ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΤΜΗΜΑ ΟΔΟΝΤΙΑΤΡΙΚΗΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΣΤΗ ΒΙΟΛΟΓΙΑ ΣΤΟΜΑΤΟΣ
|
|
|
- Αμάλθεια Δημητρίου
- 3 χρόνια πριν
- Προβολές:
Transcript
1 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΤΜΗΜΑ ΟΔΟΝΤΙΑΤΡΙΚΗΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΣΤΗ ΒΙΟΛΟΓΙΑ ΣΤΟΜΑΤΟΣ ΧΑΡΑΚΤΗΡΙΣΜΟΣ ΤΩΝ ΙΝΟΒΛΑΣΤΩΝ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΠΕΡΙΡΡΙΖΙΟΥ ΚΑΙ Η ΕΠΙΔΡΑΣΗ ΤΟΥ TGF-β ΚΑΙ ΤΗΣ 17-β ΟΙΣΤΡΑΔΙΟΛΗΣ ΣΤΟΝ ΠΟΛΛΑΠΛΑΣΙΑΣΜΟ ΤΟΥΣ. IN VITRO ΜΕΛΕΤΗ. ΑΝΝΑ Χ. ΛΑΓΟΥ ΑΘΗΝΑ 2008
2 2 ΠΕΡΙΕΧΟΜΕΝΑ ΠΡΟΛΟΓΟΣ 4 ΓΕΝΙΚΟ ΜΕΡΟΣ A.ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ KAI ΜΕΘΟΔΟΙ ΤΑΥΤΟΠΟΙΗΣΗΣ ΤΩΝ ΠΕΡΙΡΡΙΖΙΚΩΝ ΙΝΟΒΛΑΣΤΩΝ 5 ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ 5 Ινοβλάστες του περιρριζίου 5 Ινοβλάστες με οστεοβλαστικό φαινότυπο και τα αδιαφοροποίητα μεσεγχυματικά κύτταρα 9 ΜΕΘΟΔΟΙ ΤΑΥΤΟΠΟΙΗΣΗΣ ΤΩΝ ΠΕΡΙΡΡΙΖΙΚΩΝ ΙΝΟΒΛΑΣΤΩΝ 12 Ταυτοποίηση της μεσεγχυματικής προέλευσης των ινοβλαστών του περιρριζίου 12 Ταυτοποίηση του ινοβλαστικού χαρακτήρα των κυττάρων του περιρριζίου 13 Ταυτοποίηση των περιρριζικών ινοβλαστών ως προς τον οστεοβλαστικό τους φαινότυπο 17 Αμυντικά κύτταρα 21 Άλλα κύτταρα του περιρριζίου 22 B.Ο ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ ΤΩΝ ΑΥΞΗΤΙΚΩΝ ΠΑΡΑΓΟΝΤΩΝ ΣΤΑ ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ 25 Αυξητικός παράγοντας των αιμοπεταλίων (PDGF) 25 Αυξητικός παράγοντας ο προσομοιάζων με την ινσουλίνη (IGF) 30 Αυξητικός παράγοντας των ινοβλαστών (FGF) 32 Γ.Ο ΤΡΟΠΟΠΟΙΗΤΙΚΟΣ ΑΥΞΗΤΙΚΟΣ ΠΑΡΑΓΟΝΤΑΣ (TGF-β) 36 Η κυτταρική βιολογία του TGF-β 39 Η δράση τoυ TGF-β στα οστικά κύτταρα 41 Ο ρόλος του TGF-β στην οστική επανόρθωση και διατήρηση 44 Η έκφραση του TGF-β και των υποδοχέων του στα κύτταρα των περιοδοντικών ιστών 46
3 3 Η δράση του TGF-β 1 στους περιοδοντικούς ιστούς 49 Δ.Η ΒΙΟΛΟΓΙΚΗ ΚΑΙ Η ΒΙΟΧΗΜΙΚΗ ΣΥΜΠΕΡΙΦΟΡΑ ΤΗΣ ΟΙΣΤΡΑ- ΔΙΟΛΗΣ ΣΤΑ ΟΣΤΙΚΑ ΚΥΤΤΑΡΑ ΚΑΙ ΤΑ ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ 55 Οιστρογόνα 55 Οιστρογονικοί υποδοχείς 55 Ο υποδοχέας ER-β 58 ER- α ΚΑΙ ER- β knock out επίμυες 59 Μηχανισμοι δράσης των οιστρογόνων στο οστούν 61 Η δράση των οιστρογόνων στις οστεοβλάστες 62 Η δράση των οιστρογόνων στις οστεοκλάστες 64 Η δράση των οιστρογόνων στα περιρριζικά κύτταρα 65 ΕΙΔΙΚΟ ΜΕΡΟΣ Α. ΣΚΟΠΟΣ 69 Β. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 72 Υλικά 72 Διαλύματα- Όργανα 74 Μέθοδοι 76 Γ. ΣΤΑΤΙΣΤΙΚΗ ΕΠΕΞΕΡΓΑΣΙΑ ΚΑΙ ΑΠΟΤΕΛΕΣΜΑΤΑ 87 Στατιστική επεξεργασία 87 Ανοσοϊστοχημική αντίδραση 88 Η εκτίμηση της δραστηριότητας της αλκαλικής φωσφατάσης και ο ποσοτικός προσδιορισμός της οστεοκαλσίνης 91 Η εκτίμηση του πολλαπλασιασμού των κυττάρων του περιρριζίου υπό την επίδραση του TGF-β 1 και της 17-β οιστραδιόλης (Ε 2 ). 93 Δ. ΣΥΖΗΤΗΣΗ 102 Ε.ΣΥΜΠΕΡΑΣΜΑΤΑ 123 ΣΤ. ΠΕΡΙΛΗΨΗ 124 Ζ. SUMMARY 128 Η. ΒΙΒΛΙΟΓΡΑΦΙΑ 132
4 4 ΠΡΟΛΟΓΟΣ Η περιοδοντική νόσος αποτελεί μία χρόνια φλεγμονώδη κατάσταση που οδηγεί στη βαθμιαία απώλεια τόσο μαλακών όσο και σκληρών ιστών και τελικά στην απώλεια των δοντιών. Διάφορες μελέτες έχουν ασχοληθεί με τη θεραπεία της νόσου μέσω της διερεύνησης των βιολογικών μηχανισμών που συντελούν στην επούλωση και ανάπλαση των περιοδοντικών ιστών. Σημαντικό ρόλο σε αυτές τις διεργασίες παίζει το περιρρίζιο και συγκεκριμένα κάποιοι κυτταρικοί πληθυσμοί που εντοπίζονται σε αυτό και που μπορούν να οδηγήσουν στη δημιουργία νέας οστεΐνης, νέου οστού, νέων ινών του περιρριζίου και κατά συνέπεια στην ανάπλαση των περιοδοντικών ιστών. Η μελέτη των βιολογικών μηχανισμών που προάγουν τον πολλαπλασιασμό, τη χημειοταξία και τη διαφοροποίηση των περιρριζικών κυττάρων καθώς και η ανεύρεση των παραγόντων εκείνων που συμβάλλουν στην ενεργοποίηση αυτών των μηχανισμών παρουσιάζουν ιδιαίτερο ενδιαφέρον. Η ερμηνεία ωστόσο, των σχετικών ερευνών δυσχεραίνεται λόγω της ετερογένειας που παρατηρείται στο κυτταρικό φαινότυπο γεγονός που καθιστά απαραίτητο τον χαρακτηρισμό των κυττάρων που απομονώνονται από το περιρρίζιο. Διάφοροι αυξητικοί παράγοντες και ορμόνες έχουν μελετηθεί in vitro ως προς την επίδραση τους στις κυτταρικές λειτουργίες του περιρριζίου. Ένας τέτοιος παράγοντας είναι ο τροποποιητικός αυξητικός παράγοντας TGF-β 1, του οποίου όμως η δράση είναι αμφιλεγόμενη, καθώς τροποποιείται από πολλές παραμέτρους. Παράλληλα, οι ορμόνες του φύλου και συγκεκριμένα τα οιστρογόνα με κυρίαρχη την 17-β οιστραδιόλη επηρεάζουν τόσο τη παθογένεια της
5 5 περιοδοντικής νόσου όσο και τις διαδικασίες επούλωσης και ανάπλασης των περιοδοντικών ιστών. Τα οιστρογόνα επιδρούν στον πολλαπλασιασμό και στη διαφοροποίηση των περιρριζικών κυττάρων, όμως η μελέτη της δράση τους δίνει αντικρουόμενα αποτελέσματα, που οφείλονται σε διαφορές στη συγκέντρωση εφαρμογής τους, στο είδος και στην προέλευση των κυττάρων, στην κατανομή των υποδοχέων τους και στο στάδιο της κυτταρικής διαφοροποίησης, με αποτέλεσμα να απαιτείται επιπλέον διερεύνηση του ρόλου τους στις διαδικασίες επούλωσης των περιοδοντικών ιστών. Αυτοί όλοι οι προβληματισμοί αποτέλεσαν το έναυσμα για την εκπόνηση της παρούσας in vitro έρευνας σχετικά με το χαρακτηρισμό των προς μελέτη κυττάρων που απομονώθηκαν από το ανθρώπινο περιρρίζιο και την δράση του TGF-β 1 και της 17-β οιστραδιόλης στα κύτταρα αυτά. Παράλληλα έγινε μια προσπάθεια να προηγηθεί μια σύντομη αναφορά των σχετικών με όλα αυτά στοιχείων και των μέχρι τώρα ερευνητικών δεδομένων. Το πειραματικό μέρος αυτής της μελέτης, όσον αφορά στην καλλιέργεια των κυττάρων και στην επίδραση των προαναφερθέντων παραγόντων, έγινε στο Εργαστήριο της Πειραματικής Φυσιολογίας της Ιατρικής Σχολής Αθηνών ενώ, όσον αφορά στο χαρακτηρισμό των κυττάρων, πραγματοποιήθηκε στο Παθολογοανατομικό τμήμα του Γενικού Κρατικού Νοσοκομείου «Γεννηματάς». Μετά την ολοκλήρωση αυτής της διδακτορικής διατριβής θα ήθελα να αναφερθώ σε όλους εκείνους που με βοήθησαν στη διεκπεραίωση της συγκεκριμένης έρευνας. Έτσι επιθυμώ να εκφράσω τις θερμές μου ευχαριστίες: Στον Καθηγητή της Οδοντιατρικής και Διευθυντή του Εργαστηρίου της Περιοδοντολογίας κ. Ι. Α. Βρότσο για την ένδειξη εμπιστοσύνης στο άτομο
6 6 μου, την υποστήριξη του και την παρότρυνση του σε όλες τις φάσεις αυτής της μελέτης αλλά και σε όλα τα στάδια της επιστημονικής μου εξέλιξης τόσο στην Ελλάδα όσο και στις Μεταπτυχιακές μου Σπουδές στις Η.Π.Α. Στην Καθηγήτρια της Οδοντιατρικής και Αναπληρώτρια του Εργαστηρίου της Περιοδοντολογίας κ. Κ. Μαρκοπούλου την οποία και ιδιαίτερα ευχαριστώ, γιατί πέρα από την καθοδήγηση, τις πολύτιμες συμβουλές και το χρόνο που μου αφιέρωσε κατά τη διάρκεια αυτής της έρευνας, με βοήθησε και με στήριξε με τη πάντα γλυκιά και ανθρώπινη προσέγγιση της να ολοκληρώσω αυτή την διδακτορική διατριβή. Στον καθηγητή της Ιατρικής και Διευθυντή του Εργαστηρίου της Πειραματικής Φυσιολογίας κ. Μ. Κουτσιλιέρη για την ουσιαστική συμβολή και βοήθεια του στον πειραματικό σχεδιασμό και στην πειραματική τεκμηρίωση αυτής της έρευνας. Στον Μεταπτυχιακό Φοιτητή του Εργαστηρίου της Περιοδοντολογίας κ. Α. Μάμαλη για την σημαντική του βοήθεια στην εκτέλεση και ολοκλήρωση του πειραματικού τμήματος αυτής της διατριβής. Στον Καθηγητή της Ιατρικής και Διευθυντή του Παθολογοανατομικού Τμήματος του Γενικού Κρατικού Νοσοκομείου «Γεννηματάς» κ. Γ. Κοντογιώργο για την συμβολή του στην πραγματοποίηση μέρους των πειραμάτων αυτής της έρευνας. Στη στατισικολόγο κ. Ε. Δελιχά για την πρόθυμη βοήθεια της στην διεκπεραίωση της στατιστικής αξιολόγησης και τεκμηρίωσης των αποτελεσμάτων.
7 7 Στην φίλη Γραμματέα της Οδοντοστοματολογικής Ερεύνης κ. Κατερίνα Πολυχρονοπούλου για την βοήθεια της στη συγγραφή στον Υπολογιστή μεγάλων τμημάτων των κειμένων. Τελειώνοντας, θα ήθελα να ευχαριστήσω τους γονείς μου και την αδελφή μου για τη συνεχή παρότρυνση και συμπαράσταση τους σε όλα τα βήματα τόσο αυτής της διδακτορικής διατριβής όσο και της συνολικής επιστημονικής μου πορείας.
8 8 A. ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ KAI ΜΕΘΟΔΟΙ ΤΑΥΤΟΠΟΙΗΣΗΣ ΤΩΝ ΠΕΡΙΡΡΙΖΙΚΩΝ ΙΝΟΒΛΑΣΤΩΝ ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ Το περιρρίζιο αποτελεί ένα δυναμικό βιολογικό σύστημα το οποίο συμμετέχει στην ανατολή των δοντιών, στη φυσιολογική τους κινητικότητα κατά τη μάσηση καθώς και στην επούλωση και ανάπλαση των περιοδοντικών ιστών. Το περιρρίζιο προέρχεται από τον οδοντικό σάκο και είναι ιδιαίτερα πλούσιο σε κύτταρα υπεύθυνα για τις άνω λειτουργίες. Το μεγαλύτερο ποσοστό κυττάρων συνιστούν οι ινοβλάστες μεσεγχυματικής προέλευσης. Άλλοι κυτταρικοί πληθυσμοί που συναντώνται στο περιρρίζιο είναι τα αδιαφοροποίητα μεσεγχυματικά κύτταρα, τα επιθηλιακά κύτταρα, καθώς και τα αμυντικά κύτταρα. Στον περιρριζικό χώρο απαντώνται και κύτταρα τα οποία συντελούν στη διατήρηση και στην ανακατασκευή των σκληρών ιστών, που περιβάλλουν το περιρρίζιο και αυτά είναι οι οστεϊνοβλάστες, οι οστεοβλάστες, και οι οστεοκλάστες (Berkovitz και Shore 1995). ΙΝΟΒΛΑΣΤΕΣ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ Το μεγαλύτερο ποσοστό των κυττάρων του περιρριζίου είναι οι ινοβλάστες που προέρχονται από το εκτομεσέγχυμα. Συγκεκριμένα εκτιμάται ότι κατέχουν το 20% του περιρριζίου του τομέα στα πρόβατα (Berkovitz 1988), ενώ στους αρουραίους το ποσοστό αυτό ανέρχεται στο 43-55% (Beertsen και Everts 1977, Shore και συν. 1981). Οι ινοβλάστες είναι υπεύθυνες για το μεταβολισμό των συστατικών της εξωκυττάριας θεμέλιας ουσίας, καθώς επίσης και για τη διατήρηση του εύρους του περιρριζίου (Melcher 1970).
9 9 Βασική λειτουργία των ινοβλαστών είναι η σύνθεση και η δημιουργία των ινών κολλαγόνου καθώς και η απομάκρυνση αυτών (Cho και Garant 2000). Επίσης, in vitro μελέτες έδειξαν ότι οι ινοβλάστες συμμετέχουν στη φαγοκυττάρωση των ινών κολλαγόνου της εξωκυττάριας θεμέλιας ουσίας και στην αποδόμηση τους μέσα σε φαγολυσοσωμάτια. Συγκεκριμένα, φάνηκε ότι οι ινοβλάστες συμμετέχουν στη λυσοσωμική πέψη των ινών κολλαγόνου (Garant 1976). Απόδειξη αυτής της θεωρίας ήταν η εντόπιση γραμμωτών ινών κολλαγόνου και όξινης φωσφατάσης μέσα στα κοκκία των ινοβλαστών. Επιπλέον μελέτες αυτής τους της δραστηριότητας, έδειξαν ότι η κολλαγενάση δεν συμμετέχει στην ενδοκυττάρια φάση αποδόμησης του κολλαγόνου ενώ αντίθετα, υπεύθυνες γι αυτό είναι οι καθεψίνες B, L και Ν (Everts και συν. 1989, Lee και συν. 1996). Πρόσφατες μελέτες έδειξαν επίσης ότι ο ενδοκυττάριος μηχανισμός αποδόμησης του κολλαγόνου ευθύνεται για την ανακύκλωση αυτού στο περιρρίζιο (Beertsen και συν. 1997). Αντίθετα, η εξωκυττάρια οδός, με την οποία σχετίζεται η κολλαγενάση, συντελεί στην απομάκρυνση των ινών κολλαγόνου σε περιπτώσεις όπως εκείνη της φλεγμονής όπου η αποδόμηση του κολλαγόνου γίνεται με μη ελεγχόμενο τρόπο. Ινοβλάστες με οστεοβλαστικό φαινότυπο και τα αδιαφοροποίητα μεσεγχυματικά κύτταρα Οι ινοβλάστες του περιρριζίου παρουσιάζουν ετερογένεια ως προς τα λειτουργικά χαρακτηριστικά τους. Έτσι, υποπληθυσμοί αυτών είναι κύτταρα που μορφολογικά μοιάζουν στις ινοβλάστες, αλλά παρουσιάζουν οστεοβλαστική δραστηριότητα όπως αυτή χαρακτηρίζεται από: α) την
10 10 έκφραση υψηλών επιπέδων αλκαλικής φωσφατάσης, β) την παραγωγή πρωτεϊνών της εξωκυττάριας θεμέλιας ουσίας σχετιζομένων με το οστούν όπως οστεονεκτίνη, οστεοκαλσίνη, οστεοποντίνη, οστική σιαλοπρωτεΐνη, γ) την αύξηση της παραγωγής c-amp παρουσία της παραθυρεοειδούς ορμόνης (ΡΤΗ) και δ) την παραγωγή ενασβεστιωμένων οζιδίων (Somerman και συν. 1988, Piche και συν. 1989, Nohutcu και συν. 1997, Μarkopoulou 2002, Μarkopoulou και συν. 2003). Οι υποπληθυσμοί αυτοί των ινοβλαστών του περιρριζίου μπορούν να διαφοροποιηθούν σε οστεοβλάστες ή οστεϊνοβλάστες, ενώ, είναι υπεύθυνοι για την παραγωγή της ακύτταρης οστεΐνης με εξωγενείς ίνες (Cho και συν. 1992, Liu και συν.1997). Κατά την επούλωση των περιοδοντικών ιστών νέες ινοβλάστες μεταναστεύουν στην υπό επούλωση περιοχή. Αυτές προέρχονται από τα αρχέγονα κύτταρα τα οποία εντοπίζονται σε μία περιαγγειακή περιοχή σε απόσταση 5 μm από τα αγγεία (Cho και Garant 2000). Η πιθανή πηγή αυτών των αρχέγονων κυττάρων είναι ο μυελός του φατνιακού οστού όπου άλλωστε και παρατηρείται ο πολλαπλασιασμός τους. Τα νεοδιαιρούμενα κύτταρα φαίνεται να μπορούν να οδηγήσουν στο σχηματισμό τόσο νέας οστεΐνης όσο και νέων ινών του περιρριζίου. Έτσι, ενώ ακόμη υπάρχει σύγχυση ως προς την προέλευση των οστεοβλαστών και των οστεϊνοβλαστών, έχει παρατηρηθεί ότι, υποπληθυσμοί ινοβλαστών με οστεοβλαστικό χαρακτήρα προκύπτουν από τα αρχέγονα κύτταρα των περιαγγειακών χώρων του περιρριζίου, καθώς και από τα προγονικά κύτταρα του μυελού του οστού (Melcher 1970, Cho και Garant 2000).
11 11 Ιδιαίτερα σημαντικός είναι ο ρόλος των κυττάρων του περιρριζίου και ιδιαίτερα τον ινοβλαστών με οστεοβλαστική δραστηριότητα στην ανάπλαση των περιοδοντικών ιστών όπως αυτή ορίζεται από τη δημιουργία νέου οστίτη ιστού, νέας οστεΐνης και νέων, λειτουργικά προσανατολισμένων, ινών του περιρριζίου. Η ανάπλαση των ιστών του περιοδοντίου βασίζεται στην ενεργοποίηση μηχανισμών ικανών να οδηγήσουν στη χημειοταξία, στη προσκόλληση, στο πολλαπλασιασμό και στη διαφοροποίηση των περιρριζικών κυττάρων καθώς και στη παραγωγή μεσοκυττάριας θεμέλιας ουσίας (Bartold και συν. 2000). Πολλές σημαντικές μελέτες αναφέρονται στο ρόλο των περιρριζικών ινοβλαστών, στην επούλωση και ανάπλαση των περιοδοντικών ιστών. Η μελέτη των μηχανισμών αυτών, καθώς και η ερμηνεία των σχετικών ερευνών συχνά καθίστανται δύσκολες λόγω της ετερογένειας των ινοβλαστών του περιρριζίου. Για τον λόγο αυτό, κρίνεται σημαντικός ο χαρακτηρισμός των διαφόρων κυτταρικών πληθυσμών που λαμβάνονται από το περιρρίζιο προκειμένου να μελετηθεί η βιολογική τους συμπεριφορά. Διάφορες μελέτες έχουν ασχοληθεί με το χαρακτηρισμό των ινοβλαστών του περιρριζίου ως προς την μεσεγχυματική τους προέλευση και τον οστεοβλαστικό ή ινοβλαστικό τους φαινότυπο αν και μέχρι σήμερα δεν έχουν βρεθεί σαφείς συγκεκριμένοι δείκτες.
12 12 ΜΕΘΟΔΟΙ ΤΑΥΤΟΠΟΙΗΣΗΣ ΤΩΝ ΠΕΡΙΡΡΙΖΙΚΩΝ ΙΝΟΒΛΑΣΤΩΝ Ταυτοποίηση της μεσεγχυματικής προέλευσης των περιρριζικών ινοβλαστών Οι περιρριζικές ινοβλάστες προέρχονται από το εκτομεσέγχυμα. Η μεσεγχυματική τους προέλευση συχνά ταυτοποιείται με τον εντοπισμό της πρωτεΐνης βιμεντίνης στο κυτταρικό τους σκελετό. Η βιμεντίνη ανήκει στα ενδιάμεσα νημάτια που απαντώνται στο κυτταρικό σκελετό. Τα νημάτια αυτά είναι μεγέθους 11 nm και είναι μεγαλύτερα των λεπτών νηματίων της ακτίνης και μικρότερα των παχιών μυϊκών νηματίων. Πέντε είδη ενδιάμεσων νηματίων συναντώνται ανάλογα με την προέλευση των ιστών. Οι μεσεγχυματικοί ιστοί εκφράζουν την πρωτεΐνη βιμεντίνη που αποτελεί υποκατηγορία της τάξης ΙΙΙ των ενδιάμεσων νηματίων (Steinert και συν. 1985). Στους ιστούς της στοματικής κοιλότητας η βιμεντίνη εντοπίζεται στις ινοβλάστες του περιρριζίου και του πολφού, στις οδοντοβλάστες, ενώ κατά την οδοντική ανάπτυξη παρατηρείται στις ινοβλάστες της οδοντικής θηλής και του οδοντοθυλακίου (Lombardi και συν. 1992, Webb και συν. 1996, Basdra και συν. 1997). Η ανίχνευση της βιμεντίνης στηρίζεται κατά κύριο λόγο σε ανοσοϊστοχημικές μεθόδους και εντοπίζεται συνήθως γύρω από τον κυτταρικό πυρήνα ενώ η θέση της στις περιρριζικές ινοβλάστες εξαρτάται από το πειραματικό μοντέλο. Έτσι, στο περιρρίζιο των σκύλων και των αρουραίων η κατανομή της είναι διάχυτη μέσα στο κύτταρο (Cho και συν. 1988), ενώ στον άνθρωπο και στο πρόβατο, εντοπίζεται στις κυτταρικές προεξοχές (Berkovitz 1988). Οι περιρριζικές ινοβλάστες και οι οστεϊνοβλάστες, σε αντίθεση με τις ουλικές ινοβλάστες, συνεκφράζουν βιμεντίνη και κερατίνη αμέσως μετά την ανατολή των δοντιών (Webb και συν. 1994). Η έκφραση της κερατίνης, όμως, σταματά
13 13 μετά την ολοκλήρωση της ανατολής. Η βιμεντίνη εντοπίζεται πιο αδύναμα σε ηλικιωμένους ιστούς (Moxham και συν. 1998), ενώ στο νεοσχηματισμένο μετά από ιστική ανάπλαση περιρρίζιο και κοντά στη νεοσχηματισμένη οστεΐνη, εκφράζεται κατά τρόπο όμοιο με εκείνο που εκφράζεται στο άθικτο τμήμα του περιρριζίου. Το εύρημα αυτό, οδηγεί στη υπόθεση ότι τα κύτταρα που ευθύνονται για τη δημιουργία του νέου περιρριζίου είναι μεσεγχυματικής προέλευσης και προέρχονται από το ανέπαφο περιρρίζιο (Sculean και συν. 2003). Ταυτοποίηση του ινοβλαστικού χαρακτήρα των κυττάρων του περιρριζίου Γενικά, o χαρακτηρισμός των κυττάρων αυτών ως προς τον ινοβλαστικό τους φαινότυπο βασίζεται, συνήθως, στη μελέτη μορφολογικών χαρακτηριστικών και στην υψηλή περιεκτικότητα των κυττάρων αυτών σε ακτίνη. Το σχήμα των ινοβλαστών, καθορίζεται από τη συνθετική τους δραστηριότητα και την περιβάλλουσα θεμέλια ουσία. Σε in vitro καλλιέργειες σε επίπεδες επιφάνειες οι ινοβλάστες φέρουν το κλασσικό πεπλατυσμένο ατρακτοειδές σχήμα, όπου ο πυρήνας εντοπίζεται στο στενό άκρο του κυττάρου ενώ το σύστημα Golgi βρίσκεται στο ευρύτερο άκρο του. Αντίθετα, στους συνδετικούς ιστούς in vivo ή σε τρισδιάστατες καλλιέργειες (γέλες κολλαγόνου), οι ινοβλάστες εμφανίζονται με πιο πολύπλοκο σχήμα, γεγονός που αποδίδεται στην επαφή τους με τη θεμέλια ουσία (Shore και συν. 1981). Οι συνθετικά δραστήριες ινοβλάστες είναι μεγαλύτερες, πιο επιμηκυμένες και περισσότερο πολωμένες από τις ανενεργείς ινοβλάστες. Οι περιρριζικές ινοβλάστες περιέχουν έναν εμφανή πυρήνα με ένα ευδιάκριτο πυρηνίσκο και σαφώς περιγεγραμμένους πυρηνικούς πόρους (Berkovitz και
14 14 Shore 1995). Ο πυρήνας έχει σχήμα πεπλατυσμένου δίσκου διαμέτρου 10 μm (Shore και Berkovitz 1979) και κατέχει το 30% του κυτταρικού όγκου (Yamasaki και συν. 1987b). Οι προγονικές ινοβλάστες είναι μικρότερες, λιγότερο πολωμένες, περιέχουν πτωχότερο ενδοπλασματικό δίκτυο και λιγότερα κυστίδια Golgi (Gould και συν. 1983). Χαρακτηριστικό των ινοβλαστών του περιρριζίου είναι το πλούσιο αδρό ενδοπλασματικό τους δίκτυο και το πολυσύνθετο σύστημα Golgi, ενδεικτικό της υψηλής πρωτεϊνικής σύνθεσης. Διάφορες αυτοραδιογραφικές μελέτες, που βασίζονται στην ενσωμάτωση και την έκκριση ραδιοενεργών ισοτόπων διαφόρων αμινοξέων όπως της προλίνης και της γλυκίνης, αλλά και βιοχημικές μελέτες έχουν επιβεβαιώσει την αυξημένη παραγωγή και έκκριση κολλαγόνου από τις περιρριζικές ινοβλάστες (Sodek και συν. 1977, Cho και Garant 1981, Kirkham και συν. 1991). Συγκεκριμένα, οι Cho και Garant (1981) παρατήρησαν ότι, η οδός που ακολουθείται για τη σύνθεση κολλαγόνου ξεκινά από το αδρό ενδοπλασματικό δίκτυο, συνεχίζει στο σύστημα Golgi και καταλήγει μέσω των εκκριτικών κυστιδίων στην κυτταρική μεμβράνη. Για το πείραμα αυτό, χρησιμοποιήθηκε το ραδιενεργό ισότοπο του αμινοξέος προλίνη, το οποίο ενσωματώθηκε αρχικά στα πολυπεπτίδια κολλαγόνου μέσα στο αδρό ενδοπλασματικό δίκτυο. Σε χρονικό διάστημα δέκα λεπτών, νεοσχηματισμένα μόρια προκολλαγόνου εμφανίστηκαν μέσα στο σύστημα Golgi και στα επόμενα είκοσι λεπτά τα μόρια αυτά εκκρίθηκαν μέσω των εκκριτικών κοκκίων. Έτσι, σε χρόνο τριάντα λεπτών, νεοσυντεθιμένες ίνες κολλαγόνου εμφανίστηκαν στον εξωκυττάριο χώρο και σε μία ώρα οι ίνες αυτές είχαν επισημανθεί με 3 Η-προλίνη.
15 15 Προκειμένου να πραγματοποιηθεί η μετακίνηση αυτή των εκκριτικών κοκκίων από το trans-golgi δίκτυο στον εκκριτικό πόλο του κυττάρου απαιτείται ένα ανέπαφο δίκτυο μικροσωληνίσκων (Cho και Garant 1981). Οι μικροσωληνίσκοι αυτοί, είναι κύλινδροι διαμέτρου 22 nm και εμφανίζονται με τυχαία διάταξη μέσα στο κύτταρο. Πέρα από τους μικροσωληνίσκους που ο κύριος ρόλος τους είναι να διευκολύνουν την έκκριση κολλαγόνου, ο κυτταρικός σκελετός των περιρριζικών ινοβλαστών φέρει νημάτια διαφόρων μεγεθών. Έτσι, οι περιρριζικές ινοβλάστες περιέχουν στον κυτταρικό σκελετό τους ένα καλά οργανωμένο σύστημα από δέσμες μικρονηματίων ακτίνης καθώς και λείας μυϊκής ακτίνης και λείας μυϊκής μυοσίνης. Οι πρωτεΐνες αυτές, είναι χαρακτηριστικές των μυϊκών ινοβλαστών που απαντώνται στην επούλωση τραυμάτων και συμμετέχουν στο σχηματισμό κυτταροπλασματικών νηματίων συστολής ή αλλιώς νηματίων τάσης (stress fibers) (Aubin και συν. 1989, Giannopoulou και Cimasoni 1996, Serini και Gabbiani 1999). Τα νημάτια αυτά, προσανατολίζονται παράλληλα με τον επιμήκη άξονα του κυττάρου και καταλήγουν στις πλάκες πρόσφυσης με τις οποίες τα κύτταρα προσκολλώνται στο εξωκυττάριο υπόστρωμα. Το σύστημα αυτό προσκόλλησης (Fibronexus), εγκαθιστά ένα μηχανισμό έλξης (traction) των ινοβλαστών πάνω στις ίνες κολλαγόνου, υπεύθυνο τόσο για τη μετανάστευση τους όσο και για την οργάνωση της εξωκυττάριας θεμέλιας ουσίας. Η συσταλτική αυτή ικανότητα των περιρριζικών ινοβλαστών αλλά και των μυοινοβλαστών γενικότερα φαίνεται να αποδίδεται κατά κύριο λόγο στην παρουσία και έκφραση της λείας μυϊκής ακτίνης από τα συγκεκριμένα κύτταρα. Συγκεκριμένα, η αυξημένη έκφραση αυτής της πρωτεΐνης ενισχύει τη
16 16 συσταλτική δραστηριότητα των κυττάρων (Arora και McCulloch 1994, Hinz και συν. 2001), ενώ παράγοντες όπως η νικοτίνη, που αναστέλλουν την έκφραση της, οδηγούν σε μειωμένη συσταλτική ικανότητα των ινοβλαστών και κατά συνέπεια σε καθυστερημένη επούλωση (Fang και συν. 2005). Η λεία μυϊκή ακτίνη θεωρείται αξιόπιστος δείκτης διαφοροποίησης των ινοβλαστών σε μυοινοβλάστες (Gabiani 2003) ενώ συχνά χρησιμοποιείται και ως δείκτης του ινοβλαστικού χαρακτήρα των κυττάρων του περιρριζίου. Ανοσοϊστοχημικές μέθοδοι έδειξαν ότι τα κύτταρα του περιρριζίου βάφονται θετικά για τη λεία μυϊκή ακτίνη τόσο in vivo όσο και in vitro, ενώ αυξημένη βαφή παρατηρείται υπό συνθήκες στεγνής αποθήκευσης των δοντιών ( λεπτά σε στεγνό αέρα), από όπου λαμβάνονται τα κύτταρα (Giannopoulou and Cimasoni 1996, Lin και συν. 2000, Lekic και συν. 2001). Ταυτοποίηση των περιρριζικών ινοβλαστών ως προς τον οστεοβλαστικό τους φαινότυπο Ο χαρακτηρισμός των περιρριζικών ινοβλαστών ως προς τον οστεοβλαστικό τους φαινότυπο βασίζεται: α) στην έκφραση υψηλών επιπέδων αλκαλικής φωσφατάσης, β) στη παραγωγή πρωτεϊνών της εξωκυττάριας θεμέλιας ουσίας σχετιζομένων με το οστούν όπως, οστεοκαλσίνη, οστεοποντίνη, οστική σιαλοπρωτεΐνη, γ) στην αύξηση της παραγωγής c-amp παρουσία της παραθυρεοειδούς ορμόνης (ΡΤΗ) και δ) στην παραγωγή ενασβεστιωμένων οζιδίων (Somerman και συν. 1988, Piche και συν. 1989, Nohutsu και συν. 1997, Μarkopoulou και συν. 2003). Α) Μέτρηση της δραστηριότητας της αλκαλικής φωσφατάσης στα κύτταρα του περιρριζίου.
17 17 Η αλκαλική φωσφατάση είναι ένζυμο που σχετίζεται με τη διαδικασία ενασβεστίωσης. Συγκεκριμένα, η υδρόλυση των μονοφωσφορικών εστέρων από την αλκαλική φωσφατάση παράγει υψηλή συγκέντρωση φωσφορικών αλάτων (Beersten και Van den Bos 1989), οδηγώντας έτσι σε υπερκορεσμό και καθίζηση αλάτων φωσφορικού ασβεστίου στο κολλαγονώδες υπόστρωμα (Robinson 1923). Υψηλά επίπεδα δραστηριότητας της ενδογενούς αλκαλικής φωσφατάσης εντοπίζονται σε κύτταρα οστεοβλαστικού τύπου και λιγότερο στις ινοβλάστες (Robey και Termine 1985). Ανοσοϊστοχημικά, η αλκαλική φωσφατάση έχει ανιχνευθεί σε κύτταρα του περιρριζίου (Giannopoulou και Cimasoni 1996, Lin και συν. 2000, Lekic και συν. 2001). Βιοχημικές μέθοδοι έδειξαν αυξημένη δραστηριότητα της αλκαλικής φωσφατάσης στους περιρριζικούς κυτταρικούς πληθυσμούς, τόσο γενικά όσο και σε σχέση με τις ουλικές ινοβλάστες (Somerman και συν. 1988, Piche και συν. 1989, Nojima και συν. 1990, Basdra και συν.1997, Carnes και συν. 1997, Μarkopoulou και συν. 2003). Η δραστηριότητα αυτή αυξάνεται παρουσία της 1,25 διυδροξυβιταμίνης D 3 η οποία είναι ο πιο ενεργός μεταβολίτης της βιταμίνης D 3 και συμμετέχει στη ρύθμιση της ομεόστασης του ασβεστίου (Piche και συν. 1989, Basdra και συν. 1997, Carnes και συν. 1997). Τα υψηλά επίπεδα της δραστηριότητας της αλκαλικής φωσφατάσης που παρατηρούνται στα κύτταρα του περιρριζίου αποτελούν δείκτη του οστεοβλαστκού φαινοτύπου των κυττάρων αυτών. Β) Παραγωγή πρωτεϊνών της εξωκυττάριας θεμέλιας ουσίας σε σχέση με το οστούν (οστεοκαλσίνη, οστεοποντίνη, οστική σιαλοπρωτεΐνη)
18 18 Ανοσοϊστοχημικές μελέτες έχουν δείξει ότι, μέσα στους περιοδοντικούς ιστούς κατανέμονται μη-κολλαγονώδεις πρωτεΐνες οστού όπως η οστεοκαλσίνη, η οστεοποντίνη και η οστική σιαλοπρωτεΐνη (Bronckers και συν. 1994, MacNeil και συν. 1995, Ivanovski και συν. 2001). Οι πρωτεΐνες αυτές εκκρίνονται σε διαφορετικά στάδια της διαφοροποίησης των οστεογενών κυττάρων (Boskey 1996). Η οστεοκαλσίνη γνωστή και ως γ-καρβοξυ-γλουταμικού οξέος πρωτεΐνη του οστού (bone gla protein) είναι ένα 49-υπόλοιπο πολυπεπτίδιο. Στους ανθρώπους το γονίδιο της οστεοκαλσίνης εντοπίζεται στο χρωμόσωμα 1 (1q2r-q31) και ρυθμίζεται σε μεταγραφικό επίπεδο από την 1,25 διυδροξυβιταμίνη D 3. Η οστεοκαλσίνη είναι πρωτεΐνη εξαρτώμενη από τις βιταμίνες Κ και D, απαντάται σε κύτταρα ενασβεστιωμένων ιστών (ώριμες οστεοβλάστες, οδοντοβλάστες) και αποτελεί δείκτη διαφοροποίησης των οστεοβλαστών και σχηματισμού του οστού (Boskey 1996, Basdra και συν. 1997, Lee at al 2000). Ο ποσοτικός προσδιορισμός της οστεοκαλσίνης γίνεται κυρίως με μεθόδους, όπως τον ραδιοανοσοπροσδιορισμό (RIA), τον ανοσοραδιομετρικό προσδιορισμό (IRMA), τον ενζυμικό άνοσο προσδιορισμό (ELISA), τη χημειοφωταυγή ανοσομετρική μέθοδο, ενώ για τον εντοπισμό της χρησιμοποιούνται ανοσοϊστοχημικές μέθοδοι (Lee at al 2000). Η οστεοκαλσίνη, έχει ανιχνευθεί σε ανθρώπινα κύτταρα του περιρριζίου, ιδιαίτερα μετά από έκθεση σε 1,25 διυδροξυβιταμίνη D 3 και αποτελεί δείκτη του οστεοβλαστικού τους φαινοτύπου (Basdra και συν. 1997, Carnes και συν. 1997, Ivanovski και συν. 2001). Επίσης, κύτταρα του βόειου περιρριζίου σε αντίθεση με τις ουλικές ινοβλάστες φαίνεται να συνθέτουν μια πρωτεΐνη που είναι ανοσολογικά σε διασταυρούμενη αντίδραση με την βόεια οστεοκαλσίνη
19 19 (Nojima και συν. 1994). Σε αντίθεση με τα ανθρώπινα και τα βόεια κύτταρα η οστεοκαλσίνη δεν ανιχνεύθηκε στις περιρριζικές ινοβλάστες αρουραίων ενώ ανιχνεύθηκε στις οστεϊνοβλάστες και στις οδοντοβλάστες αυτών (Bronckers και συν. 1994,Takano-Yamamoto και συν. 1994). Η οστική σιαλοπρωτεΐνη είναι πρωτεΐνη πού προσκολλάται στον υδροξυαπατίτη και στα κύτταρα. Ανοσοκυτταροχημικά, η οστική σιαλοπρωτεΐνη περιορίζεται σε περιοχές ενασβεστιωμένης θεμέλιας ουσίας (Pinero και συν. 1995). Η οστεοποντίνη, παρόμοια με τη οστική σιαλοπρωτεΐνη, εκφράζεται σε διαφοροποιημένα οστικά κύτταρα, συγκεντρώνεται στη θεμέλια ουσία του οστού, συμμετέχει στην ενασβεστίωση του καθώς και στην πρόσφυση των οστεοβλαστών και των οστεοκλαστών στην εξωκυττάρια ουσία (McKee και Nanci 1995). Ανοσοϊστοχημικά, η οστεοποντίνη και η οστική σιαλοπρωτεΐνη εντοπίζονται κυρίως κοντά στην οστεΐνη κα το φατνιακό οστούν, γεγονός πού επιβεβαιώνει το ρόλο τους στην οστεϊνογένεση και την οστεοβλαστική διαφοροποίηση (Bronckers και συν. 1994, D Errico at al 1995, Lin και συν. 2000, Lekic και συν. 2001, Ivanovski και συν. 2001). Η ανίχνευση των πρωτεϊνών αυτών στα κύτταρα του περιρριζίου υποδηλώνει την ύπαρξη περιρριζικών ινοβλαστών με οστεοβλαστικό φαινότυπο. Γ) Η αύξηση της παραγωγής c-amp παρουσία της παραθυρεοειδούς ορμόνης (ΡΤΗ) Γενικά, η παραθυρεοειδής ορμόνη συμμετέχει σε εκείνες τις διεργασίες που οδηγούν στην οστική απορρόφηση. Παρόλα αυτά, η συστηματική χορήγηση της αυξάνει το σχηματισμό οστού λόγω της ικανότητας της παραθορμόνης να αυξάνει τη σύνθεση του ινσουλινικού αυξητικού παράγοντα Ι και του
20 20 υποδοχέα του στα οστεογενή κύτταρα αλλά και να διεγείρει τη κυκλική μονοφωσφορική αδενοσίνη και την πρωτεϊνική κινάση Α (Partridge και συν. 1994,Civitelli και συν. 1998). Αύξηση της παραγωγής c-amp παρουσία της παραθυρεοειδούς ορμόνης (ΡΤΗ) παρατηρήθηκε και στις περιρριζικές ινοβλάστες από έγκλειστα δόντια, υπονοώντας έτσι, την ύπαρξη οστεοβλαστικών χαρακτηριστικών στα κύτταρα αυτά (Piche και συν.1989). Αξίζει να αναφερθεί ότι την ίδια χρονική περίοδο οι Somerman και συν. (1989), χρησιμοποιώντας περιρριζικά κύτταρα από δόντια πού είχαν ανατείλει στη στοματική κοιλότητα, απέτυχαν να αποδώσουν τα ίδια αποτελέσματα πιθανά λόγω διαφορετικού πειραματικού μοντέλου. Δ) Παραγωγή ενασβεστιωμένων οζιδίων Διάφορες μελέτες έχουν δείξει ότι η καλλιέργεια περιρριζικών κύτταρων, παρουσία δεξαμεθαζόνης, για διάστημα 3-4 εβδομάδων, οδηγεί στο σχηματισμό ενασβεστιωμένων οζιδίων, τα οποία μετά από βαφή με χρωστική Van Kossa δίνουν στο μικροσκόπιο την εικόνα των αλάτων φωσφορικού ασβεστίου, ενώ η μορφολογική εικόνα της ενασβεστίωσης, προσομοιάζει εκείνη της in vivo ενδομεβρανώδους ενασβεστίωσης (Cho και συν.1992, Basdra και συν. 1997, Carnes και συν.1997). Η δυνατότητα παραγωγής ενασβεστιωμένων οζιδίων από τα κύτταρα του περιρριζίου παρουσία γλυκοκορτικοστεροειδών, υποδηλώνει, την παρουσία στο περιρρίζιο κυτταρικών πληθυσμών με οστεοβλαστικό φαινότυπο και την δυνατότητα διαφοροποίησης τους σε οστεοβλάστες (Lekic και συν. 2001). Τα τελευταία χρόνια γίνεται προσπάθεια ανεύρεσης πιο εξειδικευμένων δεικτών για τον χαρακτηρισμό των περιρριζικών ινοβλαστών και τη διάκριση τους από τις ουλικές ινοβλάστες. Δύο πρωτεΐνες που συνδέονται με το
21 21 ασβέστιο, η S100A4 και η S100A2, έχουν απομονωθεί από βόειους περιοδοντικούς ιστούς και η S100A4 έχει προταθεί ως δείκτης διάκρισης κυττάρων μεταξύ του περιρριζίου και των ούλων (Duarte και συν. 1998). Η πρωτεΐνη περιοστίνη, επίσης, έχει ειδικά εκφραστεί στο περιόστεο και στο περιρρίζιο, ενώ φαίνεται να συμμετέχει ενεργά στο σχηματισμό ενασβεστιωμένων ιστών (Horiuchi και συν. 1999). Τέλος, οι Park και συν.(2001) έδειξαν ότι ο χαρακτηρισμός των κυττάρων του περιρριζίου μπορεί να επιτευχθεί με την ταυτοποίηση ειδικών, για τα κύτταρα αυτά, γονιδίων που απουσιάζουν από τις ουλικές ινοβλάστες. ΑΜΥΝΤΙΚΑ ΚΥΤΤΑΡΑ Τα αμυντικά κύτταρα που συναντώνται στο περιρρίζιο είναι κυρίως κύτταρα της μονοκυτταρικής και μακροφαγικής σειράς. Αυτά συνήθως απαντώνται στην περιαγγειακή και περινευρική περιοχή του χαλαρού συνδετικού ιστού και σπανιότερα στον πυκνό συνδετικό ιστό των δεσμίδων κολλαγόνου. Τα μονοκύτταρα εμφανίζονται στο ηλεκτρονικό μικροσκόπιο σε ωοειδές σχήμα, με κυματοειδή επιφάνεια. Πυκνά λυσοσωματικά κοκκία απαντώνται στο κυτταρόπλασμά τους, ενώ τα ενεργοποιημένα μονοκύτταρα παρουσιάζουν πυκνό ενδοπλασματικό δίκτυο, πλούσιο σύστημα Golgi καθώς και κυτταροπλασματική πόλωση. Τα μακροφάγα που προκύπτουν από την ωρίμανση των ενεργοποιημένων μονοκύτταρων, είναι μεγαλύτερα σε μέγεθος και περιέχουν μεγάλο αριθμό φαγολυσοσωματίων. Τα μονοκύτταρα κινούνται στο περιρρίζιο μέσω χημειοτακτικών ερεθισμάτων. Σε φλεγμονή του περιρριζίου, οι κυτταροκίνες ιντερλευκίνη 1β (IL-1β) και ο παράγοντας νέκρωσης των όγκων (TNF-α), συμβάλλουν στη διείσδυση των μονοκυττάρων διεγείροντας τις ινοβλάστες να εκκρίνουν τη χημειοτακτική πρωτεΐνη 1 των
22 22 μακροφάγων. Τα ενεργοποιημένα μονοκύτταρα εκκρίνουν κολλαγενάση, ελαστάση, λυσοσωματικές υδρολάσες και τον ενεργοποιητή του πλασμινογόνου (ΡΑ). Μ αυτόν τον τρόπο συμμετέχουν στην αποδόμηση της εξωκυττάριας θεμέλιας ουσίας (Garant 2003). ΑΛΛΑ ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ Άλλοι κυτταρικοί πληθυσμοί που υπάρχουν στο περιρριζικό χώρο είναι τα επιθηλιακά κύτταρα καθώς και οι οστεοβλάστες και οι οστεϊνοβλάστες. Τα επιθηλιακά κύτταρα είναι υπολείμματα του επιθηλιακού ελύτρου του Hertwig, το οποίο, κατά τη διάπλαση των δοντιών, συμμετέχει στο καθορισμό του σχήματος της ρίζας και στη διαφοροποίηση των οδοντοβλαστών της ρίζας. Τα επιθηλιακά αυτά κύτταρα, εκκρίνουν πρωτεΐνες αμελογενίνης (enamel like proteins) στην επιφάνεια της ρίζας (Slarkin και συν. 1988). Τα επιθηλιακά κυτταρικά υπολείμματα διακρίνονται ιστολογικά από τις ινοβλάστες του περιρριζίου λόγω της πολύ στενής επαφής των κυβοειδών τους κυττάρων και την τάση τους να χρωματίζονται πολύ πιο έντονα με χρωστικές αιματοξυλίνης ηωσίνης. Περιβάλλονται πλήρως από κύτταρα συνδετικού ιστού και εντοπίζονται κοντά στην οστεΐνη. Τα κύτταρα αυτά είναι μη λειτουργικά, ενώ in vivo, δεν παρατηρείται πολλαπλασιασμός τους. Στη βιβλιογραφία είναι γνωστά και ως επιθηλιακά κύτταρα του Malassez. Οι οστεοβλάστες και οι οστεϊνοβλάστες εντοπίζονται στην περιοχή του περιρριζίου, όμως, στην πραγματικότητα, σχετίζονται με τους σκληρούς οδοντικούς ιστούς που σχηματίζουν. Οι οστεϊνοβλάστες ευθύνονται για την έκκριση της οργανικής ουσίας της οστεΐνης. Όταν αυτές είναι ενεργείς εμφανίζονται ως ένα ευδιάκριτο στρώμα
23 23 κυττάρων στην επιφάνεια της ρίζας. Συχνά τα κύτταρα αυτά διακρίνονται με δυσκολία από τις ινοβλάστες. Χαρακτηριστικό γνώρισμά τους στον άνθρωπο είναι η πληθώρα κοκκίων γλυκογόνου και ενδιάμεσων νηματίων και νηματίων ακτίνης (Yamasaki και συν. 1986). Η κυτταρική τους μεμβράνη φέρει υποδοχείς αυξητικών ορμονών και του επιδερμικού αυξητικού παράγοντα (EGF) σε μικρότερο όμως βαθμό από ότι εκείνη των ινοβλαστών (Cho και συν. 1991, Zhang και συν. 1993). Οι οστεοβλάστες του περιρριζίου εντοπίζονται στην επιφάνεια του φατνιακού οστού. Η εμφάνιση και η μορφολογία τους είναι όμοια με εκείνη των οστεοβλαστών οπουδήποτε στο σκελετό. Όταν αυτές είναι ενεργείς, δημιουργούν μία στοιβάδα κυβοειδών κυττάρων με έντονα βασεόφιλο κυτταρόπλασμα. Όπως και οι ινοβλάστες διαθέτουν έντονο αδρό ενδοπλασματικό δίκτυο και πολυάριθμα μιτοχόνδρια και κοκκία. Τα κύτταρα αυτά, ως έντονα διαφοροποιημένα κύτταρα δεν διαθέτουν υποδοχείς για τον επιδερμικό αυξητικό παράγοντα (EGF). Μεταξύ των οστεοβλαστών απαντώνται χασματικές συνδέσεις (gap junction) και δεσμοσωμάτια. Η διακυτταρική αυτή σύνδεση έχει ως αποτέλεσμα την καθήλωση των ινών του περιρριζίου που εμπλέκονται στην οστική ανακατασκευή (Puras 1996, Sodek και Mc Kee 2000). Οι οστεοβλάστες εμφανίζουν επίσης υποδοχείς οιστρογόνων και βιταμίνης D 3 στο πυρήνα τους καθώς και υποδοχείς κυτταροκινών (Baron 1999). Στην εξωκυττάρια περιοχή οι οστεοβλάστες απελευθερώνουν ινίδια κολλαγόνου και ειδικές πρωτεΐνες της θεμέλιας ουσίας και έτσι προκύπτει το
24 24 οστεοειδές, που αποτελεί μία ζώνη εύρους 10 μm, η οποία ενασβεστιώνεται στους ενήλικες με ημερήσιο ρυθμό 0.55 μm (Parfitt και συν. 1997). Μετά την ολοκλήρωση της οστικής εναπόθεσης οι οστεοβλάστες μετατρέπονται σε οστεοκύτταρα.
25 25 B. Ο ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ ΤΩΝ ΑΥΞΗΤΙΚΩΝ ΠΑΡΑΓΟΝΤΩΝ ΣΤΑ ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ Διάφοροι αυξητικοί παράγοντες επηρεάζουν τους βιολογικούς μηχανισμούς που αφορούν στη χημειοταξία, στον πολλαπλασιασμό, στη διαφοροποίηση και στη σύνθεση πρωτεϊνών των κυτταρικών πληθυσμών του περιρριζίου. Οι αυξητικοί παράγοντες ανήκουν στις κυτταροκίνες και εντοπίζονται σε πλήθος ιστών και κυττάρων κυρίως στα αιμοπετάλια, στις ινοβλάστες και στις οστεοβλάστες (Jiang και συν. 1999). Πρόκειται για πολυπεπτίδια μεγάλου μοριακού βάρους με αυτοκρινική και παρακρινική δράση όπου δρουν σε συγκεκριμένο υποδοχέα της κυτταρικής επιφάνειας. Είναι φυσικά προϊόντα κυττάρων που έχουν τη δυνατότητα να επηρεάζουν πολλές κυτταρικές δραστηριότητες, καθώς και το μεταβολισμό συγκεκριμένων κυττάρων (Mundy και συν. 1994, Weryha και συν. 1995). Ορισμένοι αυξητικοί παράγοντες, όπως ο PDGF, TGF-β, FGF, IGF-I, μόνοι τους, ή σε συνδυασμό, φαίνεται να επιδρούν στις λειτουργίες των κυττάρων του περιρριζίου και κατ επέκταση να δρούν στην επούλωση ή ανάπλαση των περιοδοντικών ιστών (Lynch και συν. 1994). Οι παράγοντες αυτοί,όπως και η δράση τους, αναφέρονται ακολούθως ενώ αναλυτικότερα περιγράφεται ο TGF-β. Αυξητικός παράγοντας των αιμοπεταλίων (PDGF) O αυξητικός παράγοντας των αιμοπεταλίων (PDGF) είναι πολυπεπτίδιο που σχηματίζεται από δύο αλυσίδες αμινοξέων οι οποίες αποτελούν το προϊόν δύο σχετιζόμενων γονιδίων των PDGF-Α και PDGF-Β. Οι αλυσίδες αυτές, μπορεί να είναι ομοδιμερείς ή ετεροδιμερείς και ο PDGF μπορεί να εκφραστεί ως
26 26 PDGF-ΑΑ, -ΒΒ ή - ΑΒ. Πρόσφατα, δύο ακόμα ισομορφές του PDGF έχουν βρεθεί, η C και η D, που όμως ελάχιστα χρησιμοποιούνται στις διάφορες μελέτες. Ο PDGF-ΒΒ είναι ο πιο δυναμικός τύπος του αυξητικού παράγοντα των αιμοπεταλίων και μελετάται ιδιαίτερα ως προς τη δράση του στα κύτταρα του οργανισμού. Ο PDGF προέρχεται κυρίως από τα αιμοπετάλια, εκφράζεται όμως και από άλλα κύτταρα όπως τις ινοβλάστες, οστεοβλάστες, και τα μακροφάγα (Centrella και συν.1992, Boyan και συν.1994, Canalis και Ornitz 2000). Οι ισομορφές του PDGF πραγματοποιούν τη βιολογική τους δράση μέσω του υποδοχέα α ο οποίος προσδένεται στις αλυσίδες PDGF- Α και Β και του υποδοχέα β ο οποίος συνδέεται μόνο με την αλυσίδα PDGF-Β. Ο PDGF με τη πρόσδεση του με τους υποδοχείς αυτούς οδηγεί στο διμερισμό τους και στην ενεργοποίηση της δραστηριότητας της τυροσίνης κινάσης με αποτέλεσμα την αυτό-φωσφορυλίωση τους (Heldin και συν. 1989). Η ενεργοποίηση αυτών των υποδοχέων διεγείρει διάφορα μεταγραφικά μονοπάτια τα οποία καθορίζονται από το τύπο των κυττάρων. Οι υποδοχείς του PDGF και συγκεκριμένα ο υποδοχέας-β έχουν εντοπισθεί ανοσοκυτταροχημικά τόσο στους ιστούς όσο και στα κύτταρα του περιοδοντίου. Η έκφραση τους παρουσιάζεται εντονότερη στους υπό ανάπλαση ιστούς από ότι στους φυσιολογικούς, γεγονός πού υποδηλώνει τη πιθανή ανάμειξη του PDGF στην επούλωση και ανάπλαση των περιοδοντικών ιστών (Parkar και συν. 2001). Χαρακτηριστική λειτουργία του PDGF είναι η μιτογόνος και χημειοτακτική δράση του στα κύτταρα μεσεγχυματικής προέλευσης συμπεριλαμβανομένων των οστεοβλαστών, των κυττάρων του περιρριζίου, και των ινοβλαστών
27 27 (Graves και συν. 1994, Boyan και συν.1994). Σε μια μελέτη σε αρουραίους, η καλλιέργεια περιρριζικών κυττάρων ινοβλαστικού χαρακτήρα παρουσία του παράγοντα PDGF είχε ως αποτέλεσμα την αύξηση της μιτογένεσης των κυττάρων αυτών (Blom και συν. 1994). Παρόμοια, η εφαρμογή 10ng/ml PDGF-ΒΒ σε κύτταρα του ανθρώπινου περιρριζίου καθώς και σε ανθρώπινες ουλικές ινοβλάστες οδήγησε σε αύξηση της μιτογόνου δραστηριότητας των κυττάρων αυτών (Markopoulou και συν. 2003). Αύξηση του πολλαπλασιασμού των κυττάρων του περιρριζίου παρατηρήθηκε μετά από εφαρμογή του PDGF-ΒΒ σε επιφάνειες οδοντίνης όπου είχε προηγηθεί απασβεστίωση με EDTA (Zaman και συν.1999). Οι Gamal και Maihlot (2000) διαπίστωσαν in vitro, ότι η εφαρμογή του PDGF-ΒΒ σε δόσεις ίσες ή μεγαλύτερες των 50ng/ml διέγειρε σημαντικά τη προσκόλληση των ανθρώπινων κυττάρων του περιρριζίου σε ριζικές επιφάνειες προσβεβλημένες από περιοδοντική νόσο. Ο PDGF-ΒΒ φαίνεται να αυξάνει τη σύνθεση του υαλουρονικού οξέος από τις ανθρώπινες ουλικές ινοβλάστες, ενώ διεγείρει τη σύνθεση κολλαγόνου από τα κύτταρα του περιρριζίου (Bartold και συν. 1993, Boltchi 1999). Η παρατήρηση αυτή, σε συνδυασμό με τη μιτογόνο δράση του PDGF-ΒΒ, δικαιολογεί την αντίληψη ότι ο παράγοντας αυτός συμμετέχει στην επούλωση και ανάπλαση των περιοδοντικών ιστών. Η συνδυαστική χρήση του PDGF με άλλους αυξητικούς παράγοντες στα κύτταρα του περιρριζίου δείχνει να ενισχύει τη δράση του παράγοντα αυτού στην επούλωση και στην ανάπλαση των περιοδοντικών ιστών. Η δράση του αυξητικού παράγοντα των αιμοπεταλίων μεμονωμένα αλλά και σε συνδυασμό με άλλους αυξητικούς παράγοντες μελετήθηκε σε κύτταρα του περιρριζίου των αρουραίων (Matsuda και συν. 1992). Η μελέτη αυτή απέδειξε και πάλι τη
28 28 μιτογόνο και χημειοτακτική δράση του PDGF-ΒΒ, καθώς και την επίδραση του στην αύξηση της σύνθεσης του κολλαγόνου. Στην ίδια μελέτη η ταυτόχρονη εφαρμογή του PDGF με τον IGF οδήγησε σε ακόμα καλύτερα αποτελέσματα. Αντίθετα, οι Nguyen και συν. (1995) παρατήρησαν ότι η συνδυαστική εφαρμογή των δύο αυτών παραγόντων δεν οδηγεί σε καλύτερα αποτελέσματα στη μιτογένεση των κυττάρων του περιρριζίου απ ότι η εφαρμογή του καθενός μεμονωμένα. Οι διαφορές αυτές στα αποτελέσματα των ερευνών μπορεί να είναι αποτέλεσμα της χρήσης διαφορετικών πειραματικών μοντέλων και συγκεντρώσεων των αυξητικών παραγόντων. Παράλληλα, η προσθήκη του TGF-β 1 στον PDGF προήγαγε την ανταπόκριση των κυττάρων του περιρριζίου στον PDGF σε επίπεδο πολλαπλασιασμού (Οates και συν. 1993). Ο συνδυασμός του TGF-β 1 με το PDGF επιδρά θετικά στο πολλαπλασιασμό των κυττάρων του περιρριζίου και μάλιστα περισσότερο από ότι στις ουλικές ινοβλάστες (Dennison και συν. 1994). Η παρατήρηση αυτή, υποδεικνύει ότι η συνδυαστική χρήση των δύο αυτών παραγόντων διεγείρει επιλεκτικά τα κύτταρα του περιρριζίου, γεγονός που είναι σημαντικό στην ανάπλαση των περιοδοντικών ιστών. Ο Lynch και συν. (1989) ήταν η πρώτη ομάδα ερευνητών που πέτυχαν περιοδοντική ανάπλαση με τη τοπική εφαρμογή του PDGF σε συνδυασμό με τον προσομοιάζοντα στην ινσουλίνη αυξητικό παράγοντα (IGF) σε περιοδοντικές βλάβες σε σκύλους Beagle. Έκτοτε, διάφοροι ερευνητές οδηγήθηκαν σε ανάλογα αποτελέσματα, χρησιμοποιώντας διαφορετικά μοντέλα πειραματόζωων (Rutherford συν.1992,1993, Giannobile και συν. 1994). Η πρώτη κλινική μελέτη σε ανθρώπους που μελέτησε την ασφάλεια και την αποτελεσματικότητα της συνδυαστικής εφαρμογής των PDGF/ IGF
29 29 πραγματοποιήθηκε από τους Howell και συν. το Τριανταοκτώ ασθενείς με περιοδοντίτιδα θεραπεύτηκαν με την εφαρμογή 150μg/ml PDGF-BB/ IGF σε μεθυλοκυτταρικό μεταφορέα, με την εφαρμογή μόνο του μεθυλοκυτταρικού μεταφορέα ή μόνο με συμβατικό χειρουργείο. Τα αποτελέσματα έδειξαν ότι με τη συνδυαστική χρήση των αυξητικών παραγόντων επιτεύχθηκε ανάπλαση της οστικής βλάβης κατά 43.2% σε σχέση με το 18.5% του συμβατικού χειρουργείου. Η δράση του PDGF-ΒΒ στην ανάπλαση του περιοδοντίου έχει μελετηθεί σε συνδυασμό με τη χρήση διάφορων μοσχευμάτων και άλλων μεταφορέων. Οι Papadopoulos και συν. (2003), μελετώντας in vitro τη δράση του PDGF-ΒΒ σε συνδυασμό με τα μοσχεύματα DFDBA και FDBA σε ανθρώπινα περιρριζικά κύτταρα, έδειξαν τον επιτυχή ρόλο των μοσχευμάτων αυτών ως μεταφορέων του PDGF-ΒΒ στην ανάπλαση του περιοδοντίου. Οι Camelo και συν. (2003) και οι Nevins και συν. (2003) έδειξαν τόσο κλινικά όσο και ιστολογικά ότι ο συνδυασμός του rh PDGF-ΒΒ με μόσχευμα DFDBA οδήγησε σε πλήρη ανάπλαση σε μεσορριζικές βλάβες ΙΙ βαθμού στους ανθρώπους. Σε μια άλλη πιο εκτεταμένη κλινική μελέτη της ασφάλειας και της αποτελεσματικότητας της χρήσης του PDGF-ΒΒ στην ανάπλαση των περιοδοντικών ιστών, η χρήση του rh PDGF-ΒΒ σε συνδυασμό με συνθετικό β-τριφωσφορικό ασβέστιο οδήγησε σε σημαντική αύξηση του επιπέδου πρόσφυσης, μείωση της υφίζησης των ούλων και βελτίωση της οστικής ανάπλασης (Nevins και συν.2005, Reynolds και Aichermann-Reidy 2005). Εφαρμογή του ίδιου συνδυασμού σε δόσεις 0.3 και 1mg/mL σε ενδοοστικές περιοδοντικές βλάβες σε ανθρώπους έδειξε ανάπλαση των περιοδοντικών ιστών ιστολογικά και κλινικά (Ridgway και συν 2008).
