ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΓΕΝΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ
|
|
|
- Ῥαχήλ Γερμανός
- 7 χρόνια πριν
- Προβολές:
Transcript
1 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΓΕΝΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ Οι ρυθµιστές της αδειοδότησης της αντιγραφής Geminin, McIdas και GemC1 στη γονιδιωµατική σταθερότητα και στη διαφοροποίηση των πολυκροσσωτών κυττάρων ΑΡΜΠΗ ΜΑΡΙΝΑ ΒΙΟΛΟΓΟΣ ΠΑΤΡΑ 2017
2 2
3 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΓΕΝΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ Οι ρυθµιστές της αδειοδότησης της αντιγραφής Geminin, McIdas και GemC1 στη γονιδιωµατική σταθερότητα και στη διαφοροποίηση των πολυκροσσωτών κυττάρων ΑΡΜΠΗ ΜΑΡΙΝΑ ΒΙΟΛΟΓΟΣ Επιβλέπουσα καθηγήτρια Λυγερού Ζωή Καθηγήτρια Τµήµα Ιατρικής Πανεπιστήµιο Πατρών ΠΑΤΡΑ
4 4
5 Τριµελής συµβουλευτική επιτροπή Λυγερού Ζωή Σταθόπουλος Γεώργιος Ταραβήρας Σταύρος Καθηγήτρια Τµήµα Ιατρικής Πανεπιστήµιο Πατρών Αναπληρωτής Καθηγητής Τµήµα Ιατρικής Πανεπιστήµιο Πατρών Καθηγητής Τµήµα Ιατρικής Πανεπιστήµιο Πατρών Επταµελής εξεταστική επιτροπή Λυγερού Ζωή Σταθόπουλος Γεώργιος Ταραβήρας Σταύρος Πέτρου-Παπαδάκη Ελένη Σανταµά Νιόβη Σπυρούλιας Γεώργιος Ταλιανίδης Ιωάννης Καθηγήτρια Τµήµα Ιατρικής Πανεπιστήµιο Πατρών Αναπληρωτής Καθηγητής Τµήµα Ιατρικής Πανεπιστήµιο Πατρών Καθηγητής Τµήµα Ιατρικής Πανεπιστήµιο Πατρών Καθηγήτρια Τµήµα Ιατρικής Πανεπιστήµιο Πατρών Αναπληρώτρια Καθηγήτρια Τµήµα Βιολογικών Επιστηµών Πανεπιστήµιο Κύπρου Καθηγητής Τµήµα Φαρµακευτικής Πανεπιστήµιο Πατρών Διευθυντής Ερευνών Διευθυντής Ινστιτούτου Μοριακής Βιολογίας και Βιοτεχνολογίας (ΙΜΒΒ) Κρήτη 5
6 6
7 ΠΕΡΙΕΧΟΜΕΝΑ ΠΕΡΙΛΗΨΗ ABSTRACT ΕΙΣΑΓΩΓΗ Συντονισμός των κύκλων διπλασιασμού και διαχωρισμού των χρωμοσωμάτων και των κεντροσωματίων Ο κύκλος διπλασιασμού των κεντροσωματίων Δομή και λειτουργία κεντροσωματίων/κεντριολίων Οι φάσεις διπλασιασμού των κεντροσωματίων κατά τη διάρκεια του κυτταρικού κύκλου Το μοριακό μονοπάτι που ελέγχει το διπλασιασμό των κεντριολίων κατά τη διάρκεια του κυτταρικού κύκλου Οι μηχανισμοί που εξασφαλίζουν το σωστό διπλασιασμό των κεντριολίων Πολυκροσσωτά κύτταρα Δομή κροσσών Προέλευση κροσσών Εντοπισμός και λειτουργία των πολυκροσσωτών κυττάρων Διαφοροποίηση των πολυκροσσωτών κυττάρων Τα μέλη της υπεροικογένειας της Geminin Δομικά χαρακτηριστικά Ο ρόλος των μελών της υπεροικογένειας της Geminin στη ρύθμιση του κυτταρικού κύκλου Ο ρόλος των μελών της υπεροικογένειας της Geminin σε διαδικασίες κυτταρικής διαφοροποίησης Σκοπός Υλικά και Μέθοδοι Μέθοδοι ανασυνδυασμένου DNA κλωνοποιήσεις Λίστα πλασμιδίων Πορεία κλωνοποίησης των πλασμιδιακών φορέων έκφρασης Μέθοδοι κυτταροκαλλιέργειας Καλλιέργεια καρκινικών κυττάρων Απομόνωση και καλλιέργεια των επιθηλιακών κυττάρων της τραχείας μυών (Mouse tracheal epithelial cell culture MTEC) Πλασμιδιακά συστήματα παραγωγής λεντιιών Μόλυνση 293Τ κυττάρων για τιτλοδότηση των παραγόμενων λεντιιών Μόλυνση των επιθηλιακών κυττάρων τραχείας μυών με λεντιιούς Δοκιμασία λουσιφεράσης Μέθοδοι απομόνωσης ιστών Απομόνωση ιστών εμβρύων και νεογέννητων μυών Απομόνωση ιστών ενήλικων μυών Προετοιμασία δειγμάτων για Real-time PCR Απομόνωση RNA από ιστούς εμβρύων μυών Απομόνωση RNA από ιστούς ενήλικων μυών Απομόνωση RNA από MTEC Απομόνωση RNA από 293T Σύνθεση cdna Real-time PCR Ταυτοποίηση γονοτύπου των μυών Απομόνωση γενωμικού DNA PCR για την ανίχνευση των WT και KO αλληλομόρφων του GemC Ανοσοφθορισμός
8 3.7.1 Πρωτόκολλο ανοσοφθορισμού σε κύτταρα Πρωτόκολλο ανοσοφθορισμού σε τομές ιστών Πρωτόκολλο ανοσοφθροσισμού σε MTEC Πίνακας αντισωμάτων Υβριδοποίηση ιn situ Παραγωγή γραμμικών πλασμιδιακών φορέων και απομόνωσή τους Παραγωγή των RNA ανιχνευτών Πρωτόκολλο υβριδοποίησης in situ Διπλασιασμός των κεντροσωματίων με τη χρήση υδροξυουρίας (HU) ΑΠΟΤΕΛΕΣΜΑΤΑ Μελέτη της έκφρασης των μελών της υπεροικογένειας της Geminin στα πολυκροσσωτά κύτταρα Μελέτη της έκφρασης των μελών της υπεροικογένειας της Geminin κατά την εμβρυϊκή ανάπτυξη και σε ενήλικους μυς Τα γονίδια GemC1 και McIdas εκφράζονται ειδικά στα πολυκροσσωτά κύτταρα του αναπνευστικού επιθηλίου Μελέτη της έκφρασης των μελών της υπεροικογένειας της Geminin κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων Η πρωτεΐνη McIdas εκφράζεται κατά τα πρώιμα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Η έκφραση της πρωτεΐνης McIdas περιορίζεται σημαντικά στα μεταγενέστερα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Μελέτη του ρόλου των μελών της υπεροικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων Η υπερέκφραση των παραγόντων McIdas και GemC1 επάγει τα πρώιμα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Η έλλειψη της πρωτεΐνης Geminin επάγει τα πρώιμα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Μεταγραφική ρύθμιση του μονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων Οι παράγοντες GemC1 και McIdas δρουν ως μεταγραφικοί ενεργοποιητές γονιδίων που εμπλέκονται στη διαφοροποίηση των πολυκροσσωτών κυττάρων Οι παράγοντες GemC1 και McIdas ενεργοποιούν μεταγραφικά το μονοπάτι της διαφοροποίησης των πολυκροσσωτών κυττάρων σε συνεργασία με το μεταγραφικό παράγοντα Ε2F Ο παράγοντας McIdas δεν ελέγχει μεταγραφικά την έκφραση του γονιδίου GemC Το γονίδιο GemC1 είναι απαραίτητο για τη διαφοροποίηση των πολυκροσσωτών κυττάρων στο αναπνευστικό επιθήλιο μυών Δημιουργία διαγονιδιακών μυών από τους οποίους απουσιάζει η πρωτεΐνη GemC Οι ελλειμματικοί μύες για το γονίδιο GemC1 εμφανίζουν καθυστέρηση στην ανάπτυξη και πεθαίνουν σύντομα μετά τη γέννηση Η πρωτεΐνη GemC1 είναι απαραίτητη για τη δημιουργία των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου Η έλλειψη του γονιδίου GemC1 εμποδίζει την έκφραση παραγόντων που εμπλέκονται σε διαφορετικά στάδια της δημιουργίας των πολυκροσσωτών κυττάρων Η έλλειψη του γονιδίου GemC1 δεν επηρεάζει τον αριθμό των βασικών διαιρούμενων κυττάρων, ενώ αυξάνει τα βλεννοπαραγωγά κύτταρα του αναπνευστικού επιθηλίου
9 4.5 Προκαταρκτική μελέτη της συμμετοχής της πρωτεΐνης McIdas στο διπλασιασμό των κεντριολίων σε διαιρούμενα κύτταρα Η πρωτεΐνη McIdas εντοπίζεται στο μητρικό κεντριόλιο των κεντροσωματίων των διαιρούμενων κυττάρων Η υπερέκφραση της πρωτεΐνης McIdas προκαλεί αύξηση του αριθμού των κεντροσωματίων ΣΥΖΗΤΗΣΗ Έκφραση των μελών της υπεροικογένειας της Geminin στα πολυκροσσωτά κύτταρα Ιστοειδική έκφραση των παραγόντων GemC1 και McIdas στα πολυκροσσωτά κύτταρα Έκφραση των παραγόντων GemC1 και McIdas κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων Η πρωτεΐνη McIdas εκφράζεται στα πρώιμα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων και όχι στα τελικώς διαφοροποιημένα κύτταρα Τα μέλη της υπεροικογένειας της Geminin ελέγχουν τη διαφοροποίηση των πολυκροσσωτών κυττάρων Μεταγραφική ρύθμιση του μονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων Οι παράγοντες GemC1 και McIdas δρουν ως μεταγραφικοί ενεργοποιητές του μονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων Οι παράγοντες GemC1 και McIdas δρουν συνεργατικά με μέλη της οικογένειας των Ε2F μεταγραφικών παραγόντων H πρωτεΐνη Geminin αναστέλλει τη μεταγραφική ενεργότητα των GemC1 και McIdas Ο in vivo ρόλος του παράγοντα GemC1 στη διαφοροποίηση των πολυκροσσωτών κυττάρων Ο παράγοντας GemC1 αποτελεί κεντρικό ρυθμιστή της διαφοροποίησης των πολυκροσσωτών κυττάρων Συσχέτιση του γονιδίου GemC1 με ασθένειες που δημιουργούνται εξαιτίας της μη σωστής διαφοροποίησης των πολυκροσσωτών κυττάρων Σύγκριση του ρόλου των GemC1 και McIdas στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Δρουν κατά πλεονασμό ή εξειδικεύονται; Συνοψίζοντας το ρόλο της υπεροικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων Οι πρωτεΐνες της οικογένειας της Geminin στη ρύθμιση του κύκλου των κεντροσωματίων Η συμμετοχή του McIdas στον πολλαπλασιασμό των κεντριολίων σε διαιρούμενα κύτταρα Τα μέλη της υπεροικογένειας της Geminin ως κοινοί ρυθμιστές των κύκλων διπλασιασμού και διαχωρισμού των χρωμοσωμάτων και των κεντροσωματίων ΒΙΒΛΙΟΓΡΑΦΙΑ ΔΗΜΟΣΙΕΥΣΕΙΣ
10 10
11 ΠΡΟΛΟΓΟΣ Η παρούσα διδακτορική διατριβή πραγµατοποιήθηκε στο εργαστήριο Γενικής Βιολογίας της Ιατρικής σχολής του Πανεπιστηµίου Πατρών. Η ανάθεση του θέµατος και η επίβλεψη της εργασίας έγιναν από την καθηγήτρια κα Λυγερού Ζωή, την οποία και θα ήθελα να ευχαριστήσω ειλικρινά για την εµπιστοσύνη που µου έδειξε από την πρώτη στιγµή που βρέθηκα στο εργαστήριό της. Η καθοδήγησή της ήταν ουσιαστική καθόλη την πορεία µου. Μου έδωσε όλα τα απαραίτητα εφόδια για µια σωστή και συγκροτηµένη επιστηµονική σκέψη που θα µε βοηθήσουν στη µελλοντική µου πορεία. Αποτελεί πρότυπο ερευνητή για εµένα και χαίροµαι γιατί δουλεύοντας στο εργαστήριό της αγάπησα ακόµα περισσότερο την επιστήµη που επέλεξα. Τέλος, θα ήθελα να την ευχαριστήσω και σε προσωπικό επίπεδο για το ενδιαφέρον της και τη βοήθειά της όποτε χρειάστηκα κάτι. Θα ήθελα επίσης να ευχαριστήσω τα άλλα δυο µέλη της τριµελούς συµβουλευτικής µου επιτροπής, τον κ. Ταραβήρα Σταύρο και τον κ. Σταθόπουλο Γεώργιο, καθώς και τα υπόλοιπα µέλη της επταµελούς εξεταστικής επιτροπής, κα Πέτρου-Παπαδάκη Ελένη, κα Σανταµά Νιόβη, κ. Σπυρούλια Γεώργιο και κ. Ταλιανίδη Ιωάννη για το χρόνο τους και την τιµή που µου έκαναν να συµµετέχουν στην ολοκλήρωση της διδακτορικής µου διατριβής. Ένα ιδιαίτερο ευχαριστώ στον κ. Ταραβήρα Σταύρο, µε τον οποίο συνεργαστήκαµε στενά σε όλη την πορεία της διδακτορικής µου διατριβής. Ένα µεγάλο µέρος των πειραµάτων µου πραγµατοποιήθηκε στο εργαστήριό του, δίνοντάς µου έτσι την ευκαιρία να αποκτήσω πολύτιµη εµπειρία στο χειρισµό των πειραµατοζώων και σε επιπλέον πειραµατικές προσεγγίσεις. Το ενδιαφέρον του ήταν µεγάλο και συνεχές σε κάθε µου πείραµα και ήταν πάντα πρόθυµος να µε συµβουλέψει όποτε χρειάστηκα κάτι. Ακόµα, θα ήθελα να ευχαριστήσω τον κ. Σταθόπουλο Γεώργιο για τις συζητήσεις µας, τις συµβουλές, το ειλικρινές ενδιαφέρον του για την πορεία µου και την ε µπιστοσύνη που έδειξε στη δουλειά µου. Ένα µεγάλο ευχαριστώ στην Dr. Monica Bettencourt-Dias που µε φιλοξένησε για σχεδόν τρεις µήνες στο εργαστήριό της Cell Cycle Regulation Lab, στη Λισαβώνα. Εκεί πραγµατοποιήθηκαν τα πρώτα πειράµατα που αφορούν το διπλασιασµό των κεντριολίων. Η επίσκεψη αυτή αποτέλεσε για εµένα µια µεγάλη ευκαιρία τόσο σε ερευνητικό όσο και σε προσωπικό επίπεδο, δίνοντάς µου πολύτιµη εµπειρία για την µετέπειτα πορεία µου. Θα ήθελα επίσης να εκφράσω ένα ιδιαίτερο ευχαριστώ στους καθηγητές και στα µέλη των εργαστηρίων Γενικής Φαρµακολογίας, Βιολογικής Χηµ είας και Φυσιολογίας της Ιατρικής Σχολής του Πανεπιστηµίου Πατρών που προσέφεραν πάντα τη βοήθειά τους σε οτιδήποτε χρειαζόµουν. Επίσης, θα ήθελα να ε υχαριστήσω τον Διευθυντή του εργαστηρίου Γενικής Βιολογίας κ. Νικόλαο Μοσχονά, την Οµότιµη καθηγήτρια κ. Αγλαΐα Αθανασιάδου, τα µέλη ΔΕΠ κ. Ζαρκάδη Ιωάννη και κα Παπαχατζοπούλου Αδαµ αντία, καθώς και τα µέλη ΕΤΕΠ κ. Καραχάλιο Παναγιώτη και κα Σάκκουλα Ελένη, για το ευχάριστο κλίµα και τις συζητήσεις µας. Δεν θα µπορούσα βέβαια να παραλείψω να ευχαριστήσω ειλικρινά όλους τους προπτυχιακούς και µεταπτυχιακούς φοιτητές, καθώς και τους υποψήφιους διδάκτορες 11
12 των εργαστηρίων Γενικής Βιολογίας και Φυσιολογίας. Πραγµατικά αποτέλεσαν για εµένα δεύτερη οικογένεια. Θα ήθελα να τους ευχαριστήσω για την αµέριστη στήριξή τους, τη συµπαράσταση, τη βοήθεια και την κατανόησή τους σε όλη µου την πορεία, καθώς και για το ευχάριστο κλίµα τόσο µέσα στο εργαστήριο όσο και εκτός αυτού. Ένα ιδιαίτερο ευχαριστώ στους Δάφνη-Ελευθερία Πεφάνη, Ιωάννα-Ελένη Συµεωνίδου, Παπαναστασίου Αναστάσιο, Νικόλαο-Νικηφόρο Γιακουµάκη και Γιαβρίδη Θεόδωρο που ήταν δίπλα µου στο ξεκίνηµά µου στο εργαστήριο. Επίσης, να ευχαριστήσω τους προπτυχιακούς φοιτητές που ανέλαβα κατά τη διάρκεια της διατριβής µου, κάθως µέσα από αυτούς µάθαινα και εξελισσόµουν και εγώ: Ιωάννη Λουκά, Μπεκιάρη Αλεξάνδρα, Γρηγοριάδη Σωκράτη, Σταυρούλα Τσαρίδου και Μαργαρίτα Σκαµνέλου. Θα ήθελα να ξεχωρίσω ιδιαιτέρως το φίλο µου Ιωάννη Λουκά, ο οποίος συνέχισε και ως µεταπτυχιακός φοιτητής και έτσι συνεργαστήκαµε στενά σε όλη τη διάρκεια της πορείας µας στο εργαστήριο. Επίσης να ευχαριστήσω τους: Μιχάλη Πετρόπουλο, Πατρούλα Ναθαναηλίδου, Στέλλα Μαξούρη, Βίκυ Πετρουλάκη, Ειρήνη Κασελίµη, Αλεξάνδρα Πατµανίδη, Ίονα Χαµπέρη και Πηνελόπη Νικολοπούλου, γιατί όλα αυτά τα χρόνια ο καθένας τους µου έδωσε κάτι ξεχωριστό. Θα ήθελα επίσης να µου επιτραπεί να ξεχωρίσω και να ευχαριστήσω ιδιαίτερα τους φίλους µου Αλεξάνδρα Κανέλλου, Νικόλαο Καραντζέλη, Χριστίνα Κυρούση, Μαρία-Ελένη Λαλιώτη, Αργυρώ Καλογεροπούλου, Ανδρέα Παναγόπουλο, Ευδοξία Τασιούδη, Κωνσταντίνα Καπλάνη και Μελέτιο Βέρρα. Ένα µεγάλο ευχαριστώ οφείλω και στον κ. Σωτήρη Τσάκα, µέλος Ε.ΔΙ.Π του τµήµατος Βιολογίας του Πανεπιστηµίου Πατρών, καθώς αποτέλεσε για εµένα τον πρώτο µου δάσκαλο στα φοιτητικά µου χρόνια και µου µετέδωσε όλη την αγάπη του για την έρευνα. Τέλος, θα ήθελα να ευχαριστήσω τους πολυαγαπηµένους µου φίλους Διονύση Αυγουστάτο, Κατερίνα Βασιλοπούλου και Αναστασία Φερλέµη π ου είναι πάντοτε δίπλα µου στα δύσκολα και κυρίως πάντοτε δίπλα µου στη χαρά µου. Ένα µεγάλο ευχαριστώ στους αθέατους αλλά ισχυρούς συνοδοιπόρους µου, στους πολυαγαπηµένους µου γονείς Ηλία και Αργυρώ, στα αδέρφια µου Ρούλα και Άλκη, στα λατρεµένα µου ανύψια Λάµπρο και Ηλία που αποτέλεσαν το αγχολυτικό µου σε όλη µου την πορεία και στον άνθρωπο της ζωής µου Θανάση, για τη συνεχή τους υποστήριξη και την αγάπη τους για εµένα... Νιώθω ειλικρινά συγκινηµένη και συνάµα περήφανη που µπορώ σήµερα στο σύντοµο αυτό γραπτό µου κείµενο να εκφράσω τα βαθύτατα συναισθήµατά µου σε όλους εσάς που υπήρξατε οι αρχιτέκτονες της επιτυχίας µου και να οµολογήσω πως µετά από όλα αυτά συναισθάνοµαι τη µεγάλη ευθύνη και γοητεία της επιστήµης που συνειδητά επέλεξα! να εύχεσαι ναναι μακρύς ο δρόμος, γεμάτος περιπέτειες, γεμάτος γνώσεις... Κ. Π. Καβάφης 12
13 ΠΕΡΙΛΗΨΗ ΠΕΡΙΛΗΨΗ Η γονιδιωµατική σταθερότητα στα κύτταρα απαιτεί την πλήρη αντιγραφή του γονιδιώµατος και τον ακριβοδίκαιο διαχωρισµό των διπλασιασµένων χρωµοσωµάτων στα θυγ ατρικά κύτταρα σε κάθε κυτταρική διαίρεση. Αυτό επιτυγχάνεται µέσω της συντονισµένης λειτουργίας των κύκλων διπλασιασµού και διαχωρισµού των χρωµοσωµάτων και των κεντροσωµατίων. Τα τελευταία χρόνια, ένας αυξανόµενος αριθµός µορίων έχει χαρακτηριστεί να συµµετέχει και στους δύο κύκλους. Ωστόσο, ο τρόπος µε τον οποίο δρουν για το συντονισµό τους παραµένει άγνωστος. Στην παρούσα διατριβή εξετάστηκαν τα µέλη της υπεροικογένειας της Geminin στον κυτταρικό κύκλο κ αι στην κυτταρική διαφοροποίηση. Τα ευρήµατα καθιστούν τις πρωτεΐνες Geminin, McIdas και GemC1 πιθανoύς κοινούς ρυθµιστές της αντιγραφής του DNA και του διπλασιασµού των κεντριολίων. Η πρωτεΐνη Geminin είναι το πρώτο µέλος της οικογένειας που χαρακτηρίστηκε στα µετάζωα και έχει µελετηθεί εκτενώς για το ρόλο της στον κυτταρικό κύκλο. Συνεισφέρει σηµαντικά στη διατήρηση της γονιδωµατικής σταθερότητας. Μέσω µια περιοχής σπειροειδούς σπειράµατος που διαθέτει αλληλεπιδρά µε τον παράγοντα αδειοδότησης της αντιγραφής Cdt1, αναστέλλοντας την πρόωρη πυροδότηση των αφετηριών της αντιγραφής. Ακόµα, δρα ανασταλτικά στον υπερδιπλασιασµό των κεντροσωµατίων. Τέλος, συµµετέχει και σε αποφάσεις κυτταρικής διαφοροποίησης. Α λληλεπιδρά µε µεταγραφικούς παράγοντες και σύµπλοκα αναδιάταξης της χρωµατίνης, ελέγχοντας τη γονιδιακή έκφραση. Τα τελευταία χρόνια ταυτοποιήθηκαν στα σπονδυλωτά τα δυο νέα µέλη της οικογένειας, GemC1 και McIdas, βάση της οµοιότητας που παρουσίαζαν µε τη Geminin στην περιοχή του σπειροειδούς σπειράµατος που διαθέτουν. Η πρωτεΐνη McIdas αλληλεπιδρά µε τη Geminin και εµποδίζει την πρόσδεσή της στον παράγοντα Cdt1, ενώ ο παράγοντας GemC1 συµβάλλει στο σχηµατισµό του προεναρκτήριου συµπλόκου της αντιγραφής. Αρχικά πειράµατα στο εργαστήριό µας έδειξαν ότι οι παράγοντες GemC1 και McIdas είναι πιθανό να εµπλέκονται, όπως και η Geminin, σε διαδικασίες κυτταρικής διαφοροποίησης. Στην παρούσα διατριβή µελετήσαµε το ρόλο τους κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Τα πολυκροσσωτά κύτταρα είναι τελικώς διαφοροποιηµένα κύτταρα, στα οποία παράγεται ένας µεγάλος αριθµός κεντριολίων που δρουν ως βασικά σωµάτια για το σχηµατισµό πολλαπλών κροσσών. Αρχικά, δείχθηκε ότι οι παράγοντες GemC1 και McIdas εκφράζονται σε υψηλά επίπεδα κατά την ανάπτυξη των µυών σε ιστούς όπου απαντώνται πολυκροσσωτά κύτταρα. Η έκφρασή τους συµπίπτει µε αυτή του παράγοντα Foxj1, που αποτελεί ένα σηµαντικό ρυθµ ιστή της µετανάστευσης των νεοσυντιθέµενων κεντριολίων στην πλασµατική µεµβράνη και της δηµιουργίας του αξονήµατος των κροσσών. Η έκφρασή τους µελετήθηκε πιο ειδικά στα στάδια της διαφοροποίησης, σε ένα ex vivo σύστηµα καλλιέργειας επιθηλιακών κυττάρων τραχείας µυών. Οι GemC1 και McIdas εκφράζονται αµέσως µετά την επαγωγή της διαφοροποίησης και µειώνονται µε την ολοκλήρωση της διαδικασίας. Τα υψηλότερα επίπεδα της έκφρασής τους ανιχνεύονται στο στάδιο του πολλαπλασιασµού των κεντριολίων. Αντίθετα, η Geminin εµφανίζει ένα ανεστραµµένο πρότυπο έκφρασης, µε υψηλότερα επίπεδα στα 13
14 ΠΕΡΙΛΗΨΗ διαιρούµενα προγονικά κύτταρα. Περαιτέρω µελέτη έδειξε ότι η πρωτεΐνη McIdas εκφράζεται ειδικά στα πρώιµα στάδια της διαφοροποίησης, ενώ η έκφρασή της εξαφανίζεται στα ώριµα πολυκροσσωτά κύτταρα. Για το χ αρακτηρισµό της λειτουργίας των µελών της υπεροικογένειας της Geminin κατά την εξειδίκευση των πολυκροσσωτών κυττάρων πραγµατοποιήθηκαν πειράµατα υπερέκφρασης και αποσιώπησης. Υπερέκφραση των GemC1 κ αι McIdas έδειξε ότι είναι ικανοί να επάγουν τα πρώιµα στάδια διαφοροποίησης. Ο ίδιος φαινότυπος παρατηρήθηκε µετά την αποσιώπηση της Geminin, ενισχύοντας την υπόθεση για ανταγωνιστική δράση των µελών της υπεροικογένειας. Οι παράγοντες GemC1 και McIdas προκαλούν τη µεταγραφική ενεργοποίηση του Foxj1. Επιπλέον, ο παράγοντας GemC1 επάγει τη µεταγραφή του McIdas, ενώ το αντίθετο δεν παρατηρήθηκε. Η λειτουργία τους αυτή ενισχύεται µέσω συνεργασίας µε το µεταγραφικό παράγοντα E2F5, ενώ η Geminin αναστέλλει τη µεταγραφική ενεργότητα των GemC1 και McIdas. Τέλος, ο McIdas δρα µέσω ενός µηχανισµού θετικής ανατροφοδότησης, καθώς είναι ικανός να επάγει τη µεταγραφή από τα ρυθµιστικά στοιχεία του γονιδίου του. Τα παραπάνω πειράµατα συνέβαλαν στο να αποσαφηνιστεί ο µοριακός µηχανισµός µέσω του οποίου τα µέλη της υπεροικογένειας της Geminin ελέγχoυν το µονοπάτι της διαφοροποίησης. Η κατασκευή διαγονιδιακών µυών ανέδειξε τον in vivo ρόλο του παράγοντα GemC1. Οι ελλειµµατικοί για το γονίδιο GemC1 µύες εµφανίζουν καθυστέρηση στην ανάπτυξη και πεθαίνουν σύντοµα µετά τη γέννα. Επίσης, αναδείχθηκε ο καθοριστικός ρόλος του στη διαφοροποίηση των πολυκροσσωτών κυττάρων, αφού αυτά απουσιάζουν εντελώς από την τραχεία των διαγονιδιακών µυών. Τέλος, µελετήθηκε η συµµετοχή της πρωτεΐνης McIdas στον πολλαπλασιασµό των κεντριολίων στα διαιρούµενα κύτταρα. Η πρωτεΐνη McIdas εκφράζεται ειδικά στο µητρικό κεντριόλιο των κεντροσωµατίων και προκαταρκτικά πειράµατα υπερέκφρασής της προκάλεσαν αύξηση στον αριθµό των κεντριολίων, υποδηλώνοντας την εµπλοκή της στο διπλασιασµό τους. Τα παραπάνω δεδοµένα µαρτυρούν ότι τα µέλη της υπεροικογένειας της Geminin αποτελούν κεντρικούς ρυθµιστές του πολλαπλασιασµού των κεντριολίων και της διαφοροποίησης των πολυκροσσωτών κυττάρων. Μέσω θετικών ή αρνητικών αλληλεπιδράσεων που αναπτύσσουν, τόσο µεταξύ τους όσο και µε άλλους παράγοντες είναι πιθανό να συνδέουν την αντιγραφή του DNA µε τον πολλαπλασιασµό των κεντριολίων και τη δηµιουργία των πολυκροσσωτών κυττάρων. 14
15 ABSTRACT ABSTRACT To maintain genome integrity cells must ensure that chromosomes are fully replicated and sister chromatids are segregated accurately to the daughter cells in each cell cycle. The chromosome and centrosome cycles in proliferating cells must be coordinated to ensure that genome and centriole duplication occur only once per cell cycle. Common regulators have been identified, but their role in linking the two cycles awaits elucidation. Geminin family members are implicated both in cell cycle and differentiation control and are likely to constitute common key regulators of genome and centrosome duplication. Geminin, the founding member of the family, was initially characterized as a negative regulator of the initiation of DNA replication in metazoa. Geminin is a coiled-coil protein that interacts and inactivates the licensing factor Cdt1, preventing the re-firing of origins until the beginning of the next cell cycle. Geminin also acts as a negative regulator of the centriole cycle, safeguarding genome stability. Geminin has also been linked to cell fate decisions, through binding to transcription factors and chromatin remodelling complexes, thereby controlling transcription of specific target genes. Recently, the vertebrate-specific GemC1 and McIdas were identified due to the significant conservation of their coiled-coils to the coiled-coil domain of Geminin. McIdas binds to Geminin and inhibits its association with Cdt1. GemC1 mediates the formation of the pre-initiation complex during DNA replication. In previous studies from our lab it has been shown that similar to Geminin, GemC1 and McIdas are implicated in differentiation decisions. In this study, we aim to understand their roles during multiciliate cell differentiation. Multiciliated cells are terminally differentiating cells, which massively amplify their centrioles, bypassing the rule for the once-per-cell-cycle centriole duplication. Hundreds of centrioles dock to the apical cell surface and generate motile cilia, whose concerted movement ensures fluid flow across epithelia. In ciliated epithelia GemC1 and McIdas are expressed at high levels during development. Their expression closely mirrors Foxj1 expression, a transcription factor specifically expressed in cells with motile cilia and implicated in multiciliogenesis. An ex vivo ciliogenesis model using mouse tracheal epithelial cells was used to assess the timing of GemC1 and McIdas expression during multiciliogenesis. Their expression is increased directly after the induction of multiciliate cell differentiation, dropping when ciliogenesis is completed. Their levels of expression are increased when multiple centriole formation is taking place. On the contrary, Geminin shows an inverted expression pattern, with high levels in cycling progenitor cells. Its expression diminishes as cells progress towards differentiation. In more detail, McIdas protein is expressed in early differentiating cells and its expression is lost as multiciliated cells mature. The ectopic expression of McIdas and GemC1 can promote the early steps of multiciliogenesis. The same phenotype was observed after the depletion of Geminin, suggesting its antagonistic role. GemC1 and McIdas activate transcriptionally the upstream regulatory elements of FOXJ1 gene. Their transcriptional activity is enhanced by the transcription factor E2F5 and inhibited by Geminin. GemC1 is able to induce transcription from the MCIDAS gene, while the opposite is not possible. 15
16 ABSTRACT Also, McIdas is able to transactivate its own promoter, creating a positive feedback loop that locks commitment toward multiciliogenesis. The in vivo role of GemC1 during multiciliate cell differentiation was delineated after the depletion of GemC1 from mice. Mice lacking GemC1 are born with airway epithelia devoid of multiciliated cells and die soon after birth. Finally, we studied the role of McIdas protein in centriole duplication, in dividing cells. McIdas is specifically localized in the distal part of the mother centriole. Overexpression of the McIdas protein affects centriole numbers in cycling cells, suggesting a possible role of McIdas during the centrosome cycle. The above data suggest that Geminin family members are central regulators of centriole duplication and multiciliate cell differentiation. Intricate positive and negative regulatory interactions among them appear important for their function. Geminin, GemC1 and McIdas may link DNA replication to centriole duplication and multiciliogenesis, acting close to the transition from proliferation to differentiation. 16
17 1. ΕΙΣΑΓΩΓΗ
18 1. ΕΙΣΑΓΩΓΗ 18
19 1. ΕΙΣΑΓΩΓΗ 1.1 Συντονισµός των κύκλων διπλασιασµού και διαχωρισµού των χρωµοσωµάτων και των κεντροσωµατίων Για να εξασφαλίζεται η γονιδιωµατική σταθερότητα στα κύτταρα απαιτείται αυστηρή χωροχρονική ρύθµ ιση του κυτταρικού κύκλου. Στα διαιρούµενα κύτταρα, ένα σύνολο µηχανισµών ελέγχει τη σωστή διέλευση του κυττάρου από την κάθε φάση του κυτταρικού κύκλου, µε στόχο τον πλήρη διπλασιασµό του γενετικού υλικού, καθώς και τον ακριβοδίκαιο διαχωρισµό του στα θυγατρικά κύτταρα. Κατά τη G1 φάση το κύτταρο ανταποκρίνεται σε σήµατα που δέχεται από το περιβάλλον του και είτε συνεχίζει στην επόµενη φάση του κύκλου διπλασιάζοντας το γενετικό του υλικό, είτε σταµατά και εξέρχεται από αυτόν παροδικά ή µόνιµα, εάν τα σήµατα που λαµβάνει είναι ανασταλτικά. Κατά τη διαφοροποίηση, απατείται η µόνιµη έξοδος των κυττάρων από τον κυτταρικό κύκλο. Μ όρια-ρυθµιστές του κυτταρικού κύκλου είναι καθοριστικά για την απόφαση µόνιµης εξόδου από τον κυτταρικό κύκλο και τη διαφοροποίηση. O κυτταρικός κύκλος περιλαµβάνει τον κύκλο διπλασιασµού και διαχωρισµού των κεντροσωµατίων και τον κύκλο διπλασιασµού και διαχωρισµού των χρωµοσωµάτων. Οι δυο κύκλοι συνδέονται άρρηκτα και βρίσκονται κάτω από αυστηρό έλεγχο, ώστε να εξασφαλίζεται ότι σε κάθε κύκλο το γενετικό υ λικό διπλασιάζεται πλήρως και οι αδελφές χρωµατίδες διαχωρίζονται σωστά στα θυγατρικά κύτταρα (Εικόνα 1.1). Η αδειοδότηση, ο διπλασιασµός και ο διαχωρισµός των χρωµοσωµάτων και των κεντροσωµατίων εξασφαλίζει ότι κάθε θυγατρικό κύτταρο κληρονοµεί ένα αντίγραφο του γονιδιώµατος και µόνο ένα κεντροσωµάτιο. Ανωµαλίες στον κύκλο των κεντροσωµατίων ή/και των χρωµοσωµάτων οδηγεί σε γονιδιωµατική αστάθεια. 19

20 1. ΕΙΣΑΓΩΓΗ Εικόνα 1.1 Οι κύκλοι διπλασιασµού και δαχωρισµού των χρωµοσωµάτων και των κεντροσωµατίων συνδέονται άρρηκτα. Η αδειοδότηση, ο διπλασιασµός και ο διαχωρισµός αποτελούν κοινά βήµατα που συντονίζουν τους δυο κύκλους. Μετά το τέλος της µίτωσης, κατά τη διάρκεια της G1 φάσης, αδειοδοτείται ένας νέος κύκ λος διπλασιασµού του γονιδιώµατος και των κεντριολίων. Ο διπλασιασµός λαµβάνει χώρα κατά τη φάση S, ενώ οι µηχανισµοί που εµποδίζουν ένα νέο κύκλο διπλασιασµού κατά τις φάσεις S και G2, εξασφαλίζουν ότι θα συµβεί µόνο µια φορά κατά τη διάρκεια του κυτταρικού κύκλου. Μετά τη µίτωση, κάθε θυγατρικό κύτταρο κληρονοµεί ένα πλήρες αντίγραφο του γονιδιώµατος και ένα κεντροσωµάτιο, αποτελούµενο από δυο κεντριόλια. Αλλαγές στο φυσιολογικό µηχανισµό αντιγραφής του DNA µπορούν να οδηγήσουν σε περίσσεια ή έλλειψη του γενετικού υλικού και σε χρωµοσωµικές ανωµαλίες. Η δηµιουργία πολυπολικών ατράκτων αποτελεί µια συχνή αιτία µη σωστού διαχωρισµού τ ων χρωµοσωµάτων κατά τη µίτωση. Ωστόσο, ο ι παραπάνω αλλαγές παρατηρούνται και φυσιολογικά και οδηγούν σε κύτταρα διαφορετικού αριθµού χρωµοσωµάτων ή κεντροσωµατίων (Εικόνα 1.2). Επαναλαµβανόµενοι κύκλοι 20
21 1. ΕΙΣΑΓΩΓΗ αντιγραφής του DNA, όπου εναλλάσσονται S-G κ ύκλοι χωρίς κυτταρική διαίρεση (ενδοαντιγραφή/ενδοµίτωση, Εικόνα 1.2 Α) ή επαναλαµβανόµενη πυροδότηση της αντιγραφής συγκεκριµένων χρωµοσωµικών περιοχών (επαναντιγραφή, Εικόνα 1.2 Β) αλλάζουν την ποσότητα του γενετικού υλικού. Oι µηχανισµοί αυτοί οδηγούν σε πολυπλοειδία ή σε αύξηση του γενετικού υλικού σε συγκεκριµένες περιοχές του γονιδιώµατος, αντίστοιχα. Οι κύκλοι αυτοί παρατηρούνται σε συγκεκριµένα αναπτυξιακά στάδια κατά τον κύκλο ζωής πρωτοζώων, εντόµων, φυτών και θηλαστικών, µε σκοπό να παραχθούν τελικώς διαφοροποιηµένα κύτταρα µε εξειδικευµένες ιδιότητες. Τα πολυταινικά χρωµοσώµατα που απαντώνται στους σιελογόνους αδένες της Drosophila, αποτελούν το πιο καλά µελετηµένο παράδειγµα επαναλαµβανόµενης αντιγραφής του γενετικού υλικού, χωρίς κυτταρική διαίρεση. Επιπρόσθετα, κατά την ανάπτυξη η επαναντιγραφή του γονιδιώµατος συνεισφέρει στην απαιτούµενη αύξηση της γονιδιακής έκφρασης σε συγκεκριµένους κυτταρικούς τύπους. Για παράδειγµα στα θυλακιακά κύτταρα στη Drosophila παρατηρείται επαναντιγραφή συγκεκριµένων γονιδίων που κωδικοποιούν δοµικές πρωτεΐνες του ωοκυττάρου. Στον άνθρωπο, τα κύτταρα της τροφοβλάστης του πλακούντα και τα µεγακαρυοκύτταρα είναι χαρακτηριστικά παραδείγµατα πολυπλοειδικών κυττάρων που έχουν προκύψει από πολλαπλούς κύκλους αντιγραφής απουσία κυτταρικής διαίρεσης. Οι µηχανισµοί που ελέγχουν τη µετάβαση από ένα φυσιολογικό κυτταρικό κύκλο σε έναν προγραµµατισµένο κύκλο επαναντιγραφής του γονιδιώµατος παραµένουν άγνωστοι. Τα πολυκροσσωτά κύτταρα αποτελούν το πιο χαρακτηριστικό παράδειγµα κυττάρων που αποκλίνουν από ένα φυσιολογικό κύκλο διπλασιασµού των κεντροσωµατίων (Εικόνα 1.2 Γ). Τα συγκεκριµένα κύτταρα εξέρχονται από τον κυτταρικό κύκλο και σχηµατίζουν εκατοντάδες κεντριόλια ανά κύτταρο, υπερβαίνοντας τους κανόνες για τη δη µιουργία ενός µόνο θυγατρικού κεντριολίου, κάθετα προς το µητρικό κεντριόλιο (ο µηχανισµός αυτός αναλύεται παρακάτω). Τα νεοσυντιθέµενα κεντριόλια µεταναστεύουν και «αγκυροβολούν» στην πλασµατική µεµβράνη του κυττάρου, δρώντας ως βασικά σωµάτια, που θα δώσουν γένεση στους κροσσούς. Κύριος ρόλος των κροσσών είναι η κίνηση του περιβάλλοντος υγρού του ιστού στον οποίο βρίσκονται, όπως για παράδειγµα η κίνηση της βλέννας στο αναπνευστικό επιθήλιο. Οι µηχανισµοί που διέπουν την απόφαση ενός κυττάρου για το σχηµατισµό πολλαπλών κεντριολίων, έναντι του ενός, παραµένουν άγνωστοι. 21
 Πολλαπλοί κύκλοι (S-G) αντιγραφής του DNA, χωρίς κυτταρική διαίρεση, οδηγούν σε πολυπλοειδία. (Β) Επαναλαµβανόµενη πυροδότηση της αντιγραφής συγκεκριµένων χρωµοσωµικών περιοχών.")
22 1. ΕΙΣΑΓΩΓΗ A Β Γ Εικόνα 1.2 Φυσιολογικές αποκλίσεις στην ποσότητα του γενετικού υλικού ή στον αριθµού των κεντροσωµατίων. (Α) Πολλαπλοί κύκλοι (S-G) αντιγραφής του DNA, χωρίς κυτταρική διαίρεση, οδηγούν σε πολυπλοειδία. (Β) Επαναλαµβανόµενη πυροδότηση της αντιγραφής συγκεκριµένων χρωµοσωµικών περιοχών. (Γ) Τα πολυκροσσωτά κύτταρα ακολουθούν έναν ιδιότυπο κύκλο κεντροσωµατίων, που οδηγεί στο σχηµατισµό πολλαπλών κεντριολίων. Τα κεντριόλια µεταναστεύουν και «αγκυροβολούν» στην πλασµατική µεµβράνη του κυττάρου, αποτελώντας κέντρα πυρήνωσης για τους κροσσούς. Πρόσφατες µελέτες έχουν αναδείξει τα µέλη της οικογένειας της Geminin ως µια οµάδα πρωτεϊνών που συµµετέχει τόσο στον κυτταρικό πολλαπλασιασµό όσο και στη διαφοροποίηση των κυττάρων. Οι πρωτεΐνες της οικογένειας της Geminin, Geminin, McIdas και GemC1, χαρακτηρίστηκαν αρχικά για το ρόλο τους κατά την αντιγραφή του DNA. Ωστόσο, τα αποτελέσµατα της παρούσας διατριβής αναδεικνύουν το ρόλο τους, κυρίως τ ων GemC1 και McIdas, κατά τον πολλαπλασιασµό των κεντριολίων και τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Στη συνέχεια αναλύονται οι µηχανισµοί που ελέγχουν το διπλασιασµό των κεντριολίων στα διαιρούµενα κύτταρα, αλλά και πώς επιτυγχάνεται ο σχηµατισµός 22
23 1. ΕΙΣΑΓΩΓΗ πολλαπλών κεντριολίων στα πολυκροσσωτά κύτταρα. Τέλος, αναλύονται τα δοµικά χαρακτηριστικά και η λειτουργία των πρωτεϊνών της οικογένειας της Geminin. 23
24 1. ΕΙΣΑΓΩΓΗ 1.2 Ο κύκλος διπλασιασµού των κεντροσωµατίων Δοµή και λειτουργία κεντροσωµατίων/κεντριολίων Το κεντροσωµάτιο είναι µια δοµή που συνήθως απαντάται κοντά στον πυρήνα των περισσότερων ζωικών κυττάρων (Conduit et al, 2015). Τα κεντροσωµάτια συµµετέχουν σε σηµαντικές λειτουργίες των κυττάρων καθώς αποτελούν το κέντρο οργάνωσης των µικροσωληνίσκων που συγκροτούν τον κυτταροσκελετό (microtubule organizing center, MTOC), οργανώνουν τη µιτωτική άτρακτο συµβάλλοντας έτσι στο διαχωρισµό των χρωµοσωµάτων κατά την κυτταρική διαίρεση και τέλος λειτουργούν ως βασικά σωµάτια για τη δηµιουργία των κροσσών. Κάθε κεντροσωµάτιο αποτελείται από δυο κεντριόλια που βρίσκονται σε ορθή γωνία το ένα ως προς το άλλο και από µια άµορφη πρωτεϊνική µάζα που συµβάλλει στην οργάνωση των µικροσωληνίσκων (PCM, pericentriolar material). Φυλογενετικές µελέτες έχουν δείξει ότι τα κεντριόλια εντοπίζονται στον τελευταίο κοινό πρόγονο των ευκαρυωτικών οργανισµών (Carvalho-Santos et al, 2011). Ωστόσο, απουσιάζουν από τα κύτταρα της ζύµης και από τους ανώτερους φυτικούς οργανισµούς, τα οποία διαθέτουν ανάλογα κέντρα οργάνωσης των µικροσωληνίσκων, χωρίς όµ ως κεντριόλια. Στα ανθρώπινα κύτταρα ένα ώριµο κεντριόλιο είναι δοµηµένο από εννέα τριάδες µικροσωληνίσκων που βρίσκονται τοποθετηµένες κυκλικά, σχηµατίζοντας έναν κύλινδρο µε ύψος 450 nm και διάµετρο 250 nm (Εικόνα 1.3) (Gonczy, 2012). Κατά µήκος του κεντριολίου διακρίνεται στη βάση του το σηµείο όπου ενώνεται µε ένα δεύτερο κεντριόλιο και ονοµάζεται εγγύς άκρο (proximal end), ενώ στο άλλο του άκρο διακρίνεται το αποµακρυσµένο τµήµα του (distal end) (Εικόνα 1.3 Α). Τα δυο κεντριόλια του κυττάρου που σχηµατίζουν ένα κεντροσωµάτιο κατά τη G1 φάση του κυτταρικού κύκλου, διακρίνονται στο θυγατρικό κεντριόλιο που έχει δηµιουργηθεί κατά τη διάρκεια του προηγούµενου κυτταρικού κύκλου και στο µητρικό που αποτελεί το πιο παλιό κεντριόλιο. Το µητρικό κεντριόλιο διαφέρει από το θυγατρικό, αφού µέσω µιας διαδικασίας ωρίµανσης που υφίσταται στον προηγούµενο κυτταρικό κύκλο έχει αποκτήσει στο αποµακρυσµένο άκρο του αποφύσεις (distal/subdistal appendages). Οι αποφύσεις αυτές συµβάλλουν στην οργάνωση των µικροσωληνίσκων, καθώς και στην «αγκυροβόληση» του µητρικού κεντριολίου στην κυτταρική µεµβράνη για την οργάνωση των κροσσών (µια διαδικασία που αναλύεται παρακάτω). Τελικά κάθε κεντριόλιο ωριµάζει ενάµιση κύκλο µετά τη δηµιουργία του. 24
 Κάθε κεντροσωµάτιο αποτελείται από ένα µητρικό και ένα θυγατρικό κεντριόλιο, που ενώνονται µεταξύ τους µε έναν χαλαρό δεσµό (G1-G2 tether).")
. (B, Γ) Κάθε κεντριόλιο είναι δοµηµένο από 9 τριάδες µικροσωληνίσκων που βρίσκονται τοποθετηµένες σε κυκλική διάταξη (modified from Gonczy, 20")

25 1. ΕΙΣΑΓΩΓΗ A Θυγατρικό κεντριόλιο Distal end Μητρικό κεντριόλιο G1-G2 tether Proximal end B Γ Εικόνα 1.3 Δοµή κεντροσωµατίου. (A) Κάθε κεντροσωµάτιο αποτελείται από ένα µητρικό και ένα θυγατρικό κεντριόλιο, που ενώνονται µεταξύ τους µε έναν χαλαρό δεσµό (G1-G2 tether). Κατά µήκος του κεντριολίου διακρίνεται το εγγύς άκρο (proximal end) µε το οποίο ενώνεται µε ένα δεύτερο κεντριόλιο και το αποµακρυσµένο τµήµα (distal end), το οποίο στο µητρικό κεντριόλιο φέρει αποφύσεις (distal/subdistal appendages). (B, Γ) Κάθε κεντριόλιο είναι δοµηµένο από 9 τριάδες µικροσωληνίσκων που βρίσκονται τοποθετηµένες σε κυκλική διάταξη (modified from Gonczy, 2012) Οι φάσεις διπλασιασµού των κεντροσωµατίων κατά τη διάρκεια του κυτταρικού κύκλου Ο κύκλος διπλασιασµού των κεντροσωµατίων συνδέεται άρρηκτα µε το διπλασιασµό του DNA και περιλαµβάνει διακριτές φάσεις καθώς εξελίσσεται ο κυτταρικός κύκλος. Η αδειοδότηση για την εκκίνηση ενός νέου κύκλου διπλασιασµού, ο διπλασιασµός και ο διαχωρισµός των κεντροσωµατίων είναι τα βασικά βήµατα που συµβάλλουν στη γονιδιωµατική σταθερότητα, αφού έτσι σε κάθε νέο κύτταρο θα κληροδοτηθεί τελικά µόνο ένα κεντροσωµάτιο. Τα παραπάνω βήµατα οµοιάζουν µε τα βήµατα που ακολουθούνται και κατά το διπλασιασµό του DNA. Κατά τη G1 φάση του κυτταρικού κύκλου κάθε κύτταρο διαθέτει ένα κεντροσωµάτιο που αποτελείται από δυο κεντριόλια, ενωµένα µεταξύ τους µε έναν χαλαρό δεσµό (G1-G2 tether) (Εικόνα 1.3 Α) (Nigg & Stearns, 2011). Ο διπλασιασµός των κεντριολίων ξεκινά κατά τη µετάβαση από τη G1 στην S φάση του κυτταρικού 25
 αρχίζει να σχηµατίζεται σε κάθετη διάταξη ένα νέο θυγατρικό κεντριόλιο, που ονοµάζεται προκεντριόλιο (κανονικό")
26 1. ΕΙΣΑΓΩΓΗ κύκλου (Εικόνα 1.4) (Firat-Karalar & Stearns, 2014; Fu et al, 2015; Gonczy, 2012). Πιο συγκεκριµένα, από το εγγύς άκρο κάθε προϋπάρχοντος κεντριολίου (µητρικό κεντριόλιο) αρχίζει να σχηµατίζεται σε κάθετη διάταξη ένα νέο θυγατρικό κεντριόλιο, που ονοµάζεται προκεντριόλιο (κανονικό κεντριολιο-εξαρτώµενο µονοπάτι, canonical centriole-dependent pathway, CD). Ανάµεσα σε κάθε µητρικό και θυγατρικό κεντριόλιο σχηµατίζεται ένας ισχυρός δεσµός που τα κρατάει ενωµένα µεταξύ τους (S-M linker). Τα νεοσυντιθέµενα κεντριόλια επιµηκύνονται κατά τη διάρκεια της S και της G2 φάσης, ενώ στη G2 φάση πραγµατοποιείται και η ωρίµανση των µητρικών κεντριολίων. Στο τέλος της G2 φάσης, σπάει ο G1-G2 χαλαρός δεσµός που υπάρχει ανάµεσα στα δυο µητρικά κεντριόλια, ώστε κατά τη µίτωση τα δυο ζεύγη των κεντριολίων να διαχωριστούν και να τοποθετηθούν αντιδιαµετρικά στους πόλους της µιτωτικής ατράκτου, συµβάλλοντας στον ακριβοδίκαιο διαχωρισµό των χρωµοσωµάτων. Μετά το τέλος της µιτωτικής διαίρεσης κάθε θυγατρικό κύτταρο κληρονοµεί από ένα κεντροσωµάτιο, αποτελούµενο από ένα µητρικό και ένα θυγατρικό κεντριόλιο. 26
27 1. ΕΙΣΑΓΩΓΗ Εικόνα 1.4 Ο κύκλος διπλασιασµού των κεντροσωµατίων. Η αδειοδότηση για την εκκίνηση ενός νέου κύκλου διπλασιασµού, ο διπλασιασµός και ο διαχωρισµός των κεντροσωµατίων είναι τα βασικά βήµατα που καθορίζουν τον κύ κλο αυτό. Κατά το τέλος της µίτωσης προς αρχή της G1 φάσης αδειοδοτείται ένας νέος κύκλος διπλασιασµού των κεντριολίων. Ο διπλασιασµός των κεντριολίων λαµβάνει χώρα κατά την S φάση, ενώ ταυτόχρονα ε µποδίζεται να συµβεί ξανά ο διπλασιασµός τους µέσα στον ίδιο κύκλο µε µηχανισµούς που ενεργοποιούνται κατά τις S και G2 φάσεις. Μετά το τέλος της µίτωσης κάθε θυγατρικό κύτταρο έχει κληροδοτήσει ένα κεντροσωµάτιο αποτελούµενο από ένα µητρικό και ένα θυγατρικό κεντριόλιο. Το µητρικό ώριµο κεντριόλιο, δρώντας ως βα σικό σωµάτιο, συµβάλλει επίσης και στο σχηµατισµό του πρωτογενούς κροσσού σε κύτταρα που βρίσκονται εκτός κυτταρικού κύκλου. Σε πολλούς κυτταρικούς τύπους, όταν το κύτταρο εξέρχεται από τον κυτταρικό κύκλο, το ώριµο µητρικό κεντριόλιο «αγκυροβολεί» στην πλασµατική µεµβράνη, δρώντας ως βασικό σωµάτιο για τη δηµιουργία ενός πρωτογενούς κροσσού (Εικόνα 1.4). Ο πρωτογενής κροσσός, που λειτουργεί ως «κεραία» λήψης σηµάτων για το κύτταρο, αποδιοργανώνεται όταν το κύτταρο εισέρχεται ξανά στον κυτταρικό κύκλο Το µοριακό µονοπάτι που ελέγχει το διπλασιασµό των κεντριολίων κατά τη διάρκεια του κυτταρικού κύκλου Κεντρικό ρόλο στο διπλασιασµό των κεντριολίων κατέχουν πέντε πρωτεΐνες που προσδιορίστηκαν αρχικά σε µελέτες που πραγµατοποιήθηκαν σ το νηµατώδη σκώληκα Caenorhabditis elegans: η κινάση ZYG-1 και οι πρωτεΐνες SPD-2, SAS-4, SAS-5 και SAS-6, που έχουν ως κοινό χαρακτηριστικό την ύπαρξη µιας περιοχής σπειροειδούς σπειράµατος στο πρωτεϊνικό τους µόριο (Brito et al, 2012; Gonczy, 2012; Nigg & Stearns, 2011). Οι παράγοντες αυτοί εµφανίζουν σηµαντική εξελικτική συντήρηση και σε άλλα είδη οργανισµών, ενώ πρόσφατες µελέτες έχουν αποσαφηνίσει τα κύρια βήµατα που ρυθµίζουν το διπλασιασµό των κεντριολίων και στα ανθρώπινα κύτταρα. Όπως προαναφέρθηκε, ο διπλασιασµός των κεντριολίων στα διαιρούµενα κύτταρα πραγµατοποιείται κατά τη διάρκεια της φάσης S του κυτταρικού κύκλου. Στους ανώτερους ευκαρυώτες, συµπεριλαµβανοµένων και των ανθρώπινων κυττάρων, στα αρχικά στάδια της διαδικασίας αυτής, ο παράγοντας Cep63 µαζί µε τον παράγοντα Cep152 συνεντοπίζονται στο εγγύς άκρο του µητρικού κεντριολίου και σχηµατίζουν µια δοµή που οµοιάζει µε δακτύλιο (Εικόνα 1.3) (Sir et al, 2011). Το σύµπλοκο αυτό στρατολογείται στο κεντροσωµάτιο από τον παράγοντα Cep192, που αποτελεί το ορθόλογο του SPD-2 στον άνθρωπο και δρα πρώτο κατά το διπλασιασµό (Brown et al, 2013). Στη συνέχεια, στο σηµείο αυτό στρατολογείται η κινάση Plk4 (ορθόλογο του ZYG-1), που σηµαίνει το σηµείο στο µητρικό κεντριόλιο από όπου θα ξεκινήσει η σύνθεση του θυγατρικού, ο παράγοντας STIL (ορθόλογο του SAS5), καθώς και οι παράγοντες SAS6 και CPAP (ορθόλογο του SAS4) που θα οδηγήσουν 27
28 1. ΕΙΣΑΓΩΓΗ στο σχηµατισµό µιας δοµ ής µε εννιακτινωτή συµµ ετρία (cartwheel), που δρα ως ικρίωµα και ελέγχει τον πολυµερισµό των µικροσωληνίσκων (Arquint & Nigg, 2016; Strnad & Gonczy, 2008) Οι µηχανισµοί που εξασφαλίζουν το σωστό διπλασιασµό των κεντριολίων Ο διπλασιασµός των κεντριολίων είναι µια διαδικασία που βρίσκεται κάτω από αυστηρό έλεγχο, ώστε να εξασφαλίζεται ότι κάθε κεντροσωµάτιο κατά τη G1 φάση του κυτταρικού κύκλου θα αποτελείται µόνο από δυο κεντριόλια, ένα µητρικό και ένα θυγατρικό. Αποκλίσεις από το φυσιολογικό αριθµό των κεντριολίων στα διαιρούµενα κύτταρα µπορούν να οδηγήσουν σε γονιδιωµατική αστάθεια και καρκινογένεση. Έτσι, σε κάθε κυτταρικό κύκλο κάθε κεντριόλιο πρέπει να διπλασιαστεί µια µόνο φορά (cell cycle control) και µόνο ένα νέο κεντριόλιο πρέπει να σχηµατίζεται δίπλα από κάθε προϋπάρχον (copy number control). Μέχρι σήµερα οι κύριοι µηχανισµοί που είναι γνωστό ότι ελέγχουν τη διαδικασία αυτή σχετίζονται µε το δεσµό που συγκρατεί τα δυο κεντριόλια, καθώς και µε τη ρύθµιση της έκφρασης της κινάσης Plk4. Κατά τη µετάβαση από την Μ στη G1 φάση του κυτταρικού κύκλου, ο ισχυρός δεσµός (S-M linker) που συγκρατεί το µητρικό και το θυγατρικό κεντριόλιο του κεντροσωµατίου χάνεται, µε αποτέλεσµα να χάνεται και η κάθετη διάταξή τους. Το γεγονός αυτό είναι ιδιαίτερα σηµαντικό καθώς επιτρέπει την αδειοδότηση του διπλασιασµού των κεντριολίων στον κυτταρικό κύκλο που έπεται (Nigg, 2007; Tsou & Stearns, 2006). Έτσι εξασφαλίζεται ότι ο διπλασιασµός τους θα συµβεί µόνο µια φορά κατά τη διάρκεια κάθε κυτταρικού κύκλου. Η διάλυση της ισχυρής σύνδεσης που έχει αναπτυχθεί ανάµεσα στο µητρικό και στο θυγατρικό κεντριόλιο εξαρτάται από τη δράση της κινάσης Plk1, καθώς και της πρωτεάσης σεπαράσης (separase) (Tsou et al, 2009). Η δράση τους κατά τη διαδικασία αυτή χαρακτηρίστηκε πρόσφατα ως εξής: Η pericentrin και ο παράγοντας Cep215, που βρίσκονται κοντά στο εγγύς άκρο του µητρικού κεντριολίου και αποτελούν συστατικά του στρώµατος των πρωτεϊνών που απαντάται εκεί, συµβάλλουν στη σύνδεση που συγκρατεί ενωµένα τα δυο κεντριόλια. Κατά το τέλος της µίτωσης, η pericentrin φωσφορυλιώνεται από την κινάση Plk1 και στη συνέχεια πρωτεολύεται από την σεπαράση. Η πρωτεόλυση της pericentrin, που έχει ως αποτέλεσµα την αποµάκρυνση του παράγοντα Cep215 από τα κεντροσωµάτια, δείχθηκε να είναι απαραίτητη αλλά και επαρκής για το διαχωρισµό των κεντριολίων (Kim et al, 2015; Lee & Rhee, 2012; Matsuo et al, 2012; Pagan et al, 2015). Επιπρόσθετα, στη σύνδεση ανάµεσα στα δύο κεντριόλια φαίνεται να συµµ ετέχουν και οι κοχεσίνες (cohesins), οι οποίες είναι γνωστό ότι κρατούν ενωµένες τις αδελφές χρωµατίδες (Sluder, 2013). Η διάσπαση των κοχεσινών από τη σεπαράση επιτρέπει όχι µόνο να ελευθερώνονται οι αδελφές χρωµατίδες, αλλά συµ βάλλει και στο διαχωρισµό των κεντριολίων, τουλάχιστον σε ορισµένους κυτταρικούς τύπους. Είναι σηµαντικό να τονιστεί από τα παραπάνω ότι ο µηχανισµός για το διαχωρισµό των κεντριολίων οµοιάζει σηµαντικά µε αυτόν του διαχωρισµού των αδελφών χρωµατίδων κατά τη κυτταρική διαίρεση, αποτελώντας µια ακόµα ένδειξη της 28
29 1. ΕΙΣΑΓΩΓΗ συνεργατικότητας ανάµεσα στον κύκλο του διπλασιασµού των κεντριολίων και του DNA, µέσω κοινών ρυθµιστικών παραγόντων. Επιπρόσθετα, ο έλεγχος του διπλασιασµού των κεντριολίων επιτυγχάνεται και µέσω της ρύθµισης των επιπέδων έκφρασης και των αλληλεπιδράσεων µεταξύ των παραγόντων Plk4-STIL-SAS6 (Arquint et al, 2015; Kratz et al, 2015; Moyer et al, 2015; Ohta et al, 2014). Ιδιαίτερα σηµαντική είναι η χωροχρονική ρύθµιση των επιπέδων έκφρασης της κινάσης Plk4 στο εγγύς άκρο του µητρικού κεντριολίου, η οποία επιτυγχάνεται µέσω ποικίλων µηχανισµών αυτορρύθµισης, όπως µέσω αυτοφωσφορυλίωσης, καθώς και ουβικιτινυλίωσης µέσω του συµ πλόκου της SCFβTrCP Ε3 λιγάσης (Cunha-Ferreira et al, 2013; Cunha-Ferreira et al, 2009; Guderian et al, 2010; Holland et al, 2012; Holland et al, 2010; Lopes et al, 2015; Nakamura et al, 2013; Rogers et al, 2009). Η κινάση Plk4 µαρκάρει τη θέση έναρξης της δηµιουργίας ενός προκεντριολίου, ώστε µόνο ένα θυγατρικό κεντριόλιο να δηµιουργείται κάθετα στο µητρικό. Είναι ιδιαίτερα σηµαντικό το γεγονός ότι η υπερέκφραση της κινάσης Plk4 επάγει την παραγωγή πολυάριθµων κεντριολίων στα κύτταρα, αφού προκαλεί τη δηµιουργία πολλών θέσεων έναρξης και εποµένως το σχηµατισµό πολλών θυγατρικών κεντριολίων γύρω από το µητρικό (Εικόνα 1.5) (Habedanck et al, 2005; Kleylein-Sohn et al, 2007; Rodrigues-Martins et al, 2007). Τα θυγατρικά κεντριόλια που προκύπτουν δηµιουργούν χαρακτηριστικές δοµές ροζέτας (rosette type) γύρω από το µητρικό κεντριόλιο. Η υπερέκφραση των παραγόντων STIL (Arquint et al, 2012; Vulprecht et al, 2012) και SAS6 (Leidel et al, 2005; Strnad et al, 2007) προκαλεί επίσης τη δηµιουργία υπεράριθµων κεντριολίων, κατά παρόµοιο τρόπο µε αυτόν της Plk4 (Εικόνα 1.5). Έχει προταθεί ότι η δηµιουργία µιας µόνο θέσης έναρξης για το σχηµατισµού του θυγατρικού κεντριολίου παρακείµενα στο µητρικό επιτυγχάνεται µε τον περιορισµό της έκφρασης της κινάσης Plk4 στο σηµείο αυτό, µέσω της στρατολόγησης των παραγόντων STIL και SAS6 στα κεντριόλια (Dzhindzhev et al, 2014; Ohta et al, 2014). 29

30 1. ΕΙΣΑΓΩΓΗ Εικόνα 1.5 Μοριακός µηχανισµός διπλασιασµού των κεντριολίων κατά τη διάρκεια του κυτταρικού κύκλου. O παράγοντας Cep192 εντοπίζεται πρώτος κατά το διπλασιασµό των κεντριολίων και στρατολογεί τους παράγοντες Cep63 και Cep152 στο εγγύς άκρο του µητρικού κεντριολίου. Στη συνέχεια στρατολογείται και η κινάση Plk4, η οποία οριοθετεί το σηµείο εκείνο στο µητρικό κεντριόλιο από όπου θα ξεκινήσει η βιοσύνθεση ενός νέου κεντριολίου, µαζί µε τους παράγοντες STIL, SAS6 και CPAP. Έτσι ξεκινά η δηµιουργία µια δοµής µε εννιακτινωτή συµµετρία, η οποία δρώντας ως ικρίωµα ελέγχει τον πολυµερισµό των µικροσωληνίσκων. Η υπερέκφραση των παραγόντων Plk4, STIL και SAS6 επάγει τον πολλαπλασιασµό των κεντριολίων, δηµιουργώντας χαρακτηριστικές δοµές ροζέτας (rosette type) γύρω από το µητρικό κεντριόλιο. Υπερπολλαπλασιασµός αλλά και επιµήκυνση των κεντριολίων έχει παρατηρηθεί και µέσω υπερέκφρασης των παραγόντων CPAP και Cep120, που επάγουν το σχηµατισµό θυγατρικών κεντριολίων καθ όλο το µήκος του µητρικού κεντριολίου (orthogonal type) (Tang, 2013). Τέλος, οι κυκλινοεξαρτώµενες κινάσες (CDKs) προσδιορίσθηκαν πολύ πρόσφατα ως ένας ακόµα µηχανισµός για τον έ λεγχο του διπλασιασµού των κεντριολίων (Zitouni et al, 2016). Πιο συγκεκριµένα, η Cdk1/cyclinB προσδένεται στην επικράτεια σπειροειδούς σπειράµατος του παράγοντα STIL και αναστέλλει έτσι το σχηµατισµό του συµπλόκου Plk4-STIL, καθώς και τη συνεπακόλουθη φωσφορυλίωση της πρωτεΐνης STIL από την κινάση Plk4. Ο µηχανισµός αυτός εµποδίζει την πρόωρη έναρξη του διπλασιασµού των κεντριολίων κατά το τέλος της µίτωσης, όπου ο ισχυρός δεσµός ανάµεσα στα δυο κεντριόλια έχει ήδη αρθεί. Μετά το τέλος της µίτωσης, αυξάνονται τα επίπεδα έκφρασης της πρωτεΐνης STIL, η οποία είναι πια ελεύθερη να προσδεθεί και να φωσφορυλιωθεί από την κινάση Plk4, επάγοντας τα επόµενα βήµατα για το διπλασιασµό των κεντριολίων. 30
31 1. ΕΙΣΑΓΩΓΗ 1.3 Πολυκροσσωτά κύτταρα Δοµή κροσσών Οι κροσσοί είναι οργανίδια που απαντώνται στην επιφάνεια ενός µεγάλου αριθµού κυττάρων και εκφύονται από µια τροποποιηµένη δοµή των κεντριολίων που ονοµάζεται βασικό σωµάτιο (basal body) (Ishikawa & Marshall, 2011) (Εικόνα 1.6). Το τµήµα του κροσσού που εκφύεται από το βασικό σωµάτιο προς το εξωκυττάριο περιβάλλον ονοµάζεται αξόνηµα. Το βασικό σωµάτιο αποτελείται από 9 τριάδες µικροσωληνίσκων, δυο εκ των οποίων συνεχίζουν και στο αξόνηµα, ενώ η τρίτη σταµατά σε µια ενδιάµεση περιοχή ανάµεσα στο βασικό σωµάτιο και το αξόνηµα που ονοµάζεται µεταβατική περιοχή (transition zone). Το αξόνηµα είναι µια αρκετά συντηρηµένη κυτταρική δοµή. Αποτελείται από 9 ζευγάρια µικροσωληνίσκων που είναι τοποθετηµένα κυκλικά και περιβάλλονται από µεµβράνη, που αποτελεί συνέχεια της πλασµατικής µεµβράνης του κυττάρου. Ορισµένα είδη κροσσών µπορεί να φέρουν επιπλέον στο κέντρο του δακτυλίου και δυο µονούς µικροσωληνίσκους. Στο βασικό σωµάτιο, όπως και στο κεντριόλιο, απουσιάζει το κεντρικό ζεύγος των µικροσωληνίσκων. Κύρια λειτουργία τόσο του βασικού σωµατίου, όσο και του κεντριολίου είναι ότι αποτελούν κέντρα εµπυρήνωσης των µικροσωληνίσκων. Οι κύριοι τύποι κροσσών που απαντώνται είναι δύο: 1) ο πρωτογενής κροσσός, ο οποίος είναι ακίνητος και συνήθως ένας ανά κύτταρο (primary cilium) και 2) οι κινητοί κροσσοί, που είναι συνήθως πολλαπλοί ανά κύτταρο (multiple motile cilia) (Ishikawa & Marshall, 2011) (Εικόνα 1.6). Μια βασική δοµική διαφορά ανάµεσα στον πρωτογενή κροσσό και στους κινητούς κροσσούς είναι ότι το αξόνηµα του πρωτογενούς κροσσού διαθέτει µόνο 9 ζευγάρια µικροσωληνίσκων (διάταξη 9+0 µικροσωληνίσκοι), ενώ οι κινητοί κροσσοί διαθέτουν επιπρόσθετα και δυο κεντρικούς µικροσωληνίσκους (διάταξη 9+2 µικροσωληνίσκοι), καθώς και πρωτεΐνες (ακτίνη, δυνεΐνη) συνδεδεµένες µε τα ζεύγη των µικροσωληνίσκων, τα οποία συµ βάλλουν στην κίνησή τους. O πρωτογενής κροσσός συνήθως είναι ένας σε αριθµό και µεγαλύτερος σε µήκος σε σύγκριση µε τους πολλαπλούς κροσσούς (Goetz & Anderson, 2010). Μπορεί να βρεθεί στην επιφάνεια των περισσότερων κυττάρων των σπονδυλωτών και κύρια λειτουργία του είναι η λήψη µηχανο-αισθητικών σηµ άτων για το κύτταρο. Πιο συγκεκριµένα, οι πρωτεΐνες της κυτταροπλασµατικής µεµβράνης γύρω από τον πρωτογενή κροσσό µεταβιβάζουν εξωκυττάρια σήµατα προς το εσωτερικό του κυττάρου, ενεργοποιώντας οδούς σηµατοδότησης και τροποποιώντας την κυτταρική δραστηριότητα. Αντίθετα, οι κινητοί κροσσοί βρίσκονται σε έναν περιορισµένο αριθµό εξειδικευµένων κυτταρικών τύπων, είναι συνήθως µερικές εκατοντάδες σε αριθµό (30-300) ανά κύτταρο και κύρια λειτουργία τους είναι η κίνηση του περιβάλλοντος υγρού του ιστού στον οποίο βρίσκονται (Spassky & Meunier, 2017). Η κίνηση των κροσσών επιτυγχάνεται µέσω της συντονισµένης λειτουργίας του εξωτερικού και του εσωτερικού βραχίονα των µορίων της δυνεΐνης που υπάρχουν στο αξόνηµα και συ µβάλλουν στην ολίσθηση των µικροσωληνίσκων (Brooks & Wallingford, 2014). Mόρια νεξίνης, που διασυνδέουν τις παρακείµενες δυάδες των µικροσωληνίσκων, καθώς και µόρια ακτίνης βοηθούν στην κάµψη των 31


32 1. ΕΙΣΑΓΩΓΗ µικροσωληνίσκων. Η κίνηση των κροσσών είναι χαρακτηριστική και συνοπτικά περιλαµβάνει δυο φάσεις: αρχίζει µε ένα ενεργό χτύπηµα και σ τη συνέχεια επανέρχεται στην αρχική του θέση µε µια κάµψη από τη βάση προς την κορυφή. Απαραίτητη είναι η υδρόλυση του ATP που αλλάζει τη διαµόρφωση της δυνεΐνης ανάµεσα στα ζεύγη των µικροσωληνίσκων και επιτρέπει την ολίσθηση. A Κινητοί κροσσοί B Πρωτογενής κροσσός 32
33 1. ΕΙΣΑΓΩΓΗ Εικόνα 1.6 Δοµικά χαρακτηριστικά κινητών κροσσών και πρωτογενούς κροσσού. Οι κροσσοί οργανώνονται από ένα τροποποιηµένο κεντριόλιο που έχει «αγκυροβολήσει» στην πλασµατική µεµβράνη του κυττάρου και ονοµάζεται βασικό σωµάτιο. Η περιοχή του βασικού σωµατίου που ενώνεται µε την πλασµατική µεµβράνη αποτελεί τη µεταβατική περιοχή και από εκεί οργανώνεται το αξόνηµα του κροσσού που εκτείνεται προς το εξωκυττάριο περιβάλλον. Το βασικό σωµάτιο αποτελείται από 9 τριάδες µικροσωληνίσκων που είναι τοποθετηµένες σε σφαιρική διάταξη, δυο εκ των οποίων συνεχίζουν και στο αξόνηµα, ενώ η τρίτη σταµατά στη µεταβατική περιοχή. Οι κινητοί κροσσοί (Α) φέρουν επιπρόσθετα και δυο κεντρικούς µικροσωληνίσκους (διάταξη 9+2 µικροσωληνίσκοι) καθώς και πρωτεΐνες ακτίνης, νεξίνης και δυνεΐνης συνδεδεµένες κατάλληλα και µε τους µικροσωληνίσκους, ώστε να είναι δυνατή η κίνησή τους. Αντίθετα, ο πρωτογενής κροσσός (Β) δεν είναι κινητός και εποµένως δεν φέρει τους κεντρικούς µικροσωληνίσκους, καθώς και τις πρωτεΐνες που συµβάλλουν στην κίνηση των πολλαπλών κροσσών (Reiter & Leroux, 2017) Προέλευση κροσσών Οι κροσσοί απαντώνται σε ένα µεγάλο αριθµό ευκαρυωτικών οργανισµών (Brooks & Wallingford, 2014). Πιο συγκεκριµένα, κινητοί κροσσοί απαντώνται τόσο σε µονοκύτταρους ευκαρυωτικούς οργανισµούς, όπως στην περίπτωση του αµοιβαδοειδούς Multicilia marina, καθώς και σε άλλα πρώτιστα, όσο και σε πολυκύτταρους οργανισµούς, από τους σπόγγους έως τον άνθρωπο. Στους µονοκύτταρους ευκαρυωτικούς οργανισµούς τα οργανίδια αυτά χρησιµεύουν για την κίνησή τους καθώς και για τη δέσµευση της τροφής τους. Στα σπονδυλωτά, τα πολυκροσσωτά κύτταρα απαντώνται σε µια ποικιλία ιστών (Brooks & Wallingford, 2014; Spassky & Meunier, 2017). Στα θηλαστικά, πολυκροσσωτά κύτταρα απαντώνται στο αναπνευστικό επιθήλιο, στις κοιλίες του εγκεφάλου, καθώς και στους ωαγωγούς θηλυκών ατόµων µε κύρια λειτουργία την κίνηση του περιβάλλοντος υγρού. Ιδιαίτερο ενδιαφέρον παρουσιάζει η εύρεση πολυκροσσωτών κυττάρων, που οµοιάζουν µε αυτά των θηλαστικών, στο δέρµα εµβρύων του βατράχου Xenopus, καθώς και στον πρόνεφρο του ψαριού zebrafish Εντοπισµός και λειτουργία των πολυκροσσωτών κυττάρων Τα πολυκροσσωτά κύτταρα ανάλογα µε τον ιστό στον οποίο εντοπίζονται επιτελούν διαφορετικές λειτουργίες ζωτικής σηµασίας για τον οργανισµό. Τα πολυκροσσωτά κύτταρα βρίσκονται κυρίως σε 3 συστήµατα του ανθρώπινου οργανισµού: στο αναπνευστικό, στο αναπαραγωγικό και στο νευρικό σύστηµα (Εικόνα 1.7) (Fliegauf et al, 2007; Spassky & Meunier, 2017). Το επιθήλιο, τόσο στο ανώτερο όσο και στο κατώτερο αναπνευστικό σύστηµα του ανθρώπου, έχει µια τέτοια κυτταρική σύσταση που του επιτρέπει να δρα προστατευτικά, ως ένας εξωτερικός µηχανισµός άµυνας, αποµακρύνοντας παθογόνους µικροοργανισµούς, νεκρά κύτταρα, σκόνη και άλλα σωµατίδια στα οποία 33
34 1. ΕΙΣΑΓΩΓΗ εκτίθεται συνεχώς ο ανθρώπινος οργανισµός (Brooks & Wallingford, 2014; Spassky & Meunier, 2017). Αυτό επιτυγχάνεται µε τη βοήθεια µιας διαδικασίας που ονοµάζεται βλεννοκροσσωτή κάθαρση και επιτελείται µέσω της λειτουργίας των πολυκροσσωτών και των βλεννοεκκριτικών κυττάρων του αναπνευστικού επιθηλίου. Πιο συγκεκριµένα, το αναπνευστικό επιθήλιο φέρει βλεννοπαραγωγά κύτταρα (goblet cells) που εκκρίνουν βλέννα (mucins), η οποία εξαπλώνεται πάνω από τους κροσσούς. Η βλέννα εγκλωβίζει σωµατίδια, τα οποία στη συνέχεια αποµακρύνονται από την αναπνευστική οδό µε τη συντονισµένη κίνηση των κροσσών του επιθηλίου. Με παρόµοιο τρόπο επιτυγχάνεται βλεννοκροσσωτή κάθαρση και στη ρινική κοιλότητα, καθώς και στην τυµπανική κοιλότητα (µέσω ους) και στην ευσταχιανή σάλπιγγα. Τέλος, ένας ιδιαίτερος τύπος πολυκροσσωτών κυττάρων εντοπίζεται και στο οσφρητικό επιθήλιο. Τα κύτταρα αυτά διαθέτουν κροσσούς το καθένα, οι οποίοι παρόλο που έχουν διάταξη «9+2» µικροσωληνίσκων, δεν διαθέτουν βραχίονες µορίων δυνεΐνης, µε αποτέλεσµα να µην κινούνται. Στο αναπαραγωγικό σύστηµα των θηλυκών ατόµων απαντώνται πολυκροσσωτά κύτταρα από την κοιλότητα της µήτρας µέχρι τους ωαγωγούς και τις ωοθήκες (Spassky & Meunier, 2017). Το ποσοστό των πολυκροσσωτών κυττάρων κατά µήκος αυτού του τµήµατος ποικίλει ανάλογα µε το σηµείο στο οποίο εντοπίζονται, καθώς και τη χρονική στιγµή του γεννητικού κύκλου. Άν και ο ρόλος των πολυκροσσωτών κυττάρων της µήτρας δεν έχει διευκρινιστεί, στους ωαγωγούς αποτελούν την κινητήρια δύναµη για τη µεταφορά των ωαρίων, ενώ ένας αριθµός µελετών δείχνει ότι µπορεί να δρούν και προστατευτικά για τη διατήρηση των σπερµατοζωαρίων που έχουν εισέλθει. Στο αρσενικό αναπαραγωγικό σύστηµα τα πολυκροσσωτά κύτταρα συµµετέχουν στη συµ πύκνωση των σπερµατοζωαρίων. Επίσης, το µαστίγιο του σπερµατοζωαρίου αποτελεί έναν εξειδικευµένο, ασυνήθιστα µεγάλου µήκους κινητό κροσσό. Στο νευρικό σύστηµα, και συγκεκριµένα στις κοιλίες του εγκεφάλου, απαντώνται τα επενδυµατικά πολυκροσσωτά κύτταρα τα οποία συµµετέχουν στην κίνηση του εγκεφαλονωτιαίου υγρού που βρίσκεται κοντά στο τοίχωµα των κοιλιών (Spassky & Meunier, 2017). Ένας ιδιαίτερος τύπος πολυκροσσωτών κυττάρων εντοπίζεται και στο χοριοειδές πλέγµα των κοιλιών του εγκεφάλου, εκεί όπου παράγεται το εγκεφαλονωτιαίο υγρό. Ωστόσο, δεν έχει ακόµα διευκρινιστεί η ακριβής λειτουργία τους, καθώς και η δοµή τους. Τέλος, πολυκροσσωτά κύτταρα µε άγνωστη λειτουργία εντοπίζονται και στο νωτιαίο µυελό. Επιπρόσθετα, στον άνθρωπο έχουν εντοπιστεί και πολυκροσσωτά κύτταρα µε άγνωστη λειτουργία ή ελάχιστα µελετηµένα στα νεφρά, υπό παθολογικές καταστάσεις, καθώς και στο επιθήλιο τ ου οισοφάγου των εµβρύων τα οποία σταδιακά µειώνονται και τελικά εξαφανίζονται µε τη γέννηση ενός ατόµου. 34
, στη βλεννοκροσσωτή κάθαρση και στη")
35 1. ΕΙΣΑΓΩΓΗ Εικόνα 1.7 Εντοπισµός των πολυκροσσωτών κυττάρων στον ανθρώπινο οργανισµό. Τα πολυκροσσωτά κύτταρα εντοπίζονται κυρίως στο νευρικό, στο αναπνευστικό κα ι στο αναπαραγωγικό σύστηµα, συµβάλλοντας στην κίνηση του εγκεφαλονωτιαίου υγρού (CSF), στη βλεννοκροσσωτή κάθαρση και στη µετακίνηση των ωαρίων, αντίστοιχα. Ωστόσο, πολυκροσσωτά κύτταρα λιγότερο µελετηµένα ως προς τη δοµή και τη λειτουργία τους απαντώνται και σε άλλα σηµεία του ανθρώπινου οργανισµού (Spassky & Meunier, 2017) Διαφοροποίηση των πολυκροσσωτών κυττάρων Τα πολυκροσσωτά κύτταρα είναι µια ειδική κατηγορία τελικώς διαφοροποιηµένων κυττάρων, τα οποία εξέρχονται από τον κυτταρικό κύκλο και µέσω ενός ιδιότυπου κύκλου κεντροσωµατίων πολλαπλασιάζουν τα κεντριόλιά τους εκατοντάδες φορές (Brooks & Wallingford, 2014) (Εικόνα 1.8). Έτσι µπορούν να παράγονται κεντριόλια ανά κύτταρο, ανάλογα µε τον κυτταρικό τύπο. Τα κύτταρα αυτά αποτελούν ένα χαρακτηριστικό παράδειγµα κυττάρων στα οποία ο διπλασιασµός των κεντριολίων αποκλίνει από τους µηχανισµούς ελέγχου που περιγράφηκαν προηγουµένως, παραβιάζοντας τους κανόνες για το σχηµατισµό ενός µόνο θυγατρικού κεντριολίου κάθετα στο µητρικό κατά τη διάρκεια του κυτταρικού κύκλου. Τα νεοσυντιθέµενα κεντριόλια µεταναστεύουν και «αγκυροβολούν» στην κορυφαία επιφάνεια της πλασµατικής µεµβράνης των κυττάρων, δρώντας ως κέντρα για την οργάνωση των µικροσωληνίσκων των κροσσών. Οι κροσσοί πάλλονται συγχρονισµένα εξασφαλίζοντας την κίνηση του περιβάλλοντος υγρού του ιστού στον οποίο βρίσκονται. 35
36 1. ΕΙΣΑΓΩΓΗ Έξοδος από τον κυτταρικό κύκλο Πολ/σμός κεντριολίων Μετανάστευση αγκυροβόληση Σχηματισμός αξονήματος Προγονικό κύτταρο Ώριμο πολυκροσσωτό κύτταρο Εικόνα 1.8 Στάδια διαφοροποίησης των πολυκροσσωτών κυττάρων. Ένα σύνολο από διακριτά στάδια χαρακτηρίζει τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Ένα πρόδροµο προγονικό κύτταρο εξέρχεται από τον κυτταρικό κύκλο και µέσω ενός ιδιόµορφου κύκλου κεντροσωµατίων δηµιουργεί εκατοντάδες κεντριόλια. Τα κεντριόλια µεταναστεύουν και «αγκυροβολούν» στην πλασµατική µεµβράνη, όπου δρώντας ως βασικά σωµάτια δίνουν γένεση στους κροσσούς. Η διαφοροποίηση των πολυκροσσωτών κυττάρων τελικά επιτυγχάνεται µέσω 3 διακριτών σταδίων που έχουν ως στόχο: να καθοριστεί αρχικά η µοίρα των πρόδροµων κυττάρων προς πολυκροσσωτά κύτταρα, την εξειδίκευσή τους στη συνέχεια προς το συγκεκριµένο κυτταρικό τύπο και τέλος την ωρίµανση των κυττάρων και τη διατήρηση της µοίρας αυτής. Ένα δίκτυο µεταγραφικών παραγόντων ελέγχει την έκφραση όλων των γονιδίων που απαιτούνται για κάθε ένα από τα παραπάνω στάδια Καθορισµός της µοίρας των πρόδροµων κυττάρων σε πολυκροσσωτά κύτταρα Η διαφοροποίηση των πολυκροσσωτών κυττάρων απαιτεί τον καθορισµό της µοίρας ενός αριθµού πρόδροµων διαιρούµενων κυττάρων προς αυτόν τον κυτταρικό τύπο, µέσω εξόδου τους από τον κυτταρικό κύκλο. Στην κορυφή του µονοπατιού αυτού βρίσκεται η σηµατοδότηση Notch η οποία σταµατά τον πολλαπλασιασµό των κυττάρων και ανάλογα µε τα επίπεδα της έκφρασής της οδηγεί τα κύτταρα στο αναπνευστικό επιθήλιο των θηλαστικών, στην επιδερµίδα εµβρύων Xenopus, καθώς και στον πρόνεφρο των ψαριών zebrafish προς συγκεκριµένους κυτταρικούς τύπους (Deblandre et al, 1999; Liu et al, 2007; Ma & Jiang, 2007; Mori et al, 2015; Morimoto et al, 2012; Tsao et al, 2009). Υψηλά επίπεδα έκφρασης του Notch αναστέλλουν τη διαφοροποίηση των κυττάρων προς τη µοίρα των πολυκροσσωτών και ευνοούν τη διαφοροποίησή τους προς εκκριτικά κύτταρα. Αντίθετα, η αναστολή του Notch µονοπατιού προάγει τη µοίρα των κυττάρων προς πολυκροσσωτά. 36
37 1. ΕΙΣΑΓΩΓΗ Στον πρόνεφρο του zebrafish, η έλλειψη είτε του συνδέτη Jagged 2, είτε του Notch3 προάγει τη διαφοροποίηση των πολυκροσσωτών κυττάρων, ενώ αντίθετα η υπερέκφραση Notch µειώνει τον αριθµό τους (Liu et al, 2007). Οµοίως, στο αναπνευστικό επιθήλιο των µυών η απενεργοποίηση του Notch οδηγεί σε µείωση των εκκριτικών Clara κυττάρων και αύξηση των πολυκροσσωτών και των νευροενδοκρινικών κυττάρων (Tsao et al, 2009). Ο έλεγχος της σηµατοδότησης Notch δείχθηκε να επιτυγχάνεται µέσω των micrornas της οικογένειας mir-449, τα οποία αναστέλλουν το Notch1 και τον συνδέτη του Delta-like 1, προάγοντας έτσι τη διαφοροποίηση των πολυκροσσωτών κυττάρων στο αναπνευστικό επιθήλιο του ανθρώπου, καθώς και στην επιδερµίδα εµβρύων βατράχων Xenopus (Marcet et al, 2011). Τέλος, εκτός από τη σηµατοδότηση Notch προκαταρκτικές µελέτες εµπλέκουν στο αρχικό αυτό στάδιο του καθορισµού της µοίρας των κυττάρων προς πολυκροσσωτά και τη BMP και HIF σηµατοδότηση (Cibois et al, 2015; Gerovac et al, 2014). Πιο συγκεκριµένα, από πειράµατα στην επιδερµίδα εµβρύων Xenopus και στο αναπνευστικό επιθήλιο των µυών, έχει προταθεί ότι η µείωση της BMP σηµατοδότησης προάγει τη διαφοροποίηση των κυττάρων προς πολυκροσσωτά, ενώ αντίθετα η αύξησή της προάγει τη διαφοροποίηση των βλεννοπαραγωγών κυττάρων (Cibois et al, 2015). Ακόµα, σε καλλιέργεια των επιθηλιακών κυττάρων που έχουν αποµονωθεί από βρόγχους ανθρώπου δείχθηκε ότι οι συνθήκες υποξίας µειώνουν τον αριθµό των πολυκροσσωτών κυττάρων. Μάλιστα, αυτό οφείλεται στη µειωµένη έκφραση των γονιδίων που απαιτούνται για τη διαφοροποίηση των πολυκροσσωτών κυττάρων (π.χ. Foxj1), αφού επάγεται η σηµατοδότηση Notch (Gerovac et al, 2014) Εξειδίκευση των πολυκροσσωτών κυττάρων Τα προγονικά κύτταρα που καθορίστηκαν να γίνουν πολυκροσσωτά εξειδικεύονται ως προς τη µοίρα αυτή µέσω της παραγωγής πολυάριθµων κεντριολίων στο κυτταρόπλασµά τους, τα ο ποία στη συνέχεια θα µεταναστεύσουν στην πλασµατική µεµβράνη του κυττάρου και θα δώσουν γ ένεση στους κροσσούς (Εικόνα 1.8) Πολλαπλασιασµός των κεντριολίων στα πολυκροσσωτά κύτταρα Κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, όπως προαναφέρθηκε, είναι απαραίτητη η δηµιουργία υπεράριθµων κεντριολίων που θα δώσουν γένεση στους κροσσούς. Έτσι, σχεδόν ταυτόχρονα, δηµ ιουργείται ένας µεγάλος αριθµός κεντριολίων, σε αντίθεση µε τον «ηµι-συντηρητικό» µηχανισµό διπλασιασµού τους στα διαιρούµενα κύτταρα, όπου κάθε µητρικό κεντριόλιο λειτουργεί ως καλούπι για το σχηµατισµό ενός θυγατρικού κεντριολίου. Ο πολλαπλασιασµός των κεντριολίων στα πολυκροσσωτά κύτταρα επιτυγχάνεται µέσω δυο διαφορετικών µονοπατιών (Tang, 2013): 37
38 1. ΕΙΣΑΓΩΓΗ 1) Σύνθεση κεντριολίων από ένα προϋπάρχον µητρικό κεντριόλιο (Mother-centrioledependent pathway): Το µονοπάτι αυτό οµοιάζει µε τον κλασσικό µηχανισµό διπλασιασµού των κεντριολίων που απαντάται στα διαιρούµενα κύτταρα. Ωστόσο, µε καλούπι το µητρικό κεντριόλιο δηµιουργούνται πολλά θυγατρικά, και 2) De novo σύνθεση κεντριολίων µέσω του σχηµατισµού δευτεροσωµατίων (Deuterosome-dependent pathway): Στο µονοπάτι αυτό δηµιουργούνται πολυάριθµα κεντριόλια, χωρίς να χρησιµοποιείται ως καλούπι ένα προϋπάρχον κεντριόλιο, τριγύρω από ακαθόριστες κυτταροπλασµατικές δοµές που ευνοούν τη βιοσύνθεσή τους και ονοµάζονται δευτεροσωµάτια. Ο µεγάλος αριθµός των κεντριολίων στα πολυκροσσωτά κύτταρα επιτυγχάνεται µέσω της αύξησης της γονιδιακής έκφρασης των παραγόντων εκείνων που συµµετέχουν στο διπλασιασµό τους στα διαιρούµενα κύτταρα (Firat-Karalar & Stearns, 2014; Vladar & Stearns, 2007). Σηµαντικό ρόλο στη διαδικασία αυτή φαίνεται να διαδραµατίζουν τα επίπεδα έκφρασης της κινάσης Plk4, η ο ποία δηµιουργεί τριγύρω από το µητρικό κεντριόλιο πολλαπλές θέσεις έναρξης για το σχηµατισµό των θυγατρικών κεντριολίων. Ο µηχανισµός της δηµιουργίας των κεντριολίων µέσω των δευτεροσωµατίων διαφέρει από το µηχανισµό δηµ ιουργίας τους µέσω ενός µητρικού κεντριολίου, ως προς τα αρχικά µόρια που επάγουν το κάθε µονοπάτι. Πρόσφατα δεδοµένα ανέδειξαν το µόριο Deup1 ως τον κεντρικό ρυθµιστή του µονοπατιού σύνθεσης κεντριολίων µέσω των δευτεροσωµατίων (Zhao et al, 2013). Φυλογενετική ανάλυση έδειξε ότι ο παράγοντας Deup1 αποτελεί παράλογο του Cep63, που εµφανίστηκε µετά το διαχωρισµό των ιχθύων κατά την εξέλιξη των σπονδυλωτών, ύστερα από το διπλασιασµό του γονιδίου Cep63. Ο παράγοντας Deup1 αλληλεπιδρά µε το Cep152 και δρα µε παρόµοιο τρόπο µε το σύµπλοκο Cep63/Cep152, για τη στρατολόγηση της κινάσης Plk4, ώστε να ενεργοποιηθεί ο σχηµατισµός των κεντριολίων. Επιπρόσθετα, ύστερα από πειράµατα στα πολυκροσσωτά κύτταρα της επιδερµίδας εµβρύων Xenopus, ταυτοποιήθηκε στα δευτεροσωµάτια ο παράγοντας Ccdc78 (Klos Dehring et al, 2013). Ο ρόλος του παράγοντα Ccdc78 κατά το διπλασιασµό των κεντριολίων δεν έχει αποσαφηνιστεί πλήρως, όµως φαίνεται να δρα σχηµατίζοντας ένα ικρίωµα για τη στρατολόγηση του παράγοντα Cep152 για τη µετέπειτα ενεργοποίηση της κινάσης Plk4. Ο πολλαπλασιασµός των κεντριολίων µέσω των δευτεροσωµατίων αποτελεί το κύριο µονοπάτι δηµιουργίας των πολυκροσσωτών κυττάρων, µιας και µπορεί να εξασφαλίσει το µεγάλο αριθµό των κεντριολίων που απαιτούνται. Παρά τις διαφορές που εντοπίζονται στα σύµπλοκα που δηµιουργούνται στα αρχικά στάδια του κ άθε µονοπατιού, φαίνεται ότι στα επόµενα βήµατα του καθενός συµµετέχουν κοινοί ρυθµιστές (Zhao et al, 2013). Ωστόσο, τα δεδοµένα για το πώς σχετίζονται τα µονοπάτια αυτά µεταξύ τους είναι ελάχιστα. Πρόσφατα, προτάθηκε ένα µοντέλο που εξηγεί το µηχανισµό της µαζικής παραγωγής των κεντριολίων κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων (Al Jord et al, 2014; Meunier & Spassky, 2016). In vivo πειράµατα τόσο στα επενδυµατικά π ολυκροσσωτά κύτταρα του εγκεφάλου, όσο και στα πολυκροσσωτά κύτταρα της τραχείας έδειξαν ότι το θυγατρικό κεντριόλιο του κεντροσωµατίου χρησιµεύει ως καλούπι για τη σύνθεση νέων κεντριολίων, υποστηρίζοντας το 38

39 1. ΕΙΣΑΓΩΓΗ σχηµατισµό των δευτεροσωµατίων. Πιο συγκεκριµένα, µέσω ενός άγνωστου µηχανισµού οι παράγοντες Deup1 και Ccdc78 συσσωρεύονται στο θυγατρικό κεντριόλιο του κεντροσωµατίου και προκαλούν τη στρατολόγηση βασικών ρυθµιστών του διπλασιασµού των κεντριολίων, όπως του παράγοντα Cep152 και της κινάσης Plk4. Αυτό έχει ως αποτέλεσµα το σχηµατισµό του πρώτου κεντριολίου (προκεντριόλιο). Η συσσώρευση των παραγόντων που βοηθούν στο διπλασιασµό των κεντριολίων συνεχίζεται στο σηµείο αυτό, ευνοώντας έτσι το σχηµατισµό και ενός δεύτερου προκεντριολίου κ.ο.κ., έως ότου το δευτεροσωµάτιο να αποκολληθεί από το τοίχωµα του θυ γατρικού κεντριολίου (Εικόνα 1.9). Η ίδια διαδικασία επαναλαµβάνεται για το σχηµατισµό ενός δεύτερου δευτεροσωµατίου που θα δώσει γένεση σε νέα προκεντριόλια. Πολλά δευτεροσωµάτια µπορεί να σχηµατίζονται ταυτόχρονα τριγύρω από το αρχικό κεντριόλιο. Τα νεοσυντιθέµενα κεντριόλια παραµένουν «υπό καταστολή» µέχρι να παραχθεί ο επιθυµητός αριθµός. Αφού παραχθεί ο απαιτούµενος αριθµός, ξεκινά ταυτόχρονα η επιµήκυνσή τους. Τέλος, όλα τα κεντριόλια, τόσο αυτά που έχουν παραχθεί στα δευτεροσωµάτια, όσο και αυτά που έχουν παραχθεί µε καλούπι το µητρικό κεντριόλιο αποσπώνται και µεταναστεύουν στην πλασµατική µεµβράνη, ώστε δρώντας ως βασικά σωµάτια να οργανώσουν το αξόνηµα των κροσσών. Από την παραπάνω µελέτη είναι πλέον φανερό ότι τα κεντροσωµάτια των κυττάρων συµµετέχουν τόσο στο διπλασιασµό των κεντριολίων στα διαιρούµενα κύτταρα, όσο και στην παραγωγή υπεράριθµων κεντριολίων στα πολυκροσσωτά κύτταρα. Εικόνα 1.9 Προτεινόµενο µοντέλο του διπλασιασµού των κεντριολίων στα πολυκροσσωτά κύτταρα. Το θυγατρικό κεντριόλιο του κεντροσωµατίου χρησιµεύει ως καλούπι για τη σύνθεση νέων κεντριολίων, υποστηρίζοντας το σχηµατισµό δευτεροσωµατίων. Η συσσώρευση των παραγόντων που χρειάζονται για το διπλασιασµό των κεντριολίων έχει ως αποτέλεσµα τη συνεχή βιοσύνθεση προκεντριολίων γύρω από ένα δευτεροσωµάτιο. Με τον τρόπο αυτό σχηµατίζονται πολλά δευτεροσωµάτια τα οποία παραµένουν σε «καταστολή», µέχρι να παραχθεί ο επιθυµητός αριθµός (modified from Meunier & Spassky, 2016). 39
40 1. ΕΙΣΑΓΩΓΗ Οι παράγοντες Ccno και cmyb κατά το διπλασιασµό των κεντριολίων Ωστόσο, ο µοριακός µηχανισµός που ελέγχει τα αρχικά βήµατα της επαγωγής του πολλαπλασιασµού των κεντριολίων, µετά την έξοδο των κυττάρων από τον κυτταρικό κύκλο και τον καθορισµό της τύχης τους προς τη µοίρα των πολυκροσσωτών, δεν έχει ακόµα αποσαφηνιστεί. Πρόσφατα, προσδιορίστηκε το γονίδιο Ccno να συµµετέχει στο στάδιο αυτό. Χωρίς να είναι σαφής ο τρόπος µε τον οποίο δρα, δείχθηκε ότι το γονίδιο Ccno εµφάνιζε µεταλλάξεις σε ασθενείς που είχαν µειωµένο αριθµό πολυκροσσωτών κυττάρων, µε αποτέλεσµα να παρουσιάζουν διαταραχή της βλεννοκροσωτής καθάρσεως του αναπνευστικού συστήµατος λόγω µειωµένης παραγωγής πολλαπλών κινητών κροσσών, µια ασθένεια που ονοµάστηκε RGMC (Reduced Generation of Multiple Motile Cilia) (Amirav et al, 2016; Wallmeier et al, 2014). Η ανάλυση των κυττάρων του αναπνευστικού συστήµατος που προέρχονταν από τους ασθενείς αυτούς έδειξε ότι ο πολλαπλασιασµός των κεντριολίων είχε µειωθεί σε µεγάλο βαθµό, ενώ τα ελάχιστα κεντριόλια που παράγονταν δεν µετανάστευαν προς την πλασµατική µεµβράνη. Επίσης, υπήρχαν ελάχιστα κύτταρα που έφεραν έναν µε δυο κινητούς κροσσούς. Ο ίδιος φαινότυπος παρατηρήθηκε στα πολυκροσσωτά κύτταρα της επιδερµίδας εµβρύων Xenopus, καθώς και του αναπνευστικού επιθηλίου στους µύες (Funk et al, 2015; Wallmeier et al, 2014). Μάλιστα, σε ex vivo καλλιέργειες των επιθηλιακών κυττάρων της τραχείας µυών παρατηρήθηκε µείωση του αριθµού των δευτεροσωµατίων που δηµιουργούνταν, µε αποτέλεσµα τη µειωµένη π αραγωγή κεντριολίων και κατά συνέπεια κροσσών. Ένας ακόµα παράγοντας που χαρακτηρίστηκε τελευταία στο στάδιο αυτό είναι ο µεταγραφικός παράγοντας cmyb. Ο παράγοντας αυτός βρέθηκε να εκφράζεται πρώιµα κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, µετά την έξοδο των κυττάρων από τον κυτταρικό κύκλο (Tan et al, 2013). Η έλλειψή του εµποδίζει τον πολλαπλασιασµό των κεντριολίων στο αναπτυσσόµενο αναπνευστικό επιθήλιο των µυών (E15.5). Επιπρόσθετα, ο παράγοντας cmyb επάγει τη δηµιουργία των πολυκροσσωτών κυττάρων µέσω µεταγραφικής ενεργοποίησης του γονιδίου Foxj1 (η δράση του αναλύεται παρακάτω στα επόµενα βήµατα της διαδικασίας) και η έκφρασή του χάνεται στα ώριµα πολυκροσσωτά κύτταρα. Ωστόσο, ο ρόλος του συγκεκριµένου µεταγραφικού παράγοντα παραµένει αδιευκρίνιστος, αφού µια ακόµα µελέτη έδειξε ότι εµπλέκεται και σε πιο πρώιµα στάδια της διαδικασίας, καθορίζοντας τη µοίρα των κυττάρων του αναπνευστικού επιθηλίου (Pan et al, 2014). Ο παράγοντας cmyb εκφράζεται σε έναν ενδιάµεσο πληθυσµό κυττάρων κατά τη διαφοροποίηση, που σε συνεργασία και µε άλλους παράγοντες µπορεί να καθοδηγήσει τη µοίρα των κυττάρων εκτός από πολυκροσσωτά και προς εκκριτικά κύτταρα. 40
41 1. ΕΙΣΑΓΩΓΗ Μετανάστευση και «αγκυροβόληση» των κεντριολίων στην πλασµατική µεµβράνη Οι µεταγραφικοί παράγοντες που έχουν χαρακτηριστεί περισσότερο στη διαφοροποίηση των πολυκροσσωτών κυττάρων εµπλέκονται κυρίως στα ύστερα στάδια της διαδικασίας, µετά τον πολλαπλασιασµό των κεντριολίων, και σχετίζονται µε τη µετανάστευση και την «αγκυροβόληση» των βασικών σωµατίων στην πλασµατική µεµβράνη, καθώς και µε το σχηµατισµό του αξονήµατος των κροσσών (Choksi et al, 2014). Οι βασικοί µεταγραφικοί παράγοντες που εµπλέκονται στα παραπάνω βήµατα είναι ο παράγοντες Foxj1 και µέλη της οικογένειας των Rfx παραγόντων. Οι παράγοντες αυτοί δρουν συνεργατικά κατά την εξειδίκευση των πολυκροσσωτών κυττάρων. Ωστόσο, ο παράγοντας Foxj1 εµπλέκεται στη δηµιουργία των κινητών κροσσών, ενώ οι Rfx έχει δειχθεί ότι συµµετέχουν τόσο στη δηµιουργία των κινητών όσο και των ακίνητων κροσσών. Foxj1 Ο µεταγραφικός παράγοντας Foxj1 µελετήθηκε κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων ύστερα από το ειδικό πρότυπο έκφρασης που δείχθηκε να έχει, τόσο σε επίπεδο mrna όσο και σε πρωτεϊνικό επίπεδο, στο χοριοειδές πλέγµα του εγκεφάλου, στο αναπνευστικό επιθήλιο, καθώς και στους ωαγωγούς και στους όρχεις των θηλαστικών (reviewed in Choksi et al, 2014). Επίσης, κατά την ανάπτυξη των µυών, ο Foxj1 εκφράζεται και στα κύτταρα του οργανωτή του εµβρύου (embryonic node) που φέρουν έναν κινητό κροσσό και είναι απαραίτητα για τη δηµιουργία ασυµµετρίας κατά την ανάπτυξη ανάµεσα στο δεξί και το αριστερό µισό του σώµατος (Left-Right asymmetry). Πράγµατι, δείχθηκε ότι οι µύες οι οποίοι είναι ελλειµµατικοί για το γονίδιο Foxj1 παρουσιάζουν ανωµαλίες ως προς την τοποθέτηση των οργάνων κατά τον άξονα συµµετρίας, µια κατάσταση που ονοµάζεται situs inversus, και επιπρόσθετα δεν διαθέτουν πολλαπλούς κροσσούς (Brody et al, 2000). Η απουσία κινητών κροσσών έχει αποτέλεσµα οι περισσότεροι µύες να πεθαίνουν νωρίς µετά τη γέννησή τους, ενώ όσοι καταφέρνουν να επιβιώσουν αναπτύσσουν υδροκεφαλία και πεθαίνουν αργότερα. Προσεκτική µελέτη µε ηλεκτρονική µικροσκοπία έδειξε ότι το γονίδιο Foxj1 επηρεάζει τους κινητούς κροσσούς, που εµφανίζουν διάταξη «9+2» µικροσωληνίσκοι, ενώ ο ακίνητος πρωτογενής κροσσός µε διάταξη «9+0» µικροσωληνίσκοι παραµένει ανεπηρέαστος. Επιπρόσθετα, δείχθηκε ότι η παραγωγή των υπεράριθµων κεντριολίων στα διαφοροποιηµένα κύτταρα πραγµατοποιείται φυσιολογικά, όµ ως τα κεντριόλια αυτά αδυνατούν να µεταναστεύσουν και να «αγκυροβολήσουν» στην πλασµατική µεµβράνη. Με παρόµοιο τ ρόπο ο παράγοντας Foxj1 ελέγχει τη διαφοροποίηση των πολυκροσσωτών κυττάρων στην επιδερµίδα εµβρύων Xenopus και στον πρόνεφρο των ψαριών zebrafish, υποδηλώνοντας την εξελικτική του συντήρηση (Stubbs et al, 2008; Yu et al, 2008). Σήµερα είναι γνωστό ότι ο Foxj1 σε µοριακό επίπεδο δρα κατά 41
42 1. ΕΙΣΑΓΩΓΗ τη διαφοροποίηση των πολυκροσσωτών κυττάρων ελέγχοντας µεταγραφικά την έκφραση µιας σειράς γονιδίων που κωδικοποιούν δοµικά συστατικά των µικροσωληνίσκων των κροσσών (Choksi et al, 2014), συµπεριλαµβανοµένου και των κεντρικών µικροσωληνίσκων, τις κινητήριες πρωτεΐνες του αξονήµατος, µόρια τουµπουλίνης, ένζυµα που τροποποιούν τις τουµπουλίνες, καθώς και άλλα γονίδια. Η δράση του αυτή εκδηλώνεται µέσω άµεσης πρόσδεσης στα ρυθµιστικά στοιχεία των γονιδίων αυτών. Rfx µεταγραφικοί παράγοντες Τα µέλη της οικογένειας των Rfx µεταγραφικών παραγόντων έχουν συνδεθεί κυρίως µε τη δηµιουργία κροσσών στα κύτταρα, τόσο κινητών όσο και µη κινητών (Choksi et al, 2014). Μέχρι σήµερα στα σπονδυλωτά έχουν αναγνωριστεί 8 µέλη της οικογένειας, µε εξαίρεση τα ψάρια στα οποία υπάρχει ένα ακόµα µέλος. Μέλη της υπεροικογένειας εντοπίζονται επίσης και στα ασπόνδυλα, όπως στη Drosophila και στο C. elegans, καθώς και στους µύκητες S. pombe και S. cerevisiae, γεγονός που υποδηλώνει την εξελικτική τους συντήρηση. Όλα τα µέλη της οικογένειας διαθέτουν µια ιδιόµορφη περιοχή πρόσδεσης στο DNA και εµφανίζουν υψηλή συντήρηση ως προς την επικράτεια που αναγνωρίζουν και προσδένονται στη χρωµατίνη, η οποία ονοµάζεται X-box (Choksi et al, 2014). Η περιοχή αυτή εντοπίζεται στους υποκινητές πολλών γονιδίων και οι Rfx παράγοντες µπορούν να προσδένονται εκεί είτε ως µονοµερή, είτε σχηµατίζοντας όµο- ή έτερο- διµερή. Στα σπονδυλωτά τα µέλη της οικογένειας που εµπλέκονται στη δηµιουργία των κροσσών είναι οι Rfx1-4, µε καλύτερα χαρακτηρισµένους τους Rfx2 και 3. Οι Rfx2 και 3 εµφανίζουν ένα σχετικά επικαλυπτόµενο πρότυπο έκφρασης και λειτουργίας. Επάγουν γονίδια που κωδικοποιούν δοµ ικά συστατικά των βασικών σωµατίων, της µεταβατικής ζώνης, του αξονήµατος και των κινητήριων πρωτεϊνών (Choksi et al, 2014). Πιο συγκεκριµένα, ο παράγοντας Rfx2 εκφράζεται στα πολυκροσσωτά κύτταρα στην επιδερµίδα του Xenopus και στον πρόνεφρο εµβρύων zebrafish. Χαρακτηρίστηκε για πρώτη φορά στο zebrafish, όπου η έλλειψή του προκαλεί µείωση στον αριθµό των µη κινητών πρωτογενών κ ροσσών που απαντώνται στον αναπτυσσόµενο νευρικό σωλήνα (Yu et al, 2008). Παρόµοια αποτελέσµατα προκαλεί και στο Xenopus επηρεάζοντας τη SHH σηµατοδότηση (Bisgrove et al, 2012; Chung et al, 2012). Και στα δυο αυτά συστήµατα επηρεάζεται το µήκος και η κινητικότητα των κροσσών. Επίσης, ο Rfx2 εκφράζεται από νωρίς κατά την εµβρυογένεση και σε υψηλά επίπεδα στον εγκέφαλο, στα νεφρά και στους όρχεις. Ο παράγοντας Rfx3 εκφράζεται στους µύες σε ιστούς που διαθέτουν κινητούς κροσσούς, όπως στον οργανωτή του εµβρύου και στον εγκέφαλο, στα επενδυµατικά κύτταρα των κοιλιών, στο φλοιό και στο χοριοειδές πλέγµα (reviewed in Choksi et al, 2014). Επίσης εκφράζεται και στα πολυκροσσωτά κύτταρα στο Xenopus (Chung et al, 2012). Η απουσία του στους µύς προκαλεί ανωµαλίες ως προς τη συµµετρία των οργάνων τους, µια κατάσταση που ονοµάζεται situs inversus, καθώς και υδροκεφαλία εξαιτίας της µη σωστής διαφοροποίησης των πολυκροσσωτών κυττάρων του 42
43 1. ΕΙΣΑΓΩΓΗ εγκεφάλου (Baas et al, 2006). Σε καλλιέργεια των πολυκροσσωτών επενδυµατικών κυττάρων του εγκεφάλου µυών δείχθηκε ότι η έλλειψή του επηρεάζει την ανάπτυξη, τον αριθµό και την κινητικότητα των κροσσών. Τέλος, ανάµεσα στους παράγοντες Foxj1 και Rfx2/3 υπάρχει συνεργατικότητα για την επιτυχή διαφοροποίηση των πολυκροσσωτών κυττάρων (Didon et al, 2013; El Zein et al, 2009). Συγκεκριµένα, έχει δειχθεί ότι ο παράγοντας Rfx3 µπορεί να επάγει την έκφραση του Foxj1, αλλά και ο Foxj1 να επάγει την έκφραση τόσο του Rfx2 όσο και του Rfx3. Μάλιστα, πρόσφατα αποκαλύφθηκε ότι ο παράγοντας Rfx2 µπορεί να δρα ως ικρίωµα για την πρόσδεση του Foxj1 στις κατάλληλες ρυθµ ιστικές περιοχές των γονιδίων που χρειάζεται να εκφραστούν για τη διαφοροποίηση των πολυκροσσωτών κυττάρων (Quigley & Kintner, 2017). Ο ρόλος του κάθε µέλους της οικογένειας των Rfx παραγόντων δεν είναι µέχρι σήµερα απόλυτα ξεκάθαρος και παρότι φαίνεται ότι δρουν κατά πλεονασµό, ωστόσο η δράση τους είναι σίγουρα και συνεργατική αφού µπορούν να φτιάχνουν ετεροδιµερή και µέσω αυτών να ελέγχουν την έκφραση µιας σειράς γονιδίων. Από όλα τα παραπάνω είναι φανερό ότι οι παράγοντες αυτοί δρούν µετά τον καθορισµό της τύχης των κυττάρων προς πολυκροσσωτά. Όµως πολύ πρόσφατα δείχθηκε ότι ο Rfx2 συµµετέχει και στη µετανάστευση των κυττάρων που πρόκειται να διαφοροποιηθούν σε πολυκροσσωτά, προς την κορυφαία πλευρά του ιστού στον οποίο βρίσκονται (Chung et al, 2014). Η λειτουργία αυτή είναι ιδιαίτερα σηµαντική για τα πολυκροσσωτά κύτταρα της επιδερµίδας εµβρύων Xenopus και του αναπνευστικού επιθηλίου των θηλαστικών, µιας και προέρχονται από έναν πληθυσµό κυττάρων p63 + που µεταναστεύει κατά τη διαφοροποίησή του. P73 Ο παράγοντας p73, µαζί µε τους p53 και p63, ανήκει στην οικογένεια των p53 µεταγραφικών παραγόντων. Ο παράγοντας p73 είναι ο λιγότερο µελετηµένος κατά τη διαδικασία της διαφοροποίησης των πολυκροσσωτών κυττάρων, µιας και τον τελευταίο χρόνο προσδιορίστηκε ότι συµµετέχει σε αυτή και µάλιστα σε πιο ύστερα στάδια της διαδικασίας (Marshall et al, 2016; Nemajerova et al, 2016). Πιο συγκεκριµένα, τα επιθηλιακά κύτταρα της τραχείας µυών από τα οποία απουσιάζει ο παράγοντας p73 εµφανίζουν φαινότυπο όµοιο µε αυτό των κυττάρων από τα οποία απουσιάζει ο παράγοντας Foxj1: ενώ δηµιουργούνται πολλαπλά κεντριόλια, αυτά δεν είναι ικανά να «αγκυροβολήσουν» στην πλασµατική µεµβράνη (Nemajerova et al, 2016). Ο παράγοντας p73 δρα µέσω άµ εσης πρόσδεσης στις ρυθµ ιστικές περιοχές διαφόρων γονιδίων, µεταξύ άλλων και των Foxj1 και Rfx2/3 (Marshall et al, 2016; Nemajerova et al, 2016). Ακόµα, παρατηρήθηκε ότι η έλλειψη του παράγοντα p73 µειώνει τον αριθµό των βασικών κυττάρων του αναπνευστικού επιθηλίου που έχουν την ικανότητα του κυτταρικού πολλαπλασιασµού, αυξάνοντας ταυτόχρονα τον αριθµό των εκκριτικών clara κυττάρων (Marshall et al, 2016). Ωστόσο, το αποτέλεσµα αυτό χρήζει περαιτέρω διερεύνησης ώστε να διαπιστωθεί εάν ο p73 συµµετέχει και σε αποφάσεις καθορισµού της µοίρας των κυττάρων άµ εσα, ή εξειδικεύεται µόνο στη διαφοροποίηση των πολυκροσσωτών κυττάρων. 43
44 1. ΕΙΣΑΓΩΓΗ Τέλος, στο στάδιο αυτό εµπλέκονται και τα micrornas της οικογένειας mir- 449, τα οποία ρυθµίζουν την έκφραση του παράγοντα CP110, αφού η πτώση των επιπέδων του CP110 προάγει την «αγκυροβόληση» των βασικών σωµατίων στη µεµβράνη του κυττάρου (Song et al, 2014) Ωρίµανση των πολυκροσσωτών κυττάρων Το τελικό σ τάδιο της διαφοροποίησης των πολυκροσσωτών κυττάρων, προκειµένου να γίνουν πλήρως λειτουργικά, περιλαµβάνει την απόκτηση της κατάλληλης πολικότητας των κροσσών τους στον ιστό που βρίσκονται, καθώς και της κατάλληλης κινητικότητάς τους. H στρατολόγηση συστατικών του σηµατοδοτικού µονοπατιού PCP (Planar Cell Polarity signaling), σε συνεργασία µε υδροδυναµικές δυνάµεις που ασκούνται στους ιστούς καθορίζουν την πολικότητα που θα αποκτήσουν οι κροσσοί στα πολυκροσσωτά κύτταρα (reviewed in Spassky & Meunier, 2017). Αξίζει να σηµειωθεί µια διαφορά ανάµεσα στα πολυκροσσωτά κύτταρα του ανθρώπινου οργανισµού ως προς την ωρίµανσή τους. Στα πολυκροσσωτά κύτταρα του αναπνευστικού επιθηλίου και των ωαγωγών οι κροσσοί καταλαµβάνουν όλη την επιφάνεια του κυττάρου και ο προσανατολισµός της κίνησής τους έχει ήδη καθοριστεί πριν ολοκληρωθεί ο σχηµατισµός τους, µε αποτέλεσµα ο προσανατολισµός τους να τελειοποιείται σε αυτό το τελικό στάδιο µέσω της κατάλληλης σηµατοδότησης (το ίδιο παρατηρείται και στα πολυκροσσωτά κύτταρα της επιδερµίδας των εµβρύων Xenopus). Αντίθετα, στα πολυκροσσωτά επενδυµατικά κύτταρα, τα βασικά σωµάτια «αγκυροβολούν» στην πλασµατική µεµβράνη µε τυχαίο τρόπο και στο τέλος µε την κατάλληλη σηµατοδότηση και µέσω υδροδυναµικών δυνάµεων προσανατολίζονται σωστά καταλαµβάνοντας ένα µέρος της επιφάνειας των κυττάρων. Τέλος, η «9+2» διάταξη των µικροσωληνίσκων εξασφαλίζει την αποτελεσµατική κίνηση των κροσσών σε συνδυασµό µε τις υδροδυναµικές δυνάµεις που ασκούνται και τον κυτταροσκελετό (reviewed in Spassky & Meunier, 2017). Ωστόσο, η συχνότητα κίνησης των κροσσών µπορεί να µεταβάλλεται από διάφορα εξωτερικά ερεθίσµατα, όπως για παράδειγµα σε περίπτωση φλεγµονής, οι ιντερλευκίνες ή διάφορες ορµόνες, όπως η προγεστερόνη, αλλάζουν τη συχνότητα µε την οποία πάλλονται οι κροσσοί του αναπνευστικού συτήµατος ή των ωαγωγών, αντίστοιχα. 44
45 1. ΕΙΣΑΓΩΓΗ 1.4 Τα µέλη της υπεροικογένειας της Geminin Οι παράγοντες Geminin, McIdas και GemC1 αποτελούν µια οµάδα πρωτεϊνών µε πολλαπλούς ρόλους τόσο κατά τον πολλαπλασιασµό, όσο και κατά τη διαφοροποίηση των κυττάρων. Η πρωτεΐνη Geminin αποτελεί το πρώτο µέλος της οικογένειας που χαρακτηρίστηκε και έκτοτε έχει µελετηθεί εκτενώς. Τα τελευταία χρόνια ταυτοποιήθηκαν και τα άλλα δυο µέλη της οικογένειας, εξαιτίας της οµοιότητας που παρουσίαζαν µε την Geminin, τόσο στην αλληλουχία όσο και στη λειτουργία τους (Εικόνα 1.8) (Balestrini et al, 2010; Pefani et al, 2011) Δοµικά χαρακτηριστικά Τα µέλη της υπεροικογένειας της Geminin εµφανίζουν µικρή οµοιότητα όσον αφορά στο σύνολο της αλληλουχίας τους. Ωστόσο, οµ οιάζουν σηµ αντικά στις επικράτειες σπειροειδούς σπειράµατος που διαθέτουν και οι τρείς πρωτεΐνες (Εικόνα 1.10 Α και Β) (Balestrini et al, 2010; Caillat et al, 2015; Caillat et al, 2013; Pefani et al, 2011). Το τµήµα µε τη µεγαλύτερη συντήρηση εντοπίζεται στο καρβοξυτελικό άκρο του σπειροειδούς σπειράµατός τους ( αα, στο πρωτεϊνικό µόριο της hgeminin). Oι πρωτεΐνες Geminin και McIdas εµφανίζουν υψηλή οµολογία σε ένα τµήµα επτά αµινοξέων που βρίσκεται στο αµινοτελικό τ µήµα του σπειροειδούς σπειράµατός τους (QYWKEVA) (Εικόνα 1.10 B, µαύρο πλαίσιο). Ιδιαίτερο ενδιαφέρον παρουσιάζει το γεγονός ότι αυτό το τµήµα συµπίπτει µε την πρώτη επικράτεια πρόσδεσης της Geminin µε τον παράγοντα αδειοδότησης της αντιγραφής Cdt1, µε τον οποίο αλληλεπιδρά εξασφαλίζοντας τη σωστή προαγωγή της αντιγραφής του DNA (Lee et al, 2004). Η µελέτη της κρυσταλλικής δοµής των πρωτεϊνών της οικογένειας της Geminin αποκάλυψε σηµαντικές πληροφορίες σχετικά µε τη διαµόρφωσή τους στο χώρο και τη λειτουργία τους. Όλα τα µέλη της οικογένειας µπορούν να φτιάχνουν οµοδιµερή, µε το οµοδιµερές της Geminin όµως να είναι το πιο σταθερό (Caillat et al, 2015; Caillat et al, 2013). Ακόµα, µέσω των σπειροειδών σπειραµάτων που διαθέτουν µπορούν να αλληλεπιδρούν µεταξύ τους σχηµατίζοντας ισχυρά συνδεδεµένα ετεροδιµερή (Caillat et al, 2015; Caillat et al, 2013), όµοια µε το οµ οδιµερές του σπειροειδούς σπειράµατος της Geminin (Thepaut et al, 2004). Ιn vitro µελέτες, αλλά και µελέτες σε κύτταρα έδειξαν ότι ο σχηµατισµός ετεροδιµερών Geminin-McIdas και Geminin-GemC1 προτιµάται, σε σχέση µε το οµοδιµερές της Geminin. Αυτό υποδηλώνει ότι τα ετεροδιµερή σχηµατίζονται αυθόρ µητα κατά την ταυτόχρονη έκφραση των πρωτεϊνών αυτών στα κύτταρα. Ωστόσο, η αλληλεπίδραση ανάµεσα στα µέλη McIdas και GemC1 δεν είναι ισχυρή (Caillat et al, 2015). Είναι σηµαντικό επίσης ότι πέραν της οµ οιότητας που παρουσιάζουν στην περιοχή του σπειροειδούς σπειράµατός τους, οι McIdas και GemC1 εµφανίζουν σηµαντική οµ οιότητα (49%) και σε µια ακόµα περιοχή που εντοπίζεται στο καρβοξυτελικό τους άκρο, η οποία όµως απουσιάζει από την Geminin (Εικόνα 1.10 Α και Γ). 45
46 1. ΕΙΣΑΓΩΓΗ Τα παραπάνω δεδοµένα σχετικά µε την οµοιότητα στην αµινοξική αλληλουχία των πρωτεϊνών της οικογένειας της Geminin, σε συνδυασµό και µε το γεγονός ότι ορθόλογα της Geminin εντοπίζονται στα µετάζωα, αλλά οι McIdas και GemC1 µόνο στα σπονδυλωτά οδηγεί στο εξής συµ πέρασµα: κατά την εξέλιξη οι McIdas και GemC1 προέκυψαν από το διπλασιασµό ενός κοινού προγονικού γονιδίου της Geminin, µετά το διαχωρισµό των ιχθύων, όπου στη συνέχεια ένας ακόµα διπλασιασµός οδήγησε στα µόρια McIdas και GemC1 (Terre et al, 2016) (Διδακτορική Διατριβή Δάφνη-Ελευθερία Πεφάνη, 2012). O McIdas διατήρησε µεγαλύτερη οµοιότητα στην περιοχή του σπειροειδούς σπειράµατος µε τη Geminin, ενώ ο GemC1 όχι, αφού δεν διαθέτει το συντηρηµένο επταπεπτίδιο (QYWKEVA) που εντοπίζεται στο αµ ινοτελικό άκρο του σπειροειδούς σπειράµατος των άλλων δυο µελών. Αυτό υποδηλώνει έναν λειτουργικό διαχωρισµό ανάµεσα στα δυο παράλογα. Επίσης, οι πρωτεΐνες McIdas και GemC1 εµφανίζουν, όπως προαναφέρθηκε, µια ακόµα περιοχή υψηλής συντήρησης στο καρβοξυτελικό τους άκρο, η οποία όµως απουσιάζει από τη Geminin. Το δεδοµένο αυτό υποδεικνύει ότι κατά την εξέλιξη των οργανισµών τα δυο νέα µέλη της οικογένειας απέκτησαν διαφορετικές λειτουργίες που διατηρήθηκαν στην εξελικτική πορεία των οργανισµών. 46

 Πολλαπλή ευθυγράµµιση της περιοχής του σπειροειδούς σπειράµατος των πρωτεϊνών Geminin, McIdas και GemC1 στον άνθρωπο, στο µύ και στο Xenopus, µε τη χρήση του προγράµµατος Clustal Omega.")
47 1. ΕΙΣΑΓΩΓΗ A B Γ Εικόνα 1.10 Τα µέλη της υπεροικογένειας της Geminin. (Α) Οι παράγοντες Geminin, McIdas και GemC1 συνιστούν την οικογένεια των πρωτεϊνών της Geminin. Όλα τα µέλη εµ φανίζουν σηµαντική οµ ολογία στην επικράτεια σπειροειδούς σπειράµατος που διαθέτουν, µέσω της οποίας µπορούν να σχηµατίζουν όµο- και έτεροδιµερή. Οι McIdas και GemC1, αλλά όχι η Geminin, διαθέτουν µια ακόµα περιοχή µε σηµαντική συντήρηση στο καρβοξυτελικό τους άκρο. (Β) Πολλαπλή ευθυγράµµιση της περιοχής του σπειροειδούς σπειράµατος των πρωτεϊνών Geminin, McIdas και GemC1 στον άνθρωπο, στο µύ και στο Xenopus, µε τη χρήση του προγράµµατος Clustal Omega. Στο µαύρο πλαίσιο περιλαµβάνεται το συντηρηµένο επταπεπτίδιο (QYWKEVA) που βρίσκεται στο αµινοτελικό τµήµα του σπειροειδούς σπειράµατος των Geminin και McIdas. (Γ) Πολλαπλή ευθυγράµµιση της συντηρηµένης περιοχής στο καρβοξυτελικό τµήµα των πρωτεϊνών McIdas και GemC1 στον άνθρωπο, στο µύ και στο Xenopus, µε τη χρήση του προγράµµατος Clustal Omega. 47
48 1. ΕΙΣΑΓΩΓΗ Ο ρόλος των µελών της υπεροικογένειας της Geminin στη ρύθµιση του κυτταρικού κύκλου Geminin Η πρωτεΐνη Geminin χαρακτηρίστηκε για πρώτη φορά ως ένας αρνητικός ρυθµιστής που δρα κατά την έναρξη της αντιγραφής του DNA στα µετάζωα (McGarry & Kirschner, 1998). Η Geminin εκφράζεται κατά τις φάσεις S και G2 του κυτταρικού κύκλου και πρωτεολύεται κατά τη διάρκεια της ανάφασης από το σύµπλοκο προαγωγής της ανάφασης/κυκλόσωµα (APC/C). Η Geminin αλληλεπιδρά και αναστέλλει τον παράγοντα αδειοδότησης της αντιγραφής Cdt1 κατά τη διάρκεια της S και G2 φάσης του κυτταρικού κύκλου, εµποδίζοντας έτσι να γίνει ξανά πρόσδεση των MCMs στη χρωµατίνη και να αδειοδοτηθεί ένα νέος κύκλος αντιγραφής µέχρι το τέλος της µίτωσης (Tada et al, 2001; Wohlschlegel et al, 2000). Ο µηχανισµός µέσω του οποίου η Geminin αναστέλλει το Cdt1 περιλαµβάνει άµεση πρόσδεση της Geminin στο Cdt1 (Lee et al, 2004), το σχηµατισµό ενός ανασταλτικού εξαµερούς συµπλόκου (2:4 Cdt1:Geminin) (De Marco et al, 2009; Lutzmann et al, 2006), καθώς και τη στρατολόγηση της Geminin στη χρωµατίνη µέσω του παράγοντα Cdt1 (Xouri et al, 2007). Πειράµατα αποσιώπησης αλλά και υπερέκφρασης της Geminin στα κύτταρα έχουν αναδείξει τη σηµαντικότητά της στη διατήρηση της γονιδιωµατικής σταθερότητας. Η αποσιώπηση της Geminin σε κύτταρα θηλαστικών προκαλεί επαναντιγραφή του γονιδιώµατος, βλάβες στο DNA και έτσι ενεργοποιείται το σηµείο ελέγχου από την G2 στην Μ φάση και εµποδίζεται η είσοδος των κυττάρων αυτών στη µίτωση (Klotz-Noack et al, 2012; Melixetian et al, 2004; Zhu et al, 2004). Ο φαινότυπος της υπερέκφρασης της Geminin στα κύτταρα έχει δειχθεί να εξαρτάται από τον κυτταρικό τύπο, µε τα κύτταρα να σταµατούν κατά τη µετάβαση από τη G1 στην S φάση του κυτταρικού κύκλου ή να παρουσιάζουν µια προβληµατική S φάση που οδηγεί τελικά τα κύτταρα σε απόπτωση (Shreeram et al, 2002). Εκτός από το ρόλο της κατά την αντιγραφή του DNA, η Geminin δείχθηκε να εµπλέκεται και στο διπλασιασµό των κεντριολίων σε διαιρούµενα κύτταρα, κατά τη διάρκεια του κυτταρικού κύκλου. Η αποσιώπηση της Geminin τόσο σε φυσιολογικά κύτταρα (ινοβλάστες ανθρώπου), όσο και σε καρκινικές κυτταρικές σειρές (U2OS και HCT116), οδηγεί σε υπερδιπλασιασµό των κεντροσωµατίων (Tachibana et al, 2005). Μάλιστα ο φαινότυπος της απουσίας της Geminin είναι ανεξάρτητος από το ρόλο της κατά το διπλασιασµό του DNA, αφού παρατηρείται χωρίς πέρασµα των κυττάρων από τη µίτωση. Ο άµ εσος ρόλος της Geminin στον κύκλο των κεντροσωµατίων ενισχύθηκε ακόµα παραπάνω από πειράµατα στα οποία η αποσιώπησή της, που πραγµατοποιήθηκε σε κύτταρα στα οποία έχει γίνει αναστολή της σύνθεσης του DNA ή της κυτταροκίνησης µε τη χρήση υδροξυουρίας (HU), προκαλούσε και πάλι υπερδιπλασιασµό των κεντροσωµατίων (Tachibana & Nigg, 2006). Επιπρόσθετα πειράµατα αργότερα έδειξαν ότι η Geminin εντοπίζεται στα κεντροσωµάτια (Lu et al, 2009). Η πρωτεΐνη Geminin µέσω του σπειροειδούς σπειράµατος που διαθέτει αλληλεπιδρά µε την υποµονάδα Arp1 του συµπλόκου της 48
49 1. ΕΙΣΑΓΩΓΗ δυνεΐνης-ακτίνης. Αυτή η αλληλεπίδραση µάλιστα είναι απαραίτητη για τον κεντροσωµατικό εντοπισµό της Geminin (Lu et al, 2009) McIdas Η πρωτεΐνη McIdas χαρακτηρίστηκε επίσης αρχικά ως ρυθµ ιστής του κυτταρικού κύκλου, αφού δείχθηκε ότι µπορεί να σχηµατίζει σύµπλοκο µε την πρωτεΐνη Geminin και να ανταγωνίζεται την πρόσδεση της Geminin στον παράγοντα αδειοδότησης της αντιγραφής Cdt1 (Εικόνα 1.11) (Pefani et al, 2011). Πειράµατα σε ολικά εκχυλίσµατα αβγών Xenopus έδειξαν ότι ο McIdas είναι ικανός να εµποδίσει την ανασταλτική δράση που εµφανίζει η Geminin κατά την αντιγραφή του DNA. Η πρωτεΐνη McIdas εντοπίζεται στον πυρήνα των κυττάρων και τα επίπεδά της ρυθµίζονται κατά τη διάρκεια του κυτταρικού κύκλου, αφού µειώνονται κατά τη διάρκεια της ανάφασης της µίτωσης και αυξάνουν ξανά κατά την είσοδο των κυττάρων στη G1 φάση. Η ρύθµιση αυτή που παρατηρήθηκε οµοιάζει µε το πρότυπο έκφρασης που παρουσιάζουν υποστρώµατα που πρωτεολύονται από το σύµπλοκο προαγωγής της ανάφασης/κυκλόσωµα (APC/C). Πράγµατι, οι Pefani et al. έδειξαν ότι η πρωτεΐνη McIdas θα µπορούσε να αποτελεί υπόστρωµα του APC/C συ µπλόκου, καθώς εντοπίστηκε στο αµινοτελικό της άκρο µια αλληλουχία RAΑL, που οµοιάζει µε τη συντηρηµένη αλληλουχία αποικοδόµησης RXXL που ε µφανίζουν οι µιτωτικές κυκλίνες που πρωτεολύονται από το σύµπλοκο προαγωγής της ανάφασης/κυκλόσωµα (Pefani et al, 2011). Η έλλειψη της πρωτεΐνης McIdas από τα κύτταρα επηρεάζει τη φυσιολογική προαγωγή του κυτταρικού κύκλου και προκαλεί συσσώρευση των κυττάρων στην S φάση. Τα κύτταρα αυτά αδυνατούν να εισέλθουν στη µίτωση και να προχωρήσουν στη G1 φάση του επόµενου κυτταρικού κύκλου (Pefani et al, 2011). Η υπερέκφραση του παράγοντα McIdas οδηγεί σε υπεραντιγραφή του γενετικού υλικού, αφού παρατηρήθηκε να αυξάνεται το µέγεθος του πυρήνα των κυττάρων, αλλά και η ποσότητα του DNA. Όπως αναφέρθηκε νωρίτερα, ο φαινότυπος αυτός έχει παρατηρηθεί και µετά την έλλειψη της πρωτεΐνης Geminin (Melixetian et al, 2004; Zhu et al, 2004). Ιδιαίτερο ενδιαφέρον παρουσιάζει ότι το αποτέλεσµα της υπερέκφρασης του McIdas διαµεσολαβείται από τη Geminin, αφού η ταυτόχρονη υπερέκφρασή τους σε U2OS κύτταρα α ναστέλλει το φαινότυπο που προκαλεί ο McIdas µόνος του (Caillat et al, 2013). Τέλος, δείχθηκε ότι η υπερέκφραση της π ρωτεΐνης McIdas έχει ως αποτέλεσµα και τη δηµ ιουργία µη φυσιολογικών πολυπολικών ατράκτων στα διαιρούµενα κύτταρα, υποδηλώνοντας την πιθανή εµπλοκή του παράγοντα McIdas και κατά τον κύκλο του διπλασιασµού των κεντροσωµατίων (Pefani et al, 2011). Το αποτέλεσµα αυτό παρουσιάζει µεγάλο ενδιαφέρον καθώς έχει ήδη αποδειχθεί ότι η Geminin εµπλέκεται στον κύκλο των κεντροσωµατίων. Συνοψίζοντας τα παραπάνω αποτελέσµατα, φαίνεται ότι η πρωτεΐνη McIdas διαδραµατίζει σηµ αντικό ρόλο κατά την έναρξη της αντιγραφής του DNA µέσω αλληλεπίδρασης µε την πρωτεΐνη Geminin και ενδέχεται να εµπλέκεται και στον 49
50 1. ΕΙΣΑΓΩΓΗ κύκλο του διπλασιασµού των κεντροσωµατίων. Ωστόσο και οι δυο αυτοί ρόλοι χρήζουν περαιτέρω διερεύνησης GemC1 Η πρωτεΐνη GemC1, όπως και τα άλλα δυο µέλη της οικογένειας, προσδιορίστηκε αρχικά για το ρόλο της κατά την αντιγραφή του DNA (Balestrini et al, 2010). Πιο συγκεκριµένα δείχθηκε ότι δρα κατά τη µετάβαση από τη G1 στην S φάση του κυτταρικού κύκλου, συµµετέχοντας στο σχηµατισµό του προεναρκτήριου συµπλόκου της αντιγραφής του DNA (pre-ic) (Εικόνα 1.11). Αντίθετα, τα άλλα δυο µέλη της οικογένειας, Geminin και McIdas, όπως αναφέρθηκε δρουν νωρίτερα κατά την αδειοδότηση της αντιγραφής του DNA. Πειράµατα σε ολικά εχκυλίσµατα αβγών βατράχου Xenopus έδειξαν ότι ο παράγοντας GemC1 αλληλεπιδρά άµ εσα µε τους παράγοντες της αντιγραφής TopBP1 και Cdc45, και ευθύνεται για τη φόρτωση του Cdc45 στο προεναρκτήριο σύµ πλοκο της αντιγραφής. Ο παράγοντας GemC1 προσδένεται ισχυρά στην κυκλίνη Ε και φωσφορυλιώνεται από το σύµπλοκο Cdk2- CyclinE (Balestrini et al, 2010). Η έλλειψη του GemC1 αναστέλλει την αντιγραφή του DNA, τόσο στο Xenopus όσο και σε κύτταρα µυός NIH3T3 (Balestrini et al, 2010). Πιο συγκεκριµένα, η έλλειψη του GemC1 εµποδίζει την πρόσδεση των παραγόντων Cdc45 και Sld5 (αποτελεί υποµονάδα του συµπλόκου GINS) στη χρωµατίνη, ενώ η πρόσδεση των TopBP1, Cdc7, ORC1-6 και MCM2-7 δεν επηρεάζεται. Η ανάλυση αυτή έδειξε ότι ο παράγοντας GemC1 είναι απαραίτητος για τη φόρτωση των Cdc45 και GINS στη χρωµατίνη και την επακόλουθη ενεργοποίηση της ελικάσης, ώστε να ξεκινήσει η αντιγραφή του DNA (Balestrini et al, 2010). 50
51 1. ΕΙΣΑΓΩΓΗ G1 S McIdas Geminin Cdc6 Cdc6 Cdt1 ORC MCMs Cdt1 ORC Cdc45 MCMs Treslin DUE-B TopBP1 GemC1 GINS RecQ4 MCM10 prerc preic Εικόνα 1.11 Τα µέλη της υπεροικογένειας της Geminin στον έλεγχο της αντιγραφής του DNA. Κατά τη διάρκεια της G1 φάσης λαµβάνει χώρα η αδειοδότηση της αντιγραφής του DNA µε το σχηµατισµό των προαντιγραφικών συµπλόκων (prerc) στις θέσεις έναρξης της αντιγραφής. Συστατικά του prerc αποτελούν το εξαµερές σύµπλοκο των ORC1-6, το Cdc6, το Cdt1 και το εξαµερές µε ενεργότητα ελικάσης MCM2-7. Σε αυτή τη φάση η πρωτεΐνη Geminin δεν εκφράζεται και έτσι ο παράγοντας Cdt1 επιτρέπει την αδειοδότηση της αντιγραφής του DNA. Κατά τη µετάβαση από τη G1 στην S φάση του κυτταρικού κύκλου η Geminin ξεκινά να εκφράζεται, προσδένεται µε τον παράγοντα Cdt1 και εµ ποδίζεται έτσι η περαιτέρω φόρτωση των MCMs στη χρωµατίνη. Ο McIdas φτιάχνει ετεροδιµερή µε την Geminin, εµ ποδίζοντας την πρόσδεση της Geminin στον παράγοντα Cdt1. Κατά την S φάση το προαντιγραφικό σύµπλοκο της αντιγραφής µετατρέπεται στο προεναρκτήριο σύµπλοκο µετά τη στρατολόγηση διαφόρων παραγόντων, συµπεριλαµβανοµένου των Cdc45, TopBP1, RecQ4, Treslin και MCM10. O παράγοντας GemC1 είναι απαραίτητος για τη στρατολόγηση του Cdc45, µέσω αλληλεπίδρασης µε το TopBP1 και το σύµπλοκο Cdk2CyclinE. Παρά τη σηµαντική οµοιότητα στην περιοχή του σπειροειδούς σπειράµατος ανάµεσα στα µέλη της υπεροικογένειας της Geminin, είναι σηµαντικό να αναφερθεί ότι οι παράγοντες GemC1 και McIdas δεν αλληλεπιδρούν µε τον παράγοντα αδειοδότησης της αντιγραφής Cdt1 (Caillat et al, 2015; Pefani et al, 2011). Όµως σχηµατίζουν σύµπλοκα µε την Geminin. Έχει προταθεί λοιπόν από τους Pefani et al. ότι οι ισορροπηµένες αλληλεπιδράσεις ανάµεσα στα µέλη της υπεροικογένειας της Geminin είναι σηµαντικές για την έναρξη της αντιγραφής του DNA (Pefani et al, 51
52 1. ΕΙΣΑΓΩΓΗ 2011) (Εικόνα 1.11). Όµως, είναι απαραίτητο να διεξαχθούν επιπρόσθετες µελέτες ώστε να διευκρινιστεί µε ακρίβεια ο ρόλος τους στον έλεγχο του κυτταρικού κύκλου Ο ρόλος των µελών της υπεροικογένειας της Geminin σε διαδικασίες κυτταρικής διαφοροποίησης Geminin Η Geminin είναι µια πολυλειτουργική πρωτεΐνη, αφού εκτός από το ρόλο της κατά την αντιγραφή του DNA, έχει µελετηθεί εκτενώς και για τη συµµετοχή της σε αποφάσεις κυτταρικής διαφοροποίησης σε µια ποικιλία οργανισµών και συστηµάτων (Patmanidi et al, 2017). Η συµµετοχή της Geminin στην κυτταρική διαφοροποίηση αποκαλύφθηκε παράλληλα µε το ρόλο της κατά την αντιγραφή του DNA (Kroll et al, 1998). Η Geminin αλληλεπιδρά µε παράγοντες αναδιάταξης της χρωµατίνης όπως ο παράγοντας Brg1 (Seo et al, 2005) ή το σύµπλοκο Polycomb (Luo et al, 2004), µε µεταγραφικούς παράγοντες όπως οι Six3 (Del Bene et al, 2004) και οι πρωτεΐνες Hox (Luo et al, 2004), ή µε άλλες πρωτεΐνες που διαθέτουν σπειροειδές σπείραµα, όπως οι ERNI και BERT (Papanayotou et al, 2008), ελέγχοντας έτσι τη διαφοροποίηση µέσω της µεταγραφής συγκεκριµένων γονιδίων-στόχων. Η απαλοιφή της Geminin από τους µύες και η in vivo µελέτη της έδωσε πιο ολοκληρωµένη εικόνα σχετικά µε το ρόλο της σε αποφάσεις καθορισµού της µοίρας των κυττάρων. Η πλήρης απουσία του γονιδίου της Geminin προκαλεί θάνατο στα έµβρυα των µυών στο στάδιο των 8 κυττάρων, καθώς αποτυγχάνουν να σχηµατίσουν την έσω κυτταρική µάζα και προκαλείται διαφοροποίηση των κυττάρων προς κύτταρα τροφοβλάστης (Gonzalez et al, 2006; Hara et al, 2006). Παράλληλα, προκαλούνται και επαναλαµβανόµενοι κύκλοι αντιγραφής του γονιδιώµατος (endoreduplication). Περαιτέρω µελέτες µετά από ιστοειδική αποσιώπηση της Geminin και σε συγκεκριµένα αναπτυξιακά στάδια, έδειξαν ότι η Geminin ρυθµίζει τις αποφάσεις διαφοροποίησης των νευρικών βλαστικών κυττάρων στον αναπτυσσόµενο εγκεφαλικό φλοιό (Spella et al, 2011), στο νευρικό σωλήνα (Patterson et al, 2014), στη νευρική ακρολοφία στη ραχιαία πλευρά του νευρικού σωλήνα (Stathopoulou et al, 2016), καθώς και των πρόδροµων κυττάρων του ανοσοποιητικού και αιµοποιητικού συστήµατος (Karamitros et al, 2010; Karamitros et al, 2015; Shinnick et al, 2010). Τέλος, η Geminin εµπλέκεται και σε διαδικασίες απόπτωσης (Roukos et al, 2007) και κυτταρικής γήρανσης (Iliou et al, 2013). Συµπερασµατικά λοιπόν η Geminin δρα προκαλώντας αναδιαµόρφωση της χρωµατίνης και ελέγχοντας µεταγραφικά την έκφραση γονιδίων που εµπλέκονται στον καθορισµό της µοίρας των πρόδροµων βλαστικών κυττάρων (Yellajoshyula et al, 2011). H Geminin έχει επίσης εµπλακεί στον καθορισµό α συµµετρίας (Left-Right asymmetry) κατά την ανάπτυξη των ψαριών zebrafish (Huang et al, 2011). Ο φαινότυπος αυτός παρατηρήθηκε εξαιτίας της µη σωστής δηµ ιουργίας κινητού κροσσού. Η έλλειψη της Geminin προκαλεί µείωση στον αριθµό των κροσσών του 52
53 1. ΕΙΣΑΓΩΓΗ οργάνου «Kupffer s vesicle», ένα όργανο που οµοιάζει µε τον οργανωτή του εµβρύου στο µύ (node), διαταράσσοντας την ορθή ανάπτυξη ασυµµετρίας κατά την ανάπτυξη McIdas Παράλληλα µε τη µελέτη της πρωτεΐνης McIdas κατά τη διάρκεια του κυτταρικού κύκλου, δείχθηκε ότι η πρωτεΐνη αυτή µπορεί να εµπλέκεται και σε διαδικασίες διαφοροποίησης, ύστερα από το ειδικό πρότυπο έκφρασης που εµφανίζει κατά τη διάρκεια της ανάπτυξης εµβρύων µυός (Pefani et al, 2011). Πιο συγκεκριµένα, δείχθηκε ότι ο McIdas εκφράζεται σε υψηλά επίπεδα, νωρίς κατά την ανάπτυξη, σε έµβρυα ηλικίας 12.5 ηµερών (Ε12.5), στο χοριοειδές πλέγµα (choroid plexus) και στην φλοιϊκή περιέλιξη (cortical hem) των πλάγιων κοιλιών του εγκεφάλου, τόσο σε επίπεδο mrna όσο και σε πρωτεϊνικό επίπεδο. Ενδιαφέρον παρουσιάζει το γεγονός ότι η έκφραση του McIdas δεν συνεντοπίζεται µε αυτή της Geminin, καθώς και του παράγοντα Pax6, υποδηλώνοντας ότι ο Μ cidas δεν εκφράζεται στα νευρικά πρόδροµα κύτταρα του εγκεφάλου, αλλά σε ένα διαφοροποιηµένο πληθυσµό κυττάρων. Η έκφραση του McIdas µειώνεται κατά τη διάρκεια της εµβρυογένεσης και τελικά περιορίζεται στο τέλος αυτής, στη βάση του χοριοειδούς πλέγµατος (Διδακτορική Διατριβή, Δάφνη-Ελευθερία Πεφάνη, 2012). Επιπρόσθετα, δείχθηκε ότι ο McIdas εντοπίζεται και στο αναπτυσσόµενο αναπνευστικό επιθήλιο των µυών. Κοινό χαρακτηριστικό των παραπάνω ιστών είναι ότι διαθέτουν πολυκροσσωτά κύτταρα, οδηγώντας στην υπόθεση ότι ο McIdas µπορεί να εµπλέκεται στη διαφοροποίηση των συγκεκριµένων κυττάρων. Πράγµατι, ένα χρόνο αργότερα, δείχθηκε ότι ο McIdas αποτελεί ένα γονίδιο που ρυθµ ίζεται από το σηµατοδοτικό µονοπάτι Notch και εµπλέκεται στη διαφοροποίηση των πολυκροσσωτών κυττάρων της επιδερµίδας εµβρύων Xenopus (Stubbs et al, 2012). Πιο συγκεκριµένα, δείχθηκε ότι τo Notch αναστέλλει την έκφραση του McIdas, ενώ όταν ο McIdas εκφράζεται απουσία το υ Notch, επάγει τη διαφοροποίηση των κυττάρων προς πολυκροσσωτά κύτταρα, στην επιδερµίδα και στο αναπτυσσόµενο νεφροστόµιο των εµβρύων του βατράχου Xenopus. Η αποσιώπηση του McIdas εµποδίζει τη διαφοροποίηση των πολυκροσσωτών κυττάρων, ενώ η υπερέκφρασή του µπορεί να προκαλέσει εκτοπικά τη δηµιουργία δοµ ών που οµ οιάζουν µε δευτεροσωµάτια, καθώς και πολυκροσσωτά κύτταρα. Μάλιστα, η δράση αυτή του McIdas φαίνεται να είναι συντηρηµένη, αφού δείχτηκε να εµπλέκεται και στη διαφοροποίηση των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου στο µυ (Stubbs et al, 2012). Η υπερέκφραση του McIdas σε ex vivo καλλιέργειες των επιθηλιακών κυττάρων της τραχείας µυών (mtecs) αυξάνει την έκφραση των παραγόντων Foxj1 και pericentrin, που εµπλέκονται στη διαφοροποίηση των πολυκροσσωτών κυττάρων. 53
54 1. ΕΙΣΑΓΩΓΗ GemC1 Παράλληλα µε τη µελέτη του προτύπου έκφρασης του παράγοντα McIdas και τη πιθανή ε µπλοκή του σε διαδικασίες κυτταρικής διαφοροποίησης, µελετήθηκε και το πρότυπο έκφρασης το υ GemC1 κατά την ανάπτυξη εµβρύου µυός (Διδακτορική Διατριβή, Δάφνη-Ελευθερία Πεφάνη, 2012). Πειράµατα in situ υβριδοποίησης έδειξαν ότι το mrna του GemC1 εντοπίζεται κατά την εµβρυογένεση, στο χοριοειδές πλέγµα όλων των εγκεφαλικών κοιλιών και στην τελεγκεφαλική φλοιϊκή περιέλιξη. Είναι ιδιαίτερα ενδιαφέρον ότι η έκφραση του παράγοντα GemC1 δεν περιορίζεται κατά το τέλος της εµβρυογένεσης, όπως παρατηρήθηκε για τον παράγοντα McIdas. Oι παράγοντες McIdas και GemC1 εµφανίζουν λοιπόν ένα επικαλυπτόµενο πρότυπο έκφρασης. Με δεδοµένο τα προκαταρκτικά αποτελέσµατα για την εµπλοκή του McIdas στη διαφοροποίηση των πολυκροσσωτών κυττάρων, είναι πιθανό ότι και ο παράγοντας GemC1 σχετίζεται µε αποφάσεις κυτταρικής διαφοροποίησης. Συνοψίζοντας, η υπεροικογένεια της Geminin αποτελείται από τις πρωτεΐνες Geminin, McIdas και GemC1, οι οποίες φαίνεται να εµπλέκονται τόσο στη σωστή προαγωγή του κυτταρικού κύκλου, όσο και σε αποφάσεις κυτταρικής διαφοροποίησης. Ιδιαίτερο ενδιαφέρον παρουσιάζει το ιστοειδικό πρότυπο έκφρασης των McIdas και GemC1, που διαφέρει από αυτό της Geminin, υποδηλώνοντας ότι τα δυο νέα µέλη της οικογένειας απέκτησαν και επιπρόσθετους ρόλους σε σχέση µε αυτούς της Geminin. 54
55 2. Σκοπός
56 2. ΣΚΟΠΟΣ 56
57 2. ΣΚΟΠΟΣ Προκειµένου να διατηρείται η γονιδιωµατική σταθερότητα στα κύτταρα, πρέπει να εξασφαλίζεται ότι σε κάθε κυτταρικό κύκλο το γενετικό υλικό διπλασιάζεται πλήρως και οι αδελφές χρωµατίδες διαχωρίζονται ακριβοδίκαια στα θυγατρικά κύτταρα. Στα διαιρούµενα κύτταρα, ο διπλασιασµός των κεντροσωµατίων συνδέεται άρρηκτα µε το διπλασιασµό των χρωµοσωµάτων. Ανωµαλίες στο διπλασιασµό και διαχωρισµό των κεντροσωµατίων ή/και των χρωµοσωµάτων µπορεί να οδηγήσει σε γονιδιωµατική αστάθεια. Ωστόσο, παρεκλίσεις από το φυσιολογικό αριθµό των κεντροσωµατίων απαντώνται σε µια ιδιαίτερη κατηγορία κυττάρων, στα πολυκροσσωτά κύτταρα. Τα πολυκροσσωτά κύτταρα είναι τελικώς διαφοροποιηµένα κύτταρα, τα οποία εξέρχονται από τον κυτταρικό κύκλο και µέσω ενός ιδιόµορφου κύκλου κεντροσωµατίων, πολλαπλασιάζουν τα κε ντριόλιά τους εκατοντάδες φορές. Τα πολυάριθµα κεντριόλια που σχηµατίζονται µεταναστεύουν στην πλασµατική µεµβράνη του κυττάρου και δρώντας ως βασικά σωµάτια αποτελούν τα κέντρα εµπυρήνωσης των κροσσών. Οι κροσσοί ωριµάζουν και κινούνται συγχρονισµένα µε σκοπό την κίνηση της βλέννας στο αναπνευστικό επιθήλιο, του εγκεφαλονωτιαίου υγρού στον εγκέφαλο, καθώς και των ωαρίων στο αναπαραγωγικό σύστηµα των θηλυκών ατόµων. Αλλαγές στον αριθµό, στη δοµή ή στη λειτουργία των κροσσών έχουν συνδεθεί µε ασθένειες στον άνθρωπο, όπως είναι χρόνιες υποτροπιάζουσες λοιµώξεις λόγω ελαττωµατικής βλεννοκροσωτής κάθαρσης του αναπνευστικού επιθηλίου, η υδροκεφαλία και η υπογονιµότητα. Οι µηχανισµοί που ελέγχουν την ισορροπία ανάµεσα στην παραγωγή ενός κεντριολίου κάθετα προς το µητρικό κεντριόλιο στα διαιρούµενα κύτταρα ή την παραγωγή υπεράριθµων κεντριολίων στα πολυκροσσωτά κύτταρα παραµένουν άγνωστοι. Στόχο της παρούσας διατριβής αποτέλεσε η µελέτη των µελών της υπεροικογένειας της Geminin -Geminin, McIdas και GemC1- στον πολλαπλασιασµό των κεντριολίων και στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Επίσης, πραγµατοποιήθηκε προκαταρκτική µελέτη ώστε να αποσαφηνιστεί η πιθανή συµµετοχή τους και κατά το διπλασιασµό των κεντριολίων στα διαιρούµενα κύτταρα. Μια σειρά από διαφορετικές µεθοδολογίες και ζωικά µοντέλα συντέλεσαν στο σκοπό να µελετηθούν τα εξής: Μελέτη της έκφρασης των Geminin, McIdas και GemC1 στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Η συγκεκριµένη µελέτη πραγµατοποιήθηκε σε ιστούς που διαθέτουν πολυκροσσωτά κύτταρα και αποµονώθηκαν από µυς κατά την ανάπτυξη και κατά την ενήλικη ζωή. Επιπρόσθετα, µελετήθηκε η έκφρασή τους κατά τα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. Αυτό κατέστει δυνατό µέσω της ex vivo καλλιέργειας επιθηλιακών κυττάρων που αποµονώνται από την τραχεία µυών, στα οποία µπορεί να επιτευχθεί ελεγχόµενη επαγωγή της διαφοροποίησής τους προς πολυκροσσωτά κύτταρα, κάτω από ειδικές συνθήκες καλλιέργειας. Τέλος, µελετήθηκε πιο στοχευµένα, η έκφραση της πρωτεΐνης McIdas σε συγκεκριµένα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. Λειτουργικός χαρακτηρισµός της δράσης των µελών της υπεροικογένειας της Geminin, µέσω πειραµάτων εκτοπικής έκφρασης ή αποσιώπησης της έκφρασής τους. 57
58 2. ΣΚΟΠΟΣ Επίσης, στόχος ήταν να αποσαφηνιστεί ο µηχανισµός µέσω του οποίου δρουν, εστιάζοντας στη µεταγραφική ρύθµιση του µονοπατιού. Μελέτη του in vivo ρόλου του γονιδίου GemC1 στη διαφοροποίηση των πολυκροσσωτών κυττάρων, µε τη χρήση διαγονιδιακών µυών στους οποίους δεν εκφραζόταν το γονίδιο GemC1. Έτσι, πραγµατοποιήθηκε φαινοτυπική µελέτη των µυών, αλλά κυρίως µελετήθηκαν τα πολυκροσσωτά κύτταρα του επιθηλίου της τραχείας των διαγονιδιακών µυών. Τέλος, σκοπός ήταν η µελέτη της πιθανής έκφρασης της πρωτεΐνης McIdas στα κεντροσωµάτια των διαιρούµενων κυττάρων. Επίσης, πραγµατοποιήθηκε προκαταρκτική µελέτη της λειτουργίας της στα κεντροσωµάτια, µέσω υπερέκφρασής της και µελέτης του φαινοτύπου που προκαλούσε στον αριθµό των κεντριολίων. 58
59 3. Υλικά και Μέθοδοι
60 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 60
61 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.1 Μέθοδοι ανασυνδυασµένου DNA κλωνοποιήσεις Λίστα πλασµιδίων Τα πλασµίδια που κατασκευάστηκαν στην παρούσα διδακτορική διατριβή και η στρατηγική κλωνοποίησης που ακολουθήθηκε για κάθε ένα από αυτά παρουσιάζονται συγκεντρωτικά στον πίνακα που ακολουθεί: Πίνακας 3.1 Στρατηγική κλωνοποίησης πλασµιδίων Ένθεµα Φορέας κλωνοποίησης Στρατηγική κλωνοποίησης GFP Το cdna του GFP έχει pentr1aminuscmr κλωνοποιηθεί στις θέσεις BamHI-XhoI GFP plvdest Πραγµατοποιήθηκε οµόλογος ανασυνδυασµός µεταξύ των θέσεων attl1/attl2 του pentr-gfp και των attr1/attr2 του plvdest. Το cdna του McIdas έχει κλωνοποιηθεί στις θέσεις GFP-mMcIdas KpnI-XhoI. Αµινοτελικά του pentr1aminuscmr McIdas, στο ίδιο πλαίσιο ανάγνωσης, έχει συντηχθεί το γονίδιο GFP. Πραγµατοποιήθηκε οµόλογος GFP-mMcIdas ανασυνδυασµός µεταξύ των plvdest θέσεων attl1/attl2 του pentr-gfp-mmcidas και των attr1/attr2 του plvdest. Το cdna του GemC1 έχει κλωνοποιηθεί στις θέσεις GFP-mGemC1 KpnI-XbaI. Αµινοτελικά του pentr1aminuscmr GemC1, στο ίδιο πλαίσιο ανάγνωσης, έχει συντηχθεί το γονίδιο GFP. Πραγµατοποιήθηκε οµόλογος GFP-mGemC1 ανασυνδυασµός µεταξύ των plvdest θέσεων attl1/attl2 του pentr-gfp-mgemc1 και των attr1/attr2 του plvdest. 1 kb (hg19, chr5: pgl3-mcidas ) του γονιδίου pgl3-basic vector McIdas έχουν κλωνοποιηθεί µεταξύ των θέσεων SacI/NheI (αναλυτικά στον πίνακα 3.4) 61
62 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ pgl3-gemc1 pgl3-foxj1 pgl3-basic vector pgl3-basic vector 2 kb (hg19, chr3: ) του γονιδίου GemC1 έχουν κλωνοποιηθεί µεταξύ των θέσεων KpnI/NheI (αναλυτικά στον πίνακα 3.4) 1,5 kb (hg19, chr17: ή hg19, chr17: ) του γονιδίου Foxj1 έχουν κλωνοποιηθεί µεταξύ των θέσεων SacI/NheI (αναλυτικά στον πίνακα 3.4) Επιπρόσθετα χρησιµοποιήθηκαν και οι εξής πλασµιδιακοί φορείς: GFP Διδακτορική διατριβή, Μάγδα pcaggs Σπέλλα, 2010 mmcidas Διδακτορική διατριβή, pcaggs Χριστίνα Κυρούση, 2014 mgemc1 Διδακτορική διατριβή, pcaggs Χριστίνα Κυρούση, 2014 hgeminin Διδακτορική διατριβή, Μάγδα pcaggs Σπέλλα, 2010 E2F1 pcmv Addgene, E2F4 pcmv Addgene, E2F5 pcmv Addgene, pcmv-neo-bam Addgene, pspax2 Addgene, pmd2.g Addgene,12259 mmcidas pflci Προπτυχιακή διπλωµατική εργασία, Χριστίνα Κυρούση, 2008 mgemc1 pbluescript KS + Δάφνη-Ελευθερία Πεφάνη, Διδακτορική διατριβή, Πορεία κλωνοποίησης των πλασµιδιακών φορέων έκφρασης Η κατασκευή των παραπάνω πλασµιδίων επιτυγχάνεται συνοπτικά µέσα από τα εξής βήµατα: o Πέψη των επιθυµητών τµηµάτων DNA (cdna και πλασµιδίου στο οποίο πρόκειται να ενσωµατωθεί) µε τις κατάλληλες περιοριστικές ενδονουκλεάσες. 62
63 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Ηλεκτροφορητική ανάλυση των προϊόντων των πέψεων και αποµόνωση των επιθυµητών ζωνών από το πήκτωµα της αγαρόζης (gel extraction). Ηλεκτροφόρηση των αποµονωµένων ζωνών για τον υπολογισµό των σχετικών τους συγκεντρώσεων. o Αντίδραση λιγάσης (ligation reaction) στους 16 o C, για 16 ώρες. o Μετασχηµατισµός δεκτικών βακτηρίων µε µια ποσότητα του προϊόντος της παραπάνω αντίδρασης. Επίστρωση των µετασχηµατισµένων βακτηρίων σε τρυβλία που περιέχουν LB-άγαρ µε το κατάλληλο αντιβιοτικό, για την ανάπτυξη µοναδιαίων αποικιών. Επώαση στους 37 o C, για 16 ώρες. o Εµβολιασµός, µερικών από τις αποικίες που αναπτύχθηκαν, σε θρεπτικό υλικό LB παρουσία του κατάλληλου ανιτβιοτικού. Ανάπτυξη των υγρών βακτηριακών καλλιεργειών (µικρής κλίµακας) στους 37 o C, για 16 ώρες. o Αποµόνωση πλασµιδιακού DNA (µικρής κλίµακας). Πέψεις και ηλεκτροφορητική ανάλυση των προϊόντων, ώστε να βρεθούν οι κλώνοι που φέρουν το επιθυµητό τµήµα DNA. 1-2 θετικοί κλώνοι επιστρώνονται σε τρυβλία LB-άγαρ παρουσία του κατάλληλου αντιβιοτικού. Επώαση στους 37 o C, για 16 ώρες. o Εµβολιασµός 1-2 αποικιών που αναπτύχθηκαν σε θρεπτικό υλικό LB, παρουσία του κατάλληλου αντιβιοτικού. Ανάπτυξη των υγρών βακτηριακών καλλιεργειών (µεσαίας κλίµακας) στους 37 o C, για 16 ώρες. o Αποµόνωση πλασµιδιακού DNA (µεσαίας κλίµακας). Πέψεις και ηλεκτροφορητική ανάλυση των προϊόντων, ώστε να επιβεβαιωθεί ότι οι κλώνοι που επιλέχθηκαν φέρουν το επιθυµητό τµήµα DNA Κατάτµηση τµηµάτων DNA Οι αντιδράσεις κατάτµησης των πλασµιδιακών φορέων πραγµατοποιούνται στους 37 o C, για 1-5 ώρες, ανάλογα µε την ποσότητα του DNA που προστίθεται στην κάθε αντίδραση. Πίνακας 3.2 Αντίδραση Κατάτµησης Τελικές συγκεντρώσεις Συστατικά αντίδρασης Πέψη µικρού Πέψη µεγάλου όγκου (20 µl) όγκου (100 µl) Ρυθµιστικό διάλυµα (10x) 1x 1x Ένζυµο περιορισµού 1U/µg DNA 1U/µg DNA BSA (όπου απαιτείται) 0,01% 0,01% DNA 1 µg 5-10 µg H 2 O (WFI) Μέχρι τελικό όγκο 20 µl Μέχρι τελικό όγκο 100 µl 63
64 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Ηλεκτροφορητική ανάλυση των προϊόντων και αποµόνωση του DNA από το πήκτωµα αγαρόζης Μετά το τέλος των πέψεων ακολουθεί ηλεκτροφορητική ανάλυση των προϊόντων σε πήκτωµα αγαρόζης. Η % σύσταση του πηκτώµατος εξαρτάται από το µέγεθος των προϊόντων της αντίδρασης. Η αποµόνωση του DNA από το πήκτωµα αγαρόζης πραγµατοποιήθηκε µε τη χρήση του NucleoSpin Gel and PCR Clean-up kit (Macherey-Nagel), σύµφωνα µε τις οδηγίες της εταιρείας Αντίδραση Λιγάσης Μετά την επιτυχή αποµόνωση του cdna και του πλασµιδίου στο οποίο πρόκειται να ενσωµατωθεί, πραγµατοποιείται η αντίδραση συνδετάσης. Για τη σύνδεση των τµηµάτων αυτών χρησιµοποιήθηκε το Takara DNA ligation kit, σύµφωνα µε τις οδηγίες της εταιρείας. Η αντίδραση πραγµατοποιείται συνήθως στους 16 o C, για 16 ώρες. Παράλληλα πραγµατοποιείται µια ακόµα αντίδραση ως αρνητικός µάρτυρας, που περιέχει µόνο τον πλασµιδιακό φορέα και το διάλυµα της Τ4 λιγάσης. Έτσι µπορεί να διαπιστωθεί πιθανή επανακυκλοποίηση του πλασµιδιακού φορέα, χωρίς δηλαδή να έχει προσλάβει το επιθυµητό τµήµα DNA Καλλιέργειες βακτηριακών κυττάρων Για τις υγρές καλλιέργειες των βακτηριακών κυττάρων χρησιµοποιείται ως θρεπτικό µέσο το LB, ενώ για τις στερεές καλλιέργειες χρησιµοποιείται LB-άγαρ. Στερεή καλλιέργεια βακτηριακών κυττάρων Στις στερεές καλλιέργειες βακτηριακών κυττάρων επιτυγχάνεται η ανάπτυξη µοναδιαίων αποικιών. o Θέρµανση του LB άγαρ µέχρι να υγροποιηθεί. o Χρήση λύχνου Bunsen για την επίτευξη στείρων συνθηκών. o Όταν η θερµοκρασία του LB-άγαρ φτάσει στους o C προστίθεται η κατάλληλη συγκέντρωση αντιβιοτικού (η α µπικιλλίνη χρησιµοποιείται σε τελική συγκέντρωση 100 µg/ml και η καναµυκίνη 30 µg/ml). o Σε κάθε τρυβλίο petri τοποθετούνται 20 ml LB-άγαρ, που αφήνεται να στερεοποιηθεί. o Επίστρωση των βακτηρίων στα τρυβλία. Τα βακτήρια µπορεί να προέρχονται είτε από τους -80 o C (glycerol stock, streaking), είτε από τη διαδικασία του µετασχηµατισµού. o Επώαση των τρυβλίων στους 37 o C, για 16 ώρες. 64
65 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Υγρή καλλιέργεια βακτηριακών κυττάρων Οι υγρές καλλιέργειες των βακτηριακών κυττάρων µικρής (3 ml) κλίµακας πραγµατοποιούνται ως εξής: o Χρήση λύχνου Bunsen για την επίτευξη στείρων συνθηκών. o Σε δοκιµαστικό σωλήνα τοποθετούνται 3 ml LB µε την κατάλληλη συγκέντρωση αντιβιοτικού (η αµ πικιλλίνη χρησιµοποιείται σε τελική συγκέντρωση 100 µg/ml και η καναµυκίνη 30 µg/ml). o Εµβολιασµός της καλλιέργειας µε µια µοναδιαία αποικία. o Επώαση των καλλιεργειών στους 37 o C για 16 ώρες, υπό ανάδευση. Οι υγρές καλλιέργειες των βακτηριακών κυττάρων µεσαίας (200 ml) κλίµακας πραγµατοποιούνται ως εξής: o Χρήση λύχνου Bunsen για την επίτευξη στείρων συνθηκών. o Σε κωνική φιάλη τοποθετούνται 200 ml LB µε την κατάλληλη συγκέντρωση αντιβιοτικού (η αµ πικιλλίνη χρησιµοποιείται σε τελική συγκέντρωση 100 µg/ml και η καναµυκίνη 30 µg/ml). o Εµβολιασµός της καλλιέργειας µε µια µοναδιαία αποικία. o Επώαση των καλλιεργειών στους 37 o C για 16 ώρες, υπό ανάδευση. Υλικά και Διαλύµατα LB 950 ml ddh 2 O 10 gr bacto -tryptone 5 gr bacto-yeast extract 10 gr NaCl Πραγµατοποιείται ρύθµιση του ph=7, ογκοµέτρηση σε τελικό όγκο 1 lt και αποστείρωση. LB άγαρ Παρασκευάζεται όπως το LB Μετά τη ρύθµιση του ph προστίθενται 15 gr άγαρ/lt LB Μετασχηµατισµός δεκτικών βακτηριακών κυττάρων DH5a µε πλασµιδιακό DNA o Χρήση λύχνου Bunsen για την επίτευξη στείρων συνθηκών. o Απόψυξη 25 µl δεκτικών βακτηρίων DH5a/αντίδραση µετασχηµατισµού σε πάγο. o Σε κάθε αντίδραση προστίθενται 1-5 µl DNA (1-10 ng DNA). o Επώαση στον πάγο για 30 λεπτά. 65
66 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Τα κύτταρα υφίστανται θερµικό πλήγµα µε επώαση στους 37 o C, για 20 δευτερόλεπτα (Προσοχή η θερµοκρασία αυτή µπορεί να διαφέρει ανάλογα µε το batch των δεκτικών κυττάρων που χρησιµοποιούνται). o Επώαση στον πάγο για 2 λεπτά. o Σε κάθε αντίδραση προστίθεται 1 ml LB. Επώαση στους 37 o C για 1 ώρα, υπό ανάδευση. o 250 µl κυττάρων επιστρώνονται σε τρυβλία µε LB-άγαρ, παρουσία του κατάλληλου αντιβιοτικού. Τα τρυβλία επωάζονται στους 37 o C, για 16 ώρες Μετασχηµατισµός δεκτικών βακτηριακών κυττάρων Stbl3 µε πλασµιδιακό DNA Τα δεκτικά βακτηριακά κύτταρα Stbl3 (Invitrogen) µετασχηµατίστηκαν µε τους λεντι-ιικούς φορείς έκφρασης: plvdest-gfp, plvdest-gfpmcidas και plvdest- GFPGemC1. Οι φορείς αυτοί διαθέτουν επαναλαµβανόµενες αλληλουχίες (LTRs) και η κλωνοποίησή τους καθίσταται λιγότερο αποδοτική. Τα Stbl3 βακτηριακά κύτταρα µειώνουν τα φαινόµενα οµ όλογου ανασυνδυασµού που παρατηρούνται µεταξύ των LTRs. o Χρήση λύχνου Bunsen για την επίτευξη στείρων συνθηκών. o Απόψυξη 30 µl δεκτικών βακτηρίων Stbl3/αντίδραση µετασχηµατισµού σε πάγο. o Σε κάθε αντίδραση προστίθενται 1-5 µl DNA (1-10 ng DNA). o Επώαση στον πάγο για 30 λεπτά. o Τα κύτταρα υφίστανται θερµικό πλήγµα µε επώαση στους 42 o C, για 45 δευτερόλεπτα. o Επώαση στον πάγο για 2 λεπτά. o Σε κάθε αντίδραση προστίθενται 250 µl SOC. Επώαση στους 37 o C για 1 ώρα, υπό ανάδευση. o 100 µl κυττάρων επιστρώνονται σε τρυβλία µε LB-άγαρ, παρουσία του κατάλληλου αντιβιοτικού. Τα τρυβλία επωάζονται στους 37 o C, για 16 ώρες Αποµόνωση πλασµιδιακού DNA µικρής κλίµακας (Minipreps) Για την αποµόνωση πλασµιδιακού DNA µικρής κλίµακας χρησιµοποιείται η µέθοδος της αλκαλικής λύσης ως εξής: o Εµβολιασµός µια µεµονωµένης αποικίας σε 3 ml LB, που περιέχει το κατάλληλο αντιβιοτικό. Επώαση στους 37 o C για 16 ώρες, υπό ανάδευση. o 1,5 ml καλλιέργειας µεταφέρονται σε σωλήνα eppendorf και φυγοκεντρούνται στις rpm, για 1 λεπτό. o Αποµάκρυνση του υπερκείµενου. o Επαναδιάλυση του ιζήµατος σε 300 µl διαλύµατος TENS, που θα προκαλέσει τη λύση των κυττάρων. 66
67 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Προσθήκη 150 µl διαλύµατος NaAc 3N (ph 5,4), για την εξουδετέρωση της αντίδρασης λύσης των κυττάρων. o Φυγοκέντρηση στις rpm για 10 λεπτά, για να ιζηµατοποιηθούν οι πρωτεΐνες και το γενωµικό DNA. o Μεταφορά του υπερκείµενου σε νέο eppendorf και προσθήκη 900 µl αιθανόλης 100% (-20 o C). Καλή ανάδευση και φυγοκέντρηση στις rpm, για 2 λεπτά. o Αποµάκρυνση του υπερκείµενου και προσθήκη 900 µl α ιθανόλης 70% (-20 o C). Καλή ανάδευση και φυγοκέντρηση στις rpm, για 2 λεπτά. o Αποµάκρυνση του υπερκείµενου και επαναδιάλυση του ιζήµατος σε 20 µl διαλύµατος TE που περιέχει RNAse (20 µg/ml). Υλικά και διαλύµατα Διάλυµα TENS Tris 10 mm EDTA 1 mm SDS 0,5% NaOH 0,1 M Διάλυµα NaAc 3M (ph 5,4) 12,3 gr NaAc Ρύθµιση του ph mε glacial acetic acid ddh 2 O µέχρι τελικού όγκου 50 ml Διάλυµα Tris EDTA, ph µl Tris 1M 100 µl EDTA 0,5M 49,4 ml ddh 2 O Αποµόνωση πλασµιδιακού DNA µεσαίας κλίµακας (Midipreps) Η αποµόνωση πλασµιδιακού DNA µεσαίας κλίµακας έγινε µε το NucleoBond Xtra Midi kit (Macherey-Nagel), σύµφωνα µε τις οδηγίες της εταιρείας. Για την αποµόνωση των λεντιιικών φορέων έκφρασης χρησιµοποιήθηκε το NucleoBond Xtra Midi EF (endotoxin free) kit. 67
68 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ LR αντίδραση ανασυνδυασµού Η τεχνολογία Gateway παρέχει τη δυνατότητα αποτελεσµατικής κλωνοποίησης τµηµάτων DNA, εκµεταλλευόµενη το σύστηµα τοπο-ειδικού ανασυνδυασµού του βακτηριοφάγου λ. Ένας αρχικός φορέας pentr1aminuscmr, φέρει τις αλληλουχίες attl1 και attl2, ενώ ο τελικός φορέας (προορισµού) plvdest, φέρει τις αλληλουχίες attr1 και attr2. Ο ανασυνδυασµός των att αλληλουχιών συµβάλλει στη µεταφορά του επιθυµητού DNA από το pentr1aminuscmr στο plvdest πλασµίδιο. Το σύστηµα αυτό χρησιµοποιήθηκε για την κατασκευή των λεντιιικών φορέων έκφρασης plvdest-gfp, plvdest-gfpmcidas και plvdest- GFPGemC1, ως εξής: Πίνακας 3.3 LR αντίδραση ανασυνδυασµού Συστατικά αντίδρασης Entry πλασµίδιο (pentry- GFP/GFPMcIdas/GFPGemC1) plvdest πλασµίδιο Πέψη µικρού όγκου 120 ng 150 ng LR clonase II enzyme mix 2 µl TE buffer, ph 8 Μέχρι τελικό όγκο 8 µl o Ξεπάγωµα του LR clonase II enzyme mix σε πάγο, για 2 λεπτά. o Ανάδευση µε σύντοµο vortex, 2 φορές από 2 δευτερόλεπτα. o Σε κάθε αντίδραση προστίθενται 2 µl LR clonase II enzyme mix. o Ανάδευση µε σύντοµο vortex, 2 φορές από 2 δευτερόλεπτα. o Σύντοµη φυγοκέντρηση. o Επώαση στους 25 0 C, για 16 ώρες. o Την επόµενη ηµέρα προστίθεται 1 µl πρωτεϊνάσης Κ/αντίδραση. o Επώαση στους 37 0 C, για 10 λεπτά. o Η αντίδραση φυλάσσεται στους C. 68
69 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.2 Μέθοδοι κυτταροκαλλιέργειας Καλλιέργεια καρκινικών κυττάρων Στην παρούσα διδακτορική διατριβή χρησιµοποιήθηκαν οι εξής κυτταρικές σειρές: 293T (CRL-3216, ATCC), πρόκειται για αθανατοποιηµένα επιθηλιακά κύτταρα, που προέρχονται από το νεφρό εµβρύου ανθρώπου. MCF7 (παραχωρήθηκαν από το εργαστήριο της Dr. Monica Bettencourt-Dias), επιθηλιακά κύτταρα αδενοκαρκινώµατος µαστού. HeLa (παραχωρήθηκαν από το εργαστήριο της Dr. Monica Bettencourt-Dias), κύτταρα από καρκίνο τραχήλου µήτρας. Τα κύτταρα καλλιεργούνται σε θρεπτικό µέσο DMEM (Gibco) εµπλουτισµένο µε 10% FBS, κάτω από άσηπτες συνθήκες και διατηρούνται σε επωαστή µε σταθερή θερµοκρασία 37 o C και 5% CO Απόψυξη κυττάρων o Σε τρυβλίο διαµέτρου 100mm, προστίθενται 10 ml προθερµασµένου (37 o C ) θρεπτικού µέσου. o Το σωληνάριο (cryovial) µε τα κύτταρα µεταφέρεται άµ εσα από το υγρό άζωτο σε υδατόλουτρο θερµοκρασίας 37 o C και επωάζεται σύντοµα, υπό ανάδευση. o Το 1ml του εναιωρήµατος των κυττάρων που περιέχεται στο cryovial, τοποθετείται στο τρυβλίο. o Όταν τα κύτταρα προσκολληθούν στο τρυβλίο πραγµατοποιείται αλλαγή του θρεπτικού µέσου, ώστε να αποµακρυνθεί το DMSO Διατήρηση της κυτταροκαλλιέργειας Τα κύτταρα πολλαπλασιάζονται σε τρυβλία (διαµέτρου 100mm), µέχρι να καλύψουν το 80-90% της επιφάνειας του τρυβλίου και κάθε δυο ηµ έρες πραγµατοποιείται αλλαγή του θρεπτικού µέσου στο οποίο αναπτύσσονται, ως εξής: o Αφαίρεση του υπερκείµενου θρεπτικού µέσου των κυττάρων. o Έκπλυση µε διάλυµα PBS (1x), 2 φορές. o Αφαίρεση του PBS και προσθήκη 10 ml φρέσκου θρεπτικού µέσου Ανακαλλιέργεια κυττάρων (από τρυβλίο διαµέτρου 100mm) o o Αφαίρεση υπερκείµενου θρεπτικού µέσου. Έκπλυση µε διάλυµα PBS (1x), 2 φορές. 69
70 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o o o o o o Προσθήκη 800 µl διαλύµατος τρυψίνης (1x) (κατάλληλη ποσότητα που να καλύπτει την επιφάνεια του τρυβλίου) και επώαση των κυττάρων στον επωαστή των 37 o C, για 1 λεπτό. Προσθήκη 9,5 ml θρεπτικού µέσου που περιέχει 10% FBS, ώστε να απενεργοποιηθεί η τρυψίνη. Καλή ανάµειξη των κυττάρων µε την πιπέτα, ώστε να ανασηκωθούν όλα τα κύτταρα και να γίνει διάσπαση τυχόν κυτταρικών συσσωµατωµάτων. Μεταφορά του κυτταρικού εναιωρήµατος σε σωλήνα φυγοκέντρησης 15 ml και φυγοκέντρηση για 5 λεπτά, σε 800 rpm (rounds per minute). Αφαίρεση του υπερκείµενου και αναδιάλυση του ιζήµατος των κυττάρων σε 10 ml θρεπτικό µέσο. Διαχωρισµός του κυτταρικού εναιωρήµατος σε νέα τρυβλία, στην επιθυµητή αναλογία, δεδοµένου και του ρυθµού πολλαπλασιασµού της κάθε κυτταρικής σειράς Ψύξη κυττάρων Η διατήρηση των κυττάρων επιτυγχάνεται µέσω της αποθήκευσής τους σε υγρό άζωτο. Προκειµένου να αποφεύγεται ο σχηµατισµός κρυστάλλων πάγου και να ελαχιστοποιείται η απώλεια νερού, η ψύξη τους γίνεται σταδιακά. Η προσθήκη του DMSO προστατεύει τα κύτταρα από την απότοµη µείωση της θερµοκρασίας κατά τη διάρκεια της ψύξης τους. o Αφαίρεση του υπερκείµενου θρεπτικού µέσου. o Έκπλυση µε διάλυµα PBS (1x), 2 φορές. o Προσθήκη 800 µl διαλύµατος τρυψίνης (1x) (κατάλληλη ποσότητα που να καλύπτει την επιφάνεια του τρυβλίου) και επώαση των κυττάρων στον επωαστή των 37 o C, για 1 λεπτό. o Προσθήκη 9,5 ml θρεπτικού µέσου που περιέχει 10% FBS, ώστε να απενεργοποιηθεί η τρυψίνη. o Καλή ανάµειξη των κυττάρων µε την πιπέτα, ώστε να ανασηκωθούν όλα τα κύτταρα και να γίνει διάσπαση τυχόν κυτταρικών συσσωµατωµάτων. o Μεταφορά του κυτταρικού εναιωρήµατος σε σωλήνα φυγοκέντρησης 15 ml και φυγοκέντρηση για 5 λεπτά, στις 800 rpm. o Αφαίρεση του υπερκείµενου και επαναιώρηση των κυττάρων σε 5 ml PBS (1x). o Φυγοκέντρηση για 5 λεπτά, στις 800 rpm. o Αφαίρεση του υπερκείµενου και επαναιώρηση του ιζήµατος σε 1 ml FBS, που περιέχει 10% DMSO. Μεταφορά των κυττάρων σε ειδικά για πάγωµα σωληνάρια (cryovials). o Τοποθέτηση των cryovials σε ειδικό δοχείο παγώµατος (-1 o C/λεπτό), σε καταψύκτη -80 o C. Μεταφορά των κυττάρων σε δοχείο υγρού αζώτου, για να διατηρηθούν για µεγάλο χρονικό διάστηµα. 70
71 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Υλικά και διαλύµατα Θρεπτικό µέσο καλλιέργειας καρκινικών κυτταρικών σειρών 450 ml DMEM (Gibco, 41966) 50 ml FBS (Fetal Bovine Serum, Gibco) Το θρεπτικό µέσο φιλτράρεται και φυλάσσεται στους 4 o C. PBS (10x) 80 gr NaCl 2 gr KCl 2,4 gr KH 2 PO 4 14,4 gr Na 2 HPO 4 Τελικός όγκος ddh 2 O: 1 lt PBS (1x) Αραίωση 1/10 του PBS 10x σε ddh 2 O, ph 7,2-7,4. Αποστείρωση και φύλαξη σε θερµοκρασία δωµατίου. Διάλυµα τρυψίνης (0,5x ή 1x) 1,5 ml από stock διάλυµα τρυψίνης 10x (Gibco) 13,5 ml PBS (1x) Φιλτράρεται και φυλάσσεται στους 4 o C Αποµόνωση και καλλιέργεια των επιθηλιακών κυττάρων της τραχείας µυών (Mouse tracheal epithelial cell culture MTEC) Η συγκεκριµένη καλλιέργεια αποτελεί ένα αξιόπιστο σύστηµα καλλιέργειας των επιθηλιακών κυττάρων που αποµονώνονται από την τραχεία είτε αγρίου τύπου, είτε γενετικά τροποποιηµένων µυών (You et al, 2002). Η αποµόνωση ενός ικανοποιητικού αριθµού κυττάρων, καθώς και η χρήση κατάλληλων θρεπτικών µέσων, είναι εξαιρετικής σηµασίας για τον επιτυχή πολλαπλασιασµό τους και τη µετέπειτα διαφοροποίησή τους σε πολυκροσσωτά κύτταρα. Τα κύτταρα αυτά αποµονώνονται και καλλιεργούνται σε ειδικές, ηµιδιαπερατές µεµβράνες, ένθετες σε πηγαδάκια πιάτου 24-θέσεων (24-well plate). Κατά το στάδιο του πολλαπλασιασµού τους απαιτείται ένα θρεπτικό µέσο ε µπλουτισµένο µε αυξητικούς παράγοντες. Το συγκεκριµένο θρεπτικό µέσο ονοµάζεται εµπλουτισµένο θρεπτικό µέσο καλλιέργειας (MTEC plus θρεπτικό µέσο) και χορηγείται στα κύτταρα τόσο στην κορυφαία, όσο και στη βασική πλευρά της µεµβράνης καλλιέργειας. Αφού τα κύτταρα πολλαπλασιαστούν και καταλάβουν όλη την επιφάνεια της µεµβράνης στην οποία καλλιεργούνται, επάγεται η διαφοροποίησή τους προς πολυκροσσωτά, µέσω της έκθεσής τους στον αέρα, µια κατάσταση που 71
72 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ ονοµάζεται ALI (Air Liquid Interface). Κατά τη φάση αυτή χορηγείται θρεπτικό υλικό µόνο στη βασική πλευρά της µεµβράνης καλλιέργειας, ενώ δεν παρέχεται θρεπτικό µέσο στην κορυφαία πλευρά της µεµβράνης. Το θρεπτικό µέσο που χορηγείται στα κύτταρα κατά το στάδιο αυτό δεν περιέχει ορό και είναι φτωχό σε αυξητικούς παράγοντες. Το συγκεκριµένο θρεπτικό µέσο θα αναφέρεται ως θρεπτικό µέσο διαφοροποίησης των MTEC (MTEC serum free θρεπτικό µέσο). Η αποµόνωση των κυττάρων γίνεται µέσω µιας διαδικασίας δύο ηµερών, αφού απαιτείται επώαση των αποµονωµένων τραχειών µε την πρωτεάση Pronase, για ώρες Επίστρωση των µεµβρανών καλλιέργειας των MTEC µε κολλαγόνο Τα επιθηλιακά κύτταρα που αποµονώνονται από την τραχεία των µυών επιστρώνονται σε ειδικές ηµι-διαπερατές µεµβράνες 0.33 cm 2, ένθετες σε πηγαδάκια πολυκυψελιδικών πιάτων (24-well plate) (Transwell filters, Corning, 3470). Προκειµένου να επιτευχθεί η προσκόλληση των κυττάρων, οι µεµβράνες επωάζονται µε διάλυµα κολλαγόνου. o Τοποθέτηση 150 µl του διαλύµατος κολλαγόνου (rat tale type I collagen), µόνο στην κορυφαία πλευρά της µεµβράνης καλλιέργειας. o Επώαση των µεµβρανών µε το διάλυµα κολλαγόνου για ώρες σε θερµοκρασία δωµατίου ή το λιγότερο για 4 ώρες στον επωαστή θερµοκρασίας 37 o C. o Αναρρόφηση του διαλύµατος κολλαγόνου από την κορυφαία πλευρά της κάθε µεµβράνης. o Οι µεµβράνες αφήνονται να στεγνώσουν για τουλάχιστον 5 λεπτά. o Έκπλυση της κορυφαίας και της βασικής πλευράς της µεµβράνης καλλιέργειας µε διάλυµα PBS (1x), 3 φορές. Στην κορυφαία πλευρά τοποθετούνται 150 µl διαλύµατος PBS (1x), ενώ στη βασική πλευρά τοποθετούνται 500 µl. o Οι µεµβράνες αφήνονται να στεγνώσουν για τουλάχιστον 5 λεπτά. o Οι µεµβράνες που έχουν επιστρωθεί µε κολλαγόνο µπορούν να χρησιµοποιηθούν άµεσα ή να διατηρηθούν (καλά κλεισµένες στο 24-well τρυβλίο µε parafilm) για πολλούς µήνες σε θερµοκρασία δωµατίου Αποµόνωση των MTEC και έναρξη της καλλιέργειας 1 η ηµέρα o Σε 2 τρυβλία (100mm) τοποθετείται παγωµένο και αποστειρωµένο θρεπτικό µέσο Ham s F-12/Pen-Strep (10 ml/τρυβλίο). Η προετοιµασία τους πραγµατοποιείται σε θάλαµο νηµατικής ροής, κάτω από άσηπτες συνθήκες. Τα τρυβλία διατηρούνται σε πάγο, καθόλη τη διάρκεια της διαδικασίας, και χρησιµοποιούνται για την τοποθέτηση των τραχειών που αποµονώνονται. 72
73 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o o o o o o o o o o o o o o Αναισθητοποίηση των ενήλικων µυών (είτε αγρίου τύπου, είτε διαγονιδιακών) µε διοξείδιο του άνθρακα. Ψεκασµό των µυών µε αιθανόλη 70%, ώστε να επιτυγχάνονται όσο το δυνατό πιο στείρες συνθήκες. Η πλήρης αναισθησία ελέγχεται µε την έλλειψη αντανακλαστικών στα κάτω άκρα. Μετά την επίτευξη της αναισθησίας πραγµατοποιείται διατοµή του δέρµατος στη θωρακική χώρα. Με ψαλίδι ξεκινώντας από το στέρνο ανοίγεται, µε δυο επιµήκεις πλάγιες τοµές, ο θώρακας, έτσι ώστε να αποκαλυφθούν τα εσωτερικά όργανα. Η τοµή εκτείνεται µέχρι το κεφάλι, έως ότου να αποκαλυφθεί ο λαιµός. Με µια ενδιάµεση τοµή αποµακρύνονται προσεκτικά οι ιστοί από την κλείδα έως τους υπογνάθιους αδένες, ώστε να αποκαλυφθεί η τραχεία. Αποµάκρυνση των παρακείµενων ιστών από την τραχεία. Αποµάκρυνση του οισοφάγου από την τραχεία. Αποµάκρυνση της τραχείας προσεκτικά από το ύψος του λάρυγγα µέχρι το σηµείο που διακλαδίζεται στους βρόγχους. Τοποθέτηση της κάθε τραχείας που αποµονώνεται σε τρυβλίο που περιέχει το παγωµένο θρεπτικό µέσο Ham s F-12/Pen-Strep. Η ίδια διαδικασία επαναλαµβάνεται για την αποµόνωση όλων των τραχειών. Καθόλη τη διάρκεια της διαδικασίας, οι αποµονωµένες τραχείες διατηρούνται στο παγωµένο θρεπτικό µέσο Ham s F-12/Pen-Strep. Μετά την αποµόνωση όλων των τραχειών πραγµατοποιείται λεπτοµερής εκκαθάριση της κάθε τραχείας από τους παρακείµενους ιστούς, µε τη χρήση στερεοσκοπίου. Αποµάκρυνση του λάρυγγα από τις τραχείες. Οι τραχείες που έχουν εκκαθαριστεί τοποθετούνται στο δεύτερο τρυβλίο που περιέχει το παγωµένο Ham s F-12/Pen-Strep. Σε κάθε τραχεία πραγµατοποιείται µια διαµήκης τοµή, ώστε να αποκαλυφθεί η κοιλότητά της. Τοποθέτηση των τραχειών σε 15 ml σωλήνα φυγοκέντρησης, που περιέχει το φρέσκο διάλυµα Pronase (πρωτεάση). Το διάλυµα της Pronase πρέπει να καλύπτει πλήρως όλες τις τραχείες (3 ml διαλύµατος επαρκούν για την κάλυψη έως 30 τραχειών). Επώαση στους 4 o C, για ώρες. 2 η ηµέρα o Τοποθέτηση του διαλύµατος της DNase σε πάγο. o Ξεπάγωµα του FBS σε υδατόλουτρο θερµοκρασίας 37 o C. o Τοποθέτηση του σωλήνα φυγοκέντρησης που περιέχει τις τραχείες σε θάλαµο νηµατικής ροής. o Ήπια ανάδευση των τραχειών, από τη µια άκρη του σωλήνα έως την άλλη (end-over-end), 5 φορές. o Επώαση των τραχειών σε θερµοκρασία δωµατίου, για 10 λεπτά. o Ήπια ανάδευση των τραχειών, από τη µια άκρη του σωλήνα έως την άλλη (end-over-end), 5 φορές. 73
74 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o o o o o o o o o o o o o o o o o o o Στο σωληνάριο µε τις τραχείες προστίθεται προθερµασµένο FBS σε τελική συγκέντρωση 10% (v/v). Ήπια ανάδευση των τραχειών, από τη µια άκρη του σωλήνα έως την άλλη (end-over-end), 20 φορές. Μεταφορά των τραχειών, µε τη βοήθεια πιπέτας Pasteur, σε ένα δεύτερο σωλήνα φυγοκέντρησης 15 ml, που περιέχει 3 ml προθερµασµένου Ham s F- 12/Pen-Strep/10% FBS. Ήπια ανάδευση των τραχειών, από τη µια άκρη του σωλήνα έως την άλλη (end-over-end), 15 φορές. Μεταφορά των τραχειών, µε τη βοήθεια πιπέτας Pasteur, σε έναν τρίτο σωλήνα φυγοκέντρησης 15 ml, που περιέχει 3 ml προθερµασµένου Ham s F- 12/Pen-Strep/10% FBS. Ήπια ανάδευση των τραχειών, από τη µια άκρη του σωλήνα έως την άλλη (end-over-end), 15 φορές. Μεταφορά των τραχειών, µε τη βοήθεια πιπέτας Pasteur, στα απόβλητα. Το θρεπτικό µέσο και από τα τρία σωληνάρια συγκεντρώνεται σε ένα από αυτά και ακολουθεί φυγοκέντρηση στις 1200rpm, στους 4 o C για 10 λεπτά. Αναρρόφηση του υπερκείµενου και επαναιώρηση των κυττάρων στο διάλυµα της DNase. Τοποθετούνται 200µl διαλύµατος DNase/τραχεία. Επώαση στον πάγο για 5 λεπτά. Φυγοκέντρηση στις 1200 rpm, στους 4 o C για 5 λεπτά. Αναρρόφηση του υπερκείµενου και επαναιώρηση στο βασικό θρεπτικό µέσο της καλλιέργειας των MTEC (MTEC basic), που περιέχει 10% FBS. Tοποθέτηση του εναιωρήµατος των κυττάρων σε τρυβλίο 100mm και επώαση στους 37 o C για 3-4 ώρες, ώστε να προσκολληθούν οι ινοβλάστες. Μετά το πέρας των 3-4 ωρών, πραγµατοποιείται ήπια ανάδευση του θρεπτικού µέσου στο τρυβλίο, το οποίο τοποθετείται σε αποστειρωµένο 50 ml σωλήνα φυγοκέντρησης. Η ανάδευση πρέπει να είναι ήπια, ώστε να µην αποκολληθούν οι ινοβλάστες. Έκπλυση του τρυβλίου, 2 φορές, µε 10 ml προθερµασµένου βασικού θρεπτικού µέσου της καλλιέργειας των MTEC, που περιέχει 10% FBS. Το θρεπτικό µέσο από κάθε έκπλυση συγκεντρώνεται σ τον 50 ml σωλήνα φυγοκέντρησης. Φυγοκέντρηση στις 1200 rpm, στους 4 o C για 5 λεπτά. Προετοιµασία του εµπλουτισµένου θρεπτικού µέσου καλλιέργειας των MTEC (MTEC plus), µε την προσθήκη διαλύµατος ρετινοϊκού οξέος Β (RA-B), σε αραίωση 1:1000. Αναρρόφηση του υπερκείµενου θρεπτικού µέσου και επαναιώρηση των κυττάρων σε µικρό όγκο ΜTEC plus θρεπτικού µέσου (που περιέχει RA-Β, σε αραίωση 1:1000). Υπολογίζονται µl/τραχεία. Κατά την επαναιώρηση των κυττάρων απαιτείται ήπια ανάδευση. Πρέπει να αποφεύγεται η προσπάθεια δηµιουργίας µοναδιαίων κυττάρων. Υπολογισµός του αριθµού των κυττάρων µε τη χρήση αιµοκυτταρόµετρου τύπου Neubauer (προστίθεται και χρωστική trypan blue για τον έλεγχο της βιωσιµότητας των κυττάρων). 74
75 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Τα κύτταρα επιστρώνονται στις ειδικές ηµι -διαπερατές µεµβράνες 0.33 cm 2 που έχουν επωαστεί µε κολλαγόνο και βρίσκονται ένθετες σε πηγαδάκια πολυκυψελιδικών πιάτων. Στην κορυφαία πλευρά της κάθε µεµβράνης επιστρώνονται 0,33 x 10 5 κύτταρα, σε τελικό όγκο 150 µl MTEC plus θρεπτικού µέσου (που περιέχει RA-Β, σε αραίωση 1:1000). Στη βασική πλευρά της µεµβράνης τοποθετούνται 500 µl MTEC plus θρεπτικού µέσου (που περιέχει RA-Β, σε αραίωση 1:1000). Εάν τα κύτταρα της καλλιέργειας πρόκειται να µολυνθούν µε λεντιιούς, για τη µεταφορά γονιδίων, τότε επιστρώνονται 0,4 x 10 5 κύτταρα/µεµβράνη. o Τα κύτταρα διατηρούνται σε επωαστή µε σταθερή θερµοκρασία 37 o C και 5% CO 2. Η ηµέρα αυτή σηµειώνεται ως preali DAY 0. o 3 ηµ έρες µετά πραγµατοποιείται η πρώτη αλλαγή θρεπτικού µέσου. Στην κορυφαία πλευρά της κάθε µεµβράνης τοποθετούνται 150 µl MTEC plus θρεπτικού µέσου, ενώ στη βασική της πλευρά τοποθετούνται 500 µl MTEC plus θρεπτικού µέσου (στο MTEC plus θρεπτικό µέσο προστίθεται κάθε φορά πριν χρησιµοποιηθεί διάλυµα RA-Β, σε αραίωση 1:1000) Πολλαπλασιασµός των MTEC Μετά την εκκίνησή της, η καλλιέργεια διατηρείται στο στάδιο του πολλαπλασιασµού για 5-7 ηµ έρες συνολικά. Μετά το πέρας των 3 ηµ ερών που πραγµατοποιείται η πρώτη αλλαγή θρεπτικού µέσου, απαιτείται αλλαγή του κάθε δυο ηµέρες Διαφοροποίηση των MTEC Αφού τα κύτταρα πολλαπλασιαστούν επιτυχώς και καταλάβουν όλη την επιφάνεια της µεµβράνης καλλιέργειας, συνήθως 7 ηµέρες µετά την έναρξη της καλλιέργειας, ξεκινά η φάση της διαφοροποίησης (Air Liquid Interface, ΑLI). Σε αυτή τη φάση, αναρροφάται το θρεπτικό µέσο από την κορυφαία πλευρά των µεµβρανών, ώστε τα κύτταρα να εκτίθενται στον αέρα. Στα κύτταρα παρέχεται θρεπτικό µέσο µόνο στη βασική πλευρά της µεµβράνης καλλιέργειας. Η πρώτη µέρα ορίζεται ως ALI DAY 0 (AD0). Η καλλιέργεια στη φάση αυτή συνεχίζεται για όσο απαιτείται, σύµφωνα µε την πειραµατική διαδικασία. Η διαφοροποίηση των κυττάρων έχει ολοκληρωθεί 21 ηµέρες µετά την επαγωγή της (ΑD21). o Αναρρόφηση του MTEC plus θρεπτικού µέσου από την κορυφαία και βασική πλευρά της µεµβράνης. o Τοποθέτηση 500 µl MTEC serum free θρεπτικού µέσου, µόνο στη βασική πλευρά της κάθε µεµβράνης καλλιέργειας. o Ανά δυο ηµέρες πραγµατοποιείται αλλαγή θρεπτικού µέσου, µόνο στη βασική πλευρά της κάθε µεµβράνης καλλιέργειας. o Η καλλιέργεια διατηρείται για όσο διάστηµα απαιτείται σύµφωνα µε την πειραµατική διαδικασία που ακολουθείται. 75
76 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Υλικά και διαλύµατα Βασικό θρεπτικό µέσο καλλιέργειας MTEC (MTEC basic θρεπτικό µέσο) DMEM/F-12 (Sigma, D8437) 15mM Hepes 4mM L-glutamine 3,6 mm NaHCO U/ml Pen 100µg/ml Strep Το MTEC basic θρεπτικό µέσο φιλτράρεται και φυλάσσεται στους 4 o C. Εµπλουτισµένο θρεπτικό µέσο καλλιέργειας MTEC (MTEC plus θρεπτικό µέσο) ΜTEC basic θρεπτικό µέσο 10 µg/ml ινσουλίνη 5 µg/ml τρανσφερίνη 0,1 µg/ml cholera toxin 25 ng/ml EGF 30 µg/ml BPE (Bovine Pituitary Extract) 5% FBS Το MTEC plus θρεπτικό µέσο φιλτράρεται και φυλάσσεται στους 4 o C. Παρασκευάζεται µια µικρή ποσότητα (50 ml) και προκειµένου να διατηρηθούν όλα τα αντιδραστήρια που έχουν προστεθεί, προθερµαίνεται κάθε φορά µόνο όση ποσότητα χρειάζεται για την καλλιέργεια των κυττάρων. Πριν χρησιµοποιηθεί στην καλλιέργεια, προστίθεται διάλυµα ρετινοϊκού οξέος (RA-B), σε αραίωση 1:1000. Θρεπτικό µέσο διαφοροποίησης MTEC (MTEC serum free θρεπτικό µέσο) ΜTEC basic θρεπτικό µέσο 5 µg/ml ινσουλίνη 5 µg/ml τρανσφερίνη 0,025 µg/ml cholera toxin 5 ng/ml EGF 30 µg/ml BPE 1 mg/ml BSA Το MTEC serum free θρεπτικό µέσο φιλτράρεται και φυλάσσεται στους 4 o C. Παρασκευάζεται µια µικρή ποσότητα (50 ml) και προκειµένου να διατηρηθούν όλα τα αντιδραστήρια που έχουν προστεθεί, προθερµαίνεται κάθε φορά µόνο όση ποσότητα χρειάζεται για την καλλιέργεια των κυττάρων. Πριν χρησιµοποιηθεί στην καλλιέργεια, προστίθεται διάλυµα ρετινοϊκού οξέος (RA-B), σε αραίωση 1:
77 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Ham s F-12/Pen-Strep Ham s F U/ml Pen 100µg/ml Strep Το Ham s F-12/Pen-Strep θρεπτικό µέσο φιλτράρεται και φυλάσσεται στους 4 o C. Διάλυµα Pronase (Roche) 0,15% w/v σε θρεπτικό µέσο Ham s F-12/Pen-Strep Φιλτράρεται και φυλάσσεται στους 4 o C. Κάθε φορά παρασκευάζεται φρέσκο. Διάλυµα DNase Ham s F-12/Pen-Strep 0,5 mg/ml crude pancreatic DNase I (DN25, Sigma) 1 mg/ml BSA Φιλτράρεται και φυλάσσεται σε µικρούς όγκους στους -20 o C. Rat tail type I collagen 50 µg/ml κολλαγόνο (Sigma) 0.02 Ν acetic acid (προετοιµάζεται σε WFI) Φιλτράρεται και φυλάσσεται στους 4 o C., έως 8 εβδοµάδες HBSS/BSA BSA διαλύεται σε HBSS σε συγκέντρωση 1 mg/ml Φιλτράρεται και φυλάσσεται στους 4 o C. Ρετινοϊκό οξύ (διάλυµα Α, RΑ-Α) 50 mg ρετινοϊκού οξέος (Sigma) 33,3 ml αιθανόλη (100%) Η προετοιµασία του διαλύµατος γίνεται σε σκοτάδι. Φιλτράρεται και φυλάσσεται σε µιρκούς όγκους στους -80 o C, στο σκοτάδι για 1 χρόνο. Ρετινοϊκό οξύ (διάλυµα Β, RΑ-Β) 1:10 αραίωση του RA-A σε διάλυµα HBSS/BSA Η προετοιµασία του διαλύµατος γίνεται σε σκοτάδι. Φιλτράρεται και φυλάσσεται σε µικρούς όγκους στους -20 o C, στο σκοτάδι για 1 µήνα. 77
78 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Ινσουλίνη 10 mg/ml Διαλύεται σε WFI, PH=2 (Η ρύθµιση του ph γίνεται µε glacial acetic acid). Φιλτράρεται και φυλάσσεται σε µικρούς όγκους στους -20 o C. Τρανσφερίνη 10 mg/ml Διαλύεται σε HBSS/BSA Φιλτράρεται και φυλάσσεται σε µικρούς όγκους στους -20 o C. EGF 20 µg/ml Διαλύεται σε HBSS/BSA Φιλτράρεται και φυλάσσεται σε µικρούς όγκους στους -20 o C. Cholera toxin 100 µg/ml Διαλύεται σε HBSS/BSA Φιλτράρεται και φυλάσσεται σε µικρούς όγκους στους -20 o C. Bovine pituitary extract Προετοιµασία διαλύµατος µε 7,5 mg συνολικής πρωτεΐνης Διαλύεται σε HBSS/BSA Φιλτράρεται και φυλάσσεται σε µικρούς όγκους στους -20 o C Πλασµιδιακά συστήµατα παραγωγής λεντιιών Οι λεντιιοί έχουν συνεισφέρει αποτελεσµατικά στη µεταφορά, στην ενσωµάτωση και στη σταθερή ή ελεγχόµενη έκφραση ενός γονιδίου, τόσο σε διαιρούµενα κύτταρα όσο και σε µη διαιρούµενα κύτταρα, τόσο in vitro όσο και in vivo. Η παραγωγή τους στηρίζεται στο συνδυασµό του RNA, των δοµικών πρωτεϊνών και των ενζύµων, καθώς και των πρωτεϊνών του περιβλήµατος των ιών. Τα συστατικά τους αυτά είναι διαχωρισµένα και µοιρασµένα σε ένα σύστηµα πλασµιδίων. Η παραγωγή τους επιτυγχάνεται µέσω της διαµόλυνσης κυττάρων 293Τ, ταυτόχρονα µε όλα τα πλασµίδια: ένα πλασµίδιο που κωδικοποιεί τις δοµικές πρωτεΐνες του ιού (pspax2, Addgene 12260), ένα πλασµίδιο που κωδικοποιεί τις πρωτεΐνες που σχηµατίζουν το έλυτρο του ιού (pmd2.g, Addgene 12259) και ένα πλασµίδιο που φέρει το γονίδιο που θέλουµε να µεταφέρουµε στα κύτταρα (plvdest-cag, παραχωρήθηκε από το εργαστήριο της Dr. Μ. Götz). 78
79 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Μέχρι σήµερα έχουν αναπτυχθεί 3 γενιές πλασµιδιακών συστηµάτων για την παραγωγή των λεντιιών. Στην παρούσα διατριβή χρησιµοποιήθηκαν τα πλασµιδιακά συστήµατα παραγωγής λεντιιών δεύτερης γενιάς, για τη µεταφορά γονιδίων στα επιθηλιακά κύτταρα που αποµονώνονται από την τραχεία των µυών. Η µόλυνση των συγκεκριµένων κυττάρων επιτυγχάνεται χωρίς τη συγκέντρωση των λεντιιών. Τα βήµατα που ακολουθούνται για την παραγωγή µη συγκεντρωµένων λεντιιών είναι η εξής: o Κύτταρα 293Τ επιστρώνονται σε τρυβλίο 100mm, ώστε την επόµενη ηµέρα να καλύπτουν το 80-90% της επιφάνειας του τρυβλίου. Τα κύτταρα καλλιεργούνται σε θρεπτικό µέσο DMEM που περιέχει 10% FBS. o Την επόµενη ηµέρα πραγµατοποιείται διαµόλυνση των κυττάρων µε το αντιδραστήριο Turbofect (κατιονικό πολυµερές, Fermentas) ως εξής: 1. Σε ένα eppendorf προετοιµάζεται το µείγµα του DNA που θα χρησιµοποιηθεί: 7,5 µg pspax2, 5 µg pmd2.g και 6,6 µg plvdest που περιέχει το επιθυµητό γονίδιο. 2. Σε ένα δεύτερο eppendorf προστίθενται 20 µl turbofect µε κατάλληλη ποσότητα Opti-MEM, ώστε ο τελικός όγκος (µετά την προσθήκη και του µείγµατος του DNA) να είναι 600µl. Ακολουθεί επώαση 5 λεπτά. 3. Στο µείγµα προστίθεται το DNA και ακολουθεί επώαση 20 λεπτών. o Αναρρόφηση του θρεπτικού µέσου από το τρυβλίο µε τα κύτταρα και προσθήκη 9,4 ml φρέσκου θρεπτικού µέσου DMEM, που περιέχει 10% FBS. o Στο τρυβλίο προστίθενται τα 600 µl του διαλύµατος διαµόλυνσης. o Μετά το πέρας 16 ωρών το θρεπτικό µέσο των κυττάρων που περιέχει και το διάλυµα διαµόλυνσης αναρροφάται και προστίθεται νέο θρεπτικό µέσο (8 ml DMEM που περιέχει 10% FBS). o Η καλλιέργεια συνεχίζεται για 48 ώρες. Κατά το διάστηµα αυτό εκφράζονται όλα τα απραίτητα συστατικά και παράγονται νέοι ιοί που απελευθερώνονται στο θρεπτικό µέσο. o Συλλογή του υπερκείµενου θρεπτικού µέσου του τρυβλίου, στο οποίο έχουν παραχθεί οι ιοί. o Φιλτράρισµα σε φίλτρο µε πόρους διαµέτρου 0,45 µm. o Οι ιοί φυλάσσονται σε µικρούς όγκους στους -80 o C και είναι δυνατό να αποψυχθούν και να χρησιµοποιηθούν µέχρι 3 φορές, ώστε να µην ελαττώνεται σε µεγάλο βαθµό η βιωσιµότητά τους Μόλυνση 293Τ κυττάρων για τιτλοδότηση των παραγόµενων λεντιιών Για την τιτλοδότηση των παραγόµενων λεντιιών, κύτταρα 293Τ µολύνονται µε ελαττούµενες ποσότητες του µη συγκεντρωµένου ιού (100 µl, 40 µl και 20 µl). Οι παραγόµενοι ιοί εκφράζουν είτε την πράσινη φθορίζουσα πρωτεΐνη (GFP) µόνη της, είτε κάποιο γονίδιο συντηγµένο µε GFP. Έτσι, µέσω ανοσοφθορισµού, επιτρέπεται η τιτλοδότηση των παραγόµενων λεντιιών. 79
80 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Σε τρυβλίο 24-well τοποθετούνται καλυπτρίδες που έχουν κατεργαστεί µε poly-l-lysine. o Σε κάθε καλυπτρίδα τοποθετούνται: 10 5 κύτταρα 293Τ, 100 µl, 40 µl ή 20 µl ιού polybrene (Hexadimethrine bromide, Sigma, H9268) θρεπτικό µέσο DMEM που περιέχει 10% FBS, σε τελικό όγκο 500 µl o Μετά από 24 ώρες πραγµατοποιείται αλλαγή του θρεπτικού µέσου και τα κύτταρα καλλιεργούνται για 48 ώρες ακόµα. o Μονιµοποίηση των κυττάρων και ανοσοφθορισµός µε αντίσωµα έναντι της πρωτεΐνης GFP, για ανίχνευση των κυττάρων που έχουν µολυνθεί. o Σε µικροσκόπιο φθορισµού µετράται ο αριθµός των αποικιών που εκφράζουν την πρωτεΐνη GFP. Για µεγαλύτερη ακρίβεια η µέτρηση πραγµατοποιείται στην καλυπτρίδα εκείνη που παρατηρούνται 20<θετικές αποικίες κυττάρων<200. o Ο αριθµός των ιών που προσβάλλει τα κύτταρα (infectious units/ml) υπολογίζεται ως εξής: (Αριθµός GFP + αποικιών/ποσότητα ιού που χρησιµοποιήθηκε) x 1000 Λεντιιοί µε infectious units/ml, συνήθως µολύνουν τα MTEC σε ποσοστό 50-60% Επίστρωση καλυπτρίδων µε poly-l-lysine Για την επίστρωση καλυπτρίδων µε poly-l-lysine ακολουθούνται τα εξής βήµατα: o Οι καλυπτρίδες (διαµέτρου 13 mm, thickness No. 1, VWR) που πρόκειται να χρησιµοποιηθούν αποστειρώνονται από την προηγούµενη ηµέρα σε ξηρή αποστείρωση. o Προετοιµασία 10 ml διαλύµατος της poly-l-lysine, ώστε να έχει τελική συγκέντρωση 100 µg/ml. o Το διάλυµα της poly-l-lysine, µαζί µε τις καλυπτρίδες που θα χρησιµοποιηθούν, τοποθετούνται σε τρυβλίο 100 mm και γίνεται επώαση στους 37 o C, για 15 λεπτά. o Έκπλυση των καλυπτρίδων µε WFI. o Ξήρανση και φύλαξη σε αποστειρωµένο τρυβλίο 100 mm. Υλικά και διαλύµατα Διάλυµα poly-l-lysine 10 µl poly-l-lysine, από stock 100 mg/ml (Sigma) 10 ml WFI 80
81 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Το διάλυµα φιλτράρεται και χρησιµοποιείται άµεσα Μόλυνση των επιθηλιακών κυττάρων τραχείας µυών µε λεντιιούς Η χρήση των λεντιιών συνεισφέρει αποτελεσµατικά στη µεταφορά γονιδίων στις ex vivo καλλιέργειες των επιθηλιακών κυττάρων της τραχείας των µυών. Τα κύτταρα µπορούν να µολυνθούν σε οποιαδήποτε φάση της καλλιέργειας. Ωστόσο, η µόλυνση είναι πιο αποτελεσµατική κατά το στάδιο του πολλαπλασιασµού τους. Η µόλυνση των κυττάρων περιλαµβάνει δυο απαραίτητα βήµατα: α) την επώαση των κυττάρων µε διάλυµα EGTA, το οποίο καταστρέφει τους συνδέσµους µεταξύ των επιθηλιακών κυττάρων, για να γίνουν πιο δεκτικά και 2) τη φυγοκέντρηση των µεµβρανών πάνω στις οποίες καλλιεργούνται τα κύτταρα, γεγονός που θα βοηθήσει την προσκόλληση των ιών πάνω στα κύτταρα. Η µόλυνση των κυττάρων πραγµατοποιείται 2 ηµέρες πριν την επαγωγή της διαφοροποίησης (AD -2). Η διαδικασία που ακολουθείται είναι η εξής: o Έκπλυση των µεµβρανών µε PBS (1x) ph 7,4, 2 φορές,. o Προσθήκη 12 mm EGTA (σε 10 mm HEPES, ph 7,4) στην κορυφαία πλευρά της κάθε µεµβράνης. Στη βασική τους πλευρά προστίθενται 500 µl MTEC plus θρεπτικού µέσου, που περιέχει RA-B, σε αραίωση 1:1000. Επώαση για 25 λεπτά, στον επωαστή θερµοκρασίας 37 o C. o Κατά τη διάρκεια της επώασης προετοιµάζεται το µείγµα των λεντιιών µε του οποίους θα µολυνθούν τα κύτταρα. Για κάθε µόλυνση απαιτούνται: 50 µl µη συγκεντρωµένου λεντιιού polybrene, τελικής συγκέντρωσης 5 µg/ml MTEC plus που περιέχει RA-B (αραίωση 1:1000), µέχρι τελικό όγκο 150 µl. o Αναρρόφηση του διαλύµατος EGTA και του θρεπτικού µέσου από την κορυφαία και τη βασική πλευρά της µεµβράνης, αντίστοιχα. Έκπλυση των µεµβρανών µε PBS (1x), 2 φορές,. o Προσθήκη 500 µl MTEC plus θρεπτικού µέσου (που περιέχει RA-B, 1:1000) στη βασική πλευρά της κάθε µεµβράνης. Προσθήκη του µείγµατος που περιέχει τον λεντιιό στην κορυφαία πλευρά της µεµβράνης. o Το τρυβλίο των 24-θέσεων που περιέχει τις µεµβράνες µε τα καλλιεργούµενα κύτταρα κλείνεται καλά µε parafilm και ακολουθεί φυγοκέντρηση (plate-spin) σε 1300 g, για 80 λεπτά. o Αφαίρεση του parafilm και επώαση των κυττάρων στον επωαστικό θάλαµο θερµοκρασίας 37 o C, για 24 ώρες. o Την επόµενη ηµέρα, το θρεπτικό µέσο αναρροφάται και από τις δύο πλευρές της µεµβράνης και προστίθεται φρέσκο MTEC plus θρεπτικό µέσο (που περιέχει RA-B, 1:1000). Στην κορυφαία πλευρά της κάθε µεµβράνης τοποθετούνται 150 µl MTEC plus θρεπτικού µέσου, ενώ στη βασική της πλευρά τοποθετούνται 500 µl. 81
82 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o o o o 48 ώρες µετά τη µόλυνση, επάγεται η διαφοροποίηση των κυττάρων. Αναρρόφηση του MTEC plus θρεπτικού µέσου από την κορυφαία και βασική πλευρά της µεµβράνης. Τοποθέτηση 500 µl MTEC serum free θρεπτικού µέσου, µόνο στη βασική πλευρά της κάθε µεµβράνης καλλιέργειας. Ανά δυο ηµέρες πραγµατοποιείται αλλαγή θρεπτικού µέσου, µόνο στη βασική πλευρά της κάθε µεµβράνης καλλιέργειας. Η καλλιέργεια διατηρείται για όσο διάστηµα απαιτείται σύµφωνα µε την πειραµατική διαδικασία που ακολουθείται. Υλικά και διαλύµατα Διάλυµα EGTA (150 µl/µεµβράνη) 12 mm EGTA (Sigma) 10 mm Hepes (Gibco) WFI (water for injection) έως τελικό όγκο 150 µl Ρύθµιση του ph=7,4 Φιλτράρεται και χρησιµοποιείται αµέσως στα κύτταρα. Polybrene (5 µg/ml) To stock polybrene συγκέντρωσης 8 mg/ml αραιώνεται 100x 9,4 µl από το αραιωµένο polybrene αναµειγνύονται µε 50 µl ιού και 90,6 µl MTEC plus θρεπτικού µέσου. 82
83 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.3 Δοκιµασία λουσιφεράσης Η δοκιµασία λουσιφεράσης (luciferase assay) χρησιµοποιείται για τη µελέτη της γονιδιακής έκφρασης. Η µέθοδος αυτή µπορεί να χρησιµοποιηθεί για το χαρακτηρισµό νέων άγνωστων υποκινητών-ρυθµιστικών στοιχείων ή την ποσοτικοποίηση της δράσης γνωστών υποκινητών ή των επιµέρους λειτουργικών στοιχείων τους. Ακόµα, µπορεί να χρησιµοποιηθεί και για τη µελέτη της ικανότητας διαφόρων παραγόντων να επάγουν τη µεταγραφή µέσω συγκεκριµένων ρυθµιστικών στοιχείων. Για το σκοπό αυτό οι υποκινητές-ρυθµιστικά στοιχεία των υπό µελέτη γονιδίων κλωνοποιούνται σε ένα πλασµίδιο ανοδικά από µια κασέτα αναφοράς, που περιέχει το γονίδιο της λουσιφεράσης. Στην παρούσα µελέτη χρησιµοποιήθηκε το Dual-Luciferase Reporter Assay Kit (Promega), που επιτρέπει την ταυτόχρονη ανίχνευση και µέτρηση δυο διαφορετικών ενζύµων αναφοράς, της Firefly και της Renilla λουσιφεράσης. Τα δυο γονίδια που κωδικοποιούν τα παραπάνω ένζυµα βρίσκονται σε δυο διαφορετικά πλασµίδια. Το γονίδιο της Firefly λουσιφεράσης χρησιµοποιείται για τη µέτρηση της µεταγραφικής ενεργότητας του ρυθµιστικού στοιχείου που είναι κλωνοποιηµένο ανοδικά αυτού. Το γονίδιο της Renilla λουσιφεράσης χρησιµοποιείται για την κανονικοποίηση των τιµών της εκπεµπόµενης βιοφωταύγειας, αφού η έκφρασή του επηρεάζεται από τεχνικές παραµέτρους του πειράµατος, όπως τον αριθµό των κυττάρων και την επιτυχία της διαµόλυνσης. Σε κάθε δείγµα µετράται η εκπεµπόµενη βιοφωταύγεια του κάθε ενζύµου. Oι µονάδες της εκπεµπόµενης βιοφωταύγειας της Firefly λουσιφεράσης κανονικοποιoύνται ως προς τις τιµές που προκύπτουν από τη βιοφωταύγεια της Renilla λουσιφεράσης (Firefly/Renilla). Έτσι, επιτρέπεται η σύγκριση της µεταγραφικής ενεργότητας ανάµεσα στις διαφορετικές συνθήκες. Τα ρυθµιστικά στοιχεία που xρησιµοποιήθηκαν στην παρούσα µελέτη και κλωνοποιήθηκαν στο πλασµίδιο pgl3 basic vector (Promega) ανοδικά του γονιδίου της λουσιφεράσης είναι τα εξής: Πίνακας 3.4 Τα ρυθµιστικά στοιχεία των γονιδίων MCIDAS, FOXJ1 και GEMC1 Νουκλεοτίδια (σε σχέση µε τη θέση Ρυθµιστικά στοιχεία έναρξης της µεταγραφής, TSS) -767 έως +233 pgl3-mcidas hg19, chr5: pgl3-foxj1 pgl3-gemc to +143 (TSS1) hg19, chr17: to +837 (TSS2) hg19, chr17: *Υπάρχουν δυο διαφορετικές πιθανές θέσεις έναρξης της µεταγραφής -535 to hg19, chr3:
84 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Οι παράγοντες που µελετήθηκαν για την ικανότητά τους να επάγουν τη µεταγραφή ήταν οι mgemc1, mmcidas και hgeminin, που είναι κλωνοποιηµένοι στο πλασµίδιο pcaggs, καθώς και οι he2f1, he2f4 και he2f5, που είναι κλωνοποιηµένοι στο πλασµίδιο pcmv (βλέπε ενότητα: Λίστα πλασµιδίων). o o Η πορεία που ακολουθήθηκε είναι η εξής: Κύτταρα 293T (3,5 x 10 4 κύτταρα) επιστρώνονται σε τρυβλίο 96-well. Την επόµενη ηµέρα πραγµατοποιείται διαµόλυνση των κυττάρων µε συνολική ποσότητα DNA 130 ng, µε τη χρήση του αντιδραστηρίου Turbofect. Οι ποσότητες του DNA που προστίθενται αναλύονται παρακάτω: pgl3 πλασµίδιο (pgl3-mcidas/pgl3-foxj1/pgl3-gemc1) Φορείς έκφρασης (pcaggs-gfp, pcaggs-gemc1, pcaggs- McIdas, pcaggs-geminin, pcmv-neo-bam, pcmv-e2f5, pcmv-e2f4, pcmv-e2f1) Renilla luciferase phrl-sv40 πλασµίδιο (Promega) pbluescript 25 ng 50 ng 5 ng Συµπληρώνεται όπου χρειάζεται µέχρι τελικής ποσότητας DNA=130 ng 1. Σε ένα eppendorf προετοιµάζεται το µείγµα του DNA που θα χρησιµοποιηθεί. 2. Σε ένα δεύτερο eppendorf προστίθενται 0,5 µl turbofect µε κατάλληλη ποσότητα Opti-MEM, ώστε ο τελικός όγκος µετά την προσθήκη και του µείγµατος του DNA να είναι 25 µl. Ακολουθεί επώαση 5 λεπτά. 3. Στο µείγµα προστίθεται το DNA και ακολουθεί επώαση 20 λεπτών. o Αναρρόφηση του θρεπτικού µέσου από το τρυβλίο µε τα κύτταρα και προσθήκη 37,5 µl φρέσκου θρεπτικού µέσου DMEM, που περιέχει 10% FBS. o Στο τρυβλίο προστίθενται τα 25 µl του διαλύµατος διαµόλυνσης. o Μετά από 5 ώρες προστίθενται επιπλέον 62,5 µl θρεπτικού µέσου (DMEM που περιέχει 10% FBS). o Την επόµενη ηµέρα γίνεται αναρρόφηση του θρεπτικού µέσου και προστίθενται 125 µl φρέσκου θρεπτικού µέσου. Η καλλιέργεια συνεχίζεται για επιπλέον 24 ώρες. o 48 ώρες συνολικά µέτα τη διαµόλυνση γίνεται η µέτρηση της βιοφωταύγειας µε τη χρήση του Dual-Luciferase Reporter Assay Kit (Promega), σύµφωνα µε τις οδηγίες της εταιρείας. Τα πειράµατα αποσιώπησης του γονιδίου McIdas παράλληλα µε τη δοκιµασία λουσιφεράσης, έγιναν µε τη χρήση του λιπόφιλου αντιδραστηρίου lipofectamine 2000 (Invitrogen). Δυο sirna oλογονουκλεοτίδια (STEALTH sirnas, Invitrogen) 84
85 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ χρησιµοποιήθηκαν για την αποσιώπηση του γονιδίου McIdas (Pefani et al, 2011): 5 - CCACCAAACGGAAGCAGACTTCAAT-3 και 5 -CCAAACAGGAACGGACATTCCCAAT-3. Ως αρνητικός µάρτυρας χρησιµοποιήθηκε το ολιγονουκλεοτίδιο: 5 - CCAAACAGGAACGGACATTCCCAAT-3. Η πορεία που ακολουθήθηκε είναι η εξής: o Κύτταρα 293T (3,5 x 10 4 κύτταρα) επιστρώνονται σε τρυβλίο 96-well. o Την επόµενη ηµέρα πραγµατοποιείται διαµόλυνση των κυττάρων µε συνολική ποσότητα DNA 130 ng. Η διαµόλυνση γίνεται µε τη χρήση του πλασµιδίου pgl3-foxj1, παράλληλα µε το φορέα έκφρασης pcaggs-gemc1 ή pcaggs- GFP, ως δείγµα-µάρτυρας. Σε κάθε δείγµα προστίθενται τα McIdas sirnas (9 nm από το καθένα) ή 18 nm από το sirna που αναγνωρίζει µια τυχαία αλληλουχία. 1. Σε ένα eppendorf προετοιµάζεται το µείγµα DNA και sirna ολιγονουκλεοτιδίων που θα χρησιµοποιηθούν. Προστίθεται Opti-MEM, ώστε ο τελικός όγκος να είναι 16 µl. Ακολουθεί επώαση 5 λεπτά. Μείγµα DNA/siRNAs Ποσότητα pgl3-foxj1 pcaggs-gfp phrl-sv40 control sirna pgl3-foxj1 pcaggs-gfp phrl-sv40 McIdas sirna pgl3-foxj1 pcaggs-gemc1 phrl-sv40 control sirna pgl3-foxj1 pcaggs-gemc1 phrl-sv40 McIdas sirna 25 ng 50 ng 5 ng 18 nm 25 ng 50 ng 5 ng 18 nm 25 ng 50 ng 5 ng 18 nm 25 ng 50 ng 5 ng 18 nm 2. Σε ένα δεύτερο eppendorf προστίθενται 0,5 µl lipofectamin 2000 µε κατάλληλη ποσότητα Opti-MEM, ώστε ο τελικός όγκος να είναι 16 µl. Ακολουθεί επώαση 5 λεπτά. 3. Τα δυο παραπάνω eppendorf ενώνονται σε ένα κοινό (master mix). Το µείγµα επωάζεται για 20 λεπτά. o Αναρρόφηση του θρεπτικού µέσου από το τρυβλίο µε τα κύτταρα και προσθήκη 32 µl φρέσκου θρεπτικού µέσου DMEM, που περιέχει 10% FBS. o Στο τρυβλίο προστίθενται τα 32 µl του διαλύµατος διαµόλυνσης. 85
86 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Μετά το πέρας 5 ωρών προστίθενται επιπλέον 61 µl θρεπτικού µέσου (DMEM που περιέχει 10% FBS). o 24 ώρες µετά πραγµατοποιείται µια ακόµα διαµόλυνση των κυττάρων µόνο µε τα sirna oλιγονουκλεοτίδια, ακολουθώντας τις ίδιες συνθήκες µε την πρώτη διαµόλυνση. o Η καλλιέργεια συνεχίζεται για επιπλέον 24 ώρες. o 24 ώρες µετά τη δεύτερη διαµόλυνση τα κύτταρα παραλαµβάνονται και πραγµατοποιείται η µέτρηση της βιοφωταύγειας µε τη χρήση του Dual- Luciferase Reporter Assay Kit (Promega), σύµφωνα µε τις οδηγίες της εταιρείας. Παράλληλα, παραλαµβάνονται δείγµατα και για qpcr, ώστε να ελεγχθεί η αποτελεσµατικότητα της αποσιώπησης του γονιδίου McIdas. 86
87 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.4 Μέθοδοι αποµόνωσης ιστών Αποµόνωση ιστών εµβρύων και νεογέννητων µυών Οι µύες που χρησιµοποιήθηκαν στην παρούσα εργασία προέρχονταν από συγχρονισµένες εγκυµοσύνες µυών Parkes και ως ηµέρα Ε 0.5 ορίζουµε την ηµέρα παρατήρησης του παχύρευστου λευκού εκκρίµατος (vaginal plug). Στην παρούσα διατριβή αποµονώθηκαν ιστοί από έµβρυα µυών αγρίου τύπου. Αποµονώθηκε το χοριοειδές πλέγµα από τις πλάγιες και την τέταρτη κοιλία του εγκεφάλου από έµβρυα µυών ηλικίας 12.5 (Ε12.5), 14.5 (Ε14.5) και 18.5 (Ε18.5) ηµερών, καθώς και οι βρόγχοι από έµβρυα ηλικίας 14.5 (Ε14.5) και 18.5 (Ε18.5) ηµερών. Οι παραπάνω ιστοί χρησιµοποιήθηκαν για την αποµόνωση RNA και τη µετέπειτα ανάλυση της έκφρασης των γονιδίων GemC1, McIdas, Geminin και Foxj1. Επίσης, από τους GemC1 WT και GemC1 ΚΟ µυς αποµονώθηκαν τραχείες είτε από έµβρυα ηλικίας 16.5 ηµερών (E18.5), είτε από νεογέννητους µυς (P0). Οι ιστοί αυτοί χρησιµοποιήθηκαν για αποµόνωση RNA και για πειράµατα ανοσοφθορισµού, αντίστοιχα. Το αρχικό στάδιο που αφορά την αναισθητοποίηση των µυών για την αποµόνωση των ιστών εµβρύων ή νεογέννητων µυών διαφέρει. Η διαδικασία που ακολουθήθηκε για την αποµόνωση των εµβρύων ήταν η εξής: o Αναισθητοποίηση των έγκυων θηλυκών µυών µε διοξείδιο του άνθρακα. o Πραγµατοποίηση αυχενικής µετατόπισης για τη θανάτωση των µυών. o Πραγµατοποίηση τοµής κατά µήκος της κοιλιακής χώρας των µυών. o Αποµάκρυνση των εµβρύων από τις ωοθήκες και τοποθέτησή τους σε κρύο διάλυµα PBS (1x). o Αφαίρεση των εµβρυϊκών µεµβρανών. Η διαδικασία που ακολουθήθηκε για την αναισθητοποίηση των νεογέννητων µυών ήταν η εξής: o Αναισθητοποίηση των νεογέννητων µυών µε ισοφλουράνιο (Isofluran, 1ml/ml, CP-Pharma). Τα βήµατα που ακολουθούνται στη συνέχεια είναι κοινά: o Με τη βοήθεια στερεοσκοπίου (στους νεογέννητους µυς δεν είναι απαραίτητη η χρήση του) πραγµατοποιείται διατοµή του δέρµατος στη θωρακική χώρα κάθε µυός. o Με δυο επιµήκεις πλάγιες τοµές ανοίγεται ο θώρακας, ώστε να αποκαλυφθούν τα εσωτερικά όργανα. o Η τοµή εκτείνεται µέχρι το κεφάλι, έως ότου να αποκαλυφθεί ο λαιµός. o Με προσοχή αποµακρύνονται οι παρακείµενοι ιστοί, ώστε να αποκαλυφθούν η τραχεία και οι βρόγχοι, οι οποίοι αποµονώνονται προσεκτικά και τοποθετούνται αµέσως σε eppendorf που βρίσκεται µέσα σε πάγο. o Για την αποµόνωση του χοριοειδούς πλέγµατος των κοιλιών του εγκεφάλου αποκεφαλίζεται το έµβρυο µε τη βοήθεια λαβίδων. 87
88 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Αποµάκρυνση του δέρµατος και του κρανίου για την αποµόνωση του εγκεφάλου. o Αποµόνωση του χοριοειδούς πλέγµατος από τις πλάγιες και την τέταρτη κοιλία του εγκεφάλου. o Άµεση τοποθέτηση των ιστών σε eppendorf που βρίσκεται µέσα σε πάγο. o Στους ιστούς που πρόκειται να πραγµατοποιηθεί αποµόνωση του RNA προστίθεται κατάλληλη ποσότητα διαλύµατος λύσης (η διαδικασία αποµόνωσης RNA περιγράφεται παρακάτω). Οι τραχείες που προέρχονται από τους GemC1 WT/WT και GemC1 ΚΟ/ΚΟ νεογέννητους µυς (P0) χρησιµοποιήθηκαν σε πειράµατα ανοσοφθορισµού και έτσι η διαδικασία συνεχίζεται ως εξής: o Έκπλυση των τραχειών µε κρύο PBS (1x), 3 φορές. o Επώαση µε µονιµοποιητικό διάλυµα PFA 4% στους 4 o C, για 16 ώρες. o Έκπλυση των τραχειών µε κρύο PBS (1x), 3 φορές. o Τοποθέτηση των τραχειών σε διάλυµα σουκρόζης 30% στους 4 o C, µέχρι να βυθιστούν. o Πάγωµα των τραχειών σε ειδικό µέσο (Tissue-Tek O.C.T) και διατήρηση στους -80 o C, µέχρι να κοπούν σε τοµές. o Οι τραχείες κόβονται σε εγκάρσιες τοµές πάχους 10 µm µε τη βοήθεια µικροτόµου, σε θερµοκρασία -24 o C. o Αποθήκευση των τοµών στους -80 o C. Υλικά και διαλύµατα PFA 4% 20 gr PFA (Sigma) PBS (1x) µέχρι τελικό όγκο 500 ml Διάλυµα σουκρόζης 30 gr σουκρόζης (Sigma) PBS (1x) µέχρι τελικό όγκο 100 ml Αποµόνωση ιστών ενήλικων µυών Από ενήλικες θηλυκούς µυς αποµονώθηκαν οι εξής ιστοί: το χοριοειδές πλέγµα από τις πλάγιες και την τέταρτη κοιλία του εγκεφάλου, οι βρόγχοι, οι ωοθήκες µαζί µε τους ωαγωγούς και το ήπαρ. Οι παραπάνω ιστοί χρησιµοποιήθηκαν για την αποµόνωση RNA και τη µετέπειτα ανάλυση της έκφρασης των γονιδίων GemC1, McIdas, Geminin και Foxj1. Η διαδικασία που ακολουθήθηκε για την αποµόνωση των ιστών ήταν η εξής: 88
89 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o o o o o o o o o o Αναισθητοποίηση των ενήλικων µυών µε διοξείδιο του άνθρακα. Ψεκασµός των µυών µε αιθανόλη 70%, ώστε να εξασφαλίζονται όσο το δυνατό πιο στείρες συνθήκες. Η πλήρης αναισθησία ελέγχεται µε την έλλειψη αντανακλαστικών στα κάτω άκρα. Μετά την επίτευξη της αναισθησίας πραγµατοποιείται τοµή του δέρµατος κατά µήκος της κοιλιακής και θωρακικής χώρας των µυών. Αποµακρύνονται οι παρακείµενοι ιστοί της κοιλιακής χώρας και αφαιρούνται οι ωοθήκες µε τους ωαγωγούς. Με ψαλίδι ξεκινώντας από το στέρνο ανοίγεται, µε δυο επιµήκεις πλάγιες τοµές, ο θώρακας, έτσι ώστε να αποκαλυφθούν τα εσωτερικά όργανα. Αποµόνωση των βρόγχων και άµεση τοποθέτηση σε eppendorf που βρίσκεται µέσα σε πάγο. Για την αποµόνωση του χοριοειδούς πλέγµατος των κοιλιών του εγκεφάλου, αποκεφαλίζεται ο µυς. Αποµάκρυνση του δέρµατος και του κρανίου για την αποµόνωση του εγκεφάλου. Αποµόνωση του χοριοειδούς πλέγµατος από τις πλάγιες και την τέταρτη κοιλία του εγκεφάλου. Άµεση τοποθέτηση των ιστών σε eppendorf που βρίσκεται µέσα σε πάγο. Στους ιστούς που πρόκειται να πραγµατοποιηθεί αποµόνωση του RNA προστίθεται κατάλληλη ποσότητα διαλύµατος λύσης (η διαδικασία αποµόνωσης RNA περιγράφεται παρακάτω). 89
90 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.5 Προετοιµασία δειγµάτων για Real-time PCR Τα βήµατα που ακολουθούνται για την προετοιµασία δειγµάτων για real-time PCR είναι τα εξής: 1. Αποµόνωση ολικού RNA από το επιθυµητό δείγµα, µε τη χρήση του κατάλληλου πρωτοκόλλου. 2. Παραγωγή cdna. 3. qpcr Αποµόνωση RNA από ιστούς εµβρύων µυών Η αποµόνωση του RNA από βρόγχους εµβρύων ηλικίας 14.5 (Ε14.5) και 18.5 (Ε18.5) ηµερών έγινε µε τη χρήση Trizol. Σε κάθε ιστό προστίθεται 1 ml Trizol (εξαρτάται από το µέγεθος του ιστού και την επιτυχή ο µογενοποίησή του). Η διαδικασία που ακολουθείται είναι η εξής: o Σε κάθε δείγµα προστίθενται 0,2 ml xλωροφόρµιου ανά 1 ml Trizol. Καλή ανάδευση για 15 δευτερόλεπτα. o Επώαση για 2-3 λεπτά, σε θερµοκρασία δωµατίου. o Φυγοκέντρηση στις rpm στους 4 o C, για 15 λεπτά. o Συλλογή της ανώτερης υδατικής φάσης που περιέχει το RNA. o Προσθήκη ισοπροπανόλης για την κατακρήµνιση του RNA (0,5 ml ισοπροπανόλης ανά 1 ml Trizol). o Επώαση -20 o C, για 16 ώρες. o Φυγοκέντρηση στις rpm στους 4 o C, για 10 λεπτά. o Αποµάκρυνση του υπερκείµενου. o Προσθήκη αιθανόλης 75% (1ml αιθανόλης ανά 1 ml Trizol). o Ανάµειξη µε vortex. o Φυγοκέντρηση στις rpm στους 4 o C, για 5 λεπτά. o Ξήρανση του ιζήµατος του RNA (στον αέρα ή υπό κενό για 5-10 λεπτά). o Επαναδιάλυση σε 20 µl RNase free H 2 O. Η αποµόνωση του RNA από το χοριοειδές πλέγµα του εγκεφάλου εµβρύων µυών πραγµατοποιήθηκε µε τη χρήση του RNeasy micro kit (Qiagen). Η αρχική λύση πραγµατοποιείται µε 350 µl διαλύµατος RLT. Η αποµόνωση συνεχίζεται σύµφωνα µε τις οδηγίες της εταιρείας. Η αποµόνωση του RNA από τις τραχείες των GemC1 WT/WT, GemC1 WT/ΚΟ και GemC1 ΚΟ/ΚΟ Ε16.5 µυών πραγµατοποιήθηκε µε το RNeasy Lipid Tissue Mini kit (Qiagen). Η αρχική λύση πραγµατοποιείται µε 100 µl Qiazol/Ε16.5 τραχεία. Η αποµόνωση συνεχίζεται σύµφωνα µε τις οδηγίες της εταιρείας. 90
91 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Αποµόνωση RNA από ιστούς ενήλικων µυών Η αποµόνωση του RNA από βρόγχους, ήπαρ και ωοθήκες/ωαγωγούς ενήλικων µυών έγινε µε τη χρήση Trizol, σύµφωνα µε το πρωτόκολλο που περιγράφεται παραπάνω (υποκεφάλαιο 3.5.1). Σε κάθε ιστό προστίθενται 1-2 ml Trizol (εξαρτάται από το µέγεθος του ιστού και την επιτυχή οµογενοποίησή του). Η αποµόνωση του RNA από το χοριοειδές πλέγµα των ενήλικων µυών έγινε µε τη χρήση του NucleoSpin RNA II kit (Macherey-Nagel). Η αρχική λύση πραγµατοποιείται µε 300 µl διαλύµατος RA1 και 3 µl β-µερκαπτοαιθανόλη (β-με) (αραίωση 1:100). Η αποµόνωση συνεχίζεται σύµφωνα µε τις οδηγίες της εταιρείας Αποµόνωση RNA από MTEC Η αποµόνωση του RNA από τα MTEC πραγµατοποιείται σε διαφορετικές χρονικές στιγµές, πριν ή µετά την επαγωγή της διαφοροποίησης. Για την αποµόνωση επαρκούς ποσότητας RNA, απαιτείται η ένωση δυο µεµβρανών για κάθε χρονική στιγµή (time point) που µελετάται. Ανεξαρτήτως του σταδίου στο οποίο βρίσκονται τα κύτταρα (κατά το στάδιο του πολλαπλασιασµού ή της διαφοροποίησης), το πρωτόκολλο που ακολουθείται είναι το εξής: o Αναρρόφηση του θρεπτικού µέσου καλλιέργειας των κυττάρων. o Στην κορυφαία επιφάνεια της µεµβράνης καλλιέργειας προστίθενται 200 µl Trizol. Καλή επαναδιάλυση των κυττάρων µε την πιπέτα, ώστε να ανασηκωθούν και να σπάσουν όλα τα κύτταρα από τη µεµβράνη. o Σε κάθε δείγµα προστίθενται 40 µl xλωροφόρµιο (στα 200 µl Trizol). Καλή ανάδευση για 15 δευτερόλεπτα. o Επώαση σε θερµοκρασία δωµατίου, για 2-3 λεπτά. o Φυγοκέντρηση στις rpm στους 4 o C, για 15 λεπτά. o Συλλογή της ανώτερης φάσης. Στο σηµείο αυτό ενώνονται τα ίδια δείγµατα (2 δείγµατα/χρονική στιγµή που εξετάζεται). o Προσθήκη ίσου όγκου αιθανόλης (70%). o Μεταφορά των δειγµάτων σε κολώνες (RNeasy MinElute spin column). Η αποµόνωση συνεχίζεται µε τη χρήση του RNeasy Micro kit (Qiagen), σύµφωνα µε τις οδηγίες της εταιρείας Αποµόνωση RNA από 293T Η αποµόνωση του RNA από τα 293Τ κύτταρα πραγµατοποιήθηκε µε τη χρήση του NucleoSpin RNA II kit (Macherey-Nagel). Η αρχική λύση πραγµατοποιείται σε 300 µl διαλύµατος RA1 και 3 µl β-µερκαπτοαιθανόλη (β-με) (αραίωση 1:100). Η αποµόνωση συνεχίζεται σύµφωνα µε τις οδηγίες της εταιρείας. 91
92 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Σύνθεση cdna Η σύνθεση του cdna γίνεται µε τη χρήση της M-MLV αντίστροφης µεταγραφάσης (Invitrogen) και ως εκκινητές χρησιµοποιούνται Oligo(dT) (Invitrogen). Κατά την προετοιµασία των δειγµάτων για τη σύνθεση του cdna, προετοιµάζεται και ένα δείγµα-µάρτυρας στο οποίο δεν προστίθεται η αντίστροφη µεταγραφάση (-RT). Το δείγµα αυτό χρησιµοποιείται στην qpcr για την εύρεση πιθανής επιµόλυνσης του δείγµατος από γενωµικό DNA. Η ποσότητα του RNA που χρησιµοποιείται είναι συνήθως 1 µg/αντίδραση. Ωστόσο, αυτή µπορεί να ποικίλει από 0,7-1 µg, ανάλογα µε τον ιστό/κύτταρα και την ποσότητα του RNA που είναι δυνατό να αποµονωθεί. Η διαδικασία που ακολουθείται για την παρασκευή του cdna είναι η εξής: o Για κάθε αντίδραση απαιτείται: Αντιδραστήρια Ποσότητα Oligo(dT) (0,5 µg/µl) 1 µl RNA 1 µg dntps (10 mm) 1 µl WFI Μέχρι τελικού όγκου 12 µl o Θέρµανση στους 65 o C, για 5 λεπτά. o Επώαση των δειγµάτων σε πάγο για 3 λεπτά. o Σε κάθε αντίδραση προστίθεται: Αντιδραστήρια Ποσότητα Ρυθµιστικό διάλυµα 5x 4 µl DTT (0,1 M) 2 µl RNase out (40 U/µl) 1 µl o Επώαση στους 37 o C, για 2 λεπτά. o Σε κάθε δείγµα προστίθεται 1 µl M-MLV. Στο δείγµα-µάρτυρας (-RT) προστίθεται 1 µl WFI. o Επώαση στους 37 o C, για 50 λεπτά. o Επώαση στους 70 o C για 15 λεπτά, ώστε να τερµατιστεί η αντίδραση. o Τα δείγµατα φυλάσσονται στους -20 o C Real-time PCR Στην παρούσα µελέτη προσδιορίστηκαν τα σχετικά επίπεδα έκφρασης µιας σειράς γονιδίων, µέσω ποσοτικής Real-Time PCR και µε τη χρήση ειδικών εκκινητών. Οι αντιδράσεις πραγµατοποιήθηκαν στο µηχάνηµα Step One, της Applied Biosystems, µε τη χρήση του αντιδραστηρίου KAPA SYBR FAST qpcr kit (KapaBiosystems), σύµφωνα µε τις οδηγίες του κατασκευαστή. Τα επίπεδα έκφρασης των γονιδίων κανονικοποιήθηκαν µε τα γονίδια HPRT (για τα δείγµατα που προέρχονταν από τους 92
93 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ µυς) ή YWHAZ (για τα δείγµατα που προέρχονταν από τα 293Τ κύτταρα). Η ανάλυση των αποτελεσµάτων έγινε µε τη χρήση του προγράµµατος REST-MCS beta. Για κάθε αντίδραση απαιτούνται τα εξής: Αντιδραστήρια Ποσότητα Kapa Master Mix (2x) 5 µl Fw εκκινητής (20 µμ) 0,15 µl Rv εκκινητής (20 µm) 0,15 µl cdna 45 ng WFI Μέχρι τελικού όγκου 10 µl Οι εκκινητές που χρησιµοποιήθηκαν συνολικά ήταν οι εξής: Πίνακας 3.4 Εκκινητές που χρησιµοποιήθηκαν κατά την qpcr mgemc1_fw 5 -TGGTCTCCTGGACAACACTG-3 mgemc1_rv 5 -TAACTCAGAGGGCGATTCCA-3 mmcidas_fw 5 -AACCGAAGCGTCTCCTAGTG-3 mmcidas_rv 5 -GGTCATCCATTGCATCTCTG-3 mfoxj1_fw 5 -GGCCACCAAGATCACTCTGT-3 mfoxj1_rv 5 -TGTTCAAGGACAGGTTGTGG-3 mgeminin_fw 5 -CTCTCTACGAAGCGCTGAAA-3 mgeminin_rv 5 -AGCTCCTGAGTCTTCCAGTT-3 mjagged 1_Fw 5 -TGGACTGGCCCCACGTGTTC-3 mjagged 1_Rv 5 -GGGCGGGCACACACACTTGAA-3 mjagged 2_Fw 5 -ACCCGGGCCTCGTCGTCAT-3 mjagged 2_Rv 5 -TGCAGGCTCTTCCAGCGGTC-3 mtap63_fw 5 -GTGCGACAAACAAGATTGAGATTAG-3 mtap63_rv 5 -TGTTCAGGAGCCCCAGGTT-3 mtap73_fw 5 -GAGCACCTGTGGAGTTCTCTAGAG-3 mtap73_rv 5 -GGTATTGGAAGGGATGACAGGCG-3 mcmyb_fw 5 -CCTCACCTCCATCTCAGCTC-3 mcmyb_rv 5 -ACATGCAATGCGACAGGATA-3 mplk4_fw 5 -GGAGAGGATCGAGGACTTTAAGG-3 mplk4_rv 5 -CCAGTGTGTATGGACTCAGCTC-3 mdeup1_fw 5 -ATATGAGAACGAAAGACTCCGA-3 mdeup1_rv 5 -TGGCTAATTCTGTCTCCACTG-3 mcdc20b_fw 5 -GAAGGAAAATCTTGCCACCA-3 mcdc20b_rv 5 -CATCTTCCCATCGATTTGCT-3 mccna1_fw 5 -TTCTGGAAGCTGACCCATTC-3 mccna1_rv 5 -GGCAAGGCACAATCTCATTT-3 mccno_fw 5 -CTGCAGCTCCCTACTCAACC-3 mccno_rv 5 -CCTTTCGGAAGTCGTAGCAG-3 mhprt_fw 5 -TCCCTGGTTAAGCAGTACAG-3 mhprt_rv 5 -GCTTTGTATTTGGCTTTTCC-3 hmcidas_fw 5 -GACGCGCTTGTTGAGAATAA-3 hmcidas_rv 5 -CACGTTCCGCTCCTTGAG-3 hywhaz_fw 5 -GATCCCCAATGCTTCACAAG-3 hywhaz_rv 5 -TGCTTGTTGTGACTGATCGAC-3 93
94 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.6 Ταυτοποίηση γονοτύπου των µυών Στην παρούσα µελέτη κατασκευάστηκαν διαγονιδιακοί µύες στους οποίους δεν εκφράζεται το γονίδιο GemC1 (trans-nih Knock-Out Mouse Project, KOMP). H εύρεση του γονοτύπου των µυών πραγµατοποιείται µε την αποµόνωση γενωµικού DNA από την ουρά τους και την ανίχνευση των WT και KO αλληλοµόρφων µε PCR Αποµόνωση γενωµικού DNA o Ένα τµήµα της ουράς των µυών τοποθετείται σε eppendorf. o Προσθήκη 500 µl tail buffer και 3 µl πρωτεϊνάσης Κ (stock 10 mg/ml, Sigma). o Επώαση στους 55 ο C, για 16 ώρες. o Προσθήκη 500 µl PCI (φαινόλη-χλωροφόρµιο-ισοαµυλική αλκοόλη σε αναλογία 25:24:1, Sigma). o Έντονη ανάδευση σε θερµοκρασία δωµατίου, στις 500 rpm για 10 λεπτά. o Φυγοκέντρηση στις rpm σε θερµοκρασία δωµατίου, για 30 λεπτά. o Η ανώτερη υδατική φάση µεταφέρεται σε νέο eppendorf. o Προσθήκη 500 µl ισοπροπανόλης. o Ήπια ανάδευση. o Επώαση στους -80 ο C, για λεπτά (εναλλακτικά η επώαση πραγµατοποιείται στους -20 ο C, για 16 ώρες). o Φυγοκέντρηση στις rpm, για 30 λεπτά στους 4 ο C. o Αφαίρεση του υπερκείµενου. o Προσθήκη 500 µl αιθανόλης 70%. o Φυγοκέντρηση στις rpm, για λεπτά στους 4 ο C. o Αφαίρεση του υπερκείµενου. o Ξήρανση του ιζήµατος. o Επαναδιάλυση του DNA σε µl WFI. Το DNA που προέρχεται από έµβρυα µυών επαναδιαλύεται σε 100 µl WFI, ενώ το DNA ενήλικων µυών επαναδιαλύεται σε 50 µl WFI. o Επώαση στους 55 ο C, για 1 ώρα. o To DNA διατηρείται στους -20 ο C. Υλικά και διαλύµατα Tail buffer 100 mm Tris-HCL ph mm EDTA 0.2 % SDS 200 mm NaCl WFI 94
95 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ PCR για την ανίχνευση των WT και KO αλληλοµόρφων του GemC1 Οι εκκινητές 546 και 548 ενισχύουν ένα τµήµα του WT αλληλοµόρφου (413 bp), ενώ οι εκκινητές 542 και 547 ενισχύουν ένα τµήµα µόνο στο KO αλληλόµορφο (285 bp) (Εικόνα 3.6.1). 546 Text Text Text Εικόνα Διαγραµµατική απεικόνιση του KO αλληλοµόρφου του GemC1. Οι εκκινητές 546 και 548 ενισχύουν το WT αλληλόµορφο, ενώ οι εκκινητές 542 και 547 ενισχύουν µόνο το KO αλληλόµορφο. Οι αλληλουχίες των εκκινητών που χρησιµοποιήθηκαν είναι οι εξής: Εκκινητής Αλληλουχία GAGATGGCGCAACGCAATTAATG CAAGAAGTATATCACTGAGGTCAATGGC TAATTCTCTGAACTGTAAGGCAGCTCG CCTTGAATGGATATGTACCTTCTTGC-3 Το πρόγραµµα που ακολουθήθηκε στη συγκεκριµένη PCR είναι το εξής: Χρόνος (Λεπτά) λεπτά Αριθµός κύκλων Θερµοκρασία ο C 10x (-1 ο C/κύκλο) 30x sec 30 sec 40 sec 15 sec 30 sec 40 sec 5 λεπτά τέλος 95
 0,5 µl 542 εκκινητής (20 µμ) 0,5 µl 546 εκκινητής (20 µμ) 0,5 µl 547 εκκινητής (20 µμ) 0,5 µl 548 εκκινητής (20 µμ) 0,5 µl Platinum Taq polymerase 0,08 µl DMSO 0,325 µl DNA 1 µl WFI 17,72 µl")
96 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ H αντίδραση που πραγµατοποιείται για την ανίχνευση των συγκεκριµένων αλληλοµόρφων είναι η εξής: Αντιδραστήρια Ποσότητα Ρυθµιστικό διάλυµα (10x) 2,5 µl MgCl (50 mμ) 0,875 µl dntps (10 mm) 0,5 µl 542 εκκινητής (20 µμ) 0,5 µl 546 εκκινητής (20 µμ) 0,5 µl 547 εκκινητής (20 µμ) 0,5 µl 548 εκκινητής (20 µμ) 0,5 µl Platinum Taq polymerase 0,08 µl DMSO 0,325 µl DNA 1 µl WFI 17,72 µl Τελικός όγκος αντίδρασης 25 µl Τα πιθανά αποτελέσµατα από την παραπάνω γονοτύπηση φαίνονται στην εικόνα WT/WT WT/KO WT/WT WT/KO KO/KO WT/WT KO/KO 500 kb Εικόνα Ανίχνευση των WT και KO αλληλοµόρφων του GemC1. 96
97 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.7 Ανοσοφθορισµός Η µέθοδος του ανοσοφθορισµού επιτρέπει τον υποκυτταρικό εντοπισµό µιας πρωτεΐνης µε τη χρήση αντισώµατος που αναγνωρίζει ειδικά έναν επίτοπο αυτής της πρωτεΐνης. Η σταθερή περιοχή του αντισώµατος αναγνωρίζεται από ένα δευτερογενές αντίσωµα που φέρει ένα φθοριόχρωµα. Αυτό διεγείρεται από κατάλληλου µήκους κύµατος πηγή µονοχρωµατικού φωτός και φθορίζει Πρωτόκολλο ανοσοφθορισµού σε κύτταρα 1 η ηµέρα o Τοποθέτηση των τρυβλίων που περιέχουν τις καλυπτρίδες µε τα κύτταρα σε πάγο. o Το υπερκείµενο θρεπτικό µέσο αφαιρείται και τα κύτταρα εκπλένονται γρήγορα µε παγωµένο PBS (1x), 2 φορές. o Μονιµοποίηση των κυττάρων µε 4% παραφορµαλδεϋδη (PFA) για 10 λεπτά, σε θερµοκρασία δωµατίου. Ανάλογα µε το αντίσωµα που χρησιµοποιείται, µπορεί να απαιτείται µονιµοποίηση των κυττάρων µε µεθανόλη (100%), για 10 λεπτά στους -20 o C. Στην περίπτωση που χρησιµοποιείται µεθανόλη παραλείπονται τα δυο επόµενα βήµατα o Έκπλυση των κυττάρων µε παγωµένο PBS (1x) για 5 λεπτά, 2 φορές. o Επώαση των κυττάρων µε διάλυµα Triton 0.3% (σε PBS 1x), για 5 λεπτά, σε θερµοκρασία δωµατίου, ώστε να γίνει διαπερατή η µεµβράνη των κυττάρων. o Έκπλυση των κυττάρων µε παγωµένο PBS (1x) για 5 λεπτά, 3 φορές o Επώαση των κυττάρων µε διάλυµα (διάλυµα blocking) που περιέχει BSA 3% (Bovine Serum Albumin, Santa Cruz), FBS 10% σε PBS 1x, για 1 ώρα, σε θερµοκρασία δωµατίου. Έτσι, επικαλύπτονται οι µη ειδικές θέσεις πρόσδεσης του αντισώµατος. o Αφαίρεση του διαλύµατος blocking. Σε κάθε καλυπτρίδα τοποθετούνται 40µl πρωτογενούς αντισώµατος στην κατάλληλη αραίωση σε διάλυµα FBS 10% σε PBS 1x, για 16 ώρες, στους 4 o C, σε συνθήκες υγρασίας. 2 η ηµέρα o Την επόµενη ηµέρα πραγµατοποιέιται έκπλυση των κυττάρων µε παγωµένο PBS-Tween 0.1%, για 5 λεπτά, 3 φορές. o Επώαση των κυττάρων µε το δευτερογενές αντίσωµα (40µl/καλυπτρίδα), αραιωµένο σε διάλυµα που περιέχει BSA 3%, FBS 10% σε PBS (1x), για 1 ώρα, σε θερµοκρασία δωµατίου, σε σκοτεινό σηµείο. Σε περίπτωση που απαιτείται χρώση των πυρήνων µε την χρωστική Draq-5 (Biostatus), αυτή προστίθεται µέσα στο διάλυµα του δευτερογενούς αντισώµατος σε αραίωση 1:1000. o Έκπλυση των κυττάρων µε παγωµένο PBS-Tween 0.1%, για 5 λεπτά, 3 φορές. 97
98 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Χρώση των πυρήνων των κυττάρων µε τη χρωστική Hoechst (1:1500, Sigma), για 2 λεπτά σε θερµοκρασία δωµατίου. o Έκπλυση των κυττάρων µε παγωµένο PBS (1x) για 5 λεπτά, 3 φορές o Εµβάπτιση των καλυπτρίδων σε ddh 2 O. o Τοποθέτηση των καλυπτρίδων σε αντικειµενοφόρο πλάκα, επάνω σε σταγόνα στερεοποιητικού διαλύµατος MOVIOL 4-88 (Calbiochem), για την προσκόλλησή τους. Αφήνονται να στεγνώσουν και παρατηρούνται σε µικροσκόπιο φθορισµού Πρωτόκολλο ανοσοφθορισµού σε τοµές ιστών 1 η ηµέρα o Ξεπάγωµα των τοµών (από τους -80 C) σε θερµοκρασία δωµατίου, για 30 λεπτά. Η διαδικασία πραγµατοποιείται σε επωαστικό θάλαµο που περιέχει νερό για να µην ξεραθούν οι τοµές. o Μονιµοποίηση των τοµών σε διάλυµα παραφορµαλδεΰδης (PFA) 4% για 10 λεπτά, σε θερµοκρασία δωµατίου. o Έκπλυση των τοµών µε PBS (1x) για 5 λεπτά, 2 φορές. o Επώαση των τοµών µε διάλυµα Triton 0.3% (σε PBS 1x), για 5 λεπτά, σε θερµοκρασία δωµατίου. o Έκπλυση των τοµών µε PBS (1x) για 5 λεπτά, 3 φορές. o Επώαση των τοµών µε διάλυµα (διάλυµα blocking) που περιέχει BSA 3% (Bovine Serum Albumin, Santa Cruz), FBS 10% σε PBS (1x), για 1 ώρα σε θερµοκρασία δωµατίου. Έτσι, επικαλύπτονται οι µη ειδικές θέσεις πρόσδεσης του αντισώµατος. o Αφαίρεση του διαλύµατος blocking. Σε κάθε αντικειµενοφόρο πλάκα που περιέχονται οι τοµές τοποθετείται επαρκής ποσότητα πρωτογενούς αντισώµατος, στην κατάλληλη αραίωση σε διάλυµα blocking, για 16 ώρες στους 4 o C, σε συνθήκες υγρασίας. 2 η ηµέρα o Την επόµενη ηµέρα πραγµατοποιέιται έκπλυση των τοµών µε PBS-Tween 0.1%, για 5 λεπτά, 3 φορές. o Επώαση των τοµών µε επαρκή ποσότητα δευτερογενούς αντισώµατος, αραιωµένο σε διάλυµα blocking, για 1 ώρα σε θερµοκρασία δωµατίου, σε σκοτεινό σηµ είο. Σε περίπτωση που απαιτείται χρώση των πυρήνων µε την χρωστική Draq-5 (Biostatus), αυτή προστίθεται µέσα στο διάλυµα του δευτερογενούς αντισώµατος σε αραίωση 1:1000. o Έκπλυση των ιστών µε PBS-Tween 0.1%, για 5 λεπτά, 3 φορές. o Χρώση των πυρήνων µε τη χρωστική Hoechst (1:1500, Sigma), για 2 λεπτά σε θερµοκρασία δωµατίου. o Έκπλυση των τοµών µε PBS (1x) για 5 λεπτά, 3 φορές 98
99 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Τοποθέτηση καλυπτρίδων στην αντικειµενοφόρο πλάκα, µε τη χρήση του στερεοποιητικού διαλύµατος MOVIOL 4-88 (Calbiochem). Αφήνονται να στεγνώσουν και παρατηρούνται σε µικροσκόπιο φθορισµού Πρωτόκολλο ανοσοφθροσισµού σε MTEC 1 η ηµέρα o Τα ένθετα µε τις µεµβράνες πάνω στις οποίες καλλιεργούνται τα MTEC παραλαµβάνονται την επιθυµητή ηµέρα κατά τη διάρκεια της καλλιέργειας (time point). o Αναρρόφηση του θρεπτικού µέσου που βρίσκεται στη βασική ή και στην κορυφαία πλευρά της µεµβράνης και έκπλυση των κυττάρων µε PBS (1x), 2 φορές. Το PBS (1x) προστίθεται κ αι στην κορυφαία και στη βασική πλευρά της µεµβράνης. o Μονιµοποίηση των κυττάρων µε 4% παραφορµαλδεΰδη (PFA) για 12 λεπτά, σε θερµοκρασία δωµατίου. Το διάλυµα της παραφορµαλδεΰδης προστίθεται και στην κορυφαία και στη βασική πλευρά της µεµβράνης. o Έκπλυση των κυττάρων µε PBS (1x), 2 φορές. Το PBS (1x) προστίθεται και στην κορυφαία και στη βασική πλευρά της µεµβράνης. o Με τη βοήθεια λαβίδας µε λεπτά άκρα αποµακρύνεται προσεκτικά η µεµβράνη επάνω από το ένθετο στο οποίο βρίσκεται. o Η µεµβράνη κόβεται προσεκτικά µε νυστέρι σε 4 ίσου µεγέθους κοµµάτια. Κάθε τµήµα της µεµβράνης σηµαδεύεται µε ένα µικρό κόψιµο (π.χ. στο επάνω δεξί άκρο της), ώστε να είναι πάντοτε γνωστή η πλευρά στην οποία βρίσκονται τα κύτταρα. o Τα τµήµατα της κάθε µεµβράνης φυλάσσονται σε PBS (1x) σ τους 4 o C, σε τρυβλία καλά κλεισµένα, για πολλούς µήνες. o Το τµήµα της µεµβράνης που πρόκειται να πραγµατοποιηθεί ο ανοσοφθορισµός τοποθετείται σε τρυβλία 96-well και γίνεται έκπλυση των µεµβρανών µε PBS-Triton 0,1%, 2 φορές. o Επώαση των µεµβρανών σε PBS-Triton 0,1%, για 5 λεπτά, σε θερµοκρασία δωµατίου. o Επώαση των µεµβρανών µε διάλυµα (διάλυµα blocking) που περιέχει PBS- Triton 0,1% και FBS 10% σε θερµοκρασία δωµατίου, για 1 ώρα. o Αφαίρεση του διαλύµατος blocking. Σε κάθε µεµβράνη τοποθετούνται 80µl πρωτογενούς αντισώµατος στην κατάλληλη αραίωση σε διάλυµα blocking για 16 ώρες, στους 4 o C. 2 η ηµέρα o Την επόµενη ηµέρα πραγµατοποιείται έκπλυση των µεµβρανών µε PBS-Triton 0,1%, 2 φορές. o Επώαση των µεµβρανών σε PBS-Triton 0,1% για 5 λεπτά, σε θερµοκρασία δωµατίου. 99
100 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Επώαση των µεµβρανών µε 80 µl δευτερογενές αντίσωµα, αραιωµένο σε διάλυµα blocking, για 1 ώρα σε θερµοκρασία δωµατίου, σε σκοτεινό σηµείο. Σε περίπτωση που απαιτείται χρώση των πυρήνων µε την χρωστική Draq-5 (Biostatus), αυτή προστίθεται µέσα στο διάλυµα τ ου δευτερογενούς αντισώµατος σε αραίωση 1:1000. o Έκπλυση των µεµβρανών µε PBS-Triton 0,1%, 2 φορές. o Επώαση των µεµβρανών σε PBS-Triton 0,1% για 5 λεπτά, σε θερµοκρασία δωµατίου. o Χρώση των πυρήνων των κυττάρων µε τη χρωστική Hoechst (1:1500, Sigma), για 2 λεπτά σε θερµοκρασία δωµατίου. o Έκπλυση των µεµβρανών µε PBS-Triton 0,1%, 2 φορές. o Εµβάπτιση των µεµβρανών σε ddh 2 O. o Τοποθέτηση του τµήµατος της µεµβράνης σε αντικειµενοφόρο πλάκα, επάνω σε σταγόνα στερεοποιητικού διαλύµατος MOVIOL 4-88 (Calbiochem), για την προσκόλλησή τους. Γίνεται έγκλειση µε καλυπτρίδες (22 x 22 mm) και αφήνονται να στεγνώσουν, ώστε να παρατηρηθούν σε µικροσκόπιο φθορισµού. Υλικά και διαλύµατα Διάλυµα Hoechst Διάλυση Hoechst (Sigma 33258) σε WFI για τελική συγκέντρωση 10mg/ml 100
101 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Πίνακας αντισωµάτων Τα πρωτογενή και δευτερογενή αντισώµατα που χρησιµοποιήθηκαν συνολικά στην παρούσα διατριβή είναι τα εξής: Αντισώµατα Αραίωση Μονιµοποίηση Προέλευση rabbit anti-hmcidas 1:500 PFA (Pefani et al, 2011) mouse anti-foxj1 1:500 PFA ebioscience rabbit anti-foxj1 1:500 PFA Sigma rabbit anti-pericentrin 1:1000 PFA Covance rabbit anti-γ-tubulin 1:1000 PFA/methanol Sigma mouse anti-acetylated 1:2000 PFA/methanol Sigma α-tubulin rat anti-e-cadherin 1:500 PFA Invitrogen rabbit anti-krt5 1:2000 PFA Covance mouse ThermoFisher 1:500 PFA anti-muc5ac Scientific mouse anti-gfp 1:500 PFA Molecular Probes rabbit anti-gfp 1:1000 PFA Molecular Probes chicken anti-gfp 1:1500 PFA/methanol 2B Scientific rat anti-cep135 1:500 methanol goat anti-cep164 1:500 rabbit anti-cp110 1:250 mouse anti-cyclina methanol methanol Dr. Monica Bettencourt-Dias lab Santa Cruz Dr. Monica Bettencourt-Dias lab 1:500 methanol Abcam 101
102 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Αντισώµατα Αραίωση Προέλευση Alexa Fluor antirabbit 488 1:1000 Molecular Probes Alexa Fluor antirabbit 568 1:1000 Molecular Probes Alexa Fluor antirabbit 647 1:1000 Molecular Probes Alexa Fluor antimouse 488 1:1000 Molecular Probes Alexa Fluor antimouse 568 1:1000 Molecular Probes Alexa Fluor antimouse 647 1:1000 Molecular Probes Alexa Fluor antimouse IgG :1000 Molecular Probes Alexa Fluor antigoat 488 1:1000 Molecular Probes Alexa Fluor antirat 488 1:1000 Molecular Probes Alexa Fluor antirat 568 1:1000 Molecular Probes Alexa Fluor antichicken 488 1:1000 Molecular Probes 102
103 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.8 Υβριδοποίηση ιn situ Η υβριδοποίηση in situ είναι µια τεχνική που βασίζεται στην επιτόπια υβριδοποίηση νουκλεϊκών οξέων µε ση µασµένο ανιχνευτικό µόριο, ώστε να εντοπιστεί η θέση συγκεκριµένου mrna σε έναν ακέραιο οργανισµό ή σε τοµή ενός ιστού. Οι ανιχνευτές RNA προκύπτουν από τη µεταγραφή κατάλληλων πλασµιδιακών φορέων, που φέρουν το cdna του γονιδίου που µελετάται. Η σήµανση των ανιχνευτών RNA γίνεται µε διγοξιγενίνη (DIG), η οποία αναγνωρίζεται από το αντίσωµα anti-dig. To anti-dig αντίσωµα είναι συνδεδεµένο µε µόρια αλκαλικής φωσφατάσης, τα οποία στο τελικό στάδιο της διαδικασίας αποφωσφορυλιώνουν τα υποστρώµατα της αλκαλικής φωσφατάσης BCIP και NBT, επιτυγχάνοντας την ανάπτυξη χρώµατος ειδικά στις περιοχές που έγινε δέσµευση του ανιχνευτή µε το mrna. To πρωτόκολλο της υβριδοποίησης in situ µπορεί να χρησιµοποιηθεί συνδυαστικά µε το πρωτόκολλο του ανοσοφθορισµού, ώστε να επιτευχθεί ταυτόχρονη ανίχνευση του mrna ενός γονιδίου και της πρωτεΐνης ενός δεύτερου γονιδίου (Jekely & Arendt, 2007). Συνοπτικά, τα βήµατα που απαιτούνται για την υβριδοποίηση in situ είναι τα εξής: 1. Παραγωγή γραµµικών πλασµιδιακών φορέων και αποµόνωσή τους 2. Παραγωγή των RNA ανιχνευτών 3. Υβριδοποίηση in situ Παραγωγή γραµµικών πλασµιδιακών φορέων και αποµόνωσή τους Στην παρούσα διατριβή τo πρωτόκολλο της υβριδοποίησης in situ χρησιµοποιήθηκε συνδυαστικά µε το πρωτόκολλο του ανοσοφθορισµού, ώστε να επιτευχθεί ταυτόχρονη ανίχνευση του mrna των γονιδίων McIdas και GemC1 και της πρωτεΐνης FoxJ1. Τα cdna των γονιδίων McIdas και GemC1 βρίσκονται κλωνοποιηµένα στα πλασµίδια pflci και pbluescript KS +, αντίστοιχα. Για να γίνουν γραµµικά τα παραπάνω πλασµίδια ακολουθείται η εξής διαδικασία: o Αντίδραση κατάτµησης (τελικού όγκου 1 µl) µε τα ένζυµα περιορισµού NotI (για τ ο πλασµίδιο που περιέχει το cdna του McIdas) και Χ hoi (για το πλασµίδιο που περιέχει το cdna του GemC1). o Την επόµενη ηµέρα, 1 µl της αντίδρασης αναλύεται ηλεκτροφορητικά σε πήκτωµα αγαρόζης 1%, ώστε να ελεγχθεί η επιτυχία της αντίδρασης. o Παράλληλα, στην αντίδραση προστίθενται επιπλέον 2 µl της περιοριστικής ενδονουκλεάσης που έχει χρησιµοποιηθεί και ακολουθεί επώαση στους 37 o C, για 2 ώρες. Για την αποµόνωση των γρα µµικών πλασµιδιακών φορέων ακολουθείται η εξής διαδικασία: o Στα 100 µl της αντίδρασης κατάτµησης προστίθενται 100 µl WFI και 200 µl PCI. 103
104 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Ισχυρή ανάδευση µε vortex. o Φυγοκέντρηση στις rpm, για 15 λεπτά σε θερµοκρασία δωµατίου. o Η ανώτερη υδατική φάση µεταφέρεται σε ένα νέο eppendorf (περίπου 200 µl). o Προστίθεται 1/10 του όγκου (20 µl) 3M οξικό νάτριο (CH 3 COONa), ph 5.3. o Ισχυρή ανάδευση µε vortex. o Προστίθενται 550 µl αιθανόλης 100% (-20 o C). o Ισχυρή ανάδευση µε vortex. o Επώαση στους -20 o C, για 16 ώρες. o Φυγοκέντρηση στις rpm, για 20 λεπτά στους 4 o C. o Αποµάκρυνση του υπερκείµενου. o Έκπλυση του ιζήµατος µε 100 µl αιθανόλης 70% (-20 o C). o Φυγοκέντρηση στις rpm, για 20 λεπτά στους 4 o C. o Αποµάκρυνση του υπερκείµενου. o Ξήρανση του ιζήµατος. o Επαναδιάλυση σε 30 µl WFI. o Φωτοµέτρηση για τον υπολογισµό της συγκέντρωσής του και ηλεκτροφορητική ανάλυση του DNA (0,5 µl) σε πήκτωµα αγαρόζης 1%. Υλικά και διαλύµατα Οξικό νάτριο (CH 3 COONa) 3M, ph 5.3 Για τελικό όγκο 1 Lt 408 gr Sodium acetate 3H 2 O Ρύθµιση του ph µε οξικό οξύ Παραγωγή των RNA ανιχνευτών Η παραγωγή των RNA ανιχνευτών γίνεται µέσω in vitro µεταγραφής. Το cdna του επιθυµητού γονιδίου που αποµονώθηκε παραπάνω χρησιµοποιείται ως µήτρα για τη µεταγραφή. Επίσης, µε βάση τον υποκινητή που διαθέτει ο φορέας έκφρασης για την παραγωγή του αντινοηµατικού RNA που θα υβριδοποιηθεί µε το RNA στόχος, χρησιµοποιείται και η κατάλληλη πολυµεράση. Στην παρούσα εργασία χρησιµοποιήθηκε η Τ3 πολυµεράση για τη παραγωγή του αντινοηµατικού RNA που θα υβριδοποιηθεί µε το mrna του γονίδιο McIdas και η Τ7 πολυµεράση για τη παραγωγή του αντινοηµατικού RNA που θα υβριδοποιηθεί µε το mrna του γονίδιο GemC1. Η αντίδραση πραγµατοποιείται ως εξής: 104
105 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Αντιδραστήρια Ποσότητα DNA (γραµµικό πλασµίδιο) 1 µg Ρυθµιστικό διάλυµα (5x) (Transcription buffer, Promega) 4 µl DTT (0,1 M) (Promega) 2 µl DIG-UTP mix 10x (Roche) 2 µl RNasin (Promega) 0,5 µl RNA πολυµεράση (10 U/µl) T3/T7 1,5 µl WFI Μέχρι τελικού όγκου 20 µl o Επώαση στους 37 o C, για 1 ώρα. o Ηλεκτροφορητική ανάλυση 1 µl της αντίδρασης σε πήκτωµα αγαρόζης 0,8 %, για έλεγχο της ποιότητας του παραγόµενου RNA. o Προσθήκη 2 µl DNase I (Roche) και επώαση στους 37 o C, για 45 λεπτά. o Προσθήκη 100 µl WFI και 10 µl LiCl (4M). o Ανάδευση και προσθήκη 300 µl αιθανόλης 100% (-20 o C). o Επώαση στους -20 o C, για 16 ώρες για την κατακρήµνιση του RNA. o Φυγοκέντρηση στις rpm, για 30 λεπτά στους 4 o C. o Αποµάκρυνση του υπερκείµενου και έκπλυση του ιζήµατος µε 150 µl αιθανόλης 70% (-20 o C). o Φυγοκέντρηση στις rpm, για 15 λεπτά στους 4 o C. o Ξήρανση του ιζήµατος σε πάγο, στον απαγωγό. o Επαναδιάλυση σε 30 µl WFI. o Ηλεκτροφορητική ανάλυση 1 µl σε πήκτωµα αγαρόζης 0,8 %, για έλεγχο της ποιότητας του παραγόµενου RNA. o Φύλαξη του RNA ανιχνευτή στους -80 o C Πρωτόκολλο υβριδοποίησης in situ 1 η ηµέρα o Ξεπάγωµα των τοµών (από τους -80 C) σε θερµοκρασία δωµατίου, για λεπτά. Η διαδικασία πραγµατοποιείται σε επωαστικό θάλαµο που περιέχει 5x SSC ή WFI για να µην ξεραθούν οι τοµές. o Μονιµοποίηση των τοµών σε διάλυµα παραφορµαλδεΰδης (PFA) 4% για 10 λεπτά, σε θερµοκρασία δωµατίου. o Έκπλυση των τοµών µε PBS (1x) για 5 λεπτά, 3 φορές. o Επώαση των τοµών σε διάλυµα ακετυλίωσης για 10 λεπτά σε θερµοκρασία δωµατίου, υπό συνεχή ανάδευση. Η ακετυλίωση των τοµών επιτυγχάνεται µε 295ml WFI στα οποία έχουν προστεθεί 4ml triethanolamine (Fluka 90278) και 525µl HCl υπό µαγνητική ανάδευση. Αφού έχουν αναµιχθεί καλά, προσθέτουµε στο παραπάνω διάλυµα 750µl οξικού ανυδρίτη (Sigma A6404) αµέσως πριν τοποθετήσουµε τις τοµές. o Έκπλυση των τοµών µε PBS (1x) για 5 λεπτά, 3 φορές. 105
106 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o o o o Επώαση των τοµών µε το διάλυµα υβριδοποίησης στον επωαστικό θάλαµο σε θερµοκρασία δωµατίου, για τουλάχιστον 3-4 ώρες. Προς το τέλος της προηγούµενης επώασης πραγµατοποιείται αναδιάλυση των ανιχνευτών RNA στο διάλυµα υβριδοποίησης, σε αναλογία 1µl ανιχνευτή RNA: 50µl διάλυµα υβριδοποίησης (η αναλογία αυτή µπορεί να αλλάξει ανάλογα µε την ποιότητα του ανιχνευτή RNA). Αποδιάταξη των ανισνευτών µε θέρµανση στους 80 C για 5 λεπτά και τοποθέτησή τους αµέσως σε πάγο. Αποµάκρυνση του διαλύµατος υβριδοποίησης και προσθήκη στις τοµές των RNA ανιχνευτών. Αφού τους προσθέσουµε οι τοµές καλύπτονται µε καλυπτρίδες και επωάζονται στους 72 C, για 16 ώρες. Είναι σηµαντικό ο επωαστικός θάλαµος να περιέχει αρκετό διάλυµα 5x SSC και να σφραγιστεί ερµητικά, ώστε οι τοµές να µην ξεραθούν κατά τη διάρκεια της επώασης. 2 η ηµέρα o Προετοιµασία των διαλυµάτων 5x SSC και 0.2x SSC, θερµοκρασίας 72 C. o Κρατώντας τα πλακάκια µε τις τοµές µε µία λαβίδα, πραγµατοποιείται αφαίρεση των καλυπτρίδων στο διάλυµα 5x SSC. o Επώαση των τοµών στο διάλυµα 0.2x SSC, για 1 ώρα στους 72 C. o Επώαση των τοµών σε διάλυµα 0.2x SSC, για 5 λεπτά σε θερµοκρασία δωµατίου. o Επώαση των τοµών στο διάλυµα Β1, για 5 λεπτά σε θερµοκρασία δωµατίου. o Μεταφορά των τοµών στον επωαστικό θάλαµο. Στις τοµές τοποθετούνται 0.5-1ml διάλυµα Β1 στο οποίο έχει προστεθεί 10% FBS. Επώαση σε θερµοκρασία δωµατίου, για 1 ώρα. o Επώαση των τοµών σε 0.5ml διάλυµα Β1 (που περιέχει FBS), στο οποίο έχει διαλυθεί το anti-dig αντίσωµα (Roche) σε αραίωση 1:5000, καθώς και το αντίσωµα έναντι της πρωτεΐνης του δεύτερου γονιδίου που µας ενδιαφέρει. Στην παρούσα εργασία, χρησιµοποιήθηκε αντίσωµα έναντι της πρωτεΐνης Foxj1, σε αραίωση 1:200. o Επώαση στους 4 C, για 16 ώρες. 3 η ηµέρα o Έκπλυση των τοµών µε διάλυµα Β1 σε θερµοκρασία δωµατίου, για 5 λεπτά, 3 φορές. o Επώαση µε διάλυµα Β3 σε θερµοκρασία δωµατίου, για 10 λεπτά. o Στις τοµές προστίθενται περίπου 100µl από το τελικό διάλυµα ανάπτυξης σήµατος (διάλυµα Β4). Επώαση στο σκοτάδι µέχρι να αναπτυχθεί το ειδικό χρώµα. Είναι σηµαντικό σε αυτό το βήµα να µην αφήσουµε να αναπτυχθεί πολύ το σήµα, καθώς µπορεί να καλύψει το σήµα τ ου φθορισµού του αντισώµατος που θα χρησιµοποιηθεί στη συνέχεια. o Η αντίδραση σταµατά µε βύθιση των τοµών σε νερό. o Έκπλυση των τοµών µε διάλυµα Β1 (που περιέχει 10% FBS) σε θερµοκρασία δωµατίου, για τουλάχιστον 1 ώρα. 106
107 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ o Επώαση των τοµών µε ένα δευτερογενές αντίσωµα το οποίο είναι αραιωµένο σε διάλυµα Β1 (που περιέχει 10% FBS) στους 4 o C, για 16 ώρες. Στην παρούσα µελέτη χρησιµοποιήθηκε το Alexa Fluor anti-rabbit 488 (Molecular Probes), σε αραίωση 1: η ηµέρα o Έκπλυση των τοµών µε διάλυµα Β1 σε θερµοκρασία δωµατίου, για 5 λεπτά, 2 φορές. o Επώαση των τοµών µε Hoechst σε διάλυµα B1 (που περιέχει 10% FBS) σε αραίωση 1:1000, σε θερµοκρασία δωµατίου για 5 λεπτά. o Τοποθετούµε καλυπτρίδες χρησιµοποιώντας ειδικό µέσο (Glycergel mounting medium, C0563 DakoCytomation), το οποίο έχουµε προθερµάνει στους 55 o C. o Παρατηρούµε τις τοµές σε συνεστιακό µικροσκόπιο φθορισµού. Υλικά και διαλύµατα Διάλυµα υβριδοποίησης 50% φορµαµίδιο (Sigma F9037) 5x SSC 5x Denhardts (Sigma D2532) 250µg/ml baker s yeast RNA (Sigma R6750) 500µg/ml herring sperm DNA (Promega D1815) WFI SSC 20x ph 7.0 Για τελικό όγκο 100ml 17,5g NaCl 8,8g Na 3 Citrate.2H 2 O Ρυθµίζουµε το ph µε HCl Διάλυµα Β1 0.1Μ TrisCl ph M NaCl Διάλυµα Β3 0.1M TrisCl ph M NaCl 50mM MgCl 2 Διάλυµα Β4 3.5µl/ml NBT (Roche ) 3.5µl/ml BCIP (Roche ) 107
108 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 0.24mg/ml levamisole (Sigma L9756) σε διάλυµα Β3 TrisCl ph M Για τελικό όγκο 500ml 60,5g TrisCl Ρυθµίζουµε το ph µε HCl NaCl 4M Για τελικό όγκο 100ml 23,4g NaCl WFI µέχρι τον τελικό όγκο MgCl 2 1M Για τελικό όγκο 1L Διαλύουµε 203,3g MgCl 2.6H 2 O Προσθέτουµε WFI µέχρι τον τελικό όγκο 108
109 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.9 Διπλασιασµός των κεντροσωµατίων µε τη χρήση υδροξυουρίας (HU) Ο κύκλος διπλασιασµού και διαχωρισµού των κεντροσωµατίων στα κύτταρα είναι άρρηκτα συνδεδεµένος µε τον κύκλο διπλασιασµού και διαχωρισµού των χρωµοσωµάτων. Προκειµένου να γίνει αποσύνδεση των δυο κύκλων για να µελετηθεί ειδικά ο διπλασιασµός των κεντριολίων, τα κύτταρα επωάζονται µε αναστολείς του διπλασιασµού του DNA, όπως είναι η υδροξυουρία (HU). Στην παρούσα µελέτη το πρωτόκολλο αυτό εφραµόστηκε, ώστε να µελετηθεί ειδικά η επίδραση της υπερέκφρασης της πρωτεΐνης McIdas στο διπλασιασµό των κεντριολίων, ανεξάρτητα από το ρόλο της κατά την αντιγραφή του DNA. Η διαδικασία που ακολουθήθηκε ήταν η εξής: o 4 x 10 5 κύτταρα επιστρώνονται σε τρυβλίο 6-well. Στο τρυβλίο έχουν τοποθετηθεί καλυπτρίδες, που έχει επιστρωθεί µε poly-l-lysine, για την καλύτερη προσκόλληση των κυττάρων. o Την επόµενη ηµέρα προστίθενται στα κύτταρα 4 mm ΗU. o Μετά από 24 ώρες από την προσθήκη της υδροξυουρίας πραγµατοποιείται διαµόλυνση των κυττάρων. Η διαµόλυνση γίνεται µε τη χρήση του πλασµιδίου plvdest-gfpmcidas γ ια την έκφραση της πρωτεΐνης Μ cidas. Παράλληλα, χρησιµοποιήθηκε και ο φορέας έκφρασης plvdest-gfp, ως δείγµα-µάρτυρας. Η διαµόλυνση έγινε µε τη χρήση του λιπόφιλου αντιδραστηρίου lipofectamine 2000, ως εξής: 1. Σε ένα eppendorf προστίθεται 1 µg DNA και κατάλληλη ποσότητα Opti- MEM, ώστε ο τελικός όγκος να είναι 250 µl. Ακολουθεί επώαση 5 λεπτά. 2. Παράλληλα σε ένα δεύτερο eppendorf προστίθενται 5 µl lipofectamin 2000 µε κατάλληλη ποσότητα Opti-MEM, ώστε ο τελικός όγκος να είναι 250 µl. Ακολουθεί επώαση 5 λεπτά. 3. Τα δυο παραπάνω eppendorf ενώνονται σε ένα κοινό (master mix). Το µείγµα επωάζεται για 20 λεπτά. o Αναρρόφηση του θρεπτικού µέσου από το τρυβλίο µε τα κύτταρα και προσθήκη 500 µl φρέσκου θρεπτικού µέσου DMEM, που περιέχει 10% FBS και 4 mm ΗU. o Στο τρυβλίο προστίθενται τα 500 µl του διαλύµατος διαµόλυνσης. o Μετά το πέρας 3 ωρών προστίθεται επιπλέον 1 ml θρεπτικού µέσου DMEM, που περιέχει 10% FBS και 4 mm ΗU. o 24 ώρες µετά το θρεπτικό µέσο αποµακρύνεται από τα κύτταρα και προστίθεται φρέσκο θρεπτικού µέσου DMEM (2 ml/well), που περιέχει 10% FBS και 4 mm ΗU. o Η καλλιέργεια συνεχίζεται για επιπλέον 24 ώρες. o 24 ώρες µέτα τη δεύτερη διαµόλυνση τα κύτταρα παραλαµβάνονται, ώστε να µονιµοποιηθούν. 109
110 3. ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 110
111 4. ΑΠΟΤΕΛΕΣΜΑΤΑ
112 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 112
113 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 4.1 Μελέτη της έκφρασης των µελών της υπεροικογένειας της Geminin στα πολυκροσσωτά κύτταρα Προηγούµενες µελέτες του εργαστηρίου µας συνέβαλαν στην εύρεση δυο νέων πρωτεϊνών που κωδικοποιούνται από το ανθρώπινο γονιδίωµα, των McIdas και GemC1. Οι νέες αυτές πρωτεΐνες εντοπίστηκαν διότι φέρουν µια περιοχή σπειροειδούς σπειράµατος µε σηµαντική οµοιότητα µε το σπειροειδές σπείραµα της πρωτεΐνης Geminin. Μελέτες έδειξαν, πέραν της δοµικής και λειτουργική οµοιότητα µε την πρωτεΐνη Geminin, αφού και οι δυο πρόσφατα χαρακτηρισµένες πρωτεΐνες ταυτοποιήθηκαν αρχικά για τη συµµ ετοχή τους στην έναρξη της αντιγραφής του DNA (Balestrini et al, 2010; Pefani et al, 2011). Είναι γνωστό ότι η πρωτεΐνη Geminin συµµετέχει στη ρύθµ ιση της αντιγραφής του DNA. Αναστέλλει τον παράγοντα αδειοδότησης της αντιγραφής Cdt1, ρυθµίζοντας µε αυτόν τον τρόπο τη σωστή πυροδότηση των αφετηριών της αντιγραφής (reviewed in Lygerou & Nurse, 2000; Tada et al, 2001; Wohlschlegel et al, 2000). Η πρωτεΐνη McIdas σχηµατίζει ετεροδιµερή µε την πρωτεΐνη Geminin και αναστέλλει την πρόσδεσή της στο Cdt1 (Pefani et al, 2011). Επιπρόσθετα, δείχτηκε ότι τόσο το mrna όσο και η πρωτεΐνη του παράγοντα McIdas εκφράζονται σε υψηλά επίπεδα στον αναπτυσσόµενο εγκέφαλο µυός. Η πρωτεΐνη GemC1 δρα κατά την έναρξη της αντιγραφής του DNA για το σχηµατισµό του προεναρκτήριου συµπλόκου της αντιγραφής (pre-ic). Α λληλεπιδρά µε τον παράγοντα TopBP1 και στρατολογεί τον παράγοντα Cdc45 στη χρωµατίνη (Balestrini et al, 2010). Εκτός από το ρόλο της Geminin κατά την αντιγραφή του DNA, µια σειρά από µελέτες σε διαφορετικούς οργανισµούς και συστήµατα χαρακτήρισαν το ρόλο της και σε διαδικασίες κυτταρικής διαφοροποίησης (Patmanidi et al, 2017). Επίσης, ενώ αυτή η εργασία βρισκόταν σε εξέλιξη, προέκυψαν δεδοµένα για την εµπλοκή του παράγοντα McIdas στη διαφοροποίηση των πολυκροσσωτών κυττάρων της επιδερµίδας ε µβρύων Xenopus, καθώς και στο αναπτυσσόµενο νεφροστόµιο των νεφρών (Stubbs et al, 2012). Πιο συγκεκριµένα, βρέθηκε ότι ο McIdas αποτελεί στόχο του µονοπατιού Notch, αφού η έκφρασή του αναστέλλεται όταν εκφράζεται το Notch. Η αποσιώπηση του McIdas εµποδίζει τη δηµιουργία των πολυκροσσωτών κυττάρων της επιδερµίδας, ενώ η υπερέκφρασή του µπορεί να προκαλέσει εκτοπικά τη δηµιουργία των συγκεκριµένων κυττάρων και µάλιστα τη δηµιουργία δοµών που µοιάζουν µε δευτεροσωµάτια. Α ργότερα δείχθηκε ότι η δράση αυτή του McIdas επιτυγχάνεται µέσω της δηµιουργίας συµπλόκων µε τους µεταγραφικούς παράγοντες E2F4/5 και TFDP1 (Ma et al, 2014). Το σύµπλοκο που σχηµατίζεται δρα επάγοντας την έκφραση γονιδίων που είναι απαραίτητα για τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Η οικογένεια των πρωτεϊνών της Geminin φαίνεται να συνδέει την αντιγραφή του DNA µε τον πολλαπλασιασµό των κεντριολίων και τη δηµιουργία των πολυκροσσωτών κυττάρων, αποτελώντας έναν κοινό ρυθµιστή των διαδικασιών αυτών. Στόχος της παρούσας διατριβής είναι να µελετηθεί η έκφραση και η λειτουργία των µελών της υπεροικογένειας της Geminin µε έµφαση στο ρόλο τους στη διαφοροποίηση των πολυκροσσωτών κυττάρων. 113
114 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Μελέτη της έκφρασης των µελών της υπεροικογένειας της Geminin κατά την εµβρυϊκή ανάπτυξη και σε ενήλικους µυς Προηγούµενα δεδοµένα του εργαστηρίου µας έδειξαν ότι τα γονίδια McIdas και GemC1 εµφανίζουν ένα επικαλυπτόµενο πρότυπο έκφρασης στο αναπτυσσόµενο κεντρικό νευρικό και αναπνευστικό σύστηµα (Διδακτορική Διατριβή, Δάφνη- Ελευθερία Πεφάνη, 2012) (Pefani et al, 2011), γεγονός που υποδηλώνει τη συµµετοχή τους σε διαδικασίες δι αφοροποίησης των κυττάρων. Κοινό χαρακτηριστικό των ιστών που εµφάνιζαν υψηλά επίπεδα έκφρασης ήταν η παρουσία κυττάρων που διέθεταν κινητούς κροσσούς. Για να διερευνηθεί περαιτέρω η έκφραση των µελών της υπεροικογένειας της Geminin κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, πραγµατοποιήθηκε αρχικά µελέτη της έκφρασής τους σε ιστούς που αποµονώθηκαν τόσο από έµβρυα, όσο και από ενήλικες µύες. Για τη µελέτη αυτή επιλέχθηκαν ιστοί που διαθέτουν πολυκροσσωτά κύτταρα, καθώς και το ήπαρ το οποίο δεν περιέχει αυτόν τον κυτταρικό τύπο. Πιο συγκεκριµένα, αποµονώθηκε το χοριοειδές πλέγµα από τις πλάγιες και την τέταρτη κοιλία του εγκεφάλου από έµβρυα µυών ηλικίας 12.5 (Ε12.5), 14.5 (Ε14.5) και 18.5 (Ε18.5) ηµερών, καθώς και οι βρόγχοι από έµβρυα ηλικίας 14.5 (Ε14.5) και 18.5 (Ε18.5) ηµερών. Από ενήλικες µυς αποµονώθηκε το χοριοειδές πλέγµα των κοιλιών του εγκεφάλου και οι βρόγχοι, καθώς και οι ωοθήκες µε τους ωαγωγούς θηλυκών µυών και το ήπαρ. Ακολούθησε αποµόνωση ολικού RNA από κάθε ιστό και σύνθεση συµ πληρωµατικού DNA (cdna). Τα επίπεδα έκφρασης του κάθε γονιδίου στους αποµονωµένους ιστούς συγκρίθηκαν µέσω ποσοτικής Real- Time PCR, µε τη χρήση ειδικών εκκινητών για την ανίχνευση των GemC1, McIdas, Geminin και Foxj1 που εκφράζεται ειδικά στα πολυκροσσωτά κύτταρα (Εικόνα 4.1.1). Παρατηρήθηκε ότι το mrna του παράγοντα GemC1 εµφανίζει υψηλή έκφραση κατά την ανάπτυξη στο χοριοειδές πλέγµα του εγκεφάλου (Εικόνα Α), η οποία µειώνεται στους ενήλικες µυς. Παρόµοιο πρότυπο έκφρασης παρατηρήθηκε και στο αναπνευστικό επιθήλιο. Η έκφρασή του εντοπίζεται και στις ωοθήκες και στους ωαγωγούς ενήλικων θηλυκών µυών. Κοινό χαρακτηριστικό όλων των παραπάνω ιστών είναι ότι διαθέτουν πολυκροσσωτά κύτταρα. Ιδιαίτερο ενδιαφέρον παρουσιάζει το γεγονός ότι στο ήπαρ ενήλικου µυός, το οποίο δεν περιέχει πολυκροσσωτά κύτταρα, δεν ανιχνεύεται το mrna του γονιδίου GemC1. Το πρότυπο έκφρασης του γονιδίου McIdas οµοιάζει µε αυτό του GemC1, καθώς παρουσιάζει επίσης αυξηµένη έκφραση κατά την ε µβρυογένεση στο επιθήλιο του χοριοειδούς πλέγµατος και στο αναπνευστικό επιθήλιο (Εικόνα Β). Ωστόσο, η έκφραση του mrna του McIdas εντοπίζεται σε όλους τους ιστούς. Ταυτόχρονα ελέγχθηκε το πρότυπο έκφρασης των γονιδίων Geminin και Foxj1. Είναι γνωστό ότι η Geminin εκφράζεται σε όλα τα διαιρούµενα κύτταρα (Spella et al, 2007), ενώ το γονίδιο Foxj1 εµφανίζει ένα ειδικό πρότυπο έκφρασης στα πολυκροσσωτά κύτταρα (Hackett et al, 1995; Lim et al, 1997; Murphy et al, 1997), συµµετέχοντας σε σηµ αντικές διαδικασίες κατά τη δηµ ιουργία τους. To πρότυπο έκφρασης του γονιδίου GemC1 οµοιάζει περισσότερο µε αυτό του παράγοντα Foxj1, ο οποίος επίσης δεν ανιχνεύεται στο ήπαρ, αλλά όπως αναµενόταν εντοπίζεται ειδικά στους ιστούς εκείνους που διαθέτουν πολυκροσσωτά κύτταρα (Εικόνα Γ). 114
115 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Τέλος, όπως φαίνεται στο διάγραµµα Δ της εικόνας 4.1.1, το γονίδιο Geminin εκφράζεται σε όλους τους ιστούς, µε υψηλότερα επίπεδα έκφρασης κατά την εµβρυογένεση του µυός. Από τα παραπάνω συµπεραίνουµε ό τι τα µέλη της υπεροικογένειας της Geminin εκφράζονται σε υψηλά επίπεδα κατά την ανάπτυξη, µε τα γονίδια McIdas και Geminin να ανιχνεύονται σε όλους τους ιστούς, ενώ το γονίδιο GemC1 να εµφανίζει ένα ειδικό πρότυπο έκφρασης µόνο στους ιστούς ε κείνους που π εριλαµβάνουν πολυκροσσωτά κύτταρα. A B Σχετικά επίπεδα έκφρασης mrna ως προς CP adult Γ * * 0 GemC1 Foxj1 Δ McIdas * Geminin 40 0 * CP E12.5 CP E14.5 CP E18.5 CP adult Bronchi E14.5 Bronchi E18.5 Bronchi adult Ovaries with OV adult Liver adult CP E12.5 CP E14.5 CP E18.5 CP adult Bronchi E14.5 Bronchi E18.5 Bronchi adult Ovaries with OV adult Liver adult Εικόνα Έκφραση των µελών της υπεροικογένειας της Geminin κατά την εµβρυογένεση και σε ενήλικες µύες. Μελέτη της έκφρασης του mrna των γονιδίων GemC1 (A), McIdas (B), Foxj1 (Γ) και Geminin (Δ) µέσω ποσοτικής real-time PCR σε ιστούς που αποµονώθηκαν από έµ βρυα και ενήλικες µύες. Οι τιµές αντιστοιχούν στα επίπεδα έκφρασης (+/- τυπική απόκλιση) του κάθε γονιδίου σε κάθε ιστό, σε σχέση µε την αντίστοιχη έκφρασή τους στο χοριοειδές πλέγµα των ενήλικων µυών (CP adult, σηµειώνεται µε *) που ορίζεται ως 1. Η κανονικοποίηση των τιµών έγινε µε τη χρήση των επιπέδων έκφρασης του HPRT γονιδίου και ακολούθησε ανάλυση των αποτελεσµάτων µέσω του προγράµµατος REST-MCS beta. Τα δεδοµένα εξήλθαν από 3 τουλάχιστον ανεξάρτητα πειράµατα. Συντοµογραφίες: CP: χοριοειδές πλέγµα, OV: ωαγωγός 115
116 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Τα γονίδια GemC1 και McIdas εκφράζονται ειδικά στα πολυκροσσωτά κύτταρα του αναπνευστικού επιθηλίου Προκειµένου να προσδιοριστεί η ειδικότητα της έκφρασης των GemC1 και McIdas στα πολυκροσσωτά κύτταρα πραγµατοποιήθηκε η µέθοδος της in situ υβριδοποίησης συνδυαστικά µε τη µέθοδο του ανοσοφθορισµού στο αναπτυσσόµενο αναπνευστικό επιθήλιο µυών. Για το σκοπό αυτό, το αναγνωστικό πλαίσιο των ορθόλογων των GemC1 και McIdas στο µυ κλωνοποιήθηκε στους φορείς έκφρασης pbluescript και pflci αντίστοιχα, οι οποίοι φέρουν υποκινητές για την Τ3 και Τ7 πολυµεράση (Προπτυχιακή Διπλωµατική Εργασία, Κυρούση Χριστίνα, 2008). Οι φορείς αυτοί χρησιµοποιήθηκαν για την παραγωγή σηµασµένων νοηµατικών και αντι-νοηµατικών ιχνηθετών RNA (sense και anti-sense RNA probes). Στη συνέχεια, σε εγκάρσιες τοµές εµβρύου µυός, ε µβρυϊκής ηλικίας 18.5 ηµερών (E18.5), ανιχνεύθηκε το πρότυπο έκφρασης του mrna των GemC1 και McIdas µέσω in situ υβριδοποίησης και συγκρίθηκε µε την έκφραση της πρωτεΐνης Foxj1, η οποία εντοπίστηκε µέσω ανοσοφθορισµού µε τη χρήση ειδικού αντισώµατος (Εικόνα 4.1.2). Η ειδικότητα του σήµατος ελέγχθηκε µε τη χρήση νοηµατικών ιχνηθετών στους οποίους δεν ανιχνεύτηκε σήµα. Παρατηρήθηκε ότι η πλειοψηφία των κυττάρων που εκφράζουν το mrna των GemC1 (Εικόνα Α-A ) και McIdas (Εικόνα B-B ) εξέφραζαν παράλληλα και την πρωτεΐνη Foxj1. Συµπεραίνουµε ότι η έκφραση των γονιδίων GemC1 και McIdas ανιχνεύεται ειδικά στα πολυκροσσωτά κύτταρα του αναπτυσσόµενου αναπνευστικού επιθηλίου. 116



 και McIdas (Β-Β ) και ακολούθησε")
.")

117 4. ΑΠΟΤΕΛΕΣΜΑΤΑ A E18.5 A GemC1/Foxj1 A B E18.5 B McIdas/Foxj1 B Εικόνα Τα γονίδια GemC1 και McIdas εκφράζονται στα πολυκροσσωτά κύτταρα του αναπτυσσόµενου αναπνευστικού επιθηλίου. Σε εγκάρσιες τοµές εµβρύου µυός ηλικίας 18.5 ηµερών (Ε18.5) πραγµατοποιήθηκε in situ υβριδοποίηση για την ανίχνευση του mrna των GemC1 (Α-Α ) και McIdas (Β-Β ) και ακολούθησε ανοσοφθορισµός µε ειδικό αντίσωµα έναντι της πρωτεΐνης Foxj1 (πράσινο). Παρατηρείται έκφραση του mrna των GemC1 και McIdas στο επίπεδο των αεραγωγών. Στην πλειοψηφία των κυττάρων που εκφράζονται τα γονίδια GemC1 και McIdas συνεκφράζεται και η πρωτεΐνη Foxj1, η οποία είναι γνωστό ότι εντοπίζεται στα πολυκροσσωτά κύτταρα του αναπνευστικού επιθηλίου. Scale bar, 10µm. 117
118 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Μελέτη της έκφρασης των µελών της υπεροικογένειας της Geminin κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων Ex vivo καλλιέργεια των επιθηλιακών κυττάρων της τραχείας ενήλικων µυών Η παρακολούθηση της διαφοροποίησης των πολυκροσσωτών κυττάρων δυσχεραίνεται από την έλλειψη κυτταρικών σειρών που να µπορούν να διαφοροποιηθούν προς αυτόν τον κυτταρικό τύπο in vitro, ενώ η µελέτη τ ης δηµιουργίας των κροσσών in vivo περιπλέκεται από την ασύγχρονη διαφοροποίηση του αναπνευστικού επιθηλίου κατά µήκος των αεραγωγών κατά την ανάπτυξη. Στην παρούσα διατριβή, µε στόχο να µελετηθεί η έκφραση και η λειτουργία των µελών της υπεροικογένειας της Geminin κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, χρησιµοποιήθηκε ένα ex vivo σύστηµα το οποίο παρέχει τη δυνατότητα καλλιέργειας επιθηλιακών κυττάρων που αποµονώνονται από την τραχεία είτε αγρίου τύπου, είτε γενετικά τροποποιηµένων µυών και τη διαφοροποίησή τους προς πολυκροσσωτά κύτταρα (You et al, 2002). Κατά την καλλιέργεια των κυττάρων αυτών µπορούν να πραγµατοποιηθούν µελέτες αποσιώπησης ή υπερέκφρασης γονιδίων, καθώς και χρήση φαρµακευτικών αναστολέων. Μιµούµενη τη φυσιολογική διαφοροποίηση του αναπνευστικού επιθηλίου, η καλλιέργεια των κυττάρων αυτών απαιτεί την παρουσία των κατάλληλων αυξητικών παραγόντων (κυρίως της ινσουλίνης, του επιδερµικού αυξητικού παράγοντα EGF και του ρετινοϊκού οξέος), καθώς και την αποµόνωση ένας ικανοποιητικού αριθµού κυττάρων που θα συµ βάλλει στον επιτυχή πολλαπλασιασµό τους (You & Brody, 2013). Μετά την αποµόνωση των κυττάρων εντοπίζονται στην καλλιέργεια τόσο διαφοροποιηµένα κύτταρα (πολυκροσσωτά και εκκριτικά Clara κύτταρα), όσο και βασικά κύτταρα που διατηρούν την ικανότητα της κυτταρικής διαίρεσης. Ωστόσο, τα τελικώς διαφοροποιηµένα κύτταρα πεθαίνουν σε διάστηµα περίπου 3 ηµερών µετά την αποµόνωσή τους, µε αποτέλεσµα ένας ορισµένος αριθµός διαιρούµενων βασικών κυττάρων να προσκολλάται τελικά στη µεµβράνη καλλιέργειας και να καταφέρνει να πολλαπλασιαστεί. Η διαφοροποίηση των κυττάρων προς πολυκροσσωτά εξαρτάται από τον επιτυχή πολλαπλασιασµό τους και την καλλιέργειά τους µε τρόπο κατά τον οποίο η κορυφαία επιφάνειά τους να εκτίθεται στον αέρα (Air Liquid Interface, ALI) (You & Brody, 2013). Πιο συγκεκριµένα, α υτό επιτυγχάνεται µέσω της καλλιέργειας των επιθηλιακών κυττάρων που αποµονώνονται από την τραχεία σε ειδικές ηµιδιαπερατές µεµβράνες, ένθετες σε πηγαδάκια πολυκυψελιδικών πιάτων, µε αποτέλεσµα να επιτρέπεται κατά το στάδιο του πολλαπλασιασµού να τους χορηγείται ένα πλούσιο σε αυξητικούς παράγοντες θρεπτικό υλικό, τόσο στην κορυφαία όσο και στη βασική πλευρά της µεµβράνης καλλιέργειας (Εικόνα 4.1.3). Η επαγωγή της διαφοροποίησης ξεκινά 5 έως 7 ηµέρες µετά από την έναρξη της καλλιέργειας, όπου τα κύτταρα έχουν πολλαπλασιαστεί αρκετά και έχουν καταλάβει όλη την επιφάνεια της µεµβράνης πάνω στην οποία αναπτύσσονται. Κατά το στάδιο αυτό το θρεπτικό υλικό αναρροφάται από την κορυφαία πλευρά της µεµβράνης και τα κύτταρα 118
119 4. ΑΠΟΤΕΛΕΣΜΑΤΑ αναπτύσσονται παρουσία θρεπτικού υλικού που χορηγείται µόνο στη βασική τους πλευρά. Η επαφή των κυττάρων µε τον αέρα, σε συνδυασµό µε την παροχή θρεπτικού υλικού φτωχού σε αυξητικούς παράγοντες και απουσία ορού, επάγει τα κύτταρα να διαφοροποιηθούν. Κορυφαία επιφάνεια καλλιέργειας Βασική επιφάνεια καλλιέργειας Ημιδιαπερατή μεμβράνη καλλιέργειας Εικόνα Ex vivo καλλιέργεια των επιθηλιακών κυττάρων της τραχείας ενήλικων µυών. Τα επιθηλιακά κύτταρα που αποµονώνονται από την τραχεία ενήλικων µυών καλλιεργούνται σε ειδικές ηµιδιαπερατές µεµβράνες µε τρόπο ώστε να τους παρέχεται θρεπτικό υλικό τόσο στην κορυφαία όσο και στη βασική πλευρά της µεµβράνης καλλιέργειας. Μετά τον επιτυχή πολλαπλασιασµό τους και αφού καταλάβουν πλήρως την επιφάνεια της µεµβράνης που καλλιεργούνται, αφαιρείται το θρεπτικό υλικό που τους παρέχεται στην κορυφαία τους πλευρά. Έτσι τα κύτταρα εκτίθενται στον αέρα (Air Liquid Interface, ALI), ενώ τους χορηγείται θρεπτικό υλικό φτωχό σε ορό και αυξητικούς παράγοντες µόνο στη βασική πλευρά. Οι συγκεκριµένες συνθήκες καλλιέργειας ευνοούν τη διαφοροποίηση των κυττάρων προς πολυκροσσωτά (You et al, 2002). Οι παραπάνω συνθήκες καλλιέργειας ευνοούν τη διαφοροποίηση των κυττάρων προς τη µοίρα των πολυκροσσωτών κυττάρων και όχι προς βλεννοπαραγωγά (goblet) ή εκκριτικά (clara) κύτταρα. Αρχικά, η καλλιέργεια απαρτίζεται κυρίως από διαιρούµενα προγονικά κύτταρα και από έναν πολύ µικρό αριθµό των προαναφερθέντων κυττάρων. Μετά την επαγωγή της διαφοροποίησης ξεκινά άµεσα ο πολλαπλασιασµός των κεντριολίων, η µετανάστευσή τους στην κορυφαία επιφάνεια των κυττάρων και η δηµιουργία κινητών κροσσών. Σε διάστηµα επτά έως δεκατεσσάρων ηµερών τα κύτταρα έχουν διαφοροποιηθεί πλήρως (You & Brody, 2013). Συνολικά, οι διαφορετικοί κυτταρικοί τύποι που απαντώνται στην καλλιέργεια πριν και µετά την επαγωγή της διαφοροποίησης είναι οι εξής (Εικόνα 4.1.4) (Vladar & Brody, 2013): Μη πολυκροσσωτά κύτταρα: Τα κύτταρα αυτά διαθέτουν δυο κεντριόλια στην κορυφαία τους πλευρά χωρίς να φέρουν πρωτογενή κροσσό. Πρόκειται για µη διαφοροποιηµένα κύτταρα που έχουν την ικανότητα κυτταρικής διαίρεσης (βασικά κύτταρα) και απαντώνται σε καλλιέργειες που δεν έχει γίνει επαγωγή της διαφοροποίησης ή σε καλλιέργειες που έγινε επαγωγή και εποµένως είτε είναι µη διαφοροποιηµένα κύτταρα ή έχουν διαφοροποιηθεί προς άλλους κυτταρικούς τύπους, όπως βλεννοπαραγωγά ή εκκριτικά. Τα κύτταρα αυτά δεν εκφράζουν τον παράγοντα Foxj1. 119
120 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Κύτταρα µε πρωτογενή κροσσό: Τα κύτταρα αυτά διαθέτουν δυο κεντριόλια στην κορυφαία τους πλευρά µε το µητρικό κεντριόλιο να δρα ως βασικό σωµάτιο από το οποίο εκφύεται ένας µικρού µήκους πρωτογενής κροσσός. Τέτοια κύτταρα απαντώνται είτε σε καλλιέργειες που είναι ακόµα στο στάδιο του πολλαπλασιασµού, είτε µετά την επαγωγή της διαφοροποίησης και πρόκειται να διαφοροποιηθούν προς πολυκροσσωτά. Τα κύτταρα αυτά αρχικά είναι αρνητικά ως προς την έκφραση του Foxj1. Στάδιο Ι: Τα κύτταρα στο στάδιο αυτό διαθέτουν δυο κεντριόλια στην κορυφαία τους πλευρά και από το µητρικό κεντριόλιο εκφύεται ένας ασυνήθιστα µεγάλου µήκους πρωτογενής κροσσός. Τα κύτταρα είναι α ρνητικά ως προς την έκφραση του παράγοντα Foxj1. Τέτοια κύτταρα ξεκινούν να ανιχνεύονται περίπου κατά τη 2 η ηµέρα από την έναρξη της καλλιέργειας, ενώ µειώνονται σταδιακά έως τη 10 η ηµέρα της καλλιέργειας. Στάδιο ΙΙ: Στο στάδιο αυτό τα κύτταρα πολλαπλασιάζουν µαζικά τα κεντριόλιά τους µε αποτέλεσµα να σχηµατίζεται σε καθένα ένας µεγάλος αριθµός από νεοσυντιθέµενα κεντριόλια. Τα κύτταρα αυτά εκφράζουν τον παράγοντα Foxj1 και παρατηρούνται αρχικά κατά την 3 η ηµέρα της διαφοροποίησης, ενώ µειώνονται σταδιακά έως τη 12 η ηµέρα. Στάδιο ΙΙΙ: Κατά το στάδιο αυτό τα πολλαπλά κεντριόλια που έχουν παραχθεί σε κάθε κύτταρο µετακινούνται προς την κορυφαία επιφάνειά τους, µε σκοπό να αγκυροβολήσουν στην πλασµατική µεµβράνη. Τα κύτταρα παρουσιάζουν υψηλή έκφραση του παράγοντα Foxj1. Τέτοια κύτταρα εντοπίζονται από την 4 η ηµέρα της διαφοροποίησης και ύστερα, ενώ είναι µειωµένα κατά τη 14 η ηµέρα. Στάδιο IV: Στο στάδιο αυτό τα κεντριόλια που έχουν µεταναστεύσει στη µεµβράνη των κυττάρων κατανέµονται οµοιόµορφα σε αυτήν και δρουν ως βασικά σωµάτια. Από κάθε βασικό σωµάτιο εκφύεται ένας κροσσός. Σε αυτό το στάδιο θα προκύψουν τελικώς διαφοροποιηµένα, ώριµα πολυκροσσωτά κύτταρα που φέρουν στην επιφάνειά τους δεκάδες κινητούς κροσσούς. Ο παράγοντας Foxj1 συνεχίζει να εκφράζεται σε πολύ υψηλά επίπεδα. Τέτοια κύτταρα ξεκινούν να δηµιουργούνται κατά την 4 η ηµέρα της διαφοροποίησης, ενώ µετά τη 14 η ηµέρα η καλλιέργεια αποτελείται κατά κύριο λόγο από κύτταρα αυτού του σταδίου. Αρχικά, οι κροσσοί που δηµιουργούνται είναι ακίνητοι και ποικίλου µήκους, ενώ από την 7 η ηµέρα και µετά γίνονται κινητοί. 120
121 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Πρωτογενής κροσσός Στάδιο I Στάδιο II Στάδιο III Πρώιμα στάδια διαφοροποίησης Στάδιο IV Ώριμο πολυκροσσωτό κύτταρο Εικόνα Σχηµατική αναπαράσταση των σταδίων της διαφοροποίησης των πολυκροσσωτών κυττάρων. Κατά το στάδιο του πολλαπλασιασµού ή αµέσως µετά την επαγωγή της διαφοροποίησης παρατηρούνται κύτταρα µε έναν µικρού µήκους πρωτογενή κροσσό, τα οποί α θα διαφοροποιηθούν σε πολυκροσσωτά. Στη συνέχεια από τα κύτταρα αυτά εκφύεται από το µητρικό κεντριόλιο ένας ασυνήθιστα µακρύς πρωτογενής κροσσός. Τα κύτταρα αυτά δεν εκφράζουν ακόµα τον παράγοντα Foxj1. Κατά το στάδιο ΙΙ ξεκινούν να πολλαπλασιάζονται µαζικά τα κεντριόλια των κυττάρων και παρατηρούνται χαµηλά επίπεδα έκφρασης του Foxj1 (αχνή µπλε πυρηνική χρώση), ενώ κατά τη µετανάστευση των κεντριολίων στη βασική µεµβράνη του κυττάρου (Στάδιο ΙΙΙ ), τα επίπεδά του ανιχνεύονται σε υψηλά επίπεδα (έντονη µπλέ πυρηνική χρώση). Τέλος, κατά το στάδιο IV προκύπτουν ώριµα πολυκροσσωτά κύτταρα από τα οποία εκφύονται δεκάδες έως εκατοντάδες κινητοί κροσσοί. Προκειµένου να χρησιµοποιηθεί το παραπάνω σύστηµα για τη µελέτη της έκφρασης των µελών της οικογένειας της Geminin, πραγµατοποιήθηκε καλλιέργεια των επιθηλιακών κυττάρων της τραχείας ενήλικων µυών και έγινε ταυτοποίηση των παραπάνω σταδίων. Τα κύτταρα αποµονώθηκαν, επιστρώθηκαν στις ειδικές µεµβράνες καλλιέργειας, πολλαπλασιάστηκαν για 7 ηµέρες και ακολούθησε επαγωγή της διαφοροποίησής τους. Στη συνέχεια, έγινε µονιµοποίηση των κυττάρων στις µεµβράνες, σε διαφορετικές χρονικές στιγµές πριν και µετά την επαγωγή της διαφοροποίησης. Η µελέτη των κυττάρων κατά τα διάφορα στάδια της διαφοροποίησης πραγµατοποιήθηκε µέσω ανοσοφθορισµού, µε τη χρήση ειδικών αντισωµάτων έναντι της ακετυλιωµένης α -τουµπουλίνης (ACT) για τη χρώση του αξονήµατος των κροσσών και της γ -τουµπουλίνης για τη χρώση των κεντριολίωνβασικών σωµατίων (Εικόνα 4.1.5). Όπως παρατηρείται στην εικόνα 4.1.5, πολύ νωρίς κατά το στάδιο του πολλαπλασιασµού (ALI DAY -5) ανιχνεύεται ένας µικρός αριθµός πολυκροσσωτών κυττάρων. Τα κύτταρα αυτά παρέµειναν στη µεµβράνη µετά την αποµόνωσή τους από την τραχεία. Αργότερα, τα διαφοροποιηµένα κύτταρα πεθαίνουν και τελικά παραµένουν κύτταρα τα οποία κατά ένα µεγάλο ποσοστό φέρουν έναν πρωτογενή κροσσό (ALI DAY 0 & ALI DAY 3). Tα πρώτα πολυκροσσωτά κύτταρα παρατηρούνται έξι ηµέρες µετά την επαγωγή της διαφοροποίησης (ALI DAY 6), τα οποία στη συνέχεια αυξάνονται σε αριθµό και ωριµάζουν. Κατά την ALI DAY 21 η διαδικασία της διαφοροποίησης έχει ολοκληρωθεί και η καλλιέργεια διαθέτει ένα µεγάλο αριθµό ώριµων πολυκροσσωτών κυττάρων. 121

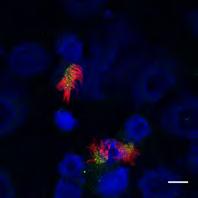

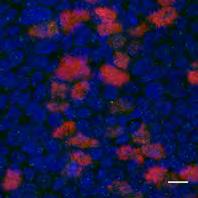
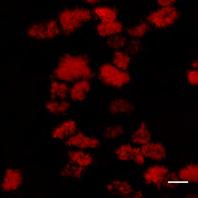
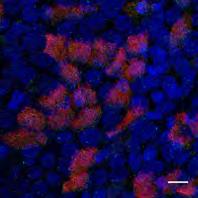
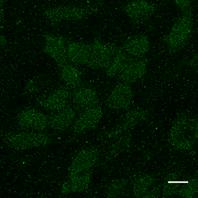
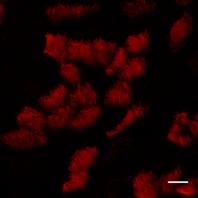
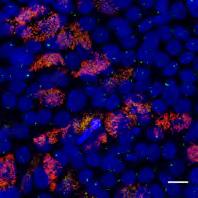
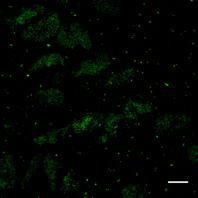
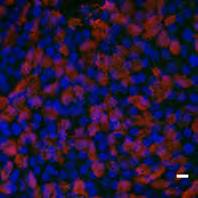
122 4. ΑΠΟΤΕΛΕΣΜΑΤΑ ACT/γ-tubulin/Draq-5 AD 21 AD 14 AD 9 AD 6 AD 3 AD 0 AD
123 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Στάδια διαφοροποίησης των πολυκροσσωτών κυττάρων κατά την ex vivo καλλιέργεια των επιθηλιακών κ υττάρων της τραχείας ενήλικων µυών. Κύτταρα αποµονώνονται από την τραχεία ενήλικων µυών και καλλιεργούνται σε ειδικές µεµβράνες. Οι συνθήκες καλλιέργειας ευνοούν αρχικά τον πολλαπλασιασµό τους, ενώ στη συνέχεια µέσω της έκθεσης των κυττάρων στον αέρα (ALI) και µε τη χρήση κατάλληλης σύστασης θρεπτικού υλικού επάγεται η διαφοροποίησή τους. Για τον εντοπισµό των πολυκροσσωτών κυττάρων χρησιµοποιήθηκαν αντισώµατα έναντι της ακετυλιωµένης α -τουµπουλίνης (ACT, κόκκινο) για τη χρώση του αξονήµατος των κροσσών και έναντι της γ -τουµπουλίνης (πράσινο) για τη χρώση των κεντριολίωνβασικών σωµατίων από τα οποία εκφύονται οι κροσσοί. Κατά την έναρξη της καλλιέργειας (ALI DAY -5) εντοπίζονται κύτταρα που φέρουν κινητούς κροσσούς, τα οποία στη συνέχεια πεθαίνουν. Κατά τα πρώτα στάδια της διαφοροποίησης ανιχνεύονται κύτταρα που διαθέτουν πρωτογενή κροσσό (ALI DAY 0&3), ενώ στη συνέχεια ένα µεγάλο ποσοστό των κυττάρων της καλλιέργειας φέρει πολλαπλούς κροσσούς που εκφύονται από πολλαπλά βασικά σωµάτια. Όλο και περισσότερα κύτταρα διαφοροποιούνται κατά την εξέλιξη της καλλιέργειας και τελικά προκύπτουν ώριµα πολυκροσσωτά κύτταρα (ALI DAY 21). Η χρώση των πυρήνων των κυττάρων έγινε µε τη χρωστική Drap-5. Scale bar, 10µm. Συντοµογραφίες: AD: ALI DAY Μελέτη της έκφρασης των µελών της υπεροικογένειας της Geminin κατά την ex vivo καλλιέργεια τ ων επιθηλιακών κυττάρων της τραχείας ενήλικων µυών Προκειµένου να µελετηθεί η έκφραση των γονιδίων GemC1, McIdas και Geminin κατά τα διάφορα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων, χρησιµοποιήθηκε το ex vivo σύστηµα καλλιέργειας των επιθηλιακών κυττάρων της τραχείας ενήλικων µυών που περιγράφηκε προηγουµένως. Για το σκοπό αυτό αποµονώθηκε ολικό RNA από κύτταρα που βρίσκονταν σε διαφορετικές χρονικές στιγµές, πριν και µετά την επαγωγή της διαφοροποίησης. Ακολούθησε σύνθεση cdna και µε τη χρήση ειδικών µορίων-εκκινητών ελέγχθηκε το πρότυπο έκφρασης των παραγόντων GemC1, McIdas, Foxj1 και Geminin µέσω ποσοτικής realtime PCR (Εικόνα 4.1.6). Όπως απεικονίζεται και στο διάγραµµα Α της εικόνας δεν ανιχνεύεται έκφραση του παράγοντα GemC1 κατά το στάδιο του πολλαπλασιασµού των κυττάρων (ALI DAY -2, ALI DAY 0). Όµως, αµέσως µετά την επαγωγή της διαφοροποίησης, η έκφρασή του αρχίζει να αυξάνεται σταδιακά, ενώ ελαττώνεται σηµαντικά κατά το τέλος της διαδικασίας, όπου έχουν πια σχηµατιστεί ώριµα πολυκροσσωτά κύτταρα (ALI DAY 21). Το πρότυπο έκφρασης του παράγοντα McIdas οµοιάζει µε αυτό του παράγοντα GemC1 (Εικόνα Β), µε τη διαφορά ότι ανιχνεύονται χαµηλά επίπεδα έκφρασής του και κατά το στάδιο του πολλαπλασιασµού. Τα γονίδια GemC1 και McIdas παρουσιάζουν επίσης παρόµοιο πρότυπο έκφρασης µε αυτό του παράγοντα 123
124 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Foxj1 (Εικόνα Γ), ο οποίος είναι γνωστό ότι διαδραµατίζει καθοριστικό ρόλο στη δηµιουργία των πολυκροσσωτών κυττάρων. Ιδιαίτερο ενδιαφέρον παρουσιάζει το ανεστραµµένο πρότυπο έκφρασης που εµφανίζει το γονίδιο της Geminin, αφού η έκφρασή του µειώνεται καθώς τα κύτταρα εξέρχονται του κυτταρικού κύκλου και διαφοροποιούνται προς πολυκροσσωτά (Εικόνα Δ). Τα παραπάνω δεδοµένα µας οδηγούν στο συµπέρασµα ότι οι παράγοντες GemC1 και McIdas εκφράζονται ειδικά κατά τη διάρκεια της διαφοροποίησης των πολυκροσσωτών κυττάρων, υποδηλώνοντας τη συµµετοχή τους στη διαδικασία αυτή, ενώ η Geminin εµφανίζει ένα αντίθετο πρότυπο έκφρασης υποδεικνύοντας έναν ανταγωνιστικό ρόλο σε σχέση µε τα άλλα δυο µέλη της υπεροικογένειας. A B Σχετικά επίπεδα έκφρασης mrna ως προς ALI DAY Γ * * GemC1 Foxj Δ McIdas * * Geminin ALI DAY ALI DAY Εικόνα Έκφραση των µελών της υπεροικογένειας της Geminin κατά την ex vivo διαφοροποίηση των πολυκροσσωτών κυττάρων της τραχείας µυών. Τα επιθηλιακά κύτταρα της τραχείας ενήλικων µυών αποµονώθηκαν και καλλιεργήθηκαν σε ειδικές µεµβράνες µε σκοπό τη διαφοροποίησή τους σε πολυκροσσωτά. Μέσω ποσοτικής real-time PCR µετρήθηκαν τα επίπεδα έκφρασής των GemC1 (A), McIdas (B), Foxj1 (Γ) και Geminin (Δ) σε διαφορετικές χρονικές στιγµές, πριν και κατά τη διάρκεια της διαφοροποίησής τους. Οι τιµές αντιστοιχούν στα επίπεδα έκφρασης (+/- τυπική απόκλιση) του κάθε γονιδίου σε σχέση µε την αντίστοιχη έκφρασή τους µια ηµέρα µετά τη διαφοροποίηση (ALI DAY 1, σηµειώνεται µε *) που ορίζεται ως τιµή 1. Η κανονικοποίηση των τιµών έγινε µε τη χρήση των επιπέδων έκφρασης του HPRT γονιδίου και ακολούθησε ανάλυση των αποτελεσµάτων µέσω του προγράµµατος REST- MCS beta. Τα δεδοµένα εξήλθαν από 3 τουλάχιστον ανεξάρτητα πειράµατα. 124
125 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Η πρωτεΐνη McIdas εκφράζεται κατά τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Προκειµένου να µελετηθεί η έκφραση της πρωτεΐνης McIdas κατά τη διάρκεια της διαφοροποίησης των πολυκροσσωτών κυττάρων σε επίπεδο µεµονωµένων κυττάρων, πραγµατοποιήθηκαν πειράµατα ανοσοφθορισµού σε αποµονωµένα επιθηλιακά κύτταρα της τραχείας µυών πριν και µετά την επαγωγή της διαφοροποίησης. Για το σκοπό αυτό χρησιµοποιήθηκε πολυκλωνικό αντίσωµα που αναγνωρίζει ειδικά την πρωτεΐνη McIdas (Pefani et al, 2011), καθώς και αντίσωµα έναντι του παράγοντα Foxj1, που εκφράζεται στα κύτταρα εκείνα που θα διαφοροποιηθούν σε πολυκροσσωτά, καθώς και σε αργότερα στάδια της διαφοροποίησής τους. Η πρωτεΐνη McIdas δεν ανιχνεύεται κατά το στάδιο του πολλαπλασιασµού των κυττάρων (Εικόνα Α&Β, ALI DAY -2 & ALI DAY 0), όµως ξεκινά να εκφράζεται σε έναν περιορισµένο αριθµό κυττάρων αµέσως µετά την επαγωγή της διαφοροποίησης (ALI DAY 1). Η πλειοψηφία των κυττάρων που εκφράζουν McIdas, εκφράζουν ταυτόχρονα και την πρωτεΐνη Foxj1. Ωστόσο, σε αυτή τη χρονική στιγµή, ανιχνεύονται και κύτταρα θετικά ως προς την έκφραση του McIdas, τα οποία όµως δεν εκφράζουν ακόµα Foxj1. Στην πορεία της διαφοροποίησης υπάρχει ένα επικαλυπτόµενο πρότυπο έκφρασης των McIdas και Foxj1 πρωτεϊνών στα κύτταρα (ALI DAY 3), ενώ σε µεταγενέστερες χρονικές στιγµές κατά τη διαφοροποίηση, η έκφραση της πρωτεΐνης McIdas περιορίζεται σηµαντικά (ALI DAY 6), µε αποτέλεσµα να εκφράζεται σε ελάχιστα κύτταρα κατά την ολοκλήρωση της διαδικασίας (ALI DAY 14). Αντίθετα, η έκφραση της πρωτεΐνης Foxj1 παραµένει σε περισσότερα κύτταρα, µιας και είναι γνωστό ότι εκφράζεται και στα διαφοροποιηµένα πολυκροσσωτά κύτταρα. Από τα παραπάνω συµπεραίνουµε ότι η πρωτεΐνη McIdas ξεκινά να εκφράζεται σε πρώιµα στάδια κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων και η έκφρασή της προηγείται της έκφρασης του Foxj1. Φαίνεται επίσης ότι η έκφραση του McIdas περιορίζεται σε µεταγενέστερα στάδια της διαφοροποίησης, όπου εντοπίζονται ώριµα πολυκροσσωτά κύτταρα. 125
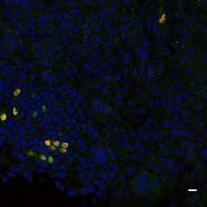
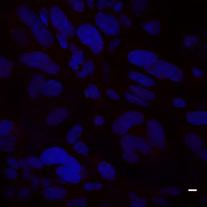
126 4. ΑΠΟΤΕΛΕΣΜΑΤΑ A McIdas/Foxj1/Draq-5 ALI DAY 14 ALI DAY 6 ALI DAY 3 ALI DAY 1 ALI DAY 0 ALI DAY
127 4. ΑΠΟΤΕΛΕΣΜΑΤΑ B Ποσοστό Idas+ ή Foxj+ κυττάρων ALI DAY -2 ALI DAY 0 ALI DAY 1 Εικόνα Η πρωτεΐνη McIdas εκφράζεται κατά τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. (Α) Κύτταρα αποµονώθηκαν από την τραχεία ενήλικων µυών και καλλιεργήθηκαν κάτω από ειδικές συνθήκες που ευνοούν τη διαφοροποίησή τους προς τη µοίρα των πολυκροσσωτών κυττάρων. Μέσω ανοσοφθορισµού και µε τη χρήση ειδικών αντισωµάτων έναντι των πρωτεϊνών McIdas (πράσινο) και Foxj1 (κόκκινο) µελετήθηκε η έκφρασή τους κατά τη διάρκεια της διαφοροποίησης. Δεν ανιχνεύεται έκφραση των πρωτεϊνών κατά το στάδιο του πολλαπλασιασµού (ALI DAY -2&0), ενώ αµέσως µετά την επαγωγή της διαφοροποίησης ξεκινoύν να εκφράζονται (ALI DAY 1), µε την έκφραση του McIdas να προηγείται αυτής του Foxj1. Τα επίπεδα έκφρασης της πρωτεΐνης McIdas µειώνονται κατά τη διάρκεια της διαφοροποίησης. Αντίθετα, η πρωτεΐνη Foxj1 εκφράζεται και σε µεταγενέστερα στάδια. Η χρώση των πυρήνων των κυττάρων έγινε µε τη χρωστική Draq-5. Τα βέλη υποδεικνύουν κύτταρα στα οποία προηγείται η έκφραση της πρωτεΐνης McIdas αυτής του Foxj1. (Β) Ποσοστό κυττάρων που εκφράζουν την πρωτεΐνη McIdas ή Foxj1 κατά τα διάφορα στάδια της διαφοροποίησης των επιθηλιακών κυττάρων της τραχείας µυών. Scale bar, 10µm. ALI DAY 3 ALI DAY 6 Idas Foxj1 ALI DAY 14 Παράλληλα µελετήθηκε και συγκρίθηκε το πρότυπο έκφρασης της πρωτεΐνης McIdas µε αυτό του µεταγραφικού παράγοντα cmyb, που συµµετέχει σηµαντικά στον καθορισµό της µοίρας και της δηµιουργίας των πολυκροσσωτών κυττάρων (Pan et al, 2014; Tan et al, 2013). Όπως φαίνεται και στην εικόνα δεν ανιχνεύονται επίπεδα έκφρασης των πρωτεϊνών McIdas και cmyb κατά το στάδιο του πολλαπλασιασµού των κυττάρων (ALI DAY -2). Όµως αµέσως µετά την επαγωγή της διαφοροποίησης ξεκινούν να εκφράζονται (ALI DAY 1) και παρατηρείται συνεντοπισµός της έκφρασής τους, µε τα επίπεδα του cmyb να περιορίζονται πιο δραστικά από αυτά του McIdas, κατά τα µεταγενέστερα στάδια της διαφοροποίησης (ALI DAY 14). 127
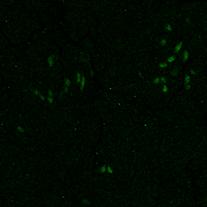
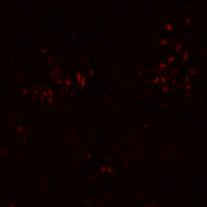

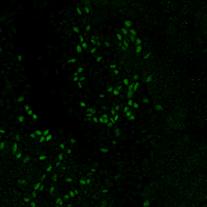
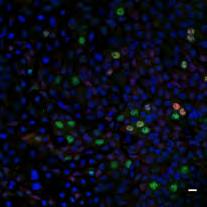
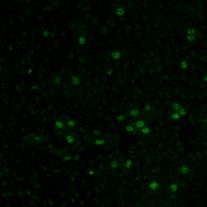
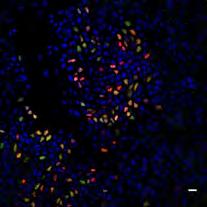
128 4. ΑΠΟΤΕΛΕΣΜΑΤΑ McIdas/cMyb/Draq-5 ALI DAY 14 ALI DAY 3 ALI DAY 1 ALI DAY 0 ALI DAY
129 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Η πρωτεΐνη McIdas συνεντοπίζεται µε το µεταγραφικό παράγοντα cmyb κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Κύτταρα αποµονώθηκαν από την τραχεία ενήλικων µυών και καλλιεργήθηκαν κάτω από ειδικές συνθήκες π ου ευνοούν τη διαφοροποίησή τους προς τη µοίρα των πολυκροσσωτών κυττάρων. Μέσω ανοσοφθορισµού και µε τη χρήση ειδικών αντισωµάτων έναντι των πρωτεϊνών McIdas (πράσινο) και cmyb (κόκκινο) µελετήθηκε η έκφρασή τους κατά τη διάρκεια της διαφοροποίησης. Δεν ανιχνεύεται έκφραση των πρωτεϊνών κατά το στάδιο του πολλαπλασιασµού των κυττάρων (ALI DAY -2&0). Αµέσως µετά την επαγωγή της διαφοροποίησης ξεκινoύν να εκφράζονται (ALI DAY 1). Η πλειοψηφία των κυττάρων που εκφράζουν την πρωτεΐνη McIdas, εκφράζουν παράλληλα και τον παράγοντα cmyb (ALI DAY 1&3). Όµως, κατά τη διάρκεια της διαφοροποίησης, τα επίπεδα έκφρασης της πρωτεΐνης cmyb µειώνονται πιο γρήγορα σε σχέση µε αυτά του McIdas. Η χρώση των πυρήνων των κυττάρων έγινε µε τη χρωστική Draq-5. Scale bar, 10µm Η έκφραση της πρωτεΐνης McIdas περιορίζεται σηµαντικά στα µεταγενέστερα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Τα παραπάνω πειράµατα έδειξαν ότι ο παράγοντας McIdas εκφράζεται νωρίς κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, ενώ η έκφρασή του περιορίζεται σηµαντικά στα ύστερα στάδια της διαδικασίας αυτής. Η µελέτη πραγµατοποιήθηκε συγκριτικά µε το πρότυπο έκφρασης των παραγόντων Foxj1 και cmyb, οι οποίοι όπως προαναφέρθηκε εκφράζονται νωρίς στα κύτταρα που θα γίνουν πολυκροσσωτά, µε τον παράγοντα Foxj1 να εντοπίζεται και αργότερα κατά τη διαφοροποίησή τους. Προκειµένου να διερευνηθεί περαιτέρω σε ποια στάδια της διαδικασίας εκφράζεται συγκεκριµένα η πρωτεΐνη McIdas, µελετήθηκε ξανά η έκφρασή της παράλληλα µε την έκφραση της ακετυλιωµένης α -τουµπουλίνης (acetylated α-tubulin, ACT), η οποία αποτελεί δείκτη των κεντριολίων και του αξονήµατος του κροσσών. Για τ ο σκοπό αυτό πραγµατοποιήθηκαν πειράµατα ανοσοφθορισµού, σε διαφορετικές χρονικές στιγµές, πριν και κατά τη διάρκεια της διαφοροποίησης (Εικόνα 4.1.9). Όπως αναµενόταν από προηγούµενα δεδοµένα, η πρωτεΐνη McIdas δεν εκφράζεται κατά το στάδιο του πολλαπλασιασµού των κυττάρων (ALI DAY -5 & ALI DAY 0), όµως ξεκινά να εκφράζεται µετά την επαγωγή της διαφοροποίησης των κυττάρων (ALI DAY 3) (Εικόνα Α&Β). Προσεκτική παρατήρηση έδειξε ότι όλα τα κύτταρα που εκφράζουν την πρωτεΐνη McIdas διαθέτουν έναν πρωτογενή κροσσό, αποτέλεσµα που καταδεικνύει ότι η έκφραση του McIdas εντοπίζεται πολύ πρώιµα, στα κύτταρα εκείνα που πρόκειται να διαφοροποιηθούν σε πολυκροσσωτά (ALI DAY 3). Αντίθετα, παρατηρήθηκε ότι σε µεταγενέστερα στάδια της διαφοροποίησης τα ώριµα πολυκροσσωτά κύτταρα δεν εκφράζουν πια την πρωτεΐνη McIdas (Εικόνα Β, ALI DAY 6, 11 & 14). Συµπεραίνουµε ότι η πρωτεΐνη McIdas εκφράζεται στα πρώιµα στάδια κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, ενώ η έκφρασή της χάνεται στα ώριµα πολυκροσσωτά κύτταρα. 129











 η έκφραση του McIdas")


.")





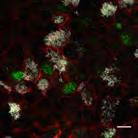
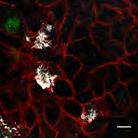
130 4. ΑΠΟΤΕΛΕΣΜΑΤΑ A AD -2 AD 0 AD 3 AD 6 AD 11 AD 14 B AD 3 AD 6 AD 11 AD 14 McIdas/ACT E-cadherin McIdas/ACT/E-cadherin Εικόνα Η πρωτεΐνη McIdas εκφράζεται νωρίς κατά τη διαφοροποίηση, ενώ η έκφρασή της χάνεται στα ώριµα πολυκροσσωτά κύτταρα. Κύτταρα αποµονώθηκαν από την τραχεία ενήλικων µυών και καλλιεργήθηκαν κάτω από ειδικές συνθήκες που ευνοούν τη διαφοροποίησή τ ους προς τη µοίρα των πολυκροσσωτών. Μέσω ανοσοφθορισµού και µε τη χρήση ειδικών αντισωµάτων έναντι των πρωτεϊνών McIdas (πράσινο), E-cadherin (κόκκινο) για τη χρώση της κυτταρικής µεµβράνης ώστε να διευκολύνεται η οριοθέτηση των κυττάρων και ακετυλιωµένης α-τουµπουλίνης (γκρι) για τη χρώση του αξονήµατος των κροσσών, µελετήθηκε η έκφρασή τους κατά τη διάρκεια της διαφοροποίησης. Κατά τα πρώιµα στάδια της διαφοροποίησης (ALI DAY 3) η έκφραση του McIdas εντοπίζεται στα κύτταρα εκείνα που διαθέτουν πρωτογενή κροσσό και θα διαφοροποιηθούν σε πολυκροσσωτά κύτταρα. Αντίθετα, δεν παρατηρούνται κύτταρα µε πολλαπλούς κροσσούς που να εκφράζουν την πρωτεΐνη McIdas (ALI DAY 6, 11 & 14). Scale bar, 10µm. Παράλληλα χρησιµοποιήθηκε αντίσωµα έναντι της πρωτεΐνης pericentrin, που αποτελεί δείκτη των νεο-συντιθέµενων κεντριολίων. Εξετάζοντας τη χρώση των κυττάρων ως προς την κατανοµή των κεντριολίων τους, έγινε κατηγοριοποίησή τους σε πρώιµα και ύστερα στάδια της διαδικασίας δηµιουργίας των κροσσών (βλέπε σχηµατική αναπαράσταση, εικόνα 4.1.4) και παράλληλα µελετήθηκε και η έκφραση της πρωτεΐνης McIdas. Τα κύτταρα που βρίσκονται στα πρώιµα στάδια της διαφοροποίησης χαρακτηρίζονται από την ύπαρξη ε νός πρωτογενούς κροσσού (στάδιο Ι) ή αναγνωρίζονται από τη µαζική παραγωγή κεντριολίων (στάδιο II & III). Αντίθετα, στο τελευταίο στάδιο της διαφοροποίησης βρίσκονται τα κύτταρα εκείνα 130
 εκφράζουν την πρωτεΐνη McIdas.")
131 4. ΑΠΟΤΕΛΕΣΜΑΤΑ στα οποία τα κεντριόλια έχουν πλέον µεταναστεύσει στη µεµβράνη και έχουν κατανεµηθεί οµοιόµορφα (στάδιο IV). Όπως παρατηρείται και στην εικόνα , 3 ηµέρες µετά την επαγωγή της διαφοροποίησης η πλειοψηφία των κυττάρων που βρίσκονται στα πρώιµα στάδια της διαφοροποίησης (Στάδιο II και III) εκφράζουν την πρωτεΐνη McIdas. Αντίθετα, ελάχιστα από τα τελικώς διαφοροποιηµένα κύτταρα, στα οποία η pericentrin εντοπίζεται κατανεµηµένη οµοιόµορφα στην επιφάνειά τους) εκφράζουν την πρωτεΐνη McIdas. Β Α ALI DAY 3 * * Idas/PCNT Ποσοστό McIdas+PCNT+/PCNT+ κυττάρων Πρώιμα στάδια (Στάδιο II & III) Μεταγενέστερα στάδια (Στάδιο IV) Εικόνα Η πρωτεΐνη McIdas εµφανίζει υψηλά επίπεδα έκφρασης κατά το στάδιο του πολλαπλασιασµού των κεντριολίων. (Α) Κύτταρα αποµονώθηκαν από την τραχεία ενήλικων µυών και καλλιεργήθηκαν κάτω από ειδικές συνθήκες που ευνοούν τη διαφοροποίησή τους προς τη µοίρα των πολυκροσσωτών κυττάρων. Μέσω ανοσοφθορισµού και µε τη χρήση ειδικών αντισωµάτων έναντι των πρωτεϊνών McIdas (πράσινο) και pericentrin (PCNT, κόκκινο) για τη χρώση των νεοσυντιθέµενων κεντριολίων, µελετήθηκε η έκφραση της πρωτεΐνης McIdas στα διάφορα στάδια της διαφοροποίησης. Αξιολογώντας το πρότυπο της κα τανοµής των κεντριολίων έγινε κατηγοριοποίηση των κυττάρων σε αυτά που βρίσκονταν στα πρώιµα στάδια της διαφοροποίησης, τα οποία έχουν πολλαπλασιάσει µαζικά τα κεντριόλιά τους, που µεταναστεύουν στην κυτταρική µεµβράνη (επισηµαίνονται µε αστερίσκο, *) και σε αυτά που βρίσκονται σε µεταγενέστερα στάδια, στα οποία τα κεντριόλια βρίσκονται οµοιόµορφα κατανεµηµένα στην κυτταρική µεµβράνη, δρώντας ως βασικά σωµάτια για το σχηµατισµό των κροσσών (επισηµαίνονται µε βέλος). Η έκφραση της πρωτεΐνης McIdas εντοπίζεται στα κύτταρα εκείνα που βρίσκονται στα πρώιµα στάδια της διαφοροποίησης σύµφωνα µε το πρότυπο χρώσης της pericentrin. (Β) Ποσοτικοποίηση του αριθµού των κυττάρων που εκφράζουν την πρωτεΐνη McIdas µε βάση την κατανοµή της πρωτεΐνης pericentrin και κατηγοριοποίησή τους σε πρώιµα και ύστερα στάδια της διαδικασία αυτής. Η πλειοψηφία των κυττάρων που εκφράζουν την πρωτεΐνη McIdas βρίσκονται στα πρώιµα στάδια της διαφοροποίησης (κύτταρα σταδίου ΙΙ και III). Scale bar, 10µm. 131
132 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 4.2 Μελέτη του ρόλου των µελών της υπεροικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων Η υπερέκφραση των παραγόντων McIdas και GemC1 επάγει τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Από τα µέχρι τώρα δεδοµένα έχει καταστεί σαφές ότι τα µέλη της υπεροικογένειας της Geminin, GemC1 και McIdas εµφανίζουν ειδική έκφραση στα πολυκροσσωτά κύτταρα του αναπνευστικού επιθηλίου. Η έκφραση των παραγόντων αυτών κατά τα πρώιµα στάδια της διαφοροποίησης µας οδήγησε στο να διερευνήσουµε στη συνέχεια πώς εµπλέκονται στη συγκεκριµένη διαδικασία. Για το σκοπό αυτό πραγµατοποιήθηκαν ex vivo πειράµατα υπερέκφρασης των παραγόντων McIdas και GemC1 στα επιθηλιακά κύτταρα της τραχείας µυών. Πιο συγκεκριµένα, αποµονώθηκαν τα επιθηλιακά κύτταρα της τραχείας µυών αγρίου τύπου τα οποία καλλιεργήθηκαν και στη συνέχεια, κατά το στάδιο του πολλαπλασιασµού, µολύνθηκαν µε λεντιιούς που εξέφραζαν είτε τα γονίδια GemC1 ή McIdas συντηγµένα µε την πράσινη φθορίζουσα πρωτεΐνη GFP, είτε την πρωτεΐνη GFP µόνη ως µάρτυρα. Δυο ηµέρες µετά, ακολούθησε επαγωγή της διαφοροποίησης προς πολυκροσσωτά (συνθήκη καλλιέργειας ALI). Το αποτέλεσµα της υπερέκφρασης αξιολογήθηκε µέσω ανοσοφθορισµού, σε διαφορετικές χρονικές στιγµές, µε τη χρήση ειδικών αντισωµάτων για την ανίχνευση παραγόντων που συµµετέχουν στη διαφοροποίηση των πολυκροσσωτών κυττάρων (Εικόνα και 4.2.2). Ο φαινότυπος των κυττάρων µετά την υπερέκφραση της πρωτεΐνης McIdas ταυτοποιήθηκε µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών pericentrin (PCNT) και Foxj1 για τη χρώση των νεοσυντιθέµενων κεντριολίων και των πολυκροσσωτών κυττάρων, αντίστοιχα (Εικόνα 4.2.1). Στα πρώιµα στάδια της διαφοροποίησης (ALI DAY 3), στα κύτταρα-µάρτυρες, τα οποία εκφράζουν την πρωτεΐνη GFP, δεν παρατηρείται ακόµα πολλαπλασιασµός των κεντριολίων. Αντίθετα, σε ποσοστό 72,9% των κυττάρων που υπερεκφράζεται η πρωτεΐνη McIdas παρατηρούνται υπεράριθµα κεντριόλια, τα οποία στη συνέχεια θα ωριµάσουν σε βασικά σωµάτια για να δώσουν γένεση στους κροσσούς (Εικόνα Α και Γ ). Οµοίως, σε αργότερα στάδια κατά τη διαφοροποίηση (ALI DAY 6), το 80,7% των κυττάρων που εκφράζουν την πρωτεΐνη McIdas εµφανίζουν αυξηµένη παραγωγή κεντριολίων, ενώ µόνο το 11,07% των κυττάρων που εκφράζουν την πρωτεΐνη GFP εµφανίζουν και πολλαπλά σήµατα της pericentrin. Ο φαινότυπος αυτός επιβεβαιώνεται και µε τη χρήση αντισώµατος έναντι του παράγοντα Foxj1 ο οποίος αποτελεί δείκτη τόσο των κυττάρων που θα γίνουν πολυκροσσωτά, όσο και των κυττάρων που έχουν ήδη διαφοροποιηθεί. Όπως φαίνεται στην εικόνα Β και στην αντίστοιχη ποσοτικοποίηση στο διάγραµµα Δ, το 81,92% των κυττάρων που εκφράζουν την πρωτεΐνη McIdas εκφράζουν και τον παράγοντα Foxj1 κατά τα πρώιµα στάδια της διαφοροποίησης (ALI DAY 3), ενώ το ποσοστό αυτό παραµένει και σε µεταγενέστερα στάδια (ALI DAY 6), σε αντίθεση µε τα κύτταρα-µάρτυρες που εκφράζουν σε αυτή τη χρονική στιγµή Foxj1 σε ποσοστό περίπου 2%. Συµπεραίνουµε λοιπόν ότι στα κύτταρα εκείνα στα οποία εκφράζεται εκτοπικά η 132









133 4. ΑΠΟΤΕΛΕΣΜΑΤΑ πρωτεΐνη McIdas παρατηρούνται πολυάριθµα κεντριόλια, καθώς και έκφραση του παράγοντα Foxj1, σε αντίθεση µε τα κύτταρα-µάρτυρες. A Ποσοστό PCNT+GFP+/GFP+ κυττάρων Γ GFP McIdas ** ** ALI DAY 6 GFP McIdas Foxj1 Foxj1 GFP/Foxj1 GFP/Foxj1 Δ Ποσοστό Foxj1+GFP+/GFP+ κυττάρων McIdas McIdas/PCNT PCNT GFP PCNT GFP/PCNT Εικόνα ALI DAY 3 Β ALI DAY 3 GFP McIdas ** *** ALI DAY 6 133
134 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Η πρωτε ΐνη McIdas επάγει τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. (Α, Β) Επιθηλιακά κύτταρα της τραχείας µυών αποµονώθηκαν και καλλιεργήθηκαν κάτω από ειδικές συνθήκες που ευ νοούν τον πολλαπλασιασµό τους. Στη συνέχεια τα κύτταρα µολύνθηκαν µε λεντιιούς που εξέφραζαν είτε το γονίδιο McIdas συντηγµένο µε GFP (McIdas), είτε την πρωτεΐνη GFP µόνο, ως µάρτυρα. Δυο µέρες αργότερα έγινε επαγωγή της διαφοροποίησής τους (ALI) και τα κύτταρα µονιµοποιήθηκαν 3 και 6 ηµέρες µετά. Ακολούθησε χρώση των κυττάρων µε αντισώµατα έναντι της πρωτεΐνης GFP (πράσινο) για τη χρώση των µολυσµένων κυττάρων, καθώς και έναντι της πρωτεΐνης pericentrin (PCNT, Α) ή Foxj1 (B) (κόκκινο). Τα βέλη υποδεικνύουν κύτταρα τα οποία εκφράζουν PCNT ή Foxj1 µετά την υπερέκφραση της πρωτεΐνης McIdas. (Γ, Δ) Ποσοτικοποίηση των κυττάρων που εκφράζουν την πρωτεΐνη PCNT ή Foxj1 µετά την υπερέκφραση της πρωτεΐνης McIdas ή GFP (δείγµα-µάρτυρας). Σε κάθε συνθήκη τα δεδοµένα αποδίδονται ως µέση τιµή (± SEM) από τουλάχιστον 10 ανεξάρτητα πεδία, ενός αντιπροσωπευτικού πειράµατος. **P < 0.01, ***P < Scale bar, 10µm. Στη συνέχεια µελετήθηκε εάν και ο παράγοντας GemC1 µπορεί να επάγει τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Στα κύτταρα στα οποία είχε υπερεκφραστεί είτε ο παράγοντας GemC1 συντηγµένος µε την πρωτεΐνη GFP, είτε η πρωτεΐνη GFP µόνη της ως µάρτυρας ακολούθησε ανάλυση µέσω ανοσοφθορισµού, µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas και Foxj1 (Εικόνα 4.2.2). Κατά τα πρώιµα στάδια της διαφοροποίησης (ALI DAY 1, 2 και 3), ενώ η διαφοροποίηση δεν έχει ξεκινήσει ακόµα στα κύτταρα εκείνα στα οποία υπερεκφράζεται η πρωτεΐνη GFP, στα κύτταρα στα οποία υπερεκφράζεται ο παράγοντας GemC1 παρατηρείται στατιστικά σηµαντική αύξηση της έκφρασης τόσο του McIdas (Εικόνα Α και Γ), όσο και του Foxj1 (Εικόνα Β και Δ). 134




, είτε την πρωτεΐνη GFP ως µάρτυρα.")

 ή Foxj1 (B)")

135 4. ΑΠΟΤΕΛΕΣΜΑΤΑ n=235 n=417 n=432 n=625 n=488 n=439 0 n=299 n=682 5 ALI DAY GemC1 Foxj1 Foxj1 P0 GFP/Foxj1 GemC1/Foxj1 P0 * *** 20 * n=322 n= n=421 n=789 *** 15 GFP GemC1 n=325 n=527 *** * Δ Ποσοστό Foxj1+GFP+/GFP+ κυττάρων ALI DAY 3 GFP GemC1 McIdas GemC1/McIdas GFP McIdas GFP/McIdas GFP GemC1 Γ Ποσοστό McIdas+GFP+/GFP+ κυττάρων B ALI DAY 2 n=280 n=505 A ALI DAY Εικόνα Η πρωτεΐνη GemC1 επάγει τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. (Α, Β) Επιθηλιακά κύτταρα της τραχείας µυών αποµονώθηκαν και καλλιεργήθηκαν κάτω από ειδικές συνθήκες που ευ νοούν τον πολλαπλασιασµό τους. Στη συνέχεια τα κύτταρα µολύνθηκαν µε λεντιιούς που εξέφραζαν είτε το γονίδιο GemC1 συντηγµένο µε GFP (GemC1), είτε την πρωτεΐνη GFP ως µάρτυρα. Δυο µέρες αργότερα έγινε επαγωγή της διαφοροποίησής τους (ALI) και τα κύτταρα µονιµοποιήθηκαν τις χρονικές στιγµές 0, 1, 2 και 3 ηµέρες µετά τη συνθήκη ALI. Ακολούθησε χρώση των κυττάρων µε αντισώµατα έναντι της πρωτεΐνης GFP (πράσινο) για τη σήµανση των κυττάρων που εξέφραζαν εκτοπικά GemC1 ή GFP, καθώς και έναντι της πρωτεΐνης McIdas (Α) ή Foxj1 (B) (κόκκινο). Τα βέλη υποδεικνύουν κύτταρα τα οποία εκφράζουν McIdas ή Foxj1 µετά την υπερέκφραση της πρωτεΐνης GemC1. (Γ, Δ) Ποσοτικοποίηση του αριθµού των κυττάρων που εκφράζουν την πρωτεΐνη McIdas ή Foxj1 µετά την υπερέκφραση της πρωτεΐνης GemC1 ή GFP ως µάρτυρα. Σε κάθε συνθήκη τα δεδοµένα αποδίδονται ως µέση τιµή (± SEM) από τουλάχιστον 10 ανεξάρτητα πεδία, που προέκυψαν από 2 ανεξάρτητα πειράµατα. *P < 0.05, ***P < Scale bar, 10µm. 135
136 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Τα παραπάνω αποτελέσµατα δείχνουν ότι οι πρωτεΐνες McIdas και GemC1 µπορούν να προάγουν τη µοίρα των κυττάρων προς πολυκροσσωτά, µέσω της επαγωγής της έκφρασης σηµαντικών παραγόντων της διαφοροποίησης αυτών, όπως των Foxj1 και pericentrin. Ιδιαίτερο ενδιαφέρον παρουσιάζει και η επαγωγή της έκφρασης του McIdas µετά την υπερέκφραση του GemC1. Καταλήγουµε λοιπόν ότι οι παράγοντες GemC1 και McIdas αποτελούν κεντρικούς ρυθµ ιστές της διαφοροποίησης των πολυκροσσωτών κυττάρων Η έλλειψη της πρωτεΐνης Geminin επάγει τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων Από προηγούµενα δεδοµένα δείχθηκε ότι τα µέλη της οικογένειας της Geminin εµφανίζουν ένα διαφορικό πρότυπο έκφρασης κατά τη διαφοροποίηση των επιθηλιακών κυττάρων της τραχείας µυών. Το γονίδιο της Geminin εκφράζεται στα διαιρούµενα προγονικά κύτταρα, σε αντίθεση µε την έκφραση των GemC1 και McIdas που εντοπίζεται σε µη ανιχνεύσιµα ή πολύ χαµηλά επίπεδα (Εικόνα 4.1.6). Μετά την επαγωγή της διαφοροποίησης των κυττάρων σε πολυκροσσωτά, οι παράγοντες GemC1 και McIdas εκφράζονται σε υψηλά επίπεδα, ενώ η έκφραση της Geminin περιορίζεται σηµαντικά, γεγονός που υποδηλώνει µια ανταγωνιστική δράση κατά τη διαδικασία αυτή. Επιπρόσθετα, δ είχθηκε ότι οι παράγοντες GemC1 και McIdas είναι ικανοί να επάγουν τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων ex vivo. Θέλοντας λοιπόν να εξετάσουµε και το ρόλο της Geminin στη διαδικασία αυτή, πραγµατοποιήθηκαν ex vivo πειράµατα καλλιέργειας των επιθηλιακών κυττάρων της τραχείας γενετικά τροποποιηµένων µυών από τους οποίους µπορούσε να γίνει ειδικά αποσιώπηση του γονιδίου της Geminin µε τη χρήση του Cre-LoxP συστήµατος (Kwan, 2002; Nagy, 2000). Πιο συγκεκριµένα, έχει κατασκευαστεί µια µεταλλαγµένη µορφή του γονιδίου της Geminin, στην οποία έχουν τοποθετηθεί 2 LoxP αλληλουχίες εκατέρωθεν των εξονίων 3 και 4 (Floxed αλληλόµορφο) (Karamitros et al, 2010; Διδακτορική Διατριβή, Πανωραία Κωταντάκη 2010). Στην παρούσα µελέτη χρησιµοποιήθηκαν µύες είτε οµόζυγοι για το Floxed αλληλόµορφο της Geminin (Geminin FL/FL ), είτε ετερόζυγοι, οι οποίοι έφεραν ένα αλληλόµορφο αγρίου τύπου και ένα Floxed αλληλόµορφο (Geminin WT/FL ) Επιπλέον, για να επιτυγχάνεται ο εντοπισµός των κυττάρων από τα οποία απουσιάζει η Geminin, οι µύες έφεραν τροποποιηµένο το γενετικό τόπο Rosa26 STOPYFP (Srinivas et al, 2001). Στους µυς αυτούς έχει ενσωµατωθεί η αλληλουχία Floxed Stop, καθώς και το cdna για τη φθορίζουσα πρωτεΐνη YFP (Yellow Fluorescence Protein) στο γενετικό τόπο Rosa26. H αλληλουχία Floxed Stop, φέρει ένα κωδικόνιο λήξης, το οποίο δεν επιτρέπει την έκφραση της YFP. Tο κωδικόνιο λήξης περιβάλλεται από δύο LoxP θέσεις, µε αποτέλεσµα σε κύτταρα που εκφράζεται η Cre ρεκοµπινάκη, η αλληλουχία Stop αποµακρύνεται και επιτρέπεται η έκφραση της πρωτεΐνης YFP. Επιθηλιακά κύτταρα της τραχείας µυών Geminin FL/FL;R26STOPYFP και Geminin WT/FL;R26STOPYFP αποµονώθηκαν, καλλιεργήθηκαν και κατά το στάδιο του 136

.")




 και 3 (ALI DAY 3) ηµέρες µετά την επαγωγή της διαφοροποίησής τους")
. Όπως φαίνεται και στην εικόνα 4.2.")

.")
137 4. ΑΠΟΤΕΛΕΣΜΑΤΑ πολλαπλασιασµού µολύνθηκαν µε λεντιιούς που εξέφραζαν την Cre ρεκοµπινάση. Δυο ηµέρες µετά ακολούθησε επαγωγή της διαφοροποίησης προς πολυκροσσωτά (συνθήκη καλλιέργειας ALI). Η δράση του ενζύµου έχει ως αποτέλεσµα στα κύτταρα που προέρχονται από τους GemininFL/FL;R26STOPYFP µύες να µην παράγεται το φυσιολογικό προϊόν του γονιδίου, σε αντίθεση µε τα επιθηλιακά κύτταρα από τους µύες µε γονότυπο GemininWT/FL;R26STOPYFP, στους οποίους παράγεται η πρωτεΐνη Geminin και χρησιµοποιήθηκαν ως κύτταρα-µάρτυρες. Παράλληλα, είναι δυνατό να εντοπιστούν τα κύτταρα στα οποία έδρασε η Cre ρεκοµπινάση αφού εκφράζουν την πρωτεΐνη YFP. Μέσω τριπλού ανοσοφθορισµού, µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών GFP, McIdas και Foxj1 αξιολογήθηκε το αποτέλεσµα της έλλειψης της Geminin, 1 (ALI DAY 1) και 3 (ALI DAY 3) ηµέρες µετά την επαγωγή της διαφοροποίησής τους (Εικόνα 4.2.3). Όπως φαίνεται και στην εικόνα Α, η έλλειψη του γονιδίου της Geminin επάγει τη διαφοροποίηση τω ν κυττάρων µέσω αύξησης της έκφρασης των παραγόντων McIdas και Foxj1 σε ποσοστό 12,84% και 16,67%, σε αντίθεση µε τα κύτταρα µάρτυρες που παρατηρείται αύξηση µόλις 4,54% και 8%, αντίστοιχα (Εικόνα Β και Γ). A McIdas ALI DAY 3 Foxj1 GFP/McIdas GFP/Foxj1 Geminin FL/FL;R26 LVCre Geminin WT/FL;R26 LVCre GFP n=1339 n= ALI DAY n=1339 n= Ποσοστό Foxj1+GFP+/GFP+ κυττάρων 12 n=159 n=1395 Ποσοστό McIdas+GFP+/GFP+ κυττάρων 14 0 GemininWT/FL;R26;LVCre GemininFL/FL;R26;LVCre Γ n=159 n=1395 GemininWT/FL;R26;LVCre GemininFL/FL;R26;LVCre B ALI DAY 137
138 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Η έλλειψη της Geminin επάγει τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. (Α) Επιθηλιακά κύτταρα της τραχείας που προέρχονταν από µύες µε γονότυπο Geminin WT/FL;R26STOPYFP ή Geminin FL/FL;R26STOPYFP αποµονώθηκαν και καλλιεργήθηκαν κάτω από ειδικές συνθήκες που ευνοούν τον πολλαπλασιασµό τους. Στη συνέχεια τα κύτταρα µολύνθηκαν µε λεντιιούς που εξέφραζαν την Cre ρεκοµπινάση. Δυο µέρες αργότερα έγινε επαγωγή της διαφοροποίησής τους (ALI) και τα κύτταρα µονιµοποιήθηκαν 1 και 3 ηµέρες µετά τη συνθήκη ALI (ALI DAY 1 και 3). Ακολούθησε χρώση των κυττάρων µε αντισώµατα έναντι της πρωτεΐνης GFP (πράσινο) για τη σήµανση των κυττάρων στα οποία είχε δράσει η Cre ρεκοµπινάση, καθώς και έναντι των πρωτεϊνών McIdas (γκρί) και Foxj1 (κόκκινο). Τα βέλη υποδεικνύουν κύτταρα στα οποία έχει δράσει η Cre ρεκοµπινάση µε αποτέλεσµα να µην εκφράζεται η πρωτεΐνη Geminin, ενώ εκφράζονται οι πρωτεΐνες McIdas και Foxj1. (Β, Γ) Ποσοτικοποίηση των κυττάρων που εκφράζουν την πρωτεΐνη McIdas (Β) ή Foxj1 (Γ) στα κύτταρα που παρατηρείται έλλειψη της πρωτεΐνης Geminin και σύγκρισή τους µε τα κύτταρα-µάρτυρες, 1 και 3 ηµέρες µετά την επαγωγή της διαφοροποίησής τους σε πολυκροσσωτά. Σε κάθε συνθήκη τα δεδοµένα αποδίδονται ως µέση τιµή από τουλάχιστον 5 ανεξάρτητα πεδία για τη συνθήκη µάρτυρα και 10 ανεξάρτητα πεδία για την πειραµατική συνθήκη και προέρχονται από ένα αντιπροσωπευτικό πείραµα. Scale bar, 10µm. 138
139 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 4.3 Μεταγραφική ρύθµιση του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων Οι παράγοντες GemC1 και McIdas δρουν ω ς µεταγραφικοί ενεργοποιητές γονιδίων που εµπλέκονται στη διαφοροποίηση των πολυκροσσωτών κυττάρων Από τα παραπάνω αποτελέσµατα είναι ε µφανές ότι τα µέλη της υπεροικογένειας της Geminin µπορούν να ρυθµίζουν τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Είναι αξιοσηµείωτο το γεγονός ότι φαίνεται να δρουν ανταγωνιστικά στη διαδικασία αυτή, µιας και η υπερέκφραση των McIdas και GemC1 στα επιθηλιακά κύτταρα της τραχείας µυών προκαλεί επαγωγή της διαφοροποίησης (Εικόνα και 4.2.2), ενώ αντίθετα κάτι τέτοιο παρατηρείται µέσω της έλλειψης της πρωτεΐνης Geminin (Εικόνα 4.2.3). Η δράση τους αυτή εκδηλώνεται ρυθµίζοντας την έκφραση παραγόντων που εµπλέκονται στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Προκειµένου να µελετήσουµε το µηχανισµό µέσω του οποίου τα µέλη της οικογένειας της Geminin δρουν στην επαγωγή του συγκεκριµένου µονοπατιού, πραγµατοποιήθηκαν πειράµατα µε τη χρήση του γονιδίου αναφοράς της λουσιφεράσης. Κ ατασκευάστηκαν πλασµίδια (pgl3-promoter πλασµίδιο) στα οποία τοποθετήθηκαν ανοδικά του γονιδίου της λουσιφεράσης τα ρυθµιστικά στοιχείαπιθανοί υποκινητές των γονιδίων MCIDAS και FOXJ1 (βλέπε υλικά και µέθοδοι). Στη συνέχεια, κύτταρα HEK293T διαµολύνθηκαν µε το αντίστοιχο πλασµίδιο, καθώς επίσης και µε πλασµίδια που εξέφραζαν τ ους παράγοντες McIdas, GemC1 και Geminin όπως φαίνεται στην εικόνα 4.3.1, ή µε πλασµίδιο που δεν έφερε κάποιο από τα παραπάνω γονίδια, ως µάρτυρα. 48 ώρες µετά τη διαµόλυνση µετρήθηκε η ενεργότητα των ενζύµων της firefly και της renilla λουσιφεράσης και ακολούθησε ανάλυση των αποτελεσµάτων. Ο παράγοντας GemC1 µπορεί να προκαλέσει επαγωγή της µεταγραφής τόσο µέσω των ρυθµιστικών στοιχείων του MCIDAS (4,2 φορές επαγωγή σε σχέση µε την επαγωγή που προκαλεί το δείγµα-µάρτυρας που ορίζεται ως 1), όσο και µέσω του FOXJ1 (5,5 φορές επαγωγή της έκφρασης σε σχέση µε το µάρτυρα) (Εικόνα Α και Β). Παρόµοια επαγωγή παρατηρείται και από την πρωτεΐνη McIdas µέσω των ρυθµιστικών στοιχείων των γονιδίων MCIDAS και FOXJ1, αλλά όχι από τη Geminin. Είναι αξιοσηµείωτο ότι η Geminin αναστέλλει σηµαντικά τη µεταγραφική ενεργότητα των GemC1 και McIdas στα ρυθµιστικά στοιχεία των γονιδίων MCIDAS και FOXJ1. Έχει ήδη δειχθεί ότι ο GemC1 προκαλεί επαγωγή της έκφρασης του McIdas στα επιθηλιακά κύτταρα της τραχείας µυός. Προκειµένου να διαπιστωθεί εάν ο GemC1 µπορεί να επάγει άµεσα την έκφραση του Foxj1 ή έµµεσα µέσω της ενεργοποίησης του McIdas, πραγµατοποιήθηκαν πειράµατα αποσιώπησης του γονιδίου McIdas µε την τεχνολογία sirna. Για το σκοπό αυτό κύτταρα HEK293T διαµολύνθηκαν παροδικά µε το πλασµίδιο που φέρει ανοδικά του γονιδίου της λουσιφεράσης τον υποκινητή του γονιδίου FOXJ1, καθώς και µε το πλασµίδιο για την έκφραση του GemC1 ή µε ένα πλασµίδιο που δεν περιέχει κανένα γονίδιο, ω ς µάρτυρα. Παράλληλα έγινε αποσιώπηση του γονιδίου McIdas µε τη χρήση sirna ολιγονουκλεοτιδίων που στοχεύουν ειδικά το mrna του. Παράλληλα, χρησιµοποιήθηκαν µη ειδικά sirna ολιγονουκλεοτίδια, ως µάρτυρας. Συνολικά έγιναν δυο διαδοχικές διαµολύνσεις µε τα sirna µε διαφορά 24 ωρών. Η πτώση των επιπέδων του mrna του McIdas (πτώση 70%) πιστοποιήθηκε µέσω ποσοτικής Real- 139
140 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Time PCR, µε τη χρήση ειδικών εκκινητών (Εικόνα Γ). 48 ώρες συνολικά µετά την πρώτη διαµόλυνση µετρήθηκε στα κύτταρα η µεταγραφική ενεργότητα του υποκινητή του γονιδίου FOXJ1 και παρατηρήθηκε ότι ο GemC1 µπορεί να προκαλέσει άµεσα µεταγραφική ενεργοποίηση µέσω του συγκεκριµένου υποκινητή, ανεξάρτητα από τον παράγοντα McIdas (Εικόνα Δ). Τα ανωτέρω ευρήµατα οδηγούν στο συµπέρασµα ότι ο GemC1 αποτελεί έναν από τους πιο πρώιµους µεταγραφικούς ενεργοποιητές του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων, ενεργοποιώντας µεταγραφικά τους παράγοντες McIdas και Foxj1. Επίσης είναι ιδιαίτερα ενδιαφέρον ότι τα µέλη της υπεροικογένειας της Geminin φαίνεται να συµµετέχουν στο µονοπάτι µέσω πολύπλοκων ρυθµιστικών αλληλεπιδράσεων µεταξύ τους. A MCIDAS ρυθμιστικά στοιχεία B FOXJ1 ρυθμιστικά στοιχεία RLF GemC1 - McIdas - Geminin - ** ** ** ** GemC1 - McIdas - Geminin - RLF ** ** ** ** Γ Δ FOXJ1 ρυθμιστικά στοιχεία Σχετικά επίπεδα έκφρασης mrna control sirna McIdas sirna RLF GemC1 - control sirna + McIdas sirna
141 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Οι παράγοντες GemC1 και McIdas ελέγχουν µεταγραφικά γονίδια που συµµετέχουν στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Κύτταρα HEK293T διαµολύνθηκαν παροδικά µε τα πιθανά ρυθµιστικά στοιχεία των γονιδίων MCIDAS (Α) ή FOXJ1 (Β, Δ), που είχαν κλωνοποιηθεί ανοδικά του γονιδίου της firefly λουσιφεράσης, καθώς και µε πλασµίδια που εξέφραζαν τους παράγοντες GemC1, McIdas, Geminin ή µε ένα πλασµίδιο χωρίς ένθεµα ως µάρτυρα, όπως φαίνεται στην εικόνα. Στο (Δ) πραγµατοποιήθηκε παράλληλα διαµόλυνση των κυττάρων και µε sirna ολιγονουκλεοτίδια που στοχεύουν ειδικά το mrna του McIdas ή µε µη ειδικά ολιγονουκλεοτίδια για µια τυχαία αλληλουχία. Τα επίπεδα έκφρασης του mrna του McIdas µετά την επίδραση του sirna, πιστοποιήθηκαν µέσω ποσοτικής real-time PCR (Γ). 48 ώρες µετά τη διαµόλυνση µετρήθηκε η ενεργότητα των ενζύµων της firefly και της renilla λουσιφεράσης. Σε κάθε δείγµα οι µονάδες της εκπεµπόµενης βιοφωταύγειας της firefly λουσιφεράσης κανονικοποίηθηκαν ως προς τις τιµές που προέκυψαν από τη βιοφωταύγεια της renilla λουσιφεράσης. Τα σχετικά επίπεδα επαγωγής κάθε δείγµατος προσδιορίστηκαν από το λόγο των κανονικοποιηµένων τιµών της βιοφωταύγειας που προέκυψαν από τα πλασµίδια που εξέφραζαν το επιθυµητό γονίδιο προς την τιµή βιοφωταύγειας που προέκυψε µετά την υπερέκφραση του πλασµιδίου-µάρτυρας. Οι τιµές αντιπροσωπεύουν το µέσο όρο των δεδοµένων από 3 τουλάχιστον ανεξάρτητα πειράµατα (± SEM). **P < Συντοµεύσεις: RLF, relative luciferase fold induction, σχετική επαγωγή έκφρασης γονιδίου λουσιφεράσης Οι παράγοντες GemC1 και McIdas ενεργοποιούν µεταγραφικά το µονοπάτι της διαφοροποίησης των πολυκροσσωτών κυττάρων σε συνεργασία µε το µεταγραφικό παράγοντα Ε2F5 Ενώ αυτή η εργασία βρισκόταν σε εξέλιξη οι Ma et al. έδειξαν ότι η πρωτεΐνη McIdas σχηµατίζει ένα τριµερές σύµ πλοκο µε τους µεταγραφικούς παράγοντες E2F4/5 και TFDP1, µέσω του οποίου επάγεται η έκφραση γονιδίων που εµπλέκονται στη διαφοροποίηση των πολυκροσσωτών κυττάρων (Ma et al, 2014). Προκειµένου να µελετηθεί εάν ο παράγοντας GemC1 δρα κατά τη διαδικασία αυτή µέσω συνεργασίας µε τους E2F µεταγραφικούς παράγοντες, πραγµατοποιήθηκαν πειράµατα µε γονίδιο αναφοράς τη λουσιφεράση και ταυτόχρονη υπερέκφραση του GemC1 µε µέλη της οικογένειας των E2F µεταγραφικών παραγόντων (Εικόνα 4.3.2). Ο µεταγραφικός παράγοντας E2F5 δεν µπορεί να προκαλέσει από µόνος του µεταγραφική ενεργοποίηση των γονιδίων MCIDAS (Α) και FOXJ1 (Β). Όµως, ύστερα από ταυτόχρονη υπερέκφραση των GemC1 και E2F5 παραγόντων παρατηρείται επαγωγή της µεταγραφής των γονιδίων MCIDAS και FOXJ1 κατά 17,73 και 10,84 φορές σε σχέση µε το δείγµα-µάρτυρας, αντίστοιχα. Ο παράγοντας McIdas δρα επίσης συνεργατικά µε τον E2F5 για τη µεταγραφική ενεργοποίηση των παραπάνω γονιδίων, όµως δεν παρατηρήθηκε το ίδιο και παρουσία της Geminin. 141
142 4. ΑΠΟΤΕΛΕΣΜΑΤΑ A MCIDAS regulatory elements B FOXJ1 regulatory elements RLF GemC1 McIdas Geminin E2F *** *** RLF GemC1 McIdas Geminin E2F *** *** Εικόνα Οι παράγοντες GemC1 και McIdas ελέγχουν τη µεταγραφή γονιδίων που συµµετέχουν στη διαφοροποίηση των πολυκροσσωτών κυττάρων σε συνεργασία µε τον µεταγραφικό παράγοντα E2F5. Κύτταρα HEK293T διαµολύνθηκαν παροδικά µε τα πιθανά ρυθµιστικά στοιχεία των γονιδίων MCIDAS (Α) ή FOXJ1 (Β) που είχαν κλωνοποιηθεί ανοδικά του γονιδίου της firefly λουσιφεράσης, καθώς και µε πλασµίδια που εξέφραζαν τους παράγοντες GemC1, McIdas, Geminin, E2F5 ή µε ένα πλασµίδιο χωρίς ένθεµα ως µάρτυρα, όπως φαίνεται στην εικόνα. 48 ώρες µετά τη διαµόλυνση µετρήθηκε η ενεργότητα των ενζύµων της firefly και της renilla λουσιφεράσης. Σε κάθε δείγµα οι µονάδες της εκπεµπόµενης βιοφωταύγειας της firefly λουσιφεράσης κανονικοποίηθηκαν ως προς τις τιµές που προέκυψαν από τη βιοφωταύγεια της renilla λουσιφεράσης. Τα σχετικά επίπεδα επαγωγής κάθε δείγµατος προσδιορίστηκαν από το λόγο των κανονικοποιηµένων τιµών της βιοφωταύγειας που προέκυψαν από τα πλασµίδια που εξέφραζαν το επιθυµητό γονίδιο προς την τιµή βιοφωταύγειας που προέκυψε µετά την υπερέκφραση του πλασµιδίου-µάρτυρας. Οι τιµές αντιπροσωπεύουν το µέσο όρο των δεδοµένων από 3 τουλάχιστον ανεξάρτητα πειράµατα (± SEM). ***P < Συντοµεύσεις: RLF, relative luciferase fold induction, σχετική επαγωγή έκφρασης γονιδίου λουσιφεράσης Παράλληλα εξετάστηκε και η πιθανή συνεργασία µε το µεταγραφικό παράγοντα E2F4. Παρατηρήθηκε ότι ο Ε2F4 προκαλεί από µόνος του υψηλά επίπεδα µεταγραφικής ενεργοποίησης µέσω των ρυθµιστικών στοιχείων των MCIDAS και FOXJ1 γονιδίων (Εικόνα Α και Β), γεγονός που δυσκόλεψε τη µελέτη της πιθανής συνεργατικής του δράσης µε τα µέλη της υπεροικογένειας της Geminin. Αντίθετα, όπως φαίνεται στην εικόνα 4.3.3, ο µεταγραφικός παράγοντας E2F1 δεν µπορεί να επάγει τη µεταγραφή των γονιδίων MCIDAS (Γ) ή FOXJ1 (Δ) σε συνεργασία µε κάποιο από τα µέλη της υπεροικογένειας της Geminin. Το αποτέλεσµα αυτό καταδεικνύει ότι οι παράγοντες GemC1 και McIdas επάγουν τη µεταγραφή γονιδίων σε συνεργασία µε συγκεκριµένα µέλη της οικογένειας των E2F µεταγραφικών παραγόντων. 142
143 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 143 A MCIDAS ρυθμιστικά στοιχεία E2F5 E2F4 E2F1 RLF B FOXJ1 ρυθμιστικά στοιχεία RLF E2F5 E2F4 E2F Γ MCIDAS ρυθμιστικά στοιχεία Geminin McIdas GemC1 E2F1 RLF ** ns ns ns Δ FOXJ1 ρυθμιστικά στοιχεία RLF Geminin McIdas GemC1 E2F1 ** ns ns ns
144 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Αξιολόγηση της συνεργατικής δράσης µελών της υπεροικογένειας της Geminin και µελών της οικογένειας των E2F µεταγραφικών παραγόντων για το µεταγραφικό έλεγχο του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων. Κύτταρα HEK293T διαµολύνθηκαν παροδικά µε τα πιθανά ρυθµιστικά στοιχεία των γονιδίων MCIDAS (Α, Γ ) ή FOXJ1 (Β, Δ ) που είχαν κλωνοποιηθεί ανοδικά του γονιδίου της firefly λουσιφεράσης, καθώς και µε πλασµίδια που εξέφραζαν τους παράγοντες GemC1, McIdas, Geminin, E2F1, E2F4, E2F5 ή µε ένα πλασµίδιο χωρίς ένθεµα ως µάρτυρα, όπως φαίνεται στην εικόνα. 48 ώρες µετά τη διαµόλυνση µετρήθηκε η ενεργότητα των ενζύµων της firefly και της renilla λουσιφεράσης. Σε κάθε δείγµα οι µονάδες της εκπεµπόµ ενης βιοφωταύγειας της firefly λουσιφεράσης κανονικοποιήθηκαν ως προς τις τιµές που προέκυψαν από τη βιοφωταύγεια της renilla λουσιφεράσης. Τα σχετικά επίπεδα επαγωγής κάθε δείγµατος προσδιορίστηκαν από το λόγο των κανονικοποιηµένων τιµών της βιοφωταύγειας που προέκυψαν από τα πλασµίδια που εξέφραζαν το επιθυµητό γονίδιο προς την τιµή βιοφωταύγειας που προέκυψε µετά την υπερέκφραση του πλασµιδίου-µάρτυρας. Ο µεταγραφικός παράγοντας E2F4 προκαλεί από µόνος του υψηλά επίπεδα µεταγραφικής ενεργοποίησης µέσω των ρυθµιστικών στοιχείων των γονιδίων MCIDAS (A) και FOXJ1 (B), σε αντίθεση µε τον E2F5 παράγοντα που δεν µπορεί µόνος του να επάγει µεταγραφή µέσω αυτών των ρυθµιστικών στοιχείων. Ο µεταγραφικός παράγοντας E2F1 επάγει τη µεταγραφή µέσω των υποκινητών των γονιδίων MCIDAS (Γ) και FOXJ1(Δ), όµ ως δεν συνεργάζεται µε τα µέλη της υπεροικογένειας της Geminin για αυτή του τη δράση. Οι τιµές αντιπροσωπεύουν το µέσο όρο των δεδοµένων από 3 τουλάχιστον ανεξάρτητα πειράµατα (± SEM). ***P < Συντοµεύσεις: RLF, relative luciferase fold induction, σχετική επαγωγή έκφρασης γονιδίου λουσιφεράσης, ns, non-significant Ο παράγοντας McIdas δεν ελέγχει µεταγραφικά την έκφραση του γονιδίου GemC1 Όπως δείχθηκε προηγουµένως ο παράγοντας GemC1 µπορεί να προκαλέσει επαγωγή της έκφρασης του McIdas ex vivo, σ τα επιθηλιακά κύτταρα της τραχείας µυός (Εικόνα Α και Γ), αλλά και ε παγωγή της µεταγραφής µέσω των ρυθµιστικών στοιχείων του είτε µόνος του, είτε συνεργατικά µε το µεταγραφικό παράγοντα E2F5 (Εικόνα Α). Προκειµένου να αποσαφηνιστεί εάν και ο McIdas µπορεί να ελέγχει την έκφραση του GemC1, πραγµατοποιήθηκαν επιπρόσθετα πειράµατα µε το γονίδιο αναφοράς της λουσιφεράσης. Τα πιθανά ρυθµιστικά στοιχεία του γονιδίου GEMC1 κλωνοποιήθηκαν ανοδικά του πλασµιδίου που φέρει το γονίδιο της λουσιφεράσης. Κύτταρα HEK293T διαµολύνθηκαν παροδικά µε το πλασµίδιο που φέρει τον πιθανό υποκινητή του γονιδίου GEMC1, καθώς και µε πλασµίδιο για την έκφραση του McIdas, Ε2F5 ή µε ένα πλασµίδιο χωρίς ένθεµα ως µάρτυρα. Δείχθηκε ότι ο παράγοντας McIdas δεν µπορεί να επάγει σηµαντικά τη µεταγραφή µέσω των πιθανών ρυθµιστικών στοιχείων του γονιδίου GEMC1 που εξετάστηκαν στην παρούσα µελέτη (Εικόνα 4.3.4). 144
145 4. ΑΠΟΤΕΛΕΣΜΑΤΑ McIdas E2F RLF GEMC1 MCIDAS 50 Ρυθμιστικά στοιχεία 45 Εικόνα Ο παράγοντας McIdas δεν ελέγχει µεταγραφικά την έκφραση του γονιδίου GemC1. Κύτταρα HEK293T διαµολύνθηκαν παροδικά µε τα πιθανά ρυθµιστικά στοιχεία του γονιδίου GEMC1 που είχαν κλωνοποιηθεί ανοδικά του γονιδίου της firefly λουσιφεράσης, καθώς και µε πλασµίδια που εξέφραζαν τους παράγοντες McIdas, E2F5 ή µε ένα πλασµίδιο χωρίς ένθεµα ως µάρτυρα, όπως φαίνεται στην εικόνα. 48 ώρες µετά τη διαµόλυνση µετρήθηκε η ενεργότητα των ενζύµων της firefly και της renilla λουσιφεράσης. Σε κάθε δείγµα οι µονάδες της εκπεµπόµενης βιοφωταύγειας της firefly λουσιφεράσης κανονικοποίηθηκαν ως προς τις τιµές που προέκυψαν από τη βιοφωταύγεια της renilla λουσιφεράσης. Τα σχετικά επίπεδα επαγωγής κάθε δείγµατος προσδιορίστηκαν από το λόγο των κανονικοποιηµένων τιµών της βιοφωταύγειας που προέκυψαν από τα πλασµίδια που εξέφραζαν το επιθυµητό γονίδιο προς την τιµή βιοφωταύγειας που προέκυψε µετά την υπερέκφραση του πλασµιδίου-µάρτυρας. Ο παράγοντας McIdas δεν ελέγχει τη µεταγραφή µέσω των πιθανών ρυθµιστικών στοιχείων του γονιδίου GEMC1, είτε µόνος του είτε συνεργατικά µε το µεταγραφικό παράγοντα E2F5. Για συγκριτικούς λόγους παρατίθεται το διάγραµµα όπου απεικονίζεται η επαγωγή της έκφρασης που µπορεί να προκαλεί ο McIdas στα ρυθµιστικά του στοιχεία σε συνεργασία µε το µεταγραφικό παράγοντα E2F5. Οι τιµές αντιπροσωπεύουν το µέσο όρο των δεδοµένων από 3 τουλάχιστον ανεξάρτητα πειράµατα (± SEM). Συντοµεύσεις: RLF, relative luciferase fold induction, σχετική επαγωγή έκφρασης γονιδίου λουσιφεράσης Συµπερασµατικά φαίνεται ότι οι GemC1 και McIdas επάγουν τη µεταγραφή των γονιδίων MCIDAS και FOXJ1 σε συνεργασία µε τον µεταγραφικό παράγοντα E2F5. Ο GemC1 προκαλεί επαγωγή της έκφρασης του McIdas, ενώ ο McIdas δεν µπορεί να ρυθµίζει την έκφραση του GemC1, αποτέλεσµα που υποδηλώνει ότι ο GemC1 αποτελεί τον κορυφαίο ρυθµ ιστή στο µονοπάτι της διαφοροποίησης των πολυκροσσωτών κυττάρων. 145
146 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 4.4 Το γονίδιο GemC1 είναι απαραίτητο για τ η διαφοροποίηση των πολυκροσσωτών κυττάρων στο αναπνευστικό επιθήλιο µυών Δηµιουργία διαγονιδιακών µυών από τους οποίους απουσιάζει η πρωτεΐνη GemC1 Τα µέχρι τώρα αποτελέσµατα οδηγούν στο συµ πέρασµα ότι ο GemC1 αποτελεί έναν από τους πιο πρώιµους και κεντρικούς ρυθµιστές της διαφοροποίησης των πολυκροσσωτών κυττάρων. Με σκοπό να διαπιστωθεί εάν ο GemC1 είναι απαραίτητος για τη διαφοροποίηση των πολυκροσσωτών κυττάρων in vivo, κατασκευάστηκαν διαγονιδιακοί µύες στους οποίους δεν εκφράζεται το γονίδιο GemC1. Η φυσιολογική έκφραση του γονιδίου GemC1 διαταράσσεται εξαιτίας της ένθεσης ενός τ µήµατος DNA που περιέχει το γονίδιο αναφοράς της β - γαλακτοσιδάσης (lacz) µαζί µε το γονίδιο που προσφέρει ανθεκτικότητα στη νεοµυκίνη (neo), ανάµεσα στα εξόνια 2 και 3 του γονιδίου GEMC1 (Εικόνα 4.4.1, GemC1 KO allele, knock-out first) (Testa et al, 2004). Μια θέση µατίσµατος ανοδικά του γονιδίου της γαλακτοσιδάσης οδηγεί σε παραγωγή ενός mrna που δεν περιέχει τα εξόνια 3-5 του γονιδίου. Επιπρόσθετα, προκειµένου να είναι δυνατή η δηµιουργία µυών για ιστοειδική ή χρονοειδική απαλοιφή του γονιδίου σε δεύτερο χρόνο, η κασέτα φέρει θέσεις loxp και FRT που αναγνωρίζονται από τις Cre και FLP ρεκοµπινάσες, αντίστοιχα. Στην παρούσα µελέτη χρησιµοποιήθηκαν φυσιολογικοί µύες που διέθεταν το αγρίου τύπου αλληλόµορφο του GemC1 είτε σε οµοζυγωτία (GemC1 WT/WT ), είτε σε ετεροζυγωτία (GemC1 ΚΟ/WT ), καθώς και µύες που ήταν οµόζυγοι για το αδρανοποιηµένο αλληλόµορφο (GemC1 ΚΟ/ΚΟ ) GemC1 WT allele FRT 2 SA lacz pa loxp neo pa FRT loxp 3 4 loxp GemC1 KO allele Εικόνα GemC1 knockout αλληλόµορφο. Σχηµατική αναπαράσταση του αγρίου τύπου (WT) και του τροποποιηµένου (KO) αλληλοµόρφου του γονιδίου GemC1. Μια αλληλουχία DNA που περιέχει το γονίδιο της β-γαλακτοσιδάσης (lacz) και το γονίδιο που προσδίδει ανθεκτικότητα στη νεοµυκίνη (neo) έχει ενσωµατωθεί ανάµεσα στα εξόνια 2 και 3 του γονιδίου GemC1. Ανοδικά του γονιδίου της β -γαλακτοσιδάσης υπάρχει µια εναλλακτική θέση µατίσµατος (splicing acceptor site) που εµ ποδίζει τη σωστή έκφραση του γονιδίου. 146
. Οι µύες αυτοί συγκρίθηκαν µε τους µύες που έφεραν το φυσιολογικό αλληλόµορφο είτε σε οµοζυγωτία (GemC1 WT/WT ), είτε σε ετεροζυγωτία (GemC1 ΚΟ/WT ).")

147 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Οι ελλειµµατικοί µύες για το γονίδιο GemC1 εµφανίζουν καθυστέρηση στην ανάπτυξη και πεθαίνουν σύντοµα µετά τη γέννηση Πραγµατοποιήθηκε φαινοτυπική µελέτη των µυών που εµφάνιζαν έλλειψη του γονιδίου GemC1 (GemC1 ΚΟ/ΚΟ ). Οι µύες αυτοί συγκρίθηκαν µε τους µύες που έφεραν το φυσιολογικό αλληλόµορφο είτε σε οµοζυγωτία (GemC1 WT/WT ), είτε σε ετεροζυγωτία (GemC1 ΚΟ/WT ). Από συνεχείς διασταυρώσεις παρατηρήθηκε ότι η συχνότητα των απογόνων που προέκυπταν ήταν η αναµενόµενη µε βάση τη Μεντελική κληρονοµικότητα. Επίσης, οι µύες που γεννιούνταν δεν εµφάνιζαν φαινοτυπικές ανωµαλίες. Ωστόσο, πολύ σύντοµα µετά τη γέννησή τους (κυρίως από την τρίτη ηµέρα και µετά P3) παρατηρήθηκε ότι οι GemC1 ΚΟ/ΚΟ µύες, παρόλο που τρέφονταν φυσιολογικά, ε µφάνιζαν καθυστέρηση στην ανάπτυξή τους µε αποτέλεσµα να είναι υποανάπτυκτοι σε σύγκριση µε τους φυσιολογικούς µύες. Όπως φαίνεται και στην εικόνα Α, το µέγεθος των GemC1 ΚΟ/ΚΟ µυών είναι πολύ µικρότερο σε σύγκριση µε αυτό των φυσιολογικών µυών και η πλειοψηφία τους αδυνατούσε να επιβιώσει ήδη από την πρώτη εβδοµάδα µετά τη γέννηση (Καµπύλη επιβίωσης, Εικόνα Β). Δ εν παρατηρήθηκαν φαινοτυπικές διαφορές ανάµεσα στους µυς που είχαν το φυσιολογικό αλληλόµορφο σε οµοζυγωτία ή σε ετεροζυγωτία, οπότε στην παρούσα µελέτη χρησιµοποιήθηκαν εξίσου ως αγρίου τύπου µύες. Α P5 GemC1 KO/WT GemC1 KO/KO Β 100 Ποσοστό επιβίωσης (%) Ημέρες μετά τη γέννηση GemC1 KO GemC1 WT Εικόνα Οι µύες µε έλλειψη του γονιδίου GemC1 εµφανίζουν καθυστέρηση στην α νάπτυξη και πεθαίνουν σύντοµα µετά τη γέννηση. (Α) Σύγκριση µυών ελλειµµατικών για το γονίδιο GemC1 (GemC1 ΚΟ/ΚΟ ) µε φυσιολογικούς µύες (GemC1 ΚΟ/WT ) της ίδιας τοκετοµάδας, κατά την πέµπτη ηµέρα µετά τη γέννηση (P5). (B) Καµπύλη επιβίωσης µυών οµ όζυγων για τ ο KO αλληλόµορφο του GemC1 (GemC1 ΚΟ, n=8) και φυσιολογικών µυών οµόζυγων ή ετερόζυγων για το αγρίου τύπου αλληλόµορφο (GemC1 WT, n=14). 147


 και της pericentrin (PCNT) για τη χρώση των κροσσών και των νεοσυντιθέµενων")


 χαρακτηρίζεται από πολλαπλούς κροσσούς, οι")


148 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Η πρωτεΐνη GemC1 είναι απαραίτητη για τη δηµιουργία των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου Με σκοπό να διερευνηθεί η συµµετοχή του γονιδίου GemC1 στη διαφοροποίηση των πολυκροσσωτών κυττάρων, έγινε αποµόνωση και µελέτη του επιθηλίου της τραχείας νεογέννητων µυών (P0) στους οποίους δεν εκφραζόταν ο παράγοντας GemC1 και παράλληλα συγκρίθηκε µε τραχείες που προέρχονταν από φυσιολογικούς µύες της ίδιας τοκετοµάδας. Σε εγκάρσιες τοµές της τραχείας των µυών πραγµατοποιήθηκε ανοσοφθορισµός µε τη χρήση αντισωµάτων έναντι της ακετυλιωµένης α-τουµπουλίνης (ACT) και της pericentrin (PCNT) για τη χρώση των κροσσών και των νεοσυντιθέµενων κεντριολίων/βασικών σωµατίων, αντίστοιχα (Εικόνα 4.4.3). Παράλληλα χρησιµοποιήθηκε αντίσωµα έναντι της E-cadherin για τη χρώση της κυτταρικής µεµβράνης, ώστε να διευκολύνεται η οριοθέτηση των κυττάρων. Από την παραπάνω ανάλυση δείχθηκε ότι το επιθήλιο της τραχείας που προερχόταν από τους GemC1 ΚΟ/ΚΟ µύες στερείται παντελώς κροσσών και πολλαπλών κεντριολίων (Εικόνα A). Αντίθετα, η τραχεία που προέρχεται από φυσιολογικούς µύες (GemC1 WT/WT ) χαρακτηρίζεται από πολλαπλούς κροσσούς, οι οποίοι εκφύονται από πολλαπλά βασικά σωµάτια που βρίσκονται κατανεµηµένα στην ε πιφάνεια των κυττάρων. Η έλλειψη υπεράριθµων κεντριολίων που δίνουν γένεση στους κροσσούς επιβεβαιώθηκε και µε τη χρήση ενός ακόµα αντισώµατος ένταντι της γ-τουµπουλίνης, που χρησιµοποιείται επίσης ως δείκτης των κεντριολίων (Εικόνα Β). Συµπεραίνουµε ότι ο παράγοντας GemC1 είναι απαραίτητος για τη δηµιουργία των κινητών κροσσών του αναπνευστικού επιθηλίου. Η έλλειψή του ε µποδίζει τη διαφοροποίηση των πολυκροσσωτών κυττάρων σε πρώιµα στάδια, αφού δεν παρατηρούνται καν τα αρχικά στάδια της διαφοροποίησης που σχετίζονται µε τον µαζικό πολλαπλασιασµό των κεντριολίων. A ACT/PCNT/E-cadherin GemC1 KO/KO GemC1 WT/WT 148
 Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν σε εγκάρσιες τοµές από τραχείες που αποµονώθηκαν από νεογέννητους (P0) µύες αγρίου τύπου (GemC1 WT/WT ) ή από µύες της ίδιας τοκετοµάδας από τους")
 για την ανίχνευση των νεοσυντιθέµενων κεντριολίων/βασικών σωµατίων και έναντι της E- cadherin (µπλέ) για τη χρώση της κυτταρικής µεµβράνης µε σκοπό την οριοθέτηση των κυττάρων.")
149 4. ΑΠΟΤΕΛΕΣΜΑΤΑ B GemC1 WT/WT GemC1 KO/KO ACT/γ-tubulin/E-cadherin Εικόνα Η έλλειψη του γονιδίου GemC1 εµποδίζει τη δηµιουργία των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου. (Α) Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν σε εγκάρσιες τοµές από τραχείες που αποµονώθηκαν από νεογέννητους (P0) µύες αγρίου τύπου (GemC1 WT/WT ) ή από µύες της ίδιας τοκετοµάδας από τους οποίους απουσίαζε το γονίδιο GemC1 (GemC1 KO/KO ), µε τη χρήση αντισωµάτων έναντι της ακετυλιωµένης α-τουµπουλίνης για τη χρώση του αξονήµατος των κροσσών (ACT, κόκκινο), της pericentrin (PCNT, πράσινο) για την ανίχνευση των νεοσυντιθέµενων κεντριολίων/βασικών σωµατίων και έναντι της E- cadherin (µπλέ) για τη χρώση της κυτταρικής µεµβράνης µε σκοπό την οριοθέτηση των κυττάρων. Οι εικόνες των τραχειών (αριστερά) προέρχονται από τη σύνθεση επιµέρους φωτογραφιών που ελήφθησαν σε συνεστιακό µικροσκόπιο φθορισµού. Στα πλαίσια απεικονίζονται οι αντίστοιχες µεγεθύνσεις της χρώσης των τραχειών των GemC1 WT/WT και GemC1 KO/KO µυών. (Β) Επιπλέον πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν µε τη χρήση αντισώµατος έναντι της γ-τουµπουλίνης (γ-tubulin, πράσινο) για την ανίχνευση των κεντριολίων. Τα βέλη δείχνουν την απουσία πολλαπλών κεντριολίων στην τραχεία των GemC1 KO/KO µυών. Scale bar, 10µm Η έλλειψη του γονιδίου GemC1 εµποδίζει την έκφραση παραγόντων που εµπλέκονται σε διαφορετικά στάδια της δηµιουργίας των πολυκροσσωτών κυττάρων Από τα παραπάνω καθίσταται σαφές ότι ο παράγοντας GemC1 είναι απαραίτητος για το σχηµατισµό των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου και έτσι στη συνέχεια εξετάστηκε ο µηχανισµός µέσω του οποίου δρα στο συγκεκριµένο µονοπάτι. Για το λόγο αυτό µελετήθηκε η επίδραση της έλλειψης του γονιδίου GemC1 σε παράγοντες που εµπλέκονται στη δηµιουργία των πολυκροσσωτών κυττάρων σε διαφορετικά βήµατα κατά τη διάρκεια της διαδικασίας αυτής. Σε εγκάρσιες τοµές της τραχείας GemC1 WT/WT και GemC1 ΚΟ/ΚΟ νεογέννητων µυών (P0) που προέρχονταν από την ίδια τοκετοµάδα, πραγµατοποιήθηκε ανοσοφθορισµός µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas, p73 και Foxj1 (Εικόνα 4.4.4). Ο παράγοντας McIdas έχει ήδη δειχθεί, τόσο στην παρούσα µελέτη όσο και από άλλες ερευνητικές οµ άδες, ότι ελέγχει τη διαφοροποίηση των 149
.")




, µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas (Α-Α, πράσινο), p73 (Β-Β, µωβ)")
150 4. ΑΠΟΤΕΛΕΣΜΑΤΑ πολυκροσσωτών κυττάρων στα πολύ αρχικά της στάδια, συµµετέχοντας στην έξοδο των κυττάρων από τον κυτταρικό κύκλο ώστε να επιτευχθεί ο πολλαπλασιασµός των κεντριολίων (Ma et al, 2014; Stubbs et al, 2012). Οι p73 (Marshall et al, 2016; Nemajerova et al, 2016) και Foxj1 (Brody et al, 2000; Stubbs et al, 2008; Yu et al, 2008) µεταγραφικοί παράγοντες συµµετέχουν αργ ότερα κατά τη διαδικασία αυτή, ελέγχοντας τη µεταφορά των κεντριολίων στην κυτταρική µεµβράνη, καθώς και άλλα µεταγενέστερα στάδια. Όπως φαίνεται στην εικόνα στην τραχεία των µυών αγρίου τύπου (GemC1 WT/WT ) ανιχνεύεται έκφραση των πρωτεϊνών McIdas (Α), p73 (Β) και Foxj1 (Γ), ενώ στα επιθηλιακά κύτταρα της τραχείας που προέρχονται από τους GemC1 ΚΟ/ΚΟ µύες, απουσιάζει εντελώς η έκφρασή τους. Συµπεραίνουµε ότι απουσία του γονιδίου GemC1 εµποδίζεται το µονοπάτι της διαφοροποίησης των πολυκροσσωτών κυττάρων, αφού δεν εκφράζονται οι παράγοντες εκείνοι που είναι απαραίτητοι για τη σωστή διεκπεραίωσή του, συµπεριλαµβανοµένου των McIdas, p73 και Foxj1. A McIdas/Draq-5 Β p73/e-cadherin Γ Foxj1/Draq-5 GemC1 KO/KO GemC1 WT/WT A Β Γ Εικόνα Η έλλειψη του GemC1 εµποδίζει τη διαφοροποίηση των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου. (Α) Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν σε εγκάρσιες τοµές από τραχείες που αποµονώθηκαν από νεογέννητους (P0) µύες αγρίου τύπου (GemC1 WT/WT, Α-Γ) ή από µύες από τους οποίους απουσίαζε το γονίδιο GemC1 (GemC1 KO/KO, Α -Γ ), µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas (Α-Α, πράσινο), p73 (Β-Β, µωβ) και Foxj1 (Γ-Γ, κόκκινο). Αντίσωµα έναντι της E-cadherin χρησιµοποιήθηκε για την οριοθέτηση των κυττάρων (Β-Β ), ενώ η χρωστική draq-5 χρησιµοποιήθηκε για τη σήµανση των πυρήνων των κυττάρων (Α-Α, Γ -Γ ). Τα βέλη υποδεικνύουν την παρουσία θετικών κυττάρων για την έκφραση των McIdas, p73 και Foxj1 στις τοµές τραχείας από τους GemC1 WT/WT µύες. Scale bar, 10µm. 150
151 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Στη συνέχεια ελέγχθηκε εάν η έλλειψη του γονιδίου GemC1 επηρεάζει τη διαφοροποίηση των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου από τα πρώιµα στάδια της διαδικασίας, και συγκεκριµένα κατά τη 16η εµβρυϊκή ηµέρα (Ε16.5). Έτσι µελετήθηκε η έκφραση µιας σειράς παραγόντων που εµπλέκονται είτε στην έξοδο των κυττάρων από τον κυτταρικό κύκλο και στο διπλασιασµό των κεντριολίων τους για τη δηµιουργία των βασικών σωµατίων, είτε στη µεταγραφική ρύθµιση του µονοπατιού. Για το σκοπό αυτό πραγµατοποιήθηκε αποµόνωση ολικού RNA από τις τραχείες φυσιολογικών µυών (GemC1 WT/WT ή GemC1 WT/ΚO ) ή µυών οµόζυγων για το µεταλλαγµένο αλληλόµορφο (GemC1 ΚΟ/ΚΟ ), από έµβρυα ηλικίας 16.5 ηµερών (Ε16.5) που προέρχονταν από την ίδια τοκετοµάδα. Το RNA αυτό χρησιµοποιήθηκε ως καλούπι για τη σύνθεση συµπληρωµατικού DNA (cdna) και στη συνέχεια χρησιµοποιήθηκαν ειδικοί εκκινητές για την α νίχνευση της έκφρασης του κάθε γονιδίου στους αποµονωµένους ιστούς µέσω ποσοτικής Real-Time PCR (Εικόνα 4.4.5). Όπως φαίνεται στο παρακάτω διάγραµµα η έλλειψη του γονιδίου GemC1 επηρεάζει την έκφραση µιας ποικιλίας παραγόντων. Όσον αφορά στους παράγοντες που εµπλέκονται στη µεταγραφική ρύθµιση του µονοπατιού παρατηρείται σηµαντική πτώση της έκφρασης του mrna των παραγόντων McIdas, p73 και Foxj1. Το αποτέλεσµα αυτό έρχεται σε συµ φωνία µε προηγούµενα δεδοµένα στους νεογέννητους µύες, όπου µετά την έλλειψη της πρωτεΐνης GemC1, χάνεται η έκφρασή τους σε πρωτεϊνικό επίπεδο. Επίσης, παρατηρείται σηµαντική πτώση της έκφρασης του µεταγραφικού παράγοντα cmyb. Ιδιαίτερο ενδιαφέρον παρουσιάζει η διαφορική ρύθµ ιση της έκφρασης των παραγόντων της οικογένειας των p53 πρωτεϊνών, αφού ο παράγοντας p63 επηρεάζεται αρκετά λιγότερο σε σύγκριση µε τον p73. Ο παράγοντας p73 χαρακτηρίστηκε πρόσφατα ότι συµµετέχει ειδικά στη διαφοροποίηση των πολυκροσσωτών κυττάρων του αναπνευστικού επιθηλίου, ενώ ο p63 εκφράζεται στα βασικά κύτταρα του επιθηλίου και επηρεάζει και άλλους κυτταρικούς πληθυσµούς (Marshall et al, 2016; Nemajerova et al, 2016). Τέλος, οι παράγοντες Jagged 1 και 2 επηρεάζονται µε διαφορετικό τρόπο µετά την έλλειψη του GemC1, αφού ο παράγοντας Jagged 1 παραµένει ανεπηρέαστος σε αντίθεση µε τον Jagged 2. Ακόµα µελετήθηκε η έκφραση παραγόντων που συνδέονται είτε µε τη ρύθµιση του κυτταρικού κύκλου, είτε µε το διπλασιασµό των κεντριολίων. Το γονίδιο Deup1, που αποτελεί το κεντρικό µόριο που ρυθµ ίζει το διπλασιασµό των κεντριολίων µέσω του σχηµατισµού των δευτεροσωµατίων, επηρεάζεται σηµαντικά, ενώ η κινάση Plk4 λιγότερο. Ο παράγοντας Ccno πoυ συµµετέχει επίσης σε αυτό το µονοπάτι µειώνεται δραστικά, σε σύγκριση µε την έκφρασή του στις τ ραχείες των φυσιολογικών µυών. Τέλος, παρατηρήθηκε ότι το mrna του γονιδίου της Geminin δεν επηρεάζεται από την έλλειψη του GemC1, ενώ τα γονίδια Cdc20b και Ccna1 επηρεάζονται σηµαντικά. Συµπεραίνουµε ότι το γονίδιο GemC1 αποτελεί έναν κορυφαίο ρυθµιστή του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων, αφού επηρεάζει την έκφραση παραγόντων που εµπλέκονται σε διάφορα στάδια κατά τη δηµιουργία των κυττάρων αυτών. 151
152 4. ΑΠΟΤΕΛΕΣΜΑΤΑ GemC1 WT Σχετικά επίπεδα έκφρασης mrna ως προς Gemc1 WT E16.5 τραχεία Jagged 1 Jagged 2 TAp63 TAp73 McIdas cmyb Foxj1 Plk4 Deup1 Geminin Cdc20b GemC1 KO Ccna1 Ccno Παράγοντες μεταγραφής Ρυθμιστές διπλασιασμού κεντριολίων Ρυθμιστές κυτταρικού κύκλου Εικόνα Η έλλειψη του γονιδίου GemC1 επηρεάζει µια οµάδα παραγόντων που εµπλέκονται στη διαφοροποίηση των πολυκροσσωτών κυττάρων του αναπτυσσόµενου αναπνευστικού επιθηλίου. Μελέτη της έκφρασης του mrna διαφόρων γονιδίων µέσω ποσοτικής real-time PCR σε τραχείες που αποµονώθηκαν από φυσιολογικούς µύες (GemC1 WT/WT ή GemC1 WT/ΚO ) ή µύες οµ όζυγους για το µεταλλαγµένο αλληλόµορφο του γονιδίου GemC1 (GemC1 KO/ΚO ). Οι τιµές αντιστοιχούν στα επίπεδα έκφρασης (+/- τυπική απόκλιση) του κάθε γονιδίου σε κάθε συνθήκη, σε σχέση µε την αντίστοιχη έκφρασή τους στις τραχείες των φυσιολογικών µυών που ορίζεται ως τιµή 1. Η κανονικοποίηση των τιµών έγινε µε τη χρήση των επιπέδων έκφρασης του HPRT γονιδίου και ακολούθησε ανάλυση των αποτελεσµάτων µέσω του προγράµµατος REST-MCS beta. Τα δεδοµένα εξήλθαν από τις τραχείες 3 εµ βρύων µυών, που ενώθηκαν και αναλύθηκαν παράλληλα εις διπλούν Η έλλειψη του γονιδίου GemC1 δεν επηρεάζει τον αριθµό των βασικών διαιρούµενων κυττάρων, ενώ αυξάνει τα βλεννοπαραγωγά κύτταρα του αναπνευστικού επιθηλίου Στη συνέχεια µελετήθηκε εάν ο παράγοντας GemC1 µπορεί να επηρεάζει και άλλους κυτταρικούς τύπους που απαντώνται στο αναπνευστικό επιθήλιο, εκτός των πολυκροσσωτών κυττάρων. Για το σκοπό αυτό πραγµατοποιήθηκαν πειράµατα ανοσοφθορισµού σε εγκάρσιες τοµές τραχείας που προέρχονταν από νεογέννητους µύες (P0) αγρίου τύπου (GemC1 WT/WT ) ή από µύες που ήταν ελλειµµατικοί για το γονίδιο GemC1 (GemC1 ΚΟ/ΚΟ ) της ίδιας τοκετοµάδας κ αι µελετήθηκαν ως προς τη σύσταση του επιθηλίου σε βασικά (basal cells) και βλεννοπαραγωγά κύτταρα (goblet cells) (Εικόνα 4.4.6). Πιο συγκεκριµένα χρησιµοποιήθηκαν αντισώµατα έναντι της κερατίνης-5 (Krt5), για τον εντοπισµό των βασικών κυττάρων του επιθηλίου που διατηρούν την ικανότητα της κυτταρικής διαίρεσης, καθώς και έναντι της βλέννης Muc5ac που αποτελεί δείκτη των βλεννοπαραγωγών κυττάρων. Παρατηρήθηκε ότι η έλλειψη του γονιδίου GemC1 δεν επηρεάζει τον αριθµό των βασικών κυττάρων που εντοπίζονται στο επιθήλιο της τραχείας σε σύγκριση µε το επιθήλιο φυσιολογικών τραχειών (Εικόνα Α). Επίσης ως προς τον αριθµό των 152








153 4. ΑΠΟΤΕΛΕΣΜΑΤΑ βλεννοπαραγωγών κυττάρων παρατηρήθηκε αύξηση αυτών απουσία του γονιδίου GemC1, σε σύγκριση µε τον αριθµό τους στο φυσιολογικό επιθήλιο (Εικόνα Β). Καταλήγουµε στο συµπέρασµα ότι το γονίδιο GemC1 δεν επηρεάζει τον πληθυσµό των βασικών κυττάρων του επιθηλίου που εκφράζουν την πρωτεΐνη κερατίνη-5. Αντίθετα, ο αριθµός των βλεννοπαραγωγών κυττάρων εµφανίζεται αυξηµένος στην τραχεία των ελλειµµατικών µυών για το γονίδιο GemC1. Το εύρηµα αυτό υποδηλώνει πιθανή εµπλοκή του GemC1 στην επιλογή κυτταρικής µοίρας στο αναπνευστικό επιθήλιο. A Krt5/ACT/Draq-5 GemC1 KO/KO GemC1 WT/WT B Muc5ac/E-cadherin/Draq-5 GemC1 KO/KO GemC1 WT/WT 153
154 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Η έλλειψη του γονιδίου GemC1 δεν επηρεάζει τα βασικά κύτταρα του επιθηλίου της τραχείας, ενώ προκαλεί αύξηση των βλεννοπαραγωγών κυττάρων. (Α) Σε εγκάρσιες τοµές από τραχείες που αποµονώθηκαν από νεογέννητους (P0) µύες αγρίου τύπου (GemC1 WT/WT ) ή από µύες από τους οποίους απουσίαζε το γονίδιο GemC1 (GemC1 KO/KO ), πραγµατοποιήθηκαν πειράµατα ανοσοφθορισµού µε τη χρήση αντισωµάτων έναντι της κερατίνης-5 (Κ5, πράσινο) και της ακετυλιωµένης α - τουµπουλίνης (ACT, κόκκινο) για την ανίχνευση των βασικών και των πολυκροσσωτών κυττάρων του επιθηλίου, αντίστοιχα. (Β) Σε εγκάρσιες τοµές από τραχείες που αποµονώθηκαν από νεογέννητους (P0) µύες αγρίου τύπου (GemC1 WT/WT ) ή από µύες από τους οποίους απουσίαζε το γονίδιο GemC1 (GemC1 KO/KO ), πραγµατοποιήθηκαν πειράµατα ανοσοφθορισµού µε τη χρήση αντισωµάτων έ ναντι των Muc5ac (κόκκινο), για τη χρώση των βλεννοπαραγωγών κυττάρων και της E-cadherin (πράσινο) για τη χρώση της κυτταρικής µεµβράνης, ώστε να επιτυγχάνεται η οριοθέτηση των κυττάρων. Οι εικόνες των τραχειών (αριστερά) προέρχονται από τη σύνθεση επιµέρους φωτογραφιών που ελήφθησαν σε συνεστιακό µικροσκόπιο φθορισµού ώστε να επιτευχθεί πλήρης απεικόνισή τους. Μεγαλύτερες µεγεθύνσεις µιας περιοχής των τραχειών απεικονίζονται δεξιά. Η χρωστική draq-5 χρησιµοποιήθηκε για τη σήµανση των πυρήνων των κυττάρων. Scale bar, 10µm. 154
155 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 4.5 Προκαταρκτική µελέτη της συµµετοχής της πρωτεΐνης McIdas στο διπλασιασµό των κεντριολίων σε διαιρούµενα κύτταρα Από τη µέχρι τώρα πορεία της παρούσας διατριβής δείχθηκε ότι τα µέλη της υπεροικογένειας της Geminin, και κυρίως οι GemC1 και McIdas, επάγουν τη µεταγραφή µιας σειράς γονιδίων που είναι απαραίτητα σε διάφορα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. Συνδυάζοντας τα αποτελέσµατα της µελέτης αυτής µε βιβλιογραφικά δεδοµένα, φαίνεται ότι οι παράγοντες GemC1 και McIdas επηρεάζουν, µεταξύ άλλων, και σηµαντικούς ρυθµ ιστές του πολλαπλασιασµού των κεντριολίων, συµπεριλαµβανοµένου των Deup1, Plk4 και STIL (Ma et al, 2014). Στα διαιρούµενα κύτταρα προκειµένου να διατηρείται η γονιδιωµατική σταθερότητα, ελέγχεται η δηµιουργία ενός µόνο θυγατρικού κεντριολίου δίπλα από κάθε µητρικό κεντριόλιο. Όµως, στα πολυκροσσωτά κύτταρα ο έλεγχος αυτός παρακάµπτεται ώστε να προκύψει τελικά ένας µεγάλος αριθµός κεντριολίων/βασικών σωµατίων. Το κύριο µοριακό µονοπάτι που ελέγχει το διπλασιασµό των κεντριολίων κατά τη διάρκεια του κυτταρικού κύκλου στα διαιρούµενα κύτταρα, ελέγχει και την παραγωγή των πολυάριθµων κεντριολίων στα διαφοροποιηµένα κύτταρα (Vladar & Stearns, 2007). Ωστόσο, δεν είναι γνωστοί οι µηχανισµοί εκείνοι που ελέγχουν αρχικά την απόφαση ενός κυττάρου ως προς τον αριθµό των κεντριολίων που θα αποκτήσει. Tα σχετικά επίπεδα έκφρασης των παραγόντων που συµµετέχουν στον πολλαπλασιασµό των κεντριολίων φαίνεται να είναι σηµαντικά για τον τελικό αριθµό τους στα κύτταρα (Firat-Karalar & Stearns, 2014; Vladar & Stearns, 2007). Οι παράγοντες GemC1 και McIdas µπορεί να ρυθµίζουν τα επίπεδα έκφρασης αυτών των παραγόντων. Επίσης, η πρωτεΐνη Geminin έχει ήδη δειχθεί να εντοπίζεται στα κεντροσωµάτια και να ελέγχει το διπλασιασµό τους (Lu et al, 2009; Tachibana et al, 2005). Στη συνέχεια της εργασίας αυτής µελετήθηκε η πιθανή έκφραση της πρωτεΐνης McIdas στα κεντριόλια και ο ρόλος της κατά τον κύκλο του διπλασιασµού των κεντροσωµατίων. Εξαιτίας της έλλειψης ενός αντισώµατος που να ανιχνεύει τα πρωτεϊνικά επίπεδα του GemC1, η µελέτη αυτή επικεντρώθηκε στην πρωτεΐνη McIdas Η πρωτεΐνη McIdas εντοπίζεται στο µητρικό κεντριόλιο των κεντροσωµατίων των διαιρούµενων κυττάρων Με δεδοµένο τη συµµετοχή των µελών της οικογένειας της Geminin στον πολλαπλασιασµό των κεντριολίων κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, θελήσαµε να διερευνήσουµε εάν η πρωτεΐνη McIdas µπορεί να συµµετέχει και στο διπλασιασµό των κεντριολίων κατά τη διάρκεια του κυτταρικού κύκλου. Για το σκοπό αυτό µελετήθηκε αρχικά η έκφρασή της στα κεντροσωµάτια. Πραγµατοποιήθηκαν πειράµατα ανοσοφθορισµού σε ασύγχρονα κύτταρα HeLa και MCF-7, µε τη χρήση αντισωµάτων έναντι της πρωτεΐνης McIdas, καθώς και έναντι των παραγόντων Cep135 και Cep164, που εντοπίζονται στα κεντριόλια και συµµετέχουν στο διπλασιασµό τους. Οι παράγοντες Cep135 και Cep164 εµφανίζουν συγκεκριµένο πρότυπο έκφρασης στα κεντριόλια των κυττάρων, ανάλογα µε τη φάση του κυτταρικού κύκλου. Ο δείκτης Cep135 εντοπίζεται στο εγγύς άκρο των κεντριολίων. Κατά τη G1 φάση του κυτταρικού κύκλου, όπου δεν παρατηρείται ακόµα διπλασιασµός των κεντριολίων, τα κύτταρα διαθέτουν ένα κεντροσωµάτιο που αποτελείται από δυο κεντριόλια συνδεδεµένα µεταξύ τους. Αυτό έχει ως αποτέλεσµα παρόλο που ο 155
.")
156 4. ΑΠΟΤΕΛΕΣΜΑΤΑ παράγοντας Cep135 εντοπίζεται και στα δυο κεντριόλια να µην είναι δυνατό να ξεχωρίσει η χρώση του από κάθε κεντριόλιο και έτσι να εµφανίζεται ως µια χρώση και από τα δυο κεντριόλια (1 εστία ανά κύτταρο). Μετά το διπλασιασµό των κεντριολίων κάθε ζεύγος έχει από ένα σήµα Cep135, και έτσι τα κύτταρα κατά τις φάσεις S, G2 και M εµφανίζουν 2 διακριτές εστίες Cep135 (2 εστίες ανά κύτταρο). Ο δείκτης Cep164 σηµαίνει το ώριµο µητρικό κεντριόλιο του κεντροσωµατίου. Έτσι κατά τις φάσεις G1 και S εµφανίζεται 1 εστία ανά κύτταρο, ενώ µετά τη G2 φάση όπου γίνεται η ωρίµανση και του δεύτερου πια µητρικού κεντριολίου εµφανίζονται 2 εστίες. Όπως φαίνεται στην εικόνα Α, η πρωτεΐνη McIdas εντοπίζεται µόνο στο ένα από τα δυο κεντριόλια του κεντροσωµατίου. Παράλληλα χρησιµοποιήθηκε και ο δείκτης Cep164, ώστε να διαπιστωθεί εάν αυτό το κεντριόλιο είναι το µητρικό ή το θυγατρικό. Ο McIdas συνεντοπίζεται µε το δείκτη Cep164, αποτέλεσµα που υποδεικνύει ότι εντοπίζεται ειδικά στο µητρικό κεντριόλιο των κεντροσωµατίων. Ιδιαίτερο ενδιαφέρον παρουσιάζει το γεγονός ότι ο McIdas φαίνεται να εντοπίζεται στο αποµακρυσµένο άκρο του µητρικού κεντριολίου, αλλά όχι τόσο αποµακρυσµένα όσο ο δείκτης Cep164, που είναι γνωστό ότι βρίσκεται στο πιο ακραίο τµήµα του µητρικού κεντριολίου. Τα αποτελέσµατα αυτά επιβεβαιώθηκαν και στην κυτταρική σειρά MCF7 (Εικόνα Β). A McIdas/Cep135/Cep164 HeLa B McIdas/Cep135/Cep164 MCF7 156
157 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Η πρωτεΐνη McIdas εντοπίζεται στο ακραίο τµήµα του µητρικού κεντριολίου των κεντροσωµατίων. Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν σε κύτταρα HeLa (A) και MCF-7 (B), µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas (κόκκινο), Cep135 (µωβ) και Cep164 (πράσινο). Η λευκή γραµµή απεικονίζει νοητά το περίγραµµα του πυρήνα του κυττάρου. Σε κάθε φωτογραφία τα ένθετα αντιστοιχούν σε µεγαλύτερη µεγέθυνση των κεντριολίων που υποδεικνύονται µε τα βέλη. Η πρωτεΐνη McIdas εντοπίζεται στο αποµακρυσµένο τµήµα του µητρικού κεντριολίου των κεντροσωµατίων. Προκειµένου να διαπιστωθεί η ειδικότητα του παραπάνω σήµατος πραγµατοποιήθηκαν πειράµατα αποσιώπησης του γονιδίου McIdas µε την τεχνολογία sirna. Για το σκοπό αυτό κύτταρα HeLa διαµολύνθηκαν παροδικά µε sirna ολιγονουκλεοτίδια που στοχεύουν ειδικά το mrna του McIdas, καθώς και µη ειδικά sirna ολιγονουκλεοτίδια για µια τυχαία αλληλουχία ως µάρτυρας. Συνολικά έγιναν δυο διαδοχικές διαµολύνσεις µε τα sirna µε διαφορά 24 ωρών. 72 ώρες µετά την πρώτη διαµόλυνση τα κύτταρα µονιµοποιήθηκαν και ακολούθησε ανοσοφθορισµός µε αντισώµατα έναντι των πρωτεϊνών McIdas και Cep135 για τη σήµανση των κεντροσωµατίων (Εικόνα 4.5.2). Ακολούθησε ανάλυση των κυττάρων µε βάση την έκφραση της πρωτεΐνης McIdas στα κεντροσωµάτια. Στα κύτταρα-µάρτυρες η πρωτεΐνη ΜcIdas εντοπίζεται στα κεντροσωµάτια στο 42% των κυττάρων, ενώ µετά την αποσιώπησή της η έκφρασή της στα κεντροσωµάτια µειώνεται στο 9,52% των κυττάρων. Συµπεραίνουµε λοιπόν ότι η έκφραση του McIdas στα κεντροσωµάτια είναι ειδική. Ποσοστό Cep135+McIdas+/Cep135+ κυττάρων Luc sirna McIdas sirna Εικόνα Η πρωτεΐνη McIdas εντοπίζεται ειδικά στα κεντροσωµάτια των διαιρούµενων κυττάρων. Κύτταρα HeLa διαµολύνθηκαν παροδικά µε sirna ολιγονουκλεοτίδια που στοχεύουν ειδικά το mrna του γονιδίου McIdas ή µε µη ειδικά ολιγονουκλεοτίδια για µια τυχαία αλληλουχία. 72 ώρες µετά τη διαµόλυνση πραγµατοποιήθηκαν πειράµατα ανοσοφθορισµού µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas και Cep135 για τη σήµανση των κεντριολίων. Το διάγραµµα απεικονίζει το ποσοστό των κεντροσωµατίων στα οποία εκφράζεται η πρωτεΐνη McIdas στα κύτταρα-µάρτυρες (42%), σε σύγκριση µε την αντίστοιχη έκφραση της πρωτεΐνης µετά την αποσιώπηση του γονιδίου της (9,52%). Τα δεδοµένα προέρχονται από ένα αντιπροσωπευτικό πείραµα και στην κάθε συνθήκη µετρήθηκαν περίπου 100 κύτταρα. Τέλος, θελήσαµε να µελετήσουµε εάν η έκφραση της πρωτεΐνης McIdas στα κεντροσωµάτια διαφέρει ανάλογα µε τη φάση του κυτταρικού κύκλου. Για το σκοπό 157



.")
158 4. ΑΠΟΤΕΛΕΣΜΑΤΑ αυτό αρχικά συγκρίθηκε η έκφραση της πρωτεΐνης McIdas µε αυτή της κυκλίνης Α, η οποία εκφράζεται κατά τις φάσεις S και G2 του κυτταρικού κύκλου. Όπως φαίνεται στην εικόνα 4.5.3, η πρωτεΐνη McIdas εντοπίζεται στα κεντροσωµάτια τόσο κυττάρων που εκφράζουν την κυκλίνη Α, όσο και κυττάρων που δεν την εκφράζουν. Eποµένως, φαίνεται ότι η πρωτεΐνη McIdas εντοπίζεται στα κεντροσωµάτια για µεγάλο διάστηµα κατά τη διάρκεια του κυτταρικού κύκλου. McIdas/Cep135/CyclinA CyclinA- κύτταρο CyclinA+ κύτταρο Εικόνα Η πρωτεΐνη McIdas εντοπίζεται τόσο σε κύτταρα που εκφράζουν την κυκλίνη Α, όσο και σε κύτταρα που δεν την εκφράζουν. Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν σε κύτταρα HeLa, µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas (κόκκινο), Cep135 (πράσινο) και κυκλίνη Α (µώβ). Η λευκή γραµµή απεικονίζει νοητά το περίγραµµα του πυρήνα του κυττάρου. Παράλληλα µελετήθηκε η έκφραση του McIdas στα κεντροσωµάτια και κατά τη διάρκεια της µίτωσης. Για το σκοπό αυτό σε κύτταρα HeLa πραγµατοποιήθηκε ανοσοφθορισµός µε τη χρήση αντισωµάτων έναντι της πρωτεΐνης McIdas και Cep135 για την ανίχνευση των κεντριολίων. Η κατάταξη των κυττάρων στα διαφορετικά στάδια της µίτωσης έγινε µε βάση τη χρώση του DNA (χρωστική Hoechst). Όπως αναµενόταν από τα παραπάνω αποτελέσµατα η πρωτεΐνη McIdas εντοπίζεται στο ένα από τα δύο κεντριόλια των κεντροσωµατίων κατά τη µεσόφαση. Όµως, δεν ανιχνεύεται έκφρασή της στα κεντροσωµάτια που οργανώνουν τη µιτωτική άτρακτο σε καµία από τις φάσεις της µίτωσης. Συµπεραίνουµε λοιπόν ότι η πρωτεΐνη McIdas δεν εντοπίζεται στα κεντροσωµάτια κατά τη διάρκεια της µίτωσης. 158


 και Cep135 (πράσινο).")



.")




159 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Μεσόφαση Πρόφαση Πρό-μετάφαση Μετάφαση Ανάφαση Τελόφαση * * * * * * * * * * McIdas/Cep135 * * * * * * * * * * * * * * * * * * * * Εικόνα Η πρωτεΐνη McIdas δεν εντοπίζεται στα κεντροσωµάτια κατά τη διάρκεια της µιτωτικής διαίρεσης. Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν σε κύτταρα HeLa, µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών McIdas (κόκκινο) και Cep135 (πράσινο). Η κατάταξη των κυττάρων στις διαφορετικές φάσεις της µίτωσης επιτελέσθηκε µε βάση τη χρώση των χρωµοσωµάτων µε τη χρωστική Hoechst. Ενώ η πρωτεΐνη McIdas εντοπίζεται στα κεντριόλια κατά τη διάρκεια της µεσόφασης, δεν εντοπίζεται σε αυτά σε κανένα στάδιο της µίτωσης Η υπερέκφραση της πρωτεΐνης McIdas προκαλεί αύξηση του αριθµού των κεντροσωµατίων Προηγούµενα πειράµατα έχουν δείξει ότι η υπερέκφραση της Geminin αναστέλλει τον πολλαπλασιασµό των κεντριολίων που επάγεται µετά την επώαση των κυττάρων µε υδροξυουρία (ΗU) και την επακόλουθη συσσώρευσή τους στη φάση S του κυτταρικού κύκλου (Tachibana & Nigg, 2006). Με δεδοµένο τον εντοπισµό της πρωτεΐνης McIdas στα κεντροσωµάτια θελήσαµε στη συνέχεια να µελετήσουµε το ρόλο της κατά το διπλασιασµό των κεντροσωµατίων στα διαιρούµενα κύτταρα. Για τη µελέτη αυτή πραγµατοποιήθηκαν πειράµατα υπερέκφρασης της πρωτεΐνης McIdas. Συγκεκριµένα, κύτταρα HeLa επωάστηκαν µε υδροξυουρία και 24 ώρες µετά διαµoλύνθηκαν παροδικά µε πλασµίδια που εξέφραζαν είτε την πρωτεΐνη McIdas συντηγµένη µε GFP, είτε µόνο την πρωτεΐνη GFP ως µάρτυρα. Τα κύτταρα µονιµοποιήθηκαν 48 ώρες µετά τη διαµόλυνση και αξιολογήθηκαν ως προς τον αριθµό των κεντριολίων που διαθέτουν. Για την ανάλυση αυτή π ραγµατοποιήθηκε ανοσοφθορισµός µε τη χρήση αντισώµατος έναντι της πράσινης φθορίζουσας πρωτεΐνης για την ανίχνευση των κυττάρων στα οποία υπερεκφράζεται είτε η πρωτεΐνη McIdas, είτε η GFP ως µάρτυρας. Παράλληλα, χρησιµοποιήθηκαν οι δείκτες Cep135 και CP110. Όπως αναφέρθηκε και προηγουµένως ο δείκτης Cep135 σηµαίνει το εγγύς άκρο των κεντριολίων, ενώ το CP110 σηµαίνει το αποµακρυσµένο άκρο των κεντριολίων. Έτσι, κατά τη G1 φάση του κυτταρικού κύκλου όπου δεν έχει γίνει ο διπλασιασµός του κεντροσωµατίου θα παρατηρούνται 2 dots (ένα από κάθε κεντριόλιο του κεντροσωµατίου), ενώ µετά το διπλασιασµό κατά τις φάσεις S, G2 και 159
.")

160 4. ΑΠΟΤΕΛΕΣΜΑΤΑ M παρατηρούνται φυσιολογικά 4 dots, αφού υπάρχουν συνολικά και 4 κεντριόλια ανά κύτταρο. Αρχικά, ταυτοποιήθηκε ο πολλαπλασιασµός των κεντριολίων στα κύτταρα µετά την επώασή τους µε υδροξυουρία και την αναστολή της αντιγραφής του DNA (Meraldi et al, 1999). Σε κύτταρα HeLa που επωάστηκαν για 72 ώρες µε υδροξυουρία, πραγµατοποιήθηκε ανοσοφθορισµός µε τη χρήση αντισωµάτων έναντι των πρωτεΐνών CP110 και Cep135. Παράλληλα, χρησιµοποιήθηκαν και κύτταρα που δεν είχαν επωαστεί µε την υδροξυουρία ως δείγµατα-µάρτυρες. Στα δείγµατα- µάρτυρες, σε ποσοστό 98%, εντοπίζονται κύτταρα που διαθέτουν είτε ένα κεντροσωµάτιο µε δυο κεντριόλια (1 Cep135 εστία µαζί µε 2 CP110 εστίες), είτε δυο κεντροσωµάτια (σε αυτή τη περίπτωση ανιχνεύονται 2 Cep135 εστίες και συνολικά 4 CP110 εστίες, 2 σε κάθε κεντροσωµάτιο) (Εικόνα Β και Δ). Αντίθετα, στα κύτταρα που έγινε επώαση µε υδροξυουρία, το 53,6% των κυττάρων ε µφανίζουν φυσιολογικό αριθµό κεντριολίων (2-4 κεντριόλια ανά κύτταρο), ενώ το 46,6% των κυττάρων διαθέτει πάνω από 4 κεντριόλια (Εικόνα Γ και Δ). Α 72h Ημέρα HU arrest IF B CP110/Cep135 -HU Γ CP110/Cep135 +HU 160
161 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Δ Ποσοστό κυττάρων ανά κατηγορία >4 0 -HU Εικόνα Η επώαση των κυττάρων HeLa µε υδροξυουρία προκαλεί αύξηση του αριθµού των κεντριολίων. (Α) Κύτταρα HeLa επιστρώθηκαν και επωάστηκαν µε υδροξυουρία για 72 ώρες. (Β, Γ) Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών CP110 (κόκκινο) και Cep135 (πράσινο). Στα κύτταρα-µάρτυρες που δεν χορηγήθηκε υδροξυουρία παρατηρούνται 1 έως 2 κεντροσωµάτια, δηλαδή 2-4 κεντριόλια ανάλογα µε τη φάση του κυτταρικού κύκλου. Αντίθετα, στα κύτταρα που χορηγήθηκε η υδροξυουρία παρατηρούνται κύτταρα µε αριθµό κεντριολίων µεγαλύτερο του 4. (Δ) Ταξινόµηση των κυττάρων µε βάση τον αριθµό των κεντριολίων που διαθέτουν απουσία ή παρουσία υδροξυουρίας. Η λευκή γραµµή απεικονίζει νοητά το περίγραµµα του πυρήνα του κυττάρου. +HU Στη συνέχεια µελετήθηκε η επίδραση της υπερέκφρασης της πρωτεΐνης McIdas στον αριθµό των κεντριολίων που προκύπτουν σύµ φωνα µε το παραπάνω πρωτόκολλο. Για το σκοπό αυτό, κύτταρα HeLa επιστρώθηκαν και καλλιεργήθηκαν απουσία ή παρουσία υδροξυουρίας. 24 ώρες µετά τα κύτταρα διαµολύνθηκαν παροδικά, παρουσία υδροξυουρίας, µε πλασµίδια που εξέφραζαν είτε την πρωτεΐνη McIdas συντηγµένη µε GFP, είτε την πρωτεΐνη GFP µόνο, ως µάρτυρα (Εικόνα Α). 48 ώρες µετά τη διαµόλυνση πραγµατοποιήθηκε ανοσοφθορισµός µε τη χρήση αντισωµάτων έναντι των πρωτεΐνών CP110 και Cep135. Στο 70,91% των κυττάρων που εκφράζουν την πρωτεΐνη GFP ανιχνεύονται 2-4 κεντριόλια, ενώ στο 29,09% εντοπίζονται περισσότερα από 4 κεντριόλια (Εικόνα Β και Δ). Αντίθετα, στα κύτταρα που υπερεκφράζεται ο παράγοντας McIdas, το 42,62% των κυττάρων διαθέτουν περισσότερα από 4 κεντριόλια (Εικόνα Γ και Δ). 161



162 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Α 72h Ημέρα HU arrest Παροδική διαμόλυνση με: GFP GFPMcIdas HU arrest IF B GFP CP110/Cep135 +HU +HU Γ GFPMcIdas CP110/Cep135 Δ Ποσοστό κυττάρων ανά κατηγορία >4 0 HU+GFP HU+GFPIdas 162
163 4. ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα Η υπερέκφραση της πρωτεΐνης McIdas προκαλεί αύξηση του αριθµού των κεντριολίων. (Α) Κύτταρα HeLa επιστρώθηκαν και επωάστηκαν µε υδροξυουρία για 24 ώρες. Στη συνέχεια διαµολύνθηκαν παροδικά µε πλασµίδια που εξέφραζαν την πρωτεΐνη McIdas συντηγµένη µε GFP, ή την πρωτεΐνη GFP µόνο ως µάρτυρα. (Β, Γ) Πειράµατα ανοσοφθορισµού πραγµατοποιήθηκαν µε τη χρήση αντισωµάτων έναντι των πρωτεϊνών GFP (πράσινο) για τον εντοπισµό των κυττάρων που διαµολύνθηκαν, CP110 (κόκκινο) και Cep135 (πράσινο). Στα κύτταρα-µάρτυρες που υπερεκφράζεται η πρωτεΐνη GFP παρατηρούνται 1 έως 2 κεντροσωµάτια, δηλαδή 2-4 κεντριόλια. Αντίθετα, στα κύτταρα που υπερεκφράζεται η πρωτεΐνη McIdas παρατηρούνται κύτταρα µε αριθµό κεντριολίων µεγαλύτερο του 4. (Δ) Ταξινόµηση των κυττάρων µε βάση τον αριθµό των κεντριολίων που διαθέτουν απουσία ή παρουσία υδροξυουρίας. Η λευκή γραµµή απεικονίζει νοητά το περίγραµµα του πυρήνα του κυττάρου. Τα παραπάνω αποτελέσµατα έδειξαν τον εντοπισµό της πρωτεΐνης McIdas στα κεντροσωµάτια και αποτελούν µια πρώτη ένδειξη για τη συµµ ετοχή της στο διπλασιασµό των κεντριολίων. Περαιτέρω πειράµατα θα αποσαφηνίσουν το ρόλο των µελών της υπεροικογένειας της Geminin στον κύκλο των κεντροσωµατίων και την πιθανή συµµετοχή τους ως µόρια που συνδέουν τον κ υτταρικό κύκλο µε το διπλασιασµό των κεντριολίων και τη διαφοροποίηση των πολυκροσσωτών κυττάρων, δρώντας µε αυτό τον τρόπο τόσο σε αποφάσεις πολλαπλασιασµού όσο και διαφοροποίησης των κυττάρων. 163
164 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 164
165 5. ΣΥΖΗΤΗΣΗ
166 5. ΣΥΖΗΤΗΣΗ 166
167 5. ΣΥΖΗΤΗΣΗ Σκοπός της παρούσας διδακτορικής διατριβής ήταν να µελετηθεί ο ρόλος των ρυθµιστών της αδειοδότησης της αντιγραφής Geminin, McIdas και GemC1 στον πολλαπλασιασµό των κεντριολίων και στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Στα διαιρούµενα κύτταρα, για να διατηρείται η γονιδιωµατική σταθερότητα απαιτείται η συντονισµένη λειτουργία των κύκλων διπλασιασµού και διαχωρισµού των χρωµοσωµάτων και των κεντροσωµατίων. Μετά την κυτταρική διαίρεση κάθε θυγατρικό κύτταρο πρέπει να διαθέτει ένα πλήρες αντίγραφο του γονιδιώµατος και ένα κεντροσωµάτιο. Αντίθετα, τα πολυκροσσωτά κύτταρα, όπως προαναφέρθηκε, αποτελούν µια ιδιαίτερη κατηγορία κυττάρων, που εξέρχονται από τον κυτταρικό κύκλο και υπερβαίνουν τους κανόνες ενός κανονικού κύκλου διπλασιασµού, σχηµατίζοντας πολυάριθµα κεντριόλια. Ωστόσο, οι µηχανισµοί που ελέγχουν το διπλασιασµό των κεντριολίων στα διαιρούµενα και στα διαφοροποιηµένα πολυκροσσωτά κύτταρα παραµένουν άγνωστοι. Οι παράγοντες Geminin, GemC1 και McIdas αποτελούν µια οικογένεια πρωτεϊνών που χαρακτηρίστηκαν αρχικά για το ρόλο τους στον κυτταρικό κύκλο, κατά την αντιγραφή του DNA. Η Geminin έχει επίσης χαρακτηριστεί ως ένας αρνητικός ρυθµ ιστής του διπλασιασµού των κεντροσωµατίων, συµ βάλλοντας έτσι στη γονιδιωµατική σταθερότητα. Παράλληλα, δείχθηκε να συµµετέχει και σε αποφάσεις κυτταρικής διαφοροποίησης. Αρχικές µελέτες στο εργαστήριό µας έδειξαν ότι και τα δυο άλλα νέα µέλη είναι πιθανό να εµπλέκονται σε αποφάσεις διαφοροποίησης των κυττάρων. Στο κεφάλαιο αυτό θα συζητηθεί το ειδικό πρότυπο έκφρασης που εµφάνισαν οι παράγοντες GemC1 και McIdas κατά την ανάπτυξη των µυών, σε ιστούς που απαντώνται πολυκροσσωτά κύτταρα. Επίσης, αναλύεται και η έκφρασή τους κατά τα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων της τραχείας των µυών, µε ιδιαίτερη έµφαση στο διαφορικό πρότυπο έκφρασης που εµφανίζουν, σε σχέση µε αυτό της πρωτεΐνης Geminin. Στη συνέχεια, αναλύονται οι λειτουργικές µελέτες που πραγµατοποιήθηκαν µε στόχο να αποσαφηνιστεί ο τρόπος δράσης τους στο συγκεκριµένο µονοπάτι, δίνοντας ιδιαίτερη έµφαση στη µεταγραφική ρύθµ ιση του µονοπατιού και στις ισορροπηµένες, θετικές ή αρνητικές, αλληλεπιδράσεις που εµφανίζουν, τόσο µεταξύ τους όσο και µε άλλους µεταγραφικούς παράγοντες. Επίσης, γίνεται αναφορά στον in vivo ρόλο του παράγοντα GemC1. Αυτό επέτρεψε τη σύγκριση του φαινοτύπου των ελλειµµατικών για το γονίδιο GemC1 µυών, µε το φαινότυπο που εµφανίζουν ασθενείς ή οργανισµοί-µοντέλα από τους οποίους απουσιάζουν άλλοι σηµ αντικοί ρυθµ ιστές της διαφοροποίησης των πολυκροσσωτών κυττάρων, συµπεριλαµβανοµένου και των ασθενών µε µεταλλάξεις στο γονίδιο McIdas. Επίσης, είναι πολύ σηµαντικό να γίνει µια συγκριτική ανάλυση των ρόλων των γονιδίων GemC1 και McIdas κατά τη διαφοροποίηση, ώστε να αποσαφηνιστεί εάν δρουν κατά πλεονασµό ή εξειδικεύονται στο συγκεκριµένο µονοπάτι. Τέλος, δεδοµένου του εντοπισµού και του ρόλο της πρωτεΐνης McIdas στο διπλασιασµό των κεντριολίων και των βιβλιογραφικών δεδοµένων για τo ρόλο της Geminin στα κεντροσωµάτια, θα αναλυθεί η πιθανή εµπλοκή τους στον κύκλο διπλασιασµού των κεντροσωµατίων. 167
168 5. ΣΥΖΗΤΗΣΗ Στην παρούσα διατριβή προτείνουµε ότι τα µέλη της υπεροικογένειας της Geminin µπορεί να αποτελούν ρυθµιστές του διπλασιασµού των κεντριολίων, τόσο στα διαιρούµενα όσο και στα διαφοροποιηµένα κύτταρα, µέσω ελέγχου της έκφρασης των παραγόντων που απαιτούνται σε αυτή τη διαδικασία. Τα σχετικά επίπεδα της έκφρασής τους και οι αλληλεπιδράσεις που αναπτύσσουν µπορεί να ελέγχουν το σχηµατισµό ενός ή περισσότερων κεντριολίων στα κύτταρα, ανάλογα µε την κατάσταση στην οποία βρίσκονται. 168
169 5. ΣΥΖΗΤΗΣΗ 5.1 Έκφραση των µελών της υπεροικογένειας της Geminin στα πολυκροσσωτά κύτταρα Ιστοειδική έκφραση των παραγόντων GemC1 και McIdas στα πολυκροσσωτά κύτταρα Στο εργαστήριό µας µελετήθηκε το πρότυπο έκφρασης των γονιδίων GemC1 και McIdas, κατά την εµβρυϊκή ανάπτυξη των µυών, προκειµένου να διαπιστωθεί εάν τα δύο νέα µέλη της υπεροικογένειας της Geminin εµπλέκονται, όπως και η Geminin, σε διαδικασίες κυτταρικής διαφοροποίησης (Pefani et al, 2011, Διδακτορική Διατριβή, Δάφνη-Ελευθερία Πεφάνη, 2012). Από πειράµατα κυρίως in situ υβριδοποίησης, δείχθηκε ότι οι παράγοντες GemC1 και McIdas εµφανίζουν καθ όλη τη διάρκεια της ανάπτυξης ένα ειδικό, επικαλυπτόµενο και σε υψηλά επίπεδα πρότυπο έκφρασης στο νευρικό σύστηµα και στο αναπνευστικό επιθήλιο των µυών. Συγκεκριµένα, ξεκινούν να εκφράζονται νωρίς κατά την ανάπτυξη στο χοριοειδές πλέγµα (choroid plexus) των κοιλιών του εγκεφάλου, καθώς και στη φλοιϊκή περιέλιξη (cortical hem). Επίσης, εµφάνισαν ειδική έκφραση και στο αναπτυσσόµενο αναπνευστικό επιθήλιο των µυών. Κοινό χαρακτηριστικό των προαναφερθέντων ιστών είναι ότι διαθέτουν πολυκροσσωτά κύτταρα, τα οποία συµβάλλουν στην κίνηση του εγκεφαλονωτιαίου υγρού και της βλέννας, αντίστοιχα. Τα πολυκροσσωτά κύτταρα διαφοροποιούνται κατά τη διάρκεια της ανάπτυξης και είτε διατηρούνται για πολλά χρόνια (π.χ. στον εγκέφαλο), είτε µπορούν να ανανεώνονται κατά τη διάρκεια της ζωής (π.χ. στο αναπνευστικό ή στο αναπαραγωγικό σύστηµα). Στην παρούσα διατριβή, µε σκοπό να διερευνηθεί η συµµετοχή των µελών της υπεροικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων, µελετήθηκε αρχικά η έκφραση του mrna τους, σε ιστούς που είναι γνωστό ότι διαθέτουν πολυκροσσωτά κύτταρα: στο επιθήλιο του χοριοειδούς πλέγµατος των κοιλιών του εγκεφάλου, στο επιθήλιο των βρόγχων του αναπνευστικού συστήµατος και στις ωοθήκες και ωαγωγούς του αναπαραγωγικού συστήµατος θηλυκών µυών (Spassky & Meunier, 2017). Οι ιστοί αυτοί αποµονώθηκαν από έµβρυα µυών που βρίσκονταν σε διαφορετικά αναπτυξιακά στάδια, αλλά και από ενήλικες µύες. Το mrna των GemC1 και McIdas ανιχνεύθηκε σε υψηλά επίπεδα κατά τη διάρκεια της εµβρυογένεσης σε όλους τους προαναφερθέντες ιστούς που εξετάστηκαν. Η έκφρασή τους εντοπίζεται και στους αντίστοιχους ιστούς που αποµονώθηκαν από τους ενήλικες µυς, σε χαµηλότερα όµως επίπεδα. Ιδιαίτερο ενδιαφέρον παρουσιάζει το διαφορικό πρότυπο έκφρασης των GemC1 και McIdas στο ήπαρ. Το ήπαρ των ενήλικων µυών χρησιµοποιήθηκε ως αρνητικό δείγµα-µάρτυρας, αφού δεν διαθέτει πολυκροσσωτά κύτταρα. Το mrna του McIdas εκφράζεται και στο ήπαρ, ενώ δεν ανιχνεύεται έκφραση του GemC1. Παράλληλα, η έκφρασή τους συγκρίθηκε µε την έκφραση των γονιδίων Foxj1 και Geminin. Το πρότυπο έκφρασης του παράγοντα Foxj1 οµοιάζει περισσότερο µε αυτό του γονιδίου GemC1, α φού παρουσιάζει υψηλά επίπεδα έκφρασης κατά την ανάπτυξη, τα οποία παραµένουν και στα ενήλικα άτοµα σε χαµηλότερα όµως επίπεδα 169
170 5. ΣΥΖΗΤΗΣΗ και δεν ανιχνεύεται η έκφρασή του στο ήπαρ των ενήλικων µυών. Ο παράγοντας Foxj1 αποτελεί έναν από τους πρώτους µεταγραφικούς παράγοντες που δείχθηκε ότι κατέχει καθοριστικό ρόλο στη διαφοροποίηση των πολυκροσσωτών κυττάρων, ελέγχοντας τη µετανάστευση και την αγκυροβόληση των κεντριολίων στην πλασµατική µεµβράνη των κυττάρων, καθώς και µετέπειτα στάδια της δηµιουργίας των κροσσών (Brody et al, 2000; Stubbs et al, 2008; Yu et al, 2008). Όσον αφορά στο πρότυπο έκφρασης του mrna της Geminin, αυτό εκφράζεται σε υψηλά επίπεδα κατά την εµβρυογένεση, αλλά και στους αποµονωµένους ιστούς από τα ενήλικα άτοµα. Το αποτέλεσµα αυτό ήταν αναµενόµενο, αφού η Geminin εκφράζεται σε διαιρούµενα κύτταρα. Το πρότυπο έκφρασης του McIdas οµοιάζει περισσότερο µε αυτό της Geminin, καθώς και τα δύο εκφράζονται και στο ήπαρ. Λόγω της πληθώρας των κυτταρικών τύπων που απαντώνται στους ιστούς που αποµονώθηκαν, εκτός των πολυκροσσωτών κυττάρων, ήταν απαραίτητο να διαπιστωθεί η ειδικότητα της έκφρασης των GemC1 και McIdas στο συγκεκριµένο κυτταρικό τύπο. Στο αναπνευστικό επιθήλιο των µυών απαντώνται, πέρα από τα πολυκροσσωτά κύτταρα, βλεννοπαραγωγά (goblet cells), εκκριτικά (clara cells), νευροενδοκρινή και βασικά διαιρούµενα κύτταρα (Rock & Hogan, 2011). Η χρήση ειδικού αντισώµατος έναντι της πρωτεΐνης Foxj1, που εντοπίζεται στα πολυκροσσωτά κύτταρα, παράλληλα µε τη χρήση ειδικών ανιχνευτών για τον εντοπισµό του mrna των GemC1 και McIdas, έδειξε ότι η πλειοψηφία των κυττάρων που εκφράζουν GemC1 ή McIdas εκφράζουν και την πρωτεΐνη Foxj1, υποδηλώνοντας την έκφρασή τους ειδικά στον πληθυσµό των πολυκροσσωτών κυττάρων. Από τα παραπάνω συµπεραίνουµε ότι τα µέλη της υπεροικογένειας της Geminin εµφανίζουν ειδική και υψηλή έκφραση κατά την ανάπτυξη, σε ιστούς που διαθέτουν πολυκροσσωτά κύτταρα. Ο παράγοντας GemC1 εµφανίζει ένα πιο ειδικό πρότυπο έκφρασης, αφού η έκφρασή του περιορίζεται αποκλειστικά στα πολυκροσσωτά κύτταρα, ενώ ο McIdas εµφανίζει ένα γενικότερο πρότυπο έκφρασης σε όλους τους ιστούς που εξετάστηκαν, κάτι που µπορεί να αποδοθεί στο ρόλο του κατά την αντιγραφή του DNA. Ωστόσο, παρουσιάζει υψηλότερα επίπεδα έκφρασης κατά τη διάρκεια της ανάπτυξης Έκφραση των παραγόντων GemC1 και McIdas κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων Προκειµένου να µελετηθεί η έκφραση των GemC1 και McIdas ειδικά κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, χρησιµοποιήθηκε το ex vivo σύστηµα καλλιέργειας των επιθηλιακών κυττάρων της τραχείας µυών, στα οποία µπορεί να γίνει ελεγχόµενη επαγωγή της διαφοροποίησής τους (Vladar & Brody, 2013; You & Brody, 2013). Το mrna των GemC1 και McIdas εκφράζεται ειδικά και αυξάνεται κατά τη διάρκεια της διαφοροποίησης των πολυκροσσωτών κυττάρων, ενώ η έκφρασή τους µειώνεται όταν πια η διαδικασία έχει ολοκληρωθεί. Είναι σηµαντικό να τονιστεί ότι το συγκεκριµένο πρότυπο έκφρασης συµ πίπτει µε αυτό που έχει παρατηρηθεί για µια πληθώρα παραγόντων που εκφράζονται κατά τη διάρκεια της διαφοροποιήσης των επιθηλιακών κυττάρων της τραχείας των µυών και 170
171 5. ΣΥΖΗΤΗΣΗ εµπλέκονται στο αρχικό στάδιο του πολλαπλασιασµού των κεντριολίων τους. Τέτοιοι παράγοντες είναι οι: Deup1, Ccno, Cep152, Plk4 κ.ά. (Funk et al, 2015; Zhao et al, 2013). Η συγκεκριµένη παρατήρηση υπογραµµίζει τη πιθανή εµπλοκή των GemC1 και McIdas στον πολλαπλασιασµό των κεντριολίων των κυττάρων που καθορίστηκαν να γίνουν πολυκροσσωτά. Επιπρόσθετα, οι GemC1 και McIdas εµφανίζουν διαφορά στην έκφρασή τους κατά το στάδιο του πολλαπλασιασµού των κυττάρων. Το mrna του GemC1 δεν ανιχνεύεται στα διαιρούµενα προγονικά κύτταρα, σε αντίθεση µε το mrna του McIdas που εντοπίζεται σε αυτά, αλλά σε χαµηλά επίπεδα. Όµως, αµέσως µετά την επαγωγή της διαφοροποίησης τα επίπεδα έκφρασης και των δυο παραγόντων αυξάνονται σηµαντικά. Το γενικότερο πρότυπο της έκφρασής τους συµπίπτει µε αυτό του µεταγραφικού παράγοντα Foxj1. Παράλληλα, µελετήθηκε και το πρότυπο έκφρασης της Geminin, η οποία δεν είχε προηγουµένως εµπλακεί στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Η Geminin εµφανίζει ένα ανεστραµµένο πρότυπο έκφρασης σε σχέση µε αυτό των άλλων δυο µελών της υπεροικογένειας. Τα επίπεδά της είναι υψηλότερα στα διαιρούµενα προγονικά κύτταρα και µειώνονται σταδιακά µετά την επαγωγή της διαφοροποίησης. Το δεδοµένο αυτό είναι ιδιαίτερα ενδιαφέρον, αφού υποδηλώνει µια ανταγωνιστική δράση ανάµεσα στη Geminin και στους GemC1 και McIdas. Τέτοιες ανταγωνιστικές σχ έσεις ανάµεσα στις πρωτεΐνες της υπεροικογένειας είναι ήδη γνωστές και από το ρόλο κατά τον κυτταρικό κύκλο. Ο McIdas δρα ανταγωνιστικά στη Geminin, αφού προσδένεται σε αυτήν και εµποδίζει τη δράση της κατά την αντιγραφή του DNA (Pefani et al, 2011). Συνοψίζοντας όλα τα αποτελέσµατα από τη µελέτη της έκφρασης των µελών της υπεροικογένειας της Geminin σε διαφορετικούς ιστούς και συστήµατα, αναδεικνύονται εκτός των οµ οιοτήτων τους και κάποιες σηµ αντικές διαφορές. Ιδιαίτερο ενδιαφέρον εµφανίζει το διαφορικό πρότυπο έκφρασης των GemC1 και McIdas σε σχέση µε αυτό της Geminin. Στο κεντρικό νευρικό σύστηµα έχει δειχθεί ότι η Geminin εκφράζεται στα πρόδροµα νευρικά κύτταρα της κοιλιακής και υποκοιλιακής ζώνης του φλοιού, νωρίς κατά τη νευρογένεση (Spella et al, 2007). Ωστόσο, δεν έχει δειχθεί έκφραση της Geminin στις δοµ ές που εντοπίζονται οι GemC1 και McIdas. Φαίνεται λοιπόν ότι τα νέα µέλη της οικογένειας απέκτησαν επιπρόσθετους ρόλους στον καθορισµό της µοίρας των πολυκροσσωτών κυττάρων, διαφοροποιώντας το ρόλο τους σε σχέση µε αυτόν της Geminin, ή προσθέτοντας νέες λειτουργίες στην υπεροικογένεια. Τέλος, παρατηρείται διαφορά στην έκφραση του GemC1, σε σχέση µε αυτή των McIdas και Geminin, αφού η έκφραση του GemC1 φαίνεται να περιορίζεται µόνο στα πολυκροσσωτά κύτταρα, ενώ οι McIdas και Geminin εµφανίζουν ένα πιο καθολικό πρότυπο έκφρασης. Ωστόσο, η παρατήρηση αυτή χρήζει περαιτέρω διερεύνησης, αφού µπορεί να υποδηλώνει µια πιο ειδική δράση του GemC1 µόνο στα πολυκροσσωτά κύτταρα, ή ότι η έκφρασή του περιορίζεται σε συγκεκριµένα διαιρούµενα κύτταρα, και εκφράζεται σε αρκετά χαµηλά επίπεδα. Άλλωστε πρέπει να λάβουµε υπόψιν και το ρόλο του GemC1 στα πολλαπλασιαζόµενα κύτταρα, κατά το 171
172 5. ΣΥΖΗΤΗΣΗ σχηµατισµό του προεναρκτήριου συµπλόκου της αντιγραφής του DNA (Balestrini et al, 2010) Η πρωτεΐνη McIdas εκφράζεται στα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων και όχι στα τελικώς διαφοροποιηµένα κύτταρα Η ύπαρξη ενός λειτουργικού πολυκλωνικού αντισώµατος έναντι της πρωτεΐνης McIdas επέτρεψε την περαιτέρω µελέτη της, σε επίπεδο µεµονωµένων κυττάρων, κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Στο ex vivo σύστηµα καλλιέργειας των επιθηλιακών κυττάρων που αποµονώνονται από την τραχεία των ενήλικων µυών, δεν ανιχνεύεται έκφραση της πρωτεΐνης McIdas στα διαιρούµενα προγονικά κύτταρα. Όµως, αµέσως µετά την επαγωγή της διαφοροποίησης, η πρωτεΐνη McIdas ξεκινά να εκφράζεται. Η έκφρασή της συγκρίθηκε µε αυτή του µεταγραφικού παράγοντα Foxj1 και δείχθηκε ότι η πλειοψηφία των κυττάρων που εκφράζουν την πρωτεΐνη McIdas, εκφράζουν και την πρωτεΐνη Foxj1. Αρχικά, εντοπίζεται ένας ελάχιστος αριθµός κυττάρων που ενώ εκφράζουν την πρωτεΐνη McIdas, δεν εκφράζουν ακόµα την πρωτεΐνη Foxj1 σε υψηλά επίπεδα. Όµως, στη συνέχεια της διαφοροποίησης η έκφραση του παράγοντα Foxj1 αυξάνεται και παραµένει και στα διαφοροποιηµένα κύτταρα, ενώ αυτή του παράγοντα McIdas µειώνεται σηµαντικά. Το αποτέλεσµα αυτό έρχεται σε συµφωνία µε βιβλιογραφικά δεδοµένα, αφού ο παράγοντας Foxj1 χρειάζεται και σε µεταγενέστερα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων (Choksi et al, 2014). Ο παράγοντας Foxj1 είτε µόνος του, είτε σ ε συνεργασία µε τους Rfx µεταγραφικούς παράγοντες, δρα ελέγχοντας τη µετανάστευση και την «αγκυροβόληση» των πολυάριθµων κεντριολίων που έχουν παραχθεί, στην πλασµατική µεµβράνη των κυττάρων. Ακόµα, ελέγχει και την έκφραση γονιδίων που χρειάζονται για τα δ οµικά συστατικά των κινητών κροσσών. Η διαφορική έκφρασή τους κατά τη διάρκεια της διαφοροποίησης των πολυκροσσωτών κυττάρων, υποδηλώνει ότι ο McIdas απαιτείται για τα πιο πρώιµα στάδια της διαδικασίας. Το παραπάνω αποτέλεσµα ενισχύθηκε και από επιπρόσθετα δεδοµένα, καθώς δείχθηκε ότι η πρωτεΐνη McIdas συνεντοπίζεται στα κύτταρα και µε το µεταγραφικό παράγοντα cmyb, που είναι γνωστό ότι εκφράζεται νωρίς κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, συµβάλλοντας στο στάδιο του πολλαπλασιασµού των κεντριολίων (Tan et al, 2013). Παράλληλα, διαπιστώθηκε ότι η πρωτεΐνη McIdas εκφράζεται πολύ νωρίς κατά τη διαφοροποίηση, στα κύτταρα εκείνα που διαθέτουν πρωτογενή κροσσό. Αντίθετα, δεν ανιχνεύεται έκφρασή της σε µεταγενέστερα στάδια, στα κύτταρα που διαθέτουν πολλαπλούς κροσσούς. Το αποτέλεσµα αυτό δείχνει ότι ο McIdas εκφράζεται κατά τα πρώιµα στάδια της διαφοροποίησης, στα κύτταρα που καθορίζονται να γίνουν πολυκροσσωτά. Τέλος, η χρήση ενός ακόµα δείκτη των κεντριολίων/βασικών σωµατίων, της pericentrin, έδειξε ότι η έκφραση του McIdas παρατηρείται πιο συγκεκριµένα στο στάδιο εκείνο που λαµβάνει χώρα ο πολλαπλασιασµός των κεντριολίων. Έτσι, η έκφρασή του εξαφανίζεται όταν τα κεντριόλια έχουν πλέον µεταναστεύσει στην πλασµατική µεµβράνη για να δώσουν γένεση στους κροσσούς. 172
173 5. ΣΥΖΗΤΗΣΗ Από όλα τα παραπάνω είναι φανερό πως ο McIdas δρα νωρίς κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων και µάλιστα φαίνεται να συµµετέχει στο στάδιο του πολλαπλασιασµού των κεντριολίων. Σε µια µελέτη π ου πραγµατοποιήθηκε παράλληλα µε την παρούσα διατριβή, κατά την οποία αποσιωπήθηκε το γονίδιο McIdas από τα κύτταρα της ακτινωτής γλοίας του εγκεφάλου µυών που διαφοροποιούνται σε πολυκροσσωτά επενδυµατικά κύτταρα, προκλήθηκε αναστολή του πολλαπλασιασµού των κεντριολίων, υποδηλώνοντας έτσι ότι ο McIdas µπορεί να επηρεάζει άµ εσα τον πολλαπλασιασµό τους (Kyrousi et al, 2015). Η δράση του αυτή φαίνεται να είναι συντηρηµένη, αφού δείχθηκε να εµπλέκεται στον πολλαπλασιασµό των κεντριολίων και κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων της επιδερµίδας εµβρύων Xenopus (Stubbs et al, 2012), καθώς και των επιθηλιακών κυττάρων της αναπνευστικής οδού του ανθρώπου (Boon et al, 2014). 173
174 5. ΣΥΖΗΤΗΣΗ 5.2 Τα µέλη της υπεροικογένειας της Geminin ελέγχουν τη διαφοροποίηση των πολυκροσσωτών κυττάρων Με δεδοµένο την ειδική έκφραση των GemC1 και McIdas κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, πραγµατοποιήθηκαν πειράµατα υπερέκφρασής τους, κατά την ex vivo καλλιέργεια των επιθηλιακών κυττάρων της τραχείας µυών, προκειµένου να χαρακτηριστεί ο ρόλος τους. Η υπερέκφραση τόσο του McIdas, όσο και του GemC1 επάγει τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. Ο McIdas επάγει τον πολλαπλασιασµό των κεντριολίων, καθώς και την έκφραση του παράγοντα Foxj1. Οµοίως, ο παράγοντας GemC1 αυξάνει την έκφραση του παράγοντα Foxj1, όµως επάγει και την έκφραση του McIdas. Τα δεδοµένα που προέκυψαν σχετικά µε την έκφραση των GemC1 και McIdas, έδειξαν ότι εµφανίζουν ένα επικαλυπτόµενο πρότυπο έκφρασης τόσο µεταξύ τους, όσο και µε τον παράγοντα Foxj1, υποδηλώνοντας ότι όλα µαζί συνεργάζονται για να ενεργοποιήσουν το µονοπάτι της διαφοροποίησης των πολυκροσσωτών κυττάρων. Πράγµατι, η υπερέκφρασή τους επάγει τον παράγοντα Foxj1 και ιδιαίτερα σηµαντική είναι και η επαγωγή της έκφρασης της πρωτεΐνης McIdas από τον GemC1. Προκειµένου να µελετηθεί και η πιθανή εµπλοκή της Geminin στη διαδικασία αυτή, εξετάσθηκε ο φαινότυπος που προκαλείται στα επιθηλιακά κύτταρα της τραχείας των µυών ύστερα από την αποσιώπησή της. Η έλλειψη της Geminin προκαλεί πρόωρη επαγωγή της διαφοροποίησης των πολυκροσσωτών κυττάρων, µέσω αύξησης της έκφρασης των πρωτεϊνών McIdas και Foxj1. Το αποτέλεσµα αυτό ενισχύει την υπόθεσή µας ότι υπάρχει µια ανταγωνιστική δράση µεταξύ της Geminin και των GemC1 και McIdas κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Όπως αναφέρθηκε στην προηγούµενη ενότητα (5.1.2), κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, το πρότυπο έκφρασης της Geminin εµφανίζεται ανεστραµµένο σε σχέση µε αυτό των άλλων δυο παραγόντων. Η Geminin µειώνεται κατά τη διάρκεια της διαφοροποίησης, ενώ οι GemC1 και McIdas αυξάνονται.η ανταγωνιστική τους δράση είναι πιθανό να επιτυγχάνεται µέσω των αλληλεπιδράσεων που είναι γνωστό ότι αναπτύσσουν οι πρωτεΐνες της οικογένειας της Geminin. Αλληλεπιδρούν µεταξύ τους µέσω των περιοχών σπειροειδούς σπειράµατος που διαθέτουν (Caillat et al, 2015; Caillat et al, 2013). Με παρόµοιο τρόπο ο McIdas αναστέλλει τη δράση της Geminin κατά την αντιγραφή του DNA (Pefani et al, 2011). Τα δεδοµένα αυτά αναδεικνύουν τη σηµαντικότητα των αλληλεπιδράσεων µεταξύ των µελών της υπεροικογένειας της Geminin, για την προαγωγή της διαφοροποίησης των πολυκροσσωτών κυττάρων. 174
175 5. ΣΥΖΗΤΗΣΗ 5.3 Μεταγραφική ρύθµιση του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων Οι παράγοντες GemC1 και McIdas δρουν ως µεταγραφικοί ενεργοποιητές του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων Από την προηγούµενη ενότητα είναι σαφές ότι τα µέλη της οικογένειας της Geminin, GemC1 και McIdas, δρουν κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων, επάγοντας την έκφραση παραγόντων που απαιτούνται για τη λειτουργική τους εξειδίκευση. Προκειµένου να διαπιστωθεί ο µηχανισµός µέσω του οποίου δρουν, πραγµατοποιήθηκαν πειράµατα µε τη χρήση του γονιδίου αναφοράς της λουσιφεράσης, όπου ελέγχθηκε εάν µπορούν να επάγουν τη µεταγραφή µέσω των πιθανών υποκινητών-ρυθµιστικών στοιχείων των γονιδίων MCIDAS και FOXJ1. Πράγµατι, δείχθηκε ότι οι παράγοντες GemC1 και McIdas µπορούν να δρουν ως µεταγραφικοί ενεργοποιητές µέσω των ρυθµιστικών στοιχείων των γονιδίων MCIDAS και FOXJ1. Ο παράγοντας GemC1 µπορεί να επάγει τη µεταγραφή µέσω των ρυθµιστικών στοιχείων του γονιδίου MCIDAS. Επίσης, οι παράγοντες GemC1 και McIdas δρουν κατά παρόµοιο τρόπο, ενεργοποιώντας τον παράγοντα Foxj1. Στην πορεία αυτής της µελέτης τέθηκε λοιπόν το ερώτηµα σχετικά µε το εάν ο GemC1 επάγει άµεσα την έκφραση του παράγοντα Foxj1 ή εάν η δράση του αυτή επιτελείται µέσω ενεργοποίησης του McIdas. Η αποσιώπηση της πρωτεΐνης McIdas από τα κύτταρα έδειξε ότι ο GemC1 είναι ικανός να επάγει από µόνος του τη µεταγραφή µέσω του υποκινητή του γονιδίου Foxj1. Το ίδιο ερώτηµα απαντήθηκε και από µια ακόµα ερευνητική οµάδα, χρησιµοποιώντας µια διαφορετική πειραµατική διάταξη. Οι Terre et al. έδειξαν ότι η αποσιώπηση του McIdas µειώνει την ικανότητα του GemC1 να επάγει τον παράγοντα Foxj1, χωρίς όµως να µηδενίζεται (Terre et al, 2016). Καταλήγουµε λοιπόν στο συµπέρασµα ότι ο παράγοντας GemC1 δρα µαζί µε τον McIdas, αλλά µπορεί να επάγει και άµεσα την έκφραση του γονιδίου Foxj Οι παράγοντες GemC1 και McIdas δρουν συνεργατικά µε µέλη της οικογένειας των Ε2F µεταγραφικών παραγόντων Ενώ αυτή η εργασία βρισκόταν σε εξέλιξη, οι Ma et al. σε µελέτες στο Xenopus έδειξαν ότι ο McIdas δρα κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων σχηµατίζοντας ένα τριµερές σύµπλοκο µε τους µεταγραφικούς παράγοντες E2F4/5 και DP1 (το σύµπλοκο αυτό ονοµάστηκε EDM σύµπλοκο) (Ma et al, 2014). Ο McIdas προσδένεται στους παράγοντες αυτούς µέσω της συντηρηµένης καρβοξυτελικής του περιοχής. Περαιτέρω ανάλυση έδειξε ότι το συγκεκριµένο µεταγραφικό σύµπλοκο ενεργοποιεί µια σειρά γονιδίων, συµπεριλαµβανοµένων 175
176 5. ΣΥΖΗΤΗΣΗ µεταγραφικών παραγόντων (Foxj1, Rfx2 και cmyb) και παραγόντων απαραίτητων για τον πολλαπλασιασµό των κεντριολίων (Ccno, Plk4, Cep152, STIL και Deup1). Στη συγκεκριµένη διατριβή ελέγχθηκε εάν οι παράγοντες GemC1 και McIdas επάγουν τη µεταγραφή µέσω του υποκινητή του γονιδίου FOXJ1, σε συνεργασία µε την οικογένεια των E2F µεταγραφικών παραγόντων. Συγκεκριµένα, εξετάσθηκαν οι παράγοντες Ε2F1, E2F4 και E2F5. Είναι γνωστό ότι οι µεταγραφικοί παράγοντες E2F4 και E2F5 σχηµατίζουν µια υποοµάδα µέσα στην οικογένεια των Ε2F παραγόντων, που χαρακτηρίζονται ως κατασταλτικοί E2F παράγοντες, αφού εµποδίζουν την έκφραση γονιδίων απαραίτητων για τον κυτταρικό πολλαπλασιασµό, προκαλώντας την έξοδο των κυττάρων από τον κυτταρικό κύκλο και την τελική διαφοροποίησή τους (reviewed in Trimarchi & Lees, 2002). Αντίθετα, ο µεταγραφικός παράγοντας E2F1 ανήκει στην υποοµάδα των µεταγραφικών ενεργοποιητών της οικογένειας, καθώς προάγουν τον πολλαπλασιασµό των κυττάρων. Τα πολυκροσσωτά κύτταρα, για να διαφοροποιηθούν, πρέπει να εξέλθουν από τον κυτταρικό κύκλο και να εκφράσουν έναν µεγάλο αριθµό γονιδίων που προάγει τη λειτουργική τους εξειδίκευση. Έτσι, δείχθηκε ότι η µεταγραφή που επάγεται από τους GemC1 και McIdas, µέσω του υποκινητή του γονιδίου FOXJ1, αυξάνεται σηµαντικά σε συνεργασία µε τον µεταγραφικό παράγοντα E2F5. Ωστόσο, δεν ήταν δυνατό να πραγµατοποιηθούν παρόµοια πειράµατα µε το µεταγραφικό παράγοντα E2F4. Ο E2F4 από µόνος του προκαλούσε µεγάλη επαγωγή της µεταγραφής, µέσω των ρυθµιστικών στοιχείων των γονιδίων MCIDAS και FOXJ1, γεγονός που δεν επέτρεπε την αξιολόγηση της συνεργατικής του δράσης µε τα µέλη της οικογένειας της Geminin. Τέλος, δείχθηκε ότι οι GemC1 και McIdas συνεργάζονται ειδικά µε τον κατασταλτικό E2F5 παράγοντα, µιας και η ταυτόχρονη υπερέκφρασή τους µε τον µεταγραφικό ενεργοποιητή E2F1 δεν άλλαζε τη µεταγραφική τους ενεργότητα. Η συνεργασία του παράγοντα GemC1 µε τους µεταγραφικούς παράγοντες Ε2F4/5 επιτυγχάνεται µέσω του σχηµατισµού ενός κοινού συµ πλόκου. Ενώ αρχικά δείχθηκε ότι o GemC1 λειτουργεί διαφορετικά από τον McIdas ως προς τα σύµπλοκα που δηµιουργεί (Zhou et al, 2015), τελικά ο GemC1, όπως και ο McIdas (Ma et al, 2014), αλληλεπιδρά µέσω της καρβοξυτελικής του περιοχής µε τ ους παράγοντες E2F4/5 και DP1 (το σύµπλοκο αυτό ονοµάστηκε EDG) (Terre et al, 2016, Μεταπτυχιακή διπλωµατική εργασία, Ιωάννης Λουκάς, 2017). Ο παράγοντας GemC1 σε σύµ πλοκο µε το µεταγραφικό παράγοντα E2F5 µπορεί να συνδέεται ειδικά στις ρυθµιστικές περιοχές των γονιδίων McIdas και Foxj1 (Arbi et al, 2016). Τέλος, παρατηρήθηκε ότι η πρωτεΐνη McIdas λειτουργεί µέσω ενός µηχανισµού θετικής ανατροφοδότησης. Μπορεί να προκαλέσει µεταγραφική ενεργοποίηση µέσω των ίδιων των ρυθµιστικών της στοιχείων. Οι πρωτεΐνες GemC1 και McIdas σχηµατίζουν µεταγραφικά σύµπλοκα µε τους E2F4/5, πετυχαίνοντας την ενεργοποίηση της κατάλληλης οµάδας γονιδίων που προάγει τον καθορισµό και την εξειδίκευση των πολυκροσσωτών κυττάρων. Παράλληλα, ο παράγοντας McIdas συµβάλλει αποτελεσµατικά σε αυτό το µονοπάτι, κλειδώνοντας την αναπτυξιακή τύχη των κυττάρων προς τη συγκεκριµένη µοίρα. 176
177 5. ΣΥΖΗΤΗΣΗ H πρωτεΐνη Geminin αναστέλλει τη µεταγραφική ενεργότητα των GemC1 και McIdas Kατά τη µελέτη των GemC1, McIdas και Geminin στη µεταγραφική ρύθµιση του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων, είναι ιδιαίτερα ενδιαφέρον ότι η Geminin αναστέλλει τη µεταγραφική ενεργότητα των GemC1 και McIdas. Το δεδοµένο αυτό σε συνδυασµό µε το αποτέλεσµα ότι η αναστολή της Geminin επάγει τη διαφοροποίηση των πολυκροσσωτών κυττάρων, οδηγεί στο συµπέρασµα ότι η Geminin αποτελεί έναν αρνητικό ρυθµιστή της διαδικασίας και δρα ανταγωνιστικά στους GemC1 και McIdas. Πειράµατα στο εργαστήριό µας έδειξαν ότι η Geminin σ τρατολογείται στα EDG σύµπλοκα (Ε2F5-DP1-GemC1), µέσω αλληλεπίδρασής της µε το σπειροειδές σπείραµα του GemC1 (Μεταπτυχιακή διπλωµατική εργασία, Ιωάννης Λουκάς, 2017). Επίσης, στο Xenopus έχει δειχθεί ότι η παρουσία της Geminin αναστέλλει τη δράση του EDM συ µπλόκου (Ε2F4/5-DP1-McIdas) κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων της επιδερµίδας των εµβρύων (Ma et al, 2014). Η Geminin λοιπόν µπορεί να δρα ανασταλτικά στη διαδικασία αυτή µέσω άµεσης πρόσδεσης στα µεταγραφικά σύµπλοκα που σχηµατίζουν οι GemC1 και McIdas, εµποδίζοντας πιθανόν την πρόσδεσή τους στα γονίδια-στόχους τους. Ωστόσο, περαιτέρω πειράµατα απαιτούνται για να διαλευκανθεί η συγκεκριµένη υπόθεση. 177
178 5. ΣΥΖΗΤΗΣΗ 5.4 Ο in vivo ρόλος του παράγοντα GemC1 στη διαφοροποίηση των πολυκροσσωτών κυττάρων Ο παράγοντας GemC1 αποτελεί κεντρικό ρυθµιστή της διαφοροποίησης των πολυκροσσωτών κυττάρων Η κατασκευή διαγονιδιακών µυών από τους οποίους απουσιάζει η έκφραση του γονιδίου GemC1 συνέβαλε αποτελεσµατικά στη µελέτη του in vivo ρόλου του. Οι ελλειµµατικοί για το γονίδιο GemC1 µύες γεννιούνταν µε την αναµενόµενη συχνότητα µε βάση τη Μεντελική κληρονοµικότητα και δεν παρουσίαζαν αναπτυξιακές ανωµαλίες. Ωστόσο, από τις πρώτες κιόλας ηµ έρες µετά τη γέννησή τους εµφάνιζαν σηµαντική καθυστέρηση στην ανάπτυξη και πέθαιναν αρκετά σύντοµα (Arbi et al, 2016). Ο ίδιος φαινότυπος παρατηρήθηκε και στους ελλειµµατικούς για το γονίδιο GemC1 µυς, που κατασκευάστηκαν παράλληλα από µια ακόµα ερευνητική οµάδα (Terre et al, 2016). Ωστόσο, οι συγκεκριµένοι διαγονιδιακοί µύες κατάφερναν να επιβιώνουν περισσότερο και έτσι παρατηρήθηκαν και µύες που ανέπτυσσαν υδροκεφαλία. Ο συγκεκριµένος φαινότυπος οµο ιάζει µε το φαινότυπο που παρατηρήθηκε για τους µυς που ήταν ελλειµµατικοί για το γονίδιο Foxj1 (Brody et al, 2000; Chen et al, 1998). Οι ελλειµµατικοί µύες για το γονίδιο GemC1 όµως δεν εµφάνιζαν ανωµαλίες ως προς την τοποθέτηση των οργάνων τους, όπως παρατηρήθηκε στους µυς που δεν εκφραζόταν το γονίδιο Foxj1. Αυτό υποδηλώνει ότι ο παράγοντας GemC1 πιθανόν να µην εµπλέκεται στη διαφοροποίηση των κροσσών που διαθέτουν τα κύτταρα του οργανωτή του εµβρύου (embryonic node). Δεν αποκλείεται βέβαια ο φαινότυπος αυτός να καλύπτεται, εξαιτίας της έκφρασης του γονιδίου GemC1 σε κάποια χαµηλά επίπεδα. Η έκφραση του παράγοντα GemC1 στα κύτταρα του οργανωτή του εµβρύου κατά την ανάπτυξη χρήζει περαιτέρω διερεύνησης. Ο παράγοντας GemC1 χαρακτηρίστηκε αρχικά για το ρόλο του κατά την έναρξη της αντιγραφής του DNA, αφού συµβάλλει στο σχηµατισµό του προεναρκτήριου συµπλόκου της αντιγραφής (Balestrini et al, 2010). Ωστόσο, η γέννηση των διαγονιδιακών µυών από τους οποίους απουσιάζει το γονίδιο GemC1, εγείρει ερωτήµατα σχετικά µε το ρόλο του κατά τον κυτταρικό πολλαπλασιασµό. Αντίστοιχα, η απαλοιφή του γονιδίου της Geminin από τους µυς προκαλεί θάνατο των εµβρύων στο στάδιο των 8 κυττάρων (Gonzalez et al, 2006; Hara et al, 2006), υποδεικνύοντας τη σπουδαιότητά της για τον κυτταρικό πολλαπλασιασµό. Παρόλο που ο φαινότυπος των µυών εµφανίζεται αντιφατικός µε ένα ρόλο του GemC1 στον πολλαπλασιασµό των κυττάρων, ο ρόλος αυτός δεν µπορεί να αποκλιστεί. Είναι γνωστό ότι η έλλειψη πολλών ρυθµιστών του κυτταρικού κ ύκλου δεν οδηγεί σε θάνατο των εµβρύων, όπως στην περίπτωση των κυκλινών E1 ή Ε2 και της κινάσης Cdk2 (Berthet et al, 2003; Geng et al, 2003; Ortega et al, 2003). Η ανάλυση των κυττάρων του αναπνευστικού επιθηλίου στους ελλειµµατικούς για το γονίδιο GemC1 µυς, ανέδειξε τον καθοριστικό του ρόλο στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Από την τραχεία των µυών απουσιάζουν εντελώς τα πολυκροσσωτά κύτταρα (Arbi et al, 2016). Επίσης, η απουσία πολλαπλών 178
179 5. ΣΥΖΗΤΗΣΗ κεντριολίων υποδηλώνει ότι η διαδικασία αυτή αναστέλλεται από τα πρώιµα κιόλας στάδια. Ο ρόλος του GemC1 στη διαφοροποίηση των πολυκροσσωτών κυττάρων είναι καθολικός, αφού η απαλοιφή του αναστέλλει τη διαφοροποίησή τους και στο χοριοειδές πλέγµα των κοιλιών του εγκεφάλου, καθώς και στο επιθήλιο των ωαγωγών των θηλυκών µυών (Terre et al, 2016). Η παρατήρηση αυτή συνάδει µε τα συµπτώµατα υδροκεφαλίας και στειρότητας που εµφανίζουν οι συγκεκριµένοι διαγονιδιακοί µύες. Επίσης, η απαλοιφή του γονιδίου GemC1 εµποδίζει το σχηµατισµό των πολυκροσσωτών κυττάρων σε έµβρυα βατράχου Xenopus, αλλά και στο zebrafish (Zhou et al, 2015) Συσχέτιση του γονιδίου GemC1 µε ασθένειες που δηµιουργούνται εξαιτίας της µη σωστής διαφοροποίησης των πολυκροσσωτών κυττάρων Μέχρι στιγµής δεν έχουν κατασκευαστεί διαγονιδιακοί µύες από τους οποίους να απουσιάζει η έκφραση του γονιδίου McIdas, ώστε να γίνει άµεση σύγκριση µε τους µυς που είναι ελλειµµατικοί για το γονίδιο GemC1. Ωστόσο, σε µια πρόσφατη µελέτη, προσδιορίστηκαν µεταλλάξεις στο γονίδιο McIdas σε ασθενείς που εµφάνιζαν µια σπάνια κληρονοµική ασθένεια που ονοµάζεται RGMC (Μειωµένη παραγωγή πολλαπλών κινητών κροσσών, Reduced Generation of Multiple Motile Cilia) (Boon et al, 2014). Τα επιθηλιακά κύτταρα της αναπνευστικής οδού των συγκεκριµένων ασθενών διαθέτουν ελάχιστους κροσσούς, οι οποίοι είναι ακίνητοι. Έτσι, παρατηρούνται ανωµαλίες στη βλεννοκροσσωτή κάθαρση του αναπνευστικού επιθηλίου, µε αποτέλεσµα να εµφανίζονται συνεχείς µολύνσεις στο ανώτερο και στο κατώτερο αναπνευστικό σύστηµα. Επίσης εµφανίζονται και συµπτώµατα υδροκεφαλίας και υπογονιµότητας στα θηλυκά άτοµα, καθώς επηρεάζεται η διαφοροποίηση των πολυκροσσωτών κυττάρων που απαντώνται και στο νευρικό και στο αναπαραγωγικό σύστηµα. Οι µεταλλάξεις που προσδιορίστηκαν στο γονίδιο McIdas στους ασθενείς µε RGMC, βρίσκονται στη συντηρηµένη καρβοξυτελική περιοχή του µορίου, µέσω της οποίας αλληλεπιδρά, όπως προαναφέρθηκε, µε τους µεταγραφικούς παράγοντες E2F4/5 και DP1 (Ma et al, 2014). Οι Ma et al. µελέτησαν την επίδραση των µεταλλάξεων αυτών και διαπίστωσαν ότι η µετάλλαξη G335D αναστέλλει το σχηµατισµό του EDM συµπλόκου (E2F4/5-DP1-McIdas), ενώ η µετάλλαξη R370H επιτρέπει το σχηµατισµό του συµπλόκου, όµως αυτό είναι λειτουργικά ανενεργό. Σε κάθε περίπτωση αναστέλλεται η µεταγραφική ενεργότητα του συµπλόκου, µε αποτέλεσµα να µην ενεργοποιούνται τα γονίδια που χρειάζονται να εκφραστούν για την επιτυχή διαφοροποίηση των πολυκροσσωτών κυττάρων. Το γονίδιο Ccno έχει επίσης δειχθεί να είναι µεταλλαγµένο σε ασθενείς µε το σύνδροµο RGMC (Amirav et al, 2016; Wallmeier et al, 2014). Στα κύτταρα του αναπνευστικού επιθηλίου των ασθενών µειώνεται ο αριθµός των κεντριολίων που σχηµατίζονται. Η µετέπειτα µετανάστευσή τους στην πλασµατική µεµβράνη των κυττάρων είναι επίσης προβληµατική. Ωστόσο, είναι δυνατόν να εντοπιστεί ένας µικρός αριθµός κυττάρων, που διαθέτουν 1-2 κινητούς κροσσούς, σε αντίθεση µε τους ελάχιστους κροσσούς που απαντώνται στους ασθενείς µε µεταλλάξεις στο 179
180 5. ΣΥΖΗΤΗΣΗ γονίδιο McIdas, οι οποίοι είναι ακίνητοι (Boon et al, 2014). Η απαλοιφή της έκφρασης του Ccno προκαλεί τον ίδιο φαινότυπο και στους µυς, καθώς και στα πολυκροσσωτά κύτταρα της επιδερµίδας εµβρύων Xenopus (Funk et al, 2015; Wallmeier et al, 2014). Οι µύες που είναι ελλειµµατικοί για το γονίδιο Ccno εµφανίζουν επίσης αυξηµένη θνησιµότητα κατά τη γέννησή τους. Οι µύες που καταφέρνουν να επιβιώσουν ε µφανίζουν καθυστερηµένη ανάπτυξη και υδροκεφαλία, µε αποτέλεσµα να πεθαίνουν τις πρώτες 6 εβδοµάδες µετά τη γέννησή τους. Τα θηλυκά και αρσενικά άτοµα που δεν επηρεάζονται τόσο πολύ είναι γόνιµα. Τέλος, οι µύες αυτοί εµφανίζουν συµφόρηση στο βλεννογόνο της ρινικής κοιλότητας, αλλά δεν παρατηρείται αναποτελεσµατική κάθαρση της αναπνευστικής οδού, όπως στους ασθενείς. Μέχρι στιγµής δεν υπάρχουν διαθέσιµα δεδοµένα που να συνδέουν τέτοιους ασθενείς µε µεταλλάξεις στο γονίδιο GemC1. Ωστόσο, τα δεδοµένα από τους µυς δείχνουν ότι προκαλείται καθυστέρηση στην ανάπτυξη και θάνατος σύντοµα µετά τη γέννηση. Οι GemC1 ελλειµµατικοί µύες που καταφέρνουν να επιβιώσουν εµφανίζουν υδροκεφαλία και στειρότητα, τόσο στα θηλυκά όσο και στα αρσενικά (Terre et al, 2016). Σε αντίθεση µε τα κύτταρα που αποµονώνονται από τους ασθενείς µε µεταλλάξεις στο γονίδιο McIdas ή από ασθενείς ή µύες µε µεταλλάξεις στο γονίδιο Ccno, από τα κύτταρα των µυών που δεν εκφράζεται το γονίδιο GemC1 απουσιάζουν παντελώς τα πολυκροσσωτά κύτταρα από την αναπνευστική οδό, από τις κοιλίες του εγκεφάλου, καθώς και από το αναπαραγωγικό σύστηµα. Έτσι, φαίνεται ότι ο παράγοντας GemC1 αποτελεί τον κορυφαίο ρυθµ ιστή της διαφοροποίησης των πολυκροσσωτών κυττάρων. Το γονίδιο GemC1 είναι πιθανό να συνδέεται µε ασθένειες του ανθρώπου που σχετίζονται µε τα πολυκροσσωτά κύτταρα. 180
181 5. ΣΥΖΗΤΗΣΗ 5.5 Σύγκριση του ρόλου των GemC1 και McIdas στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Δρουν κατά πλεονασµό ή εξειδικεύονται; Η δράση των παραγόντων GemC1 και McIdas επιτελείται µέσω της δηµιουργίας συµ πλόκων µε τους µεταγραφικούς παράγοντες E2F4/5 και DP1. Τα σύµπλοκα αυτά οµοιάζουν µεταξύ τους, τόσο δοµ ικά όσο και λειτουργικά, και συνδέονται σε ρυθµ ιστικές περιοχές-υποκινητές γονιδίων που λειτουργούν σε διάφορα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων. Παρόλο που δεν είναι ακόµα γνωστός ο µηχανισµός που πυροδοτεί το σχηµατισµό τους, γνωρίζουµε ότι και τα δυο σύµπλοκα αναστέλλονται από τη Geminin. Οι Ma et al. πραγµατοποίησαν µια RNA-seq ανάλυση στα κύτταρα της επιδερµίδας εµβρύων Xenopus, στα οποία προκάλεσαν επαγωγή της διαφοροποίησής τους προς πολυκροσσωτά κύτταρα, µετά την υπερέκφραση του McIdas. Τα κύτταρα αυτά συγκρίθηκαν µε κύτταρα στα οποία έγινε ταυτόχρονη υπερέκφραση του McIdas µε ένα µετάλλαγµα του παράγοντα Ε 2F4 (E2F4ΔCT) (Ma et al, 2014). Στο µετάλλαγµα αυτό απουσίαζε η καρβοξυτελική περιοχή του E2F4, γεγονός που εµπόδιζε την ικανότητα του McIdas να επάγει τον πολλαπλασιασµό των κεντριολίων. Η ανάλυση έδειξε ότι το EDM σύµπλοκο ενεργοποιεί µια σειρά γονιδίων, συµπεριλαµβανοµένων µεταγραφικών παραγόντων (Foxj1, Rfx2 και cmyb), καθώς και ρυθµιστών του διπλασιασµού των κεντριολίων (Ccno, Plk4, Cep152, STIL κ αι Deup1). Αντίθετα, τα γονίδια που σχετίζονται µε τον κυτταρικό κύκλο παρέµεναν ανεπηρέαστα, µε εξαίρεση την Plk1 και την κυκλίνη Ccna1, που αν και είναι άγνωστος ο ρόλος της, εκφράζεται ειδικά στα πολυκροσσωτά κύτταρα. Επιπρόσθετη ανάλυση έδειξε ότι η ικανότητα πρόσδεσης του µεταγραφικού παράγοντα E2F4 στους υποκινητές των γονιδίων που σχετίζονται µε τη διαφοροποίηση των πολυκροσσωτών κυττάρων, αυξάνεται παρουσία του παράγοντα McIdas (Ma et al, 2014). Στην παρούσα µελέτη δείχθηκε ότι η απουσία του παράγοντα GemC1 εµποδίζει τη διαφοροποίηση των πολυκροσσωτών κυττάρων, εξαιτίας της αναστολής της έκφρασης µιας σειράς µεταγραφικών παραγόντων (cmyb, p73, Foxj1, Rfx3) και ρυθµιστών του διπλασιασµού των κεντριολίων (Plk4, Deup1, Ccno). Το ίδιο αποτέλεσµα αποκαλύφθηκε ύστερα από τη σύγκριση της γονιδιακής έκφρασης των κυττάρων της τραχείας και των ωαγωγών, που προέρχονταν από αγρίου τύπου µυς και από µυς ελλειµµατικούς για το γονίδιο GemC1 (Terre et al, 2016). Ο παράγοντας GemC1 αλληλεπιδρά µε το µεταγραφικό παράγοντα E2F5 (Terre et al, 2016, Μεταπτυχιακή διπλωµατική εργασία, Ιωάννης Λουκάς, 2017), µε τον οποίο µπορεί να συνδέονται ειδικά στις ρυθµιστικές περιοχές των γονιδίων McIdas και Foxj1 (Arbi et al, 2016). Συµπεραίνουµε λοιπόν ότι οι παράγοντες GemC1 και McIdas εµφανίζουν επικάλυψη στα γονίδια που ρυθµίζουν µεταγραφικά, οδηγώντας στο συµπέρασµα ότι µπορεί να δρουν στη συγκεκριµένη διαδικασία κατά πλεονασµό. Η παραπάνω υπόθεση δεν ισχύει εάν λάβουµε υπόψιν όλα τα δεδοµένα της παρούσας εργασίας, σε συνδυασµό και µε την υπάρχουσα βιβλιογραφία. Η αποσιώπηση του γονιδίου GemC1 οδηγεί σε πλήρη αναστολή του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων (Arbi et al, 2016; Terre et al, 2016). 181
182 5. ΣΥΖΗΤΗΣΗ Επίσης, ο παράγοντας GemC1 µπορεί να επάγει µεταγραφικά τον McIdas, ενώ ο McIdas όχι (Arbi et al, 2016; Kyrousi et al, 2015) και η έκφραση του McIdas αναστέλλεται µετά την έλλειψη του GemC1 σε µυς, Xenopus και zebrafish (Arbi et al, 2016; Kyrousi et al, 2015; Terre et al, 2016; Zhou et al, 2015). Εάν δρούσαν κατά πλεονασµό ο McIdas θα έπρεπε να µπορεί να ανασυγκροτήσει πλήρως το µονοπάτι της διαφοροποίησης, µετά την έλλειψη του παράγοντα GemC1. Φαίνεται λοιπόν ότι οι παράγοντες GemC1 και McIdas λειτουργούν ιεραρχικά κατά τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Ωστόσο, απαιτούνται επιπρόσθετα πειράµατα ώστε να διευκρινιστούν όλα τα γονίδια-στόχοι τους και να αποκαλυφθούν οι διαφορές της δράσης τους κατά το µονοπάτι αυτό. Τέλος, είναι πολύ ενδιαφέρον ότι ο McIdas µπορεί να επάγει τη µεταγραφή από τα δικά του ρυθµ ιστικά στοιχεία, δηµ ιουργώντας έτσι ένα µηχανισµό θετικής ανατροφοδότησης, που συµ βάλλει αποτελεσµατικά στην ενίσχυση και εγκαθίδρυση της γονιδιακής έκφρασης. Εκατέρωθεν του γονιδίου McIdas στο γονιδίωµα εδράζονται τα γονίδια Ccno και Ccdc20b (Εικόνα 5.1). Όλα τα γονίδια της συγκεκριµένης χρωµοσωµικής περιοχής δείχθηκε να ρυθµίζονται από τον παράγοντα GemC1. Το γεγονός αυτό αναδεικνύει τη σηµ αντικότητα της συγκεκριµένης χρωµοσωµικής περιοχής στη διαφοροποίηση των πολυκροσσωτών κυττάρων, η οποία ρυθµίζεται από τον παράγοντα GemC1, µε στόχο την εκκίνηση του µονοπατιού της διαφοροποίησης των πολυκροσσωτών κυττάρων. 182
183 5. ΣΥΖΗΤΗΣΗ Chromosome 5 21 q q11.2 p MIR449C MIR449A MIR449B MCIDAS CCNO CDC20B 10 kb Εικόνα 5.1 Η περιοχή του γονιδιώµατος που εδράζεται το γονίδιο McIdas είναι καθοριστικής σηµασίας για τη διαφοροποίηση των πολυκροσσωτών κυττάρων. Το γονίδιο McIdas εδράζεται στο χρωµόσωµα 5, στην περιοχή 5q11.2, ανάµεσα στα γονίδια Ccno και Ccdc20b, που βρίσκονται 3.85 kb ανοδικά και 47 kb καθοδικά του γονιδίου McIdas, αντίστοιχα. Το γονίδιο Ccno είναι απαραίτητο για τον πολλαπλασιασµό των κεντριολίων στα πολυκροσσωτά κύτταρα, ελέγχοντας το σχηµατισµό των δευτεροσωµατίων. Από το γονίδιο Cdc20b προκύπτουν τα micrornas της οικογένειας mir449, τα οποία εκφράζονται σε υψηλά επίπεδα στα πολυκροσσωτά κύτταρα και αναστέλλουν τη σηµατοδότηση Notch, καθώς και τον παράγοντα CP
184 5. ΣΥΖΗΤΗΣΗ 5.6 Συνοψίζοντας το ρόλο της υπεροικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων Συνοψίζοντας όλα τα δεδοµένα οδηγούµαστε στο εξής µοντέλο σχετικά µε το ρόλο των πρωτεϊνών της οικογένειας της Geminin στον πολλαπλασιασµό των κεντριολίων και στη µετέπειτα διαφοροποίηση των πολυκροσσωτών κυττάρων: στα διαιρούµενα προγονικά κύτταρα, η πρωτεΐνη Geminin εκφράζεται σε υψηλά επίπεδα, ενώ οι πρωτεΐνες GemC1 και McIdas εκφράζονται σε χαµηλά επίπεδα και η λειτουργία τους αναστέλλεται µέσω πρόσδεσης µε την Geminin (Εικόνα 5.2). Καθώς γίνεται ο καθορισµός της µοίρας των κυττάρων προς πολυκροσσωτά κύτταρα, η πρωτεΐνη Geminin µειώνεται και αυξάνεται η έκφραση των GemC1 και McIdas. Ο GemC1 είναι ελεύθερος να σχηµατίσει ενεργά µεταγραφικά σύµπλοκα µε τους παράγοντες E2F4/5 και να επάγει τη µεταγραφή του McIdas, καθώς και µια πληθώρα ακόµα παραγόντων που συµµετέχουν στο διπλασιασµό των κεντριολίων (cmyb και Ccno) και στο σχηµατισµό των κροσσών (Foxj1, Rfx2/3). O McIdas σχηµατίζει επίσης µεταγραφικά σύµπλοκα µε τους παράγοντες E2F4/5 και ισχυροποιεί την ενεργοποίηση της πληθώρας των γονιδίων που απαιτούνται για τη λειτουργική εξειδίκευση των πολυκροσσωτών κυττάρων. Μέσω ενός µηχανισµού θετικής ανατροφοδότησης ο McIdas ενισχύει την έκφρασή του, «κλειδώνοντας» την τύχη των κυττάρων προς την πολυκροσσωτή µοίρα. 184
185 5. ΣΥΖΗΤΗΣΗ Cycling progenitor cell Multiciliated cell cell cycle exit E2F4/5 Geminin Cell cycle genes GemC1 Cilia-related genes Transcriptional regulatory network for: Expression levels centriole amplification cmyb CCNO cilia formation FOXJ1 RFX DP1 Cell cycle genes E2F4/5 McIdas DP1 Cilia-related genes Transcriptional regulatory network for: centriole amplification cmyb CCNO cilia formation FOXJ1 RFX Geminin GemC1 High levels Low levels McIdas Εικόνα 5.2 Οι πρωτεΐνες της οικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Το µοντέλο που προκύπτει για το ρόλο των µελών της οικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων, στηρίζεται στις ισορροπηµένες αλληλεπιδράσεις που αναπτύσσουν οι πρωτεΐνες αυτές και στα σχετικά επίπεδα της έκφρασής τους. Στα διαιρούµενα προγονικά κύτταρα, η πρωτεΐνη Geminin εκφράζεται σε υψηλά επίπεδα, ενώ οι GemC1 και McIdas βρίσκονται σε χαµηλά επίπεδα. Σε αυτή την κατάσταση, οι GemC1 και McIdas αναστέλλονται από τη Geminin, εκφράζονται γονίδια που συµµετέχουν στον κυτταρικό πολλαπλασιασµό, ενώ τα γονίδια που χρειάζονται για τη διαφοροποίηση των πολυκροσσωτών κυττάρων παραµένουν µεταγραφικά ανενεργά. Κατά τον καθορισµό της µοίρας των κυττάρων προς πολυκροσσωτά κύτταρα, τα επίπεδα της Geminin πρέπει να µειωθούν, ενώ τα επίπεδα έκφρασης των GemC1 και McIdas πρέπει να αυξηθούν. Ο παράγοντας GemC1, όταν πια δεν υφίσταται την ανασταλτική δράση της Geminin, σχηµατίζει σύµπλοκα µε τους µεταγραφικούς παράγοντες E2F4/5 και DP1 και ενεργοποιεί τη µεταγραφή του McIdas, που σχηµατίζει επίσης σύµπλοκα µε τους E2F4/5 και DP1. Τα σύµπλοκα αυτά συµβάλλουν στη λειτουργική εξειδίκευση των πολυκροσσωτών κυττάρων, µέσω ενεργοποίησης µιας σειράς γονιδίων που απαιτούνται για το διπλασιασµό των κεντριολίων και το σχηµατισµό των κροσσών 185
186 5. ΣΥΖΗΤΗΣΗ Στο µοντέλο που προτείνεται για τη δράση των µελών της υπεροικογένειας της Geminin στη διαφοροποίηση των πολυκροσσωτών κυττάρων, προκύπτουν ερωτήµατα σχετικά µε το ανoδικό µονοπάτι που ελέγχει την έκφρασή τους. Πώς ρυθµίζονται οι πρωτεΐνες της οικογένειας της Geminin, ώστε να ενεργοποιήσουν το µονοπάτι της διαφοροποίησης των πολυκροσσωτών κυττάρων την κατάλληλη στιγµή και στα κατάλληλα κύτταρα; Πώς πραγµατοποιείται η µειώση των επιπέδων έκφρασης της Geminin και η αύξηση των επιπέδων των GemC1 και McIdas; H σηµατοδότηση Notch βρίσκεται στην κορυφή του µονοπατιού. Στο αναπνευστικό επιθήλιο των µυών, τα χαµηλά επίπεδα Notch οδηγούν σε διαφοροποίηση των πρόδροµων κυττάρων προς πολυκροσσωτά και νευροενδοκρινή κύτταρα, ενώ δεν ευνοείται η διαφοροποίησή τους προς εκκριτικά Clara κύτταρα (Tsao et al, 2009). Οι GemC1 και McIdas δείχθηκε να βρίσκονται υπό τον έλεγχο της σηµατοδότησης Notch (Kyrousi et al, 2015; Stubbs et al, 2012; Zhou et al, 2015). Ο McIdas χαρακτηρίστηκε αρχικά στο Xenopus, ως ένας παράγοντας που αναστέλλεται από το σηµ ατοδοτικό µονοπάτι Notch (Stubbs et al, 2012). Αργότερα, µια άλλη µελέτη από το εργαστήριό µας έδειξε ότι το Notch δρα ανταγωνιστικά έναντι των GemC1 και McIdas κατά τη διαφοροποίηση των κυττάρων της ακτινωτής γλοίας των κοιλιών του εγκεφάλου σε επενδυµατικά πολυκροσσωτά κύτταρα (Kyrousi et al, 2015). Ο µηχανισµός αυτός φαίνεται να είναι συντηρηµένος, αφού δείχθηκε να ισχύει και στο Xenopus και στο zebrafish (Zhou et al, 2015). Ωστόσο, απαιτούνται επιπρόσθετα πειράµατα ώστε να διαπιστωθεί πώς δρα η σηµατοδότηση Notch, αλλά και η Geminin, στην έκφραση των παραγόντων GemC1 και McIdas. Ιδιαίτερα ελλιπής είναι και η γνώση σχετικά µε το εάν ο παράγοντας GemC1 µπορεί να εµπλέκεται και σε αποφάσεις καθορισµού της µοίρας των κυττάρων, µιας και φαίνεται να είναι ο πιο κορυφαίος ρυθµιστής του µονοπατιού. Η απουσία του από το αναπνευστικό επιθήλιο των µυών δεν επηρέασε τον πληθυσµό των προγονικών κυττάρων (Κ5 θετικά κύτταρα). Ωστόσο, αυξήθηκε ο αριθµός των βλεννοπαραγωγών κυττάρων (Muc5ac θετικά κύτταρα). Το αποτέλεσµα αυτό µπορεί να ερµηνευτεί είτε ως άµεση επίδραση του GemC1 στη µοίρα των βλεννοπαραγωγών κυττάρων, είτε ως έµµεση, λόγω της έλλειψης των πολυκροσσωτών κυττάρων, της συσσώρευσης βλέννης και του σχηµατισµού ενός µη φυσιολογικού αναπνευστικού επιθηλίου. Παράλληλα, απουσία του GemC1 δείχθηκε µια µικρού βαθµού µείωση της έκφρασης του µεταγραφικού παράγοντα p63 που εκφράζεται στα προγονικά βασικά κύτταρα του αναπνευστικού επιθηλίου. Σε µεγαλύτερο βαθµό επηρεάζεται και ο µεταγραφικός παράγοντας cmyb και το γονίδιο Cdc20b. Ο µεταγραφικός παράγοντας cmyb έχει ήδη εµπλακεί σε αποφάσεις καθορισµού της µοίρας των κυττάρων του επιθηλίου, αφού εκφράζεται σε έναν ενδιάµεσο πληθυσµό κυττάρων του αναπνευστικού επιθηλίου των µυών, που µπορούν να διαφοροποιηθούν είτε προς πολυκροσσωτά, ε ίτε προς εκκριτικά κύτταρα (Pan et al, 2014). Από το γονίδιο Cdc20b προκύπτουν τα micrornas της οικογένειας των mir449, τα οποία εκφράζονται σε υψηλά επίπεδα στα πολυκροσσωτά κύτταρα και αναστέλλουν τη σηµατοδότηση Notch (Marcet et al, 2011). Όλα τα παραπάνω κάνουν αναγκαία τη µελέτη του παράγοντα GemC1 και σε αποφάσεις καθορισµού της µοίρας των κυττάρων. 186
187 5. ΣΥΖΗΤΗΣΗ 5.7 Οι πρωτεΐνες της οικογένειας της Geminin στη ρύθµιση του κύκλου των κεντροσωµατίων Η συµµετοχή του McIdas στον πολλαπλασιασµό των κεντριολίων σε διαιρούµενα κύτταρα Κατά τη διάρκεια του κυτταρικού κύκλου, σε κάθε διαιρούµενο κύτταρο, ελέγχεται η δηµιουργία ενός µόνο θυγατρικού κεντριολίου κάθετα προς το µητρικό. Αντίθετα, τα πολυκροσσωτά κύτταρα εξέρχονται από τον κυτταρικό κύκλο και µέσα από έναν ιδιόµορφο κύκλο διπλασιασµού των κεντροσωµατίων, καταφέρνουν να πολλαπλασιάσουν τα κεντριόλιά τους εκατοντάδες φορές. Τα αποτελέσµατα της παρούσας διατριβής έδειξαν ότι οι παράγοντες GemC1 και McIdas κατέχουν κεντρικό ρόλο στη διαφοροποίηση των πολυκροσσωτών κυττάρων. Οι παράγοντες αυτοί δείχθηκε να επηρεάζουν, µεταξύ άλλων, και την έκφραση γονιδίων που εµπλέκονται στη βιοσύνθεση των κεντριολίων, όπως των Deup1, Plk4 και STIL. Στα πολυκροσσωτά κύτταρα ο σχηµατισµός πολλαπλών κεντριολίων επιτυγχάνεται µέσω αύξησης της έκφρασης των παραγόντων που ελέγχουν το διπλασιασµό τους και κατά τη διάρκεια ενός κανονικού κυτταρικού κύκλου (Vladar & Stearns, 2007; Zhao et al, 2013). Ωστόσο, δεν είναι ακόµα γνωστοί οι µηχανισµοί εκείνοι που ελέγχουν τα επίπεδα έκφρασης των παραπάνω ρυθµιστών, ώστε να επιτυγχάνεται ο σχηµατισµός ενός ή πολλαπλών κεντριολίων ανά κύτταρο. Είναι λοιπόν πολύ ενδιαφέρον να µελετηθεί ο πιθανός ρόλος των GemC1 και McIdas στο διπλασιασµό των κεντριολίων, τόσο κατά τη διάρκεια του κυτταρικού κύκλου, όσο και κατά τη διάρκεια της δηµιουργίας των πολυκροσσωτών κυττάρων. Προκειµένου να διερευνηθεί η συµµετοχή των GemC1 και McIdas στο διπλασιασµό των κεντριολίων στα διαιρούµενα κύτταρα, διερευνήθηκε αρχικά εάν µπορεί να εντοπίζονται στα κεντριόλια. Η µελέτη επικεντρώθηκε στην πρωτεΐνη McIdas, µιας και στο εργαστήριό µας έχει παραχθεί πολυκλωνικό αντίσωµα που την αναγνωρίζει ειδικά. Η πρωτεΐνη McIdas εντοπίζεται ειδικά µόνο στο µητρικό κεντριόλιο των διαιρούµενων κυττάρων. Η έκφρασή της µελετήθηκε σε δυο διαφορετικές κυτταρικές σειρές και συγκρίθηκε µε αυτή των πρωτεϊνών Cep135, που εντοπίζεται στο εγγύς τ µήµα και των δύο κεντριολίων, και Cep164, η οποία εντοπίζεται στις αποφύσεις που υπάρχουν στο αποµακρυσµένο τµήµα του µητρικού κεντριολίου. Η πρωτεΐνη McIdas εντοπίζεται σε ένα ενδιάµεσο τ µήµα του µητρικού κεντριολίου, αφού βρίσκεται ανάµεσα στις δυο παραπάνω πρωτεΐνες. Ωστόσο, είναι απαραίτητο να διενεργηθούν περαιτέρω πειράµατα µε τη χρήση µικροσκοπίας υψηλής ανάλυσης (super-resolution microscopy), ώστε να διαπιστωθεί ο ακριβής εντοπισµός της πρωτεΐνης και να συγκριθεί µε το πρότυπο έκφρασης που εµφανίζουν γνωστοί ρυθµιστές των κεντριολίων. Η παραγωγή ενός αντισώµατος που θα αναγνωρίζει ειδικά την πρωτεΐνη GemC1 θα συµβάλλει στο να µελετηθεί εάν και αυτό το µέλος της οικογένειας εντοπίζεται στα κεντροσωµάτια. Η µελέτη του προτύπου έκφρασης της πρωτεΐνης McIdas κατά τη διάρκεια του κυτταρικού κύκλου, µε τη χρήση µιας σταθερά διαµολυσµένης κυτταρικής 187
188 5. ΣΥΖΗΤΗΣΗ σειράς, έδειξε ότι τα επίπεδά της µειώνονται κατά τη διάρκεια της ανάφασης και αυξάνουν ξανά µε την είσοδο στη φάση G1. Προκαταρκτική µελέτη του προτύπου έκφρασης της πρωτεΐνης McIdas στα κεντροσωµάτια έδειξε ότι εντοπίζεται στα κεντροσωµάτια τόσο κυττάρων που εκφράζουν την κυκλίνη Α (S και G2 φάση), όσο και κυττάρων που δεν την εκφράζουν (G1 φ άση). Επιπλέον, η έκφραση της πρωτεΐνης χάνεται από τα κεντροσωµάτια κατά τη διάρκεια της κυτταρικής διαίρεσης. Η συγκριτική µελέτη της έκφρασης της πρωτεΐνης McIdas παράλληλα µε ρυθµιστές του κυτταρικού κύκλου (π.χ. PCNA) και των κεντριολίων (π.χ. STIL, SAS6) που εκφράζονται σε συγκεκριµένες φάσεις του κυτταρικού κύκλου, θα βοηθήσει να αποσαφηνιστεί το πρότυπο της έκφρασής της και κατά συνέπεια η λειτουργία της στα κεντροσωµάτια. Γνωρίζουµε ήδη ότι η Geminin έχει ε µπλακεί άµεσα στον κύκλο των κεντροσωµατίων (Tachibana et al, 2005; Tachibana & Nigg, 2006) και µάλιστα εντοπίζεται στα κεντροσωµάτια και αλληλεπιδρά µέσω της περιοχής του σπειροειδούς σπειράµατος που διαθέτει µε την υποµονάδα Arp1 του συµπλόκου της δυνεΐνηςακτίνης (Lu et al, 2009). Πολύ πρόσφατα δείχθηκε ότι ο µεταγραφικός παράγοντας E2F4, κατά τα πρώιµα στάδια της διαφοροποίησης των πολυκροσσωτών κυττάρων, µετατοπίζεται από τον πυρήνα στο κυτταρόπλασµα, όπου αλληλεπιδρά µε τους παράγοντες Deup1 και SAS6, συµµετέχοντας στο σχηµατισµό των δευτεροσωµατίων (Mori et al, 2017). Οι GemC1 και McIdas σχηµατίζουν σύµ πλοκα µε τους µεταγραφικούς παράγοντες E2F4/5. Τα δεδοµένα αυτά ισχυροποιούν την υπόθεση για άµεσο ρόλο των µελών της οικογένειας της Geminin στα κεντροσωµάτια. Τέλος, προκαταρκτικά λειτουργικά πειράµατα έδειξαν ότι η υπερέκφραση της πρωτεΐνης McIdas αυξάνει τον αριθµό των κεντριολίων στα κύτταρα. Ο ρόλος αυτός δεν φαίνεται να σχετίζεται µε το ρόλο του McIdas κατά την αντιγραφή του DNA, µιας και µπορούσε να προκαλέσει αύξηση στον αριθµό των κεντροσωµατίων, ενώ είχε ανασταλλεί η αντιγραφή του DNA µε τη χρήση υδροξυουρίας. Πειράµατα αποσιώπησης του γονιδίου McIdas θα αποσαφηνίσουν εάν εµπλέκεται στο διπλασιασµό των κεντριολίων Τα µέλη της υπεροικογένειας της Geminin ως κοινοί ρυθµιστές των κύκλων διπλασιασµού και διαχωρισµού των χρωµοσωµάτων και των κεντροσωµατίων Ο κύκλος διπλασιασµού και διαχωρισµού των χρωµοσωµάτων στα διαιρούµενα κύτταρα εξελίσσεται παράλληλα µε τον κύκλο διπλασιασµού και διαχωρισµού των κεντροσωµατίων. Και οι δύο κύκλοι βρίσκονται κάτω από αυστηρό έλεγχο, ώστε να εξασφαλίζεται ότι ο διπλασιασµός του γονιδιώµατος και των κεντριολίων θα συµβεί µόνο µια φορά σε κάθε κυτταρικό κύκλο. Έχουν βρεθεί µόρια που συµµετέχουν και στους δυο κύκλους, όµως παραµένει άγνωστος ο µηχανισµός µέσω του οποίου συνδέονται. Με δεδοµένο ότι τα µέλη της οικογένειας της Geminin κατέχουν πολλαπλούς ρόλους, τόσο στον κυτταρικό πολλαπλασιασµό κατά την αντιγραφή του DNA, όσο και στον πολλαπλασιασµό των κεντριολίων κατά τη 188
189 5. ΣΥΖΗΤΗΣΗ διαφοροποίηση των πολυκροσσωτών κυττάρων, θα µπορούσε να θεωρηθεί ότι αποτελούν µια οµάδα κοινών ρυθµιστών για τους δύο κύκλους (Εικόνα 5.3). Η πρωτεΐνη Geminin έχει µελετηθεί εκτενώς και γνωρίζουµε ότι συµβάλει στη γονιδιωµατική σταθερότητα, αφού αναστέλλει τον παράγοντα αδειοδότησης της αντιγραφής Cdt1 (Tada et al, 2001; Wohlschlegel et al, 2000). Επίσης, εντοπίζεται στα κεντροσωµάτια, δρώντας ως ένας αρνητικός ρυθµ ιστής του διπλασιασµού των κεντροσωµατίων (Lu et al, 2009; Tachibana et al, 2005; Tachibana & Nigg, 2006). Ακόµα, λειτουργεί ως ανασταλτικός παράγοντας και της διαφοροποίησης των πολυκροσσωτών κυττάρων. Δρα ανταγωνιστικά έναντι των GemC1 και McIdas και ίσως αυτό να αποτελεί ένα µηχανισµό που διατηρεί τα κύτταρα σε µια πρόδροµη κατάσταση, ώστε να αποφευχθεί ο πρόωρος πολλαπλασιασµός των κεντριολίων και η διαφοροποίησή τους προς πολυκροσσωτά κύτταρα. Οµοίως, οι GemC1 και McIdas χαρακτηρίστηκαν αρχικά για το ρόλο τους κατά την έναρξη της αντιγραφής του DNA (Balestrini et al, 2010; Pefani et al, 2011), µε τον παράγοντα McIdas να δρα ανταγωνιστικά στη Geminin για την πρόσδεσή της στον παράγοντα Cdt1. Από την παρούσα διατριβή είναι σαφές ότι οι GemC1 και McIdas είναι κεντρικοί ρυθµιστές του πολλαπλασιασµού των κεντριολίων και της διαφοροποίησης των πολυκροσσωτών κυττάρων. Οι µηχανισµοί που ελέγχουν τον πολλαπλασιασµό των κεντριολίων στα διαιρούµενα και στα πολυκροσσωτά κύτταρα δεν έχουν πλήρως αποσαφηνιστεί. Κατά τη διάρκεια του κυτταρικού κύκλου λαµβάνει χώρα ο κανονικός διπλασιασµός των κεντριολίων, όπου ένα θυγατρικό κεντριόλιο σχηµατίζεται κάθετα στο µητρικό. Στα πολυκροσσωτά κύτταρα, οι µηχανισµοί αυτοί ακυρώνονται και δηµ ιουργούνται πολλαπλά κεντριόλια. Η αύξηση των επιπέδων έκφρασης πάνω από έναν ουδό (threshold) και οι αλληλεπιδράσεις των Geminin, GemC1 και McIdas τόσο µεταξύ τους, όσο και µε άλλους παράγοντες φαίνεται να παίζουν καθοριστικό ρόλο στην έξοδο ενός κυττάρου από τον κυτταρικό κύκλο και στην επαγωγή του µετέπειτα µονοπατιού που ελέγχουν. 189

190 5. ΣΥΖΗΤΗΣΗ Εικόνα 5.3 Οι πρωτεΐνες της οικογένειας της Geminin στον κύκλο διπλασιασµού των χρωµοσωµάτων και των κεντροσωµατίων. Κατά τη διάρκεια του κυτταρικού κύκλου, η πρωτεΐνη Geminin εκφράζεται κατά τις φάσεις S, G2 και στην αρχή της Μ φάσης. Η Geminin συµβάλλει στη γονιδιωµατική σταθερότητα, εµ ποδίζοντας την πρόωρη αδειοδότηση ενός νέου κύκλου αντιγραφής του DNA και δρώντας ανασταλτικά στον υπερδιπλασιασµό των κεντροσωµατίων. Τα µέλη GemC1 και McIdas διαδραµατίζουν κεντρικό ρόλο στον καθορισµό της µοίρας των κυττάρων προς πολυκροσσωτά κύτταρα, επάγοντας τον πολλαπλασιασµό των κεντριολίων και ενεργοποιώντας το δίκτυο των γονιδίων που απαιτούνται για τη διαφοροποίηση των πολυκροσσωτών κυττάρων. 190
Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών
 Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών Τίτλος Ερευνητικού Έργου «Οι πρωτεΐνες της υπεροικογένειας της Geminin
Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών Τίτλος Ερευνητικού Έργου «Οι πρωτεΐνες της υπεροικογένειας της Geminin
Έλεγχος κυτταρικού κύκλου Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Έλεγχος κυτταρικού κύκλου Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Πως το κύτταρο διπλασιάζει τα συστατικά του; Πως γίνεται ο διαχωρισμός των συστατικών στα
Έλεγχος κυτταρικού κύκλου Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Πως το κύτταρο διπλασιάζει τα συστατικά του; Πως γίνεται ο διαχωρισμός των συστατικών στα
ΚΥΤΤΑΡΙΚΗ ΙΑΙΡΕΣΗ. αναπαραγωγή. αύξηση αριθµού κυττάρων ανάπτυξη
 ΚΥΤΤΑΡΙΚΗ ΙΑΙΡΕΣΗ αναπαραγωγή αύξηση αριθµού κυττάρων ανάπτυξη επιδιόρθωση ιστών Κυτταρική οργάνωση του γενετικού υλικού Γονιδίωµα: Το σύνολο του γενετικού υλικού (DNA) ενός κυττάρου Στα προκαρυωτικά κύτταρα
ΚΥΤΤΑΡΙΚΗ ΙΑΙΡΕΣΗ αναπαραγωγή αύξηση αριθµού κυττάρων ανάπτυξη επιδιόρθωση ιστών Κυτταρική οργάνωση του γενετικού υλικού Γονιδίωµα: Το σύνολο του γενετικού υλικού (DNA) ενός κυττάρου Στα προκαρυωτικά κύτταρα
ΚΕΦΑΛΑΙΟ 16. Ο κυτταρικός κύκλος. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ. Με αναφορά τόσο στους προκαρυωτικούς όσο και στους ευκαρυωτικούς οργανισμούς
 ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ Με αναφορά τόσο στους προκαρυωτικούς όσο και στους ευκαρυωτικούς οργανισμούς Λειτουργίες Γενετικού Υλικού o Αποθήκευση της γενετικής πληροφορίας. Η οργάνωση της γενετικής πληροφορίας
ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ Με αναφορά τόσο στους προκαρυωτικούς όσο και στους ευκαρυωτικούς οργανισμούς Λειτουργίες Γενετικού Υλικού o Αποθήκευση της γενετικής πληροφορίας. Η οργάνωση της γενετικής πληροφορίας
Εργασία Στο Μάθημα Της Βιολογίας. Τάξη: Γ 3 Μαθήτρια: Στίνη Αΐντα Θέμα: Κυτταρική Διαίρεση: Μίτωση
 Εργασία Στο Μάθημα Της Βιολογίας Τάξη: Γ 3 Μαθήτρια: Στίνη Αΐντα Θέμα: Κυτταρική Διαίρεση: Μίτωση ΚΥΤΤΑΡΙΚΗ ΔΙΑΙΡΕΣΗ: ΜΙΤΩΣΗ Τι είναι η κυτταρική διαίρεση; Η κυτταρική διαίρεση είναι η διαδικασία κατά
Εργασία Στο Μάθημα Της Βιολογίας Τάξη: Γ 3 Μαθήτρια: Στίνη Αΐντα Θέμα: Κυτταρική Διαίρεση: Μίτωση ΚΥΤΤΑΡΙΚΗ ΔΙΑΙΡΕΣΗ: ΜΙΤΩΣΗ Τι είναι η κυτταρική διαίρεση; Η κυτταρική διαίρεση είναι η διαδικασία κατά
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1
 Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
Δομή των μυϊκών κυττάρων.
 Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
Εργασία Στο Μάθημα Της Βιολογίας. Τάξη: Γ 3. Μαθήτρια: Στίνη Αΐντα. Θέμα: Κυτταρική Διαίρεση: Μίτωση
 Εργασία Στο Μάθημα Της Βιολογίας Τάξη: Γ 3 Μαθήτρια: Στίνη Αΐντα Θέμα: Κυτταρική Διαίρεση: Μίτωση ΚΥΤΤΑΡΙΚΗ ΔΙΑΙΡΕΣΗ: ΜΙΤΩΣΗ Τι είναι η κυτταρική διαίρεση; Η κυτταρική διαίρεση είναι η διαδικασία κατά
Εργασία Στο Μάθημα Της Βιολογίας Τάξη: Γ 3 Μαθήτρια: Στίνη Αΐντα Θέμα: Κυτταρική Διαίρεση: Μίτωση ΚΥΤΤΑΡΙΚΗ ΔΙΑΙΡΕΣΗ: ΜΙΤΩΣΗ Τι είναι η κυτταρική διαίρεση; Η κυτταρική διαίρεση είναι η διαδικασία κατά
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη
 Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ
 ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής:
ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής:
Kυτταρική Bιολογία. Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος ΔIAΛEΞΗ 20 (9/5/2017) Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.
 Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.") Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ
 ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
Μίτωση - Μείωση και φυλετικοί βιολογικοί κύκλοι Γ. Παπανικολάου MD, PhD
 Μίτωση - Μείωση και φυλετικοί βιολογικοί κύκλοι Γ. Παπανικολάου MD, PhD Ομοιότητα και διαφορά Κληρονομικότητα: η μεταβίβαση χαρακτηριστικών από τη μια γενιά στην άλλη Ποικιλία: εκτός από την ομοιότητα
Μίτωση - Μείωση και φυλετικοί βιολογικοί κύκλοι Γ. Παπανικολάου MD, PhD Ομοιότητα και διαφορά Κληρονομικότητα: η μεταβίβαση χαρακτηριστικών από τη μια γενιά στην άλλη Ποικιλία: εκτός από την ομοιότητα
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος. 09/04/ Μοριακή Βιολογία Κεφ. 1 Καθηγητής Δρ. Κ. Ε. Βοργιάς
 Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Παραγωγή, απομόνωση και καθαρισμός της φαρμακευτικής πρωτεΐνης.
 ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 1 ΙΟΥΝΙΟΥ 2004 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1o 1. δ 2. β 3. β 4. γ 5. δ ΘΕΜΑ 2o 1. Σχολικό βιβλίο, σελ.
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 1 ΙΟΥΝΙΟΥ 2004 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1o 1. δ 2. β 3. β 4. γ 5. δ ΘΕΜΑ 2o 1. Σχολικό βιβλίο, σελ.
ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ Ο.Ε.Φ.Ε. 2004 ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ Ο.Ε.Φ.Ε. 2004 ΘΕΜΑ 1 Ο Α. Να επιλέξετε την ορθή πρόταση: ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1. Το κωδικόνιο του mrna που κωδικοποιεί το αµινοξύ µεθειονίνη είναι α. 5 GUA
ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ Ο.Ε.Φ.Ε. 2004 ΘΕΜΑ 1 Ο Α. Να επιλέξετε την ορθή πρόταση: ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1. Το κωδικόνιο του mrna που κωδικοποιεί το αµινοξύ µεθειονίνη είναι α. 5 GUA
Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν
 Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν ένα δυναµικό σκελετό που χρησιµεύει στη στήριξη και την
Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο το κυτταρόπλασµα και σχηµατίζουν ένα δυναµικό σκελετό που χρησιµεύει στη στήριξη και την
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ 1ο 1. β 2. β 3. α 4. α 5. β
 ΘΕΜΑ 1ο 1. β 2. β 3. α 4. α 5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΡΙΤΗ 21 ΣΕΠΤΕΜΒΡΙΟΥ 2004 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ)
ΘΕΜΑ 1ο 1. β 2. β 3. α 4. α 5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΡΙΤΗ 21 ΣΕΠΤΕΜΒΡΙΟΥ 2004 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ)
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ
 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ: ΜΙΚΡΟΪΝΙΔΙΑ- ΕΝΔΙΑΜΕΣΑ ΙΝΙΔΙΑ- ΜΙΚΡΟΣΩΛΗΝΙΣΚΟΙ Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ: ΜΙΚΡΟΪΝΙΔΙΑ- ΕΝΔΙΑΜΕΣΑ ΙΝΙΔΙΑ- ΜΙΚΡΟΣΩΛΗΝΙΣΚΟΙ Η δοµή και η λειτουργία του κυτταροσκελετού: Ο κυτταροσκελετός είναι ένα δίκτυο από ινίδια που εκτείνονται σε όλο
Ενότητα 10: Κυτταρική Διαίρεση
 Ενότητα 10: Κυτταρική Διαίρεση Κυτταρική διαίρεση: παραγωγή γενετικά πανομοιότυπων θυγατρικών κυττάρων Κυτταρική διαίρεση Μονοκύτταροι οργανισμοί: η διαίρεση του κυττάρου συνεπάγεται αναπαραγωγή ολόκληρου
Ενότητα 10: Κυτταρική Διαίρεση Κυτταρική διαίρεση: παραγωγή γενετικά πανομοιότυπων θυγατρικών κυττάρων Κυτταρική διαίρεση Μονοκύτταροι οργανισμοί: η διαίρεση του κυττάρου συνεπάγεται αναπαραγωγή ολόκληρου
ΘΕΜΑ Α Α1. β Α2. δ Α3. α Α4. α Α5. γ
 ΘΕΜΑ Α Α1. β Α2. δ Α3. α Α4. α Α5. γ ΘΕΜΑ B B1. Η συχνότητα των ετερόζυγων ατόμων με δρεπανοκυτταρική αναιμία ή β- θαλασσαιμία είναι αυξημένη σε περιοχές όπως οι χώρες της Μεσογείου, της Δυτικής και Ανατολικής
ΘΕΜΑ Α Α1. β Α2. δ Α3. α Α4. α Α5. γ ΘΕΜΑ B B1. Η συχνότητα των ετερόζυγων ατόμων με δρεπανοκυτταρική αναιμία ή β- θαλασσαιμία είναι αυξημένη σε περιοχές όπως οι χώρες της Μεσογείου, της Δυτικής και Ανατολικής
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ: 2/12/2016 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ: ΛΑΖΑΡΑΚΗ ΝΟΤΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ: 2/12/2016 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ: ΛΑΖΑΡΑΚΗ ΝΟΤΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες
Κυτταρική διαίρεση:μίτωση
 ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΙΑΝΟΥΑΡΙΟΣ 2013 ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΑΠΟ ΤΗΣ ΜΑΘΗΤΡΙΕΣ ΤΟΥ Γ 2 ΓΕΩΡΓΙΑ ΣΤΑΜΟΥΛΗ, ΜΑΡΙΝΕΛΑ ΥΜΕΡΑΙ Κυτταρική διαίρεση:μίτωση Δύο είναι οι ανώτερες κυτταρικές λειτουργίες ενός
ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΙΑΝΟΥΑΡΙΟΣ 2013 ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΑΠΟ ΤΗΣ ΜΑΘΗΤΡΙΕΣ ΤΟΥ Γ 2 ΓΕΩΡΓΙΑ ΣΤΑΜΟΥΛΗ, ΜΑΡΙΝΕΛΑ ΥΜΕΡΑΙ Κυτταρική διαίρεση:μίτωση Δύο είναι οι ανώτερες κυτταρικές λειτουργίες ενός
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου
 Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Η αναλογία Α+G/T+C στο γενετικό υλικό ενός ιού είναι ίση με 2/3. Ο ιός μπορεί να είναι: α. ο φάγος λ. β. ο ιός της πολιομυελίτιδας. γ. φορέας κλωνοποίησης
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Η αναλογία Α+G/T+C στο γενετικό υλικό ενός ιού είναι ίση με 2/3. Ο ιός μπορεί να είναι: α. ο φάγος λ. β. ο ιός της πολιομυελίτιδας. γ. φορέας κλωνοποίησης
Καραπέτσας Θανάσης. Διπλωματική Εργασία:
 Διαπανεπιστημιακό Πρόγραμμα Μεταπτυχιακών Σπουδών «Κλινική Φαρμακολογία & Θεραπευτική» Επιβλέπων: Δρ. Αλ. Γαλάνης, Λέκτορας Μορ. Βιολογίας, «Σχεδιασμός Ειδικών Πεπτιδίων Αναστολέων της Αλληλεπίδρασης του
Διαπανεπιστημιακό Πρόγραμμα Μεταπτυχιακών Σπουδών «Κλινική Φαρμακολογία & Θεραπευτική» Επιβλέπων: Δρ. Αλ. Γαλάνης, Λέκτορας Μορ. Βιολογίας, «Σχεδιασμός Ειδικών Πεπτιδίων Αναστολέων της Αλληλεπίδρασης του
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
Moριακή Kυτταρική Bιολογία & Έλεγχος Μεταβολισμού ΔIAΛEΞΕΙΣ 4 & 5 Κυτταρική διαίρεση & Απόπτωση
 Moριακή Kυτταρική Bιολογία & Έλεγχος Μεταβολισμού ΔIAΛEΞΕΙΣ 4 & 5 Κυτταρική διαίρεση & Απόπτωση Τα κύρια σημεία της διάλεξης είναι τα παρακάτω: Ο κυτταρικός κύκλος και τα στάδια του Ρύθμιση του κυτταρικού
Moριακή Kυτταρική Bιολογία & Έλεγχος Μεταβολισμού ΔIAΛEΞΕΙΣ 4 & 5 Κυτταρική διαίρεση & Απόπτωση Τα κύρια σημεία της διάλεξης είναι τα παρακάτω: Ο κυτταρικός κύκλος και τα στάδια του Ρύθμιση του κυτταρικού
Εργασία Βιολογίας - Β Τριμήνου
 Εργασία Βιολογίας - Β Τριμήνου Γυμνάσιο Κερατέας Ονοματεπώνυμο: Σίντυ Οζοκουέρε-Αγγελική Τόλε Τάξη: Γ 5 Θέμα: Κυτταρική Διαίρεση-Μίτωση Τίτλος: Ιανουάριος 2013 ΠΕΡΙΕΧΟΜΕΝΑ 1. Διαδικασία 2. Φάσεις αναπτύξεως
Εργασία Βιολογίας - Β Τριμήνου Γυμνάσιο Κερατέας Ονοματεπώνυμο: Σίντυ Οζοκουέρε-Αγγελική Τόλε Τάξη: Γ 5 Θέμα: Κυτταρική Διαίρεση-Μίτωση Τίτλος: Ιανουάριος 2013 ΠΕΡΙΕΧΟΜΕΝΑ 1. Διαδικασία 2. Φάσεις αναπτύξεως
Κύτταρα πολυκύτταρων οργανισμών
 Μίτωση - Μείωση Τα ευκαρυωτικά κύτταρα διαιρούνται με δύο τρόπους: τη μίτωση και τη μείωση. Η Μίτωση είναι ο τύπος της κυτταρικής διαίρεσης που από ένα πατρικό κύτταρο καταλήγει σε δύο γενετικά πανομοιότυπα
Μίτωση - Μείωση Τα ευκαρυωτικά κύτταρα διαιρούνται με δύο τρόπους: τη μίτωση και τη μείωση. Η Μίτωση είναι ο τύπος της κυτταρικής διαίρεσης που από ένα πατρικό κύτταρο καταλήγει σε δύο γενετικά πανομοιότυπα
Οι φάσεις που περιλαμβάνει ο κυτταρικός κύκλος είναι:
 ΚΥΚΛΟΣ ΖΩΗΣ ΤΟΥ ΚΥΤΤΑΡΟΥ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Τι είναι ο κυτταρικός κύκλος; ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 4 ο ΚΕΦΑΛΑΙΟ «ΓΕΝΕΤΙΚΗ» 2. Οι φάσεις που περιλαμβάνει ο κυτταρικός κύκλος είναι: 3. Κατά τη
ΚΥΚΛΟΣ ΖΩΗΣ ΤΟΥ ΚΥΤΤΑΡΟΥ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Τι είναι ο κυτταρικός κύκλος; ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 4 ο ΚΕΦΑΛΑΙΟ «ΓΕΝΕΤΙΚΗ» 2. Οι φάσεις που περιλαμβάνει ο κυτταρικός κύκλος είναι: 3. Κατά τη
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 22 ΜΑΪΟΥ 2015 ΑΠΑΝΤΗΣΕΙΣ
 ΘΕΜΑ Α Α1. Β Α2. Γ Α3. Α Α4. Α5. Γ ΘΕΜΑ Β ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 22 ΜΑΪΟΥ 2015 ΑΠΑΝΤΗΣΕΙΣ B1. Α (Σωµατικά κύτταρα στην αρχή της µεσόφασης): 1, 4, 5, 6 Β (Γαµέτες): 2, 3, 7, 8 Β2. (Κάθε
ΘΕΜΑ Α Α1. Β Α2. Γ Α3. Α Α4. Α5. Γ ΘΕΜΑ Β ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 22 ΜΑΪΟΥ 2015 ΑΠΑΝΤΗΣΕΙΣ B1. Α (Σωµατικά κύτταρα στην αρχή της µεσόφασης): 1, 4, 5, 6 Β (Γαµέτες): 2, 3, 7, 8 Β2. (Κάθε
ΒΙΟΛΟΓΙΑ OΠ- Γ ΓΕΛ 13:20
 ΒΙΟΛΟΓΙΑ OΠ- Γ ΓΕΛ 13:20 Σελίδα 2 από 7 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 19/ 06 / 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: Βιολογία OΠ Γ ΓΕΛ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ
ΒΙΟΛΟΓΙΑ OΠ- Γ ΓΕΛ 13:20 Σελίδα 2 από 7 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 19/ 06 / 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: Βιολογία OΠ Γ ΓΕΛ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ
2) Ανάπτυξη βασικών αρχών της in vivo μεταφοράς γονιδίων με την βοήθεια ειδικών συστημάτων μεταφοράς (πλασμίδια, αδενοϊοί)
 Ανάπτυξη βασικών αρχών της in vivo μεταφοράς γονιδίων με την βοήθεια ειδικών συστημάτων μεταφοράς (πλασμίδια, αδενοϊοί)") ΠΕΙΡΑΜΑΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ 4 η Ενότητα: 1) Ανάπτυξη βασικών αρχών πειραματικών in vivo μεθόδων για την δημιουργία διαγονιδιακών ζώων (knock-out / knock-in) 2) Ανάπτυξη βασικών αρχών της in
ΠΕΙΡΑΜΑΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ 4 η Ενότητα: 1) Ανάπτυξη βασικών αρχών πειραματικών in vivo μεθόδων για την δημιουργία διαγονιδιακών ζώων (knock-out / knock-in) 2) Ανάπτυξη βασικών αρχών της in
Πρόλογος. Σπύρος Δ. Συγγελάκης
 ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ «Ερωτήσεις θεωρίας» «Σχόλια και Επισημάνσεις» «Ερωτήσεις πολλαπλής επιλογής» «Ερωτήσεις Σωστό - Λάθος» «Ερωτήσεις κρίσεως Συνδυαστικές» «Αναλυτική μεθοδολογία ασκήσεων» «Λυμένες ασκήσεις
ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ «Ερωτήσεις θεωρίας» «Σχόλια και Επισημάνσεις» «Ερωτήσεις πολλαπλής επιλογής» «Ερωτήσεις Σωστό - Λάθος» «Ερωτήσεις κρίσεως Συνδυαστικές» «Αναλυτική μεθοδολογία ασκήσεων» «Λυμένες ασκήσεις
ΚΥΤΤΑΡΙΚΗ ΔΙΑΙΡΕΣΗ:ΜΕΙΩΣΗ- ΓΑΜΕΤΟΓΕΝΕΣΗ. Μητρογιάννη Ευαγγελία Βαμβούνης Ιωάννης
 ΚΥΤΤΑΡΙΚΗ ΔΙΑΙΡΕΣΗ:ΜΕΙΩΣΗ- ΓΑΜΕΤΟΓΕΝΕΣΗ Μητρογιάννη Ευαγγελία Βαμβούνης Ιωάννης 5/3/2013 Η κυτταρική διαίρεση είναι η διαδικασία κατά την οποία ένα αρχικό κύτταρο διαιρείται σε δύο θυγατρικά. Στους πολυκύτταρους
ΚΥΤΤΑΡΙΚΗ ΔΙΑΙΡΕΣΗ:ΜΕΙΩΣΗ- ΓΑΜΕΤΟΓΕΝΕΣΗ Μητρογιάννη Ευαγγελία Βαμβούνης Ιωάννης 5/3/2013 Η κυτταρική διαίρεση είναι η διαδικασία κατά την οποία ένα αρχικό κύτταρο διαιρείται σε δύο θυγατρικά. Στους πολυκύτταρους
Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους.
 Τι είναι τα βλαστικά κύτταρα? Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους. Είναι σε θέση να δρουν επισκευαστικά, αναδημιουργώντας
Τι είναι τα βλαστικά κύτταρα? Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους. Είναι σε θέση να δρουν επισκευαστικά, αναδημιουργώντας
ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΕΤΑΡΤΗ 4 ΙΟΥΛΙΟΥ 2007 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ
 ΘΕΜΑ 1ο 1. α 2. γ 3. δ 4. γ 5. β 1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΕΤΑΡΤΗ 4 ΙΟΥΛΙΟΥ 2007 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 2ο 1. Σχολικό βιβλίο,
ΘΕΜΑ 1ο 1. α 2. γ 3. δ 4. γ 5. β 1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΕΤΑΡΤΗ 4 ΙΟΥΛΙΟΥ 2007 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 2ο 1. Σχολικό βιβλίο,
ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Ως φορείς κλωνοποίησης χρησιμοποιούνται:
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ / Γ ΛΥΚΕΙΟΥ ΧΕΙΜΕΡΙΝΑ ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 04/03/12 ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Ως φορείς κλωνοποίησης
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ / Γ ΛΥΚΕΙΟΥ ΧΕΙΜΕΡΙΝΑ ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 04/03/12 ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Ως φορείς κλωνοποίησης
ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ. 3. Τι γενετικές πληροφορίες μπορεί να φέρει ένα πλασμίδιο;
 ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ ΕΝΟΤΗΤΑ 1.4. Οργάνωση του γενετικού υλικού προκαρυωτικών και ευκαρυωτικών κυττάρων. 1. Ποια είναι η μορφή του DNA των προκαρυωτικών κυττάρων και ποιο είναι το μήκος τους; 2. Ποια είναι
ΕΡΩΤΗΣΕΙΣ ΘΕΩΡΙΑΣ ΕΝΟΤΗΤΑ 1.4. Οργάνωση του γενετικού υλικού προκαρυωτικών και ευκαρυωτικών κυττάρων. 1. Ποια είναι η μορφή του DNA των προκαρυωτικών κυττάρων και ποιο είναι το μήκος τους; 2. Ποια είναι
Κυτταρική Διαίρεση (Μίτωση και Μείωση) Μέρος Α Μοριακή Βιολογία και Γενετική BIOL 123 Άνοιξη 2015 Δρ. Χαρίτα Χρίστου
 Μέρος Α Μοριακή Βιολογία και Γενετική BIOL 123 Άνοιξη 2015 Δρ. Χαρίτα Χρίστου") Κυτταρική Διαίρεση (Μίτωση και Μείωση) Μέρος Α Μοριακή Βιολογία και Γενετική BIOL 123 Άνοιξη 2015 Δρ. Χαρίτα Χρίστου Παρουσιάσεις Power Point με υλικό από: Campbell και Reece (2010) ΒΙΟΛΟΓΙΑ τόμος Ι, 1
Κυτταρική Διαίρεση (Μίτωση και Μείωση) Μέρος Α Μοριακή Βιολογία και Γενετική BIOL 123 Άνοιξη 2015 Δρ. Χαρίτα Χρίστου Παρουσιάσεις Power Point με υλικό από: Campbell και Reece (2010) ΒΙΟΛΟΓΙΑ τόμος Ι, 1
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΚΕΦΑΛΑΙΟ ΤΕΤΑΡΤΟ ΑΝΑΠΑΡΑΓΩΓΗ
 ΒΙΟΛΟΓΙΑ Β ΛΥΚΕΙΟΥ ΚΕΦΑΛΑΙΟ ΤΕΤΑΡΤΟ 2016 2 Το συνώνυμο της αναπαραγωγής είναι ο πολλαπλασιασμός, η δημιουργία νέων ατόμων που έχουν παρόμοια χαρακτηριστικά με τους γονείς τους. Όλοι οι οργανισμοί κάποια
ΒΙΟΛΟΓΙΑ Β ΛΥΚΕΙΟΥ ΚΕΦΑΛΑΙΟ ΤΕΤΑΡΤΟ 2016 2 Το συνώνυμο της αναπαραγωγής είναι ο πολλαπλασιασμός, η δημιουργία νέων ατόμων που έχουν παρόμοια χαρακτηριστικά με τους γονείς τους. Όλοι οι οργανισμοί κάποια
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ. ΘΕΜΑ Β Β1. Στήλη Ι Στήλη ΙΙ 1 Α 2 Γ 3 Α 4 Β 5 Α 6 Α 7 Γ
 ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ Τρίτη 18 Ιουνίου 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ. (Ενδεικτικές Απαντήσεις)
") ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ Τρίτη 18 Ιουνίου 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Ενδεικτικές Απαντήσεις) ΘΕΜΑ Α Α1. α Α2. β Α3. γ Α4. γ Α5. β ΘΕΜΑ Β Β1. 1-ζ 2-στ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ Τρίτη 18 Ιουνίου 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Ενδεικτικές Απαντήσεις) ΘΕΜΑ Α Α1. α Α2. β Α3. γ Α4. γ Α5. β ΘΕΜΑ Β Β1. 1-ζ 2-στ
ΑΠΑΝΤΗΣΕΙΣ ΚΡΙΤΗΡΙΟΥ ΑΞΙΟΛΟΓΗΣΗΣ ΓΙΑ ΤΟ 1 Ο ΚΕΦΑΛΑΙΟ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΥΓΟΥΣΤΟΣ 2015
 ΑΠΑΝΤΗΣΕΙΣ ΚΡΙΤΗΡΙΟΥ ΑΞΙΟΛΟΓΗΣΗΣ ΓΙΑ ΤΟ 1 Ο ΚΕΦΑΛΑΙΟ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΥΓΟΥΣΤΟΣ 2015 ΘΕΜΑ 1 Ο 1. γ 2. δ 3. α 4. δ 5. δ ΘΕΜΑ 2 Ο Α) Το γενετικό υλικό των προκαρυωτικών κυττάρων είναι ένα δίκλωνο
ΑΠΑΝΤΗΣΕΙΣ ΚΡΙΤΗΡΙΟΥ ΑΞΙΟΛΟΓΗΣΗΣ ΓΙΑ ΤΟ 1 Ο ΚΕΦΑΛΑΙΟ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΥΓΟΥΣΤΟΣ 2015 ΘΕΜΑ 1 Ο 1. γ 2. δ 3. α 4. δ 5. δ ΘΕΜΑ 2 Ο Α) Το γενετικό υλικό των προκαρυωτικών κυττάρων είναι ένα δίκλωνο
Κεφάλαιο 1 ο Το γενετικό υλικό Μεθοδολογία Ασκήσεων
 Κεφάλαιο 1 ο Το γενετικό υλικό Μεθοδολογία Ασκήσεων 1. Ένα μόριο νουκλεϊκού οξέος για να χαρακτηρισθεί πλήρως θα πρέπει να γνωρίζουμε αν είναι: i. DNA ή RNA ii. iii. Μονόκλωνο ή δίκλωνο Γραμμικό ή κυκλικό
Κεφάλαιο 1 ο Το γενετικό υλικό Μεθοδολογία Ασκήσεων 1. Ένα μόριο νουκλεϊκού οξέος για να χαρακτηρισθεί πλήρως θα πρέπει να γνωρίζουμε αν είναι: i. DNA ή RNA ii. iii. Μονόκλωνο ή δίκλωνο Γραμμικό ή κυκλικό
3 ΩΡΕΣ. Σελίδα 1 από 3 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΜΑΘΗΜΑ ΙΑΡΚΕΙΑ
 ΜΑΘΗΜΑ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΙΑΡΚΕΙΑ 3 ΩΡΕΣ ΘΕΜΑ 1 Ο Στις ερωτήσεις 1-5, να γράψετε τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Από τη διασταύρωση
ΜΑΘΗΜΑ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΙΑΡΚΕΙΑ 3 ΩΡΕΣ ΘΕΜΑ 1 Ο Στις ερωτήσεις 1-5, να γράψετε τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Από τη διασταύρωση
ΕΞΩΤΕΡΙΚΕΣ ΔΙΠΛΩΜΑΤΙΚΕΣ ΕΡΓΑΣΙΕΣ Β ΚΑΤΑΝΟΜΗ ΑΚΑΔΗΜΑΪΚΟ ΕΤΟΣ
 ΕΞΩΤΕΡΙΚΕΣ ΔΙΠΛΩΜΑΤΙΚΕΣ ΕΡΓΑΣΙΕΣ Β ΚΑΤΑΝΟΜΗ ΑΚΑΔΗΜΑΪΚΟ ΕΤΟΣ 2019-2020 A/A: 6 «Επίπεδα mirnas και δείκτες παχυσαρκίας σε πληθυσμό ενηλίκων» ΠΕΡΙΛΗΨΗ ΘΕΜΑΤΟΣ: Θα μετρηθούν τα επίπεδα mirnas στο πλάσμα εθελοντών
ΕΞΩΤΕΡΙΚΕΣ ΔΙΠΛΩΜΑΤΙΚΕΣ ΕΡΓΑΣΙΕΣ Β ΚΑΤΑΝΟΜΗ ΑΚΑΔΗΜΑΪΚΟ ΕΤΟΣ 2019-2020 A/A: 6 «Επίπεδα mirnas και δείκτες παχυσαρκίας σε πληθυσμό ενηλίκων» ΠΕΡΙΛΗΨΗ ΘΕΜΑΤΟΣ: Θα μετρηθούν τα επίπεδα mirnas στο πλάσμα εθελοντών
Χρωμοσώματα & κυτταροδιαιρέσεις
 Δασική Γενετική Χρωμοσώματα & κυτταροδιαιρέσεις Χειμερινό εξάμηνο 2014-2015 Σύνοψη Το DNA αναπαράγεται, εκφράζεται και μεταλλάσσεται Το DNA είναι οργανωμένα σε χρωμοσώματα Τα ευκαρυωτικά γενώματα έχουν
Δασική Γενετική Χρωμοσώματα & κυτταροδιαιρέσεις Χειμερινό εξάμηνο 2014-2015 Σύνοψη Το DNA αναπαράγεται, εκφράζεται και μεταλλάσσεται Το DNA είναι οργανωμένα σε χρωμοσώματα Τα ευκαρυωτικά γενώματα έχουν
ιαγονιδιακή τεχνολογία G. Patrinos
 ιαγονιδιακή τεχνολογία Αντίστροφη γενετική Οργανισμός Γονιδίωμα ιαγονίδιο Γονίδιο Forward genetics Επαγόμενη Οργανισμός μεταλλαξογένεση Μεταλλαγμένος οργανισμός Εύρεση και μελέτη του υπεύθυνου γονιδίου
ιαγονιδιακή τεχνολογία Αντίστροφη γενετική Οργανισμός Γονιδίωμα ιαγονίδιο Γονίδιο Forward genetics Επαγόμενη Οργανισμός μεταλλαξογένεση Μεταλλαγμένος οργανισμός Εύρεση και μελέτη του υπεύθυνου γονιδίου
Κυτταρική Βιολογία. Ενότητα 11 : Κυτταρική διαίρεση. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής Α.Π.Θ. ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 11 : Κυτταρική διαίρεση Παναγιωτίδης Χρήστος Φαρμακευτικής Α.Π.Θ. Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 11 : Κυτταρική διαίρεση Παναγιωτίδης Χρήστος Φαρμακευτικής Α.Π.Θ. Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία
 ΣΥΓΧΡΟΝΕΣ ΜΕΘΟΔΟΛΟΓΙΕΣ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ - ΓΕΝΕΤΙΚΗΣ Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία ΠΑΠΑΝΙΚΟΛΑΟΥ ΕΛΕΝΗ, Ph.D. Λέκτορας Εργαστήριο Βιολογίας, Ιατρική Σχολή Αθηνών
ΣΥΓΧΡΟΝΕΣ ΜΕΘΟΔΟΛΟΓΙΕΣ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ - ΓΕΝΕΤΙΚΗΣ Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία ΠΑΠΑΝΙΚΟΛΑΟΥ ΕΛΕΝΗ, Ph.D. Λέκτορας Εργαστήριο Βιολογίας, Ιατρική Σχολή Αθηνών
MAPS Πρωτείνες που συνδέονται με μικροσωληνίσκους και σταθεροποιούν ή καταστρέφουν τα ινίδια
 Ταξόλη - Κολχικίνη MAPS Πρωτείνες που συνδέονται με μικροσωληνίσκους και σταθεροποιούν ή καταστρέφουν τα ινίδια H οικογένεια των πρωτεινών MAP Κυτταρική κατανομή μικροσωληνίσκων/μαps/νευροινιδίωνps/νευροινιδίων
Ταξόλη - Κολχικίνη MAPS Πρωτείνες που συνδέονται με μικροσωληνίσκους και σταθεροποιούν ή καταστρέφουν τα ινίδια H οικογένεια των πρωτεινών MAP Κυτταρική κατανομή μικροσωληνίσκων/μαps/νευροινιδίωνps/νευροινιδίων
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταροσκελετός και Κυτταρική Διαίρεση Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταροσκελετός και Κυτταρική Διαίρεση Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος
ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΙΑΓΩΝΙΣΜΑ Θέµα 1 ο 1. Τα άτοµα που είναι ετερόζυγα για τη β-θαλασσαιµία: α. Εµφανίζουν ήπια αναιµία β. Έχουν ευαισθησία στην ελονοσία γ. Συνθέτουν µεγάλη ποσότητα HbF δ.
ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΙΑΓΩΝΙΣΜΑ Θέµα 1 ο 1. Τα άτοµα που είναι ετερόζυγα για τη β-θαλασσαιµία: α. Εµφανίζουν ήπια αναιµία β. Έχουν ευαισθησία στην ελονοσία γ. Συνθέτουν µεγάλη ποσότητα HbF δ.
Φάσμα& Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι.
 σύγχρονο Φάσμα& Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο 1. 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 50.27.990 50.20.990 2. 25ης Μαρτίου 74 Π. ΠΕΤΡΟΥΠΟΛΗΣ 50.50.658 50.60.845 3. Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ
σύγχρονο Φάσμα& Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο 1. 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 50.27.990 50.20.990 2. 25ης Μαρτίου 74 Π. ΠΕΤΡΟΥΠΟΛΗΣ 50.50.658 50.60.845 3. Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ
igenetics ΜΑΘΗΜΑ 3 Το γενετικό υλικό
 igenetics ΜΑΘΗΜΑ 3 Το γενετικό υλικό ΛΕΙΤΟΥΡΓΙΕΣ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΥΛΙΚΟΥ ΑΠΟΘΗΚΕΥΣΗ Στο DNA (RNA ιών) οι πληροφορίες για τα χαρακτηριστικά ενός οργανισμού (γονίδια) ΔΙΑΤΗΡΗΣΗ Από κύτταρο σε κύτταρο και από
igenetics ΜΑΘΗΜΑ 3 Το γενετικό υλικό ΛΕΙΤΟΥΡΓΙΕΣ ΤΟΥ ΓΕΝΕΤΙΚΟΥ ΥΛΙΚΟΥ ΑΠΟΘΗΚΕΥΣΗ Στο DNA (RNA ιών) οι πληροφορίες για τα χαρακτηριστικά ενός οργανισμού (γονίδια) ΔΙΑΤΗΡΗΣΗ Από κύτταρο σε κύτταρο και από
Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης. Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010
 ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΒΙΟΛΟΓΙΑ. 1 ο ΔΙΑΓΩΝΙΣΜΑ ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ 1 o
 ΘΕΜΑ 1 o Γ ΛΥΚΕΙΟΥ-ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ 1 ο ΔΙΑΓΩΝΙΣΜΑ Α. Γιατί τα βακτήρια μπορούν να χρησιμοποιηθούν σαν «εργοστάσια παραγωγής ανθρώπινων πρωτεϊνών»; Β. Σε ένα βακτήριο εισάγεται με τη μέθοδο του ανασυνδυασμένου
ΘΕΜΑ 1 o Γ ΛΥΚΕΙΟΥ-ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ 1 ο ΔΙΑΓΩΝΙΣΜΑ Α. Γιατί τα βακτήρια μπορούν να χρησιμοποιηθούν σαν «εργοστάσια παραγωγής ανθρώπινων πρωτεϊνών»; Β. Σε ένα βακτήριο εισάγεται με τη μέθοδο του ανασυνδυασμένου
5. Η μεταγραφή σ ένα ευκαρυωτικό κύτταρο γίνεται α. στα ριβοσώματα. β. στο κυτταρόπλασμα. γ. στον πυρήνα. δ. στο κεντρομερίδιο.
 ΑΡΧΗ 1ΗΣ ΣΕΛΙ ΑΣ ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 16 ΣΕΠΤΕΜΒΡΙΟΥ 2005 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ: ΒΙΟΛΟΓΙΑ ΣΥΝΟΛΟ ΣΕΛΙ
ΑΡΧΗ 1ΗΣ ΣΕΛΙ ΑΣ ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 16 ΣΕΠΤΕΜΒΡΙΟΥ 2005 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ: ΒΙΟΛΟΓΙΑ ΣΥΝΟΛΟ ΣΕΛΙ
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη φράση που συμπληρώνει
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη φράση που συμπληρώνει
Για το χρώµα σπέρµατος επικρατής είναι η ιδιότητα κίτρινο και η υπολειπόµενη το πράσινο. Συµβολίζουµε: Κ:Κίτρινο κ: Πράσινο Κ>κ
 Απαντήσεις Βιολογία Κατεύθυνσης 2011 ΘΕΜΑ Α Α1 α Α2 δ Α3 γ Α4 β Α5 β ΘΕΜΑ Β Β1 Σελ. 13 «Το 1928 γίνεται αυτό» Β2 Σελ 101 «βλάβες στους επιδιορθωτικά ένζυµα» (Χωρίς να απαιτείται θα θεωρηθεί σωστό να γίνει
Απαντήσεις Βιολογία Κατεύθυνσης 2011 ΘΕΜΑ Α Α1 α Α2 δ Α3 γ Α4 β Α5 β ΘΕΜΑ Β Β1 Σελ. 13 «Το 1928 γίνεται αυτό» Β2 Σελ 101 «βλάβες στους επιδιορθωτικά ένζυµα» (Χωρίς να απαιτείται θα θεωρηθεί σωστό να γίνει
Ποια κύτταρα έχουν κυτταροσκελετό; Όλα τα ευκαρυωτικά!
 Ποια κύτταρα έχουν κυτταροσκελετό; Όλα τα ευκαρυωτικά! Αρχικά πιστευόταν ότι τα µικροϊνίδια είναι χαρακτηριστικό µόνο των γραµµωτών µυών. Το ΗΜ υπερ-υψηλής τάσης συνέβαλε (άθελά του) στην ανακάλυψη του
Ποια κύτταρα έχουν κυτταροσκελετό; Όλα τα ευκαρυωτικά! Αρχικά πιστευόταν ότι τα µικροϊνίδια είναι χαρακτηριστικό µόνο των γραµµωτών µυών. Το ΗΜ υπερ-υψηλής τάσης συνέβαλε (άθελά του) στην ανακάλυψη του
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΑΠΑΝΤΗΣΕΙΣ
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. α Α. β Α3. γ Α4. γ Α5. β ΘΕΜΑ Β Β. ζ στ 3 α 4 ε 5 β 6 δ Β. Κάθε πολυνουκλεοτιδική αλυσίδα αποτελείται από νουκλεοτίδια ενωμένα μεταξύ τους με ομοιοπολικό δεσμό.
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. α Α. β Α3. γ Α4. γ Α5. β ΘΕΜΑ Β Β. ζ στ 3 α 4 ε 5 β 6 δ Β. Κάθε πολυνουκλεοτιδική αλυσίδα αποτελείται από νουκλεοτίδια ενωμένα μεταξύ τους με ομοιοπολικό δεσμό.
Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού
 Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού Θέμα Α Α1) γ Α2) γ Α3) δ Α4) β Α5) β Θέμα Β Β1. Α = υδροξύλιο, Β = πρωταρχικό τμήμα, Γ = θέση έναρξης αντιγραφής, Δ = φωσφορική ομάδα, Ε = τμήμα
Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού Θέμα Α Α1) γ Α2) γ Α3) δ Α4) β Α5) β Θέμα Β Β1. Α = υδροξύλιο, Β = πρωταρχικό τμήμα, Γ = θέση έναρξης αντιγραφής, Δ = φωσφορική ομάδα, Ε = τμήμα
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΓΕΝΙΚΟ ΛΥΚΕΙΟ
 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΓΕΝΙΚΟ ΛΥΚΕΙΟ 19-6-2019 ΘΕΜΑ Α Α1. - α Α2. β Α3. γ Α4. γ Α5. β ΘΕΜΑ Β Β1. α1 αντιθρυψίνη-εμφύσημα Πρωτεΐνη επιθηλικών κυττάρων κυστική ίνωση Απαμινάση της αδενοσίνης Ανοσολογική
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΓΕΝΙΚΟ ΛΥΚΕΙΟ 19-6-2019 ΘΕΜΑ Α Α1. - α Α2. β Α3. γ Α4. γ Α5. β ΘΕΜΑ Β Β1. α1 αντιθρυψίνη-εμφύσημα Πρωτεΐνη επιθηλικών κυττάρων κυστική ίνωση Απαμινάση της αδενοσίνης Ανοσολογική
ΤΑΞΗ: Α ΛΥΚΕΙΟΥ ΗΜΕΡ. : 19/05/2016 ΜΑΘΗΜΑ: ΧΗΜΕΙΑ-ΒΙΟΛΟΓΙΑ ΣΥΝΟΛΙΚΟΣ ΧΡΟΝΟΣ: 2 ΩΡΕΣ (120 ΛΕΠΤΑ) ΟΝΟΜΑΤΕΠΩΝΥΜΟ:... ΤΜΗΜΑ:.. ΑΡ.: ΠΡΟΣΟΧΗ ΚΑΘΕ ΕΠΙΤΥΧΙΑ
 ΟΝΟΜΑΤΕΠΩΝΥΜΟ:... ΤΜΗΜΑ:.. ΑΡ.: ΠΡΟΣΟΧΗ ΚΑΘΕ ΕΠΙΤΥΧΙΑ") ΛΥΚΕΙΟ ΑΚΡΟΠΟΛΕΩΣ ΣΧΟΛΙΚΗ ΧΡΟΝΙΑ 2015-2016 ΓΡΑΠΤΕΣ ΠΡΟΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΙΟΥΝΙΟΥ 2016 ΒΑΘΜΟΣ.../35 ΟΛΟΓΡΑΦΩΣ ΥΠΟΓΡΑΦΗ:. ΤΑΞΗ: Α ΛΥΚΕΙΟΥ ΗΜΕΡ. : 19/05/2016 ΜΑΘΗΜΑ: ΧΗΜΕΙΑ-ΒΙΟΛΟΓΙΑ ΣΥΝΟΛΙΚΟΣ ΧΡΟΝΟΣ: 2 ΩΡΕΣ
ΛΥΚΕΙΟ ΑΚΡΟΠΟΛΕΩΣ ΣΧΟΛΙΚΗ ΧΡΟΝΙΑ 2015-2016 ΓΡΑΠΤΕΣ ΠΡΟΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΙΟΥΝΙΟΥ 2016 ΒΑΘΜΟΣ.../35 ΟΛΟΓΡΑΦΩΣ ΥΠΟΓΡΑΦΗ:. ΤΑΞΗ: Α ΛΥΚΕΙΟΥ ΗΜΕΡ. : 19/05/2016 ΜΑΘΗΜΑ: ΧΗΜΕΙΑ-ΒΙΟΛΟΓΙΑ ΣΥΝΟΛΙΚΟΣ ΧΡΟΝΟΣ: 2 ΩΡΕΣ
ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2015 Β ΦΑΣΗ ΑΠΑΝΤΗΣΕΙΣ ÏÅÖÅ
 ΤΑΞΗ: ΜΑΘΗΜΑ: 3 η ΤΑΞΗ ΕΠΑ.Λ. (Β ΟΜΑ Α) ΒΙΟΛΟΓΙΑ ΙΙ Ηµεροµηνία: Παρασκευή 19 Απριλίου 2015 ιάρκεια Εξέτασης: 3 ώρες ΘΕΜΑ Α Α1-β, Α2-γ, Α3-γ, Α4-α, Α5-α ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Β Β1. α. Με τη µέθοδο της επιλογής
ΤΑΞΗ: ΜΑΘΗΜΑ: 3 η ΤΑΞΗ ΕΠΑ.Λ. (Β ΟΜΑ Α) ΒΙΟΛΟΓΙΑ ΙΙ Ηµεροµηνία: Παρασκευή 19 Απριλίου 2015 ιάρκεια Εξέτασης: 3 ώρες ΘΕΜΑ Α Α1-β, Α2-γ, Α3-γ, Α4-α, Α5-α ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Β Β1. α. Με τη µέθοδο της επιλογής
Τρίτη, 27 Μαΐου 2008 Γ ΛΥΚΕΙΟΥ ΚΑΤΕΥΘΥΝΣΗΣ ΒΙΟΛΟΓΙΑ
 Τρίτη, 27 Μαΐου 2008 Γ ΛΥΚΕΙΟΥ ΚΑΤΕΥΘΥΝΣΗΣ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1o Να γράψετε στο τετράδιο σας τον αριθµό καθεµιάς από τις παρακάτω ηµιτελείς προτάσεις 1 έως 5 και δίπλα το γράµµα που αντιστοιχεί στη λέξη ή τη
Τρίτη, 27 Μαΐου 2008 Γ ΛΥΚΕΙΟΥ ΚΑΤΕΥΘΥΝΣΗΣ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1o Να γράψετε στο τετράδιο σας τον αριθµό καθεµιάς από τις παρακάτω ηµιτελείς προτάσεις 1 έως 5 και δίπλα το γράµµα που αντιστοιχεί στη λέξη ή τη
3. Σχ. Βιβλίο σελ «το βακτήριο Αgrobacterium.ξένο γονίδιο» Και σελ 133 «το βακτήριο Bacillus.Βt».
 2 ο Διαγώνισμα Βιολογίας Γ Λυκείου Θέμα Α 1. Α 2. Β 3. Β 4. Α 5. C Θέμα Β 1. Σελ 40 «τα ριβοσώματα μπορούν..πρωτεινών» Και σελ 39 «ο γενετικός κώδικας είναι σχεδόν καθολικός πρωτείνη». 2. Σελ 98 «η φαινυλκετονουρία.φαινυλαλανίνης»
2 ο Διαγώνισμα Βιολογίας Γ Λυκείου Θέμα Α 1. Α 2. Β 3. Β 4. Α 5. C Θέμα Β 1. Σελ 40 «τα ριβοσώματα μπορούν..πρωτεινών» Και σελ 39 «ο γενετικός κώδικας είναι σχεδόν καθολικός πρωτείνη». 2. Σελ 98 «η φαινυλκετονουρία.φαινυλαλανίνης»
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ
 ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
Η ζητούμενη σειρά έχει ως εξής: αδενίνη < νουκλεοτίδιο < νουκλεόσωμα < γονίδιο < χρωματίδα < χρωμόσωμα < γονιδίωμα.
 ΚΕΦ. 1 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ 1. Να κατατάξετε σε σειρά αυξανόμενου μεγέθους τις παρακάτω έννοιες που σχετίζονται με το γενετικό υλικό των οργανισμών: νουκλεόσωμα, χρωμόσωμα, αδενίνη, νουκλεοτίδιο, γονίδιο
ΚΕΦ. 1 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ 1. Να κατατάξετε σε σειρά αυξανόμενου μεγέθους τις παρακάτω έννοιες που σχετίζονται με το γενετικό υλικό των οργανισμών: νουκλεόσωμα, χρωμόσωμα, αδενίνη, νουκλεοτίδιο, γονίδιο
Γονιδιωματική. G. Patrinos
 Γονιδιωματική Η μεταγονιδιωματική εποχή... Σημαντικότερα επιτεύγματα POST GENOME ERA Ολοκλήρωση της αποκρυπτογράφησης της αλληλουχίας των γονιδιωμάτων πολλών οργανισμών. Προτύπωση μεθοδολογιών για προσδιορισμό
Γονιδιωματική Η μεταγονιδιωματική εποχή... Σημαντικότερα επιτεύγματα POST GENOME ERA Ολοκλήρωση της αποκρυπτογράφησης της αλληλουχίας των γονιδιωμάτων πολλών οργανισμών. Προτύπωση μεθοδολογιών για προσδιορισμό
Λειτουργική Περιοχή της GTP-ασης
 Λειτουργική Περιοχή της GTP-ασης Οι πρωτεΐνες πού φαίνεται να εµπλέκονται στην περιοχή είναι οι πρωτεΐνες L7/L12. Οι πρωτεΐνες αυτές φαίνεται να είναι απαραίτητες για την ενεργότητα του ριβοσώµατος και
Λειτουργική Περιοχή της GTP-ασης Οι πρωτεΐνες πού φαίνεται να εµπλέκονται στην περιοχή είναι οι πρωτεΐνες L7/L12. Οι πρωτεΐνες αυτές φαίνεται να είναι απαραίτητες για την ενεργότητα του ριβοσώµατος και
Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος
 MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞΕΙΣ 10-12 Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής
MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞΕΙΣ 10-12 Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής
Β. Σελ 60 σχολικού: «Η αποµόνωση του συνολικού έως και σελ 61 από µία cdna βιβλιοθήκη.». Γ. ι ι α α α ι α α ι α α α! " # $ % & ' ( ) ( ) ( * % + α ι α
 ( ) ( * % + α ι α") ! THΛ: 270727 222594 THΛ: 919113 949422 Απαντήσεις: " # $ % & ' 1=γ, 2=β, 3=γ, 4=β, 5=δ. " # $ % ( ' εδοµένα από την ανάλυση του ποσοστού των βάσεων σε µόρια DNA από διαφορετικούς οργανισµούς έδειχναν
! THΛ: 270727 222594 THΛ: 919113 949422 Απαντήσεις: " # $ % & ' 1=γ, 2=β, 3=γ, 4=β, 5=δ. " # $ % ( ' εδοµένα από την ανάλυση του ποσοστού των βάσεων σε µόρια DNA από διαφορετικούς οργανισµούς έδειχναν
ΚΕΦΑΛΑΙΟ 1 ο... 2 I. Το γενετικό υλικό... 2 ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ... 5 ΝΑ ΣΥΜΠΛΗΡΩΣΕΤΕ ΤΑ ΚΕΝΑ ΜΕ ΤΗΝ ΚΑΤΑΛΛΗΛΗ ΛΕΞΗ... 8 ΠΡΟΒΛΗΜΑΤΑ...
 ΚΕΦΑΛΑΙΟ 1 ο ΚΕΦΑΛΑΙΟ 1 ο... 2 I. Το γενετικό υλικό... 2 ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ... 5 ΝΑ ΣΥΜΠΛΗΡΩΣΕΤΕ ΤΑ ΚΕΝΑ ΜΕ ΤΗΝ ΚΑΤΑΛΛΗΛΗ ΛΕΞΗ.... 8 ΠΡΟΒΛΗΜΑΤΑ... 9 ΙΑΓΩΝΙΣΜΑ...12 ΚΕΦΑΛΑΙΟ 1 ο I. Το γενετικό
ΚΕΦΑΛΑΙΟ 1 ο ΚΕΦΑΛΑΙΟ 1 ο... 2 I. Το γενετικό υλικό... 2 ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ... 5 ΝΑ ΣΥΜΠΛΗΡΩΣΕΤΕ ΤΑ ΚΕΝΑ ΜΕ ΤΗΝ ΚΑΤΑΛΛΗΛΗ ΛΕΞΗ.... 8 ΠΡΟΒΛΗΜΑΤΑ... 9 ΙΑΓΩΝΙΣΜΑ...12 ΚΕΦΑΛΑΙΟ 1 ο I. Το γενετικό
Β1. Β2. ΘΕΜΑ 2ο 1. 2.
 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 20 ΣΕΠΤΕΜΒΡΙΟΥ 2002 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ) ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο Α1.
1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 20 ΣΕΠΤΕΜΒΡΙΟΥ 2002 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ) ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο Α1.
Κυτταρική Βιολογία. Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
Kυτταρική Bιολογία. Μείωση & φυλετική αναπαραγωγή ΔIAΛEΞH 21 (16/5/2016) Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.
 Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.") Kυτταρική Bιολογία ΔIAΛEΞH 21 (16/5/2016) Μείωση & φυλετική αναπαραγωγή Τι είναι φυλετική και τι αφυλετική αναπαραγωγή; Η αφυλετική αναπαραγωγή: 1. Είναι απλή και άμεση 2. Συνήθως αποδίδει απογόνους που
Kυτταρική Bιολογία ΔIAΛEΞH 21 (16/5/2016) Μείωση & φυλετική αναπαραγωγή Τι είναι φυλετική και τι αφυλετική αναπαραγωγή; Η αφυλετική αναπαραγωγή: 1. Είναι απλή και άμεση 2. Συνήθως αποδίδει απογόνους που
ΦΡΟΝΤΙΣΤΗΡΙΟ «ΕΝΑ» ΓΙΟΜΠΛΙΑΚΗΣ ΛΑΖΑΡΟΣ ΠΕΤΡΟΜΕΛΙΔΗΣ ΒΑΣΙΛΗΣ
 ΦΡΟΝΤΙΣΤΗΡΙΟ «ΕΝΑ» ΓΙΟΜΠΛΙΑΚΗΣ ΛΑΖΑΡΟΣ ΠΕΤΡΟΜΕΛΙΔΗΣ ΒΑΣΙΛΗΣ ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 ο Α. Να βάλετε σε κύκλο το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Ο µέγιστος αριθµός διαφορετικών
ΦΡΟΝΤΙΣΤΗΡΙΟ «ΕΝΑ» ΓΙΟΜΠΛΙΑΚΗΣ ΛΑΖΑΡΟΣ ΠΕΤΡΟΜΕΛΙΔΗΣ ΒΑΣΙΛΗΣ ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 ο Α. Να βάλετε σε κύκλο το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Ο µέγιστος αριθµός διαφορετικών
Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της
 ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο. 3. Το DNA των μιτοχονδρίων έχει μεγαλύτερο μήκος από αυτό των χλωροπλαστών.
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014. Απαντήσεις Θεμάτων
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2018 ΘΕΜΑ Α Α1: δ Α2: β Α3: α Α4: α Α5: β ΘΕΜΑ Β Β1. 1 γ, 2 β, 3 γ, 4 α, 5 γ, 6 γ, 7 - β B2. Η καμπύλη που αντιστοιχεί στον Lactobacillus είναι
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2018 ΘΕΜΑ Α Α1: δ Α2: β Α3: α Α4: α Α5: β ΘΕΜΑ Β Β1. 1 γ, 2 β, 3 γ, 4 α, 5 γ, 6 γ, 7 - β B2. Η καμπύλη που αντιστοιχεί στον Lactobacillus είναι
1 ο #Κεφάλαιο# 1)#Πειράματα: α)$να#περιγράψεις#το#πείραμα#των#hershey#και#chase.# Υπόδειξη:#σελ#14#σχολ.
#Πειράματα: α)$να#περιγράψεις#το#πείραμα#των#hershey#και#chase.# Υπόδειξη:#σελ#14#σχολ.") 1 ο #Κεφάλαιο# ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ 1)#Πειράματα: α)$να#περιγράψεις#το#πείραμα#των#hershey#και#chase.# Υπόδειξη:#σελ#14#σχολ. Παραλλαγή:#δίνονται##τα#παρακάτω#διαγράμματα#που#απεικονίζουν# τη#ραδιενέργεια#στο#εσωτερικό#των#βακτηρίων,#μετά#τη#μόλυνση#με#
1 ο #Κεφάλαιο# ΤΟ ΓΕΝΕΤΙΚΟ ΥΛΙΚΟ 1)#Πειράματα: α)$να#περιγράψεις#το#πείραμα#των#hershey#και#chase.# Υπόδειξη:#σελ#14#σχολ. Παραλλαγή:#δίνονται##τα#παρακάτω#διαγράμματα#που#απεικονίζουν# τη#ραδιενέργεια#στο#εσωτερικό#των#βακτηρίων,#μετά#τη#μόλυνση#με#
Κυτταρογενετική Ι Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Εργαστήριο Βιολογίας
 Κυτταρογενετική Ι 22.03.2019 Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Εργαστήριο Βιολογίας Κυτταρογενετική Η μελέτη της δομής των χρωμοσωμάτων και της κληρονόμησης τους Αντικείμενο: Χρωμοσωμικές ανωμαλίες- δομή
Κυτταρογενετική Ι 22.03.2019 Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Εργαστήριο Βιολογίας Κυτταρογενετική Η μελέτη της δομής των χρωμοσωμάτων και της κληρονόμησης τους Αντικείμενο: Χρωμοσωμικές ανωμαλίες- δομή
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014. Απαντήσεις Θεμάτων
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 19/ 06 / 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: Βιολογία OΠ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 19/ 06 / 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: Βιολογία OΠ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ ΘΕΜΑ Α Α1. Δ Α2. Β Α3. Α Α4. Α Α5. Β ΘΕΜΑ Β
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 19/ 06 / 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: Βιολογία OΠ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ ΘΕΜΑ Α Α1. Δ Α2. Β Α3. Α Α4. Α Α5. Β ΘΕΜΑ Β
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 2004
 ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 2004 ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθµό καθεµιάς από τις παρακάτω ηµιτελείς προτάσεις 1 έως 5 και δίπλα το γράµµα που αντιστοιχεί στη φράση
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 2004 ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθµό καθεµιάς από τις παρακάτω ηµιτελείς προτάσεις 1 έως 5 και δίπλα το γράµµα που αντιστοιχεί στη φράση
Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ÏÅÖÅ
 1 Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 ο Α. 1 - Γ 2 - Β 3-4 - Γ 5 - Β. 1 - Σ 2 - Λ 3 - Λ 4 - Λ 5 - Σ ΘΕΜΑ 2 ο ΑΠΑΝΤΗΣΕΙΣ 1. Κάθε είδος αντισώµατος που αναγνωρίζει έναν αντιγονικό καθοριστή παράγεται
1 Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 ο Α. 1 - Γ 2 - Β 3-4 - Γ 5 - Β. 1 - Σ 2 - Λ 3 - Λ 4 - Λ 5 - Σ ΘΕΜΑ 2 ο ΑΠΑΝΤΗΣΕΙΣ 1. Κάθε είδος αντισώµατος που αναγνωρίζει έναν αντιγονικό καθοριστή παράγεται
Η µελέτη της ρύθµισης της πρωτεινοσύνθεσης στο επίπεδο του Ριβοσώµατος εντοπίζεται σε τρία επίπεδα
 Η µελέτη της ρύθµισης της πρωτεινοσύνθεσης στο επίπεδο του Ριβοσώµατος εντοπίζεται σε τρία επίπεδα ΣτονΣτον ρόλο των διαφόρων οµάδων των ριβοσωµικών πρωτεινών. Κατά πόσο δηλαδή υπάρχει ετερογένεια στις
Η µελέτη της ρύθµισης της πρωτεινοσύνθεσης στο επίπεδο του Ριβοσώµατος εντοπίζεται σε τρία επίπεδα ΣτονΣτον ρόλο των διαφόρων οµάδων των ριβοσωµικών πρωτεινών. Κατά πόσο δηλαδή υπάρχει ετερογένεια στις
Θέματα Πανελλαδικών 2000-2013
 Θέματα Πανελλαδικών 2000-2013 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΗΜΕΡΗΣΙΩΝ ΛΥΚΕΙΩΝ ΕΣΠΕΡΙΝΩΝ ΛΥΚΕΙΩΝ ΕΠΑΝΑΛΗΠΤΙΚΕΣ Κεφάλαιο 1 ΚΕΦΑΛΑΙΟ 1 ΘΕΜΑ 1 ο Γράψτε τον αριθμό καθεμιάς από τις παρακάτω προτάσεις και δίπλα το γράμμα
Θέματα Πανελλαδικών 2000-2013 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΗΜΕΡΗΣΙΩΝ ΛΥΚΕΙΩΝ ΕΣΠΕΡΙΝΩΝ ΛΥΚΕΙΩΝ ΕΠΑΝΑΛΗΠΤΙΚΕΣ Κεφάλαιο 1 ΚΕΦΑΛΑΙΟ 1 ΘΕΜΑ 1 ο Γράψτε τον αριθμό καθεμιάς από τις παρακάτω προτάσεις και δίπλα το γράμμα
Βιολογία Γ Γενικού Λυκείου Θετικής κατεύθυνσης. Κεφάλαιο 1α Το Γενετικό Υλικό
 Βιολογία Γ Γενικού Λυκείου Θετικής κατεύθυνσης Κεφάλαιο 1α Το Γενετικό Υλικό Το DNA είναι το γενετικό υλικό Αρχικά οι επιστήμονες θεωρούσαν ότι οι πρωτεΐνες αποτελούσαν το γενετικό υλικό των οργανισμών.
Βιολογία Γ Γενικού Λυκείου Θετικής κατεύθυνσης Κεφάλαιο 1α Το Γενετικό Υλικό Το DNA είναι το γενετικό υλικό Αρχικά οι επιστήμονες θεωρούσαν ότι οι πρωτεΐνες αποτελούσαν το γενετικό υλικό των οργανισμών.
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ 1ο. 1. γ 2. γ 3. δ 4. α 5. β
 ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΠΑΝΕΛΛΑ ΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΕΠΑΛ (ΟΜΑ Α Β ) ΠΑΡΑΣΚΕΥΗ 22 ΜΑΪΟΥ 2009 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1ο 1. γ 2. γ 3.
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΠΑΝΕΛΛΑ ΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΕΠΑΛ (ΟΜΑ Α Β ) ΠΑΡΑΣΚΕΥΗ 22 ΜΑΪΟΥ 2009 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1ο 1. γ 2. γ 3.
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI