Βιοπληροφορική. Πίνακες Αντικατάστασης BLOSUM & Οπτική Σύγκριση Αλληλουχιών. Αλέξανδρος Τζάλλας
|
|
|
- Λίγεια Γαλάνη
- 8 χρόνια πριν
- Προβολές:
Transcript
1 Βιοπληροφορική Πίνακες Αντικατάστασης BLOSUM & Οπτική Σύγκριση Αλληλουχιών Αλέξανδρος Τζάλλας ΤΕΙ Ηπείρου Σχολή Τεχνολογικών Εφαρμογών Τμήμα Μηχανικών Πληροφορικής ΤΕ



2 Copyright Πανεπιστήμιο Δυτικής Μακεδονίας, Τμήμα Μηχανικών Πληροφορικής και Τηλεπικοινωνιών, Αγγελίδης Παντελής. «Βιοπληροφορική». Έκδοση: 1.0. Κοζάνη Διαθέσιμο από τη δικτυακή διεύθυνση: https: //eclass.uowm.gr/courses/icte102/ Πίνακες Αντικατάστασης BLOSUM & Οπτική Σύγκριση Αλληλουχιών
3 Σκοπός του μαθήματος Πίνακες αντικατάστασης PAM. The log odds form (the mutation data matrix) of PAM250. Πίνακες αντικατάστασης BLOSUM. Nucleic acid PAM Scoring Matrices. Οι ποινές για τα κενά. Στατιστική σημαντικότητα της στοίχισης αλληλουχιών. P-value, E-value, Z-score. Οπτική σύγκριση αλληλουχιών. Διαγράμματα πινάκων σημείων Dot plots. Θόρυβος» στα dot plots.
4 Πίνακες αντικατάστασης BLOSUM και οπτική σύγκριση αλληλουχιών
5 Γιατί σύγκριση αλληλουχιών;;; Συμπεράσματα για λειτουργία ενός γονιδίου (μίας πρωτεΐνης). Όταν δύο γονίδια έχουν μεγάλο ποσοστό ομοιότητας, τότε πιθανά κωδικοποιούν πρωτεΐνες με παρόμοια λειτουργία. Συμπεράσματα για σημαντικές περιοχές στην αλληλουχία γονιδίων/πρωτεϊνών. Όταν πρωτεΐνες με συγκεκριμένο χαρακτηριστικό μοιράζονται κοινή περιοχή, τότε αυτή η κοινή περιοχή είναι πιθανά υπεύθυνη για το συγκεκριμένο χαρακτηριστικό. Συμπεράσματα για την εξελεγκτική απόσταση μεταξύ 3 ειδών. Όταν η αλληλουχία μίας πρωτεΐνης είναι σχεδόν η ίδια μεταξύ 2 ειδών (ποντίκι και αρουραίος), τότε τα είδη αυτά είναι «κοντά».
6 Σύγκριση πρωτεϊνών - Εξαγωγή συμπερασμάτων για τη λειτουργία (1/4) Protein A expressed in colon cancer: TCGCTGCGAAGGACATTTGGGCTGTGTGTGCGACGCGGGTCGGAGGGGCAGTCGGGGGAACC GCGAAGAAGCCGAGGAGCCCGGAGCCCCGCGTGACGCTCCTCTCTCAGTCCAAAAGCGGCTTT TGGTTCGGCGCAGAGAGACCCGGGGGTCTTCAGGACAGCGATTGTCATTGCTGAAGCTTTTCCT CGAAAAGCGCCGCCCTGCCCTTGGCCCCGAGAA Protein B expressed in colon cancer : CAGACAAAGAGCACCGCAGGGCCGATCACGCTGGGGGCGCTGAGGCCGGCCATGGTCATGGA AGTGGGCACCCTGGACGCTGGAGGCCTGCGGGCGCTGCTGGGGGAGCGAGCGGCGCAATGCC TGCTGCTGGACTGCCGCTCCTTCTTCGCTTTCAACGCCGGCCACATCGCCGGCTCTGTCAACGT GCGCTTCAGCACCATCGTGCGGCGCCGGGCCAAGGGCGCCATGGGCCTGGAGCACATCGTGC CCAACGCCGAGCTCCGCGGTCAGGACAGCGATTGTCATTGCTGA Protein C expressed in colon cancer: CCGCCTGCTGGCCGGCGCCTACCACGCCGTGGTGTTGCTGGACGAGCGCAGCGCCGCCCTGG ACGGCGCCAAGCGCGACGGCACCCTGGCCCTGGCGGCCGGCGCGCTCTGCCGCGAGGCGCG CGCCGCGCAAGTCTTCTTCCTCAAAGGAGGATACGAAGCGTTTTCGGCTTCCTGCCCGGAGCTC AGGACAGCGATTGTCATTGCTGATGTGCAGCAAACAGTCGACCCCCATGGGGCTCAGCCTTCCC CTGAGTACTAGCGTCCCTGACAGCGCGGAATCTGGGTGCAGTT 7
7 Σύγκριση πρωτεϊνών - Εξαγωγή συμπερασμάτων για τη λειτουργία (2/4) Protein D expressed in colon cancer: TAACTGCCTTGATCAACGTCTCAGCCAATTGTCCCAACCATTTTGAGGGTCACTACCAGTACAAG AGCATCCCTGTGGAGGACAACCACAAGGCAGACATCAGCTCCTGGTTCAACGAGGCCATTGACT TCATAGACTCCATCAAGAATGCTGGAGGAAGGGTGTTTGTCCACTGCCAGGCAGGCATTTCCCG GTCAGCCACCATCTGCCTTGCTTACCTTATGAGGACTAATCGAGTCATCAGGACAGCGATTGTCA TTGCTGAAGCTGGACGAGGCCTTTGAGTTTGTGAAGCAGAGGC Protein E expressed in colon cancer: GAAGCATCATCTCTCCCAACTTCAGCTTCATGGGCCAGCTGCTGCAGTTTGAGTCCCAGGTGCTG GCTCCGCACTGTTCGGCAGAGGCTGGGAGCCCCGCCATGGCTGTGCTCGACCGAGGCACCTCC ACCACCACCGTGTTCAACTTATCAGGACAGCGATTGTCATTGCTGACCCCGTCTCCATCCCTGTC CACTCCACGAACAGTGCGCTGAGCTACCTTCAGAGCCCCATTACGACCTCTCCCAGCTGCTGAA AGGCCACGGGAGGTGAGGCTCTTCACATCCCATTGGGACTC Protein F with unknown function: CATGCTCCTTGAGAGGAGAAATGCAATAACTCTGGGAGGGGCTCAGGACAGCGATTGTCATTGC TGATCGAGAGGGCTGGTCCTTATTTATTTAACTTCACCCGAGTTCCTCTGGGTTTCTAAGCAGTTA TGGTGATGACTTAGCGTCAAGACATTTGCTGAACTCAGCACATTCGGGACCAATATATAGTGGGT ACATCAAGTCCATCTGACAAAATGGGGCAGAAGAGAAAGGACTCAGTGTGTGATCCGGTTTCTTT TTGCTCGCCCCTGTTTTTTGTAGAATCTCTTCATGCTTGACATACCTACCAGTATTATTCCCGACG ACACATATACATATGAGAATATACCTTATTTATTTTTGTGTAGGTGTCTGCCTTCACAAATGTCATT GTCTACTCCTAGAAGAACCAAATACCTCAATTTTTGTTTTTGAGTACTGTACTATCCTGTAAATATA TCTTAAGCAGGTTTGTTTTCA
8 Σύγκριση πρωτεϊνών - Εξαγωγή συμπερασμάτων για τη λειτουργία (3/4) Protein A expressed in colon cancer: TCGCTGCGAAGGACATTTGGGCTGTGTGTGCGACGCGGGTCGGAGGGGCAGTCGGGGGAACCGCGAAGAAGCC GAGGAGCCCGGAGCCCCGCGTGACGCTCCTCTCTCAGTCCAAAAGCGGCTTTTGGTTCGGCGCAGAGAGACCCGG GGGTCTTCAGGACAGCGATTGTCATTGCTGAAGCTTTTCCTCGAAAAGCGCCGCCCTGCCCTTGGCCCCGAGAA Protein B expressed in colon cancer : CAGACAAAGAGCACCGCAGGGCCGATCACGCTGGGGGCGCTGAGGCCGGCCATGGTCATGGAAGTGGGCACCCT GGACGCTGGAGGCCTGCGGGCGCTGCTGGGGGAGCGAGCGGCGCAATGCCTGCTGCTGGACTGCCGCTCCTTCT TCGCTTTCAACGCCGGCCACATCGCCGGCTCTGTCAACGTGCGCTTCAGCACCATCGTGCGGCGCCGGGCCAAGGG CGCCATGGGCCTGGAGCACATCGTGCCCAACGCCGAGCTCCGCGGTCAGGACAGCGATTGTCATTGCTGA Protein C expressed in colon cancer : CCGCCTGCTGGCCGGCGCCTACCACGCCGTGGTGTTGCTGGACGAGCGCAGCGCCGCCCTGGACGGCGCCAAGC GCGACGGCACCCTGGCCCTGGCGGCCGGCGCGCTCTGCCGCGAGGCGCGCGCCGCGCAAGTCTTCTTCCTCAAAG GAGGATACGAAGCGTTTTCGGCTTCCTGCCCGGAGCTCAGGACAGCGATTGTCATTGCTGATGTGCAGCAAACAG TCGACCCCCATGGGGCTCAGCCTTCCCCTGAGTACTAGCGTCCCTGACAGCGCGGAATCTGGGTGCAGTT
9 Σύγκριση πρωτεϊνών - Εξαγωγή συμπερασμάτων για τη λειτουργία (4/4) Protein D expressed in colon cancer : TAACTGCCTTGATCAACGTCTCAGCCAATTGTCCCAACCATTTTGAGGGTCACTACCAGTACAAGAGCATCCCTGTGGAGGACAACCAC AAGGCAGACATCAGCTCCTGGTTCAACGAGGCCATTGACTTCATAGACTCCATCAAGAATGCTGGAGGAAGGGTGTTTGTCCACTGCC AGGCAGGCATTTCCCGGTCAGCCACCATCTGCCTTGCTTACCTTATGAGGACTAATCGAGTCATCAGGACAGCGATTGTCATTGCTGAA GCTGGACGAGGCCTTTGAGTTTGTGAAGCAGAGGC Protein E expressed in colon cancer : GAAGCATCATCTCTCCCAACTTCAGCTTCATGGGCCAGCTGCTGCAGTTTGAGTCCCAGGTGCTGGCTCCGCACTGTTCGGCAGAGGC TGGGAGCCCCGCCATGGCTGTGCTCGACCGAGGCACCTCCACCACCACCGTGTTCAACTTATCAGGACAGCGATTGTCATTGCTGACC CCGTCTCCATCCCTGTCCACTCCACGAACAGTGCGCTGAGCTACCTTCAGAGCCCCATTACGACCTCTCCCAGCTGCTGAAAGGCCACG GGAGGTGAGGCTCTTCACATCCCATTGGGACTC Protein F with unknown function: CATGCTCCTTGAGAGGAGAAATGCAATAACTCTGGGAGGGGCTCAGGACAGCGATTGTCATTGCTGATCGAGAGGGCTGGTCCTTAT TTATTTAACTTCACCCGAGTTCCTCTGGGTTTCTAAGCAGTTATGGTGATGACTTAGCGTCAAGACATTTGCTGAACTCAGCACATTCGG GACCAATATATAGTGGGTACATCAAGTCCATCTGACAAAATGGGGCAGAAGAGAAAGGACTCAGTGTGTGATCCGGTTTCTTTTTGCTC GCCCCTGTTTTTTGTAGAATCTCTTCATGCTTGACATACCTACCAGTATTATTCCCGACGACACATATACATATGAGAATATACCTTATTTATT TTTGTGTAGGTGTCTGCCTTCACAAATGTCATTGTCTACTCCTAGAAGAACCAAATACCTCAATTTTTGTTTTTGAGTACTGTACTATCCTG TAAATATATCTTAAGCAGGTTTGTTTTCA Συμπέρασμα: Πιθανά και η πρωτεΐνη F παίζει ρόλο στον καρκίνο παχύ εντέρου 10
