ΜΕΛΕΤΗ ΤΩΝ ΠΑΡΑΓΟΝΤΩΝ ΠΟΥ ΡΥΘΜΙΖΟΥΝ ΤΗΝ ΕΠΙΒΙΩΣΗ, ΤΗ ΙΑΦΟΡΟΠΟΙΗΣΗ ΚΑΙ ΤΗΝ
|
|
|
- Ἁλκυόνη Ἀγλαΐη Αλεβίζος
- 6 χρόνια πριν
- Προβολές:
Transcript
1 AΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΚΤΗΝΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΟΜΗΣ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑΣ ΤΩΝ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ ΚΛΕΑΝΘΗ Κ. ΜΕΛΛΙΟΥ Κτηνιάτρου ΜΕΛΕΤΗ ΤΩΝ ΠΑΡΑΓΟΝΤΩΝ ΠΟΥ ΡΥΘΜΙΖΟΥΝ ΤΗΝ ΕΠΙΒΙΩΣΗ, ΤΗ ΙΑΦΟΡΟΠΟΙΗΣΗ ΚΑΙ ΤΗΝ ΑΠΟΠΤΩΣΗ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ ΚΑΙ ΜΕΤΑ ΑΠΟ ΠΕΙΡΑΜΑΤΙΚΕΣ ΒΛΑΒΕΣ Ι ΑΚΤΟΡΙΚΗ ΙΑΤΡΙΒΗ ΘΕΣΣΑΛΟΝΙΚΗ 2008
2
3 ΚΛΕΑΝΘΗ Κ. ΜΕΛΛΙΟΥ ΜΕΛΕΤΗ ΤΩΝ ΠΑΡΑΓΟΝΤΩΝ ΠΟΥ ΡΥΘΜΙΖΟΥΝ ΤΗΝ ΕΠΙΒΙΩΣΗ, ΤΗ ΙΑΦΟΡΟΠΟΙΗΣΗ ΚΑΙ ΤΗΝ ΑΠΟΠΤΩΣΗ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ ΚΑΙ ΜΕΤΑ ΑΠΟ ΠΕΙΡΑΜΑΤΙΚΕΣ ΒΛΑΒΕΣ Ι ΑΚΤΟΡΙΚΗ ΙΑΤΡΙΒΗ Υποβλήθηκε στην Κτηνιατρική Σχολή Τοµέας οµής και Λειτουργίας των Ζωικών Οργανισµών Ηµεροµηνία Προφορικής Εξέτασης: Εξεταστική Επιτροπή Επίκ. Καθηγήτρια Ι. ωρή Καθηγητής Α. Ντινόπουλος Αν. Καθηγητής Ι. Αντωνόπουλος Επιβλέπουσα Καθηγήτρια Μέλος Τριµελούς Συµβουλευτικής Επιτροπής Μέλος Τριµελούς Συµβουλευτικής Επιτροπής Καθηγήτρια Λ. Ανδριοπούλου-Οικονόµου Καθηγητής Ι. Βλέµµας Αν. Καθηγήτρια Ε. Μιχαλούδη Λέκτορας Α. Τσιγκοτζίδου Εξετάστρια Εξεταστής Εξετάστρια Εξετάστρια
4 Κλεάνθης Μέλλιος Α.Π.Θ. ΜΕΛΕΤΗ ΤΩΝ ΠΑΡΑΓΟΝΤΩΝ ΠΟΥ ΡΥΘΜΙΖΟΥΝ ΤΗΝ ΕΠΙΒΙΩΣΗ, ΤΗ ΙΑΦΟΡΟΠΟΙΗΣΗ ΚΑΙ ΤΗΝ ΑΠΟΠΤΩΣΗ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ ΚΑΙ ΜΕΤΑ ΑΠΟ ΠΕΙΡΑΜΑΤΙΚΕΣ ΒΛΑΒΕΣ ISBN «Η έγκριση της παρούσας ιδακτορικής ιατριβής από την Κτηνιατρική Σχολή του Αριστοτέλειου Πανεπιστηµίου Θεσσαλονίκης δεν υποδηλώνει αποδοχή των γνωµών του συγγραφέως». (Νόµος 5343/1932, άρθρο 202 παρ.2)
5 Στους γονείς µου
6
7 Πίνακας Περιεχοµένων ΠΙΝΑΚΑΣ ΠΕΡΙΕΧΟΜΕΝΩΝ ΠΡΟΛΟΓΟΣ 1 ΕΙΣΑΓΩΓΗ 7 Α. ΑΝΑΤΟΜΙΚΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΚΗ ΟΡΓΑΝΩΣΗ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ 7 Β. ΠΕΡΙΓΡΑΦΗ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ 8 1. ΜΟΡΦΟΛΟΓΙΚΑ ΚΑΙ ΝΕΥΡΟΧΗΜΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΜΕΣΟΜΕΓΕΘΕΙΣ ΑΚΑΝΘΩΤΟΙ ΠΡΟΒΛΗΤΙΚΟΙ ΝΕΥΡΩΝΕΣ ΙΑΜΕΣΟΙ ΝΕΥΡΩΝΕΣ Α. ΜΕΓΑΛΟΙ ΜΗ ΑΚΑΝΘΩΤΟΙ ΧΟΛΙΝΕΡΓΙΚΟΙ ΙΑΜΕΣΟΙ ΝΕΥΡΩΝΕΣ Β. ΜΕΣΟΜΕΓΕΘΕΙΣ ΜΗ ΑΚΑΝΘΩΤΟΙ GABAΕΡΓΙΚΟΙ ΙΑΜΕΣΟΙ ΝΕΥΡΩΝΕΣ ΣΥΝ ΕΣΕΙΣ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΠΡΟΣΑΓΩΓΕΣ ΙΝΕΣ Α. ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΦΛΟΙΟ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Β. ΣΥΝ ΕΣΕΙΣ ΜΕ ΥΠΟΦΛΟΙΙΚΕΣ ΠΕΡΙΟΧΕΣ Β 1 ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΘΑΛΑΜΟ Β 2 ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΜΕΣΟ ΕΓΚΕΦΑΛΟ Β 3 ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΗΝ ΩΧΡΗ ΣΦΑΙΡΑ B 4 ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΥΠΟΘΑΛΑΜΙΟ ΠΥΡΗΝΑ ΑΠΑΓΩΓΕΣ ΙΝΕΣ Ι ΙΑΙΤΕΡΟΤΗΤΕΣ ΤΗΣ ΟΡΓΑΝΩΣΗΣ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΑΝΑΠΤΥΞΗ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ 24 Γ. ΠΡΟΓΡΑΜΜΑΤΙΣΜΕΝΟΣ ΚΥΤΤΑΡΙΚΟΣ ΘΑΝΑΤΟΣ ΜΟΡΦΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ PCD ΡΥΘΜΙΣΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ 35 i
8 Πίνακας Περιεχοµένων 2.1. KYTTAΡΙΚΕΣ ΠΡΩΤΕΪΝΕΣ ΝΕΥΡΟΤΡΟΦΙΚΟΙ ΠΑΡΑΓΟΝΤΕΣ Η ΑΠΟΠΤΩΣΗ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΚΑΙ ΤΩΝ ΠΕΡΙΟΧΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ ΜΕ ΤΙΣ ΟΠΟΙΕΣ ΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΣΥΝ ΕΕΤΑΙ ΣΕ ΝΕΥΡΟΕΚΦΥΛΙΣΤΙΚΕΣ ΑΣΘΕΝΕΙΕΣ 46 ΥΛΙΚΑ ΚΑΙ ΜΕΘΟ ΟΙ ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΤΩΝ ΜΕΘΟ ΩΝ ΠΟΥ ΧΡΗΣΙΜΟΠΟΙΗΘΗΚΑΝ ΜΕΘΟ ΟΣ TUNEL ΜΕΘΟ ΟΣ ΑΒΙ ΙΝΗΣ-ΒΙΟΤΙΝΗΣ ANΟΣΟΪΣΤΟΧΗΜΕΙΑ ΠΕΙΡΑΜΑΤΙΚΟ ΥΛΙΚΟ ΚΑΙ ΜΕΘΟ ΟΙ ΠΕΙΡΑΜΑΤΙΚΟΙ ΧΕΙΡΙΣΜΟΙ Α. ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Β. ΦΑΡΜΑΚΟΛΟΓΙΚΗ ΒΛΑΒΗ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΣΥΛΛΟΓΗ ΠΕΙΡΑΜΑΤΙΚΟΥ ΥΛΙΚΟΥ ΜΕΘΟ ΟΣ TUNEL ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΘΟ ΟΣ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3, ΤΗΣ ΦΡΑΚΤΙΝΗΣ KAI TOY GABA ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΘΟ ΟΣ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ChAT ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΘΟ ΟΣ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΝΤΟΠΑΜΙΝΗΣ ΚΑΙ ΤΗΣ ΝΟΡΑ ΡΕΝΑΛΙΝΗΣ ΗΛΕΚΤΡΟΝΙΚΗ ΜΙΚΡΟΣΚΟΠΙΑ ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ 65 ΑΠΟΤΕΛΕΣΜΑΤΑ 67 ΣΧΟΛΙΑ ΓΙΑ ΤΙΣ ΤΕΧΝΙΚΕΣ ΚΑΙ ΤΟΥΣ ΠΕΙΡΑΜΑΤΙΣΜΟΥΣ ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ 70 ii
9 Πίνακας Περιεχοµένων 1.1. ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ Α. ΜΕΛΕΤΗ ΜΕ ΤΟ ΟΠΤΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Β. ΜΕΛΕΤΗ ΜΕ ΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3 ΚΑΙ ΤΗΣ ΦΡΑΚΤΙΝΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΕΠΙΜΥΩΝ ΠΟΥ ΕΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΠΕΡΙΓΡΑΦΗ ΚΑΙ ΑΞΙΟΛΟΓΗΣΗ ΤΗΣ ΒΛΑΒΗΣ ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ Α. ΜΕΛΕΤΗ ΜΕ ΤΟ ΟΠΤΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Β. ΜΕΛΕΤΗ ΜΕ ΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΕ ΕΠΙΜΥΣ ΠΟΥ ΕΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΣΤΟ ΝΕΟΓΕΝΝΗΤΟ ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΕ ΕΠΙΜΥΣ ΠΟΥ EΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩN ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΣΕ ΜΕΤΑΓΕΝΕΣΤΕΡΑ ΣΤΑ ΙΑ ΤΗΣ ΑΝΑΠΤΥΞΗΣ ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3 ΚΑΙ ΤΗΣ ΦΡΑΚΤΙΝΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΜΕΤΑ ΑΠΟ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΥΠΟΛΟΓΙΣΜΟΣ ΤΟΥ ΑΡΙΘΜΟΥ ΤΩΝ «ΖΩΝΤΑΝΩΝ» ΝΕΥΡΩΝΩΝ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΕΠΙΜΥΩΝ ΠΟΥ ΕΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΠΡΟΣΑΓΩΓΩΝ ΣΥΣΤΗΜΑΤΩΝ ΙΝΩΝ ΜΕ ΤΗ ΝΕΥΡΟΤΟΞΙΝΗ 6-OHDA ΠΕΡΙΓΡΑΦΗ ΚΑΙ ΑΞΙΟΛΟΓΗΣΗ ΤΗΣ ΒΛΑΒΗΣ ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ Α. ΜΕΛΕΤΗ ΜΕ ΤΟ ΟΠΤΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Β. ΜΕΛΕΤΗ ΜΕ ΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ 95 iii
10 Πίνακας Περιεχοµένων 3.4. ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3 ΚΑΙ ΤΗΣ ΦΡΑΚΤΙΝΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΜΕΤΑ ΑΠΟ ΒΛΑΒΗ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΜΕ 6-OHDA ΜΕΛΕΤΗ ΤΗΣ ΕΠΙ ΡΑΣΗΣ ΤΩΝ ΒΛΑΒΩΝ ΤΩΝ ΠΡΟΣΑΓΩΓΩΝ ΣΥΝ ΕΣΕΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΣΤΗΝ ΕΠΙΒΙΩΣΗ ΚΑΙ ΣΤΟΝ ΦΑΙΝΟΤΥΠΟ ΤΩΝ GABAΕΡΓΙΚΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ ΜΕΛΕΤΗ ΤΗΣ ΕΠΙ ΡΑΣΗΣ ΤΗΣ ΒΛΑΒΗΣ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΣΤΗΝ ΕΠΙΒΙΩΣΗ ΚΑΙ ΣΤΟΝ ΦΑΙΝΟΤΥΠΟ ΤΩΝ ΧΟΛΙΝΕΡΓΙΚΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ 105 ΣΥΖΗΤΗΣΗ O ΚΥΤΤΑΡΙΚΟΣ ΘΑΝΑΤΟΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ ΕΙΝΑΙ ΑΠΟΠΤΩΤΙΚΟΥ ΤΥΠΟΥ Η ΑΠΟΠΤΩΣΗ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΣΥΜΒΑΙΝΕΙ ΚΑΤΑ ΤΙΣ ΠΡΩΤΕΣ ΕΒ ΟΜΑ ΕΣ ΤΗΣ ΜΕΤΑΓΕΝΝΗΤΙΚΗΣ ΑΝΑΠΤΥΞΗΣ ΚΑΙ ΣΥΜΠΙΠΤΕΙ ΜΕ ΤΗΝ ΠΕΡΙΟ Ο ΙΑΦΟΡΟΠΟΙΗΣΗΣ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΚΑΙ ΟΡΓΑΝΩΣΗΣ ΤΩΝ ΣΥΝ ΕΣΕΩΝ ΤΟΥΣ ΟΙ ΒΛΑΒΕΣ ΤΩΝ ΣΥΝ ΕΣΕΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΠΟΥ ΠΡΑΓΜΑΤΟΠΟΙΟΥΝΤΑΙ ΣΕ ΠΡΩΙΜΑ ΣΤΑ ΙΑ ΤΗΣ ΑΝΑΠΤΥΞΗΣ ΠΡΟΚΑΛΟΥΝ ΕΛΑΤΤΩΣΗ ΤΟΥ ΑΡΙΘΜΟΥ ΤΩΝ ΝΕΥΡΩΝΩΝ ΠΟΥ ΕΠΙΒΙΩΝΟΥΝ ΚΑΙ ΕΠΗΡΕΑΖΟΥΝ ΤΟΝ ΦΑΙΝΟΤΥΠΟ ΤΟΥΣ ΓΕΝΙΚΕΣ ΠΑΡΑΤΗΡΗΣΕΙΣ ΠΟΥ ΑΦΟΡΟΥΝ ΤΟΝ ΡΟΛΟ ΤΩΝ ΠΡΟΣΑΓΩΓΩΝ ΣΥΝ ΕΣΕΩΝ ΣΤΗΝ ΕΠΙΒΙΩΣΗ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ Η ΑΦΑΙΡΕΣΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΣΤΟ ΝΕΟΓΕΝΝΗΤΟ ΠΡΟΚΑΛΕΙ ΑΥΞΗΣΗ ΤΟΥ ΚΥΤΤΑΡΙΚΟΥ ΘΑΝΑΤΟΥ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ 117 iv
11 Πίνακας Περιεχοµένων 3.3. Η ΒΛΑΒΗ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΠΡΟΣΑΓΩΓΩΝ ΙΝΩΝ ΠΡΟΚΑΛΕΙ ΑΥΞΗΣΗ ΤΟΥ ΚΥΤΤΑΡΙΚΟΥ ΘΑΝΑΤΟΥ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ H ΣΥΓΚΡΙΣΗ ΤΩΝ ΥΟ ΠΕΙΡΑΜΑΤΙΚΩΝ ΜΟΝΤΕΛΩΝ ΑΠΟΚΑΛΥΠΤΕΙ ΟΤΙ Ο ΚΥTΤΑΡΙΚΟΣ ΘΑΝΑΤΟΣ ΜΕΤΑ ΑΠΟ ΒΛΑΒΕΣ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΚΑΙ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΠΡΟΣΑΓΩΓΩΝ ΙΝΩΝ ΑΥΞΑΝΕΤΑΙ ΣΤΟΝ Ι ΙΟ ΒΑΘΜΟ ΚΑΙ ΚΟΡΥΦΩΝΕΤΑΙ ΣΤΟ Ι ΙΟ ΑΝΑΠΤΥΞΙΑΚΟ ΣΤΑ ΙΟ Ο ΚΥΤΤΑΡΙΚΟΣ ΘΑΝΑΤΟΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΜΕΤΑ ΑΠΟ ΒΛΑΒΕΣ ΤΩΝ ΣΥΝ ΕΣΕΩΝ ΤΟΥ ΕΙΝΑΙ ΑΠΟΠΤΩΤΙΚΟΥ ΤΥΠΟΥ ΟΙ ΒΛΑΒΕΣ ΤΩΝ ΣΥΝ ΕΣΕΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΠΡΟΚΑΛΟΥΝ ΑΛΛΑΓΕΣ ΟΧΙ ΜΟΝΟ ΣΤΗΝ ΕΠΙΒΙΩΣΗ ΑΛΛΑ ΚΑΙ ΣΤΟΝ ΦΑΙΝΟΤΥΠΟ ΤΩΝ GABAΕΡΓΙΚΩΝ KΑI ΤΩΝ ΧΟΛΙΝΕΡΓΙΚΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΣΥΜΠΕΡΑΣΜΑΤΑ 138 ΠΕΡΙΛΗΨΗ 141 SUMMARY 145 ΣΥΝΤΟΜΟΓΡΑΦΙΕΣ 147 ΒΙΒΛΙΟΓΡΑΦΙΑ 151 v
12
13 Πρόλογος ΠΡΟΛΟΓΟΣ Οι πυρήνες του τελικού, του διάµεσου και του µέσου εγκεφάλου που συγκροτούν τα βασικά γάγγλια αποτελούν µέρη ενός κλειστού νευρωνικού κυκλώµατος και συµµετέχουν στην εκτέλεση εξειδικευµένων κινήσεων, στην οργάνωση συνδυασµένων προσαρµογών της στάσης του σώµατος καθώς και σε ανώτερες γνωστικές λειτουργίες, όπως είναι η µάθηση, η ανταµοιβή και η κινητοποίηση. Ο µεγαλύτερος από τους πυρήνες αυτούς είναι το ραβδωτό σώµα. Η πλειονότητα των νευρώνων του ραβδωτού σώµατος χρησιµοποιεί ως χηµικό διαβιβαστή το γ-αµινοβουτυρικό οξύ (GABA), ενώ ένα µικρό ποσοστό χρησιµοποιεί την ακετυλοχολίνη (ACh). Το ραβδωτό σώµα δέχεται γλουταµινεργικές ίνες από τον φλοιό των εγκεφαλικών ηµισφαιρίων και τον θάλαµο καθώς και σηµαντικό αριθµό ντοπαµινεργικών ινών από τη συµπαγή µοίρα της µέλαινας ουσίας. Οι νευράξονες των προβλητικών νευρώνων του ραβδωτού σώµατος απολήγουν σε άλλους πυρήνες των βασικών γαγγλίων. Το ραβδωτό σώµα αποτελεί αντικείµενο εκτεταµένων µελετών, διότι η απώλεια των νευρώνων του διαδραµατίζει σηµαντικό ρόλο στην παθογένεια διαφόρων νευροεκφυλιστικών νόσων. Στη νόσο Parkinson, µια προοδευτική νευροεκφυλιστική ασθένεια που εκδηλώνεται προς το τέλος της µέσης ηλικίας, παρατηρείται απώλεια των ντοπαµινεργικών νευρώνων της µέλαινας ουσίας µε αποτέλεσµα τη µείωση της ντοπαµινεργικής εννεύρωσης και τον επακόλουθο θάνατο κυττάρων του ραβδωτού σώµατος. Στη νόσο Huntington, µια από τις συχνότερες κληρονοµικές νευροεκφυλιστικές ασθένειες, παρατηρείται µαζική απώλεια των προβλητικών νευρώνων του ραβδωτού σώµατος. Πιστεύεται ότι οι µηχανισµοί που ενεργοποιούνται στις νόσους αυτές 1
14 Πρόλογος σχετίζονται µε τους µηχανισµούς του προγραµµατισµένου κυτταρικού θανάτου κατά την ανάπτυξη. Η απόπτωση αποτελεί τον κυριότερο τύπο προγραµµατισµένου κυτταρικού θανάτου στο Νευρικό Σύστηµα (NΣ) και οφείλεται στην ενεργοποίηση µιας σειράς γονιδίων για την υλοποίηση ενός ενδογενούς προγράµµατος αυτοκαταστροφής των κυττάρων. Η επιβίωση ή η απόπτωση των νευρικών κυττάρων κατά την ανάπτυξη εξαρτάται από ενδοκυτταρικούς και εξωκυτταρικούς παράγοντες. Οι πρώτοι περιλαµβάνουν συγκεκριµένες κυτταρικές πρωτεΐνες µε αποπτωτικές ή αντι-αποπτωτικές ιδιότητες. Η µεταβολή στην ισορροπία έκφρασης των πρωτεϊνών αυτών καθορίζει την επιβίωση ή τον θάνατο των κυττάρων. Οι εξωκυτταρικοί παράγοντες περιλαµβάνουν τους νευροτροφικούς παράγοντες, οι οποίοι εκλύονται είτε από µετασυναπτικά κύτταρα σε περιοχές-στόχους και µεταφέρονται στους νευρώνες ανάδροµα µέσω των νευραξόνων τους είτε από προσαγωγές ίνες. Υποστηρίζεται ότι η αιτιοπαθογένεια πολλών νευροεκφυλιστικών νόσων βασίζεται στην απώλεια της υποστήριξης συγκεκριµένων νευρωνικών πληθυσµών από νευροτροφικούς παράγοντες. Η µελέτη των παραγόντων που επηρεάζουν την επιβίωση των νευρώνων του ραβδωτού σώµατος έχει µεγάλη σηµασία για την κατανόηση της ανάπτυξης και της λειτουργίας της περιοχής. Στην εργασία αυτή µελετήθηκε το φαινόµενο της απόπτωσης στο ραβδωτό σώµα κατά την ανάπτυξη µε τη µέθοδο TUNEL, µε την οποία επιτυγχάνεται η in situ σήµανση των αποπτωτικών κυττάρων. Η ανάλυση έγινε στο επίπεδο του οπτικού και του ηλεκτρονικού µικροσκοπίου. Ο κυτταρικός θάνατος επιβεβαιώθηκε µε τη µελέτη της έκφρασης γονιδίων που προάγουν την απόπτωση και συγκεκριµένα εκείνων που κωδικοποιούν την ενεργοποιηµένη κασπάση-3 και τη φρακτίνη. Στη συνέχεια, µε στόχο τη µελέτη των πιθανών επιπτώσεων των βλαβών σε 2
15 Πρόλογος κύριες προσαγωγές συνδέσεις του ραβδωτού σώµατος στην επιβίωση των νευρώνων του χρησιµοποιήθηκαν δύο πειραµατικά µοντέλα που περιελάµβαναν α) τη χειρουργική αφαίρεση του µεγαλύτερου µέρους του φλοιού του ενός εγκεφαλικού ηµισφαιρίου και β) τη φαρµακολογική καταστροφή των κατεχολαµινεργικών προσαγωγών συστηµάτων ινών. Οι στόχοι της εργασίας αυτής ήταν οι εξής: 1) να περιγραφεί ποιοτικά και ποσοτικά η απόπτωση στο ραβδωτό σώµα σε διάφορα στάδια της ανάπτυξης και να γίνει συσχετισµός µε αναπτυξιακά γεγονότα της περιοχής αυτής, 2) να πιστοποιηθεί µορφολογικά και ανοσοϊστοχηµικά ο αποπτωτικός κυτταρικός θάνατος, 3) να διερευνηθεί αν και κατά πόσο διαφορετικού τύπου βλάβες σε διαφορετικά προσαγωγά συστήµατα του ραβδωτού σώµατος επηρεάζουν τη διεργασία της απόπτωσης στην περιοχή αυτή και 4) να µελετηθεί η επίδραση των παραπάνω βλαβών στην επιβίωση και στον φαινότυπο των GABAεργικών και των χολινεργικών νευρώνων του ραβδωτού σώµατος. Η εργασία αυτή χωρίζεται σε τέσσερις ενότητες. Στην εισαγωγή, γίνεται η ανασκόπηση των βιβλιογραφικών δεδοµένων σχετικά µε τα µορφολογικά και νευροχηµικά χαρακτηριστικά των νευρώνων του ραβδωτού σώµατος, τις συνδέσεις τους µε τις διάφορες περιοχές του εγκεφάλου καθώς και την οργάνωση και ανάπτυξη της περιοχής αυτής. Επιπλέον, περιγράφονται τα σύγχρονα βιβλιογραφικά δεδοµένα που αφορούν τη διεργασία του προγραµµατισµένου κυτταρικού θανάτου και τους παράγοντες που συµµετέχουν και ρυθµίζουν τη διεργασία της απόπτωσης. Στην δεύτερη ενότητα, γίνεται αναφορά στις µεθόδους που χρησιµοποιήθηκαν, στο πειραµατικό υλικό και στον τρόπο ανάλυσης των αποτελεσµάτων. Στην τρίτη ενότητα, περιγράφονται αναλυτικά τα αποτελέσµατα των πειραµάτων της παρούσας εργασίας σε φυσιολογικούς επίµυς και σε επίµυς που είχαν υποστεί βλάβες των συνδέσεων του ραβδωτού σώµατος σε διάφορα στάδια της ανάπτυξης. Στην τελευταία 3
16 Πρόλογος ενότητα, γίνεται η συζήτηση των αποτελεσµάτων σε σχέση µε αποτελέσµατα άλλων εργασιών µε παρόµοιο αντικείµενο. Η εργασία αυτή εκπονήθηκε στο Εργαστήριο Ανατοµικής, Ιστολογίας και Εµβρυολογίας της Κτηνιατρικής Σχολής του Αριστοτέλειου Πανεπιστηµίου Θεσσαλονίκης. Οφείλω ένα µεγάλο ευχαριστώ στην επιβλέπουσά µου, Επίκουρη Καθηγήτρια κα Ιωάννα ωρή για την συνεχή επιστηµονική καθοδήγηση, τη γόνιµη κριτική και τη διαρκή υποστήριξη που µου προσέφερε τόσο κατά το πειραµατικό µέρος όσο και κατά τη διάρκεια της συγγραφής της παρούσας διατριβής. Ευχαριστώ ιδιαίτερα τον Καθηγητή κ. Αθανάσιο Ντινόπουλο και τον Αναπληρωτή Καθηγητή κ. Ιωάννη Αντωνόπουλο, µέλη της συµβουλευτικής επιτροπής, για την πολύτιµη βοήθεια και τις εύστοχες υποδείξεις τους. Θα ήταν παράλειψη να µην εκφράσω την ευγνωµοσύνη µου στη Λέκτορα Βιοχηµείας κα Κατερίνα Αγγελοπούλου για τη διαρκή ενθάρρυνση και συµπαράσταση στην προσπάθειά µου αυτή. Θερµές ευχαριστίες εκφράζω στον Καθηγητή Οικονοµίας Ζωικής Παραγωγής και Εφαρµοσµένης Στατιστικής κ. Χρήστο Μπάτζιο για τη συµβολή του στην στατιστική ανάλυση των αποτελεσµάτων. Τις ειλικρινείς ευχαριστίες µου εκφράζω στην Καθηγήτρια Ιατρικής κα Λουίζα Ανδριοπούλου-Οικονόµου, στον Καθηγητή Παθολογικής Ανατοµικής κ. Ιωάννη Βλέµµα καθώς και στην Αναπληρώτρια Καθηγήτρια κα Ελένη Μιχαλούδη και τη Λέκτορα κα Αναστασία Τσιγκοτζίδου, µέλη της επταµελούς εξεταστικής επιτροπής, για τις εύστοχες παρατηρήσεις τους κατά τη διόρθωση των δοκιµίων. Ένα θερµό ευχαριστώ στα µέλη του Εργαστηρίου Ανατοµικής και Ιστολογίας κα Μαρία Χιωτέλλη και κ. Χρήστο ρόσο για την πολύτιµη συνεισφορά τους σε τεχνικά και άλλα θέµατα. Ευχαριστώ τις κυρίες 4
17 Πρόλογος Άννα Τσιπηνιά και Ευαγγελία Τασούλη καθώς και τους κ.κ. Γιώργο Φαρδέλλα, Πολυχρόνη Τσουµπελίδη και Κώστα Φαρδέλλα. Εκφράζω ένα µεγάλο ευχαριστώ στις φίλες και συναδέλφους µου Θέτιδα Ζαχαράκη, Μαρία Λάτσαρη και Βιβή Σοφού για την αµέριστη βοήθεια, τη συµπαράσταση, την υποµονή τους και τη συµβολή τους κατά την εκτέλεση των πειραµάτων. Θερµά ευχαριστώ, επίσης, τους συναδέλφους Γιάννη Γρίβα, Γιάννη Λιαγκούρα και Χρύσα Μπεκιάρη. Η εκπόνηση της διατριβής αυτής δεν θα ήταν δυνατή χωρίς την αγάπη, την υποστήριξη και τη δύναµη που µου προσέφεραν όλα αυτά τα χρόνια οι γονείς µου. 5
18
19 Εισαγωγή ΕΙΣΑΓΩΓΗ Α. ΑΝΑΤΟΜΙΚΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΚΗ ΟΡΓΑΝΩΣΗ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ Το ραβδωτό σώµα είναι πυρήνας του τελικού εγκεφάλου. Η ονοµασία του οφείλεται στη ραβδωτή υφή που εµφανίζει σε εγκάρσιες τοµές λόγω της διέλευσης δεσµίδων ινών της έσω κάψας. Αποτελείται από δύο επιµέρους πυρήνες, τον κερκοφόρο και τον φακοειδή. Ο τελευταίος διακρίνεται σε µια έξω µοίρα, το κέλυφος, και µια έσω µοίρα, την ωχρή σφαίρα (Heimer και συν., 1995 Gerfen, 2004). Κατά την οντογενετική ανάπτυξη, τo τµήµα του ραβδωτού σώµατος που σχηµατίζεται αρχικά ονοµάζεται παλαιοραβδωτό σώµα και περιλαµβάνει την ωχρή σφαίρα. Το νεοραβδωτό σώµα σχηµατίζεται µεταγενέστερα και συγκροτείται από τον κερκοφόρο πυρήνα και το κέλυφος. Σήµερα, µε τη σύγχρονη ορολογία, ο όρος «ραβδωτό σώµα» θεωρείται ταυτόσηµος µε τον όρο «νεοραβδωτό σώµα» και περιλαµβάνει τον κερκοφόρο πυρήνα και το κέλυφος. Εξάλλου, σε ορισµένες µελέτες στο ραβδωτό σώµα περιλαµβάνεται και ο επικλινής πυρήνας του διαφράγµατος (Heimer και συν., 1995 Whetsell, 2002 Gerfen, 2004). Το ραβδωτό σώµα ανήκει λειτουργικά σε ένα σύνολο πυρήνων του τελικού, του διάµεσου και του µέσου εγκεφάλου που συγκροτούν τα βασικά γάγγλια. Οι πυρήνες αυτοί είναι, εκτός από το ραβδωτό σώµα, η έξω και η έσω µοίρα της ωχρής σφαίρας, ο υποθαλάµιος πυρήνας και η µέλαινα ουσία, που αποτελείται από την συµπαγή και τη δικτυωτή µοίρα (Côté και Crutcher, 1991 Gerfen, 2004). Οι πυρήνες των βασικών γαγγλίων αποτελούν µέρη ενός κλειστού νευρωνικού κυκλώµατος και συµµετέχουν στον προγραµµατισµό και στην εκτέλεση σύνθετων κινήσεων, καθώς και σε ανώτερες γνωστικές λειτουργίες, όπως είναι η 7
20 Εισαγωγή µάθηση, η κινητοποίηση και η ανταµοιβή (Parent και Hazrati, 1995). Το ραβδωτό σώµα είναι ο «πυρήνας εισόδου» των βασικών γαγγλίων καθώς δέχεται προσαγωγές ίνες κυρίως από τον φλοιό των εγκεφαλικών ηµισφαιρίων, τον θάλαµο και τη συµπαγή µοίρα της µέλαινας ουσίας. Απαγωγές ίνες από το ραβδωτό σώµα καταλήγουν στην έσω µοίρα της ωχρής σφαίρας και στη δικτυωτή µοίρα της µέλαινας ουσίας είτε απευθείας είτε µέσω της έξω µοίρας της ωχρής σφαίρας και του υποθαλάµιου πυρήνα. Η έσω µοίρα της ωχρής σφαίρας και η δικτυωτή µοίρα της µέλαινας ουσίας αποτελούν τους «πυρήνες εξόδου» των βασικών γαγγλίων και προβάλλουν στον φλοιό των εγκεφαλικών ηµισφαιρίων, στον θάλαµο, στο πρόσθιο διδύµιο και στον σκελιαιογεφυρικό πυρήνα (Côté και Crutcher, 1991 Gerfen, 2004). Άλλοι πυρήνες του τελικού εγκεφάλου, όπως είναι το προτείχισµα και οι πυρήνες της αµυγδαλής που υποστηρίζονταν ότι ανήκουν στα βασικά γάγγλια λόγω της τοπογραφικής γειτνίασής τους µε το ραβδωτό σώµα, σήµερα θεωρούνται ως ανεξάρτητοι πυρήνες, διότι συµµετέχουν σε διαφορετικές λειτουργίες από εκείνες των βασικών γαγγλίων (Heimer και συν., 1995). B. ΠΕΡΙΓΡΑΦΗ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ 1. ΜΟΡΦΟΛΟΓΙΚΑ ΚΑΙ ΝΕΥΡΟΧΗΜΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ Οι νευρώνες του ραβδωτού σώµατος διακρίνονται στους µεσοµεγέθεις ακανθωτούς προβλητικούς νευρώνες και στους διάµεσους νευρώνες (Parent και Hazrati, 1995 Gerfen, 2004). 8
21 Εισαγωγή 1.1. ΜΕΣΟΜΕΓΕΘΕΙΣ ΑΚΑΝΘΩΤΟΙ ΠΡΟΒΛΗΤΙΚΟΙ ΝΕΥΡΩΝΕΣ Η πλειονότητα των νευρώνων του ραβδωτού σώµατος είναι οι µεσοµεγέθεις ακανθωτοί προβλητικοί νευρώνες (MSN), οι οποίοι αντιπροσωπεύουν το 90-95% του συνόλου των νευρώνων του πυρήνα (Parent και Hazrati, 1995 Gerfen, 2004 Tepper και συν., 2004). Η ονοµασία των νευρώνων αυτών περιγράφει τα κύρια µορφολογικά χαρακτηριστικά τους. Οι MSN εµφανίζουν κυτταρικό σώµα διαµέτρου 20-25µm, από το οποίο εκφύονται 7-10 δενδρίτες, µετρίως διακλαδισµένοι, µε πολλές ακανθοειδείς προεξοχές. Οι δενδρίτες ενός τέτοιου µεµονωµένου νευρώνα είναι δυνατό να εκτείνονται σε περιοχή διαµέτρου 200µm περίπου (DiFiglia και συν., 1976 Wilson και Groves, 1980 Bishop και συν., 1982 Chang και συν., 1982). Οι νευράξονες των MSN απολήγουν στην έξω και στην έσω µοίρα της ωχρής σφαίρας και στη δικτυωτή µοίρα της µέλαινας ουσίας. Xορηγούν, επίσης, παράπλευρους κλάδους που παραµένουν µέσα στα όρια του ραβδωτού σώµατος και σχηµατίζουν συνάψεις µε άλλους MSN καθώς και παράπλευρους κλάδους που απολήγουν στον υποθαλάµιο πυρήνα (Kawaguchi και συν., 1990). Στους MSN απολήγουν ίνες που προέρχονται από τον φλοιό των εγκεφαλικών ηµισφαιρίων και από τον θάλαµο και χρησιµοποιούν το γλουταµινικό οξύ ως διεγερτικό χηµικό διαβιβαστή. Έχει παρατηρηθεί ότι τοπογραφικά υπάρχει σύγκλιση των φλοιόφυγων γλουταµινεργικών ινών και των νευραξόνων των διάµεσων χολινεργικών νευρώνων του ραβδωτού σώµατος στις ακανθοειδείς προεξοχές των MSN (Calabresi και συν., 2000). Επιπλέον, στις ακανθοειδείς προεξοχές των MSN συγκλίνουν ντοπαµινεργικές ίνες από τη µέλαινα ουσία µε φλοιοραβδωτές ίνες (Parent και Hazrati, 1995). Ανοσοϊστοχηµικές µελέτες καταδεικνύουν ότι όλοι οι MSN χρησιµοποιούν τον ανασταλτικό χηµικό διαβιβαστή GABA 9
22 Εισαγωγή (Τepper και συν., 2004). Με βάση τις συνδέσεις και τα νευροχηµικά χαρακτηριστικά τους οι MSN διακρίνονται σε δύο υποοµάδες. Στην πρώτη υποοµάδα περιλαµβάνονται οι περισσότεροι από τους MSN στους οποίους συνυπάρχει το GABA µε το νευροπεπτίδιο εγκεφαλίνη. Οι νευρώνες αυτοί προβάλλουν στην έξω µοίρα της ωχρής σφαίρας. Στη δεύτερη υποοµάδα ανήκουν οι MSN στους οποίους συνυπάρχει το GABA µε την ουσία P και τη δυνορφίνη. Οι νευρώνες αυτοί προβάλλουν στην έσω µοίρα της ωχρής σφαίρας και στη δικτυωτή µοίρα της µέλαινας ουσίας (Haber και Watson, 1983 Beckstead και Κersey, 1985 Gerfen και Young, 1988). Η παραπάνω διάκριση δεν φαίνεται να είναι τόσο ακριβής όσο αρχικά πιστευόταν, καθώς ένα µικρό ποσοστό MSN (3-4%) σε ενήλικους επίµυς εκφράζει ταυτόχρονα την εγκεφαλίνη και την ουσία P και προβάλλει κυρίως στη µέλαινα ουσία (Wang και συν., 2006). Ποικίλες µελέτες υποστηρίζουν την ύπαρξη δύο τύπων υποδοχέων µέσω των οποίων ρυθµίζεται η δράση του GABA στα βασικά γάγγλια: των ιοντοτρόπων GABA A και των µεταβολοτρόπων GABA B υποδοχέων (Lacey και συν., 2005). Ηλεκτροφυσιολογικές έρευνες έδειξαν ότι η δράση του GABA στο ραβδωτό σώµα ρυθµίζεται κατά κύριο λόγο από τους µετασυναπτικούς GABA A υποδοχείς, ενώ η ενεργοποίηση των προσυναπτικών GABA B υποδοχέων επηρεάζει, επίσης, τη λειτουργικότητα των νευρώνων του πυρήνα αυτού (Τepper και συν., 2004 Lacey και συν., 2005). Με τη χρήση ανοσοϊστοχηµικών µεθόδων οι GABA B υποδοχείς εντοπίσθηκαν στα τελικά κοµβία των γλουταµινεργικών ινών που προέρχονται από τον φλοιό των εγκεφαλικών ηµισφαιρίων και τον θάλαµο και απολήγουν στο ραβδωτό σώµα (Lacey και συν., 2005). 10
23 Εισαγωγή 1.2. ΙΑΜΕΣΟΙ ΝΕΥΡΩΝΕΣ Οι διάµεσοι νευρώνες αποτελούν το 5-10% του συνόλου των νευρώνων του ραβδωτού σώµατος. Με βάση τα µορφολογικά και νευροχηµικά χαρακτηριστικά τους, οι νευρώνες αυτοί διακρίνονται σε δύο κατηγορίες, στους µεγάλους µη ακανθωτούς χολινεργικούς νευρώνες (Βolam και συν., 1984 Wilson και συν., 1990 Καwaguchi, 1992, 1993) και στους µεσοµεγέθεις µη ακανθωτούς GABAεργικούς νευρώνες (Κita, 1993 Kawaguchi και συν., 1995). Οι νευράξονες των νευρώνων αυτών περιορίζονται µέσα στα όρια του ραβδωτού σώµατος. 1.2.Α. ΜΕΓΑΛΟΙ ΜΗ ΑΚΑΝΘΩΤΟΙ ΧΟΛΙΝΕΡΓΙΚΟΙ ΝΕΥΡΩΝΕΣ Οι νευρώνες αυτοί ταυτοποιήθηκαν µε ανοσοϊστοχηµικές µελέτες στις οποίες χρησιµοποιήθηκαν αντισώµατα εναντίον της χολινακετυλοτρανσφεράσης (ChAT), του βιοσυνθετικού ενζύµου της ακετυλοχολίνης. Aντιπροσωπεύουν λιγότερο από το 2% του συνολικού πληθυσµού των νευρικών κυττάρων στο ραβδωτό σώµα (Woolf και Butcher, 1981 Phelps και συν., 1985). Oι χολινεργικοί νευρώνες του ραβδωτού σώµατος εµφανίζουν µεγάλο κυτταρικό σώµα µε διάµετρο µέχρι 40µm, από το οποίο εκφύονται µεγάλοι λείοι δενδρίτες µε δευτερογενείς και τριτογενείς διακλαδώσεις (Gerfen, 2004). Είναι αξιοσηµείωτο ότι, ενώ ο αριθµός τους είναι µικρός, οι χολινεργικοί νευρώνες του ραβδωτού σώµατος παράγουν από τα υψηλότερα επίπεδα ακετυλοχολίνης που ανιχνεύονται στο Κεντρικό Νευρικό Σύστηµα (ΚΝΣ) και παίζουν σηµαντικό ρόλο στη ρύθµιση της λειτουργίας του ραβδωτού σώµατος (Calabresi και συν., 2000). Στους νευρώνες αυτούς απολήγουν γλουταµινεργικές ίνες, που προέρχονται από τον φλοιό των εγκεφαλικών ηµισφαιρίων και από τον θάλαµο (Wilson και συν., 1990 Lapper και Bolam, 1992). Οι νευράξονες των 11
24 Εισαγωγή χολινεργικών νευρώνων του ραβδωτού σώµατος σχηµατίζουν συµµετρικές συνάψεις µε τα κυτταρικά σώµατα, τους δενδρίτες και τις ακανθοειδείς προεξοχές των MSN (Calabresi και συν., 2000). Λόγω της ανατοµικής σύγκλισης των νευραξόνων των χολινεργικών νευρώνων και των γλουταµινεργικών φλοιοραβδωτών ινών στους MSN, οι χολινεργικοί νευρώνες επηρεάζουν τη ρύθµιση της ροής πληροφοριών από τον φλοιό των εγκεφαλικών ηµισφαιρίων στους προβλητικούς νευρώνες του ραβδωτού σώµατος. Επιπλέον, έχει αποδειχθεί ότι η ακετυλοχολίνη ρυθµίζει την απελευθέρωση άλλων χηµικών διαβιβαστών στο ραβδωτό σώµα µέσω προσυναπτικών µηχανισµών. Πράγµατι, µελέτες έδειξαν ότι η απελευθέρωση του GABA από τους MSN και του γλουταµινικού οξέος από τους νευρώνες του φλοιού των εγκεφαλικών ηµισφαιρίων στο ραβδωτό σώµα αναστέλλεται από την ενεργοποίηση των προσυναπτικών µουσκαρινικών υποδοχέων της ακετυλοχολίνης (Sugita και συν., 1991). 1.2.Β. MΕΣΟΜΕΓΕΘΕΙΣ ΜΗ ΑΚΑΝΘΩΤΟΙ GABAΕΡΓΙΚΟΙ ΝΕΥΡΩΝΕΣ Με βάση την ταυτόχρονη έκφραση του χηµικού διαβιβαστή GABA και πρωτεϊνών που δεσµεύουν το ασβέστιο ή διάφορων νευροπεπτιδίων, οι µεσοµεγέθεις µη ακανθωτοί GABAεργικοί νευρώνες διακρίνονται στις παρακάτω τρεις κατηγορίες: α) Μεσοµεγέθεις µη ακανθωτοί GABAεργικοί νευρώνες που εκφράζουν την πρωτεΐνη δέσµευσης του ασβεστίου παρβαλβουµίνη (Cowan και συν., 1990 Kubota και Kagawuchi, 1993). Αποτελούν το 1% του συνολικού πληθυσµού των νευρώνων του ραβδωτού σώµατος και συνιστούν τον πολυπληθέστερο τύπο των GABΑεργικών διάµεσων νευρώνων του πυρήνα αυτού στους επίµυς (Luk και Sadikot, 2001). Οι νευρώνες αυτοί δέχονται ίνες από τον φλοιό των εγκεφαλικών ηµισφαιρίων και οι νευράξονές τους σχηµατίζουν συνάψεις µε τους MSN. Κατανέµονται σε όλη την έκταση του πυρήνα µε µεγαλύτερη 12
25 Εισαγωγή πυκνότητα στην έξω και στη ραχιαία µοίρα του ραβδωτού σώµατος (Kita και συν., 1990 Kawaguchi και συν., 1995 Gerfen, 2004). β) Μεσοµεγέθεις µη ακανθωτοί GABAεργικοί νευρώνες που εκφράζουν την πρωτεΐνη δέσµευσης του ασβεστίου καλρετινίνη (Βennett και Bolam, 1993). Η καλρετινίνη συµβάλλει στη διατήρηση της οµοιόστασης του ενδοκυτταρικού ασβεστίου και πιστεύεται ότι προστατεύει τους νευρώνες από τη µαζική παραγωγή ασβεστίου που προκαλεί η υπερδιέγερση των γλουταµινικών υποδοχέων. Υποστηρίζεται ότι η µαζική αυτή παραγωγή ασβεστίου παίζει ρόλο στην αιτιοπαθογένεια της νόσου Huntington (Cicchetti και συν., 2000 Lévesque και συν., 2003). Οι µεσοµεγέθεις µη ακανθωτοί GABAεργικοί νευρώνες που εκφράζουν την καλρετινίνη βρίσκονται κυρίως στο έσω τµήµα της ραχιαίας µοίρας του ραβδωτού σώµατος και αποτελούν τον πολυπληθέστερο τύπο των GABAεργικών διάµεσων νευρώνων του ραβδωτού σώµατος στα πρωτεύοντα, σε αντίθεση µε τους επίµυς (Lévesque και συν., 2003). γ) Μεσοµεγέθεις µη ακανθωτοί GABAεργικοί νευρώνες που εκφράζουν τα πεπτίδια σωµατοστατίνη και νευροπεπτίδιο Υ και το ένζυµο NADPH-διαφοράση (Vincent και συν., 1983a, 1983b Smith και Parent, 1986 Pasik και συν., 1988 Chesselet και Robbins, 1989 Dawson και συν., 1991). Το τελευταίο είναι υπεύθυνο για τη σύνθεση του νιτρικού οξειδίου (ΝΟ), ενός νευρωνικού µεταφορέα που θεωρείται ότι παίζει ρόλο στην ανάπτυξη και στην πλαστικότητα του ΚΝΣ (Murata και Masuko, 2003). Οι διάµεσοι αυτοί νευρώνες του ραβδωτού σώµατος εντοπίζονται κυρίως στην κοιλιακή µοίρα του πυρήνα, ενώ ένα µικρό ποσοστό τους (περίπου 20%) εκφράζει ταυτόχρονα και την πρωτεΐνη καλβιδίνη (Gerfen, 2004). Η συνύπαρξη των παραπάνω ουσιών στους διάµεσους νευρώνες του ραβδωτού σώµατος αποτέλεσε αντικείµενο µελέτης µε τη χρήση 13
26 Εισαγωγή ανοσοϊστοχηµικών µεθόδων σε συνδυασµό µε τη µέθοδο του υβριδισµού in situ. Σύµφωνα µε τα αποτελέσµατα της µελέτης αυτής, το 84% των νευρώνων που περιέχουν σωµατοστατίνη εκφράζουν και το νευροπεπτίδιο Υ, ενώ όλοι οι νευρώνες που περιέχουν το νευροπεπτίδιο Υ περιέχουν και σωµατοστατίνη. Επιπλέον, όλοι οι νευρώνες που εκφράζουν τη σωµατοστατίνη σηµαίνονται θετικά για το ένζυµο NADPH-διαφοράση (Rushlow και συν., 1995). 2. ΣΥΝ ΕΣΕΙΣ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ Το ραβδωτό σώµα ανήκει σε ένα νευρωνικό κύκλωµα µε πολλούς σταθµούς, το οποίο ξεκινάει από αισθητικές και κινητικές περιοχές του φλοιού των εγκεφαλικών ηµισφαιρίων, περιλαµβάνει τους πυρήνες των βασικών γαγγλίων και καταλήγει στον κινητικό και στον προκινητικό φλοιό (Σχήµα 1). Οι συνδέσεις του ραβδωτού σώµατος έχουν αποτελέσει αντικείµενο εκτεταµένης έρευνας µε τη χρήση τεχνικών σήµανσης µε ορθόδροµους και ανάδροµους δείκτες σε συνδυασµό µε ανοσοϊστοχηµικές µεθόδους (Staines και συν., 1981 Beckstead, 1985 Gerfen και Young, 1988 Gonzales και Chesselet, 1990) ΠΡΟΣΑΓΩΓΕΣ ΙΝΕΣ 2.1.A. ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΦΛΟΙΟ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Το µεγαλύτερο µέρος των προσαγωγών ινών του ραβδωτού σώµατος προέρχεται από τον οµοπλάγιο και σε µικρότερο βαθµό από τον ετεροπλάγιο φλοιό των εγκεφαλικών ηµισφαιρίων (Σχήµα 1) (Heimer και συν., 1995). Οι ίνες αυτές αποτελούν τους νευράξονες των γλουταµινεργικών πυραµιδοειδών νευρώνων που εντοπίζονται κυρίως στη στιβάδα 5 και σπανιότερα στις στιβάδες 3 και 6 του φλοιού των εγκεφαλικών ηµισφαιρίων (Gerfen, 2004). 14
27 Εισαγωγή Σύµφωνα µε τους Parent και Hazrati (1995), το ραβδωτό σώµα διακρίνεται σε τρεις λειτουργικά ανεξάρτητες περιοχές, τη συνειρµική, την αισθητικοκινητική και τη µεταιχµιακή, στις οποίες προβάλλουν µε τοπογραφικά οργανωµένο τρόπο οι συνειρµικές, οι αισθητικοκινητικές και οι µεταιχµιακές περιοχές του φλοιού των εγκεφαλικών ηµισφαιρίων, αντίστοιχα. Η συνειρµική περιοχή του ραβδωτού σώµατος περιλαµβάνει µεγάλα τµήµατα του κελύφους, κεφαλικώς του πρόσθιου συνδέσµου και το µεγαλύτερο µέρος του κερκοφόρου πυρήνα και δέχεται προσαγωγές ίνες από συνειρµικές φλοιικές περιοχές του µετωπιαίου, του κροταφικού και του βρεγµατικού λοβού (Künzle, 1975 Goldman και Νauta, 1977 Carpenter, 1981 Preston και συν., 1981). Η αισθητικοκινητική περιοχή περιλαµβάνει το ραχιαίο και έξω τµήµα του κελύφους καθώς και την έξω και ραχιαία µοίρα του κερκοφόρου πυρήνα και δέχεται προσαγωγές ίνες κυρίως από κινητικές και αισθητικοκινητικές περιοχές του φλοιού των εγκεφαλικών ηµισφαιρίων (Künzle, 1975 DeLong και Georgopoulos, 1981 Liles και Updyke, 1985). Τέλος, η µεταιχµιακή περιοχή του ραβδωτού σώµατος αποτελείται από τον επικλινή πυρήνα του διαφράγµατος και τα κοιλιακά τµήµατα του κερκοφόρου πυρήνα και του κελύφους. έχεται ίνες από περιοχές του µεταιχµιακού συστήµατος, µεταξύ των οποίων και ο µεταιχµιακός φλοιός (Hemphill και συν., 1981 Phillipson και Griffiths, 1985 Russchen και συν., 1985 Alheid και Heimer, 1988 Ηaber και συν., 1990 Berendse και συν., 1992). Οι MSN είναι οι κύριοι µετασυναπτικοί στόχοι των φλοιόφυγων ινών στο ραβδωτό σώµα. Υποστηρίζεται ότι στους επίµυς το 90% των φλοιοραβδωτών ινών σχηµατίζει ασύµµετρες συνάψεις µε την κεφαλή των ακανθοειδών προεξοχών, το 5% µε τα δενδριτικά στελέχη και το υπόλοιπο 5% µε τα κυτταρικά σώµατα των MSN (Xu και συν., 1989). Οι συνάψεις στις ακανθοειδείς προεξοχές των MSN έχουν µελετηθεί εκτεταµένα, καθώς στα µετασυναπτικά αυτά στοιχεία απολήγουν οι 15
28 Εισαγωγή προσαγωγές ντοπαµινεργικές ίνες από τη µέλαινα ουσία και οι νευράξονες των διάµεσων χολινεργικών νευρώνων. Οι συνάψεις των φλοιοραβδωτών ινών µε τους MSN παρουσιάζουν µεγάλες συναπτικές επιφάνειες, ακανόνιστου σχήµατος. Στο 9% του συνολικού αριθµού των συνάψεων αυτών παρατηρούνται εξειδικεύσεις της µετασυναπτικής µεµβράνης, οι οποίες εµφανίζουν οπές (Kemp και Powell, 1971α Somogyi και συν., 1981 Meshul και Casey, 1989 Chen και Hillman, 1990 Ingham και συν., 1998). Η µείωση του συνολικού αριθµού των ασύµµετρων συνάψεων στο ραβδωτό σώµα µε ταυτόχρονη αύξηση εκείνων µε τις εξειδικεύσεις της µετασυναπτικής µεµβράνης έχει παρατηρηθεί σε πειραµατικά µοντέλα της νόσου Parkinson (Ingham και συν., 1998). 2.1.Β. ΣΥΝ ΕΣΕΙΣ ΜΕ ΥΠΟΦΛΟΙΙΚΕΣ ΠΕΡΙΟΧΕΣ 2.1.B 1. ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΘΑΛΑΜΟ Οι οµοπλάγιοι ενδοπετάλιοι πυρήνες του θαλάµου συνιστούν την κυριότερη πηγή των θαλαµοραβδωτών ινών (Smith και συν., 2004). To σύµπλεγµα των πρόσθιων ενδοπετάλιων πυρήνων περιλαµβάνει τον έσω κεντρικό, τον παρακεντρικό και τον έξω κοιλιακό πυρήνα, ενώ στους οπίσθιους ενδοπετάλιους πυρήνες περιλαµβάνεται ο µέσος κεντρικόςπαραδεσµικός πυρήνας (Smith και συν., 2004). Επιπλέον, το ραβδωτό σώµα δέχεται ίνες από τους πυρήνες της µέσης γραµµής του θαλάµου καθώς και από τους κοιλιακούς κινητικούς πυρήνες του θαλάµου (Beckstead, 1984 Smith και Parent, 1986 Berendse και Groenewegen, 1990 Erro και συν., 2001 McFarland και Haber, 2001 van der Werf και συν., 2002). Oι θαλαµοραβδωτές ίνες σχηµατίζουν, όπως και οι φλοιοραβδωτές, ασύµµετρες συνάψεις µε τους ΜSN. Είναι αξιοσηµείωτο ότι οι ίνες από τον µέσο κεντρικό-παραδεσµικό πυρήνα σχηµατίζουν σε 16
29 Εισαγωγή µεγαλύτερο ποσοστό συνάψεις µε τους δενδρίτες των νευρώνων αυτών, σε αντίθεση µε τις ίνες από τους πρόσθιους ενδοπετάλιους πυρήνες, που συνάπτονται κυρίως µε τις ακανθοειδείς προεξοχές των MSN (Xu και συν., 1989). Ο χηµικός διαβιβαστής που χρησιµοποιείται στις θαλαµοραβδωτές συνάψεις είναι το γλουταµινικό οξύ (Smith και συν., 2004). Η απελευθέρωση και η δράση του ρυθµίζεται από τους ιοντοτρόπους υποδοχείς ΝΜDA (Ν-µεθυλο-D-ασπαρτικού οξέος), AMPA (α-αµινο-3- υδροξυ-5-µεθυλο-4-ισοξαζολοπροπιονικού οξεός) και καϊνικού οξέος καθώς και τους µεταβολοτρόπους υποδοχείς τύπου I (Smith και συν., 2004). 2.1.Β 2. ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΜΕΣΟ ΕΓΚΕΦΑΛΟ Το ραβδωτό σώµα δέχεται πυκνή µονοαµινεργική εννεύρωση κυρίως από οµοπλάγιες περιοχές του στελέχους του εγκεφάλου (Heimer και συν., 1995 Gerfen, 2004). Ντοπαµινεργικές ίνες, που εκφύονται από νευρώνες της µέλαινας ουσίας (Σχήµα 1), του κοιλιακού καλυπτρικού πεδίου και της οπισθοερυθράς περιοχής, διανέµονται στο ραβδωτό σώµα µε τοπογραφικά οργανωµένο τρόπο. Συγκεκριµένα, νευρώνες που βρίσκονται στο κοιλιακό καλυπτρικό πεδίο, στο ραχιαίο τµήµα της συµπαγούς µοίρας της µέλαινας ουσίας και στην οπισθοερυθρά περιοχή στέλνουν τους νευράξονές τους στο υπόστρωµα του ραβδωτού σώµατος (βλ. Εισαγωγή, κεφ. Β.3). Οι νευρώνες αυτοί εκφράζουν, εκτός από τη ντοπαµίνη, και την πρωτεΐνη καλβιδίνη. Ντοπαµινεργικοί νευρώνες του κοιλιακού τµήµατος της συµπαγούς µοίρας της µέλαινας ουσίας και περιοχών της δικτυωτής µοίρας της µέλαινας ουσίας προβάλλουν στα ραβδοσωµάτια (βλ. Εισαγωγή, κεφ. Β.3) (Gerfen, 2004). Η αισθητικοκινητική περιοχή του ραβδωτού σώµατος εννευρώνεται από το έξω τµήµα της συµπαγούς µοίρας της µέλαινας 17
30 Εισαγωγή ουσίας, η συνειρµική από το έσω τµήµα της συµπαγούς µοίρας της µέλαινας ουσίας και το κοιλιακό καλυπτρικό πεδίο και η µεταιχµιακή περιοχή δέχεται ίνες κυρίως από το κοιλιακό καλυπτρικό πεδίο. Η οπισθοερυθρά περιοχή αποστέλλει ίνες και στις τρεις αυτές περιοχές του ραβδωτού σώµατος (Joel και Weiner, 2000). Οι ντοπαµινεργικές ίνες που προέρχονται από τη µέλαινα ουσία και απολήγουν στο ραβδωτό σώµα παρουσιάζουν µεγάλο κλινικό ενδιαφέρον, καθώς η εκφύλισή τους θεωρείται υπεύθυνη για την εκδήλωση των κλινικών συµπτωµάτων της νόσου Parkinson (Honig και Rosenberg, 2000 Conforti και συν., 2007). Οι ντοπαµινεργικές µελαινοραβδωτές ίνες σχηµατίζουν συµµετρικές συνάψεις µε τον αυχένα των ακανθοειδών προεξοχών των MSN (Bouyer και συν., 1984 Freund και συν., 1984 Voorn και συν., 1986 Groves και συν., 1994 Smith και συν., 1994 Antonopoulos και συν., 2002). Οι ίνες αυτές συγκλίνουν µε τις φλοιοραβδωτές ίνες στις ακανθοειδείς προεξοχές των MSN (Smith και Bolam, 1990). Τo ποσοστό των ακανθοειδών προεξοχών, στις οποίες απολήγουν ταυτόχρονα γλουταµινεργικές και ντοπαµινεργικές ίνες δεν είναι πάντα σταθερό και εξαρτάται από την περιοχή του ραβδωτού σώµατος στην οποία εντοπίζονται οι MSN και από την περιοχή στην οποία προβάλλουν (Wilson και συν., 1983 Freund και συν., 1984 Groves και συν., 1994 Ingham και συν., 1998). Υποστηρίζεται ότι, λόγω της ανατοµικής αυτής γειτνίασης, η ντοπαµίνη ρυθµίζει την διεγερτική εννεύρωση των MSN από τον φλοιό των εγκεφαλικών ηµισφαιρίων (Arbuthnott και συν., 1998). Η διαταραχή της αλληλεπίδρασης των ντοπαµινεργικών και των γλουταµινεργικών ινών στο ραβδωτό σώµα έχει µελετηθεί εκτεταµένα και πιθανόν παίζει ρόλο στην αιτιοπαθογένεια της νόσου Parkinson (Bolam και συν., 2000). Επιπρόσθετα, πιστεύεται ότι η ντοπαµίνη ρυθµίζει την έκφραση των πεπτιδίων στους MSN (Parent και Ηazrati, 1995). Βλάβη της 18
31 Εισαγωγή ντοπαµινεργικής µελαινοραβδωτής οδού προκαλεί µείωση των επιπέδων του mrna που κωδικoποιεί την ουσία Ρ και σηµαντική αύξηση των επιπέδων του mrna που κωδικοποιεί την εγκεφαλίνη (Parent και Ηazrati, 1995). Άλλες µονοαµινεργικές προσαγωγές ίνες του ραβδωτού σώµατος περιλαµβάνουν αυτές που προέρχονται από τον ραχιαίο πυρήνα της ραφής και χρησιµοποιούν τη σεροτονίνη ως χηµικό διαβιβαστή. Οι ίνες αυτές σχηµατίζουν ασύµµετρες συνάψεις µε τις ακανθοειδείς προεξοχές και τα δενδριτικά στελέχη των MSN. Υποστηρίζεται ότι οι ντοπαµινεργικές και οι σεροτονινεργικές ίνες είναι δυνατόν να ρυθµίζουν τη λειτουργία των χολινεργικών νευρώνων του ραβδωτού σώµατος λόγω της ανατοµικής γειτνίασης των απολήξεων τους στο ραβδωτό σώµα (Jackson και συν., 1988 Jacobs και Azmitia, 1992). Τέλος, στο ραβδωτό σώµα απολήγουν νοραδρενεργικές ίνες που προέρχονται από τον υποµέλανα τόπο (Parent και Hazrati, 1995 Gerfen, 2004). 2.1.Β 3. ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΗΝ ΩΧΡΗ ΣΦΑΙΡΑ Οι νευρώνες του ραβδωτού σώµατος δέχονται ίνες από GABAεργικούς νευρώνες της ωχρής σφαίρας (Staines και συν., 1981 Beckstead, 1983). Οι νευρώνες αυτοί αποτελούν σηµαντική πηγή εννεύρωσης του ραβδωτού σώµατος, καθώς κάθε νευρώνας της ωχρής σφαίρας που προβάλλει στη µέλαινα ουσία παρέχει και ένα παράπλευρο κλάδο στο ραβδωτό σώµα (Staines και Fibiger, 1984). 2.1.Β 4. ΣΥΝ ΕΣΕΙΣ ΜΕ ΤΟΝ ΥΠΟΘΑΛΑΜΙΟ ΠΥΡΗΝΑ Ο υποθαλάµιος πυρήνας αποστέλλει ίνες στο ραβδωτό σώµα. Ο αριθµός των ινών αυτών είναι σχετικά µικρός σε σύγκριση µε την πυκνότητα των ινών του πυρήνα που απολήγουν στη µέλαινα ουσία και την ωχρή σφαίρα (Kita και Kitai, 1988). 19
32 Εισαγωγή 2.2. ΑΠΑΓΩΓΕΣ ΙΝΕΣ Οι προβλητικοί νευρώνες του ραβδωτού σώµατος στέλνουν τους νευράξονές τους αποκλειστικά σε άλλους πυρήνες των βασικών γαγγλίων και συγκεκριµένα στην έξω και στην έσω µοίρα της ωχρής σφαίρας και στη δικτυωτή µοίρα της µέλαινας ουσίας (Wilson, 1998). H έσω µοίρα της ωχρής σφαίρας και η δικτυωτή µοίρα της µέλαινας ουσίας συνιστούν τους «πυρήνες εξόδου» των βασικών γαγγλίων και προβάλλουν στους έξω κοιλιακό, πρόσθιο κοιλιακό και µέσο ραχιαίο πυρήνα του θαλάµου, στον έξω πυρήνα της ηνίας, στο πρόσθιο διδύµιο και στον σκελιαιογεφυρικό πυρήνα, ενώ ίνες από την έξω µοίρα της ωχρής σφαίρας απολήγουν στον υποθαλάµιο πυρήνα (Wilson, 1998 Gerfen, 2004). Οι απαγωγές ίνες του ραβδωτού σώµατος συγκροτούν δύο διαφορετικές οδούς µέσα στο κύκλωµα των βασικών γαγγλίων. Στην «άµεση» απαγωγό οδό, οι νευράξονες των MSN απολήγουν απευθείας στην έσω µοίρα της ωχρής σφαίρας και τη δικτυωτή µοίρα της µέλαινας ουσίας και σχηµατίζουν συνάψεις µε GABAεργικούς νευρώνες των περιοχών αυτών (Σχήµα 1). Η «έµµεση» απαγωγός οδός συγκροτείται από τους νευράξονες των ΜSΝ που διανέµονται αποκλειστικά στην έξω µοίρα της ωχρής σφαίρας. Παράπλευροι κλάδοι των νευραξόνων αυτών απολήγουν στον υποθαλάµιο πυρήνα. Ίνες από τον υποθαλάµιο πυρήνα απολήγουν στην έσω µοίρα της ωχρής σφαίρας και στη µέλαινα ουσία (Σχήµα 1). Με τον τρόπο αυτό οι MSN συνδέονται µέσω του υποθαλάµιου πυρήνα µε τους «πυρήνες εξόδου» των βασικών γαγγλίων (Beckstead και Cruz, 1986 Kawaguchi και συν., 1990 Wilson, 1998). Η έξω µοίρα της ωχρής σφαίρας και ο υποθαλάµιος πυρήνας συνδέονται µεταξύ τους σχηµατίζοντας ένα πολύπλοκο δίκτυο, που είναι απαραίτητο για την εύρυθµη λειτουργία των βασικών γαγγλίων (Bevan και συν., 2002). 20
33 Εισαγωγή Ανοσοϊστοχηµικές µελέτες έδειξαν ότι ίνες που προέρχονται από το ραβδωτό σώµα και απολήγουν στην έξω µοίρα της ωχρής σφαίρας περιέχουν το νευροπεπτίδιο εγκεφαλίνη (Hökfelt και συν., 1977 Ρickel και συν., 1980 DiFiglia και συν., 1982 Haber και Watson, 1983 Beckstead, 1985), ενώ, αντίθετα, ίνες που απολήγουν στη µέλαινα ουσία περιέχουν τη δυνορφίνη και την ουσία P (Brownstein και συν., 1977 Hong και συν., 1977 Kanazawa και συν., 1977 Vincent και συν., 1982a, 1982b). Επιπρόσθετα, το mrna που κωδικοποιεί τους ντοπαµινεργικούς υποδοχείς D1 ανιχνεύθηκε εκλεκτικά στους νευρώνες του ραβδωτού σώµατος που προβάλλουν στη µέλαινα ουσία, ενώ το mrna που κωδικοποιεί τους ντοπαµινεργικούς υποδοχείς D2 ανιχνεύθηκε στους νευρώνες που προβάλλουν στην ωχρή σφαίρα (Gerfen και συν., 1990). 21
34 Εισαγωγή Σχήµα 1. Σχηµατική παράσταση του νευρωνικού κυκλώµατος που συγκροτούν οι πυρήνες των βασικών γαγγλίων. To ραβδωτό σώµα δέχεται προσαγωγές ίνες από τον φλοιό των εγκεφαλικών ηµισφαιρίων, τον θάλαµο και τη συµπαγή µοίρα της µέλαινας ουσίας. Ίνες από το ραβδωτό σώµα απολήγουν στην έσω µοίρα της ωχρής σφαίρας και τη δικτυωτή µοίρα της µέλαινας ουσίας, είτε απευθείας είτε µέσω της έξω µοίρας της ωχρής σφαίρας και του υποθαλάµιου πυρήνα. Τα κύτταρα της έσω µοίρας της ωχρής σφαίρας και της δικτυωτής µοίρας της µέλαινας ουσίας προβάλλουν µέσω του θαλάµου στον φλοιό των εγκεφαλικών ηµισφαιρίων. ΦΛΟΙΟΣ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Κινητικός φλοιός Σωµατοαισθητικός φλοιός Προκινητικός φλοιός Θάλαµος Ωχρή σφαίρα Έσω µοίρα Έξω µοίρα Υποθαλάµιος πυρήνας Συµπαγής µοίρα µέλαινας ουσίας Ραβδωτό σώµα ικτυωτή µοίρα µέλαινας ουσίας 22
35 Εισαγωγή 3. Ι ΙΑΙΤΕΡΟΤΗΤΕΣ ΤΗΣ ΟΡΓΑΝΩΣΗΣ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ Ανοσοϊστοχηµικές µελέτες απέδειξαν ότι το ραβδωτό σώµα παρουσιάζει αξιοσηµείωτη χηµική ετερογένεια. Συγκεκριµένα, µε τη χρήση αντισωµάτων εναντίον της ακετυλοχολινεστεράσης (AChE), του ενζύµου που διασπά την ACh, ταυτοποιήθηκαν στο ραβδωτό σώµα ζώνες, τα ραβδοσωµάτια, µε χαµηλή ανοσοδραστικότητα για το ένζυµο αυτό, οι οποίες διακρίνονται από το υπόστρωµα, το οποίο είναι πλούσιο σε AChE (Graybiel και Ragsdale, 1978). Τα ραβδοσωµάτια αποτελούν το 10-20% του όγκου του ραβδωτού σώµατος και σχηµατίζουν τρισδιάστατες δοµές µε τη µορφή λαβυρίνθου µέσα στον πυρήνα (Graybiel, 1990). Eµφανίζουν υψηλά επίπεδα δυνορφίνης, νευροτενσίνης, ουσίας P και οπιοειδών υποδοχέων, ενώ το υπόστρωµα παρουσιάζει υψηλά επίπεδα σωµατοστατίνης και νευροτενσίνης (Jain και συν., 2001). Τα ραβδοσωµάτια και το υπόστρωµα παρουσιάζουν, επιπλέον, διαφορές στις συνδέσεις τους (Gerfen, 1984, 1985, 1992). Συγκεκριµένα, τα ραβδοσωµάτια δέχονται ίνες από τον µεταιχµιακό φλοιό καθώς και ντοπαµινεργικές ίνες από το κοιλιακό τµήµα της συµπαγούς µοίρας και από περιοχές της δικτυωτής µοίρας της µέλαινας ουσίας. Στο υπόστρωµα απολήγουν ίνες που προέρχονται από συνειρµικές, οφθαλµοκινητικές και αισθητικοκινητικές περιοχές του φλοιού των εγκεφαλικών ηµισφαιρίων και από τον θάλαµο καθώς και ντοπαµινεργικές ίνες από το κοιλιακό καλυπτρικό πεδίο, το ραχιαίο τµήµα της συµπαγούς µοίρας της µέλαινας ουσίας και την οπισθοερυθρά περιοχή (Gerfen και συν., 1985 Parent και Hazrati, 1995 Gerfen, 2004) 23
36 Εισαγωγή 4. ΑΝΑΠΤΥΞΗ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ Η οργάνωση του ραβδωτού σώµατος στα ραβδοσωµάτια και στο υπόστρωµα γίνεται εµφανής από τα αρχικά στάδια της εµβρυογένεσης. Αυτοραδιογραφικές µελέτες που πραγµατοποιήθηκαν σε επίµυς µε τη χρήση της τριτιωµένης θυµιδίνης έδειξαν ότι οι νευρώνες που γεννιούνται µεταξύ της εµβρυϊκής ηµέρας (Ε)12 και της Ε17 εντοπίζονται κυρίως στα ραβδοσωµάτια, ενώ οι νευρώνες που γεννιούνται µεταξύ της Ε18 και της µεταγεννητικής ηµέρας (Μ)2 παρουσιάζουν εκλεκτική εντόπιση στο υπόστρωµα (Marchand και Lajoie, 1986 van der Kooy και Fishell, 1987 Fishell και van der Kooy, 1991). O πρώτοι νευρώνες που γεννιούνται στο ραβδωτό σώµα είναι οι διάµεσοι χολινεργικοί νευρώνες (Phelps και συν., 1989). Μελέτες στις οποίες χρησιµοποιήθηκαν αυτοραδιογραφικές τεχνικές µε τριτιωµένη θυµιδίνη σε συνδυασµό µε τη µέθοδο της ανοσοϊστοχηµείας µε αντισώµατα εναντίον της ChAT, έδειξαν ότι το 75% των χολινεργικών νευρώνων του ραβδωτού σώµατος γεννιέται από την Ε13 έως την Ε15 (Phelps και συν., 1989). Στην ίδια µελέτη υποστηρίχθηκε, επίσης, ότι η µιτωτική δραστηριότητα των νευρώνων αυτών ακολουθεί ένα χωροχρονικό πρότυπο. Έτσι, τα κύτταρα αυτά εµφανίζονται πρώτα στην οπίσθια και ακολούθως στην πρόσθια µοίρα του ραβδωτού σώµατος. Παρόµοιο πρότυπο ανάπτυξης παρατηρείται και στους προβλητικούς χολινεργικούς νευρώνες του βασικού πρόσθιου εγκεφάλου οι οποίοι, ωστόσο, ωριµάζουν φαινοτυπικά νωρίτερα σε σχέση µε τους χολινεργικούς νευρώνες του ραβδωτού σώµατος. Υποστηρίζεται ότι η φαινοτυπική ωρίµαση των χολινεργικών νευρώνων του ραβδωτού σώµατος ολοκληρώνεται την τέταρτη µεταγεννητική εβδοµάδα (Phelps και συν., 1989). 24
37 Εισαγωγή Εφόσον οι χολινεργικοί νευρώνες του ραβδωτού σώµατος γεννιούνται πρώτοι θα ήταν αναµενόµενο, σύµφωνα µε τις παραπάνω µελέτες, να εντοπίζονται κυρίως στα ραβδοσωµάτια. Εντούτοις, οι νευρώνες αυτοί εµφανίζουν οµοιογενή κατανοµή στο ραβδωτό σώµα των ενήλικων επιµύων (Graybiel και συν., 1986 Κubota και Kawaguchi, 1993). Οι van Vulpen και van der Kooy (1996) παρατήρησαν ότι έως την Μ7 οι νευρώνες που εκφράζουν την ChAT εντοπίζονται κυρίως στα ραβδοσωµάτια, ενώ από το στάδιο αυτό και έκτοτε εµφανίζουν εκλεκτική εντόπιση στο υπόστρωµα. Το γεγονός αυτό αποδόθηκε κυρίως στο ότι οι χολινεργικοί νευρώνες στα ραβδοσωµάτια και εκείνοι στο υπόστρωµα ωριµάζουν σε διαφορετικές χρονικές περιόδους κατά την ανάπτυξη. Έτσι, οι νευρώνες των ραβδοσωµατίων εκφράζουν την ChAT πρώτοι, ακολουθούµενοι από τους νευρώνες του υποστρώµατος. Νεότερες µελέτες έδειξαν ότι ο χρόνος γέννησης των χολινεργικών νευρώνων καθορίζει την εντόπισή τους στο ενήλικο ζώο (van Vulpen και van der Kooy, 1998). Οι νευρώνες που γεννιούνται νωρίς, την Ε13, έχουν περισσότερες πιθανότητες να καταλήξουν στην περιοχή των ραβδοσωµατίων, σε αντίθεση µε εκείνους που γεννιούνται αργότερα, την Ε17, οι οποίοι καταλήγουν κυρίως στο υπόστρωµα. H πρωιµότερη ωρίµαση των χολινεργικών νευρώνων που εντοπίζονται στα ραβδοσωµάτια θα µπορούσε να ερµηνευθεί µε βάση τον µεγαλύτερο χρόνο παραµονής στο ραβδωτό σώµα των χολινεργικών νευρώνων που γεννιούνται νωρίς και, κατά συνέπεια, τη µεγαλύτερη έκθεσή τους στους τροφικούς παράγοντες της περιοχής αυτής (van Vulpen και van der Kooy, 1998). Οι χολινεργικοί νευρώνες του ραβδωτού σώµατος εκφράζουν τον υποδοχέα υψηλής συγγένειας της κινάσης A της τυροσίνης (ΤrkA) και τον υποδοχέα χαµηλής συγγένειας p75 (p75 NTR ) (Koh και Loy, 1989 van der Zee και Ηagg, 1998), στους οποίους δεσµεύεται ο αυξητικός 25
38 Εισαγωγή παράγοντας των νεύρων (NGF). Είναι πιθανό ότι ο ΝGF σχετίζεται µε τη χρονικά διαφορετική ωρίµαση των χολινεργικών νευρώνων του ραβδωτού σώµατος, καθώς έχει βρεθεί ότι η έκφραση του TrkA αυξάνεται στο ίδιο αναπτυξιακό στάδιο που παρατηρείται η αύξηση του αριθµού των κυττάρων που εκφράζουν την ChAT στο υπόστρωµα (Holtzman και συν., 1992 Steiniger και συν., 1993). Παρότι ο NGF µπορεί να ρυθµίζει τα επίπεδα της ChAT στους χολινεργικούς νευρώνες, δεν θεωρείται απαραίτητος για την επιβίωση τους κατά τις πρώτες ηµέρες της µεταγεννητικής ζωής (van Vulpen και van der Kooy, 1996). Επιπλέον, ο νευροτροφικός παράγοντας που προέρχεται από τον εγκέφαλο (BDNF) φαίνεται ότι παίζει ρόλο στην ωρίµαση των χολινεργικών νευρώνων του ραβδωτού σώµατος κατά τη µεταγεννητική ανάπτυξη. Τα αποτελέσµατα της έρευνας των Ward και Hagg (2000), στην οποία µελετήθηκαν BDNF-/- µυς, έδειξαν ότι ο BDNF είναι απαραίτητος τόσο για την αύξηση των επιπέδων της ChAT και του p75 NTR κατά τη διάρκεια της µεταγεννητικής ανάπτυξης όσο και για την αύξηση του µεγέθους των χολινεργικών νευρώνων. Από τα στοιχεία αυτά συµπεραίνεται ότι η ανάπτυξη και η ωρίµαση των κυττάρων αυτών του ραβδωτού σώµατος εξαρτάται από έναν πολύπλοκο ρυθµιστικό µηχανισµό, στον οποίο συµµετέχουν οι νευροτροφικοί παράγοντες BDNF και NGF. Η ανάπτυξη των υπόλοιπων διάµεσων νευρώνων του ραβδωτού σώµατος έχει µελετηθεί εκτεταµένα µε τη χρήση της 5-βροµο-2- δεοξυουριδίνης (BrdU), ενός δείκτη κυτταρικού πολλαπλασιασµού, σε συνδυασµό µε τη µέθοδο της ανοσοϊστοχηµείας. Συγκεκριµένα, έχει δειχθεί ότι οι νευρώνες που εκφράζουν την παρβαλβουµίνη και εκείνοι που εκφράζουν την καλρετινίνη γεννιούνται κατά την χρονική περίοδο µεταξύ Ε13 και Ε20, παρουσιάζοντας µέγιστο µεταξύ Ε14 και Ε17 (Sadikot και Sasseville, 1997 Rymar και συν., 2004). Οι νευρώνες που 26
39 Εισαγωγή εκφράζουν τη σωµατοστατίνη γεννιούνται µεταξύ Ε15 και Ε16 (Semba και συν., 1988). Ανοσοϊστοχηµικές µελέτες έδειξαν, επίσης, ότι η έκφραση της παρβαλβουµίνης ξεκινάει από τη δεύτερη µεταγεννητική εβδοµάδα και συγκεκριµένα µετά τη Μ9, ενώ, αντίθετα, η έκφραση της καλρετινίνης παρατηρείται ήδη από την ηµέρα της γέννησης (Schlösser και συν., 1999). Επιπλέον, η ωρίµαση των διάµεσων νευρώνων του ραβδωτού σώµατος ακολουθεί διαφορετικό χωροταξικό πρότυπο. Οι νευρώνες που εκφράζουν την παρβαλβουµίνη και εκείνοι που εκφράζουν την καλρετινίνη αναπτύσσονται πρώτα στην έξω µοίρα του ραβδωτού σώµατος και µετά στην έσω, ενώ οι νευρώνες που εκφράζουν τη σωµατοστατίνη δεν ακολουθούν κανένα συγκεκριµένο πρότυπο ανάπτυξης κατά τη γένεσή τους (Rymar και συν., 2004). Οι διαφορές αυτές υποδηλώνουν ότι καθεµία από τις ανωτέρω πρωτεΐνες ίσως να παίζει συγκεκριµένο και ειδικό ρόλο στα διάφορα στάδια της ανάπτυξης και ωρίµασης των νευρώνων του ραβδωτού σώµατος. Η συµβολή των νευροτροφικών παραγόντων στην ανάπτυξη και στη φαινοτυπική διαφοροποίηση των GABAεργικών διάµεσων νευρώνων του ραβδωτού σώµατος είναι εξίσου σηµαντική. Έχει βρεθεί ότι ο BDNF προκαλεί αύξηση της πυκνότητας των νευρώνων του ραβδωτού σώµατος που εκφράζουν την παρβαλβουµίνη και την καλβιδίνη καθώς και των επιπέδων της σωµατοστατίνης και του νευροπεπτιδίου Υ (Mizuno και συν., 1994). Η γέννηση των ΜSN ακολουθεί το γενικό κανόνα που ισχύει για τους χολινεργικούς νευρώνες. Όσοι από αυτούς γεννιούνται από την Ε12 και µετά θα καταλήξουν στην περιοχή των ραβδοσωµατίων, ενώ οι νευρώνες του υποστρώµατος γεννιούνται αργότερα (Ε17-Ε22) (Bayer, 1984 Fishell και van der Kooy, 1987 van der Kooy και Fishell, 1987). Επιπλέον, από την Ε13 οι MSN εµφανίζουν ανοσοδραστικότητα για το GABA (Lauder και συν., 1986). 27
