Ταξινόμηση Πρωτεϊνικών Δομών με Χρήση Τοπολογικής, Γεωμετρικής και Βιολογικής Πληροφορίας
|
|
|
- Παραμονος Ἐφραίμ Φλέσσας
- 8 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ & ΜΗΧΑΝΙΚΩΝ ΥΠΟΛΟΓΙΣΤΩΝ ΤΟΜΕΑΣ ΗΛΕΚΤΡΟΝΙΚΗΣ & ΥΠΟΛΟΓΙΣΤΩΝ Ταξινόμηση Πρωτεϊνικών Δομών με Χρήση Τοπολογικής, Γεωμετρικής και Βιολογικής Πληροφορίας Διπλωματική εργασία του: Βασιλείου Τσατσάια Α.Ε.Μ.: 4859 Επιβλέπων: Καθ. Μιχαήλ Γ. Στρίντζης Θεσσαλονίκη 2007
2 Ευχαριστίες Ευχαριστώ πολύ τον επιβλέποντα καθηγητή κ. Μ. Γ. Στρίντζη για την εμπιστοσύνη που μου έδειξε στην εκπόνηση της παρούσης διπλωματικής εργασίας, τον ερευνητή Δ του Ινστιτούτου Πληροφορικής και Τηλεματικής κ. Πέτρο Δάρα για την στενή συνεργασία και καθοδήγησή του, επιπλέον ευχαριστώ τον υποψήφιο διδάκτορα κ. Αθανάσιο Μαδεμλή για την σημαντική βοήθεια που μου προσέφερε. Τέλος, ευχαριστώ την οικογένειά μου και τους φίλους μου για την ηθική υποστήριξη κατά την διάρκεια της εκπόνησης της εργασίας αυτής.
3 . ΕΙΣΑΓΩΓΗ ΠΑΡΟΥΣΙΑΣΗ ΤΩΝ ΣΥΓΧΡΟΝΩΝ ΜΕΘΟΔΩΝ Σύγκριση Πρωτεϊνών Βασισμένη στις Δευτερεύουσες Δομές (Δ.Δ.) Σύγκριση Πρωτεϊνικών Δομών με Γράφους απαρτισμένους από Χημικές Ομάδες Μέθοδος σύγκρισης Βιολογικών Δεδομένων με Χρήση Διμερών Γράφων Αναζήτηση ενεργών Περιοχών στην ομάδα Πρωτεϊνών G Εκτίμηση της λειτουργίας πρωτεϊνικών μορίων ταυτίζοντας 3D δομικά στοιχεία Κατηγοριοποιήση Πρωτεϊνών κάνοντας χρήση τον Μετασχηματισμό Ίχνους Άλλες Μέθοδοι Συμπεράσματα ΠΡΟΕΠΕΞΕΡΓΑΣΙΑ-ΚΑΤΑΣΚΕΥΗ ΓΡΑΦΩΝ Ανάγνωση αρχείων PDB(Protein Data Bank)[4] Δημιουργία των 3D μοντέλων-διαμέριση (segmentation) Εξαγωγή Γεωμετρικών περιγραφέων Κατασκευή Γράφων ΣΥΓΚΡΙΣΗ ΓΡΑΦΩΝ (GRAPH MATCHING) ΕΥΡΕΣΗ ΤΕΛΙΚΟΥ ΒΑΘΜΟΥ ΟΜΟΙΟΤΗΤΑΣ ΜΕΤΑΞΥ ΠΡΩΤΕΪΝΩΝ Σύγκριση Γράφων(Graph Matching) Το πρόβλημα της Σύγκρισης Γράφων Μέθοδος Σύγκρισης Γράφων Απαραίτητη θεωρητική προσέγγιση Ο Αλγόριθμος Εξαγωγή Τελικού Βαθμού Ομοιότητας ΠΕΙΡΑΜΑΤΙΚΑ ΑΠΟΤΕΛΕΣΜΑΤΑ - ΣΥΜΠΕΡΑΣΜΑΤΑ...53 ΒΙΒΛΙΟΓΡΑΦΙΑ...64
4 . Εισαγωγή Η κατανόηση των μοριακών μηχανισμών της ζωής απαιτεί την αποκωδικοποίηση των λειτουργιών που εκτελούν οι πρωτεΐνες σε έναν οργανισμό. Η αλληλουχία δεκάδων χιλιάδων πρωτεϊνών έχει μελετηθεί τα τελευταία χρόνια καθώς επίσης και η δομή χιλιάδων από αυτές. Παρ όλα αυτά οι πειραματικές μέθοδοι για την εύρεση της ακριβούς λειτουργίας πρωτεϊνών με γνωστή αλληλουχία και δομή είναι ακόμα ιδιαιτέρως δύσκολες και απαιτητικές σε χρόνο και οικονομικό κόστος. Για το λόγω αυτό επιχειρείται, με υπολογιστικές μεθόδους, να μειωθεί το κόστος της ανάλυσης πρωτεϊνικών δεδομένων. Η κατανόηση της δομής των πρωτεϊνών είναι θεμελιώδης και για το λόγω αυτό παραθέτουμε κάποια στοιχεία της δομής αυτής. Ένα μόριο πρωτεΐνης αποτελείται από μία μακριά αλυσίδα αμινοξέων, στην οποία κάθε αμινοξύ συνδέεται με το επόμενο με έναν ομοιοπολικό πεπτιδικό δεσμό[]. Η επαναληπτική αλληλουχία ατόμων κατα μήκος της αλυσίδας αποκαλείται πολυπεπτιδικός σκελετός (polypeptide backbone). Στην επαναληπτική αυτή αλυσίδα συνδέονται οι πλευρικές αλυσίδες (side chain) των διάφορων αμινοξέων: πρόκειται για τις ομάδες των μορίων των αμινοξέων που δε συμμετέχουν στο σχηματισμό του πεπτιδικού δεσμού και που προσδίδουν στο κάθε αμινοξύ τις μοναδικές του ιδιότητες. Από τις πλευρικές αυτές αλυσίδες άλλες δεν έχουν φορτίο και είναι υδρόφοβες, άλλες έχουν αρνητικό άλλες έχουν θετικό φορτίο, άλλες είναι δραστικές, άλλες μη δραστικές κ.λ.π. Οι ιδιότητες αυτές των πλευρικών αλυσίδων χαρακτηρίζουν τη μοναδικότητα του κάθε αμινοξέος και του δίνουν τη δυνατότητα να αλληλεπιδράσει με άλλα αμινοξέα και άλλα μόρια. Τα αμινοξέα αυτά που είναι μέλη μιας πολυπεπτιδικής αλυσίδας και έχουν δώσει κάποιο κομμάτι τους για τη δημιουργία των δεσμών της αλυσίδας ονομάζονται και αμινοξεϊκά κατάλοιπα ή υπόλοιπα (aminoacid residues). Η αλληλουχία αυτών των κατάλοιπων είναι που κάνει την κάθε πρωτεΐνη μοναδική. Η μακριά πρωτεϊνική αλυσίδα αναδιπλώνεται στο χώρο λαμβάνοντας μία συγκεκριμένη τρισδιάστατη μορφή, η οποία σταθεροποιείται χάρη σε
5 ασθενείς μη ομοιοπολικούς δεσμούς που δημιουργούνται τόσο από άτομα του πολυπεπτιδικού σκελετού, όσο και από άτομα των πλευρικών αλυσίδων των αμινοξέων. Οι ασθενείς αυτοί δεσμοί είναι δεσμοί υδρογόνου, ιοντικοί δεσμοί και δεσμοί van der Waals. Οι δεσμοί αυτοί μόνοι τους είναι αρκετά ασθενείς σε σχέση με τους ομοιοπολικούς δεσμούς του μορίου έτσι, είναι απαραίτητοι πολλοί μη ομοιοπολικοί δεσμοί, ώστε να συγκρατηθούν δύο περιοχές της διπλωμένης πολυπεπτιδικής αλυσίδας. Επομένως, η σταθερότητα κάθε διπλωμένης δομής επηρεάζεται από τη συνολική ισχύ πολλών τέτοιων δεσμών. Ένα τέταρτο είδος ασθενούς δύναμης παίζει σημαντικό ρόλο στον καθορισμό του σχήματος μιας πρωτεΐνης. Για την κατανόηση της συγκεκριμένης αλληλεπίδρασης πρέπει να αναφέρουμε ότι οι πλευρικές αλυσίδες των αμινοξέων κατατάσσονται σε δύο μεγάλες ομάδες τις Υδρόφοβες (μη-πολικές), που δεν διαθέτουν στο σύνολο τους κάποιο ηλεκτρικό φορίο ή διαθέτουν πολύ μικρό και χάνουν τη δομή τους, όταν βρεθούν σε επαφή με μόρια νερού, καθώς επίσης και τις Υδρόφιλες (πολικές) ομάδες τα μέλη των οποίων διαθέτουν κάποιο σημαντικό θετικό ή αρνητικό φορτίο και δεν επηρεάζονται από την παρουσία μορίων νερού στο περιβάλλον τους. Σύμφωνα με τα παραπάνω χαρακτηριστικά των πλευρικών ομάδων έχουμε την εξής διαδικασία: τα υδρόφοβα μόρια μεταξύ τους και οι μη πολικές πλευρικές αλυσίδες συγκεκριμένων αμινοξέων, όταν βρεθούν σε υδατικό περιβάλλον, συνωθούνται μαζί έτσι ώστε να ελαχιστοποιηθεί η διασπαστική δράση τους στο δίκτυο των υδρογονοδεσμών του νερού. Επομένως, ένας σημαντικός παράγοντας που κατευθύνει το δίπλωμα ή πτύχωση (folding) κάθε πρωτεΐνης είναι η κατανομή των πολικών και μη πολικών αμινοξέων της. Ο μηχανισμός είναι ο εξής: Οι μη πολικές (υδρόφοβες) πλευρικές αλυσίδες μιας πρωτεΐνης τείνουν να συναθροίζονται στο εσωτερικό του μορίου αποφεύγοντας έτσι την επαφή με το νερό, αντίθετα οι πολικές (υδρόφιλες) πλευρικές αλυσίδες τείνουν να διατάσσονται κοντά στην εξωτερική επιφάνεια του μορίου, όπου μπορεί να σχηματίσουν δεσμούς υδρογόνου με το νερό ή με άλλα πολικά μόρια. Όταν πολικά αμινοξέα βρίσκονται κρυμμένα στο εσωτερικό του μορίου συνήθως 2


6 συνδέονται με δεσμούς υδρογόνου με άλλα πολικά αμινοξέα ή με τον πολυπεπτιδικό σκελετό. Εικόνα.: α-έλικα Εικόνα.2: β-πτυχωτό φύλλο 3
7 Κατά συνέπεια, ένα πρωτεϊνικό μόριο διατάσσεται στο χώρο ακολουθώντας του νόμους των παραπάνω δεσμών δημιουργώντας μια σχετικά χαοτική δομή, μέσα στη δομή αυτή όμως παρατηρούμε και κάποια επναλαμβανόμενα μοτίβα. Τα πιο συχνά μοτίβα είναι η α-έλικα (Εικόνα.) και το β-πτυχωτό φύλλο (εικόνα.2). Οι δομές αυτές δημιουργούνται λόγω αλληλεπιδράσεων των ατόμων κυρίως του πεπτιδικού σκελετού και μας δίνουν ιδιαίτερα σταθερές δομές που συνάδουν και με το φαινόμενο της υδροφοβίας και υδροφιλίας. Μια α-έλικα σχηματίζεται όταν η ίδια πολυπεπτιδική αλυσίδα περιστρέφεται γύρω από τον εαυτό της για να σχηματίσει έναν άκαμπτο κύλινδρο. Η δομή αυτή σταθεροποιείται χάρη στη δημιουργία σύνδεσης μεταξύ ενός καρβονυλίου (C = O) ενός πεπτιδικού δεσμού με την αμινομάδα (Ν-Η) ενός άλλου πεπτιδικού δεσμού. Ένα β-πτυχωτό φύλλο μπορεί να σχηματιστεί από παρακείμενες πολυπεπτιδικές αλυσίδες που έχουν τον ίδιο προσανατολισμό, είτε από μία αλυσίδα που κάμπτεται αλλάζοντας κατεύθυνση, με συνέπεια το κάθε τμήμα της να έχει αντίθετο προσανατολισμό από τα αμέσως γειτονικά. Οι μηχανισμοί που συντηρούν τη δομή σταθερή είναι πανομοιότυποι με αυτούς της α-έλικας. Οι βιολογικές ιδιότητες μιας πρωτεΐνης εξαρτώνται από την ικανότητά τους να αλληλεπιδρούν με άλλα μόρια. Για παράδειγμα, τα αντισώματα συνδέονται με τους ιούς ή τα βακτήρια και με τον τρόπο αυτό ενεργοποιούν τους αμυντικούς μηχανισμούς του οργανισμού. Όλες οι πρωτεΐνες συνδέονται με άλλα μόρια. Σε κάθε περίπτωση η σύνδεση αυτή παρουσιάζει μεγάλη εξειδίκευση. Αυτό σημαίνει ότι κάθε πρωτεΐνη μπορεί να αλληλεπιδράσει μόνο με ένα ή έστω λίγα απο τα χιλιάδες μόρια που έρχεται σε επαφή. Η ουσία που συνδέεται με την πρωτεΐνη μπορεί να είναι είτε ένα ιόν, είτε ένα μικρό μόριο, είτε κάποιο μακρομόριο (πρωτεΐνη) και αποκαλείται συνδέτης ή προσδέτης (ligand) για τη συγκεκριμένη πρωτεΐνη. Η ικανότητα μιας πρωτεΐνης να συνδέεται εκλεκτικά και με μεγάλη συγγένεια με ένα συνδέτη οφείλεται στο σχηματισμό ενός συνόλου ασθενών, μη ομοιοπολικών δεσμών (δεσμοί υδρογόνου, ιοντικοί δεσμοί, έλξεις van der Waals) και ευνοϊκών υδρόφοβων αλληλεπιδράσεων. Από 4
8 μόνος του κάθε ένας από τους παραπάνω δεσμούς είναι ασθενής. Έτσι, προϋπόθεση για να αναπτυχθεί μία αποτελεσματική αλληλεπίδραση είναι να σχηματιστούν ταυτόχρονα πολλοί ασθενείς δεσμοί. Αυτό μπορεί να επιτευχθεί μόνο αν το επιφανειακό περίγραμμα του μορίου του συνδέτη ταιριάζει πολύ καλά με την πρωτεΐνη, όπως το «χέρι μέσα σε ένα γάντι». Η περιοχή της πρωτεΐνης που αλληλεπιδρά με το συνδέτη αποκαλείται θέση σύνδεσης (binding site). Η θέση σύνδεσης συνήθως αποτελείται από μία κοιλότητα στην επιφάνεια της πρωτεΐνης που σχηματίζεται από μία συγκεκριμένη διάταξη αμινοξέων. Τα αμινοξέα αυτά ανήκουν σε περιοχές της πολυπεπτιδικής αλυσίδας που απέχουν πολύ μεταξύ τους. Εν τούτοις, έχουν πλησιάσει πολύ κοντά χάρη στην πτύχωση της πρωτεΐνης. Επιπλέον, μία μεγάλη πρωτεΐνη μπορεί να αποτελείται από δύο ή περισσότερες πρωτεΐνες. Αυτό συμβαίνει όταν μια θέση σύνδεσης αναγνωρίζει την επιφάνεια μιας δεύτερης πρωτεΐνης, τότε η ισχυρή σύνδεση των δύο πτυχωμένων πολυπεπτιδικών αλυσίδων στη θέση αυτή, μπορεί να δημιουργήσει μια μεγαλύτερη πρωτεΐνη με μοναδική γεωμετρία. Στην απλούστερη περίπτωση, δύο (ή παραπάνω) ταυτόσημες αλυσίδες συνδέονται μεταξύ τους σχηματίζοντας ένα συμμετρικό σύμπλεγμα γνωστό ως διμερές (πολυμερές). Η ακεραιότητα του συμπλέγματος εξασφαλίζεται από τις αληλεπιδράσεις ανάμεσα σε δύο ταυτόσημες θέσεις σύνδεσης. Μελετώντας όλα τα πραπάνω στοιχεία παρατηρούμε ότι η πλήρης αναγνώριση της λειτουργίας μιας πρωτεΐνης θα μπορούσε να γίνει με δύο προσεγγίσεις. Πρώτον, θα μπορούσαμε να κατατάξουμε το πρωτεϊνικό μόριο σε μία από τις γνωστές οικογένειες πρωτεϊνών με βάση την τρισδιάστατη δίπλωση του στο χώρο κληρονομώντας του έτσι τις βιολογικές ιδιότητες της οικογένειας, η προσέγγιση αυτή γνωστή και ως κατηγοριοποίηση (classification) παρ ότι εν γένει σωστή προϋποθέτει την ύπαρξη μίας οικογένειας γνωστών πρωτεϊνών όπου μπορούμε να κατατάξουμε το υπο έλεγχο μόριο. Επίσης υπάρχουν περιπτώσεις πρωτεϊνών που γεωμετρικά ανήκουν σε μία οικογένεια, αλλά δεν έχουν τον ίδιο βιολογικό ρόλο στον οργανισμό. Έτσι υπάρχει περίπτωση να μην υπάρχει γνωστή οικογένεια 5
9 όπου μπορούμε να κατατάξουμε μία πρωτεΐνη ή ακόμα και αν υπάρχει να μη μοιράζεται τις ίδιες βιολογικές δυνατότητες με αυτήν. Δεύτερη προσέγγιση είναι η ένα-προς-ένα συγκριση πρωτεΐνης και πιθανού προσδέτη. Στην περίπτωση που μπορούμε να αναγνωρίσουμε την ικανότητα αλληλεπίδρασης πρωτεΐνης και προσδέτη μπορούμε και να εκτιμήσουμε με ιδιαίτερα μεγάλη ακρίβεια τη λειτουργικότητά της. Η προσέγγιση όμως αυτή απαιτεί σημαντική υπολογιστική ισχύ, σε σχεση με την προαναφερθείσα, καθώς εκτός των γεωμετρικών χαρακτηριστικών πρωτεΐνης-προσδέτη πρέπει να ληφθεί υπ όψιν έστω και μία στοιχειώδης προσομοίωση των χημικών χαρακτηριστικών των δύο προς σύγκριση μορίων. Γι αυτό το λόγω, έχουμε μεγαλύτερη πολυπλοκότητα από την πρώτη προσέγγιση, αλλά πιθανώς μεγαλύτερη ακρίβεια και σαφέστερη αναγνώριση της λειτουργίας του προς εξέταση μορίου. Τα τελευταία χρόνια η εύρεση ακριβούς υπολογιστικής μεθόδου για την εξέταση της λειτουργικότητας μιας πρωτεΐνης έγινε απαραίτητη, έτσι σχεδόν ολόκληρος ο κλάδος της βιοπληροφορικής στράφηκε σε αυτήν την κατεύθυνση χρησιμοποιώντας και τις δύο προαναφερθείσες προσεγγίσεις. Τα ωφέλη από την εύρεση μιας τέτοιας μεθόδου είναι ιδιαίτερα σημαντικά, καθώς θα επιταχύνει σημαντικά τη μελέτη του ανθρώπινου γονιδιώματος, κύριοι εκφραστές του οποίου είναι οι πρωτεΐνες, ανοίγοντας νέους ορίζοντες στην Ιατρική, τη Βιολογία αλλά και την κατανόηση της ζωής. Επιπλέον, η εύρεση μιας μεθόδου ικανής να εξακριβώσει την αλληλεπίδραση μεταξύ δύο πρωτεϊνικών μορίων θα έβρισκε άμεση εφαρμογή στη φαρμακολογία και συγκεκριμένα στη σχεδίαση νέων αποτελεσματικότερων και ασφαλέστερων φαρμάκων βελτιώνοντας σημαντικά την καθημερινότητα και βιωσιμότητα πολλών ανθρώπων (ενδεικτικά αναφέρουμε ότι τα νέας γενιάς αντικαρκινικά φάρμακα βασίζονται σε αυτή τη σχέση πρωτεΐνης-συνδέτη για να στοχεύσουν σε καρκινικά κύτταρα χωρίς να επηρεάσουν σημαντικά τα υγιή κύτταρα του ασθενούς). Στην εργασία αυτή θα αναφέρουμε όσο το δυνατόν πιο σφαιρικά τις υπάρχουσες μεθόδους για την ταυτοποίηση της λειτουργικότητας των πρωτεϊνών. Επιπλέον, θα αναπτύξουμε μία μέθοδο η οποία στοχεύει στην 6
10 κατηγοριοποίηση πρωτεινικών μορίων, κάνοντας χρήση της θεωρίας γράφων για την αναπαράσταση και σύγκριση των μορίων, σε οικογένειες με κοινό βιολογικό ρόλο. Η δομή της διπλωματικής εργασίας είναι ως εξής: Στο δεύτερο κεφάλαιο θα παραθέσουμε τις σημαντικότερες μεθόδους για την αναγνώριση της λειτουργίας πρωτεϊνών, ώστε να γίνει κατανοητός ο τρόπος αντιμετώπισης του προβλήματος από τις σύγχρονες μεθόδους και να αντιπαραβληθούν αυτές με την προτεινόμενη. Στο τρίτο κεφάλαιο θα παρουσιαστεί η διαδικασία προεπεξεργασίας των πρωτεϊνικών δεδομένων ώστε να δομηθούν οι γράφοι, ενώ στο τέταρτο κεφάλαιο παρουσιάζεται η μέθοδος σύγκρισης γράφων (graph matching) που χρησιμοποιείται. Στο πέμπτο κεφάλαιο θα παρατεθούν τα πειραματικά αποτελέσματα της παρούσης εργασίας συνοδευόμενα από τα εξαχθέντα συμπεράσματα. Τέλος, στο παράρτημα υπάρχει η βιβλιογραφία από την οποία αντλήθηκε το απαραίτητο γνωστικό υπόβαθρο για την εκτέλεση του παρόντος έργου. 7
11 2. Παρουσίαση των σύγχρονων μεθόδων. Στο σημείο αυτό της εργασίας παρουσιάζονται κάποιες από τις πλέον σύγχρονες υπολογιστικές μεθόδους για τον έλεγχο της λειτουργίας πρωτεϊνικών μακρομορίων. Γίνεται προσπάθεια να παρουσιαστεί η σημερινή αντιμετώπιση του προβλήματος με τη μεγαλύτερη δυνατή πληρότητα και όσο το δυνατόν σφαιρικά. Η μελέτη στις σύγχρονες μεθόδους είναι και το ερέθισμα που οδήγησε στην ανάπτυξη της παρούσης αντιμετώπισης στο προκείμενο πρόβλημα. 2. Σύγκριση Πρωτεϊνών Βασισμένη στις Δευτερεύουσες Δομές (Δ.Δ.). Μία από τις πιο διαδεδομένες προσεγγίσεις του προβλήματος είναι η εκμετάλλευση των δευτερευόντων δομών που σχηματίζει το μακρομόριο της πρωτεΐνης, όπως στην εργασία των E.Krissinel και K.Henrick[2]. Δευτερεύουσες Δομές (Δ.Δ) ονομάζουμε τις δομές α-έλικα και β-πτυχωτό φύλλο που, όπως προαναφέραμε, σχηματίζονται αυθόρμητα σε μία πρωτεΐνη. Οι συγγραφείς στοχεύουν στη σύγκριση των τρισδιάστατων δομών δύο πρωτεϊνών με απώτερο σκοπό την κατηγοριοποίηση τους (classification). Αυτό το επιχειρούν κάνοντας χρήση της ύπαρξης των Δ.Δ. και της θεωρίας γράφων. Έτσι δημιουργούν ένα γράφο στο χώρο του οποίου οι κόμβοι αναπαριστούν τις Δ.Δ. ενός πρωτεϊνικού μορίου. Κάθε κόμβος λαμβάνει μία ετικέτα { Ti, Li } όπου Ti ο τύπος του εκάστοτε κόμβου (έλικα ή φύλλο) και Li το μήκος του σε αριθμό αμινοξέων. Επιπλέον κάθε Δ.Δ. συμβολίζεται με ένα διάνυσμα r SSE, το οποίο συγκεντρώνει τα χαρακτηριστικά της Δ.Δ. σύμφωνα με την παρακάτω σχέση: όπου: r SSE = r b r e r b = (0.74r p + r p + + r p r p +3)/3.48 r e = (0.74r q -3 + r q -2 + r q r q )/3.48 για έλικες και 8
12 r b = ( r p + r p + )/2 r e = ( r q - + r q )/2 για φύλλα. Στις παραπάνω σχέσεις με r συμβολίζουμε το διάνυσμα θέσης του εκάστοτε αμινοξέος από το σημείο αναφοράς ως συντεταγμένες του αμινοξέος χρησιμοποιούμε τις συντεταγμένες του κεντρικού ατόμου άνθρακα ( Cα ), ενώ οι δείκτες p,q φανερώνουν τον αύξοντα αριθμό του πρώτου και του τελευταίου αμινοξέος της Δ.Δ. Πρέπει να σημειώσουμε ότι η μέθοδος δε λαμβάνει υπ όψιν φύλλα μικρότερα από τρία αμινοξέα και έλικες μικρότερες από πέντε αμινοξέα. Στη συνέχεια οι ακμές λαμβάνουν και αυτές μία ετικέτα της μορφής ij ij ij ij { pij, a, a2, a3, a4, N i, N, C, C j i j } όπου pij το μήκος της ακμής σε Angstrom,με a 2 ij, ij ij ij a συμβολίζονται οι γωνίες μεταξύ των κόμβων i και j ενώ με a a, 3 4 συμβολίζονται οι γωνίες στροφής των κόμβων i και j. Αφού κατασκευάζονται οι δύο προς σύγκριση γράφοι λαμβάνει χώρα η διαδικασία της σύγκρισης. Δύο κόμβοι θεωρούνται συμβατοί όταν: Ti = Tj και Li-Lj <ε L (Li+Lj)/2 + σ L Ενώ δύο ακμές e ij και e kl θεωρούνται συμβατές όταν ισχύουν τα παρακάτω: p ij p < ε ( p + p ) / 2 + σ, kl p ij kl p a ij,2 a kl,2 < δ και ij 3 a kl 3 < δ 2 a, ij kl, sign( )=sign( a 4 ) όταν a ij kl + nπ > δ s =,2, 4, a 4 πρέπει να σημειώσουμε ότι τα μεγέθη ε L, σ L, σ p, ε p, δ, δ 2, δ 3 επιλέγονται με καθαρά εμπειρικές μεθόδους για να δώσουν τα βέλτιστα αποτελέσματα. Μετά τη σύγκριση των δύο γράφων με βάση τα παραπάνω προκύπτουν ζεύγη συμβατών σημείων των δύο γράφων. Τα σημεία αυτά όμως δεν αποτελούν το σύνολο των σημείων των γράφων, αλλά μόνο τις Δ.Δ. για το λόγω αυτό με αφετηρία τα ήδη ζευγαρωμένα σημεία προχωράμε στην εύρεση της υπέρθεσης των δύο γράφων που ελαχιστοποιεί τη μέση τετραγωνική απόσταση των σημείων των γράφων με κάποια από τις γνωστές μεθόδους Ταχείας Βέλτιστης Υπέρθεσης (Fast Optimal Superposition)[3]. Έτσι, καταλήγουμε να έχουμε μία s 3, 9
13 χαρτογράφηση των δύο γράφων που απεικονίζει τα συμβατά τους σημεία. Πρέπει να σημειώσουμε ότι ως σημεία έχουμε λάβει τα άτομα άνθρακα C της κεντρικής αλυσίδας της πρωτεΐνης (επονομαζόμενα Cα). Αποτελέσματα Η μέθοδος αυτή επιταχύνει σημαντικά τη διαδικασία της χαρτογράφησης των κοινών σημείων δύο πρωτεϊνών και συνεπώς τη σύγκρισή τους, με αποτέλεσμα να επιταχύνεται η ολοκλήρωση της κατάταξης μιας πρωτεΐνης σε κάποια κλάση (classification). Αυτό δίνει τη δυνατότητα της χρήσης της μεθόδου σε εφαρμογές πραγματικού χρόνου (real time) ή για τη σάρωση ολόκληρης της πρωτεϊνικής βάσης δεδομένων (Protein Data Bank) [4] σε ρεαλιστικό χρόνο. Επιπλέον η μέθοδος αυτή συμφωνεί στα αποτελέσματά της με τις μεθόδους που χρησιμοποιούνται ευρέως σήμερα (όπως οι DALI[5], VAST[6] και CE[7]) οι οποίες έχουν αποδειχθεί ακριβείς. Παρ όλα αυτά η συγκεκριμένη μέθοδος αδυναμεί να δώσει ικανοποιητική ακρίβεια όταν αναφέρεται σε πρωτεΐνες με μικρό αριθμό Δ.Δ., καθώς βασίζεται σε αυτές. Στην περίπτωση που έχουμε πρωτεΐνη με μικρό αριθμό Δ.Δ., όχι μόνο χάνεται η ακρίβεια της μεθόδου, αλλά αυξάνεται και ο χρόνος εκτέλεσης της διαδικασίας βέλτιστης υπέρθεσης των ατόμων άνθρακα της κεντρικής αλυσίδας. 2.2 Σύγκριση Πρωτεϊνικών Δομών με Γράφους απαρτισμένους από Χημικές Ομάδες. Στη μέθοδο της αναφοράς [8] εισάγεται για άλλη μια φορά η θεωρία γράφων για τη σύγκριση δύο πρωτεϊνών. Η διαφορά με την προηγούμενη μέθοδο έγγειται στην επιλογή των κόμβων που χρησιμοποιούνται για την κατασκευή των γράφων. Αρχικά γίνεται η κατασκευή των γράφων από τα δεδομένα που παρέχονται στα αρχεία *.pdb όπως αυτά βρίσκονται στην Πρωτεϊνική Βάση Δεδομένων. Καθώς αυτή η διαδικασία απαιτεί περισσότερο χρόνο από τη σύγκριση καθαυτή τα δεδομένα των γράφων προϋπολογίζονται και αποηθηκεύονται στη βάση δεδομένων. Τέσσερα διαδοχικά βήματα ακολουθούνται για την κατασκευή του γράφου. 0
14 Αρχικά σε ατομικό επίπεδο υπολογίζεται η παράμετρος D που ονομάζεται τοπική ατομική πυκνότητα για κάθε άτομο Α. Σκοπός της παραμέτρου είναι να δώσει ένα μέτρο για την επικάλυψη του ατόμου μέσα στο μόριο. Η σχέση που δίνει την D είναι : D ( A) = u max r r max W ( r) όπου m r η μάζα των ατόμων μία σφαίρα ακτίνας r επικεντρωμένη στο άτομο Α και u max ο όγκος της σφαίρας ακτίνας r max. Mία συνάρτηση βάρους W εφαρμόζεται για να μειώσει την επιρροή των περιφερειακών ατόμων έτσι ώστε η D(A) είναι μία συνεχής συνάρτηση: W ( r) = 4( r r max ) όπου 4 συντελεστής απαραίτητος για να πάρουμε μία τιμή πυκνότητας που δε θα εξαρτάται από τη r max σε ένα θεωρητικά ομογενές μέσο. m r Μία δεύτερη παράμετρος που ονομάζεται τοπικό κέντρο μάζας (C) υπολογίζεται. Το μέγεθος C θεωρείται ως το κέντρο των ατόμων μέσα στη σφαίρα. Είναι σημαντικό να σημειώσουμε ότι το διάνυσμα CA δείχνει προς το εξωτερικό του μορίου καθώς το γεγονός αυτό μας επιτρέπει να χρησιμοποιήσουμε το διάνυσμα CA για τον προσδιορισμό του προσανατολισμού του κάθε μορίου. Το επόμενο επίπεδο οργάνωσης της αναπαράστασής μας είναι η ομαδοποίηση των ατόμων σε χημικές ομάδες. Οι χημικές ομάδες είναι χαρακτηριστικά συμπλέγματα ατόμων για το κάθε αμινοξύ. Έτσι, κάθε χημική ομάδα του μορίου αποτελείτα από ένα σύνολο ατόμων και αντιστοιχεί σε ένα συγκεκριμένο αμινοξύ. Για δεδομένη χημική ομάδα, υπολογίζονται και καταγράφονται η μέση θέση P των ατόμων, η μέση θέση C του τοπικού κάντρου μάζας και η μέση τοπική πυκνότητα D. Το βήμα αυτό απλουστεύει την αναπαράσταση του μορίου από το σύνολο των ατόμων στις χημικές ομάδες. Στην επόμενη φάση οι χημικές ομάδες χρησιμοποιούνται για τη δημιουργία τριγώνων. Μόνο τρίγωνα με ακμές μικρότερες από 8Å λαμβάνονται υπ όψιν, έτσι ώστε να λαμβάνουμε μόνο γειτονικές ομάδες στο ίδιο τρίγωνο και όχι απομακρυσμένες. Πρέπει να σημειωθεί ότι η
15 απόσταση αυτή ορίστηκε χρησιμοποιώντας εμπειρικές μεθόδους. Για κάθε τρίγωνο που δημιουργείται από τις χημικές ομάδες (P,P 2,P 3 ) έχουμε και τα αντίστοιχα τοπικά κέντρα μάζας (C,C 2,C 3 ). Οι αποστάσεις μεταξύ γειτονικών κορυφών καταγράφονται. Ο προσανατολισμός του τριγώνου προσεγγίζεται από το τριπλό βαθμωτό γινόμενο : (CP,CP 2,CP 3 ). Τέλος, η τελική αναπαράσταση του μορίου επιτυγχάνεται με τη δημιουργία του γράφου που προκύπτει αν ενώσουμε γειτονικά τρίγωνα, δηλαδή τρίγωνα που μοιράζονται ακριβώς δύο χημικές ομάδες. Η σύγκριση δύο γράφων είναι το δεύτερο κομμάτι της μεθόδου και χωρίζεται σε τρία μέρη. Αρχικά δημιουργούμε ζεύγη όμοιων τριγώνων με βάση τους εξής βασικούς γεωμετρικούς κανόνες : )ομοιότητα των ακμών μήκος-μήκος2 <2Å 2)ομοιότητα της κάλυψης δύο χημικών ομάδων πυκνότητα-πυκνότητα2 <0.08D/A 3 3)ομοιότητα πρσανατολισμόυ (τριπλό βαθμωτό γινόμενο) X-X2<00Å. Στο δεύτερο βήμα τα κοινά ζεύγη που υπολογίσαμε αποτελούν τους κόμβους ενός γράφου σύγκρισης. Για να θέσουμε μία ακμή μεταξύ δύο ζεύγων (Τ,Τ ) και (Τ2,Τ2 ) τα ζεύγη πρέπει να έχουν τις παρακάτω ιδιότητες ) Το τρίγωνο Τ πρέπει να είναι γειτονικό με το Τ2 (ίδιο μόριο) όπως επίσης και το Τ με το Τ2. 2) Η γωνία που δημιουργούν τα επίπεδα Τ,Τ2 πρέπει να είναι παρόμοια με αυτήν που δημιουργούν τα Τ και Τ2. Έτσι, καταλήγουμε σε ανεξάρτητους υπό-γράφους που αντιστοιχούν σε ζεύγη παρόμοιων περιοχών σε ολόκληρα τα μόρια. Στο σημείο αυτό τα ζεύγη τριγώνων μετατρέπονται ξανά σε ζεύγη χημικών ομάδων. Αυτά τα υποσύνολα χημικών ομάδων αποκαλούνται μπαλώματα (patches) Τελικά τα μπαλώματα αυτά τα επεξεργαζόμαστε ώστε να απορίψουμε ζεύγη χημικών ομάδων που δεν είναι συμβατά. Για το σκοπό αυτό χρησιμοποιείται μια διαδικασία Ταχείας Βέλτιστης Υπέρθεσης που ελαχιστοποιεί τη μέση Τετραγωνική Απόσταση χρησιμοποιώντας τη συνάρτηση απόστασης: 2
16 dist(g, g2) = α pos g ) pos( ) +β D g ) D( ) ( g 2 ( g 2 όπου pos g ) pos( ) είναι η ευκλείδια απόσταση μεταξύ g και g2 μετά ( g 2 από μια βέλτιστη υπέρθεση και D g ) D( ) η διαφορά των μέσων ( g 2 ατομικών πυκνοτήτων. Οι μεταβλητές α,β επιλέγονται έτσι για να σταθμίσουν τη σημαντικότητα των μεγεθών. Τέλος, ζεύγη με dist(g,g2) μεγαλύτερη από ένα ορισμένο κατώφλι απορρίπτονται, με αποτέλεσμα στο τέλος να μας μένουν μόνο μπαλώματα με τις βέλτιστα ταυτισμένες χημικές ομάδες. Συνεπώς, έχουμε εντοπίσει τις κοινές περιοχές δύο μορίων. Αποτελέσματα Τα πειραματικά αποτελέσματα της συγκεκριμένης μεθόδου κατέδειξαν ότι μπορεί με σημαντική ακρίβεια να εντοπίσει την ομοιότητα δύο πρωτεϊνικών μορίων τόσο γεωμετρικά, αλλά κυρίως ως προς τη λειτουργικότητά τους. Συγκεκριμένα, αποδείχθηκε ότι η μέθοδος αυτή δύναται να εντοπίσει δομικές ομοιότητες μεταξύ πρωτεϊνικών μορίων, που δεν έχουν καμία ομοιότητα μεταξύ τους σε ότι αφορά την αλληλουχία της κεντρικής αλυσίδας, ικανότητα που δε διαθέτει, για παράδειγμα, η ιδιαίτερα δημοφιλής εφαρμογή DALI. Επιπλέον, η μέθοδος αυτή καταφέρνει, χρησιμοποιώντας ένα ενεργό μόριο ως σημείο αναφοράς, να διαχωρίσει τα ενεργά από τα μή-ενεργα μόρια μιας οικογένειας πρωτεϊνών (λεκτίνες με ενεργές περιοχές για πρόσδεση ζακχάρων) παρότι όλα τα μέλη της οικογένειας μοιράζονται κοινά γεωμετρικά χαρακτηριστικά. Ως ενεργά μόρια αναφέρουμε μόρια της οικογένειας που έχουν πράγματι την ικανότητα να προσδέσουν ζάκχαρα, ώστε να τα διαχωρίσουμε από μέλη της οικογένειας αυτής που λόγω μεταλλάξεων έχουν χάσει την ικανότητα αυτή. Αυτό επιτυγχάνεται χάρη στην ενσωμάτωση στα δεδομένα του γράφου και των χημικών χαρακτηριστικών των χημικών ομάδων που περιέχει το μόριο, εισάγωντας έτσι την πληροφορία για τη δραστική ικανότητα του μορίου. Η μέθοδος αυτή όμως λόγω των εμπειρικών στοιχείων που χρησιμοποιεί, χάνει τη γενικότητα της και δίνει σε μερικές περιπτώσεις 3
17 λανθασμένα αποτελέσματα. Αναφορικά, αδυνατεί να εντοπίσει αλληλεπιδράσεις που βασίζονται σε δεσμούς υδρογόνου. Αυτό συμβαίνει διότι η ομαδοποίηση των ατόμων σε χημικές ομάδες δεν επιτρέπει τη διάκριση μεταξύ των ατόμων που δίνουν και δέχονται δεσμούς υδρογόνου. Τέλος, προβληματικά αποτελέσματα δίνουν, σε κάποιες περιπτώσεις, οι εμπειρικές θεωρήσεις που έχουν ληφθεί για τη συνάρτηση ατομικής πυκνότητας που αναφέραμε παραπάνω, μειώνοντας έτσι τη διακριτική ικανότητα της μεθόδου. 2.3 Μέθοδος σύγκρισης Βιολογικών Δεδομένων με Χρήση Διμερών Γράφων. Στην εργασία [9] το πρόβλημα για την ευθυγράμμιση των στοιχείων δύο πρωτεϊνών στο χώρο εντοπίζεται στην εύρεση μιας χαρτογράφησης των στοιχείων των δύο πρωτεϊνών που ορίζει ζεύγη σημείων, τα οποία ικανοποιούν κάποια συνθήκη ομοιότητας, όπως αυτή της Ελάχιστης Μέσης Τετραγωνικής Απόστασης (R.M.S.D.). Οι συγγραφείς επιχειρούν να βρούν τέτοιες αντιστοιχίες μεταξύ δύο πρωτεϊνών κάνοντας χρήση διμερών γράφων σύμφωνα με την παρακάτω λογική. Αρχικά λαμβάνονται τα δομικά στοιχεία της κάθε πρωτεΐνης από τα αρχεία των πρωτεϊνικών δεδομένων. Στη συνέχεια, διανύσματα δομικών χαρακτηριστικών υπολογίζονται και τοποθετούνται ως κόμβοι του γράφου. Η δόμηση του γράφου και η εξαγωγή των στοιχείων που τοποθετούνται στους κόμβους μπορεί να γίνει με βάση όλα τα άτομα του μορίου, τα άτομα Ca της κεντρικής αλυσίδας, ολόκληρα αμινοξεϊκά κατάλοιπα είτε δευτερεύουσες δομές του μορίου (π.χ. α-έλικες και β- πτυχωτά φύλλα). Τα στοιχεία που περιέχονται στους κόμβους αυτούς μπορούν να είναι είτε γεωμετρικά, είτε χημικά χαρακτηριστικά. Οι κόμβοι αριστερά και δεξιά του γράφου αντιπροσωπεύουν την πρωτεΐνη Α και Β αντίστοιχα. Κάθε κόμβος είναι συνδεδεμένος με κάθε κόμβο που βρίσκεται στην απέναντι πλευρά του γράφου σχηματίζοντας έτσι έναν πλήρη διμερή γράφο. Το βάρος κάθε ακμής υπολογίζεται ως η έξοδος μιας ειδικά σχεδιασμένης συνάρτησης που δέχεται ως είσοδο τα δομικά χαρακτηριστικά που περιέχονται σε κάθε κόμβο, που βρίσκεται στην αρχή και το τέλος της ακμής. 4
18 Τέλος, εφαρμόζεται κάποιος από τους γνωστούς αλγορίθμους ταυτοποίησης διμερών γράφων στον παρόν γράφο ώστε να λάβουμε το σύνολο ακμών με τη βέλτιστη ταυτοποίηση που αποτελεί και την προς έρευνα αντιστοίχιση των δύο πρωτεϊνών. Ως βέλτιστη ταυτοποίηση σε ένα διμερή γράφο εννοούμε ένα σύνολο ακμών του γράφου, έτσι ώστε να μην υπάρχει κανένας κόμβου της μίας πλευράς του γράφου, που να συνδέεται με περισσότερους από έναν κόμβους της άλλης πλευράς. Επιπλέον, αυτό το σύνολο ακμών θα πρέπει να ικανοποιεί κάποια σχέση βελτιστοποίησης όπως για παράδειγμα να έχει μέγιστο (ή ελάχιστο κατα περίπτωση) άθροισμα των βαρών των ακμών. Ο καλύτερος γνωστός αλγόριθμος, ο οποίος και απαιτεί χρόνο εκτέλεσης φραγμένο πολυωνυμικά, είναι αυτός του Kuhn[0] ο οποίος εκτελείται σε χρόνο της τάξης : O( V ( E V logv )) +. Τέτοιου είδους αλγόριθμοι μπορούν να υλοποιηθούν αποτελεσματικά και να δύνανται να επεξεργαστούν γράφους με ένα ρεαλιστικά μεγάλο αριθμό κόμβων (περίπου 00,000 κόμβους). Ειδική μνεία πρέπει να γίνει στη διατήρηση της αλληλουχίας των κόμβων κατα το ταίριασμα τους από τον αλγόριθμο έτσι ώστε να μη διαταράσσεται η αλληλουχία της πρωτεΐνης. Αυτό επιτυγχάνεται εύκολα με τη χρήση της παρακάτω συνάρτησης για τον υπολογισμό του ολικού βάρους μιας ακμής, M ij ( ) = w e ij ( i, j ) ( M, i' < i and e E) w e + max max i', j i', j ( ) max M i, j', j' < j and ei, j' E όπου M ij είναι το ολικό βάρος μία ακμής και Ε το σύνολο των ακμών του γράφου. Συμπεράσματα Με την εργασία αυτή εισάγεται ένα πλαίσιο για την σύγκριση δύο πρωτεϊνικών μορίων με χρήση διμερών γράφων. Η μέθοδος αυτή καταφέρνει να λύσει το πρόβλημα της αντιστοίχησης δύο πρωτεϊνών με έναν αλγόριθμο εύκολα υλοποιήσιμο ο οποίος δύναται να υλοποιηθεί σε 5
19 πολυωνυμικό χρόνο δίνοντας μία έξοδο που καταλήγει σε αντιστοίχηση του συνόλου των μορίων και όχι μόνο μερών αυτών. Το πλαίσιο αυτό έχει υλοποιηθεί σε C++ και Java χρησιμοποιώντας ως δομικά δεδομένα τα άτομα Ca της κεντρικής αλυσίδας δίνοντας αισιόδοξα αποτελέσματα. Στόχος είναι να μετρηθεί η αποτελεσματικότητα του στο σύνολο της βάσης δεδομένων PDB (Protein Data Bank). Επιπλέον υλοποιούνται και εκδόσεις του, που χρησιμοποιούν ως δομικά στοιχεία δευτερεύουσες δομές, καθώς επίσης και άλλες εναλακτικές επιλογές. 2.4 Αναζήτηση ενεργών Περιοχών στην ομάδα Πρωτεϊνών G. Μία ιδιαίτερη μέθοδος [] εφαρμοσμένη στη μεγαλύτερη οικογένεια κυτταρικών αποδεκτών τα G-πρωτεϊνικά συμπλέγματα (Gprotein coupled receptors GPCR) αναφέρεται στη συνέχεια. Η οικογένεια αυτή είναι στόχος περισσότερο από το 50% των φαρμάκων στην αγορά σήμερα καθώς, είναι υπεύθυνη για τη μεταφορά δραστικών ουσιών μέσα στα κύτταρα. Η μέθοδος αυτή χρησιμοποιεί μία μικτή προσέγγιση, για να ανιχνεύσει πιθανές θέσεις πρόσδεσης και τυχόν προσδέτες στα μόρια της οικογένειας GPCR συνδυάζοντας εργαστηριακά δεδομένα με υπολογιστικές μεθόδους. Οι εισηγητές της θέτουν τρεις στόχους: ) να ελέγξουν αν μπορεί να ανιχνευθεί κάποια κοιλότητα τσέπη για πρόσδεση ενός προσδέτη (ligand) χωρίς να έχουμε καμία πληροφορία για τον προσδέτη, 2) να ελεγχθεί εάν είναι δυνατή η πρόβλεψη του τρόπου ένωσης του προσδέτη με την πρωτεΐνη, εάν εντοπιστεί ενεργή περιοχή και 3) να ελεγχθεί εάν μπορεί για μία πρωτεΐνη να εντοπιστεί πιθανός προσδέτης από ένα πολυπληθές σύνολο βιολογικών ουσιών, ώστε να βρεθεί πιθανή φαρμακευτική ουσία που να συνεργάζεται με την πρωτεΐνη. Οι παραπάνω στόχοι σήμερα επιχειρείται να ικανοποιηθούν με αμιγώς πειραματικές μεθόδους στο εργαστήριο οι οποίες είναι ιδιαίτερα επίπονες και χρονοβόρες. Η νέα πρόταση, την οποία και η μέθοδος αυτή προσπαθεί να εφαρμόσει, είναι η χρήση της μηχανικής των εσωτερικών συντεταγμένων (Internal Coordinates Mechanics ICM), η οποία χρησιμοποιεί έναν εικονικό προσδέτη και επιχειρεί να τον τοποθετήσει σε 6
20 πιθανές θέσεις πρόσδεσης στην υπό διερεύνηση πρωτεΐνη της οποίας έχει κατασκευαστεί η κρυσταλλογραφία, είτε ένα ακριβές τρισδιάστατο μοντέλο με πειραματικές και υπολογιστικές μεθόδους. Η προσέγγιση αυτή δίνει τη δυνατότητα να ελεγχθεί η αλληλεπίδραση ενός μακρομορίου με εκατομμύρια πιθανούς προσδέτες. Ο πυρήνας της μεθόδου αυτής βρίσκεται στο ιδανικό ταίριασμα ενός πιθανού προσδέτη με την ενεργό περιοχή που έχει προϋπολογιστεί με βάση τα πειραματικά δεδομένα. Ο αλγόριθμος επιχειρεί να βρει την πιθανή αλληλεπίδραση προσδέτη-πρωτεΐνης τοποθετώντας τον πρώτο στην ενεργό περιοχή με διαφορετικό προσανατολισμό κάθε φορά για κάθε διαφορετικό προσανατολισμό υπολογίζεται η έξοδος μιας συνάρτησης που λαμβάνει υπ όψιν συνεχή και διακριτά ηλεκτροστατικά πεδία υδροφοβικότητες και απώλεια εντροπίας των μορίων. Η διαδικασία αυτή που ονομάζεται και Σάρωση Εικονικού προσδέτη (Virtual Ligand Screening VLS) μπορεί να ελέγξει γρήγορα και με ακρίβεια εκατοντάδες χιλιάδες πιθανούς προσδέτες από τη βάση δεδομένων τοποθετόντας κάθε προσδέτη στη θέση πρόσδεσης με τρεις διαφορετικούς προσανατολισμούς και κρατώντας αυτόν με τη βέλτιστη επίδοση σε ότι αφορά την ολική ενέργεια του συμπλέγματος όπως θα δούμε παρακάτω. Η ολική ενέργεια ενέργεια του συμπλέγματος υπολογίζεται χρησιμοποιώντας τα πειραματικά δεδομένα για τέτοιου είδους αλληλεπιδράσεις μορίων. Ενδεικτικά στοιχεία είναι οι ενέργειες των μορίων πριν και μετά τη σύνδεσή τους, τα ενεργειακά περιεχόμενα των πλευρικών αλυσίδων προσδέτη και πρωτεΐνης, καθώς επίσης και της εντροπίας του συστήματος. Η συνολική ενεργειακή στάθμη τότε εκτιμάται από μία συνάρτηση ελέγχου της αποδοχής ή απόρριψης της αλληλεπίδρασης με βάση το κριτήριο Metropolis [2]. Αποτελέσματα Η μέθοδος καταφέρνει να προσφέρει μία πολύ δυνατή προσομοίωση των εργαστηριακών πειραμάτων με μεγάλη ακρίβεια, αλλά και ταχύτητα. Δύναται να εντοπίσει τον ακριβή προσανατολισμό του προσδέτη στην ενεργό περιοχή της πρωτεΐνης βρίσκοντας την ιδανικότερη θέση από 7
21 πλευράς ελαχιστοποίησης της μέσης τετραγωνικής απόστασης των ατόμων του προσδέτη από τα άτομα της πρωτεΐνης, αλλά και από πλευράς ενεργειακού περιεχομένου του συμπλέγματος. Όλα τα παραπάνω καθιστούν τη μέθοδο αυτή σαν ένα πολύ δυνατό εργαλείο για τη μελέτη της οικογένειας GPCR και μία σημαντική προσέγγιση για την ανακάλυψη νέων φαρμάκων που αναφέρονται στην οικογένεια αυτή. Η δυναμική αυτή όμως της προκείμενης μεθόδου είναι και το κύριο μειονέκτημά της, καθώς είναι αποκλειστικά εξειδικευμένη στη μελέτη της οικογένειας GPCR με αποτέλεσμα να μην μπορεί να χρησιμοποιηθεί σε ευρύτερη και γενικότερη έρευνα για τον εντοπισμό αλληλεπιδράσεων πρωτεϊνικών μορίων με πιθανούς προσδέτες. Επιπλέον, απαιτεί γνώση εκ των προτέρων των ενεργών περιοχών της κάθε υπό έλεγχο πρωτεΐνης, κάνοντας τη μέθοδο αυτή έντονα συνδεδεμένη με τα εργαστηριακά δεδομένα, που προέρχονται από επίπονες και χρονοβόρες διαδικασίες κατά συνέπεια λόγω της αναγκαίας a priori γνώσης των ενεργών περιοχών η μέθοδος καθίσταται ανίκανη να εντοπίσει καινούργιες ενεργές περιοχές που δεν έχουν ερευνηθεί πειραματικά. 2.5 Εκτίμηση της λειτουργίας πρωτεϊνικών μορίων ταυτίζοντας 3D δομικά στοιχεία. Στόχος της μεθόδου που παρουσιάζεται στην [3] είναι ο έλεγχος της λειτουργίας μιας πρωτεΐνης με χρήση υπολογιστικών μεθόδων βασιζόμενη μόνο στη γνώση της γεωμετρίας του μορίου. Η προσέγγιση αυτή επιτάσσεται από το γεγονός ότι πρωτεΐνες με κοινή τρισδιάσταστη δομή επιτελούν παρόμοιες λειτουργίες ακόμα και αν δεν έχουν κοινή αλληλουχία αμινοξέων και επιπλέον αν χαθεί η χωρική γεωμετρία μιας πρωτεΐνης χάνεται αυτόματα και η λειτουργικότητα της. Έτσι επικεντρώνονται οι συγγραφείς στη σύγκριση τρισδιάτων δομών γνωστών (ως προς την λειτουργία τους) με πρωτεΐνες που ερευνάται η λειτουργία τους. Σύμφωνα με τα παραπάνω πιθανή ταύτιση θα σημαίνει και κοινή λειτουργία. Η μέθοδος αυτή εξελίσεται σε δύο στάδια: το στάδιο της προεπεξεργασίας των δεδομένων και το στάδιο της σύγκρισης. Στο πρώτο 8
22 στάδιο υπολογίζεται ένας πίνακας αναφοράς για κάθε αμινοξύ της πρωτεΐνης.το επίπεδο που ορίζεται από το κεντρικό άτομο άνθρακα (Ca) του αμινοξέος, το άτομο του αζώτου (N) και το άτομο ανθρακα (C) που βρίσκονται στο κεντρικό στέλεχος του αμινοξέος το ονομάζουμε πλαίσιο αναφοράς και με βάση αυτό υπολογίζεται μια ορθοκανονική βάση συντεταγμένων στην οποία μεταφράζονται όλα τα άτομα του μορίου της πρωτεΐνης και κρατούνται ως γεωμετρικά χαρακτηριστικά για το τρέχον αμινοξύ. Αυτό γίνεται για κάθε αμινοξύ, δημιουργώντας έτσι έναν πίνακα αναφοράς για κάθε αμινοξύ ο οποίος υπολογίζεται offline και αποθηκεύεται στη βάση δεδομένων πριν από οποιαδήποτε χρήση της μεθόδου. Σε δεύτερο χρόνο γίνεται η σύγκριση των προαποθηκευμένων δεδομένων με χρήση του παρακάτω αλγορίθμου hash matching. Η σύγκριση γίνεται σύμφωνα με τα παρακάτω: αρχικά επιλέγεται τυχαία ένα αμινοξύ της προς διερεύνηση πρωτεΐνης (query protein), ανακαλείται από τη μνήμη ο πίνακας αναφοράς για το συγκεκριμένο αμινοξύ της πρωτεΐνης και συγκρίνεται με τους αντίστοιχους πίνακες όλων των αμινοξέων της πρωτεΐνης αναφορας. Για κάθε ζεύγος πινάκων λαμβάνουμε ένα αριθμό στοιχείων που ταυτίζονται στους δύο πίνακες. Αυτόν τον αριθμό τον αποκαλούμε ψήφο για το συγκεκριμένο ζεύγος. Η διαδικασία αυτή επαναλαμβάνεται μέχρι να συγκριθούν όλα τα πλαίσια αναφοράς της διερευνόμενης πρωτεΐνης με όλα τα πλαίσια αναφοράς της πρωτεΐνης αναφοράς. Τα ζεύγη που ξεπερνούν έναν αριθμό ψήφων που ο χρήστης έχει θέσει ως κατώφλι αποθηκεύονται και με βάση αυτά υπολογίζονται πίνακες μετασχηματισμού που αντιστοιχίζουν τα άτομα της μιας πρωτεΐνης στα άτομα της άλλης με βάση το κριτήριο της Ελάχιστης Μέσης Τετραγωνικής Απόστασης (RMSD) Η αντιστοίχηση με το μεγαλύτερο πλήθος μορίων σε συνάρτηση με τη μικρότερη δυνατή RMSD είναι και το βέλτιστο ταίριασμα των δύο μορίων. Αν το πλήθος του βέλτιστου ταιριάσματος υπερβαίνει έναν προαποφασισμένο αριθμό συμπεραίνουμε ότι οι δύο πρωτεΐνες έχουν κοινή γεωμετρία και συνεπώς μοιράζονται και κοινές λειτουργίες. Η ίδια ακριβώς διαδικασία μπορεί να ακολουθηθεί για τη σύγκριση πρωτεϊνών με ενεργές περιοχές πρόσδεσης (binding sites), όπως αυτές 9
23 βρίσκονται στα αρχεία της PDB (Protein Data Bank) με σκοπό τον εντοπισμό ενεργών περιοχών σε πρωτεϊνικά μόρια. Στο σημείο αυτό πρέπει να αναφέρουμε ότι πιθανή χαμηλή ανάλυση της πρωτεΐνης, όπως αυτή προέρχεται από τα αρχεία της PDB, μπορεί να δώσει λανθασμένα αποτελέσματα. Επιπλέον, είναι πιθανό μόρια με κοινή γεωμετρία να μην έχουν και κοινή λειτουργία. Έτσι, για την ισχυροποίηση της μεθόδου εισάγουμε δεδομενα ομοιότητας των αμινοξέων, όπως τα λαμβάνουμε από τον πίνακα Dayhoff PAM250 [4], ο οποίος αρχικά χρησιμοποιήθηκε σε περιπτώσεις ελέγχου ομοιότητας στην αλληλουχία πρωτεϊνικών μακρομορίων. Αποτελέσματα Τα πειραματικά στοιχεία της μεθόδου αυτής δείχνουν ότι καταφέρνει να κατηγοριοποιήσει, με βάση τη λειτουργία τους, μία πειραματική βάση ελέγχου (dataset) αποτελούμενη από 43 πρωτεΐνες με ποσοστό ακριβείας 90,7%. Επιπλέον, με αντίστοιχη ακρίβεια μπορεί να εντοπίσει ενεργές περιοχές σε πρωτεΐνες. Μειονεκτήματα της μεθόδου είναι αναγκαιότητα μεγάλου όγκου μνήμης για την αποθήκευση των προεπεξεργασμένων δεδομένων που οδηγεί σε δυσκολία χειρισμού μορίων με μεγάλο αριθμό αμινοξέων. Επιπλέον, η ελλιπής δυνατότητα εισαγωγής χημικών δεδομένων στα χαρακτηριστικά της πρωτεΐνης θα οδηγήσουν, πιθανώς, σε λανθασμένα αποτελέσματα όταν έχουμε περιπτώσεις όπου η χημεία παίζει εξίσου σημαντικό ή και σημαντικότερο ρόλο από τη γεωμετρία του μορίου στην εκτέλεση της λειτουργίας του. Τέλος, η εκτέλεση της διαδικασίας σύγκρισης απαιτεί χρόνο που δεν επιτρέπει την εφαρμογή του σε μεγάλη έκταση δεδομένων, γεγονός που οι εισηγητές της μεθόδου προσπαθούν να βελτιώσουν σε συνδυασμό με την αύξηση της ακρίβειας. 2.6 Κατηγοριοποιήση Πρωτεϊνών κάνοντας χρήση τον Μετασχηματισμό Ίχνους. Ένας νέος τρόπος περιγραφής των πρωτεϊνών εισάγεται στην εργασία [5] ώστε να γίνει δυνατή η ταχύτερη σύγκριση δύο μορίων χώρις 20
24 έλλειψη ακρίβειας. Οι συγγραφείς εισάγουν το μετασχηματισμό σφαιρικού ίχνους (Spherical Trace Transform) [6], ο οποίος δύναται να περιγράψει ένα 3D μοντέλο με απόλυτη ακρίβεια και χωρίς να επηρεάζεται από την περιστροφή, δηλαδή το ίδιο μοντέλο έχει σταθερό μετασχηματισμό ακόμα και αν περιστραφεί στο χώρο. Αρχικά τα δεδομένα του pdb αρχείου όπως αυτό λαμβάνεται από την Πρωτεϊνική Βάση Δεδομένων απεικονίζεται στο χώρο ως ένα 3D μοντέλο. Το μοντέλο αυτό αποτελείται από σφαίρες, που ισοδυναμούν με τα άτομα, και οι οποίες έχουν υποστεί τριγωνοποίηση, έτσι ώστε κάθε μία σφαίρα να είναι ένα σύνολο από διακριτά σημεία συνοδευόμενο από ένα σύνολο ακμών που συνδέουν τα σημεία αυτά. Στη συνέχεια υπολογίζεται το κέντρο μάζας του μοντέλου. Όλα τα σημεία του μοντέλου, με μια διαδικασία μετατόπισης και περιστροφής, μετασχηματίζονται έτσι ώστε το κέντρο μάζας να συμπίπτει με την αρχή του συστήματος συντεταγμένων. Επιπλέον, οι αποστάσεις των σημείων από την αρχή κανονικοποιούνται, ώστε η μέγιστη απόσταση σημείου από την αρχή να είναι ίση με τη μονάδα. Τέλος, το μοντέλο βοξελοποιείται και εφαρμόζεται ο Μετασχηματισμός Σφαιρικού Ίχνους σύμφωνα με το [7]. Για την περιγραφή του μοντέλου χρησιμοποιούνται ως σφαιρικοί τελεστές ο πολικός μετασχηματισμός Fourier και οι ροπές Krawtchouk. Τελικά η παραπάνω διαδικασία μας οδηγεί στην περιγραφή του μοντέλου με τα διανύσματα περιγραφέων D FTkm (l) και D Kkm (l) για περιγραφή με χρήση μετασχηματισμού Fourier ή ροπών Krawtchouk αντίστοιχα. Μετά από πειραματικές δοκιμές τα διανύσματα : FT={FT 00, FT 0,FT 0, FT 02 } και Κ={ K 00, K 0, K 02 } αποδείχθηκε ότι παράγουν τα καλύτερα αποτελέσματα επομένως αυτά χρησιμοποιούνται στη μέθοδο αυτή. Το πλήθος των γεωμετρικών περιγραφέων ανά διάνυσμα στην περίπτωση αυτή είναι N G =960. Επιπλέον, για την ορθότερη αναπαράσταση του πρωτεϊνικού μοντέλου χρησιμοποιούνται και οι εξής περιγραφείς, που αντιπροσωπεύουν τη δομή της πρωτεΐνης ως μακρομόριο: Λόγος κάθε αμινοξέος ως προς το συνολικό πλήθος αμινοξέων που συναντώνται στην πρωτεΐνη (20 περιγραφείς), 2
25 Λόγος υδροφοβικών αμινοξέων προς συνολικό πλήθος αμινοξέων ( περιγραφέας), Αριθμός ελίκων, πτυχωτών δομών (φύλλων), στροφών που συναντώνται στη δευτεροταγή δομή της πρωτεϊνης (3 περιγραφείς). Έτσι συνολικά το κάθε μοντέλο αναπαρίσταται με N = 984 περιγραφείς. Για την κατηγοριοποίηση μιας μη ταξινομημένης πρωτεΐνης χρησιμοποιήθηκε το κριτήριο Ευκλείδειας απόστασης σύμφωνα με τα παρακάτω. Έστω D i (j) = [D i (),, D i (N)] ένα διάνυσμα περιγραφέων, όπου i =,,Ν total.ν total είναι το πλήθος των πρωτεϊνών και N είναι το πλήθος των περιγραφέων ανά διάνυσμα. Έστω, επίσης, κατηγορία C με διανύσματα περιγραφέων: D () = i Mc D () Nc D () D ( k) i D ( k) D Nc ( k) i D () i D ( S) Nc D ( S) Όπου N C είναι το πλήθος των πρωτεϊνών που ανήκουν στην κατηγορία C. Προκύπτουν τα χαρακτηριστικά διανύσματα (feature vectors) Χ C,, Χ Ck,, Χ CN, όπου C=,, N class, f Ck = [D (k),, D i (k),, D Nc (k)] T και N class ο συνολικός αριθμός των κατηγοριών. Τέλος ορίζετε U = [U(),..., U(N)] το διάνυσμα περιγραφέων μιας μη ταξινομημένης πρωτεΐνης U. Η κατηγοριοποίηση γίνεται ως εξής: Στην αρχή υπολογίζεται η απόσταση μεταξύ των διανυσμάτων περιγραφέων: M N 2 / 2 ( D, U) = [ ( D( j) U ( j)) ] j= Οπότε υπολογίζονται οι ευκλείδιες αποστάσεις Μ (X i,u), i =,2,..., N total όπου N total το πλήθος των πρωτεϊνών. Οι αποστάσεις ταξινομούνται κατά φθίνουσα σειρά και η προς κατηγοριοποίηση πρωτεΐνη ορίζεται ότι ανήκει στην κατηγορία της πρωτεΐνης με την οποία προκύπτει η μικρότερη απόσταση. 22
26 Αποτελέσματα Η μέθοδος δοκιμάστηκε σε ένα σύνολο δεδομένων αποτελούμενο από 263 πρωτεΐνες κατηγοριοποιημένες σε 27 κλάσεις σύμφωνα με το FSSP/DALI. Kάνοντας χρήση της κατηγοριοποίησης βάσει της Ευκλείδιας Απόστασης και μόνο των διανυσμάτων Κ 00, FT 02 επιτυγχάνει ακρίβεια, σε ότι αφορά την ταξινόμηση, που ανέρχεται στο 98,82%. Εάν χρησιμοποιηθούν και οι περιγραφείς που ανικατοπτρίζουν την πρωτεϊνική δομή η ακρίβεια αυξάνεται στο 99.63%. Το 00% της ακρίβειας δεν επιτυγχάνεται καθώς σε μία κατηγορία, σύμφωνα με την κατηγοριοποίηση κατα FSSP/DALI, μπορεί να ανήκουν και μακρομόρια που μοιράζονται κοινές λειτουργίες, αλλά διαθέτουν διαφορετική γεωμετρία και μερικώς διαφορετική αλληλουχία αμινοξέων, με αποτέλεσμα η μέθοδος να αδυνατεί να διακρίνει τα μέλη αυτά της κλάσης. Παρ όλα αυτά η ακρίβεια που επιτυγχάνεται είναι σημαντική και σε συνδυασμό με το μικρό χρόνο που απαιτείται για την κατηγοριοποίηση μιας πρωτεΐνης καθιστούν την μέθοδο αυτή ισχυρό εργαλείο. 2.7 Άλλες Μέθοδοι Πέραν των επικρατέστερων προαναφερθέντων μεθόδων για την κατηγοριοποίηση και τη διερεύνηση της λειτουργίας των πρωτεϊνικών μορίων, μία σειρά από άλλες μεθόδους είναι υπό ανάπτυξη και διερεύνηση. Στην [8] μία μέθοδος βασισμένη σε έναν σφαιρικό περιγραφέα αναπτύσεται. Η μέθοδος βασίζεται στην εξαγωγή χαρακτηριστικών, για κάθε αμινοξύ της αλυσίδας, περιγραφέων. Το ρόλο των περιγραφέων αυτών παίζει ο αριθμός ατόμων άνθρακα-ca που βρίσκονται μέσα σε μία σφαίρα με κέντρο το άτομο άνθρακα-ca του εκάστοτε αμινοξέος. Ο περιγραφέας αυτός μπορεί να χρησιμοποιηθεί έναντι των τύπων αμινοξεϊκών καταλοίπων σε αλγορίθμους δυναμικού προγραμματισμού, προσφέροντας ένα αποτελεσματικό τρόπο για την σύγκριση πρωτεϊνικών μορίων με στόχο την κατηγοριοποίηση. Τα πειραματικά αποτελέσματα 23
27 δείχνουν ότι η μέθοδος δύναται να ευθυγραμμίσει στο χώρο πρωτεϊνικά μόρια και να τα συγκρίνει με ταχύτητα και ακρίβεια. Στην [9] η θεωρία γράφων χρησιμοποιείται για τη σύγκριση πρωτεϊνικών μορίων που αναπαρίστανται με γράφους. Η μέθοδος αυτή δημιουργεί γράφους που συνδυάζουν δεδομένα που αφορούν τη δομή της αλληλουχίας της κάθε πρωτεΐνης με χημικές πληροφορίες. Η εφαρμογή της μεθόδου σε διαδικασίες κατηγοριοποίησης, χρησιμοποιώντας graph kernels και support vector machines, καταφέρνει να ξεπεράσει σε ακρίβεια μεθόδους που απαιτούν πληροφορίες για την δομή της επιφάνειας του μακρομορίου ώστε να δώσουν τα επιθυμητά αποτελέσματα. Στην [20] παρουσιάζεται μία μέθοδος που χρησιμοποιεί μία χαμηλής ανάλυσης αναπαράσταση για τη δομή της επιφάνειας του κάθε μακρομορίου. Η αναπαράσταση αυτή βασίζεται στη χρήση σημείων της επιφάνειας που είναι σημαντικά από χημική σκοπιά εκτελώντας κατακερματισμό hashing σε τρίγωνα με φυσικο χημικές ιδιότητες για την επίτευξη εκτενούς ανάλυσης των επιφανειών των μακρομορίων, τόσο για τοπικές, όσο και για συνολικές ομοιότητες. Η μέθοδος εφαρμόστηκε για την αναζήτηση γνωστών ενεργών περιοχών με επιφάνειες μακρομορίων για την εύρεση πιθανών ενεργών περιοχών στις τελευταίες. Επιπλέον, ελέχθηκε για τη δυνατότητα της να συγκρίνει ολόκληρη την επιφάνεια μιας πρωτεΐνης με μία σειρά από ενεργές περιοχές, ώστε να αναγνωριστούν κοινά στοιχεία. Τέλος, εξετάστηκε και η δυνατότητα ελέγχου μιας μακρομοριακής δομής για πιθανές, άγνωστες ενεργές περιοχές. Τα αποτελέσματα έδειξαν ότι η μέθοδος αυτή δίνει ικανοποιητικά αποτελέσματα σε σύγκριση με άλλες παρόμοιες μεθόδους. Τέλος όπως καταδεικνύεται στην [2] μια πλειάδα μεθόδων που χρησιμοποιούν την θεωρία του Μέγιστου Κοινού Ισομορφισμού Υπογράφων-Maximum Common Subgraph Isomorphism (MCS) για την, σύγκριση 2D και 3D χημικές δομές όπως οι πρωτεΐνες. Η θεωρία αυτή μετατρέπει το πρόβλημα της σύγκρισης των πρωτεϊνικών μορίων σε πρόβλημα αναζήτησης κοινών ισόμορφων υπό-γράφων στους γράφους με τους οποίους αναπαρίστανται τα προαναφερθέντα μόρια. Η πρακτική αυτή επιταχύνει σημαντικά τη σύγκριση των πολύπλοκων αυτών δομών και 24
28 προσφέρει σημαντική ακρίβεια. Εφαρμογές της λογικής αυτής εμφανίζονται στην κατηγοριοποίηση πρωτεϊνών, στην ανίχνευση πιθανής διάδρασης μεταξύ πρωτεΐνης και προσδέτη και γενικά στην εξακρίβωση της βιολογικής λειτουργίας κάθε πρωτεΐνης. Παρ ότι τα αποτελέσματα είναι ιδιαιτέρως ενθαρρυντικά περαιτέρω μελέτη πρέπει να λάβει χώρα για την εξέλιξη της πρακτικής αυτής. 2.8 Συμπεράσματα Ο έλεγχος της βιολογικής λειτουργίας των πρωτεϊνικών μορίων είναι σημαντική απαίτηση της σύγχρονης βιολογίας με σκοπό την πλήρη κατανόηση της λειτουργίας του ανθρώπινου οργανισμού και την ανάπτυξη νέων εργαλείων καταπολέμησης των σύγχρονων ασθενειών. Παρ ότι οι εργαστηριακές μέθοδοι δίνουν τα επιθυμητά αποτελέσματα η πολυπλοκότητα τους και ο χρόνος που απαιτούν, καθιστούν την εφαρμογή τους σε μεγάλης εμβέλειας μελέτες σχεδόν μή εφικτή. Το γεγονός αυτό επιβάλλει στον κλάδο της βιοπληροφορικής την ανάπτυξη υπολογιστικών μεθόδων που θα επιταχύνει την εξαγωγή των αποτελεσμάτων. Ένας μεγάλος αριθμός μεθόδων για τη διερεύνηση της βιολογικής λειτουργίας των πρωτεϊνικών μορίων έχει προταθεί στην παγκόσμια βιβλιογραφία. Οι μέθοδοι αυτές είτε προσπαθούν να επιτύχουν το στόχο του εκτελώντας κατηγοριοποίηση των πρωτεϊνών σε κλάσεις με κοινή λειτουργία, είτε μελετώντας τη δυνατότητα αλληλεπίδρασης των πρωτεϊνικών μορίων με άλλες, πρωτεϊνικές ή μη, χημικές δομές. Τα αποτελέσματα των μεθόδων αυτών είναι σημαντικά, όμως δεν έχει αναφερθεί μία μέθοδος που να δύναται να προσφέρει την ακρίβεια των εργαστηριακών μεθόδων εισάγοντας χαμηλή πολυπλοκότητα και χρονική απαίτηση. Στην εργασία αυτή παρουσιάζεται μία καινοτόμος μέθοδος που επιχειρεί να εφαρμόσει τη θεωρία γράφων στη σύγκριση πρωτεϊνικών μοντέλων με στόχο την κατηγοριοποίησή τους. Για κάθε 3D πρωτεϊνικό μοντέλο κατασκευάζεται η αναπαράσταση του με τη μορφή γράφου. Για την κατασκευή και σύγκριση των γράφων αυτών γίνεται συνδυασμένη χρήση τόσο τοπολογικών και γεωμετρικών χαρακτηριστικών, κάνοντας 25
29 χρήση των σφαιρικών αρμονικών [22] του κάθε 3D μοντέλου, όσο και βιοχημικών πληροφοριών χρησιμοποιώντας τα δεδομένα περί συμβατότητας των αμινοξέων όπως αυτά περιέχονται στους πίνακες PAM [4]. Ετσι η μέθοδος αυτή καταφέρνει να συνδυάσει το σύνολο των πληροφοριών που διέπουν μία πρωτεϊνική δομή. Τα αποτελέσματα της προτεινόμενης μεθόδου δείχνουν ότι είναι ικανή να κατηγοριοποιήσει πρωτεϊνικά μόρια με ακρίβεια μεγαλύτερη από τις ήδη υπάρχουσες μεθόδους. 26
30 Η Προτεινόμενη μέθοδος Στα επόμενα κεφάλαια της εργασίας παρουσιάζεται η μέθοδος που προτείνεται για την κατηγοριοποίηση πρωτεϊνικών δομών με στόχο τον έλεγχο της βιολογικής τους σημασίας. Στο τρίτο κεφάλαιο επεξηγείται η διαδικασία της προεπεξεργασίας των δεδομένων καθώς και της κατασκευής των γράφων, στο τέταρτο κεφάλαιο αναπτύσεται η μέθοδος σύγκρισης των γράφων που εφαρμόζεται και παρουσιάζεται ο τρόπος σύγκρισης δύο πρωτεϊνών που παράγει το μέτρο της συνολικής ομοιότητας των δύο μακρομορίων. Τέλος, στο πέμπτο κεφάλαιο παρουσιάζονται τα αποτελέσματα της μεθόδου και συνάγονται τα συμπεράσματα. 3. Προεπεξεργασία-κατασκευή γράφων. Βασικό κομμάτι της προτεινόμενης μεθόδου αποτελεί η προεπεξεργασία των πρωτεϊνικών δεδομένων που οδηγεί στην κατασκευή των γράφων με τους οποίους αυτά αναπαρίστανται. Η διαδικασία αυτή χωρίζεται σε τέσσερα στάδια: Ανάγνωση και εξαγωγή των πρωτεϊνικών δεδομένων από τα αρχεία pdb. Κατασκευή των 3D μοντέλων και διαμέριση κάθε πρωτεϊνικού μοντέλου (segmentation). Εξαγωγή γεωμετρικών περιγραφέων για κάθε μέρος (segment) των 3D μοντέλων. Τελική κατασκευή γράφων. 3. Ανάγνωση αρχείων PDB(Protein Data Bank)[4]. Κάθε αρχείο pdb αποτελείται από μία σειρά εγγραφών οι οποίες αποτελούνται απο 80 στήλες. Κάθε εγγραφή διαθέτει ένα κωδικό έξι χαρακτήρων, στην αρχή της, επεξηγηματικό της πληροφορίας που η εγγραφή περιέχει. Κάθε εγγραφή δύναται να καταλαμβάνει μία ή περισσότερες γραμμές, ενώ ορισμένες εγγραφές είναι προαιρετικές. Οι εγγραφές που συναντώνται σε ένα αρχείο pdb (Εικ.3.) είναι οι εξής: 27
31 HEADER OXYGEN TRANSPORT 8-SEP-92 CMY CMY 2 COMPND HEMOGLOBIN YPSILANTI (CARBON MONOXY FORM) CMY 3 SOURCE HUMAN (HOMO SAPIENS) BLOOD CMY 4 AUTHOR F.R.SMITH,E.E.LATTMAN, C.W.CARTER JUNIOR CMY 5 REVDAT 3-OCT-93 CMY 0 CMY 6 JRNL AUTH F.R.SMITH,E.E.LATTMAN,C.W.CARTER JUNIOR CMY 7 JRNL TITL THE MUTATION BETA99 ASP-TYR STABILIZES Y-A NEW, CMY 8... REMARK CMY 3 REMARK 2 CMY 4 REMARK 2 RESOLUTION. 3.0 ANGSTROMS. CMY 5 REMARK 3 CMY 6 REMARK 3 REFINEMENT. CMY 7 REMARK 3 PROGRAM PROLSQ CMY 8 REMARK 3 AUTHORS KONNERT,HENDRICKSON CMY 9... SEQRES D 46 VAL HIS LEU THR PRO GLU GLU LYS SER ALA VAL THR ALA CMY 90 SEQRES 2 D 46 LEU TRP GLY LYS VAL ASN VAL ASP GLU VAL GLY GLY GLU CMY 9 SEQRES 3 D 46 ALA LEU GLY ARG LEU LEU VAL VAL TYR PRO TRP THR GLN CMY HET HEM A PROTOPORPHYRIN IX WITH FE(II) CMY 02 HET HEM B PROTOPORPHYRIN IX WITH FE(II) CMY 03 HET HEM C PROTOPORPHYRIN IX WITH FE(II) CMY 04 HET HEM D PROTOPORPHYRIN IX WITH FE(II) CMY 05 FORMUL 5 HEM 4(C34 H32 N4 O4 FE ++) CMY 06 HELIX 23 DA THR D 4 VAL D 8 CMY 29 HELIX 24 DB ASN D 9 VAL D 34 CMY 30 HELIX 25 DC TYR D 35 PHE D 4 CMY 3... CRYST P CMY HETATM 07 FE HEM A CMY27 HETATM 072 CHA HEM A CMY28 HETATM 073 CHB HEM A CMY29... ATOM 3398 N VAL D CMY3544 ATOM 3399 CA VAL D CMY3545 ATOM 3400 C VAL D CMY3546 ATOM 340 O VAL D CMY3547 ATOM 3403 CG VAL D CMY3549 Εικόνα 3.: Αρχείο PDB. ) HEADER: Τετραψήφιος κωδικός για την αναγνώριση της εγγραφής στην (Protein Data Bank) PDB, περιέχει μια γενική ταξινόμηση του μακρομορίου καθώς και την ημερομηνία κατάθεσης της δομής στην PDB. 2) TITLE: Περιλαμβάνει συνήθως τα περιεχόμενα της εγγραφής, την πειραματική διαδικασία που χρησιμοποιήθηκε, την ύπαρξη μεταλλάξεων. Είναι το πεδίο όπου ο ερευνητής που καταθέτει τη δομή, αποδεικνύει τη σημαντικότητα της εργασίας του. 3) COMPOUND: Στο σημείο αυτό περιέχονται πληροφορίες για το μακρομόριο οι οποίες αναφέρονται στη δομή του καθώς και σε άλλα μικρότερα μόρια με τα οποία, πιθανώς, έχει αλληλεπιδράσει. 28
32 4) SOURCE: Βιολογική προέλευση του μακρομορίου. 5) KEYWDS: Χαρακτηριστικές λέξεις κλειδιά για το χαρακτηρισμό της δομής και την αναζήτηση της στην PDB. 6) EXPDATA: Πειραματική τεχνική προσδιορισμού της δομής. 7) AUTHOR: Λίστα με τα ονόματα των ερευνητών που συνετέλεσαν στην καταγραφή του μακρομορίου. 8) JRNL: Βιβλιογραφική αναφορά για τον προσδιορισμό της δομής που αναφέρει η παρούσα εγγραφή. 9) REMARK: Στη θέση αυτή, περιλαμβάνονται μια σειρά από πληροφορίες σχετικές με τη δομή που περιέχεται στο αρχείο. Περιέχονται βασικές βιβλιογραφικές αναφορές που σχετίζονται άμεσα με το προς εξέταση μακρομόριο. Στο πεδίο αυτό επιπλέον περιλαμβάνονται και στοιχεία σχετικά με την πειραματική διαδικασία που ακολουθήθηκε για την ανάλυση της δομής, όπως τα διάφορα προγράμματα που οι ερευνητές χρησιμοποίησαν, οι τιμές διάφορων δεικτών και γενικότερα πληροφορίες που δείχνουν την ορθότητα της εγγραφής. 0) SEQRES: Περιέχει την αμινοξεϊκή αλληλουχία του προκείμενου μακρομορίου. Στην περίπτωση των πρωτεϊνών ακολουθείται ο κώδικας τριών γραμμάτων για τα αμινοξέα. ) HET: Αναφορά στα μόρια (ετεροάτομα) που δεν είναι αμινοξέα ή νουκλεοτίδια. Αυτά μπορεί να είναι προσθετικές ομάδες και ιόντα για τα οποία έχουν προσδιοριστεί οι συντεταγμένες τους. Τα στοιχεία που παρουσιάζονται εδώ είναι ένας κώδικα αναγνώρισης για τη διάκριση με τα υπόλοιπα κατάλοιπα της εγγραφής, η 29
33 αρίθμηση που καταλαμβάνουν μέσα στο αρχείο των συντεταγμένων και τέλος ο αριθμός των ατόμων από τα οποία αποτελούνται. 2) HETNAM: Ονοματολογία των καταλοίπων που περιέχονται στο πεδίο HET. 3) FORMUL: Μοριακός τύπος των καταλοίπων που αναφέρονται στο πεδίο HET. 4) HELIX: Τμήματα της αλληλουχίας που έχουν ελικοειδή δομή. 5) SHEET: Τμήματα της αλληλουχίας που έχουν δομή πτυχωτού φύλλου. 6) TURN: Τμήματα της αλληλουχίας που έχουν στροφές. 7) CRYST: Στο πεδίο αυτό περιέχονται οι παράμετροι μοναδιαίας κυψελίδας και της ομάδας συμμετρίας. 8) ATOM: Εδώ περιέχονται οι συντεταγμένες των ατόμων στους άξονες Χ, Υ, Ζ. Το πεδίο περιλαμβάνει επίσης και άλλα στοιχεία όπως τα άτομα για τα οποία αναφέρονται οι συντεταγμένες και σε ποια κατάλοιπα ανήκουν. 30
34 3.2 Δημιουργία των 3D μοντέλων-διαμέριση (segmentation). Έχοντας αναγνώσει τα δεδομένα της πρωτεΐνης (P) είναι δυνατον, πλέον, να κατασκευαστεί η 3D απεικονισή της στο χώρο. Αυτό επιτυγχάνεται αναπαριστώντας κάθε άτομο Άνθρακα (C), Αζώτου (N), Οξυγόνου (O) και Θείου (S) με σφαίρες που έχουν κέντρο τις συντεταγμένες που δίνονται στο pdb αρχείο και ακτίνες ίσες με τις ατομικές ακτίνες του κάθε στοιχείου (Πίν.3.). Πρέπει να σημειωθεί, ότι στην παρούσα εφαρμογή δε γίνεται χρήση των ατόμων Υδρογόνου (H) καθώς τυχόν χρήση τους θα δημιουργούσε ιδιαίτερα μεγάλη αύξηση στην πολυπλοκότητα του 3D μοντέλου, επιπλέον τα άτομα υδρογόνου δεν έχουν ιδιαίτερη επίδραση στη συμπεριφορά του πρωτεΐνικού μορίου από γεωμετρικής απόψεως, ενώ τη χημική επίδρασή τους τη λαμβάνουμε υπ όψιν όταν μελετάμε συνολικά τη χημική συμπεριφορά του κάθε αμινοξέος. Επιπλέον, πρέπει να αναφερθεί ότι τα άτομα που βρίσκονται στο πεδίο HETNAM δε λαμβάνονται υπ όψιν. Άτομο Σύμβολο Ακτίνα(Angstrom) Άνθρακας C 0.77 Άζωτο N 0.70 Οξυγόνο O 0.66 Θείο S.04 Πίνακας 3.: Σύμβολα και ακτίνες των ατόμων που χρησιμοποιούνται για την 3D αναπαράσταση των πρωτεϊνών Η επιφάνεια του τρισδιάστατου μοντέλου της πρωτεΐνης (P) που προκύπτει από την παραπάνω διαδικασία στη συνέχεια τριγωνοποιείται. Αυτό έχει ως αποτέλεσμα να αναπαρίσταται ως ένα σύνολο κορυφών και ένα σύνολο ακμών. Στην Εικ.3.2 παρουσιάζεται το 3D μοντέλο μίας πρωτεΐνης. 3

35 Εικόνα 3.2: Το 3D μοντέλο της πρωτεΐνης 2dabA Στη συνέχεια η διαμέριση των κατασκευασμένων μοντέλων λαμβάνει χώρα ώστε να καταστεί δυνατή η εξαγωγή των γεωμετρικών περιγραφέων και η κατασκευή των γράφων. Για το διαμερισμό της πρωτεΐνης σε ένα σύνολο μερών S={S t, t=,,n} όπου N ο συνολικός αριθμός μερών της πρωτεΐνης λαμβάνονται υπ όψιν οι ακόλουθοι εμπειρικοί κανόνες: αμινοξεϊκά κατάλοιπα (residues) που ανήκουν στον ίδιο τύπο Δευτερεύουσας Δομής (έλικα ή φύλλο) σχηματίζουν ένα μέρος (segment) S t. w γειτονικά κατάλοιπα της ακολουθίας αμινοξέων, που δεν ανήκουν σε καμία Δευτερεύουσα Δομή σχηματίζουν ένα μέρος S t. Και τα δύο ήδη μερών παρουσιάζονται στην Εικ

 θα αντιστοιχεί σε ένα κόμβο γράφου. Στην Εικ.3.")
36 Εικόνα 3.3: Δύο μέρη(segments) της πρωτεΐνης 2dabA. Αριστερά μέρος που αντιστοιχεί σε ελικοειδή Δευτερεύουσα Δομή και δεξιά μέρος που αντιστοιχεί σε πέντε αμινοξέα. Σημειώνεται ότι μετά από δοκιμές αποφασίστηκε το μέγιστο μέγεθος ενός μέρους που δεν αποτελεί κάποια Δευτερεύουσα Δομή να μην ξεπερνά τα πέντε αμινοξέα. Η διαμέριση γίνεται ώστε η αναπαράσταση του μορίου να καταστεί πιο ευέλικτη ώστε να εξαχθούν τα γεωμετρικά χαρακτηριστικά (σφαιρικές αρμονικές) και να δημιουργηθούν οι γράφοι. Πρέπει να σημειώσουμε ότι κατά την κατασκευή των γράφων κάθε μέρος (segment) θα αντιστοιχεί σε ένα κόμβο γράφου. Στην Εικ.3.4 εμφανίζεται το τρισδιάστατο μοντέλο μια πρωτεΐνης μετά τη διαμέριση, όπου κάθε διαφορετικό μέρος είναι χρωματισμένο με διαφορετικό χρώμα. 33
37 Εικόνα 3.4:Το τρισδιάστατο μοντέλο της πρωτeΐνης 2dabA μετά την διαμέριση 3.3 Εξαγωγή Γεωμετρικών περιγραφέων. Στη μέθοδο που παρουσιάζεται, για τη γεωμετρική περιγραφή των τρισδιάστατων μοντέλων γίνεται χρήση των Σφαιρικών Αρμονικών (Spherical Harmonics) όπως αυτές παρουσιάζονται στην εργασία [22]. Ακολουθεί η περιγραφή των σφαιρικών αρμονικών και στην συνέχεια θα περιγραφεί η εφαρμογή τους στη μέθοδο αυτή. Οι σφαιρικές αρμονικές προτάθηκαν στην [22] ως ένας μετασχηματισμός σε περιγραφείς αντικειμένων ώστε να καταστούν ανεξάρτητοι από τυχόν περιστροφή του αντικειμένου στο χώρο. Οι σφαιρικές αρμονικές εισήχθησαν ως μία επέκταση του γνωστού μετασχηματισμού Fourier ο οποίος πλέον εφαρμόζεται σε σφαίρα. Οι αρμονικές βασίζονται στη θεώρηση ότι, μία οποιαδήποτε συνάρτηση f(θ,φ) (εκφραζόμενη σε σφαιρικές συντεταγμένες) δύναται να αναλυθεί σε μία σειρά σφαιρικών συντεταγμένων σύμφωνα με την παρακάτω σχέση: 34
38 f + l ( θ, φ ) = a Y m lm ( θ, φ ) l= 0 m= l Οι σφαιρικές αρμονικές αποτελούν τις λύσεις των εξίσωσεων Laplace εκφρασμένη σε σφαιρικές συντεταγμένες και ορίζονται ως εξής: Y m l ( θ, φ) = 2l + 4π l ( l m) ( l + m)! P! όπου P τα γενικευμένα πολυώνυμα Legandre. m l imφ ( cosθ ) e Η μέθοδος αυτή είναι εφαρμόσιμη σε περιγραφείς που ορίζονται ως ένα σύνολο σφαιρικών συναρτήσεων. Επιπλέον μπορεί να εφαρμοστεί σε περιγραφείς που είναι εκφρασμένοι ως συναρτήσεις μέσα σε τρισδιάστατο κυβικό πλέγμα (voxel grid), στην περίπτωση αυτή το πλέγμα αναλύεται σε σφαίρες, με κοινό κέντρο, ώστε να προκύψει το σύνολο των σφαιρικών συναρτήσεων. Τα στοιχεία που χρησιμοποιούνται για την περιγραφή των 3D μοντελών είναι τα διανύσματα συντελεστών : a l = l l + l l [ a a... a a ] T l l l l όπου l είναι η τάξη της αρμονικής. Στην Εικ.3.5 εμφανίζεται ανάλυση ενός αντικειμένου σε σφαιρικές αρμονικές. Το κυριότερο πλεονέκτημα αυτής της μεθόδου, που μας οδήγησε στη χρήση της, είναι ότι οι συντελεστές a l είναι ανεξάρτητοι από τον οποιοδήποτε μετασχηματισμό στροφής ενός αντικειμένου γύρω από το κέντρο μάζας του. Η ικανότητα αυτή των συντελεστών εμφανίζεται καθαρά στην Εικ.3.6, όπου το αρχικό αντικείμενο (αριστερά) μετασχηματίζεται με μία περιστροφή γύρω από το Εικόνα 3.5: Η ανάλυση ενός 3D μοντέλου σε σφαιρικές αρμονικές. 35

39 Εικόνα 3.6: Αντικείμενο πριν και μετά την εκτέλεση μετασχηματισμού στροφής. κέντρο μάζας του (δεξιά) και στις δύο περιπτώσεις οι συντελεστές των σφαιρικών αρμονικών είναι ιδιοι. Πρέπει να σημειωθεί στο σημείο αυτό ότι η ικανότητα της μεθόδου των σφαιρικών αρμονικών ορισμένες φορές θα μπορούσε να θεωρηθεί και ως μειονέκτημα, καθώς δε δύναται να ξεχωρίσει τα αντικείμενα, πριν και μετά το μετασχηματισμό, αν αυτό είναι επιθυμητό. Στην περίπτωση όμως, των πρωτεϊνικών δεδομένων, όπου θέλουμε να ταιριάξουμε αντικείμενα με κοινή γεωμετρία, αλλά περιεστραμμένα γύρω από το κέντρο μάζας τους, η ικανότητα αυτή είναι σημαντικό πλεονέκτημα. Η διαδικασία με την οποία υπολογίζονται οι συντελεστές των σφαιρικών αρμονικών είναι, εν γέννει, μια διαδικασία που εισάγει απώλεια πληροφορίας. Ο εισηγητής της (M.Kazhdan), βασιζόμενος σε πειραματικά αποτελέσματα, διατείνεται ότι η απώλεια πληροφορίας που εισάγει η μέθοδος είναι μικρότερη από τις αντίστοιχες μεθόδους που χρησιμοποιούνται σήμερα. Στην περίπτωση των πρωτεϊνικών μοντέλων η μεθοδολογία των σφαιρικών αρμονικών εφαρμόζεται ως εξής: Το κάθε μέρος του 3D μοντέλου (segment) εκφράζεται σε μορφή κυβικού πλέγματος (voxel). Σε κάθε ένα από αυτά τα βοξελοποιημένα μέρη ορίζονται Κ ομόκεντρες σφαίρες, με κέντρο το βαρύκεντρο του αντικειμένου και διαδοχικά αυξανόμενης ακτίνας. Από τις σφαίρες αυτές λαμβάνεται η τομή τους με το αντικείμενο. Οι τομές που προκύπτουν, θεωρούνται ως το σύνολο των σφαιρικών συναρτήσεων που απαιτουνται ως είσοδος για την εξαγωγή των σφαιρικών αρμονικών. 36
40 Τέλος, υπολογίζονται οι συντελεστές των l πρώτων σφαιρικών αρμονικών για κάθε μία από τις παραπάνω σφαιρικές συναρτήσεις, ώστε να κατασκευαστούν τα διανύσματα περιγραφής. Ο αριθμός Κ, των σφαιρών, λαμβάνεται έτσι ώστε να γίνεται η καλύτερη δυνατή αναπαράσταση του αντικειμένου και στην περίπτωσή μας είναι ίσος με Κ = 6. Επιπλέον για την περιγραφή κάθε μιας από τις 6 σφαιρικές συναρτήσεις που προκύπτουν υπολογίζονται οι l=34 πρώτες σφαιρικές αρμονικές. Με αυτόν τον τρόπο για κάθε μέρος (S t ) του πρωτεϊνικού μοντέλου (P) έχουμε ένα διάνυσμα : όπου D t = L = K l = 6 34 = 544 t t t [ a a a ] T 0 L το σύνολο των συντελεστών για κάθε μέρος του μοντέλου. Η παραπάνω διαδικασία εκτελέστηκε κάνοντας χρήση της εφαρμογής που ο ίδιος ο εισηγητής, της μεθόδου, προσφέρει για ελεύθερη χρήση στον παρακάτω διαδικτυακό τόπο: Κατασκευή Γράφων. Τελευταίο βήμα της προεπεξεργασίας αποτελεί η τελική δόμηση του γράφου που περιέχει όλες τις γεωμετρικές και τοπολογικές πληροφορίες που διέπουν το εκάστοτε πρωτεϊνικό μοντέλο. Σύμφωνα με τη θεωρία γράφων, ένας γράφος μαθηματικά εκφράζεται ως G = r s = { V, E, { A i } i=, { Bi} i } [23] όπου V {( u) p, p =,, n} n E Ai = το μηκενό σύνολο των κόμβων, το σύνολο των ακμών, ο πίνακας γειτονίας που φέρει την i-οστή ιδιότητα ακμής, B i η τιμή της i-οστής ιδιότητας κόμβου, r ο αριθμός των ιδιοτήτων των ακμών και s ο συνολικός αριθμός ιδιοτήτων που τοποθετούνται στους κόμβους. Με σκοπό να εφαρμόσουμε τη θεωρία αυτη, κατασκεύαζεται ένας πρωτεϊνικός γράφος αποτελούμενος από ότι N n = N κόμβους (υπενθυμίζεται είναι ο συνολικός αριθμός μερών (segments) που διαθέτει κάθε πρωτεϊνικό 3D μοντέλο). Κάθε μέρος (St) αντιστοιχίζεται σε έναν κόμβο 37
41 v t, του γράφου. Οι ακμές σχηματίζονται έτσι ώστε να ακολουθούν τον τρόπο με τον οποίο συνδέονται τα διάφορα μέρη (segments) μέσα στη πρωτεϊνική δομή. Έτσι ο τελικός γράφος για την πρωτεϊνική δομή (P) έχει τη μορφή: G P { V E A B i i= s =,,, { } }, όπου A ένας δυαδικός ορθογώνιος πίνακας γειτνίασης(adjacency matrix). Το γεγονός αυτό είναι αποτέλεσμα της παραδοχής ότι για την εφαρμογή της θεωρίας γράφων, στην προτεινόμενη μέθοδο, ολόκληρη η πληροφορία της πρωτεΐνης τοποθετείται στους κόμβους του γράφου έτσι, οι ακμές δε διαθέτουν ιδιότητες (attributes). Για λόγους ευελιξίας και ορθής λειτουργίας όμως, θεωρούμε ότι όλες οι ακμές έχουν μία ιδιότητα που για όλες έχει την ίδια τιμή, ορισμένη να είναι ίση με την μονάδα () για λόγους απλότητας. Το τελικό διάνυσμα ιδιοτήτων για κάθε κόμβο v t είναι : dt = [ D, ω] T t όπου ω είναι η γωνία που σχηματίζεται μεταξύ δύο διαδοχικών ακμών. Η τιμή της γωνίας ω χρησιμοποιείται, πολλαπλασιασμένη με βάρος 00 ώστε να ενισχυθεί η επιρροή της, με στόχο να εμπλουτιστεί ο γράφος με στοιχεία της τοπολογίας του πρωτεϊνικού μοντέλου, εκτός των γεωμετρικών στοιχείων που ήδη περιέχει. Στην Εικ.3.7 παρουσιάζεται ένας ολοκληρωμένος γράφος όπου φαίνονται οι κόμβοι και οι ακμές του, καθώς επίσης και η γωνία ω δύο διαδοχικών ακμών. 38

42 Εικόνα 3.7: Ο τελικός γράφος που αντιστοιχεί στο 3D μοντέλο της πρωτεΐνης 2dabA 39
43 4. Σύγκριση Γράφων (Graph Matching) Εύρεση τελικού βαθμού ομοιότητας μεταξύ πρωτεϊνών. Εφ όσον έχουν κατασκευαστεί οι γράφοι που αναπαριστούν τα πρωτεϊνικά μόρια, ακολουθεί η σύγκριση των γράφων με απώτερο σκοπό την εξαγωγή ενός μέτρου ομοιότητας μεταξύ δύο πρωτεϊνών. Η διαδικασία αυτή παρουσιάζεται στο παρόν κεφάλαιο ξεκινώντας με τη θεωρία και πρακτική που ακολουθείται για τη σύγκριση των γράφων και καταλήγοντας με την εύρεση του τελικού μέτρου ομοιότητας. 4. Σύγκριση Γράφων(Graph Matching). Στη μέθοδο αυτή για την επίτευξη της σύγκρισης δύο πρωτεϊνικών μορίων που απεικονίζονται ως γράφοι ακολουθείται η μέθοδος SPGM(Successive Projection Graph Matching) όπως αυτή εισάγεται στην εργασία [23]. Στην παράγραφο αυτή θα γίνει μία σύντομη εισαγωγή στο πρόβλημα της σύγκρισης των γράφων και θα αναπτυχθεί η μέθοδος που ακολουθείται στην εργασία αυτή. 4.. Το πρόβλημα της Σύγκρισης Γράφων Όπως προαναφέραμε στο προηγούμενο κεφάλαιο ένας γράφος G G = r s = { V, E, { A i } i=, { Bi} i } εκφράζεται μαθηματικά με τη μορφή. Με βάση την αναπαράσταση αυτή ορίζεται ότι : Ένας γράφος G = G = r s = { V, E, { A i } i=, { Bi} i } r s ' = { V ', E', { A' i } i=, { B' i } i } είναι ισόμορφος,με ένα γράφο, εάν υπάρχει μία, ένα προς ένα, αντιστοιχία μεταξύ των συνόλων των κόμβων V και V και μία, ένα προς ένα, αντιστοιχία των συνόλων E και E των ακμών έτσι, αν e είναι μία ακμή με άκρα τους κόμβους u και v, του γράφου G, τότε η αντίστοιχη ακμή e του γράφου G έχει τα άκρα της στους κόμβους u και v του G που αντιστοιχίζονται στα u και v. Ένα τέτοιο ζευγάρι αντιστοιχιών ονομάζεται ισομορφισμός γράφων. 40
44 Με δεδομένους δύο γράφους G = r s = { V, E, { A i } i=, { Bi} i } και G = r s ' = { V ', E', { A' i } i=, { B' i } i } Άν υπάρχει ένας n n' ορίζεται ότι : υποπίνακας P όπου T Ai = PA' i P, i =,..., r και B = PB' j =, j j,, s Τότε οι γράφοι G και G είναι ισόμορφοι και τους θεωρούμε ταιριασμένους (matched). Το πρόβλημα της σύγκρισης γράφων έγκειται στην εύρεση του πίνακα P που ικανοποιεί τις παραπάνω εξισώσεις. Πρέπει να σημειώσουμε στο σημείο αυτό ότι ως G θα αποκαλούμε τον γράφο αναφοράς και ως G το γράφο προς σύγκριση. Το πρόβλημα αυτό ονομάζεται σύγκριση πλήρων γράφων (Fullgraph matching) όταν n = n', ενώ όταν υπό-γράφων (sub-graph matching). n < n', αναφέρεται ως σύγκριση Στην εργασία αυτή χρησιμοποιούμε τη θεωρία που αντιμετωπίζει την περίπτωση της σύγκρισης υπογράφων τόσο λόγω γενικότητας της οπτικής αυτής, αλλά όσο και του γεγονότος ότι είναι δυνατόν δύο πρωτεΐνες να έχουν διαφορετικό μέγεθος, αλλά η γεωμετρία τους να ταιριάζει, μερικώς, ώστε να εκτελούν την ίδια λειτουργία μέσα στο βιολογικό τους περιβάλλον. Σε τελική ανάλυση το πρόβλημα της σύγκρισης των γράφων ανάγεται στο σύνθετο πρόβλημα βελτιστοποίησης που επιτάσσει την εύρεση ενός πίνακα μετασχηματισμού P που ανήκε στο σύνολο όλων των δυνατών πινάκων P n n' ελαχιστοποιεί την παρακάτω εξίσωση: μεταξύ των δύο, υπό σύγκριση, γράφων και r i= A i PA P i T q + s j= B j PB j q () όπου μία νόρμα πινάκων (π.χ. μέτρο πίνακα) και q σταθερά με τυπικές τιμές και 2. Λόγω της πολυπλοκότητας που παρουσιάζει το πρόβλημα 4
45 αυτό αναζητούνται προσεγγιστικές λύσεις, ώστε να μειωθεί η δυσκολία επίλυσης και ο απαραίτητος, γι αυτήν, χρόνος. Μία τέτοια λύση χρησιμοποιείται και στην παρούσα εργασία Μέθοδος Σύγκρισης Γράφων Για την επίλυση του προαναφερθέντος προβλήματος γίνεται χρήση μιας μεθόδου γνωστής ως SPGM που εισάγεται για πρώτη φορά στην [23] και βασίζεται στη θεωρία. Η μέθοδος αυτή επιχειρεί να μετασχηματίσει το σύνθετο πρόβλημα βελτιστοποίησης σε ένα πρόβλημα ελαχιστοποίησης το οποίο αντιμετωπίζεται με μία, σχετικά, απλή επαναληπτική διαδικασία Απαραίτητη θεωρητική προσέγγιση Για την επίτευξη της απλοποίησης του προβλήματος αρχικά θέτουμε ως στόχο την εύρεση ενός πίνακα μετασχηματισμού P, ο οποίος θα προσεγγίζει ικανοποιητικά τον πίνακα P που επαληθεύει την εξίσωση (). Εάν στον πίνακα P εφαρμοστεί ο, γνωστός από την γραμμική άλγεβρα, μετασχηματισμός της ανυσματοποίησης (vectorization) που συμβολίζεται με vec(.) το πρόβλημα ελαχιστοποίησης της εξ.() μετατρέπεται στο πρόβλημα της ελαχιστοποίησης της παρακάτω συνάρτησης: T T J(p) = p Xp p Ip y 2 2 T T = p (X + I)p y p 2 στη συνάρτηση αυτή p = vec(p) και ακολουθούνται οι παρακάτω περιορισμοί: T p (2) 0 P ij (3) n j= P ij =, i =,, n (4) και 42
46 n i= P ij, j =,, n (5) Στην περίπτωση αυτή P : = ( P ). Επιπλέον X είναι πίνακας που φέρει τιμές ij συμβατότητας μεταξύ των ακμών, y ένα άνυσμα που περιέχει τιμές συμβατότητας μεταξύ κόμβων και I ένας πίνακας για την αναγνώριση των διαφόρων συστατικών των γράφων (ακμές και κόμβους). Όπως T αποδεικνύεται στην [24] ο όρος p Ip, οδηγεί στην ευκολότερη 2 σύγκλιση των στοιχείων P ij σε τιμές είτε ίσες με μηδέν, είτε ίσες με τη μονάδα. Τα στοιχεία του πίνακα συμβατότητας τη συνάρτηση συμβατότητας: Χ R n n nn βρίσκονται από X kl = r g= A g l k l n, k n n n a A g l + k, n n + + a (6) k l k = l, + = + n n εκτός όταν k l ή k n = l n n n g n n όπου στην περίπτωση αυτή X = 0. Επιπλέον έχουμε : = ( ) R, ) kl g n n A g : = ( A ij R,. συμβολίζει την απόλυτη τιμή, k =,, nn, A g A ij l =,, nn και a παράμετρος που ελέγχει την ταχύτητα σύγκλισης της συνάρτησης συμβατότητας και συνήθως επιλέγεται ίση με τη μονάδα. Επιπλέον πρέπει να επισημάνουμε ότι A και αντίστοιχα g A g αντιπροσωπεύουν τους πίνακες με τις ιδιότητες των ακμών των δύο γράφων. Με παρόμοια λογική το άνυσμα συμβατότητας y βρίσκεται από τη συνάρτηση : 43
47 y k = s h= B h k k n n a B h k n + + a (7) B h h : = ( B i ) R n h n h : = ( B i ) R Όπου,, B j B j B k =,, nn. Σημειώνεται ότι και αντιστοίχως είναι τα ανύσματα που φέρουν τις ιδιότητες των κόμβων για το γράφο αναφοράς και το γράφο σύγκρισης. Όπως φαίνεται στις (3), (4) και (5) έχουν επιβληθεί στις συναρτήσεις μία σειρά από περιορισμούς ώστε να καταστούν ορθές για την περίπτωση των γράφων, αυτοί οι περιορισμοί μπορούν να διατυπωθούν με τη μορφή συνόλων ως εξής: πίνακα και : C C r = pr R = p για τις στήλες πίνακα. c C R n n n n : : pr = [p,, pn ], pi = [Pi,, Pin ], n = Pij j= 0 Pij p p i n c i= = [p = [P P ij 0 P ij j,, p,, P n nj ], ],, για τις γραμμές του Και τα δύο αυτά σύνολα αποδεικνύεται [23 παρ.7.6] ότι είναι κλειστά και κοίλα, επιπλέον οι ιδιότητες αυτές κληρονομούνται και στη μηκενή τομή τους C 0 = Cr Cc. Το γεγονός ότι η προαναφερθείσα τομή είναι κοίλη κλειστή και μη-κενή μας δίνει τη δυνατότητα να χρησιμοποιήσουμε τα αποτελέσματα της θεωρίας Προβολών σε Κοίλα 44
48 Σύνολα (Projections Onto Convex Sets, POCS) όπως παρουσιάζονται στην παράγραφο 7.2 της [23], ώστε να επιβάλουμε στον επαναληπτικό μας αλγόριθμο τους παραπάνω περιορισμούς Ο Αλγόριθμος Ο κύριος βρόγχος του αλγορίθμου που υλοποιείται στη μέθοδο αυτή προσδιορίζεται από τα παρακάτω βήματα:. Δημιουργία μιας τοπικής προσέγγισης της συνάρτησης γύρω από ένα σημείο που είναι δυνατό να προσεγγιστεί. 2. Υπολογισμός του ελάχιστου της τοπικής προσέγγισης που είναι πιθανόν να μη συμπίπτει με το σημείο που επιλέξαμε παραπάνω. 3. Χρήση επαναληπτικών διαδικασιών ώστε να βρεθεί ένα νέο σημείο όσο το δυνατό ποιο κοντά στο σημείο που υπολογίστηκε στο βήμα Έξοδος αν τα κριτήρια σύγκλισης επιτευχθούν, ειδάλλως επιστροφή στο βήμα κάνοντας χρήση το σημείο που υπολογίστηκε στο βήμα 3 ως το νέο προς προσέγγιση σημείο. Ο παραπάνω βρόγχος παρουσιάζεται στον ψευδοκώδικα που ακολουθεί: Ψευδοκώδικας :Βασικός βρόγχος του αλγορίθμου begin initialize p while( k > I and δ > ρ) or ( k < 3) ~ p p k+ k+ δ = ( p k+ k k + end while end 0 0, k =, δ > ρ. k J ( p = p γ J ( p k T ( ~ + = p ) p k ) T ( p k k k+ ) ) p k ) 45
49 T ( ~ p 0 p [,,, ] n n n 0 Το άνυσμα αρχικοποιείται ως ) αντιπροσωπεύει τη διαδικασία προβολής. Ο συμβολισμός k+ ~ k+ της τιμής p στο σύνολο C 0, που θα παρουσιαστεί παρακάτω. Λαμβάνουμε το ~ k+ p προσεγγίζοντας της εξίσωση (2) με τη σφαιρική συνάρτηση : J( ~ p ) = J(p) + J(p )( ~ p p ) + ( ~ p 2 ~ k+ T k k+ k k+ k k+ k S k = diag ( s k,, s k ) = s k I J(p k ) p k )S ( ~ p p όπου, συμβολίζει το διαφορικό άνυσμα - Xp y p της εξίσωσης (2) δοσμένο ως και παράμετρος k s k ) (8) καμπυλότητας. Το άνυσμα ~ k+ p τελικά υπολογίζεται ως το ελάχιστο της σφαιρικής συνάρτησης, το οποίο επιτυγχάνεται όταν : T J(p k ) + ( ~ p k+ p k ) T S k = 0 (9) Έτσι, καθώς S k = s k I εξάγουμε τον απλό κανόνα αναδρομής p = p k s [ J(p )] ~ k+ k k. Με σκοπό να κατασταθεί η παράμετρος καμπυλότητας ανεξάρτητη από το μέγεθος των τιμών των χαρακτηριστικών των γράφων, το διαφορικό άνυσμα κανονικοποιείται οπότε εξάγεται η παρακάτω εξίσωση: ~ p k+ = p k J(p J(p k k ) ) γ (0) Η εξίσωση (0) είναι ο κανόνας που χρησιμοποιείται τελικά στον παραπάνω αλγόριθμο. Η τιμή γ είναι παράμετρος που ορίζει το μέγεθος του βήματος, και συνήθως επιλέγεται ως μία σχετικά μεγάλη τιμή, για παράδειγμα 0, επιπλέον. συμβολίζει το Ευκλείδειο μέτρο του ανύσματος. Στην παρούσα εφαρμογή μετά από πειραματικές δοκιμές, με στόχο να λάβουμε τα καλύτερα δυνατά αποτελέσματα, επιλέχθηκε ρ = 0. 00, γ =.0 I < 6n και, σημειωτέον ότι η μεταβλητή ορίζει το μέγιστο αριθμό I 46
50 επαναλήψεων που θα λάβουν χώρα όπως φαίνεται και από τον παραπάνω ψευδοκώδικα. διαδικασία Ολοκληρώνοντας, κατά τον τερματισμό του αλγόριθμου εκτελείται η P = devec(p), ώστε να πάρουμε την τελική εκτίμηση για τον πίνακα μετασχηματισμού που οδηγεί στον ισομορφισμό των δύο γράφων σύμφωνα με τον αρχικό ορισμό του προβλήματος. Όπως είναι προφανές η διαδικασία p = vec(p). P = devec(p) συμβολίζει τη διαδικασία που αντιστρέφει την Σύμφωνα με τον παραπάνω αλγόριθμο έχοντας το άνυσμα ~ k+ p k+ p C0 στόχος είναι να βρούμε το άνυσμα έτσι ώστε : ~ k+ k+ k+ p p = min ~ p z C 0 z () Κάνοντας χρήση των αποτελεσμάτων της θεωρίας POCS η παραπάνω εξίσωση υπονοεί ότι το άνυσμα k p + ~ k+ p είναι η ορθογώνια προβολή του πάνω στο σύνολο C 0 Cr Cc = ([25],[26]). Εφαρμόζοντας την θεωρία αυτή μπορούμε να κατασκευάσουμε μία αλληλουχία που να συγκλίνει το T ( ~ p k+ εξαγόμενο σε ένα σημείο του αρκετά κοντά στο. Ο 0 αλγόριθμος για την εύρεση του ) T ( ~ p 0 k+ C 0 ) k p + περιγράφεται παρακάτω για λόγους απλότητας ο μετρητής k + και η περισπωμένη (~) δεν αναφέρονται αλλά εννοούνται. 47
51 Begin While( δ end end p k while Initialize C h p p δ 0 C c 0 > = p T T r c = (p k : k (p) (p) c h c p) + =, δ and T k (p C c h 0 > < n ) p) Ψευδοκώδικας 2: Αλγόριθμος για την Ευρεση του ~ k+ T ( p ) Στον ψευδοκώδικα αυτό με p 0 συμβολίζεται η προβολή του σημείου στο σύνολο C, ενώ με T c (p) συμβολίζεται η αντίστοιχη προβολή στο σύνολο. Αλγόριθμοι για τις προβολές αυτές παρουσιάζονται παρακάτω. Αξιοσημείωτο είναι ότι για τον υπολογισμό των προβολών αυτών γίνεται χρήση αλγόριθμων που υλοποιούν σταθερό αριθμό επαναλήψεων κάνοντας χρήση σταθερού αριθμού επαναλήψεων, καθώς μία κλειστή λύση με βάση τις γνωστές συνθήκες Kuhn-Tucker [26]. Επιπλέον ο συμβολισμός p ότι επιλέγετε κάθε j-οστό στοιχείο του ανύσματος ξεκινώντας από το i-οστο στοιχείο και καταλήγοντας στο k- οστό. Η διαδικασία r C c p(i:j:k) (p) T r στοιχείων του p σε αύξουσα σειρά, όπου και d [s, d] = sort[p] αντιστοιχεί στην ταξινόμηση των είναι το ταξινομημένο άνυσμα ένα άνυσμα που φέρει τις θέσεις των στοιχείων πριν την ταξινόμηση. s 48
52 Ψευδοκώδικας 3:Υπολογισμός της προβολής στο σύνολο C r begin for i = :n φ = sum [p( i:n:n ( n -) + i)] σ = n p = p( i:n:n ( n -) + i) [s, d] = sort [p ] for j = :n -φ s( j) s( j) + σ if s( j) < 0 s( j) = 0 φ φ - p (d( j)) σ σ - end if p (d( j)) = s( j) end for p( i:n:n end for end ( n -) + i) = p 49
53 Όταν, ο υπολογισμός του γίνεται με τον ίδιο ακριβώς τρόπο με το περίπτωση όπου. n = n' T c (p) (p) T r n < n'. Παρακάτω παραθέτουμε τον ψευδοκώδικα στην Ψευδοκώδικας 4:Υπολογισμός της προβολής στο σύνολο C c begin for j = :n φ = sum [p( n( j ) + : nj )] if φ > σ = n p = p( n( j ) + : nj ) [s, d] = sort [p ] for ι = :n -φ s( ι ) s( ι ) + σ if s( ι ) < 0 s( ι ) = 0 φ φ - p (d( ι )) σ σ - end if p (d( ι )) = s( ι ) end for p( n( j ) + : nj ) = p end for end 50
54 Σημειώνουμε ότι η εξαγωγή των προαναφερθέντων αλγορίθμων παρουσιάζεται αναλυτικά από τον εισηγητή τους στην εργασία [23] και συγκεκριμένα στην παράγραφο 7.6. Ολοκληρώνοντας την παρουσίαση της μεθόδου για τη σύγκριση των γράφων θα παρουσιαστούν κάποια στοιχεία για την πολυπλοκότητα της. Αρχικά, πρέπει να σημειώσουμε ότι οι αλγόριθμοι που παρουσιάζονται στους ψευδοκώδικες 3 και 4 χρησιμοποιούν μία διαδικασία ταξινόμησης των στοιχείων που περιέχονται στο άνυσμα εάν η διαδικασία που θα χρησιμοποιηθεί είναι αυτή της Φυσσαλίδας (Bubble Sort), όπως γίνεται στη μέθοδό μας, έχουμε να εκτελέσουμε συγκρίσεις και 2 n 2 αλλαγές θέσεων, κατά μέσο όρο και στη χειρότερη περίπτωση, όπου n το πλήθος των στοιχείων του ανύσματος υπολογισμός του απαιτεί p p 2 n 2. Συνεπώς ο 2 2 n n T r (p) n ( + ) λειτουργίες όταν 2 2 χρησιμοποιείται bubble sort και αν θεωρήσουμε την περίπτωση που n = n' υπολογισμό. Επιπλέον συγκρίσεις απαιτούνται από τον αλγόριθμο για τον του 2 n T ( ~ p 0 k+ ) συνεπώς ο υπολογισμός του θα έχει πολυπλοκότητα O( n 4 ). Καθώς και ο υπολογισμός του διαφορικού J(p k ) εισάγει πολυπλοκότητα O( n 4 ), ο συνολικός αλγόριθμος πολυπλοκότητα O( n 4 ) για κάθε επανάληψη. 4.2 Εξαγωγή Τελικού Βαθμού Ομοιότητας. Έχοντας δύο πρωτεϊνικά μόρια τα οποία αναπαρίστανται με τους γράφους και, ακολουθώντας τον φορμαλισμό που προτάθηκε στην Παρ.3..4 με αριθμό κόμβων s και G P B i = s = { V, E, A, { } i } s G s = { V, E, A, { } i } P B i αντίστοιχα, εκτελείται η διαδικασία σύγκρισης των = 5
55 γράφων. Από τη διαδικασία αυτή εξάγεται ο πίνακας P = [ p ] όπου τα ij στοιχεία p ij εκφράζουν την πιθανότητα ο i-οστός κόμβος του πρώτου γράφου να είναι όμοιος με τον j-οστό κόμβο του δεύτερου γράφου. Το τελευταίο στάδιο είναι να συνδυάσουμε τα αποτελέσματα αυτά που αντιπροσωπεύουν γεωμετρικά και τοπολογικά χαρακτηριστικά με τα βιοχημικά δεδομένα ώστε να εξαχθεί ο τελικός βαθμός ομοιότητας. Για το σκοπό αυτό, ο πίνακας P σαρώνεται, ώστε να ανιχνευθούν συνεχόμενα ζεύγη κόμβων με μεγάλο βαθμό ομοιότητας και να σχηματιστούν αλυσίδες ομοίων κόμβων. Ένα ζεύγος κόμβων για να ανήκει σε μία τέτοια αλυσίδα θα πρέπει να έχει πιθανότητα ομοιότητας μεγαλύτερη από ένα συγκεκριμένο κατώφλι (το κατώφλι αυτό μετά από πειραματικές δοκιμές ορίστηκε να είναι 0.90 για τους σκοπούς της εφαρμογής μας). Στη συνέχεια από τις αλυσίδες που δημιουργήθηκαν χρησιμοποιούνται μόνο αυτές που έχουν μήκος μεγαλύτερο από το 5% του πλήθος κόμβων που φέρει ο μικρότερος, από τους προς σύγκριση δύο, γράφος. Αυτό συμβαίνει ώστε να μη συμπεριληφθούν στην ανάλυση πολύ μικρές αλυσίδες που θα μπορούσαν να δημιουργηθούν λόγω τυχαίας τοπικής ομοιότητας μεταξύ δύο πρωτεϊνών διαφορετικής κλάσης. Στο τελευταίο στάδιο από κάθε αλυσίδα ομοιότητας που απομένει εξετάζονται ένα ένα τα ζεύγη κόμβων που αυτή περιέχει. Κάθε κόμβος όπως αναφέραμε κατά την κατασκευή του γράφου αποτελείται από μία σειρά αμινοξέων, έτσι κάθε ζεύγος όμοιων κόμβων αποτελείται από ζεύγη όμοιων αμινοξέων. Κάθε ένα τέτοιο ζεύγος αμινοξέων εξετάζεται με γνώμονα τον πίνακα PAM250 [4] και αντιστοιχίζεται σε ένα βαθμό ομοιότητας των αμινοξέων που το απαρτίζουν. Πρέπει να σημειωθεί ότι ο πίνακας PAM250 παρέχει ένα μέτρο βιολογικής ομοιότητας δύο αμινοξέων και δίνει αρνητικό, έως -8, για ανόμοια αμινοξέα και θετικό βαθμό, έως +2, για όμοια. Οι βαθμοί των αμινοξέων, κανονικοποιούνται σε τιμές μεταξύ και, αθροίζονται ώστε να αποδώσουν ένα βαθμό βιολογικής ομοιότητας των δύο κόμβων, οι βαθμοί αυτοί των κόμβων με τη σειρά τους αθροίζονται ώστε να δώσουν ένα συνολικό μέτρο για την ποιότητα προς εξέταση αλυσίδας. Τα μέτρα αυτά υπολογίζονται για κάθε μία από τις 52
56 αλυσίδες ομοιότητας και στη συνέχεια αθροίζονται παράγοντας έτσι ένα τελικό score ομοιότητας για τις δύο συγκρινόμενες πρωτεΐνες. 5. Πειραματικά αποτελέσματα - συμπεράσματα. Για τον έλεγχο της ακρίβειας της μεθόδου ως προς την κατηγοριοποίηση κατασκευάστηκε ένα σύνολο από πρωτεΐνες. Το σύνολο αυτό κατηγοριοποιήθηκε σύμφωνα με τη μέθοδο DALI/FSSP. Η μέθοδος χρησιμοποιείται ως βάση αλήθειας καθώς είναι η πλέον ακριβής, μηυπολογιστική μέθοδος, σε ό,τι αφορά την κατηγοριοποίηση. Το σύνολο ελέγχου που δημιουργήθηκε αποτελείται από 066 πρωτεΐνες χωρισμένες σε 3 κλάσεις όπως φαίνεται στον πίνακα 5.. Κατηγορίες Πληθυσμός a0ca 0 3lzt 38 3nul 5 4icb 8 6mhtA 4 aofa 7 daaa 4 ctqa 24 a6m 60 igtb 20 abwa 235 cnza 26 l Σύνολο 066 Πίνακας 5.: Σύνολο πρωτεϊνών για τον έλεγχο της μεθόδου Για τον έλεγχο της μεθόδου ακουλουθήθηκε το παρακάτω πείραμα κατηγοριοποίσης: 53
57 Οι πρωτεΐνες συγκρίθηκαν μεταξύ τους ανά ζεύγη και ο τελικός βαθμός ομοιότητας κάθε ζεύγους αποθηκεύτηκε σε έναν πίνακα ομοιοτήτων. Οι βαθμοί ομοιοτήτων ταξινομήθηκαν και οι πρωτεΐνες κατηγοριοποιήθηκαν. Για την κατηγοριοποίηση των πρωτεϊνικών δεδομένων ακολουθήθηκε η μέθοδος του κοντινότερου γείτονα (nearest neighbor approach). Σύμφωνα με την προσέγγιση αυτή κάθε πρωτεΐνη αντιστοιχίζεται στην κλάση που ανήκει η εγγύτερη σε αυτήν πρωτεΐνη. Το παραπάνω πείραμα κατέδειξε ότι η προτεινόμενη μέθοδος μπορεί αποτελεσματικά να κατηγοριοποιήσει το παραπάνω σύνολο πρωτεϊνών στην ολότητα του επιτυγχάνοντας την αξιοσημείωτη, για υπολογιστική μέθοδο, ακρίβεια 98,69%. Το αποτέλεσμα αυτό μπορεί να θεωρηθεί εξαιρετικής σημασίας, καθώς η επίδοση αυτή είναι ιδιαίτερα κοντά στη μέθοδο-πρότυπο που χρησιμοποιήθηκε ως βάση αλήθειας. Η επίδοση αυτή είναι πολύ ενθαρρυντική για τη χρήση της προτεινόμενης μεθόδου είτε αυτόνομα, είτε ως φίλτρο πριν τη χρήση άλλων πολυπλοκότερων μεθόδων. Πέραν της επίδοσης ως προς την κατηγοριοποίηση, εκτιμήθηκε και η αποτελεσματικότητα, της προτεινόμενης μεθόδου σύγκρισης πρωτεϊνών, ως προς την ανάκτηση. Για το σκοπό αυτό, κάθε πρωτεϊνικό μοντέλο της βάσης, χρησιμοποιείται ως ερώτημα (query) και τα εξαγόμενα αποτελέσματα ιεραρχούνται με βάση το βαθμό ομοιότητας τους με το μοντέλο-ερώτημα, τέλος το αντικείμενο με τη μεγαλύτερη ομοιότητα λαμβάνεται ως επιστροφή του συστήματος. Μία αναζήτηση θεωρείται σωστή όταν το αντικείμενο που επιστρέφεται ανήκει στην ίδια κατηγορία με το μοντέλο ερώτημα. Παρακάτω παρουσιάζονται ενδεικτικά μία σειρά από μοντέλα-ερωτήματα με τα μοντέλα που επιστρέφει η μέθοδος. Στις εικόνες αυτές [5.,5.2] και στην αριστερή στήλη παρουσιάζονται τα μοντέλαερωτήματα (query) που εισάγονται στη μέθοδο, ενώ στην δεξιά στήλη εμφανίζονται τα μοντέλα που επιστρέφει η μέθοδος. 54
58 Στην Εικόνα 5. παρουσιάζεται μία σειρά από τέσσερα ζευγάρια πρωτεϊνών που αποτελούν επιτυχημένες ανακλήσεις. Στα ζευγάρια αυτά είναι φανερή η γεωμετρική ομοιότητα των 3D μοντέλων, επιπλέον το γεγονός ότι τα μοντέλα αυτά δίνουν θετικό βαθμό ομοιότητας μαρτυρεί και τη βιολογική τους αντιστοιχία. Έτσι η κρίση ότι τα αντικείμενα αυτά είναι όμοια και η τοποθέτησή τους στην ίδια κατηγορία, δικαιολογείται απόλυτα. Είναι αξιοπρόσεκτο, ότι ο εντοπισμός της ομοιότητας δύο μοντέλων είναι δυνατός είτε αυτά είναι σχετικά αραιά και μικρά σε πλήθος ατόμων, ζευγάρι pdbazf-pdbat5, είτε πυκνά και μεγάλα, ζευγάρια pdba0dd pdba0db και pdbg2ua pdbgc9a. Τέλος, η αποτελεσματικότητα δεν χάνεται ούτε όταν εφαρμόζεται η μέθοδος σε συμπλέγματα με περισσότερες από μία αλυσίδες, ζευγάρι pdbe6jh pdbceh. Στην Εικόνα 5.2 εμφανίζονται μία σειρά από ανεπιτυχείς ανακλήσεις. Τα ζεύγη των μοντέλων αυτών, όπως καταδεικνύουν οι εικόνες αυτές, παρουσιάζουν τοπική γεωμετρική ομοιότητα, το γεγονός αυτό σε συνδυασμό με την εμφάνιση θετικού βαθμού ομοιότητας για τα αμινοξέα των όμοιων περιοχών οδηγεί τη μέθοδο να τα επιστρέψει ως όμοια αντικείμενα και να τα κατατάξει, λανθασμένα, στην ίδια κλάση. Η προαναφερθείσα γεωμετρική ομοιότητα είναι ευκρινής, για παράδειγμα, στο ζευγάρι υπ αριθμόν 4 (pdb05m pdb2hbda) όπου τα δύο μοντέλα είναι σχεδόν πανομοιότυπα. Ο λόγος που η μέθοδος FSSP/DALI δε θεωρεί τα παραπάνω αντικείμενα όμοια και δεν τα κατατάσσει στην ίδια κλάση είναι ότι απαιτεί τουλάχιστον το 25% των αμινοξέων δύο πρωτεϊνών να είναι απολύτως ίδια, και όχι απλά όμοια, για να τις κατατάξει στην ίδια κλάση και ότι λαμβάνει υπ όψιν πρόσθετα κριτήρια που αποδίδουν μία λεπτομερέστερη άποψη της βιολογικής συμπεριφοράς μιας πρωτεΐνης. Η χρήση των κριτηρίων αυτών καθιστούν τη μέθοδο FSSP/DALI ακριβέστερη, αλλά σε μεγάλο βαθμό πολυπλοκότερη. Το μοναδικό ζευγάρι που προκαλεί προβληματισμό είναι το υπ αριθμόν (pdbyprb pdbcm7b) τα δύο αυτά μοντέλα είναι γεωμετρικά ανόμοια. Η μέθοδος, τους αποδίδει γεωμετρική ομοιότητα διότι το μοντέλο pdbcm7b είναι πολύ μεγαλύτερο και πυκνότερο από το μοντέλο pdbyprb 55
59 με αποτέλεσμα να είναι δυνατή η ύπαρξη σημαντικής ομοιότητας του συνόλου του pdbyprb με ένα μέρος του pdbcm7b. Το «λάθος» αυτό προκαλείται από την επιλογή να απαιτηθεί μερική ομοιότητα μεταξύ των μοντέλων και όχι αυστηρά συνολική ομοιότητα. Στο σημείο αυτό πρέπει να σημειωθεί ότι τα κριτήρια που χρησιμοποιούνται για την κατηγοριοποίηση των πρωτεϊνών, τοπική ομοιότητα των 3D μοντέλων και ομοιότητα αμινοξέων, επιλέχθηκαν έτσι ώστε να μπορεί στο μέλλον να χρησιμοποιηθεί η μέθοδος και για την αναγνώριση περιοχών αλληλεπίδρασης μεταξύ δύο πρωτεϊνών. Η δυνατότητα αυτή δεν θα ήταν υπαρκτή εάν απαιτούνταν ολική ομοιότητα των 3D μοντέλων και ύπαρξη ακριβώς ίδιων αμινοξέων. Ειδικότερα η επιλογή να χρησιμοποιηθεί τοπική ομοιότητα έναντι της ολικής είναι μια επιλογή που κάνει την μέθοδο πιο ευέλικτη θυσιάζοντας ένα κομμάτι από την ακρίβεια της. Τα αποτελέσματα όμως δείχνουν (ακρίβεια κατηγοριοποίησης που αγγίζει το 98,7%) ότι το κόστος της επιλογής αυτής, το οποίο είναι ένα κομμάτι από το.3% των λανθασμένων ανακτήσεων, είναι πολύ μικρότερο από το κέρδος που προσφέρει. Το κέρδος αυτό, όπως προαναφέραμε, είναι η επεκτασιμότητα της μεθόδου ώστε, μελλοντικά, να είναι ικανή να ελέγξει τυχόν συμπληρωματική γεωμετρία μεταξύ πρωτεϊνών, που θα οδηγήσει στην αναγνώριση πρωτεϊνικών αλληλεπιδράσεων. Επιπλέον, πρέπει να υπενθυμίσουμε ότι ο προσανατολισμός των μοντέλων, όπως αυτά λαμβάνονται μετά την ανάγνωση των 3D μοντέλων, είναι εντελώς τυχαίος. Ανεξαρτήτως του γεγονότος αυτού όμως η μέθοδος είναι ιδιαίτερα αποτελεσματική. Έτσι οδηγούμαστε στην εκτίμηση ότι η επιλογή χρήσης σφαιρικών αρμονικών για την εξαγωγή γεωμετρικών περιγραφέων ήταν απολύτως ορθή καθώς καθιστά την μέθοδο ανεξάρτητη του προσανατολισμού των μοντέλων. Το γεγονός αυτό προσφέρει την δυνατότητα να αποφευχθεί η ανάγκη χρήσης μιας διαδικασίας ευθυγράμμισης των μοντέλων, πριν την εξαγωγή περιγραφέων και την εκτέλεση της σύγκρισης. Δυνατότητα την οποία δε διαθέτουν αρκετές σύγχρονες μέθοδοι με αποτέλεσμα να είναι πιο χρονοβόρες από την παρούσα. 56





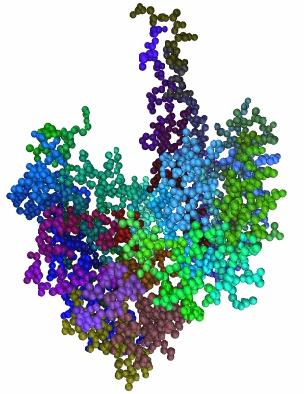
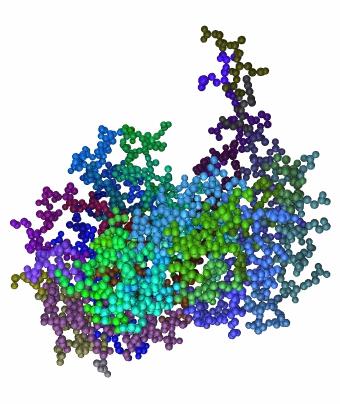
60 pdba0dd pdba0db pdbazf pdbat5 pdbe6jh pdbceh pdbg2ua Εικόνα 5.: επιτυχείς ανακλήσεις της μεθόδου pdbgc9a 57





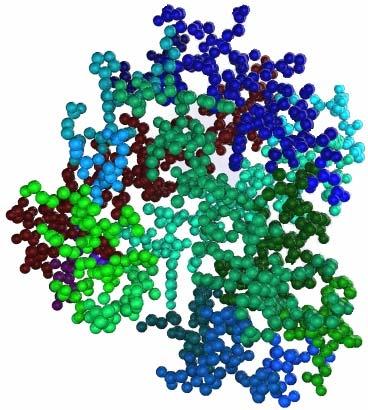

61 pdbyprb pdbcm7b pdbdctb pdbcm7b pdbdaaa pdbabwb pdb05m pdb2hbda Εικόνα 5.2: ανεπιτυχείς ανακλήσεις της μεθόδου 58
τα βιβλία των επιτυχιών
 Τα βιβλία των Εκδόσεων Πουκαμισάς συμπυκνώνουν την πολύχρονη διδακτική εμπειρία των συγγραφέων μας και αποτελούν το βασικό εκπαιδευτικό υλικό που χρησιμοποιούν οι μαθητές των φροντιστηρίων μας. Μέσα από
Τα βιβλία των Εκδόσεων Πουκαμισάς συμπυκνώνουν την πολύχρονη διδακτική εμπειρία των συγγραφέων μας και αποτελούν το βασικό εκπαιδευτικό υλικό που χρησιμοποιούν οι μαθητές των φροντιστηρίων μας. Μέσα από
ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ
 ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ ΠPΩTEINEΣ Οι πρωτεΐνες παίζουν σημαντικό ρόλο σε όλες σχεδόν τις βιολογικές διεργασίες. H σημασία τους φαίνεται στις παρακάτω περιπτώσεις: 1. Κατάλυση (πχ. ένζυμα) 2. Μεταφορά
ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ ΠPΩTEINEΣ Οι πρωτεΐνες παίζουν σημαντικό ρόλο σε όλες σχεδόν τις βιολογικές διεργασίες. H σημασία τους φαίνεται στις παρακάτω περιπτώσεις: 1. Κατάλυση (πχ. ένζυμα) 2. Μεταφορά
Άσκηση 7. Προσομοίωση 3D Δομών Βιομορίων μέσω. Ομολογίας & Threading
 Άσκηση 7 Προσομοίωση 3D Δομών Βιομορίων μέσω Ομολογίας & Threading Προσομοίωση 2ταγούς δομής πρωτεϊνών Δευτεροταγής Δομή: Η 2ταγής δομή των πρωτεϊνών είναι σταθερή τοπική διαμόρφωση της πολυπεπτιδικής
Άσκηση 7 Προσομοίωση 3D Δομών Βιομορίων μέσω Ομολογίας & Threading Προσομοίωση 2ταγούς δομής πρωτεϊνών Δευτεροταγής Δομή: Η 2ταγής δομή των πρωτεϊνών είναι σταθερή τοπική διαμόρφωση της πολυπεπτιδικής
Δευτεροταγής Δομή Πρωτεϊνών
 Δευτεροταγής Δομή Πρωτεϊνών Πρωτεϊνικό δίπλωμα Oι στόχοι μιας πρωτεΐνης όταν διπλωθεί είναι: 1. H xαμηλή ενέργεια διαμόρφωσης του κάθε αμινοξέος 2. Να επιτευχθούν υδρογονικοί δεσμοί από πολικά αμινοξέα
Δευτεροταγής Δομή Πρωτεϊνών Πρωτεϊνικό δίπλωμα Oι στόχοι μιας πρωτεΐνης όταν διπλωθεί είναι: 1. H xαμηλή ενέργεια διαμόρφωσης του κάθε αμινοξέος 2. Να επιτευχθούν υδρογονικοί δεσμοί από πολικά αμινοξέα
ΚΕΦΑΛΑΙΟ 2 Ο H XHΜΕΙΑ ΤΗΣ ΖΩΗΣ. Χημεία της ζωής 1
 ΚΕΦΑΛΑΙΟ 2 Ο H XHΜΕΙΑ ΤΗΣ ΖΩΗΣ Χημεία της ζωής 1 2.1 ΒΑΣΙΚΕΣ ΧΗΜΙΚΕΣ ΕΝΝΟΙΕΣ Η Βιολογία μπορεί να μελετηθεί μέσα από πολλά και διαφορετικά επίπεδα. Οι βιοχημικοί, για παράδειγμα, ενδιαφέρονται περισσότερο
ΚΕΦΑΛΑΙΟ 2 Ο H XHΜΕΙΑ ΤΗΣ ΖΩΗΣ Χημεία της ζωής 1 2.1 ΒΑΣΙΚΕΣ ΧΗΜΙΚΕΣ ΕΝΝΟΙΕΣ Η Βιολογία μπορεί να μελετηθεί μέσα από πολλά και διαφορετικά επίπεδα. Οι βιοχημικοί, για παράδειγμα, ενδιαφέρονται περισσότερο
αποτελούν το 96% κ.β Ποικιλία λειτουργιών
 ΧΗΜΙΚΑ ΣΤΟΙΧΕΙΑ ΠΟΥ ΣΥΝΘΕΤΟΥΝ ΤΟΥΣ ΟΡΓΑΝΙΣΜΟΥΣ 92 στοιχεία στο φλοιό της Γης 27 απαραίτητα για τη ζωή H, Ο, Ν, C αποτελούν το 96% κ.β S, Ca, P, Cl, K, Na, Mg αποτελούν το 4% κ.β. Fe, I Ιχνοστοιχεία αποτελούν
ΧΗΜΙΚΑ ΣΤΟΙΧΕΙΑ ΠΟΥ ΣΥΝΘΕΤΟΥΝ ΤΟΥΣ ΟΡΓΑΝΙΣΜΟΥΣ 92 στοιχεία στο φλοιό της Γης 27 απαραίτητα για τη ζωή H, Ο, Ν, C αποτελούν το 96% κ.β S, Ca, P, Cl, K, Na, Mg αποτελούν το 4% κ.β. Fe, I Ιχνοστοιχεία αποτελούν
ΔΟΜΗ ΠΡΩΤΕΪΝΩΝ II. Σελίδα 1 ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ. Τ. Θηραίου
 ΔΟΜΗ ΠΡΩΤΕΪΝΩΝ II Σελίδα 1 Υπολογιστικός Προσδιορισμός Δομής πειραματικός προσδιορισμός δομών κρυσταλλογραφία ακτίνων X πυρηνικός μαγνητικός συντονισμός (NMR) χρόνος / κόστος / περιορισμοί sequence - structure
ΔΟΜΗ ΠΡΩΤΕΪΝΩΝ II Σελίδα 1 Υπολογιστικός Προσδιορισμός Δομής πειραματικός προσδιορισμός δομών κρυσταλλογραφία ακτίνων X πυρηνικός μαγνητικός συντονισμός (NMR) χρόνος / κόστος / περιορισμοί sequence - structure
Ενεργειακή ανάλυση βιομορίων
 Ενεργειακή ανάλυση βιομορίων Τα βιομόρια ως φυσικά συστήματα πρωτεΐνες, DNA, πεπτίδια, μικρά μόρια (ligands, φάρμακα) Αλληλεπιδράσεις μεταξύ των ατόμων + επίδραση του περιβάλλοντος νερού σταθεροποίηση
Ενεργειακή ανάλυση βιομορίων Τα βιομόρια ως φυσικά συστήματα πρωτεΐνες, DNA, πεπτίδια, μικρά μόρια (ligands, φάρμακα) Αλληλεπιδράσεις μεταξύ των ατόμων + επίδραση του περιβάλλοντος νερού σταθεροποίηση
Στοιχεία Φυσικοχηµείας και Βιοφυσικής
 Στοιχεία Φυσικοχηµείας και Βιοφυσικής Β. Φαδούλογλου 2008 Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία
Στοιχεία Φυσικοχηµείας και Βιοφυσικής Β. Φαδούλογλου 2008 Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία
ΕΡΓΑΣΤΗΡΙΑΚΟ ΚΕΝΤΡΟ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Ν. ΜΑΓΝΗΣΙΑΣ ( Ε.Κ.Φ.Ε ) ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ
 ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ") ΕΡΓΑΣΤΗΡΙΑΚΟ ΚΕΝΤΡΟ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Ν. ΜΑΓΝΗΣΙΑΣ ( Ε.Κ.Φ.Ε ) ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ Θέμα: ΜΕΤΟΥΣΙΩΣΗ ΠΡΩΤΕΪΝΩΝ (άσκηση 7 του εργαστηριακού οδηγού) Μέσος χρόνος πειράματος: 45 λεπτά Α. ΑΝΑΛΩΣΙΜΑ Εργαλεία
ΕΡΓΑΣΤΗΡΙΑΚΟ ΚΕΝΤΡΟ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Ν. ΜΑΓΝΗΣΙΑΣ ( Ε.Κ.Φ.Ε ) ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ Θέμα: ΜΕΤΟΥΣΙΩΣΗ ΠΡΩΤΕΪΝΩΝ (άσκηση 7 του εργαστηριακού οδηγού) Μέσος χρόνος πειράματος: 45 λεπτά Α. ΑΝΑΛΩΣΙΜΑ Εργαλεία
Δομή και λειτουργία πρωτεϊνών. Το κύριο δομικό συστατικό των κυττάρων. Το κύριο λειτουργικό μόριο
 Δομή και λειτουργία πρωτεϊνών Το κύριο δομικό συστατικό των κυττάρων. Το κύριο λειτουργικό μόριο Πρωτεΐνες Κύριο συστατικό κυττάρου Δομικοί λίθοι αλλά και επιτελεστές λειτουργίας κυττάρου ένζυμα, μεταφορά,
Δομή και λειτουργία πρωτεϊνών Το κύριο δομικό συστατικό των κυττάρων. Το κύριο λειτουργικό μόριο Πρωτεΐνες Κύριο συστατικό κυττάρου Δομικοί λίθοι αλλά και επιτελεστές λειτουργίας κυττάρου ένζυμα, μεταφορά,
Μικρά αμινοξέα. Βιοχημεία Ι Β-3
 Βιοχημεία Ι Β-2 Μικρά αμινοξέα Βιοχημεία Ι Β-3 Aλειφατικά αμινοξέα Βιοχημεία Ι Β-4 Ιμινοξύ Βιοχημεία Ι Β-5 Αρωματικά αμινοξέα Βιοχημεία Ι Β-6 Βιοχημεία Ι Β-7 Η Tyr και η Trp απορροφούν στα 280nm-έτσι μετράται
Βιοχημεία Ι Β-2 Μικρά αμινοξέα Βιοχημεία Ι Β-3 Aλειφατικά αμινοξέα Βιοχημεία Ι Β-4 Ιμινοξύ Βιοχημεία Ι Β-5 Αρωματικά αμινοξέα Βιοχημεία Ι Β-6 Βιοχημεία Ι Β-7 Η Tyr και η Trp απορροφούν στα 280nm-έτσι μετράται
Ασκήσεις 3& 4. Πρωτεϊνική Αρχιτεκτονική. Πλατφόρμες Πρόβλεψης & Προσομοίωσης 2ταγούς Δομής. Μοριακή Απεικόνιση
 Ασκήσεις 3& 4 Πρωτεϊνική Αρχιτεκτονική Πλατφόρμες Πρόβλεψης & Προσομοίωσης 2ταγούς Δομής Μοριακή Απεικόνιση Πρωτεϊνική Αρχιτεκτονική Πρωτεϊνική Αρχιτεκτονική: Η τρισδιάστατη δομή μιας πρωτεΐνης και πως
Ασκήσεις 3& 4 Πρωτεϊνική Αρχιτεκτονική Πλατφόρμες Πρόβλεψης & Προσομοίωσης 2ταγούς Δομής Μοριακή Απεικόνιση Πρωτεϊνική Αρχιτεκτονική Πρωτεϊνική Αρχιτεκτονική: Η τρισδιάστατη δομή μιας πρωτεΐνης και πως
Ανόργανη Χημεία. Τμήμα Τεχνολογίας Τροφίμων. Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή. Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής
 Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ο Ομοιοπολικός Δεσμός 2 Ο δεσμός Η Η στο μόριο Η
Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ο Ομοιοπολικός Δεσμός 2 Ο δεσμός Η Η στο μόριο Η
Τα χημικά στοιχεία που είναι επικρατέστερα στους οργανισμούς είναι: i..
 ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 1 ο ΚΕΦΑΛΑΙΟ «XHMIKH ΣΥΣΤΑΣΗ ΤΟΥ ΚΥΤΤΑΡΟΥ» ΕΙΣΑΓΩΓΗ ΚΑΙ Η ΧΗΜΕΙΑ ΤΗΣ ΖΩΗΣ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Όταν αναφερόμαστε στον όρο «Χημική Σύσταση του Κυττάρου», τί νομίζετε ότι
ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 1 ο ΚΕΦΑΛΑΙΟ «XHMIKH ΣΥΣΤΑΣΗ ΤΟΥ ΚΥΤΤΑΡΟΥ» ΕΙΣΑΓΩΓΗ ΚΑΙ Η ΧΗΜΕΙΑ ΤΗΣ ΖΩΗΣ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Όταν αναφερόμαστε στον όρο «Χημική Σύσταση του Κυττάρου», τί νομίζετε ότι
Εισαγωγή στους αλγορίθμους Βιοπληροφορικής. Στοίχιση αλληλουχιών
 Στοίχιση αλληλουχιών Σύνοψη Καθολική στοίχιση Μήτρες βαθμολόγησης Τοπική στοίχιση Στοίχιση με ποινές εισαγωγής κενών Από την LCS στη στοίχιση: αλλαγές στη βαθμολόγηση Το πρόβλημα της Μεγαλύτερης Κοινής
Στοίχιση αλληλουχιών Σύνοψη Καθολική στοίχιση Μήτρες βαθμολόγησης Τοπική στοίχιση Στοίχιση με ποινές εισαγωγής κενών Από την LCS στη στοίχιση: αλλαγές στη βαθμολόγηση Το πρόβλημα της Μεγαλύτερης Κοινής
Βιοπληροφορική. Ενότητα 20: Υπολογιστικός Προσδιορισμός Δομής (2/3), 1 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου
, 1 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου") Βιοπληροφορική Ενότητα 20: Υπολογιστικός Προσδιορισμός Δομής (2/3), 1 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι κατανόηση της μεθόδου προτυποποίησης πρωτεϊνών με ομολογία. παρουσίαση
Βιοπληροφορική Ενότητα 20: Υπολογιστικός Προσδιορισμός Δομής (2/3), 1 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι κατανόηση της μεθόδου προτυποποίησης πρωτεϊνών με ομολογία. παρουσίαση
Πρόγνωση δομής πρωτεϊνών (Μέρος Ι)
") Πρόγνωση δομής πρωτεϊνών (Μέρος Ι) Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου Ταχ.Κιβ. 20537 1678, Λευκωσία ΚΥΠΡΟΣ
Πρόγνωση δομής πρωτεϊνών (Μέρος Ι) Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου Ταχ.Κιβ. 20537 1678, Λευκωσία ΚΥΠΡΟΣ
Διαλέξεις Χημείας Αγγελική Μαγκλάρα, PhD Εργαστήριο Κλινικής Χημείας Ιατρική Σχολή Πανεπιστημίου Ιωαννίνων
 Διαλέξεις Χημείας -2014 Αγγελική Μαγκλάρα, PhD Εργαστήριο Κλινικής Χημείας Ιατρική Σχολή Πανεπιστημίου Ιωαννίνων 1. Κατάταξη 2. Λειτουργίες 1. Πεπτιδικές ορμόνες 3. Πεπτιδικός δεσμός 1. Χαρακτηριστικά
Διαλέξεις Χημείας -2014 Αγγελική Μαγκλάρα, PhD Εργαστήριο Κλινικής Χημείας Ιατρική Σχολή Πανεπιστημίου Ιωαννίνων 1. Κατάταξη 2. Λειτουργίες 1. Πεπτιδικές ορμόνες 3. Πεπτιδικός δεσμός 1. Χαρακτηριστικά
Αρχιτεκτονική της τρισδιάστατης δομής πρωτεϊνών
 Αρχιτεκτονική της τρισδιάστατης δομής πρωτεϊνών Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου Ταχ.Κιβ. 20537 1678,
Αρχιτεκτονική της τρισδιάστατης δομής πρωτεϊνών Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου Ταχ.Κιβ. 20537 1678,
Αναγνώριση Προτύπων Ι
 Αναγνώριση Προτύπων Ι Ενότητα 1: Μέθοδοι Αναγνώρισης Προτύπων Αν. Καθηγητής Δερματάς Ευάγγελος Τμήμα Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Υπολογιστών Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται
Αναγνώριση Προτύπων Ι Ενότητα 1: Μέθοδοι Αναγνώρισης Προτύπων Αν. Καθηγητής Δερματάς Ευάγγελος Τμήμα Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Υπολογιστών Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται
Φυσική Στερεών στις Πρωτεΐνες
 Φυσική Στερεών στις Πρωτεΐνες Νίκος Απ. Παπανδρέου Τ.Ε.Ι. Πειραιά Φεβρουάριος 2010 Ένα ελικοϊδές μονοπάτι Χημική δομή μίας πρωτεΐνης Μήκος αλυσίδας ~30 έως ~1000 αµινοξέα Συνολικός αριθµός ατόµων έως ~
Φυσική Στερεών στις Πρωτεΐνες Νίκος Απ. Παπανδρέου Τ.Ε.Ι. Πειραιά Φεβρουάριος 2010 Ένα ελικοϊδές μονοπάτι Χημική δομή μίας πρωτεΐνης Μήκος αλυσίδας ~30 έως ~1000 αµινοξέα Συνολικός αριθµός ατόµων έως ~
ΓΩΝΙΕΣ φ, ψ ΚΑΙ ΕΠΙΤΡΕΠΤΕΣ ΔΙΑΜΟΡΦΩΣΕΙΣ ΤΗΣ ΠΟΛΥΠΕΠΤΙΔΙΚΗΣ ΑΛΥΣΙΔΑΣ
 ΓΩΝΙΕΣ φ, ψ ΚΑΙ ΕΠΙΤΡΕΠΤΕΣ ΔΙΑΜΟΡΦΩΣΕΙΣ ΤΗΣ ΠΟΛΥΠΕΠΤΙΔΙΚΗΣ ΑΛΥΣΙΔΑΣ ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ φ φ Ccarbonyl n Ccarbonyl n N Cα n Ccarbonyl n-1 Cα n N φ Ccarbonyl n-1 ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ ψ φ ψ Ccarbonyl n N (Ca
ΓΩΝΙΕΣ φ, ψ ΚΑΙ ΕΠΙΤΡΕΠΤΕΣ ΔΙΑΜΟΡΦΩΣΕΙΣ ΤΗΣ ΠΟΛΥΠΕΠΤΙΔΙΚΗΣ ΑΛΥΣΙΔΑΣ ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ φ φ Ccarbonyl n Ccarbonyl n N Cα n Ccarbonyl n-1 Cα n N φ Ccarbonyl n-1 ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ ψ φ ψ Ccarbonyl n N (Ca
Ασκήσεις 1 & 2. Βάσεις Δεδομένων. Εργαλεία Αναζήτησης ClustalW & Blast
 Ασκήσεις 1 & 2 Βάσεις Δεδομένων Εργαλεία Αναζήτησης ClustalW & Blast Μοριακή Προσομοίωση Εισαγωγή: Δομική Βάση Βιολογικών Φαινομένων Η αξιοποίηση του πλήθους των δομικών στοιχείων για την εξαγωγή βιολογικά
Ασκήσεις 1 & 2 Βάσεις Δεδομένων Εργαλεία Αναζήτησης ClustalW & Blast Μοριακή Προσομοίωση Εισαγωγή: Δομική Βάση Βιολογικών Φαινομένων Η αξιοποίηση του πλήθους των δομικών στοιχείων για την εξαγωγή βιολογικά
Εισαγωγή στη Μοριακή Προσοµοίωση
 Κεφάλαιο 6 Εισαγωγή στη Μοριακή Προσοµοίωση 6.1. Μοριακή Μηχανική 6.1.1. Εισαγωγή στη µεθοδολογία του «απ αρχής» διπλώµατος της πρωτείνης. Η ενέργεια κάθε µορίου µπορεί θεωρητικά να υπολογιστεί µε την
Κεφάλαιο 6 Εισαγωγή στη Μοριακή Προσοµοίωση 6.1. Μοριακή Μηχανική 6.1.1. Εισαγωγή στη µεθοδολογία του «απ αρχής» διπλώµατος της πρωτείνης. Η ενέργεια κάθε µορίου µπορεί θεωρητικά να υπολογιστεί µε την
ΠΡΩΤΕΪΝΕΣ: Δομή και λειτουργία. (Διδακτική προσέγγιση με πειράματα και αναλογίες)
") ΠΡΩΤΕΪΝΕΣ: Δομή και λειτουργία (Διδακτική προσέγγιση με πειράματα και αναλογίες) Η αλληλεπίδραση δομής-λειτουργίας αποτελεί βασικό θέμα της βιολογίας. Ένα σημαντικό παράδειγμα αυτής της αλληλεπίδρασης
ΠΡΩΤΕΪΝΕΣ: Δομή και λειτουργία (Διδακτική προσέγγιση με πειράματα και αναλογίες) Η αλληλεπίδραση δομής-λειτουργίας αποτελεί βασικό θέμα της βιολογίας. Ένα σημαντικό παράδειγμα αυτής της αλληλεπίδρασης
Δομές (Διαμορφώσεις) Πρωτεινικών μορίων
 Πρωτεινικών μορίων") Δομές (Διαμορφώσεις) Πρωτεινικών μορίων Πρωτοταγής δομή (αλληλουχία αμινοξέων) Δευτεροταγής δομή Η διάταξη της πεπτιδικής αλυσίδας στον χωρο αυτής καθ αυτής (χωρίς να ληφθούν υπ όψη οι ομάδες R) Τριτοταγής
Δομές (Διαμορφώσεις) Πρωτεινικών μορίων Πρωτοταγής δομή (αλληλουχία αμινοξέων) Δευτεροταγής δομή Η διάταξη της πεπτιδικής αλυσίδας στον χωρο αυτής καθ αυτής (χωρίς να ληφθούν υπ όψη οι ομάδες R) Τριτοταγής
ΦΑΡΜΑΚΩΝ WORKSHOP ΣΧΕ ΙΑΣΜΟΥ
 ΕΡΓΑΣΤΗΡΙΟΣΧΕ ΙΑΣΜΟΥ ΣΧΕ ΙΑΣΜΟΥ ΦΑΡΜΑΚΩΝ ΕΘΝΙΚΟ Ι ΡΥΜΑ ΕΡΕΥΝΩΝ ΙΝΣΤΙΤΟΥΤΟ ΟΡΓΑΝΙΚΗΣ & ΦΑΡΜΑΚΕΥΤΙΚΗΣ ΧΗΜΕΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΜΟΡΙΑΚΗΣ ΑΝΑΛΥΣΗΣ WORKSHOP ΣΧΕ ΙΑΣΜΟΥ ΦΑΡΜΑΚΩΝ Βασικές στρατηγικές Στο σχεδιασµό νέων
ΕΡΓΑΣΤΗΡΙΟΣΧΕ ΙΑΣΜΟΥ ΣΧΕ ΙΑΣΜΟΥ ΦΑΡΜΑΚΩΝ ΕΘΝΙΚΟ Ι ΡΥΜΑ ΕΡΕΥΝΩΝ ΙΝΣΤΙΤΟΥΤΟ ΟΡΓΑΝΙΚΗΣ & ΦΑΡΜΑΚΕΥΤΙΚΗΣ ΧΗΜΕΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΜΟΡΙΑΚΗΣ ΑΝΑΛΥΣΗΣ WORKSHOP ΣΧΕ ΙΑΣΜΟΥ ΦΑΡΜΑΚΩΝ Βασικές στρατηγικές Στο σχεδιασµό νέων
Οι πρωτεΐνες δομούνται από ένα σύνολο αμινοξέων. 1/10/2015 Δ.Δ. Λεωνίδας
 αμινοξέα Οι πρωτεΐνες δομούνται από ένα σύνολο αμινοξέων Λυσίνη CORN Ισομερές L Ισομερές D R = πλευρική αλυσίδα (side chain) Τα περισσότερα αμινοξέα είναι ασύμμετρα Όλα τα αμινοξέα που βρίσκονται στις
αμινοξέα Οι πρωτεΐνες δομούνται από ένα σύνολο αμινοξέων Λυσίνη CORN Ισομερές L Ισομερές D R = πλευρική αλυσίδα (side chain) Τα περισσότερα αμινοξέα είναι ασύμμετρα Όλα τα αμινοξέα που βρίσκονται στις
Δευτεροταγής Δομή Πρωτεϊνών
 Δευτεροταγής Δομή Πρωτεϊνών Πρωτεϊνικό δίπλωμα Η κινητήρια δύναμη για την αναδίπλωση μιας πρωτεΐνης είναι η επίτευξη των εξής: 1. xαμηλή ενέργεια διαμόρφωσης του κάθε αμινοξέος 2. δημιουργία μιας συμπαγούς
Δευτεροταγής Δομή Πρωτεϊνών Πρωτεϊνικό δίπλωμα Η κινητήρια δύναμη για την αναδίπλωση μιας πρωτεΐνης είναι η επίτευξη των εξής: 1. xαμηλή ενέργεια διαμόρφωσης του κάθε αμινοξέος 2. δημιουργία μιας συμπαγούς
Βιολογία Γενικής Παιδείας Β Λυκείου
 Απρίλιος Μάιος 12 Βιολογία Γενικής Παιδείας Β Λυκείου Βιολογία Γενικής Παιδείας Β Λυκείου (Ερωτήσεις που παρουσιάζουν ενδιαφέρον) 1. Τι είναι τα βιομόρια και ποια είναι τα βασικά χαρακτηριστικά τους; Βιομόρια
Απρίλιος Μάιος 12 Βιολογία Γενικής Παιδείας Β Λυκείου Βιολογία Γενικής Παιδείας Β Λυκείου (Ερωτήσεις που παρουσιάζουν ενδιαφέρον) 1. Τι είναι τα βιομόρια και ποια είναι τα βασικά χαρακτηριστικά τους; Βιομόρια
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ
 ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ") ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2017-2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1. Σε ποια μορφή της αιμοσφαιρίνης συνδέεται το 2,3, BPG, ποιο είναι το ηλεκτρικό του φορτίο, με
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2017-2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1. Σε ποια μορφή της αιμοσφαιρίνης συνδέεται το 2,3, BPG, ποιο είναι το ηλεκτρικό του φορτίο, με
Εξόρυξη Γνώσης από Βιολογικά εδομένα
 Παρουσίαση Διπλωματικής Εργασίας Εξόρυξη Γνώσης από Βιολογικά εδομένα Καρυπίδης Γεώργιος (Μ27/03) Επιβλέπων Καθηγητής: Ιωάννης Βλαχάβας MIS Πανεπιστήμιο Μακεδονίας Φεβρουάριος 2005 Εξόρυξη Γνώσης από Βιολογικά
Παρουσίαση Διπλωματικής Εργασίας Εξόρυξη Γνώσης από Βιολογικά εδομένα Καρυπίδης Γεώργιος (Μ27/03) Επιβλέπων Καθηγητής: Ιωάννης Βλαχάβας MIS Πανεπιστήμιο Μακεδονίας Φεβρουάριος 2005 Εξόρυξη Γνώσης από Βιολογικά
και χρειάζεται μέσα στο ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. ενζύμων κύτταρο τρόπους
 Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
ΠΡΩΤΕΪΝΕΣ. Φατούρος Ιωάννης Αναπληρωτής Καθηγητής
 ΠΡΩΤΕΪΝΕΣ Φατούρος Ιωάννης Αναπληρωτής Καθηγητής Θέματα Διάλεξης Δομή, αριθμός και διαχωρισμός των αμινοξέων Ένωση αμινοξέων με τον πεπτιδικό δεσμό για τη δημιουργία πρωτεΐνης Λειτουργίες των πρωτεϊνών
ΠΡΩΤΕΪΝΕΣ Φατούρος Ιωάννης Αναπληρωτής Καθηγητής Θέματα Διάλεξης Δομή, αριθμός και διαχωρισμός των αμινοξέων Ένωση αμινοξέων με τον πεπτιδικό δεσμό για τη δημιουργία πρωτεΐνης Λειτουργίες των πρωτεϊνών
Μελέτη και Υλοποίηση Αλγορίθμων για Βιολογικές Εφαρμογές σε MapReduce Περιβάλλον
 Μελέτη και Υλοποίηση Αλγορίθμων για Βιολογικές Εφαρμογές σε MapReduce Περιβάλλον Δανάη Κούτρα Eργαστήριο Συστημάτων Βάσεων Γνώσεων και Δεδομένων Εθνικό Μετσόβιο Πολυτεχνείο Θέματα Σκοπός της διπλωματικής
Μελέτη και Υλοποίηση Αλγορίθμων για Βιολογικές Εφαρμογές σε MapReduce Περιβάλλον Δανάη Κούτρα Eργαστήριο Συστημάτων Βάσεων Γνώσεων και Δεδομένων Εθνικό Μετσόβιο Πολυτεχνείο Θέματα Σκοπός της διπλωματικής
Μάθημα 10 ο. Περιγραφή Σχήματος ΤΜΗΥΠ / ΕΕΣΤ 1
 Μάθημα 10 ο Περιγραφή Σχήματος ΤΜΗΥΠ / ΕΕΣΤ 1 Εισαγωγή (1) Η περιγραφή μίας περιοχής μπορεί να γίνει: Με βάση τα εξωτερικά χαρακτηριστικά (ακμές, όρια). Αυτή η περιγραφή προτιμάται όταν μας ενδιαφέρουν
Μάθημα 10 ο Περιγραφή Σχήματος ΤΜΗΥΠ / ΕΕΣΤ 1 Εισαγωγή (1) Η περιγραφή μίας περιοχής μπορεί να γίνει: Με βάση τα εξωτερικά χαρακτηριστικά (ακμές, όρια). Αυτή η περιγραφή προτιμάται όταν μας ενδιαφέρουν
ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΠΑΤΡΩΝ ΤΜΗΜΑ ΜΗΧΑΝΟΛΟΓΩΝ ΚΑΙ ΑΕΡΟΝΑΥΠΗΓΩΝ ΜΗΧΑΝΙΚΩΝ ΕΡΓΑΣΤΗΡΙΟ ΜΗΧΑΝΙΚΗΣ ΤΩΝ ΡΕΥΣΤΩΝ ΚΑΙ ΕΦΑΡΜΟΓΩΝ ΑΥΤΗΣ
 ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΠΑΤΡΩΝ ΤΜΗΜΑ ΜΗΧΑΝΟΛΟΓΩΝ ΚΑΙ ΑΕΡΟΝΑΥΠΗΓΩΝ ΜΗΧΑΝΙΚΩΝ ΕΡΓΑΣΤΗΡΙΟ ΜΗΧΑΝΙΚΗΣ ΤΩΝ ΡΕΥΣΤΩΝ ΚΑΙ ΕΦΑΡΜΟΓΩΝ ΑΥΤΗΣ Διευθυντής: Διονύσιος-Ελευθ. Π. Μάργαρης, Αναπλ. Καθηγητής ΕΡΓΑΣΤΗΡΙΑΚΗ
ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΠΑΤΡΩΝ ΤΜΗΜΑ ΜΗΧΑΝΟΛΟΓΩΝ ΚΑΙ ΑΕΡΟΝΑΥΠΗΓΩΝ ΜΗΧΑΝΙΚΩΝ ΕΡΓΑΣΤΗΡΙΟ ΜΗΧΑΝΙΚΗΣ ΤΩΝ ΡΕΥΣΤΩΝ ΚΑΙ ΕΦΑΡΜΟΓΩΝ ΑΥΤΗΣ Διευθυντής: Διονύσιος-Ελευθ. Π. Μάργαρης, Αναπλ. Καθηγητής ΕΡΓΑΣΤΗΡΙΑΚΗ
KΕΦΑΛΑΙΟ 1ο Χημική σύσταση του κυττάρου. Να απαντήσετε σε καθεμιά από τις παρακάτω ερωτήσεις με μια πρόταση:
 KΕΦΑΛΑΙΟ 1ο Χημική σύσταση του κυττάρου Ενότητα 1.1: Χημεία της ζωής Ενότητα 2.1: Μακρομόρια Να απαντήσετε σε καθεμιά από τις παρακάτω ερωτήσεις με μια πρόταση: 1. Για ποιο λόγο θεωρείται αναγκαία η σταθερότητα
KΕΦΑΛΑΙΟ 1ο Χημική σύσταση του κυττάρου Ενότητα 1.1: Χημεία της ζωής Ενότητα 2.1: Μακρομόρια Να απαντήσετε σε καθεμιά από τις παρακάτω ερωτήσεις με μια πρόταση: 1. Για ποιο λόγο θεωρείται αναγκαία η σταθερότητα
7 ο Κεφάλαιο Οργανική Χημεία. Δ. Παπαδόπουλος, χημικός
 7 ο Κεφάλαιο Οργανική Χημεία Δ. Παπαδόπουλος, χημικός Βύρωνας, 2015 Θεωρίες ερμηνείας του ομοιοπολικού δεσμού με βάση την κβαντική θεωρία. Θεωρία δεσμού σθένους. Θεωρία των μοριακών τροχιακών. Κάθε θεωρία
7 ο Κεφάλαιο Οργανική Χημεία Δ. Παπαδόπουλος, χημικός Βύρωνας, 2015 Θεωρίες ερμηνείας του ομοιοπολικού δεσμού με βάση την κβαντική θεωρία. Θεωρία δεσμού σθένους. Θεωρία των μοριακών τροχιακών. Κάθε θεωρία
Κεφάλαιο 8. Οπτικοποίηση Απαλοιφή
 Κεφάλαιο 8. Οπτικοποίηση Απαλοιφή Oι οπτικές επιδράσεις, που μπορεί να προκαλέσει μια εικόνα στους χρήστες, αποτελούν ένα από τα σπουδαιότερα αποτελέσματα των λειτουργιών γραφικών με Η/Υ. Τον όρο της οπτικοποίησης
Κεφάλαιο 8. Οπτικοποίηση Απαλοιφή Oι οπτικές επιδράσεις, που μπορεί να προκαλέσει μια εικόνα στους χρήστες, αποτελούν ένα από τα σπουδαιότερα αποτελέσματα των λειτουργιών γραφικών με Η/Υ. Τον όρο της οπτικοποίησης
Οδηγίες σχεδίασης στο περιβάλλον Blender
 Οδηγίες σχεδίασης στο περιβάλλον Blender Στον πραγματικό κόσμο, αντιλαμβανόμαστε τα αντικείμενα σε τρεις κατευθύνσεις ή διαστάσεις. Τυπικά λέμε ότι διαθέτουν ύψος, πλάτος και βάθος. Όταν θέλουμε να αναπαραστήσουμε
Οδηγίες σχεδίασης στο περιβάλλον Blender Στον πραγματικό κόσμο, αντιλαμβανόμαστε τα αντικείμενα σε τρεις κατευθύνσεις ή διαστάσεις. Τυπικά λέμε ότι διαθέτουν ύψος, πλάτος και βάθος. Όταν θέλουμε να αναπαραστήσουμε
Τάξη. Γνωστικό αντικείµενο: Ειδικοί διδακτικοί στόχοι
 Αµινοξέα και πεπτίδια Τάξη Μάθηµα Γνωστικό αντικείµενο: ιδακτική ενότητα Απαιτούµενος χρόνος Χηµεία,Βιοχηµεία. Αµινοξέα και πεπτίδια 2 διδακτικές ώρες Ειδικοί διδακτικοί στόχοι Οι διδακτικοί στόχοι αυτών
Αµινοξέα και πεπτίδια Τάξη Μάθηµα Γνωστικό αντικείµενο: ιδακτική ενότητα Απαιτούµενος χρόνος Χηµεία,Βιοχηµεία. Αµινοξέα και πεπτίδια 2 διδακτικές ώρες Ειδικοί διδακτικοί στόχοι Οι διδακτικοί στόχοι αυτών
Οι πρωτεΐνες συμμετέχουν σε όλες τις κυτταρικές λειτουργίες
 Οι πρωτεΐνες συμμετέχουν σε όλες τις κυτταρικές λειτουργίες Γένωμα vs Πρωτέωμα Όλη η αλληλουχία βάσεων στο DNA Τι είναι δυνατόν Συγκεκριμένο Στατικό Οι πρωτεΐνες που κωδικοποιούνται από το γένωμα Τι είναι
Οι πρωτεΐνες συμμετέχουν σε όλες τις κυτταρικές λειτουργίες Γένωμα vs Πρωτέωμα Όλη η αλληλουχία βάσεων στο DNA Τι είναι δυνατόν Συγκεκριμένο Στατικό Οι πρωτεΐνες που κωδικοποιούνται από το γένωμα Τι είναι
τα Λεπτά Υμένια στις Νανοδομές και στις Νανο- & Mεγάλης κλίμακας κατασκευές.
 Από τα Λεπτά Υμένια στις Νανοδομές και στις Νανο- & Mεγάλης κλίμακας κατασκευές. Η εξέλιξη της επιστημονικής έρευνας, πέρα της ικανοποίησης της έμφυτης ανάγκης του ανθρώπου για γνώση, είχε και ως παράλληλο
Από τα Λεπτά Υμένια στις Νανοδομές και στις Νανο- & Mεγάλης κλίμακας κατασκευές. Η εξέλιξη της επιστημονικής έρευνας, πέρα της ικανοποίησης της έμφυτης ανάγκης του ανθρώπου για γνώση, είχε και ως παράλληλο
Δομικές κατηγορίες πρωτεϊνών
 3-1 Κεφάλαι ο Δομικές κατηγορίες πρωτεϊνών 3.1. α-δομές πρωτεϊνών Οι α-έλικες είναι δομικά στοιχεία που μπορούν να σχηματίσουν πολλές κατηγορίες στερεοδομών και με πολλές διαφορετικές λειτουργίες. Εκτός
3-1 Κεφάλαι ο Δομικές κατηγορίες πρωτεϊνών 3.1. α-δομές πρωτεϊνών Οι α-έλικες είναι δομικά στοιχεία που μπορούν να σχηματίσουν πολλές κατηγορίες στερεοδομών και με πολλές διαφορετικές λειτουργίες. Εκτός
Βιοπληροψορική, συσιημική βιολογία και εξατομικευμένη θεραπεία
 Βιοπληροψορική, συσιημική βιολογία και εξατομικευμένη θεραπεία Φραγκίσκος Κολίσης Καθηγητής Βιοτεχνολογίας, Σχολή Χημικών Μηχανικών ΕΜΠ, Διευθυντής Ινστιτούτου Βιολογικών Ερευνών και Βιοτεχνολογίας, EIE
Βιοπληροψορική, συσιημική βιολογία και εξατομικευμένη θεραπεία Φραγκίσκος Κολίσης Καθηγητής Βιοτεχνολογίας, Σχολή Χημικών Μηχανικών ΕΜΠ, Διευθυντής Ινστιτούτου Βιολογικών Ερευνών και Βιοτεχνολογίας, EIE
Μία μέθοδος προσομοίωσης ψηφιακών κυκλωμάτων Εξελικτικής Υπολογιστικής
 Μία μέθοδος προσομοίωσης ψηφιακών κυκλωμάτων Εξελικτικής Υπολογιστικής Βασισμένο σε μια εργασία των Καζαρλή, Καλόμοιρου, Μαστοροκώστα, Μπαλουκτσή, Καλαϊτζή, Βαλαή, Πετρίδη Εισαγωγή Η Εξελικτική Υπολογιστική
Μία μέθοδος προσομοίωσης ψηφιακών κυκλωμάτων Εξελικτικής Υπολογιστικής Βασισμένο σε μια εργασία των Καζαρλή, Καλόμοιρου, Μαστοροκώστα, Μπαλουκτσή, Καλαϊτζή, Βαλαή, Πετρίδη Εισαγωγή Η Εξελικτική Υπολογιστική
Ποσοτικές Μέθοδοι στη Διοίκηση Επιχειρήσεων ΙΙ Σύνολο- Περιεχόμενο Μαθήματος
 Ποσοτικές Μέθοδοι στη Διοίκηση Επιχειρήσεων ΙΙ Σύνολο- Περιεχόμενο Μαθήματος Χιωτίδης Γεώργιος Τμήμα Λογιστικής και Χρηματοοικονομικής Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης
Ποσοτικές Μέθοδοι στη Διοίκηση Επιχειρήσεων ΙΙ Σύνολο- Περιεχόμενο Μαθήματος Χιωτίδης Γεώργιος Τμήμα Λογιστικής και Χρηματοοικονομικής Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης
ΕΝΑΣ ΔΙΚΡΙΤΗΡΙΟΣ ΑΛΓΟΡΙΘΜΟΣ SIMPLEX
 ΚΕΦΑΛΑΙΟ 3 ΕΝΑΣ ΔΙΚΡΙΤΗΡΙΟΣ ΑΛΓΟΡΙΘΜΟΣ SIMPLEX 3.1 Εισαγωγή Ο αλγόριθμος Simplex θεωρείται πλέον ως ένας κλασικός αλγόριθμος για την επίλυση γραμμικών προβλημάτων. Η πρακτική αποτελεσματικότητά του έχει
ΚΕΦΑΛΑΙΟ 3 ΕΝΑΣ ΔΙΚΡΙΤΗΡΙΟΣ ΑΛΓΟΡΙΘΜΟΣ SIMPLEX 3.1 Εισαγωγή Ο αλγόριθμος Simplex θεωρείται πλέον ως ένας κλασικός αλγόριθμος για την επίλυση γραμμικών προβλημάτων. Η πρακτική αποτελεσματικότητά του έχει
ΤΕΧΝΟΛΟΓΙΑ & ΕΠΙΣΤΗΜΗ ΤΩΝ ΥΛΙΚΩΝ
 Τμήμα Τεχνολόγων Περιβάλλοντος Κατεύθυνσης Συντήρησης Πολιτισμικής Κληρονομιάς ΤΕΧΝΟΛΟΓΙΑ & ΕΠΙΣΤΗΜΗ ΤΩΝ ΥΛΙΚΩΝ 3 η Ενότητα ΔΕΣΜΟΙ Δημήτριος Λαμπάκης ΜΟΡΙΑΚΗ ΔΟΜΗ Μεμονωμένα άτομα: Μόνο τα ευγενή αέρια
Τμήμα Τεχνολόγων Περιβάλλοντος Κατεύθυνσης Συντήρησης Πολιτισμικής Κληρονομιάς ΤΕΧΝΟΛΟΓΙΑ & ΕΠΙΣΤΗΜΗ ΤΩΝ ΥΛΙΚΩΝ 3 η Ενότητα ΔΕΣΜΟΙ Δημήτριος Λαμπάκης ΜΟΡΙΑΚΗ ΔΟΜΗ Μεμονωμένα άτομα: Μόνο τα ευγενή αέρια
ΠΕΡΙΟΔΙΚΟΣ ΠΙΝΑΚΑΣ ΠΕΡΙΟΔΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΑΤΟΜΙΚΗ ΑΚΤΙΝΑ ΕΝΕΡΓΕΙΑ ΙΟΝΤΙΣΜΟΥ
 ΠΕΡΙΟΔΙΚΟΣ ΠΙΝΑΚΑΣ ΠΕΡΙΟΔΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΑΤΟΜΙΚΗ ΑΚΤΙΝΑ ΕΝΕΡΓΕΙΑ ΙΟΝΤΙΣΜΟΥ Ν. ΜΠΕΚΙΑΡΗΣ ΕΙΣΑΓΩΓΗ Η ηλεκτρονιακή δομή και κυρίως τα ηλεκτρόνια σθένους (τελευταία ηλεκτρόνια) προσδίδουν στο άτομο τη χημική
ΠΕΡΙΟΔΙΚΟΣ ΠΙΝΑΚΑΣ ΠΕΡΙΟΔΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΑΤΟΜΙΚΗ ΑΚΤΙΝΑ ΕΝΕΡΓΕΙΑ ΙΟΝΤΙΣΜΟΥ Ν. ΜΠΕΚΙΑΡΗΣ ΕΙΣΑΓΩΓΗ Η ηλεκτρονιακή δομή και κυρίως τα ηλεκτρόνια σθένους (τελευταία ηλεκτρόνια) προσδίδουν στο άτομο τη χημική
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ
 BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΜΟΡΙΑΚΑ ΜΟΝΤΕΛΑ 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING) 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING)
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΜΟΡΙΑΚΑ ΜΟΝΤΕΛΑ 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING) 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING)
Διδάσκων: Καθηγητής Εμμανουήλ Μ. Παπαμιχαήλ
 Τίτλος Μαθήματος: Ενζυμολογία Ενότητα: Εισαγωγή Διδάσκων: Καθηγητής Εμμανουήλ Μ. Παπαμιχαήλ Τμήμα: Χημείας 8 1. EIΣAΓΩΓH Tα ένζυμα είναι οι καταλύτες της ζώσης ύλης. Καταλύουν τις χημικές αντιδράσεις,
Τίτλος Μαθήματος: Ενζυμολογία Ενότητα: Εισαγωγή Διδάσκων: Καθηγητής Εμμανουήλ Μ. Παπαμιχαήλ Τμήμα: Χημείας 8 1. EIΣAΓΩΓH Tα ένζυμα είναι οι καταλύτες της ζώσης ύλης. Καταλύουν τις χημικές αντιδράσεις,
Κεφάλαιο 1 Χημικός δεσμός
 Κεφάλαιο 1 Χημικός δεσμός 1.1 Άτομα, Ηλεκτρόνια, και Τροχιακά Τα άτομα αποτελούνται από + Πρωτόνια φορτισμένα θετικά μάζα = 1.6726 X 10-27 kg Νετρόνια ουδέτερα μάζα = 1.6750 X 10-27 kg Ηλεκτρόνια φορτισμένα
Κεφάλαιο 1 Χημικός δεσμός 1.1 Άτομα, Ηλεκτρόνια, και Τροχιακά Τα άτομα αποτελούνται από + Πρωτόνια φορτισμένα θετικά μάζα = 1.6726 X 10-27 kg Νετρόνια ουδέτερα μάζα = 1.6750 X 10-27 kg Ηλεκτρόνια φορτισμένα
Αναγνώριση Προτύπων Ι
 Αναγνώριση Προτύπων Ι Ενότητα 2: Δομικά Συστήματα Αν. Καθηγητής Δερματάς Ευάγγελος Τμήμα Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Υπολογιστών Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
Αναγνώριση Προτύπων Ι Ενότητα 2: Δομικά Συστήματα Αν. Καθηγητής Δερματάς Ευάγγελος Τμήμα Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Υπολογιστών Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
Αριθμητική Ανάλυση και Εφαρμογές
 Αριθμητική Ανάλυση και Εφαρμογές Διδάσκων: Δημήτριος Ι. Φωτιάδης Τμήμα Μηχανικών Επιστήμης Υλικών Ιωάννινα 07-08 Αριθμητική Παραγώγιση Εισαγωγή Ορισμός 7. Αν y f x είναι μια συνάρτηση ορισμένη σε ένα διάστημα
Αριθμητική Ανάλυση και Εφαρμογές Διδάσκων: Δημήτριος Ι. Φωτιάδης Τμήμα Μηχανικών Επιστήμης Υλικών Ιωάννινα 07-08 Αριθμητική Παραγώγιση Εισαγωγή Ορισμός 7. Αν y f x είναι μια συνάρτηση ορισμένη σε ένα διάστημα
ΚΕΦΑΛΑΙΟ 3. Περιγραφή της Μεθόδου ΠΕΡΙΓΡΑΦΗ ΤΗΣ ΜΕΘΟΔΟΥ
 ΚΕΦΑΛΑΙΟ 3 Περιγραφή της Μεθόδου Το αντικείμενο αυτής της εργασίας είναι η χρήση μιας μεθόδου προσέγγισης συναρτήσεων που έχει προταθεί από τον hen-ha huang και ονομάζεται Ασαφώς Σταθμισμένη Παλινδρόμηση
ΚΕΦΑΛΑΙΟ 3 Περιγραφή της Μεθόδου Το αντικείμενο αυτής της εργασίας είναι η χρήση μιας μεθόδου προσέγγισης συναρτήσεων που έχει προταθεί από τον hen-ha huang και ονομάζεται Ασαφώς Σταθμισμένη Παλινδρόμηση
Μοριακά πρότυπα. Σε τι διαφέρουν από τα μεταλλικά συστήματα; Παραδείγματα τύπων ατόμων. Η έννοια του τύπου ατόμου
 Τεχνικές προσομοίωσης και σχεδιασμού υλικών σε ΗΥ Σε τι διαφέρουν από τα μεταλλικά συστήματα; Μοριακά πρότυπα Στα μοριακά συστήματα: Η φύση του δεσμού είναι διαφορετική (ομοιοπολικός δεσμός). Υπάρχει συγκεκριμένη
Τεχνικές προσομοίωσης και σχεδιασμού υλικών σε ΗΥ Σε τι διαφέρουν από τα μεταλλικά συστήματα; Μοριακά πρότυπα Στα μοριακά συστήματα: Η φύση του δεσμού είναι διαφορετική (ομοιοπολικός δεσμός). Υπάρχει συγκεκριμένη
Τεχνικές Μείωσης Διαστάσεων. Ειδικά θέματα ψηφιακής επεξεργασίας σήματος και εικόνας Σ. Φωτόπουλος- Α. Μακεδόνας
 Τεχνικές Μείωσης Διαστάσεων Ειδικά θέματα ψηφιακής επεξεργασίας σήματος και εικόνας Σ. Φωτόπουλος- Α. Μακεδόνας 1 Εισαγωγή Το μεγαλύτερο μέρος των δεδομένων που καλούμαστε να επεξεργαστούμε είναι πολυδιάστατα.
Τεχνικές Μείωσης Διαστάσεων Ειδικά θέματα ψηφιακής επεξεργασίας σήματος και εικόνας Σ. Φωτόπουλος- Α. Μακεδόνας 1 Εισαγωγή Το μεγαλύτερο μέρος των δεδομένων που καλούμαστε να επεξεργαστούμε είναι πολυδιάστατα.
Φυσική Β Λυκείου Γενικής
 Η ΕΝΝΟΙΑ ΠΕΔΙΟ - ΕΝΤΑΣΗ. 1.ΕΡΩΤΗΣΕΙΣ ΣΥΝΤΟΜΗΣ ΑΠΑΝΤΗΣΗΣ 1. Ποιες είναι οι διαφορές μεταξύ της θεωρίας της δράσης από απόσταση και της θεωρίας του πεδίου. Ποια η επικρατέστερη θεωρία σήμερα; 2. Ποιος είναι
Η ΕΝΝΟΙΑ ΠΕΔΙΟ - ΕΝΤΑΣΗ. 1.ΕΡΩΤΗΣΕΙΣ ΣΥΝΤΟΜΗΣ ΑΠΑΝΤΗΣΗΣ 1. Ποιες είναι οι διαφορές μεταξύ της θεωρίας της δράσης από απόσταση και της θεωρίας του πεδίου. Ποια η επικρατέστερη θεωρία σήμερα; 2. Ποιος είναι
Θέµατα ιάλεξης ΠΡΩΤΕΪΝΕΣ - ΕΝΖΥΜΑ ΠΡΩΤΕΪΝΕΣ. ιαχωρισµός Αµινοξέων
 MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΠΡΩΤΕΪΝΕΣ - ΕΝΖΥΜΑ Θέµατα ιάλεξης οµή, αριθµός και διαχωρισµός των αµινοξέων Ένωση αµινοξέων µε τον πεπτιδικό δεσµό
MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΠΡΩΤΕΪΝΕΣ - ΕΝΖΥΜΑ Θέµατα ιάλεξης οµή, αριθµός και διαχωρισµός των αµινοξέων Ένωση αµινοξέων µε τον πεπτιδικό δεσµό
Συστήματα συντεταγμένων
 Κεφάλαιο. Για να δημιουργήσουμε τρισδιάστατα αντικείμενα, που μπορούν να παρασταθούν στην οθόνη του υπολογιστή ως ένα σύνολο από γραμμές, επίπεδες πολυγωνικές επιφάνειες ή ακόμη και από ένα συνδυασμό από
Κεφάλαιο. Για να δημιουργήσουμε τρισδιάστατα αντικείμενα, που μπορούν να παρασταθούν στην οθόνη του υπολογιστή ως ένα σύνολο από γραμμές, επίπεδες πολυγωνικές επιφάνειες ή ακόμη και από ένα συνδυασμό από
Διακριτά Μαθηματικά ΙΙ Χρήστος Νομικός Τμήμα Μηχανικών Η/Υ και Πληροφορικής Πανεπιστήμιο Ιωαννίνων 2018 Χρήστος Νομικός ( Τμήμα Μηχανικών Η/Υ Διακριτά
 Διακριτά Μαθηματικά ΙΙ Χρήστος Νομικός Τμήμα Μηχανικών Η/Υ και Πληροφορικής Πανεπιστήμιο Ιωαννίνων 2018 Χρήστος Νομικός ( Τμήμα Μηχανικών Η/Υ Διακριτά και Πληροφορικής Μαθηματικά Πανεπιστήμιο ΙΙ Ιωαννίνων
Διακριτά Μαθηματικά ΙΙ Χρήστος Νομικός Τμήμα Μηχανικών Η/Υ και Πληροφορικής Πανεπιστήμιο Ιωαννίνων 2018 Χρήστος Νομικός ( Τμήμα Μηχανικών Η/Υ Διακριτά και Πληροφορικής Μαθηματικά Πανεπιστήμιο ΙΙ Ιωαννίνων
O μετασχηματισμός μιας «διαθεματικής» δραστηριότητας σε μαθηματική. Δέσποινα Πόταρη Πανεπιστήμιο Πατρών
 O μετασχηματισμός μιας «διαθεματικής» δραστηριότητας σε μαθηματική Δέσποινα Πόταρη Πανεπιστήμιο Πατρών Η έννοια της δραστηριότητας Δραστηριότητα είναι κάθε ανθρώπινη δράση που έχει ένα κίνητρο και ένα
O μετασχηματισμός μιας «διαθεματικής» δραστηριότητας σε μαθηματική Δέσποινα Πόταρη Πανεπιστήμιο Πατρών Η έννοια της δραστηριότητας Δραστηριότητα είναι κάθε ανθρώπινη δράση που έχει ένα κίνητρο και ένα
ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ
 ΜΙΝΟΠΕΤΡΟΣ ΚΩΝΣΤΑΝΤΙΝΟΣ ΦΥΣΙΚΟΣ - Ρ/Η ΚΑΘΗΓΗΤΗΣ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΥΠΕΥΘΥΝΟΣ ΣΕΦΕ 2 ου ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΠΕΡΑΜΑΤΟΣ ΕΡΓΑΣΤΗΡΙΑΚΗ ΑΣΚΗΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ
ΜΙΝΟΠΕΤΡΟΣ ΚΩΝΣΤΑΝΤΙΝΟΣ ΦΥΣΙΚΟΣ - Ρ/Η ΚΑΘΗΓΗΤΗΣ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΥΠΕΥΘΥΝΟΣ ΣΕΦΕ 2 ου ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΠΕΡΑΜΑΤΟΣ ΕΡΓΑΣΤΗΡΙΑΚΗ ΑΣΚΗΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ
Μάθημα 16 ο ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ
 Μάθημα 16 ο ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ Περιεχόμενα Παρουσίασης Βιολογικό υπόβαθρο Το κεντρικό αξίωμα Σύνοψη της Βιοπληροφορικής Ερευνητικές περιοχές Πηγές πληροφοριών Τι είναι η Βιοπληροφορική Βιο Πληροφορική μοριακή
Μάθημα 16 ο ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ Περιεχόμενα Παρουσίασης Βιολογικό υπόβαθρο Το κεντρικό αξίωμα Σύνοψη της Βιοπληροφορικής Ερευνητικές περιοχές Πηγές πληροφοριών Τι είναι η Βιοπληροφορική Βιο Πληροφορική μοριακή
Διάλεξη 7: Μοριακή Δομή
 Μεμονωμένα άτομα: Μόνο τα ευγενή αέρια Μόρια: Τα υπόλοιπα άτομα σχηματίζουν μόρια Γιατί; Διότι η ολική ενέργεια ενός ευσταθούς μορίου είναι μικρότερη από την ολική ενέργεια των μεμονωμένων ατόμων που αποτελούν
Μεμονωμένα άτομα: Μόνο τα ευγενή αέρια Μόρια: Τα υπόλοιπα άτομα σχηματίζουν μόρια Γιατί; Διότι η ολική ενέργεια ενός ευσταθούς μορίου είναι μικρότερη από την ολική ενέργεια των μεμονωμένων ατόμων που αποτελούν
ΗΥ562 Προχωρημένα Θέματα Βάσεων Δεδομένων Efficient Query Evaluation over Temporally Correlated Probabilistic Streams
 ΗΥ562 Προχωρημένα Θέματα Βάσεων Δεδομένων Efficient Query Evaluation over Temporally Correlated Probabilistic Streams Αλέκα Σεληνιωτάκη Ηράκλειο, 26/06/12 aseliniotaki@csd.uoc.gr ΑΜ: 703 1. Περίληψη Συνεισφοράς
ΗΥ562 Προχωρημένα Θέματα Βάσεων Δεδομένων Efficient Query Evaluation over Temporally Correlated Probabilistic Streams Αλέκα Σεληνιωτάκη Ηράκλειο, 26/06/12 aseliniotaki@csd.uoc.gr ΑΜ: 703 1. Περίληψη Συνεισφοράς
ΑΣΚΗΣΗ 3η Στοίχιση ακολουθιών βιολογικών µακροµορίων
 ΑΣΚΗΣΗ 3η Στοίχιση ακολουθιών βιολογικών µακροµορίων ΕΙΣΑΓΩΓΗ Ένας από τους πρωταρχικούς στόχους της σύγκρισης των ακολουθιών δύο µακροµορίων είναι η εκτίµηση της οµοιότητάς τους και η εξαγωγή συµπερασµάτων
ΑΣΚΗΣΗ 3η Στοίχιση ακολουθιών βιολογικών µακροµορίων ΕΙΣΑΓΩΓΗ Ένας από τους πρωταρχικούς στόχους της σύγκρισης των ακολουθιών δύο µακροµορίων είναι η εκτίµηση της οµοιότητάς τους και η εξαγωγή συµπερασµάτων
ΘΕΩΡΙΑ ΔΕΣΜΟΥ ΣΘΕΝΟΥΣ ΘΕΩΡΙΑ ΜΟΡΙΑΚΩΝ ΤΡΟΧΙΑΚΩΝ
 ΕΡΜΗΝΕΙΑ ΣΧΗΜΑΤΙΣΜΟΥ ΧΗΜΙΚΩΝ ΕΣΜΩΝ ΘΕΩΡΙΑ ΔΕΣΜΟΥ ΣΘΕΝΟΥΣ ΘΕΩΡΙΑ ΜΟΡΙΑΚΩΝ ΤΡΟΧΙΑΚΩΝ ΘΕΩΡΙΑ ΕΣΜΟΥ ΣΘΕΝΟΥΣ 1. Κατά την ανάπτυξη ομοιοπολικού δεσμού ανάμεσα σε δύο άτομα, τροχιακά της στιβάδας σθένους του
ΕΡΜΗΝΕΙΑ ΣΧΗΜΑΤΙΣΜΟΥ ΧΗΜΙΚΩΝ ΕΣΜΩΝ ΘΕΩΡΙΑ ΔΕΣΜΟΥ ΣΘΕΝΟΥΣ ΘΕΩΡΙΑ ΜΟΡΙΑΚΩΝ ΤΡΟΧΙΑΚΩΝ ΘΕΩΡΙΑ ΕΣΜΟΥ ΣΘΕΝΟΥΣ 1. Κατά την ανάπτυξη ομοιοπολικού δεσμού ανάμεσα σε δύο άτομα, τροχιακά της στιβάδας σθένους του
Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά
 Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά Είναι απλή πολυπεπτιδική αλυσίδα 307 αμινοξέων Είναι συμπαγής και έχει σχήμα ελλειψοειδές διαστάσεων 50 x 42 x 38 A Περιέχει περιοχές α-έλικος 38%
Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά Είναι απλή πολυπεπτιδική αλυσίδα 307 αμινοξέων Είναι συμπαγής και έχει σχήμα ελλειψοειδές διαστάσεων 50 x 42 x 38 A Περιέχει περιοχές α-έλικος 38%
Θεωρία Πληροφορίας. Διάλεξη 4: Διακριτή πηγή πληροφορίας χωρίς μνήμη. Δρ. Μιχάλης Παρασκευάς Επίκουρος Καθηγητής
 Θεωρία Πληροφορίας Διάλεξη 4: Διακριτή πηγή πληροφορίας χωρίς μνήμη Δρ. Μιχάλης Παρασκευάς Επίκουρος Καθηγητής 1 Ατζέντα Διακριτή πηγή πληροφορίας χωρίς μνήμη Ποσότητα πληροφορίας της πηγής Κωδικοποίηση
Θεωρία Πληροφορίας Διάλεξη 4: Διακριτή πηγή πληροφορίας χωρίς μνήμη Δρ. Μιχάλης Παρασκευάς Επίκουρος Καθηγητής 1 Ατζέντα Διακριτή πηγή πληροφορίας χωρίς μνήμη Ποσότητα πληροφορίας της πηγής Κωδικοποίηση
Γραφικά με υπολογιστές. Διδάσκων: Φοίβος Μυλωνάς. Διαλέξεις #11-#12
 Ιόνιο Πανεπιστήμιο Τμήμα Πληροφορικής Χειμερινό εξάμηνο Γραφικά με υπολογιστές Διδάσκων: Φοίβος Μυλωνάς fmlonas@ionio.gr Διαλέξεις #-# Σύνθεση Δ Μετασχηματισμών Ομογενείς Συντεταγμένες Παραδείγματα Μετασχηματισμών
Ιόνιο Πανεπιστήμιο Τμήμα Πληροφορικής Χειμερινό εξάμηνο Γραφικά με υπολογιστές Διδάσκων: Φοίβος Μυλωνάς fmlonas@ionio.gr Διαλέξεις #-# Σύνθεση Δ Μετασχηματισμών Ομογενείς Συντεταγμένες Παραδείγματα Μετασχηματισμών
Δομή πρωτεϊνών: Τριτοταγής διαμόρφωση της δομής
 Δομή πρωτεϊνών: Τριτοταγής διαμόρφωση της δομής - Αναφέρεται στην αναδίπλωση της πολυπεπτιδικής αλυσίδας πάνω στον εαυτό της και στο τελικό σχήμα που θα πάρει στο χώρο -Σ αυτή τη διαμόρφωση σημαντικό ρόλο
Δομή πρωτεϊνών: Τριτοταγής διαμόρφωση της δομής - Αναφέρεται στην αναδίπλωση της πολυπεπτιδικής αλυσίδας πάνω στον εαυτό της και στο τελικό σχήμα που θα πάρει στο χώρο -Σ αυτή τη διαμόρφωση σημαντικό ρόλο
Δ10. Συμπίεση Δεδομένων
 Συμπίεση Δεδομένων 203-204 Κωδικοποίηση εικονοροής (Video) Δρ. Ν. Π. Σγούρος 2 Ανάλυση Οθονών Δρ. Ν. Π. Σγούρος 3 Πρωτόκολλα μετάδοσης εικονοροών Πρωτόκολλο Ρυθμός (Hz) Φίλμ 23.976 ATSC 24 PAL,DVB-SD,DVB-HD
Συμπίεση Δεδομένων 203-204 Κωδικοποίηση εικονοροής (Video) Δρ. Ν. Π. Σγούρος 2 Ανάλυση Οθονών Δρ. Ν. Π. Σγούρος 3 Πρωτόκολλα μετάδοσης εικονοροών Πρωτόκολλο Ρυθμός (Hz) Φίλμ 23.976 ATSC 24 PAL,DVB-SD,DVB-HD
ΑΕΠΠ Ερωτήσεις θεωρίας
 ΑΕΠΠ Ερωτήσεις θεωρίας Κεφάλαιο 1 1. Τα δεδομένα μπορούν να παρέχουν πληροφορίες όταν υποβάλλονται σε 2. Το πρόβλημα μεγιστοποίησης των κερδών μιας επιχείρησης είναι πρόβλημα 3. Για την επίλυση ενός προβλήματος
ΑΕΠΠ Ερωτήσεις θεωρίας Κεφάλαιο 1 1. Τα δεδομένα μπορούν να παρέχουν πληροφορίες όταν υποβάλλονται σε 2. Το πρόβλημα μεγιστοποίησης των κερδών μιας επιχείρησης είναι πρόβλημα 3. Για την επίλυση ενός προβλήματος
Κεφάλαιο 1. Οι δομικοί λίθοι
 Κεφάλαιο 1 Οι δομικοί λίθοι Κεφάλαιο 1 Οι Δομικοί Λίθοι των Πρωτεϊνών Εικόνα 1.1 Η αμινοξική αλληλουχία μιας πρωτεϊνικής πολυπεπτιδικής αλυσίδας ονομάζεται πρωτοταγής δομή. Διαφορετικές περιοχές της αλληλουχίας
Κεφάλαιο 1 Οι δομικοί λίθοι Κεφάλαιο 1 Οι Δομικοί Λίθοι των Πρωτεϊνών Εικόνα 1.1 Η αμινοξική αλληλουχία μιας πρωτεϊνικής πολυπεπτιδικής αλυσίδας ονομάζεται πρωτοταγής δομή. Διαφορετικές περιοχές της αλληλουχίας
Βασίλειος Μαχαιράς Πολιτικός Μηχανικός Ph.D.
 Βασίλειος Μαχαιράς Πολιτικός Μηχανικός Ph.D. Μη γραμμικός προγραμματισμός: βελτιστοποίηση χωρίς περιορισμούς Πανεπιστήμιο Θεσσαλίας Σχολή Θετικών Επιστημών ΤμήμαΠληροφορικής Διάλεξη 7-8 η /2017 Τι παρουσιάστηκε
Βασίλειος Μαχαιράς Πολιτικός Μηχανικός Ph.D. Μη γραμμικός προγραμματισμός: βελτιστοποίηση χωρίς περιορισμούς Πανεπιστήμιο Θεσσαλίας Σχολή Θετικών Επιστημών ΤμήμαΠληροφορικής Διάλεξη 7-8 η /2017 Τι παρουσιάστηκε
Ενδεικτική πολυ-εργασία 1 - εφαρμογή στην υπολογιστική όραση
 Ενδεικτική πολυ-εργασία 1 - εφαρμογή στην υπολογιστική όραση Εντοπισμός ενός σήματος STOP σε μια εικόνα. Περιγράψτε τη διαδικασία με την οποία μπορώ να εντοπίσω απλά σε μια εικόνα την ύπαρξη του παρακάτω
Ενδεικτική πολυ-εργασία 1 - εφαρμογή στην υπολογιστική όραση Εντοπισμός ενός σήματος STOP σε μια εικόνα. Περιγράψτε τη διαδικασία με την οποία μπορώ να εντοπίσω απλά σε μια εικόνα την ύπαρξη του παρακάτω
Συνήθεις διαφορικές εξισώσεις προβλήματα οριακών τιμών
 Συνήθεις διαφορικές εξισώσεις προβλήματα οριακών τιμών Οι παρούσες σημειώσεις αποτελούν βοήθημα στο μάθημα Αριθμητικές Μέθοδοι του 5 ου εξαμήνου του ΤΜΜ ημήτρης Βαλουγεώργης Καθηγητής Εργαστήριο Φυσικών
Συνήθεις διαφορικές εξισώσεις προβλήματα οριακών τιμών Οι παρούσες σημειώσεις αποτελούν βοήθημα στο μάθημα Αριθμητικές Μέθοδοι του 5 ου εξαμήνου του ΤΜΜ ημήτρης Βαλουγεώργης Καθηγητής Εργαστήριο Φυσικών
Ασκήσεις. Γράψτε μια δομή Lewis για καθένα από τα παρακάτω μόρια και βρείτε τα τυπικά φορτία των ατόμων. (α) CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3
 CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3") Ασκήσεις Γράψτε μια δομή Lewis για καθένα από τα παρακάτω μόρια και βρείτε τα τυπικά φορτία των ατόμων. (α) CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3 Γεωμετρία Μορίων Θεωρία VSEPR Μοριακή γεωμετρία: είναι η διάταξη
Ασκήσεις Γράψτε μια δομή Lewis για καθένα από τα παρακάτω μόρια και βρείτε τα τυπικά φορτία των ατόμων. (α) CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3 Γεωμετρία Μορίων Θεωρία VSEPR Μοριακή γεωμετρία: είναι η διάταξη
Γενική & Ανόργανη Χημεία
 Γενική & Ανόργανη Χημεία 2017-18 1 Ηλεκτρονιακή Συγγένεια (Electron Affinity) Η πρόβλεψη ορισμένων χημικών ιδιοτήτων, προϋποθέτει τη γνώση του τρόπου μεταβολής της ενέργειας κατά την προσθήκη ηλεκτρονίων
Γενική & Ανόργανη Χημεία 2017-18 1 Ηλεκτρονιακή Συγγένεια (Electron Affinity) Η πρόβλεψη ορισμένων χημικών ιδιοτήτων, προϋποθέτει τη γνώση του τρόπου μεταβολής της ενέργειας κατά την προσθήκη ηλεκτρονίων
Ασκήσεις 5& 6. Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR. Σύγκριση & Ανάλυση Δομών Βιομορίων
 Ασκήσεις 5& 6 Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Σύγκριση & Ανάλυση Δομών Βιομορίων Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Κρυσταλλογραφία Ακτίνων-Χ & Φασματοσκοπία ΝΜR Πλεονεκτήματα -
Ασκήσεις 5& 6 Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Σύγκριση & Ανάλυση Δομών Βιομορίων Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Κρυσταλλογραφία Ακτίνων-Χ & Φασματοσκοπία ΝΜR Πλεονεκτήματα -
ΓΡΑΜΜΙΚΗ ΑΛΓΕΒΡΑ (Εξ. Ιουνίου - 02/07/08) ΕΠΙΛΕΓΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ
 ΕΠΙΛΕΓΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ") Ονοματεπώνυμο:......... Α.Μ....... Ετος... ΑΙΘΟΥΣΑ:....... I. (περί τις 55μ. = ++5++. Σωστό ή Λάθος: ΓΡΑΜΜΙΚΗ ΑΛΓΕΒΡΑ (Εξ. Ιουνίου - //8 ΕΠΙΛΕΓΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ (αʹ Αν AB = BA όπου A, B τετραγωνικά και
Ονοματεπώνυμο:......... Α.Μ....... Ετος... ΑΙΘΟΥΣΑ:....... I. (περί τις 55μ. = ++5++. Σωστό ή Λάθος: ΓΡΑΜΜΙΚΗ ΑΛΓΕΒΡΑ (Εξ. Ιουνίου - //8 ΕΠΙΛΕΓΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ (αʹ Αν AB = BA όπου A, B τετραγωνικά και
ΕΙΣΑΓΩΓΗ ΣΤΙΣ ΑΡΧΕΣ ΤΗΣ ΕΠΙΣΤΗΜΗΣ ΤΩΝ Η/Υ
 ΕΙΣΑΓΩΓΗ ΣΤΙΣ ΑΡΧΕΣ ΤΗΣ ΕΠΙΣΤΗΜΗΣ ΤΩΝ Η/Υ ΜΕΡΛΙΑΟΥΝΤΑΣ ΣΤΕΦΑΝΟΣ, ΠΕ19 ΚΕΦΑΛΑΙΟ 3 Αλγόριθμοι 3. Αλγόριθμοι 2 3. Αλγόριθμοι 3.1 Η έννοια του αλγορίθμου 3.2 Χαρακτηριστικά αλγορίθμου 3.3 Ανάλυση αλγορίθμων
ΕΙΣΑΓΩΓΗ ΣΤΙΣ ΑΡΧΕΣ ΤΗΣ ΕΠΙΣΤΗΜΗΣ ΤΩΝ Η/Υ ΜΕΡΛΙΑΟΥΝΤΑΣ ΣΤΕΦΑΝΟΣ, ΠΕ19 ΚΕΦΑΛΑΙΟ 3 Αλγόριθμοι 3. Αλγόριθμοι 2 3. Αλγόριθμοι 3.1 Η έννοια του αλγορίθμου 3.2 Χαρακτηριστικά αλγορίθμου 3.3 Ανάλυση αλγορίθμων
HY213. ΑΡΙΘΜΗΤΙΚΗ ΑΝΑΛΥΣΗ ΕΛΑΧΙΣΤΑ ΤΕΤΡΑΓΩΝΑ AΝΑΛΥΣΗ ΙΔΙΑΖΟΥΣΩΝ ΤΙΜΩΝ
 HY3. ΑΡΙΘΜΗΤΙΚΗ ΑΝΑΛΥΣΗ ΕΛΑΧΙΣΤΑ ΤΕΤΡΑΓΩΝΑ AΝΑΛΥΣΗ ΙΔΙΑΖΟΥΣΩΝ ΤΙΜΩΝ Π. ΤΣΟΜΠΑΝΟΠΟΥΛΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ & ΜΗΧΑΝΙΚΩΝ ΥΠΟΛΟΓΙΣΤΩΝ Μέθοδος ελαχίστων τετραγώνων Τα σφάλματα
HY3. ΑΡΙΘΜΗΤΙΚΗ ΑΝΑΛΥΣΗ ΕΛΑΧΙΣΤΑ ΤΕΤΡΑΓΩΝΑ AΝΑΛΥΣΗ ΙΔΙΑΖΟΥΣΩΝ ΤΙΜΩΝ Π. ΤΣΟΜΠΑΝΟΠΟΥΛΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ & ΜΗΧΑΝΙΚΩΝ ΥΠΟΛΟΓΙΣΤΩΝ Μέθοδος ελαχίστων τετραγώνων Τα σφάλματα
Ποια είναι κατά τη γνώμη σας τα 30 μικρομόρια που συνιστούν τα πρόδρομα μόρια των βιομακρομορίων; Πώς μπορούν να ταξινομηθούν;
 Ποια είναι κατά τη γνώμη σας τα 30 μικρομόρια που συνιστούν τα πρόδρομα μόρια των βιομακρομορίων; Πώς μπορούν να ταξινομηθούν; Γενικά Για να προσδιορίσουμε τα 30 πρόδρομα μόρια των βιομακρομορίων θα πρέπει
Ποια είναι κατά τη γνώμη σας τα 30 μικρομόρια που συνιστούν τα πρόδρομα μόρια των βιομακρομορίων; Πώς μπορούν να ταξινομηθούν; Γενικά Για να προσδιορίσουμε τα 30 πρόδρομα μόρια των βιομακρομορίων θα πρέπει
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ
 ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
7 ο Κεφάλαιο Οργανική Χημεία. Δ. Παπαδόπουλος, χημικός
 7 ο Κεφάλαιο Οργανική Χημεία Δ. Παπαδόπουλος, χημικός ΓΕΛ Καρέα, 2018 Θεωρίες ερμηνείας του ομοιοπολικού δεσμού με βάση την κβαντική θεωρία. Θεωρία δεσμού σθένους. Θεωρία των μοριακών τροχιακών. Κάθε θεωρία
7 ο Κεφάλαιο Οργανική Χημεία Δ. Παπαδόπουλος, χημικός ΓΕΛ Καρέα, 2018 Θεωρίες ερμηνείας του ομοιοπολικού δεσμού με βάση την κβαντική θεωρία. Θεωρία δεσμού σθένους. Θεωρία των μοριακών τροχιακών. Κάθε θεωρία
Γιατί ο σχηματισμός του CΗ 4 δεν μπορεί να ερμηνευθεί βάσει της διεγερμένης κατάστασης του ατόμου C;
 Γιατί ο σχηματισμός του CΗ 4 δεν μπορεί να ερμηνευθεί βάσει της διεγερμένης κατάστασης του ατόμου C; 1. Οι 4 ομοιοπολικοί δεσμοί στο μεθάνιο θα ήταν δύο τύπων: ένας δεσμός από την επικάλυψη του τροχιακού
Γιατί ο σχηματισμός του CΗ 4 δεν μπορεί να ερμηνευθεί βάσει της διεγερμένης κατάστασης του ατόμου C; 1. Οι 4 ομοιοπολικοί δεσμοί στο μεθάνιο θα ήταν δύο τύπων: ένας δεσμός από την επικάλυψη του τροχιακού
Ca. Να μεταφέρετε στην κόλλα σας συμπληρωμένο τον παρακάτω πίνακα που αναφέρεται στο άτομο του ασβεστίου: ΣΤΙΒΑΔΕΣ νετρόνια K L M N Ca 2
 Ερωτήσεις Ανάπτυξης 1. Δίνεται ότι: 40 20 Ca. Να μεταφέρετε στην κόλλα σας συμπληρωμένο τον παρακάτω πίνακα που αναφέρεται στο άτομο του ασβεστίου: ΣΤΙΒΑΔΕΣ νετρόνια K L M N Ca 2 2. Tι είδους δεσμός αναπτύσσεται
Ερωτήσεις Ανάπτυξης 1. Δίνεται ότι: 40 20 Ca. Να μεταφέρετε στην κόλλα σας συμπληρωμένο τον παρακάτω πίνακα που αναφέρεται στο άτομο του ασβεστίου: ΣΤΙΒΑΔΕΣ νετρόνια K L M N Ca 2 2. Tι είδους δεσμός αναπτύσσεται
Κεφάλαιο 2 Χημικοί Δεσμοί
 Κεφάλαιο 2 Χημικοί Δεσμοί Σύνοψη Παρουσιάζονται οι χημικοί δεσμοί, ιοντικός, μοριακός, ατομικός, μεταλλικός. Οι ιδιότητες των υλικών τόσο οι φυσικές όσο και οι χημικές εξαρτώνται από το είδος ή τα είδη
Κεφάλαιο 2 Χημικοί Δεσμοί Σύνοψη Παρουσιάζονται οι χημικοί δεσμοί, ιοντικός, μοριακός, ατομικός, μεταλλικός. Οι ιδιότητες των υλικών τόσο οι φυσικές όσο και οι χημικές εξαρτώνται από το είδος ή τα είδη
Στ Τάξη. Α/Α Μαθηματικό περιεχόμενο Δείκτες Επιτυχίας Ώρες Διδ. 1 ENOTHTA 1
 Ενδεικτική Οργάνωση Ενοτήτων Στ Τάξη Α/Α Μαθηματικό περιεχόμενο Δείκτες Επιτυχίας Ώρες Διδ. 1 ENOTHTA 1 15 Αρ3.1 Απαγγέλουν, διαβάζουν, γράφουν και αναγνωρίζουν ποσότητες αριθμών Επανάληψη μέχρι το 1 000
Ενδεικτική Οργάνωση Ενοτήτων Στ Τάξη Α/Α Μαθηματικό περιεχόμενο Δείκτες Επιτυχίας Ώρες Διδ. 1 ENOTHTA 1 15 Αρ3.1 Απαγγέλουν, διαβάζουν, γράφουν και αναγνωρίζουν ποσότητες αριθμών Επανάληψη μέχρι το 1 000
Γεωμετρία Μορίων Θεωρία VSEPR
 Γεωμετρία Μορίων Θεωρία VSEPR Γεωμετρία Μορίων Θεωρία VSEPR Γεωμετρία Μορίων Θεωρία VSEPR Μεθοδολογία για την πρόβλεψη της μοριακής γεωμετρία: Γράφουμε τον ηλεκτρονιακό τύπο κατά Lewis. Μετρούμε το συνολικό
Γεωμετρία Μορίων Θεωρία VSEPR Γεωμετρία Μορίων Θεωρία VSEPR Γεωμετρία Μορίων Θεωρία VSEPR Μεθοδολογία για την πρόβλεψη της μοριακής γεωμετρία: Γράφουμε τον ηλεκτρονιακό τύπο κατά Lewis. Μετρούμε το συνολικό
Βιοπληροφορική και Πολυµέσα. Ειρήνη Αυδίκου Αθήνα
 Βιοπληροφορική και Πολυµέσα Αθήνα 1.2.2009 ΠΕΡΙΕΧΟΜΕΝΑ 1. Πως σχετίζεται µε τα Πολυµέσα 2. Τι είναι η Βιοπληροφορική 3. Χρήσεις 4. Συµπεράσµατα 5. Βιβλιογραφία Βιοπληροφορική και Πολυµέσα 2 1. Τι είναι
Βιοπληροφορική και Πολυµέσα Αθήνα 1.2.2009 ΠΕΡΙΕΧΟΜΕΝΑ 1. Πως σχετίζεται µε τα Πολυµέσα 2. Τι είναι η Βιοπληροφορική 3. Χρήσεις 4. Συµπεράσµατα 5. Βιβλιογραφία Βιοπληροφορική και Πολυµέσα 2 1. Τι είναι
«ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ»
 «ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ» Τι είναι οι πρωτεΐνες; Από τι αποτελούνται; Ποιος είναι ο βιολογικός του ρόλος; Ας ρίξουμε μία ματιά σε όλα αυτά τα ερωτήματα που μας απασχολούν ΚΕΦΑΛΑΙΟ 1:
«ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ» Τι είναι οι πρωτεΐνες; Από τι αποτελούνται; Ποιος είναι ο βιολογικός του ρόλος; Ας ρίξουμε μία ματιά σε όλα αυτά τα ερωτήματα που μας απασχολούν ΚΕΦΑΛΑΙΟ 1:
Η διαδικασία Παραγωγής Συνθετικής Εικόνας (Rendering)
") Υφή Η διαδικασία Παραγωγής Συνθετικής Εικόνας (Rendering) Θέσεις αντικειμένων και φωτεινών πηγών Θέση παρατηρητή 3D Μοντέλα 3Δ Μετασχ/σμοί Μοντέλου 3Δ Μετασχ/σμός Παρατήρησης Απομάκρυνση Πίσω Επιφανειών
Υφή Η διαδικασία Παραγωγής Συνθετικής Εικόνας (Rendering) Θέσεις αντικειμένων και φωτεινών πηγών Θέση παρατηρητή 3D Μοντέλα 3Δ Μετασχ/σμοί Μοντέλου 3Δ Μετασχ/σμός Παρατήρησης Απομάκρυνση Πίσω Επιφανειών
ΕΝΟΤΗΤΑ 4 ΕΙΔΗ ΓΡΑΜΜΩΝ, ΕΙΔΗ ΤΡΙΓΩΝΩΝ, ΠΑΡΑΛΛΗΛΟΓΡΑΜΜΑ, ΜΕΤΑΣΧΗΜΑΤΙΣΜΟΙ
 ΕΝΟΤΗΤΑ 4 ΕΙΔΗ ΓΡΑΜΜΩΝ, ΕΙΔΗ ΤΡΙΓΩΝΩΝ, ΠΑΡΑΛΛΗΛΟΓΡΑΜΜΑ, ΜΕΤΑΣΧΗΜΑΤΙΣΜΟΙ ΔΕΙΚΤΕΣ ΕΠΙΤΥΧΙΑΣ ΑΡΙΘΜΟΙ Εκτίμηση και μέτρηση Μ3.6 Εκτιμούν, μετρούν, ταξινομούν και κατασκευάζουν γωνίες (με ή χωρίς τη χρήση της
ΕΝΟΤΗΤΑ 4 ΕΙΔΗ ΓΡΑΜΜΩΝ, ΕΙΔΗ ΤΡΙΓΩΝΩΝ, ΠΑΡΑΛΛΗΛΟΓΡΑΜΜΑ, ΜΕΤΑΣΧΗΜΑΤΙΣΜΟΙ ΔΕΙΚΤΕΣ ΕΠΙΤΥΧΙΑΣ ΑΡΙΘΜΟΙ Εκτίμηση και μέτρηση Μ3.6 Εκτιμούν, μετρούν, ταξινομούν και κατασκευάζουν γωνίες (με ή χωρίς τη χρήση της
5. Απλή Ταξινόμηση. ομές εδομένων. Χρήστος ουλκερίδης. Πανεπιστήμιο Πειραιώς Σχολή Τεχνολογιών Πληροφορικής και Επικοινωνιών Τμήμα Ψηφιακών Συστημάτων
 Πανεπιστήμιο Πειραιώς Σχολή Τεχνολογιών Πληροφορικής και Επικοινωνιών Τμήμα Ψηφιακών Συστημάτων 5. Απλή Ταξινόμηση 2 ομές εδομένων 4 5 Χρήστος ουλκερίδης Τμήμα Ψηφιακών Συστημάτων 11/11/2016 Εισαγωγή Η
Πανεπιστήμιο Πειραιώς Σχολή Τεχνολογιών Πληροφορικής και Επικοινωνιών Τμήμα Ψηφιακών Συστημάτων 5. Απλή Ταξινόμηση 2 ομές εδομένων 4 5 Χρήστος ουλκερίδης Τμήμα Ψηφιακών Συστημάτων 11/11/2016 Εισαγωγή Η