ΔΙΠΛΩΜΑΤΙΚΗ ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΜΟΡΙΑΚΗ ΑΝΑΛΥΣΗ ΤΟΥ ΡΕΠΕΡΤΟΡΙΟΥ ΤΟΥ Τ ΚΥΤΤΑΡΙΚΟΥ ΥΠΟΔΟΧΕΑ ΣΕ ΑΣΘΕΝΕΙΣ ΜΕ ΑΙΜΑΤΟΛΟΓΙΚΑ ΝΟΣΗΜΑΤΑ
|
|
|
- φώλος Μοσχοβάκης
- 8 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΓΝΘ «Γ. ΠΑΠΑΝΙΚΟΛΑΟΥ» ΑΙΜΑΤΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ - ΜΟΝΑΔΑ ΜΕΤΑΜΟΣΧΕΥΣΗΣ ΑΙΜΟΠΟΙΗΤΙΚΩΝ ΚΥΤΤΑΡΩΝ ΔΙΠΛΩΜΑΤΙΚΗ ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΜΟΡΙΑΚΗ ΑΝΑΛΥΣΗ ΤΟΥ ΡΕΠΕΡΤΟΡΙΟΥ ΤΟΥ Τ ΚΥΤΤΑΡΙΚΟΥ ΥΠΟΔΟΧΕΑ ΣΕ ΑΣΘΕΝΕΙΣ ΜΕ ΑΙΜΑΤΟΛΟΓΙΚΑ ΝΟΣΗΜΑΤΑ ΕΥΑΓΓΕΛΙΑ ΣΤΑΛΙΚΑ Υπότροφος ΙΚΥ Θεσσαλονίκη, Νοέμβριος 2013 [i]
2 Πίνακας Περιεχομένων Πίνακας Περιεχομένων... i Πρόλογος Εισαγωγή Εισαγωγή στο ανοσοποιητικό σύστημα Δομή του Τ κυτταρικού υποδοχέα Οργάνωση του συμπλόκου TCR-CD Βιοσύνθεση και συναρμολόγηση του συμπλόκου TCR-CD Ενεργοποίηση του συμπλόκου TCR-CD Γενετικοί τόποι των αλυσίδων του T κυτταρικού υποδοχέα Γενικά Γενετικός τόπος της β αλυσίδας του Τ κυτταρικού υποδοχέα Μηχανισμοί δημιουργίας ποικιλότητας των υποδοχέων των Τ κυττάρων Γενικά Συνδυαστική ποικιλότητα και ανασυνδυασμός V(D)J Συνδετική ποικιλότητα (ποικιλότητα συμβολών) Διαφοροποίηση των T λεμφοκυττάρων Φάση διαφοροποίησης ανεξαρτητη από το αντιγόνο Φάση διαφοροποίησης εξαρτώμενη από το αντιγόνο Επιλογή των Τ λεμφοκυττάρων στο θύμο αδένα CD4 λεμφοκύτταρα Th1 κύτταρα Th2 κύτταρα Th17 κύτταρα Treg κύτταρα Ενεργοποίηση των παρθένων βοηθητικών Τ λεμφοκυττάρων CD8 λεμφοκύτταρα [i]
3 Ενεργοποίηση των παρθένων κυτταροτοξικών Τ λεμφοκυττάρων Μνημονικά Τ λεμφοκύτταρα Απόπτωση των Τ λεμφοκυττάρων Γενικά Μηχανισμός επαγόμενης ενεργοποίησης της απόπτωσης στα Τ λεμφοκύτταρα Χρόνια Ιδιοπαθής Ουδετεροπενία Γενικά Επιδημιολογικά στοιχεία Κλινική εικόνα Εργαστηριακά ευρήματα Παθοφυσιολογία της νόσου CIN: η μελέτη της νόσου στα πλαίσια των συνδρόμων μυελικής ανεπάρκειας ο ρόλος της αυτοανοσίας Αντικείμενο της μελέτης Μεθοδολογία Ομάδα Μελέτης Υλικά και Μέθοδοι Απομόνωση μονοπύρηνων κυττάρων από δείγμα αίματος Απομόνωση RNA Σύνθεση cdna Ενίσχυση κλωνικών αναδιατάξεων της μεταβλητής περιοχής του Τ κυτταρικού υποδοχέα με αλυσιδωτή αντίδραση πολυμεράσης Καθαρισμός των προϊόντων PCR Kλωνοποίηση Aνάλυση των αποτελεσμάτων της κλωνοποίησης Αποτελέσματα Περιγραφή ασθενών Ρεπερτόριο TRBV στον ασθενή 1 _(CR6) [ii]
4 4.3. Ρεπερτόριο TRBV στον ασθενή 2_CR Ρεπερτόριο TRBV στον ασθενή 3_CIN Ρεπερτόριο TRBV στον ασθενή 4_CIN Ρεπερτόριο TRBV στον ασθενή 5_CIN Ρεπερτόριο TRBV στον ασθενή 6_CIN Μελέτη του ρεπερτορίου στο σύνολο των ασθενών Συζήτηση Χρόνια Ιδιοπαθής Ουδετεροπενία : ένα ευρύ φάσμα κλινικών εκδηλώσεων Μοριακή ανάλυση του ρεπερτορίου των γονιδίων TRB στην Χρόνια Ιδιοπαθή Ουδετεροπενία Περίληψη ΑΓΓΛΙΚΗ ΠΕΡΙΛΗΨΗ Abstract Βιβλιογραφία [iii]
5 Πρόλογος Δεδομένα από σειρά μελετών σε νοσήματα που σχετίζονται με σύνδρομα μυελικής ανεπάρκειας, όπως είναι η Απλαστική Αναιμία, η T-LGL λευχαιμία και τα μυελοδυσπλαστικά σύνδρομα, συγκλίνουν στην παρουσία αυτοάνοσων διαδικασιών μέσα στο γενικότερο πλαίσιο της ύπαρξης ολιγο/μονοκλωνικών εκπτύξεων κυτταροτοξικών Τ λεμφοκυττάρων που πιθανά στρέφονται έναντι των διαφόρων προβαθμίδων της μυελικής σειράς. Η μοριακή ανάλυση των γονιδίων της β αλυσίδας του Τ κυτταρικού υποδοχέα σε ασθενείς που εμφανίζουν υπερπλαστικές εξεργασίες των CD3+CD8+ Τ λεμφοκυττάρων συμβάλλει στην κατανόηση των υποκείμενων παθογενετικών μηχανισμών. Επίσης, προσφέρει ανοσογενετικές ενδείξεις επιλογής από αντιγόνο, υπό την έννοια ότι η έντονη αντιγονική διέγερση μπορεί αρχικά να οδηγήσει σε πολυκλωνική υπερπλασία και, υπό άγνωστες ακόμα προϋποθέσεις να αναδείξει ολιγοκλωνικούς ή μονοκλωνικούς Τ πληθυσμούς με μυελοκατασταλτικές ιδιότητες στο αίμα ή και στον μυελό των ασθενών. Στην παρούσα μελέτη παρουσιάζουμε τα αποτελέσματα από τη λεπτομερή ανάλυση έξι ασθενών με Χρόνια Ιδιοπαθή Ουδετεροπενία, ένα σύνδρομο μυελικής ανεπάρκειας χαρακτηριζόμενο από υποπλασία και αριστρεφή στροφή της κοκκιώδους σειράς στο μυελό. Η μελέτη πραγματοποιήθηκε στο Εργαστήριο Μοριακής και Κυτταρικής Βιολογίας της Αιματολογικής Κλινικής και Μονάδας Μεταμόσχευσης Αιμοποιητικών Κυττάρων του ΠΓΝΘ «Γ. Παπανικολάου» που διευθύνει ο κ. Αχιλλέας Αναγνωστόπουλος, τον οποίο ευχαριστώ θερμά για τη δυνατότητα που μου προσέφερε και την υποστήριξη που παρείχε στην εκπόνηση της παρούσας μελέτης. Θα ήθελα να ευχαριστήσω επίσης την επιβλέπουσα της διπλωματικής εργασίας κ. Αναστασία Κουβάτση, Αναπληρώτρια Καθηγήτρια του Τμήματος Βιολογίας του Αριστοτελείου Πανεπιστημίου Θεσσαλονίκης για την καθοδήγηση και τη βοήθειά της σε κάθε φάση της δημιουργίας. Η βασική εκπαίδευση, το πειραματικό μέρος και η ανάλυση των αποτελεσμάτων της μελέτης έγιναν υπό την καθοδήγηση του κ. Κώστα Σταματόπουλου, Επιμελητή της Αιματολογικής Κλινικής και Μονάδας Μεταμόσχευσης Αιμοποιητικών Κυττάρων του ΠΓΝΘ «Γ. Παπανικολάου» Ο κ. Σταματόπουλος παρακολούθησε ενεργά όλες τις φάσεις εκπόνησης της παρούσας διπλωματικής εργασίας και έδωσε λύσεις στα μεθοδολογικά και άλλα προβλήματα που ανέκυψαν στην εξέλιξη της μελέτης, για όλους τους παραπάνω λόγους τον ευχαριστώ θερμα. [1]
6 Η παρούσα διπλωματική εργασία δε θα είχε ολοκληρωθεί χωρίς τη βοήθεια των συνεργατών του Εργαστηρίου Μοριακής και Κυτταρικής Βιολογίας της Αιματολογικής Κλινικής και Μονάδας Μεταμόσχευσης Αιμοποιητικών Κυττάρων του ΠΓΝΘ «Γ. Παπανικολάου»: η βιολόγος Αναστασία Χατζηδημητρίου και η τεχνολόγος Μαρία Καρυπίδου συνέβαλαν ουσιαστικά στη διεκπεραίωση μεγάλου μέρους των πειραμάτων. Τέλος, η ολοκλήρωση της παραπάνω εργασίας έγινε στο πλαίσιο της υλοποίησης του μεταπτυχιακού προγράμματος το οποίο συγχρηματοδοτήθηκε μέσω της Πράξης «Πρόγραμμα χορήγησης υποτροφιών ΙΚΥ με διαδικασία εξατομικευμένης αξιολόγησης ακαδημαϊκού έτους » από τους πόρους του Ε.Π. «Εκαπίδευση και Δια Βίου Μάθηση» του Ευρωπαϊκού Κοινωνικού Ταμείου (ΕΚΤ) και του ΕΣΠΑ( )» Θεσσαλονίκη, Νοέμβριος 2013 [2]
7 [3] ΕΙΣΑΓΩΓΗ
8 1. Εισαγωγή 1.1. Εισαγωγή στο ανοσοποιητικό σύστημα Το ανοσοποιητικό σύστημα εξελίχθηκε για να προστατεύει τον οργανισμό από παθογόνους παράγοντες. Κάθε άνοση απάντηση περιλαμβάνει αναγνώριση του παθογόνου ή άλλου ξένου υλικού και αντίδραση εναντίον του, με σκοπό την εξάλειψή του. Γενικά, οι διάφοροι τύποι άνοσης απάντησης διακρίνονται σε δύο κατηγορίες: έμφυτες (innate) και προσαρμοστικές (adaptive). Η σημαντική διαφορά μεταξύ τους είναι ότι η προσαρμοστική άνοση απάντηση είναι πολύ πιο εξειδικευμένη για ένα συγκεκριμένο παθογόνο. Επιπλέον, η έμφυτη απάντηση δε μεταβάλλεται μετά από επανειλημμένη έκθεση σε συγκεκριμένο παθογόνο, ενώ η προσαρμοστική απάντηση γίνεται πιο αποτελεσματική σε κάθε επόμενη επαφή με το ίδιο παθογόνο. Επομένως, τα δύο κύρια χαρακτηριστικά της προσαρμοστικής άνοσης απάντησης είναι η εξειδίκευση σε συγκεκριμένο αντιγονικό επίτοπο και η ύπαρξη μνήμης που προσφέρει στο ανοσοποιητικό σύστημα την ικανότητα ν αντιδρά πολύ πιο γρήγορα και αποτελεσματικά σε επόμενη επαφής μ ένα συγκεκριμένο αντιγόνο. Στα κύτταρα του ανοσοποιητικού συστήματος ανήκουν τα λεμφοκύτταρα, τα οποία διαχωρίζονται σε Β, Τ και ΝΚ κύτταρα. Η προσαρμοστική άνοση απάντηση διακρίνεται σε χυμική (Β λεμφοκύτταρα) και κυτταρική (Τ λεμφοκύτταρα). Τα Τ λεμφοκύτταρα παράγονται στο μυελό των οστών και μεταναστεύουν στο θύμο αδένα κατά την εμβρυϊκή και νεογνική ζωή. Εκεί συμβαίνει η διαφοροποίησή τους σε ώριμα Τ-λεμφοκύτταρα, τα οποία υπάρχουν στον περιφερικό λεμφικό ιστό, στο αίμα και στη λέμφο. Κάθε T λεμφοκύτταρο φέρει στην επιφάνειά του έναν υποδοχέα (T cell receptor,tcr) που είναι εξειδικευμένος για ένα συγκεκριμένο αντιγόνο. Η τυχαία αναδιάταξη του γενετικού υλικού των T λεμφοκυττάρων είναι υπεύθυνη για τη δημιουργία διαφορετικής ειδικότητας υποδοχέα σε κάθε T λεμφοκύτταρο. Προκειμένου ν αναγνωριστεί το αντιγόνο από τα Τ λεμφοκύτταρα, πρέπει να έχει προηγηθεί η επεξεργασία και παρουσίασή του σε κατάλληλη μορφή από κάποιο αντιγονοπαρουσιαστικό κύτταρο (μακροφάγο, δενδριτικό κύτταρο, Β λεμφοκύτταρο). Τα Τ λεμφοκύτταρα διαιρούνται σε βοηθητικά/επαγωγικά (χαρακτηρίζονται από έκφραση του δείκτη CD4) και σε κατασταλτικά/κυτταροτοξικά (χαρακτηρίζονται από έκφραση του δείκτη CD8). Τα κύτταρα της πρώτης κατηγορίας ενισχύουν την άνοση απάντηση και βοηθούν στη διαφοροποίηση των Β λεμφοκυττάρων σε πλασματοκύτταρα. Τα κατασταλτικά λεμφοκύτταρα έχουν ρυθμιστικό ρόλο, περιορίζοντας την άνοση απάντηση. Τα κυτταροτοξικά λεμφοκύτταρα έχουν ως αποστολή τη λύση κυττάρων που έχουν [4]
 και")
9 προσβληθεί από μικροοργανισμούς ή έχουν υποστεί κακοήθη εξαλλαγή και αναπτύσσονται ανεξέλεγκτα Δομή του Τ κυτταρικού υποδοχέα Η α και β αλυσίδα του TCRβα και η γ και δ αλυσίδα του TCRγδ διαθέτουν : Ν-τελικό εξωκυττάριο τμήμα αποτελούμενο από μια μεταβλητή περιοχή ( αμινοξέα) και μια σταθερή περιοχή ( αμινοξέα) συνδετικό πεπτίδιο ( αμινοξέα) διαμεμβρανικό τμήμα (17-26 αμινοξέα) C-τελικό κυτταροπλασματικό ακρο (0-7 αμινοξέα) 1,2 Κάθε αλυσίδα περιλαμβάνει ένα κατάλοιπο κυστεϊνης κοντά στο διαμεμβρανικό τμήμα. Αυτό το κατάλοιπο παίζει ρόλο στο σχηματισμό δισουλφυδριλικού δεσμού ανάμεσα στις α- β και γ-δ αλυσίδες. Εξαίρεση από τον κανόνα αποτελεί η γ2 αλυσίδα του ανθρώπινου Τ κυτταρικού υποδοχέα η οποία δε διαθέτει κυστεϊνη και δε συνδέεται με δισουλφυδρυλικούς δεσμούς με τη δ αλυσίδα. Επίσης κάθε αλυσίδα συνδέεται με τη λιπιδική διπλοστοιβάδα με μια υδρόφοβη διαμεμβρανική περιοχή, της οποίας χαρακτηριστικό είναι η παρουσία ηλεκτροθετικών αμινοξέων (Εικόνα 1). Η παρουσία τέτοιων φορτισμένων καταλοίπων σε διαμεμβρανικές περιοχές δεν είναι συνήθης και συχνά επιδρά αποσταθεροποιητικά. Ωστόσο, στη συγκεκριμένη περίπτωση, αυτά τα φορτισμένα κατάλοιπα παίζουν σημαντικό ρόλο στην αλληλεπίδραση και σταθεροποίηση του συμπλόκου TCR-CD3. Εικόνα 1. Βασική δομή του Τ κυτταρικού υποδοχέα. [5]

10 Οργάνωση του συμπλόκου TCR-CD3 Οι πρωτεΐνες του συμπλόκου CD3 έχουν κυτταροπλασματικές προεξοχές που τους επιτρέπουν να αλληλεπιδρούν με πρωτεΐνες μεταβίβασης σήματος. Αντίθετα καμία από τις αλυσίδες του Τ κυτταρικού υποδοχέα δεν έχει ανάλογο κυτταροπλασματικό τμήμα που θα μπορούσε να δώσει σήμα στο κύτταρο ότι έχει ενεργοποιηθεί ο Τ κυτταρικός υποδοχέας. Το CD3 αποτελείται από πέντε πολυπεπτίδικες αλυσίδες: γ, δ, ε (υπάρχουν ως ετεροδιμερή: γ-ε και δ-ε), ζ και η (υπάρχουν είτε ως ομοδιμερή (ζ-ζ) είτε ως ετεροδιμερή (ζ-η) (εικόνα 2). Η ζ και η αλυσίδα κωδικοποιούνται από το ίδιο γονίδιο και διαφέρουν μόνο στο καρβοξυτελικό άκρο. Εικόνα 2: Δομή συμπλόκου CD3-TCR Οι αλυσίδες CD3γ, CD3δ και CD3ε αποτελούνται από ένα Ν-τελικό εξωκυττάριο τμήμα, ένα συνδετικό πεπτίδιο, μια διαμεμβρανική και μια κυτταροπλασματική περιοχή. Οι διαμεμβρανικές περιοχές χαρακτηρίζονται από την παρουσία όξινων (αρνητικά φορτισμένων) καταλοίπων που σχηματίζουν ιοντικούς δεσμούς με τα βασικά (θετικά φορτισμένα) αμινοξέα στη διαμεμβρανική περιοχή του Τ κυτταρικού υποδοχέα. Οι κυτταροπλασματικές περιοχές φέρουν χαρακτηριστικές ακολουθίες με γενικό τύπο Υxx(L/I)x6-8Yxx(L/I), γνωστές ως ΙΤΑΜ (Immunoreceptor Tyrosine-based Activation Motif) χάρη στις οποίες μπορεί να συνδέονται με κυτταροπλασματικές κινάσες τυροσίνης μετά τη διέγερση του Τ κυτταρικού υποδοχέα. Οι αλυσίδες CD3γ, CD3δ και CD3ε διαθέτουν μια ακολουθία ΙΤΑΜ, ενώ οι αλυσίδες CD3ζ και CD3η τρεις και δύο ακολουθίες, αντιστοίχως. [6]
11 Βιοσύνθεση και συναρμολόγηση του συμπλόκου TCR-CD3 Το Τ λεμφοκύτταρο δε μπορεί ν αναγνωρίσει αντιγόνο αν το σύμπλοκο TCR-CD3 δε συναρμολογηθεί κι εκφραστεί σωστά στην κυτταρική επιφάνεια 3,4,5 [7] Οι πρωτεΐνες του συμπλόκου TCR-CD3 εκφράζονται διαδοχικά στα διάφορα στάδια ωρίμανσης του Τ λεμφοκυττάρου. Οι πρωτεΐνες CD3γ, CD3δ και CD3ε συνθέτονται στα πρώιμα θυμοκύτταρα, αλλά παραμένουν μέσα στο κύτταρο. Η ολοκλήρωση της δομής του Τ κυτταρικού υποδοχέα γίνεται μέσα στο ενδοπλασματικό δίκτυο με τη βοήθεια της πρωτεΐνης CD3ω ή TRAP (T cell Receptor Associated Protein). Στα πρώιμα στάδια ωρίμανσης, το Τ λεμφοκύτταρο εκφράζει στην επιφάνεια του τον pre-t κυτταρικό υποδοχέα ο οποίος είναι ένα ετεροδιμερές της ΤCRβ αλυσίδας και της pta. Μετά τη σύνθεση της TCRα αλυσίδας και τη γλυκοζυλίωση των Ν-τελικών άκρων των αλυσίδων του συμπλόκου TCR-CD3 στη συσκευή Golgi, ολοκληρώνεται η βιοσύνθεση του Τ κυτταρικού υποδοχέα. Τα σύμπλοκα TCR-CD3 μεταφέρονται στη κυτταρική μεμβράνη, ένω οι Τ κυτταρικοί υποδοχείς οι οποίοι δε δημιούργησαν σύμπλοκο με τον CD3 μεταφέρονται στα λυσοσωμάτια όπου και αποδομούνται Ενεργοποίηση του συμπλόκου TCR-CD3 Κυτταροπλασματικές κινάσες τυροσίνης όπως η p56lck και η p59fyn συνδέονται με το σύμπλοκο TCR-CD3 μετά τη διέγερση του Τ κυτταρικού υποδοχέα και είναι υπεύθυνες για τη μεταβίβαση σήματος (Εικόνα 3). Τα μοτίβα CXCP στο κυτταροπλασματικό τμήμα των συν-υποδοχέων CD4 και CD8 προάγουν την πρόσδεσή τους στην κινάση p56lck. Η ενεργοποίηση της lck βρίσκεται υπό τον έλεγχο του CD45. Μετά την αναγνώριση του αντιγόνου από τον TCRβα ή TCRγδ, οι κινάσες p56lck και p59fyn φωσφορυλιώνουν τις αλληλουχίες ITAM των αλυσίδων CD3γ, CD3δ, CD3ε, CD3ζ και την ZAP-70 ξεκινώντας έτσι το σηματοδοτικό μονοπάτι 6,7. Η φωσφορυλίωση της ZAP-70 ενεργοποιεί επίσης τις πρωτεΐνες LAT (Linker for Activation of T cells) και SLP-76 οι οποίες επίσης συμμετέχουν στο σηματοδοτικό μονοπάτι 8. Συγκεκριμένα, οι πρωτεΐνες LAT και SLP-76 συνδέουν τον Τ κυτταρικό υποδοχέα με τις PLC-γ1, Grb-2/Sos και P13K, οδηγώντας σε ενεργοποίηση των μονοπατιών τα οποία εξαρτώνται από κινάσες ασβεστίου και κινάσες MAP (Mitogen- Activated Protein) 7,8,9 H ενεργοποίηση του Τ κυτταρικού υποδοχέα οδηγεί σε αναδιοργάνωση της κυτταρικής μεμβράνης και στο σχηματισμό των SMAC (SupraMolecular Activation Cluster), δηλαδή συνάψεων με τα αντιγονοπαρουσιαστικά κύτταρα. Το κεντρικό τμήμα των SMAC περιλαμβάνει το σύμπλοκο TCR-CD3, τους συν-υποδοχείς CD4 ή CD8, το συν-διεγερτικό

12 μόριο CD28, το CD2, τις κινάσες p56lck, p59fyn, PKC και θ (theta),ε νώ η περιφέρεια είναι εμπλουτισμένη με μόρια προσκόλλησης LFA-1 (Lymphocyte Fuction-Associated-1) και την ταλίνη, η οποία είναι πρωτεΐνη του κυτταροσκελετού 10. Αλλαγές στον κυτταροσκελετό του Τ κυτταρικού υποδοχέα πυροδοτούν και άλλα σηματοδοτικά μονοπάτια, όπως το σηματοδοτικό μονοπάτι SLP76-Vav-Nck το οποίο ενεργοποιεί τα μόρια της οικογένειας Rho 9. Εικόνα 3. Μοντέλο ενεργοποίησης του συμπλόκου TCR-CD3 για τη μεταβίβαση σήματος. Τα CD4- και CD8- Τ λεμφοκύτταρα σε συνδυασμό με την LcK, ενεργοποιούν την ZAP-70 και την φωσφολιπάση Cγ-1 (PLCγ-1) και οδηγούν το μόριο CD45 σε αποκλεισμό. Η φωσφολιπάση PLCg-1 εμπλέκεται στον μεταβολισμό του ασβεστίου, στην ενεργοποίηση της πρωτεϊνικής κινάσης C, ενώ η ZAP-70 φωσφορυλιώνει τον συνδέτη των ενεργοποιημένων Τ λεμφοκυττάρων (LAT). Το μόριο LAT σχετίζεται με πλήθος άλλων μορίων, συμπεριλαμβανομένων και των Vav, επηρεάζοντας την διαμόρφωση του κυτταροσκελετού. Συνεργατικά οι αλλαγές αυτές σε επίπεδο έκφρασης, φωσφορυλίωσης και αναδιαμόρφωσης, προκαλούν διαφορετικά πρότυπα ενεργοποίησης των ΜΑΡΚ κινασών και μετατόπιση των μεταγραφικών παραγόντων, συμπεριλαμβανομένων και του NF-κB, NF- AT, and AP-1.(Εικόνα 4) [8]
.")

13 Εικόνα 4: Μοντέλο αλλαγών που συμβαίνουν κατά την διάρκεια της σύναψης Γενετικοί τόποι των αλυσίδων του T κυτταρικού υποδοχέα Γενικά Οι γενετικοί τόποι του Τ κυτταρικού υποδοχέα περιλαμβάνουν ομάδες γονιδίων (V-D-J-C). Για τη διαμόρφωση των κωδικοποιητικών αλληλουχιών της μεταβλητής περιοχής του Τ κυτταρικού υποδοχέα, επιλέγεται τυχαία ένα γονίδιο από την κάθε ομάδα. Εκτός από τις ομάδες γονιδίων, οι γενετικοί τόποι των αλυσίδων του Τ κυτταρικού υποδοχέα περιλαμβάνουν και πλήθος ρυθμιστικών γονιδίων που καθορίζουν τη μεταγραφή ή την αποσιώπησή, όπως οι αλληλουχίες-οδηγοί (leader sequences, L) πριν από κάθε γονίδιο V. (Εικόνες 5-6) [9]
14 Εικόνα 5: Η διάταξη των γονιδίων στο γενετικό τόπο της α αλυσίδας του Τ κυτταρικού υποδοχέα. Προέλευση: LocusGenes. [10]
15 Εικόνα 6: Η διάταξη των γονιδίων στο γενετικό τόπο της β αλυσίδας του Τ κυτταρικού υποδοχέα. Προέλευση: LocusGenes Γενετικός τόπος της β αλυσίδας του Τ κυτταρικού υποδοχέα Στον άνθρωπο, ο γενετικός τόπος της β αλυσίδας του Τ κυτταρικού υποδοχέα περιλαμβάνει εκτεταμένες περιοχές γονιδίων TRBV και δύο περιοχές D-J-C. Ένα γονίδιο TRBV συνήθως περιλαμβάνει ρυθμιστικές αλληλουχίες πριν από την κωδικοποιητική αλληλουχία και μια αλληλουχία-οδηγό (leader sequence) στο 5 άκρο της, ενώ στο 3 άκρο περιλαμβάνει την αλληλουχία-σήμα του ανασυνδυασμού (recombination signal sequence, RSS) (Εικόνα 7). Στις ρυθμιστικές αλληλουχίες περιλαμβάνεται η αλληλουχία ΤΑΤΑ (ΤΑΤΑ box), μια διατηρημένη περιοχή πλούσια σε αδενίνη και θυμίνη. Η αλληλουχία-σήμα του ανασυνδυασμού (RSS) στο 3 άκρο κάθε γονιδίου TRBV αποτελείται από ένα 7μερές κι ένα 9μερές που χωρίζονται από μια διαστηματική περιοχή (spacer) μήκους 12 bp. Σύμφωνα με τον κανόνα 12-23, ένα γενετικό τμήμα μπορεί να συνδεθεί μόνο με ένα άλλο τμήμα που έχει RSS με spacer 23 bp. Έτσι διασφαλίζεται η σύνδεση των γονιδίων TRBV μόνο με τα γονίδια TRBJ και όχι με τις σταθερές περιοχές TRBC (Εικόνα 8). [11]
J.")
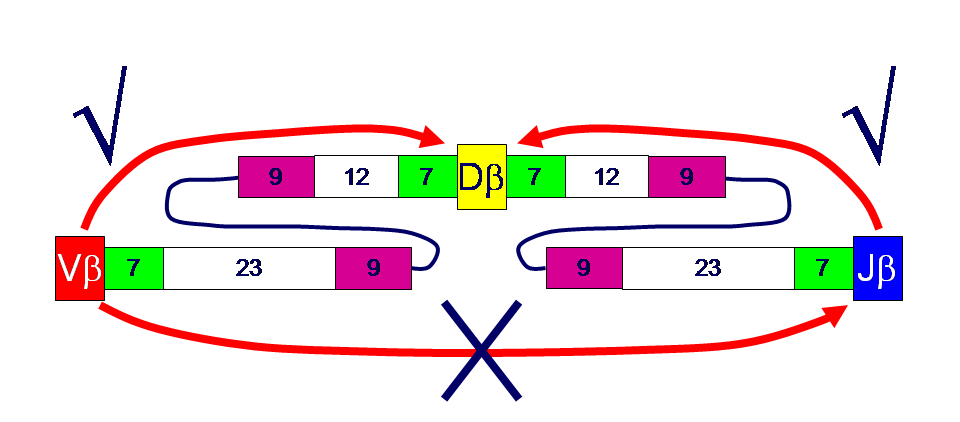
16 Εικόνα 7: Αλληλουχίες-σήματα του ανασυνδυασμού V(D)J. Προέλευση: Εικόνα 8: Ο κανόνας «12-23». [12]
17 Εικόνα 9: Ο γενετικός τόπος TRB. Προέλευση: Ο γενετικός τόπος της β αλυσίδας (TRBV) εντοπίζεται στο χρωμόσωμα 7, στη ζώνη 7q34 του μεγάλου βραχίονα του χρωμοσώματος 7 κι έχει μήκος 620 kb (Εικόνα 9). Συνολικά αποτελείται από γονίδια ανά απλοειδές γονιδίωμα, από τα οποία τα είναι λειτουργικά. Τα από αυτά ανήκουν στη συστοιχία γονιδίων TRBV και κατατάσσονται σε 32 υποομάδες. Επίσης, υπάρχουν δύο γονίδια TRBC. Τα γονίδια TRBD-J-C οργανώνονται σε δύο συστοιχίες. Η πρώτη αποτελείται από ένα γονίδιο TRBD, έξι γονίδια TRBJ και το γονίδιο TRBC1, ενώ η δεύτερη από ένα γονίδιο TRBD, οκτώ γονίδια TRBJ και το γονίδιο TRBC2 1,11,12,13. Τα δύο γονίδια της σταθερής περιοχής κωδικοποιούν πρωτεϊνες οι οποίες διαφέρουν κατά έξι αμινοξέα. Η 5-3 πολικότητα των γονιδίων TRBV υποδεικνύει τον τύπο της αναδιάταξης. Όλα τα γονίδια TRBV ανασυνδυάζονται με απαλοιφή (deletion) του DNA που παρεμβάλλεται ανάμεσα στα γονίδια TRBV και TRBJ, εκτός από το γονίδιο TRBV30 το οποίο ανασυνδυάζεται με αναστροφή (inversion), επειδή έχει αντίστροφη πολικότητα σε σχέση με τα γονίδια TRBJ-TRBC. [13]
18 1.3 Μηχανισμοί δημιουργίας ποικιλότητας των υποδοχέων των Τ κυττάρων Γενικά Το πλέον εντυπωσιακό χαρακτηριστικό του ανοσοποιητικού συστήματος των σπονδυλωτών είναι η ικανότητα αναγνώρισης εκατομμυρίων διαφορετικών παθογόνων μέσω των αντιγονοειδικών υποδοχέων των Τ κυττάρων. Τα λεμφοκύτταρα έχουν την ικανότητα να δημιουργούν μία τεράστια ποικιλότητα αντιγονικών υποδοχέων (>10 8 ). Για τη δημιουργία αυτής της τεράστιας ποικιλότητας υπεύθυνοι είναι τρεις ειδικοί μηχανισμοί: Συνδυαστική ποικιλότητα Ανασυνδυασμός V(D)J Συνδετική ποικιλότητα Συνδυαστική ποικιλότητα και ανασυνδυασμός V(D)J Τα Τ λεμφοκύτταρα αναπτύσσονται από αρχέγονα μητρικά κύτταρα του μυελού των οστών και οι πρόγονοί τους μεταναστεύουν στο θύμο αδένα όπου πολλαπλασιάζονται και διαφοροποιούνται. Ο ανασυνδυασμός V(D)J πραγματοποιείται στο θύμο αδένα κατά την πρώιμη φάση της οντογένεσης των Τ λεμφοκύτταρων. Εξελίσσεται με ιεραρχική σειρά: αρχικά συμβαίνουν αναδιατάξεις των β, γ, και δ γονιδίων. Τα κύτταρα στα οποία συμβαίνουν επιτυχημένες αναδιατάξεις των γ και δ γονιδίων, παράγουν ένα λειτουργικό γδ υποδοχέα μέσω του οποίου μεταβιβάζονται σήματα για την περαιτέρω ανάπτυξη και διαφοροποίησή του προς την γδ σειρά. Σε θυμοκύτταρα στα οποία έχει συμβεί παραγωγική αναδιάταξη των γονιδίων της β αλυσίδας, η παραγόμενη β αλυσίδα συνδέεται μ ένα υποκατάστατο πεπτίδιο της α αλυσίδας (ptα). Αυτά τα κύτταρα πολλαπλασιάζονται γρήγορα, εκφράζουν CD4 και CD8 κι έπειτα αναδιατάσσουν τα γονίδια της α αλυσίδας 14. Κάθε γονίδιο που μπορεί ν αναδιαταχθεί έχει μια αλληλουχία-σήμα του ανασυνδυασμού (RSS, recombination signal sequence) 15,16,17. Οι RSS αποτελούνται από ένα καλά διατηρημένο παλίνδρομο επτανουκλεοτίδιο (heptamer) (5 -CACTGTG-3 ), ένα καλά διατηρημένο εννεανουκλεοτίδιο (nonamer) (5 -ACAAAAACC-3 ), πλούσιο σε Α ή Τ και ανάμεσα τους μια διαστηματική αλληλουχία (spacer region) μήκους 12 ή 23 νουκλεοτιδίων (±1 νουκλεοτίδιο) με διατηρημένο μήκος αλλά χωρίς συντηρημένη αλληλουχία. Το μήκος της διαστηματικής αλληλουχίας παίζει σημαντικό ρόλο στην αντίδραση. Για να συμβεί αποτελεσματικός ανασυνδυασμός απαιτούνται δύο αλληλουχίες RSS, μια με διαστηματική αλληλουχία 12- και μια με 23-bp. Αυτός ο σχετικά απλός περιορισμός, γνωστός ως [14]
19 «κανόνας 12/23», έχει σημαντικές βιολογικές συνέπειες. Πρώτον, όλα τα γονίδια ενός συγκεκριμένου είδους περιβάλλονται από έναν τύπο αλληλουχίας RSS και όλα τα γονίδια με τα οποία θα μπορούσε ν αναδιαταχθούν φέρουν τον αντίθετο τύπο αλληλουχίας. Έτσι, διασφαλίζεται ότι ο ανασυνδυασμός οδηγεί σε λειτουργικό προϊόν. Τα γονίδια της β αλυσίδας ανασυνδυάζονται πρώτα. Αρχικά ανασυνδυάζεται ένα γονίδιο D (TRBD) μ ένα γονίδιο J (TRBJ) και, στη συνέχεια, ένα γονίδιο V ανασυνδυάζεται με το σύμπλοκο DJ. Ακολουθεί μεταγραφή του γενετικού συμπλόκου TCR VDJ, επεξεργασία και συρραφή του προ-mrna (RNA splicing), μετάφραση του ώριμου mrna σε πολυπεπτιδική αλυσίδα στα ριβοσωμάτια και μετασυνθετική τροποποίηση των αντίστοιχων πεπτιδίων (κυρίως με γλυκοζυλίωση στο ενδοπλασματικό δίκτυο). Kάθε γονίδιο V φέρει στο 5 άκρο του ένα μικρό εξόνιο, την αλληλουχία-οδηγό (leader). Αυτό συνθέτει ένα μικρό πεπτίδιοοδηγό ή πεπτίδιο-σήμα (signal ή leader L peptide). Το πεπτίδιο-οδηγός τελικά αφαιρείται από μια πεπτιδάση μετα την είσοδο της πολυπεπτιδικής αλυσίδας στο ενδοπλασματικό δίκτυο (Εικόνα 10). Εικόνα 10: Σχηματική απεικόνιση της σύνθεσης της β αλυσίδας του Τ κυτταρικού υποδοχέα. Όποτε συμβεί παραγωγική αναδιάταξη γονιδίων της β αλυσίδας του TCR, η αντίστοιχη πρωτεΐνη αρχικά εκφράζεται στο εσωτερικό του κυττάρου και στη συνέχεια στην κυτταρική επιφάνεια μαζί με μια αλυσίδα που ονομάζεται ptα 14. Η ptα αλυσίδα είναι ένα υποκατάστατο α αλυσίδας μεγέθους 33kDa. Το ετεροδιμερές β:ptα είναι ένας λειτουργικός υποδοχέας που μεταβιβάζει σήματα μέσω της κινάσης τυροσίνης Lck προκαλώντας [15]
20 γρήγορο πολλαπλασιασμό των κυττάρων, διακοπή της αναδιάταξης των γονιδίων της β αλυσίδας και έκφραση των συνυποδοχέων CD4 και CD8. Στη συνέχεια, συμβαίνει αναδιάταξη των γονιδίων της α αλυσίδας μέχρι να παραχθεί μια λειτουργική α αλυσίδα η οποία συνδυάζεται αποτελεσματικά με τη β αλυσίδα. Τελικά, με την έκφραση ενός λειτουργικού αβ TCR υποδοχέα ικανού ν αναγνωρίζει MHC μόρια, το θυμοκύτταρο είναι έτοιμο να υποβληθεί σε επιλογή Συνδετική ποικιλότητα (ποικιλότητα συμβολών) Η ποικιλότητα που δημιουργείται από την προσθαφαίρεση νουκλεοτιδίων στις συμβολές των αναδιατασσόμενων γονιδίων κατά τον ανασυνδυασμό V(D)J ονομάζεται συνδετική ποικιλότητα (junctional diversity). Επειδή σε κάθε αναδιάταξη η συμβολή των γονιδίων V(D)J περιλαμβάνεται στην περιοχή CDR3, οι διαφορετικές αλληλουχίες αμινοξέων των περιοχών CDR3 είναι περισσότερες από αυτές που μπορεί να κωδικοποιηθούν από τα τμήματα V, J και D της γαμετικής σειράς. Οι γονιδιακές αναδιατάξεις των αντιγονικών υποδοχέων είναι χαρακτηριστικές των ξεχωριστών κλώνων λεμφοκυττάρων και χάρη στη μοναδικότητά τους, αποτελούν ένα είδος «μοριακής ταυτότητας» των κλώνων. Κατά τον ανασυνδυασμό VDJ, τα ελεύθερα άκρα αναγνωρίζονται από ένα σύμπλοκο ΚU πρωτεΐνων (KU70-KU80), το οποίο με τη σειρά του ενεργοποιεί την καταλυτική δράση του συμπλόκου DNA-PKcs. Το ενεργοποιημένο σύμπλοκο DNA-PKcs φωσφορυλιώνει στόχους που ελέγχουν τον κυτταρικό κύκλο αλλά και την πρωτεΐνη Artemis 18.(5) Η πρωτεΐνη Artemis εντοπίζει τη δομή φουρκέτας στα κωδικοποιητικά άκρα και πραγματοποιεί εκτομή σ ένα από τα συμμετέχοντα νουκλεοτίδια. Αν η εκτομή δεν γίνει στην κορυφή της φουρκέτας θα προκύψουν παλίνδρομα νουκλεοτίδια, γνωστά ως P νουκλεοτίδια (palindromic nucleotides). Επίσης, στην ποικιλότητα των ανοσοσφαιρινών συνεισφέρει το ένζυμο τελική δεοξυνουκλεοτιδική τρανσφεράση (Tdt). Το ένζυμο αυτό έχει δράση παρόμοια με τη δράση της DNA πολυμεράσης, δηλαδή καταλύει την προσθήκη νουκλεοτιδίων σε μονόκλωνες αλυσίδες, με τη διαφόρα ότι η Tdt δεν απαιτεί εκμαγείο. Tα νουκλεοτίδια που προστίθενται από την Tdt είναι γνωστά ως Ν νουκλεοτίδια (non-templated nucleotides) (Εικόνα 11). H ποικιλότητα αυξάνει ακόμη περισότερο από άλλα ένζυμα και συγκεκριμένα εξωνουκλεάσες οι οποίες αφαιρούν νουκλεοτίδια πριν την ένωση των γονιδιακών τμημάτων 15. [16]

21 Εικόνα 11: Μηχανισμός δημιουργίας ποικιλότητας από την προσθήκη και αφαίρεση νουκλεοτιδίων στις συνάψεις κατά τον ανασυνδυασμό V(D)J Διαφοροποίηση των T λεμφοκυττάρων Τα Τ λεμφοκύτταρα αναπτύσσονται από πολυδύναμα, αρχέγονα αιμοποιητικά κύτταρα. Τα πρόδρομα κύτταρα των Τ λεμφοκυττάρων μεταναστεύουν από το μυελό των οστών στο θύμο αδένα, όπου και ωριμάζουν. Τα στάδια ωρίμανσης των Τ λεμφοκυττάρων διακρίνονται με βάση την έκφραση διαφορετικών μεμβρανικών πρωτεϊνών, γνωστών ως δείκτες επιφανείας (CD4, CD8 κ.α.). Η ανάπτυξη των Τ λεμφοκυττάρων παραδοσιακά διακρίνεται σε δύο φάσεις: ανεξάρτητη από το αντιγόνο, που εξελίσσεται στο μυελό των οστών, και εξαρτώμενη από το αντιγόνο, που διαδραματίζεται στα δευτερογενή λεμφικά όργανα 5,6, Φάση διαφοροποίησης ανεξαρτητη από το αντιγόνο Η ανεξάρτητη από το αντιγόνo φάση της ανάπτυξης των Τ λεμφοκυττάρων συμβαίνει στο θύμο αδένα και είναι σύνθετη και πολυσταδιακή διαδικασία. Ο θύμος είναι απαραίτητος για τη διαφοροποίηση των ανώριμων προγονικών Τ λεμφοκυττάρων σε κύτταρα που έχουν [17]
22 τα χαρακτηριστικά των ώριμων Τ λεμφοκυττάρων. Έτσι, παιδιά που γεννιούνται χωρίς θύμο αδένα (σύνδρομο DiGeorge) δεν έχουν ώριμα Τ λεμφοκύτταρα. Τα προγονικά, ανώριμα Τ λεμφοκύτταρα εισέρχονται στο φλοιό του θύμου, όπου αρχίζει ο ανασυνδυασμός των γονιδίων του T κυτταρικού υποδοχέα (προ-τ-λεμφοκύτταρα). Η ανεξάρτητη από το αντιγόνο πρώτη φάση της ανάπτυξης των Τ λεμφοκυττάρων οδηγεί στη δημιουργία ώριμων, «παρθένων» Τ λεμφοκυττάρων (naive, virgin Τ cells). H ανάπτυξη των Τ κυττάρων εξελίσσεται σε ποικίλα στάδια διαφοροποίησης. Κάθε κύτταρο φέρει στο γονιδίωμα του αλληλόμορφα για τους γενετικούς τόπους της δ, γ, β και α αλυσίδας του Τ κυτταρικού υποδοχέα. Ωστόσο κάθε Τ λεμφοκύτταρο εκφράζει μόνο έναν λειτουργικό Τ κυτταρικό υποδοχέα. Το φαινόμενο αυτό ονομάζεται αποκλεισμός αλληλομόρφου (allelic exclusion) και είναι σημαντικό για τη ρύθμιση της άνοσης απάντησης. Ο αποκλεισμός αλληλομόρφου γενικά ρυθμίζεται στο επίπεδο του ανασυνδυασμού και της έκφρασης των γονιδίων του Τ κυτταρικού υποδοχέα. Έχει διατυπωθεί η άποψη ότι ο κύριος μηχανισμός για να πραγματοποιηθεί μια τέτοια επιλογή είναι η αναστολή του ανασυνδυασμού του ενός αλληλομόρφου από το προϊόν επιτυχημένου ανασυνδυασμού του άλλου αλληλομόρφου. Μόλις εκφραστεί λειτουργικός αντιγονικός υποδοχέας στην επιφάνεια του Τ λεμφοκυττάρου, παύουν οι περαιτέρω ανασυνδυασμοί. Το μικροπεριβάλλον του θύμου αδένα είναι υπεύθυνο για τη διαφοροποίηση των αρχέγονων αιμοποιητικών κυττάρων σε ώριμα Τ λεμφοκύτταρα. Ο θύμος αδένας αποτελείται από φλοιώδες και μυελικό επιθήλιο τα οποία μαζί με το συνδετικό ιστό σχηματίζουν το θυμικό στρώμα. Σε όλα τα στάδια της διαφοροποίησης τους στο θύμο, τα Τ λεμφοκύτταρα έρχονται σε στενή επαφή με τα κύτταρα του στρώματος του θύμου (stromal cells of thymus). Τα κυριότερα από αυτά τα κύτταρα είναι: (α) επιθηλιακά κύτταρα του θύμου και (β) διαπλεκόμενα δενδριτικά κύτταρα, που είναι αντιγονοπαρουσιαστικά (antigen-presenting cells, APC) κύτταρα 5,7. Το πρώτο βήμα για την ανάπτυξη του Τ λεμφοκυττάρου είναι η δέσμευση του αρχέγονου αιμοποιητικού κυττάρου για διαφοροποίηση στον κοινό πρόδρομο των λεμφοκυττάρων (common lymphoid progenitor), κύτταρο που χαρακτηρίζεται από την ικανότητα διαφοροποίησης σε Β ή Τ λεμφοκύτταρα ή κύτταρα φυσικούς-φονείς (natural killer, NK) (Εικόνα 12). [18]
23 Εικόνα 12. Η δέσμευση του αρχέγονου αιμοποιητικού κυτταρου (HSC) και, αργότερα, του CLP προς μια κυτταρική σειρά καθορίζεται από τα επίπεδα των μεταγραφικών παραγόντων E2A και Notch-1. Η σηματοδότηση μέσω του Notch1 υποδοχέα είναι βασική για την διαφοροποίηση των Τ λεμφοκυττάρων στο θύμο αδένα. Τα στρωματικά κύτταρα του θύμου ΟP9 εκφράζουν το δείκτη Delta- like 1, ο οποίος είναι ο προσδέτης για τον Notch υποδοχέα. Αυτό προάγει τη διαφοροποίηση των αρχέγονων αιμοποιητικών κυττάρων σε ώριμα Τ λεμφοκύτταρα. Η σηματοδότηση μέσω του Notch υποδοχέα επάγει: έκφραση του c-kit υποδοχέα στην κυτταρική μεμβράνη μεταγραφή των γονιδίων στόχων Deltex1 και pre-ta μεταγραφή των γονιδίων TRBV και των ρυθμιστικών γονιδίων GATA3 και Tcf1 Στον κοινό πρόδρομο των λεμφοκυττάρων, τα γονίδια του Τ κυτταρικού υποδοχέα βρίσκονται σε διάταξη βλαστικού DNA, δηλαδή δεν έχουν αναδιαταχθεί. Τα στάδια διαφοροποίησης για την Τ λεμφική σειρά καθορίζονται από τις αναδιατάξεις των γονίδιων του Τ κυτταρικού υποδοχέα αλλά και από την έκφραση μορίων προσκόλλησης και υποδοχέων αυξητικών παραγόντων στην επιφάνεια των Τ λεμφοκυττάρων. Η διαδικασία διαφοροποίησης των Τ λεμφοκυττάρων διακρίνεται σε στάδια ανάλογα με την έκφραση των δεικτών επιφανείας CD44 (υποδοχέας υαλουρονικού οξέος ο οποίος συμμετέχει στην ενεργοποίηση των λεμφοκυττάρων, στην αιμοποίηση και στη μετάσταση των όγκων) και CD25 (α αλυσίδα του υποδοχέα της IL-2) στην κυτταρική μεμβράνη των CD4- CD8- (Double Negative) θυμοκυττάρων (εικόνα 14). Ανάλογα με την έκφραση των δεικτών επιφάνειας, τα Τ λεμφοκύτταρα διακρίνονται σε υποσύνολα χαρακτηριστικά για τα [19]
 και ολοκληρώνεται στα κύτταρα με χαρακτηριστικά")

24 διάφορα στάδια διαφοροποίησης τους. Το υποσύνολο DN1 περιλαμβάνει κύτταρα με χαρακτηριστικά CD44+ CD25-. Οι αναδιατάξεις των γονιδίων του Τ κυτταρικού υποδοχέα γίνονται ιεραρχικά. Αρχικά η αναδιάταξη γίνεται στα γονίδια που κωδικοποιούν την δ και γ αλυσίδα του Τ κυτταρικού υποδοχέα σε κύτταρα με χαρακτηριστικά CD44+ CD25+ (DN2) και ολοκληρώνεται στα κύτταρα με χαρακτηριστικά CD44- CD25+ (DN3), τα οποία διαφοροποιούνται στην Τ λεμφική σειρά. Εάν και οι δύο αναδιατάξεις είναι πετυχημένες και συντεθεί γ και δ αλυσίδα, τότε το κύτταρο θα εκφράσει TCRγδ στην κυτταρική επιφάνεια. Η έκφραση ενός λειτουργικού γδ Τ κυτταρικού υποδοχέα στην κυτταρική επιφάνεια αναστέλλει τις περαιτέρω αναδιατάξεις των γονιδίων που κωδικοποιούν την β και α αλυσίδα. Όταν η αvαδιάταξη είναι μη λειτουργική ή μη παραγωγική, το κύτταρο προχωρεί στην αναδιάταξη των γονιδίων της β αλυσίδας. Η επιτυχημένη αναδιάταξη των γονιδίων της β αλυσίδας επάγει την έκφραση του προ-tcr (pre-tcr) και την περαιτέρω διαφοροποίηση των DN3 σε CD4+CD8+ (Double Positive) θυμοκύτταρα. Η διαφοροποίηση ολοκληρώνεται με την αναδιάταξη των γονιδίων που κωδικοποιούν την α αλυσίδα του Τ κυτταρικού υποδοχέα. Τελικά, τα Τ κύτταρα εκφράζουν έναν λειτουργικό αβ Τ κυτταρικό υποδοχέα και υπόκεινται σε θετική ή αρνητική επιλογή 7 (Εικόνα 13). Εικόνα 13: Τα στάδια διαφοροποίησης των Τ λεμφοκυττάρων. Προέλευση: Nature Immunology Τα Τ λεμφοκύτταρα τα οποία κατά την ανάπτυξή τους στο θύμο παρουσιάζουν ακατάλληλες ειδικότητες υποδοχέα, δηλαδή υποδοχείς με δυνητικά επικίνδυνη ειδικότητα (π.χ. έντονη αυτοαντιδραστικότητα) εξαλείφονται με κυτταρικό θάνατο, γεγονός που αντανακλά την αυστηρή επιλογή των Τ λεμφοκυτταρικών κλώνων. Οι τύποι επιλογής των Τ λεμφοκυττάρων είναι δυο: θετική επιλογή, που εξασφαλίζει ότι όλα τα ώριμα Τ λεμφοκύτταρα είναι ικανά να αναγνωρίζουν τα μόρια του μείζονος συμπλέγματος ιστοσυμβατότητας (MHC-major histocompatibility complex) του οργανισμού, και αρνητική [20]
25 επιλογή, κατά την οποία εξαλείφονται τα Τ λεμφοκύτταρα με Τ κυτταρικό υποδοχέα που παρουσιάζει μεγάλη συγγένεια για τις πρωτεΐνες του οργανισμού (αυτοαντιδραστικά T κύτταρα, self-reactive T cells) και εξασφαλίζει την ανοχή έναντι των αυτοαντιγόνων. Μόνο ένα 3% των πρόδρομων Τ λεμφοκυττάρων που εισέρχονται στο θύμο αδένα επιβιώνει από τις διαδικασίες της θετικής και της αρνητικής επιλογής. Στη συνέχεια, αυτά τα λεμφοκύτταρα μεταναστεύουν στην περιφέρεια και είναι σε συνεχή κυκλοφορία μεταξύ των δευτερογενών λεμφοποιητικών οργάνων. Η θετική και η αρνητική επιλογή των Τ λεμφοκυττάρων αποτελούν βασικούς μηχανισμούς ρύθμισης των αυτοαντιδραστικών Τ λεμφοκυττάρων κατά την ωρίμανσή τους στο θύμο αδένα και είναι συλλογικά γνωστές ως κεντρική ανοχή (central self-tolerance). Δεδομένου όμως ότι τα αυτοαντιδρώντα Τ κύτταρα μπορεί να προκαλέσουν σημαντική βλάβη στον ίδιο τον οργανισμό, για τη ρύθμιση τους είναι απαραίτητοι και άλλοι ειδικοί ρυθμιστικοί μηχανισμοί οι οποιοι είναι συλλογικά γνωστοί ως περιφερική ανοχή (peripheral selftolerance). Κάποιοι από τους μηχανισμούς αυτούς διαμεσολαβούνται από συγκεκριμένες υποομάδες Τ λεμφοκυττάρων (Τ ρυθμιστικά λεμφοκύτταρα) και ενεργοποιούνται όταν κάποιο από τα αυτοαντιδαστικά κύτταρα διαφύγει από τους μηχανισμούς της αρνητικής επιλογής 6, Φάση διαφοροποίησης εξαρτώμενη από το αντιγόνο Τα Τ λεμφοκύτταρα τα οποία έχουν ωριμάσει στο θύμο αδένα αλλά δεν έχουν συναντήσει ακόμα αντιγόνα ονομάζονται παρθένα (naïve) λεμφοκύτταρα. Η συνάντηση Τ λεμφοκυττάρων-αντιγόνου πραγματοποιείται στα δευτερογενή λεμφικά όργανα και περιλαμβάνει αλληλεπίδραση του αντιγόνου με κατάλληλης ειδικότητας Τ κυτταρικό υποδοχέα στην επιφάνεια των Τ λεμφοκυττάρων. Η επαφή του Τ κυτταρικού υποδοχέα με το αντιγόνο αποτελεί απαραίτητη προϋπόθεση για την έναρξη των ειδικών άνοσων απαντήσεων. Ένα παρθένο Τ λεμφοκύτταρο που έρχεται σ επαφή με αντιγόνο σταματά να μεταναστεύει και πολλαπλασιάζεται. Στο αίμα, τα περισσότερα κύτταρα του ανοσοποιητικού συστήματος βρίσκονται σε κατάσταση ηρεμίας στη φάση G0 του κυτταρικού κύκλου. Μόλις ενεργοποιηθούν, τα κύτταρα περνούν σε φάση G1 κι έπειτα σε φάση S, κατά την οποία συμβαίνει η σύνθεση του DNA. Στη συνέχεια, περνούν σε φάση G2 και τέλος διαιρούνται. Μετά την ενεργοποίηση του Τ λεμφοκυττάρου, η χρωματίνη του πυρήνα του γίνεται λιγότερο πυκνή, εμφανίζονται πυρηνίσκοι, ο όγκος του κυτταροπλάσματος αυξάνει κι επάγεται η σύνθεση RNA και πρωτεϊνών. Στη συνέχεια το [21]
26 κύτταρο διαιρείται, δυο ή τέσσερις φορές κάθε 24 ώρες επί 3-5 ημέρες. Έτσι, από το αρχικό Τ λεμφοκύτταρο δημιουργείται ένας κλώνος περίπου 1000 θυγατρικών κυττάρων πανομοιότυπης ειδικότητας (κλωνική έκπτυξη, clonal expansion). Τα κύτταρα αυτά στη συνέχεια διαφοροποιούνται σε δραστικά κύτταρα ικανά να καταστρέψουν τα μολυσμένα κύτταρα ή να ενεργοποιήσουν άλλα κύτταρα του ανοσολογικού συστήματος. Μερικά από τα αντιγόνο-ειδικά κύτταρα που αναπτύσσονται από την κλωνική έκπτυξη των παρθένων Τ λεμφοκυττάρων, γνωστά ως μνημονικά Τ λεμφοκύτταρα (memory T cells), παραμένουν και μετά την απομάκρυνση του αντιγόνου και αποτελούν τη βάση της άνοσης μνήμης, η οποία εξασφαλίζει μια πιο ταχεία και αποτελεσματική απόκριση σε δεύτερη συνάντηση με το ίδιο παθογόνο και συνεπώς διασφαλιζει ανοσία στον οργανισμό Επιλογή των Τ λεμφοκυττάρων στο θύμο αδένα Το επόμενο βήμα στην ωρίμανση των Τ λεμφοκυττάρων στον θύμο, αφότου εκφράσουν στην κυτταρική μεμβράνη τους τα μόρια TCR, CD4 και CD8, είναι η επιλογή εκείνων των κυττάρων που θ αποτελέσουν το ρεπερτόριο των ώριμων Τ λεμφοκυττάρων στην περιφέρεια. Τα Τ λεμφοκύτταρα αναγνωρίζουν μόνο πεπτιδικά αντιγόνα τα οποία εκτίθενται από τα MHC μόρια της επιφανείας των αντιγονοπαρουσιαστικών κυττάρων. Τα MHC μόρια είναι μεμβρανικές πρωτεΐνες, οι οποίες διακρίνονται σε δύο τάξεις, I (MHC I) και II (MHC II): Τα MHC μόρια τάξης Ι είναι γλυκοπρωτεΐνες οι οποίες μεσολαβούν στην παρουσίαση αντιγονικών πεπτιδίων που αποτελούνται 8-9 αμινοξέα στα CD8 Τ λεμφοκύτταρα. Τα πεπτίδια προέρχονται από ενδογενείς πρωτεΐνες που έχουν συντεθεί ενδογενώς και διασπαστεί ενζυμικά στο κυτταρόπλασμα Τα MHC μόρια τάξης ΙΙ είναι γλυκοπρωτεΐνες οι οποίες μεσολαβούν στην παρουσίαση αντιγονικών πεπτιδίων που αποτελούνται από αμινοξέα στα CD4 Τ λεμφοκύτταρα. Τα πεπτίδια συνήθως προέρχονται από εξωγενείς πρωτεΐνες που έχουν διασπαστεί στο εσωτερικό του κυττάρου (εικόνα 14) [22]

27 Εικόνα 14. Παρουσίαση αντιγόνων από τα APCs, μέσω των MHC I και MHC II μορίων, στα Τ λεμφοκύτταρα. Όπως προαναφέρθηκε, η σύνδεση TCR-MHC μπορεί να διευκολύνει την ωρίμανση, επιβίωση, διαφοροποίηση ή το θάνατο των Τ-λεμφοκυττάρων. Έτσι, η επιλογή των αβ CD4 + CD8 + Τ-λεμφοκυττάρων στο θύμο χωρίζεται σε δύο φάσεις, γνωστές ως θετική επιλογή (positive selection) και αρνητική επιλογή (negative selection). Το επικουρικό μόριο που θα διατηρηθεί στα ώριμα Τ λεμφοκύτταρα (Εικόνα 15) φαίνεται ότι καθορίζεται από την τάξη του μορίου MHC που αναγνωρίζεται από τον TCR υποδοχέα των CD4 + CD8 + θυμοκυττάρων. Έτσι, το CD8 επικρατεί όταν ο TCR υποδοχέας αναγνωρίζει μόριο MHC τάξης Ι, ενώ αντίθετα, κατά τη σύνδεση του TCR με μόριο MHC τάξης ΙΙ επικρατεί το CD4. Ο μηχανισμός με τον οποίο επιτελείται η επιλογή του επικουρικού μορίου δεν έχει αποσαφηνιστεί πλήρως. Τα δύο βασικά μοντέλα είναι: 1. Το «εκπαιδευτικό» μοντέλο (instructive model). Σύμφωνα με αυτό το μοντέλο, τα δύο επικουρικά μόρια στέλνουν ξεχωριστά ενδοκυττάρια σήματα. Έτσι, το σήμα που προέρχεται από το CD4 αναστέλλει τη λειτουργία του γονιδίου για το CD8, ενώ διεγείρει τη μεταγραφή του γονιδίου για το CD4. Το αντίθετο παρατηρείται όταν αποστέλλεται σήμα από το CD8. 2. Το «στοχαστικό» μοντέλο (stochastic/selection model). Σύμφωνα με αυτό το μοντέλο, η αδρανοποίηση των γονιδίων που κωδικοποιούν το CD4 ή CD8 είναι τυχαία ή καθορίζεται από μηχανισμό που ελέγχει τη συμβατότητα του επικουρικού [23]

28 μορίου με την τάξη του μορίου MHC που αναγνωρίζεται από τον TCR υποδοχέα. Έτσι, πεθαίνουν τα κύτταρα στα οποία επικρατεί επικουρικό μόριο με ειδικότητα ασύμβατη με την τάξη του μορίου MHC που αναγνωρίζεται από τον TCR υποδοχέα. Αντίθετα, επιζούν τα κύτταρα στα οποία επικρατεί επικουρικό μόριο με ειδικότητα συμβατή με την τάξη του μορίου MHC που αναγνωρίζεται από τον TCR υποδοχέα. Στην πράξη, πιθανόν ισχύουν και τα δύο μοντέλα, τουλάχιστον εν μέρει. Επίσης, πρέπει να υπάρχουν γονίδια που καθορίζουν το αναπτυξιακό πρόγραμμα (developmental program) των κυττάρων, δηλαδή τη διαφοροποίηση των πρόδρομων κυττάρων σε συγκεκριμένες κατηγορίες δραστικών κυττάρων. Έτσι, τα γονίδια που ευθύνονται για την εκδήλωση της κυτταροτοξικής δράσης ενεργοποιούνται στα δραστικά CD8 κυτταροτοξικά Τ λεμφοκύτταρα, ενώ τα γονίδια των διαφόρων κυτταροκινών ενεργοποιούνται στα δραστικά CD4 βοηθητικά Τ λεμφοκύτταρα. Παράδειγμα γονιδίου που πιθανόν παίζει σημαντικό ρόλο στον αναπτυξιακό προγραμματισμό των Τ λεμφοκυττάρων είναι το γονίδιο Notch, το οποίο φαίνεται να επηρεάζει την ανάπτυξη πρόδρομων κυττάρων που έχουν τη δυνατότητα να ακολουθήσουν διαφορετικές οδούς διαφοροποίησης 19, 20, 21. Εικόνα 15. Επιλογή επικουρικού μορίου (CD4 ή CD8) κατά την ωρίμανση των Τ λεμφοκυττάρων. [24]
29 1.4.4 CD4 λεμφοκύτταρα Τα Τ βοηθητικά κύτταρα (Τhelper cells) αποτελούν υποσύνολο των λεμφοκυττάρων με σημαντικό ρόλο στην λειτουργία του ανοσοποιητικού συστήματος. Τα κύτταρα αυτά δεν έχουν κυτταροτοξική και φαγοκυτταρική δράση αλλά διαδραματίζουν πολύ σημαντικό ρόλο στην ενεργοποίηση των Β λεμφοκυττάρων και την ενεργοποίηση και διαφοροποίηση των κυτταροτοξικών CD8 Τ λεμφοκυττάρων. Τα ώριμα Τ βοηθητικά κύτταρα πάντα εκφράζουν το δείκτη επιφανείας CD4 παράλληλα με το σύμπλοκο TCR/CD3. Το 1986, οι Mosmann και Coffman διατύπωσαν την άποψη ότι τα δραστικά CD4 Τ λεμφοκύτταρα διακρίνονται σε δύο υποσύνολα με βάση τις κυτταροκίνες που παράγουν και τις λειτουργίες που επιτελούν. Σήμερα, τα δραστικά CD4 Τ λεμφοκύτταρα διακρίνονται σε τέσσερα υποσύνολα: Th1, Th2, Th17 και Treg κύτταρα, τα οποία επιτελούν διαφορετικές λειτουργίες, παράγουν διαφορετικά προϊόντα, ελέγχονται από διαφορετικούς μεταγραφικούς παράγοντες και χαρακτηρίζονται από διαφορετικό προφίλ έκκρισης κυτταροκινών (Εικόνα 16) 21. Εικόνα 16: τα τέσσερα υποσύνολα των CD4 T λεμφοκυττάρων: λειτουργίες, παραγόμενα προϊόντα, μεταγραφικοί παράγοντες και κυτταροκίνες που επιδρούν σε κάθε υποσύνολο. Προέλευση:Zhu Jinphang and William E.Paul.Blood 2008;112: [25]
30 1.4.5 Th1 κύτταρα Τα Th1 κύτταρα συμμετέχουν στις άνοσες απαντήσεις εναντίον ενδοκυττάριων παθογόνων κι επάγουν την εκδήλωση κάποιων αυτοάνοσων ασθενειών. Παράγουν INFγ, λεμφοτοξίνη α (LΤα) και IL-2. Η παρουσία της IFN-γ εξασφαλίζει τις συνθήκες που απαιτούνται για την επαγωγή της ενεργοποίησης των μακροφάγων από τα Τ λεμφοκύτταρα. Στη διαδικασία διαφοροποίησης των Th1 κυττάρων συμμετέχουν πολλοί μεταγραφικοί παράγοντες, οι περισσότεροι από τους οποίους επάγουν την παραγωγή της IFN-γ επιδρώντας σε διαφορετικά επίπεδα. Η δράση της IFN-γ επάγει το μεταγραφικό παράγοντα Τ-bet του οποίου η λειτουργία ελέγχεται από τον Stat1 που ρυθμίζει όλες τις διαμεσολαβούμενες από την IFN-γ διαδικασίες. Επίσης, η IL-12 επάγει το μεταγραφικό παράγοντα STAT4 ο οποίος, μαζι με τον NF-κΒ, προκαλεί την παραγωγή IFN-γ ανεξάρτητα από την ενεργοποίηση του T κυτταρικού υποδοχέα. Τέλος, οι μεταγραφικοί παράγοντες Runx3 και Hlx επάγουν την παραγωγή της INFγ και κατευθύνουν τη διαφοροποίηση των παρθένων CD4 λεμφοκυττάρων σε Th1 κύτταρα. Η διαφοροποίηση των Th1 κυττάρων περιλαμβάνει αλληλεπίδραση των αντιγονοπαρουσιαστικών κυττάρων (δενδριτικά κύτταρα) με τον Τ κυτταρικό υποδοχέα των CD4 λεμφοκυττάρων. Αυτό έχει ως αποτέλεσμα τη μεγάλη παραγωγή IL-12 από τα αντιγονοπαρουσιαστικά κύτταρα εξαιτίας της ενεργοποίησης τους. Η παραγόμενη IL-12 επιδρά σε δύο διαφορετικούς στόχους. Σε πρώτη φάση ενεργοποιεί τον παράγοντα Stat1, ο οποίος με την σειρά του ρυθμίζει την έκφραση του T-bet και στη συνέχεια επιδρά στον παράγοντα Stat4 με αποτέλεσμα την παραγωγή INFγ, η οποία επάγει τη διαφοροποίηση των CD4 λεμφοκυττάρων σε Th1 κύτταρα Th2 κύτταρα Τα Th2 κύτταρα αναγνωρίζουν εξωκυττάρια παράσιτα και συμμετέχουν σε αλλεργικές αντιδράσεις και στο άσθμα. Παράγουν IL-4, IL-5, IL-9, IL-10, IL-13 και IL-25. H ΙL-10 αναστέλει τη δράση των ενεργοποιημένων μακροφάγων και των δενδριτικών κυττάρων και επαναφέρει το σύστημα σε κατάσταση ηρεμίας μετά την απομάκρυνση της μικροβιακής λοίμωξης. Συγκεκριμένα, αναστέλει την παραγωγή της IL-12 και την έκφραση συνδιεγερτικών μορίων και μορίων MHC τάξης II από τα μακροφάγα και τα δενδριτικά κύτταρα. Η κύρια δράση της IL-4 είναι η μεταστροφή της βαριάς αλυσίδας των ανοσοσφαιρινών στον IgE ισότυπο: επομένως παίζει ρόλο στην άμυνα εναντίον παρασίτων και στις [26]
31 αλλεργικές αντιδράσεις. Επιπλέον, διεγείρει την ανάπτυξη κι επαύξηση των Th2 κυττάρων από τα παρθένα CD4+ κύτταρα και δρα ως αυτοκρινής αυξητικός παράγοντας για τα διαφοροποιημένα Th2 κύτταρα. Τέλος, αναστέλλει την ενεργοποίηση των μακροφάγων, ανταγωνιζόμενη τη δράση της IFN-γ. Η IL-5 ενεργοποιεί τα ώριμα ηωσινόφιλα και διεγείρει την αύξηση και διαφοροποίηση των κυττάρων αυτών. Τέλος διεγείρει τον πολλαπλασιασμό των Β λεμφοκυττάρων και την παραγωγή IgA αντισωμάτων. Η IL-25 παράγεται από τα επιθηλιακά κύτταρα του πνεύμονα και προάγει την παραγωγή της IL-4, IL-5 και IL-13. Επάγει τη παραγωγή χημειοκινών, όπως η RANTES (CCL5) και η ηωταξίνη, οι οποίες προσελκύουν ηωσινόφιλα. Στη διαδικασία διαφοροποίησης των Th2 κυττάρων συμμετέχει ο μεταγραφικός παράγοντας GATA-3, του οποίου η λειτουργία επάγεται από την IL-4 κι ελέγχεται από τον παράγοντα Stat6. Άλλοι παράγοντες με σημαντικό ρόλο στη διαφοροποίηση των Th2 κυττάρων είναι ο Stat5, ο οποίος επάγεται από την IL-2, ο c-maf, ο οποίος ενισχύει την παραγωγή της IL-4, ο ρυθμιστικός παράγοντας της ιντερφερόνης 4 και ο παράγοντας Gfi-1, ο οποίος επιλέγει τα GATA-3 hi κύτταρα για να διαφοροποιηθούν σε Th2 κύτταρα. Κατά τη διαφοροποίηση των Th2 κυττάρων, η IL-4 επάγει την έκφραση του GATA-3 παράλληλα με την ενεργοποίηση του παράγοντα Gfi-1 μέσω του μονοπατιού IL-4/Stat6. Έτσι επιλέγονται τα κατάλληλα κύτταρα (GATA-3 hi ) για περαιτέρω διαφοροποίηση τους σε Th2 κύτταρα 21. Συμπερασματικά, τόσο τα Τh1 όσο και τα Τh2 T λεμφοκύτταρα, προέρχονται από τα Th0 Τ λεμφοκύτταρα τα οποία παράγουν IL-2, IL-4 και INF-g. Κατά τη διάρκεια της διαφοροποίησης τους στα δύο παραπάνω υποσύνολα, η IL-12 προάγει την διαφοροποίηση σε Th1 κύτταρα μέσω ενεργοποίησης σημάτων μεταγωγής και ενεργοποίηση του μεταγραφικού παράγοντα STAT-4, ο οποίος επιδρά στην έκφραση του T-bet, του κύριου ρυθμιστή της διαφοροποίησης και ανάπτυξης των Τh1 κυττάρων. Όσον αφορά τα Th2 κύτταρα, η IL-4 προκαλεί την ενεργοποίηση του GATA-3, του κύριου ρυθμιστή της διαφοροποίησης και ανάπτυξης των Τh2 κυττάρων μέσω STAT-6. Συνεπώς τα Th1 κύτταρα χαρακτηρίζονται από την έκφραση του T-bet και την παραγωγή IFN-γ,IL-2, TNF-α, και IL-18,ενώ Th2 κύτταρα χαρακτηρίζονται από την έκφραση του GATA- 3,και την σύνθεση της IL-4,IL-5,IL-6,IL-10,και IL-13. (Εικόνα 17) [27]
32 Εικόνα 17: Πρότυπο έκφρασης ρυθμιστικών παραγόντων κατά τη διαφοροποίηση των Th0 κυττάρων στα υποσύνολα Τh1/Th2. [28]
 και οι ιντερλευκίνες IL-23 και IL-17.")
33 1.4.7 Th17 κύτταρα Ο υποπληθυσμός Th17 χαρακτηρίζεται από την παραγωγή IL-17, IL-21 και IL-22 και σχετίζεται με αυτοάνοση ιστική βλάβη, π.χ. ρευματοειδή αρθρίτιδα και ειδικές αλλεργικές αντιδράσεις. Οι κύριοι παράγοντες που πυροδοτούν την παραγωγή νέων δραστικών Th17 κυττάρων είναι ο TGF-β (Τransforming Growth Factor- β) και οι ιντερλευκίνες IL-23 και IL-17. Ο TGF-β αναστέλλει τον πολλαπλασιασμό και τη διαφοροποίηση των Τ λεμφοκυττάρων καθώς και την ενεργοποίηση των μακροφάγων. Διεγείρει επίσης την παραγωγή IgA αντισωμάτων. Έχει ποικίλες δράσεις και εκτός του ανοσοποιητικού συστήματος: επάγει τη σύνθεση πρωτεϊνών της εξωκυττάριας ουσίας (π.χ. κολλαγόνο), ενζύμων που τροποποιούν τη θεμέλια ουσία (π.χ. μεταλλοπρωτεϊνάσες) και κυτταρικών υποδοχέων για πρωτεΐνες της θεμέλιας ουσίας (π.χ. ιντεγκρίνες). Έτσι επάγει την επαναδόμηση των ιστών, αφού έχει τεθεί υπό έλεγχο η άνοση και φλεγμονώδης απάντηση. Οι κύριοι μεταγραφικοί παράγοντες που συμμετέχουν στη διαφοροποίηση των Th17 κυττάρων είναι ο RORγt, του οποίου η υπερέκφραση επάγει την παραγωγή IL-17, και ο πυρηνικός υποδοχέας RORα, που επηρεάζει ελάχιστα την παραγωγή της IL-17 αλλά μπορεί να προκαλέσει καταστολή της παραγωγής της σε συνδυασμό με απώλεια του RORγt. Τέλος, ο μεταγραφικός παράγοντας Stat3 επάγει την παραγωγή των IL-6, IL-21 και IL-23 και είναι απάραίτητος για την παραγωγή της IL-17. Στη διαφοροποίηση των Th17 κυττάρων σημαντικό ρόλο παίζει ο TGF-β παρουσία IL-6, αφού επάγει την παραγωγή IL-21 και την έκφραση του RORγt. Η IL-21 μπορεί να αντικαταστήσει την IL-6 προκαλώντας επαγωγή των RORγt και IL-17, λειτουργώντας έτσι ως ενισχυτής των κυτταροκινών που συμμετέχουν στη διαφοροποίηση των Th17 κυττάρων 21.(Εικόνα 18) Εικόνα 18: Στάδια διαφοροποίησης του Τh17 κυττάρων. [29]
, τα οποία επάγουν την καταστολή μέσω μηχανισμών στους οποίους συμμετέχουν κυτταροκίνες.")
34 1.4.8 Treg κύτταρα Τα ρυθμιστικά Τ κύτταρα (Τregs) διαχωρίζονται στα φυσικώς παραγόμενα Τregs (natural Treg, ntreg), τα οποία για να δράσουν απαιτούν άμεση κυτταρική επαφή, και στα επαγόμενα Τregs (induced Treg, itreg), τα οποία επάγουν την καταστολή μέσω μηχανισμών στους οποίους συμμετέχουν κυτταροκίνες. Τα Τregs καταστέλουν την ενεργοποίηση του ανοσοποιητικού συστήματος και βοηθούν στην ομοιόσταση της άνοσης απάντησης καθώς και στην ανοχή σε αυτοαντιγόνα. Η ανάπτυξη και διαφοροποίηση των Τ ρυθμιστικών κυττάρων ρυθμίζεται από τη δράση του μεταγραφικού παράγοντα forkhead box protein 3 (foxp3). Τα ntreg εμφανίζουν το δείκτη επιφανείας CD25 (α αλυσίδα του υποδοχέα της IL-2), ενώ τα itreg το δείκτη επιφανείας CD103. Ο δείκτης αυτός αντιστοιχεί στον υποδοχέα της ιντεγκρίνης alpha E ο οποίος συμμετέχει στην επικοινωνία και αλληλεπίδραση των κυττάρων με τους περιβάλλοντες ιστούς. Στη διαφοροποίηση των Τregs σημαντικός είναι ο ρόλος του TGF-β που ενεργοποιεί τον παράγοντα Smad3, ο οποίος με τη σειρά του επάγει την ενεργοποίηση του NFAT. Οι Smad3 και NFAT επάγουν την έκφραση του foxp3, η οποία ενισχύεται και από την ενεργοποίηση του μεταγραφικού παράγοντα Stat5 μέσω IL Προκύπτει ένα μοντέλο αμοιβαίας ανάπτυξης των Τ ρυθμιστικών και των Th17 κυττάρων το οποίο ρυθμίζεται από την έκκριση διαφορετικών παραγόντων. Πιο συγκεκριμένα, ενεργοποίηση των παρθένων Τ λεμφοκυττάρων παρουσία IL-23 δεν επάγει την ανάπτυξη των Τ ρυθμιστικών ή των Th17 κυττάρων. Παρουσία TGF-β επάγεται η διαφοροποίηση σε Foxp3+ Treg κύτταρα, ενώ η παρουσία IL-6 όχι μόνο αναστέλλει την δημιουργία τουςαλλά προκαλεί ταυτόχρονα την επαγωγή Th17 κυττάρων. (Εικόνα 19) Εικόνα 19: Μοντέλο αμοιβαίας διαφοροποίησης των T ρυθμιστικών και των Τh17 Κυττάρων. [30]
35 1.4.9 Ενεργοποίηση των παρθένων βοηθητικών Τ λεμφοκυττάρων Μετά την ωρίμανση τους, τα Τ λεμφοκύτταρα εγκαταλείπουν το θύμο αδένα και περνούν στα δευτερογενή λεμφικά όργανα (π.χ. λεμφαδένες, σπλήνας). Στο στάδιο αυτό εκφράζουν το σύμπλοκο TCR/CD3 και αναγνωρίζουν πεπτιδικά αντιγόνα τα οποία εκτίθενται από τα μόρια MHC τάξης ΙΙ της επιφανείας των αντιγονοπαρουσιαστικών κυττάρων. Η διαδικασία ενεργοποίησης περιλαμβάνει δύο στάδια τα οποία διακρίνονται από την έκφραση διαφορετικών μηνυμάτων: το στάδιο της αναγνώρισης και το στάδιο της συνδιέγερσης. Η άνοση απάντηση του Τ λεμφοκυττάρου αρχίζει με αναγνώριση του συμπλόκου ενός αντιγονικού πεπτιδίου (Ag) με αντιγόνο ιστοσυμβατότητας HLA (Ag/HLA) στην επιφάνεια του αντιγονοπαρουσιαστικού κυττάρου. Όμως, για την ενεργοποίηση του βοηθητικού Τ λεμφοκυττάρου (Τ helper) απαιτούνται δύο σήματα. Το 1ο σήμα παρέχεται από την αντίδραση TCR-Ag/HLA, ενώ το 2ο παρέχεται από τα συνδιεγερτικά μόρια. Τέτοια μόρια είναι το CD28 (στο Τ λεμφοκύτταρο) που συνδέεται με τα CD80 και CD86 (στο αντιγονοπαρουσιαστικό κύτταρο). Η σύνδεση CD28/CD80/86 επάγει μεταγραφικό σήμα για την παραγωγή ιντερλευκίνης-2 (IL-2), που είναι αυξητικός παράγοντας για το Τ λεμφοκύτταρο. Εάν δεν υπάρχει το 2ο σήμα (από το συνδιεγερτικό μόριο CD28/CD80/86), το Τ λεμφοκύτταρο θα υποστεί απόπτωση ή θα μεταπέσει σε ανεργία (anergy). Ανάλογα με τον τύπο του αντιγονοπαρουσιαστικού κυττάρου (δενδριτικό κύτταρο, μακροφάγο, Β- λεμφοκύτταρο), το είδος του αντιγόνου, τη συγγένεια σύνδεσης του αντιγoνικού πεπτιδίου με το HLA και τον τύπο των κυτταροκινών στο άμεσο περιβάλλον του, το Τ λεμφοκύτταρο διαφοροποιείται και πολλαπλασιάζεται σε ΤΗ1 κύτταρο (παράγει IFNγ), TH2 κύτταρο (παράγει IL-4, IL-5, IL-13) ή TH17 κύτταρο (παράγει IL-21 και κυτταροκίνες της οικογένειας της IL-17). Μόλις ενεργοποιηθεί, το Τ λεμφοκύτταρο εκφράζει στην επιφάνεια του το μόριο CTLA-4 (CD152) που έχει μεγαλύτερη συγγένεια σύνδεσης με τα CD80 και CD86. Η σύνδεση CTLA-4/CD80/86 μεταβιβάζει αρνητικό σήμα στο Τ λεμφοκύτταρο και αναστέλλει την ενεργοποίησή 21.(Εικόνα 20) [31]
 έχουν κυτταροτοξική και φαγοκυτταρική δράση εναντίον ιών και άλλων παθογόνων κι επάγουν το θάνατο των μολυσμένων σωματικών κυττάρων ή των")
36 Εικόνα 20: Πρότυπα έκφρασης διαφορετικών παραγόντων στα διαφορετικά υπσύνολα των Τ βοηθητικών κυττάρων CD8 λεμφοκύτταρα Τα Τ κυτταροτοξικά κύτταρα (Τ c cells) έχουν κυτταροτοξική και φαγοκυτταρική δράση εναντίον ιών και άλλων παθογόνων κι επάγουν το θάνατο των μολυσμένων σωματικών κυττάρων ή των νεοπλασματικών κυττάρων αποφεύγοντας την καταστροφή των υγιών ιστών. Τα CD8 κύτταρα αποθηκεύουν κοκκιένζυμα (granzymes) σ εκκριτικά κοκκία τα οποία απελευθερώνουν με εξωκυττάρωση όταν αναγνωρίσουν κάποιο αντιγόνο πάνω σ ένα κύτταρο-στόχο. Ένα από αυτά είναι η περφορίνη, η οποία ανήκει στην οικογένεια των σερινοπρωτεασών. Η περφορίνη καταστρέφει τα μολυσμένα κύτταρα δημιουργώντας πόρους στη μεμβράνη τους. Ένα δραστικό μόριο στην επιφάνεια των CD8 κυττάρων είναι ο Fas-L, συνδετικό μόριο για τον υποδοχέα Fas, του οποίου η ενεργοποίηση προκαλεί απόπτωση. Επίσης τα CD8 κύτταρα απελευθερώνουν INF-γ, η οποία εμποδίζει την αντιγραφή των ιών, προκαλεί αυξημένη έκφραση των μορίων MHC τάξης Ι και ενεργοποιεί τα μακροφάγα. Τα ώριμα Τ κυτταροτοξικά κύτταρα πάντα εκφράζουν το δείκτη επιφανείας CD8 παράλληλα με το σύμπλοκο TCR/CD3. Τα Τ κυτταροτοξικά κύτταρα εκφράζουν Τ κυτταρικό υποδοχέα που αναγνωρίζει συγκεκριμένο αντιγόνο το οποίο προσδένεται σε μόριο MHC τάξης Ι 21. [32]
37 Ενεργοποίηση των παρθένων κυτταροτοξικών Τ λεμφοκυττάρων Η διαφοροποίηση των παρθένων κυτταροτοξικών Τ λεμφοκυττάρων σε δραστικά και μνημονικά κύτταρα είναι πολύπλοκη διαδικασία η οποία περιλαμβάνει ενεργοποίηση πολλών σηματοδοτικών μονοπατιών, δομικές αλλαγές στην οργάνωση της μεμβράνης και του κυτταροσκελετού, αναδιαμόρφωση της χρωματίνης κι έκφραση πολλών διαφορετικών γονιδίων. Πολλές μελέτες υποστηρίζουν ότι η διαδικασία διαφοροποίησης των δραστικών σε μνημονικά CD8 Τ λεμφοκύτταρα υπακούει το γραμικό πρότυπο ανάπτυξης (παρθένα δραστικά μνημονικά). Ωστόσο, αυτό αμφισβητείται από πιο πρόσφατα δεδομένα που προτείνουν ότι τα ενεργοποιημένα CD8 Τ κύτταρα μπορεί να διαφοροποιηθούν σε μνημονικά, παρακάμπτοντας το στάδιο των δραστικών Τ λεμφοκυττάρων 19. Τα δραστικά CD8 Τ λεμφοκύτταρα αποκτούν την ικανότητα μετανάστευσης από λεμφικούς σε μη λεμφικούς ιστούς, γεγονός που συνοδεύεται από αύξηση του αριθμού των υποδοχέων χημειοκινών και άλλων κυτταρικών μορίων προσκόλλησης 20. Παρατηρήθηκε ότι γονίδια που κωδικοποιούν χημειοτακτικές πρωτεϊνες, π.χ. CCR2, CCR5, νευροπιλίνη και σεμαφορίνη 4 D, καθώς και πρωτεϊνες προσκόλλησης, π.χ. MAC-2, CD11c, CD18, CD44 και ο προσδέτης της Ρ-σελεκτίνης, εκφράζονται στα δραστικά CD8 Τ λεμφοκύτταρα, επιτρέποντας τους να μεταναστεύουν σε εστίες φλεγμονής. Η κυτταροτοξική και φαγοκυτταρική δράση των CD8 Τ λεμφοκυττάρων εναντίον ιών και άλλων παθογόνων οφείλεται στα κοκκιένζυμα τα οποία αποθηκεύονται σ εκκριτικά κοκκία και απελευθερώνονται με εξωκυττάρωση όταν αναγνωρίσουν κάποιο αντιγόνο πάνω σ ένα κύτταρο-στόχο. Μελέτες δείχνουν ότι η έκφραση της περφορίνης και των κοκκιενζύμων Α, Β, D και Κ, καθώς και των δραστικών μορίων INFγ, RANTES και Fas-ligand αυξάνεται στα δραστικά CD8 Τ κύτταρα συγκριτικά με τα παρθένα. Επίσης, πολλά γονίδια που κωδικοποιούν μόρια τα οποία συμμετέχουν σε σηματοδοτικά μεταγωγικά μονοπάτια υπερεκφράζονται στα δραστικά Τ λεμφοκύτταρα. Πιο συγκεκριμένα παρατηρήθηκε υπερέκφραση των εξής μορίων: 1) φωσφατάση CD45 και PEP και των μορίων grb-2, lck, fyn, LIME που συμμετέχουν στην ενεργοποίηση του Τ κυτταρικού υποδοχέα. 2) καλσικλίνη και καταλυτική υπομονάδα της καλσινευρίνης και της ανεξίνης Α2 και Α6 που συμμετέχουν στην ενδοκυττάρια σηματοδότηση με τη βοήθεια ιόντων Ca +2. 3) JAK1, STAT4 και SOCS-5 που συμμετέχουν στη σηματοδότηση μέσω κυτταροκινών. [33]
38 Χαρακτηριστική είναι επίσης και η υπερέκφραση των γονιδίων που ανήκουν στο σύμπλοκο ARP2/3, της ταλίνης, της φιλαμίνης και της cdc42 rho GTPάσης που ευθύνονται για την ανάπτυξη δόμων όπως τα ινοπόδια στα δραστικά κυτταροτοξικά λεμφοκύτταρα Μνημονικά Τ λεμφοκύτταρα Μερικά από τα αντιγόνο-ειδικά κύτταρα που αναπτύσσονται από την κλωνική επαύξηση των παρθένων Τ λεμφοκυττάρων, γνωστά ως μνημονικά Τ λεμφοκύτταρα (memory T cells), παραμένουν και μετά την απομάκρυνση του αντιγόνου και αποτελούν τη βάση της άνοσης μνήμης, η οποία εξασφαλίζει πιο ταχεία και αποτελεσματική απάντηση σε μεταγενέστερη συνάντηση με το ίδιο παθογόνο και συνεπώς διασφαλίζει ανοσία στον οργανισμό. Τα Τ λεμφοκύτταρα μνήμης χαρακτηρίζονται από την έκφραση γονιδίων τα οποία σχετίζονται με τη μετανάστευση και προσκόλληση των κυττάρων και είχαν κατασταλεί στα δραστικά Τ λεμφοκύτταρα. Τα γονίδια αυτά κωδικοποιούν τα μόρια CCR2, CCR5, CD44, CD18, CD11c και glycam-1. Επίσης το μόριο CXCR4 απορυθμίζεται επιλεκτικά στα μνημονικά Τ λεμφοκύτταρα σε σχέση με τα δραστικά και αυτό ίσως υποδηλώνει ότι τα μνημονικά Τ λεμφοκύτταρα έχουν την ικανότητα να μεταναστεύουν προς τα στρωματικά κύτταρα, τα οποία εκφράζουν τον παράγοντα SDF-1. Παρατηρώντας επίσης τα επίπεδα της L σελεκτίνης στα παρθένα κύτταρα και συγκρίνοντας τα με τα επίπεδα έκφρασης της στα μνημονικά και δραστικά διαπίστωνεται μείωση της έκφρασης της L σελεκτίνης η οποία είναι μεγαλύτερη στα δραστικά κύτταρα συγκρινόμενη με τα μνημονικά. Το γεγονός αυτό σχετίζεται με την ενεργοποίηση της μεταγραφικής δραστηριότητας του γενετικού τόπου που κωδικοποιεί την L σελεκτίνη και συνοδεύει τη διαφοροποίηση των δραστικών σε μνημονικά Τ λεμφοκύτταρα. Συνεχίζοντας τη σύγκριση σε επίπεδο έκφρασης γονιδίων μεταξύ παρθένων και των μνημονικών Τ λεμφοκυττάρων παρατηρούμε ότι: 1) Γονίδια που σχετίζονται με την μεταγωγή σημάτων υπερεκφράζονται στα μνημονικά κύτταρα συγκρινόμενα με τα παρθένα. 2) Μόρια τα οποία ανήκουν στο σύμπλοκο p38 και στο σηματοδοτικό μονοπάτι MKK4, καθώς επίσης και τα μόρια ler-2, junb, fos και ATF-2 επίσης υπερεκφράζονται. Η υπερέκφραση αυτών των γονιδίων μπορεί να είναι το αποτέλεσμα πρόσφατης αντιγονικής διέγερσης ή να οφείλεται στην παρουσία παραγόντων ανάπτυξης, κυτταροκινών και άλλων διεγερτών. Η έκφραση αυτών των σηματοδοτικών πρωτεϊνων αυξάνει την ευαισθησία του Τ κυτταρικού υποδοχέα και επισπεύδει την άνοση απάντηση. [34]

39 3) Επίσης παρατηρείται υπερέκφραση πολλών γονιδίων που κωδικοποιούν δραστικά μόρια όπως είναι η INF-γ, ο Fas προσδέτης, το μόριο RANTES, o ΜΙΡ-1β, η περφορίνη και τα κοκκιένζυμα Β,Κ και Μ στα μνημονικά συγκριτικά με τα παρθένα κύτταρα.(πίνακας α) Διακρίνονται οι παρακάτω υποπληθυσμοί μνημονικών Τ λεμφοκυττάρων: Κεντρικά μνημονικά κύτταρα (central memory cells, T CM ), τα οποία διαθέτουν ικανότητα αυτοανανέωσης μέσω φωσφορυλίωσης του μεταγραφικού παράγοντα STAT5. Χαρακτηρίζονται από τον εξής φαινότυπο: CD44 high, CD62L high, IL-2Rβ +, Ly- 6C high και CCR7+. Τα συγκεκριμένα κύτταρα εντοπίζονται σε δευτερογενή λεμφικά όργανα και διαφέρουν από τα δραστικά κυτταροτοξικά Τ λεμφοκύτταρα. Πιο συγκεκριμένα, εκφράζουν L-σελεκτίνη, η οποία είναι υπεύθυνη για την εγκατάσταση (homing) των λεμφοκυττάρων σε περιφερικούς λεμφαδαδένες, και υποδοχείς χημειοκινών για την πρόσδεση των ιών στα λευκοκύτταρα. Δραστικά μνημονικά κύτταρα (effector memory cells), τα οποία είναι όμοια με τα δραστικά κύτταρα αλλά παραμένουν μετά από την απομάκρυνση του αντιγόνου από τους ιστούς. Τα κύτταρα αυτά δεν εκφράζουν L-σελεκτίνη και υποδοχείς χημειοκινών αλλά παράγουν δραστικές κυτταροκίνες, όπως INFγ και IL (Εικόνα 21) Εικόνα 21: Υποσύνολα μνημονικών Τα λεμφοκυττάρων. [35]
40 1.5 Απόπτωση των Τ λεμφοκυττάρων Γενικά Η απόπτωση ή προγραμματισμένος κυτταρικός θάνατος είναι ζωτικής σημασίας για την ανάπτυξη και τη διατήρηση της ομοιόστασης των οργανισμών. Πρόκειται για αναπτυξιακά ελεγχόμενη διαδοχή γεγονότων η οποία προϋποθέτει ενεργοποίηση συγκεκριμένων γονιδίων. Απορύθμιση της απόπτωσης οδηγεί σε πλήθος παθολογικών καταστάσεων, συμπεριλαμβανομένων νεοπλασιών και αυτοάνοσων διαταραχών. Σημαντικό ρόλο στον προγραμματισμένο κυτταρικό θάνατο διαδραματίζουν οι κασπάσες, μια εξελικτικά συντηρημένη οικογένεια κυτταροπλασματικών πρωτεασών που διακρίνονται από την παρουσία κυστεϊνης στο ενεργό κέντρο και υψηλή συγγένεια σύνδεσης με κατάλοιπα ασπαρτικού οξέος στα υποστρώματα-στόχους. Διακρίνονται στις εναρκτήριες κασπάσες, ή τύπου Ι, οι οποίες περιέχουν μεγάλες αρχικές αμινοτελικές περιοχές και σε κασπάσες τελεστές, ή τύπου ΙΙ, οι οποίες δεν περιέχουν, ή περιέχουν πολύ μικρές, αμινοτελικές περιοχές. Η αμινοτελική περιοχή είναι σημαντική για τη ρύθμιση της ενεργότητας των κασπασών επειδή περιέχει μοτίβα αλληλεπίδρασης μεταξύ πρωτεϊνών. Για παράδειγμα, η κασπάση 9 περιέχει το μοτίβο CARD με το οποίο συνδέεται στο αποπτόσωμα (apoptosome) μέσω CARD-CARD αλληλεπιδράσεων με την πρωτεϊνη Apaf-1, ενώ η κασπάση 8 περιέχει το μοτίβο DED (Death Effector Domain) με το οποίο συνδέεται στο σύμπλοκο DISC μέσω DED-DED αλληλεπιδράσεων με την πρωτεϊνη FADD. Ο μηχανισμός της απόπτωσης διακρίνεται στο εξωγενές και στο ενδογενές μονοπάτι. Το ενδογενές είναι υπεύθυνο για την εξάλειψη ανεπιθύμητων κυττάρων λόγω έλλειψης ορμονών, κυτταροκινών ή μιτοχονδριακών ανωμαλιών, ένω το εξωγενές για την εξάλειψη ανεπιθύμητων κυττάρων μετά την ανάπτυξη και το σχηματισμό του ανοσοποιητικού συστήματος Μηχανισμός επαγόμενης ενεργοποίησης της απόπτωσης στα Τ λεμφοκύτταρα Η ανάπτυξη των Τ λεμφοκυττάρων ρυθμίζεται όχι μόνο από μηχανισμούς πολλαπλασιασμού και διαφοροποίησης αλλά και από τη διαδικασία της απόπτωσης 22,23,24. Θυμοκύτταρα τα οποία αποτυγχάνουν ν αναδιατάξουν τα γονίδια του Τ κυτταρικού υποδοχέα ή αναγνωρίζουν αυτοαντιγόνα εξαλείφονται μέσω της διαδικασίας απόπτωσης που είναι γνωστή ως αρνητική επιλογή 25,26,27. Στα Τ λεμφοκύτταρα, η απόπτωση επάγεται από επίμονη αντιγονική διέγερση του Τ κυτταρικού υποδοχέα, γνωστή ως διαδικασία [36]

41 επαγόμενης ενεργοποίησης της απόπτωσης (activation-induced cell death, ΑΙCD), η οποία οδηγεί στην εξάλειψη των αυτοαντιδραστικών Τ κυττάρων 28,29. Η AICD επιτυγχάνεται μέσω αλληλεπίδρασης μεταξύ Fas και Fas προσδέτη. Τα ενεργοποιημένα Τ λεμφοκύτταρα εκφράζουν ταυτόχρονα Fas και Fas προσδέτη, με αποτέλεσμα να οδηγούνται σε απόπτωση είτε μέσω αλληλεπιδράσεων στο ίδιο Τ κύτταρο είτε εξαιτίας αλληλεπιδράσεων μεταξύ δύο διαφορετικών Τ λεμφοκυττάρων 29,30,31,32,33,34. Η αλληλεπίδραση αυτή επάγει τον τριμερισμό του υποδοχέα Fas, o οποίος, μέσω αλληλεπίδρασης με τον ενδιάμεσο προσαρμοστή FADD, επιστρατεύει την προ-κασπάση 8. Η προ-κασπάση 8 (FLICE) ενεργοποιείται σε κασπάση 8 μέσω σύνδεσης με τον FADD και τις DEDs. Το σύμπλοκο Fas υποδοχέα, FADD και προ-κασπάσης 8 είναι γνωστό ως σηματοδοτικό σύμπλοκο επαγόμενο από την απόπτωση (death-induced signaling complex, DISC). Η ενεργοποιημένη κασπάση 8 απελευθερώνεται στο κυτταροδιάλυμα και πυροδοτεί την απόπτωση. Η απόπτωση των Τ λεμφοκυττάρων μέσω της διαδικασίας AICD ρυθμίζεται σε μεταγραφικό επίπεδο από την έκφραση του Fas προσδέτη. Οι υπεύθυνοι μεταγραφικοί παράγοντες που εμπλέκονται σε αυτή τη διαδικασία είναι οι Egr (early growth response genes), NF-AT, NFκB, c-myc, AP-1, SP-1 και παράγοντες οι οποίοι ελέγχονται από την ιντερφερόνη (IRFs). O NF-AT ενεργοποιείται από τη διέγερση του Τ κυτταρικού υποδοχέα και επάγει την έκφραση του Fas προσδέτη. Μελέτες αποδεικνύουν την ύπαρξη θέσης πρόσδεσης των παραγόντων NF-AT και SP-1 στον υποκινητή του προσδέτη Fas (Εικόνα 22). Εικόνα 22: Διαφορετικοί μεταγραφικοί παράγοντες επηρεάζουν την έκφραση των γονιδίων που κωδικοποιούν τον Fas προσδέτη. Προέλευση:Zhang Jian,Xu Xuemei,Liu Yong.Cellular and molecular Immunology.2004;1(3): Σημαντικός είναι επίσης και ο ρόλος κάποιων πρωτεϊνικών κινασών, όπως οι πρωτεϊνικές κινάσες της τυροσίνης (PTKs), οι πρωτεϊνικές κινάσες της οικογένειας MAPK και η ZAP-70, στην ενεργοποίηση του προσδέτη Fas και την επαγωγή της διαδικασίας AICD 35,36. Οι πρωτεϊνες FLICE περεμβαίνουν με την κασπάση 8 στο σύμπλοκο DED και FADD καταστέλλοντας την απόπτωση των Τ λεμφοκυττάρων. Στη ρύθμιση της έκφρασης των [37]
42 πρωτεϊνών FLICE συμμετέχουν οι MKK, ο NF-κΒ και η κινάση Ρ13Κ, τόσο σε φυσιολογικά όσο και σε νεοπλασματικά Τ λεμφοκύτταρα Προφανώς, οποιαδήποτε απόκλιση από τη φυσιολογική έκφραση όλων των παραπάνω ρυθμιστικών παραγόντων έχει ως αποτέλεσμα την αναστολή της απόπτωσης των Τ λεμφοκυττάρων και τη γένεση παθολογικών καταστάσεων. 1.6 Χρόνια Ιδιοπαθής Ουδετεροπενία Γενικά Η χρόνια Ιδιοπαθής ουδετεροπενία του ενηλίκου (Chronic Idiopathic Neutropenia of the Adult, CIN) είναι μια σχετικά συχνή κοκκιοκυταρική διαταραχή που ορίζεται ως η εμμένουσα και «ανεξήγητη» ελάττωση του απόλυτου αριθμού των κυκλοφορούντων ουδετεροφίλων κοκκιοκυττάρων(absolute Neutrophil Count, ANC) κάτω από τα κατώτερα όρια της φυσιολογικής διακύμανσης για μακρά χρονική περίοδο, μεγαλύτερη των τριών μηνών ή ακόμα και για ολόκληρη τη ζωή Η διαταραχή περιγράφηκε για πρώτη φορά από τους Kyle και Limman 41 και λίγα χρόνια αργότερα κα από τον Dale. Η ταξινόμηση των χρόνιων ουδετεροπενιών με βάση τον παθοφυσιολογικό μηχανισμό, είτε λόγω ελαττωμένης παραγωγής στο μυελό των οστών, μη αποδοτικής κοκκιοποίησης, αυξημένης περιφερικής καταστροφής ή διαταραχής της κατανομής στο αγγειακό χώρο, έχει αντικατασταθεί από την εμπειρική ταξινόμηση που διαχωρίζει τις χρόνιες ουδετεροπενίες σε συγγενείς και επίκτητες. Οι επίκτητες ουδετεροπενίες ταξινομούνται περαιτέρω σε δευτεροπαθείς, όταν σχετίζονται μα κάποιο υποκείμενο νόσημο, σε φαρμακευτικές όταν αποτελούν ανεπιθύμητη ενέργεια ενός φαρμακευτικού σκευάσματος, και στις αγνώστου αιτιολογίας, οι οποίες μπορεί να είναι πρωτοπαθείς ή ιδιοπαθείς Επιδημιολογικά στοιχεία Η διαταραχή είναι περισσότερο συχνή σε άτομα μέσης ηλικίας, με διάμεση ηλικία διάγνωσης τα 50 έτη, ενώ η συσχέτιση γυναικών προς άνδρες είναι περίπου 8 προς Γενετική προδιάθεση στην εμφάνιση της νόσου έχει περιγραφεί σε άτομα που φέρουν τον απλότυπο HLA DRB1* Το γεγονός αυτό θα μπορούσε ενδεχομένως να ερμηνευθεί από την παρατηρούμενη συσχέτιση του συστήματος HLA με χρόνιες φλεγμονώδεις εξεργασίες, όπως για παράδειγμα με το σακχαρώδη διαβήτη και τη φλεγμονώδη νόσο του εντέρου Crohn. Εξωγενείς παράγοντες, όπως η επαγγελματική δραστηριότητα, η χρήση φυτοφαρμάκων, η επαφή με οργανικούς διαλύτες και η συχνή χρήση μη στεροειδών αντιφλεγμονωδών φαρμάκων δεν φαίνονται να σχετίζονται με τη νόσο 45 [38]
43 1.6.3 Κλινική εικόνα Η διαφοροποίηση της χρόνιας ιδιοπαθούς ουδετεροπενίας από τις άλλες ουδετεροπενίες έγκειται στον επίκτητο χαρακτήρα της, στο γεγονός ότι εμμένει καθ όλη την διάρκεια της ζωής και στην καλή πορεία κα πρόγνωση. 46 Στο ιστορικό των πασχόντων δεν αναφέρεται έκθεση σε ακτινοβολία, χρόνια χρήση φαρμάκων ή άλλων χημικών παραγόντων που θα μπορούσαν να προκαλέσουν μυελοκαταστολή και κατά προέκταση ουδετεροπενία. Επίσης απουσιάζει κλινική ή εργαστηριακή ένδειξη συσχετιζόμενης συστηματικής νόσου. Η ουδετεροπενία μπορεί να διαρκέσει για χρόνια ή ακόμα και για την υπόλοιπη ζωή του ασθενούς με παροδική αύξηση των ουδετεροφίλων κατά τη διάρκεια βακτηριακών λοιμόξεων. 47 Αυτόματη ίαση είναι σπάνια. Οι ασθενείς που πάσχουν από CIN είναι συνήθως ασυμπτωματικοί και η διάγνωση είναι τυχαία μετά από εργαστηριακό έλεγχο ρουτίνας. Οι ασθενείς με CIN δεν παρουσιάζουν ιδιαίτερη ευαισθησία σε λοιμώξεις 48. Παρόλα αυτά οι συχνότερες λοιμώξεις αφορούν το αναπνευστικό σύστημα, το ουροποιητικό και το δέρμα Εργαστηριακά ευρήματα Η ουδετεροπενία συχνά συνοδεύεται από άλλες διαταραχές στο περιφερικό αίμα και στο μυελό των οστών. Παρατηρείται αναιμία χρόνιας νόσου στο 15% των ασθενών χωρίς όμως να απαιτούνται μεταγγίσεις. Η ελάττωση των ερυθρών αιμοσφαιρίων αποδόθηκε στην αυξημένη απόπτωση των κυτταρικών προβαθμίδων τους, σχετιζόμενη με τα υψηλά επίπεδα των προ- φλεγμονωδών κυτταροκινών TNF-α και ιντερφερόνης INF-γ στο μικροπεριβάλλον του μυελού των οστών, καθώς επίσης και στη καταστολή του υποδοχέα της ερυθροποιητίνης στις ερυθροκυτταρικές σειρές. Το 37% των ασθενών παρουσιάζει λεμφοπενία. Η μείωση αντιστοιχεί κυρίως στη ελάττωση των μνημονικών Τ λεμφοκυττάρων CD45RO+/CD4+ και CD45RO+/CD8+ κυττάρων Η αναλογία των Τ βοηθητικών CD4+ και κυτταροτοξικών CD8+ κυττάρων παρέμενε σταθερή γιατί μειωνόταν παράλληλα και οι δυο πληθυσμοί. Ο πιο πιθανός μηχανισμός που προκαλεί λεμφοπενία είναι η ενίσχυση της διαπίδυσης των λεμφοκυττάρων στον εξωαγγειακό χώρο, λόγω της ενεργοποίησης των ενδοθηλιακών κυττάρων από της φλεγμονώδεις κυτταροκίνες TNF-α και IL-1β, καθώς επίσης και εξαιτίας της επίδρασης της χημειοκίνης RANTES στα CD45RO+ κύτταρα. Περίπου το 10% των ασθενών εμφανίζουν μειωμένο αριθμό κυκλοφορούντων μονοκυττάρων καθώς και θρομβοπενία. Εικάζεται ότι η ελάττωση των μονοκυττάρων [39]
44 οφείλεται σε μείωση των προγονικών CD34+/CD33+ κυττάρων (3) ενώ η πτώση των αιμοπεταλίων σε αυξημένη απόπτωση των άωρων μεγακαρυοκυττάρων, λόγω υψηλών επιπέδων του αυξητικού παράγοντα TGF-β1 στο μυελό των ασθενών. 51 Η συγκέντρωση της πρωτεϊνης α1-αντιθρυψίνη (ΑΑΤ) εντοπίζεται σημαντικά αυξημένη, πιθανώς λόγω της μειωμένης παραγωγής της ελαστάσης από τα ουδετερόφιλα κοκκιοκύτταρα, της οποίας η ΑΑΤ αποτελεί τον φυσικό αναστολέα 52. Στην CIN η ελαττωμένη παραγωγή της ελαστάσης σχετίζεται με τον χαμηλό αριθμό των ουδετερόφιλων κοκκιοκυττάρων Παθοφυσιολογία της νόσου Ο μηχανισμός που προκαλεί την χρόνια ιδιοπαθή ουδετεροπενία δεν είναι ακόμα γνωστός. Παλαιότερες θεωρίες αναφέρουν ότι η μείωση του αριθμού των ουδετερόφιλων κοκκιοκυττάρων πιθανώς να οφείλεται σε αυξημένη περιφερική καταστροφή σε διαπίδυση των κυττάρων στους ιστούς 58-59, σε αυξημένη προσκόλληση των κοκκιοκυττάρων στα τοιχώματα των αγγείων 60 ή σε μειωμένη μυελική παραγωγή λόγω χαμηλού αριθμού προγονικών αιμοποιητικών κυττάρων Άλλη πιθανή αιτία μπορεί να είναι η δυσλειτουργία του αιμοποιητικού μικροπεριβάλλοντος στο μυελό των οστών, λόγω ελλείψεως επαγωγικών παραγόντων, όπως του αυξητικού παράγοντα G-CSF ή αυξημένης συγκέντρωσης ανασταλτικών παραγόντων, όπως του TGF-β1 και του ΤΝF-α,(50) με συνέπεια την ανεπαρκή κοκκιοποίηση. Σήμερα, όλες οι εργαστηριακές κλινικές μελέτες συγκλίνουν στην άποψη ότι ο κύριος παθογενετικός μηχανισμός της χρόνιας ιδιοπαθούς ουδετεροπενίας είναι η μειωμένη παραγωγή ουδετερόφιλων κοκκιοκυττάρων στο μυελό των πασχόντων Από τον έλεγχο της έκφρασης του αντιγόνου Fas, το οποίο ως γνωστό επάγει απόπτωση, στις αιμοποιητικές προγονικές προβαθμίδες, προέκυωε ότι η αναλογία των Fas+κυττάρων στα CD34+/CD33+, αλλά όχι και στα CD34-/CD33+ κα στα CD33-/CD15+κυτταρικά διαμερίσματα του μυελού, ήταν σημαντικά αυξημένη στους ασθενείς και σε αναλογία με τον αριθμό των αποπτωτικών κυττάρων. Επιπρόσθετα, στον ορό και στα υπερκείμενα μακρόχρονων μυελικών καλλιεργειών πασχόντων από CIN, έχουν βρεθεί αυξημένες συγκεντρώσεις προ-φλεγμονωδών και προαποπτωτικών κυτταροκινών μακροφαγικής στρωματικής προελεύσεως 67 Συγκεκριμένα παρατηρήθηκαν υψηλά επίπεδα του TNF-α, της INF-γ και του συνδέτη του Fas (Fas-ligand), κυτταροκίνες που επάγουν την έκφραση του Fas στα CD34+μυελικά κυτταρα και ευθύνονται τελικά για την αυξημένη απόπτωση των CD34+/CD33+κοκκιοκυτταρικών [40]
45 προβαθμίδων. Επιπλέον ανιχνεύθηκαν υψηλές συγκεντρώσεις IL-1β, IL-6 και του αναστολές της αιμοποίησης TGF-β1, ενώ η αντιφλεγμονώδης κυτταροκίνη IL-10 παρέμενε σε χαμηλά επίπεδα 68 Η χρήση ειδικών αντισωμάτων που αναστέλλουν τη δράση των προαναφερθέντων κυτταροκινών είχε ως αποτέλεσμα την αύξηση του αριυμού των κυτταρικών προβαθμίδων Το πιο σημαντικό ρόλο στην παθοφυσιολογία της νόσου φαίνεται να επιτελούν τα ενεργοποιημένα Τ λεμφοκύτταρα, τα οποία ανιχνεύονται σε μεγάλο βαθμό στο μυελό των ασθενών. Εκφράζουν υψηλά επίπεδα HLA-DR, CD25, CD38, CD69 και Fas στην επιφάνεια τους 71 και αποτελούν την βασική πηγή παραγωγής INF-γ και FAS-L, που επάγουν την έκφραση του αντιγόνου Fas στα CD34+κύτταρα του μυελού των ασθενών. Πρέπει να σημειωθεί τέλος, ότι στο μηχανισμό της ουδετεροπενίας, πιθανώς συμβάλλει και η διαπίδυση των ουδετερόφιλων κοκκιοκυττάρων στο εξωαγγειακό χώρο. Η IL-1β και ο ΤΝF-α προκαλούν ενεργοποίηση του ενδοθηλίου όπως αυτή εκφράζεται από τα αυξημένα επίπεδα του της διαλυτής Ε-σελεκτίνης, του διαλυτού διακυτταρικού μορίου προσκόλλησης (s ICAM ) και του διαλυτού αγγειακού μορίου προσκόλησης στον ορό των ασθενών (svcam ) Τα μόρια ICAM-1 και VCAM-1 συνδέονται με τις β2 ιντεγκρίνες των ουδετερόφιλων και κυρίως με την CD11b/CD18. Με τη δράση της IL-8, η οποία είναι χημειοκίνη ειδική για τα ουδετερόφιλα, η προσκόλληση τους ακολουθείται από διαπίδυση και υποενδοθηλιακή μετανάστευση, με αποτέλεσμα τη μείωση του αριθμού τους στην κυκλοφορία. Όλα τα ευρήματα οδηγούν στην σκέψη ότι η CIN μπορεί να οφείλεται σε κάποια χρόνια φλεγμονώδη εξεργασία χαμηλού βαθμού στον οργανισμό η οποία προκαλείται από έναν άγνωστο παράγοντα και με άγνωστο μηχανισμό. Η φλεγμονή οδηγεί σε χρόνιο αντιγονικό ερεθισμό, ο οποίος εξηγεί και τον αυξημένο σπληνικό όγκο, τα αυξημένα επίπεδα των IgG1 και IgA και τον διαλυτών ανοσοσυμπλεγμάτων στον ορό των ασθενών, καθώς και τον αυξημένο ποσοστό των πλασματοκυττάρων στο μυελό. Παράλληλα επάγει την ενεργοποίηση των μακροφάγων και των Τ λεμφοκυττάρων με αποτέλεσμα την παραγωγή προ-φλεγμονωδών και προ-αποπτωτικών κυτταροκινών κα χημειοκινών οι οποίες ενεργοποιούν τους οστεοκλάστες, τα ενδοθηλιακά κύτταρα και ελαττώνουν την παραγωγή των ουδετεροφιλιών κοκκιοκυττάρων διαμέσου της απόπτωσης των μυελικών προγονικών κυττάρων. [41]
46 1.6.6 CIN: η μελέτη της νόσου στα πλαίσια των συνδρόμων μυελικής ανεπάρκειας ο ρόλος της αυτοανοσίας Δεδομένα από σειρά μελετών που ανήκουν στα σύνδρομα μυελικής ανεπάρκειας, όπως η απλαστική αναιμία (Α.Α), η Λευχαιμία από Μεγάλα Τ Λεμφοκύτταρα με κοκκία (Τ Cell Large Granular Lymphocytic Leukemia- TLGL), τα μυελοδυσπλαστικά σύνδρομα ( MDS) και η Παροξυσμική Νυκτερινή Αιμοσφαιρινουρία (PNH), προτείνουν ως βάση της μυελικής ανεπάρκειας την παρουσία αυτοάνοσων διαδικασιών στα πλαίσια της ανάπτυξης ολιγο/μονοκλωνικών κυτταροτοξικών πληθυσμών Τ λεμφοκυττάρων. Όλες οι σχετικές μελέτες συγκλίνουν στο ρόλο των κυτταροτοξικών Τ λεμφοκυττάρων μέσα στα πλαίσια ειδικής ανοσολογικής απάντησης. Η παρουσία συγκεκριμένων κυτταροτοξικών αποκρίσεων, ή ένταση και η ειδικότητα τους ίσως να καθορίζουν την κλινική εμφάνιση ενός συνδρόμου μυελικής ανεπάρκειας δεδομένου ότι οι προβαθμίδες της μυελικής σειράς είναι δυνητικοί στόχοι μιας τέτοιας κυτταροτοξικής απόκρισης. [42]
47 ΑΝΤΙΚΕΙΜΕΝΟ ΜΕΛΕΤΗΣ [43]
48 2. Αντικείμενο της μελέτης Η ανάλυση των αναδιατάξεων των γονιδίων του Τ κυτταρικού υποδοχέα προσφέρει ενδείξεις για το ρόλο του αντιγόνου στη παθογένεια διαταραχών του αιμοποιητικού ιστού, υπό την έννοια ότι η έντονη αντιγονική διέγερση μπορεί αρχικά να οδηγήσει σε πολυκλωνική υπερπλασία και, υπό άγνωστες ακόμα προϋποθέσεις, να καταλήξει σε κλωνικές χρόνιες διαταραχές του αιμοποιητικού ιστού, όπως είναι η εκδήλωση της Χρόνιας Ιδιοπαθούς Ουδετεροπενίας. Στην παρούσα εργασία μελετήθηκε το ρεπερτόριο των γονιδίων της β αλυσίδας του Τ κυτταρικού υποδοχέα έξι ασθενών με Χρόνια Ιδιοπαθή Ουδετεροπενία. Οι βασικοί άξονες της μελέτης είχαν ως εξής: λεπτομερής μελέτη ρεπερτορίου γονιδίων TRBV, TRBD, TRDJ και ανάλυση των περιοχών CDR3 αντιπαραβολή με τις κατατεθειμένες σε βάσεις δεδομένων αλληλουχίες από άλλες οντότητες ανίχνευσή ταυτόσημων αναδιατάξεων σε κάθε ασθενή χωριστά αλλά και μεταξύ διαφορετικών ασθενών. [44]
49 ΜΕΘΟΔΟΛΟΓΙΑ [45]
50 3. Μεθοδολογία 3.1 Ομάδα Μελέτης Η ομάδα μελέτης περιλαμβάνει έξι ασθενείς με Χρόνια Ιδιοπαθή Ουδετεροπενία οι οποίοι παρακολουθούνται στην Αιματολογική Κλινική του ΓΝΘ «Γ. Παπανικολάου» και στην Αιματολογική Κλινική του ΠΕ.ΠΑ.Γ.Ν.Η Υλικά και Μέθοδοι Απομόνωση μονοπύρηνων κυττάρων από δείγμα αίματος Η απομόνωση των μονοπύρηνων κυττάρων από αίμα έγινε μετά από διαχωρισμό με φυγοκέντρηση σε βαθμίδωση πυκνότητας, με χρήση του αντιδραστηρίου φικόλλη (Ficoll- Hypaque). Διαδικασία: 1. Επιστοίβαση 35 ml ολικού αίματος σε 15 ml φικόλλης. 2. Φυγοκέντρηση στις 2100 rpm επί 20 λεπτά. 3. Συλλογή της στοιβάδας των μονοπύρηνων και μεταφορά τους σε νέο σωληνάριο. 4. Προσθήκη ανάλογης ποσότητας διαλύματος HBSS/RPMI, ώστε να προκύψει τελικός όγκος 12 ml. 5. Φυγοκέντρηση στις 2100 rpm επί 8 λεπτά. 6. Απόρριψη του υπερκείμενου και προσθήκη 10 ml διαλύματος HBSS/RPMI. Επανάληψη της φυγοκέντρησης. 7. Επαναιώρηση των κυττάρων σε 1 ml διαλύματος RPMI. 8. Μέτρηση των κυττάρων σε μικροσκόπιο με τη χρήση της πλάκας Neubauer και της χρωστικής TURK Απομόνωση RNA Η απομόμωση ολικού RNA πραγματοποιήθηκε με τη μέθοδο του 4Μ θειοκυανικού γουανιδινίου. [46]
51 Διαδικασία: 1. Ομογενοποίηση. Λύση της μεμβράνης των κυττάρων σε 1.2 ml διαλύματος φαινόλης/θειοκυανικού γουανιδινίου (Fenozol). 2. Διαχωρισμός φάσεων. Κατά τη διάρκεια της διαδικασίας, τα δείγματα είναι τοποθετημένα στον πάγο. Προσθήκη 250 ml χλωροφορμίου κι έντονη ανάδευση επί 15 λεπτά. Φυγοκέντρηση στις rpm επί 15 λεπτά στους 40 C. Mετά τη φυγοκέντρηση το δείγμα έχει διαχωριστεί στην υποκείμενη οργανική φάση που περιέχει φαινόλη-χλωροφόρμιο, στην ενδιάμεση φάση που περιέχει τις πρωτεΐνες και στην υπερκείμενη υδατική φάση που περιέχει το RNA. 3. Κατακρήμνιση του RNA. Η υδατική φάση μεταφέρεται σε νέο σωληνάριο eppendorf. Το RNA κατακρημνίζεται με προσθήκη ίσης ποσότητας ισοπροπανόλης και το δείγμα ψύχεται στους C, τουλάχιστον επί ώρα. Ακολουθεί φυγοκέντρηση στις rpm επί 15 λεπτά στους 4 0 C. Σχηματίζεται το ίζημα RNA. 4. Έκπλυση του RNA. Το υπερκείμενο απορρίπτεται και προστίθενται 850 ml αιθανόλης 70%. Ισχυρή ανακίνηση και φυγοκέντρηση στις rpm επί 15 λεπτά στους 4 0 C. Αφαίρεση του υπερκείμενου κι επανάληψη. 5. Επαναδιάλυση του RNA. Το ίζημα του RNA στεγνώνει και διαλυέται σε ποσότητα DEPC νερού ανάλογη του μεγέθους του ιζήματος. 6. Αποθήκευση στους C Σύνθεση cdna Η σύνθεση του cdna έγινε με αντίστροφη μεταγραφή (reverse transcription, RT) in vitro χρησιμοποιώντας ως εκκινητές εξανουκλεοτίδια τυχαίας αλληλουχίας (random hexamers, 500 μg/ml). Η αντίδραση έχει τελικό όγκο 40 μl (Βλέπε Πίνακα 1). Το ένζυμο που χρησιμοποιήθηκε για την αντίδραση είναι η αντίστροφη μεταγραφάση SuperScriptII (200U/μl, Ipsogen). Για την αντίδραση της αντίστροφης μεταγραφής χρησιμοποιήθηκαν επίσης RT buffer (5x), dntps (10Mm), αναστολέας RNAασών, DTT (0.1 M). Ως υπόστρωμα χρησιμοποιήθηκε ολικό κυτταρικό RNA, το οποίο αρχικά διαλύθηκε σε ddη 2 Ο. To διάλυμα RNA αρχικά θερμάνθηκε στους 70 0 C επί 10 min για την αποδιάταξη δευτεροταγών δομών του RNA. [47]
52 Πίνακας 1. Αντιδραστήρια και συγκεντρώσεις για την αντίδραση σύνθεσης cdna Αντιδραστήρια Όγκοι/συγκεντρώσεις RT buffer 5x (250 mm Tris-HCl, 375 mm KC1, 15 mm MgCl 2 ) Αναστολέας RNAασών dntps (10Mm) DTT (0.1 M) Tυχαία εξανουκλεοτίδια SuperScript II (200U/μl) MgCl 2 (50μΜ) 8 μl 1 μl 4 μl 4 μl 2 μl 1 μl (200U) 1.6 μl Υπόστρωμα RNA ddη 2 Ο 1 μg Μέχρι τελικό όγκο 18.4 μl Συνθήκες αντίδρασης: C, 10 λεπτά C, 45 λεπτά C, 3 λεπτά 4. τελική θερμοκρασία: 10 0 C Tο cdna αραιώνεται με ddh 2 0 σε τελικό όγκο 100 μl. To ενδεχόμενο ψευδώς αρνητικών αποτελεσμάτων από αδυναμία ενίσχυσης του RNA μπορεί να αποκλεισθεί με ενίσχυση αλληλουχιών που αντιστοιχούν σε μετάγραφα γονιδίων τα οποία εκφράζονται πάντοτε στον υπό ανάλυση ιστό (μετάγραφα αναφοράς ). Στην παρούσα μελέτη, ως μετάγραφο αναφοράς χρησιμοποιείται το mrna του γονιδίου RARa (retinoic acid receptor a) το οποίο κωδικοποιεί έναν από τους υποδοχείς του ρετινοϊκού οξέος. Έτσι, ως μάρτυρας για την αποτελεσματικότητα της σύνθεσης cdna, ενισχύθηκαν 2 μl του προϊόντος cdna με PCR χρησιμοποιώντας τους ειδικούς εκκινητές για τις αλληλουχίες του RARa cdna (RAR6/RAR8). [48]
53 Συνθήκες αντίδρασης: 1. Αρχική αποδιάταξη: 94 0 C, 5 min 2. Κυρίως αντίδραση: διεξάγεται σε 40 κύκλους. Κάθε κύκλος περιλαμβάνει: Φάση αποδιάταξης: 94 0 C, 1 min Φάση σύνδεσης εκκινητών: 53 0 C, 1 min Φάση επέκτασης μορίων DNA: 72 0 C, 1.5 min 3. Τελική επέκταση συντιθέμενων μορίων DNA: 72C, 10 min. Πίνακας 2. Αντιδραστήρια και συγκεντρώσεις για την αντίδραση ενίσχυσης RARa. Aντιδραστήρια Υπόστρωμα: cdna RB 10x MgCl 2 (50 mm) dntps (10 mm) RAR6 RAR8 Taq polymerase (5 units/μl) ddη 2 Ο Όγκοι/συγκεντρώσεις 2 μl 2.5 μl 1 μl (2 mm) 0.5 μl (200 μμ για καθένα) 0.75 μl 0.75 μl 0.5 μl (2.5 units) Μέχρι τελικό όγκο αντίδρασης 18 μl Ενίσχυση κλωνικών αναδιατάξεων της μεταβλητής περιοχής του Τ κυτταρικού υποδοχέα με αλυσιδωτή αντίδραση πολυμεράσης Κλωνικές αναδιατάξεις της μεταβλητής περιοχής του Τ κυτταρικού υποδοχέα ενισχύθηκαν με την τεχνική PCR. Για την ενίσχυση των συμβολών TRBV-TRBD-TRBJ χρησιμοποιήθηκαν οι εκκινητές (primers) Vβ2-24, και Jβ και Jβ2.2,Jβ 2.6, Jβ 2.7. Οι εκκινητές είναι αντιπροσωπευτικοί για καθεμιά από τις υποομάδες γονιδίων TRBV και αντίστοιχα για τα γονίδια TRBJ και 2.2, 2.6, 2.7. Στις παραπάνω αντιδράσεις το υπόστρωμα ήταν cdna και γενωμικό DNA. Πίνακας 3. Όγκοι αντιδραστηρίων για την αντίδραση ενίσχυσης των αναδιατάξεων TRBV-TRBD-TRBJ με τους εκκινητές Vβ2-Vβ24 και Jβ και Jβ 2.2, Jβ2.6, Jβ 2.7 Αντιδραστήρια Υπόστρωμα: cdna Υπόστρωμα: gdna RB 10x MgCl 2 (50 mm) dntps (10 mm) Μίγμα εκκινητών Vβ-Jβ(10 pmol/μl) Taq polymerase (5 units/μl) ddη 2 Ο Όγκοι/συγκεντρώσεις 5 μl 1μl 10 μl 3 μl(2 mm) 2 μl(200 μμ για κάθε ένα) 33μl 2.5 μl(12.5 units) 47 μl [49]
54 Συνθήκες αντίδρασης: 1. Αρχική αποδιάταξη: 94 0 C, 5 min 2. Αποδιάταξη 94 0 C, 45 sec 3. Υβριδισμός 60 0 C, 45 sec 4. Επιμήκυνση 72 0 C, 1.5 min 5. Επανάληψη βημάτων 2 4 για 39 κύκλους 6. Τελικό στάδιο επιμήκυνσης 72 0 C, 12 min Καθαρισμός των προϊόντων PCR Τα προϊόντα PCR ηλεκτροφορούνται σε πηκτή αγαρόζης χαμηλού σημείου τήξης 3% με χρώση βρωμιούχου αιθιδίου (EtBr, 10mg/ml) σε ρυθμιστικό διάλυμα 1xTAE buffer. Κάθε προϊόν PCR κόβεται και τοποθετείται σε σωληνάριο eppendorf. Για τον καθαρισμό του προϊόντος PCR από την πηκτή αγαρόζης χρησιμοποιείται το QIAquick Gel Extraction Kit (QIAGEN). Χρησιμοποιούνται στήλες με ικανότητα δέσμευσης του προϊόντος PCR σε μεμβράνη από σιλικόνη. Το DNA προσροφάται στη μεμβράνη σε συνθήκες υψηλής αλατότητας, ενώ τα υπόλοιπα συστατικά διέρχονται από τη στήλη. Το προϊόν εκλούεται με διάλυμα Tris. Διαλύματα που χρησιμοποιούνται: Διάλυμα QG :περιέχει θειοκυανικό γουανιδίνιο Διάλυμα PE :διάλυμα έκπλυσης,απαιτεί προσθήκη αιθανόλης (96-100%) Διάλυμα EB :διάλυμα έκλουσης (10mM Tris-Cl,pH 8.5) Διαδικασία : Προσθήκη 600 μl διαλύματος QG στα σωληνάρια eppendorf. Eπώαση στους 50 o C για 20 λεπτά. Vortex στη μέση και στο τέλος της επώασης. Πλήρης διαλυτοποίηση της αγαρόζης. Προσθήκη 200 μl ισοπροπανόλης και ανάδευση. Μεταφορά του διαλύματος στη στήλη. Φυγοκέντρηση ( rpm, 2 λεπτά). Απόρριψη του διηθήματος. Προσθήκη 500 μl διαλύματος QG και επανάληψη της φυγοκέντρησης. [50]
55 Απόρριψη του διηθήματος. Προσθήκη 750 μl διαλύματος PE, αναμονή 5 λεπτών και επανάληψη της φυγοκέντρησης. Απόρριψη του διηθήματος και επανάληψη της φυγοκέντρησης. Τοποθέτηση της στήλης σε σωληνάριο eppendorf. Προσθήκη 50 μl διαλύματος EB και φυγοκέντρηση ( rpm, 2 λεπτά) Kλωνοποίηση i. Ενσωμάτωση του προϊόντος PCR σε φορέα κλωνοποίησης Στην αλυσιδωτή αντίδραση πολυμεράσης χρησιμοποιήθηκε το ένζυμο Taq πολυμεράση. Η Τaq έχει την ιδιότητα να προσθέτει μια δεοξυαδενοσίνη στο 3 άκρο του προϊόντος PCR. Ο φορέας κλωνοποίησης pcr2.1 έχει κατάλοιπα δεοξυθυμιδίνης στα 3 μονόκλωνα άκρα της θέσης ένθεσης (Πίνακας 4). Αυτή η ιδιότητα επιτρέπει την αποτελεσματική ένθεση του προϊόντος PCR στο πλασμίδιο. Πίνακας 4. Χαρακτηριστικά του πλασμιδίου pcr2.1 Χαρακτηριστικά Υποκινητής γονιδίου lac Γονιδιακό τμήμα lacza Γονίδιο ανθεκτικότητας στην καναμυκίνη Γονίδιο ανθεκτικότητας στην αμπικιλλίνη Περιοχή puc Yποκινητής και θέση εκκίνησης Τ7 Θέσεις εκκίνησης Μ13 Περιοχή f1 Λειτουργία Συμβάλλει στην α-συμπληρωματικότητα και την έκφραση του γονιδιακού τμήματος lacza Κωδικοποιεί τα αρχικά 146 αμινοξέα της πρωτεΐνης β-γαλακτοζιδάσης Επιλογή και επιβίωση κυττάρων E.coli Επιλογή και επιβίωση κυττάρων E.coli Aντιγραφή, επιβίωση και υψηλά επίπεδα αντιγραφής σε κύτταρα E.coli Αποκωδικοποίηση του ενθέματος Αποκωδικοποίηση του ενθέματος Αποκωδικοποίηση μονόκλωνης αλυσίδας H αντίδραση ένθεσης γίνεται σε τελικό όγκο 10 μl. [51]
56 Στο διάλυμα της αντίδρασης περιέχονται : Αντιδραστήρια Αποστειρωμένο H 2 O 10x Ρυθμιστικό διάλυμα Πλασμίδιο pcr2.1 (25ng/μl) DNA λιγκάση Τ 4 Προϊόν PCR προς κλωνοποίηση Όγκοι 5 μl 1 μl 2 μl 1 μl 1 μl Για να επιτευχθεί μέγιστη αποτελεσματικότητα, η αντίδραση πρέπει να γίνεται την ίδια ημέρα με την αντίδραση PCR επειδή τα μονόκλωνα 3 άκρα του προϊόντος σταδιακά αποδιατάσσονται. Το διάλυμα της αντίδρασης επωάζεται στους 14 ο C τουλάχιστον επί ώρες. ii. Μετασχηματισμός βακτηριακών κυττάρων με ανασυνδυασμένα πλασμίδια Χρησιμοποιείται το στέλεχος κυττάρων E.coli TOP10F, το οποίο εκφράζει τον καταστολέα του οπερονίου της λακτόζης που αναστέλλει την μεταγραφή από τον υποκινητή lac. Διαδικασία: 1. Προσθήκη 1 μl από το διάλυμα της αντίδρασης ένθεσης σε 50 μl διαλύματος το οποίο περιέχει TOP10F κύτταρα. 2. Για την εισαγωγή των πλασμιδίων στα βακτηριακά κύτταρα εφαρμόζεται θερμικό shock, που καθιστά διαπερατή την κυτταρική μεμβράνη. Το δείγμα που περιέχει τα κύτταρα και τα πλασμίδια τοποθετείται στον πάγο επί 30 λεπτά. Tο δείγμα επωάζεται επί 30 δευτερόλεπτα στους 42 ο C και στη συνέχεια επανατοποθετείται στον πάγο. 3. Προσθήκη 250 μl διαλύματος SOC medium στο δείγμα, το οποίο έχει θερμοκρασία δωματίου. 4. Το δείγμα επωάζεται σε κλίβανο σε θερμοκρασία 37 ο C τουλάχιστον επί 1 ώρα. 5. Επίστρωση στο τρυβλίο 40 μl X-GAL (40 mg/ml) και 40 μl IPTG (100 mm). 6. Όλη η ποσότητα του δείγματος επιστρώνεται σε τρυβλίο με θρεπτικό υλικό LB άγαρ. 7. Επώαση στους 37 ο C τουλάχιστον επί 18 ώρες. Το πλασμίδιο φέρει ένα μικρό τμήμα του γονιδιώματος του E.coli το οποίο περιλαμβάνει την αλληλουχία του γονιδίου lacz που κωδικοποιεί τo α τμήμα του ενζύμου β- [52]
57 γαλακτοζιδάση. Μέσα σε αυτό το τμήμα, βρίσκεται η θέση κλωνοποίησης. Το γονίδιο lacz ανήκει στο οπερόνιο της λακτόζης. Η β-γαλακτοζιδάση καταλύει τη διάσπαση της λακτόζης σε γλυκόζη και γαλακτόζη. Απουσία λακτόζης ή κάποιου ανόλογου μορίου στο θρεπτικό υλικό, ο καταστολέας προσδένεται στον χειριστή του οπερονίου και σταματά τη μεταγραφή των γονιδίων. Τα κύτταρα TOP10F φέρουν στο γονιδίωμα τους την αλληλουχία που κωδικοποιεί το ω τμήμα του ενζύμου β-γαλακτοζιδάση. Μετά το μετασχηματισμό, προκύπτουν δύο είδη μετασχηματισμένων βακτηρίων: βακτήρια που περιέχουν πλασμίδιο το οποίο φέρει ενσωματωμένο εξωγενές DNA, βακτήρια τα οποία έχουν μετασχηματιστεί με πλασμίδιο που δε φέρει ένθετο DNA. Η διάκριση των δύο ειδών βακτηρίων γίνεται με βάση το φαινόμενο της α- συμπληρωματικότητας. Ο συνδυασμός του α τμήματος της β-γαλακτοσιδάσης που κωδικοποιείται από το πλασμίδιο με το ω τμήμα που κωδικοποιείται από τον ξενιστή έχει ως αποτέλεσμα την αποκατάσταση της ενεργότητας β-γαλακτοζιδάσης. Το φαινόμενο αυτό ονομάζεται α-συμπληρωματικότητα. Τα μετασχηματισμένα κύτταρα επιστρώνονται σε τρυβλία που περιέχουν ΙPTG και X-GAL. Το ΙPTG συνδέεται με τον καταστολέα του οπερονίου και τον απενεργοποιεί. Το X-GAL είναι ένα ανάλογο μόριο της λακτόζης, που δίνει μπλε προϊόν παρουσία β-γαλακτοσιδάσης. Τελικά το τρυβλίο επωάζεται σε κλίβανο στους 37 ο C τουλάχιστον επί 18 ώρες, ώστε να σχηματιστούν βακτηριακές αποικίες. Αν τα κύτταρα έχουν μετασχηματιστεί με πλασμίδιο χωρίς ένθετο DNA, συμβαίνει το φαινόμενο της α-συμπληρωματικότητας και οι αποικίες βάφονται μπλέ. Αν τα κύτταρα έχουν μετασχηματιστεί με πλασμίδιο που φέρει εξωγενές DNA εντός της αλληλουχίας laczα, δεν παράγεται το α τμήμα της β-γαλακτοζιδάσης με αποτέλεσμα να καταστρέφεται η συμπληρωματικότητα και να μην μεταβολίζεται το X-GAL Έτσι οι αποικίες βακτηρίων που περιέχουν πλασμίδια με ένθετο DNA έχουν λευκό χρώμα. iii. Απομόνωση βακτηριακών αποικιών Με χρήση μικροβιολογικού κρίκου συλλέγονται από το τρυβλίο λευκές αποικίες για κάθε δείγμα, οι οποίες αποτελούνται από κύτταρα που περιέχουν ανασυνδυασμένο πλασμίδιο. Κάθε αποικία μεταφέρεται σε σωληνάριο των 15 ml που περιέχει υγρό θρεπτικό υλικό LB (με συγκέντρωση 40 gr/200 ml) στο οποίο έχει προστεθεί αμπικιλλίνη σε συγκέντρωση 100 μl/ ml. Ακολουθεί επώαση του διαλύματος σε κλίβανο στους 37 ο C για ώρες. iv. Απομόνωση πλασμιδιακού DNA σε μικρή κλίμακα Η απομόνωση του πλασμιδιακού DNA έγινε με το QIAGEN Plasmid mini kit σύμφωνα με τις οδηγίες της εταιρείας. Η απομόνωση πλασμιδιακού DNA στηρίζεται στην αλκαλική λύση [53]
58 των βακτηρίων που ακολουθείται από δέσμευση του πλασμιδιακού DNA σε ρητίνηανταλλάκτη ανιόντων υπό κατάλληλες συνθήκες ph και χαμηλής συγκέντρωσης αλάτων. Πρωτεΐνες, RNA, χρωστικές και ανεπιθύμητα μόρια χαμηλού μοριακού βάρους απομακρύνονται με εφαρμογή διαλύματος πλύσης ενδιάμεσης συγκέντρωσης αλάτων. Το πλασμιδιακό DNA εκχυλίζεται σε διάλυμα υψηλής συγκέντρωσης αλάτων και έπειτα κατακρημνίζεται με προσθήκη ισοπροπανόλης. Το kit περιέχει τις κατάλληλες στήλες, το διάλυμα P1 (διάλυμα επαναδιάλυσης),το διάλυμα P2 (διάλυμα λύσης), το διάλυμα P3 (διάλυμα εξουδετέρωσης), το διάλυμα QBT (διάλυμα εξισορρόπησης), το διάλυμα QC (διάλυμα πλύσης), το διάλυμα QF (διάλυμα έκλουσης), και τα αντιδραστήρια RNάση Α και LyseBlue. Ενέργειες που πρέπει να γίνουν πριν την έναρξη της διαδικασίας: 1. Προσθήκη RNάσης Α στο διάλυμα P1 έτσι ώστε η τελική συγκέντρωση να είναι 100 mg/ml. Μετά την προσθήκη της RNάσης Α, το διάλυμα P1 φυλάσσεται στους 4 o C. 2. Τοποθέτηση του διαλύματος P3 στους 4 o C ώστε να διατηρείται παγωμένο. 3. Προσθήκη του αντιδραστηρίου LyseBlue στο διάλυμα P1. Το LyseBlue είναι δείκτης που επιτρέπει την οπτική επιβεβαίωση της σωστής ανάμιξης των αντιδραστηρίων έτσι ώστε ν αποφεύγονται τυχόν λάθη. Διαδικασία: 1. Προσθήκη 0,3 ml διαλύματος P1 στο βακτηριακό ίζημα και έντονο vortex ή πιπετάρισμα ώστε να μην παραμείνει κάποιο συσσωμάτωμα βακτηριακών κυττάρων. 2. Προσθήκη 0,3 ml διαλύματος P2, ανακίνηση και επώαση σε θερμοκρασία δωματίου για 5 λεπτά. 3. Προσθήκη 0,3 ml παγωμένου διαλύματος P3, άμεση ανακίνηση και επώαση για 5 λεπτά σε πάγο. 4. Φυγοκέντρηση (15.000g, 10min, 4 o C) 5. Προετοιμασία μιας στήλης με προσθήκη 1 ml διαλύματος QBT. Το διάλυμα μετακινείται μέσω της στήλης σε κατάλληλο σωληνάριο συλλογής λόγω της βαρύτητας. 6. Επιστοίβαση του υπερκειμένου του βήματος 4 στη στήλη. 7. Πλύση της στήλης δύο φορές με 2 ml διαλύματος QC. Το διάλυμα μετακινείται μέσω της στήλης λόγω της βαρύτητας. [54]
59 8. Τοποθέτηση της στήλης σε νέο σωληνάριο συλλογής. 9. Προσθήκη 0,8 ml διαλύματος QF στη στήλη. 10. Κατακρήμνιση του πλασμιδιακού DNA με προσθήκη 0,7 όγκου ισοπροπανόλης (0,56 ml για 0,8 ml όγκο έκλουσης), ανάδευση και κατευθείαν φυγοκέντρηση (15.000g, 30min, 4 o C). Απομάκρυνση του υπερκειμένου. 11. Πλύση του ιζήματος του DNA με 1 ml 70% αιθανόλης, φυγοκέντρηση (15000g, 10min) και απομάκρυνση του υπερκειμένου με προσοχή. 12. Τα σωληνάρια με το ίζημα αφήνονται ανοιχτά επί 5-10 λεπτά σε θερμοκρασία δωματίου για να στεγνώσει το ίζημα και κατόπιν το DNA επαναδιαλύεται σε κατάλληλο όγκο ενέσιμου νερού Aνάλυση των αποτελεσμάτων της κλωνοποίησης Μετά την απομόνωση των προϊόντων PCR από το πλασμιδιακό DNA πραγματοποιήθηκε ανάλυση της αλληλουχίας με διδεοξυριβονουκλεοτίδια (μέθοδος Sanger) σε αυτόματο αναλυτή (CEQ 8000 Genetic Analysis System)και την χρήση του εκκινητή Μ13 Forward. Οι αλληλουχίες των υποκλώνων που προέκυψαν, μελετήθηκαν και συγκρίθηκαν με τη χρήση του αλγορίθμου του προγράμματος IMGT V-QUEST. [55]
60 ΑΠΟΤΕΛΕΣΜΑΤΑ [56]
61 4. Αποτελέσματα 4.1 Περιγραφή ασθενών Μελετήθηκαν έξι ασθενείς με Χρόνια Ιδιοπαθή Ουδετεροπενία, 2 άνδρες και 4 γυναίκες, ηλικίας ετών (μέση ηλικία 55 έτη), οι οποίοι πληρούσαν τα διαγνωστικά κριτήρια της νόσου όπως αυτά έχουν περιγραφεί. Ειδικότερα, οι ασθενείς είχαν αριθμό ουδετεροφίλων <1800 χ 10 6 /L, και περίοδο παρακολούθησης μήνες (διάμεση διάρκεια παρακολούθησης 58 μήνες), δεν είχαν κανένα κλινικό, ορολογικό ή υπερηχογραφικό εύρημα υποκείμενης νόσου που συσχετίζεται με ουδετεροπενία, ούτε ιστορικό έκθεσης σε ακτινοβολία ή χρήσης χημικών ή φαρμάκων στα οποία θα μπορούσε να αποδοθεί η ουδετεροπενία, ενώ είχαν φυσιολογικό καρυότυπο μυελού και αρνητικές δοκιμασίες αντιπολυμορφοπύρηνων αντισωμάτων. Περιπτώσεις κυκλικής και οικογενούς ουδετεροπενίας αποκλείσθηκαν μετά από επαναλαμβανόμενες μετρήσεις των πολυμορφοπύρηνων στους ασθενείς καθώς και στο οικογενειακό περιβάλλον. Κανένας από τους ασθενείς δεν ανέφερε ενεργό λοίμωξη κατά τη διάρκεια της μελέτης, αλλά ούτε και το τελευταίο τρίμηνο. Η βασική διαφορά μεταξύ των ασθενών αυτών αναδείχθηκε κατά το μοριακό έλεγχο και αφορούσε στα πρότυπα αναδιατάξεων των γονιδίων TCRβ. Οι αναδιατάξεις των γονιδίων της β αλυσίδας του Τ κυτταρικού υποδοχέα ενισχύθηκαν με RT-PCR και τα προϊόντα υποβλήθηκαν σε κλασική ανάλυση υποκλωνοποίησης (subcloning) η οποία περιλάμβανε ενσωμάτωση στο πλασμίδιο pcr 2.1 και μετασχηματισμό κυττάρων Ε.Coli/TOP10F. Ο αριθμός των υποκλώνων που αναλύθηκαν ανά ασθενή ήταν (Δm: 31 κλώνοι/ ασθενή). Η βιοπληροφορική επεξεργασία των αποτελεσμάτων πραγματοποιήθηκε με το εργαλείο V-QUEST/IMGT. [57]
62 4.2. Ρεπερτόριο TRBV στον ασθενή 1 _(CR6) Στον ασθενή 1 αναλύθηκαν συνολικά 39 κλώνοι ( 35 παραγωγικές αναδιατάξεις και 5 μη παραγωγικές αναδιατάξεις) και προέκυψαν τρία διαφορετικά υποσύνολα αναδιατάξεων σε πολυκλωνικό υπόστρωμα.(εικόνα 23) Εικόνα 23: Σχηματική αναπαράσταση των αναδιατάξεων TRBV-TRBJ στον ασθενή 1 Πιο συγκεκριμένα ο ασθενής 1 εκφράζει διάφορα TRBV και TRBJ γονίδια (Εικόνες 24 και 25) Εικόνα 24: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBV στον ασθενή 1 [58]
63 Εικόνα 25: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBJ στον ασθενή 1 Τα γονίδια TRBD1 και TRBD2 ανιχνεύθηκαν σε ποσοστό 74,8% και 11,5%, αντιστοίχως. Το μήκος της περιοχής CDR3 κυμαινόταν από 10 εώς 16 νουκλεοτίδια. Οι περισσότερες αναδιατάξεις (16/35 περιπτώσεις, 45,71%) εμφάνιζαν CDR3 περιοχή 13 νουκλεοτιδίων Ρεπερτόριο TRBV στον ασθενή 2_CR7 Στον ασθενή 2 αναλύθηκαν συνολικά 19 κλώνοι, όλες οι αναδιατάξεις ήταν παραγωγικές και προέκυψαν δύο διαφορετικά υποσύνολα αναδιατάξεων σε πολυκλωνικό υπόστρωμα. (Εικόνα26) Εικόνα 26: Σχηματική αναπαράσταση των αναδιατάξεων TRBV-TRBJ στον ασθενή 2 [59]
64 Πιο συγκεκριμένα ο ασθενής 2 εκφράζει διάφορα γονίδια TRBV και TRBJ, με την συντριπτική πλειοψηφία των αναδιατάξεων να χρησιμοποίει το TRBV19 γονίδιο και το TRBJ2-7 γονίδιο.(εικόνες 27 και 28) Εικόνα 27: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBV στον ασθενή 2. Εικόνα 28: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBJ στον ασθενή 2 Τα γονίδια TRBD1 και TRBD2 ανιχνεύθηκαν σε ποσοστό 26,31 % και 73,69%, αντιστοίχως. Το μήκος της περιοχής CDR3 κυμαινόταν από 13 έως 17 νουκλεοτίδια. Οι 18/19 παραγωγικές αναδιατάξεις (94,73%) είχαν μήκος CDR3 13 νουκλεοτιδίων. [60]
65 4.4. Ρεπερτόριο TRBV στον ασθενή 3_CIN32 Στον ασθενή 3 αναλύθηκαν συνολικά 29 κλώνοι ( 27 παραγωγικές αναδιατάξεις και 2 μη παραγωγικές αναδιατάξεις) και προέκυψαν πέντε διαφορετικά υποσύνολα αναδιατάξεων σε πολυκλωνικό υπόστρωμα.(εικόνα 29) Εικόνα 29: Σχηματική αναπαράσταση των αναδιατάξεων TRBV-TRBJ στον ασθενή 3 Πιο συγκεκριμένα ο ασθενής 3 εμφανίζει διάφορα TRBV και TRBJ γονίδια (εικόνα 30 και 31) Εικόνα 30: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBV στον ασθενή 3. [61]
66 Εικόνα 31: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBJ στον ασθενή 3. Τα γονίδια TRBD1 και TRBD2 ανιχνεύθηκαν σε ποσοστό 55,17 % και 20,6%, αντιστοίχως. Επίσης 7/29 αλληλουχίες δεν χρησιμοποιούσαν TRBD γονίδιο. Το μήκος της περιοχής CDR3 κυμαινόταν από 9 έως 19 νουκλεοτίδια. Οι 7/27 παραγωγικές αναδιατάξεις (26 %) είχαν μήκος CDR3 13 νουκλεοτιδίων Ρεπερτόριο TRBV στον ασθενή 4_CIN34 Στον ασθενή 4 αναλύθηκαν συνολικά 38 κλώνοι ( 37 παραγωγικές αναδιατάξεις και 1 μη παραγωγική αναδιάταξη) και προέκυψαν τρία διαφορετικά υποσύνολα αναδιατάξεων σε πολυκλωνικό υπόστρωμα.(εικόνα 32) Εικόνα 32: Σχηματική αναπαράσταση των αναδιατάξεων TRBV-TRBJ στον ασθενή 4 [62]
67 Πιο συγκεκριμένα ο ασθενής 3 εμφανίζει διάφορα TRBV και TRBJ γονίδια (εικόνα 33 και 34) Εικόνα 33: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBV στον ασθενή 4. Εικόνα 34: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBVJστον ασθενή 4. Τα γονίδια TRBD1 και TRBD2 ανιχνεύθηκαν σε ποσοστό 43,24 % και 48,64%, αντιστοίχως. Επίσης 3/37 αλληλουχίες δεν χρησιμοποιούσαν TRBD γονίδιο. Το μήκος της περιοχής CDR3 κυμαινόταν από 9 έως 16 νουκλεοτίδια. Οι 14/37 παραγωγικές αναδιατάξεις (37,9 %) είχαν μήκος CDR3 14 νουκλεοτιδίων. [63]
68 4.6. Ρεπερτόριο TRBV στον ασθενή 5_CIN38 Στον ασθενή 5 αναλύθηκαν συνολικά 28 κλώνοι,όλοι οδηγούσαν σε παραγωγικές αναδιατάξεις και προέκυψαν έξι διαφορετικά υποσύνολα αναδιατάξεων σε πολυκλωνικό υπόστρωμα.(εικόνα 35) Εικόνα 35: Σχηματική αναπαράσταση των αναδιατάξεων TRBV-TRBJ στον ασθενή 5 Πιο συγκεκριμένα ο ασθενής 5 εμφανίζει διάφορα TRBV και TRBJ γονίδια (εικόνα 36 και 37) Εικόνα 36: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBV στον ασθενή 5. [64]
69 Εικόνα 37: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBJ στον ασθενή 5. Τα γονίδια TRBD1 και TRBD2 ανιχνεύθηκαν σε ποσοστό 67,9 % και 32,1%, αντιστοίχως. Το μήκος της περιοχής CDR3 κυμαινόταν από 10 έως 14 νουκλεοτίδια. Οι 12/28 παραγωγικές αναδιατάξεις (42,9 %) είχαν μήκος CDR3 12 νουκλεοτιδίων Ρεπερτόριο TRBV στον ασθενή 6_CIN21 Στον ασθενή 6 αναλύθηκαν συνολικά 31 κλώνοι (29 παραγωγικές αναδιατάξεις και 2 μη παραγωγικές αναδιατάξεις) και προέκυψαν τρία διαφορετικά υποσύνολα αναδιατάξεων σε πολυκλωνικό υπόστρωμα.(εικόνα 38) Εικόνα 38: Σχηματική αναπαράσταση των αναδιατάξεων TRBV-TRBJ στον ασθενή 5 [65]
70 Πιο συγκεκριμένα ο ασθενής 6 εμφανίζει διάφορα TRBV και TRBJ γονίδια (εικόνα 39 και 40) Εικόνα 39: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBV στον ασθενή 6. Εικόνα 40: Σχηματική αναπαράσταση του ρεπερτορίου των γονιδίων TRBJ στον ασθενή 6. Τα γονίδια TRBD1 και TRBD2 ανιχνεύθηκαν σε ποσοστό 62,1 % και 37,9 %, αντιστοίχως. Το μήκος της περιοχής CDR3 κυμαινόταν από 8 έως 16 νουκλεοτίδια. Οι 8/29 παραγωγικές αναδιατάξεις (25,6 %) είχαν μήκος CDR3 11 νουκλεοτιδίων. [66]

 Εικόνα 41: Σχηματική αναπαράσταση των επτά πιο συχνά χρησιμοποιούμενων γονιδίων στο σύνολο των παραγωγικών αναδιατάξεων όλων των ασθενών.")
71 4.8 Μελέτη του ρεπερτορίου στο σύνολο των ασθενών Αναγνωρίστηκαν δεκαπέντε γονίδια τα οποία συμμετείχαν στην δημιουργία των κύριων κλωνοτύπων στο σύνολο των ασθενών. Έξι κοινά γονίδια, τα TRBV2, TRBV19, TRBV27, TRBV7-8, TRBV10-2 και TRBV28 χρησιμοποιούνται από όλους τους ασθενείς για την δημιουργία των κλωνοτύπων. Αν μελετήσουμε το ρεπερτόριο των TRBV γονιδίων στο σύνολο των ασθενών παρατηρούμε ότι μόλις επτά γονίδια αντιπροσωπεύουν το 67,56% όλων των αλληλουχιών. (εικόνα 41) Εικόνα 41: Σχηματική αναπαράσταση των επτά πιο συχνά χρησιμοποιούμενων γονιδίων στο σύνολο των παραγωγικών αναδιατάξεων όλων των ασθενών. Σε επίπεδο ασθενή παρατηρήθηκε επίσης επιλεκτικότητα των γονιδίων TRBV. Οι ασθενείς χρησιμοποιούσαν τα ίδια TRBV γονίδια αλλά με διαφορετική συχνότητα σε κάθε περίπτωση. (Εικόνα 42) Εικόνα 42: Επιλεκτικότητα ρεπερτορίου σε επίπεδο ασθενούς. [67]
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς. Οι φάσεις των Τ κυτταρικών απαντήσεων
 ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα
 Β λεμφοκύτταρα") Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα φυσική ή μη ειδική ανοσία δεν απαιτεί προηγούμενη έκθεση στο παθογόνο και δεν διαθέτει μνήμη. σε επίκτητη ή ειδική ανοσία χυμική ανοσία με παραγωγή
Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα φυσική ή μη ειδική ανοσία δεν απαιτεί προηγούμενη έκθεση στο παθογόνο και δεν διαθέτει μνήμη. σε επίκτητη ή ειδική ανοσία χυμική ανοσία με παραγωγή
Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 14/10/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Γενικά στοιχεία
Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 14/10/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Γενικά στοιχεία
ΑΝΟΣΟΛΟΓΙΑ. 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ
 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ") ΑΝΟΣΟΛΟΓΙΑ 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ ΑΝΤΙΓΟΝΟ 3. Η πρόσληψη του αντιγόνου και η παρουσίασή του στα λεμφοκύτταρα 4. Η αναγνώριση του αντιγόνου. Αντιγονικοί
ΑΝΟΣΟΛΟΓΙΑ 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ ΑΝΤΙΓΟΝΟ 3. Η πρόσληψη του αντιγόνου και η παρουσίασή του στα λεμφοκύτταρα 4. Η αναγνώριση του αντιγόνου. Αντιγονικοί
ΥΠΟΠΛΗΘΥΣΜΟΙ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ. Αλεξάνδρα Φλέβα Ph.D Βιολόγος Τμήμα Ανοσολογίας-Ιστοσυμβατότητας Γ.Ν. «Παπαγεωργίου»
 ΥΠΟΠΛΗΘΥΣΜΟΙ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ Αλεξάνδρα Φλέβα Ph.D Βιολόγος Τμήμα Ανοσολογίας-Ιστοσυμβατότητας Γ.Ν. «Παπαγεωργίου» Η αξιοποίηση του εύρους των δυνατοτήτων της κυτταρομετρίας ροής στην Ανοσολογία προσφέρει
ΥΠΟΠΛΗΘΥΣΜΟΙ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ Αλεξάνδρα Φλέβα Ph.D Βιολόγος Τμήμα Ανοσολογίας-Ιστοσυμβατότητας Γ.Ν. «Παπαγεωργίου» Η αξιοποίηση του εύρους των δυνατοτήτων της κυτταρομετρίας ροής στην Ανοσολογία προσφέρει
Μοριακή κυτταρική βιοχημεία Ανοσοποιητικό σύστημα
 Μοριακή κυτταρική βιοχημεία Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY-335 Χειμερινό εξάμηνο 2014 / 2015 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα
Μοριακή κυτταρική βιοχημεία Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY-335 Χειμερινό εξάμηνο 2014 / 2015 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα
οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά
 οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά Ανοσολογικό σύστηµα Βασικό σύστηµα του οργανισµού Λειτουργικές µονάδες του ανοσολογικού συστήµατος Οργανωµένος λεµφικός
οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά Ανοσολογικό σύστηµα Βασικό σύστηµα του οργανισµού Λειτουργικές µονάδες του ανοσολογικού συστήµατος Οργανωµένος λεµφικός
Ανοσιακή απάντηση (immune response)
") Ανοσιακή απάντηση (immune response) Το σύνολο των μηχανισμών που καθιστούν τον οργανισμό ικανό να αναγνωρίζει,να εξουδετερώνει και να απομακρύνει κάθε ξένη ουσία προς αυτόν χαρακτηρίζεται ως ανοσία. Αποτέλεσμα
Ανοσιακή απάντηση (immune response) Το σύνολο των μηχανισμών που καθιστούν τον οργανισμό ικανό να αναγνωρίζει,να εξουδετερώνει και να απομακρύνει κάθε ξένη ουσία προς αυτόν χαρακτηρίζεται ως ανοσία. Αποτέλεσμα
ΚΕΦΑΛΑΙΟ 7 Χυμικές ανοσοαπαντήσεις Ενεργοποίηση των Β λεμφοκυττάρων και παραγωγή των αντισωμάτων
 ΚΕΦΑΛΑΙΟ 7 Χυμικές ανοσοαπαντήσεις Ενεργοποίηση των Β λεμφοκυττάρων και παραγωγή των αντισωμάτων Φάσεις και τύποι των χυμικών ανοσοαπαντήσεων Διέγερση των Β λεμφοκυττάρων από το αντιγόνο Μεταβίβαση σήματος
ΚΕΦΑΛΑΙΟ 7 Χυμικές ανοσοαπαντήσεις Ενεργοποίηση των Β λεμφοκυττάρων και παραγωγή των αντισωμάτων Φάσεις και τύποι των χυμικών ανοσοαπαντήσεων Διέγερση των Β λεμφοκυττάρων από το αντιγόνο Μεταβίβαση σήματος
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ. ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ
 ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
όλοι αναπνευστική οδός στομάχι στόμα
 κεράτινη στιβάδα περιέχει σμήγμα λιπαρά οξέα Μηχανισμοί που παρεμποδίζουν την είσοδο Δέρμα περιέχει ιδρώτας φυσιολογική μικροχλωρίδα λυσοζύμη γαλακτικό οξύ μικροοργανισμών Βλεννογόνοι όλοι αναπνευστική
κεράτινη στιβάδα περιέχει σμήγμα λιπαρά οξέα Μηχανισμοί που παρεμποδίζουν την είσοδο Δέρμα περιέχει ιδρώτας φυσιολογική μικροχλωρίδα λυσοζύμη γαλακτικό οξύ μικροοργανισμών Βλεννογόνοι όλοι αναπνευστική
ΦΡΟΝΤΙΣΤΗΡΙΑ ΠΡΟΟΠΤΙΚΗ
 Απαντήσεις του κριτηρίου αξιολόγησης στη βιολογία γενικής παιδείας 1 ο ΚΕΦΑΛΑΙΟ ΘΕΜΑ 1 ο Να γράψετε τον αριθμό καθεμίας από τις ημιτελείς προτάσεις 1 έως και 5, και δίπλα σε αυτόν το γράμμα που αντιστοιχεί
Απαντήσεις του κριτηρίου αξιολόγησης στη βιολογία γενικής παιδείας 1 ο ΚΕΦΑΛΑΙΟ ΘΕΜΑ 1 ο Να γράψετε τον αριθμό καθεμίας από τις ημιτελείς προτάσεις 1 έως και 5, και δίπλα σε αυτόν το γράμμα που αντιστοιχεί
ΑΝΟΣΟΒΙΟΛΟΓΙΑ. Εξεταστική Ιανουαρίου 2010
 Εξεταστική Ιανουαρίου 2010 Ποιες είναι οι διαφορές μιας πρωτογενούς από μια δευτερογενή χυμική ανοσολογική απόκριση; Περιγράψετε τους μηχανισμούς ενεργοποίησης στις δυο περιπτώσεις. ΘΕΜΑ 2 (1 μονάδα) Περιγράψετε
Εξεταστική Ιανουαρίου 2010 Ποιες είναι οι διαφορές μιας πρωτογενούς από μια δευτερογενή χυμική ανοσολογική απόκριση; Περιγράψετε τους μηχανισμούς ενεργοποίησης στις δυο περιπτώσεις. ΘΕΜΑ 2 (1 μονάδα) Περιγράψετε
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ ΜΟΝΟΚΛΩΝΙΚΑ ΑΝΤΙΣΩΜΑΤΑ ΕΜΒΟΛΙΑ. Εργαστήριο Γενετικής, ΓΠΑ
 ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ ΜΟΝΟΚΛΩΝΙΚΑ ΑΝΤΙΣΩΜΑΤΑ ΕΜΒΟΛΙΑ Στάδια μικροβιακής λοίμωξης δημιουργία αποικίας σε εξωτερική επιφάνεια διείσδυση στον οργανισμό τοπική μόλυνση συστηματική (γενικευμένη) μόλυνση H σημασία
ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ ΜΟΝΟΚΛΩΝΙΚΑ ΑΝΤΙΣΩΜΑΤΑ ΕΜΒΟΛΙΑ Στάδια μικροβιακής λοίμωξης δημιουργία αποικίας σε εξωτερική επιφάνεια διείσδυση στον οργανισμό τοπική μόλυνση συστηματική (γενικευμένη) μόλυνση H σημασία
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ. Παθοφυσιολογία Ι
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Παθοφυσιολογία Ι Ανοσολογία - Ρευματολογία Υπεύθυνος μαθήματος: Καθηγητής Παθολογίας/Ρευματολογίας, Αλέξανδρος Α. Δρόσος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Παθοφυσιολογία Ι Ανοσολογία - Ρευματολογία Υπεύθυνος μαθήματος: Καθηγητής Παθολογίας/Ρευματολογίας, Αλέξανδρος Α. Δρόσος Άδειες Χρήσης Το παρόν εκπαιδευτικό
7. Χυµικές ανοσοαπαντήσεις. Ενεργοποίηση των Β λεµφοκυττάρων και παραγωγή αντισωµάτων
 7. Χυµικές ανοσοαπαντήσεις. Ενεργοποίηση των Β λεµφοκυττάρων και παραγωγή αντισωµάτων Οι φάσεις και οι τύποι των χυµικών ανοσοαπαντήσεων Η διέγερση των Β λεµφοκυττάρων από το αντιγόνο Ο ρόλος των βοηθητικών
7. Χυµικές ανοσοαπαντήσεις. Ενεργοποίηση των Β λεµφοκυττάρων και παραγωγή αντισωµάτων Οι φάσεις και οι τύποι των χυµικών ανοσοαπαντήσεων Η διέγερση των Β λεµφοκυττάρων από το αντιγόνο Ο ρόλος των βοηθητικών
Αυτοφαγία & Ανοσολογικό Σύστημα. Χαράλαμπος Μ. Μουτσόπουλος
 Αυτοφαγία & Ανοσολογικό Σύστημα Χαράλαμπος Μ. Μουτσόπουλος Απρίλιος 2018 Αυτοφαγία & Ανοσολογικό Σύστημα Ευκαρυωτικά κύτταρα: Δυο μηχανισμούς αποδόμησης και ανακύκλωσης κυτταροπλασματικών αποβλήτων Πρωτεάσες
Αυτοφαγία & Ανοσολογικό Σύστημα Χαράλαμπος Μ. Μουτσόπουλος Απρίλιος 2018 Αυτοφαγία & Ανοσολογικό Σύστημα Ευκαρυωτικά κύτταρα: Δυο μηχανισμούς αποδόμησης και ανακύκλωσης κυτταροπλασματικών αποβλήτων Πρωτεάσες
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΑΝΟΣΟΓΕΝΕΤΙΚΗ ΑΝΑΛΥΣΗ ΤΟΥ ΡΕΠΕΡΤΟΡΙΟΥ ΤΟΥ Τ ΚΥΤΤΑΡΙΚΟΥ ΥΠΟΔΟΧΕΑ ΤΩΝ ΚΥΤΤΑΡΟΤΟΞΙΚΩΝ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΜΕ ΑΛΛΗΛΟΥΧΗΣΗ ΥΨΗΛΗΣ ΑΠΟΔΟΣΗΣ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ-ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ Α ΠΑΘΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ ΔΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ ΠΑΝΤΕΛΗΣ ΖΕΜΠΕΚΑΚΗΣ ΠΑΝΕΠ. ΕΤΟΣ 2018-2019 Αριθμ. Διατριβής: 4107
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ-ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ Α ΠΑΘΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ ΔΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ ΠΑΝΤΕΛΗΣ ΖΕΜΠΕΚΑΚΗΣ ΠΑΝΕΠ. ΕΤΟΣ 2018-2019 Αριθμ. Διατριβής: 4107
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ
 ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ Η πρώτη γραπτή αναφορά στο φαινόμενο της ανοσίας μπορεί να αναζητηθεί στον Θουκυδίδη, τον μεγάλο ιστορικό του Πελοποννησιακού Πολέμου Ανάπτυξη και επιβίωση o
ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ Η πρώτη γραπτή αναφορά στο φαινόμενο της ανοσίας μπορεί να αναζητηθεί στον Θουκυδίδη, τον μεγάλο ιστορικό του Πελοποννησιακού Πολέμου Ανάπτυξη και επιβίωση o
ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ. Εξειδίκευση: προϊόντα (κύτταρα ή αντισώματα) ειδικά για το αντιγόνο. Μνήμη: κύτταρα
 ειδικά για το αντιγόνο. Μνήμη: κύτταρα") ΕΙΔΙΚΗ ΑΜΥΝΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ Εξειδίκευση: προϊόντα (κύτταρα ή αντισώματα) ειδικά για το αντιγόνο Μνήμη: κύτταρα ΑΝΤΙΓΟΝΟ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΣ ΤΜΗΜΑ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΤΟΞΙΚΕΣ ΟΥΣΙΕΣ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΓΥΡΗ, ΦΑΡΜΑΚΕΥΤΙΚΕΣ
ΕΙΔΙΚΗ ΑΜΥΝΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ Εξειδίκευση: προϊόντα (κύτταρα ή αντισώματα) ειδικά για το αντιγόνο Μνήμη: κύτταρα ΑΝΤΙΓΟΝΟ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΣ ΤΜΗΜΑ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΤΟΞΙΚΕΣ ΟΥΣΙΕΣ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΓΥΡΗ, ΦΑΡΜΑΚΕΥΤΙΚΕΣ
Ανοσιακή απάντηση Αικατερίνη Ταράση
 Ανοσιακή απάντηση Αικατερίνη Ταράση ιευθύντρια Τµ. Ανοσολογίας- Ιστοσυµβατότητας Γ.Ν.Α.. " Ο Ευαγγελισµός" ανοσιακή απάντηση είναι το σύνολο των πολύπλοκων διεργασιών µε τις οποίες ο οργανισµός (ξενιστής)
Ανοσιακή απάντηση Αικατερίνη Ταράση ιευθύντρια Τµ. Ανοσολογίας- Ιστοσυµβατότητας Γ.Ν.Α.. " Ο Ευαγγελισµός" ανοσιακή απάντηση είναι το σύνολο των πολύπλοκων διεργασιών µε τις οποίες ο οργανισµός (ξενιστής)
Κυτταρική Βιολογία. Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; ΘΩΜΑΣ ΑΠΑΝΤΗΣΗ. 2. Ποιες είναι οι κατηγορίες γονιδίων με κριτήριο το προϊόν της μεταγραφής τους;
 Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
Ανοσοποιητικό σύστημα
 Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY- 335 Χειμερινό εξάμηνο 2016 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα Η ανοσία είναι η κατάσταση προστασίας
Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY- 335 Χειμερινό εξάμηνο 2016 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα Η ανοσία είναι η κατάσταση προστασίας
Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών
 Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
Ερωτήµατα προς απάντηση
 Κεφ. 9. Ανοσιακή ανοχή και αυτοανοσία: Η διάκριση εαυτού-ξένου και οι διαταραχές της Ερωτήµατα προς απάντηση Πως διατηρεί το ανοσοποιητικό σύστηµα τηµη απαντητικότητα σε εαυτά αντιγόνα (=ανοχή); Ποιοι
Κεφ. 9. Ανοσιακή ανοχή και αυτοανοσία: Η διάκριση εαυτού-ξένου και οι διαταραχές της Ερωτήµατα προς απάντηση Πως διατηρεί το ανοσοποιητικό σύστηµα τηµη απαντητικότητα σε εαυτά αντιγόνα (=ανοχή); Ποιοι
ΑΝΟΣΟΛΟΓΙΚΟ ΣΥΣΤΗΜΑ-ΕΙΣΑΓΩΓΗ. Στην έμφυτη ανοσία, η οποία δραστηριοποιείται άμεσα, σε λίγα λεπτά, περιλαμβάνονται:
 ΑΝΟΣΟΛΟΓΙΚΟ ΣΥΣΤΗΜΑ-ΕΙΣΑΓΩΓΗ Περικλής Γ. Φούκας Β`Εργαστήριο Παθολογικής Ανατομικής Ιατρικής Σχολής Ε.Κ.Π.Α. Πανεπιστημιακό Γενικό Νοσοκομείο «Αττικόν» pfoukas@med.uoa.gr Το ανοσολογικό ή αμυντικό σύστημα
ΑΝΟΣΟΛΟΓΙΚΟ ΣΥΣΤΗΜΑ-ΕΙΣΑΓΩΓΗ Περικλής Γ. Φούκας Β`Εργαστήριο Παθολογικής Ανατομικής Ιατρικής Σχολής Ε.Κ.Π.Α. Πανεπιστημιακό Γενικό Νοσοκομείο «Αττικόν» pfoukas@med.uoa.gr Το ανοσολογικό ή αμυντικό σύστημα
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
Μεταβολικές ανάγκες ανοσοκυττάρων
 Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ Διακρίνονται σε: - Πρωτογενή και - Δευτερογενή Πρωτογενή είναι τα όργανα στα οποία γίνεται η ωρίμανση των κυττάρων του ανοσοποιητικού: - Θύμος
ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ Διακρίνονται σε: - Πρωτογενή και - Δευτερογενή Πρωτογενή είναι τα όργανα στα οποία γίνεται η ωρίμανση των κυττάρων του ανοσοποιητικού: - Θύμος
Ανοσιακή Αποκατάσταση μετά την Μεταμόσχευση Αρχέγονων Αιμοποιητικών Κυττάρων
 Ανοσιακή Αποκατάσταση μετά την Μεταμόσχευση Αρχέγονων Αιμοποιητικών Κυττάρων Μαριάννα Τζανουδάκη Τμήμα Ανοσολογίας & Ιστοσυμβατότητας ΓΝ Παίδων Αθηνών «Η ΑγίαΣοφία» ΜΑΚ Βασική θεραπευτική επιλογή σε πολλές
Ανοσιακή Αποκατάσταση μετά την Μεταμόσχευση Αρχέγονων Αιμοποιητικών Κυττάρων Μαριάννα Τζανουδάκη Τμήμα Ανοσολογίας & Ιστοσυμβατότητας ΓΝ Παίδων Αθηνών «Η ΑγίαΣοφία» ΜΑΚ Βασική θεραπευτική επιλογή σε πολλές
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ
 14.1 ΕΙΣΑΓΩΓΗ") 1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
ΘΕΜΑ Α Α1. γ Α2. γ Α3. α Α4. β Α5. β ΘΕΜΑ B B1. B2.
 ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
να ταράξουν την λειτουργία των ιστών και των οργάνων του; α. τη θέση τους στο ανθρώπινο σώμα β. την γενικευμένη ή εξειδικευμένη δράση
 Ερωτήσεις κατανόησης της θεωρίας του 1 ο κεφαλαίου (συνέχεια) 1. Από τι εξαρτάται η επιβίωση του ανθρώπου και ποιοι εξωτερικοί παράγοντες θα μπορούσαν να ταράξουν την λειτουργία των ιστών και των οργάνων
Ερωτήσεις κατανόησης της θεωρίας του 1 ο κεφαλαίου (συνέχεια) 1. Από τι εξαρτάται η επιβίωση του ανθρώπου και ποιοι εξωτερικοί παράγοντες θα μπορούσαν να ταράξουν την λειτουργία των ιστών και των οργάνων
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ. Παθοφυσιολογία Ι
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Παθοφυσιολογία Ι Ανοσολογία - Ρευματολογία Υπεύθυνος μαθήματος: Καθηγητής Παθολογίας/Ρευματολογίας, Αλέξανδρος Α. Δρόσος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Παθοφυσιολογία Ι Ανοσολογία - Ρευματολογία Υπεύθυνος μαθήματος: Καθηγητής Παθολογίας/Ρευματολογίας, Αλέξανδρος Α. Δρόσος Άδειες Χρήσης Το παρόν εκπαιδευτικό
Βιολογία γενικής παιδείας τάξη Γ
 Βιολογία γενικής παιδείας τάξη Γ Παραδόσεις του μαθήματος Επιμέλεια: Γιάννης Αργύρης Βιολόγος M.Sc. Καθηγητής 3 ου Γεν. Λυκ. Ηλιούπολης Κεφάλαιο 1ο Άνθρωπος και υγεία 2. Μηχανισμοί Άμυνας του Ανθρώπινου
Βιολογία γενικής παιδείας τάξη Γ Παραδόσεις του μαθήματος Επιμέλεια: Γιάννης Αργύρης Βιολόγος M.Sc. Καθηγητής 3 ου Γεν. Λυκ. Ηλιούπολης Κεφάλαιο 1ο Άνθρωπος και υγεία 2. Μηχανισμοί Άμυνας του Ανθρώπινου
ΑΝΟΣΟΛΟΓΙΑ. 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία
 2. Φυσική ανοσία") ΑΝΟΣΟΛΟΓΙΑ 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ ΑΝΤΙΓΟΝΟ και ΥΠΟ ΟΧΕΙΣ ΑΝΤΙΓΟΝΟΥ 3. Η πρόσληψη του αντιγόνου και η παρουσίασή του στα λεµφοκύτταρα 4. Η αναγνώριση
ΑΝΟΣΟΛΟΓΙΑ 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ ΑΝΤΙΓΟΝΟ και ΥΠΟ ΟΧΕΙΣ ΑΝΤΙΓΟΝΟΥ 3. Η πρόσληψη του αντιγόνου και η παρουσίασή του στα λεµφοκύτταρα 4. Η αναγνώριση
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
3. Η ΠΡΟΣΛΗΨΗΤΟΥ ΑΝΤΙΓΟΝΟΥ ΚΑΙ Η ΠΑΡΟΥΣΙΑΣΗ ΤΟΥ ΣΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ
 3. Η ΠΡΟΣΛΗΨΗΤΟΥ ΑΝΤΙΓΟΝΟΥ ΚΑΙ Η ΠΑΡΟΥΣΙΑΣΗ ΤΟΥ ΣΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ ΤΙ ΒΛΕΠΟΥΝ ΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ Τα αντιγόνα που αναγνωρίζονται από τα Τ λεµφοκύτταρα Πρόσληψη των πρωτεϊνικών αντιγόνων από τα αντιγονοπαρουσιαστικά
3. Η ΠΡΟΣΛΗΨΗΤΟΥ ΑΝΤΙΓΟΝΟΥ ΚΑΙ Η ΠΑΡΟΥΣΙΑΣΗ ΤΟΥ ΣΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ ΤΙ ΒΛΕΠΟΥΝ ΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ Τα αντιγόνα που αναγνωρίζονται από τα Τ λεµφοκύτταρα Πρόσληψη των πρωτεϊνικών αντιγόνων από τα αντιγονοπαρουσιαστικά
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: 16 / 06 / 2017 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1: δ Α2:
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: 16 / 06 / 2017 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1: δ Α2:
ΧΡΗΣΤΟΣ ΚΑΚΑΒΑΣ 1 ΚΑΘΗΓΗΤΗΣ ΒΙΟΛΟΓΟΣ Μ.Δ.Ε
 ΚΕΦΑΛΑΙΟ 2 ον. ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ ΤΙ ΠΡΕΠΕΙ ΝΑ ΞΕΡΩ. 1. Τη δομή της δίκλωνης έλικας πάρα πολύ καλά. 2. Τους δεσμούς υδρογόνου μεταξύ των συμπληρωματικών βάσεων και την επίπτωσή
ΚΕΦΑΛΑΙΟ 2 ον. ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ ΤΙ ΠΡΕΠΕΙ ΝΑ ΞΕΡΩ. 1. Τη δομή της δίκλωνης έλικας πάρα πολύ καλά. 2. Τους δεσμούς υδρογόνου μεταξύ των συμπληρωματικών βάσεων και την επίπτωσή
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Χρόνια φλεγμονή. Βαλεντίνη Τζιούφα-Ασημακοπούλου. Νοέμβριος 2018
 Χρόνια φλεγμονή Βαλεντίνη Τζιούφα-Ασημακοπούλου Νοέμβριος 2018 Οξεία φλεγμονή Ταχεία εισβολή και λύση Εξιδρωματικά στοιχεία Πολυμορφοπύρηνα Χρόνια φλεγμονή Ύπουλη εισβολήπαρατεταμένη πορείαβραδεία λύση
Χρόνια φλεγμονή Βαλεντίνη Τζιούφα-Ασημακοπούλου Νοέμβριος 2018 Οξεία φλεγμονή Ταχεία εισβολή και λύση Εξιδρωματικά στοιχεία Πολυμορφοπύρηνα Χρόνια φλεγμονή Ύπουλη εισβολήπαρατεταμένη πορείαβραδεία λύση
6. ΡΑΣΤΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΑΝΟΣΙΑΣ: Εξάλειψη των ενδοκυττάριων µικροοργανισµών
 6. ΡΑΣΤΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΑΝΟΣΙΑΣ: Εξάλειψη των ενδοκυττάριων µικροοργανισµών Οι δύο τύποι της κυτταρικής ανοσίας Η µετανάστευση των δραστικών Τ κυττάρων στις εστίες της λοίµωξης (εντόπιση
6. ΡΑΣΤΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΑΝΟΣΙΑΣ: Εξάλειψη των ενδοκυττάριων µικροοργανισµών Οι δύο τύποι της κυτταρικής ανοσίας Η µετανάστευση των δραστικών Τ κυττάρων στις εστίες της λοίµωξης (εντόπιση
ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ
 ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ 16/06/2017 ΕΠΙΜΕΛΕΙΑ: ΑΝΔΡΙΑΝΗ ΠΑΡΑΧΗ ΘΕΜΑ Α A1. Δ Α2. Δ Α3. Β Α4. Γ Α5.Α ΘΕΜΑ Β Β1) I A II E III Σ IV Β V Ζ VI Γ VII Δ Β2) ε προκαρυωτικό γιατί
ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ 16/06/2017 ΕΠΙΜΕΛΕΙΑ: ΑΝΔΡΙΑΝΗ ΠΑΡΑΧΗ ΘΕΜΑ Α A1. Δ Α2. Δ Α3. Β Α4. Γ Α5.Α ΘΕΜΑ Β Β1) I A II E III Σ IV Β V Ζ VI Γ VII Δ Β2) ε προκαρυωτικό γιατί
Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της
 ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΚΕΦΑΛΑΙΟ 15. Κυτταρική ρύθμιση. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
Αντιγόνα & Ανοσοσφαιρίνες
 Αντιγόνα & Ανοσοσφαιρίνες Ανοσολογικά χαρακτηριστικά των αντιγόνων I Τα αντιγόνα είναι ουσίες ικανές να επάγουν ειδική ανοσιακή απάντηση (ανοσογόνα) Ανοσογονικότητα (immunogenicity) ικανότητα επαγωγής
Αντιγόνα & Ανοσοσφαιρίνες Ανοσολογικά χαρακτηριστικά των αντιγόνων I Τα αντιγόνα είναι ουσίες ικανές να επάγουν ειδική ανοσιακή απάντηση (ανοσογόνα) Ανοσογονικότητα (immunogenicity) ικανότητα επαγωγής
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β
 ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
Το μονοπάτι της κινάσης MAP- ERK
 Το μονοπάτι της κινάσης MAP- ERK 1 Σηματοδότηση μέσω μικρών GTPασών Η οικογένεια μορίων Ras (Rat Sarcoma virus) Ρύθμιση των πρωτεϊνών Ras Οι πρωτεΐνες Ras μετατρέπονται από την ανενεργή μορφή τους, που
Το μονοπάτι της κινάσης MAP- ERK 1 Σηματοδότηση μέσω μικρών GTPασών Η οικογένεια μορίων Ras (Rat Sarcoma virus) Ρύθμιση των πρωτεϊνών Ras Οι πρωτεΐνες Ras μετατρέπονται από την ανενεργή μορφή τους, που
ΣΤΗΛΗ Α Αντιβιοτικό Αντισώματα ιντερφερόνες Τ- Τ- (αντιγόνα) κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera
 κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera") Α1. 1. β Βιολογία ΘΕΜΑ Α γενιικής παιιδείίας 2. γ 3. γ 4. γ 5. δ Α2. ΣΤΗΛΗ Α Αντιβιοτικό Αντισώματα ιντερφερόνες Τ- Τ- (αντιγόνα) κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera Ηπατίτιδα C + Candida albicans
Α1. 1. β Βιολογία ΘΕΜΑ Α γενιικής παιιδείίας 2. γ 3. γ 4. γ 5. δ Α2. ΣΤΗΛΗ Α Αντιβιοτικό Αντισώματα ιντερφερόνες Τ- Τ- (αντιγόνα) κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera Ηπατίτιδα C + Candida albicans
ΘΕΜΑ Α ΘΕΜΑ Β. Α1 δ. Α2 δ. Α3 β. Α4 γ. Α5 α. Β1 Ι Α ( φωσφορική ομάδα) ΙΙ Ε (υδροξυλομάδα) ΙΙΙ ΣΤ (αμινομάδα) IV B (mrna) V Z (RNA πολυμεράση)
 ΙΙ Ε (υδροξυλομάδα) ΙΙΙ ΣΤ (αμινομάδα) IV B (mrna) V Z (RNA πολυμεράση)") ΘΕΜΑ Α Α1 δ Α2 δ Α3 β Α4 γ Α5 α ΘΕΜΑ Β Β1 Ι Α ( φωσφορική ομάδα) ΙΙ Ε (υδροξυλομάδα) ΙΙΙ ΣΤ (αμινομάδα) IV B (mrna) V Z (RNA πολυμεράση) VI Γ (μεταγραφόμενη αλυσίδα) VII Δ (κωδική αλυσίδα) Β2 Αντιστοιχεί
ΘΕΜΑ Α Α1 δ Α2 δ Α3 β Α4 γ Α5 α ΘΕΜΑ Β Β1 Ι Α ( φωσφορική ομάδα) ΙΙ Ε (υδροξυλομάδα) ΙΙΙ ΣΤ (αμινομάδα) IV B (mrna) V Z (RNA πολυμεράση) VI Γ (μεταγραφόμενη αλυσίδα) VII Δ (κωδική αλυσίδα) Β2 Αντιστοιχεί
Η πρωτογενής ανοσοβιολογική απόκριση ενεργοποιείται κατά την πρώτη επαφή του οργανισμού με ένα αντιγόνο. Περιλαμβάνει τα εξής στάδια:
 Εικόνα 1.24: α) Σύνδεση αντισώματος - αντιγόνου, β) συμπληρωματικότητα αντισώματος - αντιγόνου, γ) ένα αντίσωμα συνδέεται με περισσότερα από ένα αντιγόνα. Στάδια ανοσοβιολογικής απόκρισης Η αντίδραση του
Εικόνα 1.24: α) Σύνδεση αντισώματος - αντιγόνου, β) συμπληρωματικότητα αντισώματος - αντιγόνου, γ) ένα αντίσωμα συνδέεται με περισσότερα από ένα αντιγόνα. Στάδια ανοσοβιολογικής απόκρισης Η αντίδραση του
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
ΘΕΜΑ 1 Ο ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 10/11/2013
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 10/11/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Οι αποικοδομητές είναι:
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 10/11/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Οι αποικοδομητές είναι:
ΑΝΟΣΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΡΑΣΗΣ ΕΜΒΟΛΙΩΝ. Μαρία Χατζηστυλιανού Σιδηροπούλου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Τμήματος Ιατρικής, ΣΕΥ, ΑΠΘ
 ΑΝΟΣΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΡΑΣΗΣ ΕΜΒΟΛΙΩΝ. Μαρία Χατζηστυλιανού Σιδηροπούλου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Τμήματος Ιατρικής, ΣΕΥ, ΑΠΘ ΑΝΟΣΟΠΟΙΗΣΗ ΕΝΕΡΓΗΤΙΚΗ ΠΑΘΗΤΙΚΗ ΦΥΣΙΚΗ (ΝΟΣΗΣΗ) ΤΕΧΝΗΤΗ
ΑΝΟΣΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΡΑΣΗΣ ΕΜΒΟΛΙΩΝ. Μαρία Χατζηστυλιανού Σιδηροπούλου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Τμήματος Ιατρικής, ΣΕΥ, ΑΠΘ ΑΝΟΣΟΠΟΙΗΣΗ ΕΝΕΡΓΗΤΙΚΗ ΠΑΘΗΤΙΚΗ ΦΥΣΙΚΗ (ΝΟΣΗΣΗ) ΤΕΧΝΗΤΗ
Ενδεικτικές απαντήσεις
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ
Κεφάλαιο 4: Ανασυνδυασμένο DNA
 Κεφάλαιο 4: Ανασυνδυασμένο DNA 1. Η ανάπτυξη της γενετικής μηχανικής επέτρεψε: α. την κατανόηση των μηχανισμών αντιγραφής του γενετικού υλικού β. την απομόνωση των πλασμιδίων από τα βακτήρια γ. την πραγματοποίηση
Κεφάλαιο 4: Ανασυνδυασμένο DNA 1. Η ανάπτυξη της γενετικής μηχανικής επέτρεψε: α. την κατανόηση των μηχανισμών αντιγραφής του γενετικού υλικού β. την απομόνωση των πλασμιδίων από τα βακτήρια γ. την πραγματοποίηση
Μονοπάτια ενεργοποίησης κινασών MAP σε κύτταρα θηλαστικών
 Μονοπάτια ενεργοποίησης κινασών MAP σε κύτταρα θηλαστικών Εκτός από την ERK, τα κύτταρα των θηλαστικών διαθέτουν τις κινάσες MAP JNK και p38. Η ενεργοποίηση των κινασών JNK και p38 προκαλείται από μέλη
Μονοπάτια ενεργοποίησης κινασών MAP σε κύτταρα θηλαστικών Εκτός από την ERK, τα κύτταρα των θηλαστικών διαθέτουν τις κινάσες MAP JNK και p38. Η ενεργοποίηση των κινασών JNK και p38 προκαλείται από μέλη
ΕΡΩΤΗΣΕΙΣ ΣΩΣΤΟΥ - ΛΑΘΟΥΣ. ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ
 ΕΡΩΤΗΣΕΙΣ ΣΩΣΤΟΥ - ΛΑΘΟΥΣ. ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ 1. Η πήξη του αίματος συμβάλλει στην άμυνα του οργανισμού 2. Η φαγοκυττάρωση είναι αποτελεσματική μόνο έναντι των βακτηρίων 3. Οι ιντερφερόνες δρουν άμεσα
ΕΡΩΤΗΣΕΙΣ ΣΩΣΤΟΥ - ΛΑΘΟΥΣ. ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ 1. Η πήξη του αίματος συμβάλλει στην άμυνα του οργανισμού 2. Η φαγοκυττάρωση είναι αποτελεσματική μόνο έναντι των βακτηρίων 3. Οι ιντερφερόνες δρουν άμεσα
Συστήματα επικοινωνίας Ανθρωπίνου σώματος. ενδοκρινολογικό νευρικό σύστημα
 Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Επιλεγόµενο µάθηµα:λευχαιµίες. Γεωργόπουλος Χρήστος Καρκατσούλης Μάριος Μπρίκος Νικήτας
 Επιλεγόµενο µάθηµα:λευχαιµίες Γεωργόπουλος Χρήστος Καρκατσούλης Μάριος Μπρίκος Νικήτας Ανοσολογία και καρκίνος Γενετικές και κυτταρικές αλλαγές που συµβαίνουν στον καρκίνο προσφέρουν στο ανοσιακό σύστηµα
Επιλεγόµενο µάθηµα:λευχαιµίες Γεωργόπουλος Χρήστος Καρκατσούλης Μάριος Μπρίκος Νικήτας Ανοσολογία και καρκίνος Γενετικές και κυτταρικές αλλαγές που συµβαίνουν στον καρκίνο προσφέρουν στο ανοσιακό σύστηµα
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη
 Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ
 ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής
 Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
ΦΡΟΝΤΙΣΤΗΡΙΑ ΜΗΤΑΛΑΣ ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ
 ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ ΚΕΦΑΛΑΙΟ 1.3 : ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΑΝΟΣΙΑΣ ΘΕΜΑ 1 Χαρακτηρίστε τις προτάσεις ως σωστές ή λανθασµένες. 1. Οι βλεννογόνοι
ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ ΚΕΦΑΛΑΙΟ 1.3 : ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΑΝΟΣΙΑΣ ΘΕΜΑ 1 Χαρακτηρίστε τις προτάσεις ως σωστές ή λανθασµένες. 1. Οι βλεννογόνοι
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. :
 ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. : 2010336 ΕΡΓΑΣΙΑ ΓΙΑ ΤΟ Μ.Π.Σ. ΜΕΤΑΒΟΛΙΚΑ ΝΟΣΗΜΑΤΑ ΟΣΤΩΝ ΚΥΤΟΚΙΝΕΣ ΠΑΡΑΓΟΜΕΝΕΣ ΑΠΟ ΤΑ Τ-ΚΥΤΤΑΡΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΠΟΥ ΕΜΠΛΕΚΟΝΤΑΙ ΣΤΗΝ ΟΣΤΕΟΚΛΑΣΤΟΓΕΝΕΣΗ
ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. : 2010336 ΕΡΓΑΣΙΑ ΓΙΑ ΤΟ Μ.Π.Σ. ΜΕΤΑΒΟΛΙΚΑ ΝΟΣΗΜΑΤΑ ΟΣΤΩΝ ΚΥΤΟΚΙΝΕΣ ΠΑΡΑΓΟΜΕΝΕΣ ΑΠΟ ΤΑ Τ-ΚΥΤΤΑΡΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΠΟΥ ΕΜΠΛΕΚΟΝΤΑΙ ΣΤΗΝ ΟΣΤΕΟΚΛΑΣΤΟΓΕΝΕΣΗ
Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά;
 ΒΙΟΛΟΓΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ 12 26/10/2016 Κεφάλαιο 3 Α μέρος Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά; Ποια είναι η δομή
ΒΙΟΛΟΓΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ 12 26/10/2016 Κεφάλαιο 3 Α μέρος Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά; Ποια είναι η δομή
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Linking Pes*cide Exposure with Pediatric Leukemia: Poten*al Underlying Mechanisms
 Πανεπιστήμιο Ιωαννίνων Τμήμα Ιατρικής Λευχαιμίες Linking Pes*cide Exposure with Pediatric Leukemia: Poten*al Underlying Mechanisms Ιωάννης- Μάριος Ρόκας Χρήστος Σταφυλίδης Λευχαιμία παιδιών Το συχνότερο
Πανεπιστήμιο Ιωαννίνων Τμήμα Ιατρικής Λευχαιμίες Linking Pes*cide Exposure with Pediatric Leukemia: Poten*al Underlying Mechanisms Ιωάννης- Μάριος Ρόκας Χρήστος Σταφυλίδης Λευχαιμία παιδιών Το συχνότερο
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY
 ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Γενικά χαρακτηριστικά Απεικόνιση των κυτταρικών πληθυσμών με βάση: το μέγεθος την κοκκίωση το φθορισμό ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Υδροδυναμική ροή Πρόσθιος
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Γενικά χαρακτηριστικά Απεικόνιση των κυτταρικών πληθυσμών με βάση: το μέγεθος την κοκκίωση το φθορισμό ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Υδροδυναμική ροή Πρόσθιος
ΚΕΦΑΛΑΙΟ 16. Ο κυτταρικός κύκλος. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
2 Ο ΜΑΘΗΜΑ ΣΥΣΤΗΜΑ ΣΥΜΠΛΗΡΩΜΑΤΟΣ ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ
 2 Ο ΜΑΘΗΜΑ ΣΥΣΤΗΜΑ ΣΥΜΠΛΗΡΩΜΑΤΟΣ ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις μεμβράνες 2. Φυσιολογικοί φραγμοί
2 Ο ΜΑΘΗΜΑ ΣΥΣΤΗΜΑ ΣΥΜΠΛΗΡΩΜΑΤΟΣ ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις μεμβράνες 2. Φυσιολογικοί φραγμοί
Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ
 Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΝΤΙΓΡΑΦΗ ΜΕΤΑΓΡΑΦΗ Γίνεται σύνθεση DNA. Γίνεται σύνθεση RNA. Εξασφαλίζεται η διαιώνιση της γενετικής
Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΝΤΙΓΡΑΦΗ ΜΕΤΑΓΡΑΦΗ Γίνεται σύνθεση DNA. Γίνεται σύνθεση RNA. Εξασφαλίζεται η διαιώνιση της γενετικής
Βασικοί μηχανισμοί προσαρμογής
 ΣΥΓΚΡΙΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 23-24, 18/4/2016 Π.Παπαζαφείρη Βασικοί μηχανισμοί προσαρμογής Προσαρμογή σε μοριακό και γονιδιακό επίπεδο Επίπεδα ελέγχου 1. Πρωτεïνική δράση 2. Πρωτεïνοσύνθεση 3. Ρύθμιση της
ΣΥΓΚΡΙΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 23-24, 18/4/2016 Π.Παπαζαφείρη Βασικοί μηχανισμοί προσαρμογής Προσαρμογή σε μοριακό και γονιδιακό επίπεδο Επίπεδα ελέγχου 1. Πρωτεïνική δράση 2. Πρωτεïνοσύνθεση 3. Ρύθμιση της
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ. ΘΕΜΑ Β Β1. Στήλη Ι Στήλη ΙΙ 1 Α 2 Γ 3 Α 4 Β 5 Α 6 Α 7 Γ
 ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1.
1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1.
ANOΣΟΓΗΡΑΝΣΗ. Ιωάννα Οικονοµίδου. Αναπληρώτρια Καθηγήτρια Ιατρικής Πανεπιστηµίου Αθηνών
 ANOΣΟΓΗΡΑΝΣΗ Ιωάννα Οικονοµίδου Αναπληρώτρια Καθηγήτρια Ιατρικής Πανεπιστηµίου Αθηνών Το ανοσιακό σύστηµα θεωρείται αποφασιστικός παράγοντας για την διατήρηση της υγείας και την επιβίωση στους ηλικιωµένους
ANOΣΟΓΗΡΑΝΣΗ Ιωάννα Οικονοµίδου Αναπληρώτρια Καθηγήτρια Ιατρικής Πανεπιστηµίου Αθηνών Το ανοσιακό σύστηµα θεωρείται αποφασιστικός παράγοντας για την διατήρηση της υγείας και την επιβίωση στους ηλικιωµένους
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ. Παθοφυσιολογία Ι
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Παθοφυσιολογία Ι Ανοσολογία - Ρευματολογία Υπεύθυνος μαθήματος: Καθηγητής Παθολογίας/Ρευματολογίας, Αλέξανδρος Α. Δρόσος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Παθοφυσιολογία Ι Ανοσολογία - Ρευματολογία Υπεύθυνος μαθήματος: Καθηγητής Παθολογίας/Ρευματολογίας, Αλέξανδρος Α. Δρόσος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΚΕΦΑΛΑΙΟ 11. Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα
 ΚΕΦΑΛΑΙΟ 11 Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα Τα ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΑ Μέρος Ε ΤΑ ΒΑΣΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΩΝ - Περιέχουν ένζυµα για ποικίλες µεταβολικές
ΚΕΦΑΛΑΙΟ 11 Βιοενεργητική & Μεταβολισµός: Μιτοχόνδρια, Χλωροπλάστες & Υπεροξειδιοσώµατα Τα ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΑ Μέρος Ε ΤΑ ΒΑΣΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΥΠΕΡΟΞΕΙΔΙΟΣΩΜΑΤΩΝ - Περιέχουν ένζυµα για ποικίλες µεταβολικές
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ. Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη
 Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY
 ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY Κυτταρομετρία Ροής Η Κυτταρομετρία Ροής είναι μια τεχνική που εκμεταλλεύεται την μέτρηση ορισμένων βιοχημικών και βιοφυσικών παραμέτρων των κυττάρων μεμονωμένα, όταν αυτά
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY Κυτταρομετρία Ροής Η Κυτταρομετρία Ροής είναι μια τεχνική που εκμεταλλεύεται την μέτρηση ορισμένων βιοχημικών και βιοφυσικών παραμέτρων των κυττάρων μεμονωμένα, όταν αυτά
Τριαδικό σύστημα του Salthe (1985) επίπεδα σημειωτικών διεργασιών.
 επίπεδα σημειωτικών διεργασιών.") Τριαδικό σύστημα του Salthe (1985) επίπεδα σημειωτικών διεργασιών. Υψηλότερο επίπεδο-εκλεκτικό σημειωτικό περιβάλλον ή πλαίσιο-όροι ορίου-συνοριακές συνθήκες-εκλεκτικός ρυθμιστικός ρόλος-μακροσημειωτικό
Τριαδικό σύστημα του Salthe (1985) επίπεδα σημειωτικών διεργασιών. Υψηλότερο επίπεδο-εκλεκτικό σημειωτικό περιβάλλον ή πλαίσιο-όροι ορίου-συνοριακές συνθήκες-εκλεκτικός ρυθμιστικός ρόλος-μακροσημειωτικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ. Ανοσολογία. Ο Τ κυτταρικός υποδοχέας (TCR) Διδάσκων: Αναπληρωτής Καθηγητής Γεώργιος Θυφρονίτης
 Διδάσκων: Αναπληρωτής Καθηγητής Γεώργιος Θυφρονίτης") ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ανοσολογία Ο Τ κυτταρικός υποδοχέας (TCR) Διδάσκων: Αναπληρωτής Καθηγητής Γεώργιος Θυφρονίτης Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Ανοσολογία Ο Τ κυτταρικός υποδοχέας (TCR) Διδάσκων: Αναπληρωτής Καθηγητής Γεώργιος Θυφρονίτης Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4
 ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
Ιοί & HPV. Ευστάθιος Α. Ράλλης. Επικ. Καθηγητής Δερματολογίας - Αφροδισιολογίας
 Ιοί & HPV Ευστάθιος Α. Ράλλης Επικ. Καθηγητής Δερματολογίας - Αφροδισιολογίας Περί ΙΩΝ Ορισμός Οι ιοί είναι ατελείς, λοιμώδεις οργανισμοί μικρότεροι σε μέγεθος από τα μικρόβια και ορατοί μόνο με το ηλεκτρονικό
Ιοί & HPV Ευστάθιος Α. Ράλλης Επικ. Καθηγητής Δερματολογίας - Αφροδισιολογίας Περί ΙΩΝ Ορισμός Οι ιοί είναι ατελείς, λοιμώδεις οργανισμοί μικρότεροι σε μέγεθος από τα μικρόβια και ορατοί μόνο με το ηλεκτρονικό
Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό
 Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
ITP Ιδιοπαθής/ αυτοάνοση θρομβοπενική πορφύρα
 ITP Ιδιοπαθής/ αυτοάνοση θρομβοπενική πορφύρα Αθανασία Μουζάκη, Καθηγήτρια Εργαστηριακής Αιματολογίας-Αιμοδοσίας, Αιματολογικό Τμήμα, Παθολογική Κλινική, Τμήμα Ιατρικής, Παν/ο Πατρών Τα είδη της ITP είναι:
ITP Ιδιοπαθής/ αυτοάνοση θρομβοπενική πορφύρα Αθανασία Μουζάκη, Καθηγήτρια Εργαστηριακής Αιματολογίας-Αιμοδοσίας, Αιματολογικό Τμήμα, Παθολογική Κλινική, Τμήμα Ιατρικής, Παν/ο Πατρών Τα είδη της ITP είναι:
ΑΝΟΣΙΑΚΗ ΑΠΑΝΤΗΣΗ ΣΤΑ ΕΜΒΟΛΙΑ-ΑΝΟΣΙΑΚΗ ΜΝΗΜΗ
 ΑΝΟΣΙΑΚΗ ΑΠΑΝΤΗΣΗ ΣΤΑ ΕΜΒΟΛΙΑ-ΑΝΟΣΙΑΚΗ ΜΝΗΜΗ Φ. Κανακούδη Τσακαλίδου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Ιατρικής Σχολής ΑΠΘ ΠΕΡΙΓΡΑΜΜΑ ΟΜΙΛΙΑΣ Σύνδεση βασικών γνώσεων Ανοσολογίας ( Immunology)
ΑΝΟΣΙΑΚΗ ΑΠΑΝΤΗΣΗ ΣΤΑ ΕΜΒΟΛΙΑ-ΑΝΟΣΙΑΚΗ ΜΝΗΜΗ Φ. Κανακούδη Τσακαλίδου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Ιατρικής Σχολής ΑΠΘ ΠΕΡΙΓΡΑΜΜΑ ΟΜΙΛΙΑΣ Σύνδεση βασικών γνώσεων Ανοσολογίας ( Immunology)
ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΠΑΝΕΛΛΑΔΙΚΩΝ ΕΞΕΤΑΣΕΩΝ 2107 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Γ ΛΥΚΕΙΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ Β Β1. Ι: κωδική αλυσίδα (Δ) ΙΙ: μεταγραφόμενη αλυσίδα (Γ) ΙΙΙ: αμινομάδα (ΣΤ) ΙV: mrna (Β) V: RNA πολυμεράση (Ζ) VI: φωσφορική
ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ Β Β1. Ι: κωδική αλυσίδα (Δ) ΙΙ: μεταγραφόμενη αλυσίδα (Γ) ΙΙΙ: αμινομάδα (ΣΤ) ΙV: mrna (Β) V: RNA πολυμεράση (Ζ) VI: φωσφορική