10 Σύγκριση πρωτεϊνών - Εξαγωγή συμπερασμάτων για λειτουργικό τμήμα πρωτεΐνης (1/2) Protein A with membrane localization: TCGCTGCGAAGGACATTTGGGCTGTGTGTGCGACGCGGGTCGGAGGGGCAGTCGGGGGAACCGCGAAGAAGCCGAGGAGCCCGGAGCCCCGCGTGACG CTCCTCTCTCAGTCCAAAAGCGGCTTTTGGTTCGGCGCAGAGAGACCCGGGGGTCTTCAGGACAGCGATTGTCATTGCTGAAGCTTTTCCTCGAAAAGCGCC GCCCTGCCCTTGGCCCCGAGAACAGACAAAGAGCACCGCAGGGCCGATCACGCTGGGGGCGCTGAGGCCGGCCATGGTCATGGAAGTGGGCACCCTGGAC GCTGGAGGCCTGCGGGCGCTGCTGGGGGAGCGAGCGGCGCAATGCCTGCTGCTGGACTGCCGCTCCTTCTTCGCTTTCAACGCCGGCCACATCGCCGGCTC TGTCAACGTGCGCTTCAGCACCATCGTGCGGCGCCGGGCCAAGGGCGCCATGGGCCTGGAGCACATCGTGCCCAACGCCGAGCTCCGCGGTCAGGACAGC GATTGTCATTGCTGACCGCCTGCTGGCCGGCGCCTACCACGCCGTGGTGTTGCTGGACGAGCGCAGCGCCGCCCTGGACGGCGCCAAGCGCGACGGCACCC TGGCCCTGGCGGCCGGCGCGCTCTGCCGCGAGGCGCGCGCCGCGCAAGTCTTCTTCCTCAAAGGAGGATACGAAGCGTTTTCGGCTTCCTGCCCGGAGCTC AGGACAGCGATTGTCATTGCTGATGTGCAGCAAACAGTCGACCCCCATGGGGCTCAGCCTTCCCCTGAGTACTAGCGTCCCTGACAGCGCGGAATCTGGGTG CAGTTCCTGCAGTACCCCACTCTACGATCAGGGTGGCCCGGTGGAAATCCTGCCCTTTCTGTACCTGGGCAGTGCGTATCACGCTTCCCGCAAGGACATGCTG GATGCCTTGGGCA Protein B with membrane localization : TAACTGCCTTGATCAACGTCTCAGCCAATTGTCCCAACCATTTTGAGGGTCACTACCAGTACAAGAGCATCCCTGTGGAGGACAACCACAAGGCAGACATCA GCTCCTGGTTCAACGAGGCCATTGACTTCATAGACTCCATCAAGAATGCTGGAGGAAGGGTGTTTGTCCACTGCCAGGCAGGCATTTCCCGGTCAGCCACCA TCTGCCTTGCTTACCTTATGAGGACTAATCGAGTCATCAGGACAGCGATTGTCATTGCTGAAGCTGGACGAGGCCTTTGAGTTTGTGAAGCAGAGGCGAAGC ATCATCTCTCCCAACTTCAGCTTCATGGGCCAGCTGCTGCAGTTTGAGTCCCAGGTGCTGGCTCCGCACTGTTCGGCAGAGGCTGGGAGCCCCGCCATGGCT GTGCTCGACCGAGGCACCTCCACCACCACCGTGTTCAACTTATCAGGACAGCGATTGTCATTGCTGACCCCGTCTCCATCCCTGTCCACTCCACGAACAGTGC GCTGAGCTACCTTCAGAGCCCCATTACGACCTCTCCCAGCTGCTGAAAGGCCACGGGAGGTGAGGCTCTTCACATCCCATTGGGACTCCATGCTCCTTGAGA GGAGAAATGCAATAACTCTGGGAGGGGCTCAGGACAGCGATTGTCATTGCTGATCGAGAGGGCTGGTCCTTATTTATTTAACTTCACCCGAGTTCCTCTGGG TTTCTAAGCAGTTATGGTGATGACTTAGCGTCAAGACATTTGCTGAACTCAGCACATTCGGGACCAATATATAGTGGGTACATCAAGTCCATCTGACAAAATG GGGCAGAAGAGAAAGGACTCAGTGTGTGATCCGGTTTCTTTTTGCTCGCCCCTGTTTTTTGTAGAATCTCTTCATGCTTGACATACCTACCAGTATTATTCCCG ACGACACATATACATATGAGAATATACCTTATTTATTTTTGTGTAGGTGTCTGCCTTCACAAATGTCATTGTCTACTCCTAGAAGAACCAAATACCTCAATTTTT GTTTTTGAGTACTGTACTATCCTGTAAATATATCTTAAGCAGGTTTGTTTTCA Συμπέρασμα: Πιθανά το κοινό τμήμα να είναι υπεύθυνο για τη μεμβρανική εντόπιση των πρωτεϊνών
11 Σύγκριση πρωτεϊνών - Εξαγωγή συμπερασμάτων για λειτουργικό τμήμα πρωτεΐνης (2/2) Protein A with membrane localization: TCGCTGCGAAGGACATTTGGGCTGTGTGTGCGACGCGGGTCGGAGGGGCAGTCGGGGGAACCGCGAAGAAGCCGAGGAGCCCGGAGCCCCGCGTGACGCTCCTC TCTCAGTCCAAAAGCGGCTTTTGGTTCGGCGCAGAGAGACCCGGGGGTCTTCAGGACAGCGATTGTCATTGCTGAAGCTTTTCCTCGAAAAGCGCCGCCCTGCCCTT GGCCCCGAGAACAGACAAAGAGCACCGCAGGGCCGATCACGCTGGGGGCGCTGAGGCCGGCCATGGTCATGGAAGTGGGCACCCTGGACGCTGGAGGCCTGCGG GCGCTGCTGGGGGAGCGAGCGGCGCAATGCCTGCTGCTGGACTGCCGCTCCTTCTTCGCTTTCAACGCCGGCCACATCGCCGGCTCTGTCAACGTGCGCTTCAGCAC CATCGTGCGGCGCCGGGCCAAGGGCGCCATGGGCCTGGAGCACATCGTGCCCAACGCCGAGCTCCGCGGTCAGGACAGCGATTGTCATTGCTGACCGCCTGCTGG CCGGCGCCTACCACGCCGTGGTGTTGCTGGACGAGCGCAGCGCCGCCCTGGACGGCGCCAAGCGCGACGGCACCCTGGCCCTGGCGGCCGGCGCGCTCTGCCGCG AGGCGCGCGCCGCGCAAGTCTTCTTCCTCAAAGGAGGATACGAAGCGTTTTCGGCTTCCTGCCCGGAGCTCAGGACAGCGATTGTCATTGCTGATGTGCAGCAAAC AGTCGACCCCCATGGGGCTCAGCCTTCCCCTGAGTACTAGCGTCCCTGACAGCGCGGAATCTGGGTGCAGTTCCTGCAGTACCCCACTCTACGATCAGGGTGGCCCG GTGGAAATCCTGCCCTTTCTGTACCTGGGCAGTGCGTATCACGCTTCCCGCAAGGACATGCTGGATGCCTTGGGCA Protein B with membrane localization : TAACTGCCTTGATCAACGTCTCAGCCAATTGTCCCAACCATTTTGAGGGTCACTACCAGTACAAGAGCATCCCTGTGGAGGACAACCACAAGGCAGACATCAGCTCCT GGTTCAACGAGGCCATTGACTTCATAGACTCCATCAAGAATGCTGGAGGAAGGGTGTTTGTCCACTGCCAGGCAGGCATTTCCCGGTCAGCCACCATCTGCCTTGCTT ACCTTATGAGGACTAATCGAGTCATCAGGACAGCGATTGTCATTGCTGAAGCTGGACGAGGCCTTTGAGTTTGTGAAGCAGAGGCGAAGCATCATCTCTCCCAACTT CAGCTTCATGGGCCAGCTGCTGCAGTTTGAGTCCCAGGTGCTGGCTCCGCACTGTTCGGCAGAGGCTGGGAGCCCCGCCATGGCTGTGCTCGACCGAGGCACCTCC ACCACCACCGTGTTCAACTTATCAGGACAGCGATTGTCATTGCTGACCCCGTCTCCATCCCTGTCCACTCCACGAACAGTGCGCTGAGCTACCTTCAGAGCCCCATTA CGACCTCTCCCAGCTGCTGAAAGGCCACGGGAGGTGAGGCTCTTCACATCCCATTGGGACTCCATGCTCCTTGAGAGGAGAAATGCAATAACTCTGGGAGGGGCTC AGGACAGCGATTGTCATTGCTGATCGAGAGGGCTGGTCCTTATTTATTTAACTTCACCCGAGTTCCTCTGGGTTTCTAAGCAGTTATGGTGATGACTTAGCGTCAAGA CATTTGCTGAACTCAGCACATTCGGGACCAATATATAGTGGGTACATCAAGTCCATCTGACAAAATGGGGCAGAAGAGAAAGGACTCAGTGTGTGATCCGGTTTCTT TTTGCTCGCCCCTGTTTTTTGTAGAATCTCTTCATGCTTGACATACCTACCAGTATTATTCCCGACGACACATATACATATGAGAATATACCTTATTTATTTTTGTGTAGG TGTCTGCCTTCACAAATGTCATTGTCTACTCCTAGAAGAACCAAATACCTCAATTTTTGTTTTTGAGTACTGTACTATCCTGTAAATATATCTTAAGCAGGTTTGTTTTCA Συμπέρασμα: Πιθανά το κοινό τμήμα να είναι υπεύθυνο για τη μεμβρανική εντόπιση των πρωτεϊνών 12
12 Σύστημα βαθμονόμησης στοίχισης - DNA & RNA Sequence 1: ATCGGATCT Sequence 2: ACGGACT ΠΡΟΣΟΧΗ: Συγκρίνουμε νουκλεοτίδια, μόνο 4 πιθανές βάσεις A T C G G A T - C T A - C - G G - A C T A T C G G A T C T A - C G G A - C T Πιθανό σύστημα βαθμονόμησης: όμοιο κατάλοιπο: +2 διαφορετικό κατάλοιπο: -1 κενό: 2 Alignment 1: 5 x 2 1(1) 4(2) = = 1 Alignment 2: 7 x 2 0(1) 2 (2) = = 10 13
13 Στοίχιση πρωτεϊνών;;; A S K T M P I I I?? I I I A S R H M P I A S K T M P I I I?? I I A S Y H M P I Πώς στοιχίζουμε πρωτεΐνες. Αμινοξέα με παρόμοιες ιδιότητες. Αμινοξέα με διαφορετικές ιδιότητες.
14 Πίνακες βαθμονόμησης (scoring matrices) ή πίνακες αντικατάστασης Για την αντικατάσταση αμινοξέων: PAM. BLOSUM. Για την αντικατάσταση νουκλεοτιδίων (DNA): Το DNA είναι λιγότερο συντηρημένο από τις πρωτεΐνες. Δεν είναι αποτελεσματικό να συγκρίνουμε κωδικοποιούσες περιοχές, από τις οποίες προκύπτουν οι πρωτεΐνες, σε επίπεδο νουκελοτιδίων.
15 The log odds form (the mutation data matrix) of PAM250 Tryptophan: W Cysteine: C
16 Πίνακες αντικατάστασης PAM (Percent Accepted Mutation - PAM or Dayhoff Matrices) Μελετήθηκαν από την Margaret Dayhoff. Αξιολογούν - βαθμολογούν την αντικατάσταση ενός αμινοξέος από ένα άλλο. Αξιολογήθηκαν: 1572 αμινοξικές αντικαταστάσεις από, 71 πρωτεϊνικά γκρουπ με, 85% ομοιότητα τουλάχιστον, Η κατασκευή τους βασίζεται στις: στοιχίσεις πολλών όμοιων πρωτεϊνών, μικρής εξελεγκτικής απόστασης αποδεκτές σημειακές μεταλλάξεις - accepted mutations (αντικατάσταση αμινοξέος από κάποιο άλλο που δεν επηρεάζει τη λειτουργία της πρωτεΐνης).
17 Πίνακες αντικατάστασης BLOSUM (BLOcks amino acid SUbstitution Matrices) Αναπτύχθηκαν από τους Stephen και Georgia Henikoff, Βασίζονται στην απαρίθμηση αντικατάστασης αμινοξέων από άλλα αμινοξέα σε συντηρημένες περιοχές αλληλουχιών. μικρής ομοιότητας, και απομακρυσμένης εξελεγκτικής σχέσης. Εξετάστηκαν πάνω από 2000 συντηρημένες περιοχές (blocks) που ανήκουν σε περισσότερες από 500 οικογένειες συσχετιζόμενων πρωτεϊνών. Οι περιοχές (blocks) δεν περιέχουν κενά.
18 BLOSUM MATRICES 2000 α.α. : Συντηρημένη περιοχή Οι περιοχές βρέθηκαν σε πρωτεϊνικές βάσεις δεδομένων και αντιπροσωπεύουν περισσότερες από 500 οικογένειες πρωτεϊνών. Οι πίνακες BLOSUM βασίζονται σε ανάλυση διαφορετικού τύπου αλληλουχιών και μεγαλύτερου αριθμού δεδομένων από τους πίνακες PAM. Βάση δεδομένων: PROSITE. Prosite: Database with protein domains. It provides list of proteins that are in the same family because they have a similar biochemical action. For each family, a pattern of amino acids that are characteristic of that function is provided.
19 Πίνακες αντικατάστασης BLOSUM (BLOcks SUbstitution Matrices) BLOSUMn: Το n δείχνει το ποσοστό ομοιότητας των αλληλουχιών που χρησιμοποιήθηκαν για να προκύψει ο συγκεκριμένος πίνακας. Αλληλουχίες με ομοιότητα τουλάχιστον 62% δίνουν τον πίνακα αντικατάστασης BLOSUM62. BLOSUM με μεγάλο n: αλληλουχίες με μεγάλη ομοιότητα. BLOSUM με μεγάλο n -> PAM με μικρό n. Γενικά, οι πίνακες BLOSUM είναι καλύτεροι για την εύρεση τοπικών στοιχίσεων.
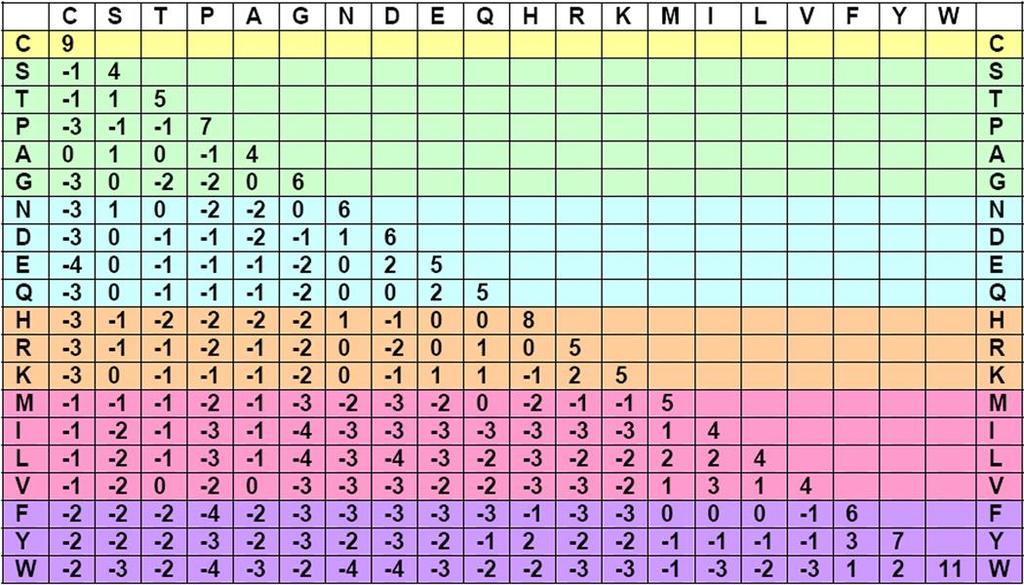
20 Παράδειγμα BLOSUM62
21 Ερμηνεία τιμής του πίνακα BLOSUM (1/4) Η τιμή κάθε ζεύγους αμινοξέος δείχνει την πιθανότητα να εμφανιστεί η συγκεκριμένη αντικατάσταση κατά την εξέλιξη. Δίνεται από το λόγο: λ = συχνότητα εμφάνισης του ζεύγους / αναμενόμενη συχνότητα εμφάνισης του ζεύγους. λ = q ij /e ij, q: expected, and e: observed frequency of appearance.
22 Ερμηνεία τιμής του πίνακα BLOSUM (2/4) A A A A S A A A A A Pairs Frequency of appearance, 1 st Frequency of appearance, 2 nd AA 9 9 AS 9 1 SS 1 1 Pair frequency c ij, for each pair i and j, for each column k using: For like comparisons c n 2 ( k) i ii For unlike comparisons ( k) ij c n i n j
23 Ερμηνεία τιμής του πίνακα BLOSUM (3/4) A A A A S A A A A A c c c AA SS AS Pairs Frequency of appearance, 1 st Frequency of appearance, 2 nd AA 9 9 AS 9 1 SS (9 1) 2 1 1(1 1) ( k) ni cii 2 ( k) c n n ij c ij A S A 36 S 9 0 i j
24 Ερμηνεία τιμής του πίνακα BLOSUM - Κανονικοποίηση A A A A S A A A A A c ij A S A 36 S 9 0 n( n 1) T cij w 2 w : columns n : sequences Normalization factor: 10(10 1) T Observed pair frequency, q ij : q AA q AS q SS q ij A S A 0.8 S 0.2 0
25 Ερμηνεία τιμής του πίνακα BLOSUM (4/4) Η τιμή κάθε ζεύγους αμινοξέος δείχνει την πιθανότητα να εμφανιστεί η συγκεκριμένη αντικατάσταση κατά την εξέλιξη. Δίνεται από το λόγο: λ = συχνότητα εμφάνισης του ζεύγους / αναμενόμενη συχνότητα εμφάνισης του ζεύγους. λ = q ij /e ij, q: observed, and e: expected frequency of appearance.
26 Ερμηνεία τιμής του πίνακα BLOSUM Expected pair frequency, eij (1/3) 2 eii p i, for like comparisons e 2p ij i p j, for unlike comparisons where, p i : expected probability of occurrence of the ith residue p i q ii ji q ij 2
27 Ερμηνεία τιμής του πίνακα BLOSUM Expected pair frequency, eij (2/3) e 2p p i ij q ii i p j ji q 2 ii p i e ij 2 p p A S q q AA SS Για κάθε κατάλοιπο q 2 q 2 AS AS Για κάθε ζεύγος q ij A S A 0.8 S e e e AA SS AS p p 2 A 2 S 2 p A p S
28 Ερμηνεία τιμής του πίνακα BLOSUM Expected pair frequency, eij (3/3) Observed frequency q ij A S A 0.8 S Expected frequency e e e AA SS AS p p 2 A 2 S 2 p A p S Entry : S S S AA AS SS Entry: S = 2log 2 (observed/expected) 2log 2log 2log observed exp ected observed exp ected observed exp ected 2log 2log 2log ? Δεν παρέχονται πληροφορίες για το ζεύγος SS
29 Παράδειγμα - Pair frequency cij Sequence 1 A A I Sequence 2 S A L Sequence 3 T A L Sequence 4 T A V Sequence 5 A A L c n 2 ( k) i ii ( k) ij c n i n j Pairs n i n j AT 2 2 IL 1 3 LL 3 3 AA 2 2 AA 5 5 Pair frequency c ij, for each pair i and j c ij A I L S T V A 10+1 I 0 0 L S T V
30 Παράδειγμα - Normalization Sequence 1 A A I Sequence 2 S A L Sequence 3 T A L Sequence 4 T A V Sequence 5 A A L n( n 1) T cij w 2 w : columns n : sequences Normalization factor, T: T 5(5 1) Pair frequency cij, for each pair i and j c ij A I L S T V A 10+1 I 0 0 L S T V cij / 30 = q ij q ij A I L S T V A I 0 0 L S T V
31 Παράδειγμα - Observed frequency qij Sequence 1 A A I Sequence 2 S A L Sequence 3 T A L Sequence 4 T A V Sequence 5 A A L Observed normalized pair frequency c ij, for each pair i and j c ij A I L S T V A I 0 0 L S T V
32 Παράδειγμα - Expected frequency of ij pair, eij 2 eii p i e 2p ij i p j p i q ii ji q ij 2 Αναμενόμενη συχνότητα εμφάνισης ζεύγους Πιθανότητα εμφάνισης ith καταλοίπου q ij A I L S T V A I 0 0 L S T V Πιθανότητα εμφάνισης ith καταλοίπου p A ( )/2=0.466 p I 0+( )/2=0.066 p L 0.1+( )/2=0.2 p S 0+( )/2=0.066 p T ( )/2=0.133 p V 0+( )/2=0.066
33 Expected frequency of ij pair, eij 2 eii p i e 2p ij i p j e AA =p A2 = =0.217 p i q ii ji q ij 2 e AI =2p A p I = =0.062 e AI =2p A p L = =0.186 e II =p I2 = =0.217 Expected frequency of ij pair, e ij p Πιθανότητα εμφάνισης ith καταλοίπου i p A ( )/2=0.466 p I 0+( )/2=0.066 p L 0.1+( )/2=0.2 p S 0+( )/2=0.066 p T ( )/2=0.133 p V 0+( )/2=0.066 e ij A I L S T V A I L S T V
34 Entry in BLOSUM matrix, Sij Observed normalized pair frequency cij, for each pair i and j A I L S T V A I 0 0 L S T V Expected frequency of ij pair, e ij e ij A I L S T V A I L S T V Entry : S S S ij AA SA 2log 2log 2log observed exp ected Finally S ij A I L S T V A 2 I?? L? 4 3 S 0??? T 0?? 4 2 V? 4 4???
35 Comparison PAM vs. BLOSUM PAM is based on an evolutionary model using phylogenetic trees (85% similarity). BLOSUM assumes no evolutionary model, but rather conserved blocks of proteins.
36 Πίνακες αντικατάστασης - Ισοδυναμία PAM100 PAM120 PAM160 PAM200 PAM250 Blosum90 Blosum80 Blosum62 Blosum52 Blosum45
37 Nucleic acid PAM Scoring Matrices (1/2) PAM: Point accepted mutations. Για τον DNA PAM1 πίνακα θεωρούμε ότι το 99% των αλληλουχιών συντηρείται ενώ το 1% μεταλλάσσεται. Υποθέτουμε ότι και οι 4 βάσεις, A-C-G-T, μεταλλάσσονται το ένα στο άλλο με την ίδια πιθανότητα. Υποθέτουμε ότι και τα 4 βάσεις έχουν την ίδια συχνότητα εμφάνισης.
38 Nucleic acid PAM Scoring Matrices (2/2) Θεωρούμε ότι είναι 3 φορές πιο πιθανό οι πυριμιδίνες να μεταλλαχθούν σε πυριμιδίνες (Τ C) και οι πουρίνες σε πουρίνες (A G) σε σχέση με τη μετάλλη των πυριμιδών σε πουρίνες και το αντίστροφο. 4 αζωτούχες βάσεις: Θυμίνη (Τ), Κυτοσίνη (C), Αδενίνη (Α) και Γουανίνη (G)
39 Οι ποινές για τα κενά (1/2) Η εισαγωγή κενών είναι απαραίτητη έτσι ώστε να έχουμε την καλύτερη στοίχιση των αλληλουχιών. Συσχετισμένη ή αφινική ποινή (affine gap penalty). Πιο πιθανό 1 κενό μήκους Κ, παρά Κ κενά μήκους 1. Αφαίρεση μιας τιμής για την εισαγωγή ενός κενού και πρόσθετες αφαιρέσεις για επόμενες επεκτάσεις του κενού. Το σύστημα βαθμονόμησης θα πρέπει να «τιμωρεί» περισσότερο τα καινούργια κενά.
40 Παράδειγμα - PAM250 C - K H V F C R V C I C K K C - F C - K C V PAM250 Εισαγωγή κενού: -10 Score = x10 Score = 19
41 Οι ποινές για τα κενά (2/2) p = - A - B (n-1): όπου Α: ποινή έναρξης του κενού (gap opening penalty). Β: ποινή επέκτασης (gap extension penalty). n: μήκος του κενού. Συνήθως: Α=12 και Β=4, οπότε p = (n-1) Πολύ μικρή ποινή: Εισαγωγή πολλών κενών. Πολύ μεγάλη ποινή: Μη εισαγωγή κενών. Γενικά: Η ποινή για ένα κενό n καταλοίπων θα πρέπει να είναι μικρότερη από τη συνολική ποινή για n κενά του 1 καταλοίπου.
42 Typical score matrix DNA: Match = +1. Mismatch = -3. Gap penalty = -5. Gap extension penalty = -2. Protein sequences: Blossum62 matrix. Gap open penalty = -11. Gap extension = -1.
43 Ποινές κενών - Παράδειγμα
44 Εξετάζοντας τη σημαντικότητα της στοίχισης αλληλουχιών Για αλληλουχίες που ανήκουν στην ίδια οικογένεια και είναι προφανές, κάτι τέτοιο δεν είναι απαραίτητο. Για αλληλουχίες που δεν είναι ξεκάθαρο ότι είναι παρόμοιες, αυτό είναι αναγκαίο. Συμπέρασμα: Είναι πράγματι όμοιες ή το αποτέλεσμα της σύγκρισης είναι τυχαίο, δηλαδή το ίδιο με την περίπτωση που οι δύο αλληλουχίες δεν έχουν καμία σχέση. Ο έλεγχος θα γίνει σε όλες τις στοιχίσεις έτσι ώστε να επιλεχτεί εκείνη που είναι στατιστικά η σημαντικότερη.
45 Gumbel extreme value distribution Σκοπός: Να υπολογιστεί η πιθανότητα το σκορ της στοίχισης μεταξύ δύο τυχαίων αλληλουχιών να φτάσει το σκορ της στοίχισης δύο σχετικών αλληλουχιών. Όσο μικρότερη είναι αυτή η πιθανότητα, τόσο στατιστικά σημαντικό είναι το σκορ στοίχισης των 2 πραγματικών αλληλουχιών. The distribution of alignment scores between random sequences follows the extreme value distribution (A), not the normal distribution (B)
46 Βαθμολόγηση στοίχισης αλληλουχιών Score: A number used to assess the biological relevance of a finding. In the context of sequence alignment, a score is a numerical value that describes the overall quality of an alignment. Higher numbers correspond to higher similarity. The score scale depends on the scoring system used (matrix, gap penalty). 1: 27 matches x 5 = 135. No gap penalty 2: 26 matches x 5 = 130, 7 mismatch x (-4) = -28, 7 gaps x (-5) =-35 TOTAL= =67 3: 10 matches x 5 = 50
47 Converting to Bit-scores The bit-score S is a normalized score expressed in bits. Lets you estimate the magnitude of the search space you would have to look through before you would expect to find a score as good as or better that this one by chance. S S' ln( K) ln2 where S: raw score. Parameters λ and K depend on the substitution matrix and on the gap penalty The bit-score is a rescaled version of the raw alignment score that is independent of the size of the search space
48 Δείκτες στατιστικής σημαντικότητας (1/4) P-value: Probability that an event occurs by chance The P-value associated to a score S is the probability to obtain by chance a score x at least equal to S Low P-value significant alingment P( S' x) 1 exp e x e x
49 Δείκτες στατιστικής σημαντικότητας (2/4) E-value: Correlation of the p-value for multiple testing. Expected number of sequences that will produce same or better score by chance. The lower the E value, the more significant the score is Low E-value significant alingment. Αναμενόμενος αριθμός στοιχίσεων με σκορ τουλάχιστον S: E = K m n e -λs m,n: Μήκος αλληλουχιών Κ, λ: Στατιστικές σταθερές που εξαρτώνται από το σύστημα βαθμονόμησης και τη συχνότητα εμφάνισης των καταλοίπων. π.χ. για τον πίνακα PAM250, Κ=0.1 και λ=0.229
50 Z-score: Δείκτες στατιστικής σημαντικότητας (3/4) Measures how much standard deviations above the mean of the score distribution.
 http://www.")
51 Δείκτες στατιστικής σημαντικότητας (4/4)

52 Alignment - Output
53 Μέθοδοι σύγκρισης αλληλουχιών 1. Οπτικοί. 2. Με αλγόριθμους δυναμικού προγραμματισμού. 3. Με ευρετικούς αλγόριθμους, που βασίζονται στην έννοια των «λέξεων». Πρέπει να λαμβάνεται υπόψη ο λόγος για τον οποίο γίνεται η στοίχιση αλληλουχιών. Ανάλογα επιλέγεται και η μέθοδος σύγκρισης. Ομοιότητα της τάξης το 90%: Εύκολη κατασκευή της στοίχισης με όλους τους αλγόριθμους. Ομοιότητα της τάξης του 25% (ζώνη του λυκόφωτος, «twilight zone of sequence alignment»): Οριακή για ασφαλή εξαγωγή συμπερασμάτων.
54 Διαγράμματα πινάκων σημείων - Dot plots Αποτελούν μία από τις απλούστερες μεθόδους οπτικοποίησης της ομοιότητας μεταξύ δύο αλληλουχιών. Μέθοδος: Σε ένα δισδιάστατο ορθογώνιο διάγραμμα ο κάθε άξονας αντιστοιχεί και σε μία αλληλουχία. Αλληλουχία 1: A C A G C G C G A G C Αλληλουχία 2: A C A C G A G G Στο πλέγμα που προκύπτει:
55 Dot plots - Μέθοδος (1/3) Σε ένα δισδιάστατο ορθογώνιο διάγραμμα ο κάθε άξονας αντιστοιχεί και σε μία αλληλουχία A C A C G A G G A C A G C G C G A G C
56 Dot plots - Μέθοδος (2/3) Τα τετράγωνα που αντιστοιχούν σε ταυτόσημα κατάλοιπα σημειώνονται (χρώμα) A C A C G A G G A C A G C G C G A G C
57 Dot plots - Μέθοδος (3/3) Όταν οι αλληλουχίες είναι όμοιες κατά μήκος μιας περιοχής, σχηματίζεται μία διαγώνιο A C A C G A G G γραμμή με κατεύθυνση από πάνω αριστερά κάτω δεξιά A C A G C G C G A G C

58 Dot plots - Παράδειγμα (1/2)

59 Dot plots - Παράδειγμα (2/2)
60 «Θόρυβος» στα dot plots 1 από τις 4 βάσεις έχει την πιθανότητα να ταιριάζει τυχαία. Μέθοδος φιλτραρίσματος: μία περιοχή σκιάζεται αν υπερβαίνει κάποιον συγκεκριμένο αριθμό (φίλτρο) όμοιων καταλοίπων.
61 Μείωση θορύβου στα dot plots (1/3) A C A C G A G G A C A G C G C G A G C

62 Μείωση θορύβου στα dot plots (2/3) A C A C G A G G A C A G C G C G A G C
63 Μείωση θορύβου στα dot plots (3/3) Window: 2, Stringency: 1 Window size: Number of nucleotides compared each time Stringency: The minimum number of nucleotides in the window must be match, so the dot can be placed


64 Μείωση θορύβου στα dot plots (Dotlet)
65 Πληροφορίες από τα dot plots 1. Περιοχές ομοιότητας: Διαγώνιος. 2. Προσθήκες (insertions) / εξαλείψεις (deletions). 3. Επαναλαμβανόμενες περιοχές: Ορθές. Αντιστρέψιμες (αντίστροφη φορά).

66 Check out the «mutated» intersequence comparison below:

67 Easy to visualize with dot matrix

68 Now consider the more complicated «mutation» in the following comparison:
 Ολική παλινδρόμηση. d) Μερική παλινδρόμηση.")
 Επαναλαμβανόμενες περιοχές και στις δύο αλληλουχίες. g) Διακοπές - Κοινός πρόγονος.")
69 Dot plots - Παράδειγμα a) Απόλυτη ομοιότητα. b) Επαναλαμβανόμενες περιοχές. c) Ολική παλινδρόμηση. d) Μερική παλινδρόμηση. e) Επαναλαμβανόμενο σύμβολο (αμινοξύ ή νουκλεοτίδιο) και στις δύο αλληλουχίες. f) Επαναλαμβανόμενες περιοχές και στις δύο αλληλουχίες. g) Διακοπές - Κοινός πρόγονος. h) Προσθήκη στην αλληλουχία 1 ή εξάλειψη στην αλληλουχία 2.
70 Size of the search space (1/2) Size of the search space ~ query sequence length (n) the sum of the lengths of the sequences in the database (m), N=n m. Size of the search space = N K, where K: Altschul coefficient. Example: Calculate the search space for a protein of 235a.a. length against a protein database of size m= a.a. The value of K is given equal to Size of the search space = 0.13*235*12,496,420=0.38 billion. In this case, a bit score of 30 (which correspond to a space of 2 30 = 1 billion) may have occurred by chance alone.

71 Converting to Bit - scores 'ln2 'ln 2 ln ln ln 2 ' ln 2 ln ' S S e K S S K K S S K S S S e n m K E ' ln2 S ' 2 S S S n m e e e n m E


72 Μείωση θορύβου στα Dot Plots (1/2)
73 Μείωση θορύβου στα Dot Plots (2/2)
74 Ερμηνεία τιμής του πίνακα BLOSUM - Παράδειγμα A A A A S A A A AA pairs: =36 possible AA pairs, F AA. AS pairs: 9 possible AS pairs, f AS. Η συχνότητα εμφάνισης του ζεύγους ΑΑ: q AA =f AA / f AA +f AS = 36/(36+9)=0.8. Η συχνότητα εμφάνισης του ζεύγους AS: q AS =f AS / f AA +f AS = 9/(36+9)=0.2. Η αναμενόμενη συχνότητα το Α να βρίσκεται σε ζεύγος: p A =q AA +q AS /2= /2=0.9. Η αναμενόμενη συχνότητα το S να βρίσκεται σε ζεύγος: p S =q AS /2=0.2/2=0.1. Η αναμενόμενη συχνότητα εμφάνισης του ζεύγους ΑΑ: e AA = p A x p A = 0.9x0.9=0.81. Η αναμενόμενη συχνότητα εμφάνισης του ζεύγους ΑS: e AS = 2xp A x p S =0.18. Η τιμή στον πίνακα για το ζεύγος ΑΑ: e AA/ q AA =0.8/0.81= Η τιμή στον πίνακα για το ζεύγος ΑS: e AS/ q AS =0.18/0.2= Οι τιμές μετατρέπονται σε λογαρίθμους με βάση το 2 και πολλαπλασιάζονται με το 2.
75 Size of the search space (2/2) Bit-score S : A log-scaled version of a score, log 2 (score)=s Sequence: m Database of protein sequences: Total length n. Size of the search space ~ m*n. Size of the search space =K m*n, K: Example: Protein database with sequences of total length: a.a. Sequence: 235 a.a. length For protein database, K=0.13 Size of the search space:
76 Significance of a local alignment Altschul and Gish (1996) have provided estimations of K=0.09 and λ=0,229 for PAM250 matrix, for a typical amino acid distribution and for an alignment score based on using a very high gap penalty. S' S ln( Kmn) S' ln( ) 8.09bits P( S' 8.55) e ***Note that the calculated S of 8.09 bits in the previous step is approximately the same as the 9 bits calculated by the simpler method***
77 Relative Entropy Indicates power of scoring scheme to distinguish from background noise (i.e., randomness). H i j q ij s ij Can use H to compare different scoring matrices. Relative entropy of a random alignment should be negative. Scores can be related to biology: negative=dissimilarity, zero=indifference, positive=similar.
78 Στατιστική σημαντικότητα στοίχισης αλληλουχιών χωρίς κενά The raw score of an alignment is determined as the sum of the log-odds scores and the gap penalties and is given in the units of log to the base 2 (log 2 ) or bits. Bit-score S : A log-scaled version of a score, log 2 (score)=s. Αν μετατρέψουμε τις τιμές σε log 2 x έχουμε bits πληροφορίας. Όταν χρησιμοποιούμε bit score system, το κατώφλι της στατιστικής σημαντικότητας δίνεται από τη σχέση: log 2 (m n).
79 Στατιστική σημαντικότητα στοίχισης αλληλουχιών χωρίς κενά - Παράδειγμα (1/2) 2 sequences, each 250 a.a. long Significance cut off: log 2 (250*250)=16 bit F W L E V E G N S M T A P T G F W L D V Q G D S M T A P A G Using PAM250 the score is calculated: S= =73 S=73
80 Στατιστική σημαντικότητα στοίχισης αλληλουχιών χωρίς κενά - Παράδειγμα (2/2) S is in 10*log 10 x. Convert S to a bit score, i.e. calculate log 2 x. S 10log log log log log S' x x 1 3 x S x S 10 S log 10 1 S 3 log x S 10
81 Στατιστική σημαντικότητα στοίχισης αλληλουχιών χωρίς κενά - Παράδειγμα Significance cut-offs (log 2 (m n)) = 16 bits. 1 1 S' S bits 3 3 Το S είναι 9 bits μεγαλύτερο από το κατώφλι της στατιστικής σημαντικότητας, επομένως η στοίχιση στο συγκεκριμένο σημείο των αλληλουχιών είναι στατιστικά σημαντική.

82 Μείωση θορύβου στα dot plots (1/2) Window: 5, Stringency: 3 Window size: Number of nucleotides compared each time Stringency: The minimum number of nucleotides in the window must be match, so the dot can be placed
 Window: 5, Stringency:")
83 Μείωση θορύβου στα dot plots (2/2) Window: 5, Stringency: 3
84 Τέλος 7 ης Διάλεξης: Πίνακες Αντικατάστασης BLOSUM & Οπτική Σύγκριση Αλληλουχιών tzallas@teiep.gr
Πανεπιστήμιο Δυτικής Μακεδονίας. Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών. Βιοπληροφορική
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 5: Πίνακες αντικατάστασης BLOSUM και οπτική σύγκριση αλληλουχιών Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 5: Πίνακες αντικατάστασης BLOSUM και οπτική σύγκριση αλληλουχιών Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ
Βιοπληροφορική. Ενότητα 6: Στοίχιση ακολουθιών ανά ζεύγη Σύστημα βαθμολόγησης, 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.
 Βιοπληροφορική Ενότητα 6: Στοίχιση ακολουθιών ανά ζεύγη Σύστημα βαθμολόγησης, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της σημασίας του συστήματος βαθμολόγησης
Βιοπληροφορική Ενότητα 6: Στοίχιση ακολουθιών ανά ζεύγη Σύστημα βαθμολόγησης, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της σημασίας του συστήματος βαθμολόγησης
Βιοπληροφορική. Ενότητα 6: Στοίχιση ακολουθιών ανά ζεύγη Σύστημα βαθμολόγησης, 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.
 Βιοπληροφορική Ενότητα 6: Στοίχιση ακολουθιών ανά ζεύγη Σύστημα βαθμολόγησης, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της σημασίας του συστήματος βαθμολόγησης
Βιοπληροφορική Ενότητα 6: Στοίχιση ακολουθιών ανά ζεύγη Σύστημα βαθμολόγησης, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της σημασίας του συστήματος βαθμολόγησης
LALING/PLALING :
 1. Άρθρα- δημοσιεύσεις Scopus DBLP Pubmed Google Scholar 2. Αναζήτηση νουκλεοτιδίου- πρωτεΐνης Entrez : http://www.ncbi.nlm.nih.gov/nuccore/ Uniprot (πρωτεΐνης): http://www.uniprot.org/ Blast : http://blast.ncbi.nlm.nih.gov/blast.cgi
1. Άρθρα- δημοσιεύσεις Scopus DBLP Pubmed Google Scholar 2. Αναζήτηση νουκλεοτιδίου- πρωτεΐνης Entrez : http://www.ncbi.nlm.nih.gov/nuccore/ Uniprot (πρωτεΐνης): http://www.uniprot.org/ Blast : http://blast.ncbi.nlm.nih.gov/blast.cgi
ΑΣΚΗΣΗ 3η Στοίχιση ακολουθιών βιολογικών µακροµορίων
 ΑΣΚΗΣΗ 3η Στοίχιση ακολουθιών βιολογικών µακροµορίων ΕΙΣΑΓΩΓΗ Ένας από τους πρωταρχικούς στόχους της σύγκρισης των ακολουθιών δύο µακροµορίων είναι η εκτίµηση της οµοιότητάς τους και η εξαγωγή συµπερασµάτων
ΑΣΚΗΣΗ 3η Στοίχιση ακολουθιών βιολογικών µακροµορίων ΕΙΣΑΓΩΓΗ Ένας από τους πρωταρχικούς στόχους της σύγκρισης των ακολουθιών δύο µακροµορίων είναι η εκτίµηση της οµοιότητάς τους και η εξαγωγή συµπερασµάτων
ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ ΑΝΑ ΖΕΥΓΗ
 ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ ΑΝΑ ΖΕΥΓΗ Σελίδα 1 Ομολογία Σελίδα 2 Ομολογία Ομολογία κοινή εξελικτική καταγωγή Ορθόλογα γονίδια ειδογένεση συνήθως, ίδια βιολογική λειτουργία Παράλογα γονίδια γονιδιακός διπλασιασμός
ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ ΑΝΑ ΖΕΥΓΗ Σελίδα 1 Ομολογία Σελίδα 2 Ομολογία Ομολογία κοινή εξελικτική καταγωγή Ορθόλογα γονίδια ειδογένεση συνήθως, ίδια βιολογική λειτουργία Παράλογα γονίδια γονιδιακός διπλασιασμός
Στοίχιση Ακολουθιών. Μέθοδοι σύγκρισης ακολουθιών. Είδος στοίχισης. match. gap. mismatch
 Οµολογία ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ ΑΝΑ ΖΕΥΓΗ Σελίδα 1 Σελίδα 2 Οµολογία Οµολογία Οµολογία κοινή εξελικτική καταγωγή Ορθόλογα γονίδια ειδογένεση συνήθως, ίδια βιολογική λειτουργία Παράλογα γονίδια γονιδιακός
Οµολογία ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ ΑΝΑ ΖΕΥΓΗ Σελίδα 1 Σελίδα 2 Οµολογία Οµολογία Οµολογία κοινή εξελικτική καταγωγή Ορθόλογα γονίδια ειδογένεση συνήθως, ίδια βιολογική λειτουργία Παράλογα γονίδια γονιδιακός
Κατα ζέυγη στοίχιση και στατιστική σημαντικότητα αυτής
 ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ ΙΙ Κατα ζέυγη στοίχιση και στατιστική σημαντικότητα αυτής Παντελής Μπάγκος 1 Διάλεξη 2 Αναζήτηση ομοιότητας και κατά ζεύγη στοίχιση ακολουθιών 2 Κατά ζεύγη στοίχιση ακολουθιών Από τα πιο
ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ ΙΙ Κατα ζέυγη στοίχιση και στατιστική σημαντικότητα αυτής Παντελής Μπάγκος 1 Διάλεξη 2 Αναζήτηση ομοιότητας και κατά ζεύγη στοίχιση ακολουθιών 2 Κατά ζεύγη στοίχιση ακολουθιών Από τα πιο
ΑΣΚΗΣΗ 4η Αναζήτηση οµοιοτήτων σε βάσεις δεδοµένων ακολουθιών
 ΑΣΚΗΣΗ 4η Αναζήτηση οµοιοτήτων σε βάσεις δεδοµένων ακολουθιών ΕΙΣΑΓΩΓΗ Η αναζήτηση οµοιοτήτων σε βάσεις δεδοµένων ακολουθιών (database similarity searching) αποτελεί µια από τις συχνότερα χρησιµοποιούµενες
ΑΣΚΗΣΗ 4η Αναζήτηση οµοιοτήτων σε βάσεις δεδοµένων ακολουθιών ΕΙΣΑΓΩΓΗ Η αναζήτηση οµοιοτήτων σε βάσεις δεδοµένων ακολουθιών (database similarity searching) αποτελεί µια από τις συχνότερα χρησιµοποιούµενες
Βιοπληροφορική. Ενότητα 9: Αναζήτηση Ομοιοτήτων σε ΒΔ Ακολουθιών - Στατιστική Σημαντικότητα, 1 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.
 Βιοπληροφορική Ενότητα 9: Αναζήτηση Ομοιοτήτων σε ΒΔ Ακολουθιών - Στατιστική Σημαντικότητα, 1 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Παρουσίαση των εφαρμογών της αναζήτησης
Βιοπληροφορική Ενότητα 9: Αναζήτηση Ομοιοτήτων σε ΒΔ Ακολουθιών - Στατιστική Σημαντικότητα, 1 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Παρουσίαση των εφαρμογών της αναζήτησης
Βιοπληροφορική. Ενότητα 5: Στοίχιση ακολουθιών ανά ζεύγη, 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου
 Βιοπληροφορική Ενότητα 5: Στοίχιση ακολουθιών ανά ζεύγη, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της συσχέτισης ομολογίας ομοιότητας. Παρουσίαση των πληροφοριών
Βιοπληροφορική Ενότητα 5: Στοίχιση ακολουθιών ανά ζεύγη, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της συσχέτισης ομολογίας ομοιότητας. Παρουσίαση των πληροφοριών
Πανεπιστήμιο Δυτικής Μακεδονίας. Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών. Βιοπληροφορική
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 12: Αναζήτηση ομοιοτήτων έναντι βάσεων δεδομένων με τη χρήση ευρετικών αλγορίθμων Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 12: Αναζήτηση ομοιοτήτων έναντι βάσεων δεδομένων με τη χρήση ευρετικών αλγορίθμων Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr
ΕΠΑΝΑΛΗΨΗ. Σελίδα 1 ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ. Τ. Θηραίου
 ΕΠΑΝΑΛΗΨΗ Σελίδα 1 τεχνική σύγκρισης ακολουθιών υπολογισµός ενός µέτρου οµοιότητας αναζήτηση ομολογίας S-S match S1 HFCGGSLINEQWVVSAGHC HFCG S NE AGHC S2 HFCGASIYNENYA-TAGHC gap mismatch Σελίδα 2 ολική
ΕΠΑΝΑΛΗΨΗ Σελίδα 1 τεχνική σύγκρισης ακολουθιών υπολογισµός ενός µέτρου οµοιότητας αναζήτηση ομολογίας S-S match S1 HFCGGSLINEQWVVSAGHC HFCG S NE AGHC S2 HFCGASIYNENYA-TAGHC gap mismatch Σελίδα 2 ολική
Πρόβλημα. Σύνολο γνωστών αλληλουχιών
 BLAST Πρόβλημα Άγνωστη αλληλουχία Σύνολο γνωστών αλληλουχιών Η χρήση ενός υπολογιστή κι ενός αλγόριθμου είναι απαραίτητη για την ανακάλυψη της σχέσης μιας αλληλουχίας με τις γνωστές υπάρχουσες Τί είναι
BLAST Πρόβλημα Άγνωστη αλληλουχία Σύνολο γνωστών αλληλουχιών Η χρήση ενός υπολογιστή κι ενός αλγόριθμου είναι απαραίτητη για την ανακάλυψη της σχέσης μιας αλληλουχίας με τις γνωστές υπάρχουσες Τί είναι
Homework 3 Solutions
 Homework 3 Solutions Igor Yanovsky (Math 151A TA) Problem 1: Compute the absolute error and relative error in approximations of p by p. (Use calculator!) a) p π, p 22/7; b) p π, p 3.141. Solution: For
Homework 3 Solutions Igor Yanovsky (Math 151A TA) Problem 1: Compute the absolute error and relative error in approximations of p by p. (Use calculator!) a) p π, p 22/7; b) p π, p 3.141. Solution: For
Πίνακες αντικατάστασης PAM και BLOSUM και εναλλακτικές προσεγγίσεις
 Πίνακες αντικατάστασης PAM και BLOSUM και εναλλακτικές προσεγγίσεις Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου
Πίνακες αντικατάστασης PAM και BLOSUM και εναλλακτικές προσεγγίσεις Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 19/5/2007
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
Κεφάλαιο 5 ο : Αλγόριθµοι Σύγκρισης Ακολουθιών Βιολογικών εδοµένων
 Κεφάλαιο 5 ο : Αλγόριθµοι Σύγκρισης Ακολουθιών Βιολογικών εδοµένων Σε αυτό το κεφάλαιο παρουσιάζουµε 2 βασικούς αλγορίθµους σύγκρισης ακολουθιών Βιολογικών εδοµένων τους BLAST & FASTA. Οι δυο αλγόριθµοι
Κεφάλαιο 5 ο : Αλγόριθµοι Σύγκρισης Ακολουθιών Βιολογικών εδοµένων Σε αυτό το κεφάλαιο παρουσιάζουµε 2 βασικούς αλγορίθµους σύγκρισης ακολουθιών Βιολογικών εδοµένων τους BLAST & FASTA. Οι δυο αλγόριθµοι
5.4 The Poisson Distribution.
 The worst thing you can do about a situation is nothing. Sr. O Shea Jackson 5.4 The Poisson Distribution. Description of the Poisson Distribution Discrete probability distribution. The random variable
The worst thing you can do about a situation is nothing. Sr. O Shea Jackson 5.4 The Poisson Distribution. Description of the Poisson Distribution Discrete probability distribution. The random variable
Εφαρμοσμένη Βιοτεχνολογία Εργαστηριακή Άσκηση Εισαγωγή στην Βιοπληροφορική
 Εφαρμοσμένη Βιοτεχνολογία Εργαστηριακή Άσκηση Εισαγωγή στην Βιοπληροφορική Δραστηριότητες 1. Εύρεση γονιδίων/πρωτεϊνών από βάσεις δεδομένων 2. Ευθυγράμμιση και σύγκριση γονιδίων/πρωτεϊνών 3. Δημιουργία
Εφαρμοσμένη Βιοτεχνολογία Εργαστηριακή Άσκηση Εισαγωγή στην Βιοπληροφορική Δραστηριότητες 1. Εύρεση γονιδίων/πρωτεϊνών από βάσεις δεδομένων 2. Ευθυγράμμιση και σύγκριση γονιδίων/πρωτεϊνών 3. Δημιουργία
Πανεπιστήμιο Δυτικής Μακεδονίας. Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών. Βιοπληροφορική. Ενότητα 7: Σύγκριση αλληλουχιών Part II
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 7: Σύγκριση αλληλουχιών Part II Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία e-mail: sbellou@uowm.gr
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 7: Σύγκριση αλληλουχιών Part II Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία e-mail: sbellou@uowm.gr
Αλγόριθµοι Εύρεσης Οµοιοτήτων Ακολουθιών Μέρος ΙΙΙ: Έλεγχος στατιστικής σηµαντικότητας. Πίνακες αντικατάστασης για σύγκριση ακολουθιών
 Αλγόριθµοι Εύρεσης Οµοιοτήτων Ακολουθιών Μέρος ΙΙΙ: Έλεγχος στατιστικής σηµαντικότητας Πίνακες αντικατάστασης για σύγκριση ακολουθιών Vasilis Promponas Bioinformatics Research Laboratory Department of
Αλγόριθµοι Εύρεσης Οµοιοτήτων Ακολουθιών Μέρος ΙΙΙ: Έλεγχος στατιστικής σηµαντικότητας Πίνακες αντικατάστασης για σύγκριση ακολουθιών Vasilis Promponas Bioinformatics Research Laboratory Department of
Εισαγωγή στους αλγορίθμους Βιοπληροφορικής. Στοίχιση αλληλουχιών
 Στοίχιση αλληλουχιών Σύνοψη Καθολική στοίχιση Μήτρες βαθμολόγησης Τοπική στοίχιση Στοίχιση με ποινές εισαγωγής κενών Από την LCS στη στοίχιση: αλλαγές στη βαθμολόγηση Το πρόβλημα της Μεγαλύτερης Κοινής
Στοίχιση αλληλουχιών Σύνοψη Καθολική στοίχιση Μήτρες βαθμολόγησης Τοπική στοίχιση Στοίχιση με ποινές εισαγωγής κενών Από την LCS στη στοίχιση: αλλαγές στη βαθμολόγηση Το πρόβλημα της Μεγαλύτερης Κοινής
ΒΙΟ230 - Εισαγωγή στην Υπολογιστική Βιολογία Πρακτικό Εργαστήριο: Basic Local Alignment Search Tool BLAST
 ΒΙΟ230 - Εισαγωγή στην Υπολογιστική Βιολογία Πρακτικό Εργαστήριο: Basic Local Alignment Search Tool BLAST Στέλλα Ταμανά, Βασίλης Προμπονάς Λευκωσία 2016-2018 Περίληψη (Overview) Κατά τη διάρκεια αυτού
ΒΙΟ230 - Εισαγωγή στην Υπολογιστική Βιολογία Πρακτικό Εργαστήριο: Basic Local Alignment Search Tool BLAST Στέλλα Ταμανά, Βασίλης Προμπονάς Λευκωσία 2016-2018 Περίληψη (Overview) Κατά τη διάρκεια αυτού
ΕΙΣΑΓΩΓΗ ΣΤΗ ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ
 ΕΙΣΑΓΩΓΗ ΣΤΗ ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ ΕΛΕΝΑ ΦΛΟΚΑ Επίκουρος Καθηγήτρια Τµήµα Φυσικής, Τοµέας Φυσικής Περιβάλλοντος- Μετεωρολογίας ΓΕΝΙΚΟΙ ΟΡΙΣΜΟΙ Πληθυσµός Σύνολο ατόµων ή αντικειµένων στα οποία αναφέρονται
ΕΙΣΑΓΩΓΗ ΣΤΗ ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ ΕΛΕΝΑ ΦΛΟΚΑ Επίκουρος Καθηγήτρια Τµήµα Φυσικής, Τοµέας Φυσικής Περιβάλλοντος- Μετεωρολογίας ΓΕΝΙΚΟΙ ΟΡΙΣΜΟΙ Πληθυσµός Σύνολο ατόµων ή αντικειµένων στα οποία αναφέρονται
ΑΝΑΖΗΤΗΣΗ ΟΜΟΙΟΤΗΤΩΝ ΣΕ ΒΑΣΕΙΣ Ε ΟΜΕΝΩΝ ΑΚΟΛΟΥΘΙΩΝ
 Αναζήτηση οµοιοτήτων ΑΝΑΖΗΤΗΣΗ ΟΜΟΙΟΤΗΤΩΝ ΣΕ ΒΑΣΕΙΣ Ε ΟΜΕΝΩΝ ΑΚΟΛΟΥΘΙΩΝ Σελίδα 1 εδοµένα Ακολουθία επερώτησης (query sequence) Ακολουθίες στη Βάση εδοµένων (subject sequences) Αναζήτηση Μέθοδοι δυναµικού
Αναζήτηση οµοιοτήτων ΑΝΑΖΗΤΗΣΗ ΟΜΟΙΟΤΗΤΩΝ ΣΕ ΒΑΣΕΙΣ Ε ΟΜΕΝΩΝ ΑΚΟΛΟΥΘΙΩΝ Σελίδα 1 εδοµένα Ακολουθία επερώτησης (query sequence) Ακολουθίες στη Βάση εδοµένων (subject sequences) Αναζήτηση Μέθοδοι δυναµικού
Numerical Analysis FMN011
 Numerical Analysis FMN011 Carmen Arévalo Lund University carmen@maths.lth.se Lecture 12 Periodic data A function g has period P if g(x + P ) = g(x) Model: Trigonometric polynomial of order M T M (x) =
Numerical Analysis FMN011 Carmen Arévalo Lund University carmen@maths.lth.se Lecture 12 Periodic data A function g has period P if g(x + P ) = g(x) Model: Trigonometric polynomial of order M T M (x) =
Πρόβλημα 1: Αναζήτηση Ελάχιστης/Μέγιστης Τιμής
 Πρόβλημα 1: Αναζήτηση Ελάχιστης/Μέγιστης Τιμής Να γραφεί πρόγραμμα το οποίο δέχεται ως είσοδο μια ακολουθία S από n (n 40) ακέραιους αριθμούς και επιστρέφει ως έξοδο δύο ακολουθίες από θετικούς ακέραιους
Πρόβλημα 1: Αναζήτηση Ελάχιστης/Μέγιστης Τιμής Να γραφεί πρόγραμμα το οποίο δέχεται ως είσοδο μια ακολουθία S από n (n 40) ακέραιους αριθμούς και επιστρέφει ως έξοδο δύο ακολουθίες από θετικούς ακέραιους
Βιοπληροφορική. Πίνακες Αντικατάστασης & Οπτική Σύγκριση Αλληλουχιών. Αλέξανδρος Τζάλλας
 Βιοπληροφορική Πίνακες Αντικατάστασης & Οπτική Σύγκριση Αλληλουχιών Αλέξανδρος Τζάλλας e-mail: tzallas@teiep.gr ΤΕΙ Ηπείρου Σχολή Τεχνολογικών Εφαρμογών Τμήμα Μηχανικών Πληροφορικής ΤΕ Copyright Πανεπιστήμιο
Βιοπληροφορική Πίνακες Αντικατάστασης & Οπτική Σύγκριση Αλληλουχιών Αλέξανδρος Τζάλλας e-mail: tzallas@teiep.gr ΤΕΙ Ηπείρου Σχολή Τεχνολογικών Εφαρμογών Τμήμα Μηχανικών Πληροφορικής ΤΕ Copyright Πανεπιστήμιο
CHAPTER 25 SOLVING EQUATIONS BY ITERATIVE METHODS
 CHAPTER 5 SOLVING EQUATIONS BY ITERATIVE METHODS EXERCISE 104 Page 8 1. Find the positive root of the equation x + 3x 5 = 0, correct to 3 significant figures, using the method of bisection. Let f(x) =
CHAPTER 5 SOLVING EQUATIONS BY ITERATIVE METHODS EXERCISE 104 Page 8 1. Find the positive root of the equation x + 3x 5 = 0, correct to 3 significant figures, using the method of bisection. Let f(x) =
HOMEWORK 4 = G. In order to plot the stress versus the stretch we define a normalized stretch:
 HOMEWORK 4 Problem a For the fast loading case, we want to derive the relationship between P zz and λ z. We know that the nominal stress is expressed as: P zz = ψ λ z where λ z = λ λ z. Therefore, applying
HOMEWORK 4 Problem a For the fast loading case, we want to derive the relationship between P zz and λ z. We know that the nominal stress is expressed as: P zz = ψ λ z where λ z = λ λ z. Therefore, applying
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 6/5/2006
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Ολοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα είναι μικρότεροι το 1000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Διάρκεια: 3,5 ώρες Καλή
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Ολοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα είναι μικρότεροι το 1000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Διάρκεια: 3,5 ώρες Καλή
Αλγόριθμοι Εύρεσης Ομοιοτήτων Ακολουθιών Μέρος ΙΙΙ: Έλεγχος στατιστικής σημαντικότητας
 Αλγόριθμοι Εύρεσης Ομοιοτήτων Ακολουθιών Μέρος ΙΙΙ: Έλεγχος στατιστικής σημαντικότητας Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161
Αλγόριθμοι Εύρεσης Ομοιοτήτων Ακολουθιών Μέρος ΙΙΙ: Έλεγχος στατιστικής σημαντικότητας Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161
ΑΝΑΖΗΤΗΣΗ ΟΜΟΙΟΤΗΤΩΝ ΣΕ ΒΑΣΕΙΣ ΔΕΔΟΜΕΝΩΝ ΑΚΟΛΟΥΘΙΩΝ
 ΑΝΑΖΗΤΗΣΗ ΟΜΟΙΟΤΗΤΩΝ ΣΕ ΒΑΣΕΙΣ ΔΕΔΟΜΕΝΩΝ ΑΚΟΛΟΥΘΙΩΝ Σελίδα 1 Αναζήτηση ομοιοτήτων Δεδομένα Ακολουθία επερώτησης (query sequence) Ακολουθίες στη Βάση Δεδομένων (subject sequences) Αναζήτηση Μέθοδοι δυναμικού
ΑΝΑΖΗΤΗΣΗ ΟΜΟΙΟΤΗΤΩΝ ΣΕ ΒΑΣΕΙΣ ΔΕΔΟΜΕΝΩΝ ΑΚΟΛΟΥΘΙΩΝ Σελίδα 1 Αναζήτηση ομοιοτήτων Δεδομένα Ακολουθία επερώτησης (query sequence) Ακολουθίες στη Βάση Δεδομένων (subject sequences) Αναζήτηση Μέθοδοι δυναμικού
Μηχανική Μάθηση Hypothesis Testing
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Μηχανική Μάθηση Hypothesis Testing Γιώργος Μπορμπουδάκης Τμήμα Επιστήμης Υπολογιστών Procedure 1. Form the null (H 0 ) and alternative (H 1 ) hypothesis 2. Consider
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Μηχανική Μάθηση Hypothesis Testing Γιώργος Μπορμπουδάκης Τμήμα Επιστήμης Υπολογιστών Procedure 1. Form the null (H 0 ) and alternative (H 1 ) hypothesis 2. Consider
Βιοπληροφορική. Blast/PSI-Blast 3o εργαστήριο
 Βιοπληροφορική Blast/PSI-Blast 3o εργαστήριο Αναζήτηση οµόλογων ακολουθιών σε βάσεις δεδοµένων (i) Οµόλογες ακολουθίες πιθανόν να έχουν παρόµοιες λειτουργίες. Ακολουθία επερώτησης (query sequence) Υποκείµενες
Βιοπληροφορική Blast/PSI-Blast 3o εργαστήριο Αναζήτηση οµόλογων ακολουθιών σε βάσεις δεδοµένων (i) Οµόλογες ακολουθίες πιθανόν να έχουν παρόµοιες λειτουργίες. Ακολουθία επερώτησης (query sequence) Υποκείµενες
Στοίχιση κατά ζεύγη. Στοίχιση ακολουθιών κατά ζεύγη (Pairwise alignment)
") Στοίχιση ακολουθιών κατά ζεύγη (Pairwise alignment) Στοίχιση κατά ζεύγη: Τι είναι Αντιστοίχιση των νουκλεοτιδίων/αµινοξέων δυο ακολουθιών, ώστε να εντοπιστούν οι οµοιότητες και οι διαφορές τους. Χρησιµοποιείται
Στοίχιση ακολουθιών κατά ζεύγη (Pairwise alignment) Στοίχιση κατά ζεύγη: Τι είναι Αντιστοίχιση των νουκλεοτιδίων/αµινοξέων δυο ακολουθιών, ώστε να εντοπιστούν οι οµοιότητες και οι διαφορές τους. Χρησιµοποιείται
Βιοπληροφορική Ι. Παντελής Μπάγκος Αναπληρωτής Καθηγητής. Πανεπιστήμιο Θεσσαλίας Λαμία, 2015
 Βιοπληροφορική Ι Παντελής Μπάγκος Αναπληρωτής Καθηγητής Πανεπιστήμιο Θεσσαλίας Λαμία, 2015 1 Στοίχιση αλληλουχιών 2 Τρόποι μελέτης των ακολουθιών Global information Η ακολουθία αναπαρίσταται από ένα διάνυσμα
Βιοπληροφορική Ι Παντελής Μπάγκος Αναπληρωτής Καθηγητής Πανεπιστήμιο Θεσσαλίας Λαμία, 2015 1 Στοίχιση αλληλουχιών 2 Τρόποι μελέτης των ακολουθιών Global information Η ακολουθία αναπαρίσταται από ένα διάνυσμα
Approximation of distance between locations on earth given by latitude and longitude
 Approximation of distance between locations on earth given by latitude and longitude Jan Behrens 2012-12-31 In this paper we shall provide a method to approximate distances between two points on earth
Approximation of distance between locations on earth given by latitude and longitude Jan Behrens 2012-12-31 In this paper we shall provide a method to approximate distances between two points on earth
Other Test Constructions: Likelihood Ratio & Bayes Tests
 Other Test Constructions: Likelihood Ratio & Bayes Tests Side-Note: So far we have seen a few approaches for creating tests such as Neyman-Pearson Lemma ( most powerful tests of H 0 : θ = θ 0 vs H 1 :
Other Test Constructions: Likelihood Ratio & Bayes Tests Side-Note: So far we have seen a few approaches for creating tests such as Neyman-Pearson Lemma ( most powerful tests of H 0 : θ = θ 0 vs H 1 :
Πανεπιστήμιο Δυτικής Μακεδονίας. Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών. Βιοπληροφορική
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 4: Πίνακες αντικατάστασης & οπτική σύγκριση αλληλουχιών Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 4: Πίνακες αντικατάστασης & οπτική σύγκριση αλληλουχιών Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία
Μέθοδοι μελέτης εξέλιξης
 H διερεύνηση της μοριακής βάσης της εξέλιξης βασίζεται σε μεγάλο βαθμό στη διευκρίνιση της διαδικασίας με την οποία μετασχηματίσθηκαν στη διάρκεια της εξέλιξης πρωτεϊνες, άλλα μόρια και βιοχημικές πορείες
H διερεύνηση της μοριακής βάσης της εξέλιξης βασίζεται σε μεγάλο βαθμό στη διευκρίνιση της διαδικασίας με την οποία μετασχηματίσθηκαν στη διάρκεια της εξέλιξης πρωτεϊνες, άλλα μόρια και βιοχημικές πορείες
Concrete Mathematics Exercises from 30 September 2016
 Concrete Mathematics Exercises from 30 September 2016 Silvio Capobianco Exercise 1.7 Let H(n) = J(n + 1) J(n). Equation (1.8) tells us that H(2n) = 2, and H(2n+1) = J(2n+2) J(2n+1) = (2J(n+1) 1) (2J(n)+1)
Concrete Mathematics Exercises from 30 September 2016 Silvio Capobianco Exercise 1.7 Let H(n) = J(n + 1) J(n). Equation (1.8) tells us that H(2n) = 2, and H(2n+1) = J(2n+2) J(2n+1) = (2J(n+1) 1) (2J(n)+1)
Εργαστήριο Ανάπτυξης Εφαρμογών Βάσεων Δεδομένων. Εξάμηνο 7 ο
 Εργαστήριο Ανάπτυξης Εφαρμογών Βάσεων Δεδομένων Εξάμηνο 7 ο Procedures and Functions Stored procedures and functions are named blocks of code that enable you to group and organize a series of SQL and PL/SQL
Εργαστήριο Ανάπτυξης Εφαρμογών Βάσεων Δεδομένων Εξάμηνο 7 ο Procedures and Functions Stored procedures and functions are named blocks of code that enable you to group and organize a series of SQL and PL/SQL
6.1. Dirac Equation. Hamiltonian. Dirac Eq.
 6.1. Dirac Equation Ref: M.Kaku, Quantum Field Theory, Oxford Univ Press (1993) η μν = η μν = diag(1, -1, -1, -1) p 0 = p 0 p = p i = -p i p μ p μ = p 0 p 0 + p i p i = E c 2 - p 2 = (m c) 2 H = c p 2
6.1. Dirac Equation Ref: M.Kaku, Quantum Field Theory, Oxford Univ Press (1993) η μν = η μν = diag(1, -1, -1, -1) p 0 = p 0 p = p i = -p i p μ p μ = p 0 p 0 + p i p i = E c 2 - p 2 = (m c) 2 H = c p 2
Βιοπληροφορική. Ενότητα 7: Στοίχιση ακολουθιών ανά ζεύγη Τεχνικές Στοίχισης Ακολουθιών,(2/2) 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.
 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.") Βιοπληροφορική Ενότητα 7: Στοίχιση ακολουθιών ανά ζεύγη Τεχνικές Στοίχισης Ακολουθιών,(2/2) 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Παρουσίαση της μεθόδου κατασκευής και
Βιοπληροφορική Ενότητα 7: Στοίχιση ακολουθιών ανά ζεύγη Τεχνικές Στοίχισης Ακολουθιών,(2/2) 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Παρουσίαση της μεθόδου κατασκευής και
ΚΥΠΡΙΑΚΟΣ ΣΥΝΔΕΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY 21 ος ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ Δεύτερος Γύρος - 30 Μαρτίου 2011
 Διάρκεια Διαγωνισμού: 3 ώρες Απαντήστε όλες τις ερωτήσεις Μέγιστο Βάρος (20 Μονάδες) Δίνεται ένα σύνολο από N σφαιρίδια τα οποία δεν έχουν όλα το ίδιο βάρος μεταξύ τους και ένα κουτί που αντέχει μέχρι
Διάρκεια Διαγωνισμού: 3 ώρες Απαντήστε όλες τις ερωτήσεις Μέγιστο Βάρος (20 Μονάδες) Δίνεται ένα σύνολο από N σφαιρίδια τα οποία δεν έχουν όλα το ίδιο βάρος μεταξύ τους και ένα κουτί που αντέχει μέχρι
Matrices and vectors. Matrix and vector. a 11 a 12 a 1n a 21 a 22 a 2n A = b 1 b 2. b m. R m n, b = = ( a ij. a m1 a m2 a mn. def
 Matrices and vectors Matrix and vector a 11 a 12 a 1n a 21 a 22 a 2n A = a m1 a m2 a mn def = ( a ij ) R m n, b = b 1 b 2 b m Rm Matrix and vectors in linear equations: example E 1 : x 1 + x 2 + 3x 4 =
Matrices and vectors Matrix and vector a 11 a 12 a 1n a 21 a 22 a 2n A = a m1 a m2 a mn def = ( a ij ) R m n, b = b 1 b 2 b m Rm Matrix and vectors in linear equations: example E 1 : x 1 + x 2 + 3x 4 =
Phys460.nb Solution for the t-dependent Schrodinger s equation How did we find the solution? (not required)
") Phys460.nb 81 ψ n (t) is still the (same) eigenstate of H But for tdependent H. The answer is NO. 5.5.5. Solution for the tdependent Schrodinger s equation If we assume that at time t 0, the electron starts
Phys460.nb 81 ψ n (t) is still the (same) eigenstate of H But for tdependent H. The answer is NO. 5.5.5. Solution for the tdependent Schrodinger s equation If we assume that at time t 0, the electron starts
derivation of the Laplacian from rectangular to spherical coordinates
 derivation of the Laplacian from rectangular to spherical coordinates swapnizzle 03-03- :5:43 We begin by recognizing the familiar conversion from rectangular to spherical coordinates (note that φ is used
derivation of the Laplacian from rectangular to spherical coordinates swapnizzle 03-03- :5:43 We begin by recognizing the familiar conversion from rectangular to spherical coordinates (note that φ is used
Βιοπληροφορική. Ενότητα 7: Στοίχιση ακολουθιών ανά ζεύγη Τεχνικές Στοίχισης Ακολουθιών, (1/2) 1ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.
 1ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.") Βιοπληροφορική Ενότητα 7: Στοίχιση ακολουθιών ανά ζεύγη Τεχνικές Στοίχισης Ακολουθιών, (1/2) 1ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Παρουσίαση της μεθόδου κατασκευής και
Βιοπληροφορική Ενότητα 7: Στοίχιση ακολουθιών ανά ζεύγη Τεχνικές Στοίχισης Ακολουθιών, (1/2) 1ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Παρουσίαση της μεθόδου κατασκευής και
BMI/CS 776 Lecture #14: Multiple Alignment - MUSCLE. Colin Dewey
 BMI/CS 776 Lecture #14: Multiple Alignment - MUSCLE Colin Dewey 2007.03.08 1 Importance of protein multiple alignment Phylogenetic tree estimation Prediction of protein secondary structure Critical residue
BMI/CS 776 Lecture #14: Multiple Alignment - MUSCLE Colin Dewey 2007.03.08 1 Importance of protein multiple alignment Phylogenetic tree estimation Prediction of protein secondary structure Critical residue
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ. Ψηφιακή Οικονομία. Διάλεξη 7η: Consumer Behavior Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Ψηφιακή Οικονομία Διάλεξη 7η: Consumer Behavior Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών Τέλος Ενότητας Χρηματοδότηση Το παρόν εκπαιδευτικό υλικό έχει αναπτυχθεί
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Ψηφιακή Οικονομία Διάλεξη 7η: Consumer Behavior Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών Τέλος Ενότητας Χρηματοδότηση Το παρόν εκπαιδευτικό υλικό έχει αναπτυχθεί
Solution Series 9. i=1 x i and i=1 x i.
 Lecturer: Prof. Dr. Mete SONER Coordinator: Yilin WANG Solution Series 9 Q1. Let α, β >, the p.d.f. of a beta distribution with parameters α and β is { Γ(α+β) Γ(α)Γ(β) f(x α, β) xα 1 (1 x) β 1 for < x
Lecturer: Prof. Dr. Mete SONER Coordinator: Yilin WANG Solution Series 9 Q1. Let α, β >, the p.d.f. of a beta distribution with parameters α and β is { Γ(α+β) Γ(α)Γ(β) f(x α, β) xα 1 (1 x) β 1 for < x
Math 6 SL Probability Distributions Practice Test Mark Scheme
 Math 6 SL Probability Distributions Practice Test Mark Scheme. (a) Note: Award A for vertical line to right of mean, A for shading to right of their vertical line. AA N (b) evidence of recognizing symmetry
Math 6 SL Probability Distributions Practice Test Mark Scheme. (a) Note: Award A for vertical line to right of mean, A for shading to right of their vertical line. AA N (b) evidence of recognizing symmetry
2 Composition. Invertible Mappings
 Arkansas Tech University MATH 4033: Elementary Modern Algebra Dr. Marcel B. Finan Composition. Invertible Mappings In this section we discuss two procedures for creating new mappings from old ones, namely,
Arkansas Tech University MATH 4033: Elementary Modern Algebra Dr. Marcel B. Finan Composition. Invertible Mappings In this section we discuss two procedures for creating new mappings from old ones, namely,
Exercises 10. Find a fundamental matrix of the given system of equations. Also find the fundamental matrix Φ(t) satisfying Φ(0) = I. 1.
 satisfying Φ(0) = I. 1.") Exercises 0 More exercises are available in Elementary Differential Equations. If you have a problem to solve any of them, feel free to come to office hour. Problem Find a fundamental matrix of the given
Exercises 0 More exercises are available in Elementary Differential Equations. If you have a problem to solve any of them, feel free to come to office hour. Problem Find a fundamental matrix of the given
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 24/3/2007
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Όλοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα μικρότεροι του 10000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Αν κάπου κάνετε κάποιες υποθέσεις
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Όλοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα μικρότεροι του 10000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Αν κάπου κάνετε κάποιες υποθέσεις
Exercises to Statistics of Material Fatigue No. 5
 Prof. Dr. Christine Müller Dipl.-Math. Christoph Kustosz Eercises to Statistics of Material Fatigue No. 5 E. 9 (5 a Show, that a Fisher information matri for a two dimensional parameter θ (θ,θ 2 R 2, can
Prof. Dr. Christine Müller Dipl.-Math. Christoph Kustosz Eercises to Statistics of Material Fatigue No. 5 E. 9 (5 a Show, that a Fisher information matri for a two dimensional parameter θ (θ,θ 2 R 2, can
Math221: HW# 1 solutions
 Math: HW# solutions Andy Royston October, 5 7.5.7, 3 rd Ed. We have a n = b n = a = fxdx = xdx =, x cos nxdx = x sin nx n sin nxdx n = cos nx n = n n, x sin nxdx = x cos nx n + cos nxdx n cos n = + sin
Math: HW# solutions Andy Royston October, 5 7.5.7, 3 rd Ed. We have a n = b n = a = fxdx = xdx =, x cos nxdx = x sin nx n sin nxdx n = cos nx n = n n, x sin nxdx = x cos nx n + cos nxdx n cos n = + sin
ΑΛΓΟΡΙΘΜΟΙ Άνοιξη I. ΜΗΛΗΣ
 ΑΛΓΟΡΙΘΜΟΙ http://eclass.aueb.gr/courses/inf161/ Άνοιξη 216 - I. ΜΗΛΗΣ ΔΥΝΑΜΙΚΟΣ ΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ ΑΛΓΟΡΙΘΜΟΙ - ΑΝΟΙΞΗ 216 - Ι. ΜΗΛΗΣ 9 DP II 1 Dynamic Programming ΓΕΝΙΚΗ ΙΔΕΑ 1. Ορισμός υπο-προβλήματος/ων
ΑΛΓΟΡΙΘΜΟΙ http://eclass.aueb.gr/courses/inf161/ Άνοιξη 216 - I. ΜΗΛΗΣ ΔΥΝΑΜΙΚΟΣ ΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ ΑΛΓΟΡΙΘΜΟΙ - ΑΝΟΙΞΗ 216 - Ι. ΜΗΛΗΣ 9 DP II 1 Dynamic Programming ΓΕΝΙΚΗ ΙΔΕΑ 1. Ορισμός υπο-προβλήματος/ων
Lecture 2: Dirac notation and a review of linear algebra Read Sakurai chapter 1, Baym chatper 3
 Lecture 2: Dirac notation and a review of linear algebra Read Sakurai chapter 1, Baym chatper 3 1 State vector space and the dual space Space of wavefunctions The space of wavefunctions is the set of all
Lecture 2: Dirac notation and a review of linear algebra Read Sakurai chapter 1, Baym chatper 3 1 State vector space and the dual space Space of wavefunctions The space of wavefunctions is the set of all
( ) 2 and compare to M.
 2 and compare to M.") Problems and Solutions for Section 4.2 4.9 through 4.33) 4.9 Calculate the square root of the matrix 3!0 M!0 8 Hint: Let M / 2 a!b ; calculate M / 2!b c ) 2 and compare to M. Solution: Given: 3!0 M!0 8
Problems and Solutions for Section 4.2 4.9 through 4.33) 4.9 Calculate the square root of the matrix 3!0 M!0 8 Hint: Let M / 2 a!b ; calculate M / 2!b c ) 2 and compare to M. Solution: Given: 3!0 M!0 8
Bayesian statistics. DS GA 1002 Probability and Statistics for Data Science.
 Bayesian statistics DS GA 1002 Probability and Statistics for Data Science http://www.cims.nyu.edu/~cfgranda/pages/dsga1002_fall17 Carlos Fernandez-Granda Frequentist vs Bayesian statistics In frequentist
Bayesian statistics DS GA 1002 Probability and Statistics for Data Science http://www.cims.nyu.edu/~cfgranda/pages/dsga1002_fall17 Carlos Fernandez-Granda Frequentist vs Bayesian statistics In frequentist
SOLUTIONS TO MATH38181 EXTREME VALUES AND FINANCIAL RISK EXAM
 SOLUTIONS TO MATH38181 EXTREME VALUES AND FINANCIAL RISK EXAM Solutions to Question 1 a) The cumulative distribution function of T conditional on N n is Pr T t N n) Pr max X 1,..., X N ) t N n) Pr max
SOLUTIONS TO MATH38181 EXTREME VALUES AND FINANCIAL RISK EXAM Solutions to Question 1 a) The cumulative distribution function of T conditional on N n is Pr T t N n) Pr max X 1,..., X N ) t N n) Pr max
ST5224: Advanced Statistical Theory II
 ST5224: Advanced Statistical Theory II 2014/2015: Semester II Tutorial 7 1. Let X be a sample from a population P and consider testing hypotheses H 0 : P = P 0 versus H 1 : P = P 1, where P j is a known
ST5224: Advanced Statistical Theory II 2014/2015: Semester II Tutorial 7 1. Let X be a sample from a population P and consider testing hypotheses H 0 : P = P 0 versus H 1 : P = P 1, where P j is a known
EPL 603 TOPICS IN SOFTWARE ENGINEERING. Lab 5: Component Adaptation Environment (COPE)
") EPL 603 TOPICS IN SOFTWARE ENGINEERING Lab 5: Component Adaptation Environment (COPE) Performing Static Analysis 1 Class Name: The fully qualified name of the specific class Type: The type of the class
EPL 603 TOPICS IN SOFTWARE ENGINEERING Lab 5: Component Adaptation Environment (COPE) Performing Static Analysis 1 Class Name: The fully qualified name of the specific class Type: The type of the class
ANSWERSHEET (TOPIC = DIFFERENTIAL CALCULUS) COLLECTION #2. h 0 h h 0 h h 0 ( ) g k = g 0 + g 1 + g g 2009 =?
 COLLECTION #2. h 0 h h 0 h h 0 ( ) g k = g 0 + g 1 + g g 2009 =?") Teko Classes IITJEE/AIEEE Maths by SUHAAG SIR, Bhopal, Ph (0755) 3 00 000 www.tekoclasses.com ANSWERSHEET (TOPIC DIFFERENTIAL CALCULUS) COLLECTION # Question Type A.Single Correct Type Q. (A) Sol least
Teko Classes IITJEE/AIEEE Maths by SUHAAG SIR, Bhopal, Ph (0755) 3 00 000 www.tekoclasses.com ANSWERSHEET (TOPIC DIFFERENTIAL CALCULUS) COLLECTION # Question Type A.Single Correct Type Q. (A) Sol least
ΠΟΛΛΑΠΛΗ ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ I
 ΠΟΛΛΑΠΛΗ ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ I Σελίδα 1 Πολλαπλή στοίχιση αποκαλύπτει συντηρημένες περιοχές αντιστοίχιση καταλοίπων με κριτήρια ομοιότητας σε επίπεδο δομής εξέλιξης λειτουργίας ακολουθίας Σελίδα 2 Πολλαπλή
ΠΟΛΛΑΠΛΗ ΣΤΟΙΧΙΣΗ ΑΚΟΛΟΥΘΙΩΝ I Σελίδα 1 Πολλαπλή στοίχιση αποκαλύπτει συντηρημένες περιοχές αντιστοίχιση καταλοίπων με κριτήρια ομοιότητας σε επίπεδο δομής εξέλιξης λειτουργίας ακολουθίας Σελίδα 2 Πολλαπλή
Βιοπληροφορική. Ενότητα 8: Αναζήτηση Ομοιοτήτων σε Βάσεις Δεδομένων Ακολουθιών, 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.
 Βιοπληροφορική Ενότητα 8: Αναζήτηση Ομοιοτήτων σε Βάσεις Δεδομένων Ακολουθιών, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της αναγκαιότητας των ευριστικών αλγορίθμων
Βιοπληροφορική Ενότητα 8: Αναζήτηση Ομοιοτήτων σε Βάσεις Δεδομένων Ακολουθιών, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Κατανόηση της αναγκαιότητας των ευριστικών αλγορίθμων
Matrices and Determinants
 Matrices and Determinants SUBJECTIVE PROBLEMS: Q 1. For what value of k do the following system of equations possess a non-trivial (i.e., not all zero) solution over the set of rationals Q? x + ky + 3z
Matrices and Determinants SUBJECTIVE PROBLEMS: Q 1. For what value of k do the following system of equations possess a non-trivial (i.e., not all zero) solution over the set of rationals Q? x + ky + 3z
Statistical Inference I Locally most powerful tests
 Statistical Inference I Locally most powerful tests Shirsendu Mukherjee Department of Statistics, Asutosh College, Kolkata, India. shirsendu st@yahoo.co.in So far we have treated the testing of one-sided
Statistical Inference I Locally most powerful tests Shirsendu Mukherjee Department of Statistics, Asutosh College, Kolkata, India. shirsendu st@yahoo.co.in So far we have treated the testing of one-sided
A Bonus-Malus System as a Markov Set-Chain. Małgorzata Niemiec Warsaw School of Economics Institute of Econometrics
 A Bonus-Malus System as a Markov Set-Chain Małgorzata Niemiec Warsaw School of Economics Institute of Econometrics Contents 1. Markov set-chain 2. Model of bonus-malus system 3. Example 4. Conclusions
A Bonus-Malus System as a Markov Set-Chain Małgorzata Niemiec Warsaw School of Economics Institute of Econometrics Contents 1. Markov set-chain 2. Model of bonus-malus system 3. Example 4. Conclusions
Problem Set 3: Solutions
 CMPSCI 69GG Applied Information Theory Fall 006 Problem Set 3: Solutions. [Cover and Thomas 7.] a Define the following notation, C I p xx; Y max X; Y C I p xx; Ỹ max I X; Ỹ We would like to show that C
CMPSCI 69GG Applied Information Theory Fall 006 Problem Set 3: Solutions. [Cover and Thomas 7.] a Define the following notation, C I p xx; Y max X; Y C I p xx; Ỹ max I X; Ỹ We would like to show that C
1. Ηλεκτρικό μαύρο κουτί: Αισθητήρας μετατόπισης με βάση τη χωρητικότητα
 IPHO_42_2011_EXP1.DO Experimental ompetition: 14 July 2011 Problem 1 Page 1 of 5 1. Ηλεκτρικό μαύρο κουτί: Αισθητήρας μετατόπισης με βάση τη χωρητικότητα Για ένα πυκνωτή χωρητικότητας ο οποίος είναι μέρος
IPHO_42_2011_EXP1.DO Experimental ompetition: 14 July 2011 Problem 1 Page 1 of 5 1. Ηλεκτρικό μαύρο κουτί: Αισθητήρας μετατόπισης με βάση τη χωρητικότητα Για ένα πυκνωτή χωρητικότητας ο οποίος είναι μέρος
Reminders: linear functions
 Reminders: linear functions Let U and V be vector spaces over the same field F. Definition A function f : U V is linear if for every u 1, u 2 U, f (u 1 + u 2 ) = f (u 1 ) + f (u 2 ), and for every u U
Reminders: linear functions Let U and V be vector spaces over the same field F. Definition A function f : U V is linear if for every u 1, u 2 U, f (u 1 + u 2 ) = f (u 1 ) + f (u 2 ), and for every u U
Πανεπιστήμιο Δυτικής Μακεδονίας. Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών. Βιοπληροφορική. Ενότητα 6: Σύγκριση αλληλουχιών Part I
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 6: Σύγκριση αλληλουχιών Part I Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία e-mail: sbellou@uowm.gr
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 6: Σύγκριση αλληλουχιών Part I Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία e-mail: sbellou@uowm.gr
Calculating the propagation delay of coaxial cable
 Your source for quality GNSS Networking Solutions and Design Services! Page 1 of 5 Calculating the propagation delay of coaxial cable The delay of a cable or velocity factor is determined by the dielectric
Your source for quality GNSS Networking Solutions and Design Services! Page 1 of 5 Calculating the propagation delay of coaxial cable The delay of a cable or velocity factor is determined by the dielectric
Βιοπληροφορική. Ενότητα 10: Αναζήτηση Ομοιοτήτων σε ΒΔ Ακολουθιών - Blast, (1/2) 1ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.
 1ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ.") Βιοπληροφορική Ενότητα 10: Αναζήτηση Ομοιοτήτων σε ΒΔ Ακολουθιών - Blast, (1/2) 1ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Αναφορά στις παραλλαγές του BLAST. Εξοικείωση με τη
Βιοπληροφορική Ενότητα 10: Αναζήτηση Ομοιοτήτων σε ΒΔ Ακολουθιών - Blast, (1/2) 1ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Αναφορά στις παραλλαγές του BLAST. Εξοικείωση με τη
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ. Ψηφιακή Οικονομία. Διάλεξη 9η: Basics of Game Theory Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Ψηφιακή Οικονομία Διάλεξη 9η: Basics of Game Theory Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών Course Outline Part II: Mathematical Tools Firms - Basics of Industrial
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Ψηφιακή Οικονομία Διάλεξη 9η: Basics of Game Theory Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών Course Outline Part II: Mathematical Tools Firms - Basics of Industrial
Areas and Lengths in Polar Coordinates
 Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
Correction Table for an Alcoholometer Calibrated at 20 o C
 An alcoholometer is a device that measures the concentration of ethanol in a water-ethanol mixture (often in units of %abv percent alcohol by volume). The depth to which an alcoholometer sinks in a water-ethanol
An alcoholometer is a device that measures the concentration of ethanol in a water-ethanol mixture (often in units of %abv percent alcohol by volume). The depth to which an alcoholometer sinks in a water-ethanol
Περιοχές με ακραία σύσταση / χαμηλή πολυπλοκότητα
 Περιοχές με ακραία σύσταση / χαμηλή πολυπλοκότητα Vasilis Promponas Bioinformatics Research Laboratory Department of Biological Sciences University of Cyprus Σύνοψη Βασικές έννοιες XNU SEG LCRs και αναζητήσεις
Περιοχές με ακραία σύσταση / χαμηλή πολυπλοκότητα Vasilis Promponas Bioinformatics Research Laboratory Department of Biological Sciences University of Cyprus Σύνοψη Βασικές έννοιες XNU SEG LCRs και αναζητήσεις
4.6 Autoregressive Moving Average Model ARMA(1,1)
") 84 CHAPTER 4. STATIONARY TS MODELS 4.6 Autoregressive Moving Average Model ARMA(,) This section is an introduction to a wide class of models ARMA(p,q) which we will consider in more detail later in this
84 CHAPTER 4. STATIONARY TS MODELS 4.6 Autoregressive Moving Average Model ARMA(,) This section is an introduction to a wide class of models ARMA(p,q) which we will consider in more detail later in this
3.4 SUM AND DIFFERENCE FORMULAS. NOTE: cos(α+β) cos α + cos β cos(α-β) cos α -cos β
 cos α + cos β cos(α-β) cos α -cos β") 3.4 SUM AND DIFFERENCE FORMULAS Page Theorem cos(αβ cos α cos β -sin α cos(α-β cos α cos β sin α NOTE: cos(αβ cos α cos β cos(α-β cos α -cos β Proof of cos(α-β cos α cos β sin α Let s use a unit circle
3.4 SUM AND DIFFERENCE FORMULAS Page Theorem cos(αβ cos α cos β -sin α cos(α-β cos α cos β sin α NOTE: cos(αβ cos α cos β cos(α-β cos α -cos β Proof of cos(α-β cos α cos β sin α Let s use a unit circle
Econ 2110: Fall 2008 Suggested Solutions to Problem Set 8 questions or comments to Dan Fetter 1
 Eon : Fall 8 Suggested Solutions to Problem Set 8 Email questions or omments to Dan Fetter Problem. Let X be a salar with density f(x, θ) (θx + θ) [ x ] with θ. (a) Find the most powerful level α test
Eon : Fall 8 Suggested Solutions to Problem Set 8 Email questions or omments to Dan Fetter Problem. Let X be a salar with density f(x, θ) (θx + θ) [ x ] with θ. (a) Find the most powerful level α test
Instruction Execution Times
 1 C Execution Times InThisAppendix... Introduction DL330 Execution Times DL330P Execution Times DL340 Execution Times C-2 Execution Times Introduction Data Registers This appendix contains several tables
1 C Execution Times InThisAppendix... Introduction DL330 Execution Times DL330P Execution Times DL340 Execution Times C-2 Execution Times Introduction Data Registers This appendix contains several tables
k A = [k, k]( )[a 1, a 2 ] = [ka 1,ka 2 ] 4For the division of two intervals of confidence in R +
[a 1, a 2 ] = [ka 1,ka 2 ] 4For the division of two intervals of confidence in R +](/thumbs/73/69566903.jpg "k A = [k, k]( )[a 1, a 2 ] = [ka 1,ka 2 ] 4For the division of two intervals of confidence in R +") Chapter 3. Fuzzy Arithmetic 3- Fuzzy arithmetic: ~Addition(+) and subtraction (-): Let A = [a and B = [b, b in R If x [a and y [b, b than x+y [a +b +b Symbolically,we write A(+)B = [a (+)[b, b = [a +b
Chapter 3. Fuzzy Arithmetic 3- Fuzzy arithmetic: ~Addition(+) and subtraction (-): Let A = [a and B = [b, b in R If x [a and y [b, b than x+y [a +b +b Symbolically,we write A(+)B = [a (+)[b, b = [a +b
DESIGN OF MACHINERY SOLUTION MANUAL h in h 4 0.
 DESIGN OF MACHINERY SOLUTION MANUAL -7-1! PROBLEM -7 Statement: Design a double-dwell cam to move a follower from to 25 6, dwell for 12, fall 25 and dwell for the remader The total cycle must take 4 sec
DESIGN OF MACHINERY SOLUTION MANUAL -7-1! PROBLEM -7 Statement: Design a double-dwell cam to move a follower from to 25 6, dwell for 12, fall 25 and dwell for the remader The total cycle must take 4 sec
Statistics 104: Quantitative Methods for Economics Formula and Theorem Review
 Harvard College Statistics 104: Quantitative Methods for Economics Formula and Theorem Review Tommy MacWilliam, 13 tmacwilliam@college.harvard.edu March 10, 2011 Contents 1 Introduction to Data 5 1.1 Sample
Harvard College Statistics 104: Quantitative Methods for Economics Formula and Theorem Review Tommy MacWilliam, 13 tmacwilliam@college.harvard.edu March 10, 2011 Contents 1 Introduction to Data 5 1.1 Sample
Areas and Lengths in Polar Coordinates
 Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
w o = R 1 p. (1) R = p =. = 1
 R = p =. = 1") Πανεπιστήµιο Κρήτης - Τµήµα Επιστήµης Υπολογιστών ΗΥ-570: Στατιστική Επεξεργασία Σήµατος 205 ιδάσκων : Α. Μουχτάρης Τριτη Σειρά Ασκήσεων Λύσεις Ασκηση 3. 5.2 (a) From the Wiener-Hopf equation we have:
Πανεπιστήµιο Κρήτης - Τµήµα Επιστήµης Υπολογιστών ΗΥ-570: Στατιστική Επεξεργασία Σήµατος 205 ιδάσκων : Α. Μουχτάρης Τριτη Σειρά Ασκήσεων Λύσεις Ασκηση 3. 5.2 (a) From the Wiener-Hopf equation we have:
ΑΡΧΕΣ ΒΙΟΛΟΓΙΚΗΣ ΜΗΧΑΝΙΚΗΣ
 ΑΡΧΕΣ ΒΙΟΛΟΓΙΚΗΣ ΜΗΧΑΝΙΚΗΣ Εργαστήριο Βιοπληροφορικής 7 ο εξάμηνο Σχολή Μηχανολόγων Μηχανικών ΕΜΠ Διδάσκων: Λεωνίδας Αλεξόπουλος Fritz Kahn (1888 1968) 1 Περιεχόμενα Ομοιότητα πρωτεϊνών Σύγκριση αλληλουχιών
ΑΡΧΕΣ ΒΙΟΛΟΓΙΚΗΣ ΜΗΧΑΝΙΚΗΣ Εργαστήριο Βιοπληροφορικής 7 ο εξάμηνο Σχολή Μηχανολόγων Μηχανικών ΕΜΠ Διδάσκων: Λεωνίδας Αλεξόπουλος Fritz Kahn (1888 1968) 1 Περιεχόμενα Ομοιότητα πρωτεϊνών Σύγκριση αλληλουχιών
The Probabilistic Method - Probabilistic Techniques. Lecture 7: The Janson Inequality
 The Probabilistic Method - Probabilistic Techniques Lecture 7: The Janson Inequality Sotiris Nikoletseas Associate Professor Computer Engineering and Informatics Department 2014-2015 Sotiris Nikoletseas,
The Probabilistic Method - Probabilistic Techniques Lecture 7: The Janson Inequality Sotiris Nikoletseas Associate Professor Computer Engineering and Informatics Department 2014-2015 Sotiris Nikoletseas,
Block Ciphers Modes. Ramki Thurimella
 Block Ciphers Modes Ramki Thurimella Only Encryption I.e. messages could be modified Should not assume that nonsensical messages do no harm Always must be combined with authentication 2 Padding Must be
Block Ciphers Modes Ramki Thurimella Only Encryption I.e. messages could be modified Should not assume that nonsensical messages do no harm Always must be combined with authentication 2 Padding Must be
the total number of electrons passing through the lamp.
 1. A 12 V 36 W lamp is lit to normal brightness using a 12 V car battery of negligible internal resistance. The lamp is switched on for one hour (3600 s). For the time of 1 hour, calculate (i) the energy
1. A 12 V 36 W lamp is lit to normal brightness using a 12 V car battery of negligible internal resistance. The lamp is switched on for one hour (3600 s). For the time of 1 hour, calculate (i) the energy
Mean bond enthalpy Standard enthalpy of formation Bond N H N N N N H O O O
 Q1. (a) Explain the meaning of the terms mean bond enthalpy and standard enthalpy of formation. Mean bond enthalpy... Standard enthalpy of formation... (5) (b) Some mean bond enthalpies are given below.
Q1. (a) Explain the meaning of the terms mean bond enthalpy and standard enthalpy of formation. Mean bond enthalpy... Standard enthalpy of formation... (5) (b) Some mean bond enthalpies are given below.
ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ. Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων
 ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων Φυλογένεση Η φυλογένεσης αφορά την ανεύρεση των συνδετικών εκείνων κρίκων που συνδέουν τα διάφορα είδη µεταξύ τους εξελικτικά, σε µονοφυλετικές
ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων Φυλογένεση Η φυλογένεσης αφορά την ανεύρεση των συνδετικών εκείνων κρίκων που συνδέουν τα διάφορα είδη µεταξύ τους εξελικτικά, σε µονοφυλετικές
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ. Ψηφιακή Οικονομία. Διάλεξη 10η: Basics of Game Theory part 2 Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Ψηφιακή Οικονομία Διάλεξη 0η: Basics of Game Theory part 2 Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών Best Response Curves Used to solve for equilibria in games
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ Ψηφιακή Οικονομία Διάλεξη 0η: Basics of Game Theory part 2 Mαρίνα Μπιτσάκη Τμήμα Επιστήμης Υπολογιστών Best Response Curves Used to solve for equilibria in games
Αλγόριθμοι Εύρεσης Ομοιοτήτων Ακολουθιών
 Αλγόριθμοι Εύρεσης Ομοιοτήτων Ακολουθιών Μέρος Ι: Στοιχίσεις ακολουθιών κατά ζεύγη Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο
Αλγόριθμοι Εύρεσης Ομοιοτήτων Ακολουθιών Μέρος Ι: Στοιχίσεις ακολουθιών κατά ζεύγη Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο