«Έκφραση, απομόνωση και μελέτη της Σχέσης Δομής-Δραστικότητας πρωτεϊνών Φαρμακευτικού ενδιαφέροντος, μέσω Φασματοσκοπίας NMR»
|
|
|
- Βασιλική Βενιζέλος
- 6 χρόνια πριν
- Προβολές:
Transcript
1 ΑΛΕΞΑΝΔΡΟΠΟΥΛΟΣ ΙΩΑΝΝΗΣ ΧΗΜΙΚΟΣ «Έκφραση, απομόνωση και μελέτη της Σχέσης Δομής-Δραστικότητας πρωτεϊνών Φαρμακευτικού ενδιαφέροντος, μέσω Φασματοσκοπίας NMR» ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΥΠΟΒΛΗΘΗΚΕ ΣΤΟ ΦΑΡΜΑΚΕΥΤΙΚΟ ΤΜΗΜΑ ΤΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΠΑΤΡΩΝ ΠΑΤΡΑ 2016 i
2 ΑΛΕΞΑΝΔΡΟΠΟΥΛΟΣ ΙΩΑΝΝΗΣ ΧΗΜΙΚΟΣ «Έκφραση, απομόνωση και μελέτη της Σχέσης Δομής-Δραστικότητας πρωτεϊνών Φαρμακευτικού ενδιαφέροντος, μέσω Φασματοσκοπίας NMR» «Μελέτη της H-NOX C139A επικράτειας του οργανισμού Nostoc sp.μέσω Φασματοσκοπίας NMR» ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Ε Ξ Ε Τ Α Σ Τ Ι Κ Η Ε Π Ι Τ Ρ Ο Π Η ΓΕΩΡΓΙΟΣ Α. ΣΠΥΡΟΥΛΙΑΣ ΚΑΘΗΓΗΤΗΣ, ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΕΠΙΒΛΕΠΩΝ / ΠΡΟΕΔΡΟΣ ΕΞΕΤΑΣΤΙΚΗΣ ΕΠΙΤΡΟΠΗΣ ΕΜΜΑΝΟΥΗΛ ΦΟΥΣΤΕΡΗΣ ΕΠΙΚΟΥΡΟΣ ΚΑΘΗΓΗΤΗΣ, ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΓΕΩΡΓΙΟΣ ΠΑΪΡΑΣ ΑΝΑΠΛΗΡΩΤΗΣ ΚΑΘΗΓΗΤΗΣ, ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ii
3 1. Περιεχόμενα 2. ΕΙΣΑΓΩΓΗ ΝΟ σηματοδοτικό μονοπάτι: Βιοχημεία/Ασθένειες/Φαρμακολογία Διαλυτή γουανυλική κυκλάση Αρχιτεκτονική του μορίου της sgc Επικράτεια H-NOX Επικράτειες Per/Arnt/Sim (PAS) και σπειρωμένου σπειράματος Καταλυτική περιοχή Απομόνωση της sgc Ισομορφές της sgc Ομόλογα της sgc Άτυπες μορφές της sgc σε ευκαρυωτικούς οργανισμούς H-NOX πρωτεΐνες σε προκαρυωτικούς οργανισμούς Σημαντικά αμινοξέα στην πρόσδεση των υποκαταστατών της H-NOX Ανόργανη χημεία του συμπλόκου αίμη-h-nox-υποκαταστάτης Το μόριο της αίμης Η σημασία του μορίου της αίμης στο σύμπλοκο της πρωτεΐνης Αμινοξέα ικανά να προσδεθούν με το μόριο της αίμης Οξειδωτικές καταστάσεις του σιδήρου Ενεργειακός διαχωρισμός των d τροχιακών Σύμπλοκα υψηλού και χαμηλού spin Ενεργοποίηση της sgc από πλευράς ανόργανης χημείας Φασματοσκοπία Πυρηνικού Μαγνητικού Συντονισμού (Nuclear Magnetic Resonance, NMR) Εισαγωγή Θεωρητική προσέγγιση Μεταπτωτική κίνηση των πυρήνων Επίδραση του παλμού ραδιοσυχνότητας στις ενεργειακές καταστάσεις του πυρήνα Μηχανισμοί αποδιέγερσης (relaxation) στο NMR Ανατομία ενός NMR πειράματος Η έννοια της χημικής μετατόπισης i
4 2.8.8 Η σταθερά σύζευξης J Ένταση και εύρος κορυφής Μονοδιάστατο (1D) φάσμα NMR NMR φασματοσκοπία δύο διαστάσεων (2D) Τρισδιάστατη NMR φασματοσκοπία Επίλυση τρισδιάστατων δομών βιομορίων μέσω φασματοσκοπίας NMR Συλλογή NMR δεδομένων Ανάλυση NMR φασμάτων Μετατροπή της έντασης των NOE σημάτων σε ανώτερα όρια διαπρωτονιακών αποστάσεων Υπολογισμός της τριτοταγούς δομής Τελική οικογένεια δομών Πρωτεϊνική δυναμική Υπολογιστικά εργαλεία Το πρόγραμμα TALOS (Torsion Angle Likelihood Obtained from chemical shift and sequence Similarity) Το πρόγραμμα DYANA ΣΚΟΠΟΣ ΕΡΓΑΣΙΑΣ ΠΕΙΡΑΜΑΤΙΚΟ ΜΕΡΟΣ NMR πειράματα Επεξεργασία των NMR φασμάτων Ανάλυση των NMR φασμάτων Δομικοί υπολογισμοί Μελέτη πρωτεϊνικής δυναμικής ΑΠΟΤΕΛΕΣΜΑΤΑ Η H-NOX επικράτεια του οργανισμού Nostoc sp Ανάλυση NMR φασμάτων και δομικοί υπολογισμοί ΣΥΖΗΤΗΣΗ Επιλογή της κατάλληλης επικράτειας H-NOX για δομική μελέτη μέσω NMR φασματοσκοπίας Ανάλυση των δυσχερειών κατά τη διάρκεια της επεξεργασίας των πειραματικών δεδομένων Aνάλυση των στοιχείων δευτεροταγούς δομής της Nostoc sp. H-NOX C139A και σύγκριση με τα αντίστοιχα της κρυσταλλικής δομής ii
5 5.4 Σύγκριση της κρυσταλλογραφικής δομής με την τριτοταγή διαμόρφωση που προέκυψε από τους δομικούς υπολογισμούς βάσει των NMR πειραματικών δεδομένων Μελλοντικοί στόχοι ΒΙΒΛΙΟΓΡΑΦΙΑ ΠΕΡΙΛΗΨΗ ΑBSTRACT ΠΑΡΑΡΤΗΜΑ Πίνακες Χημικών Μετατοπίσεων iii
6 2. ΕΙΣΑΓΩΓΗ 1
7 2.1 ΝΟ/cGMP σηματοδοτικό μονοπάτι: Βιοχημεία/Ασθένειες/Φαρμακολογία Το μόριο του NO αποτελεί ένα πολύ σημαντικό σηματοδοτικό μόριο στα βιολογικά συστήματα. Στα θηλαστικά, το διατομικό αυτό αέριο είναι καθοριστικής σημασίας για το μονοπάτι NO/cGMP (κυκλική μονοφωσφορική γουανοσίνη) καθώς λειτουργεί ως ενεργοποιητής της διαλυτής γουανυλικής κυκλάσης (sgc). Τα κλάσματα του κυτοσολίου καθώς και κάποια κλάσματα ιστού θηλαστικών εμφανίζουν δράση γουανυλικής κυκλάσης. Συγκεκριμένα, η σωματιδιακή γουανυλική κυκλάση (pgc), η οποία εντοπίζεται στο μη διαλυτό μέρος των κυττάρων, βρίσκεται δεσμευμένη στην κυτταρική μεμβράνη και ενεργοποιείται από νατριουρητικά πεπτίδια [6,7]. Tα παράγωγα του κυτοσολίου περιέχουν διαλυτή γουανυλική κυκλάση, η οποία ενεργοποιείται από το NO. Η διαδικασία ενεργοποίησης του σηματοδοτικού μονοπατιού NO/cGMP ξεκινά με απελευθέρωση ασβεστίου. Ακολουθεί δέσμευση του συμπλέγματος ασβεστίου/καλμοδουλίνης στο ένζυμο συνθάση του νιτρικού οξειδίου (NOS). Στην NOS συντίθεται το NO από L-αργινίνη και οξυγόνο (O2) [8] συμβάλλοντας έτσι στην ενεργοποίηση της sgc. Το παραγόμενο NO διαχέεται στα κύτταρα στόχους, όπου προσδένεται στο μόριο αίμης της sgc (Εικόνα 1). Το μόριο αίμης είναι συνδεδεμένο με ένα κατάλοιπο ιστιδίνης του πρωτεϊνικού μορίου. Αποτέλεσμα της πρόσδεσης του ΝΟ με την αιμοπρωτεΐνη είναι η ραγδαία αύξηση σύνθεσης cgmp. Εικόνα 1: Σηματοδοτικό μονοπάτι NO/cGMP [8]. Μετά το σχηματισμό της cgmp, ακολουθεί η πρόσδεση και η ενεργοποίηση φωσφοδιεστερασών (PDEs), ιοντικών καναλιών και cgmp-εξαρτώμενων 2
8 πρωτεϊνικών κινασών (cgks) συμβάλλοντας έτσι στη ρύθμιση διαφόρων φυσιολογικών λειτουργιών του κυττάρου, όπως είναι η αγγειοδιαστολή [9] και η επακόλουθη ρύθμιση της καρδιακής ομοιόστασης. Η διαδικασία επιτυγχάνεται μέσω της χαλάρωσης των λείων μυϊκών κυττάρων. Επίσης, αναστέλλεται η συσσώρευση αιμοπεταλίων [10] στα αγγεία με αποτέλεσμα τη ρύθμιση της αρτηριακής πίεσης. Ο ρόλος που διαδραματίζει η cgmp είναι καθοριστικός και στη διαδικασία της νευροδιαβίβασης [11]. Επίσης, συμμετέχει και σε άλλες λειτουργίες, όπως στη ρύθμιση διαφόρων σταδίων του κύκλου ζωής του κυττάρου (πολλαπλασιασμός, διαφοροποίηση, ανάπτυξη και απόπτωση), στην ανοσολογική απόκριση, στην έκκριση απαραίτητων ουσιών του γαστρεντερικού συστήματος και σε πλήθος άλλων λειτουργιών. Το 1998, το βραβείο Νόμπελ στη Φυσιολογία και Ιατρική απονεμήθηκε στους Robert F. Furchgott, Louis J. Ignarro, και Ferid Murad, για τα επιτεύγματά τους σχετικά με την ανακάλυψη του σηματοδοτικού μονοπατιού NO/cGMP. Ιδιαίτερο ενδιαφέρον αποτελεί η έρευνα με σκοπό την ανακάλυψη φαρμάκων, τα οποία θα συμβάλουν στην καταπολέμηση πολλών ασθενειών, όπως είναι οι καρδιαγγειακές και νευροεκφυλιστικές νόσοι, και θα στοχεύουν κάποιο από τα μόρια του ΝΟσηματοδοτικού μονοπατιού. 2.2 Διαλυτή γουανυλική κυκλάση Η διαλυτή γουανυλική κυκλάση (sgc) αποτελεί μια πρωτεΐνη-αισθητήρα του νιτρικού οξειδίου (NO) [1,2], η οποία για πρώτη φορά απομονώθηκε το Παρουσία του NO, η sgc ενεργοποιείται και συμμετέχει στο NO-σηματοδοτικό μονοπάτι, το οποίο διαδραματίζει καθοριστικό ρόλο στις φυσιολογικές λειτουργίες του οργανισμού. Μελέτες σχετικά με την sgc, έχουν δείξει ότι φέρει ένα μόριο αίμης [3,4], στο οποίο το NO προσδένεται εκλεκτικά επιτυγχάνοντας έτσι την ενεργοποίησή της. Αυτή η εκλεκτικότητα γίνεται εύκολα αντιληπτή από το γεγονός ότι, παρά τις μεγάλες ποσότητες διατομικού οξυγόνου (O2) στα ευκαρυωτικά κύτταρα, πραγματοποιείται πρόσδεση με το NO το οποίο συναντάται σε πολύ μικρότερες ποσότητες. Συγκεκριμένα, η πρόσδεση του NO [5] γίνεται στο σίδηρο (Fe) της αίμης. 3
9 2.3 Αρχιτεκτονική του μορίου της sgc Η sgc αποτελείται από δύο υπομονάδες, μια μεγάλη, την α και μια μικρότερη, τη β, οι οποίες συνδέονται μεταξύ τους και σχηματίζουν ένα ετεροδιμερές. Οι υπομονάδες α1 και β1, οι οποίες απομονώθηκαν από sgc ποντικών φέρουν 690 και 619 αμινοξέα, αντίστοιχα. Επίσης, οι υπομονάδες α2 και β2 εντοπίστηκαν σε εμβρυικό εγκέφαλο ανθρώπου και σε νεφρούς αρουραίου. Οι συγκεκριμένες υπομονάδες εμφανίζουν μοριακά βάρη 82 kda και 76 kda, αντίστοιχα [31,32]. Οι πρωτεΐνες αυτές αποτελούν μέρος της μεγάλης οικογένειας των sgc υπομονάδων, οι οποίες είναι συντηρημένες στους ευκαρυωτικούς οργανισμούς. Κάθε υπομονάδα μπορεί να χωριστεί σε τρία επιμέρους τμήματα: (α) το N-τελικό τμήμα, το οποίο καλείται ρυθμιστική περιοχή, (β) το κεντρικό τμήμα, το οποίο συμβάλλει στη δημιουργία του ετεροδιμερούς και (γ) το C-τελικό άκρο το οποίο είναι υπεύθυνο για την καταλυτική δράση του ενζύμου [6] (Εικόνα 2). (Α) (Β) Εικόνα 2: Αρχιτεκτονική επικράτειας της sgc. (α) Η επικράτεια πρόσδεσης αίμης-νιτρικού οξειδίου/οξυγόνου (Heme-nitric oxide/oxygen binding, H-NOX) απεικονίζεται με κίτρινο χρώμα. (β) Χαρακτηριστικά τμήματα πρόσδεσης αίμης και GTP. Η αίμη αναπαριστάται από το κόκκινο παραλληλόγραμμο [8]. 4
10 2.3.1 Επικράτεια H-NOX Πρόκειται για μια περιοχή συντηρημένη στους οργανισμούς, η οποία αποτελείται περίπου από τα 200 πρώτα αμινοξέα της β-αλυσίδας, ενώ εντοπίζεται και στην α- αλυσίδα [6]. Προκειμένου να εξακριβωθεί η δράση της H-NOX επικράτειας (Heme- Nitric Oxide/Oxygen Binding Domain) και η ακριβής θέση πρόσδεσης της αίμης, ήταν αναγκαίο να πραγματοποιηθούν τοποκατευθυνόμενες μεταλλάξεις καθώς και δημιουργία μικρότερων τμημάτων της sgc. Μεταλλάξεις των συντηρημένων ιστιδινών [36] στο μόριο της sgc, έκφραση διαφόρων τμημάτων της σε συστήματα έκφρασης όπως E.coli [26] και διαγραφή του N-τελικού άκρου της β1 υπομονάδας [37], έδειξαν πως η αίμη είναι προσδεδεμένη στο N-τελικό άκρο της β1 υπομονάδας και πρότειναν ως εγγύ προσδέτη την ιστιδίνη 105 [6]. Συγκεκριμένα, η επικράτεια πρόσδεσης της αίμης εντοπίστηκε στα αμινοξικά κατάλοιπα 1 έως περίπου 194 της β1 υπομονάδας [25] της. Το μόριο της sgc και συγκεκριμένα η επικράτεια πρόσδεσης της αίμης συνδέεται με NO και CO, αλλά όχι με O2. Το Ν-τελικό άκρο της α1 υπομονάδας παρουσιάζει ομολογία με αυτό της β1 [27]. Το Ν-τελικό άκρο της sgc αποτελεί μέρος μιας συντηρημένης οικογένειας πρωτεϊνών οι οποίες απαντώνται τόσο στους προκαρυωτικούς όσο και στους ευκαρυωτικούς οργανισμούς [38]. Αυτή η οικογένεια πρωτεΐνών ονομάστηκε H-NOX [39] (Εικόνα 4). Όσες πρωτεΐνες H-NOX έχουν χαρακτηριστεί μέχρι σήμερα, προσδένονται με το NO και το CO [40] τόσο καλά όσο προσδένονται και με την αίμη. Κάποιες επικράτειες H-NOX δεν προσδένονται με το O2, ενώ άλλες δημιουργούν με αυτό πολύ σταθερά σύμπλοκα. Στους ευκαρυωτικούς οργανισμούς, οι H-NOX πρωτεΐνες έχουν βρεθεί μόνο με τη γνωστή αρχιτεκτονική επικράτειας που έχουν και στο μόριο της sgc. Στα βακτήρια, απαντώνται ως πρωτεΐνες με μήκος περίπου 200 αμινοξέων με μια μοναδική προβλεπόμενη λειτουργία ή ως επικράτειες μέσα σε μεγαλύτερα πρωτεϊνικά μόρια. Αξιοσημείωτο είναι ότι τα βακτηριακά γονίδια, τα οποία κωδικοποιούν H-NOX πρωτεΐνες, παρουσιάζουν ομοιότητες με εκείνα που κωδικοποιούν ιστιδινικές κινάσες και διγουανυλικές κυκλάσες Επικράτειες Per/Arnt/Sim (PAS) και σπειρωμένου σπειράματος (coiled coil) Στην κεντρική περιοχή της sgc περιλαμβάνονται δύο επικράτειες των οποίων η λειτουργία δεν έχει ακόμα αποσαφηνιστεί. Η επικράτεια, η οποία βρίσκεται πλησιέστερα στην Ν-τελική περιοχή, εμφανίζει αναδίπλωση επικράτειας PAS. Οι επικράτειες PAS, κατά κύριο λόγο, μεσολαβούν στις αλληλεπιδράσεις μεταξύ 5
11 πρωτεϊνικών μορίων, ενώ έχουν βρεθεί προσδεδεμένες με αίμη, με φλαβίνη ή με ένα νουκλεοτίδιο [65]. Η άλλη επικράτεια, γνωστή ως σπειρωμένο σπείραμα, φαίνεται να είναι μοναδική στο μόριο της sgc και μάλιστα δεν παρουσιάζει σημαντική ομολογία με πρωτεΐνες οι οποίες είναι κατατεθειμένες στη βάση δεδομένων NCBI. Συγκεκριμένα, η επικράτεια σπειρωμένο σπείραμα, η οποία απομονώθηκε από την β1 υπομονάδα ποντικού, ήταν ένα τετραμερές του οποίου η δομή επιλύθηκε με Κρυσταλλογραφία Ακτίνων Χ [28]. Η παραπάνω δομική μελέτη, καθώς και πειράματα τα οποία περιελάμβαναν μεταλλάξεις σε αμινοξέα της α1 υπομονάδας [86] και δομικές αναλύσεις ομόλογων PAS επικρατειών της sgc [87], απέδειξαν πως η κεντρική περιοχή και των δύο υπομονάδων της sgc συμβάλλει στη δημιουργία ενός λειτουργικού ετεροδιμερούς Καταλυτική περιοχή Οι C-τελικές περιοχές των α1 και β1 πρωτεϊνών παρουσιάζουν μεγάλη ομολογία με την καταλυτική περιοχή της αδενυλικής και γουανυλικής κυκλάσης. Τη δεκαετία του 90, από ομόλογα μοντέλα βασισμένα στην κρυσταλλική δομή των ίδιων περιοχών της αδενυλικής κυκλάσης [41,42], προέκυψαν κάποιες ιδέες σχετικά με τη δομή των καταλυτικών περιοχών των sgc πρωτεϊνών. Αυτά τα μοντέλα χαρακτηρίζουν καταλυτικά αμινοξέα «κλειδιά», συμπεριλαμβανομένων δύο συντηρημένων καταλοίπων ασπαραγινικού στην υπομονάδα α1 (D485 και D529, σύμφωνα με την αρίθμηση σε sgc ποντικών), τα οποία προβλέπεται ότι συνδέονται με δύο ιόντα Mg 2+[43]. Ο ρόλος των αμινοξέων αυτών είναι πολύ σημαντικός στην κατάλυση, γιατί αφενός διεγείρουν την ενεργοποίηση του ΝΟ μονοπατιού και αφετέρου σταθεροποιούν το προϊόν cgmp και το υπόστρωμα ATP. Επιπλέον, το αμινοξύ N548 της β1 υπομονάδας φαίνεται να προσανατολίζει το δακτύλιο της ριβόζης κατάλληλα ώστε να πραγματοποιηθεί η αντίδραση, ενώ άλλα όπως το R573 και R552 των α1 και β1 υπομονάδων, αντίστοιχα, φαίνεται να αλληλεπιδρούν με το τριφωσφορικό νουκλεοτίδιο [43]. Με την εκλεκτικότητα ως προς το υπόστρωμα φαίνεται να σχετίζονται τα κατάλοιπα της β1 υπομονάδας E473 και C541. Μετά την ταυτοποίηση αυτών των σημαντικών για την αντίδραση αμινοξέων, κατέστη δυνατό να γίνουν προβλέψεις για τη δράση της γουανυλικής κυκλάσης με τη βοήθεια της ανάλυσης της αμινοξικής ακολουθίας. Αυτή η ανάλυση οδήγησε στο συμπέρασμα ότι η β2 ισομορφή μπορεί να είναι δραστική ως ομοδιμερές, αλλά οι β1, α1 και α2 είναι λειτουργικές μόνο ως ετεροδιμερή [6]. 6
.")
12 Οι καταλυτικές περιοχές έχουν πλέον εντοπιστεί στη C-τελική περιοχή των υπομονάδων α1 και β1 και περιλαμβάνουν τα αμινοξέα και , αντίστοιχα. Αυτές οι καταλυτικές περιοχές σχηματίζουν ένα ετεροδιμερές για να συμβάλουν στη σύνθεση cgmp (Εικόνα 4). Στο μόριο της sgc η καταλυτική ικανότητα του μορίου εξαρτάται από την κατάσταση πρόσδεσης της αίμης στην H- NOX περιοχή. Η sgc παρουσιάζει μεγάλη εκλεκτικότητα ως προς το υπόστρωμα GTP. Εικόνα 3: Ετεροδιμερής καταλυτική (CAT) επικράτεια [44]. 2.3 Απομόνωση της sgc Παρά το γεγονός ότι η sgc μελετάται αρκετά χρόνια, δεν είχε βρεθεί μία μέθοδος απομόνωσής της με χαμηλό κόστος. Βέβαια, με την πάροδο των χρόνων αναπτύχθηκαν διάφορες μέθοδοι αλλά απέδιδαν μικρές ποσότητες (μερικά μικρογραμμάρια) πρωτεΐνης. Αρχικά, ο χαρακτηρισμός της sgc πραγματοποιήθηκε σε ιστούς ποντικών και βοειδών. Από το 1980, οι μελέτες πραγματοποιούνταν με μόρια της sgc η οποία απομονώθηκε από τον πνεύμονα [12] ή το ήπαρ [13] ποντικού καθώς και από ιστούς βόειου πνεύνομα [14,15]. Οι παραπάνω μελέτες έδειξαν ότι η sgc είναι ετεροδιμερές μόριο. Είναι σημαντικό να αναφερθεί ότι η sgc μπορούσε να 7
13 απομονωθεί με ή χωρίς τον συμπαράγοντα αίμης. Επίσης, αξιοσημείωτο είναι ότι η χρήση διαλυτοποιητικών παραγόντων καθώς και η κατακρήμνισή της με θειικό αμμώνιο συνέβαλαν σε μια μορφή μερικώς αναδιπλωμένης απο-πρωτεΐνης. Μέχρι σήμερα, η διαδικασία απομόνωσης από βόειο πνεύμονα είναι η πιο αποτελεσματική καθώς αποδίδει 1 mg πρωτεΐνης ανά κιλό (kg) πνεύμονα [89,90]. Η ανάπτυξη ετερόλογων συστημάτων έκφρασης για την έκφραση ανασυνδυασμένης sgc προσέφερε νέες δυνατότητες στη μελέτη της sgc. Το πρώτο επιτυχές σύστημα έκφρασης τελειοποιήθηκε σε ευκαρυωτικά κύτταρα COS-7 [16]. Η συγκεκριμένη κυτταρική σειρά, απομονώθηκε από ηπατικό ιστό πιθήκου. Παρ όλο που τα συγκεκριμένα κύτταρα δεν παρείχαν αρκετή ποσότητα πρωτεΐνης προς απομόνωση, η διαδικασία ήταν ζωτικής σημασίας ώστε να αποδειχθεί ότι η sgc σχηματίζει ένα ετεροδιμερές αποτελούμενο από τις υπομονάδες α1 και β1 [17]. Επιπροσθέτως, η κυτταρική σειρά COS-7 χρησιμοποιήθηκε για τη δημιουργία μεταλλάξεων και απαλοιφών περιοχών στο μόριο της sgc [18,19]. Η υπερέκφραση της sgc προερχόμενης από ποντικούς επετεύχθη σε σύστημα έκφρασης Sf9/βακυλοϊού [18,20,21]. Αρχικά, η έκφραση της sgc σε κύτταρα πραγματοποιήθηκε επιτυχώς χωρίς επίτοπο [18]. Ωστόσο, κάποια νεότερα πρωτόκολλα έκφρασης περιελάμβαναν μια ιστιδινική επίτοπο προς διευκόλυνση της διαδικασίας [22,23,24]. Παρά τα υψηλά επίπεδα έκφρασης, η μεγαλύτερη ποσότητα πρωτεΐνης (>90%) εντοπιζόταν στο μη διαλυτό μέρος των κυττάρων. Η μέθοδος αυτή απέδωσε mg καθαρής διαλυτής πρωτεΐνης ανά λίτρο καλλιέργειας και αποτέλεσε την κοινώς χρησιμοποιούμενη διαδικασία απομόνωσης της sgc με τις ακολουθίες της πρωτεΐνης από άνθρωπο και ποντικό. Ένα ξεκάθαρο πλεονέκτημα του συγκεκριμένου συστήματος έκφρασης είναι ο in vitro χαρακτηρισμός του, ο βιοχημικός χαρακτηρισμός του διμερισμού των υπομονάδων και η δημιουργία κατευθυνόμενων μεταλλάξεων. Μέχρι σήμερα, δεν έχει απομονωθεί ολόκληρο το ετεροδιμερές μόριο α1β1 των θηλαστικών από βακτηριακό σύστημα έκφρασης. Ωστόσο, μικρότερα τμήματα της πρωτεΐνης έχουν εκφραστεί σε βακτηριακά κύτταρα Escherichia coli. Οι πρωτεΐνες που προκύπτουν περιλαμβάνουν τμήματα των υπομονάδων α1, β1 και β2 του Ν- τελικού άκρου (H-NOX επικράτειες) [25,26,27], τμήματα των υπομονάδων α1 και β1 του C-τελικού άκρου (CAT επικράτειες) [28,29] και μία επικράτεια στο κέντρο της β1(pas) 8
14 (Εικόνα 2) [28]. Αυτά τα τμήματα απομονώνονται με απόδοση η οποία κυμαίνεται από 0.5 έως 5 mg καθαρής πρωτεΐνης ανά λίτρο καλλιέργειας. Βακτηριακά στελέχη E.coli, χρησιμοποιήθηκαν στην προσπάθεια έκφρασης του ετεροδιμερούς α1β1 του οργανισμού Manduca sexta [30]. Με τη συγκεκριμένη μέθοδο τόσο η απόδοση (0.5-1 mg/l) όσο και η καθαρότητα της πρωτεΐνης ήταν χαμηλή. Ωστόσο, η απόδοση αυξανόταν σημαντικά (1-2 mg/l) μετά την έκφραση των α1 και β1 υπομονάδων χωρίς το C-τελικό άκρο [30]. Οι ετεροδιμερείς πρωτεΐνες που προέκυπταν στερούνταν της δυνατότητας να κυκλοποιήσουν την GTP. Με βάση τα παραπάνω, καθίσταται αναγκαία η βελτιστοποίηση των συνθηκών έκφρασης και απομόνωσης της sgc από E.coli. 2.4 Ισομορφές της sgc Το ετεροδιμερές μόριο της sgc αποτελείται, όπως προαναφέρθηκε, από δύο όμοιες υπομονάδες α και β. Η ισομορφή, η οποία απαντάται πιο συχνά στους ιστούς, είναι η ετεροδιμερής πρωτεΐνη α1β1. Ωστόσο, έχουν ενταθεί οι έρευνες όσον αφορά τις υπομονάδες α2 και β2 [31,32]. Αυτές οι πρωτεΐνες αρχικά εντοπίστηκαν στα θηλαστικά, σε έντομα όπως η Drosophila melanogaster και η M. sexta καθώς επίσης και στα ψάρια. Μεταξύ αυτών των οργανισμών, οι α-υπομονάδες εμφανίζουν υψηλή ομολογία μεταξύ τους, η οποία κυμαίνεται σε ποσοστό περίπου 48% και οι β- υπομονάδες, αντίστοιχα, με ομολογία περίπου 41%. Κάθε υπομονάδα ανιχνεύτηκε σε ιστούς θηλαστικών, συμπεριλαμβανομένου του ανθρώπου, του ποντικού και της αγελάδας. Οι α1 και οι β1 υπομονάδες εκφράζονται σε διάφορους ιστούς και είναι, πλέον, κοινώς αποδεκτό ότι σχηματίζουν ένα λειτουργικό ετεροδιμερές [33]. Επιπροσθέτως, έχει παρατηρηθεί ότι η α2 υπομονάδα εντοπίζεται σε λιγότερους ιστούς συκγριτικά με τις ισομορφές α1 και β1 αλλά είναι υπερεκφρασμένη στον εγκέφαλο, στον πνεύμονα, στο παχύ έντερο, στην καρδιά, στον σπλήνα, στη μήτρα και στον πλακούντα [33]. Ακόμα, έχει αποδειχθεί ότι τα ετεροδιμερή α2β1 εμφανίζουν όμοια χαρακτηριστικά πρόσδεσης με τα αντίστοιχα του ετεροδιμερούς α1β1 [34,35]. Παρά τις παρόμοιες βιοχημικές ιδιότητες των δύο αυτών ετεροδιμερών της sgc, καθένα από αυτά διαδραματίζει ξεχωριστό ρόλο στη σηματοδότηση της cgmp, γεγονός το οποίο μπορεί να αποδοθεί στο ότι εντοπίζονται σε διαφορετικά σημεία του κυττάρου. Πιο συγκεκριμένα, το ετεροδιμερές α2β1 9
15 συνδέεται με την κυτταρική μεμβράνη σε διάφορους ιστούς, και κατά συνέπεια, αποκρίνεται διαφορετικά από το ετεροδιμερές α1β1, το οποίο εντοπίζεται στο κυτοσόλιο [34]. 2.6 Ομόλογα της sgc Άτυπες μορφές της sgc σε ευκαρυωτικούς οργανισμούς Είναι γνωστό πως υπάρχει μεγάλος αριθμός ευκαρυωτικών οργανισμών, οι οποίοι περιλαμβάνουν στο γονιδίωμα τους γονίδια που εκφράζουν sgc πρωτεΐνες. Κάποια διμερή της sgc των ευκαρυωτικών οργανισμών παρουσιάζουν ομοιότητες με τις α1 και β1 υπομονάδες των ποντικών ενώ άλλες διαφέρουν σημαντικά. Οι τελευταίες ονομάστηκαν «άτυπες» sgc. Η ονομασία τους προέκυψε λόγω των ξεχωριστών ιδιοτήτων ενεργοποίησης και διμερισμού τους. Επίσης, κάποιες «άτυπες» sgc εμφανίζουν απόκριση στα διατομικά αέρια ΝΟ, CO και O2 ενώ άλλες αναστέλλονται παρουσία αυτών των μορίων H-NOX πρωτεΐνες σε προκαρυωτικούς οργανισμούς Είναι σημαντικό να αναφερθεί ότι οι πρωτεΐνες H-NOX που έχουν απομονωθεί από δυνητικά αερόβιους οργανισμούς παρουσιάζουν όμοιες ιδιότητες πρόσδεσης με αυτές της sgc, δηλαδή προσδένονται με ΝΟ και CO, ενώ φαίνεται να μην εμφανίζουν συγγένεια με το O2. Οι οργανισμοί αυτοί κωδικοποιούν μια μοναδική επικράτεια με ιδιότητες αυτόνομης πρωτεΐνης, και εμπεριέχουν γονίδια τα οποία κωδικοποιούν είτε κινάσες ιστιδίνης είτε διγουανυλικές κυκλάσες. Το γεγονός αυτό υποδηλώνει ότι η σηματοδότηση των H-NOX πρωτεΐνών μεσολαβείται από δύο μόρια. Προκειμένου να επιβεβαιωθεί αυτή η υπόθεση, απομονώθηκε μία περιοχή H- NOX και μια ιστιδινική κινάση από το βακτήριο Shewanella oneidensis και φάνηκε να αλληλεπιδρούν in vitro. Επιπροσθέτως, η αλληλεπίδραση της H-NOX και της κινάσης πραγματοποιήθηκε με τη μεσολάβηση του NO [46]. Στους υποχρεωτικά αναερόβιους οργανισμούς, η H-NOX επικράτεια είναι συγχωνευμένη με το C-τελικό άκρο μιας μεθυλ-χημειοεπαγώμενης πρωτεΐνης (methyl-accepting chemotaxis protein), γεγονός που υποδηλώνει τη συμμετοχή της σε κάποια σηματοδοτική λειτουργία. Η επικράτεια H-NOX φαίνεται να σχηματίζει ένα πολύ σταθερό σύμπλοκο αίμης-ο2 [47], υποδεικνύοντας έτσι τη μοριακή διάκριση που υπάρχει με την αντίστοιχη περιοχή των αερόβιων οργανισμών. Η απομόνωση και ο 10
16 χαρακτηρισμός της H-NOX από τον οργανισμό Τ.tengcongensis επηρέασε σημαντικά την μέχρι τότε κατανόηση της δράσης της sgc καθώς η H-NOX επικράτεια ήταν η πρώτη η οποία χαρακτηρίστηκε δομικά και κρυσταλλώθηκε μαζί με το μόριο του O2 [39,48]. Μέχρι σήμερα, αυτές οι πρωτεΐνες χρησιμοποιήθηκαν ως «εργαλεία» διερεύνησης της δομής και της ρύθμισης της sgc. Ωστόσο, ιδιαίτερα σημαντικές είναι οι μελέτες σε μικροβιακά συστήματα, καθώς οι μεταβλητές ιδιότητες πρόσδεσης των H-NOX πιθανά να έχουν συνέπειες στην ικανότητά τους να αποκρίνονται σε διαφορετικά αέρια. Έτσι θα αποσαφηνιστεί ο λόγος για τον οποίο κάποιες πρωτεΐνες προτιμούν το O2 έναντι των άλλων αερίων Σημαντικά αμινοξέα στην πρόσδεση των υποκαταστατών της H-NOX Κρυσταλλική δομή Τ.tengcongensis Η πρώτη κρυσταλλική δομή που επιλύθηκε ήταν το σύμπλοκο της H-NOX με Ο2 του αναερόβιου οργανισμού Τ.tengcongensis [39,48]. Το μόριο αναδιπλώνεται με τέτοιο τρόπο ώστε να σχηματίζονται επτά α-έλικες και 4 αντιπαράλληλα β-φύλλα (Εικόνα 4). Σημαντικό εύρημα για τη δομή της συγκεκριμένης επικράτειας, είναι η παρουσία ενός δικτύου δεσμών υδρογόνου γύρω από το προσδεμένο μόριο Ο2. Πιο συγκεκριμένα, η Y140 σχηματίζει δεσμό υδρογόνου με το Ο2 και τα αμινοξέα N74 και W9 συμμετέχουν σε δεσμούς υδρογόνου με το φαινολικό οξυγόνο της Y140 (Εικόνα 4). Όσον αφορά το μόριο της αίμης είναι συνδεδεμένο στο αμινοξικό κατάλοιπο H
 της Y140 με τα Ο2, N74 και W9. Απεικονίζεται και ο εγγύς προσδέτης της αίμης H102 [39].")

17 Εικόνα 4: Αριστερά: Κρυσταλλική δομή επικράτειας H-NOX του Τ. tengcongensis. Δεξιά: Περιβάλλον της αίμης. Απεικονίζονται οι δεσμοί υδρογόνου (κόκκινη διακεκομμένη γραμμή) της Y140 με τα Ο2, N74 και W9. Απεικονίζεται και ο εγγύς προσδέτης της αίμης H102 [39]. Είναι αξιοσημείωτο, ότι η σημασία των αμινοξέων ιστιδίνης, αργινίνης και τυροσίνης εξακριβώθηκε ύστερα από μεταλλάξεις που πραγματοποιήθηκαν στην β1 υπομονάδα sgc προερχόμενης από βοειδή και αρουραίους [49,50]. Παρ όλα αυτά, ο ρόλος των συγκεκριμένων αμινοξέων αποσαφηνίστηκε ύστερα από την επίλυση της δομής της H-NOX επικράτειας του οργανισμού Τ. tengcongensis Κρυσταλλική δομή H-NOX του Nostoc sp. Οι δομές του μορίου της Ns Η-ΝΟΧ, οι οποίες επιλύθηκαν μέσω κρυσταλλογραφίας ακτίνων Χ, έδειξαν ότι ο σίδηρος της αίμης έχει αριθμό ένταξης 5 απουσία υποκαταστάτη και αριθμό ένταξης 6 παρουσία υποκαταστατών των μορίων ΝΟ και CO. Επίσης, η θέση πρόσδεσης του μορίου της αίμης είναι η Η105. Αξιοσημείωτο είναι ότι τα φάσματα ηλεκτρονικής απορρόφησης της H-NOX παρουσιάζουν ομοιότητες με αυτά της sgc, ενώ παράλληλα εμφανίζουν σημαντικές διαφορές με αυτά της Τt H-NOX.. Ένα πολύ σημαντικό χαρακτηριστικό της Ns H- NOX, είναι ότι παρουσιάζει πολύ υψηλή ακολουθιακή ομολογία (33%) με την H- NOX επικράτεια της β1 υπομονάδας της ανθρώπινης sgc. Επίσης, το 63% των αμινοξέων που βρίσκονται στην κοιλότητα της αίμης είναι ακριβώς ίδια στα δύο μόρια. Οι αλληλεπιδράσεις της αίμης με τα αμινοξέα της πρωτεΐνης, εκτός της H105, είναι κυρίως υδροφοβικές, ενώ οι αλληλεπιδράσεις των προπιονικών ομάδων του πορφυρινικού δακτυλίου της εμπλέκουν το άζωτο της κύριας αλυσίδας του αμινοξέος Y2 και τη συντηρημένη αλληλουχία Y134-X-S136-X-R138. Επίσης, σημαντικός 12
18 κρίνεται ο ρόλος του αρωματικού αμινοξέος W74, το οποίο προσανατολίζεται κάθετα στον πορφυρινικό δακτύλιο της αίμης (Εικόνα 5). Η δέσμευση του Ο2 σε ενώσεις μοντέλων αίμης, όπως το Fe(II)PP(1-Melm), είναι αρκετά πιο ασθενής συγκριτικά με τη δέσμευση των αερίων ΝΟ και CO [74] και, συνεπώς, οι αιμοπρωτεΐνες απαιτούν επιπλέον ηλεκτροστατικές αλληλεπιδράσεις προκειμένου να δεσμευτεί το Ο2 [79]. Η απουσία, λοιπόν, τέτοιων πολικών αλληλεπιδράσεων παρέχει την εξήγηση για την απουσία της δέσμευσης του Ο2 στο μόριο της Νs Η-ΝΟΧ. Όσον αφορά το μόριο της αίμης στο πρωτεϊνικό μόριο της Νs H-NOX, η πρώτη είναι σχετικά κεκαμένη με ήπιες πτυχώσεις και παραμορφώσεις σε σχήμα θόλου. Η μέτρηση φασμάτων ηλεκτρονικής απορρόφησης (UV-Visible) που πραγματοποιήθηκε προκειμένου να διαπιστωθούν οι δομικές αλλαγές που επέρχονται στο μόριο της Νs HNOX κατά τη δέσμευση των μορίων ΝΟ και CO απέδειξε πως το σύμπλοκο ΝΟ-Νs ΗΝΟΧ έχει κυρίως αριθμό ένταξης 6 με ένα μικρό ποσοστό συμπλόκου να κατέχει αριθμό συναρμογής 5. Όπως αναμενόταν, η πρόσδεση του CO οδηγεί στην κατάσταση του μορίου της Νs HNOX με αριθμό ένταξης 6. Ενδιαφέρον παρουσιάζει το γεγονός πως οι κρυσταλλικές δομές των συμπλόκων ΝΟ-Νs H-NOX και CO-Ns Η-ΝΟΧ έδειξαν ότι τα μόρια ΝΟ και CO βρίσκονται τοποθετημένα σε μακρινή απόσταση από την αίμη. Επίσης, και οι δύο προσδέτες τοποθετούνται εντός ενός υδροφοβικού πυρήνα, ο οποίος απαρτίζεται από τα αμινοξέα της V5, F70, W74, M144 και L148. Διαμορφωτικές αλλαγές κατά τη δέσμευση των ΝΟ και CO στην Νs Η-ΝΟΧ Η υπέρθεση των δομών της Νs H-NOX χωρίς προσδέτη, με προσδέτη το ΝΟ και με προσδέτη το CO αποκαλύπτει μία εντυπωσιακή περιστροφική μετατόπιση του Fe της αίμης περίπου 0,2-0,3 Å στις δομές με το ΝΟ και 0,8-0,9 Å στις δομές με το CO. Oι άξονες της αίμης, ενώ συγκρατούν τις προπιονικές ομάδες τους προσαρτημένες στον πυρρολικό δακτύλιο D (PD) (Εικόνα 5Β), κατά τόπους χρησιμεύουν ως σημεία περιστροφής. Στις δομές με το CO, παρατηρούνται μετατοπίσεις μεγαλύτερες του ενός 1 Å για τα άτομα της αίμης που βρίσκονται μακριά από το σημείο περιστροφής. Το κέντρο του σιδήρου της αίμης στρέφεται μακριά από το αμινοξύ W74, το οποίο φαίνεται πως είναι απαραίτητο στην παροχή χώρου ώστε τα μόρια ΝΟ και CO να δεσμευτούν στην απώτερη περιοχή της αίμης. Η μεγαλύτερη μετατόπιση που 13
![υφίσταται ο σίδηρος κατά την πρόσδεση του CO ίσως οφείλεται στη μεγαλύτερη ατομική ακτίνα του ατόμου του άνθρακα αλλά και της προτίμησης του CO να δεσμεύεται κάθετα προς το μόριο της αίμης [83]](/docs-images/74/69937456/images/19-0.jpg "συγκριτικά με τη δέσμευση του μορίου του ΝΟ, το οποίο προσδένεται υπό γωνία. Η μετατόπιση της αίμης συνοδεύεται από τη μετατόπιση της πλευρικής αλυσίδας της Η105 κατά μήκος της αίμης.")
19 υφίσταται ο σίδηρος κατά την πρόσδεση του CO ίσως οφείλεται στη μεγαλύτερη ατομική ακτίνα του ατόμου του άνθρακα αλλά και της προτίμησης του CO να δεσμεύεται κάθετα προς το μόριο της αίμης [83] συγκριτικά με τη δέσμευση του μορίου του ΝΟ, το οποίο προσδένεται υπό γωνία. Η μετατόπιση της αίμης συνοδεύεται από τη μετατόπιση της πλευρικής αλυσίδας της Η105 κατά μήκος της αίμης. Η πρόσδεση του ΝΟ στο μόριο της Νs HNOX προκαλεί μικρές δομικές αλλαγές, εν αντιθέσει με τις μεγάλες δομικές αλλαγές που επάγονται κατά την πρόσδεση του CO. Κάποιες από αυτές είναι οι εξής: το αμινοξύ Η105 της έλικας αf, μαζί με τα γειτονικά αμινοξέα, μετατοπίζονται μαζί με την αίμη. Δεύτερον, η περιοχή της έλικας αα γύρω από το αμινοξύ Υ2, του οποίου η κύρια αλυσίδα αλληλεπιδρά με την PA προπιονική ομάδα της αίμης (Εικόνα 5Β), μετατοπίζεται μαζί με την αίμη περίπου 0,5 Å. Τρίτον, η περιοχή του βρόγχου β1 κοντά στο F112 που αλληλεπιδρά με την αf καθώς και το C- τελικό άκρο της έλικας αf, διαταράσσονται σημαντικά εμφανίζοντας πτωχή ηλεκτρονιακή πυκνότητα και αυξάνοντας τους παράγοντες θερμοκρασίας από 45 Å 2 που είναι στην ελεύθερη μορφή, σε 75 Å 2 που εμφανίζεται στη δομή με το CO. Η αυξημένη αυτή ευλυγισία προκαλείται από τη μετατόπιση της αίμης οδηγώντας σε μειωμένες αλληλεπιδράσεις Van der Waals μεταξύ της αίμης και των πλευρικών αλυσίδων των F112 και V108. Aυτές οι διαμορφωτικές αλλαγές που προκύπτουν κατά την πρόσδεση του CO πιθανά παρέχουν τη μοριακή βάση για την τετραπλάσια ενεργοποίηση της sgc από το CO [84,85]. Εικόνα 5: (A) περιοχή πρόσδεσης της αίμης στην Ns H-NOX. Υπέρθεση των δομών χωρίς υποκαταστάτη και με υποκαταστάτες NO και CO. (B) το μόριο της αίμης. Οι πυρρολικοί δακτύλιοι συμβολίζονται με τα γράμματα A-D, ενώ οι προπιονικές ομάδες PA και PD [92]. 14
20 NMR δομή Shewanella oneidensis H-NOX με υποκαταστάτη το μόριο του CO Η αγρίου τύπου μορφή της H-NOX του οργανισμού S. oneidensis ρυθμίζει τη δράση μιας κινάσης ιστιδίνης μ έναν τρόπο εξαρτώμενο από κάποιον υποκαταστάτη. Συγκεκριμένα, η δράση της κινάσης αναστέλλεται από την H-NOX που φέρει ως υποκαταστάτη το ΝΟ, αλλά όχι από την ελεύθερη μορφή της πρωτεΐνης [46]. Επίσης, η μετάλλαξη της ιστιδίνης 103 σε γλυκίνη (H103G) του συμπλόκου H-NOX-CO εμφανίζει δράση αναστολέα της κινάσης όμοια με αυτή που εμφανίζει η αγρίου τύπου πρωτεΐνη με υποκαταστάτη το NO. Επειδή η H103 αποτελεί τον προσδέτη της αίμης στην S. oneidensis H-NOX, η μετάλλαξη αυτού του αμινοξικού καταλοίπου σε γλυκίνη οδηγεί σε απομόνωση της απο-μορφής της πρωτεΐνης. Βέβαια, η πρόσδεση της αίμης στο μόριο με τη μετάλλαξη H103G μπορεί να επιτευχθεί με τη χρήση ιμιδαζολίου από την αρχική διαδικασία έκφρασης της πρωτεΐνης μέχρι και την απομόνωσή της [50,52]. Έχοντας πραγματοποιηθεί αυτή η μετάλλαξη, η α-έλικα F είναι ελεύθερη να υιοθετήσει στο χώρο μια διαμόρφωση όμοια με αυτή που διαθέτει όταν το μόριο σχηματίζει σύμπλοκο με το μόριο του ΝΟ. Η δομή του μορίου με την H103G μετάλλαξη καθώς και η αγρίου τύπου με υποκαταστάτη το CO (η οποία παρουσιάζει χαμηλή δραστικότητα), έχουν επιλυθεί με τη χρήση φασματοσκοπίας NMR. Διαμορφωτικές αλλαγές μεταξύ Fe(II)-CO WT και Fe(II)-CO H103G. Προκειμένου να ανακτηθεί μια αρχική εκτίμηση των περιοχών με διαμορφωτικές διαφορές μεταξύ των Fe(II)-CO WT και Fe(II)-CO H103G, ποσοτικοποιήθηκαν οι διαφορές των χημικών μετατοπίσεων των 15 Ν πυρήνων των ατόμων του αζώτου του πεπτιδικού σκελετού και οι χημικές μετατοπίσεις των μεθυλίων των πλευρικών αλυσίδων. Τα αμινοξέα που έχουν επηρεαστεί βρίσκονται κυρίως στις περιοχές των δευτεροταγών δομών και συνορεύουν άμεσα με την εγγύτατη περιοχή του συμπαράγοντα της αίμης. Οι μεγαλύτερες αλλαγές εντοπίζονται μεταξύ των αμινοξέων , τα οποία απαρτίζουν την αf έλικα και τη στροφή που συνδέει την αf έλικα με το β1 φύλλο. Μικρότερες αλλαγές παρατηρούνται στις πιο απομακρυσμένες περιοχές της αίμης, όπως στα αμινοξέα D86 και Κ87 της αε έλικας και του αμινοξέος Ι118 του β1 φύλλου. Τα μεγέθη και οι αλλαγές των χημικών μετατοπίσεων μεταξύ Fe(II)-CO WT και Fe(II)-CO H103G αναμένεται να αντικατοπτρίζουν μια αλλαγή στη θέση της εγγούς αf έλικας ως προς το συμπαράγοντα της αίμης στην επικράτεια της ΗΝΟΧ. Επίσης, συσχετίσεις των 15
21 αλλαγών της ενεργότητας που πραγματοποιήθηκαν μεταξύ CO-HNOX και CO- HNOX(H103G) αποκάλυψαν πως οι κυριότερες αλλαγές εντοπίζονταν στις υποεπικράτειες των αντίθετων πλευρών της αίμης. Το συμπέρασμα αυτό επιβεβαιώθηκε εξετάζοντας την υπέρθεση των υπο-επικρατειών. Πιο συγκεκριμένα το rmsd (root mean square deviation) της εγγούς επικράτειας του Fe(II)-CO WT (αμινοξέα και ) ήταν 0,48 Å ενώ για το σύνολο των δομών του Fe(II)-CO H103G ήταν 0.58 Å. Η τιμή του rmsd καθορίζει την ακρίβεια των υπολογισμένων δομών, καθώς αντιπροσωπεύει τη μέση τυπική απόκλιση των θέσεων των ατόμων κάθε μέλους μιας οικογένειας δομών από τη μέση δομή. Επομένως, οι παραπάνω τιμές αυτές υποδηλώνουν τον αρκετά καλό χαρακτηρισμό των δομών μέσω Φασματοσκοπίας ΝΜR. Συγκρίνοντας τις εγγείς επικράτειες των δομών WT με τις αντίστοιχες των Η103G παρατηρείται μικρή αύξηση του rmsd (0,72 Å ), γεγονός που υποδηλώνει τη μικρή δομική διαφορά μεταξύ αυτών των περιοχών. Ωστόσο, όταν πραγματοποιηθεί απευθείας σύγκριση μεταξύ των μακρινών αμινοξέων της οικογένειας δομών WT ως προς την οικογένεια Η103G, το rmsd αυξάνεται αισθητά (1,35 Å). 2.7 Ανόργανη χημεία του συμπλόκου αίμη-h-nox-υποκαταστάτης Το μόριο της αίμης Όλες οι αιμο-πρωτεΐνες περιλαμβάνουν ένα τετραπυρρολικό δακτύλιο, όπως για παράδειγμα την πρωτοπορφυρίνη IX [53]. Οι δομές αυτού του είδους ονομάζονται πορφυρίνες και οι δακτύλιοί τους είναι εκτεταμένα π-συζυγιακά συστήματα με επίπεδες δομές. 16
22 Εικόνα 6: Το μόριο της αίμης. Μεθυλικές ομάδες: 1,3,5,8. Βινυλικές ομάδες: 2,4. Προπιονικές ομάδες: 6,7. Πυρρολικοί δακτύλιοι: A-D [53]. Οι πρωτοπορφυρίνες αποτελούνται από τέσσερις πυρρολικούς δακτυλίους που συνδέονται µεταξύ τους µε µεθενυλικές γέφυρες (=CH-), ώστε να σχηµατίζεται ένας πολυκυκλικός δακτύλιος (τετραπυρρολικός δακτύλιος). Περιφερειακά στο δακτύλιο συνδέονται τέσσερις µεθυλικές (-CH3), δυο βινυλικές (-CH=CH2) και δυο προπιονικές (-CH2-CH2-COOH) οµάδες (Εικόνα 6). Από αυτές, οι βινυλικές και οι µεθυλικές οµάδες είναι µη πολικές, ενώ οι προπιονικές είναι πολικές. Οι οµάδες αυτές είναι δυνατόν να διαταχθούν µε 15 διαφορετικούς τρόπους, ώστε να προκύπτουν τα αντίστοιχα ισοµερή. Στα βιολογικά συστήµατα, απαντάται, εκτός των άλλων, και η πρωτοπορφυρίνη ΙΧ. Στο µόριο της αίµης, τα τέσσερα άτοµα αζώτου στις κορυφές των πυρρολικών δακτυλίων της πρωτοπορφυρίνης ΙΧ ενώνονται µε ομοιοπολικούς δεσµούς µε ένα άτοµο δισθενούς σιδήρου. Ο συνολικός αριθμός των δεσμών γύρω από το μόριο του σιδήρου ονομάζεται «αριθμός συναρμογής ή ένταξης». Οι άλλοι δύο δεσµοί συναρµογής του σιδήρου, ο πέμπτος και ο έκτος, παραµένουν ελεύθεροι και κατευθύνονται κάθετα, του επιπέδου του µορίου της αίµης. Οτιδήποτε σχηματίζει δεσμό με το κεντρικό μεταλλικό ιόν ονομάζεται «υποκαταστάτης» (ligand) και η ένωση που προκύπτει ονομάζεται «ένωση 17
23 συναρμογής». Επίσης, αν ένας υποκαταστάτης μπορεί να σχηματίσει περισσότερους του ενός δεσμού με ένα μεταλλικό ιόν, όπως η πορφυρίνη, ονομάζεται «χειλικός». Στη συνέχεια, κατηγοριοποιείται ανάλογα με τον αριθμό των δεσμών που σχηματίζει με το μεταλλοϊόν. Έτσι, οι πορφυρίνες αποτελούν τετραδοντικούς χειλικούς υποκαταστάτες για το ιόν του σιδήρου. Ο δισθενής σίδηρος (Fe 2+ ), είναι δυνατόν να οξειδωθεί και να µετατραπεί σε τρισθενή µορφή (Fe 3+ ), χάνοντας ένα ηλεκτρόνιο. Το µόριο που φέρει τρισθενή σίδηρο ονοµάζεται αιµατίνη. Η αιµατίνη µε τη σειρά της είναι δυνατόν να αναχθεί και να µετατραπεί σε αίµη. Γενικά, στα βιολογικά συστήματα ο σίδηρος απαντάται στην οξειδωτική κατάσταση (+2) η οποία ονομάζεται σιδηρο- (ferrous), και στην οξειδωτική κατάσταση (+3) η οποία ονομάζεται σιδηρι- (ferric). Η πρώτη αποτελεί την «ανηγμένη» κατάσταση και η δεύτερη την «οξειδωμένη» κατάσταση του Fe Η σημασία του μορίου της αίμης στο σύμπλοκο της πρωτεΐνης Η αίμη λειτουργεί ως προσθετική ομάδα σε πολλά πρωτεϊνικά μόρια που συχνά απαντώνται στη φύση. Μερικά από αυτά είναι οι πρωτεΐνες σπονδυλωτών και ασπόνδυλων, τα κυτοχρώματα P450, οι αιμογλοβίνες και οι μυογλοβίνες των φυτών καθώς και αιμο-πρωτεΐνες που λειτουργούν ως αισθητήρες για NO, CO και οξειδοαναγωγικά μόρια. Είναι αξιοσημείωτο, ότι στο μόριο της αίμης σε ελεύθερη μορφή το ιόν του σιδήρου βρίσκεται στο επίπεδο του πορφυρινικού δακτυλίου. Ωστόσο, όταν η αίμη είναι υποκαταστάτης κάποιου πρωτεϊνικού μορίου το ιόν παρεκλίνει σημαντικά από το επίπεδο του δακτυλίου. Στα σύμπλοκα των αίμοπρωτεϊνών, όταν το μόριο της αίμης απομακρυνθεί από το μόριο τότε η πρωτεΐνη είναι μερικώς αναδιπλωμένη και μη λειτουργική [54]. Σε αυτή την περίπτωση το πρωτεϊνικό μόριο ονομάζεται «απο-πρωτεΐνη». Παρ όλα αυτά, το φαινόμενο απομάκρυνσης μορίων αίμης από σύμπλοκα με πρωτεϊνικά μόρια συναντάται πολύ συχνά στη φύση Αμινοξέα ικανά να προσδεθούν με το μόριο της αίμης Όπως προαναφέρθηκε το μεταλλοϊόν του μορίου της αίμης είναι ικανό να δημιουργήσει δεσμούς συναρμογής με άλλους υποκαταστάτες, εκτός του πορφυρινικού δακτυλίου. Επομένως, το μόριο της αίμης μπορεί να συνδεθεί με κάποια πολυπεπτιδική αλυσίδα σχηματίζοντας μία αιμο-πρωτεΐνη. Παρ όλα αυτά, ο 18
24 δεσμός μεταξύ του μεταλλοϊόντος και του υποκαταστάτη είναι ασθενής, κάτι που προκύπτει από την ευκολία με την οποία ένα μόριο αίμης απομακρύνεται από ένα σύμπλοκο. Το σύμπλοκο αίμη-πρωτεΐνη σχηματίζεται με τη δημιουργία ενός δεσμού συναρμογής του μετάλλου της αίμης με την πλευρική αλυσίδα κάποιου αμινοξικού καταλοίπου. Ωστόσο, για να επιτευχθεί αυτό θα πρέπει το αμινοξύ στην πλευρική του αλυσίδα να διαθέτει άτομα-δότες ηλεκτρονίων (Ν,Ο,S). Το γεγονός αυτό, περιορίζει των αριθμό των αμινοξέων που μπορούν να συναρμοστούν με την αίμη. Συγκεκριμένα, τα αμινοξέα αυτά είναι τα εξής: Arg, Lys, His, Thr, Ser, Met και Trp [54] Οξειδωτικές καταστάσεις του σιδήρου Τα μεταβατικά μέταλλα, στα οποία ανήκει και ο σίδηρος, απαντώνται σε διάφορες οξειδωτικές καταστάσεις. Οι πιο κοινές οξειδωτικές καταστάσεις του σιδήρου είναι +2, +3 και σε πιο σπάνιες περιπτώσεις +4. Ειδικότερα, στις αιμο-πρωτεΐνες το ιόν του σιδήρου βρίσκεται στις οξειδωτικές καταστάσεις +2 ή +3. Έτσι, ο σίδηρος μπορεί να δημιουργήσει σύμπλοκα με μέγιστο αριθμό ένταξης 6 (δηλαδή να δημιουργήσει δεσμούς συναρμογής με έξι υποκαταστάτες) δημιουργώντας σύμπλοκα με οκταεδρική γεωμετρία Ενεργειακός διαχωρισμός των d τροχιακών Όσον αφορά μόρια της αίμης, τα οποία βρίσκονται δεσμευμένα με κάποια πρωτεΐνη ή είναι ελεύθερα σε διάλυμα, υπάρχει άρρηκτη σύνδεση μεταξύ του αριθμού ένταξης του ατόμου του σιδήρου, της ηλεκτρονικής δομής της αίμης και των μαγνητικών της ιδιοτήτων. Είναι γνωστό, ότι η χημεία των μεταβατικών μετάλλων είναι η χημεία των d τροχιακών. Ο σίδηρος είναι ένα μεταβατικό μέταλλο το οποίο στη θεμελιώδη κατάσταση, όπως και στην οξειδωτική κατάσταση +2 διαθέτει 6 ηλεκτρόνια σε d τροχιακά, ενώ στην κατάσταση +3 διαθέτει 5 d ηλεκτρόνια. Τα d ηλεκτρόνια κατανέμονται στα d τροχιακά τα οποία είναι: dxy, dxz, dyz, dx 2 -y 2, dz 2[55]. Κάτω από την επίδραση του κρυσταλλικού πεδίου των υποκαταστατών, οι οποίοι βρίσκονται γύρω από το ιόν του σιδήρου, τα d τροχιακά χωρίζονται σε δύο μη εκφυλισμένες ομάδες (Εικόνα 7). Τα τροχιακά της πρώτης ομάδας (dx 2, dx 2 -y 2 ) έχουν υψηλότερη ενέργεια από τα τροχιακά της δεύτερης ομάδας (dxy, dyz, dxz). Η ενεργειακή 19
25 διαφορά μεταξύ των δύο ομάδων d τροχιακών ενός κεντρικού μεταλλικού ιόντος η οποία οφείλεται στην αλληλεπίδραση των τροχιακών με το ηλεκτρικό πεδίο των υποκαταστατών ονομάζεται διαχωρισμός κρυσταλλικού πεδίου. Εικόνα 7: Ενεργειακός διαχωρισμός των d τροχιακών [55] Σύμπλοκα υψηλού και χαμηλού spin Γνωρίζοντας τα ενεργειακά επίπεδα των d τροχιακών ενός οκταεδρικού συμπλόκου, είναι δυνατόν να καταλήξουμε σε συμπεράσματα για τον τρόπο κατανομής των d ηλεκτρονίων του μεταλλικού ιόντος σε αυτά. Αυτή η κατανομή επιτρέπει την πρόβλεψη των μαγνητικών ιδιοτήτων του συμπλόκου. Τα ιόντα των μεταβατικών μετάλλων της μορφής d 4, d 5, d 6 και d 7 είναι δυνατόν να συμμετάσχουν σε σύμπλοκα, στα οποία ανάλογα με το είδος των υποκαταστατών, τον αριθμό και τη γεωμετρία συναρμογής έχουν σαν αποτέλεσμα το σχηματισμό συμπλόκων υψηλού ή χαμηλού spin. Σύμπλοκο υψηλού spin (Εικόνα 8Α) είναι ένα σύμπλοκο, στο οποίο παρατηρείται η ελάχιστη σύζευξη ηλεκτρονίων στα d τροχιακά του μεταλλικού ατόμου. Σύμπλοκο χαμηλού spin (Εικόνα 8Β) είναι ένα σύμπλοκο, στο οποίο υπάρχει μεγαλύτερη σύζευξη ηλεκτρονίων στα d τροχιακά του μεταλλικού ατόμου απ ότι σε ένα αντίστοιχο σύμπλοκο υψηλού spin [55] και ο μικρότερος δυνατός αριθμός ασύζευκτων ηλεκτρονίων στα τροχιακά του. 20
26 Α Β Εικόνα 8: Κατάληψη των 3d τροχιακών σε σύμπλοκα του Fe 2+ : A) υψηλού spin. B) χαμηλού spin [55]. Ο τρόπος με τον οποίο κατανέμονται τα ηλεκτρόνια στα d τροχιακά εξαρτάται από δύο θεμελιώδεις αντιτιθέμενες δυνάμεις. Η μία εκ των δύο δυνάμεων είναι η ενέργεια, η οποία απαιτείται ώστε δύο ηλεκτρόνια να συζευχθούν σε ένα τροχιακό (ενέργεια σύζευξης). Η άλλη δύναμη είναι η ενέργεια, η οποία απαιτείται για να μεταβεί ένα ηλεκτρόνιο σε τροχιακό υψηλότερης ενέργειας, αντί να συζευχθεί με κάποιο άλλο σε τροχιακό χαμηλότερης ενέργειας. Είναι αξιοσημείωτο, ότι όταν ο αριθμός ένταξης του συμπλόκου είναι 5 σχηματίζεται σύμπλοκο υψηλού spin, ενώ όταν είναι 6 σχηματίζεται σύμπλοκο χαμηλού spin. Το παραπάνω ισχύει για σύμπλοκα στα οποία ο σίδηρος βρίσκεται στις οξειδωτικές καταστάσεις +2 και +3. Όλα τα σύμπλοκα του σιδήρου με αριθμό οξείδωσης +2 και +3 έχουν μονήρη ηλεκτρόνια σε d τροχιακά, έκτος από αυτά στα οποία ο σίδηρος εμφανίζει αριθμό ένταξης 6 και βρίσκεται στην οξειδωτική κατάσταση +2. Τα σύμπλοκα, στα οποία ο σίδηρος έχει ασύζευκτα ηλεκτρόνια είναι παραμαγνητικά, δηλαδή έλκονται από εξωτερικά εφαρμοζόμενο μαγνητικό πεδίο. Μάλιστα, όσο μεγαλύτερος είναι ο αριθμός των ασύζευκτων ηλεκτρονίων τόσο περισσότερο παραμαγνητικό είναι το μόριο. Για παράδειγμα, τα υψηλού spin σύμπλοκα της αίμης με αριθμό ένταξης 5 και το σίδηρο στην οξειδωτική κατάσταση +3 εμφανίζουν τις πιο έντονες παραμαγνητικές ιδιότητες. Αντίθετα, τα σύμπλοκα στα οποία ο σίδηρος δεν έχει μονήρη ηλεκτρόνια είναι διαμαγνητικά, δηλαδή δεν έλκονται ή και απωθούνται 21
![ελαφρώς από το μαγνητικό πεδίο. Σε αυτό το σημείο μπορεί να γίνει κατανοητή η σχέση που υπάρχει μεταξύ του αριθμού ένταξης και των μαγνητικών ιδιοτήτων [55].](/docs-images/74/69937456/images/27-1.jpg "Παρ όλα αυτά, υπάρχει μια πιο λεπτή σχέση μεταξύ των παραπάνω ιδιοτήτων και της πραγματικής δομής της αίμης.")
27 ελαφρώς από το μαγνητικό πεδίο. Σε αυτό το σημείο μπορεί να γίνει κατανοητή η σχέση που υπάρχει μεταξύ του αριθμού ένταξης και των μαγνητικών ιδιοτήτων [55]. Παρ όλα αυτά, υπάρχει μια πιο λεπτή σχέση μεταξύ των παραπάνω ιδιοτήτων και της πραγματικής δομής της αίμης. Συγκεκριμένα, η δομή του μορίου της αίμης είναι πραγματικά επίπεδη μόνο όταν το μεταλλικό ιόν του σιδήρου εμφανίζει αριθμό ένταξης 6 (Εικόνα 9Α). Όταν ο αριθμός συναρμογής είναι 5, τότε το ιόν του σιδήρου μετακινείται έξω από το χηλικό δακτύλιο της πορφυρίνης προς τον πέμπτο υποκαταστάτη (Εικόνα 9Β). Επομένως, σε σύμπλοκο υψηλού spin με αριθμό συναρμογής 5, το ιόν σιδήρου βρίσκεται πάνω από το επίπεδο της αίμης και η πορφυρίνη δημιουργεί ένα θόλο προς την κατεύθυνση του ιόντος [54]. Α Β XO Εικόνα 9: Γεωμετρία του μορίου της αίμης. Α) Σύμπλοκο με αριθμό ένταξης 6. Με ΧΟ συμβολίζονται τα διατομικά αέρια ΝΟ, CO και Ο2. Β) Σύμπλοκο με αριθμό ένταξης 5 [54] Ενεργοποίηση της sgc από πλευράς ανόργανης χημείας Το μόριο του ΝΟ λειτουργεί ως παράγοντας ενεργοποίησης για το μόριο της sgc. Ένας πιθανός λόγος, για τον οποίο η φύση έχει επιλέξει ένα ασταθές και πολύ δραστικό μόριο, όπως είναι το ΝΟ, να επιτελεί σημαντικές σηματοδοτικές λειτουργίες είναι οι ιδιότητες που εμφανίζει το μόριο του ΝΟ ως υποκαταστάτης μεταβατικών μετάλλων. Μεταξύ των κοινών διατομικών αερίων που απαντώνται στα βιολογικά συστήματα, τα οποία είναι ΝΟ, CO και Ο2, το μόριο του ΝΟ διαφέρει σημαντικά. Γενικά, οι υποκαταστάτες της μορφής ΧΟ μπορούν να προσδεθούν στο ιόν σιδήρου της αίμης δίνοντας στο τελικό σύμπλοκο γραμμική ή κεκαμένη διαμόρφωση ( Εικόνα 10). 22
28 Εικόνα 10: Διάγραμμα μοριακών τροχιακών το οποίο απεικονίζει τον τρόπο με τον οποίο οι υποκαταστάτες αλληλεπιδρούν με το ιόν σιδήρου της αίμης. α) κεκαμένη διαμόρφωση. β) γραμμική διαμόρφωση. Η διαμόρφωση η οποία ευνοείται, εξαρτάται από το αν ο υποκαταστάτης διαθέτει ή όχι ηλεκτρόνια στα π* τροχιακά. Τόσο το Ο2 όσο και το ΝΟ, έχουν ηλεκτρόνια στα π* τροχιακά. Αύτο έχει σαν αποτέλεσμα να ευνοείται η κεκαμένη γεωμετρία καθώς με τον τρόπο αυτό μεγιστοποιείται η επικάλυψη των π* τροχιακών με τα dz 2 τροχιακά του Fe [1]. Ωστόσο, το πότε ευνοείται η γραμμική ή η κεκαμένη διαμόρφωση εξαρτάται από το πλήθος των ηλεκτρονίων του υποκαταστάτη ΧΟ, τα οποία βρίσκονται στα π* τροχιακά. Συγκεκριμένα, το μόριο του CO έχει κενά π* τροχιακά. Το γεγονός αυτό έχει σαν αποτέλεσμα, η ροή των ηλεκτρονίων από τα dπ τροχιακά του σιδήρου προς τα κενά π* τροχιακά του CO, να είναι μέγιστη κατά την γραμμική διαμόρφωση. Όσον αφορά στα υπόλοιπα διατομικά αέρια, το Ο2 διαθέτει δύο ηλεκτρόνια στα π* τροχιακά, ενώ το μόριο του ΝΟ διαθέτει μόνο ένα ηλεκτρόνιο. Στις δύο αυτές περιπτώσεις, η μέγιστη σταθεροποίηση επιτυγχάνεται με την επικάλυψη των π* τροχιακών του ΧΟ με τα dz 2 τροχιακά Fe, σύμφωνα με την κεκαμένη διαμόρφωση. Η διαφορά που εμφανίζει το ΝΟ συγκρινόμενο με τους υπόλοιπους υποκαταστάτες, είναι ότι λόγω ισχυρού trans φαινομένου η ηλεκτρονική πυκνότητα απεντοπίζεται στα τροχιακά dz 2 τα οποία βρίσκονται κοντά στην ιστιδίνη, αποδυναμώνοντας έτσι το δεσμό His-Fe(II) [56]. Στην ανόργανη χημεία το φαινόμενο trans είναι η δραστικότητα των υποκαταστατών, οι οποίοι βρίσκονται σε θέση trans ως προς άλλους συγκεκριμένους υποκαταστάτες και ως εκ τούτου θεωρούνται trans κατευθυντήριοι υποκαταστάτες. Το φαινόμενο αυτό οφείλεται στα ηλεκτρονιακά φαινόμενα που λαμβάνουν χώρα σε επίπεδα τετραεδρικά και οκταεδρικά σύμπλοκα. 23
29 Αυτή η ιδιότητα του ΝΟ να αποδυναμώνει το δεσμό His-Fe(II), παρέχει ένα μέσο για τη συμπλοκοποίησή του με το ιόν σιδήρου ύστερα από λειτουργικά σημαντικές διαμορφωτικές αλλαγές στο πρωτεϊνικό μόριο. Συγκεκριμένα, είναι πιθανό ότι η ενεργοποίηση της sgc πραγματοποιείται από την λύση του δεσμού His-Fe(II), η οποία ωθείται από την πρόσδεση του μορίου του ΝΟ στο ιόν σίδηρου. Προφανώς, η απομάκρυνση της ιστιδίνης, ως αποτέλεσμα της ρήξης του δεσμού His-Fe(II), σχετίζεται με δομικές αλλαγές στο πρωτεϊνικό μόριο, οι οποίες εντοπίζονται στην καταλυτική υπομονάδα. Στην περιοχή αυτή, όπως προαναφέρθηκε, πραγματοποιείται η μετατροπή της GTP σε cgmp. Τέλος, με βάση τα παραπάνω, γίνεται αντιληπτός ο λόγος για τον οποίο το ΝΟ αποτελεί τον καταλληλότερο υποκαταστάτη για το μόριο της αίμης, καθώς και ο τρόπος με τον οποίο η φύση έχει εκμεταλλευτεί τις μοναδικές ιδιότητες κάποιων ανόργανων μορίων σε εξειδικευμένες σηματοδοτικές λειτουργίες [1]. 2.8 Φασματοσκοπία Πυρηνικού Μαγνητικού Συντονισμού (Nuclear Magnetic Resonance, NMR) Εισαγωγή Η φασματοσκοπία NMR βασίζεται στη μέτρηση της απορρόφησης ηλεκτρομαγνητικής ακτινοβολίας στην περιοχή των ραδιοσυχνοτήτων, δηλαδή 4 έως 900 MHz. Στη συγκεκριμένη φασματοσκοπική τεχνική, στη διαδικασία απορρόφησης μετέχουν οι πυρήνες των ατόμων. Επιπλέον, για τη δημιουργία των πυρηνικών ενεργειακών καταστάσεων, που απαιτούνται για να υπάρξει απορρόφηση, είναι απαραίτητη η τοποθέτηση του αναλύτη εντός ισχυρού μαγνητικού πεδίου. Η NMR φασματοσκοπία αποτελεί ένα πολύ χρήσιμο εργαλείο για την ανάλυση της δομής τόσο απλών χημικών ενώσεων όσο και μακρομορίων [57]. Η θεωρητική βάση του NMR παρουσιάστηκε πρώτη φορά από τον W. Pauli to 1924, o οποίος πρότεινε ότι ορισμένοι ατομικοί πυρήνες πρέπει να έχουν ιδιότητες αυτοστροφορμής (spin) και μαγνητικής ροπής και συνεπώς η έκθεση τους σε μαγνητικό πεδίο θα οδηγούσε σε διαχωρισμό των ενεργειακών τους επιπέδων. Οι παραπάνω υποθέσεις επιβεβαιώθηκαν πειραματικά το 1946 από τους Block και Purcell, οι οποίοι εργαζόμενοι ανεξάρτητα απέδειξαν ότι οι πυρήνες απορροφούν ηλεκτρομαγνητική ακτινοβολία, εντός ισχυρού μαγνητικού πεδίου ως αποτέλεσμα του μαγνητικώς επαγόμενου διαχωρισμού των ενεργειακών επιπέδων [58]. 24
30 Η ανακάλυψη του NMR έφερε επανάσταση στο χώρο της Χημείας, Βιοχημείας και Δομκής Βιολογίας, καθώς συντέλεσε στην επίλυση της δομής μικρών μορίων αλλά και μακρομορίων καθώς επίσης και στη μελέτη της δυναμικής τους. Το πρώτο βιολογικό μακρομόριο, του οποίου η δομή επιλύθηκε με χρήση της φασματοσκοπίας NMR, ήταν ο αναστολέας πρωτεϊνάσης ΙΙΑ [90]. Πρόκειται, για μια πρωτεΐνη μήκους 57 αμινοξέων, η οποία απομονώθηκε από ταύρο. Από τότε μέχρι σήμερα, στη βάση δεδομένων Protein Data Bank (PDB) ο αριθμός των δομών μακρομορίων και νουκλεϊκών οξέων που έχουν κατατεθεί, έχει ξεπεράσει τις Η Φασματοσκοπία NMR και η Κρυσταλλογραφία ακτίνων Χ αποτελούν τις δύο βασικές πειραματικές μεθόδους που έχουν αναπτυχθεί προς επίλυση της δομής βιομορίων. Καθεμιά από τις δύο αυτές τεχνικές παρουσιάζει τους δικούς της περιορισμούς. Ένα βασικό μειονέκτημα της φασματοσκοπίας NMR είναι ότι περιορίζεται στο μέγεθος των μορίων που μπορούν να αναλυθούν. Αυτό συμβαίνει διότι με την αύξηση του μεγέθους του μορίου, η αλληλεπικάλυψη των κορυφών καθώς και η διεύρυνση των σημάτων είναι τέτοια, που πολλές φορές δεν είναι ανιχνεύσιμα. Μέχρι σήμερα, έχουν αναλυθεί μόρια με μοριακό βάρος kda. Η επίτευξη της ανάλυσης μορίων μεγαλύτερου μεγέθους αποτελεί πρόκληση για τους ερευνητές που ασχολούνται με την ανάπτυξη νέων NMR μεθοδολογιών. Ένα ακόμα στοιχείο στο οποίο μειονεκτεί η φασματοσκοπία NMR, είναι ότι η μέθοδος παρουσιάζει χαμηλή ευαισθησία. Αυτό έχει σαν αποτέλεσμα, να απαιτούνται συγκεντρώσεις πρωτεΐνης της τάξης του 0,3-1mM. Επιπλέον, σε αυτές τις συγκεντρώσεις τα πρωτεϊνικά μόρια δεν θα πρέπει να σχηματίζουν συσσωματώματα. Παράλληλα, κρίνεται απαραίτητος ο εμπλουτισμός των δειγμάτων με ενεργούς για το NMR πυρήνες, συνήθως 15 Ν και 13 C. Ακόμα, για πειράματα NMR πρωτονίων, η τιμή του ph πρέπει να ρυθμιστεί περίπου στο 7. Και αυτό γιατί, σε υψηλότερες τιμές ph συμβαίνει γρήγορη ανταλλαγή των αμιδικών υδρογόνων με τα πρωτόνια του διαλύτη. Ο προσδιορισμός της δομής κάποιου μακρομορίου με τη φασματοσκοπία NMR, είναι μια διαδικασία λιγότερο αυτοματοποιημένη, από αυτή της κρυσταλογραφίας ακτίνων Χ. Τέλος, αξίζει να σημειωθεί ότι η δομικοί περιορισμοί στη φασματοσκοπία NMR εξάγονται βάσει των διαπρωτονιακών αποστάσεων. Το μεγαλύτερο ίσως πλεονέκτημα, που παρουσιάζει η φασματοσκοπία NMR έναντι της κρυσταλλογραφίας ακτίνων Χ, είναι η δυνατότητα μελέτης δυναμικών διεργασιών που συμβαίνουν μέσα στο διάλυμα. Αυτό, στην κρυσταλλογραφία ακτίνων Χ δεν 25
31 είναι εφικτό, διότι τα άτομα στον κρύσταλλο είναι διατεταγμένα σε συγκεκριμένες θέσεις και η κινητικότητά τους είναι σχεδόν μηδαμινή [59]. Από την άλλη πλευρά, η κρυσταλλογραφία ακτίνων Χ, παρέχει τη δυνατότητα δομικού χαρακτηρισμού, μεγάλων πρωτεϊνικών μορίων καθώς και μακρομοριακών συμπλόκων. Η τεχνική είναι κατάλληλη κυρίως για δυσδιάλυτες πρωτεΐνες, καθώς απαιτείται κρυστάλλωση του πρωτεϊνικού μορίου. Στα αρχεία PDB των κρυσταλλογραφικών δομών εισάγονται πληροφορίες σχετικές με τα βαρέα άτομα και όχι με τα υδρογόνα. Είναι σημαντικό να αναφερθεί, πως η Χ-ray δομή δεν αντιπροσωπεύει απαραιτήτως τη δομή που κυριαρχεί στο διάλυμα, η οποία θεωρητικά, είναι πλησιέστερη στην πραγματική δομή της πρωτεΐνης. Αυτό, μπορεί να οδηγήσει σε εσφαλμένα συμπεράσματα τα οποία σχετίζονται με τη λειτουργία της πρωτεΐνης. Χαρακτηριστικό παράδειγμα, αποτελεί η δομή της πρωτεΐνης Ηpr του οργανισμού Enterococcus faecalis. Η δομή αρχικά είχε επιλυθεί με κρυσταλλογραφία ακτίνων Χ, και οι ερευνητές υποστίρηζαν πως η διαμόρφωση μιας θηλιάς συμμετέχει στην κατάλυση. Αργότερα όμως, η επίλυση της ίδιας δομής με τη φασματοσκοπία NMR, έδειξε ότι η συγκεκριμένη διαμόρφωση βρισκόταν σε πολύ μικρό ποσοστό στο διάλυμα [60]. Συμπερασματικά, μπορούμε να πούμε ότι οι δύο παραπάνω μέθοδοι δομικού χαρακτηρισμού μακρομορίων, πρέπει να θεωρούνται συμπληρωματικές. Από τη μία πλευρά, η κρυσταλλογραφία ακτίνων Χ παραμένει η μόνη μέθοδος για την επίλυση της δομής ενός μεγάλου πρωτεϊνικού μορίου. Από την άλλη, η φασματοσκοπία NMR παρέχει τη δυνατότητα μελέτης της δυναμικής μικρότερων βιομορίων σε διάλυμα. Οι δύο τεχνικές μπορούν να χρησιμοποιηθούν συνδυαστικά ώστε να δώσουν πιο εμπεριστατωμένα αποτελέσματα Θεωρητική προσέγγιση Για να ερμηνευθούν κάποιες ιδιότητες των πυρήνων πρέπει να υποτεθεί ότι εκτελούν περιστροφική κίνηση γύρω από ένα άξονα και επομένως εμφανίζουν την ιδιότητα της αυτοστροφορμής (spin). Οι πυρήνες αυτοί, παρουσιάζουν στροφορμή p. Ο μέγιστος αριθμός των τιμών της στροφορμής για ένα συγκεκριμένο πυρήνα, είναι ο κβαντικός αριθμός του spin Ι. Σύμφωνα με τα παραπάνω, ο πυρήνας θα εμφανίζει 2Ι+1 ενεργειακές καταστάσεις. Η ενέργεια των ενεργειακών αυτών καταστάσεων, απουσία εξωτερικού πεδίου, είναι η ίδια. 26
32 Οι πυρήνες που είναι ενεργοί στη φασματοσκοπία NMR, έχουν συνισταμένη στροφορμή περιττό πολλαπλάσιο του ½. Αυτοί που κυρίως χρησιμοποιούνται στη φασματοσκοπία NMR είναι οι 1 Η, 13 C, 19 F και 31 P. Ένας περιστρεφόμενος πυρήνας δημιουργεί μαγνητικό πεδίο ίδιο ακριβώς με το πεδίο που δημιουργείται, όταν ένα πηνίο διαρρέεται από ρεύμα. Η προσπίπτουσα μαγνητική ροπή μ προσανατολίζεται κατά τη διεύθυνση του άξονα περιστροφής και είναι ανάλογη προς τη στροφορμή p. Επομένως, προκύπτει: μ = γ*p όπου γ είναι μια σταθερά αναλογίας, γνωστή ως γυρομαγνητικός λόγος (magnetogyric ratio). Η τιμή του γυρομαγνητικού λόγου είναι χαρακτηριστική για κάθε είδος πυρήνα. Όταν ένα πυρήνας με κβαντικό αριθμό spin 1/2 εισέλθει σε εξωτερικό μαγνητικό πεδίο Β0, η μαγνητική ροπή του προσανατολίζεται σε μια από τις δύο δυνατές διευθύνσεις σε σχέση με το πεδίο, οι οποίες εξαρτώνται από τη μαγνητική κβαντική του κατάσταση. Η ενεργειακή διαφορά μεταξύ των δύο κβαντικών καταστάσεων δίνεται από τη σχέση: ΔΕ = γh/2π*β0 Όπου Β0 η ένταση του εφαρμοζόμενου μαγνητικού πεδίου και h η σταθερά του Planck. Η μετάπτωση μεταξύ των ενεργειακών καταστάσεων επιτυγχάνεται με απορρόφηση ηλεκτρομαγνητικής ακτινοβολίας συχνότητας v0, η οποία υπολογίζεται από τη σχέση: V0 = γβ0/2π Μεταπτωτική κίνηση των πυρήνων Το εξωτερικό εφαρμοζόμενο μαγνητικό πεδίο, έκτος από το διαχωρισμό των ενεργειακών καταστάσεων του πυρήνα, ευθύνεται και για τη μεταπτωτική κίνηση του. Η κίνηση αυτή προσανατολίζεται προς τη διεύθυνση του πεδίου, και προσομοιάζει την περιστροφή ενός γυροσκοπίου υπο την επίδραση του βαρυτικού πεδίου της γης (Εικόνα 11). Η συχνότητα με την οποία εκτελείται η μεταπτωτική 27
33 κίνηση του πυρήνα, η οποία είναι γνωστή ως συχνότητα Larmor δίνεται από τη σχέση: V0= -γb0/2π Εικόνα 11: Μεταπτωτική κίνηση περιστρεφόμενου σωματιδίου σε ένα μαγνητικό πεδίο [58] Επίδραση του παλμού ραδιοσυχνότητας στις ενεργειακές καταστάσεις του πυρήνα Κατά την εφαρμογή παλμού ραδιοσυχνότητας σε κάποιους πυρήνες, πραγματοποιούνται μεταπτώσεις ανάμεσα στις επιτρεπόμενες ενεργειακές καταστάσεις. Οι μεταπτώσεις από τη χαμηλότερη προς την υψηλότερη ενεργειακή στάθμη είναι περισσότερες, λόγω του γεγονότος ότι οι περισσότεροι πυρήνες βρίσκονται στο χαμηλότερο ενεργειακό επίπεδο. Από την ανακατανομή των πυρήνων στις δύο ενεργειακές καταστάσεις προκαλείται μείωση ή και αναστροφή του διανύσματος Μz. Επιπροσθέτως, οι μεταπτωτικές κινήσεις των πυρήνων στις διάφορες ενεργειακές στάθμες, οδηγούν στο συγχρονισμό της ραδιοσυχνότητας με τους πυρήνες. Αυτό έχει σαν αποτέλεσμα, η ολική μαγνήτιση στο επίπεδο χψ να αποκτά κάποια ορισμένη τιμή Mχψ διαφορετική του μηδενός και να περιστρέφεται με συχνότητα Larmor. Απώτερος στόχος της φασματοσκοπίας NMR είναι η ανίχνευση της συνιστώσας Μχψ. 28
34 Τέλος, είναι κατανοητό, πως η εφαρμογή παλμού ηλεκτρομαγνητικής ακτινοβολίας στην περιοχή των ραδιοσυχνοτήτων μειώνει ή αναστρέφει τη μαγνητική συνιστώσα Μz και συντελεί στην λήψη κάποιας τιμής διάφορης του μηδενός από τη συνιστώσα Μxy. Η συνισταμένη των δύο μαγνητικών συνιστωσών δίνει τη μαγνήτιση Μ (Εικόνα 12). Εικόνα 12: Επίδραση του παλμού ραδιοσυχνοτήτων στη συνισταμένη μαγνήτιση [58]. Η συνισταμένη μαγνήτιση Μ μπορεί να θεωρηθεί και ως απόκλιση της συνιστώσας Μz από τον άξονα z. Όσο μεγαλύτερη είναι η διάρκεια και η ένταση του παλμού ραδιοσυχνότητας τόσο μεγαλύτερη και η τιμή της Μ [58,61] Μηχανισμοί αποδιέγερσης (relaxation) στο NMR Ένας πυρήνας, όταν εκτεθεί σε ηλεκτρομαγνητική ακτινοβολία κατάλληλης συχνότητας, απορροφάται ενέργεια εξαιτίας της πολύ μικρής περίσσειας πυρήνων χαμηλότερης ενεργειακής κατάστασης, οι οποίοι εκτίθενται στο ισχυρό μαγνητικό πεδίο. Λόγω αυτής της απορρόφησης υπάρχει κίνδυνος εξίσωσης των πυρήνων στις δύο ενεργειακές καταστάσεις, η οποία έχει σαν αποτέλεσμα τη μείωση της έντασης του σήματος απορρόφησης. Για να αποφευχθεί αυτό, η ταχύτητα αποδιέγερσης των πυρήνων στη χαμηλότερη ενεργειακή κατάσταση, θα πρέπει να είναι μεγαλύτερη από την ταχύτητα με την οποία τα spin απορροφούν ενέργεια. Αυτό μπορεί να επιτευχθεί, με εκπομπή ηλεκτρομαγνητικής ακτινοβολίας με συχνότητα αντίστοιχη προς τη διαφορά των δύο ενεργειακών καταστάσεων, η οποία οδηγεί σε εκπομπή φθορισμού. Σύμφωνα με τον Block, η αποδιέγερση πραγματοποιείται εκθετικά. Πιο συγκεκριμένα, η συνιστώσα Μz (Εικόνα 13), η οποία είναι η συνολική μαγνήτιση που προέρχεται από τη μικρή περίσσεια των spin στην μικρότερη ενεργειακή κατάσταση 29

35 και έχει τη διεύθυνση του μαγνητικού πεδίου, αυξάνει εκθετικά και λαμβάνει την αρχική της τιμή με μια διαδικασία γνωστή ως διαμήκης αποδιέγερση. Η διαδικασία αυτή πραγματοποιείται σε χρόνο Τα1. Επιπρόσθετα, η συνιστώσα Μχψ, η οποία είναι η ολική μαγνήτιση στο επίπεδο χψ και περιστρέφεται με συχνότητα Larmor, ελαττώνεται εκθετικά και λαμβάνει την τιμή μηδέν, με μια διαδικασία γνωστή ως εγκάρσια αποδιέγερση. Η συγκεκριμένη διαδικασία πραγματοποιείται σε χρόνο Τ2. Εικόνα 13: Συνιστώσες Μz και Μχψ. Σε κατάσταση ισορροπίας η συνιστώσα Μz είναι μέγιστη ενώ η Μχψ ισούται με μηδέν [61]. Τέλος, αξίζει να αναφερθεί, πως η πιθανότητα να πραγματοποιηθεί κάποια μετάπτωση από μια ενεργειακή κατάσταση σε άλλη, είναι ανάλογη προς την τρίτη δύναμη της ενέργειας μετάπτωσης. Επομένως, προκύπτει ότι η αυθόρμητη αποδιέγερση καθίσταται σημαντική μόνο για σχετικά υψηλές ενέργειες. Για μικρές ενεργειακές μεταβολές, όπως αυτές που μελετώνται με τη φασματοσκοπία NMR, η αποδιέγερση δεν μπορεί να πραγματοποιηθεί αυθόρμητα. Οι δύο σημαντικοί μηχανισμοί αποδιέγερσης στη Φασματοσκοπία NMR είναι η αποδιέγερση spin- πλέγματος ή διαμήκης αποδιέγερση, Τ1 και η αποδιέγερση spin- spin ή εγκάρσια αποδιέγερση, Τ2. z 2 x 2 -y Αποδιέγερση spin-πλέγματος Η ευθύγραμμη και κυκλική κίνηση των μορίων ενός NMR δείγματος, συντελεί στη μεταφορά φορτίου μέσω πυρήνων οι οποίοι εκτελούν μεταπτωτική κίνηση. Συνεπώς, κάθε μόριο μπορεί να συμβάλλει στη διακύμανση του μαγνητικού πεδίου. Μέσα σε αυτό το μαγνητικό πλέγμα, κάποιες συνιστώσες εμφανίζουν συχνότητα ίση 30
36 με τη συχνότητα Larmor συγκεκριμένου πυρήνα και αλληλεπιδρούν με αυτόν. Αποτέλεσμα της αλληλεπίδρασης αυτής είναι η μετάπτωση από υψηλότερη σε χαμηλότερη ενεργειακή κατάσταση spin. Η αποδιέγερση spin-πλέγματος χαρακτηρίζεται από το χρόνο αποδιέγερσης Τ1, ο οποίος αποτελεί το χρόνο παραμονής των πυρήνων στην υψηλότερη ενεργειακή κατάσταση. Ο χρόνος Τ1 εξαρτάται από το γυρομαγνητικό λόγο των πυρήνων καθώς και από το ιξώδες του διαλύματος. Πιο συγκεκριμένα, όσο μεγαλύτερο είναι το ιξώδες του διαλύματος, και επομένως η κινητικότητα είναι μικρή, τόσο μεγαλύτερος είναι και ο χρόνος Τ1. Αντίθετα, όταν η κινητικότητα είναι μεγάλη, αυξάνεται η πιθανότητα επίτευξης κατάλληλης μαγνητικής διακύμανσης ικανής να προκαλέσει αποδιέγερση, και η τιμή του Τ1 μειώνεται. Συμπερασματικά, τα μόρια με μέσους χρόνους κίνησης, εμφανίζουν μικρότερο Τ1 και αποδιεγείρονται πιο αποτελεσματικά μέσω του μηχανισμού spin-πλέγματος Αποδιέγερση spin-spin Όταν δύο γειτονικοί πυρήνες, εμφανίζουν ίδιες ταχύτητες αποδιέγερσης και βρίσκονται σε διαφορετικές μαγνητικές κβαντικές καταστάσεις, προκαλείται αλληλεπίδραση των μαγνητικών τους πεδίων και ανταλλαγή ενεργειακών καταστάσεων μεταξύ τους. Αυτό, έχει σαν αποτέλεσμα ο πυρήνας ο οποίος βρίσκεται στη χαμηλότερη ενεργειακή κατάσταση να διεγερθεί, ενώ αυτός ο οποίος βρίσκεται στη διεγερμένη κατάσταση να επιστρέψει στο χαμηλότερο ενεργειακό επίπεδο. Με τον τρόπο αυτό, οι πληθυσμοί των ενεργειακών καταστάσεων παραμένουν αμετάβλητοι. Ωστόσο, ο πληθυσμός των πυρήνων που εκτελούν ταυτόχρονη μεταπτωτική κίνηση μειώνεται και κατά συνέπεια και η συνιστώσα Μχψ. Επομένως, ενώ η ανταλλαγή spin συνεισφέρει στην εγκάρσια αποδιέγερση, δεν επηρεάζει το χρόνο Τ1. Ο μηχανισμός ανταλλαγής spin- spin αποτελεί κύριο μέσο αποδιέγερσης για μόρια χαμηλής κινητικότητας. Πράγματι, ο χρόνος Τ2 στα στερεά εμφανίζει χαμηλότερες τιμές. Οι χρόνοι αποδιέγερσης Τ1 και Τ2 συμβάλουν σημαντικά στη μελέτη της κινητικότητας των μορίων. Ο Block απέδειξε ότι το εύρος ενός NMR σήματος είναι ανάλογο του λόγου 1/Τ2. Μέσω αυτής της προσέγγισης εξηγείται γιατί τα μόρια τα 31
37 οποία κινούνται αργά, και επομένως έχουν μικρό Τ2, εμφανίζουν ευρείες κορυφές σε ένα φάσμα NMR [58,61] Ανατομία ενός NMR πειράματος Σύμφωνα με αυτά που προαναφέρθηκαν, ένα NMR πείραμα λειτουργεί ως εξής: ένας πυρήνας με στροφορμή p, μπορεί να λάβει 2Ι+1 ενεργειακούς προσανατολισμούς οι οποίοι έχουν μεταξύ τους την ίδια ενέργεια. Όμως η εφαρμογή εξωτερικού μαγνητικού πεδίου Β0, οδηγεί στη διαφοροποίηση των ενεργειακών τους καταστάσεων. Όταν το σύστημα βρίσκεται σε κατάσταση ισορροπίας, οι πληθυσμοί των πυρήνων στις επιτρεπόμενες ενεργειακές στάθμες δεν είναι ίσοι. Μάλιστα, ο αριθμός των πυρήνων στην χαμηλότερη ενεργειακή κατάσταση είναι ελαφρώς μεγαλύτερος. Λόγω της εφαρμογής του μαγνητικού πεδίου, κάθε πυρήνας εκτελεί μεταπτωτική κίνηση με χαρακτηριστική συχνότητα Larmor. Επίσης, προκαλείται επαγωγή μαγνήτισης Μz. Οι πληθυσμοί των πυρήνων στα διάφορα ενεργειακά επίπεδα διαταράσσονται, λόγω ακτινοβόλησης τους με κατάλληλη ραδιοσυχνότητα. Η εφαρμογή ραδιοσυχνότητας έχει σαν αποτέλεσμα το διάνυσμα της πυρηνικής μαγνήτισης να αλλάζει διεύθυνση περιστροφής από τον άξονα z προς το επίπεδο χψ. Στη συνέχεια, η μαγνήτιση Μχψ γίνεται μηδέν και προκαλείται αποδιέγερση. Η συνάρτηση απόσβεσης της μαγνήτισης του πυρήνα ως προς το χρόνο απόσβεσης ονομάζεται «ελεύθερη επαγόμενη απόσβεση» (Free Induction Decay, FID). Το κλασσικό μονοδιάστατο NMR φάσμα προκύπτει από το μετασχηματισμό Fourier της FID. Στο συγκεκριμένο φάσμα, κάθε κορυφή αντιστοιχεί σε διαφορετικό πυρήνα με χαρακτηριστικό Τ2 και επομένως χαρακτηριστικό εύρος σήματος Η έννοια της χημικής μετατόπισης Μέσα σε ένα διάλυμα, το οποίο βρίσκεται υπό την επίδραση εξωτερικού μαγνητικού πεδίου, η συχνότητα Larmor ενός πυρήνα επηρεάζεται έντονα από το χημικό του περιβάλλον, με αποτέλεσμα να διαφοροποιείται η συχνότητα συντονισμού των διαφόρων πυρήνων. Ως χημικό περιβάλλον του πυρήνα αναφέρονται τα γειτονικά ηλεκτρόνια και πυρήνες. Η διαφοροποίηση αυτή στη συχνότητα συντονισμού καλείται χημική μετατόπιση. Πρόκειται για αδιάστατη ποσότητα η οποία εκφράζεται σε ppm (parts per million) και συμβολίζεται με δ. Ο υπολογισμός της χημικής μετατόπισης πραγματοποιείται με τη βοήθεια μιας ουσίας αναφοράς, της οποίας η χημική μετατόπιση δεν μεταβάλλεται. Συνήθως χρησιμοποιούμενο εσωτερικό 32
38 πρότυπο στη φασματοσκοπία NMR, είναι το τετρα-μέθυλ-σιλάνιο (ΤΜS) και το σουλφονικό οξύ (DSS). Η σχέση από την οποία εξάγεται η χημική μετατόπιση είναι η εξής: δ = [(vsignal-vreference)/vreference]*10 6 όπου vsignal η συχνότητα συντονισμού του πυρήνα και vreference η συχνότητα συντονισμού του εσωτερικού προτύπου Η σταθερά σύζευξης J Η μαγνητική σύζευξη γειτονικών πυρήνων, οι οποίοι συνδέονται μεταξύ τους με ομοιοπολικούς δεσμούς, οδηγεί στην πολλαπλότητα των κορυφών χημικής μετατόπισης. Πιο συγκεκριμένα, το μαγνητικό πεδίο το οποίο δημιουργείται από έναν περιστρεφόμενο πυρήνα επηρεάζει την κατανομή των ηλεκτρονίων στους δεσμούς του με τους άλλους πυρήνες, με αποτέλεσμα να προκαλούνται πολλαπλές ενεργειακές μεταπτώσεις. Η πολλαπλότητα των μεταπτώσεων από τη σχάση των ενεργειακών επιπέδων των πυρήνων. Ως σταθερά σύζευξης J ορίζεται η απόσταση σε Hz μεταξύ των συνιστωσών μιας πολλαπλής κορυφής Ένταση και εύρος κορυφής Το εμβαδόν του χωρίου που περικλείεται από ένα σήμα συντονισμού είναι ανάλογο του αριθμού των πυρήνων που συνεισφέρουν στη δημιουργία του. Το εύρος του σήματος είναι χαρακτηριστικό της κινητικότητας του βιομορίου, και μετράται σε Hz. Η διεύρυνση μιας κορυφής οφείλεται στην ύπαρξη δυναμικών διεργασιών στο διάλυμα [61] Μονοδιάστατο (1D) φάσμα NMR Το πρώτο πείραμα το οποίο πραγματοποιείται κατά τη μελέτη της δομής μιας πρωτεΐνης είναι το 1 Η-1D (Εικόνα 14). Στο φάσμα που εξάγεται, κάθε κορυφή αντιστοιχεί σε ένα πρωτόνιο. Το φάσμα αυτό παρέχει τις πρώτες ενδείξεις σχετικά με την αναδίπλωση αλλά και τη δυναμική του πρωτεϊνικού μορίου. Η αρχική πληροφορία προέρχεται από την αμιδική περιοχή (6-11 ppm), η οποία επικαλύπτεται με την περιοχή των αρωματικών υδρογόνων. Πιο συγκεκριμένα, μια καλά αναδιπλωμένη πρωτεΐνη εμφανίζει μεγάλη διασπορά σημάτων στο φάσμα 1D στην περιοχή των αμιδικών πρωτονίων. Επίσης, η θωράκιση των αρωματικών υδρογόνων, 33
39 σε ένα καλά αναδιπλωμένο μόριο, οδηγεί σε μετατόπιση των μεθυλικών ομάδων σε τιμές 0,5-1 ppm. Τέλος, χαρακτηριστικά σήματα στο συγκεκριμένο φάσμα, είναι οι πλευρικές αμιδικές ομάδες των αμινοξικών καταλοίπων της τρυπτοφάνης. Αντιθέτως, βασική ένδειξη για μια μη αναδιπλωμένη πρωτεΐνη είναι η φτωχή διασπορά των σημάτων του 1D φάσματος [62]. Εικόνα 14: Τυπικό 1 H-1D NMR φάσμα μιας καλά αναδιπλωμένης πρωτεΐνης [63] NMR φασματοσκοπία δύο διαστάσεων (2D) Ένα μεγάλο μειονέκτημα του 1D φάσματος είναι η μεγάλη αλληλεπικάλυψη των σημάτων, ιδιαίτερα σε μεγάλα πρωτεϊνικά μόρια. Αυτό συμβαίνει, καθώς όσο αυξάνεται το μοριακό βάρος της πρωτεΐνης τόσο αυξάνεται και ο συνολικός αριθμός των σημάτων, ενώ το εύρος των χημικών μετατοπίσεων παραμένει το ίδιο. Επίσης, ένας ακόμα λόγος είναι ότι τα μεγάλα μόρια κινούνται πιο αργά με αποτέλεσμα να μεγάλωνει το εύρος κορυφής. Η αλληλεπικάλυψη των κορυφών δυσχεραίνει την ταυτοποίηση των συχνοτήτων συντονισμού των πυρήνων. Η λύση στο πρόβλημα της αλληλεπικάλυψης των σημάτων δόθηκε από ομοπυρηνικά πειράματα δύο διαστάσεων Φασματοσκοπίας NMR. Μερικά γνωστά δυσδιάστατα πειράματα είναι τα COSY, TOCSY και NOESY, η ανακάλυψη των οποίων χάρισε στον Richard R. Ernst και στον Kurt Wütrich το βραβείο Nobel Χημείας το 1991 και το 2002, αντίστοιχα. Τα συγκεκριμένα πειράματα 34
40 περιγράφονται από κάποια βασικά στάδια. Αρχικά, άλλοι πυρήνες λαμβάνους τυχαίους προσανατολισμούς αυτοστροφορμών ενώ άλλοι βρίσκονται σε κατάσταση θερμικής ισορροπίας. Στη συνέχεια, ακολουθεί η περίοδος ανάπτυξης t1, κατά την οποία οι πυρήνες εκτελούν μεταπτωτική κίνηση με χαρακτηριστική συχνότητα Larmor. Με την εφαρμογή κατάλληλων παλμών ραδιοσυχνότητας, η μαγνήτιση μεταφέρεται μεταξύ δύο πυρήνων Α και Β. Η περίοδος κατά την οποία οι πυρήνες αλληλεπιδρούν μεταξύ τους, ονομάζεται περίοδος μίξης ΤΑΒ. Τέλος, ακολουθεί η περίοδος ανίχνευσης t2, με την οποία ολοκληρώνεται το πείραμα. Η παραπάνω διαδικασία διεργασιών, αποτελεί μια παλμική ακολουθία. Όταν το πείραμα ολοκληρωθεί, εξάγεται το επιθυμητό 2D φάσμα έπειτα από μετασχηματισμό Fourier [64]. Οι κορυφές του 2D φάσματος παριστάνουν τη συσχέτιση μεταξύ δύο διαφορετικών πυρήνων. Χαρακτηριστικό δισδιάστατο, ετεροπυρηνικό φάσμα είναι το 15 Ν-HSQC, στο οποίο κάθε κορυφή αποτελεί και μια συσχέτιση 1 H- 15 N. Σε ένα τυπικό 15 Ν-HSQC φάσμα, μιας καλά αναδιπλωμένης πρωτεΐνης, εμφανίζονται: α) μια κορυφή για κάθε αμιδική ομάδα (Ν-Η) του σκελετού, β) μια κορυφή για κάθε πλευρική αμινομάδα των Trp και γ) ζεύγη κορυφών για τις πλευρικές αμινομάδες των Asn και Gln. Το 15 Ν- HSQC χαρακτηρίζεται ως το «δακτυλικό αποτύπωμα» της πρωτεΐνης [65], καθώς περιλαμβάνει τόσες κορυφές όσες και τα αμινοξικά κατάλοιπα της ακολουθίας, με εξαίρεση τις Pro. Οι Pro δεν εμφανίζονται στο φάσμα καθώς δεν διαθέτουν 1 Η, και επομένως δεν υφίσταται συσχέτιση με το 15 Ν. Κάποια αμινοξέα κατέχουν συγκεκριμένη θέση στο φάσμα όπως παραδείγματος χάριν οι Gly, οι πλευρικές αμινομάδες των Asn/Glu καθώς και τα πλευρικά ινδολικά ΗΝ των Τrp (Εικόνα 15). 35
41 Εικόνα 15: Τυπικό 15 Ν-HSQC φάσμα. Αποτελεί το «δακτυλικό αποτύπωμα» της πρωτεΐνης καθώς συγκεκριμένα αμινοξέα εμφανίζονται σε συγκεκριμένες περιοχές του φάσματος. Για μια καλά αναδιπλωμένη πρωτεΐνη το 15 Ν-HSQC φάσμα εμφανίζει μεγάλη διασπορά σημάτων. Η μεγάλη διασπορά των σημάτων είναι αποτέλεσμα του διαφορετικού χημικού, αλλά κυρίως του μαγνητικού περιβάλλοντος των αμιδικών πρωτονίων. Αντίθετα, όταν το μαγνητικό περιβάλλον των αμιδικών πρωτονίων είναι παρόμοιο, η πρωτεΐνη δεν είναι καλά αναδιπλωμένη και το φάσμα εμφανίζει μικρή διασπορά. Επίσης, πρωτεΐνες πλούσιες σε β- φύλλα εμφανίζουν μεγαλύτερη διασπορά σημάτων σε σχέση με τις α-έλικες. Σε ένα φάσμα 15 Ν-HSQC, είναι εμφανής και η ανταλλαγή διαμορφωτικών καταστάσεων στο διάλυμα. Πιο συγκεκριμένα, όταν η ανταλλαγή μεταξύ δύο διαμορφώσεων είναι αργή, εμφανίζονται στο φάσμα δύο σήματα για το ίδιο αμινοξύ, καθένα από τα οποία αντιστοιχεί σε μια διαφορετική διαμορφωτική κατάσταση. Με αύξηση του ρυθμού ανταλλαγής δημιουργείται μια οξεία κορυφή, η οποία αποτελεί το μέσο όρο των διαμορφώσεων (Εικόνα 16). 36
42 Εικόνα 16: Η αλληλομετατροπή των διαμορφωτικών καταστάσεων της πρωτεΐνης. Στο πλαίσιο της διερεύνησης της δυναμικής πρωτεϊνικού μορίου σε διάλυμα, είναι δυνατόν, να εξαχθούν οι χρόνοι αποδιέγερσης T1 και Τ2 από μια σειρά φασμάτων 15 Ν- HSQC. Μέσα από αυτή τη διαδικασία, είναι εφικτό να γίνει αποτίμηση της κινητικότητας του μορίου. Τέλος, από τη μελέτη ενός 15 Ν-HSQC φάσματος, μπορούν να εξαχθούν ασφαλή συμπεράσματα για τη σταθερότητα της πρωτεΐνης. Συγκεκριμένα, αν η εικόνα του φάσματος μεταβάλλεται με την πάροδο του χρόνου σημαίνει πως το μόριο στερείται σταθερότητας στο διάλυμα. Στην περίπτωση αυτή, στο κάτω δεξιά τμήμα του φάσματος εμφανίζονται κάποια νέα σήματα, τα οποία υποδηλώνουν πρωτεόλυση ή αποδιάταξη της πρωτεΐνης. Από όσα προαναφέρθηκαν εξηγείται ο λόγος για τον οποίο το 15 Ν-HSQC είναι το πρώτο ετεροπυρηνικό πείραμα που πραγματοποιείται σε ένα πρωτεϊνικό δείγμα. Τα εξαγόμενα συμπεράσματα από το φάσμα αυτό βοηθούν τον ερευνητή να αποφασίσει αν θα προχωρήσει στη συλλογή των υπόλοιπων ετεροπυρηνικών πειραμάτων τα οποία είναι αναγκαία για το δομικό χαρακτηρισμό της πρωτεΐνης ή αν θα πρέπει πρώτα να βελτιστοποιήσει τις συνθήκες έκφρασης και απομόνωσης του μορίου. 37
43 Τρισδιάστατη NMR φασματοσκοπία Παρά τη βελτίωση του προβλήματος της αλληλεπικάλυψης των κορυφών, κρίθηκε αναγκαία η εισαγωγή και τρίτης διάστασης στα NMR πειράματα. Η εισαγωγή μιας παραπάνω διάστασης αύξησε κατά πολύ τη διακριτική ικανότητα των φασμάτων. Στα τρισδιάστατα NMR πειράματα υπάρχουν δύο επιπρόσθετοι χρόνοι ανάπτυξης t1 και t2, οι οποίοι διαχωρίζονται από δύο περιόδους μίξης. Το τρισδιάστατο φάσμα παρίσταται ως ένας κύβος με ακμές τις συχνότητες συντονισμού των σχετιζόμενων πυρήνων, και προκύπτει έπειτα από μετασχηματισμό Fourier καθενός από τους τρεις ορθογώνιους άξονες. Η ανάλυση των τρισδιάστατων φασμάτων γίνεται με τη βοήθεια παράλληλων τομών προς τον ένα από τους τρείς άξονες. Το επίπεδο που ορίζεται από τους δύο πυρήνες διατηρείται σταθερό και η ταυτοποίηση των συσχετίσεων με τον τρίτο πυρήνα πραγματοποιείται στον κάθετο άξονα (Εικόνα 17). Εικόνα 17: Ανάλυση τρισδιάστατου φάσματος Επίλυση τρισδιάστατων δομών βιομορίων μέσω φασματοσκοπίας NMR Τα στάδια που ακολουθούνται κατά την επίλυση δομών βιομορίων (Εικόνα 18) είναι τα εξής: 1. Συλλογή δεδομένων 38

44 2. Ανάλυση NMR φασμάτων, η οποία περιλαμβάνει ταυτοποίηση των σημάτων του πολυπεπτιδικού σκελετού και των πλευρικών αλυσίδων των αμινοξέων, καθώς και αποτίμηση των διαπρωτονιακών σημάτων. 3. Μετατροπή του όγκου των NOE σημάτων σε ανώτατα όρια διατομικών αποστάσεων. 4. Εξαγωγή γεωμετρικών πληροφοριών 5. Δομικοί υπολογισμοί με χρήση κατάλληλου υπολογιστικού λογισμικού 6. Βελτιστοποίηση των τελικών δομών μέσω μοριακής δυναμικής Εικόνα 18: Διαδικασία επίλυσης τρισδιάστατων δομών βιομορίων με τη χρήση φασματοσκοπίας NMR [65] Συλλογή NMR δεδομένων Ζητούμενο για κάθε ερευνητή σε αυτό το στάδιο, είναι η απόκτηση δεδομένων στις καλύτερες συνθήκες ευαισθησίας και ανάλυσης της μεθόδου. Για να επιτευχθεί αυτό, καθίσταται αναγκαίο να επιλεγούν η κατάλληλη παλμική ακολουθία καθώς και οι πειραματικές παράμετροι. Ο σχετικός χρόνος περιστροφής του μορίου (correlation time, τc), είναι καθοριστικός για την επιλογή των πειραματικών συνθηκών. Ο τc, ορίζεται ως ο χρόνος που απαιτείται για την περιστροφή του μορίου κατά ένα ακτίνιο. 39
45 Τα μικρά μόρια περιστρέφονται ταχύτερα, πράγμα το οποίο σημαίνει ότι έχουν μικρότερο τc και μεγαλύτερους χρόνους αποδιέγερσης Τ1. Για τα μεγάλα μόρια ισχύει το ακριβώς αντίστροφο. Με βάση τα παραπάνω εξάγεται ο ακριβής παλμός 90⁰, ο οποίος είναι καθοριστικός για την ανίχνευση του σήματος στη μέγιστη του ένταση. Η διαδικασία συλλογής των NMR δεδομένων, τα οποία είναι αναγκαία για τον υπολογισμό της δομής του βιομορίου, απαιτεί μερικές εβδομάδες NMR μετρήσεων. Παρ όλα αυτά, σήμερα, με τη βελτιστοποίηση των τεχνικών και ιδιαίτερα με τη χρήση κρυογονικά ψυχόμενων υποδοχέων δείγματος (cryoprobes) o χρόνος που απαιτείται για τη συλλογή των δεδομένων έχει μειωθεί αισθητά. Τα φάσματα μπορούν να ταξινομηθούν σε αυτά που χρησιμοποιούνται για την ταυτοποίηση των συχνοτήτων συντονισμού των πυρήνων και σε εκείνα που παρέχουν πληροφορίες σχετικές με τις διαπρωτονιακές αλληλεπιδράσεις μέσω χώρου [66]. Όλα τα πειράματα, εκτός του 15 Ν-HSQC είναι τρισδιάστατα Ανάλυση NMR φασμάτων Ταυτοποίηση σημάτων πολυπεπτιδικού σκελετού Η ταυτοποίηση των σημάτων συντονισμού των πυρήνων του πολυπεπτιδικού σκελετού είναι το πρώτο στάδιο που απαιτείται για τον υπολογισμό της τρισδιάστατης δομής του μορίου. Τα ζεύγη πειραμάτων τα οποία χρησιμοποιούνται για το σκοπό αυτό είναι: HNCA [67,68,69] και HN(CO)CA [69,70], HNCO και HN(CA)CO, CBCANH [71] και CBCA(CO)NH [72]. Τα φάσματα που προκύπτουν παρέχουν πληροφορίες για τις συσχετίσεις των πρωτονίων δύο διαδοχικών αμινοξέων στην ακολουθία, καθώς και για τις ενδοαμινοξικές συσχετίσεις. Ο άξονας x παριστάνει το ΗΝ, o y το Ν και ο z ένα άτομο C (Cα, Cβ ή Co). Για την επίλυση των φασμάτων ακολουθείται η ίδια στρατηγική. Ως παράδειγμα, αναφέρεται το πείραμα HNCA του οποίου οι διαστάσεις είναι HN, N και Cα. 40
 ώστε στο φάσμα να αποτυπώνεται η συσχέτιση μεταξύ του αμιδικού πρωτονίου με τον α- άνθρακα του ίδιου και του προηγούμενου")

46 Εικόνα 19: Η μεταφορά μαγνήτισης στο πείραμα HNCA. [72] Στο πείραμα HNCA, η μαγνήτιση μεταφέρεται με τέτοιο τρόπο (Εικόνα 19) ώστε στο φάσμα να αποτυπώνεται η συσχέτιση μεταξύ του αμιδικού πρωτονίου με τον α- άνθρακα του ίδιου και του προηγούμενου στην ακολουθία αμινοξέος. Η διάκριση μεταξύ ενδοαμινοξικών και διαμινοξικών σημάτων γίνεται με τη βοήθεια του πειράματος HN(CO)CA. Στο συγκεκριμένο πείραμα, συσχετίζεται το αμιδικό πρωτόνιο με τον α-άνθρακα του προηγούμενου, στην ακολουθία, αμινοξέος. Οι παρατηρούμενες κορυφές αντιστοιχούν μόνο σε διαμινοξικές συζεύξεις, καθώς η μαγνήτιση μεταφέρεται μέσω του καρβονυλικού άνθρακα (Εικόνα 20). Εικόνα 20: Η μεταφορά μαγνήτισης μέσω του καρβονιλικού άνθρακα (γαλάζιο) στο πείραμα ΗΝ(CO)CA [72]. Παρομοίως, τo πείραμα ΗΝCO συσχετίζει το πρωτόνιο της αμινομάδας με τον άνθρακα του καρβονυλίου του ιδίου και του αμέσως προηγούμενου αμινοξέος, με βάση την αμινοξική ακολουθία του πρωτεϊνικού μορίου, ενώ το πείραμα ΗΝ(CA)CO συσχετίζει το αμιδικό πρωτόνιο μόνο με τον καρβονυλικό άνθρακα του ιδίου αμινοξέος. Η παραπάνω διαδικασία παρουσιάζει από ένα σημαντικό μειονέκτημα: λόγω του σχετικά μικρού εύρους των Cα σημάτων υπάρχει μεγάλη πιθανότητα 41

47 συντονισμού Cα ατόμων διαφορετικών αμινοξέων στην ίδια συχνότητα ιδιαίτερα στην περιοχή των 55ppm. Επομένως, η χρήση μόνο των συγκεκριμένων φασμάτων που περιεγράφηκαν ανωτέρω εγείρει πολλές αμφιβολίες κατά τη διαδικασία της αποτίμησης των φασμάτων συντονισμού. Για να αποφευχθούν τυχόν λάθη, επινοήθηκαν και άλλα πειράματα, στα οποία η μαγνήτιση μεταφέρεται κατά ένα επιπλέον βήμα ώστε να συμπεριληφθεί και ο Cβ (Εικόνα 21A). Πιο συγκεκριμένα, το πείραμα CBCANH συσχετίζει το άζωτο της αμινομάδας τόσο με τους Cα και Cβ του ίδιου αμινοξέος όσο και με τους αντίστοιχους άνθρακες του προηγούμενου αμινοξέος. Αντιθέτως, στο πείραμα CBCA(CO)NH παρατηρούνται μόνο διαμινοξικές συσχετίσεις μεταξύ του αμιδικού αζώτου και των Cα και Cβ του προηγούμενου αμινοξέος (Εικόνα 21Β), με βάση την ακολουθία. Εικόνα 21: Η μεταφορά της μαγνήτισης στα πειράματα Α) CBCANH και Β) CBCA(CO)NH. Με γαλάζιο χρώμα απεικονίζονται τα άτομα που διευκολύνουν τη μεταφορά της μαγνήτισης αλλά δεν ανιχνεύονται [72]. Τέλος, το πείραμα CBCANH, εκτός από την εύρεση των Cβ χημικών μετατοπίσεων και την επιβεβαίωση, των ήδη ταυτοποιημένων ατόμων Cα μέσω HNCA και HN(CO)CA πειραμάτων, εξυπηρετεί στην αναδημιουργία της πρωτεϊνικής ακολουθίας. Προκειμένου να γίνει αυτό, επιλέγεται ένα αμινοξύ τέτοιο ώστε είτε να είναι μοναδικό στην ακολουθία του πρωτεϊνικού μορίου είτε να εμπεριέχει χαρακτηριστικές τιμές Cα και Cβ (π.χ Ser, Gly, Thr) και αναζητάται το προηγούμενό του. Είναι προφανές πως οι χημικές μετατοπίσεις των ανθράκων Cα και Cβ του προηγούμενου αμινοξέος θα πρέπει να είναι ίδιες με τις αντίστοιχες των Cα(i-1) και Cβ(i-1) του συγκεκριμένου αμινοξέος. Η παραπάνω διαδικασία, είναι γνωστή και ως «μέθοδος των διαδοχικών αναθέσεων» (Εικόνα 22). 42

NH [75], το οποίο συσχετίζει πρωτόνιο της αμινομάδας με τα")
48 Εικόνα 22: Μέθοδος των «διαδοχικών αναθέσεων» Ταυτοποίηση ατόμων πλευρικών αλυσίδων Εκτός από τις συχνότητες συντονισμού των Cα, Cβ και Cο είναι πολύ σημαντικό να γνωστοποιηθούν και οι συχνότητες συντονισμού των υδρογόνων των Cα και Cβ (Ηα και Ηβ αντιστοίχως). Αυτό καθίσταται εφικτό με τη χρήση του τρισδιάστατου πειράματος ΗΒΗΑ(CO)NH [75], το οποίο συσχετίζει πρωτόνιο της αμινομάδας με τα πρωτόνια Ηα και Ηβ του προηγούμενου αμινοξέος (Εικόνα 23). Εικόνα 23: Η μεταφορά της μαγνήτισης στο πείραμα ΗΒΗΑ(CO)NH [72]. 43

49 Ο προσδιορισμός των Cα, Cβ, Ηα και Ηβ χημικών μετατοπίσεων είναι απαραίτητος προκειμένου να γίνει εφικτή η ταυτοποίηση των σημάτων του φάσματος, που αντιστοιχούν στους άνθρακες των πλευρικών ομάδων των αμινοξέων και στα αντίστοιχα πρωτόνια. Η ταυτοποίηση των σημάτων αυτών πραγματοποιείται μέσω του φάσματος HCCH-TOCSY [76]. Εικόνα 24: Η πορεία της μαγνήτισης στο πείραμα HCCH-TOCSY. Η μαγνήτιση μεταφέρεται από το πλευρικό 1 Η στον 13 C με τον οποίο συνδέεται ομοιοπολικά. Στη συνέχεια, η μαγνήτιση επιστρέφει στο πλευρικό 1 Η για ανίχνευση [72]. Συνοψίζοντας, το πρώτο στάδιο του υπολογισμού της τρισδιάστατης δομής ενός πρωτεϊνικού μορίου περιλαμβάνει την εύρεση και ταυτοποίηση των πυρήνων του σκελετού οι οποίοι είναι οι 1 Η, 13 C, 15 Ν καθώς και των πυρήνων Ηβ/Ηγ/Cβ/Cγ και μπορεί να συνοψιστεί στα ακόλουθα βήματα: 1. Kαταγραφή τρισδιάστατων πειραµάτων HNCA - HN(CO)CA, HNCO - HN(CA)CO, CBCANH - CBCA(CO)NH, ΗΒΗΑ(CO)NH και ταυτοποίηση των συχνοτήτων συντονισµού των ατόμων Cο, Cα, Cβ, Ηα, Ηβ. 2. Αναγνώριση των αµινοξέων της πολυπεπτιδικής αλυσίδας µε χρήση του φάσµατος CBCANH. 3. Ταυτοποίηση των πυρήνων των πλευρικών αλυσίδων µέσω του πειράματος HCCH-TOCSY Αναγνώριση ακολουθιακών συζεύξεων και αποτίµηση διαπρωτονιακών σηµάτων Οι περισσότερες δομικές πληροφορίες ώστε να υπολογιστεί η τρισδιάστατη δομή ενός πρωτεϊνικού μορίου προέρχονται από τα φάσματα 15 Ν-ΝΟΕSY και 13 C-NOESY (Εικόνα 26). Η ανάλυση των εν λόγω φασμάτων απαιτεί κυρίως την κρίση και την 44

50 παρέμβαση του αναλυτή. Τα φάσματα αυτά αποτελούνται από δύο άξονες 1 Η και έναν ετεροπυρηνικό άξονα ο οποίος είναι είτε 15 Ν ή 13 C. Τα συγκεκριμένα φάσματα περιέχουν μεγάλο όγκο πληροφοριών με εκατοντάδες και πλέον κορυφές διασταύρωσης ακόμα και όταν πρόκειται για μικρού μοριακού βάρους πρωτεϊνικά μόρια. Στηρίζονται στο καλούμενο Πυρηνικό Φαινόμενο Οverhauser (Νuclear Overhauser Effect, NOE) το οποίο ορίζει πως τα πρωτόνια που βρίσκονται σε απόσταση μεταξύ τους έως 6 Å, αλληλεπιδρούν μέσω χώρου και εκπέμπουν σήμα. Η ένταση του φαινομένου ΝΟΕ, όπως φαίνεται και από την ακόλουθη εξίσωση, εξαρτάται σε μεγάλο βαθμό από την απόσταση των σχετιζόμενων πρωτονίων. ΝΟΕ=(1/r 6 )f(τ c) όπου r είναι η απόσταση μεταξύ των πυρήνων που προκαλούν το φαινόμενο και τc ο χρόνος περιστροφής του μορίου [77]. Εικόνα 25: Η πορεία της μαγνήτισης στα πειράματα NOESY. A) Στο 1 Η- 15 Ν NOESY η μαγνήτιση ανταλλάσεται μεταξύ όλων των υδρογόνων που συμμετέχουν στο φαινόμενο NOE. Στη συνέχεια μεταφέρεται στο γειτονικό 15 Ν για να καταλήξει στο 1 Η για ανίχνευση. Β) Στο 1 Η- 13 C NOESY, η μαγνήτιση ακολουθεί παρόμοια πορεία με αυτή του 1 Η- 15 Ν NOESY μεταξύ των υδρογόνων. Έπειτα, μεταφέρεται στον 13 C και τέλος στο 1 Η για ανίχνευση [72]. H ανάλυση των φασµάτων ΝΟΕSY, ξεκινά µε την ταυτοποίηση των ακολουθιακών συζεύξεων, µικρής και µέσης απόστασης. Ως συζεύξεις µέσης απόστασης χαρακτηρίζονται οι αλληλεπίδρασης πρωτονίων µέσω χώρου, τα οποία απέχουν μέχρι και πέντε θέσεις στην πολυπεπτιδική αλυσίδα. Κάποιες από τις ακολουθιακές συζεύξεις είναι χαρακτηριστικές για τη δευτεροταγή δοµή του μορίου. Πιο συγκεκριµένα, όταν ο πολυπεπτιδικός σκελετός αναδιπλώνεται σε δοµή α-έλικας, παρατηρούνται συζεύξεις του τύπου Ηα(i) -NH(i+1,2,3,4), NH(i)-NH(i±1) και Hα(i)-Ηβ(i+3). Χαρακτηριστικές συζεύξεις για β-πτύχωση, είναι ισχυρής έντασης Ηα(i)-NH(i+1) και ασθενούς έντασης ΝΗ(i)-ΝΗ(i±1). 45
51 Αξίζει να σημειωθεί πως τα φάσματα ΝΟΕSY συνεισφέρουν και βοηθούν αρκετά στη διαδικασία επίλυσης μιας πρωτεϊνικής δομής μέσω των 1 Η- 1 Η συζεύξεων μακράς απόστασης. Ο όρος μακράς απόστασης συζεύξεις αναφέρεται στις συζεύξεις μεταξύ πρωτονίων που απέχουν μεταξύ τους στην πολυπεπτιδική αλυσίδα περισσότερο από πέντε θέσεις. Ο αρχικός σκοπός είναι η σωστή ταυτοποίηση ενός αριθμού σημάτων ΝΟΕ μακράς απόστασης τα οποία δε χωρούν αμφιβολίας. Ο συγκεκριμένος πυρήνας ΝΟΕ συσχετίσεων καθορίζει την αναδίπλωση του πρωτεϊνικού μορίου και αποτελεί την πρώτη ύλη για την επίλυση της τρισδιάστατης δομής. Κατά τη διάρκεια της ανάλυσης αυτών των φασμάτων είναι αρκετά σημαντικός ο διεξοδικός έλεγχος των συζεύξεων που μπορεί να αντιστοιχούν σε περισσότερα του ενός πρωτόνια έτσι ώστε να οδηγηθούμε σε εκείνα τα δύο πρωτόνια στα οποία οφείλεται, αποκλειστικά, η κορυφή διασταύρωσης. Με το πέρας του υπολογισμού της πρώτης οικογένειας δομών, προστίθενται και άλλα ΝΟΕ σήματα, τα οποία δεν αντιπαρατίθενται με τα αρχικά εξαγόμενα μοντέλα. Σε αυτό το στάδιο των υπολογισμών η επιλεκτική επισήμανση μεθυλικών ομάδων αλλά και πλευρικών αλυσίδων είναι δυνατό να μειώσει σημαντικά την πολυπλοκότητα των φασμάτων. Οι υπολογισμοί πραγματοποιούνται μέχρις ότου καταλήξουμε σε έναν ικανοποιητικό αριθμό συζεύξεων ΝΟΕ οι οποίες συμφωνούν μεταξύ τους και δεν εισάγουν στατιστικά σημαντικό σφάλμα. Στην αρχή της διαδικασίας του υπολογισμού εμφανίζονται λανθασμένα σήματα ΝΟΕ τα οποία, όμως, είναι εφικτό να αναγνωριστούν εύκολα λόγω ασυμφωνίας με το αρκετά πολύπλοκo δίκτυο των ΝΜR δομικών περιορισμών Μετατροπή της έντασης των NOE σημάτων σε ανώτερα όρια διαπρωτονιακών αποστάσεων Όπως προαναφέρθηκε, η ένταση του παραγόμενου σήματος από την αλληλεπίδραση μεταξύ δύο πρωτονίων μέσω χώρου, εξαρτάται από τη μεταξύ τους απόσταση. Το σήματα NOE, τα οποία έχουν ταυτοποιηθεί στο φάσμα, ολοκληρώνονται, και στη συνέχεια ο όγκος του σήματος μεταφράζεται σε απόσταση. Συγκεκριμένα, τα σήματα βαθμονομούνται με βάση τις αποστάσεις πρωτονίων του πολυπεπτιδικού σκελετού γνωστής δευτεροταγούς δομής, οι οποίες είναι γνωστές και σταθερές. Στη συνέχεια ταξινομούνται σε πέντε τάξεις και ισάριθμες υποκατηγορίες. Η 46
52 βαθμονόμηση της έντασης των σημάτων αλλά και η μετατροπή τους σε ανώτατα όρια αποστάσεων συμφωνούν με την εξίσωση: V=C/b n Όπου V η ένταση της κορυφής, C μια σταθερά και b το ανώτατο όριο απόστασης με 2.4Å<b<5.5Å και n=6 [77] Υπολογισμός της τριτοταγούς δομής Για τον υπολογισμό της τρισδιάστατης δομής ενός πρωτεϊνικού μορίου, μέσω κατάλληλων λογισμικών σε ηλεκτρονικό υπολογιστή, είναι αναγκαία η καταχώρηση των δίεδρων γωνιών και άλλων δομικών περιορισμών στο χώρο καθώς και η απεικόνιση των διαπρωτονιακών αποστάσεων, οι οποίες αποτελούν τον κύριο όγκο των γεωμετρικών όρων βάση των οποίων θα υπολογισθεί η δομή του βιομορίου. Εκτός όμως των πειραματικών δεδομένων, καθίσταται αναγκαία η χρήση εμπειρικών δεδομένων όπως είναι μήκη και γωνίες δεσμών καθώς και η γνώση διαφόρων ατομικών ιδιοτήτων των ομοιοπολικά συνδεδεμένων ατόμων. Τα παραπάνω δεδομένα καταχωρούνται σε ειδικές βιβλιοθήκες, γνωστές ως «Force Field Parameters». Έπειτα, το κατάλληλο υπολογιστικό πρόγραμμα δίνει αναδίπλωση στην αρχική δομή με τέτοιο τρόπο ώστε να ικανοποιούνται οι δομικοί περιορισμοί αλλά και η συνολική ενέργεια να είναι η ελάχιστη δυνατή. Από τα πιο γνωστά χρησιμοποιούμενα υπολογιστικά προγράμματα είναι το πρόγραμμα DYANA [78]. Το συγκεκριμένο πρόγραμμα, λειτουργεί με βάση το πρωτόκολλο «προσομοίωσης ανόπτησης» (simulated annealing) [77]. Σύμφωνα με το συγκεκριμένο πρωτόκολλο, τα πρώτα στάδια των υπολογισμών πραγματοποιούνται σε υψηλή θερμοκρασία. Από την έκθεση τους σε υψηλή θερμοκρασία τα άτομα αποκτούν υψηλή κινητική ενέργεια, και λόγω έντονης θερμικής κίνησης, καταλαμβάνουν το μέγιστο δυνατό χώρο. Με τον τρόπο αυτό αποφεύγεται ο εγκλωβισμός της πρωτεΐνης σε κάποιο ενεργειακό ελάχιστο. Οι υπολογισμοί συνεχίζονται με την ψύξη του συστήματος. Έτσι η δομή αναπτύσσεται προς μια ενεργειακά προτιμητέα διαμόρφωση, η οποία αντιστοιχεί στο ολικό ελάχιστο. Η διαμόρφωση υπόκειται στα όρια του πεδίου δυνάμεων και των πειραματικών διαπρωτονιακών αποστάσεων. 47
53 Τελική οικογένεια δομών Μετά την ολοκλήρωση των δομικών υπολογισμών, δεν προκύπτει ένα μοντέλο αντιπροσωπευτικό για την πρωτεΐνη. Αντίθετα, επιλέγεται ένας αριθμός δομών, συνήθως 10 έως 40, με τη χαμηλότερη τιμή συνάρτησης, γνωστή ως target function (tf). Η συνάρτηση tf αποτελεί μέτρο της ορθότητας των εξαγόμενων αποτελεσμάτων, καθώς περιγράφει την απόκλιση των υπολογιζόμενων δομών από την πραγματική. Είναι αξιοσημείωτο, ότι όλα τα μέλη της οικογένειας δομών συμφωνούν με τους δομικούς περιορισμούς και συγκλίνουν προς μια διαμόρφωση. Αυτό συμβαίνει, καθώς οι δομικοί περιορισμοί έχουν ένα εύρος επιτρεπόμενων τιμών. Ο λόγος για τον οποίο καθίσταται αναγκαία η χρήση εύρους τιμών, εντοπίζεται στο ότι τα σήματα των NOE συσχετίσεων αντιστοιχούν σε όλες τις δυνατές διαμορφώσεις τις οποίες μπορεί να υιοθετήσει η πολυπεπτιδική αλυσίδα στο διάλυμα στην κλίμακα των μs. Επομένως, κάθε δομική πληροφορία που εξάγεται από τα πειραματικά δεδομένα, αντιπροσωπεύει το μέσο όρο των τιμών της υπολογιζόμενης δομικής παραμέτρου. Η ακρίβεια των τελικών δομών χαρακτηρίζεται με βάση τη μέση τυπική απόκλιση των συντεταγμένων κάθε μέλους της οικογένειας δομών από τη μέση δομή και περιγράφεται από τη μαθηματική σχέση: Όπου xi, ψi, zi οι ατομικές συντεταγμένες και Ν ο αριθμός των συγκρινόμενων ατόμων. Η τιμή του RMSD ανα αμινοξύ παρουσιάζει έντονη διακύμανση. Πιο συγκεκριμένα, τα αμινοξέα τα οποία βρίσκονται σε ευκίνητα τμήματα εμφανίζουν υψηλές τιμές RMSD, ενώ τα αμινοξέα σε τμήματα με καθορισμένη δευτεροταγή δομή παρουσιάζουν χαμηλές τιμές. Η τιμή του RMSD, υπολογίζεται πάνω στα βαρέα άτομα όλης της πρωτεΐνης ή του πρωτεϊνικού σκελετού. Η ποιότητα και η ακρίβεια των τελικών δομών δεν εξαρτάται μόνο από τον υπολογισμό του RMSD. Για την απόλυτη μέτρηση της ακρίβειας μιας NMR δομής χρειάζονται δεδομένα για την πραγματική δομή. Για το σκοπό αυτό υπάρχουν 48
54 κατάλληλα λογισμικά, τα οποία υπολογίζουν διάφορους στερεοχημικούς παράγοντες, όπως μήκη και γωνίες δεσμών. Επίσης καθορίζουν τη διαμόρφωση των δίεδρων γωνιών φ και ψ και των άλλων στοιχείων δευτεροταγούς δομής Πρωτεϊνική δυναμική Τα βιολογικά μόρια, όπως είναι οι πρωτεΐνες, όταν βρίσκονται σε διάλυμα δε λαμβάνουν μια μοναδική διαμόρφωση αλλά ένα ευρύ φάσμα διαμορφωτικών καταστάσεων. Η αλλαγή διαμόρφωσης, σε επίπεδο δίεδρων γωνιών, ισοδυναμεί με τη μετάβαση από μια επιτρεπόμενη ενεργειακή κατάσταση σε μια άλλη. Επομένως, γίνεται εύκολα αντιληπτό πως για να επιτευχθεί αλληλομετατροπή διαμορφώσεων απαιτείται μια ορισμένη ενέργεια ενεργοποίησης. Αυτή η ενέργεια εξαρτάται από την ταχύτητα της κινητικής διεργασίας. Συγκεκριμένα, όσο μεγαλύτερη είναι η ενέργεια που χρειάζεται το σύστημα, τόσο βραδύτερη η κίνηση. Εικόνα 26: Η χρονική κλίμακα των κινήσεων που εκτελεί το μόριο στο διάλυμα. H κλίμακα των χρόνων των διαφόρων κινήσεων που εκτελεί ένα πρωτεϊνικό μόριο σε διάλυμα κυμαίνεται από ps έως αρκετά s (Εικόνα 26). Ως διαχωριστική γραμμή για τα διάφορα είδη κίνησης των πρωτεϊνικών μορίων έχει οριστεί ο χρόνος περιστροφής τc. 49
55 Τα πειράματα τα οποία χρησιμοποιούνται για τη μελέτη της πρωτεϊνικής δυναμικής είναι τύπου HSQC. Από τα συγκεκριμένα πειράματα εξάγονται οι χρόνοι αποδιέγερσης των ατόμων 15 N. Συγκεκριμένα, πρόκειται για μια σειρά 15 N-HSQC πειραμάτων, τα οποία διαφέρουν μεταξύ τους στο χρονικό διάστημα που μεσολαβεί μεταξύ δυο διαδοχικών παλμικών ακολουθιών. Σε ένα τυπικό NMR πείραμα, για να εξασφαλιστεί ικανοποιητικός λόγος σήματος προς θόρυβο, η εκάστοτε παλμική ακολουθία επαναλαμβάνεται αρκετές φορές. Το μεσοδιάστημα μεταξύ δυο διαδοχικών παλμικών ακολουθιών ονομάζεται χρόνος υστέρησης (delay time) και ακολουθεί του χρόνου ανίχνευσης του NMR σήματος. Όσο μεγαλύτερος είναι ο χρόνος υστέρησης τόσο περισσότερα spin έχουν επιστρέψει στην θεμελιώδη ενεργειακή τους κατάσταση με άμεση συνέπεια την εξασθένηση του NMR σήματος. Οι χρόνοι αποδιέγερσης Τ1 και Τ2, υπολογίζονται με βάση τη μεταβολή της έντασης του σήματος. H τιμή του ρυθμού αποδιέγερσης R1 είναι ενδεικτική για κινήσεις της τάξης των ps-ns, ενώ η τιμή του R2 παρέχει πληροφορίες για διεργασίες που πραγματοποιούνται στη χρονική κλίμακα των μs-ms [80]. Τέλος, από το λόγο των ρυθμών αποδιέγερσης για όλα τα αμινοξέα ενός πρωτεϊνικού μορίου είναι εφικτό να εξαχθούν ασφαλή συμπεράσματα σχετικά με το εάν η πρωτεΐνη πολυμερίζεται ή όχι. 2.9 Υπολογιστικά εργαλεία Το πρόγραμμα TALOS (Torsion Angle Likelihood Obtained from chemical shift and sequence Similarity) Σε ένα πρωτεϊνικό μόριο, υπάρχει άμεση συσχέτιση των χημικών μετατοπίσεων των ατόμων του πολυπεπτιδικού σκελετού με τη δευτεροταγή διαμόρφωση του μορίου. Στις περιοχές με καθορισμένη δευτεροταγή δομή, οι ομόλογες πρωτεΐνες εμφανίζουν παρόμοιες χημικές μετατοπίσεις. Το υπολογιστικό πρόγραμμα TALOS [82] εκμεταλλευόμενο την ιδιότητα αυτή, παρέχει τη δυνατότητα πρόβλεψης των τιμών των δίεδρων γωνιών φ και ψ του πρωτεϊνικού σκελετού. Στο TALOS είναι καταχωρημένες σε μια βάση δεδομένων, οι χημικές μετατοπίσεις των ατόμων Cα, Cβ, CΟ, Hα και Ν διακοσίων πρωτεϊνών, των οποίων οι δομές έχουν επιλυθεί με κρυσταλλογραφία ακτίνων Χ. Το πρόγραμμα, με βάση την ομοιότητα της αμινοξικής ακολουθίας και των χημικών μετατοπίσεων, αναζητά το σημείο στο οποίο η κάθε τριπλέτα της πρωτεΐνης ταιριάζει καλύτερα. Σε περίπτωση κατά την οποία τα 50
56 κεντρικά κατάλοιπα των δέκα δομών με τη μεγαλύτερη ταύτιση, εμφανίζουν παρόμοιες γωνίες φ και ψ, τότε οι τιμές αυτές χρησιμοποιούνται ως γεωμετρικοί περιορισμοί στους δoμικoύς υπoλoγισμoύς. Το πρόγραμμα TALOS αποτελεί ένα αξιόπιστο υπολογιστικό λογισμικό για την πρόβλεψη της δευτεροταγούς δομής διάφορων πρωτεϊνών [83], καθώς το σφάλμα της μεθόδου είναι της τάξης του 3% Το πρόγραμμα DYANA Το υπολογιστικό πρόγραμμα DYANA, μέσω προσομοίωσης μοριακής δυναμικής και αξιοποίησης της συστροφής των δίεδρων γωνιών, δύναται να υπολογίσει τις τρισδιάστατες δομές των πρωτεϊνικών μορίων. Με τη βοήθεια του συγκεκριμένου προγράμματος η μεταβλητότητα του συστήματος ελαττώνεται σημαντικά, καθώς οι παράμετροι που καθορίζουν τη δομή του μορίου διατηρούνται στις βέλτιστες τιμές τους. Εξαιτίας της περιορισμένης διαμορφωτικής ελευθερίας, τα βήματα του υπολογισμού διαρκούν περισσότερο, με αποτέλεσμα ο δομικός υπολογισμός να είναι αποτελεσματικότερος. Η προσομοίωση μοριακής δυναμικής με το πρόγραμμα DYANA, πραγματοποιείται με επίλυση των κινητικών εξισώσεων Lagrange. H συγκεκριμένη προσέγγιση απαιτεί επίλυση n γραμμικών εξισώσεων σε κάθε βήμα υπολογισμού, κάτι που συνεπάγεται πως ο υπολογιστικός χρόνος για κάθε υπολογισμό θα είναι της τάξης του n 3. Το πρόγραμμα DYANA, με τη χρήση ενός γρήγορου αλγορίθμου, κατάφερε να ξεπεράσει αυτό το εμπόδιο και παρέχει τη δυνατότητα γρήγορων υπολογισμών. 51
57 ΣΚΟΠΟΣ ΕΡΓΑΣΙΑΣ 52
58 Η διαλυτή γουανυλική κυκλάση (sgc) αποτελεί μια πρωτεΐνη-αισθητήρα για το διατομικό μόριο του νιτρικού οξειδίου (ΝΟ). Το μόριο του ΝΟ παράγεται από τη συνθάση του νιτρικού οξειδίου (ΝΟs) και στη συνέχεια προσδένεται στο μόριο της sgc. Μετά την πρόσδεση η sgc ενεργοποιείται και καταλύει τη μετατροπή της 5- τριφωσφορικής γουανοσίνης (GTP) σε κυκλική 3,5- μονοφωσφορική γουανοσίνη (cgmp). Το μόριο της sgc αποτελείται από δύο υπομονάδες, α1 και β1, οι οποίες σχηματίζουν μεταξύ τους ένα ετεροδιμερές. Η β1 υπομονάδα περιλαμβάνει τρεις επικράτειες στο Ν-τελικό άκρο της μεταξύ των οποίων και η H-NOX (Heme Nitric Oxygen/Oxide). Η επικράτεια H-NOX φέρει ένα μόριο αίμης επάνω στο οποίο, και συγκεκριμένα στο ιόν σιδήρου, προσδένεται το μόριο του ΝΟ επιτυγχάνοντας έτσι την ενεργοποίηση της sgc. Επομένως, ο ρόλος της συγκεκριμένης επικράτειας κρίνεται καθοριστικός στην ενεργοποίηση του μονοπατιού NO/cGMP. Το συγκεκριμένο μονοπάτι εμπλέκεται σε ένα πλήθος φυσιολογικών λειτουργιών του οργανισμού κυρίως του νευρικού και του καρδιαγγειακού συστήματος και έτσι το μόριο της sgc αποτελεί φαρμακευτικό στόχο για ασθένειες όπως η καρδιακή ανεπάρκεια, η υπέρταση και η στυτική δυσλειτουργία. Η επίλυση και μελέτη της δομής διαφόρων H-NOX επικρατειών αναμένεται να δώσει πληροφορίες για το ρόλο του μορίου της sgc σε διάφορες ασθένειες και να συνεισφέρει στο σχεδιασμό φαρμακευτικών ενώσεων. Σκοπός της συγκεκριμένης εργασίας είναι η μελέτη της διαμόρφωσης του μορίου της H-NOX επικράτειας του οργανισμού Nostoc sp. με μετάλλαξη της κυστεΐνης στη θέση 139 σε αλανίνη (C139A) σε διάλυμα. H συγκεκριμένη H-NOX εμφανίζει 33,86% ακολουθιακή ομολογία με την αντίστοιχη ανθρώπινη. Αν και η ίδια δομή έχει επιλυθεί με τη χρήση κρυσταλλογραφίας ακτίνων-χ, η ανάλυσή της με φασματοσκοπία NMR αφενός θα δώσει πληροφορίες για τη διαμόρφωση της σε διάλυμα, κατάσταση η οποία προσομοιάζει με τις φυσιολογικές κυτταρικές συνθήκες, αφετέρου θα μελετηθεί η δυναμική του μορίου. Για τον προσδιορισμό της δομής θα αναλυθούν 2D και 3D ομο/ετερο-πυρηνικά NMR φάσματα, ενώ για την επίλυση της τρισδιάστατης δομής θα χρησιμοποιηθούν διάφορα υπολογιστικά εργαλεία. 53
59 3. ΠΕΙΡΑΜΑΤΙΚΟ ΜΕΡΟΣ 54
60 3.1 NMR πειράματα Η έκφραση και η απομόνωση της επικράτειας H-NOX C139A του Nostoc sp πραγματοποιήθηκε με τεχνικές ανασυνδυασμένου DNA. Πιο συγκεκριμένα, το γονίδιο που έφερε τη μετάλλαξη εισήχθη σε βέκτορα έκφρασης pet-22b(+). Η παραπάνω διαδικασία πραγματοποιήθηκε από τον Dr Focco van Akker (Case Western Reserve University, USA), ο οποίος και μας παραχώρησε το ανασυνδυασμένο γονίδιο. Η H-NOX πρωτεΐνη, περιελάμβανε τα αμινοξέα της αντίστοιχης επικράτειας του Nostoc sp. Η έκφραση πραγματοποιήθηκε με τη χρήση βακτηριακών στελεχών BL21 (DE3) Star Escherichia coli. Οι βακτηριακές καλλιέργειες αναπτύχθηκαν σε θρεπτικό υλικό M9 εμπλουτισμένο με 0,3 mm δ- αμινολεβουνιλικό οξύ (δ-ala), ενώ η επαγωγή της έκφρασης πραγματοποιήθηκε με τη χρήση 0,15 mm IPTG. Η ανασυνδυασμένη πρωτεΐνη απομονώθηκε με χρωματογραφία ιονανταλλαγής ακολουθούμενη από μια διαδικασία χρωματογραφίας αποκλεισμού μεγέθους. Τα πρωτεϊνικά δείγματα για τα NMR πειράματα παρασκευάστηκαν σε διαλύτη 90% H20 και 10% D2O. Ως ουσία αναφοράς χρησιμοποιήθηκε DSS (4,4-διμεθυλ-4-σιλαπενταν-1-σουλφονικό οξύ). Η συγκέντρωση της πρωτεΐνης στα δείγματα ήταν 0,6 mm για πρωτεΐνη επισημασμένη με 15 Ν και 1,0 mm για διπλά επισημασμένη πρωτεΐνη 13 C/ 15 N. Όσον αφορά την επιλεκτική επισήμανση οι τιμές συγκεντρώσεων ήταν 0,8 mm στα δείγματα Αλανινή/Τυροσίνη και Λευκίνη/Φαινυλαλανίνη, και 0,25 mm στο δείγμα Ισολευκίνη/Βαλίνη. Οι διαδικασίες έκφρασης και απομόνωσης της ανασυνδυασμένης πρωτεΐνης πραγματοποιήθηκαν από την υποψήφια διδάκτορα Κα Κατερίνα Αργυρίου. Τα πειράματα NMR πραγματοποιήθηκαν σε θερμοκρασία Τ=298Κ, και καταγράφηκαν από φασματογράφο Βruker Avance III HD 700MHz εξοπλισμένο με κρυογονικά ψυχόμενη probe για καλύτερη ανίχνευση των πυρήνων 1 Η, 13 C και 15 Ν. Τα πρωτεϊνικά δείγματα τα οποία χρησιμοποιήθηκαν, παρασκευάστηκαν με τεχνικές ανασυνδυασμένου DNA. Για τη μελέτη μέσω φασματοσκοπίας NMR, τα πρωτεϊνικά δείγματα επισημάνθηκαν κατάλληλα με ενεργούς για το NMR πυρήνες, 13 C και 15 Ν. Ο τελικός όγκος κάθε δείγματος ήταν 500 μl. Από αυτά, τα 450 μl ήταν ρυθμιστικό διάλυμα φωσφορικών και 50 μl δευτεριωμένο νερό (D2O). Η προσθήκη του D2O πραγματοποιείται για την αποφυγή διακυμάνσεων του μαγνητικού πεδίου. Ως εσωτερικό πρότυπο χρησιμοποιήθηκε η ουσία DSS. Η συμπίεση του σήματος 55
61 συμπιεσμού του νερού, πραγματοποιήθηκε με την τεχνική του προκορεσμού (presaturation). Η συγκεκριμένη διαδικασία περιλαμβάνει την εφαρμογή παλμού ραδιοσυχνότητας μεγάλης διάρκειας και χαμηλής ισχύος, ο οποίος ακολουθείται από έναν παλμό RF 90⁰ για τη διέγερση των επιθυμητών πυρήνων. Η ταυτοποίηση των σημάτων συντονισμού των πυρήνων, 1 Η, 15 Ν και 13 C τόσο στον πολυπεπτιδικό σκελετό όσο και στην πλευρικές αλυσίδες των αμινοξέων, πραγματοποιήθηκε με τη βοήθεια των παρακάτω πειραμάτων: 15 N- HSQC, HNCO και HN(CA)CO, HNCA και HN(CO)CA, HNCACB και HN(CO)CACB, HBHA(CBCACO)NH, HNHA, HCCH-TOCSY. Με τη βοήθεια των τρισδιάστατων φασμάτων, 15 Ν- και 13 C- NOESY, προέκυψαν οι περιορισμοί διαπρωτονιακών αποστάσεων. 3.2 Επεξεργασία των NMR φασμάτων Η επεξεργασία των προς ανάλυση φασμάτων πραγματοποιήθηκε με τη βοήθεια του λογισμικού TOPSPIN 3.2 της Bruker. Επίσης, χρησιμοποιήθηκαν διάφορες υπορουτίνες για το χειρισμό των ακατέργαστων NMR δεδομένων. 3.3 Ανάλυση των NMR φασμάτων Η ταυτοποίηση των σημάτων συντονισμού των πυρήνων πραγματοποιήθηκε με το πρόγραμμα CARA, ενώ η ολοκλήρωση του όγκου των κορυφών συσχέτισης με το πρόγραμμα NEASY. 3.4 Δομικοί υπολογισμοί Οι δομικοί υπολογισμοί πραγματοποιήθηκαν με τη χρήση του προγράμματος DYANA. Οι όγκοι των σημάτων μετατράπηκαν σε διαπρωτονιακές αποστάσεις μέσω της ρουτίνας Caliba και με τον τρόπο αυτόν προέκυψαν οι δομικοί περιορισμοί υπό τη μορφή ανώτατων ορίων διατομικών αποστάσεων. Κατά τους δομικούς υπολογισμούς, χρησιμοποιήθηκαν και γεωμετρικοί περιορισμοί όπως προέκυψαν από το πρόγραμμα TALOS. Στους υπολογισμούς συμπεριλήφθηκαν και δεσμοί υδρογόνου, οι οποίοι προβλέφθηκαν από το πρόγραμμα DYANA. 56
62 3.5 Μελέτη πρωτεϊνικής δυναμικής Οι ρυθμοί αποδιέγερσης R1 και R2 υπολογίστηκαν μετά από καταγραφή μιας σειράς δισδιάστατων 15 N-HSQC πειραμάτων με διαφορετικούς χρόνους υστέρησης. 57
63 4. ΑΠΟΤΕΛΕΣΜΑΤΑ 58
64 4.1 Η H-NOX επικράτεια του οργανισμού Nostoc sp Ανάλυση NMR φασμάτων και δομικοί υπολογισμοί Η επικράτεια Η-ΝΟX του οργανισμού Nostoc sp. αποτελείται από 183 αμινοξέα. Στη συγκεκριμένη επικράτεια, πραγματοποιήθηκε σημειακή μετάλλαξη στη θέση 139. Συγκεκριμένα, το αμινοξύ κυστεΐνη στη συγκεκριμένη θέση αντικαταστάθηκε από αλανίνη (C139A). Παρακάτω φαίνονται τα αμινοξικά κατάλοιπα, απο τα οποία απαρτίζεται η πολυπεπτιδική αλυσίδα, καθώς και η θέση τους σε αυτή. Ο αριθμός των φορτισμένων αμινοξέων της πολυπεπτιδικής αλυσίδας είναι 14 θετικά (5 Arg+, 9 Lys+) και 30 αρνητικά (11 Asp-, 19 Glu-). Για την παρασκευή των πρωτεϊνικών δειγμάτων ήταν αναγκαία η εύρεση των βέλτιστων συνθηκών έκφρασης και απομόνωσης του πρωτεϊνικού μορίου. Η εύρεση των κατάλληλων συνθηκών εξασφαλίζει τη λήψη φασμάτων με την καλύτερη δυνατή ποιότητα, καθώς και τη μέγιστη δυνατή σταθερότητα του πρωτεϊνικού μορίου στις συχνότητες συντονισμού των πυρήνων 1 Η και 15 Ν. Το αρχικό συμπέρασμα που εξάγεται από το 1 H- 15 N HSQC (Εικόνα 27) φάσμα είναι ότι η προς μελέτη πρωτεΐνη είναι αναδιπλωμένη. Αυτό προκύπτει από τη διασπορά των σημάτων σε μεγάλο εύρος χημικών μετατοπίσεων. Στο φάσμα επίσης φαίνεται ο αριθμός των σημάτων να είναι μεγαλύτερος από τον αριθμό των αμινοξέων στην ακολουθία. Αυτό μπορεί να οφείλεται σε διάφορους παράγοντες όπως είναι η παρουσία δύο μορφών του ίδιου αμινοξέος. 59
65 Εικονα 27: 1 H- 15 N HSQC της επικράτειας H-NOX C139A του Nostoc sp. Η ταυτοποίηση των σημάτων του πολυπεπτιδικού σκελετού πραγματοποιήθηκε με τη χρήση της στρατηγικής που περιεγράφηκε στην ενότητα έπειτα από ανάλυση των αντίστοιχων φασμάτων. Επίσης για την ταυτοποίηση των αμινοξικών καταλοίπων χρησιμοποιήθηκαν και φάσματα επιλεκτικής επισήμανσης. Για την εξαγωγή των φασμάτων αυτών παρασκευάστηκαν 3 δείγματα στα οποία είχαν επισημανθεί επιλεκτικά τα αμινοξέα Leu/Phe, Ile/Val και Ala/Tyr. 60
66 Εικόνα 28: Φάσμα HN(CA)CO της H-NOX C139A επικράτειας του Nostoc sp. Οι συσχετίσεις οι οποίες απεικονίζονται αντιστοιχούν στην περιοχή His16-Asp20. Εικόνα 29: Φάσμα HNCACB της H-NOX C139A επικράτειας του Nostoc sp. Οι συσχετίσεις οι οποίες απεικονίζονται αντιστοιχούν στην περιοχή Gln27-Leu31. 61
67 62
68 Εικόνα 30: 1 H- 15 N HSQC φάσματα με επιλεκτική επισήμανση. Α) Ala/Tyr, B) Ile/Val και Γ) Leu/Phe. Για την ταυτοποίηση των αμινοξέων της ακολουθίας της πρωτεΐνης χρησιμοποιήθηκαν τα φάσματα 1 H- 15 N HSQC, HNCO, HN(CA)CO, HN(CO)CA, HNCA, HN(CO)CACB και HNCACB. Τα συγκεκριμένα φάσματα αναλύθηκαν σε συνδυασμό με τα 1 H- 15 N HSQC με επιλεκτική επισήμανση αμινοξέων (Εικόνα 30), συμβάλλοντας έτσι στην ορθή ταυτοποίηση των αμινοξικών καταλοίπων. Από την ανάλυση των φασμάτων ταυτοποιήθηκε το 82% των αμινοξέων της ακολουθίας της πρωτεΐνης. Λόγω της παρουσίας έξι προλινών (Pro62, Pro95, Pro113, Pro117, Pro118, Pro143) στην αμινοξική ακολουθία, ο μέγιστος αριθμός αμινοξικών καταλοίπων προς ταυτοποίηση μόνο με τη χρήση των παραπάνω φασμάτων ήταν 177 από τα 183. Αυτό συμβαίνει επειδή οι προλίνες στερούνται αμιδικού υδρογόνου και ως εκ τούτου δεν εμφανίζονται στο φάσμα 1 Η- 15 Ν ΗSQC. Στον παρακάτω πίνακα συγκεντρώνονται τα αμινοξέα τα οποία δεν ήταν εφικτό να ταυτοποιηθούν. 63
69 Πίνακας 1 Μη ταυτοποιημένα αμινοξέα στο φάσμα 1 Η- 15 Ν HSQC (34) Met1, Tyr2, Gly3, Ile4, Val5, Asn6, Asp45, Pro62 Trp74, Val75, Thr76, Tyr77, Thr78, Leu87, Pro95, Leu101, Asp102, Asn103, Leu104, His105, Ala106, Arg107, Val108, Gly109, Pro113, Arg116, Pro117, Pro118, Ala119, Phe120, His124, Ser127, Pro143, Met144 Παρ όλα αυτά, επιτεύχθηκε ο προσδιορισμός των χημικών μετατοπίσεων Cα, Cβ, Hα και Ηβ για τις προλίνες Pro62, Pro95 και Pro113 από τα αμινοξέα τα οποία βρίσκονται μπροστά από τις προλίνες στην αμινοξική ακολουθία. Για το σκοπό αυτό αξιοποιήθηκαν δεδομένα τα οποία προέκυψαν από τα φάσματα HN(CO)CACB και HBHA(CBCACO)NH. Μετά την παραπάνω διαδικασία ακολούθησε η ταυτοποίηση των χημικών μετατοπίσεων των ατόμων των πλευρικών ομάδων έπειτα από ανάλυση του φάσματος hcch-tocsy. Το συγκεκριμένο φάσμα παρέχει τη δυνατότητα ταυτοποίησης των πλευρικών αλυσιδών των αμινοξέων τα οποία δεν εμφανίζονται στο φάσμα 1 Η- 15 Ν HSQC. Έπειτα από ανάλυση όλων των διαθέσιμων φασμάτων και αξιοποίηση όλων των NMR δεδομένων, το συνολικό ποσοστό των ταυτοποιημένων συχνοτήτων συντονισμού έφτασε το 64,5%. Το ποσοστό δεν εμφανίζεται τόσο υψηλό καθώς η ταυτοποίηση κάποιων αμινοξέων δεν κατέστη δυνατή. Πίνακας 2 Αμινοξέα τα οποία δεν ταυτοποιήθηκαν έπειτα από αξιοποίηση όλων των NMR δεδομένων (23) Met1, Tyr2, Gly3, Ile4, Val5, Trp74, Val75, Thr76, Tyr77, Leu101, Asp102, Asn103, Leu104, His105, Ala106, Arg107, Val108, Arg116, Pro117, Pro118, Ala119, Phe120, Pro143 64
70 Για την ταυτοποίηση της διαμόρφωσης του δεσμού Χaa-Pro (Xaa: τυχαίο αμινοξύ), χρησιμοποιήθηκαν οι χημικές μετατοπίσεις 13 Cγ και 13 Cβ των ταυτοποιημένων αμινοξικών καταλοίπων Pro. Συγκεκριμένα, η διαφορά δ[ 13 Cβ] δ[ 13 Cγ] μπορεί να χρησιμοποιηθεί ως δείκτης για cis/trans διαμόρφωση. Η trans διαμόρφωση συναντάται συχνότερα από τη cis, σύμφωνα με δομικές και υπολογιστικές αναλύσεις [91]. Πίνακας 3 Δβγ Pro trans Δβγ Pro cis Μέση τιμή Δ( 13 Cβ- 13 Cγ) (ppm) Τυπική απόκλιση (ppm) Πίνακας 3: Αντιστοιχία της διαφοράς Δ( 13 Cβ- 13 Cγ) με τη διαμόρφωση του δεσμού Xaa-Pro. Οι τιμές της αντίστοιχης διαφοράς για την H-NOX C139A επικράτεια του Nostoc sp. έχουν ως εξής: Pro62: 4.88, Pro95: 2.00, Pro113: Οι τιμές αυτές, σύμφωνα με το Πίνακα 3, δείχνουν πως πρόκειται για προλίνες με trans διαμόρφωση. Όσον αφορά τις υπόλοιπες προλίνες της αλληλουχίας, δεν κατέστη δυνατός ο προσδιορισμός της διαμόρφωσής τους, εξαιτίας της έλλειψης των χημικών τους μετατοπίσεων. 65

71 Εικόνα 31: 1 H- 15 N HSQC της επικράτειας H-NOX C139A του Nostoc sp έπειτα από ταυτοποίηση των αμινοξέων. Μετά την ταυτοποίηση των πυρήνων των αμινοξέων στο φάσμα 1 Η- 15 Ν HSQC, ακολουθεί η πρόβλεψη δευτεροταγούς διαμόρφωσης από το λογισμικό TALOS. Το συγκεκριμένο πρόγραμμα παρέχει τη δυνατότητα πρόβλεψης δευτεροταγούς δομής της πρωτεΐνης με τη βοήθεια στατιστικής ανάλυσης και λαμβάνοντας υπόψη τις δίεδρες γωνίες. 66

72 Εικόνα 32: Πάνω: Δευτεροταγής δομή της επικράτειας H-NOX C139A του Nostoc sp. σύμφωνα με την αντίστοιχη κρυσταλλική δομή (PDB: 4IAM). Κάτω: Πρόβλεψη δευτεροταγούς δομής από το TALOS+. Οι στήλες με μπλε χρώμα υποδεικνύουν την ύπαρξη α-έλικας ενώ οι κόκκινες την παρουσία β-φύλλων. Σε γκρι πλαίσιο βρίσκονται οι περιοχές οι οποίες δεν ταυτοποιήθηκαν με συνέπεια να μην είναι δυνατή η πρόβλεψη δευτεροταγούς διαμόρφωσης από το TALOS για τις περιοχές αυτές. Τα εξαγόμενα αποτελέσματα από το λογισμικό TALOS+ για τη δευτεροταγή διαμόρφωση της πρωτεΐνης στο διάλυμα παρουσιάζουν μεγάλη ομοιότητα με τα αντίστοιχα της κρυσταλλικής δομής της επικράτειας H-NOX C139A του Nostoc sp. Το γεγονός αυτό επιβεβαιώνει την ορθή ταυτοποίηση των αμινοξέων από την ανάλυση των NMR δεδομένων. Το λογισμικό δεν εμφανίζει πρόβλεψη για την περιοχή L101-G109 όπου στην κρυσταλλική δομή υπάρχει μια α-έλικα. Αυτό συμβαίνει καθώς τα αμινοξικά κατάλοιπα της συγκεκριμένης περιοχής δεν έχουν ταυτοποιηθεί και συνεπώς το TALOS δεν διαθέτει τις απαραίτητες πληροφορίες ώστε να πραγματοποιήσει την πρόβλεψη. Επίσης, το TALOS+ προέβλεψε την ύπαρξη μιας επιπλέον μικρής β-πτυχωτής επιφάνειας η οποία έπεται της πρώτης α-έλικας στο N- τελικό άκρο. Έπειτα από την πρόβλεψη δευτεροταγούς δομής της πρωτεΐνης ακολούθησε η ανάλυση των φασμάτων 15 Ν-ΝOESY και 13 C-NOESY με σκοπό την εύρεση διαπρωτονιακών αποστάσεων. Η ανάλυση πραγματοποιήθηκε λαμβάνοντας υπόψη και τα εξαγόμενα αποτελέσματα από το TALOS. 67

 1 H- 1 H- 15 N NOESY φάσμα της επικράτειας σε προβολή 2D.")
 1 H- 1 H- 13 C NOESY φάσμα της επικράτειας")
 1 H- 1 H- 13 C NOESY φάσμα της επικράτειας σε προβολή 2D.")
73 Εικόνα 33: A) 1 H- 1 H- 15 N NOESY φάσμα της επικράτειας H-NOX C139A Nostoc sp. F1: δ 1 Η (ppm) F2: δ 15 Ν (ppm) F3: δ 1 Η (ppm). B) 1 H- 1 H- 15 N NOESY φάσμα της επικράτειας H-NOX C139A Nostoc sp. σε προβολή 2D. F1: 1 H (ppm) F3: 1 H (ppm). Εικόνα 34: A) 1 H- 1 H- 13 C NOESY φάσμα της επικράτειας H-NOX C139A Nostoc sp. F1: δ 1 Η (ppm) F2: δ 13 C (ppm) F3: δ 1 Η (ppm). B) 1 H- 1 H- 13 C NOESY φάσμα της επικράτειας H-NOX C139A Nostoc sp. σε προβολή 2D. F1: 1 H (ppm) F3: 1 H (ppm). 68
.")
74 Τα στοιχεία δευτεροταγούς δομής επιβεβαιώθηκαν και μέσω NOE συζεύξεων, οι οποίες λειτουργούν ως διαγνωστικά στοιχεία για την ύπαρξη α-έλικας και β-φύλλου. Χαρακτηριστικό διαγνωστικό στοιχείο για την ύπαρξη α-έλικας αποτελεί η σύζευξη του τύπου dαν(i, i+4). Ο συμβολισμός αυτός υποδηλώνει ότι το αμιδικό υδρογόνο ενός αμινοξέος το οποίο εντοπίζεται στην α-έλικα συσχετίζεται μέσω χώρου με το ΗΑ υδρογόνο του αμινοξέος το οποίο βρίσκεται τέσσερις θέσεις πριν στην αλληλουχία. Ομοίως, συζεύξεις του τύπου dnn(i, i+1), dnn(i, i+2), dαn(i, i+3) και dαβ(i, i+3) είναι χαρακτηριστικές για την ύπαρξη α-έλικας ενώ ισχυρής έντασης σύζευξη dαn(i, i+1) και ασθενούς έντασης dnn(i, i+1) χαρακτηρίζουν τα β-φύλλα. Όλες οι παραπάνω συσχετίσεις συγκεντρώνονται στο διάγραμμα ακολουθιακών συζεύξεων. Εικόνα 35: 15 N NOESY φάσμα της επικράτειας H-NOX C139A Nostoc sp. Απεικονίζονται συσχετίσεις του τύπου dnn(i, i+1) και dnn(i, i+2) στην περιοχή Αla24-Lys26. Τα συγκεκριμένα αμινοξέα αποτελούν τμήμα α- έλικας. 69
75 Εικόνα 36: 15 N NOESY φάσμα της επικράτειας H-NOX C139A Nostoc sp. Απεικονίζονται συσχετίσεις του τύπου dαn(i, i+3) και dαn(i, i+4) οι οποίες αποτελούν διαγνωστικά στοιχεία για την ύπαρξη α-έλικας. Εικόνα 37: 15 N NOESY φάσμα της επικράτειας H-NOX C139A Nostoc sp. Ισχυρής έντασης σύζευξη dαn(i, i+1) πιστοποιεί την ύπαρξη β-πτυχωτής επιφάνειας. 70
76 Εικόνα 38: Διάγραμμα ακολουθιακών συζεύξεων. Τα πειραματικά δεδομένα αποστάσεων επιβεβαιώνουν την ύπαρξη των στοιχείων δευτεροταγούς δομής, τα οποία προέβλεψε το λογισμικό TALOS. Σε μαύρο πλαίσιο περικλείονται οι περιοχές των αμινοξέων τα οποία δεν έχουν ταυτοποιηθεί. Η ανάλυση των φασμάτων 15 Ν/ 13 C-NOESY και η αναγνώριση των διαφόρων ΝΟΕ συζεύξεων, εκτός από την επαλήθευση των στοιχείων δευτεροταγούς δομής, συντελούν και στην διαμόρφωση της τριτοταγούς δομής του πρωτεϊνικού μορίου. Η διαμόρφωση της τρισδιάστατης δομής ενός πρωτεϊνικού μορίου καθορίζεται από το πλήθος των ΝΟΕ συζεύξεων. Για τον υπολογισμό μιας καλά διαμορφωμένης πρωτεϊνικής δομής απαιτείται η εύρεση και ο χαρακτηρισμός ενός ικανοποιητικού αριθμού ΝΟΕ συζεύξεων μακράς απόστασης ([i-j] > 5). Αυτές οι ΝΟΕ συσχετίσεις εμπεριέχονται στα φάσματα 1 Η- 15 Ν NOESY και 1 Η- 13 C NOESY και αναπτύσσονται ανάμεσα σε αμινοξέα, τα οποία 71
, η οποία προέκυψε από ανάλυση μέσω κρυσταλλογραφίας ακτίνων-χ, αναζητήθηκαν και ταυτοποιήθηκαν NOE συζεύξεις μακράς απόστασης (Εικόνα 40).")
77 βρίσκονται μακριά στην αλληλουχία, αλλά συσχετίζονται μέσω χώρου λόγω εγγύτητας των ατόμων τους. Τα άτομα των αμινοξέων αυτών προσεγγίζουν μεταξύ τους στο χώρο εξαιτίας της πρωτεϊνικής διαμόρφωσης. Χρησιμοποιώντας ως μοντέλο την τρισδιάστατη δομή (Εικόνα 39), η οποία προέκυψε από ανάλυση μέσω κρυσταλλογραφίας ακτίνων-χ, αναζητήθηκαν και ταυτοποιήθηκαν NOE συζεύξεις μακράς απόστασης (Εικόνα 40). Εικόνα 39: Τρισδιάστατη δομή του μορίου της Ns H-NOX η οποία προέκυψε με ανάλυση κρυσταλλογραφίας ακτίνων-χ. οι α-έλικες συμβολίζονται αα-αf και οι β-επιφάνειες β1-β4. 72




78 Εικόνα 40: Εκδεικτικές περιοχές στις οποίες αναπτύσσονται ΝΟΕ συζεύξεις μακράς απόστασης. Πάνω: διαπρωτονιακές συζεύξεις μεταξύ ατόμων των α-ελίκων αα και αβ. Κάτω: διαπρωτονιακές συζεύξεις μεταξύ ατόμων των β-πτυχωτών επιφανειών β3 και β4. 73

79 Εικόνα 41: Διάγραμμα NOE συσχετίσεων ανα αμινοξύ. Μαύρο: NOE μακράς απόστασης, σκούρο γκρί: NOE μέσης απόστασης, γκρί: διαδοχικές συσχετίσεις, λευκό: ενδοαμινοξικές συσχετίσεις. Όσο μεγαλύτερος είναι ο αριθμός των ΝΟΕ συσχετίσεων, και ιδιαίτερα των μέσης και μακράς απόστασης, τόσο καλύτερη είναι και η διαμόρφωση της δομής του πρωτεϊνικού μορίου. Έπειτα από ανάλυση των φασμάτων 1 Η- 15 Ν NOESY και 1 Η- 13 C NOESY για την περιοχή των αλειφατικών πρωτονίων προέκυψαν οι συσχετίσεις των διαπρωτονιακών αποστάσεων. Οι συσχετίσεις αυτές εισήχθησαν στο πρόγραμμα DYANA για την πραγματοποίηση των δομικών υπολογισμών. Κατά τη διάρκεια των υπολογισμών πραγματοποιείται επαναπροσδιορισμός ή και προσθήκη νέων συσχετίσεων. Επίσης, συσχετίσεις οι οποίες φαίνονται να είναι λανθασμένες αφαιρούνται. Οι αλλαγές αυτές κατά τη διάρκεια των δομικών υπολογισμών έχουν ως σκοπό τη δημιουργία μιας οικογένειας 20 δομών στην οποία τα υπολογισθέντα μοντέλα θα συγκλίνουν προς μια μέση διαμόρφωση (χαμηλή τιμή rmsd). Για να επιτευχθεί αυτό θα πρέπει να υπάρχει όσο το δυνατόν μεγαλύτερη συμφωνία των γεωμετρικών περιορισμών στα μοντέλα αυτά. 74
80 Στο υπολογιστικό πρόγραμμα DYANA, εκτός από τους περιορισμούς των διαπρωτονιακών αποστάσεων, εισήχθησαν και περιορισμοί στρέψης των δίεδρων γωνιών φ και ψ όπως προέκυψαν από το πρόγραμμα TALOS+. Εικόνα 42: Διαγραμματική απεικόνιση των ΝΟΕ διαπρωτονιακών συζεύξεων σε συνάρτηση με το εύρος της σύζευξης.. 75
81 5. ΣΥΖΗΤΗΣΗ 76

82 5.1 Επιλογή της κατάλληλης επικράτειας H-NOX για δομική μελέτη μέσω NMR φασματοσκοπίας Οι επικράτειες H-NOX είναι γενικά συντηρημένες στα διάφορα είδη των οργανισμών. Αποτελούνται από περίπου 190 αμινοξέα και παρουσιάζουν μεταξύ τους ακολουθιακή ομολογία από 10 έως 40%. Επίσης, υποστηρίζεται ότι οι βακτηριακές H-NOX εμφανίζουν παρόμοιες ιδιότητες πρόσδεσης με την sgc. Λόγω αδυναμίας επίλυσης της δομής της ανθρώπινης H-NOX επικράτειας κατέστη αναγκαία η αποσαφήνιση του μηχανισμού ενεργοποίησης από τα μόρια ΝΟ και CO μέσα από τη μελέτη βακτηριακών ομολόγων. Για το σκοπό αυτό πολλές δομές βακτηριακών H-NOX επικρατειών έχουν επιλυθεί μέσω κρυσταλλογραφίας ακτίνων-χ και φασματοσκοπίας NMR. Η Nostoc sp. H-NOX εμφανίζει περίπου 34% (Εικόνα 43) ακολουθιακή ομολογία με την αντίστοιχη επικράτεια της sgc. Επίσης, 17 από τα 27 αμινοξέα τα οποία βρίσκονται στην κοιλότητα της αίμης είναι ακριβώς ίδια στην Ns H-NOX και την sgc. Η δομή της Νs H-NOX έχει επιλυθεί μέσω κρυσταλλογραφίας ακτίνων-χ τόσο στην ελεύθερη μορφή της όσο και με προσδέτες τα δυατομικά αέρια ΝΟ και CO. Ως εκ τούτου, η Ns H-NOX αποτελεί πιθανά ένα αντιπροσωπευτικό μοντέλο πρωτεΐνης το οποίο χρήζει μελέτης μέσω φασματοσκοπίας NMR. Με τη συγκεκριμένη φασματοσκοπική τεχνική παρέχεται η δυνατότητα μελέτης της δυναμικής του μορίου και χαρακτηρισμού των διαμορφωτικών μεταβάσεων του μορίου ύστερα από αλληλεπίδραση με σηματοδοτικά αέρια ή άλλες βιοδραστικές ενώσεις. Εικόνα 43: Σύγκριση των αμινοξικών ακολουθιών της Ns H-NOX και της επικράτειας H-NOX της β1 υπομονάδας του μορίου της sgc. Με κόκκινο χρώμα απεικονίζονται τα ομόλογα αμινοξέα. Σε μαύρο πλαίσιο εμφανίζονται τα αμινοξέα στην κοιλότητα της αίμης τα οποία είναι ακριβώς ίδια και στις δύο επικράτειες. 77

83 5.2 Ανάλυση των δυσχερειών κατά τη διάρκεια της επεξεργασίας των πειραματικών δεδομένων Στο φάσμα 15 Ν-HSQC εμφανίζονταν περισσότερα σήματα από τον αριθμό των αμινοξέων της πρωτεΐνης. Επομένως, αναμέναμε πως θα επιτευχθεί η ταυτοποίηση όλων των αμινοξικών καταλοίπων τα οποία απαρτίζουν το πρωτεϊνικό μόριο. Προς μεγάλη μας έκπληξη όμως κάτι τέτοιο δεν συνέβη. Ύστερα από την ανάλυση των απαραίτητων φασμάτων διαπιστώθηκε πως κάποια αμινοξέα δεν ήταν δυνατό να αναγνωριστούν λόγω έλλειψης φασματικών δεδομένων, δηλαδή απουσία του αναμενόμενου αριθμού ενδοαμινοξικών σημάτων συντονισμού 13 C και 1 Η, καθώς και των αντίστοιχων συζεύξεων μεταξύ των σημάτων αυτών. Για το λόγο αυτό αναζητήθηκαν τα αίτια τα οποία προκάλεσαν αυτή την απώλεια των σημάτων. Η χαρτογράφηση των μη ταυτοποιημένων αμινοξέων στην ακολουθία και την κρυσταλλική δομή έκανε εμφανές ότι τα αμινοξέα αυτά δεν βρίσκονται διάσπαρτα στην πρωτεΐνη αλλά εντοπίζονται σε συγκεκριμένες περιοχές και συγκεκριμένα στοιχεία β-ταγούς δομής της H-NOX. Συγκεκριμένα, αυτά τα αμινοξέα εντοπίζονται σε δύο α-έλικες οι οποίες βρίσκονται πάνω και κάτω από το μόριο της αίμης (Εικόνα 44). Εικόνα 44: Κρυσταλλογραφική δομή Ns H-NOX C139A. Με κόκκινο χρώμα απεικονίζονται τα αμινοξέα των ελίκων που βρίσκονται πάνω και κάτω από το μόριο της αίμης και δεν κατέστει δυνατό να ταυτοποιηθούν. 78
84 Μια πιθανή αιτία απώλειας αυτών των περιοχών είναι η ύπαρξη δύο ή περισσότερων διαμορφωτικών καταστάσεων αυτών όπως παρατηρήθηκε και στις κρυσταλλικές δομές της επικράτειας H-NOX του οργανισμού Shewanella oneidensis. Σε περιπτώσεις διαμορφωτικής ανταλλαγής τα εκπεμπόμενα σήματα των πυρήνων στα NMR φάσματα διευρύνονται και αλληλεπικαλύπτονται με άλλα με αποτέλεσμα να μην είναι εύκολα ανιχνεύσιμα. Η διαμορφωτική ανταλλαγή αυτών των τμημάτων υποστηρίζεται σθεναρά και από το γεγονός ότι η α-έλικα στην περιοχή Leu101-Val108, γνωστή και ως «έλικα σηματοδότησης», εντοπίζεται παραμορφωμένη στην κρυσταλλική δομή Shewanella oneidensis Η-NOX. Αυτό συμβαίνει καθώς η λύση του δεσμού His-Fe επιτρέπει την στρέψη της «έλικας σηματοδότησης» κατά 45⁰ γύρω από έναν άξονα ο οποίος έχει το κέντρο του κατά μήκος της ελικοειδούς διαμόρφωσης. Αύτη η δυνατότητα στρέψης προκαλεί δομικές αλλαγές στην κοιλότητα της αίμης. Αυτή η διαμορφωτική ευκαμψία η οποία παρατηρείται στην κρυσταλλική δομή, μπορεί να λαμβάνει χώρα και στο διάλυμα με αποτέλεσμα να ασκείται επιρροή στην περιοχή της κοιλότητας της αίμης. Οι αλλαγές αυτές είναι ικανές να προκαλέσουν διεύρυνση σημάτων στα NMR φάσματα και συνεπώς αδυναμία ταυτοποίησής τους. Πάντως, σύμφωνα με μετρήσεις ηλεκτρονιακής απορρόφησης UV-Visible δεν παρατηρείται καμία αλλαγή στην ηλεκτρονιακή δομή του σιδήρου (ο σίδηρος παραμένει Fe 2+ ) και στον αριθμό συναρμογής του (αριθμός ένταξης:5) κατά τη διάρκεια λήψης των NMR φασμάτων. Εικόνα 45: Φάσμα ατομικής απορρόφησης UV του ατόμου σιδήρου στο μόριο της αίμης της H-NOX επικράτειας του οργανισμού Nostoc sp. Μπλέ: Fe(II) Κόκκινο: Fe(III). 79
85 Επιπλέον, η επίδραση του ρεύματος δακτυλίου του εκτεταμένου π-συστήματος του μορίου της πορφυρίνης, μπορεί να περιπλέξει την ταυτοποίηση των συντονισμών που επηρεάζονται από αυτό το φαινόμενο. Το ρεύμα δακτυλίου είναι ένα φαινόμενο το οποίο παρατηρείται σε μόρια με αρωματικό χαρακτήρα, δηλαδή με συζυγιακό σύστημα και ο πορφυρινικός δακτύλιος αποτελεί ένα τετραπυρρολικό, εκτεταμένο π- συζυγιακό σύστημα. Σύμφωνα με αυτό, εάν ένα μαγνητικό πεδίο κατευθύνεται κάθετα προς το επίπεδο του αρωματικού συστήματος ένα ρεύμα δακτυλίου προκαλείται στα απεντοπισμένα π ηλεκτρόνια. Δεδομένου ότι τα ηλεκτρόνια αυτά είναι ελεύθερα να κυκλοφορούν, αντί να είναι εντοπισμένα στους δεσμούς, ανταποκρίνονται καλύτερα στο μαγνητικό πεδίο, δημιουργώντας ένα τοπικό μαγνητικό πεδίο, το οποίο επιδρά στους πυρήνες του μορίου. Συνήθως, οι πυρήνες οι οποίοι βρίσκονται πάνω και κάτω από το επίπεδο του δακτυλίου μετατοπίζονται σε χαμηλότερες τιμές ppm ενώ οι πυρήνες οι οποίοι βρίσκονται στο επίπεδο του αρωματικού δακτυλίου μετατοπίζονται σε μεγαλύτερες τιμές της κλίμακας ppm. Η τελευταία υπόθεση υποστηρίζεται έντονα από την αυξημένη χημική μετατόπιση κάποιων αμιδικών πρωτονίων των ταυτοποιημένων αμινοξέων της H-NOX τα οποία σύμφωνα με την κρυσταλλογραφική δομή βρίσκονται πάνω και κάτω από το μόριο της αίμης (Val145-Leu149, Ser111, Phe112, Ser79-Glu81). Ωστόσο, η πιθανότητα ύπαρξης παραμαγνητικού μεταλλικού κέντρου Fe(III) απορρίπτεται σύμφωνα με τα δεδομένα από τα φάσματα ηλεκτρονιακής απορρόφησης UV-Visible (Εικόνα 45). Η καμπύλη, η οποία παρουσιάζει μέγιστο στα 425 nm και είναι χαρακτηριστική για μεταλλικό ιόν Fe(II) με αριθμό συναρμογής 5 σύμφωνα με τη βιβλιογραφία, προέκυψε ύστερα από μετρήσεις που πραγματοποιήθηκαν στο πρωτεϊνικό μόριο κατά τη διάρκεια λήψης των NMR φασμάτων. Η οξείδωση του μεταλλικού κέντρου, ώστε να προκύψει η καμπύλη απορρόφησης στα 407 nm, η οποία είναι χαρακτηριστική για οξειδωτική κατάσταση Fe(III), πραγματοποιήθηκε σύμφωνα με τα πρότυπα πειραματικά πρωτόκολλα για την επικράτεια H-NOX του Nostoc sp. 80
86 5.3 Aνάλυση των στοιχείων δευτεροταγούς δομής της Nostoc sp. H-NOX C139A και σύγκριση με τα αντίστοιχα της κρυσταλλικής δομής Για την εύρεση των στοιχείων δευτεροταγούς δομής του πρωτεϊνικού μορίου χρησιμοποιήθηκαν συνδυαστικά οι NOE συσχετίσεις, οι οποίες προσδιορίστηκαν ύστερα από την ανάλυση των φασμάτων 15 Ν- και 13 C-NOESY, και η πρόβλεψη της δευτεροταγούς διαμόρφωσης του μορίου όπως προέκυψε από το TALOS+. Από την ανάλυση των φασμάτων και την ολοκλήρωση των όγκων των σημάτων προέκυψαν περίπου 2500 NOE διαπρωτονιακές συσχετίσεις, οι οποίες μετατράπηκαν σε ανώτατα όρια αποστάσεων με σκοπό τη χρήση τους ως γεωμετρικών περιορισμών στη διαδικασία των δομικών υπολογισμών. Επίσης, για τους δομικούς υπολογισμούς χρησιμοποιήθηκαν και περιορισμοί στρέψης δίεδρων γωνιών όπως προέκυψαν από το πρόγραμμα TALOS+, το οποίο ανέλυσε τα δεδομένα των χημικών μετατοπίσεων των πυρήνων και τα συσχέτισε με τα στοιχεία της δευτεροταγούς δομής. Οι α-έλικες και οι β-πτυχωτές επιφάνειες καθορίστηκαν από την αναγνώριση των απαραίτητων διαγνωστικών στοιχείων στα φάσματα. Μέσω των δομικών υπολογισμών προέκυψαν τα στοιχεία δευτεροταγούς δομής της Nostoc sp. H-NOX τα οποία είναι 6 α-έλικες και 4 αντιπαράλληλα β-φύλλα. Οι α-έλικες εντοπίζονται στις εξής περιοχές: αα (Asn6-Gly18), αβ (Glu19-Ala29), αc (44-60), αd (Ala63-Glu72), αε (Glu85-Ala90) και αf (Ala142-Arg155). Στις συγκεκριμένες περιοχές ο σχηματισμός των α-ελίκων προήλθε έπειτα από την ταυτοποίηση και χρήση των απαραίτητων συσχετίσεων(διαγνωστικά στοιχεία) στα φάσματα NOESY. Όσον αφορά τα β-φύλλα, τέτοιες διαμορφώσεις του πολυπεπτιδικού σκελετού εντοπίστηκαν στις εξής περιοχές: β1 (Gln123-Ser126), β2 (Ser129-Glu131), β3 (Val160-Gln164) και β4 (Ser178-Glu182). Οι β-πτυχωτές επιφάνειες, ομοίως με τις α-έλικες, σχηματίστηκαν σε αυτές τις περιοχές εξαιτίας της ύπαρξης συσχετίσεων μεταξύ των αμινοξέων που τις απαρτίζουν, οι οποίες αποτελούν διαγνωστικά στοιχεία για την ύπαρξη β-φύλλου. Συγκρίνοντας τα στοιχεία δευτεροταγούς δομής, τα οποία προέκυψαν από την NMR ανάλυση και την πρόβλεψη του TALOS+, με αυτά της κρυσταλλογραφικής δομής παρατηρείται ότι εμφανίζουν μεγάλη ομοιότητα μεταξύ τους (Εικόνα 46). 81
87 Συγκεκριμένα, φαίνεται να έχουν τα ίδια στοιχεία δευτεροταγούς διαμόρφωσης στις ίδιες περιοχές. Η μόνη διαφορά εντοπίζεται στην περιοχή Asn100-Leu110, όπου στην κρυσταλλική δομή υπάρχει μια α-έλικα. Η αδυναμία ταυτοποίησης των συγκεκριμένων αμινοξέων, λόγω απουσίας πειραματικών δεδομένων, δεν επέτρεψε τον καθορισμό α-έλικας στην περιοχή αυτή. 82
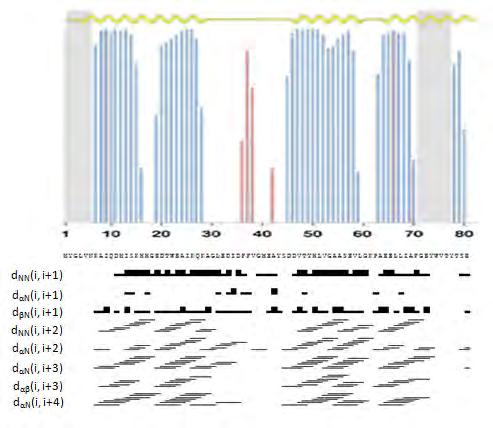
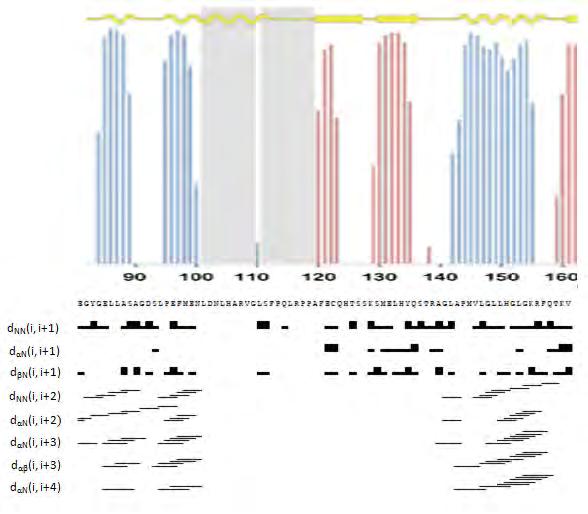
88 83

89 Εικόνα 46: Στο επάνω μέρος με κίτρινο χρώμα εμφανίζονται τα στοιχεία δευτεροταγούς της Nostoc sp. H-NOX C139A της κρυσταλλικής δομής (PDB:4IAM). Ακολουθεί η πρόβλεψη δευτεροταγούς δομής του μορίου όπως προέκυψε από το TALOS+. Τέλος απεικονίζεται το διάγραμμα ακολουθιακών συζεύξεων. 5.4 Σύγκριση της κρυσταλλογραφικής δομής με την τριτοταγή διαμόρφωση που προέκυψε από τους δομικούς υπολογισμούς βάσει των NMR πειραματικών δεδομένων Αναλύοντας τμηματικά τις τριτοταγείς διαμορφώσεις του μορίου παρατηρείται ότι συγκλίνουν σημαντικά μεταξύ τους. Πιο συγκεκριμένα οι έλικες αα και αβ φαίνεται να βρίσκονται κοντά στο χώρο. Στην NMR διαμόρφωση το γεγονός αυτό οφείλεται στην ύπαρξη και ανίχνευση NOE συσχετίσεων μεταξύ αμινοξέων τα οποία συμμετέχουν στις δύο α-έλικες. Συγκεκριμένα, στα φάσματα εμφανίζονται 84

90 συσχετίσεις μεταξύ των αμινοξέων Ιle13 και Ser14 της αα με τα αμινοξέα Trp22 και Ile25 της αβ.(εικόνα 47), τόσο στην κρυσταλλική δομή, όσο και στη δομή στο διάλυμα. Εικόνα 47: A: Κρυσταλλική δομή. Β: NMR μοντέλο. Γ: Χαρακτηριστικές NOE συζεύξεις για την συγκεκριμένη περιοχή. Άλλα τμήματα του μορίου τα οποία φαίνεται να βρίσκονται κοντά στο χώρο είναι οι έλικες αβ και αc. Οι συγκεκριμένες περιοχές προσεγγίζουν στο χώρο και ως αποτέλεσμα της εγγύτητας τους αναπτύσσονται NOE συζεύξεις μεταξύ των αμινοξέων Ala24 και Lys28 της αβ και Ala54 και Val58 της αc (Εικόνα 48), κατά τρόπο ανάλογο με αυτόν στην κρυσταλλική δομή. 85

91 Εικόνα 48: A: Κρυσταλλική δομή. Β: NMR μοντέλο. Γ: Χαρακτηριστικές NOE συζεύξεις για την συγκεκριμένη περιοχή. NOE συσχετίσεις μακράς απόστασης παρατηρούνται και μεταξύ των α-ελίκων αd και αf. Πιο συγκεκριμένα, στα NOESY φάσματα εντοπίζονται συζεύξεις μεταξύ των αμινοξέων Ile68 και Glu72 της αd και Gly147 και His150 της αf (Εικόνα 49). 86

92 Εικόνα 49: A: Κρυσταλλική δομή. Β: NMR μοντέλο. Γ: Χαρακτηριστικές NOE συζεύξεις για την συγκεκριμένη περιοχή. Στην περιοχή των β-κλώνων αναπτύσσονται συσχετίσεις μακράς απόστασης μεταξύ των αμινοξέων Met130 του β2 και Ile179 του β4 με αποτέλεσμα οι δύο κλώνοι να προσεγγίζουν στο χώρο (Εικόνα 50). 87

93 Εικόνα 50: : A: Κρυσταλλική δομή. Β: NMR μοντέλο. Γ: Χαρακτηριστικές NOE συζεύξεις για την συγκεκριμένη περιοχή. Όλες αυτές οι NOE συσχετίσεις μακράς απόστασης, οι οποίες αναπτύσσονται μεταξύ των διαφόρων στοιχείων δευτεροταγούς δομής του μορίου της Nostoc sp. H- NOX C139A, καθορίζουν την τριτοταγή διαμόρφωση του μορίου στο χώρο. Πριν ακόμα ολοκληρωθούν οι δομικοί υπολογισμοί και προκύψει η τελική οικογένεια δομών είναι εμφανές ότι η διαμόρφωση του μορίου, το οποίο μελετήθηκε μέσω φασματοσκοπίας NMR, παρουσιάζει όλα εκείνα τα χαρακτηριστικά της αναδίπλωσης και τους πακεταρίσματος των στοιχείων της δευτεροταγούς δομής με την κρυσταλλική δομή, υιοθετώντας διευθέτηση στο χώρο η οποία είναι εξαιρετικά παρόμοια με αυτή που αποκαλύφθηκε από την κρυσταλλογραφική ανάλυση της δομής της Η-ΝΟΧ.. 88
Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών
 Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών Τίτλος Ερευνητικού Έργου «Διαφώτιση των διαμορφωτικών αλλαγών της
Περιγραφή Χρηματοδοτούμενων Ερευνητικών Έργων 1η Προκήρυξη Ερευνητικών Έργων ΕΛ.ΙΔ.Ε.Κ. για την ενίσχυση Μεταδιδακτόρων Ερευνητών/Τριών Τίτλος Ερευνητικού Έργου «Διαφώτιση των διαμορφωτικών αλλαγών της
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ
 ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ") ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2015-2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1)Πώς το φαινόμενο Bohr επηρεάζει την πρόσδεση οξυγόνου στην αιμοσφαιρίνη; Που συνδέονται τα ιόντα
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2015-2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1)Πώς το φαινόμενο Bohr επηρεάζει την πρόσδεση οξυγόνου στην αιμοσφαιρίνη; Που συνδέονται τα ιόντα
Ασκήσεις 5& 6. Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR. Σύγκριση & Ανάλυση Δομών Βιομορίων
 Ασκήσεις 5& 6 Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Σύγκριση & Ανάλυση Δομών Βιομορίων Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Κρυσταλλογραφία Ακτίνων-Χ & Φασματοσκοπία ΝΜR Πλεονεκτήματα -
Ασκήσεις 5& 6 Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Σύγκριση & Ανάλυση Δομών Βιομορίων Διαμόρφωση Βιομορίων μέσω Φασματοσκοπίας NMR Κρυσταλλογραφία Ακτίνων-Χ & Φασματοσκοπία ΝΜR Πλεονεκτήματα -
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ
 ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
ΦΥΣΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗΝ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ ΦΥΣΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗΝ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ Φασματοσκοπία Mossbauer ΠΕΡΙΚΛΗΣ ΑΚΡΙΒΟΣ Τμήμα Χημείας Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ ΦΥΣΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗΝ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ Φασματοσκοπία Mossbauer ΠΕΡΙΚΛΗΣ ΑΚΡΙΒΟΣ Τμήμα Χημείας Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
Βιοφυσική. ΦΥΣ 415 Διδάσκων Σ. Σκούρτης (χειμερινό εξάμηνο ) 3 η Διάλεξη
 3 η Διάλεξη") Βιοφυσική ΦΥΣ 415 Διδάσκων Σ. Σκούρτης (χειμερινό εξάμηνο 2009-10) 3 η Διάλεξη Από την προηγούμενη διάλεξη: Οι πρωτεΐνες εκτελούν τις περισσότερες βιολογικές λειτουργίες π.χ Ενζυμική κατάλυση (επιτάχυνση
Βιοφυσική ΦΥΣ 415 Διδάσκων Σ. Σκούρτης (χειμερινό εξάμηνο 2009-10) 3 η Διάλεξη Από την προηγούμενη διάλεξη: Οι πρωτεΐνες εκτελούν τις περισσότερες βιολογικές λειτουργίες π.χ Ενζυμική κατάλυση (επιτάχυνση
ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ
 ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ ΠPΩTEINEΣ Οι πρωτεΐνες παίζουν σημαντικό ρόλο σε όλες σχεδόν τις βιολογικές διεργασίες. H σημασία τους φαίνεται στις παρακάτω περιπτώσεις: 1. Κατάλυση (πχ. ένζυμα) 2. Μεταφορά
ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ ΠPΩTEINEΣ Οι πρωτεΐνες παίζουν σημαντικό ρόλο σε όλες σχεδόν τις βιολογικές διεργασίες. H σημασία τους φαίνεται στις παρακάτω περιπτώσεις: 1. Κατάλυση (πχ. ένζυμα) 2. Μεταφορά
Διδάσκων: Καθηγητής Εμμανουήλ Μ. Παπαμιχαήλ
 Τίτλος Μαθήματος: Ενζυμολογία Ενότητα: Εισαγωγή Διδάσκων: Καθηγητής Εμμανουήλ Μ. Παπαμιχαήλ Τμήμα: Χημείας 8 1. EIΣAΓΩΓH Tα ένζυμα είναι οι καταλύτες της ζώσης ύλης. Καταλύουν τις χημικές αντιδράσεις,
Τίτλος Μαθήματος: Ενζυμολογία Ενότητα: Εισαγωγή Διδάσκων: Καθηγητής Εμμανουήλ Μ. Παπαμιχαήλ Τμήμα: Χημείας 8 1. EIΣAΓΩΓH Tα ένζυμα είναι οι καταλύτες της ζώσης ύλης. Καταλύουν τις χημικές αντιδράσεις,
ΑΙΜΟΣΦΑΙΡΙΝΗ (ΑΜΦ) ΑΙΜΟΣΦΑΙΡΙΝΗ: Hb, είναι τετραμερής πρωτείνη. ΜΕΤΑΠΤΩΣΗ ΑΠΟ Τ <=> R
 ΑΙΜΟΣΦΑΙΡΙΝΗ: Hb, είναι τετραμερής πρωτείνη. ΜΕΤΑΠΤΩΣΗ ΑΠΟ Τ <=> R") ΑΙΜΟΣΦΑΙΡΙΝΗ (ΑΜΦ) ΑΙΜΟΣΦΑΙΡΙΝΗ: Hb, είναι τετραμερής πρωτείνη. ΜΕΤΑΠΤΩΣΗ ΑΠΟ Τ R ΔΕΟΞΥΑΙΜΟΣΦΑΙΡΙΝΗ ΟΞΥΑΙΜΟΣΦΑΙΡΙΝΗ (Σταθερότητα, χαμηλή συγγένεια για Ο2Εύκαμπτη, υψηλή συγγένεια για Ο2) Λόγο των
ΑΙΜΟΣΦΑΙΡΙΝΗ (ΑΜΦ) ΑΙΜΟΣΦΑΙΡΙΝΗ: Hb, είναι τετραμερής πρωτείνη. ΜΕΤΑΠΤΩΣΗ ΑΠΟ Τ R ΔΕΟΞΥΑΙΜΟΣΦΑΙΡΙΝΗ ΟΞΥΑΙΜΟΣΦΑΙΡΙΝΗ (Σταθερότητα, χαμηλή συγγένεια για Ο2Εύκαμπτη, υψηλή συγγένεια για Ο2) Λόγο των
ΓΩΝΙΕΣ φ, ψ ΚΑΙ ΕΠΙΤΡΕΠΤΕΣ ΔΙΑΜΟΡΦΩΣΕΙΣ ΤΗΣ ΠΟΛΥΠΕΠΤΙΔΙΚΗΣ ΑΛΥΣΙΔΑΣ
 ΓΩΝΙΕΣ φ, ψ ΚΑΙ ΕΠΙΤΡΕΠΤΕΣ ΔΙΑΜΟΡΦΩΣΕΙΣ ΤΗΣ ΠΟΛΥΠΕΠΤΙΔΙΚΗΣ ΑΛΥΣΙΔΑΣ ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ φ φ Ccarbonyl n Ccarbonyl n N Cα n Ccarbonyl n-1 Cα n N φ Ccarbonyl n-1 ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ ψ φ ψ Ccarbonyl n N (Ca
ΓΩΝΙΕΣ φ, ψ ΚΑΙ ΕΠΙΤΡΕΠΤΕΣ ΔΙΑΜΟΡΦΩΣΕΙΣ ΤΗΣ ΠΟΛΥΠΕΠΤΙΔΙΚΗΣ ΑΛΥΣΙΔΑΣ ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ φ φ Ccarbonyl n Ccarbonyl n N Cα n Ccarbonyl n-1 Cα n N φ Ccarbonyl n-1 ΜΕΤΡΗΣΗ ΤΗΣ ΓΩΝΙΑΣ ψ φ ψ Ccarbonyl n N (Ca
Χαρακτηριστικά της δομής της Μυοσφαιρίνης
 Χαρακτηριστικά της δομής της Μυοσφαιρίνης Η μυοσφαιρίνη είναι ενα εξαιρετικά συμπαγές μόριο.οι διαστάσεις είναι 45Χ35Χ25 Α και υπαρχει πολύ λίγος αδειος χώρος στο εσωτερικό τού μορίου Γυρω στα 75% της
Χαρακτηριστικά της δομής της Μυοσφαιρίνης Η μυοσφαιρίνη είναι ενα εξαιρετικά συμπαγές μόριο.οι διαστάσεις είναι 45Χ35Χ25 Α και υπαρχει πολύ λίγος αδειος χώρος στο εσωτερικό τού μορίου Γυρω στα 75% της
Οργανική Χημεία. Κεφάλαια 12 &13: Φασματοσκοπία μαζών και υπερύθρου
 Οργανική Χημεία Κεφάλαια 12 &13: Φασματοσκοπία μαζών και υπερύθρου 1. Γενικά Δυνατότητα προσδιορισμού δομών με σαφήνεια χρησιμοποιώντας τεχνικές φασματοσκοπίας Φασματοσκοπία μαζών Μέγεθος, μοριακός τύπος
Οργανική Χημεία Κεφάλαια 12 &13: Φασματοσκοπία μαζών και υπερύθρου 1. Γενικά Δυνατότητα προσδιορισμού δομών με σαφήνεια χρησιμοποιώντας τεχνικές φασματοσκοπίας Φασματοσκοπία μαζών Μέγεθος, μοριακός τύπος
ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΠΥΡΗΝΙΚΟΥ ΜΑΓΝΗΤΙΚΟΥ ΣΥΝΤΟΝΙΣΜΟΥ ΦΑΣΜΑΤΑ 1 H-NMR. Επίκουρος καθηγητής Ν. Αλιγιάννης
 ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΠΥΡΗΝΙΚΟΥ ΜΑΓΝΗΤΙΚΟΥ ΣΥΝΤΟΝΙΣΜΟΥ ΦΑΣΜΑΤΑ 1 -NMR Επίκουρος καθηγητής Ν. Αλιγιάννης Εισαγωγή Η φασματοσκοπία πυρηνικού μαγνητικού συντονισμού NMR (Nuclear Magnetic Resonance) αποτελεί ένα είδος
ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΠΥΡΗΝΙΚΟΥ ΜΑΓΝΗΤΙΚΟΥ ΣΥΝΤΟΝΙΣΜΟΥ ΦΑΣΜΑΤΑ 1 -NMR Επίκουρος καθηγητής Ν. Αλιγιάννης Εισαγωγή Η φασματοσκοπία πυρηνικού μαγνητικού συντονισμού NMR (Nuclear Magnetic Resonance) αποτελεί ένα είδος
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ
 ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ") ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2017-2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1. Σε ποια μορφή της αιμοσφαιρίνης συνδέεται το 2,3, BPG, ποιο είναι το ηλεκτρικό του φορτίο, με
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2017-2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1. Σε ποια μορφή της αιμοσφαιρίνης συνδέεται το 2,3, BPG, ποιο είναι το ηλεκτρικό του φορτίο, με
Άσκηση 7. Προσομοίωση 3D Δομών Βιομορίων μέσω. Ομολογίας & Threading
 Άσκηση 7 Προσομοίωση 3D Δομών Βιομορίων μέσω Ομολογίας & Threading Προσομοίωση 2ταγούς δομής πρωτεϊνών Δευτεροταγής Δομή: Η 2ταγής δομή των πρωτεϊνών είναι σταθερή τοπική διαμόρφωση της πολυπεπτιδικής
Άσκηση 7 Προσομοίωση 3D Δομών Βιομορίων μέσω Ομολογίας & Threading Προσομοίωση 2ταγούς δομής πρωτεϊνών Δευτεροταγής Δομή: Η 2ταγής δομή των πρωτεϊνών είναι σταθερή τοπική διαμόρφωση της πολυπεπτιδικής
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ
 ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ") ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΘΕΜΑΤΑ 1.Πώς οι κινητικές παράμετροι Κ m και K cat χρησιμεύουν για να συγκριθεί η ανακύκλωση διαφορετικών
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2014-2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ ΘΕΜΑΤΑ 1.Πώς οι κινητικές παράμετροι Κ m και K cat χρησιμεύουν για να συγκριθεί η ανακύκλωση διαφορετικών
τα βιβλία των επιτυχιών
 Τα βιβλία των Εκδόσεων Πουκαμισάς συμπυκνώνουν την πολύχρονη διδακτική εμπειρία των συγγραφέων μας και αποτελούν το βασικό εκπαιδευτικό υλικό που χρησιμοποιούν οι μαθητές των φροντιστηρίων μας. Μέσα από
Τα βιβλία των Εκδόσεων Πουκαμισάς συμπυκνώνουν την πολύχρονη διδακτική εμπειρία των συγγραφέων μας και αποτελούν το βασικό εκπαιδευτικό υλικό που χρησιμοποιούν οι μαθητές των φροντιστηρίων μας. Μέσα από
ΠΡΩΤΕΪΝΕΣ. Φατούρος Ιωάννης Αναπληρωτής Καθηγητής
 ΠΡΩΤΕΪΝΕΣ Φατούρος Ιωάννης Αναπληρωτής Καθηγητής Θέματα Διάλεξης Δομή, αριθμός και διαχωρισμός των αμινοξέων Ένωση αμινοξέων με τον πεπτιδικό δεσμό για τη δημιουργία πρωτεΐνης Λειτουργίες των πρωτεϊνών
ΠΡΩΤΕΪΝΕΣ Φατούρος Ιωάννης Αναπληρωτής Καθηγητής Θέματα Διάλεξης Δομή, αριθμός και διαχωρισμός των αμινοξέων Ένωση αμινοξέων με τον πεπτιδικό δεσμό για τη δημιουργία πρωτεΐνης Λειτουργίες των πρωτεϊνών
Φυσική Στερεών στις Πρωτεΐνες
 Φυσική Στερεών στις Πρωτεΐνες Νίκος Απ. Παπανδρέου Τ.Ε.Ι. Πειραιά Φεβρουάριος 2010 Ένα ελικοϊδές μονοπάτι Χημική δομή μίας πρωτεΐνης Μήκος αλυσίδας ~30 έως ~1000 αµινοξέα Συνολικός αριθµός ατόµων έως ~
Φυσική Στερεών στις Πρωτεΐνες Νίκος Απ. Παπανδρέου Τ.Ε.Ι. Πειραιά Φεβρουάριος 2010 Ένα ελικοϊδές μονοπάτι Χημική δομή μίας πρωτεΐνης Μήκος αλυσίδας ~30 έως ~1000 αµινοξέα Συνολικός αριθµός ατόµων έως ~
Περίληψη 1 ου Κεφαλαίου
 Περίληψη 1 ου Κεφαλαίου Άτοµο: θετικά φορτισµένος πυρήνας περικυκλωµένος από αρνητικά φορτισµένα ηλεκτρόνια Ηλεκτρονική δοµή ατόµου περιγράφεται από κυµατοσυνάρτηση Ηλεκτρόνια καταλαµβάνουν τροχιακά γύρω
Περίληψη 1 ου Κεφαλαίου Άτοµο: θετικά φορτισµένος πυρήνας περικυκλωµένος από αρνητικά φορτισµένα ηλεκτρόνια Ηλεκτρονική δοµή ατόµου περιγράφεται από κυµατοσυνάρτηση Ηλεκτρόνια καταλαµβάνουν τροχιακά γύρω
αποτελούν το 96% κ.β Ποικιλία λειτουργιών
 ΧΗΜΙΚΑ ΣΤΟΙΧΕΙΑ ΠΟΥ ΣΥΝΘΕΤΟΥΝ ΤΟΥΣ ΟΡΓΑΝΙΣΜΟΥΣ 92 στοιχεία στο φλοιό της Γης 27 απαραίτητα για τη ζωή H, Ο, Ν, C αποτελούν το 96% κ.β S, Ca, P, Cl, K, Na, Mg αποτελούν το 4% κ.β. Fe, I Ιχνοστοιχεία αποτελούν
ΧΗΜΙΚΑ ΣΤΟΙΧΕΙΑ ΠΟΥ ΣΥΝΘΕΤΟΥΝ ΤΟΥΣ ΟΡΓΑΝΙΣΜΟΥΣ 92 στοιχεία στο φλοιό της Γης 27 απαραίτητα για τη ζωή H, Ο, Ν, C αποτελούν το 96% κ.β S, Ca, P, Cl, K, Na, Mg αποτελούν το 4% κ.β. Fe, I Ιχνοστοιχεία αποτελούν
ΚΕΦΑΛΑΙΟ 10. Στρατηγικές ρύθμισης
 ΚΕΦΑΛΑΙΟ 10. Στρατηγικές ρύθμισης Oι μεταβολικές πορείες, όπως και η κυκλοφοριακή κίνηση ρυθμίζονται από σήματα. Η CTP, το τελικό προϊόν μιας πορείας πολλών βημάτων, ελέγχει τη ροή των αντιδράσεων σύνθεσής
ΚΕΦΑΛΑΙΟ 10. Στρατηγικές ρύθμισης Oι μεταβολικές πορείες, όπως και η κυκλοφοριακή κίνηση ρυθμίζονται από σήματα. Η CTP, το τελικό προϊόν μιας πορείας πολλών βημάτων, ελέγχει τη ροή των αντιδράσεων σύνθεσής
ΣΥΝΟΨΗ ΠΑΡΑΓΩΓΗΣ ΕΝΕΡΓΕΙΑΣ
 ΣΥΝΟΨΗ ΠΑΡΑΓΩΓΗΣ ΕΝΕΡΓΕΙΑΣ ΤΡΟΦΗ Λίπη Πολυσακχαρίτες Γλυκόζη κι άλλα σάκχαρα Πρωτεΐνες Αμινοξέα Λιπαρά Οξέα Γλυκόλυση Πυροσταφυλικό Οξύ Ακέτυλο-CoA Αναπνευστική Αλυσίδα μεταφοράς ηλεκτρονίων / Οξειδωτική
ΣΥΝΟΨΗ ΠΑΡΑΓΩΓΗΣ ΕΝΕΡΓΕΙΑΣ ΤΡΟΦΗ Λίπη Πολυσακχαρίτες Γλυκόζη κι άλλα σάκχαρα Πρωτεΐνες Αμινοξέα Λιπαρά Οξέα Γλυκόλυση Πυροσταφυλικό Οξύ Ακέτυλο-CoA Αναπνευστική Αλυσίδα μεταφοράς ηλεκτρονίων / Οξειδωτική
NMR - πορφυρινών v=(γ/2π)(1-σ).ηο σ=σταθερά προστασίας
(1-σ).ηο σ=σταθερά προστασίας") NMR - πορφυρινών B r Σ v=(γ/2π)(1-σ).ηο σ=σταθερά προστασίας θ i μ H Κύριο χαρακτηριστικό του πορφυρινικού δακτυλίου είναι η μεγάλη μαγνητική ανισοτροπία που προκαλείται από το π ηλεκτρονιακό νέφος του
NMR - πορφυρινών B r Σ v=(γ/2π)(1-σ).ηο σ=σταθερά προστασίας θ i μ H Κύριο χαρακτηριστικό του πορφυρινικού δακτυλίου είναι η μεγάλη μαγνητική ανισοτροπία που προκαλείται από το π ηλεκτρονιακό νέφος του
ΦΥΣΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗΝ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ ΦΥΣΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗΝ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ Φωτοηλεκτρονιακή φασματοσκοπία ΠΕΡΙΚΛΗΣ ΑΚΡΙΒΟΣ Τμήμα Χημείας Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ ΦΥΣΙΚΕΣ ΜΕΘΟΔΟΙ ΣΤΗΝ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ Φωτοηλεκτρονιακή φασματοσκοπία ΠΕΡΙΚΛΗΣ ΑΚΡΙΒΟΣ Τμήμα Χημείας Άδειες Χρήσης Το παρόν εκπαιδευτικό
Ενόργανη Ανάλυση Εργαστήριο. Φασματοσκοπία πυρηνικού μαγνητικού συντονισμού Nuclear Magnetic Resonance spectroscopy, NMR. Πέτρος Α.
 Ενόργανη Ανάλυση Εργαστήριο Φασματοσκοπία πυρηνικού μαγνητικού συντονισμού Πέτρος Α. Ταραντίλης 1 Βασικές αρχές Που βασίζεται; Στη μέτρηση της απορρόφησης της ακτινοβολίας στην περιοχή των ραδιοσυχνοτήτων
Ενόργανη Ανάλυση Εργαστήριο Φασματοσκοπία πυρηνικού μαγνητικού συντονισμού Πέτρος Α. Ταραντίλης 1 Βασικές αρχές Που βασίζεται; Στη μέτρηση της απορρόφησης της ακτινοβολίας στην περιοχή των ραδιοσυχνοτήτων
ΕΞΕΤΑΣΕΙΣ ΣΤΗ ΓΕΝΙΚΗ ΧΗΜΕΙΑ ΘΕΜΑΤΑ
 ΕΞΕΤΑΣΕΙΣ ΣΤΗ ΓΕΝΙΚΗ ΧΗΜΕΙΑ ΘΕΜΑΤΑ 1. Ο άργυρος εμφανίζεται στη φύση υπό τη μορφή δύο ισοτόπων τα οποία έχουν ατομικές μάζες 106,905 amu και 108,905 amu. (α) Γράψτε το σύμβολο για καθένα ισότοπο του αργύρου
ΕΞΕΤΑΣΕΙΣ ΣΤΗ ΓΕΝΙΚΗ ΧΗΜΕΙΑ ΘΕΜΑΤΑ 1. Ο άργυρος εμφανίζεται στη φύση υπό τη μορφή δύο ισοτόπων τα οποία έχουν ατομικές μάζες 106,905 amu και 108,905 amu. (α) Γράψτε το σύμβολο για καθένα ισότοπο του αργύρου
Θεώρημα Jahn Teller: Μια Απλουστευμένη Προσέγγιση
 Θεώρημα Jahn Teller: Μια Απλουστευμένη Προσέγγιση 1. Τι λέει το Θεώρημα Jahn Teller; Το φαινόμενο Jahn Teller, γνωστό και ως παραμόρφωση Jahn Teller, περιγράφει τη γεωμετρική παραμόρφωση που υφίστανται
Θεώρημα Jahn Teller: Μια Απλουστευμένη Προσέγγιση 1. Τι λέει το Θεώρημα Jahn Teller; Το φαινόμενο Jahn Teller, γνωστό και ως παραμόρφωση Jahn Teller, περιγράφει τη γεωμετρική παραμόρφωση που υφίστανται
Φασματοσκοπίας UV/ορατού Φασματοσκοπίας υπερύθρου Φασματοσκοπίας άπω υπερύθρου / μικροκυμάτων Φασματοσκοπίας φθορισμού Φασματοσκοπίας NMR
 Φασματοσκοπία Ερμηνεία & εφαρμογές : Φασματοσκοπίας UV/ορατού Φασματοσκοπίας υπερύθρου Φασματοσκοπίας άπω υπερύθρου / μικροκυμάτων Φασματοσκοπίας φθορισμού Φασματοσκοπίας NMR Ποια φαινόμενα παράγουν τα
Φασματοσκοπία Ερμηνεία & εφαρμογές : Φασματοσκοπίας UV/ορατού Φασματοσκοπίας υπερύθρου Φασματοσκοπίας άπω υπερύθρου / μικροκυμάτων Φασματοσκοπίας φθορισμού Φασματοσκοπίας NMR Ποια φαινόμενα παράγουν τα
BIOΛ154 ΒΙΟΧΗΜΕΙΑ. ΒΙΟΧΗΜΕΙΑ (Lubert Stryer)
") BIOΛ154 ΒΙΟΧΗΜΕΙΑ ΒΙΟΧΗΜΕΙΑ (Lubert Stryer) ΠΕΡΙΓΡΑΜΜΑ 18.1 H οξειδωτική φωσφορυλίωση στους ευκαρυωτικούς οργανισμούς λαμβάνει χώρα στα μιτοχόνδρια. 18.2 H οξειδωτική φωσφορυλίωση εξαρτάται από τη μεταφορά
BIOΛ154 ΒΙΟΧΗΜΕΙΑ ΒΙΟΧΗΜΕΙΑ (Lubert Stryer) ΠΕΡΙΓΡΑΜΜΑ 18.1 H οξειδωτική φωσφορυλίωση στους ευκαρυωτικούς οργανισμούς λαμβάνει χώρα στα μιτοχόνδρια. 18.2 H οξειδωτική φωσφορυλίωση εξαρτάται από τη μεταφορά
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής
 Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
και χρειάζεται μέσα στο ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. ενζύμων κύτταρο τρόπους
 Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
 Η πεπτιδυλοτρανσφεράση είναι τό ενζυμο το οποίο καταλύει τον σχηματισμό του πεπτιδικού δεσμού.το ενζυμο διερευνάται εντατικά τα τελευταία 30 χρόνια και εχουν αναπτυχθεί ποικίλες απόψεις οσον αφορά την
Η πεπτιδυλοτρανσφεράση είναι τό ενζυμο το οποίο καταλύει τον σχηματισμό του πεπτιδικού δεσμού.το ενζυμο διερευνάται εντατικά τα τελευταία 30 χρόνια και εχουν αναπτυχθεί ποικίλες απόψεις οσον αφορά την
Εξερευνώντας την Εξέλιξη Κεφάλαιο 7
 Εξερευνώντας την Εξέλιξη Κεφάλαιο 7 Εξερευνώντας την Εξέλιξη Σχέση μεταξύ αλληλουχίας αμινοξέων, δομής και λειτουργίας πρωτεϊνών Καταγωγή από έναν κοινό πρόγονο Εξελικτική Συγγένεια/Προέλευση Δύο ομάδες
Εξερευνώντας την Εξέλιξη Κεφάλαιο 7 Εξερευνώντας την Εξέλιξη Σχέση μεταξύ αλληλουχίας αμινοξέων, δομής και λειτουργίας πρωτεϊνών Καταγωγή από έναν κοινό πρόγονο Εξελικτική Συγγένεια/Προέλευση Δύο ομάδες
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Επίδραση και άλλων παραγόντων στην Αλλοστερική συμπεριφορά της Αιμοσφαιρίνης
 Επίδραση και άλλων παραγόντων στην Αλλοστερική συμπεριφορά της Αιμοσφαιρίνης Καθώς το οξυγόνο χρησιμοποιείται στους ιστούς παράγεται CO2 το οποίο πρέπει να μεταφερθεί πίσω στους πνεύμονες ή τα βράγχια
Επίδραση και άλλων παραγόντων στην Αλλοστερική συμπεριφορά της Αιμοσφαιρίνης Καθώς το οξυγόνο χρησιμοποιείται στους ιστούς παράγεται CO2 το οποίο πρέπει να μεταφερθεί πίσω στους πνεύμονες ή τα βράγχια
οµή και Αναδίπλωση πρωτεϊνών
 οµή και Αναδίπλωση πρωτεϊνών Νηφόρου Κατερίνα Μεταδιδακτορική Ερευνήτρια, Οµάδα Μοριακής Καρκινογένεσης, Εργ/ριο Ιστολογίας-Εµβρυολογίας, Ιατρική Σχολή Αθηνών Σηµασία των πρωτεϊνών Ενζυµική κατάλυση Μεταφορά
οµή και Αναδίπλωση πρωτεϊνών Νηφόρου Κατερίνα Μεταδιδακτορική Ερευνήτρια, Οµάδα Μοριακής Καρκινογένεσης, Εργ/ριο Ιστολογίας-Εµβρυολογίας, Ιατρική Σχολή Αθηνών Σηµασία των πρωτεϊνών Ενζυµική κατάλυση Μεταφορά
Θεωρία του δεσμού σθένους
 ΣΚΟΠΟΣ Ο σκοπός αυτού του κεφαλαίου είναι να γνωρίσουμε μια αρκετά απλή θεωρία, τη θεωρία του δεσμού σθένους, με την οποία θα μπορούμε να εξηγούμε με αρκετή επιτυχία τη γεωμετρία των συμπλόκων, καθώς και
ΣΚΟΠΟΣ Ο σκοπός αυτού του κεφαλαίου είναι να γνωρίσουμε μια αρκετά απλή θεωρία, τη θεωρία του δεσμού σθένους, με την οποία θα μπορούμε να εξηγούμε με αρκετή επιτυχία τη γεωμετρία των συμπλόκων, καθώς και
Εθνικόν και Καποδιστριακόν Πανεπιστήμιον Αθηνών ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ Τομέας Φαρμακευτικής Χημείας. Ιωάννης Ντότσικας. Επικ.
 Εθνικόν και Καποδιστριακόν Πανεπιστήμιον Αθηνών ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ Τομέας Φαρμακευτικής Χημείας Ιωάννης Ντότσικας Επικ. Καθηγητής 1 Οι κυκλοδεξτρίνες (Cyclodextrins, CDs) είναι κυκλικοί ολιγοσακχαρίτες
Εθνικόν και Καποδιστριακόν Πανεπιστήμιον Αθηνών ΤΜΗΜΑ ΦΑΡΜΑΚΕΥΤΙΚΗΣ Τομέας Φαρμακευτικής Χημείας Ιωάννης Ντότσικας Επικ. Καθηγητής 1 Οι κυκλοδεξτρίνες (Cyclodextrins, CDs) είναι κυκλικοί ολιγοσακχαρίτες
Μικρά αμινοξέα. Βιοχημεία Ι Β-3
 Βιοχημεία Ι Β-2 Μικρά αμινοξέα Βιοχημεία Ι Β-3 Aλειφατικά αμινοξέα Βιοχημεία Ι Β-4 Ιμινοξύ Βιοχημεία Ι Β-5 Αρωματικά αμινοξέα Βιοχημεία Ι Β-6 Βιοχημεία Ι Β-7 Η Tyr και η Trp απορροφούν στα 280nm-έτσι μετράται
Βιοχημεία Ι Β-2 Μικρά αμινοξέα Βιοχημεία Ι Β-3 Aλειφατικά αμινοξέα Βιοχημεία Ι Β-4 Ιμινοξύ Βιοχημεία Ι Β-5 Αρωματικά αμινοξέα Βιοχημεία Ι Β-6 Βιοχημεία Ι Β-7 Η Tyr και η Trp απορροφούν στα 280nm-έτσι μετράται
Εργασία Βιολογίας. Β. Γιώργος. Εισαγωγή 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ. Μεταφορά ενέργειας στα κύτταρα
 Εργασία Βιολογίας Β. Γιώργος Εισαγωγή Η ενεργεια εχει πολυ μεγαλη σημασια για εναν οργανισμο, γιατι για να κανει οτιδηποτε ενας οργανισμος ειναι απαραιτητη. Ειναι απαραιτητη ακομη και οταν δεν κανουμε
Εργασία Βιολογίας Β. Γιώργος Εισαγωγή Η ενεργεια εχει πολυ μεγαλη σημασια για εναν οργανισμο, γιατι για να κανει οτιδηποτε ενας οργανισμος ειναι απαραιτητη. Ειναι απαραιτητη ακομη και οταν δεν κανουμε
Βασικές αρχές της Φασµατοσκοπίας NMR
 Βασικές αρχές της Φασµατοσκοπίας NMR Φώτης Νταής Καθηγητής Πανεπιστηµίου Κρήτης, Τµήµα Χηµείας Φασµατοσκοπία NMR Ο Πυρηνικός µαγνητικός Συντονισµός (NMR) είναι ένα φαινόµενο που συµβαίνει όταν πυρήνες
Βασικές αρχές της Φασµατοσκοπίας NMR Φώτης Νταής Καθηγητής Πανεπιστηµίου Κρήτης, Τµήµα Χηµείας Φασµατοσκοπία NMR Ο Πυρηνικός µαγνητικός Συντονισµός (NMR) είναι ένα φαινόµενο που συµβαίνει όταν πυρήνες
Δομή και λειτουργία πρωτεϊνών. Το κύριο δομικό συστατικό των κυττάρων. Το κύριο λειτουργικό μόριο
 Δομή και λειτουργία πρωτεϊνών Το κύριο δομικό συστατικό των κυττάρων. Το κύριο λειτουργικό μόριο Πρωτεΐνες Κύριο συστατικό κυττάρου Δομικοί λίθοι αλλά και επιτελεστές λειτουργίας κυττάρου ένζυμα, μεταφορά,
Δομή και λειτουργία πρωτεϊνών Το κύριο δομικό συστατικό των κυττάρων. Το κύριο λειτουργικό μόριο Πρωτεΐνες Κύριο συστατικό κυττάρου Δομικοί λίθοι αλλά και επιτελεστές λειτουργίας κυττάρου ένζυμα, μεταφορά,
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ
 ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ") ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2016-2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1) Δίνονται τα αμινοξέα σερίνη, αλανίνη, γλουταμικό, κυστεΐνη, λυσίνη, αργινίνη, διαχωρίστε τα
ΙΑΤΡΙΚΗ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΑΘΗΝΩΝ (ΕΚΠΑ) ΚΑΤΑΤΑΚΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΑΚ.ΕΤΟΥΣ 2016-2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΧΗΜΕΙΑ 1) Δίνονται τα αμινοξέα σερίνη, αλανίνη, γλουταμικό, κυστεΐνη, λυσίνη, αργινίνη, διαχωρίστε τα
BIOΛ154 ΒΙΟΧΗΜΕΙΑ Ι. ΒΙΟΧΗΜΕΙΑ (Lubert Stryer)
") BIOΛ154 ΒΙΟΧΗΜΕΙΑ Ι ΒΙΟΧΗΜΕΙΑ (Lubert Stryer) ΠΕΡΙΓΡΑΜΜΑ 17.1 Η πυροσταφυλική αφυδρογονάση συνδέει τη γλυκόλυση με τον κύκλο του κιτρικού οξέος 17.2 O κύκλος του κιτρικού οξέος οξειδώνει μονάδες δύο ατόμων
BIOΛ154 ΒΙΟΧΗΜΕΙΑ Ι ΒΙΟΧΗΜΕΙΑ (Lubert Stryer) ΠΕΡΙΓΡΑΜΜΑ 17.1 Η πυροσταφυλική αφυδρογονάση συνδέει τη γλυκόλυση με τον κύκλο του κιτρικού οξέος 17.2 O κύκλος του κιτρικού οξέος οξειδώνει μονάδες δύο ατόμων
Ca. Να μεταφέρετε στην κόλλα σας συμπληρωμένο τον παρακάτω πίνακα που αναφέρεται στο άτομο του ασβεστίου: ΣΤΙΒΑΔΕΣ νετρόνια K L M N Ca 2
 Ερωτήσεις Ανάπτυξης 1. Δίνεται ότι: 40 20 Ca. Να μεταφέρετε στην κόλλα σας συμπληρωμένο τον παρακάτω πίνακα που αναφέρεται στο άτομο του ασβεστίου: ΣΤΙΒΑΔΕΣ νετρόνια K L M N Ca 2 2. Tι είδους δεσμός αναπτύσσεται
Ερωτήσεις Ανάπτυξης 1. Δίνεται ότι: 40 20 Ca. Να μεταφέρετε στην κόλλα σας συμπληρωμένο τον παρακάτω πίνακα που αναφέρεται στο άτομο του ασβεστίου: ΣΤΙΒΑΔΕΣ νετρόνια K L M N Ca 2 2. Tι είδους δεσμός αναπτύσσεται
KΕΦΑΛΑΙΟ 1ο Χημική σύσταση του κυττάρου. Να απαντήσετε σε καθεμιά από τις παρακάτω ερωτήσεις με μια πρόταση:
 KΕΦΑΛΑΙΟ 1ο Χημική σύσταση του κυττάρου Ενότητα 1.1: Χημεία της ζωής Ενότητα 2.1: Μακρομόρια Να απαντήσετε σε καθεμιά από τις παρακάτω ερωτήσεις με μια πρόταση: 1. Για ποιο λόγο θεωρείται αναγκαία η σταθερότητα
KΕΦΑΛΑΙΟ 1ο Χημική σύσταση του κυττάρου Ενότητα 1.1: Χημεία της ζωής Ενότητα 2.1: Μακρομόρια Να απαντήσετε σε καθεμιά από τις παρακάτω ερωτήσεις με μια πρόταση: 1. Για ποιο λόγο θεωρείται αναγκαία η σταθερότητα
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ ΤΟΜΕΑΣ ΑΝΟΡΓΑΝΗΣ ΧΗΜΕΙΑΣ ΜΕΝΕΛΑΟΥ ΜΕΛΙΤΑ. Διπλωματούχου Χημικού Μηχανικού
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ ΤΟΜΕΑΣ ΑΝΟΡΓΑΝΗΣ ΧΗΜΕΙΑΣ ΜΕΝΕΛΑΟΥ ΜΕΛΙΤΑ Διπλωματούχου Χημικού Μηχανικού Μελέτη της δομικής ειδογένεσης και των φθορισμομετρικών ιδιοτήτων
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΧΗΜΙΚΩΝ ΜΗΧΑΝΙΚΩΝ ΤΟΜΕΑΣ ΑΝΟΡΓΑΝΗΣ ΧΗΜΕΙΑΣ ΜΕΝΕΛΑΟΥ ΜΕΛΙΤΑ Διπλωματούχου Χημικού Μηχανικού Μελέτη της δομικής ειδογένεσης και των φθορισμομετρικών ιδιοτήτων
Ανόργανη Χημεία. Τμήμα Τεχνολογίας Τροφίμων. Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή. Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής
 Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ο Ομοιοπολικός Δεσμός 2 Ο δεσμός Η Η στο μόριο Η
Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 5 η : Ομοιοπολικοί δεσμοί & μοριακή δομή Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Ο Ομοιοπολικός Δεσμός 2 Ο δεσμός Η Η στο μόριο Η
ΦΑΣΜΑΤΟΣΚΟΠΙΑ IR/NMR
 ΦΑΣΜΑΤΟΣΚΟΠΙΑ IR/NMR ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΤΥΠΟΙ Φασματοσκοπία Μάζας (Ms) προσδιορισμός μεγέθους και μοριακού βάρους Φασματοσκοπία Υπερύθρου (UV) προσδιορισμός π συζυγιακού συστήματος Φασματοσκοπία Υπεριώδους
ΦΑΣΜΑΤΟΣΚΟΠΙΑ IR/NMR ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΤΥΠΟΙ Φασματοσκοπία Μάζας (Ms) προσδιορισμός μεγέθους και μοριακού βάρους Φασματοσκοπία Υπερύθρου (UV) προσδιορισμός π συζυγιακού συστήματος Φασματοσκοπία Υπεριώδους
Δελτίο μαθήματος (Syllabus): ΓΕΝΙΚΗ ΚΑΙ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ
: ΓΕΝΙΚΗ ΚΑΙ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ") Δελτίο μαθήματος (Syllabus): ΓΕΝΙΚΗ ΚΑΙ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ Κωδικός μαθήματος: ΝΠ-01 Κύκλος/Επίπεδο σπουδών: Προπτυχιακό Εξάμηνο σπουδών: Πρώτο (1 ο ) Τύπος μαθήματος Χ Υποβάθρου / Γενικών Γνώσεων Επιστημονικής
Δελτίο μαθήματος (Syllabus): ΓΕΝΙΚΗ ΚΑΙ ΑΝΟΡΓΑΝΗ ΧΗΜΕΙΑ Κωδικός μαθήματος: ΝΠ-01 Κύκλος/Επίπεδο σπουδών: Προπτυχιακό Εξάμηνο σπουδών: Πρώτο (1 ο ) Τύπος μαθήματος Χ Υποβάθρου / Γενικών Γνώσεων Επιστημονικής
Τα χημικά στοιχεία που είναι επικρατέστερα στους οργανισμούς είναι: i..
 ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 1 ο ΚΕΦΑΛΑΙΟ «XHMIKH ΣΥΣΤΑΣΗ ΤΟΥ ΚΥΤΤΑΡΟΥ» ΕΙΣΑΓΩΓΗ ΚΑΙ Η ΧΗΜΕΙΑ ΤΗΣ ΖΩΗΣ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Όταν αναφερόμαστε στον όρο «Χημική Σύσταση του Κυττάρου», τί νομίζετε ότι
ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 1 ο ΚΕΦΑΛΑΙΟ «XHMIKH ΣΥΣΤΑΣΗ ΤΟΥ ΚΥΤΤΑΡΟΥ» ΕΙΣΑΓΩΓΗ ΚΑΙ Η ΧΗΜΕΙΑ ΤΗΣ ΖΩΗΣ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Όταν αναφερόμαστε στον όρο «Χημική Σύσταση του Κυττάρου», τί νομίζετε ότι
ΣΗΜΕΙΩΣΕΙΣ ΚΑΘΗΓΗΤΗ κ. ΚΟΥΠΠΑΡΗ
 ΣΗΜΕΙΩΣΕΙΣ ΚΑΘΗΓΗΤΗ κ. ΚΟΥΠΠΑΡΗ 1 2 3 4 5 6 7 8 9 10 11 Σημειώσεις από τα μαθήματα Φαρμακευτικής Ανάλυσης του καθηγητή κ. Ιωάννη Κουντουρέλλη ΑΝΙΣΟΤΡΟΠΙΚΑ ΦΑΙΝΟΜΕΝΑ 12 13 Nuclear Magnetic Resonance Spectroscopy
ΣΗΜΕΙΩΣΕΙΣ ΚΑΘΗΓΗΤΗ κ. ΚΟΥΠΠΑΡΗ 1 2 3 4 5 6 7 8 9 10 11 Σημειώσεις από τα μαθήματα Φαρμακευτικής Ανάλυσης του καθηγητή κ. Ιωάννη Κουντουρέλλη ΑΝΙΣΟΤΡΟΠΙΚΑ ΦΑΙΝΟΜΕΝΑ 12 13 Nuclear Magnetic Resonance Spectroscopy
Κεφάλαιο 3 ΜΕΤΑΒΟΛΙΣΜΟΣ
 Κεφάλαιο 3 ΜΕΤΑΒΟΛΙΣΜΟΣ 3.1 Ενέργεια και οργανισμοί Όλοι οι οργανισμοί, εκτός από αυτούς από αυτούς που έχουν την ικανότητα να φωτοσυνθέτουν, εξασφαλίζουν ενέργεια διασπώντας τις θρεπτικές ουσιές που περιέχονται
Κεφάλαιο 3 ΜΕΤΑΒΟΛΙΣΜΟΣ 3.1 Ενέργεια και οργανισμοί Όλοι οι οργανισμοί, εκτός από αυτούς από αυτούς που έχουν την ικανότητα να φωτοσυνθέτουν, εξασφαλίζουν ενέργεια διασπώντας τις θρεπτικές ουσιές που περιέχονται
ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ. Μαντώ Κυριακού 2015
 ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ Μαντώ Κυριακού 2015 Ενεργειακό Στα βιολογικά συστήματα η διατήρηση της ενέργειας συμπεριλαμβάνει οξειδοαναγωγικές αντιδράσεις παραγωγή ATP Οξείδωση: απομάκρυνση e από ένα υπόστρωμα
ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ Μαντώ Κυριακού 2015 Ενεργειακό Στα βιολογικά συστήματα η διατήρηση της ενέργειας συμπεριλαμβάνει οξειδοαναγωγικές αντιδράσεις παραγωγή ATP Οξείδωση: απομάκρυνση e από ένα υπόστρωμα
Καραπέτσας Θανάσης. Διπλωματική Εργασία:
 Διαπανεπιστημιακό Πρόγραμμα Μεταπτυχιακών Σπουδών «Κλινική Φαρμακολογία & Θεραπευτική» Επιβλέπων: Δρ. Αλ. Γαλάνης, Λέκτορας Μορ. Βιολογίας, «Σχεδιασμός Ειδικών Πεπτιδίων Αναστολέων της Αλληλεπίδρασης του
Διαπανεπιστημιακό Πρόγραμμα Μεταπτυχιακών Σπουδών «Κλινική Φαρμακολογία & Θεραπευτική» Επιβλέπων: Δρ. Αλ. Γαλάνης, Λέκτορας Μορ. Βιολογίας, «Σχεδιασμός Ειδικών Πεπτιδίων Αναστολέων της Αλληλεπίδρασης του
Κεντρικό δόγμα της βιολογίας
 Κεντρικό δόγμα της βιολογίας DNA RNA Πρωτεΐνη Μεταγραφή Σύνθεση (μονόκλωνου) RNA από ένα δίκλωνο μόριο DNA κυρίως με τη βοήθεια του ενζύμου RNA πολυμεράση Το προϊόν της μεταγραφής ονομάζεται πρωτογενές
Κεντρικό δόγμα της βιολογίας DNA RNA Πρωτεΐνη Μεταγραφή Σύνθεση (μονόκλωνου) RNA από ένα δίκλωνο μόριο DNA κυρίως με τη βοήθεια του ενζύμου RNA πολυμεράση Το προϊόν της μεταγραφής ονομάζεται πρωτογενές
Αρχιτεκτονική της τρισδιάστατης δομής πρωτεϊνών
 Αρχιτεκτονική της τρισδιάστατης δομής πρωτεϊνών Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου Ταχ.Κιβ. 20537 1678,
Αρχιτεκτονική της τρισδιάστατης δομής πρωτεϊνών Βασίλης Προμπονάς, PhD Ερευνητικό Εργαστήριο Βιοπληροφορικής Τμήμα Βιολογικών Επιστημών Νέα Παν/πολη, Γραφείο B161 Πανεπιστήμιο Κύπρου Ταχ.Κιβ. 20537 1678,
Γιατί ο σχηματισμός του CΗ 4 δεν μπορεί να ερμηνευθεί βάσει της διεγερμένης κατάστασης του ατόμου C;
 Γιατί ο σχηματισμός του CΗ 4 δεν μπορεί να ερμηνευθεί βάσει της διεγερμένης κατάστασης του ατόμου C; 1. Οι 4 ομοιοπολικοί δεσμοί στο μεθάνιο θα ήταν δύο τύπων: ένας δεσμός από την επικάλυψη του τροχιακού
Γιατί ο σχηματισμός του CΗ 4 δεν μπορεί να ερμηνευθεί βάσει της διεγερμένης κατάστασης του ατόμου C; 1. Οι 4 ομοιοπολικοί δεσμοί στο μεθάνιο θα ήταν δύο τύπων: ένας δεσμός από την επικάλυψη του τροχιακού
3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ
 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Όλοι οι οργανισμοί προκειμένου να επιβιώσουν και να επιτελέσουν τις λειτουργίες τους χρειάζονται ενέργεια. Οι φυτικοί οργανισμοί μετατρέπουν την ηλιακή ενέργεια με τη διαδικασία
3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Όλοι οι οργανισμοί προκειμένου να επιβιώσουν και να επιτελέσουν τις λειτουργίες τους χρειάζονται ενέργεια. Οι φυτικοί οργανισμοί μετατρέπουν την ηλιακή ενέργεια με τη διαδικασία
Χημεία Ενώσεων Συναρμογής
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Χημεία Ενώσεων Συναρμογής Ενότητα 2: Δεσμοί στις Ενώσεις Συναρμογής Περικλής Ακρίβος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Χημεία Ενώσεων Συναρμογής Ενότητα 2: Δεσμοί στις Ενώσεις Συναρμογής Περικλής Ακρίβος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται
Βιολογία Β Λυκείου θέματα
 Ι. Οι υδατάνθρακες διακρίνονται σε μονοσακχαρίτες, δισακχαρίτες και πολυσακχαρίτες. α) Να αναφέρετε από δύο παραδείγματα μονοσακχαριτών, δισακχαριτών και πολυσακχαριτών. (6μ) β) Σε ένα κύτταρο συναντώνται
Ι. Οι υδατάνθρακες διακρίνονται σε μονοσακχαρίτες, δισακχαρίτες και πολυσακχαρίτες. α) Να αναφέρετε από δύο παραδείγματα μονοσακχαριτών, δισακχαριτών και πολυσακχαριτών. (6μ) β) Σε ένα κύτταρο συναντώνται
Μάθημα 20 ο. Το σχήμα των μορίων
 Μάθημα 20 ο Το σχήμα των μορίων Tα μόρια Μπορεί να είναι μη πολικά έστω και άν οι δεσμοί μεταξύ των ατόμων τους είναι πολωμένοι Δεν είναι επίπεδα (έχουν τρισδιάστατη διάταξη στο χώρο) Γενική και Ανόργανη
Μάθημα 20 ο Το σχήμα των μορίων Tα μόρια Μπορεί να είναι μη πολικά έστω και άν οι δεσμοί μεταξύ των ατόμων τους είναι πολωμένοι Δεν είναι επίπεδα (έχουν τρισδιάστατη διάταξη στο χώρο) Γενική και Ανόργανη
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
Προσδιορισμός της Δομής Οργανικών Μορίων
 Προσδιορισμός της Δομής Οργανικών Μορίων Φασματοσκοπία - Φασματοσκοπικές τεχνικές ανάλυσης Πέτρος Ταραντίλης Αναπληρωτής Καθηγητής Φασματοσκοπικές τεχνικές. Υπεριώδους-ορατού (Ultraviolet-Visible UV-Vis)
Προσδιορισμός της Δομής Οργανικών Μορίων Φασματοσκοπία - Φασματοσκοπικές τεχνικές ανάλυσης Πέτρος Ταραντίλης Αναπληρωτής Καθηγητής Φασματοσκοπικές τεχνικές. Υπεριώδους-ορατού (Ultraviolet-Visible UV-Vis)
Συνδυάζοντας το πρώτο και το δεύτερο θερμοδυναμικό αξίωμα προκύπτει ότι:
 Συνδυάζοντας το πρώτο και το δεύτερο θερμοδυναμικό αξίωμα προκύπτει ότι: Για να είναι μια αντίδραση αυθόρμητη, πρέπει η μεταβολή της ελεύθερης ενέργειας της αντίδρασης να είναι αρνητική. Η μεταβολή της
Συνδυάζοντας το πρώτο και το δεύτερο θερμοδυναμικό αξίωμα προκύπτει ότι: Για να είναι μια αντίδραση αυθόρμητη, πρέπει η μεταβολή της ελεύθερης ενέργειας της αντίδρασης να είναι αρνητική. Η μεταβολή της
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
Ανόργανη Χημεία. Τμήμα Τεχνολογίας Τροφίμων. Ενότητα 4 η : Ιοντικοί Δεσμοί Χημεία Κύριων Ομάδων. Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής
 Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 4 η : Ιοντικοί Δεσμοί Χημεία Κύριων Ομάδων Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Δόμηση Ηλεκτρονίων στα Ιόντα 2 Για τα στοιχεία
Τμήμα Τεχνολογίας Τροφίμων Ανόργανη Χημεία Ενότητα 4 η : Ιοντικοί Δεσμοί Χημεία Κύριων Ομάδων Οκτώβριος 2018 Δρ. Δημήτρης Π. Μακρής Αναπληρωτής Καθηγητής Δόμηση Ηλεκτρονίων στα Ιόντα 2 Για τα στοιχεία
Εξερευνώντας τα Βιομόρια Ένζυμα: Βασικές Αρχές και Κινητική
 Εξερευνώντας τα Βιομόρια Ένζυμα: Βασικές Αρχές και Κινητική Βιοχημεία Βιομορίων Αθήνα 2015 Γενικές Ιδιότητες Ένζυμα : Βιολογικοί Καταλύτες Τα ένζυμα είναι πρωτεϊνικά μόρια Μικρή ομάδα καταλυτικών RNA H
Εξερευνώντας τα Βιομόρια Ένζυμα: Βασικές Αρχές και Κινητική Βιοχημεία Βιομορίων Αθήνα 2015 Γενικές Ιδιότητες Ένζυμα : Βιολογικοί Καταλύτες Τα ένζυμα είναι πρωτεϊνικά μόρια Μικρή ομάδα καταλυτικών RNA H
ΦΑΣΜΑΤΑ ΕΚΠΟΜΠΗΣ ΑΠΟΡΡΟΦΗΣΗΣ
 ΚΒΑΝΤΙΚΗ ΦΥΣΙΚΗ: Τα άτομα έχουν διακριτές ενεργειακές στάθμες Τα άτομα και μόρια, βρίσκονται σε διακριτές ενεργειακές στάθμες και Υφίστανται μεταβάσεις μεταξύ αυτών των ενεργειακών σταθμών όταν αλληλεπιδρούν
ΚΒΑΝΤΙΚΗ ΦΥΣΙΚΗ: Τα άτομα έχουν διακριτές ενεργειακές στάθμες Τα άτομα και μόρια, βρίσκονται σε διακριτές ενεργειακές στάθμες και Υφίστανται μεταβάσεις μεταξύ αυτών των ενεργειακών σταθμών όταν αλληλεπιδρούν
Από τα δεδομένα της πρωτοδιάταξης σε συνδυασμό με τα δεδομένα που λαμβάνονται από ανοσοχημικές
 Εν αντιθέσει με αυτά που έχουν βρεθεί για τους προκαρυωτικούς οργανισμούς, για τους ευκαρυωτικούς υπάρχουν διαθέσιμες μόνο λίγες ακολουθίες ριβοσωμικών πρωτεϊνών. Από τα δεδομένα της πρωτοδιάταξης σε συνδυασμό
Εν αντιθέσει με αυτά που έχουν βρεθεί για τους προκαρυωτικούς οργανισμούς, για τους ευκαρυωτικούς υπάρχουν διαθέσιμες μόνο λίγες ακολουθίες ριβοσωμικών πρωτεϊνών. Από τα δεδομένα της πρωτοδιάταξης σε συνδυασμό
πρωτεΐνες πολυμερείς ουσίες δομούν λειτουργούν λευκώματα 1.Απλές πρωτεΐνες 2.Σύνθετες πρωτεΐνες πρωτεΐδια μη πρωτεϊνικό μεταλλοπρωτεΐνες
 ΠΡΩΤΕΙΝΕΣ Οι πρωτεΐνες είναι πολυμερείς ουσίες με κυρίαρχο και πρωταρχικό ρόλο στη ζωή. Πρωτεΐνες είναι οι ουσίες που κυρίως δομούν και λειτουργούν τους οργανισμούς. Λέγονται και λευκώματα λόγω του λευκού
ΠΡΩΤΕΙΝΕΣ Οι πρωτεΐνες είναι πολυμερείς ουσίες με κυρίαρχο και πρωταρχικό ρόλο στη ζωή. Πρωτεΐνες είναι οι ουσίες που κυρίως δομούν και λειτουργούν τους οργανισμούς. Λέγονται και λευκώματα λόγω του λευκού
1. Να οξειδωθούν και να παράγουν ενέργεια. (ΚΑΤΑΒΟΛΙΣΜΟΣ)
") Θάνος Α. Β1 ΠΕΡΙΛΗΨΗ ΚΕΦΑΛΑΙΟ ΤΡΙΤΟ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Όλοι οι οργανισμοί προκειμένου να επιβιώσουν και να επιτελέσουν τις λειτουργίες τους χρειάζονται ενέργεια. Οι φυτικοί οργανισμοί μετατρέπουν
Θάνος Α. Β1 ΠΕΡΙΛΗΨΗ ΚΕΦΑΛΑΙΟ ΤΡΙΤΟ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Όλοι οι οργανισμοί προκειμένου να επιβιώσουν και να επιτελέσουν τις λειτουργίες τους χρειάζονται ενέργεια. Οι φυτικοί οργανισμοί μετατρέπουν
Κεφάλαιο 13 Φασματοσκοπία
 Κεφάλαιο 13 Φασματοσκοπία Φασματοσκοπία υπερύθρου Φασματοσκοπία ορατού-υπεριώδους Φασματοσκοπία πυρηνικού μαγνητικού συντονισμού Φασματοσκοπία μάζας 13.1 Οι αρχές της μοριακής φασματοσκοπίας: Ηλεκτρομαγνητική
Κεφάλαιο 13 Φασματοσκοπία Φασματοσκοπία υπερύθρου Φασματοσκοπία ορατού-υπεριώδους Φασματοσκοπία πυρηνικού μαγνητικού συντονισμού Φασματοσκοπία μάζας 13.1 Οι αρχές της μοριακής φασματοσκοπίας: Ηλεκτρομαγνητική
Οι πρωτεΐνες συμμετέχουν σε όλες τις κυτταρικές λειτουργίες
 Οι πρωτεΐνες συμμετέχουν σε όλες τις κυτταρικές λειτουργίες Γένωμα vs Πρωτέωμα Όλη η αλληλουχία βάσεων στο DNA Τι είναι δυνατόν Συγκεκριμένο Στατικό Οι πρωτεΐνες που κωδικοποιούνται από το γένωμα Τι είναι
Οι πρωτεΐνες συμμετέχουν σε όλες τις κυτταρικές λειτουργίες Γένωμα vs Πρωτέωμα Όλη η αλληλουχία βάσεων στο DNA Τι είναι δυνατόν Συγκεκριμένο Στατικό Οι πρωτεΐνες που κωδικοποιούνται από το γένωμα Τι είναι
Δομές (Διαμορφώσεις) Πρωτεινικών μορίων
 Πρωτεινικών μορίων") Δομές (Διαμορφώσεις) Πρωτεινικών μορίων Πρωτοταγής δομή (αλληλουχία αμινοξέων) Δευτεροταγής δομή Η διάταξη της πεπτιδικής αλυσίδας στον χωρο αυτής καθ αυτής (χωρίς να ληφθούν υπ όψη οι ομάδες R) Τριτοταγής
Δομές (Διαμορφώσεις) Πρωτεινικών μορίων Πρωτοταγής δομή (αλληλουχία αμινοξέων) Δευτεροταγής δομή Η διάταξη της πεπτιδικής αλυσίδας στον χωρο αυτής καθ αυτής (χωρίς να ληφθούν υπ όψη οι ομάδες R) Τριτοταγής
ΧΗΜΙΚΟΣ ΕΣΜΟΣ ΙΙ : ΚΒΑΝΤΟΜΗΧΑΝΙΚΗ ΘΕΩΡΗΣΗ ΤΟΥ ΕΣΜΟΥ
 ΧΗΜΙΚΟΣ ΕΣΜΟΣ ΙΙ : ΚΒΑΝΤΟΜΗΧΑΝΙΚΗ ΘΕΩΡΗΣΗ ΤΟΥ ΟΜΟΙΟΠΟΛΙΚΟΥ Ή ΟΜΟΣΘΕΝΟΥΣ ΕΣΜΟΥ Παππάς Χρήστος Επίκουρος Καθηγητής Το μόριο του Η 2 Σύμφωνα με τη θεωρία του Lewis στο μόριο του Η 2 τα άτομα συγκρατούνται
ΧΗΜΙΚΟΣ ΕΣΜΟΣ ΙΙ : ΚΒΑΝΤΟΜΗΧΑΝΙΚΗ ΘΕΩΡΗΣΗ ΤΟΥ ΟΜΟΙΟΠΟΛΙΚΟΥ Ή ΟΜΟΣΘΕΝΟΥΣ ΕΣΜΟΥ Παππάς Χρήστος Επίκουρος Καθηγητής Το μόριο του Η 2 Σύμφωνα με τη θεωρία του Lewis στο μόριο του Η 2 τα άτομα συγκρατούνται
Λειτουργική Περιοχή της GTP-ασης
 Λειτουργική Περιοχή της GTP-ασης Οι πρωτεΐνες πού φαίνεται να εµπλέκονται στην περιοχή είναι οι πρωτεΐνες L7/L12. Οι πρωτεΐνες αυτές φαίνεται να είναι απαραίτητες για την ενεργότητα του ριβοσώµατος και
Λειτουργική Περιοχή της GTP-ασης Οι πρωτεΐνες πού φαίνεται να εµπλέκονται στην περιοχή είναι οι πρωτεΐνες L7/L12. Οι πρωτεΐνες αυτές φαίνεται να είναι απαραίτητες για την ενεργότητα του ριβοσώµατος και
ΜΕΤΑΦΟΡΑ ΑΕΡΙΩΝ ΠΡΟΣ ΚΑΙ ΑΠΟ ΤΟΥΣ ΣΤΟΥΣ ΙΣΤΟΥΣ
 ΜΕΤΑΦΟΡΑ ΑΕΡΙΩΝ ΠΡΟΣ ΚΑΙ ΑΠΟ ΤΟΥΣ ΣΤΟΥΣ ΙΣΤΟΥΣ Μεταφορά οξυγόνου (Ο 2 ) από τον αέρα μέσω κυψελίδων στο αίμα και ιστούς Μεταφορά διοξειδίου άνθρακα (CO 2 ) από ιστούς σε κυψελίδες Οι κλίσεις των μερικών
ΜΕΤΑΦΟΡΑ ΑΕΡΙΩΝ ΠΡΟΣ ΚΑΙ ΑΠΟ ΤΟΥΣ ΣΤΟΥΣ ΙΣΤΟΥΣ Μεταφορά οξυγόνου (Ο 2 ) από τον αέρα μέσω κυψελίδων στο αίμα και ιστούς Μεταφορά διοξειδίου άνθρακα (CO 2 ) από ιστούς σε κυψελίδες Οι κλίσεις των μερικών
Ασκήσεις. Γράψτε μια δομή Lewis για καθένα από τα παρακάτω μόρια και βρείτε τα τυπικά φορτία των ατόμων. (α) CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3
 CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3") Ασκήσεις Γράψτε μια δομή Lewis για καθένα από τα παρακάτω μόρια και βρείτε τα τυπικά φορτία των ατόμων. (α) CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3 Γεωμετρία Μορίων Θεωρία VSEPR Μοριακή γεωμετρία: είναι η διάταξη
Ασκήσεις Γράψτε μια δομή Lewis για καθένα από τα παρακάτω μόρια και βρείτε τα τυπικά φορτία των ατόμων. (α) CΟ (β) ΗΝO 3 (γ) ClΟ 3 (δ) ΡΟCl 3 Γεωμετρία Μορίων Θεωρία VSEPR Μοριακή γεωμετρία: είναι η διάταξη
Κεφάλαιο 2 Χημικοί Δεσμοί
 Κεφάλαιο 2 Χημικοί Δεσμοί Σύνοψη Παρουσιάζονται οι χημικοί δεσμοί, ιοντικός, μοριακός, ατομικός, μεταλλικός. Οι ιδιότητες των υλικών τόσο οι φυσικές όσο και οι χημικές εξαρτώνται από το είδος ή τα είδη
Κεφάλαιο 2 Χημικοί Δεσμοί Σύνοψη Παρουσιάζονται οι χημικοί δεσμοί, ιοντικός, μοριακός, ατομικός, μεταλλικός. Οι ιδιότητες των υλικών τόσο οι φυσικές όσο και οι χημικές εξαρτώνται από το είδος ή τα είδη
ΘΕΜΑΤΑ ΑΠΟ ΠΜΔΧ ΣΧΕΤΙΚΑ ΜΕ ΤΟ 1 ΚΕΦΑΛΑΙΟ ΤΗΣ Γ ΛΥΚΕΙΟΥ
 ΘΕΜΑΤΑ ΑΠΟ ΠΜΔΧ ΣΧΕΤΙΚΑ ΜΕ ΤΟ ΚΕΦΑΛΑΙΟ ΤΗΣ Γ ΛΥΚΕΙΟΥ 27 ος ΠΜΔΧ Γ ΛΥΚΕΙΟΥ 30 03 203. Στοιχείο Μ το οποίο ανήκει στην πρώτη σειρά στοιχείων μετάπτωσης, σχηματίζει ιόν Μ 3+, που έχει 3 ηλεκτρόνια στην υποστιβάδα
ΘΕΜΑΤΑ ΑΠΟ ΠΜΔΧ ΣΧΕΤΙΚΑ ΜΕ ΤΟ ΚΕΦΑΛΑΙΟ ΤΗΣ Γ ΛΥΚΕΙΟΥ 27 ος ΠΜΔΧ Γ ΛΥΚΕΙΟΥ 30 03 203. Στοιχείο Μ το οποίο ανήκει στην πρώτη σειρά στοιχείων μετάπτωσης, σχηματίζει ιόν Μ 3+, που έχει 3 ηλεκτρόνια στην υποστιβάδα
Στοιχεία Φυσικοχηµείας και Βιοφυσικής
 Στοιχεία Φυσικοχηµείας και Βιοφυσικής Β. Φαδούλογλου 2008 Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία
Στοιχεία Φυσικοχηµείας και Βιοφυσικής Β. Φαδούλογλου 2008 Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία Φυσικοχηµείας και Βιοφυσικής Εργαστήρια Βιβλίο Εξετάσεις Ύλη Στοιχεία
ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΟΡΓΑΝΙΚΩΝ ΕΝΩΣΕΩΝ
 ΑΘ.Π.ΒΑΛΑΒΑΝΙΔΗΣ ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΟΡΓΑΝΙΚΩΝ ΕΝΩΣΕΩΝ ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΥΠΕΡΥΘΡΟΥ ΠΥΡΗΝΙΚΟΥ ΜΑΓΝΗΤΙΚΟΥ ΣΥΝΤΟΝΙΣΜΟΥ ΦΑΣΜΑΤΟΜΕΤΡΙΑ ΜΑΖΩΝ ΥΠΕΡΙΩΔΟΥΣ-ΟΡΑΤΟΥ, RΑΜΑΝ, ΗΛΕΚΤΡΟΝΙΚΟΥ ΠΑΡΑΜΑΓΝΗΤΙΚΟΥ ΣΥΝΤΟΝΙΣΜΟΥ Τμήμα Χημείας
ΑΘ.Π.ΒΑΛΑΒΑΝΙΔΗΣ ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΟΡΓΑΝΙΚΩΝ ΕΝΩΣΕΩΝ ΦΑΣΜΑΤΟΣΚΟΠΙΑ ΥΠΕΡΥΘΡΟΥ ΠΥΡΗΝΙΚΟΥ ΜΑΓΝΗΤΙΚΟΥ ΣΥΝΤΟΝΙΣΜΟΥ ΦΑΣΜΑΤΟΜΕΤΡΙΑ ΜΑΖΩΝ ΥΠΕΡΙΩΔΟΥΣ-ΟΡΑΤΟΥ, RΑΜΑΝ, ΗΛΕΚΤΡΟΝΙΚΟΥ ΠΑΡΑΜΑΓΝΗΤΙΚΟΥ ΣΥΝΤΟΝΙΣΜΟΥ Τμήμα Χημείας
«ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ»
 «ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ» Τι είναι οι πρωτεΐνες; Από τι αποτελούνται; Ποιος είναι ο βιολογικός του ρόλος; Ας ρίξουμε μία ματιά σε όλα αυτά τα ερωτήματα που μας απασχολούν ΚΕΦΑΛΑΙΟ 1:
«ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ» Τι είναι οι πρωτεΐνες; Από τι αποτελούνται; Ποιος είναι ο βιολογικός του ρόλος; Ας ρίξουμε μία ματιά σε όλα αυτά τα ερωτήματα που μας απασχολούν ΚΕΦΑΛΑΙΟ 1:
ΘΕΜΑ Α Α1. γ Α2. γ Α3. α Α4. β Α5. β ΘΕΜΑ B B1. B2.
 ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΗ ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ
 ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΗ ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΕΝΟΤΗΤΑ: ΕΝΖΥΜΑ ΚΑΘΗΓΗΤΗΣ: ΠΑΤΗΡ ΑΝΑΣΤΑΣΙΟΣ ΙΣΑΑΚ 1. Να εξηγήσετε γιατί πολλές βιταμίνες, παρά τη μικρή συγκέντρωσή τους στον οργανισμό, είναι πολύ σημαντικές για
ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΗ ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΕΝΟΤΗΤΑ: ΕΝΖΥΜΑ ΚΑΘΗΓΗΤΗΣ: ΠΑΤΗΡ ΑΝΑΣΤΑΣΙΟΣ ΙΣΑΑΚ 1. Να εξηγήσετε γιατί πολλές βιταμίνες, παρά τη μικρή συγκέντρωσή τους στον οργανισμό, είναι πολύ σημαντικές για
ΒΙΟΛΟΓΙΑ Β ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΠΕΡΙΛΗΨΗ ΚΕΦΑΛΑΙΟΥ 3
 ΒΙΟΛΟΓΙΑ Β ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΠΕΡΙΛΗΨΗ ΚΕΦΑΛΑΙΟΥ 3 Το θέμα που απασχολεί το κεφάλαιο σε όλη του την έκταση είναι ο μεταβολισμός και χωρίζεται σε τέσσερις υποκατηγορίες: 3.1)Ενέργεια και οργανισμοί,
ΒΙΟΛΟΓΙΑ Β ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΠΕΡΙΛΗΨΗ ΚΕΦΑΛΑΙΟΥ 3 Το θέμα που απασχολεί το κεφάλαιο σε όλη του την έκταση είναι ο μεταβολισμός και χωρίζεται σε τέσσερις υποκατηγορίες: 3.1)Ενέργεια και οργανισμοί,
ΒΙΟΧΗΜΕΙΑ Ι. ΚΕΦΑΛΑΙΟ 2 ο Βιοχημική εξέλιξη
 ΒΙΟΧΗΜΕΙΑ Ι ΚΕΦΑΛΑΙΟ 2 ο Βιοχημική εξέλιξη ΣΥΝΔΕΣΗ ΜΕ ΤΑ ΠΡΟΗΓΟΥΜΕΝΑ Τι είναι ΒΙΟΧΗΜΕΙΑ DNA ΠΡΩΤΕΙΝΕΣ ΑΛΛΑ ΣΥΝΔΕΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΧΗΜΕΙΑΣ (Δεσμοί, ενέργεια, δομή) ΕΞΕΛΙΞΗ ΤΗΣ ΖΩΗΣ Υπάρχει μια συνεχή εξελικτική
ΒΙΟΧΗΜΕΙΑ Ι ΚΕΦΑΛΑΙΟ 2 ο Βιοχημική εξέλιξη ΣΥΝΔΕΣΗ ΜΕ ΤΑ ΠΡΟΗΓΟΥΜΕΝΑ Τι είναι ΒΙΟΧΗΜΕΙΑ DNA ΠΡΩΤΕΙΝΕΣ ΑΛΛΑ ΣΥΝΔΕΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΧΗΜΕΙΑΣ (Δεσμοί, ενέργεια, δομή) ΕΞΕΛΙΞΗ ΤΗΣ ΖΩΗΣ Υπάρχει μια συνεχή εξελικτική
Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά
 Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά Είναι απλή πολυπεπτιδική αλυσίδα 307 αμινοξέων Είναι συμπαγής και έχει σχήμα ελλειψοειδές διαστάσεων 50 x 42 x 38 A Περιέχει περιοχές α-έλικος 38%
Το ένζυμο Καρβοξυπεπτιδάση Α έχει τα εξής χαρακτηριστικά Είναι απλή πολυπεπτιδική αλυσίδα 307 αμινοξέων Είναι συμπαγής και έχει σχήμα ελλειψοειδές διαστάσεων 50 x 42 x 38 A Περιέχει περιοχές α-έλικος 38%
igenetics Mια Μεντελική προσέγγιση
 igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 19 Ρύθμιση της γονιδιακής έκφρασης σε βακτήρια και βακτηριοφάγους Ο καταστολέας του οπερονίου lac προσδεδεμένος στο DNA. 2 Ιδιοστατικά γονίδια Ρυθμιζόμενα γονίδια
igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 19 Ρύθμιση της γονιδιακής έκφρασης σε βακτήρια και βακτηριοφάγους Ο καταστολέας του οπερονίου lac προσδεδεμένος στο DNA. 2 Ιδιοστατικά γονίδια Ρυθμιζόμενα γονίδια
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
Κ Ε Φ Α Λ Α Ι Ο 25 : Το καταλυτικό RNA
 Κ Ε Φ Α Λ Α Ι Ο 25 : Το καταλυτικό RNA Εικόνα 25.1 Το αυτο-µάτισµα του πρώιµου rrna 35S της Tetrahymena thermophila µπορεί να µελετηθεί µε ηλεκτροφόρηση σε πήκτωµα. Το αποδεσµευµένο ιντρόνιο σχηµατίζει
Κ Ε Φ Α Λ Α Ι Ο 25 : Το καταλυτικό RNA Εικόνα 25.1 Το αυτο-µάτισµα του πρώιµου rrna 35S της Tetrahymena thermophila µπορεί να µελετηθεί µε ηλεκτροφόρηση σε πήκτωµα. Το αποδεσµευµένο ιντρόνιο σχηµατίζει
πρωτεϊνες νουκλεϊκά οξέα Βιολογικά Μακρομόρια υδατάνθρακες λιπίδια
 πρωτεϊνες νουκλεϊκά οξέα Βιολογικά Μακρομόρια υδατάνθρακες λιπίδια Περιγραφή μαθήματος Επανάληψη σημαντικών εννοιών από την Οργανική Χημεία Χημική σύσταση των κυττάρων Μονοσακχαρίτες Αμινοξέα Νουκλεοτίδια
πρωτεϊνες νουκλεϊκά οξέα Βιολογικά Μακρομόρια υδατάνθρακες λιπίδια Περιγραφή μαθήματος Επανάληψη σημαντικών εννοιών από την Οργανική Χημεία Χημική σύσταση των κυττάρων Μονοσακχαρίτες Αμινοξέα Νουκλεοτίδια
3.1 Ενέργεια και οργανισμοί..σελίδα 2 3.2 Ένζυμα βιολογικοί καταλύτες...σελίδα 4 3.3 Φωτοσύνθεση..σελίδα 5 3.4 Κυτταρική αναπνοή.
 5ο ΓΕΛ ΧΑΛΑΝΔΡΙΟΥ Μ. ΚΡΥΣΤΑΛΛΙΑ 2/4/2014 Β 2 ΚΕΦΑΛΑΙΟ 3 ΒΙΟΛΟΓΙΑΣ 3.1 Ενέργεια και οργανισμοί..σελίδα 2 3.2 Ένζυμα βιολογικοί καταλύτες...σελίδα 4 3.3 Φωτοσύνθεση..σελίδα 5 3.4 Κυτταρική
5ο ΓΕΛ ΧΑΛΑΝΔΡΙΟΥ Μ. ΚΡΥΣΤΑΛΛΙΑ 2/4/2014 Β 2 ΚΕΦΑΛΑΙΟ 3 ΒΙΟΛΟΓΙΑΣ 3.1 Ενέργεια και οργανισμοί..σελίδα 2 3.2 Ένζυμα βιολογικοί καταλύτες...σελίδα 4 3.3 Φωτοσύνθεση..σελίδα 5 3.4 Κυτταρική
Μέθοδοι έρευνας ορυκτών και πετρωμάτων
 Μέθοδοι έρευνας ορυκτών και πετρωμάτων Μάθημα 9 ο Φασματοσκοπία Raman Διδάσκων Δρ. Αδαμαντία Χατζηαποστόλου Τμήμα Γεωλογίας Πανεπιστημίου Πατρών Ακαδημαϊκό Έτος 2017-2018 Ύλη 9 ου μαθήματος Αρχές λειτουργίας
Μέθοδοι έρευνας ορυκτών και πετρωμάτων Μάθημα 9 ο Φασματοσκοπία Raman Διδάσκων Δρ. Αδαμαντία Χατζηαποστόλου Τμήμα Γεωλογίας Πανεπιστημίου Πατρών Ακαδημαϊκό Έτος 2017-2018 Ύλη 9 ου μαθήματος Αρχές λειτουργίας
Δομικές κατηγορίες πρωτεϊνών
 3-1 Κεφάλαι ο Δομικές κατηγορίες πρωτεϊνών 3.1. α-δομές πρωτεϊνών Οι α-έλικες είναι δομικά στοιχεία που μπορούν να σχηματίσουν πολλές κατηγορίες στερεοδομών και με πολλές διαφορετικές λειτουργίες. Εκτός
3-1 Κεφάλαι ο Δομικές κατηγορίες πρωτεϊνών 3.1. α-δομές πρωτεϊνών Οι α-έλικες είναι δομικά στοιχεία που μπορούν να σχηματίσουν πολλές κατηγορίες στερεοδομών και με πολλές διαφορετικές λειτουργίες. Εκτός
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών
 Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.