40 Εισαγωγή Ανατοµικές και ηλεκτροφυσιολογικές µελέτες απέδειξαν ότι οι MSN παρουσιάζουν παρατεταµένη µεταγεννητική ωρίµαση, η οποία ολοκληρώνεται αρκετές εβδοµάδες µετά τη γέννηση. Οι µορφολογικές αλλαγές των MSN, οι οποίες αφορούν κυρίως τη µορφολογία των δενδριτών τους, µελετήθηκαν µε in vivo ενδοκυτταρική σήµανση µε τη χρήση βιοκυτίνης (Tepper και συν., 1998). Έχει βρεθεί ότι κατά τη διάρκεια των δύο πρώτων µεταγεννητικών εβδοµάδων οι δενδρίτες των MSN παρουσιάζουν κυστίδια και δεν εµφανίζουν ακανθοειδείς προεξοχές (Trent και Tepper, 1993), ενώ µέχρι το τέλος της τέταρτης µεταγεννητικής εβδοµάδας οι δενδρίτες εµφανίζουν ακανθοειδείς προεξοχές, ο αριθµός των οποίων είναι µικρότερος σε σύγκριση µε το ενήλικο (Sharpe και Tepper, 1998). Στους επίµυς, σε αντίθεση µε άλλα είδη ζώων όπως είναι η γάτα, ο αριθµός των δενδριτών καθώς και η ακτίνα της περιοχής που αυτοί καταλαµβάνουν, δεν αλλάζουν σηµαντικά µετά το τέλος της πρώτης µεταγεννητικής εβδοµάδας (Tepper και συν., 1998). Η ανάπτυξη των GABAεργικών MSN εξαρτάται, επίσης, από την παρουσία των νευροτροφικών παραγόντων και κυρίως του BDNF, όπως συµβαίνει και µε τους διάµεσους νευρώνες. Παρατηρήθηκε ότι ο BDNF προκαλεί αύξηση των κυτταρικών επιπέδων του GABA σε καλλιέργειες αναπτυσσόµενων νευρώνων του ραβδωτού σώµατος. ιαπιστώθηκε, επίσης, in vivo αύξηση του GABA στο ραβδωτό σώµα νεογέννητων επιµύων, στους οποίους χορηγήθηκε η νευροτροφίνη αυτή διαµέσου των κοιλιών του εγκεφάλου (Mizuno και συν., 1994). Οι συνάψεις που σχηµατίζονται στο ραβδωτό σώµα κατά τη µεταγεννητική ανάπτυξη των επιµύων µελετήθηκαν για πρώτη φορά από τους Ηattori και McGeer (1973). Σύµφωνα µε τη µελέτη αυτή, η µεγαλύτερη αύξηση στην πυκνότητα των συνάψεων παρατηρείται µεταξύ Μ13 και Μ17. ιαπιστώθηκε, επίσης, ότι η πυκνότητα των συµµετρικών 28
41 Εισαγωγή συνάψεων παρουσιάζει τη µέγιστη τιµή της τη Μ17 σε αντίθεση µε την πυκνότητα των ασύµµετρων συνάψεων που αυξάνεται µέχρι την ενηλικίωση. Μεταγενέστερες υπερµικροσκοπικές µελέτες απέδειξαν ότι η µεγαλύτερη αύξηση στην πυκνότητα των ασύµµετρων συνάψεων που σχηµατίζουν οι προσαγωγές ίνες από τον φλοιό των εγκεφαλικών ηµισφαιρίων µε τους MSN παρατηρείται κατά τη διάρκεια της τρίτης µεταγεννητικής εβδοµάδας και συγκεκριµένα µεταξύ Μ15 και Μ21 (Sharpe και Τepper, 1998 Tepper και συν., 1998). Nεότερες µελέτες σήµανσης µε τη χρήση ορθόδροµα µεταφερόµενης βιοκυτίνης απέδειξαν ότι οι φλοιοραβδωτές ίνες εντοπίζονται στο ραβδωτό σώµα από την ηµέρα της γέννησης (Christensen και συν., 1999). Υποστηρίζεται ότι οι ντοπαµινεργικοί νευρώνες της µέλαινας ουσίας στέλνουν τους νευράξονές τους στο ραβδωτό σώµα ήδη από την Ε14 (Voorn και συν., 1988). Την Ε18 εντοπίζονται ντοπαµινεργικές ίνες στην έξω κοιλιακή µοίρα του ραβδωτού σώµατος, ενώ µέχρι τη Μ4 ολοκληρώνεται η ντοπαµινεργική εννεύρωση του πυρήνα (Kalsbeek και συν., 1992). Οι ντοπαµινεργικοί νευρώνες αρχίζουν να εκφράζουν το ένζυµο υδροξυλάση της τυροσίνης (ΤΗ), που είναι απαραίτητο για τη σύνθεση της ντοπαµίνης, αµέσως µετά το τέλος της τελευταίας µιτωτικής τους διαίρεσης, κατά τη διάρκεια της µετανάστευσής τους στις τελικές τους θέσεις (Vitalis και συν., 2005). Nεότερες ανοσοκυτταροχηµικές µελέτες µε ταυτόχρονη χρήση της BrdU και της ΤΗ απέδειξαν ότι η πλειοψηφία των ντοπαµινεργικών νευρώνων της µέλαινας ουσίας πιθανώς γεννιέται νωρίτερα, την Ε12, αλλάζοντας µε αυτόν τον τρόπο τα δεδοµένα στις µελέτες που χρησιµοποιούν πρόδροµες µορφές νευρικών κυττάρων σε πειραµατικά µοντέλα της νόσου Parkinson (Gates και συν., 2006). Είναι χαρακτηριστικό ότι οι ντοπαµινεργικές µελαινοραβδωτές απολήξεις εµφανίζουν οµοιογενή κατανοµή στο ραβδωτό σώµα την Ε14, κατά το τέλος της εµβρυϊκής ζωής σχηµατίζουν παροδικά 29
42 Εισαγωγή συσσωµατώµατα και ανακατανέµονται οµοιογενώς µέχρι το τέλος της πρώτης µεταγεννητικής εβδοµάδας (Moon Edley και Herkenham, 1984 Murrin και Ferrier, 1984 van der Kooy, 1984). Η ανάπτυξη των ντοπαµινεργικών προσαγωγών συστηµάτων του ραβδωτού σώµατος συµπίπτει χρονικά µε µορφολογικές µεταβολές των MSN και πιθανόν να επηρεάζει την ωρίµασή τους (Antonopoulos και συν., 2002). Κατά τη διάρκεια των δύο πρώτων µεταγεννητικών εβδοµάδων οι ντοπαµινεργικές ίνες συνάπτονται στην πλειονότητά τους µε τους δενδρίτες και τα κυτταρικά σώµατα των MSN, ενώ από την τρίτη µεταγεννητική εβδοµάδα και µετέπειτα, περίοδο κατά την οποία παρατηρείται η µεγαλύτερη πυκνότητα των ακανθοειδών προεξοχών των MSN, αυξάνεται ο αριθµός των ντοπαµινεργικών συνάψεων σε ακανθοειδείς προεξοχές. Επιπρόσθετα, οι ντοπαµινεργικές συνάψεις στο ραβδωτό σώµα εµφανίζουν µεγαλύτερη πυκνότητα κατά τις πρώτες ηµέρες της πρώτης µεταγεννητικής εβδοµάδας σε αντίθεση µε τις λοιπές συνάψεις που σχηµατίζονται στον πυρήνα, η πυκνότητα των οποίων κορυφώνεται κατά την τρίτη µεταγεννητική εβδοµάδα. Τα ευρήµατα αυτά υποδηλώνουν ότι η ντοπαµίνη πιθανόν να παίζει ρόλο στην ανάπτυξη του ραβδωτού σώµατος (Tepper και συν., 1998 Αntonopoulos και συν., 2002). Γ. ΠΡΟΓΡΑΜΜΑΤΙΣΜΕΝΟΣ ΚΥΤΤΑΡΙΚΟΣ ΘΑΝΑΤΟΣ Η ανάπτυξη του ΝΣ χαρακτηρίζεται από µια σειρά διεργασιών που περιλαµβάνουν τη γέννηση των νευρικών κυττάρων, τη µετανάστευσή τους στις τελικές τους θέσεις, τη φαινοτυπική διαφοροποίηση καθώς και την εδραίωση των λειτουργικών συνδέσεών τους (Burek και Oppenheim, 1996). Στις διεργασίες αυτές, θεµελιώδη ρόλο παίζει ο προγραµµατισµένος κυτταρικός θάνατος (PCD). 30
43 Εισαγωγή Σύµφωνα µε την θεωρία της «ποσοτικής αντιστοίχισης», κατά τα αρχικά στάδια της ανάπτυξης οι νευρώνες παράγονται σε µεγάλους αριθµούς και ανταγωνίζονται για τη δέσµευση τροφικών παραγόντων σε ποσότητες ικανές να εξασφαλίσουν την επιβίωσή τους. Οι νευρώνες που προσλαµβάνουν επαρκείς ποσότητες τροφικών παραγόντων επιβιώνουν, ενώ οι υπόλοιποι αποµακρύνονται µε τη διεργασία του PCD (Burek και Oppenheim, 1996). Μελέτες µε στόχο τον προσδιορισµό των αλληλεπιδράσεων µεταξύ των κυττάρων και την κατανόηση των µοριακών µηχανισµών που ρυθµίζουν την επιβίωση και το θάνατο των νευρώνων βρίσκονται στο επίκεντρο της αναπτυξιακής νευροβιολογίας (Burek και Oppenheim, 1996 Leist και Jäätelä, 2001 Krantic και συν., 2005). Με βάση το στάδιο της ανάπτυξης και τα κύτταρα στα οποία εκδηλώνονται, διακρίνονται δύο διαφορετικές κατηγορίες PCD. Η πρώτη κατηγορία είναι ο «πολλαπλασιαστικός» κυτταρικός θάνατος, ο οποίος αφορά κύτταρα που διαιρούνται µιτωτικά (Blaschke και συν., 1996 Thomaidou και συν., 1997 Burek και Oppenheim, 1999). Έτσι, ενώ αρχικά υποστηρίζονταν ότι οι νευροβλάστες, µετά από κάθε κυτταρική διαίρεση, είτε συνέχιζαν τον πολλαπλασιασµό τους είτε διαφοροποιούνταν, νεότερες έρευνες έδειξαν ότι µπορεί να ακολουθήσουν διαφορετική πορεία και να υποστούν PCD (Blaschke και συν., 1996, 1998). Οι µοριακοί και κυτταρικοί µηχανισµοί που ρυθµίζουν τον πολλαπλασιαστικό κυτταρικό θάνατο δεν έχουν διευκρινιστεί πλήρως, θεωρείται όµως ότι η κατηγορία αυτή PCD είναι απαραίτητη για την αποµάκρυνση πρόδροµων µορφών νευρικών κυττάρων που φέρουν µεταλλάξεις ή κυττάρων που εκφράζουν λανθασµένο φαινότυπο (Blaschke και συν.,1996, 1998 Burek και Oppenheim, 1999). Η δεύτερη κατηγορία PCD παρατηρείται σε πληθυσµούς µεταµιτωτικών νευρώνων και έχει περιγραφεί σε όλες σχεδόν τις 31
44 Εισαγωγή περιοχές του ΝΣ. Η κατηγορία αυτή του PCD παρατηρείται σε νευρωνικούς πληθυσµούς κατά την περίοδο που εδραιώνουν τις συνδέσεις τους (Burek και Oppenheim, 1999). Υπάρχουν ισχυρές ενδείξεις ότι η επιβίωση των νευρώνων εξαρτάται από την αλληλεπίδρασή τους µε κύτταρα στις περιοχές µε τις οποίες συνδέονται, καθώς έχει αποδειχθεί ότι πειραµατική µείωση του µεγέθους µιας περιοχής-στόχου οδηγεί σε αύξηση του αριθµού των προσυναπτικών κυττάρων που πεθαίνουν και, κατά συνέπεια, σε µείωση των νευρώνων που επιβιώνουν και διαφοροποιούνται (Oppenheim, 1991 Burek και Oppenheim, 1996). Νεότερα δεδοµένα δείχνουν ότι η επιβίωση των νευρώνων κατά την ανάπτυξη εξαρτάται και από τις προσαγωγές ίνες, οι οποίες µεταφέρουν ορθόδροµα σήµατα που προάγουν την επιβίωσή τους (Oppenheim, 1991 Linden, 1994). Πιστεύεται ότι η κατηγορία αυτή του PCD παίζει ρόλο στην τοπογραφική οργάνωση των νευρωνικών δικτύων, καθώς έχει δειχθεί ότι συµµετέχει στην αποµάκρυνση κάθε προβλητικού νευρώνα που εµφανίζει ανωµαλίες στις θέσεις σύνδεσης µε την περιοχήστόχο του (Clarke, 1985 Catsicas και συν., 1987). 1. ΜΟΡΦΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ PCD Αν και δεν υπάρχει οµοφωνία µεταξύ των ερευνητών, θεωρείται ότι η πιο ακριβής ταξινόµηση του PCD είναι εκείνη που βασίζεται στις µορφολογικές αλλαγές που συντελούνται στον πυρήνα των κυττάρων που πεθαίνουν. Σύµφωνα µε την ταξινόµηση αυτή, ο PCD διακρίνεται σε τρεις τύπους: α) στην απόπτωση, β) στον παρόµοιο µε την απόπτωση PCD και γ) στον παρόµοιο µε την νέκρωση PCD (Krantic και συν., 2005). α) Απόπτωση. Βασικό χαρακτηριστικό της απόπτωσης είναι η ενεργοποίηση µιας σειράς κυτταροπλασµατικών πρωτεολυτικών ενζύµων, των κασπασών. Οι κασπάσες ενεργοποιούν ένζυµα του πυρήνα, 32
45 Εισαγωγή όπως είναι οι DNAσες, οι οποίες µε τη σειρά τους καταλύουν τη διάσπαση του πυρηνικού DNA (Krantic και συν., 2005). Το αποτέλεσµα των αντιδράσεων αυτών είναι ο κατακερµατισµός του χρωµοσωµικού DNA σε διαµεσονουκλεοσωµατικά θραύσµατα της τάξεως των ζευγών βάσεων. Ο τεµαχισµός του DNA των αποπτωτικών κυττάρων µπορεί να ανιχνευθεί µε την ηλεκτροφόρησή του σε γέλη αγαρόζης, κατά την οποία τα θραύσµατα σχηµατίζουν χαρακτηριστικές «βαθµίδες» (Ηäcker, 2000). Μορφολογικά, ο τύπος αυτός του PCD χαρακτηρίζεται από την συµπύκνωση και τον κατακερµατισµό της χρωµατίνης σε οµοιογενείς σφαιρικές µάζες. Ο πυρήνας και το κυτταρόπλασµα συρρικνώνονται, ενώ το κύτταρο αποκολλάται από το περιβάλλον νευροπίληµα. Τα κυτταρικά οργανίδια των κυττάρων παραµένουν άθικτα και η κυτταρική µεµβράνη ακέραιη. Σε µεταγενέστερα στάδια, ο πυρήνας κατακερµατίζεται σε δύο ή τρία θραύσµατα ενωµένα µεταξύ τους. Κατά τα τελικά στάδια της απόπτωσης, σχηµατίζονται τα αποπτωτικά σωµάτια, τα οποία είναι κυτταρικά θραύσµατα που περιβάλλονται από κυτταρική µεµβράνη και περιέχουν οποιοδήποτε τµήµα του κυττάρου. Τα αποπτωτικά σωµάτια αποµακρύνονται στο τέλος της αποπτωτικής διεργασίας µε κυτταροφαγία από µικρονευρογλοιακά κύτταρα (Stadelmann και Lassmann, 2000). Η απόπτωση θεωρείται ως ο επικρατέστερος τύπος PCD κατά την ανάπτυξη, καθώς αποτελεί τον πιο αποτελεσµατικό µηχανισµό που διαθέτει το ΝΣ προκειµένου να εξασφαλιστεί το τελικό µέγεθος των νευρωνικών οµάδων (Βurek και Oppenheim, 1996). O αποπτωτικός µηχανισµός ενεργοποιείται και µετά από βλάβη του ΝΣ. Συγκεκριµένα, έχει αποδειχθεί ότι η απόπτωση ενεργοποιείται σε τραυµατικές (Ng και συν., 1997 Yakovlev και συν., 1997 Conti και συν., 1998 Newcomb και συν., 1999 Clark και συν., 1999, 2000), φαρµακολογικές (Marti και συν., 1997, 2002) και ισχαιµικές βλάβες του εγκεφάλου (Ruan και συν., 2003), 33
46 Εισαγωγή µετά από διατοµή των νευραξόνων (Elliott και Snider, 1999), σε τοξίκωση µε διεγερτικά αµινοξέα (Macaya και συν., 1994) καθώς και σε νευροεκφυλιστικές ασθένειες, όπως είναι οι νόσοι Alzheimer, Parkinson και Huntington (Portera-Cailliau και συν., 1995 Jenner και Olanow, 1998 Shimohama, 2000). β) Παρόµοιος µε την απόπτωση PCD. Ο παρόµοιος µε την απόπτωση PCD χαρακτηρίζεται από τη µερική συµπύκνωση της χρωµατίνης του πυρήνα και τον κατακερµατισµό της σε µεγαλύτερα σε σχέση µε την απόπτωση τµήµατα, µεγέθους ζευγών βάσεων. Στον τύπο αυτό του PCD, δεν ενεργοποιούνται οι κασπάσες αλλά ένα άλλο πρωτεϊνικό µόριο, ο παράγοντας που διεγείρει την απόπτωση (AIF), που βρίσκεται στα µιτοχόνδρια του κυττάρου (Krantic και συν., 2005). Ο AIF, µε τη σειρά του, µέσω της ενεργοποίησης άλλων νουκλεασών, προκαλεί κατάτµηση του DNA (Susin και συν., 1999). γ) Παρόµοιος µε τη νέκρωση PCD. Ο παρόµοιος µε τη νέκρωση PCD ενεργοποιείται από διαφορετικές πρωτεάσες, όπως είναι οι καθεψίνες και οι καλπαΐνες, που απελευθερώνονται από τα λυσοσωµάτια και από το ενδοπλασµατικό δικτυωτό του κυττάρου, αντίστοιχα (Leist και Jäätelä, 2001 Jäätelä και Tschopp, 2003 Lockshin και Zakeri, 2004). Οι πρωτεάσες αυτές δεν προκαλούν τεµαχισµό του DNA. Οι µοριακοί µηχανισµοί που καθορίζουν τον τύπο αυτό του PCD δεν είναι πλήρως γνωστοί. Εντούτοις, έχει υποστηριχθεί η ύπαρξη δύο υποκατηγοριών αυτού του PCD: η παράπτωση που χαρακτηρίζεται από τον σχηµατισµό µεγάλων, κενών κυστιδίων στο κυτταρόπλασµα (Sperandio και συν., 2004) και ο αυτοφαγικός θάνατος, στον οποίο σχηµατίζονται κυτταροπλασµατικά κυστίδια που προέρχονται από τα λυσοσωµάτια και περιβάλλονται από διπλή µεµβράνη (Chi και συν., 1999). Οι βιοχηµικές και µορφολογικές µεταβολές που περιγράφηκαν χρησιµεύουν στη διάκριση µεταξύ του PCD, που αποτελεί ενεργητική 34
47 Εισαγωγή διεργασία, και της νέκρωσης που αποτελεί παθητική µορφή κυτταρικού θανάτου (Leist και Jäätelä, 2001). Η νέκρωση προκαλείται συνήθως µετά από οξέα ισχαιµικά, τραυµατικά ή τοξικά ερεθίσµατα (Friedlander, 2003). Κατά τον κυτταρικό αυτό θάνατο, παρατηρείται τεµαχισµός του DNA σε τυχαίες θέσεις µέσω της ενεργοποίησης λυσοσωµικών DNΑσών (Stadelmann και Lassmann, 2000). Τα νεκρωτικά κύτταρα παρουσιάζουν διόγκωση του πυρήνα και των µιτοχονδρίων και τελικά διάλυση των οργανιδίων τους που οδηγεί στη λύση των κυττάρων και την πρόκληση φλεγµονής στον περιβάλλοντα ιστό (Friedlander, 2003). H απόπτωση και η νέκρωση είναι δυνατόν να συνυπάρχουν σε έναν κυτταρικό πληθυσµό (Formigli και συν., 2000). Μελέτες σε νεογέννητους επίµυς απέδειξαν ότι ο κυτταρικός θάνατος που παρατηρείται σε νευρώνες µετά τη χορήγηση διεγερτικών αµινοξέων παρουσιάζει µορφολογικά χαρακτηριστικά τόσο της απόπτωσης όσο και της νέκρωσης καθώς και ενδιάµεσες µορφές (Portera-Cailliau και συν., 1997). Έχει αποδειχθεί, επίσης, ότι η αναστολή του νεκρωτικού κυτταρικού θανάτου µπορεί να προκαλέσει απώλεια των κυττάρων διαµέσου της αποπτωτικής διεργασίας (Pohl και συν., 1999). 2. ΡΥΘΜΙΣΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ 2.1. ΚΥΤΤΑΡΙΚΕΣ ΠΡΩΤΕΪΝΕΣ Τις τελευταίες δεκαετίες µοριακές και βιοχηµικές έρευνες οδήγησαν στην ταυτοποίηση πολλών γονιδίων, καθώς και των προϊόντων τους, που ρυθµίζουν την απόπτωση (Abrams, 1999 Wang, 2001 Horvitz, 2003). Ανάµεσα στις κυτταρικές πρωτεΐνες που ανακαλύφθηκαν, τον σηµαντικότερο ρόλο παίζουν, όπως προαναφέρθηκε, οι κασπάσες. Μέχρι σήµερα, έχουν περιγραφεί 14 διαφορετικά µέλη της οικογένειας των κασπασών (Zimmermann και συν., 2001). 35
48 Εισαγωγή Φυλογενετικές αναλύσεις έδειξαν ότι η οικογένεια των κασπασών διακρίνεται σε δύο υποοµάδες. Στην πρώτη ανήκουν οι κασπάσες-1, -4, -5, -11,-12 και -13, που παίζουν ρόλο στην ωρίµαση και ενεργοποίηση προ-φλεγµονωδών κυτοκινών, όπως είναι οι προ-ιντερλευκίνες-1ß και -18 (Denault και Salvesen, 2002) και στη δεύτερη οι κασπάσες-2, -3, -6, -7, -8, -9 και -10, που συµµετέχουν στη διεργασία του αποπτωτικού θανάτου (Yakovlev και Faden, 2004). Οι κασπάσες-2, -8, -9 και -10 είναι οι «εναρκτήριες» κασπάσες, οι οποίες διεγείρουν τον µηχανισµό της απόπτωσης, ενώ οι κασπάσες-3, -6 και -7 είναι οι «δραστικές» κασπάσες, οι οποίες ευθύνονται για τις µορφολογικές και βιοχηµικές αλλαγές που χαρακτηρίζουν τον αποπτωτικό κυτταρικό θάνατο (Yakovlev και Faden, 2004). Οι κασπάσες, αρχικά, συντίθενται ως αδρανή ζυµογόνα ένζυµα (προκασπάσες) και ενεργοποιούνται µέσω της πρωτεολυτικής τους διάσπασης που προκαλείται είτε από τις ίδιες είτε από άλλες κασπάσες (Shiozaki και Shi, 2004). Υποστηρίζεται ότι υπάρχουν δύο διαφορετικές οδοί διαµέσου των οποίων είναι δυνατόν να ενεργοποιηθεί και να ολοκληρωθεί η απόπτωση. Η «εξωτερική» οδός χαρακτηρίζεται από την ενεργοποίηση των υποδοχέων της οικογένειας του παράγοντα νέκρωσης των όγκων (TNF), που βρίσκονται στην επιφάνεια του κυττάρου, από αποπτωτικά σηµατοδοτικά µόρια, όπως είναι οι κυτοκίνες TNF-α, ο προσδέτης Fas και ο σχετιζόµενος µε τον TNF προσδέτης που επάγει την απόπτωση (TRAIL) (Choi και Benveniste, 2004). Η σύνδεση των υποδοχέων αυτών, που συχνά αναφέρονται και ως υποδοχείς θανάτου, µε τα σηµατοδοτικά µόρια οδηγεί στο σχηµατισµό ειδικού συµπλόκου (σηµατοδοτικό σύµπλοκο που επάγει θάνατο, DISC), το οποίο µε τη σειρά του ενεργοποιεί τις «εναρκτήριες» κασπάσες-2, -8, -9 και -10 (Yakovlev και Faden, 2004). 36
49 Εισαγωγή Στην «εσωτερική» οδό, το αποπτωτικό ερέθισµα προκαλεί την απελευθέρωση πρωτεϊνών από τα µιτοχόνδρια του κυττάρου στο κυτταρόπλασµα, όπως είναι το κυτόχρωµα c και ο παράγοντας Smac/DIABLO (Shiozaki και Shi, 2004). Ειδικότερα, το κυτόχρωµα c προσδένεται και ενεργοποιεί την πρωτεΐνη Apaf-1 στο κυτταρόπλασµα (Li και συν., 1997) σχηµατίζοντας ένα σύµπλοκο, το «αποπτώσωµα», το οποίο µε τη σειρά του ενεργοποιεί την προκασπάση-9 (Rodriguez και Lazebnik, 1999), ενώ ο Smac/DIABLO δεσµεύεται και απενεργοποιεί τις πρωτεΐνες-αναστολείς της απόπτωσης (ΙΑΡs) (Zimmermann και συν., 2001). Στη συνέχεια, οι δύο παραπάνω οδοί συγκλίνουν στην ενεργοποίηση των «δραστικών» κασπασών, µεταξύ των οποίων η κασπάση-3 παίζει τον σηµαντικότερο ρόλο, καθώς ενεργοποιεί άλλα ένζυµα που καταλύουν την κατάτµηση του χρωµοσωµικού DNA (Zimmermann και συν., 2001). Η ενεργοποίηση της κασπάσης-3 θεωρείται ότι χαρακτηρίζει τον αποπτωτικό κυτταρικό θάνατο, καθώς πολλά από τα µορφολογικά και βιοχηµικά χαρακτηριστικά της απόπτωσης οφείλονται στην αποδόµηση βασικών για την επιβίωση του κυττάρου πρωτεϊνών από την κασπάση αυτή. Συγκεκριµένα, έχει βρεθεί ότι η κασπάση-3 απενεργοποιεί τις πρωτεΐνες που είναι υπεύθυνες για την επιδιόρθωση του DNA (Davoli και συν., 2002) και καταστρέφει δοµικές πρωτεΐνες του κυτταροσκελετού, επηρεάζοντας την ακεραιότητα των κυττάρων (Wang και συν., 1998 Yang και συν., 1998). Μια πρωτεΐνη του κυτταροσκελετού είναι η ακτίνη, η αποδόµηση της οποίας από την κασπάση-3 κατά τα τελικά στάδια της απόπτωσης οδηγεί στο σχηµατισµό ενός πρωτεϊνικού µορίου, της φρακτίνης. Ανοσοϊστοχηµικές µελέτες απέδειξαν ότι τα κύτταρα που είναι ανοσοδραστικά για τη φρακτίνη εµφανίζουν τα τυπικά µορφολογικά χαρακτηριστικά της απόπτωσης, όπως είναι η συρρίκνωση του πυρήνα και η συµπύκνωση της 37
50 Εισαγωγή χρωµατίνης, καθιστώντας την πρωτεΐνη αυτή αξιόπιστο δείκτη του αποπτωτικού κυτταρικού θανάτου (Suurmeijer και συν., 1999 Jackson- Lewis και συν., 2000 Οo και συν., 2002). Ο ρόλος της κασπάσης-3 στην απόπτωση έχει δειχθεί τόσο κατά τη διάρκεια της ανάπτυξης του ΝΣ όσο και σε παθολογικές καταστάσεις. Έρευνες σε κασπάση-3 -/- µυς έδειξαν σηµαντικές ανωµαλίες κατά την ανάπτυξη του εγκεφάλου (Kuida και συν., 1996), ενώ σε µελέτες σε πειραµατικά µοντέλα πρόκλησης κυτταρικού θανάτου µετά από αξονότµηση (Kermer και συν., 1998), καταστροφή της περιοχής-στόχου (Sophou και συν., 2006) και τραυµατισµό του εγκεφάλου λόγω µηχανικής βλάβης (Yakovlev και συν., 1997) ή αγγειακού εγκεφαλικού επεισοδίου (Chen και συν., 1998 Namura και συν., 1998) παρατηρήθηκε ενεργοποίηση της κασπάσης-3. Η έκφραση της κασπάσης-3 έχει µελετηθεί εκτεταµένα στους νευρώνες της µέλαινας ουσίας κατά την ανάπτυξη (Jeon και συν., 1999) καθώς και σε πειραµατικά µοντέλα βλάβης του ραβδωτού σώµατος είτε µετά από τοξίκωση µε διεγερτικά αµινοξέα (Macaya και συν., 1994) είτε µετά τη χορήγηση της νευροτοξίνης 6-υδροξυντοπαµίνης (6-OHDA) (Marti και συν., 1997). Πιστεύεται ότι η ενεργοποίηση της κασπάσης-3 πιθανόν να παίζει ρόλο στους µηχανισµούς εκφύλισης των ντοπαµινεργικών νευρώνων της µέλαινας ουσίας που χαρακτηρίζουν τη νόσο Parkinson (El-Khodor και Βurke, 2002). Σηµαντικό ρόλο στη ρύθµιση του αποπτωτικού µηχανισµού και κυρίως της «εσωτερικής» οδού παίζουν, επίσης, οι πρωτεΐνες της οικογένειας Bcl. Τα µέλη της οικογένειας αυτής διακρίνονται σε δύο κατηγορίες, ανάλογα µε το αν αναστέλλουν ή αν προάγουν την απόπτωση (Lindsten και συν., 2005). Στην πρώτη κατηγορία ανήκουν οι πρωτεΐνες Bcl-2 και Βcl-x L, οι οποίες πιστεύεται ότι εντοπίζονται στην εξωτερική µεµβράνη των µιτοχονδρίων, συντελώντας στη διατήρηση της 38
51 Εισαγωγή ακεραιότητάς της (Lindsten και συν., 2005). Αντίθετα, οι δύο σηµαντικότερες πρωτεΐνες που προάγουν την απόπτωση, Bax και Βak, προκαλούν µεταβολές στη µιτοχονδριακή µεµβράνη, µε αποτέλεσµα την απελευθέρωση του κυτοχρώµατος c στο κυτταρόπλασµα, την ενεργοποίηση των κασπασών και τον επακόλουθο θάνατο του κυττάρου (Lindsten και συν., 2005). Υπάρχουν, επίσης, ενδείξεις ότι οι Bax και Βak είναι δυνατόν να επιδράσουν στο ενδοπλασµατικό δικτυωτό του κυττάρου προκαλώντας διαταραχές των ενδοκυτταρικών επιπέδων του Ca ++ και ενεργοποίηση της κασπάσης-12 (Scorrano και συν., 2003 Ζong και συν., 2003). Η οικογένεια Bcl-2 περιλαµβάνει επίσης ένα µεγάλο αριθµό από τις ΒΗ3 πρωτεΐνες που προάγουν την απόπτωση, όπως είναι οι Bid, Bad, Bik, Bim, Bmf, Noxa, Puma και Βnip3 (Cory και Adams, 2002 Lindsten και συν., 2005). Κάτω από φυσιολογικές συνθήκες, οι Bcl-2 και Βcl-x L προστατεύουν τη µιτοχονδριακή µεµβράνη από τη δράση των Bax και Βad. Παρουσία, όµως, ενός αποπτωτικού ερεθίσµατος προκαλείται έκφραση µιας ΒΗ3 πρωτεΐνης, η οποία προσδένεται στις Bcl-2 και Βcl-x L απελευθερώνοντας, µε αυτόν τον τρόπο, τη δράση των Bax και Βad µε στόχο την ενεργοποίηση του αποπτωτικού µηχανισµού (Lindsten και συν., 2005). Η έκφραση των αντιαποπτωτικών πρωτεϊνών Bcl-2 και Βcl-x L ρυθµίζεται από τον πυρηνικό παράγοντα kappa Β (NF-κΒ), ο οποίος περιγράφεται συχνά ως αντιαποπτωτικός µεταγραφικός παράγοντας (Heckman και συν., 2002 Karin και Lin, 2002), παρόλο που κάτω από ορισµένες συνθήκες µπορεί να προκαλέσει διέγερση του αποπτωτικού µηχανισµού (Grimm και συν., 1996). Επίσης, ο ΝF-κΒ φαίνεται ότι παίζει ρόλο στην ενεργοποίηση και άλλων αντιαποπτωτικών γονιδίων, όπως εκείνων που είναι υπεύθυνα για την έκφραση των ΙΑPs (Karin και Lin, 2002). Τα κυριότερα µέλη των ΙΑPs είναι οι πρωτεΐνες c-iap1, c-iap2, NAIP και XAIP, οι οποίες προσδένονται και απενεργοποιούν τις 39
52 Εισαγωγή κασπάσες-3 και -7 και εµποδίζουν την ενεργοποίηση των προκασπασών -6 και -9 (Deveraux και συν., 1998 Salvesen και Duckett, 2002). Τέλος, πρόσφατες έρευνες απέδειξαν ότι η απόπτωση ρυθµίζεται και από τα µέλη της οικογένειας της p53 πρωτεΐνης, p53, p63 και p73, και τις µεταξύ τους αλληλεπιδράσεις (Bradley Jacobs και συν., 2006). Μελέτες που πραγµατοποιήθηκαν σε p53 -/- µυς έδειξαν ότι ο αριθµός των αποπτωτικών νευρώνων εµφανίζεται σηµαντικά µειωµένος (Martin και συν., 2001). Πιστεύεται ότι η p53 συµµετέχει στην ενεργοποίηση των πρωτεϊνών Bax και Βak που προάγουν την απόπτωση µε αποτέλεσµα την πρόκληση µεταβολών στη διαπερατότητα της µιτοχονδριακής µεµβράνης και τη διέγερση της «εσωτερικής» οδού της απόπτωσης (Bradley Jacobs και συν., 2006) ΝΕΥΡΟΤΡΟΦΙΚΟΙ ΠΑΡΑΓΟΝΤΕΣ Με τον όρο νευροτροφικοί παράγοντες περιγράφεται µια οµάδα πολυπεπτιδίων µε πολλαπλούς ρόλους τόσο στο αναπτυσσόµενο όσο και στο ενήλικο ΝΣ (Chao και συν., 2006). Κατά τα αρχικά στάδια της ανάπτυξης, οι παράγοντες αυτοί συµµετέχουν στη ρύθµιση της γέννησης των νευρώνων, στην επιβίωση και στη διαφοροποίησή τους καθώς και στην εδραίωση των συνδέσεών τους (Caleo και Cenni, 2004), ενώ σε µεταγενέστερα στάδια, πιστεύεται ότι ρυθµίζουν την πλαστικότητα των νευρωνικών δικτύων (Poo, 2001 Caleo και Μaffei, 2002). O πρώτος νευροτροφικός παράγοντας που αποµονώθηκε και χαρακτηρίστηκε από τη Levi-Montalcini και τους συνεργάτες της (1968) ήταν ο NGF. Σήµερα, είναι γνωστό ότι ο NGF είναι µέλος µιας οικογένειας νευροτροφικών παραγόντων, των νευροτροφινών, που περιλαµβάνει επίσης τον BDNF, τις νευροτροφίνες ΝΤ-3, ΝΤ-4/5 και ΝΤ-6 (Μaisonpierre και συν., 1990 Bothwell, 1991 Ibanez και συν., 1992 Ιp και Yancopoulos, 1992). Οι νευροτροφίνες δρουν στα κύτταρα- 40
53 Εισαγωγή στόχους µέσω της δέσµευσής τους σε δύο διαφορετικούς τύπους διαµεµβρανικών υποδοχέων, τους Trk υποδοχείς και τον υποδοχέα p75 ΝΤR (Patapoutian και Reichardt, 2001 Dechant και Barde, 2002 Hempstead, 2002). Εκτός, όµως, από τις νευροτροφίνες έχει δειχθεί ότι και άλλοι νευροτροφικοί παράγοντες, όπως είναι ο νευροτροφικός παράγοντας που προέρχεται από τα νευρογλοιακά κύτταρα (GDNF) (Lin και συν., 1993), ο ακτινοειδής νευροτροφικός παράγοντας (CNTF) (Ιp και Yancopoulos, 1992 Lindsay και συν., 1994) και, τέλος, η οικογένεια του αυξητικού παράγοντα των ινοβλαστών (FGF) (Walicke, 1988) προάγουν την επιβίωση διάφορων νευρωνικών πληθυσµών (Mufson και συν., 1999). Παραδείγµατος χάριν, ο GDNF προάγει την επιβίωση των ντοπαµινεργικών νευρώνων της µέλαινας ουσίας και των νοραδρενεργικών νευρώνων του υποµέλανα τόπου. Επιπλέον, ο GDNF και ο CNTF προάγουν την επιβίωση των κινητικών νευρώνων που εκφυλίζονται στην πλάγια µυατροφική σκλήρυνση (Sendtner και συν., 1992 Mitsumoto και συν., 1994 Lapchak, 1996 Lapchak και συν., 1996). Ο βασικός αυξητικός παράγοντας των ινοβλαστών (bfgf) διεγείρει τη νευρογένεση µετά από τραυµατικές βλάβες του εγκεφάλου (Monfils και συν., 2005) και συµβάλλει στην αποκατάσταση διαταραχών κίνησης και συµπεριφοράς (Li και Stephenson, 2002). Νευροτροφικές ιδιότητες εµφανίζει και ο αιµοπεταλιακός αυξητικός παράγοντας (PDGF- BB), καθώς εκφράζεται σε υψηλά επίπεδα κατά την περίοδο πολλαπλασιασµού και µετανάστευσης των νευρικών κυττάρων (Mohapel και συν., 2005). Σύµφωνα µε την κλασική «νευροτροφική θεωρία», οι µεταµιτωτικοί νευρώνες κατά την ανάπτυξή τους ανταγωνίζονται για τη δέσµευση και ανάδροµη µεταφορά των περιορισµένων σε διαθεσιµότητα νευροτροφικών παραγόντων από τις περιοχές στις οποίες οι νευρώνες 41
54 Εισαγωγή προβάλλουν, ενώ όσοι από αυτούς αποτύχουν, πεθαίνουν µέσω της διεργασίας του PCD (Burek και Oppenheim, 1996 Yuan και Yankner, 2000). Οι νευροτροφικοί αυτοί παράγοντες εκλύονται από τα µετασυναπτικά κύτταρα στις περιοχές-στόχους, δεσµεύονται µέσω ειδικών υποδοχέων στις απολήξεις των προσυναπτικών νευρώνων και µεταφέρονται ανάδροµα κατά µήκος των νευραξόνων µέχρι το σώµα των νευρικών κυττάρων (Μiller και Kaplan, 2002). Πρόσφατες έρευνες έδειξαν ότι κάποιοι νευροτροφικοί παράγοντες είναι δυνατό να δρουν και µέσω παρακρινών ή αυτοκρινών µηχανισµών (Caleo και Μaffei, 2002), ενώ είναι πλέον αποδεδειγµένο ότι µπορούν να µεταφερθούν και ορθόδροµα κατά µήκος των νευραξόνων στα µετασυναπτικά κύτταραστόχους (Linden, 1994 von Bartheld και συν., 2001). Χαρακτηριστικά παραδείγµατα νευροτροφικών παραγόντων που µεταφέρονται ορθόδροµα είναι ο BDNF και η ΝΤ-3 (Altar και συν., 1997 Μufson και συν., 1999 Caleo και Μaffei, 2002). O NGF και το mrna του νευροτροφικού αυτού παράγοντα ανιχνεύονται στο ραβδωτό σώµα (Dreyfus, 1989). Η ενδογενής αυτή παραγωγή του NGF πιστεύεται ότι συµβάλλει στη φαινοτυπική διαφοροποίηση των χολινεργικών νευρώνων του ραβδωτού σώµατος (van Vulpen και van der Kooy, 1996). Ο NGF ανιχνεύεται και σε περιοχές του εγκεφάλου που προβάλλουν στο ραβδωτό σώµα, όπως είναι ο φλοιός των εγκεφαλικών ηµισφαιρίων, ωστόσο δεν είναι δυνατή η ορθόδροµη µεταφορά του µέσω των φλοιοραβδωτών ινών (Large και συν., 1986 Μufson και συν., 1999). Υποστηρίζεται ότι από τους νευροτροφικούς παράγοντες, ο BDNF παίζει τον σηµαντικότερο ρόλο στην επιβίωση των νευρώνων του ραβδωτού σώµατος (Mizuno και συν., 1994 Νakao και συν., 1995 Ventimiglia και συν., 1995). Ο παράγοντας αυτός δρα µέσω της ενεργοποίησης του υποδοχέα υψηλής συγγένειας ΤrkB 42
55 Εισαγωγή (Bibel και Barde, 2000 Huang και Reichardt, 2001). Ιn vivo µελέτες έδειξαν ότι ο BDNF προστατεύει τους GABAεργικούς νευρώνες του ραβδωτού σώµατος µετά από τοξίκωση µε διεγερτικά αµινοξέα (Nakao και συν., 1995), ενώ είναι δυνατό να αναστείλει την απόπτωση σε γενετικά τροποποιηµένα κύτταρα που εκφράζουν την µεταλλαγµένη πρωτεΐνη χαντινγκτίνη, η οποία σχετίζεται µε την παθογένεια της νόσου Huntington (Saudou και συν., 1998). Έχει διαπιστωθεί ότι οι νευρώνες του φλοιού των εγκεφαλικών ηµισφαιρίων καθώς και οι ντοπαµινεργικοί νευρώνες της µέλαινας ουσίας έχουν την ικανότητα να παράγουν BDNF σε ποσότητες που ποικίλουν κατά τα διάφορα στάδια της ανάπτυξης (Friedman και συν., 1991 Aliaga και συν., 2000 Rite και συν., 2003). Η παρατήρηση αυτή σε συνδυασµό µε το εύρηµα ότι στο ραβδωτό σώµα ανιχνεύεται mrna για τον υποδοχέα TrkB αλλά όχι για τον BDNF, οδήγησε στο συµπέρασµα ότι ο ΒDNF µεταφέρεται στους MSN κυρίως ορθόδροµα µέσω των προσαγωγών ινών από τον φλοιό των εγκεφαλικών ηµισφαιρίων και σε µικρότερο βαθµό από τη µέλαινα ουσία και προάγει την επιβίωσή τους (Altar και συν., 1997 Altar και DiStefano, 1998 Mufson και συν., 1999 Caleo και συν., 2000 Fawcett και συν., 2000 von Bartheld και συν., 2001). Οι συνέπειες της µακροπρόθεσµης έλλειψης του BDNF που προέρχεται από τον φλοιό των εγκεφαλικών ηµισφαιρίων στο ραβδωτό σώµα εκτιµήθηκαν σε µελέτες που πραγµατοποιήθηκαν σε BDNF -/- µυς (Βaquet και συν., 2004). Τα πειραµατόζωα αυτά παρουσίασαν σηµαντικές µεταβολές στη µορφολογία και στον αριθµό των MSN, οι οποίες ήταν εντονότερες στο ενήλικο (Βaquet και συν., 2004). Επιπρόσθετα, η έλλειψη του BDNF λόγω µετάλλαξης των υπεύθυνων για την έκφρασή του γονιδίων ή λόγω βλάβης του φλοιού των εγκεφαλικών ηµισφαιρίων είχε σαν αποτέλεσµα την ελάττωση της έκφρασης της παρβαλβουµίνης και της καλβιδίνης σε νευρώνες του ραβδωτού σώµατος (Altar και συν., 1997 Ivkovic και Ehrlich, 1999). 43
56 Εισαγωγή Έχει αποδειχθεί, επίσης, ότι ο BDNF διεγείρει τη µετανάστευση και διαφοροποίηση πρόδροµων µορφών νευρικών κυττάρων στο ραβδωτό σώµα είτε µετά από έγχυσή του στις κοιλίες του εγκεφάλου (Pencea και συν., 2001) είτε µετά από έκφρασή του µέσω της µεταφοράς του υπεύθυνου γονιδίου σε αδενοϊό (Benraiss και συν., 2001). Επιπλέον, έχει βρεθεί ότι ο BDNF και ο PDGF-BB διεγείρουν τη νευρογένεση στο ραβδωτό σώµα µετά από βλάβες µε τη νευροτοξίνη 6-OHDA (Mohapel και συν., 2005). Ο GDNF, ένας άλλος νευροτροφικός παράγοντας, έχει βρεθεί ότι ασκεί ευεργετική δράση στην επιβίωση των νευρώνων του ραβδωτού σώµατος σε πειραµατικά µοντέλα της νόσου Huntington (Pérez-Navarro και συν., 1996 Araujo και Hilt, 1997). Επιπλέον, πιστεύεται ότι ο ρόλος του GDNF είναι σηµαντικός για την επιβίωση των ντοπαµινεργικών µελαινοραβδωτών νευρώνων, η εκφύλιση των οποίων θεωρείται υπεύθυνη για την αιτιοπαθογένεια της νόσου Parkinson (Mattson, 2000). Ο GDNF θεωρείται πιθανός θεραπευτικός παράγοντας για τη νόσο αυτή (Choi-Lundberg και συν., 1997 Βjörklund και συν., 2003 Gill και συν., 2003 Κordower, 2003). Τέλος, in vitro µελέτες απέδειξαν ότι οι νευροτροφίνες NT-3 και ΝΤ-4/5 συµβάλλουν στην επιβίωση και διαφοροποίηση των νευρώνων του ραβδωτού σώµατος (Ardelt και συν., 1994 Widmer και Hefti, 1994 Ventimiglia και συν., 1995). Η ΝΤ-3 µεταφέρεται στο ραβδωτό σώµα ορθόδροµα από τον φλοιό των εγκεφαλικών ηµισφαιρίων, καθώς τα επίπεδα του mrna που ανιχνεύονται στον πυρήνα αυτό είναι χαµηλά (Friedman και συν., 1991 Mufson και συν., 1999). H αποτελεσµατικότητα της ΝΤ-3 σε σύγκριση µε τον BDNF και την ΝΤ- 4/5 στην αποτροπή νευροεκφυλιστικών αλλαγών σε µοντέλα της νόσου Huntington, την καθιστούν πιθανό θεραπευτικό εργαλείο για την προστασία των προβλητικών νευρώνων του ραβδωτού σώµατος από την 44
57 Εισαγωγή ατροφία και την απώλεια χηµικών διαβιβαστών που χαρακτηρίζουν τη νόσο (Pérez-Navarro και συν., 1999). Αυτό πιστεύεται ότι οφείλεται στο γεγονός ότι τα επίπεδα του mrna για τους υποδοχείς ΤrkC, µε τους οποίους συνδέεται η ΝΤ-3, είναι υψηλότερα στο ραβδωτό σώµα από εκείνα για τους ΤrkB (Altar και συν., 1994), µε αποτέλεσµα µεγαλύτερος αριθµός νευρώνων να εκφράζει τους ΤrkC και να αποκρίνεται στη δράση της ΝΤ-3. Επιπλέον, η µεγαλύτερη αποτελεσµατικότητα της ΝΤ-3 µπορεί να σχετίζεται µε την ικανότητά της να ενεργοποιεί όχι µόνο τους ΤrkC αλλά, αν και σε µικρότερο βαθµό, και τους ΤrkB υποδοχείς (Barbacid, 1994). Οι MSN διαθέτουν και τις δύο κατηγορίες υποδοχέων, εποµένως, η δράση της NT-3 µπορεί να αποδοθεί στην ταυτόχρονη ενεργοποίησή τους (Μerlio και συν., 1992). Εκτός από τους κλασικούς νευροτροφικούς παράγοντες, νευροτροφικές ιδιότητες κατά την ανάπτυξη του NΣ εµφανίζουν, επίσης, ορισµένοι χηµικοί διαβιβαστές. Σε καλλιέργειες εµβρυϊκών κυττάρων, έχει δειχθεί ότι το GABA παίζει ρυθµιστικό ρόλο κατά τη διαφοροποίηση και µετανάστευση των σεροτονινεργικών και των νοραδρενεργικών νευρώνων (Lauder και συν., 1998). Επιπλέον, υποστηρίζεται ότι οι µονοαµινεργικές ίνες επηρεάζουν την ανάπτυξη και ωρίµαση των περιοχών στις οποίες απολήγουν (Levitt και συν., 1997). Έχει αποδειχθεί ότι σε καλλιέργειες νευρικών κυττάρων η σεροτονίνη διεγείρει την ανάπτυξη του νευροπιλήµατος και την µυελίνωση των νευραξόνων και επηρεάζει το µέγεθος του κυτταρικού σώµατος (Emerit και συν., 1992). Τέλος, in vitro µελέτες έδειξαν ότι η σεροτονίνη προάγει την επιβίωση και τη διαφοροποίηση και αναστέλλει τον κυτταρικό θάνατο των γλουταµινεργικών νευρώνων στον φλοιό των εγκεφαλικών ηµισφαιρίων (Dooley και συν., 1997 Lavdas και συν., 1997). 45
58 Εισαγωγή 3. Η ΑΠΟΠΤΩΣΗ ΤΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΚΑΙ ΤΩΝ ΠΕΡΙΟΧΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ ΜΕ ΤΙΣ ΟΠΟΙΕΣ ΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΣΥΝ ΕΕΤΑΙ ΣΕ ΝΕΥΡΟΕΚΦΥΛΙΣΤΙΚΕΣ ΑΣΘΕΝΕΙΕΣ Η πληθώρα δεδοµένων που υπάρχουν στη βιβλιογραφία σχετικά µε το ρόλο της απόπτωσης στις νευροεκφυλιστικές ασθένειες προέρχεται κυρίως από πειραµατικά µοντέλα ζώων. Είναι πρακτικά δύσκολο να επιβεβαιωθεί ο αποπτωτικός κυτταρικός θάνατος σε ανθρώπινο υλικό, διότι η απόπτωση είναι φαινόµενο που εξελίσσεται σχετικά γρήγορα µε αποτέλεσµα να δυσχεραίνεται η ταυτοποίηση των αποπτωτικών κυττάρων σε κάθε χρονική στιγµή (Μattson, 2000). H νόσος Huntington είναι κληρονοµική νευροεκφυλιστική ασθένεια µε σοβαρές κινητικές και γνωστικές διαταραχές. Χαρακτηρίζεται από σηµαντική απώλεια των MSN και επιβίωση των µεγάλων µη ακανθωτών διάµεσων νευρώνων του ραβδωτού σώµατος (Reiner και συν., 1988 Vonsattel και DiFiglia, 1998 Ross και συν., 1999). Μηχανισµοί του κυτταρικού θανάτου που πιθανόν να εµπλέκονται στην παθογένεια της νόσου περιλαµβάνουν το οξειδωτικό στρες, τη µιτοχονδριακή δυσλειτουργία, την τοξίκωση από διεγερτικά αµινοξέα και την ενεργοποίηση πρωτεασών (Bizat και συν., 2003). Έχει αποδειχθεί µε τη χρήση της µέθοδου TUNEL ότι η απώλεια των MSN σε ασθενείς που πάσχουν από τη νόσο Huntington οφείλεται σε αποπτωτικό κυτταρικό θάνατο (Dragunow και συν., 1995 Τhomas και συν., 1995 Ferrer και συν., 2000). Σε πειραµατικά µοντέλα µυών της νόσου Huntington διαπιστώθηκε ενεργοποίηση των κασπασών-3, -8 και -9 και απελευθέρωση του κυτοχρώµατος c (Sanchez και συν., 1999 Chen και συν., 2000 Kiechle και συν., 2002). Ασθενείς που πάσχουν από τη νόσο Parkinson εµφανίζουν κινητικές διαταραχές που χαρακτηρίζονται από βραδυκινησία, τρόµο, ακαµψία και απώλεια της σταθερότητας της στάσης. Oι διαταραχές αυτές 46
59 Εισαγωγή οφείλονται στην απώλεια των ντοπαµινεργικών νευρώνων της µέλαινας ουσίας που προβάλλουν στο ραβδωτό σώµα (Honig και Rosenberg, 2000 Conforti και συν., 2007). Πειραµατική έγχυση της νευροτοξίνης 6-OHDA στο ραβδωτό σώµα νεογέννητων (Marti και συν., 1997) και ενήλικων επιµύων (Marti και συν., 2002) προκάλεσε την απόπτωση των ντοπαµινεργικών νευρώνων της µέλαινας ουσίας ενισχύοντας, έτσι, την άποψη ότι η απώλεια ντοπαµινεργικών νευρώνων στη νόσο Parkinson οφείλεται σε αποπτωτικό κυτταρικό θάνατο. Επιπλέον, η ανάλυση εγκεφάλων από ασθενείς έδειξε κατάτµηση του DNA µε τον χαρακτηριστικό τρόπο που παρατηρείται κατά την απόπτωση και ενεργοποίηση γονιδίων που χαρακτηρίζουν τον κυτταρικό αυτό θάνατο (Jenner και Olanow, 1998). 47
60
61 Υλικά και Μέθοδοι ΥΛΙΚΑ ΚΑΙ ΜΕΘΟ ΟΙ H απόπτωση στο ραβδωτό σώµα µελετήθηκε ποιοτικά και ποσοτικά κατά την ανάπτυξη και µετά από καταστροφή συγκεκριµένων προσαγωγών συστηµάτων ινών. Χρησιµοποιήθηκε ένα τροποποιηµένο πρωτόκολλο της µεθόδου ΤUNEL (Terminal deoxynucleotidyl transferase-mediated dutp-biotin Nick End Labeling) (Gavrieli και συν., 1992) για την ταυτοποίηση των αποπτωτικών κυττάρων, ενώ η επιβεβαίωση των αποπτωτικών µορφολογικών χαρακτηριστικών των κυττάρων έγινε µε τη χρήση του ηλεκτρονικού µικροσκοπίου. Επιπλέον, για την πιστοποίηση του αποπτωτικού κυτταρικού θανάτου χρησιµοποιήθηκε η µέθοδος της ανοσοϊστοχηµείας µε αντισώµατα εναντίον της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης, που θεωρούνται αξιόπιστοι αποπτωτικοί δείκτες. Τέλος, για τη µελέτη των επιπτώσεων της καταστροφής των προσαγωγών ινών στην επιβίωση και στον φαινότυπο των GABAεργικών και των χολινεργικών νευρώνων του ραβδωτού σώµατος χρησιµοποιήθηκε η µέθοδος της ανοσοϊστοχηµείας µε αντισώµατα εναντίον του GABA και της ChAT, αντίστοιχα. 1. ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΤΩΝ ΜΕΘΟ ΩΝ ΠΟΥ ΧΡΗΣΙΜΟΠΟΙΗΘΗΚΑΝ 1.1. ΜΕΘΟ ΟΣ ΤUNEL Όπως προαναφέρθηκε (βλ. Εισαγωγή, κεφ. Γ.1α), χαρακτηριστικό γνώρισµα του αποπτωτικού κυτταρικού θανάτου είναι η συµπύκνωση της χρωµατίνης και ο κατακερµατισµός του χρωµοσωµικού DNA σε διαµεσονουκλεοσωµατικά θραύσµατα της τάξεως των ζευγών βάσεων (Kerr και συν., 1972 Wyllie, 1980 Wyllie και συν., 1980 Arends και συν. 1990). Με τη µέθοδο ΤUNEL επιτυγχάνεται η σήµανση των 49
62 Υλικά και Μέθοδοι θραυσµάτων αυτών in situ. Η µέθοδος αυτή θεωρείται αξιόπιστη όταν χρησιµοποιείται σε συνδυασµό µε την ανάλυση των µορφολογικών χαρακτηριστικών των κυττάρων µε το οπτικό και το ηλεκτρονικό µικροσκόπιο και µε τη µελέτη της έκφρασης των γονιδίων που προάγουν ή αναστέλλουν την απόπτωση (Ishimaru και συν., 1999 Stadelmann και Lassmann, 2000). Η µέθοδος TUNEL βασίζεται στην ιδιότητα του ενζύµου τελική δεοξυνουκλεοτιδιλική τρανσφεράση (TdT) να µεταφέρει και να ενσωµατώνει νουκλεοτίδια στα ελεύθερα 3 -ΟΗ άκρα που προκύπτουν από τον κατακερµατισµό του DNA. Η ανίχνευση των αποπτωτικών κυττάρων περιλαµβάνει τρία βασικά στάδια: α) την πρωτεόλυση των ιστονών, των πρωτεϊνών που συνδέονται µε το DNA για τον σχηµατισµό των νουκλεοσωµάτων, µε στόχο την αποκάλυψη των θέσεων κατάτµησης του DNA, β) την ενσωµάτωση, µε τη δράση του ενζύµου TdT, του συνδεδεµένου µε βιοτίνη νουκλεοτιδίου δεοξυουριδίνη (b-dutp) στις ελεύθερες 3 -ΟΗ θέσεις κατάτµησης του DNA και γ) την ανίχνευση του b-dutp, που στη συγκεκριµένη µελέτη έγινε µε τη µέθοδο της Αβιδίνης- Βιοτίνης ΜΕΘΟ ΟΣ ΑΒΙ ΙΝΗΣ-ΒΙΟΤΙΝΗΣ Στη µέθοδο αυτή χρησιµοποιείται το σύµπλεγµα Αβιδίνης- Βιοτίνης-Υπεροξειδάσης (ABC). Το σύµπλεγµα αυτό αποτελείται από τη γλυκοπρωτεΐνη αβιδίνη, µε την οποία συνδέεται η µικρού µοριακού βάρους βιταµίνη βιοτίνη ενωµένη µε µεγάλο αριθµό µορίων υπεροξειδάσης. Η συγγένεια της αβιδίνης µε τη βιοτίνη είναι 10 6 φορές µεγαλύτερη από εκείνη µεταξύ αντιγόνου και αντισώµατος και εποµένως είναι µη αντιστρεπτή (Wilchek και Βayer, 1990). Η αβιδίνη έχει 4 θέσεις δέσµευσης για τη βιοτίνη. Μία από αυτές είναι κενή. Όταν η µέθοδος της Αβιδίνης-Βιοτίνης συνδυάζεται µε τη µέθοδο TUNEL, η θέση αυτή 50
63 Υλικά και Μέθοδοι µπορεί να καλυφθεί από τη βιοτίνη που είναι ενωµένη µε το νουκλεοτίδιο dutp, µε αποτέλεσµα την έµµεση δέσµευση του συµπλέγµατος ABC στα ελεύθερα 3 -ΟΗ άκρα του κατακερµατισµένου DNA. Η ανίχνευση του συµπλέγµατος γίνεται µε τη χρήση της αµίνης 3,3 -διαµινοβενζιδίνης (DAB), η οποία οξειδώνεται όταν έρθει σε επαφή µε το υπεροξείδιο του υδρογόνου (Η 2 Ο 2 ) παρουσία της υπεροξειδάσης του ABC. Η οξειδωµένη µορφή της DAB είναι ένα αδιάλυτο, µόνιµο πολυµερές, καστανού χρώµατος, το οποίο είναι ορατό µε το οπτικό µικροσκόπιο, υποδηλώνοντας έµµεσα τις θέσεις κατάτµησης του DNA και κατ επέκταση τους πυρήνες των αποπτωτικών κυττάρων ΑΝΟΣΟΪΣΤΟΧΗΜΕΙΑ Με τη µέθοδο της ανοσοϊστοχηµείας επιτυγχάνεται η ανίχνευση in situ ουσιών που θεωρούνται ως αντιγόνα στους ιστούς χρησιµοποιώντας τα κατάλληλα σεσηµασµένα αντισώµατα. Η µέθοδος στηρίζεται στην ειδική ανοσολογική αντίδραση που αναπτύσσεται µεταξύ αντιγόνουαντισώµατος. Στη συγκεκριµένη µελέτη χρησιµοποιήθηκε η µέθοδος της έµµεσης ανοσοϊστοχηµείας, η οποία περιλαµβάνει 3 στάδια: 1) η προς ανίχνευση ουσία αναγνωρίζεται και δεσµεύεται από το πρώτο αντίσωµα, 2) χρησιµοποιείται δεύτερο αντίσωµα, το οποίο είναι ειδικό εναντίον των ανοσοσφαιρινών G (IgG) του ζώου που παρήγαγε το πρώτο αντίσωµα. Το αντίσωµα αυτό είναι συνδεδεµένο µε βιοτίνη και δεσµεύεται στο σύµπλεγµα αντιγόνου-πρώτου αντισώµατος, 3) γίνεται ανίχνευση του σεσηµασµένου δεύτερου αντισώµατος µε τη µέθοδο Αβιδίνης-Βιοτίνης. Πριν από την ανοσοϊστοχηµική µέθοδο εφαρµόστηκε η τεχνική αποκάλυψης των αντιγονικών επιτόπων (AR). H τεχνική αυτή χρησιµοποιείται ευρέως σε τοµές ιστών που έχουν µονιµοποιηθεί και έχουν εγκλειστεί σε παραφίνη, στις οποίες ενδεχοµένως να έχει µειωθεί η αντιγονικότητα λόγω της πρόκλησης µορφολογικών µεταβολών των 51
64 Υλικά και Μέθοδοι πρωτεϊνών στους ιστούς (Shi και συν., 2001). Η AR γίνεται µε ενζυµικές ή χηµικές µεθόδους ή µε θέρµανση των τοµών σε ρυθµιστικά διαλύµατα. Με την τεχνική αυτή επιτυγχάνεται η υδρόλυση των χηµικών δεσµών που δηµιουργούνται κατά την µονιµοποίηση και η επαναφορά των πρωτεϊνών στην αρχική τους δοµή (Shi και συν., 2001). Στη συγκεκριµένη µελέτη, για την ανοσοϊστοχηµική ανίχνευση της ενεργοποιηµένης κασπάσης-3, της φρακτίνης και του GABA εφαρµόστηκε η θέρµανση των τοµών σε κιτρικό ρυθµιστικό διάλυµα µε τη χρήση συσκευής µικροκυµάτων. 2. ΠΕΙΡΑΜΑΤΙΚΟ ΥΛΙΚΟ ΚΑΙ ΜΕΘΟ ΟΙ Για τη διενέργεια των πειραµάτων χρησιµοποιήθηκε ο απολύτως αναγκαίος αριθµός πειραµατοζώων. Επίσης, εφαρµόστηκαν όλα τα απαραίτητα µέτρα ώστε να διασφαλιστεί η ευζωία των πειραµατοζώων, σύµφωνα µε την οδηγία της Ε.Ε. για την προστασία τους (24 th November 1986; 86/609/EEC). Στην εργασία αυτή χρησιµοποιήθηκαν συνολικά 111 επίµυες της φυλής Wistar, οι οποίοι χωρίστηκαν σε τρεις οµάδες: Η Α οµάδα περιελάµβανε 38 πειραµατόζωα, στα οποία µελετήθηκε η απόπτωση στο ραβδωτό σώµα κατά την ανάπτυξη. Στη Β οµάδα περιλαµβάνονταν 56 πειραµατόζωα, στα οποία έγινε αφαίρεση του φλοιού του δεξιού εγκεφαλικού ηµισφαιρίου και µελέτη του αποπτωτικού θανάτου στο οµοπλάγιο ραβδωτό σώµα. Οι επίµυες της οµάδας αυτής κατατάχθηκαν σε τρεις υποοµάδες: Στη Β1 υποοµάδα (n=32), η βλάβη πραγµατοποιήθηκε σε νεογέννητους επίµυς (Μ0), στη Β2 (n=12) σε επίµυς 7 ηµερών και στη Β3 (n=12) η βλάβη έγινε σε επίµυς 14 ηµερών. Η Γ οµάδα, τέλος, περιελάµβανε 17 επίµυες, στους οποίους προκλήθηκε βλάβη των κατεχολαµινεργικών συστηµάτων µε την 6-OHDA. 52
65 Υλικά και Μέθοδοι Για τον εντοπισµό και την καταµέτρηση των αποπτωτικών κυττάρων του ραβδωτού σώµατος µε τη µέθοδο TUNEL, χρησιµοποιήθηκαν συνολικά 84 επίµυες από όλες τις οµάδες. Εξετάστηκαν 3 πειραµατόζωα για κάθε ηλικία. Από την Α οµάδα χρησιµοποιήθηκαν 30 επίµυες των παρακάτω ηλικιών: Μ0, Μ1, Μ3, Μ5, Μ7, Μ10, Μ14, Μ21, Μ28 και ενήλικοι επίµυες (Μ60). Από τους επίµυς των ηλικιών Μ0, Μ1, Μ5 και Μ7 χρησιµοποιήθηκαν τοµές για την ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης µε τη µέθοδο της ανοσοϊστοχηµείας. Σε 18 επίµυς της Β1 υποοµάδας οι ηλικίες που εξετάστηκαν ήταν η πρώτη ηµέρα µετά τη βλάβη (ΜΒ1), η ΜΒ3, η ΜΒ7, η ΜΒ14, η ΜΒ21 και η ΜΒ60. Για την ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης χρησιµοποιήθηκαν τοµές από τους επίµυς των ηλικιών ΜΒ1 και ΜΒ7, ενώ εξετάστηκαν επιπλέον και οι ηλικίες ΜΒ0 (βλάβη σε νεογέννητους επίµυς) και ΜΒ5 (n=3 ζώα/ηλικία). Στους 24 συνολικά επίµυς των Β2 και Β3 υποοµάδων οι ηλικίες που εξετάστηκαν ήταν η ΜΒ1, η ΜΒ7, η ΜΒ14 και η ΜΒ60. Σε 12 επίµυς της Γ οµάδας εξετάστηκαν οι ηλικίες: Μ3, Μ7, Μ21 και Μ60. Από τους επίµυς των ηλικιών Μ3 και Μ7 χρησιµοποιήθηκαν τοµές για την ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης. Τέλος, από τους επίµυς ηλικίας Μ3 χρησιµοποιήθηκαν τοµές για την ανοσοϊστοχηµική ανίχνευση της ντοπαµίνης και της νοραδρεναλίνης. Για τη µελέτη των GABAεργικών νευρώνων του ραβδωτού σώµατος χρησιµοποιήθηκαν συνολικά 9 ενήλικοι επίµυες από τις οµάδες Α, Β1 και Γ (n=3 ζώα/οµάδα). Για τη µελέτη των χολινεργικών νευρώνων του ραβδωτού σώµατος χρησιµοποιήθηκαν 6 ενήλικα πειραµατόζωα από τις οµάδες Α και Β1 (n=3 ζώα/οµάδα). Τέλος, για τη µελέτη των αποπτωτικών κυττάρων µε το ηλεκτρονικό µικροσκόπιο χρησιµοποιήθηκαν συνολικά 6 επίµυες των οµάδων Α, Β1 και Γ, ηλικιών Μ7, Β7 και Μ7, αντίστοιχα (n=2 ζώα/οµάδα). 53
66 Υλικά και Μέθοδοι Οι οµάδες και υποοµάδες των πειραµατοζώων που χρησιµοποιήθηκαν στις διάφορες πειραµατικές µεθόδους φαίνονται στον Πίνακα ΠΕΙΡΑΜΑΤΙΚΟΙ ΧΕΙΡΙΣΜΟΙ 2.1.A. ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Όλες οι επεµβάσεις γίνονταν κάτω από γενική αναισθησία. Για τα νεογέννητα ζώα χρησιµοποιήθηκε η µέθοδος της υποθερµίας µε τοποθέτησή τους σε πάγο, ενώ για ζώα µεγαλύτερης ηλικίας η αναισθησία γινόταν µε τη χορήγηση αιθυλικού αιθέρα. Κατόπιν, γινόταν τοµή του δέρµατος κατά µήκος της µέσης γραµµής του κρανίου και αφαίρεση τµήµατος του κρανίου ώστε να αποκαλυφθεί το δεξιό εγκεφαλικό ηµισφαίριο. Με ειδική αναρροφητική αντλία γινόταν αφαίρεση του µεγαλύτερου τµήµατος του φλοιού και ακολουθούσε η συρραφή του δέρµατος. Οι επίµυες παρακολουθούνταν µέχρι την πλήρη ανάνηψή τους και, κατόπιν, επέστρεφαν στον κλωβό τους. 2.1.Β. ΦΑΡΜΑΚΟΛΟΓΙΚΗ ΒΛΑΒΗ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ Για την καταστροφή των κατεχολαµινεργικών συστηµάτων του εγκεφάλου η κατεχολαµινεργική νευροτοξίνη 6-OHDA (Sigma) χορηγούνταν ενδοπεριτοναϊκά στα πειραµατόζωα σε 4 δόσεις (1 δόση ανά ηµέρα) από τη Μ0 έως και τη Μ3. Η 6-OHDA ήταν διαλυµένη σε φυσιολογικό ορό χλωριούχου νατρίου 0,9% που περιείχε 1mg/ml ασκορβικού οξέος. Η πρώτη έγχυση (50mg/g σωµατικού βάρους) γίνονταν περίπου 6 ώρες µετά τη γέννηση, ενώ οι υπόλοιπες (100mg/g σωµατικού βάρους) γίνονταν σε εικοσιτετράωρα διαστήµατα µετά την πρώτη χορήγηση, σύµφωνα µε το πρωτόκολλο που προτάθηκε από τους Blue και Parnavelas (1982). 54
67 Υλικά και Μέθοδοι ΠΕΙΡΑΜΑΤΙΚΕΣ ΟΜΑ ΕΣ ΚΑΙ ΥΠΟΟΜΑ ΕΣ Πίνακας 1. Οµάδες και υποοµάδες των πειραµατοζώων που χρησιµοποιήθηκαν στις διάφορες πειραµατικές µεθόδους Μέθοδος TUNEL Aνοσοϊστοχηµική µέθοδος για την ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης ΠΕΙΡΑΜΑΤΙΚΕΣ ΜΕΘΟ ΟΙ Aνοσοϊστοχηµική µέθοδος για την ανίχνευση του GABA Aνοσοϊστοχηµική µέθοδος για την ανίχνευση της ChAT Aνοσοϊστοχηµική µέθοδος για την ανίχνευση της ντοπαµίνης και της νοραδρεναλίνης Hλεκτρονικό µικροσκόπιο (Α) Αναπτυσσόµενα πειραµατόζωα (n=38) Μ Μ Μ Μ Μ Μ Μ Μ Μ (Β1) Βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 (n=32) ΜΒ ΜΒ ΜΒ ΜΒ ΜΒ ΜΒ ΜΒ ΜΒ (Β2) Βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ7 (n=12) ΜΒ ΜΒ ΜΒ ΜΒ (Β3) Βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ14 (n=12) ΜΒ ΜΒ ΜΒ ΜΒ (Γ) Βλάβη των κατεχολαµινεργικών προσαγωγών συστηµάτων µε την 6-OHDA (n=17) Μ Μ Μ Μ
68 Υλικά και Μέθοδοι 2.2. ΣΥΛΛΟΓΗ ΠΕΙΡΑΜΑΤΙΚΟΥ ΥΛΙΚΟΥ Στα αναισθητοποιηµένα µε χορήγηση αιθυλικού αιθέρα ζώα γίνονταν έγχυση, διαµέσου της καρδιάς και µε τη βοήθεια ειδικής αντλίας, φυσιολογικού ορού χλωριούχου νατρίου 0,9% στην αρχή και κατόπιν µονιµοποιητικού διαλύµατος παραφορµαλδεΰδης 4% σε ρυθµιστικό διάλυµα φωσφορικού νατρίου (ΡΒ) 0,1Μ, ph 7,4. Στους ενήλικες επίµυς η έγχυση αυτή γινόταν διαµέσου της πρόσθιας αορτής, ενώ η οπίσθια αορτή κλείνονταν µε τη βοήθεια αιµοστατικής λαβίδας. Η ποσότητα του µονιµοποιητικού διαλύµατος που χρησιµοποιούνταν κυµαινόταν ανάλογα µε την ηλικία των πειραµατοζώων από ml. Μετά τη µονιµοποίηση, οι εγκέφαλοι αφαιρούνταν, τοποθετούνταν στο ίδιο µονιµοποιητικό διάλυµα για 2 ώρες στους 4ºC και στη συνέχεια µεταφέρονταν σε ΡΒ για 24 ώρες στην ίδια θερµοκρασία. Για τους εγκεφάλους που προορίζονταν να εξεταστούν µε τη µέθοδο ΤUNEL και µε την ανοσοϊστοχηµική µέθοδο για την ανίχνευση της ενεργοποιηµένης κασπάσης-3, της φρακτίνης και του GABA, ακολουθούσε αφυδάτωση σε µια σειρά διαλυµάτων αιθυλικής αλκοόλης αυξανόµενης πυκνότητας, από 50% έως 100% κ.ο. Στη συνέχεια, οι εγκέφαλοι τοποθετούνταν σε χλωροφόρµιο, σε ισοµερές µείγµα χλωροφορµίου και υγρής παραφίνης και τέλος, σε υγρή παραφίνη σε κλίβανο θερµοκρασίας 58ºC. Ο χρόνος παραµονής των εγκεφάλων στα παραπάνω διαλύµατα διέφερε ανάλογα µε την ηλικία των πειραµατοζώων, όπως φαίνεται στον Πίνακα 2. Μετά την έγκλεισή τους σε παραφίνη οι εγκέφαλοι διατηρούνταν σε θερµοκρασία 4ºC. Στη συνέχεια, οι εγκέφαλοι κόβονταν µε µικροτόµο παραφίνης σε διαδοχικές εγκάρσιες τοµές πάχους 10µm. Μία τοµή κάθε 200µm χρωµατίζονταν µε τη µέθοδο Nissl για να προσδιοριστούν τα όρια του ραβδωτού σώµατος. 56
69 Υλικά και Μέθοδοι Ηλικία Πίνακας 2. Χρόνος παραµονής των εγκεφάλων στα διάφορα διαλύµατα κατά την αφυδάτωσή τους ιαλύµατα αιθυλικής αλκοόλης 50% 70% 95% 100% 100% Χλωροφόρµιο Μείγµα χλωροφορµίουπαραφίνης Παραφίνη Μ0-24ω 1ω 1ω 45λ 45λ 2x15λ 15λ 3ω Μ5 Μ7-24ω 1ω 1ω 45λ 1ω 2x20λ 15λ 3,5ω Μ10 Μ14 1ω 1ω 1ω 45λ 1ω 2x30λ 20λ 4ω Μ21 1ω 15ω 1ω 50λ 1ω30λ 2x30λ 20λ 4ω Μ28 1ω 15ω 1ω 1ω15λ 1ω45λ 2x30λ 20λ 4ω Μ60 1ω 15ω 1ω 1ω30λ 1ω45λ 2x30λ 20λ 4ω ω=ώρα(-ες), λ=λεπτά Οι εγκέφαλοι που προορίζονταν για την ανοσοϊστοχηµική ανίχνευση της ChAT, της ντοπαµίνης και της νοραδρεναλίνης, µετά τη µονιµοποίησή τους και την έκπλυση µε ΡΒ, κόβονταν σε συνεχείς εγκάρσιες τοµές πάχους 50µm µε µικροτόµο δονήσεως και συλλέγονταν σε κρύο ΡΒ. Για τη συλλογή των εγκεφάλων που θα εξετάζονταν µε το ηλεκτρονικό µικροσκόπιο χρησιµοποιήθηκε µονιµοποιητικό διάλυµα παραφορµαλδεΰδης 4%, που περιείχε γλουταραλδεΰδη 0,2%, σε ΡΒ. Μετά την παραµονή in situ στους 4ºC για 1 ώρα, οι εγκέφαλοι αφαιρούνταν, τοποθετούνταν σε µονιµοποιητικό διάλυµα παραφορµαλδεΰδης 4% για 2 ώρες στους 4ºC και µετά µεταφέρονταν σε διάλυµα ΡΒ για 24 ώρες στην ίδια θερµοκρασία. Κατόπιν, οι εγκέφαλοι, κόβονταν σε εγκάρσιες τοµές πάχους 50µm µε µικροτόµο δονήσεως. Οι τοµές αυτές συλλέγονταν σε κρύο ΡΒ. 57
70 Υλικά και Μέθοδοι 2.3. ΜΕΘΟ ΟΣ ΤUNEL Οι τοµές συλλέγονταν σε αντικειµενοφόρους πλάκες καλυµµένες µε ζελατίνη και ακολουθούσε η αποπαραφίνωσή τους µε εµβάπτιση σε ξυλόλη (για 30 λεπτά) και κατόπιν η ενυδάτωσή τους σε διαλύµατα αιθυλικής αλκοόλης µειούµενης πυκνότητας, από 100% έως 50% κ.ο. Οι τοµές, στη συνέχεια, πλένονταν σε απεσταγµένο νερό για 5 λεπτά και µετά τοποθετούνταν σε διάλυµα που περιείχε 26mM CaCl 2 σε 76mM Tris/HCl, ph 7,6 για 5 λεπτά. Το επόµενο στάδιο περιελάµβανε την πρωτεόλυση των ιστονών του DNA, η οποία γίνονταν µε επώαση των τοµών σε πρωτεϊνάση Κ (Sigma), συγκέντρωσης 2µg/ml για 15 λεπτά. Μετά την πλύση τους µε απεσταγµένο νερό και την νέα επώασή τους για 15 λεπτά σε ρυθµιστικό διάλυµα TdT (διάλυµα 25mM Tris/HCl µε ph 6,6 που περιείχε 200mM κακοδυλικό νάτριο και 0,25mg/ml αλβουµίνη ορού βοός), οι τοµές επωάζονταν στο διάλυµα TUNEL (Roche) σε κλίβανο θερµοκρασίας 37ºC για 1½ ώρες. Το διάλυµα αυτό περιείχε 1,25mM χλωριούχο κοβάλτιο, 18µΜ b-dutp και 1600U/ml ανασυνδυασµένο ένζυµο TdT. Κατόπιν, οι τοµές πλένονταν εκ νέου σε απεσταγµένο νερό και σε ρυθµιστικό διάλυµα PB που περιείχε NaCl 0,9%, ph 7,4 (ΡΒS) για 10 λεπτά και επωάζονταν σε φυσιολογικό ορό αίγας (NGS) 1% για 15 λεπτά, ώστε να καλυφθούν οι µη ειδικές αντιγονικές θέσεις. Στη συνέχεια, τοποθετούνταν για 2 ώρες στο σκοτάδι στο σύµπλεγµα ABC (Vector Laboratories), 1:100 σε PBS. Μετά την έκπλυση σε PBS και σε ρυθµιστικό διάλυµα Tris 0,1M, ph 7,4 (ΤΒ) για 15 λεπτά οι τοµές µεταφέρονταν σε διάλυµα DAB (0,075% σε ΤΒ) για 5 λεπτά και κατόπιν σε όµοιο διάλυµα DAB, το οποίο περιείχε 0,01% Η 2 Ο 2 για 15 λεπτά. Ακολουθούσε πλύσιµο των τοµών σε διαλύµατα TΒ, PB και σε απεσταγµένο νερό για 30 λεπτά συνολικά. Οι τοµές αντιχρωµατίζονταν µε 10% κυανό της τολουιδίνης για 3 δευτερόλεπτα. Τέλος, οι τοµές αφυδατώνονταν σε διαλύµατα αιθυλικής αλκοόλης 58
71 Υλικά και Μέθοδοι αυξανόµενης πυκνότητας, από 50% έως 100% κ.ο. (5 λεπτά σε κάθε διάλυµα) και κατόπιν γινόταν η διαύγαση σε ξυλόλη (10 λεπτά), η έγκλειση σε DPX και η παρατήρηση µε το οπτικό µικροσκόπιο. Για τον έλεγχο της ειδικότητας της µεθόδου TUNEL εφαρµόστηκαν δύο δοκιµές: α) επώαση των τοµών µε το ένζυµο δεοξυριβονουκλεάση ΙΙ (DNAase II) 5%, το οποίο προκαλεί τη διάσπαση του DNA σε 3 ολιγονουκλεοτίδια (θετικοί µάρτυρες), β) επώαση των τοµών µε το διάλυµα TUNEL, στο οποίο παραλείφθηκε η προσθήκη του ενζύµου TdT (αρνητικοί µάρτυρες) ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΘΟ ΟΣ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3, ΤΗΣ ΦΡΑΚΤΙΝΗΣ ΚΑΙ ΤΟΥ GABA Οι τοµές συλλέγονταν σε αντικειµενοφόρους πλάκες καλυµµένες µε ζελατίνη. Ακολουθούσε η αποπαραφίνωσή τους µε εµβάπτιση σε ξυλόλη (για 30 λεπτά) και κατόπιν η ενυδάτωσή τους σε µια σειρά διαλυµάτων αιθυλικής αλκοόλης µειούµενης πυκνότητας, από 100% έως 50% κ.ο. Στη συνέχεια, πλένονταν σε απεσταγµένο νερό για 5 λεπτά και ακολουθούσε η µέθοδος αποκάλυψης των αντιγονικών επιτόπων µε τη χρήση µικροκυµάτων. Συγκεκριµένα, οι τοµές εµβαπτίζονταν σε κιτρικό ρυθµιστικό διάλυµα 10mM, ph 6 (CB). Tο διάλυµα αυτό αποτελούνταν από 1,2% διαλύµατος κιτρικού οξέος 0,1Μ και 8,8% διαλύµατος κιτρικού νατρίου 0,1Μ. Η µέθοδος περιελάµβανε 3 κύκλους θέρµανσης των τοµών. Στον πρώτο κύκλο οι τοµές θερµαίνονταν για 5 λεπτά σε 500W και παρέµεναν σε θερµοκρασία δωµατίου για άλλα 5 λεπτά. Στον δεύτερο και στον τρίτο κύκλο οι τοµές θερµαίνονταν µέχρι βρασµού του ρυθµιστικού διαλύµατος, ενώ µετά τον τρίτο κύκλο θερµαίνονταν για επιπλέον 2½ λεπτά σε 90W. Τέλος, οι τοµές παρέµεναν σε θερµοκρασία δωµατίου για 30 λεπτά. 59
72 Υλικά και Μέθοδοι Στη συνέχεια, οι τοµές πλένονταν σε PBS για 15 λεπτά και επωάζονταν σε NGS 10% για 15 λεπτά µε σκοπό να καλυφθούν οι µη ειδικές αντιγονικές θέσεις. Ακολουθούσε η επώασή τους σε διάλυµα που περιείχε το πρώτο πολυκλωνικό αντίσωµα, είτε εναντίον της ενεργοποιηµένης κασπάσης-3 (Chemicon Int.) σε συγκέντρωση 1:50 είτε εναντίον της φρακτίνης (Chemicon Int.) σε συγκέντρωση 1:800 ή εναντίον του GABA (Sigma) σε συγκέντρωση 1:500, σε PBS µε 0,5% Τriton-Χ (TX) για 1 ώρα σε θερµοκρασία δωµατίου, για 24 ώρες σε θερµοκρασία 4ºC και για 1 ώρα επιπλέον σε θερµοκρασία δωµατίου. Οι τοµές πλένονταν σε PBS για 15 λεπτά και κατόπιν επωάζονταν στο συνδεδεµένο µε βιοτίνη δεύτερο αντίσωµα. Το δεύτερο αντίσωµα ήταν ειδικό εναντίον πολυκλωνικών αντισωµάτων (Vector Laboratories) και χρησιµοποιήθηκε σε συγκέντρωση 1:200 σε PBS µε 0,5% TX για 2 ώρες. Κατόπιν, οι τοµές πλένονταν σε PBS για 15 λεπτά και επωάζονταν στο σύµπλεγµα ABC για 1½ ώρα. Μετά το πλύσιµό τους σε PBS και σε ΤΒ για 15 λεπτά οι τοµές µεταφέρονταν σε διάλυµα DAB (0,075% σε ΤΒ) για 5 λεπτά και κατόπιν σε όµοιο διάλυµα DAB που περιείχε 0,01% Η 2 Ο 2 για 15 λεπτά. Ακολουθούσε πλύσιµό τους σε ΤΒ και PB για 15 λεπτά. Τέλος, γίνονταν η αφυδάτωση των τοµών σε µια σειρά διαλυµάτων αιθυλικής αλκοόλης αυξανόµενης πυκνότητας, από 50% έως 100% κ.ο., η διαύγαση σε ξυλόλη (2x5 λεπτά), η έγκλειση σε DPX και η παρατήρηση µε το οπτικό µικροσκόπιο. Για τον έλεγχο της ανοσοϊστοχηµικής αντίδρασης παραλείφθηκε το συνδεδεµένο µε τη βιοτίνη αντίσωµα ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΘΟ ΟΣ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ChAT Οι τοµές µετά την πλύση τους σε κρύο ΡΒ (3x10 λεπτά) µεταφέρονταν σε φιαλίδια που περιείχαν το πολυκλωνικό αντίσωµα εναντίον της ChAT (Chemicon Int.) σε συγκέντρωση 1:500 σε PBS µε 60
73 Υλικά και Μέθοδοι 0,5% ΤΧ. Επωάζονταν στο αντίσωµα αυτό για 1 ώρα σε θερµοκρασία δωµατίου, για 24 ώρες σε θερµοκρασία 4ºC και για 1 ώρα ακόµη σε θερµοκρασία δωµατίου. Κατόπιν, πλένονταν σε PBS για 15 λεπτά. Ακολουθούσε η επώασή τους για 2 ώρες στο συνδεδεµένο µε βιοτίνη δεύτερο αντίσωµα (Vector Laboratories) σε συγκέντρωση 1:200 σε PBS. Στη συνέχεια, οι τοµές πλένονταν σε PBS για 15 λεπτά και ακολουθούσε επώασή τους στο σύµπλεγµα ABC για 1 ώρα. Μετά την πλύση τους σε PBS και ΤΒ για 15 λεπτά οι τοµές µεταφέρονταν σε διάλυµα DAB (0,05% σε ΤΒ) για 5 λεπτά και σε όµοιο διάλυµα DAB που περιείχε 0,01% Η 2 Ο 2 για 10 λεπτά. Στη συνέχεια, τοποθετούνταν σε αντικειµενοφόρους πλάκες καλυµµένες µε ζελατίνη µέχρι να στεγνώσουν. Τέλος, ακολουθούσε η αφυδάτωση των τοµών σε διαλύµατα αιθυλικής αλκοόλης αυξανόµενης πυκνότητας, από 50% έως 100% κ.ο., η διαύγαση σε ξυλόλη, η έγκλειση σε DPX και η παρατήρηση µε το οπτικό µικροσκόπιο. Ο έλεγχος της ανοσοϊστοχηµικής αντίδρασης έγινε όπως αναφέρθηκε παραπάνω (βλ. Υλικά και Μέθοδοι, κεφ. 2.4) ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΘΟ ΟΣ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΝΤΟΠΑΜΙΝΗΣ ΚΑΙ ΤΗΣ ΝΟΡΑ ΡΕΝΑΛΙΝΗΣ Οι τοµές συλλέγονταν σε κρύο ρυθµιστικό διάλυµα ΤB που περιείχε NaCl 0,9%, ph 7,6 (ΤΒS) και στη συνέχεια επωάζονταν σε NGS 5% σε ΤΒS για 30 λεπτά. Κατόπιν, µεταφέρονταν σε φιαλίδια τα οποία περιείχαν το πρώτο πολυκλωνικό αντίσωµα. Το αντίσωµα εναντίον της ντοπαµίνης (προσφορά του Dr. R.M. Buijs, Brain Research Institute, Amsterdam, The Netherlands) χρησιµοποιήθηκε σε συγκέντρωση 1:1.500 σε ΤΒS µε 1% µεταδιθειώδες νάτριο και 0,5% ΤΧ. Για την ανίχνευση της νοραδρεναλίνης χρησιµοποιήθηκε πολυκλωνικό αντίσωµα εναντίον της υδροξυλάσης β της ντοπαµίνης (DBH) (Incstar, USA), του ενζύµου που καταλύει τη µετατροπή της ντοπαµίνης σε νοραδρεναλίνη, σε 61
74 Υλικά και Μέθοδοι συγκέντρωση 1:2.000 σε TBS µε 0,3% ΤΧ. Οι τοµές επωάζονταν για 1 ώρα σε θερµοκρασία δωµατίου, για ώρες σε θερµοκρασία 4ºC και για 1 ώρα ακόµη σε θερµοκρασία δωµατίου. Ακολουθούσε πλύσιµο σε TBS για 30 λεπτά και επώαση στο συνδεδεµένο µε βιοτίνη δεύτερο αντίσωµα (Vector Laboratories) (1:400 σε TBS µε 0,3% ΤΧ) για 2 ώρες σε θερµοκρασία δωµατίου. Στη συνέχεια, οι τοµές πλένονταν σε ΤBS για 30 λεπτά και επωάζονταν στο σύµπλεγµα ABC για 1 ώρα. Μετά την πλύση τους σε ΤΒS για 30 λεπτά οι τοµές µεταφέρονταν σε διάλυµα DAB (0,025% σε ΤΒS) για 10 λεπτά και σε όµοιο διάλυµα DAB που περιείχε 0,01% Η 2 Ο 2 για 10 λεπτά. Kατόπιν, τοποθετούνταν σε αντικειµενοφόρους πλάκες καλυµµένες µε ζελατίνη µέχρι να στεγνώσουν. Ακολουθούσε η αφυδάτωση των τοµών σε διαλύµατα αιθυλικής αλκοόλης αυξανόµενης πυκνότητας, από 50% έως 100% κ.ο., η διαύγαση σε ξυλόλη, η έγκλειση σε DPX και η παρατήρηση µε το οπτικό µικροσκόπιο ΗΛΕΚΤΡΟΝΙΚΗ ΜΙΚΡΟΣΚΟΠΙΑ Οι τοµές που προορίζονταν να µελετηθούν µε το ηλεκτρονικό µικροσκόπιο, µετά την επεξεργασία τους µε τη µέθοδο TUNEL, όπως περιγράφηκε παραπάνω (βλ. Υλικά και Μέθοδοι, κεφ. 2.3), µεταφέρονταν σε φιαλίδια που περιείχαν διάλυµα τετροξειδίου του οσµίου 1% για 30 λεπτά και στη συνέχεια πλένονταν σε διάλυµα οξικού νατρίου 0,1Μ (2x5 λεπτά). Κατόπιν, τοποθετούνταν σε υδατικό διάλυµα οξικού ουρανυλίου συγκέντρωσης 1% για 30 λεπτά, πλένονταν σε διάλυµα οξικού νατρίου (2x5 λεπτά) και στη συνέχεια αφυδατώνονταν σε σειρά διαλυµάτων αιθυλικής αλκοόλης αυξανόµενης πυκνότητας, από 25% έως 100% κ.ο. Ακολουθούσε εµβάπτιση των τοµών σε οξείδιο του προπυλενίου για 5 λεπτά και εµποτισµός τους σε µείγµα 1:1 οξειδίου του προπυλενίου και εποξειδικής ρητίνης για 12 ώρες. Οι τοµές µεταφέρονταν σε νέα ρητίνη 62
75 Υλικά και Μέθοδοι (2 αλλαγές), τοποθετούνταν σε αντικειµενοφόρους πλάκες και παρέµεναν σε κλίβανο θερµοκρασίας 60ºC για 48 ώρες. Στη συνέχεια, οι τοµές εξετάζονταν µε το οπτικό µικροσκόπιο και επιλέγονταν περιοχές του ραβδωτού σώµατος, που περιείχαν κύτταρα σεσηµασµένα µε τη µέθοδο TUNEL. Οι επιλεγµένες αυτές περιοχές κόβονταν κάτω από στερεοσκόπιο και επικολλούνταν στην κορυφή µικρών κυλίνδρων πολυµερισµένης ρητίνης. Στη συνέχεια, µε υπερµικροτόµο λαµβάνονταν ηµίλεπτες τοµές πάχους 1µm, οι οποίες χρωµατίζονταν µε κυανό της τολουιδίνης και εξετάζονταν στο οπτικό µικροσκόπιο µε σκοπό τον εντοπισµό TUNEL+ κυττάρων. Στη συνέχεια, κόβονταν υπέρλεπτες τοµές πάχους περίπου 90nm, οι οποίες µετά την επώασή τους σε υδατικό διάλυµα οξικού ουρανυλίου και κιτρικού µολύβδου εξετάζονταν µε ηλεκτρονικό µικροσκόπιο διέλευσης και φωτογραφίζονταν. 3. ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ Tα µορφολογικά χαρακτηριστικά των κυττάρων µε πυκνωτικό πυρήνα µελετήθηκαν εκτεταµένα µε το οπτικό και το ηλεκτρονικό µικροσκόπιο κατά την ανάπτυξη και µετά από βλάβες των κύριων προσαγωγών συνδέσεων του ραβδωτού σώµατος. ιαπιστώθηκε ότι τα κύτταρα αυτά παρουσίαζαν τα χαρακτηριστικά µορφολογικά γνωρίσµατα των αποπτωτικών κυττάρων. Επιπλέον, µε τη µέθοδο της ανοσοϊστοχηµείας, µελετήθηκε η έκφραση ειδικών αποπτωτικών δεικτών, όπως είναι η ενεργοποιηµένη κασπάση-3 και η φρακτίνη. Τέλος, µελετήθηκαν ανοσοϊστοχηµικά οι GABAεργικοί νευρώνες στο ραβδωτό σώµα ενήλικων επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο και ενήλικων επιµύων που είχαν υποστεί βλάβη µε 6-OHDA κατά τις πρώτες ηµέρες της 63
76 Υλικά και Μέθοδοι µεταγεννητικής ανάπτυξης καθώς και οι χολινεργικοί νευρώνες σε επίµυς που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο και εξετάστηκαν κατά την ενηλικίωση. 4. ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ Για την ποσοτική ανάλυση των αποπτωτικών κυττάρων έγινε καταµέτρηση των σεσηµασµένων µε τη µέθοδο TUNEL πυρήνων (TUNEL+) σε τοµές του ραβδωτού σώµατος που προέρχονταν από εγκεφάλους επιµύων κατά την ανάπτυξη και µετά από βλάβες των προσαγωγών συνδέσεών του. Επιλέχθηκαν 4 αντιπροσωπευτικές τοµές εντός των ορίων του πυρήνα από κάθε ζώο (οι τοµές απείχαν µεταξύ τους µm ανάλογα µε την ηλικία και το µέγεθος του εγκεφάλου). Για κάθε ηλικία χρησιµοποιήθηκαν 3 ζώα. Για τον προσδιορισµό και τη χαρτογράφηση των ορίων του ραβδωτού σώµατος χρησιµοποιήθηκαν τοµές χρωµατισµένες µε τη µέθοδο Nissl καθώς και άτλαντες του εγκεφάλου του αναπτυσσόµενου και του ενήλικου επίµυος (Paxinos και Watson, 1986 Paxinos και συν., 1991). Με ειδική συσκευή σχεδίασης (Camera lucida) προσαρτηµένης στο οπτικό µικροσκόπιο γίνονταν σχηµατογραφήµατα, µε τη χρήση του αντικειµενικού φακού Χ25, που απεικόνιζαν τα όρια του πυρήνα και την κατανοµή των σεσηµασµένων κυττάρων. Η πυκνότητα των αποπτωτικών κυττάρων υπολογίστηκε σε πεδία εµβαδού 0,1mm 2 και εκφράστηκε ως ο µέσος όρος του αριθµού των TUNEL+ κυττάρων ανά τοµή (TC/sect). Στις ίδιες τοµές και στο ίδιο εµβαδόν, η πυκνότητα των αποπτωτικών κυττάρων εκφράστηκε, επίσης, και ως ο µέσος όρος των λόγων των TUNEL+ κυττάρων ανά «ζωντανά κύτταρα» (TC/LC). Ως «ζωντανά» θεωρούνταν τα µη σεσηµασµένα κύτταρα, τα οποία εµφάνιζαν ευδιάκριτο πυρηνίσκο ή πυρήνα (TUNEL ). Για το σκοπό αυτό, σε σταθερό εµβαδόν τοµής 64
77 Υλικά και Μέθοδοι 0,1mm 2, έγινε καταµέτρηση των «ζωντανών» κυττάρων και προσδιορίστηκε µε αναγωγή η πυκνότητα των αποπτωτικών κυττάρων στο ραβδωτό σώµα που αντιστοιχεί σε «ζωντανά» κύτταρα του πυρήνα. Πραγµατοποιήθηκαν, επίσης, µετρήσεις του αριθµού των «ζωντανών» νευρώνων σε 12 τοµές από το ραβδωτό σώµα ενήλικων επιµύων της οµάδας Α και σε 12 τοµές από το ραβδωτό σώµα ενήλικων επιµύων της υποοµάδας Β1, σε σταθερό εµβαδόν τοµής µεγέθους 0,1 mm 2. Για την ποσοτική ανάλυση της πυκνότητας των GABAεργικών νευρώνων του ραβδωτού σώµατος καθώς και των σεσηµασµένων µε την ενεργοποιηµένη κασπάση-3 κυττάρων (κασπάση-3+) κατά την ανάπτυξη και µετά από βλάβες των προσαγωγών συστηµάτων ινών επιλέχθηκαν 4 τοµές εντός των ορίων του ραβδωτού σώµατος για κάθε ένα από τα 3 ζώα που χρησιµοποιήθηκαν από τις οµάδες Α, Β1 και Γ. Για την ποσοτική ανάλυση της πυκνότητας των χολινεργικών νευρώνων του ραβδωτού σώµατος επιλέχθηκαν, επίσης, 4 τοµές για κάθε ένα από τα 3 ζώα που χρησιµοποιήθηκαν από τις οµάδες Α και Β1. Η πυκνότητα των GABAεργικών και χολινεργικών νευρώνων υπολογίστηκε σε πεδία εµβαδού 0,1mm 2. Η πυκνότητα των κασπάση-3+ κυττάρων ήταν πολύ µικρή σε πεδία εµβαδού 0,1mm 2. Για το λόγο αυτό, έγινε αναγωγή των τιµών που υπολογίστηκαν σε πεδία εµβαδού 1mm 2. Όλες οι µετρήσεις πραγµατοποιήθηκαν στην ίδια περιοχή του ραβδωτού σώµατος (βλ. Αποτελέσµατα, Εικ. 2). 5. ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ Για τη στατιστική ανάλυση των αποτελεσµάτων χρησιµοποιήθηκαν παραµετρικές και µη παραµετρικές µέθοδοι. Η 65
78 Υλικά και Μέθοδοι οµοιογένεια των διακυµάνσεων εξετάστηκε µε τον έλεγχο του Levene. Για τον έλεγχο της κανονικότητας των διακυµάνσεων έγινε µετατροπή των αποτελεσµάτων σε log 10 και sqrt. Η µεθοδολογία της ανάλυσης των διακυµάνσεων µιας κατεύθυνσης (One-way ANOVA) και η µέθοδος Duncan χρησιµοποιήθηκαν για την εκτίµηση των διαφορών της πυκνότητας των ΤUNEL+ κυττάρων µεταξύ διαφορετικών ηλικιών, των διαφορών του αριθµού των «ζωντανών» νευρώνων µεταξύ των πειραµατοζώων µε βλάβες και των µαρτύρων καθώς και για τη σύγκριση της πυκνότητας των κασπάση-3+ κυττάρων µεταξύ διαφορετικών ηλικιών και µεταξύ πειραµατοζώων µε βλάβες και µαρτύρων αντίστοιχων ηλικιών. Η παραµετρική µέθοδος t-test χρησιµοποιήθηκε για τη σύγκριση των χολινεργικών κυττάρων µεταξύ των διαφορετικών οµάδων. Οι µη παραµετρικές µέθοδοι Kruskal-Wallis και Mann-Whitney U-test χρησιµοποιήθηκαν για την εκτίµηση των διαφορών της πυκνότητας των ΤUNEL+ κυττάρων µεταξύ των πειραµατοζώων µε βλάβες και των µαρτύρων αντίστοιχων ηλικιών, µεταξύ πειραµατοζώων µε διαφορετικές βλάβες σε αντίστοιχες ηλικίες και για τη σύγκριση της πυκνότητας των GABAεργικών κυττάρων µεταξύ των διαφορετικών οµάδων. Οι µέθοδοι αυτοί χρησιµοποιήθηκαν επίσης, για την εκτίµηση των διαφορών της πυκνότητας των TUNEL+ κυττάρων µεταξύ διαφορετικών ηλικιών και των κασπάση-3+ κυττάρων µεταξύ διαφορετικών ηλικιών και µεταξύ των πειραµατοζώων µε βλάβες και των µαρτύρων αντίστοιχων ηλικιών, σε περίπτωση µη οµοιογένειας. Όλες οι αναλύσεις των αποτελεσµάτων έγιναν µε το στατιστικό πρόγραµµα SPSS (v. 11.5) και όλοι οι έλεγχοι έγιναν σε επίπεδο πιθανότητας P 0,05. 66
79 Αποτελέσµατα ΑΠΟΤΕΛΕΣΜΑΤΑ ΣΧΟΛΙΑ ΓΙΑ ΤΙΣ ΤΕΧΝΙΚΕΣ ΚΑΙ ΤΟΥΣ ΠΕΙΡΑΜΑΤΙΣΜΟΥΣ Για τη µελέτη της απόπτωσης στο ραβδωτό σώµα χρησιµοποιήθηκε η µέθοδος TUNEL, µε την οποία επιτυγχάνεται η in situ σήµανση διαµεσονουκλεοσωµατικών θραυσµάτων στο DNA κυττάρων που βρίσκονται σε διαδικασία κυτταρικού θανάτου (Gavrieli και συν., 1992). Έχει διαπιστωθεί ότι µε τη µέθοδο αυτή µπορούν να σηµανθούν όχι µόνο τα αποπτωτικά αλλά και τα νεκρωτικά κύτταρα (Stadelmann και Lassmann, 2000). Η αδιαµφισβήτητη πιστοποίηση του αποπτωτικού τύπου κυτταρικού θανάτου επιτυγχάνεται µε τη χρήση της TUNEL σε συνδυασµό µε την ανάλυση των µορφολογικών χαρακτηριστικών των κυττάρων µε το οπτικό και το ηλεκτρονικό µικροσκόπιο καθώς και µε τη µελέτη των ειδικών γονιδίων που εκφράζονται κατά τον αποπτωτικό θάνατο (Ishimaru και συν., 1999 Stadelmann και Lassmann, 2000). Ανάµεσα στα προϊόντα έκφρασης των γονιδίων που σχετίζονται µε την απόπτωση, πρωταρχικό ρόλο παίζουν οι κασπάσες, οι οποίες θεωρούνται αξιόπιστοι αποπτωτικοί δείκτες, διότι ενεργοποιούνται κατά τον απoπτωτικό κυτταρικό θάνατο (Bratton και συν., 2000). Στην απόπτωση των νευρικών κυττάρων η κασπάση-3 θεωρείται ότι παίζει κεντρικό ρόλο, διότι όλες οι αποπτωτικοί οδοί συγκλίνουν στην ενεργοποίησή της (Zimmermann και συν., 2001). Η ενεργοποίηση της κασπάσης-3 κατά τα τελικά στάδια της απόπτωσης οδηγεί στο σχηµατισµό ενός πρωτεϊνικού µορίου, της φρακτίνης, η ανίχνευση της οποίας, επίσης, χρησιµοποιείται για την πιστοποίηση του αποπτωτικού κυτταρικού θανάτου (Suurmeijer και συν., 1999 Jackson- Lewis και συν., 2000 Oo και συν., 2002). 67
80 Αποτελέσµατα Τα TUNEL+ κύτταρα που παρατηρήθηκαν στο ραβδωτό σώµα κατά την ανάπτυξη και µετά από τις βλάβες προσαγωγών συνδέσεών του, παρουσίαζαν τα τυπικά µορφολογικά χαρακτηριστικά των αποπτωτικών κυττάρων, όπως αυτά έχουν περιγραφεί σε άλλες µελέτες (Kerr και συν., 1972 Wyllie και συν., 1980 Häcker, 2000 Stadelmann και Lassmann, 2000). Επιπλέον, η υπερµικροσκοπική µελέτη απέδειξε ότι τα κύτταρα που εµφάνιζαν πυκνωτικό πυρήνα στο οπτικό µικροσκόπιο ήταν αποπτωτικά (Janec και Burke, 1993 Μacaya και συν., 1994 Jackson- Lewis και συν., 2000 Marti και συν., 2002). Η ανοσοϊστοχηµική ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης επιβεβαίωσε ότι ο κυτταρικός θάνατος στο ραβδωτό σώµα κατά την ανάπτυξη και µετά από βλάβες των συνδέσεών του είναι αποπτωτικού τύπου (Jeon και συν., 1999 El-Khodor και Burke, 2000 Jackson-Lewis και συν., 2000 Oo και συν., 2002 Sophou και συν., 2006). Για τον λόγο αυτό, στην παρούσα µελέτη οι όροι TUNEL+ και αποπτωτικά κύτταρα χρησιµοποιούνται ως ταυτόσηµοι. Για τη µελέτη των επιπτώσεων της καταστροφής των κύριων προσαγωγών συστηµάτων στην επιβίωση των νευρώνων του ραβδωτού σώµατος πραγµατοποιήθηκαν δύο σειρές πειραµάτων. Στην πρώτη, πραγµατοποιήθηκε καταστροφή των φλοιοραβδωτών συστηµάτων ινών µε χειρουργική αφαίρεση του µεγαλύτερου τµήµατος του φλοιού του δεξιού εγκεφαλικού ηµισφαιρίου. Η βλάβη αυτή πραγµατοποιήθηκε σε επίµυς κατά τη γέννηση και σε συγκεκριµένα στάδια της µεταγεννητικής ανάπτυξης, στα οποία ολοκληρώνονται σηµαντικές αναπτυξιακές διεργασίες στο ραβδωτό σώµα, όπως είναι η διαφοροποίηση των νευρώνων του και η εδραίωση των συνδέσεών τους. Οι διεργασίες αυτές συνοδεύονται από προοδευτική αύξηση της συγκέντρωσης των νευροτροφικών παραγόντων στον φλοιό των εγκεφαλικών ηµισφαιρίων. Στη δεύτερη σειρά πειραµάτων, χορηγήθηκε ενδοπεριτοναϊκά η 68
81 Αποτελέσµατα νευροτοξίνη 6-OHDA σε επίµυς κατά τις 4 πρώτες ηµέρες της µεταγεννητικής ανάπτυξης. Με τον τρόπο αυτό προκλήθηκε καταστροφή όλων των κατεχολαµινεργικών συστηµάτων του εγκεφάλου, τα οποία περιλαµβάνουν και τις προσαγωγές κατεχολαµινεργικές ίνες του ραβδωτού σώµατος. Για να διαπιστωθεί εάν µε τη µέθοδο TUNEL εκτιµήθηκε σωστά το φαινόµενο της απόπτωσης έγινε καταµέτρηση των «ζωντανών» νευρώνων του ραβδωτού σώµατος σε ενήλικα πειραµατόζωα που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο και σύγκρισή τους µε τον αριθµό των «ζωντανών» νευρώνων σε µάρτυρες. Επίσης, µελετήθηκε ανοσοϊστοχηµικά η επίδραση των βλάβης του φλοιού των εγκεφαλικών ηµισφαιρίων και της βλάβης των κατεχολαµινεργικών συστηµάτων του ραβδωτού σώµατος στην επιβίωση και στον φαινότυπο των GABAεργικών νευρώνων, οι οποίοι συνιστούν την πλειονότητα των νευρώνων του ραβδωτού σώµατος. Τέλος, εκτιµήθηκαν οι επιπτώσεις της βλάβης του φλοιού των εγκεφαλικών ηµισφαιρίων στον αριθµό και στο φαινότυπο των χολινεργικών νευρώνων, οι οποίοι αποτελούν τον πολυπληθέστερο τύπο των διάµεσων νευρώνων του ραβδωτού σώµατος. Κατά τον έλεγχο της ειδικότητας της µεθόδου TUNEL, στις τοµές που χρησιµοποιήθηκαν ως θετικοί µάρτυρες (βλ. Υλικά και Μέθοδοι, κεφ. 2.3) όλα τα κύτταρα ήταν σεσηµασµένα, ενώ στις τοµές που χρησιµοποιήθηκαν ως αρνητικοί µάρτυρες (βλ. Υλικά και Μέθοδοι, κεφ. 2.3) δεν βρέθηκαν σεσηµασµένα κύτταρα. Κατά τον έλεγχο των ανοσοϊστοχηµικών αντιδράσεων που χρησιµοποιήθηκαν στην παρούσα µελέτη, στον οποίο παραλείφθηκε το συνδεδεµένο µε τη βιοτίνη αντίσωµα, δεν παρατηρήθηκε ανοσοδραστικότητα. 69
82 Αποτελέσµατα 1. ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ 1.1. ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ 1.1.Α. ΜΕΛΕΤΗ ΜΕ ΤΟ ΟΠΤΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Η ταυτοποίηση των TUNEL+ κυττάρων στο επίπεδο του οπτικού µικροσκοπίου έγινε µε βάση την έντονη και επιλεκτική σήµανση του πυρήνα τους. Πιο συγκεκριµένα, οι πυρήνες των αποπτωτικών κυττάρων ήταν πυκνωτικοί, καστανού χρώµατος και εµφάνιζαν µορφολογικά χαρακτηριστικά, τα οποία υποστηρίζεται ότι αντιστοιχούν σε διαφορετικά στάδια της απόπτωσης (Häcker, 2000). Ειδικότερα, το προϊόν σήµανσης σε αρχικά στάδια της απόπτωσης είχε δρεπανοειδή ή δακτυλιοειδή µορφή στην περιφέρεια του πυρήνα, ενώ σε µεταγενέστερα στάδια παρουσίαζε οµοιογενή διάχυτη κατανοµή στον πυρήνα. Σε τελικά στάδια της απόπτωσης, οι σεσηµασµένοι πυρήνες παρουσίαζαν δορυφορικές προεκβολές της χρωµατίνης ή εµφανίζονταν κατακερµατισµένοι σε δύο ή τρία θραύσµατα ενωµένα µεταξύ τους. Το κυτταρόπλασµα των αποπτωτικών κυττάρων είχε συνήθως τη µορφή λεπτότατου αντιχρωµατισµένου δακτυλίου που περιέβαλε τον πυρήνα. Σε πολλές περιπτώσεις τα σεσηµασµένα κύτταρα ήταν συρρικνωµένα και αποκολληµένα από το περιβάλλον νευροπίληµα. Τα αποπτωτικά κύτταρα ήταν διάσπαρτα ανάµεσα σε µεγάλο αριθµό µη σεσηµασµένων «ζωντανών» κυττάρων. Τα τελευταία ήταν αντιχρωµατισµένα µε τολουιδίνη (Εικόνα 1). Κατά τα τελικά στάδια της απόπτωσης, παρατηρήθηκαν, επίσης, αποπτωτικά σωµάτια, τα οποία συνιστούν κυτταρικά θραύσµατα που περιβάλλονται από κυτταρική µεµβράνη. Στις ηλικίες στις οποίες βρέθηκαν TUNEL+ κύτταρα, η κατανοµή τους ήταν οµοιογενής σε όλη την έκταση του ραβδωτού σώµατος (Εικόνα 2). 70
 και στο άλλο κύτταρο ο πυρήνας εµφανίζει δορυφορικές προεκβολές της χρωµατίνης (µαύρο βέλος).")
, Μ1 (Β), Μ3 (Γ), Μ5 ( ), Μ7 (Ε), και Μ10 (ΣΤ). Ράβδος=100µm.")
83 Αποτελέσµατα Α Β Εικόνα 1. Μικροφωτογραφίες στις οποίες φαίνονται TUNEL+ κύτταρα (βέλη) στο ραβδωτό σώµα του επίµυος τη Μ5. Στην Α ο πυρήνας του αποπτωτικού κυττάρου έχει οµοιογενή διάχυτο χρωµατισµό. Στη Β στο ένα κύτταρο το προϊόν σήµανσης έχει δακτυλιοειδή µορφή στην περιφέρεια του πυρήνα (λευκό βέλος) και στο άλλο κύτταρο ο πυρήνας εµφανίζει δορυφορικές προεκβολές της χρωµατίνης (µαύρο βέλος). Τα αποπτωτικά κύτταρα είναι διάσπαρτα ανάµεσα σε µεγάλο αριθµό «ζωντανών» αντιχρωµατισµένων κυττάρων. Ράβδος=20µm. Εικόνα 2. Μικροφωτογραφίες στις οποίες φαίνεται η τοπογραφική κατανοµή των TUNEL+ κυττάρων στο ραβδωτό σώµα του επίµυος στις ηλικίες Μ0 (Α), Μ1 (Β), Μ3 (Γ), Μ5 ( ), Μ7 (Ε), και Μ10 (ΣΤ). Ράβδος=100µm. Στο σχηµατογράφηµα Ζ της επιµήκους τοµής του εγκεφάλου επίµυος φαίνεται το εγκάρσιο επίπεδο από το οποίο συλλέχθηκαν οι τοµές του ραβδωτού σώµατος, οι οποίες απεικονίζονται στις µικροφωτογραφίες Α-ΣΤ. Στο σχηµατογράφηµα Η της εγκάρσιας τοµής του εγκεφάλου, η περιοχή που οριοθετείται από το διακεκοµµένο ορθογώνιο πλαίσιο απεικονίζει την περιοχή του ραβδωτού σώµατος στην οποία έγιναν οι µετρήσεις των TUNEL+ κυττάρων. Το γραµµοσκιασµένο τµήµα του ορθογωνίου αυτού απεικονίζει την περιοχή του ραβδωτού σώµατος από την οποία λήφθηκαν οι µικροφωτογραφίες Α-ΣΤ. 71
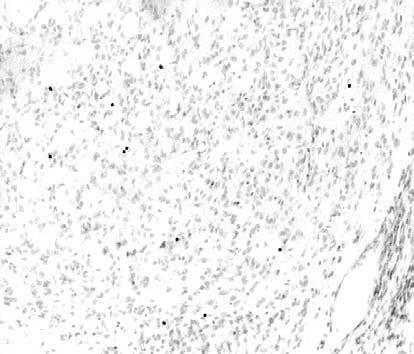
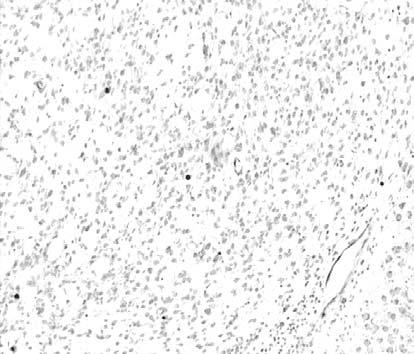
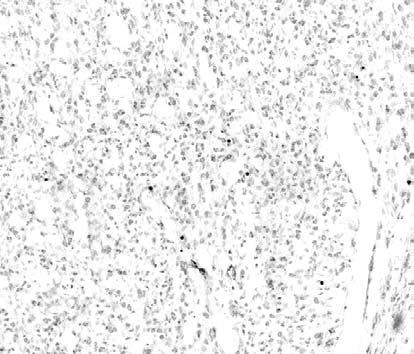
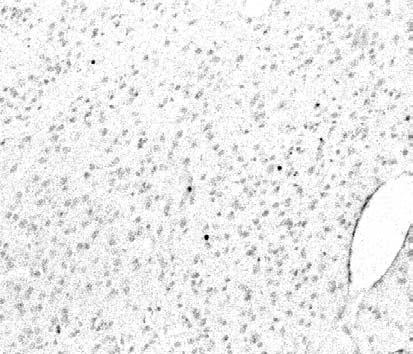
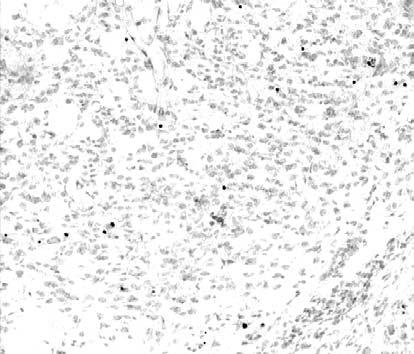
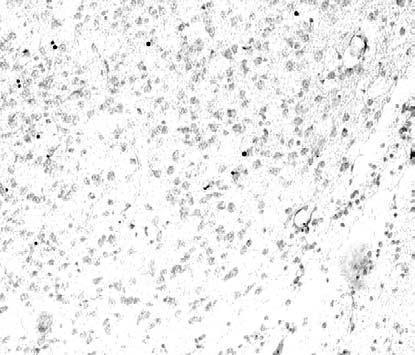
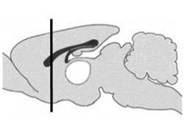
84 Αποτελέσµατα A Β Γ Ε ΣΤ Ζ Η 72
85 Αποτελέσµατα 1.1.B. ΜΕΛΕΤΗ ΜΕ ΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ H εξέταση µε το ηλεκτρονικό µικροσκόπιο πραγµατοποιήθηκε για να διερευνηθεί ο τύπος του κυτταρικού θανάτου σε κύτταρα του ραβδωτού σώµατος που ήταν σεσηµασµένα µε τη µέθοδο TUNEL. Για τη µελέτη αυτή η συλλογή του υλικού έγινε από επίµυς ηλικίας Μ7, διότι στα ζώα αυτής της ηλικίας εντοπίστηκε µεγάλος αριθµός ΤUNEL+ κυττάρων. Κατά την εξέταση του υλικού παρατηρήθηκαν κύτταρα µε συρρίκνωση του κυτταροπλάσµατος και πύκνωση του πυρήνα, ο οποίος συχνά εµφάνιζε οµοιογενή κατανοµή της χρωµατίνης. Τα µιτοχόνδρια εµφανίζονταν φυσιολογικά σε µέγεθος και η κυτταρική µεµβράνη διατηρούσε την ακεραιότητά της (Εικόνες 3Α, 3Β). Παρατηρήθηκαν, επίσης, αποπτωτικά σωµάτια, δηλαδή συµπυκνωµένες κυτταροπλασµατικές µάζες µικρού µεγέθους, οι οποίες περιβάλλονταν από άθικτη κυτταρική µεµβράνη. Σε µερικές περιπτώσεις, τα αποπτωτικά σωµάτια ήταν εγκολπωµένα από νευρογλοιακά κύτταρα. Τα παραπάνω γνωρίσµατα αποτελούν τυπικά χαρακτηριστικά αποπτωτικών κυττάρων (Häcker, 2000 Stadelmann και Lassmann, 2000). Σε περιπτώσεις, στις οποίες ήταν εφικτή η διάκριση µεταξύ νευρώνων και νευρογλοιακών κυττάρων διαπιστώθηκε ότι η πλειοψηφία των αποπτωτικών κυττάρων ήταν νευρώνες. Μορφολογικά χαρακτηριστικά της νέκρωσης, όπως είναι η διόγκωση των µιτοχονδρίων, η διακοπή της συνέχειας της κυτταρικής µεµβράνης ή η λύση του κυττάρου δεν παρατηρήθηκαν στο υλικό που εξετάστηκε (Κerr και Harmon, 1991 Kerr και συν., 1995). 73

.")

 βρίσκεται σε επαφή µε το αποπτωτικό κύτταρο.")
86 Αποτελέσµατα Ν Ν ΝΓ Α Β Γ Εικόνα 3. Μικροφωτογραφίες από ηλεκτρονικό µικροσκόπιο στις οποίες φαίνονται κύτταρα µε αποπτωτική µορφολογία στο ραβδωτό σώµα επίµυος τη Μ7 (Α, Β) και επίµυος που είχε υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 και θυσιάστηκε τη ΜΒ7 (Γ). Τα κύτταρα εµφανίζονται συρρικνωµένα και µερικώς αποκολληµένα από το νευροπίληµα. Οι πυρήνες είναι πυκνωτικοί και εµφανίζουν οµοιογενή κατανοµή της χρωµατίνης. Στην Α και Β διακρίνονται οι πυρήνες «ζωντανών» νευρώνων (Ν) δίπλα στα αποπτωτικά κύτταρα. Στη Β ένα νευρογλοιακό κύτταρο (ΝΓ) βρίσκεται σε επαφή µε το αποπτωτικό κύτταρο. Στη απεικονίζεται ένα αποπτωτικό σωµάτιο στο ραβδωτό σώµα επίµυος που είχε υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων και θυσιάστηκε τη Μ7. Ράβδος=1,15µm (Α, Β, Γ), 0,6µm ( ). 74
87 Αποτελέσµατα 1.2. ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ Aποπτωτικά κύτταρα στο ραβδωτό σώµα παρατηρήθηκαν κατά τις τέσσερις πρώτες µεταγεννητικές εβδοµάδες. TUNEL+ κύτταρα παρατηρήθηκαν για πρώτη φορά στο νεογέννητο και η πυκνότητά τους αυξήθηκε σηµαντικά τη Μ1. Ακολούθησε µείωση της πυκνότητάς τους τη Μ3, ενώ τη Μ5 η πυκνότητα των TUNEL+ κυττάρων αυξήθηκε σηµαντικά και παρουσίασε τη µέγιστη τιµή της. Από το τέλος της πρώτης µεταγεννητικής εβδοµάδας η πυκνότητα των αποπτωτικών κυττάρων παρουσίασε σηµαντική και συνεχή µείωση και µηδενίστηκε µέχρι το τέλος της τέταρτης µεταγεννητικής εβδοµάδας. Αποπτωτικά κύτταρα δεν παρατηρήθηκαν σε ενήλικα πειραµατόζωα. Η ποσοτική ανάλυση της πυκνότητας των αποπτωτικών κυττάρων στο ραβδωτό σώµα κατά την ανάπτυξη φαίνεται στον Πίνακα 3 και στο ιάγραµµα 1. 75
88 Αποτελέσµατα Πίνακας 3. Πυκνότητα των αποπτωτικών κυττάρων στο ραβδωτό σώµα επιµύων κατά την ανάπτυξη Ηλικία Πυκνότητα των TUNEL+ κυττάρων TC/sect TC/LC M0 4,75±0,25 α 5,03±0,23 α M1 6,00±0,43 β 9,67±0,31 β M3 5,00±0,21 α 8,26±0,40 β M5 9,75±0,80 γ 16,61±1,81 δ M7 6,41±0,39 β 12,70±1,11 γ M10 1,83±0,11 δ 4,55±0,35 α M14 1,00±0,00 δ,ε 2,74±0,19 α M21 0,50±0,15 ε 2,50±0,79 α M28 0,08±0,08 ε 0,37±0,37 ε Μ Μ: µεταγεννητική ηµέρα TC/sect: µέσος όρος του αριθµού των ΤUNEL+ κυττάρων ανά τοµή, ανά 0,1mm 2. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι της ίδιας στήλης µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). 76
89 Αποτελέσµατα ιάγραµµα 1 ΠYKNOTHTΑ ΤΩΝ TUNEL+ ΚΥΤΤΑΡΩΝ * TC/sect TC/LC M0 M1 M3 M5 M7 M10 M14 M21 M28 ΗΛΙΚΙΑ Η πυκνότητα των ΤUNEL+ κυττάρων στο ραβδωτό σώµα κατά την ανάπτυξη εµφανίζει µέγιστη τιµή τη Μ5. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. TC/sect: µέσος όρος του αριθµού των ΤUNEL+ κυττάρων ανά τοµή, ανά 0,1mm 2 (µαύρες στήλες), TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2 (λευκές στήλες). Η στατιστική ανάλυση του διαγράµµατος φαίνεται στον Πίνακα 3 (*Ρ 0,05). 77
90 Αποτελέσµατα 1.3. ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3 ΚΑΙ ΤΗΣ ΦΡΑΚΤΙΝΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ Η ανοσοϊστοχηµική µελέτη για την ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης κατά την ανάπτυξη έγινε τη Μ0, Μ1, Μ5 και Μ7, ηλικίες στις οποίες παρατηρήθηκε κυρίως το φαινόµενο της απόπτωσης µε τη µέθοδο TUNEL (βλ. Αποτελέσµατα, κεφ. 1.2). Η εξέταση των τοµών µε το οπτικό µικροσκόπιο έδειξε ότι το προϊόν της ανοσοαντίδρασης για την ενεργοποιηµένη κασπάση-3 εντοπίζονταν στον πυρήνα και στο κυτταρόπλασµα των κυττάρων. Το προϊόν της ανοσοαντίδρασης για την φρακτίνη εντοπίζονταν στο κυτταρόπλασµα των κυττάρων. Τα σεσηµασµένα κύτταρα εµφάνιζαν τα τυπικά µορφολογικά χαρακτηριστικά της απόπτωσης (Kerr και συν., 1972 Wyllie και συν., 1980 Häcker, 2000 Stadelmann και Lassmann, 2000). Τα κύτταρα αυτά ήταν συρρικνωµένα και είχαν πυκνωτικό πυρήνα (Εικόνα 4). Η ποσοτική ανάλυση έδειξε ότι η πυκνότητα των κασπάση-3+ κυττάρων ήταν µεγάλη τη Μ0 και αυξήθηκε σηµαντικά τη Μ1. Από τη Μ5 παρουσίασε σηµαντική µείωση. Η ποσοτική ανάλυση της πυκνότητας των κασπάση-3+ κυττάρων στο ραβδωτό σώµα κατά την ανάπτυξη φαίνεται αναλυτικά στον Πίνακα 4 και στο ιάγραµµα 2. Όπως προανάφερθηκε (βλ. Εισαγωγή, κεφ. Γ.2.1), ο σχηµατισµός της φρακτίνης πραγµατοποιείται κατά τα τελικά στάδια της απόπτωσης. Εποµένως, η πυκνότητα των σεσηµασµένων για τη φρακτίνη (φρακτίνη+) κυττάρων αντιπροσωπεύει ένα µικρό ποσοστό του συνόλου των αποπτωτικών κυττάρων, σε µια δεδοµένη χρονική στιγµή κατά το διάστηµα Μ0-Μ7. Πράγµατι, διαπιστώθηκε ότι η πυκνότητα των φρακτίνη+ κυττάρων ήταν µικρότερη από εκείνη των κασπάση-3+ 78


 και τη Μ1 (Β) και της")

91 Αποτελέσµατα κυττάρων σε πεδία αντίστοιχου εµβαδού. Για το λόγο αυτό, τα φρακτίνη+ κύτταρα µελετήθηκαν µόνο ποιοτικά. A B Γ Εικόνα 4. Ανοσοϊστοχηµική έκφραση της ενεργοποιηµένης κασπάσης-3 τη Μ0 (Α) και τη Μ1 (Β) και της φρακτίνης τη Μ0 (Γ) και τη Μ5 ( ) στο ραβδωτό σώµα του αναπτυσσόµενου επίµυος. Τα σεσηµασµένα κύτταρα βρίσκονται ανάµεσα σε αντιχρωµατισµένα «ζωντανά» κύτταρα. Ράβδος=20µm. 79
92 Αποτελέσµατα Πίνακας 4. Πυκνότητα των κασπάση-3+ κυττάρων στο ραβδωτό σώµα επιµύων κατά την ανάπτυξη Ηλικία M0 M1 M5 M7 Πυκνότητα των κασπάση-3+ κυττάρων 7,25±0,21 α 11,50±0,38 β 4,17±0,24 γ 0,67±0,59 δ Μ: µεταγεννητική ηµέρα Οι τιµές στη δεύτερη στήλη αντιπροσωπεύουν τον µέσο όρο του αριθµού των κασπάση-3+ κυττάρων, ανά τοµή, ανά 1mm 2 ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). ιάγραµµα 2 ΠYKNOTHTΑ ΤΩΝ ΚΑΣΠΑΣΗ-3+ ΚΥΤΤΑΡΩΝ * Μ0 Μ1 Μ5 Μ7 HΛΙΚΙΑ Η πυκνότητα των κασπάση-3+ κυττάρων στο ραβδωτό σώµα κατά την ανάπτυξη εµφανίζει µέγιστη τιµή τη Μ1. Οι τιµές αντιπροσωπεύουν τον µέσο όρο του αριθµού των κασπάση-3+ κυττάρων, ανά τοµή, ανά 1mm 2 ± τυπικό σφάλµα του µέσου. Η στατιστική ανάλυση του διαγράµµατος φαίνεται στον Πίνακα 4 (*Ρ 0,05). 80

93 Αποτελέσµατα 2. ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΕΠΙΜΥΩΝ ΠΟΥ ΕΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ 2.1. ΠΕΡΙΓΡΑΦΗ ΚΑΙ ΑΞΙΟΛΟΓΗΣΗ ΤΗΣ ΒΛΑΒΗΣ Η βλάβη πραγµατοποιήθηκε µε αναρρόφηση του µεγαλύτερου µέρους του φλοιού του δεξιού εγκεφαλικού ηµισφαιρίου και περιελάµβανε την αφαίρεση σηµαντικού τµήµατος του οπτικού, του σωµατοαισθητικού και του κινητικού φλοιού καθώς και τµηµάτων του ακουστικού και του προµετωπιαίου φλοιού. Αφαιρούνταν όλες οι στιβάδες του φλοιού, η λευκή ουσία και το µεγαλύτερο τµήµα του ιπποκάµπου (Σχήµα 2). Σχήµα 2. (Α) Σχηµατογράφηµα του εγκεφάλου επίµυος στο οποίο φαίνεται η έκταση της βλάβης στον φλοιό του δεξιού εγκεφαλικού ηµισφαιρίου (σκιασµένη περιοχή). (Β) Σχηµατογράφηµα µιας επιµήκους τοµής του εγκεφάλου, στο οποίο φαίνεται το επίπεδο της τοµής (µαύρη γραµµή) η οποία απεικονίζεται στη Γ. (Γ) Μικροφωτογραφία εγκάρσιας τοµής, χρωµατισµένης µε τη µέθοδο Nissl, στην οποία φαίνεται η έκταση και το βάθος της βλάβης. Η βλάβη έγινε τη Μ0 και το πειραµατόζωο θυσιάστηκε τη ΜΒ3. Η ράβδος (=600µm) αντιστοιχεί στο Γ. 81
94 Αποτελέσµατα Η ακριβής έκταση της βλάβης εκτιµήθηκε σε τοµές χρωµατισµένες µε τη µέθοδο Νissl. Οι τοµές εξετάζονταν προσεκτικά ώστε να επιλεγούν για ανάλυση µόνο οι εγκέφαλοι εκείνοι που είχαν βλάβες συγκρίσιµες ως προς την έκταση και το βάθος τους. Με τη βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων προκαλούνταν διατοµή των νευραξόνων των νευρώνων του φλοιού που προβάλλουν στο ραβδωτό σώµα, µε αποτέλεσµα την αποστέρηση των νευρώνων του ραβδωτού σώµατος από το µεγαλύτερο µέρος των προσαγωγών ινών τους και τους νευροτροφικούς παράγοντες που µεταφέρονται µέσω αυτών. Η βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων πραγµατοποιήθηκε σε 3 διαφορετικά στάδια της ανάπτυξης 2.2. ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ 2.2.Α. ΜΕΛΕΤΗ ΜΕ ΤΟ ΟΠΤΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Μετά από αφαίρεση του φλοιού του δεξιού εγκεφαλικού ηµισφαιρίου τη Μ0, τα TUNEL+ κύτταρα στο οµοπλάγιο ραβδωτό σώµα ταυτοποιήθηκαν από την έντονη και επιλεκτική σήµανση του πυρήνα τους (Εικόνα 5). Tα κύτταρα αυτά παρουσίαζαν οµοιογενή κατανοµή στο ραβδωτό σώµα και εµφάνιζαν τα τυπικά µορφολογικά χαρακτηριστικά των αποπτωτικών κυττάρων (Kerr και συν., 1972 Wyllie και συν., 1980 Häcker, 2000 Stadelmann και Lassmann, 2000). 2.2.Β. ΜΕΛΕΤΗ ΜΕ ΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Η εξέταση µε το ηλεκτρονικό µικροσκόπιο επιβεβαίωσε ότι στο ραβδωτό σώµα τα TUNEL+ κύτταρα παρουσίαζαν τα τυπικά µορφολογικά χαρακτηριστικά της απόπτωσης (Häcker, 2000 Stadelmann και Lassmann, 2000) (Εικόνα 3Γ). Στις περιπτώσεις που ήταν εφικτή η διάκριση, διαπιστώθηκε ότι η πλειονότητα των κυττάρων παρουσίαζε τα µορφολογικά χαρακτηριστικά νευρώνων. Η ανάλυση 82



, ΜΒ3 (Β),")
95 Αποτελέσµατα έγινε στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 και θυσιάστηκαν τη ΜΒ7, στάδιο κατά το οποίο παρατηρείται η µεγαλύτερη πυκνότητα των ΤUNEL+ κυττάρων. Κύτταρα µε νεκρωτική µορφολογία δεν παρατηρήθηκαν στο υλικό που εξετάστηκε (Κerr και Harmon, 1991 Kerr και συν., 1995). Α Β Γ Εικόνα 5. Μικροφωτογραφίες στις οποίες φαίνεται η τοπογραφική κατανοµή των TUNEL+ κυττάρων στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 και θανατώθηκαν στις ηλικίες ΜΒ1 (Α), ΜΒ3 (Β), ΜΒ7 (Γ) και ΜΒ14 ( ). Ράβδος=100µm. 83
96 Αποτελέσµατα 2.3. ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ MΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΕ ΕΠΙΜΥΣ ΠΟΥ ΕΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΣΤΟ ΝΕΟΓΕΝΝΗΤΟ Η µελέτη της απόπτωσης στο ραβδωτό σώµα σε πειραµατόζωα που είχαν υποστεί βλάβη των εγκεφαλικών ηµισφαιρίων τη Μ0 έγινε σε διάφορα στάδια της ανάπτυξης και στο ενήλικο. Τη ΜΒ1 παρατηρήθηκαν αποπτωτικά κύτταρα, η πυκνότητα των οποίων αυξήθηκε σηµαντικά τη ΜΒ3 και παρουσίασε µέγιστο τη ΜΒ7. Κατά τη δεύτερη και τρίτη µεταγεννητική εβδοµάδα η πυκνότητα των αποπτωτικών κυττάρων παρουσίασε συνεχή και σηµαντική µείωση. εν παρατηρήθηκαν αποπτωτικά κύτταρα στα ενήλικα πειραµατόζωα. Η ποσοτική ανάλυση της πυκνότητας των TUNEL+ κυττάρων στους επίµυς που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο φαίνεται στον Πίνακα 5. Πίνακας 5. Πυκνότητα των αποπτωτικών κυττάρων στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητo Ηλικία Πυκνότητα των TUNEL+ κυττάρων TC/sect TC/LC ΜΒ1 4,91±0,31 α 10,66±0,63 α ΜΒ3 6,50±0,35 β 13,80±0,92 β ΜΒ7 8,50±0,54 γ 24,42±1,31 γ ΜΒ14 1,83±0,16 δ 6,38±0,61 δ ΜΒ21 1,08±0,14 ε 4,09±0,57 ε ΜΒ ΜΒ: ηµέρα µετά τη βλάβη TC/sect: µέσος όρος του αριθµού των ΤUNEL+ κυττάρων ανά τοµή, ανά 0,1mm 2. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι της ίδιας στήλης µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). 84
97 Αποτελέσµατα Όπως φαίνεται στο ιάγραµµα 3, η σύγκριση των τιµών σε πειραµατόζωα που είχαν υποστεί βλάβη µε εκείνες των µαρτύρων απέδειξε ότι η βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο προκαλεί αύξηση της πυκνότητας των αποπτωτικών κυττάρων στο ραβδωτό σώµα σε όλες τις ηλικίες που εξετάστηκαν. Η αύξηση αυτή είναι στατιστικά σηµαντική τη ΜΒ3, ΜΒ7 και ΜΒ14. ιάγραµµα 3 ΠYKNOTHTA TΩΝ TUNEL+ ΚΥΤΤΑΡΩΝ (TC/LC) Μάρτυρες * Βλάβη του φλοιού τη Μ0 * * M1/MB1 M3/MB3 M7/MB7 M14/MB14 M21/MB21 ΗΛΙΚΙΑ Σύγκριση της πυκνότητας των ΤUNEL+ κυττάρων µεταξύ των φυσιολογικών επιµύων (λευκές στήλες) και επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 (µαύρες στήλες) και εξετάστηκαν σε διάφορες ηλικίες µετά τη βλάβη. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2. Τη ΜΒ3, ΜΒ7 και ΜΒ14 η αύξηση είναι στατιστικά σηµαντική (*Ρ 0,05). 85
98 Αποτελέσµατα MΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΕ ΕΠΙΜΥΣ ΠΟΥ ΕΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΣΕ ΜΕΤΑΓΕΝΕΣΤΕΡΑ ΣΤΑ ΙΑ ΤΗΣ ΑΝΑΠΤΥΞΗΣ Σε πειραµατόζωα που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ7 παρατηρήθηκαν TUNEL+ κύτταρα τη ΜΒ1 και η πυκνότητά τους παρουσίασε σηµαντική µείωση τη ΜΒ7 και τη ΜΒ14. Αποπτωτικά κύτταρα δεν εντοπίστηκαν στα ενήλικα πειραµατόζωα (Πίνακας 6). Πίνακας 6. Πυκνότητα των αποπτωτικών κυττάρων στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων σε µεταγενέστερα στάδια της ανάπτυξης Ηλικία Πυκνότητα των TUNEL+ κυττάρων (TC/LC) Βλάβη του φλοιού τη Μ7 Βλάβη του φλοιού τη Μ14 ΜΒ1 15,11±1,22 α 3,39±0,71 α ΜΒ7 5,51±0,71 β 3,31±0,71 α ΜΒ14 4,72±1,02 β 0,96±0,65 β ΜΒ ΜΒ: ηµέρα µετά τη βλάβη TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2.Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). Η σύγκριση της πυκνότητας των αποπτωτικών κυττάρων στα παραπάνω πειραµατόζωα µε την πυκνότητά τους στους µάρτυρες καταδεικνύει ότι δεν υπάρχουν στατιστικά σηµαντικές διαφορές µεταξύ τους στις αντίστοιχες ηλικίες ( ιάγραµµα 4). 86
99 Αποτελέσµατα ιάγραµµα 4 ΠYKNOTHTA ΤΩΝ TUNEL+ ΚΥΤΤΑΡΩΝ (TC/LC) Mάρτυρες Βλάβη του φλοιού τη Μ7 M7/MB1 M14/MB7 M21/MB14 ΗΛΙΚΙΑ Σύγκριση της πυκνότητας των ΤUNEL+ κυττάρων µεταξύ των φυσιολογικών επιµύων (λευκές στήλες) και επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ7 (µαύρες στήλες) και εξετάστηκαν σε διάφορες ηλικίες µετά τη βλάβη. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2. εν παρατηρείται στατιστικά σηµαντική διαφορά µεταξύ των δύο οµάδων (Ρ>0,05). Σε πειραµατόζωα που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ14 παρατηρήθηκε µικρός αριθµός αποπτωτικών κυττάρων τη ΜΒ1, ο οποίος ελαττώθηκε και σχεδόν µηδενίστηκε µέχρι τη ΜΒ14 (Πίνακας 6). ΤUNEL+ κύτταρα δεν παρατηρήθηκαν σε ενήλικους επίµυς. Η σύγκριση της πυκνότητας των αποπτωτικών κυττάρων στα παραπάνω πειραµατόζωα µε την πυκνότητά τους στους µάρτυρες έδειξε ότι δεν υπάρχουν στατιστικά σηµαντικές διαφορές µεταξύ τους στις αντίστοιχες ηλικίες ( ιάγραµµα 5). 87
100 Αποτελέσµατα ιάγραµµα 5 ΠYKNOTHΤΑ ΤΩΝ TUNEL+ ΚΥΤΤΑΡΩΝ (TC/LC) Μάρτυρες Βλάβη του φλοιού τη Μ14 M14/MB1 M21/MB7 M28/MB14 HΛΙΚΙΑ Σύγκριση της πυκνότητας των ΤUNEL+ κυττάρων µεταξύ των φυσιολογικών επιµύων (λευκές στήλες) και επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ14 (µαύρες στήλες) και εξετάστηκαν σε διάφορες ηλικίες µετά τη βλάβη. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2. εν παρατηρείται στατιστικά σηµαντική διαφορά µεταξύ των δυο οµάδων (Ρ>0,05) ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3 ΚΑΙ ΤΗΣ ΦΡΑΚΤΙΝΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΜΕΤΑ ΑΠΟ ΒΛΑΒΗ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Η ανοσοϊστοχηµική µελέτη για την ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης µετά από βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων έγινε σε πειραµατόζωα, τα οποία είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 και θυσιάστηκαν στις ηλικίες ΜΒ0, ΜΒ1, ΜΒ5 και ΜΒ7. Την περίοδο αυτή παρατηρείται, όπως προαναφέρθηκε (βλ. Αποτελέσµατα, κεφ ), αύξηση των TUNEL+ κυττάρων στο ραβδωτό σώµα ως συνέπεια της βλάβης αυτής. Το προϊόν της ανοσοαντίδρασης για την ενεργοποιηµένη 88


. Α Β Γ Εικόνα 6.")
 στο ραβδωτό σώµα επίµυος που είχε υποστεί βλάβη του φλοιού των εγκεφαλικών")
101 Αποτελέσµατα κασπάση-3 εντοπίζονταν στον πυρήνα και στο κυτταρόπλασµα των κυττάρων. Το προϊόν της ανοσοαντίδρασης για τη φρακτίνη εντοπίζονταν στο κυτταρόπλασµα των κυττάρων. Ο πυρήνας των σεσηµασµένων κυττάρων ήταν πυκνωτικός και συρρικνωµένος, ενώ το κυτταρόπλασµα είχε τη µορφή λεπτότατου δακτυλίου που περιέβαλλε τον πυρήνα (Εικόνα 6). Α Β Γ Εικόνα 6. Ανοσοϊστοχηµική έκφραση της ενεργοποιηµένης κασπάσης-3 (Α, Β) και της φρακτίνης (Γ, ) στο ραβδωτό σώµα επίµυος που είχε υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 και θυσιάστηκε τη ΜΒ1 (Α, Γ) και τη ΜΒ5 (Β, ). Τα σεσηµασµένα κύτταρα βρίσκονται ανάµεσα σε αντιχρωµατισµένα «ζωντανά» κύτταρα. Ράβδος=20µm. 89
102 Αποτελέσµατα Η ποσοτική ανάλυση των κασπάση-3+ κυττάρων έδειξε µεγάλο αριθµό σεσηµασµένων κυττάρων τη ΜΒ0. Η πυκνότητά τους αυξήθηκε σηµαντικά τη ΜΒ1 και παρουσίασε σταδιακή µείωση τη ΜΒ5 και τη ΜΒ7 (Πίνακας 7). Πίνακας 7. Πυκνότητα των κασπάση-3+ κυττάρων στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητo Ηλικία MΒ0 MΒ1 MΒ5 MΒ7 Πυκνότητα των κασπάση-3+ κυττάρων 10,33±0,52 α 26,25±1,11 β 11,58±0,38 α 2,08±0,19 γ ΜΒ: ηµέρα µετά τη βλάβη Οι τιµές στη δεύτερη στήλη αντιπροσωπεύουν τον µέσο όρο του αριθµού των κασπάση-3+ κυττάρων, ανά τοµή, ανά 1mm 2 ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). Όπως φαίνεται στο ιάγραµµα 6, η σύγκριση των τιµών σε πειραµατόζωα που είχαν υποστεί βλάβη µε εκείνες των µαρτύρων απέδειξε ότι η βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο προκαλεί στατιστικά σηµαντική αύξηση της πυκνότητας των κασπάση-3+ κυττάρων στο ραβδωτό σώµα σε όλες τις ηλικίες που εξετάστηκαν. 90
103 Αποτελέσµατα ιάγραµµα 6 ΠYKNOTHTA ΤΩΝ ΚΑΣΠΑΣΗ-3+ ΚΥΤΤΑΡΩΝ * Μάρτυρες Βλάβη του φλοιού τη Μ0 * * * Μ0/ΜΒ0 Μ1/ΜΒ1 Μ5/ΜΒ5 Μ7/ΜΒ7 ΗΛΙΚΙΑ Σύγκριση της πυκνότητας των κασπάση-3+ κυττάρων µεταξύ των φυσιολογικών επιµύων (γαλάζιες στήλες) και επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 (µαύρες στήλες) και εξετάστηκαν σε διάφορες ηλικίες µετά τη βλάβη. Οι τιµές αντιπροσωπεύουν τους µέσους όρους του αριθµού των κυττάρων, ανά τοµή, ανά 1mm 2 ± τυπικό σφάλµα του µέσου. Η αύξηση είναι στατιστικά σηµαντική σε όλες τις ηλικίες που εξετάστηκαν (*Ρ 0,05) YΠΟΛΟΓΙΣΜΟΣ ΤΟΥ ΑΡΙΘΜΟΥ ΤΩΝ «ΖΩΝΤΑΝΩΝ» ΝΕΥΡΩΝΩΝ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ Λαµβάνοντας υπ όψιν ότι η απόπτωση είναι φαινόµενο που εξελίσσεται σε διάστηµα λίγων ωρών (Thomaidou και συν., 1997) και για να επιβεβαιώσουµε ότι µε τη µέθοδο TUNEL εκτιµήσαµε σωστά το φαινόµενο της απόπτωσης, πραγµατοποιήσαµε µετρήσεις του αριθµού των «ζωντανών» νευρώνων (LC) στο ραβδωτό σώµα σε ενήλικους επίµυς που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0. Ως µάρτυρες χρησιµοποιήθηκαν φυσιολογικοί ενήλικοι επίµυες. Η στατιστική ανάλυση των αποτελεσµάτων απέδειξε ότι η πυκνότητα των «ζωντανών» νευρώνων στα πειραµατόζωα που 91
104 Αποτελέσµατα είχαν υποστεί τη βλάβη ήταν µικρότερη από εκείνη των µαρτύρων και η διαφορά αυτή ήταν στατιστικά σηµαντική (Πίνακας 8). Πίνακας 8. Πυκνότητα των «ζωντανών» νευρώνων στο ραβδωτό σώµα ενήλικων φυσιολογικών επιµύων και επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0. Πειραµατική οµάδα Μάρτυρες Βλάβη τη Μ0 LC 203,08±2,78 α 152,83±2,72 β Μ: µεταγεννητική µέρα LC: µέσος όρος των «ζωντανών» νευρώνων ανά τοµή, ανά 0,1mm 2. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). 92
105 Αποτελέσµατα 3. ΜΕΛΕΤΗ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΕΠΙΜΥΩΝ ΠΟΥ ΕΙΧΑΝ ΥΠΟΣΤΕΙ ΒΛΑΒΗ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΠΡΟΣΑΓΩΓΩΝ ΣΥΣΤΗΜΑΤΩΝ ΙΝΩΝ ΜΕ ΤΗ ΝΕΥΡΟΤΟΞΙΝΗ 6-OHDA 3.1. ΠΕΡΙΓΡΑΦΗ ΚΑΙ ΑΞΙΟΛΟΓΗΣΗ ΤΗΣ ΒΛΑΒΗΣ H πρόκληση της βλάβης πραγµατοποιήθηκε µε ενδοπεριτοναϊκές εγχύσεις της νευροτοξίνης 6-OHDA σε επίµυς κατά τις 4 πρώτες ηµέρες της µεταγεννητικής ανάπτυξης. Σύµφωνα µε αποτελέσµατα προηγούµενων µελετών, µε τον τρόπο αυτό προκαλείται καταστροφή των κατεχολαµινεργικών συστηµάτων του εγκεφάλου (Jonsson και Sachs, 1976 Jonsson και Ponzio, 1978 Blue και Parnavelas, 1982). Τα συστήµατα αυτά (βλ. Εισαγωγή, κεφ. Β.2.1.β 2 ) περιλαµβάνουν τις προσαγωγές κατεχολαµινεργικές ίνες του ραβδωτού σώµατος και συγκεκριµένα τις ντοπαµινεργικές ίνες, που προέρχονται από τη µέλαινα ουσία, το κοιλιακό καλυπτρικό πεδίο και την οπισθοερυθρά περιοχή και τις νοραδρενεργικές ίνες, που προέρχονται από τον υποµέλανα τόπο. Η ολοκληρωτική καταστροφή των κατεχολαµινεργικών συστηµάτων του εγκεφάλου µετά τη χορήγηση της 6-OHDA επιβεβαιώθηκε στην παρούσα µελέτη µε τη χρήση της µεθόδου της ανοσοϊστοχηµείας µε αντισώµατα εναντίον της ντοπαµίνης και της νοραδρεναλίνης. Οι εγκέφαλοι των επιµύων που είχαν υποστεί βλάβες µε 6-OHDA εξετάστηκαν 12 ώρες µετά την τελευταία έγχυση της 6-ΟHDA, τη Μ3. Στην ηλικία αυτή οι παραπάνω εγκέφαλοι δεν εµφάνιζαν ανοσοδραστικότητα για τη ντοπαµίνη και τη νοραδρεναλίνη, σε αντίθεση µε τους µάρτυρες, στους οποίους τα αποτελέσµατα της ανοσοϊστοχηµικής ανάλυσης ήταν σύµφωνα µε προηγούµενες µελέτες των κατεχολαµινεργικών συστηµάτων του εγκεφάλου (Antonopoulos και συν., 2002 Latsari και συν., 2002, 2004). 93


 και ότι είχαν οµοιογενή κατανοµή στο")

106 Αποτελέσµατα 3.2. ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ 3.2.A. ΜΕΛΕΤΗ ΜΕ ΤΟ ΟΠΤΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Μετά τη βλάβη των κατεχολαµινεργικών συστηµάτων µε 6-OHDA τα TUNEL+ κύτταρα ταυτοποιήθηκαν από την έντονη και επιλεκτική σήµανση του πυρήνα τους. ιαπιστώθηκε ότι τα TUNEL+ κύτταρα παρουσίαζαν τα τυπικά µορφολογικά χαρακτηριστικά των αποπτωτικών κυττάρων (Kerr και συν., 1972 Wyllie και συν., 1980 Häcker, 2000 Stadelmann και Lassmann, 2000) και ότι είχαν οµοιογενή κατανοµή στο ραβδωτό σώµα (Εικόνα 7). Α Β Γ Εικόνα 7. Μικροφωτογραφίες στις οποίες φαίνεται η τοπογραφική κατανοµή των TUNEL+ κυττάρων στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων και θυσιάστηκαν στις ηλικίες Μ3 (A), Μ7 (B, Γ) και Μ21 ( ). Ράβδος=100µm. 94
107 Αποτελέσµατα 3.2.B. ΜΕΛΕΤΗ ΜΕ ΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ Για τη µελέτη υλικού µε το ηλεκτρονικό µικροσκόπιο χρησιµοποιήθηκαν οι εγκέφαλοι επιµύων που είχαν υποστεί βλάβη των κατεχολαµινεργικών συστηµάτων µε 6-OHDA και θανατώθηκαν τη Μ7, ηλικία στην οποία παρατηρήθηκε ο µεγαλύτερος αριθµός TUNEL+ κυττάρων. Η µελέτη σε υπερµικροσκοπικό επίπεδο επιβεβαίωσε ότι τα TUNEL+ κύτταρα στο ραβδωτό σώµα παρουσίαζαν τα τυπικά µορφολογικά χαρακτηριστικά της απόπτωσης, όπως είναι η συρρίκνωση του κυτταροπλάσµατος και η πύκνωση του πυρήνα (Häcker, 2000 Stadelmann και Lassmann, 2000). Παρατηρήθηκε, επίσης, µεγάλος αριθµός αποπτωτικών σωµατίων (Εικόνα 3 ). Κύτταρα µε µορφολογικά χαρακτηριστικά της νέκρωσης δεν βρέθηκαν (Κerr και Harmon, 1991 Kerr και συν., 1995) ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ Για τη µελέτη της απόπτωσης στα πειραµατόζωα που είχαν υποστεί βλάβη µε 6-OHDA πραγµατοποιήθηκε ποσοτική ανάλυση σε τρία στάδια της ανάπτυξης, τη Μ3 (12 ώρες µετά την τελευταία έγχυση της 6-OHDA), τη Μ7 και τη Μ21. Τη Μ3 παρατηρήθηκαν πολλά TUNEL+ κύτταρα στο ραβδωτό σώµα, η πυκνότητά τους παρουσίασε µέγιστο τη Μ7 και µειώθηκε σηµαντικά µέχρι το τέλος της τρίτης µεταγεννητικής εβδοµάδας. ΤUNEL+ κύτταρα δεν παρατηρήθηκαν σε ενήλικα πειραµατόζωα. Η ποσοτική ανάλυση της πυκνότητας των αποπτωτικών κυττάρων στο ραβδωτό σώµα των πειραµατόζωων που είχαν υποστεί βλάβη των κατεχολαµινεργικών συστηµάτων φαίνεται στον Πίνακα 9. 95
108 Αποτελέσµατα Πίνακας 9. Πυκνότητα των αποπτωτικών κυττάρων στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων Ηλικία Πυκνότητα των TUNEL+ κυττάρων TC/sect TC/LC Μ3 6,41±0,19 α 14,73±0,45 α Μ7 9,91±0,41 β 25,58±1,52 β Μ21 1,25±0,13 γ 5,56±0,57 γ M Μ: µεταγεννητική ηµέρα TC/sect: µέσος όρος του αριθµού των ΤUNEL+ κυττάρων ανά τοµή, ανά 0,1mm 2. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2.Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι της ίδιας στήλης µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). Όπως φαίνεται στο ιάγραµµα 7, η βλάβη των κατεχολαµινεργικών συστηµάτων µε 6-OHDA προκαλεί σηµαντική αύξηση της πυκνότητας των αποπτωτικών κυττάρων στο ραβδωτό σώµα σε σύγκριση µε τους µάρτυρες, σε όλες τις αντίστοιχες ηλικίες που εξετάστηκαν. Η συγκριτική µελέτη των πυκνοτήτων των αποπτωτικών κυττάρων στο ραβδωτό σώµα πειραµατοζώων που είχαν υποστεί βλάβη µε 6-OHDA και πειραµατοζώων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων σε αντίστοιχες ηλικίες έδειξε ότι δεν υπάρχει στατιστικά σηµαντική διαφορά µεταξύ τους. Η πυκνότητα των αποπτωτικών κυττάρων στο ραβδωτό σώµα εµφανίζει µέγιστο τη Μ7 και στις δύο οµάδες πειραµατοζώων ( ιάγραµµα 8). 96
109 Αποτελέσµατα ιάγραµµα 7 ΠYKNOTHTA ΤΩΝ TUNEL+ ΚΥΤΤΑΡΩΝ (TC/LC) * Μάρτυρες Βλάβη µε 6-OHDA M3 M7 M21 ΗΛΙΚΙΑ Σύγκριση της πυκνότητας των ΤUNEL+ κυττάρων µεταξύ των φυσιολογικών επιµύων (λευκές στήλες) και επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων (γκρι στήλες). Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2. Η αύξηση είναι στατιστικά σηµαντική σε όλες τις ηλικίες που εξετάστηκαν (*Ρ 0,05). * * ιάγραµµα 8 ΠYKNOTHTA TΩΝ TUNEL+ ΚΥΤΤΑΡΩΝ (TC/LC) Βλάβη του φλοιού τη Μ0 Βλάβη µε 6-OHDA MB3/M3 MB7/M7 MB21/M21 ΗΛΙΚΙΑ Σύγκριση της πυκνότητας των ΤUNEL+ κυττάρων µεταξύ των επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 (µαύρες στήλες) και επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων (γκρι στήλες). Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. TC/LC: µέσος όρος των λόγων των ΤUNEL+ κυττάρων ανά «ζωντανά» κύτταρα, ανά τοµή, ανά 0,1mm 2. εν παρατηρείται στατιστικά σηµαντική διαφορά µεταξύ των δυο οµάδων (Ρ>0,05). 97
110 Αποτελέσµατα 3.4. ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΜΕΝΗΣ ΚΑΣΠΑΣΗΣ-3 ΚΑΙ ΤΗΣ ΦΡΑΚΤΙΝΗΣ ΣΤΟ ΡΑΒ ΩΤΟ ΣΩΜΑ ΜΕΤΑ ΑΠΟ ΒΛΑΒΗ ΤΩΝ ΚΑΤΕΧΟΛΑΜΙΝΕΡΓΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΜΕ 6-OHDA Η ανοσοϊστοχηµική µελέτη για την ανίχνευση της ενεργοποιηµένης κασπάσης-3 και της φρακτίνης µετά από βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων έγινε στις ηλικίες Μ3 και Μ7, στις οποίες παρατηρήθηκε η µεγαλύτερη πυκνότητα των TUNEL+ κυττάρων (βλ. Αποτελέσµατα, κεφ. 3.3). Στα κύτταρα που παρουσίαζαν ανοσοδραστικότητα για το αντίσωµα εναντίον της ενεργοποιηµένης κασπάσης-3 το προϊόν σήµανσης ήταν εντοπισµένο στον πυρήνα και στο κυτταρόπλασµα. Στα κύτταρα που ήταν σεσηµασµένα µε το αντίσωµα εναντίον της φρακτίνης, το προϊόν σήµανσης εντοπίζονταν στο κυτταρόπλασµα. Όλα τα σεσηµασµένα κύτταρα παρουσίαζαν τυπικά µορφολογικά χαρακτηριστικά της απόπτωσης (Kerr και συν., 1972 Wyllie και συν., 1980 Häcker, 2000 Stadelmann και Lassmann, 2000) (Εικόνα 8). Κασπάση-3+ κύτταρα εντοπίστηκαν στο ραβδωτό σώµα και στις δυο ηλικίες που εξετάστηκαν, µε µεγαλύτερη πυκνότητα τη Μ3 (Πίνακας 10). Πίνακας 10. Πυκνότητα των κασπάση-3+ κυττάρων στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων Ηλικία M3 M7 Πυκνότητα των κασπάση-3+ κυττάρων 18,00±0,53 α 15,25±0,39 β Μ: µεταγεννητική ηµέρα Οι τιµές στη δεύτερη στήλη αντιπροσωπεύουν τον µέσο όρο του αριθµού των κασπάση-3+ κυττάρων, ανά τοµή, ανά 1mm 2 ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). 98




111 Αποτελέσµατα Α Β Γ Εικόνα 8. Ανοσοϊστοχηµική έκφραση της ενεργοποιηµένης κασπάσης-3 (Α, Β) και της φρακτίνης (Γ, ) στο ραβδωτό σώµα επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων και θυσιάστηκαν στις ηλικίες Μ3 (Α, Γ) και Μ7 (Β, ). Τα σεσηµασµένα κύτταρα βρίσκονται ανάµεσα σε αντιχρωµατισµένα «ζωντανά» κύτταρα. Ράβδος=20µm. 99
112 Αποτελέσµατα 4. ΜΕΛΕΤΗ ΤΗΣ ΕΠΙ ΡΑΣΗΣ ΤΩΝ ΒΛΑΒΩΝ ΤΩΝ ΠΡΟΣΑΓΩΓΩΝ ΣΥΝ ΕΣΕΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ ΣΤΗΝ ΕΠΙΒΙΩΣΗ ΚΑΙ ΣΤΟΝ ΦΑΙΝΟΤΥΠΟ ΤΩΝ GABAΕΡΓΙΚΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ 4.1. ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ Οι GABAεργικοί νευρώνες συνιστούν τη συντριπτική πλειοψηφία των νευρώνων του ραβδωτού σώµατος, καθώς το GABA χρησιµοποιείται ως χηµικός διαβιβαστής από τους MSN και από τους µεσοµεγέθεις µη ακανθωτούς διάµεσους νευρώνες του πυρήνα αυτού. Στους GABAεργικούς αυτούς νευρώνες απολήγουν γλουταµινεργικές ίνες από τον φλοιό των εγκεφαλικών ηµισφαιρίων και κατεχολαµινεργικές ίνες από περιοχές του µέσου εγκεφάλου (βλ. Εισαγωγή, κεφ. Β.2.1.α, Β.2.1.β 2 ). Με βάση τα παραπάνω και τα αποτελέσµατά µας µε τη µέθοδο ΤUNEL, τα οποία καταδεικνύουν ότι οι βλάβες του φλοιού των εγκεφαλικών ηµισφαιρίων και των κατεχολαµινεργικών συστηµάτων µε 6-OHDA προκαλούν σηµαντική αύξηση της πυκνότητας των αποπτωτικών κυττάρων στο ραβδωτό σώµα, διερευνήσαµε την επίδραση των βλαβών αυτών στην επιβίωση και στον φαινότυπο των GABAεργικών νευρώνων του ραβδωτού σώµατος. Η µελέτη πραγµατοποιήθηκε σε επίµυς που υπέστησαν βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο και σε επίµυς, στους οποίους προκλήθηκε καταστροφή των κατεχολαµινεργικών συστηµάτων µε 6-OHDA και εξετάστηκαν κατά την ενηλικίωσή τους (τη Μ60). Τα αποτελέσµατα της ανοσοϊστοχηµικής µελέτης έδειξαν ότι οι ανοσοδραστικοί για το αντίσωµα εναντίον του GABA νευρώνες (GABA+) εµφάνιζαν συσσώρευση του προϊόντος αντίδρασης στο κυτταρόπλασµα και στους πρωτοταγείς δενδρίτες. Στους µάρτυρες, οι GABAεργικοί νευρώνες εµφάνιζαν πολυγωνικό ή ωοειδές σχήµα µε έναν 100
113 Αποτελέσµατα ή περισσότερους δενδρίτες, οι οποίοι εκφύονταν από τους πόλους του περικαρύου. Σε αντίθεση µε τους µάρτυρες, τόσο στα πειραµατόζωα που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων όσο και στα πειραµατόζωα που είχαν υποστεί βλάβη των κατεχολαµινεργικών συστηµάτων οι GABAεργικοί νευρώνες ήταν ατροφικοί και παρουσίαζαν ασθενέστερη σήµανση, ενώ οι δενδρίτες τους δεν ήταν συνήθως διακριτοί (Εικόνα 9) ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ Η ποσοτική ανάλυση απέδειξε ότι η πυκνότητα των GABAεργικών νευρώνων στο ραβδωτό σώµα µειώθηκε σηµαντικά τόσο µετά τη βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 όσο και µετά τη βλάβη των κατεχολαµινεργικών συστηµάτων µε 6-OHDA. Η πυκνότητα των GABAεργικών νευρώνων σε σχέση µε τους µάρτυρες µειώθηκε κατά 48,8% στα ζώα που υπέστησαν βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 και κατά 47,8% στα ζώα που υπέστησαν βλάβη µε 6-OHDA (Πίνακας 11, ιάγραµµα 9). 101
114 Αποτελέσµατα Πίνακας 11. Πυκνότητα των GABA+ νευρώνων στο ραβδωτό σώµα φυσιολογικών επιµύων, επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων στο νεογέννητο και επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων και εξετάστηκαν κατά την ενηλικίωση Πειραµατική οµάδα Μάρτυρες Βλάβη του φλοιού τη Μ0 Βλάβη µε 6-OHDA Πυκνότητα των GABA+νευρώνων 17,75±0,66 α 9,08±0,40 β 9,25±0,32 β Μ: µεταγεννητική µέρα GABA+: µέσος όρος των GABAεργικών νευρώνων ανά τοµή, ανά 0,1mm 2. Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. Οι µέσοι όροι µε διαφορετικό εκθέτη παρουσιάζουν στατιστικά σηµαντική διαφορά µεταξύ τους (Ρ 0,05). ιάγραµµα 9 ΠΥΚΝΟΤΗΤΑ ΤΩΝ GABA+ ΝΕΥΡΩΝΩΝ Mάρτυρες * * Βλάβη του φλοιού τη Μ0 Bλάβη µε 6- OHDA Πυκνότητα των GABA+ νευρώνων: µέσος όρος των GABA+ νευρώνων ανά τοµή, ανά 0,1mm 2.Οι τιµές αντιπροσωπεύουν τους µέσους όρους ± τυπικό σφάλµα του µέσου. Η µείωση στην πυκνότητα των GABAεργικών νευρώνων σε ενήλικους επίµυς µετά από βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 και µετά από βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων του ραβδωτού σώµατος είναι στατιστικά σηµαντική σε σύγκριση µε τους µάρτυρες (*Ρ 0,05). 102






115 Αποτελέσµατα A Β Γ Ε ΣΤ Εικόνα 9. Μικροφωτογραφίες στις οποίες φαίνονται τα µορφολογικά χαρακτηριστικά των GABAεργικών νευρώνων στο ραβδωτό σώµα φυσιολογικών επιµύων (Α, Β), επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 (Γ, ) και επιµύων που είχαν υποστεί βλάβη µε 6-OHDA των κατεχολαµινεργικών συστηµάτων (Ε, ΣΤ). Στα πλαίσια των Α, Γ και Ε οι περιοχές του ραβδωτού σώµατος που περιέχουν GABAεργικούς νευρώνες απεικονίζονται σε µεγαλύτερη µεγέθυνση στις Β, και ΣΤ, αντίστοιχα. Ράβδος=50µm (Α, Γ, Ε), 20µm ( Β,, ΣΤ). 103
116 Αποτελέσµατα 5. ΜΕΛΕΤΗ ΤΗΣ ΕΠΙ ΡΑΣΗΣ ΤΗΣ ΒΛΑΒΗΣ ΤΟΥ ΦΛΟΙΟΥ ΤΩΝ ΕΓΚΕΦΑΛΙΚΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΣΤΗΝ ΕΠΙΒΙΩΣΗ ΚΑΙ ΣΤΟΝ ΦΑΙΝΟΤΥΠΟ ΤΩΝ ΧΟΛΙΝΕΡΓΙΚΩΝ ΝΕΥΡΩΝΩΝ ΤΟΥ ΡΑΒ ΩΤΟΥ ΣΩΜΑΤΟΣ 5.1. ΠΟΙΟΤΙΚΗ ΑΝΑΛΥΣΗ Όπως προαναφέρθηκε (βλ. Εισαγωγή, κεφ. Β.1.2.α), παρόλο που ο πληθυσµός των διάµεσων χολινεργικών νευρώνων στο ραβδωτό σώµα είναι µικρός, τα επίπεδα της ακετυλοχολίνης που παράγονται από αυτούς είναι από τα υψηλότερα στο ΚΝΣ. Στους χολινεργικούς νευρώνες του ραβδωτού σώµατος απολήγουν γλουταµινεργικές ίνες που προέρχονται από τον φλοιό των εγκεφαλικών ηµισφαιρίων. Με βάση τα παραπάνω, ελέγξαµε την επίδραση της βλάβης του φλοιού των εγκεφαλικών ηµισφαιρίων στην επιβίωση και στη φαινοτυπική έκφραση των χολινεργικών νευρώνων του ραβδωτού σώµατος. Η µελέτη πραγµατοποιήθηκε σε επίµυς που είχαν υποστεί βλάβη του φλοιού στο νεογέννητο και εξετάστηκαν τη Μ60. Οι νευρώνες, οι οποίοι παρουσίαζαν ανοσοδραστικότητα για το αντίσωµα εναντίον της ChAT (ChAT+) αναγνωρίστηκαν από τη συσσώρευση του προϊόντος αντίδρασης στο κυτταρόπλασµα και στους πρωτοταγείς δενδρίτες τους. Στους µάρτυρες παρατηρήθηκαν χολινεργικοί νευρώνες, οι οποίοι είχαν τριγωνικό σχήµα και δενδρίτες που εκφύονταν από τους πόλους του περικαρύου ή ωοειδές σχήµα µε δύο δενδρίτες. Σε αντίθεση µε τους µάρτυρες, στα πειραµατόζωα που είχαν υποστεί βλάβη του φλοιού στο νεογέννητο, οι χολινεργικοί νευρώνες παρουσίαζαν ασθενέστερη σήµανση, είχαν ατροφική εµφάνιση και εµφάνιζαν λιγότερους και βραχύτερους δενδρίτες (Εικόνα 10). 104

.")


117 Αποτελέσµατα A B Γ Εικόνα 10. Μικροφωτογραφίες στις οποίες φαίνονται τα µορφολογικά χαρακτηριστικά των χολινεργικών νευρώνων στο ραβδωτό σώµα φυσιολογικών επιµύων (Α, Β) και επιµύων που είχαν υποστεί βλάβη του φλοιού των εγκεφαλικών ηµισφαιρίων τη Μ0 (Γ, ). Στα πλαίσια της Α και Γ οι περιοχές του ραβδωτού σώµατος, οι οποίες περιέχουν χολινεργικούς νευρώνες απεικονίζονται σε µεγαλύτερη µεγέθυνση στις Β και, αντίστοιχα. Ράβδος=100µm (Α, Γ), 50µm (Β, ) ΠΟΣΟΤΙΚΗ ΑΝΑΛΥΣΗ Η ποσοτική ανάλυση απέδειξε ότι η πυκνότητα των χολινεργικών νευρώνων στο ραβδωτό σώµα στα ζώα που υπέστησαν βλάβη του φλοιού στο νεογέννητο µειώθηκε σηµαντικά (κατά 54%) σε σύγκριση µε τους µάρτυρες (Πίνακας 12, ιάγραµµα 10). 105
1. Ανάπτυξη του νοραδρενεργικού συστήµατος στον VC και στον MC
 ΑΠΟΤΕΛΕΣΜΑΤΑ ιάφορα αντισώµατα, µεταξύ των οποίων και αντισώµατα εναντίον της DBH, έχουν χρησιµοποιηθεί για την in situ σήµανση και ανίχνευση των νοραδρενεργικών ινών. Μεγάλος αριθµός µελετών, ενδεικτικά
ΑΠΟΤΕΛΕΣΜΑΤΑ ιάφορα αντισώµατα, µεταξύ των οποίων και αντισώµατα εναντίον της DBH, έχουν χρησιµοποιηθεί για την in situ σήµανση και ανίχνευση των νοραδρενεργικών ινών. Μεγάλος αριθµός µελετών, ενδεικτικά
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ Ορίζουμε ως διαβιβαστή μια ουσία που απελευθερώνεται από έναν νευρώνα σε μια σύναψη και που επηρεάζει ένα άλλο κύτταρο, είτε έναν νευρώνα είτε ένα κύτταρο
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ Ορίζουμε ως διαβιβαστή μια ουσία που απελευθερώνεται από έναν νευρώνα σε μια σύναψη και που επηρεάζει ένα άλλο κύτταρο, είτε έναν νευρώνα είτε ένα κύτταρο
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων
 Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Διάµεσος Εγκέφαλος (Θάλαµος) Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Διάµεσος Εγκέφαλος (Θάλαµος) Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά
Βασικά γάγγλια. Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας
 Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί η πρόθεση δράσης
Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί η πρόθεση δράσης
Οι Κυριότερες Νευρικές Οδοί
 Οι Κυριότερες Νευρικές Οδοί Κατιόντα (φυγόκεντρα) δεµάτια Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή φυσιολογικά δεµάτια (κατά τον επιµήκη άξονα) έχουν κοινή έκφυση πορεία απόληξη λειτουργία
Οι Κυριότερες Νευρικές Οδοί Κατιόντα (φυγόκεντρα) δεµάτια Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή φυσιολογικά δεµάτια (κατά τον επιµήκη άξονα) έχουν κοινή έκφυση πορεία απόληξη λειτουργία
Βασικά γάγγλια. Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017
 Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί
Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί
Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός. Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας
 Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Θάλαμος Ο θάλαμος: «πύλη προς τον εγκέφαλο» Είναι μια μεγάλη συλλογή νευρώνων στο διεγκέφαλο Παίρνει
Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Θάλαμος Ο θάλαμος: «πύλη προς τον εγκέφαλο» Είναι μια μεγάλη συλλογή νευρώνων στο διεγκέφαλο Παίρνει
Εγκέφαλος και Έλεγχος της Κίνησης. 4 διακριτά υποσυστήματα που αλληλεπιδρούν
 Τα Βασικά Γάγγλια Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης
Τα Βασικά Γάγγλια Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης
Γνωστική-Πειραµατική Ψυχολογία
 Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Γνωστική λειτουργία & φλοιός. Γνωστική λειτουργία & φλοιός. Γνωστικές λειτουργίες à επεξεργασία πληροφοριών από διαφορετικές περιοχές (µεγαλύτερη ( αποκλειστική)
Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Γνωστική λειτουργία & φλοιός. Γνωστική λειτουργία & φλοιός. Γνωστικές λειτουργίες à επεξεργασία πληροφοριών από διαφορετικές περιοχές (µεγαλύτερη ( αποκλειστική)
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ
 ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Εγκέφαλος Μεγάλη αιµάτωση, πολύ σηµαντική για την λειτουργία του Επικοινωνία µε το περιβάλλον Χρησιµοποιεί το 20% του Ο 2 και ως πηγή ενέργειας γλυκόζη Στις χειρουργικές επεµβάσεις
ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Εγκέφαλος Μεγάλη αιµάτωση, πολύ σηµαντική για την λειτουργία του Επικοινωνία µε το περιβάλλον Χρησιµοποιεί το 20% του Ο 2 και ως πηγή ενέργειας γλυκόζη Στις χειρουργικές επεµβάσεις
ΜΑΘΗΜΑ 7ο ΜΕΡΟΣ Α Η ΔΟΜΗ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ
 ΜΑΘΗΜΑ 7ο ΜΕΡΟΣ Α Η ΔΟΜΗ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Η ΛΕΥΚΗ ΟΥΣΙΑ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ Η λευκή ουσία συντίθεται από εμύελες νευρικές ίνες διαφόρων διαμέτρων και νευρογλοία Οι νευρικές ίνες κατατάσσονται
ΜΑΘΗΜΑ 7ο ΜΕΡΟΣ Α Η ΔΟΜΗ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Η ΛΕΥΚΗ ΟΥΣΙΑ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ Η λευκή ουσία συντίθεται από εμύελες νευρικές ίνες διαφόρων διαμέτρων και νευρογλοία Οι νευρικές ίνες κατατάσσονται
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ
 Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΜΕΛΕΤΗ ΤΩΝ ΥΠΟΜΟΝΑΔΩΝ ΤΩΝ ΥΠΟΔΟΧΕΩΝ ΔΙΕΓΕΡΤΙΚΩΝ ΚΑΙ ΑΝΑΣΤΑΛΤΙΚΩΝ ΑΜΙΝΟΞΕΩΝ ΣΤΟΝ ΕΓΚΕΦΑΛΟ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΜΕΛΕΤΗ ΤΩΝ ΥΠΟΜΟΝΑΔΩΝ ΤΩΝ ΥΠΟΔΟΧΕΩΝ ΔΙΕΓΕΡΤΙΚΩΝ ΚΑΙ ΑΝΑΣΤΑΛΤΙΚΩΝ ΑΜΙΝΟΞΕΩΝ ΣΤΟΝ ΕΓΚΕΦΑΛΟ
λκλλκλκλλκκκκ Βασικά Γάγγλια Εισαγωγή Μορφολογία & Τοπογραφική Ανατοµική Ραβδωτό Σώµα Προτείχισµα Αµυδαλοειδές Σώµα Ανώνυµη Ουσία Επικλινής Πυρήνας
 9 λκλλκλκλλκκκκ Βασικά Γάγγλια Εισαγωγή Μορφολογία & Τοπογραφική Ανατοµική Ραβδωτό Σώµα Προτείχισµα Αµυδαλοειδές Σώµα Ανώνυµη Ουσία Επικλινής Πυρήνας Συνδέσεις του Ραβδωτου Σώµατος Προσαγωγές Ίνες Απαγωγές
9 λκλλκλκλλκκκκ Βασικά Γάγγλια Εισαγωγή Μορφολογία & Τοπογραφική Ανατοµική Ραβδωτό Σώµα Προτείχισµα Αµυδαλοειδές Σώµα Ανώνυµη Ουσία Επικλινής Πυρήνας Συνδέσεις του Ραβδωτου Σώµατος Προσαγωγές Ίνες Απαγωγές
ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ
 ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ Δρ. Κυριακή Σιδηροπούλου Λέκτορας Νευροφυσιολογίας Γραφείο: Γ316δ ΤΗΛ: 28103940871 (γραφείο) E- MAIL: sidirop@imbb.forth.gr Εισαγωγή Σιδηροπούλου - Νευροβιολογία 1 Δομή μαθήματος
ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ Δρ. Κυριακή Σιδηροπούλου Λέκτορας Νευροφυσιολογίας Γραφείο: Γ316δ ΤΗΛ: 28103940871 (γραφείο) E- MAIL: sidirop@imbb.forth.gr Εισαγωγή Σιδηροπούλου - Νευροβιολογία 1 Δομή μαθήματος
Κυτταροαρχιτεκτονική Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή
 Κυτταροαρχιτεκτονική Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή Τελικός Εγκ Εγκεφαλικά ηµισφαίρια Διάµεσος εγκ & Βασικά γάγγλια Διαίρεση του ΚΝΣ Στέλεχος του εγκέφαλου Μέσος εγκ Γέφυρα Προµήκης
Κυτταροαρχιτεκτονική Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή Τελικός Εγκ Εγκεφαλικά ηµισφαίρια Διάµεσος εγκ & Βασικά γάγγλια Διαίρεση του ΚΝΣ Στέλεχος του εγκέφαλου Μέσος εγκ Γέφυρα Προµήκης
Σύναψη µεταξύ της απόληξης του νευράξονα ενός νευρώνα και του δενδρίτη ενός άλλου νευρώνα.
 ΟΙ ΝΕΥΡΩΝΕΣ ΕΠΙΚΟΙΝΩΝΟΥΝ ΜΕΣΩ ΤΗΣ ΣΥΝΑΨΗΣ Άντα Μητσάκου Αναπληρώτρια Καθηγήτρια, Ιατρική Σχολή, Πανεπιστήµιο Πατρών Γνωρίζουµε ότι είµαστε ικανοί να εκτελούµε σύνθετες νοητικές διεργασίες εξαιτίας της
ΟΙ ΝΕΥΡΩΝΕΣ ΕΠΙΚΟΙΝΩΝΟΥΝ ΜΕΣΩ ΤΗΣ ΣΥΝΑΨΗΣ Άντα Μητσάκου Αναπληρώτρια Καθηγήτρια, Ιατρική Σχολή, Πανεπιστήµιο Πατρών Γνωρίζουµε ότι είµαστε ικανοί να εκτελούµε σύνθετες νοητικές διεργασίες εξαιτίας της
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ. Κάντε κλικ για να επεξεργαστείτε τον υπότιτλο του υποδείγματος
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Κάντε κλικ για να επεξεργαστείτε τον υπότιτλο του υποδείγματος ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα θέτει σε επικοινωνία τον οργανισμό μας με τον έξω κόσμο. Μοιάζει με τηλεφωνικό δίκτυο,
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Κάντε κλικ για να επεξεργαστείτε τον υπότιτλο του υποδείγματος ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα θέτει σε επικοινωνία τον οργανισμό μας με τον έξω κόσμο. Μοιάζει με τηλεφωνικό δίκτυο,
Φυσιολογία ΙΙ Ενότητα 1:
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 1: Το Νευρικό Σύστημα Ανωγειανάκις Γεώργιος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 1: Το Νευρικό Σύστημα Ανωγειανάκις Γεώργιος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
M.Sc. Bioinformatics and Neuroinformatics
 M.Sc. Bioinformatics and Neuroinformatics Recording and Processing Brain Signals Μαρία Σαγιαδινού Ο ανθρώπινος εγκέφαλος Πιο πολύπλοκο δημιούργημα της φύσης Προιόν βιολογικής εξέλιξης εκατομμυρίων ετών
M.Sc. Bioinformatics and Neuroinformatics Recording and Processing Brain Signals Μαρία Σαγιαδινού Ο ανθρώπινος εγκέφαλος Πιο πολύπλοκο δημιούργημα της φύσης Προιόν βιολογικής εξέλιξης εκατομμυρίων ετών
Φυσιολογία ΙΙ Ενότητα 1:
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 1: Το Νευρικό Σύστημα Ανωγειανάκις Γεώργιος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ενότητα 1: Το Νευρικό Σύστημα Ανωγειανάκις Γεώργιος Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
2
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΝΕΥΡΟΕΠΙΣΤΗΜΩΝ Α ΝΕΥΡΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ Η ΚΥΤΤΑΡΟΑΡΧΙΤΕΚΤΟΝΙΚΗ ΚΑΙ ΟΙ ΣΥΝ ΕΣΕΙΣ ΤΟΥ ΚΕΡΚΟΦΟΡΟΥ ΠΥΡΗΝΑ ΣΤΟΝ ΑΝΘΡΩΠΟ ΚΑΙ ΟΙ ΑΛΛΟΙΩΣΕΙΣ ΤΟΥΣ ΣΤΗ ΝΟΣΟ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΝΕΥΡΟΕΠΙΣΤΗΜΩΝ Α ΝΕΥΡΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ Η ΚΥΤΤΑΡΟΑΡΧΙΤΕΚΤΟΝΙΚΗ ΚΑΙ ΟΙ ΣΥΝ ΕΣΕΙΣ ΤΟΥ ΚΕΡΚΟΦΟΡΟΥ ΠΥΡΗΝΑ ΣΤΟΝ ΑΝΘΡΩΠΟ ΚΑΙ ΟΙ ΑΛΛΟΙΩΣΕΙΣ ΤΟΥΣ ΣΤΗ ΝΟΣΟ
Συστήματα αισθήσεων. Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
 Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας 2018 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας 2018 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Κινητικό σύστηµα. Κινητικός φλοιός
 Κινητικό σύστηµα Κινητικός φλοιός Κινητικός φλοιός Όλες οι εκούσιες κινήσεις ελέγχονται από τον εγκέφαλο Μια από τις περιοχές του εγκεφάλου που εµπλέκονται στον έλεγχο των εκούσιων κινήσεων είναι ο κινητικός
Κινητικό σύστηµα Κινητικός φλοιός Κινητικός φλοιός Όλες οι εκούσιες κινήσεις ελέγχονται από τον εγκέφαλο Μια από τις περιοχές του εγκεφάλου που εµπλέκονται στον έλεγχο των εκούσιων κινήσεων είναι ο κινητικός
Βιολογία Α Λυκείου Κεφ. 9. Νευρικό Σύστημα. Δομή και λειτουργία των νευρικών κυττάρων
 Βιολογία Α Λυκείου Κεφ. 9 Νευρικό Σύστημα Δομή και λειτουργία των νευρικών κυττάρων Νευρικό Σύστημα Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων φροντίζουν να διατηρείται σταθερό το εσωτερικό
Βιολογία Α Λυκείου Κεφ. 9 Νευρικό Σύστημα Δομή και λειτουργία των νευρικών κυττάρων Νευρικό Σύστημα Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων φροντίζουν να διατηρείται σταθερό το εσωτερικό
ΜΕΛΕΤΗ ΤΟΥ ΦΑΙΝΟΜΕΝΟΥ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟΝ ΕΞΩ ΓΟΝΑΤΩ Η ΠΥΡΗΝΑ ΤΟΥ ΕΠΙΜΥΟΣ ΚΑΤΑ ΤΗΝ ΑΝΑΠΤΥΞΗ ΚΑΙ ΜΕΤΑ ΑΠΟ ΒΛΑΒΕΣ ΠΡΟΣΑΓΩΓΩΝ ΚΑΙ ΑΠΑΓΩΓΩΝ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΚΤΗΝΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΟΜΗΣ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑΣ ΤΩΝ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ ΘΕΤΙ ΑΣ Γ. ΖΑΧΑΡΑΚΗ Κτηνιάτρου ΜΕΛΕΤΗ ΤΟΥ ΦΑΙΝΟΜΕΝΟΥ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟΝ ΕΞΩ ΓΟΝΑΤΩ Η ΠΥΡΗΝΑ ΤΟΥ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΚΤΗΝΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΟΜΗΣ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑΣ ΤΩΝ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ ΘΕΤΙ ΑΣ Γ. ΖΑΧΑΡΑΚΗ Κτηνιάτρου ΜΕΛΕΤΗ ΤΟΥ ΦΑΙΝΟΜΕΝΟΥ ΤΗΣ ΑΠΟΠΤΩΣΗΣ ΣΤΟΝ ΕΞΩ ΓΟΝΑΤΩ Η ΠΥΡΗΝΑ ΤΟΥ
Νευροδιαβιβαστές και συμπεριφορά
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Νευροδιαβιβαστές και συμπεριφορά Νευροβιολογία της λήψης τροφής Διδάσκοντες: Καθ. Α. Ψαρροπούλου, Λεκ. Χ. Λαμπρακάκης Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Νευροδιαβιβαστές και συμπεριφορά Νευροβιολογία της λήψης τροφής Διδάσκοντες: Καθ. Α. Ψαρροπούλου, Λεκ. Χ. Λαμπρακάκης Άδειες Χρήσης Το παρόν εκπαιδευτικό
Ερωτήσεις Εξετάσεων από το βιβλίο του Γ.Χ. Παπαδόπουλου. «Λειτουργική Οργάνωση του ΚΝΣ» Κεφάλαιο 1
 Ερωτήσεις Εξετάσεων από το βιβλίο του Γ.Χ. Παπαδόπουλου «Λειτουργική Οργάνωση του ΚΝΣ» Κεφάλαιο 1 1. Τα νευρικά κύτταρα µπορούν να επηρεάζουν τη λειτουργία των νευρικών κυττάρων, των... κυττάρων και των...
Ερωτήσεις Εξετάσεων από το βιβλίο του Γ.Χ. Παπαδόπουλου «Λειτουργική Οργάνωση του ΚΝΣ» Κεφάλαιο 1 1. Τα νευρικά κύτταρα µπορούν να επηρεάζουν τη λειτουργία των νευρικών κυττάρων, των... κυττάρων και των...
ΕΙΣΑΓΩΓΗ ΣΤΗ ΝΕΥΡΟΠΑΘΟΛΟΓΙA Γεώργιος Καρκαβέλας Καθηγητής Παθολογικής Ανατοµικής ΑΠΘ
 ΕΙΣΑΓΩΓΗ ΣΤΗ ΝΕΥΡΟΠΑΘΟΛΟΓΙA Γεώργιος Καρκαβέλας Καθηγητής Παθολογικής Ανατοµικής ΑΠΘ ΚΝΣ: πολυσύνθετο σύστηµα πολλές από τις λειτουργίες του αδιευκρίνιστες Πρώτες ανατοµικές µελέτες Αριστοτέλης και Γαληνός
ΕΙΣΑΓΩΓΗ ΣΤΗ ΝΕΥΡΟΠΑΘΟΛΟΓΙA Γεώργιος Καρκαβέλας Καθηγητής Παθολογικής Ανατοµικής ΑΠΘ ΚΝΣ: πολυσύνθετο σύστηµα πολλές από τις λειτουργίες του αδιευκρίνιστες Πρώτες ανατοµικές µελέτες Αριστοτέλης και Γαληνός
Ανατοµία του Εγκεφάλου
 Ανατοµία του Εγκεφάλου Κύριες Σχισµές & Αύλακες: 1. Η επιµήκης σχισµή 2. Η εγκάρσια σχισµή 3. Η πλάγια σχισµή (του Sylvius) πρόσθιο οριζόντιο κλαδο πρόσθιο ανιόντα κλάδο οπίσθιο κλάδο 4. Κεντρική αύλακα
Ανατοµία του Εγκεφάλου Κύριες Σχισµές & Αύλακες: 1. Η επιµήκης σχισµή 2. Η εγκάρσια σχισµή 3. Η πλάγια σχισµή (του Sylvius) πρόσθιο οριζόντιο κλαδο πρόσθιο ανιόντα κλάδο οπίσθιο κλάδο 4. Κεντρική αύλακα
K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ
 K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ κατάσταση ετοιμότητος του μυός ενός βαθμού μόνιμης σύσπασης που διατηρούν οι μύες στην ηρεμία αποτελεί
K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ κατάσταση ετοιμότητος του μυός ενός βαθμού μόνιμης σύσπασης που διατηρούν οι μύες στην ηρεμία αποτελεί
Γνωστική Ψυχολογία Ι (ΨΧ32)
") Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 3 Η φυσιολογία των γνωστικών διεργασιών Πέτρος Ρούσσος Η νευροψυχολογική βάση των γνωστικών διεργασιών Γνωστική νευροεπιστήμη: μελετάει τους τρόπους με τους οποίους
Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 3 Η φυσιολογία των γνωστικών διεργασιών Πέτρος Ρούσσος Η νευροψυχολογική βάση των γνωστικών διεργασιών Γνωστική νευροεπιστήμη: μελετάει τους τρόπους με τους οποίους
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α. Ο ηλεκτρονικός υπολογιστής του οργανισμού μας
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
Συστήματα αισθήσεων. Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
 Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΝΕΥΡΙΚΩΝ. Νευρώνες
 9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος (ομοιόσταση), ελέγχοντας και συντονίζοντας τις λειτουργίες των
9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος (ομοιόσταση), ελέγχοντας και συντονίζοντας τις λειτουργίες των
ΜΑΘΗΜΑ 6ο ΜΕΡΟΣ Β ΤΑ ΕΓΚΕΦΑΛΙΚΑ ΗΜΙΣΦΑΙΡΙΑ
 ΜΑΘΗΜΑ 6ο ΜΕΡΟΣ Β ΤΑ ΕΓΚΕΦΑΛΙΚΑ ΗΜΙΣΦΑΙΡΙΑ ΤΑ ΕΓΚΕΦΑΛΙΚΑ ΗΜΙΣΦΑΙΡΙΑ Τα εγκεφαλικά ημισφαίρια διακρίνονται σε δεξιό και αριστερό Διαχωρίζονται μεταξύ τους με μια βαθιά σχισμή, την επιμήκη σχισμή Εντός
ΜΑΘΗΜΑ 6ο ΜΕΡΟΣ Β ΤΑ ΕΓΚΕΦΑΛΙΚΑ ΗΜΙΣΦΑΙΡΙΑ ΤΑ ΕΓΚΕΦΑΛΙΚΑ ΗΜΙΣΦΑΙΡΙΑ Τα εγκεφαλικά ημισφαίρια διακρίνονται σε δεξιό και αριστερό Διαχωρίζονται μεταξύ τους με μια βαθιά σχισμή, την επιμήκη σχισμή Εντός
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων. Μεταιχµιακό Σύστηµα
 Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Μεταιχµιακό Σύστηµα Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά Γάγγλια µεταιχµιακό (στεφανιαίο) σύστηµα διάµεσος εγκέφαλος
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Μεταιχµιακό Σύστηµα Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά Γάγγλια µεταιχµιακό (στεφανιαίο) σύστηµα διάµεσος εγκέφαλος
Η Λευκή Ουσία του Νωτιαίου Μυελού
 Η Λευκή Ουσία του Νωτιαίου Μυελού λκλλκλκλλκκκκ Εισαγωγή Ανιόντα Δεµάτια του Νωτιαίου Μυελού Ανιόντα Δεµάτια της Πρόσθιας Δέσµης Ανιόντα Δεµάτια της Πλάγιας Δέσµης Ανιόντα Δεµάτια της Οπίσθιας Δέσµης Κατιόντα
Η Λευκή Ουσία του Νωτιαίου Μυελού λκλλκλκλλκκκκ Εισαγωγή Ανιόντα Δεµάτια του Νωτιαίου Μυελού Ανιόντα Δεµάτια της Πρόσθιας Δέσµης Ανιόντα Δεµάτια της Πλάγιας Δέσµης Ανιόντα Δεµάτια της Οπίσθιας Δέσµης Κατιόντα
ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 Page1 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Μαθητές: Ρουμπάνης Γιάννης και Οικονομίδης Αριστείδης Τάξη: Γ γυμνασίου Κερατέας Τμήμα: Γ 4 Οκτώβριος 2013 Page2 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί
Page1 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Μαθητές: Ρουμπάνης Γιάννης και Οικονομίδης Αριστείδης Τάξη: Γ γυμνασίου Κερατέας Τμήμα: Γ 4 Οκτώβριος 2013 Page2 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί
Υποφλοιϊκοί και στελεχιαίοι πυρήνες
 Υποφλοιϊκοί και στελεχιαίοι πυρήνες Σχεδιασµός κινήσεων Παρακολούθηση της κίνησης ηµιουργία κινητικών προγραµµάτων Εκµάθηση επιδέξιων κινήσεων Βασικά γάγγλια κέλυφος Κερκοφόρος πυρήνας Ωχρά σφαίρα Ωχρά
Υποφλοιϊκοί και στελεχιαίοι πυρήνες Σχεδιασµός κινήσεων Παρακολούθηση της κίνησης ηµιουργία κινητικών προγραµµάτων Εκµάθηση επιδέξιων κινήσεων Βασικά γάγγλια κέλυφος Κερκοφόρος πυρήνας Ωχρά σφαίρα Ωχρά
Απ. Χατζηευθυμίου Αν Καθηγήτρια Ιατρικής Φυσιολογίας
 Απ. Χατζηευθυμίου Αν Καθηγήτρια Ιατρικής Φυσιολογίας Το 80% περίπου της γεύσης του φαγητού παρέχεται στην πραγματικότητα από την αίσθηση της όσφρησης. Η μυρωδιά μιας ουσίας σχετίζεται άμεσα με τη χημική
Απ. Χατζηευθυμίου Αν Καθηγήτρια Ιατρικής Φυσιολογίας Το 80% περίπου της γεύσης του φαγητού παρέχεται στην πραγματικότητα από την αίσθηση της όσφρησης. Η μυρωδιά μιας ουσίας σχετίζεται άμεσα με τη χημική
Σκοπός του μαθήματος είναι ο συνδυασμός των θεωρητικών και ποσοτικών τεχνικών με τις αντίστοιχες περιγραφικές. Κεφάλαιο 1: περιγράφονται οι βασικές
 Εισαγωγή Ασχολείται με τη μελέτη των ηλεκτρικών, η λ ε κ τ ρ ο μ α γ ν η τ ι κ ώ ν κ α ι μ α γ ν η τ ι κ ώ ν φαινομένων που εμφανίζονται στους βιολογικούς ιστούς. Το αντικείμενο του εμβιοηλεκτρομαγνητισμού
Εισαγωγή Ασχολείται με τη μελέτη των ηλεκτρικών, η λ ε κ τ ρ ο μ α γ ν η τ ι κ ώ ν κ α ι μ α γ ν η τ ι κ ώ ν φαινομένων που εμφανίζονται στους βιολογικούς ιστούς. Το αντικείμενο του εμβιοηλεκτρομαγνητισμού
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ
 ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΑΠΕΛΕΥΘΕΡΩΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ ΥΠΟΔΟΧΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ 1 Ηλεκτρικές και Χημικές Συνάψεις 2 Ηλεκτρικές
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΑΠΕΛΕΥΘΕΡΩΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ ΥΠΟΔΟΧΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ 1 Ηλεκτρικές και Χημικές Συνάψεις 2 Ηλεκτρικές
ΑΣΚΗΣΗ: MΕΤΡΗΣΗ ΜΕΤΑΣΥΝΑΠΤΙΚΩΝ ΔΥΝΑΜΙΚΩΝ ΣΕ ΤΟΜΕΣ ΙΠΠΟΚΑΜΠΟΥ ΑΡΟΥΡΑΙΟΥ.
 ΑΣΚΗΣΗ: MΕΤΡΗΣΗ ΜΕΤΑΣΥΝΑΠΤΙΚΩΝ ΔΥΝΑΜΙΚΩΝ ΣΕ ΤΟΜΕΣ ΙΠΠΟΚΑΜΠΟΥ ΑΡΟΥΡΑΙΟΥ. Η μετάδοση σημάτων μεταξύ των νευρώνων επιτυγχάνεται μέσω χημικών συνάψεων. Κατά τη χημική συναπτική διαβίβαση απελευθερώνεται από
ΑΣΚΗΣΗ: MΕΤΡΗΣΗ ΜΕΤΑΣΥΝΑΠΤΙΚΩΝ ΔΥΝΑΜΙΚΩΝ ΣΕ ΤΟΜΕΣ ΙΠΠΟΚΑΜΠΟΥ ΑΡΟΥΡΑΙΟΥ. Η μετάδοση σημάτων μεταξύ των νευρώνων επιτυγχάνεται μέσω χημικών συνάψεων. Κατά τη χημική συναπτική διαβίβαση απελευθερώνεται από
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ
 ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
Εγκέφαλος και Έλεγχος της Κίνησης
 Παρεγκεφαλίδα Εγκέφαλος και Έλεγχος της Κίνησης 3 4 Γενικά Cerebellum = Little brain 10% όγκου εγκεφάλου, >50% του συνόλου των νευρώνων του εγκεφάλου Κανονικότητα στη διάταξη των νευρώνων και τις διασυνδέσεις
Παρεγκεφαλίδα Εγκέφαλος και Έλεγχος της Κίνησης 3 4 Γενικά Cerebellum = Little brain 10% όγκου εγκεφάλου, >50% του συνόλου των νευρώνων του εγκεφάλου Κανονικότητα στη διάταξη των νευρώνων και τις διασυνδέσεις
To «πρόσωπο του γιγαντιαίου πάντα» και το. «πρόσωπο του μικροσκοπικού πάντα» στη νόσο του Wilson
 To «πρόσωπο του γιγαντιαίου πάντα» και το «πρόσωπο του μικροσκοπικού πάντα» στη νόσο του Wilson Ζήκου Αναστασία, Μούκα Βασιλική, Μούκα Γεωργία, Κώστα Παρασκευή, Ξύδης Βασίλειος, Αργυροπούλου Ι. Μαρία.
To «πρόσωπο του γιγαντιαίου πάντα» και το «πρόσωπο του μικροσκοπικού πάντα» στη νόσο του Wilson Ζήκου Αναστασία, Μούκα Βασιλική, Μούκα Γεωργία, Κώστα Παρασκευή, Ξύδης Βασίλειος, Αργυροπούλου Ι. Μαρία.
Οι Κυριότερες Νευρικές Οδοί
 Οι Κυριότερες Νευρικές Οδοί λκλλκλκλλκκκκ Εισαγωγή Κινητικότητα Αισθητικότητα Φυγόκεντρες Νευρικές Οδοί Οργάνωση Κεντροµόλες Νευρικές Οδοί Οργάνωση Λειτουργική Ανατοµική Θέσεων των Οδών Εισαγωγή Οι µακρές
Οι Κυριότερες Νευρικές Οδοί λκλλκλκλλκκκκ Εισαγωγή Κινητικότητα Αισθητικότητα Φυγόκεντρες Νευρικές Οδοί Οργάνωση Κεντροµόλες Νευρικές Οδοί Οργάνωση Λειτουργική Ανατοµική Θέσεων των Οδών Εισαγωγή Οι µακρές
ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ
 ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Νευρικό Σύστημα 2 Ανατομία ΝΣ ΝΣ ΚΝΣ (εγκέφαλος + ΝΜ) ΠΝΣ Προσαγωγό (αισθητικό) Απαγωγό Κινητικοί νευρώνες ΑΝΣ Συμπαθητικό Παρασυμπαθητικό 3 Οι διαταραχές της λειτουργίας
ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Νευρικό Σύστημα 2 Ανατομία ΝΣ ΝΣ ΚΝΣ (εγκέφαλος + ΝΜ) ΠΝΣ Προσαγωγό (αισθητικό) Απαγωγό Κινητικοί νευρώνες ΑΝΣ Συμπαθητικό Παρασυμπαθητικό 3 Οι διαταραχές της λειτουργίας
Νωτιαίος Μυελός. Ντελής Κων/νος MD, PhD Ρευματολόγος
 Νωτιαίος Μυελός Ντελής Κων/νος MD, PhD Ρευματολόγος Νωτιαίος Μυελός (Spinal Cord) Επίμηκες μόρφωμα που βρίσκεται μέσα στον σπονδυλικό σωλήνα και μεταφέρει προσαγωγά ερεθίσματα (πληροφορίες) από το σώμα
Νωτιαίος Μυελός Ντελής Κων/νος MD, PhD Ρευματολόγος Νωτιαίος Μυελός (Spinal Cord) Επίμηκες μόρφωμα που βρίσκεται μέσα στον σπονδυλικό σωλήνα και μεταφέρει προσαγωγά ερεθίσματα (πληροφορίες) από το σώμα
Αμυγδαλή (ΑΜΥ)* Ι. Εισαγωγή ΙΙ. Ανατομική οργάνωση
* Ι. Εισαγωγή ΙΙ. Ανατομική οργάνωση") Αμυγδαλή (ΑΜΥ)* Ι. Εισαγωγή Η αμυγδαλή είναι μια σύνθετη δομή που εμπλέκεται σε μια πληθώρα φυσιολογικών συμπεριφορικών λειτουργιών αλλά και ψυχιατρικών καταστάσεων. Για πολύ καιρό η αμυγδαλή αποτελούσε
Αμυγδαλή (ΑΜΥ)* Ι. Εισαγωγή Η αμυγδαλή είναι μια σύνθετη δομή που εμπλέκεται σε μια πληθώρα φυσιολογικών συμπεριφορικών λειτουργιών αλλά και ψυχιατρικών καταστάσεων. Για πολύ καιρό η αμυγδαλή αποτελούσε
K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ
 K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ κατάσταση ετοιμότητος του μυός ενός βαθμού μόνιμης σύσπασης που διατηρούν οι μύες στην ηρεμία αποτελεί
K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ κατάσταση ετοιμότητος του μυός ενός βαθμού μόνιμης σύσπασης που διατηρούν οι μύες στην ηρεμία αποτελεί
ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ. Βιολογία A λυκείου. Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος:
 ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ Βιολογία A λυκείου Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος: 2013-2014 Ένα αισθητικό σύστημα στα σπονδυλωτά αποτελείται από τρία βασικά μέρη: 1. Τους αισθητικούς υποδοχείς,
ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ Βιολογία A λυκείου Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος: 2013-2014 Ένα αισθητικό σύστημα στα σπονδυλωτά αποτελείται από τρία βασικά μέρη: 1. Τους αισθητικούς υποδοχείς,
Αισθητικά συστήματα Σωματοαισθητικό σύστημα
 Αισθητικά συστήματα Σωματοαισθητικό σύστημα Κεφάλαιο 18, 20 Σιδηροπούλου - Νευροβιολογία 44 Υποδοχείς σωματοαισθητικού συστήματος Μηχανοϋποδοχείς Ταχέως προσαρμοσμένοι υποδοχείς Υποδοχείς θυλάκων των τριχών
Αισθητικά συστήματα Σωματοαισθητικό σύστημα Κεφάλαιο 18, 20 Σιδηροπούλου - Νευροβιολογία 44 Υποδοχείς σωματοαισθητικού συστήματος Μηχανοϋποδοχείς Ταχέως προσαρμοσμένοι υποδοχείς Υποδοχείς θυλάκων των τριχών
ΕΡΩΤΗΣΕΙΣ Επιλέξτε τη σωστή απάντηση στις παρακάτω προτάσεις: 1) Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά
 Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά") ΕΡΩΤΗΣΕΙΣ Επιλέξτε τη σωστή απάντηση στις παρακάτω προτάσεις: 1) Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά συστατικά και να απομακρύνουν τις άχρηστες ουσίες. β. Να
ΕΡΩΤΗΣΕΙΣ Επιλέξτε τη σωστή απάντηση στις παρακάτω προτάσεις: 1) Τα νευρογλοιακά κύτταρα δεν μπορούν: α. Να προμηθεύουν τους νευρώνες με θρεπτικά συστατικά και να απομακρύνουν τις άχρηστες ουσίες. β. Να
Γνωστική-Πειραµατική Ψυχολογία
 Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Μηχανισµοί της ΣΔ à Βάση διεργασιών όπως η αντίληψη, µάθηση, εκούσια κίνηση. 10.000 à Μέσος νευρώνας à 1000 (1011 1014). 2 θεµελιώδεις µηχανισµοί ΣΔς:
Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Μηχανισµοί της ΣΔ à Βάση διεργασιών όπως η αντίληψη, µάθηση, εκούσια κίνηση. 10.000 à Μέσος νευρώνας à 1000 (1011 1014). 2 θεµελιώδεις µηχανισµοί ΣΔς:
ΑΝΑΠΤΥΞΗ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΑΝΑΠΤΥΞΗ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Η τρίτη εβδομάδα (έναρξη) Περιλαμβάνει το σχηματισμό: της αρχικής γραμμής της νωτιαίας χορδής των τριών βλαστικών στιβάδων ή βλαστικά δέρματα (γαστριδίωση) Η τρίτη εβδομάδα
ΑΝΑΠΤΥΞΗ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Η τρίτη εβδομάδα (έναρξη) Περιλαμβάνει το σχηματισμό: της αρχικής γραμμής της νωτιαίας χορδής των τριών βλαστικών στιβάδων ή βλαστικά δέρματα (γαστριδίωση) Η τρίτη εβδομάδα
ΕΙΣΑΓΩΓΗ ΣΤΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ
 ΕΙΣΑΓΩΓΗ ΣΤΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Σχηματική Λειτουργία Νευρικού Συστήματος Ποιος είναι ο ταχύτερος τρόπος αντίδρασης στα ερεθίσματα; Α) Το ηλεκτρικό ρεύμα Β) Βιοχημικές αντιδράσεις. Κίνηση των μεταβολιτών
ΕΙΣΑΓΩΓΗ ΣΤΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Σχηματική Λειτουργία Νευρικού Συστήματος Ποιος είναι ο ταχύτερος τρόπος αντίδρασης στα ερεθίσματα; Α) Το ηλεκτρικό ρεύμα Β) Βιοχημικές αντιδράσεις. Κίνηση των μεταβολιτών
5o Μάθηµα. Αισθητικά Συστήµατα
 5o Μάθηµα Αισθητικά Συστήµατα Γραφή Μπράιγ Σωµαταισθητικό Σύστηµα Αισθήσεις:Αφής,Ιδιοδεκτικότητας Πόνου,Θερµοκρασίας Οργάνωση:Ιεραρχική-Παράλληλη Τρείς Νευρώνες:Πρωτοταγής ευτεροταγής Τριτοταγής Ραχιαίο
5o Μάθηµα Αισθητικά Συστήµατα Γραφή Μπράιγ Σωµαταισθητικό Σύστηµα Αισθήσεις:Αφής,Ιδιοδεκτικότητας Πόνου,Θερµοκρασίας Οργάνωση:Ιεραρχική-Παράλληλη Τρείς Νευρώνες:Πρωτοταγής ευτεροταγής Τριτοταγής Ραχιαίο
3. Να συμπληρώσετε κατάλληλα τα μέρη από τα οποία αποτελείται ένας νευρώνας.
 ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 9 ο ΚΕΦΑΛΑΙΟ «ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ» ΜΕΡΟΣ Α: ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Να συμπληρώσετε το παρακάτω διάγραμμα. 2. Ποιος είναι ο ρόλος του
ΦΥΛΛΟ ΕΡΓΑΣΙΑΣ ΣΤΟ 9 ο ΚΕΦΑΛΑΙΟ «ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ» ΜΕΡΟΣ Α: ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ Α. ΔΡΑΣΤΗΡΙΟΤΗΤΕΣ ΜΕΣΑ ΣΤΗΝ ΤΑΞΗ 1. Να συμπληρώσετε το παρακάτω διάγραμμα. 2. Ποιος είναι ο ρόλος του
IΣTOΛOΓIA. Tα δείγµατα του βιολογικού υλικού λαµβάνονται µε > βελόνες ενδοσκοπικούς σωλήνες εύκαµπτους καθετήρες
 IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
Δοµή και Λειτουργία του Νευρώνα και του Εγκεφάλου
 Δοµή και Λειτουργία του Νευρώνα και του Εγκεφάλου Επισκόπηση o Νευρικό σύστηµα o Κύτταρα του νευρικού συστήµατος o Εγκέφαλος o Επικοινωνία εντός νευρώνα (νευρική αγωγιµότητα) o Επικοινωνία µεταξύ νευρώνων
Δοµή και Λειτουργία του Νευρώνα και του Εγκεφάλου Επισκόπηση o Νευρικό σύστηµα o Κύτταρα του νευρικού συστήµατος o Εγκέφαλος o Επικοινωνία εντός νευρώνα (νευρική αγωγιµότητα) o Επικοινωνία µεταξύ νευρώνων
Οπτική οδός. Έξω γονατώδες σώµα. Οπτική ακτινοβολία
 Όραση Γ Όραση Οπτική οδός Έξω γονατώδες σώµα Οπτική ακτινοβολία Οπτικό χίασµα: Οι ίνες από το ρινικό ηµιµόριο περνούν στην αντίπλευρη οπτική οδό ενώ τα κροταφικά ηµιµόρια δεν χιάζονται. Εποµένως κάθε οπτική
Όραση Γ Όραση Οπτική οδός Έξω γονατώδες σώµα Οπτική ακτινοβολία Οπτικό χίασµα: Οι ίνες από το ρινικό ηµιµόριο περνούν στην αντίπλευρη οπτική οδό ενώ τα κροταφικά ηµιµόρια δεν χιάζονται. Εποµένως κάθε οπτική
Β. Να επιλέξετε την ορθή απάντηση αναγράφοντας στον πίνακα της ακόλουθης
 Ονοματεπώνυμο:.. Βαθμός: Ωριαία γραπτή εξέταση Α Τετραμήνου στη Βιολογία [Κεφ. 9 ο, σελ. 153-158] Α. Να χαρακτηρίσετε τις ακόλουθες προτάσεις με το γράμμα Ο, εφόσον είναι ορθές, ή με το γράμμα Λ, αν είναι
Ονοματεπώνυμο:.. Βαθμός: Ωριαία γραπτή εξέταση Α Τετραμήνου στη Βιολογία [Κεφ. 9 ο, σελ. 153-158] Α. Να χαρακτηρίσετε τις ακόλουθες προτάσεις με το γράμμα Ο, εφόσον είναι ορθές, ή με το γράμμα Λ, αν είναι
Εργαστηριακη διάγνωση Νευροεκφυλιστικων νοσημάτων. Χρυσούλα Νικολάου
 Εργαστηριακη διάγνωση Νευροεκφυλιστικων νοσημάτων Χρυσούλα Νικολάου Νευροεκφυλιστικά Νοσήματα Νοσολογικές οντότητες, συχνά κληρονομικής αρχής, με προσβολή συγκεκριμένων ανατομικών δομών του Ν.Σ, (επηρεάζουν
Εργαστηριακη διάγνωση Νευροεκφυλιστικων νοσημάτων Χρυσούλα Νικολάου Νευροεκφυλιστικά Νοσήματα Νοσολογικές οντότητες, συχνά κληρονομικής αρχής, με προσβολή συγκεκριμένων ανατομικών δομών του Ν.Σ, (επηρεάζουν
Αλκοόλ, Εθεβεία & Εγκέθαλορ. Γιώργος Παναγής Πανεπιστήμιο Κρήτης Τμήμα Ψυχολογίας Εργαστήριο Νευροεπιστημών & Συμπεριφοράς
 Αλκοόλ, Εθεβεία & Εγκέθαλορ Γιώργος Παναγής Πανεπιστήμιο Κρήτης Τμήμα Ψυχολογίας Εργαστήριο Νευροεπιστημών & Συμπεριφοράς Κατανάλωση οινοπνευματωδών στους Έλληνες μαθητές (2011) Στην Ελλάδα, τα αγόρια
Αλκοόλ, Εθεβεία & Εγκέθαλορ Γιώργος Παναγής Πανεπιστήμιο Κρήτης Τμήμα Ψυχολογίας Εργαστήριο Νευροεπιστημών & Συμπεριφοράς Κατανάλωση οινοπνευματωδών στους Έλληνες μαθητές (2011) Στην Ελλάδα, τα αγόρια
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ & ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ. Πτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ & ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή ΚΙΝΗΤΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΑΝΤΙΔΡΑΣΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΑΝΑΓΩΓΗΣ ΤΩΝ ΟΞΕΙΔΙΩΝ ΤΟΥ ΑΖΩΤΟΥ (NO X
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΕΠΙΣΤΗΜΗΣ & ΤΕΧΝΟΛΟΓΙΑΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Πτυχιακή διατριβή ΚΙΝΗΤΙΚΗ ΜΕΛΕΤΗ ΤΗΣ ΑΝΤΙΔΡΑΣΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΑΝΑΓΩΓΗΣ ΤΩΝ ΟΞΕΙΔΙΩΝ ΤΟΥ ΑΖΩΤΟΥ (NO X
Τι θα προτιμούσατε; Γνωστική Ψυχολογία Ι (ΨΧ32) 25/4/2012. Διάλεξη 5 Όραση και οπτική αντίληψη. Πέτρος Ρούσσος. Να περιγράψετε τι βλέπετε στην εικόνα;
 25/4/2012. Διάλεξη 5 Όραση και οπτική αντίληψη. Πέτρος Ρούσσος. Να περιγράψετε τι βλέπετε στην εικόνα;") Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 5 Όραση και οπτική αντίληψη Πέτρος Ρούσσος Να περιγράψετε τι βλέπετε στην εικόνα; Τι θα προτιμούσατε; Ή να αντιμετωπίσετε τον Γκάρι Κασπάροβ σε μια παρτίδα σκάκι; 1
Γνωστική Ψυχολογία Ι (ΨΧ32) Διάλεξη 5 Όραση και οπτική αντίληψη Πέτρος Ρούσσος Να περιγράψετε τι βλέπετε στην εικόνα; Τι θα προτιμούσατε; Ή να αντιμετωπίσετε τον Γκάρι Κασπάροβ σε μια παρτίδα σκάκι; 1
1. Κεντρικό Νευρικό Σύστημα
 1. Κεντρικό Νευρικό Σύστημα 1.1. Νευρικό Σύστημα 1.1.1. Ανατομία του Νευρικού Συστήματος: Το νευρικό σύστημα αποτελείται από ένα κεντρικό και ένα περιφερικό τμήμα (πίνακας 1, σχήμα 1). (α) Το κεντρικό
1. Κεντρικό Νευρικό Σύστημα 1.1. Νευρικό Σύστημα 1.1.1. Ανατομία του Νευρικού Συστήματος: Το νευρικό σύστημα αποτελείται από ένα κεντρικό και ένα περιφερικό τμήμα (πίνακας 1, σχήμα 1). (α) Το κεντρικό
ΠΡΩΤΟΚΟΛΛΟ ΕΡΕΥΝΑΣ ΜΥΟΚΑΡ ΙΑΚΗ ΑΝΑΓΕΝΝΗΣΗ ΜΕΤΑ ΤΟ ΕΜΦΡΑΓΜΑ ΕΡΕΥΝΗΤΙΚΗ ΟΜΑ Α
 ΠΡΩΤΟΚΟΛΛΟ ΕΡΕΥΝΑΣ ΜΥΟΚΑΡ ΙΑΚΗ ΑΝΑΓΕΝΝΗΣΗ ΜΕΤΑ ΤΟ ΕΜΦΡΑΓΜΑ ΕΡΕΥΝΗΤΙΚΗ ΟΜΑ Α ΘΕΟΦΙΛΟΣ ΚΩΛΕΤΤΗΣ ΑΠΟΣΤΟΛΟΣ ΠΑΠΑΛΟΗΣ ΓΙΑΝΝΗΣ ΜΠΑΛΤΟΓΙΑΝΝΗΣ ΗΜΗΤΡΙΟΣ ΟΙΚΟΝΟΜΙ ΗΣ ΜΑΡΙΑ ΑΓΓΕΛΑΚΗ ΑΝΤΩΝΙΟΣ ΒΛΑΧΟΣ ΑΓΑΘΟΚΛΕΙΑ ΜΗΤΣΗ
ΠΡΩΤΟΚΟΛΛΟ ΕΡΕΥΝΑΣ ΜΥΟΚΑΡ ΙΑΚΗ ΑΝΑΓΕΝΝΗΣΗ ΜΕΤΑ ΤΟ ΕΜΦΡΑΓΜΑ ΕΡΕΥΝΗΤΙΚΗ ΟΜΑ Α ΘΕΟΦΙΛΟΣ ΚΩΛΕΤΤΗΣ ΑΠΟΣΤΟΛΟΣ ΠΑΠΑΛΟΗΣ ΓΙΑΝΝΗΣ ΜΠΑΛΤΟΓΙΑΝΝΗΣ ΗΜΗΤΡΙΟΣ ΟΙΚΟΝΟΜΙ ΗΣ ΜΑΡΙΑ ΑΓΓΕΛΑΚΗ ΑΝΤΩΝΙΟΣ ΒΛΑΧΟΣ ΑΓΑΘΟΚΛΕΙΑ ΜΗΤΣΗ
 ΦυσιολογίαΝευρικούΣυστήµατος 1 0 Μάθηµα Στοιχεία Ανατοµίας Λειτουργίας του ΝΣ Τεχνικές Λειτουργικής Απεικόνισης Λειτουργία της Γλώσσας ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Το ΝΣ συντονίζει τη λειτουργία
ΦυσιολογίαΝευρικούΣυστήµατος 1 0 Μάθηµα Στοιχεία Ανατοµίας Λειτουργίας του ΝΣ Τεχνικές Λειτουργικής Απεικόνισης Λειτουργία της Γλώσσας ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Το ΝΣ συντονίζει τη λειτουργία
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ. Δομή και λειτουργία των νευρικών κυττάρων. Ηλιάνα Καρβουντζή Βιολόγος
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Δομή και λειτουργία των νευρικών κυττάρων Ρόλος του νευρικού συστήματος Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Δομή και λειτουργία των νευρικών κυττάρων Ρόλος του νευρικού συστήματος Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος
ΜΑΘΗΜΑ 4ο ΜΕΡΟΣ Α ΝΩΤΙΑΙΟΣ ΜΥΕΛΟΣ
 ΜΑΘΗΜΑ 4ο ΜΕΡΟΣ Α ΝΩΤΙΑΙΟΣ ΜΥΕΛΟΣ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ (ΚΝΣ) ΕΓΚΕΦΑΛΟΣ ΝΩΤΙΑΙΟΣ ΜΥΕΛΟΣ ΝΩΤΙΑΙΟΣ ΜΥΕΛΟΣ Είναι το πιο ουραίο τμήμα του Κ.Ν.Σ. Εκτείνεται από τη βάση του κρανίου μέχρι τον 1 ο οσφυϊκό
ΜΑΘΗΜΑ 4ο ΜΕΡΟΣ Α ΝΩΤΙΑΙΟΣ ΜΥΕΛΟΣ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ (ΚΝΣ) ΕΓΚΕΦΑΛΟΣ ΝΩΤΙΑΙΟΣ ΜΥΕΛΟΣ ΝΩΤΙΑΙΟΣ ΜΥΕΛΟΣ Είναι το πιο ουραίο τμήμα του Κ.Ν.Σ. Εκτείνεται από τη βάση του κρανίου μέχρι τον 1 ο οσφυϊκό
ΥΠΟΔΟΧΕΙΣ ΣΩΜΑΤΙΚΕΣ ΑΙΣΘΗΣΕΙΣ
 ΥΠΟΔΟΧΕΙΣ ΣΩΜΑΤΙΚΕΣ ΑΙΣΘΗΣΕΙΣ ΑΙΣΘΗΤΙΚΟΙ ΥΠΟΔΟΧΕΙΣ (συγκεντρωμένοι ή διάσπαρτοι) ΝΕΥΡΙΚΕΣ ΟΔΟΙ ΕΓΚΕΦΑΛΙΚΟΣ ΦΛΟΙΟΣ Ειδικά κύτταρα - υποδοχείς, ευαίσθητα στις αλλαγές αυτές, είναι τα κύρια μέσα συλλογής
ΥΠΟΔΟΧΕΙΣ ΣΩΜΑΤΙΚΕΣ ΑΙΣΘΗΣΕΙΣ ΑΙΣΘΗΤΙΚΟΙ ΥΠΟΔΟΧΕΙΣ (συγκεντρωμένοι ή διάσπαρτοι) ΝΕΥΡΙΚΕΣ ΟΔΟΙ ΕΓΚΕΦΑΛΙΚΟΣ ΦΛΟΙΟΣ Ειδικά κύτταρα - υποδοχείς, ευαίσθητα στις αλλαγές αυτές, είναι τα κύρια μέσα συλλογής
ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ
 ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής:
ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ I ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιμος Π. Βανδώρος ΒΑΣΙΚΕΣ ΔΟΜΕΣ - ΤΟ ΚΥΤΤΑΡΟ Οι βασικές δομές που εξετάζουμε στην ανατομία μπορούν ιεραρχικά να ταξινομηθούν ως εξής:
Kυτταρική Bιολογία. Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος ΔIAΛEΞΗ 20 (9/5/2017) Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.
 Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.") Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
ράσεις Οιστρογόνων στον Εγκέφαλο
 ράσεις Οιστρογόνων στον Εγκέφαλο S2 Γενικά για τα Οιστρογόνα ιαφυλικές διαφορές σε γνωστικές λειτουργίες Χωρικές και ποσοτικές δοκιµασίες Λεκτικές ικανότητες, ταχύτητα αντίληψης Μεταβολές ς γνωστικών λειτουργιών
ράσεις Οιστρογόνων στον Εγκέφαλο S2 Γενικά για τα Οιστρογόνα ιαφυλικές διαφορές σε γνωστικές λειτουργίες Χωρικές και ποσοτικές δοκιµασίες Λεκτικές ικανότητες, ταχύτητα αντίληψης Μεταβολές ς γνωστικών λειτουργιών
χρόνιου πόνου κι των συναισθημάτων. Μάλιστα, μεγάλο μέρος αυτού
 Το μαιτεχμιακό σύστημα συνδέεται με τμήματα του μετωπιαίου κι κροταφικού λοβού ( τμήματα των εγκεφαλικών ημισφαιρίων,ονομασμένα σύμφωνα με το κρανιακό οστό που τα καλύπτει). Το ίδιο σχετίζεται με τον έλεγχο
Το μαιτεχμιακό σύστημα συνδέεται με τμήματα του μετωπιαίου κι κροταφικού λοβού ( τμήματα των εγκεφαλικών ημισφαιρίων,ονομασμένα σύμφωνα με το κρανιακό οστό που τα καλύπτει). Το ίδιο σχετίζεται με τον έλεγχο
Δυνάμεις Starling. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 03/10/2017
 Δυνάμεις Starling Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 03/10/2017 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2017-2018 Πιέσεις σε όλο το μήκος της συστημικής
Δυνάμεις Starling Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 03/10/2017 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2017-2018 Πιέσεις σε όλο το μήκος της συστημικής
ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ
 ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 5-6/29-02-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ 1. ΕΙΣΑΓΩΓΗ 2. ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 3. ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 4. ΚΕΝΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΡΥΘΜΙΣΗ 1. ΕΙΣΑΓΩΓΗ διαβίβαση στο νευρικό
ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 5-6/29-02-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ 1. ΕΙΣΑΓΩΓΗ 2. ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 3. ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 4. ΚΕΝΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΡΥΘΜΙΣΗ 1. ΕΙΣΑΓΩΓΗ διαβίβαση στο νευρικό
ΑΥΤΟΝΟΜΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ (ΑΝΣ) ΠΑΥΛΟΣ Γ. ΚΑΤΩΝΗΣ ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΙΑΤΡΙΚΗΣ ΣΧΟΛΗΣ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΡΗΤΗΣ
 ΠΑΥΛΟΣ Γ. ΚΑΤΩΝΗΣ ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΙΑΤΡΙΚΗΣ ΣΧΟΛΗΣ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΡΗΤΗΣ") ΑΥΤΟΝΟΜΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ (ΑΝΣ) ΠΑΥΛΟΣ Γ. ΚΑΤΩΝΗΣ ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΙΑΤΡΙΚΗΣ ΣΧΟΛΗΣ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΡΗΤΗΣ ΑΥΤΟΝΟΜΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ (ΑΝΣ) ΑΝΣ ΚΙΝΗΤΙΚΑ ΝΕΥΡΑ (λείοι μύες, καρδιακός μυς, αδένες) (Σπλαχνικά Νεύρα)
ΑΥΤΟΝΟΜΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ (ΑΝΣ) ΠΑΥΛΟΣ Γ. ΚΑΤΩΝΗΣ ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΙΑΤΡΙΚΗΣ ΣΧΟΛΗΣ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΚΡΗΤΗΣ ΑΥΤΟΝΟΜΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ (ΑΝΣ) ΑΝΣ ΚΙΝΗΤΙΚΑ ΝΕΥΡΑ (λείοι μύες, καρδιακός μυς, αδένες) (Σπλαχνικά Νεύρα)
ΜΑΘΗΜΑ 5ο ΜΕΡΟΣ Β ΠΑΡΕΓΚΕΦΑΛΙΔΑ
 ΜΑΘΗΜΑ 5ο ΜΕΡΟΣ Β ΠΑΡΕΓΚΕΦΑΛΙΔΑ ΠΑΡΕΓΚΕΦΑΛΙΔΑ Η παρεγκεφαλίδα βρίσκεται στον οπίσθιο κρανιακό βόθρο, πίσω από τη γέφυρα και τον προμήκη μυελό Αποτελείται από δύο ημισφαίρια που συνδέονται μεταξύ τους με
ΜΑΘΗΜΑ 5ο ΜΕΡΟΣ Β ΠΑΡΕΓΚΕΦΑΛΙΔΑ ΠΑΡΕΓΚΕΦΑΛΙΔΑ Η παρεγκεφαλίδα βρίσκεται στον οπίσθιο κρανιακό βόθρο, πίσω από τη γέφυρα και τον προμήκη μυελό Αποτελείται από δύο ημισφαίρια που συνδέονται μεταξύ τους με
94 Η χρήση των νευρωνικών µοντέλων για την κατανόηση της δοµής και λειτουργίας τού εγκεφάλου. = l b. K + + I b. K - = α n
 Nευροφυσιολογία Η μονάδα λειτουργίας του εγκεφάλου είναι ένας εξειδικευμένος τύπος κυττάρου που στη γλώσσα της Νευροφυσιολογίας ονομάζεται νευρώνας. Το ηλεκτρονικό μικροσκόπιο αποκαλύπτει ότι ο ειδικός
Nευροφυσιολογία Η μονάδα λειτουργίας του εγκεφάλου είναι ένας εξειδικευμένος τύπος κυττάρου που στη γλώσσα της Νευροφυσιολογίας ονομάζεται νευρώνας. Το ηλεκτρονικό μικροσκόπιο αποκαλύπτει ότι ο ειδικός
Το Παρασυµπαθητικό Νευρικό Σύστηµα λκλλκλκλλκκκκ
 Το Παρασυµπαθητικό Νευρικό Σύστηµα λκλλκλκλλκκκκ Εισαγωγή Παρασυµπαθητική Φυγόκεντρος Οδός Κεντρική Μοίρα (Εγκεφαλικό Σκέλος) Ιερή Μοίρα (Ιερό Σκέλος) Προγαγγλιακές Ίνες Τα Παρασυµπαθητικά Γάγγλια και
Το Παρασυµπαθητικό Νευρικό Σύστηµα λκλλκλκλλκκκκ Εισαγωγή Παρασυµπαθητική Φυγόκεντρος Οδός Κεντρική Μοίρα (Εγκεφαλικό Σκέλος) Ιερή Μοίρα (Ιερό Σκέλος) Προγαγγλιακές Ίνες Τα Παρασυµπαθητικά Γάγγλια και
Συστήματα επικοινωνίας Ανθρωπίνου σώματος. ενδοκρινολογικό νευρικό σύστημα
 Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ. Οι ρυθμιστές του οργανισμού
 ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Οι ρυθμιστές του οργανισμού Είδη αδένων στον άνθρωπο o Εξωκρινείς αδένες: εκκρίνουν το προϊόν τους μέσω εκφορητικού πόρου είτε στην επιφάνεια του σώματος (π.χ. ιδρωτοποιοί και σμηγματογόνοι
ΕΝΔΟΚΡΙΝΕΙΣ ΑΔΕΝΕΣ Οι ρυθμιστές του οργανισμού Είδη αδένων στον άνθρωπο o Εξωκρινείς αδένες: εκκρίνουν το προϊόν τους μέσω εκφορητικού πόρου είτε στην επιφάνεια του σώματος (π.χ. ιδρωτοποιοί και σμηγματογόνοι
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ. ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΣΥΣΠΑΣΗΣ
 ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
Μεταιχμιακό Σύστημα του Εγκεφάλου
 Μεταιχμιακό Σύστημα του Εγκεφάλου Άρθρο του ΧΑΡΑΛΑΜΠΟY ΤΙΓΓΙΝΑΓΚΑ, MT, CST, MNT Το μεταιχμιακό σύστημα ελέγχει το κύκλωμα του χρόνιου πόνου και των συναισθημάτων, ενώ συνδέεται με τα βαθύτερα τμήματα του
Μεταιχμιακό Σύστημα του Εγκεφάλου Άρθρο του ΧΑΡΑΛΑΜΠΟY ΤΙΓΓΙΝΑΓΚΑ, MT, CST, MNT Το μεταιχμιακό σύστημα ελέγχει το κύκλωμα του χρόνιου πόνου και των συναισθημάτων, ενώ συνδέεται με τα βαθύτερα τμήματα του
Μεταπτυχιακή διατριβή
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΜΕΛΕΤΗ ΤΗΣ ΕΚΛΕΚΤΙΚΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΟΞΕΙΔΩΣΗΣ ΤΟΥ ΜΕΘΑΝΙΟΥ ΠΡΟΣ ΔΙΟΞΕΙΔΙΟ ΤΟΥ ΑΝΘΡΑΚΑ Βασιλική
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΓΕΩΤΕΧΝΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΚΑΙ ΔΙΑΧΕΙΡΙΣΗΣ ΠΕΡΙΒΑΛΛΟΝΤΟΣ Μεταπτυχιακή διατριβή ΜΕΛΕΤΗ ΤΗΣ ΕΚΛΕΚΤΙΚΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΟΞΕΙΔΩΣΗΣ ΤΟΥ ΜΕΘΑΝΙΟΥ ΠΡΟΣ ΔΙΟΞΕΙΔΙΟ ΤΟΥ ΑΝΘΡΑΚΑ Βασιλική
Εγκέφαλος-Αισθητήρια Όργανα και Ορμόνες. Μαγδαληνή Γκέιτς Α Τάξη Γυμνάσιο Αμυγδαλεώνα
 Εγκέφαλος-Αισθητήρια Όργανα και Ορμόνες O εγκέφαλος Ο εγκέφαλος είναι το κέντρο ελέγχου του σώματος μας και ελέγχει όλες τις ακούσιες και εκούσιες δραστηριότητες που γίνονται μέσα σε αυτό. Αποτελεί το
Εγκέφαλος-Αισθητήρια Όργανα και Ορμόνες O εγκέφαλος Ο εγκέφαλος είναι το κέντρο ελέγχου του σώματος μας και ελέγχει όλες τις ακούσιες και εκούσιες δραστηριότητες που γίνονται μέσα σε αυτό. Αποτελεί το
ΛΕΙΤΟΥΡΓΙΚΗ ΑΝΑΤΟΜΙΚΗ ΠΑΡΕΓΚΕΦΑΛΙΔΟΣ
 ΛΕΙΤΟΥΡΓΙΚΗ ΑΝΑΤΟΜΙΚΗ ΠΑΡΕΓΚΕΦΑΛΙΔΟΣ ΦΑΙΔΩΝΑΣ ΛΙΑΚΟΣ ΝΕΥΡΟΧΕΙΡΟΥΡΓΟΣ ΕΠΙΣΤΗΜΟΝΙΚΟΣ ΣΥΝΕΡΓΑΤΗΣ ΠΕΙΡΑΜΑΤΙΚΟΥ ΕΡΓΑΣΤΗΡΙΟΥ ΝΕΥΡΟΧΕΙΡΟΥΡΓΙΚΗΣ ΑΝΑΤΟΜΙΑΣ ΛΕΙΤΟΥΡΓΙΚΗ ΑΝΑΤΟΜΙΚΗ ΠΑΡΕΓΚΕΦΑΛΙΔΟΣ ΕΜΒΡΥΟΛΟΓΙΑ ΧΕΙΡΟΥΡΓΙΚΗ
ΛΕΙΤΟΥΡΓΙΚΗ ΑΝΑΤΟΜΙΚΗ ΠΑΡΕΓΚΕΦΑΛΙΔΟΣ ΦΑΙΔΩΝΑΣ ΛΙΑΚΟΣ ΝΕΥΡΟΧΕΙΡΟΥΡΓΟΣ ΕΠΙΣΤΗΜΟΝΙΚΟΣ ΣΥΝΕΡΓΑΤΗΣ ΠΕΙΡΑΜΑΤΙΚΟΥ ΕΡΓΑΣΤΗΡΙΟΥ ΝΕΥΡΟΧΕΙΡΟΥΡΓΙΚΗΣ ΑΝΑΤΟΜΙΑΣ ΛΕΙΤΟΥΡΓΙΚΗ ΑΝΑΤΟΜΙΚΗ ΠΑΡΕΓΚΕΦΑΛΙΔΟΣ ΕΜΒΡΥΟΛΟΓΙΑ ΧΕΙΡΟΥΡΓΙΚΗ
Συστήµατα Αισθήσεων Σωµατικές Αισθήσεις
 Συστήµατα Αισθήσεων Σωµατικές Αισθήσεις ΟΡΓΑΝΩΣΗ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Φλοιός (Ανώτερος Εγκέφαλος) Κατώτερος Εγκέφαλος Ειδικές Αισθήσεις Εν τω Βάθει Αισθητικότητα Επί πολλής Αισθητικότητα Χυµικά Ερεθίσµατα
Συστήµατα Αισθήσεων Σωµατικές Αισθήσεις ΟΡΓΑΝΩΣΗ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Φλοιός (Ανώτερος Εγκέφαλος) Κατώτερος Εγκέφαλος Ειδικές Αισθήσεις Εν τω Βάθει Αισθητικότητα Επί πολλής Αισθητικότητα Χυµικά Ερεθίσµατα
Το μυϊκό σύστημα αποτελείται από τους μύες. Ο αριθμός των μυών του μυϊκού συστήματος ανέρχεται στους 637. Οι μύες είναι όργανα για τη σωματική
 Μύες Το μυϊκό σύστημα αποτελείται από τους μύες. Ο αριθμός των μυών του μυϊκού συστήματος ανέρχεται στους 637. Οι μύες είναι όργανα για τη σωματική κινητικότητα, την σπλαχνική κινητικότητα και τη κυκλοφορία
Μύες Το μυϊκό σύστημα αποτελείται από τους μύες. Ο αριθμός των μυών του μυϊκού συστήματος ανέρχεται στους 637. Οι μύες είναι όργανα για τη σωματική κινητικότητα, την σπλαχνική κινητικότητα και τη κυκλοφορία
ΜΑΘΗΜΑ 8ο ΜΕΡΟΣ Α ΑΙΜΑΤΟ-ΕΓΚΕΦΑΛΙΚΟΣ ΦΡΑΓΜΟΣ
 ΜΑΘΗΜΑ 8ο ΜΕΡΟΣ Α ΑΙΜΑΤΟ-ΕΓΚΕΦΑΛΙΚΟΣ ΦΡΑΓΜΟΣ ΑΙΜΑΤΟ-ΕΓΚΕΦΑΛΙΚΟΣ ΦΡΑΓΜΟΣ ΚΑΙ ΦΡΑΓΜΟΣ ΑΙΜΑΤΟΣΕΓΚΕΦΑΛΟΝΩΤΙΑΙΟΥ ΥΓΡΟΥ Το ΚΝΣ για να λειτουργεί φυσιολογικά χρειάζεται πολύ σταθερό περιβάλλον Η σταθερότητα αυτή
ΜΑΘΗΜΑ 8ο ΜΕΡΟΣ Α ΑΙΜΑΤΟ-ΕΓΚΕΦΑΛΙΚΟΣ ΦΡΑΓΜΟΣ ΑΙΜΑΤΟ-ΕΓΚΕΦΑΛΙΚΟΣ ΦΡΑΓΜΟΣ ΚΑΙ ΦΡΑΓΜΟΣ ΑΙΜΑΤΟΣΕΓΚΕΦΑΛΟΝΩΤΙΑΙΟΥ ΥΓΡΟΥ Το ΚΝΣ για να λειτουργεί φυσιολογικά χρειάζεται πολύ σταθερό περιβάλλον Η σταθερότητα αυτή
«Η ομορφιά εξαρτάται από τα μάτια εκείνου που τη βλέπει»
 «Η ομορφιά εξαρτάται από τα μάτια εκείνου που τη βλέπει» Γνωστική Νευροεπιστήμη Πώς γίνεται αντιληπτή η αισθητική πληροφορία; Πώς σχηματίζονται οι μνήμες; Πώς μετασχηματίζονται σε λόγο οι αντιλήψεις και
«Η ομορφιά εξαρτάται από τα μάτια εκείνου που τη βλέπει» Γνωστική Νευροεπιστήμη Πώς γίνεται αντιληπτή η αισθητική πληροφορία; Πώς σχηματίζονται οι μνήμες; Πώς μετασχηματίζονται σε λόγο οι αντιλήψεις και
ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΓΙΑ ΤΙΣ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ HELLENIC SOCIETY FOR NEUROSCIENCE
 Τ.Θ. 5, Πανε ιστήµιο Πατρών, 650 Πάτρα, P.O. Box 5, University of Patras, 6 50 Patra ΠΑΓΚΟΣΜΙΑ ΕΒ ΟΜΑ Α ΕΝΗΜΕΡΩΣΗΣ ΓΙΑ ΤΟΝ ΕΓΚΕΦΑΛΟ ΕΓΚΕΦΑΛΟΣ ΚΑΙ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ: ΤΙ ΓΝΩΡΙΖΟΥΝ ΟΙ ΤΕΛΕΙΟΦΟΙΤΟΙ ΤΗΣ ΥΠΟΧΡΕΩΤΙΚΗΣ
Τ.Θ. 5, Πανε ιστήµιο Πατρών, 650 Πάτρα, P.O. Box 5, University of Patras, 6 50 Patra ΠΑΓΚΟΣΜΙΑ ΕΒ ΟΜΑ Α ΕΝΗΜΕΡΩΣΗΣ ΓΙΑ ΤΟΝ ΕΓΚΕΦΑΛΟ ΕΓΚΕΦΑΛΟΣ ΚΑΙ ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ: ΤΙ ΓΝΩΡΙΖΟΥΝ ΟΙ ΤΕΛΕΙΟΦΟΙΤΟΙ ΤΗΣ ΥΠΟΧΡΕΩΤΙΚΗΣ
ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I)
") ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν