές πρωτεΐνες που συμμετέχουν στη σύνθεση του DNA, στη μεταγραφή ή στην τροποποίηση της χρωματίνης, παρόλο που η σημασία αυτών των αλληλεπιδράσεων μόλις έχει αρχίσει να κατανοείται. ΕΙΚΟΝΑ πυρηνική Λαμίνες 9.5: λάμινα. Η Η εσωτερική πυρηνική μεμβράνη περιέχει αρκετές διαμεμβρανικές πρωτεΐνες, όπως είναι η εμερίνη και ο υποδοχέας της λαμίνης Β (LBR, Lamin B Receptor), που αλληλεπιδρούν με τις λαμίνες του πυρήνα. Οι λαμίνες και οι πρωτεΐνες που συνδέονται με αυτές αλληλεπιδρούν επίσης με τη χρωματίνη. 14
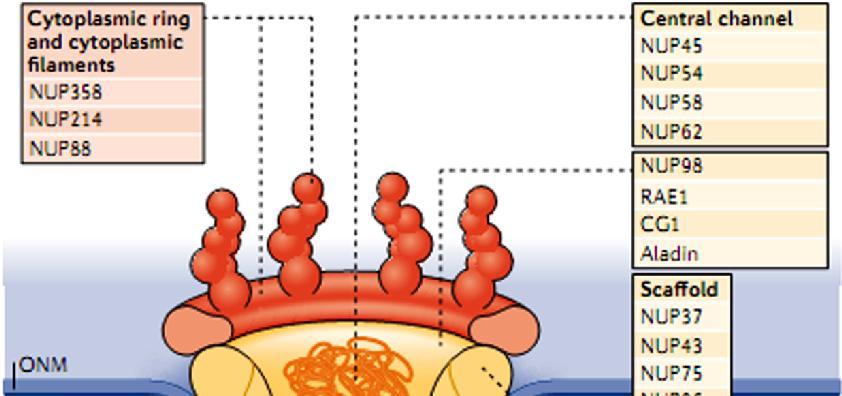
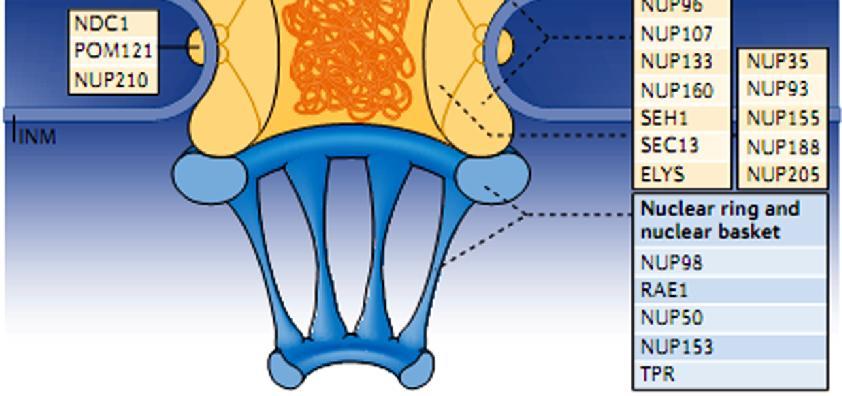
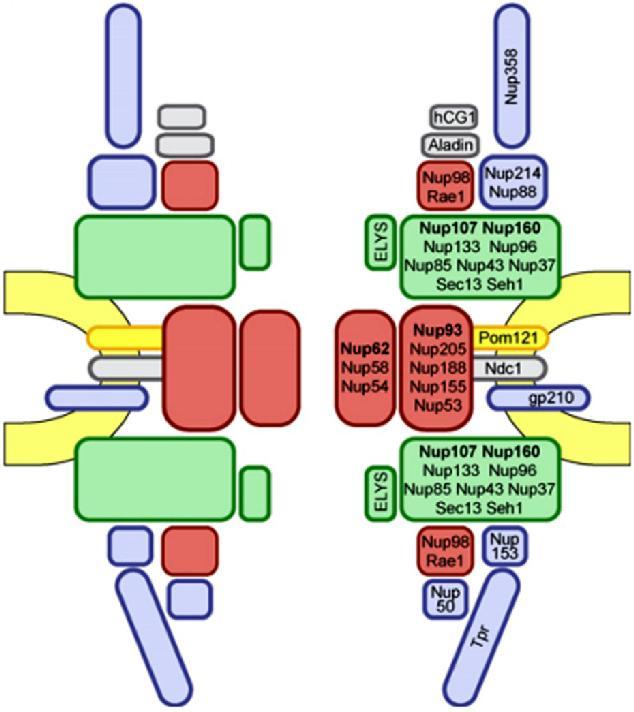
. Το NPC αποτελείται από ~30 διαφορετικές πρωτεΐνες, τις νουκλεοπορίνες.")
15 Τα σύμπλοκα των πυρηνικών πόρων (NPC) είναι οι μόνοι δίαυλοι που επιτρέπουν τη μεταφορά μικρών πολικών μορίων, ιόντων και μακρομορίων (πρωτεϊνών & RNA) ανάμεσα στον πυρήνα και το κυτταρόπλασμα. Είναι ευμεγέθεις δομές με διάμετρο ~120 nm και ΜΒ~125x103 kda (30 φορές > από μέγεθος ριβοσώματος). Το NPC αποτελείται από ~30 διαφορετικές πρωτεΐνες, τις νουκλεοπορίνες. Μικρά μόρια και πρωτεΐνες MB<20-40 kd Πρωτεΐνες MB>40 kd και μόρια RNA ΕΙΚΟΝΑ 9.6: Κυκλοφορία μορίων διαμέσου των συμπλόκων των πυρηνικών πόρων (NPC). Τα μικρά μόρια μπορούν να διέλθουν ταχύτατα με παθητική διάχυση, από ανοικτούς διαύλους του NPC. Αντίθετα, τα μακρομόρια (πρωτεΐνες & RNA) μεταφέρονται με έναν επιλεκτικό μηχανισμό που απαιτεί κατανάλωση ενέργειας. 15
16 Τα σύμπλοκα των πυρηνικών πόρων (nuclear pore complexes; NPC) είναι οι μόνοι δίαυλοι που επιτρέπουν τη μεταφορά μικρών πολικών μορίων, ιόντων και μακρομορίων (πρωτεϊνών & RNA) ανάμεσα στον πυρήνα και το κυτταρόπλασμα. Είναι ευμεγέθεις δομές με διάμετρο ~120 nm και ΜΒ~125x103 kda (μέγεθος 30 φορές > από μέγεθος ριβοσώματος). Το NPC αποτελείται από ~30 διαφορετικές πρωτεΐνες, τις νουκλεοπορίνες. Τα μόρια RNA που συντίθενται στον πυρήνα εξάγονται στο κυτταρόπλασμα, προκειμένου να λάβουν μέρος στην πρωτεϊνοσύνθεση. Αντίθετα, οι πρωτεΐνες που απαιτούνται για τη λειτουργία του πυρήνα (π.χ. μεταγραφικοί παράγοντες) θα πρέπει να μεταφερθούν από το κυτταρόπλασμα, όπου γίνεται η σύνθεση τους, στον πυρήνα. Επιπρόσθετα, πολλές πρωτεΐνες μετακινούνται συνεχώς δηλαδή παλινδρομούν (shuttle) ανάμεσα στον πυρήνα και το κυτταρόπλασμα. Η μεταφορά μορίων μέσω του NPC διεξάγεται με 2 διαφορετικούς μηχανισμούς, ανάλογα με το μέγεθος των μορίων: 1. Τα μικρά μόρια και πρωτεΐνες MB<20-40 kda διέρχονται ελεύθερα από τον πόρο προς οποιαδήποτε από τις 2 κατευθύνσεις: από κυτταρόπλασμα προς πυρήνα ή αντίθετα, με παθητική διάχυση από το υδατικό μέσο ανοικτών διαύλων του NPC, που έχουν διάμετρο ~9 nm. 2. Όμως, οι περισσότερες πρωτεΐνες και τα περισσότερα μόρια RNA δεν μπορούν να περάσουν από αυτούς τους ανοικτούς διαύλους. Τα μεγαλύτερα αυτά μακρομόρια περνούν από τον κεντρικό δίαυλο του NPC, που έχει διάμετρο ~10-40 nm, με ενεργή διεργασία κατά την οποία κατάλληλες πρωτεΐνες και RNA αναγνωρίζονται και μεταφέρονται επιλεκτικά προς μία συγκεκριμένη κατεύθυνση. 16


17 Η απεικόνιση των συμπλοκών των πυρηνικών πόρων με ΗΜ αποκαλύπτει μια δομή με οκταπλή συμμετρία, οργανωμένη γύρω από έναν μεγάλο κεντρικό δίαυλο. Ο κεντρικός αυτός δίαυλος αποτελεί την οδό διαμέσου της οποίας διασχίζουν τον πυρηνικό φάκελο οι πρωτεΐνες και τα RNA. Κεντρικός δίαυλος Δομές με οκταπλή συμμετρία ΕΙΚΟΝΑ 9.7: Φωτογραφία ΗΜ που δείχνει σύμπλοκα πυρηνικών πόρων. Στην κάτοψη αυτή διακρίνονται μεμονωμένα σύμπλοκα πυρηνικών πόρων, που αποτελούνται από 8 δομικές υπομονάδες που περιβάλλουν έναν κεντρικό δίαυλο. 17


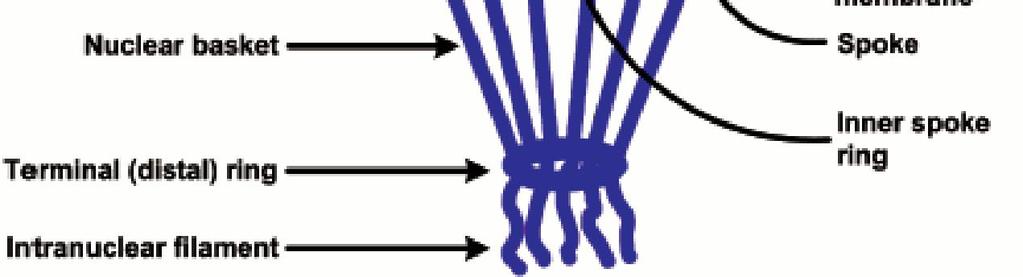
18 Το NPC αποτελείται από 8 ακτίνες διατεταγμένες γύρω από έναν κεντρικό δίαυλο. Οι 8 ακτίνες συνδέονται με δακτυλίους στην πυρηνική και στην κυτταροπλασματική επιφάνεια, συγκροτώντας μια δομή που βρίσκεται αγκυροβολημένη στο εσωτερικό του πυρηνικού φακέλου σε θέσεις όπου συνενώνονται η εσωτερική και η εξωτερική πυρηνική μεμβράνη. Ινίδια πρωτεϊνών εκτείνονται τόσο από τον κυτταροπλασματικό όσο και από τον πυρηνικό δακτύλιο, σχηματίζοντας μια διακριτή καλαθοειδή δομή στην πλευρά του πυρήνα. ΕΙΚΟΝΑ 9.8: Μοντέλο του συμπλόκου του πυρηνικού πόρου. Το σύμπλοκο: αποτελείται από ένα συγκρότημα 8 ακτίνων προσδεδεμένων σε δακτυλίους στην κυτταροπλασματική και στην πυρηνική πλευρά του πυρηνικού φακέλου. Το συγκρότημα δακτυλίωνακτίνων περιβάλλει έναν κεντρικό δίαυλο. Από τον κυτταροπλασματικό δακτύλιο εκτείνονται κυτταροπλασματικά ινίδια, ενώ από τον πυρηνικό δακτύλιο εκτείνονται ινίδια που σχηματίζουν τον πυρηνικό κάλαθο. 18

19 19
20 20
21 ΜΕΤΑΦΟΡΑ ΠΡΩΤΕΙΝΩΝ ΑΠΟ ΚΑΙ ΠΡΟΣ ΤΟΝ ΠΥΡΗΝΑ 21
, που αναγνωρίζονται από υποδοχείς πυρηνικής μεταφοράς (nuclear transport receptors) και κατευθύνουν τη μεταφορά των πρωτεϊνών διαμέσου του NPC.")
22 Επιλεκτική μεταφορά πρωτεϊνών από και προς τον πυρήνα Η στόχευση πρωτεϊνών στον πυρήνα γίνεται διαμέσου ειδικών αλληλουχιών αμινοξέων που ονομάζονται σήματα πυρηνικού εντοπισμού (NLS; Nuclear Localisation Signals), που αναγνωρίζονται από υποδοχείς πυρηνικής μεταφοράς (nuclear transport receptors) και κατευθύνουν τη μεταφορά των πρωτεϊνών διαμέσου του NPC. Σήματα NLS έχουν ταυτοποιηθεί σε πρωτεΐνες και είναι συχνά πλούσια σε βασικά αμινοξέα (Lys & Arg). Δύο τύποι κλασικών NLS: 1. Μονοπαραγοντικά σήματα NLS: Διαδοχική (ενιαία) αλληλουχία αμινοξέων στα σήματα NLS: π.χ. η αλληλουχία του αντιγόνου-τ. 2. Διπαραγοντικά (bipartite) σήματα NLS: Tα αμινοξέα που σχηματίζουν το σήμα NLS βρίσκονται κοντά το ένα στο άλλο, αλλά δεν είναι διαδοχικά μεταξύ τους. π.χ. το σήμα NLS της νουκλεοπλασμίνης (συμμετέχει στη συγκρότηση της χρωματίνης) αποτελείται από 2 μέρη που χωρίζονται από ένα συνδετικό μεσοδιάστημα 10 αμινοξέων. Και οι δύο αυτές ξεχωριστές αλληλουχίες είναι απαραίτητες για την πυρηνική στόχευση. Τα 10 αμινοξέα που μεσολαβούν ανάμεσα σε αυτές τις αλληλουχίες μπορούν να αντικατασταθούν με μεταλλαξιγένεση χωρίς να επηρεαστεί ο πυρηνικός εντοπισμός. Τα διπαραγοντικά κλασσικά σήματα NLS είναι συχνότερα, άλλα υπάρχουν και σήματα NLS που εμφανίζουν μεγάλη ποικιλία αμινοξικής αλληλουχίας και δομής. PKKKRKV ζεύγος Lys-Arg συνδετικό μεσοδιάστημα 10 αμινοξέων 4 κατάλοιπα Lys ΕΙΚΟΝΑ 9.9: Σήματα πυρηνικού εντοπισμού (NLS). Το NLS του αντιγόνου Τ αντιστοιχεί σε μια ενιαία αλληλουχία αμινοξέων. Αντίθετα, το NLS της νουκλεοπλασμίνης είναι διπαραγοντικό, καθώς αποτελείται από 2 αλληλουχίες, μία Lys-Arg και μία Lys-Lys-LysLys, μεταξύ των οποίων μεσολαβεί ένα μεσοδιάστημα 10 αμινοξέων. 22

23 ΠΕΙΡΑΜΑ-ΣΤΑΘΜΟΣ: Η ταυτοποίηση των σημάτων πυρηνικού εντοπισμού Το ευρύτερο πλαίσιο Η διατήρηση του πυρήνα ως διακριτού βιοχημικού διαμερίσματος απαιτεί έναν μηχανισμό διαχωρισμού μεταξύ πρωτεϊνών του πυρήνα και κυτταροπλάσματος. Μελέτες τη δεκαετία του 1970 έδειξαν ότι μικρά μόρια διαχέονται ταχύτατα διαμέσου του πυρηνικού φακέλου, αλλά οι περισσότερες πρωτεΐνες δεν μπορούν να διέλθουν με αυτόν τον τρόπο. Επομένως, φαινόταν πιθανό ότι οι πυρηνικές πρωτεΐνες αναγνωρίζονται με κάποιον ειδικό τρόπο από τις θέσεις σύνθεσης τους στο ριβοσώματα του κυτταροπλάσματος και μεταφέρονται επιλεκτικά στον πυρήνα. Προηγούμενα πειράματα του Günter Blobel απέδειξαν ότι η στόχευση πρωτεϊνών στο ΕΔ γίνεται μέσω σηματοδοτικών αλληλουχιών μικρού μήκους. Ο Alan Smith (1984) ταυτοποίησε μια μικρή αλληλουχία αμινοξέων που λειτουργεί ως σήμα πυρηνικού εντοπισμού (NLS) στο T αντιγόνο του DNA ιού SV40. Alan Smith (1984) 23
 ανήκει στην")

24 DNA ιός SV40 Ο DNA ιός Simian Virus 40 (SV40) ανήκει στην οικογένεια των ιών πολυώμα, και φέρει κυκλικό γονιδίωμα μεγέθους 5 kb 24
 εισήχθησαν σε κύτταρα με μικροένεση. Στη συνέχεια, προσδιορίστηκε ο κυτταρικός εντοπισμός των υβριδικών πρωτεϊνών με μικροσκοπία ανοσοφθορισμού.")
25 ΠΕΙΡΑΜΑ-ΣΤΑΘΜΟΣ: Η ταυτοποίηση των σημάτων πυρηνικού εντοπισμού Πλασμιδιακά DNA που κωδικοποιούν για χιμαιρικές πρωτεΐνες που φέρουν αλληλουχίες του SV40 Tag σε σύντηξη με την κινάση του πυροσταφυλικού (εντοπίζεται στο κυτοσόλιο) εισήχθησαν σε κύτταρα με μικροένεση. Στη συνέχεια, προσδιορίστηκε ο κυτταρικός εντοπισμός των υβριδικών πρωτεϊνών με μικροσκοπία ανοσοφθορισμού. (Α) Η υβριδική πρωτεΐνη περιέχει ένα ακέραιο NLS του ιού SV40 (αλληλουχία ). (Β) Το NLS έχει απενεργοποιηθεί με απαλοιφή των καταλοίπων των αμινοξέων 131 και 132. Μετάλλαξη του καταλοίπου Lys-128 προς Thr ή Asn εμπόδισε τη συσσώρευση του αντιγόνου Τ στον πυρήνα κυττάρων πιθήκου και τρωκτικών. PKKKRKV PKTKRKV ή PKNKRKV Το SV40 TAg αναγνωρίζεται από την ιμπορτίνη-α Ο αντίκτυπος: Διατυπώθηκε η άποψη ότι το NLS του αντιγόνου Τ του ιού SV40 αντιπροσωπεύει μια πρότυπη μορφή παρόμοιων αλληλουχιών-σημάτων που συναντώνται και σε άλλες πυρηνικές πρωτεΐνε, και υπαγορεύουν τη στόχευση πρωτεϊνών για εισαγωγή στον πυρήνα και επομένως έχουν βασικό ρόλο στον καθορισμό της βιοχημικής ταυτότητας του πυρήνα και στη διατήρηση της θεμελιώδους οργάνωσης του ευκαρυωτικού κυττάρου σε διαμέρισμα πυρήνα και διαμέρισμα κυτταροπλάσματος. Τα NLS είναι γνωστό ότι αναγνωρίζονται από κυτταροπλασματικούς υποδοχείς που μεταφέρουν τις πρωτεΐνες-υποστρώματά τους διαμέσου του NPC. Η ταυτοποίηση των NLS αποτέλεσε καθοριστικής σημασίας για την κατανόηση της εισαγωγής πρωτεϊνών στον πυρήνα 25
 Μόρια μικρότερα από 40 kda (και με διάμετρο 4-5 nm) περνούν διαμέσου του συμπλόκου του πυρηνικού πόρου (NPC) χωρίς να απαιτούν πρωτεΐνες-μεταφορείς, αλλά με απλή διάχυση.")
 δεσμεύονται σε υποδοχείς πυρηνικής μεταφοράς (transport proteins) που αναγνωρίζουν αλληλουχίες-σήματα πυρηνικού εντοπισμού (NLS) και")
26 Μηχανισμός πυρηνικής μεταφοράς. (a) Μόρια μικρότερα από 40 kda (και με διάμετρο 4-5 nm) περνούν διαμέσου του συμπλόκου του πυρηνικού πόρου (NPC) χωρίς να απαιτούν πρωτεΐνες-μεταφορείς, αλλά με απλή διάχυση. Η επιλεκτική διαπεροτότητα του NPC παρεμποδίζει την απλή διάχυση των μεγαλυτέρων μορίων. Μεγαλύτερα μόρια όπως οι πρωτεΐνες ΜΒ>40 kda και RNAs (cargo) δεσμεύονται σε υποδοχείς πυρηνικής μεταφοράς (transport proteins) που αναγνωρίζουν αλληλουχίες-σήματα πυρηνικού εντοπισμού (NLS) και αλληλουχίες-σήματα πυρηνικής εξαγωγής. Η διέλευση μακρομορίων από τον πυρηνικό πόρο ρυθμίζεται από την πρωτεΐνη Ran. Πρόκειται για έναν τύπο μικρών πρωτεϊνών πρόσδεσης GTP, η διαμόρφωση και η ενεργότητα της οποίας ρυθμίζεται από την πρόσδεση και την υδρόλυση GTP. Παραδείγματα μικρών πρωτεϊνών πρόσδεσης GTP είναι οι πρωτεΐνες Ras, αρκετοί από τους μεταφραστικούς παράγοντες που συμμετέχουν στην πρωτεϊνοσύνθεση, οι πρωτεΐνες Arf και Rab, και οι πρωτεΐνες Rac, Rho και Cdc42. 26


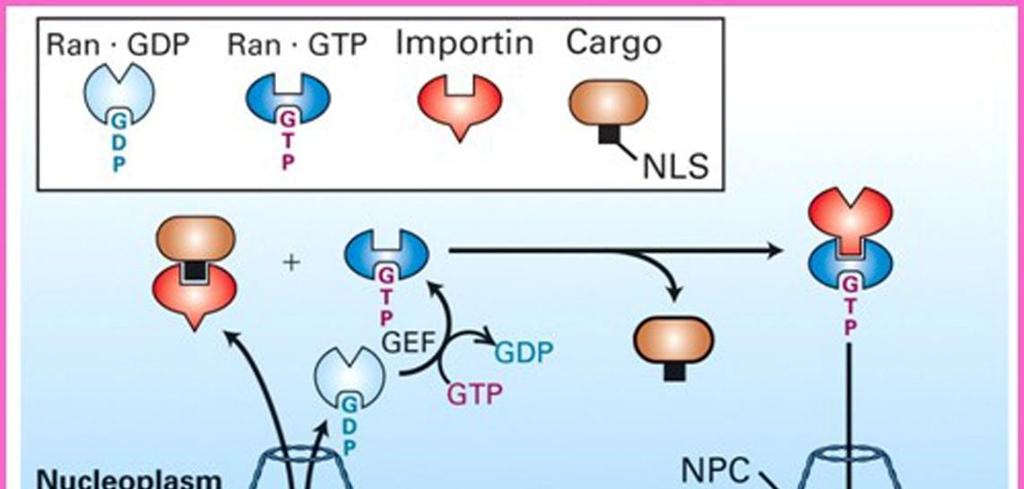
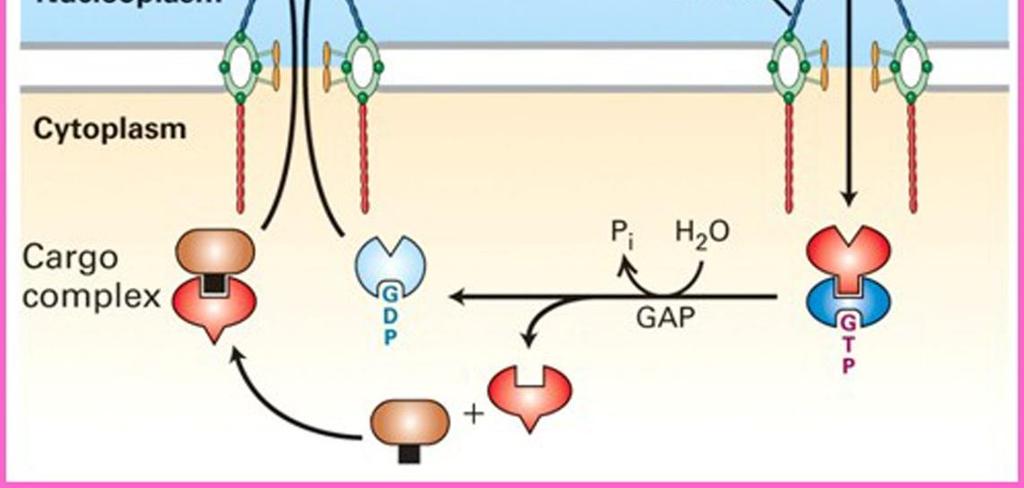
27 Τα NLS αναγνωρίζονται από υποδοχείς πυρηνικής μεταφοράς τις ιμπορτίνες (importins). Η διέλευση μακρομορίων από τον πυρηνικό πόρο ρυθμίζεται από την πρωτεΐνη Ran που δεσμεύει GTP, με αποτέλεσμα η διαμόρφωση και η ενεργότητα της να ρυθμίζεται από την πρόσδεση και την υδρόλυση GTP. Τα ένζυμα που ενεργοποιούν την υδρόλυση της GTP προς GDP εντοπίζονται στην κυτταροπλασματική πλευρά του πυρηνικού φακέλου, ενώ τα ένζυμα που ενεργοποιούν την ανταλλαγή GDP με GTP εντοπίζονται στην πυρηνική πλευρά. Επομένως, δεν υπάρχει ισόποση κατανομή των Ran/GTP από τις δύο πλευρές του πυρηνικού πόρου. Στο εσωτερικό του πυρήνα διατηρείται υψηλότερη συγκέντρωση μορίων Ran/GTP, που καθορίζει την κατεύθυνση της πυρηνικής μεταφοράς των πρωτεϊνών-φορτίων που αναγνωρίζουν οι ιμπορτίνες. ΕΙΚΟΝΑ των δύο Η άνιση κατανομή της Ran/GTP μεταξύ των Ran/GDP > Ran/GTP 2 πλευρών του πυρηνικού φακέλου διατηρείται χάρη στον εντοπισμό της πρωτεΐνης ενεργοποίησης της GTPάσης Ran (RanGAP, Ran GTPase-Αctivating Protein) στο κυτταρόπλασμα και του παράγοντα ανταλλαγής νουκλεοτιδίων γουανίνης Ran (RanGEF, Ran Guanine nucleotide Exchange Factor ή RCC1) στον πυρήνα. Στο κυτταρόπλασμα, η RanGAΡ (που προσδένεται στα κυτταροπλασματικά ινίδια NPC) ενεργοποιεί την υδρόλυση της GTP που είναι προσδεδεμένη στη Ran, οδηγώντας σε μετατροπή της Ran/GTP σε Ran/GDP. Στον πυρήνα, η Ran-GEF (RCC1) ενεργοποιεί την ανταλλαγή της GDP που Ran/GTP > Ran/GDP είναι προσδεδεμένη στη Ran με GTP, οδηγώντας σε μετατροπή του Ran/GDP σε Ran/GTP. Κατά συνέπεια, το εσωτερικό του 9.10: Κατανομή της Ran/GTP μεταξύ πυρήνα διατηρεί μια υψηλή συγκέντρωση Ran/GTP. πλευρών του πυρηνικού φακέλου. 27
28 Η πρωτεΐνη Ran ρυθμίζει τη διέλευση πρωτεϊνών από τον πυρηνικό πόρο ελέγχοντας την ενεργότητα των υποδοχέων πυρηνικής μεταφοράς. Η εισαγωγή πρωτεϊνών διαμέσου του συμπλόκου του πυρηνικού πόρου αρχίζει όταν μια συγκεκριμένη ιμπορτίνη προσδεθεί στο σήμα πυρηνικού εντοπισμού μιας πρωτεΐνης προς μεταφορά στο κυτταρόπλασμα. Κατόπιν, αυτό το σύμπλοκο ιμπορτίνης/φορτίου προσδένεται σε πρωτεΐνες των κυτταροπλασματικών ινιδίων του συμπλόκου του πυρηνικού πόρου και η μεταφορά προχωρεί με διαδοχική πρόσδεση σε ειδικές πρωτεΐνες του πυρηνικού πόρου που βρίσκονται πλησιέστερα στην πυρηνική πλευρά του συμπλόκου του πόρου. Ιδιαίτερα σημαντικές για τη διεργασία αυτή είναι ορισμένες πρωτεΐνες (νουκλεοπορίνες) που επενδύουν τον κεντρικό δίαυλο του πόρου, που φέρουν πολλαπλές επαναλήψεις Phe-Gly και ονομάζονται πρωτεΐνες FG. Ακολούθως, στην πυρηνική πλευρά του πόρου, το σύμπλοκο ιμπορτίνης/φορτίου διίσταται καθώς προσδένεται σε αυτό η RanGTP. Η πρόσδεση αυτή προκαλεί μια αλλαγή στη διαμόρφωση της ιμπορτίνης, που εκτοπίζει από το σύμπλοκο την πρωτεΐνη-φορτίο και την απελευθερώνει στο εσωτερικό του πυρήνα. Στη συνέχεια, το σύμπλοκο ιμπορτίνης-ran/gtp εξάγεται από τον πυρήνα διαμέσου του συμπλόκου του πυρηνικού πόρου και επιστρέφει στο κυτταρόπλασμα, όπου η GTP υδρολύεται σε GDP από την RanGAP. Με αυτόν τον τρόπο αποδεσμεύεται η ιμπορτίνη, που μπορεί πλέον να προσδεθεί σε μια νέα πρωτεΐνη-φορτίο στο κυτταρόπλασμα και να συμμετάσχει σε έναν νέο κύκλο πυρηνικής μεταφοράς. Η Ran/GDP που παράγεται στο κυτταρόπλασμα μεταφέρεται πίσω στον πυρήνα διαμέσου του δικού της μεταφορέα (που ονομάζεται NTF2), όπου και αναγεννάται η Ran/GTP. 28
: Μια οικογένεια πρωτεϊνών, οι ιμπορτίνες (καρυοφορίνες) είναι υπεύθυνες για τη")

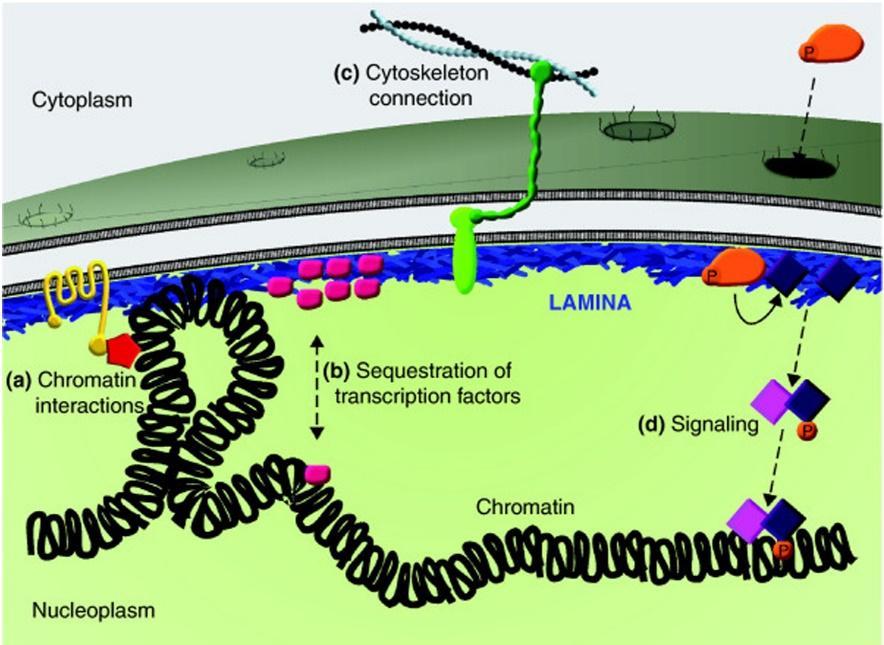
29 ΕΙΚ. 9.11: Μεταφορά πρωτεϊνών διαμέσου του συμπλόκου του πυρηνικού πόρου Μηχανισμός μεταφοράς προς τον πυρήνα (Nuclear import): Μια οικογένεια πρωτεϊνών, οι ιμπορτίνες (καρυοφορίνες) είναι υπεύθυνες για τη μεταφορά των πρωτεϊνών στον πυρήνα. Οι ιμπορτίνες δεσμεύουν στο κυτταρόπλασμα τις πρωτεΐνες που φέρουν ένα σήμα NLS, και μετακινούνται διαμέσου του κεντρικού διαύλου του NPC καθώς αλληλοεπιδρούν με τις νουκλεοπρωτεΐνες του NPC (FG-NUPs). Αφού εισέλθουν μέσα στον πυρήνα, δεσμεύονται σε μια πρωτεΐνη Ran GTP, που προκαλεί δομικές μεταβολές και την απελευθέρωση της μεταφερόμενης πρωτεΐνης. Το σύμπλοκο ιμπορτίνηςran GTP που προκύπτει εξάγεται πίσω στο κυτταρόπλασμα διαμέσου του NPC και η πρωτεΐνη ενεργοποίησης της GTPάσης (RanGAP) υδρολύει τη GTP, που φέρει η Ran, προς GDP, προκαλώντας απελευθέρωση της ιμπορτίνης στο κυτταρόπλασμα. Η Ran/GDP που παράγεται στο κυτταρόπλασμα μεταφέρεται πίσω στον πυρήνα διαμέσου του δικού της μεταφορέα NTF2, όπου και αναγεννάτε η Ran/GTP. 29
30 27
31 Μηχανισμός πυρηνικής μεταφοράς - Μεταφορά προς τον πυρήνα (Nuclear import) Η μεταφορά μιας πρωτεΐνης διαμέσου του NPC αρχίζει με την αναγνώριση της αλληλουχίας πυρηνικού εντοπισμού της (NLS) από έναν υποδοχέα πυρηνικής μεταφοράς (ιμπορτίνη). Το σύμπλοκο της πρωτεΐνης-φορτίου με την ιμπορτίνη προσδένεται σε ειδικές πρωτεΐνες του πυρηνικού πόρου στα κυτταροπλασματικά ινίδια, λόγω της ικανότητας των ιμπορτίνων να αλληλοεπιδρούν μ αυτές. Στη συνέχεια, μεταφέρεται διαμέσου του πυρηνικού πόρου (κεντρικού διαύλου), καθώς η ιμπορτίνη προσδένεται διαδοχικά σε πρωτεΐνες που βρίσκονται βαθύτερα στο εσωτερικό του, ιδιαίτερα νουκλεοπορίνες που επενδύουν τον κεντρικό δίαυλο του πόρου, που φέρουν πολλαπλές επαναλήψεις Phe-Gly και ονομάζονται πρωτεΐνες FG. Στην πυρηνική πλευρά του πόρου, το σύμπλοκο πρωτεΐνης-φορτίου/ιμπορτίνης διίσταται λόγω της πρόσδεσης Ran/GTP στην ιμπορτίνη. Η αλλαγή διαμόρφωσης της ιμπορτίνης εκτοπίζει την πρωτεΐνη-φορτίο και την απελευθερώνει στο εσωτερικό του πυρήνα. Το σύμπλοκο ιμπορτίνης-ran/gtp εξάγεται και πάλι από τον πυρήνα μέσω του πυρηνικού πόρου και η πρωτεΐνη ενεργοποίησης της GTPάσης (RanGAP) υδρολύει τη GTP, που φέρει η Ran, προς GDP, προκαλώντας απελευθέρωση της ιμπορτίνης στο κυτταρόπλασμα. Η Ran/GDP που παράγεται στο κυτταρόπλασμα μεταφέρεται πίσω στον πυρήνα μέσω του δικού της μεταφορέα (που ονομάζεται NTF2), όπου και αναγεννάται η Ran/GTP. 31
. ΠΙΝΑΚΑΣ 9.")

32 Η πυρηνική εισαγωγή ή εξαγωγή πρωτεϊνών από και προς τον πυρήνα απαιτεί ειδικές πρωτεΐνες υποδοχείς, τις ιμπορτίνες και τις εξπορτίνες, αντίστοιχα, που ανήκουν σε μια οικογένεια υποδοχέων πυρηνικής μεταφοράς, τις καρυοφερίνες (karyopherins). ΠΙΝΑΚΑΣ 9.1: Καρυοφερίνες με γνωστά υποστρώματα Αναγνωρίζουν σήματα NLS Ιμπορτίνη (α/β) Αναγνωρίζουν σήματα NES Η καρυοφερίνη Kapα / Kapβ1 ή ιμπορτίνη είναι ένα ετεροδιμερές που αποτελείται από 2 υπομονάδες, την ιμπορτίνη-α που αναγνωρίζει και δεσμεύεται σε ένα σήμα NLS μιας πρωτεΐνης, και από την ιμπορτίνη-β που αλληλεπιδρά με νουκλεοπορίνες και FGNUPs καθώς το σύμπλοκο μετακινείται διαμέσου του κεντρικού διαύλου του NPC. Η εξπορτίνη Crm1 αναγνωρίζει σήματα NES πλούσια σε leu. 32
33 Η εξαγωγή πρωτεϊνών από τον πυρήνα υπαγορεύεται από ειδικές αλληλουχίες αμινοξέων, που ονομάζονται σήματα πυρηνικής εξαγωγής (NES; Nuclear Export Signals). Τα NES, όπως και τα NLS, αναγνωρίζονται από υποδοχείς μέσα στον πυρήνα. Οι υποδοχείς των NES, οι εξπορτίνες (nuclear export proteins ή exportins), βρίσκονται μέσα στον πυρήνα και κατευθύνουν τη μεταφορά των πρωτεϊνών από τον πυρήνα προς το κυτταρόπλασμα μέσω του NPC. Πολλές από τις εξπορτίνες, όπως και οι ιμπορτίνες, ανήκουν σε μια οικογένεια υποδοχέων πυρηνικής μεταφοράς, τις καρυοφερίνες (karyopherins). Ένα σήμα πυρηνικής εξαγωγής είναι μια μικρή αλληλουχία υδρόφοβων αμινοξέων σε μια πρωτεΐνη η οποία θα πρέπει να μεταφερθεί από τον πυρήνα στο κυτταρόπλασμα διαμέσου του NPC, με έναν από τους μηχανισμούς της πυρηνικής μεταφοράς. Τα σήματα NES αναγνωρίζονται από υποδοχείς, τις εξπορτίνες. Τα πιο συχνά αλλά και εξελικτικά διατηρημένα σήματα NES αποτελούνται από την αλληλουχία αμινοξέων: LxxxLxxLxL όπου L είναι ένα υδρόφοβο κατάλοιπο, συνήθως λευκίνη (leu), και x οποιοδήποτε άλλο αμινοξύ. Η καρυοφερίνη CRM1 έχει ταυτοποιηθεί ως ο υποδοχές πυρηνικής εξαγωγής πρωτεϊνών που φέρουν σήματα NES πλούσια σε leu. Η πυρηνική εξαγωγή διαμεσολαβούμενη από την CRM1 μπορεί να παρεμποδιστεί αποτελεσματικά με τη χρήση του μυκητοκτόνου λεπτομυκίνης Β (LMB), καταδεικνύοντας την ύπαρξη αυτού του μηχανισμού πυρηνικής εξαγωγής. 33
, εξπορτινών και Ran/GTP.")


34 Μηχανισμός μεταφοράς από τον πυρήνα (Nuclear export): ΕΙΚΟΝΑ 9.12: Εξαγωγή πρωτεϊνών από τον πυρήνα. Στον πυρήνα σχηματίζονται σύμπλοκα μεταξύ πρωτεϊνών-φορτίων που φέρουν σήματα πυρηνικής εξαγωγής (NES), εξπορτινών και Ran/GTP. Μετά τη διέλευση από το σύμπλοκο του πυρηνικού πόρου, η RanGAP ενεργοποιεί την υδρόλυση της προσδεδεμένης GTP, οδηγώντας σε σχηματισμό Ran/GDP και απελευθέρωση της πρωτεΐνης-φορτίου και της εξπορτίνης στο κυτταρόπλασμα. H Ran GDP που προκύπτει αλληλoεπιδρά με τον πυρηνικό παράγοντα μεταφοράς NTF2 και μετατοπίζεται πίσω στον πυρήνα διαμέσου του NPC όπου ο παράγοντας ανταλλαγής νουκλεοτιδίων γουανίνης Ran GEF, RCC1, αποκαθιστά τα επίπεδα της Ran GTP. Οι εξπορτίνες ανακυκλώνονται και επιστρέφουν στον πυρήνα διαμέσου του NPC, ώστε να μπορούν να χρησιμοποιηθούν εκ νέου. 34
35 Μεταφορά από τον πυρήνα προς το κυτταρόπλασμα ή Πυρηνική εξαγωγή (Nuclear protein export). Πυρηνικές πρωτεΐνες εξαγωγής (Nuclear export proteins), οι εξπορτίνες δεσμεύονται στον πυρήνα στην πρωτεΐνη προς εξαγωγή καθώς επίσης και στην RAN/GTP και μετακινούνται διαμέσου του NPC προς το κυτταρόπλασμα. Ενώ η πρόσδεση της Ran/GTP στις ιμπορτίνες προκαλεί διάσπαση των συμπλοκών μεταξύ των ιμπορτινών και των πρωτεϊνών-φορτίων τους, η πρόσδεση της στις εξπορτίνες συμβάλλει στον σχηματισμό σταθερών συμπλοκών μεταξύ των εξπορτινών και των πρωτεϊνών-φορτίων. Επομένως, η πρόσδεση της Ran/GTP στις εξπορτίνες καθορίζει τη μετακίνηση των πρωτεϊνών που περιέχουν σήματα NES προς το κυτταρόπλασμα. Έτσι, στο εσωτερικό του πυρήνα σχηματίζονται σταθερά σύμπλοκα μεταξύ των εξπορτινών και των πρωτεϊνών-φορτίων τους, στα οποία συμβάλλει και η πρόσδεση της Ran/GTP. Μετά τη μεταφορά αυτών των συμπλοκών στην κυτταροπλασματική πλευρά του πυρηνικού φακέλου, η ενεργοποιούσα RAN GTPάση, RANGAP, προκαλεί την υδρόλυση της GTP από την RAN σε συνεργασία με δύο πρωτεΐνες που αλληλεπιδρούν με την RAN GTP (RAN GTPbinding proteins RANBP1 & RANBP2; δεν απεικονίζονται) στην κυτταροπλασματική πλευρά του με αποτέλεσμα αποδέσμευση της πρωτεΐνης-φορτίου, που απελευθερώνεται στο κυτταρόπλασμα. Η συγκέντρωση της RAN GTP στο κυτταρόπλασμα είναι χαμηλή. Το σύμπλοκο RAN GDP που προκύπτει αλληλεπιδρά με τον πυρηνικό παράγοντα μεταφοράς NTF2 και μετατοπίζεται πίσω στον πυρήνα μέσω του πυρηνικού πόρου όπου ο παράγοντας ανταλλαγής νουκλεοτιδίων γουανίνης RAN GEF (Guanine nucleotide ecchange factor), RCC1, αποκαθιστά τα επίπεδα της RAN GTP. Στη συνέχεια, οι εξπορτίνες ανακυκλώνονται και επιστρέφουν στον πυρήνα μέσω του πυρηνικού πόρου, ώστε να μπορούν να χρησιμοποιηθούν εκ νέου. 35
36 Ρύθμιση της εισαγωγής πρωτεϊνών στον πυρήνα Η μεταφορά πρωτεϊνών στον πυρήνα είναι ένα ακόμη επίπεδο στο οποίο μπορούν να ελεγχθούν οι ενεργότητες των πυρηνικών πρωτεϊνών. Οι μεταγραφικοί παράγοντες είναι λειτουργικοί μόνο όταν βρίσκονται μέσα στον πυρήνα και επομένως η ρύθμιση της εισαγωγής και της εξαγωγής τους προς και από τον πυρήνα συνιστά έναν νέο τρόπο ελέγχου της γονιδιακής έκφρασης. Η ρυθμιζόμενη εισαγωγή στον πυρήνα μεταγραφικών παραγόντων και πρωτεϊνικών κινασών έχει σημαντικό ρόλο στον έλεγχο της κυτταρικής συμπεριφοράς, καθώς παρέχει έναν μηχανισμό διαμέσου του οποίου σήματα που λαμβάνονται στην επιφάνεια του κυττάρου μπορούν να μεταδοθούν στον πυρήνα. Η σημασία της ρυθμιζόμενης εισαγωγής στον πυρήνα αποδεικνύεται από το ότι η αλλαγή στη συγγένεια της αλληλεπίδρασης δύο μόλις πρωτεϊνών του συμπλόκου του πυρηνικού πόρου προς τον υποδοχέα πυρηνικής μεταφοράς έχει συμβάλει στον εξελικτικό διαχωρισμό των ειδών Drosophila melanogaster και Drosophila simulans. Σε έναν συγκεκριμένο μηχανισμό ρύθμισης, οι μεταγραφικοί παράγοντες (ή άλλες πρωτεΐνες) συνδέονται με κυτταροπλασματικές πρωτεΐνες που καλύπτουν τα σήματα πυρηνικού εντοπισμού τους και οδηγούν σε παραμονή των παραγόντων αυτών στο κυτταρόπλασμα, εφόσον τα σήματα πυρηνικού εντοπισμού δεν είναι πλέον αναγνωρίσιμα. Χαρακτηριστικό παράδειγμα αποτελεί ο μεταγραφικός παράγοντας NFκB, που ενεργοποιείται σε κύτταρα των θηλαστικών από πολλά διαφορετικά εξωκυτταρικά σήματα. Στο κυτταρόπλασμα μη διεγερμένων κυττάρων, ο NF-κB σχηματίζει ένα ανενεργό σύμπλοκο με μια πρωτεΐνη-αναστολέα (την ΙκΒ). Η πρόσδεση της ΙκΒ καλύπτει το σήμα πυρηνικού εντοπισμού του NF-κB, εμποδίζοντας τη μεταφορά του στον πυρήνα. Σε διεγερμένα κύτταρα, η πρωτεΐνη ΙκΒ φωσφορυλιώνεται και αποικοδομείται πρωτεολυτικά διαμέσου της ουβικιτίνης, επιτρέποντας στον NF-κB να εισέλθει στον πυρήνα και να ενεργοποιήσει τη μεταγραφή των γονιδίων-στόχων του. 36
.")
37 ΕΙΚΟΝΑ 9.13: Ρύθμιση της εισαγωγής μεταγραφικών παραγόντων στον πυρήνα. Ο μεταγραφικός παράγοντας NF-κΒ διατηρείται στο κυτταρόπλασμα υπό μορφή ανενεργού συμπλόκου με την πρωτεΐνη ΙκΒ, που καλύπτει την αλληλουχία πυρηνικού εντοπισμού του (NLS). Όταν δεχτεί τα κατάλληλα εξωκυτταρικά σήματα, η ΙκΒ φωσφορυλιώνεται και αποικοδομείται μέσω πρωτεόλυσης, επιτρέποντας την εισαγωγή του NF-κΒ στον πυρήνα. Ο μεταγραφικός παράγοντας Pho4 του σακχαρομύκητα ρυθμίζεται άμεσα διαμέσου της φωσφορυλίωσης του και όχι διαμέσου σύνδεσης με πρωτεΐνες-αναστολείς. Ο Pho4 διατηρείται στο κυτταρόπλασμα λόγω της φωσφορυλίωσης ενός καταλοίπου σερίνης που βρίσκεται κοντά στην αλληλουχία πυρηνικού εντοπισμού του. Υπό κατάλληλες συνθήκες, όμως, η ρυθμιζόμενη αποφωσφορυλίωση του καταλοίπου αυτού οδηγεί σε έκθεση του NLS και επιτρέπει την μετατόπιση του Pho4 στον πυρήνα και την ενεργοποίηση του. 37

38 Η ρύθμιση της νουκλεοκυτραοπλασματικής μεταφοράς είναι δυναμική και διαφορετική Η πυρηνική εισαγωγή του ετεροδιμερούς p65-p50 του μεταγραφικού παράγοντα ΝF-κB ρυθμίζεται από μια διαμοριακή κάλυψη του NLS. Ο αναστολέας του NF-κB, IκBα παρεμποδίζει άμεσα το NLS του p65-p50, με αποτέλεσμα τον κυτταροπλασματικό εντοπισμό του ετεροδιμερούς. Όμως όταν ένα κύτταρο δεχθεί προφλεγμονώδη ερεθίσματα, η ΙκΒα φωσφορυλιώνεται και αποδομείται από το πρωτεόσωμα, και επομένως το ετεροδιμερές του NF-κΒ, p65-p50 απελευθερώνεται και μετατοπίζεται (μεταφέρεται) στον πυρήνα με το μηχανισμό των ιμπορτινών α/β διαμέσου του NPC. Η ογκοκατασταλτική πρωτεΐνη p53 παλινδρομεί μεταξύ του πυρήνα και του κυτταροπλάσματος, και αυτή η μετακίνηση εξαρτάται από τη φάση του κυτταρικού κύκλου. Υπό συνθήκες κυτταρικού στρες, η p53 συγκρατείται στον πυρήνα με έναν μηχανισμό κάλυψης του σήματος NES. Η p53 σχηματίζει ομοτετραμερή με αποτέλεσμα το σήμα NES να αποκρύπτεται και να παρεμποδίζεται μ αυτόν τον τρόπο η αλληλεπίδραση της με την εξπορτίνη Crm1/kapβ. Η νουκλεοκυτταροπλασματική μεταφορά της p53 ισχυροποιείται επίσης λόγω της επαγόμενης από στρες Διαμοριακές και ενδομοριακές πολύ-adp-ριβοσυλίωση της, η οποία παρεμποδίζει την αλληλεπιδράσεις (NF-κB ή p53, αλληλοεπίδραση της με την Crm1. Ελαττώματα που αντίστοιχα) ρυθμίζουν τη μεταφορά παρεμποδίζουν την πυρηνική διατήρηση της p53 σχετίζονται με έναν αριθμό νεοπλασμάτων, μιας πρωτεΐνης-φορτίου. αναδεικνύοντας τη σημασία του κατάλληλου κυτταρικού εντοπισμού μιας πρωτεΐνης. 38
39 ΜΕΤΑΦΟΡΑ RNAs ΑΠΟ ΚΑΙ ΠΡΟΣ ΤΟΝ ΠΥΡΗΝΑ 39
40 Η ΠΥΡΗΝΙΚΗ ΕΙΣΑΓΩΓΗ / ΕΞΑΓΩΓΗ ΕΙΝΑΙ ΒΑΣΙΚΕΣ ΔΙΑΔΙΚΑΣΙΕΣ ΠΟΥ ΕΜΠΛΕΚΟΝΤΑΙ ΣΤΗ ΡΥΘΜΙΣΗ ΤΗΣ ΓΟΝΙΔΙΑΚΗΣ ΕΚΦΡΑΣΗΣ ΣΤΑ ΕΥΚΑΡΥΩΤΙΚΑ ΚΑΙ ΟΙ ΟΠΟΙΕΣ ΥΠΟΚΕΙΝΤΑΙ ΣΕ ΡΥΘΜΙΣΗ Γονιδιακή έκφραση στους Προκαρυώτες: DNA RNA Πρωτεΐνη Γονιδιακή έκφραση στους Ευκαρυώτες: ΠΥΡΗΝΙΚΟΣ ΦΑΚΕΛΟΣ DNA ΠΥΡΗΝΑΣ RNAN εξαγωγή RNAC Πρωτεΐνη ΚΥΤΤΑΡΟΠΛΑΣΜΑ Πυρηνικές πρωτεΐνες Εισαγωγή Εξαγωγή Πρωτεΐνες με παλινδρομική κίνηση Ενώ πολλές πρωτεΐνες μεταφέρονται επιλεκτικά από το κυτταρόπλασμα στον πυρήνα, τα περισσότερα μόρια RNA εξάγονται από τον πυρήνα στο κυτταρόπλασμα. Εφόσον οι πρωτεΐνες συντίθενται στο κυτταρόπλασμα, η εξαγωγή από τον πυρήνα μορίων mrna, rrna, trna και mirna αποτελεί ένα καθοριστικό βήμα της γονιδιακής έκφρασης στα ευκαρυωτικά κύτταρα. Η εξαγωγή αυτών των μορίων RNA μέσω του συμπλόκου του πυρηνικού πόρου είναι, όπως και η εισαγωγή πρωτεϊνών, μια ενεργή, εξαρτώμενη από ενέργεια, διεργασία που απαιτεί αλληλεπίδραση των υποδοχέων μεταφοράς με το σύμπλοκο του πυρηνικού πόρου. 40
, όπως οι ιμπορτίνες και οι εξπορτίνες και Ran/GTP.")

41 Μεταφορά των μορίων RNA Συγκρότηση σε RNPs Καρυοφερίνες Ran/GTP Μικρά RNAs όπως τα trnas και mirnas εξάγονται με σχετικά απλές πορείες, με τη δέσμευση τους σε υποδοχείς εξαγωγείς (καρυοφορίνες), όπως οι ιμπορτίνες και οι εξπορτίνες και Ran/GTP. Τα μεγαλύτερα μόρια όπως τα rrnas και τα snrnas συγκροτούνται σε πολύπλοκα ριβονουκλεοπρωτεϊνικά σωμάτια (Ribonucleoproteins; RNP) και στρατολογούν τις καρυοφερίνες διαμέσου μιας κατηγορίας ειδικών προσαρμοστικών πρωτεϊνών και χρησιμοποιούν Ran/GTP. Η εξαγωγή των mrnas είναι μοναδική καθώς εκτός από τη συγκρότηση σε RNPs, είναι συζευγμένη με τη μεταγραφή (σακχαρομύκητες) και με το μάτισμα (μετάζωα), και δεν χρησιμοποιούν Ran/GTP, αλλά διεξάγεται διαμέσου ενός συμπλόκου δύο πρωτεϊνών, γνωστού με το όνομα «εξαγωγέας mrnas». 41
42 Οι κύριες πορείες πυρηνικής εξαγωγής μορίων RNA: trna, microrna (mirna), snrna, mrna, rrna. Σε κάθε περίπτωση απεικονίζεται το μετάγραφο RNA μετά την επεξεργασία, την ωρίμανση, και την συγκρότηση τους σε σύμπλοκα με παράγοντες εξαγωγής [οι προσαρμοστικές πρωτεΐνες εξαγωγής (export adaptors) εμφανίζονται μπλε, οι πρωτεϊνικοί υποδοχείς μεταφοράς (export receptors) με κίτρινο]. Απεικονίζονται επίσης τα επικρατή δομικά χαρακτηριστικά στα προ-rnas (μονόκλωνες/δίκλωνες περιοχές RNA, βρόγχοι, εξόνια και ιντρόνια, 5' cap και 3' πολυ(a) ουρά). Στην πορεία εξαγωγής mrna, απεικονίζεται η ονομασία των πρωτεϊνών των μεταζώων και σακχαρομυκήτων, και τα mrnas μαζί με επιπλέον προσαρμοστικές πρωτεΐνες και παράγοντες δέσμευσης RNA (RNA-binding factors) με οβαλ πορτοκαλί. Στην πορεία του rrna, απεικονίζονται ο γενικός εξαγωγέας στα ευκαρυωτικά, CRM1, και δύο βοηθητικές πρωτεΐνες εξαγωγής, Mex67 Mtr2 και Arx1, που έχουν μελετηθεί στον σακχαρομύκητα. CBC, Σύμπλοκο δέσμευσης cap (Cap-binding complex); Exp, Εξπορτίνη (exportin). 42

43 Μεταφορά μορίων trna και mirna Τα trnas εξάγονται από την εξπορτίνη-t που δρα ως υποδοχέας τους. Η μεταφορά trna απαιτεί Ran/GTP. Η εξαγωγή trna μπορεί να επηρεαστεί από τροποποιήσεις του trnas. Προεξοχή 2 nt αναγνωρίζεται από Exp-5 Τα trnas μπορούν να επαναεισαχθούν στον πυρήνα. Τα πρωτογενή μετάγραφα mirnas (προ-mirnas) εξάγονται από τον πυρήνα με τη δράση της εξπορτίνης-5 και η επεξεργασία τους διεξάγεται στο κυτταρόπλασμα 43
44 Πυρηνική εξαγωγή μορίων trna και mirna (a) Η μεταγραφή ενός γονιδίου trna από την RNA πολυμεράση III (Pol III) παράγει ένα πρωτογενές μετάγραφο, που μερικές φορές περιέχει ένα ιντρόνιο, με επιμηκύνσεις των 5 και 3 άκρων. Μετά την επεξεργασία των 5 και 3 άκρων, τροποποιήσεις βάσεων (κόκκινοι κύκλοι) και την προσθήκη των νουκλεοτιδίων CCA στο 3 άκρο, τα παραγόμενα trnas μπορούν να ακολουθήσουν διαφορετικές πορείες εξαγωγής. Τα trna που περιέχουν ένα ιντρόνιο ή ελεύθερα ιντρονίου trnas (με ένα αμινοξύ ή όχι) μεταφέρονται με την πορεία που εξαρτάται από την εξπορτίνη-t/los1, μεταξύ άλλων την αλληλουχία CAA. Μετά την εξαγωγή στο κυτταρόπλασμα και τη διάσταση του trna από τον υποδοχέα, το trna που περιέχει ένα ιντρόνιο υφίσταται μάτισμα από τις πρωτεΐνες ματίσματος που εντοπίζονται στην εξωτερική μιτοχονδριακή μεμβράνη, στους σακχαρομύκητες. Το ώριμο trna διοχετεύεται στη συνέχεια στα ριβοσώματα από παράγοντες αποδέσμευσης όπως Cex1 και eef1. (b) Για τα mirna, απεικονίζεται μόνον η παραγωγή του πρωτογενούς mrna (προ-mrnas) που μεταγράφεται από την Pol II. Τα mirnas μπορεί να κωδικοποιούνται από ένα γονίδιο mirna που εντοπίζεται στο ιντρόνιο ενός γονιδίου που κωδικοποιεί μια πρωτεΐνη (αριστερός κλάδος) ή από ένα ξεχωριστό γονίδιο mirna (δεξιός κλάδος). Και στις 2 περιπτώσεις, το πρωτογενές μετάγραφο διασπάται από το σύμπλοκο Drosha DGCR8 σχηματίζοντας ένα προ-mrna που φέρει μια δομή μίσχου-βρόγχου (stem loop structure). Επιπλέον, παράγεται ένα mirna που έχει υποστεί μάτισμα μετά από την αποκοπή ενός ιντρονικού mrna. Το προ-mirna που φέρει μια προεξοχή 2 νουκλεοτιδίων στο 3' άκρο του αναγνωρίζεται ειδικά από την εξπορτίνη-5 και μεταφέρεται στο κυτταρόπλασμα, όπου διίσταται από τον υποδοχέα του μετά την υδρόλυση του RanGTP. Μετά την αποδέσμευση του από την εξπορτίνη-5, το Dicer διασπά επιπλέον το προ-mirna και παράγει ένα μονόκλωνο mirna, το οποίο συγκροτείται σε ένα επαγόμενο σύμπλοκο αποσιώπησης RNA (RNA-induced silencing complex; RISC). Με μπλε χρώμα φαίνεται η περιοχή του RNA που αντιστοιχεί σε ένα ώριμο mirna. 44
 συγκροτούνται σε ριβονουκλεοπρωτεϊνικά σωματίδια τα προ-mrnp (pre-mrnp) ή ετερογενή πυρηνικά mrnp ή hnrnp. Protein RNA Προ-mRNA ΕΙΚΟΝΑ 9.")
, που περιέχουν 35-40 kb RNA και έχουν συνολική μάζα περίπου 30 εκατομμύρια Da.")
45 Μεταφορά των μορίων mrna, rrna και snrna Τα μόρια mrna, rrna και snrna μεταφέρονται διαμέσου του πυρηνικού πόρου υπό μορφή συμπλοκών ριβονουκλεοπρωτεΐνης (RNP; Ribonucleoprotein complexes). Στον πυρήνα, τα προ-mrna (pre-mrna) συγκροτούνται σε ριβονουκλεοπρωτεϊνικά σωματίδια τα προ-mrnp (pre-mrnp) ή ετερογενή πυρηνικά mrnp ή hnrnp. Protein RNA Προ-mRNA ΕΙΚΟΝΑ 9.14: Μεταφορά ενός Σωμάτιο RNP συμπλόκου ριβονουκλεοπρωτεΐνης. Τα κύτταρα των σιελογόνων αδένων των εντόμων παράγουν ευμεγέθη σύμπλοκα ριβονουκλεοπρωτεΐνης (RNΡ), που περιέχουν kb RNA και έχουν συνολική μάζα περίπου 30 εκατομμύρια Da. Σε αυτή τη σειρά φωτογραφιών ΗΜ παρουσιάζονται (Α) η πρόσδεση ενός τέτοιου RNΡ σε ένα σύμπλοκο πυρηνικού πόρου και (Β-Δ) η αποδίπλωση του RNA κατά τη μεταφορά του στο κυτταρόπλασμα. 45
, υπόκεινται σε εναλλακτικό μάτισμα και υφίστανται επεξεργασία και προσθήκη της 5 πολύ-α ουράς.")

46 Πυρηνική εξαγωγή των μορίων mrna Τα πρόδρομα μόρια προ-mrna συνθέτονται από την RNA πολυμεράση (Pol)-II, αποκτούν την 5 καλύπτρα 7-μεθυλογουανοσίνης (Cap) όπου δεσμεύονται οι πρωτεΐνες CBC (Cap Binding Protein Complex), υπόκεινται σε εναλλακτικό μάτισμα και υφίστανται επεξεργασία και προσθήκη της 5 πολύ-α ουράς. Κατά τη συγκρότηση αυτών των επεξεργασμένων μορίων mrna σε ένα ώριμο σύμπλοκο ριβονουκλεοπρωτεϊνών (mrnps) συνδέονται με ένα σύνολο τουλάχιστον 20 πρωτεϊνών. Η μεταφορά του mrnp από τον πυρήνα στο κυτταρόπλασμα επιτυγχάνεται μέσω του συμπλόκου του εξαγωγέα mrna, που στρατολογείται στο επεξεργασμένο mrna. Εξαγωγή mrna συζευγμένη με το μάτισμα. Στα μετάζωα, η εξαγωγή mrna είναι συζευγμένη με το μάτισμα και εξαρτάται από την παρουσία της 5 cap που αλληλεπιδρά με το σύμπλοκο TREX (THO υποσύμπλοκο) και στη συνέχεια στρατολογείται στο mrnp. Στη συνέχεια στρατολογείται ο υποδοχέας εξαγωγής mrna, TAP p15 (κίτρινο), και προσαρμοστικές πρωτεΐνες (μπλε). Πολλά mrna εξάγονται με το μηχανισμό Crm1Ran/GTP με τον οποίο το σύμπλοκο Crm1-Ran/GTP προσδένει στις πρωτεΐνες CBC, διαμέσου μιας προσαρμοστικής πρωτεΐνης HuR. 46


47 Ο υποδοχέας εξαγωγής Crm1 δεσμεύεται παρουσία της RanGTP στην αλληλουχία NES της προσαρμοστικής πρωτεΐνης Nmd3 NPC RanGAP Μεταφορά των μορίων rrna: Τα ριβοσωμικά RNA συνδέονται αρχικά με ριβοσωμικές πρωτεΐνες και με ειδικές πρωτεΐνες επεξεργασίας RNA μέσα στον πυρηνίσκο, δημιουργώντας τις δύο ριβοσωμικές υπομονάδες 60S και 40S. Οι υπομονάδες αυτές στη συνέχεια μεταφέρονται στο κυτταρόπλασμα μέσω ενός μηχανισμού στον οποίο συμμετέχει η καρυοφερίνη Crm1. Η εξαγωγή τους από τον πυρήνα επιτυγχάνεται διαμέσου της αναγνώρισης σημάτων NES που περιέχονται σε πρωτεΐνες του συμπλόκου κάθε ριβοσωμικής υπομονάδας, ιδιαίτερα στην προσαρμοστική πρωτεΐνη Nmd3. Οι άλλοι υποδοχείς εξαγωγής Mex67 Mtr2 και Arx1 δεσμεύονται άμεσα στην υπομονάδα pre-60s. 47
48 Πυρηνική εξαγωγή των ριβοσωμικών υπομονάδων. Η εξαγωγή της ριβοσωμικής υπομονάδς pre-60s στο σακχαρομύκητα αρχίζει με τη συγκρότηση της στον πυρηνίσκο με μια πολυσταδιακή και πολύπλοκη διαδικασία. Μόλις το σωμάτιο pre-60s εξάγεται στο νουκλεόπλασμα στρατολογεί διάφορους υποδοχείς εξαγωγής. Ο υποδοχέας εξαγωγής Crm1 δεσμεύεται παρουσία της RanGTP στην αλληλουχία πυρηνικής εξαγωγής (NES) της προσαρμοστικής πρωτεΐνης Nmd3. Οι άλλοι υποδοχείς εξαγωγής Mex67 Mtr2 και Arx1 δεσμεύονται άμεσα στην υπομονάδα pre-60s. Μετά τη μετατόπιση του διαμέσου του NPC, απομακρύνονται στο κυτταρόπλασμα οι παράγοντες εξαγωγής από σωμάτιο pre-60s. Η Crm1 διίσταται από την Nmd3 με την υδρόλυση της RanGTPάση (RanGAP). Η Nmd3 απελευθερώνεται από την Lsg1 GTPάση και αντικαθίσταται από τη ριβοσωμική πρωτεΐνη and Rpl10. Η Arx1 αποδεσμεύεται από τον παράγοντα απελευθέρωσης Rei1. Δεξιά απεικονίζεται ο δίαυλος του NPC και οι υποδοχείς εξαγωγής που αλληλεπιδρούν με το σωμάτιο pre-60s particle και τις νουκλεοπορίνες FG. Οι υποδοχείς εξαγωγής εμφανίζονται με κίτρινο, οι προσαρμοστικές πρωτεΐνες με μπλε και οι παράγοντες αποδέσμευσης/απελευθέρωσης με ροζ. 48


 εξάγονται αρχικά από τον πυρήνα στο κυτταρόπλασμα μέσω μιας εξπορτίνης (Crm1) που αναγνωρίζει την καλύπτρα 7-μεθυλογουανοσίνης του 5 άκρου.")
49 Μεταφορά των μορίων snrna: Σε αντιδιαστολή με τα μόρια mrna, trna και rrna, που ασκούν τη λειτουργία τους στο κυτταρόπλασμα, πολλά μικρά μόρια RNA (snrna και snorna) λειτουργούν μέσα στον πυρήνα ως συστατικά του μηχανισμού επεξεργασίας RNA. Τα snrna μεταφέρονται αρχικά από τον πυρήνα στο κυτταρόπλασμα, όπου συνδέονται με πρωτεΐνες για να σχηματίσουν λειτουργικά snrnp και στη συνέχεια επαναφέρονται στον πυρήνα. Στην εξαγωγή των μορίων snrna στο κυτταρόπλασμα συμμετέχουν η πρωτεΐνη Crm1 και άλλοι υποδοχείς μεταφοράς που προσδένονται στις 5' καλύπτρες 7μεθυλογουανοσίνης των snrna, ενώ για τη μεταφορά των snrnp από το κυτταρόπλασμα στον πυρήνα ευθύνονται αλληλουχίες που φέρουν οι πρωτεΐνες snrnp. ΕΙΚΟΝΑ 9.15: Μεταφορά των snrna μεταξύ πυρήνα και κυτταροπλάσματος. Τα μικρά πυρηνικά RNA (snrna) εξάγονται αρχικά από τον πυρήνα στο κυτταρόπλασμα μέσω μιας εξπορτίνης (Crm1) που αναγνωρίζει την καλύπτρα 7-μεθυλογουανοσίνης του 5 άκρου. Στο κυτταρόπλασμα, τα snrna συνδέονται με πρωτεΐνες και σχηματίζουν snrnp, τα οποία στη συνέχεια μεταφέρονται και πάλι στον πυρήνα. 49
50 Εσωτερική οργάνωση του πυρήνα 50
51 Εσωτερική οργάνωση του πυρήνα Ο πυρήνας διαθέτει μια εσωτερική δομή που εξασφαλίζει την οργάνωση του γενετικού υλικού και τον καταμερισμό των πυρηνικών λειτουργιών. Στα ζωικά κύτταρα, ένα χαλαρά οργανωμένο πλέγμα πυρηνικών λαμινών εκτείνεται από την πυρηνική λάμινα ως το εσωτερικό του πυρήνα. Αυτές οι λαμίνες χρησιμεύουν ως θέσεις πρόσδεσης της χρωματίνης και οργανώνουν άλλες πρωτεΐνες σε δομές λειτουργικών πυρηνικών σωματίων. Μέσα στον πυρήνα, η χρωματίνη είναι οργανωμένη σε μεγάλους βρόχους DNA, συγκεκριμένες περιοχές των οποίων προσδένονται στο στρώμα των λαμινών μέσω πρωτεϊνών πρόσδεσης λαμινών που βρίσκονται στη χρωματίνη. Πολλές πυρηνικές πρωτεΐνες σχηματίζουν σύμπλοκα εξαρτώμενα από λαμίνες, που με τη σειρά τους σχηματίζουν πυρηνικά σωμάτια που συμμετέχουν στην επιδιόρθωση του DNA, στην οργάνωση της χρωματίνης, στη γονιδιακή ρύθμιση και στη μεταγωγή σήματος. Αυτός ο ρόλος της πυρηνικής λάμινας και των λαμινών στον καταμερισμό των λειτουργιών της επιδιόρθωσης του DNA και της μεταγραφής γονιδίων αποτελεί τη βάση πολλών διαφορετικών γενετικών ασθενειών που σχετίζονται με τις λαμίνες. 51


52 Εσωτερική οργάνωση του πυρήνα Απεικόνιση των αλληλεπιδράσεων των λαμινών με πρωτεΐνες της εσωτερικής πυρηνικής μεμβράνης, του πυρηνικού πόρου και πυρηνοπλάσματος. Οι λαμίνες βρίσκονται συγκεντρωμένες στην πυρηνική λάμινα (παχιές κόκκινες γραμμές) καθώς επίσης και διάσπαρτες στο πυρηνόπλασμα (λεπτές κόκκινες γραμμές). 52

, αλλά και με άλλες πυρηνικές πρωτεΐνες")
53 Εσωτερική οργάνωση του πυρήνα Μοντέλο των πυρηνικών λαμίνων και των αλληλεπιδράσεων τους με γειτονικές πρωτεΐνες. Οι λαμίνες αλληλεπιδρούν με συναφείς πρωτεΐνες (lamina-associated proteins (LBR, LAP2, emerin, MAN1, nesprins-1 & -2), αλλά και με άλλες πυρηνικές πρωτεΐνες (BAF, Rb, SREBP1), ιστόνες και DNA (χρωματίνη). 53
54 Εσωτερική οργάνωση του πυρήνα Η σύνδεση των λαμινών με την εσωτερική πυρηνική μεμβράνη διευκολύνεται από τη μεταμεταφραστική προσθήκη λιπιδίων - πρενυλίωση των καρβοξυτελικών καταλοίπων κυστεΐνης. Οι λαμίνες προσδένονται σε πρωτεΐνες της εσωτερική πυρηνική μεμβράνη, όπως η εμερίνη και ο LBR, μεσολαβώντας στην πρόσδεση τους στον πυρηνικό φάκελο. Η πυρηνική λάμινα προσδένεται στη χρωματίνη μέσω των ιστονών Η2Α και Η2Β, και άμεσα στο DNA. Οι λαμίνες σχηματίζουν ένα χαλαρό δίχτυ που εκτείνεται σε ολόκληρο το εσωτερικό του πυρήνα. Στις λαμίνες προσδένονται πολλές πυρηνικές πρωτεΐνες που συμμετέχουν στη σύνθεση του DNA, στη μεταγραφή ή στην τροποποίηση της χρωματίνης, παρόλο που η σημασία αυτών των αλληλεπιδράσεων μόλις έχει αρχίσει να κατανοείται. Η πυρηνική λάμινα, ένα ινώδες δίκτυο που αποτελείται κυρίως από λαμίνες, επενδύει την εσωτερική πυρηνική μεμβράνη. Σ' αυτήν την περιοχή διεξάγονται αλληλεπιδράσεις πρωτεΐνης-πρωτεΐνης οι οποίες συνδέουν την χρωματίνη με την πυρηνική περιφέρεια, την πυρηνική λάμινα με τα σύμπλοκα NPC και τις πρωτεΐνες της εσωτερικής πυρηνικής μεμβράνης. Είναι επομένως φανερό ότι αυτές οι πρωτεϊνικές αλληλεπιδράσεις διασυνδέουν την περιφέρεια του νουκλεοπλάσματος με τον κυτταροσκελετό. Ιδιαίτερα τα αυλικά τμήματα της οικογένειας των πρωτεϊνών SUN που εντοπίζονται στην εσωτερική πυρηνική μεμβράνη, αλληλεπιδρούν με τις πρωτεΐνες Nesprins, που ανήκουν στην οικογένεια των πρωτεϊνών KASH και εντοπίζονται στην εξωτερική πυρηνική μεμβράνη. Στη συνέχεια κυτταροπλασματικές περιοχές (επικράτειες) των νεσπρίνες (nesprins) αλληλεπιδρούν με τα ινίδια ακτίνης, τους μικροσωληνίσκους και τα ενδιάμεσα ινίδια του κυτταροσκελετού. Συνολικά αυτό το δίκτυο των πρωτεϊνικών αλληλεπιδράσεων ονομάζεται συνδετικό πρωτεϊνικό σύμπλεγμα του πυρηνικού σκελετού και του κυτταροσκελετού (σύμπλεγμα LINC). Οι πρωτεΐνες του συμπλέγματος LINC προσδένουν επίσης και στα κεντροσωμάτια, τα οποία εντοπίζονται κοντά στην εξωτερική πυρηνική μεμβράνη. 54

55 Χρωμοσώματα και υψηλότερη οργάνωση δομής της χρωματίνης Η χρωματίνη συμπυκνώνεται σε υψηλό βαθμό κατά τη μίτωση, σχηματίζοντας τα συμπαγή μεταφασικά χρωμοσώματα που κατανέμονται στους θυγατρικούς πυρήνες. Κατά τη μεσόφαση, το DNA βρίσκεται σε 2 μορφές που αντανακλούν τη μεταγραφική ενεργότητα του κυττάρου. Η ετεροχρωματίνη (κλειστή χρωματίνη) παραμένει ιδιαίτερα συμπυκνωμένη και μεταγραφικά ανενεργή. Η υπόλοιπη χρωματίνη ή ευχρωματϊνη (ανοικτή χρωματίνη) είναι αποσυμπυκνωμένη, κατανέμεται σε ολόκληρο τον πυρήνα, και επικρατεί στα κύτταρα που είναι μεταγραφικά ενεργά. Η ετεροχρωματίνη έχει διάφορες λειτουργίες: Αποσιώπιση έκφρασης γονιδίων Δομική ακεραιότητα του γονιδιώματος Πυρηνίσκος ΕΙΚΟΝΑ 9.16: Ετεροχρωματίνη σε μεσοφασικούς πυρήνες. Η ευχρωματίνη είναι κατανεμημένη σε ολόκληρο τον πυρήνα. Η ετεροχρωματίνη υποδεικνύεται με τρίγωνα και ο πυρηνίσκος με βέλος. Η ετεροχρωματίνη απεικονίζεται ως μικρά, μαύρα ακανόνιστα σωματίδια διάσπαρτα στον πυρήνα και συναθροισμένα στον πυρηνικό φάκελο. 55

56 Η ετεροχρωματίνη αλληλεπιδρά με την πυρηνική λάμινα αλλά δεν εντοπίζεται στο σύμπλοκο των πυρηνικών πόρων. Επομένως, οι περιοχές στις οποίες αποκλείεται η χρωματίνη περιέχουν μεταγραφική ενεργά ευχρωματίνη. 56
 Στα θηλαστικά, η νουκλεοπορίνη TPR απαιτείται για να δημιουργήσει μια αποκλειστική ζώνη ελεύθερης ετεροχρωματίνης κάτω από το σύμπλοκο NPC (+TPR).")
57 Η περιφερειακή ετεροχρωματίνη αποκλείεται από το σύμπλοκο NPC. (a) Σχηματικό διάγραμμα της κατανομής της ετεροχρωματίνης κάτω από τον πυρηνικό φάκελο που απεικονίζει τα κενά στα σύμπλοκα των NPC. (b) Στα θηλαστικά, η νουκλεοπορίνη TPR απαιτείται για να δημιουργήσει μια αποκλειστική ζώνη ελεύθερης ετεροχρωματίνης κάτω από το σύμπλοκο NPC (+TPR). Η απώλεια της TRP μέσω παρεμβολής RNAi οδηγεί στην κάλυψη της περιοχής του NPC με ετεροχρωματίνη (-TPR). 57
58 Τα κύτταρα περιέχουν 2 τύπους ετεροχρωματίνης: την ιδιοστατική (constitutive) ετεροχρωματίνη, που περιέχει αλληλουχίες DNA που δε μεταγράφονται, όπως είναι οι δορυφορικές αλληλουχίες στα κεντρομερή και τελομερή, και σε τμήματα φυλετικών χρωμοσωμάτων όπως το Υ. τη δυνητική (facultative) ετεροχρωματίνη, που περιέχει αλληλουχίες που δε μεταγράφονται στο υπό εξέταση κύτταρο αλλά μεταγράφονται σε άλλους τύπους κυττάρων. Επομένως, μπορεί να αποσυμπυκνωθεί ως απόκριση σε ένα ερέθισμα και να γίνει μεταγραφικά ενεργή. Κατά συνέπεια, η ποσότητα της δυνητικής ετεροχρωματίνης ποικίλλει ανάλογα με τη μεταγραφική ενεργότητα του κυττάρου, μια δεδομένη χρονική περίοδο. Για παράδειγμα, το γονίδιο της αιμοσφαιρίνης σε ένα νευρώνα, και σε ένα ερυθροκύτταρο. 58
 Molecular Cell")
59 Η δυνητική χρωματίνη (Facultative heterochromatin) μπορεί να ενεργοποιηθεί ως απόκριση σε διάφορα αναπτυξιακά ή εξωτερικά ερεθίσματα, με τη στρατολόγηση διαφόρων πρωτεϊνικών συμπλεγμάτων ενεργοποίησης της μεταγραφής, απώλεια της ιστόνης Η1 και με ιστονικές τροποποιήσεις, όπως ακετυλίωση, Trojer & Reinberg (2007) Molecular Cell 28:

 και τα τελομερή να προσδένονται σε αντίθετες πλευρές του πυρηνικού φακέλου. ΕΙΚΟΝΑ 9.17: Οργάνωση των χρωμοσωμάτων.")
60 Αν και η μεσοφασική χρωματίνη φαίνεται να είναι ομοιόμορφα κατανεμημένη, στην πραγματικότητα τα χρωμοσώματα είναι διατεταγμένα με οργανωμένο τρόπο και χωρίζονται σε διακριτές λειτουργικές επικράτειες που έχουν σημαντικό ρόλο στη ρύθμιση της γονιδιακής έκφρασης. Τη μη τυχαία κατανομή της χρωματίνης στον μεσοφασικό πυρήνα εισηγήθηκε το 1885 ο Carl Rabl, διατυπώνοντας ότι κάθε χρωμόσωμα καταλαμβάνει μια διακριτή επικράτεια, με τα κεντρομερή (επαναλαμβανόμενες δορυφορικού αλληλουχίες μεθυλιωμένου DNAκεντρομερικών πρωτεϊνών CEN) και τα τελομερή να προσδένονται σε αντίθετες πλευρές του πυρηνικού φακέλου. ΕΙΚΟΝΑ 9.17: Οργάνωση των χρωμοσωμάτων. Αναπαραγωγή χειρόγραφων σχεδίων χρωμοσωμάτων σε κύτταρα σαλαμάνδρας. (Α) Πλήρη χρωμοσώματα. (Β) Μόνο τα τελομερή (που εντοπίζονται στην πυρηνική μεμβράνη). (Rabl C, Morphologisches Jahrbuch 10: 214.) 60

61 Αυτό το βασικό μοντέλο οργάνωσης της χρωματίνης επιβεβαιώθηκε το 1984 με λεπτομερείς μελέτες των πολυταινικών χρωμοσωμάτων των σιελογόνων αδένων της Drosophila. Τα χρωμοσώματα δεν τυλίγονται τυχαία το ένα γύρω από το άλλο, αλλά κάθε χρωμόσωμα καταλαμβάνει μια διακριτή περιοχή του πυρήνα. Τα χρωμοσώματα συνδέονται στενά με τον πυρηνικό φάκελο σε πολλές θέσεις. Πολλές από αυτές τις συνδέσεις οδηγούν σε καταστολή της γονιδιακής έκφρασης, κάποιες άλλες όμως, όπως οι συνδέσεις με τα σύμπλοκα του πυρηνικού πόρου, προάγουν τη μεταγραφή συγκεκριμένων γονιδίων. Οι χρωμοσωμικές επικράτειες πλούσιες σε γονίδια βρίσκονται στο εσωτερικό του πυρήνα, ενώ εκείνες οι περιοχές που είναι φτωχές σε γονίδια εντοπίζονται στην περιφέρεια του πυρήνα. Αυτές οι χρωμοσωμικές επικράτειες είναι δυναμικές δομές και ο εντοπισμός τους μπορεί να μεταβληθεί, για παράδειγμα, ανάλογα με τη φάση του κυτταρικού κύκλου. ΕΙΚΟΝΑ 9.18: Οργάνωση των χρωμοσωμάτων της Drosophila (Α) Μοντέλο του πυρήνα, στο οποίο παρουσιάζονται με διαφορετικά χρώματα οι βραχίονες των 5 χρωμοσωμάτων. Υποδεικνύονται oι θέσεις των τελομερών και των κεντρομερών. (Β) Παρουσιάζονται οι δύο βραχίονες του χρωμοσώματος 3, προκειμένου να φανεί ο τοπολογικός διαχωρισμός των χρωμοσωμάτων. (Matdog D, et al Nature 308:414.) 61


62 Μεμονωμένα χρωμοσώματα καταλαμβάνουν διακριτές επικράτειες στους πυρήνες των κυττάρων θηλαστικών. Τα ενεργώς μεταγραφόμενα γονίδια εντοπίζονται στην περιφέρεια αυτών των επικρατειών, δίπλα σε «διαύλους» που χωρίζουν τα χρωμοσώματα μεταξύ τους. Θεωρείται ότι τα νεοσυντιθέμενα μόρια RNA απελευθερώνονται μέσα σε αυτούς τους «διαύλους» και εκεί λαμβάνει χώρα η επεξεργασία του RNA. Μεγάλο μέρος της ετεροχρωματίνης εντοπίζεται στην περιφέρεια του πυρήνα, καθώς πρωτεΐνες που συνδέονται με την ετεροχρωματίνη προσδένονται στο πλέγμα της πυρηνικής λάμινας. Εφόσον διαφορετικοί τύποι κυττάρων εκφράζουν διαφορετικά γονίδια, η δυνητική ετεροχρωματίνη, καθώς και οι περιοχές των χρωμοσωμάτων που αλληλεπιδρούν με την πυρηνική λάμινα διαφέρουν μεταξύ διαφορετικών κυττάρων και ιστών. Σε μερικά κύτταρα, τα κεντρομερή και τα τελομερή είναι συγκεντρωμένα σε αντίθετους πόλους, ενώ σε άλλα κύτταρα τα χρωμοσώματα διατάσσονται ακτινωτά. Οι θέσεις των χρωμοσωμάτων μέσα στον πυρήνα διαφέρουν επίσης ανάλογα με τον οργανισμό και τον τύπο του ιστού. Εξάλλου, η χρωματίνη στο εσωτερικό του πυρήνα αναδιοργανώνεται κατά την κυτταρική διαφοροποίηση και σε συντονισμό με αλλαγές στη γονιδιακή έκφραση. Αν και αυτή η δυναμική αναδιοργάνωση της χρωματίνης είναι πολύπλοκη και δεν έχει κατανοηθεί πλήρως, είναι γνωστό ότι βασίζεται, τουλάχιστον εν μέρει, στην ακτίνη και στη μυοσίνη του πυρήνα - πρωτεΐνες που χαρακτηρίστηκαν για πρώτη φορά στον κυτταροσκελετό. ΕΙΚΟΝΑ 9.19: Οργάνωση των χρωμοσωμάτων στον πυρήνα των θηλαστικών. (Α) Ανιχνευτές επαναλαμβανόμενων αλληλουχιών του χρωμοσώματος 4 χρησιμοποιήθηκαν για πείραμα υβριδισμού σε ένα ανθρώπινο κύτταρο. Τα δύο αντίγραφα του χρωμοσώματος 4, που ταυτοποιείται με τον φθορισμό κίτρινου χρώματος, καταλαμβάνουν διακριτές επικράτειες μέσα στον πυρήνα. (Β) Μοντέλο οργάνωσης των χρωμοσωμάτων. Τα χρωμοσώματα καταλαμβάνουν διακριτές επικράτειες, που χωρίζονται μεταξύ τους με διαχρωμοσωμικές περιοχές, όπου θεωρείται ότι λαμβάνουν χώρα η επεξεργασία και η μεταφορά του RNA. (Lamond AI & Earnshaw, WC. 1999, Science 280: 547) 62
:167 Χρωμοσωμικές επικράτειες σε ινοβλάστες κοτόπουλου. (a) Μεταφασικά χρωμοσώματα ινοβλαστών κοτόπουλου μετά από χρώση.")
. (c) Τα χρώματα της φθορίζουσας ουσίας που χρησιμοποιήθηκε για τη σήμανση κάθε χρωμοσώματος.")
63 Τα χρωμοσώματα καταλαμβάνουν διακριτές περιοχές στον πυρήνα που ονομάζονται χρωμοσωμικές επικράτειες. Misteli, T. (2008) Chromosome territories: The arrangement of chromosomes in the nucleus. Nature Education 1(1):167 Χρωμοσωμικές επικράτειες σε ινοβλάστες κοτόπουλου. (a) Μεταφασικά χρωμοσώματα ινοβλαστών κοτόπουλου μετά από χρώση. (b) Τα ίδια μεταφασικά χρωμοσώματα μετά από υβριδισμό ανιχνευτών που έχουν σημανθεί με μια φθορίζουσα χρωστική διαφορετικού χρώματος [Φθορίζων υβριδισμός πολλαπλών χρωμάτων in situ (mfish; multicolour Fluorescence in situ hybridisation). (c) Τα χρώματα της φθορίζουσας ουσίας που χρησιμοποιήθηκε για τη σήμανση κάθε χρωμοσώματος. Χρωμόσωμα 1 κόκκινο; Chr2 - πράσινο; Chr3 - μπλε; Chr4 - πορτοκαλί; Chr5 - μωβ; Chr6 - ροζ; ChrZ - μπλε ανοικτό, κλπ. (d) Οπτική τομή διαμέσου του πυρήνα μιας ινοβλάστη κοτόπουλου που απεικονίζει με διαφορετικό χρώμα τις διαφορετικές χρωμοσωμικές επικράτειες, κάθε μια από τις οποίες αντιστοιχεί σε ένα διαφορετικό χρωμόσωμα. Κάθε χρωμόσωμα εντοπίζεται σε διαφορετική θέση στον πυρήνα, και τα ομόλογα χρωμοσώματα έχουν επίσης διαφορετική θέση εντοπισμού μέσα στον πυρήνα (π.χ. χρωμοσώματα 2 και 5). 63


64 Όπως και το DNA στα μεταφασικά χρωμοσώματα, η χρωματίνη των μεσοφασικών πυρήνων είναι οργανωμένη σε περιοχές βρόχων που περιέχουν ~ kb DNA. Παράδειγμα αυτής της οργάνωσης είναι τα εντόνως μεταγραφόμενα χρωμοσώματα των ωοκυττάρων των αμφιβίων, όπου οι ενεργώς μεταγραφόμενες περιοχές DNA είναι ορατές στο μικροσκόπιο ως μεγάλοι βρόχοι αποδιπλωμένης χρωματίνης. Αυτές οι περιοχές φαίνεται ότι αντιστοιχούν σε διακριτές λειτουργικές μονάδες, οι οποίες ρυθμίζουν ανεξάρτητα τη γονιδιακή έκφραση. ΕΙΚΟΝΑ 9.20: Φωτογραφία φωτονικού μικροσκοπίου που δείχνει ένα χρωμόσωμα ωοκυττάρου αμφιβίου. Φαίνονται οι αποσυμπυκνωμένοι βρόχοι ενεργώς μεταγραφόμενης χρωματίνης που εκτείνονται από έναν άξονα μη μεταγραφόμενης χρωματίνης υψηλής συμπύκνωσης. 64

65 Υποδιαμερίσματα μέσα στον πυρήνα Η εσωτερική οργάνωση του πυρήνα καταδεικνύεται περαιτέρω από τον καταμερισμό των περισσότερων πυρηνικών διεργασιών σε συγκεκριμένες, διακριτές περιοχές του πυρήνα. Πολλά σημαντικά ένζυμα και πρωτεΐνες του πυρήνα εντοπίζονται σε διακριτά πυρηνικά σωμάτια με σπογγοειδή δομή χαμηλής πυκνότητας, που επιτρέπει την είσοδο και έξοδο μακρομορίων από τις υπόλοιπες περιοχές του πυρήνα. Πυρηνικά στίγματα (nuclear speckles): Συστατικά της συσκευής ματίσματος του RNA Σωμάτια PML: Αλληλεπιδρούν με τη χρωματίνη και αποτελούν θέσεις συσσώρευσης μεταγραφικών παραγόντων και πρωτεϊνών τροποποίησης της χρωματίνης Σωμάτια Cajal: Περιέχουν τη χαρακτηριστική πρωτεΐνη κοϊλίνη (coilin) και πλήθος μικρών RNP. Τα σωμάτια αυτά θεωρείται ότι λειτουργούν ως θέσεις συναρμολόγησης και επεξεργασίας των RNP. Πυρηνίσκος (nucleolus): Αποτελεί τη θέση μεταγραφής και επεξεργασίας του rrna, καθώς και τη θέση έναρξης της συναρμολόγησης των ριβοσωμάτων 65

66 Οι πυρήνες των κυττάρων των θηλαστικών περιέχουν ομαδοποιημένες θέσεις (εστίες) αντιγραφής DNA όπου διεξάγεται η αντιγραφή πολλαπλών μορίων DNA. Αυτές οι διακριτές θέσεις αντιγραφής του DNA προσδιορίστηκαν με πειράματα απεικόνισης του νεοσυντιθέμενου DNA στο εσωτερικό των πυρήνων έπειτα από σήμανση των κυττάρων με βρωμοδεοξυουριδίνη, ένα ανάλογο θυμιδίνης που ενσωματώνεται στο DNA και μπορεί να ανιχνευθεί μέσω χρώσης με φθορίζοντα αντισώματα. Κατά την έναρξη της σύνθεσης DNA, το DNA που μόλις είχε αντιγραφεί ανιχνεύθηκε σε ~20 εστίες που ήταν συγκεντρωμένες γύρω από τον πυρηνίσκο και συνδεδεμένες με πυρηνικές λαμίνες. Στη συνέχεια, η διαδικασία της σύνθεσης του DNA επεκτάθηκε σε εκατοντάδες εστίες διάσπαρτες σε ολόκληρο τον πυρήνα. Εφόσον σε ένα διπλοειδές κύτταρο θηλαστικού μπορούν να είναι ενεργές ανά πάσα στιγμή ~4.000 θέσεις έναρξης της αντιγραφής, καθεμία από αυτές τις εστίες αντιγραφής θα πρέπει να περιέχει πολλαπλές διχάλες αντιγραφής του DNA. Επομένως, η αντιγραφή του DNA διεξάγεται σε μεγάλες δομές που περιέχουν πολυάριθμα σύμπλοκα αντιγραφής οργανωμένα σε διακριτά λειτουργικά σωμάτια. Τα σωμάτια αυτά έχουν ονομαστεί εργοστάσια αντιγραφής. ΕΙΚΟΝΑ 9.21: Ομαδοποιημένες θέσεις αντιγραφής του DNA. DNA που έχει μόλις αντιγραφεί σημάνθηκε με σύντομη έκθεση των κυττάρων σε βρομοδεοξυουριδίνη, η οποία ενσωματώνεται στο DNA στη θέση της θυμιδίνης. Αυτή η αντικατάσταση επιτρέπει την ανίχνευση του νεοσυντιθέμενου DNA μέσω ανοσοφθορισμού μετά από χρώση με ένα αντίσωμα που αναγνωρίζει τη βρομοδεοξυουριδίνη. Παρατηρήστε ότι το νεοσυντιθέμενο DNA συναντάται σε διακριτές εστίες που κατανέμονται σε ολόκληρο τον πυρήνα. Οι δύο φωτογραφίες δείχνουν την κατανομή στην αρχή και στο τέλος της σύνθεσης του DNA αντίστοιχα. (Kennedy BK, et al., Genes Dev. 14: 2855.) 66
. Τα πυρηνικά στίγματα έχουν είναι δομές με ακανόνιστα σχήματα και μέγεθος 0.8-1.")

67 Πυρηνικά στίγματα: Ενώ τα ενεργώς μεταγραφόμενα γονίδια εμφανίζονται κατανεμημένα σε ολόκληρο τον πυρήνα, τα συστατικά της συσκευής ματίσματος του RNA και RNPs είναι συγκεντρωμένα σε διακριτά πυρηνικά σωμάτια, τα πυρηνικά στίγματα (nuclear speckles). Τα πυρηνικά στίγματα έχουν είναι δομές με ακανόνιστα σχήματα και μέγεθος μm, και ένας τυπικός πυρήνας περιέχει τέτοιες υποπυρηνικές δομές που δεν κατανέμονται ομοιόμορφα σε ολόκληρο τον πυρήνα. Χρώσεις ανοσοφθορισμού με χρήση αντισωμάτων κατέδειξαν ότι στα πυρηνικά στίγματα εντοπίζονται τα συστατικά της συσκευής ματίσματος του RNA συμπεριλαμβανομένων των μικρών πυρηνικών ριβονουκλεοπρωτεϊνικών σωματίων (snrnps) και μη-snrnp πρωτεϊνικών συμπλόκων ματίσματος (π.χ. SC35). Τα πυρηνικά στίγματα είναι θέσεις αποθήκευσης συστατικών της συσκευής ματίσματος (π.χ. οι πρωτεΐνες SR), τα οποία στη συνέχεια στρατολογούνται σε ενεργώς μεταγραφόμενα γονίδια κατά την επεξεργασία του προ-mrna. ΕΙΚΟΝΑ 9.22: Εντοπισμός συστατικών της συσκευής ματίσματος. Η χρώση με αντισώματα ανοσοφθορισμού δείχνει ότι οι παράγοντες ματίσματος συγκεντρώνονται σε διακριτά σωμάτια στο εσωτερικό του πυρήνα, τα οποία ονομάζονται πυρηνικά στίγματα. 67

.")
, που ενδέχεται να στοχεύονται στα σωμάτια PML από το μικρό πολυπεπτίδιο SUMO.")
68 Οι πυρήνες περιέχουν και άλλους τύπους διακριτών δομών, όπως τα σωμάτια PML και τα σωμάτια Cajal. Τα σωμάτια PML (έχουν μέγεθος μm και απαντώνται περίπου 5-20 σε έναν τυπικό πυρήνα. Τα σωμάτια PML ταυτοποιήθηκαν ως διακριτές θέσεις εντοπισμού μιας πρωτεΐνης ρύθμισης της μεταγραφής που συνδέεται με την οξεία προμυελοκυτταρική λευχαιμία (PML; Promyelocytic Leukemia). Τα σωμάτια PML αλληλεπιδρούν με τη χρωματίνη και αποτελούν θέσεις συσσώρευσης μεταγραφικών παραγόντων και πρωτεϊνών τροποποίησης της χρωματίνης (όπως είναι οι απακετυλάσες των ιστονών), που ενδέχεται να στοχεύονται στα σωμάτια PML από το μικρό πολυπεπτίδιο SUMO. Τα σωμάτια PML εντοπίζονται σε θέσεις πλούσιες σε γονίδια και μεταγραφικά ενεργές περιοχές της χρωματίνης. Αντίσωμα PML; Χρώση: Hoescht ΕΙΚΟΝΑ 9.23: Ένα σωμάτιο PML. Το σωμάτιο PML (βέλος) περιβάλλεται από χρωματίνη. 68

 Εικόνα του πυρήνα ενός κυττάρου HeLa σε μικροσκοπία αντίθεσης-διαφορικής συμβολής. Τα βέλη υποδεικνύουν τα δύο σωμάτια Cajal.")
69 Τα σωμάτια Cajal μεγέθους (έως 2) μm περιέχουν τη χαρακτηριστική πρωτεΐνη κοϊλίνη (p80 coilin) και πλήθος μικρών RNP. Τα σωμάτια αυτά φαίνονται ως σπειρωμένα νημάτια και θεωρείται ότι λειτουργούν ως θέσεις συναρμολόγησης και επεξεργασίας των RNP, ιδιαίτερα των μετα-μεταγραφικών τροποποιήσεων των μικρών πυρηνισκικών RNAs (snrnps & snornps). Υπάρχουν περίπου 6 σωμάτια Cajal ανά κύτταρο. Η κοϊλίνη είναι σημαντική για την επεξεργασία των snrnps αλλά επίσης διασυνδέει τα σωμάτια Cajal με τον πυρηνίσκο. ΕΙΚΟΝΑ 9.24: Σωμάτια Cajal στον πυρήνα. (Α) Εικόνα του πυρήνα ενός κυττάρου HeLa σε μικροσκοπία αντίθεσης-διαφορικής συμβολής. Τα βέλη υποδεικνύουν τα δύο σωμάτια Cajal. (Β) Σήμανση του ίδιου πυρήνα με ανοσοφθορισμό, χρησιμοποιώντας αντισώματα που αναγνωρίζουν τις πρωτεΐνες κοϊλίνη (πράσινο) και φιμπριλλαρίνη (κόκκινο). Η φιμπριλλαρίνη συναντάται τόσο στις πυκνές ινώδεις ζώνες των πυρηνίσκων όσο και στα σωμάτια Cajal. Η κοϊλίνη ανιχνεύεται μόνο στα σωμάτια Cajal. 69
 Κύτταρα HeLa επωάστηκαν με βρωμοδεοξυουριδίνη (Br-UTP) και στη συνέχεια μονιμοποιήθηκαν και επωάστηκαν με ένα μονοκλωνικό αντίσωμα κοϊλίνης.")
 (άσπρα βέλη) και σε πολυάριθμες πυρηνοπλασματικές εστίες (κεφαλές βελών).")
70 Η κοϊλίνη (p80; ΜΒ~80 kda) δεν εντοπίζεται μαζί τις θέσεις (εστίες) μεταγραφής DNA. (A) Κύτταρα HeLa επωάστηκαν με βρωμοδεοξυουριδίνη (Br-UTP) και στη συνέχεια μονιμοποιήθηκαν και επωάστηκαν με ένα μονοκλωνικό αντίσωμα κοϊλίνης. Οι θέσεις ενσωμάτωσης της βρωμοδεοξυουριδίνης απεικονίζονται πράσινες, ενώ οι θέσεις που περιέχουν κοϊλίνη εμφανίζονται κόκκινες. Η κοϊλίνη εντοπίζεται σε σπειρωμένα σωμάτια (νημάτια) (άσπρα βέλη) και σε πολυάριθμες πυρηνοπλασματικές εστίες (κεφαλές βελών). (B) Σε πυρήνες κυττάρων HeLa έγινε μικροέγχυση αντισωμάτων για την κοϊλίνη, και 24 h μετά τα κύτταρα επωάστηκαν με Br-UTP. Οι μικροεστίες κοϊλίνης (κόκκινες) δεν συνεντοπίζονται με τις θέσεις (εστίες) έναρξης της μεταγραφής (πράσινες). Bar, 10 μm. Almeida F et al. (1998). Microinjection of anti-coilin antibodies affects the structure of coiled bodies. J Cell Sci 142:899 70
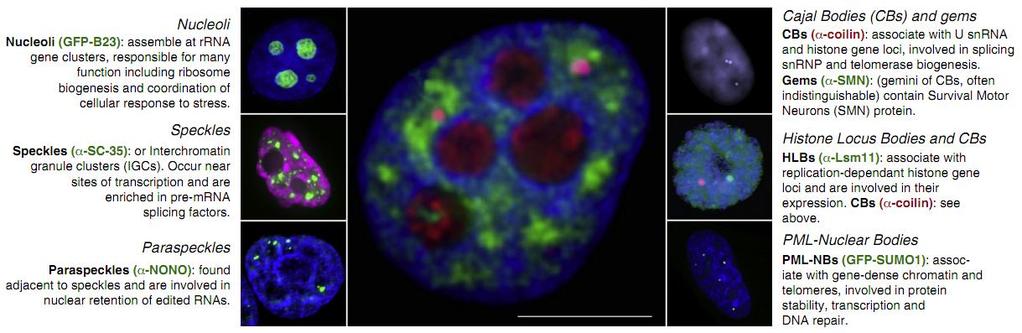
71 71

72 Ο πυρηνίσκος και η επεξεργασία του rrna Το εμφανέστερο από τα πυρηνικά σωμάτια είναι ο πυρηνίσκος (nucleolus), που αποτελεί τη θέση μεταγραφής και επεξεργασίας του rrna, καθώς και τη θέση έναρξης της συναρμολόγησης των ριβοσωμάτων. Τα κύτταρα απαιτούν μεγάλο αριθμό ριβοσωμάτων σε συγκεκριμένες χρονικές στιγμές ώστε να ικανοποιήσουν τις ανάγκες τους για πρωτεϊνοσύνθεση. Τα ενεργώς αυξανόμενα κύτταρα θηλαστικών περιέχουν 5-10 εκατομμύρια ριβοσώματα, τα οποία πρέπει να συντεθούν κάθε φορά που διαιρείται το κύτταρο. Ο πυρηνίσκος είναι ένα εργοστάσιο παραγωγής ριβοσωμάτων που πληροί την ανάγκη για ρυθμιζόμενη και αποδοτική παραγωγή RRNA και για συναρμολόγηση των ριβοσωμικών υπομονάδων. Οι πυρηνίσκοι έχουν επίσης έναν πιο γενικό ρόλο στην τροποποίηση του RNA και αρκετοί τύποι μορίων RNA κινούνται από και προς τον πυρηνίσκο σε συγκεκριμένα στάδια της επεξεργασίας τους. 72

73 Ευκαρυωτικό ριβόσωμα και Γονίδια rrna "S" σημαίνει "Svedberg", μια μονάδα σχετικού ρυθμού καθίζησης σε διαβαθμισμένες στήλες CsCl. Προκειμένου να ανταποκριθούν στην ανάγκη για μεγάλο αριθμό μορίων rrna, όλα τα κύτταρα περιέχουν πολυάριθμα αντίγραφα των γονιδίων rrna. Το γονιδίωμα του ανθρώπου περιέχει ~200 αντίγραφα του γονιδίου που κωδικοποιεί τα 5.8S,18S και 28S rrna και ~2.000 αντίγραφα του γονιδίου που κωδικοποιεί το 5S rrna. Τα γονίδια των 5.8S, 18S και 28S rrna ομαδοποιούνται σε συστοιχίες διαδοχικών επαναλήψεων σε 5 διαφορετικά ανθρώπινα χρωμοσώματα (Chr 13, 14, 15, 21 και 22), ενώ τα γονίδια του 5S rrna εντοπίζονται σε μία μόνο συστοιχία διαδοχικών επαναλήψεων στο Chr 1. Venkatraman Ramakrishnan, Thomas A Steitz και Ada E Yonath, Nobel Χημείας (2009) για τις μελέτες τους στη δομή και τη λειτουργία του ριβοσώματος 73
, 5.8S (160 bp), 18S (1874 bp) και 28S (4718 bp) rrna. Τα γονίδια 5.")
74 Γονίδια του ριβοσωμικού rrna και οργάνωση του πυρηνίσκου Ο πυρηνίσκος, που δεν περιβάλλεται από μεμβράνη, συνδέεται με χρωμοσωμικές περιοχές όπου περιέχονται τα γονίδια 5.8S, 18S και 28S rrna. Τα ριβοσώματα των ανώτερων ευκαρυωτών περιέχουν 4 τύπους RNA: 5S (120 bp), 5.8S (160 bp), 18S (1874 bp) και 28S (4718 bp) rrna. Τα γονίδια 5.8S, 18S και 28S rrna, που εντοπίζονται ως συστοιχία, μεταγράφονται ως ενιαία μονάδα μέσα στον πυρηνίσκο από την RNA πολυμεράση I, που παράγει ένα πρόδρομο rrna, το 45S. Το 45S προ-rrna υπόκειται σε επεξεργασία, παράγοντας το 18S rrna της μικρής ριβοσωμικής υπομονάδας (40S) και τα 5.8S και 28S rrna της μεγάλης ριβοσωμικής υπομονάδας (60S). Η μεταγραφή του 5S rrna, που επίσης συναντάται στη μεγάλη ριβοσωμική υπομονάδα 60S, διεξάγεται εκτός του πυρηνίσκου, (διεξάγεται στον πυρήνα) στους ανώτερους ευκαρυώτες και καταλύεται από την RNA πολυμεράση III. Γενετικός τόπος rdna στον πυρηνίσκο RNA Pol I 18S rrna (40S) και 5.8S & 28S (60S) ΕΙΚΟΝΑ 9.25: Τα γονίδια του ριβοσωμικού RNA. Κάθε γονίδιο rrna αποτελεί μια μονάδα μεταγραφής που περιέχει τα 18S, 5,8S και 28S rrna, καθώς και μεσοδιαστήματα μεταγραφόμενων αλληλουχιών. Τα γονίδια rrna οργανώνονται σε συστοιχίες διαδοχικών επαναλήψεων, οι οποίες χωρίζονται μεταξύ τους από μεσοδιαστήματα μη μεταγραφόμενου DNA. Πολλά από τα ανθρώπινα γονίδια 45S rrna είναι ψευδογονίδια. 74

75 Γονίδιο 5S rrna Σε αντίθεση με τα γονίδια 5.8S, 18S και 28S rrna που εντοπίζονται στον πυρηνίσκο όπου και μεταγράφονται από την RNA πολυμεράση I, τα γονίδια 5S rrna εντοπίζονται στον πυρήνα ως συστοιχίες διαδοχικών επαναλαμβανόμενων αλληλουχιών, και μεταγράφονται από την RNA πολυμεράση ΙΙΙ. Οργάνωση των γονιδίων 5S rrna. Τα γονίδια 5S rrna εντοπίζονται σε πολλαπλά αντίγραφα, καθένα από τα οποία φέρει μια κωδική περιοχή 200 bp από το κωδικόνιο έναρξης της μεταγραφής στο κωδικόνιο λήξης της μεταγραφής που μετά από επεξεργασία το ώριμο 5S rrna που ενσωματώνεται στην μεγάλη ριβοσωμική υπομονάδα 60S, έχει μέγεθος 120 bp. Τα γονίδια 5S rrna διαχωρίζονται μεταξύ τους με αλληλουχίες μεσοδιαστήματος (NTS; non transcribed DNA segments) διαφορετικού μεγέθους. 75


76 Η σημασία της παραγωγής ριβοσωμάτων είναι ιδιαίτερα εμφανής στα ωοκύτταρα, όπου τα γονίδια rrna ενισχύονται ώστε να υποστηριχτεί η σύνθεση του μεγάλου αριθμού ριβοσωμάτων που απαιτούνται για τα πρώιμα στάδια ανάπτυξης των εμβρύων. Στα ωοκύτταρα του βατράχου Xenopus, τα γονίδια rrna πολλαπλασιάζονται ~2.000 φορές, με αποτέλεσμα να υπάρχουν ~106 αντίγραφα τους ανά κύτταρο. Αυτά τα πολλαπλασιασμένα γονίδια rrna κατανέμονται σε χιλιάδες πυρηνίσκους, που υποστηρίζουν τη συσσώρευση σχεδόν 1012 ριβοσωμάτων ανά ωοκύτταρο. ΕΙΚΟΝΑ 9.26: Πυρηνίσκοι σε ωοκύτταρα αμφιβίων. Τα πολλαπλασιασμένα γονίδια rrna ωοκυττάρων Xenopus συγκεντρώνονται σε πολυάριθμους πυρηνίσκους (σκουρόχρωμες κηλίδες). (Brown DD and Dawid IB Science 160: 272.) 76

 το πυκνό ινώδες συστατικό (DFC,")

77 ΕΙΚΟΝΑ 9.27: Δομή του πυρηνίσκου. Φωτογραφία ηλεκτρονικού μικροσκοπίου που δείχνει το ινώδες κέντρο (FC, Fibrillar Center) το πυκνό ινώδες συστατικό (DFC, Dense Fibrillar Component) και το κοκκιώδες συστατικό (G, Granular component) ενός πυρηνίσκου. 77
78 Μορφολογικά, ο πυρηνίσκος διακρίνεται σε 3 περιοχές: το ινώδες κέντρο, το πυκνό ινώδες συστατικό και το κοκκιώδες συστατικό, που αντιπροσωπεύουν τις θέσεις όπου επιτελούνται τα διαδοχικά στάδια της μεταγραφής rrna, της επεξεργασίας rrna και της συναρμολόγησης των ριβοσωμάτων αντίστοιχα. Η τροποποίηση άλλων μικρών μορίων RNA, όπως αυτών του σωματίου αναγνώρισης σήματος, διεξάγεται σε άλλες περιοχές του πυρηνίσκου. Έπειτα από κάθε κυτταρική διαίρεση, οι πυρηνίσκοι συνδέονται με τις χρωμοσωμικές περιοχές που περιέχουν τα γονίδια των 5.8S, 18S και 28S rrna, που γι' αυτόν τον λόγο ονομάζονται περιοχές οργανωτή του πυρηνίσκου (nucleolar organizing regions). Ο σχηματισμός πυρηνίσκων απαιτεί τη μεταγραφή του 45S pre-mrna, που οδηγεί στη σύντηξη μικρών πρόδρομων σωματίων που περιέχουν παράγοντες επεξεργασίας και άλλα συστατικά του πυρηνίσκου. Στα περισσότερα κύτταρα, πυρηνίσκοι που αρχικά είναι μεμονωμένοι συντήκονται, σχηματίζοντας έναν ενιαίο, μεγαλύτερο πυρηνίσκο. Το μέγεθος του πυρηνίσκου εξαρτάται από τη μεταβολική δραστηριότητα του κυττάρου, δηλαδή μεγάλοι πυρηνίσκοι συναντώνται σε κύτταρα με έντονη πρωτεϊνοσυνθετική ενεργότητα. Αυτή η διακύμανση μεγέθους οφείλεται κυρίως σε διαφορές στο μέγεθος του κοκκιώδους συστατικού του πυρηνίσκου, που αντανακλούν τα διαφορετικά επίπεδα συναρμολόγησης των ριβοσωμάτων. 78

79 Μεταγραφή και επεξεργασία του rrna Κάθε περιοχή οργανωτή του πυρηνίσκου περιέχει μια συστοιχία διαδοχικά επαναλαμβανόμενων γονιδίων rrna, που χωρίζονται μεταξύ τους από μεσοδιαστήματα μη μεταγραφόμενου DNA. Τα γονίδια αυτά μεταγράφονται πολύ ενεργά από την RNA πολυμεράση I και είναι ορατά με ευχέρεια σε ηλεκτρονική μικροσκοπία. Στις φωτογραφίες ΗΜ, καθένα από τα γονίδια rrna της συστοιχίας φαίνεται να περιβάλλεται από αυξανόμενες αλυσίδες RNA πυκνής διάταξης, που σχηματίζουν μια δομή που θυμίζει χριστουγεννιάτικο δέντρο. Η υψηλή πυκνότητα των αυξανόμενων αλυσίδων RNA αντανακλά την υψηλή πυκνότητα των μορίων RNA πολυμεράσης, που συναντώνται με μέγιστη πυκνότητα ~1 μορίου πολυμεράσης ανά 100 ζεύγη βάσεων (bp) της μήτρας DNA. Κατεύθυνση μεταγραφής Μονάδα μεταγραφής Μονάδα μεταγραφής Μονάδα μεταγραφής Νεοσυντιθέμενο προ-rnp Μη μεταγραφόμενη συνδετική περιοχή χρωματίνης στον πυρηνίσκο Μη μεταγραφόμενη συνδετική περιοχή χρωματίνης στον πυρηνίσκο ΕΙΚΟΝΑ 9.28: Μεταγραφή γονιδίων rrna. Φωτογραφία ηλεκτρονικού μικροσκοπίου που δείχνει τη χρωματίνη του πυρηνίσκου. Φαίνονται τρία γονίδια rrna χωρισμένα μεταξύ τους από μεσοδιαστήματα μη μεταγραφόμενου DNA. Κάθε γονίδιο rrna περιβάλλεται από μια συστοιχία αυξανόμενων αλυσίδων RNA, σχηματίζοντας μια δομή που θυμίζει χριστουγεννιάτικο δέντρο. 79
80 Στους ανώτερους ευκαρυώτες, το πρωτογενές μετάγραφο των γονιδίων rrna είναι το μεγάλο pre-rrna 45S, που περιέχει τα 18S, 5.8S και 28S rrna, καθώς και μεταγραφόμενες περιοχές μεσοδιαστήματος. Τόσο τα 5' άκρα όσο και τα 3' άκρα των pre-rrna περιέχουν εξωτερικά μεσοδιαστήματα μεταγραφόμενης αλληλουχίας, ενώ μεταξύ των αλληλουχιών των 18S, 5.8S και 28S rrna υπάρχουν επίσης δύο εσωτερικά μεσοδιαστήματα μεταγραφόμενης αλληλουχίας. Τα αρχικά βήματα επεξεργασία του μεταγράφου περιλαμβάνουν τομές στο εξωτερικό μεσοδιάστημα μεταγραφόμενης αλληλουχίας του 5' άκρου του pre-rrna και απομάκρυνση του εξωτερικού μεσοδιαστήματος μεταγραφόμενης αλληλουχίας του 3' άκρου. Ακολουθούν επιπλέον τομές, που οδηγούν στον σχηματισμό των ώριμων μορίων rrna και τροποποιήσεις βάσεων (π.χ. μεθυλίωση). Επίσης, snorna συμμετέχουν στην επεξεργασία του pre-rrna που σχηματίζουν σύμπλοκα με πρωτεΐνες (8-10), παράγοντας snornp. ΕΙΚΟΝΑ 9.29: Επεξεργασία του pre-rrna. Το μετάγραφο 45S pre-rrna των ανώτερων ευκαρυωτών περιέχει εξωτερικά μεσοδιαστήματα μεταγραφόμενης αλληλουχίας (ETS, External Transcribed Spacers) στα δύο άκρα του και εσωτερικά μεσοδιαστήματα μεταγραφόμενης αλληλουχίας (ΙTS, Internal Transcribed Spacers) ανάμεσα στις αλληλουχίες των 18S, 5,8S και 28S rrna. Το pre-rrna υπόκειται σε επεξεργασία μέσω μιας σειράς τομών, από τις οποίες παράγονται τα ώριμα rrna. 80
81 Εκτός από τομές, η επεξεργασία του pre-rrna περιλαμβάνει σε σημαντικό βαθμό τροποποίηση βάσεων, που προκύπτει τόσο από την προσθήκη μεθυλομάδων σε συγκεκριμένες βάσεις και κατάλοιπα ριβόζης όσο και από τη μετατροπή ουριδίνης σε ψευδοουριδίνη. Στα ζωικά κύτταρα, η επεξεργασία του pre-rrna περιλαμβάνει μεθυλίωση ~100 καταλοίπων ριβόζης και 10 αζωτούχων βάσεων, καθώς και σχηματισμό ~100 ψευδοουριδινών. Οι περισσότερες από αυτές τις τροποποιήσεις συμβαίνουν κατά τη διάρκεια ή λίγο μετά την ολοκλήρωση της σύνθεσης του pre-rrna αν και ορισμένες από αυτές λαμβάνουν χώρα σε μετέπειτα στάδια της επεξεργασίας του pre-rrna. Η επεξεργασία του pre-rrna προϋποθέτει τη δράση τόσο πρωτεϊνών όσο και μορίων RNA που εδράζονται στον πυρηνίσκο. Είναι γνωστή η συμμετοχή μικρών πυρηνικών RNA (snrna) στο μάτισμα του pre-mrna. Οι πυρηνίσκοι περιέχουν πάνω από 300 πρωτεΐνες και πολλά (~200) μικρά πυρηνισκικά RNA (snorna; small nucleolar RNA), που συμμετέχουν στην επεξεργασία του pre-rrna. Τα snorna, όπως και τα snrna του σωματίου ματίσματος, σχηματίζουν σύμπλοκα με πρωτεΐνες, δίνοντας snornp. Κάθε snornp αποτελείται από ένα μόριο snorna συνδεδεμένο με 8-10 πρωτεΐνες. Στη συνέχεια, τα snornp συγκεντρώνονται πάνω στο pre-rrna και σχηματίζουν σύμπλοκα επεξεργασίας κατά τρόπο ανάλογο με τον σχηματισμό των σωματίων ματίσματος pre-mrna. Ορισμένα από τα snorna είναι υπεύθυνα για τις τομές του pre-rrna, απ όπου προκύπτουν προϊόντα 18S, 5,8S και 28S. Για παράδειγμα, το snorna U3, που συναντάται σε ~ αντίγραφα ανά κύτταρο και είναι αυτό που υπάρχει σε μεγαλύτερη αφθονία στον πυρηνίσκο, απαιτείται για την τομή του pre-rrna στα 5' εξωτερικά μεσοδιαστήματα μεταγραφόμενης αλληλουχίας. Ομοίως, το snorna U8 απαιτείται για την τομή του pre-rrna ώστε να προκύψουν τα 5,8S και 28S rrna, ενώ το snorna U12 απαιτείται για την τομή του pre-rrna ώστε να προκύψει το 18S rrna. 81


82 Τα περισσότερα snorna συμμετέχουν στη σύνθεση rrna ως RNA-οδηγοί που κατευθύνουν τις ειδικές τροποποιήσεις βάσεων του pre-rrna, όπως είναι η μεθυλίωση συγκεκριμένων καταλοίπων ριβόζης και ο σχηματισμός ψευδοουριδινών. Τα περισσότερα snorna περιέχουν αλληλουχίες μικρού μήκους, ~15 νουκλεοτιδίων, συμπληρωματικές προς τις αλληλουχίες των 18S ή 28S rrna που περιλαμβάνουν τις θέσεις τροποποίησης βάσεων στο rrna. Σχηματίζοντας ομόλογα ζεύγη βάσεων με τις συγκεκριμένες περιοχές του pre-rrna, τα snorna λειτουργούν ως RNA-οδηγοί που καθοδηγούν τα ένζυμα της μεθυλίωσης της ριβόζης ή της ψευδοουριδυλίωσης στις σωστές θέσεις του μορίου pre-rrna. Η τροποποίηση βάσεων είναι απαραίτητη και σε άλλους τύπους RNA εκτός του rrna. Ένα σχετικό παράδειγμα είναι το σωμάτιο αναγνώρισης σήματος RNA. Ο εντοπισμός των snornp στον πυρηνίσκο θεωρείται ότι αποτελεί τη βάση του γενικότερου ρόλου που έχει ο πυρηνίσκος στις διεργασίες τροποποίησης RNA. ΕΙΚΟΝΑ 9.30: Ο ρόλος των snorna στην τροποποίηση των βάσεων του pre-rrna. Τα snorna περιέχουν μικρές αλληλουχίες συμπληρωματικές με rrna. Το ζευγάρωμα βάσεων μεταξύ snorna και pre-rrna οδηγεί σε στόχευση των ενζύμων που καταλύουν τη χημική τροποποίηση αζωτούχων βάσεων (π.χ. μεθυλίωση) στις κατάλληλες θέσεις του pre-rrna. 82

83 Η συναρμολόγηση των ριβοσωμάτων Ο σχηματισμός των ριβοσωμάτων περιλαμβάνει τη συναρμολόγηση του πρόδρομου ριβοσωμικού RNA με ριβοσωμικές πρωτεΐνες και με το 5S rrna. Τα γονίδια που κωδικοποιούν τις ριβοσωμικές πρωτεΐνες μεταγράφονται έξω από τον πυρηνίσκο, από την RNA πολυμεράση II, παράγοντας μόρια mrna που μεταφράζονται στα ριβοσώματα του κυτταροπλάσματος. Στη συνέχεια, οι ριβοσωμικές πρωτεΐνες μεταφέρονται από το κυτταρόπλασμα στον πυρηνίσκο, όπου συναρμολογούνται μαζί με τα rrna για να σχηματίσουν προριβοσωμικά σωμάτια. Αν και τα γονίδια του 5S rrna μεταγράφονται επίσης έξω από τον πυρηνίσκο, από την RNA πολυμεράση III, τα 5S rrna μεταφέρονται και αυτά στο εσωτερικό του πυρηνίσκου, όπου σχηματίζουν προριβοσωμικά σωμάτια. ΕΙΚΟΝΑ 9.31: Συναρμολόγηση του ριβοσώματος. Οι ριβοσωμικές πρωτεΐνες εισάγονται από το κυτταρόπλασμα στον πυρηνίσκο και αρχίζουν να συναρμολογούνται πάνω στο pre-rrna πριν ακόμη αρχίσει η επεξεργασία του. Κατά την επεξεργασία του pre-rrna συναρμολογούνται επιπλέον ριβοσωμικές πρωτεΐνες και το 5S rrna (που συντίθεται σε άλλη περιοχή του πυρήνα), ώστε να σχηματιστούν προριβοσωμικά σωμάτια. Τα τελικά στάδια ωρίμανσης λαμβάνουν χώρα μετά την εξαγωγή των προριβοσωμικών σωματίων στο κυτταρόπλασμα και οδηγούν στον σχηματισμό των ριβοσωμικών υπομονάδων 40S και 60S. 83

 και μικρής")

84 Βιογένεση ευκαρυωτικού ριβοσώματος. Σχηματισμός της μεγάλης (60S) και μικρής ριβοσωμικής (40S) υπομονάδας από τις οποίες προκύπτει το ευκαρυωτικό ριβόσωμα 80S. 84
85 Η συναρμολόγηση των ριβοσωμάτων Η σύνδεση ριβοσωμικών πρωτεϊνών με το rrna ξεκινά ενώ η σύνθεση του pre-rrna βρίσκεται ακόμη σε εξέλιξη και περισσότερες από τις μισές ριβοσωμικές πρωτεΐνες σχηματίζουν σύμπλοκο με το pre-rrna πριν αρχίσει η τομή του. Οι υπόλοιπες ριβοσωμικές πρωτεΐνες καθώς και το 5S rrna ενσωματώνονται στα προριβοσωμικά σωμάτια προοδευτικά κατά την τομή του pre-rrna. Από τα αρχικά στάδια συναρμολόγησης του ριβοσώματος, η επεξεργασία των δύο νεοσχηματιζόμενων ριβοσωμικών υπομονάδων ακολουθεί διαφορετική πορεία. Η επεξεργασία της μικρότερης υπομονάδας (40S), που περιέχει μόνο το 18S rrna, είναι απλούστερη και περιλαμβάνει μόνο 4 ενδονουκλεολυτικές τομές. Στους ανώτερους ευκαρυώτες, η επεξεργασία αυτή ολοκληρώνεται μέσα στον πυρήνα. Η επεξεργασία της μεγαλύτερης υπομονάδας (60S), που περιέχει τα 28S, 5,8S και 5S rrna, περιλαμβάνει εκτενείς νουκλεολυτικές τομές και ολοκληρώνεται μέσα στον πυρηνίσκο. Επομένως, τα περισσότερα προριβοσωμικά σωμάτια στον πυρηνίσκο αντιστοιχούν σε πρόδρομα σύμπλοκα της μεγάλης υπομονάδας (60S). Τα τελικά στάδια ωρίμανσης των ριβοσωμικών υπομονάδων ολοκληρώνονται μετά την εξαγωγή των προριβοσωμικών σωματίων στο κυτταρόπλασμα και οδηγούν στον σχηματισμό των ενεργών υπομονάδων 40S και 60S των ευκαρυωτικών ριβοσωμάτων. 85
86 ΜΟΡΙΑΚΗ ΙΑΤΡΙΚΗ: Ασθένειες που συνδέονται με την πυρηνική λάμινα 1. Μυϊκής δυστροφία Emery-Dreifuss: Μορφή φιλοσύνδετης μυϊκής δυστροφίας που συνδέεται με το χρωμόσωμα Χ λόγω μεταλλάξεων της διαμεμβρανικής πρωτεΐνης του πυρηνικού φακέλου, εμερίνης [Alan Emery και Fritz E. Dreifuss (1996)] Κατά τα πρώτα στάδια της ασθένειας εμφανίζεται ακαμψία στους αγκώνες, λαιμό και φτέρνες των ασθενών και συχνά υπάρχει αποκλεισμός αγωγής στην καρδιά. Τα συμπτώματα αυτά παρατηρούνται μέχρι την ηλικία των 10 ετών και συμπεριλαμβάνουν «ακροποδητί βάδιση» λόγω ακαμψίας του αχίλλειου τένοντα στις φτέρνες και δυσκολία κάμψης των αγκώνων. Από την ηλικία των 20 ετών οι ασθενείς αναπτύσσουν καρδιακά προβλήματα και πολλές φορές κρίνεται απαραίτητη η χρήση βηματοδότη. Παρουσιάζεται σταδιακός εκφυλισμός και εξασθένηση των μυών του ώμου και του βραχίονα, καθώς και των μυών της κνήμης, κάτι που όμως συμβαίνει με αργό ρυθμό και συχνά δεν αποτελεί πρόβλημα προτού το άτομο φτάσει σε προχωρημένη ηλικία. Το εύρημα ήταν απροσδόκητο, καθώς φαινόταν ότι μεταλλάξεις μιας πρωτεΐνης του πυρηνικού φακέλου που εκφράζεται σε όλα τα κύτταρα προκαλούν μια ιστοειδική ασθένεια. Αν και η πρωτεΐνη απουσίαζε από όλα τα κύτταρα του σώματος των ασθενών, η παθολογία εκδηλωνόταν μόνο στον μυϊκό ιστό. 2. Η ίδια μορφή δυστροφίας μπορεί να κληρονομηθεί επίσης με μη φυλοσύνδετο τρόπο. Οι οικογένειες που εμφανιζόταν η μη φυλοσύνδετη μορφή της μυϊκής δυστροφίας Emery-Dreifuss έφεραν μεταλλάξεις στο γονίδιο LMNA, το γονίδιο που κωδικοποιεί τις πυρηνικές λαμίνες Α και C. Μεταλλάξεις επομένως σε 2 διαφορετικά γονίδια - το γονίδιο μιας πρωτεΐνης της εσωτερικής πυρηνικής μεμβράνης και το γονίδιο μιας από τις κυριότερες πυρηνικές λαμίνες - προκαλούν μυϊκή δυστροφία με την ίδια κλινική εικόνα. 3. Παράλληλες μελέτες συνέδεσαν διαφορετικές μεταλλάξεις του γονιδίου LMNA με διαφορετικές ασθένειες: - Μερική λιποδυστροφία τύπου Dunnigan - Διαταραχή Charcot-Marie-Tooth τύπου 2Β1 - Σύνδρομο προγηρίας Hutchinson-Gilford, μια ασθένεια που προκαλεί πρόωρη γήρανση. 4. Διαταραχή Peiger-Huët που οφείλεται σε μεταλλάξεις μιας άλλης πρωτεΐνης της εσωτερικής πυρηνικής μεμβράνης, του υποδοχέα της λαμίνης Β (LBR). 86
 Παιδί που πάσχει από προγηρία Hutchinson-Gilford. Το σύνδρομο Hutchinson-Gilford μεταλλάξεις του γονιδίου LMNA.")
87 ΣΥΝΔΡΟΜΟ ΠΡΟΓΗΡΙΑΣ Hutchinson-Gilford (HGPS) Συχνότητα 1: νεογέννητα παγκοσμίως Τα παιδιά με HGPS γεννιούνται φυσιολογικά και τα συμπτώματα εμφανίζονται πριν την ηλικία των 2 ετών. Τα παιδιά με HGPS πεθαίνουν με μέση ηλικία 13 ετών κυρίως από καρδιακές επιπλοκές. (Α) Παιδί που πάσχει από προγηρία Hutchinson-Gilford. Το σύνδρομο Hutchinson-Gilford μεταλλάξεις του γονιδίου LMNA. οφείλεται σε Έχουν ταυτοποιηθεί >250 μεταλλάξεις στο γονίδιο LMNA που προκαλούν διάφορες ασθένειες, και 21 απ αυτές προκαλούν σύνδρομα προγηρίας. 87


88 (B) Διάγραμμα της δομής ιντρονίων-εξονίων του γονιδίου LMNA και της πρωτεΐνης της λαμίνης Α. Οι σφαιρικές επικράτειες υποδεικνύονται με κόκκινο χρώμα και οι ραβδοειδείς επικράτειες με κίτρινο. Στο μεταλλαγμένο γονίδιο του διαγράμματος υπάρχει μια έλλειψη 150 bp (μαύρο χρώμα) στο εξόνιο 11 88

.")
89 Ανοσοφθορισμός με χρήση αντισώματος για τη λαμίνη Α σε ινοβλάστες δέρματος ενός φυσιολογικού ατόμου (αριστερά) και ενός ασθενή με HGPS (δεξιά) που καταδεικνύει nuclear blebbing. Οι περισσότεροι ασθενείε με HGPS φέρουν τη μετάλλαξη 1824CT (G608G), που ενεργοποιεί μια κρυφή θέση ματίσματος που οδηγεί στην έκφραση μιας μορφής λαμίνης Α στην οποία υπάρχει μια απάλειψη 50 αμινοξέων κοντά στο C-τελικό άκρο της πρωτεΐνης, που αναφέρεται ως LA50/προγερίνη (progerin). Σαν αποτέλεσμα η LA50/προγερίνη διατηρείται μόνιμα φαρνεσυλιωμένη και η συνάθροιση της στα κύτταρα των ασθενών σχετίζεται με την απώλεια της ετεροχρωματίνης και με μεταβολές στη μεθυλίωση των ιστονών. 89
90 90
91 Μοριακή και κυτταρική βάση: Θεωρήθηκε ότι οι μεταλλάξεις των λαμινών θα προκαλούσαν γενικευμένα ελαττώματα στην αρχιτεκτονική του πυρήνα και σοβαρά προβλήματα στα ταχέως διαιρούμενα κύτταρα. Ωστόσο, η δομή του πυρήνα των κυττάρων των ασθενών δεν εμφανίζει παρά ελάχιστες αποκλίσεις από το φυσιολογικό. Το ερώτημα λοιπόν είναι με ποιον τρόπο οι μεταλλάξεις των πυρηνικών λαμινών ή των πρωτεϊνών που προσδένονται σε αυτές προκαλούν διαφορετικές ιστοειδικές ασθένειες. Δύο είναι οι επικρατέστερες υποθέσεις: Η πρώτη είναι η υπόθεση της «γονιδιακής έκφρασης». Σύμφωνα με αυτήν, η ενδεδειγμένη αλληλεπίδραση των δύο λαμινών, Α και C με τον πυρηνικό φάκελο είναι απαραίτητη για την κανονική ιστοειδική έκφραση ορισμένων γονιδίων. Μεταγραφικώς ανενεργά γονίδια εντοπίζονται κατά προτίμηση στην περιφέρεια του πυρήνα, ενώ γονίδια που εκφράζονται συγκεντρώνονται στο κέντρο του πυρήνα, με πρότυπο κατανομής που εξαρτάται από τον κυτταρικό τύπο. Επομένως, η βάση των παραπάνω ασθενειών θα μπορούσε να είναι μια αλλαγή στη γονιδιακή έκφραση λόγω ελαττωματικών αλληλεπιδράσεων μεταξύ πρωτεϊνών. Σύμφωνα με μια δεύτερη υπόθεση, την υπόθεση του «μηχανικού στρες», οι μεταλλαγές του πυρηνικού συμπλόκου λαμινών-εμερίνης θεωρείται ότι εξασθενούν τη δομική ακεραιότητα ενός ενιαίου κυτταροσκελετικού δικτύου. Σε όλα τα κύτταρα, η λάμινα, η εσωτερική πυρηνική μεμβράνη και τα σύμπλοκα των πυρηνικών πόρων είναι στενά συνδεδεμένα μεταξύ τους. Σύμφωνα με την υπόθεση του «μηχανικού στρες», η λάμινα θα μπορούσε, μέσω ινιδίων που προσδένονται στο σύμπλοκο του πυρηνικού πόρου, να συνδέεται έμμεσα με τον κυτταροσκελετό των μυϊκών κυττάρων. Μια τέτοια ερμηνεία αρμόζει περισσότερο στην περίπτωση των μυϊκών δυστροφιών. Πρόληψη και θεραπεία: Η ανακάλυψη ότι μεταλλαγές σε κοινά εκφραζόμενες πρωτεΐνες του συμπλέγματος της πυρηνικής λάμινας προκαλούν διαφορετικές κληρονομούμενες ιστοειδικές ασθένειες άλλαξε τον γενικό τρόπο θεώρησης των πυρηνικών λαμινών. Περαιτέρω ερευνητικές προσπάθειες απαιτούνται για να κατανοηθεί κατά πόσο η κλινική εικόνα για καθεμία από αυτές τις ασθένειες βασίζεται σε μειωμένη μηχανική σταθερότητα του πυρηνικού φακέλου ή σε απορρύθμιση της γονιδιακής έκφρασης. Ωστόσο, η γνώση της μοριακής φύσης των ασθενειών απλοποιεί σημαντικά τη διάγνωση τους και καθιστά πιθανή την εύρεση της κατάλληλης θεραπευτικής αντιμετώπισης. Ένα πρώτο βήμα προς αυτήν την κατεύθυνση αποτελεί η πρόσφατη ανάπτυξη ενός πειραματικού μοντέλου του γονιδίου LMNA (knockout). Συμπτώματα μυϊκής δυστροφίας Emery-Dreifuss εμφανίζονται στα ποντίκια αυτά κατά την εμβρυϊκή ανάπτυξη. Τέλος, γνωρίζουμε πλέον ότι στις «λαμινοπάθειες» του πυρήνα είναι πιθανόν να συμπεριλαμβάνονται αρκετές συγγενείς ασθένειες βραδείας ανάπτυξης. 91
, με αποτέλεσμα ο φαινότυπος τέτοιων διαγονιδιακών ζώων να είναι όμοιος με μια ανωμαλία στον άνθρωπο που ονομάζεται")
.")
92 Διαταραχή Pelger-Huët που οφείλεται σε μεταλλάξεις μιας άλλης πρωτεΐνης της εσωτερικής πυρηνικής μεμβράνης, του υποδοχέα της λαμίνης Β (LBR) Μεταλλάξεις στο γενετικό τόπου ιχθύωσης (ichthyosis) του ποντικού εντοπίζονται εντός του γονιδίου του υποδοχέα της λαμίνης Β (LBR), με αποτέλεσμα ο φαινότυπος τέτοιων διαγονιδιακών ζώων να είναι όμοιος με μια ανωμαλία στον άνθρωπο που ονομάζεται διαταραχή Pelger Huët (PHA). Το γονίδιο του LBR εντοπίζεται στο ανθρώπινο χρωμόσωμα 1. Ραδιογραφία εμπρόσθιων και ποντικών ηλικίας 5 εβδομάδων. πίσω άκρων Τομές δέρματος ενός φυσιολογικού και ενός ζώου με ιχθύωση. Shultz LD, et al. (2002). Mutations at the mouse ichthyosis locus are within the lamin B receptor gene: a single gene model for human Pelger Huët anomaly. Hum. Mol. Genet. 12:

>(b)>(c) Hoffmann et al. (2002).")
93 Διαταραχή Pelger-Huët Ασθενής με PHA ο οποίος έχει κοντύτερα το τρίτο και το πέμπτο metacarpals στο αριστερό του χέρι και από το τρίτο στο πέμπτο metacarpals στο δεξί του χέρι. Έκφραση του LBR με χρήση αντισώματος σε λεμφοκύτταρα ενός: (a) φυσιολογικού ατόμου, (b) ετεροζυγώτη, και (c) ομοζυγώτη που καταδεικνύει τη μειωμένη έκφραση του LBR στα (a)>(b)>(c) Hoffmann et al. (2002). Mutations in the gene encoding the lamin B receptor produce an altered nuclear morphology in granulocytes (Pelger-Huet anomaly). Nature Genetics 93
ΚΕΦΑΛΑΙΟ 9. Ο πυρήνας. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 9 Ο πυρήνας Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 9.1 Ο πυρηνικός φάκελος. (Α) Φωτογραφία ηλεκτρονικού μικροσκοπίου που δείχνει έναν πυρήνα. Η εσωτερική και η εξωτερική
ΚΕΦΑΛΑΙΟ 9 Ο πυρήνας Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 9.1 Ο πυρηνικός φάκελος. (Α) Φωτογραφία ηλεκτρονικού μικροσκοπίου που δείχνει έναν πυρήνα. Η εσωτερική και η εξωτερική
ΜΕΡΟΣ ΙII: ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΚΥΤΤΑΡΩΝ ΚΕΦΑΛΑΙΟ 9
 Ευάγγελος Κωλέττας, B.Sc (HONS), Ph.D.(LON) Αναπληρωτής Καθηγητής, Εργαστήριο Βιολογίας Ιατρική Σχολή, Παν/μιο Ιωαννίνων ΜΕΡΟΣ ΙII: ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΚΥΤΤΑΡΩΝ ΚΕΦΑΛΑΙΟ 9 Ο Πυρήνας ΕΙΣΑΓΩΓΗ Η παρουσία
Ευάγγελος Κωλέττας, B.Sc (HONS), Ph.D.(LON) Αναπληρωτής Καθηγητής, Εργαστήριο Βιολογίας Ιατρική Σχολή, Παν/μιο Ιωαννίνων ΜΕΡΟΣ ΙII: ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΚΥΤΤΑΡΩΝ ΚΕΦΑΛΑΙΟ 9 Ο Πυρήνας ΕΙΣΑΓΩΓΗ Η παρουσία
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη
 Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10
 Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ-ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής Βιολογίας Εργαστήριο Φαρμακολογίας, Τομέας Φαρμακογνωσίας/Φαρμακολογίας
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ-ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής Βιολογίας Εργαστήριο Φαρμακολογίας, Τομέας Φαρμακογνωσίας/Φαρμακολογίας
Κυτταρική Βιολογία. Ενότητα 08 : Βιολογικές μεμβράνες, μεμβρανικά διαμερίσματα, μεταφορά πρωτεϊνών Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 08 : Βιολογικές μεμβράνες, μεμβρανικά διαμερίσματα, μεταφορά πρωτεϊνών Παναγιωτίδης Χρήστος Φαρμακευτικής ΑΠΘ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 08 : Βιολογικές μεμβράνες, μεμβρανικά διαμερίσματα, μεταφορά πρωτεϊνών Παναγιωτίδης Χρήστος Φαρμακευτικής ΑΠΘ
ΚΕΦΑΛΑΙΟ 16. Ο κυτταρικός κύκλος. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; ΘΩΜΑΣ ΑΠΑΝΤΗΣΗ. 2. Ποιες είναι οι κατηγορίες γονιδίων με κριτήριο το προϊόν της μεταγραφής τους;
 Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
Κεντρικό δόγμα της βιολογίας
 Κεντρικό δόγμα της βιολογίας DNA RNA Πρωτεΐνη Μεταγραφή Σύνθεση (μονόκλωνου) RNA από ένα δίκλωνο μόριο DNA κυρίως με τη βοήθεια του ενζύμου RNA πολυμεράση Το προϊόν της μεταγραφής ονομάζεται πρωτογενές
Κεντρικό δόγμα της βιολογίας DNA RNA Πρωτεΐνη Μεταγραφή Σύνθεση (μονόκλωνου) RNA από ένα δίκλωνο μόριο DNA κυρίως με τη βοήθεια του ενζύμου RNA πολυμεράση Το προϊόν της μεταγραφής ονομάζεται πρωτογενές
Η κυτταρική µετατόπιση των πρωτεϊνών
 9-1 Κεφάλαιο 9 Η κυτταρική µετατόπιση των πρωτεϊνών Εισαγωγή Στο κύτταρο η έκφραση των πρωτεϊνών γίνεται από µόνο ένα τύπο ριβοσώµατος (εκτός των µιτοχονδριακών και των χλωροπλαστικών που µοιάζουν µε αυτά
9-1 Κεφάλαιο 9 Η κυτταρική µετατόπιση των πρωτεϊνών Εισαγωγή Στο κύτταρο η έκφραση των πρωτεϊνών γίνεται από µόνο ένα τύπο ριβοσώµατος (εκτός των µιτοχονδριακών και των χλωροπλαστικών που µοιάζουν µε αυτά
ΚΕΦΑΛΑΙΟ 5. Η οργάνωση και οι αλληλουχίες των κυτταρικών γονιδιωμάτων
 ΚΕΦΑΛΑΙΟ 5 Η οργάνωση και οι αλληλουχίες των κυτταρικών γονιδιωμάτων Όπου δεν αναφέρεται ρητά, οι αριθμημένες εικόνες, οι πίνακες και το αντίστοιχο κείμενό τους προέρχονται από το βιβλίο: Το κύτταρο-μια
ΚΕΦΑΛΑΙΟ 5 Η οργάνωση και οι αλληλουχίες των κυτταρικών γονιδιωμάτων Όπου δεν αναφέρεται ρητά, οι αριθμημένες εικόνες, οι πίνακες και το αντίστοιχο κείμενό τους προέρχονται από το βιβλίο: Το κύτταρο-μια
Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της
 ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής
 Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΕΙΣ 4 & 5 (3/3 & 6/3/2017)
") Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (3/3 & 6/3/2017) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (3/3 & 6/3/2017) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
Κυτταρικά οργανίδια Μορφολογία-Σύσταση-Λειτουργία. Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Κυτταρικά οργανίδια Μορφολογία-Σύσταση-Λειτουργία Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικά οργανίδια-ενδοκυττάρια διαμερίσματα-διαλογή πρωτεϊνών -(23/10)
Κυτταρικά οργανίδια Μορφολογία-Σύσταση-Λειτουργία Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικά οργανίδια-ενδοκυττάρια διαμερίσματα-διαλογή πρωτεϊνών -(23/10)
Μοριακή Βιολογία. Ενότητα # (5): Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής
: Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής") ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (5): Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (5): Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν
ΚΕΦΑΛΑΙΟ 19. Τα χρωμοσώματα
 ΚΕΦΑΛΑΙΟ 19 Τα χρωμοσώματα Παράγραφοι 19.1-19.4 (σελ. 771-780) Παράγραφοι 19.7-19.12 (σελ. 784-793) Παράγραφοι 19.16-19.17 (σελ. 797-799) Παράγραφοι 19.19-19.20 (σελ. 801-804) Το μήκος του νουκλεϊκού οξέος
ΚΕΦΑΛΑΙΟ 19 Τα χρωμοσώματα Παράγραφοι 19.1-19.4 (σελ. 771-780) Παράγραφοι 19.7-19.12 (σελ. 784-793) Παράγραφοι 19.16-19.17 (σελ. 797-799) Παράγραφοι 19.19-19.20 (σελ. 801-804) Το μήκος του νουκλεϊκού οξέος
Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν
 Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν ένα δυναµικό σκελετό που χρησιµεύει στη στήριξη και την
Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν ένα δυναµικό σκελετό που χρησιµεύει στη στήριξη και την
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ. ΘΕΜΑ Β Β1. Στήλη Ι Στήλη ΙΙ 1 Α 2 Γ 3 Α 4 Β 5 Α 6 Α 7 Γ
 ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ
 14.1 ΕΙΣΑΓΩΓΗ") 1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ )
") Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016)
") Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
ΘΕΜΑ Α Α1. γ Α2. γ Α3. α Α4. β Α5. β ΘΕΜΑ B B1. B2.
 ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ
 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ: ΜΙΚΡΟΪΝΙΔΙΑ- ΕΝΔΙΑΜΕΣΑ ΙΝΙΔΙΑ- ΜΙΚΡΟΣΩΛΗΝΙΣΚΟΙ Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ: ΜΙΚΡΟΪΝΙΔΙΑ- ΕΝΔΙΑΜΕΣΑ ΙΝΙΔΙΑ- ΜΙΚΡΟΣΩΛΗΝΙΣΚΟΙ Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
Εργασία στην Βιολογία
 Εργασία στην Βιολογία Μεμβράνη του Πυρήνα του Κυττάρου Χρονιά: 2013-2014 Επιμέλεια: Σταμάτης Ορφανός, Μάριος Παναγιωτόπουλος [1] Πυρήνας του Κυττάρου Ο πυρήνας είναι το πιο μεγάλο και ευδιάκριτο οργανίδιο
Εργασία στην Βιολογία Μεμβράνη του Πυρήνα του Κυττάρου Χρονιά: 2013-2014 Επιμέλεια: Σταμάτης Ορφανός, Μάριος Παναγιωτόπουλος [1] Πυρήνας του Κυττάρου Ο πυρήνας είναι το πιο μεγάλο και ευδιάκριτο οργανίδιο
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος. 09/04/ Μοριακή Βιολογία Κεφ. 1 Καθηγητής Δρ. Κ. Ε. Βοργιάς
 Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Κ Ε Φ Α Λ Α Ι Ο 21 : Υποκινητές και Ενισχυτές
 Κ Ε Φ Α Λ Α Ι Ο 21 : Υποκινητές και Ενισχυτές Εικόνα 21.1 Ένα τυπικό γονίδιο που µεταγράφεται από την RNA πολυµεράση ΙΙ έχει έναν υποκινητή ο οποίος εκτείνεται ανοδικά από τη θέση έναρξης της µεταγραφής.
Κ Ε Φ Α Λ Α Ι Ο 21 : Υποκινητές και Ενισχυτές Εικόνα 21.1 Ένα τυπικό γονίδιο που µεταγράφεται από την RNA πολυµεράση ΙΙ έχει έναν υποκινητή ο οποίος εκτείνεται ανοδικά από τη θέση έναρξης της µεταγραφής.
Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA
 Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA Εικόνα 24.1 Το hnrna απαντάται ως ριβονουκλεοπρωτεϊνικό σύµπλοκο µε µορφή µιας σειράς από χάντρες. Εικόνα 24.2 Το RNA τροποποιείται στον πυρήνα
Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA Εικόνα 24.1 Το hnrna απαντάται ως ριβονουκλεοπρωτεϊνικό σύµπλοκο µε µορφή µιας σειράς από χάντρες. Εικόνα 24.2 Το RNA τροποποιείται στον πυρήνα
γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A 1. Να επιλέξετε τη σωστή απάντηση: Α1. Το γονίδιο
γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A 1. Να επιλέξετε τη σωστή απάντηση: Α1. Το γονίδιο
Δοµή και ιδιότητες του DNA
 Δοµή και ιδιότητες του DNA Βακτηριακό χρωµόσωµα ευκαρυωτικό χρωµόσωµα και χρωµατίνη 28/02/2014 1 Tο βακτηριακό γονιδίωµα περιέχεται σε ένα κυκλικό DNA µήκους 1300 µm εντός του βακτηριακού κυττάρου που
Δοµή και ιδιότητες του DNA Βακτηριακό χρωµόσωµα ευκαρυωτικό χρωµόσωµα και χρωµατίνη 28/02/2014 1 Tο βακτηριακό γονιδίωµα περιέχεται σε ένα κυκλικό DNA µήκους 1300 µm εντός του βακτηριακού κυττάρου που
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών
 Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
ΚΥΤΤΑΡΙΚΟΣ ΚΥΤΚΛΟΣ. abee/biobk/biobookmeiosis.html. pdf
 ΚΥΤΤΑΡΙΚΟΣ ΚΥΤΚΛΟΣ http://www2.estrellamountain.edu/faculty/far abee/biobk/biobookmeiosis.html pdf https://www.youtube.com/watch?v=wy3n5nczbhq http://www.youtube.com/watch?v=lf9rcqifx34&feature=related
ΚΥΤΤΑΡΙΚΟΣ ΚΥΤΚΛΟΣ http://www2.estrellamountain.edu/faculty/far abee/biobk/biobookmeiosis.html pdf https://www.youtube.com/watch?v=wy3n5nczbhq http://www.youtube.com/watch?v=lf9rcqifx34&feature=related
Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014
 Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014 ΘΕΜΑ Α Α1. β Α2. β Α3. β Α4. β Α5. β ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ B B1. Ο όρος γονιδιακή έκφραση αναφέρεται συνήθως σε όλη τη διαδικασία με την οποία
Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014 ΘΕΜΑ Α Α1. β Α2. β Α3. β Α4. β Α5. β ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ B B1. Ο όρος γονιδιακή έκφραση αναφέρεται συνήθως σε όλη τη διαδικασία με την οποία
Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Σε ένα
ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Σε ένα
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
Κυτταρική Βιολογία. Ενότητα 07 : Κυτταροσκελετός. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 07 : Κυτταροσκελετός Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 07 : Κυτταροσκελετός Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
ΑΠΑΝΤΗΣΕΙΣ. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
 ΒΙΟΛΟΓΙΑΣ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
ΒΙΟΛΟΓΙΑΣ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
Ρύθμιση της μετάφρασης. Η μετάφραση του mrna της φερριτίνης ρυθμίζεται από τη διαθεσιμότητα σιδήρου
 Ρύθμιση της μετάφρασης Η μετάφραση του mrna της φερριτίνης ρυθμίζεται από τη διαθεσιμότητα σιδήρου Ρύθμιση της μετάφρασης 2. Πρόσδεση πρωτεϊνικών καταστολέων σε αλληλουχίες της 3 περιοχής, στη μη-μεταφραζόμενη
Ρύθμιση της μετάφρασης Η μετάφραση του mrna της φερριτίνης ρυθμίζεται από τη διαθεσιμότητα σιδήρου Ρύθμιση της μετάφρασης 2. Πρόσδεση πρωτεϊνικών καταστολέων σε αλληλουχίες της 3 περιοχής, στη μη-μεταφραζόμενη
Δομή των μυϊκών κυττάρων.
 Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
ΚΕΦΑΛΑΙΟ 15. Κυτταρική ρύθμιση. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ)
") ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ) ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό κάθε μιας από τις παρακάτω ημιτελείς προτάσεις 1-5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ) ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό κάθε μιας από τις παρακάτω ημιτελείς προτάσεις 1-5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα
 Εργασία στη Βιολογία ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Η ροή της πληροφορίας για το σχηματισμό των πρωτεϊνών, προϋποθέτει τη μεταφορά της από το DNA στο RNA (ΜΕΤΑΓΡΑΦΗ).
Εργασία στη Βιολογία ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Η ροή της πληροφορίας για το σχηματισμό των πρωτεϊνών, προϋποθέτει τη μεταφορά της από το DNA στο RNA (ΜΕΤΑΓΡΑΦΗ).
3. Σε ένα σωματικό κύτταρο ανθρώπου που βρίσκεται στη μεσόφαση πριν την αντιγραφή υπάρχουν:
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Στο οπερόνιο της λακτόζης: Α. Η πρωτεΐνη καταστολέας
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Στο οπερόνιο της λακτόζης: Α. Η πρωτεΐνη καταστολέας
ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ & ΕΠΑΛ (ΟΜΑΔΑ Β ) ΝΕΟ & ΠΑΛΑΙΟ ΣΥΣΤΗΜΑ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 27/05/2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΟΠ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1:
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ & ΕΠΑΛ (ΟΜΑΔΑ Β ) ΝΕΟ & ΠΑΛΑΙΟ ΣΥΣΤΗΜΑ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 27/05/2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΟΠ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1:
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Βασικοί μηχανισμοί προσαρμογής
 ΣΥΓΚΡΙΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 23-24, 18/4/2016 Π.Παπαζαφείρη Βασικοί μηχανισμοί προσαρμογής Προσαρμογή σε μοριακό και γονιδιακό επίπεδο Επίπεδα ελέγχου 1. Πρωτεïνική δράση 2. Πρωτεïνοσύνθεση 3. Ρύθμιση της
ΣΥΓΚΡΙΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 23-24, 18/4/2016 Π.Παπαζαφείρη Βασικοί μηχανισμοί προσαρμογής Προσαρμογή σε μοριακό και γονιδιακό επίπεδο Επίπεδα ελέγχου 1. Πρωτεïνική δράση 2. Πρωτεïνοσύνθεση 3. Ρύθμιση της
Βιολογία Προσανατολισμού Γ Λυκείου
 Β ΓΕΛ 05/ 05/ 2019 Βιολογία Προσανατολισμού Γ Λυκείου ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Εσώνια δεν υπάρχουν: Α. Στο DNA των ιών που προσβάλλουν
Β ΓΕΛ 05/ 05/ 2019 Βιολογία Προσανατολισμού Γ Λυκείου ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Εσώνια δεν υπάρχουν: Α. Στο DNA των ιών που προσβάλλουν
ΚΕΦΑΛΑΙΟ 29 Σύνθεση πρωτεϊνών
 ΚΕΦΑΛΑΙΟ 29 Σύνθεση πρωτεϊνών Αφού είδαμε πως το DNA αντιγράφεται και μεταγράφεται, τώρα θα εξετάσουμε τη διαδικασία με την παράγονται οι πρωτεϊνες Στην ουσία θα πρέπει να συνδυαστεί ο κώδικας δύο βιβλιοθηκών,
ΚΕΦΑΛΑΙΟ 29 Σύνθεση πρωτεϊνών Αφού είδαμε πως το DNA αντιγράφεται και μεταγράφεται, τώρα θα εξετάσουμε τη διαδικασία με την παράγονται οι πρωτεϊνες Στην ουσία θα πρέπει να συνδυαστεί ο κώδικας δύο βιβλιοθηκών,
Σύνθεση πρωτεϊνών και σημειακές μεταλλάξεις Γ. Παπανικολαόυ MD, PhD
 Σύνθεση πρωτεϊνών και σημειακές μεταλλάξεις Γ. Παπανικολαόυ MD, PhD Κεντρικό δόγμα DNA Μεταγραφή RNA Αντιγραφή Αντίστροφη μεταγραφάση Μετάφραση mrna Πρωτείνη RNA γονίδια trna rrna SnRNA microrna κα Ροή
Σύνθεση πρωτεϊνών και σημειακές μεταλλάξεις Γ. Παπανικολαόυ MD, PhD Κεντρικό δόγμα DNA Μεταγραφή RNA Αντιγραφή Αντίστροφη μεταγραφάση Μετάφραση mrna Πρωτείνη RNA γονίδια trna rrna SnRNA microrna κα Ροή
ΑΝΤΙΓΡΑΦΗ, ΕΚΦΡΑΣΗ ΚΑΙ ΡΥΘΜΙΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ. Πώς από το DNA φτάνουμε στις πρωτεΐνες
 ΑΝΤΙΓΡΑΦΗ, ΕΚΦΡΑΣΗ ΚΑΙ ΡΥΘΜΙΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ Πώς από το DNA φτάνουμε στις πρωτεΐνες Αντιγραφή του DNA o Ο μηχανισμός αντιγραφής του DNA ονομάζεται ημισυντηρητικός διότι κατά την αντιγραφή του
ΑΝΤΙΓΡΑΦΗ, ΕΚΦΡΑΣΗ ΚΑΙ ΡΥΘΜΙΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ Πώς από το DNA φτάνουμε στις πρωτεΐνες Αντιγραφή του DNA o Ο μηχανισμός αντιγραφής του DNA ονομάζεται ημισυντηρητικός διότι κατά την αντιγραφή του
ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 02/12/2012 ΑΠΑΝΤΗΣΕΙΣ
 ΤΣΙΜΙΣΚΗ &ΚΑΡΟΛΟΥ ΝΤΗΛ ΓΩΝΙΑ THΛ: 270727 222594 ΑΡΤΑΚΗΣ 12 - Κ. ΤΟΥΜΠΑ THΛ: 919113 949422 ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 02/12/2012 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1 ο Α. Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη
ΤΣΙΜΙΣΚΗ &ΚΑΡΟΛΟΥ ΝΤΗΛ ΓΩΝΙΑ THΛ: 270727 222594 ΑΡΤΑΚΗΣ 12 - Κ. ΤΟΥΜΠΑ THΛ: 919113 949422 ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 02/12/2012 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1 ο Α. Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη
Μάθηµα: Κίνηση πρωτεινών
 Μάθηµα: Κίνηση πρωτεινών ιάλεξη 1:Σύνθεση πρωτεινών- Ριβόσωµα Κώστας Τοκατλίδης Η σύνθεση πρωτεινών απαιτεί την µετάφραση αλληλουχίας νουκλεοτιδίων σε αλληλουχία αµινοξέων Οι συνθετάσες των αµινοακυλο-trna
Μάθηµα: Κίνηση πρωτεινών ιάλεξη 1:Σύνθεση πρωτεινών- Ριβόσωµα Κώστας Τοκατλίδης Η σύνθεση πρωτεινών απαιτεί την µετάφραση αλληλουχίας νουκλεοτιδίων σε αλληλουχία αµινοξέων Οι συνθετάσες των αµινοακυλο-trna
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1
 Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
Κοζάνογλου Αικατερίνη Βιολόγος Συνεργάτης ΕΚΦΕ
 ΜΟΝΤΕΛΑ DNA ΚΑΙ ΝΟΥΚΛΕΟΣΩΜΑΤΩΝ κοπός 1. Να παρατηρήσουν τη δομή του DNA στο χώρο. 2. Να διαπιστώσουν από ποια επιμέρους τμήματα αποτελούνται οι αλυσίδες του DNA και πως συνδέονται μεταξύ τους. 3. Να παρατηρήσουν
ΜΟΝΤΕΛΑ DNA ΚΑΙ ΝΟΥΚΛΕΟΣΩΜΑΤΩΝ κοπός 1. Να παρατηρήσουν τη δομή του DNA στο χώρο. 2. Να διαπιστώσουν από ποια επιμέρους τμήματα αποτελούνται οι αλυσίδες του DNA και πως συνδέονται μεταξύ τους. 3. Να παρατηρήσουν
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΗ 4 (6/3/2013)
") Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ
 ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ") ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ Θέματα 1. Ποια είναι η δομή και τα βασικά χαρακτηριστικά των λιπαρών οξέων 2. Πώς απομακρύνονται
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ Θέματα 1. Ποια είναι η δομή και τα βασικά χαρακτηριστικά των λιπαρών οξέων 2. Πώς απομακρύνονται
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1 ο Α. Ερωτήσεις πολλαπλής επιλογής 1. δ 2. β 3. γ 4. γ 5. β Β. Ερωτήσεις σωστού λάθους 1. Λάθος 2. Σωστό 3. Λάθος 4. Λάθος 5. Σωστό ΘΕΜΑ
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1 ο Α. Ερωτήσεις πολλαπλής επιλογής 1. δ 2. β 3. γ 4. γ 5. β Β. Ερωτήσεις σωστού λάθους 1. Λάθος 2. Σωστό 3. Λάθος 4. Λάθος 5. Σωστό ΘΕΜΑ
Μηχανισμοί Μεταμεταγραφικού Ελέγχου
 Μηχανισμοί Μεταμεταγραφικού Ελέγχου Δημήτρης Λ. Κοντογιάννης, PhD Ερευνητής Β, Ινστιτούτο Ανοσολογίας ΕΕΠΑ Αθήνα 6-2-2010 Crowne-Plaza Μεταμεταγραφικοί Μηχανισμοί Χρήσης- Τα Βασικά... ΕΕΠΑ Αθήνα 6-2-2010
Μηχανισμοί Μεταμεταγραφικού Ελέγχου Δημήτρης Λ. Κοντογιάννης, PhD Ερευνητής Β, Ινστιτούτο Ανοσολογίας ΕΕΠΑ Αθήνα 6-2-2010 Crowne-Plaza Μεταμεταγραφικοί Μηχανισμοί Χρήσης- Τα Βασικά... ΕΕΠΑ Αθήνα 6-2-2010
) 4 x 10 5 ) 2 x 10 5
 4 x 10 5 ) 2 x 10 5") 1 & ( ) 27 2016 - : ( ) ( ) : (5) 1 5,,,. 1.. DNA. DNA. DNA. RNA. 2.. 46... DNA 1,5 x 10 9. 3. Dolly. DNA.. 1. DNA. 4. (ADA),. AIDS.... 1 5 2 & 5. Ti. Agrobacterium tumefaciens. T 2. DNA.. 1. N,,,. 1.
1 & ( ) 27 2016 - : ( ) ( ) : (5) 1 5,,,. 1.. DNA. DNA. DNA. RNA. 2.. 46... DNA 1,5 x 10 9. 3. Dolly. DNA.. 1. DNA. 4. (ADA),. AIDS.... 1 5 2 & 5. Ti. Agrobacterium tumefaciens. T 2. DNA.. 1. N,,,. 1.
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ
 BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΜΕΡΟΣ ΠΡΩΤΟ - ΠΕΡΙΓΡΑΜΜΑ 5.1 ΈΝΑ ΝΟΥΚΛΕΙΝΙΚΟ ΟΞΥ ΑΠΟΤΕΛΕΙΤΑΙ ΑΠΌ ΤΕΣΣΕΡΑ ΕΙΔΗ ΒΑΣΕΩΝ, ΠΟΥ ΠΡΟΣΔΕΝΟΝΤΑΙ ΣΕ ΈΝΑ ΚΟΡΜΟ ΣΑΚΧΑΡΩΝ ΦΩΣΦΟΡΙΚΩΝ 5.2 ΈΝΑ ΖΕΥΓΟΣ
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΜΕΡΟΣ ΠΡΩΤΟ - ΠΕΡΙΓΡΑΜΜΑ 5.1 ΈΝΑ ΝΟΥΚΛΕΙΝΙΚΟ ΟΞΥ ΑΠΟΤΕΛΕΙΤΑΙ ΑΠΌ ΤΕΣΣΕΡΑ ΕΙΔΗ ΒΑΣΕΩΝ, ΠΟΥ ΠΡΟΣΔΕΝΟΝΤΑΙ ΣΕ ΈΝΑ ΚΟΡΜΟ ΣΑΚΧΑΡΩΝ ΦΩΣΦΟΡΙΚΩΝ 5.2 ΈΝΑ ΖΕΥΓΟΣ
ΧΡΗΣΤΟΣ ΚΑΚΑΒΑΣ 1 ΚΑΘΗΓΗΤΗΣ ΒΙΟΛΟΓΟΣ Μ.Δ.Ε
 ΚΕΦΑΛΑΙΟ 2 ον. ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ ΤΙ ΠΡΕΠΕΙ ΝΑ ΞΕΡΩ. 1. Τη δομή της δίκλωνης έλικας πάρα πολύ καλά. 2. Τους δεσμούς υδρογόνου μεταξύ των συμπληρωματικών βάσεων και την επίπτωσή
ΚΕΦΑΛΑΙΟ 2 ον. ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ ΤΙ ΠΡΕΠΕΙ ΝΑ ΞΕΡΩ. 1. Τη δομή της δίκλωνης έλικας πάρα πολύ καλά. 2. Τους δεσμούς υδρογόνου μεταξύ των συμπληρωματικών βάσεων και την επίπτωσή
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ
 ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ
 Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΝΤΙΓΡΑΦΗ ΜΕΤΑΓΡΑΦΗ Γίνεται σύνθεση DNA. Γίνεται σύνθεση RNA. Εξασφαλίζεται η διαιώνιση της γενετικής
Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΝΤΙΓΡΑΦΗ ΜΕΤΑΓΡΑΦΗ Γίνεται σύνθεση DNA. Γίνεται σύνθεση RNA. Εξασφαλίζεται η διαιώνιση της γενετικής
Ημερομηνία: Κυριακή 29 Οκτωβρίου 2017 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ
 ΤΑΞΗ: ΜΑΘΗΜΑ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ημερομηνία: Κυριακή 29 Οκτωβρίου 2017 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ Στις ημιτελείς προτάσεις Α1 Α4 να γράψετε στο τετράδιό σας τον αριθμό
ΤΑΞΗ: ΜΑΘΗΜΑ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ημερομηνία: Κυριακή 29 Οκτωβρίου 2017 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ Στις ημιτελείς προτάσεις Α1 Α4 να γράψετε στο τετράδιό σας τον αριθμό
Θέματα πριν τις εξετάσεις. Καλό διάβασμα Καλή επιτυχία
 Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
Έλεγχος κυτταρικού κύκλου Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Έλεγχος κυτταρικού κύκλου Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Πως το κύτταρο διπλασιάζει τα συστατικά του; Πως γίνεται ο διαχωρισμός των συστατικών στα
Έλεγχος κυτταρικού κύκλου Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Πως το κύτταρο διπλασιάζει τα συστατικά του; Πως γίνεται ο διαχωρισμός των συστατικών στα
ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΟΜΑΔΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Γ ΛΥΚΕΙΟΥ
 ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΟΜΑΔΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Γ ΛΥΚΕΙΟΥ Επιμέλεια: Βουδούρη Καλλιρρόη Ριζηνίας 69 & Λασαίας 21 τηλ.2810313170 www.kmathisi.com ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΟΝΟΜΑ:..
ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΟΜΑΔΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Γ ΛΥΚΕΙΟΥ Επιμέλεια: Βουδούρη Καλλιρρόη Ριζηνίας 69 & Λασαίας 21 τηλ.2810313170 www.kmathisi.com ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΟΝΟΜΑ:..
ΚΥΤΤΑΡΙΚΗ ΙΑΙΡΕΣΗ. αναπαραγωγή. αύξηση αριθµού κυττάρων ανάπτυξη
 ΚΥΤΤΑΡΙΚΗ ΙΑΙΡΕΣΗ αναπαραγωγή αύξηση αριθµού κυττάρων ανάπτυξη επιδιόρθωση ιστών Κυτταρική οργάνωση του γενετικού υλικού Γονιδίωµα: Το σύνολο του γενετικού υλικού (DNA) ενός κυττάρου Στα προκαρυωτικά κύτταρα
ΚΥΤΤΑΡΙΚΗ ΙΑΙΡΕΣΗ αναπαραγωγή αύξηση αριθµού κυττάρων ανάπτυξη επιδιόρθωση ιστών Κυτταρική οργάνωση του γενετικού υλικού Γονιδίωµα: Το σύνολο του γενετικού υλικού (DNA) ενός κυττάρου Στα προκαρυωτικά κύτταρα
Β. Σελ 60 σχολικού: «Η αποµόνωση του συνολικού έως και σελ 61 από µία cdna βιβλιοθήκη.». Γ. ι ι α α α ι α α ι α α α! " # $ % & ' ( ) ( ) ( * % + α ι α
 ( ) ( * % + α ι α") ! THΛ: 270727 222594 THΛ: 919113 949422 Απαντήσεις: " # $ % & ' 1=γ, 2=β, 3=γ, 4=β, 5=δ. " # $ % ( ' εδοµένα από την ανάλυση του ποσοστού των βάσεων σε µόρια DNA από διαφορετικούς οργανισµούς έδειχναν
! THΛ: 270727 222594 THΛ: 919113 949422 Απαντήσεις: " # $ % & ' 1=γ, 2=β, 3=γ, 4=β, 5=δ. " # $ % ( ' εδοµένα από την ανάλυση του ποσοστού των βάσεων σε µόρια DNA από διαφορετικούς οργανισµούς έδειχναν
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς
 Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο. 3. Το DNA των μιτοχονδρίων έχει μεγαλύτερο μήκος από αυτό των χλωροπλαστών.
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
Ενδεικτικές απαντήσεις
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
Βιολογία προσανατολισμού
 Βιολογία προσανατολισμού ΘΕΜΑ Α Στις προτάσεις από Α1-Α5 να βρείτε την σωστή απάντηση. Α1. Ένας ερευνητής απομόνωσε ένα ασυνεχές γονίδιο από το γονιδίωμα ανθρώπινων κυττάρων. Το γονίδιο συνδέθηκε με βακτηριακό
Βιολογία προσανατολισμού ΘΕΜΑ Α Στις προτάσεις από Α1-Α5 να βρείτε την σωστή απάντηση. Α1. Ένας ερευνητής απομόνωσε ένα ασυνεχές γονίδιο από το γονιδίωμα ανθρώπινων κυττάρων. Το γονίδιο συνδέθηκε με βακτηριακό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
Τριαδικό σύστημα του Salthe (1985) επίπεδα σημειωτικών διεργασιών.
 επίπεδα σημειωτικών διεργασιών.") Τριαδικό σύστημα του Salthe (1985) επίπεδα σημειωτικών διεργασιών. Υψηλότερο επίπεδο-εκλεκτικό σημειωτικό περιβάλλον ή πλαίσιο-όροι ορίου-συνοριακές συνθήκες-εκλεκτικός ρυθμιστικός ρόλος-μακροσημειωτικό
Τριαδικό σύστημα του Salthe (1985) επίπεδα σημειωτικών διεργασιών. Υψηλότερο επίπεδο-εκλεκτικό σημειωτικό περιβάλλον ή πλαίσιο-όροι ορίου-συνοριακές συνθήκες-εκλεκτικός ρυθμιστικός ρόλος-μακροσημειωτικό
4 DNA ελικάση Δ Περιέχουν πανομοιότυπο DNA. 6 Πριμόσωμα ΣΤ Σταθερότητα κατά μήκος της κάθε πολυνουκλεοτιδικής αλυσίδας
 Θέμα 1 ο 1. Αντιστοιχείστε όλες τις έννοιες της στήλης 1 με όλες τις φράσεις της στήλης 2 (Οι αντιστοιχίσεις να γραφούν στην κόλλα απαντήσεων σας & όχι στη φωτοτυπία των θεμάτων) 1 2 1 Αδελφές χρωματίδες
Θέμα 1 ο 1. Αντιστοιχείστε όλες τις έννοιες της στήλης 1 με όλες τις φράσεις της στήλης 2 (Οι αντιστοιχίσεις να γραφούν στην κόλλα απαντήσεων σας & όχι στη φωτοτυπία των θεμάτων) 1 2 1 Αδελφές χρωματίδες
ΚΕΦΑΛΑΙΟ 11. Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα
 ΚΕΦΑΛΑΙΟ 11 Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα Τα ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΑ Μέρος Ε ΤΑ ΒΑΣΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΩΝ - Περιέχουν ένζυµα για ποικίλες µεταβολικές
ΚΕΦΑΛΑΙΟ 11 Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα Τα ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΑ Μέρος Ε ΤΑ ΒΑΣΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΩΝ - Περιέχουν ένζυµα για ποικίλες µεταβολικές
Μετάφραση - Translation Μετα-μεταφραστικές τροποποιήσεις
 Μετάφραση Πρωτεϊνοσύνθεση Μετάφραση - Translation Διαδικασία κατά την οποία η γενετική πληροφορία μετατρέπεται σε λειτουργικά μόρια, τις πρωτεΐνες. Μετα-μεταφραστικές τροποποιήσεις Post-translational modifications
Μετάφραση Πρωτεϊνοσύνθεση Μετάφραση - Translation Διαδικασία κατά την οποία η γενετική πληροφορία μετατρέπεται σε λειτουργικά μόρια, τις πρωτεΐνες. Μετα-μεταφραστικές τροποποιήσεις Post-translational modifications
Βιολογία Θετικής Κατεύθυνσης. Κεφάλαιο 2 ο Αντιγραφή, έκφραση & ρύθμιση της γενετικής πληροφορίας
 Βιολογία Θετικής Κατεύθυνσης Κεφάλαιο 2 ο Αντιγραφή, έκφραση & ρύθμιση της γενετικής πληροφορίας Αντιγραφή του DNA Οι Watson & Crick το 1953 μαζί με το μοντέλο της διπλής έλικας, πρότειναν και έναν τρόπο
Βιολογία Θετικής Κατεύθυνσης Κεφάλαιο 2 ο Αντιγραφή, έκφραση & ρύθμιση της γενετικής πληροφορίας Αντιγραφή του DNA Οι Watson & Crick το 1953 μαζί με το μοντέλο της διπλής έλικας, πρότειναν και έναν τρόπο
Λειτουργική Περιοχή της GTP-ασης
 Λειτουργική Περιοχή της GTP-ασης Οι πρωτεΐνες πού φαίνεται να εµπλέκονται στην περιοχή είναι οι πρωτεΐνες L7/L12. Οι πρωτεΐνες αυτές φαίνεται να είναι απαραίτητες για την ενεργότητα του ριβοσώµατος και
Λειτουργική Περιοχή της GTP-ασης Οι πρωτεΐνες πού φαίνεται να εµπλέκονται στην περιοχή είναι οι πρωτεΐνες L7/L12. Οι πρωτεΐνες αυτές φαίνεται να είναι απαραίτητες για την ενεργότητα του ριβοσώµατος και
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΤΡΗΝΑ ΣΟΤ ΚΤΣΣΑΡΟΤ: ΔΟΜΗ, ΛΕΙΣΟΤΡΓΙΑ, ΔΙΑΚΙΝΗΗ ΤΣΑΣΙΚΩΝ ΔΙΑΜΕΟΤ ΣΗ ΠΤΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ
 ΠΤΡΗΝΑ ΣΟΤ ΚΤΣΣΑΡΟΤ: ΔΟΜΗ, ΛΕΙΣΟΤΡΓΙΑ, ΔΙΑΚΙΝΗΗ ΤΣΑΣΙΚΩΝ ΔΙΑΜΕΟΤ ΣΗ ΠΤΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ ΓΤΜΝΑΙΟ ΚΕΡΑΣΕΑ ΦΟΛ. ΕΣΟ 2011-12 ΣΜΗΜΑ Γ3 ΟΡΥΑΝΙΔΗ ΓΙΩΡΓΟ ΙΔΕΡΗ ΒΑΓΓΕΛΗ Σο κύτταρο είναι η μικρότερη ζωντανή μονάδα,
ΠΤΡΗΝΑ ΣΟΤ ΚΤΣΣΑΡΟΤ: ΔΟΜΗ, ΛΕΙΣΟΤΡΓΙΑ, ΔΙΑΚΙΝΗΗ ΤΣΑΣΙΚΩΝ ΔΙΑΜΕΟΤ ΣΗ ΠΤΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ ΓΤΜΝΑΙΟ ΚΕΡΑΣΕΑ ΦΟΛ. ΕΣΟ 2011-12 ΣΜΗΜΑ Γ3 ΟΡΥΑΝΙΔΗ ΓΙΩΡΓΟ ΙΔΕΡΗ ΒΑΓΓΕΛΗ Σο κύτταρο είναι η μικρότερη ζωντανή μονάδα,
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης
 Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Κυτταρική Βιολογία. Ενότητα 05 : Η μεταγραφή του DNA και η ρύθμισή της. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 05 : Η μεταγραφή του DNA και η ρύθμισή της Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 05 : Η μεταγραφή του DNA και η ρύθμισή της Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού
 Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού Θέμα Α Α1) γ Α2) γ Α3) δ Α4) β Α5) β Θέμα Β Β1. Α = υδροξύλιο, Β = πρωταρχικό τμήμα, Γ = θέση έναρξης αντιγραφής, Δ = φωσφορική ομάδα, Ε = τμήμα
Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού Θέμα Α Α1) γ Α2) γ Α3) δ Α4) β Α5) β Θέμα Β Β1. Α = υδροξύλιο, Β = πρωταρχικό τμήμα, Γ = θέση έναρξης αντιγραφής, Δ = φωσφορική ομάδα, Ε = τμήμα
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΒΙΟΛΟΓΙΑ. 1 ο ΔΙΑΓΩΝΙΣΜΑ ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ 1 o
 ΘΕΜΑ 1 o Γ ΛΥΚΕΙΟΥ-ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ 1 ο ΔΙΑΓΩΝΙΣΜΑ Α. Γιατί τα βακτήρια μπορούν να χρησιμοποιηθούν σαν «εργοστάσια παραγωγής ανθρώπινων πρωτεϊνών»; Β. Σε ένα βακτήριο εισάγεται με τη μέθοδο του ανασυνδυασμένου
ΘΕΜΑ 1 o Γ ΛΥΚΕΙΟΥ-ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ 1 ο ΔΙΑΓΩΝΙΣΜΑ Α. Γιατί τα βακτήρια μπορούν να χρησιμοποιηθούν σαν «εργοστάσια παραγωγής ανθρώπινων πρωτεϊνών»; Β. Σε ένα βακτήριο εισάγεται με τη μέθοδο του ανασυνδυασμένου
 Σύμφωνα με τις απόψεις που σήμερα επικρατούν η διαδικασία της έναρξης μπορεί να διαιρεθεί σε πέντε στάδια Διάσταση του 80S Ριβοσώματος στη 60S και την 40S υπομοναδα Φόρτιση του 40S S eif-3 συμπλέγματος
Σύμφωνα με τις απόψεις που σήμερα επικρατούν η διαδικασία της έναρξης μπορεί να διαιρεθεί σε πέντε στάδια Διάσταση του 80S Ριβοσώματος στη 60S και την 40S υπομοναδα Φόρτιση του 40S S eif-3 συμπλέγματος
ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ. Με αναφορά τόσο στους προκαρυωτικούς όσο και στους ευκαρυωτικούς οργανισμούς
 ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ Με αναφορά τόσο στους προκαρυωτικούς όσο και στους ευκαρυωτικούς οργανισμούς Λειτουργίες Γενετικού Υλικού o Αποθήκευση της γενετικής πληροφορίας. Η οργάνωση της γενετικής πληροφορίας
ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ Με αναφορά τόσο στους προκαρυωτικούς όσο και στους ευκαρυωτικούς οργανισμούς Λειτουργίες Γενετικού Υλικού o Αποθήκευση της γενετικής πληροφορίας. Η οργάνωση της γενετικής πληροφορίας
Το μονοπάτι της κινάσης MAP- ERK
 Το μονοπάτι της κινάσης MAP- ERK 1 Σηματοδότηση μέσω μικρών GTPασών Η οικογένεια μορίων Ras (Rat Sarcoma virus) Ρύθμιση των πρωτεϊνών Ras Οι πρωτεΐνες Ras μετατρέπονται από την ανενεργή μορφή τους, που
Το μονοπάτι της κινάσης MAP- ERK 1 Σηματοδότηση μέσω μικρών GTPασών Η οικογένεια μορίων Ras (Rat Sarcoma virus) Ρύθμιση των πρωτεϊνών Ras Οι πρωτεΐνες Ras μετατρέπονται από την ανενεργή μορφή τους, που
ΓENIKA ΣTOIXEIA. Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό.
 1 ΓENIKA ΣTOIXEIA Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό. ΤΑ ΚΥΤΤΑΡΑ Η βασική ζώσα μονάδα του σώματος είναι
1 ΓENIKA ΣTOIXEIA Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό. ΤΑ ΚΥΤΤΑΡΑ Η βασική ζώσα μονάδα του σώματος είναι
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β
 ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
ΑΠΑΝΤΗΣΕΙΣ. Β2. Η εικόνα αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς οργανισμούς το mrna αρχίζει να μεταφράζεται σε πρωτεΐνη πριν ακόμη
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 6 ΙΟΥΝΙΟΥ 207 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. δ Α2. δ Α3. β Α4. γ Α5.
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 6 ΙΟΥΝΙΟΥ 207 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. δ Α2. δ Α3. β Α4. γ Α5.
Βιολογία Προσανατολισμού Γ Λυκείου Κεφάλαιο: Κεφάλαια 1,2,4 Ονοματεπώνυμο Μαθητή: Ημερομηνία: 08/12/2018 Επιδιωκόμενος Στόχος: 75/100
 Μάθημα/Τάξη: Βιολογία Προσανατολισμού Γ Λυκείου Κεφάλαιο: Κεφάλαια 1,2,4 Ονοματεπώνυμο Μαθητή: Ημερομηνία: 08/12/2018 Επιδιωκόμενος Στόχος: 75/100 ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς
Μάθημα/Τάξη: Βιολογία Προσανατολισμού Γ Λυκείου Κεφάλαιο: Κεφάλαια 1,2,4 Ονοματεπώνυμο Μαθητή: Ημερομηνία: 08/12/2018 Επιδιωκόμενος Στόχος: 75/100 ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς