Η επίδραση των βαρέων μετάλλων (Cd, Ni, Pb) στη δράση των μυκορριζών.
|
|
|
- Θεοφάνης Παπαδάκης
- 8 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΓΕΩΠΟΝΙΚΗ ΣΧΟΛΗ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΕΔΑΦΟΛΟΓΙΑΣ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΕΔΑΦΙΚΩΝ ΠΟΡΩΝ Η επίδραση των βαρέων μετάλλων (Cd, Ni, Pb) στη δράση των μυκορριζών. ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΑΤΡΙΒΗ Πηνελόπης Μπαλτζώη Υπότροφος ΙΚΥ Επιβλέπουσα Καθηγήτρια Παυλάτου-Βε Αθηνά Αναπλ. Καθηγήτρια Θεσσαλονίκη 2006
2 ΠΕΡΙΕΧΟΜΕΝΑ 1 ΕΙΣΑΓΩΓΗ 1 2 ΑΝΑΣΚΟΠΗΣΗ ΒΙΒΛΙΟΓΡΑΦΙΑΣ Μυκόρριζες Τύποι μυκορριζών Η βιολογία του μύκητα που σχηματίζει VAM Τα καλλιεργητικά πλεονεκτήματα της VAM μυκόρριζας Ο εμβολιασμός με μύκητα που σχηματίζει VAM Η επίδραση των βαρέων μετάλλων στο μικροβιακό πληθυσμό του εδάφους Η επίδραση των βαρέων μετάλλων στη λειτουργία του μύκητα που σχηματίζει VAM 16 και στη συμβιωτική σχέση φυτού - μύκητα 3 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ ΕΔΑΦΗ Αναλύσεις χαρακτηρισμού των εδαφών Εμπλουτισμός των εδαφών με μέταλλα Απολύμανση εδάφους και σπόρων Μικροβιακή βιομάζα C και Ν Σταθερότητα των συσσωματωμάτων στη διαβροχή Διασπορά της αργίλου ΦΥΤΟ Ο εμβολιασμός με μυκόρριζα Glomus mosseae 26 4 ΑΠΟΤΕΛΕΣΜΑΤΑ ΚΑΙ ΣΥΖΗΤΗΣΗ Εδάφη Μικροβιακή βιομάζα C και Ν Φυτό Σταθερότητα των εδαφών Διασπορά της αργίλου Βιομάζα φυτού Πρόσληψη των μετάλλων από το φυτό Συγκεντρώσεις των μετάλλων στο ριζικό σύστημα του φυτού Παρατήρηση των χρωματισμένων ριζών στο μικροσκόπιο 51 5 ΣΥΜΠΕΡΑΣΜΑΤΑ 60 RESULTS 62 ΒΙΒΛΙΟΓΡΑΦΙΑ 63 ΠΑΡΑΡΤΗΜΑ 68
3
4 ΠΡΟΛΟΓΟΣ Για την ολοκλήρωση της μεταπτυχιακής μου αυτής διατριβής θα ήθελα να εκφράσω τις θερμές ευχαριστίες μου στην επιβλέπουσα της εργασίας αυτής Αναπλ. Καθηγήτρια Εδαφολογίας κ. Παυλάτου-Βε Αθηνά τόσο για την επιλογή του θέματος όσο και για την αμέριστη επιστημονική και ηθική υποστήριξη που μου παρείχε κατά τη διεξαγωγή και συγγραφή αυτής της διατριβής. Θερμότατες ευχαριστίες επίσης εκφράζονται: στα δύο υπόλοιπα μέλη της Εξεταστικής επιτροπής, τον κ. Δημήτρη Αλιφραγκή Αναπλ. Καθηγητή της Δασολογικής και Φυσικού Περιβάλλοντος Σχολής και την κ. Κωστοπούλου Σοφία Επ. Καθηγήτρια Εδαφολογίας για τις ουσιαστικές παρατηρήσεις και διορθώσεις τους. Επιπλέον ευχαριστώ θερμά τον κ. Δημήτρη Αλιφραγκή για τη παραχώρηση του εργαστηρίου του κατά τη διενέργεια των εργαστηριακών αναλύσεων, τον διδάσκοντα του N. 407 της Δασολογικής και Φυσικού Περιβάλλοντος Σχολής κ. Μιχάλη Ορφανουδάκη για τις πολύτιμες πρακτικές και επιστημονικές του γνώσεις που προσέφερε κατά τη διεξαγωγή του πειράματος, καθώς και όλους τους καθηγητές και το προσωπικό του εργαστηρίου Εδαφολογίας της Γεωπονικής Σχολής του Α.Π.Θ. για τις πολύτιμες γνώσεις και τη βοήθεια που μου παρείχαν. Τέλος, θα ήθελα να ευχαριστήσω το Ίδρυμα Κρατικών Υποτροφιών για την οικονομική συμπαράσταση κατά τη διάρκεια των μεταπτυχιακών μου σπουδών. Μπαλτζώη Πηνελόπη
5 ΚΕΦΑΛΑΙΟ 1 ΕΙΣΑΓΩΓΗ Η μόλυνση της βιόσφαιρας με τοξικά μέταλλα εξαιτίας ανθρωπογενών δραστηριοτήτων αποτελεί μεγάλο πρόβλημα τόσο για το ίδιο το περιβάλλον όσο και για την υγεία του ανθρώπου. Οι υψηλές συγκεντρώσεις των μετάλλων στο έδαφος είναι τοξικές, εκτός των άλλων, και στους μικροοργανισμούς του εδάφους (βακτήρια και μύκητες). Τόσο η πρόσληψη των βαρέων μετάλλων όσο και η ανθεκτικότητα των φυτών σε αυτά, εξαρτάται από φυτικούς και εδαφικούς παράγοντες, συμπεριλαμβανομένου και των μικροοργανισμών του εδάφους. Στο πλαίσιο εύρεσης μεθόδου απομάκρυνσης των μετάλλων από τα εδάφη έχει διερευνηθεί η αντοχή των μικροοργανισμών του εδάφους στη τοξικότητα των μετάλλων. Μεταξύ των πιο σημαντικών φαινόμενων ωφέλιμης συμβίωσης στη φύση, είναι και η μυκόρριζα. Οι μυκόρριζες αποτελούν αναπόσπαστα λειτουργικά μέρη των ριζών των φυτών και ο μύκητας που εμπλέκεται παρέχει άμεση σύνδεση του ριζικού συστήματος των φυτών με το έδαφος. Έχει βρεθεί ότι η μυκόρριζα συμβάλλει σημαντικά στην αποκατάσταση ρυπασμένων από μέταλλα εδαφών και η χρήση της αποτελεί μια από τις πιο ελπιδοφόρες μεθόδους για τον αποικισμό φυτών σε υποβαθμισμένα από μέταλλα εδάφη. Για τη μελέτη των αλληλεπιδράσεων μεταξύ μυκόρριζας και βαρέων μετάλλων θα πρέπει να διερευνηθούν δύο πλευρές: 1) η επίδραση των μετάλλων στους πληθυσμούς των μυκορριζικών μυκήτων καθώς και η αντοχή των μυκήτων αυτών στα μέταλλα και 2) η επίδραση των μυκορριζικών μυκήτων στη διαθεσιμότητα και μεταφορά των μετάλλων στα φυτά. Στη παρούσα εργασία διερευνήθηκε η επίδραση των Cd, Ni και Pb σε φυτά του είδους Lollium perrene (γκαζόν) και Allium cepa (κρεμμύδι) τα οποία σπάρθηκαν σε δύο διαφορετικά εδάφη. Ένας αριθμός φυτών εμβολιάστηκε με το μύκητα Glomus mosseae και διερευνήθηκε, τόσο η επίδραση αυτού του μύκητα στη πρόσληψη των μετάλλων από τα φυτά όσο και η επίδραση των αυτοχθόνων μυκήτων των φυτών. Επίσης εξετάστηκε και η επίδραση των τριών υπό μελέτη μετάλλων στο μικροβιακό πληθυσμό των δύο εδαφών που χρησιμοποιήθηκαν. Διερευνήθηκε η επίδραση τόσο των τριών μετάλλων όσο και των μυκορριζών στη δομή των δύο εδαφών. Τέλος έγινε παρατήρηση με το μικροσκόπιο των μυκορριζικών σχηματισμών στο ριζικό σύστημα των φυτών, αφού προηγήθηκε ειδική διαδικασία χρωματισμού των ριζών των φυτών. 1
6 ΚΕΦΑΛΑΙΟ 2 ΑΝΑΣΚΟΠΗΣΗ ΤΗΣ ΒΙΒΛΙΟΓΡΑΦΙΑΣ 2.1 Μυκόρριζες Η ύπαρξη των μυκόρριζων είναι γνωστή από τις αρχές του Παλαιοντολογικά ευρήματα δείχνουν ότι μυκόρριζες σχημάτιζαν και τα πρώτα φυτικά είδη τα οποία εμφανίσθηκαν στη Γη. Με τον όρο μυκόρριζα καλείται η σχέση μεταξύ των ριζών των περισσοτέρων φυτών με το συμβιωτικό μύκητα και ο μορφολογικός μετασχηματισμός αυτών των ριζών το οποίο και καλείται μυκόρριζα ( μύκητας ρίζες ) (Tate, L.R., 1987). Αυτού του είδους η συμβίωση αναμφισβήτητα αποτελεί ένα από τα πιο διαδεδομένα και ενδιαφέροντα παραδείγματα παρασιτισμού που συμβαίνει στη φύση. Ο Allen, (1991) περιέγραψε την αλληλεπίδραση μύκητα φυτού (μυκόρριζα) σαν μία αμοιβαία συμβίωση μεταξύ φυτού και μύκητα η οποία λαμβάνει χώρα μέσα στη ρίζα του φυτού και στην οποία η ενέργεια κινείται κυρίως από το φυτό στο μύκητα και ανόργανα συστατικά κινούνται από το μύκητα προς το φυτό. Σε όλα τα εδαφικά συστήματα κάθε αναπτυσσόμενο φυτό σχηματίζει κάποιου είδους μικροβιακής αλληλεπίδρασης στη περιοχή της ριζοσφαιράς του. Η συντριπτική πλειοψηφία των ανώτερων φυτών σχηματίζουν μυκόρριζες και οι μύκητες αυτοί συντελούν στην αύξηση της διαθεσιμότητας των θρεπτικών στοιχείων για τα φυτά, σε οικοσυστήματα όπου παρατηρείται έλλειψη αυτών. Οι περισσότερες έρευνες που έχουν γίνει έχουν δείξει βελτίωση της μεταφοράς των θρεπτικών στους φυτικούς ιστούς μέσω της αύξησης της επιφάνειας απορρόφησης των ριζών λόγω της επέκτασης του μυκηλίου του μύκητα σε σημεία αρκετά πιο μακριά από τη ριζόσφαιρα. Οι πιο συνήθεις παρατηρήσεις που έχουν γίνει είναι αυτές της αύξησης της μεταφοράς Ρ, Ν και νερού. Οι μύκητες που σχηματίζουν μυκόρριζες όχι μόνο αυξάνουν τη μεταφορά των θρεπτικών από το έδαφος στο φυτό, αλλά μπορούν επίσης να συντελέσουν στην αύξηση της μεταφοράς των θρεπτικών μεταξύ των φυτών. Αυτή η γέφυρα υφών δημιουργείται μεταξύ φυτών τόσο του ιδίου όσο και διαφορετικού είδους. Με αυτό τον τρόπο φυτά που βρίσκονται σε μη ευνοϊκά σημεία (π.χ. σκιερά μέρη) μπορούν να επωφεληθούν από τη φωτοσυνθετική δραστηριότητα άλλων φυτών καθώς και να προσλάβουν θρεπτικά από το σύστημα σύνδεσης των υφών. Ένα ακόμη πλεονέκτημα για τη φυτική βιομάζα αποτελεί και η ικανότητα των μυκόρριζων να μειώνουν ή ακόμη και να αποτρέπουν την ανάπτυξη ασθενειών. Η σύνδεση του μύκητα με τον ευπρόσβλητο ιστό της ρίζας μπορεί να μειώσει την ευπάθεια σε παθογενή τροποποιώντας τη σύνθεση και τη συγκέντρωση των εκκρίσεων, διεγείροντας το αμυντικό 2
7 σύστημα του φυτού μέσω της δικής του προσβολής του ριζικού ιστού, εμποδίζοντας άμεσα άλλους πληθυσμούς μικροβίων μέσω της σύνθεσης αντιβιοτικών και μειώνοντας τη πρόσβαση του παθογόνου μέσα στο φυτικό ιστό μέσω της φυσικής κατάληψης της επιφάνειας της ρίζας (Tate, L.R., 1987) Τύποι μυκορριζών Σήμερα είναι γνωστοί δύο τύποι μυκόρριζας: οι εκτομυκόρριζες και οι ενδομυκόρριζες. Οι εκτομυκόρριζες χαρακτηρίζονται από μια εξωκυτταρική ανάπτυξη του μύκητα στο φλοιό της ρίζας. Εμφανίζονται κυρίως στα δάση εύκρατων και βόρειων περιοχών και απαρτίζονται από 5000 είδη, μαζί με τους βασιδιομύκητες (Quilambo, 2003). Ωστόσο πολλά τροπικά είδη δέντρων έχει βρεθεί ότι σχηματίζουν εκτομυκόρριζες. Οι ενδομυκόρριζες χαρακτηρίζονται από μία μέσοκαι ενδοκυτταρική ανάπτυξη του μύκητα στο φλοιό της ρίζας, σχηματίζοντας ειδικά όργανα γνωστά ως κυστίδια (vesicles) και θύσανοι (arbuscules). Αυτή η χαρακτηριστική ανάπτυξη δίνει στην ενδομυκόρριζα και το εναλλακτικό όνομα vesicular arbuscular mychorrhiza (VAM) (θυσανώδης μυκόρριζα). Είναι η πιο ευρέως διαδεδομένη σχέση των φυτών. Περίπου το 80% των χερσαίων φυτικών ειδών σχηματίζουν αυτού του είδους τη συμβίωση και το 95% των τραχειόφυτων ανήκουν σε οικογένειες οι οποίες είναι χαρακτηριστικά μυκορριζικές (Quilambo, 2003). Στις ενδομυκόρριζες ανήκουν και οι ερικοειδείς μυκόρριζες οι οποίες απαντώνται στα φυτά της τάξεως Ericales καθώς και η μυκόρριζα των φυτών που ανήκουν στην οικογένεια Orchidaceae. Στο Πίνακα 1 παρουσιάζονται συγκεντρωτικά τα σπουδαιότερα χαρακτηριστικά των κύριων τύπων μυκόρριζας. Όλοι οι μύκητες του εδάφους που σχηματίζουν θυσάνους σε συνεργασία με τα φυτά, ανήκουν στη τάξη Glomeales ( Ζυγομύκητες ). Αυτή η τάξη που σχηματίζει κυστίδια ( Acaulospora, Entrophospora, Glomus και Sclerocystis ) ανήκει στην υπόταξη Glominae και εκείνοι που σχηματίζουν βοηθητικά κύτταρα και όχι κυστίδια ( Gigaspora και Scutellispora ) ανήκουν στην υπόταξη Gigasporinae. Η ταξινόμηση των μυκορριζικών μυκήτων VAM βασίζεται στη μορφολογία των χλαμυδοσπορίων και των ζυγοσπορίων. Έτσι η θυσανώδης μυκόρριζα ταξινομικά ανήκει στη τάξη Glomeales, η οποία αποτελείται από έξι γένη: 3
8 Τάξη Glomeales Glomaceae Acaulosporaceae Gigasporace Glomus Acaulospora Gigaspora Sclerocystis Entrophospora Scutelospora 4
9 Πίνακας 1. Τα χαρακτηριστικά των τριών κύριων τύπων μυκόρριζας Τύπος Μυκόρριζας Φυτά Ξενιστές Μυκητολογική Ταξινόμηση Arbuscular Αγγειόσπερμα Ζυγομύκητες Mychorrhza Μερικά Γυμνόσπερμα (ΑΜ) Πτεριδόφυτα Μερικά κατώτερα φυτά Μορφολογικά χαρακτηριστικά Ισχυρά διακλαδωμένοι θύσανοι στα φλοιώδη κύτταρα Εκτεταμένες υφές που αναπτύσσονται από την επιφάνεια της ρίζας στο έδαφος Λειτουργικά χαρακτηριστικά Οι υφές αυξάνουν την επιφάνεια απορρόφησης θρεπτικών Εκτομυκόρριζα (ECM) Ξυλώδη δένδρα και θάμνοι Μερικά ποώδη φυτά Βασιδιομύκητες Ασκομύκητες Ζυγομύκητες Περίβλημα από σφιχτά συμπιεσμένες υφές γύρω από τις κοντές ρίζες Ισχυρά διακλαδωμένο δίκτυο υφών Hartig net μεταξύ των επιδερμικών και/ή των φλοιωδών κυττάρων Οι υφές αυξάνουν την επιφάνεια απορρόφησης θρεπτικών Μερικοί μύκητες εκκρίνουν ορισμένες πρωτεϊνάσες και φωσφατάσες οι οποίες περιέχουν οργανικό Ν και Ρ Μερικοί μύκητες παράγουν ένζυμα τα οποία αλλοιώνουν το κυτταρικό τοίχωμα τα οποία διευκολύνουν τη πρόσβαση στον οργανικό Ν και Ρ Ερικοείδής Μυκόρριζα (ΕRM) Ericales Ασκομύκητες Εκτεταμένες σπειροειδείς υφές στα επιδερμικά κύτταρα Μικρή η ανάπτυξη των υφών από την επιφάνεια των ριζών στο έδαφος Οι μύκητες εκκρίνουν ορισμένες πρωτεϊνάσες και φωσφατάσες οι οποίες περιέχουν οργανικό Ν και Ρ Οι μύκητες παράγουν ένζυμα τα οποία αλλοιώνουν το κυτταρικό τοίχωμα και φαινολ-οξειδωτικά ένζυμα τα οποία διευκολύνουν τη πρόσβαση στο οργανικό Ν και Ρ του χούμου 5
10 Τρεις κύριες συνιστώσες συνθέτουν τη μυκορριζική σχέση: 1) Το έδαφος 2) ο μύκητας και 3) το φυτό. Η επίδραση του μύκητα αφορά τη παρουσία του στα κύτταρα της ρίζας και την επέκταση του μυκηλίου του στο έδαφος. Το μυκήλιο του μύκητα μπορεί να είναι αρκετά εκτεταμένο κάτω από ορισμένες συνθήκες, αλλά δε σχηματίζει κάποια φυτική δομή. Η κύρια λειτουργία του είναι η απορρόφηση των θρεπτικών του εδάφους. Η μεγάλη αποδοτικότητα των μυκορριζικών ριζών, σε αντίθεση με τις μη μυκορριζικές, οφείλεται στη δραστική πρόσληψη και μεταφορά των θρεπτικών στοιχείων από τις μυκόρριζες. Οι μυκόρριζες βελτιώνουν τη πρόσληψη πολλών θρεπτικών όπως P, N, K, Mg, Cu, Zn, Ca, Fe, Cd, Ni (Quilambo, 2003). Οι εκτομυκόρριζες αναπτύσσονται εξωκυτταρικά και αντικαθιστούν μερικώς τα μεσαία κύτταρα του φλοιού των ριζικών τριχιδίων. Οι μύκητες αυτοί σχηματίζουν ένα πυκνό μυκηλιακό δίκτυο γύρω και ανάμεσα από τα φυτικά κύτταρα, το οποίο ονομάζεται δίκτυο Hartig. Οι εκτομυκόρριζες χαρακτηρίζονται επίσης και από ένα πυκνό και γενικά συνεχές δίκτυο υφών πάνω από την επιφάνεια των ριζικών τριχιδίων το οποίο ονομάζεται μανδύας. Οι ενδομυκόρριζες διακρίνονται από το γεγονός ότι ο μύκητας διαπερνά τα φλοιώδη κύτταρα των ριζικών τριχιδίων και μπορεί να σχηματίσει μεγάλα κυστίδια και θυσάνους. Αυτοί οι μύκητες δε σχηματίζουν πυκνούς μυκηλιακούς μανδύες, αλλά σχηματίζουν μια χαλαρή και ασυνεχή μυκηλιακή διάταξη στην επιφάνεια της ρίζας. Σε σύγκριση με τις VAM μυκόρριζες των ετησίων φυτών, οι γνώσεις μας για τη συνεισφορά της εκτομυκόρριζας στην ικανότητα πρόσληψης θρεπτικών στοιχείων από τα δασικά είδη, είναι ελλιπείς. Υπάρχουν τουλάχιστον δύο κύριες διαφορές από τις VAM μυκόρριζες: (1) οι υφές της εκτομυκόρριζας μπορεί να περιβάλουν εντελώς τις πλευρικές ρίζες και αυτό εμποδίζει την απ ευθείας πρόσληψη θρεπτικών από τις ρίζες αυτές, και (2) το εξωτερικό μυκήλιο ορισμένων τουλάχιστον ειδών κινητοποιεί τα ανόργανα θρεπτικά από σημεία που δεν είναι προσβάσιμα για τις ρίζες του φυτού. (Read et al., 1992). Η μόλυνση από μυκόρριζες μπορεί να μεταβάλλει τόσο τη λειτουργία και το ρυθμό ανάπτυξης της ρίζας όσο και την εκκριτική της δραστηριότητα, μέσω του εκτεταμένου μυκηλίου τους. Οι υφές της VAM μυκόρριζας μπορεί να επεκτείνονται αρκετά εκατοστά από την επιφάνεια της ρίζας και με αυτό το τρόπο αυξάνουν τη πρόσληψη των θρεπτικών συστατικών (Read et al., 1992). Επίσης, διαδραματίζουν σημαντικό ρόλο στη δημιουργία, τη λειτουργία και την αναπαραγωγή των μυκορριζών σε φυσικά αλλά και διαταραγμένα οικοσυστήματα. Οι υφές που βρίσκονται στο έδαφος και προέρχονται είτε από υπάρχον μυκηλιακό δίκτυο είτε από άλλου είδους πολλαπλασιαστικό υλικό ( σπόρια, κυστίδια και τμήματα ριζών ), οδηγούν στη μόλυνση και το περαιτέρω αποικισμό των ριζών. Η διανομή των υφών στο έδαφος και η σχέση τους με το σχηματισμό σπορίων επηρεάζει τη θέση των κυστιδίων σε σχέση με τις νεοσχηματισθέντες ρίζες. Ο ρόλος των υφών στη σταθερότητα του εδάφους εξαρτάται από τη συνεισφορά τους στην 6
11 "επικάλυψη των εδαφικών τεμαχιδίων"σε σχέση με την επιφάνεια της ρίζας. Οι εξωτερικές υφές της VAM μυκόρριζας έχουν την ικανότητα να ικανοποιούν τις ανάγκες των φυτών τόσο για Ρ όσο και για Zn και Cu. Αντίθετα, η ικανότητα της πρόσληψης και μεταφοράς K, Ca και Mn φαίνεται να είναι πολύ χαμηλή έως ανύπαρκτη. Αυτό μπορεί να έχει ως αποτέλεσμα τη μείωση της συγκέντρωσης K και Mn μέσα στο φυτό όταν η παρουσία μυκόρριζας μειώνει την επιφάνεια απορρόφησης της ρίζας ή την έκκριση της ρίζας (Read et al., 1992). Υπάρχουν αρκετές ενδείξεις για σημαντική ικανότητα απορρόφησης NH 4 -N των εξωτερικών υφών της μυκόρριζας και μεταφοράς στο φυτό ξενιστή. Όπως στη ριζόσφαιρα έτσι και στην υφόσφαιρα η πρόσληψη NH 4 - N εξαρτάται από το βαθμό οξίνισης του υποστρώματος. 2.2 Η βιολογία του μύκητα που σχηματίζει VAM Όλες σχεδόν οι οικογένειες των φυτών περιλαμβάνουν είδη που σχηματίζουν VAM μυκόρριζες. Εξαίρεση αποτελούν οι οικογένειες Chenopodiaceae, Brassicaceae, Caryophyllaceae, και μερικές άλλες. Ακόμη και οι παραπάνω οικογένειες περιλαμβάνουν είδη στις ρίζες των οποίων έχουν βρεθεί κυστίδια και υφές μυκήτων που σχηματίζουν μυκόρριζες. Οι τοξίνες και η φυσιολογία των ριζών των παραπάνω φυτών ίσως είναι η αιτία μη σχηματισμού μυκόρριζας. Υπάρχουν όμως ορισμένες αποδείξεις που εξηγούν ότι γενετικοί παράγοντες του φυτού ξενιστή είναι αυτοί που είτε προάγουν είτε αναστέλλουν το σχηματισμό μυκόρριζας (Jarstfer and Sylvia, 1993). Όταν οι εδαφικές συνθήκες ευνοούν το γρήγορο φύτρωμα των σπόρων τότε ευνοείται παράλληλα και η βλάστηση των σπορίων του VAM μύκητα. Τα σπόρια του μύκητα μπορεί να απαιτούν μία περίοδο λήθαργου πριν βλαστήσουν. Παρόλα αυτά έχει παρατηρηθεί ότι η βλάστηση των σπορίων επιταχύνεται σε περιόδους με χαμηλή θερμοκρασία, καθώς και σε συνθήκες ξηρασίας στα φυλλοβόλα δάση και στις ξερές εύκρατες περιοχές. Μετά τη βλάστηση των σπορίων, οι εδαφικές συνθήκες που ευνοούν την ανάπτυξη της ρίζας, ευνοούν επίσης και την ανάπτυξη των υφών του μύκητα. Θερμοκρασιακό ή υδατικό στρες μπορούν να μειώσουν την ανάπτυξη των υφών και να καθυστερήσουν την εγκατάσταση του μύκητα. Η ανάπτυξη των υφών στη περιοχή της ριζόσφαιρας και η διείσδυσή τους μέσα στη ρίζα επηρεάζεται από τη συγκέντρωση του Ρ στους φυτικούς ιστούς, από τις εκκρίσεις της ρίζας και από το CO 2 (Jarstfer and Sylvia, 1993). Ο μύκητας που σχηματίζει VAM διαπερνά την επιδερμίδα της ρίζας τουλάχιστον κατά ένα μέρος με μηχανική πίεση, αλλά στη διαδικασία αυτή λαμβάνει χώρα και κάποια ενζυματική δραστηριότητα. 7
12 Η εξωτερική εξάπλωση του μύκητα από ένα μόλυσμα γίνεται όταν οι εσωτερικές υφές αναπτύσσονται από το φλοιό διαμέσου της επιδερμίδας. Αυτές οι εξωτερικές υφές αναπτύσσονται μέσα στο έδαφος εωσότου αντιμετωπίσουν δυσμενείς εδαφικές συνθήκες ή συναντήσουν κάποια δεκτική ρίζα την οποία και μολύνουν. Κατά τη δεύτερη περίπτωση λαμβάνει χώρα μια δευτερογενής διείσδυση με διαδικασίες όμοιες με αυτές που συνέβησαν κατά την αρχική εγκατάσταση του μύκητα από το σπερματικό σωλήνα. Για το σχηματισμό των σπορίων απαιτούνται μεγάλες ποσότητες C από τα προϊόντα της φωτοσύνθεσης (Jarstfer and Sylvia, 1993). Οι εξωτερικές υφές λειτουργούν και σαν όργανα πολλαπλασιασμού για τους περισσότερους μύκητες. Οι εξωτερικές υφές μεταφέρουν επίσης C και άλλα θρεπτικά στοιχεία στα αναπτυσσόμενα σπόρια. Ο ρόλος των εξωτερικών υφών είναι πιθανόν πιο σημαντικός για τη προσρόφηση των θρεπτικών από το έδαφος. Η εσωτερική ανάπτυξη του μύκητα ξεκινά όταν η υφή του μύκητα περνά μέσω της επιδερμίδας στο φλοιό. Ακολουθεί μια ταχεία μεσοκυτταρική εξάπλωση με ταυτόχρονη ενδοκυτταρική εισχώρηση στα φλοιώδη κύτταρα. Στη συνέχεια σχηματίζονται οι θύσανοι, που έχουν δενδρώδη μορφή, από τις μεσοκυττάριες υφές. Μετά από 8-20 μέρες οι θύσανοι γηράσκουν και αφήνουν ταννίνες. Σε αυτά τα φυτικά κύτταρα μπορεί να ξαναεισχωρήσουν ενδοκυτταρικά κυστίδια, τα οποία μπορεί να πάρουν το ορθογώνιο σχήμα των κυτταρικών τοιχωμάτων. Τα κυστίδια σχηματίζονται από τη διόγκωση της άκρης των υφών ένδο- ή μεσοκυττάρια. Αυτή η ακολουθία των (1) γραμμική επέκταση των υφών, (2) η δημιουργία και ο γηρασμός των θυσάνων, και (3) σχηματισμός κυστιδίων, φαίνεται ότι συμβαίνει σε συνεχή ρυθμό και είναι άμεσα συνδεδεμένη με το ρυθμό επέκτασης των ριζών (Jarstfer and Sylvia, 1993). Με την αύξηση του μήκους της ρίζας εμφανίζεται και μια εκθετική αύξηση της αποίκισης του μύκητα κατά μήκος της ρίζας. O μύκητας που σχηματίζει VAM που βρίσκεται σε ένα σημείο της ρίζας, μπορεί να περάσει απευθείας σε μια άλλη διακλάδωση της ίδιας ρίζας. Είναι όμως άγνωστο το αν τα περισσότερα ριζικά τριχίδια μολύνονται απευθείας από εξωτερικές υφές γειτονικών ριζών ή από εσωτερικές υφές. Το ποσοστό του μήκους της ρίζας που καταλαμβάνεται από το μύκητα εξαρτάται από μια αλληλεπίδραση μεταξύ της φυσιολογικής κατάστασης των ριζών τη στιγμή που συναντά το πολλαπλασιαστικό υλικό του μύκητα και από τη δυναμική ανάπτυξης αυτών των ριζών μετά το σχηματισμό της μυκόρριζας. Η αύξηση της ανάπτυξης της ρίζας λόγω αύξησης της διαθεσιμότητας των θρεπτικών δε σημαίνει απαραίτητα και μείωση του ποσοστού του μήκους της ρίζας που αποικείται από το μύκητα. Παρόλα αυτά η εξάπλωση του μύκητα μπορεί να καθυστερήσει κατά την ανάπτυξη της ρίζας. Στο αγγειακό σύστημα του φυτού ποτέ δεν εισχωρούν οι υφές του μύκητα. Όταν ο φλοιός παύει να είναι ενεργός οι θύσανοι σταματούν να 8
13 αναπτύσσονται και να λειτουργούν (Jarstfer and Sylvia, 1993). Όταν η ρίζα φελλοποιείται δε μπορεί πια να σχηματίσει μυκόρριζα. Η μολυσματικότητα της VAM μυκόρριζας στο έδαφος μπορεί να μειωθεί αρκετά με διατάραξη του εδάφους. Αυτός ο βαθμός μείωσης εξαρτάται από το είδος του μύκητα και το στάδιο ανάπτυξης του (Read et al., 1992) Τα καλλιεργητικά πλεονεκτήματα της VAM μυκόρριζας Ο μύκητας που σχηματίζει VAM μπορεί να παίξει αποφασιστικό ρόλο στα χαμηλών εισροών συστήματα καλλιέργειας ή στην οργανική γεωργία, λόγω της συμμετοχής του στη σύνδεση φυτικών και εδαφικών διεργασιών. Ο μύκητας προμηθεύεται υδρογονάνθρακες από το φυτό-ξενιστή και με τη σειρά του το φυτό προμηθεύεται θρεπτικά στοιχεία που μπορούν να προσληφθούν από το έδαφος με τις μυκηλιακές υφές του μύκητα. Επίσης, εκτός από τα οφέλη που προκύπτουν για το φυτό μέσο της θρέψης του, έχει αποδειχθεί ότι τα φυτά που έχουν μολυνθεί από μύκητες που σχηματίζουν μυκόρριζες παρουσιάζουν βελτιωμένη αναπαραγωγική ικανότητα, αυξημένη ικανότητα προσαρμογής σε συνθήκες έλλειψης νερού και βελτιωμένη φυτική υγεία απέναντι στις επιδράσεις διαφόρων παθογενών οργανισμών (Eason et al., 1999). Το μυκηλιακό δίκτυο που σχηματίζει ο μύκητας της VΑΜ, συνεισφέρει σημαντικά στη βελτίωση της δομής του εδάφους και στις υδατικές σχέσεις, με ιδιαίτερα οφέλη στη συσσωμάτωση του εδάφους (Harrier and Watson, 2003). Όμως το σχετικό μέγεθος της συνεισφοράς του φυτού ή του ενδόφυτου (μύκητα) στη σταθερότητα των συσσωματωμάτων δεν έχει μετρηθεί άμεσα. Επίσης, οι διάφοροι τύποι ριζών μπορεί να διαφέρουν στις σχέσεις τους με τους VAM μύκητες (Thomas et al., 1993). H επίδραση του VΑΜ μύκητα στη συσσωμάτωση μπορεί να περιγραφεί ως τρεις ξεχωριστές αλλά ταυτόχρονες διαδικασίες: (1) ανάπτυξη των υφών στο περίβλημα των εδαφικών τεμαχιδίων δημιουργώντας τη σκελετική δομή η οποία εμπλέκει φυσικά τα κύρια εδαφικά τεμαχίδια, (2) ρίζες και υφές δημιουργούν συνθήκες που καθιστούν δυνατό το σχηματισμό των μικροσυσσωματωμάτων μέσα στο έδαφος, (3) ρίζες και υφές ενώνουν τα μικροσυσσωματώματα και τα μικρά μακροσυσσωματώματα σε μεγαλύτερα συσσωματώματα (Bearden and Petersen, 2000). Η αποτελεσματικότητα των υφών του μύκητα καθορίζεται από το μήκος των υφών και την ειδική τους επιφάνεια καθώς επίσης και από τις εκκρίσεις τους σε πολυσακχαρίτες (Bearden and Petersen, 2000). 9
14 Έχει διαπιστωθεί ότι η VAM μυκόρριζα αυξάνει τη παραγωγικότητα εδαφών χαμηλής γονιμότητας και είναι ιδιαίτερα σημαντική για την αύξηση της πρόσληψης δυσδιάλυτων ιόντων όπως τα PO 3-4, δυσκίνητων θρεπτικών όπως Ρ, Zn και Cu καθώς και άλλων στοιχείων όπως το Cd (Quilambo, 2003). Κάτω από συνθήκες ξηρασίας, η πρόσληψη των ευκίνητων θρεπτικών - όπως τα ΝΟ 3 μπορεί επίσης να βελτιωθεί με την ύπαρξη μυκόρριζας. Οι μύκητες που σχηματίζουν μυκόρριζες μπορούν επίσης να βελτιώσουν τη πρόσληψη του Ν που προέρχεται από την ανόργανη λίπανση (Quilambo, 2003). Η αύξηση της προσρόφησης του εδαφικού Ρ είναι το πιο φανερό όφελος της ύπαρξης VAM μυκόρριζας. Οι υφές του μύκητα επιτρέπουν στο ριζικό σύστημα να εκμεταλλευτεί μεγαλύτερο όγκο εδάφους με το 1) να επεκτείνονται μέχρι και 8 cm πιο μακριά από τις ρίζες, 2) να χρησιμοποιούν μικρότερους εδαφικούς πόρους, καθώς οι υφές του μύκητα έχουν διάμετρο μικρότερη από το 20% της διαμέτρου των ριζικών τριχιδίων και 3) αυξάνουν την ενεργό επιφάνεια απορρόφησης (Jarstfer and Sylvia, 1993). Τα πλεονεκτήματα της αυξημένης προσρόφησης Ρ είναι πιο εμφανή σε εδάφη με χαμηλή περιεκτικότητα σε Ρ, αλλά παρατηρούνται και σε εδάφη με χαμηλή περιεκτικότητα σε ευδιάλυτο Ρ. Η μεταφορά του εδαφικού Ρ δια μέσου των μυκηλιακών υφών μπορεί να θεωρηθεί ως αποτέλεσμα τριών βημάτων: πρόσληψη μέσω των υφών, αναδιανομή μέσα στις υφές και μεταφορά δια μέσου της συμβιωτικής αλληλεπίδρασης (Read et al., 1992). Η προσρόφηση των μικροθρεπτικών αυξάνεται με τον ίδιο μηχανισμό όπως συμβαίνει και με το Ρ. Έτσι η παρουσία VAM μυκόρριζας αυξάνει τη προσρόφηση του Cu και του Zn (Jarstfer and Sylvia, 1993).. Η ύπαρξη της VAM μπορεί να προστατέψει τα φυτά ξενιστές από τις επιβλαβείς συνέπειες της ξηρασίας (Quilambo, 2003). Διάφοροι μηχανισμοί έχουν προταθεί για να εξηγήσουν τη προστατευτική δράση της VAM όπως, αλλαγές των φυτικών ορμονών, αυξημένη ανταλλαγή αερίων από τα φύλλα και αυξημένος ρυθμός φωτοσύνθεσης, άμεση πρόσληψη του εδαφικού νερού από τις υφές του μύκητα και μεταφορά του στο φυτό ξενιστή, αυξημένη δραστηριότητα των ενζύμων που σχετίζονται με την αντιοξειδωτική άμυνα, αφομοίωση των νιτρικών, αυξημένη πρόσληψη νερού μέσω βελτιωμένης υδραυλικής αγωγιμότητας και αυξημένης φυλλικής αγωγιμότητας και φωτοσυνθετικής δραστηριότητας, οσμωτική ρύθμιση και μεταβολές στην ελαστικότητα του κυτταρικού τοιχώματος (Quilambo, 2003). Η VAM μπορεί επίσης να μειώσει τις επιπτώσεις και άλλων δυσμενών συνθηκών, όπως της αλατότητας του εδάφους (Quilambo, 2003). Οι παθογενείς μύκητες και οι νηματώδεις αλληλεπιδρούν με τη VAM μυκόρριζα στις ρίζες των φυτών. Στις περισσότερες περιπτώσεις, η κατάληψη του φλοιού της ρίζας από το μύκητα της μυκόρριζας ήταν ικανή να εμποδίσει ή και να μειώσει την εγκατάσταση του παθογενή μύκητα. Είναι άγνωστο αν το κατά πόσο ο μύκητας που σχηματίζει μυκόρριζα 10
15 προκαλεί το σχηματισμό συγκεκριμένων αντιμικροβιακών ουσιών, όπως οι φυτοαλεξίνες. Είναι γνωστό ότι οι φυσιολογικές μεταβολές που προκαλεί ο μύκητας της μυκόρριζας και ιδιαίτερα η αυξημένη προσρόφηση του Ρ, μειώνουν το ποσοστό προσβολής της ρίζας από παθογενείς μύκητες. Αυτή η αυξημένη αντοχή που δημιουργείται μπορεί να οφείλεται στην αύξηση του ρυθμού ανάπτυξης των ριζών λόγω αύξησης της προσρόφησης Ρ. Η αυξημένη θρέψη με Ρ επιτρέπει επίσης στο φυτό να αντικαταστήσει τις κατεστραμμένες από το παθογενή ρίζες. Σε άλλες περιπτώσεις, ο σχηματισμός μυκόρριζας μπορεί έμμεσα να μειώσει το ρυθμό προσβολής των ριζών μέσο των υψηλών επιπέδων Ρ και N. Από την άλλη, οι μυκόρριζες μπορεί να μην έχουν τα παραπάνω αποτελέσματα σε περιπτώσεις ραγδαίας μόλυνσης των ριζών από είδη όπως Pythium και Phytophthora. Σε γενικές γραμμές, η αντοχή σε παθογενείς μύκητες είναι πιο πιθανό να εμφανιστεί όταν γίνεται εμβολιασμός με μυκόρριζες, σε υγιή σπορόφυτα, πριν αυτά μεταφυτευτούν. Τα φυτά που σχηματίζουν μυκόρριζες μπορούν να παρουσιάσουν αντοχή και σε νηματώδεις (Jarstfer and Sylvia, 1993). 2.3 Ο εμβολιασμός με μύκητα που σχηματίζει VAM Τα τελευταία χρόνια παρουσιάζεται έντονο ενδιαφέρον για τη χρήση VAM μυκόρριζας στη γεωργική παραγωγή. Τα οφέλη αυτής της συμβίωσης επικεντρώνονται κυρίως στην αύξηση της πρόσληψης των θρεπτικών στοιχείων από το έδαφος καθώς και στην αύξηση της αντοχής των φυτών τόσο στη ξηρασία όσο και σε διάφορα παθογόνα (Vosátka, 1995). Ορισμένες μελέτες έχουν αποκαλύψει μία αναζωογονητική επίδραση της μυκόρριζας στην ανάπτυξη και τη θρέψη φυτών κρεμμυδιού τα οποία καλλιεργήθηκαν τόσο σε γλαστράκια όσο και σε συνθήκες αγρού (Vosátka, 1995). Η ανάπτυξη των φυτών μπορεί να αυξηθεί σημαντικά με τον εμβολιασμό τους με το κατάλληλο είδος μύκητα που δημιουργεί μυκόρριζα, ιδιαίτερα αν τα επίπεδα του διαθέσιμου εδαφικού Ρ είναι χαμηλά (Vosátka, 1995). Στη πράξη όμως οι καλλιέργειες τις περισσότερες φορές υπερλιπαίνονται, επειδή το κόστος της λίπανσης με φώσφορο είναι ασήμαντο σε σχέση με τα υψηλά επίπεδα κέρδους που προκύπτουν από τη καλλιέργεια. Ο εμβολιασμός των φυτών με μύκητα που σχηματίζει VAM μπορεί να πραγματοποιηθεί είτε με τη χρήση μολυσμένων ριζών είτε με πολλαπλασιαστικά όργανα του μύκητα, όπως υφές και σπόρια. Η εισαγωγή του εμβολίου, για τη μόλυνση των φυτών, γίνεται τόσο προφυτρωτικά όσο και μετάφυτρωτικά. Επίσης είναι δυνατός και ο εμβολιασμός σπορόφυτων, πριν τη μεταφύτευσή τους. Σε περιπτώσεις όπου το καλλιεργούμενο έδαφος έχει απολυμανθεί, και οι περισσότεροι ιθαγενείς μύκητες έχουν εξολοθρευτεί, ο εμβολιασμός με μυκόρριζα είναι συχνά επιτυχής 11
16 (Vosátka, 1995). Η καλύτερη μέθοδος για την εξαφάνιση του ιθαγενούς πληθυσμού μυκήτων φαίνεται να είναι η εφαρμογή ζεστού ατμού παρά η απολύμανση με καπνό. Παρόλα αυτά έχει επίσης διαπιστωθεί ότι η χρήση ατμού θα μπορούσε να εμποδίσει τη μελλοντική μόλυνση με μυκόρριζες των μεταφυτευμένων και εμβολιασμένων σποροφύτων (Vosátka, 1995). Επίσης, και σε μη απολυμασμένα εδάφη, ο εμβολιασμός με μυκόρριζα μπορεί να είναι επιτυχής αν χρησιμοποιηθεί αποτελεσματικό είδος μύκητα ή αν έχουμε ταχύτερο ρυθμό μόλυνσης. Τα σπορόφυτα μπορούν να εμβολιαστούν με μύκητες που δημιουργούν, μυκόρριζα οι οποίοι μπορεί να είναι πιο ανταγωνιστικοί από τον ιθαγενή πληθυσμό μυκήτων που περιέχονται στα καλλιεργούμενα εδάφη (Vosátka, 1995). Ο ρυθμός ανάπτυξης μπορεί να εξαρτάται από τη ταχεία εγκατάσταση του μύκητα. Η πυκνότητα του πολλαπλασιαστικού υλικού, η βιωσιμότητά του, και η τοποθεσία όπου βρίσκεται είναι παράγοντες που μπορούν να συμβάλουν στη ταχεία εγκατάσταση του μύκητα. Ως πυκνότητα του πολλαπλασιαστικού υλικού ορίζεται η συγκέντρωση των μολυσματικών μονάδων των μυκήτων μέσα στο εμβόλιο ή στο αναπτυσσόμενο καλλιεργητικό μέσο. Η ταχέως αναπτυσσόμενη μυκόρριζα μπορεί να εγκατασταθεί σε δεκτικές ρίζες σποροφύτων μέσα σε διάστημα δύο ημερών, ενώ η εγκατάσταση από σπόρια μπορεί να διαρκέσει πάνω από δέκα μέρες, μετά τη βλάστησή τους (Jarstfer and Sylvia, 1993). Η εξάπλωση του μύκητα εξαρτάται από το βαθμό ανάπτυξης των ριζών. Όταν υπάρχουν περισσότεροι από ένας μύκητες που σχηματίζουν VAM μέσα στο καλλιεργητικό μέσο, η κατάληψη των ριζών εξαρτάται από το σημείο τοποθέτησης του εμβολίου και την επιθετικότητα του κάθε μύκητα. Μία ρίζα μπορεί να καταληφθεί από περισσότερους του ενός μύκητες VAM. Σε μεγάλα δοχεία καλλιέργειας ή σε συνθήκες αγρού η ανάπτυξη της μυκόρριζας εξαρτάται από το σημείο τοποθέτησης του εμβολίου επειδή ένα τμήμα των ριζών δε θα συναντήσει το πολλαπλασιαστικό υλικό, αν αυτό δε τοποθετηθεί σωστά. Αντίθετα, στη περίπτωση καλλιέργειας σε δοχεία, δε παίζει ρόλο το σημείο τοποθέτησης του εμβολίου, αλλά μόνο η ύπαρξη δεκτικών ριζών (Jarstfer and Sylvia, 1993). Ο μύκητας που σχηματίζει VAM μπορεί να παραμένει στο έδαφος με τη μορφή σπορίων, μολυσμένων ριζών, ή σαν δίκτυο υφών. Για την εισαγωγή ενός τέτοιου μύκητα που θέλουμε να πραγματοποιεί συνεχή εγκατάσταση, θα πρέπει αυτός συνεχώς να καταλαμβάνει νέες ρίζες ή να σχηματίζει ανθεκτικά όργανα ( σπόρια ή κυστίδια ) με τα οποία θα επιβιώνει κατά την απουσία ριζών. Στα περισσότερα καλλιεργητικά συστήματα, η χρησιμοποίηση των μυκηλιακών υφών ως πολλαπλασιαστικό υλικό, για συνεχή εγκατάσταση, είναι μη αποτελεσματική λόγω της συχνής διατάραξης του εδάφους. Η χρήση μυκορριζών θα μπορούσε να αποτελέσει μια εναλλακτική λύση για συμπληρωματική θρέψη των φυτών ακόμη και κάτω από συνθήκες ξηρασίας, αλλά περισσότερο 12
17 χρήσιμη αποδεικνύεται για τη βιολογική γεωργία όπου απαιτείται μείωση της χρήσης λιπασμάτων (Vosátka, 1995). 2.4 Η επίδραση των βαρέων μετάλλων στο μικροβιακό πληθυσμό του εδάφους. Πολλά μέταλλα και παίζουν ένα ιδιαίτερο ρόλο στις λειτουργίες πολλών ζωντανών μικροοργανισμών, ως μικροστοιχεία (π.χ. Fe, Mn, Mg, Ni, Zn, Cu, κ.τ.λ.), χρησιμεύοντας ως συστατικό ενζύμων, δομικών πρωτεϊνών και διατηρώντας την ιονική ισορροπία και το οσμωτικό δυναμικό των κυττάρων (Kosolapov et al., 2004). Παρόλα αυτά, οι υψηλές συγκεντρώσεις των μετάλλων μπορούν να αποτελέσουν μια σημαντική απειλή για τη χλωρίδα και πανίδα αλλά και για την ανθρώπινη υγεία. Στο εδαφικό περιβάλλον, τα μέταλλα μπορούν να διαλυθούν, να συγκρατηθούν από διάφορα ανόργανα συστατικά του εδάφους μέσω απορρόφησης και ιονικής ανταλλαγής, να σχηματίσουν σύμπλοκα με διάφορα οργανικά συστατικά του εδάφους, ή να καταβυθιστούν ως απλά ή σύνθετα στερεά (Fomina et al., 2005). Σε αντίθεση με τους διασπάσιμους οργανικούς ρύπους, οι οποίοι γίνονται λιγότερο τοξικοί με το πέρασμα του χρόνου, τα μέταλλα δεν αποσυντίθενται στο περιβάλλον. Μπορούν όμως να μετασχηματιστούν σε περισσότερο (ή λιγότερο) ευκίνητες μορφές μέσω τροποποίησης των φυσικών και χημικών τους χαρακτηριστικών (Kosolapov et al., 2004). Η τοξικότητα των μετάλλων στο έδαφος εξαρτάται από τη βιοδιαθεσιμότητά τους, η οποία ορίζεται ως η ικανότητά τους να μπορούν να μεταφερθούν από ένα τμήμα του εδάφους σε ένα ζωντανό οργανισμό (Leyval et al., 1997). Η βιοδιαθεσιμότητα των μετάλλων είναι συνάρτηση όχι μόνο της συνολικής τους συγκέντρωσης αλλά και φυσικοχημικών (π.χ. ph, οργανική ουσία, περιεκτικότητα σε άργιλο)και βιολογικών παραγόντων (βιοπροσρόφηση, βιοσυσσώρευση, διαλυτοποίηση) (Leyval et al., 1997). Οι υψηλές συγκεντρώσεις βαρέων μετάλλων στο έδαφος έχουν αρνητική επίδραση στους μικροοργανισμούς και στις βιοχημικές διεργασίες. Οι πρώτες παρατηρήσεις της επίδρασης των βαρέων μετάλλων στους μικροοργανισμούς έγιναν στις αρχές του προηγούμενου αιώνα. Μετά από έρευνες έχει προκύψει η διαπίστωση ότι οι μικροοργανισμοί είναι πολύ πιο ευαίσθητοι στα βαριά μέταλλα από τα ανώτερα φυτά. Οι επιδράσεις της ρύπανσης από βαρέα μέταλλα στη μικροβιακή κοινότητα είναι πολύ δύσκολο να ποσοτικοποιηθούν (McGrath, 1994). Η ρύπανση 13
18 από μέταλλα μπορεί να μειώσει τη συνολική μικροβιακή βιομάζα ενός εδάφους ή μπορεί να οδηγήσει σε φυσική επιλογή και επικράτηση των ειδών που παρουσιάζουν αντοχή (McGrath, 1994). Οι μικροοργανισμοί διαφέρουν στο βαθμό αντοχής τους στη τοξικότητα των μετάλλων και μια έκθεση σε υψηλές συγκεντρώσεις μετάλλων θα έχει ως αποτέλεσμα την άμεση καταστροφή των κυττάρων, λόγω της απότομης ρήξης ζωτικών λειτουργιών, και επίσης σταδιακές αλλαγές στα μεγέθη των πληθυσμών λόγω των αλλαγών της βιωσιμότητας ή της ικανότητας ανταγωνισμού (McGrath, 1994). Οι μικροοργανισμοί που εκτείθονται σε μεγάλες συγκεντρώσεις μετάλλων για μεγάλο χρονικό διάστημα, δεν είναι ικανοί να διατηρήσουν την ίδια ολική βιομάζα όπως συμβαίνει σε μη ρυπασμένα εδάφη. Συγκεκριμένα, έχει παρατηρηθεί ότι υπό συνθήκες ρύπανσης με μέταλλα, παράλληλα με τη μείωση της μικροβιακής βιομάζας πραγματοποιείται και μεταβολή της δομής της μικροβιακής κοινότητας (McGrath, 1994). Τα στοιχεία για τη τοξικότητα των μετάλλων στις μικροβιακές διεργασίες του εδάφους και στο πληθυσμό των μικροοργανισμών, παρουσιάζουν μια τεράστια μεταβλητότητα (Giller et al., 1998) το οποίο οφείλεται σε: (1) παράγοντες που διαφοροποιούν τη τοξικότητα των μετάλλων και (2) διαφορές στην ευαισθησία των μικροοργανισμών ή των μικροβιακών διεργασιών. Οι επιδράσεις των μετάλλων στο μικροβιακό πληθυσμό του εδάφους επηρεάζονται από ιδιότητες του εδάφους όπως, το ph, το ποσοστό της οργανικής ουσίας, το ποσοστό της αργίλου και τα οξείδια Fe (Giller et al., 1998). Από τους παραπάνω παράγοντες, το ph φαίνεται να προκαλεί τη μεγαλύτερη μεταβολή, λόγω της μεγάλης επίπτωσης που έχει στη διαλυτότητα των μετάλλων, τόσο στο σύνολο του εδάφους όσο και στο εδαφικό διάλυμα. Έχει βρεθεί ότι για κάθε βαθμό μείωσης του ph προκύπτει διπλασιασμός της συγκέντρωσης μετάλλων όπως του Zn, του Ni και του Cd στο εδαφικό διάλυμα (Giller et al., 1998). Τα μέταλλα που βρίσκονται στο εδαφικό διάλυμα δεν είναι όλα βιοδιαθέσιμα καθώς σχηματίζουν σύμπλοκα με οργανικά μόρια και μετατρέπονται σε χημικές μορφές οι οποίες δεν είναι άμεσα διαθέσιμες. Επίσης, οι ριζικές εκκρίσεις των φυτών επηρεάζουν τη διαθεσιμότητα των μετάλλων τόσο άμεσα (π.χ.fe 3+ ) ή μέσω της επίδρασης που έχουν οι εκκρίσεις αυτές στη μικροβιακή δραστηριότητα (Giller et al., 1998). Ακόμη και οι ίδιοι οι μικροοργανισμοί μπορούν να μεταβάλλουν τη διαθεσιμότητα των μετάλλων γύρω από τη περιοχή στην οποία βρίσκονται λόγω της τοπικής οξίνισης που προκαλούν ή λόγω της παραγωγής ουσιών οι οποίες σχηματίζουν σύμπλοκα με τα μέταλλα (Giller et al., 1998). Το Cd θεωρείται ότι είναι το μέταλλο με τις πιο αρνητικές επιδράσεις στη μικροβιακή βιομάζα και στη μικροβιακή δραστηριότητα, σε εδάφη ρυπασμένα με βαριά μέταλλα (Renella et al., 2004), ενώ δεν απαιτείται για κάποια γνωστή βιολογική λειτουργία. Είναι επίσης πολύ ευκίνητο λόγω της μικρής έλξης από τα κολλοειδή του εδάφους (Renella et al., 2004). Η 14
19 συσσώρευση του Cd στους φυτικούς ιστούς εξαρτάται από τα επίπεδα του διαθέσιμου Cd στο έδαφος όπου αναπτύσσονται τα φυτά (Qadir et al., 2000). Ακόμη και κάτω από συνθήκες ήπιας ρύπανσης με Cd, οι συγκεντρώσεις του Cd στα φυτά είναι γενικά μικρότερες από τα επίπεδα φυτοτοξικότητας, για τις περισσότερες καλλιέργειες (Qadir et al., 2000). Για το λόγο αυτό, πολλές καλλιέργειες μπορούν να αναπτυχθούν σε εδάφη ρυπασμένα με Cd χωρίς να παρουσιάζουν μείωση της παραγωγής τους. Το Cd μεταφέρεται στα φύλλα και τους καρπούς όλων των λαχανικών. Μία τιμή της τάξης των 0.3 mg Cd kg 1 ξηρού φυτικού βάρους θεωρείται γενικά κρίσιμη (Qadir et al., 2000) αν και παρουσιάζονται μεγάλες γενοτυπικές διαφορές μεταξύ των φυτών κατά τη πρόσληψη Cd. Θα πρέπει να σημειωθεί ότι μικρής διάρκειας εργαστηριακές επωάσεις δεν αντικατοπτρίζουν τις τοξικές επιδράσεις μετά από χρόνια έκθεση της μικροβιακής μικροχλωρίδας σε βαριά μέταλλα (Giller et al., 1998). Ο προσδιορισμός της ρύπανσης του εδάφους με βαριά μέταλλα πραγματοποιείται κυρίως με βιοδείκτες. Τόσο η αναπνοή του εδάφους όσο και η εδαφική μικροβιακή βιομάζα επηρεάζονται από τη παρουσία μετάλλων, αλλά η ολική αναπνοή δε φαίνεται να επηρεάζεται καθώς μειώνεται μόνο σε πολύ υψηλές συγκεντρώσεις μετάλλων (Ross, 1994). Ο ρυθμός της ανοργανοποίησης του οργανικού C του εδάφους (γενικώς μετρούμενη ως ο ρυθμός παραγωγής CO 2 κάτω από πρότυπες συνθήκες και αναφέρεται ως βασική αναπνοή) έχει χρησιμοποιηθεί εκτενώς από τους Giller et al., 1998 ως δείκτης της τοξικότητας των μετάλλων. Σύμφωνα με αυτούς, εκτός του ότι απεικονίζει το ρυθμό ανοργανοποίησης του οργανικού άνθρακα του εδάφους, εκφράζει και την αναπνευστική δραστηριότητα των μικροοργανισμών. Εκφρασμένος ανά μονάδα μικροβιακής βιομάζας εδάφους ( ρυθμός σχετικής αναπνοής ή qco 2 ) προτάθηκε να είναι ένα φυσιολογικό χαρακτηριστικό της μικροβιακής κοινότητας. Σε περιπτώσεις μακροχρόνιας ρύπανσης εδαφών από βαριά μέταλλα παρατηρείται ένα υψηλό qco 2 και έχει προταθεί ως ένας χρήσιμος δείκτης της ρύπανσης εδαφών από μέταλλα. Το γεγονός ότι σε ορισμένα εδάφη ρυπασμένα με βαριά μέταλλα έχει παρατηρηθεί χαμηλό qco 2 μπορεί να οφείλεται στο ότι οι μικροοργανισμοί χρησιμοποιούν διαφορετικά υποστρώματα. Η μικροβιακή βιομάζα είναι πολύ πιο ευαίσθητη και η ποσοστιαία αναλογία του συνολικού εδαφικού άνθρακα η οποία εκφράζεται σαν ζωντανή βιομάζα αποτελεί έναν καλό δείκτη του βαθμού ρύπανσης από μέταλλα (McGrath, 1994). Έχουν διερευνηθεί οι εδαφικές ενζυμικές δραστηριότητες σε εδάφη ρυπασμένα με βαριά μέταλλα αλλά τα αποτελέσματα αυτά είναι πολύ δύσκολο να ερμηνευτούν, σε σχέση με τη μέτρηση των ίδιων των μικροοργανισμών ή των δραστηριοτήτων τους. Ο λόγος του C της βιομάζας προς τον οργανικό C του εδάφους (Cmic:Corg) αντικατοπτρίζει τη συμβολή της μικροβιακής βιομάζας στον οργανικό C του εδάφους (Moscatelli et al., 2005). 15
20 Έχει αποδειχθεί ότι τα βαριά μέταλλα προκαλούν μείωση της αναλογίας του C της μικροβιακής βιομάζας στη συνολική οργανική ουσία του εδάφους (Dai et al., 2004) και ο λόγος του εδαφικού μικροβιακού C προς τον οργανικό εδαφικό C έχει προταθεί ως ένα χρήσιμο μέτρο της εδαφικής ρύπανσης από βαριά μέταλλα (Dai et al., 2004). Από άλλες μελέτες έχει παρατηρηθεί μια μείωση αυτού του λόγου ως αποτέλεσμα της ρύπανσης από μέταλλα (Dai et al., 2004). Οι συσχετισμοί για το Ν της βιομάζας με τη περιεκτικότητα σε μέταλλα ήταν χαμηλότερο από αυτούς που παρατηρήθηκαν με τον C της βιομάζας. Αυτό είχε σαν αποτέλεσμα, ο λόγος C:Ν της βιομάζας να διαφέρει έντονα στα διάφορα επίπεδα ρύπανσης. Αυτός ο λόγος ήταν περίπου 3,5 στα ελαφρώς ρυπασμένα εδάφη και αυξανόταν προοδευτικά μέχρι περίπου τη τιμή 10 στα πολύ ρυπασμένα εδάφη. Σύμφωνα με προηγούμενες έρευνες οι μύκητες φάνηκαν να είναι πιο ανθεκτικοί στα βαριά μέταλλα από ότι τα βακτήρια και οι ακτινομύκητες (Dai et al., 2004, Jordan and Lechevallier, 1975, Hiroki, 1992). Αναφέρεται πως στα βακτήρια ο λόγος C:Ν είναι χαμηλός, της τάξης των 3,5:1 και στους μύκητες στο 10 έως 15:1(Paul and Clear, 1996) και έχει προταθεί από τους Joergensen et al. (1995) ότι οι υψηλοί λόγοι C:Ν της μικροβιακής βιομάζας προκαλούνται από το αυξημένο ποσοστό μυκήτων στους λόγους της μικροβιακής βιομάζας. 2.5 Η επίδραση των βαρέων μετάλλων στη λειτουργία του μύκητα που σχηματίζει VAM και στη συμβιωτική σχέση φυτού - μύκητα Μεταξύ των μικροοργανισμών του εδάφους, οι μύκητες που σχηματίζουν VAM είναι οι μόνοι που παρέχουν άμεση σύνδεση των ριζών με το έδαφος και γι αυτό έχουν μεγάλη σπουδαιότητα όσον αφορά τη διαθεσιμότητα των βαρέων μετάλλων και τη τοξικότητά τους στα φυτά (Leyval et al., 1997). Η επίδραση των μυκορριζών στη θρέψη των φυτών θεωρείται να είναι μεγαλύτερη για στοιχεία με περιορισμένες ζώνες διάχυσης γύρω από τις ρίζες των φυτών, όπως ο Ρ και τα περισσότερα βαριά μέταλλα (Leyval et al., 1997). Από τις αρχές του 1980 πολλοί ερευνητές απέδειξαν ότι ο σχηματισμός μυκόρριζας θα μπορούσε να έχει επίδραση στην αφομοίωση των βαρέων μετάλλων από τα φυτά (Leyval et al., 1997). Σε εδάφη ρυπασμένα με βαριά μέταλλα έχει παρατηρηθεί μια επιβλαβής επίδραση των μετάλλων στους μύκητες που σχηματίζουν VAM (Giller et al., 1998). Έχει διαπιστωθεί ότι το μήκος των μυκηλιακών υφών του μύκητα που σχηματίζει VAM αποτελεί ένα καλό δείκτη αξιολόγησης των αρνητικών επιδράσεων που δέχεται το έδαφος (Vivas et al., 2005). Η ανάπτυξη των υφών παρεμποδίζεται από τα βαριά μέταλλα, το οποίο μπορεί να καθυστερήσει σημαντικά τον αποικισμό των ριζών σε ρυπασμένα εδάφη (Weissenhorn et al., 1993, Leyval et al., 1994). Από πειράματα σε φυτοδοχεία, έχει βρεθεί ότι τα βαριά μέταλλα μειώνουν ή και σταματούν τον 16
21 αποικισμό των ριζών από το μύκητα VΑΜ, ακόμη και σε συγκεντρώσεις μετάλλων όπου δε παρατηρείται φυτοτοξικότητα (Weissenhorn et al., 1993, Weissenhorn and Leyval, 1995, Mikuš et al., 2005). Θα πρέπει να σημειωθεί ότι τα μέταλλα διαφέρουν στις επιδράσεις τους στη βλάστηση των σπορίων και την ανάπτυξη του μυκηλίου του μύκητα. Από πειράματα, βρέθηκε ότι το Cd μειώνει τη βλάστηση των σπορίων, του μύκητα Glomus mosseae, περισσότερο από ότι την επέκταση των υφών, οι οποίες θα προκύψουν αφού βλαστήσει ένα σπόριο (Weissenhorn et al., 1993). Η τοξικότητα των μετάλλων απέναντι στους μύκητες γενικά αυξάνεται με την αύξηση της ηλεκτραρνητικότητας του στοιχείου: ο Zn βρέθηκε να είναι διαλλακτικά τοξικός, ο Cu και ο Pb τοξικοί και το Cd πολύ τοξικό (Fomina et al., 2005). Παρόλα αυτά, οι προσαρμοσμένοι μύκητες που σχηματίζουν VAM είναι ικανοί να επιβιώνουν σε περιοχές όπου έχουν μολυνθεί από βαρέα μέταλλα και επίσης μπορούν να αυξήσουν την αντοχή των φυτών στα μέταλλα (Vivas et al., 2005). Έχει βρεθεί ότι μυκόρριζες μπορούν να σχηματίσουν και τα φυτά τα οποία αναπτύσσονται σε εδάφη που είναι ρυπασμένα με βαριά μέταλλα (Weissenhorn et al., 1993; Weissenhorn and Leyval, 1995) πράγμα που αποδεικνύει ότι αυτοί οι μύκητες έχουν αναπτύξει αντοχή στα βαριά μέταλλα και θα μπορούσαν να παίξουν καθοριστικό ρόλο στη διαχείριση αυτών των εδαφών. Η αντοχή στα μέταλλα που επιδυκνείουν τα φυτά με μυκόρριζα βασίζεται σε μια σειρά μηχανισμών και συμβιωτικών επιδράσεων όπως η ικανότητα δέσμευσης και ακινητοποίησης των μετάλλων από το μυκήλιο του μύκητα της μυκόρριζας, τόσο στη περιοχή της ριζόσφαιρας όσο και μέσα στις ρίζες (Fomina et al., 2005; Vivas et al., 2005). Έχει διαπιστωθεί ότι, οι μύκητες που σχηματίζουν VAM μπορούν να προσαρμοστούν σε μια ποικιλία συνθηκών και η επιτυχής ανάπτυξη των φυτών σε ρυπασμένα εδάφη εξαρτάται πάρα πολύ από τη δραστηριότητα του μικροβιακού πληθυσμού (Vivas et al., 2005). Η κίνηση των μετάλλων προς το φυτό πραγματοποιείται με τις υφές του μύκητα. Σε υψηλές συγκεντρώσεις μετάλλων, τα στοιχεία τα οποία ανεμπόδιστα φτάνουν στο εσωτερικό των ριζών συγκεντρώνονται στα εσωτερικά παρεγχυματικά κύτταρα της ρίζας, εκεί όπου κυρίως βρίσκονται οι εσωτερικές δομές του μύκητα (arbuscules, κυστίδια, εσωτερικές υφές) (Ouziad et al., 2005). Οι διαφορές της συμπεριφοράς κίνησης των μετάλλων μεταξύ των μυκήτων που σχηματίζουν VAM και η αποτελεσματικότητα τους σχετίζονται με τη καταγωγή του μύκητα. Διαφορετικοί μύκητες που σχηματίζουν VAM έχουν διαφορετικές δυνατότητες στο να ακινητοποιούν τα μέταλλα, μέσα ή κοντά στη ρίζα, και να μειώνουν τη μετατόπισή τους στο βλαστό (Vivas et al., 2005). Γενικά, οι μύκητες που σχηματίζουν VAM που έχουν απομονωθεί από ρυπασμένα εδάφη έχουν την ικανότητα να αντέχουν τις υψηλές συγκεντρώσεις βαρέων μετάλλων σε σύγκριση με τα είδη που απομονώθηκαν από μη μολυσμένα εδάφη (Vivas et al., 2005). 17
22 Οι μύκητες που σχηματίζουν VAM βελτιώνουν τις δυσμενείς συνέπειες της τοξικότητας των μετάλλων για τα φυτά. Καταρχήν μειώνουν τη συγκέντρωση των μετάλλων στο υπέργειο τμήμα των φυτών (Read et al., 1992). Έχει βρεθεί ότι τα περισσότερα γένη μυκήτων είναι πιο ανθεκτικά στα βαριά μέταλλα από ότι τα ανώτερα φυτά με τα οποία συμβιούν (Read et al., 1992). Από μελέτες που έγιναν διαπιστώθηκε ότι φυτά γκαζόν με μυκόρριζα μπορούσαν να ευδοκιμήσουν πιο εύκολα σε εδάφη με υψηλές συγκεντρώσεις μετάλλων σε αντίθεση με τα φυτά χωρίς μυκόρριζα (Leyval et al., 1997). Αυτό υποδηλώνει ότι η αντοχή στα βαριά μέταλλα καθώς και άλλα ευεργετικά αποτελέσματα αποδίδονται στη συμβίωση με μυκόρριζα. Η ύπαρξη μυκόρριζας αυξάνει την επιφάνεια απορρόφησης των φυτών λόγω του εκτεταμένου μυκηλιακού δικτύου στη περιοχή της ριζόσφαιρας, το οποίο συμβάλει και στην αύξηση του ρυθμού πρόσληψης νερού και θρεπτικών (Khan et al., 2000). Αυτό οδηγεί σε αύξηση της παραγωγής βιομάζας η οποία αποτελεί απαραίτητη προϋπόθεση για τη προστασία των φυτών από τα βαριά μέταλλα. Μύκητας που σχηματίζει μυκόρριζα Πρόσληψη Μεταφορά Διαθέσιμα βαριά μέταλλα στο έδαφος Φυτό Τοξικότητα Αντοχή Είναι επίσης γνωστό πως οι μυκόρριζες παράγουν αυξητικές ουσίες, για το φυτό και έτσι προάγουν την ανόργανη θρέψη και αυξάνουν το ρυθμό ανάπτυξης και τη βιομάζα του φυτού, που όπως αναφέρθηκε, είναι απαραίτητη για τη προστασία του φυτού από τα βαριά μέταλλα. (Khan et al., 2000) Η επίδραση του μύκητα που σχηματίζει VAM στη πρόσληψη των μετάλλων από τα φυτά δεν είναι πάντα συγκεκριμένη. Σε γενικές γραμμές, τα αποτελέσματα των ερευνών δείχνουν ότι η επίδραση του μύκητα, στη πρόσληψη των βαρέων μετάλλων από τα φυτά, είναι σχετική με το μέταλλο και εξαρτάται από τη συγκέντρωσή του. Επίσης η επίδραση της VΑΜ στη πρόσληψη των μετάλλων από τα φυτά εξαρτάται και από το ph του εδάφους (Leyval et al., 1997). Έχει διαπιστωθεί ότι η αποίκιση των ριζών από μυκόρριζα VAM προκαλεί αύξηση της πρόσληψης των μεταλλικών ιόντων από τα φυτά, όταν αυτά τα ιόντα βρίσκονται σε χαμηλές συγκεντρώσεις 18
23 (Dehn and Schüepp, 1989, Weissenhorn et al., 1995, Weissenhorn and Leyval, 1995, Leyval et al., 1997, Joner et al., 2000, Ouziad et al., 2005, Giasson et al., 2005). Αντίθετα, η παρουσία μυκόρριζας στις ρίζες φυτών που βρίσκονται σε εδάφη μολυσμένα με υψηλές συγκεντρώσεις βαριών μετάλλων, προκαλεί μείωση της συγκέντρωσης των μετάλλων αυτών στα φυτά σε σχέση με τα μη μυκορριζικά φυτά (Bi et al., 2003; Ouziad et al., 2005). Έχει διαπιστωθεί και το αντίθετο από τους Killham and Firestone, 1983, οι οποίοι βρήκαν ότι, σε φυτά γρασιδιού, μερικά από τα οποία εμβολιάστηκαν με μύκητα του γένους Glomus fasciculatum, και σε όξινες συνθήκες υπό τη παρουσία μεγάλων συγκεντρώσεων βαρέων μετάλλων, οι συγκεντρώσεις των μετάλλων στις ρίζες και τους βλαστούς των φυτών με μυκόρριζα ήταν μεγαλύτερες από εκείνες των φυτών χωρίς μυκόρριζα. Η ανάπτυξη των φυτών με μυκόρριζα μειώθηκε, σε σύγκριση με τα μη τα άλλα φυτά, γεγονός το οποίο αποδόθηκε στην αυξημένη πρόσληψη των βαρέων μετάλλων λόγω της παρουσίας του μύκητα της μυκόρριζας. Έχει αναφερθεί μείωση της αρνητικής επίδρασης του Zn στην ανάπτυξη δύο ειδών γρασιδιού, λόγω της παρουσίας του μύκητα που σχηματίζει VΑΜ, αν και η πρόσληψη του Zn και η μεταφορά του στους βλαστούς δεν εμποδίστηκε (Weissenhorn et al., 1995). Από πειράματα σε φυτοδοχεία των Weissenhorn et al., (1995), βρέθηκε ότι το ξηρό βάρος των βλαστών και των ριζών φυτών καλαμποκιού που είχαν σχηματίσει μυκόρριζα ήταν σημαντικά μεγαλύτερο από εκείνο των φυτών χωρίς μυκόρριζα. Οι συγκεντρώσεις στους ιστούς και η συνολική πρόσληψη Cd από τα φυτά ήταν σημαντικά χαμηλότερη για τα φυτά με μυκόρριζα, ενώ η συνολική τους περιεκτικότητα δε διέφερε μεταξύ των φυτών με και χωρίς μυκόρριζα. Αντίθετα, για το Pb, δε παρατηρήθηκαν διαφορές στις συγκεντρώσεις των φυτικών ιστών και η συνολική πρόσληψη Pb έτεινε να είναι υψηλότερη για τα φυτά με μυκόρριζα αν και δεν ήταν στατιστικά σημαντική. Ο αποικισμός από το μύκητα VΑΜ δεν απέτρεψε τις συγκεντρώσεις των μετάλλων να φτάσουν πολύ πιο πάνω από τις κανονικές τιμές. Από την άλλη, ο μύκητας VΑΜ δεν αύξησε τη πρόσληψη των μετάλλων από τα φυτά σε επίπεδο ανησυχητικό, σε σύγκριση με τα φυτά χωρίς μυκόρριζα. Οι Giasson et al., (2005), βρήκαν ότι ο σχηματισμός μυκόρριζας αύξησε τη συσσώρευση των βαρέων μετάλλων στις ρίζες του μαρουλιού αλλά όχι στους βλαστούς. Παρόμοια αποτελέσματα βρήκαν και με το γρασίδι. Επίσης έχουν διαπιστώσει ότι τα βαριά μέταλλα απομονώνονται και μεταφέρονται στις ρίζες των φυτών μέσω των εξωτερικών υφών της VΑΜ. Όσο για το Cd, εισέρχεται μέσα στις υφές μέσω των συστημάτων μεταφοράς των μαγγανίου και ψευδαργύρου και συσσωρεύεται σε υψηλές συγκεντρώσεις στα φυτά. Σύμφωνα με τους ιδίους, κατά τη παρουσία βαρέων μετάλλων σε μη διαθέσιμες μορφές για τα φυτά, οι υφές της VAM μπορούν να μεταβάλουν το μέταλλο από ανθρακικό σε είδος διαλυτό στο νερό. Οι υφές εκκρίνουν οργανικά οξέα όπως τα κιτρικά και τα οξαλικά τα οποία καθιστούν τα βαριά μέταλλα 19
24 διαλυτά. Αυτό το φαινόμενο είναι πιο εμφανές με ένα μη ουσιώδες μέταλλο (Cd) παρά με ένα ουσιώδες (Zn). Οι Joner et al., (2000), βρήκαν ότι το μυκήλιο του μύκητα της VΑΜ έχει μια υψηλή ικανότητα ρόφησης μετάλλων σε σχέση με τους άλλους μικροοργανισμούς και μια και μια CEC συγκρίσιμη με άλλους μύκητες. Οι Weissenhorn and Leyval, 1995, ανέφεραν αυξημένη προσρόφηση Cd από φυτά που είχαν σχηματίσει VAM ενώ ταυτόχρονα, βρήκαν ότι τα φυτά χωρίς VAM είχαν υψηλότερη βιομάζα από τα αντίστοιχα με VAM φυτά. Η μείωση της βιομάζας των φυτών που είχαν σχηματίσει VAM μπορεί να σχετίζεται με τις απαιτήσεις σε C του μύκητα, οι οποίες είναι πολύ δύσκολο να καλυφθούν από το φυτό λόγω του stress που έχει υποστεί από την υψηλή συγκέντρωση Cd. Από πειράματα σε φυτοδοχεία των Bi, et al., (2003), σε φυτά τριφυλλιού στα οποία έγινε εμβολιασμός με το μύκητα Glomus mosseae και αναπτυχθήκαν σε έδαφος εμπλουτισμένο με μέταλλα, παρατηρήθηκε, 40 ημέρες μετά τη σπορά, ότι οι συγκεντρώσεις των μετάλλων ήταν πολύ μεγαλύτερες στις ρίζες σε σχέση με τις συγκεντρώσεις στους βλαστούς. Σκοπός της εργασίας Σκοπός της παρούσας εργασίας ήταν η διερεύνηση της επίδρασης τριών βαρέων μετάλλων (Cd, Ni, Pb) στη δράση της μυκόρριζας στα φυτά Lollium perrene και Allium cepa, τα οποία σπάρθηκαν σε δύο εδάφη, από τη περιοχή Γαλάτιστας και Κερασιάς, φυσικά και εμπλουτισμένα με τα υπό μελέτη μέταλλα. Επιπλέον εξετάστηκε η επίδραση των τριών μετάλλων στη μικροβιακή βιομάζα C και Ν των δύο εδαφών και διερευνήθηκε η επίδραση τόσο της μυκόρριζας όσο και των μετάλλων, στη σταθερότητα της δομής και στη διασπορά της αργίλου των εδαφών. 20
25 ΚΕΦΑΛΑΙΟ 3 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3.1 ΕΔΑΦΗ Για τη διεξαγωγή της μελέτης αυτής χρησιμοποιήθηκαν δύο επιφανειακά εδάφη με διαφορετικές ιδιότητες (Πίν.3.1). Το ένα λήφθηκε από τη περιοχή της Κερασιάς, του νομού Θεσσαλονίκης και το άλλο από τη περιοχή της Γαλάτιστας του νομού Χαλκιδικής. Πίνακας 3.1: Φυσικοχημικά χαρακτηριστικά των εδαφών ΙΔΙΟΤΗΤΕΣ ΓΑΛΑΤΙΣΤΑ ΚΕΡΑΣΙΑ ph 5,51 6,87 Μηχανική σύσταση Αμμοαργιλοπηλώδες Αμμοαργιλοπηλώδες Οργανική ουσία % 1,87 1,56 NO - 3 ppm 2,67 2,44 P ppm 5,9 14,5 EC μs/cm Ολικό Ν % 0,09 0,09 ΣΣ % 71,2 40,32 CEC (me/100 g) 11,3 17,39 21
26 3.1.1 Αναλύσεις χαρακτηρισμού των εδαφών Μετά τη συλλογή τους, τα εδάφη αυτά αεροξηράθηκαν, λειοτριβήθηκαν και περάστηκαν από κόσκινο με οπές 2 mm. Οι αναλύσεις χαρακτηρισμού τους έγιναν ως εξής: η μηχανική σύσταση με τη Διεθνή μέθοδο του σιφωνίου, το ph σε αναλογία 1:2,5 CaCl 2, η οργανική ουσία με τη μέθοδο της υγρής οξείδωσης κατά Walkey and Black, ο αφομοιώσιμος Ρ προσδιορίσθηκε - με τη μέθοδο κατά Olsen, τα ΝΟ 3 με εκχύλιση με 1Μ KCl (Norman et al., 1985), τα ανταλλάξιμα με τη μέθοδο του CH 3 COONH 4, η ικανότητα ανταλλαγής κατιόντων (Ι.Α.Κ.) με τη μέθοδο του CH 3 COONa ph 8,2 και ο προσδιορισμός των βαρέων μετάλλων έγινε με τη μέθοδο DTPA (Lindsay and Norvell, 1978) Εμπλουτισμός των εδαφών με μέταλλα Για τη διεξαγωγή της μελέτης χρειάστηκε να εμπλουτιστούν τα εδάφη με μεγαλύτερη συγκέντρωση των υπό μελέτη μετάλλων. Σε μία ποσότητα 6 kg από τα υπό μελέτη εδάφη προστέθηκαν τα μέταλλα Cd, Pb και Ni με τη μορφή διαλύματος που περιείχε 15 μg Cd/g εδ, (Cd(NO 3 ) 2 ), 25 μgpb/g εδ, (Pb (NO 3 ) 2 ) και 20 μgni/g εδ (NiCl 2 6H 2 O) (Νικολογιάννης, 2005). Τα εδάφη αναμίχθηκαν και επωάσθηκαν για ένα μήνα Απολύμανση εδάφους και σπόρων Μία ποσότητα εδάφους των 800 g τοποθετείται στο πυραντήριο σε θέρμανση στους 105 C για 24 h. Κατόπιν τοποθετείτε σε γλαστράκια για τη σπορά. Έγινε και απολύμανση των σπόρων (στις μεταχειρίσεις με το απολυμασμένο έδαφος) με εμβάπτιση των σπόρων σε διάλυμα νερού : χλωρίνης (10:1) Μικροβιακή βιομάζα C και Ν Δείγματα εδάφους των 20 και 25 g τοποθετούνται σε επωαστήριο στους 25 0 C για 2 εβδομάδες αφού προστέθηκε σε αυτά νερό μέχρι το σημείο της υδατοχωρητικότητας Το επωαστήριο αεριζόταν κάθε μέρα και ελεγχόταν η εδαφική υγρασία, να διατηρείται σταθερή. Για κάθε μεταχείριση χρησιμοποιούνταν τέσσερα δείγματα 22
27 Η μικροβιακή βιομάζα -C προσδιορίσθηκε όπως προτείνουν οι Jenkinson and Powlson, (1976), και Vance et al., (1987). Δύο δείγματα από τα τέσσερα απολυμαίνονται με χλωροφόρμιο σε αντλία κενού για 24 h. Τα τέσσερα δείγματα εδάφους των 20 g, δύο απολυμασμένα με χλωροφόρμιο και δύο μη, ανακινούνται με 80 ml 0,5 M K 2 SO 4 για 30 min. Μετά την ανακίνηση το αιώρημα φυγοκεντρείται και διηθείται. Στο εκχύλισμα προσδιορίζεται ο διαλυτός C με την μέθοδο COD (οξείδωση με διχρωμικό κάλιο). Συγκεκριμένα, λαμβάνονται από το εκχύλισμα 1-10 ml και τοποθετούνται σε κωνικές φιάλες των 100 ml. Κατόπιν προστίθενται 10 ml K 2 Cr 2 O 7 0,25 N, 30 ml πυκνό H 2 SO 4 και μικρή ποσότητα (περίπου 70 mg) HgO. Το μίγμα αυτό τοποθετείται για βρασμό (150 C) στην εστία για 2 h. Μετά το τέλος του βρασμού αφήνεται να κρυώσει σε θερμοκρασία περιβάλλοντος. Κατόπιν προστίθεται 2 ml διφαινυλαμίνη και ογκομετρείται με 0,25 Ν (NH 4 ) 2 SO 4 FeSO 4 6H 2 O. Η μικροβιακή βιομάζα -C υπολογίζεται από τον τύπο: BC=2,64*EC Όπου EC η διαφορά μεταξύ του Μέσου Όρου (ΜΟ) του εκχυλιζόμενου C από το απολυμασμένο και του μη εδάφους. Η μικροβιακή βιομάζα -N προσδιορίσθηκε όπως προτείνει ο Rowell, (1994). Δύο δείγματα απολυμαίνονται με χλωροφόρμιο σε αντλία κενού για 24 h. Τα τέσσερα δείγματα εδάφους των 25 g, δύο απολυμασμένα με χλωροφόρμιο και δύο μη, ανακινούνται με 100 ml 2 M KCl για 30 min. Μετά την ανακίνηση το αιώρημα φυγοκεντρείται και διηθείται. Στο εκχύλισμα προσδιορίζεται το διαλυτό Ν με την μέθοδο της nindhydrin. Συγκεκριμένα, από το εκχύλισμα παίρνονται 2 ml και τοποθετούνται σε δοκιμαστικούς σωλήνες των 50 ml, για το προσδιορισμό του Ν, και προστίθενται αργά και με ανακίνηση 1 ml διαλύματος ninhydrin. Οι δοκιμαστικοί σωλήνες τοποθετούνται σε υδρόλουτρο (100 C) για 25 min. Αφήνονται να κρυώσουν σε θερμοκρασία δωματίου και στη συνέχεια προστίθενται αργά 20 ml διαλύματος νερό με αιθανόλη (1:1). Το διάλυμα χρωματίζεται μωβ και μετράται η προσροφητικότητά του στο σπεκτοφωτόμετρο σε μήκος κύματος 570 nm. Για τον υπολογισμό της συγκέντρωσης Ν σε ppm, χρησιμοποιείται η καμπύλη αναφοράς η οποία κατασκευάζεται με γνωστές συγκεντρώσεις Ν. Η μικροβιακή βιομάζα -N υπολογίζεται από τον τύπο: BN=4.6*EN Όπου EN η διαφορά μεταξύ του (ΜΟ) του εκχυλιζόμενου N από το απολυμασμένο και του μη εδάφους. 23
28 Από τη μικροβιακή βιομάζα-c και από τη μικροβιακή βιομάζα-ν υπολογίσθηκε ο μικροβιακός λόγος C/N των δύο εδαφών τόσο στο φυσικό έδαφος όσο και στο εμπλουτισμένο με τα βαρέα μέταλλα Σταθερότητα των συσσωματωμάτων στη διαβροχή Η ΣΣ προσδιορίστηκε σε συσσωματώματα μεγέθους 2-1 mm, με τη μέθοδο του υγρού κοσκινίσματος (Angers and Mehuys, 1993). Χρησιμοποιήθηκε ένα κόσκινο με διάμετρο οπών 250 μm. Το κόσκινο με τα κορεσμένα συσσωματώματα ανακινείτο παλινδρομικά κατά τη κατακόρυφη διεύθυνση μέσα σε νερό, επί 3 min με συχνότητα 60 παλινδρομήσεις/min. Εν συνεχεία προσδιορίζονταν με ξήρανση στους 105 C η μάζα των σταθερών συσσωματωμάτων. Κατόπιν, τα σταθερά συσσωματώματα, διαμερίζονταν και προσδιορίζονταν η χονδρή άμμος (> 250 μm). Η ΣΣ υπολογίζονταν από το τύπο: μάζα σταθερών σε νερό συσσωματωμάτων μάζα άμμου (> 250 μm) / αρχική μάζα δείγματος μάζα άμμου (> 250 μm) Διασπορά της αργίλου Για το προσδιορισμό της μηχανικά διασπειρόμενης αργίλου, ποσότητα εδάφους ίση με 1 g και με διάμετρο συσσωματωμάτων μικρότερη των 2 mm, τοποθετείται σε πλαστικό σωλήνα φυγοκέντρησης των 50 ml. Προστίθενται 30 ml νερό, ο σωλήνας πωματίζεται και ανακινείται οριζόντια για 30 min με συχνότητα 100 στρ/min. Μετά την ανακίνηση ακολουθεί ακινησία για 2 h. Στη συνέχεια, με τη βοήθεια ενός σιφωνίου των 5 ml λαμβάνεται μία ποσότητα από το αιώρημα που περιέχεται στο σωλήνα, από βάθος 2 cm. Σε αυτή τη ποσότητα μετριέται στο σπεκτοφωτόμετρο η % διαπερατότητα (Τ %) σε μήκος κύματος 640 nm (Curtin et al., 1994). Για το προσδιορισμό της αυθόρμητα διασπειρόμενης αργίλου, ποσότητα εδάφους ίση με 1 g και με διάμετρο συσσωματωμάτων μικρότερη των 2 mm, τοποθετείται σε πλαστικό σωλήνα φυγοκέντρησης των 50 ml. Προστίθονται 0,5 ml νερό και παραμείνουν έτσι για κάποιες ώρες. Στη συνέχεια προστίθονται με αργό ρυθμό 30 ml νερό, ο σωλήνας πωματίζεται και παραμένει σε ηρεμία για 30 min. Κατόπιν πραγματοποιούνται τρεις ήπιες αναστροφές με το χέρι και ακολουθεί ακινησία για 2 h. Στη συνέχεια, με τη βοήθεια ενός σιφωνίου των 5 ml λαμβάνεται μία ποσότητα από το αιώρημα που περιέχεται στο σωλήνα, από βάθος 2 cm. Σε αυτή τη ποσότητα μετριέται στο σπεκτοφωτόμετρο η % διαπερατότητα (Τ %) σε μήκος κύματος 640 nm (Rengasamy et al., 1984). 24
29 3.2 ΦΥΤΟ Σε πλαστικά δοχεία του 1 kg τοποθετήθηκαν 800 g εδάφους του οποίου τα συσσωματώματα είχαν μέγεθος μικρότερο των 6 mm. Η βάση του δοχείου ήταν διάτρητη και τα δοχεία τοποθετήθηκαν μέσα σε πλαστικούς υποδοχείς. Για τη σπορά χρησιμοποιήθηκαν δύο είδη φυτών: το Lollium perrene (γκαζόν) και Allium cepa (κρεμμύδι). Για τη σπορά του κρεμμυδιού κάθε δοχείο σπάρθηκε με περίπου 50 σπόρους του Allium cepa και για τη σπορά του γκαζόν κάθε δοχείο σπάρθηκε με 0,5 g σπόρου Lollium perrene. Εφαρμόστηκαν έξι διαφορετικές μεταχειρίσεις για κάθε έδαφος: έδαφος φυσικό, έδαφος απολυμασμένο (μάρτυρας), έδαφος εμπλουτισμένο, έδαφος εμπλουτισμένο απολυμασμένο (μάρτυρας), έδαφος απολυμασμένο με μυκόρριζα του γένους Glomus, έδαφος εμπλουτισμένο απολυμασμένο με μυκόρριζα του γένους Glomus. Για το Allium cepa πραγματοποιήθηκαν τρεις επαναλήψεις και για το Lollium perrene πραγματοποιήθηκαν δύο επαναλήψεις. Ένα μήνα περίπου από τη σπορά, εφαρμόστηκε λίπανση των στοιχείων Κ και Ν. Εφαρμόστηκαν 10 ml διαλύματος/δοχείο με διάλυμα όγκου 1000 ml που περιείχε 0,932 g KNO 3 και 2,546 g ΝΗ 4 ΝΟ 3 (Terman and al., 1974). Για το Lollium perrene πραγματοποιήθηκαν συνολικά τρεις κοπές του υπέργειου τμήματος (μία κοπή/μήνα). Για το Allium cepa πραγματοποιήθηκε μια κοπή του υπέργειου τμήματος στο τέλος της καλλιέργειας. Η φυτομάζα ξηράθηκε στους 60 C και έγινε προσδιορισμός των μετάλλων μετά από καύση της ξηρής φυτομάζας (Benton-Jones and Case,1990). Στο τέλος της καλλιέργειας έγινε και παραλαβή του ριζικού συστήματος των φυτών. Το ριζικό σύστημα κάθε φυτού απομακρύνονταν με το χέρι και το πολύ έδαφος εκκαθαρίζονταν με τη βοήθεια συσκευής υπερήχων. Ακολούθησε ο χρωματισμός ενός μέρους των ριζών με τη χρωστική Trypan blue, για να είναι δυνατή η παρατήρηση, μέσω μικροσκοπίου, της παρουσίας ή όχι μυκορριζικών μυκήτων, καθώς και ο προσδιορισμός της συγκέντρωσης των υπό μελέτη βαρέων μετάλλων στο ριζικό σύστημα των φυτών. Για τη περίπτωση του Lollium perrene, λόγω της μεγάλης ποσότητας των ριζών, ο χρωματισμός πραγματοποιήθηκε στο 30 % του ριζικού συστήματος των φυτών και το υπόλοιπο ξηράθηκε στους 60 C και έγινε προσδιορισμός της συγκέντρωσης των μετάλλων μετά από καύση τω ξηρών ριζών (Benton-Jones and Case,1990). Στη περίπτωση του Allium cepa είχαμε μικρή ποσότητα ριζικού συστήματος και πραγματοποιήθηκε ο χρωματισμός σε όλο το ριζικό σύστημα των φυτών. Για το λόγο αυτό δεν ήταν δυνατός ο προσδιορισμός της συγκέντρωσης των υπό μελέτη βαρέων μετάλλων στο ριζικό σύστημα του Allium cepa. Ο χρωματισμός των ριζών έγινε σύμφωνα με τη μέθοδο που περιγράφουν οι R.E. Koske and J. N. Gemma (1989). Σύμφωνα με τη μέθοδο αυτή, οι ρίζες, αφού καθαριστούν προσεκτικά από τα υπολείμματα εδάφους, τεμαχίζονται σε πολύ μικρά 25
30 κομμάτια. Στη συνέχεια εμβαπτίζονται σε διάλυμα ΚΟΗ 10 % το οποίο είναι πάνω σε θερμαινόμενη εστία στους C για min. Κατόπιν ξεπλένονται αρκετές φορές με νερό και εμβαπτίζονται σε διάλυμα το οποίο περιέχει 20 ml NH 4 OH και 45 ml Η 2 Ο 2 και έχει συμπληρωθεί μέχρι το 1 L με Η 2 Ο, το οποίο βρίσκεται πάνω σε θερμαινόμενη εστία στους 90 C. Οι ρίζες παραμένουν σε αυτό το διάλυμα για λίγα λεπτά και στη συνέχεια ξεπλένονται αρκετές φορές με νερό. Έπειτα εμβαπτίζονται σε διάλυμα HCl 1 % και μετά πραγματοποιείται ξέπλυμα με νερό κατευθείαν. Κατόπιν εμβαπτίζονται στο διάλυμα της χρωστικής το οποίο περιέχει 500 ml glycerol, 450 ml Η 2 Ο, 50 ml HCl 1 % και 0,5 g/l Trypan blue. Οι ρίζες παραμένουν στο διάλυμα της χρωστικής για 1 h. Στη συνέχεια, οι ρίζες ξεπλένονται με το παραπάνω διάλυμα στο οποίο δεν έχουμε προσθέσει τη χρωστική. Θα πρέπει να σημειωθεί ότι με αυτό το διάλυμα, χωρίς τη χρωστική, οι ρίζες μπορούν να αποθηκευτούν εωσότου γίνει η παρατήρηση στο μικροσκόπιο. Μετά τη χρώση των ριζών είναι δυνατή η παρατήρηση στο μικροσκόπιο της ύπαρξης ή όχι μυκήτων που σχημάτισαν μυκόρριζα καθώς τα όργανα του μύκητα (υφές, σπόρια κ.τ.λ.) αποκτούν ένα έντονο μπλε χρώμα. Επίσης, με τον τρόπο αυτό μπορούμε να διακρίνουμε εάν η πυκνότητα αποικισμού του μύκητα στις ρίζες είναι μεγάλη ή μικρή Ο εμβολιασμός με μυκόρριζα Glomus mosseae Ο εμβολιασμός με μυκόρριζα πραγματοποιήθηκε ένα μήνα μετά τη σπορά. Χρησιμοποιήθηκε το είδος Glomus mosseae (BEG 12) υπό μορφή σπορίων. Μία ποσότητα εδάφους που περιείχε τα σπόρια του μύκητα αναμείχθηκε με λίγο νερό και εφαρμόστηκε στην επιφάνεια της γλάστρας. 26
31 ΚΕΦΑΛΑΙΟ 4 ΑΠΟΤΕΛΕΣΜΑΤΑ ΚΑΙ ΣΥΖΗΤΗΣΗ 4.1 Εδάφη Τα δύο εδάφη, της Γαλάτιστας και της Κερασιάς, που χρησιμοποιήθηκαν για την μελέτη αυτή ανήκουν στα Alfisol. Η επιλογή τους, έγινε αρχικά, γιατί είχαν διαφορετικό ph, το έδαφος της Κερασιάς είχε ph = 6,87 και το έδαφος της Γαλάτιστας είχε ph = 5,51. Είχαν όμως την ίδια μηχανική σύσταση, η άργιλος τους περιείχε πολλά οξείδια σιδήρου και αργιλίου και είχαν το ίδιο ποσοστό οργανικής ουσίας. Διέφεραν όμως ως προς την ΙΑΚ, που για το έδαφος της Κερασιάς ήταν 17,39 me/100g και για το έδαφος της Γαλάτιστας 11,3 me/100g, ως προς τη σταθερότητα δομής, το έδαφος της Κερασιάς είχε σταθερότητα 40,32 % και το έδαφος της Γαλάτιστας 71,2 % (Πίν. 3.1) και ως προς τη συγκέντρωση των βαρέων μετάλλων Cd, Ni και Pb που για το έδαφος της Κερασιάς ήταν 0,346, 2,8 και 2,25 ppm αντίστοιχα και για το έδαφος της Γαλάτιστας 0,053, 1,27 και 3,23 ppm αντίστοιχα. Τα εδάφη αυτά για τις ανάγκες της μελέτης αυτής, εμπλουτίστηκαν με βαριά μέταλλα. Οι συγκεντρώσεις των βαρέων μετάλλων μετά τον εμπλουτισμό τους ήταν 14,7, 24,1 και 14,93 ppm για τη Κερασιά και 2,63, 34,8 και 35,8 για τη Γαλάτιστα για Cd, Ni και Pb αντίστοιχα.(πίν. 4.1). Πίνακας 4.1 Συγκεντρώσεις των μετάλλων Cd, Ni και Pb σε ppm πριν και μετά τον εμπλουτισμό των εδαφών της Γαλάτιστας και της Κερασιάς Στοιχείο Γαλάτιστα Κερασιά πριν μετά πριν μετά Cd ppm 0,053a 2,63b 0,346c 14,7d Pb ppm 3,23e 35,8f 2,25e 14,93g Ni ppm 1,27k 34,8m 2,8n 24,1q Οι τιμές διαφέρουν στατιστικά μεταξύ τους σε επίπεδο p<0,05 (a,b,c συμβολίζονται οι σημαντικές διαφορές μεταξύ των τιμών, ενώ ίδια γράμματα μεταξύ των τιμών συμβολίζουν μη σημαντικές διαφορές) 27
32 4.2 Μικροβιακή βιομάζα C και Ν Η μικροβιακή βιομάζα θεωρείται ένας πολύ ευαίσθητος δείκτης της μεταβολής των εδαφικών συνθηκών και ιδίως της ρύπανσης των εδαφών με βαριά μέταλλα. Έχει αποδειχθεί ότι η παρουσία μεγάλης συγκέντρωσης βαρέων μετάλλων προκαλεί μείωση του C και του Ν της μικροβιακής βιομάζας (Brookes and McGrath 1984, Dai et al., 2004, Murrieta et al., 2006). Η μικροβιακή βιομάζα - C (ΒC) για τα εδάφη της Κερασιάς και της Γαλάτιστας ήταν 229,79 και 66,29 mgc/kg εδάφους αντίστοιχα και η μικροβιακή βιομάζα - Ν (ΒΝ) ήταν 14,47 και 3,096 mgν/kg εδάφους αντίστοιχα (Πίν. 4.2). Η ΒC συμφωνεί με τα ευρήματα άλλων ερευνητών (Dai et al., 2004, Kautz, 2004, Murrieta et al., 2006, κ.α.) αλλά η ΒΝ ήταν μικρότερη από αυτή που αναφέρεται στη βιβλιογραφία. Αυτό πιθανόν να οφείλεται στη διαφορετική μέθοδο προσδιορισμού της μικροβιακής μάζας C και Ν που χρησιμοποιήθηκε. Αύξηση των μετάλλων στα εδάφη, είχε σαν αποτέλεσμα για το έδαφος της Κερασιάς τη μείωση των μικροβιακών παραμέτρων ΒC και BN (Πίν. 4.2) από 229,79 σε 194,44 και 14,47 σε 7,649 αντίστοιχα. Η ίδια τάση όμως δεν παρατηρήθηκε στο έδαφος της Γαλάτιστας όπου σημειώθηκε αύξηση των μικροβιακών παραμέτρων ΒC, BN (Πίν. 4.2) από 66,29 σε 185,603 και 3,096 σε 13,75 αντίστοιχα. Το ίδιο βρήκαν και άλλοι ερευνητές οι οποίοι πρότειναν οι μικροβιακές αυτές παράμετροι να μπορούν να χρησιμοποιηθούν σαν δείκτες ρύπανσης των εδαφών από βαριά μέταλλα (Brookes and McGrath 1984, Lavelle and Spain 2001, Yuangen et al., 2006, Murrieta et al., 2006, κ.α.). Εκτός από τους μικροβιακούς παραμέτρους ΒC, BN ως δείκτες της εδαφικής ρύπανσης από βαριά μέταλλα, πολλοί ερευνητές έχουν προτείνει να χρησιμοποιείται ο λόγος του εδαφικού μικροβιακού C ως προς τον C της οργανικής ουσίας (Brookes and McGrath 1984, Giller et al. 1998, Murrieta et al 2006). Ένας δείκτης που δείχνει την κατανομή της δομής των μικροοργανισμών στο έδαφος είναι ο λόγος του C προς το Ν της μικροβιακής βιομάζας. Ο μικροβιακός λόγος C/N της βιομάζας για τα βακτήρια έχει βρεθεί ότι έχει τιμή 3,5-10 ενώ οι μύκητες χαρακτηρίζονται συνήθως από υψηλότερους λόγους (10-15:1) (Breland, 1997, Dai et al., 2004). O μέσος μικροβιακός λόγος C/N σε ένα υγιές έδαφος είναι 5,3-10 (Bremer and Kessel, 1992). Ο μικροβιακός λόγος C/N για το έδαφος της Κερασιάς ήταν 15,8 και για το έδαφος της Γαλάτιστας 21,46. Από αυτά τα δεδομένα φαίνεται ότι στο πληθυσμό των μικροοργανισμών και των δύο εδαφών υπερτερούσαν οι μύκητες, με μεγαλύτερη κατανομή δομής τους μύκητες στο έδαφος της Γαλάτιστας (Joergensen et al., 1995). Αυτό πιθανόν μπορεί να οφείλεται στο χαμηλό ph που είχε το έδαφος της Γαλάτιστας, που ως γνωστόν οι μύκητες αντέχουν σε όλο το φάσμα του ph εν αντιθέσει με τα βακτήρια και τους ακτινομύκητες, που είναι ευαίσθητοι στα χαμηλά ph. 28
33 Πίνακας 4.2: Η μικροβιακή βιομάζα - C (BC) και - N (BN) σε mg/kg εδάφους και ο μικροβιακός λόγος C/N των εδαφών Κερασιάς και Γαλάτιστας ως έχει (Φ) και με βαριά μέταλλα (Μ). Κερασιά Γαλάτιστα Φ Μ Φ Μ BC mgc/kg εδ. 229,79a 194,44b 66,29c 185,603d BN mgn/kg εδ. 14,47e 7,649f 3,096m 13,75n BC/Co 2,5 2,26 0,6 2,99 C/N 15,8 25,42 21,46 13,50 Οργ. Ουσία % 1,56a 1,348a 1,878a 1,062b Οι τιμές διαφέρουν στατιστικά μεταξύ τους σε επίπεδο p<0,05 (a,b,c,d συμβολίζονται οι σημαντικές διαφορές μεταξύ των τιμών, ενώ ίδια γράμματα μεταξύ των τιμών συμβολίζουν μη σημαντικές διαφορές) O λόγος C/N της βιομάζας διαφέρει σημαντικά στα διάφορα επίπεδα ρύπανσης με βαριά μέταλλα. Οι Dai et al., 2004 αναφέρουν ότι ο λόγος αυτός κυμαίνεται από 10,3 μέχρι 3,5 από πολύ ρυπασμένα μέχρι ελαφρώς ρυπασμένα εδάφη, ενώ αύξηση του λόγου αυτού κατά τους Joergensen et al., (1995) οφείλεται σε αύξηση του ποσοστού των μυκήτων στο έδαφος. Σύμφωνα με πολλούς ερευνητές, οι μύκητες είναι πιο ανθεκτικοί στα βαριά μέταλλα από ότι τα βακτήρια και οι ακτινομύκητες (Giller et al Dai et al., 2004). Για το έδαφος της Κερασιάς ο μικροβιακός λόγος C/N αυξήθηκε από 15,8 σε 25,4 με αύξηση της συγκέντρωσης των βαρέων μετάλλων (Πίν. 4.2). Αυτό πιθανόν να οφείλεται στο ότι, η αύξηση της συγκέντρωσης των βαρέων μετάλλων κατέστρεψε ένα μεγάλο πληθυσμό ακτινομυκήτων και βακτηρίων που ήταν ευαίσθητα στις μεγάλες συγκεντρώσεις βαρέων μετάλλων στο έδαφος με παράλληλη αύξηση του πληθυσμού των μυκήτων που αντέχουν στη παρουσία τους (Hiroki, 1992, McGrath, 1994, Giller et al. 1998, Dai et al., 2004, Yuangen et al. 2006). Για το έδαφος της Γαλάτιστας, ο μικροβιακός λόγος C/N μειώθηκε από 21,46 σε 13,3 με αύξηση της συγκέντρωσης των βαρέων μετάλλων (Πίν. 4.2). H μείωση του μικροβιακού λόγου C/N, στο έδαφος της Γαλάτιστας, οφείλεται στην μικρότερη συγκέντρωση του Cd που είχε το έδαφος αυτό μετά τον εμπλουτισμό του με βαριά μέταλλα. Αυτή η συγκέντρωση του Cd (2,63 ppm) φαίνεται ότι δεν επηρεάζει την 29
34 ανάπτυξη των μικροοργανισμών στο έδαφος και αυτό φαίνεται από την οργανική ουσία του εδάφους αυτού που μειώθηκε από 1,878 σε 1,062 % κατά την επώαση εμπλουτισμού των εδαφών ενώ η οργανική ουσία στο έδαφος της Κερασιάς παρέμεινε σταθερή (πιν 4.2). Η μείωση της οργανικής ουσίας συνεπάγει και αύξηση του πληθυσμού των μικροοργανισμών όχι μόνο των μυκήτων και επομένως ελάττωση του μικροβιακού λόγου C/N. Στο κεφάλαιο αυτό εξετάσθηκε η επίδραση της αύξησης των βαρέων μετάλλων στο έδαφος στη βιομάζα των μικροοργανισμών του εδάφους γιατί ως γνωστόν οι μυκόρριζες αποτελούν ένα μέρος των μυκήτων του εδάφους. 4.3 Φυτό Για τη μελέτη της επίδρασης των βαρέων μετάλλων στις μυκόρριζες, φυτά Lollium perrene (γκαζόν) και Allium cepa (κρεμμύδι) σπάρθηκαν α)σε εδάφη ως έχουν (φυσικά), β) σε εδάφη που είχαν απολυμανθεί, γ) σε εδάφη που είχαν εμπλουτισθεί με βαριά μέταλλα δ) σε εμπλουτισμένα με μέταλλα εδάφη που είχαν απολυμανθεί τόσο τα εδάφη όσο και οι σπόροι των φυτών και ε) σε απολυμασμένα εδάφη φυσικά και εμπλουτισμένα με μέταλλα που είχαν μολυνθεί με μυκόρριζα Glomus mosseae. Η επιλογή των δύο αυτών φυτών έγινε γιατί αυτά σχηματίζουν μυκόρριζες, χωρίς να είναι απαραίτητος ο εμβολιασμός τους με μυκορριζικό μύκητα. Επίσης το Lollium perrene παρουσιάζει ανθεκτικότητα στις υψηλές συγκεντρώσεις βαρέων μετάλλων καθώς επίσης σχηματίζει και πλούσιο ριζικό σύστημα. Κατά τη διάρκεια της ανάπτυξης των φυτών στις γλάστρες παρατηρήθηκε η δημιουργία επιφανειακής κρούστας η οποία εμπόδισε το φύτρωμα των σπόρων και το πότισμα των γλαστρών (Παράρτημα φωτογραφιών). Μάλιστα, λόγω της ύπαρξης της επιφανειακής αυτής κρούστας, δεν ήταν δυνατή η ανάπτυξη των φυτών του Allium cepa στο έδαφος της Γαλάτιστας. Αν και η βλάστηση των σπόρων πραγματοποιήθηκε κανονικά, δεν ήταν δυνατή η περαιτέρω ανάπτυξη των φυτών καθώς παρατηρήθηκε ξήρανση λόγο έλλειψης νερού των μικρών φυτών περίπου μια εβδομάδα μετά τη βλάστησή τους. Για το λόγο αυτό θεωρήθηκε σκόπιμο να μελετηθεί η σταθερότητα των εδαφών αυτών και η διασπορά της αργίλου σε όλες τις μεταχειρίσεις, καθώς αυτές οι παράμετροι καθορίζουν το σχηματισμό της επιφανειακής κρούστας. 30
35 4.3.1 Σταθερότητα των εδαφών Για τα δύο εδάφη πραγματοποιήθηκε προσδιορισμός της σταθερότητας (ΣΣ %) των συσσωματωμάτων (2-1 mm) στη διαβροχή, τόσο πριν όσο και μετά τη σπορά, κατά την ανάπτυξη των φυτών. Επίσης προσδιορίστηκε η οργανική ουσία για τις διάφορες μεταχειρίσεις πριν τη σπορά, επειδή: α) αποτελεί παράγοντα σταθερότητας της δομής και β) το ποσοστό της μπορεί να μεταβληθεί με την επώαση. Τα αποτελέσματα παρουσιάζονται στο Πίνακα 4.3. Τα αποτελέσματα του Πίνακα 4.3 παριστάνονται γραφικά και για τα δύο εδάφη στο Σχήμα 4.1. Πίνακας 4.3: Σταθερότητα των συσσωματωμάτων στη διαβροχή (ΣΣ %) για τα δύο εδάφη, πριν και μετά τη σπορά. ΜΕΤΑΧΕΙΡΙΣΕΙΣ %Οργανική ουσία Πρίν τη σπορά ΣΣ % Πριν τη σπορά ΣΣ % Lollium perrene ΣΣ % Allium cepa Κερασιά φυσικό 1,56 41,21a 37,96a 42,89a Κερασιά απολυμασμένο 1,479-45,91a 48,80a Κερασιά εμπλουτισμένο 1,348 56,02b 39,52a 54,81a Κερασιά εμπλουτισμένο απολυμασμένο 1,477 37,04a 39,16a 41,00a Κερασιά απολυμασμένο (Glomus) - 41,76a 46,76a Κερασιά εμπλουτισμένο απολυμασμένο (Glomus) - 31,09a 42,67a Γαλάτιστα φυσικό 1,878 68,12c 56,16a - Γαλάτιστα απολυμασμένο 1,445 75,47c 36,19b - Γαλάτιστα εμπλουτισμένο 1,062 37,59d 41,98c - Γαλάτιστα εμπλουτισμένο απολυμασμένο 1,126 42,79d 37,91d - Γαλάτιστα απολυμασμένο (Glomus) - 33,73e - Γαλάτιστα εμπλουτισμένο απολυμασμένο (Glomus) - 31,79e - Οι τιμές διαφέρουν στατιστικά μεταξύ τους σε επίπεδο p<0,05 Η σύγκριση έχει πραγματοποιηθεί μεταξύ των τιμών της ίδιας στήλης και χωριστά για κάθε έδαφος (a,b,c,d κ.τ.λ. συμβολίζονται οι σημαντικές διαφορές μεταξύ των τιμών, ενώ ίδια γράμματα μεταξύ των τιμών συμβολίζουν μη σημαντικές διαφορές.) 31
36 ΣΣ % πριν τη σπορά Lollium perrene Κερασιά Allium cepa Allium cepa(g) Lollium perrene (G) Φ Απ. Εμ. Εμ-Απ ΣΣ % πριν τη σπορά Γαλάτιστα Lollium perrene Lollium perrene (G) Φ Απ Εμ Εμ-Απ Σχήμα 4.1: Γραφική αναπαράσταση της σταθερότητας των συσσωματωμάτων στη διαβροχή Για το έδαφος της Κερασιάς πριν τη σπορά, ο εμπλουτισμός με βαριά μέταλλα προκαλεί σημαντική αύξηση της ΣΣ %, σε σχέση με το φυσικό έδαφος. Επειδή δε παρατηρήθηκε μεταβολή της οργανικής ουσίας, αυτή η αύξηση της ΣΣ % πρέπει να οφείλεται στη προσρόφηση των μετάλλων στο έδαφος, όπως φαίνεται από το λόγο του αθροίσματος των βαρέων μετάλλων στο φυσικό προς το επωασμένο έδαφος (1:10) και στο ρόλο τους ως συνδετικών παραγόντων σε αυτά τα εδάφη. Τα πολυσθενή μεταλλικά κατιόντα αποτελούν γέφυρες που συνδέουν την οργανική ουσία με τα τεμαχίδια της αργίλου και συμβάλλουν στη αύξηση της σταθερότητας (Jiovannini and Sequi, 1976). Δεδομένου αυτού, και για να διερευνηθεί ο ρόλος των βαρέων μετάλλων ως συνδετικών παραγόντων, συσχετίστηκε η ΣΣ % των εμπλουτισμένων με βαριά μέταλλα επεμβάσεων με το άθροισμα των βαρέων μετάλλων στη στερεή φάση του εδάφους (Πίν. 4.4). Η σχέση ήταν γραμμική και σημαντική (p < 0,001) (Σχ. 4.2) και δείχνει ότι σε αυτό το έδαφος η τιμή της ΣΣ % καθορίζεται κυρίως από το ρόλο των πολυσθενών μετάλλων ως γεφυρών μεταξύ των ανόργανων και των οργανικών ενεργών επιφανειών του εδάφους. Σε παρόμοια αποτελέσματα κατέληξαν και οι Román et al., (2003). Μετά τη σπορά, οι τιμές της ΣΣ % στο έδαφος της Κερασιάς δεν διέφεραν στατιστικά μεταξύ των μεταχειρίσεων, για κανένα από τα δύο φυτά (Πίν. 4.3), αλλά μειώθηκαν σε σύγκριση με το έδαφος πριν τη σπορά. Ο εμβολιασμός με Glomus δε μετέβαλε τη ΣΣ% σε σύγκριση με τις αντίστοιχες μεταχειρίσεις χωρίς Glomus και στα δύο φυτά. 32
37 Πίνακας 4.4: Το άθροισμά των τριών βαρέων μετάλλων και η ΣΣ % για το επωασμένο έδαφος της Κερασιάς των φυτοδοχείων, στα δύο φυτά. ΜΕΤΑΧΕΙΡΙΣΗ Βαρέων μετάλλων ΣΣ % (μg/g) Κερασιά εμπλουτισμένο (γκαζόν) 31,58 39,52 Κερασιά εμπλουτισμένο απολυμασμένο (γκαζόν) 38,25 39,16 Κερασιά εμπλουτισμένο απολυμασμένο-glomus- (γκαζόν) 22,19 31,09 Κερασιά εμπλουτισμένο (κρεμμύδι) 39,63 54,81 Κερασιά εμπλουτισμένο απολυμασμένο (κρεμμύδι) 32,94 41 Κερασιά εμπλουτισμένο απολυμασμένο-glomus-(κρεμμύδι) 34,34 42,67 Κερασιά εμπλουτισμένο (πριν τη σπορά) 53,7 56,02 Στο έδαφος της Γαλάτιστας, πριν τη σπορά, στα εμπλουτισμένα με βαριά μέταλλα δείγματα, παρατηρήθηκε αξιοσημείωτη μείωση τόσο της ΣΣ % όσο και της οργανικής ουσίας (Πιν. 4.3). Η απώλεια της οργανικής ουσίας κατά τη διάρκεια της επώασης μετά τον εμπλουτισμό πρέπει να είναι μία αιτία μείωσης της ΣΣ %. Για να διερευνηθεί αυτό, πραγματοποιήθηκε επώαση του εδάφους απουσία βαρέων μετάλλων και προσδιορίστηκε η οργανική ουσία (0,41 %) και η ΣΣ (27 %). Σε αυτή τη περίπτωση η μείωση της οργανικής ουσίας και της ΣΣ % ήταν μεγαλύτερη από αυτή που παρατηρήθηκε κατά την επώαση με μέταλλα. Φαίνεται λοιπόν y = 0,8142x + 14,084 R 2 = 0,7644 ΣΣ % Σύνολο βαρέων μετάλλων (μg/g) Σχήμα 4.2: Η γραφική απεικόνιση της μεταβολής της ΣΣ % με το σύνολο των βαριών μετάλλων στο εμπλουτισμένο έδαφος της Κερασιάς ότι τα μέταλλα δρουν τοξικά στους μικροοργανισμούς με ανάλογα αποτελέσματα στο ποσοστό της οργανικής ουσίας και κατά συνέπεια στη τιμή της ΣΣ %. Η συσχέτιση της ΣΣ % με το ποσοστό της οργανικής ουσίας ήταν γραμμική (p < 0,05) (Σχ. 4.3) και έδειχνε ότι σε αυτό το έδαφος η τιμή της ΣΣ % πριν τη σπορά επηρεαζόταν από τη μεταβολή της οργανικής ουσίας. Μια άλλη πιθανή αιτία μείωσης της ΣΣ % μετά τον εμπλουτισμό με βαριά μέταλλα ήταν η 33
38 μείωση που παρατηρήθηκε στη συμμετοχή των μυκήτων στο μικροβιακό πληθυσμό. Αυτό επιβεβαιώνεται y = 33,36x + 10,63 R 2 = 0,7622 ΣΣ % ,5 1 1,5 2 οργανική ουσία % Σχήμα 4.3: Η γραφική απεικόνιση του ποσοστού της οργανικής ουσίας σε σχέση με τη ΣΣ%, για κάθε μεταχείριση, στο έδαφος της Γαλάτιστας από τη μείωση του μικροβιακού λόγου C/N, που προκάλεσε ο εμπλουτισμός του εδάφους με βαριά μέταλλα, από 21,46 σε 13,50 (Πιν. 4.2). Όπως αναφέρθηκε και στα προηγούμενα, αυτή η μείωση του μικροβιακού λόγου C/N υποδηλώνει μεταβολή της δομής της μικροβιακής κοινότητας του εδάφους της Γαλάτιστας, με μείωση του πληθυσμού των μυκήτων (Yuangen et al. 2006). Την ένδειξη της συμβολής των μυκήτων στη σταθερότητα της δομής αυτού του εδάφους ενισχύει το γεγονός ότι και η καταστροφή των μικροοργανισμών με απολύμανση προκάλεσε επίσης σημαντική μείωση της ΣΣ % σε σχέση με τις αντίστοιχες μεταχειρίσεις χωρίς απολύμανση. Η ανάπτυξη του φυτού σε αυτό το έδαφος επηρέασε τη τιμή της ΣΣ %. Κατά την ανάπτυξη του Lollium perrene γενικά παρατηρήθηκε μείωση της ΣΣ % σε σχέση με το έδαφος πριν τη σπορά. Αυτό πιθανώς οφείλεται στην εκκριτική δραστηριότητα των ριζών κατά τα πρώτα στάδια ανάπτυξης του φυτού (Oades, 1984). Αντίθετα από το αναμενόμενο, ο εμβολιασμός των απολυμασμένων εδαφών με Glomus δε συνετέλεσε σε αύξηση της ΣΣ %. Από τα παραπάνω φαίνεται ότι η ΣΣ %, στα εδάφη πριν από την ανάπτυξη των φυτών, επηρεάζεται από τις μεταβολές των παραγόντων συσσωμάτωσης (βαριά μέταλλα Κερασιά, οργανική ουσία Γαλάτιστα). Μετά την ανάπτυξη φυτού, η τιμή της ΣΣ % διαμορφώνεται περισσότερο από την άμεση επίδραση των ριζών των φυτών παρά από την ύπαρξη του μυκορριζικού μύκητα (Thomas et al., 1986). Αυτό πιθανώς οφείλεται και στη μικρή χρονική διάρκεια του πειράματος. 34
39 4.3.2 Διασπορά της αργίλου Πραγματοποιήθηκε προσδιορισμός της αυθόρμητα και μηχανικά διασπειρόμενης αργίλου για το έδαφος των φυτοδοχείων, μετά το τέλος της καλλιέργειας. Επειδή η διασπορά της αργίλου προσδιορίσθηκε ως εκατοστιαίο ποσοστό της διαπερατότητας στο φως (Τ %) ενός υδατικού αιωρήματος αργίλου, όσο αυξάνει η τιμή της Τ τόσο μειώνεται το ποσοστό της εν διασπορά αργίλου. Οι τιμές της διαπερατότητας για την αυθόρμητα (Τα %) και μηχανικά (Τμ %) διασπειρόμενη άργιλο παρουσιάζονται στο Πίνακα 4.5 και παριστάνονται γραφικά στο Σχήμα 4.4. Πίνακας 4.5: Οι τιμές της διαπερατότητας για την αυθόρμητα (Τα %) και μηχανικά (Τμ %) διασπειρόμενη άργιλο του εδάφους των φυτοδοχείων. Τα % Τμ % ΜΕΤΑΧΕΙΡΙΣΕΙΣ Lollium perrene Allium cepa Lollium perrene Allium cepa Κερασιά φυσικό 53,75b 72,10ab 16,6abc 29,15a Κερασιά απολυμασμένο 38,05a 73,95ab 12,75a 17,40b Κερασιά εμπλουτισμένο 54,2b 77,95bc 20,9bc 32a Κερασιά εμπλουτισμένο απολυμασμένο Κερασιά απολυμασμένο (G) Κερασιά εμπλουτισμένο απολυμασμένο (G) 35,3a 79,07c 49,3b 76,10bc 80,75c 68,2a 15,5ab 19bc 39,25d 22,50b 32,3a 48,9b Γαλάτιστα φυσικό 38,80a 51,65a 9,85ab 8,10a Γαλάτιστα απολυμασμένο 22,55b 61,75a 4,15a 12,85a Γαλάτιστα εμπλουτισμένο 41,45a 52,7a 12,30bc 12,55a Γαλάτιστα εμπλουτισμένο απολυμασμένο Γαλάτιστα απολυμασμένο (G) Γαλάτιστα εμπλουτισμένο απολυμασμένο (G) 31,85c 31,55c 40,25d 48,45a 62,4a 54,85a 5,50a 19,90d 20,95d 9,15a 17,70a 11,40a Οι τιμές διαφέρουν στατιστικά μεταξύ τους σε επίπεδο p<0,05 Η σύγκριση έχει πραγματοποιηθεί μεταξύ των τιμών της ίδιας στήλης και χωριστά για κάθε έδαφο.ς (a,b,c,d κ.τ.λ. συμβολίζονται οι σημαντικές διαφορές μεταξύ των τιμών, ενώ ίδια γράμματα μεταξύ των τιμών συμβολίζουν μη σημαντικές διαφορές ) 35
40 Τα, % Lollium perrene Allium cepa Κερασιά Allium cepa (G) Lollium perrene (G) Φ Απ. Εμ. Ε-Α Τα, % Lollium perrene Allium cepa Γαλάτιστα Allium cepa (G) Lollium perrene (G) Φ Απ. Εμ. Ε-Α Τμ, % Lollium perrene Allium cepa Κερασιά Allium cepa (G) Lollium perrene (G) Φ Απ. Εμ. Ε-Α Τμ, % Lollium perrene Allium cepa Γαλάτιστα Σχήμα 4.4: Οι τιμές των Τα % και Τμ % για τα εδάφη των φυτοδοχείων Allium cepa (G) Lollium perrene (G) Φ Απ. Εμ. Ε-Α Από το Πίνακα 4.5 και το Σχήμα 4.4 φαίνεται ότι ο εμπλουτισμός δε προκάλεσε μεταβολή στο ποσοστό της Τα και στα δύο εδάφη. Αντίθετα η απολύμανση αύξησε σημαντικά το ποσοστό της Τα και στα δύο εδάφη, όταν αναπτύσσεται το Lollium perrene. Αυτό οφείλεται αφ ενός στην απώλεια της συνδετικής δράσης της μικροβιακής βιομάζας και αφ ετέρου στη δραστηριότητα του ριζικού συστήματος του φυτού η οποία έχει βρεθεί ότι διεγείρει τη δημιουργία διασπειρόμενης αργίλου (Oades, 1984). Ο εμβολιασμός με το μύκητα Glomus mosseae μείωσε σημαντικά το ποσοστό της αυθόρμητα διασπειρόμενης αργίλου, στις περιπτώσεις όπου αναπτύχθηκε φυτό. Θα μπορούσε να σημειωθεί ότι η μείωση της διασποράς που προκάλεσε ο μύκητας επισκίαζε την αρνητική επίδραση του Lollium perrene στη διασπορά της αργίλου. Οι Thomas και συν. (1993) βρήκαν επίσης ότι η δράση της μυκόρριζας επιβράδυνε τη διασπορά της αργίλου. Στο έδαφος της Γαλάτιστας, όπου δε μπόρεσε να αναπτυχθεί το Allium cepa, δε παρατηρήθηκε μεταβολή της Τα μετά τον εμβολιασμό με το μύκητα Glomus mosseae. Πιθανώς, λόγω της απουσίας φυτού δεν επέζησε και ο μύκητας. Οι μεταβολές της μηχανικά διασπειρόμενης αργίλου στις διάφορες επεμβάσεις ήταν παρόμοιες με αυτές που παρατηρήθηκαν και για την Τα (Πίν. 4.5) και οφείλονται στους ίδιους 36
41 λόγους όπως αναφέρθηκε παραπάνω. Έτσι ο εμπλουτισμός με βαριά μέταλλα δεν επηρέασε το ποσοστό της Τμ. Η απολύμανση αντίθετα, προκάλεσε μια τάση αύξησης της Τμ η οποία όμως ήταν σημαντική μόνο σε μερικές περιπτώσεις. Η παρουσία του μύκητα Glomus mosseae μείωσε σημαντικά το ποσοστό της Τμ, εκτός από τη περίπτωση του Allium cepa στο έδαφος της Γαλάτιστας όπου δε παρατηρήθηκε κάποια μεταβολή της Τμ. Η διασπορά της αργίλου και στα δύο εδάφη προκάλεσε επιφανειακή κρούστα η οποία δυσχέραινε το πότισμα των φυτοδοχείων. Έτσι, το μηδενικό ποσοστό φύτρωσης των σπόρωντου Allium cepa στο έδαφος της Γαλάτιστας μπορεί να δικαιολογηθεί από τη δημιουργία επιφανειακής κρούστας στα φυτοδοχεία, λίγες μέρες μετά τη σπορά. Η αυξημένη διασπορά της αργίλου και η ανάπτυξη επιφανειακής κρούστας παρατηρήθηκε επίσης και στη περίπτωση του Lollium perrene και στα δύο εδάφη, η οποία όμως δεν εμπόδισε το φύτρωμα και την ανάπτυξη αυτού του φυτού. Αυτό πιθανώς να οφείλεται στο είδος του φυτού και στη διαφορετική αντοχή μεταξύ τους, δεδομένου ότι στη περίπτωση του Allium cepa αν και είχαμε τη φύτρωση των σπόρων, ακολουθούσε μαρασμός των φυτών, λίγες μέρες μετά, λόγω της ύπαρξης επιφανειακής κρούστας. 37
42 4.4 Βιομάζα φυτού Αφού συλλέχθηκε, η ποσότητα της φυτομάζας κάθε μεταχείρισης, ξηράθηκε και ζυγίστηκε (ξηρό βάρος). Οι ποσότητες που προέκυψαν φαίνονται στο Πίνακα 4.6 και παριστάνονται γραφικά στο Σχήμα 4.5. Πίνακας 4.6: Το ξηρό βάρος (g) της φυτομάζας για κάθε μεταχείριση ΜΕΤΑΧΕΙΡΙΣΗ Ξ.Β. Lollium perrene φυτομ. (g) Allium cepa Κερασιά φυσικό 0,906a 0,399ab Κερασιά απολυμασμένο 2,683b 0,5006a Κερασιά εμπλουτισμένο 0,923a 0,124cd Κερασιά εμπλουτισμένο απολυμασμένο 3,094cb 0,244ec Κερασιά απολυμασμένο (Glomus) 1,509c 0,369be Κερασιά εμπλουτισμένο απολυμασμένο (Glomus) 2,28b 0,089d Γαλάτιστα φυσικό 1,104a - Γαλάτιστα απολυμασμένο 2,060b - Γαλάτιστα εμπλουτισμένο 0,784c - Γαλάτιστα εμπλουτισμένο απολυμασμένο 2,218d - Γαλάτιστα απολυμασμένο (Glomus) 1,664e - Γαλάτιστα εμπλουτισμένο απολυμασμένο (Glomus) 1,383e - Οι τιμές διαφέρουν στατιστικά μεταξύ τους σε επίπεδο p<0,05 Η σύγκριση έχει πραγματοποιηθεί μεταξύ των τιμών της ίδιας στήλης και χωριστά για κάθε έδαφος (a,b,c,d κ.τ.λ. συμβολίζονται οι σημαντικές διαφορές μεταξύ των τιμών, ενώ ίδια γράμματα μεταξύ των τιμών συμβολίζουν μη σημαντικές διαφορές) 38
43 Ξηρό βάρος (g) 3,5 3 2,5 2 1,5 1 0,5 0 Lollium perrene (Κερασιά) Φ Απ Εμ Εμ-Απ Απ(G) Eμ-Απ(G) Mεταχειρίσεις Ξηρό βάρος (g) 2,5 2 1,5 1 0,5 0 Lollium perrene (Γαλάτιστα) Φ Απ Εμ Εμ-Απ Απ(G) Eμ-Απ(G) Μεταχειρίσεις Ξηρό βάρος (g) 0,6 0,5 0,4 0,3 0,2 0,1 0 Αllium cepa (Κερασιά) Φ Απ Εμ Εμ-Απ Απ(G) Eμ-Απ(G) Μεταχειρίσεις Σχήμα 4.5: Γραφική αναπαράσταση του ξηρού βάρους της φυτομάζας όλων των μεταχειρίσεων. Cd, Ni mg/kg φυτ y = -109,42x + 53,785 R 2 = 0,754 Cd Νi y = -169,2x + 77,645 R 2 = 0, ,1 0,2 0,3 0,4 0,5 0,6 Φυτομάζα Allium cepa g Σχήμα 4.6: Απεικόνιση της σχέσης των συγκεντρώσεων των Cd, Ni και Pb στο υπέργειο τμήμα του Allium cepa με τη φυτομάζα του 39
44 Παρατηρείται ότι στους μάρτυρες (απολυμασμένα και εμπλουτισμένα απολυμασμένα εδάφη) η αύξηση της συγκέντρωσης των τριών βαρέων μετάλλων δεν επηρέασε τη παραγόμενη φυτομάζα του Lollium perrene στο έδαφος της Κερασιάς (από 2,683 σε 3,094), ή παρατηρήθηκε οριακή αύξηση στο έδαφος της Γαλάτιστας, (από 2,06 g σε 2,218 g). Αυτό συμβαίνει γιατί το Lollium perrene, παρουσιάζει ανθεκτικότητα στις υψηλές συγκεντρώσεις των βαρέων μετάλλων. Αυτό φαίνεται και από τα πειραματικά δεδομένα όπου δεν βρέθηκε καμία στατιστική σχέση με τις συγκεντρώσεις των τριών βαρέων μετάλλων στη φυτομάζα. Ενώ για το Allium cepa στο έδαφος της Κερασιάς παρατηρείται σημαντική μείωση της παραγόμενης φυτομάζας από 0,500 σε 0,244 g και αυτό συμφωνεί με τη στενή αρνητική σχέση που βρέθηκε μεταξύ των συγκεντρώσεων των δύο βαρέων μετάλλων και της φυτομάζας του Allium cepa (Cd: r = - 0,907*** και Ni: r = -0,869*) (Σχ. 4.6). Στα φυσικά εδάφη (εμπλουτισμένα και μη) ως γνωστόν υπάρχουν αυτόχθονες μύκητες και επομένως μυκόρριζες. Μεταξύ των φυσικών εδαφών (εμπλουτισμένα και μη) με αύξηση της συγκέντρωσης των τριών βαρέων μετάλλων, παρατηρείται σημαντική μείωση της παραγόμενης φυτομάζας του Lollium perrene από 1,104 σε 0,784 g στο έδαφος της Γαλάτιστας και μια τάση μείωση στο έδαφος της Κερασιάς. Για το Allium cepa, παρατηρήθηκε σημαντική μείωση της παραγόμενης φυτομάζας με αύξηση της συγκέντρωσης των τριών βαρέων μετάλλων από 0,399 σε 0,124 g στο έδαφος της Κερασιάς. Η μείωση αυτή συμφωνεί και με τα ευρήματα άλλων ερευνητών (Bethlenfalvay and Franson, 1989, Díaz et al., 1996) σύμφωνα με τους οποίους οι υψηλές συγκεντρώσεις των βαρέων μετάλλων προκαλούν μείωση της ανάπτυξης του φυτού και της παραγωγής φυτομάζας. Μεταξύ των μαρτύρων και των φυσικών εδαφών, παρατηρείται σημαντική μείωση της παραγόμενης φυτομάζας για το Lollium perrene και για τα δύο εδάφη. Από 2,683 σε 0,906 g και από 3,094 σε 0,923 g για τα μη και εμπλουτισμένα εδάφη αντίστοιχα στο έδαφος της Κερασιάς και από 2,06 σε 1,104 g και από 2,218 σε 0,784 g για τα μη και εμπλουτισμένα εδάφη αντίστοιχα στο έδαφος της Γαλάτιστας. Για το Allium cepa παρατηρείται μια τάση μείωση της παραγόμενης φυτομάζας. Ο εμβολιασμός των φυτών με το μυκορριζικό μύκητα Glomus mosseae προκάλεσε μια στατιστικά σημαντική μείωση της παραγόμενης φυτομάζας και στα δύο εδάφη και στα δύο φυτά τόσο στις εμπλουτισμένες όσο και στις μη εμπλουτισμένες μεταχειρίσεις. Για το έδαφος της Κερασιάς και για το Lollium perrene από 2,683 σε 1,506 g και από 3,094 σε 2,28 g για τα μη και εμπλουτισμένα εδάφη αντίστοιχα ενώ για το Allium cepa από 0,5 σε 0,369 g και από 0,244 σε 0,089 g για τα μη και εμπλουτισμένα εδάφη αντίστοιχα. Για το έδαφος της Γαλάτιστας και για το Lollium perrene από 2,06 σε 1,664 g και από 2,218 σε 1,383 g για τα μη και εμπλουτισμένα εδάφη αντίστοιχα Αυτή η μείωση της φυτομάζας συμφωνεί και με τα ευρήματα των J.P. Liao et al., (2003) σύμφωνα με τους οποίους μία από τις αιτίες είναι το 40
45 γεγονός ότι η μυκόρριζα δεν προσλαμβάνει δυσκίνητα θρεπτικά συστατικά από το έδαφος για την ανάπτυξη της αλλά καταναλώνει μερικά από τα φωτοσυνθετικά προϊόντα των φυτών μειώνοντας έτσι τη παραγωγή της φυτομάζας του φυτού. Για το λόγο αυτό δικαιολογείται και η μεγαλύτερη ποσότητα φυτομάζας στις απολυμασμένες μεταχειρίσεις, απουσία μικροοργανισμών. Στα ίδια αποτελέσματα έχουν καταλήξει και οι Weissenhorn and Leyval, (1995), σύμφωνα με τους οποίους η μειωμένη φυτομάζα των μυκορριζικών φυτών, στο εμπλουτισμένο έδαφος, μπορεί να σχετίζεται με τις απαιτήσεις σε άνθρακα του μυκορριζικού μύκητα, οι οποίες είναι πολύ δύσκολο να καλυφθούν από το φυτό λόγω του stress που έχει υποστεί από τις υψηλές συγκεντρώσεις των μετάλλων. Τέλος, με τα παραπάνω συμφωνούν και τα ευρήματα των Killham and Firestone (1983), οι οποίοι βρήκαν μειωμένη ανάπτυξη των μυκορριζικών φυτών σε εδάφη ρυπασμένα από μέταλλα, την οποία απέδωσαν στην αυξημένη πρόσληψη των μετάλλων από τα φυτά. 41
46 4.5 Πρόσληψη των μετάλλων από το φυτό Προσδιορίστηκαν οι συγκεντρώσεις των υπό μελέτη βαρέων μετάλλων, Cd, Ni και Pb, που συγκεντρώθηκαν στη φυτομάζα του υπέργειου τμήματος του Lollium perrene και του Allium cepa. Τα αποτελέσματα παρουσιάζονται στο Πίνακα 4.7. Στο Σχήμα 4.8 παριστάνονται γραφικά οι συγκεντρώσεις των Cd, Ni και Pb για κάθε μεταχείριση. Πίνακας 4.7: Η συγκέντρωση των μετάλλων Cd, Ni και Pb στη φυτομάζα του υπέργειου τμήματος των φυτών Lollium perrene ΜΕΤΑΧΕΙΡΙΣΕΙΣ Cd(ppm) Ni(ppm) Pb(ppm) Κερασιά φυσικό 6,6a 14,256a 24,295a Κερασιά απολυμασμένο 4,195a 27,251a 13,267b Κερασιά εμπλουτισμένο 55,206b 71,894b 34,882ac Κερασιά εμπλουτισμένο απολυμασμένο 46,089b 97,494c 14,759ab Κερασιά απολυμασμένο (Glomus) 13,544a 77,32bc 45,519abc Κερασιά εμπλουτισμένο απολυμασμένο (Glomus) 37,673b 99,438dc 39,235c Γαλάτιστα φυσικό 4,63a 11,604a 29,145a Γαλάτιστα απολυμασμένο 3,587a 12,738a 18,781a Γαλάτιστα εμπλουτισμένο 49,121bc 283,225bcd 58,716ab Γαλάτιστα εμπλουτισμένο απολυμασμένο 36,269c 155,75b 16,441a Γαλάτιστα απολυμασμένο (Glomus) 13,688ab 61,76c 72,026b Γαλάτιστα εμπλουτισμένο απολυμασμένο (Glomus) 28,996cb 111,48d 63,974ab Allium cepa Κερασιά φυσικό Κερασιά απολυμασμένο 2,206a 1,076b 8,826a 6,101b Κερασιά εμπλουτισμένο 67,615c 56,939c - Κερασιά εμπλουτισμένο απολυμασμένο 51,699d 16,288d - Κερασιά απολυμασμένο (Glomus) 2,34a 8,426a - Κερασιά εμπλουτισμένο απολυμασμένο (Glomus) 49,065d 37,383e - Οι τιμές διαφέρουν στατιστικά μεταξύ τους σε επίπεδο p<0,05 Η σύγκριση έχει πραγματοποιηθεί μεταξύ των τιμών της ίδιας στήλης και χωριστά για κάθε έδαφος (a,b,c,d κ.τ.λ. συμβολίζονται οι σημαντικές διαφορές μεταξύ των τιμών, ενώ ίδια γράμματα μεταξύ των τιμών συμβολίζουν μη σημαντικές διαφορές) - - Από τα πειραματικά δεδομένα ( Πίν. 4.7 και Σχ. 4.8), για το Lollium perrene, στους μάρτυρες (απολυμασμένα και εμπλουτισμένα απολυμασμένα εδάφη) παρατηρήθηκε ότι στη φυτομάζα του υπέργειου τμήματος η συγκέντρωση του Ni ήταν η μεγαλύτερη, ακλουθούσε η 42
47 συγκέντρωση του Pb και κατόπιν η συγκέντρωση του Cd, (27,25 και 97,49 ppm Ni, 13,266 και 14,75 ppm Pb, 4,19 και 46,09 ppm Cd για το έδαφος της Κερασιάς, για τα μη και εμπλουτισμένα εδάφη αντίστοιχα και 12,738 και 155,75 ppm Ni, 18,781 και 16,441 ppm Pb, 3,587 και 36,269 ppm Cd για το έδαφος της Γαλάτιστας για τα μη και εμπλουτισμένα εδάφη αντίστοιχα ). Οι συγκεντρώσεις των μετάλλων Cd, Ni στο υπέργειο τμήμα του φυτού Allium cepa ακολουθούσαν την ίδια τάση με το φυτό Lollium perrene αλλά η υπέργεια φυτομάζα δεν περιείχε καθόλου Pb. Αυτό συμφωνεί με τον Alloway (1995), που αναφέρει ότι το Cd και το Ni μεταφέρονται ευκολότερα στο υπέργειο τμήμα του φυτού από ότι ο Pb που ως δυσκίνητο στοιχείο παραμένει στη ρίζα του φυτού και μετακινείται με πολύ αργό ρυθμό στο υπέργειο τμήμα του φυτού. Ακόμη παρατηρήθηκε ότι οι συγκεντρώσεις των Cd, Ni και Pb στο υπέργειο τμήμα των φυτών είχαν στενή στατιστική σχέση με τις αντίστοιχες συγκεντρώσεις των στο έδαφος. Για το Cd ήταν: r = 0,936* στα φυσικά και εμπλουτισμένα εδάφη και r = 0,945* στα απολυμασμένα φυσικά και απολυμασμένα εμπλουτισμένα εδάφη, για το Ni ήταν: r = 0,949* στα φυσικά και εμπλουτισμένα εδάφη και r = 0,994** στα απολυμασμένα φυσικά και απολυμασμένα εμπλουτισμένα εδάφη και για τον Pb ήταν: r = 0,986* στα φυσικά και εμπλουτισμένα εδάφη στα απολυμασμένα φυσικά και απολυμασμένα εμπλουτισμένα εδάφη δεν βρέθηκε καμία σχέση με τη συγκέντρωση του στο έδαφος. Από τα πειραματικά δεδομένα (Πίν. 4.7 και Σχ. 4.8), παρατηρήθηκε ότι οι συγκεντρώσεις των τριών βαρέων μετάλλων στη φυτομάζα και των δύο φυτών, στα φυσικά και εμπλουτισμένα φυσικά εδάφη, ή ήταν σημαντικά μεγαλύτερες από τις αντίστοιχες συγκεντρώσεις των μαρτύρων ή είχαν τάση αύξησης από τις αντίστοιχες συγκεντρώσεις των μαρτύρων. Μεταξύ των μαρτύρων και φυσικών απολυμασμένων εμβολιασμένων με το μύκητα Glomus εδαφών, προκαλείται μια στατιστικά σημαντική αύξηση της συγκέντρωσης των τριών μετάλλων, στο υπέργειο φυτικό τμήμα. Για το Lollium perrene στο έδαφος της Κερασιάς παρατηρήθηκε αύξηση του Cd από 4,195 σε 13,544 ppm, του Ni από 27,251 σε 77,32 ppm και του Pb από 13,267 σε 45,519 ppm. Στο έδαφος της Γαλάτιστας παρατηρήθηκε αύξηση του Cd από 3,587 σε 13,688 ppm, του Ni από 12,738 σε 61,76 ppm και του Pb από 18,781 σε 72,026 ppm. Ίδια τάση παρατηρήθηκε και για στο φυτό Allium cepa. Μεταξύ των εμπλουτισμένων απολυμασμένων μεταχειρίσεων, ο εμβολιασμός με το μυκορριζικό μύκητα Glomus mosseae προκαλεί μια τάση αύξησης της συγκέντρωσης των μετάλλων στο υπέργειο τμήμα των φυτών. Για το Lollium perrene στο έδαφος της Κερασιάς παρατηρείται μια τάση αύξησης του Ni από 97,494 σε 99,438 ppm ενώ του Pb από 14,759 σε 39,235 ppm και μια μείωση του Cd από 46,089 σε 37,673 ppm. Στο έδαφος της Γαλάτιστας παρατηρείται μια αύξηση του Pb από 16,441 σε 63,974 ppm.και μία μείωση του Ni από 155,75 σε 111,48 ppm και του Cd από 36,269 σε 28,996 43
48 Για το Allium cepa σημειώθηκε αύξηση του Ni από 16,288 σε 37,383 ppm και μία μείωση του Cd από 51,699 σε 49,065. Αυτό βρήκαν οι Killham and Firestone, (1983), σε φυτά γρασιδιού, μερικά από τα οποία εμβολιάστηκαν με μυκορριζικό μύκητα του γένους Νι μg/g φυτ y = 10,262e 0,0906x R 2 = 0,9862 y = 4,0075x + 10,218 R 2 = 0,9876 Φυσικο απ ολυμ Νι mg/kg εδαφ Cd μg/gr φυτου y = 10,361Ln(x) + 29,773 R 2 = 0,8772 y = 8,4827Ln(x) + 23,264 R 2 = 0,8945 Φυσικό απολυμασμένο Cd mg/kg Εδαφ y = 8,1198x - 18,859 R 2 = 0,9738 Pb μg/g φυτου y = 49,743Ln(x) - 29,47 R 2 = 0,9516 φυσικο απολυμ Pb mg/kg εδαφ. Σχήμα 4.7: Απεικόνιση της σχέσης των συγκεντρώσεων των Cd, Ni και Pb στο υπέργειο τμήμα του Lollium perrene με τις αντίστοιχες συγκεντρώσεις των στο έδαφος 44
49 Glomus σε όξινες συνθήκες, υπό τη παρουσία μεγάλων συγκεντρώσεων βαρέων μετάλλων (Cu, Ni, Pb, Zn, Fe, και Co), διαπιστώθηκε ότι οι συγκεντρώσεις των μετάλλων στις ρίζες και τους βλαστούς των μυκορριζικών φυτών ήταν μεγαλύτερες από εκείνες των μη μυκορριζικών φυτών. Τα παραπάνω υποδηλώνουν ότι η παρουσία μυκορριζικού μύκητα αυξάνει τη πρόσληψη των τριών υπό μελέτη μετάλλων από τα φυτά, τόσο εάν ο μυκορριζικός μύκητας είναι αυτόχθων, φυσικά εδάφη, όσο και αν είναι ο Glomus mosseae. Αυτή η διαπίστωση συμφωνεί και με τα ευρήματα των Killham and Firestone (1983), Weissenhorn et al., (1995), Weissenhorn and Leyval (1995), Díaz et al. (1996), Liao et al. (2003), Bi et al. (2003), Giasson et al. (2005). Ακόμη παρατηρήθηκε ότι η εφαρμογή του Glomus mosseae σε απολυμασμένα εδάφη εμπλουτισμένα με Cd μείωσε την πρόσληψη του Cd στα φυτά. Σύμφωνα με τους Liao et al., (2003), η αύξηση της πρόσληψης των μετάλλων από τα μυκορριζικά φυτά πραγματοποιείται μαζί με την αύξηση της πρόσληψης του Ρ ενώ παράλληλα η παρουσία του μυκορριζικού μύκητα βελτιώνει τη μεταφορά των μετάλλων από τις ρίζες στους βλαστούς των φυτών. Οι δε Giasson et al., (2005) αναφέρουν ότι η μεταφορά των βαρέων μετάλλων στα φυτά πραγματοποιείται μέσω των υφών της μυκόρριζας. Τα ουσιώδη και μη ουσιώδη βαριά μέταλλα απομονώνονται και μεταφέρονται στις ρίζες των φυτών μέσω των εξωτερικών υφών της ΑΜ. Όσο για το Cd, εισέρχεται μέσα στις υφές μέσω των συστημάτων μεταφοράς των μαγγανίου και ψευδαργύρου και συσσωρεύεται σε υψηλές συγκεντρώσεις στα φυτά. Επίσης, κατά τη παρουσία βαρέων μετάλλων σε μη διαθέσιμες μορφές για τα φυτά, οι υφές του AM μπορούν να μεταβάλουν τα μέταλλα από ανθρακικά, σε μορφές διαλυτές στο νερό. Οι υφές εκκρίνουν οργανικά οξέα όπως τα κιτρικά και τα οξαλικά τα οποία καθιστούν τα βαριά μέταλλα διαλυτά. Αυτό το φαινόμενο είναι πιο εμφανές με ένα μη ουσιώδες μέταλλο (Cd) παρά με ένα ουσιώδες (Zn) (Giasson et al., 2005). Οι Bi et al., (2003) διαπίστωσαν επίσης μια αυξημένη προσρόφηση των μετάλλων από τα μυκορριζικά φυτά, όταν τα μέταλλα βρίσκονται σε χαμηλές συγκεντρώσεις στο έδαφος ενώ αναφέρουν μια ικανότητα του μυκηλίου διαφόρων μυκήτων του γένους Glomus spp. να προσροφούν με ταχύ ρυθμό τα βαρέα μέταλλα. Θα πρέπει να αναφερθεί η διαπίστωση των Weissenhorn et al., (1995), σύμφωνα με τους οποίους, η ΑΜ μυκόρριζα, υπό τη παρουσία υψηλών συγκεντρώσεων βαρέων μετάλλων, δεν αποτρέπει τις συγκεντρώσεις των μετάλλων μέσα στο φυτό να ξεπεράσουν τις κανονικές τιμές αλλά δεν αυξάνει τη πρόσληψη των μετάλλων από τα φυτά, σε επίπεδο ανησυχητικό, σε σύγκριση με τα μη μυκορριζικά φυτά. Τα παραπάνω δικαιολογούν και τη μείωση της πρόσληψης των μετάλλων που παρατηρήθηκε κατά την απολύμανση του εδάφους, καθώς η απολύμανση εξαφανίζει και τους αυτόχθονες μύκητες που σχηματίζουν μυκόρριζα οι οποίοι θα μπορούσαν να αυξήσουν τη πρόσληψη των μετάλλων αυτών. 45
50 Κερασιά (Lollium perrene) 150 μg/g Φ Απ Εμ Ε-Α Α(G) E-A(G) Cd Ni Pb Mεταχειρίσεις Γαλάτιστα (Lollium perrene) 300 μg/g Φ Απ Εμ Ε-Α Α(G) E-A(G) Cd Ni Pb Μεταχειρίσεις Κερασιά (Allium cepa) μg/g Φ Απ Εμ Ε-Α Α(G) E-A(G) Mεταχειρίσεις Cd Ni Σχήμα 4.8: Γραφική αναπαράσταση της συγκεντρώσεως των Cd, Ni και Pb στη φυτομάζα, για κάθε μεταχείριση 46
51 4.6 Συγκεντρώσεις των μετάλλων στο ριζικό σύστημα του φυτού Προσδιορίστηκε η συγκέντρωση σε ppm των υπό μελέτη βαρέων μετάλλων στο ριζικό σύστημα του Lollium perrene. Οι τιμές των συγκεντρώσεων φαίνονται στο Πίνακα 4.8 και η γραφική αναπαράσταση αυτών των τιμών παρουσιάζεται στο Σχήμα Δεν έγινε προσδιορισμός των μετάλλων στη ρίζα του Allium cepa. γιατί η ποσότητα του ριζικού συστήματος ήταν μικρή και χρησιμοποιήθηκε για τον προσδιορισμό των μυκορρίζων. Πίνακας 4.8: Οι συγκεντρώσεις σε ppm των Cd, Ni και Pb στο ριζικό σύστημα του Lollium perrene. Lollium perrene ΜΕΤΑΧΕΙΡΙΣΕΙΣ Cd(ppm) Ni(ppm) Pb(ppm) Κερασιά φυσικό 41,095a 171,232a 68,493a Κερασιά απολυμασμένο 12,012b 61,561b 15,015b Κερασιά εμπλουτισμένο 200c 208,333c 83,333c Κερασιά εμπλουτισμένο απολυμασμένο 130d 110d 60d Κερασιά απολυμασμένο (Glomus) 3,368e 36,490e 14,326b Κερασιά εμπλουτισμένο απολυμασμένο (Glomus) 46,813a 72,081f 14,71b Γαλάτιστα φυσικό 21,367a 66,239a 64,102a Γαλάτιστα απολυμασμένο 2,192b 28,508b 10,964b Γαλάτιστα εμπλουτισμένο 104,477c 339,552c 149,253c Γαλάτιστα εμπλουτισμένο απολυμασμένο 88,235d 137,647d 70,588a Γαλάτιστα απολυμασμένο (Glomus) 1,872b 16,1e 9,362b Γαλάτιστα εμπλουτισμένο απολυμασμένο (Glomus) 20,93e 44,651f 23,255d Οι τιμές διαφέρουν στατιστικά μεταξύ τους σε επίπεδο p<0,05 Η σύγκριση έχει πραγματοποιηθεί μεταξύ των τιμών της ίδιας στήλης και χωριστά για κάθε έδαφος (a,b,c,d κ.τ.λ. συμβολίζονται οι σημαντικές διαφορές μεταξύ των τιμών, ενώ ίδια γράμματα μεταξύ των τιμών συμβολίζουν μη σημαντικές διαφορές) Παρατηρείται πως στο σύνολό τους, οι συγκεντρώσεις των τριών μετάλλων στη ρίζα του Lollium perrene είναι πιο μεγάλες από τις αντίστοιχες της φυτομάζας. Από το Σχήμα 4.9 φαίνεται ότι και οι τρείς συγκεντρώσεις των μετάλλων σε όλες τις μεταχειρίσεις, είχαν στενή σχέση με τις αντίστοιχες συγκεντρώσεις στο έδαφος. Επίσης από τον Πίνακα 4.8 και το γράφημα 4.10 φαίνεται ότι και στα δύο εδάφη, μεταξύ των μαρτύρων η αύξηση των συγκεντρώσεων των βαρέων μετάλλων στο έδαφος προκάλεσε στατιστικά σημαντική αύξηση της συγκέντρωσης των τριών μετάλλων στο ριζικό σύστημα. Στο έδαφος της Κερασιάς, για το Cd από 12,012 σε 130 ppm, για το Ni από 61,56 σε 110 ppm και για το Pb από 15,015 σε 60 47
52 ppm. Στο έδαφος της Γαλάτιστας για το Cd από 2,192 σε 88,235 ppm, για το Ni από 28,508 σε 137,647 ppm και για το Pb από 10,96 σε 70,588 ppm.. Αύξηση των συγκεντρώσεων στη ρίζα παρατηρήθηκε και στα φυσικά και φυσικά εμπλουτισμένα εδάφη. Η αύξηση των συγκεντρώσεων συμφωνεί με τις παρατηρήσεις που έγιναν, για τη συγκέντρωση των μετάλλων στο υπέργειο τμήμα των φυτών, αφού η απολύμανση εξαφάνισε τους αυτόχθονες μύκητες, που σχηματίζουν VAM, των εδαφών οι οποίοι, όπως αναφέρθηκε, αυξάνουν τη προσρόφηση των μετάλλων. Μια ερμηνεία της συμπεριφοράς αυτής των φυτών με VAM δίδεται από τους Dehn και Schüepp, (1989), σύμφωνα με τους οποίους η συγκράτηση των βαρέων μετάλλων στο ριζικό σύστημα αυτών των φυτών μπορεί να ερμηνευτεί με το σχηματισμό επιφανειακών συμπλόκων μεταξύ των μετάλλων και των υφών του μύκητα μέσα ή έξω από τις ρίζες, πιθανόν σε ειδικές θέσεις. Η ίδια διαπίστωση έχει γίνει και από τους Bi et al., (2003). Παρατηρείται επίσης ότι στις εμβολιασμένες με το μύκητα Glomus mosseae μεταχειρίσεις, η συγκέντρωση των υπό μελέτη μετάλλων είναι πολύ χαμηλότερη σε σύγκριση με τις αντίστοιχες μη εμβολιασμένες μεταχειρίσεις (Πιν. 9). Για το έδαφος της Κερασιάς, μεταξύ των μη εμπλουτισμένων μεταχειρίσεων, το Cd μειώθηκε από 12,012 σε 3,368 ppm και το Ni από 61,561 σε 36,490 ppm.και ο Pb μη σημαντική μείωση από 15,015 σε 14,32 ppm. Για το έδαφος της Γαλάτιστας, το Ni μειώθηκε από 28,508 σε 16,1 ppm ενώ και στα άλλα δύο μέταλλα παρατηρείται μια τάση μείωσης. Όμοια και στις εμπλουτισμένες μεταχειρίσεις στο έδαφος της Κερασιάς το Cd μειώθηκε από 130 σε 46,813 ppm, το Ni από 110 σε 72,081 ppm και ο Pb από 60 σε 14,71 ppm και στο έδαφος της Γαλάτιστας το Cd από 88,235 σε 20,93 ppm, το Ni από 137,647 σε 44, 651 ppm και ο Pb από 70,588 σε 23,255 ppm. Αυτή η μείωση πιθανόν να οφείλεται στην ικανότητα του μύκητα Glomus mosseae να προσροφά και να ακινητοποιεί τις ποσότητες των μετάλλων στο μυκήλιό του (Bi et al., 2003). Ενώ οι Audet and Charest, (2006), διαπίστωσαν, από πειράματα σε φυτά καπνού, χαμηλότερη συγκέντρωση Cd στις ρίζες όπου αποικούσε μύκητας του γένους Glomus, σε σύγκριση με τις ρίζες χωρίς μυκόρριζα. 48
53 Pb μg/g φυτ.ριζα y = 2,4884x + 56,327 R 2 = 0,9656 φυσ. απολ y = 22,957Ln(x) - 8,2924 R 2 = 0, Pb mg/kg εδ. Cd μg/g φυτ.ριζα y = 31,521Ln(x) + 94,443 R 2 = 0,916 Φυσ απολ y = 24,352Ln(x) + 60,204 R 2 = 0, Cd mg/kg εδ. Ni μg/g φυτ.ριζα y = 6,2186x + 98,434 R 2 = 0,8198 φυσ. απολ y = 29,76Ln(x) + 24,906 R 2 = 0, Ni mg/kg εδ. Σχήμα 4.9: Απεικόνιση της σχέσης των συγκεντρώσεων των Cd, Ni και Pb στη ρίζα του Lollium perrene με τις αντίστοιχες συγκεντρώσεις των στο έδαφος Από πειράματα των Joner et al., (2000) διαπιστώθηκε η μεγάλη ικανότητα του μυκηλίου του μύκητα Glomus mosseae να προσροφά μέταλλα από το εδαφικό διάλυμα. Η αντοχή στα 49
54 μέταλλα που επιδεικνύουν τα φυτά με μυκόρριζα βασίζεται σε μια σειρά μηχανισμών και συμβιωτικών επιδράσεων όπως η ικανότητα δέσμευσης των μετάλλων από το μυκήλιο του μύκητα τόσο στη περιοχή της ριζόσφαιρας όσο και μέσα στις ρίζες (Vivas et al., 2005). Επίσης, οι Weissenhorn et al., (1995), ανέφεραν μείωση της αρνητικής επίδρασης του Zn στην ανάπτυξη δύο ειδών γρασιδιού, λόγω της παρουσίας του μύκητα που σχηματίζει VAM, αν και η πρόσληψη του Zn από τα φυτά ήταν μεγάλη. Γαλάτιστα ppm Α Φ G (1=Cd 2=Ni 3=Pb), Εμπλουτισμένο (4=Cd 5=Ni 6=Pb) Κερασιά ppm Α Φ G (1=Cd 2=Ni 3=Pb), Εμπλουτισμένο (4=Cd 5=Ni 6=Pb) Σχήμα 4.10: Γραφική αναπαράσταση των συγκεντρώσεων σε ppm των Cd, Ni και Pb στο ριζικό σύστημα του Lollium perrene. 50
55 4.7 Παρατήρηση των χρωματισμένων ριζών στο μικροσκόπιο Αφού πραγματοποιήθηκε ο χρωματισμός του ριζικού συστήματος των φυτών, ακολούθησε η παρατήρηση των χρωματισμένων δειγμάτων στο μικροσκόπιο. Με τον τρόπο αυτό ήταν δυνατή η εξακρίβωση της ύπαρξης μυκορριζικών σχηματισμών καθώς και η δυνατότητα παρατήρησης της πυκνότητας αποικισμού των ριζών από μύκητες που σχηματίζουν μυκόρριζα. Για τη περίπτωση του Lollium perrene έγιναν οι παρακάτω παρατηρήσεις: Για τη περίπτωση του Lollium perrene, οι μεταχειρίσεις του εδάφους της Γαλάτιστας είχαν πολύ μεγαλύτερο ποσοστό αποικισμού από μύκητες που σχηματίζουν μυκόρριζα σε σύγκριση με το έδαφος της Κερασιάς. Συγκεκριμένα, οι μεταχειρίσεις "Κερασιά εμπλουτισμένο" και "Κερασιά εμπλουτισμένο απολυμασμένο" δεν είχαν καθόλου μυκορριζικούς σχηματισμούς, δηλαδή δεν είχαμε αποικισμό των ριζών των φυτών από μύκητα που σχηματίζει μυκόρριζα. Η μεταχείριση "Κερασιά απολυμασμένο" είχε ελάχιστες υφές, ενώ μόνο η μεταχείριση "Κερασιά φυσικό" είχε ικανοποιητικό ποσοστό αποικισμού από μύκητα που σχηματίζει μυκόρριζα. Μπορεί επομένως να ειπωθεί ότι για τη περίπτωση του εδάφους της Κερασιάς, με το Lollium perrene, η παρουσία των υψηλών συγκεντρώσεων των βαρέων μετάλλων έδρασε ανασταλτικά στη δημιουργία μυκόρριζας. Όσον αφορά τις μεταχειρίσεις του εδάφους της Κερασιάς όπου έγινε εμβολιασμός με το μύκητα Glomus mosseae, είχαμε ικανοποιητικό αποικισμό των ριζών από το μύκητα αυτό. Μάλιστα το ποσοστό αποικισμού ήταν πολύ μεγαλύτερο στη μεταχείριση "Κερασιά απολυμασμένο" με Glomus σε σύγκριση με τη μεταχείριση "Κερασιά εμπλουτισμένο απολυμασμένο με Glomus", γεγονός που υποδηλώνει την ανασταλτική δράση των μετάλλων στο σχηματισμό μυκόρριζας. Παρακάτω παρατίθενται φωτογραφίες από τους μυκορριζικούς σχηματισμούς αυτών των μεταχειρίσεων για το ριζικό σύστημα του Lollium perrene. 51



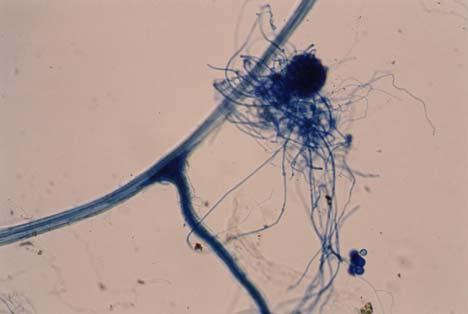

56 ΚΕΡΑΣΙΑ ΦΥΣΙΚΟ Φωτ. 1: Θύσανος Φωτ. 2: Σπόριο Φωτ. 3: Σπόριο ΚΕΡΑΣΙΑ ΑΠΟΛΥΜΑΣΜΕΝΟ Φωτ. 4: Υφές Φωτ. 5: Υφές 52

57 Φωτ. 6: Υφές Όσον αφορά το έδαφος της Γαλάτιστας, στη περίπτωση του Lollium perrene, ο αποικισμός από μυκόρριζες ήταν πολύ μεγαλύτερος. Συγκεκριμένα, σε όλες τις μεταχειρίσεις είχαμε τη παρουσία μυκήτων που σχηματίζουν μυκόρριζες με εξαίρεση τη μεταχείριση "Γαλάτιστα απολυμασμένο" όπου παρατηρήθηκαν ελάχιστες υφές. Αυτό πιθανόν να οφείλεται εξολοκλήρου στην απολύμανση του εδάφους. Το μεγαλύτερο ποσοστό αποικισμού παρατηρήθηκε στη μεταχείριση Γαλάτιστα εμπλουτισμένο απολυμασμένο καθώς επίσης και στις μεταχειρίσεις όπου έγινε εισαγωγή του μύκητα Glomus mosseae ("Γαλάτιστα απολυμασμένο με Glomus", "Γαλάτιστα εμπλουτισμένο απολυμασμένο με Glomus"). Θα πρέπει να αναφερθεί ότι τα μαύρα στίγματα που φαίνονται στις φωτογραφίες, ιδίως στα εμπλουτισμένα εδάφη, είναι συσσωρεύσεις μετάλλων στις υφές του μύκητα. Παρακάτω παρατίθενται φωτογραφίες από τους μυκορριζικούς σχηματισμούς αυτών των μεταχειρίσεων. 53





58 ΓΑΛΑΤΙΣΤΑ ΦΥΣΙΚΟ Φωτ. 7: Υφές Φωτ. 8: Υφές Φωτ. 9: Σπόριο πάνω σε υφές Φωτ. 10: Σπόριο και υφή Φωτ. 11: Σπόρια μέσα στη ρίζα Φωτ. 12: Κύστη 54
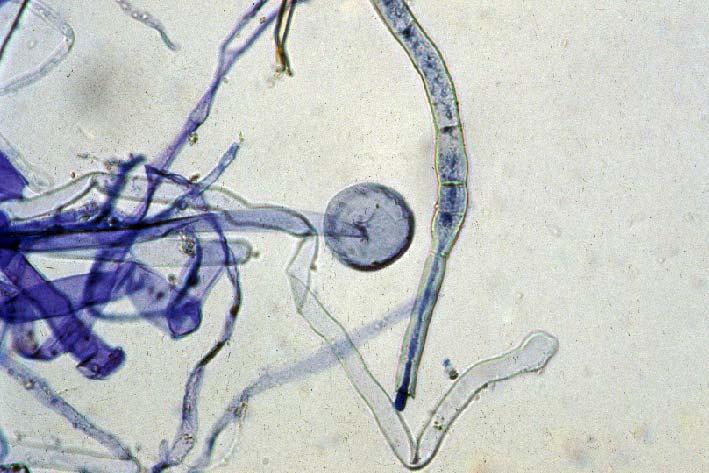
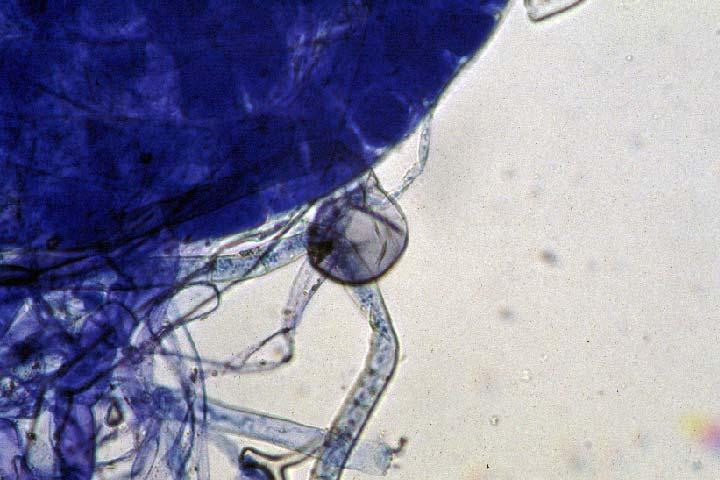

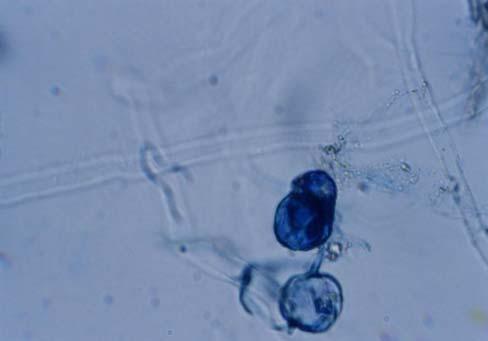
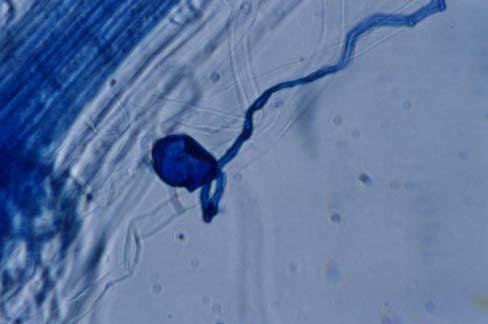

59 ΓΑΛΑΤΙΣΤΑ ΑΠΟΛΥΜΑΣΜΕΝΟ Φωτ. 13: Σπόριο Φωτ. 14: Σπόριο Φωτ. 15: Σπόριο Φωτ. 16: Σπόριο Φωτ. 17: Σπόριο Φωτ. 18: Απρεσόριο και κύστες 55
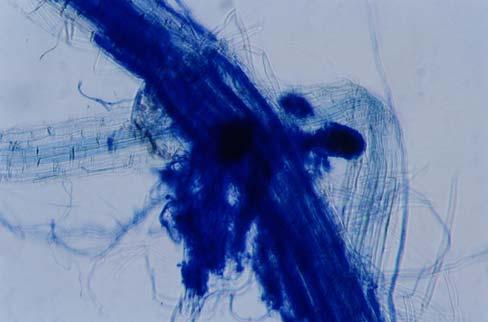
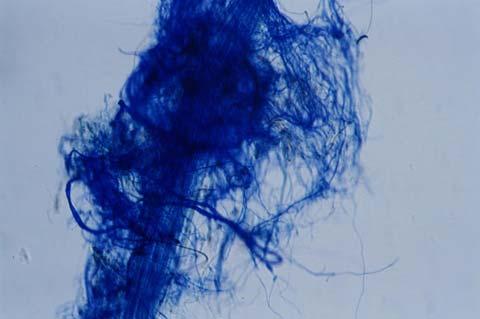


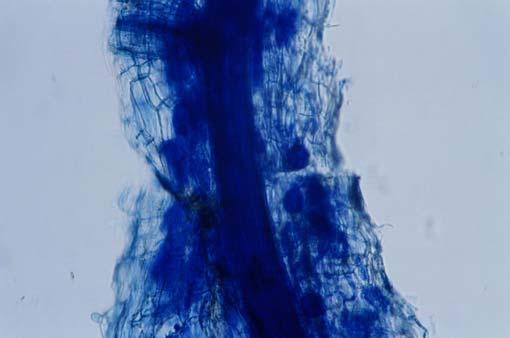

60 ΓΑΛΑΤΙΣΤΑ ΕΜΠΛΟΥΤΙΣΜΕΝΟ Φωτ. 19: Υφές Φωτ. 20: Υφές Φωτ. 21: Υφές Φωτ. 22: Υφές Φωτ. 23: Υφές Φωτ. 24: Υφές 56


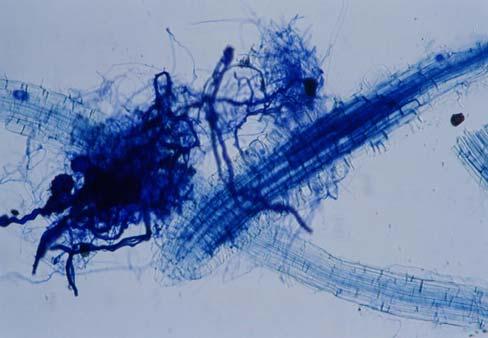

61 Φωτ. 25: Υφές Φωτ. 26: Υφές Φωτ. 27: Υφές Φωτ. 28: Υφές 57
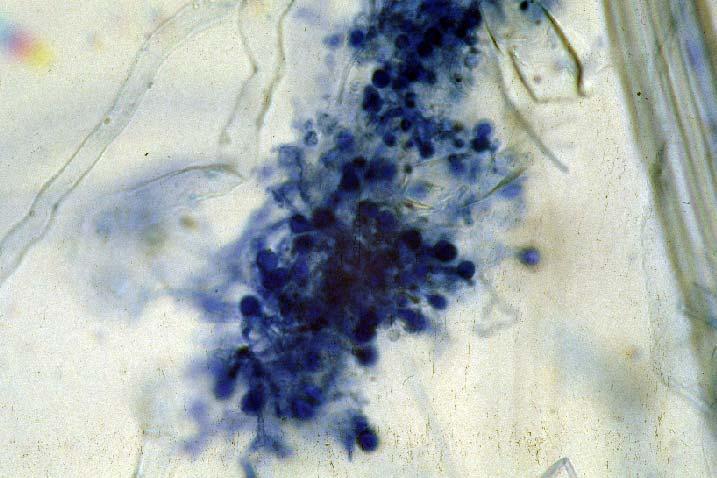
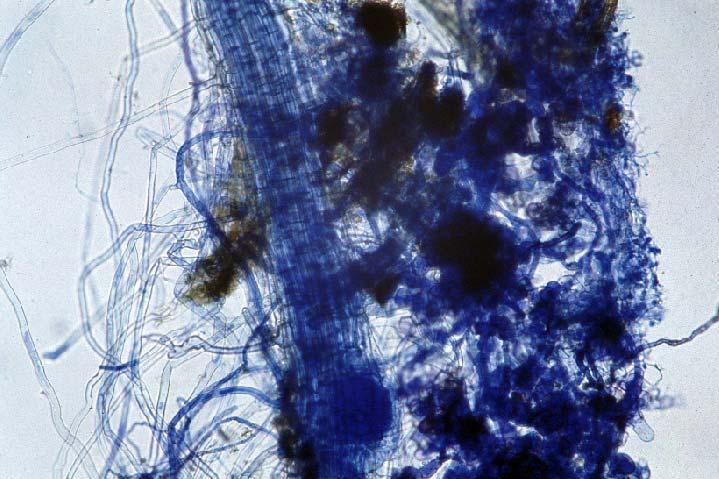
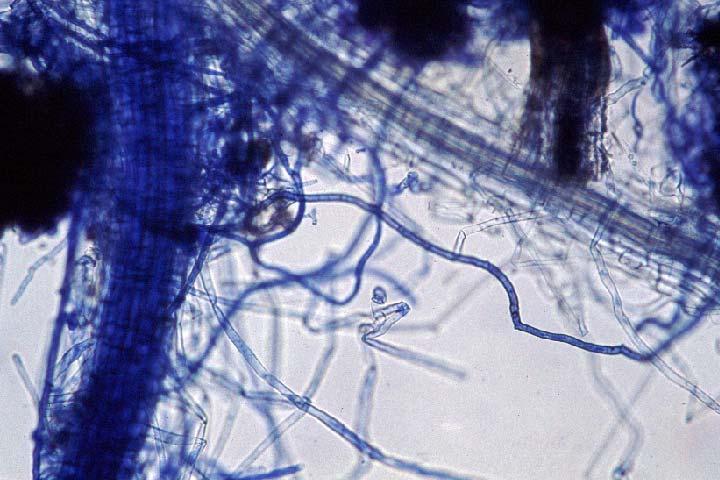
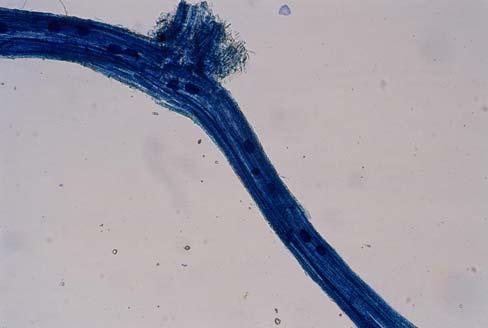
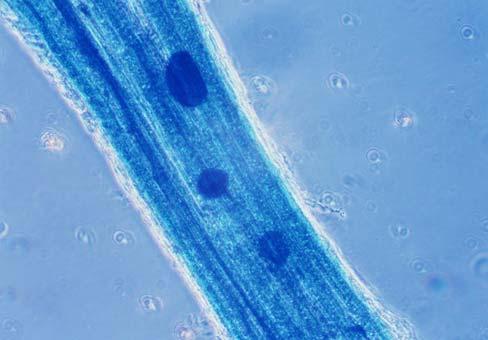
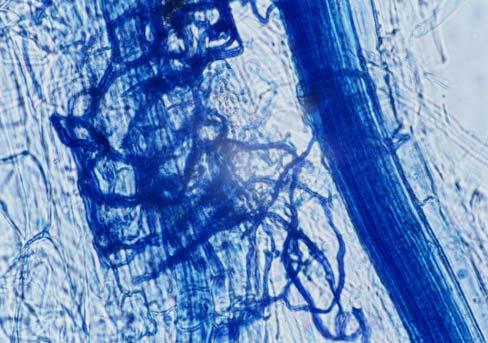
62 ΓΑΛΑΤΙΣΤΑ ΕΜΠΛΟΥΤΙΣΜΕΝΟ ΑΠΟΛΥΜΑΣΜΕΝΟ Φωτ. 29: Υφές Φωτ. 30: Υφές Φωτ. 31: Υφές Φωτ. 32: Κύστες και σπόρια Φωτ. 33: Κύστες και σπόριο Φωτ. 34: Υφές 58


63 Φωτ. 35: Υφές και σπόρια Φωτ. 36: Υφές 59
64 ΚΕΦΑΛΑΙΟ 5 ΣΥΜΠΕΡΑΣΜΑΤΑ Από τη διεξαγωγή του πειράματος και τη συζήτηση που έγινε παραπάνω μπορούμε να καταλήξουμε στα παρακάτω συμπεράσματα: 1. Όσον αφορά την επίδραση των τριών βαρειών μετάλλων στη μικροβιακή βιομάζα των δύο εδαφών, προέκυψε ότι στο έδαφος της Κερασιάς η προσθήκη των μετάλλων προκάλεσε μείωση της μικροβιακής βιομάζας C και Ν. Αντίθετα, στο έδαφος της Γαλάτιστας, ο εμπλουτισμός με μέταλλα αύξησε τη μικροβιακή βιομάζα C και Ν. Από το προσδιορισμό του μικροβιακού λόγου C/Ν προέκυψε ότι στο έδαφος της Γαλάτιστας υπερτερούσαν οι μύκητες. Η παρουσία των βαρειών μετάλλων προκάλεσε αύξηση του λόγου C/Ν στο έδαφος της Κερασιάς και μείωση του λόγου αυτού στο έδαφος της Γαλάτιστας. 2. Όσον αφορά τη ΣΣ % των δύο εδαφών, πριν τη σπορά των φυτών, προέκυψε ότι για το έδαφος της Κερασιάς, ο εμπλουτισμός με τα τρία βαριά μέταλλα προκάλεσε σημαντική αύξηση της ΣΣ %, ενώ στο έδαφος της Γαλάτιστας προκάλεσε σημαντική μείωση της ΣΣ %. Η ανάπτυξη των δύο φυτών στο έδαφος της Κερασιάς προκάλεσε μείωση της ΣΣ %. Όμοια και στο έδαφος της Γαλάτιστας η ανάπτυξη του Lollium perrene μείωσε τη ΣΣ % του εδάφους. Και στα δύο εδάφη, ο εμβολιασμός των φυτών με το μύκητα Glomus mosseae δε μετέβαλλε τη ΣΣ % των δύο εδαφών. Όσον αφορά τη διασπορά της αργίλου, ο εμπλουτισμός των εδαφών με τα τρία υπό μελέτη μέταλλα δε προκάλεσε μεταβολή της Τα και Τμ στα δύο εδάφη. Η απολύμανση όμως των εδαφών προκάλεσε σημαντική αύξηση της Τα στις μεταχειρίσεις όπου είχαμε την ανάπτυξη του Lollium perrene. Η παρουσία του μύκητα Glomus mosseae μείωσε τα ποσοστά της Τα και Τμ στις μεταχειρίσεις όπου είχαμε ανάπτυξη φυτού. 3. Όσον αφορά τη βιομάζα των φυτών, η προσθήκη των βαρειών μετάλλων μείωσε σημαντικά τη φυτομάζα του Allium cepa στο έδαφος της Κερασιάς. Το Lollium perrene αποδείχτηκε πιο ανθεκτικό στις υψηλές συγκεντρώσεις βαρειών μετάλλων και δε παρατηρήθηκε σημαντική μείωση της παραγόμενης φυτομάζας του. Στις μεταχειρίσεις των μαρτύρων είχαμε αύξηση της φυτομάζας και των δύο φυτών. Η παρουσία του μύκητα Glomus mosseae προκάλεσε μια στατιστικά σημαντική μείωση της παραγόμενης φυτομάζας των δύο φυτών και στα δύο εδάφη. 4. Όσον αφορά τις συγκεντρώσεις των βαρειών μετάλλων στο υπέργειο τμήμα των φυτών, προέκυψε ότι στο Lollium perrene, το μέταλλο με τη μεγαλύτερη συγκέντρωση ήταν το Ni, ακολούθησε ο Pb και τελευταίο το Cd. Στη φυτομάζα του Allium cepa το μέταλλο με την υψηλότερη συγκέντρωση ήταν το Ni και στη συνέχεια το Cd ενώ δε βρέθηκαν 60
65 ποσότητες Pb. Βρέθηκε ότι οι συγκεντρώσεις Cd, Ni και Pb στο υπέργειο τμήμα των φυτών είχαν στενή στατιστική σχέση με τις αντίστοιχες συγκεντρώσεις τους στο έδαφος. Διαπιστώθηκε επίσης ότι οι μεταχειρίσεις των μαρτύρων είχαν μικρότερες συγκεντρώσεις βαρειών μετάλλων στη φυτομάζα από τις αντίστοιχες μεταχειρίσεις των φυσικών εδαφών. Επιπλέον, η παρουσία μυκορριζικού μύκητα αύξησε τη πρόσληψη των τριών μετάλλων από τα φυτά ενώ η εφαρμογή του Glomus mosseae σε απολυμασμένα εδάφη εμπλουτισμένα με Cd, μείωσε τη πρόσληψη του Cd στα φυτά. Τέλος όσον αφορά τις συγκεντρώσεις των τριών μετάλλων στο ριζικό σύστημα των φυτών προέκυψε ότι αυτές στο σύνολό τους ήταν μεγαλύτερες από τις αντίστοιχες συγκεντρώσεις της φυτομάζας. Η αύξηση των συγκεντρώσεων των βαρειών μετάλλων στο έδαφος προκάλεσε στατιστικά σημαντική αύξηση της συγκέντρωσης των τριών μετάλλων στο ριζικό σύστημα. Αυτή η αύξηση των συγκεντρώσεων συμφωνεί με τις παρατηρήσεις που έγιναν για τη συγκέντρωση των μετάλλων στο υπέργειο τμήμα των φυτών. Παρατηρήθηκε επίσης ότι στις εμβολιασμένες με το μύκητα Glomus mosseae μεταχειρίσεις, η συγκέντρωση των υπό μελέτη μετάλλων ήταν πολύ χαμηλότερη σε σύγκριση με τις αντίστοιχες μη εμβολιασμένες μεταχειρίσεις. 61
66 RESULTS The microbial biomass C (BC) and N (BN) determine in two Alfisols from N. Greece, soil named Kerasia and Galatista. A mass part of each soil was incubated with heavy metals. For the Kerasia soil region, the BC and BN decreased after incubation. In reverse, for the Galatista soil the BC and BN increased after incubation. At the Kerasia soil the ratio C/N increased after the incubation, and decreased to the Galatista soil. Also, from the value of the ratio C/N it was concluded that the population of fungi was higher at the soil Galatista. The soil aggregate stability was determined for the two soils, before and after the seeding of the two plants species. Before the seeding, the incubation increased the aggregate stability of the Kerasia soil whereas it decreased the aggregate stability of the Galatista soil. At the Kerasia soil, the growth of both plants caused reduction of the aggregate stability. The same reduction appeared to the Galatista soil, due to the growth of Lollium perrene. In both soils, the inoculation of the plants with the mychorrhizal fungi G. mosseae had no effect on the aggregate stability of the soils. In both soils, the enrichment with heavy metals caused no change at the clay dispersion. Though, the sterilization of both soils increased the values of spontaneously dispersive clay, in the case of Lollium perrene. The inoculation with the mychorrhizal fungi G. mosseae decreased clay dispersion in both soils. At the Kerasia soil, the enrichment with the three heavy metals decreased significantly the shoot dry weight production of Allium cepa but Lollium perrene appeared to be more tolerant to the toxic concentrations of metals. In control plants, the shoot dry matter production of both plants was increased. The inoculation with the fungi G. mosseae caused a statistically significant decrease of the plant biomass, for both plants at the two soils. The metal with the highest concentration, in shoots of Lollium perrene, was Ni, followed by Pb and last by Cd. In shoots of Allium cepa, the metal with the highest concentration was Ni followed by Cd whereas no Pb was found. The concentrations of the three metals at the biomass of the two plants were statistically correlated with the concentrations of these metals in the two soils. The metal concentrations were lower in the biomass of control plants, in relation to the rest treatments. Also, inoculation with the mychorrhizal fungi G. mosseae increased the absorption of the Ni and Pb by the two plants, except for the Cd which was in low concentration due to the presence of fungus G. mosseae. The concentrations of the three metals were higher in roots of the two plants, than in shoots. The inoculated plants had lower metal concentrations in roots than non-inoculated plants. 62
67 ΒΙΒΛΙΟΓΡΑΦΙΑ Alloway B.J Heavy metals in soils. Second ed. Blackie Academic and Professional, an imprint of Chapman and Hall. Glasgow G 64 2NZ, UK. Angers, D. A., and G. R. Mehuys Aggregate stability to water. In soil sampling and methods of analysis. Ed. by Carter, M. R., pp Can. Soc. Soil Sci. Audet Patrick, Christiane Charest Effects of AM colonization on wild tobacco plants grown in zinc-contaminated soil. Mycorrhiza 16: Bearden Birgitte Neergaard and Leif Petersen Influence of arbuscular mycorrhizal fungi on soil structure and aggregate stability of a vertisol. Plant and Soil 218: Benton J. and V. W. Case Sampling, handling and analyzing plant tissue samples. P: In: R. L. Westerman (ed). Soil Testing and Plant Analysis Third edition SSSA Madison Wisconsin USA. Bethenfalvay, G.J., R.L. Franson Manganes toxicity alleviated by mycorrhizae in soybeans. J. Plant Nutr. 12: Bi Y.L., X.L. Li, P. Christie Influence of early stages of arbuscular mycorrhiza on uptake of zinc and phosphorus by red clover from a low-phosphorus soil amended with zinc and phosphorus. Chemosphere 50: Breland T.A Modelling mineralisation of plant residue on soil: effect of physical protection. Biol. Fertil. Soils 25: Bremer E. & Van Kessel C Seasonal microbial biomass dynamics after addition of lentil and wheat residues. Soil Sci. Soc. Am. J 56: Brookes, P.C., McGrath S.P Effects of metal toxicity on the size of the soil microbial biomass. J. Soil Sci. 95: D. Curtin, H. Stepphuhn, and F. Selles Clay Dispersion in Relation to Sodicity, Electrolyte Concentration, and Mechanical Effects. Soil Sci. Soc. Am. J. 58: Dai Jun, Thierry Becquer, James Henri Rouiller, Georges Reversat, France Bernhard- Reversat, Patrick Lavelle Influence of heavy metals on C and N mineralisation and microbial biomass in Zn-, Pb-, Cu-, and Cd- contaminated soils. Applied Soil Ecology 25: Dehn Barbara and Hannes Schüepp Influence of VA mycorrhizae on the uptake and distribution of heavy metals in plants. Agr., Ecos. and Env. 29: Díaz G., C. Azcón-Aguilar, M. Honrubia Influence of arbuscular mycorrhizae on heavy metal (Zn and Pb) uptake and growth of Lygeum spartum and Anthyllis cytisoides. Plant and Soil 180:
68 Eason W.R., J. Scullion, E.P. Scott Soil parameters and plant responses associated with arbuscular mychorrhizas from contrasting grassland management regimes. Agr. Ecos. and Envir. 73: Fomina M.A., I.J. Alexander, J.V. Colpaert, G.M. Gadd Solubilization of toxic metal minerals and metal tolerance of mychorrhizal fungi. Soil Biol. & Bioch. 37: Giasson Philippe, Alfred Jaouich, Serge Gagné, Peter Moutoglis Phytoremediation of zinc and cadmium: a study of arbuscular mychorrhizal hyphae. Remediation Autumn 2005 Giasson Philippe, Alfred Jaouich, Serge Gagné, Peter Moutoglis Arbuscular mychorrhizal fungi involvement in zinc and cadmium speciation change and phytoaccumulation. Remediation Spring 2005 Giller Ken E., Ernst Witter, Steve P. McGrath Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. Vol. 30. pp G. Giovannini and P. Sequi Iron and aluminium as cementing substances of soil aggregates. II. Changes in stability of soil aggregates following extraction of iron and aluminium by acetylacetone in a non-polar solvent. J. of Soil Sci. 27: Harrier L. A. and C.A. Watson Cropping systems. ADV AGRON 79: Hiroki M Effects of heavy metal contamination on soil microbial population. Soil Sci. Plant Nutr. 38, Jarstfer Amiel G. and David M. Sylvia Inoculum Production and Inoculation Strategies for Vesicular Arbuscular Mycorrhizal Fungi. In: Soil Microbial Ecology, Florida Agricultural Experiment Station Journal Series No R-01157, pp Jenkinson and Powlson The effects of biocidal treatment on metabolism in soil V. A method for measuring soil biomass. Soil Biol. and Bioch. 8: Joergensen, R.G., Anderson, T.H., Wolters, V Carbon and nitrogen relationships in the microbial biomass in soils of beech (Fagus Sylvatica L.) forest. Biol. Fertil. Soils 19, Joner Erik J., Roberto Briones, Corinne Leyval Metal-binding capacity of arbuscular mycorrhizal mycelium. Plant and Soil 226: Jordan, M.J., Lechevallier, M.P Effects of zinc-smelter emissions on forest soil microflora. Can. J. Microbiol. 21, Kautz Timo, Stephan Wirth, Frank Ellmer Microbial activity in a sandy arable soil is governed by the fertilization regime. European J. of Soil Biol. 40: Khan A.G., Kuck C., Chauhdry T.M., Khoo C.S., W.J. Hayes Role of plants, mychorrhizae and phytochelators in heavy metal contaminated land remediation. Chemosphere 41:
69 Killham K. and M. K. Firestone Vesicular arbuscular mycorrhizal mediation of grass response to acidic and heavy metal depositions. Plant and Soil 72, Koske R.E., J.N. Gemma A modified procedure for staining roots to detect VA mychorrhizas. Mycol. Res. 92 (4): Kosolapov D.B., P. Kuschk, M.B. Vainshtein, A.V. Vatsourina, A. Wieβner, R.A. Müller Microbial processes of heavy metal removal from carbon-deficient effluents in constructed wetlands. Eng. Life Sci. 4 No. 5 Lavelle P., Spain A.V Soil Ecology. Kluwer Academic Publishers, Dordrecht, pp Leyval Corinne, Bal Ram Singh, Erik J. Joner Occurrence and infectivity of arbuscular mycorrhizal fungi in some Norwegian soils influenced by heavy metals and soil properties. Water, Air and Soil Pollution 84: Leyval C., K. Turnau, K. Haselwandter Effect of heavy metal pollution on mychorrhizal colonization and function: physiological, ecological and applied aspects. Mychorrhiza 7: Liao J.P., X.G. Lin, Z.H. Cao, Y.Q. Shi, M.H. Wong Interactions between arbuscular mycorrhizae and heavy metals under sand culture experiment. Chemosphere 50: Lindsay, W.L. and W.A. Norvell Development of a DTPA soil test for zinc, iron, manganese and copper. Soil Sci. Soc. Am. J. 42: McGrath Steve P., Effects of Heavy Metals from Sewage Sludge on Soil Microbes in Agricultural Ecosystems. In: Toxic Metals in Soil Plant Systems. John Wiley and Sons Ltd pp Moscatelli M.C., A. Lagomarsino, S. Marinari, P. De Angelis, S. Grego Soil microbial indices as bioindicators of environmental changes in a poplar plantation. Ecological Indicators 5: Vasquez-Murrieta M. S., I. Migueles-Garduno, O. Franco-Jereandez, B. Govaerts and L. Dendooven C and N mineralization and microbial biomass in heavy-metal contaminated soil. European J. of Soil Biol. 42: Norman, R.J., J.C. Edberg and J.W. Stucki Determination of nitrate in soilextracts by dual-wavelength ultraviolet spectophotometry. Soil Sci. Soc. Am. J. 49: Νικολογιάννης Φώτιος Μεταπτυχιακή Διατριβή. Εργαστήριο Εδαφολογίας, Σχολή Γεωπονίας, ΑΠΘ Oades J. M Soil organic matter and structural stability: mechanisms and implications for management. Plant and Soil 76:
70 Paul, E.A., Clear, F.E Soil Microbiology and Biochemistry. Second ed. Academic Press, London, pp Qadir M., A. Ghafoor, G. Murtaza Cadmium concentration in vegetables grown on urban soils irrigated with untreated municipal sewage. Environment, Development and Sustainability 2: Quilambo Orlando António The vesicular-arbuscular mycorrhizal symbiosis. African Journal of Biotechnology Vol. 2 (12), pp D.J. Read, D.H. Lewis. A.H. Fitter, I.J. Alexander Mychorrhizas in Ecosystems. C A B International, Printed and Bound in the UK at the University Press, Cambridge. Renella G., M. Mench, D. van der Lelie, G. Pietramellara, J. Ascher, M.T. Ceccherini, L. Landi, P. Nannipieri Hydrolase activity, microbial biomass and community structure in long term Cd-contaminated soils. Soil Biol. & Bioch. 36: Rengasamy, P., R. S. B. Greene, G. W. Ford, and A. H. Mehanni Identification of dispersive behavior and the management of red-brown earths. Aust. J. Soil Res. 22: R. Román, C. Fortún, M. E. García López De Sá, G. Almendros Successful Soil Remediation and Reforestation of a Calcic Regosol Amended with Composted Urban Waste. Arid Land Research and Management 17: Ross Sheila M Κεφάλαιο 5. In: Toxic Metals in Soil Plant Systems. John Wiley and Sons Ltd pp, Rowell. D. L Soil Science. Methods and Applications. Addison Wesley Longman Limited. Edinburg Gate, Harlow, England. Tate, R.L Soil Organic matter. Biological and Ecological Effectors. Wiley (Interscience), New York. Terman, G.L. S.E. Allenn and L.B. Clements Greenhouse techniques for soil-plant fertilizer research. National Fertilizer Development Center. T.V.A. Techn. Rep. No 3 Robert L. Tate III Soil Microbiology. John Wiley and Sons, ING. Thomas R. S., R. L. Franson, and G. J. Bethlenfalvay Separation of Vesicular Arbuscular Mycorrhizal Fungus and Root Effects on Soil Aggregation. Soil Sci. Am. J. 57: Thomas R. S., S. Dakessian, R. N. Ames, M. S. Brown, and G. J. Bethlenfalvay, Aggregation of a Silty Clay Loam Soil by Mycorrhizal Onion Roots. Soil Sci. Soc. Am. J. 50: Vance E.D., Brookes P.C., Jenkinson D.S An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 19, Vivas A., Barea J.M., R. Azcón Interactive effect of Brevibacillus brevis and Glomus mosseae, both isolated from Cd contaminated soil, on plant growth, physiological mychorrhizal 66
71 fungal characteristics and soil enzymatic activities in Cd polluted soil. Environ. Pollut. 134: Vosátka Miroslav Influence of inoculation with arbuscular mycorrhizal fungi on the growth and mycorrhizal infection of transplanted onion. Agr. Ecos. and Envir. 53: Vogel-Mikuš Katarina, Damjana Drobne, Marjana Regvar Zn, Cd and Pb accumulation and arbuscular mycorrhizal colonisation of pennycress Thlaspi praecox Wulf. (Brassicaceae) from the vicinity of a lead mine and smelter in Slovenia. Environmental Pollution 133: Weissenhorn I., C. Leyval, G. Belgy, J. Berthelin Arbuscular mycorrhizal contribution to heavy metal uptake by maize (Zea mays L.) in pot culture with contaminated soil. Mycorrhiza 5: Weissenhorn I., C. Leyval, J. Berthelin Cd-tolerant arbuscular mycorrhizal (AM) fungi from heavy-metal polluted soils. Plant and Soil 157: Weissenhorn I. and C. Leyval Root colonization of maize by a Cd-sensitive and a Cdtolerant Glomus mosseae and cadmium uptake in sand culture. Plant and Soil 175: Ouziada Fouad, Ulrich Hildebrandta, Elmon Schmelzerb, Hermann Bothe Differential gene expressions in arbuscular mycorrhizal-colonized tomato grown under heavy metal stress. Journal of Plant Physiology 162 (2005) Yuangen Yang, C.D. Campbell, L. Clark, C.M. Cameron, E. Paterson Microbial indicators of heavy metal contamination in urban and rural soils. Chemosphere 63:

")


72 ΠΑΡΑΡΤΗΜΑ ΦΩΤΟΓΡΑΦΙΩΝ Κερασιά φυσικό (Allium cepa) Κερασιά απολυμασμένο (Allium cepa) Κερασιά εμπλουτισμένο (Allium cepa) 68

")


73 Κερασιά εμπλουτισμένο απολυμασμένo (Allium cepa) Κερασιά απολυμασμένο Glomus (Allium cepa) Κερασιά εμπλουτισμένο απολυμασμένο Glomus (Allium cepa) 69



74 Κερασιά απολυμασμένο Glomus (Lollium perrene) Κερασιά εμπλουτισμένο απολυμασμένο Glomus (Lollium perrene) Γαλάτιστα φυσικό (Allium cepa) 70

")



75 Γαλάτιστα απολυμασμένο (Allium cepa) Γαλάτιστα εμπλουτισμένο (Allium cepa) Γαλάτιστα εμπλουτισμένο απολυμασμένο (Allium cepa) 71
")


76 Γαλάτιστα απολυμασμένο Glomus (Allium cepa) Γαλάτιστα εμπλουτισμένο απολυμασμένο Glomus (Allium cepa) Γαλάτιστα απολυμασμένο Glomus (Lollium perrene) 72

77 Γαλάτιστα εμπλουτισμένο απολυμασμένο Glomus (Lollium perrene) 73
Ησυμβιωτικήσχέσημε ενδομυκορριζικούς μύκητες ως στρατηγική αφομοίωσης ανοργάνων θρεπτικών από τα φυτά
 Ησυμβιωτικήσχέσημε ενδομυκορριζικούς μύκητες ως στρατηγική αφομοίωσης ανοργάνων θρεπτικών από τα φυτά 1 Η ικανότητα των φυτών να προσλαμβάνουν νερό και ανόργανα θρεπτικά από το έδαφος σχετίζεται και με
Ησυμβιωτικήσχέσημε ενδομυκορριζικούς μύκητες ως στρατηγική αφομοίωσης ανοργάνων θρεπτικών από τα φυτά 1 Η ικανότητα των φυτών να προσλαμβάνουν νερό και ανόργανα θρεπτικά από το έδαφος σχετίζεται και με
Η ανόργανη θρέψη των φυτών
 Η ανόργανη θρέψη των φυτών Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Η μελέτη της πρόσληψης και αξιοποίησης των ανόργανων θρεπτικών από τα φυτά H αξιοποίηση περιλαμβάνει τη: διακίνηση
Η ανόργανη θρέψη των φυτών Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Η μελέτη της πρόσληψης και αξιοποίησης των ανόργανων θρεπτικών από τα φυτά H αξιοποίηση περιλαμβάνει τη: διακίνηση
Εδαφοκλιματικό Σύστημα και Άμπελος
 Εδαφοκλιματικό Σύστημα και Άμπελος Δολαπτσόγλου Χριστίνα ΤΕΙ ΑΝΑΤΟΛΙΚΗΣ ΜΑΚΕΔΟΝΙΑΣ ΚΑΙ ΘΡΑΚΗΣ ΤΜΗΜΑ ΟΙΝΟΛΟΓΙΑΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΟΤΩΝ ΔΡΑΜΑ 2019 Chr. Dolaptsoglou Οργανική ουσία είναι όλα τα οργανικά υπολείμματα
Εδαφοκλιματικό Σύστημα και Άμπελος Δολαπτσόγλου Χριστίνα ΤΕΙ ΑΝΑΤΟΛΙΚΗΣ ΜΑΚΕΔΟΝΙΑΣ ΚΑΙ ΘΡΑΚΗΣ ΤΜΗΜΑ ΟΙΝΟΛΟΓΙΑΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΠΟΤΩΝ ΔΡΑΜΑ 2019 Chr. Dolaptsoglou Οργανική ουσία είναι όλα τα οργανικά υπολείμματα
ΒΙΟΛΟΓΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΔΑΣΙΚΩΝ
 ΒΙΟΛΟΓΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΔΑΣΙΚΩΝ ΕΔΑΦΩΝ Οργανική ουσία Αποτελείται από πολύπλοκες ενώσεις οι οποίες παράγονται από τα υπολείμματα των φυτικών και ζωικών οργανισμών, με την επίδραση βιολογικών, χημικών
ΒΙΟΛΟΓΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΔΑΣΙΚΩΝ ΕΔΑΦΩΝ Οργανική ουσία Αποτελείται από πολύπλοκες ενώσεις οι οποίες παράγονται από τα υπολείμματα των φυτικών και ζωικών οργανισμών, με την επίδραση βιολογικών, χημικών
ΕΞΕΤΑΣΕΙΣ ΣΤΟ ΜΑΘΗΜΑ ΕΔΑΦΟΛΟΓΙΑ ΛΙΠΑΣΜΑΤΟΛΟΓΙΑ ΤΟΥ 4 ΟΥ ΕΞΑΜΗΝΟΥ ΑΠΡΙΛΙΟΣ 2012
 ΕΞΕΤΑΣΕΙΣ ΣΤΟ ΜΑΘΗΜΑ ΕΔΑΦΟΛΟΓΙΑ ΛΙΠΑΣΜΑΤΟΛΟΓΙΑ ΤΟΥ 4 ΟΥ ΕΞΑΜΗΝΟΥ ΑΠΡΙΛΙΟΣ 2012 ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: 1. Οι εδαφικές ιδιότητες μεταβάλλονται: Α. Κατά μήκος των τριών αξόνων (x, y, z) Β. Με το πέρασμα του
ΕΞΕΤΑΣΕΙΣ ΣΤΟ ΜΑΘΗΜΑ ΕΔΑΦΟΛΟΓΙΑ ΛΙΠΑΣΜΑΤΟΛΟΓΙΑ ΤΟΥ 4 ΟΥ ΕΞΑΜΗΝΟΥ ΑΠΡΙΛΙΟΣ 2012 ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: 1. Οι εδαφικές ιδιότητες μεταβάλλονται: Α. Κατά μήκος των τριών αξόνων (x, y, z) Β. Με το πέρασμα του
1η Διάλεξη ΚΟΛΛΟΕΙΔΕΣ ΣΥΜΠΛΟΚΟ ΕΔΑΦΟΥΣ ΙΚΑΝΟΤΗΤΑ ΑΝΤΑΛΛΑΓΗΣ ΚΑΤΙΟΝΤΩΝ ΤΕΙ ΠΕΛΟΠΟΝΝΗΣΟΥ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ
 1η Διάλεξη ΚΟΛΛΟΕΙΔΕΣ ΣΥΜΠΛΟΚΟ ΕΔΑΦΟΥΣ ΙΚΑΝΟΤΗΤΑ ΑΝΤΑΛΛΑΓΗΣ ΚΑΤΙΟΝΤΩΝ Τροφοδότηση του εδαφικού διαλύματος Απορρόφηση Ρίζας Οργανική ουσία Ανταλλαγή κατιόντων Εδαφικό διάλυμα Μικροοργανισμοί εδάφους Προσρόφηση
1η Διάλεξη ΚΟΛΛΟΕΙΔΕΣ ΣΥΜΠΛΟΚΟ ΕΔΑΦΟΥΣ ΙΚΑΝΟΤΗΤΑ ΑΝΤΑΛΛΑΓΗΣ ΚΑΤΙΟΝΤΩΝ Τροφοδότηση του εδαφικού διαλύματος Απορρόφηση Ρίζας Οργανική ουσία Ανταλλαγή κατιόντων Εδαφικό διάλυμα Μικροοργανισμοί εδάφους Προσρόφηση
Τίτλος Διάλεξης: Ο ρόλος του ανταγωνισμού των θρεπτικών στοιχείωνστηνανάπτυξηκαιτην. Χ. Λύκας
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Πρόγραμμα Μεταπτυχιακών Σπουδών «ΑΕΙΦΟΡΟΣ ΑΓΡΟΤΙΚΗ ΠΑΡΑΓΩΓΗ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗ ΠΕΡΙΒΑΛΛΟΝΤΟΣ» Μάθημα: «Επίδραση του αβιοτικού και βιοτικού περιβάλλοντος στην παραγωγή των φυτών» Τίτλος
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Πρόγραμμα Μεταπτυχιακών Σπουδών «ΑΕΙΦΟΡΟΣ ΑΓΡΟΤΙΚΗ ΠΑΡΑΓΩΓΗ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗ ΠΕΡΙΒΑΛΛΟΝΤΟΣ» Μάθημα: «Επίδραση του αβιοτικού και βιοτικού περιβάλλοντος στην παραγωγή των φυτών» Τίτλος
Η διεργασία της αποσύνθεσης γίνεται με την βοήθεια των οργανισμών που ζουν στο έδαφος και σχηματίζεται ουσία με βαθύ φαιό χρώμα Οργανική ουσία
 Δρ. Γεώργιος Ζαΐμης Αποτελείται από πολύπλοκες ενώσεις οι οποίες παράγονται από τα υπολείμματα των φυτικών και ζωικών οργανισμών, με την επίδραση βιολογικών, χημικών και φυσικών παραγόντων. Η οργανική
Δρ. Γεώργιος Ζαΐμης Αποτελείται από πολύπλοκες ενώσεις οι οποίες παράγονται από τα υπολείμματα των φυτικών και ζωικών οργανισμών, με την επίδραση βιολογικών, χημικών και φυσικών παραγόντων. Η οργανική
Δασική Εδαφολογία. Χημικές ιδιότητες του εδάφους
 Δασική Εδαφολογία Χημικές ιδιότητες του εδάφους Χημικές ιδιότητες εδάφους Εδαφικά κολλοειδή Ηλεκτρικά φορτία των ανόργανων κολλοειδών Εναλλακτική ικανότητα του εδάφους Βαθμός κορεσμού με βάσεις Ανταλλαγή
Δασική Εδαφολογία Χημικές ιδιότητες του εδάφους Χημικές ιδιότητες εδάφους Εδαφικά κολλοειδή Ηλεκτρικά φορτία των ανόργανων κολλοειδών Εναλλακτική ικανότητα του εδάφους Βαθμός κορεσμού με βάσεις Ανταλλαγή
ΕΡΓΑΣΤΗΡΙΟ 11 ο ΜΥΚΟΡΡΙΖΕΣ
 ΕΡΓΑΣΤΗΡΙΟ 11 ο ΜΥΚΟΡΡΙΖΕΣ 1 ΜΥΚΟΡΡΙΖΕΣ 2 ΜΥΚΟΡΡΙΖΕΣ Μυκορριζική συμβίωση: Η συμβιωτική σχέση μεταξύ ριζών μυκήτων / εκδηλώνεται με σχηματισμό ξεχωριστών οργάνων =μυκόρριζες Κατηγορίες μυκόρριζων ενδομυκόρριζες
ΕΡΓΑΣΤΗΡΙΟ 11 ο ΜΥΚΟΡΡΙΖΕΣ 1 ΜΥΚΟΡΡΙΖΕΣ 2 ΜΥΚΟΡΡΙΖΕΣ Μυκορριζική συμβίωση: Η συμβιωτική σχέση μεταξύ ριζών μυκήτων / εκδηλώνεται με σχηματισμό ξεχωριστών οργάνων =μυκόρριζες Κατηγορίες μυκόρριζων ενδομυκόρριζες
Δ. Μείωση του αριθμού των μικροοργανισμών 4. Να αντιστοιχίσετε τα συστατικά της στήλης Ι με το ρόλο τους στη στήλη ΙΙ
 Κεφάλαιο 7: Εφαρμογές της Βιοτεχνολογίας 1. Η βιοτεχνολογία άρχισε να εφαρμόζεται α. μετά τη βιομηχανική επανάσταση (18ος αιώνας) β. μετά την ανακάλυψη της δομής του μορίου του DNA από τους Watson και
Κεφάλαιο 7: Εφαρμογές της Βιοτεχνολογίας 1. Η βιοτεχνολογία άρχισε να εφαρμόζεται α. μετά τη βιομηχανική επανάσταση (18ος αιώνας) β. μετά την ανακάλυψη της δομής του μορίου του DNA από τους Watson και
ΤΕΙ ΙΟΝΙΩΝ ΝΗΣΩΝ ΤΜΗΜΑ ΤΕΧΝΟΛΟΓΙΑΣ ΒΙΟΛΟΓΙΚΗΣ ΓΕΩΡΓΙΑΣ & ΤΡΟΦΙΜΩΝ ΓΟΝΙΜΟΤΗΤΑ ΕΔΑΦΩΝ/ΘΡΕΨΗ ΦΥΤΩΝ ΘΕΩΡΙΑ ΕΡΩΤΗΣΕΙΣ ΕΠΙ ΤΗΣ ΕΞΕΤΑΣΤΕΑΣ ΥΛΗΣ
 ΤΕΙ ΙΟΝΙΩΝ ΝΗΣΩΝ ΤΜΗΜΑ ΤΕΧΝΟΛΟΓΙΑΣ ΒΙΟΛΟΓΙΚΗΣ ΓΕΩΡΓΙΑΣ & ΤΡΟΦΙΜΩΝ ΓΟΝΙΜΟΤΗΤΑ ΕΔΑΦΩΝ/ΘΡΕΨΗ ΦΥΤΩΝ ΘΕΩΡΙΑ ΕΡΩΤΗΣΕΙΣ ΕΠΙ ΤΗΣ ΕΞΕΤΑΣΤΕΑΣ ΥΛΗΣ 1. Ποιές πληροφορίες σχετικά µε τη θρέψη των φυτών βρίσκουµε στους
ΤΕΙ ΙΟΝΙΩΝ ΝΗΣΩΝ ΤΜΗΜΑ ΤΕΧΝΟΛΟΓΙΑΣ ΒΙΟΛΟΓΙΚΗΣ ΓΕΩΡΓΙΑΣ & ΤΡΟΦΙΜΩΝ ΓΟΝΙΜΟΤΗΤΑ ΕΔΑΦΩΝ/ΘΡΕΨΗ ΦΥΤΩΝ ΘΕΩΡΙΑ ΕΡΩΤΗΣΕΙΣ ΕΠΙ ΤΗΣ ΕΞΕΤΑΣΤΕΑΣ ΥΛΗΣ 1. Ποιές πληροφορίες σχετικά µε τη θρέψη των φυτών βρίσκουµε στους
Η αποβολή άνθρακα στη ριζόσφαιρα. 30 60% του καθαρού φωτοσυνθετικού άνθρακα κατανέµεται στη ρίζα
 Ριζοαπόθεση Η αποβολή άνθρακα στη ριζόσφαιρα 30 60% του καθαρού φωτοσυνθετικού άνθρακα κατανέµεται στη ρίζα 5 70% τουκατανεµηθέντοςστηρίζα αποβάλλεται στη ριζόσφαιρα αποβολή γίνεται από τα αυξανόµενα µέρη
Ριζοαπόθεση Η αποβολή άνθρακα στη ριζόσφαιρα 30 60% του καθαρού φωτοσυνθετικού άνθρακα κατανέµεται στη ρίζα 5 70% τουκατανεµηθέντοςστηρίζα αποβάλλεται στη ριζόσφαιρα αποβολή γίνεται από τα αυξανόµενα µέρη
Ο μικρόκοσμος του εδάφους συμβολή του στη γεωργία και παραγωγή τροφίμων
 Ο μικρόκοσμος του εδάφους συμβολή του στη γεωργία και παραγωγή τροφίμων Δρ. Αθανάσιος Γκέρτσης Καθηγητής - Επικεφαλής Τμήματος ΔΙΑΧΕΙΡΙΣΗΣ ΑΓΡΟ-ΠΕΡΙΒΑΛΛΟΝΤΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ [Agro-environmental Systems Management]
Ο μικρόκοσμος του εδάφους συμβολή του στη γεωργία και παραγωγή τροφίμων Δρ. Αθανάσιος Γκέρτσης Καθηγητής - Επικεφαλής Τμήματος ΔΙΑΧΕΙΡΙΣΗΣ ΑΓΡΟ-ΠΕΡΙΒΑΛΛΟΝΤΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ [Agro-environmental Systems Management]
Μπορεί η διαχείριση των εδαφικών πόρων να συμβάλλει στη μείωση των αερίων του θερμοκηπίου;
 Μπορεί η διαχείριση των εδαφικών πόρων να συμβάλλει στη μείωση των αερίων του θερμοκηπίου; Δημ. Αλιφραγκής Καθηγητής Εργαστήριο Δασικής Εδαφολογίας ΑΠΘ Αύξηση του ρυθμού δέσμευσης του διοξειδίου του άνθρακα
Μπορεί η διαχείριση των εδαφικών πόρων να συμβάλλει στη μείωση των αερίων του θερμοκηπίου; Δημ. Αλιφραγκής Καθηγητής Εργαστήριο Δασικής Εδαφολογίας ΑΠΘ Αύξηση του ρυθμού δέσμευσης του διοξειδίου του άνθρακα
Βραβευμένο σύστημα τροφής φυτών!
 Βραβευμένο σύστημα τροφής φυτών! Powder Feeding Τα οργανικά και ορυκτά λιπάσματα χρησιμοποιούνται στη γεωργία εδώ και χιλιάδες χρόνια, ωστόσο, οι γεωργικές τεχνικές και τα χρησιμοποιούμενα μέσα δεν έχουν
Βραβευμένο σύστημα τροφής φυτών! Powder Feeding Τα οργανικά και ορυκτά λιπάσματα χρησιμοποιούνται στη γεωργία εδώ και χιλιάδες χρόνια, ωστόσο, οι γεωργικές τεχνικές και τα χρησιμοποιούμενα μέσα δεν έχουν
Κεφάλαιο 1: Εισαγωγή. Κεφάλαιο 2: Η Βιολογία των Ιών
 Κεφάλαιο 1: Εισαγωγή 1.1 Μικροοργανισμοί, Μικροβιολογία και Μικροβιολόγοι... 19 1.1.1 Μικροοργανισμοί... 19 1.1.2 Μικροβιολογία... 20 1.1.3 Μικροβιολόγοι... 21 1.2 Σύντομη Ιστορική Εξέλιξη της Μικροβιολογίας...
Κεφάλαιο 1: Εισαγωγή 1.1 Μικροοργανισμοί, Μικροβιολογία και Μικροβιολόγοι... 19 1.1.1 Μικροοργανισμοί... 19 1.1.2 Μικροβιολογία... 20 1.1.3 Μικροβιολόγοι... 21 1.2 Σύντομη Ιστορική Εξέλιξη της Μικροβιολογίας...
Τύποι Φυτών. Ετήσια Διετή Πολυετή. Ποώδη. Ξυλώδη
 Τύποι Φυτών Ετήσια Διετή Πολυετή Ποώδη Ξυλώδη Δένδρα Θάμνοι Ανατομική των αγγειωδών φυτών Κύτταρο Ιστός Όργανο Φυτό Υπόγειο μέρος Υπέργειο μέρος Ρίζα Βλαστοί ή στελέχη Ιστοί του βλαστού Εφυμενίδα Επιδερμίδα
Τύποι Φυτών Ετήσια Διετή Πολυετή Ποώδη Ξυλώδη Δένδρα Θάμνοι Ανατομική των αγγειωδών φυτών Κύτταρο Ιστός Όργανο Φυτό Υπόγειο μέρος Υπέργειο μέρος Ρίζα Βλαστοί ή στελέχη Ιστοί του βλαστού Εφυμενίδα Επιδερμίδα
Περιεχομενα ΚΕΦΑΛΑΙΟ ΚΕΦΑΛΑΙΟ ΚΕΦΑΛΑΙΟ
 Περιεχομενα ΚΕΦΑΛΑΙΟ 1...1 1.1 Βασικά εδαφικά συστατικά...1 1.2 Στερεή εδαφική φάση:...2 1.3 Υγρή εδαφική φάση...2 1.4 Αέρια εδαφική φάση:...3 1.5 Σχηματική απεικόνιση των τριών φάσεων εδάφους...3 1.6
Περιεχομενα ΚΕΦΑΛΑΙΟ 1...1 1.1 Βασικά εδαφικά συστατικά...1 1.2 Στερεή εδαφική φάση:...2 1.3 Υγρή εδαφική φάση...2 1.4 Αέρια εδαφική φάση:...3 1.5 Σχηματική απεικόνιση των τριών φάσεων εδάφους...3 1.6
7. Βιοτεχνολογία. α) η διαθεσιμότητα θρεπτικών συστατικών στο θρεπτικό υλικό, β) το ph, γ) το Ο 2 και δ) η θερμοκρασία.
 η διαθεσιμότητα θρεπτικών συστατικών στο θρεπτικό υλικό, β) το ph, γ) το Ο 2 και δ) η θερμοκρασία.") 7. Βιοτεχνολογία Εισαγωγή Τι είναι η Βιοτεχνολογία; Η Βιοτεχνολογία αποτελεί συνδυασμό επιστήμης και τεχνολογίας. Ειδικότερα εφαρμόζει τις γνώσεις που έχουν αποκτηθεί για τις βιολογικές λειτουργίες των
7. Βιοτεχνολογία Εισαγωγή Τι είναι η Βιοτεχνολογία; Η Βιοτεχνολογία αποτελεί συνδυασμό επιστήμης και τεχνολογίας. Ειδικότερα εφαρμόζει τις γνώσεις που έχουν αποκτηθεί για τις βιολογικές λειτουργίες των
Τύποι Φυτών. Ετήσια Διετή Πολυετή. Ποώδη. Ξυλώδη
 Τύποι Φυτών Ετήσια Διετή Πολυετή Ποώδη Ξυλώδη Δένδρα Θάμνοι Ανατομική των αγγειωδών φυτών Κύτταρο Ιστός Όργανο Φυτό Υπόγειο μέρος Υπέργειο μέρος Ρίζα Βλαστοί ή στελέχη Ιστοί του βλαστού Εφυμενίδα Επιδερμίδα
Τύποι Φυτών Ετήσια Διετή Πολυετή Ποώδη Ξυλώδη Δένδρα Θάμνοι Ανατομική των αγγειωδών φυτών Κύτταρο Ιστός Όργανο Φυτό Υπόγειο μέρος Υπέργειο μέρος Ρίζα Βλαστοί ή στελέχη Ιστοί του βλαστού Εφυμενίδα Επιδερμίδα
Ανακύκλωση & διατήρηση Θρεπτικών
 Ανακύκλωση & διατήρηση Θρεπτικών 30-12-2014 EVA PAPASTERGIADOU Ανακύκλωση των Θρεπτικών είναι η χρησιμοποίηση, ο μετασχηματισμός, η διακίνηση & η επαναχρησιμοποίηση των θρεπτικών στοιχείων στα οικοσυστήματα
Ανακύκλωση & διατήρηση Θρεπτικών 30-12-2014 EVA PAPASTERGIADOU Ανακύκλωση των Θρεπτικών είναι η χρησιμοποίηση, ο μετασχηματισμός, η διακίνηση & η επαναχρησιμοποίηση των θρεπτικών στοιχείων στα οικοσυστήματα
ΓΕΝΕΤΙΚΗ ΒΕΛΤΙΩΣΗ. 9η ΙΑΛΕΞΗ ΒΕΛΤΙΩΣΗ ΓΙΑ ΑΝΘΕΚΤΙΚΟΤΗΤΑ ΣΤΙΣ ΑΣΘΕΝΕΙΕΣ
 ΓΕΝΕΤΙΚΗ ΒΕΛΤΙΩΣΗ ΦΥΤΩΝ 9η ΙΑΛΕΞΗ ΒΕΛΤΙΩΣΗ ΓΙΑ ΑΝΘΕΚΤΙΚΟΤΗΤΑ ΣΤΙΣ ΑΣΘΕΝΕΙΕΣ Ανθεκτικότητα Γενικά Η εξέλιξη των καλλιεργούµενων φυτών είναι το αποτέλεσµα φυσικής και τεχνητής επιλογής Η επιλογή για αυξηµένες
ΓΕΝΕΤΙΚΗ ΒΕΛΤΙΩΣΗ ΦΥΤΩΝ 9η ΙΑΛΕΞΗ ΒΕΛΤΙΩΣΗ ΓΙΑ ΑΝΘΕΚΤΙΚΟΤΗΤΑ ΣΤΙΣ ΑΣΘΕΝΕΙΕΣ Ανθεκτικότητα Γενικά Η εξέλιξη των καλλιεργούµενων φυτών είναι το αποτέλεσµα φυσικής και τεχνητής επιλογής Η επιλογή για αυξηµένες
ΓΕΩΡΓΙΑ ΑΚΡΙΒΕΙΑΣ ΛΙΠΑΝΣΗ ΚΑΛΛΕΙΕΡΓΕΙΩΝ
 ΓΕΩΡΓΙΑ ΑΚΡΙΒΕΙΑΣ ΛΙΠΑΝΣΗ ΚΑΛΛΕΙΕΡΓΕΙΩΝ ΠΡΟΣ ΙΟΡΙΣΜΟΣ ΤΩΝ ΑΝΑΓΚΩΝ ΤΩΝ ΦΥΤΩΝ ΣΕ ΘΡΕΠΤΙΚΑ ΣΤΟΙΧΕΙΑ ΧΡΙΣΤΟΣ ΤΣΑΝΤΗΛΑΣ ΙΝΣΤΙΤΟΥΤΟ ΧΑΡΤΟΓΡΑΦΗΣΗΣ ΚΑΙ ΤΑΞΙΝΟΜΗΣΗΣ ΕΔΑΦΩΝ Web: http://www.ismc.gr/ ΠΡΟΓΡΑΜΜΑ ΕΚΠΑΙ
ΓΕΩΡΓΙΑ ΑΚΡΙΒΕΙΑΣ ΛΙΠΑΝΣΗ ΚΑΛΛΕΙΕΡΓΕΙΩΝ ΠΡΟΣ ΙΟΡΙΣΜΟΣ ΤΩΝ ΑΝΑΓΚΩΝ ΤΩΝ ΦΥΤΩΝ ΣΕ ΘΡΕΠΤΙΚΑ ΣΤΟΙΧΕΙΑ ΧΡΙΣΤΟΣ ΤΣΑΝΤΗΛΑΣ ΙΝΣΤΙΤΟΥΤΟ ΧΑΡΤΟΓΡΑΦΗΣΗΣ ΚΑΙ ΤΑΞΙΝΟΜΗΣΗΣ ΕΔΑΦΩΝ Web: http://www.ismc.gr/ ΠΡΟΓΡΑΜΜΑ ΕΚΠΑΙ
ΦΡΟΝΤΙΣΤΗΡΙΑ ΟΡΟΣΗΜΟ ΘΑΛΗΣ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ. Α1. γ Α2. α Α3. β Α4. β Α5. δ
 ΦΡΟΝΤΙΣΤΗΡΙΑ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ Α Α1. γ Α2. α Α3. β Α4. β Α5. δ ΘΕΜΑ Β Β1. 1 Β 2 Α 3 Α 4 Β 5 Β 6 Α 7 Α 8 Β Β2. Σελ. 18 Σχολικού βιβλίου Το γενετικό
ΦΡΟΝΤΙΣΤΗΡΙΑ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ Α Α1. γ Α2. α Α3. β Α4. β Α5. δ ΘΕΜΑ Β Β1. 1 Β 2 Α 3 Α 4 Β 5 Β 6 Α 7 Α 8 Β Β2. Σελ. 18 Σχολικού βιβλίου Το γενετικό
Ε ΑΦΟΣ. Έδαφος: ανόργανα οργανικά συστατικά
 Ε ΑΦΟΣ Έδαφος: ανόργανα οργανικά συστατικά ρ. Ε. Λυκούδη Αθήνα 2005 Έδαφος Το έδαφος σχηµατίζεται από τα προϊόντα της αποσάθρωσης των πετρωµάτων του υποβάθρου (µητρικό πέτρωµα) ή των πετρωµάτων τω γειτονικών
Ε ΑΦΟΣ Έδαφος: ανόργανα οργανικά συστατικά ρ. Ε. Λυκούδη Αθήνα 2005 Έδαφος Το έδαφος σχηµατίζεται από τα προϊόντα της αποσάθρωσης των πετρωµάτων του υποβάθρου (µητρικό πέτρωµα) ή των πετρωµάτων τω γειτονικών
ΠΡΟΤΥΠΟ ΠΕΙΡΑΜΑΤΙΚΟ ΛΥΚΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΠΑΤΡΩΝ 2013-14
 ΘΕΜΑΤΑ ΟΙΚΟΛΟΓΙΑΣ Μπορεί να λειτουργήσει ένα οικοσύστημα α) με παραγωγούς και καταναλωτές; β) με παραγωγούς και αποικοδομητές; γ)με καταναλωτές και αποικοδομητές; Η διατήρηση των οικοσυστημάτων προϋποθέτει
ΘΕΜΑΤΑ ΟΙΚΟΛΟΓΙΑΣ Μπορεί να λειτουργήσει ένα οικοσύστημα α) με παραγωγούς και καταναλωτές; β) με παραγωγούς και αποικοδομητές; γ)με καταναλωτές και αποικοδομητές; Η διατήρηση των οικοσυστημάτων προϋποθέτει
Η θρέψη και η λίπανση της βιομηχανικής τομάτας
 Η θρέψη και η λίπανση της βιομηχανικής τομάτας Αθανάσιος Κουκουνάρας Λέκτορας Εργαστήριο Λαχανοκομίας Τμήμα Γεωπονίας ΑΠΘ thankou@agro.auth.gr 9 Μαρτίου 2015, Λάρισα Κύρια σημεία Η ανάγκη για λίπανση Οργανική
Η θρέψη και η λίπανση της βιομηχανικής τομάτας Αθανάσιος Κουκουνάρας Λέκτορας Εργαστήριο Λαχανοκομίας Τμήμα Γεωπονίας ΑΠΘ thankou@agro.auth.gr 9 Μαρτίου 2015, Λάρισα Κύρια σημεία Η ανάγκη για λίπανση Οργανική
Τ.Ε.Ι. Ηπείρου Σχολή Τεχνολογίας Γεωπονίας Τμήμα Φυτικής Παραγωγής ΚΑΛΛΙΕΡΓΕΙΕΣ ΚΕΦΑΛΑΙΟ 1 Ο. Εισαγωγικές Έννοιες. Δούμα Δήμητρα Άρτα, 2013
 Τ.Ε.Ι. Ηπείρου Σχολή Τεχνολογίας Γεωπονίας Τμήμα Φυτικής Παραγωγής ΚΑΛΛΙΕΡΓΕΙΕΣ IN VITRO ΚΕΦΑΛΑΙΟ 1 Ο Εισαγωγικές Έννοιες Δούμα Δήμητρα Άρτα, 2013 Καλλιέργεια in vitro (= μέσα σε γυαλί): η καλλιέργεια
Τ.Ε.Ι. Ηπείρου Σχολή Τεχνολογίας Γεωπονίας Τμήμα Φυτικής Παραγωγής ΚΑΛΛΙΕΡΓΕΙΕΣ IN VITRO ΚΕΦΑΛΑΙΟ 1 Ο Εισαγωγικές Έννοιες Δούμα Δήμητρα Άρτα, 2013 Καλλιέργεια in vitro (= μέσα σε γυαλί): η καλλιέργεια
ΧΗΜΙΚΕΣ ΙΔΙΟΤΗΤΕΣ ΤΩΝ ΕΔΑΦΩΝ
 Εδαφικά κολλοειδή Ανόργανα ορυκτά (άργιλος) ή οργανική ουσία (χούμος) με διάμετρο μικρότερη από 0,001 mm ή 1μ ανήκουν στα κολλοειδή. Ηάργιλος(
Εδαφικά κολλοειδή Ανόργανα ορυκτά (άργιλος) ή οργανική ουσία (χούμος) με διάμετρο μικρότερη από 0,001 mm ή 1μ ανήκουν στα κολλοειδή. Ηάργιλος(
ΔΠΘ - Τμήμα Δασολογίας & Διαχείρισης Περιβάλλοντος & Φυσικών Πόρων ΦΥΣΙΟΛΟΓΙΑ ΦΥΤΩΝ ΠΡΟΣΛΗΨΗ ΚΑΙ ΜΕΤΑΦΟΡΑ ΤΟΥ ΝΕΡΟΥ ΣΤΑ ΦΥΤΑ
 ΔΠΘ - Τμήμα Δασολογίας & Διαχείρισης Περιβάλλοντος & Φυσικών Πόρων ΦΥΣΙΟΛΟΓΙΑ ΦΥΤΩΝ ΠΡΟΣΛΗΨΗ ΚΑΙ ΜΕΤΑΦΟΡΑ ΤΟΥ ΝΕΡΟΥ ΣΤΑ ΦΥΤΑ Θερινό εξάμηνο 2011 Ο ρόλος του νερού στο φυτό Βασικότερο συστατικό των ιστών
ΔΠΘ - Τμήμα Δασολογίας & Διαχείρισης Περιβάλλοντος & Φυσικών Πόρων ΦΥΣΙΟΛΟΓΙΑ ΦΥΤΩΝ ΠΡΟΣΛΗΨΗ ΚΑΙ ΜΕΤΑΦΟΡΑ ΤΟΥ ΝΕΡΟΥ ΣΤΑ ΦΥΤΑ Θερινό εξάμηνο 2011 Ο ρόλος του νερού στο φυτό Βασικότερο συστατικό των ιστών
ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ. Το σύνολο των μετασχηματισμών βιολογικής ή χημικής φύσης που λαμβάνουν χώρα κατά την ανακύκλωση ορισμένων στοιχείων
 ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ Το σύνολο των μετασχηματισμών βιολογικής ή χημικής φύσης που λαμβάνουν χώρα κατά την ανακύκλωση ορισμένων στοιχείων Επιβίωση οργανισμών Ύλη o Η ύλη που υπάρχει διαθέσιμη στη βιόσφαιρα
ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ Το σύνολο των μετασχηματισμών βιολογικής ή χημικής φύσης που λαμβάνουν χώρα κατά την ανακύκλωση ορισμένων στοιχείων Επιβίωση οργανισμών Ύλη o Η ύλη που υπάρχει διαθέσιμη στη βιόσφαιρα
Newsletter ΣΥΣΤΗΜΑ ΔΙΑΧΕΙΡΙΣΗΣ CONDENSE: ΠΑΡΑΓΩΓΗ ΚΑΙΝΟΤΟΜΩΝ ΛΙΠΑΣΜΑΤΩΝ ΑΠΟ ΚΟΠΡΙΑ ΚΑΙ ΚΑΤΣΙΓΑΡΟ ΣΚΟΠΟΣ ΤΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ ΓΕΩΓΡΑΦΙΚΗ ΘΕΣΗ ΤΗΣ
 Newsletter 12 η Έκδοση ΣΥΣΤΗΜΑ ΔΙΑΧΕΙΡΙΣΗΣ CONDENSE: ΠΑΡΑΓΩΓΗ ΚΑΙΝΟΤΟΜΩΝ ΛΙΠΑΣΜΑΤΩΝ ΑΠΟ ΚΟΠΡΙΑ ΚΑΙ ΚΑΤΣΙΓΑΡΟ ΣΚΟΠΟΣ ΤΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ ΓΕΩΓΡΑΦΙΚΗ ΘΕΣΗ ΤΗΣ ΠΕΡΙΟΧΗΣ ΕΠΙΔΕΙΞΗΣ Ο οργανισμός ανάπτυξης Δυτικής
Newsletter 12 η Έκδοση ΣΥΣΤΗΜΑ ΔΙΑΧΕΙΡΙΣΗΣ CONDENSE: ΠΑΡΑΓΩΓΗ ΚΑΙΝΟΤΟΜΩΝ ΛΙΠΑΣΜΑΤΩΝ ΑΠΟ ΚΟΠΡΙΑ ΚΑΙ ΚΑΤΣΙΓΑΡΟ ΣΚΟΠΟΣ ΤΟΥ ΠΡΟΓΡΑΜΜΑΤΟΣ ΓΕΩΓΡΑΦΙΚΗ ΘΕΣΗ ΤΗΣ ΠΕΡΙΟΧΗΣ ΕΠΙΔΕΙΞΗΣ Ο οργανισμός ανάπτυξης Δυτικής
ΑΛΕΞΑΝΔΡΕΙΟ ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΕΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΤΕΧΝΟΛΟΓΙΑΣ ΓΕΩΠΟΝΙΑΣ ΤΜΗΜΑ ΦΥΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΤΟΜΕΑΣ ΟΠΩΡΟΚΗΠΕΥΤΙΚΩΝ-ΒΟΤΑΝΙΚΗΣ
 ΑΛΕΞΑΝΔΡΕΙΟ ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΕΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΤΕΧΝΟΛΟΓΙΑΣ ΓΕΩΠΟΝΙΑΣ ΤΜΗΜΑ ΦΥΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΤΟΜΕΑΣ ΟΠΩΡΟΚΗΠΕΥΤΙΚΩΝ-ΒΟΤΑΝΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Αξιοποίηση της οργανικής ουσίας για τη βελτίωση
ΑΛΕΞΑΝΔΡΕΙΟ ΤΕΧΝΟΛΟΓΙΚΟ ΕΚΠΕΔΕΥΤΙΚΟ ΙΔΡΥΜΑ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΤΕΧΝΟΛΟΓΙΑΣ ΓΕΩΠΟΝΙΑΣ ΤΜΗΜΑ ΦΥΤΙΚΗΣ ΠΑΡΑΓΩΓΗΣ ΤΟΜΕΑΣ ΟΠΩΡΟΚΗΠΕΥΤΙΚΩΝ-ΒΟΤΑΝΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Αξιοποίηση της οργανικής ουσίας για τη βελτίωση
Έδαφος. Οι ιδιότητες και η σημασία του
 Έδαφος Οι ιδιότητες και η σημασία του ΕΔΑΦΟΣ : Είναι το χαλαρό επιφανειακό στρώμα του στερεού φλοιού της γης. ΕΔΑΦΟΓΕΝΕΣΗ: Το έδαφος σχηματίζεται από την αποσάθρωση των μητρικών πετρωμάτων με την επίδραση
Έδαφος Οι ιδιότητες και η σημασία του ΕΔΑΦΟΣ : Είναι το χαλαρό επιφανειακό στρώμα του στερεού φλοιού της γης. ΕΔΑΦΟΓΕΝΕΣΗ: Το έδαφος σχηματίζεται από την αποσάθρωση των μητρικών πετρωμάτων με την επίδραση
Η ανόργανη θρέψη των φυτών
 Η ανόργανη θρέψη των φυτών Οργανικά θρεπτικά στοιχεία σάκχαρα που προέρχονται από τη διαδικασία της φωτοσύνθεσης με τις επακόλουθες μετατροπές Ανόργανα θρεπτικά στοιχεία προέρχονται από το έδαφος, με τη
Η ανόργανη θρέψη των φυτών Οργανικά θρεπτικά στοιχεία σάκχαρα που προέρχονται από τη διαδικασία της φωτοσύνθεσης με τις επακόλουθες μετατροπές Ανόργανα θρεπτικά στοιχεία προέρχονται από το έδαφος, με τη
AMINEMAX και ΤΡΟΠΟΣ ΠΑΡΑΣΚΕΥΗΣ
 AMINEMAX και ΤΡΟΠΟΣ ΠΑΡΑΣΚΕΥΗΣ Το AMINΕMAX παραλαμβάνεται από κανονική ενζυματική υδρόλυση με πρώτες ύλες από σπόρους σιτηρών και καλαμποκιού σε ισορροπία με μείγμα από ειδικά ένζυμα ( Έξω πρωτει πρωτεΐνάσες-
AMINEMAX και ΤΡΟΠΟΣ ΠΑΡΑΣΚΕΥΗΣ Το AMINΕMAX παραλαμβάνεται από κανονική ενζυματική υδρόλυση με πρώτες ύλες από σπόρους σιτηρών και καλαμποκιού σε ισορροπία με μείγμα από ειδικά ένζυμα ( Έξω πρωτει πρωτεΐνάσες-
Θρέψη Φυτών. Ενότητα 15 η Ριζόσφαιρα. Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής
 Θρέψη Φυτών Ενότητα 15 η Ριζόσφαιρα Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής Στόχοι ενότητας 1. Περιγραφή της διαδικασίας της ριζοαπόθεσης. 2. Ποια
Θρέψη Φυτών Ενότητα 15 η Ριζόσφαιρα Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής Στόχοι ενότητας 1. Περιγραφή της διαδικασίας της ριζοαπόθεσης. 2. Ποια
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΕΠΑΝΑΛΗΠΤΙΚΟ ΔΙΑΓΩΝΙΣΜΑ ( )
") ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΕΠΑΝΑΛΗΠΤΙΚΟ ΔΙΑΓΩΝΙΣΜΑ (8-4-2012) ΘΕΜΑ 1: Να επιλέξετε τη φράση που συμπληρώνει σωστά την πρόταση: (Μονάδες 25) 1. Ο πυρετός: α. Δημιουργείται σε περίπτωση τοπικής
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΕΠΑΝΑΛΗΠΤΙΚΟ ΔΙΑΓΩΝΙΣΜΑ (8-4-2012) ΘΕΜΑ 1: Να επιλέξετε τη φράση που συμπληρώνει σωστά την πρόταση: (Μονάδες 25) 1. Ο πυρετός: α. Δημιουργείται σε περίπτωση τοπικής
E. Καµπουράκης. Τηλ.. 281 0 245851, Fax.. 281 0 245873 ekab@nagref-her.gr
 ΛίπανσηστηνΒιολογικήΓεωργία E. Καµπουράκης Εθνικό Ίδρυµα Αγροτικής Έρευνας (ΕΘΙΑΓΕ) Ταχ. Θυρ.. 2229, 71003 Ηράκλειο, Κρήτη Ελλάδα Τηλ.. 281 0 245851, Fax.. 281 0 245873 ekab@nagref-her.gr Λίπανσηστηνβιολογικήγεωργία
ΛίπανσηστηνΒιολογικήΓεωργία E. Καµπουράκης Εθνικό Ίδρυµα Αγροτικής Έρευνας (ΕΘΙΑΓΕ) Ταχ. Θυρ.. 2229, 71003 Ηράκλειο, Κρήτη Ελλάδα Τηλ.. 281 0 245851, Fax.. 281 0 245873 ekab@nagref-her.gr Λίπανσηστηνβιολογικήγεωργία
ΘΕΜΑ Γ Ένας άνθρωπος μολύνεται από ιό. Το παρακάτω διάγραμμα απεικονίζει τις συγκεντρώσεις των αντιγόνων και των αντισωμάτων σε συνάρτηση με το χρόνο.
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμίας από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και, δίπλα, το γράμμα που αντιστοιχεί στη λέξη ή στη φράση η οποία συμπληρώνει
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμίας από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και, δίπλα, το γράμμα που αντιστοιχεί στη λέξη ή στη φράση η οποία συμπληρώνει
ΕΡΓΑΣΙΑ ΒΙΟΛΟΓΙΑΣ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ
 ΕΡΓΑΣΙΑ ΒΙΟΛΟΓΙΑΣ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Οι οργανισμοί εξασφαλίζουν ενέργεια, για τις διάφορες λειτουργίες τους, διασπώντας θρεπτικές ουσίες που περιέχονται στην τροφή τους. Όμως οι φωτοσυνθετικοί
ΕΡΓΑΣΙΑ ΒΙΟΛΟΓΙΑΣ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Οι οργανισμοί εξασφαλίζουν ενέργεια, για τις διάφορες λειτουργίες τους, διασπώντας θρεπτικές ουσίες που περιέχονται στην τροφή τους. Όμως οι φωτοσυνθετικοί
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ 2015 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ. Επιμέλεια: Aβραμίδου Δέσποινα
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ 2015 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ Επιμέλεια: Aβραμίδου Δέσποινα ΘΕΜΑ Α 1. γ 2. α 3. β 4. β 5. δ ΘΕΜΑ Β Β1. 1. Β 2. Α 3. Α 4. Β 5. Β 6. Α 7. Α 8. Β Β2. Η απάντηση βρίσκεται στη σελίδα
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ 2015 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ Επιμέλεια: Aβραμίδου Δέσποινα ΘΕΜΑ Α 1. γ 2. α 3. β 4. β 5. δ ΘΕΜΑ Β Β1. 1. Β 2. Α 3. Α 4. Β 5. Β 6. Α 7. Α 8. Β Β2. Η απάντηση βρίσκεται στη σελίδα
ΒΙΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΥΠΟΒΑΘΜΙΣΗΣ ΤΡΟΦΙΜΩΝ - 2
 31-7-14 ΒΙΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΥΠΟΒΑΘΜΙΣΗΣ ΤΡΟΦΙΜΩΝ - 2 Στο σχήμα 1 του άρθρου που δημοσιεύσαμε την προηγούμενη φορά φαίνεται η καθοριστικός ρόλος των μικροοργανισμών για την ύπαρξη της ζωής, αφού χωρίς
31-7-14 ΒΙΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΥΠΟΒΑΘΜΙΣΗΣ ΤΡΟΦΙΜΩΝ - 2 Στο σχήμα 1 του άρθρου που δημοσιεύσαμε την προηγούμενη φορά φαίνεται η καθοριστικός ρόλος των μικροοργανισμών για την ύπαρξη της ζωής, αφού χωρίς
ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ
 ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Οι μικροοργανισμοί είναι αναπόσπαστο τμήμα τόσο της ιστορίας του κόσμου μας όσο και της κοινωνικής εξέλιξης του ανθρώπου Βιοτεχνολογία o Ο όρος Βιοτεχνολογία χρησιμοποιήθηκε
ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Οι μικροοργανισμοί είναι αναπόσπαστο τμήμα τόσο της ιστορίας του κόσμου μας όσο και της κοινωνικής εξέλιξης του ανθρώπου Βιοτεχνολογία o Ο όρος Βιοτεχνολογία χρησιμοποιήθηκε
ΘΕΜΑ Α 1 δ 2 β 3 γ 4 β 5 α ΘΕΜΑ Β
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 30 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α 1 δ 2 β 3 γ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 30 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α 1 δ 2 β 3 γ
Soil Fertility & Plant Nutrition
 Soil Fertility & Plant Nutrition E. Katsalirou Spring 2015 Γονιμότητα Εδάφους & Θρέψη Φυτών Πηγές& Πρόσληψη Θρεπτικών Στοιχείων Φωτοσύνθεση Φως 6 CO 2 +12 H 2 O C 6 H 12 O 6 +6O 2 +6H 2 O Χλωροφύλλη Ο
Soil Fertility & Plant Nutrition E. Katsalirou Spring 2015 Γονιμότητα Εδάφους & Θρέψη Φυτών Πηγές& Πρόσληψη Θρεπτικών Στοιχείων Φωτοσύνθεση Φως 6 CO 2 +12 H 2 O C 6 H 12 O 6 +6O 2 +6H 2 O Χλωροφύλλη Ο
Πιο ενεργά συστατικά κολλοειδή κλασματα Διάμετρο μικρότερη από 0,001 mm ή 1μ ανήκουν στα κολλοειδή.
 Δρ. Γεώργιος Ζαΐμης Πιο ενεργά συστατικά κολλοειδή κλασματα Διάμετρο μικρότερη από 0,001 mm ή 1μ ανήκουν στα κολλοειδή. Ανόργανα ορυκτά (άργιλος) οργανική ουσία (χούμος) Η άργιλος (< 2μ) των εδαφών, διαμέτρου
Δρ. Γεώργιος Ζαΐμης Πιο ενεργά συστατικά κολλοειδή κλασματα Διάμετρο μικρότερη από 0,001 mm ή 1μ ανήκουν στα κολλοειδή. Ανόργανα ορυκτά (άργιλος) οργανική ουσία (χούμος) Η άργιλος (< 2μ) των εδαφών, διαμέτρου
Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000
 Ζήτηµα 1ο Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι ιοί είναι :
Ζήτηµα 1ο Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι ιοί είναι :
Δασική Εδαφολογία. Εδαφογένεση
 Δασική Εδαφολογία Εδαφογένεση Σχηματισμός της στερεάς φάσης του εδάφους Η στερεά φάση του εδάφους σχηματίζεται από τα προϊόντα της αποσύνθεσης των φυτικών και ζωικών υπολειμμάτων μαζί με τα προϊόντα της
Δασική Εδαφολογία Εδαφογένεση Σχηματισμός της στερεάς φάσης του εδάφους Η στερεά φάση του εδάφους σχηματίζεται από τα προϊόντα της αποσύνθεσης των φυτικών και ζωικών υπολειμμάτων μαζί με τα προϊόντα της
Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000
 Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 ΕΚΦΩΝΗΣΕΙΣ Ζήτηµα 1ο Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι
Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 ΕΚΦΩΝΗΣΕΙΣ Ζήτηµα 1ο Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι
ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ:Κ.Κεραμάρης ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ
 ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ:Κ.Κεραμάρης ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Κωνσταντίνος Ρίζος Γιάννης Ρουμπάνης Βιοτεχνολογία με την ευρεία έννοια είναι η χρήση ζωντανών
ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ:Κ.Κεραμάρης ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Κωνσταντίνος Ρίζος Γιάννης Ρουμπάνης Βιοτεχνολογία με την ευρεία έννοια είναι η χρήση ζωντανών
Ολοκληρωμένη διαχείριση ζιζανίων
 Οι περισσότερες στρατηγικές αντιμετώπισης των ζιζανίων έχουν σαν μέσω τους την χρήση ζιζανιοκτόνων καθώς και την κατεργασία του εδάφους ώστε να ελαχιστοποιήσουν τις απώλειες της παραγωγής λόγω των ζιζανίων.
Οι περισσότερες στρατηγικές αντιμετώπισης των ζιζανίων έχουν σαν μέσω τους την χρήση ζιζανιοκτόνων καθώς και την κατεργασία του εδάφους ώστε να ελαχιστοποιήσουν τις απώλειες της παραγωγής λόγω των ζιζανίων.
ΘΕΜΑ 1 ο Στις ερωτήσεις 1-5, να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση.
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 ο Στις ερωτήσεις 1-5, να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Τα κύτταρα που παράγουν
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 ο Στις ερωτήσεις 1-5, να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Τα κύτταρα που παράγουν
ΡΥΠΑΝΣΗ. Ρύποι. Αντίδραση βιολογικών συστημάτων σε παράγοντες αύξησης
 ΡΥΠΑΝΣΗ 91 είναι η άμεση ή έμμεση διοχέτευση από τον άνθρωπο στο υδάτινο περιβάλλον ύλης ή ενέργειας με επιβλαβή αποτελέσματα για τους οργανισμούς ( ο ορισμός της ρύπανσης από τον ΟΗΕ ) Ρύποι Φυσικοί (εκρήξεις
ΡΥΠΑΝΣΗ 91 είναι η άμεση ή έμμεση διοχέτευση από τον άνθρωπο στο υδάτινο περιβάλλον ύλης ή ενέργειας με επιβλαβή αποτελέσματα για τους οργανισμούς ( ο ορισμός της ρύπανσης από τον ΟΗΕ ) Ρύποι Φυσικοί (εκρήξεις
Γενικές εξετάσεις 2015 Βιολογία Γ λυκείου γενικής παιδείας
 Φροντιστήρια δυαδικό 1 ΦΡΟΝΤΙΣΤΗΡΙΑ δυαδικό Τα θέματα επεξεργάστηκαν οι καθηγητές των Φροντιστηρίων «δυαδικό» Μελλίδης Κ. Πασσιά Α. Θέμα Α Γενικές εξετάσεις 2015 Βιολογία Γ λυκείου γενικής παιδείας Να
Φροντιστήρια δυαδικό 1 ΦΡΟΝΤΙΣΤΗΡΙΑ δυαδικό Τα θέματα επεξεργάστηκαν οι καθηγητές των Φροντιστηρίων «δυαδικό» Μελλίδης Κ. Πασσιά Α. Θέμα Α Γενικές εξετάσεις 2015 Βιολογία Γ λυκείου γενικής παιδείας Να
ΑΖΩΤΟΥΧΟΣ ΛΙΠΑΝΣΗ ΚΑΙ ΟΡΘΗ ΓΕΩΡΓΙΚΗ ΠΡΑΚΤΙΚΗ. Δρ. Γιάννης Ασημακόπουλος Πρώην Καθηγητής Γεωπονικού Παν/μίου Αθηνών
 ΑΖΩΤΟΥΧΟΣ ΛΙΠΑΝΣΗ ΚΑΙ ΟΡΘΗ ΓΕΩΡΓΙΚΗ ΠΡΑΚΤΙΚΗ Δρ. Γιάννης Ασημακόπουλος Πρώην Καθηγητής Γεωπονικού Παν/μίου Αθηνών ΟΡΙΣΜΟΣ ΤΗΣ ΕΝΝΟΙΑΣ ΤΟΥ ΛΙΠΑΣΜΑΤΟΣ. Με την κλασσική έννοια, ως λίπασμα ορίζεται κάθε ουσία
ΑΖΩΤΟΥΧΟΣ ΛΙΠΑΝΣΗ ΚΑΙ ΟΡΘΗ ΓΕΩΡΓΙΚΗ ΠΡΑΚΤΙΚΗ Δρ. Γιάννης Ασημακόπουλος Πρώην Καθηγητής Γεωπονικού Παν/μίου Αθηνών ΟΡΙΣΜΟΣ ΤΗΣ ΕΝΝΟΙΑΣ ΤΟΥ ΛΙΠΑΣΜΑΤΟΣ. Με την κλασσική έννοια, ως λίπασμα ορίζεται κάθε ουσία
ΠΡΟΣΛΗΨΗ - ΜΕΤΑΦΟΡΑ ΘΡΕΠΤΙΚΩΝ ΣΤΟΙΧΕΙΩΝ ΣΤΟ ΥΠΕΡΓΕΙΟ ΤΜΗΜΑ ΤΟΥ ΦΥΤΟΥ
 ΠΡΟΣΛΗΨΗ - ΜΕΤΑΦΟΡΑ ΘΡΕΠΤΙΚΩΝ ΣΤΟΙΧΕΙΩΝ ΣΤΟ ΥΠΕΡΓΕΙΟ ΤΜΗΜΑ ΤΟΥ ΦΥΤΟΥ Κίνηση στα αγγεία του ξύλου Κίνηση μέσω του φλοιού Τρόποι μετακίνησης των θρεπτικών στοιχείων στο εσωτερικό της ρίζας: Κίνηση με διάχυση
ΠΡΟΣΛΗΨΗ - ΜΕΤΑΦΟΡΑ ΘΡΕΠΤΙΚΩΝ ΣΤΟΙΧΕΙΩΝ ΣΤΟ ΥΠΕΡΓΕΙΟ ΤΜΗΜΑ ΤΟΥ ΦΥΤΟΥ Κίνηση στα αγγεία του ξύλου Κίνηση μέσω του φλοιού Τρόποι μετακίνησης των θρεπτικών στοιχείων στο εσωτερικό της ρίζας: Κίνηση με διάχυση
Τα οφέλη χρήσης του Νιτρικού Καλίου έναντι του Χλωριούχου Καλίου και του Θειικού Καλίου
 Τα οφέλη χρήσης του Νιτρικού Καλίου έναντι του Χλωριούχου Καλίου και του Θειικού Καλίου Τα οφέλη του καλίου, γενικά Προάγει την φωτοσύνθεση Επιταχύνει την μεταφορά των προϊόντων μεταβολισμού Ενισχύει την
Τα οφέλη χρήσης του Νιτρικού Καλίου έναντι του Χλωριούχου Καλίου και του Θειικού Καλίου Τα οφέλη του καλίου, γενικά Προάγει την φωτοσύνθεση Επιταχύνει την μεταφορά των προϊόντων μεταβολισμού Ενισχύει την
ΗΜΕΡΙΔΑ ELQA. Καθαρισμός ύδατος από βαρέα μέταλλα με καινοτόμα τεχνολογία. Ερευνητικό εργαστήριο Food InnovaLab 1
 Καθαρισμός ύδατος από βαρέα μέταλλα με καινοτόμα τεχνολογία Σταύρος Λαλάς*, Βασίλειος Αθανασιάδης και Όλγα Γκορτζή Τμήμα Τεχνολογίας Τροφίμων Τ.Ε.Ι. ΘΕΣΣΑΛΙΑΣ Ερευνητικό εργαστήριο Food InnovaLab 1 Βαρέα
Καθαρισμός ύδατος από βαρέα μέταλλα με καινοτόμα τεχνολογία Σταύρος Λαλάς*, Βασίλειος Αθανασιάδης και Όλγα Γκορτζή Τμήμα Τεχνολογίας Τροφίμων Τ.Ε.Ι. ΘΕΣΣΑΛΙΑΣ Ερευνητικό εργαστήριο Food InnovaLab 1 Βαρέα
ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ "ΔΟΜΗ ΞΥΛΟΥ"
 "ΔΟΜΗ ΞΥΛΟΥ" ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ Καθ. Γεώργιος Μαντάνης Εργαστήριο Επιστήμης & Τεχνολογίας Ξύλου Τμήμα Σχεδιασμού & Τεχνολογίας Ξύλου & Επίπλου www.teilar.gr/~mantanis ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ ΣΥΣΤΑΣΗ
"ΔΟΜΗ ΞΥΛΟΥ" ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ Καθ. Γεώργιος Μαντάνης Εργαστήριο Επιστήμης & Τεχνολογίας Ξύλου Τμήμα Σχεδιασμού & Τεχνολογίας Ξύλου & Επίπλου www.teilar.gr/~mantanis ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ ΣΥΣΤΑΣΗ
Μετακίνηση. Απώλλεια 95-97% του νερού που απορροφάται ενώ ~2% υποστηρίζει την αύξηση ~1% συμμετέχει σε φωτοσύνθεση & άλλες μεταβολικές διεργασίες
 Το νερό στο φυτό Άριστος διαλύτης συμμετέχει στη διαμόρφωση της λειτουργικής στερεοδομής των μακρομορίων παίζει ενεργό ρόλο στο μεταβολισμό αποτελεί τον πρωταρχικό δότη e - κατά τη φωτοσυνθετική ροή e
Το νερό στο φυτό Άριστος διαλύτης συμμετέχει στη διαμόρφωση της λειτουργικής στερεοδομής των μακρομορίων παίζει ενεργό ρόλο στο μεταβολισμό αποτελεί τον πρωταρχικό δότη e - κατά τη φωτοσυνθετική ροή e
Σήµερα οι εξελίξεις στην Επιστήµη και στην Τεχνολογία δίνουν τη
 ΚΕΦΑΛΑΙΟ 7ο: ΑΡΧΕΣ & ΜΕΘΟ ΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Συνδυασµός ΤΕΧΝΟΛΟΓΙΑΣ & ΕΠΙΣΤΗΜΗΣ Προσφέρει τη δυνατότητα χρησιµοποίησης των ζωντανών οργανισµών για την παραγωγή χρήσιµων προϊόντων 1 Οι ζωντανοί οργανισµοί
ΚΕΦΑΛΑΙΟ 7ο: ΑΡΧΕΣ & ΜΕΘΟ ΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Συνδυασµός ΤΕΧΝΟΛΟΓΙΑΣ & ΕΠΙΣΤΗΜΗΣ Προσφέρει τη δυνατότητα χρησιµοποίησης των ζωντανών οργανισµών για την παραγωγή χρήσιµων προϊόντων 1 Οι ζωντανοί οργανισµοί
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή στη φράση, η οποία
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή στη φράση, η οποία
Αποσάθρωση. Κεφάλαιο 2 ο. ΣΧΗΜΑΤΙΣΜΟΣ ΕΔΑΦΩΝ
 Κεφάλαιο 2 ο. ΣΧΗΜΑΤΙΣΜΟΣ ΕΔΑΦΩΝ Αποσάθρωση Ονομάζουμε τις μεταβολές στο μέγεθος, σχήμα και την εσωτερική δομή και χημική σύσταση τις οποίες δέχεται η στερεά φάση του εδάφους με την επίδραση των παραγόντων
Κεφάλαιο 2 ο. ΣΧΗΜΑΤΙΣΜΟΣ ΕΔΑΦΩΝ Αποσάθρωση Ονομάζουμε τις μεταβολές στο μέγεθος, σχήμα και την εσωτερική δομή και χημική σύσταση τις οποίες δέχεται η στερεά φάση του εδάφους με την επίδραση των παραγόντων
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ
 10 Ιουνίου 2013 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Απαντήσεις Θεμάτων Επαναληπτικών Πανελληνίων Εξετάσεων Ημερησίων Γενικών Λυκείων Περιεχόμενα ΘΕΜΑ Α.... 2 Α1.... 2 Α3.... 2 Α5.... 2 ΘΕΜΑ B.... 2 Β1.... 2 Β2....
10 Ιουνίου 2013 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Απαντήσεις Θεμάτων Επαναληπτικών Πανελληνίων Εξετάσεων Ημερησίων Γενικών Λυκείων Περιεχόμενα ΘΕΜΑ Α.... 2 Α1.... 2 Α3.... 2 Α5.... 2 ΘΕΜΑ B.... 2 Β1.... 2 Β2....
Μέρη Δένδρου. Υπόγειο. Επίγειο
 Μέρη Δένδρου Υπόγειο Επίγειο Μέρη Δένδρου Ακραίο μερίστωμα βλαστού Οφθαλμός Καταβολή φύλλου Φύλλο Λαιμός Ριζικά τριχίδια Ακραίο μερίστωμα ρίζας Καλύπτρα Ρίζα Ο ώριμος σπόρος (έμβρυο) έχει ριζικές καταβολές
Μέρη Δένδρου Υπόγειο Επίγειο Μέρη Δένδρου Ακραίο μερίστωμα βλαστού Οφθαλμός Καταβολή φύλλου Φύλλο Λαιμός Ριζικά τριχίδια Ακραίο μερίστωμα ρίζας Καλύπτρα Ρίζα Ο ώριμος σπόρος (έμβρυο) έχει ριζικές καταβολές
«ΕΙΣΑΓΩΓΗ ΣΤΗ ΔΟΜΗ ΞΥΛΟΥ» ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ. Δρ. Γεώργιος Μαντάνης Εργαστήριο Τεχνολογίας Ξύλου Τμήμα Σχεδιασμού & Τεχνολογίας Ξύλου & Επίπλου
 «ΕΙΣΑΓΩΓΗ ΣΤΗ ΔΟΜΗ ΞΥΛΟΥ» ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ Δρ. Γεώργιος Μαντάνης Εργαστήριο Τεχνολογίας Ξύλου Τμήμα Σχεδιασμού & Τεχνολογίας Ξύλου & Επίπλου ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ ΣΥΣΤΑΣΗ ΞΥΛΟΥ ΣΕ ΔΟΜΙΚΑ ΣΥΣΤΑΤΙΚΑ
«ΕΙΣΑΓΩΓΗ ΣΤΗ ΔΟΜΗ ΞΥΛΟΥ» ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ Δρ. Γεώργιος Μαντάνης Εργαστήριο Τεχνολογίας Ξύλου Τμήμα Σχεδιασμού & Τεχνολογίας Ξύλου & Επίπλου ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΟΥ ΞΥΛΟΥ ΣΥΣΤΑΣΗ ΞΥΛΟΥ ΣΕ ΔΟΜΙΚΑ ΣΥΣΤΑΤΙΚΑ
Περιβαλλοντικοί παράγοντες. και η επίδρασή τους στους ζωντανούς οργανισμούς
 Περιβαλλοντικοί παράγοντες και η επίδρασή τους στους ζωντανούς οργανισμούς Νερό Ολοι οι οργανισμοί αποτελούνται κατά μεγάλο ποσοστό από νερό ανεξάρτητα από το εάν ζουν σε χερσαία ή υδατικά περιβάλλοντα
Περιβαλλοντικοί παράγοντες και η επίδρασή τους στους ζωντανούς οργανισμούς Νερό Ολοι οι οργανισμοί αποτελούνται κατά μεγάλο ποσοστό από νερό ανεξάρτητα από το εάν ζουν σε χερσαία ή υδατικά περιβάλλοντα
Τ.Ε.Ι. Ηπείρου Σχολή Τεχνολογίας Γεωπονίας Τμήμα Φυτικής Παραγωγής ΚΑΛΛΙΕΡΓΕΙΕΣ. Εργαστήριο 2 ο. Υλικό Καλλιέργειας. Δούμα Δήμητρα Άρτα, 2013
 Τ.Ε.Ι. Ηπείρου Σχολή Τεχνολογίας Γεωπονίας Τμήμα Φυτικής Παραγωγής ΚΑΛΛΙΕΡΓΕΙΕΣ IN VITRO Εργαστήριο 2 ο Υλικό Καλλιέργειας Δούμα Δήμητρα Άρτα, 2013 ΥΛΙΚΟ ΚΑΛΛΙΕΡΓΕΙΑΣ o Ο ζυγώτης εμφανίζει ολοδυναμικότητα
Τ.Ε.Ι. Ηπείρου Σχολή Τεχνολογίας Γεωπονίας Τμήμα Φυτικής Παραγωγής ΚΑΛΛΙΕΡΓΕΙΕΣ IN VITRO Εργαστήριο 2 ο Υλικό Καλλιέργειας Δούμα Δήμητρα Άρτα, 2013 ΥΛΙΚΟ ΚΑΛΛΙΕΡΓΕΙΑΣ o Ο ζυγώτης εμφανίζει ολοδυναμικότητα
μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους
 Η ΕΠΙΣΤΗΜΗ ΤΗΣ ΟΙΚΟΛΟΓΙΑΣ μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους Οι οργανισμοί αλληλεπιδρούν με το περιβάλλον τους σε πολλά επίπεδα στα πλαίσια ενός οικοσυστήματος Οι φυσικές
Η ΕΠΙΣΤΗΜΗ ΤΗΣ ΟΙΚΟΛΟΓΙΑΣ μελετά τις σχέσεις μεταξύ των οργανισμών και με το περιβάλλον τους Οι οργανισμοί αλληλεπιδρούν με το περιβάλλον τους σε πολλά επίπεδα στα πλαίσια ενός οικοσυστήματος Οι φυσικές
ΠΑΝΕΠΙΣΤΗΜΙΑΚΑ ΦΡΟΝΤΙΣΤΗΡΙΑ ΚΟΛΛΙΝΤΖΑ
 Κ Kάνιγγος ΠΑΝΕΠΙΣΤΗΜΙΑΚΑ ΦΡΟΝΤΙΣΤΗΡΙΑ ΚΟΛΛΙΝΤΖΑ ΠΑΝΕΠΙΣΤΗΜΙΑΚΑ ΦΡΟΝΤΙΣΤΗΡΙΑ ΟΛΛΙΝΤΖΑ 10, (5ος όροφ. Τηλ: 210-3300296-7. www.kollintzas.gr OΙΚΟΛΟΓΙΑ 1. Όσο το ποσό της ενέργειας: α) μειώνεται προς τα ανώτερα
Κ Kάνιγγος ΠΑΝΕΠΙΣΤΗΜΙΑΚΑ ΦΡΟΝΤΙΣΤΗΡΙΑ ΚΟΛΛΙΝΤΖΑ ΠΑΝΕΠΙΣΤΗΜΙΑΚΑ ΦΡΟΝΤΙΣΤΗΡΙΑ ΟΛΛΙΝΤΖΑ 10, (5ος όροφ. Τηλ: 210-3300296-7. www.kollintzas.gr OΙΚΟΛΟΓΙΑ 1. Όσο το ποσό της ενέργειας: α) μειώνεται προς τα ανώτερα
ΕΝΔΕΙΚΤΙΚΑ ΘΕΜΑΤΑ ΕΔΑΦΟΛΟΓΙΑΣ
 ΕΝΔΕΙΚΤΙΚΑ ΘΕΜΑΤΑ ΕΔΑΦΟΛΟΓΙΑΣ Κεφάλαιο 1 ο ΟΡΥΚΤΟΛΟΓΙΚΗ ΣΥΣΤΑΣΗ ΕΔΑΦΟΥΣ 1. Κυριότερες ιδιότητες της αργίλου 2. Ποια είναι τα ποιο κοινά ορυκτά της αργίλου. Ποιο θεωρείτε σημαντικότερο. 3. Κατατάξτε τα
ΕΝΔΕΙΚΤΙΚΑ ΘΕΜΑΤΑ ΕΔΑΦΟΛΟΓΙΑΣ Κεφάλαιο 1 ο ΟΡΥΚΤΟΛΟΓΙΚΗ ΣΥΣΤΑΣΗ ΕΔΑΦΟΥΣ 1. Κυριότερες ιδιότητες της αργίλου 2. Ποια είναι τα ποιο κοινά ορυκτά της αργίλου. Ποιο θεωρείτε σημαντικότερο. 3. Κατατάξτε τα
Ανοικτά Ακαδημαϊκά Μαθήματα στο ΤΕΙ Ιονίων Νήσων Τμήμα Τεχνολόγων Περιβάλλοντος Κατεύθυνση Τεχνολογιών Φυσικού Περιβάλλοντος. ΜΑΘΗΜΑ: Γενική Οικολογία
 Ανοικτά Ακαδημαϊκά Μαθήματα στο ΤΕΙ Ιονίων Νήσων Τμήμα Τεχνολόγων Περιβάλλοντος Κατεύθυνση Τεχνολογιών Φυσικού Περιβάλλοντος ΜΑΘΗΜΑ: Γενική Οικολογία 11 η Ενότητα Επίδραση περιβαλλοντικών παραγόντων στη
Ανοικτά Ακαδημαϊκά Μαθήματα στο ΤΕΙ Ιονίων Νήσων Τμήμα Τεχνολόγων Περιβάλλοντος Κατεύθυνση Τεχνολογιών Φυσικού Περιβάλλοντος ΜΑΘΗΜΑ: Γενική Οικολογία 11 η Ενότητα Επίδραση περιβαλλοντικών παραγόντων στη
Βιολογία Γενικής Παιδείας 20-5-2015
 ΘΕΜΑ Α Α1. γ Α2. α Α3. β Α4. β Α5. δ ΘΕΜΑ Β Βιολογία Γενικής Παιδείας 20-5-2015 Β1. 1 Β, 2 Α, 3 Α, 4 Β,5 Β, 6 Α, 7 Α, 8 Β Β2. Σχολικό βιβλίο σελ. 18 «Το γενετικό υλικό ενός ιού τον πολλαπλασιασμό του»
ΘΕΜΑ Α Α1. γ Α2. α Α3. β Α4. β Α5. δ ΘΕΜΑ Β Βιολογία Γενικής Παιδείας 20-5-2015 Β1. 1 Β, 2 Α, 3 Α, 4 Β,5 Β, 6 Α, 7 Α, 8 Β Β2. Σχολικό βιβλίο σελ. 18 «Το γενετικό υλικό ενός ιού τον πολλαπλασιασμό του»
ΚΕΦΑΛΑΙΟ 7 Τι είναι οι καλλιέργειες μικροοργανισμών; Τι είναι το θρεπτικό υλικό; Ποια είναι τα είδη του θρεπτικού υλικού και τι είναι το καθένα;
 ΚΕΦΑΛΑΙΟ 7 Τι είναι οι καλλιέργειες μικροοργανισμών; Καλλιέργεια είναι η διαδικασία ανάπτυξης μικροοργανισμών με διάφορους τεχνητούς τρόπους στο εργαστήριο ή σε βιομηχανικό επίπεδο. Με τη δημιουργία καλλιεργειών
ΚΕΦΑΛΑΙΟ 7 Τι είναι οι καλλιέργειες μικροοργανισμών; Καλλιέργεια είναι η διαδικασία ανάπτυξης μικροοργανισμών με διάφορους τεχνητούς τρόπους στο εργαστήριο ή σε βιομηχανικό επίπεδο. Με τη δημιουργία καλλιεργειών
Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Γενικής Παιδείας των Ημερησίων Γενικών Λυκείων και ΕΠΑΛ (Ομάδας Β ).
.") Αθήνα, 30/5/2014 ΠΑΝΕΛΛΗΝΙΑ ΕΝΩΣΗ ΒΙΟΕΠΙΣΤΗΜΟΝΩΝ Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Γενικής Παιδείας των Ημερησίων Γενικών Λυκείων και ΕΠΑΛ (Ομάδας Β ). Η
Αθήνα, 30/5/2014 ΠΑΝΕΛΛΗΝΙΑ ΕΝΩΣΗ ΒΙΟΕΠΙΣΤΗΜΟΝΩΝ Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Γενικής Παιδείας των Ημερησίων Γενικών Λυκείων και ΕΠΑΛ (Ομάδας Β ). Η
Γενική Μικροβιολογία. (Μύκητες)
") Γενική Μικροβιολογία (Μύκητες) Χαρακτηριστικά των μυκήτων Ευκαρυωτικοί μικροοργανισμοί Δεν φωτοσυνθέτουν, είναι χημειοοργανότροφοι. Σημαντικοί αποικοδομητές! Μύκητες και τρόφιμα (Αλλοιώσεις τροφίμων) Μύκητες
Γενική Μικροβιολογία (Μύκητες) Χαρακτηριστικά των μυκήτων Ευκαρυωτικοί μικροοργανισμοί Δεν φωτοσυνθέτουν, είναι χημειοοργανότροφοι. Σημαντικοί αποικοδομητές! Μύκητες και τρόφιμα (Αλλοιώσεις τροφίμων) Μύκητες
Η ανόργανη θρέψη των φυτών
 Η ανόργανη θρέψη των φυτών Οργανικά θρεπτικά στοιχεία σάκχαρα που προέρχονται από τη διαδικασία της φωτοσύνθεσης με τις επακόλουθες μετατροπές Ανόργανα θρεπτικά στοιχεία προέρχονται από το έδαφος, με τη
Η ανόργανη θρέψη των φυτών Οργανικά θρεπτικά στοιχεία σάκχαρα που προέρχονται από τη διαδικασία της φωτοσύνθεσης με τις επακόλουθες μετατροπές Ανόργανα θρεπτικά στοιχεία προέρχονται από το έδαφος, με τη
ειδική φυτοπαθολογία οπωροκηπευτικών και φυτών μεγάλης καλλιέργειας 5. Rhizoctonia spp. Sclerotium spp. Sclerotinia spp.
 ειδική φυτοπαθολογία οπωροκηπευτικών και φυτών μεγάλης καλλιέργειας 5. Rhizoctonia spp. Sclerotium spp. Sclerotinia spp. Rhizoctonia spp. R. solani τέλεια μορφή Βασιδιομύκητας Thanatephorus cucumeris ασθένεια
ειδική φυτοπαθολογία οπωροκηπευτικών και φυτών μεγάλης καλλιέργειας 5. Rhizoctonia spp. Sclerotium spp. Sclerotinia spp. Rhizoctonia spp. R. solani τέλεια μορφή Βασιδιομύκητας Thanatephorus cucumeris ασθένεια
Θρέψη Φυτών. Ενότητα 1 η Εισαγωγή (μέρος α) Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής
 Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής") Θρέψη Φυτών Ενότητα 1 η Εισαγωγή (μέρος α) Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής Στόχοι ενότητας 1. Περιγραφή της ύλης του μαθήματος. 2. Κατανόηση
Θρέψη Φυτών Ενότητα 1 η Εισαγωγή (μέρος α) Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής Στόχοι ενότητας 1. Περιγραφή της ύλης του μαθήματος. 2. Κατανόηση
ΦΩΤΟΣΥΝΘΕΣΗ. Αυτότροφοι και ετερότροφοι οργανισμοί. Καρβουντζή Ηλιάνα Βιολόγος
 ΦΩΤΟΣΥΝΘΕΣΗ Αυτότροφοι και ετερότροφοι οργανισμοί Η ζωή στον πλανήτη μας στηρίζεται στην ενέργεια του ήλιου. Η ενέργεια αυτή εκπέμπεται με τη μορφή ακτινοβολίας. Ένα πολύ μικρό μέρος αυτής της ακτινοβολίας
ΦΩΤΟΣΥΝΘΕΣΗ Αυτότροφοι και ετερότροφοι οργανισμοί Η ζωή στον πλανήτη μας στηρίζεται στην ενέργεια του ήλιου. Η ενέργεια αυτή εκπέμπεται με τη μορφή ακτινοβολίας. Ένα πολύ μικρό μέρος αυτής της ακτινοβολίας
προσλαμβάνουν από το έδαφος
 Τα φυτά με αρχική πηγή τους υδατάνθρακες που παράγουν κατά τη φωτοσύνθεση είναι ικανά να συνθέσουν όλες τις απαραίτητες οργανικές ενώσεις, χρησιμοποιώντας και άλλα ανόργανα θρεπτικά στοιχεία τα οποία μπορούν
Τα φυτά με αρχική πηγή τους υδατάνθρακες που παράγουν κατά τη φωτοσύνθεση είναι ικανά να συνθέσουν όλες τις απαραίτητες οργανικές ενώσεις, χρησιμοποιώντας και άλλα ανόργανα θρεπτικά στοιχεία τα οποία μπορούν
Bιολογία γενικής παιδείας
 Bιολογία γενικής παιδείας Α1. 1. δ 2. α 3. β 4. δ ΘΕΜΑ Α Α2. ΟΛΑ ΚΑΠΟΙΑ Τοξίνες + Πλασματική μεμβράνη + Κυτταρικό τοίχωμα + Αποικίες + Κάψα + Πλασμίδια + Μαστίγια + Ριβοσώματα + Πυρηνοειδές + Ενδοσπόρια
Bιολογία γενικής παιδείας Α1. 1. δ 2. α 3. β 4. δ ΘΕΜΑ Α Α2. ΟΛΑ ΚΑΠΟΙΑ Τοξίνες + Πλασματική μεμβράνη + Κυτταρικό τοίχωμα + Αποικίες + Κάψα + Πλασμίδια + Μαστίγια + Ριβοσώματα + Πυρηνοειδές + Ενδοσπόρια
Η ΕΝΝΟΙΑ ΤΟΥ ΟΙΚΟΣΥΣΤΗΜΑΤΟΣ. Η έννοια του οικοσυστήματος αποτελεί θεμελιώδη έννοια για την Οικολογία
 Η ΕΝΝΟΙΑ ΤΟΥ ΟΙΚΟΣΥΣΤΗΜΑΤΟΣ Η έννοια του οικοσυστήματος αποτελεί θεμελιώδη έννοια για την Οικολογία Οικολογία Οικολογία είναι η επιστήμη που μελετά τις σχέσεις των οργανισμών (συνεπώς και του ανθρώπου)
Η ΕΝΝΟΙΑ ΤΟΥ ΟΙΚΟΣΥΣΤΗΜΑΤΟΣ Η έννοια του οικοσυστήματος αποτελεί θεμελιώδη έννοια για την Οικολογία Οικολογία Οικολογία είναι η επιστήμη που μελετά τις σχέσεις των οργανισμών (συνεπώς και του ανθρώπου)
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Γενικής Παιδείας, Ημ/νία: 20 Μαΐου 2015. Απαντήσεις Θεμάτων
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Γενικής Παιδείας, Ημ/νία: 20 Μαΐου 2015 Απαντήσεις Θεμάτων ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμίας από τις παρακάτω
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Γενικής Παιδείας, Ημ/νία: 20 Μαΐου 2015 Απαντήσεις Θεμάτων ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμίας από τις παρακάτω
All from a Single Source
 All from a Single Source Το PeKacid TM είναι μια νέα, καινοτόμος λύση για τον φώσφορο σε ασβεστούχα εδάφη και νερά με μεγάλη σκληρότητα. Στερεό φωσφορικό οξύ σε σάκους Μονοκρυσταλλικό, με χαμηλό ph (2.2)
All from a Single Source Το PeKacid TM είναι μια νέα, καινοτόμος λύση για τον φώσφορο σε ασβεστούχα εδάφη και νερά με μεγάλη σκληρότητα. Στερεό φωσφορικό οξύ σε σάκους Μονοκρυσταλλικό, με χαμηλό ph (2.2)
ΦΡΟΝΤΙΣΤΗΡΙΑ «ΟΜΟΚΕΝΤΡΟ» Α. ΦΛΩΡΟΠΟΥΛΟΥ
 Τ λεµφοκύτταρα µνήµης (βοηθητικά Τ λεµφοκύτταρα µνήµης και κυτταροτοξικά Τ λεµφοκύτταρα µνήµης). Ο τερµατισµός της δευτερογενούς ανοσοβιολογικής απόκρισης γίνεται µε την βοήθεια των κατασταλτικών Τ λεµφοκυττάρων.
Τ λεµφοκύτταρα µνήµης (βοηθητικά Τ λεµφοκύτταρα µνήµης και κυτταροτοξικά Τ λεµφοκύτταρα µνήµης). Ο τερµατισµός της δευτερογενούς ανοσοβιολογικής απόκρισης γίνεται µε την βοήθεια των κατασταλτικών Τ λεµφοκυττάρων.
ΕΔΑΦΟΣ ΕΝΕΡΓΗΤΙΚΗ ΣΥΣΤΑΣΗ. Άργιλος Οργανική ουσία Ενεργοί μικροοργανισμοί. Προκαλούν δομή (μικρά μεγάλα συνενωμένα συσσωματώματα)
") ΕΔΑΦΟΣ ΕΝΕΡΓΗΤΙΚΗ ΣΥΣΤΑΣΗ Άργιλος Οργανική ουσία Ενεργοί μικροοργανισμοί. Προκαλούν δομή (μικρά μεγάλα συνενωμένα συσσωματώματα) Α. Η άργιλος (ηλεκτρικό φορτίο) συγκρατεί κατιόντα (+) και τα ανταλλάσει
ΕΔΑΦΟΣ ΕΝΕΡΓΗΤΙΚΗ ΣΥΣΤΑΣΗ Άργιλος Οργανική ουσία Ενεργοί μικροοργανισμοί. Προκαλούν δομή (μικρά μεγάλα συνενωμένα συσσωματώματα) Α. Η άργιλος (ηλεκτρικό φορτίο) συγκρατεί κατιόντα (+) και τα ανταλλάσει
Η συμβολή του Καλίου στην ανάπτυξη των φυτών
 Η συμβολή του Καλίου στην ανάπτυξη των φυτών 1. Τα φυτά δεσμεύουν την ηλιακή ενέργεια E H 2 O CO 2 χλωροφύλλη σάκχαρα Ηηλιακήενέργεια μετατρέπεται σε χημική ενέργεια μέσω της φωτοσύνθεσης. Αν και όλα τα
Η συμβολή του Καλίου στην ανάπτυξη των φυτών 1. Τα φυτά δεσμεύουν την ηλιακή ενέργεια E H 2 O CO 2 χλωροφύλλη σάκχαρα Ηηλιακήενέργεια μετατρέπεται σε χημική ενέργεια μέσω της φωτοσύνθεσης. Αν και όλα τα
ΔΟΜΗ ΚΑΙ ΠΑΡΑΓΩΓΙΚΟΤΗΤΑ ΤΩΝ ΛΙΒΑΔΙΩΝ
 ΔΟΜΗ ΚΑΙ ΠΑΡΑΓΩΓΙΚΟΤΗΤΑ ΤΩΝ ΛΙΒΑΔΙΩΝ ΛΕΙΤΟΥΡΓΙΚΕΣ ΟΜΑΔΕΣ Είναι ομάδες φυτών, από διαφορετικές βοτανικές οικογένειες αλλά που έχουν κοινά βιολογικά χαρακτηριστικά και κοινή συμπεριφορά σε σχέση με το περιβάλλον.
ΔΟΜΗ ΚΑΙ ΠΑΡΑΓΩΓΙΚΟΤΗΤΑ ΤΩΝ ΛΙΒΑΔΙΩΝ ΛΕΙΤΟΥΡΓΙΚΕΣ ΟΜΑΔΕΣ Είναι ομάδες φυτών, από διαφορετικές βοτανικές οικογένειες αλλά που έχουν κοινά βιολογικά χαρακτηριστικά και κοινή συμπεριφορά σε σχέση με το περιβάλλον.
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑΤΑ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και, δίπλα, το γράμμα που αντιστοιχεί στη λέξη ή στη φράση η οποία
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑΤΑ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και, δίπλα, το γράμμα που αντιστοιχεί στη λέξη ή στη φράση η οποία
Δασική Εδαφολογία. Γεωχημικός, Βιοχημικός, Υδρολογικός κύκλος
 Δασική Εδαφολογία Γεωχημικός, Βιοχημικός, Υδρολογικός κύκλος Μέρος 1 ο ΥΔΡΟΛΟΓΙΚΟΣ ΚΥΚΛΟΣ ΥΔΡΟΛΟΓΙΚΟΣ ΚΥΚΛΟΣ Η μεταφορά του νερού από την ατμόσφαιρα στην επιφάνεια της γης, η κίνησή του σ αυτή και η επιστροφή
Δασική Εδαφολογία Γεωχημικός, Βιοχημικός, Υδρολογικός κύκλος Μέρος 1 ο ΥΔΡΟΛΟΓΙΚΟΣ ΚΥΚΛΟΣ ΥΔΡΟΛΟΓΙΚΟΣ ΚΥΚΛΟΣ Η μεταφορά του νερού από την ατμόσφαιρα στην επιφάνεια της γης, η κίνησή του σ αυτή και η επιστροφή
ÃËÕÖÁÄÁ - ÅËËÇÍÉÊÏ ÓÕÃ ÑÏÍÏ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΓΕΝΙΚΗΣ ΠΑΙ ΕΊΑΣ Γ ΛΥΚΕΙΟΥ 20 ΜΑΪΟΥ 2015 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α ΘΕΜΑ Β B1.
 ΘΕΜΑ Α Α1. Γ Α2. Α Α3. Β Α4. Β Α5. ΘΕΜΑ Β B1. ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΓΕΝΙΚΗΣ ΠΑΙ ΕΊΑΣ Γ ΛΥΚΕΙΟΥ 20 ΜΑΪΟΥ 2015 ΑΠΑΝΤΗΣΕΙΣ Α (Λυσοζύµη): 2, 3, 6, 7 Β (Πενικιλίνη): 1, 4, 5, 8 Β2. Το γενετικό υλικό ενός ιού µπορεί
ΘΕΜΑ Α Α1. Γ Α2. Α Α3. Β Α4. Β Α5. ΘΕΜΑ Β B1. ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΓΕΝΙΚΗΣ ΠΑΙ ΕΊΑΣ Γ ΛΥΚΕΙΟΥ 20 ΜΑΪΟΥ 2015 ΑΠΑΝΤΗΣΕΙΣ Α (Λυσοζύµη): 2, 3, 6, 7 Β (Πενικιλίνη): 1, 4, 5, 8 Β2. Το γενετικό υλικό ενός ιού µπορεί
Τρόπος Δράσης. Ιδιότητες. Κυριότερα Πλεονεκτήματα
 Η αύξηση των αποδόσεων των καλλιεργειών και η υψηλή ποιότητα των παραγόμενων προϊόντων εξαρτάται από την αρμονική και προσαρμοσμένη στις απαιτήσεις των φυτών θρέψη. Για την εξασφάλιση της ιδανικής θρεπτικής
Η αύξηση των αποδόσεων των καλλιεργειών και η υψηλή ποιότητα των παραγόμενων προϊόντων εξαρτάται από την αρμονική και προσαρμοσμένη στις απαιτήσεις των φυτών θρέψη. Για την εξασφάλιση της ιδανικής θρεπτικής
που χάνεται κατά την καλλιέργεια και του Ν στην ατμόσφαιρα συνεισφέρει στο φαινόμενο του θερμοκηπίου,, στην τρύπα του όζοντος και στην όξινη βροχή.
 Επίδραση του ανθρώπου Γεωργία και επίδραση στον κύκλο του Ν Το Ν είναι το κύριο θρεπτικό που χάνεται κατά την καλλιέργεια και απελευθερώνεται ως οξείδια του Ν στην ατμόσφαιρα συνεισφέρει στο φαινόμενο
Επίδραση του ανθρώπου Γεωργία και επίδραση στον κύκλο του Ν Το Ν είναι το κύριο θρεπτικό που χάνεται κατά την καλλιέργεια και απελευθερώνεται ως οξείδια του Ν στην ατμόσφαιρα συνεισφέρει στο φαινόμενο
ΦΡΟΝΤΙΣΤΗΡΙΑ ΠΡΟΟΠΤΙΚΗ
 Απαντήσεις του κριτηρίου αξιολόγησης στη βιολογία γενικής παιδείας 1 ο ΚΕΦΑΛΑΙΟ ΘΕΜΑ 1 ο Να γράψετε τον αριθμό καθεμίας από τις ημιτελείς προτάσεις 1 έως και 5, και δίπλα σε αυτόν το γράμμα που αντιστοιχεί
Απαντήσεις του κριτηρίου αξιολόγησης στη βιολογία γενικής παιδείας 1 ο ΚΕΦΑΛΑΙΟ ΘΕΜΑ 1 ο Να γράψετε τον αριθμό καθεμίας από τις ημιτελείς προτάσεις 1 έως και 5, και δίπλα σε αυτόν το γράμμα που αντιστοιχεί
ΑΝΟΡΓΑΝΟΙ ΡΥΠΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΚΥΡΙΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΣΤΟ ΕΔΑΦΟΣ
 ΑΝΟΡΓΑΝΟΙ ΡΥΠΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΚΥΡΙΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΣΤΟ ΕΔΑΦΟΣ ΑΠΟΚΑΤΑΣΤΑΣΗ ΡΥΠΑΣΜΕΝΩΝ ΕΔΑΦΩΝ 2γ-1 ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΣΤΟ ΕΔΑΦΟΣ Μη ειδική προσρόφηση (ανταλλαγή ιόντων) Ειδική προσρόφηση
ΑΝΟΡΓΑΝΟΙ ΡΥΠΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΚΥΡΙΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΣΤΟ ΕΔΑΦΟΣ ΑΠΟΚΑΤΑΣΤΑΣΗ ΡΥΠΑΣΜΕΝΩΝ ΕΔΑΦΩΝ 2γ-1 ΜΗΧΑΝΙΣΜΟΙ ΔΕΣΜΕΥΣΗΣ ΣΤΟ ΕΔΑΦΟΣ Μη ειδική προσρόφηση (ανταλλαγή ιόντων) Ειδική προσρόφηση
Θρέψη Φυτών. Ενότητα 14 η Τοξικά μέταλλα. Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής
 Θρέψη Φυτών Ενότητα 14 η Τοξικά μέταλλα Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής Στόχοι ενότητας 1. Ποια είναι τα τοξικά μέταλλα και ποιοι οι προστατευτικοί
Θρέψη Φυτών Ενότητα 14 η Τοξικά μέταλλα Όνομα καθηγητή: Δ. Μπουράνης Όνομα καθηγητή: Σ. Χωριανοπούλου Τμήμα: Επιστήμης Φυτικής Παραγωγής Στόχοι ενότητας 1. Ποια είναι τα τοξικά μέταλλα και ποιοι οι προστατευτικοί