30 30 Τελευταία, έχει μελετηθεί η εφαρμογή της PDGF γονιδιακής θεραπείας στην ανάπλαση του περιοδοντίου. Οι σχετικές έρευνες έχουν δείξει ότι αυτού του είδους η θεραπεία σε περιοδοντικές βλάβες αρουραίων αυξάνει τον πολλαπλασιασμό των κυττάρων του περιοδοντίου και διεγείρει την οστεϊνοβλαστική δραστηριότητα, καθώς και την ανάπλαση της οστεΐνης και του φατνιακού οστού (Giannobile και συν.2001, Jin και συν. 2004). Αυξητικός παράγοντας ο προσομοιάζων με την ινσουλινη (IGF) Οι αυξητικοί παράγοντες IGFs είναι μια οικογένεια μιτογόνων πρωτεϊνών που συμμετέχουν στην ανάπτυξη, στη διαφοροποίηση και στη διατήρηση διαφόρων ιστών. Η οικογένεια αυτή αποτελείται από τρεις συνδέτες (ινσουλίνη, IGF-I και ΙGF-II), τρεις υποδοχείς της κυτταρικής επιφάνειας (τον ινσουλινικό, τον IGF-I και τον IGF-II / φωσφορική μαννόζη 6 υποδοχέα) και από τουλάχιστον έξι πρωτεΐνες που συνδέονται με υψηλή χημική συγγένεια με τους IGFs και τροποποιούν τη βιολογική τους δράση (Werner και Katz 2004). Αρχικά οι IGFs ταυτοποιήθηκαν στο ήπαρ όπου μεσολαβούν στη δράση της αυξητικής ορμόνης στην ανάπτυξη των μακρών οστών (Salmon και Daughaday 1957). Αργότερα, διαπιστώθηκε ότι οι IGFs παράγονται και από άλλους ιστούς γεγονός, πού οδήγησε στην πεποίθηση ότι δρούν στην ανάπτυξη του οργανισμού ενδοκρινικά, αυτοκρινικά και παρακρινικά (D Ercole 1980). O IGF-I συντίθεται από διάφορα κύτταρα συμπεριλαμβανομένων των οστεοβλαστών και των χονδροβλαστών.ο IGF-I διεγείρει το πολλαπλασιασμό των οστεοβλαστών και τη σύνθεση του κολλαγόνου τύπου Ι και γενικά θεωρείται σημαντικός ρυθμιστής του μεταβολισμού των οστικών κυττάρων (Graves και συν. 1994, Giannobile και συν. 1997). Επίσης έχει μιτογόνο και
31 31 χημειοτακτική δράση στις ινοβλάστες (Matsuda και συν. 1992, Cochran και Wozney 1999). Οι Ivanovski και συν. (2001) έδειξαν ότι ο IGF-I προωθεί τη σύνθεση της εξωκυττάριας θεμέλιας ουσίας και τον πολλαπλασιασμό των ουλικών ινοβλαστών, των κυττάρων του περιρριζίου καθώς και των ινοβλαστών που προέρχονται από τους αναπλασμένους ιστούς του περιοδοντίου. Ο ρυθμός του πολλαπλασιασμού φάνηκε να είναι πιο αργός για τα κύτταρα του περιρριζίου. Σημαντική είναι η δράση των IGFs στην απόπτωση των κυττάρων. Συγκεκριμένα, αυτοί οι αυξητικοί παράγοντες παρεμποδίζουν την απόπτωση των κυττάρων και η δράση τους καθορίζεται από τον αριθμό των IGF υποδοχέων που φέρουν στην κυτταρική τους επιφάνεια (Werner και Katz 2004). Η δράση αυτή του IGF-I ισχύει για τα κύτταρα του περιρριζίου αφού η θεραπεία των περιρριζικών κυττάρων με τον αυξητικό παράγοντα αναστέλλει τη σύνθεση του DNA των κυττάρων (Han και Amar 2003a). Οι φυσιολογικοί περιοδοντικοί ιστοί δεν εκφράζουν υποδοχείς των IGFs (Parkar και συν 2001). Τέτοιοι υποδοχείς ανιχνεύθηκαν ανοσοϊστοχημικά στα κύτταρα του περιρριζίου και στις οστεϊνοβλάστες ανθρώπινων νεογιλών δοντιών (Götz και συν. 2001). Ο IGF-I μόνος του δεν επιδρά ουσιαστικά στην επανόρθωση και ανάπλαση των περιοδοντικών ιστών. Αντίθετα, η εφαρμογή του σε συνδυασμό με άλλους αυξητικούς παράγοντες δίνει αισιόδοξα αποτελέσματα. Συγκεκριμένα, η εφαρμογή του σε συνδυασμό με τον PDGF σε σκυλιά Beagle με περιοδοντική νόσο και σε πιθήκους με προκλητή απώλεια πρόσφυσης οδήγησε σε αύξηση της νέας πρόσφυσης κατά 51% και 64% αντίστοιχα, σε σχέση με 8% και 34%
32 32 αντίστοιχα, που προέκυψε μετά από θεραπεία με χειρουργείο και placebo (Giannobile και συν. 1994). Στους ανθρώπους, η συνδυαστική εφαρμογή αυτών των παραγόντων σε περιοδοντικές οστικές βλάβες, οδήγησε σε σημαντική διέγερση της ανάπλασης των περιοδοντικών ιστών (Howell και συν. 1997). Συμπερασματικά, ο IGF-I σε συνδυασμό με τον PDGF φαίνεται να παίζουν σημαντικό ρόλο στην ανάπλαση των περιοδοντικών ιστών. Αυξητικός παράγοντας των ινοβλαστών (FGF) Η οικογένεια των FGFs αποτελείται από 18 γονίδια που κωδικοποιούν πρωτεΐνες οι οποίες είναι δομικά ομόλογες σε επίπεδο αμινοξέων κατά 30%- 80%. Παρ όλο που στην οικογένεια αυτή υπάρχουν διάφορα μέλη (FGF1- FGF18), στους ιστούς των ενηλίκων, τα συνηθέστερα όπως και αυτά πού χρησιμοποιούνται στις διάφορες μελέτες, είναι ο όξινος αυξητικός παράγοντας των ινοβλαστών (FGF1), ο βασικός αυξητικός παράγοντας των ινοβλαστών (FGF2) και ο κερατινοκυτταρικός παράγοντας των ινοβλαστών (FGF7) (Baird και Ornitz 2000). Επίσης, υπάρχουν τέσσερα ξεχωριστά γονίδια τα οποία κωδικοποιούν τους υποδοχείς των FGFs, FGFR1, FGFR2, FGFR3, FGFR4. Οι υποδοχείς αυτοί έχουν κοινή δομή και ανήκουν στην προσομοιάζουσα στην ανοσοσφαιρίνη οικογένεια της τυροσίνης κινάσης. Η αλληλεπίδραση του αυξητικού παράγοντα με τον υποδοχέα του επιπλέκεται από το ρόλο των πρωτεογλυκανών στην δράση των FGFs. Συγκεκριμένα, οι FGFs αλληλεπιδρούν με γλυκοζαμινογλυκάνες τύπου ηπαρίνης. Η αλληλεπίδραση αυτή αποτελεί σημαντική προϋπόθεση στη σύνδεση του παράγοντα με τον υποδοχέα του. Ο εξωκυττάριος FGF προσδένεται στις πρωτεογλυκάνες της θειικής ηπαρίνης στην εξωκυττάρια θεμέλια ουσία και στην κυτταρική επιφάνεια και η πρόσδεση αυτή προστατεύει τον FGF και
33 33 λειτουργεί ως απόθεμα διαθέσιμου FGF σε περίπτωση τραύματος ή όποιας παθοφυσιολογίας που μπορεί να απαιτεί την ταχεία κινητοποίηση της δράσης του αυξητικού παράγοντα (Gospodarowicz και συν. 1986, Vlodavsky και συν. 1991, Prestrelski και συν. 1992). Οι FGFs δρουν κυρίως στα κύτταρα που προέρχονται από το μεσόδερμα όμως η δράση τους εκτείνεται και στα κύτταρα του εκτοδέρματος και του ενδοδέρματος. Η δράση αυτή αφορά στη χημειοταξία, στη διαφοροποίηση και στον πολλαπλασιασμό των κυττάρων και τροποποιείται ανάλογα με τον τύπο των κυττάρων, τις συνθήκες καλλιέργειας και την παρουσία άλλων τοπικών παραγόντων. Παρότι οι αυξητικοί αυτοί παράγοντες ανακαλύφθηκαν από τη δράση τους στις ινοβλάστες (εξόν και το όνομα τους), οι λειτουργίες τους αφορούν μεγάλη ποικιλία κυττάρων (Baird και Klagsburn 1991). Γενικά, οι FGFs έχουν μιτογόνο και χημειοτακτική δράση στα κύτταρα του ενδοθηλίου, στις ινοβλάστες, στα χονδροκύτταρα, στις οστεοβλάστες και στα κύτταρα του περιρριζίου (Terranova και συν. 1989, Howell και συν. 1997). Οι FGFs θεωρούνται σήμερα σημαντικοί αγγειογενετικοί παράγοντες γεγονός που τους προσδίδει ιδιαίτερο ρόλο στην επούλωση των ιστών συμπεριλαμβανομένου και του περιοδοντίου. Υποδοχείς των FGFs εντοπίσθηκαν στα κύτταρα του περιρριζίου αλλά η πυκνότητα τους διαφέρει ανάλογα με την κυτταρική καλλιέργεια (Takayama και συν. 1998). Οι Parkar και συν. (2001) εντόπισαν ανοσοκυτταροχημικά την παρουσία του υποδοχέα FGFR1 μόνο στα κύτταρα των αναπλασμένων περιοδοντικών ιστών και καθόλου στους φυσιολογικούς ιστούς. Οι ίδιοι ερευνητές υπέθεσαν ότι ο FGF μπορεί να δρα στα κύτταρα του περιρριζίου μέσω άλλων FGFs υποδοχέων.
34 34 Οι FGFs παράγονται κυρίως από τις ινοβλάστες και τα ενδοθηλιακά κύτταρα του ανθρώπινου περιρριζίου ενώ παρουσία χρόνιας περιοδοντικής φλεγμονής η συγκέντρωση αυτών μειώνεται (Gao και συν. 1996). Ο βασικός FGF διεγείρει τον πολλαπλασιασμό των περιρριζικών κυττάρων και αναστέλλει τη διαφοροποίηση τους (Okamoto και συν. 1997, Murakami και συν. 1999). Ο βασικός FGF (FGF-2), παρότι μειώνει την έκφραση του mrna των πρωτεϊνών των σχετιζόμενων με το οστούν, φαίνεται να αυξάνει την έκφραση της οστεοποντίνης στα περιρριζικά κύτταρα των ποντικών τόσο σε επίπεδο mrna όσο και σε πρωτεϊνικό επίπεδο (Terashima και συν. 2008). Ο ίδιος αυξητικός παράγοντας καταστέλλει τα μεταγραφικά επίπεδα του κολλαγόνου τύπου Ι και της τροποελαστίνης των ανθρώπινων περιρριζικών κυττάρων, καθώς και την έκφραση του mrna της δεκορίνης με τρόπο εξαρτώμενο από τη δόση εφαρμογής του παράγοντα ενώ αυξάνει την έκφραση του mrna της λαμινίνης, ενός σημαντικού για την αγγειογένεση παράγοντα (Murakami και συν 1999, Palmon και συν. 2001, Wang και συν. 2008). Οι λειτουργίες του FGF-2 στα κύτταρα του περιρριζίου δίνουν το έναυσμα για την περαιτέρω διερεύνηση του ρόλου του στην ανάπλαση των περιοδοντικών ιστών. Σε μια ιστομετρική μελέτη σε σκυλιά η χρήση του βασικού FGF σε μεσορριζικές βλάβες ΙΙΙ βαθμού βελτίωσε την ανάπλαση των περιοδοντικών ιστών (Rossa και συν. 2000). Παρόμοια αποτελέσματα έδωσαν και άλλες μελέτες σε ζώα (πιθήκους και σκυλιά) σε μεσορριζικές βλάβες ΙΙ βαθμού (Murakami και συν. 1999, 2003, Takayama και συν 2001). Ο συνδυασμός του FGF-2 με τους TGF-β 1 και IGF-2 σε οστικές βλάβες στο περιοδόντιο των σκύλων οδήγησε σε μικρότερου βαθμού οστικό σχηματισμό απ ότι στην ομάδα μαρτύρων (Selvig και συν.1994). Παρόλα αυτά ταυτόχρονη χορήγηση
35 35 του FGF-2 με την μορφογενετική πρωτεΐνη του οστού-2 σε μυ αρουραίου οδήγησε σε αυξημένο οστικό σχηματισμό (Fujimura και συν. 2002). Συμπερασματικά, η δράση του FGF-2 στα κύτταρα του περιρριζίου δείχνει θετικά αποτελέσματα στον πολλαπλασιασμό αυτών ενώ ο ρόλος του είναι ασήμαντος στη διαφοροποίηση αυτών των κυττάρων.
36 36 Γ. ΤΡΟΠΟΠΟΙΗΤΙΚΟΣ ΑΥΞΗΤΙΚΟΣ ΠΑΡΑΓΟΝΤΑΣ (TGF-β) Oι TGF-βs είναι πολυλειτουργικά πεπτίδια τα οποία εκφράζονται στα θηλαστικά ως τρεις ομόλογες ισομορφές που καλούνται TGF-β 1, β 2 και β 3. Οι TGF-β 1, β 2 και β 3 εντοπίζονται σε τρία διαφορετικά χρωμοσώματα (ανθρώπινο χρωμόσωμα 19q13, 1q41 και 14q24, αντίστοιχα) και μεταγράφονται ως mrnas, χαρακτηρισμένα από μακριές 3 και 5 - αμετάφραστες περιοχές, γεγονός που συνεπάγεται μεταγραφική ρύθμιση. Ο προαγωγός (promoter) του TGF-β 1 διαφοροποιείται από εκείνους των TGF-β 2 και TGF-β 3 από την απουσία του TATAA box αλλά και από την παρουσία πολλαπλών ρυθμιστικών περιοχών που ενεργοποιούνται μέσω άμεσων πρώιμων γονιδίων όπως c-jun, c-fos και egr-1, αλλά και μέσω διαφόρων ογκογονιδίων όπως abl, fos, jun, ras και src (Roberts και Sporn 1992). Ο προαγωγός διαθέτει διάφορες GC ακολουθίες κοντά στην περιοχή που ξεκινά η μεταγραφή, οι οποίες συμμετέχουν στη ρύθμιση αυτής μέσω των Sp1 και Zf9/CPBP. Η Zf9/CPBP αποτελεί πρωτεΐνη που συμμετέχει στην ανοδική ρύθμιση της έκφρασης του TGF-β 1 και των υποδοχέων του ως απάντηση στο τραύμα (Kim και συν. 1998). Άλλες μεταγραφικές ρυθμιστικές περιοχές περιλαμβάνουν την ΑP-1 περιοχή, η οποία συμμετέχει εμφανώς στην αυτεπαγωγή του TGF-β 1 καθώς και στην ογκογενή επαγωγή του από ενεργοποιημένα jun και fos γονίδια (Kim και συν. 1990) και Egr-1 πλούσιες σε GC περιοχές (Kim και συν. 1994). Αξιοσημείωτο είναι ότι, οι ίδιες περιοχές
37 37 προκαλούν τη δράση μιας ποικιλίας ιογενών τρανσενεργοποιών πρωτεϊνών στην μεταγραφή του TGF-β 1. Τα χαρακτηριστικά αυτά του προαγωγού του TGF-β 1 παρέχουν τη βάση εκείνη που οδηγεί στην επιλεκτική υπερέκφραση αυτή της ισομορφής όπως αυτή παρατηρείται σε περίπτωση αποκατάστασης τραύματος, σε απάντηση στο stress, στη παθογένεια νόσων ιογενούς προέλευσης, καθώς και στη καρκινογένεση. Οι προαγωγοί των TGF-β 2 και 3 διαφέρουν από αυτούς του TGF-β 1 ενώ είναι παρόμοιοι μεταξύ τους. Ο καθένας χαρακτηρίζεται από την παρουσία του ΤΑΤΑ κουτιού. Όσον αφορά στο TGF-β 3, έχει περιγραφεί ένας νέος μηχανισμός ρύθμισης της μεταγραφής του μέσω της 17-β οιστραδιόλης και ενός συνθετικού τροποποιητή της ραλοξιφένης (Yang και συν. 1996). Αυτή η ενεργοποίηση φαίνεται να συμβάλει στη δράση των οιστρογόνων στη διατήρηση του οστού. Αυξητική ρύθμιση των διαφόρων ισομορφών του TGF-β γίνεται και από τα ρετινοειδή, τη Vitamin D και το αντιοιστρογόνο ταμοξιφένη. Η ρύθμιση αυτή γίνεται μέσω μεταγραφικών μηχανισμών (Roberts και συν. 1992). Τα πολυπεπτίδια αυτά παράγονται από διάφορους τύπους κυττάρων, κυρίως από τα αιμοπετάλια, τις οστεοβλάστες και τα μακροφάγα. Οι TGF-β βρίσκονται σε λανθάνουσα μορφή στα κύτταρα και ενεργοποιούνται μέσω διαφόρων μηχανισμών (Schultz-Cherry και συν.1995, Munger και συν. 1997, 1999). Η ενεργοποίηση της λανθάνουσας μορφής του TGF-β έχει μελετηθεί εκτενώς in vitro και λιγότερο in vivo. Οι περισσότερες in vitro μελέτες έχουν εστιάσει στη μη ενζυματική ενεργοποίηση μέσω έντονων αλλαγών του ph, της
38 38 θερμοκρασίας, ή μέσω χαοτροπικών παραγόντων, οι οποίοι διακόπτουν τη σύνδεση μεταξύ του LAP και του ώριμου TGF-β, καθώς και μέσω πρωτεολυτικών μηχανισμών. Το τελευταίο, έχει ιδιαίτερα μελετηθεί σε ενεργοποιημένα μακροφάγα καλλιεργημένα in vitro καθώς και σε συγκαλλιέργειες (co-culture) ενδοθηλιακών και λείων μυϊκών κυττάρων (Munger και συν. 1997). Ένας άλλος μηχανισμός ενεργοποίησης του TGF-β αφορά στη σύνδεση του με τον υποδοχέα της ανβ6 ιντεγκρίνης μέσω μιας RGD αλληλουχίας (Munger και συν. 1999). Επίσης, η ενεργοποίηση του TGF-β φαίνεται να πραγματοποιείται μέσω μίας, ανεξάρτητης από πρωτεάση, αλλαγής του λανθάνοντος συμπλέγματος που προκύπτει από τη σύνδεση του στη θρομβοποντίνη, ενός στοιχείου των α- κοκκίων των αιμοπεταλίων και της εξωκυττάριας θεμέλιας ουσίας (Schultz- Cherry και συν. 1995). Η μοριακή βάση του τρόπου δράσης των TGF-β στα κύτταρα φαίνεται να έχει γίνει πλέον κατανοητή μέσω των υποδοχέων και των μονοπατιών μεταβίβασης του εξωκυττάριου σήματος. Έτσι, μετά την ενεργοποίηση των λανθανόντων μορφών του TGF-β, το ώριμο και ενεργό διμερές TGF-β συνδέεται στην κυτταρική επιφάνεια με τους TGF-β υποδοχείς για να προκαλέσει διάφορες κυτταρικές απαντήσεις όπως αλλαγές στη κυτταρική διαφοροποίηση, στον πολλαπλασιασμό και στη γονιδιακή έκφραση. Ο TGF-β υποδοχέας είναι ένα ετεροτετραμερές σύμπλεγμα αποτελούμενο από δύο τύπου ΙΙ και δύο τύπου Ι υποδοχείς (Derynck και Feng 1997, Heldin και συν. 1997, Massagué 1998). Οι τύπου ΙΙ και Ι υποδοχείς είναι δομικά σχετιζόμενες διαμεμβρανικές κινάσες σερίνης/θρεονίνης, οι οποίες υφίστανται
39 39 ως ομοδιμερή πριν τη σύνδεση τους με το συνδέτη (ligand). H ετερομερής αυτή διασύνδεση μεταξύ των διμερών υποδοχέων τύπου Ι και ΙΙ σταθεροποιείται με τη σύνδεση του TGF-β δημιουργώντας έτσι ένα λειτουργικό ετεροτετραμερές σύμπλεγμα. Συγκεκριμένα, ένας άλλος υποδοχέας, ο υποδοχέας τύπου ΙΙΙ μεσολαβεί για τη πρόσδεση του TGF-β στον υποδοχέα τύπουιι (Franzen και συν.1995). Στη συνέχεια, ο υποδοχέας τύπου ΙΙ φωσφορυλιώνεται ενεργοποιώντας έτσι τον υποδοχέα τύπου Ι, ο οποίος στη συνέχεια φωσφορυλιώνει άλλους ειδικούς παράγοντες (effectors). Σε αντίθεση με τον υποδοχέα ΙΙ όπου έχει ταυτοποιηθεί μόνο ένας, διάφοροι υποδοχείς τύπου Ι έχουν βρεθεί. Οι πιο συνήθεις είναι ο TβRΙ ή αλλιώς ALK-5 (activin receptor-like kinase) ενώ άλλοι τέτοιοι υποδοχείς είναι ο TSR-1/ALK-1 και ο Tsk7L/ActRI/ALK-2 (Derynck και συν. 1998). Παρόλο που διάφοροι επιπρόσθετοι μηχανισμοί έχουν αναφερθεί για τη μεταβίβαση του σήματος πιο γνωστοί διαμεσολαβητές της σηματοδότησης του TGF-β είναι οι Smads. Οι Smads είναι ενδοκυττάριες πρωτεΐνες που ενεργοποιούν τη μεταγραφή (Derynck και συν. 1997, Heldin και συν. 1997, Whitman 1998, Massagué 1998). Η κυτταρική βιολογία του TGF-β Γενικά η δράση του TGF-β στον οργανισμό μπορεί να είναι αυτοκρινική, παρακρινική και ενδοκρινική. Ο TGF-β παρουσιάζει μοναδική κυτταρική επίδραση, εμποδίζοντας την ανάπτυξη των περισσοτέρων επιθηλιακών, ενδοθηλιακών και αιμοποιητικών κυττάρων ή ανταγωνίζοντας τη δράση άλλων αυξητικών παραγόντων όπως του EGF ή του PDGF (Roberts και Sporn 1990).
40 40 Αξίζει να σημειωθεί, ωστόσο, ότι ενώ η μοριακή βάση του τρόπου δράσης του TGF-β, των υποδοχέων και μονοπατιών μεταβίβασης του εξωκυττάριου σήματος είναι κατανοητή, το εξωκυττάριο έναυσμα δεν οδηγεί στη συγκεκριμένη, ίδια πάντα, κυτταρική απάντηση σε όλα τα κύτταρα. Γενικά η δράση του TGF-β αφορά κυρίως στη χημειοταξία, στην κυτταρική ανάπτυξη, διαφοροποίηση και λειτουργία (Roberts και Sporn 1990). Ο TGF-β διεγείρει τη χημειοταξία πολλών κυτταρικών τύπων όπως ινοβλαστών, μακροφάγων, λεμφοκυττάρων και ουδετερόφιλων. Το αποτέλεσμα αυτό δικαιολογεί τη δράση του TGF-β στην επανόρθωση των ιστών τόσο στη φλεγμονή όσο και στην ίνωση (McCartney-Francis και συν. 1994, Roberts 1995). Η επίδραση του TGF-β στα μεσεγχυματικά κύτταρα αφορά στη ρύθμιση της δραστηριότητας των γονιδίων στόχων παρά στην αυτή καθαυτή ανάπτυξη τους. Τόσο στην επανόρθωση των ιστών, όσο και στην ίνωση, πολλές από τις δράσεις του TGF-β αφορούν στη ρύθμιση της παραγωγής της εξωκυττάριας ουσίας από τα κύτταρα της θεμέλιας ουσίας (stromal cells). Η ρύθμιση αυτή σχετίζεται με: α) την αύξηση της έκφρασης διαφόρων πρωτεϊνών όπως της φιμπρονεκτίνης, διαφόρων τύπων κολλαγόνου και των πρωτεϊνών δεκορίνης και διγλυκάνης, β) τη μείωση της έκφρασης και της δραστηριότητας του ενεργοποιητή του πλασμινογόνου, της κολλαγενάσης, της ελαστάσης και της στρομολυσίνης, γ) τη διέγερση της έκφρασης των αναστολέων των μεταλλοπρωτεασών όπως του ΡΑΙ-Ι και TIMP-I και δ) τη ρύθμισης της έκφρασης της ιντεγκρίνης και της ικανότητας των κυττάρων να αλληλοεπιδρούν με ειδικές πρωτεΐνες της θεμέλιας ουσίας (Border και συν. 1994, Roberts 1995).
41 41 Η δράση τoυ TGF-β στα οστικά κύτταρα Στο οστούν, οι TGF-β εντοπίζονται στα οστικά κύτταρα και ιστούς, συμπεριλαμβανομένων των οστεοβλαστών, του περιχονδρίου και των περιοχών σχηματισμού ενδομεμβρανώδους οστού. Επίσης, έχει αναφερθεί ότι και οι οστεοκλάστες εκφράζουν TGF-β (κυρίως TGF-β 1 ) γεγονός που υποθέτει την επίδραση του TGF-β στην οστεοκλαστική δραστηριότητα (Sandberg και συν. 1988). Οι υποδοχείς των TGF-β εντοπίζονται, επίσης, σε περιοχές ενδομεμβρανώδους οστικού σχηματισμού και στο περιόστεο και ενδόστεο των αναπτυσσόμενων ενδοχονδρικών οστών, γεγονός που υποδηλώνει τον αυτοκρινικό ρόλο του TGF-β στις προοστεοβλάστες και οστεοβλάστες (Horner και συν. 1998, Kabasawa και συν. 1998). Ο TGF- β λειτουργεί ως χημειοτακτικός παράγοντας για τα οστικά κύτταρα, αυξάνει την παραγωγή της θεμέλιας ουσίας και αυξάνει ή μειώνει τον πολλαπλασιασμό τους ανάλογα με το στάδιο διαφοροποίησης των κυττάρων, τη συγκέντρωση του αυξητικού παράγοντα και τις συνθήκες καλλιέργειας (Cochran και Wozney 1999). Όσον αφορά στη διαφοροποίηση των προ-οστεοβλαστών και οστεοβλαστών υπό την επίδραση του TGF-β, το αποτέλεσμα της δράσης του παράγοντα εξαρτάται από τις συνθήκες καλλιέργειας και το βαθμό διαφοροποίησης των κυττάρων. Σε καλλιέργειες μυελού των οστών, ο TGF-β οδηγεί σε αύξηση της αλκαλικής φωσφατάσης, οστεονεκτίνης και οστεοκαλσίνης, γεγονός που υποδηλώνει τη διεγερτική δράση του TGF-β στην οστεοβλαστική διαφοροποίηση (Long και συν. 1995). Παρόλα αυτά, πολλές μελέτες
42 42 αναφέρονται στον TGF-β ως ανασταλτικού παράγοντα στη διαφοροποίηση των οστεοβλαστών (Centrella και συν. 1987, Centrella και συν. 1988, Rosen και συν. 1988, Guenther HL και συν. 1988, Ten Dijke P και συν. 1990). Επίσης, ενώ ο TGF-β αναστέλλει τη δημιουργία και ενασβεστίωση οζιδίων (nodule), διεγείρει την έκφραση κολλαγόνου Ι, οστεοποντίνης, οστεονεκτίνης, βασικών πρωτεϊνών της εξωκυττάριας θεμέλιας ουσίας που συνθέτουν την οστική θεμέλια ουσία (Centrella και συν. 1987, Noda και συν. 1987, Centrella και συν. 1988, Wrana και συν. 1988, Rosen και συν. 1988, Noda και συν. 1988). Συμπερασματικά, παρατηρείται ότι ο TGF-β διεγείρει τον κυτταρικό πολλαπλασιασμό και τα πρώτα στάδια διαφοροποίησης των οστεοβλαστών ενώ αναστέλλει τα πιο προχωρημένα στάδια διαφοροποίησης. Ο TGF-β φαίνεται να λειτουργεί ρυθμιστικά και στην οστεοκλαστική λειτουργία (Centrella και συν. 1994). Οι οστεοκλάστες είναι κύτταρα που προέρχονται από τη μονοκύτταρη φαγοκυτταρική σειρά σε αντίθεση με της μεσεγχυματικής προέλευσης οστεοβλάστες. Τα κύτταρα αυτά, συμμετέχουν στην οστική απορρόφηση εκκρίνοντας διάφορα λυσοσωμικά ένζυμα. Το όξινο μικροπεριβάλλον που δημιουργείται έχει ως αποτέλεσμα την ενεργοποίηση του TGF-β που βρίσκεται σε λανθάνουσα μορφή στην οστική θεμέλια ουσία (Oreffo και συν. 1989, Oursler 1994). Οι έρευνες γύρω από τη δράση του TGF-β στην οστεοκλαστική διαφοροποίηση δείχνουν τόσο διεγερτικά όσο και ανασταλτικά φαινόμενα. Η δράση του παράγοντα αυτού τροποποιείται από τη δόση εφαρμογής του (Tashjian και συν. 1985, Pfeilschifter και συν. 1988, Shinar και συν. 1990). Σε χαμηλά επίπεδα ο TGF-β διεγείρει την οστεοκλαστική διαφοροποίηση
43 43 γεγονός που συσχετίζεται και με την παρατήρηση ότι χαμηλές δόσεις TGF-β διεγείρουν την παραγωγή προσταγλανδινών, πιθανότατα μέσω των μεσεγχυματικών κυττάρων. Με αυτόν τον τρόπο, οι προσταγλανδίνες επάγουν την οστεοκλαστική διαφοροποίηση. Έτσι, η διεγερτική δράση του TGF-β σε χαμηλές δόσεις μπορεί να οφείλεται στην επαγωγή των προσταγλανδινών από τα μεσεγχυματικά κύτταρα, χωρίς να αναστέλλεται η μιτογένεση και η διαφοροποίηση των αιμοποιητικών πρόδρομων κυττάρων (Tashjian και συν. 1985, Shinar και συν. 1990, Marcelli και συν. 1990, Dieudonné και συν. 1991). Αντίθετα, ο TGF-β σε υψηλές δόσεις μπορεί άμεσα να αναστείλλει τον πολλαπλασιασμό και τη διαφοροποίηση των πρόδρομων οστεοκλαστών (Tashjian και συν. 1985, Pfeilschifter και συν. 1988, Shinar και συν. 1990). Η παρουσία των οστεοβλαστών ή των μεσεγχυματικών κυττάρων και η απάντησή τους στο TGF-β λειτουργούν καθοριστικά στη δράση του TGF-β στη διαφοροποίηση των οστεοκλαστών (Hattersley και συν. 1991). Περισσότερες πληροφορίες γύρω από το μηχανισμό της κυτταρικής διασταύρωσης (crosstalk) μεταξύ οστεοβλαστών και οστεοκλαστών αλλά και της ρύθμισης αυτών από το TGF-β δόθηκαν με την ταυτοποίηση μιας νέας διαμεμβρανικής κυτταροκίνης και του υποδοχέα της (Fivaroff και συν. 1998, Suda και συν. 1999). Η κυτταροκίνη αυτή, που έχει ονομαστεί σύνδεσμος του υποδοχέα ενεργοποίησης του NFkβ (RANKL), είναι μέλος της οικογένειας των TNF των διαμεμβρανικών συνδέσμων και επάγει την οστεοκλαστική διαφοροποίηση και δράση (Yasuda και συν. 1998, Lacey και συν. 1998). Διάφορες μελέτες ανακάλυψαν κάποια σχέση μεταξύ του συνδέσμου του υποδοχέα ενεργοποίησης του NFkβ (RANKL) και της ικανότητας του TGF-β
44 44 να αναστέλλει την οστεοκλαστική διαφοροποίηση. Ο RANKL εκφράζεται στις οστεοβλάστες και τα κύτταρα της θεμέλιας ουσίας και συνδέεται και ενεργοποιεί έναν υποδοχέα, το RANK, ο οποίος εκφράζεται στα πρόδρομα οστεοκλαστικά κύτταρα και στις οστεοκλάστες (Suda και συν. 1999). Η ενεργοποίηση του RANK από το RANKL εξηγεί το φαινόμενο της διαφοροποίησης των οστεοκλαστών σε συγκαλλιέργειες οστεοβλαστώνοστεοκλαστών. Η δράση του RANKL ρυθμίζεται επιπλέον από την οστεοπροτεγερίνη (OPG) ή παράγοντα αναστολής οστεοκλαστών (OCIF) (Simonet και συν. 1997, Yasuda και συν. 1998). Η OPG είναι ένας διαλυτός υποδοχέας των οστεοβλαστών ο οποίος επίσης συνδέεται στο RANKL και με αυτόν τον τρόπο εμποδίζει την ένωση RANK-RANKL και κατά συνέπεια την οστεοκλαστική διαφοροποίηση. Συγκεκριμένα, ο TGF-β φάνηκε να επάγει την έκφραση και έκκριση της OPG στις οστεοβλάστες και στα κύτταρα του μυελού των οστών (Takai και συν. 1998, Murakami και συν. 1998) και να αναστέλλει την έκφραση του RANKL (Takai και συν. 1998). Η διπλή αυτή δράση του TGF-β μπορεί να εξηγήσει την ανασταλτική δράση του TGF-β στην διαφοροποίηση και δράση των οστεοκλαστών. Ο ρόλος του TGF-β στην οστική ανάπλαση και διατήρηση Ο TGF-β λειτουργεί αναβολικά στο σχηματισμό του οστού. Έγχυση TGF-β 1 ή TGF-β 2 υποπεριοστικά στο κρανίο ποντικών διέγειρε το σχηματισμό οστού από το περιόστεο και αύξησε το οστικό πάχος (Noda και συν. 1989, Marcelli και συν. 1990). Επιπλέον συστηματική χορήγηση TGF-β, για μικρό διάστημα αύξησε το ρυθμό οστικού σχηματισμού και ενασβεστίωσης σε αρουραίους (Rosen και συν. 1994). Προφανώς, ο TGF-β προάγει τον οστικό σχηματισμό αυξάνοντας την πηγή των πρόδρομων οστικών κυττάρων, όπως και τον
45 45 αριθμό των διαφοροποιημένων οστεοβλαστών (Mundy και συν. 1995). Με την ίδια λογική, έγχυση του TGF-β σε χονδρογενή περιοχή οδηγεί σε σχηματισμό χόνδρου. Ο TGF-β σε αντίθεση με τις BMPs 2 και 4 αδυνατεί να επάγει στο σχηματισμό εξωγενούς χόνδρου ή οστίτη ιστού (Centrella και συν. 1994), γεγονός που καθιστά αναγκαία την ύπαρξη χονδρογενούς ή οστεογενούς περιβάλλοντος. Τα αποτελέσματα της τοπικής εφαρμογής του TGF-β δίδουν πληροφορίες για το ρόλο του ενδογενούς TGF-β στην οστική επανόρθωση. Έτσι, σε περιοχές κατάγματος αυξάνεται η έκφραση του TGF-β 1 και ενεργοποιούνται οι ανενεργές μορφές του (Sandberg και συν. 1993), ενώ επιπλέον TGF-β ελευθερώνεται από τα αιμοπετάλια στην περιοχή του κατάγματος (Bolander 1992). Γενικά, ο TGF-β φαίνεται ότι επιταχύνει την οστική επανόρθωση (Beck και συν. 1991, Nielsen και συν. 1994). Οι διαγονιδιακές μελέτες που βασίζονται στην υπερέκφραση του TGF- β 2 στις οστεοβλάστες ή στην υπερέκφραση ενός TGF-β υποδοχέα ο οποίος επιδρά με επικρατή αρνητικό τρόπο στην ενδογενή σηματοδότηση και των τριών ισομορφών TGF-β, αλλά και η μελέτη του οστικού φαινοτύπου ποντικών χωρίς TGF- β 1, αποκάλυψαν το ρόλο του TGF-β στον οστικό μεταβολισμό. Συγκεκριμένα, ο TGF-β που ελευθερώνεται από τις οστεοβλάστες δρα ρυθμιστικά, όχι μόνο σε αυτά τα κύτταρα, αλλά και στις οστεοκλάστες. Η οστεοκλαστική δραστηριότητα καθορίζεται σε ένα βαθμό από την οστεοβλαστική απάντηση στο TGF-β, υποθέτοντας έτσι ότι ο TGF-β λειτουργεί ως παράγοντας σύζευξης μεταξύ των δύο κυτταρικών τύπων. Λαμβάνοντας υπόψη τη σημασία της ισορροπίας μεταξύ οστικής εναπόθεσης απορρόφησης, γίνεται κατανοητή η σημασία του TGF-β στην αναδόμηση
46 46 του οστού. Μικρές αλλαγές στο επίπεδο ενεργού TGF-β ή στην οστεοβλαστική απάντηση στο TGF-β μπορεί να διαταράξει αυτή την ισορροπία και να οδηγήσει σε φαινοτυπικές αλλαγές που μπορεί να εκδηλωθούν ως οστεοπορωτικές ή οστεοπετρωτικές (Erlebacher και συν. 1996, Filvaroff και συν. 1999). Ο ρόλος του TGF-β στις μεταβολικές νόσους ακόμα διερευνάται. In vitro μελέτες δείχνουν ότι η έκφραση του TGF-β από τις οστεοβλάστες επάγεται από τα οιστρογόνα, ενώ ο TGF-β λειτουργεί ως μεσολαβητής στην αποπτωτική δράση των οιστρογόνων στις οστεοκλάστες. Άλλες μελέτες αναφέρουν μείωση του TGF-β στην οστική μάζα αρουραίων με ωοθηκεκτομή ή μεταεμμηνοπαυσιακών γυναικών. Σε πολλές από αυτές τις έρευνες, απώλεια του TGF-β συσχετίσθηκε με μείωση της οστικής μάζας, ενώ χορήγηση οιστρογόνων ανέτρεψε αυτή την εικόνα και αύξησε τα επίπεδα του TGF-β (Robinson και συν. 1996, Yang και συν. 1996, Hughes και συν. 1996). Παρόλο που αυτές οι έρευνες προτείνουν κάποιον πιθανό ρόλο του TGF-β στην οστεοπόρωση, περισσότερη έρευνα απαιτείται για να αποδειχθεί άμεση σχέση μεταξύ TGF-β και οστεοπόρωσης. Η έκφραση του TGF-β και των υποδοχέων του στα κύτταρα των περιοδοντικών ιστών Τα τελευταία χρόνια, πολλές in vitro και in vivo μελέτες έχουν ασχοληθεί με τη συμβολή των αυξητικών παραγόντων γενικά αλλά και του TGF- β στην επούλωση και ανάπλαση των περιοδοντικών ιστών (Lynch και συν. 1991, Oates και συν. 1993, Caffesse και Quinones 1993, Takayama και συν. 1997). Ο TGF- β, όπως αναφέρθηκε, σχετίζεται με διάφορες λειτουργίες των κυττάρων που περιλαμβάνουν τη ρύθμιση της κυτταρικής λειτουργίας και την
47 47 τροποποίηση της εξωκυττάριας ουσίας (Roberts 1995). Το γεγονός αυτό, οδήγησε στη μελέτη της δράσης του TGF- β στα κύτταρα του περιοδοντίου και στην επούλωση και ανάπλαση των περιοδοντικών ιστών. Σε μελέτη σε αρουραίους Lewis διαφορετικής ηλικίας ο TGF- β 1 βρέθηκε να εκφράζεται σε όλα τα τμήματα των περιοδοντικών ιστών (φατνιακό οστούν, περιρρίζιο, οστεΐνη) στα διάφορα στάδια ανάπτυξης που ερευνήθηκαν. Επίσης, mrna του TGF-β 1 εκφράστηκε από τις οστεοβλάστες, μερικά κύτταρα του μυελού των οστών και τις ινοβλάστες (Gao και συν. 1998). Το εύρημα αυτό, πιθανά δηλώνει το ρόλο του TGF-β 1 στην ανάπτυξη του περιοδοντίου. Οι Gao J και συν. (1999) μελετώντας το περιοδόντιο των αρουραίων εντόπισαν την ύπαρξη των υποδοχέων τύπου ΙΙ και ΙΙΙ του TGF-β στην κυτταρική μεμβράνη και το κυτταρόπλασμα των περιρριζικών ινοβλαστών, των οστεϊνοβλαστών και των οστεοβλαστών. Συγκεκριμένα, ο υποδοχέας τύπου ΙΙ εντοπίσθηκε στα κύτταρα του περιρριζίου σε μεγαλύτερο ποσοστό από τον υποδοχέα τύπου ΙΙΙ ο οποίος φάνηκε να επικρατεί στην εξωκυττάρια θεμέλια ουσία. Οι Parkar και συν. (2001), διαπίστωσαν ανοσοκυτταροχημικά ότι ο υποδοχέας τύπου Ι εκφράζεται στο 50% των κυττάρων του περιρριζίου και των ουλικών ινοβλαστών, η έκφραση αυτή όμως ήταν εντονότερη για τις ουλικές ινοβλάστες. Αντίθετα, παρόλο που ο υποδοχέας τύπου ΙΙ εκφράσθηκε στον ίδιο βαθμό και στους δύο τύπους κυττάρων, η έκφραση του εντοπίσθηκε μόνο στο 18% των κυττάρων του περιρριζίου σε σχέση με το 50% των ουλικών ινοβλαστών. Επιπλέον, οι TGFβ υποδοχείς εκφράσθηκαν αδύναμα στους φυσιολογικούς περιοδοντικούς ιστούς ενώ η έκφραση τους ήταν πολύ πιο έντονη στους αναπλασμένους περιοδοντικούς ιστούς. Η παρουσία των υποδοχέων Ι και ΙΙ στα περιρριζικά
48 48 κύτταρα και στις ουλικές ινοβλάστες επιβεβαιώθηκε και από μία μεταγενέστερη μελέτη όπου όμως το ποσοστό των κυττάρων που έφεραν τέτοιους υποδοχείς ήταν σαφώς μικρότερο (Yashiro και συν. 2006). Τα m-rna των TGF-β 1 και TGF-β 2 αλλά όχι του TGF-β 3 καθώς και τα m-rna των τριών υποδοχέων του TGF-β εκφράζονται στα κύτταρα του στοματικού και του προσπεφυκότος επιθηλίου. Η έκφραση του TGF-β 1 καθώς και του υποδοχέα τύπου ΙΙΙ είναι παρόμοια και στους δύο τύπους του επιθηλίου ενώ ο TGF-β2 και ο υποδοχέας τύπου Ι εκφράζεται περισσότερο στο στοματικό επιθήλιο. Στα κύτταρα του προσπεφυκότος επιθηλίου εκφράσθηκε ο υποδοχέας τύπου ΙΙ περισσότερο από ότι στα κύτταρα του στοματικού επιθηλίου (Lu και συν. 1997). Ο TGF-β 1 δεν ανιχνεύεται στα επιθηλιακά κύτταρα των φλεγμονωδών ούλων. Αντίθετα ο TGF-β 3 είναι παρόν σε όλες τις περιοχές των φλεγμονωδών κυττάρων ενώ η συγκέντρωση του TGF-β 2 μειώνεται σταδιακά από το στοματικό επιθήλιο προς το επιθήλιο του περιοδοντικού θυλάκου. Διαφορετική μεταξύ τους παρουσιάζεται και η κατανομή των τριών υποδοχέων του TGF-β στα επιθηλιακά κύτταρα των φλεγμονωδών περιοδοντικών ιστών (Ye και συν. 2003). Η έκφραση των τριών ισομορφών του TGF-β καθώς και των υποδοχέων του τροποποιείται ανάλογα με το στάδιο διαφοροποίησης των κυττάρων του περιρριζίου των αρουραίων. Συγκεκριμένα, η έκφραση του mrna του TGF-β 1 αυξάνεται κατά τη διάρκεια της εξέλιξης της διαφοροποίησης των κυττάρων του περιρριζίου, του TGF-β 3 μειώνεται ελαφρώς ενώ ο TGF-β 2 δεν εκφράζεται σε κανένα στάδιο (Chien και συν. 1999). Η έκφραση του mrna του υποδοχέα τύπου Ι μειώνεται με την εξέλιξη της κυτταρικής διαφοροποίησης, του τύπου ΙΙΙ αυξάνεται ενώ του τύπου ΙΙ παραμένει σταθερά υψηλή σε όλη τη διάρκεια
49 49 της εξέλιξης της διαφοροποίησης των περιρριζικών κυττάρων. Ο TGF-β 1 έχει ανιχνευθεί στο ουλικό υγρό ενώ φάνηκε, ότι η έκφρασή του, αυξάνεται μετά από τη χειρουργική του περιοδοντίου (Buduneli και συν. 2001, Kuru και συν. 2004). Η δράση του TGF-β 1 στους περιοδοντικούς ιστούς Σε in vitro μελέτες, ο TGF-β 1 φαίνεται να δρα στον πολλαπλασιασμό των κυττάρων του περιρριζίου (Dennison και συν. 1994, Markopoulou και συν. 2003, Fujita και συν. 2004). Ο παράγοντας αυτός, μόνος του, ή σε συνδυασμό με το PDGF, επιδρά θετικά στον πολλαπλασιασμό των κυττάρων του περιρριζίου και μάλιστα περισσότερο από ότι στις ουλικές ινοβλάστες (Dennison και συν. 1994). Παρόμοια, σε άλλη in vitro μελέτη η εφαρμογή 10ng/ml TGF-β 1 μόνου του ή σε συνδυασμό με τους PDGF-BB και IGF-1 σε κύτταρα του ανθρώπινου περιρριζίου διέγειρε τον πολλαπλασιασμό τους ενώ ο συνδυασμός των παραγόντων αύξησε και την προσκόλληση τους σε περιοδοντικά προσβεβλημένες επιφάνειες ριζών που είχαν υποβληθεί σε ριζική απόξεση και ταυτόχρονη εφαρμογή διαλύματος κιτρικού οξέος και τετρακυκλίνης (Sant Ana και συν. 2007). Παρόμοια, η χορήγηση του TGF-β 1 στα κύτταρα του ανθρώπινου περιρριζίου σε δόση 1ng/ml για 48 ώρες διέγειρε τον πολλαπλασιασμό τους (Oates και συν. 1993). Οι Silverio-Ruiz και συν. (2007) αναφέρουν αύξηση του αριθμού των περιρριζικών κυττάρων ύστερα από εφαρμογή 1ng/ml TGF-β 1 και 10ng/ml TGF-β 1 για 24 και 48 ώρες με τρόπο που εξαρτάται από το χρόνο και τη δόση χορήγησης του παράγοντα. Η μιτογόνος δράση του TGF-β 1 δεν παρατηρείται σε όλες τις περιπτώσεις, ενώ ο βαθμός διέγερσης των κυττάρων από τον παράγοντα αυτό ποικίλλει
50 50 (Markopoulou και συν. 2003). Στα πλαίσια της ανάπλασης του περιρριζίου βρέθηκε ότι παράγωγα της αμελογενίνης (EMD) σε δόση 50 mg/ml, διεγείρουν την έκφραση TGF-β 1 και IGF-1 στα κύτταρα του περιρριζίου σε επίπεδο πρωτεΐνης και mrna. Συγκεκριμένα, καλλιέργεια των κυττάρων αυτών για 48 ώρες με EMD (στην αυτή συγκέντρωση) οδήγησε σε αύξηση της έκκρισης TGF-β 1 από 110,7 σε 123,4 pg/ml. Επίσης, στην ίδια μελέτη παρατηρήθηκε αύξηση της σύνθεσης του DNA με την εξωγενή προσθήκη TGF-β 1 με τρόπο εξαρτώμενο από τη δόση εφαρμογής του παράγοντα. Το γεγονός αυτό έδειξε ότι η διεγερτική δράση του EMD στο πολλαπλασιασμό των κυττάρων μπορεί να πραγματοποιηθεί, σε ένα βαθμό, μέσω της αυξημένης από τα κύτταρα παραγωγής TGF-β 1 και IGF I (Οkubo και συν. 2003). Γενικά, η διέγερση του πολλαπλασιασμού των ανθρώπινων περιρριζικών κυττάρων από τον TGF-β 1 επηρεάζεται από το χρόνο και τη δόση εφαρμογής, ενώ μελέτη σε περιρρίζιο αρουραίων έδειξε αναστολή της μιτογένεσης (Matsuda και συν. 1992). Ο TGF-β 1 φαίνεται να αναστέλλει τη δραστηριότητα της αλκαλικής φωσφατάσης στα κύτταρα του περιρριζίου όπως και στα ανθρώπινα πολφικά κύτταρα (Brady και συν. 1998, Shiba και συν. 1998, Fujita και συν. 2004). Στις οστεϊνοβλάστες, ο TGF-β 1 διεγείρει τη σύνθεση του DNA, αναστέλλει την έκφραση της οστεοκαλσίνης και της οστικής σιαλοπρωτεΐνης ενώ η συνεχής χορήγηση του στα κύτταρα αναστέλλει την διαδικασία της ενασβεστίωσης (Saugin και συν.2000). Τόσο στην επανόρθωση των ιστών, όσο και στην ίνωση, πολλές από τις δράσεις του TGF-β αφορούν στη ρύθμιση της παραγωγής της εξωκυττάριας ουσίας από τα κύτταρα της θεμέλιας ουσίας (stromal cells). Η ρύθμιση αυτή
51 51 σχετίζεται με: α) την αύξηση της έκφρασης διαφόρων πρωτεϊνών όπως της φιμπρονεκτίνης, διαφόρων τύπων κολλαγόνου κυρίως του τύπου Ι και των πρωτεϊνών δεκορίνης και διγλυκάνης, β) τη μείωση της έκφρασης και της δραστηριότητας του ενεργοποιητή του πλασμινογόνου, της κολλαγενάσης, της ελαστάσης και της στρομολυσίνης, γ) τη διέγερση της έκφρασης των αναστολέων των μεταλλοπρωτεϊνασών όπως του ΡΑΙ-Ι και TIMP-I και δ) τη ρύθμισης της έκφρασης της ιντεγκρίνης και έτσι της ικανότητας των κυττάρων να αλληλοδρούν με ειδικές πρωτεΐνες της θεμέλιας ουσίας (Border και συν. 1994, Roberts 1995). Στα κύτταρα του ανθρώπινου περιρριζίου ο TGF-β αυξάνει την έκφραση του mrna για το κολλαγόνο τύπου Ι και ΙΙΙ και για τους αναστολείς των μεταλλοπρωτεϊνασών (TIMPs) ενώ αναστέλλει την έκφραση του mrna για τις μεταλλοπρωτεϊνάσες-1 και - 2 (Silverio-Ruiz και συν. 2007). Ο TGF-β 1 αυξάνει, επίσης, τα επίπεδα της SPARK και της φιμπρονεκτίνης στα κύτταρα του περιρριζίου (Fujita και συν.2004). Η SPARK είναι πρωτεΐνη που εντοπίζεται στο περιρρίζιο και ρυθμίζει το πολλαπλασιασμό και τη σύνθεση πρωτεϊνών σχετιζόμενων με το οστούν σε καλλιέργεια περιρριζικών κυττάρων (Fujita και συν. 2002α). Η πρωτεΐνη αυτή, κατέχει διάφορους ρόλους στην εκκίνηση της ενασβεστίωσης σε διάφορα πειραματικά μοντέλα και σχετίζεται με την ανάπλαση των περιοδοντικών ιστών (Takano-Yamamoto και συν. 1994, Matsura και συν. 1995). Η φιμπρονεκτίνη, από την άλλη, είναι ένα από τα βασικά στοιχεία της εξωκυττάριας ουσίας και εκφράζεται στο περιρρίζιο των σκύλων κατά την περιοδοντική ανάπλαση (Matsura και συν. 1995, Amar 1996).
52 52 Ο TGF-β1 εμπλέκεται στην παθογένεια της κληρονομικής ινωμάτωσης των ούλων και της υπερτροφίας αυτών από τη χρήση φαρμάκων. Η χορήγηση της κυκλοσπορίνης Α, η οποία ενοχοποιείται για την πρόκληση υπερτροφίας των ούλων, στα κύτταρα ανθρώπινων ούλων και ούλων αρουραίων αυξάνει την έκφραση του TGF-β 1 τόσο σε επίπεδο mrna όσο και σε πρωτεϊνικό επίπεδο. Η παρατήρηση αυτή οδηγεί στο συμπέρασμα ότι η δράση της κυκλοσπορίνης Α στα ούλα μπορεί να πραγματοποιείται μέσω της ρύθμισης της έκφρασης του TGF-β1 (Gagliano και συν. 2004, Chen και συν. 2005). Είναι γνωστό, ότι η μηχανική παραμόρφωση και οι δυνάμεις τάσης σηματοδοτούν μια διαδικασία που οδηγεί σε οστεογένεση. Ο TGF-β υποστηρίζει την οστεογένεση μέσω της συνδυαστικής του δράσης στις οστεοβλάστες και στις οστεοκλάστες. Η γονιδιακή έκφραση των TGF-β 1 και TGF-β 3 από τα κύτταρα του ανθρώπινου περιρριζίου αυξάνεται υπό την εφαρμογή μηχανικής παραμόρφωσης (Pinkerton και συν. 2008). Η δράση του TGF-β 1 στην ανάπλαση των περιοδοντικών ιστών έχει μελετηθεί με την εφαρμογή του παράγοντα στις περιοδοντικές βλάβες μεμονωμένα, ή σε συνδυασμό, με άλλους παράγοντες και αναπλαστικά υλικά. Ο συνδυασμός του TGF-β 1 με κατευθυνόμενη ιστική ανάπλαση σε περιοδοντικές βλάβες σκυλιών Beagle και με μεταφορέα ανθρακικό ασβέστιο, δεν απέδωσε διαφορές συγκριτικά με τους μάρτυρες που είχαν θεραπευθεί με κατευθυνόμενη ιστική ανάπλαση ή μόνο με το μεταφορέα (Wikesjo και συν. 1998). Στα πρόβατα η εφαρμογή του TGF-β 1 μεμονωμένα ή σε συνδυασμό με κατευθυνόμενη ιστική ανάπλαση σε μεσορριζικές βλάβες ΙΙ βαθμού ενίσχυσε την ανάπλαση των περιοδοντικών ιστών (Mohammed και συν. 1998). Σε οστικές βλάβες σκύλων η αργή απελευθέρωση του παράγοντα σε σπόγγο
53 53 κολλαγόνου ενίσχυσε τον οστικό σχηματισμό (Shigeno και συν. 2002). Αντίθετα, σε άλλες πειραματικές μελέτες ο TGF-β 1 φαίνεται να μειώνει τη δράση των BMPs κατά την οστική δημιουργία ενώ εμφανίζει αμφιλεγόμενη δράση όσον αφορά στην ανάπλαση του φατνιακού οστού και της οστεΐνης (Heckman και συν. 1999, Tatakis και συν. 2000).
54 54 Δ. Η ΒΙΟΛΟΓΙΚΗ ΚΑΙ Η ΒΙΟΧΗΜΙΚΗ ΣΥΜΠΕΡΙΦΟΡΑ ΤΗΣ ΟΙΣΤΡΑΔΙΟΛΗΣ ΣΤΑ ΟΣΤΙΚΑ ΚΥΤΤΑΡΑ ΚΑΙ ΤΑ ΚΥΤΤΑΡΑ ΤΟΥ ΠΕΡΙΡΡΙΖΙΟΥ Ένα μεγάλο μέρος της έρευνας, τελευταία, έχει στραφεί στη δράση των οιστρογόνων στους περιοδοντικούς ιστούς. Γενικά τα οιστρογόνα είναι στεροειδείς ωοθηκικές ορμόνες που παίζουν σημαντικό ρόλο στη διατήρηση της σκελετικής ακεραιότητας συμπεριλαμβανομένου του φατνιακού οστού και στο μεταβολισμό της ανόργανης μάζας του οστού. Ευθύνονται για τις φυσιολογικές αλλαγές που συμβαίνουν κατά τη διάρκεια της ζωής των γυναικών ενώ η βιολογική τους δράση μπορεί να επηρεάσει διάφορα όργανα του σώματος καθώς και τους ιστούς της στοματικής κοιλότητας (Mariotti 1994). Οι ορμόνες αυτές φαίνεται να εμπλέκονται στη παθογένεια της περιοδοντικής νόσου αλλά και στην επούλωση των περιοδοντικών ιστών. Τα οιστρογόνα και συγκεκριμένα η 17-β οιστραδιόλη (Ε2) ασκούν στα οστεοβλαστικά κύτταρα τόσο άμεση δράση, μέσω συγκεκριμένων υποδοχέων, όσο και έμμεση μέσω της ικανότητας τους να ρυθμίζουν την έκφραση και τη δράση πολλών αυξητικών παραγόντων στους κυτταρικούς πληθυσμούς (Eriksen και συν.1988, Rickard και συν. 1993, Cheng και συν.2002, Rubin και συν. 1995). Πρόσφατα, η δράση των οιστρογόνων ερευνάται στα κύτταρα του περιρριζίου. Το γεγονός ότι τα κύτταρα αυτά έχουν οστεοβλαστικό φαινότυπο οδήγησε στην αναζήτηση υποδοχέων οιστραδιόλης ανάλογων με αυτών που υπάρχουν στα οστικά κύτταρα. Πολλές μελέτες επιβεβαιώνουν την παρουσία τέτοιων υποδοχέων στα περιρριζικά κύτταρα (Lekwo και συν.1986, Parkar και συν.1996, Morishita και συν.1999, Jönsson και συν.2004) καθώς επίσης και
55 55 την άμεση δράση της οιστραδιόλης τόσο στον πολλαπλασιασμό όσο και στη διαφοροποίηση των κυττάρων (Lekwo και συν.1986, Nanba και συν. 1989, Morishita και συν.1998). Το γεγονός αυτό, οδηγεί στην υπόθεση ότι τα οιστρογόνα, πέρα από την εμπλοκή τους στη περιοδοντική νόσο, πιθανά συμμετέχουν και στις διαδικασίες ανάπλασης των περιοδοντικών ιστών με διάφορους μηχανισμούς που ακόμα ερευνώνται. Οιστρογόνα Τα οιστρογόνα εκπροσωπούνται από ένα μεγάλο αριθμό μορίων στεροειδούς και μη στεροειδούς φύσεως. Τα ενδογενή οιστρογόνα των θηλαστικών έχουν τη δομή τεσσάρων δακτυλίων αποτελούμενων από δεκαοκτώ άτομα άνθρακα (Wilson και συν.1997). Στο ανθρώπινο είδος τα ενδογενή οιστρογόνα είναι κατά κύριο λόγο η οιστρόνη (Ε1), η 17β- οιστραδιόλη (Ε2) και η οιστριόλη (Ε3). Οιστρογονική δραστηριότητα παρουσιάζουν και οι οιστρογονικοί μεταβολίτες καθώς και τα οιστρογόνα πού είναι κορεσμένα με Β-δακτύλιο. (M.O Connell και συν 1995). Ως οιστρογόνα, επίσης, χαρακτηρίζονται και οιστρογονικές ουσίες που προέρχονται από φυτικές πηγές (φυτοοιστρογόνα), τα συνθετικά οιστρογόνα (διαιθυλστιβεστρόλη) και μια σχετικά μεγάλη ομάδα ξενοβιοτικών (DDT, διφαινόλες). Τέλος, υπάρχει μια ομάδα μορίων που παλαιότερα ήταν γνωστά ως αντιοιστρογόνα και που σήμερα αναφέρονται ως επιλεκτικοί ρυθμιστές των οιστρογονικών υποδοχέων (SERMs) (Bryant και συν. 1998). Οιστρογονικοί υποδοχείς Τα οιστρογόνα εξασκούν τη δράση τους μέσω των οιστρογονικών υποδοχέων οι οποίοι είναι πρωτεΐνες που εντοπίζονται στον πυρήνα των κυττάρων. Αυτοί
56 56 διακρίνονται στον οιστρογονικό υποδοχέα α (ER-α) και στον οιστρογονικό υποδοχέα-β (ER-β) (Kuiper και συν. 1996). Οι οιστρογονικοί υποδοχείς ανήκουν σε μια μεγάλη οικογένεια υποδοχέων που δρουν στον κυτταρικό πυρήνα και εκπροσωπούνται από μέλη που συνδέονται με τις στεροειδείς ορμόνες. Δομικά, η ομάδα αυτή υποδοχέων διαχωρίζεται σε περιοχές με διαφορετικές λειτουργίες οι οποίες χαρακτηρίζονται ως A, B, C, D, E, και F. Το κοινό χαρακτηριστικό των πυρηνικών υποδοχέων είναι ένα τμήμα δυο δακτυλίων ψευδαργύρου (περιοχή C) που προσδένεται με το DNA (DNA binding domain). Οι άλλες περιοχές είναι η περιοχή πρόσδεσης του συνδέτη (περιοχή D, E, F), η περιοχή που εντοπίζεται στο πυρήνα (D) και η περιοχή αρμός (hinge). Επιπρόσθετα δύο αλληλεπιδρούσες περιοχές, οι AF-1και AF-2 είναι εγκατεστημένες στα αμινο- (περιοχή Α/Β) και καρβοξυλικά άκρα των υποδοχέων αντίστοιχα (Tzukerman και συν.1994). Ο μηχανισμός μέσω του οποίου η κάθε πληροφορία μεταφέρεται από το συνδέτη (οιστρογόνο) μέσω του υποδοχέα, έχει μελετηθεί αρκετά τις τελευταίες δεκαετίες. Είναι πλέον ξεκάθαρο, ότι η πρόσδεση του συνδέτη στον οιστρογονικό υποδοχέα δίνει την εκκίνηση σε ένα αριθμό διαδικασιών. Συγκεκριμένα η πρόσδεση αυτή προκαλεί, αρχικά, μια αλλαγή στο δομικό σχηματισμό του υποδοχέα. Οι δύο προσδεμένοι με το συνδέτη υποδοχείς διμερίζονται, τροποποιούνται βιοχημικά ( π.χ. φωσφορυλίωση ή ακετυλιώση) και τελικά προσδένονται σε ειδικές DNA ακολουθίες. Με αυτό το απλό μοντέλο το ενεργοποιημένο σύμπλεγμα του οιστρογονικού υποδοχέα μπορεί να ενεργοποιήσει ή να καταστείλει τη μεταγραφική γονιδιακή δραστηριότητα (Grody και συν 1982, Kumar και συν. 1988, Auricchio 1989).
57 57 Η ρύθμιση της μεταγραφής των γονιδίων μέσω των οιστρογονικών υποδοχέων επηρεάζεται και από την ύπαρξη κάποιων πρωτεϊνών οι οποίες αλληλοεπιδρούν με τον οιστρογονικό υποδοχέα και αναφέρονται ως συνρυθμιστές ή συν-τροποποιητές. Οι πρωτεΐνες αυτές λειτουργούν είτε ως συνενεργοποιητές είτε ως συν καταστολείς (McKenna και συν. 1999, Clement- Lacroix και συν.1999). Έχει πλέον βρεθεί ότι οι οιστρογονικοί υποδοχείς, πέρα από την ενεργοποίηση τους μέσω της πρόσδεσης τους με το συνδέτη οιστρογόνο, λειτουργούν και μέσω άλλων μηχανισμών προκειμένου να επηρεάσουν διάφορες φυσιολογικές λειτουργίες με τρόπο εξαρτώμενο ή μη από τον συνδέτη. Έτσι, αυτοί μπορούν να ενεργοποιηθούν από αυξητικούς παράγοντες μέσω των πρωτεϊνικών κινασών Α και C ή ακόμα και σε συνέργεια με αυτές τις κινάσες (Curtis, Katzenellenbogen και συν. 1990). Οι κινάσες φωσφορυλιώνουν τον υποδοχέα στη σερίνη 118. Η φωσφορυλίωση αυτή είναι αρκετή να ενεργοποιήσει τον υποδοχέα ο οποίος συντάσσει τους κατάλληλους συνενεργοποιητές, προσδένεται στο DNA και εμπλουτίζει τη μεταγραφική δραστηριότητα. Η λειτουργία αυτή, φαίνεται να συμβαίνει σε κάποια πειραματικά μοντέλα όπως στο ανθρώπινο μοντέλο και στα καρκινικά μαστικά κύτταρα. Στα μοντέλα αυτά η θεραπεία με IGF-I, EGF και άλλους ενεργοποιητές των πρωτεϊνικών κινασών A και C ενεργοποιεί τους οιστρογονικούς υποδοχείς των κυττάρων (Katzenellenbogen και συν. 1990). Παρόλο που οι αυξητικοί παράγοντες φαίνεται να ενεργοποιούν τους οιστρογονικούς υποδοχείς, τα οιστρογόνα μπορούν να αναστείλουν τη δράση κάποιων ιντερλευκινών και ειδικά των IL-1, TNF-α και IL-6. Αυτό καθιστά πάρα πολύ σημαντική τη δράση των οιστρογόνων στο σκελετό. Η IL-6 και ο
58 58 TNF-α λειτουργούν μέσω της ενεργοποίησης του μεταγραφικού παράγοντα NF-kβ και της ακόλουθης αλληλοδράσης αυτού του συμπλέγματος με ειδικές DNA ακολουθίες. Η δράση αυτή των κυτταροκινών στο σκελετό είναι αρνητική (αυξάνουν την απορρόφηση οδηγώντας σε απώλεια της οστικής μάζας). Οι οιστρογονικοί υποδοχείς αλληλοδρούν με το NF-kβ με τρόπο εξαρτώμενο από το συνδέτη αναστέλλοντας έτσι τη δράση του (Manolagas 1998, Bellido και συν. 1998). Οι οιστρογονικοί υποδοχείς πέρα από αυτή την ανασταλτική δράση πού έχουν στον παράγοντα NF-kβ όταν είναι προσδεμένοι στον συνδέτηοιστρογόνο, επηρεάζουν και τους μηχανισμούς άλλων μεταγραφικών συμπλεγμάτων όπως του AP1, Sp1, SF-1 (Webb και συν. 1995, Qin και συν. 1999, Sadovsky και συν. 1998). Τέλος, τα οιστρογόνα φαίνεται να επιδρούν μέσω διαδικασιών όπου δε χρησιμοποιούνται πυρηνικοί υποδοχείς αλλά υποδοχείς της πλασματικής μεμβράνης (Fiorelli και 1996, Cooke και 1998). Αυτό δικαιολογεί φυσιολογικές απαντήσεις του οργανισμού που συμβαίνουν γρήγορα και οι οποίες θα θεωρείτο απίθανο να προκύψουν ως αποτέλεσμα μεταγραφικών δραστηριοτήτων εξαρτώμενων από πυρηνικούς υποδοχείς. Ένα τέτοιο παράδειγμα, αποτελεί η αύξηση του c-amp παρουσία 17-β οιστραδιόλης. Ο Υποδοχέας ER-β Το 1996 η θεωρία περί ενός οιστρογονικού υποδοχέα, ανατράπηκε με την ταυτοποίηση ενός δεύτερου πρωτεϊνικού υποδοχέα που παρουσιάζει υψηλή συγγένεια σύνδεσης με τα οιστρογόνα. Ο υποδοχέας αυτός ονομάσθηκε ER-β ενώ ο αρχικός οιστρογονικός υποδοχέας μετονομάσθηκε σε ER-α (Kuiper και συν. 1996, Mosselman και συν.1996). Ποικίλες διαφορές υπάρχουν μεταξύ
59 59 των δύο οιστρογονικών υποδοχέων. Ο ER-β εντοπίζεται στο χρωμόσωμα 14 ενώ ο ER-α στο χρωμόσωμα 6. Ο ER-β αποτελεί πρωτεΐνη 530 αμινοξέων ενώ ο ER-α είναι πρωτεΐνη 590 αμινοξέων. Επιπλέον, η κατανομή των δύο υποδοχέων στους ιστούς όπως στον εγκέφαλο, στη μήτρα, στις ωοθήκες και στο προστάτη διαφέρει. In vitro μελέτες έδειξαν ότι ο ER-β όπως και ο ER-α διμερίζονται και συνδέονται με το DNA. Υπό κατάλληλες συνθήκες ο ER- β ετεροδιμερίζεται με τον ER-α δημιουργώντας ένα σύμπλεγμα που συνδέεται με το DNA περισσότερο από ότι ο ομοδιμερής ER- β. Παρόλα αυτά, η μεταγραφική δραστηριότητα του ετεροδιμερούς είναι ίδια με εκείνη του ομοδιμερούς ER- β αλλά διαφέρει από του ομοδιμερούς ER- α (Cowley και συν. 1997). Τέλος μια άλλη χαρακτηριστική διαφορά τους είναι η διαφορετική χημική συγγένεια προς το συνδέτη-οιστρογόνο. ER- α και ER- β knock out επίμυες Προκειμένου να αποσαφηνισθεί ο φυσιολογικός ρόλος των υποδοχέων ER- α και ER-β, χρησιμοποιήθηκαν τα πειραματικά μοντέλα των knock out ποντικών. Τόσο οι ER-α, όσο και οι ER-β knock out επίμυες, δεν εμφάνισαν έντονες αλλαγές στο σκελετικό τους φαινότυπο γεγονός που υποδηλώνει ότι η παρουσία ενός από τους δύο υποδοχείς είναι αρκετή για να διατηρηθεί η ανταπόκριση του σκελετού στα οιστρογόνα. Οι Couse και συν. (1999) έδειξαν ότι οι ER-α knock out επίμυες εμφανίζουν μια μείωση στο μήκος των οστών και στα δυο φύλα. Κάτι τέτοιο, δεν παρατηρήθηκε στους ER-β knock out επίμυες. Ελάχιστη μείωση της ανόργανης οστικής πυκνότητας παρατηρήθηκε και στα δύο μοντέλα.
60 60 Στους ανθρώπους υπάρχει μία αναφορά σε έναν άρενα που έπασχε από απενεργοποίηση του ER-α. Το άτομο αυτό, παρουσίαζε μειωμένη οστική πυκνότητα ενώ η ανάπτυξη των μακρών οστών δεν είχε ολοκληρωθεί. Ο σκελετικός φαινότυπος αυτού του ανθρώπου δείχνει να είναι διαφορετικός από εκείνο του ER-α knock out ποντικού γεγονός που υποδηλώνει τη διαφορετική συμπεριφορά των ανθρώπινων και ζωικών πειραματικών μοντέλων. Οι ER-α knock out επίμυες εμφανίζουν ατροφική μήτρα, ωοθηκική δυσλειτουργία και υψηλά επίπεδα οιστρογόνων στο αίμα. Οι όρχεις επίσης είναι μη φυσιολογικοί όσον αφορά στο βάρος, στο σχήμα και στη λειτουργία τους (Komm και συν. 2001). Οι Sims και συν. (2002) έδειξαν ότι η έλλειψη του υποδοχέα ER-α οδηγεί σε μειωμένη οστική αναδόμηση και αύξηση του όγκου του σπογγώδους οστού τόσο στους αρσενικούς όσο και στους θηλυκούς επίμυες. Οι αλλαγές αυτές οφείλονται στα αυξημένα επίπεδα οιστρογόνων και τεστοστερόνης στη κυκλοφορία των θηλυκών ζώων καθώς και τεστοστερόνης στη κυκλοφορία των αρσενικών ζώων. Πιο συγκεκριμένα, στους θηλυκούς ER-α knock out επίμυες, η οστική αναδόμηση καταστέλλεται και ο όγκος του σπογγώδους οστού αυξάνεται λόγω των υψηλών επιπέδων οιστραδιόλης που διεγείρουν τους ER-β υποδοχείς και των υψηλών επιπέδων τεστοστερόνης που διεγείρουν τους υποδοχείς των ανδρογόνων. Στους αρσενικούς παρατηρείται το ίδιο φαινόμενο ως αποτέλεσμα των υψηλών επιπέδων τεστοστερόνης που διεγείρουν τους υποδοχείς ανδρογόνων. Οι Sims και συν. (2002), όπως και οι προηγούμενοι ερευνητές, παρατήρησαν επίσης μείωση του πάχους και της πυκνότητας του συμπαγούς οστού.
61 61 Οι ER-β knock out επίμυες παρουσιάζουν αλλαγές στις ωοθήκες όμως εμφανίζουν φυσιολογική μήτρα και επίπεδα οιστρογόνων στο αίμα. Η ιστολογική εικόνα, καθώς και η λειτουργία των όρχεων είναι φυσιολογικές παρόλο που με τη πάροδο της ηλικίας αναφέρεται υπερπλασία του προστάτη (Komm και συν. 2001). Οι ER-β knock out αρσενικοί επίμυες δεν εμφανίζουν προβλήματα στην οστική αναδόμηση και στην οστική μάζα, ενώ αντίθετα οι αντίστοιχοι θηλυκοί εκδηλώνουν μείωση της οστικής αναδόμησης και αύξηση του όγκου του σπογγώδους οστού. Τα ευρήματα αυτά, υποδηλώνουν ότι οι υποδοχείς ER-β δεν έχουν έντονη επίδραση στα οστά του αρσενικού ενώ στα θηλυκά μπορεί να ανταγωνίζονται τη δράση των ER-α υποδοχέων σε φυσιολογικές συνθήκες αλλά μπορεί και να τους αντικαθιστούν όταν αυτοί εκλείπουν (Syed και Khosla 2004). Στους ER-α-ER-β knock out αρσενικούς επίμυες παρατηρείται καταστολή της οστικής απορρόφησης και αύξηση του όγκου του σπογγώδούς οστού γεγονός που αποδίδεται, στα υψηλά επίπεδα τεστοστερόνης στο αίμα και στη δράση τους στα οστά μέσω των υποδοχέων των ανδρογόνων. Αντιθέτως, τα επίπεδα αυτά δεν εμφανίσθηκαν αυξημένα στους αντίστοιχους θηλυκούς και έτσι η απώλεια και των δύο οιστρογονικών υποδοχέων συντέλεσε σε μείωση του όγκου του σπογγώδους οστού η οποία σχετίζεται με τη μείωση του οστικού σχηματισμού που παρατηρείται παρουσία φυσιολογικών επιπέδων οστικής απορρόφησης ( Syed και συν. 2004). Μηχανισμοί δράσης των οιστρογόνων στο οστούν Τα οιστρογόνα αποτελούν σημαντικούς ρυθμιστές της σκελετικής ανάπτυξης και ομοιοστασίας (Turner και συν. 1994). Στις γυναίκες, η έλλειψη οιστρογόνων λόγω εμμηνόπαυσης σχετίζεται με τη ταχεία απώλεια του
62 62 σπογγώδους οστού (Riggs και συν. 2002). Η έλλειψη οιστρογόνων προκαλεί απώλεια και του συμπαγούς οστού αλλά με πιο αργό ρυθμό. Η απώλεια αυτή του οστού χαρακτηρίζεται από αυξημένους ρυθμούς οστικής αναδόμησης με αύξηση τόσο της οστικής απορρόφησης όσο και του οστικού σχηματισμού. Η υπεροχή ωστόσο της απορρόφησης σε σχέση με τον οστικό σχηματισμό οδηγεί τελικά σε καθαρή οστική απώλεια. Οι παρατηρήσεις αυτές, υπογραμμίζουν τις βασικές λειτουργίες των οιστρογόνων στο σκελετό, που είναι οι ακόλουθες: α) αναστολή της οστικής αναδόμησης, β) καταστολή της οστικής απορρόφησης και γ) διέγερση του οστικού σχηματισμού. Η δράση των οιστρογόνων στο οστούν μπορεί να είναι άμεση ή έμμεση. Σήμερα είναι γνωστό, ότι οι οστεοβλάστες και οι οστεοκλάστες φέρουν τους οιστρογονικούς υποδοχείς ER-α και ER-β. Επίσης, η παραγωγή των αυξητικών παραγόντων και των κυτταροκινών από αυτά τα κύτταρα φαίνεται να ρυθμίζεται σε ένα βαθμό από τα οιστρογόνα, γεγονός που υποδεικνύει ένα έμμεσο τρόπο δράσης των οιστρογόνων στο σκελετό με τη μεσολάβηση των αυξητικών παραγόντων και των κυτταροκινών (Turner και συν. 1994, Prince και συν. 1994). Η δράση των οιστρογόνων στις οστεοβλάστες Τα οιστρογόνα δρουν άμεσα στο πολλαπλασιασμό των οστεοβλαστών αλλά και στην έκφραση των γονιδίων που κωδικοποιούν ένζυμα, πρωτεΐνες της θεμέλιας ουσίας του οστού, ορμονικούς υποδοχείς, μεταγραφικούς παράγοντες καθώς και αυξητικούς παράγοντες και κυτταροκίνες (Harris και συν. 1996, Robinson και συν.1997, Ourlser 1998, Turner 1999). Τα αποτελέσματα από τις in vitro μελέτες της γενετικής και μεταβολικής αλλαγής που προκύπτουν από τη δράση των οιστρογόνων στις οστεοβλάστες φαίνεται να είναι αντικρουόμενα. Αυτό, μπορεί να οφείλεται στις διαφορές που
63 63 υπάρχουν στη συνολική συγκέντρωση υποδοχέων στις διάφορες οστεοβλαστικές σειρές, στη συγκέντρωση ERα και ERβ στα διαφορετικά πειραματικά μοντέλα, στις διαφορετικές περιοχές του σκελετού από όπου προέρχονται τα κύτταρα καθώς και στο στάδιο διαφοροποίησης των κυττάρων. Έτσι, τα οιστρογόνα, ενώ γενικώς αναστέλλουν τον πολλαπλασιασμό σε καλλιέργειες των ώριμων ανθρώπινων οστεοβλαστών, μπορούν υπό συγκεκριμένες συνθήκες να προκαλέσουν διέγερση του πολλαπλασιασμού των οστεοβλαστών. Τα οιστρογόνα διεγείρουν τη σύνθεση των αυξητικών παραγόντων TGF- β1, IGF-1 και των πρωτεϊνών που συνδέονται με τον IGF-1 ενώ αναστέλλουν τη σύνθεση των ιντερλευκινών IL-1, IL-6, IL-11 (Ourlser 1998, Jilka 1998, Martin και συν. 1998, Khosla και συν. 1999). Η έλλειψη οιστρογόνων αυξάνει τη παραγωγή της ιντερλευκίνης 1 και του παράγοντα διέγερσης αποικιών των μακροφάγων (M-CSF) στα κύτταρα του στρώματος του μυελού των οστών. Πρόσφατα, παρατηρήθηκε ότι τα οιστρογόνα ρυθμίζουν τη σύνθεση των πρωτεϊνών της οστικής θεμέλιας ουσίας. Η επίδραση των οιστρογόνων στην έκφραση των πρωτεϊνών αυτών φαίνεται να τροποποιείται ανάλογα με το στάδιο διαφοροποίησης των οστεοβλαστών (π.χ. στάδιο σύνθεσης της θεμέλιας ουσίας, πρώιμο/καθυστερημένο στάδιο ενασβεστίωσης) αλλά και τον υπό μελέτη οιστρογονικό υποδοχέα (Bodine και συν. 1998, Waters και συν. 2001). Τέλος βρέθηκε ότι τα οιστρογόνα ρυθμίζουν τη σύνθεση των μορφογενετικών πρωτεϊνών (BMP-6), της οστεοπροτεγερίνης και των μεταγραφικών παραγόντων όπως των TIEG ( TGF inducible early gene), NF-kβ, c/ebp-β και c-fos στις ανθρώπινες οστεοβλάστες (Stein και Yang 1995, Ourlser 1998,
64 64 Jilka 1998, Martin και συν. 1998, Tau και συν. 1998, Khosla και συν. 1999, Hofbauer και συν. 1998, Rickard και συν. 1998). Η δράση των οιστρογόνων στις οστεοκλάστες Τα οιστρογόνα, πέρα από τη μείωση της ροής των προγονικών οστεοβλαστών και οστεοκλαστών από τις αιμοποιητικές και μεσεγχυματικές κυτταρικές σειρές, δρουν και στην ανάπτυξη, δραστηριότητα και απόπτωση των οστεοκλαστών. Το σημαντικότερο μόριο στην οστεοκλαστική ανάπτυξη είναι ο υποδοχέας ενεργοποίησης του συνδέτη NF-kβ (RANKL) ο οποίος εκφράζεται στην επιφάνεια των πρόδρομων οστεοβλαστικών κυττάρων, των Τ- κυττάρων και των Β κυττάρων (Eghbali-Fatourechi και συν. 2003). Ο RANKL συνδέεται με τον υποδοχέα RANK στην οστεοκλαστική κυτταρική σειρά και αδρανοποιείται από την οστεοπροτεγερίνη η οποία επίσης παράγεται από τα κύτταρα της οστεοβλαστικής σειράς. Συνδυαστικές in vitro και in vivo μελέτες έχουν δείξει ότι τα οιστρογόνα διεγείρουν την παραγωγή της οστεοπροτεγερίνης (Hofbauer και συν. 1998, Khosla και συν. 2001). Τα οιστρογόνα, πέρα από αυτή τους τη δράση, λειτουργούν ρυθμιστικά στην παραγωγή κυτταροκινών από τις οστεοβλάστες τροποποιώντας την οστεοκλαστική δραστηριότητα παρακρινικά (Manolagas και συν. 1995). Όπως αναφέρθηκε και προηγούμενα, οι κυτταροκίνες όπως η IL-1 και η IL-6, ο TNF-α, ο M-CSF και οι προσταγλανδίνες αποτελούν σημαντικούς παράγοντες στην απώλεια οστού που ακολουθεί την έλλειψη οιστρογόνων. Ουσιαστικά οι παράγοντες αυτοί αυξάνονται ελλείψει οιστρογόνων (Pacifici και συν. 1991, Tanaka και συν. 1993). Τα οιστρογόνα προάγουν, επίσης, την απόπτωση των οστεοκλαστών και καταστέλλουν την οστεοκλαστική διαφοροποίηση που προάγεται από τον
65 65 RANKL. Επιπλέον, οι ορμόνες αυτές αναστέλλουν τη δραστηριότητα των οστεοκλαστών με μηχανισμούς άμεσους που λειτουργούν μέσω υποδοχέων (Ourlser και συν. 1994, Pederson και συν. 1999). Η δράση των οιστρογόνων στα περιρριζικά κύτταρα Τα οιστρογόνα θεωρούνται σημαντικοί τροποποιητικοί παράγοντες που επηρεάζουν τη παθογένεια της περιοδοντικής νόσου αλλά και την επούλωση των περιοδοντικών ιστών (Marrioti και συν. 1994, Hofbauer και Henfelder 2001). Οι αλλαγές των επιπέδων των οιστρογόνων κατά την εφηβεία, την εγκυμοσύνη καθώς και την εμμηνόπαυση φαίνεται να σχετίζονται άμεσα με αλλαγές στην περιοδοντική κατάσταση των ασθενών. Οι μηχανισμοί με τους οποίους τα οιστρογόνα δρουν στους περιοδοντικούς ιστούς είναι ακόμα ασαφείς. Όπως αναφέρθηκε σε προηγούμενη ενότητα τα οιστρογόνα δρουν στους ιστούς μέσω των οιστρογονικών υποδοχέων. Τέτοιοι υποδοχείς έχουν αναζητηθεί και στα κύτταρα του περιρριζίου. Οι σχετικές έρευνες δίδουν ποίκιλα αποτελέσματα. Ο Lewko και συν (1986) αναφέρουν ότι τα ανθρώπινα κύτταρα του περιρριζίου παρουσιάζουν υψηλή χημική συγγένεια στα οιστρογόνα. Οι Parkar και συν. (1996) ισχυρίζονται την απουσία των οιστρογονικών υποδοχέων στα κύτταρα του περιρριζίου με τη μέθοδο RT-PCR ενώ οι Morishita και συν. (1999) χρησιμοποιώντας την ίδια μέθοδο αναφέρουν την παρουσία χαμηλών επιπέδων ER-α και καθόλου ER-β στα ανθρώπινα περιρριζικά κύτταρα. Σε αντίθετα αποτελέσματα οδηγήθηκαν οι Jöhnsson και συν. (2004, 2005, 2007) οι οποίοι έδειξαν ανοσοϊστοχημικά την παρουσία των ER-β αλλά όχι των ER-α υποδοχέων στα ανθρώπινα περιρριζικά κύτταρα. Ενδιαφέρον παρουσιάζει το γεγονός ότι οι υποδοχείς αυτοί εντοπίσθηκαν πέρα από τον πυρήνα και στα μιτοχόνδρια των κυττάρων
66 66 του περιρριζίου γεγονός που υποδηλώνει τη δράση των οιστρογόνων στη λειτουργία των μιτοχονδρίων και στο μεταβολισμό των περιρριζικών κυττάρων. Συγκεκριμένα, η 17-β οιστραδιόλη μειώνει την έκφραση της οξειδάσης του κυτοχρώματος c η οποία συμμετέχει στην οξειδωτική φωσφορυλίωση (Jöhnsson και συν. 2007). Οι Liang και συν. (2008) επιβεβαίωσαν την παρουσία των υποδοχέων ER-β στα κύτταρα του περιρριζίου ενώ οι Cao και συν. (2007) χρησιμοποιώντας τις μεθόδους RT- PCR και Western- blot έδειξαν την παρουσία και των δύο υποδοχέων (ER-α και ER-β) στα περιρριζικά κύτταρα τόσο των αρουραίων όσο και των ανθρώπων. Τα διαφορετικά αποτελέσματα που προκύπτουν από τις ανωτέρω μελέτες οφείλονται πιθανά στη διαφορετική μεθοδολογία και στα υλικά που χρησιμοποιούνται στην εκάστοτε έρευνα. Εξίσου αντικρουόμενα είναι και τα αποτελέσματα των ερευνών που αφορούν στη επίδραση των οιστρογόνων στη σύνθεση του DNA και στον πολλαπλασιασμό των περιρριζικών κυττάρων. Οι Jöhnsson και συν. (2007) υποστηρίζουν ότι η θεραπεία με φυσιολογική συγκέντρωση 17-β οιστραδιόλης για 24 ώρες διεγείρει τη σύνθεση των καρκινικών κυττάρων του μαστού MCF7 όμως δεν έχει κανένα αποτέλεσμα στα ανθρώπινα περιρριζικά κύτταρα. Αντίθετα άλλες μελέτες αναφέρουν αύξηση του ρυθμού ανάπτυξης και της σύνθεσης DNA των ανθρώπινων περιρριζικών κυττάρων παρουσία 17-β οιστραδιόλης (Nanba και συν. 1989, Kakai και συν. 1992). Τέλος, έχει αναφερθεί ότι η εφαρμογή της 17-β οιστραδιόλης σε δόσεις 10-9 Μ και 10-8 Μ οδηγεί σε μείωση της σύνθεσης του DNA των ανθρώπινων περιρριζικών κυττάρων κατά 62% και 38% αντίστοιχα (Lewko και συν. 1986). Οι διαφορές στη δοσολογία και στο χρόνο εφαρμογής της οιστραδιόλης στα κύτταρα του
67 67 περιρριζίου αλλά και οι διαφορετικές μεθοδολογίες είναι πιθανά αίτια των διαφορών που παρατηρούνται στα αποτελέσματα των προηγούμενων μελετών. Σε αντίθεση με τον πολλαπλασιασμό των κυττάρων οι περισσότερες έρευνες φαίνεται να συμφωνούν ως προς τη επίδραση της 17-β οιστραδιόλης στην οστεοβλαστική διαφοροποίηση των κυττάρων του περιρριζίου. In vitro, η οιστραδιόλη αυξάνει τη δραστηριότητα της αλκαλικής φωσφατάσης, την παραγωγή της οστεοκαλσίνης με τρόπο που εξαρτάται από τη δόση και το χρόνο εφαρμογής της στα περιρριζικά κύτταρα ενώ προάγει τη διαφοροποίηση των κυττάρων του περιρριζίου όπως προκύπτει από τον σχηματισμό, υπό την επίδραση της, ενασβεστιωμένων οζιδίων (Morishita και συν. 1998, Cao και συν. 2007, Liang και συν. 2008). Τα οιστρογόνα αναστέλλουν την απορρόφηση του φατνιακού οστού ρυθμίζοντας την έκφραση του υποδοχέα RANKL και της οστεοπροτεγερίνης κατά τη διάρκεια της οστεοβλαστικής διαφοροποίησης των κυττάρων του περιρριζίου. Συγκεκριμένα η εφαρμογή της οιστραδιόλης στα κύτταρα του περιρριζίου σε δόσεις Μ και 10-7 Μ για 24, 48 και 72 ώρες οδήγησε σε μείωση των επιπέδων RANKL m RNA με τρόπο εξαρτώμενο από το χρόνο και τη δόση εφαρμογής της ενώ αύξηση παρατηρήθηκε στα επίπεδα του mrna της οστεοπροτεγερίνης τα οποία έφθασαν στο μέγιστο στις 72 ώρες (Tang και συν. 2007). Τέλος οι Shu και συν. (2008) έδειξαν ότι τα οιστρογόνα μπορούν να τροποποιήσουν τη συμπεριφορά των περιοδοντικών ιστών ως προς το λιποσαγχαρίδιο E.Coli το οποίο, ως γνωστό, αυξάνει την παραγωγή των προφλεγμονωδών κυτταροκινών από τα κύτταρα του περιρριζίου τόσο σε
68 68 επίπεδο m RNA όσο και σε πρωτεϊνικό επίπεδο. Το εύρημα αυτό προτείνει τη πιθανότητα δράσης των οιστρογόνων στους περιοδοντικούς ιστούς μέσω της αλλαγής της έκφρασης των φλεγμονωδών κυτταροκινών από τα περιρριζικά κύτταρα.
69 69 Α. ΣΚΟΠΟΣ Τις τελευταίες δεκαετίες ιδιαίτερο ενδιαφέρον στη θεραπευτική των νόσων του περιοδοντίου παρουσιάζει η ανάπλαση των περιοδοντικών ιστών όπως αυτή ορίζεται από τη δημιουργία νέου οστίτη ιστού, νέας οστεΐνης και νέων, λειτουργικά προσανατολισμένων, ινών του περιρριζίου. Σημαντικό ρόλο στη διαδικασία επούλωσης και ανάπλασης των περιοδοντικών ιστών φαίνεται να κατέχουν τα κύτταρα του περιρριζίου (Melcher 1976). Αυτά αφορούν σε ένα μικτό ετερογενή πληθυσμό κυττάρων όπου υποπληθυσμοί αυτών είναι κύτταρα που, ενώ μορφολογικά μοιάζουν στις ινοβλάστες, παρουσιάζουν οστεοβλαστική δραστηριότητα. Η ανάπλαση των ιστών του περιοδοντίου βασίζεται στην ενεργοποίηση μηχανισμών ικανών να οδηγήσουν στη χημειοταξία, στην προσκόλληση, στον πολλαπλασιασμό και στη διαφοροποίηση αυτών των περιρριζικών κυττάρων καθώς και στην παραγωγή μεσοκυττάριας θεμέλιας ουσίας (Bartold και συν.2000). Παρόλα αυτά, η φαινοτυπική ετερογένεια των κυττάρων του περιρριζίου καθιστά δύσκολη την κατανόηση των διαδικασιών επούλωσης και ανάπλασης των περιοδοντικών ιστών. Για το λόγο αυτό κρίνεται σημαντικός ο χαρακτηρισμός των διαφόρων κυτταρικών πληθυσμών που απομονώνονται από το περιρρίζιο πριν από τη χρήση τους σε οποιαδήποτε έρευνα. Διάφοροι αυξητικοί παράγοντες και ορμόνες συμμετέχουν και επηρεάζουν αυτούς τους βιολογικούς μηχανισμούς. Ένας τέτοιος αυξητικός παράγοντας, ο TGF-β έχει μελετηθεί αρκετά όσον αφορά στη δράση του στον πολλαπλασιασμό και στη διαφοροποίηση τόσο των οστεοβλαστών όσο και των κυττάρων του περιρριζίου. Ο TGF-β είναι πολυπεπτίδιο μοριακού βάρους 25 kda που παράγεται κυρίως από το οστούν και τα αιμοπετάλια (Bonewald
70 70 και συν. 1966, Franzen και συν. 1995,). Ο παράγοντας αυτός επηρεάζει τη λειτουργία πολλών κυτταρικών τύπων, αυξάνει τη παραγωγή της θεμέλιας ουσίας και επηρεάζει τα οστικά κύτταρα ενώ έχει ανιχνευθεί και στο ουλικό υγρό περιοδοντικών ασθενών (Buduneli 2001). Η δράση του TGF-β1 στα κύτταρα του περιρριζίου διαφοροποιείται από παράγοντες που αφορούν στη δόση και στο χρόνο εφαρμογής του στα κύτταρα, στο είδος των υποδοχέων, στο βαθμό διαφοροποίησης των κυττάρων, στη προέλευση τους αλλά και στη παρουσία άλλων παραγόντων γεγονός που δίνει το έναυσμα για περισσότερη μελέτη της δράσης του αυξητικού αυτού παράγοντα στα κύτταρα του ανθρώπινου περιρριζίου. Πέρα από τους αυξητικούς παράγοντες, ένα μεγάλο μέρος της έρευνας, τελευταία, έχει στραφεί στη δράση των οιστρογόνων στους περιοδοντικούς ιστούς. Οι ορμόνες αυτές έχουν σχετισθεί με τη παθογένεια της περιοδοντικής νόσου, όπου δρούν με ποικίλους τρόπους (Mascarenhas και συν. 2003), ενώ ιδιαίτερο ενδιαφέρον παρουσιάζει η πιθανή δράση τους στην επούλωση των περιοδοντικών ιστών. Τα οιστρογόνα και συγκεκριμένα η 17-β οιστραδιόλη (Ε2) φαίνεται να δρουν απευθείας στα οστεοβλαστικά κύτταρα μέσω συγκεκριμένων υποδοχέων (Eriksen και συν. 1988). Η δράση τους αυτή σχετίζεται τόσο με τον πολλαπλασιασμό όσο και με την οστεοβλαστική δραστηριότητα των κυττάρων και εξαρτάται από την ενεργοποίηση του συγκεκριμένου υποδοχέα, τη ποσότητα οιστραδιόλης που χρησιμοποιείται καθώς και από το στάδιο διαφοροποίησης των κυττάρων (Bodine και συν. 1998, Qu και συν.1998, Waters και συν. 2001, Chen και συν. 2002). Πολύ πρόσφατη παρουσιάζεται η έρευνα της δράσης των οιστρογόνων στα κύτταρα του περιρριζίου. Το γεγονός ότι τα κύτταρα αυτά διαθέτουν και οστεοβλαστικό
71 71 φαινότυπο οδήγησε στην αναζήτηση υποδοχέων οιστραδιόλης ανάλογων με αυτών που υπάρχουν στα οστικά κύτταρα. Οι περισσότερες μελέτες επιβεβαιώνουν την παρουσία τέτοιων υποδοχέων στα περιρριζικά κύτταρα (Lekwo και συν.1986, Parkar και συν. 1996, Morishita και συν. 1999, Jonsson και συν. 2004, Cao και συν. 2007, Liang και συν. 2008) καθώς επίσης και την άμεση δράση της οιστραδιόλης τόσο στον πολλαπλασιασμό όσο και στη διαφοροποίηση των κυττάρων (Lekwo και συν.1986, Nanba και συν. 1989, Morishita και συν. 1998). Παρόλα αυτά η δράση του TGF-β1 και της οιστραδιόλης στον πολλαπλασιασμό των κυττάρων του περιρριζίου φαίνεται να είναι αμφιλεγόμενη γεγονός που μπορεί να είναι αποτέλεσμα μελέτης διαφορετικής δόσης και χρόνου εφαρμογής των παραγόντων αυτών στα κύτταρα καθώς και διαφορετικών κυτταρικών υποπληθυσμών του περιρριζίου με διαφορετικό βαθμό διαφοροποίησης ανάλογα με τον φαινότυπο και την προέλευση τους. Απαραίτητη καθίσταται η περαιτέρω έρευνα του χαρακτηρισμού των περιρριζικών κυττάρων καθώς και της βιολογικής δράση της οιστραδιόλης και του TGF-β1 στα κύτταρα του περιρριζίου αλλά και γενικότερα στην επούλωση και μεταβολισμό των περιοδοντικών ιστών. Για το λόγο αυτό, η έρευνα που ακολουθεί έχει ως σκοπό α) το χαρακτηρισμό των κυττάρων ως προς τη μεσεγχυματική τους προέλευση, τον ινοβλαστικό και τον οστεοβλαστικό τους φαινότυπο και β) την in vitro μελέτη της δράσης της17-β οιστραδιόλης και του αυξητικού παράγοντα TGF-β1 στον πολλαπλασιασμό των συγκεκριμένων κυττάρων.
72 72 Β. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Στην παρούσα μελέτη σε καλλιέργειες περιρριζικών κυττάρων τα οποία απομονώθηκαν από ανθρώπινα δόντια, έγινε χαρακτηρισμός των κυττάρων αυτών ως προς τη μεσεγχυματική τους προέλευση, τον ινοβλαστικό και οστεοβλαστικό τους χαρακτήρα. Στη συνέχεια, μελετήθηκε η επίδραση του αυξητικού παράγοντα TGF-β 1 και της 17-β οιστραδιόλης στον πολλαπλασιασμό των κυττάρων του ανθρώπινου περιρριζίου. Η επίδραση των παραγόντων αυτών μελετήθηκε με την χρήση διαφόρων δόσεων και την εφαρμογή τους στα κύτταρα σε διάφορους χρόνους. Ακολούθως αναφέρονται τα υλικά και οι μέθοδοι που χρησιμοποιήθηκαν. ΥΛΙΚΑ 1. Tρεις προγόμφιοι από 3 ενήλικα άτομα (2 άνδρες και μία γυναίκα) ετών, υγιή χωρίς περιοδοντική νόσο και τερηδόνα, όπου τα δόντια εξήχθησαν για ορθοδοντικούς λόγους και από τα οποία έγινε η απομόνωση των κυττάρων για την ανάπτυξη των κυτταροκαλλιεργειών. Για την λήψη και τη χρήση των δοντιών είχε δοθεί έγκριση από την Επιτροπή Έρευνας και Δεοντολογίας της Οδοντιατρικής Σχολής του Πανεπιστημίου Αθηνών σύμφωνα με τη Δήλωση του Ελσίνκι του Οι ασθενείς προήλθαν από την Οδοντιατρική Σχολή Αθηνών και από ιδιωτικά ιατρεία.
73 73 2. Τα αντίσωματα για τη βιμεντίνη και τη λεία μυϊκή ακτίνη (μονοκλωνικά αντισώματα ποντικού από το εργαστήριο του Παθολογοανατομικού Τμήματος του Γενικού Κρατικού Νοσοκομείου «Γεννηματάς»). 3. Ο ανθρώπινος τροποποιητικός αυξητικός παράγοντας TGF-β 1 (rhtgf-β 1 : recombinant human TGF β 1 ) από την R&D Systems (England) (Εικόνα 1). Η σειρά του DNA που κωδικoποιεί τη πρόδρομη ανθρώπινη πρωτεΐνη TGF β 1 εκφράστηκε στα ωοθηκικά κύτταρα των κινέζικων χάμστερ (CHO) (Derynck και συν.1985). Η ώριμη μορφή TGF β 1 απομονώθηκε από το οξεοποιημένο μέσο της κυτταρικής καλλιέργειας. Σύμφωνα με τις προδιαγραφές της εταιρείας διάθεσης της, η ώριμη μορφή του TGF β 1 είναι μια ομοδιμερής πρωτεΐνη που αποτελείται από δύο αλυσίδες των 112 αμινοξέων συνενωμένες με δισουλφιδικούς δεσμούς. Το μοριακό της βάρος είναι περίπου 25kDa. Η καθαρότητα της είναι >97% και το επίπεδο ενδοτοξίνης <1 EU για κάθε 1μg της κυτταροκίνης. Η δραστικότητα του TGF β 1 μετράται από την ικανότητα του να αναστέλει τον εξαρτώμενο από την IL-4 πολλαπλασιασμό των HT-2 κυττάρων των ποντικών. Η διάμεση αποτελεσματική δόση (ED 50 ) γι αυτή τη δράση είναι τυπικά ng/ml. O TGF β 1 διατίθεται σε λυοφιλοποιημένη μορφή από 0.2μm διαλύματος φιλτραρισμένου σε 35% ακετονιτριλίου και 0.1% τριφθοροξικού οξέος (TFA) το οποίο περιέχει 50μg αλβουμίνης βόειου ορρού ανά 1μg κυτταροκίνης. 4. Η 17-β οιστραδιόλη (β-estradiol 17- acetate. Product #46551) της εταιρείας Sigma-Aldrich Canada Inc., Ontario, Canada.
74 74 ΔΙΑΛΥΜΑΤΑ- ΟΡΓΑΝΑ Τα κυριότερα διαλύματα και όργανα πού χρησιμοποιήθηκαν για την ανάπτυξη και αξιολόγηση των κυτταροκαλλιεργειών ήταν τα ακόλουθα: ΔΙΑΛΥΜΑΤΑ Θρεπτικό υλικό DMEM (Dulbecco s Modified Eagle s Medium), πενικιλίνη G, στρεπτομυκίνη και fungizone, εμβρυϊκός βόειος ορός (FBS), ρυθμιστικό διάλυμα φωσφορικών (PBS), διάλυμα κολλαγενάσης Ι, διάλυμα θρυψίνης με EDTA και διάλυμα διμεθυλσουλφοξειδίου (DMSO) (Sigma-Aldrich Canada Inc., Ontario, Canada). Καθαρή αιθανόλη, φορμαλδεΰδη 10%, παραφορμαλδεύδη 2.5%, διάλυμα Sorensen s (Εργαστήριο του Παθολογοανατομικού Τμήματος του Γενικού Κρατικού Νοσοκομείου «Γεννηματάς»). Χρωμογόνο διαμινοβενζιδίνη (DAB) (Vector laboratories Inc. Burlingame CA, USA). Άχρωμο p-νιτροφαινυλοφωσφορικό οξύ (PNPP), διάλυμα διαμινοδιαιθυλπροπανόλης με θειικό ψευδάργυρο, άλας μαγνησίου και υδροξυαιθυλδιαμινο τριοξικό οξύ (ΗEDTA) (Abbott Laboratories,IL). Διάλυμα χρωμογόνου Trypan blue για τη βαφή των νεκρών κυττάρων (Sigma-Aldrich Canada Inc., Ontario, Canada.). ΟΡΓΑΝΑ Κλίβανος Forma Scientific CO 2 Incubator (Anti Σελ, Ελλάδα) για την επώαση των κυτταροκαλλιεργειών. Μικροσκόπιο Leica για τις κυτταρικές παρατηρήσεις. Μηχάνημα φυγοκέντρισης διαλύματος κυττάρων ALC International SRL PK131R.


75 75 Εικόνα 1.Το πιστοποιητικό της εταιρείας RD Systems που συνοδεύει τη συσκευασία του rh- TGF-β1
76 76 Εστία νηματικής ροής Microflow Biological safety Cabinet (Anti Σελ, Ελλάδα) (για τις άσηπτες συνθήκες όπου πραγματοποιήθηκε η μελέτη). Αιμοκυτταρόμετρο Neubauer (Sigma-Aldrich Canada Inc., Ontario, Canada). Οπτικό μικροσκόπιο Nikon ECLIPSE ME 600 της Nikon, Kogagu, Tokyo, Japan. ΜΕΘΟΔΟΙ Μέθοδος λήψης των κυττάρων Προκειμένου να γίνει η απομόνωση των κυττάρων από τα δόντια, όλα τα προς χρήση εργαλεία (λαβίδες, ψαλίδια, πορσελάνινα τρυβλία) αποστειρώθηκαν στο εργαστήριο σε αυτόκαστο ενώ οι επιφάνειες εργασίας απολυμαίνονταν με φλόγα και αιθυλική αλκοόλη 75% ύστερα από κάθε χρήση. Τα δόντια τοποθετήθηκαν στα αποστειρωμένα πορσελάνινα δοχεία που περιείχαν διάλυμα με 5ml DMEM (Dulbecco s Modified Eagle s Medium) εμπλουτισμένο με 5mg κολλαγενάση Ι. Ακολούθησε αφαίρεση των ιστών εντός του διαλύματος από το μέσο τριτημόριο της ρίζας με νυστέρι Νο15 κατά τις γνωστές μεθόδους (Somerman και συν. 1988, Markopoulou και συν. 2003). Τα ξέσματα των ιστών που αφαιρέθηκαν τοποθετήθηκαν σε διάλυμα DMEM εμπλουτισμένο με 10% εμβρυϊκό βόειο ορό (FBS) και ακολούθησε η καλλιέργεια των κυττάρων. Μέθοδος ανάπτυξης καλλιεργειών Καλλιέργεια κυττάρων Προκειμένου να διαχωριστούν τα κύτταρα από τα ξέσματα των ιστών ακολούθησε επώαση των ξεσμάτων με κολλαγενάση Ι σε θερμοκρασία 37ºC
77 77 για 30 λεπτά και φυγοκέντριση τους για άλλα 10 λεπτά σε 2000rpm. Το ίζημα του διαλύματος, το οποίο περιείχε και τα κύτταρα, επαναδιαλύθηκε σε φλάσκες 25cm 2 σε διάλυμα το οποίο συνίστατο από DMEM εμπλουτισμένο με 10% εμβρυϊκό βόειο ορό (FBS), 5000U/ml πενικιλίνη G, 5mg/ml στρεπτομυκίνη και 850 mg/ml fungizone και τοποθετήθηκε σε επωαστικό κλίβανο στους 37ºC σε περιβάλλον υγρασίας 95% και παρουσία CO 2 5%. Η ανάπτυξη των κυττάρων στο DMEM ολοκληρώθηκε σε 7-10 μέρες όπου τα κύτταρα κάλυψαν το 100% της επιφάνειας του τρυβλίου. Τρεις κυτταρικές σειρές PDL 1,PDL 2 και PDL 3 καλλιεργήθηκαν, μια από το κάθε δόντι. Ανακαλλιέργεια έγινε τη δέκατη μέρα. Συγκεκριμένα αφού αναρροφήθηκε το θρεπτικό υλικό και ξεπλύθηκε η καλλιέργεια σε 5ml ρυθμιστικό διάλυμα φωσφορικών (PBS), προστέθηκε 1ml θρυψίνης με EDTA. Ακολούθησε επώαση των κυττάρων για 5 λεπτά σε θερμοκρασία 37 C και παρουσία CO 2 5% και παρατήρηση στο μικροσκόπιο για τη πλήρη αποκόλληση τους. Στη συνέχεια τα κύτταρα τοποθετήθηκαν σε 20ml DMEM στο οποίο αιωρούνταν. Το εναιώρημα των κυττάρων χωρίστηκε σε ίσα μέρη όπου τα κύτταρα αναπτυχθήκαν και πάλι έως την επόμενη ανακαλλιέργεια. Καλλιέργεια και συγχρονισμός κυττάρων Η μελέτη της επίδρασης στα κύτταρα εξωγενών παραγόντων προϋποθέτει το συγχρονισμό των κυττάρων σε φάση ηρεμίας (GO). Για το λόγο αυτό, κύτταρα σε αραιές καλλιέργειες διατηρήθηκαν σε στέρηση ορού έτσι ώστε να συγχρονίζονται τα κύτταρα μιας υποπλήρους μονοστοιβάδας υπό την επίδραση χαμηλών συγκεντρώσεων ορρού (Tobbey και συν. 1988).
78 78 Η προετοιμασία των καλλιεργειών έγινε ως εξής: Τα κύτταρα αποκολλήθηκαν κατά τα γνωστά, μετρήθηκαν τα συνολικά κύτταρα και αφού έγινε η κατάλληλη αραίωση τα κύτταρα επιστρώθηκαν σε πλαστικά τρυβλία. Καταρχάς, δημιουργήθηκαν διάσπαρτες καλλιέργειες οι οποίες στη συνέχεια έγιναν πλήρεις. Παρατήρηση των καλλιεργειών στο μικροσκόπιο αναδεικνύει την ατρακτοειδή μορφολογία των κυττάρων αυτών, χαρακτηριστική των ινοβλαστών. Μέτρηση κυττάρων Οι μετρήσεις των κυττάρων έγιναν με το αιμοκυτταρόμετρο και αφορούσαν τη μέτρηση του συνολικού αριθμού των κυττάρων. Συγκεκριμένα, τα κύτταρα αποκολλήθηκαν από το τρυβλίο όπου αιωρούνταν σε διάλυμα DMEM εμπλουτισμένο με 10% FBS με τη μέθοδο της θρυψινοποίησης και τοποθετήθηκαν σε πλακίδιο στο αιμοκυτταρόμετρο. Οι μετρήσεις του αριθμού των κυττάρων εξέφραζαν το μέσο όρο του αριθμού των αντιστοίχων δειγμάτων πού είχαν αναπτυχθεί παράλληλα και στις ίδιες συνθήκες. Αποθήκευση των κυττάρων Προκειμένου τα κύτταρα να αποθηκευτούν, έγινε φυγοκέντριση για 10 λεπτά στις 2500 στροφές το λεπτό, αφαίρεση του υπερκείμενου διαλύματος και επαναδιάλυση του ιζήματος σε 10% DMSO. Μετά τη πλήρη διάλυση του ιζήματος σε 1ml DMSO τα κύτταρα αποθηκεύτηκαν σε φιαλίδιο cryovial. Ακολούθησε αρχική ψύξη στους -80ºC για 24 ώρες και τελικά σε υγρό άζωτο στους -180 ºC.
79 79 Μέθοδοι ταυτοποίησης των κυττάρων Ταυτοποίηση των κυττάρων του περιρριζίου Τα περιρριζικά κύτταρα είναι ένας μικτός πληθυσμός κυττάρων. Το μεγαλύτερο ποσοστό αυτών είναι ινοβλάστες μεσεγχυματικής προέλευσης. Οι ινοβλάστες του περιρριζίου παρουσιάζουν ετερογένεια ως προς τα λειτουργικά χαρακτηριστικά τους. Έτσι υποπληθυσμοί αυτών είναι κύτταρα που μορφολογικά μοιάζουν στις ινοβλάστες αλλά παρουσιάζουν οστεοβλαστική δραστηριότητα. Η ποικιλομορφία αυτή καθιστά πολύπλοκη και δυσνόητη τη διαδικασία επούλωσης των περιοδοντικών ιστών. Για το λόγο αυτό κρίνεται απαραίτητη η ταυτοποίηση των κυττάρων αυτών. Συγκεκριμένοι δείκτες για το χαρακτηρισμό του ινοβλαστικού ή οστεοβλαστικού φαινοτύπου των ινοβλαστών του περιρριζίου δεν αναφέρονται σαφώς στη βιβλιογραφία. Γενικά τα κύτταρα αυτά έχει βρεθεί ότι εκφράζουν υψηλά επίπεδα αλκαλικής φωσφατάσης, παράγουν πρωτεΐνες της εξωκυττάριας θεμέλιας ουσίας σχετιζόμενες με το οστούν όπως η οστεονεκτίνη, η οστική πρωτεογλυκάνη Ι, η οστική σιαλοπρωτεΐνη Ι, η οστεοποντίνη, η οστεοκαλσίνη, αντιδρούν παρουσία της παραθυρεοειδούς ορμόνης (PTH) με αύξηση της παραγωγής του camp και μπορούν να παράγουν ενασβεστιωμένα οζίδια in vitro και είναι πλούσια σε ακτίνη (Somerman και συν. 1988, Piche και συν. 1989, Nohutsu και συν. 1997). Η ταυτοποίηση των κυττάρων έγινε α) ανοσοϊστοχημικά με τη χρήση αντισωμάτων βιμεντίνης για την πιστοποίηση της μεσεγχυματικής τους προέλευσης και λείας μυϊκής ακτίνης, η οποία εντοπίζεται σε μεγάλες ποσότητες στις ινοβλάστες, για τη πιστοποίηση του ινοβλαστικού τους χαρακτήρα και β) με τη μέτρηση της δραστηριότητας της αλκαλικής
80 80 φωσφατάσης και γ) του ποσοτικού προσδιορισμού της οστεοκαλσίνης για την πιστοποίηση του οστεοβλαστικού τους χαρακτήρα. Kύτταρα που βρίσκονταν στη τρίτη γενιά (passage) χρησιμοποιήθηκαν σε όλα τα πειράματα. Ανοσοϊστοχημεία Τα κύτταρα αφαιρέθηκαν από το υγρό άζωτο και τοποθετήθηκαν σε κλίβανο στους 37 ºC μέχρι να ξεπαγώσουν. Στη συνέχεια τοποθετήθηκαν σε καινούργια φλάσκα με 10ml DMEM όπου παρέμειναν μέχρι να προσκολληθούν πλήρως. Ακολούθησε αφαίρεση του θρεπτικού υλικού, ξέπλυμα με ρυθμιστικό διάλυμα φωσφορικών (PBS), αφαίρεση του PBS και προσθήκη 1ml θρυψίνης και EDTA. Τα κύτταρα μεταφέρθηκαν σε ειδικό σωλήνα και φυγοκεντρήθηκαν στα 12G για 15 λεπτά. Ακολούθησε η διαδικασία της ταυτοποίησης η οποία έγινε σύμφωνα με το πρωτόκολλο του Παθολογοανατομικού Τμήματος του Γενικού Κρατικού Νοσοκομείου «Γεννηματάς». Συγκεκριμένα μετά τη φυγοκέντριση κρατήθηκε το ίζημα το οποίο με πιπέτα μικρής διαμέτρου μεταφέρθηκε στα πλακίδια πού προορίζονταν για ανοσοϊστοχημεία. Σε κάθε πλακίδιο τοποθετήθηκε υλικό όσο η κεφαλή της καρφίτσας και απλώθηκε με λοξές κινήσεις για αποφυγή σύνθλιψης των κυττάρων. Για να επιτευχθεί η καλύτερη επικόλληση και ακινητοποίηση των κυττάρων χρησιμοποιήθηκαν τρεις διαφορετικοί παράγοντες: καθαρή αιθανόλη, φορμαλδεΰδη 10% και παραφορμαλδεΰδη 2.5% σε διάλυμα Sorensen s και ph=7,4. Η αλκοόλη τοποθετήθηκε αμέσως στα πλακίδια ενώ οι άλλοι δύο παράγοντες τοποθετήθηκαν μετά το οριστικό στέγνωμα των κυττάρων. Για το πείραμα αυτό και για κάθε κυτταρική σειρά (PDL 1, PDL 2, PDL 3 ) χρησιμοποιήθηκαν 18 πλακίδια (Menzel-Glaser Co,
81 81 Super Frost Plus), 6 για κάθε υγρό ακινητοποίησης. Τρία πλακίδια από τη κάθε ομάδα δέχτηκαν αντίσωμα βιμεντίνης (μονοκλωνικό αντίσωμα ποντικού) σε διάλυση 1:50, 1:100 και 1:200 και τα υπόλοιπα αντίσωμα α-λείας μυϊκής ακτίνης (μονοκλωνικό αντίσωμα ποντικού) σε αντίστοιχες διαλυτότητες. Το πείραμα επαναλήφθηκε εις τριπλούν και για τις τρεις κυτταρικές σειρές για το υγρό που παρουσίασε την καλύτερη ακινητοποίηση και για τις συγκεντρώσεις των αντισωμάτων που κατέδειξαν την ισχυρότερη αντίδραση. Η ανοσοϊστοχημική μέθοδος που εφαρμόσθηκε είναι η μέθοδος συμπλέγματος αβιδίνης βιοτίνης-υπεροξειδάσης (ABC) (Hsu και συν.1981, Hsu και συν. 1981, Rifas και συν. 1999). Πρόκειται για ανοσοενζυμική μέθοδο και θεωρείται πιο ευαίσθητη από τη μέθοδο υπεροξειδάσηςαντιυπεροξειδάσης (PAP), λόγω της μεγάλης δεσμευτικής ικανότητας μεταξύ βιοτίνης και αβιδίνης. Η αβιδίνη είναι γλυκοπρωτεΐνη, προέρχεται από το λευκό του αυγού και έχει μεγάλη τάση σύνδεσης με τη βιοτίνη. Η βιοτίνη είναι βιταμίνη με χαμηλό μοριακό βάρος και βρίσκεται στη λέκιθο του αυγού. Η βιοτίνη συνδέεται με το δεύτερο αντιορό με τον ίδιο τρόπο που συνδέεται η υπεροξειδάση. Στη συνέχεια ο τρίτος αντιορός που αποτελείται από ένα σύμπλεγμα αβιδίνης και ενζύμου συνδεδεμένου με αβιδίνη συνδέεται μέσω μερικών ελεύθερων θέσεων αβιδίνης του ενζύμου με τη βιοτίνη του δεύτερου αντισώματος. Για την ανάδειξη της αντίδρασης χρησιμοποιήθηκε το Vectastain Elite ABC kit (Vector laboratories Inc. Burlingame, CA). Οι επωάσεις με το κύριο αντίσωμα έγιναν ολονύκτια (16 ώρες) σε θερμοκρασία 4 ºC ενώ για την αναγνώριση των αντιγόνων με το κοινό μικροσκόπιο χρησιμοποιήθηκε το χρωμογόνο διαμινοβενζιδίνη (DAB). Η διαμινοβενζιδίνη πολυμερίζεται με τη παρουσία υπεροξειδάσης και Η 2 Ο 2 σχηματίζοντας
82 82 αδιάλυτο πολυμερές που εντοπίζεται στις θέσεις αντιγόνου αντισώματος δίνοντας ένα καφέ χρώμα ορατό στο κοινό μικροσκόπιο. Οι μοριακές μεταβολές οδηγούν στο σχηματισμό σταθερού χρώματος που δεν είναι διαλυτό σε οργανικούς διαλύτες. Η παρατήρηση των πλακιδίων έγινε σε οπτικό μικροσκόπιο (Nikon ECLIPSE ME 600 της Nikon, Kogagu, Tokyo, Japan) από όπου λήφθηκαν φωτογραφίες με ψηφιακή κάμερα SONY Cyber shot. Ποσοτικός προσδιορισμός της δραστηριότητας της αλκαλικής φωσφατάσης Η αλκαλική φωσφατάση είναι ένζυμο που έχει τη δυνατότητα να υδρολύει τις οργανικές ενώσεις του φωσφόρου στην επιφάνεια της κυτταρικής μεμβράνης και χρησιμοποιείται ως δείκτης του ρυθμού απορρόφησης και εναπόθεσης οστίτη ιστού ( Nojima και συν ). Η αυξημένη δραστηριότητα της αλκαλικής φωσφατάσης αποτελεί το κυριότερο δείκτη του οστεοβλαστικού χαρακτήρα των κυττάρων του περιρριζίου ανάλογα με το δυναμικό τους φαινότυπο (Somerman και συν. 1988, Piche και συν. 1989, Hoang και συν ) και αποτελεί σημαντικότατο δείκτη κατά τη διαφοροποίηση τους. Η μέθοδος που ακολούθησε, αφορά στη περιοδική κατανομή της ενζυματικής δραστηριότητας της αλκαλικής φωσφατάσης και στηρίζεται στην υδρόλυση του άχρωμου p-νιτροφαινυλοφωσφορικού οξέος (Pnpp) όπου ελευθερώνεται p-νιτροφαινόλη της οποίας η απορρόφηση μετράται στα 404 nm και είναι ανάλογος της δραστηριότητας της αλκαλικής φωσφατάσης στο δείγμα. Η p- νιτροφαινόλη στο Ph (αλκαλικό) του assay παίρνει τη μορφή κίτρινου φαινοξειδίου (Βessey και συν. 1946, Tietz 1990).
83 83 Για τη μελέτη της δραστηριότητας της αλκαλικής φωσφατάσης αναρροφήθηκαν 5ml θρεπτικού υλικού από τις καλλιέργειες της κάθε κυτταρικής σειράς PDL 1, PDL 2, PDL 3 αφού είχε προηγηθεί φυγοκέντριση για λήψη του υπερκείμενου. Η εκτίμηση της αλκαλικής δραστηριότητας έγινε την τέταρτη μέρα σε όλες τις κυτταρικές σειρές. Η δράση του ένζυμου υπολογίστηκε σε units όπου κάθε unit αντιστοιχεί σε 1μmol/L p-νιτροφαινόλης που απελευθερώνεται ανά λεπτό. Τρία δείγματα από κάθε κυτταρική καλλιέργεια χρησιμοποιήθηκαν για το προσδιορισμό της δραστηριότητας της αλκαλικής φωσφατάσης. Για την ανάδειξη της αντίδρασης χρησιμοποιήθηκε το AEROSET SYSTEM (Abbott Laboratories,IL). Ποσοτικός προσδιορισμός της οστεοκαλσίνης Η οστεοκαλσίνη είναι η πιο συνήθης μη κολλαγενής πρωτεΐνη του οστού. Είναι πρωτεΐνη χαμηλού μοριακού βάρους πού περιέχει τρία υπολείμματα α- καρβοξυλ-γλουταμικά οξέα ανά μόριο. Η βιταμίνη Κ έχει ιδιαίτερη σημασία στη σύνθεση των υπολειμματικών α-καρβοξυγλουταμινικών οξέων. Η οστεοκαλσίνη παίζει σημαντικό ρόλο στην ενασβεστίωση του οστού. Αυτή παράγεται από οστεοβλαστικά κύτταρα και η σύνθεση της αυξάνεται κατά την ενασβεστίωση του οστού και με την οστεοβλαστική διαφοροποίηση (Garant 2003). Ο ποσοτικός προσδιορισμός της οστεοκαλσίνης έγινε με το σύστημα Nichols Advantage (Nichols Institute Diagnostics, CA). Το σύστημα αυτό βασίζεται στην χημειοφωταύγεια και στην ανοσομετρική μέθοδο. Η χημειοφωταύγεια πραγματοποιείται με τη χρήση χημειοφωταυγών εστέρων ακριδινίου οι οποίοι εκπέμπουν φως κατόπιν θεραπείας με υπεροξείδιο του υδρογόνου και ένα αλκαλικό διάλυμα. Το σύστημα περιέχει δύο διαλύματα. To Trigger 1 διάλυμα
84 84 που περιέχει υπεροξείδιο του υδρογόνου σε διαλυμένο οξύ και το Trigger2 διάλυμα που περιέχει διαλυμένο υδροξείδιο του νατρίου. Το χημειοφωταυγές σύστημα πραγματοποιεί αυτόματη έγχυση των δύο διαλυμάτων στα wells με αποτέλεσμα την οξείδωση του ακριδινικού εστέρα. Το οξειδωμένο προϊόν βρίσκεται σε φάση διέγερσης με αποτέλεσμα η επαναφορά του στο αρχικό στάδιο να οδηγεί σε εκπομπή φωτός. Το φως αυτό προσδιορίζεται ποσοτικά σε 2 δευτερόλεπτα και εκφράζεται σε σχετικές μονάδες φωτός (RLU: Relative light units) από το φωτομετρητή του συστήματος, Η ανοσομετρική μέθοδος, από την άλλη, χρησιμοποιεί δύο μονοκλωνικά αντισώματα ειδικά για την ακέραιη (intact) ανθρώπινη οστεοκαλσίνη (OC 1-49). Το ένα αντίσωμα, ειδικό για τα αμινοξέα 7-19 του πεπτιδίου, συνδέεται με τη βιοτίνη και χρησιμοποιείται για να συγκρατήσει την οστεοκαλσίνη. Το άλλο αντίσωμα, ειδικό για τα αμινοξέα του πεπτιδίου, σημαίνεται με ένα εστέρα ακριδινίου για ανίχνευση. Το δείγμα που περιέχει την οστεοκαλσίνη εγκιβωτίζεται στο σύστημα ταυτόχρονα με το ενωμένο με τη βιοτίνη αντίσωμα και το σημασμένο με ακριδίνιο αντίσωμα στους 37ºC. Στο πρώτο εγκιβωτισμό η οστεοκαλσίνη βρίσκεται ανάμεσα στα δύο αντισώματα με τη μορφή σάντουϊτς. Μετά την πρώτη περίοδο εγκιβωτισμού μαγνητικά σωμάτια καλλυμένα με σεπταβιδίνη προστίθενται στην αντίδραση για να ακολουθήσει μια δεύτερη περίοδος εγκιβωτισμού. Το σύμπλεγμα σάντουϊτς συνδέεται με τα μαγνητικά σωμάτια εξαιτίας της υψηλής συγγένειας μεταξύ βιοτίνης και σεπταβιδίνης. Ακολουθεί αναρρόφηση και πλύση του αντιδραστικού μίγματος με αποτέλεσμα το διαχωρισμό του ελεύθερου σημασμένου αντισώματος από εκείνο που είναι συνδεδεμένο με τα μαγνητικά σωμάτια. Τα δισκία (wells) που περιέχουν τα πλυμένα μαγνητικά σωμάτια μεταφέρονται στο φωτομετρητή
85 85 όπoυ γίνεται αυτόματη έγχυση του Trigger 1 και Trigger2 για να ξεκινήσει η χημειοφωταυγής αντίδραση. Το φως προσδιορίζεται ποσοτικά με το φωτομετρητή και εκφράζεται ως RLU (Relative light units). Το ποσό του συνδεδεμένου σημασμένου αντισώματος είναι ανάλογο της συγκέντρωσης της οστεοκαλσίνης στο δείγμα. Τρία δείγματα από κάθε κυτταρική καλλιέργεια χρησιμοποιήθηκαν για τον προσδιορισμό της οστεοκαλσίνης ενώ ο ποσοτικός προσδιορισμός της οστεοκαλσίνης πραγματοποιήθηκε την τέταρτη μέρα για όλες τις κυτταρικές καλλιέργειες. Μέθοδος εκτίμησης του πολλαπλασιασμού των κυττάρων της καλλιέργειας υπό την επίδραση του TGF-β 1 και της 17-β οιστραδιόλης(ε 2 ) Η επίδραση του TGF-β 1 και της 17-β οιστραδιόλης (Ε2) εφαρμόσθηκε στη κυτταρική σειρά PDL 1 λόγω του οστεοβλαστικού της φαινοτύπου όπως αυτός προέκυψε από τα προηγούμενα πειράματα. Ο rhtgf-β 1 (RD Systems, London,UK) εφαρμόσθηκε σε συγκεντρώσεις 1ng/ml και 25ng/ml ενώ η Ε 2 (Sigma-Aldrich Canada Inc., Ontario, Canada) σε συγκεντρώσεις 1nM και 100nM. Η επίδραση των παραγόντων έγινε για 48 και 72 ώρες. Ακολούθησε η μέτρηση των ζωντανών κυττάρων στους δύο αυτούς χρόνους στο αιμοκυτταρόμετρο Neubauer κατόπιν χρώσης τους με το χρωμοφόρο Trypan blue. Το Trypan blue είναι μια ζωτική χρωστική που χρησιμοποιείται προκειμένου να εκτιμηθεί η αναλογία των ζωντανών κυττάρων σε μια καλλιέργεια (Phillips και συν.1957). Η αντίδραση στη χρωστική βασίζεται στο ότι το χρωμοφόρο δεν αντιδρά με το κύτταρο παρά μόνο εάν έχει καταστραφεί η κυτταρική τους μεμβράνη. Ο χρωματισμός των κυττάρων διευκολύνει την παρατήρηση της μορφολογίας. Τα ζωντανά κύτταρα δεν αντιδρούν στη
86 86 χρωστική ενώ αντίθετα τα νεκρά κύτταρα χρωματίζονται. Ακολουθεί η μέτρηση των κυττάρων με τη χρήση του αιμοκυτταρομέτρου που φέρει τις χαρακώσεις μέτρησης Neubauer. Πιο συγκεκριμένα, τα κύτταρα αφαιρέθηκαν από το υγρό άζωτο και τοποθετήθηκαν σε κλίβανο στους 37 ºC για 2 λεπτά για να ξεπαγώσουν. Στη συνέχεια τοποθετηθήκαν σε καινούργια φλάσκα με 5ml DMEM εμπλουτισμένου με 10% FBS ελεύθερου οιστρογόνων (Phenol-red free), με 5000U/ml πενικιλίνη G και με 5000 U/ml στρεπτομυκίνη. Η φλάσκα τοποθετήθηκε σε επωαστικό κλίβανο στους 37ºC σε περιβάλλον υγρασίας 95% και παρουσία CO 2 5% μέχρι τη πλήρωση του 100% της επιφάνειας της με κύτταρα. Ακολούθησε αφαίρεση του θρεπτικού υλικού, ξέπλυμα με ρυθμιστικό διάλυμα φωσφορικών (PBS), αφαίρεση του PBS και προσθήκη 1ml θρυψίνης και EDTA για την πλήρη αποκόλληση των κυττάρων όπως αυτή παρατηρήθηκε με οπτική παρατήρηση στο ηλεκτρονικό μικροσκόπιο. Στη συνέχεια μεταφέρθηκε ίσος αριθμός κυττάρων ( ) σε 2 τρυβλία 24 θέσεων όπου προστέθηκαν ο TGF-β1 και η 17-β οιστραδιόλη (Ε2). Συγκεκριμένα μελετήθηκαν πέντε ομάδες κυττάρων που αποτελούνταν από τα κύτταρα μάρτυρες, τα κύτταρα με 1ng/ml TGF-β1, τα κύτταρα με 25ng/ml TGF-β1, τα κύτταρα με 1nM Ε 2, και τα κύτταρα με 100nM Ε 2. Τα τρυβλία τοποθετήθηκαν σε επωαστικό κλίβανο στους 37ºC σε περιβάλλον υγρασίας 95% και παρουσία CO 2 5%. Μετά την ολοκλήρωση της επώασης στις 48 και 72 ώρες και αφού προστέθηκε 1ml θρυψίνης και EDTA για να ξεκολλήσουν τα κύτταρα προστέθηκε 1ml Trypan blue για κάθε ml ξεκολλημένων κυττάρων. Ακολούθησε επώαση στον κλίβανο για 5 λεπτά και μέτρηση του αριθμού των κυττάρων στο αιμοκυτταρόμετρο Neubauer.
87 87 Γ. ΣΤΑΤΙΣΤΙΚΗ ΕΠΕΞΕΡΓΑΣΙΑ ΚΑΙ ΑΠΟΤΕΛΕΣΜΑΤΑ Τα αποτελέσματα που ακολουθούν αφορούν: Α) Στο χαρακτηρισμό των κυττάρων του περιρριζίου ως προς τη μεσεγχυματική τους προέλευση και τον ινοβλαστικό τους χαρακτήρα μέσω της ανοσοϊστοχημικής τους αντίδρασης στα αντισώματα της βιμεντίνης και της λείας μυϊκής ακτίνης αντίστοιχα. Β) Στο χαρακτηρισμό των κυττάρων του περιρριζίου ως προς τον οστεοβλαστικό τους χαρακτήρα με τη μέτρηση της δραστηριότητας της αλκαλικής φωσφατάσης και το ποσοτικό προσδιορισμό της οστεοκαλσίνης. Γ) Στην επίδραση της 17-β οιστραδιόλης και του αυξητικού παράγοντα TGFβ 1 στον πολλαπλασιασμό των κυττάρων του περιρριζίου όπως αυτός εκτιμήθηκε από τη μέτρηση του αριθμού των ζωντανών κυττάρων μετά την εφαρμογή διαφορετικών δόσεων των δύο παραγόντων και για διαφορετικά χρονικά διαστήματα. Στατιστική επεξεργασία Η στατιστική επεξεργασία χρησιμοποιήθηκε για την ανάλυση και σύγκριση των μετρήσεων που αφορούσαν στο πολλαπλασιασμό των κυττάρων του περιρριζίου στις 48 και 72 ώρες υπό την επίδραση 1ng/mL και 25ng/mL του αυξητικού παράγοντα TGF-β 1 καθώς και 1nM και 100nM της 17-β οιστραδιόλης. Για τη σύγκριση του αριθμού των κυττάρων της κάθε ομάδας μεταξύ των δύο χρονικών στιγμών (48 έναντι 72 ωρών) εφαρμόσθηκε το Student s t-test για ανεξάρτητα δείγματα. Ως κριτήριο για τη «στατιστική σημαντικότητα» χρησιμοποιήθηκε το όριο πιθανότητας p<0.05.
88 88 Για τη σύγκριση του αριθμού των κυττάρων μεταξύ των ομάδων στη κάθε χρονική στιγμή εφαρμόσθηκε η στατιστική μέθοδος one way ANOVA. Ως κριτήριο για τη «στατιστική σημαντικότητα» χρησιμοποιήθηκε το όριο πιθανότητας p<0.05. ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΑΝΤΙΔΡΑΣΗ Για το πείραμα αυτό χρησιμοποιήθηκαν 18 πλακίδια (Menzel-Glaser Co, Super Frost Plus) για κάθε κυτταρική σειρά PDL 1, PDL 2, PDL 3 τα οποία χωρίστηκαν ανά 6 για την εφαρμογή των τριών διαφορετικών υγρών ακινητοποίησης (καθαρή αιθανόλη, φορμαλδεΰδη 10%, παραφορμαλδεΰδη 25%). Τρία πλακίδια από τη κάθε ομάδα δέχτηκαν αντίσωμα βιμεντίνης (μονοκλωνικό αντίσωμα ποντικού) σε διάλυση 1:50, 1:100 και 1:200 και τα υπόλοιπα αντίσωμα α-λείας μυϊκής ακτίνης (μονοκλωνικό αντίσωμα ποντικού) στις ίδιες διαλυτότητες. Από το πρώτο πείραμα φάνηκε ότι τα κύτταρα ακινητοποιούνται στη φορμαλδεΰδη 10% και καθόλου στα άλλα μέσα ακινητοποίησης (Εικόνα 1 και 2). Η βιμεντίνη σε διάλυση 1:50 και 1:100 έδωσε αδύναμη ανοσοϊστοχημική αντίδραση (Εικόνα 3). Το ίδιο ίσχυσε και για την ακτίνη σε διαλυτότητες 1:100 και 1:200 (Εικόνα 4). Η ισχυρότερη αντίδραση παρατηρήθηκε για τη βιμεντίνη σε διάλυση 1:200 (Εικόνες 5-7). Στην εικόνα 7 διακρίνεται η ανίχνευση της βιμεντίνης (καφέ χρώμα) γύρω από τον κυτταρικό πυρήνα μέσα στο κυτταρικό σκελετό και στις κυτταρικές προεξοχές. Η α-sma παρουσίασε την ισχυρότερη ανοσοϊστοχημική αντίδραση σε διάλυση 1:50 (Εικόνες 8 και 9). Επανάληψη του πειράματος και στις τρεις κυτταρικές σειρές με τη παρατήρηση δύο νέων πλακιδίων σε φορμαλδεΰδη 10% και στις διαλυτότητες που βρέθηκαν από το προπείραμα επιβεβαίωσε τη σύνδεση των αντισωμάτων.





89 89 Εικόνα 1. Βιμεντίνη 1:200 σε διάλυμα παραφορμαλδεΰδης 2.5% σε μεγέθυνση x 100. Εικόνα 2. Βιμεντίνη 1:200 σε διάλυμα φορμαλδεΰδης 10% σε μεγέθυνση x 100. Εικόνα 3. Βιμεντίνη 1:100 σε διάλυμα φορμαλδεΰδης 10% σε μεγέθυνση x 5.



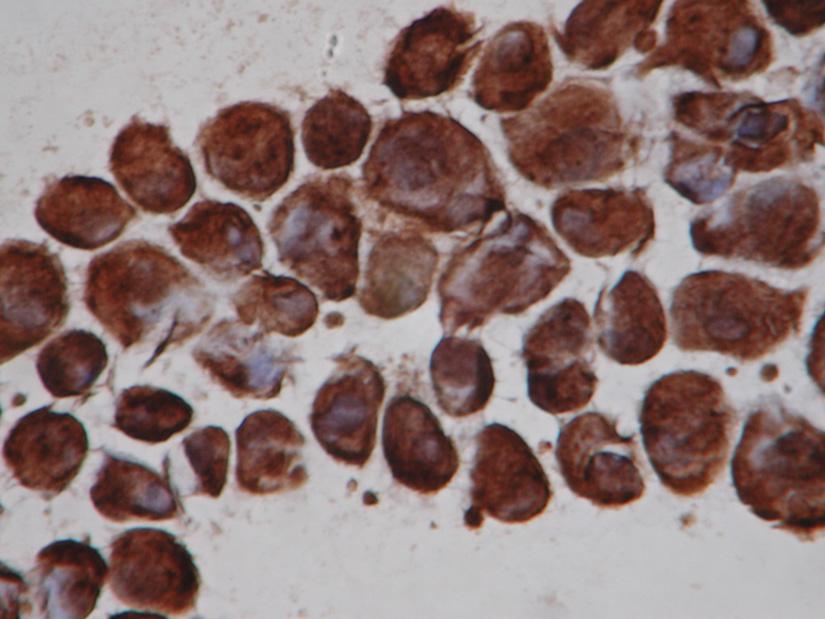
90 90 Εικόνα 4. Ακτίνη 1:100 σε διάλυμα φορμαλδεΰδης 10% σε μεγέθυνση x 100. Εικόνες 5 και 6. Βιμεντίνη 1:200 σε διάλυμα φορμαλδεΰδης 10% σε μεγέθυνση x 20 και x 50. Εικόνες 7. Βιμεντίνη 1:200 σε διάλυμα φορμαλδεΰδης 10% σε μεγέθυνση x 50.

.")

. Ο ποσοτικός προσδιορισμός της οστεοκαλσίνης πραγματοποιήθηκε με την εφαρμογή της χημειοφωταυγούς ανοσολογικής μεθόδου.")
91 91 Εικόνα 8 και 9. Ακτίνη 1:50 σε διάλυμα φορμαλδεΰδης 10% σε μεγέθυνση x 10 και x 50. Η ΕΚΤΙΜΗΣΗ ΤΗΣ ΔΡΑΣΤΗΡΙΟΤΗΤΑΣ ΤΗΣ ΑΛΚΑΛΙΚΗΣ ΦΩΣΦΑΤΑΣΗΣ ΚΑΙ Ο ΠΟΣΟΤΙΚΟΣ ΠΡΟΣΔΙΟΡΙΣΜΟΣ ΤΗΣ ΟΣΤΕΟΚΑΛΣΙΝΗΣ Η μέθοδος που χρησιμοποιήθηκε για τη μέτρηση της δραστηριότητας της αλκαλικής φωσφατάσης αφορά στη περιοδική κατανομή της ενζυματικής δραστηριότητας της αλκαλικής φωσφατάσης και στηρίζεται στην υδρόλυση του άχρωμου p- νιτροφαινυλοφωσφορικού οξέος (Pnpp). Η δράση του ένζυμου υπολογίστηκε σε units όπου κάθε unit αντιστοιχεί σε 1μmol/L p- νιτροφαινόλης που απελευθερώνεται ανά λεπτό. Και οι τρεις κυτταρικές σειρές εκφράσανε δραστηριότητα αλκαλικής φωσφατάσης. Η μέση τιμή της δραστηριότητας αυτής ήταν για την κάθε κυτταρική σειρά (PDL 1, PDL 2, PDL 3 ) 23, 20 και 11 μmol-p-nitrophenol/min αντίστοιχα (Πίνακας 1). Ο ποσοτικός προσδιορισμός της οστεοκαλσίνης πραγματοποιήθηκε με την εφαρμογή της χημειοφωταυγούς ανοσολογικής μεθόδου. Το φως που εκπέμπεται από τη χημειοφωταυγή αντίδραση προσδιορίζεται ποσοτικά με το φωτομετρητή και εκφράζεται ως RLU (Relative light units) ενώ το ποσό του συνδεδεμένου σημασμένου αντισώματος είναι ανάλογο της συγκέντρωσης της
Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης. Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010
 ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΚΕΦΑΛΑΙΟ 15. Κυτταρική ρύθμιση. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ. ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ
 ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία. Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ
 Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Συστήματα επικοινωνίας Ανθρωπίνου σώματος. ενδοκρινολογικό νευρικό σύστημα
 Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Σοφία Χαβάκη Λέκτορας Εργαστήριο Ιστολογίας-Εμβρυολογίας
 Οστίτης Ιστός Σοφία Χαβάκη Λέκτορας Εργαστήριο Ιστολογίας-Εμβρυολογίας λ ί Οστά εξειδικευμένος στηρικτικός-συνδετικός ιστός χαρακτηριστικά: σκληρή σύσταση και ακαμψία, λόγω παρουσίας αλάτων ασβεστίου στην
Οστίτης Ιστός Σοφία Χαβάκη Λέκτορας Εργαστήριο Ιστολογίας-Εμβρυολογίας λ ί Οστά εξειδικευμένος στηρικτικός-συνδετικός ιστός χαρακτηριστικά: σκληρή σύσταση και ακαμψία, λόγω παρουσίας αλάτων ασβεστίου στην
Η ΕΠΙΔΡΑΗ ΣΗ ΕΓΚΤΜΟΤΝΗ ΣΗΝ ΚΑΣΑΣΑΗ ΣΩΝ ΠΕΡΙΟΔΟΝΣΙΚΩΝ ΙΣΩΝ
 10 ο σνέδριο Φοιηηηών Οδονηιαηρικής τολής Πανεπιζηημίοσ Αθηνών ΓΝΩΣΙΚΟ ΑΝΣΙΚΕΙΜΕΝΟ ΠΕΡΙΟΔΟΝΣΟΛΟΓΙΑ Η ΕΠΙΔΡΑΗ ΣΗ ΕΓΚΤΜΟΤΝΗ ΣΗΝ ΚΑΣΑΣΑΗ ΣΩΝ ΠΕΡΙΟΔΟΝΣΙΚΩΝ ΙΣΩΝ σμμεηέτονηες: Λαμπροπούλοσ Ν. Οσρανία* Παπαγεωργίοσ
10 ο σνέδριο Φοιηηηών Οδονηιαηρικής τολής Πανεπιζηημίοσ Αθηνών ΓΝΩΣΙΚΟ ΑΝΣΙΚΕΙΜΕΝΟ ΠΕΡΙΟΔΟΝΣΟΛΟΓΙΑ Η ΕΠΙΔΡΑΗ ΣΗ ΕΓΚΤΜΟΤΝΗ ΣΗΝ ΚΑΣΑΣΑΗ ΣΩΝ ΠΕΡΙΟΔΟΝΣΙΚΩΝ ΙΣΩΝ σμμεηέτονηες: Λαμπροπούλοσ Ν. Οσρανία* Παπαγεωργίοσ
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών
 Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
ΣTHPIKTIKA KYTTAPA - EΞΩKYTTAPIA ΘEMEΛIA OYΣIA
 ΣTHPIKTIKA KYTTAPA - EΞΩKYTTAPIA ΘEMEΛIA OYΣIA µηχανική στήριξη Στηρικτικά κυτ. > παράγουν συστατικά εξωκυτ. θεµέλιας ουσίας για & οργάνωση τους χαρακτηριστικά τους > προέρχονται από το µεσέγχυµα παράγουν
ΣTHPIKTIKA KYTTAPA - EΞΩKYTTAPIA ΘEMEΛIA OYΣIA µηχανική στήριξη Στηρικτικά κυτ. > παράγουν συστατικά εξωκυτ. θεµέλιας ουσίας για & οργάνωση τους χαρακτηριστικά τους > προέρχονται από το µεσέγχυµα παράγουν
 Οστίτης Ιστός Dr. med. Λουκάς Κωνσταντίνου Ορθοπεδικός Χειρουργός Στήριξη Κίνηση Οστά εξειδικευµένος στηρικτικός-συνδετικός ιστός χαρακτηριστικά: σκληρήσύστασηκαι ακαµψία, λόγω παρουσίαςαλάτων ασβεστίουστην
Οστίτης Ιστός Dr. med. Λουκάς Κωνσταντίνου Ορθοπεδικός Χειρουργός Στήριξη Κίνηση Οστά εξειδικευµένος στηρικτικός-συνδετικός ιστός χαρακτηριστικά: σκληρήσύστασηκαι ακαµψία, λόγω παρουσίαςαλάτων ασβεστίουστην
Ατυπία Υπερπλασία- Δυσπλασία. Κίττυ Παυλάκη
 Ατυπία Υπερπλασία- Δυσπλασία Κίττυ Παυλάκη Jeanne Calment Κάπνιζε µέχρι τα 117 Πέθανε στα 122 Η σωστή λειτουργία των οργανισµών απαιτεί τη δυνατότητα προσαρµογής των κυττάρων και κατά συνέπεια και των
Ατυπία Υπερπλασία- Δυσπλασία Κίττυ Παυλάκη Jeanne Calment Κάπνιζε µέχρι τα 117 Πέθανε στα 122 Η σωστή λειτουργία των οργανισµών απαιτεί τη δυνατότητα προσαρµογής των κυττάρων και κατά συνέπεια και των
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία. Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ
 Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
ΠΕΤΡΟΣ ΔΕΔΕΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΣΙ ΠΕΡΙΛΗΨΗ
 ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΣΙ ΠΕΡΙΛΗΨΗ Το οστό αποτελεί ένα ιδανικό περιβάλλον για μετάσταση, καθώς η συνεχής και δυναμική ανάπλασή τον παρέχει μια γόνιμη βάση για την παλιννόστηση και τον πολλαπλασιασμό των καρκινικών
ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΣΙ ΠΕΡΙΛΗΨΗ Το οστό αποτελεί ένα ιδανικό περιβάλλον για μετάσταση, καθώς η συνεχής και δυναμική ανάπλασή τον παρέχει μια γόνιμη βάση για την παλιννόστηση και τον πολλαπλασιασμό των καρκινικών
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ ΤΟ ΚΥΤΤΑΡΟ Δ.ΑΡΕΘΑ
 ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ ΤΟ ΚΥΤΤΑΡΟ Δ.ΑΡΕΘΑ ΚΥΤΤΑΡΟ 2 Κατά την Βιολογία, κύτταρο ονομάζεται η βασική δομική και λειτουργική μονάδα που εκδηλώνει το φαινόμενο της ζωής. Έτσι, ως κύτταρο νοείται
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ ΤΟ ΚΥΤΤΑΡΟ Δ.ΑΡΕΘΑ ΚΥΤΤΑΡΟ 2 Κατά την Βιολογία, κύτταρο ονομάζεται η βασική δομική και λειτουργική μονάδα που εκδηλώνει το φαινόμενο της ζωής. Έτσι, ως κύτταρο νοείται
ΜΕΣΟΘΕΡΑΠΕΙΑ ΕΝΕΣΙΜΗ ΤΟΠΙΚΗ ΘΕΡΑΠΕΙΑ
 ΜΕΣΟΘΕΡΑΠΕΙΑ ΕΝΕΣΙΜΗ ΤΟΠΙΚΗ ΘΕΡΑΠΕΙΑ ΜΕΣΟΘΕΡΑΠΕΙΑ Αυτό σημαίνει ότι χρησιμοποιούμε μόνο ενέσιμα φάρμακα και μόνο στο σημείο που πάσχει. ΜΕΣΟΘΕΡΑΠΕΙΑ Ξεκίνησε στη λογική του γιατί να μη χορηγήσω ένα αντιφλεγμονώδες
ΜΕΣΟΘΕΡΑΠΕΙΑ ΕΝΕΣΙΜΗ ΤΟΠΙΚΗ ΘΕΡΑΠΕΙΑ ΜΕΣΟΘΕΡΑΠΕΙΑ Αυτό σημαίνει ότι χρησιμοποιούμε μόνο ενέσιμα φάρμακα και μόνο στο σημείο που πάσχει. ΜΕΣΟΘΕΡΑΠΕΙΑ Ξεκίνησε στη λογική του γιατί να μη χορηγήσω ένα αντιφλεγμονώδες
Ρύθµιση κυτταρικής λειτουργίας. Μεταγωγή σήµατος
 Ρύθµιση κυτταρικής λειτουργίας Μεταγωγή σήµατος 1 Εισαγωγή Η διαδικασία εξέλιξης των πολυκύτταρων οργανισµών (πρίν 2.5 δις χρόνια) άρχισε πολύ πιο αργά από την ύπαρξη των µονοκύτταρων οργανισµών (πρίν
Ρύθµιση κυτταρικής λειτουργίας Μεταγωγή σήµατος 1 Εισαγωγή Η διαδικασία εξέλιξης των πολυκύτταρων οργανισµών (πρίν 2.5 δις χρόνια) άρχισε πολύ πιο αργά από την ύπαρξη των µονοκύτταρων οργανισµών (πρίν
Παθοφυσιολογία της επούλωσης των ελκών στο διαβήτη και αιτίες αποτυχίας
 Παθοφυσιολογία της επούλωσης των ελκών στο διαβήτη και αιτίες αποτυχίας Ιωάννα Ελευθεριάδου Επιστημονικός Συνεργάτης Διαβητολογικού Κέντρου ΓΝΑ Λαϊκό Σακχαρώδης διαβήτης και έλκη κάτω άκρων 25% των ασθενών
Παθοφυσιολογία της επούλωσης των ελκών στο διαβήτη και αιτίες αποτυχίας Ιωάννα Ελευθεριάδου Επιστημονικός Συνεργάτης Διαβητολογικού Κέντρου ΓΝΑ Λαϊκό Σακχαρώδης διαβήτης και έλκη κάτω άκρων 25% των ασθενών
Φλεγμονή. Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ
 Φλεγμονή Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ Μη ειδική ανοσολογική άμυνα ΑΝΑΤΟΜΙΚΟΙ ΦΡΑΓΜΟΙ Φυσικοί: δέρμα, βλεννογόνοι, βλέννα, βήχας Χημικοί: λυσοζύμη, αντιμικροβιακά
Φλεγμονή Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ Μη ειδική ανοσολογική άμυνα ΑΝΑΤΟΜΙΚΟΙ ΦΡΑΓΜΟΙ Φυσικοί: δέρμα, βλεννογόνοι, βλέννα, βήχας Χημικοί: λυσοζύμη, αντιμικροβιακά
ΕΠΕΚΤΑΣΗ ΚΑΚΟΗΘΩΝ. Ι. ελλαδέτσιµα
 ΕΠΕΚΤΑΣΗ ΚΑΚΟΗΘΩΝ ΝΕΟΠΛΑΣΜΑΤΩΝ Ι. ελλαδέτσιµα Χαρακτηριστικά φυσιολογικών ιστών Κυτταρική συνοχή και επικοινωνία µέσω µορίων προσκόλλησης (καντχερίνες, σελεκτίνες, ιντεγκρίνες) Εξ επαφής αναστολή κυτταρικής
ΕΠΕΚΤΑΣΗ ΚΑΚΟΗΘΩΝ ΝΕΟΠΛΑΣΜΑΤΩΝ Ι. ελλαδέτσιµα Χαρακτηριστικά φυσιολογικών ιστών Κυτταρική συνοχή και επικοινωνία µέσω µορίων προσκόλλησης (καντχερίνες, σελεκτίνες, ιντεγκρίνες) Εξ επαφής αναστολή κυτταρικής
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Βιολογία Εμφυτευμάτων
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 5: Οστική Επούλωση Γεώργιος Μακρής Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons. Για
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 5: Οστική Επούλωση Γεώργιος Μακρής Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons. Για
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1
 Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
Πανεπιστημιο Θεσσαλιας Ιατρικη Σχολη
 Εισαγωγη στην Ορθοπαιδικη Κωνσταντινος Ν. ΜΑΛΙΖΟΣ Πανεπιστημιο Θεσσαλιας Ιατρικη Σχολη Εισαγωγη στην Ορθοπαιδικη Ορθοπαιδικη ή Ορθοπεδικη «παιδο-ορθωτικη» (Γ. Μπαμπινιωτης) Πανεπιστημιο Θεσσαλιας Ιατρικη
Εισαγωγη στην Ορθοπαιδικη Κωνσταντινος Ν. ΜΑΛΙΖΟΣ Πανεπιστημιο Θεσσαλιας Ιατρικη Σχολη Εισαγωγη στην Ορθοπαιδικη Ορθοπαιδικη ή Ορθοπεδικη «παιδο-ορθωτικη» (Γ. Μπαμπινιωτης) Πανεπιστημιο Θεσσαλιας Ιατρικη
ΕΡΓΑΣΤΗΡΙΑΚΟ ΚΕΝΤΡΟ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Ν. ΜΑΓΝΗΣΙΑΣ ( Ε.Κ.Φ.Ε ) ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ
 ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ") ΕΡΓΑΣΤΗΡΙΑΚΟ ΚΕΝΤΡΟ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Ν. ΜΑΓΝΗΣΙΑΣ ( Ε.Κ.Φ.Ε ) ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ Θέμα: ΜΙΚΡΟΣΚΟΠΙΚΗ ΠΑΡΑΤΗΡΗΣΗ ΜΟΝΙΜΩΝ ΠΑΡΑΣΚΕΥΑΣΜΑΤΩΝ ΚΥΤΤΑΡΩΝ ΚΑΙ ΙΣΤΩΝ Μέσος χρόνος πειράματος: 45 λεπτά Α. ΑΝΑΛΩΣΙΜΑ
ΕΡΓΑΣΤΗΡΙΑΚΟ ΚΕΝΤΡΟ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Ν. ΜΑΓΝΗΣΙΑΣ ( Ε.Κ.Φ.Ε ) ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ Θέμα: ΜΙΚΡΟΣΚΟΠΙΚΗ ΠΑΡΑΤΗΡΗΣΗ ΜΟΝΙΜΩΝ ΠΑΡΑΣΚΕΥΑΣΜΑΤΩΝ ΚΥΤΤΑΡΩΝ ΚΑΙ ΙΣΤΩΝ Μέσος χρόνος πειράματος: 45 λεπτά Α. ΑΝΑΛΩΣΙΜΑ
ΚΥΤΤΑΡΑ. Καρβουντζή Ηλιάνα (Βιολόγος) 1
 1") ΚΥΤΤΑΡΑ ΖΥΓΩΤΟ: Το πρώτο κύτταρο του οργανισμού από το οποίο με συνεχείς κυτταρικές διαιρέσεις προκύπτουν όλα. ΔΙΑΦΟΡΟΠΟΙΗΣΗ: Διαδικασία με την οποία τα κύτταρα αποκτούν διαφορετικά μορφολογικά και λειτουργικά
ΚΥΤΤΑΡΑ ΖΥΓΩΤΟ: Το πρώτο κύτταρο του οργανισμού από το οποίο με συνεχείς κυτταρικές διαιρέσεις προκύπτουν όλα. ΔΙΑΦΟΡΟΠΟΙΗΣΗ: Διαδικασία με την οποία τα κύτταρα αποκτούν διαφορετικά μορφολογικά και λειτουργικά
Λείος μυς. Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017
 Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
Ανθρώπινο σώμα: 1200 gr Ca. 99% στα οστά και τα δόντια Το υπόλοιπο βρίσκεται στους ιστούς
 ΡΥΘΜΙΣΗ ΑΣΒΕΣΤΙΟΥ Ανθρώπινο σώμα: 1200 gr Ca 99% στα οστά και τα δόντια Το υπόλοιπο βρίσκεται στους ιστούς ΡΟΛΟΣ ΕΞΩΚΥΤΤΑΡΙΟΥ Ca ++ Μυϊκή συστολή Συναπτική διαβίβαση Συσσώρευση αιμοπεταλίων Πήξη του αίματος
ΡΥΘΜΙΣΗ ΑΣΒΕΣΤΙΟΥ Ανθρώπινο σώμα: 1200 gr Ca 99% στα οστά και τα δόντια Το υπόλοιπο βρίσκεται στους ιστούς ΡΟΛΟΣ ΕΞΩΚΥΤΤΑΡΙΟΥ Ca ++ Μυϊκή συστολή Συναπτική διαβίβαση Συσσώρευση αιμοπεταλίων Πήξη του αίματος
Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν
 Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν ένα δυναµικό σκελετό που χρησιµεύει στη στήριξη και την
Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν ένα δυναµικό σκελετό που χρησιµεύει στη στήριξη και την
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
ΚΕΦΑΛΑΙΟ 1 Από το κύτταρο στον οργανισμό
 ΚΕΦΑΛΑΙΟ 1 Από το κύτταρο στον οργανισμό 1o ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11207, 2o ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11208 ΘΕΜΑ Δ Το ανθρώπινο σώμα, όπως και το σώμα κάθε πολυκύτταρου οργανισμού αποτελείται από πολλά
ΚΕΦΑΛΑΙΟ 1 Από το κύτταρο στον οργανισμό 1o ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11207, 2o ΔΙΑΓΩΝΙΣΜΑ - ΓΗ_Α_ΒΙΟ_0_11208 ΘΕΜΑ Δ Το ανθρώπινο σώμα, όπως και το σώμα κάθε πολυκύτταρου οργανισμού αποτελείται από πολλά
ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. :
 ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. : 2010336 ΕΡΓΑΣΙΑ ΓΙΑ ΤΟ Μ.Π.Σ. ΜΕΤΑΒΟΛΙΚΑ ΝΟΣΗΜΑΤΑ ΟΣΤΩΝ ΚΥΤΟΚΙΝΕΣ ΠΑΡΑΓΟΜΕΝΕΣ ΑΠΟ ΤΑ Τ-ΚΥΤΤΑΡΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΠΟΥ ΕΜΠΛΕΚΟΝΤΑΙ ΣΤΗΝ ΟΣΤΕΟΚΛΑΣΤΟΓΕΝΕΣΗ
ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. : 2010336 ΕΡΓΑΣΙΑ ΓΙΑ ΤΟ Μ.Π.Σ. ΜΕΤΑΒΟΛΙΚΑ ΝΟΣΗΜΑΤΑ ΟΣΤΩΝ ΚΥΤΟΚΙΝΕΣ ΠΑΡΑΓΟΜΕΝΕΣ ΑΠΟ ΤΑ Τ-ΚΥΤΤΑΡΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΠΟΥ ΕΜΠΛΕΚΟΝΤΑΙ ΣΤΗΝ ΟΣΤΕΟΚΛΑΣΤΟΓΕΝΕΣΗ
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ
 ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Γενετικοί δείκτες χαμηλής οστικής πυκνότητας σε άτομα με κυστική ίνωση
 Γενετική Γενετικοί δείκτες χαμηλής οστικής πυκνότητας σε άτομα με κυστική ίνωση Δρ Aleksandra Norek Βοηθός Τμήμα Ιατρικής Γενετικής Ινστιτούτο Υγείας Μητέρας και Παιδιού Βαρσοβία, Πολωνία Το μέσο προσδόκιμο
Γενετική Γενετικοί δείκτες χαμηλής οστικής πυκνότητας σε άτομα με κυστική ίνωση Δρ Aleksandra Norek Βοηθός Τμήμα Ιατρικής Γενετικής Ινστιτούτο Υγείας Μητέρας και Παιδιού Βαρσοβία, Πολωνία Το μέσο προσδόκιμο
Φλοιοτρόπος ορμόνη ή Κορτικοτροπίνη (ACTH) και συγγενή πεπτίδια
 και συγγενή πεπτίδια") ΕΠΙΝΕΦΡΙΔΙΑ Φλοιοτρόπος ορμόνη ή Κορτικοτροπίνη (ACTH) και συγγενή πεπτίδια 39 αμινοξέα Μ.Β. 4500 προοπιομελανοκορτίνη(pomc) 1. κορτικοτροπίνη (ACTH), 2. β λιποτροφίνη (β LPH), 3. γ λιποτροφίνη (γ LPH),
ΕΠΙΝΕΦΡΙΔΙΑ Φλοιοτρόπος ορμόνη ή Κορτικοτροπίνη (ACTH) και συγγενή πεπτίδια 39 αμινοξέα Μ.Β. 4500 προοπιομελανοκορτίνη(pomc) 1. κορτικοτροπίνη (ACTH), 2. β λιποτροφίνη (β LPH), 3. γ λιποτροφίνη (γ LPH),
ΕΡΓΑΣΤΗΡΙΟ ΙΣΤΟΛΟΓΙΑΣ Μ. ΠΑΥΛΙ ΗΣ
 ΕΡΓΑΣΤΗΡΙΟ ΙΣΤΟΛΟΓΙΑΣ Μ. ΠΑΥΛΙ ΗΣ Hράκλειο, εκέμβριος 2011 ΤΥΠΟΙ ΙΣΤΩΝ 1. Eπιθηλιακός Πολυεδρικά κύτταρα που είναι πάρα πολύ στενά συνδεδεμένα και φέρουν ελάχιστη μεσοκυττάρια ουσία 2. Συνδετικός Κύτταρα
ΕΡΓΑΣΤΗΡΙΟ ΙΣΤΟΛΟΓΙΑΣ Μ. ΠΑΥΛΙ ΗΣ Hράκλειο, εκέμβριος 2011 ΤΥΠΟΙ ΙΣΤΩΝ 1. Eπιθηλιακός Πολυεδρικά κύτταρα που είναι πάρα πολύ στενά συνδεδεμένα και φέρουν ελάχιστη μεσοκυττάρια ουσία 2. Συνδετικός Κύτταρα
ΣYΣTAΛTA KYTTAPA. Tα συσταλτά κύτταρα παράγουν > δυνάµεις κίνησης µε την αλληλεπίδραση > ακτίνης. & µυοσίνης
 ΣYΣTAΛTA KYTTAPA Tα συσταλτά κύτταρα παράγουν > δυνάµεις κίνησης µε την αλληλεπίδραση > ακτίνης µυοσίνης υπάρχουν 4 κατηγορίες κυττάρων > µυϊκά > που σχηµατίζουν > γραµµωτό µυϊκό ιστό καρδιακό µυϊκό ιστό
ΣYΣTAΛTA KYTTAPA Tα συσταλτά κύτταρα παράγουν > δυνάµεις κίνησης µε την αλληλεπίδραση > ακτίνης µυοσίνης υπάρχουν 4 κατηγορίες κυττάρων > µυϊκά > που σχηµατίζουν > γραµµωτό µυϊκό ιστό καρδιακό µυϊκό ιστό
ΙΣΤΟΙ Ως προς τη µορφή και τη λειτουργία τους. Κυτταρική διαφοροποίηση.
 ΙΣΤΟΙ 1. Τα κύτταρα που αποτελούν τον οργανισµό µας, διακρίνονται σε διάφορους τύπους, παρά το γεγονός ότι όλα, τελικώς, προέρχονται από το ζυγωτό, δηλαδή το πρώτο κύτταρο µε το οποίο ξεκίνησε η ζωή µας.
ΙΣΤΟΙ 1. Τα κύτταρα που αποτελούν τον οργανισµό µας, διακρίνονται σε διάφορους τύπους, παρά το γεγονός ότι όλα, τελικώς, προέρχονται από το ζυγωτό, δηλαδή το πρώτο κύτταρο µε το οποίο ξεκίνησε η ζωή µας.
314 ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗ ΜΥΪΚΗ ΥΠΕΡΤΡΟΦΙΑ. ΦΑΤΟΥΡΟΣ Γ. ΙΩΑΝΝΗΣ, Ph.D. Επίκουρος Καθηγητής Τ.Ε.Φ.Α.Α. Δ.Π.Θ.
 314 ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗ ΜΥΪΚΗ ΥΠΕΡΤΡΟΦΙΑ ΦΑΤΟΥΡΟΣ Γ. ΙΩΑΝΝΗΣ, Ph.D. Επίκουρος Καθηγητής Τ.Ε.Φ.Α.Α. Δ.Π.Θ. ΜΗΧΑΝΙΣΜΟΙ ΜΥΪΚΗΣ ΥΠΕΡΤΡΟΦΙΑΣ ΠΡΟΣΩΡΙΝΗ ΥΠΕΡΤΡΟΦΙΑ ΧΡΟΝΙΑ Ή ΜΟΝΙΜΗ ΥΠΕΡΤΡΟΦΙΑ ΥΠΕΡΤΡΟΦΙΑ ΜΥΪΚΩΝ
314 ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗ ΜΥΪΚΗ ΥΠΕΡΤΡΟΦΙΑ ΦΑΤΟΥΡΟΣ Γ. ΙΩΑΝΝΗΣ, Ph.D. Επίκουρος Καθηγητής Τ.Ε.Φ.Α.Α. Δ.Π.Θ. ΜΗΧΑΝΙΣΜΟΙ ΜΥΪΚΗΣ ΥΠΕΡΤΡΟΦΙΑΣ ΠΡΟΣΩΡΙΝΗ ΥΠΕΡΤΡΟΦΙΑ ΧΡΟΝΙΑ Ή ΜΟΝΙΜΗ ΥΠΕΡΤΡΟΦΙΑ ΥΠΕΡΤΡΟΦΙΑ ΜΥΪΚΩΝ
Μοριακή Βιολογία. Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής
: Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής") ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς. Οι φάσεις των Τ κυτταρικών απαντήσεων
 ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΠΕΙΡΑΜΑΤΙΚΗΣ ΦΑΡΜΑΚΟΛΟΓΙΑΣ
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΠΕΙΡΑΜΑΤΙΚΗΣ ΦΑΡΜΑΚΟΛΟΓΙΑΣ ΠΕΡΙΛΗΨΗ ΔΙΔΑΚΤΟΡΙΚΗΣ ΔΙΑΤΡΙΒΗΣ ΑΘΗΝΑΣ ΓΙΑΓΚΙΝΗ ΧΗΜΙΚΟΥ Μελέτη των ιστολογικών και βιοχημικών αλλοιώσεων
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΠΕΙΡΑΜΑΤΙΚΗΣ ΦΑΡΜΑΚΟΛΟΓΙΑΣ ΠΕΡΙΛΗΨΗ ΔΙΔΑΚΤΟΡΙΚΗΣ ΔΙΑΤΡΙΒΗΣ ΑΘΗΝΑΣ ΓΙΑΓΚΙΝΗ ΧΗΜΙΚΟΥ Μελέτη των ιστολογικών και βιοχημικών αλλοιώσεων
Η κυτταρική µετατόπιση των πρωτεϊνών
 9-1 Κεφάλαιο 9 Η κυτταρική µετατόπιση των πρωτεϊνών Εισαγωγή Στο κύτταρο η έκφραση των πρωτεϊνών γίνεται από µόνο ένα τύπο ριβοσώµατος (εκτός των µιτοχονδριακών και των χλωροπλαστικών που µοιάζουν µε αυτά
9-1 Κεφάλαιο 9 Η κυτταρική µετατόπιση των πρωτεϊνών Εισαγωγή Στο κύτταρο η έκφραση των πρωτεϊνών γίνεται από µόνο ένα τύπο ριβοσώµατος (εκτός των µιτοχονδριακών και των χλωροπλαστικών που µοιάζουν µε αυτά
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ
 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ: ΜΙΚΡΟΪΝΙΔΙΑ- ΕΝΔΙΑΜΕΣΑ ΙΝΙΔΙΑ- ΜΙΚΡΟΣΩΛΗΝΙΣΚΟΙ Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ: ΜΙΚΡΟΪΝΙΔΙΑ- ΕΝΔΙΑΜΕΣΑ ΙΝΙΔΙΑ- ΜΙΚΡΟΣΩΛΗΝΙΣΚΟΙ Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο
Εργασία στην Βιολογία
 Εργασία στην Βιολογία Μεμβράνη του Πυρήνα του Κυττάρου Χρονιά: 2013-2014 Επιμέλεια: Σταμάτης Ορφανός, Μάριος Παναγιωτόπουλος [1] Πυρήνας του Κυττάρου Ο πυρήνας είναι το πιο μεγάλο και ευδιάκριτο οργανίδιο
Εργασία στην Βιολογία Μεμβράνη του Πυρήνα του Κυττάρου Χρονιά: 2013-2014 Επιμέλεια: Σταμάτης Ορφανός, Μάριος Παναγιωτόπουλος [1] Πυρήνας του Κυττάρου Ο πυρήνας είναι το πιο μεγάλο και ευδιάκριτο οργανίδιο
ΟΙ ΠΑΡΑΘΥΡΕΟΕΙΔΕΙΣ ΑΔΕΝΕΣ ΚΑΙ Η ΡΥΘΜΙΣΗ ΤΟΥ ΑΣΒΕΣΤΙΟΥ
 ΟΙ ΠΑΡΑΘΥΡΕΟΕΙΔΕΙΣ ΑΔΕΝΕΣ ΚΑΙ Η ΡΥΘΜΙΣΗ ΤΟΥ ΑΣΒΕΣΤΙΟΥ ΔΗΜΗΤΡΗΣ Ε. ΚΟΥΤΣΟΝΙΚΟΛΑΣ ΑΝΑΠΛΗΡΩΤΗΣ ΚΑΘΗΓΗΤΗΣ ΦΥΣΙΟΛΟΓΙΑΣ(ΧΕΙΡΟΥΡΓΟΣ ΠΑΙΔΩΝ) ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α.Π.Θ ΟΙ ΠΑΡΑΘΥΡΕΟΕΙΔΕΙΣ ΑΔΕΝΕΣ ΑΝΑΤΟΜΙΚΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΑ
ΟΙ ΠΑΡΑΘΥΡΕΟΕΙΔΕΙΣ ΑΔΕΝΕΣ ΚΑΙ Η ΡΥΘΜΙΣΗ ΤΟΥ ΑΣΒΕΣΤΙΟΥ ΔΗΜΗΤΡΗΣ Ε. ΚΟΥΤΣΟΝΙΚΟΛΑΣ ΑΝΑΠΛΗΡΩΤΗΣ ΚΑΘΗΓΗΤΗΣ ΦΥΣΙΟΛΟΓΙΑΣ(ΧΕΙΡΟΥΡΓΟΣ ΠΑΙΔΩΝ) ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α.Π.Θ ΟΙ ΠΑΡΑΘΥΡΕΟΕΙΔΕΙΣ ΑΔΕΝΕΣ ΑΝΑΤΟΜΙΚΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΑ
Kυτταρική$Bιολογία$ Πολυκυτταρική+οργάνωση+και+ καρκίνος+ ΔIAΛEΞΕΙΣ*15*&*16! (18!&!21/5/2012)! Δρ.$Xρήστος$Παναγιωτίδης,$Τμήμα$Φαρμακευτικής$Α.Π.Θ.
! Δρ.$Xρήστος$Παναγιωτίδης,$Τμήμα$Φαρμακευτικής$Α.Π.Θ.") Kυτταρική$Bιολογία$ ΔIAΛEΞΕΙΣ*15*&*16! (18!&!21/5/2012)! Πολυκυτταρική+οργάνωση+και+ καρκίνος+ Ας+ξαναθυμηθούμε+για+λίγο+ τη+κυτταρική+θεωρία+ Η*κυτταρική*θεωρία*! OΛOI!OI!ZΩNTANOI!OPΓANIΣMOI! AΠOTEΛOYNTAI!AΠO!KYTTAPA!H!AΠO!
Kυτταρική$Bιολογία$ ΔIAΛEΞΕΙΣ*15*&*16! (18!&!21/5/2012)! Πολυκυτταρική+οργάνωση+και+ καρκίνος+ Ας+ξαναθυμηθούμε+για+λίγο+ τη+κυτταρική+θεωρία+ Η*κυτταρική*θεωρία*! OΛOI!OI!ZΩNTANOI!OPΓANIΣMOI! AΠOTEΛOYNTAI!AΠO!KYTTAPA!H!AΠO!
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΓENIKA ΣTOIXEIA. Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό.
 1 ΓENIKA ΣTOIXEIA Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό. ΤΑ ΚΥΤΤΑΡΑ Η βασική ζώσα μονάδα του σώματος είναι
1 ΓENIKA ΣTOIXEIA Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό. ΤΑ ΚΥΤΤΑΡΑ Η βασική ζώσα μονάδα του σώματος είναι
314 ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗ ΜΥΪΚΗ ΥΠΕΡΤΡΟΦΙΑ. ΦΑΤΟΥΡΟΣ Γ. ΙΩΑΝΝΗΣ, Ph.D. Επίκουρος Καθηγητής Τ.Ε.Φ.Α.Α. Δ.Π.Θ.
 314 ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗ ΜΥΪΚΗ ΥΠΕΡΤΡΟΦΙΑ ΦΑΤΟΥΡΟΣ Γ. ΙΩΑΝΝΗΣ, Ph.D. Επίκουρος Καθηγητής Τ.Ε.Φ.Α.Α. Δ.Π.Θ. Μέρος Πρώτο Νέες πληροφορίες σχετικά με τον μηχανισμό αύξησης της μυϊκής μάζας και του ρόλου των
314 ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗ ΜΥΪΚΗ ΥΠΕΡΤΡΟΦΙΑ ΦΑΤΟΥΡΟΣ Γ. ΙΩΑΝΝΗΣ, Ph.D. Επίκουρος Καθηγητής Τ.Ε.Φ.Α.Α. Δ.Π.Θ. Μέρος Πρώτο Νέες πληροφορίες σχετικά με τον μηχανισμό αύξησης της μυϊκής μάζας και του ρόλου των
Οδοντογένεση - Εµβρυολογία του δοντιού
 Οδοντογένεση - Εµβρυολογία του δοντιού Εισαγωγή Στον άνθρωπο, 20 νεογιλά και 32 µόνιµα δόντια αναπτύσσονται κατά την εµβρυϊκή περίοδο από την αλληλεπίδραση των στοµατικών επιθηλιακών κυττάρων και των υποκείµενων
Οδοντογένεση - Εµβρυολογία του δοντιού Εισαγωγή Στον άνθρωπο, 20 νεογιλά και 32 µόνιµα δόντια αναπτύσσονται κατά την εµβρυϊκή περίοδο από την αλληλεπίδραση των στοµατικών επιθηλιακών κυττάρων και των υποκείµενων
Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό
 Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
Kυτταρική$Bιολογία$ Πολυκυτταρική+οργάνωση+και+ καρκίνος+ ΔIAΛEΞΕΙΣ*19*&*20! (21!&!28/5/2014)! Δρ.$Xρήστος$Παναγιωτίδης,$Τμήμα$Φαρμακευτικής$Α.Π.Θ.
! Δρ.$Xρήστος$Παναγιωτίδης,$Τμήμα$Φαρμακευτικής$Α.Π.Θ.") Kυτταρική$Bιολογία$ ΔIAΛEΞΕΙΣ*19*&*20! (21!&!28/5/2014)! Πολυκυτταρική+οργάνωση+και+ καρκίνος+ Ας+ξαναθυμηθούμε+για+λίγο+ τη+κυτταρική+θεωρία+ Η*κυτταρική*θεωρία*! OΛOI!OI!ZΩNTANOI!OPΓANIΣMOI! AΠOTEΛOYNTAI!AΠO!KYTTAPA!H!AΠO!
Kυτταρική$Bιολογία$ ΔIAΛEΞΕΙΣ*19*&*20! (21!&!28/5/2014)! Πολυκυτταρική+οργάνωση+και+ καρκίνος+ Ας+ξαναθυμηθούμε+για+λίγο+ τη+κυτταρική+θεωρία+ Η*κυτταρική*θεωρία*! OΛOI!OI!ZΩNTANOI!OPΓANIΣMOI! AΠOTEΛOYNTAI!AΠO!KYTTAPA!H!AΠO!
Χόνδρος Αρθρώσεις. Σοφία Χαβάκη Λέκτορας Εργαστήριο Ιστολογίας-Εμβρυολογίας
 Χόνδρος Αρθρώσεις Σοφία Χαβάκη Λέκτορας Εργαστήριο Ιστολογίας-Εμβρυολογίας Χόνδρος συνδετικός-στηρικτικός ς ιστός συμπαγής αλλά εύκαμπτος Λειτουργίες Χόνδρου υποστήριξη μαλακών ιστών απορρόφηση κραδασμών
Χόνδρος Αρθρώσεις Σοφία Χαβάκη Λέκτορας Εργαστήριο Ιστολογίας-Εμβρυολογίας Χόνδρος συνδετικός-στηρικτικός ς ιστός συμπαγής αλλά εύκαμπτος Λειτουργίες Χόνδρου υποστήριξη μαλακών ιστών απορρόφηση κραδασμών
IΣTOΛOΓIA. Tα δείγµατα του βιολογικού υλικού λαµβάνονται µε > βελόνες ενδοσκοπικούς σωλήνες εύκαµπτους καθετήρες
 IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
Το μυϊκό σύστημα αποτελείται από τους μύες. Ο αριθμός των μυών του μυϊκού συστήματος ανέρχεται στους 637. Οι μύες είναι όργανα για τη σωματική
 Μύες Το μυϊκό σύστημα αποτελείται από τους μύες. Ο αριθμός των μυών του μυϊκού συστήματος ανέρχεται στους 637. Οι μύες είναι όργανα για τη σωματική κινητικότητα, την σπλαχνική κινητικότητα και τη κυκλοφορία
Μύες Το μυϊκό σύστημα αποτελείται από τους μύες. Ο αριθμός των μυών του μυϊκού συστήματος ανέρχεται στους 637. Οι μύες είναι όργανα για τη σωματική κινητικότητα, την σπλαχνική κινητικότητα και τη κυκλοφορία
ΚΕΦΑΛΑΙΟ 11. Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα
 ΚΕΦΑΛΑΙΟ 11 Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα Τα ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΑ Μέρος Ε ΤΑ ΒΑΣΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΩΝ - Περιέχουν ένζυµα για ποικίλες µεταβολικές
ΚΕΦΑΛΑΙΟ 11 Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα Τα ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΑ Μέρος Ε ΤΑ ΒΑΣΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΩΝ - Περιέχουν ένζυµα για ποικίλες µεταβολικές
1. ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ
 ΚΕΦΑΛΑΙΟ 1 1. ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ ΚΥΤΤΑΡΑ ΚΑΙ ΙΣΤΟΙ Ο ανθρώπινος οργανισμός συνίσταται α- πό τρισεκατομμύρια κύτταρα. Τα κύτταρα αυτά εμφανίζουν σημαντική ποικιλομορφία, που αφορά το μέγεθος,
ΚΕΦΑΛΑΙΟ 1 1. ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ ΚΥΤΤΑΡΑ ΚΑΙ ΙΣΤΟΙ Ο ανθρώπινος οργανισμός συνίσταται α- πό τρισεκατομμύρια κύτταρα. Τα κύτταρα αυτά εμφανίζουν σημαντική ποικιλομορφία, που αφορά το μέγεθος,
Κυτταρική Βιολογία. Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
Ο ρόλος της ΕΘΟ. στην αναγέννηση. & την επανόρθωση
 Ο ρόλος της ΕΘΟ στην αναγέννηση & την επανόρθωση Νοvo E & Parola M. Fibrogenesis & Tissue Repair 2008, 1:5 Χρόνια παγκρεατίτιδα Ιστολογία παγκρεατικού καρκινώµατος Αδενοκαρκίνωµα εξ εκφορητικών
Ο ρόλος της ΕΘΟ στην αναγέννηση & την επανόρθωση Νοvo E & Parola M. Fibrogenesis & Tissue Repair 2008, 1:5 Χρόνια παγκρεατίτιδα Ιστολογία παγκρεατικού καρκινώµατος Αδενοκαρκίνωµα εξ εκφορητικών
ΛΕΜΦΟΚΥΤΤΑΡΑ. Τ λεµφοκύτταρα:
 ΛΕΜΦΟΚΥΤΤΑΡΑ Προέλευση: µυελός των οστών. Μερικά µεταναστεύουν στο θύµο, όπου παραµένουν για ποικίλες περιόδους πριν διασκορπισθούν στο σώµα. Βίος: η ζωή τους ποικίλει. Τα µνηµοκύτταρα ζουν για πολλά χρόνια
ΛΕΜΦΟΚΥΤΤΑΡΑ Προέλευση: µυελός των οστών. Μερικά µεταναστεύουν στο θύµο, όπου παραµένουν για ποικίλες περιόδους πριν διασκορπισθούν στο σώµα. Βίος: η ζωή τους ποικίλει. Τα µνηµοκύτταρα ζουν για πολλά χρόνια
11. ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ
 11. ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Στον ανθρώπινο οργανισμό υπάρχουν δύο είδη αδένων, οι εξωκρινείς και οι ενδοκρινείς. Οι εξωκρινείς (ιδρωτοποιοί αδένες, σμηγματογόνοι αδένες κ.ά.) εκκρίνουν το προϊόν τους στην επιφάνεια
11. ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Στον ανθρώπινο οργανισμό υπάρχουν δύο είδη αδένων, οι εξωκρινείς και οι ενδοκρινείς. Οι εξωκρινείς (ιδρωτοποιοί αδένες, σμηγματογόνοι αδένες κ.ά.) εκκρίνουν το προϊόν τους στην επιφάνεια
Κυριακή 15/02/2015 Ημερομηνία
 Διαγώνισμα 2014-15 Ενδεικτικές απαντήσεις Κυριακή 15/02/2015 Ημερομηνία Βιολογία Κατεύθυνσης Εξεταζόμενο μάθημα Γ Λυκείου Τάξη Θέμα 1 ο : 1 α, 2 γ, 3 ε, 4 α, 5 ε Θέμα 2 ο : Α. Η απεικόνιση των μεταφασικών
Διαγώνισμα 2014-15 Ενδεικτικές απαντήσεις Κυριακή 15/02/2015 Ημερομηνία Βιολογία Κατεύθυνσης Εξεταζόμενο μάθημα Γ Λυκείου Τάξη Θέμα 1 ο : 1 α, 2 γ, 3 ε, 4 α, 5 ε Θέμα 2 ο : Α. Η απεικόνιση των μεταφασικών
Σακχαρώδης Διαβήτης και Περιοδοντίτιδα. Μια αμφίδρομη σχέση. ΜΑΡΑΓΚΟΣ ΠΑΝΑΓΙΩΤΗΣ ΧΕΙΡΟΥΡΓΟΣ ΟΔΟΝΤΙΑΤΡΟΣ
 Σακχαρώδης Διαβήτης και Περιοδοντίτιδα. Μια αμφίδρομη σχέση. ΜΑΡΑΓΚΟΣ ΠΑΝΑΓΙΩΤΗΣ ΧΕΙΡΟΥΡΓΟΣ ΟΔΟΝΤΙΑΤΡΟΣ Από την παρούσα εισήγηση δεν προκύπτει σύγκρουση συμφερόντων. ΠΕΡΙΟΔΟΝΤΙΚΗ ΝΟΣΟΣ Η Περιοδοντική
Σακχαρώδης Διαβήτης και Περιοδοντίτιδα. Μια αμφίδρομη σχέση. ΜΑΡΑΓΚΟΣ ΠΑΝΑΓΙΩΤΗΣ ΧΕΙΡΟΥΡΓΟΣ ΟΔΟΝΤΙΑΤΡΟΣ Από την παρούσα εισήγηση δεν προκύπτει σύγκρουση συμφερόντων. ΠΕΡΙΟΔΟΝΤΙΚΗ ΝΟΣΟΣ Η Περιοδοντική
ΕΠΙΝΕΦΡΙΔΙΑ ΚΟΡΤΙΖΟΛΗ
 ΕΠΙΝΕΦΡΙΔΙΑ ΚΟΡΤΙΖΟΛΗ Μεταβολισμός της κορτιζόλης Η κορτιζόλη μεταβολίζεται στο ήπαρ. Στην συνέχεια οι μεταβολίτες συζευγνύνται με γλυκουρονιδικές και θειικές ομάδες, γίνονται υδατοδιαλυτά, εισέρχονται
ΕΠΙΝΕΦΡΙΔΙΑ ΚΟΡΤΙΖΟΛΗ Μεταβολισμός της κορτιζόλης Η κορτιζόλη μεταβολίζεται στο ήπαρ. Στην συνέχεια οι μεταβολίτες συζευγνύνται με γλυκουρονιδικές και θειικές ομάδες, γίνονται υδατοδιαλυτά, εισέρχονται
Δομή των μυϊκών κυττάρων.
 Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
gr
 Η επίδραση της περίσσειας θυρεοειδικών ορµονών στα οστά ήταν γνωστή πριν από την εµφάνιση των αντιθυρεοειδικών φαρµάκων. Μία από τις πρώτες αναφορές καταγράφηκε το 1891από τον von Recklinghausen και αναφέρει
Η επίδραση της περίσσειας θυρεοειδικών ορµονών στα οστά ήταν γνωστή πριν από την εµφάνιση των αντιθυρεοειδικών φαρµάκων. Μία από τις πρώτες αναφορές καταγράφηκε το 1891από τον von Recklinghausen και αναφέρει
Χρόνια φλεγμονή. Βαλεντίνη Τζιούφα-Ασημακοπούλου. Νοέμβριος 2018
 Χρόνια φλεγμονή Βαλεντίνη Τζιούφα-Ασημακοπούλου Νοέμβριος 2018 Οξεία φλεγμονή Ταχεία εισβολή και λύση Εξιδρωματικά στοιχεία Πολυμορφοπύρηνα Χρόνια φλεγμονή Ύπουλη εισβολήπαρατεταμένη πορείαβραδεία λύση
Χρόνια φλεγμονή Βαλεντίνη Τζιούφα-Ασημακοπούλου Νοέμβριος 2018 Οξεία φλεγμονή Ταχεία εισβολή και λύση Εξιδρωματικά στοιχεία Πολυμορφοπύρηνα Χρόνια φλεγμονή Ύπουλη εισβολήπαρατεταμένη πορείαβραδεία λύση
Ν. Κατσίκη[1], Α. Γκοτζαμάνη-Ψαρράκου[2], Φ. Ηλιάδης[1], Τρ. Διδάγγελος[1], Ι. Γιώβος[3], Δ. Καραμήτσος[1]
![Ν. Κατσίκη[1], Α. Γκοτζαμάνη-Ψαρράκου[2], Φ. Ηλιάδης[1], Τρ. Διδάγγελος[1], Ι. Γιώβος[3], Δ. Καραμήτσος[1]](/thumbs/20/414553.jpg "Ν. Κατσίκη[1], Α. Γκοτζαμάνη-Ψαρράκου[2], Φ. Ηλιάδης[1], Τρ. Διδάγγελος[1], Ι. Γιώβος[3], Δ. Καραμήτσος[1]") Ολόγοςλεπτίνης/αδιπονεκτίνης ως ανεξάρτητος προγνωστικός παράγοντας 10ετούς καρδιαγγειακού κινδύνου σε ινσουλινοθεραπευόμενους ασθενείς με διαβήτη τύπου 2 Ν. Κατσίκη[1], Α. Γκοτζαμάνη-Ψαρράκου[2], Φ. Ηλιάδης[1],
Ολόγοςλεπτίνης/αδιπονεκτίνης ως ανεξάρτητος προγνωστικός παράγοντας 10ετούς καρδιαγγειακού κινδύνου σε ινσουλινοθεραπευόμενους ασθενείς με διαβήτη τύπου 2 Ν. Κατσίκη[1], Α. Γκοτζαμάνη-Ψαρράκου[2], Φ. Ηλιάδης[1],
Από το κύτταρο στον οργανισμό. Κεφάλαιο 1ο
 Από το κύτταρο στον οργανισμό Κεφάλαιο 1ο Ι. Ανάμεσα στους μεικτούς αδένες του ανθρώπινου οργανισμού περιλαμβάνεται και το πάγκρεας. α) Ποια είναι τα προϊόντα που παράγει το πάγκρεας (παγκρεατικό υγρό,
Από το κύτταρο στον οργανισμό Κεφάλαιο 1ο Ι. Ανάμεσα στους μεικτούς αδένες του ανθρώπινου οργανισμού περιλαμβάνεται και το πάγκρεας. α) Ποια είναι τα προϊόντα που παράγει το πάγκρεας (παγκρεατικό υγρό,
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης
 Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
ΚΡΑΝΙΟΣΤΟΜΑΤΟΓΝΑΘΙΚΟ ΣΥΣΤΗΜΑ
 ΚΡΑΝΙΟΣΤΟΜΑΤΟΓΝΑΘΙΚΟ ΣΥΣΤΗΜΑ Α. Εμβρυολογία, Διάπλαση & Ανάπτυξη του κρανιοπροσωπικού συμπλέγματος 1. Εξέλιξη και διάπλαση του Κρανιοπροσωπικού συμπλέγματος (ΚΠΣ). 2. Παράγοντες που επηρεάζουν τη διάπλαση
ΚΡΑΝΙΟΣΤΟΜΑΤΟΓΝΑΘΙΚΟ ΣΥΣΤΗΜΑ Α. Εμβρυολογία, Διάπλαση & Ανάπτυξη του κρανιοπροσωπικού συμπλέγματος 1. Εξέλιξη και διάπλαση του Κρανιοπροσωπικού συμπλέγματος (ΚΠΣ). 2. Παράγοντες που επηρεάζουν τη διάπλαση
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ 15/9/2014. ΚΕΦΑΛΑΙΟ 1 ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ Κύτταρα και ιστοί Όργανα και συστήματα οργάνων
 Κύτταρα και ιστοί: Πλήθος κυττάρων ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ Ο ανθρώπινος οργανισμός αποτελείται από τρισεκατομμύρια κύτταρα. ΚΕΦΑΛΑΙΟ 1 ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ Κύτταρα και ιστοί Όργανα και συστήματα οργάνων
Κύτταρα και ιστοί: Πλήθος κυττάρων ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ Ο ανθρώπινος οργανισμός αποτελείται από τρισεκατομμύρια κύτταρα. ΚΕΦΑΛΑΙΟ 1 ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ Κύτταρα και ιστοί Όργανα και συστήματα οργάνων
Πρόκειται για 4 μικρούς αδένες στο μέγεθος "φακής" που βρίσκονται πίσω από το θυρεοειδή αδένα. Οι αδένες αυτοί παράγουν μια ορμόνη που λέγεται
 Πρόκειται για 4 μικρούς αδένες στο μέγεθος "φακής" που βρίσκονται πίσω από το θυρεοειδή αδένα. Οι αδένες αυτοί παράγουν μια ορμόνη που λέγεται παραθορμόνη και ρυθμίζει τα επίπεδα ασβεστίου στο αίμα. Ορμόνες
Πρόκειται για 4 μικρούς αδένες στο μέγεθος "φακής" που βρίσκονται πίσω από το θυρεοειδή αδένα. Οι αδένες αυτοί παράγουν μια ορμόνη που λέγεται παραθορμόνη και ρυθμίζει τα επίπεδα ασβεστίου στο αίμα. Ορμόνες
ΠΡΟΕΚΒΟΛΕΣ ΤΗΣ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ ΚΑΙ ΚΥΤΤΑΡΙΚΗ ΚΙΝΗΣΗ
 ΠΡΟΕΚΒΟΛΕΣ ΤΗΣ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ ΚΑΙ ΚΥΤΤΑΡΙΚΗ ΚΙΝΗΣΗ Φωτογραφία ηλεκτρονικού μικροσκοπίου που δείχνει μικρολάχνες. Οι μικρολάχνες των επιθηλιακών κυττάρων του εντέρου είναι δακτυλοειδείς προεκβολές
ΠΡΟΕΚΒΟΛΕΣ ΤΗΣ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ ΚΑΙ ΚΥΤΤΑΡΙΚΗ ΚΙΝΗΣΗ Φωτογραφία ηλεκτρονικού μικροσκοπίου που δείχνει μικρολάχνες. Οι μικρολάχνες των επιθηλιακών κυττάρων του εντέρου είναι δακτυλοειδείς προεκβολές
1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; ΘΩΜΑΣ ΑΠΑΝΤΗΣΗ. 2. Ποιες είναι οι κατηγορίες γονιδίων με κριτήριο το προϊόν της μεταγραφής τους;
 Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ ΜΥΙΚΗ ΥΠΕΡΤΡΟΦΙΑ. κωδ. μαθήματος Ν314. Τίτλος 11ης Διάλεξης. Εισήγηση: Χατζηνικολάου Α.,Επίκουρος Καθηγητής
 ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ κωδ. μαθήματος Ν314 Τίτλος 11ης Διάλεξης ΜΥΙΚΗ ΥΠΕΡΤΡΟΦΙΑ Εισήγηση: Χατζηνικολάου Α.,Επίκουρος Καθηγητής Μέρος Πρώτο Νέες πληροφορίες σχετικά με τον μηχανισμό αύξησης της μυϊκής μάζας
ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ κωδ. μαθήματος Ν314 Τίτλος 11ης Διάλεξης ΜΥΙΚΗ ΥΠΕΡΤΡΟΦΙΑ Εισήγηση: Χατζηνικολάου Α.,Επίκουρος Καθηγητής Μέρος Πρώτο Νέες πληροφορίες σχετικά με τον μηχανισμό αύξησης της μυϊκής μάζας
ΣΧΗΜΑΤΙΣΜΟΣ ΕΠΑΓΩΜΕΝΩΝ ΠΟΛΥΔΥΝΑΜΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΙΚΟΣ ΕΠΑΝΑΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ
 ΣΧΗΜΑΤΙΣΜΟΣ ΕΠΑΓΩΜΕΝΩΝ ΠΟΛΥΔΥΝΑΜΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΙΚΟΣ ΕΠΑΝΑΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ Υπάρχουν Βλαστικά κύτταρα με διαφορετικές ιδιότητες: Τα Πολυδύναμα - Pluripotent Εμβρυονικά Βλαστικά κύτταρα - Embryonic Stem
ΣΧΗΜΑΤΙΣΜΟΣ ΕΠΑΓΩΜΕΝΩΝ ΠΟΛΥΔΥΝΑΜΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΙΚΟΣ ΕΠΑΝΑΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ Υπάρχουν Βλαστικά κύτταρα με διαφορετικές ιδιότητες: Τα Πολυδύναμα - Pluripotent Εμβρυονικά Βλαστικά κύτταρα - Embryonic Stem
ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ. Οι ρυθμιστές του οργανισμού
 ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Οι ρυθμιστές του οργανισμού Είδη αδένων στον άνθρωπο o Εξωκρινείς αδένες: εκκρίνουν το προϊόν τους μέσω εκφορητικού πόρου είτε στην επιφάνεια του σώματος (π.χ. ιδρωτοποιοί και σμηγματογόνοι
ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Οι ρυθμιστές του οργανισμού Είδη αδένων στον άνθρωπο o Εξωκρινείς αδένες: εκκρίνουν το προϊόν τους μέσω εκφορητικού πόρου είτε στην επιφάνεια του σώματος (π.χ. ιδρωτοποιοί και σμηγματογόνοι
ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ ΕΠΑΝΑΛΗΨΗ
 ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ ΕΠΑΝΑΛΗΨΗ ΘΕΜΑ 1ο 1. Κυτταρική διαφοροποίηση ονομάζουμε: α. Την δομική κυρίως εξειδίκευση των συστημάτων β. Την δομική και λειτουργική εξειδίκευση των κυττάρων γ. Την λειτουργική εξειδίκευση
ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ ΕΠΑΝΑΛΗΨΗ ΘΕΜΑ 1ο 1. Κυτταρική διαφοροποίηση ονομάζουμε: α. Την δομική κυρίως εξειδίκευση των συστημάτων β. Την δομική και λειτουργική εξειδίκευση των κυττάρων γ. Την λειτουργική εξειδίκευση
Osteogenesis Imperfecta (Ατελής Οστεογένεση ) Ομάδα: Πατρασκάκη Μυρτώ Τσιτσικλή Μαγδαληνή
 Ομάδα: Πατρασκάκη Μυρτώ Τσιτσικλή Μαγδαληνή") Osteogenesis Imperfecta (Ατελής Οστεογένεση ) Ομάδα: Πατρασκάκη Μυρτώ Τσιτσικλή Μαγδαληνή Osteogenesis imperfecta Μενδελικό Νόσημα Συχνότητα στον πληθυσμό: 1:20.000 80-95% αυτοσωμικό επικρατές 10-15% αυτοσωμικό
Osteogenesis Imperfecta (Ατελής Οστεογένεση ) Ομάδα: Πατρασκάκη Μυρτώ Τσιτσικλή Μαγδαληνή Osteogenesis imperfecta Μενδελικό Νόσημα Συχνότητα στον πληθυσμό: 1:20.000 80-95% αυτοσωμικό επικρατές 10-15% αυτοσωμικό
Αυτοφαγία & Ανοσολογικό Σύστημα. Χαράλαμπος Μ. Μουτσόπουλος
 Αυτοφαγία & Ανοσολογικό Σύστημα Χαράλαμπος Μ. Μουτσόπουλος Απρίλιος 2018 Αυτοφαγία & Ανοσολογικό Σύστημα Ευκαρυωτικά κύτταρα: Δυο μηχανισμούς αποδόμησης και ανακύκλωσης κυτταροπλασματικών αποβλήτων Πρωτεάσες
Αυτοφαγία & Ανοσολογικό Σύστημα Χαράλαμπος Μ. Μουτσόπουλος Απρίλιος 2018 Αυτοφαγία & Ανοσολογικό Σύστημα Ευκαρυωτικά κύτταρα: Δυο μηχανισμούς αποδόμησης και ανακύκλωσης κυτταροπλασματικών αποβλήτων Πρωτεάσες
Θέματα πριν τις εξετάσεις. Καλό διάβασμα Καλή επιτυχία
 Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016)
") Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
ΔΙΑΓΝΩΣΤΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗΝ ΟΡΘΟΠΑΙΔΙΚΗ ΠΑΥΛΟΣ Γ. ΚΑΤΩΝΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΡΗΤΗΣ
 ΠΑΥΛΟΣ Γ. ΚΑΤΩΝΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΡΗΤΗΣ ΙΣΤΟΡΙΚΟ + ΚΛΙΝΙΚΗ ΕΞΕΤΑΣΗ = Α & Ω ΣΤΗΝ! ΑΠΛΕΣ ΑΚΤΙΝΟΓΡΑΦΙΕΣ (Βασική απεικονιστική μέθοδος) ΠΡΟΣΘΙΟΠΙΣΘΙΑ ΠΛΑΓΙΑ ΛΟΞΗ ΕΙΔΙΚΗ ΑΠΛΕΣ ΑΚΤΙΝΟΓΡΑΦΙΕΣ (Βασική
ΠΑΥΛΟΣ Γ. ΚΑΤΩΝΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΡΗΤΗΣ ΙΣΤΟΡΙΚΟ + ΚΛΙΝΙΚΗ ΕΞΕΤΑΣΗ = Α & Ω ΣΤΗΝ! ΑΠΛΕΣ ΑΚΤΙΝΟΓΡΑΦΙΕΣ (Βασική απεικονιστική μέθοδος) ΠΡΟΣΘΙΟΠΙΣΘΙΑ ΠΛΑΓΙΑ ΛΟΞΗ ΕΙΔΙΚΗ ΑΠΛΕΣ ΑΚΤΙΝΟΓΡΑΦΙΕΣ (Βασική
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΗ 4 (6/3/2013)
") Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
ΚΑΤΗΓΟΡΙΕΣ. 1. Μυϊκά. 2. Μυοεπιθηλιακά. 3. Περικύτταρα. 4. Μυοϊνοβλάστες
 ΣΥΣΤΑΛΤΑ ΚΥΤΤΑΡΑ ΚΑΤΗΓΟΡΙΕΣ 1. Μυϊκά 2. Μυοεπιθηλιακά 3. Περικύτταρα 4. Μυοϊνοβλάστες ΜΥΙΚΑ 1. Γραμμωτά μυϊκά κύτταρα 2. Καρδιακά μυϊκά κύτταρα 3. Λεία μυϊκά κύτταρα ΓΡΑΜΜΩΤΟΙ ΜΥΣ 1. Μύς 2. Μυϊκή δέσμη
ΣΥΣΤΑΛΤΑ ΚΥΤΤΑΡΑ ΚΑΤΗΓΟΡΙΕΣ 1. Μυϊκά 2. Μυοεπιθηλιακά 3. Περικύτταρα 4. Μυοϊνοβλάστες ΜΥΙΚΑ 1. Γραμμωτά μυϊκά κύτταρα 2. Καρδιακά μυϊκά κύτταρα 3. Λεία μυϊκά κύτταρα ΓΡΑΜΜΩΤΟΙ ΜΥΣ 1. Μύς 2. Μυϊκή δέσμη
ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΘΝΙΚΟΥ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ ΜΟΝΑΔΑ ΕΡΕΥΝΑΣ Β'ΠΡΟΠΑΙΔΕΥΤΙΚΗΣ ΠΑΘΟΛΟΓΙΚΗΣ ΚΛΙΝΙΚΗΣ ΚΑΘΗΓΗΤΗΣ Θ.
 ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΘΝΙΚΟΥ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ ΜΟΝΑΔΑ ΕΡΕΥΝΑΣ Β'ΠΡΟΠΑΙΔΕΥΤΙΚΗΣ ΠΑΘΟΛΟΓΙΚΗΣ ΚΛΙΝΙΚΗΣ ΚΑΘΗΓΗΤΗΣ Θ. ΟΙΚΟΝΟΜΟΠΟΥΛΟΣ 1 ΙΕΡΙΛΗΨΗ ΔΙΔΑΚΤΟΡΙΚΗΣ ΔΙΑΤΡΙΒΗΣ ΒΙΟΛΟΓΟΣ: ΕΡΙΦΥΛΗ ΚΥΡΙΑΖΗ
ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΘΝΙΚΟΥ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ ΜΟΝΑΔΑ ΕΡΕΥΝΑΣ Β'ΠΡΟΠΑΙΔΕΥΤΙΚΗΣ ΠΑΘΟΛΟΓΙΚΗΣ ΚΛΙΝΙΚΗΣ ΚΑΘΗΓΗΤΗΣ Θ. ΟΙΚΟΝΟΜΟΠΟΥΛΟΣ 1 ΙΕΡΙΛΗΨΗ ΔΙΔΑΚΤΟΡΙΚΗΣ ΔΙΑΤΡΙΒΗΣ ΒΙΟΛΟΓΟΣ: ΕΡΙΦΥΛΗ ΚΥΡΙΑΖΗ
ΤΑ ΜΟΡΙΑ ΤΗΣ ΖΩΗΣ. Τι γνωρίζετε για τους υδατάνθρακες;
 1 ΤΑ ΜΟΡΙΑ ΤΗΣ ΖΩΗΣ Το κύτταρο αποτελείται από χηµικές ενώσεις, στις οποίες περιλαµβάνονται τα µικρά βιολογικά µόρια και τα βιολογικά µακροµόρια. Στα µικρά βιολογικά µόρια ανήκουν, τα ανόργανα στοιχεία
1 ΤΑ ΜΟΡΙΑ ΤΗΣ ΖΩΗΣ Το κύτταρο αποτελείται από χηµικές ενώσεις, στις οποίες περιλαµβάνονται τα µικρά βιολογικά µόρια και τα βιολογικά µακροµόρια. Στα µικρά βιολογικά µόρια ανήκουν, τα ανόργανα στοιχεία
ΒΙΟΧΗΜΕΙΑ ΤΟΥ ΜΕΤΑΒΟΛΙΣΜΟΥ
 ΒΙΟΧΗΜΕΙΑ ΤΟΥ ΜΕΤΑΒΟΛΙΣΜΟΥ ΠΑΡΟΥΣΙΑΣΗ 6 Η ΚΑΤΑΝΑΛΩΣΗ ΥΔΑΤΑΝΘΡΑΚΩΝ ΚΑΙ ΡΥΘΜΙΣΗ ΤΗΣ ΓΛΥΚΑΙΜΙΑΣ 1 Έλεγχος της ενέργειας Τα πραγματικά «Βιοκαύσιμα» 2 Υδατανθρακούχα τρόφιμα 3 Σημασία της ρύθμισης κατανάλωσης
ΒΙΟΧΗΜΕΙΑ ΤΟΥ ΜΕΤΑΒΟΛΙΣΜΟΥ ΠΑΡΟΥΣΙΑΣΗ 6 Η ΚΑΤΑΝΑΛΩΣΗ ΥΔΑΤΑΝΘΡΑΚΩΝ ΚΑΙ ΡΥΘΜΙΣΗ ΤΗΣ ΓΛΥΚΑΙΜΙΑΣ 1 Έλεγχος της ενέργειας Τα πραγματικά «Βιοκαύσιμα» 2 Υδατανθρακούχα τρόφιμα 3 Σημασία της ρύθμισης κατανάλωσης
Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ
 ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ") Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ Βιοενεργητική είναι ο κλάδος της Βιολογίας που μελετά τον τρόπο με τον οποίο οι οργανισμοί χρησιμοποιούν ενέργεια για να επιβιώσουν και να υλοποιήσουν τις
Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ Βιοενεργητική είναι ο κλάδος της Βιολογίας που μελετά τον τρόπο με τον οποίο οι οργανισμοί χρησιμοποιούν ενέργεια για να επιβιώσουν και να υλοποιήσουν τις
Ερειστικό Σύστημα. Γεωργιάδου Ελευθερία και Μηλιάδου Αθανασία.
 Ερειστικό Σύστημα Μια εργασία στο μάθημα της Βιολογίας από της μαθήτριες Γεωργιάδου Ελευθερία και Μηλιάδου Αθανασία. Υπεύθυνη Καθηγήτρια: Ζαρφτζιάν Μαριλένα Περιεχόμενα: Εισαγωγή Οστά Σύσταση του οστίτη
Ερειστικό Σύστημα Μια εργασία στο μάθημα της Βιολογίας από της μαθήτριες Γεωργιάδου Ελευθερία και Μηλιάδου Αθανασία. Υπεύθυνη Καθηγήτρια: Ζαρφτζιάν Μαριλένα Περιεχόμενα: Εισαγωγή Οστά Σύσταση του οστίτη
Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους.
 Τι είναι τα βλαστικά κύτταρα? Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους. Είναι σε θέση να δρουν επισκευαστικά, αναδημιουργώντας
Τι είναι τα βλαστικά κύτταρα? Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους. Είναι σε θέση να δρουν επισκευαστικά, αναδημιουργώντας
ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ
 ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής:
ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής: