ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ
|
|
|
- Λάμεχ Ζυγομαλάς
- 7 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΔΙΕΡΕΥΝΗΣΗ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΜΕΛΕΤΕΣ ΣΤΟ ΠΕΔΙΟ (ΘΕΡΜΑΪΚΟΣ ΣΤΡΥΜΟΝΙΚΟΣ ΚΟΛΠΟΣ) ΚΑΙ ΣΤΟ ΕΡΓΑΣΤΗΡΙΟ ΜΕΤΑ ΑΠΟ ΕΚΘΕΣΗ ΣΕ ΡΥΠΟΥΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΚΟΥΚΟΥΖΙΚΑ ΝΙΚΟΛΕΤΑ ΘΕΣΣΑΛΟΝΙΚΗ 2009
2 ΕΙΣΑΓΩΓΗ ΠΕΡΙΕΧΟΜΕΝΑ Α. ΧΡΗΣΙΜΟΠΟΙΗΣΗ ΤΟΥ ΜΥΔΙΟΥ MYTILUS GALLOPROVINCIALIS ΓΙΑ ΤΗΝ ΕΚΤΙΜΗΣΗ ΤΗΣ ΡΥΠΑΝΣΗΣ ΤΟΥ ΘΑΛΑΣΣΙΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ 1. Βιοπαρακολούθηση της θαλάσσιας ρύπανσης 2. Βιομάρτυρες 3. Καταλληλότητα των μυδιών ως οργανισμών βιοενδεικτών σε προγράμματα βιοπαρακολούθησης 4. Χαρακτηριστικά γενικής βιολογίας του είδους Mytilus galloprovincialis α) Κατανομή, συστηματική κατάταξη β) Διατροφή και πέψη Πεπτικός αδένας γ) Αιμοκύτταρα δ) Συσσώρευση βαρέων μετάλλων και οργανικών ρύπων σε ιστούς του μυδιού Mytilus galloprovincialis ε) Γενετική και μοριακή σκοπιά του στρες B. ΠΕΡΙΓΡΑΦΗ ΤΗΣ ΠΕΡΙΟΧΗΣ ΜΕΛΕΤΗΣ. ΘΕΡΜΑΪΚΟΣ ΚΑΙ ΣΤΡΥΜΟΝΙΚΟΣ ΚΟΛΠΟΣ Γ. ΒΙΟΜΑΡΤΥΡΕΣ ΠΟΥ ΕΦΑΡΜΟΣΘΗΚΑΝ ΣΤΗ ΣΥΓΚΕΚΡΙΜΕΝΗ ΜΕΛΕΤΗ 1. Βιομάρτυρες που σχετίζονται με τα λυσοσώματα α) Η εκτίμηση της διαπερατότητας των λυσοσωμικών μεμβρανών ως βιομάρτυρας ρύπανσης i. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» σε κρυοτομές ii. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» σε ζωντανά κύτταρα (κατακράτηση της χρωστικής ουδέτερο ερυθρό) β) Τα δομικά χαρακτηριστικά των λυσοσωμάτων ως βιομάρτυρες θαλάσσιας ρύπανσης γ) Η συσσώρευση λιπιδιακών υλικών και η συσσώρευση λιποφουσκινών ως βιομάρτυρες θαλάσσιας ρύπανσης 2. Ο προσδιορισμός των πυρηνικών ανωμαλιών ως βιομάρτυρας θαλάσσιας ρύπανσης α) Βιομάρτυρας «συχνότητα μικροπυρήνων» β) Βιομάρτυρας «συχνότητα συνολικών πυρηνικών ανωμαλιών» 3. Βιομάρτυρας «εκτίμησης στρες» Δ. ΕΠΟΧΙΚΗ ΔΙΑΚΥΜΑΝΣΗ ΤΩΝ ΛΥΣΟΣΩΜΑΤΩΝ, ΤΩΝ ΛΙΠΙΔΙΩΝ ΚΑΙ ΤΩΝ ΛΙΠΟΦΟΥΣΚΙΝΩΝ ΣΤΟΝ ΠΕΠΤΙΚΟ ΑΔΕΝΑ ΤΟΥ ΜΥΔΙΟΥ MYTILUS GALLOPROVINCIALIS Ε. ΣΚΟΠΟΣ ΤΗΣ ΜΕΛΕΤΗΣ
3 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Α. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΜΕΛΕΤΗ ΠΕΔΙΟΥ ΣΕ ΕΛΛΗΝΙΚΕΣ ΑΚΤΕΣ 1. Δειγματοληψίες μυδιών από το πεδίο 2. Βιομάρτυρες που εξετάσθηκαν Β. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΕΡΓΑΣΤΗΡΙΑΚΗ ΜΕΛΕΤΗ ΜΕΤΑ ΑΠΟ ΕΚΘΕΣΗ ΣΤΑ ΒΑΡΕΑ ΜΕΤΑΛΛΑ Cu,Cd, Hg KAI ΣΤΟΝ ΟΡΓΑΝΙΚΟ ΡΥΠΟ ΦΑΙΝΑΝΘΡΕΝΙΟ 1. Δειγματοληψία για επίδραση ρύπων στο εργαστήριο 2. Επίδραση ρύπων 3. Βιομάρτυρες που εξετάσθηκαν Γ. ΜΕΛΕΤΗ ΤΗΣ ΕΠΟΧΙΚΗΣ ΔΙΑΚΥΜΑΝΣΗΣ ΤΩΝ ΛΥΣΟΣΩΜΑΤΩΝ, ΤΩΝ ΛΙΠΙΔΙΩΝ ΚΑΙ ΤΩΝ ΛΙΠΟΦΟΥΣΚΙΝΩΝ ΣΤΟΝ ΠΕΠΤΙΚΟ ΑΔΕΝΑ ΤΟΥ ΜΥΔΙΟΥ MYTILUS GALLOPROVINCIALIS. 1. Δειγματοληψίες μυδιών από το πεδίο. 2. Παράμετροι που εξετάσθηκαν Δ. ΠΕΡΙΓΡΑΦΗ ΒΙΟΜΑΡΤΥΡΩΝ 1. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» 2. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» 3. Ο προσδιορισμός των πυρηνικών ανωμαλιών ως βιομάρτυρας θαλάσσιας ρύπανσης α) Βιομάρτυρας «συχνότητα μικροπυρήνων» β) Βιομάρτυρας «συχνότητα συχνότητα συνολικών πυρηνικών ανωμαλιών» 4. Βιομάρτυρας «εκτίμησης στρες» 5. Ιστοχημεία λυσοσωμάτων, ουδέτερων λιπιδίων, ολικών λιπιδίων και λιποφουσκινών στον πεπτικό αδένα του μυδιού Mytilus galloprovincialis Προετοιμασία ιστών α) Ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακέτυλο-β-εξοζαμινιδάση β) Ιστοχημεία ουδέτερων λιπιδίων γ) Ιστοχημεία ολικών λιπιδίων δ) Ιστοχημεία λιποφουσκινών Στατιστική ανάλυση δεδομένων ΑΠΟΤΕΛΕΣΜΑΤΑ Α. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΜΕΛΕΤΗ ΠΕΔΙΟΥ ΣΕ ΕΛΛΗΝΙΚΕΣ ΑΚΤΕΣ 54
4 1. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» 2. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» 3. Βιομάρτυρας «συχνότητα μικροπυρήνων» 4. Βιομάρτυρας «εκτίμησης στρες» 5. Μορφομετρικές και μορφολογικές αποκρίσεις των λυσοσωμάτων του πεπτικού αδένα 6. Συσχέτιση μεταξύ των αποτελεσμάτων βιομαρτύρων στη μελέτη πεδίου 7. Ομαδοποίηση των σταθμών δειγματοληψίας σύμφωνα με τα αποτελέσματα των βιομαρτύρων Β. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΕΡΓΑΣΤΗΡΙΑΚΗ ΜΕΛΕΤΗ ΜΕΤΑ ΑΠΟ ΕΚΘΕΣΗ ΣΤΑ ΒΑΡΕΑ ΜΕΤΑΛΛΑ Cu, Cd, Hg KAI ΣΤΟΝ ΟΡΓΑΝΙΚΟ ΡΥΠΟ ΦΑΙΝΑΝΘΡΕΝΙΟ 1. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» 2. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» 3. Πυρηνικές ανωμαλίες στα αιμοκύτταρα μυδιών Mytilus gallorpovincialis μετά από εργαστηριακή έκθεση στα βαρέα μέταλλα Cu, Cd, Hg και στον οργανικό ρύπο φαινανθρένιο. α) Βιομάρτυρας «συχνότητα μικροπυρήνων» β) Βιομάρτυρας «συχνότητα συνολικών πυρηνικών ανωμαλιών» 4. Μορφομετρικές και μορφολογικές αποκρίσεις των λυσοσωμάτων του πεπτικού αδένα μυδιών Mytilus galloprovincialis μετά από εργαστηριακή επίδραση διαφορετικών ρύπων 5. Μορφομετρικές και μορφολογικές αποκρίσεις των ουδέτερων λιπιδίων του πεπτικού αδένα μυδιών Mytilus galloprovincialis μετά από εργαστηριακή επίδραση διαφορετικών ρύπων 6. Συσχέτιση μεταξύ των αποτελεσμάτων βιομαρτύρων στην εργαστηριακή μελέτη 7. Ομαδοποίηση των ρύπων βάσει των επιπτώσεών τους στα μύδια Mytilus galloprovincialis και διερεύνηση της συγγένειας των υπό εξέταση βιομαρτύρων Γ. ΜΕΛΕΤΗ ΛΥΣΟΣΩΜΙΚΩΝ, ΛΙΠΙΔΙΑΚΩΝ ΠΑΡΑΜΕΤΡΩΝ ΚΑΙ ΛΙΠΟΦΟΥΣΚΙΝΩΝ ΣΤΟΝ ΠΕΠΤΙΚΟ ΑΔΕΝΑ ΜΥΔΙΩΝ MYTILUS GALLOPROVINCIALIS 1. Εποχική διακύμανση των λιπιδίων, των λυσοσωμάτων και των λιποφουσκινών στον πεπτικό αδένα του μυδιού Mytilus galloprovincialis α) Χαρακτηριστικά των πεπτικών σωληναρίων κατά την αρχή του καλοκαιριού β) Χαρακτηριστικά των πεπτικών σωληναρίων κατά την αρχή του χειμώνα γ) Χαρακτηριστικά των πεπτικών σωληναρίων την άνοιξη δ) Μορφομετρία λιποφουσκινών, ουδέτερων λιπιδίων και λυσοσωμάτων στον πεπτικό αδένα μυδιών που συλλέχθηκαν στην αρχή του καλοκαιριού, στην αρχή του χειμώνα και την άνοιξη
5 2. Αλληλεπίδραση ανάμεσα στο λιπιδιακό και το ενζυμικό περιεχόμενο των λυσοσωμάτων 3. Ιστοχημικός εντοπισμός λιποφουσκινών, λιπιδίων και λυσοσωμάτων στον πεπτικό αδένα μυδιών Mytilus galloprovincialis ΣΥΖΗΤΗΣΗ Α. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΜΕΛΕΤΗ ΠΕΔΙΟΥ ΣΕ ΕΛΛΗΝΙΚΕΣ ΑΚΤΕΣ 1. Εφαρμογή του βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» στο πεδίο 2. Εφαρμογή του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» στο πεδίο 3. Εφαρμογή του βιομάρτυρα «συχνότητα μικροπυρήνων» στο πεδίο 4. Εφαρμογή του βιομάρτυρα «εκτίμηση στρες» στο πεδίο 5. Αποκρίσεις των λυσοσωμάτων του πεπτικού αδένα κατά τη μελέτη πεδίου 6. Συσχέτιση μεταξύ των αποτελεσμάτων βιομαρτύρων στη μελέτη πεδίου Β. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΕΡΓΑΣΤΗΡΙΑΚΗ ΜΕΛΕΤΗ ΜΕΤΑ ΑΠΟ ΕΚΘΕΣΗ ΣΤΑ ΒΑΡΕΑ ΜΕΤΑΛΛΑ Cu, Cd, Hg KAI ΣΤΟΝ ΟΡΓΑΝΙΚΟ ΡΥΠΟ ΦΑΙΝΑΝΘΡΕΝΙΟ 1. Πυρηνικές ανωμαλίες στα αιμοκύτταρα μυδιών Mytilus gallorpovincialis μετά από εργαστηριακή έκθεση στα βαρέα μέταλλα Cu, Cd, Hg και στον οργανικό ρύπο φαινανθρένιο. α) Βιομάρτυρας «συχνότητα μικροπυρήνων» β) Βιομάρτυρας «συχνότητα συνολικών πυρηνικών ανωμαλιών» 2. Αποκρίσεις λιπιδιακών παραμέτρων του πεπτικού αδένα των μυδιών μετά από έκθεση σε διαφορετικούς ρύπους 3. Αποκρίσεις λυσοσωμικών παραμέτρων του πεπτικού αδένα μυδιών μετά από έκθεση σε διαφορετικούς ρύπους 4. Συσχέτιση μεταξύ των αποτελεσμάτων βιομαρτύρων στην εργαστηριακή μελέτη Γ. ΜΕΛΕΤΗ ΛΥΣΟΣΩΜΙΚΩΝ, ΛΙΠΙΔΙΑΚΩΝ ΠΑΡΑΜΕΤΡΩΝ ΚΑΙ ΛΙΠΟΦΟΥΣΚΙΝΩΝ ΣΤΟΝ ΠΕΠΤΙΚΟ ΑΔΕΝΑ ΜΥΔΙΩΝ MYTILUS GALLOPROVINCIALIS 1. Εποχική διακύμανση των λιπιδίων, των λυσοσωμάτων και των λιποφουσκινών στον πεπτικό αδένα του μυδιού Mytilus galloprovincialis 2. Αλληλεπίδραση ανάμεσα στο λιπιδιακό και το ενζυμικό περιεχόμενο των λυσοσωμάτων 3. Ιστοχημικός εντοπισμός λιποφουσκινών, λιπιδίων και λυσοσωμάτων στον πεπτικό αδένα μυδιών Mytilus galloprovincialis
6 ΣΥΜΠΕΡΑΣΜΑΤΑ CONCLUSIONS ΠΕΡΙΛΗΨΗ SUMMARY ΒΙΒΛΙΟΓΡΑΦΙΑ
7 ΕΙΣΑΓΩΓΗ
8 ΕΙΣΑΓΩΓΗ Α. ΧΡΗΣΙΜΟΠΟΙΗΣΗ ΤΟΥ ΜΥΔΙΟΥ MYTILUS GALLOPROVINCIALIS ΓΙΑ ΤΗΝ ΕΚΤΙΜΗΣΗ ΤΗΣ ΡΥΠΑΝΣΗΣ ΤΟΥ ΘΑΛΑΣΣΙΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ 1. Βιοπαρακολούθηση της θαλάσσιας ρύπανσης Με τον όρο θαλάσσια ρύπανση εννοούμε την άμεση ή έμμεση απελευθέρωση ρυπαντικών ουσιών στο θαλάσσιο περιβάλλον, η οποία οφείλεται στην ανθρώπινη δραστηριότητα και προκαλεί ή είναι δυνατόν να προκαλέσει κίνδυνο στην ανθρώπινη υγεία, βλάβη στους ζωντανούς οργανισμούς, υποβιβασμό των θαλασσίων οικοσυστημάτων αλλά και των φυσικών πόρων ή παρακώλυση νόμιμων δραστηριοτήτων που σχετίζονται με τη θάλασσα (OSPAR Commission 2000). Τα τελευταία χρόνια, τα επίπεδα των περιβαλλοντικών ρύπων έχουν αυξηθεί πολύ στο θαλάσσιο περιβάλλον ως συνέπεια των δραστηριοτήτων του ανθρώπου (ΕΕΑ 1999). Κύριες πηγές θαλάσσιας ρύπανσης αποτελούν τα αστικά και βιομηχανικά απόβλητα, η έκπλυση γεωργικών εδαφών, τα λύματα υπονόμων, οι διαδικασίες εξαγωγής άμμου και χαλικιού, οι βιομηχανίες εξόρυξης πετρελαίου, φυσικού αερίου και ορυκτών, η τοποθέτηση καλωδίων και σωληνώσεων, ο τουρισμός, ποικίλες ψυχαγωγικές δραστηριότητες, οι υδατοκαλλιέργειες, η αλιεία, οι θαλάσσιες μεταφορές, τα ατυχήματα διαρροής πετρελαίου κ.ά. (ΕΕΑ 1999, GESAMP 2001). Οι ρύποι που απελευθερώνονται στο θαλάσσιο περιβάλλον διακρίνονται σε βασικές κατηγορίες: αλογονωμένοι υδρογονάθρακες, μη αλογονωμένοι υδρογονάνθρακες, βαρέα μέταλλα, ανιόντα κ.ά. (Lawrence & Elliot 2003, Δημητριάδης κ.ά. 2006). Θαλάσσιοι ρύποι έχουν ανιχνευθεί όχι μόνο κοντά σε βιομηχανικές περιοχές, αλλά και σε απομακρυσμένα σημεία του πλανήτη, όπως είναι η Αρκτική και η Ανταρκτική (Wu 1999). Σε παγκόσμια κλίμακα, οι περισσότερες από τις παράκτιες περιοχές που μελετήθηκαν εμφανίζουν μικρότερα ή μεγαλύτερα επίπεδα ρύπανσης ( Πολλοί οργανικοί ρύποι που χρησιμοποιήθηκαν από τον άνθρωπο κατά το παρελθόν είναι ιδιαίτερα ανθεκτικοί, με αποτέλεσμα αυτοί ή τα παράγωγά τους να εντοπίζονται στο θαλάσσιο περιβάλλον, ακόμα και πολλά χρόνια μετά από την απαγόρευση της χρήσης τους από τον άνθρωπο. Καθίσταται λοιπόν αναγκαία η λήψη μέτρων για την προστασία του υδάτινου περιβάλλοντος από τη ρύπανση, ιδίως τη ρύπανση που προκαλείται από ορισμένες ανθεκτικές, τοξικές και βιοσυσσωρεύσιμες ουσίες, όπως ορίζεται και στην αντίστοιχη οδηγία του Ευρωπαϊκού Κοινοβουλίου (ΕC 11/2006). 1
9 ΕΙΣΑΓΩΓΗ Η παρουσία ρύπων στα θαλάσσια οικοσυστήματα μπορεί να επιφέρει πλήθος ανεπιθύμητων αποτελεσμάτων σε όλα τα επίπεδα οργάνωσης της ζωής. Στις αρνητικές συνέπειες που προκαλεί η θαλάσσια ρύπανση περιλαμβάνονται επιδράσεις σε επίπεδα όπως το μοριακό ή κυτταρικό, έως και αλλαγές στη δομή και τη λειτουργία των βιοκοινοτήτων, μέσω της διατάραξης της αναπαραγωγικής διαδικασίας και της αλλαγής στη συμπεριφορά των οργανισμών (GESAMP 2001). Η έντονη επίδραση των ρύπων στα θαλάσσια οικοσυστήματα καθιστά επιτακτική την ανάγκη ανάπτυξης μεθόδων για την ταυτοποίηση, μέτρηση, συγκριτική εκτίμηση και διαχείριση των επιπτώσεων που προκύπτουν από την απελευθέρωση χημικών ρύπων στο θαλάσσιο περιβάλλον (Cajaraville et al. 2000, Broeg et al. 2005). Αρχικά η εκτίμηση της θαλάσσιας ρύπανσης πραγματοποιούνταν με τον εντοπισμό χημικών ουσιών στο νερό, στα ιζήματα και στους ζωντανούς οργανισμούς. Η χημική ανάλυση μπορεί να αποκαλύψει την παρουσία ρύπων, χωρίς όμως να προσδιορίσει τις επιπτώσεις τους στη χλωρίδα και την πανίδα, ούτε επίσης και το αποτέλεσμα που μπορεί να επιφέρει ο συνδυασμός ποικίλων ρύπων (συνεργατικά, ανταγωνιστικά ή προσθετικά φαινόμενα), ούτε το διαφορετικό αποτέλεσμα που μπορεί να προκαλέσει ένας ρύπος σε έναν οργανισμό κάτω από την επίδραση διαφορετικών περιβαλλοντικών συνθηκών. Επιπλέον, πολλοί οργανικοί ρύποι (Aas et al. 2000) μεταβολίζονται γρήγορα μετά από την απελευθέρωσή τους στο περιβάλλον, δημιουργώντας μεταβολίτες αρκετά επικίνδυνους για τους ζωντανούς οργανισμούς (Heidelberger 1973). Σε μια τέτοια περίπτωση, η χημική ανάλυση δεν αποκαλύπτει την παρουσία των πρωτογενών ρύπων, μολονότι οι ζωντανοί οργανισμοί δέχονται τη δυσμενή επίδραση των παραγώγων τους. Επομένως, αν και η χημική ανάλυση συνιστά θεμελιώδη τεχνική για την παρακολούθηση της ρύπανσης θαλασσίων οικοσυστημάτων (chemical monitoring), η μέθοδος αυτή από μόνη της δεν αποτελεί αποτελεσματικό εργαλείο για τον προσδιορισμό των επιπτώσεων της ρύπανσης (UNEP/STAP 2003). Για το λόγο αυτό από τις αρχές της δεκαετίας του 70 άρχισε να εφαρμόζεται και η παρακολούθηση των αποτελεσμάτων της ρύπανσης σε ζωντανούς οργανισμούς ή βιοπαρακολούθηση (biomonitoring) της ρύπανσης. Δηλαδή, εκτός από τις τεχνικές για τη μέτρηση της συγκέντρωσης οργανικών ή ανόργανων ρύπων σε ιστούς ζωντανών οργανισμών, αναπτύχθηκαν και μέθοδοι για την εκτίμηση και καταγραφή των βιολογικών επιπτώσεων της ρύπανσης. Το πεδίο της βιοπαρακολούθησης είναι ιδιαίτερα ευρύ και περιλαμβάνει μια πληθώρα τεχνικών (για μια συνοπτική παρουσίαση αυτών των τεχνικών βλ. Kramer 2
10 ΕΙΣΑΓΩΓΗ 1994). Στην «παθητική βιοπαρακολούθηση» χρησιμοποιούνται οργανισμοί, οι οποίοι υπάρχουν ήδη στο φυσικό περιβάλλον της περιοχής μελέτης. Αντίθετα, στην «ενεργητική βιοπαρακολούθηση» πραγματοποιείται εισαγωγή και έκθεση δεικτώνοργανισμών στην περιοχή μελέτης για συγκεκριμένο χρόνο και σε καθορισμένες συνθήκες (Wittig 1993). Η καταγραφή αλλαγών της φυσιολογίας ή/και της συμπεριφοράς οργανισμών με τη χρήση αυτόματων συστημάτων αποτελεί άλλο ένα σημαντικό εργαλείο στον τομέα της βιοπαρακολούθησης. Επίσης, βιολογικοί προσδιορισμοί στο πεδίο ή στο εργαστήριο αξιοποιούνται για την εκτίμηση της τοξικότητας του νερού, ενώ και η χρήση ανοσολογικών μεθόδων εφαρμόζεται ως ένα γρήγορο μέσο για τη μέτρηση μεμονωμένων ή ομάδων χημικών στοιχείων. Επιπρόσθετα, η εφαρμογή «βιοτικών δεικτών», οι οποίοι χρησιμοποιούν συγκεκριμένους οργανισμούς με διαφορερικό βαθμό ανθεκτικότητας στη ρύπανση, παρέχει χρήσιμες πληροφορίες σχετικά με την καθαρότητα του νερού (NRC 1980, NCMR 1997, Δημητριάδης κ.ά. 2006). Μάλιστα, ο συνδυασμός της εφαρμογής «βιοτικών δεικτών» και της μέτρησης διάφορων φυσικοχημικών και υδρομορφολογικών παραμέτρων, όπως απαιτεί και η οδηγία του Ευρωπαϊκού Κοινοβουλίου για τα νερά (EC 60/2000), μας δίνει μια πιο ολοκληρωμένη εικόνα για την κατάσταση της ποιότητας των υδάτων. 2. Βιομάρτυρες Τα τελευταία χρόνια ιδιαίτερη σημασία έχει δοθεί στις βιολογικές παραμέτρους που χαρακτηρίζονται ως δείκτες στρες ή βιομάρτυρες για τη βιοπαρακολούθηση της θαλάσσιας ρύπανσης (Bayne 1989, Gray 1992). Ως βιομάρτυρας χαρακτηρίζεται κάθε μεταβολή που μπορεί να παρατηρηθεί ή και να μετρηθεί σε βιοχημικό και κυτταρικό επίπεδο, καθώς και μεταβολές σε επίπεδο φυσιολογίας και συμπεριφοράς του οργανισμού, οι οποίες αποκαλύπτουν την έκθεση του ατόμου στο παρόν ή στο παρελθόν σε τουλάχιστο μία χημική ουσία με χαρακτηριστικά ρύπου (Adams 1990, Roy & Hanninen 1993, Fishelson 1995, Lagadic et al. 1997, Bresler et al. 1999, Fishelson et al. 1999). Βιολογικές αποκρίσεις σε ανώτερα επίπεδα οργάνωσης (πληθυσμού, βιοκοινότητας και οικοσυστήματος) ορίζονται ως βιοδείκτες (Δημητριάδης κ.ά. 2006). Κάθε βιομάρτυρας πρέπει να συγκεντρώνει ορισμένα χαρακτηριστικά όπως η επαναληψιμότητα, η ευαισθησία, η εξειδίκευση, ο σύντομος χρόνος απόκρισης, η 3
11 ΕΙΣΑΓΩΓΗ αναστρεψιμότητα, το μεγάλο εύρος ειδών στα οποία εφαρμόζεται, η ευκολία εφαρμογής και το χαμηλό κόστος του σε σύγκριση με την αποτελεσματικότητά του (Gosling 1994). Οι μοριακοί και κυτταρικοί βιομάρτυρες αποτελούν εργαλεία έγκαιρης προειδοποίησης (early warning tools) για τις μακροπρόθεσμες επιπτώσεις της ρύπανσης στο επίπεδο του οργανισμού, του πληθυσμού ή της βιοκοινότητας, εξαιτίας της πολύ γρήγορης επίδρασης των τοξικών ουσιών στο μοριακό και κυτταρικό επίπεδο (McCarthy & Shugart 1990, Huggert et al. 1992). Οι βιομάρτυρες που χρησιμοποιούνται στη βιοπαρακολούθηση της θαλάσσιας ρύπανσης διακρίνονται σε δύο βασικές κατηγορίες: σε βιομάρτυρες έκθεσης και βιομάρτυρες τοξικής επίπτωσης. Οι βιομάρτυρες έκθεσης φανερώνουν την απόκριση ενός οργανισμού σε χημικές ενώσεις, χωρίς να προσδιορίζουν το μέγεθος του επιβλαβούς αποτελέσματος που προκαλεί η έκθεση στους ρύπους. Αντίθετα, οι βιομάρτυρες τοξικής επίπτωσης παρέχουν πληροφορίες και είναι ικανοί να εκτιμήσουν την έκταση του επιβλαβούς αποτελέσματος της ρύπανσης στους οργανισμούς (Δημητριάδης κ.ά. 2006). Το εύρος εξειδίκευσης των βιομαρτύρων είναι μεγάλο: η εξειδίκευση κυμαίνεται από εξαιρετικά υψηλή έως και χαμηλή. Οι βιομάρτυρες με υψηλή εξειδίκευση σχετίζονται με την έκθεση των οργανισμών σε συγκεκριμένους ρύπους ή ομάδες ρύπων. Αντίθετα, οι μη ειδικοί βιομάρτυρες περιλαμβάνουν αποκρίσεις που μπορεί να προκληθούν από μια μεγάλη ποικιλία ρύπων (Δημητριάδης κ.ά. 2006). Ωστόσο, για μια περισσότερο αποτελεσματική εκτίμηση των επιπτώσεων των διαφόρων ρύπων στους οργανισμούς και τη διαμόρφωση μιας σαφέστερης εικόνας της κατάστασης του περιβάλλοντος από άποψη ρύπανσης, είναι απαραίτητη η συνεξέταση βιομαρτύρων με υψηλή ειδίκευση και μη ειδικών βιομαρτύρων, λαμβάνοντας υπόψη τα χαρακτηριστικά της φυσιολογίας του εκάστοτε οργανισμού και του οικοσυστήματός του. 3. Καταλληλότητα των μυδιών ως οργανισμών βιοενδεικτών σε προγράμματα βιοπαρακολούθησης Οι βιομάρτυρες ρύπανσης (biomarkers ή stress indices) εφαρμόζονται σε ζωντανούς οργανισμούς, οι οποίοι χαρακτηρίζονται ως βιοενδείκτες (sentinel species ή bioindicators). Σπονδυλωτά (ψάρια), καρκινοειδή και μαλάκια αποτελούν τους περισσότερο διαδεδομένους οργανισμούς για την εκτίμηση της κατάστασης του 4
12 ΕΙΣΑΓΩΓΗ θαλάσσιου περιβάλλοντος. Τα ψάρια, λόγω του ότι εμφανίζουν δυνατότητα μετακίνησης, δεν μπορούν να καταδείξουν τη ρύπανση μιας συγκεκριμένης θαλάσσιας περιοχής. Ως εκ τούτου, όταν χρησιμοποιούνται τα ψάρια ως οργανισμοί βιοενδείκτες σε μελέτες βιοπαρακολούθησης θαλασσίων οικοσυστημάτων, ακολουθείται συνήθως η διαδικασία διατήρησής τους μέσα σε κλωβούς με σκοπό να περιορισθεί η κινητικότητά τους. Το παραπάνω γεγονός σε συνδυασμό με το υψηλό κόστος των ψαριών έχει ως αποτέλεσμα να μη χρησιμοποιούνται οι οργανισμοί αυτοί σε μελέτες βιοπαρακολούθησης της θαλάσσιας ρύπανσης σε κλίμακα ανάλογη με αυτή των μαλακίων. Αντίθετα, τα στρείδια, οι πεταλίδες και περισσότερο τα μύδια χρησιμοποιούνται εκτεταμένα σε προγράμματα βιοπαρακολούθησης (Cajaraville et al. 1990, Livingstone et al. 1990, Smolders et al. 2003, Marigomez et al. 2006, ICES 2006). Τα χαρακτηριστικά των διθύρων και ιδιαίτερα των μυδιών που τα καθιστούν ιδανικούς οργανισμούς βιοενδείκτες σε μελέτες βιοπαρακολούθησης είναι (σύμφωνα με τους NCR 1980, Philips 1980, Widdows 1985, Farrington et al. 1987, Δημητριάδης κ.ά. 2006): Τα μύδια κυριαρχούν στην παράκτια ζώνη, στις κοινωνίες των φωτόφιλων φυκών (Χιντήρογλου, προσωπική επικοινωνία) και παρουσιάζουν ευρεία γεωγραφική κατανομή. Είναι προσκολλημένα είδη και άρα περισσότερο κατάλληλοι οργανισμοί από άλλους, οι οποίοι έχουν τη δυνατότητα μετανάστευσης, για την πρόσληψη χημικών ρύπων από μια συγκεκριμένη περιοχή. Είναι αρκετά ανθεκτικά σε ένα ευρύ φάσμα περιβαλλοντικών συνθηκών και παρουσιάζουν μεγάλη αντοχή μετά από επίδραση σχετικά υψηλής συγκέντρωσης ρύπων. Είναι διηθηματοφάγα και λόγω της ικανότητάς τους να φιλτράρουν σημαντικό όγκο νερού, γίνονται αποδέκτες μεγάλων ποσοτήτων ρυπογόνων ουσιών, οι οποίες μπορούν να συσσωρευτούν στους ιστούς τους σε πολύ υψηλότερες συγκεντρώσεις από το θαλασσινό νερό. Επομένως, τοξικές ουσίες που εμφανίζονται σε ίχνη στο θαλασσινό νερό, μπορούν να προσδιορισθούν ευκολότερα στους ιστούς των οργανισμών αυτών. Τα δίθυρα, σε σύγκριση με τα αρθρόποδα και τα ψάρια, εμφανίζουν χαμηλό επίπεδο δραστικότητας των ενζυμικών συστημάτων, τα οποία είναι υπεύθυνα για το μεταβολισμό οργανικών τοξικών ουσιών. Επομένως, οι συγκεντρώσεις 5
13 ΕΙΣΑΓΩΓΗ ρύπων στους ιστούς τους αντικατοπτρίζουν με μεγαλύτερη ακρίβεια την έκταση της περιβαλλοντικής ρύπανσης. Η μέτρηση των τοξικών ουσιών στους ιστούς των διθύρων παρέχει πρόσθετες πληροφορίες για τη βιολογική διαθεσιμότητά τους, η οποία δεν μπορεί να αξιολογηθεί όταν οι μετρήσεις πραγματοποιούνται στο νερό, στα αιωρούμενα σωματίδια ή στα ιζήματα. Τα μύδια εμφανίζουν σχετικά σταθερούς πληθυσμούς, οι οποίοι χαρακτηρίζονται από μεγάλο αριθμό ατόμων. Το γεγονός αυτό επιτρέπει την πραγματοποίηση επανειλημμένων δειγματοληψιών για τη μακροχρόνια παρακολούθηση του υδάτινου περιβάλλοντος. Είναι εύκολη η μεταφορά και η διατήρησή τους μέσα σε κλωβούς σε περιοχές ενδιαφέροντος. Είναι δυνατή η διατήρησή τους κάτω από ελεγχόμενες εργαστηριακές συνθήκες, γεγονός που επιτρέπει πειραματισμό στο εργαστήριο. Υπάρχει εκτεταμένη βασική γνώση για τις βασικές λειτουργίες τους, με αποτέλεσμα τη δυνατότητα εξαγωγής συμπερασμάτων με οικοτοξικολογική συνάφεια. Τα δίθυρα μαλάκια χρησιμοποιούνται, λοιπόν, ευρέως ως βιοενδείκτες της θαλάσσιας ρύπανσης. Ειδικότερα, είδη του γένους Mytilus χρησιμοποιούνται σε μεγάλη κλίμακα, τόσο σε εργαστηριακό επίπεδο, όσο και στο πεδίο (Mussel Watch Programmes) για την εκτίμηση των επιπτώσεων που προκαλούν οι διάφοροι ρύποι (Goldberg 1986, Bayne 1989). Ποικίλοι περιβαλλοντικοί παράγοντες μπορούν, ωστόσο, να επιδράσουν στη φυσιολογία και τη συμπεριφορά των δίθυρων μαλακίων, επηρεάζοντας το ρυθμό αύξησης και πρόσληψης τροφής, τον αναπαραγωγικό κύκλο και την ικανότητά τους να αποκρίνονται στη ρύπανση. Η αυξημένη αλατότητα και η έντονη μεταβολή στο ph του θαλασσινού νερού αποτελούν αβιοτικούς παράγοντες οι οποίοι μπορούν να προκαλέσουν μείωση του χρόνου ανοίγματος του κελύφους, ώστε να επέλθει ομαλή εξισορρόπηση του εσωτερικού περιβάλλοντος του οργανισμού με το υδάτινο μέσο. Στην περίπτωση αυτή διατηρείται ευκολότερα η ομοιόσταση των κυττάρων, ενώ η απαιτούμενη ενέργεια προκύπτει μέσα από αναερόβιες διαδικασίες. Η έκθεση των ατόμων στην ατμόσφαιρα εξαιτίας της παλίρροιας είναι ένας άλλος παράγοντας που προκαλεί αύξηση της αναερόβιας αναπνοής επηρεάζοντας το μεταβολισμό των 6
14 ΕΙΣΑΓΩΓΗ οργανισμών, τη φυσιολογική τους ομοιόσταση και τη δράση των ενζύμων στο εσωτερικό των κυττάρων. Επίσης, οι συνθήκες έντονου συνωστισμού περιορίζουν την ικανότητα πρόσληψης τροφής, με συνέπεια τη μείωση της βιωσιμότητας των ατόμων, την ελάττωση του μεγέθους τους και τη μεταβολή του αναπαραγωγικού τους κύκλου. Ο αυξημένος ανταγωνισμός για εύρεση τροφής, ο οποίος προκαλείται λόγω του συνωστισμού, αναγκάζει τους οργανισμούς να φιλτράρουν μεγαλύτερες ποσότητες νερού, με αποτέλεσμα τη συσσώρευση αυξημένης ποσότητας ρύπων στο εσωτερικό των ιστών τους (Philips 1976a,b, Cossa et al. 1979, Davies & Pirie 1980). Τέλος, η μόλυνση από παράσιτα έχει επίπτωση στη φυσιολογική κατάσταση των μυδιών (Rayyan et al. 2004), ενώ διάφοροι βιοτικοί παράγοντες όπως η ηλικία, το βάρος του μαλακού ιστού, το μέγεθος των ατόμων αλλά και η περίοδος αναπαραγωγής φαίνεται να επηρεάζουν το φυσιολογικό ρυθμό ανάπτυξης και κατά συνέπεια τη δυνατότητα των μυδιών να αποκρίνονται στη ρύπανση (Philips 1976a,b, Cossa et al. 1979, Davies & Pirie 1980). 4. Χαρακτηριστικά γενικής βιολογίας του είδους Mytilus galloprovincialis α) Κατανομή, συστηματική κατάταξη Το γένος Mytilus παρουσιάζει ευρεία κατανομή τόσο στο βόρειο όσο και στο νότιο ημισφαίριο και περιλαμβάνει τέσσερα είδη: M. edulis, M. galloprovincialis, M. trossulus, M. californianus. Η εμφάνιση του γένους τοποθετείται πριν από 1-2 εκατομμύρια χρόνια (Seed 1976). Τα στοιχεία στα οποία στηρίζεται η κατάταξη των ειδών του γένους Mytilus, καθώς και στοιχεία σχετικά με την οικολογία, τη φυσιολογία, τη γενετική, την εκτροφή ή τη σχέση των μυδιών με τη δημόσια υγεία εμπεριέχονται στο βιβλίο της Gosling (1992). Στην παρούσα μελέτη χρησιμοποιήθηκε ως βιοενδείκτης το μύδι Mytilus galloprovincialis. Η συστηματική κατάταξη του είδους αυτού είναι η ακόλουθη: Βασίλειο: Animalia Φύλο: Mollusca Κλάση: Bivalvia Υποκλάση: Pteriomorphia Τάξη: Lamellibranchia Οικογένεια: Mytilidae Γένος: Mytilus Είδος: galloprovincialis 7
15 ΕΙΣΑΓΩΓΗ β) Διατροφή και πέψη Τα μύδια είναι διηθηματοφάγοι οργανισμοί. Τα σωματίδια της τροφής εισέρχονται με το νερό μέσω ενός προσαγωγού σιφωνίου (Bayne et al. 1976) και στη συνέχεια συνδυασμένες κινήσεις των βλεφαρίδων των βραγχίων και των χειλικών προσακτρίδων της μανδυακής κοιλότητας τα μεταφέρουν προς τον γαστρεντερικό σωλήνα. Στα δίθυρα μαλάκια ο γαστρεντερικός σωλήνας αποτελείται από έναν απλό και κοντό οισοφάγο, έναν πολύπλοκο στόμαχο και το έντερο. Το οπίσθιο τμήμα του στομάχου επικοινωνεί με μια δομή, γνωστή ως σάκος, που περιέχει το στύλο. Ο στύλος είναι κρυσταλλικής πρωτεϊνικής φύσεως και βοηθάει στη διαδικασία της πέψης (Morton 1983). Συγκεκριμένα, καθώς περιστρέφεται αργά στο στομάχι αναδεύει την τροφή, ενώ ταυτόχρονα συγκεντρώνει άπεπτα τεμάχια τροφής και τα διατηρεί στο κέντρο του, με σκοπό τη χρησιμοποίησή τους σε περιόδους κατά τις οποίες υπάρχει περιορισμένη δυνατότητα πρόσληψης τροφής. Τέλος, το όργανο αυτό ελευθερώνει κατάλληλα ένζυμα για την πέψη αμυλούχων τροφών (Morton 1983). Από τον στόμαχο ξεκινούν μικροί αγωγοί που οδηγούν στον πεπτικό αδένα, το όργανο που είναι υπεύθυνο για την απορρόφηση και την ενδοκυτταρική πέψη. Ο πεπτικός αδένας συνίσταται από μικρότερους αδένες, που ονομάζονται πεπτικά σωληνάρια (Morton 1983). Αρχικά η πέψη είναι εξωκυτταρική και πραγματοποιείται στο στομάχι. Τα απαραίτητα ένζυμα για τη διαδικασία αυτή προέρχονται από τη διάλυση του πρωτεϊνικού στύλου, που επιτυγχάνεται με τη δράση πρωτεϊνών που εκκρίνονται από τα βασεόφιλα κύτταρα του πεπτικού αδένα ή αντίστοιχων ενζύμων που περιέχονται στα σφαιρίδια θρυμματισμού (Morton 1983). Τα τελευταία αποβάλλονται από το κορυφαίο τμήμα των πεπτικών κυττάρων στον αγωγό των πεπτικών σωληναρίων, μεταφέρονται αρχικά στο στομάχι και στη συνέχεια στο έντερο. Αποτελούνται από τμήμα του κυτταροπλάσματος των πεπτικών κυττάρων και υπολείμματα της ενδοκυτταρικής πέψης και περιέχουν υδρολυτικά ένζυμα, όπως πρωτεάσες σε όξινο ph οι οποίες συμβάλλουν στη διάλυση του στύλου και κατά συνέπεια στην εξωκυτταρική πέψη. Στη συνέχεια πραγματοποιείται ενδοκυτταρική πέψη στα λυσοσώματα των πεπτικών κυττάρων του πεπτικού αδένα (Morton 1983). 8
16 ΕΙΣΑΓΩΓΗ Πεπτικός αδένας Ο πεπτικός αδένας των δίθυρων μαλακίων αποτελείται από πολυάριθμους τυφλούς σωλήνες, τα πεπτικά σωληνάρια, τα οποία επικοινωνούν με το στομάχι διαμέσου ενός συστήματος διακλαδισμένων βλεφαριδοφόρων αγωγών (Nakazima 1956). Οι κύριοι αγωγοί διακλαδίζονται και σχηματίζουν δευτερεύοντες αγωγούς (Owen 1955), που αποτελούνται από απλό επιθήλιο με μικρολάχνες. Οι δευτερεύοντες αγωγοί διακλαδίζονται ακόμα περισσότερο και σχηματίζουν τα πεπτικά σωληνάρια. Οι αγωγοί είναι υπεύθυνοι για τη μεταφορά υλικών από και προς τα πεπτικά σωληνάρια (Owen 1972, Morton 1983), για το μεταβολισμό των λιπιδίων (Owen 1973), καθώς και για την απορρόφηση θρεπτικών συστατικών (Morton 1983). Τα πεπτικά σωληνάρια των δίθυρων μαλακίων διαθέτουν επιθήλιο που συνίσταται από δύο τουλάχιστον κυτταρικούς τύπους, τα πεπτικά και τα βασεόφιλα κύτταρα (Εικ. 1). Τα πεπτικά κύτταρα είναι ο πιο συχνός κυτταρικός τύπος στο επιθήλιο των πεπτικών σωληναρίων. Τα κύτταρα αυτά είναι κιονοειδή και διαθέτουν ένα καλά ανεπτυγμένο λυσοσωμικό σύστημα (De Duve & Wattiaux, 1966) στο οποίο πραγματοποιείται η ενδοκυτταρική πέψη των θρεπτικών μορίων (Owen 1972, Morton 1983). Στο εσωτερικό των πεπτικών κυττάρων εντοπίζονται αρκετά καλυμμένα και μη-καλυμμένα κυστίδια, ενδοσώματα με ακανόνιστο σχήμα, μεγάλα ετερολυσοσώματα και υπολειπόμενα σωμάτια που χαρακτηρίζονται από μορφολογική ετερογένεια (Cajaraville et al. 1995b). Τα ετερολυσοσώματα έχουν πολύ μεγάλο μέγεθος με διάμετρο που μπορεί να φθάνει τα 10 nm και εντοπίζονται κυρίως στο κορυφαίο ή κεντρικό τμήμα των πεπτικών κυττάρων. Κάποια από τα οργανίδια αυτής της κατηγορίας περιέχουν λεπτοκοκκώδες υλικό μέτριας ηλεκτρονικής πυκνότητας, ενώ ορισμένα εμφανίζουν μάζες ηλεκτρονικά πυκνού υλικού στο εσωτερικό τους. Τα υπολειπόμενα σωμάτια είναι μικρότερα λυσοσώματα με διάμετρο που φθάνει έως τα 3 nm. Εσωτερικά της μεμβράνης τους διακρίνεται ένα φωτεινό «στεφάνι», ενώ στο εσωτερικό τους εντοπίζεται μια μήτρα με κοκκώδες υλικό υψηλής ηλεκτρονικής πυκνότητας, που σχηματίζει σφαιρικές δομές στο κέντρο ή στην περιφέρεια του οργανιδίου. Τα ετερολυσοσώματα και τα υπολειπόμενα σωμάτια που συμμετέχουν στην ενδοκυτταρική πέψη θρεπτικών μορίων μπορούν να εντοπισθούν σε διαφορετικά τμήματα του κυττάρου ανάλογα με το στάδιο της πεπτικής διεργασίας (Owen 1972, Morton 1983, Cajaraville et al. 1995b). Τα βασεόφιλα κύτταρα του πεπτικού αδένα έχουν πυραμιδοειδή μορφή και διαθέτουν ένα καλά ανεπτυγμένο αδρό ενδοπλασματικό δίκτυο και πολυάριθμα 9

.")
17 ΕΙΣΑΓΩΓΗ μιτοχόνδρια (Pugh 1963). Στο κορυφαίο τμήμα του κυτταροπλάσματός τους εμφανίζονται συμπλέγματα Golgi, κυπελοειδούς μορφής, τα οποία παράγουν κυστίδια που εντοπίζονται στην κορυφή των κυττάρων και συνιστούν πιθανότατα εκκριτικά κοκκία (Summer 1966, McQuiston 1969, Owen 1972, Owen 1973). Τα βασεόφιλα κύτταρα είναι λοιπόν υπεύθυνα για τη σύνθεση και την έκκριση πρωτεϊνών ενζυμικής φύσεως, που είναι απαραίτητες για την εξωκυτταρική πέψη των τροφών (Owen 1972). Σε ορισμένα δίθυρα εντοπίζεται κι ένας άλλος τύπος βασεόφιλων κυττάρων, τα βλεφαριδοφόρα βασεόφιλα κύτταρα. Ο τύπος αυτός χαρακτηρίζεται από πολυάριθμα ελεύθερα ριβοσώματα, αλλά δεν εμφανίζει ανεπτυγμένο αδρό ενδοπλασματικό δίκτυο και εκκριτικά κοκκία (Owen 1970, Pal et al. 1990, Weinstein 1995). Τα κύτταρα αυτά πιθανότατα ευθύνονται για την αναγέννηση των πεπτικών κυττάρων (Owen 1970). Σύμφωνα όμως με μια δεύτερη άποψη, υπάρχει μόνο ένας τύπος βασεόφιλων τα οποία μπορεί να εμφανίζονται με διαφορετικά χαρακτηριστικά (είτε ως πυραμιδοειδή, είτε ως βλεφαριδοφόρα), ανάλογα με τη φάση της πεπτικής διεργασίας στην οποία βρίσκονται τα πεπτικά σωληνάρια (Owen 1970). m en hp m sv hl ger Gb mi rb p li p α β Εικόνα 1. Σχηματική αναπαράσταση των κύριων κυτταρικών τύπων που απαντώνται στο επιθήλιο των πεπτικών σωληναρίων των δίθυρων μαλακίων, δηλαδή των πεπτικών κυττάρων (α) και των βασεόφιλων κυττάρων (β). rb: υπολειπόμενo σωμάτιo, en: ενδόσωμα, Gb: συσκευή Golgi, ger: αδρό ενδοπλασματικό δίκτυο, hl: ετερολυσόσωμα, hp: ετεροφαγόσωμα, li: λιπίδιο, m: μικρολάχνες, mi: μιτοχόνδριο, p: υπεροξειδιόσωμα, sv: εκκριτικό κοκκίο (από Owen 1973). Figure 1. Schematic view of the main cell types in the epithelium of the digestive tubules, the digestive cell (α) and the basophilic cell (β). rb: residual body, en: endosome, Gb: Golgi complex, ger: granular endoplasmic reticulum, hl: heterolysosome, hp: heterophagosome, li: lipid, m: microvilli, mi: mitochondrion, p: peroxisome, sv: secretory vesicle (from Owen 1973). 10
.")
18 ΕΙΣΑΓΩΓΗ Τα σωληνάρια (ή κυψελίδες) του πεπτικού αδένα των δίθυρων μαλακίων υφίστανται κυκλικές μεταβολές σε φυσιολογικές συνθήκες, ανάλογα με το στάδιο στο οποίο βρίσκονται οι πεπτικές διεργασίες στα πεπτικά κύτταρα (Εικ. 2). Τα στάδια δραστηριότητας των πεπτικών σωληναρίων είναι: α) η φάση ηρεμίας, β) η φάση απορρόφησης, κατά την οποία τα θρεπτικά μόρια εισέρχονται στα πεπτικά κύτταρα με συνέπεια να αυξάνεται το ύψος των κυττάρων αυτών, γ) η φάση κατακερματισμού, κατά την οποία έχει ολοκληρωθεί η ενδοκυτταρική πέψη και συμβαίνει αποδιοργάνωση των πεπτικών κυττάρων και αποβολή του κορυφαίου τμήματος του κυτταροπλάσματός τους και δ), η φάση ανασχηματισμού κατά την οποία τα πεπτικά κύτταρα ανακτούν το ύψος που είχαν στη φάση ηρεμίας (Owen 1966,1972, Langton 1975). Εικόνα 2. Διαφορετική μορφολογία του πεπτικού επιθηλίου κατά τη διάρκεια της πεπτικής διαδικασίας. Α) φάση ηρεμίας Β) φάση απορρόφησης C) φάση κατακερματισμού D) φάση ανασχηματισμού (από Langton 1975) 1300x Figure 2. Different morphology of the digestive epithelium through the digestion process A) holding phase B) absorpting phase C) disintegrating phase D) reconstituting phase (from Langton 1975). 1300x 11
19 ΕΙΣΑΓΩΓΗ Σύμφωνα με τα παραπάνω, ο κύκλος των πεπτικών σωληναρίων και η ανανέωση των κυττάρων θα έπρεπε να συμβαίνει συγχρονισμένα με αποτέλεσμα να παρατηρείται μαζική αντικατάσταση κυττάρων μέσα σε λίγες ώρες. Ωστόσο, δεν έχει παρατηρηθεί μαζική διαίρεση επιθηλιακών κυττάρων στο πεπτικό επιθήλιο (Zaldibar 2006). H τρισδιάστατη αναπαράσταση του πεπτικού αδένα μυδιών M. galloprovincialis οδήγησε στο συμπέρασμα ότι ο πεπτικός αδένας δεν αποτελείται από απλά πεπτικά σωληνάρια, αλλά από σύνθετες πεπτικές κυψελίδες, όπου οι φάσεις της πεπτικής διαδικασίας ακολουθούν ένα κυματοειδές πρότυπο από τις πιο απομακρυσμένες στις πιο κεντρικές περιοχές των κυψελίδων (Marigomez et al. 1995a,b). Συγκεκριμένα, η φάση που χαρακτηρίσθηκε προηγουμένως ως φάση ηρεμίας εμφανίζεται κυρίως σε απομακρυσμένες ζώνες, οι φάσεις απορρόφησης και κατακερματισμού εναλλάσσονται καθώς μεταβαίνουμε προς την κεντρική περιοχή, ενώ η φάση ανασχηματισμού είναι κυρίαρχη κοντά στον αγωγό των κυψελίδων (Quincoces 1995). Επιπρόσθετα, η βιολογική σημασία των φάσεων της ενδοκυτταρικής πέψης έχει επαναπροσδιορισθεί με βάση μορφομετρικές μετρήσεις και ιστοχημικά δεδομένα (Quincoces 1995, Marigomez 2007). Σύμφωνα με τη νέα προσέγγιση, η φάση ανασχηματισμού αντιστοιχεί σε μια φάση ηρεμίας κατά την οποία μερικά λυσοσώματα εμφανίζονται στα πεπτικά κύτταρα. Η κατάσταση αυτή είναι ιδιαίτερα συχνή σε μύδια που εκτίθενται στον αέρα για μεγάλο χρονικό διάστημα (Cajaraville et al. 1991, 1992). Η φάση ηρεμίας αντιστοιχεί σε ένα τελικό στάδιο της φάσης απορρόφησης ή σε ένα προχωρημένο στάδιο ενδοκυτταρικής πέψης (Marigomez 2007). Η νέα αυτή άποψη δεν λαμβάνει υπόψη την αναγέννηση των πεπτικών κυττάρων, η οποία δεν φαίνεται να συμβαίνει μαζικώς σε κάθε παλιρροιακό ή πεπτικό κύκλο (Zaldibar 2006, Zaldibar et al. 2007). Σε κάθε περίπτωση, δύο είναι τα είδη πέψης που έχουν περιγραφεί, ανάλογα με τη διαφορετική μορφολογία του επιθηλίου των πεπτικών κυψελίδων, καθώς εξελίσσεται η διαδικασία της πέψης. Στη μονοφασική πέψη, ένας τύπος σωληναρίων είναι κυρίαρχος κάθε στιγμή κατά τη διάρκεια της πέψης. Σε κάθε παλιρροιακό κύκλο, σωληνάρια που βρίσκονται στη φάση ηρεμίας περνούνε στη φάση απορρόφησης και έπειτα στη φάση κατακερματισμού, όπως έχει περιγραφεί στα είδη Dreissena polymorpha, Cardium edule, Ostrea edulis και Scrobicularia plana (Morton 1969, 1970, 1971, Owen 1972). Στη διφασική πέψη, δύο τύποι σωληναρίων κυριαρχούν κάθε στιγμή της πεπτικής διαδικασίας. Η αναλογία των σωληναρίων που βρίσκονται στη φάση κατακερματισμού είναι σταθερή, ενώ τα σωληνάρια που 12
20 ΕΙΣΑΓΩΓΗ βρίσκονται στη φάση ηρεμίας εναλλάσσονται με αυτά που βρίσκονται σε φάση απορρόφησης κατά τη διάρκεια του παλιρροιακού κύκλου (McQuiston 1969), όπως φαίνεται να συμβαίνει στα είδη Lasaea rubra, Cardium edule και Ostrea edulis (McQuiston 1969, Owen 1972, Robinson & Langton 1980, Robinson 1983). Ωστόσο, η αναλογία στην οποία εντοπίζονται οι διάφορες φάσεις των πεπτικών σωληναρίων μπορεί επίσης να διαφέρει ανάλογα με την εποχή (Usheva & Frolova 2006). Συνοψίζοντας, ο ρόλος του πεπτικού αδένα είναι η απορρόφηση των θρεπτικών μορίων που προήλθαν από την εξωκυτταρική πέψη των τροφών και οι οποίες εισέρχονται κυρίως με ενδοκύτωση στα πεπτικά κύτταρα, καθώς και η απέκκριση των άχρηστων προϊόντων της ενδοκυτταρικής πέψης. Επίσης, ο πεπτικός αδένας είναι υπεύθυνος για τη σύνθεση ενζύμων που χρησιμοποιούνται στην εξωκυτταρική πέψη (Yonge 1926, Pugh 1963, Owen 1966, 1972, Thomson et al. 1974, Dimitriadis et al. 2004). Εκτός όμως από τη συμμετοχή του στη διαδικασία της πέψης, το όργανο αυτό συγκεντρώνει ρύπους ποικίλης φύσεως και συμβάλλει ενεργά στις διαδικασίες αποτοξικοποίησής τους. Για τον παραπάνω λόγο αποτελεί καίριο όργανο στις έρευνες βιοπαρακολούθησης για την εκτίμηση της περιβαλλοντικής ρύπανσης. (Marigomez et al. 2002, Moore & Allen 2002). γ) Αιμοκύτταρα Τα αιμοκύτταρα του μυδιού Mytilus galloprovincialis διακρίνονται μορφολογικά σε δύο διαφορετικούς τύπους, τα κοκκώδη κύτταρα και τα υαλώδη κύτταρα (Εικ. 3). Τα κοκκώδη κύτταρα έχουν αυξημένο μέγεθος και σχηματίζουν μεγάλου μεγέθους αλλά και μικρότερα, επιμήκη ψευδοπόδια. Χαρακτηρίζονται από αυξημένο αριθμό κυτταροπλασματικών κοκκίων με ποικίλο μέγεθος και μεγάλη ηλεκτρονική πυκνότητα, τα οποία καταλαμβάνουν το μεγαλύτερο όγκο του κυττάρου. Μερικά από τα κοκκία αυτά είναι πιθανότατα λυσοσώματα, εφόσον αντιδρούν θετικά στην ανίχνευση λυσοσωμικών ενζύμων (Huffman & Tripp 1982, Moore & Gelder 1985, Pipe 1990, Pal & Cajaraville 1994). Επίσης, περιέχουν καλυμμένα κυστίδια μικρότερης ηλεκτρονικής πυκνότητας, ενδοκυτικά και φαγοκυτικά κυστίδια, πολυκυστιδιακά σωμάτια, πολλά υπεροξειδιοσώματα και μιτοχόνδρια. Το ενδοπλασματικό δίκτυο και το σύμπλεγμα Golgi δεν είναι ιδιαίτερα ανεπτυγμένα. Στα ώριμα κύτταρα, που είναι γεμάτα από κοκκία, ο πυρήνας είναι σφαιρικός, αλλά στα μικρότερα ανώριμα κύτταρα ο πυρήνας μπορεί να είναι επιμήκης. Χαρακτηρίζονται 13

21 ΕΙΣΑΓΩΓΗ από μια εξωτερική περιφερειακή ζώνη στο κυτταρόπλασμα, που φαίνεται να είναι απαλλαγμένη από οργανίδια. Τέλος, στα κύτταρα αυτά εντοπίζονται ορισμένα λιπιδιακά υλικά και ποικίλη ποσότητα γλυκογόνου. α β Εικόνα 3. Φωτογραφίες ηλεκτρονικού μικροσκοπίου που παρουσιάζουν τη γενική μορφολογία των αιμοκυττάρων του μυδιού M. galloprovincialis. α) Ώριμο κοκκώδες κύτταρο που εμφανίζει πολλά κυτταροπλασματικά κοκκία μεγάλης ηλεκτρονικής πυκνότητας (G), ποικίλου μεγέθους και σχήματος. Ο πυρήνας (Ν) έχει σφαιρικό σχήμα και είναι περιφερειακά τοποθετημένος. Διακρίνεται μια περιφερειακή ζώνη στο κυτταρόπλασμα (EC) χωρίς οργανίδια, που εμφανίζει ποσότητα γλυκογόνου (GL). Ενδοσωμικά κυστίδια χαμηλής ηλεκτρονικής πυκνότητας (Ε) εντοπίζονται επίσης περιφερειακά. Ράβδος: 1 μm (από Cajaraville & Pal 1995) β) Υαλώδες κύτταρο που παρουσιάζει μεγάλο πυρήνα (Ν), εμφανή πυρηνίσκο (ΝL) και λίγα μικρά λυσοσώματα (L), περιφερειακά τοποθετημένα ενδοσωμικά κυστίδια (Ε) και αρκετά μιτοχόνδρια (Μ). Ράβδος: 0.5 μm (από Cajaraville & Pal 1995) Figure 3. Low power electron micrographs displaying the general morphology of haemocytes of mussels M. galloprovincialis. α) Mature granulocyte showing high numbers of cytoplasmic electron-dense specific granules (G) of various size and shapes. The nucleus (Ν) is spherical and eccentrically placed. The organelle-free peripheral ectoplasm (EC) contains several electron lucent glycogen fields (GL). Electron lucent endosomal vesicles (E) are also placed cortically. Bar: 1μm (from Cajaraville & Pal 1995) β) Hyalinocyte illustrating various ultrastructural features including the large nucleus (N) with a prominent eccentric nucleolus (NL), few small lysosomes (L), cortically placed endocytic vesicles (E) and numerous mitochondria (M). Bar: 0.5 μm (from Cajaraville & Pal 1995). Τα μη κοκκώδη, υαλώδη αιμοκύτταρα χαρακτηρίζονται από αρκετά λεπτά και μακριά ψευδοπόδια, μεγάλο πυρήνα με μάζες πυκνής χρωματίνης, περιορισμένο κυτταρόπλασμα, καλά σχηματισμένο σύμπλεγμα Golgi, ελάχιστα λυσοσώματα, 14
22 ΕΙΣΑΓΩΓΗ αρκετά μιτοχόνδρια, σημαντική ποσότητα αδρού και λείου ενδοπλασματικού δικτύου και άφθονα ριβοσώματα. Τα κύτταρα αυτά περιέχουν ελάχιστα ή καθόλου κοκκία κι έχουν θεωρηθεί είτε πρόδρομες μορφές των κοκκωδών κυττάρων (Mix 1976) είτε διαφορετικός τύπος ώριμων κυττάρων που ανήκουν σε μια ξεχωριστή κυτταρική σειρά. (Cheng 1981, 1984, Auffret 1989). Τα αιμοκύτταρα των δίθυρων μαλακίων συμμετέχουν σε ποικίλες φυσιολογικές λειτουργίες όπως η επούλωση τραυμάτων, ο σχηματισμός και επιδιόρθωση του κελύφους, η πέψη θρεπτικών συστατικών, η μεταφορά και η απέκκριση των τελικών προϊόντων της πέψης (Cheng 1981, 1984, Bayne 1990, Cajaraville & Pal 1995). Ιδιαίτερα σημαντικός όμως είναι ο ρόλος των αιμοκυττάρων στην εσωτερική άμυνα. Η φαγοκυττάρωση συνιστά τον κύριο αμυντικό μηχανισμό κατά των άγνωστων ουσιών ή μικροοργανισμών που εισβάλλουν στον οργανισμό (Bayne 1990, Sima & Vetvicka 1993), ενώ ταυτόχρονα συμμετέχει και στην εξουδετέρωση κατεστραμμένων ή νεκρωτικών κυττάρων (Bubel et al. 1977, Cheng 1981, Rodrick & Ulrich 1984, Le Gall et al. 1988, Bayne 1990, Sima & Vetvicka 1993). Συγκεκριμένα, τα κοκκώδη αιμοκύτταρα φαίνεται να αποτελούν τον κύριο φαγοκυτικό τύπο κυττάρων στα δίθυρα μαλάκια (Foley & Cheng 1975, Rodrick & Ulrich 1984, Cajaraville & Pal 1995). Επίσης, τα κοκκώδη κύτταρα είναι η κύρια κατηγορία κυττάρων που σχετίζεται με την παραγωγή ελεύθερων ριζών οξυγόνου, καθώς και αντιμικροβιακών ουσιών που βοηθούν στην εξουδετέρωση των τοξικών εισβολέων (Cheng 1992, Pipe 1992, Cajaraville et al. 1994, Bachere et al. 1995, Oliver & Fisher 1999). Στα υαλώδη κύτταρα δεν έχει αποδοθεί συγκεκριμένη λειτουργία, παρόλο που πρόσφατα έχει συσχετισθεί η δράση τους με την αιμόσταση και υπάρχουν στοιχεία για τη συμμετοχή τους στη διαδικασία επούλωσης τραυμάτων (Suzuki et al. 1991, Suzuki & Funakoshi 1992). Τα αιμοκύτταρα δέχονται την άμεση επίδραση περιβαλλοντικών ρύπων λόγω του φυσιολογικού ρόλου τους στη μεταφορά ενδογενών και εξωγενών παραγόντων και της συμμετοχής τους στη διαδικασία της άμυνας (Mersh et al. 1996). Επίσης, η εύκολη απομόνωσή τους από το σώμα των μυδιών ελαχιστοποιεί τον κίνδυνο καταστροφής τους κατά τη χρησιμοποίησή τους στην εφαρμογή βιομαρτύρων, καθιστώντας τα πολύ εύχρηστο βιολογικό υλικό σε μελέτες βιοπαρακολούθησης. 15
23 ΕΙΣΑΓΩΓΗ δ) Συσσώρευση βαρέων μετάλλων και οργανικών ρύπων σε ιστούς του μυδιού Mytilus galloprovincialis Τα μέταλλα είναι μια μεγάλη ομάδα χημικών στοιχείων που ενώ υπάρχουν στη φύση, συνήθως θεωρούνται ρύποι. Τα παραδείγματα παραγωγής ρύπων από φυσικές διεργασίες είναι περιορισμένα. Αντίθετα, τα μέταλλα μετατρέπονται σε ρύπους κυρίως από ανθρωπογενείς δραστηριότητες, όπως κατά την εξόρυξη και την επακόλουθη απελευθέρωσή τους από τα πετρώματα στα οποία είχαν εναποτεθεί κατά τη διάρκεια της ηφαιστειακής δράσης, με αποτέλεσμα την οξείδωσή τους και τη μετατροπή τους σε μορφές που μπορεί να προκαλέσουν βιολογικές βλάβες (Δημητριάδης κ.ά. 2006). Ο όρος «βαρέα μέταλλα» έχει χρησιμοποιηθεί στο παρελθόν για να περιγράψει μέταλλα που αποτελούν περιβαλλοντικούς ρύπους. Παλαιότερα ως «βαρύ» χαρακτηριζόταν κάθε μέταλλο που είχε σχετική πυκνότητα ως προς το νερό μεγαλύτερη του πέντε. Τα τελευταία χρόνια, ωστόσο, η κατηγοριοποίηση των βαρέων μετάλλων δεν γίνεται με βάση την πυκνότητά τους, αλλά κυρίως λαμβάνοντας υπόψη τις χημικές τους ιδιότητες (Δημητριάδης κ.ά. 2006). Τα μέταλλα διακρίνονται σε δύο κατηγορίες ανάλογα με τη βιολογική τους σημασία, σε απαραίτητα και μη απαραίτητα. Τα απαραίτητα μέταλλα συμμετέχουν στην ενεργοποίηση μεταλλοενζύμων και πρωτεϊνών στρες, βοηθούν στη μεταφορά οξυγόνου, ενώ σημαντικός είναι ο ρόλος τους σε αναγωγικές και πολλές άλλες βιολογικές διαδικασίες. Παρά τη μεγάλη σημασία τους στη φυσιολογική λειτουργία των κυττάρων, μικρές μόνο συγκεντρώσεις τους απαιτούνται από τους ζωντανούς οργανισμούς, οι οποίοι εμφανίζουν ειδικούς μηχανισμούς για την πρόσληψη και μεταφορά των στοιχείων αυτών στους ιστούς. Τα μέταλλα της κατηγορίας αυτής μπορεί να εμφανίσουν τοξική δράση στην περίπτωση που η συγκέντρωσή τους ξεπεράσει κάποιο όριο (Nicaise et al. 1989, Nott 1991). Τα μη απαραίτητα μέταλλα δεν έχουν κάποιο γνωστό βιολογικό ρόλο και εμφανίζουν μεγάλη τοξικότητα, ιδιαίτερα όταν η συγκέντρωσή τους στους ιστούς υπερβεί μια οριακή τιμή (Rainbow 1993). Συγκεκριμένα, μπορούν να επιφέρουν σειρά δυσμενών επιδράσεων στους ζωντανούς οργανισμούς, όπως νευροφυσιολογικές διαταραχές, γενετικές αλλοιώσεις των κυττάρων (μεταλλάξεις), επιδράσεις στην ενζυμική και ορμονική δραστηριότητα, προβλήματα στην αναπαραγωγή, τερατογενέσεις και καρκινογενέσεις. Μάλιστα, λόγω συναγωνισμού με τα 16
24 ΕΙΣΑΓΩΓΗ απαραίτητα, μπορούν να καταλάβουν τις θέσεις σύνδεσης των τελευταίων σε ειδικά μόρια και δομές, με συνέπεια την εμφάνιση ανωμαλιών στο μεταβολισμό (Hartwig 1998). Γενικότερα, η τοξικότητα των μετάλλων αποτελεί συνδυασμό ποικίλων παραγόντων όπως η συγκέντρωσή τους, το είδος του μετάλλου, η παρουσία και η συνεργατική δράση άλλων μετάλλων καθώς και το είδος του οργανισμού. Αυξημένες συγκεντρώσεις μετάλλων εμφανίζονται σε νερά και ιζήματα παράκτιων και εκβολικών περιοχών, κυρίως σε περιοχές όπως είναι τα δέλτα ποταμών και οι χώροι απελευθέρωσης βιομηχανικών αποβλήτων. Η εισαγωγή των μετάλλων σε ιστούς μαλακίων μπορεί να πραγματοποιηθεί παθητικά, με διάχυση από την επιφάνεια του σώματος ή μέσω της τροφής (Carpene & George 1981, Depledge & Rainbow, 1990). Ορισμένα μέταλλα, όπως ο χαλκός και ο ψευδάργυρος εισάγονται στο κύτταρο μέσω πρωτεϊνών της πλασματικής μεμβράνης, με τη διαδικασία της υποβοηθούμενης διάχυσης (Simkiss & Taylor 1989, Viarengo 1989), ενώ μεγαλύτερα ιόντα χρησιμοποιούν διαμεμβρανικές πρωτεΐνες ενεργητικής μεταφοράς για την είσοδό τους στα κύτταρα (Rainbow & Dallinger 1993). Τέλος η πρόσληψη των μετάλλων μπορεί να πραγματοποιηθεί με ενδοκύτωση και πινοκύτωση οργανομεταλλικών συμπλόκων (Depledge & Rainbow 1990, Endo et al. 1998, 2000). Οι ιστοί των θαλάσσιων μαλακίων αποτελούν σημεία συσσώρευσης πολλών διαφορετικών μετάλλων (Goldberg 1980, Philips 1980, Goldberg 1986, Rainbow 1992, 1993). Η συσσώρευση μεταλλικών ιόντων στους ιστούς των μυδιών δεν επηρεάζεται μόνο από τη ρύπανση, αλλά φαίνεται να εξαρτάται και από ποικίλους φυσικούς παράγοντες όπως το μέγεθος των ατόμων, η ηλικία, το φύλο, ο ρυθμός αύξησης, η αναπαραγωγική δραστηριότητα, η εποχικότητα, το ύψος της υδάτινης στήλης, η αλατότητα, η ταυτόχρονη παρουσία ποικίλων ρύπων, ο ρυθμός έκκρισης των ρύπων από τα διάφορα όργανα (Boyden 1977, Cossa et al. 1980, Cossa, 1989, Depledge & Rainbow 1990). Ιστοί όπως ο πεπτικός αδένας, τα βράγχια και τα νεφρίδια αποτελούν τις κύριες θέσεις συσσώρευσης και αποτοξικοποίησης για τα περισσότερα μέταλλα στα θαλάσσια μαλάκια (Bryan 1979, Amiard et al. 1986, Cossa 1989, Truchet et al. 1990, Regoli & Orlando 1994, Bigas et al. 1997). Με τον τρόπο αυτό επιτυγχάνεται η διατήρηση χαμηλών τιμών ρύπων στους υπόλοιπους ιστούς του οργανισμού. Τα κύτταρα για να αντιμετωπίσουν τις τοξικές επιπτώσεις των μετάλλων έχουν αναπτύξει διάφορους προστατευτικούς μηχανισμούς, όπως η συσσώρευσή τους σε μη τοξικές μορφές, είτε με τη σύνδεσή τους με οργανικές χειλικές ενώσεις, είτε με 17
25 ΕΙΣΑΓΩΓΗ σχηματισμό συμπλόκων με ανόργανα υλικά σε μορφή ιζημάτων (Simkiss & Taylor 1989, Viarengo 1989, Depledge & Rainbow 1990). Οι περισσότεροι οργανικοί ρύποι είναι λιπόφιλες ενώσεις (Cossarini-Dunier et al. 1987) που παρουσιάζουν υψηλό ρυθμό συσσώρευσης στους ιστούς των ζωντανών οργανισμών (La Rocca et al. 1991). Εμφανίστηκαν στο φυσικό περιβάλλον ως αποτέλεσμα της βιομηχανικής ανάπτυξης κατά τις τελευταίες μόνο δεκαετίες. Το σύντομο διάστημα παρουσίας τους στη φύση δεν επέτρεψε την ανάπτυξη ειδικών προστατευτικών μηχανισμών για την εξουδετέρωση των τοξικών αποτελεσμάτων τους. Η αδυναμία βιοδιάσπασης και απομάκρυνσής τους από τους ιστούς, σε συνδυασμό με τη μικρή υδατοδιαλυτότητα που εμφανίζουν, συνιστούν κύριους παράγοντες που αυξάνουν την τοξικότητά τους στους οργανισμούς. Τα άτομα του είδους M. galloprovincialis παρουσιάζουν αυξημένη δυνατότητα συσσώρευσης οργανικών ρύπων στους ιστούς τους (Shugart et al. 1992, Stien et al. 1998, Cajaraville et al. 2000, Viarengo et al. 2000) με συνέπεια να αποτελούν αντικείμενο πολλών περιβαλλοντικών μελετών (Contardi et al. 1979, Kilikidis et al. 1981, Satsmadjis & Gabrielides 1983, Satsmadjis & Voutsinou 1983, Picer & Picer 1994, Granby & Spliid 1995). ε) Γενετική και μοριακή σκοπιά του στρες Η απόκριση των μαλακίων σε διάφορους χημικούς παράγοντες (βαρέα μέταλλα, ραδιενεργά απόβλητα κ.α.) και στο θερμικό πλήγμα αποτέλεσε αντικείμενο πολλών μελετών (Porte et al. 2001, Piano et al. 2004, Singer et al. 2005, Farcy et al. 2007, Gao et al. 2008). Οι πρωτεϊνες θερμικού πλήγματος προστατεύουν τα κύτταρα από τις πιθανές επιβλαβείς συνέπειες παραγόντων στρες. Η επαγωγή αυτών των πρωτεϊνών μπορεί να προκληθεί από την αύξηση της θερμοκρασίας, το ψυχρό πλήγμα, την υποξία, την αλατότητα, τα ιόντα βαρέων μετάλλων, οργανικές ουσίες, ιώσεις, μικροβιακές μολύνσεις, δηλητηριάσεις, καρκίνο, αναπτυξιακούς παράγοντες κ.α. (Feder & Hofmann 1999). Σε μοριακό επίπεδο έχουν περιγραφεί μέλη της οικογένειας γονιδίων HSP70 για αρκετά μαλάκια (Piano et al. 2004, Farcy et al. 2007). Για το είδος Μ. galloprovincialis, έχουν απομονωθεί τόσο θερμοεπαγόμενα όσο και συστατικά εκφραζόμενα γονίδια (Kourtidis & Scouras 2005, Kourtidis et al. 2006α,β). Έχει προταθεί ότι οι πρωτεϊνες θερμικού πλήγματος θα μπορούσαν να 18
26 ΕΙΣΑΓΩΓΗ χρησιμοποιηθούν ως βιομάρτυρες (Sole 2000, Hamer et al. 2004), εκφράζοντας το βαθμό του κυτταρικού στρες των θαλάσσιων οργανισμών, ακόμα και να παρέχουν πληροφορίες για τη φύση του παράγοντα στρες (Sanders 1988). Β. ΠΕΡΙΓΡΑΦΗ ΤΗΣ ΠΕΡΙΟΧΗΣ ΜΕΛΕΤΗΣ. ΘΕΡΜΑΪΚΟΣ ΚΑΙ ΣΤΡΥΜΟΝΙΚΟΣ ΚΟΛΠΟΣ Ο Θερμαϊκός κόλπος, που βρίσκεται στο βορειοδυτικό άκρο του Αιγαίου πελάγους, αποτελεί ένα σύνθετο και ενδιαφέρον οικοσύστημα. Είναι σχετικά ρηχός κόλπος και το μέσο βάθος του δεν ξεπερνά τα 50 m (Karageorgis et al. 2005). Αποτελεί τελικό αποδέκτη πολλών ρύπων μια και εδώ καταλήγουν τα αστικά λύματα της Θεσσαλονίκης (πάνω από κατοίκους), τα οποία μάλιστα μέχρι πρόσφατα ελευθερώνονταν στο εσωτερικό τμήμα του κόλπου, χωρίς να υποστούν κάποια προηγούμενη επεξεργασία. Επιπλέον, βιομηχανικά απόβλητα και η λειτουργία του λιμανιού της Θεσσαλονίκης επιβαρύνουν περισσότερο την κατάσταση των νερών του Θερμαϊκού. Ο κόλπος αυτός είναι αποδέκτης των νερών του Αξιού, του Λουδία και του Αλιάκμονα. O Αξιός ποταμός επιβαρύνεται με αστικά λύματα, βιομηχανικά απόβλητα που καταλήγουν ανεπεξέργαστα στον ποταμό, ενώ σε όλο σχεδόν το μήκος του η απόπλυση γεωργικών εκτάσεων έχει ως αποτέλεσμα λιπάσματα και φυτοφάρμακα να επηρεάζουν την ποιότητα του νερού του (Kampa et al. 2000, Chatzinikolaou et al. 2006, Konstantinou et al. 2006, Milovanovic 2007). Ο Λουδίας εμφανίζει αυξημένες συγκεντρώσεις ρύπων (Kontstantinou et al. 2006), ενώ ο Αλιάκμονας παρουσιάζει χαμηλή ποιότητα υδάτων κατάντη της Τάφρου 66 (Lazaridou-Dimitriadou et al. 2000). Σύμφωνα με προηγούμενες μελέτες, αυξημένες τιμές βαρέων μετάλλων ανιχνεύθηκαν σε ιζήματα από το Θερμαϊκό κόλπο, κυρίως στη βιομηχανική περιοχή, στο λιμάνι της Θεσσαλονίκης και την έξοδο του αγωγού των αστικών λυμάτων (Voutsinou-Taliadouri et al. 1998). Οι συγκεντρώσεις βαρέων μετάλλων σε ιστούς μυδιών που συλλέχθηκαν από διάφορες περιοχές του Θερμαϊκού κόλπου είναι αρκετά αυξημένες σε περιοχές της δυτικής πλευράς, κοντά στη βιομηχανική ζώνη της Θεσσαλονίκης, στην περιοχή του λιμανιού, καθώς και στις περιοχές που εκβάλλουν οι μεγάλοι ποταμοί της περιοχής (Αναγνώστου κ.ά. 1997, Voutsinou Taliadouri et al. 19
27 ΕΙΣΑΓΩΓΗ 1998, Catsiki et al. 2001). Αντίθετα με τα μέταλλα, υψηλότερες τιμές πολυκυκλικών αρωματικών υδρογονανθράκων (PAHs) και οργανοχλωριωμένων παραγώγων εντοπίσθηκαν στο ανατολικό τμήμα του Θερμαϊκού κόλπου (περιοχή Περαίας) (Catsiki et al. 2001). Ο Στρυμονικός κόλπος εκτείνεται μεταξύ της χερσονήσου της Χαλκιδικής και της χερσονήσου που σχηματίζει το όρος Παγγαίο καθώς καταλήγει στο Αιγαίο Πέλαγος. Τα ισοβαθή του Στρυμονικού κόλπου αυξάνονται απότομα καθώς απομακρυνόμαστε από την ακτή, φθάνοντας τα 40 m σε απόσταση 2 km από την ακτή, μέχρι ένα μέγιστο βάθος 90 m στο σχετικά επίπεδο κεντρικό τμήμα του κόλπου. Ο ποταμός Στρυμόνας που πηγάζει από τη Βουλγαρία και ο μικρότερος Ρήχιος ποταμός στα δυτικά αποτελούν τις κύριες πηγές γλυκού νερού, θρεπτικών ουσιών και ρύπων (οικιακής και γεωργικής προέλευσης) του Στρυμονικού κόλπου (Stamatis et al. 2002, Kalliantonis et al. 2004). Ο κόλπος αυτός αποτελεί μια από τις πιο σημαντικές περιοχές για τον πολλαπλασιασμό και την αλιεία πελαγικών ειδών στο Βόρειο Αιγαίο (Sylaios et al. 1999). Κατά το παρελθόν στην περιοχή δεν ανιχνεύονταν υψηλά επίπεδα ρύπων (Vasilikiotis et al. 1983), ωστόσο τα τελευταία χρόνια ο κόλπος δέχεται την επίδραση αυξημένης ποσότητας λιπασμάτων και εντομοκτόνων από τις καλλιέργειες γειτονικών εκτάσεων, συγκεντρώνει λύματα από τους γύρω οικισμούς και βιομηχανικά απόβλητα από τις χημικές βιομηχανίες που αναπτύσσονται στην περιοχή (Dassenakis 2000, Lazaridou-Dimitriadou et al. 2004). Παρόλα αυτά, συγκριτικές μελέτες στο νερό, στα ιζήματα και στους ιστούς μυδιών καταδεικνύουν χαμηλότερα επίπεδα ρύπανσης στο Στρυμονικό, σε σχέση με το Θερμαϊκό κόλπο (ΔΕΠΠΠΕΘ 2004). H θαλάσσια περιοχή του Θερμαϊκού κόλπου αποτελεί ιδανικό περιβάλλον για την ανάπτυξη φυσικών πληθυσμών μυδιών M. galloprovincialis (Arsenoudi et al. 2003). Οι ακτές του Θερμαϊκού κόλπου φιλοξενούν τις πιο εκτεταμένες και παραγωγικές μονάδες καλλιέργειας μυδιών στην Ελλάδα. Σύμφωνα με τα στοιχεία του Υπουργείου Γεωργίας (2002) το 70% της συνολικής Ελληνικής παραγωγής προέρχεται από τη συγκεκριμένη περιοχή, ενώ μονάδες μυδοκαλλιέργειας έχουν αναπτυχθεί και στο Στρυμονικό κόλπο. 20
28 ΕΙΣΑΓΩΓΗ Γ. ΒΙΟΜΑΡΤΥΡΕΣ ΠΟΥ ΕΦΑΡΜΟΣΘΗΚΑΝ ΣΤΗ ΣΥΓΚΕΚΡΙΜΕΝΗ ΜΕΛΕΤΗ Οι βιομάρτυρες ρύπανσης που εξετάσθηκαν στη συγκεκριμένη μελέτη αναφέρονται σε διάφορες κυτταρικές και φυσιολογικές αποκρίσεις των μυδιών. Οι βιομάρτυρες αυτοί αποτελούν εργαλεία έγκαιρης προειδοποίησης (early warning tools) για την εκτίμηση της θαλάσσιας ρύπανσης, μια και δίνουν τη δυνατότητα έγκαιρης πρόβλεψης των μακροπρόθεσμων επιπτώσεων της ρύπανσης στο επίπεδο του πληθυσμού ή της βιοκοινότητας. Η σημασία των βιομαρτύρων αυτών είναι πολύ μεγάλη ειδικά για εδώδιμα είδη, με την προϋπόθεση ότι οι βιομάρτυρες δεν χρησιμοποιούνται μεμονωμένα, αλλά εφαρμόζονται πολλοί μαζί και ο καθένας τους παρέχει καινούρια πληροφορία για τις επιπτώσεις της ρύπανσης. 1. Βιομάρτυρες που σχετίζονται με τα λυσοσώματα Τα λυσοσώματα αποτελούν υποκυτταρικές δομές που περιέχουν μεγάλο αριθμό υδρολυτικών ενζύμων, όπως νουκλεάσες, φωσφατάσες και υπεροξειδάσες, και συμμετέχουν στην αποικοδόμηση ποικίλων μακρομορίων ενδογενούς κι εξωγενούς προέλευσης (Klionsky & Emr 2000, Moore et al. 2006b, Robledo et al. 2006, Moore et al. 2007). Εκτός όμως από τον κύριο ρόλο τους, που είναι η ενδοκυτταρική πέψη, τα οργανίδια αυτά συμβάλλουν στη συγκέντρωση και αδρανοποίηση μετάλλων, αλλά και πολλών οργανικών ρύπων (Domouhtsidou & Dimitriadis 2001, Soto et al. 2002, Marigomez & Baybay-Villacorta 2003, Moore et al. 2004, Einsporn et al. 2005). Συγκεκριμένα, τα λυσοσώματα του πεπτικού αδένα των μυδιών αποτελούν την κύρια ενδοκυτταρική θέση συσσώρευσης ρύπων (Moore 1985, 1988, Sarasquete et al. 1992). Για το λόγο αυτό, οι λυσοσωμικές αποκρίσεις στους ρύπους έχουν αξιοποιηθεί σε μεγάλο βαθμό ως κυτταρικοί βιομάρτυρες για τον προσδιορισμό των επιπτώσεων της ρύπανσης και είναι αποδεκτές από διεθνείς οργανισμούς όπως ο WHO (World Health Organization) και ο FAO (Food Agruculture Organization) για την εκτίμηση της ποιότητας του περιβάλλοντος (Group of Experts on the Scientific Aspects of Marine Protection GESAMP 1980). Η διαταραχή στις λυσοσωμικές λειτουργίες προκαλεί πρόβλημα στη διαδικασία της ενδοκυτταρικής πέψης, με συνέπεια σοβαρές διαταραχές στο διατροφικό καθεστώς του κυττάρου αλλά και ολόκληρου του οργανισμού και αποτελεί ένδειξη διαταραγμένης υγείας. Συνοπτικά, οι λυσοσωμικές αποκρίσεις στο περιβαλλοντικό 21
29 ΕΙΣΑΓΩΓΗ στρες διακρίνονται σε τρεις κατηγορίες: μεταβολές στη διαπερατότητα της λυσοσωμικής μεμβράνης, αλλαγές στο μέγεθος των λυσοσωμάτων και διαφοροποιήσεις στο περιεχόμενό τους, όπως είναι η συσσώρευση ακόρεστων ουδέτερων λιπιδίων και λιποφουσκινών (Lowe 1988, Moore 1988, Marigomez & Baybay Villacorta 2003). Οι αλλαγές στα χαρακτηριστικά των λυσοσωμάτων έχουν αξιοποιηθεί ως βιομάρτυρες ρύπανσης σε πολλά είδη θαλασσίων διθύρων, σε μαλάκια του γλυκού νερού ή ακόμα και σε ψάρια με σκοπό να εκτιμηθεί η κυτταρική υγεία (Kohler 1991, Romeo et al. 2000, Alvarado et al. 2005, Broeg et al. 2005, Bilbao et al. 2006, Champeau & Narbonne 2006, Guerlet et al. 2006, Schiedek et al. 2006, Zaldibar et al. 2006, Zorita 2006, Guerlet et al. 2007). Οι λυσοσωμικές αποκρίσεις θαλασσίων διθύρων έχουν χρησιμοποιηθεί σε μελέτες πεδίου ως βιομάρτυρες έκθεσης σε περιβαλλοντικούς ρύπους, ενώ σύμφωνα και με εργαστηριακές μελέτες οι αποκρίσεις των λυσοσωμάτων των πεπτικών κυττάρων των μυδιών αποτελούν ευαίσθητους βιομάρτυρες οργανικής και ανόργανης ρύπανσης (Moore et al. 1982, Moore 1988, Regoli 1992, Krishnakumar et al. 1994, Etxeberria et al. 1995, Marigomez et al. 1996a). α) Η εκτίμηση της διαπερατότητας των λυσοσωμικών μεμβρανών ως βιομάρτυρας ρύπανσης i. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» σε κρυοτομές Η επίδραση της ρύπανσης στη διαπερατότητα της μεμβράνης των λυσοσωμάτων μπορεί να εκτιμηθεί με την εφαρμογή της «σταθερότητας της λυσοσωμικής μεμβράνης» (lysosomal membrane stability / LMS) στα πεπτικά κύτταρα. Η τεχνική αυτή έχει καθιερωθεί ως ο πλέον ευαίσθητος και αξιόπιστος από τους προτεινόμενους βιομάρτυρες για την εκτίμηση της βιολογικής απόκρισης σε μια πληθώρα οργανικών ή ανόργανων ρύπων και συνεπώς θεωρείται κατάλληλη μέθοδος για την εκτίμηση της ποιότητας του περιβάλλοντος (United Nations Environment Programme UNEP 1997, ICES 2004, Dagnino et al. 2007). Η αποσταθεροποίηση της λυσοσωμικής μεμβράνης των πεπτικών κυττάρων συνιστά, επομένως, ένα πρώτης τάξεως μέγεθος που μπορεί να δώσει ποσοτικά αποτελέσματα σε σχέση με την απόκριση των οργανισμών σε ειδικό ή γενικό στρες (Δημητριάδης κ.ά. 2006). 22
30 ΕΙΣΑΓΩΓΗ Ο βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» θεωρείται χρήσιμο και ευαίσθητο εργαλείο για την εκτίμηση των αποτελεσμάτων επιβλαβών παραγόντων που βρίσκονται ακόμα και σε πολύ μικρή συγκέντρωση στο εσωτερικό των κυττάρων (UNEP/ RAMOGE 1999). Αποσταθεροποίηση της λυσοσωμικής μεμβράνης έχει αναφερθεί μετά από έκθεση σε οργανικούς ρύπους (Moore 1988, Krishnakumar et al. 1994), μέταλλα (Viarengo et al. 1981, Harisson & Berger 1982, Regoli 1992) και άλλες πηγές περιβαλλοντικού στρες (Moore 1976, Bayne et al. 1981, Tremblay & Pellerin-Massicotte 1997). Η μέθοδος αυτή έχει χρησιμοποιηθεί σε εργαστηριακές έρευνες (Viarengo et al. 1981, Harrison & Berger 1982, Viarengo et al. 1987), αλλά και σε μελέτες πεδίου (Moore et al. 1982, 1986, 1987, Viarengo et al. 1987, Moore 1988, Regoli 1992, Krishnakumar et al. 1994) και είναι εξίσου αποτελεσματική τόσο σε προγράμματα παθητικής, όσο και ενεργητικής βιοπαρακολούθησης (Viarengo & Canessi 1991, Regoli & Orlando 1993, Stien et al. 1998). Μολονότι η «σταθερότητα της λυσοσωμικής μεμβράνης» συνιστά έναν καθιερωμένο βιομάρτυρα για την εκτίμηση των επιπτώσεων που προκαλούν οι ρύποι, ο συσχετισμός της παραμέτρου αυτής με άλλους βιομάρτυρες ρύπανσης δεν έχει αποσαφηνισθεί πλήρως. Η σύγκριση των αποτελεσμάτων της τεχνικής αυτής με τα αποτελέσματα άλλων βιομαρτύρων ρύπανσης, στο πεδίο και σε εργαστηριακές συνθήκες, θα παράσχει, λοιπόν, χρήσιμες πληροφορίες που θα μπορούσαν να αξιοποιηθούν σε προγράμματα βιοπαρακολούθησης θαλασσίων οικοσυστημάτων. ii. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» σε ζωντανά κύτταρα (κατακράτηση της χρωστικής ουδέτερο ερυθρό) Ο βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (neutral red retention assay/ NRR) συνιστά μια πολλά υποσχόμενη μέθοδο για την καταγραφή αλλαγών στη διαπερατότητα της λυσοσωμικής μεμβράνης των αιμοκυττάρων, που προκαλείται από την επίδραση ρύπων. Ο χρόνος που απαιτείται για να εισχωρήσει η χρωστική μέσα στα λυσοσώματα των αιμοκυττάρων και να τα καταστρέψει χαρακτηρίζει την ποιότητα της θαλάσσιας περιοχής από την οποία έχουν εξαχθεί τα μύδια. Μικρότεροι χρόνοι αφορούν σε μύδια των οποίων οι μεμβράνες των λυσοσωμάτων έχουν αποσταθεροποιηθεί και επιτρέπουν την εύκολη εισχώρηση της χρωστικής στα λυσοσώματα και άρα μαρτυρούν μεγαλύτερη ρύπανση των 23
31 ΕΙΣΑΓΩΓΗ θαλασσίων υδάτων. Αντίστοιχα, μεγαλύτεροι χρόνοι αφορούν σε μύδια που μεγαλώνουν σε καθαρότερες περιοχές (Δημητριάδης κ.ά. 2006). Ο βιομάρτυρας αυτός έχει εφαρμοσθεί στα αιμοκύτταρα μυδιών για την εκτίμηση των επιπτώσεων οργανικών ρύπων (Lowe & Pipe 1994, Fernley et al. 2000) ή βαρέων μετάλλων (Svendsen & Weeks 1995), τόσο σε μελέτες πεδίου (Fernley et al. 2000), όσο και σε εργαστηριακές έρευνες (Lowe et al. 1995b). Ο χρόνος κατακράτησης της χρωστικής στα λυσοσώματα επηρεάζεται κυρίως από την παρουσία ρύπων, ενώ φυσικοί παραγόντες, όπως η θερμοκρασία και η αλατότητα, έχουν μικρότερη επίδραση σε αυτή την παράμετρο (Ringwood et al. 1998). Επομένως, ο υπολογισμός του χρόνου διατήρησης της χρωστικής σε μελέτες πεδίου έχει θεωρηθεί πολύ ευαίσθητος βιομάρτυρας στρες προκαλούμενου κυρίως από ανθρώπινες δραστηριότητες. Η μέθοδος έχει εφαρμοσθεί τόσο σε προγράμματα ενεργητικής, όσο και παθητικής βιοπαρακολούθησης (Ringwood et al. 1988, Lowe et al. 1995a). Ο βιομάρτυρας αυτός έχει δοκιμασθεί επιτυχώς σε αιμοκύτταρα, καθώς και σε πεπτικά κύτταρα θαλασσίων οργανισμών, γεγονός που φανερώνει ότι τα αποτελέσματα της τεχνικής δεν επηρεάζονται από το είδος του κυττάρου, αλλά από τις επιπτώσεις των ρύπων στα λυσοσώματα (Lowe et al. 1995b). Η εφαρμογή της μεθόδου «κατακράτηση της χρωστικής ουδέτερο ερυθρό» στο πεδίο, αλλά και σε εργαστηριακές συνθήκες, θα συμβάλλει στην περαιτέρω αξιολόγηση της αποτελεσματικότητάς της να καταδεικνύει τις επιπτώσεις της ρύπανσης, ενώ η παράλληλη εφαρμογή της «κατακράτησης της χρωστικής ουδέτερο ερυθρό» και άλλων βιομαρτύρων ρύπανσης θα οδηγήσει στην εξαγωγή χρήσιμων συμπερασμάτων σχετικά με την εμφάνιση συσχέτισης ανάμεσά τους. β) Τα δομικά χαρακτηριστικά των λυσοσωμάτων ως βιομάρτυρες θαλάσσιας ρύπανσης Οι αλλαγές στο μέγεθος και στον αριθμό των λυσοσωμάτων έχουν παραδοσιακά αξιοποιηθεί ως διαγνωστικές παράμετροι σε βιοϊατρικές έρευνες και πιο πρόσφατα σε οικοτοξικολογικές μελέτες που χρησιμοποιούν μύδια και ψάρια (Dingle & Fell 1973, Alvarado et al. 2005, Marigomez et al. 2005b, 2006). Διαφορετικές αιτίες περιβαλλοντικού στρες (χημική ρύπανση, μεταβολές στην αλατότητα, αυξημένη θερμοκρασία, υποσιτισμός, αναπαραγωγικό στρες) φαίνεται να προκαλούν διαφοροποιήσεις στο μέγεθος των λυσοσωμάτων των πεπτικών κυττάρων (Lowe et 24
32 ΕΙΣΑΓΩΓΗ al. 1981, Moore et al. 1987, Lowe 1988, Cajaraville et al. 1989, Marigomez et al. 1989, Marigomez et al. 1991, Regoli 1992, Etxeberria et al. 1994, Cajaraville et al. 1995b, Etxeberria et al. 1995, Domouhtsidou & Dimitriadis 2001). Η αλλαγή στις λυσοσωμικές δομικές παραμέτρους έχει εφαρμοσθεί ως βιομάρτυρας ρύπανσης σε πολλά είδη μαλακίων και ψαριών μετά από την επίδραση διαφορετικού είδους στρες (Alvarado et al. 2005, Bilbao et al. 2006, Chanpeau & Narbonne 2006, Guerlet et al. 2006, Marigomez et al. 2006). Η μεταβολή στα δομικά χαρακτηριστικά των λυσοσωμάτων μπορεί να προσδιορισθεί σε επίπεδο ηλεκτρονικού μικροσκοπίου (Domouhtsidou & Dimitriadis 2001), αλλά συνήθως εφαρμόζεται η διαδικασία ανάλυσης εικόνας στο οπτικό μικροσκόπιο, έπειτα από χρώση για λυσοσωμικά ένζυμα (Cajaraville et al. 1991, 1995a, 1995b). Η διόγκωση που προκαλείται στα λυσοσώματα των πεπτικών κυττάρων αποτελεί την πιο κοινή αντίδραση μετά από έκθεση των ατόμων σε περιβαλλοντικούς ρύπους (Lowe et al. 1981, Lowe 1988, Moore 1988, Marigomez et al. 1989, Kohler 1991, Regoli 1992, Etxeberria et al. 1994, Regoli et al. 1998). Ωστόσο, σε περιπτώσεις μακροχρόνιας επίδρασης ρύπων έχει παρατηρηθεί μείωση στο μέγεθος των πεπτικών λυσοσωμάτων (Cajaraville et al. 1995a, Marigomez et al. 1996b). Η έκθεση σε υδρογονάνθρακες κάτω από εργαστηριακές συνθήκες προκαλεί μια σύνθετη απόκριση του ενδολυσοσωμικού συστήματος των πεπτικών κυττάρων μυδιών, αφού τα λυσοσώματα φαίνεται να διέρχονται από διαφορετικές φάσεις: (α) μεταβατική λυσοσωμική διόγκωση (β) μεταβατική μείωση στο μέγεθος των λυσοσωμάτων (γ) λυσοσωμική διόγκωση έπειτα από μακροχρόνια έκθεση (Marigomez & Baybay-Villacorta 2003). Αύξηση στο μέγεθος των πεπτικών λυσοσωμάτων και αποσταθεροποίηση της μεμβράνης τους έχει παρατηρηθεί ως συνέπεια της συγκέντρωσης μετάλλων στα λυσοσώματα κατά την προσπάθεια αποτοξικοποίησης των κυττάρων (Viarengo et al. 1981, Moore 1990, Regoli et al. 1998, Marigomez et al. 2005b). Όσον αφορά στις μεταβολές στις λυσοσωμικές δομικές παραμέτρους, οι μελέτες στο πεδίο δίνουν αντιφατικά αποτελέσματα (Moore 1988, Regoli 1992, Etxeberia et al. 1995, Marigomez et al. 1996b), που έχουν αποδοθεί σε γενετικές αλλαγές, αλληλεπίδραση με περιβαλλοντικούς παράγοντες ή συνεργατικά αποτελέσματα ρύπων που βρίσκονται ταυτόχρονα στο περιβάλλον (Moore 1988, Ward 1990, Marigomez et al. 1996b, Da Ros et al. 2000, 2002, Marigomez et al. 2005a). 25
33 ΕΙΣΑΓΩΓΗ Τα αντικρουόμενα αποτελέσματα σχετικά με την επίδραση των ρύπων στο μέγεθος και τον αριθμό των λυσοσωμάτων (Marigomez et al. 1996b, Marigomez & Baybay-Villacorta 2003, Zorita et al. 2007) καθιστούν επιτακτική την αποσαφήνιση των επιπτώσεων της ρύπανσης στα δομικά χαρακτηριστικά των οργανιδίων αυτών. Η μελέτη των αλλαγών που παρατηρούνται στον αριθμό και το μέγεθος των λυσοσώματων του πεπτικού αδένα των μυδιών λόγω της επίδρασης ρύπων και η συσχέτιση των δομικών αυτών παραμέτρων με τα αποτελέσματα άλλων βιομαρτύρων ρύπανσης θα οδηγήσουν σε χρήσιμα συμπεράσματα, τα οποία θα μπορούσαν να αξιοποιηθούν σε μελέτες βιοπαρακολούθησης. γ) Η συσσώρευση λιπιδιακών υλικών και η συσσώρευση λιποφουσκινών ως βιομάρτυρες θαλάσσιας ρύπανσης Εκτός από τις μεταβολές στη σταθερότητα της λυσοσωμικής μεμβράνης και στα δομικά χαρακτηριστικά των λυσοσωμάτων, και οι αλλαγές στο περιεχόμενο των πεπτικών λυσοσωμάτων συνιστούν αποκρίσεις στη ρύπανση. Η σύσταση του περιεχομένου και το ενζυμικό φορτίο του λυσοσωμικού συστήματος μπορεί να διαφέρει, μια και εξαρτάται από τη φύση και την ποσότητα τόσο των υλικών που πρόκειται να διασπασθούν, όσο και των προϊόντων της διάσπασης. Συσσώρευση λιποφουσκινών, μετάλλων και ουδέτερων λιπιδίων είναι οι καλύτερα παρατηρούμενες αλλαγές στο λυσοσωμικό περιεχόμενο ως αποκρίσεις στην έκθεση σε ρύπους. Για το λόγο αυτό, οι ιστοχημικές τεχνικές προσδιορισμού ουδέτερων λιπιδίων και λιποφουσκινών έχουν αναγνωρισθεί ως ευαίσθητοι δείκτες της κυτταρικής υγείας. Οι λιποφουσκίνες αποτελούν το τελικό προϊόν της υπεροξείδωσης λιπιδίων (Moore 1990, Yin 1996, Terman et al. 1999, Brunk & Terman 2002). Αποτελούνται κυρίως από τριγλυκερίδια, ελεύθερα λιπαρά οξέα, χοληστερόλη και φωσφολιπίδια, ποικίλη ποσότητα αμινοξέων, υδρογονάνθρακες και μπορεί να περιέχουν μέταλλα όπως Fe, Cu, Al, Zn, Ca, Mn. Η ακριβής σύσταση των λιποφουσκινών, όμως, φαίνεται να ποικίλλει ανάμεσα στους διάφορους κυτταρικούς τύπους (Terman & Brunk 2004). Κατά τη διάρκεια της φυσιολογικής λειτουργίας του πεπτικού αδένα, τα πεπτικά κύτταρα των μυδιών εξωκυτώνουν υπολειπόμενα σωμάτια που περιέχουν υπολείμματα της πέψης στη μορφή λιποφουσκινών (Moore 1990, Moore & Willows 26
34 ΕΙΣΑΓΩΓΗ 1998). Ωστόσο, η επίδραση ρύπων προκαλεί συσσώρευση λιποφουσκινών στα πεπτικά κύτταρα (Viarengo et al. 1990, Regoli 1992). Αυξημένη συγκέντρωση λιποφουσκίνης έχει παρατηρηθεί σε πεπτικά κύτταρα μυδιών που εκτέθηκαν σε αρωματικούς υδρογονάνθρακες (PAHs) κάτω από εργαστηριακές συνθήκες, αλλά και σε μελέτες στο πεδίο (Krishnakumar et al. 1994, 1997) καθώς και σε κύτταρα ψαριών που εκτέθηκαν σε οργανικούς ρύπους στο εργαστήριο (Biagianti-Risbourg & Bastide 1995) ή συλλέχθηκαν από περιοχές επιβαρυμένες με μέταλλα και οργανικά (Kohler et al. 1992). Η υπερβολική συσσώρευση λιποφουσκινών μέσα στα υπολειπόμενα σωμάτια του λυσοσωμικού συστήματος έχει θεωρηθεί υπεύθυνη για τη διαταραχή της λειτουργίας του συστήματος αυτού (Moore 1990). Οι λιποφουσκίνες προκαλούν δυσλειτουργία των λυσοσωμάτων, μια και δεσμεύουν τα λυσοσωμικά ένζυμα, περιορίζοντας την ικανότητα διάσπασης των οργανιδίων αυτών (Brunk & Terman 2002, Moore et al. 2006b). Ουδέτερα λιπίδια, λιπαρά οξέα, στεροειδή και προσταγλανδίνες είναι οι κύριες κατηγορίες λιπιδίων που έχουν αναφερθεί ως συστατικά των ιστών μαλακίων (Gosling 1992). Τα ουδέτερα λιπίδια αποτελούν αποθήκες ενέργειας, καταναλώνονται κατά τη γαμετογένεση και συνιστούν την κύρια μορφή αποθηκευμένης ενέργειας των ωοκυττάρων για την κάλυψη των ενεργειακών αναγκών των εμβρύων και των πρώτων προνυμφικών σταδίων (Holland 1978). Οι στεροειδικές ορμόνες συμμετέχουν κατά κύριο λόγο στη ρύθμιση της αύξησης και της αναπαραγωγής, ενώ ο ρόλος των προσταγλανδινών είναι κυρίως η ρύθμιση της απελευθέρωσης γαμετών (Osada et al. 1989). Ο κυριότερος λόγος για τον οποίο γίνεται αναφορά στα λιπίδια κατά την παρούσα έρευνα είναι η συσχέτισή τους με τη ρύπανση, καθώς η συσσώρευση ενδοκυτταρικών λιπιδίων συνιστά άλλο ένα κυτταρικό στοιχείο, που συνδέεται με την παρουσία περιβαλλοντικών ρύπων. Τα συστατικά αυτά συγκεντρώνονται είτε με τη μορφή λιπιδιακών εγκλείστων στο κυτταρόπλασμα, είτε στο εσωτερικό κυτταρικών οργανιδίων, όπως είναι τα λυσοσώματα. Η επίδραση ρύπων προκαλεί την αποσταθεροποίηση του κυτταρικού μηχανισμού παραγωγής και αποθήκευσης των ουδέτερων λιπιδίων. Ως αποτέλεσμα εμφανίζεται αυξημένη παρουσία των λιπιδιακών αυτών συστατικών σε περιοχές επιβαρυμένες από ρύπους. H ενδοκυτταρική συσσώρευση ουδέτερων λιπιδίων φαίνεται να σχετίζεται περισσότερο με οργανική χημική ρύπανση (Wolf et al. 1981, Pipe & Moore 1986, Moore 1988, Lowe & Clarke 27
35 ΕΙΣΑΓΩΓΗ 1989, Cajaraville 1991, Domouhtsidou & Dimitriadis 2001, Marigomez & Baybay- Villacorta 2003) όπως έχει παρατηρηθεί σε εργαστηριακές έρευνες (Lowe & Clarke 1989, Moore 1990, Fahraeus Van-Ree & Spurrell 2000, Marigomez & Baybay Villacorta 2003, Moore et al. 2006a,) και σε μελέτες πεδίου (Kohler et al. 1992, Regoli 1992, Simpson 1992, Domouhtsidou & Dimitriadis 2001, Kohler et al. 2002). Η μορφομετρική ανάλυση των ουδέτερων λιπιδίων στον πεπτικό αδένα μυδιών M. galloprovincialis, έπειτα από την επίδραση οργανικών και ανόργανων ρύπων, θα δώσει πρόσθετες πληροφορίες για τη μεταβολή που προκαλείται στο συγκεκριμένο κυτταρικό συστατικό από διαφορετικά είδη ρύπανσης. Επίσης η ταυτόχρονη εφαρμογή της μορφομετρίας των ουδέτερων λιπιδίων και άλλων βιομαρτύρων ρύπανσης θα καταδείξει την ύπαρξη συσχέτισης ανάμεσά τους, στοιχείο χρήσιμο σε μελέτες βιοπαρακολούθησης. Το αυξημένο ποσό λιπιδίων και λιποφουσκινών έχει θεωρηθεί υπεύθυνο για την πρόκληση μορφολογικών αλλοιώσεων και λειτουργικών διαταραχών στα λυσοσώματα (Dimitriadis et al. 2004, Marigomez & Baybay- Villacorta 2003). Επομένως, η ιστοχημεία λιπιδίων ταυτόχρονα με την ιστοχημεία λυσοσωμικών ενζύμων σε κρυοτομές από τα ίδια πεπτικά σωληνάρια θα δώσει περαιτέρω στοιχεία σχετικά με την αλληλεπίδραση των λιπιδίων και του λυσοσωμικού ενζυμικού περιεχομένου και θα συμβάλλει στην ορθότερη ερμηνεία των αποτελεσμάτων που προκύπτουν από την εφαρμογή ιστοχημικών τεχνικών για την εκτίμηση της ποιότητας του περιβάλλοντος. 2. Ο προσδιορισμός των πυρηνικών ανωμαλιών ως βιομάρτυρας θαλάσσιας ρύπανσης Η παρουσία ρύπων στο εσωτερικό των κυττάρων μπορεί να προκαλέσει αλλοιώσεις στο γενετικό υλικό. Μεταλλαξιγόνες διεργασίες, που αφορούν σε μη κανονικό αναδιπλασιασμό και μεταγραφή του DNA, καθώς και μεταφραστικές διεργασίες του RNA, συχνά αντιπροσωπεύουν την πρώτη απόκριση του πυρηνικού υλικού σε ρυπογόνες ουσίες (Δημητριάδης κ.ά. 2006). Τα βαρέα μέταλλα, σύμφωνα με in vitro μελέτες, επηρεάζουν την ικανότητα των νουκλεϊνικών οξέων να συνδέονται συμπληρωματικά και ως επακόλουθο διευκολύνουν την εξέλιξη των προηγούμενων διεργασιών. Στα ευκαρυωτικά κύτταρα, οι οργανικοί ρύποι μετατρέπονται, με τη δράση του ενζύμου του κυττοχρώματος Ρ-450, σε διάφορους 28
36 ΕΙΣΑΓΩΓΗ μεταλλαξιγόνους και καρκινογόνους μεταβολίτες, οι οποίοι συνιστούν πολύ δραστικά παραπροϊόντα που παρουσιάζουν μεγάλη ικανότητα σύνδεσης με πυρηνικά μακρομόρια, όπως το DNA. Η επίδραση αυτών των ενεργών ουσιών στο DNA μπορεί να οδηγήσει στη δημιουργία γενετικών βλαβών. Εξειδικευμένα πυρηνικά ένζυμα είναι ικανά να αποκαταστήσουν αυτές τις βλάβες κι έτσι να συντελέσουν σε κανονική έκφραση του γενώματος. Πολλές φορές όμως τα ένζυμα αυτά απενεργοποιούνται από ορισμένες κατηγορίες ρύπων, όπως τα βαρέα μέταλλα. Η ανεπαρκής επιδιόρθωση στις βλάβες που υφίσταται το γενετικό υλικό αποτελεί ένα από τα πρώτα στάδια στη διαδικασία δημιουργίας καρκινογενέσεων. Αυτό μπορεί να έχει σοβαρές επιπλοκές αν τέτοιες διεργασίες ανιχνευθούν στους αναπαραγωγικούς ιστούς, με επακόλουθη αρνητική επίδραση στην αναπαραγωγή του είδους (Δημητριάδης κ.ά. 2006). Πρόσφατα, η επιτυχής χρησιμοποίηση συγκεκριμένων μεθόδων για την αποτίμηση των γενοτοξικών βλαβών κατέστησε δυνατή την πιστοποίηση μεταλλαξιγενέσεων σε θαλάσσια ασπόνδυλα και έδειξε άμεση συσχέτιση ανάμεσα στη βιοσυσσώρευση ρύπων (βαρέα μέταλλα και PAHs) και σε διάφορες δυσμενείς καταστάσεις που σχετίζονται με το γενετικό υλικό, όπως χρωμοσωμικές ανωμαλίες ή αναστολές πυρηνικών ενζύμων (Δημητριάδης κ.ά. 2006). α) Βιομάρτυρας «συχνότητα μικροπυρήνων» Οι μικροπυρήνες είναι μικρές, κυτταροπλασματικές μάζες χρωματίνης που προκύπτουν από θραύση των χρωμοσωμάτων ή μη σωστό διαχωρισμό του γενετικού υλικού στα θυγατρικά κύτταρα κατά τη διάρκεια της κυτταρικής διαίρεσης (Bolognesi et al. 2004). Αυτές οι μικρές δομές χρωματίνης περιβάλλονται από μεμβράνη, εντοπίζονται στο κυτταρόπλασμα και δεν εμφανίζουν σημεία σύνδεσης με τον πυρήνα (Heddle 1973, Heddle et al. 1991). Ο βιομάρτυρας «συχνότητα μικροπυρήνων» (micronucleus frequency / ΜΝ), που βασίζεται στην καταμέτρηση των μικροπυρήνων σε ενεργώς διαιρούμενους πληθυσμούς κυττάρων, έχει χρησιμοποιηθεί ως δείκτης κυτογενετικής βλάβης για περισσότερα από 20 χρόνια (Ayllon & Garcia-Vazquez 2001). H κυτογενετική αυτή μέθοδος θεωρείται ικανή να εκτιμήσει το αποτέλεσμα της δράσης παραγόντων που προκαλούν θραύση στο γενετικό υλικό σε διάφορους κυτταρικούς τύπους και οργανισμούς (Tates et al. 1980, Ma et al. 1984, Das & Nanda 1986, Jaylet et al. 1986, Majone et al. 1987). Αναπτύχθηκε αρχικά σε κύτταρα θηλαστικών για τον εντοπισμό κυτογενετικών βλαβών (Boller & Schmid
37 ΕΙΣΑΓΩΓΗ Heddle 1973) και αργότερα εφαρμόσθηκε και σε άλλους οργανισμούς, μεταξύ των οποίων θαλάσσια μύδια από το γένος Mytilus, για την εκτίμηση της ποιότητας του περιβάλλοντος (Majone et al. 1987, 1988, 1990, Venier et al. 1997, Bolognesi et al. 1999, Barsiene et al. 2004, Bolognesi et al. 2004). Σε μελέτες βιοπαρακολούθησης διαφορετικοί ιστοί μπορεί να χρησιμοποιηθούν για την εφαρμογή της τεχνικής των μικροπυρήνων. Οι μικροπυρήνες σχηματίζονται στα αιμοκύτταρα και στα κύτταρα βραγχίων διάφορων μαλακίων (Mersch et al. 1996, Izquierdo et al. 2003, Bolognesi et al. 2004, Barsiene et al. 2006a, 2006b, 2006c, Villela et al. 2007) καθώς και στα ερυθροκύτταρα ποικίλων ειδών ψαριών (Ayllon & Garcia-Vasquez 2000, 2001, Gravato & Santos 2002, Pacheco et al. 2005). Η χρήση των αιμοκυττάρων συνιστάται για την εφαρμογή της συγκεκριμένης τεχνικής στα μύδια, μια και τα κύτταρα αυτά είναι εύκολο να απομονωθούν από τον προσαγωγό μυ, γεγονός που αποτρέπει πιθανό σπάσιμο των κλώνων του DNA που θα μπορούσε να προκληθεί από μια περισσότερο σύνθετη διαδικασία εξαγωγής και επεξεργασίας του ιστού (Accomando et al. 1999). Επίσης, ο φυσιολογικός ρόλος αυτών των κυττάρων στη μεταφορά τοξικών ουσιών και η συμμετοχή τους σε διάφορους αμυντικούς μηχανισμούς τα καθιστά πρώτης τάξεως υλικό για την εκτίμηση των επιπτώσεων περιβαλλοντικών ρύπων (Villela et al. 2007). Ο υπολογισμός της «συχνότητας των μικροπυρήνων» έχει εφαρμοσθεί μετά από έκθεση σε ποικίλους οργανικούς (Scarpato et al. 1990, Mersch et al. 1992, Brieger & Hunter 1993, Marvin et al. 1994, Becker-van Slooten & Tarradellas 1995) και ανόργανους ρύπους (Majone et al. 1987, Mersch & Pihan 1993, Camusso et al. 1994, Bolognesi et al. 1999) στο εργαστήριο, καθώς και σε πολλές μελέτες στο πεδίο (Brunetti et al. 1988, Mersch & Beauvais 1997). Στις ημέρες μας η ανίχνευση μικροπυρήνων αντιπροσωπεύει μια ευρέως διαδεδομένη μέθοδο, ικανή να προσδιορίσει τη θαλάσσια ρύπανση και για το λόγο αυτό έχει προταθεί ως εργαλείο βιοπαρακολούθησης για την εκτίμηση της γενοτοξικότητας στο περιβάλλον (Hooftman & Raat 1982, Manna et al. 1985). Η εφαρμογή του βιομάρτυρα «συχνότητα μικροπυρήνων» μαζί με άλλους βιομάρτυρες ρύπανσης στο πεδίο και στο εργαστήριο θα διερευνήσει την ύπαρξη συσχέτισης ανάμεσά τους. Τα στοιχεία που θα προκύψουν θα δώσουν επιπρόσθετες πληροφορίες σχετικά με τη δυνατότητα του βιομάρτυρα «συχνότητα μικροπυρήνων» να προσδιορίζει έγκαιρα τις συνέπειες γενοτοξικών παραγόντων, αξιολογώντας την αποτελεσματικότητά του σε μελέτες βιοπαρακολούθησης. 30
38 ΕΙΣΑΓΩΓΗ β) Βιομάρτυρας «συχνότητα συνολικών πυρηνικών ανωμαλιών» Στην προσπάθεια εκτίμησης της γενοτοξικότητας στο περιβάλλον, προκύπτουν ορισμένα μεθοδολογικά προβλήματα. Συγκεκριμένα, δεν έχει αποσαφηνισθεί ποιος τύπος πυρηνικών ανωμαλιών θα πρέπει να λαμβάνεται υπόψη, ως ακριβέστερη ένδειξη γενοτοξικής βλάβης. Πολλοί ερευνητές εφαρμόζουν την καταμέτρηση των κυττάρων που εμφανίζουν ένα μόνο μικροπυρήνα για την εκτίμηση της γενοτοξικότητας στο περιβάλλον (Kirsch-Volders et al. 2000, Villela et al. 2007, Binelli et al. 2008). Ωστόσο, έχει διατυπωθεί η άποψη ότι οι συνολικές αλλοιώσεις στη μορφολογία του πυρήνα έχουν, επίσης, γενοτοξική προέλευση (Pacheco & Santos 1996, 1997, 1998, Sanchez-Galan et al. 1998). Στην κατηγορία των συνολικών πυρηνικών ανωμαλιών περιλαμβάνονται, εκτός από την ύπαρξη κυττάρων με έναν μικροπυρήνα, η εμφάνιση κυττάρων που περιέχουν ταυτόχρονα δύο μικροπυρήνες, δύο ξεχωριστούς πυρήνες, πυρήνες σχήματος οκτώ ή πυρήνες που εμφανίζουν εξογκώματα ή εγκολπώσεις. Σύμφωνα λοιπόν με ορισμένες μελέτες, οι συνολικές πυρηνικές ανωμαλίες (total nuclear abnormalities / ΤΝΑ) θα πρέπει να λαμβάνονται υπόψη ως ένδειξη κυτογενετικής βλάβης για την πιο ακριβή εκτίμηση της δράσης γενοτοξικών παραγόντων (Ayllon & Garcia-Vazquez 2000, 2001, Gravato & Santos 2002, Pacheco et al. 2005, Barsiene et al. 2006a, b, c). Οι Carrasco et al. (1990) δεν βρήκαν σημαντική συσχέτιση ανάμεσα στις συνολικές πυρηνικές ανωμαλίες (συμπεριλαμβανομένων των μικροπυρήνων) και τα υψηλά επίπεδα χημικής ρύπανσης στα ιζήματα ή στους ιστούς ψαριών, καταλήγοντας στο συμπέρασμα ότι η εμφάνισή τους δεν σχετίζεται άμεσα με τη δράση γενοτοξικών ουσιών. Αντίθετα, η καταγραφή των συνολικών πυρηνικών ανωμαλιών θεωρήθηκε αξιόπιστη προσέγγιση στην εκτίμηση της γενοτοξικότητας σε μαλάκια (Venier et al. 1997, Dailianis et al. 2003). Ο υπολογισμός της συχνότητας των συνολικών πυρηνικών ανωμαλιών και η συσχέτιση αυτής της παραμέτρου με το ποσοστό των μικροπυρήνων και με τα αποτελέσματα άλλων βιομαρτύρων ρύπανσης, θα συμβάλλουν στην αξιολόγηση της τεχνικής προσδιορισμού των συνολικών πυρηνικών ανωμαλιών ως αξιόπιστο δείκτη κυτογενετικής βλάβης σε προγράμματα βιοπαρακολούθησης. 31
39 ΕΙΣΑΓΩΓΗ 3. Βιομάρτυρας «εκτίμησης στρες» Ο βιομάρτυρας εκτίμησης στρες (stress on stress / SOS) περιλαμβάνεται στους βιομάρτυρες χαμηλής εξειδίκευσης (Livingstone 1982) και στηρίζεται στη μέτρηση του χρονικού διαστήματος κατά το οποίο ορισμένοι υδρόβιοι οργανισμοί θα επιβιώσουν όταν απομακρυνθούν από το νερό. Σύμφωνα με αυτή την μέθοδο, η έκθεση στον αέρα, που αντιπροσωπεύει ένα φυσικό στρεσογόνο παράγοντα, επιβαρύνει τους οργανισμούς που έχουν ήδη δεχθεί την επίδραση ρυπογόνων ουσιών. Η θνησιμότητα στον αέρα φαίνεται, λοιπόν, να συμβαίνει γρηγορότερα σε άτομα που έχουν ήδη υποστεί στρες λόγω των ρύπων, σε σύγκριση με αυτά από περιοχές με μικρότερη ρύπανση (Viarengo et al. 1995). Επομένως, οργανισμοί που έχουν δεχθεί την επίδραση ρύπανσης και κατά συνέπεια χαρακτηρίζονται από αυξημένο φορτίο ρύπων στους ιστούς τους, είναι λιγότερο ανθεκτικοί στην επιπρόσθετη πίεση που προκαλεί η έκθεσή τους στον αέρα (Viarengo et al. 1995). Η αξιοποίηση αυτής της φυσιολογικής αντίδρασης σε μελέτες βιοπαρακολούθησης είναι γρήγορη, εύκολα υπολογίσιμη και δεν απαιτεί ιδιαίτερο οικονομικό κόστος. Στις περισσότερες έρευνες έχουν χρησιμοποιηθεί δίθυρα μαλάκια, με τα μύδια να αποτελούν τους περισσότερο διαδεδομένους οργανισμούς για την εφαρμογή της τεχνικής αυτής. Η δυνατότητα επιβίωσης στον αέρα, δηλαδή η δυνατότητα επιβίωσης των διθύρων κάτω από συνθήκες έλλειψης οξυγόνου, έχει αξιοποιηθεί σε πολλές περιβαλλοντικές μελέτες κατά τα τελευταία χρόνια για την εκτίμηση των επιπτώσεων της θαλάσσιας ρύπανσης (de Zwaan & Eertman 1996). Η τεχνική έχει εφαρμοσθεί σε εργαστηριακές μελέτες (Wang et al. 1992, Viarengo et al. 1995) και σε έρευνες στο πεδίο (Smaal et al. 1991, Veldhuizen-Tsoerkan et al. 1991, Thomas et al. 1999a) κι έχει θεωρηθεί αξιόπιστος δείκτης έγκαιρης προειδοποίησης για την εκτίμηση της πίεσης που προκαλούν οι περιβαλλοντικοί ρύποι. Εκτός όμως από την παρουσία οργανικών και ανόργανων ρύπων, φυσικοί παράγοντες του περιβάλλοντος όπως η αλατότητα, η θερμοκρασία, η περιεκτικότητα του νερού σε οξυγόνο, η διαθεσιμότητα της τροφής, η περίοδος αναπαραγωγής φαίνεται να επηρεάζουν τη δυνατότητα του βιομάρτυρα αυτού να καταδεικνύει τα αποτελέσματα της ρύπανσης (Da Ros et al. 1985, Eertman et al. 1993, Thomas et al. 1999b, Nesto et al. 2004, Pampanin et al. 2005). Η ταυτόχρονη εφαρμογή του βιομάρτυρα «εκτίμηση στρες» με άλλους βιομάρτυρες ρύπανσης παρουσιάζει ενδιαφέρον στις μελέτες βιοπαρακολούθησης. Η προσέγγιση αυτή θα συμβάλλει στη διερεύνηση της 32
40 ΕΙΣΑΓΩΓΗ συσχέτισης που ενδεχομένως εμφανίζεται ανάμεσα στους υπό εξέταση βιομάρτυρες, με αποτέλεσμα την εξαγωγή χρήσιμων συμπερασμάτων σχετικά με την αξιολόγηση της μεθόδου έκθεσης των μυδιών στον ατμοσφαιρικό αέρα ως αξιόπιστου εργαλείου για την έγκαιρη προειδοποίηση των επιπτώσεων της ρύπανσης. Δ. ΕΠΟΧΙΚΗ ΔΙΑΚΥΜΑΝΣΗ ΤΩΝ ΛΥΣΟΣΩΜΑΤΩΝ, ΤΩΝ ΛΙΠΙΔΙΩΝ ΚΑΙ ΤΩΝ ΛΙΠΟΦΟΥΣΚΙΝΩΝ ΣΤΟΝ ΠΕΠΤΙΚΟ ΑΔΕΝΑ ΤΟΥ ΜΥΔΙΟΥ MYTILUS GALLOPROVINCIALIS Τα λυσοσώματα των πεπτικών κυττάρων στα δίθυρα μαλάκια, εκτός από τον πρωταρχικό ρόλο τους στην ενδοκυτταρική πέψη (Owen 1972, Robledo et al. 1997), αποτελούν κύρια σημεία συγκέντρωσης και αποτοξικοποίησης μετάλλων και οργανικών ρύπων (Moore 1985, Viarengo et al. 1987). Συνεπώς, οι αλλαγές των δομικών λυσοσωμικών παραμέτρων των μυδιών θεωρούνται χρήσιμα εργαλεία για την έγκαιρη εκτίμηση των επιπτώσεων της ρύπανσης (Cajaraville et al. 1995b, Domouhtsidou & Dimitriadis 2001, Marigomez & Baybay-Villacorta 2003, Giamberine & Cajaraville 2005, Zorita et al. 2007). Επίσης, η αλλαγή στην ποσότητα των λιπιδίων του πεπτικού αδένα των μυδιών συνιστά μια άλλη παράμετρο, ικανή να καταδείξει περιβαλλοντική ρύπανση (Viarengo & Canesi 1991, Regoli 1992, Kagley et al. 2003). Τέλος, η συσσώρευση λιποφουσκίνης αντιπροσωπεύει έναν ακόμη παράγοντα που θεωρείται ενδεικτικός της έκθεσης σε ρύπους (Kagley et al. 2003), αποτελώντας μια γενική απόκριση σε διαφορετικά είδη ρύπανσης (Marigomez & Baybay Villacorta 2003). Σύμφωνα με προηγούμενες έρευνες, οι βιολογικές παράμετροι που αναφέρθηκαν προηγουμένως εμφανίζουν σημαντικές εποχικές μεταβολές (Marigomez et al. 1996b, Cancio et al. 1999, Petrovic et al. 2004, Guerlet et al. 2007) που θα μπορούσαν να αποκρύψουν τις επιπτώσεις της ρύπανσης. Είναι, λοιπόν, πολλές φορές δύσκολο να διαχωριστούν οι αποκρίσεις του οργανισμού που οφείλονται στην παρουσία ρύπων, από τις αλλαγές που οφείλονται σε εποχικές διακυμάνσεις (Kagley et al. 2003). Επομένως, η εφαρμογή βιομαρτύρων που σχετίζονται με τις παραπάνω παραμέτρους, σε κάποιες εποχές του έτους, μπορεί να οδηγήσει σε λάθος εκτίμηση των αποτελεσμάτων, καθώς αλλαγές στα 33
41 ΕΙΣΑΓΩΓΗ χαρακτηριστικά του πεπτικού αδένα που προκαλούνται από εποχικούς παράγοντες, θα μπορούσαν εσφαλμένα να αποδοθούν στην παρουσία ρύπων. Από τα παραπάνω γίνεται φανερό πως είναι σημαντικό να προσδιορισθεί το εύρος των εποχικών μεταβολών των ιστολογικών παραμέτρων του πεπτικού αδένα, ώστε να γίνεται καλύτερη αποτίμηση των δεδομένων που συγκεντρώνονται σε προγράμματα βιοπαρακολούθησης. Η εποχική διακύμανση των λυσοσωμάτων, των λιπιδίων και των λιποφουσκινών αποτελεί αντικείμενο διαφόρων μελετών που πραγματοποιήθηκαν κατά το παρελθόν (Marigomez et al. 1996b, Cancio et al. 1999, Petrovic et al. 2004). Ωστόσο, δεν υπάρχουν πολλές πληροφορίες σχετικά με την ταυτόχρονη μεταβολή στα χαρακτηριστικά των παραπάνω οργανιδίων και συστατικών, μετά από την εξέταση των ίδιων πεπτικών σωληναρίων κατά τη διάρκεια διαφορετικών εποχών του έτους. Η συνεξέταση των παραμέτρων αυτών θα παράσχει περαιτέρω πληροφορίες για την παράλληλη μεταβολή τους κατά τη διάρκεια του έτους, ενώ τα αποτελέσματα που θα προκύψουν θα συμβάλλουν στον προσδιορισμό της περισσότερο ευνοϊκής περιόδου για την πραγματοποίηση δειγματοληψιών σε μελέτες βιοπαρακολούθησης, που αφορούν στον πεπτικό αδένα των μυδιών. 34
42 ΕΙΣΑΓΩΓΗ Ε. ΣΚΟΠΟΣ ΤΗΣ ΜΕΛΕΤΗΣ Ο σκοπός της παρούσας μελέτης είναι η εξέταση της αξιοπιστίας βιομαρτύρων ρύπανσης σε μύδια Mytilus galloprovincialis από επιλεγμένους δειγματοληπτικούς σταθμούς του Θερμαϊκού και του Στρυμονικού κόλπου σε δύο διαφορετικές εποχές του έτους. Η ταυτόχρονη εφαρμογή των βιομαρτύρων «σταθερότητα της λυσοσωμικής μεμβράνης» σε πεπτικά κύτταρα του πεπτικού αδένα, «κατακράτηση της χρωστικής ουδέτερο ερυθρό» σε αιμοκύτταρα, «συχνότητα μικροπυρήνων» σε αιμοκύτταρα, «εκτίμηση στρες», αλλά και η εφαρμογή μορφομετρίας στα λυσοσώματα του πεπτικού αδένα θα καταδείξει την εμφάνιση στατιστικά σημαντικών συσχετίσεων μεταξύ των βιομαρτύρων στο πεδίο με τελικό στόχο την ενίσχυση του κάθε βιομάρτυρα για την εφαρμογή του σε προγράμματα βιοπαρακολούθησης υδάτινων οικοσυστημάτων. Εκτός όμως από την εκτίμηση της αξιοπιστίας των προαναφερθέντων βιομαρτύρων στο πεδίο, σκοπός αυτής της μελέτης είναι και η διερεύνηση της αποτελεσματικότητας συγκεκριμένων βιομαρτύρων ρύπανσης κάτω από εργαστηριακές συνθήκες. Ο προσδιορισμός της «συχνότητας των μικροπυρήνων», της «συχνότητας των συνολικών πυρηνικών ανωμαλιών», καθώς και η εφαρμογή μορφομετρίας για τα λυσοσώματα και τα λιπίδια του πεπτικού αδένα των μυδιών, μετά από εργαστηριακή επίδραση ενός οργανικού ρύπου (φενανθρένιο) και τριών βαρέων μετάλλων (Cu, Cd και Hg) θα καταδείξει τη δυνατότητα των παραπάνω βιομαρτύρων να φανερώνουν τις επιπτώσεις συγκεκριμένων περιβαλλοντικών ρύπων κάτω από εργαστηριακές συνθήκες. Επίσης, η ταυτόχρονη εφαρμογή των παραπάνω βιομαρτύρων μαζί με καθιερωμένους βιομάρτυρες ρύπανσης, όπως η «σταθερότητα της λυσοσωμικής μεμβράνης» και η «κατακράτηση της χρωστικής ουδέτερο ερυθρό», κάτω από εργαστηριακές συνθήκες, θα διερευνήσει τη συσχέτιση μεταξύ των αποτελεσμάτων τους, με σκοπό την ενίσχυση του ρόλου κάθε βιομαρτύρα στη βιοπαρακολούθηση της ρύπανσης. Ένας άλλος στόχος των διερευνήσεών μας είναι η μελέτη της εποχικής διακύμανσης λιπιδιακών και λυσοσωμικών παραμέτρων στον πεπτικό αδένα των μυδιών M. galloprovincialis. Ο ιστοχημικός εντοπισμός των λιποφουσκινών, των λιπιδίων και των λυσοσωμάτων, καθώς και η πραγματοποίηση μορφομετρικών μετρήσεων για τα προαναφερθέντα κυτταρικά οργανίδια και συστατικά σε τρεις 35
43 ΕΙΣΑΓΩΓΗ διαφορετικές εποχές του έτους θα οδηγήσει στην εξαγωγή χρήσιμων συμπερασμάτων σχετικά με τις επιπτώσεις εποχικών παραγόντων στα μορφολογικά και λειτουργικά χαρακτηριστικά του πεπτικού αδένα. Η παρούσα εργασία θεωρούμε ότι θα ενισχύσει τις γνώσεις πάνω σε βασικά θέματα της λειτουργίας του πεπτικού αδένα του κοινού μυδιού σε σχέση με εποχικούς περιβαλλοντικούς παράγοντες. Η μελέτη της εποχικότητας των λιπιδίων, των λυσοσωμάτων και των λιποφουσκινών του πεπτικού αδένα θα συμβάλλει, επίσης, στον προσδιορισμό της καταλληλότερης περιόδου για την πραγματοποίηση δειγματοληψιών σε μελέτες βιοπαρακολούθησης, στις οποίες αξιοποιούνται οι παραπάνω παράμετροι για την εκτίμηση των επιπτώσεων της ρύπανσης. Τέλος, ο ταυτόχρονος ιστοχημικός εντοπισμός λιπιδίων και λυσοσωμικών ενζύμων σε κρυοτομές πεπτικών κυττάρων θα παράσχει χρήσιμες πληροφορίες για την αλληλεπίδρασή τους, με σκοπό την ορθότερη αξιολόγηση ιστοχημικών αποτελεσμάτων για την εκτίμηση της ποιότητας του περιβάλλοντος. 36
44 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ
45 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Α. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΜΕΛΕΤΗ ΠΕΔΙΟΥ ΣΕ ΕΛΛΗΝΙΚΕΣ ΑΚΤΕΣ 1. Δειγματοληψίες μυδιών από το πεδίο Μύδια M. galloprovincialis συλλέχθηκαν από πέντε δειγματοληπτικές περιοχές κατά μήκος του Θερμαϊκού κόλπου κι ένα δειγματοληπτικό σταθμό στο Στρυμονικό κόλπο (Εικ. 4) στις αρχές Ιουνίου και στις αρχές Δεκεμβρίου του έτους Οι δειγματοληψίες σε κάθε δειγματοληπτική εποχή πραγματοποιήθηκαν μέσα σε διάστημα δύο ημερών. Τα μύδια συλλέχθηκαν από τις ίδιες ακριβώς περιοχές σε κάθε δειγματοληψία με βάρκα από καλλιεργούμενους πληθυσμούς που όπως ήταν φυσικό ήταν συνεχώς εμβαπτισμένοι στο θαλασσινό νερό. Οι φυσικοί πληθυσμοί συλλέχθηκαν με αποκόλληση από βράχια κοντά στις ακτές. Οι δειγματοληψίες περιέλαβαν μύδια από την Ολυμπιάδα (καλλιεργούμενος πληθυσμός, 40 o N, 23 o E) στο Στρυμονικό κόλπο και από το Αγγελοχώρι (καλλιεργούμενος πληθυσμός 40 o N, 22 o 48 8 E), την Περαία (φυσικός πληθυσμός 40 o N, 22 o 55 6 E), το Καλοχώρι (φυσικός πληθυσμός κοντά στην πρώην έξοδο του αγωγού του βιολογικού καθαρισμού του Καλοχωρίου, 40 o 34 8 N, 22 o E), τη Χαλάστρα (καλλιεργούμενος πληθυσμός 40 o 33 2 N, 22 o 45 7 E) και τα Κύμινα (καλλιεργούμενος πληθυσμός, 40 o N, 22 o E) στο Θερμαϊκό κόλπο (Βλέπε Εικ. 4, περιοχές 1-6 αντίστοιχα). Τα μύδια που συλλέχθηκαν από φυσικούς πληθυσμούς ήταν μικρότερα σε μέγεθος από τα μύδια των καλλιεργούμενων πληθυσμών, αφού τα πρώτα μεγάλωσαν σε συνθήκες συνωστισμού και ανταγωνισμού, ενώ τα δεύτερα υπέστησαν αραίωση από τους καλλιεργητές τουλάχιστο μια φορά κατά την περίοδο ανάπτυξης. Το μήκος του κελύφους των μυδιών κατά τη διάρκεια των δειγματοληψιών κυμαίνονταν ως εξής: Αγγελοχώρι 5,87 ± 0,34 cm τον Ιούνιο, 6,02 ± 0,17 cm το Δεκέμβριο, Περαία: 5,22 ± 0,42 cm τον Ιούνιο, 5,04 ± 0,3 cm το Δεκέμβριο, Καλοχώρι: 4,78 ±0,34 cm τον Ιούνιο, 5,44 ± 0,48 cm το Δεκέμβριο, Χαλάστρα: 6,56 ± 0,58 cm τον Ιούνιο, 6,76 ± 0,48 το Δεκέμβριο, Κύμινα: 7,02 ± 0,31 cm τον Ιούνιο, 6,68 ± 0,26 cm το Δεκέμβριο, Ολυμπιάδα: 6,24 ± 0,73 cm τον Ιούνιο, 6,22 ± 0,35 cm το Δεκέμβριο. Η κύρια αναπαραγωγική περίοδος των μυδιών M. galloprovincialis από το Θερμαϊκό κόλπο διαρκεί από το Φεβρουάριο έως τον Απρίλιο και από τον Οκτώβριο έως το 37

46 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Νοέμβριο. Ωστόσο, σύντομα και μικρής έντασης φαινόμενα απελευθέρωσης γαμετών φαίνεται να συμβαίνουν σε όλη τη διάρκεια του έτους (προσωπικές παρατηρήσεις) ΣΤΡΥΜΟΝΙΚΟΣ ΚΟΛΠΟΣ ΘΕΡΜΑΪΚΟΣ ΚΟΛΠΟΣ Εικόνα 4. Χάρτης της περιοχής του Θερμαϊκού και του Στρυμονικού κόλπου όπου σημειώνονται οι περιοχές από τις οποίες πραγματοποιήθηκαν οι δειγματοληψίες των μυδιών. 1. Ολυμπιάδα (καλλιεργούμενος πληθυσμός), 2. Αγγελοχώρι (καλλιεργούμενος πληθυσμός), 3. Περαία (φυσικός πληθυσμός), 4. Καλοχώρι (φυσικός πληθυσμός κοντά στην πρώην έξοδο του αγωγού του βιολογικού καθαρισμού), 5. Χαλάστρα (καλλιεργούμενος πληθυσμός), 6. Κύμινα (καλλιεργούμενος πληθυσμός). Figure 4. Map of Thermaikos and Strymonikos Gulf showing the sampling stations. 1. Olympiada (cultivated population), 2. Aggelohori (cultivated population), 3. Peraia (natural population), 4. Halastra (cultivated population), 5. Kalohori (natural population close to the past wastewater treatment plant tube), 6. Kymina (cultivated population). Ο δειγματοληπτικός σταθμός της Ολυμπιάδας επιλέχθηκε ως σταθμός ελέγχου, σύμφωνα τόσο με αποτελέσματα βιοπαρακολούθησης του εργαστηρίου μας, όσο και με βιβλιογραφικές αναφορές σχετικές με τη χημική ανάλυση των νερών του Θερμαϊκού και του Στρυμονικού κόλπου (Zahariadis & Stratis 1995, Καμαριανός κ.ά. 2002, ΔΕΠΠΠΕΘ 2004). Σύμφωνα με προηγούμενες αναφορές η δυτική πλευρά του Θερμαϊκού (σταθμοί 4, 5, 6) εμφάνισε υψηλότερο βαθμό περιβαλλοντικής επιβάρυνσης σε σχέση με την ανατολική (Σταθμοί 2, 3) (ΔΕΠΠΠΕΘ 2004). Η δυτική πλευρά αποτελεί αποδέκτη των ανεξέλεγκτων λυμάτων κάποιων βιομηχανικών μονάδων της βιομηχανικής περιοχής, του αγωγού του Βιολογικού καθαρισμού αλλά 38
47 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ και των δέλτα του Αξιού, του Λουδία και του Αλιάκμονα και θεωρείται περισσότερο επιβαρυμένη από την ανατολική πλευρά. 2. Βιομάρτυρες που εξετάσθηκαν «Σταθερότητας της λυσοσωμικής μεμβράνης» (lysosomal membrane stability LMS) «Κατακράτησης της χρωστικής ουδέτερο ερυθρό» (neutral red retention assay - NRR) «Συχνότητα μικροπυρήνων» (micronucleus frequency - MN) «Εκτίμησης στρες» (stress on stress - SOS) «Μεταβολές των λυσοσωμικών δομικών παραμέτρων στον πεπτικό αδένα μυδιών M. galloprovincialis» (lysosomal structural changes - LSC) Β. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΕΡΓΑΣΤΗΡΙΑΚΗ ΜΕΛΕΤΗ ΜΕΤΑ ΑΠΟ ΕΚΘΕΣΗ ΣΤΑ ΒΑΡΕΑ ΜΕΤΑΛΛΑ Cu,Cd, Hg KAI ΣΤΟΝ ΟΡΓΑΝΙΚΟ ΡΥΠΟ ΦΑΙΝΑΝΘΡΕΝΙΟ 1. Δειγματοληψία για επίδραση ρύπων στο εργαστήριο Μύδια M. galloprovincialis, με μέσο μέγεθος κελύφους 6.06 ± 0.56 cm, συλλέχθηκαν το Μάιο 2002 από το δειγματοληπτικό σταθμό της Χαλάστρας (καλλιεργούμενος πληθυσμός) και μεταφέρθηκαν στο εργαστήριο μέσα σε θερμός με παγοκύστεις. Εκεί τα μύδια καθαρίσθηκαν και τοποθετήθηκαν σε πλαστικά ενυδρεία χωρητικότητας 10 L (30 άτομα/ενυδρείο), μέσα σε θαλασσινό νερό στους 18 ο C. Το νερό ανανεωνόταν κάθε τρεις ημέρες επί μία εβδομάδα, χωρίς την προσθήκη τροφής, ώστε να επιτευχθεί εγκλιματισμός των ατόμων στις συνθήκες εργαστηρίου. Το θαλασσινό νερό συλλέγονταν από την περιοχή του Μικρού Καράμπουρνου δύο φορές την εβδομάδα και αφού φιλτραριζόταν διαμέσου υδρόφιλου βάμβακος και Whatman διηθητικού χαρτιού, φυλάσσονταν στους 18 ο C, σε πλαστικά δοχεία με συνεχή αερισμό έως ότου χρησιμοποιηθεί για τα πειράματα. 39
48 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 2. Επίδραση ρύπων Μετά από εγλιματισμό για μια εβδομάδα στις εργαστηριακές συνθήκες, τα μύδια εκτέθηκαν για 15 ημέρες σε διάφορους ρύπους. Η επιλογή των ρύπων έγινε με βάση το γεγονός ότι ανιχνεύονται στο θαλασσινό νερό του Θερμαϊκού κόλπου (Manoli & Samara 1999, Haritonidis & Malea 1999, ΔΕΠΠΠΕΘ 2004). Συγκεκριμένα, τα μύδια εκτέθηκαν επί 15 ημέρες σε 0.1 mg L -1 της ουσίας φαινανθρένιο (phenanthrene) διαλυμένης σε ακετόνη (συγκέντρωση της ακετόνης στο νερό 0,002%), σε 0.1 mg L -1 Cu που χορηγήθηκε ως CuCl 2, 0.1 mg L -1 Cd που χορηγήθηκε ως CdCl 2, 0.1 mg L -1 Hg που χορηγήθηκε ως HgCl 2. Οι ρύποι χορηγήθηκαν μεμονωμένα σε κάθε ομάδα μυδιών. Μια ομάδα μυδιών που διατηρήθηκε χωρίς την προσθήκη κάποιας χημικής ουσίας στο νερό χρησιμοποιήθηκε ως ομάδα ελέγχου για την επίδραση μετάλλων, ενώ άτομα που εκτέθηκαν σε ακετόνη (συγκέντρωση της ακετόνης στο νερό 0.002%) χρησιμοποιήθηκαν ως ομάδα ελέγχου για τα μύδια που δέχθηκαν την επίδραση του φαινανθρένιου. Η δόση των ρύπων που χορηγήθηκαν στο θαλασσινό νερό επιλέχθηκε με βάση στοιχεία της βιβλιογραφίας, που αναφέρουν ότι παρόμοιες δόσεις σε μύδια του ίδιου είδους και μεγέθους/ηλικίας είναι υποθανάτιες, μπορούν να προκαλούν συσσώρευση των ρύπων στους ιστούς και ευθύνονται για μια πληθώρα βιολογικών αποκρίσεων (Davenport 1977, Nelson et al. 1988, Salanki & Balogh 1989). Το νερό των δοχείων αεριζόταν συνεχώς με τη χρήση αντλιών αέρα. Τα μύδια τρέφονταν με κατεψυγμένο φυτοπλαγκτόν (30 mg/άτομο) κάθε δύο ημέρες. Το νερό και η δοσολογία των μετάλλων ανανεώνονταν κάθε τρίτη μέρα. Καθημερινά ελέγχονταν η θερμοκρασία και ο αερισμός και απομακρύνονταν ακαθαρσίες και νεκρά ζώα. Μετά την πάροδο 15 ημερών από την έναρξη της επίδρασης των ρύπων, από κάθε πειραματική ομάδα επιλέχθηκαν 15 μύδια. Η καλή κατάσταση των μυδιών, που καθορίζονταν από την καλή προσκόλλησή τους στα τοιχώματα του ενυδρείου με βύσσο και το φυσιολογικό ρυθμό ανοίγματος και κλεισίματος του κελύφους τους (Borcherding 1992), αποτέλεσε απαραίτητο κριτήριο για τη χρησιμοποίησή τους. Ακολούθησε εξαγωγή του πεπτικού αδένα πέντε μυδιών και αφαίρεση αιμόλεμφου από δέκα μύδια για την εφαρμογή βιομαρτύρων ρύπανσης. 40
49 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 3. Βιομάρτυρες που εξετάσθηκαν «Σταθερότητα της λυσοσωμικής μεμβράνης» (lysosomal membrane stability LMS) «Κατακράτηση της χρωστικής ουδέτερο ερυθρό» (neutral red retention assay - NRR) «Συχνότητα μικροπυρήνων» (micronucleus frequency - MN) «Συχνότητα συνολικών πυρηνικών ανωμαλιών» (frequency of total nuclear abnormalities TNA) «Μεταβολές των λυσοσωμικών δομικών παραμέτρων στον πεπτικό αδένα μυδιών M. galloprovincialis» (lysosomal structural changes - LSC) «Δομικές μεταβολές των ουδέτερων λιπιδίων στον πεπτικό αδένα μυδιών Mytilus galloprovincialis» (neutral lipid structural changes - NLSC) Γ. ΜΕΛΕΤΗ ΤΗΣ ΕΠΟΧΙΚΗΣ ΔΙΑΚΥΜΑΝΣΗΣ ΤΩΝ ΛΥΣΟΣΩΜΑΤΩΝ, ΤΩΝ ΛΙΠΙΔΙΩΝ ΚΑΙ ΤΩΝ ΛΙΠΟΦΟΥΣΚΙΝΩΝ ΣΤΟΝ ΠΕΠΤΙΚΟ ΑΔΕΝΑ ΤΟΥ ΜΥΔΙΟΥ MYTILUS GALLOPROVINCIALIS 1. Δειγματοληψίες μυδιών από το πεδίο Μύδια Mytilus galloprovincialis συλλέχθηκαν από τέσσερις δειγματοληπτικές περιοχές κατά μήκος του Θερμαϊκού κόλπου κι ένα δειγματοληπτικό σταθμό στο Στρυμονικό κόλπο. Οι δειγματοληψίες περιέλαβαν μύδια από την Ολυμπιάδα (καλλιεργούμενος πληθυσμός, 40 o N, 23 o E) στο Στρυμονικό κόλπο και από το Αγγελοχώρι (καλλιεργούμενος πληθυσμός 40 o N, 22 o 48 8 E, την Περαία (φυσικός πληθυσμός 40 o N, 22 o 55 6 E), το Καλοχώρι (φυσικός πληθυσμός κοντά στην πρώην έξοδο του αγωγού του βιολογικού καθαρισμού του Καλοχωρίου, 40 o 34 8 N, 22 o E), τη Χαλάστρα (καλλιεργούμενος πληθυσμός 40 o 33 2 N, 22 o 45 7 E) στο Θερμαϊκό κόλπο (Βλέπε Εικ. 4, περιοχές 1-5 αντίστοιχα). Πραγματοποιήθηκαν τρεις κύκλοι δειγματοληψιών. Ο πρώτος κύκλος έγινε κατά τη διάρκεια της άνοιξης και συγκεκριμένα τον μήνα Μάρτιο 2003, ο δεύτερος κύκλος κατά την αρχή του καλοκαιριού και συγκεκριμένα από το τέλος του Μαϊου έως τις αρχές Ιουνίου 2003 και ο τρίτος κύκλος στις αρχές του χειμώνα και συγκεκριμένα την περίοδο από το τέλος Νοεμβρίου έως τις αρχές Δεκεμβρίου
50 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Ο δειγματοληπτικός σταθμός της Ολυμπιάδας επιλέχθηκε ως σταθμός ελέγχου. Το μήκος του κελύφους των μυδιών κατά τη διάρκεια των δειγματοληψιών αναγράφεται στον πίνακα 1: Πίνακας 1. Μήκος του κελύφους των μυδιών που συλλέχθηκαν από τις πέντε περιοχές δειγματοληψίας την άνοιξη, στην αρχή του καλοκαιριού και στην αρχή του χειμώνα του έτους Table 1. Shell length of the mussels, sampled from the five sampling stations in spring, early summer and early winter Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα Άνοιξη ±0.27 cm 6.06±0.28 cm 4.91±0.34 cm 4.79±0.29 cm 6.71±0.4 cm Αρχή καλοκαιριού 2003 Αρχή χειμώνα ± 0.49 cm 6.1 ± 0.32 cm 6.42±0.51 cm 4.61±0.21 cm 6.76±0.2 cm 6.26±0.15 cm 6.12±0,25 cm 4.46±0.56 cm 5.48±0.29 cm 6.8±0.28 cm Τα μύδια συλλέγονταν από τις ίδιες ακριβώς περιοχές σε κάθε δειγματοληψία με βάρκα από πληθυσμούς οι οποία ήταν συνεχώς εμβαπτισμένοι στο νερό. Τα άτομα των φυσικών πληθυσμών συλλέγονταν με αποκόλληση από βράχια κοντά στις ακτές. Τα μύδια μεταφέρονταν στο εργαστήριο ζωντανά μέσα σε πλαστικά δοχεία με παγοκύστεις. 2. Παράμετροι που εξετάσθηκαν Εποχική μεταβολή στα χαρακτηριστικά του λυσοσωμικού συστήματος των πεπτικών κυττάρων έπειτα από ιστοχημική αντίδραση για το λυσοσωμικό ένζυμο Ν-ακέτυλο-β-εξοζαμινιδάση. Εποχική διακύμανση των ουδέτερων λιπιδίων, των ολικών λιπιδίων και των λιποφουσκινών του πεπτικού αδένα, μετά από ιστοχημικό εντοπισμό των παραπάνω κυτταρικών συστατικών. Υπολογισμός της πυκνότητας όγκου των λυσοσωμάτων, των ουδέτερων λιπιδίων και των λιποφουσκινών του πεπτικού αδένα στις τρεις περιόδους δειγματοληψίας. 42
51 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Δ. ΠΕΡΙΓΡΑΦΗ ΒΙΟΜΑΡΤΥΡΩΝ 1. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» Για την εκτίμηση της «σταθερότητας της λυσοσωμικής μεμβράνης» (lysosomal membrane stability / LMS) πραγματοποιήθηκε φυσική μονιμοποίηση με ψύξη του πεπτικού αδένα μυδιών που είχαν μεταφερθεί ζωντανά στο εργαστήριο σε θερμός με παγοκύστεις και ακολούθησε ιστοχημεία για το λυσοσωμικό ένζυμο N- ακέτυλο-β-εξοζαμινιδάση, σύμφωνα με τη διαδικασία που προτείνεται από τους Moore (1976) και UNEP (1997) και περιλαμβάνει τα εξής στάδια: 1. Γρήγορη εξαγωγή του πεπτικού αδένα από 5 μύδια και τοποθέτησή του σε γυάλινη επιφάνεια που βρισκόταν πάνω σε πάγο. 2. Λήψη μικρών τεμαχίων του πεπτικού αδένα, περίπου 5 mm 3. Τοποθέτηση των κομματιών σε ειδικές βάσεις από αλουμίνιο (aluminium cryostat chucks), έτσι ώστε σε κάθε βάση να υπάρχουν 5 τεμάχια, ένα από καθένα από τα 5 ζώα και από αντίστοιχες περιοχές του αδένα, σε μια ευθεία γραμμή που περνάει από το κέντρο της βάσης. 3. Φυσική μονιμοποίηση του ιστού με ψύξη με εμβάπτιση της βάσης με τους 5 ιστούς από τα 5 ζώα για 40 sec, μέσα σε υγρό n-hexane το οποίο είχε προηγουμένως ψυχθεί στους -70 ο C με βύθιση σε υγρό άζωτο (θ=-273 ο C). 4. Τύλιγμα των βάσεων σε parafilm και αλουμινόχαρτο, αναγραφή της περιοχής και εποχής δειγματοληψίας και αποθήκευση στους -80 ο C έως την ιστοχημική ανίχνευση. 5. Λήψη κρυοτομών πάχους 9-10 nm από τον πεπτικό αδένα 5 μυδιών ανά δειγματοληπτική περιοχή σε κρυοστάτη τύπου Reichert-Jung Cryocut 1800 (Leica) σε θερμοκρασία -27 ο C. 6. Μεταφορά συνεχόμενων τομών σε γυάλινες αντικειμενοφόρους, παραμονή σε θερμοκρασία δωματίου για 1-2 λεπτά έως ότου ξηρανθούν. Αποθήκευση αντικειμενοφόρων στον κρυοστάτη έως ότου χρησιμοποιηθούν για την ιστοχημεία, ποτέ όμως περισσότερο από 3-4 h. 7. Τοποθέτηση των τομών σε θερμοκρασία δωματίου, σε διάλυμα κατεργασίας. Το διάλυμα κατεργασίας αποτελείται από ρυθμιστικό διάλυμα κιτρικού οξέος 0.1 Μ (ph 4.5) που περιέχει 2.5% NaCl (w:v). Η κατεργασία περιέλαβε παραμονή των αντικειμενοφόρων στο προηγούμενο διάλυμα για χρονικό διάστημα από 0 έως 40 43
52 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ min. Πιο συγκεκριμένα, 10 σετ διπλών αντικειμενοφόρων υποβάλλονταν σε κατεργασία για 0, 3, 6, 9, 12, 15, 20, 25, 30 και 40 min. 8. Μεταφορά των αντικειμενοφόρων στο μέσο επώασης. Για την παρασκευή του μέσου επώασης 20 mg του υποστρώματος napthol AS-BI-N-acetyl-β-D glucosaminide (Sigma) διαλύθηκαν σε 2.5 ml 2-methoxyethanol και προστέθηκαν 47.5 ml ρυθμιστικού διαλύματος κιτρικού οξέος (0.1 Μ - ph 4.5) που περιείχε 2.5% NaCl (w:v) και 3.5 g ενός πολυπεπτιδίου χαμηλής πυκνότητας (Sigma POLYPEP, P5115) για τη σταθεροποίηση των τομών. Επώαση των τομών για 20 min στους 37 ο C σε υδατόλουτρο με συνεχή ανάδευση. 9. Πλύση με 3% NaCl στους 37 ο C για 2 min. 10. Χρώση των τομών σε 1 mg/ml Fast Violet B (Sigma) σε 0.1 Μ ρυθμιστικό φωσφορικών (ph 7.4) σε θερμοκρασία δωματίου για 10 min. 11. Πλύση με τρεχούμενο νερό βρύσης για 5 min. 12. Στερέωση σε calcium formal (30% paraformaldehyde, 2% calcium acetate) για 10 min στους 4 ο C. 13. Πλύση με ddh 2 O. 14. Τοποθέτηση υδατικού μέσου έγκλεισης (Glycergel Daco) και καλυπτρίδων. 15. Παρατήρηση στο οπτικό μικροσκόπιο για τον προσδιορισμό του χρόνου που αντιπροσωπεύει την τιμή της «σταθερότητας της λυσοσωμικής μεμβράνης». Λόγω της επεξεργασίας που είχαν υποστεί οι τομές, βάφονται με διαφορετική ένταση, ανάλογα με το χρόνο επίδρασης με το ρυθμιστικό διάλυμα κιτρικού οξέος (στάδιο 7). Η τιμή «σταθερότητας της λυσοσωμικής μεμβράνης» αντιστοιχούσε στο χρόνο στον οποίο εμφανιζόταν η εντονότερη χρώση (στο χρόνο επίδρασης με το διάλυμα κιτρικού οξέος). Στο χρόνο στον οποίο είχε παρατηρηθεί η μέγιστη ένταση χρώσης, το υδρολυτικό ένζυμο του ιστού είχε αντιδράσει πλήρως με το υπόστρωμα που προσθέτονταν (Εικ. 5). Τέσσερις τιμές λαμβάνονταν από κάθε ζώο, μετά το νοητό χωρισμό της κάθε τομής σε τέσσερα τεταρτημόρια και τον υπολογισμό του χρόνου που αντιστοιχούσε στην τιμή της «σταθερότητας της λυσοσωμικής μεμβράνης» σε καθένα από τα αντίστοιχα σετ τεταρτημορίων. Η μέση τιμή των τεσσάρων τεταρτημορίων ενός ζώου αντιστοιχούσε στην τιμή της «σταθερότητας της λυσοσωμικής μεμβράνης» για το συγκεκριμένο ζώο. Ο χρόνος (σε min) που αντιπροσώπευσε την τιμή της τεχνικής της «σταθερότητας της λυσοσωμικής μεμβράνης» για μια συγκεκριμένη δειγματοληπτική περιοχή ήταν η μέση τιμή για τα πέντε ζώα που χρησιμοποιήθηκαν. 44
 12 min, (β) 15 min και (γ) 20 min σε ρυθμιστικό διάλυμα κιτρικού.")
53 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Εικόνα 5. Διαδοχικές κρυοτομές πεπτικού αδένα μυδιού M. galloprovincialis στις οποίες εφαρμόσθηκε ιστοχημεία για το λυσοσωμικό ένζυμο N-ακέτυλο-β-εξοζαμινιδάση, μετά από κατεργασία για: (α) 12 min, (β) 15 min και (γ) 20 min σε ρυθμιστικό διάλυμα κιτρικού. Ο χρόνος στον οποίο εμφανίζεται η εντονότερη χρώση (15 min) χαρακτηρίζει την ποιότητα των υδάτων της εκάστοτε θαλάσσιας περιοχής. 1500x Figure 5. Serial cryostat sections of a mussel M. galloprovincialis showing lysosomal N-acetyl-βhexosaminidase histochemistry after: (α) 12 min; (β) 15 min and (γ) 20 min preatreatment in citrate buffer. The time showing the maximum staining intensity (15 min) is indicative of the water quality in each marine area. 1500x 45
54 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 2. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» Ο βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (neutral red retention assay / NRR) εφαρμόσθηκε σε απομονωμένα αιμοκύτταρα 10 μυδιών από κάθε περιοχή δειγματοληψίας, για τον προσδιορισμό του χρόνου αποσταθεροποίησης της λυσοσωμικής μεμβράνης των κοκκωδών αιμοκυττάρων. Η διαδικασία που ακολουθήθηκε έχει προταθεί από τους Lowe και Pipe (1994) και περιλαμβάνει τα εξής στάδια: 1. Αφαίρεση μικρής ποσότητας αιμόλεμφου με αποστειρωμένη σύριγγα από τον προσαγωγό μυ 10 μυδιών (Εικ. 6) και ανάμιξή της με φυσιολογικό διάλυμα (4.77 g/l HEPES, g/l NaCl, 13.06g/l MgSO 4, 0.75 g/l KCl, 1.47g/l CaCl 2, ph=7.3) ούτως ώστε να ληφθεί εναιώρημα κυττάρων/φυσιολογικού διαλύματος σε αναλογία 1:1 v/v. 2. Τοποθέτηση 40μl του εναιωρήματος αιμόλεμφου-φυσιολογικού διαλύματος από κάθε μύδι σε αντικειμενοφόρο πλάκα για 15 λεπτά, μέσα σε ειδικό σκοτεινό κυτίο, διαστάσεων 25cm x 35cm x 16cm, όπου υπάρχουν συνθήκες αυξημένης υγρασίας και χαμηλής θερμοκρασίας, για να επικαθίσουν τα κύτταρα. 3. Αφαίρεση της σταγόνας, ώστε να μείνουν τα κύτταρα. 4. Διάλυση 10 μl διαλύματος stock (28.8 g ουδέτερου ερυθρού σε 100 ml DMSO) σε 5 ml φυσιολογικού διαλύματος. 5. Προσθήκη 40 μl από το διάλυμα της χρωστικής ουδέτερο ερυθρό στο στρώμα των κυττάρων και κάλυψή τους με καλυπτρίδα. 6. Επώαση των κυττάρων για 15 λεπτά στο σκοτάδι, για να απορροφηθεί η χρωστική. 7. Παρατήρηση των αντικειμενοφόρων στο οπτικό μικροσκόπιο, ανά χρονικά διαστήματα 15 λεπτών. Στο μικροσκόπιο παρατηρείται η απώλεια της χρωστικής από τα λυσοσώματα προς το κυτταρόπλασμα ή άλλες παραμορφώσεις των εξετασθέντων κοκκωδών κυττάρων της αιμόλεμφου (Εικ. 6). Η χρονική περίοδος ανάμεσα στην εφαρμογή της χρωστικής ουδέτερο ερυθρό και την εμφάνιση της πρώτης ένδειξης απώλειας της χρωστικής από τα λυσοσώματα στο κυτταρόπλασμα ή η εμφάνιση άλλων ανωμαλιών, όπως η κυτταρική λύση, στο 50% των υπό εξέταση κυττάρων που ανήκουν στα κοκκώδη αιμοκύτταρα, αντιπροσωπεύει το χρόνο κατακράτησης της χρωστικής για τα μύδια. Ο 46

55 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ μέσος όρος, από το χρόνο κατακράτησης της χρωστικής από 10 μύδια, αντιστοιχεί στο χρόνο κατακράτησης της χρωστικής για την κάθε ομάδα μυδιών. Μεγαλύτεροι χρόνοι αντιστοιχούν σε καθαρότερες περιοχές, ενώ μικρότεροι χρόνοι σε περιοχές με αυξημένη ρύπανση. Εικόνα 6. Εικόνες οπτικού μικροσκοπίου (α-δ) από τα διάφορα στάδια των αιμοκυττάρων μυδιών M. galloprovincialis μετά από την επίδραση της χρωστικής ουδέτερο ερυθρό. α) άθικτα αιμοκύτταρα β) αιμοκύτταρα που εμφανίζουν διογκωμένα λυσοσώματα λόγω εισχώρησης της χρωστικής στο εσωτερικό των λυσοσωμάτων γ) αιμοκύτταρα στα οποία έχει συμβεί διάχυση της χρωστικής στο κυτταρόπλασμα δ) λύση τν αιμοκυττάρων. Οι τιμές του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» αντιστοιχούν στο χρόνο που το 50% των αιμοκυττάρων βρίσκεται στα στάδια γ και δ. 1000x Figure 6. Light microscopic views (α δ) of the various stages of haemocytes from mussels M. galloprovincialis after the influence of the neutral red staining. α) intact haemocytes β) haemocytes displaying enlarged lysosomes due to the penetration of neutral red into the lysosomes γ) haemocytes displaying dye loss from the lysosomes to the cytosol δ) lysis of the haemocytes. Neutral red retention values represent the mean time when 50% of the haemocytes are in stages γ and δ. 1000x 3. Ο προσδιορισμός πυρηνικών ανωμαλιών ως βιομάρτυρας θαλάσσιας ρύπανσης α) Βιομάρτυρας «συχνότητα μικροπυρήνων» Η συχνότητα εμφάνισης κυττάρων με κυτταροπλασματικούς πυρήνες μειωμένου μεγέθους, γνωστούς ως μικροπυρήνες, χρησιμοποιείται ως ένδειξη κυτογενετικής βλάβης που μπορεί να οφείλεται στη θραύση της χρωματίνης ή στο μη φυσιολογικό διαχωρισμό του γενετικού υλικού στα θυγατρικά κύτταρα κατά την κυτταρική διαίρεση. Ο βιομάρτυρας «συχνότητα μικροπυρήνων» (micronucleus frequency / MN) εφαρμόσθηκε σύμφωνα με τη διαδικασία που προτάθηκε από τη UNEP/RAMOGE (1999) και περιλαμβάνει τα εξής στάδια: 47
56 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 1. Εξαγωγή μικρής ποσότητας αιμόλεμφου με αποστειρωμένη σύριγγα από τον προσαγωγό μύ 10 μυδιών και ανάμιξή της με φυσιολογικό διάλυμα (4.77 g/l HEPES, g/l NaCl, 13.06g/l MgSO 4, 0.75 g/l KCl, 1.47g/l CaCl 2, ph=7.3) ούτως ώστε να ληφθεί εναιώρημα κυττάρων/φυσιολογικού διαλύματος σε αναλογία 1:1 v/v. 2. Τοποθέτηση 40μl του εναιωρήματος αιμόλεμφου-φυσιολογικού διαλύματος από κάθε μύδι σε αντικειμενοφόρο πλάκα για 15 λεπτά, μέσα σε ειδικό σκοτεινό κυτίο, διαστάσεων 25cm x 35cm x 16cm, όπου υπάρχουν συνθήκες αυξημένης υγρασίας και χαμηλής θερμοκρασίας, για να επικαθίσουν τα κύτταρα. 3. Aφαίρεση της σταγόνας, ώστε να μείνουν μόνο τα κύτταρα. 4. Στερεοποίηση των κυττάρων σε διάλυμα methanol/acetic acid (3/1). 5. Χρώση με Giemsa. 6. Μονιμοποίηση με Canada balsam και τοποθέτηση καλυπτρίδων πάνω στα κύτταρα. 7. Παρατήρηση των κυττάρων στο οπτικό μικροσκόπιο. Από κάθε αντικειμενοφόρο εξετάσθηκαν 1000 αιμοκύτταρα. Οι μικροπυρήνες καταμετρήθηκαν σε άθικτα κύτταρα με καλά διατηρημένο κυτταρόπλασμα. Ως μικροπυρήνας θεωρήθηκε κάθε κυτταροπλασματικός πυρήνας μειωμένου μεγέθους που εμφανίζει σφαιρικό σχήμα, ξεκάθαρο περίγραμμα, διάμετρο που κυμαίνεται ανάμεσα στο ένα δέκατο και το ένα τρίτο της διαμέτρου του πυρήνα και μοιάζει με τον κύριο πυρήνα στη σύσταση και το χρώμα. β) Βιομάρτυρας «συχνότητα συνολικών πυρηνικών ανωμαλιών» Με σκοπό την εκτίμηση της «συχνότητας των συνολικών πυρηνικών ανωμαλιών» (frequency of total nuclear abnormalities / TNA) στα αιμοκύτταρα μυδιών Mytilus galloprovincialis, που δέχθηκαν την εργαστηριακή επίδραση του φαινανθρένιου (phenanthrene) και τριών βαρέων μετάλλων (Cu, Cd, Hg), χρησιμοποιήθηκε αιμόλεμφος 10 μυδιών από κάθε ομάδα ατόμων. Ακολουθήθηκε η διαδικασία που περιγράφεται στην παράγραφο Δ.3.α στα «Υλικά και Μέθοδοι», με τη διαφορά πως για την καταμέτρηση των συνολικών πυρηνικών αλλοιώσεων (TNA) στα αιμοκύτταρα δεν λήφθηκε υπόψη μόνο η συχνότητα εμφάνισης κυττάρων με ένα μικροπυρήνα, αλλά υπολογίσθηκε η συχνότητα εμφάνισης κυττάρων με έναν 48
 (Εικ. 7). α β γ δ Εικόνα 7. Αιμοκύτταρα μυδιών M.")
. 1000x Figure 7. Haemocytes from M.")


57 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ μικροπυρήνα, καθώς και κυττάρων που εμφάνισαν ταυτόχρονα δύο μικροπυρήνες, κυττάρων με δύο ξεχωριστούς πυρήνες και κυττάρων με πυρήνες σχήματος οκτώ (eight-shaped nucleus) (Εικ. 7). α β γ δ Εικόνα 7. Αιμοκύτταρα μυδιών M. galloprovincialis που εμφανίζουν ένα μικροπυρήνα (α βέλος), δύο μικροπυρήνες ταυτόχρονα (β - βέλη), δύο ξεχωριστούς πυρήνες (γ - βέλη) και πυρήνα σχήματος οκτώ (δ - βέλος). 1000x Figure 7. Haemocytes from M. galloprovincialis bearing one micronucleus (α - arrow), two micronuclei (β - arrows), two separate nuclei (γ - arrows) and eight-shaped nucleus (δ - arrow). 1000x 4. Βιομάρτυρας «εκτίμησης στρες» Κατά την άφιξή τους στο εργαστήριο, 30 μύδια από κάθε σταθμό δειγματοληψίας τοποθετήθηκαν σε δίσκους και εκτέθηκαν στον αέρα σε θερμοκρασία 18 o C. Η θνησιμότητα ελέγχονταν κάθε μέρα. Τα νεκρά άτομα καταγράφηκαν και αφαιρέθηκαν έως ότου επιτεύχθηκε η θνησιμότητα 100%. Τα μύδια θεωρούνταν νεκρά όταν δεν αντιδρούσαν μετά από πίεση του κελύφους τους, όταν οι θυρίδες τους ήταν ανοιχτές ή δεν ανταποκρίνονταν μετά από την τοποθέτησή τους σε θαλασσινό νερό. Στη συνέχεια υπολογίσθηκε με τη χρήση του Kaplan Mayer test ο χρόνος στον οποίο πεθαίνει το 50 % των ατόμων από κάθε ομάδα μυδιών (LT 50 ). 49
58 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ 5. Ιστοχημεία λυσοσωμάτων, ουδέτερων λιπιδίων, ολικών λιπιδίων, και λιποφουσκινών στον πεπτικό αδένα του μυδιού Mytilus galloprovincialis Προετοιμασία ιστών Για την ιστοχημική ανίχνευση των ολικών λιπιδίων, των ουδέτερων λιπιδίων και των λιποφουσκινών καθώς και για την ενζυμική αντίδραση για το λυσοσωμικό ένζυμο N-ακέτυλο-β-εξοζαμινιδάση πραγματοποιήθηκε φυσική μονιμοποίηση με ψύξη του πεπτικού αδένα μυδιών που είχαν μεταφερθεί ζωντανά στο εργαστήριο μέσα σε θερμός με παγοκύστεις. Για τη μονιμοποίηση των ιστών ακολουθήθηκε η διαδικασία που προτάθηκε από τους Moore (1976) και UNEP (1997) και περιγράφεται στα στάδια 1-4 της παραγράφου Δ.1 στα «Υλικά και Μέθοδοι». α) Ιστοχημεία για το λυσοσωμικό ένζυμο N-ακέτυλο-β-εξοζαμινδάση Για την εφαρμογή ιστοχημείας για το λυσοσωμικό ένζυμο N-ακέτυλο-βεξοζαμινιδάση εφαρμόσθηκε η διαδικασία που περιγράφεται στα στάδια 5-14 της παραγράφου Δ.1 στα «Υλικά και Μέθοδοι», με τη μόνη διαφορά πως παραλήφθηκε το στάδιο 7, δηλαδή η κατεργασία των τομών με το διάλυμα κιτρικού οξέος. Ακολούθησε παρατήρηση των τομών στο οπτικό μικροσκόπιο και προσδιορισμός της πυκνότητας όγκου των λυσοσωμάτων (Volume Density / VD) και της αριθμητικής πυκνότητας των λυσοσωμάτων (Numerical Density / ND). Για τον υπολογισμό των παραπάνω παραμέτρων χρησιμοποιήθηκε σύστημα ανάλυσης εικόνας που αποτελούνταν από μία JVC βιντεοκάμερα προσαρμοσμένη σε ένα οπτικό μικροσκόπιο τύπου Olympus CX41. Η εικόνα προβαλλόταν στην οθόνη ηλεκτρονικού υπολογιστή και επεξεργαζόταν με τη βοήθεια του προγράμματος Adobe Premier 5. Στις επιλεγμένες εικόνες τα λυσοσώματα διαχωρίζονταν από το κυτταρόπλασμα με τη βοήθεια κατάλληλου προγράμματος (EIKONA program). Ο αγωγός του πεπτικού αδένα καθώς και τμήματα του συνδετικού ιστού που περιέβαλαν τα πεπτικά σωληνάρια δεν λαμβάνονταν υπόψη κατά την ανάλυση. Πραγματοποιήθηκαν 50 μετρήσεις για την κάθε ομάδα μυδιών. 50
59 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ β) Ιστοχημεία ουδέτερων λιπιδίων 1. Λήψη τομών πάχους 9-10 nm σε κρυοστάτη τύπου Reichert-Jung Cryocut 1800 (Leica) σε θερμοκρασία -27 ο C. Οι τομές μεταφέρθηκαν σε γυάλινες αντικειμενοφόρες πλάκες και παρέμειναν σε θερμοκρασία δωματίου για 1-2 λεπτά έως ότου ξηρανθούν. 2. Μονιμοποίηση των ιστών με calcium formal για 10 min στο ψυγείο (4 ο C) 3. Πλύση με ddh2o. 4. Προσθήκη 60% triethylphosphate για 5 min. 5. Προσθήκη oil red 1% για 5 min. (Oil Red: 1gr oil red Ο σε 100 ml triethylphosphate 60%). 6. Πλύση με ddη2ο. 7. Μονιμοποίηση με glycergel 8. Παρατήρηση των τομών στο οπτικό μικροσκόπιο. 9. Προσδιορισμός της πυκνότητας όγκου (volume density / VD NL ) και της αριθμητικής πυκνότητας (numerical density / ND NL ) των ουδέτερων λιπιδίων, όπως περιγράφεται στην παράγραφο Δ.5.α στα «Υλικά και Μέθοδοι». γ) Ιστοχημεία ολικών λιπιδίων Με σκοπό τον εντοπισμό των ολικών λιπιδίων στα κύτταρα του πεπτικού αδένα, πραγματοποιήθηκε χρώση κρυοτομών των πεπτικών σωληναρίων με τη χρωστική Sudan black. Η διαδικασία που ακολουθήθηκε ήταν η εξής: 1. Λήψη τομών πάχους 9-10 nm σε κρυοστάτη τύπου Reichert-Jung Cryocut 1800 (Leica) σε θερμοκρασία -27 ο C. Οι τομές μεταφέρθηκαν σε γυάλινες αντικειμενοφόρες πλάκες και παρέμειναν σε θερμοκρασία δωματίου για 1-2 λεπτά έως ότου ξηρανθούν. 2. Μονιμοποίηση των ιστών με calcium formal για 10 min στο ψυγείο (4 ο C). 3. Πλύση με ddη2ο. 4. Προσθήκη sudan black για 5 min. 5. Πλύση με ddη2ο. 6. Μονιμοποίηση με glycergel. 7. Παρατήρηση των τομών στο οπτικό μικροσκόπιο. 51
60 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ δ) Ιστοχημεία λιποφουσκινών Με σκοπό τον εντοπισμό λιποφουσκινών στον πεπτικό αδένα των μυδιών που συλλέχθηκαν από διαφορετικές περιοχές δειγματοληψίας στις τρεις διαφορετικές δειγματοληπτικές περιόδους, πραγματοποιήθηκε η αντίδραση Schmorl, όπως έχει προταθεί από τον Moore (1988). Η διαδικασία που ακολουθήθηκε ήταν η εξής: 1. Λήψη τομών πάχους 9-10 nm σε κρυοστάτη τύπου Reichert-Jung Cryocut 1800 (Leica) σε θερμοκρασία -27 ο C. Οι τομές μεταφέρθηκαν σε γυάλινες αντικειμενοφόρες πλάκες και παρέμειναν σε θερμοκρασία δωματίου για 1-2 λεπτά έως ότου ξηρανθούν. 2. Μονιμοποίηση των ιστών με calcium formal για 10 min στο ψυγείο (4 ο C) 3. Πλύση με ddη2ο. 4. Προσθήκη 1% ferric chloride και 1% potassium ferricyanide (3.5:0.5) για 5 min. 5. Πλύση με acetic acid 1% για 1 min. 6. Πλύση με ddη2ο. 7. Μονιμοποίηση με glycergel 8. Εξέταση των τομών σε οπτικό μικροσκόπιο Olympus BH-2 9. Υπολογισμός της πυκνότητας όγκου των λιποφουσκινών (volume density / VD LF ), όπως περιγράφεται στην παράγραφο Δ.5.α στα «Υλικά και Μέθοδοι». Στατιστική ανάλυση δεδομένων Η εξαγωγή των στατιστικών διαφορών και οι συσχετίσεις των αποτελεσμάτων των βιομαρτύρων πραγματοποιήθηκαν με χρησιμοποίηση του στατιστικού πακέτου STATISTICA (STATISTICA statistical package, Microsoft Co. Auth, Greece). Τα αποτελέσματα του βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» επεξεργάστηκαν στατιστικά με τη χρήση μη παραμετρικών δοκιμασιών. Συγκεκριμένα, πραγματοποιήθηδε σύγκριση των αποτελεσμάτων με την εφαρμογή του Mann-Whitney-U-test (P<0.05). Τα δεδομένα που προέκυψαν από τους βιομάρτυρες «κατακράτηση της χρωστικής ουδέτερο ερυθρό», «συχνότητα μικροπυρήνων», «συχνότητα ολικών πυρηνικών ανωμαλιών», «εκτίμησης στρες», καθώς και τα αποτελέσματα της μορφομετρικής ανάλυσης των λιπιδίων, των λυσοσωμάτων και των λιποφουσκινών υποβλήθηκαν σε διαδικασία Komolvorov- Smirnov για να διαπιστωθεί αν ακολουθούσαν κανονική κατανομή. 52
61 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟΔΟΙ Πραγματοποιήθηκε λογαριθμική μετατροπή όλων των παραμέτρων που δεν ακολουθούσαν κανονική κατανομή και εφαρμογή παραμετρικών δοκιμασιών. Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών προσδιορίσθηκαν σε επίπεδο σημαντικότητας P<0.05 με τη χρήση του Duncan test. Οι συσχετίσεις (θετικές και αρνητικές) μεταξύ των βιολογικών παραμέτρων πραγματοποιήθηκαν με τη χρήση της στατιστικής μεθόδου απλής γραμμικής συσχέτισης (Correlation matrices, Pearson test, P<0.05), ενώ για την ομαδοποίηση των παραμέτρων και των περιοχών, ανάλογα με το βαθμό συγγένειας, χρησιμοποιήθηκε ανάλυση ομαδοποίησης (Cluster analysis, P<0.05), χρησιμοποιώντας την Ευκλείδεια απόσταση. 53
62 ΑΠΟΤΕΛΕΣΜΑΤΑ
63 ΑΠΟΤΕΛΕΣΜΑΤΑ Α. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΜΕΛΕΤΗ ΠΕΔΙΟΥ ΣΕ ΕΛΛΗΝΙΚΕΣ ΑΚΤΕΣ 1. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» Με σκοπό την εκτίμηση του βαθμού ρύπανσης διαφορετικών δειγματοληπτικών περιοχών του Θερμαϊκού κόλπου και μιας δειγματοληπτικής περιοχής του Στρυμονικού κόλπου (σταθμός ελέγχου), εφαρμόσθηκε η κυτοχημική τεχνική της «σταθερότητας της λυσοσωμικής μεμβράνης» (lysosomal membrane stability / LMS) στα λυσοσώματα των πεπτικών κυττάρων μυδιών M. galloprovincialis. Η παραπάνω τεχνική, στηρίζεται στη μελέτη της διαπερατότητας των λυσοσωμικών μεμβρανών, που μπορεί να διαταραχθεί λόγω της επίδρασης ρυπογόνων παραγόντων. Η μέθοδος αυτή, η οποία εκτιμάται σε min, συνιστά έναν διεθνώς καθιερωμένο, ευαίσθητο και αξιόπιστο βιομάρτυρα στρες τόσο για οργανικές, όσο και για ανόργανες ουσίες. Η εφαρμογή του βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» αποκάλυψε μια έντονη διαφοροποίηση στους δειγματοληπτικούς σταθμούς που εξετάσθηκαν. Κατά τη δειγματοληψία που πραγματοποιήθηκε τον Ιούνιο του 2000 παρατηρήθηκαν αυξημένες τιμές της τεχνικής της αυτής στον πεπτικό αδένα των μυδιών που συλλέχθηκαν από την περιοχή της Ολυμπιάδας (σταθμός ελέγχου), συγκριτικά με τα αντίστοιχα αποτελέσματα της τεχνικής στον πεπτικό αδένα των μυδιών από τις άλλες περιοχές δειγματοληψίας (Διαγρ. 1). Ειδικότερα, στον πεπτικό αδένα ατόμων που συλλέχθηκαν από την περιοχή της Ολυμπιάδας (σταθμός ελέγχου), η τιμή του συγκεκριμένου βιομάρτυρα βρέθηκε αρκετά αυξημένη (33 ± 3,769 min), συγκριτικά με τις τιμές που μετρήθηκαν στις περιοχές του Θερμαϊκού κόλπου και ιδιαίτερα στην περιοχή της Χαλάστρας (4,8 ± 1,507 min). Ανάλογα με τα αποτελέσματα του Ιουνίου, κυμάνθηκαν και οι τιμές του βιομάρτυρα κατά τη δειγματοληπτική περίοδο του Δεκεμβρίου του 2000 (Διαγρ. 1). Συγκεκριμένα, τα άτομα από την περιοχή της Ολυμπιάδας (σταθμός ελέγχου) εμφάνισαν την υψηλότερη τιμή για το συγκεκριμένο βιομάρτυρα (16 ± 2,676 min), με τη διαφορά πως η χαμηλότερη τιμή εμφανίσθηκε τώρα στην περιοχή του Καλοχωρίου (4,4 ± 1,957 min) και ακολούθησε η περιοχή της Χαλάστρας (4,87 ± 1,5 min). 54
64 ΑΠΟΤΕΛΕΣΜΑΤΑ Η εποχική εκτίμηση των αποτελεσμάτων κατέδειξε πως οι τιμές της «σταθερότητας της λυσοσωμικής μεμβράνης» ήταν χαμηλότερες το Δεκέμβριο σε όλους του σταθμούς δειγματοληψίας, σε σύγκριση με τις αντίστοιχες τιμές του Ιουνίου, με εξαίρεση μόνο το δειγματοληπτικό σταθμό της Χαλάστρας, όπου και στις δύο εποχές δειγματοληψίας μετρήθηκε παρόμοια, χαμηλή τιμή για το συγκεκριμένο βιομάρτυρα (Διαγρ. 1). Σταθερότητα της λυσοσωμικής μεμβράνης (LMS) (min) * * * * * * * * * * * * * * Ιούνιος 2000 Δεκέμβριος 2000 Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα Κύμινα Διάγραμμα 1. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) στον πεπτικό αδένα μυδιών M. galloprovincialis από πέντε σταθμούς δειγματοληψίας του Θερμαϊκού κι ένα σταθμό του Στρυμονικού κόλπου (σταθμός ελέγχου), τον Ιούνιο και το Δεκέμβριο του Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Mann-Whitney U test. Diagram 1. Lysosomal membrane stability (LMS) biomarker in the digestive gland cells of mussels M. galloprovincialis from five stations in Thermaikos and one station in Strymonikos Gulf (reference station), in June and December Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0,05). Statistical signification is based on Mann-Whitney U test. 2. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» Με σκοπό την εκτίμηση του βαθμού ρύπανσης διαφορετικών περιοχών του Θερμαϊκού και του Στρυμονικού κόλπου, εφαρμόσθηκε ο βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (neutral red retention assay / NRR) στα αιμοκυττάρα μυδιών M. galloprovincialis. Η παραπάνω τεχνική, η οποία εκτιμάται σε min, θεωρείται ευαίσθητος βιομάρτυρας γενικού στρες που προκαλείται 55
65 ΑΠΟΤΕΛΕΣΜΑΤΑ τόσο από οργανικές, όσο κι από ανόργανες ουσίες. Μικρότεροι χρόνοι της τεχνικής αφορούν σε μύδια των οποίων τα λυσοσώματα έχουν αποσταθεροποιηθεί, κι άρα μαρτυρούν μεγαλύτερη ρύπανση των θαλασσίων υδάτων. Αντίστοιχα, μεγαλύτεροι χρόνοι αφορούν μύδια που προέρχονται από καθαρότερες περιοχές. Κατακράτηση της χρωστικής ουδέτερο ερυθρό (NRR) (min) * * * * * * * * * * * * * * * * * Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα Κύμινα Ιούνιος 2000 Δεκέμβριος 2000 Διάγραμμα 2. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) στα αιμοκύτταρα μυδιών M. galloprovincialis από πέντε σταθμούς δειγματοληψίας του Θερμαϊκού κι ένα σταθμό του Στρυμονικού κόλπου (σταθμός ελέγχου), τον Ιούνιο και το Δεκέμβριο του Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 2. Neutral red retention assay (NRR) applied in the haemocytes of mussels M. galloprovincialis from five stations in Thermaikos and one station in Strymonikos Gulf (reference station), in June and December Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (p<0,05). Statistical signification is based on Duncan s test. Σύμφωνα με τα αποτελέσματα, η δυνατότητα κατακράτησης της χρωστικής ουδέτερο ερυθρό από τα λυσοσώματα αιμοκυττάρων μυδιών M. galloprovicialis παρουσίασε διακυμάνσεις ανάμεσα στις διάφορες περιοχές δειγματοληψίας. Συγκεκριμένα, η περιοχή της Ολυμπιάδας (σταθμός ελέγχου) εμφάνισε τη μεγαλύτερη τιμή για το βιομάρτυρα τόσο τον Ιούνιο (64,5 ± 10,124 min), όσο και το Δεκέμβριο (51 ± 8,125 min), σε σύγκριση με τους υπόλοιπους σταθμούς (Διαγρ. 2). Αντίθετα, τα μύδια από την περιοχή της Χαλάστρας παρουσίασαν τη μικρότερη δυνατότητα κατακράτησης της χρωστικής, εμφανίζοντας την ίδια τιμή της τεχνικής NRR (16,66 ± 5 min) και το Δεκέμβριο, αλλά και τον Ιούνιο (Διαγρ. 2). Και στις δύο εποχές δειγματοληψίας, η περιοχή αναφοράς (Ολυμπιάδα) και οι σταθμοί 56
66 ΑΠΟΤΕΛΕΣΜΑΤΑ δειγματοληψίας στις ανατολικές ακτές του Θερμαϊκού εμφάνισαν μεγαλύτερες τιμές αυτής της παραμέτρου σε σύγκριση με τους σταθμούς δειγματοληψίας στη δυτική πλευρά του κόλπου (Διαγρ. 2). 3. Βιομάρτυρας «συχνότητα μικροπυρήνων» Με σκοπό την ανίχνευση διαφορών μεταξύ των διαφορετικών δειγματοληπτικών σταθμών του Θερμαϊκού και του Στρυμονικού κόλπου που θα μπορούσαν να αποδοθούν στο διαφορετικό βαθμό ρύπανσης, μελετήθηκε το ποσοστό εμφάνισης αιμοκυττάρων με μικροπυρήνες (micronucleus frequency / MN) σε άτομα μυδιών M. galloprovincialis. Συχνότητα μικροπυρήνων (ΜΝ) ποσοστό μικροπυρήνων ( ) * * * * Ιούνιος 2000 Δεκέμβριος 2000 * Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα Κύμινα Διάγραμμα 3. Βιομάρτυρας «συχνότητα μικροπυρήνων» (MN) στα αιμοκύτταρα μυδιών M. galloprovincialis από πέντε σταθμούς δειγματοληψίας του Θερμαϊκού κι ένα σταθμό του Στρυμονικού κόλπου (σταθμός ελέγχου), τον Ιούνιο και το Δεκέμβριο του Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 3. Micronuclei frequency (MN) in the haemocytes of mussels M. galloprovincialis from five stations in Thermaikos and one station in Strymonikos Gulf (reference station), in June and December Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0,05). Statistical signification is based on Duncan s test. Η συχνότητα εμφάνισης κυττάρων με μικροπυρήνες στα άτομα μυδιών M. galloprovicialis διέφερε ανάμεσα στις περιοχές δειγματοληψίας. Το Δεκέμβριο, η 57
67 ΑΠΟΤΕΛΕΣΜΑΤΑ δημιουργία μικροπυρήνων ήταν μεγαλύτερη στα μύδια από την περιοχή του Καλοχωρίου (4,20 ± 2,553 ), ενώ η μικρότερη συχνότητα μικροπυρήνων καταγράφηκε στα μύδια από την Περαία (1,42 ± 0,767 ). Τον Ιούνιο, τα μύδια από την Περαία (6,48 ± 5,155 ) και το Αγγελοχώρι (4,04 ± 2,109 ) εμφάνισαν αυξημένη συχνότητα κυττάρων με μικροπυρήνες, σε αντίθεση με τα μύδια από την Ολυμπιάδα (σταθμός ελέγχου) (2,59 ± 2,582 ) που σημείωσαν τη χαμηλότερη τιμή αυτής της παραμέτρου (Διαγρ. 3). Το πρότυπο που καταγράφηκε από την εφαρμογή του βιομάρτυρα «συχνότητα μικροπυρήνων» τον Ιούνιο του 2000, δεν επαναλήφθηκε το Δεκέμβριο του Ωστόσο, παρατηρήθηκε μείωση στο σχηματισμό μικροπυρήνων κατά τη διάρκεια του χειμώνα στις περισσότερες περιοχές δειγματοληψίας (Διαγρ. 3). 4. Βιομάρτυρας «εκτίμησης stress» Με σκοπό την ανίχνευση διαφορών μεταξύ μυδιών διαφορετικών δειγματοληπτικών σταθμών του Θερμαϊκού και του Στρυμονικού κόλπου που θα μπορούσαν να αποδοθούν στο διαφορετικό βαθμό ρύπανσης των υδάτων εφαρμόσθηκε η τεχνική «εκτίμησης stress» (stress on stress / SOS). Πιο συγκεκριμένα, η τεχνική αυτή, της οποίας η τιμή αφορά την επιβίωση σε βάθος ημερών, καταγράφει το χρονικό διάστημα (LT 50 ) που μπορεί να επιβιώσει το 50% των μυδιών κατά την έκθεσή τους στον αέρα. Κατά τη δειγματοληψία του Ιουνίου τα μύδια από την περιοχή των Κυμίνων εμφάνισαν σημαντικά μειωμένες τιμές LT 50 (4 ημέρες), σε σύγκριση με τους υπόλοιπους σταθμούς δειγματοληψίας. Αντίθετα, τα μύδια από την Περαία κατέδειξαν τις υψηλότερες τιμές αυτής της παραμέτρου (LT 50 =10 ημέρες) (Διαγρ. 4). Το Δεκέμβριο, τα μύδια από την Ολυμπιάδα (σταθμός ελέγχου) παρουσίασαν τη μικρότερη αντοχή στην έκθεσή τους στον αέρα (LT 50 = 3 ημέρες) σε σύγκριση με τα άτομα από τις άλλες περιοχές, ενώ τα άτομα που προήλθαν από τη Χαλάστρα (LT 50 =9 ημέρες) και το Καλοχώρι (LT 50 =8 ημέρες) εμφάνισαν αυξημένη δυνατότητα επιβίωσης στον αέρα (Διαγρ. 4). 58
68 ΑΠΟΤΕΛΕΣΜΑΤΑ Βιομάρτυρας "εκτίμησης stress" (SOS) *** * * * * * * * * Ολυμπιάδα Αγγελοχώρι * Περαία Καλοχώρι Χαλάστρα Κύμινα 8 LT Ιούνιος 2000 Δεκέμβριος 2000 Διάγραμμα 4. Βιομάρτυρας «εκτίμησης stress» (SOS) σε μύδια M. galloprovincialis από πέντε σταθμούς δειγματοληψίας του Θερμαϊκού κι ένα σταθμό του Στρυμονικού κόλπου (σταθμός ελέγχου), τον Ιούνιο και τον Δεκέμβριο του Οι τιμές της τεχνικής αυτής αφορούν στο χρονικό διάστημα, εκφραζόμενο σε ημέρες (LT 50 ), που μπορεί να επιβιώσει το 50% των μυδιών, μετά από την έκθεσή τους στον αέρα. Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων, όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 4. Stress on stress technique (SOS) in mussels M. galloprovincialis collected from five stations in Thermaikos and one station in Strymonikos Gulf (reference station), in June and December LT 50 values, expressed in days, represent the time required to reach 50% mortality in mussels after aerial exposure. Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0,05). Statistical signification is based on Duncan s test. Τα αποτελέσματα της προαναφερθείσας τεχνικής φανερώνουν πως υπάρχει μείωση στις τιμές LT 50 το Δεκέμβριο, σε σύγκριση με τις αντίστοιχες τιμές του Ιουνίου, σε όλους τους σταθμούς δειγματοληψίας, με εξαίρεση τις περιοχές της Χαλάστρας και των Κυμίνων (Διαγρ. 4). 5. Μορφομετρικές και μορφολογικές αποκρίσεις των λυσοσωμάτων του πεπτικού αδένα Με σκοπό την εύρεση διαφορών μεταξύ μυδιών M. galloprovincialis από πέντε δειγματοληπτικούς σταθμούς του Θερμαϊκού κόλπου και ένα δειγματοληπτικό σταθμό του Στρυμονικού κόλπου, οι οποίες θα μπορούσαν να αποδοθούν στο 59
69 ΑΠΟΤΕΛΕΣΜΑΤΑ διαφορετικό βαθμό ρύπανσης και στην επίδραση εποχικών παραγόντων, πραγματοποιήθηκε μελέτη της δομής του πεπτικού αδένα μυδιών με το οπτικό μικροσκόπιο και εφαρμόσθηκε μορφομετρία σε παραμέτρους των λυσοσωμάτων, στις δύο διαφορετικές εποχές δειγματοληψίας. Πιο συγκεκριμένα, σε κρυοτομές πεπτικού αδένα που είχαν υποστεί χρώση για το ένζυμο N-ακετυλο-β-εξοζαμινιδάση, μετρήθηκαν η πυκνότητα όγκου (volume density / VD) και η αριθμητική πυκνότητα (numerical density / ND) των λυσοσωμάτων των πεπτικών κυττάρων, στο επίπεδο του οπτικού μικροσκοπίου. Τα αποτελέσματα της παρούσης μελέτης κατέδειξαν πως η χρώση για το λυσοσωμικό ένζυμο Ν-ακετυλο-β-εξοζαμινιδάση δεν περιορίστηκε μόνο σε περιοχές γύρω από τη λυσοσωμική μεμβράνη, αλλά τα λυσοσώματα εμφάνισαν ομοιόμορφη αντίδραση σε όλη τους την έκταση. Το χαρακτηριστικό αυτό επέτρεψε τον προσδιορισμό παραμέτρων που αφορούν τα δομικά χαρακτηριστικά των λυσοσωμάτων, όπως η πυκνότητα όγκου και η αριθμητική πυκνότητα των λυσοσωμάτων, μετά από επεξεργασία κρυοτομών για την ανίχνευση του συγκεκριμένου λυσοσωμικού ενζύμου. Ο υπολογισμός της πυκνότητας όγκου των λυσοσωμάτων των πεπτικών κυττάρων μυδιών M. galloprovincialis, μέγεθος που εκτιμά τον όγκο των λυσοσωμάτων ανά μονάδα όγκου του κυττάρου, αποκάλυψε ότι η πυκνότητα όγκου των οργανιδίων αυτών ήταν αυξημένη σε όλους τους σταθμούς δειγματοληψίας τον Ιούνιο, σε σχέση με το Δεκέμβριο. Μοναδική εξαίρεση αποτέλεσαν τα μύδια από την περιοχή των Κυμίνων, που εμφάνισαν ελαφρώς μεγαλύτερη τιμή αυτής της παραμέτρου το Δεκέμβριο (Διαγρ. 5α). Η εκτίμηση της αριθμητικής πυκνότητας των λυσοσωμάτων, μέγεθος που αντιπροσωπεύει τον αριθμό των λυσοσωμάτων ανά μονάδα όγκου του κυττάρου, αποκάλυψε ότι η τιμή της παραμέτρου αυτής σε όλους τους σταθμούς δειγματοληψίας ήταν μικρότερη τον Ιούνιο σε σχέση με το Δεκέμβριο (Διαγρ. 5β). Τα παραπάνω μορφομετρικά αποτελέσματα επιβεβαίωσαν τα συμπεράσματα από τις παρατηρήσεις κρυοτομών του πεπτικού αδένα που υπέστησαν χρώση για το ένζυμο N-ακέτυλο-β-εξοζαμινιδάση, σύμφωνα με τα οποία κατά την αρχή του καλοκαιριού τα μύδια εμφάνισαν πεπτικά σωληνάρια με μεγαλύτερα σε μέγεθος και λιγότερα σε αριθμό λυσοσώματα, σε σύγκριση με τα άτομα που συλλέχθηκαν το Δεκέμβριο (Εικ. 8, 9, 10), σε όλες τις περιοχές δειγματοληψίας. 60
70 ΑΠΟΤΕΛΕΣΜΑΤΑ Επίσης, η εξέταση εγκάρσιων τομών πεπτικών σωληναρίων από μύδια, τα οποία συλλέχθηκαν στην αρχή του καλοκαιρού, έδειξε ότι εμφανίζεται διαφοροποίηση στο μέγεθος των λυσοσωμάτων ανάμεσα στα μύδια που συλλέχθηκαν από διαφορετικές περιοχές δειγματοληψίας. Συγκεκριμένα, τα λυσοσώματα των πεπτικών κυττάρων μυδιών από την περιοχή της Χαλάστρας και των Κυμίνων ήταν μικρότερα σε μέγεθος, σε σύγκριση με τα μεγαλύτερα λυσοσώματα των μυδιών από τις υπόλοιπες περιοχές που εξετάσθηκαν (Εικ. 8, 11). 61
71 ΑΠΟΤΕΛΕΣΜΑΤΑ Πυκνότητα όγκου λυσοσωμάτων (VD) VD (μm 3 /μm 3 ) α 0,3 0,25 0,2 0,15 * * * * * * * * * * * * ** * * * Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα Κύμινα 0,1 0,05 0 Ιούνιος 2000 Δεκέμβριος 2000 Αριθμητική πυκνότητα λυσοσωμάτων (ΝD) β ΝD (αριθμός/μm 3 ) 0 0,01 0,02 0,03 * * * * * * * * * * * * * Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα Κύμινα Ιούνιος 2000 Δεκέμβριος 2000 Διάγραμμα 5. Μορφομετρία λυσοσωμικών παραμέτρων του πεπτικού αδένα μυδιών M. galloprovincialis από πέντε σταθμούς δειγματοληψίας του Θερμαϊκού κι ένα σταθμό του Στρυμονικού κόλπου (σταθμός ελέγχου) τον Ιούνιο και το Δεκέμβριο του α. Πυκνότητα όγκου λυσοσωμάτων (VD) β. Αριθμητική πυκνότητα λυσοσωμάτων (ND) Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 5. Results on the morphometry of lysosomal parameters of the digestive lysosomes of mussels M. galloprovincialis collected from five stations in Thermaikos and one station in Strymonikos Gulf (reference station) in June and December α. Volume density of lysosomes (VD) β. Numerical density of lysosomes (ND) Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0,05). Statistical signification is based on Duncan s test. 62



")
, το")
 και τα Κύμινα (φ ω)")









 Figure 8.")


, Peraia (ι μ),")
 and Kymina (φ ω) in June")
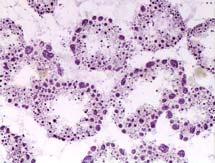
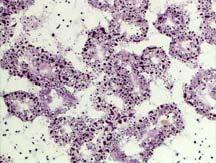
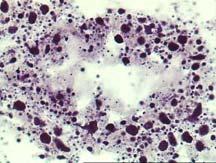
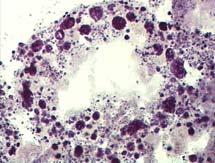
72 Ιούνιος 2000 Ολυμπιάδα Αγγελοχώρι α β ε ζ γ δ η θ Περαία Καλοχώρι ι κ ν ξ λ μ ο π Χαλάστρα Κύμινα ρ σ φ χ τ υ Εικόνα 8. Πεπτικά σωληνάρια μυδιών M. galloprovincialis από την Ολυμπιάδα (σταθμός ελέγχου) (α δ), το Αγγελοχώρι (ε θ), την Περαία (ι μ), το Καλοχώρι (ν π), τη Χαλάστρα (ρ υ) και τα Κύμινα (φ ω) τον Ιούνιο του Τα λυσοσώματα των πεπτικών κυττάρων γίνονται ορατά μετά από χρώση για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση σε κρυοτομές του πεπτικού αδένα. Κατά την περίοδο αυτή, τα πεπτικά κύτταρα των μυδιών όλων των περιοχών εμφανίζουν, σε γενικές γραμμές, μεγάλου μεγέθους λυσοσώματα που χαρακτηρίζονται από ισχυρή αντίδραση για το λυσοσωμικό ένζυμο. (α, β, ε, ζ, ι, κ, ν, ξ, ρ, σ, φ, χ : 550x γ, δ, η, θ, λ, μ, ο, π, τ, υ, ψ, ω: 1100x) Figure 8. Digestive tubules of mussels M. galloprovincialis from Olympiada (reference station) (α δ), Aggelohori (ε θ), Peraia (ι μ), Kalohori (ν π), Halastra (ρ υ) and Kymina (φ ω) in June Digestive cell lysosomes have been visualized using N-acetyl-β-hexosaminidase histochemistry on digestive gland cryosections. Digestive cells of mussels, collected from all sampling stations, display during this period enlarged lysosomes which strongly react on the lysosomal enzyme (α, β, ε, ζ, ι, κ, ν, ξ, ρ, σ, φ, χ : 550x γ, δ, η, θ, λ, μ, ο, π, τ, υ, ψ, ω: 1100x) ψ ω 63




 (α δ), το")
, το")
 και τα")











 (α δ),")
, Kalohori (ν π),")
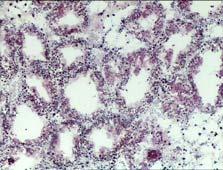
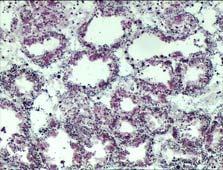
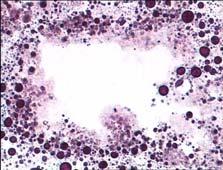
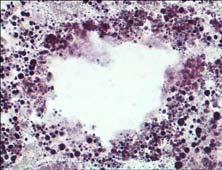
73 Δεκέμβριος 2000 Ολυμπιάδα Αγγελοχώρι α β ε ζ γ δ η θ Περαία Καλοχώρι ι κ ν ξ λ μ ο π Χαλάστρα Κύμινα ρ σ φ χ τ υ ψ ω Εικόνα 9. Πεπτικά σωληνάρια μυδιών M. galloprovincialis από την Ολυμπιάδα (σταθμός ελέγχου) (α δ), το Αγγελοχώρι (ε θ), την Περαία (ι μ), το Καλοχώρι (ν π), τη Χαλάστρα (ρ υ) και τα Κύμινα (φ ω) το Δεκέμβριο του Τα λυσοσώματα των πεπτικών κυττάρων γίνονται ορατά μετά από χρώση για το λυσοσωμικό ένζυμο Ν-ακετυλ-βεξοζαμινιδάση σε κρυοτομές του πεπτικού αδένα. Τα πεπτικά κύτταρα των μυδιών όλων των περιοχών εμφανίζουν, σε γενικές γραμμές, μικρά σε μέγεθος λυσοσώματα. (α, β, ε, ζ, ι, κ, ν, ξ, ρ, σ, φ, χ : 550x γ, δ, η, θ, λ, μ, ο, π, τ, υ, ψ, ω: 1100x) Figure 9. Digestive tubules of mussels M. galloprovincialis from Olympiada (reference station) (α δ), Aggelohori (ε θ), Peraia (ι μ), Kalohori (ν π), Halastra (ρ υ) and Kymina (φ ω) in early winter. Digestive cell lysosomes have been visualized using N-acetyl-β-hexosaminidase histochemistry on digestive gland cryosections. Digestive cells of mussels, collected from all sampling stations, display small sized lysosomes. (α, β, ε, ζ, ι, κ, ν, ξ, ρ, σ, φ, χ : 550x γ, δ, η, θ, λ, μ, ο, π, τ, υ, ψ, ω: 1100x) 64

")
 με μικρότερο μέγεθος (β).")

74 ΑΠΟΤΕΛΕΣΜΑΤΑ Ιούνιος 2000 Δεκέμβριος 2000 α β γ δ Εικόνα 10. Ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση σε κρυοτομές πεπτικού αδένα μυδιών M. galloprovincialis από την περιοχή της Περαίας τον Ιούνιο και το Δεκέμβριο του Τον Ιούνιο τα πεπτικά κύτταρα των μυδιών εμφανίζουν ένα ιδιαίτερα ενεργό λυσοσωμικό σύστημα, που χαρακτηρίζεται από λυσοσώματα (βέλος) αυξημένου μεγέθους (α). Στη δειγματοληψία του Δεκεμβρίου τα πεπτικά κύτταρα χαρακτηρίζονται από λυσοσώματα (βέλος) με μικρότερο μέγεθος (β). Οι εικόνες γ και δ αποτελούν μεγαλύτερες μεγεθύνσεις των α και β, αντίστοιχα. α, β : 900x γ, δ : 1800x Figure 10. N-acetyl-β-hexosaminidase histochemistry on digestive gland cryosections from mussels M. galloprovincialis, collected from Peraia in June and December In June the digestive cells of mussels display a very active lysosomal system, presenting increased lysosomal size (arrow) (α). In December the digestive cells are characterized by small-sized lysosomes (arrow) (β). Figures γ and δ represent higher magnification of α and β, respectively. α, β : 900x γ, δ : 1800x 65

 στα μύδια από την περιοχή της Ολυμπιάδας (α) σε σχέση με τα")


75 ΑΠΟΤΕΛΕΣΜΑΤΑ Ολυμπιάδα Χαλάστρα α β γ δ Εικόνα 11. Ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση σε κρυοτομές πεπτικού αδένα μυδιών M. galloprovincialis τον Ιούνιο του 2000 από την περιοχή της Ολυμπιάδας και της Χαλάστρας. Διακρίνεται το μεγαλύτερο μέγεθος των λυσοσωμάτων (βέλος) στα μύδια από την περιοχή της Ολυμπιάδας (α) σε σχέση με τα μικρότερα λυσοσώματα (βέλος) που εμφανίζονται στα μύδια από την περιοχή της Χαλάστρας (β). Οι φωτογραφίες γ και δ αποτελούν μεγαλύτερες μεγεθύνσεις των α και β, αντίστοιχα. α, β : 900x γ, δ : 1800x Figure 11. N-acetyl-β-hexosaminidase histochemistry on digestive gland cryosections from mussels M. galloprovincialis, collected in June 2000 from Olympiada and Halastra. Mussels from Olympiada (α) display increased lysosomal size (arrow) compared to mussels from Halastra (arrow) (β). Figures γ and δ represent higher magnification of α and β, respectively. α, β : 900x γ, δ : 1800x 66
76 ΑΠΟΤΕΛΕΣΜΑΤΑ 6. Συσχέτιση μεταξύ των αποτελεσμάτων βιομαρτύρων στη μελέτη πεδίου Με δεδομένη την ευαισθησία και ακρίβεια του βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) και με σκοπό την εκτίμηση της ευαισθησίας των υπόλοιπων βιομαρτύρων που χρησιμοποιήθηκαν, πραγματοποιήθηκε ανάλυση συσχέτισης (correlation analysis) μεταξύ των βιολογικών παραμέτρων. Τα αποτελέσματα της συσχέτισης των βιολογικών παραμέτρων που εξετάστηκαν (Pearson s test P<0,05) αναγράφονται στον πίνακα 2. Τα κύρια σημεία της συσχέτισης αυτής αναφέρονται παρακάτω: Σημαντική συσχέτιση (θετική) διαπιστώθηκε μεταξύ των αποτελεσμάτων του βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) και του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR), τόσο κατά τη περίοδο του Ιουνίου (r=0,83) (Διαγρ. 6), όσο και κατά την περίοδο του Δεκεμβρίου (r=0,89) (Διαγρ. 7). Υψηλή συσχέτιση (αρνητική) καταγράφηκε μεταξύ των αποτελεσμάτων των βιομαρτύρων «εκτίμησης stress» (SOS) και «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS) (r=-0,94) κατά τη δειγματοληψία του χειμώνα (Διαγρ. 8). Υψηλή συσχέτιση (αρνητική) καταγράφηκε μεταξύ των αποτελεσμάτων των βιομαρτύρων «εκτίμησης stress» (SOS) και «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) (r=-0,97) κατά τη δειγματοληψία του χειμώνα (Διαγρ. 9). Σημαντική συσχέτιση (θετική) εμφανίσθηκε ανάμεσα στις τιμές που προέκυψαν το Δεκέμβριο και τον Ιούνιο, όσον αφορά το βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) (r=+0,86) (Διαγρ. 10). Σημαντική συσχέτιση (θετική) εμφανίσθηκε ανάμεσα στις τιμές που προέκυψαν το Δεκέμβριο και τον Ιούνιο, όσον αφορά το βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) (r=+0,97) (Διαγρ. 11). 67
77 ΑΠΟΤΕΛΕΣΜΑΤΑ Πίνακας 2. Συντελεστές γραμμικής συσχέτισης μεταξύ των βιολογικών παραμέτρων που εξετάστηκαν. Με κόκκινα νούμερα συμβολίζονται οι στατιστικά σημαντικές συσχετίσεις (Pearson test, P<0,05). Τable 2. Correlation coefficients between the variables under test. Values in red indicate a statistically significant correlation (Pearson test, P<0,05). LMS S NRR S MN S SOS S VD S ND S LMS W NRR W MN W SOS W VD W ND W LMS S 1 0,84-0,33-0,60 0,02-0,02 0,86 0,8 0,07-0,82-0,24-0,86 NRR S 0,84 1 0,21-0,24 0,27-0,19 0,95 0,97-0,12-0,96-0,47-0,64 MN S -0,33 0,21 1 0,44 0,48-0,42 0,03 0,29-0,41-0,22-0,25 0,35 SOS S -0,6-0,24 0,44 1 0,26 0,11-0,26-0,36 0,06 0,37-0,42 0,72 VD S 0,02 0,27 0,48 0,26 1-0,55-0,04 0,27 0,37-0,07-0,54-0,07 ND S -0,02-0,19-0,42 0,11-0,55 1-0,03-0,35-0,45 0,15-0,31-0,18 LMS W 0,86 0,95 0,03-0,26-0,04-0,33 1 0,89-0,16-0,95-0,33-0,58 NRR W 0,8 0,97 0,29-0,36 0,27-0,35 0,89 1-0,16-0,97 0,29-0,61 MN W 0,07-0,12-0,41 0,06 0,37-0,45-0,16-0,16 1 0,3 0,12 0,12 SOS W -0,82-0,96-0,22 0,37-0,07 0,15-0,95-0,97 0,3 1 0,3 0,64 VD W -0,24-0,47-0,25-0,42-0,54 0,31-0,33-0,29 0,12 0,3 1 0,32 ND W -0,86-0,64 0,35 0,72-0,07-0,18-0,58-0,61 0,12 0,64 0,32 1 LMS S: Σταθερότητα της λυσοσωμικής μεμβράνης το καλοκαίρι, NRR S: Κατακράτηση του ουδέτερου ερυθρού το καλοκαίρι, ΜΝ S: Συχνότητα μικροπυρήνων το καλοκαίρι, SOS S: τεχνική εκτίμησης stress το καλοκαίρι, VD S: πυκνότητα όγκου λυσοσωμάτων το καλοκαίρι, ND S: αριθμητική πυκνότητα το καλοκαίρι, LMS W: Σταθερότητα της λυσοσωμικής μεμβράνης το χειμώνα, NRR W: Κατακράτηση του ουδέτερου ερυθρού το χειμώνα, ΜΝ W: Συχνότητα μικροπυρήνων το χειμώνα, SOS W: τεχνική εκτίμησης stress το χειμώνα, VD W: πυκνότητα όγκου λυσοσωμάτων το χειμώνα, ND W: αριθμητική πυκνότητα λυσοσωμάτων το χειμώνα LMS S : Lysosomal membrane stability test in summer, NRR S:Neutral red retention assay in summer, ΜΝ S: Micronucleus frequency in summer, VD S:Volume density of lysosomes in summer, ND S: Numerical density of lysosomes in summer, LMS W : Lysosomal membrane stability test in winter, NRR W:Neutral red retention assay in winter, ΜΝ W: Micronucleus frequency in winter, VD W:Volume density of lysosomes in winter, ND W: Numerical density of lysosomes in winter, 70 LMS vs. NRR (Ιούνιος) NRR = 16, ,4816 * LMS Συντελεστής συσχέτισης: r =, NRR LMS Διάγραμμα 6. Θετική συσχέτιση των τιμών του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) και των τιμών της «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS), στα μύδια που συλλέχθηκαν τον Ιούνιο του Diagram 6. Possitive correlation between the neutral red retention assay (NRR) results and the lysosomal membrane stability values (LMS), in mussels collected in June
78 ΑΠΟΤΕΛΕΣΜΑΤΑ 55 LMS vs. NRR (Δεκέμβριος) NRR = 12, ,4488 * LMS Συντελεστής συσχέτισης: r =, NRR LMS Διάγραμμα 7. Θετική συσχέτιση των τιμών των βιομαρτύρων «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) και «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS), στα μύδια που συλλέχθηκαν το Δεκέμβριο του Diagram 7. Possitive correlation between the neutral red retention assay (NRR) results and the lysosomal membrane stability values (LMS), in mussels collected in December LMS vs.sos (Δεκέμβριος) SOS = 10,434 -,4317 * LMS Συντελεστής συσχέτισης: r = -, SOS LMS Διάγραμμα 8. Αρνητική συσχέτιση των τιμών του βιομάρτυρα «εκτίμησης stress» (SOS) και των τιμών του βιομάρτυρα «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS), στα μύδια που συλλέχθηκαν το Δεκέμβριο του Diagram 8. Negative correlation between the values obtained from the application of the technique stress on stress (SOS) and the lysosomal membrane stability values (LMS), in mussels collected in December
79 LMS Ιουνίου vs. LMS MΔεκεμβρίουLSΔεκΣεμυβντρίελοευσ=τή3,ς3σ0υ4+χέτ8σ0,ισ9*ς:r3ηL=+MS0,8ΙουνίουΑΠΟΤΕΛΕΣΜΑΤΑ 10 NRR vs.sos (Δεκέμβριος) SOS = 12,177 -,1614 * NRR Συντελεστής συσχέτισης: r = -, SOS NRR Διάγραμμα 9. Αρνητική συσχέτιση των τιμών του βιομάρτυρα «εκτίμησης stress» (SOS) και των τιμών του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (ΝRR), στα μύδια που συλλέχθηκαν το Δεκέμβριο του Diagram 9. Negative correlation between the values obtained from the application of the biomarkers stress on stress (SOS) and neurtal red retention (NRR), in mussels collected in December LMS Δεκεμβρίου LMS Ιουνίου Διάγραμμα 10. Θετική συσχέτιση των τιμών του βιομάρτυρα «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS), στα μύδια που συλλέχθηκαν το Δεκέμβριο και στα μύδια που συλλέχθηκαν τον Ιούνιο του Diagram 10. Possitive correlation of the lysosomal membrane stability values (LMS), obtained from the mussels collected in December and the mussels collected in June
80 ΑΠΟΤΕΛΕΣΜΑΤΑ 60 NRR Δεκεμβρίου vs. NRR Ιουνίου NRR Δεκεμβρίου=0, ,6984*NRR Ιουνίου Συντελεστής συσχέτισης; r = +0, NRR Δεκεμβρίου NRR Ιουνίου Διάγραμμα 11. Θετική συσχέτιση των τιμών του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (ΝRR), στα μύδια που συλλέχθηκαν το Δεκέμβριο και στα μύδια που συλλέχθηκαν τον Ιούνιο του Diagram 11. Possitive correlation of the neurtal red retention values (NRR), obtained from the mussels collected in December and the mussels collected in June Ομαδοποίηση των σταθμών δειγματοληψίας σύμφωνα με τα αποτελέσματα των βιομαρτύρων Η ανάλυση των τιμών των βιομαρτύρων οδηγεί σε σημαντικά στοιχεία σχετικά με τη διαφοροποίηση των περιοχών δειγματοληψίας. Για την εξαγωγή των αποτελεσμάτων χρησιμοποιήθηκε στατιστική ανάλυση με την οποία διαπιστώνεται η συγγένεια των σταθμών δειγματοληψίας με βάση τις μετρήσιμες τιμές κάθε παραμέτρου (Cluster Analysis, Statistica package, Microsoft Co., Greece), χρησιμοποιώντας την Ευκλείδεια απόσταση. Συγκεκριμένα, από τα εξαγόμενα αποτελέσματα διαπιστώθηκε σημαντική διαφοροποίηση της Ολυμπιάδας (σταθμός ελέγχου) σε σχέση με τους σταθμούς του Θερμαϊκού κόλπου, βάσει των τιμών της «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS) στον πεπτικό αδένα των ατόμων που συλλέχθηκαν κατά τη διάρκεια των δειγματοληψιών στις δύο διαφορετικές εποχές του έτους (Διαγρ. 12). Από τις περιοχές του Θερμαϊκού κόλπου μεγαλύτερη συγγένεια εμφάνισε η Χαλάστρα με το Καλοχώρι, καθώς και η Περαία με το Αγγελοχώρι (Διαγρ. 12). 71
81 ΑΠΟΤΕΛΕΣΜΑΤΑ 04 LMS καλοκαίρι - χειμώνας Χαλάστρα Καλοχώρι Κύμινα Περαία Αγγελοχώρι Ολυμπιάδα Διάγραμμα 12. Διαφοροποίηση των σταθμών σύμφωνα με τα αποτελέσματα του βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) στον πεπτικό αδένα των μυδιών που συλλέχθηκαν και στις δύο εποχές δειγματοληψίας (Δεκέμβριο και Ιούνιο). Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 12. Relationships between stations concerning lysosomal membrane stability (LMS) values in the digestive gland of mussels, sampled in December and June, as occurred after cluster analysis (Statistica software package), using Euclidean distance. Από την μελέτη των αποτελεσμάτων της τεχνικής «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) στις δύο εποχές δειγματοληψίας, διαπιστώθηκε σαφής διαφοροποίηση των ανατολικών περιοχών του κόλπου από τους σταθμούς δειγματοληψίας που βρίσκονται στη δυτική πλευρά. Συγκεκριμένα, εμφανίζεται ομαδοποίηση της Περαίας και του Αγγελοχωρίου, περιοχές που εμφανίζουν μεγάλη συγγένεια και με την περιοχή της Ολυμπιάδας (σταθμός ελέγχου). Επίσης, οι δυτικές περιοχές του Θερμαϊκού παρουσιάζουν ομαδοποίηση, με μεγαλύτερη συγγένεια να παρατηρείται μεταξύ των Κυμίνων και του Καλοχωρίου (Διαγρ. 13). 72
82 ΑΠΟΤΕΛΕΣΜΑΤΑ 4,0 NRR καλοκαίρι - χειμώνας 3,5 3,0 2,5 2,0 1,5 1,0 0,5 0,0 Χαλάστρα Κύμινα Καλοχώρι Περαία Αγγελοχώρι Ολυμπιάδα Διάγραμμα 13. Διαφοροποίηση των σταθμών σύμφωνα με τα αποτελέσματα του βιομάρτυρα «κατακράτηση του ουδέτερου ερυθρού» (NRR) στα αιμοκύτταρα των μυδιών που συλλέχθηκαν και στις δύο εποχές δειγματοληψίας (Δεκέμβριο και Ιούνιο). Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 13. Relationships between stations concerning neutral red retention (NRR) assay s results as occurred after cluster analysis (Statistica software package). The technique was performed on mussel haemocytes, in December and June, using Euclidean distance. Από τον υπολογισμό του ποσοστού των μικροπυρήνων σε όλους τους σταθμούς δειγματοληψίας κατά τις δύο διαφορετικές εποχές, προέκυψε σημαντική διαφοροποίηση της Περαίας και του Καλοχωρίου σε σχέση με τις υπόλοιπες περιοχές, ενώ μεγαλύτερη συγγένεια εμφανίσθηκε ανάμεσα στα Κύμινα και τη Χαλάστρα (Διαγρ. 14). Επίσης, τα αποτελέσματα του βιομάρτυρα «εκτίμηση stress» κατέδειξαν μεγάλη συγγένεια ανάμεσα στο Καλοχώρι και τη Χαλάστρα, καθώς και στο Αγγελοχώρι με την Ολυμπιάδα (σταθμός ελέγχου) (Διαγρ. 15). Η Περαία εμφανίζει μεγαλύτερη συγγένεια με την ομάδα του Αγγελοχωρίου και της Ολυμπιάδας, ενώ τα Κύμινα είναι αυτά που διαφοροποιούνται περισσότερο από όλες τις υπόλοιπες περιοχές (Διαγρ. 15). Τα αποτελέσματα των μορφομετρικών μετρήσεων δεν κατέδειξαν κάποιο σαφές πρότυπο ομαδοποίησης των περιοχών που εξετάσθηκαν (Διαγρ. 16, 17). Τέλος, από την προσπάθεια ομαδοποίησης των βιομαρτύρων που εξετάσθηκαν προέκυψε μεγαλύτερη συγγένεια ανάμεσα στις τεχνικές LMS και NRR, οι οποίες διαφοροποιούνται αρκετά από τις υπόλοιπες τεχνικές που εφαρμόσθηκαν (Διαγρ. 18). 73
83 ΑΠΟΤΕΛΕΣΜΑΤΑ 4,0 ΜΝ καλοκαίρι - χειμώνας 3,5 3,0 2,5 2,0 1,5 1,0 0,5 0,0 Περαία Αγγελοχώρι Καλοχώρι Κύμινα Χαλάστρα Ολυμπιάδα Διάγραμμα 14. Διαφοροποίηση των σταθμών σύμφωνα με τα αποτελέσματα του βιομάρτυρα «συχνότητα μικροπυρήνων» (ΜΝ) στα αιμοκύτταρα των μυδιών που συλλέχθηκαν και στις δύο εποχές δειγματοληψίας (Δεκέμβριο και Ιούνιο). Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 14. Relationships between stations concerning micronucleus frequency (MN) technique as occurred after cluster analysis (Statistica software package). The technique was performed on mussel haemocytes, in December and June, using Euclidean distance. 3,5 SOS καλοκαίρι - χειμώνας 3,0 2,5 2,0 1,5 1,0 0,5 0,0 Κύμινα Χαλάστρα Καλοχώρι Περαία Αγγελοχώρι Ολυμπιάδα Διάγραμμα 15. Διαφοροποίηση των σταθμών σύμφωνα με τα αποτελέσματα του βιομάρτυρα «εκτίμησης stress» (SOS) σε μύδια που συλλέχθηκαν και στις δύο εποχές δειγματοληψίας (Δεκέμβριο και Ιούνιο). Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package) χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 15. Relationships between stations concerning stress on stress (SOS) technique as occurred after cluster analysis (Statistica software package). The technique was performed on mussels sampled in December and June, using Euklidean distance. 74
84 ΑΠΟΤΕΛΕΣΜΑΤΑ 3,0 VD καλοκαίρι - χειμώνας 2,5 2,0 1,5 1,0 0,5 Χαλάστρα Κύμινα Αγγελοχώρι Καλοχώρι Περαία Ολυμπιάδα Διάγραμμα 16. Διαφοροποίηση των σταθμών σύμφωνα με τα αποτελέσματα του υπολογισμού της «πυκνότητας όγκου» (VD) των λυσοσωμάτων σε μύδια που συλλέχθηκαν και στις δύο εποχές δειγματοληψίας (Δεκέμβριο και Ιούνιο). Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 16. Relationships between stations concerning volume density (VD) of lysosomes as occurred after cluster analysis (Statistica software package). The technique was performed on the digestive gland of mussels sampled in December and June, using Euclidean distance. 3,0 ND καλοκαίρι - χειμώνας 2,5 2,0 1,5 1,0 0,5 0,0 Κύμινα Περαία Καλοχώρι Χαλάστρα Αγγελοχώρι Ολυμπιάδα Διάγραμμα 17. Διαφοροποίηση των σταθμών σύμφωνα με τα αποτελέσματα του υπολογισμού της «αριθμητικής πυκνότητας» (ND) των λυσοσωμάτων σε μύδια που συλλέχθηκαν και στις δύο εποχές δειγματοληψίας (Δεκέμβριο και Ιούνιο). Για την εξαγωγή των αποτελεσμάτων εφαρφμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 17. Relationships between stations concerning numerical density (ND) of lysosomes as occurred after cluster analysis (Statistica software package). The technique was performed on the digestive gland of mussels sampled in December and June, using Euclidean distance. 75
85 ΑΠΟΤΕΛΕΣΜΑΤΑ 4,5 Ομαδοποίηση βιομαρτύρων 4,0 3,5 3,0 2,5 2,0 1,5 1,0 0,5 0,0 VD W SOS W VD S MN S NRR W LMS W ND W MN W SOS S ND S NRR S LMS S Διάγραμμα 18. Διαφοροποίηση των βιομαρτύρων σε μύδια που συλλέχθηκαν από το Θερμαϊκό και το Στρυμονικό κόλπο και στις δύο εποχές δειγματοληψίας (Δεκέμβριο και Ιούνιο). Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 18. Relationships between biomarkers applied on mussels sampled from Thermaikos and Strymonikos Gulf in both sampling seasons (December and June). The results were occurred after cluster analysis (Statistica software package), using Euclidean distance. 76
86 ΑΠΟΤΕΛΕΣΜΑΤΑ Β. ΔΙΕΡΕΥΝΗΣΗ ΤΗΣ ΑΞΙΟΠΙΣΤΙΑΣ ΒΙΟΜΑΡΤΥΡΩΝ ΡΥΠΑΝΣΗΣ ΣΤΟ ΜΥΔΙ MYTILUS GALLOPROVINCIALIS. ΕΡΓΑΣΤΗΡΙΑΚΗ ΜΕΛΕΤΗ ΜΕΤΑ ΑΠΟ ΕΚΘΕΣΗ ΣΤΑ ΒΑΡΕΑ ΜΕΤΑΛΛΑ Cu, Cd, Hg KAI ΣΤΟΝ ΟΡΓΑΝΙΚΟ ΡΥΠΟ ΦΑΙΝΑΝΘΡΕΝΙΟ 1. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» Ο βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» (lysosomal membrane stability / LMS) εφαρμόσθηκε σε κρυοτομές πεπτικού αδένα μυδιών και κατέδειξε πως υπάρχει μείωση στις τιμές της τεχνικής που αφορούν στην αποσταθεροποίηση της λυσοσωμικής μεμβράνης, μετά από την έκθεση των ατόμων στον οργανικό ρύπο και στα βαρέα μέταλλα. χρόνος (min) Σταθερότητα της λυσοσωμικής μεμβράνης (LMS) * * ** * * 0 Control Ακετόνη Cu Φαινανθρένιο Cd Hg Διάγραμμα 19. Βιομάρτυρας «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) στον πεπτικό αδένα μυδιών M. galloprovincialis μετά από έκθεση σε 0.1mg L -1 ρύπων για 15 ημέρες. Οι στατιστικώς σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0.05) και προσδιορίσθηκαν με τη χρήση του Mann-Whitney U test. Diagram 19. Lysosomal membrane stability (LMS) biomarker in the digestive gland cells of mussels M. galloprovincialis exposed to 0.1mg L -1 of various pollutants for 15 days. Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0.05). Statistical signification is based on Mann-Whitney U test. 77
87 ΑΠΟΤΕΛΕΣΜΑΤΑ Συγκεκριμένα, η στατιστική επεξεργασία των αποτελεσμάτων (Mann Whitney U-test, P<0.05) φανέρωσε πως στατιστικώς σημαντικά χαμηλότερες τιμές του συγκεκριμένου βιομάρτυρα εμφανίζονται στα μύδια που δέχθηκαν την επίδραση Cu (9.166 ± min), Cd (6 ± 1.3 min) και Hg (11,5 ± 4.27 min), σε σύγκριση με τα μύδια ελέγχου (17,5 ± min) (Διαγρ. 19). Επίσης, τα μύδια που δέχθηκαν την επίδραση της ουσίας φαινανθρένιο παρουσίασαν μείωση στην τιμή της συγκεκριμένης τεχνικής (13.5 ± 2.85 min) σε σχέση με τα άτομα ελέγχου, δηλαδή αυτά που δέχθηκαν την επίδραση ακετόνης (16.25 ± 3.75 min), χωρίς όμως η μείωση αυτή να αποτελεί στατιστικώς σημαντική διαφορά (Διάγρ. 19). 2. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» Η αποσταθεροποίηση της λυσοσωμικής μεμβράνης των αιμοκυττάρων μυδιών M. galloprovincialisιs εκτιμήθηκε με την τεχνική «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (neutral red retention assay / NRR), μετά από έκθεση των ατόμων στον οργανικό ρύπο φαινανθρένιο (phenanthrene) και σε τρία βαρέα μέταλλα (Cu, Cd, Hg). Όπως προαναφέρθηκε, μικρότεροι χρόνοι της τεχνικής αφορούν σε μύδια των οποίων τα λυσοσώματα έχουν αποσταθεροποιηθεί και άρα μαρτυρούν μεγαλύτερη περιβαλλοντική επιβάρυνση. Αντίθετα, μεγαλύτεροι χρόνοι αφορούν μύδια που διαβιούν σε περιβάλλον με μικρότερη ρύπανση. Σύμφωνα με τα αποτελέσματα, οι χρόνοι κατακράτησης της χρωστικής ουδέτερο ερυθρό εμφανίζονται μειωμένοι μετά από την επίδραση του οργανικού ρύπου και των βαρέων μετάλλων. Η στατιστική ανάλυση (Duncan s test, P<0,05) φανέρωσε πως στατιστικώς σημαντικά χαμηλότερες τιμές του συγκεκριμένου βιομάρτυρα εμφανίζονται στα μύδια που δέχθηκαν την επίδραση Cu (30 ± min) και Cd ( ± min), σε σύγκριση με τα μύδια ελέγχου ( ± 18.1 min) (Διαγρ. 20). Αντίθετα, δεν υπάρχει στατιστικώς σημαντική διαφορά ανάμεσα στα μύδια που εκτέθηκαν στον Hg (34.5 ± min) και τα μύδια ελέγχου, καθώς και μεταξύ των ατόμων που δέχθηκαν την επίδραση του φαινανθρένιου ( ± min) και των μυδιών που υπέστησαν την επίδραση ακετόνης (45 ± min) (Διαγρ. 20). 78
88 ΑΠΟΤΕΛΕΣΜΑΤΑ Κατακράτηση της χρωστικής ουδέτερο ερυθρό (NRR) * * ** 50 * (min) Control Ακετόνη Cu Φαινανθρένιο Cd Hg Διάγραμμα 20. Βιομάρτυρας «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) στα αιμοκύτταρα μυδιών M. galloprovincialis μετά από έκθεση σε 0.1mg L -1 ρύπων για 15 ημέρες. Οι στατιστικώς σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0.05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 20. Neutral red retention (NRR) assay applied in the haemocytes of mussels M. galloprovincialis exposed to 0.1mg L -1 of various pollutants for 15 days. Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0.05). Statistical signification is based on Duncan s test. 3. Πυρηνικές ανωμαλίες στα αιμοκύτταρα μυδιών Mytilus galloprovincialis μετά από εργαστηριακή έκθεση στα βαρέα μέταλλα Cu, Cd και Hg και στον οργανικό ρύπο φαινανθρένιο Με σκοπό τον προσδιορισμό γενοτοξικών βλαβών σε αιμοκύτταρα μυδιών M. galloprovincialis, που δέχθηκαν εργαστηριακή επίδραση των βαρέων μετάλλων Cu, Cd και Hg και του οργανικού ρύπου φαινανθρένιο, εφαρμόσθηκαν οι βιομάρτυρες «συχνότητα μικροπυρήνων» και «συχνότητα συνολικών πυρηνικών ανωμαλιών». Οι παραπάνω τεχνικές βασίζονται στην καταμέτρηση του ποσοστού των πυρηνικών ανωμαλιών στα αιμοκύτταρα των μυδιών. Οι μετρήσεις αυτές αποτελούν μια έγκαιρη προειδοποίηση της έκθεσης σε ρύπους και των δυνατών συνεπειών των γενοτοξικών 79
89 ΑΠΟΤΕΛΕΣΜΑΤΑ παραγόντων και για το λόγο αυτό έχουν προταθεί ως εργαλείο βιοπαρακολούθησης του θαλάσσιου περιβάλλοντος. α) Βιομάρτυρας «συχνότητα μικροπυρήνων» Μετά από έκθεση μυδιών M. galloprovincialis για 15 ημέρες σε συγκέντρωση 0.1 mg L -1 του οργανικού ρύπου φαινανθρένιο (phenanthrene) και τριών βαρέων μετάλλων (Cu, Cd και Hg), υπολογίσθηκε το ποσοστό εμφάνισης κυττάρων με μικροπυρήνες (micronucleus frequency / MN) στην αιμόλεμφο των ατόμων. Ως μύδια ελέγχου για την έκθεση στα μέταλλα χρησιμοποιήθηκαν άτομα που δεν δέχθηκαν την επίδραση κανενός ρύπου (control), ενώ ως μύδια ελέγχου για την οργανική ουσία φαινανθρένιο χρησιμοποιήθηκαν άτομα που δέχθηκαν την επίδραση της ουσίας ακετόνης (acetone). Συχνότητα μικροπυρήνων (ΜΝ) ποσοστό μικροπυρήνων ( ) * * * * Acetone Control Cu Phenanthrene Cd Hg Διάγραμμα 21. Βιομάρτυρας «συχνότητα μικροπυρήνων» (ΜΝ) στα αιμοκύτταρα μυδιών M. galloprovincialis μετά από έκθεση σε 0,1mg L -1 ρύπων για 15 ημέρες. Οι στατιστικώς σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 21. Micronuclei frequency (ΜΝ) technique in the haemocytes of mussels M. galloprovincialis exposed to 0,1mg L -1 of various pollutants for 15 days. Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0,05). Statistical signification is based on Duncan s test. 80
90 ΑΠΟΤΕΛΕΣΜΑΤΑ Σύμφωνα με τα αποτελέσματα προέκυψε στατιστικώς σημαντική αύξηση του ποσοστού εμφάνισης αιμοκυττάρων με μικροπυρήνες στα άτομα που δέχθηκαν την επίδραση Hg (7.25 ± ), σε σχέση με τα μύδια που δεν δέχθηκαν καμιά επίδραση (3.62 ± ) και ανάμεσα στα μύδια που δέχθηκαν την επίδραση φαινανθρένιου (9.71 ± 1.38 ) σε σχέση με αυτά που δέχθηκαν την επίδραση ακετόνης (3.7 ± ). Αντίθετα, τα μύδια που υπέστησαν την επίδραση Cu και Cd δεν εμφάνισαν στατιστικώς σημαντική αύξηση στο ποσοστό των κυττάρων με μικροπυρήνες σε σύγκριση με τα αντίστοιχα μύδια ελέγχου (Διαγρ. 21). β) Βιομάρτυρας «συχνότητα συνολικών πυρηνικών ανωμαλιών» Με σκοπό την εκτίμηση των συνολικών πυρηνικών ανωμαλιών στα αιμοκύτταρα μυδιών M. galloprovincialis, που δέχθηκαν την εργαστηριακή επίδραση του φαινανθρένιου (phenanthrene) και τριών βαρέων μετάλλων (Cu, Cd, Hg) καταμετρήθηκε το ποσοστό των συνολικών πυρηνικών αλλοιώσεων (total nuclear abnormalities frequency / TNA). Συγκεκριμένα, υπολογίσθηκε το ποσοστό που προέκυψε από το άθροισμα κυττάρων με έναν μικροπυρήνα, κυττάρων που εμφάνισαν ταυτόχρονα δύο μικροπυρήνες, κυττάρων με δύο ξεχωριστούς πυρήνες και κυττάρων με δύο πυρήνες ενωμένους μεταξύ τους. Η επεξεργασία των αποτελεσμάτων κατέδειξε στατιστικώς σημαντική διαφορά στο ποσοστό συνολικών πυρηνικών αλλοιώσεων ανάμεσα στα μύδια που είχαν δεχθεί την επίδραση Hg (12 ± 2 ) και Cu (12.43 ± ) σε σχέση με τα μύδια ελέγχου (7.13 ± ), καθώς και στα μύδια που δέχθηκαν την επίδραση φαινανθρένιου (15.14 ± ) σε σχέση με τα αντίστοιχα μύδια ελέγχου τους, δηλαδή αυτά που δέχθηκαν την επίδραση ακετόνης (8.1 ± ) (Διαγρ. 22). 81
91 ΑΠΟΤΕΛΕΣΜΑΤΑ ποσοστό συνολικών πυρηνικών ανωμαλιών ( ) Συχνότητα συνολικών πυρηνικών ανωμαλιών (ΤΝΑ) * * * * Control Acetone Cu Phenanthrene Cd Hg Διάγραμμα 22. Βιομάρτυρας «συχνότητα συνολικών πυρηνικών ανωμαλιών» (TNA) στα αιμοκύτταρα μυδιών M. galloprovincialis μετά από έκθεση σε 0.1mg L -1 ρύπων για 15 ημέρες. Οι στατιστικώς σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0.05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 22. Τotal nuclear abnormalities frequency (TNA) in the haemocytes of mussels M. galloprovincialis exposed to 0.1mg L -1 of various pollutants for 15 days. Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0.05). Statistical signification is based on Duncan s test. 4. Μορφομετρικές και μορφολογικές αποκρίσεις των λυσοσωμάτων του πεπτικού αδένα μυδιών Mytilus galloprovincialis μετά από εργαστηριακή επίδραση διαφορετικών ρύπων Με σκοπό την εύρεση μορφολογικών διαφορών μεταξύ των λυσοσωμάτων μυδιών που δέχθηκαν την επίδραση ρύπων στο εργαστήριο και μυδιών ελέγχου πραγματοποιήθηκε μελέτη της δομής του πεπτικού αδένα σε κρυοτομές πεπτικών σωληναρίων μετά από χρώση για το λυσοσωμικό ένζυμο Ν-ακετυλο-β-εξοζαμινιδάση, σε επίπεδο οπτικού μικροσκοπίου. Επίσης, εφαρμόσθηκε μορφομετρία λυσοσωμικών παραμέτρων στις κρυοτομές του πεπτικού αδένα για την εκτίμηση της πυκνότητας 82
92 ΑΠΟΤΕΛΕΣΜΑΤΑ όγκου (volume density / VD lys ) και της αριθμητικής πυκνότητας (numerical density / ND lys ) των λυσοσωμάτων του πεπτικού αδένα. Σύμφωνα με τη στατιστική επεξεργασία των αποτελεσμάτων (Anova, Duncan s test, P<0.05), η πυκνότητα όγκου των λυσοσωμάτων ήταν στατιστικώς χαμηλότερη στα μύδια που εκτέθηκαν σε Cd (VD lys = ± μm 3 /μm 3 ) και σε Hg (VD lys = ± μm 3 /μm 3 ) σε σχέση με τα μύδια ελέγχου (VD lys = ± μm 3 /μm 3 ), αλλά όχι και στα μύδια που δέχθηκαν την επίδραση του Cu (VD lys = ± μm 3 /μm 3 ) (Διαγρ. 23α). Επίσης, τα άτομα που δέχθηκαν την επίδραση του φαινανθρένιου εμφάνισαν στατιστικώς σημαντική μείωση στην παράμετρο αυτή (VD lys = ± μm 3 /μm 3 ), σε σύγκριση τα μύδια που δέχθηκαν την επίδραση της ακετόνης (VD lys =0.3 ± μm 3 /μm 3 ) (Διαγρ. 23α). Όσον αφορά την αριθμητική πυκνότητα των λυσοσωμάτων, τα αποτελέσματα της παρούσης μελέτης φανέρωσαν πως υπάρχει στατιστικώς σημαντική αύξηση της παραμέτρου αυτής στην περίπτωση των ατόμων που δέχθηκαν την επίδραση Cd (ND lys = ± number/μm 3 ), Hg (ND lys = ± number/μm 3 ) και Cu (ND lys = ± number/μm 3 ), σε σχέση με τα άτομα ελέγχου (ΝD lys = ± number/μm 3 ), όπως και στα μύδια που είχαν υποστεί επίδραση φαινανθρένιου (ND lys = ± number/μm 3 ), σε σύγκριση με τα μύδια που δέχθηκαν την επίδραση της ακετόνης (ND lys = ± number/μm 3 ) (Διαγρ. 23β). 83
93 ΑΠΟΤΕΛΕΣΜΑΤΑ μm 3 /μm 3 α 0,4 0,35 0,3 0,25 0,2 0,15 0,1 0,05 0 Πυκνότητα όγκου λυσοσωμάτων (VD lys ) * * * * * * κ Cu Phenanhtrene Acetone Control Cd Hg αριθμός/μm 3 β Αριθμητική πυκνότητα λυσοσωμάτων (ND lys ) 0,04 0,035 0,03 0,025 0,02 0,015 0,01 0,005 0 * * * * * ** ** * * Cu Phenanhtrene Acetone Control Cd Hg Διάγραμμα 23. Μορφομετρία λυσοσωμικών παραμέτρων του πεπτικού αδένα μυδιών M. galloprovincialis μετά από έκθεση σε 0.1mg L -1 ρύπων για 15 ημέρες. α. Πυκνότητα όγκου λυσοσωμάτων (VD lys ) β. Αριθμητική πυκνότητα λυσοσωμάτων (ND lys ) Οι στατιστικώς σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0.05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 23. Results on the morphometry of lysosomal parameters of the digestive lysosomes of mussels M. galloprovincialis exposed to 0.1mg L -1 of various pollutants for 15 days. α. Volume density of lysosomes (VD lys ) β. Numerical density of lysosomes (ND lys ) Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0.05). Statistical signification is based on Duncan s test. 84



 α β γ δ Phenanthrene ε ζ η θ Εικόνα 12.")
 (α - δ) και ατόμων που δέχθηκαν την")

. 1200x Figure 12.")
, as well as of animals exposed to 0,1mg L")
94 ΑΠΟΤΕΛΕΣΜΑΤΑ Σύμφωνα με τα παραπάνω αποτελέσματα, στις συγκεκριμένες εργαστηριακές συνθήκες η πυκνότητα όγκου των λυσοσωμάτων μειώνεται μετά από την επίδραση των περισσότερων ρύπων, ενώ η αριθμητική πυκνότητα παρουσιάζει αύξηση. Συνεπώς μικραίνει το μέγεθος και αυξάνεται ο αριθμός των λυσοσωμάτων. Η ποιοτική εξέταση των κρυοτομών στο οπτικό μικροσκόπιο επιβεβαίωσε τα παραπάνω αποτελέσματα καταδεικνύοντας περισσότερα σε αριθμό και μικρότερα σε μέγεθος λυσοσώματα στις περισσότερες περιπτώσεις μυδιών που δέχθηκαν την επίδραση ρύπων, σε σχέση με τα μύδια ελέγχου (Εικ. 12, 13). Acetone (Μύδια ελέγχου οργανικού) α β γ δ Phenanthrene ε ζ η θ Εικόνα 12. Ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση σε κρυοτομές του πεπτικού αδένα μυδιών που δέχθηκαν την επίδραση ακετόνης (acetone) (α - δ) και ατόμων που δέχθηκαν την επίδραση 0,1mg L -1 της ουσίας φαινανθρένιο (Phenanthrene) (ε - θ), διαλυμένης σε ακετόνη, για 15 ημέρες. Τα άτομα που εκτέθηκαν στο φαινανθρένιο (ε - θ) εμφάνισαν περισσότερα και μικρότερα σε μέγεθος λυσοσώματα σε σχέση με τα αντίστοιχα μύδια ελέγχου (α - δ). 1200x Figure 12. Histochemistry for the lysosomal enzyme N-acetyl-β-hexosaminidase in cryosections of the digestive gland of mussels exposed to acetone (α - δ), as well as of animals exposed to 0,1mg L -1 phenanthrene (ε - θ), dissolved in acetone, for 15 days. Transversely cut digestive tubules of animals exposed to phenanthrene (ε - θ) display smaller sized and increased in number lysosomes, compared to controls (α -δ). 1200x 85





, Cd")

 και στον Hg (ν-π)")

.")




 and Hg (ν-π) for 15")

95 ΑΠΟΤΕΛΕΣΜΑΤΑ Μύδια ελέγχου α β γ δ Χαλκός ε ζ η θ Κάδμιο ι κ λ μ Υδράργυρος ν ξ ο π Εικόνα 13. Ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση σε κρυοτομές του πεπτικού αδένα μυδιών ελέγχου (α-δ) και ατόμων που δέχθηκαν την επίδραση 0.1mg L -1 Cu (ε-θ), Cd (ι-μ) και Hg (ν-π) για 15 ημέρες. Τα άτομα που εκτέθηκαν στο Cd (ιμ) και στον Hg (ν-π) εμφάνισαν περισσότερα και μικρότερα σε μέγεθος λυσοσώματα σε σχέση με τα μύδια ελέγχου (α-δ). 1200x Figure 13. Histochemistry for the lysosomal enzyme N-acetyl-β-hexosaminidase in cryosections of the digestive gland of control mussels (αδ), as well as of animals exposed to 0.1mg L -1 of Cu (ε-θ), Cd (ι-μ) and Hg (ν-π) for 15 days. Transversely cut digestive tubules of animals exposed to Cd (ι-μ) and Hg (ν-π) display smaller sized and increased in number lysosomes, compared to controls (α-δ). 1200x 86
96 ΑΠΟΤΕΛΕΣΜΑΤΑ 5. Μορφομετρικές και μορφολογικές αποκρίσεις των ουδέτερων λιπιδίων του πεπτικού αδένα μυδιών Mytilus galloprovincialis μετά από εργαστηριακή επίδραση διαφορετικών ρύπων Με σκοπό την ανίχνευση διαφορών μεταξύ του πεπτικού αδένα μυδιών που δέχθηκαν την επίδραση ρύπων στο εργαστήριο και μυδιών ελέγχου πραγματοποιήθηκε ιστοχημεία ουδέτερων λιπιδίων σε κρυοτομές πεπτικών σωληναρίων και παρατήρησή τους, σε επίπεδο οπτικού μικροσκοπίου. Επίσης, εφαρμόσθηκε μορφομετρία λιπιδιακών παραμέτρων για την εκτίμηση της πυκνότητας όγκου (volume density / VD NL ) και της αριθμητικής πυκνότητας (numerical density / ND NL ) των ουδέτερων λιπιδίων του πεπτικού αδένα. Σύμφωνα με τη στατιστική επεξεργασία των αποτελεσμάτων (Anova, Duncan s test, P<0.05), η πυκνότητα όγκου των λιπιδιακών εγκλείστων ήταν στατιστικώς σημαντικά χαμηλότερη στα μύδια που εκτέθηκαν σε Cd (VD NL = ± μm 3 /μm 3 ), σε Hg (VD NL = ± μm 3 /μm 3 ) και σε Cu (VD NL = ± μm 3 /μm 3 ), σε σχέση με τα μύδια ελέγχου (VD NL = ± μm 3 /μm 3 ) (Διαγρ. 24α). Επίσης, τα μύδια που δέχθηκαν την επίδραση φαινανθρένιου εμφάνισαν στατιστικώς σημαντικά μικρότερη πυκνότητα όγκου ουδέτερων λιπιδίων (VD NL = ± μm 3 /μm 3 ), σε σύγκριση αντίστοιχα άτομα ελέγχου, δηλαδή τα μύδια που δέχθηκαν την επίδραση της ακετόνης (VD NL = ± μm 3 /μm 3 ) (Διαγρ. 24α). Όσον αφορά την αριθμητική πυκνότητα των λιπιδιακών εγκλείστων, τα αποτελέσματα της παρούσης μελέτης φανέρωσαν πως υπάρχει στατιστικώς σημαντική αύξηση της παραμέτρου αυτής στην περίπτωση των ατόμων που δέχθηκαν την επίδραση Cd (ND NL = ± number/μm 3 ), Hg (ND NL = ± number/μm 3 ) και Cu (ND NL = ± number/μm 3 ), σε σχέση με τα άτομα ελέγχου (ΝD NL = ± number/μm 3 ), όπως και στα μύδια που είχαν υποστεί επίδραση φαινανθρένιου (ND NL = ± number/μm 3 ), σε σύγκριση με τα μύδια που δέχθηκαν την επίδραση της ακετόνης (ND NL = ± number/μm 3 ) (Διαγρ. 24β). 87
97 ΑΠΟΤΕΛΕΣΜΑΤΑ α μm 3 /μm 3 0,3 0,25 0,2 0,15 0,1 Πυκνότητα όγκου λιπιδίων (VD NL ) * * * * * 0,05 0 Cu Phenanhtrene Acetone Control Cd Hg β αριθμός/μm 3 0,03 0,025 0,02 0,015 0,01 0,005 0 Αριθμητική πυκνότητα λιπιδίων (ND NL ) * * * * * Cu Phenanhtrene Acetone Control Cd Hg Διάγραμμα 24. Μορφομετρία ουδέτερων λιπιδίων του πεπτικού αδένα μυδιών M. galloprovincialis μετά από έκθεση σε 0.1mg L -1 ρύπων για 15 ημέρες. α. Πυκνότητα όγκου ουδέτερων λιπιδίων (VD NL ) β. Αριθμητική πυκνότητα ουδέτερων λιπιδίων (ND NL ) Οι στατιστικώς σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0.05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 24. Results on the morphometry of neutral lipid parameters of the digestive lysosomes of mussels M. galloprovincialis exposed to 0.1mg L -1 of various pollutants for 15 days. α. Volume density of neutral lipids (VD NL ) β. Numerical density of neutral lipids (ND NL ) Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0.05). Statistical signification is based on Duncan s test. 88



. Acetone (Μύδια ελέγχου οργανικού) α β γ δ Phenanthrene ε ζ η θ Εικόνα 14.")
 (α - δ) και ατόμων που δέχθηκαν την επίδραση 0.")
 εμφάνισαν περισσότερα και μικρότερα σε")

, as well as of animals exposed to 0.")
98 ΑΠΟΤΕΛΕΣΜΑΤΑ Σύμφωνα με τα παραπάνω αποτελέσματα, στις συγκεκριμένες εργαστηριακές συνθήκες, η πυκνότητα όγκου των ουδέτερων λιπιδίων μειώνεται μετά από την επίδραση των περισσότερων ρύπων, ενώ η αριθμητική πυκνότητα παρουσιάζει αύξηση. Συνεπώς μικραίνει το μέγεθος και αυξάνεται ο αριθμός των κοκκίων των ουδέτερων λιπιδίων. Η ποιοτική εξέταση των κρυοτομών στο οπτικό μικροσκόπιο επιβεβαίωσε τα παραπάνω αποτελέσματα καταδεικνύοντας αύξηση στον αριθμό και ταυτόχρονη μείωση στην ολική ποσότητα των ουδέτερων λιπιδίων στα μύδια που δέχθηκαν την επίδραση ρύπων, σε σχέση με τα μύδια ελέγχου (Εικ. 14, 15). Acetone (Μύδια ελέγχου οργανικού) α β γ δ Phenanthrene ε ζ η θ Εικόνα 14. Χρώση με Oil Red σε κρυοτομές πεπτικού αδένα μυδιών που δέχθηκαν την επίδραση ακετόνης (acetone) (α - δ) και ατόμων που δέχθηκαν την επίδραση 0.1mg L -1 του φαινανθρένιου (phenanthrene) (ε - θ), διαλυμένου σε ακετόνη, για 15 ημέρες. Τα άτομα που εκτέθηκαν στο φαινανθρένιο (ε - θ) εμφάνισαν περισσότερα και μικρότερα σε μέγεθος κοκκία ουδέτερων λιπιδίων, σε σχέση με τα αντίστοιχα μύδια ελέγχου (α -δ). 1200x Figure 14. Oil Red staining on cryosections of the digestive gland of mussels exposed to acetone (α - δ), as well as of animals exposed to 0.1mg L -1 phenanthrene (ε - θ), dissolved in acetone, for 15 days. Transversely cut digestive tubules of animals exposed to phenanthrene (ε - θ) display smaller sized and increased in number neutral lipid inclusions, compared to controls (α - δ). 1200x 89




, Cd")

, στο Cd")


.")



 and Hg (ν-π) for 15")

, Cd (ι-μ) and Hg")
99 ΑΠΟΤΕΛΕΣΜΑΤΑ Μύδια ελέγχου α β γ δ Χαλκός ε ζ η θ Κάδμιο ι κ λ μ Υδράργυρος ν ξ ο π Εικόνα 15. Χρώση με Oil Red σε κρυοτομές πεπτικού αδένα μυδιών ελέγχου (α-δ) και ατόμων που δέχθηκαν την επίδραση 0.1mg L -1 Cu (ε-θ), Cd (ι-μ) και Hg (ν-π) για 15 ημέρες. Τα άτομα που εκτέθηκαν στο Cu (ε-θ), στο Cd (ι-μ) και στον Hg (ν-π) εμφάνισαν περισσότερα και μικρότερα σε μέγεθος κοκκία ουδέτερων λιπιδίων, σε σχέση με τα μύδια ελέγχου (α-δ). 1200x Figure 15. Oil Red staining on cryosections of the digestive gland of control mussels (α-δ), as well as of animals exposed to 0.1mg L -1 Cu (ε-θ), Cd (ι-μ) and Hg (ν-π) for 15 days. Transversely cut digestive tubules of animals exposed to Cu (ε-θ), Cd (ι-μ) and Hg (ν-π) display smaller sized and increased in number neutral lipid inclusions, compared to controls (α-δ). 1200x 90
100 ΑΠΟΤΕΛΕΣΜΑΤΑ 6. Συσχέτιση μεταξύ των αποτελεσμάτων βιομαρτύρων στην εργαστηριακή μελέτη Με δεδομένη την ευαισθησία και ακρίβεια του βιομάρτυρα «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS) και με σκοπό την εκτίμηση της ευαισθησίας των υπόλοιπων βιομαρτύρων που χρησιμοποιήθηκαν, πραγματοποιήθηκε ανάλυση συσχέτισης (correlation analysis) μεταξύ των βιολογικών παραμέτρων. Τα αποτελέσματα της συσχέτισης των βιολογικών παραμέτρων που εξετάστηκαν (Pearson s test P<0.05) αναγράφονται στον πίνακα 3. Τα κύρια σημεία της συσχέτισης αυτής αναφέρονται παρακάτω: Σημαντική συσχέτιση (θετική) (r=0.98) υπήρξε μεταξύ των τιμών του βιομάρτυρα «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) και «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (ΝRR) (Διαγρ. 25), καθώς και σημαντική συσχέτιση (αρνητική) (r=-0.86) μεταξύ των τιμών της LMS και της αριθμητικής πυκνότητας των λυσοσωμάτων (ND lys ). Εμφανίσθηκε σημαντική συσχέτιση (αρνητική) (r=-0.90) μεταξύ των τιμών της «κατακράτησης της χρωστικής ουδέτερο ερυθρό» (NRR) και της αριθμητικής πυκνότητας των λυσοσωμάτων (ND lys ). Σημαντική συσχέτιση (θετική) (r=0.82) εμφανίσθηκε μεταξύ του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) και της πυκνότητας όγκου των λιπιδίων (VD NL ). Ο υπολογισμός του ποσοστού κυττάρων με μικροπυρήνες (MN) αποκάλυψε ότι υπάρχει σημαντική θετική (r=0.87) συσχέτιση ανάμεσα σε αυτήν την παράμετρο και στο ποσοστό των κυττάρων με συνολικές πυρηνικές αλλοιώσεις (TNA) (Διαγρ. 26). Το ποσοστό εμφάνισης κυττάρων με μικροπυρήνες (MN) παρουσιάζει σημαντική συσχέτιση (αρνητική) (r=-0.89) με την πυκνότητα όγκου των λυσοσωμάτων (VD lys ). H μορφομετρική εξέταση κρυοτομών του πεπτικού αδένα κατέδειξε σημαντική συσχέτιση (αρνητική) (r=-0.95) μεταξύ της πυκνότητας όγκου των λιπιδίων (VD NL ) και της αριθμητικής πυκνότητας των λιπιδίων (ND NL ) (Διαγρ. 27). Σημαντική συσχέτιση (αρνητική) (r=-0.82) εμφανίσθηκε μεταξύ του ποσοστού των κυττάρων που εμφανίζουν πυρηνικές αλλοιώσεις (TNA) και της αριθμητικής πυκνότητας των λιπιδίων (ND NL ). 91
101 ΑΠΟΤΕΛΕΣΜΑΤΑ Πίνακας 3. Συντελεστές γραμμικής συσχέτισης μεταξύ των βιολογικών παραμέτρων που εξετάστηκαν. Με κόκκινα νούμερα συμβολίζονται οι στατιστικώς σημαντικές συσχετίσεις (Pearson test, P<0.05). Τable 3. Correlation coefficients between the variables under test. Values in red indicate a statistically significant correlation (Pearson test, P<0.05). LMS NRR MN TNA VD lys ND lys VD NL ND NL LMS 1,00 0,98-0,03-0,31 0,20-0,86 0,77-0,69 NRR 0,98 1,00-0,08-0,31 0,25-0,90 0,82-0,73 MN -0,03-0,08 1,00 0,87-0,89 0,18-0,62 0,71 TNA -0,31-0,31 0,87 1,00-0,66 0,34-0,67 0,82 VD lys 0,20 0,25-0,89-0,66 1,00-0,32 0,73-0,70 ND lys -0,86-0,90 0,18 0,34-0,32 1,00-0,80 0,79 VD NL 0,77 0,82-0,62-0,67 0,73-0,80 1,00-0,95 ND NL -0,69-0,73 0,71 0,82-0,70 0,79-0,95 1,00 LMS: Σταθερότητα της λυσοσωμικής μεμβράνης, NRR: Κατακράτηση της χρωστικής ουδέτερο ερυθρό, ΜΝ: Συχνότητα μικροπυρήνων, ΤΝΑ: Συχνότητα συνολικών πυρηνικών ανωμαλιών, VD lys : πυκνότητα όγκου λυσοσωμάτων, ND lys : αριθμητική πυκνότητα λυσοσωμάτων, VD NL : πυκνότητα όγκου λιπιδίων, ND LP : αριθμητική πυκνότητα λιπιδίων. LMS: Lysosomal membrane stability test, NRR:Neutral red retention assay, ΜΝ: Micronucleus frequency, ΤΝΑ: Total nuclear abnormalities frequency, VD lys :Volume density of lysosomes, ND lys : Numerical density of lysosomes, VD NL : Volume density of lipids, ND NL : Numerical density of lipids. 50 LMS vs.nrr NRR = 9,6724+2,0516*LMS Συντελεστής συχέτισης: r = 0, NRR LMS Διάγραμμα 25. Θετική συσχέτιση των τιμών του βιομάρτυρα «κατακράτηση της χρωστικής ουδέτερο ερυθρό» (NRR) και των τιμών του βιομάρτυρα «σταθερότητας της λυσοσωμικής μεμβράνης» (LMS), μυδιών M. galloprovincialis που δέχθηκαν την επίδραση τριών βαρέων μετάλλων κι ενός οργανικού ρύπου. Diagram 25. Possitive correlation between the neutral red retention assay (NRR) results and the lysosomal membrane stability values (LMS), in mussels M. galloprovincialis, after heavy metal and organic pollutant exposure. 92
102 ΑΠΟΤΕΛΕΣΜΑΤΑ 0,016 TNA vs. MN TNA= 0,0048+1,0678*MN Συντελεστής συσχέτισης: r = 0,87 0,014 0,012 TNA 0,010 0,008 0,006 0,003 0,004 0,005 0,006 0,007 0,008 0,009 0,010 Διάγραμμα 26. Θετική συσχέτιση των τιμών του βιομάρτυρα «συχνότητα μικροπυρήνων» (ΜΝ) και των τιμών του βιομάρτυρα «συχνότητα συνολικών πυρηνικών ανωμαλιών» (ΤΝΑ), μυδιών M. galloprovincialis που δέχθηκαν την επίδραση τριών βαρέων μετάλλων κι ενός οργανικού ρύπου. Diagram 26. Possitive correlation between the micronucleus frequency (MN) results and the total nuclear abnormalities frequency (TNA), in mussels M. galloprovincialis, after heavy metal and organic pollutant exposure. MN NDNL vs. VDNL 0,024 ND NL = 0,0518-0,1618*VD NL Συντελεστής συσχέτισης: r = - 0,95 0,022 0,020 ND NL 0,018 0,016 0,014 0,012 0,18 0,19 0,20 0,21 0,22 0,23 0,24 0,25 VD NL Διάγραμμα 27. Συσχέτιση (αρνητική) της αριθμητικής πυκνότητας των λιπιδίων (ND NL ) και της πυκνότητας όγκου των λιπιδίων (VD NL ), μυδιών M. galloprovincialis που δέχθηκαν την επίδραση τριών βαρέων μετάλλων κι ενός οργανικού ρύπου. Diagram 27. Correlation (negative) between the numerical density (ND NL ) and the volume density (VD NL ) of lipids, in mussels M. galloprovincialis, after heavy metal and organic pollutant exposure. 93
103 ΑΠΟΤΕΛΕΣΜΑΤΑ 7. Ομαδοποίηση των ρύπων βάσει των επιπτώσεών τους στα μύδια Mytilus galloprovincialis και διερεύνηση της συγγένειας των υπό εξέταση βιομαρτύρων Από την ανάλυση των τιμών των βιομαρτύρων που μετρήθηκαν στους ιστούς των ατόμων μετά από εργαστηριακή επίδραση ρύπων για 15 ημέρες, προκύπτουν σημαντικά στοιχεία που σχετίζονται με τη διαφοροποίηση των ατόμων. Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε στατιστική ανάλυση με την οποία διαπιστώθηκε η συγγένεια των αποτελεσμάτων των ρύπων, με βάση τις μετρήσιμες τιμές κάθε βιομάρτυρα (Cluster Analysis, Statistica package, Microsoft Co., Greece), χρησιμοποιώντας την Ευκλείδεια απόσταση. Συγκεκριμένα, από τα εξαγόμενα αποτελέσματα διαπιστώθηκε σημαντική διαφοροποίηση των ατόμων ελέγχου (control) από τα μύδια που δέχθηκαν την επίδραση των τριών βαρέων μετάλλων. Επίσης, τα μύδια που δέχθηκαν την επίδραση της ακετόνης (control οργανικού ρύπου) διαφοροποιήθηκαν αρκετά από τα μύδια που δέχθηκαν την επίδραση της ουσίας φαινανθρένιο (Διαγρ. 28). Μεγαλύτερη συγγένεια εμφανίζουν τα άτομα ελέγχου με τα άτομα που δέχθηκαν την επίδραση ακετόνης (Διαγρ. 28). 6 LMS-NRR-MN-TNA-VD lys -ND lys -VD NL - ND NL Cd Cu Hg Acetone Phenanthrene control Διάγραμμα 28. Διαφοροποίηση των μυδιών που δέχθηκαν την επίδραση ρύπων, σύμφωνα με τα αποτελέσματα όλων των βιομαρτύρων που εξετάσθηκαν στους ιστούς των ατόμων. Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 28. Relationships between mussels exposed to pollutants concerning all the biomarkers applied in animal s tissues as occurred after cluster analysis (Statistica software package), using Euclidean distance. 94
104 ΑΠΟΤΕΛΕΣΜΑΤΑ Τέλος, από την προσπάθεια ομαδοποίησης των βιομαρτύρων που εξετάσθηκαν και λαμβάνοντας υπόψη τις επιπτώσεις όλων των ρύπων στους ιστούς των μυδιών προέκυψε μεγάλη συγγένεια ανάμεσα στους βιομάρτυρες «σταθερότητα της λυσοσωμικής μεμβράνης» (LMS) και «κατακράτηση του ουδέτερου ερυθρού» (NRR). Επίσης, σαφής ομαδοποίηση εμφανίσθηκε μεταξύ των τεχνικών «συχνότητα συνολικών πυρηνικών ανωμαλιών» (ΤΝΑ) και «συχνότητα μικροπυρήνων» (ΜΝ) (Διαγρ. 29) 4,5 Control- Acetone -Phenanthrene -Cu-Cd-Hg 4,0 3,5 3,0 2,5 2,0 1,5 1,0 0,5 0,0 VDlys VDLP NRR LMS NDLP NDlys TNA MN Διάγραμμα 29. Διαφοροποίηση των βιομαρτύρων σε μύδια που δέχθηκαν την εργαστηριακή επίδραση διάφορων ρύπων για 15 ημέρες. Για την εξαγωγή των αποτελεσμάτων εφαρμόσθηκε ανάλυση δενδρογράμματος (cluster analysis, Statistica software package), χρησιμοποιώντας την Ευκλείδεια απόσταση. Diagram 29. Relationships between biomarkers applied on mussels after laboratory exposure to various pollutants for 15 days. The results were occurred after cluster analysis (Statistica software package), using Euclidean distance. 95
105 ΑΠΟΤΕΛΕΣΜΑΤΑ Γ. ΜΕΛΕΤΗ ΛΥΣΟΣΩΜΙΚΩΝ, ΛΙΠΙΔΙΑΚΩΝ ΠΑΡΑΜΕΤΡΩΝ ΚΑΙ ΛΙΠΟΦΟΥΣΚΙΝΩΝ ΣΤΟΝ ΠΕΠΤΙΚΟ ΑΔΕΝΑ ΜΥΔΙΩΝ MYTILUS GALLOPROVINCIALIS 1. Εποχική διακύμανση των λιπιδίων, των λυσοσωμάτων και των λιποφουσκινών στον πεπτικό αδένα του μυδιού M. galloprovincialis Σκοπός της συγκεκριμένης μελέτης ήταν η εποχική εξέταση των μορφολογικών χαρακτηριστικών του πεπτικού αδένα μυδιών Mytilus galloprovincialis από περιοχές με αυξημένη και μικρότερη ρύπανση. Οι περιοχές από τις οποίες ελήφθησαν τα μύδια ήταν η Περαία, το Αγγελοχώρι, το Καλοχώρι και η Χαλάστρα στο Θερμαϊκό κόλπο, καθώς και η Ολυμπιάδα στο Στρυμονικό κόλπο, που επιλέχθηκε ως περιοχή ελέγχου. Τρεις διαφορετικοί κύκλοι δειγματοληψιών έγιναν σε χρονικό διάστημα ενός έτους. Η πρώτη δειγματοληψία πραγματοποιήθηκε την άνοιξη (μέσα Μαρτίου), η δεύτερη στην αρχή του καλοκαιριού (τέλος Μαϊου αρχή Ιουνίου) και η τρίτη στην αρχή του χειμώνα (τέλος Νοεμβρίου - αρχή Δεκεμβρίου) του Η μικροσκοπική παρατήρηση των κρυοτομών του πεπτικού αδένα αποκάλυψε την ύπαρξη διαφορών ανάμεσα στα πεπτικά σωληνάρια μυδιών που συλλέχθηκαν σε διαφορετικές εποχές δειγματοληψίας. Οι διαφοροποιήσεις αυτές αφορούσαν στην παρουσία λιποφουσκινών, στο ποσό των λιπιδίων και στην ιστοχημική αντίδραση για το λυσοσωμικό ένζυμο N-ακέτυλo-β-εξοζαμινιδάση. Επίσης, εποχικές διαφορές παρατηρήθηκαν και όταν εξετάσθηκαν τα γενικότερα μορφολογικά χαρακτηριστικά του λυσοσωμικού συστήματος του πεπτικού αδένα των μυδιών. Το πρότυπο που αποκαλύφθηκε και αφορά στις εποχικές μεταβολές των παραπάνω συστατικών και οργανιδίων ήταν, σε γενικές γραμμές, παρόμοιο για όλους τους πληθυσμούς μυδιών από όλες τις περιοχές δειγματοληψίας. Δηλαδή, οι εποχικές μεταβολές των λιποφουσκινών, των λιπιδίων και των λυσοσωμάτων παρατηρήθηκαν και στις περιοχές με αυξημένη, αλλά και στις περιοχές με μικρότερη ρύπανση. Μικρές διαφορές που καταγράφηκαν σε ορισμένα άτομα δεν προκάλεσαν απόκλιση από την γενική τάση αλλαγής των παραμέτρων αυτών λόγω εποχικών παραγόντων. 96
106 ΑΠΟΤΕΛΕΣΜΑΤΑ α) Χαρακτηριστικά των πεπτικών σωληναρίων κατά την αρχή του καλοκαιριού Σύμφωνα με τα αποτελέσματα, τα μύδια που συλλέχθηκαν στην αρχή του καλοκαιριού, εμφάνισαν εξαιρετικά μειωμένο ποσό λιποφουσκινών στα πεπτικά τους σωληνάρια (Εικ. 16, 19). Το αποτέλεσμα αυτό χαρακτηρίζει τόσο τα μύδια από την περιοχή ελέγχου, όσο και τα άτομα από τους σταθμούς δειγματοληψίας του Θερμαϊκού κόλπου, που θεωρούνται ιδιαίτερα επιβαρυμένοι με οργανικούς και ανόργανους ρύπους. Αντίθετα, οι χρώσεις Sudan black και Oil red κατέδειξαν την ύπαρξη μέτριας ποσότητας λιπιδιακού υλικού στα πεπτικά σωληνάρια των μυδιών από τις περισσότερες περιοχές δειγματοληψίας, ενώ τα λυσοσώματα εμφάνισαν έντονη θετική αντίδραση για το ένζυμο N-ακέτυλo-β-εξοζαμινιδάση. Κατά την εποχή αυτή, το λυσοσωμικό σύστημα χαρακτηριζόταν από ιδιαίτερα αυξημένο αριθμό και μέγεθος λυσοσωμάτων, σε όλες τις περιοχές δειγματοληψίας. Συμπερασματικά, τα πεπτικά σωληνάρια εμφάνισαν έντονη λειτουργία στην αρχή του καλοκαιριού, μια και χαρακτηρίζονταν από ένα ιδιαίτερα ενεργό λυσοσωμικό σύστημα, μειωμένο ποσό λιποφουσκινών και μέτρια ποσότητα λιπιδίων (Εικ. 16, 19). β) Χαρακτηριστικά των πεπτικών σωληναρίων κατά την αρχή του χειμώνα Κατά τη δειγματοληψία του χειμώνα η εικόνα των πεπτικών σωληναρίων ήταν αρκετά διαφορετική από αυτή που παρατηρήθηκε στην αρχή του καλοκαιριού (Εικ. 17, 19). Συγκεκριμένα, τα μύδια εμφάνισαν αυξημένο ποσό λιποφουσκινών στον πεπτικό τους αδένα σε σχέση με την προηγούμενη δειγματοληπτική περίοδο. Μάλιστα, την εποχή αυτή, περισσότερες λιποφουσκίνες εντοπίστηκαν στα μύδια από τις περιοχές του Θερμαϊκού, σε σύγκριση με την Ολυμπιάδα του Στρυμονικού κόλπου, που αποτελεί το σταθμό ελέγχου. Σε αντίθεση με την αύξηση στην ποσότητα των λιποφουσκινών, μειωμένο ποσό ουδέτερων και ολικών λιπιδίων ανιχνεύθηκε στον πεπτικό αδένα των ατόμων. Τα λυσοσώματα κατά την περίοδο αυτή ήταν μειωμένα σε αριθμό, είχαν ιδιαίτερα μικρό μέγεθος, ενώ παρουσίασαν εξαιρετικά ασθενή αντίδραση μετά από χρώση για το λυσοσωμικό ένζυμο Ν-ακετυλο-βεξοζαμινιδάση. Συμπερασματικά, στις αρχές του χειμώνα η λειτουργία των πεπτικών σωληναρίων έδειξε να καταστέλλεται, αφού το λυσοσωμικό σύστημα φάνηκε να 97
107 ΑΠΟΤΕΛΕΣΜΑΤΑ υπολειτουργεί, οι λιποφουσκίνες αυξήθηκαν αρκετά και στον πεπτικό αδένα εμφανίσθηκαν ιδιαίτερα μειωμένα ποσά λιπιδίων (Εικ. 17, 19). γ) Χαρακτηριστικά των πεπτικών σωληναρίων την άνοιξη Σύμφωνα με τα αποτελέσματα της παρούσης μελέτης στη δειγματοληψία της άνοιξης εμφανίσθηκε εξαιρετικά μεγάλη ποσότητα λιποφουσκινών στις κυψελίδες του πεπτικού αδένα στα μύδια από όλες τις περιοχές δειγματοληψίας (Εικ. 18, 19). Οι χρώσεις Sudan black και Oil Red κατέδειξαν την ύπαρξη μεγαλύτερης ποσότητας λιπιδίων σε σχέση με τα μύδια που συλλέχθηκαν το χειμώνα. Επίσης, εντοπίσθηκαν περισσότερα και μεγαλύτερα λυσοσώματα σε σύγκριση με την προηγούμενη δειγματοληπτική περίοδο, που εμφάνισαν μάλιστα θετική αντίδραση για το ένζυμο Ν- ακέτυλο-β-εξοζαμινιδάση. Συμπερασματικά, στα μέσα της άνοιξης, τα πεπτικά σωληνάρια επανεμφάνισαν σημάδια ενεργοποίησης, όπως κατέδειξε η επαναλειτουργία του λυσοσωμικού συστήματος και η μικρή αύξηση των λιπιδίων σε σχέση με το χειμώνα (Εικ. 18, 19). 98




, χρώση με Sudan black")
, χρώση")


.")






, Sudan black (β,")
 staining for the")



108 Αρχή καλοκαιριού Ολυμπιάδα α β γ δ Αγγελοχώρι ε ζ η θ Περαία ι κ λ μ Καλοχώρι ν ξ ο π Χαλάστρα ρ σ τ υ Εικόνα 16. Πεπτικά σωληνάρια μυδιών M. galloprovincialis, κατά την αρχή του καλοκαιριού, μετά από αντίδραση Schmrol για τον εντοπισμό λιποφουσκινών (α, ε, ι, ν, ρ), χρώση με Sudan black για τον εντοπισμό των ολικών λιπιδίων (β, ζ, κ, ξ, σ), χρώση με Oil red για τον εντοπισμό των ουδέτερων λιπιδίων (γ, η, λ, ο, τ) και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση (δ, θ, μ, π, υ). Στην περίοδο αυτή τα πεπτικά σωληνάρια χαρακτηρίζονται από ιδιαίτερα μειωμένο ποσό λιποφουσκινών, μέτρια ποσότητα λιπιδίων και αυξημένα σε μέγεθος και αριθμό λυσοσώματα, που αντιδρούν έντονα για το λυσοσωμικό ένζυμο. 1200x Figure 16. Tranversely cut digestive tubules of mussels M. galloprovincialis in early summer, after Schmrol reaction for the detection of lipofuscins (α, ε, ι, ν, ρ), Sudan black (β, ζ, κ, ξ, σ) and Oil red (γ, η, λ, ο, τ) staining for the detection of total lipids and neutral lipids, respectively, as well as after N-acetyl-β-hexosaminidase histochemistry (δ, θ, μ, π, υ). During this period digestive tubules present decreased amount of lipofuscins, considerable amount of lipids, as well as increased size and number of lysosomes, which strongly react for the lysosomal enzyme. 1200x 99




, χρώση με Sudan black")
, χρώση")


.")






 and")




109 Αρχή χειμώνα Ολυμπιάδα α β γ δ Αγγελοχώρι ε ζ η θ Περαία ι κ λ μ Καλοχώρι ν ξ ο π Χαλάστρα ρ σ τ υ Εικόνα 17. Πεπτικά σωληνάρια μυδιών M. galloprovincialis, κατά την αρχή του χειμώνα, μετά από αντίδραση Schmrol για τον εντοπισμό λιποφουσκινών (α, ε, ι, ν, ρ), χρώση με Sudan black για τον εντοπισμό των ολικών λιπιδίων (β, ζ, κ, ξ, σ), χρώση με Oil red για τον εντοπισμό των ουδέτερων λιπιδίων (γ, η, λ, ο, τ) και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση (δ, θ, μ, π, υ). Στην περίοδο αυτή εντοπίζονται αρκετές λιποφουσκίνες, περιορισμένο ποσό λιπιδίων, ενώ τα λίγα και μικρά σε μέγεθος λυσοσώματα παρουσιάζουν ασθενή αντίδραση για το λυσοσωμικό ένζυμο. 1200x Figure 17. Transversely cut digestive tubules of mussels M. galloprovincialis, in early winter, after Schmrol reaction for the detection of lipofuscins (α, ε, ι, ω, ρ), Sudan black (β, ζ, κ, ξ, σ) and Oil red (γ, η, λ, ο, τ) staining for the detection of total lipids and neutral lipids, respectively, as well as after N-acetyl-β-hexosaminidase histochemistry (δ, θ, μ, π, υ). During this period increased amount of lipofuscin and restricted lipid content are detected, while the lysosomal system presents few and small sized lysosomes, which hardly react for the lysosomal enzyme. 1200x 100



, χρώση με Sudan black")
, χρώση με")
")








, Sudan black (β, ζ, κ, ξ, σ)")


.")


110 Άνοιξη Ολυμπιάδα α β γ δ Αγγελοχώρι ε ζ η θ Περαία ι κ λ μ Καλοχώρι ν ξ ο π Χαλάστρα ρ σ τ υ Εικόνα 18. Πεπτικά σωληνάρια μυδιών M. galloprovincialis, στα μέσα της άνοιξης, μετά από αντίδραση Schmrol για τον εντοπισμό λιποφουσκινών (α, ε, ι, ν, ρ), χρώση με Sudan black για τον εντοπισμό των ολικών λιπιδίων (β, ζ, κ, ξ, σ), χρώση με Oil red για τον εντοπισμό των ουδέτερων λιπιδίων (γ, η, λ, ο, τ) και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση (δ, θ, μ, π, υ). Στην περίοδο αυτή οι λιποφουσκίνες είναι ιδιαίτερα αυξημένες, μέτρια ποσότητα λιπιδίων εντοπίζεται στα πεπτικά σωληνάρια, ενώ το λυσοσωμικό σύστημα εμφανίζει σημάδια ενεργοποίησης μια και επανεμφανίζονται λυσοσώματα που αντιδρούν θετικά για το λυσοσωμικό ένζυμο. 1200x Figure 18. Transversely cut digestive tubules of mussels M. galloprovincialis,sampled in spring, after staining for the detection of lipofuscins (α, ε, ι, ν, ρ), Sudan black (β, ζ, κ, ξ, σ) and Oil red (γ, η, λ, ο, τ) staining for the detection of total lipids and neutral lipids, respectively, as well as after N-acetyl-β-hexosaminidase histochemistry (δ, θ, μ, π, ο). During this period lipofuscins are extremely increased, moderate amount of lipids are detected in the digestive tubules and the lysosomal system presents signs of reactivation, since lysosomes, which react positive for the lysosomal enzyme, can be detected in the digestive tubules. 1200x 101


, μέτρια ποσότητα λιπιδίων (β, γ - βέλη) και αυξημένα σε μέγεθος και αριθμό λυσοσώματα,")
.")
, μειωμένο ποσό λιπιδίων (ζ, η) και σχετικά ανενεργό λυσοσωμικό")

 και το λυσοσωμικό σύστημα δείχνει να ενεργοποιείται και πάλι μια και χαρακτηρίζεται")


.")
, reduced amount of lipids (ζ, η) and rather inactive lysosomal system, characterized")
.")
111 ΑΠΟΤΕΛΕΣΜΑΤΑ Αρχή καλοκαιριού α β γ δ Αρχή χειμώνα ε ζ η θ Άνοιξη ι κ λ μ Εικόνα 19. Πεπτικά σωληνάρια μυδιών M. galloprovincialis, που συλλέχθηκαν από την περιοχή της Ολυμπιάδας στην αρχή του καλοκαιριού, στην αρχή του χειμώνα και στα μέσα της άνοιξης. Στην αρχή του καλοκαιριού τα πεπτικά σωληνάρια των μυδιών εμφανίζουν έλλειψη λιποφουσκινών (α), μέτρια ποσότητα λιπιδίων (β, γ - βέλη) και αυξημένα σε μέγεθος και αριθμό λυσοσώματα, που παρουσιάζουν ισχυρή αντίδραση για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση (δ - βέλος). Στις αρχές του χειμώνα τα μύδια εμφανίζουν πεπτικά σωληνάρια με αυξημένη ποσότητα λιποφουσκινών (ε - βέλος), μειωμένο ποσό λιπιδίων (ζ, η) και σχετικά ανενεργό λυσοσωμικό σύστημα με μικρά σε μέγεθος και λίγα σε αριθμό λυσοσώματα, που παρουσιάζουν ασθενή αντίδραση για το λυσοσωμικό ένζυμο (θ - βέλος). Στα μέσα της άνοιξης υπάρχει περαιτέρω αύξηση των λιποφουσκινών (ι - βέλος), τα λιπίδια τείνουν να αυξηθούν σε σχέση με το χειμώνα (κ, λ - βέλη) και το λυσοσωμικό σύστημα δείχνει να ενεργοποιείται και πάλι μια και χαρακτηρίζεται από περισσότερα σε αριθμό και αυξημένα σε μέγεθος λυσοσώματα, σε σύγκριση με την προηγούμενη περίοδο (μ - βέλος). 1200x Figure 19. Transversely cut digestive tubules of mussels M. galloprovincialis, collected from Olympiada at early summer, early winter and early spring. In early summer the digestive gland tubules of mussels display absence of lipofuscin inclusions (α), considerable amount of lipid material (β, γ - arrows) and increased lysosomal size, number and enzymatic content (δ - arrow). In early winter mussels show digestive tubules with increased amount of lipofuscins (ε - arrow), reduced amount of lipids (ζ, η) and rather inactive lysosomal system, characterized by reduced in size and number lysosomes, which hardly react for the marker enzyme N-acetyl-β-hexosaminidase (θ - arrow). In early spring, there is extreme accumulation of lipofuscins (ι - arrow), lipids (κ, λ - arrows) show a tendency to be increased in respect to winter and the lysosomal system presents signs of reactivation, as there are more numerous and increased in size lysosomes (μ - arrow) compared to the previous sampling period. 1200x 102
112 ΑΠΟΤΕΛΕΣΜΑΤΑ δ) Μορφομετρία λιποφουσκινών, ουδέτερων λιπιδίων και λυσοσωμάτων στον πεπτικό αδένα μυδιών που συλλέχθηκαν στην αρχή του καλοκαιριού, στην αρχή του χειμώνα και την άνοιξη Με σκοπό την επιβεβαίωση των μορφολογικών παρατηρήσεων σχετικά με την εποχική διακύμανση του ποσού των λιποφουσκινών, των ουδέτερων λιπιδίων και των λυσοσωμάτων, εφαρμόσθηκε μορφομετρία σε κρυοτομές πεπτικών σωληναρίων, σε επίπεδο οπτικού μικροσκοπίου. Συγκεκριμένα, πραγματοποιήθηκε υπολογισμός της πυκνότητας όγκου (volume density / VD) των λιποφουσκινών, των ουδέτερων λιπιδίων και των λυσοσωμάτων του πεπτικού αδένα μυδιών που συλλέχθηκαν στην αρχή του καλοκαιριού, την άνοιξη και στην αρχή του χειμώνα του έτους Το πρότυπο που αποκαλύφθηκε και αφορά στις εποχικές μεταβολές της πυκνότητας όγκου των παραπάνω κυτταρικών συστατικών και οργανιδίων ήταν παρόμοιο για όλους τους πληθυσμούς μυδιών από όλες τις περιοχές δειγματοληψίας (Διαγρ. 30, 31). Τα μορφομετρικά αποτελέσματα επιβεβαίωσαν τις μορφολογικές παρατηρήσεις που καταδεικνύουν την ύπαρξη ενεργών πεπτικών σωληναρίων κατά την αρχή του καλοκαιριού. Συγκεκριμένα, στον πεπτικό αδένα μυδιών που συλλέχθηκαν στην αρχή του καλοκαιριού και από τους πέντε σταθμούς δειγματοληψίας, μετρήθηκε εξαιρετικά μικρή πυκνότητα όγκου λιποφουσκινών. Η μικρότερη τιμή αυτής της παραμέτρου (0,004 ± 0,001 μm 3 /μm 3 ) σημειώθηκε στα μύδια από την Ολυμπιάδα (σταθμός ελέγχου) και η υψηλότερη (0.016 ± μm 3 /μm 3 ) στα μύδια από τη Χαλάστρα. Επίσης, μέτριες τιμές πυκνότητας όγκου ουδέτερων λιπιδίων καταγράφηκαν σε όλους τους σταθμούς κατά την περίοδο αυτή. Ειδικότερα, η πυκνότητα όγκου των ουδέτερων λιπιδίων κυμάνθηκε από 0.08 ± 0.03 μm 3 /μm 3 στα μύδια από τη Χαλάστρα, έως ± 0.05 μm 3 /μm 3 στα μύδια από την Ολυμπιάδα. Τέλος, αυξημένες τιμές υπολογίσθηκαν για την πυκνότητα όγκου των λυσοσωμάτων στην αρχή του καλοκαιριού σε όλους σταθμούς δειγματοληψίας. Η χαμηλότερη τιμή καταγράφηκε στα μύδια από τη Χαλάστρα (0.141 ± 0.04 μm 3 /μm 3 ), ενώ η μεγαλύτερη πυκνότητα όγκου λυσοσωμάτων μετρήθηκε στα μύδια από την Περαία (0.194 ± 0.06 μm 3 /μm 3 ) (Διαγρ. 30α). 103
113 ΑΠΟΤΕΛΕΣΜΑΤΑ Η περιορισμένη δραστηριότητα των πεπτικών σωληναρίων στην αρχή του χειμώνα, που αποκαλύφθηκε από τις μορφολογικές παρατηρήσεις, επιβεβαιώθηκε και από τα αποτελέσματα των μορφομετρικών μετρήσεων. Κατά την περίοδο αυτή, τα μύδια που συλλέχθηκαν από τους πέντε σταθμούς δειγματοληψίας εμφάνισαν αυξημένη πυκνότητα όγκου λιποφουσκινών. Η υψηλότερη τιμή καταγράφηκε στα μύδια από την Περαία (0.229 ± 0.07 μm 3 /μm 3 ), ενώ η χαμηλότερη τιμή αυτής της παραμέτρου σημειώθηκε στα μύδια από την Ολυμπιάδα (0.046 ± μm 3 /μm 3 ). Επίσης, στην αρχή του χειμώνα μικρή πυκνότητα όγκου ουδέτερων λιπιδίων καταγράφηκε στα μύδια που συλλέχθηκαν και από τους πέντε σταθμούς δειγματοληψίας. Οι τιμές για αυτή την παράμετρο κυμάνθηκαν ανάμεσα στα ± μm 3 /μm 3 στα μύδια από την Ολυμπιάδα και τα ± μm 3 /μm 3 στα μύδια από το Αγγελοχώρι. Τέλος, στην περίοδο αυτή η πυκνότητα όγκου των λυσοσωμάτων ήταν, σε γενικές γραμμές, χαμηλή, παρουσιάζοντας τη μικρότερη τιμή στα μύδια από το Αγγελοχώρι (0.048 ± μm 3 /μm 3 ) και τη μεγαλύτερη στα μύδια από το Καλοχώρι (0.048 ± μm 3 /μm 3 ) (Διαγρ. 30β). Κατά τη δειγματοληψία της άνοιξης, σύμφωνα με τα μορφομετρικά αποτελέσματα, τα μύδια από τους πέντε σταθμούς δειγματοληψίας εμφάνισαν εξαιρετικά υψηλή πυκνότητα όγκου λιποφουσκινών. Η υψηλότερη τιμή αυτής της παραμέτρου καταγράφηκε στα μύδια από το Καλοχώρι (0.302 ± μm 3 /μm 3 ), ενώ η χαμηλότερη στα μύδια από την Ολυμπιάδα (0.119 ± 0.06 μm 3 /μm 3 ). Ωστόσο, η μικρή αύξηση που καταγράφηκε στην πυκνότητα όγκου των λιπιδίων και των λυσοσωμάτων την περίοδο αυτή, σε σύγκριση με τις αντίστοιχες τιμές που ελήφθησαν το χειμώνα, φανέρωσε ενεργοποίηση των πεπτικών σωληναρίων. Η πυκνότητα όγκου των ουδέτερων λιπιδίων κυμάνθηκε μεταξύ των τιμών ± μm 3 /μm 3 στα μύδια από την Ολυμπιάδα και ± μm 3 /μm 3 στα μύδια από την Περαία. Σχετικά με την πυκνότητα όγκου των λυσοσωμάτων, η χαμηλότερη τιμή καταγράφηκε στα μύδια από το Καλοχώρι (0.073 ± 0.02 μm 3 /μm 3 ), ενώ η υψηλότερη στα μύδια από την Περαία (0,134 ± 0.053) (Διαγρ. 30γ). 104
114 ΑΠΟΤΕΛΕΣΜΑΤΑ VD μm 3 / μm 3 α 0,35 0,3 0,25 0,2 0,15 0,1 αρχή καλοκαιριού Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα 0,05 0 Λιποφουσκίνες Ουδέτερα λιπίδια Λυσοσώματα β 0,35 0,3 0,25 αρχή χειμώνα Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα VD μm3 / μm 3 0,2 0,15 0,1 0,05 0 Λιποφουσκίνες Ουδέτερα λιπίδια Λυσοσώματα γ 0,35 0,3 άνοιξη Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα VD μm 3 / μm 3 0,25 0,2 0,15 0,1 0,05 0 Λιποφουσκίνες Ουδέτερα λιπίδια Λυσοσώματα Διάγραμμα 30. Πυκνότητα όγκου λιποφουσκινών, ουδέτερων λιπιδίων και λυσοσωμάτων του πεπτικού αδένα μυδιών M. galloprovincialis που συλλέχθηκαν από πέντε σταθμούς δειγματοληψίας κατά στην αρχή του καλοκαιριού (α), στην αρχή του χειμώνα (β) και στην άνοιξη (γ). Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 30. Volume density of lipofuscins, neutral lipids and lysosomes in the digestive gland of mussels M. galloprovincialis from five different sampling stations in early summer (α), early winter (β) and spring (γ). Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0,05). Statistical signification is based on Duncan s test. 105
115 ΑΠΟΤΕΛΕΣΜΑΤΑ α 0,4 0,35 0,3 Πυκνότητα όγκου λιποφουσκινών αρχή καλοκαιριού αρχή χειμών α άν οιξη μm 3 / μm 3 0,25 0,2 0,15 0,1 0,05 0 Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα β 0,3 0,25 0,2 Πυκνότητα όγκου ουδέτερων λιπιδίων αρχή καλοκαιριού αρχή χειμών α άν οιξη 0,15 0,1 0,05 0 Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα γ 0,3 0,25 Πυκνότητα όγκου λυσοσωμάτων αρχή καλοκαιριού αρχή χειμώνα άν οιξη μm 3 / μm 3 0,2 0,15 0,1 0,05 0 Ολυμπιάδα Αγγελοχώρι Περαία Καλοχώρι Χαλάστρα Διάγραμμα 31. Πυκνότητα όγκου λιποφουσκινών (α), ουδέτερων λιπιδίων (β) και λυσοσωμάτων (γ) του πεπτικού αδένα μυδιών M. galloprovincialis που συλλέχθηκαν από πέντε σταθμούς δειγματοληψίας στην αρχή του καλοκαιριού, την αρχή του χειμώνα και την άνοιξη. Οι στατιστικά σημαντικές διαφορές μεταξύ ζευγών τιμών παρουσιάζονται στα τρίγωνα υπεράνω των ραβδογραμμάτων όπου και συμβολίζονται με αστερίσκους (P<0,05) και προσδιορίσθηκαν με τη χρήση του Duncan s test. Diagram 31. Volume density of lipofuscins (α), neutral lipids (β) and lysosomes (γ) in the digestive gland of mussels M. galloprovincialis from five different sampling stations in early summer, early winter and spring. Significant differences between pairs of mean values are indicated in the upper triangular matrix by asterisks (P<0,05). Statistical signification is based on Duncan s test. 106
116 ΑΠΟΤΕΛΕΣΜΑΤΑ 2. Αλληλεπίδραση ανάμεσα στο λιπιδιακό και το ενζυμικό περιεχόμενο των λυσοσωμάτων Κατά την παρούσα έρευνα μελετήθηκε, επίσης, η αλληλεπίδραση των ενδολυσοσωμικών λιπιδίων με την ενζυμική αντίδραση στο εσωτερικό των λυσοσωμάτων στον πεπτικό αδένα μυδιών Mytilus galloprovincialis. Για το σκοπό αυτό σε κρυοτομές πεπτικών σωληναρίων εξετάστηκαν παράλληλα, το ποσό των ολικών και των ουδέτερων λιπιδίων, καθώς και η αντίδραση στο ένζυμο N-ακέτυλoβ-εξοζαμινιδάση μέσα στα λυσοσώματα των πεπτικών κυττάρων. Η μικροσκοπική παρατήρηση των κρυοτομών των πεπτικών σωληναρίων κατέδειξε την ύπαρξη τριών διαφορετικών κατηγοριών μυδιών. Τα άτομα της πρώτης ομάδας εμφάνισαν πεπτικά σωληνάρια που χαρακτηρίζονταν από μειωμένο ποσό λιπιδιακού υλικού μέσα στα λυσοσώματα (Εικ. 20 α, β, δ, ε). Στην περίπτωση αυτή παρουσιάστηκε έντονα θετική αντίδραση για το λυσοσωμικό ένζυμο N-ακέτυλo-β-εξοζαμινιδάση (Εικ. 20 γ, ζ). Το πρότυπο αυτό ήταν και το πιο συχνό στις περιοχές που εξετάστηκαν. Η δεύτερη ομάδα περιελάμβανε άτομα που χαρακτηρίζονταν από αυξημένη ποσότητα λιπιδίων μέσα στα λυσοσώματα των πεπτικών σωληναρίων (Εικ 21 α, β, δ, ε). Σε αυτές τις περιπτώσεις το λιπιδιακό υλικό έδειχνε να έχει κατακλύσει τα λυσοσώματα αποτρέποντας τη χρώση για το λυσοσωμικό ένζυμο N-ακέτυλo-β-εξοζαμινιδάση (Εικ.21 γ, ζ). Τέλος, μια ενδιάμεση κατάσταση έχει επίσης παρατηρηθεί, όπου τα λυσοσώματα των πεπτικών σωληναρίων είναι μερικώς γεμάτα από λιπίδια, εμφανίζοντας ταυτόχρονα και χρώση για το προαναφερθέν ένζυμο. Σύμφωνα με τα αποτελέσματα, αυτές οι διαφοροποιήσεις δεν ήταν ενδεικτικές της ρύπανσης κάθε σταθμού δειγματοληψίας, αλλά πιθανόν να οφειλόταν σε συνδυασμό ρύπων και περιβαλλοντικών παραγόντων, που δημιουργεί ιδιαίτερες συνθήκες για την κάθε περιοχή. Τα αποτελέσματα που αναφέρθηκαν προηγουμένως στηρίχθηκαν σε παρατηρήσεις κρυοτομών του πεπτικού αδένα μυδιών που συλλέχθηκαν την άνοιξη και το καλοκαίρι. Το χειμώνα δεν μπορούσαν να εξαχθούν συμπεράσματα για το περιεχόμενο των λυσοσωμάτων, μια και τα άτομα κατά την εποχή αυτή παρουσιάσαν ανενεργό λυσοσωμικό σύστημα, όπως αναφέρθηκε προηγουμένως. 107
 και Oil red (β, ε)")

.")

 and N-acetyl-β-hexosaminidase histochemistry (γ, ζ).")
, a strong positive reaction for the lysosomal enzyme")
117 ΑΠΟΤΕΛΕΣΜΑΤΑ α β γ δ ε ζ Εικόνα 20. Κρυοτομές πεπτικών σωληναρίων μυδιών M. galloprovincialis μετά από χρώση με Sudan black (α, δ) και Oil red (β, ε) και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλο-β-εξοζαμινιδάση (γ, ζ). Όταν η ποσότητα των ενδολυσοσωμικών λιπιδίων στον πεπτικό αδένα του μυδιού είναι περιορισμένη (α, β, δ, ε βέλη), εμφανίζεται ισχυρή θετική αντίδραση για το λυσοσωμικό ένζυμο Ν-ακετυλ-β-εξοζαμινιδάση (γ, ζ - βέλη). α,β,γ: 1500x δ, ε, ζ: 4000x Figure 20. Cryosections from the digestive gland tubules of mussels M. galloprovincialis after the application of Sudan black (α, δ) and Oil red stainings (β, ε) and N-acetyl-β-hexosaminidase histochemistry (γ, ζ). When there is a limited accumulation of lipids into the lysosomes (α, β, δ, ε - arrows), a strong positive reaction for the lysosomal enzyme N-acetyl-β-hexosaminidase occurs (γ, ζ - arrows). α,β,γ: 1500x δ, ε, ζ: 4000x 108

.")
, εμφανίζεται")

 and Oil red")

118 ΑΠΟΤΕΛΕΣΜΑΤΑ α β γ δ ε ζ Εικόνα 21. Κρυοτομές πεπτικών σωληναρίων μυδιών M. galloprovincialis μετά από χρώση με Sudan black (α, δ), Oil red (β, ε) και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλο-β-εξοζαμινιδάση (γ, ζ). Στις περιπτώσεις όπου ο πεπτικός αδένας των μυδιών παρουσιάζει αυξημένη ποσότητα λιπιδίων μέσα στα λυσοσώματα (α, β, δ, ε - βέλη), εμφανίζεται μειωμένη αντίδραση για τα λυσοσωμικά ένζυμα (γ, ζ - βέλη). α,β,γ: 1500x δ, ε, ζ: 4000x Figure 21. Cryosections from the digestive gland tubules of mussels M. galloprovincialis after the application of Sudan black (α, δ) and Oil red stains (β, ε) and N-acetyl-β-hexosaminidase histochemistry (γ, ζ). Using mussels displaying digestive tubules full of lipid contents (α, β, δ, ε - arrows), the lysosomal structures show a restricted N-acetyl-β-hexosaminidase reaction (γ, ζ - arrows). α,β,γ: 1500x δ, ε, ζ: 4000x 109

, ενώ η χρώση των λιποφουσκινών με Οil red ήταν λιγότερο έντονη (Εικ. 23 γ, η).")
119 ΑΠΟΤΕΛΕΣΜΑΤΑ 3. Ιστοχημικός εντοπισμός λιποφουσκινών, λιπιδίων και λυσοσωμάτων στον πεπτικό αδένα μυδιών Mytilus galloprovincialis Η εφαρμογή της αντίδρασης Schmrol, σε κρυοτομές του πεπτικού αδένα μυδιών, έδειξε ότι οι κόκκοι λιποφουσκίνης εντοπίζονται κυρίως στο κυτταρόπλασμα (Εικ. 22). Σε ορισμένους ιστούς οι λιποφουσκίνες φάνηκε να καταλαμβάνουν ολόκληρες κυτταροπλασματικές περιοχές (Εικ. 23 α, ε). Σε κάποιες περιπτώσεις οι λιποφουσκίνες εντοπίσθηκαν και με χρώση με Sudan black (Εικ 23 β, ζ), ενώ η χρώση των λιποφουσκινών με Οil red ήταν λιγότερο έντονη (Εικ. 23 γ, η). Στον πεπτικό αδένα των μυδιών τα λιπίδια εντοπίσθηκαν κυρίως μέσα στα λυσοσώματα μετά από χρώση με Sudan black και Oil red. Οι χρωστικές αυτές έδωσαν το ίδιο πρότυπο όταν εφαρμόσθηκαν σε κρυοτομές των πεπτικών σωληναρίων (Εικ. 24). Τέλος, η παρούσα μελέτη έδειξε πως το λυσοσωμικό σύστημα στον πεπτικό αδένα των μυδιών εμφάνισε διαφοροποιήσεις από τη γενική μορφολογία του. Σε μια τέτοια περίπτωση η συνηθισμένη μορφολογία των λυσοσωμάτων αντικαταστάθηκε από πολλά μικρού μεγέθους λυσοσώματα. Τα ιδιαιτέρως μικρά αυτά λυσοσώματα παρουσίασαν ασθενή χρώση για το ένζυμο Ν-ακέτυλ-β-εξοζαμινιδάση, αλλά και περιορισμένη αντίδραση μετά από χρώση με Sudan black και Oil red, για τον εντοπισμό των ολικών και των ουδέτερων λιπιδίων, αντίστοιχα (Εικ. 25). α β Εικόνα 22. Μικρότερη (α) και μεγαλύτερη (β) μεγέθυνση κρυοτομών πεπτικών σωληναρίων μυδιών M. galloprovincialis, μετά από χρώση για τον εντοπισμό λιποφουσκινών. Οι κόκκοι λιποφουσκίνης (βέλη) ανιχνεύονται στο κυτταρόπλασμα. α: 1500x, β: 4000x Figure 22. Cryosections from the digestive gland tubules of mussels M. galloprovincialis after staining for the detection of lipofuscins, in lower (α) and higher (β) magnification. Lipofuscins (arrows) can be detected into the cytoplasm. α: 1500x, β: 4000x 110

.")
 και ασθενέστερη με τη χρωστική Oil red (γ, η")

 and Oil red (γ, η), as well as after")
,")

 σε κρυοτομές πεπτικών σωληναρίων μυδιών M.")
 μπορούν να εντοπισθούν τόσο μετά από χρώση με")

120 ΑΠΟΤΕΛΕΣΜΑΤΑ α β γ δ ε ζ η θ Εικόνα 23. Πεπτικά σωληνάρια μυδιών M. galloprovincialis μετά από αντίδραση Schmrol για τον εντοπισμό λιποφουσκινών (α, ε - βέλη), χρώση με Sudan black (β, ζ), χρώση με Oil red (γ, η), και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-βεξοζαμινιδάση (δ, θ). Τα κοκκία λιποφουσκίνης παρουσιάζουν έντονη χρώση μετά από επεξεργασία με τη χρωστική Sudan black (β, ζ - βέλη) και ασθενέστερη με τη χρωστική Oil red (γ, η βέλη). α-δ:1200x, ε-θ:3500x Figure 23. Transversely cut digestive tubules of mussels Mytilus galloprovincialis, after Schmrol reaction for the detection of lipofuscins (α, ε - arrows), staining with Sudan black (β, ζ) and Oil red (γ, η), as well as after N-acetyl-βhexosaminidase histochemistry (δ, θ). Lipofuscin inclusions can be also detected with Sudan black (β, ζ - arrows), while there is a weaker reaction after Oil red staining (γ, η - arrows). α-δ:1200x, ε-θ:3500x α β γ Εικόνα 24. Χρώση Sudan black (α), Oil red (β) και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλο-β-εξοζαμινιδάση (γ) σε κρυοτομές πεπτικών σωληναρίων μυδιών M. galloprovincialis. Τα ενδολυσοσωμικά λιπίδια (βέλη) μπορούν να εντοπισθούν τόσο μετά από χρώση με Sudan black, όσο και μετά από χρώση με Oil red στον πεπτικό αδένα των μυδιών. 1400x Figure 24. Cryosections from the digestive gland tubules of mussels M. galloprovincialis after the application of Sudan black (α) and oil red (β) stains and N-acetyl-β-hexosaminidase histochemistry (γ). Intralysosomal lipids (arrows) can be detected by Sudan black, as well as Oil red stainings in mussel s digestive gland. 1400x 111
, χρώση με Sudan black")
 που παρουσιάζουν ασθενή αντίδραση")

 staining, as well as after N-acetyl-β-hexosaminidase histochemistry (δ).")
121 ΑΠΟΤΕΛΕΣΜΑΤΑ α β γ δ Εικόνα 25. Πεπτικά σωληνάρια μυδιών M. galloprovincialis μετά από αντίδραση Schmrol για τον εντοπισμό λιποφουσκινών (α), χρώση με Sudan black (β), χρώση με Oil red (γ), και ιστοχημεία για το λυσοσωμικό ένζυμο Ν-ακετυλ-βεξοζαμινιδάση (δ). Στον πεπτικό αδένα των μυδιών εμφανίζονται μόνο μικρά λυσοσώματα (βέλη) που παρουσιάζουν ασθενή αντίδραση μετά από την εφαρμογή και των τεσσάρων ιστοχημικών τεχνικών. 3500x Figure 25. Transversely cut digestive tubules of mussels M. galloprovincialis, after Schmrol reaction for the detection of lipofuscins (α), Sudan black (β) and Oil red (γ) staining, as well as after N-acetyl-β-hexosaminidase histochemistry (δ). The digestive epithelium display small lysosomes (arrows), presenting weak reaction after the application of all the cytochemical techniques mentioned above. 3500x 112
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ, ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ, ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΡΑΦΤΟΠΟΥΛΟΥ ΕΛΕΝΗ ΒΙΟΛΟΓΟΣ ΙΕΡΕΥΝΗΣΗ ΚΥΤΤΑΡΙΚΩΝ ΠΑΡΑΜΕΤΡΩΝ ΣΕ ΙΣΤΟΥΣ ΤΟΥ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ, ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΡΑΦΤΟΠΟΥΛΟΥ ΕΛΕΝΗ ΒΙΟΛΟΓΟΣ ΙΕΡΕΥΝΗΣΗ ΚΥΤΤΑΡΙΚΩΝ ΠΑΡΑΜΕΤΡΩΝ ΣΕ ΙΣΤΟΥΣ ΤΟΥ
Τι είναι άμεση ρύπανση?
 ΡΥΠΑΝΣΗ ΝΕΡΟΥ Τι είναι ρύπανση; Ρύπανση μπορεί να θεωρηθεί η δυσμενής μεταβολή των φυσικοχημικών ή βιολογικών συνθηκών ενός συγκεκριμένου περιβάλλοντος ή/και η βραχυπρόθεσμη ή μακροπρόθεσμη βλάβη στην
ΡΥΠΑΝΣΗ ΝΕΡΟΥ Τι είναι ρύπανση; Ρύπανση μπορεί να θεωρηθεί η δυσμενής μεταβολή των φυσικοχημικών ή βιολογικών συνθηκών ενός συγκεκριμένου περιβάλλοντος ή/και η βραχυπρόθεσμη ή μακροπρόθεσμη βλάβη στην
ΠΤΡΗΝΑ ΣΟΤ ΚΤΣΣΑΡΟΤ: ΔΟΜΗ, ΛΕΙΣΟΤΡΓΙΑ, ΔΙΑΚΙΝΗΗ ΤΣΑΣΙΚΩΝ ΔΙΑΜΕΟΤ ΣΗ ΠΤΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ
 ΠΤΡΗΝΑ ΣΟΤ ΚΤΣΣΑΡΟΤ: ΔΟΜΗ, ΛΕΙΣΟΤΡΓΙΑ, ΔΙΑΚΙΝΗΗ ΤΣΑΣΙΚΩΝ ΔΙΑΜΕΟΤ ΣΗ ΠΤΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ ΓΤΜΝΑΙΟ ΚΕΡΑΣΕΑ ΦΟΛ. ΕΣΟ 2011-12 ΣΜΗΜΑ Γ3 ΟΡΥΑΝΙΔΗ ΓΙΩΡΓΟ ΙΔΕΡΗ ΒΑΓΓΕΛΗ Σο κύτταρο είναι η μικρότερη ζωντανή μονάδα,
ΠΤΡΗΝΑ ΣΟΤ ΚΤΣΣΑΡΟΤ: ΔΟΜΗ, ΛΕΙΣΟΤΡΓΙΑ, ΔΙΑΚΙΝΗΗ ΤΣΑΣΙΚΩΝ ΔΙΑΜΕΟΤ ΣΗ ΠΤΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ ΓΤΜΝΑΙΟ ΚΕΡΑΣΕΑ ΦΟΛ. ΕΣΟ 2011-12 ΣΜΗΜΑ Γ3 ΟΡΥΑΝΙΔΗ ΓΙΩΡΓΟ ΙΔΕΡΗ ΒΑΓΓΕΛΗ Σο κύτταρο είναι η μικρότερη ζωντανή μονάδα,
Β. ΚΑΜΙΝΕΛΛΗΣ ΒΙΟΛΟΓΙΑ. Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα).
.") ΒΙΟΛΟΓΙΑ Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα). Είδη οργανισμών Υπάρχουν δύο είδη οργανισμών: 1. Οι μονοκύτταροι, που ονομάζονται μικροοργανισμοί
ΒΙΟΛΟΓΙΑ Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα). Είδη οργανισμών Υπάρχουν δύο είδη οργανισμών: 1. Οι μονοκύτταροι, που ονομάζονται μικροοργανισμοί
ΘΑΛΑΣΣΙΑ ΡΥΠΑΝΣΗ ΣΤΟΝ ΚΟΛΠΟ ΤΗΣ ΕΛΕΥΣΙΝΑΣ. Μ.Δασενάκης ΣΥΛΛΟΓΟΣ ΕΛΛΗΝΩΝ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ, ΤΜΗΜΑ ΧΗΜΕΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΧΗΜΕΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΘΑΛΑΣΣΙΑ ΡΥΠΑΝΣΗ ΣΤΟΝ ΚΟΛΠΟ ΤΗΣ ΕΛΕΥΣΙΝΑΣ Μ.Δασενάκης ΣΥΛΛΟΓΟΣ ΕΛΛΗΝΩΝ Ο ΣΑΡΩΝΙΚΟΣ ΚΟΛΠΟΣ Επιφάνεια: 2600 km 2 Μέγιστο βάθος: 450 m
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ, ΤΜΗΜΑ ΧΗΜΕΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΧΗΜΕΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΘΑΛΑΣΣΙΑ ΡΥΠΑΝΣΗ ΣΤΟΝ ΚΟΛΠΟ ΤΗΣ ΕΛΕΥΣΙΝΑΣ Μ.Δασενάκης ΣΥΛΛΟΓΟΣ ΕΛΛΗΝΩΝ Ο ΣΑΡΩΝΙΚΟΣ ΚΟΛΠΟΣ Επιφάνεια: 2600 km 2 Μέγιστο βάθος: 450 m
ΓENIKA ΣTOIXEIA. Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό.
 1 ΓENIKA ΣTOIXEIA Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό. ΤΑ ΚΥΤΤΑΡΑ Η βασική ζώσα μονάδα του σώματος είναι
1 ΓENIKA ΣTOIXEIA Η φυσιολογία του ανθρώπου μελετά τα χαρακτηριστικά και τους λειτουργικούς μηχανισμούς που κάνουν το ανθρώπινο σώμα ζωντανό οργανισμό. ΤΑ ΚΥΤΤΑΡΑ Η βασική ζώσα μονάδα του σώματος είναι
ΡΥΠΑΝΣΗ. Ρύποι. Αντίδραση βιολογικών συστημάτων σε παράγοντες αύξησης
 ΡΥΠΑΝΣΗ 91 είναι η άμεση ή έμμεση διοχέτευση από τον άνθρωπο στο υδάτινο περιβάλλον ύλης ή ενέργειας με επιβλαβή αποτελέσματα για τους οργανισμούς ( ο ορισμός της ρύπανσης από τον ΟΗΕ ) Ρύποι Φυσικοί (εκρήξεις
ΡΥΠΑΝΣΗ 91 είναι η άμεση ή έμμεση διοχέτευση από τον άνθρωπο στο υδάτινο περιβάλλον ύλης ή ενέργειας με επιβλαβή αποτελέσματα για τους οργανισμούς ( ο ορισμός της ρύπανσης από τον ΟΗΕ ) Ρύποι Φυσικοί (εκρήξεις
ΒΟΤΑΝΙΚΗ ΜΟΡΦΟΛΟΓΙΑ ΚΑΙ ΑΝΑΤΟΜΙΑ ΦΥΤΟΥ
 ΒΟΤΑΝΙΚΗ ΜΟΡΦΟΛΟΓΙΑ ΚΑΙ ΑΝΑΤΟΜΙΑ ΦΥΤΟΥ Κύτταρο Θεμελιώδης δομική και λειτουργική μονάδα των ζωντανών οργανισμών Φυτικό κύτταρο Χαρακτηριστικά των κυττάρων των πολυκύτταρων φυτών: 1. Μερική ανεξαρτησία
ΒΟΤΑΝΙΚΗ ΜΟΡΦΟΛΟΓΙΑ ΚΑΙ ΑΝΑΤΟΜΙΑ ΦΥΤΟΥ Κύτταρο Θεμελιώδης δομική και λειτουργική μονάδα των ζωντανών οργανισμών Φυτικό κύτταρο Χαρακτηριστικά των κυττάρων των πολυκύτταρων φυτών: 1. Μερική ανεξαρτησία
ΚΕΦΑΛΑΙΟ 1. Οργάνωση της ζωής βιολογικά συστήματα
 ΚΕΦΑΛΑΙΟ 1 Οργάνωση της ζωής βιολογικά συστήματα 1.2 Κύτταρο: η μονάδα της ζωής Ιστορικά 1665: Ο Ρ.Χουκ μιλά για κύτταρα. Σύγχρονη κυτταρική θεωρία: Το κύτταρο είναι η θεμελιώδης δομική και λειτουργική
ΚΕΦΑΛΑΙΟ 1 Οργάνωση της ζωής βιολογικά συστήματα 1.2 Κύτταρο: η μονάδα της ζωής Ιστορικά 1665: Ο Ρ.Χουκ μιλά για κύτταρα. Σύγχρονη κυτταρική θεωρία: Το κύτταρο είναι η θεμελιώδης δομική και λειτουργική
ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ I ΕΡΓΑΣΤΗΡΙΟ ΜΑΛΑΚΙΑ ΑΝΑΤΟΜΙΑ ΚΑΙ ΣΥΣΤΗΜΑΤΙΚΗ. Διδάσκων: Σ. Νταϊλιάνης
 ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ I ΕΡΓΑΣΤΗΡΙΟ ΜΑΛΑΚΙΑ ΑΝΑΤΟΜΙΑ ΚΑΙ ΣΥΣΤΗΜΑΤΙΚΗ Διδάσκων: Σ. Νταϊλιάνης Γενικά χαρακτηριστικά - Πλευρικά πεπιεσμένοι οργανισμοί. - Φέρουν 2 θυρίδες (μήκος 1-2 mm μέχρι 1 m, π.χ.
ΕΡΓΑΣΤΗΡΙΟ ΒΙΟΛΟΓΙΑΣ ΖΩΩΝ I ΕΡΓΑΣΤΗΡΙΟ ΜΑΛΑΚΙΑ ΑΝΑΤΟΜΙΑ ΚΑΙ ΣΥΣΤΗΜΑΤΙΚΗ Διδάσκων: Σ. Νταϊλιάνης Γενικά χαρακτηριστικά - Πλευρικά πεπιεσμένοι οργανισμοί. - Φέρουν 2 θυρίδες (μήκος 1-2 mm μέχρι 1 m, π.χ.
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ ΤΟ ΚΥΤΤΑΡΟ Δ.ΑΡΕΘΑ
 ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ ΤΟ ΚΥΤΤΑΡΟ Δ.ΑΡΕΘΑ ΚΥΤΤΑΡΟ 2 Κατά την Βιολογία, κύτταρο ονομάζεται η βασική δομική και λειτουργική μονάδα που εκδηλώνει το φαινόμενο της ζωής. Έτσι, ως κύτταρο νοείται
ΕΙΣΑΓΩΓΗ ΣΤΗ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ ΤΟ ΚΥΤΤΑΡΟ Δ.ΑΡΕΘΑ ΚΥΤΤΑΡΟ 2 Κατά την Βιολογία, κύτταρο ονομάζεται η βασική δομική και λειτουργική μονάδα που εκδηλώνει το φαινόμενο της ζωής. Έτσι, ως κύτταρο νοείται
Φοιτητες: Σαμακός Φώτιος Παναγιώτης 7442 Ζάπρης Αδαμάντης 7458
 Φοιτητες: Σαμακός Φώτιος Παναγιώτης 7442 Ζάπρης Αδαμάντης 7458 1.ΕΙΣΑΓΩΓΗ 2.ΣΤΟΙΧΕΙΑΡΥΠΑΝΣΗΣ 2.1 ΠΑΘΟΦΟΝΟΙ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΙ 2.1.1 ΒΑΚΤΗΡΙΑ 2.1.2 ΙΟΙ 2.1.3 ΠΡΩΤΟΖΩΑ 2.2 ΑΝΟΡΓΑΝΕΣ ΚΑΙ ΟΡΓΑΝΙΚΕΣ ΔΙΑΛΥΤΕΣ ΣΤΟ
Φοιτητες: Σαμακός Φώτιος Παναγιώτης 7442 Ζάπρης Αδαμάντης 7458 1.ΕΙΣΑΓΩΓΗ 2.ΣΤΟΙΧΕΙΑΡΥΠΑΝΣΗΣ 2.1 ΠΑΘΟΦΟΝΟΙ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΙ 2.1.1 ΒΑΚΤΗΡΙΑ 2.1.2 ΙΟΙ 2.1.3 ΠΡΩΤΟΖΩΑ 2.2 ΑΝΟΡΓΑΝΕΣ ΚΑΙ ΟΡΓΑΝΙΚΕΣ ΔΙΑΛΥΤΕΣ ΣΤΟ
Εργασία στο μάθημα: ΟΙΚΟΛΟΓΙΑ ΓΙΑ ΜΗΧΑΝΙΚΟΥΣ. Θέμα: ΕΥΤΡΟΦΙΣΜΟΣ
 ΔΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΠΟΛΙΤΙΚΩΝ ΜΗΧΑΝΙΚΩΝ Εργασία στο μάθημα: ΟΙΚΟΛΟΓΙΑ ΓΙΑ ΜΗΧΑΝΙΚΟΥΣ Θέμα: ΕΥΤΡΟΦΙΣΜΟΣ 1 Ονομ/μο φοιτήτριας: Κουκουλιάντα Στυλιανή Αριθμός μητρώου: 7533 Υπεύθυνος καθηγητής:
ΔΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΠΟΛΙΤΙΚΩΝ ΜΗΧΑΝΙΚΩΝ Εργασία στο μάθημα: ΟΙΚΟΛΟΓΙΑ ΓΙΑ ΜΗΧΑΝΙΚΟΥΣ Θέμα: ΕΥΤΡΟΦΙΣΜΟΣ 1 Ονομ/μο φοιτήτριας: Κουκουλιάντα Στυλιανή Αριθμός μητρώου: 7533 Υπεύθυνος καθηγητής:
Η ρύπανση του εδάφους αφορά στη συγκέντρωση σ αυτό ρυπογόνων ουσιών σε ποσότητες που αλλοιώνουν τη σύσταση του και συνεπώς προκαλούν βλάβες στους
 ΡΥΠΑΝΣΗ ΕΔΑΦΟΥΣ Η ρύπανση του εδάφους αφορά στη συγκέντρωση σ αυτό ρυπογόνων ουσιών σε ποσότητες που αλλοιώνουν τη σύσταση του και συνεπώς προκαλούν βλάβες στους οργανισμούς και διαταραχές στα οικοσυστήματα.
ΡΥΠΑΝΣΗ ΕΔΑΦΟΥΣ Η ρύπανση του εδάφους αφορά στη συγκέντρωση σ αυτό ρυπογόνων ουσιών σε ποσότητες που αλλοιώνουν τη σύσταση του και συνεπώς προκαλούν βλάβες στους οργανισμούς και διαταραχές στα οικοσυστήματα.
ΒΑΡΙΑ ΜΕΤΑΛΛΑ ΚΑΙ ΡΥΠΑΝΣΗ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ (Λύσεις και αντιμετώπιση της ρύπανσης από βαριά μέταλλα) ΠΕΡΙΕΧΟΜΕΝΑ. ΒΙΒΛΙΟΓΡΑΦΙΑ σ.
 ΠΕΡΙΕΧΟΜΕΝΑ. ΒΙΒΛΙΟΓΡΑΦΙΑ σ.") ΒΑΡΙΑ ΜΕΤΑΛΛΑ ΚΑΙ ΡΥΠΑΝΣΗ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ (Λύσεις και αντιμετώπιση της ρύπανσης από βαριά μέταλλα) ΠΕΡΙΕΧΟΜΕΝΑ ΒΙΒΛΙΟΓΡΑΦΙΑ σ. 2 ΤΟ ΠΡΟΒΛΗΜΑ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ.σ.3 Η ΡΥΠΑΝΣΗ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΑΠΟ ΤΑ
ΒΑΡΙΑ ΜΕΤΑΛΛΑ ΚΑΙ ΡΥΠΑΝΣΗ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ (Λύσεις και αντιμετώπιση της ρύπανσης από βαριά μέταλλα) ΠΕΡΙΕΧΟΜΕΝΑ ΒΙΒΛΙΟΓΡΑΦΙΑ σ. 2 ΤΟ ΠΡΟΒΛΗΜΑ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ.σ.3 Η ΡΥΠΑΝΣΗ ΤΟΥ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΑΠΟ ΤΑ
Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ
 ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ") Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ Βιοενεργητική είναι ο κλάδος της Βιολογίας που μελετά τον τρόπο με τον οποίο οι οργανισμοί χρησιμοποιούν ενέργεια για να επιβιώσουν και να υλοποιήσουν τις
Κωνσταντίνος Π. (Β 2 ) ΚΕΦΑΛΑΙΟ 3: ΜΕΤΑΒΟΛΙΣΜΟΣ Βιοενεργητική είναι ο κλάδος της Βιολογίας που μελετά τον τρόπο με τον οποίο οι οργανισμοί χρησιμοποιούν ενέργεια για να επιβιώσουν και να υλοποιήσουν τις
ΥΔΑΤΙΝΗ ΡΥΠΑΝΣΗ ΥΔΑΤΙΝΗ ΡΥΠΑΝΣΗ-ΟΡΙΣΜΟΣ
 Τι είναι ρύπανση: Ρύπανση μπορεί να θεωρηθεί η δυσμενής μεταβολή των φυσικοχημικών ή βιολογικών συνθηκών ενός συγκεκριμένου περιβάλλοντος ή/και η βραχυπρόθεσμη ή μακροπρόθεσμη βλάβη στην ευζωία, την ποιότητα
Τι είναι ρύπανση: Ρύπανση μπορεί να θεωρηθεί η δυσμενής μεταβολή των φυσικοχημικών ή βιολογικών συνθηκών ενός συγκεκριμένου περιβάλλοντος ή/και η βραχυπρόθεσμη ή μακροπρόθεσμη βλάβη στην ευζωία, την ποιότητα
2.4 Ρύπανση του νερού
 1 Η θεωρία του μαθήματος με ερωτήσεις 2.4 Ρύπανση του νερού 4-1. Ποια ονομάζονται λύματα; Έτσι ονομάζονται τα υγρά απόβλητα από τις κατοικίες, τις βιομηχανίες, τις βιοτεχνίες και τους αγρούς. 4-2. Ποιοι
1 Η θεωρία του μαθήματος με ερωτήσεις 2.4 Ρύπανση του νερού 4-1. Ποια ονομάζονται λύματα; Έτσι ονομάζονται τα υγρά απόβλητα από τις κατοικίες, τις βιομηχανίες, τις βιοτεχνίες και τους αγρούς. 4-2. Ποιοι
Διαταραχές των βιογεωχημικών κύκλων των στοιχείων από την απελευθέρωση χημικών ουσιών στο περιβάλλον
 Διαταραχές των βιογεωχημικών κύκλων των στοιχείων από την απελευθέρωση χημικών ουσιών στο περιβάλλον Διεύθυνση Ενεργειακών, Βιομηχανικών και Χημικών Προϊόντων ΓΧΚ Δρ. Χ. Νακοπούλου Βιογεωχημικοί κύκλοι
Διαταραχές των βιογεωχημικών κύκλων των στοιχείων από την απελευθέρωση χημικών ουσιών στο περιβάλλον Διεύθυνση Ενεργειακών, Βιομηχανικών και Χημικών Προϊόντων ΓΧΚ Δρ. Χ. Νακοπούλου Βιογεωχημικοί κύκλοι
Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000
 Ζήτηµα 1ο Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι ιοί είναι :
Ζήτηµα 1ο Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι ιοί είναι :
Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000
 Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 ΕΚΦΩΝΗΣΕΙΣ Ζήτηµα 1ο Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι
Θέµατα Βιολογίας Γενική Παιδεία Γ Λυκείου 2000 ΕΚΦΩΝΗΣΕΙΣ Ζήτηµα 1ο Στις ερωτήσεις 1-5 να γράψετε στο τετράδιο σας τον αριθµό της ερώτησης και δίπλα το γράµµα που αντιστοιχεί στη σωστή απάντηση. 1. Οι
ΕΦΑΡΜΟΣΜΕΝΗ ΟΙΚΟΛΟΓΙΑ. 1. Ποια από τις παρακάτω ενώσεις αποτελεί πρωτογενή ρύπο; α. το DDT β. το νιτρικό υπεροξυακετύλιο γ. το όζον δ.
 1 ΕΦΑΡΜΟΣΜΕΝΗ ΟΙΚΟΛΟΓΙΑ 1. Ποια από τις παρακάτω ενώσεις αποτελεί πρωτογενή ρύπο; α. το DDT β. το νιτρικό υπεροξυακετύλιο γ. το όζον δ. το βενζοπυρένιο 2. Τα οξείδια του αζώτου: α. αντιδρούν με το οξυγόνο
1 ΕΦΑΡΜΟΣΜΕΝΗ ΟΙΚΟΛΟΓΙΑ 1. Ποια από τις παρακάτω ενώσεις αποτελεί πρωτογενή ρύπο; α. το DDT β. το νιτρικό υπεροξυακετύλιο γ. το όζον δ. το βενζοπυρένιο 2. Τα οξείδια του αζώτου: α. αντιδρούν με το οξυγόνο
ΤΡΑΠΕΖΑ ΘΕΜΑΤΩΝ ΚΕΦΑΛΑΙΟ 2 ΚΥΤΤΑΡΟ: Η ΘΕΜΕΛΙΩΔΗΣ ΜΟΝΑΔΑ ΤΗΣ ΖΩΗΣ ΘΕΜΑ Β 1. Η εικόνα απεικονίζει τμήμα μιας δομής του κυττάρου.
 ΤΡΑΠΕΖΑ ΘΕΜΑΤΩΝ ΚΕΦΑΛΑΙΟ 2 ΚΥΤΤΑΡΟ: Η ΘΕΜΕΛΙΩΔΗΣ ΜΟΝΑΔΑ ΤΗΣ ΖΩΗΣ ΘΕΜΑ Β 1. Η εικόνα απεικονίζει τμήμα μιας δομής του κυττάρου. I. Πώς ονομάζεται η κυτταρική δομή που απεικονίζεται στην εικόνα; Οι αριθμοί:
ΤΡΑΠΕΖΑ ΘΕΜΑΤΩΝ ΚΕΦΑΛΑΙΟ 2 ΚΥΤΤΑΡΟ: Η ΘΕΜΕΛΙΩΔΗΣ ΜΟΝΑΔΑ ΤΗΣ ΖΩΗΣ ΘΕΜΑ Β 1. Η εικόνα απεικονίζει τμήμα μιας δομής του κυττάρου. I. Πώς ονομάζεται η κυτταρική δομή που απεικονίζεται στην εικόνα; Οι αριθμοί:
ΡΥΠΑΝΣΗ. Ρύπανση : η επιβάρυνση του περιβάλλοντος με κάθε παράγοντα ( ρύπο ) που έχει βλαπτικές επιδράσεις στους οργανισμούς ΡΥΠΟΙ
 που έχει βλαπτικές επιδράσεις στους οργανισμούς ΡΥΠΟΙ") ΡΥΠΑΝΣΗ Ρύπανση : η επιβάρυνση του περιβάλλοντος με κάθε παράγοντα ( ρύπο ) που έχει βλαπτικές επιδράσεις στους οργανισμούς ΡΥΠΟΙ χημικές ουσίες μορφές ενέργειας ακτινοβολίες ήχοι θερμότητα ΕΠΙΚΥΝΔΥΝΟΤΗΤΑ
ΡΥΠΑΝΣΗ Ρύπανση : η επιβάρυνση του περιβάλλοντος με κάθε παράγοντα ( ρύπο ) που έχει βλαπτικές επιδράσεις στους οργανισμούς ΡΥΠΟΙ χημικές ουσίες μορφές ενέργειας ακτινοβολίες ήχοι θερμότητα ΕΠΙΚΥΝΔΥΝΟΤΗΤΑ
ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ
 ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής:
ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής:
ΕΡΓΑΣΙΑ ΟΙΚΙΑΚΗΣ ΟΙΚΟΝΟΜΙΑΣ ΘΕΜΑ ΕΠΙΛΟΓΗΣ: ΠΕΡΙΒΑΛΛΟΝΤΙΚΑ ΠΡΟΒΛΗΜΑΤΑ ΤΗΣ ΠΟΛΗΣ ΜΟΥ ΤΟΥ ΜΑΘΗΤΗ: ΑΣΚΟΡΔΑΛΑΚΗ ΜΑΝΟΥ ΕΤΟΣ
 ΕΡΓΑΣΙΑ ΟΙΚΙΑΚΗΣ ΟΙΚΟΝΟΜΙΑΣ ΘΕΜΑ ΕΠΙΛΟΓΗΣ: ΠΕΡΙΒΑΛΛΟΝΤΙΚΑ ΠΡΟΒΛΗΜΑΤΑ ΤΗΣ ΠΟΛΗΣ ΜΟΥ ΤΟΥ ΜΑΘΗΤΗ: ΑΣΚΟΡΔΑΛΑΚΗ ΜΑΝΟΥ ΕΤΟΣ 2013-2014 ΤΑΞΗ:B ΤΜΗΜΑ: Β1 ΡΥΠΑΝΣΗ- ΕΠΙΠΤΩΣΕΙΣ Η καθαριότητα και η λειτουργικότητα
ΕΡΓΑΣΙΑ ΟΙΚΙΑΚΗΣ ΟΙΚΟΝΟΜΙΑΣ ΘΕΜΑ ΕΠΙΛΟΓΗΣ: ΠΕΡΙΒΑΛΛΟΝΤΙΚΑ ΠΡΟΒΛΗΜΑΤΑ ΤΗΣ ΠΟΛΗΣ ΜΟΥ ΤΟΥ ΜΑΘΗΤΗ: ΑΣΚΟΡΔΑΛΑΚΗ ΜΑΝΟΥ ΕΤΟΣ 2013-2014 ΤΑΞΗ:B ΤΜΗΜΑ: Β1 ΡΥΠΑΝΣΗ- ΕΠΙΠΤΩΣΕΙΣ Η καθαριότητα και η λειτουργικότητα
Προστατευόμενες θαλάσσιες περιοχές φυσικής κληρονομιάς
 Προστατευόμενες θαλάσσιες περιοχές φυσικής κληρονομιάς Habitat: κυρίαρχη μορφή, γύρω από την οποία αναπτύσσεται ένας οικότοπος Χλωρίδα (π.χ. φυτό-φύκος) Πανίδα (π.χ. ύφαλος διθύρων) Γεωλογική μορφή (π.χ.
Προστατευόμενες θαλάσσιες περιοχές φυσικής κληρονομιάς Habitat: κυρίαρχη μορφή, γύρω από την οποία αναπτύσσεται ένας οικότοπος Χλωρίδα (π.χ. φυτό-φύκος) Πανίδα (π.χ. ύφαλος διθύρων) Γεωλογική μορφή (π.χ.
Εργασία για το μάθημα της Βιολογίας. Περίληψη πάνω στο κεφάλαιο 3 του σχολικού βιβλίου
 Εργασία για το μάθημα της Βιολογίας Περίληψη πάνω στο κεφάλαιο 3 του σχολικού βιβλίου Στο 3 ο κεφάλαιο του βιβλίου η συγγραφική ομάδα πραγματεύεται την ενέργεια και την σχέση που έχει αυτή με τους οργανισμούς
Εργασία για το μάθημα της Βιολογίας Περίληψη πάνω στο κεφάλαιο 3 του σχολικού βιβλίου Στο 3 ο κεφάλαιο του βιβλίου η συγγραφική ομάδα πραγματεύεται την ενέργεια και την σχέση που έχει αυτή με τους οργανισμούς
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ 3 Ο ΔΙΑΓΩΝΙΣΜΑ (2 Ο κεφάλαιο) ΘΕΜΑΤΑ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και δίπλα το γράμμα που αντιστοιχεί
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ 3 Ο ΔΙΑΓΩΝΙΣΜΑ (2 Ο κεφάλαιο) ΘΕΜΑΤΑ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και δίπλα το γράμμα που αντιστοιχεί
Βιολογία Γενικής Παιδείας Γ Λυκείου. Άνθρωπος και Περιβάλλον (Κεφ.2)
") Μάθημα/Τάξη: Κεφάλαιο: Βιολογία Γενικής Παιδείας Γ Λυκείου Άνθρωπος και Περιβάλλον (Κεφ.2) Ονοματεπώνυμο Μαθητή: Ημερομηνία: 13-02-2017 Επιδιωκόμενος Στόχος: 70/100 ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον
Μάθημα/Τάξη: Κεφάλαιο: Βιολογία Γενικής Παιδείας Γ Λυκείου Άνθρωπος και Περιβάλλον (Κεφ.2) Ονοματεπώνυμο Μαθητή: Ημερομηνία: 13-02-2017 Επιδιωκόμενος Στόχος: 70/100 ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον
ΤΟ ΚΥΤΤΑΡΟ ΠΥΡΗΝΑ ΚΥΤΤΑΡΟΠΛΑΣΜΑ ΟΡΓΑΝΥΛΛΙΑ ΥΑΛΟΠΛΑΣΜΑ ΠΥΡΗΝΙΟ ΠΥΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ ΠΥΡΗΝΙΚΟΣ ΦΛΟΙΟΣ
 ΤΟ ΚΥΤΤΑΡΟ ΚΥΤΤΑΡΟΠΛΑΣΜΑ ΠΥΡΗΝΑ ΟΡΓΑΝΥΛΛΙΑ ΥΑΛΟΠΛΑΣΜΑ ΠΥΡΗΝΙΟ ΠΥΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ ΠΥΡΗΝΙΚΟΣ ΦΛΟΙΟΣ ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΑΝΟΡΓΑΝΕΣ και ΟΡΓΑΝΙΚΕΣ ενώσεις ΑΝΟΡΓΑΝΕΣ C, O, N, H, P, S, Cl,, J, Fe, Cu Άλατα των παραπάνω
ΤΟ ΚΥΤΤΑΡΟ ΚΥΤΤΑΡΟΠΛΑΣΜΑ ΠΥΡΗΝΑ ΟΡΓΑΝΥΛΛΙΑ ΥΑΛΟΠΛΑΣΜΑ ΠΥΡΗΝΙΟ ΠΥΡΗΝΙΚΗ ΜΕΜΒΡΑΝΗ ΠΥΡΗΝΙΚΟΣ ΦΛΟΙΟΣ ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΑΝΟΡΓΑΝΕΣ και ΟΡΓΑΝΙΚΕΣ ενώσεις ΑΝΟΡΓΑΝΕΣ C, O, N, H, P, S, Cl,, J, Fe, Cu Άλατα των παραπάνω
gr ΜΟΥΓΙΟΣ Β.
 1ο Πανελλήνιο Συνέδριο ΙΒΙΣ «Βιολογία-Ιατρική Συστημάτων & Στρες» Άσκηση και μεταβολικόοξιδωτικό στρες Βασίλης Μούγιος Τμήμα Επιστήμης Φυσικής Αγωγής και Αθλητισμού με έδρα τη Θεσσαλονίκη Αριστοτέλειο
1ο Πανελλήνιο Συνέδριο ΙΒΙΣ «Βιολογία-Ιατρική Συστημάτων & Στρες» Άσκηση και μεταβολικόοξιδωτικό στρες Βασίλης Μούγιος Τμήμα Επιστήμης Φυσικής Αγωγής και Αθλητισμού με έδρα τη Θεσσαλονίκη Αριστοτέλειο
Η οδηγία για τα νερά κολύμβησης και η επίδραση της μυδοκαλλιέργειας στην ποιότητα νερών του Θερμαϊκού κόλπου (Βόρειο. Αιγαίο)
") Η οδηγία για τα νερά κολύμβησης και η επίδραση της μυδοκαλλιέργειας στην ποιότητα νερών του Θερμαϊκού κόλπου (Βόρειο Αιγαίο) Δρ. Σοφία Γαληνού-Μητσούδη Αλεξάνδρειο ΤΕΙ Θεσσαλονίκης Τμήμα Τεχνολογίας Αλιείας
Η οδηγία για τα νερά κολύμβησης και η επίδραση της μυδοκαλλιέργειας στην ποιότητα νερών του Θερμαϊκού κόλπου (Βόρειο Αιγαίο) Δρ. Σοφία Γαληνού-Μητσούδη Αλεξάνδρειο ΤΕΙ Θεσσαλονίκης Τμήμα Τεχνολογίας Αλιείας
Πεπτικός σωλήνας Κύρια λειτουργία του είναι η εξασφάλιση του διαρκούς ανεφοδιασμού του οργανισμού με νερό, ηλεκτρολύτες και θρεπτικά συστατικά.
 Πεπτικός σωλήνας Κύρια λειτουργία του είναι η εξασφάλιση του διαρκούς ανεφοδιασμού του οργανισμού με νερό, ηλεκτρολύτες και θρεπτικά συστατικά. Στον πεπτικό σωλήνα πραγματοποιείται ο τεμαχισμός της τροφής
Πεπτικός σωλήνας Κύρια λειτουργία του είναι η εξασφάλιση του διαρκούς ανεφοδιασμού του οργανισμού με νερό, ηλεκτρολύτες και θρεπτικά συστατικά. Στον πεπτικό σωλήνα πραγματοποιείται ο τεμαχισμός της τροφής
Προσδιορισμός φυσικοχημικών παραμέτρων υγρών αποβλήτων και υδάτων
 Προσδιορισμός φυσικοχημικών παραμέτρων υγρών αποβλήτων και υδάτων (DO - BOD - COD - TOC) Χ. Βασιλάτος Οργανική ύλη Αποξυγόνωση επιφανειακών και υπογείων υδάτων Οι οργανικές ύλες αποτελούν πολύ σοβαρό ρύπο,
Προσδιορισμός φυσικοχημικών παραμέτρων υγρών αποβλήτων και υδάτων (DO - BOD - COD - TOC) Χ. Βασιλάτος Οργανική ύλη Αποξυγόνωση επιφανειακών και υπογείων υδάτων Οι οργανικές ύλες αποτελούν πολύ σοβαρό ρύπο,
Πεπτικό σύστημα Περιγραφή
 Πεπτικό Σύστημα Πεπτικό σύστημα Περιγραφή Το γαστρεντερικό σύστημα (ΓΕΣ) αποτελείται από τα κοίλα όργανα που εκτείνονται από το στόμα έως τον πρωκτό και τους επικουρικούς αδένες που ευθύνονται για την
Πεπτικό Σύστημα Πεπτικό σύστημα Περιγραφή Το γαστρεντερικό σύστημα (ΓΕΣ) αποτελείται από τα κοίλα όργανα που εκτείνονται από το στόμα έως τον πρωκτό και τους επικουρικούς αδένες που ευθύνονται για την
ΕΠΙΔΡΑΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΚΑΙ ΕΙΔΙΚΟΤΕΡΑ ΤΟΥ ΜΟΛΥΒΔΟΥ, ΣΤΗΝ ΥΓΕΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ. ΔΙΕΡΓΑΣΙΕΣ ΑΠΟΜΑΚΡΥΝΣΗΣ ΤΟΥ ΜΕ ΤΗΝ ΜΕΘΟΔΟ ΤΗΣ ΦΥΤΟΑΠΟΚΑΤΑΣΤΑΣΗΣ.
 ΕΠΙΔΡΑΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΚΑΙ ΕΙΔΙΚΟΤΕΡΑ ΤΟΥ ΜΟΛΥΒΔΟΥ, ΣΤΗΝ ΥΓΕΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ. ΔΙΕΡΓΑΣΙΕΣ ΑΠΟΜΑΚΡΥΝΣΗΣ ΤΟΥ ΜΕ ΤΗΝ ΜΕΘΟΔΟ ΤΗΣ ΦΥΤΟΑΠΟΚΑΤΑΣΤΑΣΗΣ. (ΜΗΔΙΚΗΣ). Σαχινίδης Συμεών ΟΝΟΜΑ ΕΠΙΒΛΕΠΟΝΤΑ ΚΑΘΗΓΗΤΗ
ΕΠΙΔΡΑΣΗ ΤΩΝ ΒΑΡΕΩΝ ΜΕΤΑΛΛΩΝ ΚΑΙ ΕΙΔΙΚΟΤΕΡΑ ΤΟΥ ΜΟΛΥΒΔΟΥ, ΣΤΗΝ ΥΓΕΙΑ ΤΟΥ ΑΝΘΡΩΠΟΥ. ΔΙΕΡΓΑΣΙΕΣ ΑΠΟΜΑΚΡΥΝΣΗΣ ΤΟΥ ΜΕ ΤΗΝ ΜΕΘΟΔΟ ΤΗΣ ΦΥΤΟΑΠΟΚΑΤΑΣΤΑΣΗΣ. (ΜΗΔΙΚΗΣ). Σαχινίδης Συμεών ΟΝΟΜΑ ΕΠΙΒΛΕΠΟΝΤΑ ΚΑΘΗΓΗΤΗ
ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ:Κ.Κεραμάρης ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ
 ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ:Κ.Κεραμάρης ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Κωνσταντίνος Ρίζος Γιάννης Ρουμπάνης Βιοτεχνολογία με την ευρεία έννοια είναι η χρήση ζωντανών
ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ:Κ.Κεραμάρης ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Κωνσταντίνος Ρίζος Γιάννης Ρουμπάνης Βιοτεχνολογία με την ευρεία έννοια είναι η χρήση ζωντανών
Κεφάλαιο 2ο ΔΟΜΗ ΤΟΥ ΚΥΤΤΑΡΟΥ
 Κεφάλαιο 2ο ΔΟΜΗ ΤΟΥ ΚΥΤΤΑΡΟΥ 1. Κυτταρική μεμβράνη μοντέλο ρευστού μωσαϊκού κατά Singer και Nicolson Αποτελείται από διπλό στρώμα φωσφολιπιδίων με διάσπαρτα μόρια στεροειδών (χοληστερόλης) και μεγάλα
Κεφάλαιο 2ο ΔΟΜΗ ΤΟΥ ΚΥΤΤΑΡΟΥ 1. Κυτταρική μεμβράνη μοντέλο ρευστού μωσαϊκού κατά Singer και Nicolson Αποτελείται από διπλό στρώμα φωσφολιπιδίων με διάσπαρτα μόρια στεροειδών (χοληστερόλης) και μεγάλα
Περιβαλλοντικά Προβλήματα της πόλης μου
 Οικιακή Οικονομία Περιβαλλοντικά Προβλήματα της πόλης μου Κουτσάκη Μαρία Χρυσή Β 4 Η ρύπανση του περιβάλλοντος: το σύγχρονο πρόβλημα του 21ου αιώνα Η ρύπανση του περιβάλλοντος είναι μια ευρεία έννοια,
Οικιακή Οικονομία Περιβαλλοντικά Προβλήματα της πόλης μου Κουτσάκη Μαρία Χρυσή Β 4 Η ρύπανση του περιβάλλοντος: το σύγχρονο πρόβλημα του 21ου αιώνα Η ρύπανση του περιβάλλοντος είναι μια ευρεία έννοια,
Υδατικοί Πόροι -Ρύπανση
 Υδατικοί Πόροι -Ρύπανση Γήινη επιφάνεια Κατανομή υδάτων Υδάτινο στοιχείο 71% Ωκεανοί αλμυρό νερό 97% Γλυκό νερό 3% Εκμεταλλεύσιμο νερό 0,01% Γλυκό νερό 3% Παγόβουνα Υπόγεια ύδατα 2,99% Εκμεταλλεύσιμο νερό
Υδατικοί Πόροι -Ρύπανση Γήινη επιφάνεια Κατανομή υδάτων Υδάτινο στοιχείο 71% Ωκεανοί αλμυρό νερό 97% Γλυκό νερό 3% Εκμεταλλεύσιμο νερό 0,01% Γλυκό νερό 3% Παγόβουνα Υπόγεια ύδατα 2,99% Εκμεταλλεύσιμο νερό
Από τον Δρ. Φρ. Γαΐτη* για το foodbites.eu
 Από τον Δρ. Φρ. Γαΐτη* για το foodbites.eu Η μικροβιακή αύξηση μπορεί να επηρεάζεται από διάφορους ενδογενείς (εσωτερικούς) και εξωγενείς (εξωτερικούς) παράγοντες. Η αξιολόγηση αυτών των παραγόντων είναι
Από τον Δρ. Φρ. Γαΐτη* για το foodbites.eu Η μικροβιακή αύξηση μπορεί να επηρεάζεται από διάφορους ενδογενείς (εσωτερικούς) και εξωγενείς (εξωτερικούς) παράγοντες. Η αξιολόγηση αυτών των παραγόντων είναι
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α. Ο ηλεκτρονικός υπολογιστής του οργανισμού μας
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
Σήµερα οι εξελίξεις στην Επιστήµη και στην Τεχνολογία δίνουν τη
 ΚΕΦΑΛΑΙΟ 7ο: ΑΡΧΕΣ & ΜΕΘΟ ΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Συνδυασµός ΤΕΧΝΟΛΟΓΙΑΣ & ΕΠΙΣΤΗΜΗΣ Προσφέρει τη δυνατότητα χρησιµοποίησης των ζωντανών οργανισµών για την παραγωγή χρήσιµων προϊόντων 1 Οι ζωντανοί οργανισµοί
ΚΕΦΑΛΑΙΟ 7ο: ΑΡΧΕΣ & ΜΕΘΟ ΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Συνδυασµός ΤΕΧΝΟΛΟΓΙΑΣ & ΕΠΙΣΤΗΜΗΣ Προσφέρει τη δυνατότητα χρησιµοποίησης των ζωντανών οργανισµών για την παραγωγή χρήσιµων προϊόντων 1 Οι ζωντανοί οργανισµοί
Τα ποτάμια και οι λίμνες της Ελλάδας. Λάγιος Βασίλειος, Εκπαιδευτικός
 Τα ποτάμια και οι λίμνες της Ελλάδας Λάγιος Βασίλειος, Εκπαιδευτικός Ποτάμι είναι το ρεύμα γλυκού νερού που κινείται από τα ψηλότερα (πηγές) προς τα χαμηλότερα μέρη της επιφάνειας της Γης (πεδινά) και
Τα ποτάμια και οι λίμνες της Ελλάδας Λάγιος Βασίλειος, Εκπαιδευτικός Ποτάμι είναι το ρεύμα γλυκού νερού που κινείται από τα ψηλότερα (πηγές) προς τα χαμηλότερα μέρη της επιφάνειας της Γης (πεδινά) και
ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ Βιογεωχημικός κύκλος
 ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ Βιογεωχημικός κύκλος ενός στοιχείου είναι, η επαναλαμβανόμενη κυκλική πορεία του στοιχείου στο οικοσύστημα. Οι βιογεωχημικοί κύκλοι, πραγματοποιούνται με την βοήθεια, βιολογικών, γεωλογικών
ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ Βιογεωχημικός κύκλος ενός στοιχείου είναι, η επαναλαμβανόμενη κυκλική πορεία του στοιχείου στο οικοσύστημα. Οι βιογεωχημικοί κύκλοι, πραγματοποιούνται με την βοήθεια, βιολογικών, γεωλογικών
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΖΩΟΛΟΓΙΑΣ, ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΖΩΩΝ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΖΩΟΛΟΓΙΑΣ, ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΖΩΩΝ ΘΕΣΣΑΛΟΝΙΚΗ 2005 ΜΕΛΕΤΗ ΒΙΟΧΗΜΙΚΩΝ ΠΑΡΑΜΕΤΡΩΝ ΚΑΙ ΜΗΧΑΝΙΣΜΩΝ ΠΟΥ ΣΧΕΤΙΖΟΝΤΑΙ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΖΩΟΛΟΓΙΑΣ, ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΖΩΩΝ ΘΕΣΣΑΛΟΝΙΚΗ 2005 ΜΕΛΕΤΗ ΒΙΟΧΗΜΙΚΩΝ ΠΑΡΑΜΕΤΡΩΝ ΚΑΙ ΜΗΧΑΝΙΣΜΩΝ ΠΟΥ ΣΧΕΤΙΖΟΝΤΑΙ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 18 ΙΟΥΝΙΟΥ 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΕΠΙΛΟΓΗΣ ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 18 ΙΟΥΝΙΟΥ 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΕΠΙΛΟΓΗΣ ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α A1. β Α2. γ Α3. δ Α4. α Α5. γ ΘΕΜΑ Β Β1. α. 9 β. 8 γ.
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 18 ΙΟΥΝΙΟΥ 2019 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΕΠΙΛΟΓΗΣ ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α A1. β Α2. γ Α3. δ Α4. α Α5. γ ΘΕΜΑ Β Β1. α. 9 β. 8 γ.
ΑΠΑΝΤΗΣΕΙΣ ΤΩΝ ΕΡΩΤΗΣΕΩΝ-ΑΣΚΗΣΕΩΝ ΚΑΙ ΠΡΟΒΛΗΜΑΤΩΝ ΤΟΥ ΒΙΒΛΙΟΥ ΤΟΥ ΜΑΘΗΤΗ
 ΑΠΑΝΤΗΣΕΙΣ ΤΩΝ ΕΡΩΤΗΣΕΩΝ-ΑΣΚΗΣΕΩΝ ΚΑΙ ΠΡΟΒΛΗΜΑΤΩΝ ΤΟΥ ΒΙΒΛΙΟΥ ΤΟΥ ΜΑΘΗΤΗ 1. Περιγράψτε τη δομή της πλασματικής μεμβράνης. Στην απάντησή σας να α- ναφερθείτε στη διευθέτηση των χημικών μορίων που συνθέτουν
ΑΠΑΝΤΗΣΕΙΣ ΤΩΝ ΕΡΩΤΗΣΕΩΝ-ΑΣΚΗΣΕΩΝ ΚΑΙ ΠΡΟΒΛΗΜΑΤΩΝ ΤΟΥ ΒΙΒΛΙΟΥ ΤΟΥ ΜΑΘΗΤΗ 1. Περιγράψτε τη δομή της πλασματικής μεμβράνης. Στην απάντησή σας να α- ναφερθείτε στη διευθέτηση των χημικών μορίων που συνθέτουν
ΕΡΓΑΣΙΑ ΒΙΟΛΟΓΙΑΣ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ
 ΕΡΓΑΣΙΑ ΒΙΟΛΟΓΙΑΣ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Οι οργανισμοί εξασφαλίζουν ενέργεια, για τις διάφορες λειτουργίες τους, διασπώντας θρεπτικές ουσίες που περιέχονται στην τροφή τους. Όμως οι φωτοσυνθετικοί
ΕΡΓΑΣΙΑ ΒΙΟΛΟΓΙΑΣ 3.1 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Οι οργανισμοί εξασφαλίζουν ενέργεια, για τις διάφορες λειτουργίες τους, διασπώντας θρεπτικές ουσίες που περιέχονται στην τροφή τους. Όμως οι φωτοσυνθετικοί
Ενεργοποίησηκινδύνουγιαεπιβλαβήμικροφύκη: Το παράδειγμα του Μαλιακού κόλπου
 Ενεργοποίησηκινδύνουγιαεπιβλαβήμικροφύκη: Το παράδειγμα του Μαλιακού κόλπου Κων/νοςΚουκάρας 1, ΝικόλαοςΚαλογεράκης 2 & ΔημήτριοςΡίζος 3 1. ΤμήμαΈρευνας& Ανάπτυξης, ΝΕΑΡΧΟΣ Ο.Ε. 2. Εργαστήριο Βιοχημικής
Ενεργοποίησηκινδύνουγιαεπιβλαβήμικροφύκη: Το παράδειγμα του Μαλιακού κόλπου Κων/νοςΚουκάρας 1, ΝικόλαοςΚαλογεράκης 2 & ΔημήτριοςΡίζος 3 1. ΤμήμαΈρευνας& Ανάπτυξης, ΝΕΑΡΧΟΣ Ο.Ε. 2. Εργαστήριο Βιοχημικής
ΠΡΟΤΥΠΟ ΠΕΙΡΑΜΑΤΙΚΟ ΛΥΚΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΠΑΤΡΩΝ 2013-14
 ΘΕΜΑΤΑ ΟΙΚΟΛΟΓΙΑΣ Μπορεί να λειτουργήσει ένα οικοσύστημα α) με παραγωγούς και καταναλωτές; β) με παραγωγούς και αποικοδομητές; γ)με καταναλωτές και αποικοδομητές; Η διατήρηση των οικοσυστημάτων προϋποθέτει
ΘΕΜΑΤΑ ΟΙΚΟΛΟΓΙΑΣ Μπορεί να λειτουργήσει ένα οικοσύστημα α) με παραγωγούς και καταναλωτές; β) με παραγωγούς και αποικοδομητές; γ)με καταναλωτές και αποικοδομητές; Η διατήρηση των οικοσυστημάτων προϋποθέτει
Ορισμός το. φλψ Στάδια επεξεργασίας λυμάτων ΘΕΜΑ: ΒΙΟΛΟΓΙΚΟΣ ΚΑΘΑΡΙΣΜΟΣ ΣΤΗΝ ΚΩ ΤΙ ΕΙΝΑΙ Ο ΒΙΟΛΟΓΙΚΟΣ ΚΑΘΑΡΙΣΜΟΣ?
 ΘΕΜΑ: ΒΙΟΛΟΓΙΚΟΣ ΚΑΘΑΡΙΣΜΟΣ ΣΤΗΝ ΚΩ ΤΙ ΕΙΝΑΙ Ο ΒΙΟΛΟΓΙΚΟΣ ΚΑΘΑΡΙΣΜΟΣ? Ο βιολογικος καθαρισμος αφορα την επεξεργασια λυματων, δηλαδη τη διαδικασια μεσω της οποιας διαχωριζονται οι μολυσματικες ουσιες από
ΘΕΜΑ: ΒΙΟΛΟΓΙΚΟΣ ΚΑΘΑΡΙΣΜΟΣ ΣΤΗΝ ΚΩ ΤΙ ΕΙΝΑΙ Ο ΒΙΟΛΟΓΙΚΟΣ ΚΑΘΑΡΙΣΜΟΣ? Ο βιολογικος καθαρισμος αφορα την επεξεργασια λυματων, δηλαδη τη διαδικασια μεσω της οποιας διαχωριζονται οι μολυσματικες ουσιες από
Επιστημονικά Δεδομένα για τη βιοχημική δράση της αντιοξειδωτικής Βιταμίνης C.
 Βιταμίνη C - Ενισχύει το ανοσοποιητικό με 20 διαφορετικούς τρόπους - ΚΑΛΑΜΠΑΚΑ CITY KALAMP Επιστημονικά Δεδομένα για τη βιοχημική δράση της αντιοξειδωτικής Βιταμίνης C. Η βιταμίνη C, γνωστή και ως ασκορβικό
Βιταμίνη C - Ενισχύει το ανοσοποιητικό με 20 διαφορετικούς τρόπους - ΚΑΛΑΜΠΑΚΑ CITY KALAMP Επιστημονικά Δεδομένα για τη βιοχημική δράση της αντιοξειδωτικής Βιταμίνης C. Η βιταμίνη C, γνωστή και ως ασκορβικό
Πηγή: ΑΠΟΛΥΜΑΝΣΗ ΤΟΥ ΠΟΣΙΜΟΥ ΝΕΡΟΥ : ΠΡΟΧΩΡΗΜΕΝΕΣ ΜΕΘΟΔΟΙ ΕΝΑΛΛΑΚΤΙΚΕΣ ΤΟΥ ΧΛΩΡΙΟΥ, ΘΕΟΔΩΡΑΤΟΥ ΑΓΓΕΛΙΚΗ, ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ, ΜΥΤΙΛΗΝΗ 2005
 Πηγή: ΑΠΟΛΥΜΑΝΣΗ ΤΟΥ ΠΟΣΙΜΟΥ ΝΕΡΟΥ : ΠΡΟΧΩΡΗΜΕΝΕΣ ΜΕΘΟΔΟΙ ΕΝΑΛΛΑΚΤΙΚΕΣ ΤΟΥ ΧΛΩΡΙΟΥ, ΘΕΟΔΩΡΑΤΟΥ ΑΓΓΕΛΙΚΗ, ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ, ΜΥΤΙΛΗΝΗ 2005 ΠΡΟΧΩΡΗΜΕΝΕΣ ΤΕΧΝΙΚΕΣ ΟΞΕΙΔΩΣΗΣ 1 ΕΙΣΑΓΩΓΗ Οι προχωρημένες τεχνικές
Πηγή: ΑΠΟΛΥΜΑΝΣΗ ΤΟΥ ΠΟΣΙΜΟΥ ΝΕΡΟΥ : ΠΡΟΧΩΡΗΜΕΝΕΣ ΜΕΘΟΔΟΙ ΕΝΑΛΛΑΚΤΙΚΕΣ ΤΟΥ ΧΛΩΡΙΟΥ, ΘΕΟΔΩΡΑΤΟΥ ΑΓΓΕΛΙΚΗ, ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΙΓΑΙΟΥ, ΜΥΤΙΛΗΝΗ 2005 ΠΡΟΧΩΡΗΜΕΝΕΣ ΤΕΧΝΙΚΕΣ ΟΞΕΙΔΩΣΗΣ 1 ΕΙΣΑΓΩΓΗ Οι προχωρημένες τεχνικές
Γ. ΜΕΤΑ ΤΟ ΤΕΛΟΣ ΤΟΥ ΜΑΘΗΜΑΤΟΣ ΘΑ ΠΡΕΠΕΙ ΝΑ ΕΙΣΑΙ ΣΕ ΘΕΣΗ ΝΑ:
 ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 2 ο ΚΕΦΑΛΑΙΟ «ΚΥΤΤΑΡΟ: Η ΘΕΜΕΛΙΩΔΗΣ ΜΟΝΑΔΑ ΤΗΣ ΖΩΗΣ» ΕΙΣΑΓΩΓΗ ΚΑΙ ΤΟ ΠΟΡΤΡΕΤΟ ΤΟΥ ΕΥΚΑΡΥΩΤΙΚΟΥ ΚΥΤΤΑΡΟΥ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Τα κύτταρα με κριτήριο την ύπαρξη ή όχι μεμβράνης
ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 2 ο ΚΕΦΑΛΑΙΟ «ΚΥΤΤΑΡΟ: Η ΘΕΜΕΛΙΩΔΗΣ ΜΟΝΑΔΑ ΤΗΣ ΖΩΗΣ» ΕΙΣΑΓΩΓΗ ΚΑΙ ΤΟ ΠΟΡΤΡΕΤΟ ΤΟΥ ΕΥΚΑΡΥΩΤΙΚΟΥ ΚΥΤΤΑΡΟΥ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Τα κύτταρα με κριτήριο την ύπαρξη ή όχι μεμβράνης
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ. Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη
 Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΗ 4 (6/3/2013)
") Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
Επιδράση των υδατοκαλλιεργειών στο περιβάλλον
 Επιδράση των υδατοκαλλιεργειών στο περιβάλλον Παύλος Μακρίδης, επίκουρος καθηγητής Τμήμα Βιολογίας, Τομέας Βιολογίας Ζώων Πανεπιστήμιο Πατρών Τι είναι υδατοκαλλιέργειες; Η καλλιέργεια υδρόβιων οργανισμών,
Επιδράση των υδατοκαλλιεργειών στο περιβάλλον Παύλος Μακρίδης, επίκουρος καθηγητής Τμήμα Βιολογίας, Τομέας Βιολογίας Ζώων Πανεπιστήμιο Πατρών Τι είναι υδατοκαλλιέργειες; Η καλλιέργεια υδρόβιων οργανισμών,
ΓΕΝΕΤΙΚΑ ΤΡΟΠΟΠΟΙΗΜΕΝΑ ΦΥΤΑ (ΑΝΤΟΧΗ ΣΕ ΕΝΤΟΜΑ-ΙΟΥΣ)
") ΓΕΝΕΤΙΚΑ ΤΡΟΠΟΠΟΙΗΜΕΝΑ ΦΥΤΑ (ΑΝΤΟΧΗ ΣΕ ΕΝΤΟΜΑ-ΙΟΥΣ) 1 ΔΗΜΙΟΥΡΓΙΑ ΦΥΤΩΝ ΜΕ ΑΝΤΟΧΗ ΣΕ ΕΝΤΟΜΑ 19 Παράγοντες που συμβάλλουν σε αύξηση των εντόμων 1. Μονοκαλλιέργειες 2. Βελτίωση με κριτήριο αποκλειστικά την
ΓΕΝΕΤΙΚΑ ΤΡΟΠΟΠΟΙΗΜΕΝΑ ΦΥΤΑ (ΑΝΤΟΧΗ ΣΕ ΕΝΤΟΜΑ-ΙΟΥΣ) 1 ΔΗΜΙΟΥΡΓΙΑ ΦΥΤΩΝ ΜΕ ΑΝΤΟΧΗ ΣΕ ΕΝΤΟΜΑ 19 Παράγοντες που συμβάλλουν σε αύξηση των εντόμων 1. Μονοκαλλιέργειες 2. Βελτίωση με κριτήριο αποκλειστικά την
CAMPBELL REECE, ΒΙΟΛΟΓΙΑ ΤΟΜΟΣ Ι, ΠΕΚ 2010
 CAMPBELL REECE, ΒΙΟΛΟΓΙΑ ΤΟΜΟΣ Ι, ΠΕΚ 2010 CAMPBELL REECE, ΒΙΟΛΟΓΙΑ ΤΟΜΟΣ Ι, ΠΕΚ 2010 CAMPBELL REECE, ΒΙΟΛΟΓΙΑ ΤΟΜΟΣ Ι, ΠΕΚ 2010 Μεγεθυντική ικανότητα και διακριτική ικανότητα ή ανάλυση Μέγιστη ανάλυση
CAMPBELL REECE, ΒΙΟΛΟΓΙΑ ΤΟΜΟΣ Ι, ΠΕΚ 2010 CAMPBELL REECE, ΒΙΟΛΟΓΙΑ ΤΟΜΟΣ Ι, ΠΕΚ 2010 CAMPBELL REECE, ΒΙΟΛΟΓΙΑ ΤΟΜΟΣ Ι, ΠΕΚ 2010 Μεγεθυντική ικανότητα και διακριτική ικανότητα ή ανάλυση Μέγιστη ανάλυση
Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ ΤΩΝ ΖΩΩΝ. Αρχιτομία. Αγενής αναπαραγωγή. Παρατομία. Εκβλάστηση. Εγγενής αναπαραγωγή Διπλοφασικός κύκλος.
 Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ ΤΩΝ ΖΩΩΝ Αρχιτομία Αγενής αναπαραγωγή Παρατομία Εκβλάστηση Εγγενής αναπαραγωγή Απλοφασικός κύκλος Διπλοφασικός κύκλος Ισογαμία Ανισογαμία Ωογαμία Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ
Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ ΤΩΝ ΖΩΩΝ Αρχιτομία Αγενής αναπαραγωγή Παρατομία Εκβλάστηση Εγγενής αναπαραγωγή Απλοφασικός κύκλος Διπλοφασικός κύκλος Ισογαμία Ανισογαμία Ωογαμία Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ
Τα Αίτια Των Κλιματικών Αλλαγών
 Τα Αίτια Των Κλιματικών Αλλαγών Το Φαινόμενο του θερμοκηπίου Η τρύπα του όζοντος Η μόλυνση της ατμόσφαιρας Η μόλυνση του νερού Η μόλυνση του εδάφους Όξινη βροχή Ρύπανση του περιβάλλοντος Ραδιενεργός ρύπανση
Τα Αίτια Των Κλιματικών Αλλαγών Το Φαινόμενο του θερμοκηπίου Η τρύπα του όζοντος Η μόλυνση της ατμόσφαιρας Η μόλυνση του νερού Η μόλυνση του εδάφους Όξινη βροχή Ρύπανση του περιβάλλοντος Ραδιενεργός ρύπανση
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ
 ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
ΚΕΦΑΛΑΙΟ 3. Κύκλοι Βαρέων Μετάλλων. Βαρέα Μέταλλα στα Παράκτια Συστήματα
 ΚΕΦΑΛΑΙΟ 3 Κύκλοι Βαρέων Μετάλλων Βαρέα Μέταλλα στα Παράκτια Συστήματα Ο όρος βαρέα μέταλλα (heavy metals, trace metals, toxic metals, trace elements) χρησιμοποιείται συχνά για να περιγράψει τη παρουσία
ΚΕΦΑΛΑΙΟ 3 Κύκλοι Βαρέων Μετάλλων Βαρέα Μέταλλα στα Παράκτια Συστήματα Ο όρος βαρέα μέταλλα (heavy metals, trace metals, toxic metals, trace elements) χρησιμοποιείται συχνά για να περιγράψει τη παρουσία
ρευστότητα (εξασφαλίζεται µε τα φωσφολιπίδια)
") Λειτουργίες Πλασµατική µεµβράνη οριοθέτηση του κυττάρου εκλεκτική διαπερατότητα ή ηµιπερατότητα αναγνώριση και υποδοχή µηνυµάτων πρόσληψη και αποβολή ουσιών Πλασµατική µεµβράνη Ιδιότητες σταθερότητα ρευστότητα
Λειτουργίες Πλασµατική µεµβράνη οριοθέτηση του κυττάρου εκλεκτική διαπερατότητα ή ηµιπερατότητα αναγνώριση και υποδοχή µηνυµάτων πρόσληψη και αποβολή ουσιών Πλασµατική µεµβράνη Ιδιότητες σταθερότητα ρευστότητα
ΜΗΧΑΝΙΣΜΟΙ ΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΦΟΡΑΣ ΚΥΤΤΑΡΙΚΗ ΔΙΑΠΕΡΑΤΟΤΗΤΑ
 ΜΗΧΑΝΙΣΜΟΙ ΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΦΟΡΑΣ ΚΥΤΤΑΡΙΚΗ ΔΙΑΠΕΡΑΤΟΤΗΤΑ Προσοµοίωση Είναι γνωστό ότι η εξάσκηση των φοιτητών σε επίπεδο εργαστηριακών ασκήσεων, µε χρήση των κατάλληλων πειραµατοζώων, οργάνων και αναλωσίµων
ΜΗΧΑΝΙΣΜΟΙ ΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΦΟΡΑΣ ΚΥΤΤΑΡΙΚΗ ΔΙΑΠΕΡΑΤΟΤΗΤΑ Προσοµοίωση Είναι γνωστό ότι η εξάσκηση των φοιτητών σε επίπεδο εργαστηριακών ασκήσεων, µε χρήση των κατάλληλων πειραµατοζώων, οργάνων και αναλωσίµων
ΧΡΗΣΙΜΟΤΗΤΑ ΤΟΥ ΕΝΖΥΜΟΥ. Ο μεταβολισμός είναι μία πολύ σημαντική λειτουργία των μονογαστρικών ζώων και επιτυγχάνεται με τη δράση φυσικών
 ΧΡΗΣΙΜΟΤΗΤΑ ΤΟΥ ΕΝΖΥΜΟΥ Ο μεταβολισμός είναι μία πολύ σημαντική λειτουργία των μονογαστρικών ζώων και επιτυγχάνεται με τη δράση φυσικών Ενζύμων που βρίσκονται κατά μήκος του πεπτικού τους σωλήνα. Τα ένζυμα
ΧΡΗΣΙΜΟΤΗΤΑ ΤΟΥ ΕΝΖΥΜΟΥ Ο μεταβολισμός είναι μία πολύ σημαντική λειτουργία των μονογαστρικών ζώων και επιτυγχάνεται με τη δράση φυσικών Ενζύμων που βρίσκονται κατά μήκος του πεπτικού τους σωλήνα. Τα ένζυμα
Κεφάλαιο 7 - Ένζυμα, οι μηχανισμοί της ζωής
 1 Απόσπασμα από το βιβλίο «Πως να ζήσετε 150 χρόνια» του Dr. Δημήτρη Τσουκαλά Κεφάλαιο 7 - Ένζυμα, οι μηχανισμοί της ζωής Μικροσκοπικοί οργανισμοί που ρυθμίζουν τη λειτουργία του ανθρώπινου σώματος σε
1 Απόσπασμα από το βιβλίο «Πως να ζήσετε 150 χρόνια» του Dr. Δημήτρη Τσουκαλά Κεφάλαιο 7 - Ένζυμα, οι μηχανισμοί της ζωής Μικροσκοπικοί οργανισμοί που ρυθμίζουν τη λειτουργία του ανθρώπινου σώματος σε
Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Γενικής Παιδείας των Ημερησίων Γενικών Λυκείων και ΕΠΑΛ (Ομάδας Β ).
.") Αθήνα, 30/5/2014 ΠΑΝΕΛΛΗΝΙΑ ΕΝΩΣΗ ΒΙΟΕΠΙΣΤΗΜΟΝΩΝ Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Γενικής Παιδείας των Ημερησίων Γενικών Λυκείων και ΕΠΑΛ (Ομάδας Β ). Η
Αθήνα, 30/5/2014 ΠΑΝΕΛΛΗΝΙΑ ΕΝΩΣΗ ΒΙΟΕΠΙΣΤΗΜΟΝΩΝ Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Γενικής Παιδείας των Ημερησίων Γενικών Λυκείων και ΕΠΑΛ (Ομάδας Β ). Η
ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ Δ ΕΣΠΕΡΙΝΩΝ
 ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ Δ ΕΣΠΕΡΙΝΩΝ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 18 ΙΟΥΝΙΟΥ 2019 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΣΥΝΟΛΟ ΣΕΛΙΔΩΝ: ΠΕΝΤΕ (5) ΘΕΜΑ Α Να γράψετε στο
ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ Δ ΕΣΠΕΡΙΝΩΝ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 18 ΙΟΥΝΙΟΥ 2019 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΣΥΝΟΛΟ ΣΕΛΙΔΩΝ: ΠΕΝΤΕ (5) ΘΕΜΑ Α Να γράψετε στο
όλοι αναπνευστική οδός στομάχι στόμα
 κεράτινη στιβάδα περιέχει σμήγμα λιπαρά οξέα Μηχανισμοί που παρεμποδίζουν την είσοδο Δέρμα περιέχει ιδρώτας φυσιολογική μικροχλωρίδα λυσοζύμη γαλακτικό οξύ μικροοργανισμών Βλεννογόνοι όλοι αναπνευστική
κεράτινη στιβάδα περιέχει σμήγμα λιπαρά οξέα Μηχανισμοί που παρεμποδίζουν την είσοδο Δέρμα περιέχει ιδρώτας φυσιολογική μικροχλωρίδα λυσοζύμη γαλακτικό οξύ μικροοργανισμών Βλεννογόνοι όλοι αναπνευστική
Αποσάθρωση. Κεφάλαιο 2 ο. ΣΧΗΜΑΤΙΣΜΟΣ ΕΔΑΦΩΝ
 Κεφάλαιο 2 ο. ΣΧΗΜΑΤΙΣΜΟΣ ΕΔΑΦΩΝ Αποσάθρωση Ονομάζουμε τις μεταβολές στο μέγεθος, σχήμα και την εσωτερική δομή και χημική σύσταση τις οποίες δέχεται η στερεά φάση του εδάφους με την επίδραση των παραγόντων
Κεφάλαιο 2 ο. ΣΧΗΜΑΤΙΣΜΟΣ ΕΔΑΦΩΝ Αποσάθρωση Ονομάζουμε τις μεταβολές στο μέγεθος, σχήμα και την εσωτερική δομή και χημική σύσταση τις οποίες δέχεται η στερεά φάση του εδάφους με την επίδραση των παραγόντων
ΠΑΡΑΡΤΗΜΑ. της. Οδηγίας της Επιτροπής
 ΕΥΡΩΠΑΪΚΗ ΕΠΙΤΡΟΠΗ Βρυξέλλες, 17.5.2017 C(2017) 2842 final ANNEX 1 ΠΑΡΑΡΤΗΜΑ της Οδηγίας της Επιτροπής για την τροποποίηση της οδηγίας 2008/56/ΕΚ του Ευρωπαϊκού Κοινοβουλίου και του Συμβουλίου όσον αφορά
ΕΥΡΩΠΑΪΚΗ ΕΠΙΤΡΟΠΗ Βρυξέλλες, 17.5.2017 C(2017) 2842 final ANNEX 1 ΠΑΡΑΡΤΗΜΑ της Οδηγίας της Επιτροπής για την τροποποίηση της οδηγίας 2008/56/ΕΚ του Ευρωπαϊκού Κοινοβουλίου και του Συμβουλίου όσον αφορά
ΛΥΚΕΙΟ ΠΑΡΑΛΙΜΝΙΟΥ ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΗ ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ. ΖΗΤΗΜΑ Α Το σχεδιάγραμμα δείχνει τμήμα κυτταρικής μεμβράνης.
 ΛΥΚΕΙΟ ΠΑΡΑΛΙΜΝΙΟΥ ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΗ ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΕΝΟΤΗΤΑ: ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ ΚΑΘΗΓΗΤΗΣ: ΠΑΤΗΡ ΑΝΑΣΤΑΣΙΟΣ ΙΣΑΑΚ ΖΗΤΗΜΑ Α Το σχεδιάγραμμα δείχνει τμήμα κυτταρικής μεμβράνης. (α) Να ονομάσετε τα
ΛΥΚΕΙΟ ΠΑΡΑΛΙΜΝΙΟΥ ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΗ ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ ΕΝΟΤΗΤΑ: ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ ΚΑΘΗΓΗΤΗΣ: ΠΑΤΗΡ ΑΝΑΣΤΑΣΙΟΣ ΙΣΑΑΚ ΖΗΤΗΜΑ Α Το σχεδιάγραμμα δείχνει τμήμα κυτταρικής μεμβράνης. (α) Να ονομάσετε τα
Φλεγμονή. Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ
 Φλεγμονή Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ Μη ειδική ανοσολογική άμυνα ΑΝΑΤΟΜΙΚΟΙ ΦΡΑΓΜΟΙ Φυσικοί: δέρμα, βλεννογόνοι, βλέννα, βήχας Χημικοί: λυσοζύμη, αντιμικροβιακά
Φλεγμονή Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ Μη ειδική ανοσολογική άμυνα ΑΝΑΤΟΜΙΚΟΙ ΦΡΑΓΜΟΙ Φυσικοί: δέρμα, βλεννογόνοι, βλέννα, βήχας Χημικοί: λυσοζύμη, αντιμικροβιακά
7. Βιοτεχνολογία. α) η διαθεσιμότητα θρεπτικών συστατικών στο θρεπτικό υλικό, β) το ph, γ) το Ο 2 και δ) η θερμοκρασία.
 η διαθεσιμότητα θρεπτικών συστατικών στο θρεπτικό υλικό, β) το ph, γ) το Ο 2 και δ) η θερμοκρασία.") 7. Βιοτεχνολογία Εισαγωγή Τι είναι η Βιοτεχνολογία; Η Βιοτεχνολογία αποτελεί συνδυασμό επιστήμης και τεχνολογίας. Ειδικότερα εφαρμόζει τις γνώσεις που έχουν αποκτηθεί για τις βιολογικές λειτουργίες των
7. Βιοτεχνολογία Εισαγωγή Τι είναι η Βιοτεχνολογία; Η Βιοτεχνολογία αποτελεί συνδυασμό επιστήμης και τεχνολογίας. Ειδικότερα εφαρμόζει τις γνώσεις που έχουν αποκτηθεί για τις βιολογικές λειτουργίες των
ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ. Το σύνολο των μετασχηματισμών βιολογικής ή χημικής φύσης που λαμβάνουν χώρα κατά την ανακύκλωση ορισμένων στοιχείων
 ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ Το σύνολο των μετασχηματισμών βιολογικής ή χημικής φύσης που λαμβάνουν χώρα κατά την ανακύκλωση ορισμένων στοιχείων Επιβίωση οργανισμών Ύλη o Η ύλη που υπάρχει διαθέσιμη στη βιόσφαιρα
ΒΙΟΓΕΩΧΗΜΙΚΟΙ ΚΥΚΛΟΙ Το σύνολο των μετασχηματισμών βιολογικής ή χημικής φύσης που λαμβάνουν χώρα κατά την ανακύκλωση ορισμένων στοιχείων Επιβίωση οργανισμών Ύλη o Η ύλη που υπάρχει διαθέσιμη στη βιόσφαιρα
1. ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ
 ΚΕΦΑΛΑΙΟ 1 1. ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ ΚΥΤΤΑΡΑ ΚΑΙ ΙΣΤΟΙ Ο ανθρώπινος οργανισμός συνίσταται α- πό τρισεκατομμύρια κύτταρα. Τα κύτταρα αυτά εμφανίζουν σημαντική ποικιλομορφία, που αφορά το μέγεθος,
ΚΕΦΑΛΑΙΟ 1 1. ΑΠΟ ΤΟ ΚΥΤΤΑΡΟ ΣΤΟΝ ΟΡΓΑΝΙΣΜΟ ΚΥΤΤΑΡΑ ΚΑΙ ΙΣΤΟΙ Ο ανθρώπινος οργανισμός συνίσταται α- πό τρισεκατομμύρια κύτταρα. Τα κύτταρα αυτά εμφανίζουν σημαντική ποικιλομορφία, που αφορά το μέγεθος,
Ενότητα 3: : Ασφάλεια Βιολογικών Τροφίμων
 Ενότητα 3: : Ασφάλεια Βιολογικών Τροφίμων Διάλεξη 3.2 : Ποιοτικός έλεγχος & ασφάλεια στην αλυσίδα παραγωγής βιολογικών προϊόντων Εργαστήριο Πληροφορικής Γεωπονικό Πανεπιστήμιο Αθηνών http://infolab.aua.gr
Ενότητα 3: : Ασφάλεια Βιολογικών Τροφίμων Διάλεξη 3.2 : Ποιοτικός έλεγχος & ασφάλεια στην αλυσίδα παραγωγής βιολογικών προϊόντων Εργαστήριο Πληροφορικής Γεωπονικό Πανεπιστήμιο Αθηνών http://infolab.aua.gr
4. Ως αυτότροφοι οργανισμοί χαρακτηρίζονται α. οι καταναλωτές Α τάξης. β. οι παραγωγοί. γ. οι αποικοδομητές. δ. οι καταναλωτές Β τάξης.
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή στη φράση, η οποία
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή στη φράση, η οποία
Αρχές Βιοτεχνολογίας Τροφίμων
 Αρχές Βιοτεχνολογίας Τροφίμων Ενότητα 3: Εφαρμογές Βιομηχανικής Βιοτεχνολογίας(1/3), 2ΔΩ Τμήμα: Επιστήμης και Τεχνολογίας Τροφίμων Διδάσκων: Δρ. Σεραφείμ Παπανικολαου Μαθησιακοί Στόχοι Βιοτεχνολογικά Προϊόντα
Αρχές Βιοτεχνολογίας Τροφίμων Ενότητα 3: Εφαρμογές Βιομηχανικής Βιοτεχνολογίας(1/3), 2ΔΩ Τμήμα: Επιστήμης και Τεχνολογίας Τροφίμων Διδάσκων: Δρ. Σεραφείμ Παπανικολαου Μαθησιακοί Στόχοι Βιοτεχνολογικά Προϊόντα
ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ
 ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Οι μικροοργανισμοί είναι αναπόσπαστο τμήμα τόσο της ιστορίας του κόσμου μας όσο και της κοινωνικής εξέλιξης του ανθρώπου Βιοτεχνολογία o Ο όρος Βιοτεχνολογία χρησιμοποιήθηκε
ΑΡΧΕΣ ΚΑΙ ΜΕΘΟΔΟΛΟΓΙΑ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Οι μικροοργανισμοί είναι αναπόσπαστο τμήμα τόσο της ιστορίας του κόσμου μας όσο και της κοινωνικής εξέλιξης του ανθρώπου Βιοτεχνολογία o Ο όρος Βιοτεχνολογία χρησιμοποιήθηκε
Φαρμακοκινητική. Χρυσάνθη Σαρδέλη
 Φαρμακοκινητική Χρυσάνθη Σαρδέλη Αναπληρώτρια Καθηγήτρια Κλινικής Φαρμακολογίας Εργαστήριο Κλινικής Φαρμακολογίας Τμήμα Ιατρικής, Σχολή Επιστημών Υγείας, ΑΠΘ Φαρμακοκινητική Η Φαρμακοκινητική είναι η επιστήμη
Φαρμακοκινητική Χρυσάνθη Σαρδέλη Αναπληρώτρια Καθηγήτρια Κλινικής Φαρμακολογίας Εργαστήριο Κλινικής Φαρμακολογίας Τμήμα Ιατρικής, Σχολή Επιστημών Υγείας, ΑΠΘ Φαρμακοκινητική Η Φαρμακοκινητική είναι η επιστήμη
Επίκουρος Καθηγητής Π. Μελίδης
 Χαρακτηριστικά υγρών αποβλήτων Επίκουρος Καθηγητής Π. Μελίδης Τμήμα Μηχανικών Περιβάλλοντος Εργαστήριο Διαχείρισης και Τεχνολογίας Υγρών Αποβλήτων Τα υγρά απόβλητα μπορεί να προέλθουν από : Ανθρώπινα απόβλητα
Χαρακτηριστικά υγρών αποβλήτων Επίκουρος Καθηγητής Π. Μελίδης Τμήμα Μηχανικών Περιβάλλοντος Εργαστήριο Διαχείρισης και Τεχνολογίας Υγρών Αποβλήτων Τα υγρά απόβλητα μπορεί να προέλθουν από : Ανθρώπινα απόβλητα
Εφαρμοσμένη Διατροφική Ιατρική
 Γλωσσάρι για το Μάθημα της Διατροφικής Ιατρικής Λιπαρά οξέα: περιέχουν μακριές αλυσίδες μορίων που αποτελούν σχεδόν όλο το σύμπλεγμα λιπιδίων τόσο για τα ζωικά όσο και για τα φυτικά λίπη. Αν αποκοπούν
Γλωσσάρι για το Μάθημα της Διατροφικής Ιατρικής Λιπαρά οξέα: περιέχουν μακριές αλυσίδες μορίων που αποτελούν σχεδόν όλο το σύμπλεγμα λιπιδίων τόσο για τα ζωικά όσο και για τα φυτικά λίπη. Αν αποκοπούν
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4
 ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
ΕΠΑΝ II, KOYΠΟΝΙΑ ΚΑΙΝΟΤΟΜΙΑΣ ΓΙΑ ΜΙΚΡΟΜΕΣΑΙΕΣ ΕΠΙΧΕΙΡΗΣΕΙΣ Κωδικός Αριθμός Κουπονιού:
 ΕΛΛΗΝΙΚΟ ΚΕΝΤΡΟ ΘΑΛΑΣΣΙΩΝ ΕΡΕΥΝΩΝ ΙΝΣΤΙΤΟΥΤΟ ΘΑΛΑΣΣΙΩΝ ΒΙΟΛΟΓΙΚΩΝ ΠΟΡΩΝ ΙΝΣΤΙΤΟΥΤΟ ΩΚΕΑΝΟΓΡΑΦΙΑΣ ΣΥΛΛΟΓΗ ΠΕΡΙΒΑΛΛΟΝΤΙΚΩΝ, ΟΙΚΟΛΟΓΙΚΩΝ, ΩΚΕΑΝΟΓΡΑΦΙΚΩΝ ΚΑΙ ΑΛΙΕΥΤΙΚΩΝ ΔΕΔΟΜΕΝΩΝ ΓΙΑ ΤΗΝ ΠΕΡΙΟΧΗ ΤΟΥ ΑΡΓΟΛΙΚΟΥ
ΕΛΛΗΝΙΚΟ ΚΕΝΤΡΟ ΘΑΛΑΣΣΙΩΝ ΕΡΕΥΝΩΝ ΙΝΣΤΙΤΟΥΤΟ ΘΑΛΑΣΣΙΩΝ ΒΙΟΛΟΓΙΚΩΝ ΠΟΡΩΝ ΙΝΣΤΙΤΟΥΤΟ ΩΚΕΑΝΟΓΡΑΦΙΑΣ ΣΥΛΛΟΓΗ ΠΕΡΙΒΑΛΛΟΝΤΙΚΩΝ, ΟΙΚΟΛΟΓΙΚΩΝ, ΩΚΕΑΝΟΓΡΑΦΙΚΩΝ ΚΑΙ ΑΛΙΕΥΤΙΚΩΝ ΔΕΔΟΜΕΝΩΝ ΓΙΑ ΤΗΝ ΠΕΡΙΟΧΗ ΤΟΥ ΑΡΓΟΛΙΚΟΥ
ΗΜΕΡΙΔΑ ELQA. Καθαρισμός ύδατος από βαρέα μέταλλα με καινοτόμα τεχνολογία. Ερευνητικό εργαστήριο Food InnovaLab 1
 Καθαρισμός ύδατος από βαρέα μέταλλα με καινοτόμα τεχνολογία Σταύρος Λαλάς*, Βασίλειος Αθανασιάδης και Όλγα Γκορτζή Τμήμα Τεχνολογίας Τροφίμων Τ.Ε.Ι. ΘΕΣΣΑΛΙΑΣ Ερευνητικό εργαστήριο Food InnovaLab 1 Βαρέα
Καθαρισμός ύδατος από βαρέα μέταλλα με καινοτόμα τεχνολογία Σταύρος Λαλάς*, Βασίλειος Αθανασιάδης και Όλγα Γκορτζή Τμήμα Τεχνολογίας Τροφίμων Τ.Ε.Ι. ΘΕΣΣΑΛΙΑΣ Ερευνητικό εργαστήριο Food InnovaLab 1 Βαρέα
Διαχείριση Αποβλήτων
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Διαχείριση Αποβλήτων Ενότητα 11 : Βιομηχανικά Στερεά και Υγρά Απόβλητα Δρ. Σταυρούλα Τσιτσιφλή Τμήμα Μηχανικών Χωροταξίας, Πολεοδομίας και Περιφερειακής Ανάπτυξης Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Διαχείριση Αποβλήτων Ενότητα 11 : Βιομηχανικά Στερεά και Υγρά Απόβλητα Δρ. Σταυρούλα Τσιτσιφλή Τμήμα Μηχανικών Χωροταξίας, Πολεοδομίας και Περιφερειακής Ανάπτυξης Άδειες Χρήσης Το
ΡΑΔΙΟΧΗΜΕΙΑ 1. ΦΥΣΙΚΗ ΡΑΔΙΕΝΕΡΓΕΙΑ ΚΕΦΑΛΑΙΟ 5. ΔΙΑΧΕΙΡΙΣΗ ΡΑΔΙΕΝΕΡΓΩΝ ΑΠΟΒΛΗΤΩΝ ΤΟΞΙΚΟΤΗΤΑ ΡΑΔΙΕΝΕΡΓΩΝ ΙΣΟΤΟΠΩΝ Τμήμα Χημικών Μηχανικών
 ΡΑΔΙΟΧΗΜΕΙΑ ΔΙΑΧΕΙΡΙΣΗ ΡΑΔΙΕΝΕΡΓΩΝ ΑΠΟΒΛΗΤΩΝ ΤΟΞΙΚΟΤΗΤΑ ΡΑΔΙΕΝΕΡΓΩΝ ΙΣΟΤΟΠΩΝ Τμήμα Χημικών Μηχανικών ΚΕΦΑΛΑΙΟ 5. ΤΟΞΙΚΟΤΗΤΑ ΤΩΝ ΡΑΔΙΕΝΕΡΓΩΝ ΣΤΟΙΧΕΙΩΝ 1. ΦΥΣΙΚΗ ΡΑΔΙΕΝΕΡΓΕΙΑ Ιωάννα Δ. Αναστασοπούλου Βασιλική
ΡΑΔΙΟΧΗΜΕΙΑ ΔΙΑΧΕΙΡΙΣΗ ΡΑΔΙΕΝΕΡΓΩΝ ΑΠΟΒΛΗΤΩΝ ΤΟΞΙΚΟΤΗΤΑ ΡΑΔΙΕΝΕΡΓΩΝ ΙΣΟΤΟΠΩΝ Τμήμα Χημικών Μηχανικών ΚΕΦΑΛΑΙΟ 5. ΤΟΞΙΚΟΤΗΤΑ ΤΩΝ ΡΑΔΙΕΝΕΡΓΩΝ ΣΤΟΙΧΕΙΩΝ 1. ΦΥΣΙΚΗ ΡΑΔΙΕΝΕΡΓΕΙΑ Ιωάννα Δ. Αναστασοπούλου Βασιλική
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ
 ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Περιγραφή της περίπτωσης Άνδρας 43 ετών εισάγεται σε κλινική λόγω επιγαστραλγίας. Μετά από έλεγχο ετέθη η διάγνωση του πεπτικού
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Περιγραφή της περίπτωσης Άνδρας 43 ετών εισάγεται σε κλινική λόγω επιγαστραλγίας. Μετά από έλεγχο ετέθη η διάγνωση του πεπτικού
Σκελετικό σύστημα. Λειτουργίες: 1. Στηρικτικό πλαίσιο του σώματος των ζώων 2. Κινητική ποικιλομορφία. 2. Σκληροί σκελετοί
 Σκελετικό σύστημα Λειτουργίες: 1. Στηρικτικό πλαίσιο του σώματος των ζώων 2. Κινητική ποικιλομορφία Τύποι: 1. Υδροστατικοί σκελετοί 2. Σκληροί σκελετοί Ενδοσκελετός Εξωσκελετός Σκελετικό σύστημα υδροστατικοί
Σκελετικό σύστημα Λειτουργίες: 1. Στηρικτικό πλαίσιο του σώματος των ζώων 2. Κινητική ποικιλομορφία Τύποι: 1. Υδροστατικοί σκελετοί 2. Σκληροί σκελετοί Ενδοσκελετός Εξωσκελετός Σκελετικό σύστημα υδροστατικοί
ΘΕΜΑ Α 1 δ 2 β 3 γ 4 β 5 α ΘΕΜΑ Β
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 30 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α 1 δ 2 β 3 γ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 30 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α 1 δ 2 β 3 γ
ΕΘΝΙΚΟ ΜΕΤΣΟΒΙΟ ΠΟΛΥΤΕΧΝΕΙΟ ΙΕΠΙΣΤΗΜΟΝΙΚΟ ΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥ ΩΝ (.Π.Μ.Σ.) «ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ»
 «ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ»") ΕΘΝΙΚΟ ΜΕΤΣΟΒΙΟ ΠΟΛΥΤΕΧΝΕΙΟ ΙΕΠΙΣΤΗΜΟΝΙΚΟ ΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥ ΩΝ (.Π.Μ.Σ.) «ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ» ΜΑΘΗΜΑ ΚΟΡΜΟΥ «Υ ΑΤΙΚΟ ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ» ΡΥΠΑΝΣΗ ΝΕΡΟΥ ΜΕΡΟΣ B ΙΑΦΑΝΕΙΕΣ.
ΕΘΝΙΚΟ ΜΕΤΣΟΒΙΟ ΠΟΛΥΤΕΧΝΕΙΟ ΙΕΠΙΣΤΗΜΟΝΙΚΟ ΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥ ΩΝ (.Π.Μ.Σ.) «ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ» ΜΑΘΗΜΑ ΚΟΡΜΟΥ «Υ ΑΤΙΚΟ ΠΕΡΙΒΑΛΛΟΝ ΚΑΙ ΑΝΑΠΤΥΞΗ» ΡΥΠΑΝΣΗ ΝΕΡΟΥ ΜΕΡΟΣ B ΙΑΦΑΝΕΙΕΣ.
Εισαγωγή στην Επιστήμη του Μηχανικού Περιβάλλοντος Δ Ι Δ Α Σ Κ Ο Υ Σ Α Κ Ρ Ε Σ Τ Ο Υ Α Θ Η Ν Α Δ Ρ. Χ Η Μ Ι Κ Ο Σ Μ Η Χ Α Ν Ι Κ Ο Σ
 Εισαγωγή στην Επιστήμη του Μηχανικού Περιβάλλοντος Δ Ι Δ Α Σ Κ Ο Υ Σ Α Κ Ρ Ε Σ Τ Ο Υ Α Θ Η Ν Α Δ Ρ. Χ Η Μ Ι Κ Ο Σ Μ Η Χ Α Ν Ι Κ Ο Σ Εισαγωγή στην Επιστήμη του Μηχανικού Περιβάλλοντος 1 ΜΑΘΗΜΑ 2 Ο & 3 O
Εισαγωγή στην Επιστήμη του Μηχανικού Περιβάλλοντος Δ Ι Δ Α Σ Κ Ο Υ Σ Α Κ Ρ Ε Σ Τ Ο Υ Α Θ Η Ν Α Δ Ρ. Χ Η Μ Ι Κ Ο Σ Μ Η Χ Α Ν Ι Κ Ο Σ Εισαγωγή στην Επιστήμη του Μηχανικού Περιβάλλοντος 1 ΜΑΘΗΜΑ 2 Ο & 3 O
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΕΡΓΑΣΤΗΡΙΟ ΧΗΜΕΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ ΣΤΟΝ ΜΑΛΙΑΚΟ ΚΟΛΠΟ. Αν. Καθηγητης Μ.Δασενακης. Δρ Θ.Καστριτης Ε.Ρουσελάκη
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΕΡΓΑΣΤΗΡΙΟ ΧΗΜΕΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Ο ΚΙΝΔΥΝΟΣ ΤΟΥ ΕΥΤΡΟΦΙΣΜΟΥ ΣΤΟΝ ΜΑΛΙΑΚΟ ΚΟΛΠΟ Αν. Καθηγητης Μ.Δασενακης Δρ Θ.Καστριτης Ε.Ρουσελάκη Φ.Σάλτα Κύκλος αζώτου Κύκλος φωσφόρου Kύκλος πυριτίου
ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΕΡΓΑΣΤΗΡΙΟ ΧΗΜΕΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Ο ΚΙΝΔΥΝΟΣ ΤΟΥ ΕΥΤΡΟΦΙΣΜΟΥ ΣΤΟΝ ΜΑΛΙΑΚΟ ΚΟΛΠΟ Αν. Καθηγητης Μ.Δασενακης Δρ Θ.Καστριτης Ε.Ρουσελάκη Φ.Σάλτα Κύκλος αζώτου Κύκλος φωσφόρου Kύκλος πυριτίου
Περίληψη Βιολογίας Κεφάλαιο 3
 Περίληψη Βιολογίας Κεφάλαιο 3 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Η σημασία της ενέργειας στους οργανισμούς. Η ενέργεια είναι ένας παράγοντας σημαντικός για τους οργανισμούς γιατί όλες οι λειτουργίες τους απαιτούν
Περίληψη Βιολογίας Κεφάλαιο 3 ΕΝΕΡΓΕΙΑ ΚΑΙ ΟΡΓΑΝΙΣΜΟΙ Η σημασία της ενέργειας στους οργανισμούς. Η ενέργεια είναι ένας παράγοντας σημαντικός για τους οργανισμούς γιατί όλες οι λειτουργίες τους απαιτούν
Θέματα πριν τις εξετάσεις. Καλό διάβασμα Καλή επιτυχία
 Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
ΠΑΝΕΛΛΗΝΙΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΒΙΟΛΟΓΙΑΣ 2006
 ΠΑΝΕΛΛΗΝΙΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΒΙΟΛΟΓΙΑΣ 2006 Β φάση 18-3-2006 1. Ένα φυτό έχει ποτιστεί με νερό σημασμένο ραδιενεργά, του οποίου το οξυγόνο είναι το ισότοπο ( 18 Ο). Η ραδιενέργεια ανιχνεύεται μετά πάροδο ορισμένων
ΠΑΝΕΛΛΗΝΙΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΒΙΟΛΟΓΙΑΣ 2006 Β φάση 18-3-2006 1. Ένα φυτό έχει ποτιστεί με νερό σημασμένο ραδιενεργά, του οποίου το οξυγόνο είναι το ισότοπο ( 18 Ο). Η ραδιενέργεια ανιχνεύεται μετά πάροδο ορισμένων