Φυλογεωγραφία των ενδημικών ειδών του γένους
|
|
|
- Όφελος Κοντολέων
- 7 χρόνια πριν
- Προβολές:
Transcript
1 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ ΕΡΓΑΣΤΗΡΙΟ ΖΩΟΛΟΓΙΑΣ Φυλογεωγραφία των ενδημικών ειδών του γένους Trachelipus (Isopoda, Oniscidea) στην Ελλάδα ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ ΔΙΑΧΕΙΡΙΣΗ ΚΑΙ ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΔΙΑΤΡΙΒΗ ΜΕΤΑΠΤΥΧΙΑΚΟΥ ΔΙΠΛΩΜΑΤΟΣ ΕΙΔΙΚΕΥΣΗΣ ΜΑΡΙΑ ΚΑΜΗΛΑΡΗ Βιολόγος Α.Μ. 401 ΠΑΤΡΑ, 2010
 στην")

2 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ ΕΡΓΑΣΤΗΡΙΟ ΖΩΟΛΟΓΙΑΣ Φυλογεωγραφία των ενδημικών ειδών του γένους Trachelipus (Isopoda, Oniscidea) στην Ελλάδα ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ ΔΙΑΧΕΙΡΙΣΗ ΚΑΙ ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΔΙΑΤΡΙΒΗ ΜΕΤΑΠΤΥΧΙΑΚΟΥ ΔΙΠΛΩΜΑΤΟΣ ΕΙΔΙΚΕΥΣΗΣ ΜΑΡΙΑ ΚΑΜΗΛΑΡΗ Βιολόγος Α.Μ. 401 ΠΑΤΡΑ, 2010
3 Μέλη της τριμελούς εξεταστικής επιτροπής Γιώργος Κίλιας, Αναπληρωτής Καθηγητής, Τομέας Γενετικής Βιολογίας Κυττάρου και ανάπτυξης Έλενα Κλώσσα Κίλια, Επίκουρη Καθηγήτρια, Τομέας Βιολογίας Ζώων Σπύρος Σφενδουράκης, Επίκουρος Καθηγητής, Τομέας Βιολογίας Ζώων Επιβλέπουσα Καθηγήτρια Έλενα Κλώσσα Κίλια
4 Ευχαριστίες Η παρούσα Διατριβή αποτελεί την ερευνητική μου εργασία στο πλαίσιο του Μεταπτυχιακού Διπλώματος Ειδίκευσης του τμήματος Βιολογίας στην κατεύθυνση Οικολογία Διαχείριση και Προστασία Φυσικού Περιβάλλοντος. Θα ήθελα να ευχαριστήσω θερμά τα μέλη της τριμελούς επιτροπής, κα Έλενα Κλώσσα-Κίλια, κ. Γιώργο Κίλια και κ. Σπύρο Σφενδουράκη για την εμπιστοσύνη που μου έδειξαν και την ελευθερία με την οποία μου επέτρεψαν να εργάζομαι στο εργαστήριο, να οργανώσω το πρόγραμμά μου και να φέρω εις πέρας το θέμα της διατριβής μου. Ήταν πάντα εκεί για να απαντήσουν τις όποιες απορίες ή να ακούσουν την «γκρίνια» μου για όποιες ατυχίες στα πειράματά μου. Ιδιαίτερο ευχαριστώ οφείλω στο Σπύρο Σφενδουράκη με τον οποίο συνεργάζομαι τα τελευταία 6 χρόνια. Οι εξορμήσεις μαζί του στο πεδίο είναι πάντα ένα πολύ καλύτερο και ποιοτικότερο μάθημα από αυτό μέσα στις άχρωμες αίθουσες του Πανεπιστημίου. Τον ευχαριστώ θερμά και για το ερευνητικό πρόγραμμα «Καραθεοδωρή» στο οποίο με συμπεριέλαβε ως κύρια ερευνήτρια και από το οποίο χρηματοδοτήθηκε ένα μέρος αυτής της διατριβής. Στο πλαίσιο της ερευνητικής μου εργασίας είχα την τύχη να εργάζομαι τόσο στον ερευνητικό χώρο του κ. Σφενδουράκη στο εργαστήριο Ζωολογίας, όσο και στον ερευνητικό χώρο του κ. Κίλια στο εργαστήριο Γενετικής. Ως εκ τούτου είχα την ευκαιρία να συνεργαστώ με πολλούς ανθρώπους διαφορετικών αντικειμένων, να μάθω πολλά από αυτούς και να περάσουν πολύ ευχάριστα τα τρία αυτά χρόνια. Ελπίζοντας λοιπόν να μην ξεχνώ κανέναν, ευχαριστώ για το ευχάριστο κλίμα, τα αμέτρητα γέλια, τα ομαδικά ξενύχτια στο εργαστήριο και όλες τις στιγμές που περάσαμε μαζί τη Χριστίνα Κασσάρα, την Εβίτα Πίττα, Ονούφριο Μεττούρη, το Μιχάλη Ροβάτσο και τη Σπυριδούλα Κράιτσεκ.
5 Ξεχωριστά οφείλω να ευχαριστήσω το Στέφανο Μαρτιμιανάκη για την καθοδήγησή του στα πρώτα μου πειράματα στο πάγκο και την εκμάθηση των εργαστηριακών τεχνικών. Για μένα ο εργαστηριακός πάγκος της γενετικής ήταν κάτι καινούριο και στην αρχή ομολογώ «αγχωτικό». Ωστόσο ήταν πάντα με πολλή υπομονή δίπλα μου δίνοντας περισσότερο βάση στο επιστημονικό «γιατί» παρά στο τεχνικό «πρέπει» της όποιας πειραματικής διαδικασίας. Επίσης θα ήθελα να ευχαριστήσω θερμά το Δρ. Γιώργο Τρυφωνόπουλο για τη μεγάλη του βοήθεια στην εκμάθηση και κατανόηση των υπολογιστικών προγραμμάτων που χρησιμοποίησα στην παρούσα μελέτη. Μιας άχαρης διαδικασίας για πολλούς αλλά απολύτως απαραίτητης για όλους. Με τη Χαρά Παπαϊωάννου μοιραστήκαμε τις ίδιες καρέκλες, τις ίδιες πιπέτες και τις ίδιες «επιστημονικές ανησυχίες» στο εργαστήριο της Γενετικής. Ταιριάξαμε στον τρόπο δουλειάς και συνεννόησης αλλά ευτυχώς και στην «ευχάριστη διάθεση» που μας διακρίνει. Χαίρομαι πολύ που θα συνεχίσουμε να δουλεύουμε μαζί. Δε θα μπορούσα να παραλείψω τους γονείς μου Θανάση και Ευαγγελία, την αδερφή μου Ελένη και φυσικά τη γιαγιά μου Μαίρη που τόσα χρόνια με στηρίζουν ηθικά, ψυχολογικά και οικονομικά. Ξέρω πως κάνουν τα θέλω μου θέλω τους ακόμα και όταν δεν το πολυθέλουν Τέλος θέλω να ευχαριστήσω το Δημήτρη μου που με βοήθησε τόσο στις δειγματοληψίες όσο και στη συγγραφή της διατριβής μου με εύστοχες διορθώσεις και προτάσεις. Ο καλός του λόγος και η ανοιχτή του αγκαλιά ήταν το αγχολυτικό μου.
6 Στην γιαγιά μου Μαίρη, Την πιο φανατική μου θαυμάστρια
7 ΠΕΡΙΕΧΟΜΕΝΑ ΠΕΡΙΛΗΨΗ ABSTRACT A. ΕΙΣΑΓΩΓΗ ΓΕΝΙΚΑ ΣΤΟΙΧΕΙΑ ΓΙΑ ΤΗΝ ΤΑΞΗ ΤΩΝ ΙΣΟΠΟΔΩΝ ΙΣΤΟΡΙΚΑ ΣΤΟΙΧΕΙΑ ΚΑΙ ΣΤΟΙΧΕΙΑ ΕΞΕΛΙΞΗΣ ΤΑ ΧΕΡΣΑΙΑ ΙΣΟΠΟΔΑ Εξωτερική μορφολογία ΓΕΝΙΚΑ ΠΕΡΙ ΟΙΚΟΛΟΓΙΑΣ ΚΑΙ ΒΙΟΛΟΓΙΑΣ ΤΟΥ ΓΕΝΟΥΣ TRACHELIPUS BUDDE- LUND, ΟΙΚΟΛΟΓΙΑ ΑΝΑΠΑΡΑΓΩΓΗ ΑΝΑΠΤΥΞΗ ΤΑΞΙΝΟΜΗΣΗ - ΓΕΩΓΡΑΦΙΚΗ ΚΑΤΑΝΟΜΗ ΔΙΑΓΝΩΣΤΙΚΟΙ ΧΑΡΑΚΤΗΡΕΣ ΤΟΥ ΓΕΝΟΥΣ ΠΑΛΑΙΟΓΕΩΓΡΑΦΙΚΑ ΣΤΟΙΧΕΙΑ ΤΟΥ ΕΛΛΑΔΙΚΟΥ ΧΩΡΟΥ ΦΥΛΟΓΕΩΓΡΑΦΙΑ ΕΡΓΑΛΕIΑ ΣΤΗΝ ΥΠΗΡΕΣΙΑ ΤΗΣ ΦΥΛΟΓΕΩΓΡΑΦΙΑΣ Μοριακοί χαρακτήρες «εναντίον» μορφολογικών ΧΡΗΣΗ ΤΟΥ MTDNA ΣΤΗ ΜΕΛΕΤΗ ΤΩΝ ΠΛΗΘΥΣΜΩΝ Προέλευση του mt DNA Ιδιότητες του mtdna ΕΡΕΥΝΗΤΙΚΟ ΠΡΟΒΛΗΜΑ Β. ΜΕΘΟΔΟΛΟΓΙΑ ΚΑΙ ΥΛΙΚΑ ΣΥΛΛΟΓΗ ΔΕΙΓΜΑΤΩΝ
8 2. ΤΑ ΜΙΤΟΧΟΝΔΡΙΑΚΑ ΓΟΝΙΔΙΑ ΤΗΣ ΜΕΛΕΤΗΣ ΕΡΓΑΣΤΗΡΙΑΚΕΣ ΑΝΑΛΥΣΕΙΣ ΠΕΙΡΑΜΑΤΙΚΗ ΠΟΡΕΙΑ Εξαγωγή Ολικού Γενωμικού DNA Ηλεκτροφόρηση Ολικού Γενωμικού DNA Πολλαπλασιασμός συγκεκριμένων τμημάτων DNA με τη μέθοδο της αλυσιδωτής αντίδρασης πολυμεράσης (PCR) Καθαρισμός του πολλαπλασιασμένου τμήματος των γονιδίων COI και 16S rrna Καθορισμός της αλληλουχίας νουκλεοτιδίων των επιλεγμένων τμημάτων των μιτοχονδριακών γονιδίων ΣΤΑΤΙΣΤΙΚΗ ΕΠΕΞΕΡΓΑΣΙΑ ΤΩΝ ΔΕΔΟΜΕΝΩΝ THΣ ΑΝΑΛΥΣΗΣ ΑΛΛΗΛΟΥΧΙΑΣ ΣΤΟΙΧΙΣΗ ΑΛΛΗΛΟΥΧΙΩΝ ΕΛΕΓΧΟΣ ΔΕΔΟΜΕΝΩΝ ΓΙΑ ΟΜΟΠΛΑΣΙΑ ΚΑΙ ΚΟΡΕΣΜΟ ΕΠΕΞΕΡΓΑΣΙΑ ΤΩΝ ΣΤΟΙΧΙΣΜΕΝΩΝ ΑΛΛΗΛΟΥΧΙΩΝ Εκτίμηση γενετικών αποστάσεων Επιλογή του καταλληλότερου μοντέλου νουκλεοτιδικής υποκατάστασης ΑΝΑΛΥΣΗ ΦΑΙΝΕΤΙΚΩΝ ΚΑΙ ΦΥΛΟΓΕΝΕΤΙΚΩΝ ΣΧΕΣΕΩΝ Μέθοδος Σύνδεσης Γειτόνων (Neighbor-Joining) Η μέθοδος της μέγιστης φειδωλότητας (Maximum Parsimony) Μπεϊεσιανή συμπερασματολογία ΑΝΕΞΑΡΤΗΤΗ Η ΣΥΝΔΥΑΣΜΕΝΗ ΑΝΑΛΥΣΗ ΑΝΑΛΥΣΗ ΜΟΡΙΑΚΗΣ ΔΙΑΚΥΜΑΝΣΗΣ ANALYSIS OF MOLECULAR VARIANCE (AMOVA) Γ. ΑΠΟΤΕΛΕΣΜΑΤΑ ΑΝΑΛΥΣΗ ΑΛΛΗΛΟΥΧΙΑΣ ΜΙΤΟΧΟΝΔΡΙΑΚΩΝ ΓΟΝΙΔΙΩΝ ΑΝΑΛΥΣΗ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΤΟΠΟΥ 16S RRNA Νουκλεοτιδική σύσταση και Γενετική ποικιλότητα των μελετηθέντων πληθυσμών Έλεγχος Ομοπλασίας
9 Εκτίμηση γενετικών αποστάσεων Φαινετικές και φυλογενετικές σχέσεις των μελετηθέντων πληθυσμών Ανάλυση Μοριακής Διακύμανσης (AMOVA) Δίκτυο Ελαχίστων Αποστάσεων (MSN) Ομαδοποίηση βάσει γεωγραφικής προέλευσης Ομαδοποίηση βάσει φυλογενετικής συγγένειας ΑΝΑΛΥΣΗ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΤΟΠΟΥ COI Νουκλεοτιδική σύσταση και γενετική ποικιλότητα των μελετηθέντων πληθυσμών Έλεγχος Ομοπλασίας Εκτίμηση γενετικών αποστάσεων Φαινετικές και φυλογενετικές σχέσεις των μελετηθέντων πληθυσμών Ανάλυση Μοριακής Διακύμανσης (AMOVA) Ομαδοποίηση βάσει γεωγραφικής προέλευσης Ομαδοποίηση βάσει φυλογενετικής συγγένειας ΣΥΝΔΥΑΣΜΕΝΗ ΑΝΑΛΥΣΗ Νουκλεοτιδική σύσταση και γενετική ποικιλότητα Έλεγχος Ομοπλασίας Εκτίμηση γενετικών αποστάσεων Φαινετικές και φυλογενετικές σχέσεις των μελετηθέντων πληθυσμών Ανάλυση Μοριακής Διακύμανσης (AMOVA) Ομαδοποίηση βάσει γεωγραφικής προέλευσης Ομαδοποίηση βάσει φυλογενετικής συγγένειας Δ. ΣΥΖΗΤΗΣΗ ΠΛΗΘΟΣ ΑΠΛΟΤΥΠΩΝ ΚΟΡΕΣΜΟΣ ΚΑΙ ΠΙΣΤΟΠΟΙΗΣΗ ΤΩΝ ΜΙΤΟΧΟΝΔΡΙΑΚΩΝ ΑΛΛΗΛΟΥΧΙΩΝ ΔΙΑΦΟΡΟΠΟΙΗΣΗ ΑΛΛΗΛΟΥΧΙΩΝ ΔΙΑΦΟΡΟΠΟΙΗΣΗ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΤΟΠΟΥ 16S RRNA ΓΕΝΕΤΙΚΟΣ ΤΟΠΟΣ COI MTDNA
10 3.3. ΣΥΝΔΥΑΣΜΕΝΗ ΑΝΑΛΥΣΗ ΦΥΛΟΓΕΝΕΤΙΚΗ ΑΝΑΛΥΣΗ ΓΕΝΕΤΙΚΟΣ ΤΟΠΟΣ 16S RRNA ΓΕΝΕΤΙΚΟΣ ΤΟΠΟΣ COI ΣΥΝΔΥΑΣΜENΗ ΑΝAΛΥΣΗ Η περίπτωση του T. nsp Ο μεικτός ηπειρωτικός κλάδος Η περίπτωση της λίμνης Ζηρού Ο κλάδος του Αιγαίου Η περίπτωση της Επιδαύρου Η περίπτωση της Εύβοιας Η περίπτωση Βουραϊκού Πλανητέρου ΚΡΥΠΤΙΚΑ ΕΙΔΗ ΕΤΕΡΟΠΛΑΣΜΙΑ ΓΕΝΙΚΑ ΣΥΜΠΕΡΑΣΜΑΤΑ Ε. ΒΙΒΛΙΟΓΡΑΦΙΑ REFERENCES Ζ. ΠΑΡΑΡΤΗΜΑ
11 ΠΕΡΙΛΗΨΗ Φυλογεωγραφία των ενδημικών ειδών του γένους Trachelipus (Isopoda, Oniscidea) στην Ελλάδα. Το γένος Trachelipus περιλαμβάνει οργανισμούς σχετικά στενόοικους οι οποίοι ζουν είτε στην παρόχθια βλάστηση ρεμάτων και ποταμών είτε σε υγρά δάση. Στη χώρα μας έχουν καταγραφεί 8 από τα 50 είδη του γένους, 4 από τα οποία είναι ενδημικά της Ελλάδας. Το ένα από αυτά εξαπλώνεται από την Κρήτη μέχρι την Ήπειρο, ένα στα νησιά του κεντρικού Αιγαίου, ένα στην Κρήτη και ένα στο νότιο Ευβοϊκό. Η κατανομή κάθε είδους είναι ασυνεχής, είτε λόγω γεωγραφικών (νησιωτικοί πληθυσμοί κλπ) είτε λόγω ενδιαιτηματικών παραγόντων. Η διάκριση μεταξύ των ειδών έχει γίνει βάσει περιορισμένου αριθμού μορφολογικών χαρακτήρων και δεν είναι βέβαιο ότι αντανακλά τις πραγματικές φυλογενετικές σχέσεις τους. Από τα αποτελέσματα προηγούμενης μελέτης διαπιστώθηκε έντονη απόκλιση μεταξύ των προτύπων της γενετικής ποικιλότητας και εκείνης της τρέχουσας ταξινόμησης σε ορισμένες ομάδες πληθυσμών του γένους αυτού. Επιπλέον, φάνηκε σημαντικός βαθμός γενετικής απομόνωσης μεταξύ των πληθυσμών ενός είδους, ενισχύοντας την άποψη περί ισχυρής μεταπληθυσμιακής συγκρότησής τους. Στην παρούσα μελέτη, συλλέχθηκαν 47 πληθυσμοί στην ηπειρωτική Ελλάδα, οι οποίοι στη μεγάλη τους πλειοψηφία ανήκαν στο είδος Trachelipus kytherensis (σύμφωνα με την ισχύουσα ταξινόμηση). Σε αυτούς προστέθηκαν και τα δεδομένα των Parmakelis et al 2008 (16 πληθυσμοί) έτσι ώστε να είναι πιο ολοκληρωμένη η μελέτη και η εξαγωγή συμπερασμάτων για το γένος Trachelipus. 5
12 Συνολικά μελετήθηκαν γενετικά 63 πληθυσμοί του γένους, χρησιμοποιώντας ως μοριακούς δείκτες τα μιτοχονδριακά γονίδια 16S rrna και COI. Έπειτα από απομόνωση του DNA και τον πολλαπλασιασμό των συγκεκριμένων τμημάτων με PCR προσδιορίστηκε η αλληλουχία των βάσεων, και υπολογίστηκε η γενετική διαφοροποίηση εντός και μεταξύ των πληθυσμών. Για την ανάλυση των φυλογενετικών σχέσεων μεταξύ των πληθυσμών ή/και των ειδών χρησιμοποιήθηκαν οι μέθοδοι της Σύνδεσης Γειτόνων (Neighbor Joining-NJ), της Μέγιστης Φειδωλότητας (Maximum Parsimony-MP) και της Μπεϊεσιανής Συμπερασματολογίας (Bayesian Inference-BI). Το τελικό μήκος των αλληλουχιών μετά την επεξεργασία ήταν 386 θέσεις για το γενετικό τόπο 16S rrna και 512 θέσεις για το γενετικό τόπο COI. Με τα δεδομένα αυτά δεδομένα πραγματοποιήθηκε τόσο ανεξάρτητη όσο και συνδυασμένη ανάλυση. Από τα αποτελέσματα φαίνονται πληθυσμοί οι οποίοι παρα το ότι είναι πολύ κοντινοί γεωγραφικά, και μέχρι σήμερα θεωρείται πως ανήκουν στο ίδιο είδος (Trachelipus kytherensis), εμφανίζουν μεγάλες γενετικές αποστάσεις μεταξύ τους και ομαδοποιούνται σε διαφορετικούς και αρκετά απομακρυσμένους κλάδους των δένδρων σε όλες τις αναλύσεις (NJ, MP, BI). Η τοπολογία των κλάδων, καθώς και η απουσία σαφούς γεωγραφικού προτύπου στην ομαδοποίηση των πληθυσμών του T. kytherensis, καταδεικνύει ότι πιθανότατα δεν έχουμε να κάνουμε με ένα μόνο είδος, αλλά με περισσότερα που είναι δύσκολο να διακριθούν μορφολογικά, τουλάχιστον με τους μέχρι σήμερα χρησιμοποιούμενους ταξινομικούς διαγνωστικούς χαρακτήρες. Αυτό ενισχύεται και από τις γενετικές αποστάσεις που καταγράφηκαν στην παρούσα μελέτη και εμφανίζονται ιδιαίτερα αυξημένες (μέγιστες παρατηρηθείσες γενετικές απόστασεις: 27,3% COI, 17,6% 16S rrna) ακόμα και σε σχέση με αυτές που έχουν αναφερθεί σε άλλες έρευνες για τη διάκριση ειδών ισοπόδων. Επισημαίνεται η ιδιαίτερα μεγάλη γενετική διαφοροποίηση μεταξύ των αντιπροσώπων του γένους. Επιπλέον καταδεικνύεται πως η Πελοπόννησος φιλοξενεί τα είδη Trachelipus kytherensis και T. aegaeus (τουλάχιστον στη 6
13 χερσόνησο της Αργολίδας) αλλά και πιθανόν μια τρίτη μορφή στα βόρεια (νέο είδος;) η οποία εμφανίζεται ευρύτερα στην ηπειρωτική Ελλάδα. Θα είχε ιδιαίτερο ενδιαφέρον λοιπόν να μελετηθούν αυτές οι πιθανές «ζώνες επαφής» ως προς τη γονιδιακή τους ροή, ώστε να εκτιμηθεί το ποσοστό απομόνωσης των πληθυσμών και, κατ επέκταση, του κάθε είδους. 7
14 ABSTRACT Phylogeography of Greek endemic Trachelipus species (Isopoda, Oniscidea). The phylogenetic relationships among terrestrial isopod species are still largely unknown because robust analyses have started to appear only relatively recently. Species-level taxonomy has been based mainly on a few secondary sexual characters of males, although recent analyses based on molecular markers have indicated that species definitions based on morphology may underestimate the true levels of divergence among populations. Furthermore, within several genera or species groups, morphological characters do not provide clear-cut taxonomic resolution, so that many changes in the interpretation of nominal species have appeared in the literature. The genus Trachelipus comprises of relatively stenoecious animals living in habitats generally threatened by human activities, such as humid forest sites and riparian habitats. It includes some 50 species distributed around the Palaearctic, with 8 species recorded from Greece, 4 endemic to the country. The distribution of species is discontinuous due to the increasing fragmentation of its habitats and the expansion of agricultural land and dry woodland. Projected climatic change will restrict further gene flow between Trachelipus populations, as dry habitats are expected to expand in Greece. Species level taxonomy has been based on a few morphological characters, mainly the secondary sexual characters of males, exhibiting significant variation, and is controversial. Very high intraspecific genetic divergence among several populations has been documented. 8
15 In this study we attempt a phylogeographic analysis among the Greek endemic species of the genus. We sampled 47 populations from several sites in mainland Greece. In our analyses we incorporated data from previous work (16 populations) in order to better estimate possible geographic structure in the patterns of divergence among populations, and to throw new light in the systematics of the species. Overall, 63 populations were considered. After total DNA extraction, we sequenced the two PCR amplified mtdna gene fragments, namely 16S rrna and cytochrome oxidase subunit I (COI), and calculated the genetic divergence within and among the populations studied, as well as their phylogenetic relationships. The methods for phylogenetic reconstruction used were Neighbor Joining (NJ), Maximum Parsimony (MP) and Bayesian Inference (BI) for each mtdna sequence data and the concatenated dataset. The phylogenetic trees obtained from the molecular data from all three phylogenetic methods (NJ, MP, BI) - produced trees with quite congruent topologies. Some populations that are considered conspecific exhibit large genetic distances and cluster in different clades. The highly-structured phylogenetic tree and the lack of an overall geographic pattern in the clustering of Trachelipus populations indicates that very probably we are not dealing with a single species, but rather with a number of cryptic species, hardly distinguished by means of currently used morphological characters. This is further corroborated by the genetic distances separating the clades hosting nominal T. kytherensis populations (max_d COI =27.3% and max_d 16S rrna =17.6%). In general, it can be argued that the genetic distances recorded in the present study are quite large compared with those reported for different species and even genera in other studies of terrestrial isopods. Furthermore, it is evident than there are two species present in the Peloponnese, i.e. Trachelipus kytherensis and T. aegaeus (in Argolis peninsula). In northern Peloponnese, a third form is also present (new species?) that occurs throughout the central and 9
16 northern part of mainland Greece. These contact zones should be further investigated in terms of genetic flow and isolation of the populations and/or species. Both the phylogeny presented here and the genetic distances separating populations appear to justify the necessity of further investigation into the phylogeny of the Greek Trachelipus species using a population by population approach. It is likely that morphology inadequately describes real variation inside and among species; hence, diagnoses based on the morphological characters used so far for the delineation of Trachelipus species should be reconsidered under the light of more extensive molecular phylogenetic analyses. 10
17 A. ΕΙΣΑΓΩΓΗ 1. ΓΕΝΙΚΑ ΣΤΟΙΧΕΙΑ ΓΙΑ ΤΗΝ ΤΑΞΗ ΤΩΝ ΙΣΟΠΟΔΩΝ 1.1. ΙΣΤΟΡΙΚΑ ΣΤΟΙΧΕΙΑ ΚΑΙ ΣΤΟΙΧΕΙΑ ΕΞΕΛΙΞΗΣ Η εμφάνιση των ισοπόδων τοποθετείται στο Δεβόνιο, ενώ το αρχείο των απολιθωμάτων διαθέτει μορφές που τοποθετούνται στον Παλαιοζωικό αιώνα (ανώτερο Λιθανθρακοφόρο -περί τα 300 Μyα). Απολιθωμένα Oniscidea συναντώνται για πρώτη φορά στο Ηώκαινο, δηλαδή περίπου πενήντα εκατομμύρια χρόνια πριν. Τα παλαιότερα απολιθώματα ανήκουν στην υπόταξη Phreatoicidea που θεωρείται και ο παλαιότερος κλάδος των ισοπόδων. Τα ισόποδα έχουν αποικίσει σχεδόν το σύνολο των ενδιαιτημάτων. Απαντώνται από τις ακτές έως τις αβύσσους των θαλασσών, τα γλυκά νερά, καθώς και στη χέρσο (Schmalfuss 1983). Ο εποικισμός της χέρσου συνέβη, σύμφωνα με εκτιμήσεις, περίπου εκατό εκατομμύρια χρόνια πριν και έγινε σταδιακά από βενθικούς προγόνους που μετακινήθηκαν στην παραλιακή ζώνη και μετά ένα στάδιο κατά το οποίο ζούσαν ως αμφίβιοι, κατάφεραν να υπερπηδήσουν τις δυσκολίες της χερσαίας διαβίωσης και να παραμείνουν μόνιμα στη χέρσο. Η ονομασία Ισόποδα προέρχεται από τη δομική ομοιομορφία των επτά ζευγών θωρακικών άκρων που χαρακτηρίζει αυτήν την παλαιά και διαφοροποιημένη ομάδα των Καρκινοειδών (Wilson & Edgecombe 2003). Η τάξη των Ισοπόδων είναι η πλέον διαφοροποιημένη και πλούσια σε είδη ομάδα Καρκινοειδών της υπέρταξης Peracarida. Απαριθμούν περί τα περιγεγραμμένα είδη τα οποία ταξινομούνται σε δέκα τάξεις. Το μέγεθός τους κυμαίνεται από 1 έως 500 mm (Bathynomus giganteus).
18 1.2. ΤΑ ΧΕΡΣΑΙΑ ΙΣΟΠΟΔΑ Τα χερσαία ισόποδα (Oniscidea) είναι μια ευρέως εξαπλωμένη ομάδα οργανισμών με ένα σημαντικό ιδιαίτερο χαρακτηριστικό: είναι η καλύτερα προσαρμοσμένη ομάδα καρκινοειδών στο χερσαίο περιβάλλον. Η μετάβαση των ισοπόδων στη χέρσο συνοδεύτηκε από μια σειρά μορφολογικών και φυσιολογικών προσαρμογών καθώς και αλλαγών στη συμπεριφορά ώστε να αντιμετωπιστούν τα προβλήματα που προέκυψαν από αυτόν το νέο τύπο διαβίωσης. Οι σημαντικότερες από αυτές αφορούσαν την επαρκή στήριξη του σώματος στις νέες συνθήκες, ένα νέο τρόπο αναπνοής και κυρίως την προστασία από την αφυδάτωση, το σημαντικότερο πρόβλημα της χερσαίας διαβίωσης. Τα χερσαία ισόποδα αντιμετώπισαν επιτυχώς όλες αυτές τις δυσκολίες. Το γεγονός αυτό αποδεικνύεται εξάλλου από την ευρεία τους εξάπλωση σε όλα σχεδόν τα χερσαία οικοσυστήματα πλην αυτών των αρκτικών περιοχών και των πολύ μεγάλων υψομέτρων (Σφενδουράκης 1994). Η υπόταξη Oniscidea περιλαμβάνει σχεδόν αποκλειστικά χερσόβια είδη, για το λόγο αυτό και αποκαλείται ως «η χερσαία ομάδα» των ισοπόδων. Λίγα είναι γνωστά σχετικά με αυτή την ομάδα: ο Λινναίος αναγνώρισε μόλις ένα είδος το 1758 ενώ ο Bouffon επτά είδη 34 χρόνια αργότερα (Michael Salzat & Bouchon 2000). Επόμενες μελέτες αύξησαν τον αριθμό των ειδών αυτής της ομάδας σε 2000 περί το 1943 ενώ ο Schmalfuss (2004)προσδιόρισε τον αριθμό των χερσαίων ισοπόδων ανά τον κόσμο στα 3637 είδη περίπου. Αργότερα προτάθηκε η πρώτη φαινετική ταξινόμηση της υπόταξης των Ονισκίδων με βάση κυρίως τη μορφολογία του αρσενικού γενετικού συστήματος. Το 1946, καθορίστηκαν δύο υπερτάξεις, τα Tylomorpha και τα Ligiamorpha. Η πρώτη υπέρταξη περιελάμβανε την οικογένεια Tylidae, που αποτελείτο από 25 είδη του γένους Tylos και ένα μόνο είδος του γένους Helleria (Michael Salzat & Bouchon 2000). Η υπέρταξη Ligiamorpha υποδιαιρέθηκε σε τρία τμήματα: 12
19 1. τα Diplocheta, με περίπου 80 είδη τα οποία ανήκουν στις οικογένειες Ligiidae και Mesoniscidae 2. τα Synocheta,με περίπου 200 είδη τα οποία ανήκουν στις υπεροικογένειες Styloniscidea και Trichoniscidea 3. τα Crinocheta, που περιλαμβάνουν τις υπεροικογένειες Armadilloidea Oniscoidea. Πιο πρόσφατα η οικογένεια Mesoniscidae τοποθετήθηκε σε ένα νέο τμήμα τα Microcheta (Schmalfuss 1989). Αυτή η κατάταξη είναι αποδεκτή επί σειρά ετών με μερικές μικρές τροποποιήσεις. Ωστόσο αυτή η κατάταξη δεν είναι δεν βασίζεται σε φυλογενετικά δεδομένα με αποτέλεσμα νέες μελέτες αναφορικά με τη φυλογένεια αυτών των οργανισμών να προτείνουν και αλλαγές στις ισχύουσα ταξινόμηση. Στην Ελλάδα απαντώνται 19 οικογένειες, 47 γένη (τα 8 ενδημικά) και 232 είδη (153 ενδημικά - τα 37 εξ αυτών τρωγλόβια) Εξωτερική μορφολογία Το σώμα τους είναι νωτοκοιλιακώς πεπλατυσμένο, με εμφανή τη μεταμέρεια εξωτερικά αφού δεν υπάρχει θυρεός. Το μέγεθός τους δεν ξεπερνά συνήθως τα 5cm. Όπως και στα περισσότερα Καρκινοειδή, το σώμα των ισοπόδων είναι διαχωρισμένο σε τρεις διακριτές περιοχές: Κεφάλι Θώρακας (περαίιον) Κοιλιά (πλέον) Στο κεφάλι φέρουν σύνθετα, άμισχα μάτια αποτελούμενα από ποικίλο αριθμό ομματιδίων, δύο ζεύγη μονοσκελών κεραιών, δύο ζεύγη γνάθων και ένα ζεύγος γναθοποδίων. Το πρώτο ζεύγος κεραιών έχει χημειοδεκτική λειτουργία, είναι υποπλασμένο και δυσδιάκριτο, ενώ το δεύτερο είναι ανεπτυγμένο και αντιλαμβάνεται απτικά ερεθίσματα. Το πρώτο θωρακικό μεταμερές είναι συντηγμένο με το κεφάλι 13

 αποτελείται από έξι μεταμερή εκ των οποίων τα πέντε είναι ελεύθερα ενώ το έκτο έχει συντηχθεί με το μη μεταμερικό τέλσον σχηματίζοντας το πλεοτέλσον.")
20 και τα εξαρτήματά του έχουν διαφοροποιηθεί σε γναθοπόδια. Τα υπόλοιπα επτά θωρακικά μεταμερή συναποτελούν το περαίιον και φέρουν, πρωτογενώς, από ένα ζεύγος μονοσκελών αρθρωτών εξαρτημάτων, τα περαιιοπόδια. Τα περαιιοπόδια είναι παρόμοιας μορφολογίας και διαφοροποιημένα για βάδιση (Εικ.A-1α). Η κοιλιακή περιοχή (Εικ.A-1β) αποτελείται από έξι μεταμερή εκ των οποίων τα πέντε είναι ελεύθερα ενώ το έκτο έχει συντηχθεί με το μη μεταμερικό τέλσον σχηματίζοντας το πλεοτέλσον. Καθένα από αυτά τα μεταμερή φέρει ένα ζεύγος δισκελών, αρθρωτών εξαρτημάτων, τα πλεοπόδια (ονομάζονται ουροπόδια εκείνα του συντηγμένου με το τέλσον μεταμερούς) και αποτελούνται από τον ενδοποδίτη (εσωτερικά), τον εξωποδίτη (εξωτερικά) και το βασιποδίτη στη βάση. Στους εξωποδίτες των πλεοποδίων των περισσότερων χερσαίων ισοπόδων, ιδίως εκείνων που εξελίχθηκαν με τρόπο ώστε να εποικίσουν ξηρά περιβάλλοντα, έχουν αναπτυχθεί ειδικές αναπνευστικές δομές, κοιλότητες που ονομάζονται τραχειακοί πνεύμονες ή ψευδοτραχείες. Στα χερσαία ισόποδα που δεν διαθέτουν τέτοιους πνεύμονες, χρησιμοποιείται για την ανταλλαγή των αερίων ολόκληρη η επιφάνεια των εξωποδιτών, η οποία και πρέπει να διατηρείται διαρκώς υγρή (Sutton 1972, Σφενδουράκης 1994). κεραία μαστίγιο κεραίας γναθοπόδι τεργίτης επιμερές πλέον τέλσον περαιιοπό δ πλεοπόδ α ουροπόδια β Εικόνα A-1: Εξωτερική μορφολογία χερσόβιου ισοπόδου. α. ραχιαία άποψη, β. κοιλιακή άποψη (τροποποιημένες από Sutton 1972). 14

21 2. ΓΕΝΙΚΑ ΠΕΡΙ ΟΙΚΟΛΟΓΙΑΣ ΚΑΙ ΒΙΟΛΟΓΙΑΣ ΤΟΥ ΓΕΝΟΥΣ TRACHELIPUS BUDDE-LUND, ΟΙΚΟΛΟΓΙΑ Οι αντιπρόσωποι του γένους Trachelipus είναι υγρόφιλα είδη που ζουν κοντά σε τρεχούμενο νερό (στις όχθες ρεμάτων) ή, σπανιότερα, στάσιμο νερό, αν και μπορούν να βρεθούν και σε καλλιέργειες, μακκία βλάστηση ή δάση, σε σημεία που υπάρχει πολλή υγρασία. Συνήθως εντοπίζονται κάτω από πέτρες αλλά και πολλές φορές και μέσα στη φυλλοστρωμνή, όταν αυτή είναι πλούσια και υγρή. Όπως όλα τα χερσαία ισόποδα, είναι φυτοσαπροφάγα, (Σφενδουράκης 1994) ωστόσο δεν είναι γνωστό επακριβώς με τι τρέφονται. Στην Εικ.A-2 απεικονίζονται οι τροφικές σχέσεις των χερσόβιων ισοπόδων εν γένει. Εικόνα A-2: Τροφικές σχέσεις χερσόβιων ισοπόδων. Τα βέλη καταδεικνύουν τη ροή της οργανικής ύλης (τροποποιημένες από Sutton 1972) 15
.")
22 2.2. ΑΝΑΠΑΡΑΓΩΓΗ ΑΝΑΠΤΥΞΗ Όπως όλα τα χερσαία ισόποδα, έτσι και τα είδη του γένους Trachelipus είναι γονοχωριστικά, με τα θηλυκά άτομα να είναι συνήθως μεγαλύτερα σε μέγεθος από τα αρσενικά, αν και αυτό δεν είναι απόλυτο. Η γονιμοποίηση είναι εσωτερική και η ανάπτυξη άμεση χωρίς προνυμφικά στάδια. Στα αρσενικά άτομα υπάρχουν και εξωτερικά δευτερογενή όργανα αναπαραγωγής (Εικ.A-3). Συγκεκριμένα, τα δύο πρώτα ζεύγη πλεοποδίων των αρσενικών, είναι διαφοροποιημένα για τη μεταφορά σπέρματος και αποτελούν διαγνωστικούς χαρακτήρες, όπως και στα περισσότερα ισόποδα. Πέος Εικόνα A-3: Εξωτερικά γεννητικά όργανα αρσενικού ατόμου (Τροποποιημένη από Sutton 1972). Οι εξωποδίτες είναι λιγότερο διαφοροποιημένοι από τους ενδοποδίτες, οι οποίοι επιμηκύνονται προς τα πίσω σχηματίζοντας στύλο με τον οποίο το σπέρμα μεταφέρεται στο γενετικό πόρο του θηλυκού. Το γονιμοποιημένο θηλυκό αναπτύσσει ειδικό σχηματισμό γεμάτο με υγρό ανάμεσα στα θωρακικά πόδια, το μάρσιπο, όπου και αποθηκεύει τα αυγά. Μετά την εκκόλαψή τους τα νεαρά άτομα παραμένουν στο μάρσιπο για μικρό χρονικό διάστημα. Από εκεί εξέρχονται ως ανήλικα (mancas) των 16
23 οποίων τα μεταμερή υπολείπονται σε αριθμό σε σχέση με τα ενήλικα άτομα (Sutton 1972). Το φύλο, μπορεί να καθοριστεί γενετικά, κυτταροπλασματικά ή από περιβαλλοντικούς παράγοντες. Στην πρώτη περίπτωση, ο φυλοκαθορισμός γίνεται από δυο τύπους φυλετικών χρωμοσωμάτων, X και Y, ή Z και W. Στη δεύτερη περίπτωση το φύλο καθορίζεται από κυτταροπλασματικά μεταβιβαζόμενους μικροοργανισμούς. Στην τρίτη περίπτωση, ανάλογα με το είδος, ο προκαθορισμός εξαρτάται από διάφορους παράγοντες, όπως π.χ. τη θερμοκρασία, τη φωτοπερίοδο, τη συναθροιστικότητα κλπ (Thierry et al. 1997, Hidding et al. 2003). Η σύζευξη των χερσαίων ισοπόδων συμβαίνει στο απόλυτο σκοτάδι και μπορεί να διακοπεί από την ελάχιστη παρουσία φωτός. Στα χερσαία ισόποδα, εμφανίζεται τόσο η μονοτοκία (αναπαραγωγή μόνο μία φορά σε όλη τους τη ζωή) όσο και η πολυτοκία (περισσότερες από μια αναπαραγωγικές περίοδοι στη διάρκεια της ζωής), η οποία συχνά διαπιστώνεται από μελέτη της δομής των ωοθηκών. Στα μονότοκα είδη οι ωοθήκες περιέχουν μία ή δύο σειρές ισομεγεθών ωοκυττάρων ενώ στα πολύτοκα υπάρχουν μεγάλα, ώριμα ωοκύτταρα που θα δώσουν ζυγωτά στην τρέχουσα αναπαραγωγική περίοδο και μικρότερα που θα ωριμάσουν σε επόμενες αναπαραγωγικές περιόδους. Ως προς την αναπαραγωγική περίοδο τα ισόποδα διακρίνονται σε δυο κατηγορίες: εκείνα που αναπαράγονται την άνοιξη ή στις αρχές του καλοκαιριού και σε εκείνα που αναπαράγονται το φθινόπωρο. Τα είδη γεννούν περισσότερες από μία φορές κατά τη διάρκεια μιας ευρύτερης περιόδου όπως π.χ. από την άνοιξη ως το φθινόπωρο, ανάλογα με τις περιβαλλοντικές συνθήκες. Η ενεργοποίηση της αναπαραγωγικής περιόδου καθορίζεται από παράγοντες όπως: i) η φωτοπερίοδος, ii) η θερμοκρασία, iii) βιοτικοί παράγοντες, όπως το είδος της τροφής, και iv) άλλοι, όπως π.χ. τα βαρέα μέταλλα που επιδρούν αρνητικά σε ορισμένες συνθήκες (Sutton 1972). 17
24 3.ΤΑΞΙΝΟΜΗΣΗ - ΓΕΩΓΡΑΦΙΚΗ ΚΑΤΑΝΟΜΗ Όπως γράφηκε παραπάνω, τα χερσαία ισόποδα έχουν ευρεία εξάπλωση σε ολόκληρο τον κόσμο και ο αριθμός των ειδών τους ανέρχεται στα Σύμφωνα με την τρέχουσα ταξινόμηση, το γένος Trachelipus περιλαμβάνει 50 είδη που κατανέμονται σε όλη Φύλο: Αρθρόποδα Υπόφυλο: Καρκινοειδή Ομοταξία: Μαλακόστρακα Υφομοταξία: Ευμαλακόστρακα Υπέρταξη: Peracarida Τάξη: Isopoda Υπόταξη: Oniscidea Τμήμα: Crinocheta Οικογένεια: Trachelipodidae Γένος: Trachelipus Budde-Lund, 1908 την Παλαιαρκτική (Εικ.A-4), με 9 είδη στον ελλαδικό χώρο και 5 από αυτά ενδημικά της Ελλάδας (Εικ.A-5). Πιο συγκεκριμένα, τα είδη που απαντώνται στην Ελλάδα είναι τα: 1. Trachelipus arcuatus (Budde-Lund, 1885), με εξάπλωση στη ΒΔ Ελλάδα. 2. T. buddelundi (Strouhal, 1937), με μία καταγραφή στη Χίο. 3. T. camerani (Tua, 1900), με εξάπλωση στη ΒΔ Ελλάδα. 4. T. cavaticus (Schmalfuss, Paragamian & Sfenthourakis, 2004), εντοπισμένο στη δυτική Κρήτη. 5. T. razzautii (Arcangeli, 1933), με μία καταγραφή στη Λέσβο 6. T. squamuliger (Verhoeff, 1907), που εξαπλώνεται στην ανατολική Ελλάδα και τη Μυτιλήνη. 7. T. aegaeus (Verhoeff, 1907), με εξάπλωση στα νησιά του Αιγαίου και τα αιγιακά παράλια της ηπειρωτικής Ελλάδας. 8. T. kytherensis (Strouhal, 1929), με εξάπλωση την ηπειρωτική Ελλάδα και τα Κύθηρα. 18

.")
25 9. T. n.sp. (υπό περιγραφή), που εντοπίστηκε στα Στουρονήσια (νησίδες μεταξύ Αττικής και Εύβοιας). Από αυτά, τα T. kytherensis, T. aegaeus, T. buddelundi, T. cavaticus και T. n.sp. είναι ελληνικά ενδημικά, με το T. kytherensis να εμφανίζει τη μεγαλύτερη εξάπλωση. Το T. buddelundi έχει καταγραφεί μία και μοναδική φορά στο νησί της Χίου, οπότε και περιγράφηκε, και η εγκυρότητά του είναι υπό αμφισβήτηση. Αντιστοίχως, το T. cavaticus περιορίζεται μόνο στην Κρήτη όπου και ζει κατά κύριο λόγο σε σπήλαια. Τα άλλα είδη έχουν καταγραφεί σε πολύ λίγες περιοχές της βορειοδυτικής Ελλάδας (2 επίσης είδη καταγράφηκαν στο νησί της Λέσβου στο βορειοανατολικό Αιγαίο). Η εγκυρότητα αυτών των μορφολογικά προσδιορισμένων ειδών είναι υπό αμφισβήτηση. Για τον προσδιορισμό τους είναι ιδιαίτερα χρήσιμη η ενδελεχής μελέτη αυτών των πληθυσμών με τη χρήση μοριακών μεθόδων. Εικόνα A-4: Η παγκόσμια κατανομή του γένους Trachelipus. 19


26 Υπόμνημα T. aegaeus T. kytherensis T. cavaticus T. squamuliger T. buddelundi T. camerani T. razzauti T. n.sp. T. arcuatus Εικόνα A-5: Η κατανομή των ειδών του γένους Trachelipus στον ελλαδικό χώρο. 20
27 3.1. ΔΙΑΓΝΩΣΤΙΚΟΙ ΧΑΡΑΚΤΗΡΕΣ ΤΟΥ ΓΕΝΟΥΣ (Σύμφωνα με Schmidt 1997) Σχήμα σώματος ελλειψοειδές (οβάλ), χωρίς διακοπή μεταξύ του κεφαλοθώρακα και της κοιλιάς. Νωτοκοιλιακά πεπλατυσμένο χωρίς τη δυνατότητα σφαιροποίησης. Οι τεργίτες φέρουν φύματα. Στο κεφάλι υπάρχει καλά αναπτυγμένη μετωπιαία γραμμή, η οποία είναι πλευρικά επιμηκυμένη προκειμένου να σχηματιστούν οι πλευρικοί λοβοί. Ο μεσαίος κεφαλικός λοβός μπορεί να είναι παρών ή να απουσιάζει. Τα επιμερή στο πλέον είναι πολύ καλά ανεπτυγμένα και κυρτώνουν με κατεύθυνση προς το πλεοτέλσον, το οποίο είναι οξύληκτο. Τα σύνθετα μάτια αποτελούνται από ομματίδια. Ο μέγιστος αριθμός που έχει καταγραφεί σε αντιπρόσωπο του γένους είναι τα 31 ομματίδια. Τα τρωγλόβια είδη δε φέρουν πάνω από 2-4 υποπλασμένα ομματίδια. 21
 αποτελούν τα τόξα του Παναμά και του δυτικού Ειρηνικού, καθώς και το Ελληνικό Τόξο.")
28 4. ΠΑΛΑΙΟΓΕΩΓΡΑΦΙΚΑ ΣΤΟΙΧΕΙΑ ΤΟΥ ΕΛΛΑΔΙΚΟΥ ΧΩΡΟΥ Είναι γνωστό πως στο γεωλογικό χρόνο νησιά βυθίζονται ή/και αναδύονται ως αποτέλεσμα των κινήσεων των τεκτονικών πλακών, ιδίως σε περιοχές όπου συγκρούονται οι πλάκες και όπου εμφανίζονται «νησιωτικά τόξα». Τέτοια παραδείγματα κατά το Τριτογενές (65-2 Mya) αποτελούν τα τόξα του Παναμά και του δυτικού Ειρηνικού, καθώς και το Ελληνικό Τόξο. Εικόνα A-6: H σχετική κίνηση των τεκτονικών πλακών στην περιοχή της ανατολικής Μεσογείου (τροποποιημένη από Η λιθόσφαιρα κάτω από την περιοχή της ανατολικής Μεσογείου αποτελεί οριακή περιοχή μεταξύ τριών βασικών τεκτονικών πλακών: της Ευρασιατικής, της Αφρικανικής και της πλάκας της Ανατολίας (Εικ.A-6). Οι πλάκες στην περιοχή αυτή αλληλεπιδρούν μεταξύ τους. Η Αιγαιική πλάκα υφίσταται πιέσεις από την Ευρασιατική, η οποία κινείται με ταχύτητα 30 mm/έτος. Τα όρια μεταξύ αυτών των δύο πλακών στην περιοχή της κεντρικής και βόρειας Ελλάδας δεν είναι επακριβώς 22
 από τη δυτική Πελοπόννησο έως την Κρήτη, τη Ρόδο και τη δυτική Τουρκία, με ρυθμό περίπου 40 mm/έτος. Εικόνα A-7: Απεικόνιση του ηφαιστειακού τόξου και του τόξου ιζηματογενών πετρωμάτων.")
29 καθορισμένα. Από την άλλη, η Αφρικανική πλάκα βυθίζεται κάτω από την Αιγαιική κατά μήκος του τόξου των Ελληνίδων οροσειρών (Εικ.A-7) από τη δυτική Πελοπόννησο έως την Κρήτη, τη Ρόδο και τη δυτική Τουρκία, με ρυθμό περίπου 40 mm/έτος. Εικόνα A-7: Απεικόνιση του ηφαιστειακού τόξου και του τόξου ιζηματογενών πετρωμάτων. Η μεταξύ τους απόσταση ανέρχεται στα 120 km. Ηφαιστειακό τόξο. Τόξο ιζηματογενών πετρωμάτων (Ελληνίδες οροσειρές, Ιόνια νησιά, Κρήτη, Ρόδος, ΝΔ Τουρκία) 23
.")

30 Εικόνες A-8-13: Η κατανομή ξηράς-θάλασσας στον ελλαδικό χώρο στις περιόδους του Τριτογενούς και του Τεταρτογενούς (τροποποιημένο από Dermitzakis & Papanikolaou 1981 και Parmakelis et al. 2005). Η ιστορία του ελλαδικού χώρου ξεκινά κατά το Ολιγόκαινο (30 Mya). Την περίοδο αυτή το Αιγαίο αποτελούσε ενιαία ηπειρωτική περιοχή, γνωστή ως Αιγαιίδα, η oποία καταλάμβανε τη Μικρά Ασία, τη σημερινή ηπειρωτική Ελλάδα, τα νησιά του Αιγαίου και του Ιονίου, και εκτεινόταν νότια περιλαμβάνοντας και την Κρήτη. Στο 24
31 βορρά ο ελλαδικός χώρος χωριζόταν από την κεντρική Ευρώπη με τη θάλασσα της Παρατηθύος. Κατά το Βουρδιγάλιο (17 Mya), η επικοινωνία της Μεσογείου με τον Ινδικό ωκεανό διακόπηκε με αποτέλεσμα το κλίμα να γίνει υγρό και ξηρό (Εικ.A-8). Στη συνέχεια, στο Σερραβάλιο (14,5-12 Mya), ο ενιαίος όγκος ξηράς στην περιοχή του Αιγαίου άρχισε να κατακερματίζεται (Εικ.A-9). Ακολούθησε ο σχηματισμός λιμνών με γλυκά και υφάλμυρα νερά, και στο βορρά συνδέθηκε η ηπειρωτική Ελλάδα με την κεντρική Ευρώπη. Στο Τορτόνιο (10,5-7,5 Mya), η λεκάνη του Κρητικού Πελάγους καθιζάνει με αποτέλεσμα την εισροή θαλάσσιων υδάτων και την απαρχή της δημιουργίας του Αιγαίου Πελάγους. Παρατηρείται βάθυνση του Κρητικού Πελάγους αλλά και ανύψωση ορεινών όγκων (Ίδη). Η εισροή των γλυφών ή/και θαλάσσιων νερών οδηγεί στον κατακερματισμό της Κρήτης και την αποκοπή της από την ενιαία μάζα ξηράς (Εικ.A-10). Εν συνεχεία, βορειότερα, η θάλασσα που διεισδύει χωρίζει τις Κυκλάδες από τα νησιά του Αν. Αιγαίου και την Πελοπόννησο. Ωστόσο οι σύνδεση των Κυκλάδων διατηρείται τόσο με την Αττική όσο και με τη νότιο Εύβοια. Κατά το Μεσσήνιο (6,5-5,5 Mya), λόγω σύγκρουσης της βορειοδυτικής Αφρικής με την Ιβηρική χερσόνησο, τα στενά του Γιβραλτάρ έκλεισαν με αποτέλεσμα η επικοινωνία της Μεσογείου με τον Ατλαντικό να ωκεανό να διακοπεί. Το γεγονός αυτό οδήγησε στη αποστράγγιση της περιοχής και στη γνωστή «κρίση αλατότητας». Χαρακτηριστικά της κρίσης ήταν η ανύψωση πολλών περιοχών και οι εκτεταμένες γέφυρες ξηράς που αποτέλεσαν διαδρόμους επικοινωνίας και μετακίνησης πολλών πανιδικών και χλωριδικών στοιχείων, καθώς και η δημιουργία μεγάλων και αλμυρών λιμνών και κλειστών θαλασσών στην περιοχή του Αιγαίου, οι οποίες όμως θα πρέπει να ήταν «νεκρές» εξαιτίας της υψηλής αλατότητας (Dermitzakis 1990). Κατά το Μεσσήνιο το κλίμα δεν άλλαξε σημαντικά, όμως η κρίση αλατότητας οδήγησε σε πιο άνυδρο κλίμα (Εικ.A-11). Η Πικερμική πανίδα 25
32 που βρέθηκε στις περιοχές της Σάμου, της Κω, της Εύβοιας και της Αττικής, αποτελεί απόδειξη της μετακίνησης ζώων της Αφρικής προς τον ελλαδικό χώρο. Η κρίση αλατότητας έλαβε τέλος κατά το κατώτερο Πλειόκαινο και συγκεκριμένα στο Ζάγκλιο (5,5-4,5 Mya), μετά την αποκατάσταση της επικοινωνίας της Μεσογείου με τον Ατλαντικό (στενά Γιβραλτάρ) και την άνοδο της στάθμης της θάλασσας. Σε αυτήν την περίοδο το Αιγαίο πέλαγος αρχίζει να μορφοποιείται. Η Κρήτη αποχωρίζεται από την ηπειρωτική Ελλλάδα (Poulakakis & Sfenthourakis 2008, Dermitzaks & Papanikoloaou 1981). Η ανατολική Κρήτη ενώνεται με τη δυτική και παρατηρείται ο σχηματισμός των κόλπων Ηρακλείου και Μεσσαράς, οι νότιες και κεντρικές Κυκλάδες «απομονώνονται» αποτελώντας ενιαία ξηρά, και οι βόρειες Κυκλάδες ενώνονται μέσω της Εύβοιας με την ηπειρωτική Ελλάδα. Ακολουθεί αύξηση του ρυθμού ανύψωσης των τεκτονικών πλακών κατά το Πλακέντιο (ανώτερο Πλειόκαινο 4,5-3,5 Mya) που οδηγεί σε αλλαγές κυρίως στο σχήμα των νησιών και των ηπειρωτικών περιοχών (Εικ.A-12). Για παράδειγμα, στην Κρήτη σημειώνεται ανύψωση ορεινών όγκων άνω των 2000m με ταυτόχρονη υποχώρηση του θαλάσσιου βυθού. Οι νότιες περιοχές της Κρήτης καταβυθίζονται (καταβύθιση Γαύδου, Παξιμαδιών που οδηγεί σε σημαντική μείωση του μεγέθους τους). Κατά αυτή την περίοδο δημιουργείται ο νότιος Ευβοϊκός, ο οποίος χωρίζει τη νότιο Εύβοια από την Αττική. Με την έναρξη του Πλειστοκαίνου (2 Mya) ξεκινούν οι παγετώδεις και οι μεσοπαγετώδεις περίοδοι που χαρακτηρίζονται από έντονο ευστατισμό 1 αλλά και από ισοστατικές 2 κινήσεις. Οι παγετώνες κατά το Πλειστόκαινο προκάλεσαν μείωση της Ευστατικές αλλαγές: παγκόσμιες αυξομειώσεις της στάθμης της θάλασσας εξαιτίας της τήξης και πήξης των παγετώνων. 2 Ισοστατικές αλλαγές: συμβαίνουν όταν τμήματα του φλοιού της Γης αναδύονται ή βυθίζονται προκαλώντας τοπικές ή σχετικές αλλαγές στη στάθμη της θάλασσας (μέχρι και 300 μέτρα), ακόμη και όταν η παγκόσμια στάθμη δεν έχει αλλάξει. 26
33 στάθμης της θάλασσας από 100 έως 180 μέτρα σε σχέση με σήμερα. Οι μεταβολές της θαλάσσιας στάθμης καθ όλη τη διάρκεια της περιόδου αυτής οδήγησαν άλλες φορές σε συνένωση περιοχών και άλλες φορές σε αποχωρισμό τους. Αυτό είχε ως αποτέλεσμα την δημιουργία διόδων, για τη μετακίνηση και διακίνηση των πληθυσμών και τη μετέπειτα καταστροφή τους. Κατά το μέσο Πλειστόκαινο (Εικ.A-13) οι Κυκλάδες αποχωρίζονται από την Εύβοια και κατακερματίζονται. Τα νησιά του Αν. Αιγαίου παραμένουν ενωμένα με τη Μικρά Ασία (με εξαίρεση τη Νίσυρο) μέχρι και το Ολόκαινο (Σφενδουράκης 1994, Welter-Schultes 2000, Poulakakis et al. 2005a, Parmakelis et al. 2006) ενώ τα Κύθηρα είναι ακόμα συνδεδεμένα με την νότια Πελοπόννησο (Poulakakis & Sfenthourakis 2008). Την ίδια περίοδο σχηματίστηκε και ο βόρειος Ευβοϊκός κόλπος, διαχωρίζοντας έτσι την Εύβοια από τη Στερεά Ελλάδα. Σημαντικό στοιχείο για τη μετακίνηση των ειδών αποτελεί η κατά περιόδους ανάδυση νησιών ως αποτέλεσμα της πτώσης της στάθμης της θάλασσας και της αποκάλυψης υποθαλάσσιων ορέων. Τα νησιά αυτά λειτουργούσαν ως ενδιάμεσοι σταθμοί. Το Αιγαίο πέλαγος χαρακτηρίζεται από έντονες ορογενετικές κινήσεις και ως εκ τούτου είναι ελάχιστες οι περιπτώσεις που υπήρξαν νησιά πλήρως απομονωμένα. Η πλειονότητα των νησιών είχαν ενωθεί μεταξύ τους ή/και με κάποια ηπειρωτική περιοχή. Οι μεταβολές αυτές του Αιγαίου κατά το Πλειστόκαινο, συμπληρώνονται και περιπλέκονται από τις πολύ μεγάλες σε αριθμό και ένταση τεκτονικές κινήσεις. Οι τεκτονικές διεργασίες, παρά το ότι ήταν περιορισμένες σε γεωγραφική έκταση, έδρασαν καθοριστικά στην παλαιογεωγραφική εξέλιξη πολλών νησιών και δορυφορικών νησίδων (Van Andel & Shakleton 1982). Οι μελέτες αναφορικά με τις ανακατατάξεις στεριάς και θάλασσας στη διάρκεια του ανώτερου Τεταρτογενούς (ανώτερο Πλειστόκαινο Ολόκαινο) καταδεικνύουν πως περί τα ya η βόρεια Ελλάδα και η βόρεια Αδριατική σχημάτιζαν μεγάλες παραθαλάσσιες πεδιάδες που τις διέσχιζαν πολλά ποτάμια. Ευρείες πεδιάδες υπήρχαν στον Κορινθιακό Κόλπο. Πολλά νησιά, όπως η Εύβοια, η 27
34 Κέρκυρα και οι Β. Σποράδες, συνδέονταν με την ηπειρωτική Ελλάδα, και τα περισσότερα νησιά των Κυκλάδων ήταν ενωμένα σχηματίζοντας την λεγόμενη «Κυκλαδική ημι-χερσόνησο» (Van Andel & Shakleton 1982). Τα πλέον εντυπωσιακά γεωλογικά γεγονότα της Παλαιολιθικής και Μεσολιθικής περιόδου ήταν αυτά της ένωσης της Πελοποννήσου και της Εύβοιας με την υπόλοιπη ηπειρωτική Ελλάδα, η κυκλαδική ημι-χερσόνησος και η ύπαρξη αρκετών μεγάλων λιμνών. Περί τα ya (Εικ.A-14) μια ευμεγέθης ενιαία πεδιάδα, στη θέση της σημερινής Αδριατικής, συνέδεε την Ιταλία με την πρώην Γιουγκοσλαβία. Η πεδιάδα αυτή συνέχιζε από τα δυτικά παράλια της Αλβανίας και έφτανε μέχρι τα νοτιοδυτικά παράλια της Πελοποννήσου και τον Κορινθιακό Κόλπο, διακοπτόμενη μόνο από μια ορεινή ζώνη βόρεια της Κέρκυρας. Η Κεφαλονιά και η Ζάκυνθος ήταν ενωμένες σχηματίζοντας ένα ενιαίο νησί που απείχε μόλις km από την ηπειρωτική Ελλάδα. Στη διάρκεια της τελευταίας παγετώδους περιόδου η Πελοπόννησος ήταν ενωμένη με την κεντρική Ελλάδα τόσο στα βορειοδυτικά της όσο και στα βορειοανατολικά μέσω του Ισθμού της Κορίνθου ενώ ο Κορινθιακός Κόλπος σχημάτιζε λίμνη. Μεταξύ της Αργολίδας και της Αττικής εκτεινόταν μια πεδιάδα στην οποία τα σημερινά νησιά πρόβαλαν ως απομονωμένοι λόφοι. Τα στενά μεταξύ Εύβοιας και ηπειρωτικής περιοχής ήταν ξηρά. Πολλά από τα νησιά του Αιγαίου ήταν ενωμένα μεταξύ τους. Οι Σποράδες, με εξαίρεση τη Σκύρο, σχημάτιζαν μια μεγάλη χερσόνησο, η οποία ενδεχομένως διακοπτόταν με δύο πολύ ρηχά στενά που απείχαν μόλις μερικά χιλιόμετρα. Πολλά από τα κυκλαδίτικα νησιά είχαν και αυτά ενωθεί σχηματίζοντας ενιαία περιοχή, η οποία σύμφωνα με τις περισσότερες απόψεις χωριζόταν από την ηπειρωτική Ελλάδα με μια ρηχή «τάφρο» πλάτους 10 km αλλά ενδέχεται να ήταν και ενωμένη με αυτή. Οι θαλάσσιες περιοχές μεταξύ Μικράς Ασίας και Ελλάδας, αλλά και μεταξύ της Κρήτης και της ευρωπαϊκής και ασιατικής ενδοχώρας, είχαν κατά πολύ μειωθεί σε εύρος. 28
35 Εικόνα A-14: Παλαιογεωγραφικός χάρτης του ελλαδικού χώρου ya. Απεικονίζονται τα όρια της ηπειρωτικής περιοχής και των νήσων κατά τη διάρκεια της χαμηλότερης στάθμης της θάλασσας στο τέλος της τελευταίας παγετώδους περιόδου. Η ακτογραμμή έχει χαραχτεί στην Συνολικά, θα μπορούσαμε να θεωρήσουμε πως η γεωγραφία στα τέλη της Παλαιολιθικής περιόδου χαρακτηρίζεται από εκτεταμένες παραλιακές πεδιάδες ιδιαίτερα στα βορειοδυτικά και βορειοανατολικά της χώρας, μια μεγάλη κυκλαδική χερσόνησο και μια συνολική μεγέθυνση της ελλαδικής ηπειρωτικής περιοχής με ταυτόχρονη μείωση του πλάτους του Αιγαίου. Θα μπορούσαμε να πούμε πως η ελληνική χερσόνησος είχε πολύ πιο «λεία» ακτογραμμή από τη σημερινή και πως το μέγεθός της ήταν αρκετά μεγαλύτερο. Η ανύψωση της στάθμης της θάλασσας που ξεκίνησε περί τα ya, με την έναρξη της μεταπαγετώδους περιόδου, αποκατέστησε γύρω στα ya την ακτογραμμή του ελλαδικού χώρου περίπου όπως είναι διαμορφωμένη και σήμερα. Μετά τα ya (Εικ.A-15), η στάθμη της θάλασσας ανέβηκε κατά ακόμα 30m και η εναπόθεση υλικών και ιζημάτων σχημάτισε τα δέλτα ποταμών και παραλιακές πεδιάδες. Σε γενικές γραμμές, τα όρια των ακτών 29
36 μεταβλήθηκαν πολύ λίγο (Van Andel & Shakleton 1982, Shakleton et al. 1984, Demitzakis 1990, Westaway 1996). Εικόνα A-15: Παλαιογεωγραφικός χάρτης του ελλαδικού χώρου ya. Απεικονίζονται τα όρια της ηπειρωτικής περιοχής και των νήσων περί την περίοδο της ταχείας ανόδου της στάθμης της θάλασσας κατά την τελευταία μεσοπαγετώδη περίοδο. Η ακτογραμμή έχει χαραχτεί βάσει της ισοβαθούς των 130m. 30
37 5. ΦΥΛΟΓΕΩΓΡΑΦΙΑ Η φυλογεωγραφία είναι το πεδίο έρευνας που ασχολείται με τις αρχές και τις διαδικασίες που καθορίζουν τα γεωγραφικά πρότυπα κατανομής των γενεαλογικών γραμμών εντός του είδους καθώς και μεταξύ στενά συγγενικών ειδών. Λαμβάνει υπόψη το σύνολο των ιστορικών και φυλογενετικών παραγόντων της χωρικής κατανομής των γονιδιακών γενεαλογικών γραμμών (Avise 2000). Το 2007 ήταν τα 20ά γενέθλια της φυλογεωγραφίας. Ο όρος «φυλογεωγραφία» ήταν σύλληψη των Avise et al. (1987) προκειμένου να περιγράψουν μια αρχή με τεχνικές και θεωρητικές ρίζες που συνδέονται με το αρχόμενο πεδίο της μοριακής γενετικής στη δεκαετία του Ωστόσο, ο de Candolle το 1820 ήταν ο πρώτος που πρότεινε πως η τρέχουσα γεωγραφική εξάπλωση των ζωντανών οργανισμών οφείλεται τόσο σε οικολογικούς όσο και σε ιστορικούς παράγοντες. Η προσέγγιση της επιρροής των ιστορικών παραμέτρων πραγματοποιείται μέσω των ερευνών της ιστορικής βιογεωγραφίας (Wiley 1988α, 1988b, Morrone & Crisci 1995, Edwards & Beerli 2000). Οι περισσότερες μελέτες τέτοιου τύπου συγκρίνουν τη γεωγραφική εξάπλωση τάξων στο επίπεδο του είδους ή πάνω από αυτό. Με την ανάπτυξη των μοριακών μεθόδων, είναι πλέον εφικτό να διερευνηθούν πρότυπα γεωγραφικής ποικιλότητας με τη χρήση μοριακών δεικτών και να συναχθούν ενδοειδικές φυλογεωγραφικές δομές. Υπ αυτήν την έννοια, είναι δελεαστικό να εφαρμοστούν οι μέθοδοι της ιστορικής βιογεωγραφίας και κάτω από το επίπεδο του είδους, συγκρίνοντας έτσι ενδοειδικά φυλογεωγραφικά πρότυπα διαφόρων τάξων που εξαπλώνονται στην ίδια περιοχή. Έως τώρα πολύ λίγες μελέτες έχουν πραγματοποιηθεί σε αυτό το πεδίο. Ο κύριος στόχος αυτών των μελετών ήταν η διερεύνηση σύμφωνων προτύπων γεωγραφικής εξάπλωσης μεταξύ γενεαλογικών γραμμών ανάμεσα σε διαφορετικά είδη, τα οποία (γεωγραφικά πρότυπα) θα καταδείκνυαν την επιρροή ενός κοινού ιστορικού παράγοντα (Avise et al. 1987, Taberlet 1998). 31
38 Τα περισσότερα είδη εμφανίζουν σε ένα βαθμό πληθυσμιακή δομή που μπορεί να ερμηνευτεί με γεωγραφικά και χρονολογικά πρότυπα. Κύριοι στόχοι της φυλογεωγραφίας είναι η αποκρυπτογράφηση των χωρικών και χρονικών συνιστωσών της δομής των πληθυσμών και η ερμηνεία των εξελικτικών και οικολογικών διαδικασιών που ευθύνονται για την εκάστοτε δομή (Beheregaray 2008). Η φυλογεωγραφία είναι ένα συνδυαστικό επιστημονικό πεδίο που χρησιμοποιεί τη γενετική πληροφορία προκειμένου να ερευνήσει τη γεωγραφική κατανομή των γενεαλογικών γραμμών, ειδικά εκείνων μέσα στο ίδιο είδος (Avise 2000). Με βάση την προσεκτική δειγματοληψία σε άτομα και την προσεκτική επιλογή των γενετικών τόπων, οι φυλογεωγράφοι μπορούν να ελέγξουν βιογεωγραφικές υποθέσεις, να περιγράψουν την εξέλιξη αναπαραγωγικά απομονωμένων πληθυσμών και να καταλήξουν σε συμπεράσματα αναφορικά με την προέλευση, την κατανομή και τη διατήρηση της βιοποικιλότητας. Δεδομένου ότι η δομή των γενεαλογιών των πληθυσμών επηρεάζεται από τη δημογραφική τους ιστορία, οι φυλογεωγράφοι μπορούν να διατυπώσουν υποθέσεις /πιθανά σενάρια για τις αλλαγές που συντελούνται συν τω χρόνω στο φυσικό περιβάλλον των πληθυσμών, χρησιμοποιώντας τη σημερινή γενετική τους πληροφορία (Beheregaray 2008). Για όλους αυτούς τους λόγους, η φυλογεωγραφία έχει συνεισφέρει σημαντικά σε πολλά πεδία της βιολογίας και των γεωεπιστημών, όπως στην ειδογένεση (Avise et al. 2000, Moritz et al. 2000, 2006, Hewitt 2001, Kohn 2005), την ιστορική βιογεωγραφία (Avise 2000, Riddle & Hafner 2006), την ανθρώπινη εξέλιξη (Torroni et al. 1996, Beaumont 2004, Olivieri 2006, Templeton 2007), την εξελικτική βιολογία (Avise & Ball 1990, O Brien & Mayr 1991, Riddle 1996, Moritz & Faith 1998, Richards et al. 1998, Eizirik et al. 2001, Fraser & Bernatchez 2001), τη μελέτη της βιοποικιλότητας και την ταξινόμηση (Avise & Ball 1990, Tamberlet 1998, Beheregaray & Caccone 2007), την παλαιοοικολογία (Cruzan & Templeton 2000), την παλαιοκλιματολογία (Hewitt 2000) και την ηφαιστειολογία (Emerson 2002, Beheregaray 2003). 32
39 Οι φυλογεωγραφικές μελέτες έρχονται αντιμέτωπες με δύο βασικές δυσκολίες: την τυχαιότητα των γενετικών διαδικασιών και την ενδεχομένως πολύπλοκη ιστορία των μελετώμενων τάξων (Knowles & Maddison 2002, Knowles 2004). Η ανάλυση και η επεξήγηση της κατανομής των γενεαλογιών απαιτούν συνήθως εκτενή δεδομένα από τη γεωλογία, τη μοριακή και την πληθυσμιακή γενετική, την ηθολογία, τη φυλογένεση, τη δημογραφία, την παλαιοντολογία, τη γεωλογία και την ιστορική γεωγραφία (Avise 2000). Γίνεται κατανοητό, λοιπόν, πως η φυλογεωγραφία εδράζεται σε ένα κρίσιμο σταυροδρόμι μεταξύ μικροεξελικτικών και μακροεξελικτικών αρχών, τις οποίες και προσπαθεί να συγκεράσει (Εικ. 16). Οι φυλογεωγραφικές μελέτες προσπαθούν να ερμηνεύσουν τον τρόπο και το βαθμό στον οποίο οι διάφορες ιστορικές διαδικασίες που σχετίζονται με τη δημογραφία των πληθυσμών, έχουν αφήσει τα εξελικτικά τους αποτυπώματα στη σύγχρονη γεωγραφική κατανομή των γονιδιακών γραμμών των οργανισμών (Εικ.16). (Avise 2000). Η φυλογεωγραφία καλύπτει ποικίλα εναλλακτικά σενάρια σχετικά με την ερμηνεία των χωρικών διευθετήσεων των οργανισμών και των ιδιαίτερων χαρακτηριστικών τους. Παραδείγματος χάριν, ο βικαριανισμός και η διασπορά είναι συχνά ανταγωνιστικοί παράγοντες που καθορίζουν, ο καθένας με το δικό του τρόπο, την προέλευση του χωρικού διαχωρισμού των ταξινομικών μονάδων. Υπό βικαριανιστικές συνθήκες, οι πληθυσμοί ή άλλες ανώτερες ταξινομικές μονάδες, διαχωρίζονται όταν η λίγο έως πολύ συνεχής κατανομή της προγονικής μορφής κατακερματίζεται από περιβαλλοντικούς παράγοντες, όπως φαινόμενα ορογένεσης, αποχωρισμός ηπειρωτικών μαζών, ευστατικές ή ισοστατικές αλλαγές κλπ (Croizat 1982, Nelson & Ladiges 1991, Avise 2000). Κατά τη διασπορά, μια ταξινομική μονάδα αποκτά τη σημερινή της κατανομή μέσω ενεργητικής ή παθητικής διασποράς από ένα ή περισσότερα προγονικά κέντρα προέλευσης (Briggs 1989, 1992, 2000). Ο βικαριανισμός και η διασπορά είναι ιστορικά φαινόμενα, ο σχετικός ρόλος των 33

 όπως αναφέρθηκε είναι ένα ταχέως αναπτυσσόμενο επιστημονικό πεδίο που αναλύει τη")
40 οποίων στις διάφορες περιπτώσεις μπορεί να σταθμιστεί στο πλαίσιο της φυλογενετικής ανάλυσης. Εικόνα A-16: Η φυλογεωγραφία εδράζεται σε ένα κρίσιμο σταυροδρόμι μεταξύ πολύ καλά προσδιορισμένων ερευνητικών πεδίων (τροποποιημένο από Avise 2000) ΕΡΓΑΛΕIΑ ΣΤΗΝ ΥΠΗΡΕΣΙΑ ΤΗΣ ΦΥΛΟΓΕΩΓΡΑΦΙΑΣ Η φυλογεωγραφία (Avise 2000) όπως αναφέρθηκε είναι ένα ταχέως αναπτυσσόμενο επιστημονικό πεδίο που αναλύει τη γεωγραφική εξάπλωση των γενεαλογικών γραμμών. Σταθμός στην ανάπτυξή της υπήρξε η μεγάλη ανάπτυξη νέων τεχνικών κατά τη δεκαετία του 1980, η οποία κατέστησε δυνατό τον προσδιορισμό της ποικιλότητας του DNA μεταξύ ατόμων ενός είδους, και με αυτόν 34
41 τον τρόπο επέτρεψε την ανασύσταση της γενεαλογίας τους. Οι χωρικές σχέσεις τέτοιων γενεαλογιών μπορούν να απεικονιστούν γεωγραφικά και να αναλυθούν προκειμένου να εξαχθεί η εξελικτική ιστορία πληθυσμών, υποειδών και ειδών. Συγκεκριμένα, η δυνατότητα ανάλυση των μιτοχονδριακών αλληλουχιών τροφοδότησε αυτές τις νέες έρευνες. Σήμερα το εύρος των τεχνικών DNA, σε συνδυασμό με άλλες αναλυτικές μεθόδους καθώς και παλαιοκλιματολογικές και γεωλογικές μελέτες, παρέχουν σημαντικές πληροφορίες για τη γενετική ποικιλότητα και την εξέλιξης των πληθυσμών (Hewitt 2001, Emerson & Hewitt 2005). Οι φυλογεωγραφικές μελέτες που αφορούν ζωικούς οργανισμούς κυριαρχούνται από τη μελέτη του μιτοχονδιακού DNA, ενώ αντίστοιχα οι μελέτες που αφορούν φυτικούς οργανισμούς από το DNA του χλωροπλάστη. Και στις δύο περιπτώσεις, ωστόσο, υπάρχει πληθώρα άλλων διαθέσιμων μοριακών δεικτών. Φυλοεξαρτώμενοι δείκτες μπορούν να ανιχνεύσουν τις ιστορικές διαφορές στη δημογραφία μεταξύ των φύλων, πράγμα που, για παράδειγμα, υπήρξε ιδιαίτερα πληροφοριακό στη μελέτη της ανθρώπινης φυλογεωγραφίας. Για ανίχνευση βαθύτερων φυλογεωγραφικών σχέσεων χρειάζονται πιο αργοί μοριακοί δείκτες ενώ από την άλλη για πρόσφατα γεγονότα, πιθανότατα μόλις λίγων δεκάδων χιλιάδων ετών, απαιτούνται αλληλουχίες πιο μεταβλητές ή πιο γρήγορα εξελισσόμενες (Emerson & Hewitt 2005). Στην τελευταία περίπτωση, η χρήση τόσο των μικροδορυφόρων όσο και των πολυμορφισμών μήκους ενισχυμένων θραυσμάτων του DNA (AFLPs) μπορεί να επεκταθεί και σε μελέτες πέραν της πληθυσμιακής διαφοροποίησης, και με αυτό τον τρόπο να ανασυσταθούν γενεαλογίες. Πρόσφατες μελέτες (Mendelson & Shaw 2005) συνδύασαν μιτοχονδριακό και πυρηνικό DNA με τa AFLPs προκειμένου να ερευνηθεί η φυλογεωγραφία των γρύλων στα νησιά της Χαβάης, αποκαλύπτοντας τον υψηλότερο ρυθμό ειδογένεσης που έχει έως τώρα καταγραφεί σε αρθρόποδα. Κάθε αλληλουχία DNA έχει τη δική της γενεαλογία και η εξελικτική ιστορία του κάθε οργανισμού είναι το σύνολο των διαφορετικών γονιδιακών γενεαλογιών. Η 35
42 συνδυαστική χρήση τόσο μιτοχονδριακών όσο και πυρηνικών δεικτών έχει αναδείξει τα πιθανά παραπλανητικά συμπεράσματα στα οποία μπορεί να καταλήξει μια έρευνα όταν χρησιμοποιεί μόνο ένα δείκτη. Επιπροσθέτως η κάθε αναλυτική μέθοδος προάγει διαφορετικές οπτικές γωνίες της μοριακής και χωρικής ιστορίας των μελετώμενων τάξων. Ως εκ τούτου, στην προσπάθεια να ανασυσταθεί η φυλογεωγραφική ιστορία ενός είδους, θα ήταν ιδανικό να χρησιμοποιήσει κάποιος πληθώρα αλληλουχιών που θα περιελάμβαναν μιτοχονδριακές, πυρηνικές, φυλοεπηρεαζόμενες, αυτοσωματικές, συντηρητικές, καθώς και ουδέτερες, χρησιμοποιώντας δείκτες με υψηλούς αλλά και χαμηλούς ρυθμούς μεταλλακτικότητας. Φυλογεωγραφικές μελέτες που χρησιμοποιούν γενεαλογικά δεδομένα από διαφορετικούς και ανεξάρτητους μεταξύ τους τόπους θα έχουν ως αποτέλεσμα μια πιο πλήρη και αξιόπιστη ιστορία του είδους (Posada & Crandall 2001, Zink 2002, Gillespie 2004, Hewitt 2004, Emerson & Hewitt 2005) Μοριακοί χαρακτήρες «εναντίον» μορφολογικών Ένα από τα πιο χρήσιμα εργαλεία ση διερεύνηση των φυλογενετικών σχέσεων που διέπουν τα τάξα, έχει αποδειχθεί πως είναι η σύγκριση τμήματος ή ολόκληρης της νουκλεοτιδικής αλληλουχίας συγκεκριμένων γενετικών τόπων, η οποία εξετάζει τις σχέσεις των τάξων μέσα από συναπωμορφίες και όχι με βάση τη συνολική ομοιότητα, όπως ισχύει σε φαινετικές προσεγγίσεις που στηρίζονται σε ποσοτικούς χαρακτήρες. Σε γενικές γραμμές, η χρήση των μοριακών δεδομένων στη φυλογένεση έχει τύχει πολύ μεγάλης αποδοχής ακόμα και από πολύ φανατικούς οπαδούς της μορφολογίας, ιδιαίτερα στις περιπτώσεις όπου οι μορφολογικοί χαρακτήρες δεν καταφέρνουν να δώσουν ξεκάθαρη εικόνα για την εξελικτική ιστορία των μελετώμενων τάξων (Lenk et al. 1999, Michel-Salzat & Bouchon 2000, Wetzer 2002). Από πολλούς θεωρείται πως τα μοριακά δεδομένα είναι καταλληλότερα για εξελικτικές μελέτες συγκρινόμενα με δεδομένα της μορφολογίας, της φυσιολογίας 36
43 κλπ (Graur & Li 2000). Φυσικά δεν έλειψε η διαμάχη μεταξύ των επιστημόνων πάνω σε αυτό το ζήτημα (Kluge 1989, Patterson et al. 1993). Τόσο τα μορφομετρικά όσο και τα μοριακά και άλλα δεδομένα έχουν τα επιμέρους πλεονεκτήματά τους, και η βέλτιστη εκτίμηση προκύπτει μετά τη συνδυαστική χρήση τους ΧΡΗΣΗ ΤΟΥ MTDNA ΣΤΗ ΜΕΛΕΤΗ ΤΩΝ ΠΛΗΘΥΣΜΩΝ Προέλευση του mt DNA Φυλογενετικές μελέτες αντιπροσώπων από διάφορα Βασίλεια υποστηρίζουν τη μονοφυλετική προέλευση του mtdna μέσω δύο εναλλακτικών μοντέλων ενδοσυμβίωσης. Το κύτταρο με μιτοχόνδρια και mtdna προήλθε, κατά το πρώτο μοντέλο, από την ενδοσυμβίωση ενός αρχαιοβακτηρίου και ενός πρωτεοβακτηρίου σε ένα εξελικτικό βήμα, ενώ, κατά το δεύτερο μοντέλο, από την ενδοσυμβίωση αρχικά ενός αρχαιοβακτηρίου και ενός πρωτεοβακτηρίου, και στη συνέχεια και ενός α- Εικόνα A-17: Τα μοντέλα προέλευσης του μορίου του mtdna. (Τροποποιημένη από Τρυφωνόπουλος 2004). πρωτεοβακτηρίου σε δύο εξελικτικά βήματα (Εικ.A-17), με το πρώτο μοντέλο να κερδίζει έδαφος. Κατά τη διάρκεια της ενδοσυμβίωσης πολλά αρχικά γονίδια χάθηκαν ή μετανάστευσαν από το πρωτεοβακτήριο στον πυρήνα του ευκαρυώτη. Την τύχη αυτή ακολούθησαν γονίδια των οποίων είτε η λειτουργία συμπληρωνόταν από 37
44 πυρηνικά ομόλογα είτε υπήρχαν σε περισσότερα αντίγραφα και έτσι ήταν «διαθέσιμα» για γενετικές αλλαγές (Gray et al. 1999, Τρυφωνόπουλος 2004). Περαιτέρω συγκριτικές μελέτες με διάφορους οργανισμούς έδειξαν ότι το mtdna κατατάσσεται σε γενικές γραμμές σε δύο τύπους: α) τον αρχαίο τύπο που απαντάται στα φυτά και χαρακτηρίζεται από μεγάλο μέγεθος μορίου, περισσότερα γονίδια, ψευδογονίδια, εσώνια, εσωνικά ORF, URF, τμήματα ξένου DNA (cdna: χλωροπλαστικό DNA και ndna: πυρηνικό DNA) και τον τυπικό γενετικό κώδικα, και β) τον παράγωγο τύπο που απαντάται στα ζώα και στους περισσότερους μύκητες και χαρακτηρίζεται από μικρότερο μέγεθος μορίου και αριθμό γονιδίων, αλλαγές στη δομή των γονιδίων, μεγαλύτερο βαθμό νουκλεοτιδικής διαφοροποίησης και εκτεταμένες αλλαγές στο γενετικό κώδικα (Gray et al. 1999, Lang et al. 1999) Ιδιότητες του mtdna Ο μοριακός δείκτης που χρησιμοποιείται κυρίως στις φυλογεωγραφικές μελέτες είναι το μιτοχονδριακό DNA (Avise 2000). Την τελευταία εικοσαετία έχει χρησιμοποιηθεί εκτενώς σε μελέτες που αφορούν τη δομή των πληθυσμών και τη γονιδιακή ροή, τον υβριδισμό, τη βιογεωγραφία και τις φυλογενετικές σχέσεις (Moritz et al. 1987, Avise 2000), αφού διαθέτει μια σειρά από χαρακτηριστικά που το καθιστούν ιδιαίτερα χρήσιμο σύστημα για την ανίχνευση γενετικών διαφορών σε διαειδικό και ενδοειδικό επίπεδο. To mtdna των ζώων αποτελεί το καλύτερα μελετημένο τμήμα ευκαρυωτικού DNA. Αυτό οφείλεται στην ευκολία απομόνωσής του σε σχέση με οποιοδήποτε τμήμα του πυρηνικού DNA. Η ευκολία αυτή έγκειται στην ασυνήθιστα μικρή πυκνότητά του σε κλίση CsCl, στο μεγάλο αριθμό αντιγράφων του και στην παρουσία του σ ένα οργανίδιο διαφορετικό του πυρήνα. Επιπλέον, εξαιτίας του μικρού μεγέθους του και της απουσίας των πολύπλοκων χαρακτηριστικών του ndna, όπως τα εσώνια, οι επαναλαμβανόμενες 38
45 αλληλουχίες, τα μεταθέσιμα στοιχεία, είναι εύκολος ο χαρακτηρισμός του (εύρεση γονιδίων και σειράς αυτών, αλληλούχιση). Aπό τη μεριά της συγκριτικής ανάλυσης, το mtdna έχει το πλεονέκτημα να υπάρχει σ όλο το ζωικό βασίλειο και να έχει εξαιρετικά παρόμοιο γονιδιακό περιεχόμενο (όλα τα μετάζωα και ορισμένα πρωτόζωα έχουν την ίδια ομάδα 37 γονιδίων) (Wilson et al. 1985) (Εικ.A-18). Το περιεχόμενο και το μέγεθος του μορίου έμεινε σχετικά σταθερό για περισσότερο από 800 εκατομμύρια χρόνια, γεγονός που υποδηλώνει ότι τα χαρακτηριστικά αυτά αποτελούν επιλέξιμο εξελικτικό πλεονέκτημα. Όμως, αναλύσεις έχουν αποκαλύψει πλαστικότητα στην οργάνωση αλλά και στους τρόπους αλληλεπίδρασης με το συνεξελισσόμενο ndna, μέσω της μεγάλης ποικιλομορφίας των κύριων μη κωδικοποιουσών και άλλων ρυθμιστικών περιοχών (Saccone et al. 1999, 2000, 2002). Οι εξελικτικές μεταλλακτικές αλλαγές του ζωικού mtdna είναι σχετικά απλές και είναι κυρίως υποκαταστάσεις βάσεων και μεταλλάξεις μήκους, με τις τελευταίες να συγκεντρώνονται κυρίως στις μικρές μη κωδικοποιούσες περιοχές. To μόριο του mtdna αποτελεί το πιθανότερο εύρημα σε οργανικά κατάλοιπα ζωικών ειδών που έχουν εξαφανιστεί. Ο μεγάλος αριθμός μορίων ανά κύτταρο σε σχέση με οποιοδήποτε τμήμα του ndna αυξάνουν την πιθανότητα εύρεσης τμημάτων του ικανά για ανάλυση αλληλουχίας (Wilson et al. 1985). Το μιτοχονδριακό DNA (mtdna) εμφανίζει πολλά πλεονεκτήματα τόσο από εξελικτική όσο και από τεχνική άποψη (Hurst & Jiggins 2005), τα οποία μπορούν να συνοψιστούν στα ακόλουθα: 1. Το mtdna εμφανίζεται σε μεγάλο αριθμό αντιγράφων στα κύτταρα (Futuyma 1991), είναι πλήρως χαρακτηρισμένο, ενώ ο μητρικός, κυρίως, τρόπος κληρονομικότητας (Zhao et al. 2004) δεν αφήνει περιθώρια για ανασυνδυασμό, με ελάχιστες εξαιρέσεις (Zouros et al. 1994, Ladoukakis & Zouros 2001a,b, Rokas et al. 2003) και έτσι οι διαφοροποιήσεις που 39
46 παρατηρούνται είναι καθαρά αποτέλεσμα μεταλλαγών (Hagelberg et al. 1994, 1999). 2. είναι πολύ καλά χαρακτηρισμένο μόριο σε σχέση με το πυρηνικό γονιδίωμα (Hagelberg et al. 1994, 1999) 3. είναι απλοειδές, οπότε δεν απαιτείται κλωνοποίηση πριν από τον προσδιορισμό της αλληλουχίας (Hurst & Jiggins 2005). 4. τα ψευδογονίδια και τα εσώνια απουσιάζουν από το μιτοχονδριακό DNA (Avise et al. 1987). 5. ο γρήγορος ρυθμός εξέλιξης του mtdna το καθιστά πολύτιμο εργαλείο για εξελικτικές και φυλογενετικές μελέτες τόσο στο διαειδικό όσο και στο ενδοειδικό επίπεδο (Avise et al. 1987). Βέβαια ο ρυθμός εξέλιξης δεν είναι σταθερός για όλες τις ομάδες οργανισμών. Μέχρι σήμερα έχουν διατυπωθεί διάφορες υποθέσεις για την ερμηνεία των διαφορών που παρατηρούνται στο ρυθμό αντικατάστασης μεταξύ των διαφόρων εξελικτικών γραμμών. Οι παράγοντες που συχνά επιστρατεύονται για το σκοπό αυτό, διακρίνονται σε 2 κατηγορίες: τους εξαρτώμενους από τη γενεά, όπως ο χρόνος γενεάς, η αδυναμία επιδιόρθωσης λαθών κατά την αντιγραφή (που οδηγεί στη συσσώρευση μεταλλάξεων) και οι ανεξάρτητοι από τη γενεά, όπως ο μεταβολικός ρυθμός (τα εξώθερμα έχουν χαμηλότερο ρυθμό από τα ενδόθερμα) και το μέγεθος του σώματος. Η εξαγωγή φυλογεωγραφικών συμπερασμάτων βασίζεται στη μελέτη της αναδομημένης γενεαλογικής ιστορίας των γονιδίων (γονιδιακών δένδρων) από διαφορετικούς πληθυσμούς. Επειδή όμως πολλά γεγονότα έχουν λάβει χώρα στο παρελθόν (π.χ. επέκταση πληθυσμών, φαινόμενα στενωπού, μετανάστευση) και πολλές γενεαλογικές γραμμές ενδέχεται να έχουν χαθεί τυχαία, η ιστορία των ειδών πιθανότατα να μην ταυτίζεται με την ιστορία του γονιδιακού δένδρου. Για το λόγο 40

47 αυτόν, και για να αποφευχθεί η εξαγωγή λανθασμένων συμπερασμάτων, απαιτείται ο προσδιορισμός των φυλογενετικών σχέσεων των υπό εξέταση ταξινομικών μονάδων να βασίζεται, όχι σε ένα, αλλά σε περισσότερα γονίδια. Εικόνα A-18: Σχηματική απεικόνιση του κυκλικού μορίου του συντηρητικού μιτοχονδριακού γονιδιώματος. Απεικονίζονται η διάταξη των γονιδίων και τα μόρια που κωδικοποιούν. (τροποποιημένο από Chen & Zhao 2009). 41
48 6. ΕΡΕΥΝΗΤΙΚΟ ΠΡΟΒΛΗΜΑ Οι αντιπρόσωποι του γένους Trachelipus στην Ελλάδα παρουσιάζουν μεγάλη μορφολογική και γενετική ποικιλότητα (Parmakelis et al. 2008). Η ποικιλότητα αυτή εμφανίζεται να είναι μάλιστα και ενδοειδική στην περίπτωση του καλύτερα μελετημένου είδους, στο πλαίσιο της παρούσας μελέτης, του Trachelipus kytherensis. Ο ελλαδικός χώρος αποτελεί ένα εργαστήρι της φύσης όπου ποικίλα πειράματα έλαβαν χώρα καθοδηγούμενα από την παλαιογεωγραφία και την παλαιοοικολογία της περιοχής, οι οποίες λόγω των έντονων ανακατατάξεων θάλασσας ξηράς, επηρέασαν καθοριστικά τόσο την κατανομή όσο και τη διαφοροποίηση πολλών τάξων από διάφορες ταξινομικές βαθμίδες. Τόσο οι ανακατατάξεις κατά τους προϊστορικούς χρόνους όσο και οι πλέον πρόσφατες των τελευταίων ετών, οι οποίες διαμόρφωσαν τόσο την έκταση της σημερινής ηπειρωτικής περιοχής όσο και τις συνδέσεις των επιμέρους γεωγραφικών διαμερισμάτων, χρήζουν ιδιαίτερης προσοχής και μελέτης προκειμένου να διερευνηθεί το αποτύπωμα που ακόμα και τόσο νωρίς - στον εξελικτικό χρόνο - ενδέχεται να έχουν αφήσει στο γονιδίωμα ή/και το φαινότυπο των οργανισμών. Ο ελλαδικός χώρος παρουσιάζει ιδιαίτερο ενδιαφέρον για την ιστορική και την οικολογική βιογεωγραφία, αφού από τη μια πλευρά ο πρόσφατος διαχωρισμός των νησιών του Αιγαίου έχει οδηγήσει πολλά τάξα σε μεγάλη διαφοροποίηση και από την άλλη η περιοχή περιλαμβάνει πλήθος νησιών διαφόρων μεγεθών προσφέροντας έτσι πλούσιο έδαφος για εφαρμογή των αρχών της νησιωτικής βιογεωγραφίας (MacArthur & Wilson 1967). H επιλογή του συγκεκριμένου γένους ως πειραματικού υλικού για την παρούσα εργασία έγινε με βάση το ότι εμφανίζει 4 (ίσως και 5) ενδημικά είδη στα νησιά και την ηπειρωτική περιοχή της ελληνικής επικράτειας, τα οποία εμφανίζουν ευρεία εξάπλωση (από το επίπεδο της θάλασσας έως τις κορυφές των βουνών), σε 42
49 συνδυασμό με τις ιδιαιτερότητες των περιβαλλόντων όπου ζουν. Πρόκειται για υγρά ενδιαιτήματα, άμεσα απειλούμενα από την ανθρώπινη δραστηριότητα (παρόχθια βλάστηση ρεμάτων και ποταμών, υγρά δάση κλπ.). Ως εκ τούτου, τα ξηρά περιβάλλοντα αποτελούν φράγματα στη διασπορά των συγκεκριμένων οργανισμών. Ο συνεχώς αυξανόμενος κατακερματισμός των βιοτόπων οδηγεί σε αύξηση της απομόνωσης των φυσικών πληθυσμών και, συνεπώς, σε μείωση της γενετικής ροής μεταξύ τους. Από προηγούμενη μελέτη του εργαστηρίου μας (Parmakelis et al. 2008) στο γένος Trachelipus παρατηρήθηκαν: έντονη απόκλιση μεταξύ των προτύπων της γενετικής ποικιλότητας και εκείνης της ισχύουσας ταξινόμησης σε ορισμένες ομάδες πληθυσμών τους γένους αυτού σημαντικός βαθμός γενετικής απομόνωσης μεταξύ πληθυσμών του Trachelipus kytherensis που ενισχύουν την άποψη περί ισχυρής μεταπληθυσμιακής συγκρότησής τους,. και αναντιστοιχία μεταξύ μορφολογικής και γενετικής διαφοροποίησης Όπως προαναφέρθηκε, η περιγραφή του είδους στηρίζεται σε περιορισμένο αριθμό μορφολογικών χαρακτήρων και δεν είναι βέβαιο ότι αντανακλά την πραγματική φυλογένεση. Η παρούσα εργασία έχει ως αντικείμενα μελέτης: 1. Τον προσδιορισμό της γενετικής δομής των πληθυσμών και των ειδών 2. Την εκτίμηση του βαθμού της γενετικής διαφοροποίησης / απόστασης μεταξύ των πληθυσμών του ίδιου είδους σε συγκεκριμένη γεωγραφική περιοχή 3. Τον προσδιορισμό της φυλογενετικής ιστορίας των αντίστοιχων πληθυσμών ώστε να αναπλασθεί το ιστορικό της απομόνωσής τους 4. Το συνδυασμό των παραπάνω αποτελεσμάτων ώστε να εκτιμηθεί η επίδραση των ιστορικών (παλαιογεωγραφικών, οικολογικών κλπ) παραγόντων στη διαμόρφωση της σύγχρονης κατάστασης. 43
50 Το κεντρικό ερώτημα λοιπόν που τίθεται είναι πώς σχετίζεται ο βαθμός διαφοροποίησης μεταξύ των πληθυσμών με την κατανομή τους στο χώρο. Συνοψίζοντας όλα τα παραπάνω, στόχος της παρούσας διατριβής είναι η διερεύνηση των διεργασιών διαφοροποίησης των πληθυσμών των ενδημικών ειδών του γένους Trachelipus στην Ελλάδα, ώστε να εντοπισθεί η επίδραση της απομόνωσης των πληθυσμών στη γενετική τους σύσταση και τους μορφολογικούς τους χαρακτήρες. 44
51 Β. ΜΕΘΟΔΟΛΟΓΙΑ ΚΑΙ ΥΛΙΚΑ 1. ΣΥΛΛΟΓΗ ΔΕΙΓΜΑΤΩΝ Η συλλογή των δειγμάτων πραγματοποιήθηκε κατά την περίοδο Για την πραγματοποίηση της παρούσας εργασίας συλλέχθηκαν 150 άτομα του γένους Trachelipus. Η συλλογή έγινε σε περιοχές περιλαμβάνουν περιοχές κυρίως της δυτικής και βορειοδυτικής ηπειρωτικής Ελλάδας και της Εύβοιας (Εικ.B-1). Οι τοποθεσίες αυτές και ο αριθμός των ατόμων που συλλέχθηκαν φαίνονται στον Πίνακα Β-1. Η συλλογή πραγματοποιήθηκε με το χέρι και τα άτομα θανατώνονταν άμεσα σε αιθανόλη 95%. Κατόπιν μεταφέρονταν στο εργαστήριο, όπου ταυτοποιούνταν μορφολογικά και αποθηκεύονταν μέχρι τη χρήση τους. Η ταυτοποίηση των οργανισμών στο επίπεδο του είδους έγινε με βάση τα μορφολογικά τους χαρακτηριστικά, όπως αυτά περιγράφονται στην κλείδα του Schmidt 1997, και προσδιορίστηκαν ως αντιπρόσωποι του είδους T. kytherensis. Επιπροσθέτως, στις αναλύσεις συμπεριλήφθησαν, για συγκριτικούς λόγους, και οι 77 αλληλουχίες του γονιδίου 16S rrna και οι 75 αλληλουχίες της COI ατόμων Trachelipus από 16 τοποθεσίες (Πιν. Β-2) που χρησιμοποιήθηκαν στην εργασία των Parmakelis et al. 2008, οι οποίες ανακτήθηκαν από τη βάση δεδομένων GenBank ( Από την ίδια βάση δεδομένων ελήφθησαν και 4 αλληλουχίες ατόμων για το γενετικό τόπο 16S rrna [Porcelio laevis, Armadillidium lobocurvum (EF027622), Porcelionides pruinosus (AJ275211)] και 1 αλληλουχία για το γενετικό τόπο της υπομονάδας Ι της κυτοχρωμικής οξειδάσης (COI) [Armadillidium lobocurvum (AJ275211]) από άλλα είδη ισοπόδων (βλ. Πιν. Β-1), προκειμένου να 45
52 χρησιμοποιηθούν ως εξωτερική ομάδα αναφοράς στη στατιστική επεξεργασία των αποτελεσμάτων. Συνολικά στις αναλύσεις συμπεριελήφθησαν 231 άτομα για το γενετικό τόπο 16s rrna και 186 άτομα για το γενετικό τόπο COI. Πίνακας Β-1: Οι πληθυσμοί που συλλέχτηκαν στο πλαίσιο της παρούσας μελέτης, το είδος στο οποίο ανήκουν και η συντόμευση της κάθε περιοχής, όπως φαίνεται στο χάρτη της Εικ. Β-1. Για τα άτομα Armadillidium lobocurvum και Porcelionides pruinosus παρατίθεται σε παρένθεση ο αριθμός πρόσβασης στη GenBank. Κωδικός χάρτη ΤΟΠΟΘΕΣΙΑ ΑΡΙΘΜΟΣ ΑΤΟΜΩΝ (Ν) 16S COI AIN Αίνος 4 4 ALF Αλφειός 6 6 AMG Αμυγδαλιές 4 4 ATR Αγία Τριάδα 4 0 BEL Βέλλα (πηγές) 4 1 CHb Χελμός β 3 3 DRM Δρυμώνας 3 1 CML Χρυσομηλιά 3 0 ELTa Ελάτη α 4 4 ELTb Ελάτη β 4 3 FEN Φενεός 3 3 FLA Φλαμπουρίτσα 1 0 FNR Φανάρι 2 1 FRX Δάσος Φράξου 4 4 GAB Γάβρος 0 2 GRP Γκρόπα 4 2 KAR Καρυές 1 0 KAT Κατάρα 4 2 KLM Καλαμάς 4 2 KON Κόνιτσα 2 1 ΕΙΔΟΣ Trachelipus kytherensis 46
53 KRN Κρανιά 2 1 KRT Κουρούτα 4 4 LAG Λαγκάδια 1 1 LIB Λιμποβίσι 5 5 LOU Λούσιος (ποταμός) 1 0 MAIN Μαίναλο 1 0 MKL Μακελαριά 1 0 MNS Μανεσαίικος 2 2 NED Νέδα (ποταμός) 3 0 PAP Παπάδες 3 2 PER Περτούλι 2 2 PMS Πάμισσος 3 4 PRR Παρόρι 4 4 PRSM Μικρή Πρέσπα 1 0 SFK Σοφικό 3 0 SOL Σόλος 3 3 SSN Σωσσινίτσα 4 3 TAY Ταΰγετος 8 10 TPT Τριπόταμα 3 3 Trachelipus kytherensis VLA Βλασία 3 3 VLK Βελίκα (παραλία) 3 2 VOI Βοϊδομάτης 4 2 VTS Βίτσα 3 3 ZIR Ζήρια 3 2 ZRL Λίμνη Ζηρού 9 4 STE Στενή 4 4 EPD Επίδαυρος 3 3 Trachelipus aegaeus AKYT Αντικύθηρα 2 - Porcelio laevis PAN_N6 ATHE Παναχαϊκό Αθήνα 1 (EF027622) 1 (AJ275211) 1 (EF027706) Σύνολο Armadillidium lobocurvum - Porcelionides pruinosus 47
54 Πίνακας Β-2: Οι πληθυσμοί από Parmakelis et al που ενσωματώθηκαν στις αναλύσεις της παρούσας μελέτης. ΑΡΙΘΜΟΣ Κωδικός Κωδικός πρόσβασης Κωδικός πρόσβασης Τοποθεσία ΑΤΟΜΩΝ (Ν) ΕΙΔΟΣ χάρτη GenBank COI GenBank 16S rrna COI 16S VEL Βελούχι 2 2 EF EF (2) EF EF (2) PROU Προυσσός 5 5 EF EF (5) EF EF (5) THEO Θεόκτιστο 4 4 EF EF (4) EF027515, EF EF (4) PLAN Πλανητέρο 6 6 EF EF (6) EF EF (6) PAN Παναχαικό 9 9 EF EF (2) EF (1) EF EF (6) EF EF (2) EF (1) EF EF (6) Trachelipus kytherensis MIK Μικρουλέικα 6 6 EF EF (6) EF EF (6) LAMB Λάμπεια 6 6 EF EF (6) EF EF (6) KYT Κύθηρα 4 4 EF EF (4) EF EF (4) CRE Κρήτη 4 4 EF EF (4) EF EF (4) KAL Καλέτζι 3 3 EF EF (3) EF EF (3) CH Χελμός 6 6 EF EF (6) EF EF (6) BOU Βουραϊκός EF EF (12) EF EF (12) STN Στουρονήσι 2 2 EF EF (2) EF EF (2) AGA Άγιος Ανδρέας 2 2 Trachelipus n.sp. EF EF (2) EF EF (2) IKAR Ικαρία EF EF (2) NAX Νάξος 3 3 Trachelipus aegaeus EF EF (3) EF EF (3) STO Στουρονήσι 1 1 Σύνολο EF (1) EF (1) 48

55 49

, 13=Τριπόταμα (TPT), 14=Δάσος Φράξου (FRX), 15=Λίμνη Ζηρού (ZRL), 16=Αγ.")
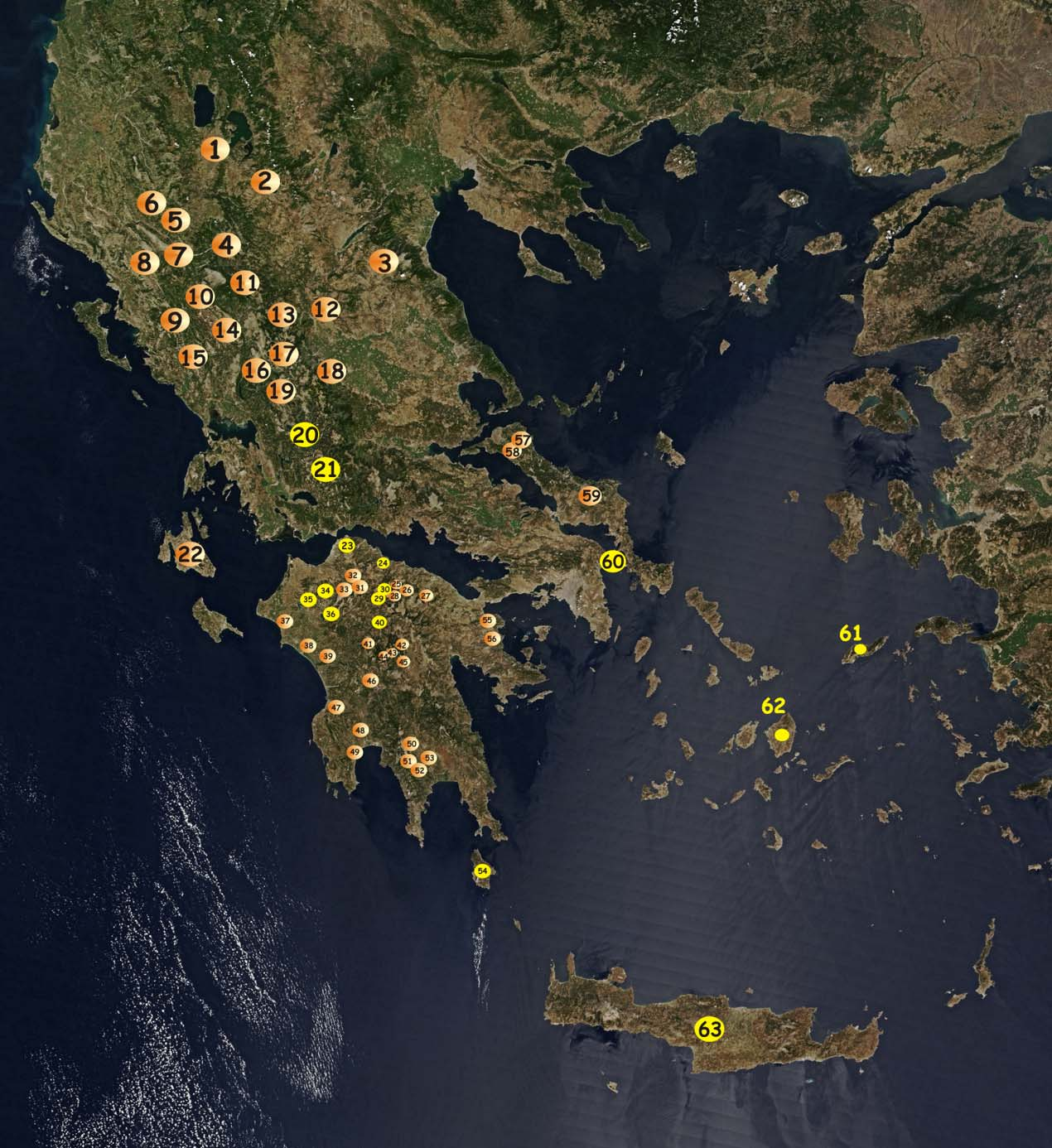
56 Εικόνα Β-1: Οι περιοχές δειγματοληψίας των πληθυσμών που αναλύθηκαν στην παρούσα μελέτη. Οι κίτρινοι κύκλοι αντιστοιχούν στους πληθυσμούς των Parmakelis et al που ενσωματώθηκαν στις αναλύσεις της παρούσας μελέτης. 1= Πρέσπες (PRSM), 2=Γαύρος (GAV), 3=Καρυές (KAR), 4=Κρανιά (KRN), 5=Κόνιτσα (KON), 6=Σωσσινίτσα (SSN), 7=Βίτσα (VTS), 8=Βοϊδομάτης (VOI), 9=Καλαμάς (KLM), 10=Βέλλα (BEL), 11=Κατάρα (KAT), 12=Χρυσομηλιά (CHM), 13=Τριπόταμα (TPT), 14=Δάσος Φράξου (FRX), 15=Λίμνη Ζηρού (ZRL), 16=Αγ. Τριάδα (ATR), 17=Περτούλι (PER), 18=Γκρόπα (GRP), 19=Φανάρι (FNR), 20=Βελούχι (VEL), 21=Προυσσός (PROU), 22=Αίνος (AIN), 23 Παναχαϊκό (PAN), 24= Βουραϊκός (VOU), 25=Σόλος (SOL), 26=Φεναιός (FEN), 27=Ζήρεια (ZIR), 28=Χελμός β (CHb), 29=Πλανητέρο (PLAN), 30=Χελμός (CH), 31=Μανεσαίικος (MNS), 32=Μακελαριά (MKL), 33=Βλασσία (VLA), 34=Καλέτζι (KAL), 35=Μικρουλέικα (MIK), 36=Λάμπεια (LAMB), 37=Κουρούτα (KRT), 38=Αλφειός (ALF), 39=Αμυγδαλιές (ΑMG), 40=Θεόκτιστο (THEO), 41=Λαγκάδια (LAG), 42=Μαίναλο (MAIN), 43=Ελάτη α (ELTa), 44=Ελάτη β (ELTb), 45=Λιμποβίσι (LIB), 46=Λούσιος, 47=Νέδα (NED), 48=Πάμισος (PMS), 49=Βελίκα (VLK), 50=Παρόρι (PRR), 51=Ταΰγετος α (TAYa), 52=Ταΰγετος β (TAYb), 53= Ταΰγετος γ (TAYc), 54=Κύθηρα (KYT), 55=Σοφικό (SFK), 56=Επίδαυρος (EPD) 57=Παπάδες (PAP), 58=Δρυμώνας (DRM), 59=Στενή (STN), 60=Στουρονήσι, Αγ.Ανδρεας (STO,STN,AGA), 61=Ικαρία (IKAR), 62=Νάξος (NAX), 63=Κρήτη (CRE) 50
57 2. ΤΑ ΜΙΤΟΧΟΝΔΡΙΑΚΑ ΓΟΝΙΔΙΑ ΤΗΣ ΜΕΛΕΤΗΣ Για τη διερεύνηση του φυλογεωγραφικού προτύπου των ενδημικών ειδών του γένους Trachelipus που μελετήθηκαν στην παρούσα εργασία (T. kytherensis, Τ. nsp. και T. aegaeus) χρησιμοποιήθηκαν δύο μιτοχονδριακά γονίδια: η υπομονάδα Ι της κυτοχρωμικής οξειδάσης (cytochrome oxidase subunit I COI) και το 16S rrna. Η υπομονάδα Ι της κυτοχρωμικής οξειδάσης αποτελεί ένα ταχέως εξελισσόμενο γονίδιο του μιτοχονδριακού γονιδιώματος, είναι ένας πολύ καλός μοριακός δείκτης και έχει χρησιμοποιηθεί ευρέως σε μελέτες της γενετικής διαφοροποίησης και της εύρεσης φυλογεωγραφικών προτύπων τόσο σε ενδοειδικό επίπεδο όσο και μεταξύ συγγενικών ειδών (Ros & Breeuwer 2007, Lewis et al. 2005, Avise 2000, Maslov et al. 1999), καθώς και από το εργαστήριό μας σε προηγούμενες μελέτες ισοπόδων (Klossa-Kilia et al. 2005, 2006, Parmakelis et al. 2008). Σύμφωνα μάλιστα με ερευνητές, υποστηρικτές του Barcoding, το γονίδιο αυτό φέρει μεγαλύτερο εύρος φυλογενετικού σήματος σε σύγκριση με τα υπόλοιπα μιτοχονδριακά γονίδια με τη βοήθεια αυτού του γονιδίου και μπορεί να ταυτοποιήσει στο επίπεδο του είδους το 98% των ζωικών ειδών. (Hebert et al. 2003, Kerr et al. 2007) Ο γενετικός τόπος 16S rrna είναι ένας από τους πιο ευρέως χρησιμοποιούμενους σε φυλογενέσεις, ανεξάρτητα από ζωική ομάδα ή ταξινομική βαθμίδα, κυρίως γιατί φέρει περιοχές που εξελίσσονται ταχύτατα και εξυπηρετεί τη διερεύνηση των φυλογενετικών σχέσεων μεταξύ πρόσφατα διαχωρισμένων ειδών, αλλά παράλληλα φέρει και περιοχές αρκετά συντηρημένες που εξυπηρετούν στη διερεύνηση σχέσεων μεταξύ τάξων που έχουν διαχωριστεί εδώ και 200 εκ. χρόνια (DeSalle 1987, 1992). Το συγκεκριμένο γονίδιο δεν κωδικοποιεί πρωτεΐνη και έτσι παίρνουμε χρήσιμες πληροφορίες για τις σχέσεις μεταξύ των πληθυσμών από τις ενθέσεις ή/και τις ελλείψεις νουκλεοτιδίων που μπορεί να παρατηρούνται σε διάφορες θέσεις μεταξύ των εξεταζόμενων αλληλουχιών. 51
58 3. ΕΡΓΑΣΤΗΡΙΑΚΕΣ ΑΝΑΛΥΣΕΙΣ 3.1. ΠΕΙΡΑΜΑΤΙΚΗ ΠΟΡΕΙΑ Εξαγωγή Ολικού Γενωμικού DNA Στη μελέτη αυτή, η απομόνωση του ολικού DNA πραγματοποιήθηκε με τη βοήθεια του kit απομόνωσης Nucleospin Tissue της εταιρείας Macherey-Nagel. Το kit περιλαμβάνει το αναλυτικό πρωτόκολλο απομόνωσης, στο οποίο περιγράφεται σταδιακά η διαδικασία εξαγωγής του DNA, καθώς και τα απαραίτητα διαλύματα και αντιδραστήρια. Φυγοκέντρηση Εικόνα Β-2: Σχηματική απεικόνιση της διαδικασίας απομόνωσης ολικού DNA από ιστό. 52
 του ιστού για 24 ώρες με διάλυμα ΤΕ9 σε ανακινούμενο υδατόλουτρο και σε θερμοκρασία 57 ο C προκειμένου να απομακρυνθεί στο μέγιστο δυνατό βαθμό η εναπομείνασα αλκοόλη")
59 Στις περιπτώσεις που το παραλαμβανόμενο DNA ήταν κακής ποιότητας αλλά και γενικότερα πολλές φορές για τη βελτιστοποίηση του Για την παρασκευή του διαλύματος TE9 απαιτούνται εξαγόμενου DNA, προηγήθηκε επώαση και ξεπλύματα (2-3 φορές) του ιστού για 24 ώρες με διάλυμα ΤΕ9 σε ανακινούμενο υδατόλουτρο και σε θερμοκρασία 57 ο C προκειμένου να απομακρυνθεί στο μέγιστο δυνατό βαθμό η εναπομείνασα αλκοόλη όπου ήταν συντηρημένα τα δείγματα. 500mM TrisHCl 20mM EDTA 10mM NaCl ph= Ηλεκτροφόρηση Ολικού Γενωμικού DNA Ο έλεγχος της επιτυχίας της απομόνωσης, της ποσότητας και ποιότητας του εξαγόμενου DNA γινόταν με ηλεκτροφόρηση ζώνης. Η παρουσία DNA σε ικανοποιητική συγκέντρωση, χωρίς προσμίξεις RNA και πρωτεϊνών, δηλωνόταν με την παρουσία μιας μόνο ευκρινούς ζώνης. Η ηλεκτροφόρηση πραγματοποιείτο σε πήκτωμα αγαρόζης 1,5% που παρασκευαζόταν διαλύοντας με βρασμό την αγαρόζη σε διάλυμα ΤΒΕ (διάλυμα Trizma Base, Η 3 ΒΟ 3, και EDTA). Το πήκτωμα περιείχε και 5-7% βρωμιούχο αιθίδιο (EtBr) το οποίο δεσμεύεται ανάμεσα σε διαδοχικές βάσεις του ίδιου κλώνου των νουκλεϊκών οξέων, απορροφά υπεριώδη ακτινοβολία (UV) και εκπέμπει ακτινοβολία στο ορατό για τον άνθρωπο φάσμα. Εικόνα Β-3: Το πρότυπο ζωνών από απομόνωση ολικού γενωμικού DNA. Παρατηρείται μια φωτεινή ζώνη η οποία αντιστοιχεί στο ολικό DNA. Η εμφάνιση των πηκτωμάτων γίνεται με την επίδραση υπεριώδους φωτός. 53
 που χρησιμοποιήθηκε για να εκτιμηθεί το μέγεθος, η ποσότητα και η ποιότητα των προϊόντων.")
60 Εικόνα Β-4: Το πρότυπο ζωνών των προϊόντων μιας αντίδρασης PCR. Στο πρώτο πηγαδακι του πηκτώματος διακρίνεται ο μάρτυρας (gelpilot low range ladder) που χρησιμοποιήθηκε για να εκτιμηθεί το μέγεθος, η ποσότητα και η ποιότητα των προϊόντων. Η εμφάνιση των πηκτωμάτων γίνεται με την επίδραση υπεριώδους φωτός Πολλαπλασιασμός συγκεκριμένων τμημάτων DNA με τη μέθοδο της αλυσιδωτής αντίδρασης πολυμεράσης (PCR) Για τον πολλαπλασιασμό κάθε γενετικού τόπου χρησιμοποιήθηκε ένα ζεύγος εκκινητών (primers) που με τη μεγαλύτερη πιστότητα πολλαπλασίαζε το ζητούμενο κάθε φορά γονίδιο. Οι αλληλουχίες των εκκινητών και το μέγεθος του προϊόντος που παράγουν δίνονται στον πίνακα B-3. Τα πρωτόκολλα παρασκευής των διαλυμάτων της PCR παρατίθενται στους πίνακες B-4 και B-5. Πίνακας B-3: Τα γονίδια που χρησιμοποιήθηκαν στην παρούσα μελέτη, οι ακολουθία των εκκινητών που χρησιμοποιήθηκαν, το μέγεθος των προϊόντων τους και η πηγή προέλευσής Γονίδιο Όνομα Εκκινητές COI 16S rrna COIF (forward) COIR-int (reverse) 16Sar-int-sf (forward) 16Sbr-sf (reverse) τους. Μέγεθος προϊόντος Αναφορά 5 -CCG GTT TGA ACT CAG ATC ATG-3 Folmer et al bp 5 -GCY CCY GCY AAW ACA GGK ARD-3 Parmakelis et al GCC GCA GTA THC TRA CTG TGC T-3 Parmakelis et al bp 5 -CCG GTT TGA ACT CAG ATC ATG-3 Palumbi
61 Πίνακας B4: Πρωτόκολλο παρασκευής διαλύματος PCR για το μιτοχονδριακό γονίδιο της COI 1λ από το διάλυμα ολικού DNA 5λ ρυθμιστικού διαλύματος αντίδρασης 10x (PCR buffer) 4λ διαλύματος 10mM MgCl 2 0,5λ διαλύματος 20pmol/lt για κάθε ολιγονουκλεοτίδιο εκκινητή 5λ διαλύματος 10mM dntp που περιέχει σε ίσες επιμέρους συγκεντρώσεις τα τέσσερα dntps 2λ BSA (αλβουμιίνη ορού βοός) 0,5λ διαλύματος 5u/λ Taq πολυμεράσης 31,5λ απεσταγμένο και αποστειρωμένο Η 2 Ο, ώστε η τελική ποσότητα κάθε δείγματος να είναι 50λ. Πίνακας B5: Πρωτόκολλο παρασκευής διαλύματος PCR για το μιτοχονδριακό γονίδιο 16S rrna 1λ από το διάλυμα ολικού DNA 5λ ρυθμιστικού διαλύματος αντίδρασης 10x (PCR buffer) 3,5λ διαλύματος 10mM MgCl 2 0,6λ διαλύματος 20pmol/lt για τον ολιγονουκλεοτίδιο εκκινητή 16Sar-int-sf 0,4λ διαλύματος 10μΜ για τον ολιγονουκλεοτίδιο εκκινητή 16Sbr-sf 5λ διαλύματος 10mM dntp που περιέχει σε ίσες επιμέρους συγκεντρώσεις τα τέσσερα dntps 0,5λ διαλύματος 5u/λ Taq πολυμεράσης 34λ απεσταγμένο και αποστειρωμένο Η 2 Ο, ώστε η τελική ποσότητα κάθε δείγματος να είναι 50λ. 55
62 Η διαδικασία που ακολουθήθηκε για τον πολλαπλασιασμό των μιτοχονδριακών τμημάτων COI και 16S rrna ήταν κατά βάση η ίδια με εκείνη της εργασίας των Klossa-Kilia et al. (2006), τροποποιημένη κατάλληλα ώστε να έχουμε το καλύτερο δυνατό αποτέλεσμα. Η διαδικασία για καθένα από τα γονίδια περιγράφεται στον ακόλουθο πίνακα (Β-6) Πίνακας B-6: Οι συνθήκες της PCR που χρησιμοποιήθηκαν στον Προγραμματιζόμενο Θερμικό Κυκλοποιητή (PTC) για τον πολλαπλασιασμό του κάθε γονιδίου. Γονίδιο Κύκλοι Χρόνος o C Στάδιο (Σχόλια) o C Χρόνος Κύκλοι Γονίδιο 1 3 min 94 ο C Denaturation (αρχικό στάδιο αποδιάταξης/προεπώαση) 94 ο C 3 min 1 30 sec 94 ο C Denaturation (στάδιο αποδιάταξης) 94 ο C 15 sec COI Χ 37 1 min 55 ο C 1 min 72 ο C Annealing (στάδιο υβριδισμού) Extension (στάδιο επιμήκυνσης) 55 ο C 30 sec 72 ο C 1 min X 35 16S rrna 1 10 min 72 ο C Extension (τελικό στάδιο επιμήκυνσης) 72 ο C 10 min 1 4 ο C End (στάδιο ολοκλήρωσης της αντίδρασης και διατήρησης των δειγμάτων) 4 ο C Τα προϊόντα της PCR ελέγχονταν ηλεκτροφορητικά όπως και στην περίπτωση του απομονωμένου ολικού DNA που περιγράφηκε προηγουμένως (βλ Γ.2.2.). Τα δείγματα-προϊόντα της PCR διατηρούνταν σε θερμοκρασία -20 o C. 56
63 Καθαρισμός του πολλαπλασιασμένου τμήματος των γονιδίων COI και 16S rrna Η διαδικασία καθαρισμού πραγματοποιήθηκε για την απομάκρυνση των κατάλοιπων ουσιών, όπως ένζυμα, απορρυπαντικά, άλατα ή ολιγονουκλεοτίδια που παραμένουν μέσα στο διάλυμα των νουκλεικών οξέων μετά το πέρας της απομόνωσής τους ή/και της PCR. Το τελικό προϊόν πρέπει να καθαρίζεται από τα κατάλοιπα αυτά, τα οποία μειώνουν τη συγκέντρωση του DNA στο διάλυμα και είναι πηγή θορύβου σε πολλές μεθόδους ανάλυσης του. Στην περίπτωση που ένα τμήμα γενετικού υλικού πολλαπλασιάζεται με σκοπό την ανάλυση της αλληλουχίας του, ο καθαρισμός του προϊόντος της PCR είναι απαραίτητος εξαιτίας της μεγάλης ευαισθησίας που έχουν οι αναλυτές αλληλουχίας στις προσμίξεις. Η αρχή της μεθόδου βασίζεται στη χρήση μεμβράνης με βασικό συστατικό το πυρίτιο, ή άλλων πηκτωμάτων που έχουν σχεδιαστεί ειδικά ώστε να προσδένουν και να συγκρατούν τα νουκλεϊκά οξέα. Η πρόσδεση των νουκλεϊκών οξέων στη μεμβράνη γίνεται παρουσία διαλύματος υψηλής αλατότητας ενώ τα κατάλοιπα που υπήρχαν στο αρχικό προϊόν διαπερνούν τη μεμβράνη και ξεπλένονται από το αρχικό διάλυμα. Τα νουκλεϊκά οξέα επαναδιαλύονται με διάλυμα Tris-HCl ή διπλά απεσταγμένο νερό. Στο τελικό διάλυμα παίρνουμε 90-95% της αρχικής ποσότητας νουκλεϊκών οξέων, μεγάλης καθαρότητας. Στην παρούσα μελέτη ο καθαρισμός πραγματοποιήθηκε με τη χρήση του kit Nucleospin Extract II της εταιρίας Macherey Nagel, βάσει του πρωτοκόλλου της εταιρίας. Η έκλουση του DNA (Εικ. B-5) γίνεται σε τελικό όγκο 20λ, και διατηρείται στους -20 C. 57
64 Φυγοκέντρηση Εικόνα Β-5: Σχηματική απεικόνιση της διαδικασίας καθαρισμού των προϊόντων της PCR Καθορισμός της αλληλουχίας νουκλεοτιδίων των επιλεγμένων τμημάτων των μιτοχονδριακών γονιδίων Η ανάγκη για τη γνώση της αλληλουχίας του DNA έγινε απαραίτητη για τη βασική βιολογική έρευνα. Έτσι αναπτύχθηκαν μέθοδοι, με τη βοήθεια των οποίων μπορεί κανείς να «διαβάσει» πλέον οποιαδήποτε αλληλουχία DNA. Η κλασική μέθοδος απαιτεί ένα μονόκλωνο μόριο DNA ως μήτρα, έναν DNA εκκινητή, μια DNA πολυμεράση, ραδιενεργά σημασμένα ή σημασμένα με χρωστική νουκλεοτίδια και τροποποιημένα νουκλεοτίδια τα οποία θα τερματίσουν την επιμήκυνση της αλυσίδας του DNA. Το δείγμα του DNA διαχωρίζεται σε τέσσερις ξεχωριστές αντιδράσεις αλληλούχισης (Εικ.B-6), στις οποίες περιλαμβάνονται τα τέσσερα βασικά dntps και η DNA πολυμεράση. 58

65 Εικόνα Β-6: Σχηματική απεικόνιση της μεθόδου Sanger. Απεικονίζονται τα συστατικά των τεσσάρων ξεχωριστών αντιδράσεων (A, B, C, D) και η τελική τους εμφάνιση στο πήκτωμα. Κάθε ζώνη αποτελεί τμήμα DNA διαφορετικού μεγέθους. Οι σχετικές θέσεις των διαφορετικών ζωνών μεταξύ των τεσσάρων γραμμών, χρησιμοποιούνται για το διάβασμα της αλληλουχίας με φορά από κάτω προς τα πάνω. Η αλληλούχιση των δειγμάτων στη συγκεκριμένη εργασία πραγματοποιήθηκε από την εταιρεία VBC Biotech με έδρα τη Βιέννη. Μετά από αποστολή των δειγμάτων, καθώς και μιας ποσότητας του ενός εκ των δύο εκκινητών που χρησιμοποιήθηκαν για την PCR, η εταιρεία πραγματοποιεί την αλληλούχιση χρησιμοποιώντας το μοντέλο ABI Prism 3100 Capillary Sequencer, ενώ τα αποτελέσματα αποστέλλονται μέσω ηλεκτρονικού ταχυδρομείου. 59

66 4. ΣΤΑΤΙΣΤΙΚΗ ΕΠΕΞΕΡΓΑΣΙΑ ΤΩΝ ΔΕΔΟΜΕΝΩΝ THΣ ΑΝΑΛΥΣΗΣ ΑΛΛΗΛΟΥΧΙΑΣ 4.1. ΣΤΟΙΧΙΣΗ ΑΛΛΗΛΟΥΧΙΩΝ Η στοίχιση αποσκοπεί στον προσδιορισμό των ομόλογων θέσεων των αλληλουχιών, αφού βάσει αυτών των διαφορών υπολογίζονται οι εξελικτικές σχέσεις των υπό μελέτη αλληλουχιών. Συνεπώς, η στοίχιση των αλληλουχιών (Εικ.Β-7) είναι μια διαδικασία ζωτικής σημασίας για την εύρεση των πραγματικών φυλογενετικών σχέσεων των υπό εξέταση ταξινομικών μονάδων (Gatesy et al. 1993, Collins et al. 1994a, Wheeler et al. 1995, Baker & DeSalle 1997, Simmons & Ochoterenam 2000). Εικόνα Β-7: Απεικόνιση της διαδικασίας στοίχισης των αλληλουχιών στο πρόγραμμα BioEdit Sequence Alignment Editor, version (Hall 1999). Τα τμήματα των δύο μιτοχονδριακών γονιδίων (COI και 16S rrna) τα οποία πολλαπλασιάστηκαν για κάθε πειραματόζωο της παρούσας μελέτης δεν είχε τον ίδιο αριθμό βάσεων σε όλα τα άτομα. Το γεγονός αυτό οφείλεται σε παράγοντες όπως η μικρή συγκέντρωση του DNA σε ορισμένα δείγματα που δυσχεραίνει την ανάγνωση των βάσεων κατά την εφαρμογή της μεθόδου των Sanger et al. (1977), και η 60
67 δέσμευση των ολιγονουκλεοτιδίων εκκινητών σε συμπληρωματικές αλληλουχίες που βρίσκονται σε διαφορετικές θέσεις κατά μήκος του τμήματος της περιοχής ελέγχου. Ενθέσεις ή ελλείψεις βάσεων μεταβάλλουν το μήκος των τμημάτων που μελετούνται. Για την αποκάλυψη αυτών των πολυμορφισμών οι αλληλουχίες στοιχίζονται. H στοίχιση των αλληλουχιών που εξετάστηκαν στην παρούσα μελέτη, για τα άτομαtrachelipus και τα άτομα της εξωομάδας, έγινε με το πρόγραμμα CLUSTAL W [version 1.4 (Thompson et al. 1994)], που περιλαμβάνεται στο πρόγραμμα BioEdit Sequence Alignment Editor, version (Hall 1999). Η διαδικασία της στοίχισης γίνεται αρχικά με βάση την αντιστοιχία όσο το δυνατόν περισσότερων ομόλογων περιοχών και εν συνεχεία γίνεται και η στοίχιση των νουκλεοτιδικών θέσεων που δεν χαρακτηρίζονται από την ίδια αλληλουχία βάσεων. Η διαδικασία που αφορά στη στοίχιση τμημάτων DNA περισσότερων από δύο δειγμάτων, ονομάζεται πολλαπλή στοίχιση (multiple alignment) και γίνεται προοδευτικά, αρχίζοντας με την στοίχιση ανά ζεύγος αλληλουχιών. Αρχικά ομαδοποιούνται οι αλληλουχίες που μοιράζονται τις περισσότερες ομολογίες και σταδιακά προστίθενται όσες είναι περισσότερο διαφοροποιημένες. Σε κάθε στάδιο εξετάζεται η ορθότητα της στοίχισης μέσω της σύγκρισης πινάκων που δημιουργεί το πρόγραμμα και καταγράφουν τον αριθμό των νουκλεοτιδίων που ταίριαξαν, όσων διαφοροποιήθηκαν και το βαθμό των ενθέσεων και ελλείψεων που παρουσιάζει η στοίχιση. Η διαδικασία συνεχίζεται μέχρι να μεγιστοποιηθεί ο αριθμός των νουκλεοτιδίων που είναι ομόλογα και ταυτόχρονα να ελαχιστοποιηθούν οι τιμές των άλλων τριών παραμέτρων (Feng & Doolittle 1987, Thompson et al. 1994). 61
68 4.2. ΕΛΕΓΧΟΣ ΔΕΔΟΜΕΝΩΝ ΓΙΑ ΟΜΟΠΛΑΣΙΑ ΚΑΙ ΚΟΡΕΣΜΟ Ο αριθμός των αλλαγών σε μια νουκλεοτιδική θέση είναι πεπερασμένος. Όσο μεγαλύτερη είναι η γενετική απόσταση δύο τάξων (όσο πιο απομακρυσμένα εξελικτικά είναι δύο είδη) τόσο μεγαλύτερη είναι και η πιθανότητα να συμβεί αλλαγή σε μια θέση στην οποία είχε ήδη συμβεί μια αλλαγή. Όταν συμβεί κάτι τέτοιο, τότε υπάρχει κορεσμός των νουκλεοτιδικών υποκαταστάσεων (substitution saturation), οπότε σταματά να ισχύει η γραμμική σχέση του χρόνου και της απόστασης δύο αλληλουχιών, με αποτέλεσμα να αλλοιώνεται ποιοτικά η φυλογενετική πληροφορία που θα έδιναν οι αλληλουχίες (Tamura & Nei 1993, Page & Holmes 1998). Ένας από τους τρόπους με τους οποίους μπορεί να ελεγχθεί η ομοπλασία των δεδομένων, είναι ο έλεγχος του Xia που χρησιμοποιεί το δείκτη κορεσμού υποκατάστασης (Xia et al. 2003) και παρατίθεται στο πρόγραμμα Dambee (Xia & Xie 2001). Η μεταβλητότητα σε κάθε νουκλεοτιδική θέση i μετράται από την εντροπία με την εξίσωση: 4 p j log 2 p j όπου, j 1 j= 1,2,3,4 αντιστοιχούν στα νουκλεοτίδια A, T, G και C αντίστοιχα και p j = η αναλογία του νουκλεοτιδίου j στη θέση i. Αν όλα τα νουκλεοτίδια στη θέση i είναι ίδια τότε προφανώς H i = 0. Οι υποκαταστάσεις οδηγούν σε πολυμορφικές θέσεις στις οποίες η τιμή H θα είναι μεγαλύτερη του μηδενός. Ο δείκτης των Xia et al. (2003) αποδίδεται από τη σχέση 62
69 I H / όπου, SS H FSS H = η παρατηρούμενη τιμή της εντροπίας H FSS = η τιμή της εντροπίας για πλήρη κορεσμό υποκαταστάσεων Σημειώνεται ότι FSS = Full substitution saturation (πλήρης κορεσμός υποκαταστάσεων) Θεωρητικά, ο έλεγχος του κορεσμού υποκατάστασης μπορεί να πραγματοποιηθεί απλά ελέγχοντας αν η παρατηρούμενη τιμή του H είναι στατιστικώς σημαντικά μικρότερη από την τιμή H FSS. Αν η H δεν είναι στατιστικώς σημαντικά μικρότερη από την H FSS, τότε οι αλληλουχίες παρουσιάζουν κορεσμό. Αυτός ο συλλογισμός όμως είναι καθαρά θεωρητικός καθώς οι αλληλουχίες αποτυγχάνουν να αποδώσουν την πραγματική φυλογένεια πολύ πριν φτάσουν στον πλήρη κορεσμό οπότε και θα ισχύει I 1 δηλαδή H = H FSS. Γι αυτό SS το λόγο προσδιορίζεται η κρίσιμη τιμή I SS. C του δείκτη I SS, στην οποία οι αλληλουχίες ξεκινούν και αποτυγχάνουν να ανασυστήσουν στο πραγματικό φυλογενετικό δέντρο. Όταν η τιμή του I SS. C είναι γνωστή, μπορεί εύκολα να υπολογιστεί και η I SS και να γίνει η μεταξύ τους σύγκριση. Στο πρόγραμμα Dambee (Xia & Xie 2001) η σύγκριση των δύο τιμών πραγματοποιείται με two tailed t test. Αν η I SS δεν είναι μικρότερη (στατιστικώς σημαντικά) τότε συμπεραίνεται πως οι αλληλουχίες είναι κορεσμένες και δεν θα πρέπει να χρησιμοποιηθούν για φυλογενετικές αναλύσεις. 63
70 4.3. ΕΠΕΞΕΡΓΑΣΙΑ ΤΩΝ ΣΤΟΙΧΙΣΜΕΝΩΝ ΑΛΛΗΛΟΥΧΙΩΝ Με το πρόγραμμα BioEdit, version (Hall 1999), εκτός από τη στοίχιση των αλληλουχιών, γίνονται εκτιμήσεις σχετικά με τις συντηρητικές περιοχές των αλληλουχιών, όπως για τον αριθμό τους, το μήκος τους και την αλληλουχία τους. Για τις στοιχισμένες αλληλουχίες κατασκευάστηκε το διάγραμμα εντροπίας (entropy diagram) το οποίο απεικονίζει την πιθανότητα που έχει μια αλληλουχία να φέρει σε συγκεκριμένη θέση της μια συγκεκριμένη νουκλεοτιδική βάση, η οποία είναι η βάση που φέρουν στη θέση αυτή οι περισσότερες από τις αλληλουχίες που εξετάζονται (Hall 1999). Ο προσδιορισμός των πολυμορφισμών που εμφανίζονται μεταξύ των εξεταζόμενων αλληλουχιών έγινε με το πρόγραμμα Mega v4.0 (Kumar et al. 2004). Mε το πρόγραμμα αυτό υπολογίστηκε η νουκλεοτιδική σύσταση (nucleotide composition), δηλαδή τα σχετικά ποσοστά εμφάνισης καθεμιάς από τις τέσσερις βάσεις στο σύνολο των αλληλουχιών, καθώς και η ενδοπληθυσμιακή, η διαπληθυσμιακή και καθαρή διαπληθυσμιακή νουκλεοτιδική απόκλιση (intrapopulational, interpopulational και net interpopulational nucleoside diversity, αντίστοιχα - βλ παρακάτω). Με το ίδιο πρόγραμμα έγινε και ο υπολογισμός των βάσεων που διαφοροποιούνταν και προσδιορίστηκαν και οι πληροφοριακές θέσεις φειδωλότητας σε κάθε σύνολο αλληλουχιών που αναλυόταν κάθε φορά. Τέλος η νουκλεοτιδική ποικιλότητα (nucleotide diversity, π), καθώς και η απλοτυπική ποικιλότητα (haplotype diversity, h) υπολογίστηκαν από το στατιστικό πρόγραμμα DnaSP v4.10 (Rozas et al. 2003). 64
71 Εκτίμηση γενετικών αποστάσεων Η απλούστερη μέτρηση της απόστασης δύο νουκλεοτιδικών αλληλουχιών είναι η καταμέτρηση του αριθμού των νουκλεοτιδικών θέσεων στις οποίες οι δύο αυτές αλληλουχίες διαφέρουν (απόσταση d), η οποία εκφράζεται ως ποσοστό p των νουκλεοτιδικών θέσεων στις οποίες οι δύο αλληλουχίες είναι διαφορετικές. Όμως στις περιπτώσεις που ο εξελικτικός ρυθμός είναι μεγάλος η απόσταση d υποεκτιμά ουσιαστικά την απόσταση μεταξύ των δύο αλληλουχιών (Page & Holmes 1998). Αυτό οφείλεται στο γεγονός ότι όσο αυξάνεται ο βαθμός διαφοροποίησης μεταξύ των αλληλουχιών τόσο αυξάνει και η πιθανότητα η ίδια νουκλεοτιδική θέση να έχει υποστεί πολλαπλές μεταλλαγές και να έχει επανέλθει στην αρχική της κατάσταση. Με αυτόν τον τρόπο έχει μείνει κρυφός ο πραγματικός αριθμός αλλαγών στην συγκεκριμένη νουκλεοτιδική θέση άρα και o πραγματικός βαθμός διαφοροποίησης της εν λόγω αλληλουχίας (Kumar et al. 2004). Αυτό οδήγησε στο να αναπτυχθούν μέθοδοι γνωστές ως «τεχνικές διόρθωσης» των αποστάσεων, που έχουν ως στόχο να ανάγουν τις παρατηρούμενες αποστάσεις σε πραγματικούς εκτιμητές των εξελικτικών αλλαγών. Αυτό πραγματοποιείται με την εκτίμηση του ποσοστού της γενετικής διαφοροποίησης που έχει καλυφθεί λόγω πολλαπλών αλλαγών σε μια νουκλεοτιδική θέση. Ο υπολογισμός των νουκλεοτιδικών αποστάσεων έγινε με βάση το μοντέλο 2 παραμέτρων του Kimura (1980). Σύμφωνα μ αυτό, οι συχνότητες υποκατάστασης των μεταπτώσεων διαφέρουν από εκείνες των μεταστροφών, γεγονός που ισχύει στις αλληλουχίες του mtdna των ζώων. Οι νουκλεοτιδικές αποστάσεις υπολογίζονται από την εξίσωση: 65
72 d log 1 2P Q 1 2Q 2 log e e όπου, 4 P=n s /n και Q= n v /n, με n s και n v τον αριθμό των διαφορών μεταστροφών και μεταπτώσεων μεταξύ δυο αλληλουχιών. Για τον υπολογισμό των γενετικών αποστάσεων χρησιμοποιήθηκε το πρόγραμμα MEGA version 4 (Kumar et al. 2004) Επιλογή του καταλληλότερου μοντέλου νουκλεοτιδικής υποκατάστασης Το αποτέλεσμα των διαφόρων φυλογενετικών μεθόδων που χρησιμοποιούνται, προκειμένου να αποκωδικοποιηθούν οι σχέσεις μεταξύ των μελετώμενων κάθε φορά τάξων, εξαρτάται άμεσα από το μοντέλο που αυτές οι μέθοδοι χρησιμοποιούν για την εκτίμηση των εξελικτικών αποστάσεων μεταξύ των υπό μελέτη αλληλουχιών. Αυτό συμβαίνει καθώς κάθε φορά αυτές οι μέθοδοι κάνουν και διαφορετικές παραδοχές σε ό,τι αφορά τις νουκλεοτιδικές υποκαταστάσεις των αλληλουχιών (Felsestein 1988a, 1988b). Για να υπάρχει λοιπόν αξιοπιστία στα συμπεράσματα των φυλογενετικών αναλύσεων, είναι αναγκαία πρωτίστως και η αξιοπιστία του μοντέλου νουκλεοτιδικής υποκατάστασης (Goldman 1993). Γι αυτό το λόγο οι φυλογενετικές μέθοδοι που βασίζονται σε μοντέλα νουκλεοτιδικής υποκατάστασης θα πρέπει να εξεταστούν προκειμένου να επιλεγεί το μοντέλο που ταιριάζει καλύτερα στο σύνολο δεδομένων που αναλύεται κάθε φορά ώστε και να δικαιολογείται και η χρήση του (Huelsenbeck & Crandall 1997, Posada & Crandall 1998,2001). Στην παρούσα μελέτη για την επιλογή του καταλληλότερου μοντέλου νουκλεοτιδικής υποκατάστασης εφαρμόστηκε η δοκιμασία του λόγου πιθοφανειών 66
73 (Likelihood Rαtio Test LTR) και το κριτήριο του Akaike - AIC (Akaike Information Criterion, Akaike 1974) με τη βοήθεια του υπολογιστικού προγράμματος MODELTEST V3.7. (Posada & Crandall 1998) και PAUP (v4.0.beta10, Swofford 2002), το οποίο έχει τη δυνατότητα να επιλέξει ανάμεσα σε 56 διαφορετικά μοντέλα πιο είναι το βέλτιστο αναφορικά με τις αλληλουχίες που αναλύονται. Μεταξύ των στατιστικών ελέγχων που έχουν προταθεί για την επιλογή του καλύτερου μοντέλου νουκλεοτιδικής υποκατάστασης, το κριτήριο του Akaike (1974, Akaike Information Criterion AIC) θεωρείται το πλέον αξιόπιστο καθώς υπολογίζει την πιθανότητα των προτεινόμενων μοντέλων επιβάλλοντας αρνητική βαθμολόγηση βασισμένη στον αριθμό των παραμέτρων που χρησιμοποιήθηκαν από τα ίδια τα μοντέλα (Bos & Posada 2004, Bozdogan 2000, Akaike 1981) ΑΝΑΛΥΣΗ ΦΑΙΝΕΤΙΚΩΝ ΚΑΙ ΦΥΛΟΓΕΝΕΤΙΚΩΝ ΣΧΕΣΕΩΝ Οι νουκλεοτιδικές αποκλίσεις των αλληλουχιών των ατόμων των μελετηθέντων πληθυσμών χρησιμοποιήθηκαν για την κατασκευή δεντρογραμμάτων που απεικονίζουν τις σχέσεις συγγένειας των πληθυσμών αυτών. Οι μέθοδοι που χρησιμοποιήθηκαν για την κατασκευή των δεντρογραμμάτων ήταν: α) μέθοδοι φαινετικής ανάλυσης που στηρίζονται στην ομαδοποίηση ανάλογα με το βαθμό της γενετικής απόστασης, και β) μέθοδοι κλαδιστικής ανάλυσης που στηρίζονται στην ομαδοποίηση με βάση τη φυλογενετική συγγένεια. Η πρώτη κατηγορία μεθόδων βασίζεται σε έναν απλό αλγόριθμο που προκύπτει από πίνακες γενετικών αποστάσεων μεταξύ των αλληλουχιών. Αυτός ο αλγόριθμος είναι που οδηγεί στον καθορισμό του δέντρου. Οι απλές αλγοριθμικές μέθοδοι υπολογιστικά είναι πολύ ταχύτερες διότι προχωρούν άμεσα στην τελική 67
74 λύση, χωρίς να καθυστερούν συγκρίνοντας εναλλακτικά ανταγωνιστικά σενάρια. Ωστόσο δε θεωρούνται πολύ αξιόπιστες, καθώς χάνεται μεγάλο ποσό εξελικτικής πληροφορίας κατά τον υπολογισμό των γενετικών αποστάσεων Στην κατηγορία αυτή ανήκουν η μέθοδος της UPGMA (Sneath & Sokal 1973), της Neighbor-Joining (Saitou & Nei 1986, 1987) κ.α. Στις μεθόδους κλαδιστικής ανάλυσης, η διαδικασία που ακολουθείται χαρακτηρίζεται από δυο βήματα. Αρχικά ορίζεται ένα κριτήριο βελτιστοποίησης (optimality criterion) που θα χρησιμοποιηθεί για την αποτίμηση ενός συγκεκριμένου δένδρου. Συνήθως, αφού οριστεί το κριτήριο, σε κάθε δένδρο αποδίδεται μια τιμή (score) και πλέον τα εναλλακτικά δένδρα συγκρίνονται ως προς αυτή την τιμή. Στο δεύτερο βήμα χρησιμοποιείται συγκεκριμένος αλγόριθμος για τον υπολογισμό της τιμής του κριτηρίου και για την ανεύρεση των δένδρων που έχουν τη βέλτιστη τιμή για αυτό το κριτήριο. Έτσι αναζητούνται ανάμεσα σε όλα τα πιθανά δέντρα, εκείνα με τη μέγιστη τιμή του κριτηρίου βελτιστοποίησης. Σε αυτή την κατηγορία μεθόδων εντάσσεται η μέθοδος της μέγιστης φειδωλότητας (Maximum Parsimony) η Μέγιστη Πιθανοφάνεια (maximum likelihood) και η Μπεϊεσιανή Συμπερασματολογία (Bayesian inference) Μέθοδος Σύνδεσης Γειτόνων (Neighbor-Joining) Η μέθοδος αυτή ανήκει στις φαινετικές μεθόδους. Χρησιμοποιεί τον αλγόριθμο των Saitou & Nei (1987) για να υπολογίσει το βραχύτερο εξελικτικά δένδρο. Τα δεδομένα που χρησιμοποιεί αυτή η μέθοδος είναι ένας πίνακας γενετικών αποστάσεων και το αρχικό δένδρο που δημιουργεί είναι ένα ακτινωτό δέντρο (star tree) στο οποίο όλα τα εσωτερικά κλαδιά έχουν μηδενικό μήκος. Το δέντρο κατασκευάζεται με την ένωση των κόμβων με τη μικρότερη απόσταση (ο υπολογισμός γίνεται μεταξύ όλων των δυνατών ζευγών) σε αυτόν τον τροποποιημένο πίνακα. Όταν βρει τις αλληλουχίες με τη μικρότερη απόσταση τις 68
75 ενώνει και πλέον τις αντιμετωπίζει ως μια αλληλουχία. Στη συνέχεια εξετάζει ποια είναι η αλληλουχία που είναι κοντινότερη προς το προηγούμενο ζεύγος για να την ενώσει με αυτές κατά τον ίδιο τρόπο. Η διαδικασία ολοκληρώνεται όταν απομένουν δύο κόμβοι, οι οποίοι διαχωρίζονται από έναν κλάδο (Saitou & Nei 1987) Η μέθοδος της μέγιστης φειδωλότητας (Maximum Parsimony) Η εν λόγω μέθοδος πραγματοποιεί την ανασυγκρότηση των δέντρων χρησιμοποιώντας νουκλεοτιδικές αλληλουχίες και αξιοποιώντας μόνο εκείνες τις νουκλεοτιδικές θέσεις που διαφοροποιούνται μεταξύ των αλληλουχιών. Αυτές οι θέσεις χαρακτηρίζονται ως πληροφοριακές θέσεις φειδωλότητας (parsimony informative sites). Όσο πιο πολλές είναι αυτές οι θέσεις τόσο πιο αξιόπιστο αναμένεται να είναι το αντίστοιχο δένδρο που θα προκύψει. Ωστόσο, όταν ο βαθμός ομοπλασίας (παράλληλες και οπισθοδρομίζουσες αντικαταστάσεις) είναι μεγάλος, αυτό αναιρείται. Η μέθοδος υπολογίζει τον ελάχιστο αριθμό νουκλεοτιδικών υποκαταστάσεων που απαιτούνται προκειμένου να εξηγηθεί το παρατηρούμενο πρότυπο στην τοπολογία του δέντρου. Ο αριθμός των αλλαγών μεταξύ των θέσεων αθροίζεται και υπολογίζεται μια τιμή (τιμή φειδωλότητας) για κάθε τοπολογία δέντρου. Η τοπολογία που φέρει την ελάχιστη τιμή (δηλαδή φέρει τις λιγότερες διαφορές) είναι και ο εκτιμητής της φυλογένειας γνωστή και ως το πλέον φειδωλό δέντρο (Felsenstein, 1978, Collins et al. 1994b, Edwards 1996). Η μέθοδος στηρίζεται στη φιλοσοφική ιδέα που διατυπώθηκε από τον William of Occam πως «δοθείσες δύο πιθανές εξηγήσεις των δεδομένων, με όλους τους άλλους παράγοντες ίδιους, η πιο απλή εξήγηση είναι προτιμητέα» (Blumer et al. 1987). Η μέθοδος της μέγιστη φειδωλότητας, είχε αρχικά προταθεί από τους Edwards and Cavalli-Sforza (1965) και Cavalli-Sforza & Edwards (1967) για ανάλυση δεδομένων γονιδιακών συχνοτήτων και από τους Camin και Sokal (1965) για την ανάλυση μορφολογικών χαρακτήρων. Η εφαρμογή της μεθόδου για μοριακά 69
76 δεδομένα αποδίδεται στους Zuckerkandl and Pauling (1965) ενώ τον αλγόριθμο της μεθόδου ανέπτυξαν οι Fitch (1971) και Hartigan (1973). Υπήρξαν πολλές αντικρουόμενες απόψεις αναφορικά με τη μεθοδολογία που ακολουθεί η μέθοδος (Farris 1973, Felsenstein 1988a, 1988b), ως προς το ότι δεν κάνει σαφείς υποθέσεις σχετικά με το μηχανισμό εξέλιξης που ακολουθεί. Έτσι η τοπολογία του πιο φειδωλού δένδρου ενδέχεται να είναι λανθασμένη όταν ο εξελικτικός ρυθμός διαφέρει από κλάδο σε κλάδο (Felsenstein 1978) ακόμα και στην περίπτωση που μελετώνται αλληλουχίες πολύ μεγάλου μήκους. Από την άλλη είχε προταθεί ότι η μέθοδος θα ήταν προτιμότερο να μην κάνει καμία υπόθεση σχετικά με το ρυθμό εξέλιξης των αλληλουχιών και να αφήνεται «ελεύθερη» να υπολογίσει το φειδωλότερο δέντρο με το δεδομένα των αλληλουχιών που υπόκεινται σε ανάλυση. (Yang 1996). Στην παρούσα μελέτη η κατασκευή του δέντρου με τη μέθοδο της μέγιστης φειδωλότητας πραγματοποιήθηκε με τη βοήθεια του προγράμματος PAUP* v4.0 beta10 (Swofford D.L. 2002) Μπεϊεσιανή συμπερασματολογία Το όνομα της μεθόδου προέρχεται από το στατιστικό του 18 ου αιώνα Thomas Bayes ( ) που εισήγαγε το Μπεϊεσιανό θεώρημα στο οποίο και βασίζονται κάποιες από τις βασικές έννοιες της μεθόδου. Η Μπεϊεσιανή συμπερασματολογία είναι μια στατιστική συμπερασματολογία στην οποία στοιχεία ή παρατηρήσεις χρησιμοποιούνται για την ανανέωση ή για την εκ νέου εξαγωγή συμπερασμάτων σχετικά με το τι είναι ήδη γνωστό για βασικές παραμέτρους ή υποθέσεις (Dempster 1968). Η Μπεϊεσιανή συμπερασματολογία είναι μια προσέγγιση που χρησιμοποιείται στη φυλογένεση προκειμένου να απαντήσει στο ερώτημα: «Με δεδομένη μια μήτρα 70
77 αποστάσεων και ένα μοντέλο εξελικτικών μεταβολών ποια είναι η πιθανότητα να είναι σωστό ένα δέντρο;» (Rubin 1978, Wade 2000, Huelsenbeck et al. 2002). Η Μπεϊεσιανή πιθανότητα ερμηνεύει την έννοια της πιθανότητας ως ένα «μέτρο της κατάστασης της γνώσης» και όχι ως τη συχνότητα ή μια άλλη φυσική ιδιότητα ενός μελετώμενου συστήματος. Πρόκειται για μια από τις πλέον δημοφιλείς ερμηνείες της έννοιας της πιθανότητας. Η εν λόγω ερμηνεία μπορεί να θεωρηθεί μια επέκταση της λογικής σκέψης που υποστηρίζει την επιχειρηματολογία στηριζόμενη σε αβέβαιους ισχυρισμούς. Προκειμένου να εκτιμηθεί η πιθανότητα μιας υπόθεσης, η μέθοδος προσδιορίζει μια εκ των προτέρων πιθανότητα, που ολοένα και ανανεώνεται με την εισροή νέων δεδομένων. Περιλαμβάνει τη συλλογή αποδείξεων, οι οποίες είναι είτε σύμφωνες είτε ασύμφωνες με μια δεδομένη υπόθεση. Καθώς συγκεντρώνονται οι αποδείξεις, ο βαθμός στον οποίο πιστεύουμε στην αρχική υπόθεση αλλάζει. Με αρκετές αποδείξεις αυτή η αρχική υπόθεση θα πρέπει είτε να μειωθεί αρκετά είτε να αυξηθεί (Huelsenbeck & Ronquist 2001, Ronquist & Huelsenbeck 2003, Nylander 2004). Η μέθοδος αξιολογεί δέντρα ξεκινώντας από κάποιες τυχαίες τοπολογίες τις οποίες και βελτιστοποιεί σταδιακά, και επιλέγει τις πλέον πιθανές, δηλαδή εκείνες που δεν επιδέχονται περαιτέρω βελτίωση. Θεωρείται μια ιδιαιτέρως αξιόπιστη μέθοδος για τα μοριακά δεδομένα. Το μοντέλο των μεταβολών απαιτεί προσδιορισμό της σχετικής συχνότητας των μεταστροφών και των μεταπτώσεων και έναν εξελικτικό ρυθμό. Πρόκειται για μια γρήγορη μέθοδο που παρέχει ταυτόχρονα και τη στατιστική υποστήριξη των κλάδων στο δέντρο. Η εφαρμογή της σε μορφολογικούς χαρακτήρες δεν έχει μελετηθεί ικανοποιητικά (Lewis 2001). Η μέθοδος χρησιμοποιεί μια σημαντική αρχή η οποία ονομάζεται εκ των υστέρων πιθανότητα (posterior probability) και στηρίζεται στο θεώρημα του Bayes (Bayes & Price 1763), σύμφωνα με το οποίο : 71
78 P( D H ) P( H ) P( H D) P( D) όπου, Η = η υπόθεση D = τα δεδομένα P(H) = η εκ των προτέρων πιθανότητα του Η, δηλ. η πιθανότητα το Η να είναι αληθής προτού αναλυθούν τα δεδομένα P(D) = η οριακή πιθανότητα του D P(D/H) = η υπό όρους πιθανότητα, δηλ η πιθανότητα επιβεβαίωσης του D υπό την προϋπόθεση ότι η H είναι αληθής. P(H/D) = η εκ των υστέρων πιθανότητα, δηλ η πιθανότητα ότι η Η είναι αληθής δεδομένης τόσο της προηγούμενης εκτίμησης για την υπόθεση όσο και των υπό ανάλυση δεδομένων (Huelsenbeck & Ronquist 2001). Η Μπεϊεσιανή Συμπερασματολογία χρησιμοποιεί τον αλγόριθμο που είναι γνωστός ως Αλυσίδα Μαρκόφ Μόντε Κάρλο (Markov Chain Monte Carlo MCMC). Ο αλγόριθμος χρησιμοποιεί την τυχαιότητα προκειμένου να βρει τυχαίους εκτιμητές των παραμέτρων, όπως γίνεται με τα τυχερά παιχνίδια στα καζίνο, από όπου πήρε και το όνομά του. Καθορίζει μια περιοχή στην οποία κάνει δειγματοληψίες ομοιογενώς και καθώς κάθε δείγμα λαμβάνεται με την ίδια πιθανότητα, το ολοκλήρωμα είναι ο μέσος όρος των δειγμάτων. Το κάθε δείγμα λαμβάνεται ανεξάρτητα. Ο αλγόριθμος παράγει στατιστικά συνεπή δείγματα από την κατανομήστόχο χωρίς να χρησιμοποιεί άλλες κατανομές και χωρίς να απορρίπτει αυθαίρετα δείγματα. Το αποτέλεσμα υπολογίζεται με δειγματοληψίες που χρησιμοποιούνται από περιοχές με υψηλότερη πυκνότητα δεδομένων. Η διαδικασία ξεκινά επιλέγοντας ένα τυχαίο σημείο εκκίνησης. Καθώς οι νέες τιμές γίνονται αποδεκτές από την ανάλυση, τα δείγματα που προκύπτουν γίνονται ανεξάρτητα του αρχικού σημείου εκκίνησης. Προκειμένου να αποφευχθεί η όποια μεροληψία που θα οφείλεται στο σημείο 72
79 εκκίνησης, μετά το πέρας της ανάλυσης, ένας συγκεκριμένος αριθμός των αρχικών επαναλήψεων απορρίπτεται με τη διαδικασία burnin (Larget & Simon 1999, Huelsenbeck & Ronquist 2001, Huelsenbeck et al. 2002). Ο αριθμός των επαναλήψεων που απορρίπτονται υπολογίζεται ως το 25% των δειγμάτων που τελικά έχουν συλλεγεί. Αν είχε επιλεγεί η δειγματοληψία των δειγμάτων (δέντρων) να πραγματοποιείται κάθε 100 γενιές σε σύνολο π.χ γενεών, τότε στο τέλος προκύπτουν δέντρα. Οπότε με τη διαδικασία burnin θα απορριφθούν οι επαναλήψεις (δέντρα) (Ronquist et al MrBayes 3.1 Manual, ΑΝΕΞΑΡΤΗΤΗ Η ΣΥΝΔΥΑΣΜΕΝΗ ΑΝΑΛΥΣΗ Στις φυλογενετικές αναλύσεις εν γένει, η ασυμφωνία μεταξύ των δεδομένων ή ακόμα και μεταξύ τμημάτων των δεδομένων μπορούν να εξηγηθούν είτε από πραγματικές διαφορές στην εξελικτική ιστορία αλλά και από παραπλανητικά φυλογενετικά σήματα (θόρυβος ή ομοπλασία). Αυτά τα «λανθασμένα» σήματα μπορεί να υπερκεράσουν το έγκυρο φυλογενετικό σήμα. Κάτι τέτοιο ενδέχεται να είναι αποτέλεσμα ανεπαρκούς δειγματοληψίας, πειραματικού λάθους ή της επίδρασης του διαχωρισμού των δεδομένων σε υποσύνολα. Η ανίχνευση λοιπόν σημαντικής ασυμφωνίας μεταξύ των δεδομένων υπό ανάλυση είναι ένα πολύ σημαντικό βήμα στις φυλογενετικές αναλύσεις με δεδομένα γενωμικού DNA (Planet & Sarkar 2005) Όταν λοιπόν έχουμε πολλαπλά σύνολα δεδομένων, όπως στην παρούσα μελέτη αλληλουχίες COI και 16S, ανακύπτει το ερώτημα αν αυτά τα δεδομένα μπορούν να συνδυαστούν σε μια ενιαία ανάλυση ή αν πρέπει τα δεδομένα αυτά να αναλυθούν μόνο ανεξάρτητα μεταξύ τους. Οι απόψεις των ερευνητών ως προς αυτό το ζήτημα διίστανται. Υπάρχει η «προσέγγιση όλων των δεδομένων» (total evidence approach) του Kluge (1989), ο οποίος υποστηρίζει ότι: 73
80 τα δεδομένα πρέπει πάντοτε να συνδυάζονται δεν υπάρχουν «κακά» δεδομένα και το συνδυαστικό σήμα που λαμβάνεται αποκαλύπτει λύσεις και σχέσεις που τα ανεξάρτητα δεδομένα δεν μπορούν μόνα τους να αποδώσουν. Άλλοι ερευνητές όμως υποστηρίζουν πως αυτή η διαδικασία μπορεί να οδηγήσει σε λανθασμένες τοπολογίες (Bull et al. 1993), και προτείνουν πως τα δεδομένα πρέπει να αναλύονται ανεξάρτητα και τα παραγόμενα δέντρα να συνδυάζονται σε συναινετικά δέντρα (consensus trees, (Miyamoto and Fitch 1995). Μια τρίτη προσέγγιση είναι αυτή του «συνδυασμού υπό προϋποθέσεις» (conditional combination, Bull et al. 1993, Huelsenbeck et al. 1996a, 1996b, Cunningham (1997b)). Η προσέγγιση αυτή προτείνει ότι: τα δεδομένα πρέπει να εξετάζονται ως προς τη συμφωνία μεταξύ τους να συνδυάζονται εάν συμφωνούν στατιστικώς σημαντικά η μικρή ασυμφωνία δεν αποτελεί σημαντικό πρόβλημα και η έντονη ασυμφωνία των δεδομένων χρήζει περαιτέρω διερεύνησης Σήμερα υπάρχει μια πληθώρα στατιστικών ελέγχων που έχουν αναπτυχθεί προκειμένου να ανιχνεύουν ασυμφωνίες φυλογενετικών χαρακτήρων που είναι «εξελικτικά» σημαντικές. Ένας από τους πλέον χρησιμοποιούμενους τέτοιους ελέγχους είναι αυτός της «ασυμφωνίας διαφοράς μήκους» των μελετώμενων αλληλουχιών (Incongruence Length Difference ILD, Μickevich & Farris 1981, Farris et al. 1994, Barker & Lutzoni 2002) ο οποίος περιγράφεται από τη σχέση: ILD L C i n 1 L i όπου L C = ο αριθμός των βημάτων στο πιο φειδωλό δέντρο που προκύπτει όταν όλα τα υποσύνολα των δεδομένων συνδυάζονται σε μια ενιαία ανάλυση. L i = ο αριθμός των βημάτων του πιο φειδωλού δέντρου που βρέθηκε στο υποσύνολο δεδομένων i μεταξύ του n αριθμού υποσυνόλων. 74
81 Στο δ ILD αποδίδεται μια τιμή p συγκρίνοντάς το με την κατανομή τυχαίων τιμών δ ILD που υπολογίζονται από υποσύνολα -ίσα σε μέγεθος με τα πραγματικά τα οποία προκύπτουν από τυχαία επαναλαμβανόμενη δειγματοληψία χαρακτήρων (δηλ, στήλες ενός φυλογενετικού πίνακα) μεταξύ όλων των υποσυνόλων. Ο ILD καθορίζει αν ο βαθμός της ασυμφωνίας μεταξύ των δεδομένων υπάρχει απλώς και μόνο εξαιτίας της διαίρεσής αυτών σε υποσύνολα για ανεξάρτητη ανάλυση, καταδεικνύοντας πότε αυτή η ασυμφωνία δεν είναι στατιστικά σημαντικότερη από αυτή που παρατηρείται μέσα σε κάθε μεμονωμένο σύνολο δεδομένων. Ο Cunningham (1997a) δοκίμασε τρία τεστ ως προς την ικανότητά τους να προβλέπουν πότε τα δεδομένα μπορούν να συνδυαστούν σε μια ανάλυση. Επιπλέον απομάκρυνε τους μη πληροφοριακούς χαρακτήρες πριν πραγματοποιήσει τη συνδυαστική ανάλυση καθώς υπογραμμίζει ότι είναι πολύ σημαντικό το αν τα σύνολα των δεδομένων έχουν διαφορετικό αριθμό φυλογενετικό πληροφοριακών χαρακτήρων. Συγκρίνοντας εμπειρικά τον ILD με άλλους δείκτες (Templeton, Rodrigo) κατέληξε στο ότι ο συνδυασμός των γονιδίων που δεν συμφωνούν, αύξησε την ακρίβεια σε κάθε περίπτωση και πως ο ILD είναι ο καλύτερος έλεγχος στο να ανιχνεύει σημαντική ασυμφωνία μεταξύ των δεδομένων. Υπογραμμίζει δε πως μόνο όταν ο ILD < η συνδυαστική ανάλυση δεδομένων μείωνε την ακρίβεια. Στην παρούσα μελέτη ελέγχτηκε η σημαντικότητα του ILD με τον έλεγχο ομοιογένειας των δεδομένων (partition homogeneity test) στο πρόγραμμα PAUP* v4.0 beta10 (Swofford DL 2002). 75
82 4.6. ΑΝΑΛΥΣΗ ΜΟΡΙΑΚΗΣ ΔΙΑΚΥΜΑΝΣΗΣ ANALYSIS OF MOLECULAR VARIANCE (AMOVA) Προκειμένου να διερευνηθεί η ενδοπληθυσμιακή και η διαπληθυσμιακή ποικιλότητα των υπό μελέτη πληθυσμών, πραγματοποιήθηκε Ανάλυση της Μοριακής Διακύμανσης (AMOVA) με τη βοήθεια του προγράμματος Arlequin v.3 (Excoffier et al. 2005). Για κάθε σύνολο δεδομένων (16S rrna, COI mtdna, κλπ) χρησιμοποιήθηκαν οι απλότυποι των πληθυσμών (αφού χαρακτήριζαν πλήρως τις αλληλουχίες των ατόμων Trachelipus sp. καθώς και των εξωομάδων) και ομαδοποιήθηκαν τόσο σύμφωνα με τη γεωγραφική τους προέλευση όσο και μετά την ομαδοποίησή τους σύμφωνα με τη φυλογενετική τους συγγένεια - όπως αυτή προέκυψε από την ανάλυση των φυλογενετικών σχέσεων ( B.4.4.). Τέλος, με το ίδιο πρόγραμμα σχεδιάστηκε και το Δίκτυο Ελάχιστων Αποστάσεων (Minimum Spanning Network, MSN), ένα δίκτυο των σχέσεων και των αποστάσεων μεταξύ των εξεταζόμενων απλοτύπων. Το πρόγραμμα δημιουργεί έναν πίνακα αποστάσεων για όλα τα ζεύγη μεταξύ των απλοτύπων, με βάση το οποίο υπολογίζεται ο αριθμός μεταλλακτικών διαφορών, δηλαδή ο μέγιστος αριθμός βημάτων που συνδέουν φειδωλά δύο απλοτύπους. Με βάση τη συχνότητα του κάθε απλοτύπου στο παραγόμενο δίκτυο υπολογίζεται η ρίζα του δικτύου, δηλαδή ο απλότυπος που πιθανώς είναι ο προγονικός. Ωστόσο, ενδέχεται να υπάρχουν και πολλαπλές συνδέσεις μεταξύ των απλοτύπων οι οποίες πολλές φορές δυσκολεύουν την ερμηνεία των μεταξύ τους συνδέσεων. Όταν δεν υπάρχουν τέτοιες πολλαπλές συνδέσεις, τότε το δίκτυο ελάχιστων αποστάσεων (Minimum Spanning Network) μετατρέπεται σε δέντρο ελάχιστων αποστάσεων (Minimum Spanning Tree). 76
83 Γ. ΑΠΟΤΕΛΕΣΜΑΤΑ Σημειώνεται πως εφεξής τα άτομα που συλλέχτηκαν στο πλαίσιο της παρούσας μελέτης (Πιν. Β-1) και τα άτομα των Parmakelis et al (Πιν. Β-2), των οποίων οι αλληλουχίες ανακτήθηκαν από την GenBank ( θα αναφέρονται ως «δεδομένα της παρούσας μελέτης» συνολικά χωρίς να διαχωρίζονται, προκειμένου να είναι πιο ολοκληρωμένη η μελέτη και η εξαγωγή συμπερασμάτων για το γένος Trachelipus. 1. ΑΝΑΛΥΣΗ ΑΛΛΗΛΟΥΧΙΑΣ ΜΙΤΟΧΟΝΔΡΙΑΚΩΝ ΓΟΝΙΔΙΩΝ 1.1. ΑΝΑΛΥΣΗ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΤΟΠΟΥ 16S RRNA Νουκλεοτιδική σύσταση και Γενετική ποικιλότητα των μελετηθέντων πληθυσμών Η Αλυσιδωτή Αντίδραση Πολυμεράσης (PCR) και η επακόλουθη αλληλούχιση του πολλαπλασιασμένου τμήματος του γενετικού τόπου 16S rrnα, έδωσαν επιτυχώς νουκλεοτιδικές αλληλουχίες για τα 150 άτομα Trachelipus sp. και τα 2 άτομα Porcelio laevis που χρησιμοποιήθηκαν ως εξωμάδα. Το τελικό σύνολο των στοιχισμένων νουκλεοτιδίων που χρησιμοποιήθηκαν στις περαιτέρω αναλύσεις και αφού εξαιρέθηκε ένα μέρος του αρχικού και τελικού τμήματος εξαιτίας του διαφορετικού μήκους των τμημάτων που οφείλεται στα διαφορετικά σημεία έναρξης και λήξης των τμημάτων κατά τη διαδικασία αλληλούχισης (βλ B. Μεθοδολογία Υλικά 4.1.)- είχε μήκος 386 βάσεων. Σημειώνεται πως αντιστοίχως επεξεργάστηκαν οι αλληλουχίες των ατόμων Armadillidium lobocurvum (EF027622) και Porcellionides pruinosus (AJ275211) που ανακτήθηκαν από την 77

84 παγκόσμια βάση δεδομένων GenBank ώστε να έχει το ίδιο μήκος (386 βάσεις) με τις υπόλοιπες αλληλουχίες. Συνολικά υπολογίζοντας και τα 77 άτομα από Parmakelis et al. (2008) βλ Πιν. Β-2-υποβλήθηκαν σε ανάλυση 227 αλληλουχίες ατόμων Trachelipus sp. και 4 αλληλουχίες ατόμων που χρησιμοποιήθηκαν ως εξωομάδες. Με βάση το διάγραμμα της εντροπίας (Εικ.Γ-1), η πλέον μεταβλητή περιοχή βρίσκεται από τη θέση 120 έως και τη θέση 220, με αρκετές ακόμα μεταβλητές περιοχές των νουκλεοτιδίων να βρίσκονται σχεδόν διάσπαρτες σε όλο σχεδόν το μήκος των 386 βάσεων που μελετήθηκαν. Εικόνα Γ-1: Το διάγραμμα της εντροπίας σε σχέση με τη θέση κάθε νουκλεοτιδίου για το γονίδιο 16S rrna. Από την ανάλυση με το πρόγραμμα Mega v4.0 (Kumar et al. 2004), προκύπτει ότι, στο σύνολο των αλληλουχιών, οι 199 από τις 386 βάσεις (51,55%) διαφοροποιούνταν μεταξύ των αλληλουχιών και οι 141 (36,53%) ήταν πληροφοριακές θέσεις φειδωλότητας (parsimony-informative sites) 3. Σε ό,τι αφορά μόνο τις αλληλουχίες των αντιπροσώπων του γένους Trachelipus, οι αντίστοιχοι αριθμοί είναι 144 (37,31%) και 120 (31,09%). Τα ποσοστά των βάσεων στις Ως Πληροφοριακές Θέσεις Φειδωλότητας προσμετρούνται οι θέσεις στις οποίες εμφανίζονται τουλάχιστον δύο διαφορετικά νουκλεοτίδια και το καθένα απαντάται τουλάχιστον δύο φορές στο σύνολο των αλληλουχιών που εξετάζονται. 78
85 μιτοχονδριακές αλληλουχίες κατά μέσο όρο ήταν 37% Α, 33,6% Τ, 14,7% C και 14,6% G (Πιν Γ-1). Από τις 231 αλληλουχίες ατόμων (Trachelipus sp και εξωομάδες) που λήφθηκαν για το 16S rrna, οι 70 (Πιν. Γ-2) αντιστοιχούσαν σε διαφορετικούς απλότυπους σύμφωνα με το πρόγραμμα DNAsp v.4 (Rozas et al. 2003). Πίνακας Γ-1: Τα ποσοστά των βάσεων των απλοτύπων, καθώς και ο μέσος όρος αυτών για κάθε απλότυπο. Hap# %T %C %A %G Hap 1 33,4 14,5 38,1 14,0 Hap 2 33,9 14,2 37,7 14,2 Hap 3 32,6 14,0 39,6 13,7 Hap 4 35,6 13,5 35,9 14,9 Hap 5 32,7 15,8 36,5 15,0 Hap 6 31,9 15,7 37,6 14,8 Hap 7 33,0 15,1 37,6 14,3 Hap 8 33,2 15,2 37,4 14,2 Hap 9 34,9 14,6 36,1 14,4 Hap 10 35,3 14,1 35,6 15,0 Hap 11 35,2 13,7 36,9 14,2 Hap 12 35,2 13,9 36,6 14,2 Hap 13 34,3 14,4 36,2 15,2 Hap 14 34,7 14,1 36,0 15,2 Hap 15 34,0 14,6 37,4 14,1 Hap 16 34,3 14,4 36,2 15,2 Hap 17 34,3 14,4 36,2 15,2 Hap 18 34,3 14,4 36,2 15,2 Hap 19 34,4 14,2 36,9 14,5 Hap 20 31,4 16,1 38,1 14,5 Hap 21 33,0 15,2 37,8 14,1 Hap 22 33,1 15,2 37,6 14,1 Hap 23 33,1 15,2 37,3 14,4 Hap 24 35,3 13,6 35,6 15,6 Hap 25 34,7 13,9 35,3 16,1 Hap 26 35,2 13,8 35,8 15,3 Hap 27 31,8 15,8 38,0 14,4 Hap 28 31,7 15,7 38,1 14,4 Hap 29 31,6 16,4 37,3 14,7 79
86 Hap 30 34,0 14,3 37,1 14,6 Hap 31 31,3 16,0 38,0 14,7 Hap 32 33,7 14,9 37,1 14,3 Hap 33 34,0 14,4 36,7 14,9 Hap 34 34,4 14,9 35,7 14,9 Hap 35 33,9 14,7 35,7 15,7 Hap 36 36,2 13,6 35,9 14,2 Hap 37 33,4 15,4 36,9 14,3 Hap 38 33,1 15,3 37,4 14,2 Hap 39 35,3 13,8 35,9 14,9 Hap 40 35,6 13,2 36,5 14,7 Hap 41 34,0 14,6 37,6 13,8 Hap 42 33,0 14,9 37,8 14,4 Hap 43 32,7 15,0 37,8 14,5 Hap 44 34,5 13,7 36,4 15,4 Hap 45 35,2 13,5 36,4 15,0 Hap 46 34,6 14,1 35,9 15,4 Hap 47 32,5 15,9 37,4 14,2 Hap 48 32,7 15,5 37,5 14,2 Hap 49 32,7 15,5 37,3 14,5 Hap 50 34,8 13,7 37,6 14,0 Hap 51 34,0 14,1 37,0 14,9 Hap 52 34,6 14,1 36,2 15,1 Hap 53 31,7 15,9 36,8 15,6 Hap 54 32,6 13,8 39,5 14,1 Hap 55 31,8 15,9 36,9 15,4 Hap 56 32,0 15,9 36,8 15,3 Hap 57 32,1 15,5 38,0 14,4 Hap 58 33,5 14,6 37,8 14,1 Hap 59 34,6 14,4 37,1 13,9 Hap 60 34,8 13,6 38,0 13,6 Hap 61 34,2 14,5 36,9 14,5 Hap 62 34,0 14,6 36,4 15,1 Hap 63 34,6 14,0 36,0 15,4 Hap 64 33,4 15,4 36,9 14,3 Hap 65 32,8 15,5 37,4 14,4 Hap 66 34,3 14,4 36,4 14,9 Hap 67 32,2 16,2 36,8 14,9 Hap 68 32,0 15,5 38,4 14,1 Hap 69 34,8 13,6 39,3 12,3 Hap 70 30,3 15,9 38,6 15,3 Μέσος όρος 33,6 14,7 37,0 14,6 80
87 Πίνακας Γ-2: Γενετικός τόπος 16s rrna. Παρατίθενται τα ονόματα των απλοτύπων, ο αριθμός ατόμων σε κάθε απλότυπο, ο πληθυσμός προέλευσης των ατόμων και το είδος στο οποίο ανήκουν. ΑΠΛΟΤΥΠΟΣ ΑΡΙΘΜΟΣ ΑΤΟΜΩΝ (Ν)/ΑΠΛΟΤΥΠΟ ΑΤΟΜΟ Hap_1 2 AIN 1,4 Hap_2 27 AIN2-3, ALF1-5,7, CH1,3-4, KRN1-2, KRT1-4, LAMB1-6, ZRL6-8 Hap_4 11 AMG 1-3, MIK 2,4,6, PLAN1-2,4-5 Hap_5 4 ATR1-4 Hap_6 9 BEL1-4, KLM1-4, ZRL9 Hap_7 14 BOU1-12, CH2,6 Hap_8 4 CH5, CHb2-3 Hap_9 2 CML1,4 Hap_10 1 CML2 Hap_11 2 DRM1-2 Hap_12 4 DRM3, PAP1-3 Hap_13 6 ELTA1-4, LOU1, MIK6 Hap_14 4 ELTb1-4 Hap_15 3 EPD1-3 Hap_16 2 FEN1,3 Hap_17 2 FEN2, SFK2 Hap_18 5 FLA1, LIB3, MAIN1, SFK1, ZIR1 Hap_19 2 FNR2-3 Hap_20 1 FRX1 Hap_21 3 FRX2-4 Hap_22 3 GRP1-2,4 Hap_23 1 GRP3 Hap_24 1 KAL1 Hap_25 2 KAL2-3 Hap_26 1 KAR1 Hap_27 1 KAT1 Hap_28 2 KAT2,4 Hap_29 1 KAT3 Hap_30 2 KON3-4 Hap_31 4 CRE3-6 Hap_32 4 KYT1-4 ΕΙΔΟΣ Trachelipus kytherensis Hap_33 1 LAG1 Hap_34 3 LIB1-2,4 Hap_35 1 LIB5 81
88 Hap_36 3 MIK1,3,5 Hap_37 5 MKL1, VLA1-3, VOI1 Hap_38 4 MNS1-2, SOL1,3 Hap_39 19 NED1,4, PMS1-3, PRR1-4, TAY1-8, VLK2-3 Hap_40 1 NED2 Hap_41 6 PAN1-6 Hap_42 3 PAN7-9 Hap_43 2 PER1-2 Hap_44 2 PLAN3, THEO1 Hap_45 1 PRSM1 Hap_46 1 SFK3 Hap_47 1 SOL2 Hap_48 3 SSN1-3 Hap_49 1 SSN4 Hap_50 4 STE 1-4 Hap_51 4 THEO2-4,6 Hap_52 1 THEO5 Hap_53 2 PROU1,3 Hap_54 1 PROU2 Hap_55 1 PROU4 Hap_56 1 PROU5 Hap_57 2 TPT1,3 Hap_58 1 TPT2 Hap_61 1 VEL1 Hap_62 2 VEL2 Hap_63 1 VLK1 Hap_64 3 VOI1-4 Hap_65 3 VTS2-4 Hap_66 1 ZIR2 Hap_67 4 ZRL1-4 Hap_68 1 ZRL5 Hap_59 2 IKAR 1-2 Hap_60 8 NAX1-3, STO1 AGA2-3, STN1-2 Hap_3 2 AKYT Porcelio laevis Hap_69 1 PAN_N6 Armadillidium lobocurvum Hap_70 1 ATHE Porcellionides pruinosus Trachelipus kutherensis T. aegaeus T. n.sp Εξωομάδες (Outgroups) Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εύβοια Εξωομάδες (outgroups) 82
89 Ο απλότυπος που εμφανίζει τη μεγαλύτερη συχνότητα είναι ο Hap 2, ο οποίος περιλαμβάνει 27 άτομα από 7 διαφορετικούς πληθυσμούς. Από αυτούς οι 6 πληθυσμοί είναι της Πελοποννήσου ενώ ο ένας είναι από τη λίμνη Ζηρού που ανήκει στους νομούς Πρεβέζης Άρτας. Ιδιαίτερο ενδιαφέρον παρουσιάζει ο απλότυπος Hap60 ο οποίος αντιπροσωπεύει άτομα από 2 διαφορετικά είδη (T. aegaeus και T. n.sp.), καταδεικνύοντας ενδεχομένως τη συντηρητικότητα του συγκεκριμένου γενετικού τόπου στην προκειμένη περίπτωση. Ωστόσο αξίζει να σημειωθεί ότι 53 από τους 70 απλοτύπους είναι μοναδικοί είτε για έναν ολόκληρο πληθυσμό είτε μοναδικοί για κάποια άτομα ενός πληθυσμού. Κάτι τέτοιο είναι ενδεικτικό της μεγάλης ποικιλότητας μεταξύ των ατόμων ακόμα και μέσα στον ίδιο τον πληθυσμό αναφορικά με το γενετικό τόπο 16S. Η νουκλεοτιδική ποικιλότητα (π) και η απλοτυπική ποικιλότητα (h) στις μελετηθείσες αλληλουχίες ήταν π=0,0967 και h=0,9675. Ειδικά για τα άτομα Trachelipus sp. οι τιμές τις νουκλεοτιδικής και απλοτυπικής ποικιλότητας παρατίθενται στον πίνακα Γ-3. Πίνακας Γ-3: Οι τιμές τις νουκλεοτιδικής και απλοτυπικής ποικιλότητας για τους αντιπροσώπους του γένους Trachelipus. Τάξον Νουκλεοτιδική ποικιλότητα (π) Απλοτυπική ποικιλότητα (h) Αριθμός Απλοτύπων Trachelipus kytherensis 0,0931 0, Trachelipus aegaeus 0,0500 0, Trachelipus n.sp. 0,0014 0, Trachelipus sp. 0,0949 0, Για λόγους ευκολίας, η κατασκευή των φυλογενετικών δέντρων πραγματοποιήθηκε με τη χρήση των απλοτύπων που προέκυψαν. 83
90 Έλεγχος Ομοπλασίας Οι πραγματικές εξελικτικές σχέσεις των υπό εξέταση αλληλουχιών ενδέχεται να μην αποκαλυφθούν εάν οι νουκλεοτιδικές θέσεις έχουν κορεστεί από πολλαπλές υποκαταστάσεις (Swofford et al. 2001). Προκειμένου να ελεγχθεί ο βαθμός κορεσμού των αλληλουχιών σε νουκλεοτιδικές υποκαταστάσεις, πραγματοποιήθηκε ο έλεγχος του Xia (βλ Β. Μεθοδολογία Υλικά 4.2.) που χρησιμοποιεί το δείκτη κορεσμού υποκατάστασης (Xia et al. 2003) και περιλαμβάνεται στο πρόγραμμα Dambee (Xia & Xie 2001). Από τον έλεγχο προέκυψε πως I SS < SS C I. και μάλιστα στατιστικώς σημαντικά, οπότε οι αλληλουχίες μπορούν να χρησιμοποιηθούν για φυλογενετικές αναλύσεις. Ακολούθως παρατίθενται αναλυτικά τα αποτελέσματα του εν λόγω ελέγχου (Πιν. Γ-4). Πίνακας Γ-4: Τα αποτελέσματα του ελέγχου των Xia et al. 2003) για διερεύνηση του κορεσμού υποκατάστασης των βάσεων στις μελετώμενες αλληλουχίες του γενετικού τόπου 16S r RNA. NumOTU Iss Iss.c DF P 4 0,187 0, <0, ,178 0, <0, ,178 0, <0, ,180 0, <0,001 Εφόσον οι αλληλουχίες των απλοτύπων δεν εμφάνισαν κορεσμό, ακολούθησε με τη χρήση αυτών η κατασκευή των φυλογενετικών δέντρων με τις μεθόδους Σύνδεσης Γειτόνων (NJ), Μέγιστης Φειδωλότητας (MP) και Μπεϊεσιανής Συμπερασματολογίας (BI). 84
91 Εκτίμηση γενετικών αποστάσεων Οι γενετικές αποστάσεις μεταξύ των υπό εξέταση αλληλουχιών εκτιμήθηκαν με το διπαραμετρικό μοντέλο Kimura (K2P). Πρόκειται για ένα απλό μοντέλο, το οποίο επιτρέπει διαφορετικό ρυθμό εμφάνισης των μεταπτώσεων και των μεταστροφών (Kumar et al. 2004). Ο υπολογισμός των αποστάσεων πραγματοποιήθηκε με το πρόγραμμα MEGA v.4 (Kumar et al. 2004). Στον πίνακα Γ-5 που ακολουθεί, παρατίθενται οι νουκλεοτιδικές αποκλίσεις μεταξύ των απλοτύπων που εξετάστηκαν για τους αντιπροσώπους του γένους Trachelipus, μετά την ομαδοποίησή τους ανά είδος. Οι αντιπρόσωποι του T. n.sp. ομαδοποιήθηκαν σε έναν κοινό απλότυπο (Hap 60) με τα άτομα του T. aegaeus από τη Νάξο και το Στουρονήσι (STO), οπότε και ενοποιήθηκαν για τη συγκεκριμένη ανάλυση. Πίνακας Γ-5: Οι μέσες νουκλεοτιδικές αποκλίσεις (σε παρενθέσεις οι καθαρές μέσες νουκλεοτιδικές αποστάσεις) βάσει των απλοτύπων μεταξύ των ειδών Trachelipus και της εξωομάδας (K2P) για το γενετικό τόπο 16S rrna. Είδος T. kytherensis T. aegaeus + T. n.sp. εξωομάδες T. aegaeus + T. n.sp. 0,124 (0,029) - εξωομάδες 0,297 (0,085) 0,258 (0,060) - Επιπλέον, υπολογίστηκαν οι νουκλεοτιδικές αποκλίσεις μεταξύ των γεωγραφικών ομάδων των πληθυσμών (Πιν. Γ-6). Σε αυτήν την περίπτωση πραγματοποιήθηκε ομαδοποίηση των 63 μελετούμενων πληθυσμών του γένους, ανάλογα με τη γεωγραφική τους προέλευση. Έτσι, διακρίθηκαν οι ακόλουθες ομάδες: Πελοπόννησος 85
92 Λοιπή ηπειρωτική Ελλάδα (πλην Πελοποννήσου) Νησιωτικές περιοχές Αιγαίου Εύβοια Πίνακας Γ-6: Οι μέσες νουκλεοτιδικές αποκλίσεις (σε παρενθέσεις οι καθαρές μέσες νουκλεοτιδικές αποκλίσεις) μεταξύ των γεωγραφικών ομάδων των αντιπροσώπων Trachelipus και της εξωομάδας (K2P) για το γενετικό τόπο 16S rrna. ΓΕΩΓΡΑΦΙΚΗ ΟΜΑΔΑ Εύβοια Λοιπή ηπειρωτική Ελλάδα Πελοπόννησος Αιγαίο εξωομάδες Λοιπή ηπειρωτική Ελλάδα 0,131 (0,051) - Πελοπόννησος 0,113 (0,043) 0,122 (0,040) - Αιγαίο 0,099 (0,055) 0,136 (0,079) 0,120 (0,074) - εξωομάδες 0,276 (0,116) 0,298 (0,125) 0,296 (0,134) 0,243 (0,105) - Στον πίνακα Π-1 του παραρτήματος παρουσιάζονται οι γενετικές αποστάσεις K2P μεταξύ όλων των μελετούμενων απλοτύπων ανά ζεύγη και στον Π-2, οι γενετικές αποστάσεις K2P μεταξύ όλων των μελετούμενων πληθυσμών ανά ζεύγη για το γενετικό τόπο 16S rrna. Η γενετική απόσταση, d, με βάση το μοντέλο του Kimura (1980) κυμαινόταν από 0 (BEL KLM) ως 0,176 (EPD-BEL, EPD-KLM, EPD-VTS). Οι γενετικές αποστάσεις που χωρίζουν τα είδη του γένους Trachelipus μεταξύ τους, σύμφωνα πάντα με την ισχύουσα ταξινόμηση, κυμαίνονταν: μεταξύ 0,037 (T. aegaeus - EPD) και 0,155 (T. aegaeus - ATR) και μεταξύ 0,011 (T. n.sp. - EPD) και 0,162 (T. n.sp. - VTS). 86
93 Οι αποστάσεις μεταξύ των αντιπροσώπων του γένους και των ατόμων από τις εξωομάδες κυμαίνονταν από 0,215 (Porcelio laevis-κυτ) μέχρι 0,337 (Porcellionides pruinosus-atr). Τέλος, στον πίνακα Γ-7 παρουσιάζεται η μέση νουκλεοτιδική διαφοροποίηση των πληθυσμών μελέτης. Πίνακας Γ-7. Η μέση ενδοπληθυσμιακή γενετική διαφοροποίηση (απόσταση Kimura, 1980) των πληθυσμών για το γενετικό τόπο 16S rrna. Ενδοπλ/κή Ενδοπλ/κή Ενδοπλ/κή Ενδοπλ/κή Πλ/σμός διαφ/ση Πλ/σμός διαφ/ση Πλ/σμός διαφ/ση Πλ/σμός διαφ/ση AIN 0,010 FEN 0,005 KRT 0,000 PAP 0,000 ALF 0,000 FLA n/c KYT 0,000 PER 0,000 AMG 0,000 FNR 0,000 LAG n/c PLAN 0,002 ATR 0,000 FRX 0,066 LAMB 0,000 PMS 0,000 BEL 0,000 GRP 0,002 LIB 0,005 PRR 0,000 BOU 0,000 KAL 0,010 LOU n/c PRSM n/c CH 0,063 KAR n/c MAIN n/c SFK 0,007 CML 0,015 KAT 0,046 MIK 0,002 SOL 0,002 DRM 0,002 KLM 0,000 MKL n/c SSN 0,002 ELT 0,006 KON 0,000 MNS 0,000 STE 0,000 EPD 0,000 CRE 0,000 NED 0,007 TAY 0,000 FEN 0,005 KRN 0,000 PAN 0,006 THEO 0,010 PROU 0,063 VLA 0,000 VTS 0,000 T.aegaeus 0,041 TPT 0,086 VLK 0,007 ZIR 0,010 T.n.sp. 0,000 VEL 0,026 VOI 0,069 ZRL 0,091 87
94 Φαινετικές και φυλογενετικές σχέσεις των μελετηθέντων πληθυσμών Τα δένδρα που προέκυψαν από τις διάφορες φυλογενετικές αναλύσεις παρουσιάζονται παρακάτω. Αρχικά παρουσιάζονται τα αποτελέσματα της μεθόδου Σύνδεσης Γειτόνων, ακολουθούν αυτά της Μέγιστης Φειδωλότητας και εν συνεχεία αυτά της Μπεϊεσιανής Συμπερασματολογίας. Το δένδρο με τη μέθοδο της Σύνδεσης Γειτόνων (NJ) (Εικ. Γ-2) κατασκευάστηκε με το πρόγραμμα Mega 4.0 (Kumar et al. 2004) βάσει του διπαραμετρικού μοντέλου - K2P - του Kimura (1980). Η αξιοπιστία των κλάδων ελέγχθηκε με την πραγματοποίηση 1000 ψευδοεπαναλήψεων (bootstrap). Την ίδια τοπολογία κλάδων εμφάνιζε και η μέθοδος UPGMA. Το δένδρο που παρουσιάζεται στην Εικ. Γ-3 είναι αυτό που προέκυψε από μια σταθμισμένη ανάλυση Μέγιστης Φειδωλότητας (weighted maximum parsimony). Συγκεκριμένα, είναι το 50% πλειοψηφικής αποδοχής δένδρο, το οποίο προέκυψε χρησιμοποιώντας την ευρετική μέθοδο αναζήτησης του πιο φειδωλού δένδρου στο πρόγραμμα PAUP (v4.0.beta10, Swofford 2002). Τέλος, στην Εικ. Γ-4 παρατίθεται το δένδρο που προέκυψε από την ανάλυση Μπεϊεσιανής Συμπερασματολογίας (BI) με τη βοήθεια του προγράμματος MrBayes v3.1.2 (Huelsenbeck & Ronquist 2001). Το εξελικτικό μοντέλο που χρησιμοποιήθηκε, όπως αυτό προέκυψε από το πρόγραμμα Modeltest, ήταν το TVM+I+G (με -lnl= ), με ποσοστό αμετάβλητων θέσεων 0,3724 και παράμετρο για τη μεταβολή γάμμα των μεταβλητών θέσεων α=0,6597. Οι συχνότητες των νουκλεοτιδίων ήταν: Α= 0,3959, C= 0,1422, G= 0,0932 και T= 0,3686. Ο αριθμός παραμέτρων που λήφθηκαν υπόψην ήταν Κ=7 με εξι διαφορετικούς ρυθμούς μεταπτώσεων / μεταστροφών (Πιν. Γ-8) 88
95 Ο συνολικός αριθμός γενεών ήταν 10 6, τα δένδρα σώζονταν ανά 100 γενεές, από τα οποία τελικά απορρίφθηκε το αρχικό 25% ως burn-in. Πίνακας Γ-8: Ρυθμός υποκαταστάσεων μεταξύ των νουκλεοτιδίων. A C G T C 1,000 - G 6,994 1,000 - T 1,000 7,521 1,000-89




96 Εικόνα Γ-2: Γενετικός τόπος 16S rrna Δένδρο NJ βάσει του διπαραμετρικού μοντέλου του Kimura. Οι αριθμοί στους κλάδους αντιστοιχούν στις τιμές bootstrap (1000 ψευδοεπαναλήψεις) που υπερβαίνουν το 50%. Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ-2. Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εύβοια Εξωομάδες 90


.")
97 Εικόνα Γ-3: Κλαδόγραμμα Μέγιστης Φειδωλότητας (ΜΡ) με βάση στοιχισμένες αλληλουχίες μήκους 386bp του γονιδίου 16S rrna που ομαδοποιήθηκαν σε 70 απλοτύπους. Οι τιμές στους κλάδους αναφέρονται σε % ποσοστά ελέγχου 1000 bootstrap. Παρουσιάζονται μόνο τιμές bootstrap μεγαλύτερες του 50%. (Tree length=483, CI=0,580, RI=0,863, HI=0,420). Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ-2. Πελοπόννησος Νησιωτικές περιοχές (Αιγαίο) Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Εξωομάδες Εύβοια 91




98 Εικόνα Γ-4: Κλαδόγραμμα Μπεϊεσιανής Συμπερασματολογίας (B.I.) ) με βάση στοιχισμένες αλληλουχίες μήκους 386bp του γονιδίου 16S rrna που ομαδοποιήθηκαν σε 70 απλοτύπους Οι αριθμοί στους κλάδους δηλώνουν τις εκ των υστέρων πιθανότητες. Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ-2. Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εύβοια Εξωομάδες 92
99 Ανάλυση Μοριακής Διακύμανσης (AMOVA) Δίκτυο Ελαχίστων Αποστάσεων (MSN) Ομαδοποίηση βάσει γεωγραφικής προέλευσης Σε αυτήν την περίπτωση πραγματοποιήθηκε ομαδοποίηση των 63 μελετούμενων πληθυσμών του γένους ανάλογα με τη γεωγραφική τους προέλευση. Έτσι διακρίθηκαν οι ακόλουθες ομάδες: Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (πλην Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο+Εύβοια) Αν εξαιρεθούν λοιπόν τα άτομα που χρησιμοποιήθηκαν ως εξωομάδα και εμφανίζουν την αναμενόμενη γενετική απόκλιση από τους πληθυσμούς του γένους Trachelipus, αυτό που απομένει είναι να ελεγχθούν οι διαπληθυσμιακές και ενδοπληθυσμιακές αποκλίσεις των πληθυσμών του εν λόγω γένους. Πίνακας Γ-9: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για το γενετικό τόπο 16S μεταξύ όλων των πληθυσμών του γένους Trachelipus της παρούσας μελέτης. Πηγή διακύμανσης Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 35,05% διαπληθυσμιακά, εντός της ίδιας ομάδας 49,52% ενδοπληθυσμιακά 15,43% Προκειμένου να διερευνηθούν τα ποσοστά μοριακής διακύμανσης αποκλειστικά μεταξύ των πληθυσμών του Trachelipus kytherensis που αποτέλεσε το κυρίαρχο είδος στο πλαίσιο αυτής της μελέτης, πραγματοποιήθηκε AMOVA μεταξύ των ίδιων γεωγραφικών ομάδων η οποία όμως περιλάμβανε μόνο τους αντιπροσώπους του είδους αυτού. 93
100 Πίνακας Γ-10: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για το γενετικό τόπο 16S μεταξύ όλων των πληθυσμών του είδους Trachelipus kytherensis της παρούσας μελέτης. Πηγή διακύμανσης Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 33,98% διαπληθυσμιακά, εντός της ίδιας ομάδας 48,72% ενδοπληθυσμιακά 17,30%. Η AMOVA έδειξε πως η διακύμανση είναι μεγαλύτερη μεταξύ των πληθυσμών της ίδιας ομάδας παρά μεταξύ των διαφορετικών γεωγραφικών ομάδων, καταδεικνύοντας έτσι τη μεγάλη ποικιλότητα που εμφανίζουν οι αντιπρόσωποι τόσο του γένους Trachelipus όσο και οι αντιπρόσωποι του είδους Trachelipus kytherensis. Τέλος, αναφέρεται πως όταν η ανάλυση επαναλήφθηκε θεωρώντας την Εύβοια ως ξεχωριστή γεωγραφική ενότητα, δεν παρατηρήθηκαν διαφοροποιήσεις στα ποσοστά της μοριακής διακύμανσης καθώς αυτά μεταβλήθηκαν μειούμενα μόλις κατά 0,5%. Ακολούθως παρατίθεται το Δίκτυο Ελαχίστων Αποστάσεων (MSN) όπως προέκυψε από την Ανάλυση Μοριακής Διακύμανσης και με χρήση των αποστάσεων K2P μεταξύ των απλοτύπων (Εικ. Γ-5). Το δίκτυο αυτό απεικονίζει τόσο τις σχέσεις όσο και τις αποστάσεις μεταξύ των απλοτύπων. Το μήκος κάθε γραμμής σύνδεσης είναι αναλογικό του αριθμού νουκλεοτιδικών αλλαγών που απαιτούνται για τη μετάβαση από τον ένα απλότυπο στον άλλο. Στην ανάλυση συμπεριλήφθησαν και τα άτομα της εξωομάδας. Σημειώνεται πως τα δίκτυα ελαχίστων αποστάσεων (MSN) στις υπόλοιπες αναλύσεις της παρούσας διατριβής εμπεριείχαν αρκετές πολλαπλές συνδέσεις, οι οποίες υποδηλώνουν αμφισβητούμενες σχέσεις μεταξύ των μελετούμενων απλοτύπων. Για το λόγο αυτόν, παρατίθεται μόνο το MSN για το γενετικό τόπο 94
101 16S. Τα αντίστοιχα δίκτυα τόσο για την υπομονάδα Ι της κυτοχρωμικής οξειδάσης (COI) όσο και για τη συνδυασμένη ανάλυση ήταν πολύ πιο πολύπλοκα και δύσκολο να αναπαρασταθούν γραφικά με τρόπο που να φαίνονται όλες οι συνδέσεις. 95
 μεταξύ των 70 απλοτύπων για το γενετικό τόπο 16S rrna.")

102 Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Εύβοια Νησιωτικές περιοχές (Αιγαίο) Εξωομάδες Εικόνα Γ-5: Το δίκτυο ελαχίστων αποστάσεων (MSN) μεταξύ των 70 απλοτύπων για το γενετικό τόπο 16S rrna. Από το όνομα του κάθε απλοτύπου έχει αποκοπεί το πρόθεμα «Hap» προκειμένου να γίνει πιο εύληπτη η εικόνα. Το μήκος των γραμμών σύνδεσης είναι αναλογικό του αριθμού νουκλεοτιδικών αλλαγών που απαιτούνται για τη μετάβαση από 96 τον έναν απλότυπο στον άλλο. Οι απλότυποι αντιστοιχούν στα άτομα του Πιν.
103 Ομαδοποίηση βάσει φυλογενετικής συγγένειας Η ομαδοποίηση στην προκειμένη περίπτωση έγινε με βάση τις ομάδες που προέκυψαν από τη φυλογενετική ανάλυση για το γενετικό τόπο 16S rrna. Οι ομάδες αυτές διαμορφώθηκαν ως εξής: PEL mix = απλότυποι από όλη την ηπειρωτική Ελλάδα αλλά στην πλειονότητά τους από την Πελοπόννησο PEL = απλότυποι μόνο από περιοχές της Πελοποννήσου AEGEAN = οι απλότυποι κυρίως από περιοχές του Αιγαίου + ο απλότυπος των ατόμων της Επιδαύρου + ο απλότυπος της Κ. Εύβοιας Ν.EUBOIA = οι απλότυποι της Β. Εύβοιας MAINLAND = μόνο απλότυποι της ηπειρωτικής περιοχής (πλην της Πελ/σου) MAINLAND_mix = απλότυποι από όλη την ηπειρωτική Ελλάδα αλλά στην πλειονότητά τους απλότυποι της ηπειρωτικής περιοχής (πλην της Πελ/σου). Πίνακας Γ-11: Οι απλότυποι που αντιστοιχούν σε κάθε φυλογενετική ομάδα όπως αυτές διαμορφώθηκαν από τη φυλογενετική ανάλυση του γενετικού τόπου 16S. Ομάδα PEL_mix PEL AEGEAN Απλότυπος Hap1 Hap2 Hap8 Hap21 Hap37 Hap41 Hap42 Hap54 Hap58 Hap68 Hap4 Hap13 Hap14 Hap16 Hap17 Hap18 Hap24 Hap25 Hap33 Hap34 Hap35 Hap36 Hap39 Hap40 Hap44 Hap46 Hap51 Hap52 Hap63 Hap66 Hap31 Hap32 Hap50 Hap59 Hap60 Hap15 Ν. EUBOIA Hap11 Hap12 MAINLAND Hap9 Hap10 Hap19 Hap26 Hap45 Hap61 Hap62 Hap5 Hap6 Hap7 Hap20 Hap22 Hap23 Hap27 Hap28 Hap29 Hap30 Hap38 MAINLAND_mix Hap43 Hap47 Hap48 Hap49 Hap53 Hap55 Hap56 Hap57 Hap64 Hap65 Hap67 97
104 Αν εξαιρεθούν λοιπόν τα άτομα που χρησιμοποιήθηκαν ως εξωομάδα και εμφανίζουν την αναμενόμενη γενετική απόκλιση από τους πληθυσμούς του γένους Trachelipus, αυτό που απομένει είναι να ελεγχθούν οι διαπληθυσμιακές και ενδοπληθυσμιακές αποκλίσεις των προαναφερθέντων ομάδων του εν λόγω γένους. Πίνακας Γ-12: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για το γενετικό τόπο 16S μεταξύ των φυλογενετικών του γένους Trachelipus της παρούσας μελέτης. Πηγή διακύμανσης Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 67,05 διαπληθυσμιακά, εντός της ίδιας ομάδας 3,27 ενδοπληθυσμιακά 29,68 Το συμπέρασμα που εξάγεται από αυτήν την ανάλυση είναι λίγο πολύ αναμενόμενο: η μεγάλη γενετική απόκλιση που παρατηρείται προέρχεται κυρίως από τις διαφορές των πληθυσμών μεταξύ των διαφορετικών κλάδων (ομάδων) όπως αυτοί προέκυψαν από τη φυλογενετική ανάλυση, και σε πολύ μικρό βαθμό από τις διαπληθυσμιακές διαφορές εντός των κλάδων (ομάδων) ή τις ενδοπληθυσμιακές διαφορές. Αξίζει να σημειωθεί βέβαια η αρκετά μεγάλη τιμή που λαμβάνει η ενδοπληθυσμιακή διακύμανση - ήτοι η διακύμανση μέσα στους απλότυπους του ίδιου κλάδου - θυμίζοντας πως οι απλότυποι που ομαδοποιήθηκαν στις φυλογενετικές αναλύσεις μέσα στον ίδιο κλάδο πολλές φορές αντιπροσώπευαν άτομα ή και πληθυσμούς που είναι αρκετά απομονωμένοι μεταξύ τους και που θεωρητικά θα ήταν αδύνατο με φυσικό τρόπο να βρεθούν μαζί. Τέτοια παραδείγματα αποτελούν οι απλότυποι Hap2 και Hap37, στους οποίους ομαδοποιούνται άτομα από την Πελοπόννησο μαζί με άτομα από περιοχές της βόρειας και κεντρικής Ελλάδας (βλ Πιν. Γ-2). 98

105 1.2. ΑΝΑΛΥΣΗ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΤΟΠΟΥ COI Νουκλεοτιδική σύσταση και γενετική ποικιλότητα των μελετηθέντων πληθυσμών Η Αλυσιδωτή Αντίδραση Πολυμεράσης (PCR) και η επακόλουθη αλληλούχιση του πολλαπλασιασμένου τμήματος του γενετικού τόπου COI, έδωσαν επιτυχώς νουκλεοτιδική αλληλουχία για τα 110 άτομα Trachelipus sp. Το τελικό σύνολο των στοιχισμένων αλληλουχιών που χρησιμοποιήθηκε στις περαιτέρω αναλύσεις και αφού εξαιρέθηκε ένα μέρος του αρχικού και τελικού τμήματος εξαιτίας του διαφορετικού μήκους των τμημάτων που οφείλεται στα διαφορετικά σημεία έναρξης και λήξης των τμημάτων κατά τη διαδικασία αλληλούχισης (βλ Β. Μεθοδολογία Υλικά 4.1.) - είχε μήκος 512 βάσεων. Αντίστοιχα, έτσι ώστε να έχει το ίδιο μήκος (512 βάσεις) με τις υπόλοιπες αλληλουχίες, έτυχε επεξεργασίας και η αλληλουχία του ατόμου Armadillidium lobocurvum (EF027706) που ανακτήθηκε από την παγκόσμια βάση δεδομένων GenBank για να χρησιμοποιηθεί ως εξωομάδα στις αναλύσεις. Εικόνα Γ-6: Το διάγραμμα της εντροπίας σε σχέση με τη θέση κάθε νουκλεοτιδίου για το γονίδιο 99
106 Με βάση το διάγραμμα της εντροπίας (Εικ. Γ-6), η πλέον μεταβλητή περιοχή βρίσκεται από τη θέση 310 έως και τη θέση 440. Από την ανάλυση με το πρόγραμμα Mega v4.0 (Kumar et al. 2004), προκύπτει ότι, στο σύνολο των αλληλουχιών, οι 210 από τις 512 βάσεις (41%) διαφοροποιούνταν μεταξύ των αλληλουχιών και οι 164 (32%) ήταν πληροφοριακές θέσεις φειδωλότητας. Σε ό,τι αφορά μόνο τις αλληλουχίες των αντιπροσώπων του γένους Trachelipus, οι αντίστοιχοι αριθμοί είναι 197 (38,48%) και 159 (31,05%). Τα ποσοστά των βάσεων στις αλληλουχίες κατά μέσο όρο ήταν 24,4% Α, 35,7% Τ, 15,3% C και 24,6% G (Πιν. Γ-12). Από τις 186 αλληλουχίες ατόμων που λήφθηκαν για το γονίδιο της COI, οι 47 (Πιν. Γ-13) αντιστοιχούσαν σε διαφορετικούς απλοτύπους, σύμφωνα με το πρόγραμμα DNAsp v. 4 (Rozas et al. 2003). Πίνακας Γ-13: Τα ποσοστά των βάσεων των απλοτύπων, καθώς και ο μέσος όρος αυτών για κάθε απλότυπο. Hap # %T %C %A %G Hap 1 36,3 14,0 25,9 23,8 Hap 2 36,7 13,6 25,9 23,8 Hap 3 36,6 14,0 25,0 24,4 Hap 4 35,1 15,5 25,5 23,9 Hap 5 35,3 15,3 25,5 23,9 Hap 6 33,5 16,8 22,5 27,2 Hap 7 36,0 15,5 24,4 24,2 Hap 8 34,5 15,5 25,0 25,0 Hap 9 34,5 15,5 25,4 24,6 Hap 10 34,3 16,3 24,3 25,1 Hap 11 34,8 15,0 25,5 24,7 Hap 12 36,8 13,7 22,3 27,2 Hap 13 37,3 14,8 23,7 24,1 Hap 14 37,7 13,8 25,6 22,9 Hap 15 36,5 13,0 26,2 24,3 Hap 16 36,3 15,3 25,2 23,3 Hap 17 36,5 15,1 25,4 23,1 100
107 Hap 18 35,5 16,1 24,4 24,0 Hap 19 34,8 15,1 22,1 28,0 Hap 20 36,9 12,8 26,2 24,1 Hap 21 36,7 13,9 25,3 24,1 Hap 22 35,5 15,9 24,4 24,2 Hap 23 35,3 15,3 25,5 23,9 Hap 24 34,3 17,1 22,9 25,7 Hap 25 35,5 14,4 26,2 23,9 Hap 26 34,4 16,7 23,1 25,8 Hap 27 33,9 17,8 23,0 25,3 Hap 28 32,3 18,5 22,4 26,7 Hap 29 34,2 15,5 22,5 27,8 Hap 30 32,6 12,8 26,7 27,8 Hap 31 37,7 15,0 24,2 23,0 Hap 32 37,2 14,4 25,7 22,8 Hap 33 34,2 16,8 22,2 26,8 Hap 34 36,4 14,8 24,3 24,5 Hap 35 36,2 14,8 24,5 24,5 Hap 36 37,6 13,6 24,4 24,4 Hap 37 35,4 15,2 24,6 24,8 Hap 38 36,6 14,3 25,6 23,5 Hap 39 36,0 15,6 23,5 24,9 Hap 40 36,0 15,6 23,7 24,7 Hap 41 35,1 16,4 22,4 26,2 Hap 42 36,5 14,8 24,0 24,8 Hap 43 35,0 15,9 25,2 23,9 Hap 44 34,9 16,9 23,2 24,9 Hap 45 36,7 14,8 25,1 23,4 Hap 46 36,5 15,2 24,8 23,4 Hap 47 36,9 16,7 23,4 23,0 Μέσος όρος 35,7 15,3 24,4 24,6 101
108 Πίνακας Γ-14: Γενετικός τόπος COI. Παρατίθενται τα ονόματα των απλοτύπων, ο αριθμός ατόμων σε κάθε απλότυπο, ο πληθυσμός προέλευσης των ατόμων και το είδος στο οποίο ανήκουν. ΑΠΛΟΤΥΠΟΣ ΑΡΙΘΜΟΣ ΑΤΟΜΩΝ (Ν)/ΑΠΛΟΤΥΠΟ ΑΤΟΜΟ ΕΙΔΟΣ Hap_1 2 AIN 1,4 Hap_2 2 AIN2-3 Hap_3 25 ALF1-5,7, CHb2-4, VLA1-3,PROU2, PAN1-6,10-11, CH1,3-5 Hap_4 3 AMG1b,c,cA Hap_5 1 AMG1a Hap_6 9 BEL1, FRX1, KLM3-4, VTS1,3-4, ZRL3,7 Hap_7 3 DRM1 PAP1,3 Hap_8 10 ELTa1,-4, FEN1,3, ZIR3,, THEO1, PLAN3-4, Hap_9 3 ELTb1,3-4 Hap_11 14 FEN2, LAG1, LIB1,2,4,5, VLK1, ZIR2, THEO4,6, PLAN2,5, MIK2,6, Hap_12 1 FLA2 Hap_13 1 FNR1 Hap_14 1 FNR3 Hap_15 3 FRX2-4 Hap_16 1 GAB1 Hap_17 1 GAB2 Hap_18 2 GRP1,4 Hap_19 4 KON4 SSN1,3,4 Hap_20 3 KRN2, VOI1 ZRL6 Hap_21 4 KRT1-4 Hap_22 7 KAT1,2, PER1-2 TPT1-3 Hap_23 1 LIB3 Trachelipus kytherensis 102
109 Hap_24 2 MNS1-2 Hap_25 18 PMS1-4, RR1-4 TAYa1-3, TAYb1-3 TAYc1,3,4, VLA2 Hap_26 3 SOL1-3 Hap_27 4 STE1-4 Hap_28 2 TAYc2 Hap_29 1 VOI3 Hap_30 1 ZRL5 Hap_31 1 VEL2 Hap_32 1 VEL1 Hap_33 4 PROU1,3-5, Hap_34 1 THEO5 Hap_35 3 PLAN1,6 MIK4 Hap_36 1 PAN8 Hap_37 3 MIK1,3,5 Hap_38 6 LAMB1-6 Hap_42 2 KAL2-3 Hap_43 1 KAL1 Hap_44 14 CH2,6 BOU1-12 Hap_39 1 KYT4 Hap_40 3 KYT1-3 Hap_41 4 CRE3-6 Hap_10 6 EPD, NAX Hap_45 1 STO1 Hap_46 4 STN1,2 AGA2,3 Hap_47 1 Trachelipus kytherensis T. aegaeus T.n.sp. Armadillidium lobocurvum Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εύβοια Εξωομάδα Σημειώνεται πως μετά από τις πρώτες προκαταρκτικές αναλύσεις, οι απλότυποι Hap 12,13,28 (επισημαίνονται με κόκκινο χρώμα) εξαιρέθηκαν από το σύνολο δεδομένων καθώς παρουσίαζαν ιδιαίτερα προβλήματα τόσο στη στοίχιση όσο και στις γενετικές αποστάσεις που εμφάνιζαν με τα υπόλοιπα δείγματα. 103
110 Η νουκλεοτιδική ποικιλότητα (π) και η απλοτυπική ποικιλότητα (h) στις μελετηθείσες αλληλουχίες ήταν π=0,126 και h=0,951. Ειδικά για τα άτομα Trachelipus sp. οι τιμές τις νουκλεοτιδικής και απλοτυπικής ποικιλότητας παρατίθενται στον πίνακα Γ-15. Πίνακας Γ-15: Οι τιμές τις νουκλεοτιδικής και απλοτυπικής ποικιλότητας για τους αντιπροσώπους του γένους Trachelipus. Τάξον Νουκλεοτιδική ποικιλότητα (π) Απλοτυπική ποικιλότητα (h) Αριθμός Απλοτύπων Trachelipus sp. 0,125 0, Trachelipus kytherensis 0,122 0, Trachelipus aegaeus ,500 2 Trachelipus n.sp , Έλεγχος Ομοπλασίας Όπως και στην περίπτωση του γονιδίου 16S rrna (βλ Γ ), προκειμένου να διασφαλιστεί η ορθολογική χρήση των αλληλουχικών δεδομένων της υπομονάδας Ι της κυτοχρωμικής οξειδάσης, πραγματοποιήθηκε ο έλεγχος κορεσμού με το πρόγραμμα Dambee (Xia & Xie 2001) που χρησιμοποιεί τον έλεγχο του δείκτη κορεσμού υποκατάστασης (Xia et al. 2003). Από τον έλεγχο προέκυψε πως I < SS C SS I. και μάλιστα στατιστικώς σημαντικά, οπότε και οι αλληλουχίες μπορούν να χρησιμοποιηθούν για φυλογενετικές αναλύσεις. Ακολούθως παρατίθενται αναλυτικά τα αποτελέσματα του εν λόγω ελέγχου (Πιν. Γ-16). 104
111 Πίνακας Γ-16: Τα αποτελέσματα του ελέγχου των Xia et al. 2003) για τη διερεύνηση του κορεσμού υποκατάστασης των βάσεων στις μελετώμενες αλληλουχίες του γενετικού τόπου COI mtdna. NumOTU Iss Iss.c DF P 4 0,153 0, <0, ,162 0, <0, ,161 0, <0, ,168 0, <0, Εκτίμηση γενετικών αποστάσεων Οι γενετικές αποστάσεις μεταξύ των υπό εξέταση αλληλουχιών εκτιμήθηκαν με το διπαραμετρικό μοντέλο Kimura (K2P). Ο υπολογισμός των αποστάσεων πραγματοποιήθηκε με το πρόγραμμα MEGA v.4 (Kumar et al. 2004). Στους πίνακες που ακολουθούν, παρατίθενται οι νουκλεοτιδικές αποκλίσεις μεταξύ των απλοτύπων που εξετάστηκαν για τους αντιπροσώπους του γένους Trachelipus της παρούσας μελέτης. Οι γενετικές αποστάσεις υπολογίστηκαν τόσο μεταξύ των διαφορετικών ειδών (Πιν. Γ-17) όσο και μεταξύ των διαφορετικών γεωγραφικών ενοτήτων (Πιν. Γ-18) από όπου συλλέχθηκαν τα δείγματα. Στη δεύτερη περίπτωση, οι μελετώμενοι πληθυσμοί ομαδοποιήθηκαν ανάλογα με τη γεωγραφική τους προέλευση στις ακόλουθες ομάδες: Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (πλην Πελοποννήσου) Νησιωτικές περιοχές Αιγαίου Εύβοια 105
112 Πίνακας Γ-17: Οι μέσες νουκλεοτιδικές αποκλίσεις (σε παρενθέσεις οι καθαρές μέσες νιουκλεοτιδικές αποκλίσεις) μεταξύ των τριών ειδών Trachelipus και της εξωομάδας (K2P) T. aegaeus T. n.sp. εξωομάδα για το γενετικό τόπο COI. SP T. kytherensis T. aegaeus T. n.sp. εξωομάδα 0,175 (0,064) 0,174 (0,103) 0,261 (0,191) - 0,200 (0,159) 0,279 (0,238) - 0,226 (0,226) - Πίνακας Γ-18: Οι μέσες νουκλεοτιδικές αποκλίσεις (σε παρενθέσεις οι καθαρές μέσες νουκλεοτιδικές αποκλίσεις) μεταξύ των γεωγραφικών ομάδων των αντιπροσώπων Trachelipus και της εξωομάδας (K2P) για το γενετικό τόπο COI. ΓΕΩΓΡΑΦΙΚΗ ΟΜΑΔΑ Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα Εύβοια Αιγαίο εξωομάδα Λοιπή ηπειρωτική Ελλάδα 0,158 (0,034) - Εύβοια 0,198 (0,082) 0,213 (0,085) - Αιγαίο 0,177 (0,044) 0,184 (0,039) 0,188 (0,051) - εξωομάδα 0,256 (0,119) 0,273 (0,206) 0,296 (0,236) 0,253 (0,176) - Οι πίνακες γενετικών αποστάσεων μεταξύ όλων των μελετούμενων απλοτύπων (Π-3) και μεταξύ όλων των πληθυσμών ανά ζεύγη (Π-4), βάσει του μοντέλου δύο παραμέτρων του Kimura (1980), παρατίθενται στο παράρτημα. Η γενετική απόσταση, d, με βάση το μοντέλο του Kimura (1980) κυμαινόταν από 0 (DRM-PAP) ως 0,273 (DRM,PAP-MSN, DRM,PAP-SOL, DRM,PAP-VOU). Οι 106
113 γενετικές αποστάσεις που χωρίζουν τα είδη του γένους Trachelipus μεταξύ τους, σύμφωνα πάντα με την ισχύουσα ταξινόμηση, κυμαίνονταν: μεταξύ 0,080 (T. aegaeus - KYT) και 0,209 (T. aegaeus - SOL) και μεταξύ 0,135 [T. n.sp. - πληθυσμοί της βόρειας Εύβοιας (PAP, DRM)] και 0,225 [(T. n.sp. - πληθυσμοί της Ηπείρου (KAL, BEL, VTS)]. Οι αποστάσεις μεταξύ των αντιπροσώπων του γένους και της εξωομάδας κυμαίνονταν από 0,212 (A. lobocurvum-gav) μέχρι 0,313 (A. lobocurvum KLM). Τέλος, στον ακόλουθο πίνακα Γ-19 παρουσιάζεται η μέση νουκλεοτιδική διαφοροποίηση των πληθυσμών μελέτης. Πίνακας Γ-19. Η μέση ενδοπληθυσμιακή γενετική διαφοροποίηση (απόσταση Kimura,1980) των πληθυσμών για το γενετικό τόπο COI mtdna. Πλ/σμός Ενδοπλ/κή διαφ/ση Πλ/σμός Ενδοπλ/κή διαφ/ση Πλ/σμός Ενδοπλ/κή διαφ/ση Πλ/σμός Ενδοπλ/κή διαφ/ση AIN 0,010 KLM 0,000 PMS 0,000 VTS 0,000 ALF 0,000 KON n/c PRR 0,000 ZIR 0,007 AMG 0,004 KRN n/c SOL 0,000 ZRL 0,129 BEL n/c KRT 0,000 SSN 0,000 VEL 0,045 CH 0,082 KTR 0,000 STE 0,000 PROU 0,085 DRM n/c LAG n/c TAY 0,043 THEO 0,007 ELT 0,008 LIB 0,006 TPT 0,000 PLAN 0,008 EPD 0,000 MNS 0,000 VLA 0,000 PAN 0,002 FEN 0,005 PAP 0,000 VLK 0,015 T.aegaeus 0,083 FNR n/c PER 0,000 VOI 0,154 T. n.sp. 0,000 KYT 0,004 KAL 0,015 MIK 0,020 CRE 0,000 BOU 0,000 LAMB 0,
114 Φαινετικές και φυλογενετικές σχέσεις των μελετηθέντων πληθυσμών Για λόγους ευκολίας, η κατασκευή των φυλογενετικών δένδρων πραγματοποιήθηκε με τη χρήση των απλοτύπων που εξήχθησαν. Τα δένδρα που προέκυψαν από τις διάφορες φυλογενετικές αναλύσεις παρουσιάζονται παρακάτω. Αρχικά παρουσιάζονται τα αποτελέσματα της μεθόδου Σύνδεσης Γειτόνων, ακολουθούν αυτά της Μέγιστης Φειδωλότητας και εν συνεχεία αυτά της Μπεϊεσιανής Συμπερασματολογίας. Το δένδρο με τη μέθοδο της Σύνδεσης Γειτόνων (NJ) (Εικ. Γ-7) κατασκευάστηκε με το πρόγραμμα Mega 4.0 (Kumar et al. 2004) βάσει του διπαραμετρικού μοντέλου K2P του Kimura (1980). Η αξιοπιστία των κλάδων ελέγχθηκε με πραγματοποίηση 1000 ψευδοεπαναλήψεων (bootstrap). Την ίδια τοπολογία κλάδων εμφάνιζε και η μέθοδος UPGMA. Το δένδρο που παρουσιάζεται στην Εικόνα Γ-8 είναι αυτό που προέκυψε από μια σταθμισμένη ανάλυση Μέγιστης Φειδωλότητας. Συγκεκριμένα, είναι το 50% πλειοψηφικής αποδοχής δένδρο Μέγιστης Φειδωλότητας, το οποίο προέκυψε χρησιμοποιώντας την ευρετική μέθοδο αναζήτησης του πιο φειδωλού δένδρου στο πρόγραμμα PAUP (v4.0.beta10, Swofford 2002). Τέλος, στην Εικόνα Γ-9, παρατίθεται το δέντρο όπως προέκυψε από την ανάλυση της Μπεϊεσιανής Συμπερασματολογίας (BI) με τη βοήθεια του προγράμματος MrBayes v3.1.2 (Huelsenbeck & Ronquist 2001). Το εξελικτικό μοντέλο που χρησιμοποιήθηκε, όπως αυτό προέκυψε από το πρόγραμμα Modeltest, ήταν το HKY+I+G (με -lnl = ), με ποσοστό αμετάβλητων θέσεων 0,475 και παράμετρο για την μεταβολή γάμμα των μεταβλητών θέσεων α=0,662. Οι συχνότητες των νουκλεοτιδίων ήταν: Α= 0,2470, C= 0,1429, G= 0,2219 και T= 0,
115 Ο αριθμός παραμέτρων που λήφθηκαν υπόψη ήταν Κ=6 με λόγο μεταστροφών/μεταπτώσεων (Ti/tv ratio) = 6,1032. Ο συνολικός αριθμός γενεών ήταν 10 6, τα δέντρα σώζονταν ανά 100 γενεές, από τα οποία τελικά απορρίφθηκε το αρχικό 25% ως burn-in. 109




116 Εικονα Γ-7: Γενετικός τόπος COI. Δένδρο NJ βάσει του διπαραμετρικού μοντέλου του Kimura. Οι αριθμοί στους κλάδους αντιστοιχούν στις τιμές bootstrap (1000 επαναλήψεις) ) που υπερβαίνουν το 50%. Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ- 14. Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εύβοια Εξωομάδα 110


117 Εικόνα Γ-8: Κλαδόγραμμα Μέγιστης Φειδωλότητας (ΜΡ) με βάση στοιχισμένες αλληλουχίες μήκους 512bp του γονιδίου COI mtdna που ομαδοποιήθηκαν σε 44 απλοτύπους. Οι τιμές στους κλάδους αναφέρονται σε % ποσοστά ελέγχου 1000 bootstrap. Παρουσιάζονται μόνο τιμές bootstrap μεγαλύτερες του 50%. (Tree length=758, CI=0,443,RI=0,726, HI=0,557). Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ-14. Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) 111 Εξωομάδα Εύβοια
 ) με βάση στοιχισμένες αλληλουχίες μήκους 512bp του γονιδίου COI mtdna που ομαδοποιήθηκαν σε 44")

118 Εικόνα Γ-9: Κλαδόγραμμα Μπεϊεσιανής συμπερασματολογίας (B.I.) ) με βάση στοιχισμένες αλληλουχίες μήκους 512bp του γονιδίου COI mtdna που ομαδοποιήθηκαν σε 44 απλοτύπους Οι αριθμοί στους κλάδους δηλώνουν τις εκ των υστέρων πιθανότητες. Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ-14. Πελοπόννησος Εύβοια Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εξωομάδα 112
119 Ανάλυση Μοριακής Διακύμανσης (AMOVA) Ομαδοποίηση βάσει γεωγραφικής προέλευσης Πραγματοποιήθηκε ομαδοποίηση των 63 μελετούμενων πληθυσμών του γένους ανάλογα με τη γεωγραφική τους προέλευση. Έτσι διακρίθηκαν οι ακόλουθες ομάδες: Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (πλην Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο+Εύβοια) Αν εξαιρεθούν λοιπόν τα άτομα που χρησιμοποιήθηκαν ως εξωομάδα και εμφανίζουν την αναμενόμενη γενετική απόκλιση από τους πληθυσμούς του γένους Trachelipus, αυτό που απομένει είναι να ελεγχθούν οι διαπληθυσμιακές και ενδοπληθυσμιακές αποκλίσεις των πληθυσμών του εν λόγω γένους. Πίνακας Γ-20: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για το γενετικό τόπο COI μεταξύ όλων των πληθυσμών του γένους Trachelipus της παρούσας μελέτης. Πηγή διακύμανσης με εξωομάδες Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 22,40% διαπληθυσμιακά, εντός της ίδιας ομάδας 64,78% ενδοπληθυσμιακά 12,82% Προκειμένου να διερευνηθούν τα ποσοστά μοριακής διακύμανσης αποκλειστικά μεταξύ των πληθυσμών του Trachelipus kytherensis που αποτέλεσε το κυρίαρχο (αριθμητικά) είδος στο πλαίσιο αυτής της μελέτης, πραγματοποιήθηκε AMOVA μεταξύ των ίδιων γεωγραφικών ομάδων, η οποία όμως περιλάμβανε μόνο τους αντιπροσώπους του είδους αυτού. 113
120 Πίνακας Γ-21: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για το γενετικό τόπο COI μεταξύ όλων των πληθυσμών του είδους Trachelipus kytherensis της παρούσας μελέτης. Πηγή διακύμανσης με εξωομάδες Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 22,60% διαπληθυσμιακά, εντός της ίδιας ομάδας 63,23% ενδοπληθυσμιακά 14,17%. Η AMOVA έδειξε πως η διακύμανση είναι μεγαλύτερη μεταξύ των πληθυσμών της ίδιας ομάδας παρά μεταξύ των διαφορετικών γεωγραφικών ομάδων, καταδεικνύοντας έτσι τη μεγάλη ποικιλομορφία που εμφανίζουν οι αντιπρόσωποι τόσο συνολικά του γένους Trachelipus όσο και οι αντιπρόσωποι του είδους Trachelipus kytherensis. Όπως και στην περίπτωση του γενετικού τόπου 16S rrna, και εδώ παρατηρήθηκε πως όταν η ανάλυση επαναλήφθηκε θεωρώντας την Εύβοια ως ξεχωριστή γεωγραφική ενότητα δεν σημειώθηκαν αξιόλογες διαφοροποιήσεις στα ποσοστά της μοριακής διακύμανσης, καθώς αυτά μεταβλήθηκαν μειούμενα μόλις κατά 0,5% Ομαδοποίηση βάσει φυλογενετικής συγγένειας Η ομαδοποίηση στην προκειμένη περίπτωση έγινε με βάση τις ομάδες που προέκυψαν από τη φυλογενετική ανάλυση για το γενετικό τόπο COI mtdna. Οι ομάδες αυτές διαμορφώθηκαν ως εξής: PEL mix = απλότυποι από όλη την ηπειρωτική Ελλάδα αλλά στην πλειονότητά τους από την Πελοπόννησο 114
121 PEL = απλότυποι μόνο από περιοχές της Πελοποννήσου AEGEAN = οι απλότυποι κυρίως από περιοχές του Αιγαίου + ο απλότυπος των ατόμων της Επιδαύρου + ο απλότυπος της Κ. Εύβοιας EUBOIA = οι απλότυποι της Β. Εύβοιας MAINLAND = μόνο απλότυποι της ηπειρωτικής περιοχής (πλην της Πελ/σου) MAINLAND_mix = απλότυποι από όλη την ηπειρωτική Ελλάδα αλλά στην πλειονότητά τους απλότυποι της ηπειρωτικής περιοχής (πλην της Πελ/σου). Πίνακας Γ-22: Οι απλότυποι που αντιστοιχούν σε κάθε φυλογενετική ομάδα όπως αυτές διαμορφώθηκαν από τη φυλογενετική ανάλυση του γενετικού τόπου COI. Ομάδα PEL_mix PEL AEGEAN Απλότυπος Hap1 Hap2 Hap3 Hap13 Hap18 Hap19 Hap27 Hap33 Hap35 Hap4 Hap5 Hap8 Hap9 Hap11 Hap21 Hap23 Hap31 Hap32 Hap34 Hap39 Hap40 Hap10 Hap25 Hap36 Hap37 Hap38 Hap42 N. EUBOIA Hap7 Hap43 MAINLAND Hap12 Hap14 Hap15 Hap28 Hap 29 MAINLAND_mix Hap6 Hap16 Hap17 Hap20 Hap22 Hap24 Hap26 Hap30 Hap41 Αν εξαιρεθούν λοιπόν τα άτομα που χρησιμοποιήθηκαν ως εξωομάδα και εμφανίζουν την αναμενόμενη γενετική απόκλιση από τους πληθυσμούς του γένους Trachelipus, αυτό που απομένει είναι να ελεγχθούν οι διαπληθυσμιακές και ενδοπληθυσμιακές αποκλίσεις των προαναφερθέντων ομάδων του εν λόγω γένους. 115
122 Πίνακας Γ-23: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για το γενετικό τόπο COI μεταξύ των φυλογενετικών ομάδων του γένους Trachelipus της παρούσας μελέτης. Πηγή διακύμανσης Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 38,96 διαπληθυσμιακά, εντός της ίδιας ομάδας 19,35 ενδοπληθυσμιακά 41,69 Το συμπέρασμα που εξάγεται από αυτήν την ανάλυση είναι ότι η μεγάλη γενετική απόκλιση που παρατηρείται προέρχεται κυρίως από τις ενδοπληθυσμιακές διαφορές και από τις διαφορές των πληθυσμών μεταξύ των διαφορετικών κλάδων (ομάδων) όπως αυτοί προέκυψαν από τη φυλογενετική ανάλυση, και σε πολύ μικρό βαθμό από τις διαπληθυσμιακές διαφορές εντός των κλάδων (ομάδων). Αναφορικά με τη διακύμανση μέσα στους απλότυπους του ίδιου κλάδου (ενδοποληθυσμιακή διακύμανση), η τιμή θα μπορούσε να δικαιολογηθεί λόγω του ότι οι απλότυποι που ομαδοποιήθηκαν στις φυλογενετικές αναλύσεις μέσα στον ίδιο κλάδο πολλές φορές αντιπροσώπευαν άτομα ή και πληθυσμούς που είναι αρκετά απομονωμένοι μεταξύ τους και που θεωρητικά θα ήταν αδύνατο με φυσικό τρόπο να βρεθούν μαζί. Τέτοιο παράδειγμα αποτελεί ο απλότυπος Hap3 στον οποίο ομαδοποιούνται άτομα από την Πελοπόννησο μαζί με ένα άτομο από τον πληθυσμό του Προυσσού (PROU) από τη Στερεά Ελλάδα. Παραμένει, πάντως, σημαντική, υποδηλώνοντας πολύ μεγάλη διαφοροποίηση μεταξύ των πληθυσμών, παρά τη γενεαλογική τους συγγένεια. Για τη διαπληθυσμιακή διακύμανση μπορούμε να πούμε πως η υψηλή της τιμή καταδεικνύει μεγάλη ποικιλπότητα στους αντιπροσώπους του Trachelipus ακόμα και μεταξύ πολύ κοντινών γεωγραφικών περιοχών. Εξάλλου η ομαδοποίηση αυτή καθαυτή πραγματοποιήθηκε βάση των κλάδων όπως αυτή προέκυψε από τη ανάλυση 116
123 της φυλογένεσης, η οποία κατέδειξε πως δεν υπάρχει γεωγραφική ομαδοποίηση των μελετούμενων πληθυσμών. Ωστόσο εμφανίζει ενδιαφέρον πως η τιμή στην περίπτωση του γενετικού τόπου COI (19,35%) είναι κατά πολύ μεγαλύτερη από την αντίστοιχη διαπληθυσμιακή τιμή διακύμανσης του γενετικού τόπου 16S rrna (3,27%). Κάτι τέτοιο μπορεί να οφείλεται στον ταχύτερο ρυθμό εξέλιξης του γονιδίου COI, οπότε και είναι ανιχνεύσιμες ακόμα και πολύ πρόσφατες αλλαγές στις αλληλουχίες που μελετώνται, έτσι ώστε αυτές να απεικονίζονται στην παρούσα ανάλυση με τη μεγαλύτερη τιμή της μοριακής διακύμανσης. 117
124 1.3. ΣΥΝΔΥΑΣΜΕΝΗ ΑΝΑΛΥΣΗ Προκειμένου να εξεταστεί αν τα δύο μιτοχονδριακά γονίδια μπορούν να συνδυαστούν για τις περαιτέρω αναλύσεις, χρησιμοποιήθηκε ο έλεγχος της «ασυμφωνίας διαφοράς μήκους» των μελετούμενων αλληλουχιών (Incongruence Length Difference ILD, Μickevich & Farris 1981, Farris et al. 1994) (βλ Μεθοδολογία Υλικά). Στην παρούσα μελέτη ελέγχτηκε η σημαντικότητα του ILD με τον έλεγχο ομοιογένειας των δεδομένων (partition homogeneity test) στο πρόγραμμα PAUP* v4.0 beta10 (Swofford 2002), με 100 επαναλήψεις. Το τελικό αποτέλεσμα του ελέγχου ήταν P=0,01, τιμή η οποία δεν υπερβαίνει το όριο σημαντικότητας P=0,05. Ωστόσο οι Sullivan 1996 και Cunningham (1997a) υποστηρίζουν πως η τιμή του P=0,05 είναι πολύ συντηρητική για το ILD. Ο Cunningham (1997a), συγκρίνοντας τρεις διαφορετικούς δείκτες ως προς την αξιοπιστία τους για το συνδυασμό ή μη δεδομένων, υποστηρίζει πως μόνο όταν ο ILD ανίχνευε τιμές P<0,001 τα δεδομένα δεν έπρεπε να συνδυάζονται. Ακολουθώντας αυτή την παραδοχή, η τιμή (P=0,01) του έλεγχου ομοιογένειας των δεδομένων (partition homogeneity test) θεωρήθηκε ικανοποιητική, προκειμένου να πραγματοποιηθεί συνδυασμένη ανάλυση Νουκλεοτιδική σύσταση και γενετική ποικιλότητα Η συνδυασμένη ανάλυση περιελάμβανε τα 176 κοινά άτομα από τους πληθυσμούς της παρούσας μελέτης, για τα οποία προσδιορίστηκε επιτυχώς η αλληλουχία και για τα δύο μιτοχονδριακά γονίδια. Το τελικό αρχείο με το σύνολο των δεδομένων από τα δύο μιτοχονδριακά γονίδια δημιουργήθηκε με χρήση των προγραμμάτων Mega v4.0 (Kumar et al. 2004) και DNAsp v4. (Rozas et al. 2003), 118

125 προσέχοντας η σειρά και ο αριθμός των αλληλουχιών να είναι κοινά και για τους δύο γενετικούς τόπους. Το τελικό μήκος των στοιχισμένων αλληλουχιών που χρησιμοποιήθηκε στις περαιτέρω αναλύσεις, είχε μήκος 898 βάσεων. Εικόνα Γ-10: Το διάγραμμα της εντροπίας σε σχέση με τη θέση κάθε νουκλεοτιδίου για τη συνδυασμένη ανάλυση των δύο γενετικών τόπων. Προηγείται ο γενετικός τόπος COI και ακολουθεί το 16S rrna. Από τις 898 θέσεις, οι 508 ήταν συντηρημένες (56,57%), οι 394 διαφοροποιούνταν μεταξύ των αλληλουχιών (43,88%), ενώ οι θέσεις που έδιναν πληροφορίες για τη φειδωλότητα ήταν 308 (34,3%). Σε ό,τι αφορά μόνο τις αλληλουχίες των αντιπροσώπων του γένους Trachelipus, οι αντίστοιχοι αριθμοί είναι 549 (61,14%), 353 (39,31%) και 298 (33,18%). Τα ποσοστά των βάσεων στις αλληλουχίες κατά μέσο όρο ήταν 29,9% Α, 34,7% Τ, 15,1% C και 20,4% G (Πιν. Γ- 24). Μετά τις απαραίτητες διορθώσεις και τη στοίχιση, ακολούθησαν οι περαιτέρω αναλύσεις όπως και στην περίπτωση της ανεξάρτητης ανάλυσης. 119
126 Από τις 176 αλληλουχίες ατόμων που λήφθηκαν για το σύνολο των δύο γονιδίων, οι 70 (Πιν. Γ-25) αντιστοιχούσαν σε διαφορετικούς απλοτύπους, σύμφωνα με το πρόγραμμα DNAsp (Rozas et al. 2003). Πίνακας Γ-24: Τα ποσοστά των βάσεων των απλοτύπων, καθώς και Ο μέσος όρος αυτών για κάθε απλότυπο Hap# %T %C %A %G Hap 1 35,1 14,2 31,0 19,7 Hap 2 35,5 13,9 30,9 19,8 Hap 3 35,5 14,1 30,3 20,1 Hap 4 35,4 14,6 29,8 20,2 Hap 5 35,3 14,7 29,8 20,2 Hap 6 32,8 16,3 28,9 21,9 Hap 7 33,8 16,5 29,5 20,3 Hap 8 35,2 14,6 31,0 19,1 Hap 9 35,7 14,7 29,6 20,0 Hap 10 34,4 15,0 29,9 20,7 Hap 11 34,5 14,9 30,1 20,4 Hap 12 34,1 15,6 30,0 20,3 Hap 13 34,9 14,5 30,0 20,5 Hap 14 34,6 14,7 30,2 20,5 Hap 15 36,3 14,0 30,4 19,3 Hap 16 32,8 16,1 29,4 21,8 Hap 17 35,0 13,9 31,2 19,9 Hap 18 34,4 15,7 30,3 19,5 Hap 19 35,1 14,9 29,5 20,4 Hap 20 35,2 14,7 29,0 21,1 Hap 21 34,5 14,7 28,2 22,6 Hap 22 33,1 16,6 30,0 20,3 Hap 23 36,0 13,3 30,9 19,9 Hap 24 35,5 14,1 30,4 20,0 Hap 25 34,9 15,3 29,3 20,5 Hap 26 35,1 15,1 29,4 20,4 Hap 27 34,0 15,3 30,3 20,4 Hap 28 35,6 13,9 31,1 19,4 Hap 29 35,1 15,1 29,5 20,3 Hap 30 34,9 14,9 30,1 20,2 Hap 31 34,8 14,9 29,7 20,6 Hap 32 35,8 14,8 29,0 20,3 Hap 33 35,6 15,0 29,2 20,2 120
127 Hap 34 35,4 15,0 29,6 20,1 Hap 35 33,8 16,4 29,1 20,8 Hap 36 35,7 14,3 30,2 19,8 Hap 37 35,5 14,2 30,4 20,0 Hap 38 35,7 14,8 29,4 20,0 Hap 39 33,6 16,0 31,3 19,1 Hap 40 35,6 14,4 29,2 20,8 Hap 41 35,0 14,9 30,2 20,0 Hap 42 35,0 14,2 30,4 20,4 Hap 43 33,7 16,2 29,2 20,8 Hap 44 33,6 16,3 29,2 20,8 Hap 45 33,4 15,7 28,8 22,0 Hap 46 33,4 15,7 28,7 22,1 Hap 47 34,2 16,1 29,1 20,6 Hap 48 35,6 14,7 29,2 20,5 Hap 49 35,7 14,5 29,2 20,6 Hap 50 33,1 16,5 29,0 21,4 Hap 51 35,1 14,0 31,4 19,5 Hap 52 33,1 16,6 29,0 21,3 Hap 53 33,3 16,4 28,3 22,0 Hap 54 33,7 16,0 30,1 20,2 Hap 55 34,3 15,6 30,0 20,1 Hap 56 34,6 15,5 29,9 20,0 Hap 57 35,7 14,3 30,6 19,3 Hap 58 35,6 14,6 30,5 19,3 Hap 59 35,9 14,4 30,5 19,2 Hap 60 36,1 14,8 29,3 19,7 Hap 61 34,6 15,5 31,3 18,6 Hap 62 35,0 14,5 29,9 20,7 Hap 63 35,4 13,8 30,8 20,0 Hap 64 33,9 15,4 28,7 22,0 Hap 65 33,4 15,8 28,8 22,0 Hap 66 34,8 15,0 29,9 20,4 Hap 67 33,1 16,0 28,8 22,0 Hap 68 32,2 14,6 34,5 18,7 Hap 69 33,9 15,4 29,1 21,7 Hap 70 36,0 15,4 30,1 18,5 Μέσος όρος 34,7 15,1 29,9 20,4 121
128 Πίνακας Γ-25: Συνδυασμένη ανάλυση. Οι απλότυποι, όπως προέκυψαν για τη συνδυασμένη ανάλυση των δύο γενετικών τόπων. Παρατίθενται ο αριθμός ατόμων σε κάθε απλότυπο, ο πληθυσμός προέλευσης των ατόμων και το είδος στο οποίο ανήκουν. ΑΠΛΟΤΥΠΟΣ ΑΡΙΘΜΟΣ ΑΤΟΜΩΝ (Ν)/ΑΠΛΟΤΥΠΟ ΑΤΟΜΟ Hap_1 2 AIN 1,4 Hap_2 2 AIN2-3 Hap_3 10 ALF1,2,4,6 CH1,3,4 PAN10,11 Hap_4 1 AMG1a Hap_5 2 AMG1b,c Hap_6 3 BEL1 KLM3,4 Hap_7 14 BOU1-12 CH2,6 Hap_8 4 CH5 CHb2,3,4 Hap_9 1 DRM1 Hap_10 5 ELTa1-4 ZIR3 Hap_11 3 ELTb1,3,4 Hap_12 3 EPD1,2,3 Hap_13 2 FEN1,3 Hap_14 1 FEN2 Hap_15 1 FNR3 Hap_16 1 FRX1 Hap_17 3 FRX2,3,4 Hap_18 2 GRP1,4 Hap_19 1 KAL1 Hap_20 2 KAL2,3 Hap_21 1 KON4 Hap_22 4 CRE3,4,5,6 Hap_23 2 KRN2 ZRL6 Hap_24 4 KRT1-4 Hap_25 3 KYT1-3 Hap_26 1 KYT4 Hap_27 1 LAG1 Hap_28 6 LAMB1-6 Hap_29 3 LIB1,2,4 Hap_30 1 LIB3 Hap_31 1 LIB5 Hap_32 3 MIK1,3,5 Hap_33 4 MIK2,6 PLAN2,5 ΕΙΔΟΣ Trachelipus kytherensis 122
129 Hap_34 3 MIK4 PLAN1,6 Hap_35 2 MNS1,2 Hap_36 6 PAN1-6 Hap_37 1 PAN8 Hap_38 2 PAP1,3 Hap_39 2 PER1,2 Hap_40 2 PLAN3 THEO1 Hap_41 1 PLAN4 Hap_42 16 PMS1-3, PRR1-4 TAYa1-3, TAYb1-3 TAYC1,3, VLK2 Hap_43 2 SOL1,3 Hap_44 1 SOL2 Hap_45 2 SSN1,3 Hap_46 1 SSN4 Hap_47 4 STE1-4 Hap_48 2 THEO4,6 Hap_49 1 THEO5 Hap_50 2 PROU1,3 Hap_51 1 PROU2 Hap_52 1 PROU4 Hap_53 1 PROU5 Hap_54 2 TPT1,3 Hap_55 1 TPT2 Hap_56 3 NAX1,2,4 Hap_57 1 STO1 T. aegaeus Hap_58 4 AGA2,3 STN1,2 T. n.sp. Hap_59 1 VEL1 Hap_60 1 VEL2 Hap_61 3 VLA1,2,3 Hap_62 1 VLK1 Hap_63 1 VOI1 Hap_64 1 VOI3 Trachelipus kytherensis Hap_65 2 VTS3,4 Hap_66 1 ZIR2 Hap_67 1 ZRL3 Hap_68 1 ZRL5 Hap_69 1 ZRL7 Hap_70 1 PAN_N6 A. lobocurvum Trachelipus kytherensis Πελοπόννησος Εύβοια Νησιωτικές περιοχές (Αιγαίο) Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Εξωομάδες 123
130 Η νουκλεοτιδική ποικιλότητα (π) και η απλοτυπική ποικιλότητα (h) στις μελετηθείσες αλληλουχίες ήταν π=0,1023 και h=0,9754. Ειδικά για τα άτομα Trachelipus sp. οι τιμές τις νουκλεοτιδικής και απλοτυπικής ποικιλότητας παρατίθενται στον πίνακα Γ-26. Πίνακας Γ-26: Οι τιμές τις νουκλεοτιδικής και απλοτυπικής ποικιλότητας για τους αντιπροσώπους του γένους Trachelipus. Τάξον Νουκλεοτιδική ποικιλότητα (π) Απλοτυπική ποικιλότητα (h) Αριθμός Απλοτύπων Trachelipus kytherensis 0,0985 0, Trachelipus aegaeus 0,0340 0, Trachelipus n.sp. 0,0006 0, Trachelipus sp. 0,1007 0, Για λόγους ευκολίας, η κατασκευή των φυλογενετικών δένδρων πραγματοποιήθηκε με τη χρήση των απλοτύπων που προέκυψαν Έλεγχος Ομοπλασίας Οι πραγματικές εξελικτικές σχέσεις των υπό εξέταση αλληλουχιών ενδέχεται να μην αποκαλυφθούν εάν οι νουκλεοτιδικές θέσεις έχουν κορεστεί από πολλαπλές υποκαταστάσεις (Swofford et al. 2001). Προκειμένου να ελεγχθεί ο βαθμός κορεσμού των αλληλουχιών σε νουκλεοτιδικές υποκαταστάσεις πραγματοποιήθηκε ο έλεγχος του Xia (βλ. Μεθοδολογία Υλικά 4.2.) που χρησιμοποιεί το δείκτη κορεσμού υποκατάστασης (Xia et al. 2003) και περιλαμβάνεται στο πρόγραμμα Dambee (Xia & Xie 2001). Από τον έλεγχο προέκυψε πως I SS < SS C I. και μάλιστα στατιστικώς σημαντικά, οπότε και οι αλληλουχίες μπορούν να χρησιμοποιηθούν για 124
131 φυλογενετικές αναλύσεις. Ακολούθως παρατίθενται αναλυτικά τα αποτελέσματα του εν λόγω ελέγχου (Πιν. Γ-27). Πίνακας Γ-27: Τα αποτελέσματα του ελέγχου των Xia et al. 2003) για διερεύνηση του κορεσμού υποκατάστασης των βάσεων στις μελετώμενες αλληλουχίες για τη συνδυασμένη ανάλυση των δύο γενετικών τόπων. NumOTU Iss Iss.c DF P 4 0,286 0, <0, ,289 0, <0, ,309 0, <0, ,328 0, <0,001 Εφόσον οι αλληλουχίες των απλοτύπων δεν εμφάνισαν κορεσμό, ακολούθησε η κατασκευή των φυλογενετικών δένδρων με τις τρεις μεθόδους. Η χρήση των απλοτύπων πραγματοποιήθηκε και για λόγους ευκολίας στην κατασκευή των φυλογενετικών δένδρων Εκτίμηση γενετικών αποστάσεων Όπως και στην ανεξάρτητη ανάλυση, έτσι και εδώ οι γενετικές αποστάσεις μεταξύ των υπό εξέταση αλληλουχιών εκτιμήθηκαν με το διπαραμετρικό μοντέλο Kimura (K2P). Στους πίνακες Γ-28 και Γ-29 παρατίθενται οι νουκλεοτιδικές αποκλίσεις μεταξύ των απλοτύπων που εξετάστηκαν για τους αντιπροσώπους του γένους Trachelipus που συλλέχθηκαν στο πλαίσιο της παρούσας μελέτης. Οι γενετικές αποστάσεις υπολογίστηκαν τόσο μεταξύ των διαφορετικών ειδών (Πιν. Γ-28) όσο και μεταξύ των διαφορετικών γεωγραφικών ενοτήτων (Πιν. Γ-29) από όπου 125
132 συλλέχθηκαν τα δείγματα. Στη δεύτερη περίπτωση, οι μελετώμενοι πληθυσμοί ομαδοποιήθηκαν ανάλογα με τη γεωγραφική τους προέλευση στις ακόλουθες ομάδες: Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (πλην Πελοποννήσου) Νησιωτικές περιοχές Αιγαίου Εύβοια Πίνακας Γ-28: Οι νουκλεοτιδικές αποκλίσεις (σε παρενθέσεις οι καθαρές μέσες νουκλεοτιδικές αποκλίσεις) μεταξύ των τριών ειδών Trachelipus και της εξωομάδας (K2P) για το σύνολο των αλληλουχιών της συνδυασμένης ανάλυσης. SP T. kytherensis T. aegaeus T. n.sp. εξωομάδα T. aegaeus 0,138 (0,070) - T. n.sp. 0,137 (0,082) 0,060 (0,048) - εξωομάδα 0,294 (0,239) 0,268 (0,256) 0,251 (0,251) - 126
133 Πίνακας Γ-29: Οι μέσες νουκλεοτιδικές αποκλίσεις (σε παρενθέσεις οι καθαρές μέσες νουκλεοτιδικές αποκλίσεις) μεταξύ των γεωγραφικών ομάδων των αντιπροσώπων Trachelipus και της εξωομάδας (K2P) για τη συνδυασμένη ανάλυση. ΓΕΩΓΡΑΦΙΚΗ ΟΜΑΔΑ Εύβοια Λοιπή ηπειρωτική Ελλάδα Πελοπόννησος Αιγαίο εξωομάδες Λοιπή ηπειρωτική Ελλάδα 0,157 (0,032) - Πελοπόννησος 0,141 (0,063) 0,130 (0,065) - Αιγαίο 0,132 (0,054) 0,155 (0,057) 0,141 (0,054) - εξωομάδες 0,293 (0,252) 0,300 (0,245) 0,294 (0,227) 0,262 (0,249) - Οι πίνακες γενετικών αποστάσεων (d) μεταξύ όλων των μελετούμενων απλοτύπων και μεταξύ όλων των πληθυσμών, βάσει του μοντέλου δύο παραμέτρων του Kimura (1980) παρατίθενται ως παραρτήματα (πίνακες Π-5 και Π-6, αντίστοιχα). Η γενετική απόσταση, d, με βάση το μοντέλο του Kimura (1980) κυμαινόταν από 0 (BEL-KLM) ως 0,198 (BOU-ELT, BOU-LIB). Οι γενετικές αποστάσεις που χωρίζουν τα είδη του γένους Trachelipus μεταξύ τους, σύμφωνα πάντα με την ισχύουσα ταξινόμηση, κυμαίνονταν: μεταξύ 0,028 (T. aegaeus - KYT) και 0,179 (T. aegaeus - SOL) και μεταξύ 0,060 (T. n.sp. - T. aegaeus και 0,186 [(T. n.sp. - πληθυσμοί της Ηπείρου (KAL. BEL VTS)] Τέλος, οι αποστάσεις μεταξύ των αντιπροσώπων του γένους και της εξωομάδας κυμαίνονταν από 0,251 (A. lobocurvum- T. nsp.) μέχρι 0,318 (A. lobocurvum-ssn). 127
134 Τέλος, στον πίνακα Γ-30 παρουσιάζεται η μέση νουκλεοτιδική διαφοροποίηση των πληθυσμών μελέτης για το συνδυασμένο σύνολο δεδομένων. Πίνακας Γ-30. Η μέση ενδοπληθυσμιακή γενετική διαφοροποίηση (απόσταση Kimura, 1980) των πληθυσμών για τη συνδυασμένη ανάλυση. Πλ/σμός Ενδοπλ/κή διαφ/ση Πλ/σμός Ενδοπλ/κή διαφ/ση Πλ/σμός Ενδοπλ/κή διαφ/ση AIN 0,010 KYT 0,001 PROU 0,072 ALF 0,000 LAG n/c TPT 0,058 AMG 0,002 LAMB 0,000 VEL 0,035 BEL n/c LIB 0,005 VLA 0,000 BOU 0,000 MIK 0,008 VLK 0,012 CH 0,071 MNS 0,000 VOI 0,147 ELT 0,007 PAN 0,003 VTS 0,000 FEN 0,005 PER 0,000 ZIR 0,012 FNR n/c PLAN 0,004 ZRL 0,115 FRX 0,078 PMS 0,000 PAP 0,000 GRP 0,000 PRR 0,000 DRM n/c KAL 0,011 SOL 0,002 STE 0,000 KLM 0,000 SSN 0,002 EPD 0,000 KON n/c TAY 0,000 CRE 0,000 KRT 0,002 THEO 0,012 T. n.sp. 0,000 T. aegaeus
135 Φαινετικές και φυλογενετικές σχέσεις των μελετηθέντων πληθυσμών Το δένδρο που παρουσιάζεται στην Εικ. Γ-12 είναι αυτό που προέκυψε από μια σταθμισμένη ανάλυση Μέγιστης Φειδωλότητας. Συγκεκριμένα, είναι το 50% πλειοψηφικής αποδοχής δένδρο Μέγιστης Φειδωλότητας, το οποίο προέκυψε χρησιμοποιώντας την ευρετική μέθοδο αναζήτησης του πιο φειδωλού δένδρου στο πρόγραμμα PAUP (v4.0.beta10, Swofford 2002). Τέλος στην Εικ. Γ-13 παρατίθεται το δέντρο όπως προέκυψε από την ανάλυση της Μπεϊεσιανής Συμπερασματολογίας (BI) με τη βοήθεια του προγράμματος MrBayes v3.1.2 (Huelsenbeck & Ronquist 2001). Το εξελικτικό μοντέλο που χρησιμοποιήθηκε, όπως αυτό προέκυψε από το πρόγραμμα Modeltest, ήταν το GTR+I+G (με lnl= ), με ποσοστό αμετάβλητων θέσεων 0,4731 και παράμετρο για τη μεταβολή γάμμα των μεταβλητών θέσεων α= 0,7813. Οι συχνότητες των νουκλεοτιδίων ήταν: Α= 0,3109, C= 0,1449, G= 0,0932 και T= 0,3678. Ο αριθμός παραμέτρων που λήφθηκαν υπόψη ήταν Κ=6 με λόγο μεταστροφών/μεταπτώσεων (Ti/tv ratio) = 3,8636. Ο συνολικός αριθμός γενεών ήταν 10 6, τα δέντρα σώζονταν ανά 100 γενεές, από τα οποία τελικά απορρίφθηκε το αρχικό 25% ως burn-in. 129


136 Εικονα Γ-11: Δέντρο NJ βάσει του διπαραμετρικού μοντέλου του Kimura για τη συνδυασμένη ανάλυση των δύο γενετικών τόπων. Οι αριθμοί στους κλάδους αντιστοιχούν στις τιμές bootstrap (1000 επαναλήψεις) που υπερβαίνουν το 50%. Οι απλότυποι αντιστοιχούν στα άτομα του Πιν Γ-25. Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εύβοια Εξωομάδα 130




137 Εικόνα Γ-12: Κλαδόγραμμα μέγιστης φειδωλότητας (ΜΡ) με βάση στοιχισμένες αλληλουχίες μήκους 898bp του συνολικού πακέτου δεδομένων που ομαδοποιήθηκαν σε 70 απλότυπους. Οι τιμές στους κλάδους αναφέρονται σε % ποσοστά ελέγχου 1000 bootstrap. Παρουσιάζονται μόνο τιμές bootstrap μεγαλύτερες του 50%. (Tree length=1358, CI=0,443,RI=0,819, HI=0,557). Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ- 25. Πελοπόννησος Νησιωτικές περιοχές (Αιγαίο) Εύβοια Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) 131 Εξωομάδα




138 Εικόνα Γ-13: Κλαδόγραμμα Μπεϊεσιανής Συμπερασματολογίας (B.I.) ) με βάση στοιχισμένες αλληλουχίες μήκους 898bp των δύο γενετικών τόπων που ομαδοποιήθηκαν σε 70 απλότυπους Οι αριθμοί στους κλάδους δηλώνουν τις εκ των υστέρων πιθανότητες. Οι απλότυποι αντιστοιχούν στα άτομα του Πιν. Γ-25. Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (εκτός Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο) Εύβοια Εξωομάδα 132
139 Ανάλυση Μοριακής Διακύμανσης (AMOVA) Ομαδοποίηση βάσει γεωγραφικής προέλευσης Πραγματοποιήθηκε ομαδοποίηση των 63 μελετούμενων πληθυσμών του γένους ανάλογα με τη γεωγραφική τους προέλευση. Έτσι διακρίθηκαν οι ακόλουθες ομάδες: Πελοπόννησος Λοιπή ηπειρωτική Ελλάδα (πλην Πελοποννήσου) Νησιωτικές περιοχές (Αιγαίο+Εύβοια) Αν εξαιρεθούν, λοιπόν, τα άτομα που χρησιμοποιήθηκαν ως εξωομάδα και εμφανίζουν την αναμενόμενη γενετική απόκλιση από τους πληθυσμούς του γένους Trachelipus, αυτό που απομένει είναι να ελεγχθούν οι διαπληθυσμιακές και ενδοπληθυσμιακές αποκλίσεις των πληθυσμών του εν λόγω γένους. Πίνακας Γ-31: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για τη συνδυασμένη ανάλυση μεταξύ όλων των πληθυσμών του γένους Trachelipus της παρούσας μελέτης. Πηγή διακύμανσης με εξωομάδες Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 25,69% διαπληθυσμιακά εντός της ίδιας ομάδας 61,61% ενδοπληθυσμιακά 12,69% Προκειμένου να διερευνηθούν τα ποσοστά μοριακής διακύμανσης αποκλειστικά μεταξύ των πληθυσμών του Trachelipus kytherensis που αποτέλεσε το κυρίαρχο είδος στο πλαίσιο αυτής της μελέτης, πραγματοποιήθηκε AMOVA 133
140 μεταξύ των ίδιων γεωγραφικών ομάδων, η οποία όμως περιλάμβανε μόνο τους αντιπροσώπους του είδους αυτού. Πίνακας Γ-32: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για το γενετικό τόπο COI μεταξύ όλων των πληθυσμών του είδους Trachelipus kytherensis της παρούσας μελέτης. Πηγή διακύμανσης με εξωομάδες Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 23,82% διαπληθυσμιακά εντός της ίδιας ομάδας 63,12% ενδοπληθυσμιακά 13,06%. Η AMOVA έδειξε πως η διακύμανση είναι μεγαλύτερη μεταξύ των πληθυσμών της ίδιας ομάδας παρά μεταξύ των διαφορετικών γεωγραφικών ομάδων, καταδεικνύοντας έτσι τη μεγάλη ποικιλομορφία που εμφανίζουν οι αντιπρόσωποι τόσο συνολικά του γένους Trachelipus όσο και οι αντιπρόσωποι του είδους Trachelipus kytherensis. Όπως και στην περίπτωση της ανεξάρτητης ανάλυσης, έτσι και εδώ παρατηρήθηκε πως όταν η ανάλυση επαναλήφθηκε θεωρώντας την Εύβοια ως ξεχωριστή γεωγραφική ενότητα δεν σημειώθηκαν αξιόλογες διαφοροποιήσεις στα ποσοστά της μοριακής διακύμανσης, καθώς αυτά μεταβλήθηκαν μειούμενα μόλις κατά 0,5% Ομαδοποίηση βάσει φυλογενετικής συγγένειας Η ομαδοποίηση στην προκειμένη περίπτωση έγινε με βάση τις ομάδες που προέκυψαν από τη φυλογενετική ανάλυση. Οι ομάδες αυτές διαμορφώθηκαν ως εξής: 134
141 PEL mix = απλότυποι από όλη την ηπειρωτική Ελλάδα αλλά στην πλειονότητά τους από την Πελοπόννησο PEL = απλότυποι μόνο από περιοχές της Πελοποννήσου AEGEAN = οι απλότυποι κυρίως από περιοχές του Αιγαίου + ο απλότυπος των ατόμων της Επιδαύρου + ο απλότυπος της Κ. Εύβοιας EUBOIA = οι απλότυποι της Β. Εύβοιας MAINLAND = μόνο απλότυποι της ηπειρωτικής περιοχής (πλην της Πελ/σου MAINLAND_mix = απλότυποι από όλη την ηπειρωτική Ελλάδα αλλά στην πλειονότητά τους απλότυποι της ηπειρωτικής περιοχής (πλην της Πελ/σου). (MAN, SOL BOU CH) STE = ο απλότυπος του πληθυσμού της Κ. Εύβοιας που δεν ομαδοποιούταν με κανέναν άλλο οπότε και αντιμετωπίστηκε ως «διαφορετική» ομάδα. Πίνακας Γ-33: Οι απλότυποι που αντιστοιχούν σε κάθε φυλογενετική ομάδα όπως αυτές διαμορφώθηκαν από τη φυλογενετική ανάλυση του γενετικού τόπου COI. Ομάδα PEL_mix PEL AEGEAN Απλότυπος Hap1 Hap2 Hap3 Hap8 Hap17 Hap23 Hap24 Hap28 Hap36 Hap37 Hap51 Hap61 Hap63 Hap68 Hap4 Hap5 Hap10 Hap11 Hap13 Hap14 Hap19 Hap20 Hap27 Hap29 Hap30 Hap31 Hap32 Hap33 Hap34 Hap40 Hap41 Hap42 Hap48 Hap49 Hap62 Hap66 Hap22 Hap25 Hap26 Hap56 Hap58 Hap12 N. EUBOIA Hap9 Hap38 MAINLAND MAINLAND_mix Hap15 Hap59 Hap60 Hap6 Hap7 Hap16 Hap18 Hap21 Hap35 Hap39 Hap43 Hap44 Hap45 Hap46 Hap50 Hap52 Hap53 Hap54 Hap55 Hap64 Hap65 Hap67 Hap69 135
142 Πίνακας Γ-34: Αποτελέσματα Ανάλυσης Μοριακής Διακύμανσης (AMOVA) για τη συνδυασμένη ανάλυση των δύο γενετικών τόπων, μεταξύ των φυλογενετικών ομάδων του γένους Trachelipus της παρούσας μελέτης. Πηγή διακύμανσης Ποσοστό (%) διακύμανσης μεταξύ των ομάδων 64,35 διαπληθυσμιακά εντός της ίδιας ομάδας 2,60 ενδοπληθυσμιακά 33,05 Το συμπέρασμα που εξάγεται από αυτή την ανάλυση είναι λίγο πολύ αναμενόμενο: η μεγάλη γενετική απόκλιση που παρατηρείται προέρχεται κυρίως από τις διαφορές των πληθυσμών μεταξύ των διαφορετικών κλάδων (ομάδων) όπως αυτοί προέκυψαν από τη φυλογενετική ανάλυση, και σε πολύ μικρό βαθμό από τις διαπληθυσμιακές διαφορές εντός των κλάδων (ομάδων). Αξίζει να σημειωθεί βέβαια η αρκετά μεγάλη τιμή που λαμβάνει η ενδοπληθυσμιακή διακύμανση, ήτοι η διακύμανση μέσα στους απλότυπους του ίδιου κλάδου, η οποία και εδώ είναι ένδειξη της μεγάλης ποικιλομορφίας που εμφανίζουν τα άτομα ακόμα και μέσα στον ίδιο πληθυσμό ή ακόμα και μέσα στον ίδιο απλότυπο. Στην περίπτωση της συνδυασμένης ανάλυσης των δεδομένων παρατηρείται πως η τιμή της διαπληθυσμιακής μοριακής διακύμανσης είναι αρκετά χαμηλή, γεγονός που συνάδει με την καλή υποστήριξη των κλάδων όπως αυτοί διαμορφώθηκαν στη φυλογενετική ανάλυση. Υπενθυμίζεται πως η τιμή αυτή ήταν χαμηλή και στην περίπτωση του 16S rrna ενώ αντίθετα στην περίπτωση του γενετικού τόπου COI ήταν πολύ μεγαλύτερη. Κάτι τέτοιο μπορεί να σημαίνει πως η αλληλουχία νουκλεοτιδίων του 16S rrna έχει πολύ μεγαλύτερη συμβολή στην εξαγωγή των αποτελεσμάτων. 136
143 Δ. ΣΥΖΗΤΗΣΗ 1. ΠΛΗΘΟΣ ΑΠΛΟΤΥΠΩΝ Από το σύνολο των 227 ατόμων του γένους Trachelipus των οποίων προσδιορίστηκε η πρωτοταγής αλληλουχία για το γενετικό τόπο 16S rrna, προέκυψαν 67 διαφορετικοί απλότυποι (Πίν. Γ-2). Οι περισσότεροι πληθυσμοί από τους οποίους χρησιμοποιήθηκαν περισσότερα από ένα άτομα, έδειξαν ότι περιλαμβάνουν περισσότερους από έναν απλοτύπους. Αυτό είναι ενδεικτικό για τη μεγάλη ενδοπληθυσμιακή μιτοχονδριακή ποικιλότητα που χαρακτηρίζει το γένος Trachelipus. Το ίδιο φαινόμενο παρατηρείται και στην περίπτωση της υπομονάδας Ι της κυτοχρωμικής οξειδάσης (COI). Από τα 185 άτομα του γένους Trachelipus προέκυψαν 43 διαφορετικοί απλότυποι (Πιν. Γ-14), και ακόμα περισσότεροι πληθυσμοί έδειξαν ότι περιλαμβάνουν παραπάνω από έναν απλοτύπους. Μια άλλη παρατήρηση, η οποία είναι αρκετά ενδιαφέρουσα, είναι το γεγονός ότι υπάρχει πληθώρα πληθυσμών, οι οποίοι μοιράζονται κοινούς απλοτύπους, πράγμα που αναδεικνύει την περίπλοκη σχέση και σύνδεση που εμφανίζουν οι πληθυσμοί μεταξύ τους ακόμα και όταν βρίσκονται σε γεωγραφική απόσταση. Μάλιστα, στην περίπτωση της COI mtdna, ένας απλότυπος (Hap11) εμφανίζεται σε άτομα από 8 διαφορετικούς πληθυσμούς (Πιν Γ-14). Τέλος, ενδιαφέρον παρουσιάζει η ομαδοποίηση ατόμων που ανήκουν σε διαφορετικά είδη σύμφωνα με την ισχύουσα ταξινόμηση. Συγκεκριμένα, παρατηρείται σταθερά ομαδοποίηση των ατόμων/απλοτύπων των Κυθήρων και της Κρήτης μαζί με τα άτομα/απλοτύπους των νησιών του Αιγαίου. Το γεγονός αυτό προκαλεί έκπληξη, καθώς σύμφωνα με την ισχύουσα ταξινόμηση, στην Κρήτη και τα Κύθηρα 137
144 εξαπλώνεται το είδος T. kytherensis ενώ στα νησιά του Αιγαίου το είδος T. aegaeus. Μια ακόμη ενδιαφέρουσα περίπτωση είναι αυτή του γενετικού τόπου 16S rrna, όπου άτομα του νέου υπό περιγραφή είδους (Parmakelis et al. 2008) από την περιοχή των Στουρονησίων (AGA,STN) και άτομα T. aegaeus από τα νησιά της Νάξου (NAX) και από τα Στουρονήσια (STO), εμφανίζουν τον ίδιο απλότυπο (Hap60, Πιν Γ-2). Αντίστοιχα, στην περίπτωση της COI, ομαδοποιούνται στον ίδιο απλότυπο (Hap10, Πιν. Γ-14) τα άτομα της Επιδαύρου (που ανήκουν στο είδος T. kytherensis) με τα άτομα της Νάξου (που ανήκουν στο T. aegaeus). 2. ΚΟΡΕΣΜΟΣ ΚΑΙ ΠΙΣΤΟΠΟΙΗΣΗ ΤΩΝ ΜΙΤΟΧΟΝΔΡΙΑΚΩΝ ΑΛΛΗΛΟΥΧΙΩΝ Οι μεταπτώσεις (Ts) συνήθως είναι περισσότερες από τις μεταστροφές (Tv) στο μιτοχονδριακό γονιδίωμα των ζώων (Brown et al. 1979, Wilson et al. 1985). Όπως αναλύθηκε στο κεφάλαιο Β (Μεθοδολογία-Υλικά, 4.2), τα αποτελέσματα για όλα τα σύνολα δεδομένων (16S rrna, COI και συνδυασμένη ανάλυση), έδειξαν πως οι αλληλουχίες δεν έχουν υποστεί κορεσμό και μπορούν να χρησιμοποιηθούν για την εξαγωγή φυλογενετικών συμπερασμάτων. Άλλωστε, δεδομένα από τάξα σπονδυλοζώων έχουν δείξει ότι στο 16S rrna ο κορεσμός στις μεταπτώσεις εκδηλώνεται έπειτα από χρόνους απόκλισης αλληλουχιών που φτάνουν ως τα 300 εκ. χρόνια (DeSalle et al. 1987, DeSalle 1992). Συγκεκριμένα, ο τελευταίος συγγραφέας ανέφερε ότι το 16S rrna έδειξε πολύ μικρό βαθμό κορεσμού έπειτα από 200 εκ. χρόνια απόκλισης, όταν σύγκρινε αλληλουχίες από είδη Drosophila με αυτή του κουνουπιού Aedes albopictus. 138
145 Η πιστοποίηση των μιτοχονδριακών αλληλουχιών πραγματοποιήθηκε με τον έλεγχο BLAST (Basic Local Alignment Search Tool, Altschul et al. 1990), ο οποίος αφορά τη σύγκριση κάθε αλληλουχίας με ήδη κατατεθειμένες αλληλουχίες στη διαδικτυακή βάση δεδομένων Genbank του NCBI (Benson et al. 2007). Το BLAST επιβεβαίωσε πως οι αλληλουχίες της παρούσας μελέτης προέρχονταν από το ίδιο γένος και απορρίφθηκε η πιθανότητα χρήσης λανθασμένων αλληλουχιών κατά την ανάλυση, ή μόλυνσης των δειγμάτων με ξένο γενετικό υλικό. Επιπροσθέτως, αυτή η διαδικασία μειώνει την πιθανότητα οι αλληλουχίες που λήφθηκαν να είναι πυρηνικά αντίγραφα των μιτοχονδριακών τμημάτων (nuclear copies of mtdna genes: numts) τα γνωστά ψευδογονίδια. Τα numts, έχει αναφερθεί ότι διαφοροποιούνται υπό πιο χαλαρούς εξελικτικούς περιορισμούς (Bensasson et al. 2001) και, συνεπώς, θα μπορούσαν να δικαιολογήσουν τα υψηλά επίπεδα διαφοροποίησης που απαντώνται στις αλληλουχίες της παρούσας μελέτης. Ωστόσο, εκτός της γενικότερης συμφωνίας των αλληλουχιών της παρούσας μελέτης με ήδη κατατεθειμένες αλληλουχίες του γένους Trachelipus, η χρήση των ποδιών μυϊκού ιστού μειώνει την πιθανότητα οι αλληλουχίες που προέκυψαν να είναι numts ή αλληλουχίες ενδοσυμβιωτικών βακτηρίων κλπ., σύμφωνα με τα συμπεράσματα των Greenwood & Pääbo (1999). Επιπρόσθετα, στα χρωματογραφήματα που λήφθηκαν, δε διαπιστώθηκε αυξημένη παρουσία διπλών κορυφών, οι οποίες είναι ενδεικτικές μόλυνσης του προϊόντος της PCR από numts (Kimura et al. 2002). 139
146 3. ΔΙΑΦΟΡΟΠΟΙΗΣΗ ΑΛΛΗΛΟΥΧΙΩΝ 3.1. ΔΙΑΦΟΡΟΠΟΙΗΣΗ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΤΟΠΟΥ 16S RRNA Από τις 386 βάσεις του γενετικού τόπου 16S rrna που εξετάστηκαν, το 52,33% διαφοροποιείται μεταξύ του συνόλου των αλληλουχιών. Η παρατήρηση αυτή καταδεικνύει αρκετά υψηλό βαθμό διαφοροποίησης του εν λόγω γενετικού τόπου. Πράγματι, με βάση τις γενετικές αποστάσεις (Kimura 2P) που καταγράφηκαν, είναι φανερή η ύπαρξη μεγάλης γενετικής διαφοροποίησης στο μιτοχονδριακό γονιδίωμα του γένους Trachelipus ακόμα και σε ενδοπληθυσμιακό επίπεδο. Συγκεκριμένα, οι πληθυσμοί του γένους απέχουν μεταξύ τους ως και 17,6% (Επίδαυρος-Βέλλα, Επίδαυρος-Καλαμάς, Επίδαυρος-Βίτσα) και η μέγιστη απόσταση μεταξύ δύο απλοτύπων φτάνει το 18,3% (Hap34,35-Hap53,55) (Πιν. Π-1, Παράρτημα). Ιδιαίτερα εντυπωσιακό είναι το γεγονός ότι η γενετική απόσταση μεταξύ δυο ατόμων του ίδιου είδους ανέρχεται στο 18,4% (Προυσσός Λιμποβίσι) ενώ ακόμα και μεταξύ αντιπροσώπων του ίδιου πληθυσμού η γενετική απόσταση υπερβαίνει σε πολλές περιπτώσεις το 10% (16,1% στο Χελμό, 15,5% στον Προυσσό, 12,9% στα Τριπόταμα, 13,8% στο Βοϊδομάτη και 15,1% στη λίμνη Ζηρού). Την ίδια στιγμή οι μέγιστες γενετικές αποστάσεις που καταγράφονται μεταξύ ατόμων των τριών ειδών που περιλαμβάνονται στην παρούσα μελέτη, ανέρχονται στο 16,6% μεταξύ των ειδών T. aegaeus και T. kytherensis (ATR και PROU) και στο 16,6% μεταξύ του T. nsp και του T. kytherensis (ΑTR και PROU). Η μέγιστη απόσταση μεταξύ ατόμων των T. nsp και T. aegaeus (IKAR) είναι μόλις 7,8%. Υπενθυμίζεται, όμως, ότι στην ανάλυση αυτού του γενετικού τόπου, άτομα της Νάξου (NAX) και των Στουρονησίων (STO) (T. aegaeus) και του νέου υπό περιγραφή είδους (AGA, STN) αντιπροσωπεύονταν από τον ίδιο απλότυπο (Hap60). Στην περίπτωση των γενετικών αποστάσεων μεταξύ των απλοτύπων, επισημαίνεται η περίπτωση του Hap15 [που αντιστοιχεί στα άτομα της Επιδαύρου 140
147 (Πιν. Γ-2)], ο οποίος παρουσιάζει κατά μέσο όρο γενετική απόσταση 14% από όλους τους πληθυσμούς της ηπειρωτικής Ελλάδας. Αντίθετα η γενετική απόσταση του ίδιου απλοτύπου από τους νησιωτικούς πληθυσμούς της Κρήτης (Hap31) και της Ικαρίας (Hap59) είναι μικρότερη του 10%, ενώ στην περίπτωση των Κυθήρων (Hap32) και των νησιών του κεντρικού Αιγαίου (Hap60) μειώνεται εντυπωσιακά στο 1,5% και 1,1%, αντίστοιχα. Οι Poulakakis & Sfenthourakis (2008) στην εργασία για το γένος Orthometopon, υποστηρίζουν πως τιμές της τάξης του 1,1%-1,3% είναι ενδεικτικές για αντιπροσώπους που ανήκουν στο ίδιο είδος, χρησιμοποιώντας στην ανάλυσή τους τις καθαρές μέσες νουκλεοτιδικές αποστάσεις μεταξύ των μελετώμενων τάξων. Λαμβάνοντας υπόψη πως οι καθαρές μέσες νουκλεοτιδικές αποστάσεις είναι κατά κανόνα πιο μικρές, εξάγεται το συμπέρασμα πως οι μέσες νουκλεοτιδικές αποκλίσεις, όπως προκύπτουν στην παρούσα εργασία, είναι ιδιαίτερα χαμηλές και μπορούν να χρησιμοποιηθούν ως ενδεικτικές των σχέσεων μεταξύ πληθυσμών ή/και ειδών ΓΕΝΕΤΙΚΟΣ ΤΟΠΟΣ COI MTDNA Από τις 512 βάσεις του γενετικού τόπου COI mtdna που εξετάστηκαν, το 41% διαφοροποιούνταν μεταξύ του συνόλου των αλληλουχιών. Η παρατήρηση αυτή καταδεικνύει αυξημένη διαφοροποίηση και αυτού του γενετικού τόπου. Με βάση τις γενετικές αποστάσεις (Kimura 2P) που υπολογίστηκαν, προκύπτει έντονη γενετική διαφοροποίηση τόσο σε διαπληθυσμιακό όσο και σε ενδοπληθυσμιακό επίπεδο. Συγκεκριμένα, οι πληθυσμοί του γένους απέχουν μεταξύ τους ως και 27,3% (Πιν. Π-4, παράρτημα) και η μέγιστη απόσταση μεταξύ δύο απλοτύπων φτάνει το 27,4% (Πιν. Π-3, Παράρτημα). Η γενετική απόσταση μεταξύ δυο ατόμων του ίδιου - με την έως τώρα ισχύουσα ταξινόμηση είδους ανέρχεται στο 27,3% (μεταξύ ατόμων από τους 141
148 πληθυσμούς Δρυμώνας και Παπάδες από τη βόρεια Εύβοια και Μανεσαίικος και Σόλος από την Πελοπόννησο). Μεταξύ αντιπροσώπων του ίδιου πληθυσμού, και συγκεκριμένα στους πληθυσμούς του Προυσσού και του Χελμού, η μέγιστη γενετική απόσταση που παρατηρήθηκε ήταν 21%. Οι μέγιστες γενετικές αποστάσεις μεταξύ αντιπροσώπων των T. aegaeus και T. kytherensis ανέρχονταν στο 25,1%. Αντίστοιχα, μεταξύ του T. nsp και πληθυσμών του T. kytherensis [που όμως εντοπίζονται μόνο στη κεντροδυτική (Φράξος, Λίμνη Ζηρού) και βόρεια Ελλάδα (πηγές Βέλλας, Καλαμάς ποταμός, Βίτσα)], η μέγιστη γενετική απόσταση ήταν 22,5%, Για λόγους σύγκρισης με τα προαναφερθέντα για το γενετικό τόπο 16S rrna, επισημαίνεται πως ο απλότυπος Hap10 που αντιπροσωπεύει τα άτομα της Επιδαύρου σε αυτή την ανάλυση, εμφανίζεται και σε άτομα από τη Νάξο, υποδηλώνοντας τη μεγάλη ομοιότητα μεταξύ των εν λόγω πληθυσμών. Σε αυτή την ανάλυση, οι γενετικές αποστάσεις μεταξύ των ατόμων της Επιδαύρου και της Νάξου είναι 0%, ενώ η μικρότερη γενετική απόσταση των ατόμων της Επιδαύρου εντοπίζεται με το άτομο T. aegaeus από τα Στουρονήσια, και είναι 4,3%. Ακολουθούν οι πολύ μικρές γενετικές αποστάσεις με του απλοτύπους των Κυθήρων (5,5%), ενώ με όλους τους άλλους απλοτύπους του γένους από την ηπειρωτική Ελλάδα και την Εύβοια, εμφανίζει ιδιαίτερα αυξημένες νουκλεοτιδικές αποκλίσεις (15,4%-20,5%) ΣΥΝΔΥΑΣΜΕΝΗ ΑΝΑΛΥΣΗ Από τις 898 βάσεις της συνδυασμένης ανάλυσης των δύο γενετικών τόπων (COI και 16S rrna) που εξετάστηκαν, το 44,2% διαφοροποιούνταν μεταξύ του συνόλου των αλληλουχιών. 142
149 Με βάση τις γενετικές αποστάσεις (Kimura 2P) που καταγράφηκαν, οι πληθυσμοί του γένους απέχουν μεταξύ τους από 0% (BEL-KLM) ως και 19,8% (BOU-ELT,LIB βλ. Πιν. Π-6, Παράρτημα), ενώ η μέγιστη απόσταση μεταξύ δύο απλοτύπων φτάνει το 19,9% σε αρκετές περιπτώσει (π.χ. Hap7-Hap29, Hap10- Hap50 κ.α. βλ. Πιν. Π-5, Παράρτημα). Η γενετική απόσταση μεταξύ δυο ατόμων του ίδιου με την έως τώρα ισχύουσα ταξινόμηση είδους αλλά από διαφορετικό πληθυσμό, ανέρχεται στο 20,4% σε δύο περιπτώσεις (Προυσσός-Αμυγδαλιές και Προυσσός-Καλέτζι). Μεταξύ ατόμων του ίδιου πληθυσμού (περίπτωση Χελμού), η μέγιστη γενετική απόσταση που παρατηρήθηκε ήταν 18,1%. Οι μέγιστες γενετικές αποστάσεις μεταξύ αντιπροσώπων των T. aegaeus και T. kytherensis ανέρχονταν στο 18,7% με πληθυσμούς που εντοπίζονται τόσο στην Πελοπόννησο (Σόλος) όσο και στην Κεντρική (Προυσσός) και Β. Ελλάδα (ποταμός Καλαμάς). Αντίστοιχα, μεταξύ του T. nsp και του T. kytherensis η μέγιστη γενετική απόσταση ήταν 18,8% με αντιπροσώπους από τον πληθυσμό του Προυσσού. Για λόγους σύγκρισης (με την ανεξάρτητη ανάλυση των δύο γενετικών τόπων) επισημαίνεται πως ο απλότυπος Hap12 που αντιπροσωπεύει τα άτομα της Επιδαύρου, και σε αυτή την ανάλυση παρουσιάζει τις μικρότερες γενετικές αποστάσεις με τον απλότυπο της Νάξου (0,7%), έπειτα με τους απλοτύπους των Κυθήρων (2,7-2,9%) και, τέλος, με τους απλοτύπους που αντιπροσωπεύουν τα άτομα από τα Στουρονήσια (5,8% με T. aegaeus και 6,8% μεt. nsp). Αξίζει δε να επισημανθεί ότι με τον απλότυπο της Κρήτης παρουσιάζεται ιδιαίτερα αυξημένη γενετική απόσταση, της τάξης του 12,5%. Ο απλότυπος της Επιδαύρου (Hap12), και σε αυτή την ανάλυση, με όλους τους άλλους απλοτύπους του γένους από την ηπειρωτική Ελλάδα και την Εύβοια, εμφανίζει ιδιαίτερα αυξημένες νουκλεοτιδικές αποκλίσεις (12,3%-18,6%). 143
150 4.ΦΥΛΟΓΕΝΕΤΙΚΗ ΑΝΑΛΥΣΗ Εξετάζοντας τους δείκτες που χαρακτηρίζουν τα δένδρα της ανάλυσης μέγιστης φειδωλότητας, αυτό που μπορεί να ειπωθεί είναι ότι ο δείκτης ομοπλασίας (HI) σε όλες τις αναλύσεις (16S rrna=0,420, COI mtdna=0,557 και συνδυασμένη ανάλυση=0,557) είναι ικανοποιητικός, με το 16S rrna να εμφανίζει την πιο χαμηλή τιμή. Παρακάτω παρουσιάζονται αναλυτικά οι ομάδες που δημιουργούνται για κάθε γενετικό τόπο και οι επιμέρους σχέσεις τους. Οι τιμές στατιστικής υποστήριξης που αναφέρονται αφορούν τη μεγαλύτερη τιμή που καταγράφηκε σε οποιαδήποτε από τις αναλύσεις (NJ, MP, ΒΙ) που έγιναν, εκτός και αν δηλώνεται αλλιώς. 4.1.ΓΕΝΕΤΙΚΟΣ ΤΟΠΟΣ 16S RRNA Όπως φαίνεται από τα φυλογενετικά δένδρα που παρουσιάζονται στις εικόνες Γ-2 (NJ), Γ-3 (MP) και Γ-4 (ΒΙ) και αφορούν το γενετικό τόπο 16S rrna, ανεξάρτητα από τη μέθοδο ανάλυσης, οι περισσότεροι από τους απλοτύπους συγκροτούν τις ίδιες ομάδες, η μονοφυλετικότητα των οποίων έχει ικανοποιητική στατιστική υποστήριξη (μεγαλύτερη του 70%). Ωστόσο, η σειρά διαδοχής των κλάδων των μονοφυλετικών ομάδων μπορεί να ποικίλλει. Ανάλογα με τη μέθοδο ανάλυσης, προκύπτει διαφορετική σειρά σύνδεσης των κλάδων. Πιο συγκεκριμένα, η πρώτη ομάδα του γένους Trachelipus που διαμορφώνεται είναι αυτή της Β. Εύβοιας (PAP, DRM) η οποία υποστηρίζεται πάρα πολύ καλά (posterior probability=0,92 - ΒΙ). Ακολουθεί ένας κλάδος που περιλαμβάνει πληθυσμούς αποκλειστικά της ηπειρωτικής Ελλάδας, συγκεκριμένα, της Στερεάς Ελλάδας και της Ηπείρου. «Παραφωνία» σε αυτό το πρότυπο αποτελεί η παρουσία ατόμων από το Βουραϊκό 144
151 (BOU), το Χελμό (CH), το Μανεσαίικο (MNS) και το Σόλο (SOL) που προέρχονται από τη Β. Πελοπόννησο. Ο επόμενος κλάδος που διαμορφώνεται είναι αυτός των νησιωτικών περιοχών από το Αιγαίο (IKAR, NAX) μαζί με την Κρήτη (CRE), τα Κύθηρα (KYT), τα Στρουρονήσια (STN,STO,AGA) και τα άτομα της Επιδαύρου (EPD), ο οποίος παρουσιάζει ιδιαίτερο ενδιαφέρον αφού περιλαμβάνει άτομα και από τα τρία είδη του γένους Trachelipus (T. kytherensis, T aegaeus και T. nsp). Ακολουθεί ο Πελοποννησιακός κλάδος που απαρτίζεται αποκλειστικά από πληθυσμούς της Πελοποννήσου και έχει ισχυρή στατιστική υποστήριξη (98% - MP, 1,00 - BI). Έπειτα, υπάρχει ο «μεικτός» κλάδος που φέρει, σχεδόν ισόποσα, πληθυσμούς από την Πελοπόννησο και την υπόλοιπη ηπειρωτική Ελλάδα και ενσωματώνει και τον πληθυσμό από την Κεφαλλονιά (ΑΙΝ). Τέλος, τα άτομα της Κ. Εύβοιας (STE) και ένα άτομο της λίμνης Ζηρού (ZRL5) αποκλίνουν, το καθένα ξεχωριστά, χωρίς να εντάσσονται σε κάποιον από τους προαναφερθέντες κλάδους ΓΕΝΕΤΙΚΟΣ ΤΟΠΟΣ COI Στα φυλογενετικά δέντρα των εικόνων Γ-7, Γ-8 και Γ-9 που αφορούν το γενετικό τόπο της υπομονάδας Ι της κυτοχρωμικής οξειδάσης (COI mtdna), η εικόνα που παρατηρείται είναι λίγο πολύ η ίδια ως προς τους βασικούς κλάδους που δημιουργούνται. Εδώ υπενθυμίζεται ότι τα άτομα και κατ επέκταση και οι πληθυσμοί - σε αυτή την ανάλυση ήταν λιγότερα, λόγω ανεπιτυχούς αλληλούχισης. Η στατιστική υποστήριξη των κλάδων είναι και εδώ ισχυρή, με εξαίρεση τους βαθύτερους κλάδους της ανάλυσης. Συγκεκριμένα, στην περίπτωση του δέντρου, όπως αυτό προέκυψε από την Μπεϊσιανή Συμπερασματολογία, βλέπουμε ότι η 145
152 υποστήριξη του πρώτου κλάδου είναι πολύ χαμηλή. Αυτό όμως ενδεχομένως οφείλεται στην ύπαρξη πολυτομίας στη βάση του δέντρου. Ο επόμενος κλάδος που αποκλίνει περιλαμβάνει αποκλειστικά πληθυσμούς της κεντρικής και βόρειας Ελλάδας (Βελούχι, Φανάρι, Γαύρος). Αυτός ο κλάδος στις αναλύσεις του 16S rrna είναι ένας υποκλάδος (στον οποίο ενσωματώνονταν και απλότυποι από 3 επιπλέον πληθυσμούς, των οποίων όμως δεν ήταν δυνατή η αλληλούχισή τους για το γενετικό τόπο COI) μέσα στην ευρύτερη ομάδα που δημιουργείται με την πλειονότητα των πληθυσμών από αυτές τις περιοχές μαζί με άτομα από τη βόρεια Πελοπόννησο (BOU, CH, MNS, SOL). Ωστόσο, στην ανάλυση για το γενετικό τόπο COI, ο κλάδος αυτός αποκλίνει πολύ «βαθύτερα» και βρίσκεται αρκετά απομακρυσμένος από τον ευρύτερο κλάδο της Κεντρικής και Β. Ελλάδας. Ο κλάδος που αποκλίνει στη συνέχεια είναι αυτός που φέρει τους πληθυσμούς του Αιγαίου, της Κ. Εύβοιας και για άλλη μια φορά της Επιδαύρου. Ακολουθεί ο κλάδος με πληθυσμούς της Κεντρικής και Βόρειας Ελλάδας μαζί με έναν εσωτερικό υποκλάδο με πληθυσμούς από τη βόρεια Πελοπόννησο (BOU, CH, MNS, SOL). Ο κλάδος που αποκλίνει στη συνέχεια είναι ο αμιγώς Πελοποννησιακός και, τέλος, υπάρχει και σε αυτόν το γενετικό τόπο ο «μεικτός» κλάδος που (όπως και στην περίπτωση του 16S rrna) ενσωματώνει στον ίδιο βαθμό πληθυσμούς από τη Β. και Κ. Ελλάδα και την Πελοπόννησο ΣΥΝΔΥΑΣΜENΗ ΑΝAΛΥΣΗ Στα φυλογενετικά δέντρα των εικόνων Γ-11, Γ-12 και Γ-13 που αφορούν τη συνδυασμένη ανάλυση των δεδομένων από τα δύο γονίδια, η εικόνα που παρατηρείται είναι λίγο πολύ η ίδια ως προς του βασικούς κλάδους που δημιουργούνται. Οι τιμές 146
153 της στατιστικής υποστήριξης των κλάδων είναι υψηλές και φέρονται ενδιάμεσες των τιμών που παρατηρούνται στις ανεξάρτητες αναλύσεις των δύο γονιδίων. Πρώτος αποκλίνει ένας κλάδος με λίγους απλοτύπους από τη Β. και Κ. Ελλάδα. Εν συνεχεία, αποκλίνει ο κλάδος που ομαδοποιεί τους πληθυσμούς του Trachelipus aegaeus μαζί με τους πληθυσμούς της Κρήτης, των Κυθήρων και της Επιδαύρου αλλά και του T. nsp. Έπειτα ακολουθεί ο απλότυπος της Κ. Εύβοιας που αποκλίνει μόνος του. Ο συγκεκριμένος είναι και ο μόνος κλάδος που φέρει χαμηλή στατιστική στήριξη (ΒΙ=0,58). Αυτό θα μπορούσε να δικαιολογηθεί ανατρέχοντας στις ανεξάρτητες αναλύσεις των δύο γονιδίων. Υπενθυμίζεται πως στην περίπτωση της COI ο απλότυπος της Στενής (STE) ενσωματωνόταν στον κλάδο του Αιγαίου και στην περίπτωση του 16S ο ίδιος απλότυπος δημιουργούσε πολυτομία με τον Αιγαιακό κλάδο. Συνυπολογίζοντας λοιπόν τα αποτελέσματα και από τις τρεις αναλύσεις, γίνεται αντιληπτό ότι αυτός ο πληθυσμός δείχνει μάλλον μεγαλύτερη εγγύτητα με τους πληθυσμούς του Αιγαίου παρά με τους υπόλοιπους ηπειρωτικούς πληθυσμούς του γένους Trachelipus. Στη συνέχεια, διαμορφώνεται και πάλι ένας κλάδος που περιλαμβάνει μόνο τους πληθυσμούς της Β. Εύβοιας. Οι επόμενοι κλάδοι περιλαμβάνουν και την πλειονότητα των ατόμων της παρούσας μελέτης. Πρώτος αποκλίνει ο κλάδος που φέρει αποκλειστικά πληθυσμούς από την Πελοπόννησο. Ακολουθεί ο κλάδος με πληθυσμούς της Κεντρικής και Βόρειας Ελλάδας μαζί με έναν εσωτερικό υποκλάδο με πληθυσμούς από τη βόρεια Πελοπόννησο (BOU, CH, MNS, SOL) και, τέλος, ο «μεικτός» υποκλάδος που φέρει πληθυσμούς τόσο από την Β. και Κ. ηπειρωτική Ελλάδα όσο και από την Πελοπόννησο. 147
154 Ακολούθως σχολιάζονται χωριστά ορισμένοι κλάδοι και ομάδες που δημιουργούνται από τις αναλύσεις και εμφανίζουν ιδιαίτερο ενδιαφέρον. Η περίπτωση του T. nsp Σημαντική διαφορά μεταξύ των αναλύσεων παρουσιάζει η τοπολογία του T. nsp: στην περίπτωση της COI, οι αντιπρόσωποί του αποκλίνουν σχηματίζοντας έναν ανεξάρτητο κλάδο ενώ στο συνδυασμένο σύνολο δεδομένων και το 16S rrna ομαδοποιούνται στον Αιγαιακό κλάδο. Σε κάθε περίπτωση, ωστόσο, οι γενετικές αποστάσεις εμφανίζονται αυξημένες τόσο με τους αντιπροσώπους του T. kytherensis όσο και τουt. aegaeus. Στην εργασία των Parmakelis et al υπήρχε σαφής διαχωρισμός των ατόμων από τη συγκεκριμένη περιοχή τόσο βάσει γενετικών αποστάσεων όσο και βάσει φυλογενετικών αναλύσεων. Από αυτά τα αποτελέσματα, σε συνδυασμό και με τη σαφή μορφολογική διαφοροποίηση των εν λόγω ατόμων αλλά και τη συντοπική εμφάνισή τους με άτομα του Τ. aegaeus στο Στουρονήσι, προτάθηκε η παρουσία στις εν λόγω βραχονησίδες ενός νέου είδους. Στην παρούσα εργασία, τα συμπεράσματα για τα άτομα αυτά είναι αντικρουόμενα. Στην περίπτωση της υπομονάδας Ι της κυτοχρωμικής οξειδάσης αποκλίνουν ανεξάρτητα σε έναν κλάδο, ενώ στην περίπτωση του 16S rrna τα άτομα αυτά ομαδοποιούνται στον κλάδο «Αιγαίου- Κρήτης-Κυθήρων-Επιδαύρου», σε κοινό μάλιστα απλότυπο με τα άτομα της Νάξου. Ωστόσο οι Parmakelis et al χρησιμοποίησαν εκτός από την COI mtdna και 16S rrna, και το γενετικό τόπο 12S rrna που σίγουρα έδωσε επιπρόσθετες πληροφορίες για τις γενετικές και φυλογενετικές σχέσεις των πληθυσμών. Επιπλέον, στην παρούσα εργασία χρησιμοποιήθηκαν 47 νέοι πληθυσμοί από την ηπειρωτική Ελλάδα στους οποίους ενσωματώθηκαν και οι αλληλουχίες των ατόμων των Parmakelis et al Η προσθήκη του μεγάλου αριθμού νέων πληθυσμών είναι αναμενόμενο σε ένα βαθμό να μεταβάλει τις σχέσεις των 148
155 πληθυσμών της προηγούμενης εργασίας και, σε συνδυασμό με τη μελέτη ενός λιγότερου γενετικού τόπου, να διαφοροποιήσει τα πρότυπα και τις σχέσεις μεταξύ των ατόμων, πληθυσμών και απλοτύπων που μελετώνται. Ο μεικτός ηπειρωτικός κλάδος Παρατηρούμε πως ο μεικτός ηπειρωτικός κλάδος, εκτός από περιοχές της Στερεάς και της Βόρειας Ελλάδας, περιλαμβάνει και πληθυσμούς της Πελοποννήσου που γεωγραφικά εντοπίζονται αποκλειστικά στα βόρεια και δυτικά του εν λόγω γεωγραφικού διαμερίσματος. Συγκεκριμένα, στο γενετικό τόπο 16S rrna, αυτοί οι πληθυσμοί περιλαμβάνουν τους απλοτύπους Hap 2 (Αλφειός, Χελμός, Κουρούτα, Λάμπεια, Παναχαϊκό), Hap41 (Παναχαϊκό), Hap 8 (Χελμός), Hap37 (Βλασσία, Μακελαριά), και Hap42 (Παναχαϊκό). Επιπλέον, στον υποκλάδο όπου περιλαμβάνονται κατά μεγάλη πλειονότητα πληθυσμοί από την ηπειρωτική Ελλάδα πλην της Πελοποννήσου (πορτοκαλί χρώμα στα δέντρα), ενσωματώνονται σταθερά σε όλες τις αναλύσεις οι αντιπρόσωποι από το Βουραϊκό, το Χελμό, το Μανεσαίικο και το Σόλο (Hap 7, Hap38, Hap47), οι οποίοι και πάλι εντοπίζονται στις βόρειες περιοχές της Πελοποννήσου. Αξίζει να σημειωθεί ότι τα αποτελέσματα έρχονται να ενισχύσουν τη μελέτη των Parmakelis et al. (2008) σε ό,τι αφορά τον προαναφερθέντα υποκλάδο (καθώς οι πληθυσμοί εκείνης της εργασίας ήταν λιγότεροι από αυτούς της παρούσης). Ένα αντίστοιχο πρότυπο είχε διαπιστωθεί και σε προηγούμενη μελέτη του εργαστηρίου μας με πειραματόζωο το χερσόβιο ισόποδο του γένους Ligidium, τόσο με RFLP ανάλυση (Klossa-Kilia et al όσο και με την ανάλυση τριών γενετικών τόπων (12S rrna, 16S rrna και COI) του μιτοχονδριακού DNA (Klossa-Kilia et al. 2006). Και στις δύο περιπτώσεις, είχε διαπιστωθεί η παρουσία ενός μεικτού κλάδου που ενσωμάτωνε πληθυσμούς της Κ. Ελλάδας με πληθυσμούς της Β. Πελοποννήσου. 149
156 Η επαναλαμβανόμενη αυτή εικόνα για διαφορετικά τάξα που μελετώνται κάθε φορά, υποδηλώνει πιθανή ιστορική σύνδεση μεταξύ της βόρειας Πελοποννήσου και της Κεντρικής Ελλάδας, καθώς δεν είναι εμφανής κάποια άλλη οικολογική ή άλλη διαφοροποίηση που θα μπορούσε να τη δικαιολογήσει. Σε κάθε περίπτωση, υπάρχει ανάγκη εκτενέστερης δειγματοληψίας στις περιοχές της κεντρικής Ελλάδας, προκειμένου να αποτυπωθεί η εξάπλωση των ειδών του γένους Trachelipus στην περιοχή. Η Πελοπόννησος μέχρι και μόλις χρόνια πριν από σήμερα, ήταν ενωμένη με τη Στερεά Ελλάδα με χαμηλές πεδιάδες ενώ τα βαθύτερα τμήματα του Κορινθιακού Κόλπου σχημάτιζαν λίμνη (Van Andel & Shakleton 1982). Ενδεχομένως, λοιπόν, να υπήρχαν τα απαιτούμενα υγρά περιβάλλοντα που ευνόησαν τη μετακίνηση πληθυσμών από τις βορειότερες ηπειρωτικές περιοχές του Προυσσού και του Τυμφρηστού (Βελούχι) προς την Πελοπόννησο. Ερχόμενοι οι πληθυσμοί αυτοί, εγκαταστάθηκαν στις ορεινές περιοχές τις Πελοποννήσου (Χελμός, Παναχαϊκό, Ερύμανθος). Αντιστρόφως, και αντιστοίχως, θα μπορούσε κανείς να υποθέσει πως η διαφοροποίηση είχε προηγηθεί στις περιοχές μεγαλύτερου υψομέτρου της Β. Πελοποννήσου (ενδεχομένως παραπάτρια ειδογένεση;) και αυτοί οι διαφοροποιημένοι πληθυσμοί προχώρησαν βορειότερα. Σε κάθε περίπτωση ενδιαφέρον παρουσιάζει το γεγονός ότι οι γενετικές αποστάσεις των πληθυσμών από το Βουραϊκό, το Χελμό, το Μανεσαίικο και το Σόλο παρουσιάζουν μεγαλύτερες τιμές από τους πληθυσμούς που ομαδοποιούνται στον αμιγώς πελοποννησιακό κλάδο σε σύγκριση με τις αποστάσεις που εμφανίζουν οι υπόλοιποι πληθυσμοί της Β. και Κ. Ελλάδας του υποκλάδου από τον ίδιο κλάδο (της Πελοποννήσου). Τέλος, η ομαδοποίηση της Κεφαλονιάς (ΑΙΝ) με την Κ. και Β. Ελλάδα είναι απολύτως λογική, καθώς μέχρι και το πρόσφατο γεωλογικό παραλθόν (Μέσο Πλειόκαινο) ήταν ενωμένη με την Στερεά Ελλάδα (Dermitzakis & Papanikolaou 1981). 150
157 Η περίπτωση της λίμνης Ζηρού Στις αναλύσεις αναδείχτηκαν κάποια πολύ ενδιαφέροντα στοιχεία αναφορικά με τον πληθυσμό της λίμνης Ζηρού. Από αυτήν την περιοχή συλλέχτηκαν 11 άτομα και αλληλουχήθηκαν επιτυχώς τα 9 (για το γενετικό τόπο 16S rrna). Από τη φυλογενετική ανάλυση του εν λόγω γενετικού τόπου, προέκυψε πως ο πληθυσμός αντιπροσωπευόταν από 4 απλοτύπους (Hap2, Hap6, Hap67, Hap68 / Πιν. Γ-2), οι οποίοι μάλιστα δεν ομαδοποιούνται σε έναν κοινό κλάδο αλλά, αντιθέτως, εντοπίζονται σε πολύ απομακρυσμένες θέσεις μεταξύ τους και στις τρεις περιπτώσεις αναλύσεων (NJ, MP, MrBayes). Οι απλότυποι Hap6 και Hap67 ομαδοποιούνται με τον κλάδο των ηπειρωτικών περιοχών (που περιλαμβάνει τους πληθυσμούς Βουραϊκού, Χελμού, Μανεσαίικου και Σόλου), ο απλότυπος Hap2 με το μεικτό ηπειρωτικό κλάδο και ο απλότυπος Hap68 αποκλίνει μόνος του από τους υπόλοιπους κλάδους. Ο πληθυσμός της λίμνης Ζηρού εμφανίζει τη μεγαλύτερη μέση τιμή ενδοπληθυσμιακής διαφοροποίησης (9,1% Πιν Γ-7) και υπενθυμίζεται επίσης πως 2 άτομα από αυτόν τον πληθυσμό είχαν γενετική απόσταση 15,1%, τιμή που αγγίζει τα όρια της διαφοροποίησης μεταξύ ειδών. Για την περίπτωση της λίμνης Ζηρού ως προς την COI και πάλι παρατηρήθηκε έντονη ποικιλομορφία. Από τα 11 άτομα αλληλουχήθηκαν επιτυχώς μόνο τα 4. Και πάλι όμως τα άτομα αυτά αντιπροσωπεύονταν από 3 διαφορετικούς απλοτύπους (Hap6, Hap20, Hap30 / Πιν Γ-14) οι οποίοι εντοπίζονται σε διαφορετικούς κλάδους και μάλιστα αρκετά απομακρυσμένους μεταξύ τους. Τέτοιοι πληθυσμοί θα μπορούσαν στην ουσία να αποτελούνται από υποπληθυσμούς κρυπτικών ειδών (βλ παρακάτω) και κάτι τέτοιο θα δικαιολογούσε τον υψηλό βαθμό ενδοπληθυσμιακής διαφοροποίησης που παρατηρείται στους πληθυσμούς. Η τιμή ενδοπληθυσμιακής διαφοροποίησης του πληθυσμού είναι 13% (Πιν. Γ-19). Ωστόσο η 151
158 γενετική απόσταση μεταξύ των ατόμων αυτού του πληθυσμού φτάνει το 19,2%, τιμή ιδιαίτερα μεγάλη που είναι ενδεικτική διαφοροποίησης μεταξύ ειδών. Και στην περίπτωση της συνδυασμένης ανάλυσης, τα 4 άτομα ομαδοποιούνται σε ξεχωριστό απλότυπο το καθένα (Hap23, Hap67, Hap68, Hap69). Οι τρεις από αυτούς εντοπίζονται σε διαφορετικούς κλάδους των αναλύσεων και μάλιστα με ισχυρή στατιστική υποστήριξη. Τόσο η τιμή ενδοπληθυσμιακής διαφοροποίησης 11,5%, όσο και η 16,8% διαφοροποίηση μεταξύ ατόμων του πληθυσμού αυτού, και σε αυτήν την περίπτωση, θεωρείται οριακή για διαφοροποίηση μεταξύ ειδών (Πιν Γ-30) (Poulakakis & Sfenthourakis 2008). Ο κλάδος του Αιγαίου Με βάση την πλειονότητα των μεθόδων ανάλυσης, τόσο για τα δύο γονίδια ανεξάρτητα όσο και για τη συνδυασμένη ανάλυση, παρατηρείται ομαδοποίηση των πληθυσμών του Αιγαίου με τα άτομα της Κρήτης και των Κυθήρων. Το ενδιαφέρον έγκειται στο γεγονός πως τα άτομα της Κρήτης και των Κυθήρων (από εδώ μάλιστα έγινε και η περιγραφή του είδους) ανήκουν στο είδος T. kytherensis, ενώ οι υπόλοιποι πληθυσμοί των νησιών που αναλύθηκαν ανήκουν στο είδος T. aegaeus. Προκειμένου, λοιπόν, να διατηρηθεί η μονοφυλετικότητα του κλάδου, οι πληθυσμοί των Κυθήρων και της Κρήτης θα έπρεπε να μεταφερθούν στο είδος T. aegaeus. Κάτι τέτοιο όμως θα οδηγούσε στην κατάργηση του ονόματος «Τ. kytherensis» για τους ηπειρωτικούς πληθυσμούς μιας και η περιγραφή, όπως προαναφέρθηκε, είχε γίνει από τα Κύθηρα και πλέον δε θα ισχύε (το T. aegaeus είχε περιγραφεί νωρίτερα οπότε και προηγείται). Έτσι, θα έπρεπε να επανέρθει σε ισχύ το όνομα T. palustris (Strouhal, 1936) που είχε δοθεί στα δείγματα από το Παναχαϊκό και εν συνεχεία έγινε συνώνυμο του T. kytherensis από τον Schmalfuss 2003, 2004). Ωστόσο, αξίζει να αναφερθεί πως στην ανάλυση της σύνδεσης γειτόνων (NJ) και για όλα τα σύνολα δεδομένων, ο πληθυσμός της Κρήτης σχημάτιζε ξεχωριστό 152
159 υποκλάδο από τους υπόλοιπους πληθυσμούς του Αιγαίου, που μάλιστα στην περίπτωση του COI συνδεόταν με πληθυσμούς της Β. και Κ. Ελλάδας. Κάτι τέτοιο θα μπορούσε να είναι ένδειξη μιας ιδιαιτερότητας που εμφανίζουν οι πληθυσμοί της Κρήτης και προκειμένου να διαλευκανθεί θα πρέπει να πραγματοποιηθούν δειγματοληψίες από περισσότερες περιοχές της αλλά και από άλλες νησιωτικές περιοχές του Αιγαίου. Με αυτόν τον τρόπο θα μπορέσουν να εκτιμηθούν τα επίπεδα γενετικής ή/και μορφολογικής ποικιλότητας μέσα στους πληθυσμούς της Κρήτης και να προσδιοριστούν με περισσότερη σαφήνεια οι φυλογενετικές της σχέσεις με άλλες περιοχές του ελλαδικού χώρου. Η περίπτωση της Επιδαύρου Ο πληθυσμός της Επιδαύρου, σύμφωνα με όλες τις αναλύσεις όλων των συνόλων δεδομένων της παρούσας μελέτης φαίνεται ότι διαφοροποιείται από το είδος T. kytherensis και ανήκει στο είδος T. aegaeus. Αυτό προκύπτει και από τις γενετικές αποκλίσεις που σε κάθε περίπτωση (COI, 16S rrna και συνδυασμένη ανάλυση) είναι πολύ μικρές σε σχέση με τα άτομα T. aegaeus από τη Νάξο (0%, 1,5% και 0,7% αντίστοιχα). Τέτοιες γενετικές αποστάσεις είναι ενδεικτικές για αντιπροσώπους που ανήκουν στο ίδιο είδος (Poulakakis & Sfenthourakis 2008). Επιπλέον, στις φυλογενετικές αναλύσεις τα άτομα/απλότυποι του συγκεκριμένου πληθυσμού περιλαμβάνονται πάντοτε στον κλάδο του Αιγαίου. Από τα παλαιογεωγραφικά στοιχεία της περιοχής είναι γνωστό πως μεταξύ της Αργολίδας και της Αττικής εκτεινόταν μια πεδιάδα στην οποία τα σημερινά νησιά πρόβαλλαν ως απομονωμένοι λόφοι (για λεπτομέρειες βλ Εισαγωγή Α.4). Αυτή ενδεχομένως να υπήρξε η δίοδος μέσω της οποίας άτομα του είδους T. aegaeus εποίκισαν από τα νησιά του Αιγαίου, μέσω της Αττικής, την περιοχή της Πελοποννήσου. 153
160 Το ερώτημα που ανακύπτει από τα νέα δεδομένα, καθώς η δειγματοληψία στη χερσόνησο της Αργολίδας και ευρύτερα στην νοτιοανατολική περιοχή (Πάρνωνας, Μονεμβάσια κλπ) της Πελοποννήσου είναι ανεπαρκής, είναι: σε ποιες ακριβώς άλλες περιοχές της Πελοποννήσου περιορίζεται η εξάπλωση του εν λόγω είδους; Η μελέτη αυτής της περιοχής ως προς τη ζωογεωγραφία των ειδών του γένους Trachelipus, θα μπορούσε να ρίξει φως τόσο στα όρια εξάπλωσης του μέχρι τώρα γνωστού ως «νησιωτικού» είδους (T. aegaeus) όσο και του T. kytherensis, αποκαλύπτοντας ενδεχόμενες παραπάτριες ή συμπάτριες κατανομές και να προτείνει πρόσθετες εναλλακτικές διόδους εποίκισης της Πελοποννήσου από το T. aegaeus (πχ από τα Κύθηρα). Η περίπτωση της Εύβοιας Οι δειγματοληψίες που πραγματοποιήθηκαν στη Β. Εύβοια περιελάμβαναν 2 τοποθεσίες που κάλυπταν τόσο την πλευρά προς τα αιγαιακά παράλια (Παπάδες - PAP) όσο και τη πλευρά προς τον Ευβοϊκό Κόλπο (Δρυμώνας - DRM). Αυτό έγινε προκείμενου να ελεγχθεί και η ενδεχόμενη διττή εποίκιση της περιοχής, αφενός από τα νησιά του Αιγαίου και αφετέρου από την ηπειρωτική περιοχή με την οποία η Εύβοια ήταν μέχρι πολύ πρόσφατα ενωμένη (Van Andel & Shakleton 1982). Μια τέτοια διαφοροποίηση δεν υποστηρίζεται από τα αποτελέσματα της παρούσας μελέτης, καθώς τα άτομα των δύο πληθυσμών ομαδοποιούνται πάντα μαζί και μάλιστα στην περίπτωση της COI αντιπροσωπεύονται από τον ίδιο απλότυπο. Αυτό, όμως, που προκύπτει είναι η σαφής διαφοροποίηση συνολικά αυτής της περιοχής τόσο από την κεντρική Εύβοια όσο και από τους υπόλοιπους πληθυσμούς της ηπειρωτικής και νησιωτικής Ελλάδας. Η κεντρική Εύβοια σε κάθε περίπτωση δείχνει να ομαδοποιείται με τους πληθυσμούς του Αιγαίου (T. aegaeus) παρά με τους υπόλοιπους ηπειρωτικούς πληθυσμούς του γένους Trachelipus. 154
161 Σε προηγούμενη μελέτη του εργαστηρίου μας για το γένος Ligidium (Klossa Kilia et al. 2006) και πάλι η φυλογενετική θέση των αντιπροσώπων της Εύβοιας εμφανιζόταν να ξεχωρίζει από τις υπόλοιπες ομάδες τόσο της νησιωτικής όσο και της ηπειρωτικής Ελλάδας. Από τα αποτελέσματα προκύπτει η ανάγκη περαιτέρω δειγματοληψίας τόσο στην περιοχή της Αττικής όσο και στην ανατολική Στερεά Ελλάδα και Θεσσαλία προκειμένου να προσδιοριστούν με μεγαλύτερη λεπτομέρεια και ακρίβεια οι σχέσεις της βόρειας Εύβοιας με την υπόλοιπη ηπειρωτική Ελλάδα και να διευκρινιστεί αν υπάρχει πράγματι μια τοπική διαφοροποίηση των πληθυσμών στην συγκεκριμένη περιοχή. Η περίπτωση Βουραϊκού Πλανητέρου Ιδιαίτερο ενδιαφέρον παρουσιάζει η περίπτωση των πληθυσμών Πλανητέρου και Βουραϊκού. Οι ανά ζεύγη γενετικές αποστάσεις μεταξύ όλων των ατόμων των δύο πληθυσμών ήταν 15,7% (για το γενετικό τόπο 16S rrna) και 25,3%-26,5% (γενετικός τόπος COI). Οι τιμές αυτές είναι ιδιαίτερα μεγάλες αν ληφθεί υπόψη πως αυτοί οι δύο πληθυσμοί βρίσκονται πολύ κοντά μεταξύ τους στο όρος Χελμός της Πελοποννήσου και δεν εμφανίζουν εμφανή οικολογική, ηθολογική η μορφολογική διαφοροποίηση τουλάχιστον στους έως τώρα χρησιμοποιούμενους διαγνωστικούς χαρακτήρες. Ωστόσο, σε όλες τις αναλύσεις, οι δύο πληθυσμοί τοποθετούνται σε διαφορετικούς κλάδους, οι οποίοι είναι αρκετά απομακρυσμένοι. Ως εκ τούτου, η εγκυρότητα των ταξινομικών χαρακτήρων που χρησιμοποιούνται ως τώρα για τους πληθυσμούς αυτούς φαίνεται να τίθεται υπό αμφισβήτηση. Η περίπτωση αυτή θα μπορούσε να αποτελέσει ένα καλό παράδειγμα για την υποστήριξη της ύπαρξης κρυπτικών ειδών (βλ παρακάτω). Όμως, αυτή η εκδοχή διερευνήθηκε και απορρίφθηκε - τουλάχιστον για την περίπτωση των δύο συγκεκριμένων περιοχών - σύμφωνα με τα αποτελέσματα της 155
162 πτυχιακής εργασίας που πραγματοποιήθηκε στο εργαστήριό μας από τον προπτυχιακό φοιτητή Γιάννη Νούρο (2010, αδημοσίευτα δεδομένα). Στην εργασία αυτή χρησιμοποιήθηκαν δύο πληθυσμοί του χερσόβιου ισοπόδου Trachelipus kytherensis από τις εν λόγω περιοχές, με σκοπό να μελετηθούν γενικά στοιχεία της αναπαραγωγής τους. Ένας επιπλέον σκοπός της μελέτης, επομένως, ήταν να βοηθήσει στον έλεγχο της εγκυρότητας της ισχύουσας ταξινόμησης των δύο πληθυσμών, προσφέροντας στοιχεία για το βαθμό της αναπαραγωγικής απομόνωσης μεταξύ τους. Τα αποτελέσματα της εν λόγω εργασίας έδειξαν ότι οι αντιπρόσωποι των δύο περιοχών διασταυρώνονται ελεύθερα μεταξύ τους και, μάλιστα, η αναπαραγωγική τους επιτυχία δεν διαφοροποιείται από εκείνη μεταξύ αμιγών διασταυρώσεων μεταξύ ατόμων καθενός από τους δύο πληθυσμούς. Ωστόσο, λόγω τεχνικών περιορισμών, ο πειραματικός σχεδιασμός δεν προχώρησε και σε εκτίμηση της γονιμότητας των απογόνων της F 2 γενεάς με περαιτέρω διασταυρώσεις. Έτσι, μπορεί να γίνει εκτίμηση για τους προσυζευκτικούς μηχανισμούς απομόνωσης των εν λόγω πληθυσμών. Οι μηχανισμοί αυτοί αφορούν στην αδυναμία συνεύρεσης των πληθυσμών λόγω της διαφορετικής ανατομίας των γεννητικών τους οργάνων, της διαφορετικής αναπαραγωγικής περίοδου, του διαφορετικού χώρου κατοικίας ή της διαφορετικής προτίμησης συντρόφου για σύζευξη (Dobzhansky 1970, Mayr 2002, Provine 2004, de Queiroz 2005). Από τα αποτελέσματα της αναπαραγωγικής επιτυχίας γίνεται αντιληπτό πως τέτοιοι παράγοντες δεν έχουν δράσει στους μελετηθέντες πληθυσμούς, καθώς παρατηρήθηκαν επιτυχείς συνευρέσεις. Ωστόσο δεν μπορούν να γίνουν συνολικές εκτιμήσεις αναφορικά με τους μετασυζευκτικούς μηχανισμούς απομόνωσης (μηχανισμοί που δεν επιτρέπουν τη βιωσιμότητα ή τη γονιμότητα των απογόνων), καθώς δεν επιβίωσαν οι απόγονοι της πρώτης θυγατρικής γενιάς, τόσο των ετεροτυπικών όσο και των ομοτυπικών διασταυρώσεων, κατά πάσα πιθανότητα λόγω των πειραματικών συνθηκών. 156
163 Μια υπόθεση που θα συνδύαζε τα αποτελέσματα της αυξημένης γενετικής απόκλισης με αυτά της επιτυχούς σύζευξης των ατόμων των δύο πληθυσμών θα μπορούσε να είναι πως οι εν λόγω πληθυσμοί, του Βουραϊκού και του Πλανητέρου, βρίσκονται στη φάση της ανάπτυξης μετασυζευτικών αναπαραγωγικών μηχανισμών. Προκειμένου να διερευνηθεί κάτι τέτοιο θα πρέπει να πραγματοποιηθούν πειραματικές διασταυρώσεις μεταξύ των πληθυσμών, κρατώντας τους απογόνους για περισσότερες γενεές και υποβάλλοντάς τους σε περαιτέρω διασταυρώσεις, έτσι ώστε να γίνει δυνατή η εκτίμηση της γονιμότητας και της βιωσιμότητας των υβριδίων αλλά και των επόμενων γενεών. Σημειώνεται δε πως σύμφωνα με την κλασσική θεωρία της ειδογένεσης, οι μηχανισμοί που αναπτύσσονται πρώτοι στην πορεία της απομόνωσης δύο πληθυσμών είναι οι μετασυζευκτικοί και έπονται οι προσυζευκτικοί. Η παρουσία κρυπτικών ειδών, σε κάθε περίπτωση, δεν αποκλείεται για άλλες περιοχές της ηπειρωτικής Ελλάδας (πχ. Λίμνη Ζηρού- βλ. παραπάνω) που εμφανίζουν ενδιαφέρουσες αποκλίσεις, μεγάλη γενετική διαφοροποίηση μεταξύ των ατόμων τους και ομαδοποιήσεις που δεν φαίνεται να σχετίζονται με την οικολογία, τη μορφολογία ή την παλαιογεωγραφία της περιοχής. 5. ΚΡΥΠΤΙΚΑ ΕΙΔΗ Μεγάλες γενετικές αποστάσεις εντός παραδοσιακά αναγνωρισμένων ειδών, συχνά σε συνδυασμό με μορφολογικές, γεωγραφικές και άλλες μικρές διαφορές, έχουν αποκαλύψει την ύπαρξη κρυπτικών ειδών στους περισσότερους τύπους οργανισμών ανεξάρτητα από τα περιβάλλοντα όπου ζουν, από τα μύδια του ανοιχτού πελάγους (Vrijenhoek et al. 1994) μέχρι τα ψάρια του γλυκού νερού (Feulner et al. 2006), τις τροπικές πεταλούδες (Hebert et al. 2004), τα αρκτικά φυτά (Grundt et 157
164 al. 2006), τις καμηλοπαρδάλεις (Brown et al. 2007) και τους βάτραχους του Αμαζονίου (Elmer et al. 2007a,b,c). Η εμφάνιση κρυπτικών ειδών μπορεί να είναι τόσο πρόσφατη ώστε τα μορφολογικά και διαγνωστικά χαρακτηριστικά των οργανισμών να μην έχουν προλάβει ακόμα να αλλάξουν (Saez & Lozano 2005). Τα κρυπτικά είδη που έχουν ανακαλυφθεί τα τελευταία χρόνια, παρουσιάζουν δυνατότητες για τη μελέτη σημαντικών μηχανισμών ειδογένεσης. Ιδιαίτερα υποσχόμενες είναι οι έρευνες που προσανατολίζονται στην εύρεση χαρακτήρων πάνω στους οποίους δρα η φυσική επιλογή και η μελέτη μη μορφολογικών χαρακτήρων που θα μπορούσαν να χρησιμοποιηθούν για το διαχωρισμό των ειδών (Bickford et al. 2006, Beheregaray & Caccone 2007). Οι γενετικοί τόποι 16S rrna και COI mtdna έχουν χρησιμοποιηθεί αρκετά για τη μελέτη πιθανών κρυπτικών ειδών (Held 2000, Held & Wägele 2005, Finston et al. 2007, 2009) και έχουν θέσει κάποια σημαντικά κριτήρια προς αυτήν την κατεύθυνση (Raupach & Wägele 2006). 6. ΕΤΕΡΟΠΛΑΣΜΙΑ Μια άλλη εξήγηση που θα μπορούσε να οδηγήσει σε υψηλά επίπεδα διαφοροποίησης των μιτοχονδριακών αλληλουχιών είναι η αυξημένη πιθανότητα μιτοχονδριακού συνδυασμού, η οποία οδηγεί σε ετεροπλασμία. Ετεροπλασμία είναι η κατάσταση κατά την οποία μέσα σε ένα κύτταρο ή έναν οργανισμό υπάρχουν μιτοχόνδρια διαφορετικής γενετικής σύστασης. Γενικά, τα μιτοχόνδρια είναι μητρικής προέλευσης, καθώς ο οργανισμός τα κληρονομεί μέσω του ωαρίου, χωρίς ο αρσενικός γαμέτης να έχει συμμετοχή. Γι αυτόν το λόγο και όλα τα μιτοχόνδρια είναι πανομοιότυπα μεταξύ τους. Ωστόσο, σε ορισμένα είδη έχει 158
165 παρατηρηθεί «διαρροή» μιτοχονδριακού γενετικού υλικού και από τα αρσενικά άτομα. Οι Doublet et al. (2008) προτείνουν πως, στην περίπτωση των ισοπόδων, η ετεροπλασμία δεν ακολουθεί απαραίτητα τον παραπάνω «κλασικό» ορισμό. Στα πειράματά τους με το Armadillidium vulgare παρατήρησαν ετεροπλασμία σε όλους τους απογόνους και για τρεις γενεές, από μια αρχική μητρική γραμμή. Συγκεκριμένα, υποστηρίζουν ότι η ετεροπλασμία δύναται να υπάρχει με αποκλειστική κληρονόμηση των μιτοχονδρίων από το θηλυκό άτομο. Χρονολογούν το φαινόμενο αυτό στα χερσαία ισόποδα περί τα 30 εκατομμύρια χρόνια πριν από σήμερα. Επισημαίνουν πως παρατηρήθηκε ετεροπλασμία και στο ισόποδο Trichoniscus pusillus που αναπαράγεται με παρθενογένεση. Προτείνουν πως κληρονομείται σταθερά μέσα στις γενεές, αφού απαντάται στα μιτοχόνδρια, και με αυτόν τον τρόπο ξεφεύγει από το φαινόμενο της μιτοχονδριακής στενωπού, η οποία συνήθως εξαλείφει την ετεροπλασμία μέσα σε λίγες γενιές. Τόσο στο Armadillidium vulgare όσο και στο Porcellionides pruinosus έχει επιβεβαιωθεί ετεροπλασμία. Τα είδη αυτά διαθέτουν ένα α-τυπικό τριμερές μιτοχονδριακό γονιδίωμα, το οποίο διαμορφώνεται από ένα κυκλικό διμερές 28kb που αποτελείται από 2 μονομερή των 14kb, τα οποία συντήκονται με αντίθετη πολικότητα, και από ένα γραμμικό μονομερές των 14kb (Raimond et al. 1999). Παρατηρείται ένα μόνο ετεροπλασματικό σημείο, το οποίο αφορά έναν πολυμορφισμό στο DNA που μπορεί να αλλάζει το αντικωδικόνιο για το trna Ala στο αντίστοιχο του trna Val. Το σημείο αυτό βρίσκεται στην ίδια θέση αλλά σε διαφορετικά μονομερή, διασφαλίζοντας με αυτόν τον τρόπο την παρουσία και των δύο απαραίτητων trna στο μιτοχόνδριο (Marcadé et al. 2007). Αυτή η τριμερής δομή θα μπορούσε να παρέχει το κατάλληλο υπόστρωμα για την εμφάνιση ενδομιτοχονδριακής ετεροπλασμίας. Οι Doublet et al. (2008) καταλήγουν πως η επιμονή ενός τέτοιου μιτοχονδριακού πολυμορφισμού σε τόσο βαθιά εξελικτική κλίμακα υποδηλώνει δράση σταθεροποιούσας επιλογής. 159
166 Με αυτόν τον τρόπο, θα μπορούσε ενδεχομένως να εξηγηθεί η αυξημένη νουκλεοτιδική ποικιλότητα που εμφανίζουν τα δεδομένα της παρούσας μελέτης, καθώς η ενδεχόμενη παρουσία διαφορετικών μιτοχονδριακών αλληλουχιών θα περνούσε απαρατήρητη και δε θα λαμβάνεται υπόψη, οπότε θα μπορούσε να οδηγήσει σε στρεβλά συμπεράσματα για τις «παράξενα» μεγάλες γενετικές αποστάσεις μεταξύ πολύ κοντινών πληθυσμών ή ακόμα και μεταξύ ατόμων του ίδιου πληθυσμού, ή/και τις κατά πρώτη ματιά μη αναμενόμενες ομαδοποιήσεις στις φυλογενετικές μας αναλύσεις. Ωστόσο, πρέπει να επισημανθεί πως στην παρούσα εργασία δεν μελετήθηκε περαιτέρω το συγκεκριμένο ενδεχόμενο, παρότι υπήρξαν περιπτώσεις στις οποίες παρατηρήθηκαν διπλές κορυφές στα χρωματογραφήματα, οι οποίες θεωρούνται μια πρώτη ένδειξη ετεροπλασμίας (Doublet et al. 2008). Για τον έλεγχο της υπόθεσης αυτής, συνεπώς, πρέπει να γίνει εξειδικευμένη έρευνα. 160
167 7. ΓΕΝΙΚΑ ΣΥΜΠΕΡΑΣΜΑΤΑ Στον ελλαδικό χώρο έχουν πραγματοποιηθεί διάφορες μελέτες οι οποίες σχετίζονται με τις βιογεωγραφικές συγγένειες οργανισμών (Apostololidis 1997, Bittkau & Comes 2005, Chatzimanolis et al. 2003, Kasapidis et al. 2005a, Kasapidis et al. 2005b, Poulakakis et al. 2003, 2005a 2005b, Poulakakis & Sfenthourakis 2008, Douris et al. 2007, Trichas et al. 2008, Kornilios et al. 2009, Papadopoulou 2009), οι οποίες συνφωνούν είτε με την παλαιογεωγραφία της περιοχής και συνεπώς εμφανίζουν σαφές γεωγραφικό πρότυπο είτε όχι (Giokas 2000, Kamilari & Sfendourakis 2009, Parmakelis 2005). Αναφορικά με τα χερσαία ισόποδα, υπάρχουν μελέτες που σε κάποιο βαθμό συμφωνούν με την παλαιογεωγραφία ή τη σύγχρονη γεωγραφία της περιοχής (Poulakakis & Sfenthourakis 2008) και άλλες που αλλά παρουσιάζουν αρκετές «αποκλίσεις» από αυτή (Klossa-Kilia et al. 2006). Στην παρούσα μελέτη, παρατηρούνται πληθυσμοί πολύ κοντινοί γεωγραφικά, οι οποίοι μέχρι σήμερα θεωρείται πως ανήκουν στο ίδιο είδος (Trachelipus kytherensis), να εμφανίζουν μεγάλες γενετικές αποστάσεις μεταξύ τους και να ομαδοποιούνται σε διαφορετικούς και αρκετά απομακρυσμένους κλάδους των δένδρων. Τέτοια παραδείγματα είναι ο πληθυσμός του Βουραϊκού (BOU) και του Πλανητέρου (PLAN) μεταξύ τους απόσταση 15,7% (16S rrna) - και οι πληθυσμοί από τη Βλασσία (VLA), το Μανεσαίικο ποταμό (ΜNS) και τη Μακελαριά (MKL) μεταξύ τους απόσταση 15,6% (16S rrna). Η τοπολογία των κλάδων, καθώς και η απουσία σαφούς γεωγραφικού προτύπου στην ομαδοποίηση των πληθυσμών του T. kytherensis, καταδεικνύει ότι πιθανότατα δεν έχουμε να κάνουμε με ένα μόνο είδος, αλλά μάλλον με περισσότερα που είναι δύσκολο να διακριθούν μορφολογικά, τουλάχιστον με τους μέχρι σήμερα χρησιμοποιούμενους ταξινομικούς διαγνωστικούς χαρακτήρες. 161
168 Αυτό ενισχύεται και από τις γενετικές αποστάσεις που καταγράφηκαν στην παρούσα μελέτη και εμφανίζονται αυξημένες ακόμα και σε σχέση με αυτές που έχουν αναφερθεί για τη διάκριση ειδών ισοπόδων σε άλλες έρευνες που χρησιμοποιούνται μεταξύ άλλων- οι γενετικοί τόποι 16S rrna και COI. Για παράδειγμα, έχουν παρατηρηθεί νουκλεοτιδικές αποκλίσεις με εύρος από 10,7% - 12,6% μεταξύ του είδους Armadillidium pelagicum Arcangeli, 1955 και του πολύ καλά διαχωρισμένου είδους Armadillidium album Dollfus, 1887, σε μελέτη του γενετικού τόπου 16S rrna (Charfi-Cheikhroua 2003). Άλλα καλά διαχωρισμένα είδη ισοπόδων αναφέρεται ότι διαχωρίζονται με γενετικές αποστάσεις της τάξης του 13% - 28% - για το γενετικό τόπο COI- (Wetzer 2001, Rivera et al. 2002, Baratti et al. 2004, 2005, 2010, McGaughran et al. 2006). Συνεπώς, οι νουκλεοτιδικές αποκλίσεις που υπολογίστηκαν μεταξύ των δύο ειδών που μελετήθηκαν βρίσκονται εντός των προαναφερθέντων ορίων και σε πλήρη συμφωνία με τις γενετικές αποστάσεις της προηγούμενης μελέτης των Parmakelis et al. (2008). Πολλοί μελετητές επιχειρηματολογούν ενάντια στη χρήση των γενετικών αποστάσεων ως μέτρο για να διαχωρίζονται ή να ενοποιούνται είδη, κυρίως γιατί δεν είναι όμοιες οι συγκρίσεις ως προς τις γενετικές αποκλίσεις μεταξύ διαφορετικών τάξων (Avise & Aquadro 1982, Avise & Johns 1999). Παραδείγματος χάριν, οι France & Kocher (1996) κατέγραψαν γενετικές αποστάσεις 4-5% μεταξύ πολύ καλά προσδιορισμένων ειδών αμφιπόδων. Γι αυτόν το λόγο και οι συγκρίσεις στην παρούσα εργασία περιορίστηκαν μεταξύ τάξων της ίδιας ταξινομικής ομάδας (της τάξης των Ισοπόδων) έτσι ώστε συγκρίσεις με τάξα από άλλες ομάδες Καρκινοειδών να μην οδηγήσουν σε παραπλανητικά συμπεράσματα. Σε κάθε περίπτωση, από τα αποτελέσματα της παρούσας μελέτης επισημαίνεται ιδιαίτερα μεγάλη γενετική ποικιλότητα μεταξύ των αντιπροσώπων του γένους. Επιπλέον καταδεικνύεται πως η Πελοπόννησος φιλοξενεί τα είδη Trachelipus kytherensis και T. aegaeus (τουλάχιστον στη χερσόνησο της Αργολίδας) αλλά και πιθανόν μια τρίτη μορφή στα βόρεια (νέο είδος;) η οποία 162
169 εμφανίζεται ευρύτερα στην ηπειρωτική Ελλάδα. Θα είχε ιδιαίτερο ενδιαφέρον λοιπόν να μελετηθούν αυτές οι πιθανές «ζώνες επαφής» ως προς τη γονιδιακή τους ροή, ώστε να εκτιμηθεί το ποσοστό απομόνωσης των πληθυσμών και, κατ επέκταση, του κάθε είδους. Επιπλέον, εξάγονται συμπεράσματα σχετικά με τις περιοχές των Κυθήρων, της Κρήτης και της Επιδαύρου, όπου όπως ήδη επισημάνθηκε θα πρέπει να θεωρηθεί ότι απαντάται το είδος Trachelipus aegaeus, έτσι ώστε να διατηρηθεί η μονοφυλετικότητα του κλάδου, όπως αυτός προκύπτει από τις αναλύσεις όλων των γενετικών τόπων. Υπενθυμίζεται πως σε αυτήν την περίπτωση, θα πρέπει να γίνει αλλαγή της ονοματολογίας και να επανέλθει σε ισχύ το όνομα T. palustris (Strouhal, 1936) που είχε δοθεί στα δείγματα από το Παναχαϊκό και, εν συνεχεία, θεωρήθηκε συνώνυμο του T. kytherensis από τον Schmalfuss, 2003, 2004). Κυρίως, όμως, η μελέτη αυτή θέτει μια σειρά ερωτημάτων βάσει όλων των προαναφερθέντων, τόσο σχετικά με την παρουσία ή μη ενός νέου είδους στην ηπειρωτική Ελλάδα όσο και με την εξήγηση του γεωγραφικού προτύπου, όπως αυτό αποτυπώνεται από τις αναλύσεις που άλλοτε συνάδει με την παλαιογεωγραφία του ελλαδικού χώρου και άλλοτε όχι. Σημειώνεται πως, παρά τις προσπάθειες που καταβλήθηκαν κατά τη διάρκεια των δειγματοληψιών, δεν συλλέχτηκαν αντιπρόσωποι από τα είδη T. camerani, T. buddelundi και T. squamuliger, τα οποία εξαπλώνονται στην ηπειρωτική Ελλάδα. Αντίθετα, συλλέχτηκαν άτομα τα οποία μορφολογικά αναγνωρίστηκαν ως αντιπρόσωποι του είδους T. kytherensis και, εν συνεχεία, οι γενετικές αναλύσεις τα διαχώρισαν σε 2 διαφορετικές «μορφές». Μια εξήγηση για την αυξημένη διαφοροποίηση που παρατηρείται ενδέχεται να είναι πως η γενετική ποικιλότητα, τουλάχιστον στο T. kytherensis, είναι πολύ μεγαλύτερη από αυτήν που καταδεικνύει η μορφολογία. Γι αυτόν το λόγο, απαιτείται η ενδελεχής μορφολογική εξέταση των αντιπροσώπων των πληθυσμών που εξετάστηκαν μοριακά στην παρούσα μελέτη, προκειμένου να βρεθούν πρόσθετοι 163
.")

170 μορφολογικοί χαρακτήρες που είτε θα διαχωρίζουν νέες μορφές είτε θα περιγράφουν καλύτερα το είδος αυτό. Η έντονη γενετική ποικιλομορφία που παρατηρείται στην παρούσα μελέτη θα μπορούσε να είναι αποτέλεσμα «αρχής του ιδρυτή» (Εικ. Δ-1). Στην πληθυσμιακή Εικόνα Δ-1: Σχηματική απεικόνιση της Αρχής του Ιδρυτή. Δεξιά ο αρχικός πληθυσμός. Αριστερά τρεις πιθανοί ιδρυτικοί πληθυσμοί. γενετική, η αρχή του ιδρυτή αφορά στην απώλεια της γενετικής ποικιλότητας, η οποία λαμβάνει χώρα όταν ένας νέος πληθυσμός ιδρύεται από πολύ μικρό αριθμό ατόμων ενός μεγαλύτερου πληθυσμού (Provine 2004). Πρόκειται, δηλαδή, για ακραία περίπτωση γενετικής παρέκκλισης. Ως αποτέλεσμα της απώλειας της γενετικής ποικιλότητας, ο νέος πληθυσμός μπορεί να είναι ευδιάκριτα διαφορετικός, τόσο γενετικά όσο και φαινοτυπικά, από τον πληθυσμό από τον οποίον προέρχεται. Σε ακραίες περιπτώσεις, πολλαπλές επαναλήψεις ιδρυτικών φαινομένων μπορούν να οδηγήσουν σε ειδογένεση και ακολούθως στην εξέλιξη νέων ειδών (Avise 2000). Μια άλλη εξήγηση για την αυξημένη γενετική ποικιλομορφία θα μπορούσε να είναι αυτή της ανεπαρκούς διαλογής γενεαλογικών γραμμών (inadequate lineage sorting) του είδους λόγω συνεχών φαινομένων αποχωρισμού και επανένωσης περιοχών. Οι αντιπρόσωποι του γένους Trachelipus έχουν περιορισμένη δυνατότητα μετακίνησης, εξαιτίας της εξάρτησής τους από τα υγρά περιβάλλοντα. Αυτές οι οικολογικές απαιτήσεις αποτελούν σημαντικά φράγματα στη γονιδιακή ροή. Στη διάρκεια ξηρών περιόδων, θα μπορούσαν να παρατηρηθούν φαινόμενα πληθυσμιακής στενωπού ή/και εξαφάνισης. Κάτι τέτοιο θα οδηγούσε σε μείωση της γενετικής 164
171 ποικιλότητας και, τελικά, σε γενετική παρέκκλιση. Όταν θα επικρατούσαν και πάλι ευνοϊκές συνθήκες, πιθανότατα θα ακολουθούσε επανεποίκιση των άδειων περιοχών, πιθανώς από διαφορετικές γενεαλογικές γραμμές ή/και από επέκταση γειτονικών πληθυσμών. Ένα τέτοιο σενάριο (Εικ. Δ-2) θα μπορούσε να εξηγεί το γεγονός ότι πολύ κοντινοί γεωγραφικά πληθυσμοί εμφανίζουν σήμερα τόσο μεγάλη γενετική απόκλιση. Όλα τα προηγούμενα μένει να διερευνηθούν μελλοντικά με τη χρήση επιπρόσθετων μοριακών εργαλείων, π.χ. με τη χρήση μικροδορυφορικού DNA, που θα εξετάσουν τα επίπεδα γονιδιακής ροής μεταξύ των πληθυσμών, ή/και πυρηνικών γονιδίων προκειμένου να προσεγγιστεί η φυλογενετική ιστορία και οι εξελικτικές σχέσεις και συνδέσεις των πληθυσμών και των ειδών του γένους Trachelipus στην Ελλάδα. Όλα αυτά, μαζί με τη συνδρομή της λεπτομερειακής μορφολογικής εξέτασης των αντιπροσώπων και τη χρήση μορφομετρικών μεθόδων, θα βοηθήσουν να προσδιοριστεί με μεγαλύτερη ακρίβεια, λεπτομέρεια και πιστότητα η φυλογεωγραφία του τάξου αυτού στον ελλαδικό χώρο. 165

172 Εικόνα Δ-2: Υποθετικό φυλογεωγραφικό σενάριο για την σημερινή εξάπλωση των αντιπροσώπων του γένους Trachelipus που μελετήθηκαν στο πλαίσιο της παρούσας εργασίας. 166
ΜΑΘΗΜΑ 1 ΑΣΚΗΣΕΙΣ ΜΑΘΗΜΑ Να γνωρίζεις τις έννοιες γεωγραφικό πλάτος, γεωγραφικό μήκος και πως αυτές εκφράζονται
 ΜΑΘΗΜΑ 1 Π. Γ Κ Ι Ν Η Σ 1. Να γνωρίζεις τις έννοιες γεωγραφικό πλάτος, γεωγραφικό μήκος και πως αυτές εκφράζονται 2. Να μπορείς να δώσεις την σχετική γεωγραφική θέση ενός τόπου χρησιμοποιώντας τους όρους
ΜΑΘΗΜΑ 1 Π. Γ Κ Ι Ν Η Σ 1. Να γνωρίζεις τις έννοιες γεωγραφικό πλάτος, γεωγραφικό μήκος και πως αυτές εκφράζονται 2. Να μπορείς να δώσεις την σχετική γεωγραφική θέση ενός τόπου χρησιμοποιώντας τους όρους
ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ. Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων
 ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων Φυλογένεση Η φυλογένεσης αφορά την ανεύρεση των συνδετικών εκείνων κρίκων που συνδέουν τα διάφορα είδη µεταξύ τους εξελικτικά, σε µονοφυλετικές
ΦΥΣΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ Πρωτεύοντα ΙΙΙ Χρήση µοριακών δεδοµένων Φυλογένεση Η φυλογένεσης αφορά την ανεύρεση των συνδετικών εκείνων κρίκων που συνδέουν τα διάφορα είδη µεταξύ τους εξελικτικά, σε µονοφυλετικές
Συστηματική και φυλογεωγραφία του συμπλέγματος ειδών Ablepharus kitaibelii στη περιοχή της ανατολικής Μεσογείου
 Συστηματική και φυλογεωγραφία του συμπλέγματος ειδών Ablepharus kitaibelii στη περιοχή της ανατολικής Μεσογείου Νίκος Πουλακάκης, Cetin Ilgaz, Yusuf Kumlutaş, Aziz Avcı, Ahmadzadeh Faraham, Jelca Crnobrnja
Συστηματική και φυλογεωγραφία του συμπλέγματος ειδών Ablepharus kitaibelii στη περιοχή της ανατολικής Μεσογείου Νίκος Πουλακάκης, Cetin Ilgaz, Yusuf Kumlutaş, Aziz Avcı, Ahmadzadeh Faraham, Jelca Crnobrnja
ΕΛΛΗΝΙΚΟ ΤΟΞΟ. Γεωλογική εξέλιξη της Ελλάδας Το Ελληνικό τόξο
 ΕΛΛΗΝΙΚΟ ΤΟΞΟ Γεωλογική εξέλιξη της Ελλάδας Το Ελληνικό τόξο ρ. Ε. Λυκούδη Αθήνα 2005 Γεωλογική εξέλιξη της Ελλάδας Ο Ελλαδικός χώρος µε την ευρεία γεωγραφική έννοια του όρου, έχει µια σύνθετη γεωλογικοτεκτονική
ΕΛΛΗΝΙΚΟ ΤΟΞΟ Γεωλογική εξέλιξη της Ελλάδας Το Ελληνικό τόξο ρ. Ε. Λυκούδη Αθήνα 2005 Γεωλογική εξέλιξη της Ελλάδας Ο Ελλαδικός χώρος µε την ευρεία γεωγραφική έννοια του όρου, έχει µια σύνθετη γεωλογικοτεκτονική
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ. Γεωγραφική διαφοροποίηση και. γενετική ποικιλότητα πληθυσμών χερσόβιων ισοπόδων.
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ Γεωγραφική διαφοροποίηση και γενετική ποικιλότητα πληθυσμών χερσόβιων ισοπόδων. ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ Μαρία Καμηλάρη Βιολόγος, M.Sc. ΠΑΤΡΑ, ΣΕΠΤΕΜΒΡΙΟΣ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ Γεωγραφική διαφοροποίηση και γενετική ποικιλότητα πληθυσμών χερσόβιων ισοπόδων. ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ Μαρία Καμηλάρη Βιολόγος, M.Sc. ΠΑΤΡΑ, ΣΕΠΤΕΜΒΡΙΟΣ
Ευρασιατική, Αφρικανική και Αραβική
 Έχει διαπιστωθεί διεθνώς ότι τα περιθώρια τεκτονικών πλακών σε ηπειρωτικές περιοχές είναι πολύ ευρύτερα από τις ωκεάνιες (Ευρασία: π.χ. Ελλάδα, Κίνα), αναφορικά με την κατανομή των σεισμικών εστιών. Στην
Έχει διαπιστωθεί διεθνώς ότι τα περιθώρια τεκτονικών πλακών σε ηπειρωτικές περιοχές είναι πολύ ευρύτερα από τις ωκεάνιες (Ευρασία: π.χ. Ελλάδα, Κίνα), αναφορικά με την κατανομή των σεισμικών εστιών. Στην
Κωνσταντίνος Στεφανίδης
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης Οικολογική
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης Οικολογική
Η ιστορική πατρότητα του όρου «Μεσόγειος θάλασσα» ανήκει στους Λατίνους και μάλιστα περί τα μέσα του 3ου αιώνα που πρώτος ο Σολίνος τη ονομάζει
 Η ιστορική πατρότητα του όρου «Μεσόγειος θάλασσα» ανήκει στους Λατίνους και μάλιστα περί τα μέσα του 3ου αιώνα που πρώτος ο Σολίνος τη ονομάζει χαρακτηριστικά «Mare Mediterraneum» ως μεταξύ δύο ηπείρων
Η ιστορική πατρότητα του όρου «Μεσόγειος θάλασσα» ανήκει στους Λατίνους και μάλιστα περί τα μέσα του 3ου αιώνα που πρώτος ο Σολίνος τη ονομάζει χαρακτηριστικά «Mare Mediterraneum» ως μεταξύ δύο ηπείρων
ENOTHTA 1: ΧΑΡΤΕΣ ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ
 1 ENOTHTA 1: ΧΑΡΤΕΣ ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ Μάθημα 1: Οι έννοιες και θέση 1. Τι ονομάζεται σχετική θέση ενός τόπου; Να δοθεί ένα παράδειγμα. Πότε ο προσδιορισμός της σχετικής θέσης
1 ENOTHTA 1: ΧΑΡΤΕΣ ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ Μάθημα 1: Οι έννοιες και θέση 1. Τι ονομάζεται σχετική θέση ενός τόπου; Να δοθεί ένα παράδειγμα. Πότε ο προσδιορισμός της σχετικής θέσης
Εξελικτική πορεία της ελληνικής χλωρίδας παράδειγμα τα νησιά του Αιγαίου
 Εξελικτική πορεία της ελληνικής χλωρίδας παράδειγμα τα νησιά του Αιγαίου Καθηγητής Δημήτριος Χριστοδουλάκης Τμήμα Βιολογίας Τομέας Βιολογίας Φυτών E-mail: dkchrist@upatras.gr Τηλ.: 2610 997277 20 Αιγηίδα:
Εξελικτική πορεία της ελληνικής χλωρίδας παράδειγμα τα νησιά του Αιγαίου Καθηγητής Δημήτριος Χριστοδουλάκης Τμήμα Βιολογίας Τομέας Βιολογίας Φυτών E-mail: dkchrist@upatras.gr Τηλ.: 2610 997277 20 Αιγηίδα:
2. ΓΕΩΓΡΑΦΙΑ ΤΗΣ Υ ΡΟΣΦΑΙΡΑΣ
 2. ΓΕΩΓΡΑΦΙΑ ΤΗΣ Υ ΡΟΣΦΑΙΡΑΣ 2.1 Ωκεανοί και Θάλασσες. Σύµφωνα µε τη ιεθνή Υδρογραφική Υπηρεσία (International Hydrographic Bureau, 1953) ως το 1999 θεωρούντο µόνο τρεις ωκεανοί: Ο Ατλαντικός, ο Ειρηνικός
2. ΓΕΩΓΡΑΦΙΑ ΤΗΣ Υ ΡΟΣΦΑΙΡΑΣ 2.1 Ωκεανοί και Θάλασσες. Σύµφωνα µε τη ιεθνή Υδρογραφική Υπηρεσία (International Hydrographic Bureau, 1953) ως το 1999 θεωρούντο µόνο τρεις ωκεανοί: Ο Ατλαντικός, ο Ειρηνικός
ΓΕΩΓΡΑΦΙΑ ΘΕΜΑΤΑ. ήταν ο κάθε ένας από αυτούς και σε ποιον από αυτούς σχηματίστηκε η Ελλάδα;
 ΓΕΩΓΡΑΦΙΑ ΘΕΜΑΤΑ ΘΕΜΑ 1 ο (Μονάδες 3,3) 1. Ποια είναι η διοικητική ιεραρχία των πόλεων στην Ελλάδα; Πως λέγεται ο διοικητής του κάθε διοικητικού τομέα; 2. Ποιους γεωλογικούς αιώνες περιλαμβάνει η γεωλογική
ΓΕΩΓΡΑΦΙΑ ΘΕΜΑΤΑ ΘΕΜΑ 1 ο (Μονάδες 3,3) 1. Ποια είναι η διοικητική ιεραρχία των πόλεων στην Ελλάδα; Πως λέγεται ο διοικητής του κάθε διοικητικού τομέα; 2. Ποιους γεωλογικούς αιώνες περιλαμβάνει η γεωλογική
Τελική Αναφορά της Κατάστασης Διατήρησης της Μεσογειακής Φώκιας Monachus monachus στη Νήσο Γυάρο Περίληψη
 Τελική Αναφορά της Κατάστασης Διατήρησης της Μεσογειακής Φώκιας Monachus monachus στη Νήσο Γυάρο Περίληψη ΙΟΥΝΙΟΣ 2018 Τελική Έκθεση Για την Κατάσταση της Μεσογειακής φώκιας στη νήσο Γυάρο Περίληψη Σελίδα
Τελική Αναφορά της Κατάστασης Διατήρησης της Μεσογειακής Φώκιας Monachus monachus στη Νήσο Γυάρο Περίληψη ΙΟΥΝΙΟΣ 2018 Τελική Έκθεση Για την Κατάσταση της Μεσογειακής φώκιας στη νήσο Γυάρο Περίληψη Σελίδα
Department of Biological Sciences, Texas Tech University, MS 43131, Lubbock,Texas 79409-3131. 3
 1 Τμήμα Βιολογίας, Πανεπιστήμιο Κρήτης, Ηράκλειο Κρήτης. 2 Department of Biological Sciences, Texas Tech University, MS 43131, Lubbock,Texas 79409-3131. 3 Department of Biology, Faculty of Science, Dokuz
1 Τμήμα Βιολογίας, Πανεπιστήμιο Κρήτης, Ηράκλειο Κρήτης. 2 Department of Biological Sciences, Texas Tech University, MS 43131, Lubbock,Texas 79409-3131. 3 Department of Biology, Faculty of Science, Dokuz
ρ. Ε. Λυκούδη Αθήνα 2005 ΩΚΕΑΝΟΙ Ωκεανοί Ωκεάνιες λεκάνες
 ρ. Ε. Λυκούδη Αθήνα 2005 ΩΚΕΑΝΟΙ Ωκεανοί Ωκεάνιες λεκάνες Ωκεανοί Το νερό καλύπτει τα δύο τρίτα της γης και το 97% όλου του κόσµου υ και είναι κατοικία εκατοµµυρίων γοητευτικών πλασµάτων. Οι ωκεανοί δηµιουργήθηκαν
ρ. Ε. Λυκούδη Αθήνα 2005 ΩΚΕΑΝΟΙ Ωκεανοί Ωκεάνιες λεκάνες Ωκεανοί Το νερό καλύπτει τα δύο τρίτα της γης και το 97% όλου του κόσµου υ και είναι κατοικία εκατοµµυρίων γοητευτικών πλασµάτων. Οι ωκεανοί δηµιουργήθηκαν
Περιεχόμενο Μαθημάτων
 Περιεχόμενο Μαθημάτων Χερσαία και Θαλάσσια Οικοσυστήματα Συντονιστές I. Καρακάσης M. Μυλωνάς Σύνολο ωρών 100 Διδάσκοντες Χ. Αρβανιτίδης, Ι. Καρακάσης, Κ. Κοτζαμπάσης, Ν. Λαμπαδαρίου, Μ. Μυλωνάς, Π. Πήττα
Περιεχόμενο Μαθημάτων Χερσαία και Θαλάσσια Οικοσυστήματα Συντονιστές I. Καρακάσης M. Μυλωνάς Σύνολο ωρών 100 Διδάσκοντες Χ. Αρβανιτίδης, Ι. Καρακάσης, Κ. Κοτζαμπάσης, Ν. Λαμπαδαρίου, Μ. Μυλωνάς, Π. Πήττα
Η Αφρική είναι η τρίτη σε μέγεθος ήπειρος του πλανήτη μας, μετά την Ασία και την Αμερική. Η έκτασή της είναι, χωρίς τα νησιά, 29,2 εκατομμύρια τετρ. χ
 Β. Π. Γ. Π. Η Αφρική είναι η τρίτη σε μέγεθος ήπειρος του πλανήτη μας, μετά την Ασία και την Αμερική. Η έκτασή της είναι, χωρίς τα νησιά, 29,2 εκατομμύρια τετρ. χιλιόμετρα, ενώ με τα νησιά φτάνει τα 30,2
Β. Π. Γ. Π. Η Αφρική είναι η τρίτη σε μέγεθος ήπειρος του πλανήτη μας, μετά την Ασία και την Αμερική. Η έκτασή της είναι, χωρίς τα νησιά, 29,2 εκατομμύρια τετρ. χιλιόμετρα, ενώ με τα νησιά φτάνει τα 30,2
Για να περιγράψουμε την ατμοσφαιρική κατάσταση, χρησιμοποιούμε τις έννοιες: ΚΑΙΡΟΣ. και ΚΛΙΜΑ
 Το κλίμα της Ευρώπης Το κλίμα της Ευρώπης Για να περιγράψουμε την ατμοσφαιρική κατάσταση, χρησιμοποιούμε τις έννοιες: ΚΑΙΡΟΣ και ΚΛΙΜΑ Καιρός: Οι ατμοσφαιρικές συνθήκες που επικρατούν σε μια περιοχή, σε
Το κλίμα της Ευρώπης Το κλίμα της Ευρώπης Για να περιγράψουμε την ατμοσφαιρική κατάσταση, χρησιμοποιούμε τις έννοιες: ΚΑΙΡΟΣ και ΚΛΙΜΑ Καιρός: Οι ατμοσφαιρικές συνθήκες που επικρατούν σε μια περιοχή, σε
Σκορπιός Χειλόποδο Διπλόποδο Ορθόπτερο Ημίπτερο Υμενόπτερο Κολεόπτερο. Φυλογενετικό δέντρο
 Σχολές Συστηματικής Σκορπιός Χειλόποδο Διπλόποδο Ορθόπτερο Ημίπτερο Υμενόπτερο Κολεόπτερο Φυλογενετικό δέντρο Σκορπιός Χειλόποδο Διπλόποδο Ορθόπτερο Ημίπτερο Υμενόπτερο Κολεόπτερο 14 12 11 7 Φαινόγραμμα
Σχολές Συστηματικής Σκορπιός Χειλόποδο Διπλόποδο Ορθόπτερο Ημίπτερο Υμενόπτερο Κολεόπτερο Φυλογενετικό δέντρο Σκορπιός Χειλόποδο Διπλόποδο Ορθόπτερο Ημίπτερο Υμενόπτερο Κολεόπτερο 14 12 11 7 Φαινόγραμμα
4. γεωγραφικό/γεωλογικό πλαίσιο
 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο /Ελληνικός χώρος Τα ελληνικά βουνά (και γενικότερα οι ορεινοί όγκοι της
4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο 4. ΜΕΛΛΟΝΤΙΚΟ γεωγραφικό/γεωλογικό πλαίσιο /Ελληνικός χώρος Τα ελληνικά βουνά (και γενικότερα οι ορεινοί όγκοι της
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή εργασία
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΟΡΙΑΚΗ ΚΑΙ ΦΑΙΝΟΤΥΠΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΚΥΠΡΙΑΚΟΥ ΠΛΗΘΥΣΜΟΥ ΣΚΛΗΡΟΥ ΣΙΤΑΡΙΟΥ ΠΟΥ ΑΠΟΚΤΗΘΗΚΕ ΑΠΟ ΤΡΑΠΕΖΑ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή εργασία ΜΟΡΙΑΚΗ ΚΑΙ ΦΑΙΝΟΤΥΠΙΚΗ ΑΝΑΛΥΣΗ ΕΝΟΣ ΚΥΠΡΙΑΚΟΥ ΠΛΗΘΥΣΜΟΥ ΣΚΛΗΡΟΥ ΣΙΤΑΡΙΟΥ ΠΟΥ ΑΠΟΚΤΗΘΗΚΕ ΑΠΟ ΤΡΑΠΕΖΑ
iv. Παράκτια Γεωμορφολογία
 iv. Παράκτια Γεωμορφολογία Η παράκτια ζώνη περιλαμβάνει, τόσο το υποθαλάσσιο τμήμα της ακτής, μέχρι το βάθος όπου τα ιζήματα υπόκεινται σε περιορισμένη μεταφορά εξαιτίας της δράσης των κυμάτων, όσο και
iv. Παράκτια Γεωμορφολογία Η παράκτια ζώνη περιλαμβάνει, τόσο το υποθαλάσσιο τμήμα της ακτής, μέχρι το βάθος όπου τα ιζήματα υπόκεινται σε περιορισμένη μεταφορά εξαιτίας της δράσης των κυμάτων, όσο και
ΚΛΙΜΑΤΙΚΗ ΤΑΞΙΝΟΝΗΣΗ ΕΛΛΑΔΑΣ
 ΚΛΙΜΑΤΙΚΗ ΤΑΞΙΝΟΝΗΣΗ ΕΛΛΑΔΑΣ ΚΡΙΤΗΡΙΑ ΤΑΞΙΝΟΜΗΣΗΣ Το κλίμα μιας γεωγραφικής περιοχής διαμορφώνεται κατά κύριο λόγο από τους 3 παρακάτω παράγοντες: 1)το γεωγραφικό πλάτος 2)την αναλογία ξηράς/θάλασσας 3)το
ΚΛΙΜΑΤΙΚΗ ΤΑΞΙΝΟΝΗΣΗ ΕΛΛΑΔΑΣ ΚΡΙΤΗΡΙΑ ΤΑΞΙΝΟΜΗΣΗΣ Το κλίμα μιας γεωγραφικής περιοχής διαμορφώνεται κατά κύριο λόγο από τους 3 παρακάτω παράγοντες: 1)το γεωγραφικό πλάτος 2)την αναλογία ξηράς/θάλασσας 3)το
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΤΟΓΕΩΓΡΑΦΙΑ
 ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΤΟΓΕΩΓΡΑΦΙΑ ΑΝΤΙΚΕΙΜΕΝΑ ΤΗΣ ΦΥΤΟΓΕΩΓΡΑΦΙΑΣ Χωρολογία των φυτών Εξελικτική ή ιστορική φυτογεωγραφία Φυτοκοινωνιολογία Οικολογική φυτογεωγραφία Η ΠΕΡΙΟΧΗ ΕΞΑΠΛΩΣΗΣ 1) Η ερμηνεία της περιοχής
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΤΟΓΕΩΓΡΑΦΙΑ ΑΝΤΙΚΕΙΜΕΝΑ ΤΗΣ ΦΥΤΟΓΕΩΓΡΑΦΙΑΣ Χωρολογία των φυτών Εξελικτική ή ιστορική φυτογεωγραφία Φυτοκοινωνιολογία Οικολογική φυτογεωγραφία Η ΠΕΡΙΟΧΗ ΕΞΑΠΛΩΣΗΣ 1) Η ερμηνεία της περιοχής
Νεοφυτικός αιώνας (περίοδος των Αγγειοσπέρμων)
") Νεοφυτικός αιώνας (περίοδος των Αγγειοσπέρμων) Η κυριαρχία των Αγγειοσπέρμων αρχίζει από το Mέσο Kρητιδικό (πριν 100 εκ. χρόνια) και συνεχίζεται μέχρι σήμερα. Υπάρχουν περίπου 250.000 είδη Αγγειοσπέρμων.
Νεοφυτικός αιώνας (περίοδος των Αγγειοσπέρμων) Η κυριαρχία των Αγγειοσπέρμων αρχίζει από το Mέσο Kρητιδικό (πριν 100 εκ. χρόνια) και συνεχίζεται μέχρι σήμερα. Υπάρχουν περίπου 250.000 είδη Αγγειοσπέρμων.
Η ΣΤΑΘΜΗ ΤΗΣ ΘΑΛΑΣΣΑΣ ΧΘΕΣ, ΣΗΜΕΡΑ, ΑΥΡΙΟ
 ΕΛΛΗΝΙΚΗ ΣΠΗΛΑΙΟΛΟΠΚΗ ΕΤΑΙΡΕΙΑ Σίνα 32, Αθήνα 106 72, τηλ.210-3617824, φαξ 210-3643476, e- mails: ellspe@otenet.gr & info@speleologicalsociety.gr website: www.speleologicalsociety.gr ΕΚΠΑΙΔΕΥΤΙΚΟ ΠΡΟΓΡΑΜΜΑ
ΕΛΛΗΝΙΚΗ ΣΠΗΛΑΙΟΛΟΠΚΗ ΕΤΑΙΡΕΙΑ Σίνα 32, Αθήνα 106 72, τηλ.210-3617824, φαξ 210-3643476, e- mails: ellspe@otenet.gr & info@speleologicalsociety.gr website: www.speleologicalsociety.gr ΕΚΠΑΙΔΕΥΤΙΚΟ ΠΡΟΓΡΑΜΜΑ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ - ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΦΥΤΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΚΑΤΕΥΘΥΝΣΗ: ΟΙΚΟΛΟΓΙΑ-ΔΙΑΧΕΙΡΙΣΗ & ΠΡΟΣΤΑΣΙΑ ΦΥΣΙΚΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Διατριβή Μεταπτυχιακού Διπλώματος Ειδίκευσης
Νησιωτική Βιογεωγραφία
 Νησιωτική Βιογεωγραφία Κατερίνα Βαρδινογιάννη Μουσείο Φυσικής Ιστορίας Κρήτης - Πανεπιστήμιο Κρήτης Ηράκλειο 2018 Δυναμική ισορροπία MacArthur & Wilson 1967 Αποίκιση και εξαφάνιση σε δυναμική ισορροπία
Νησιωτική Βιογεωγραφία Κατερίνα Βαρδινογιάννη Μουσείο Φυσικής Ιστορίας Κρήτης - Πανεπιστήμιο Κρήτης Ηράκλειο 2018 Δυναμική ισορροπία MacArthur & Wilson 1967 Αποίκιση και εξαφάνιση σε δυναμική ισορροπία
Γεωλογία - Γεωγραφία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ. Τ μαθητ : Σχολικό Έτος:
 Γεωλογία - Γεωγραφία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ Τ μαθητ : Σχολικό Έτος: 1 ΜΑΘΗΜΑ 1, Οι έννοιες «γεωγραφική» και «σχετική» θέση 1. Με τη βοήθεια του χάρτη στη σελ.12, σημειώστε τις παρακάτω πόλεις στην
Γεωλογία - Γεωγραφία Β Γυμνασίου ΦΥΛΛΑΔΙΟ ΑΣΚΗΣΕΩΝ Τ μαθητ : Σχολικό Έτος: 1 ΜΑΘΗΜΑ 1, Οι έννοιες «γεωγραφική» και «σχετική» θέση 1. Με τη βοήθεια του χάρτη στη σελ.12, σημειώστε τις παρακάτω πόλεις στην
5. κλίμα. Οι στέπες είναι ξηροί λειμώνες με ετήσιο εύρος θερμοκρασιών το καλοκαίρι μέχρι 40 C και το χειμώνα κάτω από -40 C
 5. κλίμα 5. κλίμα Οι στέπες είναι ξηροί λειμώνες με ετήσιο εύρος θερμοκρασιών το καλοκαίρι μέχρι 40 C και το χειμώνα κάτω από -40 C 5. κλίμα 5. κλίμα Οι μεσογειακές περιοχές βρίσκονται μεταξύ 30 0 και
5. κλίμα 5. κλίμα Οι στέπες είναι ξηροί λειμώνες με ετήσιο εύρος θερμοκρασιών το καλοκαίρι μέχρι 40 C και το χειμώνα κάτω από -40 C 5. κλίμα 5. κλίμα Οι μεσογειακές περιοχές βρίσκονται μεταξύ 30 0 και
Εξελικτική Οικολογία. Σίνος Γκιώκας Πανεπιστήμιο Πάτρας Τμήμα Βιολογίας 2014
 Εξελικτική Οικολογία Σίνος Γκιώκας Πανεπιστήμιο Πάτρας Τμήμα Βιολογίας 2014 Εξελικτική Οικολογία - Σίνος Γκιώκας - Παν. Πατρών - Τμ. Βιολογίας - 2014 1 Πού συναντιούνται τα δύο πεδία; Οι κιχλίδες των Μεγάλων
Εξελικτική Οικολογία Σίνος Γκιώκας Πανεπιστήμιο Πάτρας Τμήμα Βιολογίας 2014 Εξελικτική Οικολογία - Σίνος Γκιώκας - Παν. Πατρών - Τμ. Βιολογίας - 2014 1 Πού συναντιούνται τα δύο πεδία; Οι κιχλίδες των Μεγάλων
Στρωματογραφία-Ιστορική γεωλογία. Ιστορική γεωλογία Δρ. Ηλιόπουλος Γεώργιος Σχολή Θετικών Επιστημών Τμήμα Γεωλογίας
 Στρωματογραφία-Ιστορική γεωλογία Ιστορική γεωλογία Δρ. Ηλιόπουλος Γεώργιος Σχολή Θετικών Επιστημών Τμήμα Γεωλογίας Σκοποί ενότητας Σκοπός της ενότητας είναι η εξοικείωση με τους κλάδους της ιστορικής γεωλογίας.
Στρωματογραφία-Ιστορική γεωλογία Ιστορική γεωλογία Δρ. Ηλιόπουλος Γεώργιος Σχολή Θετικών Επιστημών Τμήμα Γεωλογίας Σκοποί ενότητας Σκοπός της ενότητας είναι η εξοικείωση με τους κλάδους της ιστορικής γεωλογίας.
ΦΥΣΙΚΗ -ΚΛΙΜΑΤΙΚΗ ΑΛΛΑΓΗ ΚΑΙ ΓΕΩΡΓΙΑ
 Γιάννης Λ. Τσιρογιάννης Γεωργικός Μηχανικός M.Sc., PhD Επίκουρος Καθηγητής ΤΕΙ Ηπείρου Τμ. Τεχνολόγων Γεωπόνων Κατ. Ανθοκομίας Αρχιτεκτονικής Τοπίου ΦΥΣΙΚΗ -ΚΛΙΜΑΤΙΚΗ ΑΛΛΑΓΗ ΚΑΙ ΓΕΩΡΓΙΑ Κλιματική αλλαγή
Γιάννης Λ. Τσιρογιάννης Γεωργικός Μηχανικός M.Sc., PhD Επίκουρος Καθηγητής ΤΕΙ Ηπείρου Τμ. Τεχνολόγων Γεωπόνων Κατ. Ανθοκομίας Αρχιτεκτονικής Τοπίου ΦΥΣΙΚΗ -ΚΛΙΜΑΤΙΚΗ ΑΛΛΑΓΗ ΚΑΙ ΓΕΩΡΓΙΑ Κλιματική αλλαγή
ΟΙ ΥΔΡΙΤΕΣ ΚΑΙ Η ΣΗΜΑΣΙΑ ΤΟΥΣ ΩΣ ΚΑΥΣΙΜΗ ΥΛΗ ΤΟΥ ΜΕΛΛΟΝΤΟΣ. ΤΟ ΕΡΕΥΝΗΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ANAXIMANDER. Από Δρ. Κωνσταντίνο Περισοράτη
 ΟΙ ΥΔΡΙΤΕΣ ΚΑΙ Η ΣΗΜΑΣΙΑ ΤΟΥΣ ΩΣ ΚΑΥΣΙΜΗ ΥΛΗ ΤΟΥ ΜΕΛΛΟΝΤΟΣ. ΤΟ ΕΡΕΥΝΗΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ANAXIMANDER Από Δρ. Κωνσταντίνο Περισοράτη Οι υδρίτες (εικ. 1) είναι χημικές ενώσεις που ανήκουν στους κλειθρίτες, δηλαδή
ΟΙ ΥΔΡΙΤΕΣ ΚΑΙ Η ΣΗΜΑΣΙΑ ΤΟΥΣ ΩΣ ΚΑΥΣΙΜΗ ΥΛΗ ΤΟΥ ΜΕΛΛΟΝΤΟΣ. ΤΟ ΕΡΕΥΝΗΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ANAXIMANDER Από Δρ. Κωνσταντίνο Περισοράτη Οι υδρίτες (εικ. 1) είναι χημικές ενώσεις που ανήκουν στους κλειθρίτες, δηλαδή
ΚΕΦΑΛΑΙΟ 1. Η ΠΡΟΕΛΕΥΣΗ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΣΚΕΨΗΣ
 ΚΕΦΑΛΑΙΟ 1. Η ΠΡΟΕΛΕΥΣΗ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΣΚΕΨΗΣ Οι αρχές της εξελικτικής σκέψης Η προέλευση των ειδών Ορθές και λανθασµένες αντιλήψεις σχετικά µε τη θεωρία της εξέλιξης Η θεωρία της εξέλιξης
ΚΕΦΑΛΑΙΟ 1. Η ΠΡΟΕΛΕΥΣΗ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΣΚΕΨΗΣ Οι αρχές της εξελικτικής σκέψης Η προέλευση των ειδών Ορθές και λανθασµένες αντιλήψεις σχετικά µε τη θεωρία της εξέλιξης Η θεωρία της εξέλιξης
0,5 1,1 2,2 4,5 20,8 8,5 3,1 6,0 14,9 22,5 15,0 0,9
 ΩΚΕΑΝΟΓΡΑΦΙΑ E ΕΞΑΜΗΝΟ Ακαδημαϊκό Έτος 2016-2017 ΥΠΟΘΑΛΑΣΣΙΑ ΓΕΩΜΟΡΦΟΛΟΓΙΑ (Ο ΩΚΕΑΝΙΟΣ ΠΥΘΜΕΝΑΣ) Βασίλης ΚΑΨΙΜΑΛΗΣ Γεωλόγος-Ωκεανογράφος Κύριος Ερευνητής, ΕΛ.ΚΕ.Θ.Ε. Τηλ. Γραφείου: 22910 76378 Κιν.: 6944
ΩΚΕΑΝΟΓΡΑΦΙΑ E ΕΞΑΜΗΝΟ Ακαδημαϊκό Έτος 2016-2017 ΥΠΟΘΑΛΑΣΣΙΑ ΓΕΩΜΟΡΦΟΛΟΓΙΑ (Ο ΩΚΕΑΝΙΟΣ ΠΥΘΜΕΝΑΣ) Βασίλης ΚΑΨΙΜΑΛΗΣ Γεωλόγος-Ωκεανογράφος Κύριος Ερευνητής, ΕΛ.ΚΕ.Θ.Ε. Τηλ. Γραφείου: 22910 76378 Κιν.: 6944
ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Κεντούλλα Πέτρου Αριθμός Φοιτητικής Ταυτότητας 2008761539 Κύπρος
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΜΗΤΡΙΚΟΣ ΘΗΛΑΣΜΟΣ ΚΑΙ ΓΝΩΣΤΙΚΗ ΑΝΑΠΤΥΞΗ ΜΕΧΡΙ ΚΑΙ 10 ΧΡΟΝΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Ονοματεπώνυμο Κεντούλλα Πέτρου Αριθμός Φοιτητικής Ταυτότητας 2008761539 Κύπρος
Θέμα μας το κλίμα. Και οι παράγοντες που το επηρεάζουν.
 Θέμα μας το κλίμα. Και οι παράγοντες που το επηρεάζουν. 1 Που συμβαίνουν οι περισσότερες βροχοπτώσεις; Κυρίως στη θάλασσα. Και μάλιστα στο Ισημερινό. Είδαμε γιατί στο προηγούμενο μάθημα. Ρίξε μία ματιά.
Θέμα μας το κλίμα. Και οι παράγοντες που το επηρεάζουν. 1 Που συμβαίνουν οι περισσότερες βροχοπτώσεις; Κυρίως στη θάλασσα. Και μάλιστα στο Ισημερινό. Είδαμε γιατί στο προηγούμενο μάθημα. Ρίξε μία ματιά.
Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) Εισαγωγή. Δρ. Γεωργία Τοουλάκου
 Εισαγωγή. Δρ. Γεωργία Τοουλάκου") Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) 2018-2019 Εισαγωγή Δρ. Γεωργία Τοουλάκου Η Συστηματική Βοτανική είναι κλάδος της Βοτανικής. δηλαδή της επιστήμης η οποία ασχολείται
Εργαστήριο Συστηματικής Βοτανικής / Ζιζανιολογίας 2 ο Εξάμηνο (Εαρινό) 2018-2019 Εισαγωγή Δρ. Γεωργία Τοουλάκου Η Συστηματική Βοτανική είναι κλάδος της Βοτανικής. δηλαδή της επιστήμης η οποία ασχολείται
Εξελικτική Οικολογία. Ασκήσεις Βιογεωγραφίας ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ. Σίνος Γκιώκας 2014
 Εξελικτική Οικολογία Ασκήσεις Βιογεωγραφίας ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ Σίνος Γκιώκας 24 Στόχοι Οι ασκήσεις που συνδέονται με την ιστορική και κυρίως την οικολογική βιογεωγραφία.
Εξελικτική Οικολογία Ασκήσεις Βιογεωγραφίας ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ Σίνος Γκιώκας 24 Στόχοι Οι ασκήσεις που συνδέονται με την ιστορική και κυρίως την οικολογική βιογεωγραφία.
μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους
 Η ΕΠΙΣΤΗΜΗ ΤΗΣ ΟΙΚΟΛΟΓΙΑΣ μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους Οι οργανισμοί αλληλεπιδρούν με το περιβάλλον τους σε πολλά επίπεδα στα πλαίσια ενός οικοσυστήματος Οι φυσικές
Η ΕΠΙΣΤΗΜΗ ΤΗΣ ΟΙΚΟΛΟΓΙΑΣ μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους Οι οργανισμοί αλληλεπιδρούν με το περιβάλλον τους σε πολλά επίπεδα στα πλαίσια ενός οικοσυστήματος Οι φυσικές
Πολλαπλασιασμός των ειδών - Ειδογένεση. Αλλοπάτρια, παραπάτρια και συμπάτρια ειδογένεση. Μηχανισμοί αναπαραγωγικής απομόνωσης.
 Πολλαπλασιασμός των ειδών - Ειδογένεση. Αλλοπάτρια, παραπάτρια και συμπάτρια ειδογένεση. Μηχανισμοί αναπαραγωγικής απομόνωσης. Εισηγητής: Ν. Πουλακάκης Ειδογένεση H ειδογένεση αναφέρεται στο σχηματισμό
Πολλαπλασιασμός των ειδών - Ειδογένεση. Αλλοπάτρια, παραπάτρια και συμπάτρια ειδογένεση. Μηχανισμοί αναπαραγωγικής απομόνωσης. Εισηγητής: Ν. Πουλακάκης Ειδογένεση H ειδογένεση αναφέρεται στο σχηματισμό
Δημογραφία. Ενότητα 11.1: Παράδειγμα - Περιφερειακές διαφοροποιήσεις και ανισότητες του προσδόκιμου ζωής στη γέννηση
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Δημογραφία Ενότητα 11.1: Παράδειγμα - Περιφερειακές διαφοροποιήσεις και ανισότητες του προσδόκιμου ζωής στη γέννηση Μιχάλης Αγοραστάκης Τμήμα Μηχανικών Χωροταξίας, Πολεοδομίας &
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Δημογραφία Ενότητα 11.1: Παράδειγμα - Περιφερειακές διαφοροποιήσεις και ανισότητες του προσδόκιμου ζωής στη γέννηση Μιχάλης Αγοραστάκης Τμήμα Μηχανικών Χωροταξίας, Πολεοδομίας &
ΤΟ ΦΥΣΙΚΟ ΠΕΡΙΒΑΛΛΟΝ ΤΗΣ ΑΦΡΙΚΗΣ. Φύλλο εργασίας 1 Το φυσικό περιβάλλον της Αφρικής. Ονοματεπώνυμο Τάξη... Ημερομηνία.
 Ενότητα Ηπειροι (στιγμιότυπα): Η Αφρική Φύλλο εργασίας 1 Το φυσικό περιβάλλον της Αφρικής Γεωλογία - Γεωγραφία A Γυμνασίου Ονοματεπώνυμο Τάξη... Ημερομηνία. Πρώτη εργασία : Η θέση της Αφρικής στον παγκόσμιο
Ενότητα Ηπειροι (στιγμιότυπα): Η Αφρική Φύλλο εργασίας 1 Το φυσικό περιβάλλον της Αφρικής Γεωλογία - Γεωγραφία A Γυμνασίου Ονοματεπώνυμο Τάξη... Ημερομηνία. Πρώτη εργασία : Η θέση της Αφρικής στον παγκόσμιο
Νησιωτική Βιογεωγραφία
 Νησιωτική Βιογεωγραφία Κατερίνα Βαρδινογιάννη Μουσείο Φυσικής Ιστορίας Κρήτης - Πανεπιστήμιο Κρήτης Ηράκλειο 2018 Τι είναι νησί Τύποι νησιών Νησιά Πραγματικά νησιά (γεωγραφικά νησιά) Ωκεάνια νησιά Ηπειρωτικά
Νησιωτική Βιογεωγραφία Κατερίνα Βαρδινογιάννη Μουσείο Φυσικής Ιστορίας Κρήτης - Πανεπιστήμιο Κρήτης Ηράκλειο 2018 Τι είναι νησί Τύποι νησιών Νησιά Πραγματικά νησιά (γεωγραφικά νησιά) Ωκεάνια νησιά Ηπειρωτικά
Πληθυσμιακή και Εξελικτική Γενετική
 Οικολογία και Προστασία Δασικών Οικοσυστημάτων Πληθυσμιακή και Εξελικτική Γενετική & πληθυσμιακή δομή Εργαστήριο Δασικής Γενετικής Αριστοτέλης Χ. Παπαγεωργίου Πολλοί πληθυσμοί Εξετάσαμε τους εξελικτικούς
Οικολογία και Προστασία Δασικών Οικοσυστημάτων Πληθυσμιακή και Εξελικτική Γενετική & πληθυσμιακή δομή Εργαστήριο Δασικής Γενετικής Αριστοτέλης Χ. Παπαγεωργίου Πολλοί πληθυσμοί Εξετάσαμε τους εξελικτικούς
ΚΑΘΟΡΙΣΜΟΣ ΤΟΥ ΠΕΔΙΟΥ ΤΩΝ ΤΑΣΕΩΝ
 ΚΑΘΟΡΙΣΜΟΣ ΤΟΥ ΠΕΔΙΟΥ ΤΩΝ ΤΑΣΕΩΝ Εισαγωγή: Η σεισμικότητα μιας περιοχής χρησιμοποιείται συχνά για την εξαγωγή συμπερασμάτων σχετικών με τις τεκτονικές διαδικασίες που λαμβάνουν χώρα εκεί. Από τα τέλη του
ΚΑΘΟΡΙΣΜΟΣ ΤΟΥ ΠΕΔΙΟΥ ΤΩΝ ΤΑΣΕΩΝ Εισαγωγή: Η σεισμικότητα μιας περιοχής χρησιμοποιείται συχνά για την εξαγωγή συμπερασμάτων σχετικών με τις τεκτονικές διαδικασίες που λαμβάνουν χώρα εκεί. Από τα τέλη του
Ταξινόµιση οργανισµών
 Ταξινόµιση οργανισµών Ιεραρχική κατηγοριοποίηση/ οµαδοποίηση οργανισµών. Linnaeus (1707-1778) οµαδοποίησε οργανισµούς µε βάση κοινούς χαρακτήρες. Αργότερα, η ταξινόµιση προσαρµόστηκε στην εξελικτική θεωρία
Ταξινόµιση οργανισµών Ιεραρχική κατηγοριοποίηση/ οµαδοποίηση οργανισµών. Linnaeus (1707-1778) οµαδοποίησε οργανισµούς µε βάση κοινούς χαρακτήρες. Αργότερα, η ταξινόµιση προσαρµόστηκε στην εξελικτική θεωρία
Η πορεία της βιοποικιλότητας στο χρόνο
 Τμήμα Δασολογίας & Διαχείρισης Περιβάλλοντος & Φυσικών Πόρων / ΔΠΘ Η πορεία της βιοποικιλότητας στο χρόνο ΕΞΕΛΙΚΤΙΚΗ ΒΙΟΛΟΓΙΑ & ΒΙΟΠΟΙΚΙΛΟΤΗΤΑ Αριστοτέλης Χ. Παπαγεωργίου Ιδανικός πλανήτης για τη ζωή Θερμοκρασία
Τμήμα Δασολογίας & Διαχείρισης Περιβάλλοντος & Φυσικών Πόρων / ΔΠΘ Η πορεία της βιοποικιλότητας στο χρόνο ΕΞΕΛΙΚΤΙΚΗ ΒΙΟΛΟΓΙΑ & ΒΙΟΠΟΙΚΙΛΟΤΗΤΑ Αριστοτέλης Χ. Παπαγεωργίου Ιδανικός πλανήτης για τη ζωή Θερμοκρασία
ΚΛΙΜΑΤΙΚΗ ΚΑΤΑΤΑΞΗ ΤΗΣ ΕΛΛΑΔΑΣ ΚΑΤΑ KOPPEN Το κλίμα μιας γεωγραφικής περιοχής διαμορφώνεται κατά κύριο λόγο από τους 3 παρακάτω παράγοντες: 1) το
 το") ΚΛΙΜΑΤΙΚΗ ΚΑΤΑΤΑΞΗ ΤΗΣ ΕΛΛΑΔΑΣ ΚΑΤΑ KOPPEN Το κλίμα μιας γεωγραφικής περιοχής διαμορφώνεται κατά κύριο λόγο από τους 3 παρακάτω παράγοντες: 1) το γεωγραφικό πλάτος 2) την αναλογία ξηράς/θάλασσας 3) το
ΚΛΙΜΑΤΙΚΗ ΚΑΤΑΤΑΞΗ ΤΗΣ ΕΛΛΑΔΑΣ ΚΑΤΑ KOPPEN Το κλίμα μιας γεωγραφικής περιοχής διαμορφώνεται κατά κύριο λόγο από τους 3 παρακάτω παράγοντες: 1) το γεωγραφικό πλάτος 2) την αναλογία ξηράς/θάλασσας 3) το
ΤΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΥΜΒΑΛΕΙ ΟΥΣΙΑΣΤΙΚΑ ΣΤΗΝ ΑΝΤΙΜΕΤΩΠΙΣΗ ΤΗΣ ΠΑΡΑΝΟΜΗΣ ΘΗΡΕΥΣΗΣ ΤΩΝ ΑΜΠΕΛΟΠΟΥΛΙΩΝ
 ΑΝΑΚΟΙΝΩΣΗ ΤΥΠΟΥ Επικοινωνία: Γραφείο Επικοινωνίας Τομέας Προώθησης και Προβολής, Πανεπιστήμιο Κύπρου Τηλ. 22894304 ηλ. διεύθυνση: prinfo@ucy.ac.cy, ιστοσελίδα: www.ucy.ac.cy/pr ΠΡΟΣ ΑΜΕΣΗ ΔΗΜΟΣΙΕΥΣΗ Λευκωσία,
ΑΝΑΚΟΙΝΩΣΗ ΤΥΠΟΥ Επικοινωνία: Γραφείο Επικοινωνίας Τομέας Προώθησης και Προβολής, Πανεπιστήμιο Κύπρου Τηλ. 22894304 ηλ. διεύθυνση: prinfo@ucy.ac.cy, ιστοσελίδα: www.ucy.ac.cy/pr ΠΡΟΣ ΑΜΕΣΗ ΔΗΜΟΣΙΕΥΣΗ Λευκωσία,
PROJECT 2017 ΟΜΑΔΑ: ΑΝΕΣΤΗΣ ΠΑΠΑΖΟΓΛΟΥ ΧΡΥΣΟΥΛΑ ΜΙΧΟΠΟΥΛΟΥ ΚΑΤΕΡΙΝΑ ΝΤΙΝΗ ΓΙΑΝΝΗΣ ΜΟΣΧΟΦΙΔΗΣ
 PROJECT 2017 ΟΜΑΔΑ: ΑΝΕΣΤΗΣ ΠΑΠΑΖΟΓΛΟΥ ΧΡΥΣΟΥΛΑ ΜΙΧΟΠΟΥΛΟΥ ΚΑΤΕΡΙΝΑ ΝΤΙΝΗ ΓΙΑΝΝΗΣ ΜΟΣΧΟΦΙΔΗΣ ΑΦΡΙΚΗ Η Αφρική είναι η δεύτερη σε έκταση και πληθυσμό ήπειρος της γης με πληθυσμό περίπου 1 δισ κατοίκους.
PROJECT 2017 ΟΜΑΔΑ: ΑΝΕΣΤΗΣ ΠΑΠΑΖΟΓΛΟΥ ΧΡΥΣΟΥΛΑ ΜΙΧΟΠΟΥΛΟΥ ΚΑΤΕΡΙΝΑ ΝΤΙΝΗ ΓΙΑΝΝΗΣ ΜΟΣΧΟΦΙΔΗΣ ΑΦΡΙΚΗ Η Αφρική είναι η δεύτερη σε έκταση και πληθυσμό ήπειρος της γης με πληθυσμό περίπου 1 δισ κατοίκους.
γεωγραφικό γλωσσάρι για την πέμπτη τάξη (από το βιβλίο «Μαθαίνω την Ελλάδα» του ΟΕΔΒ)
") γεωγραφικό γλωσσάρι για την πέμπτη τάξη (από το βιβλίο «Μαθαίνω την Ελλάδα» του ΟΕΔΒ) Α Κεφ. αβιοτικό κάθε στοιχείο που δεν έχει ζωή 4 αιολική διάβρωση Η διάβρωση που οφείλεται στον άνεμο 5 ακρωτήριο ακτογραμμή
γεωγραφικό γλωσσάρι για την πέμπτη τάξη (από το βιβλίο «Μαθαίνω την Ελλάδα» του ΟΕΔΒ) Α Κεφ. αβιοτικό κάθε στοιχείο που δεν έχει ζωή 4 αιολική διάβρωση Η διάβρωση που οφείλεται στον άνεμο 5 ακρωτήριο ακτογραμμή
μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους
 Η ΕΠΙΣΤΗΜΗ ΤΗΣ ΟΙΚΟΛΟΓΙΑΣ μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους Οι οργανισμοί αλληλεπιδρούν με το περιβάλλον τους σε πολλά επίπεδα στα πλαίσια ενός οικοσυστήματος Οι φυσικές
Η ΕΠΙΣΤΗΜΗ ΤΗΣ ΟΙΚΟΛΟΓΙΑΣ μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους Οι οργανισμοί αλληλεπιδρούν με το περιβάλλον τους σε πολλά επίπεδα στα πλαίσια ενός οικοσυστήματος Οι φυσικές
Η ΧΛΩΡΙΔΑ ΚΑΙ Η ΠΑΝΙΔΑ ΣΤΗΝ ΧΩΡΑ ΜΑΣ. ΟΜΑΔΑ 1 Κορμπάκη Δέσποινα Κολακλίδη Ναταλία Ζαχαροπούλου Φιλιππούλα Θανοπούλου Ιωαννά
 Η ΧΛΩΡΙΔΑ ΚΑΙ Η ΠΑΝΙΔΑ ΣΤΗΝ ΧΩΡΑ ΜΑΣ ΟΜΑΔΑ 1 Κορμπάκη Δέσποινα Κολακλίδη Ναταλία Ζαχαροπούλου Φιλιππούλα Θανοπούλου Ιωαννά ΓΙΑΤΙ ΤΟ ΕΠΙΛΕΞΑΜΕ: Η χλωρίδα και η πανίδα στην χώρα μας είναι ένα πολύ σημαντικό
Η ΧΛΩΡΙΔΑ ΚΑΙ Η ΠΑΝΙΔΑ ΣΤΗΝ ΧΩΡΑ ΜΑΣ ΟΜΑΔΑ 1 Κορμπάκη Δέσποινα Κολακλίδη Ναταλία Ζαχαροπούλου Φιλιππούλα Θανοπούλου Ιωαννά ΓΙΑΤΙ ΤΟ ΕΠΙΛΕΞΑΜΕ: Η χλωρίδα και η πανίδα στην χώρα μας είναι ένα πολύ σημαντικό
ΧΕΡΣΑΙΑ ΟΙΚΟΣΥΣΤΗΜΑΤΑ. Δομή και σύσταση οικοσυστημάτων - αβιοτικοί και βιοτικοί παράγοντες
 ΕΝΟΤΗΤΕΣ ΧΕΡΣΑΙΑ ΟΙΚΟΣΥΣΤΗΜΑΤΑ Η δομή του πλανήτη Η οργάνωσης τα ζωής Φυσικοί πόροι νερό έδαφος - αέρας Δομή και σύσταση οικοσυστημάτων - αβιοτικοί και βιοτικοί παράγοντες Μεγαδιαπλάσεις Μεσογειακά Οικοσυστήματα
ΕΝΟΤΗΤΕΣ ΧΕΡΣΑΙΑ ΟΙΚΟΣΥΣΤΗΜΑΤΑ Η δομή του πλανήτη Η οργάνωσης τα ζωής Φυσικοί πόροι νερό έδαφος - αέρας Δομή και σύσταση οικοσυστημάτων - αβιοτικοί και βιοτικοί παράγοντες Μεγαδιαπλάσεις Μεσογειακά Οικοσυστήματα
πληθυσμών του αρουραίου (Rattus rattus)σε συμπλέγματα νησιών και βραχονησίδων των ελληνικών θαλασσών
σε συμπλέγματα νησιών και βραχονησίδων των ελληνικών θαλασσών") Γενετική δομή και πρότυπα διαφοροποίησης των πληθυσμών του αρουραίου (Rattus rattus)σε συμπλέγματα νησιών και βραχονησίδων των ελληνικών θαλασσών Δημήτρης Τσαπάρης Jacob Fric Αθήνα, Οκτώβριος 2012 Jon
Γενετική δομή και πρότυπα διαφοροποίησης των πληθυσμών του αρουραίου (Rattus rattus)σε συμπλέγματα νησιών και βραχονησίδων των ελληνικών θαλασσών Δημήτρης Τσαπάρης Jacob Fric Αθήνα, Οκτώβριος 2012 Jon
2ο φύλο του ζωικού βασιλείου ( ~100.000 είδη) Μεγαλύτερο ασπόνδυλο
 Μεγαλύτερο ασπόνδυλο") 2ο φύλο του ζωικού βασιλείου ( ~100.000 είδη) Μεγαλύτερο ασπόνδυλο Απολιθώματα πολλά και σε όλο το γεωλογικό χρόνο Οικοσυστήματα Μέγεθος Διάρκεια ζωής Θηρευτές χρήσεις από τον άνθρωπο κόσμηματα τέχνη
2ο φύλο του ζωικού βασιλείου ( ~100.000 είδη) Μεγαλύτερο ασπόνδυλο Απολιθώματα πολλά και σε όλο το γεωλογικό χρόνο Οικοσυστήματα Μέγεθος Διάρκεια ζωής Θηρευτές χρήσεις από τον άνθρωπο κόσμηματα τέχνη
Πρόλογος...11. 1. Οργανισμοί...15
 Περιεχόμενα Πρόλογος...11 1. Οργανισμοί...15 1.1 Οργανισμοί και είδη...15 1.1.1 Ιδιότητες των οργανισμών...15 1.1.2 Φαινότυπος, γονότυπος, οικότυπος...17 1.1.3 Η έννοια του είδους και ο αριθμός των ειδών...19
Περιεχόμενα Πρόλογος...11 1. Οργανισμοί...15 1.1 Οργανισμοί και είδη...15 1.1.1 Ιδιότητες των οργανισμών...15 1.1.2 Φαινότυπος, γονότυπος, οικότυπος...17 1.1.3 Η έννοια του είδους και ο αριθμός των ειδών...19
Φυλογένεση. 5o εργαστήριο
 Φυλογένεση 5o εργαστήριο Φυλογένεση οργανισµών Δείχνει την εξελικτική πορεία µιας οµάδας οργανισµών. Οι κόµβοι (nodes) στο δένδρο απεικονίζουν γεγονότα ειδογένεσης. H φυλογένεση µπορεί να γίνει από µια
Φυλογένεση 5o εργαστήριο Φυλογένεση οργανισµών Δείχνει την εξελικτική πορεία µιας οµάδας οργανισµών. Οι κόµβοι (nodes) στο δένδρο απεικονίζουν γεγονότα ειδογένεσης. H φυλογένεση µπορεί να γίνει από µια
ΑΣΚΗΣΗ: ΚΑΡΚΙΝΟΕΙΔΗ ΑΝΑΤΟΜΙΑ ΓΑΡΙΔΑΣ
 ΑΣΚΗΣΗ: ΚΑΡΚΙΝΟΕΙΔΗ Ομοταξία: Βραγχιόποδα Κλαδοκεραιωτά (Daphnia μικροσκοπικό παρασκεύασμα) Ανόστρακα (Artemia, ενήλικα αυγά και ναύπλιοι σε διάφορα στάδια ανάπτυξης σε μικροσκοπικό παρασκεύασμα και ζωντανό
ΑΣΚΗΣΗ: ΚΑΡΚΙΝΟΕΙΔΗ Ομοταξία: Βραγχιόποδα Κλαδοκεραιωτά (Daphnia μικροσκοπικό παρασκεύασμα) Ανόστρακα (Artemia, ενήλικα αυγά και ναύπλιοι σε διάφορα στάδια ανάπτυξης σε μικροσκοπικό παρασκεύασμα και ζωντανό
ΟΙΚΟΛΟΓΙΑ ΤΟΠΙΟΥ. Χειμερινό εξάμηνο
 ΟΙΚΟΛΟΓΙΑ ΤΟΠΙΟΥ Χειμερινό εξάμηνο 2009 2010 Κ. Ποϊραζίδης Ανάλυση ψηφίδων ΕΙΣΗΓΗΣΗ 4 Οικολογία Τοπίου 22 Νοεμβρίου 2009 Ανάλυση ψηφίδων Το μέγεθος τους (Patch size / perimeter / edges ) Έκταση Περίμετρος
ΟΙΚΟΛΟΓΙΑ ΤΟΠΙΟΥ Χειμερινό εξάμηνο 2009 2010 Κ. Ποϊραζίδης Ανάλυση ψηφίδων ΕΙΣΗΓΗΣΗ 4 Οικολογία Τοπίου 22 Νοεμβρίου 2009 Ανάλυση ψηφίδων Το μέγεθος τους (Patch size / perimeter / edges ) Έκταση Περίμετρος
Η ελληνική βιοποικιλότητα Ενας κρυμμένος θησαυρός. Μανώλης Μιτάκης Φαρμακοποιός Αντιπρόεδρος Ελληνικής Εταιρείας Εθνοφαρμακολογίας
 Η ελληνική βιοποικιλότητα Ενας κρυμμένος θησαυρός Μανώλης Μιτάκης Φαρμακοποιός Αντιπρόεδρος Ελληνικής Εταιρείας Εθνοφαρμακολογίας Ο μεγάλος αριθμός και η ποικιλομορφία των σύγχρονων μορφών ζωής στη Γη
Η ελληνική βιοποικιλότητα Ενας κρυμμένος θησαυρός Μανώλης Μιτάκης Φαρμακοποιός Αντιπρόεδρος Ελληνικής Εταιρείας Εθνοφαρμακολογίας Ο μεγάλος αριθμός και η ποικιλομορφία των σύγχρονων μορφών ζωής στη Γη
Τρίκαλα, 27/12/2011. Συνεντεύξεις. «Μεγαλύτερες σε διάρκεια ξηρασίες»
 Τρίκαλα, 27/12/2011 Συνεντεύξεις «Μεγαλύτερες σε διάρκεια ξηρασίες» Τι επισημαίνει στην ΕΡΕΥΝΑ για την περιοχή μας ο κ. Σοφοκλής Ε. Δρίτσας, ερευνητής στο Εργαστήριο Δημογραφικών και Κοινωνικών Αναλύσεων
Τρίκαλα, 27/12/2011 Συνεντεύξεις «Μεγαλύτερες σε διάρκεια ξηρασίες» Τι επισημαίνει στην ΕΡΕΥΝΑ για την περιοχή μας ο κ. Σοφοκλής Ε. Δρίτσας, ερευνητής στο Εργαστήριο Δημογραφικών και Κοινωνικών Αναλύσεων
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων Καθηγητής: Δρ. Νίκος Μίτλεττον Η ΣΧΕΣΗ ΤΟΥ ΜΗΤΡΙΚΟΥ ΘΗΛΑΣΜΟΥ ΜΕ ΤΗΝ ΕΜΦΑΝΙΣΗ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 2 ΣΤΗΝ ΠΑΙΔΙΚΗ ΗΛΙΚΙΑ Ονοματεπώνυμο: Ιωσηφίνα
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ Επιβλέπων Καθηγητής: Δρ. Νίκος Μίτλεττον Η ΣΧΕΣΗ ΤΟΥ ΜΗΤΡΙΚΟΥ ΘΗΛΑΣΜΟΥ ΜΕ ΤΗΝ ΕΜΦΑΝΙΣΗ ΣΑΚΧΑΡΩΔΗ ΔΙΑΒΗΤΗ ΤΥΠΟΥ 2 ΣΤΗΝ ΠΑΙΔΙΚΗ ΗΛΙΚΙΑ Ονοματεπώνυμο: Ιωσηφίνα
ΤΑ ΝΗΣΙΑ ΤΩΝ ΚΥΚΛΑΔΩΝ
 ΤΑ ΝΗΣΙΑ ΤΩΝ ΚΥΚΛΑΔΩΝ Η Σύρος είναι νησί των Κυκλάδων. Πρωτεύουσά της είναι η Ερμούπολη, η οποία είναι πρωτεύουσα της Περιφέριας Νότιου Αιγαίου αλλά και του πρώην Νομού Κυκλάδων. Η Σύρος αναπτύχθηκε ιδιαίτερα
ΤΑ ΝΗΣΙΑ ΤΩΝ ΚΥΚΛΑΔΩΝ Η Σύρος είναι νησί των Κυκλάδων. Πρωτεύουσά της είναι η Ερμούπολη, η οποία είναι πρωτεύουσα της Περιφέριας Νότιου Αιγαίου αλλά και του πρώην Νομού Κυκλάδων. Η Σύρος αναπτύχθηκε ιδιαίτερα
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ ΣΧΟΛΗ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΤΜΗΜΑ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μυτιλήνη, 24.03.2017 Αρ.Πρωτ.: 397 Θέμα: Συγκρότηση Εκλεκτορικού Σώματος για την εκλογή καθηγητή του Τμήματος Περιβάλλοντος στο γνωστικό αντικείμενο
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ ΣΧΟΛΗ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΤΜΗΜΑ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μυτιλήνη, 24.03.2017 Αρ.Πρωτ.: 397 Θέμα: Συγκρότηση Εκλεκτορικού Σώματος για την εκλογή καθηγητή του Τμήματος Περιβάλλοντος στο γνωστικό αντικείμενο
ΓΕΩΛΟΓΙΑ - ΓΕΩΓΡΑΦΙΑ Β ΓΥΜΝΑΣΙΟΥ ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ
 ΓΕΩΛΟΓΙΑ - ΓΕΩΓΡΑΦΙΑ Β ΓΥΜΝΑΣΙΟΥ ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ 1. Η πόλη Λ βρίσκεται: α. βόρεια της πόλης Κ. β. νότια της πόλης Κ. γ. ΒΑ της πόλης Κ. δ. ΒΔ της πόλης Κ. 2. Η πόλη Λ βρίσκεται: α. βόρεια
ΓΕΩΛΟΓΙΑ - ΓΕΩΓΡΑΦΙΑ Β ΓΥΜΝΑΣΙΟΥ ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ 1. Η πόλη Λ βρίσκεται: α. βόρεια της πόλης Κ. β. νότια της πόλης Κ. γ. ΒΑ της πόλης Κ. δ. ΒΔ της πόλης Κ. 2. Η πόλη Λ βρίσκεται: α. βόρεια
ΜΕΤΑΝΑΣΤΕΥΣΗ ΚΑΙ ΙΑΣΠΟΡΑ ΤΩΝ ΕΝΤΟΜΩΝ ΣΕ ΜΕΣΟΓΕΙΑΚΟΥ ΤΥΠΟΥ ΟΙΚΟΣΥΣΤΗΜΑΤΑ
 ΜΕΤΑΝΑΣΤΕΥΣΗ ΚΑΙ ΙΑΣΠΟΡΑ ΤΩΝ ΕΝΤΟΜΩΝ ΣΕ ΜΕΣΟΓΕΙΑΚΟΥ ΤΥΠΟΥ ΟΙΚΟΣΥΣΤΗΜΑΤΑ Α. Λεγάκις* & Π.Β. Πετράκης** * Ζωολογικό Μουσείο Πανεπιστημίου Αθηνών ** ΕΘΙΑΓΕ, Ινστιτούτο Μεσογειακών Οικοσυστημάτων 11 ο Πανελλήνιο
ΜΕΤΑΝΑΣΤΕΥΣΗ ΚΑΙ ΙΑΣΠΟΡΑ ΤΩΝ ΕΝΤΟΜΩΝ ΣΕ ΜΕΣΟΓΕΙΑΚΟΥ ΤΥΠΟΥ ΟΙΚΟΣΥΣΤΗΜΑΤΑ Α. Λεγάκις* & Π.Β. Πετράκης** * Ζωολογικό Μουσείο Πανεπιστημίου Αθηνών ** ΕΘΙΑΓΕ, Ινστιτούτο Μεσογειακών Οικοσυστημάτων 11 ο Πανελλήνιο
«Οι επιπτώσεις της ανθρωπογενούς κλιματικής αλλαγής στο περιβάλλον, την κοινωνία και την οικονομία της Ελλάδος»
 «Οι επιπτώσεις της ανθρωπογενούς κλιματικής αλλαγής στο περιβάλλον, την κοινωνία και την οικονομία της Ελλάδος» Χρήστος Σ. Ζερεφός Ακαδημαïκός Ακαδημία Αθηνών 28 Νοεμβρίου 2017 Χωρίς την παρουσία των αερίων
«Οι επιπτώσεις της ανθρωπογενούς κλιματικής αλλαγής στο περιβάλλον, την κοινωνία και την οικονομία της Ελλάδος» Χρήστος Σ. Ζερεφός Ακαδημαïκός Ακαδημία Αθηνών 28 Νοεμβρίου 2017 Χωρίς την παρουσία των αερίων
Ε.Μ. Σκορδύλης Καθηγητής Σεισμολογίας Τομέας Γεωφυσικής, Α.Π.Θ.
 Ε.Μ. Σκορδύλης Καθηγητής Σεισμολογίας Τομέας Γεωφυσικής, Α.Π.Θ. 223 Μa 200 Μa 135 Μa 35 Μa Present 2 Σχετικές Κινήσεις Λιθοσφαιρικών Πλακών 1. Απόκλισεις λιθοσφαιρικών πλακών (μεσο-ωκεάνιες ράχες) 2. Εφαπτομενικές
Ε.Μ. Σκορδύλης Καθηγητής Σεισμολογίας Τομέας Γεωφυσικής, Α.Π.Θ. 223 Μa 200 Μa 135 Μa 35 Μa Present 2 Σχετικές Κινήσεις Λιθοσφαιρικών Πλακών 1. Απόκλισεις λιθοσφαιρικών πλακών (μεσο-ωκεάνιες ράχες) 2. Εφαπτομενικές
ΦΥΣΙΚΗ ΧΗΜΙΚΗ ΓΕΩΛΟΓΙΚΗ ΒΙΟΛΟΓΙΚΗ ΜΑΘΗΜΑΤΙΚΗ
 ΦΥΣΙΚΗ ΩΚΕΑΝΟΓΡΑΦΙΑ Αρχές και έννοιες της Ωκεανογραφίας, με ιδιαίτερη έμφαση στις φυσικές διεργασίες των ωκεάνιων συστημάτων. Φυσικές ιδιότητες και οι φυσικές παράμετροι του θαλασσινού νερού, και χωροχρονικές
ΦΥΣΙΚΗ ΩΚΕΑΝΟΓΡΑΦΙΑ Αρχές και έννοιες της Ωκεανογραφίας, με ιδιαίτερη έμφαση στις φυσικές διεργασίες των ωκεάνιων συστημάτων. Φυσικές ιδιότητες και οι φυσικές παράμετροι του θαλασσινού νερού, και χωροχρονικές
ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ
 Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ Άντρια Πολυκάρπου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
Σχολή Επιστημών Υγείας Πτυχιακή εργασία ΕΠΑΝΑΛΗΨΗ ΨΕΥΔΟΛΕΞΕΩΝ ΑΠΟ ΠΑΙΔΙΑ ΜΕ ΕΙΔΙΚΗ ΓΛΩΣΣΙΚΗ ΔΙΑΤΑΡΑΧΗ ΚΑΙ ΠΑΙΔΙΑ ΤΥΠΙΚΗΣ ΑΝΑΠΤΥΞΗΣ Άντρια Πολυκάρπου Λεμεσός, Μάιος 2017 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ
TreeTOPS. ένα εισαγωγικό παιχνίδι για τα φυλογενετικά δέντρα. Teacher s Guide. ELLS European Learning Laboratory for the Life Sciences
 TreeTOPS ένα εισαγωγικό παιχνίδι για τα φυλογενετικά δέντρα Teacher s Guide ELLS European Learning Laboratory for the Life Sciences 1 Γενικός σκοπός Το συγκεκριμένο παιχνίδι έχει ως στόχο να εισάγει τους
TreeTOPS ένα εισαγωγικό παιχνίδι για τα φυλογενετικά δέντρα Teacher s Guide ELLS European Learning Laboratory for the Life Sciences 1 Γενικός σκοπός Το συγκεκριμένο παιχνίδι έχει ως στόχο να εισάγει τους
Τμήμα Βιολογίας Πανεπιστήμιο Κρήτης. Μορφολογία και Ανατομία Δεκάποδου Καρκινοειδούς
 Τμήμα Βιολογίας Πανεπιστήμιο Κρήτης Μορφολογία και Ανατομία Δεκάποδου Καρκινοειδούς Iάσμη Στάθη Δέσποινα Δοκιανάκη Ηράκλειο, Σπονδυλωτά Ασπόνδυλα Καρκινοειδή ΦΥΛΟ: Αρθρόποδα Χαρακτηριστικά Μεταμέρεια
Τμήμα Βιολογίας Πανεπιστήμιο Κρήτης Μορφολογία και Ανατομία Δεκάποδου Καρκινοειδούς Iάσμη Στάθη Δέσποινα Δοκιανάκη Ηράκλειο, Σπονδυλωτά Ασπόνδυλα Καρκινοειδή ΦΥΛΟ: Αρθρόποδα Χαρακτηριστικά Μεταμέρεια
Γιατί είναι σημαντική η Βιοποικιλότητα;
 Γενετική Διαχείριση Ο όρος βιοποικιλότητα αναφέρεται σε όλους τους διαφορετικούς οργανισμούς του πλανήτη μας και περιλαμβάνει τόσο την ποικιλότητα σε επίπεδο ειδών, όσο και τη γενετική ποικιλότητα. Γιατί
Γενετική Διαχείριση Ο όρος βιοποικιλότητα αναφέρεται σε όλους τους διαφορετικούς οργανισμούς του πλανήτη μας και περιλαμβάνει τόσο την ποικιλότητα σε επίπεδο ειδών, όσο και τη γενετική ποικιλότητα. Γιατί
Τ Α Η Φ Α Ι Σ Τ Ε Ι Α
 ΤΑ ΗΦΑΙΣΤΕΙΑ ΤΑ ΗΦΑΙΣΤΕΙΑ ΠΑΝΩ ΣΤΗ ΓΗ ΒΡΙΣΚΟΝΤΑΙ ΣΥΝΗΘΩΣ ΕΚΕΙ ΟΠΟΥ ΔΥΟ Η ΤΡΕΙΣ ΤΕΚΤΟΝΙΚΕΣ ΠΛΑΚΕΣ ΣΥΓΚΡΟΥΟΝΤΑΙ Η ΑΠΟΜΑΚΡΥΝΟΝΤΑΙ: ΜΙΑ ΜΕΣΟΩΚΕΑΝΕΙΑ ΡΑΧΗ Η ΟΡΟΣΕΙΡΑ, ΟΠΩΣ ΣΤΟ ΜΕΣΟ ΤΟΥ ΑΤΛΑΝΤΙΚΟΥ ΩΚΕΑΝΟΥ, ΔΙΝΕΙ
ΤΑ ΗΦΑΙΣΤΕΙΑ ΤΑ ΗΦΑΙΣΤΕΙΑ ΠΑΝΩ ΣΤΗ ΓΗ ΒΡΙΣΚΟΝΤΑΙ ΣΥΝΗΘΩΣ ΕΚΕΙ ΟΠΟΥ ΔΥΟ Η ΤΡΕΙΣ ΤΕΚΤΟΝΙΚΕΣ ΠΛΑΚΕΣ ΣΥΓΚΡΟΥΟΝΤΑΙ Η ΑΠΟΜΑΚΡΥΝΟΝΤΑΙ: ΜΙΑ ΜΕΣΟΩΚΕΑΝΕΙΑ ΡΑΧΗ Η ΟΡΟΣΕΙΡΑ, ΟΠΩΣ ΣΤΟ ΜΕΣΟ ΤΟΥ ΑΤΛΑΝΤΙΚΟΥ ΩΚΕΑΝΟΥ, ΔΙΝΕΙ
Δασική Γενετική Εισαγωγή: Βασικές έννοιες
 Δασική Γενετική Εισαγωγή: Βασικές έννοιες Χειμερινό εξάμηνο 2014-2015 Γενετική Πειραματική επιστήμη της κληρονομικότητας Προέκυψε από την ανάγκη κατανόησης της κληρονόμησης οικονομικά σημαντικών χαρακτηριστικών
Δασική Γενετική Εισαγωγή: Βασικές έννοιες Χειμερινό εξάμηνο 2014-2015 Γενετική Πειραματική επιστήμη της κληρονομικότητας Προέκυψε από την ανάγκη κατανόησης της κληρονόμησης οικονομικά σημαντικών χαρακτηριστικών
Μέθοδοι Φυλογένεσης. Μέθοδοι που βασίζονται σε αποστάσεις UPGMA Κοντινότερης γειτονίας (Neighbor joining) Fitch-Margoliash Ελάχιστης εξέλιξης
 Fitch-Margoliash Ελάχιστης εξέλιξης") Μέθοδοι Φυλογένεσης Μέθοδοι που βασίζονται σε αποστάσεις UPGMA Κοντινότερης γειτονίας (Neighbor joining) Fitch-Margoliash Ελάχιστης εξέλιξης Μέθοδοι που βασίζονται σε χαρακτήρες Μέγιστη φειδωλότητα (Maximum
Μέθοδοι Φυλογένεσης Μέθοδοι που βασίζονται σε αποστάσεις UPGMA Κοντινότερης γειτονίας (Neighbor joining) Fitch-Margoliash Ελάχιστης εξέλιξης Μέθοδοι που βασίζονται σε χαρακτήρες Μέγιστη φειδωλότητα (Maximum
Προσδιορισμός Σημαντικών Χαρακτηριστικών της Αυθόρμητης Δραστηριότητας Απομονωμένου Εγκεφαλικού Φλοιού in vitro
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΠΛΗΡΟΦΟΡΙΚΗΣ ΚΑΙ ΤΗΛΕΠΙΚΟΙΝΩΝΙΩΝ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ "ΤΕΧΝΟΛΟΓΙΕΣ ΠΛΗΡΟΦΟΡΙΚΗΣ ΣΤΗΝ ΙΑΤΡΙΚΗ ΚΑΙ ΤΗ ΒΙΟΛΟΓΙΑ"
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΠΛΗΡΟΦΟΡΙΚΗΣ ΚΑΙ ΤΗΛΕΠΙΚΟΙΝΩΝΙΩΝ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ "ΤΕΧΝΟΛΟΓΙΕΣ ΠΛΗΡΟΦΟΡΙΚΗΣ ΣΤΗΝ ΙΑΤΡΙΚΗ ΚΑΙ ΤΗ ΒΙΟΛΟΓΙΑ"
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ. Πτυχιακή εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΟΥ ΚΛΙΜΑΤΟΣ ΑΣΦΑΛΕΙΑΣ ΤΩΝ ΑΣΘΕΝΩΝ ΣΤΟ ΝΟΣΟΚΟΜΕΙΟ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΟΥ ΚΛΙΜΑΤΟΣ ΑΣΦΑΛΕΙΑΣ ΤΩΝ ΑΣΘΕΝΩΝ ΣΤΟ ΝΟΣΟΚΟΜΕΙΟ ΑΝΔΡΕΑΣ ΛΕΩΝΙΔΟΥ Λεμεσός, 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ Πτυχιακή εργασία ΔΙΕΡΕΥΝΗΣΗ ΤΟΥ ΚΛΙΜΑΤΟΣ ΑΣΦΑΛΕΙΑΣ ΤΩΝ ΑΣΘΕΝΩΝ ΣΤΟ ΝΟΣΟΚΟΜΕΙΟ ΑΝΔΡΕΑΣ ΛΕΩΝΙΔΟΥ Λεμεσός, 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ
ΟΙ ΥΔΡΟΒΙΟΤΟΠΟΙ ΚΑΙ Η ΣΗΜΑΣΙΑ ΤΟΥΣ
 ΟΙ ΥΔΡΟΒΙΟΤΟΠΟΙ ΚΑΙ Η ΣΗΜΑΣΙΑ ΤΟΥΣ Υδατικά οικοσυστήματα Στη βιόσφαιρα υπάρχουν δύο είδη οικοσυστημάτων: τα υδάτινα και τα χερσαία. Tα υδάτινα οικοσυστήματα διαχωρίζονται ανάλογα με την αλατότητα του νερού
ΟΙ ΥΔΡΟΒΙΟΤΟΠΟΙ ΚΑΙ Η ΣΗΜΑΣΙΑ ΤΟΥΣ Υδατικά οικοσυστήματα Στη βιόσφαιρα υπάρχουν δύο είδη οικοσυστημάτων: τα υδάτινα και τα χερσαία. Tα υδάτινα οικοσυστήματα διαχωρίζονται ανάλογα με την αλατότητα του νερού
2. ΓΕΩΛΟΓΙΑ - ΝΕΟΤΕΚΤΟΝΙΚΗ
 2. 2.1 ΓΕΩΛΟΓΙΑ ΤΗΣ ΕΥΡΥΤΕΡΗΣ ΠΕΡΙΟΧΗΣ Στο κεφάλαιο αυτό παρουσιάζεται συνοπτικά το Γεωλογικό-Σεισμοτεκτονικό περιβάλλον της ευρύτερης περιοχής του Π.Σ. Βόλου - Ν.Ιωνίας. Η ευρύτερη περιοχή της πόλης του
2. 2.1 ΓΕΩΛΟΓΙΑ ΤΗΣ ΕΥΡΥΤΕΡΗΣ ΠΕΡΙΟΧΗΣ Στο κεφάλαιο αυτό παρουσιάζεται συνοπτικά το Γεωλογικό-Σεισμοτεκτονικό περιβάλλον της ευρύτερης περιοχής του Π.Σ. Βόλου - Ν.Ιωνίας. Η ευρύτερη περιοχή της πόλης του
ΠΑΡΑΡΤΗΜΑ. της. Οδηγίας της Επιτροπής
 ΕΥΡΩΠΑΪΚΗ ΕΠΙΤΡΟΠΗ Βρυξέλλες, 17.5.2017 C(2017) 2842 final ANNEX 1 ΠΑΡΑΡΤΗΜΑ της Οδηγίας της Επιτροπής για την τροποποίηση της οδηγίας 2008/56/ΕΚ του Ευρωπαϊκού Κοινοβουλίου και του Συμβουλίου όσον αφορά
ΕΥΡΩΠΑΪΚΗ ΕΠΙΤΡΟΠΗ Βρυξέλλες, 17.5.2017 C(2017) 2842 final ANNEX 1 ΠΑΡΑΡΤΗΜΑ της Οδηγίας της Επιτροπής για την τροποποίηση της οδηγίας 2008/56/ΕΚ του Ευρωπαϊκού Κοινοβουλίου και του Συμβουλίου όσον αφορά
CYCLADES Life: Integrated monk seal conservation of Northern Cyclades
 Α.Καραμανλίδης/MOm 2 η Ε τ ή σ ι α Α ν α φ ο ρ ά Της κ α τ ά σ τ α σ η ς τ ο υ π λ η θ υ σ μ ο ύ τ η ς μ ε σ ο γ ε ι α κ ή ς φ ώ κ ι α ς σ τ η ν ή σ ο Γ υ ά ρ ο ( Π ε ρ ί λ η ψ η ) Ιούλιος 2015 ΠΕΡΙΕΧΟΜΕΝΑ
Α.Καραμανλίδης/MOm 2 η Ε τ ή σ ι α Α ν α φ ο ρ ά Της κ α τ ά σ τ α σ η ς τ ο υ π λ η θ υ σ μ ο ύ τ η ς μ ε σ ο γ ε ι α κ ή ς φ ώ κ ι α ς σ τ η ν ή σ ο Γ υ ά ρ ο ( Π ε ρ ί λ η ψ η ) Ιούλιος 2015 ΠΕΡΙΕΧΟΜΕΝΑ
Περιεχόμενα. 1 Η ιστορία της εξελικτικής βιολογίας: Εξέλιξη και Γενετική 2 Η Προέλευση της Μοριακής Βιολογίας 3 Αποδείξεις για την εξέλιξη 89
 Περιεχόμενα Οι Συγγραφείς Πρόλογος της Ελληνικής Έκδοσης Πρόλογος της Αμερικανικής Έκδοσης Σκοπός και Αντικείμενο του Βιβλίου ΜΕΡΟΣ Ι ΜΙΑ ΕΠΙΣΚΟΠΗΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ 1 Η ιστορία της εξελικτικής
Περιεχόμενα Οι Συγγραφείς Πρόλογος της Ελληνικής Έκδοσης Πρόλογος της Αμερικανικής Έκδοσης Σκοπός και Αντικείμενο του Βιβλίου ΜΕΡΟΣ Ι ΜΙΑ ΕΠΙΣΚΟΠΗΣΗ ΤΗΣ ΕΞΕΛΙΚΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ 1 Η ιστορία της εξελικτικής
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η ΣΥΓΚΕΝΤΡΩΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΣΤΟ ΕΔΑΦΟΣ ΚΑΙ ΜΕΘΟΔΟΙ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΤΟΥΣ Μιχαήλ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή Η ΣΥΓΚΕΝΤΡΩΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΣΤΟ ΕΔΑΦΟΣ ΚΑΙ ΜΕΘΟΔΟΙ ΠΡΟΣΔΙΟΡΙΣΜΟΥ ΤΟΥΣ Μιχαήλ
Ορισμός επιδημιολογίας
 Ορισμός επιδημιολογίας Επιδημιολογία είναι η μελέτη της κατανομής των παραγόντων που επηρεάζουν τη συχνότητα των νοσημάτων στον άνθρωπο (MacMahon, 1970) Επιδημιολογία είναι η μελέτη της κατανομής των παραγόντων
Ορισμός επιδημιολογίας Επιδημιολογία είναι η μελέτη της κατανομής των παραγόντων που επηρεάζουν τη συχνότητα των νοσημάτων στον άνθρωπο (MacMahon, 1970) Επιδημιολογία είναι η μελέτη της κατανομής των παραγόντων
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ ΤΜΗΜΑ ΓΕΩΓΡΑΦΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ: «ΓΕΩΓΡΑΦΙΑ ΚΑΙ ΕΦΑΡΜΟΣΜΕΝΗ ΓΕΩΠΛΗΡΟΦΟΡΙΚΗ»
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ ΤΜΗΜΑ ΓΕΩΓΡΑΦΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ: «ΓΕΩΓΡΑΦΙΑ ΚΑΙ ΕΦΑΡΜΟΣΜΕΝΗ ΓΕΩΠΛΗΡΟΦΟΡΙΚΗ» «ΠΑΛΑΙΟΓΕΩΓΡΑΦΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΠΕΡΙΟΧΗΣ ΛΑΨΑΡΝΩΝ ΛΕΣΒΟΥ ΜΕ ΒΑΣΗ ΤΑ ΠΑΛΑΙΟΝΤΟΛΟΓΙΚΑ ΕΥΡΗΜΑΤΑ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ ΤΜΗΜΑ ΓΕΩΓΡΑΦΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ: «ΓΕΩΓΡΑΦΙΑ ΚΑΙ ΕΦΑΡΜΟΣΜΕΝΗ ΓΕΩΠΛΗΡΟΦΟΡΙΚΗ» «ΠΑΛΑΙΟΓΕΩΓΡΑΦΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΠΕΡΙΟΧΗΣ ΛΑΨΑΡΝΩΝ ΛΕΣΒΟΥ ΜΕ ΒΑΣΗ ΤΑ ΠΑΛΑΙΟΝΤΟΛΟΓΙΚΑ ΕΥΡΗΜΑΤΑ
Ειδικά Θέματα Δημογραφίας: Χωρικές Διαστάσεις Δημογραφικών Δεδομένων
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Ειδικά Θέματα Δημογραφίας: Χωρικές Διαστάσεις Δημογραφικών Δεδομένων Ενότητα 4.3: Οι δημογραφικές δομές και ο δημογραφικός δυναμισμός των ελληνικών δήμων (1999-2009) Μαρί-Νοέλ Ντυκέν,
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Ειδικά Θέματα Δημογραφίας: Χωρικές Διαστάσεις Δημογραφικών Δεδομένων Ενότητα 4.3: Οι δημογραφικές δομές και ο δημογραφικός δυναμισμός των ελληνικών δήμων (1999-2009) Μαρί-Νοέλ Ντυκέν,
ΓΕΩΧΗΜΕΙΑ Δ ΕΞΑΜΗΝΟ ΕΙΣΑΓΩΓΗ- ΠΕΡΙΕΧΟΜΕΝΑ ΜΑΘΗΜΑΤΟΣ. Χριστίνα Στουραϊτη
 1 ΓΕΩΧΗΜΕΙΑ Δ ΕΞΑΜΗΝΟ ΕΙΣΑΓΩΓΗ- ΠΕΡΙΕΧΟΜΕΝΑ ΜΑΘΗΜΑΤΟΣ Χριστίνα Στουραϊτη ΠΡΟΓΡΑΜΜΑ ΜΑΘΗΜΑΤΩΝ ΓΕΩΧΗΜΕΙΑΣ Δ ΕΞΑΜΗΝΟ ΑΚΑΔΗΜΑΪΚΟ ΕΤΟΣ 2016-2017 ΕΒΔΟΜΑΔΑ ΗΜ/ΝΙΑ ΠΕΡΙΕΧΟΜΕΝΟ ΔΙΔΑΣΚΟΥΣΑ 1 η Τετ 22/2/17 Εισαγωγή-
1 ΓΕΩΧΗΜΕΙΑ Δ ΕΞΑΜΗΝΟ ΕΙΣΑΓΩΓΗ- ΠΕΡΙΕΧΟΜΕΝΑ ΜΑΘΗΜΑΤΟΣ Χριστίνα Στουραϊτη ΠΡΟΓΡΑΜΜΑ ΜΑΘΗΜΑΤΩΝ ΓΕΩΧΗΜΕΙΑΣ Δ ΕΞΑΜΗΝΟ ΑΚΑΔΗΜΑΪΚΟ ΕΤΟΣ 2016-2017 ΕΒΔΟΜΑΔΑ ΗΜ/ΝΙΑ ΠΕΡΙΕΧΟΜΕΝΟ ΔΙΔΑΣΚΟΥΣΑ 1 η Τετ 22/2/17 Εισαγωγή-
ΕΡΓΑΣΤΗΡΙΟ 1: ΕΚΤΙΜΗΣΗ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ
 ΕΡΓΑΣΤΗΡΙΟ 1: ΕΚΤΙΜΗΣΗ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ Α. Θεωρητικό υπόβαθρο Τι είναι βιοποικιλότητα; Η «βιολογική ποικιλότητα» ή «βιοποικιλότητα» αναφέρεται στην ποικιλία όλων των ζωντανών οργανισμών και των οικοσυστημάτων
ΕΡΓΑΣΤΗΡΙΟ 1: ΕΚΤΙΜΗΣΗ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ Α. Θεωρητικό υπόβαθρο Τι είναι βιοποικιλότητα; Η «βιολογική ποικιλότητα» ή «βιοποικιλότητα» αναφέρεται στην ποικιλία όλων των ζωντανών οργανισμών και των οικοσυστημάτων
ΓΕΩΛΟΓΙΑ ΓΕΩΜΟΡΦΟΛΟΓΙΑ
 ΕΡΩΤΗΜΑΤΟΛΟΓΙΟ Για τη διευκόλυνση των σπουδαστών στη μελέτη τους και την καλύτερη κατανόηση των κεφαλαίων που περιλαμβάνονται στο βιβλίο ΓΕΝΙΚΗ ΓΕΩΛΟΓΙΑ ΓΕΩΜΟΡΦΟΛΟΓΙΑ Σημείωση: Το βιβλίο καλύπτει την ύλη
ΕΡΩΤΗΜΑΤΟΛΟΓΙΟ Για τη διευκόλυνση των σπουδαστών στη μελέτη τους και την καλύτερη κατανόηση των κεφαλαίων που περιλαμβάνονται στο βιβλίο ΓΕΝΙΚΗ ΓΕΩΛΟΓΙΑ ΓΕΩΜΟΡΦΟΛΟΓΙΑ Σημείωση: Το βιβλίο καλύπτει την ύλη
Βιοπληροφορική. Ενότητα 16: Μεθοδολογίες (Ανα-) Κατασκευής, 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου
 Κατασκευής, 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου") Βιοπληροφορική Ενότητα 16: Μεθοδολογίες (Ανα-) Κατασκευής, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Επεξήγηση των μεθόδων (ανα-)κατασκευής φυλογενετικών δέντρων. Παρουσίαση
Βιοπληροφορική Ενότητα 16: Μεθοδολογίες (Ανα-) Κατασκευής, 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι Επεξήγηση των μεθόδων (ανα-)κατασκευής φυλογενετικών δέντρων. Παρουσίαση
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου
 Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Η αναλογία Α+G/T+C στο γενετικό υλικό ενός ιού είναι ίση με 2/3. Ο ιός μπορεί να είναι: α. ο φάγος λ. β. ο ιός της πολιομυελίτιδας. γ. φορέας κλωνοποίησης
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Η αναλογία Α+G/T+C στο γενετικό υλικό ενός ιού είναι ίση με 2/3. Ο ιός μπορεί να είναι: α. ο φάγος λ. β. ο ιός της πολιομυελίτιδας. γ. φορέας κλωνοποίησης
Η ΣΗΜΑΣΙΑ ΤΩΝ ΠΑΡΑΠΟΤΑΜΙΩΝ ΟΙΚΟΣΥΣΤΗΜΑΤΩΝ ΤΟΥ ΠΗΝΕΙΟΥ ΓΙΑ ΤΗ ΔΙΑΤΗΡΗΣΗ ΤΗΣ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ ΣΤΟ ΘΕΣΣΑΛΙΚΟ ΑΓΡΟΤΙΚΟ ΤΟΠΙΟ
 Η ΣΗΜΑΣΙΑ ΤΩΝ ΠΑΡΑΠΟΤΑΜΙΩΝ ΟΙΚΟΣΥΣΤΗΜΑΤΩΝ ΤΟΥ ΠΗΝΕΙΟΥ ΓΙΑ ΤΗ ΔΙΑΤΗΡΗΣΗ ΤΗΣ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ ΣΤΟ ΘΕΣΣΑΛΙΚΟ ΑΓΡΟΤΙΚΟ ΤΟΠΙΟ Μακρή Μαρία, Βιολόγος, Υπ. Διδάκτορας Βλαχόπουλος Κωνσταντίνος, Περιβαλλοντολόγος,
Η ΣΗΜΑΣΙΑ ΤΩΝ ΠΑΡΑΠΟΤΑΜΙΩΝ ΟΙΚΟΣΥΣΤΗΜΑΤΩΝ ΤΟΥ ΠΗΝΕΙΟΥ ΓΙΑ ΤΗ ΔΙΑΤΗΡΗΣΗ ΤΗΣ ΒΙΟΠΟΙΚΙΛΟΤΗΤΑΣ ΣΤΟ ΘΕΣΣΑΛΙΚΟ ΑΓΡΟΤΙΚΟ ΤΟΠΙΟ Μακρή Μαρία, Βιολόγος, Υπ. Διδάκτορας Βλαχόπουλος Κωνσταντίνος, Περιβαλλοντολόγος,
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 19/5/2007
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
Λίγη εξέλιξη: οµολογία
 Φυλογένεση Η εκτίµηση της εξελικτικής ιστορίας γονιδίων/πρωτεϊνών ή οργανισµών. Η απεικόνιση αυτής της ιστορίας γίνεται µε φυλογράµµατα/ κλαδογράµµατα Λίγη εξέλιξη: οµολογία Οµόλογα γονίδια: κοινός εξελικτικός
Φυλογένεση Η εκτίµηση της εξελικτικής ιστορίας γονιδίων/πρωτεϊνών ή οργανισµών. Η απεικόνιση αυτής της ιστορίας γίνεται µε φυλογράµµατα/ κλαδογράµµατα Λίγη εξέλιξη: οµολογία Οµόλογα γονίδια: κοινός εξελικτικός
ΔΕΛΤΙΟ ΑΠΟΤΕΛΕΣΜΑΤΩΝ ΔΕΙΚΤΗ SET15: ΕΞΩΤΕΡΙΚΟ ΕΜΠΟΡΙΟ
 ΔΕΛΤΙΟ ΑΠΟΤΕΛΕΣΜΑΤΩΝ ΔΕΙΚΤΗ ΟΡΙΣΜΟΣ - ΣΚΟΠΙΜΟΤΗΤΑ Ο δείκτης εκτιμά στην αξία των εισαγωγών και εξαγωγών ανά Περιφέρεια από και προς τις χώρες της ΕΕ ή τρίτες χώρες, καθώς και τη σχέση των εξαγωγών ως προς
ΔΕΛΤΙΟ ΑΠΟΤΕΛΕΣΜΑΤΩΝ ΔΕΙΚΤΗ ΟΡΙΣΜΟΣ - ΣΚΟΠΙΜΟΤΗΤΑ Ο δείκτης εκτιμά στην αξία των εισαγωγών και εξαγωγών ανά Περιφέρεια από και προς τις χώρες της ΕΕ ή τρίτες χώρες, καθώς και τη σχέση των εξαγωγών ως προς
Σχέδιο Μαθήματος Φύλλο Εργασίας Τα ηφαίστεια στην Ελλάδα
 Σχέδιο Μαθήματος Φύλλο Εργασίας Τα ηφαίστεια στην Ελλάδα Εισαγωγή: Η σύγχρονη - σε γωολογικούς όρους ηφαιστειακή δραστηριότητα στην Ελληνική Επικράτεια πηγαίνει πίσω περίπου 40 εκατομμύρια χρόνια και συνεχίζει
Σχέδιο Μαθήματος Φύλλο Εργασίας Τα ηφαίστεια στην Ελλάδα Εισαγωγή: Η σύγχρονη - σε γωολογικούς όρους ηφαιστειακή δραστηριότητα στην Ελληνική Επικράτεια πηγαίνει πίσω περίπου 40 εκατομμύρια χρόνια και συνεχίζει