SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV
|
|
|
- Αντίγονος Ζάχος
- 7 χρόνια πριν
- Προβολές:
Transcript


1 SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV
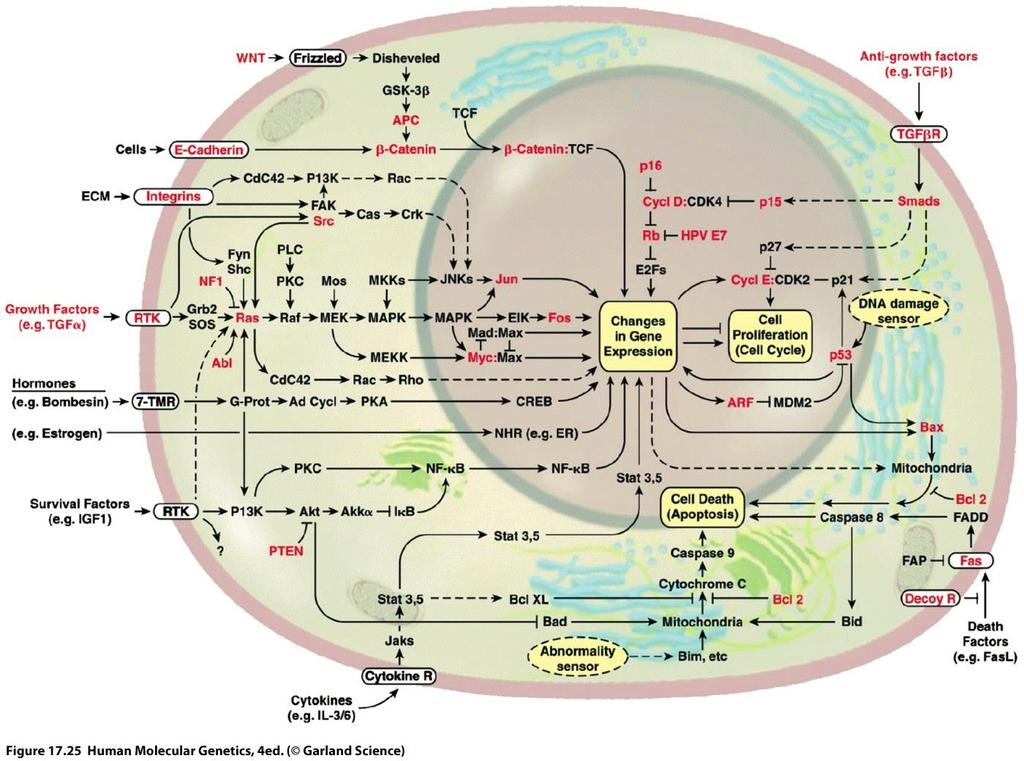
2
3 Literatura za temo Signalne poti, ki regulirajo izražanje genov: Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION 16. poglavje 15. poglavje (GPCR) 7. poglavje (jedrni receptorji, od 323. strani dalje) Copyright 2013 by W. H. Freeman and Company
4 Prenos signala Signalne molekule Receptorji Molekule za prenos informacij Efektorski proteini
5 Prenos signala Signalna molekula se veže na receptor: Hitri odziv: sprememba v aktivnosti ali funkciji celičnih proteinov Počasnejši odziv: spremembe v količini proteinov (preko transkripcisjkih faktorjev)
6 Prenos signala - receptorji GPCR 900: Kratkotrajne spremembe v celični funkciji (metabolizem, gibanje ) Drugi receptorji: Spremembe v izražanju genov (diferenciacija, delitev celic, dolgotrajni vpliv)

7 Prenos signala preko štirih tipičnih receptorjev celične membrane
8 Signalne kaskade GPCR/CREB Protein-tirozin-kinaze Serinska kinaza/smad Ubikvitinacija (Wnt, Hh, NT- B) Proteoliza (Notch/Delta, SREBP) Jedrni receptorji

9 Delovanje signalnih molekul na receptorske celice

10 Vezava rastnega hormona na receptor 8 od 28 ak, ki so v stiku z receptorjem Esencialna 2 triptofana Dimerizacija receptorja po vezavi rastnega faktorja
11 Regulacija aktivnosti proteinov s kinazami/fosfatazami 600 P na Tyr ali Ser/Thr 100
12 Prenos signala preko receptorja in kinaze do tarčnega proteina Kinaza je lahko del receptorja ali prosta molekula.

13 GTPase so pogosto stikala
14 Ojačanje signala

15 1. camp in PKA preko CREB aktivirata transkripcijo Z G-proteini povezani receptorji (800)

16 Splošen mehanizem aktivacije preko z G-proteini povezanimi receptorji
 aktivirajo adenil ciklazo (G-protein je konstantno aktiven,gtp ne more v GDP).")
17 Bakterije (V. cholera, E. coli) aktivirajo adenil ciklazo (G-protein je konstantno aktiven,gtp ne more v GDP). Visok nivo znotrajceličnega camp vodi do izločanja elektrolitov in vode v lumen prebavil posledica driska.

18 Hormoni v maščobnih celicah lahko aktivirajo ali inhibirajo adenilil ciklazo
19 Adenilil ciklaza sentetizira camp


20 Adenilil ciklaza se veže na Gαs podenoto, sintetizira se camp, ki aktivira protein-kinazo A (PKA)

21

22 Aktivacija CREB transkripcijskega faktorja
 Ko se veže na koaktivator CBP, se zvije v 2")
23 CREB CREB je aktiviran s fosforilacijo Ser 123 Aktivacijska domena nima strukture (random coil) Ko se veže na koaktivator CBP, se zvije v 2 heliksa
 - regulacija PDE camp")

24 Subcelularna lokalizacija PKA na jedrno membrano z AKAP (A-kinase-associated protein) - regulacija PDE camp fosfodiesteraza nizka koncetracija camp Aktivacija receptorja s hormonom PKA s fosforilacijo aktivira PDE hidroliza camp, inaktivacija PKA in PDE
25 2. signalna pot preko protein-tirozin-kinaze
26 Receptorji, ki aktivirajo protein-tirozin-kinazo 90 PTK pri človeku Fosforilacija tirozinov v tarčnih proteinih Regulacija proliferacije, diferenciacije, preživetja in metabolizma celic Dve vrsti receptorjev: - tirozin kinaza je del receptorja (RTK tirozin-kinazni receptorji) - receptorji citokinov; samostojna gena za receptor in kinazo, proteina sta močno vezana), JAK kinaza
27 Signalne poti preko RTK in citokinskih receptorjev Direktna aktivacija STAT TF Pot z Ras/MAP kinazo Pot s PLC, Ca 2+ sekundarni prenašalec Pot s PI-3 kinazo, PKB
 živčni (NGF), PDGI (platelet derived GF), FGH (fibroblast")
28 Receptorji tirozin-kinaz Ligandi so topni ali vezani na membrano. Hormoni (rastni faktorji) živčni (NGF), PDGI (platelet derived GF), FGH (fibroblast GF), epidermalni (EGF),
 Transforming growth factor α (TGF-α)")
29 Dimerizacija HER1, človeškega receptorja za epidermalni rastni faktor (EGF) Transforming growth factor α (TGF-α) HER1
")
30 Aktivacija EGF receptorja z EGF Konformacijska sprememba aktivira (receiver) tirozin kinazo
31 Družina človeških HER in njihovih ligandov 4 homo in heterodimeri HER2 ne veže liganda HER2 je vedno heterodimer HER3 je šibko katalitično aktiven HB vezan heparan NRG neuregulin iz EGF družine TGF-α - Transforming growth factor alpha
32 HER in rak dojke Več kopij gena za HER2 Prekomerno izražanje HER2 Pri 25 % pacientk z rakom na dojki, rak na pljučih Tumorske celice občutljive že na šibko stimulacijo z rastnimi faktorji Zdravilo: mab proti HER2, inhibitorji TK, mab, ki zavirajo angiogenezo HB vezan heparan NRG neuregulin iz EGF družine
 - interlevkini (proliferacija T in B celice) - interferoni (virusna infekcija-aktivacija encimov na sosednjih celicah, da so bolj")
33 Citokini preko citokinskih receptorjev aktivirajo tirozin-kinaze Male molekule, 160 aminokislin 4 dolge -vijačnice Kontrola rasti in diferenciacije celic Primeri: - prolaktin (z diferenciacijo epitelijskih celic te proizvajajo mlečne proteine v mlečnih žlezah) - interlevkini (proliferacija T in B celice) - interferoni (virusna infekcija-aktivacija encimov na sosednjih celicah, da so bolj odporne na viruse) - citokini, ki inducirajo nastanek krvnih celic iz predniških celic (G-CSF, granulocyte colony stimulating factor, eritropoetin)

34 Razvoj eritrocitov z eritropoetinom Znižanje kisika v krvi preko TF HIF-1 aktivira sintezo eritropoetina v ledvičnih celicah. V nekaj dneh iz 1 predniške celice nastane ~ 50 eritrocitov. Zdravljenje anemije pri ledvičnih bolnikih. Del terapije pri rakavih obolenjih.

35 Struktura eritropoetina z receptorjem

36 Aktivacija citokinskih receptorjev Receptor je dimer Na citosolni del sta vezani 2 protein-tirozin-kinazi Vezava liganda približa obe JAK, ki postaneta aktivni Sledi fosforilacija tirozinov na C-koncu receptorja Na P-Tyr se vežejo proteini s specifičnimi domenami (SH2, PTB)

37 Signalni proteini se vežejo na P-Tyr v RTK in citokinskih receptorjih ali na umestitvene proteine (npr. IRS-1) IRS insulin receptor substrate PTB phosphotyrosine binding

38 Proteini z SH2 domeno se vežejo na fosforiliran receptor v citoplazmi Hidrofobni žep je specifičen. SH2 Src homology 2 domain
39 Aktivacija STAT transkripcijskih faktorjev STAT proteini imajo: - N-konec, ki se veže na DNA - SH2 domeno - C-konec s tirozinom Fosforilacija Tyr 2 prosta STAT proteina dimerizirata, konformacijske spremembe N C NLS se izpostavi, STAT potuje v jedro Veže se na ojačevalna zaporedja

40 Struktura STAT proteina
41 Aktivacija transkripcije s STAT Določen STAT protein aktivira transkripcijo različnih genov v različnih celicah. V celicah so različni TF, drugačne epigenetske modifikacije, geni, pripravljeni za transkripcijo s STAT. Primer: aktivacija STAT5 z eritropoetinom v eritroidnih predniških celicah in s prolaktinom v mlečnih žlezah. Različni ligandi receptorji celice enaka signalna molekula aktivira druge gene!
42 Znižanje učinka signalne poti z RTK in s citokini Endocitoza receptorjev: HER1 ima brez EGF razpolovni čas ur, internalizacija receptorja vsakih 30 min (v endosome in nazaj v celično membrano) Ob prisotnsoti EGF se internalizacija 10x poveča, malo receptorjev se vrne v membrano, večina se razgradi v lizosomih Razgradnja v lizosomu: Singnal za razgradnjo (ubikvitinacija) ali recikliranje Če ni internalizacije, ima HER1 konstantni odgovor na EGF, kar vodi v nastanek tumorjev. Fosfotirozin fosfataze SOCS proteini
 in inaktivira JAK2 (dokler se na receptor ne veže nov citokin).")
43 Regulacija prenosa signala na eritropoetinskem receptorju Fosfotirozin fosfataza SHP1 SHP1 se veže na P-Tyr na EpoR receptorju (z SH2 domeno) in inaktivira JAK2 (dokler se na receptor ne veže nov citokin).

44 Prekinitev prenosa signala na eritropoetinskem receptorju SOCS proteini (Suppressor of cytokine signaling) TF STAT (negativna regulacija) Kompetitivna vezava z SH2 domeno na receptor SOCS1 se veže na JAK2, inaktivacija Z SOCS box domeno veže E3 ubikvitin ligazo Poliubikvitinilacija JAK2 in razgradnja v proteasomu

45 Signalna pot z Ras/MAP kinazo Receptorji: RTK in citokinski receptorji Ras GTPaza Študija na Drosophili, C. elegans, proteini ohranjeni Ras stikalo

46 Signalna pot z Ras/MAP kinazo Receptorji: RTK in citokinski receptorji Ras GTPaza Kaskada kinaz se konča z MAP kinazo (Mitogen-Activated Protein Kinase) MAP kinaza aktivira TF, ki vplivajo na celični cikel in diferenciacijo celic. Mutacije v RTK, Ras in downstream proteinih so našli v skoraj vseh vrstah človeških tumorjev. Ras se na tirozin-kinazo veže preko vmesnih proteinov, Sos je GEF GRB2 growth factor receptor-bound protein 2, pomožni protein Sos Son of Sevenless, GEF

47 Specifične interakcije SH3 domene s prolin bogato regijo npr. Sos npr. GBR2

48 Strukturne spremembe Ras, ko je vezan na GDP, Sos in GTP Ko se GTP veže v žep, se Sos sprosti iz kompleksa. Strukturne spremembe omogočijo interakcije s proteini iz kaskade kinaz.
49 Signalna pot preko Ras/MAP kinaze Veže se na P-Ser Ser/Thr kinaza Ser/Thr in Tyr kinaza Kompleksnost sistema: P-Thr in P-Tyr 3 Ras 3 Raf 2 MEK (MAPK/ERK Kinase) 2 Erk (Extracellular signal-regulated Kinases) Ser/Thr kinaza
50 Mutacije v B-Raf povzročijo melanom 40 % primerov melanoma je mutacija v genu za B-Raf 90 % mutacij je Glu 600 Val Stimulacija MEK brez rastnega faktorja Inhibitorji B-Raf kinaze v kliničnih študijah

51 Aktivna in neaktivna oblika MAP kinaze Fosforilacija povzroči aktivacijo in dimerizacijo (transport v jedro).
 Transkripcijski faktorji (c-fos, c-jun) SRE serum response element SRF serum response factor TCF ternary complex factor")
52 Aktivacija transkripcije z MAP kinazo Aktivacija 100 genov (early response genes, ki inducirajo izražanje proteinov za celični cikel) Transkripcijski faktorji (c-fos, c-jun) SRE serum response element SRF serum response factor TCF ternary complex factor

53 Scaffold proteini ločijo različne signalne poti preko MAP kinaz Figure Yeast MAP kinase cascades in the mating and osmoregulatory pathways.
54 Signalna pot preko fosfoinozitidov
 RTK, citokinski receptorji - PLC (SH2 domena) - DAG in IP")
55 Sinteza sekundarnih prenašalcev DAG in IP 3 s PLC GPCR - PLC - DAG in IP 3 (Ca 2+ ) RTK, citokinski receptorji - PLC (SH2 domena) - DAG in IP 3
, protein-kinaza C")
56 Fosfatidil inozitol 3,4,5-trisfosfat RTK, citokinski receptorji PI-3 kinaza (SH2 domena), 9 človeških homologov PI-3 kinaza se veže na receptor na citosolni strani približa se substratu Pot sproži celično delitev, prepreči apoptozo, inducira spremembe v metabolizmu Dolgotrajni efekt na izražanje genov Protein-kinaza B (PKB), protein-kinaza C (PKC)

57 Aktivacija PKB v signalni poti preko PI-3 kinaze ZUNAJCELIČNI SIGNAL PH domena veže se na 3-fosfat Thr Ser Ser/Thr kinaza PH domena - Pleckstrin homology domain 3-phosphoinositide dependent protein kinase
58 Vloga PKB Fosforilacija ter inaktivacija pro-apoptotičnih proteinov Primer: fosforilacija Forkhead TF FOXO3a prepreči njegovo indukcijo izražanja proapoptotičnih proteinov Rastni faktorji PKB fosforilacija FOXO3a nanj se veže prenos v citosol Rak, diabetes Insulin RTK PKB poraba in skladiščenje glukoze (ena signalna pot kontrolira različne celične funkcije v različnih celicah)
59 Negativna regulacija PI-3 kinaze s PTEN fosfatazo Fosforilacija s PI-3 kinazo je reverzibilna PTEN (Phosphatase and tensin homolog), tumor-supresorski gen Široka specifičnost (Tyr, Ser, Thr) Poglavitna vlog: defosforilacija 3-fosfata iz PI 3,4,5-trisfosfata apoptoza Delecije gena v naprednih rakih (PKB ni inhibirana, prekomerna celična rast) Miške z izbitim genom za PTEN imajo večje možgane in večje število nevronov (kontrola normalnega razvoja)
60 3. signalna pot preko receptorja serinske kinaze
 V matriksu v neaktivni obliki Hitra aktivacija s proteolizo ali odcepitvijo inhibitorja 7 Cys 3 disulfidne vezi, 1 Cys za dimerizacijo (homodimer,")
61 Signalna molekula TGF- TGF- (transforming growth factor beta) zunajcelična signalna molekula TGF- 1,2,3 normalna vloga: inhibicija celičnega cikla (zgodnji raki, metastaze), BMP (bone morphogenic protein) V matriksu v neaktivni obliki Hitra aktivacija s proteolizo ali odcepitvijo inhibitorja 7 Cys 3 disulfidne vezi, 1 Cys za dimerizacijo (homodimer, heterodimer)
62 TGF- /Smad signalna pot Receptor serinske kinaze Kompleks 3 TGF- receptorjev RIII je največ, -glikan proteoglikan (GAG-protein) RI in RII sta dimera RII konstitutivno aktiven RII in RIII vežeta TGF- Kompleks RIII, 2x RII, 2xRI (ko je veliko TGF- ) RII fosforilira RI Fosforilirajo Smad TF
63 TGF-b/Smad signalna pot Smad TF Neaktivni v citosolu (R-, co-, i) Imajo 2 domeni: MH1 (veže na DNA, NLS) in MH2 domeni Fosforilacija aktivira Smad 2/3 Kompleks dveh Smad2/3, Smad 4 in dveh importinov potuje v jedro Smad 2-4 ali Smad3-4 se vežeta na druge TF Defosforilacija Smad2/3, recikliranje v citoplazmo Smad vežejo različne TF v različnih celicah drugačen celični odgovor Smad regulirajo rast in diferenciacijo
64 TGF- /Smad signalna pot Smad vežejo različne TF v različnih celicah drugačen celični odgovor Primer: v epitelijskih celicah in fibroblastih TGF- inducirajo izražanje proteinov zunajceličnega matriksa (fibronektin, kolagene) in proteine, ki inhibirajo serinske proteaze (cepijo zgornje proteine). S tem se stabilizira matriks, posledično tkivo. Inhibitor serinskih proteaz je PAI- 1 (plasminogen activator inhibitor 1). Preprečijo proliferacijo celic Brez TGF- pride do zgodnjega razvoja rakov (mutacije). Pankreatični raki imajo delecijo v genu za Smad4. Zdravljenje z rekombinantnimi proteini TGF-.
65 Negativna regulacija TGF- signalizacije Inhibicija transkripcije, ki jo povzročijo Smad proteini S Smad7 (i Smad) S Ski in SnoN proteinoma
 Znižano izražanje manjša rast pankretičnega tumorja Negativna zanka: po TGF-")
66 Zmanjšanje signalizacije preko Smad TF s Ski ali SnoN proteinoma Inducirata deacetilacijo histonov. Izražanje SnoN in Ski vodi v nenormalno proliferacijo celic Prekomerno izražena v rakih (melanoma, rak na prsih) Znižano izražanje manjša rast pankretičnega tumorja Negativna zanka: po TGF- stimulaciji se SnoN in Ski razgradita. Po nekaj urah se poveča izražanje SnoN (Smad2-4 se veže na njegov promotor) izniči se dolgotrajni efekt prenosa signalov zaradi neprestanega delovanja TGF-.
67 4. signalne poti, kontrolirane z ubikvitinacijo Večinoma ireverzibilen proces Wnt in Hedgehog poti podobni (TF-U, razgradnja TF) NF-κB (U-inhibitor TF)
68 Wnt signalna pot Wnt: wingless (Drosophila) + int (integracija retrovirusa pri miškah) Wint1 (miške) je protooknogen Kontrola razvoja (možgani, udi, organogeneza), nastanka osteoblastov, kontrola matičnih celic, proliferacija, preživetje, polarnost, migracija Wnt so zunajčelični glikoproteini (19) Modifikacija na N-koncu hidrofoben palmitat za vezavo na membrano celic (bližnje celice, so lokalno omejeni) 2 receptorja Fz (Frizzled - GPCR) in LRP

69 APC adenomatous polyposis coli GSK3 glycogen synthase kinase 3 CK1 casein kinase 1 TCF - transcription factor Gro Groucho, repressor Wnt signalna pot
70 Hedgehog signalna pot Hedgehog je sekretorni protein, avtoproteoliza Morfogen: koncentracijsko odvisna usoda celic Potuje do 20 celic daleč Modifikacije za vezavo na celično membrano 3 hh geni (sonic, desert, indian), 2 ptc, 3 Gli TF Nepravilna aktivacija Hh signalne poti raki (tumor v cerebelumu, mišicah)

71 Hedgehog signalna pot
72 NF- B signalna pot NF- B (Nuclear Factor kappa-light chain enhancer of activated B cells) je inhibiran v citosolu Aktivacija stres, Toll-podobni receptorji, vnetni citokini (TNFα, IL-1) Nastane proteinski kompleks blizu celične membrane, kjer se inhibitor NF- B ubikvitinira in razgradi Negativna povratna zveza: aktivacija transkripcije inhibitorja z NF- B. Inhibitor v jerdu veže NF- B in ga vrne v citoplazmo.

73 NF- B signalna pot

74 Aktivacija NF- B signalne poti z IL-1 E3 ligaza kinaza Poli U kot ogrodje
75 5. signalne poti, kontrolirane s proteolizo Receptor se cepi 2x: 1. metaloproteaze v matriksu cepijo zunajcelični del receptorja 2. druge proteaze cepijo v celični membrani Citosolna domena se sprosti in deluje kot transkripcijski faktor Notch/Delta Cepitev prekurzorjev rastnih faktorjev SREBP
76 Notch/Delta signalna pot Notch se sintetizira kot monomer v ER, v Golgiju se cepi na 2 verigi. ADAM a disintegrin and metalloprotease
77 Lateralna inhibicija Različna diferenciacija sosednjih celic Med enakovrednimi celicami ena celica določi, da imajo sosednje drugačno usodo (Notch stimulira izražanje Notcha). Različna usoda celic pri razvoju Notch/Delta signalna pot
78 Cepitev signalnih molekul z matriks-metaloproteazami Podobna cepitev kot pri Notch 19 metaloproteaz iz družine ADAM Rastni faktorji se sintetizirajo kot transmembranske molekule Matriks-metaloproteaze cepijo prekurzor, sprosti se topna molekula rastnega faktorja. Odcepljeni protein je signalna molekula. Nepravilnosti vodijo v nenormalno celično proliferacijo. Cepitev prekurzorjev EGF družine (EGP, HB-EGF, TGF-α, NRG1,2)
79 Posledice nepravilnega delovanja ADAM metaloproteaz Povečano delovanje ADAM proteinov (opazili pri rakih): - Visoka koncentracija EGF zunaj celice (avtokrina, parakrina stimulacija) nepravilna proliferacija. - S proteolizo uničijo zunajcelični matriks, kar vodi v nastanek metastaz. ADAM so vpletene v bolezni srca: epinefrin stimulira delovanje β- andrenergičnega receptorja glikogenoliza in povečano krčenje srčne mišice. Pri daljšem zdravljenju z epinefrinom se aktivira ADAM9, cepi HB-EGF prekurzor, ta se veže na receptor v celicah srčne mišice nepravilna rast (povečano, a šibko srce).
 GRASP (γ")
80 Proteolitčna cepitev APP z ADAM pri Alzheimerjevi bolezni Kopičenje Aβ 42 peptida v plakih smrt nevronov Nepravilna cepitev amiloid-prekurzor-proteina (funkcija neznana) 1 zunajcelična in 1 membranska cepitev Genetske napake v APP na mestih, kjer cepijo sekretaze in v proteazah, kar favorizira nastanek amiloida Aβ 42 Sekretaze niso dobra tarča za zdravila (Notch/Delta) GRASP (γ secretase activating protein) poveča količino amiloida Aβ 42, ker interagira z ββ, inhibitor tega proteina potencialno, dovolj specifično zdravilo.
81 Proteoliza intramembranskega proteina SREBP Kontrola količine membranskih lipidov (pravo razmerje med fosfolipidi in holesterolom) poteka z regulacijo sinteze holesterola in njegovega vnosa v celico. Sinteza holesterola (iz manjših prekurzorjev s HMG-CoA reduktazo) in LDL receptorja (vnos v celico) je znižana, če je dovolj holesterola v celici (regulacija transkripcije). SREBP (SRE binding protein) je TF, odvisen od holesterola Znotraj celice Geni, katerih izražanje je regulirano s steroli imajo mesto SRE (sterol regulatory element, 10 bp).
.")

82 Aktivacija SREBP s holesterolom ER membrana < 5% vseh lipidov v ER membrani N C Na membrano vezikla za transport iz ER v Golgi je vezan protein COPII. sterol-sensing domain SREBP cleavageactivating protein C Ko je sterol-sensing domain v SCAP vezan na holesterol, je protein vezan tudi na insig-1(2). Vezan insig-1(2) prepreči vezavo na Sec24 (protein na COPII veziklu, ki potujejo v Golgi) ko je holesterola v ER membrani več kot 5% vseh membranskih lipidov. Če insig ni vezan na SCAP, se nanj veže Sec24, SCAP-SREBP kompleks gre v Golgi. V Golgiju se SREBP cepi z S1P in S2P. nsrebp gre v jedro, veže se na SRE in sproži transkripcijo (npr. genov za LDL receptor, HMG-CoA reduktazo). nsrebp se hitro razgradi, a hitro nastane hiter celični odgovor.
83 Regulacija lipidov v celici Koordinirana regulacija: vseh lipidov (rast membran) Diferencialna regulacija: potreba po nekaterih lipidih (npr. holesterolu za sintezo steroidnih hormonov) Diferencialna regulacija s 3 SREBP SREBP1a in 1c (alternativni splicing) imata večji vpliv na metabolizem maščobnih kislin kot na holesterol SREBP2 ima večji vpliv na metabolizem holesterola Ateroskleroza preveč LDL v krvi, zato naj bi zmanjšali LDL in povečali HDL S statini se inhibira HMG-CoA reduktaza (količina holesterola v jetrih). Odgovor je aktivacija SREBP in s tem izražanje HMG-CoA reduktaze ter LDL receptorja (več receptorjev v jetrih-zmanjša se količina LDL v krvi).
84 6. regulacija transkripcije z malimi hormoni Extracelularna signalizacija Signalne molekule so mali hidrofobni hormoni Hormoni so topni v lipidih. Difundirajo skozi celično in jedrno membrano. Vežejo se na proteine iz naddružine jedrnih receptorjev. Primeri: Steroidni hormoni, retinoidi, tiroidni hormoni

85 Jedrni receptorji So transkripcijski faktorji, regulirani z lipidotopnimi hormoni, ki jih dobimo z dieto in preko okolja. > 50 človeških transkripcijskih faktorjev S Cys4 cinkovim prstom se vežejo na DNA. Homodimer ali heterodimer Vežejo se na DNA ponovitve.

86 Ohranjena struktura domen transkripcijskih faktorjev iz naddružine jedrnih receptorjev Ena ali več aktivacijskih domen Cinkov prst Od hormona odvisna aktivacijska domena

87 C-terminalni del receptorja: hormon-vezavna domena Receptor estrogena Z zelenim heliksom nastane hidrofobni žep za hormon Pri vezavi antagonista se zeleni heliks pomakne v mesto za koaktivator - represija

88 Odzivni elementi jedrnih receptorjev Glukokortikoid Estrogen obratne ponovitve l receptor je homodimer

89 Odzivni elementi jedrnih receptorjev Vitamin D3 Tiroidni hormon Specifičnost vezave receptor je heterodimer Retinojska ksl

90 Vezava liganda na homodimerni jedrni receptor povzroči prenos kompleksa v jedro Glukokortikoidni hormon deksametazon


91 Hormon se na heterodimerni jedrni receptor veže v jedru HDAC h n r HAT h n r Brez hormona se heterodimerni receptor veže na DNA odzivni element SRE. Na receptor se veže tudi HDAC (histone deacetylases). Transkripcija je blokirana. Ko v jedro difundira hormon, se HDAC sprosti, veže pa se histon-acetilaza. Transkripcija je aktivirana.
92 7. regulacija transkripcije med elongacijo Hsp Ustvljena elongacija Pol II 200bp od transkripcijskega začetka Aktivacija HSTF s toplotnim šokom, elongacija s Pol II se nadaljuje hiter odgovor Myc kontrola elongacije s Pol II Regulacija celične rasti in delitve Povečano izražanje v rakih Glavni TF pri reprogramiranju somatski celic v pluripotenetne matične celice

93 8. regulacija transkripcije s Ca 2+ ioni GPCR in receptorji s tirozin kinazo (RTK in receptorji citokinov)
94 Regulacija transkripcije s Ca 2+ ioni PLC katalizira nastanek 2 sekundarnih prenašalcev DAG in IP 3 IP 3 je odgovoren za kratkotrajno sproščanje Ca 2+ ionov iz lumna ER (ni povečanja ionov zunaj celice) Ca 2+ ioni pomagajo pri aktivaciji PKC z DAG. PKC vplivajo na celično rast in metabolizem: npr. na metabolizem glikogena v jetrih. Regulirajo transkripcijo s fosforilacijo TF.
 Sproščanje gladkih")
95 Aktivacija PLC s citosolnim Ca 2+ ioni Poleg prenosa PKC k membrani, se Ca 2+ ioni vežejo na kalmodulin (citosolno stikalo) kooperativno. Vloga Ca 2+ - kalmodulin kompleksa: - krčenje mišic z aktivacijo miozina - degradacija camp fina regulacija - aktivacija TF (aktivacija kinaz in fosfataz) Sproščanje gladkih mišic v žilah (NO) Regulacija glikogenolize

. Tudi TF C/EBP se inducira med diferenciacijo adepocitov.")
96 Regulacija diferenciacije adipocitov z interakcijo različnih signalnih poti PPAR - peroxisome proliferator-activated receptors Beli adepociti za skladiščenje maščob v trigliceridnih globulah So endokrine celice Iz mezenhimalnih matičnih celic lahko nastanejo adipociti, osteoblasti in celice, ki proizvajajo hrustanec TF PPAR je glavni transkripcijski regulator za diferenciacijo adepocitov (jedrni receptor) Aktivni PPAR verjetno potrebuje vezan ligand (oksidiran derivat maščobne kisline). Tudi TF C/EBP se inducira med diferenciacijo adepocitov. Skupaj inducirata izražanje vseh proteinov, potrebnih za nastanek maščobnih celic Inzulin +, Wnt -, TGF- -
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks'
 Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Κυτταρική Επικοινωνία Cell Communication
 Κυτταρική Επικοινωνία Cell Communication Η μοίρα του κυττάρου ρυθμίζεται από εξωτερικά σήματα (από το περιβάλλον και άλλα κύτταρα) survival division differentiation apoptosis Είδη κυτταρικής επικοινωνίας
Κυτταρική Επικοινωνία Cell Communication Η μοίρα του κυττάρου ρυθμίζεται από εξωτερικά σήματα (από το περιβάλλον και άλλα κύτταρα) survival division differentiation apoptosis Είδη κυτταρικής επικοινωνίας
PROTEINI POVEZANI S PRENOSOM SIGNALOV. Funkcije proteinov
 PROTEINI POVEZANI S PRENOSOM SIGNALOV Funkcije proteinov Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni proteini) Oporna funkcija (strukturni proteini,
PROTEINI POVEZANI S PRENOSOM SIGNALOV Funkcije proteinov Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni proteini) Oporna funkcija (strukturni proteini,
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda
![[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda](/thumbs/89/99439684.jpg "[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda") kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Ο ΡΟΛΟΣ ΤΩΝ ΚΑΝΝΑΒΙΝΟΕΙ ΩΝ ΚΑΤΑ ΤΗΝ ΕΜΒΡΥΪΚΗ ΑΝΑΠΤΥΞΗ
 ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ Πρόγραµµα Μεταπτυχιακών Σπουδών Εφαρµογές στις Βασικές Ιατρικές Επιστήµες Εργαστήριο Ανατοµικής-Ιστολογίας-Εµβρυολογίας ΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Ο ΡΟΛΟΣ ΤΩΝ ΚΑΝΝΑΒΙΝΟΕΙ
ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ Πρόγραµµα Μεταπτυχιακών Σπουδών Εφαρµογές στις Βασικές Ιατρικές Επιστήµες Εργαστήριο Ανατοµικής-Ιστολογίας-Εµβρυολογίας ΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Ο ΡΟΛΟΣ ΤΩΝ ΚΑΝΝΑΒΙΝΟΕΙ
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Κυτταρική Επικοινωνία Cell Communication
 Κυτταρική Επικοινωνία Cell Communication Κυτταρική Επικοινωνία Είναι η ικανότητα του κυττάρου να αποκρίνεται σε εξωτερικά σήματα (σήματα από το περιβάλλον του ή άλλα κύτταρα) Τα σήματα αυτά μπορεί να είναι
Κυτταρική Επικοινωνία Cell Communication Κυτταρική Επικοινωνία Είναι η ικανότητα του κυττάρου να αποκρίνεται σε εξωτερικά σήματα (σήματα από το περιβάλλον του ή άλλα κύτταρα) Τα σήματα αυτά μπορεί να είναι
PROVOĐENJE SIGNALA BORIS MILDNER. Tri primjera prijenosa signala
 PROVOĐENJE SIGNALA BORIS MILDNER Tri primjera prijenosa signala (adrenalin) Vezanjem signalne molekule za specifičan receptor započinju važni fiziološki procesi u stanici. 1 Osnovni princip prijenosa (provođenja)
PROVOĐENJE SIGNALA BORIS MILDNER Tri primjera prijenosa signala (adrenalin) Vezanjem signalne molekule za specifičan receptor započinju važni fiziološki procesi u stanici. 1 Osnovni princip prijenosa (provođenja)
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
IONOTROPNI in METABOTROPNI receptorji. Klara Kropivšek
 IONOTROPNI in METABOTROPNI receptorji Klara Kropivšek Membranski receptorji Specializirani integralni membranski proteini. Vezava ekstracelularnih mediatorjev - ligandov (hormoni, nevrotransmiterji, citokini
IONOTROPNI in METABOTROPNI receptorji Klara Kropivšek Membranski receptorji Specializirani integralni membranski proteini. Vezava ekstracelularnih mediatorjev - ligandov (hormoni, nevrotransmiterji, citokini
Encimi v zunajceličnem matriksu
 Encimi v zunajceličnem matriksu - Proteaze - metaloproteaze - serinske proteaze - cisteinske proteaze - aspartatne proteaze - Glikozidaze - Lizil-oksidaze - Superoksid dismutaza Metaloproteaze Sterchi
Encimi v zunajceličnem matriksu - Proteaze - metaloproteaze - serinske proteaze - cisteinske proteaze - aspartatne proteaze - Glikozidaze - Lizil-oksidaze - Superoksid dismutaza Metaloproteaze Sterchi
Razgradnja maščobnih kislin. Ketonska telesa H + NAD+ NADH. Pregled metabolizma MK. lipoprotein-lipaza. maščobne kisline.
 Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
Το μονοπάτι της κινάσης MAP- ERK
 Το μονοπάτι της κινάσης MAP- ERK 1 Σηματοδότηση μέσω μικρών GTPασών Η οικογένεια μορίων Ras (Rat Sarcoma virus) Ρύθμιση των πρωτεϊνών Ras Οι πρωτεΐνες Ras μετατρέπονται από την ανενεργή μορφή τους, που
Το μονοπάτι της κινάσης MAP- ERK 1 Σηματοδότηση μέσω μικρών GTPασών Η οικογένεια μορίων Ras (Rat Sarcoma virus) Ρύθμιση των πρωτεϊνών Ras Οι πρωτεΐνες Ras μετατρέπονται από την ανενεργή μορφή τους, που
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Provođenje signala. Boris Mildner
 Seminar 10 Provođenje signala Boris Mildner Rješenja zadaće 9. 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. C A B B A 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. C C B A C A C A A A 1 1. Membranski receptorski
Seminar 10 Provođenje signala Boris Mildner Rješenja zadaće 9. 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. C A B B A 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. C C B A C A C A A A 1 1. Membranski receptorski
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Energetski metabolizem
 Energetski metabolizem Metabolne povezave med jetri in mišicami Corijev ciklus Glukoza-alaninski ciklus Možgani potrebe po glukozi: 120 g/dan (t.j. ca 60% ob počitku) za vzdrževanje membranskih potencialov
Energetski metabolizem Metabolne povezave med jetri in mišicami Corijev ciklus Glukoza-alaninski ciklus Možgani potrebe po glukozi: 120 g/dan (t.j. ca 60% ob počitku) za vzdrževanje membranskih potencialov
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
Nekatere interakcije v lipidnem metabolizmu
 Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
Ćelijska signalizacija
 Ćelijska signalizacija Interakcije organizma i sredine posredovane su signalima Primanje signala iz okoline, odgovor na signale i odašiljanje signala u okolinu osobinu živih organizama Posebno važno i
Ćelijska signalizacija Interakcije organizma i sredine posredovane su signalima Primanje signala iz okoline, odgovor na signale i odašiljanje signala u okolinu osobinu živih organizama Posebno važno i
SABiosciences PCR Array Catalog #: PAHS-021 SA+ SCF 4h stimulaton AVG(Ct) Position Unigene Refseq Symbol Description shcontr shcontr-4h shgskβ A01
 Position Unigene Refseq Symbol Description shcontr shcontr-4h shgskβ A01") SABiosciences PCR Array Catalog #: PAHS-021 SA+ SCF 4h stimulaton AVG(Ct) Position Unigene Refseq Symbol Description shcontr shcontr-4h shgskβ A01 Hs.1274 NM_006129 BMP1 Bone morphogenetic protein 1 25.25
SABiosciences PCR Array Catalog #: PAHS-021 SA+ SCF 4h stimulaton AVG(Ct) Position Unigene Refseq Symbol Description shcontr shcontr-4h shgskβ A01 Hs.1274 NM_006129 BMP1 Bone morphogenetic protein 1 25.25
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
Najpomembnejši človeški Hb
 Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
ΥΠΕΡΙΝΣΟΥΛΙΝΑΙΜΙΑ ΚΑΙ ΑΝΤΙΣΤΑΣΗ ΣΤΗΝ ΙΝΣΟΥΛΙΝΗ ΣΕ ΠΑΧΥΣΑΡΚΑ ΠΑΙΔΙΑ ΚΑΙ ΕΦΗΒΟΥΣ ΜΕ ΠΡΩΙΜΗ ΑΔΡΕΝΑΡΧΗ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΠΡΟΛΗΠΤΙΚΗΣ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΧΗΜΕΙΑΣ ΔΙΕΥΘΥΝΤΡΙΑ: Η ΚΑΘΗΓΗΤΡΙΑ Ν. ΒΑΒΑΤΣΗ-ΧΡΙΣΤΑΚΗ ΠΑΝΕΠ. ΕΤΟΣ 2009-2010 Αριθμ.2456
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΠΡΟΛΗΠΤΙΚΗΣ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΧΗΜΕΙΑΣ ΔΙΕΥΘΥΝΤΡΙΑ: Η ΚΑΘΗΓΗΤΡΙΑ Ν. ΒΑΒΑΤΣΗ-ΧΡΙΣΤΑΚΗ ΠΑΝΕΠ. ΕΤΟΣ 2009-2010 Αριθμ.2456
Μονοπάτια ενεργοποίησης κινασών MAP σε κύτταρα θηλαστικών
 Μονοπάτια ενεργοποίησης κινασών MAP σε κύτταρα θηλαστικών Εκτός από την ERK, τα κύτταρα των θηλαστικών διαθέτουν τις κινάσες MAP JNK και p38. Η ενεργοποίηση των κινασών JNK και p38 προκαλείται από μέλη
Μονοπάτια ενεργοποίησης κινασών MAP σε κύτταρα θηλαστικών Εκτός από την ERK, τα κύτταρα των θηλαστικών διαθέτουν τις κινάσες MAP JNK και p38. Η ενεργοποίηση των κινασών JNK και p38 προκαλείται από μέλη
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
Rang & Dale's Pharmacology (7th) IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod)
 IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod)") Rang & Dale's Pharmacology (7th) IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod) Domen Sever, študent 3. letnika medicine Kazalo 2. POGLAVJE: MEHANIZMI DELOVANJA ZDRAVIL... 3 3. POGLAVJE: MEHANIZMI
Rang & Dale's Pharmacology (7th) IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod) Domen Sever, študent 3. letnika medicine Kazalo 2. POGLAVJE: MEHANIZMI DELOVANJA ZDRAVIL... 3 3. POGLAVJE: MEHANIZMI
Metabolizem in energija
 Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Hepatic Stellate Cells: Multifunctional mesenchymal cells of the Liver
 Nagoya Med. J., 137 Hepatic Stellate Cells: Multifunctional mesenchymal cells of the Liver KAZUO IKEDA Department of Anatomy and Cell Biology, Graduate School of Medical Sciences, Nagoya City University
Nagoya Med. J., 137 Hepatic Stellate Cells: Multifunctional mesenchymal cells of the Liver KAZUO IKEDA Department of Anatomy and Cell Biology, Graduate School of Medical Sciences, Nagoya City University
Funkcije proteinov (pogojene s strukturo)
") Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Najpogostejše hemoglobinopatije
 Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina
Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina
Neavtorizirani povzetki izbranih predavanj iz biokemije. UN študij Okolje Politehnika Nova Gorica. doc. dr. Marko Dolinar
 Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Debashish Sahay. To cite this version: HAL Id: tel
 Identification of genes activated and biological markers involved in lysophosphatidic acid (LPA)-induced breast cancer metastasis through its receptor LPA1 Debashish Sahay To cite this version: Debashish
Identification of genes activated and biological markers involved in lysophosphatidic acid (LPA)-induced breast cancer metastasis through its receptor LPA1 Debashish Sahay To cite this version: Debashish
Μελέτη της αντιμεταλλαξιγόνου δράσης φλαβονοειδών του φυτού Lotus Edulis με τη μέθοδο Ames test
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΒΙΟΧΗΜΕΙΑΣ & ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Μελέτη της αντιμεταλλαξιγόνου δράσης φλαβονοειδών του φυτού Lotus Edulis με τη μέθοδο Ames test Επιμέλεια
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΒΙΟΧΗΜΕΙΑΣ & ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Μελέτη της αντιμεταλλαξιγόνου δράσης φλαβονοειδών του φυτού Lotus Edulis με τη μέθοδο Ames test Επιμέλεια
Η ΦΛΕΓΜΟΝΩ ΗΣ ΑΝΤΙ ΡΑΣΗ ΤΟΥ ΓΑΣΤΡΙΚΟΥ ΒΛΕΝΝΟΓΟΝΟΥ ΣΤΗ ΛΟΙΜΩΞΗ ΜΕ ΕΛΙΚΟΒΑΚΤΗΡΙ ΙΟ ΤΟΥ ΠΥΛΩΡΟΥ ΠΡΙΝ ΚΑΙ ΜΕΤΑ ΤΗ ΘΕΡΑΠΕΙΑ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΤΟΜΕΑΣ ΥΓΕΙΑΣ ΕΡΓΑΣΤΗΡΙΑΚΟΣ ΙΕΥΘΥΝΤΗΣ: Ο ΚΑΘΗΓΗΤΗΣ ΓΕΩΡΓΙΟΣ ΚΑΡΚΑΒΕΛΑΣ ΠΑΝΕΠ. ΕΤΟΣ 2008-2009 Αριθµ. 2084 Η ΦΛΕΓΜΟΝΩ ΗΣ ΑΝΤΙ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΤΟΜΕΑΣ ΥΓΕΙΑΣ ΕΡΓΑΣΤΗΡΙΑΚΟΣ ΙΕΥΘΥΝΤΗΣ: Ο ΚΑΘΗΓΗΤΗΣ ΓΕΩΡΓΙΟΣ ΚΑΡΚΑΒΕΛΑΣ ΠΑΝΕΠ. ΕΤΟΣ 2008-2009 Αριθµ. 2084 Η ΦΛΕΓΜΟΝΩ ΗΣ ΑΝΤΙ
Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner
 Boris Mildner") Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner Čestice glikogena u hepatocitima. Glikogen, čestice za pohranjivanje glukoze, u hepatocitima se nalazi u citoplazmi blizu tubula endoplazmatske
Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner Čestice glikogena u hepatocitima. Glikogen, čestice za pohranjivanje glukoze, u hepatocitima se nalazi u citoplazmi blizu tubula endoplazmatske
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Metabolizem oz. presnova
 Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Hormoni ščitnice. Anatomija in histologija. Transport jodida (J - ) (1) Nastanek ščitnice
 (1) Nastanek ščitnice") Anatomija in histologija Hormoni ščitnice Dva povezana lobusa Bogata prekrvitev Kuboidne epitelijske celice tvorijo folikle V foliklih se skladiščijo hormoni v obliki koloida Parafolikularne C celice (kalcitonin)
Anatomija in histologija Hormoni ščitnice Dva povezana lobusa Bogata prekrvitev Kuboidne epitelijske celice tvorijo folikle V foliklih se skladiščijo hormoni v obliki koloida Parafolikularne C celice (kalcitonin)
Združevanje celic v tkiva. Lodish 4: 22. poglavje
 Združevanje celic v tkiva Lodish 4: 22. poglavje Razdelitev poglavja Pritrjevanje celic in komuniciranje med njimi Pritrjevanje celic na medceličnino Kolageni v medceličnini Nekolagenske sestavine medceličnine
Združevanje celic v tkiva Lodish 4: 22. poglavje Razdelitev poglavja Pritrjevanje celic in komuniciranje med njimi Pritrjevanje celic na medceličnino Kolageni v medceličnini Nekolagenske sestavine medceličnine
http / / cjbmb. bjmu. edu. cn Chinese Journal of Biochemistry and Molecular Biology Gab2 SHP2 / Ras / ERK PI3K / AKT Gab2 Signaling in Breast Cancer
 ISSN 1007-7626 CN 11-3870 / Q http / / cjbmb bjmu edu cn Chinese Journal of Biochemistry and Molecular Biology 2011 4 27 4 300 ~ 304 Gab2 * 310058 Gab2 Gabs Gab2 SH2 SHP2 / Ras / ERK PI3K / AKT GAB2 ErbB2
ISSN 1007-7626 CN 11-3870 / Q http / / cjbmb bjmu edu cn Chinese Journal of Biochemistry and Molecular Biology 2011 4 27 4 300 ~ 304 Gab2 * 310058 Gab2 Gabs Gab2 SH2 SHP2 / Ras / ERK PI3K / AKT GAB2 ErbB2
ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΙΣ «ΚΛΙΝΙΚΕΣ ΚΑΙ ΚΛΙΝΙΚΟΕΡΓΑΣΤΗΡΙΑΚΕΣ ΙΑΤΡΙΚΕΣ ΕΙΔΙΚΟΤΗΤΕΣ»
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΙΣ «ΚΛΙΝΙΚΕΣ ΚΑΙ ΚΛΙΝΙΚΟΕΡΓΑΣΤΗΡΙΑΚΕΣ ΙΑΤΡΙΚΕΣ ΕΙΔΙΚΟΤΗΤΕΣ» ΑΡΝΗΤΙΚΗ ΡΥΘΜΙΣΗ ΤΗΣ ΜΕΤΑΒΙΒΑΣΗΣ ΤΟΥ ΣΗΜΑΤΟΣ ΤΗΣ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΙΣ «ΚΛΙΝΙΚΕΣ ΚΑΙ ΚΛΙΝΙΚΟΕΡΓΑΣΤΗΡΙΑΚΕΣ ΙΑΤΡΙΚΕΣ ΕΙΔΙΚΟΤΗΤΕΣ» ΑΡΝΗΤΙΚΗ ΡΥΘΜΙΣΗ ΤΗΣ ΜΕΤΑΒΙΒΑΣΗΣ ΤΟΥ ΣΗΜΑΤΟΣ ΤΗΣ
ΕΝΔΟΚΥΤΤΑΡΙΑ ΣΗΜΑΤΑ Δεύτεροι αγγελιοφόροι BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ
 ΕΝΔΟΚΥΤΤΑΡΙΑ ΣΗΜΑΤΑ Δεύτεροι αγγελιοφόροι BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ Ενδοκυττάρια σήματα Κυτοσολικά camp cgmp Φωσφοϊνοσιτίδια Ca 2+ Μεμβρανικά DAG (Διακυλογλυκερόλη) Φωσφατιδυλοϊνοσιτόλες
ΕΝΔΟΚΥΤΤΑΡΙΑ ΣΗΜΑΤΑ Δεύτεροι αγγελιοφόροι BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ Ενδοκυττάρια σήματα Κυτοσολικά camp cgmp Φωσφοϊνοσιτίδια Ca 2+ Μεμβρανικά DAG (Διακυλογλυκερόλη) Φωσφατιδυλοϊνοσιτόλες
Ο ΡΟΛΟΣ ΤΗΣ ΑΓΓΕΙΟΓΕΝΕΣΗΣ ΣΤΙΣ ΝΟΣΟΥΣ ΤΟΥ ΝΕΦΡΙΚΟΥ ΠΑΡΕΓΧΥΜΑΤΟΣ
 ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΡΟΓΡΑΜΜΑ ΒΙΕ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΤΟΜΙΑΣ ΚΑΙ ΙΣΤΟΛΟΓΙΑΣ ΜΠΕΛΛΑΣ ΑΘΑΝΑΣΙΟΣ ΑΜ: 456 ΚΑΘΗΓΗΤΗΣ: ΒΑΡΑΚΗΣ ΙΩΑΝΝΗΣ ΥΠΕΥΘΥΝΗ ΚΑΘΗΓΗΤΡΙΑ: ΠΑΠΑΔΑΚΗ-ΠΕΤΡΟΥ ΕΛΕΝΗ Ο ΡΟΛΟΣ ΤΗΣ ΑΓΓΕΙΟΓΕΝΕΣΗΣ ΣΤΙΣ ΝΟΣΟΥΣ ΤΟΥ
ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΡΟΓΡΑΜΜΑ ΒΙΕ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΤΟΜΙΑΣ ΚΑΙ ΙΣΤΟΛΟΓΙΑΣ ΜΠΕΛΛΑΣ ΑΘΑΝΑΣΙΟΣ ΑΜ: 456 ΚΑΘΗΓΗΤΗΣ: ΒΑΡΑΚΗΣ ΙΩΑΝΝΗΣ ΥΠΕΥΘΥΝΗ ΚΑΘΗΓΗΤΡΙΑ: ΠΑΠΑΔΑΚΗ-ΠΕΤΡΟΥ ΕΛΕΝΗ Ο ΡΟΛΟΣ ΤΗΣ ΑΓΓΕΙΟΓΕΝΕΣΗΣ ΣΤΙΣ ΝΟΣΟΥΣ ΤΟΥ
Viri. Nukleinske kisline. Program predavanj maj-junij DNA, RNA struktura, podvajanje, transkripcija, translacija
 Program predavanj maj-junij 2013 Datum izvajanja Vsebina predavanja / vaj 8. 5. 2013 P2 11-15 DNA, replikacija, RNA, transkripcija, translacija 15. 5. 2013 P2 11-15 Mutacije, variabilnost genoma PBL- naloge
Program predavanj maj-junij 2013 Datum izvajanja Vsebina predavanja / vaj 8. 5. 2013 P2 11-15 DNA, replikacija, RNA, transkripcija, translacija 15. 5. 2013 P2 11-15 Mutacije, variabilnost genoma PBL- naloge
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
BIOLOGIJA CELICE. Uredil Primož Pirih Društvo študentov biologije
 Društvo študentov biologije Stran 1/44 http://dsb.biologija.org BIOLOGIJA CELICE Zapiski s predavanj pri predmetu Biologija celice za študente biologije. Predavatelji Jezernik, Strle, Pšeničnik. Zadnja
Društvo študentov biologije Stran 1/44 http://dsb.biologija.org BIOLOGIJA CELICE Zapiski s predavanj pri predmetu Biologija celice za študente biologije. Predavatelji Jezernik, Strle, Pšeničnik. Zadnja
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
I. OSNOVNI STRUKTURNI PRINCIPI
 I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
ΙΩΑΝΝΗΣ ΚΛΑΓΚΑΣ. Χημικός, MSc ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΥΠΟΒΛΗΘΗΚΕ ΣΤΗΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΥ ΑΡΙΣΤΟΤΕΛΕΙΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΘΕΣΣΑΛΟΝΙΚΗΣ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΦΥΣΙΟΛΟΓΙΑΣ ΦΑΡΜΑΚΟΛΟΓΙΑΣ B ΕΡΓΑΣΤΗΡΙΟ ΦΑΡΜΑΚΟΛΟΓΙΑΣ ΔΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ ΓΙΩΡΓΟΣ ΚΑΡΑΚΙΟΥΛΑΚΗΣ ΠΑΝΕΠ. ΕΤΟΣ 2008-2009 ΑΡΙΘΜ. 2376 Η ΕΠΙΔΡΑΣΗ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΦΥΣΙΟΛΟΓΙΑΣ ΦΑΡΜΑΚΟΛΟΓΙΑΣ B ΕΡΓΑΣΤΗΡΙΟ ΦΑΡΜΑΚΟΛΟΓΙΑΣ ΔΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ ΓΙΩΡΓΟΣ ΚΑΡΑΚΙΟΥΛΑΚΗΣ ΠΑΝΕΠ. ΕΤΟΣ 2008-2009 ΑΡΙΘΜ. 2376 Η ΕΠΙΔΡΑΣΗ
Družina globinov pri človeku in bolezni.
 Družina globinov pri človeku in bolezni www.muhlenberg.edu/ Mioglobin in hemoglobin spadata v družino globinov Globinsko zvitje Mb β podenota Hb Podobnost aminokislinskega zaporedja Podobnost 3D strukture
Družina globinov pri človeku in bolezni www.muhlenberg.edu/ Mioglobin in hemoglobin spadata v družino globinov Globinsko zvitje Mb β podenota Hb Podobnost aminokislinskega zaporedja Podobnost 3D strukture
Υποδοχείς κινάσες τυροσίνης 1. Δομή και λειτουργία των υποδοχέων κινάσες τυροσίνης
 9 o Υποδοχείς κινάσες τυροσίνης 1. Δομή και λειτουργία των υποδοχέων κινάσες τυροσίνης Γενική δομή και ταξινόμηση Σύνδεση του προσδέτη και ολιγομερισμός του υποδοχέα Σχηματισμός ετεροδιμερών Η ενεργοποίηση
9 o Υποδοχείς κινάσες τυροσίνης 1. Δομή και λειτουργία των υποδοχέων κινάσες τυροσίνης Γενική δομή και ταξινόμηση Σύνδεση του προσδέτη και ολιγομερισμός του υποδοχέα Σχηματισμός ετεροδιμερών Η ενεργοποίηση
TOLL-U PODOBNI RECEPTORJI
 UNIVERZA V LJUBLJANI FAKULTETA ZA KEMIJO IN KEMIJSKO TEHNOLOGIJO TOLL-U PODOBNI RECEPTORJI Seminarska naloga pri predmetu Biološke membrane Mentor: prof. dr. Igor Križaj Avtorja: Angelika Vižintin in Urban
UNIVERZA V LJUBLJANI FAKULTETA ZA KEMIJO IN KEMIJSKO TEHNOLOGIJO TOLL-U PODOBNI RECEPTORJI Seminarska naloga pri predmetu Biološke membrane Mentor: prof. dr. Igor Križaj Avtorja: Angelika Vižintin in Urban
ΚΕΦΑΛΑΙΟ 15. Κυτταρική ρύθμιση. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
Biološke membrane in transport Model tekočega mozaika, asimetričnost, omogočeno lateralno gibanje lipidov in proteinov, oligosahardine verige lipidov
 Biološke membrane in transport Model tekočega mozaika, asimetričnost, omogočeno lateralno gibanje lipidov in proteinov, oligosahardine verige lipidov in proteinov na zunanji strani Sestava in struktura
Biološke membrane in transport Model tekočega mozaika, asimetričnost, omogočeno lateralno gibanje lipidov in proteinov, oligosahardine verige lipidov in proteinov na zunanji strani Sestava in struktura
Sinteza RNA - transkripcija
 Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL
 PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
Uravnavanje encimske aktivnosti
 Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
ΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΓΟΝΙ ΙΑΚΗ ΕΚΦΡΑΣΗ ΚΑΙ ΣΗΜΑΤΟ ΟΤΗΣΗ
 ΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΓΟΝΙ ΙΑΚΗ ΕΚΦΡΑΣΗ ΚΑΙ ΣΗΜΑΤΟ ΟΤΗΣΗ ρ. Α. ΓΑΛΑΝΗΣ agalanis@mbg.duth.gr Figure 6.1 The Biology of Cancer ( Garland Science 2007) Figure
ΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΓΟΝΙ ΙΑΚΗ ΕΚΦΡΑΣΗ ΚΑΙ ΣΗΜΑΤΟ ΟΤΗΣΗ ρ. Α. ΓΑΛΑΝΗΣ agalanis@mbg.duth.gr Figure 6.1 The Biology of Cancer ( Garland Science 2007) Figure
CM707. GR Οδηγός χρήσης... 2-7. SLO Uporabniški priročnik... 8-13. CR Korisnički priručnik... 14-19. TR Kullanım Kılavuzu... 20-25
 1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
Nevrodegeneracija. Boris Rogelj
 Nevrodegeneracija Boris Rogelj boris.rogelj@ijs.si Pregled predavanja Osnove in skupne lastnosti Alzheimerjava bolezen (AB) Parkinsonova bolezen (PB) Huntingtonova bolezen (HB) Frontotemporalna demenca
Nevrodegeneracija Boris Rogelj boris.rogelj@ijs.si Pregled predavanja Osnove in skupne lastnosti Alzheimerjava bolezen (AB) Parkinsonova bolezen (PB) Huntingtonova bolezen (HB) Frontotemporalna demenca
Βιολογικές Μεμβράνες και Μεταγωγή Σήματος
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογικές Μεμβράνες και Μεταγωγή Σήματος Ενδοκυττάρια σήματα Διδάσκουσα: Καθ. Μαρία - Ελένη Ε. Λέκκα Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογικές Μεμβράνες και Μεταγωγή Σήματος Ενδοκυττάρια σήματα Διδάσκουσα: Καθ. Μαρία - Ελένη Ε. Λέκκα Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Statistična analiza. doc. dr. Mitja Kos, mag. farm. Katedra za socialno farmacijo Univerza v Ljubljani- Fakulteta za farmacijo
 Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Biologija rakave celice
 Predavanje 12.1.2009 M Robert Zorec 1 1. Opredelitev 2. Lastnosti rakave celice 3. Lastnosti maligne novotvorbe 4. Nekateri sistemski znaki maligne neoplazme (patofiziologija neoplazme) 5. Strategije za
Predavanje 12.1.2009 M Robert Zorec 1 1. Opredelitev 2. Lastnosti rakave celice 3. Lastnosti maligne novotvorbe 4. Nekateri sistemski znaki maligne neoplazme (patofiziologija neoplazme) 5. Strategije za
Research progress in inducing stem cells to differentiate toward the β-like cells of pancreatic islet
 19 5 2007 10 Chinese Bulletin of Life Sciences Vol. 19, No. 5 Oct., 2007 1004-0374(2007)05-0526-05 β ( 510632) β 1 β Q813 R587.1 Research progress in inducing stem cells to differentiate toward the β-like
19 5 2007 10 Chinese Bulletin of Life Sciences Vol. 19, No. 5 Oct., 2007 1004-0374(2007)05-0526-05 β ( 510632) β 1 β Q813 R587.1 Research progress in inducing stem cells to differentiate toward the β-like
13o Μεμβρανικοί υποδοχείς με εσωτερική δραστικότητα κινάσης Ser/Thr 1. Σηματοδότηση μέσω TGFβ
 13 o TGF-β Μεμβρανικοί υποδοχείς με εσωτερική δραστικότητα κινάσης Ser/Thr 1. Σηματοδότηση μέσω TGFβ Ωρίμανση του μορίου TGFβ Ενεργοποίηση των υποδοχέων TGFβ Οι μεταγραφικοί παράγοντες Smads Η ρύθμιση
13 o TGF-β Μεμβρανικοί υποδοχείς με εσωτερική δραστικότητα κινάσης Ser/Thr 1. Σηματοδότηση μέσω TGFβ Ωρίμανση του μορίου TGFβ Ενεργοποίηση των υποδοχέων TGFβ Οι μεταγραφικοί παράγοντες Smads Η ρύθμιση
UPORABA POLYHIPE-BIOKOMPATIBILNIH
 UPORABA POLYHIPE-BIOKOMPATIBILNIH AKRILNIH POLIMEROV V TKIVNEM INŽENIRSTVU KOSTNIH NADOMESTKOV PoliMaT Academy TalentCamp 2012 Naloga za samostojno delo; področje C: Polimeri v medicini ddr. Matjaž DEŽELAK,
UPORABA POLYHIPE-BIOKOMPATIBILNIH AKRILNIH POLIMEROV V TKIVNEM INŽENIRSTVU KOSTNIH NADOMESTKOV PoliMaT Academy TalentCamp 2012 Naloga za samostojno delo; področje C: Polimeri v medicini ddr. Matjaž DEŽELAK,
Chinese Bulletin of Life Sciences. NTAL/LAB: immuno-regulatory role in lymphocyte development and function. WANG Ying
 17 3 2005 6 Chinese Bulletin of Life Sciences Vol. 17, No. 3 Jun., 2005 1004-0374(2005)03-0251-05 NTAL/LAB 200025 NTAL/LAB B FcγRI FcεRI NTAL/LAB NTAL/LAB ; NTAL/LAB; Q25; Q51 A NTAL/LAB: immuno-regulatory
17 3 2005 6 Chinese Bulletin of Life Sciences Vol. 17, No. 3 Jun., 2005 1004-0374(2005)03-0251-05 NTAL/LAB 200025 NTAL/LAB B FcγRI FcεRI NTAL/LAB NTAL/LAB ; NTAL/LAB; Q25; Q51 A NTAL/LAB: immuno-regulatory
Η παθογένεια της απώλειας οστικής μάζας στις φλεγμονώδεις αρθροπάθειες
 Ανασκόπηση EΛΛΗΝΙΚΗ ΡΕΥΜΑΤΟΛΟΓΙΑ 2007,18(1):60-75 Η παθογένεια της απώλειας οστικής μάζας στις φλεγμονώδεις αρθροπάθειες Ν.Γ. ΓΑΛΑΝΟΠΟΥΛΟΣ Ι.Α. ΠΑΠΑΔΟΠΟΥΛΟΣ Γ.Π. ΚΑΜΠΑΚΗΣ K. ΜΠΛΑΜΗΣ ΠΕΡΙΛΗΨΗ Οι φλεγμονώδεις
Ανασκόπηση EΛΛΗΝΙΚΗ ΡΕΥΜΑΤΟΛΟΓΙΑ 2007,18(1):60-75 Η παθογένεια της απώλειας οστικής μάζας στις φλεγμονώδεις αρθροπάθειες Ν.Γ. ΓΑΛΑΝΟΠΟΥΛΟΣ Ι.Α. ΠΑΠΑΔΟΠΟΥΛΟΣ Γ.Π. ΚΑΜΠΑΚΗΣ K. ΜΠΛΑΜΗΣ ΠΕΡΙΛΗΨΗ Οι φλεγμονώδεις
p 1 ENTROPIJSKI ZAKON
 ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
OSNOVE IMUNOLOGIJE PREGLED SNOVI
 ONOVE IMUNOLOGIJE PREGLED NOVI Po knjigi Vozelj, M.: Temelji imunologije DZ d.d., Ljubljana, 2000 Berete kratek pregled snovi iz predmeta Osnove imunologije. To pomeni, da sem izpustil par poglavij in
ONOVE IMUNOLOGIJE PREGLED NOVI Po knjigi Vozelj, M.: Temelji imunologije DZ d.d., Ljubljana, 2000 Berete kratek pregled snovi iz predmeta Osnove imunologije. To pomeni, da sem izpustil par poglavij in
Στεροειδή σηματοδοτικά μόρια δεσμεύονται σε πυρηνικούς υποδοχείς
 1 Στεροειδή σηματοδοτικά μόρια δεσμεύονται σε πυρηνικούς υποδοχείς στεροειδών (Οι πυρηνικοί υποδοχείς είναι μεταγραφικοί παράγοντες) Το στεροειδές προκαλεί δομικές αλλαγές στον πυρηνικό υποδοχέα, και μπορεί
1 Στεροειδή σηματοδοτικά μόρια δεσμεύονται σε πυρηνικούς υποδοχείς στεροειδών (Οι πυρηνικοί υποδοχείς είναι μεταγραφικοί παράγοντες) Το στεροειδές προκαλεί δομικές αλλαγές στον πυρηνικό υποδοχέα, και μπορεί
Ιωάννης Α. Βουτσαδάκης
 Πανεπιστήμιο Θεσσαλίας Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Τομέας Παθολογίας Πανεπιστημιακή Παθολογική Κλινική Διευθυντής: Καθ. Ν. Σταθάκης Παθογένεση του κολο-ορθικού καρκίνου: Ο ρόλος των αναστολέων
Πανεπιστήμιο Θεσσαλίας Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Τομέας Παθολογίας Πανεπιστημιακή Παθολογική Κλινική Διευθυντής: Καθ. Ν. Σταθάκης Παθογένεση του κολο-ορθικού καρκίνου: Ο ρόλος των αναστολέων
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Contents Part I Psychoneuroimmunology and Systems Biology Mechanisms 1 From Psychoneuroimmunology to Personalized, Systems, and Dynamical Medicine
 Contents Part I Psychoneuroimmunology and Systems Biology Mechanisms 1 From Psychoneuroimmunology to Personalized, Systems, and Dynamical Medicine... 3 1.1 Psychoneuroimmunology (PNI) and Systems Biology...
Contents Part I Psychoneuroimmunology and Systems Biology Mechanisms 1 From Psychoneuroimmunology to Personalized, Systems, and Dynamical Medicine... 3 1.1 Psychoneuroimmunology (PNI) and Systems Biology...
Imunofluorescenčna mikroskopska preiskava
 Imunofluorescenčna mikroskopska preiskava Imunofluorescenčna mikroskopska preiskava Obvezna dopolnilna preiskava pri ledvični biopsiji (imunohistokemija imunoglobulinov in komponent komplementa na zmrznjenih
Imunofluorescenčna mikroskopska preiskava Imunofluorescenčna mikroskopska preiskava Obvezna dopolnilna preiskava pri ledvični biopsiji (imunohistokemija imunoglobulinov in komponent komplementa na zmrznjenih
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Τι το νεότερο στη φαρμακευτική θεραπεία; Παραμένουν οι στατίνες ως φάρμακα επιλογής;
 Τι το νεότερο στη φαρμακευτική θεραπεία; Παραμένουν οι στατίνες ως φάρμακα επιλογής; Άννα Ταυρίδου,, PhD Εργαστήριο Φαρμακολογίας, Τμήμα Ιατρικής, ημοκρίτειο Πανεπιστήμιο Θράκης Υπάρχει ανάγκη για καινούργια
Τι το νεότερο στη φαρμακευτική θεραπεία; Παραμένουν οι στατίνες ως φάρμακα επιλογής; Άννα Ταυρίδου,, PhD Εργαστήριο Φαρμακολογίας, Τμήμα Ιατρικής, ημοκρίτειο Πανεπιστήμιο Θράκης Υπάρχει ανάγκη για καινούργια
Μοριακή Βιολογία. Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής
: Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής") ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν εκπαιδευτικό
Supplemental Table 1. Oligonucleotides used to identify H-RAS, K-RAS and N-RAS mutations and PTEN gene expression.
 Supplemental Table 1. Oligonucleotides used to identify H-RAS, K-RAS and N-RAS mutations and PTEN gene expression. Gene Exon Forward primer Reverse primer Temp ( C) HRAS 2-3 ATGACGGAATATAAGCTGGT ATGGCAAACACACACAGGAA
Supplemental Table 1. Oligonucleotides used to identify H-RAS, K-RAS and N-RAS mutations and PTEN gene expression. Gene Exon Forward primer Reverse primer Temp ( C) HRAS 2-3 ATGACGGAATATAAGCTGGT ATGGCAAACACACACAGGAA
LIPIDI IN PREHRANA ŠPORTNIKA. Žiga Drobnič, Filip Zupančič, 1.b
 LIPIDI IN PREHRANA ŠPORTNIKA Žiga Drobnič, Filip Zupančič, 1.b Lipide delimo na: maščobe (masti,olja) - kombinacija molekule glicerola s tremi dolgoverižnimi organskimi kislinami - maščobnimi kislinami
LIPIDI IN PREHRANA ŠPORTNIKA Žiga Drobnič, Filip Zupančič, 1.b Lipide delimo na: maščobe (masti,olja) - kombinacija molekule glicerola s tremi dolgoverižnimi organskimi kislinami - maščobnimi kislinami
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,