Σχολή Επιστημών Υγείας, Τμήμα Ιατρικής ΠΜΣ Βασικές Ιατρικές Επιστήμες ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΤΟΜΙΑΣ ΕΠΙΒΛΕΠΟΥΣΑ: ΕΠ. ΚΑΘΗΓΗΤΡΙΑ ΜΠΡΑΒΟΥ ΒΑΣΙΛΙΚΗ
|
|
|
- Τηθύς Βονόρτας
- 9 χρόνια πριν
- Προβολές:
Transcript
1 Σχολή Επιστημών Υγείας, Τμήμα Ιατρικής ΠΜΣ Βασικές Ιατρικές Επιστήμες ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΤΟΜΙΑΣ ΕΠΙΒΛΕΠΟΥΣΑ: ΕΠ. ΚΑΘΗΓΗΤΡΙΑ ΜΠΡΑΒΟΥ ΒΑΣΙΛΙΚΗ Μελέτη της έκφρασης ρυθμιστών της δυναμικής του κυτταροσκελετού στο καρκίνο του παχέος εντέρου στον άνθρωπο. ΑΓΓΕΛΟΥ ΕΛΕΝΗ Ιατρός ΠΑΤΡΑ 2014

2 ΤΡΙΜΕΛΗΣ ΕΠΙΤΡΟΠΗ ΕΛΕΝΗ ΠΕΤΡΟΥ ΠΑΠΑΔΑΚΗ. Καθηγήτρια ΒΑΣΙΛΙΚΗ ΜΠΡΑΒΟΥ. Επίκουρη Καθηγήτρια (επιβλέπουσα) ΣΠΥΡΙΔΩΝ ΣΥΓΓΕΛΟΣ. Λέκτορας Ένα μεγάλο ευχαριστώ στην επιβλέπουσα μου, επίκουρη καθηγήτρια κ Μπράβου για την ευκαιρία που μου έδωσε να δουλέψω μαζί της, την υπομονή και την αμέριστη συμπαράστασή της. 2

3 Αφιερωμένο στους Γονείς μου. Ένα μεγάλο ευχαριστώ στην οικογένεια μου που με στήριξαν υλικά και ηθικά όλα αυτά τα χρόνια των σπουδών μου. 3

4 ΠΕΡΙΕΧΟΜΕΝΑ Εισαγωγή...7 ΚΕΦΑΛΑΙΟ 1 1.1Μακροσκοπική Μικροσκοπική και ανατομική Παχέoς Εντέρου Εμβρυική Ανάπτυξη Επιδημιολογία Κλινική εικόνα Αιτιολογία-παθογένεια Περιβαλλοντικοί παράγοντες Γενετική προδιάθεση Προδιαθεσιακοί παράγοντες-προκαρκινικές αλλοιώσεις Αδενώματα...21 ΚΕΦΑΛΑΙΟ 2 2.0Καρκίνωμα παχέος εντέρου Μοριακή παθογένεια Μικροδορυφορική αστάθεια (MSI) Χρωμοσωμική Αστάθεια (CIN, chromosomal instability) Μοριακοί μηχανισμοί που ενεργοποιούνται από την χρωμοσωμική αστάθεια στον καρκίνο του παχέος εντέρου Ο ρόλος του μικροπεριβάλοντος...37 ΚΕΦΑΛΑΙΟ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ 3.0Παθολογοανατομικά χαρακτηριστικά Μακροσκοπικά Χαρακτηριστικά Ιστοπαθολογικά Χαρακτηριστικά Διαφοροποίηση Grading Σταδιοποίηση Προγνωστική Παράγοντες Θεραπεία Παρακολούθηση...48 ΚΕΦΑΛΑΙΟ Κυτταροσκελετός ακτίνης WASP ( W iskot- A ldrich S yndrome)

5 4.3 LIM ΚΙΝΑΣΕΣ (LIMK1 και LIMK2)...66 ΚΕΦΑΛΑΙΟ 5 ΕΙΔΙΚΟ ΜΕΡΟΣ Αντικείμενο παρούσας μελέτης Υλικά και μέθοδοι Ανοσοϊστοχημική χρώση Εκτίμηση ανοσοϊστοχημικής χρώσης Στατιστική Ανάλυση ΑΠΟΤΕΛΕΣΜΑΤΑ ΣΥΖΗΤΗΣΗ SUMMARY ΠΕΡΙΛΗΨΗ ΒΙΒΛΙΟΓΡΑΦΙΑ

6 6
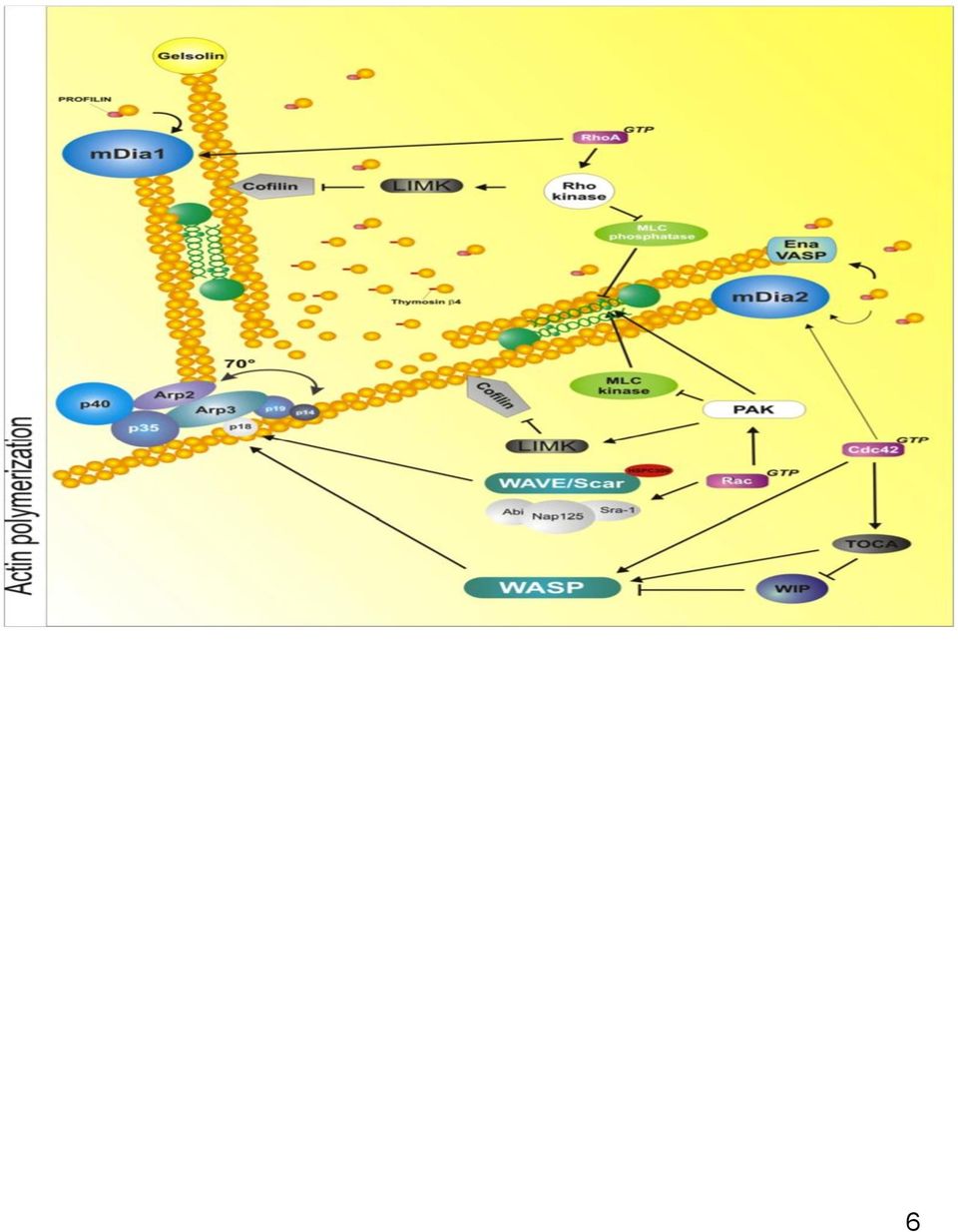
7 ΕΙΣΑΓΩΓΗ Ο καρκίνος του παχέος εντέρου αποτελεί σημαντική αιτία νοσηρότητας και θνητότητας παγκοσμίως. Ευθύνεται για πάνω από 9% όλων των κρουσμάτων καρκίνου. Πρόκειται για τον 3 0 συχνότερο καρκίνο στους άνδρες και 2 0 στις γυναίκες παγκοσμίως. Η επιβίωση των ασθενών με καρκίνο παχέος εντέρου εξαρτάται σημαντικά από το στάδιο της νόσου κατά την διάγνωση και κυμαίνεται από 90% 5ετή επιβίωση για περιπτώσεις με τοπική εντόπιση της νόσου, 70% για περιπτώσεις με τοπική επέκταση και 10% στις περιπτώσεις με απομακρυσμένες μεταστάσεις. Ο συχνότερος ιστολογικός τύπος καρκίνου του παχέος εντέρου είναι το αδενοκαρκίνωμα. Όπως και με άλλους τύπους νεοπλασματικής εξεργασίας ο καρκίνος του παχέος εντέρου είναι αποτέλεσμα συσσώρευσης γενετικών και επιγενετικών αλλαγών που οδηγούν την κακοήθη εξαλλαγή του φυσιολογικού επιθηλιακού κυττάρου. Βασικός μοριακός παθογενετικός μηχανισμός είναι η γονιδιωματική αστάθεια. Έχουν περιγραφεί δύο βασικοί τύποι γονιδιωματικής αστάθειας:1) η μικροδορυφορική αστάθεια (microsatellite instability, MSI) και 2) η χρωμοσωμική αστάθεια ( chromosomal instability,cin). Πρόκειται για μια πολυσταδιακή διαδικασία που χαρακτηρίζεται από αδρανοποίηση ογκοκατασταλτικών γονιδίων π.χ ( APC, DCC, p53) και γονιδίων επιδιόρθωσης DNA πχ (hmsh, hmslh1) και ταυτόχρονη ενεργοποίηση πρώτο- ογκογονιδίων ( πχ K-ras, N-ras, c-myc). Καθώς η διήθηση και η μετάσταση αποτελούν την κύρια αιτία νοσηρότητας και θνητότητας από τον καρκίνο του παχέος εντέρου, η καλύτερη κατανόηση των μηχανισμών που ελέγχουν την διηθητική και μεταστατική ικανότητα των καρκινικών κυττάρων είναι μέγιστης σημασίας. Ένα από τα χαρακτηριστικά γνωρίσματα των καρκινικών κυττάρων είναι η αυξημένη ικανότητα κυτταρικής μετανάστευσης διήθησης και μετάστασης. H απώλεια των κυτταρικών συνδέσεων μεταξύ των επιθηλιακών κυττάρων και η απόκτηση ιδιοτήτων μεσεγχυματικών κυττάρων, με κύρια την αυξημένη ικανότητα κυτταρικής κίνησης, δίνει την δυνατότητα στα καρκινικά κύτταρα να διηθούν τους γύρω ιστούς, να εισβάλλουν σε αγγεία και να δώσουν τελικά απομακρυσμένες μεταστάσεις. Αυτή η διαδικασία είναι γνωστή ως επιθήλιομεσεγχυματική μετατροπή (epithelial mesenhymal transition, ΕΜΤ) και πιστεύεται ότι παίζει σημαντικό ρόλο στην εξέλιξη του όγκου, στην διηθητική και μεταστατική ικανότητα των καρκίνων επιθηλιακής αρχής (καρκινωμάτων). Ο κυτταροσκελετός ακτίνης αποτελείται από νημάτια F-ακτίνης, διαμέτρου 6 nm, τα οποία με την σειρά τους σχηματίζονται από τον πολυμερισμό μορίων σφαιρικής πρωτεΐνης ακτίνης. Ο ρόλος του κυτταροσκελετού της ακτίνης φαίνεται να είναι κρίσιμος σε διαδικασίες όπως: η δημιουργία κυτταρικών προσεκβολών κίνησης (ελασματοπόδια, νηματοπόδια) και συνεπώς η κυτταρική κίνηση-μετανάστευση, η επιθήλιο-μεσεγχηματική μετατροπή (ΕΜΤ), η πολικότητα των επιθηλιακών κύτταρων, η κυτταροκίνηση (η δημιουργία του συσταλτού δακτυλίου κατά την μίτωση και μείωση), η ενδοκυττάρωση, η φαγοκυττάρωση, η απόπτωση και η μεταγωγή σήματος. Τα καρκινικά κύτταρα χρησιμοποιούν εξειδικευμένες κυτταρικές δομές πού έχουν ως κύριο δομικό συστατικό τον κυτταροσκελετό ακτίνης, όπως τα πόδια διήθησης (invadopodia), για να μεταναστεύσουν, να εισβάλουν σε γειτονικές δομές,στα αγγεία και τελικά να δώσουν μετάσταση σε απομακρυσμένες θέσεις. Σηματοδότηση μέσω διαφορετικών οδών μπορεί να οδηγήσει στον σχηματισμό διαφορετικών δομών ακτίνης πχ (νηματοπόδια, ελασματοπόδια, ίνες στρες κ.α.). Η 7

8 δυναμική του κυτταροσκελετού της ακτίνης (πολυμερισμός-αποπολυμερισμός) ρυθμίζεται από μια μεγάλη ομάδα πρωτεϊνών (ABPs, actin binding proteins), οι οποίες ταξινομούνται σε πέντε κατηγορίες 1) πρωτεΐνες που ελέγχουν την δεξαμενή μη πολυμερισμένης ακτίνης (πχ προφιλίνη) 2) πρωτεΐνες που επάγουν τον πολυμερισμό 3) πρωτεΐνες που ελέγχουν την κατεύθυνση της ανάπτυξης (capping proteins) 4) πρωτεΐνες που κόβουν νημάτια ακτίνης και δημιουργούν ελεύθερα άκρα( severing proteins). Πολλές από τις πρωτεΐνες αυτές που ελέγχουν την δυναμική του κυτταροσκελετού ακτίνης όπως η πρωτεΐνης N- WASP ( Wiskott Aldrich Syndrome Protein), η οικογένεια πρωτεϊνών Arp2/3, οι LIM κινάσες (LIMK),η κοφιλίνη και μονοπάτια που ελέγχονται από την cortactin, έχουν μελετηθεί εκτενώς εξαιτίας της σημασίας τους στην κυτταρική μετανάστευση και έχει δειχθεί ότι υπερκεράζονται σε διάφορους τύπους καρκίνου. Η οικογένεια LIM κινασών αποτελεί μια νέα υποκατηγορία κινασών σερίνης/θρεονίνης και περιλαμβάνει δύο πρωτεΐνες: LIMK1 και LIMK2. Οι κινάσες αυτές εμπλέκονται σημαντικά στην ρύθμιση της δυναμικής του κυτταροσκελετού ακτίνης και ρυθμίζουν πολύ σημαντικές κυτταρικές λειτουργίες όπως η μορφολογία των κυττάρων, η κυτταρική κίνηση, η μετανάστευση των κυττάρων, ο κυτταρικός κύκλος, η μεταγραφή γονιδίων και η νευρωνική διαφοροποίηση. Λόγω των σημαντικών τους λειτουργιών εμπλέκονται σε παθήσεις του ανθρώπου όπως νευροαναπτυξιακές και νευροεκφυλιστικές διαταραχές, πνευμονική υπέρταση καθώς και στην εξέλιξη του καρκίνου στον άνθρωπο. Η λειτουργία τους διαμεσολαβείτε μέσω της ρύθμισης μιας σημαντικής ABP πρωτεΐνης της κοφιλίνης. Η Κοφιλίνη ρυθμίζει τον πολυμερισμόαποπολυμερισμό της ακτίνης και φαίνεται να παίζει σημαντικό ρόλο στη διατήρηση και επέκταση δομών όπως τα ελασματοπόδια (lamellipodial) που βρίσκονται στο προπορευόμενο-οδηγό άκρο των μεταναστευτικών κυττάρων. Οι LΙΜΚ1 και 2 αδρανοποιούν την κοφιλίνη μέσω της φωσφορυλίωσης της στην Ser-3 (Arber et al,1998;yang et al, 1998;Toshima et al,2001). Αυτή η φωσφορυλίωση αναστέλλει την αλληλεπίδραση της κοφιλίνης με τα νημάτια και τα μονομερή ακτίνης. Ο πολυμερισμός της ακτίνης επάγεται όχι μόνο από τις πρωτεΐνες (ADF) / κοφιλίνη αλλά και από την νευρωνική πρωτεΐνη Ν- WASP (Wiskott Aldrich Syndrome protein), η οποία ανήκει στην οικογένεια των πρωτεϊνών WASP που επάγουν τον πολυμερισμό της ακτίνης με την ενεργοποίηση του συμπλόκου Arp2/3, το οποίο ενεργοποιεί της έναρξης του πολυμερισμού της ακτίνης (nucleation) και της δημιουργίας διακλαδώσεων. Παρά τα δεδομένα που υποστηρίζουν ότι η πρωτεΐνη N-WASP, η οικογένεια πρωτεϊνών Arp2/3, οι LΙΜ κινάσες και η κοφιλίνη αποτελούν βασικούς ρυθμιστές του κυτταροσκελετού της ακτίνης και εμπλέκονται στην καρκινογένεση, λίγα είναι γνωστά για το ρόλο τους στην εξέλιξη του καρκίνου του παχέος εντέρου. Σκοπός της παρούσας μελέτης ήταν η διερεύνηση της έκφρασης και του ρόλου των ρυθμιστών της δυναμικής του κυτταροσκελετού ακτίνης LIMK1, LIMK2 και Ν- WASP στον καρκίνο του παχέος εντέρου στον άνθρωπο. Για το σκοπό αυτό εξετάσαμε, με ανοσοϊστοχημεία την έκφραση των LIMK1, LIMK2 και Ν- WASP σε 101 ορθοκολικά καρκινώματα σε σχέση με παθολογοανατομικές παραμέτρους, όπως η διήθηση του υποβλεννογόνιου, το βάθος διήθησης, η παρουσία λεμφαδενικών μεταστάσεων, ο βαθμός διαφοροποίησης και το στάδιο της νόσου. Επίσης ελέγξαμε την πιθανή τους συσχέτιση με σηματοδοτικές οδούς που παίζουν ρόλο στην ΕΜΤ, την διήθηση και μετάσταση καθώς είναι γνωστό ότι εμπλέκονται στην εξέλιξη του καρκίνου του παχέος εντέρου, όπως η Integrin-linked kinase, p-akt, E-cadherin και Wnt/β-catenin. 8
, η οικογένεια πρωτεϊνών Arp2/3, οι LIM κινάσες")
9 ΚΕΦΑΛΑΙΟ 1 1.1Μακροσκοπική Μικροσκοπική και Μοριακή ανατομική Παχέoς Εντέρου Το παχύ έντερο αποτελεί το τελικό μέρος του πεπτικού συστήματος. Αποτελείται από το τυφλό, την σκωληκοειδή απόφυση, το κόλον, το ορθό και τον πρωκτικό σωλήνα. Η πρωταρχική του λειτουργία είναι η επαναρρόφηση ύδατος και ανόργανων αλάτων, η αποβολή της άπεπτης τροφής και των άχρηστων προϊόντων. Μακροσκοπικά το κόλον έχει διακριτά μορφολογικά χαρακτηριστικά σε σύγκριση με το λεπτό έντερο. Με εξαίρεση το ορθό, τον πρωκτικό σωλήνα και την σκωληκοειδή απόφυση η έξω επιμήκης στιβάδα του μυϊκού χιτώνα εμφανίζει τρείς παχυσμένες, ισαπέχουσες κολικές ταινίες. Η εξωτερική επιφάνεια του τυφλού και του κόλουν εμφανίζει σακοειδής προεκβολές γνωστές ως κολίκες κυψέλες που είναι ορατές μεταξύ των ταινιών. Η λεία επιφάνεια του βλεννογόνου εμφανίζει ημισεληνοειδής πτυχές που σχηματίζονται ως αποτέλεσμα συστολών του μυϊκού χιτώνα. Μικρές λιπώδεις προεκβολές του οργάνου γνωστές ως επιπλοικές αποφύσεις είναι ορατές στην έξω επιφάνεια του εντέρου. ΒΛΕΝΝΟΓΟΝΟΣ Ο βλεννογόνος του παχέος εντέρου δεν εμφανίζει προεκβολές όπως πτυχές ή λάχνες. Αποτελείται από: επιθήλιο, χόριο και βλεννογόνιο μυϊκή στιβάδα. Το επιθήλιο εμφανίζει καταδύσεις στο χόριο, αδένες ή κρύπτες του Lieberkuhn. Τα επιφανειακά κύτταρα είναι κυρίως απορροφητικά και καλυκοειδή κύτταρα goblet. Κατά μήκος των αδένων τα απορροφητικά κύτταρα λιγοστεύουν σε αριθμό ενώ τα Goblet αυξάνουν. Η κύρια λειτουργία των απορροφητικών κυττάρων είναι η επαναρρόφηση ύδατος και ηλεκτρολυτών. Τα κυλινδρικά απορροφητικά κύτταρα είναι ο πολυπληθέστερος τύπος επιθηλιακών κυττάρων (4%) συγκριτικά με τα goblet, η αναλογία μειώνεται 1:1 κοντά στο ορθό όπου αυξάνεται ο αριθμός των καλυκοειδών κυττάρων. Τα επιθηλιακά κύτταρα του βλεννογόνου του παχέος εντέρου προκύπτουν από αρχέγονα κύτταρα που εντοπίζονται στον πυθμένα του εντερικού αδένα. Το κατώτερο τμήμα του αδένα αποτελεί την φυσιολογική ζώνη πολλαπλασιασμού όπου νεοδημιουργηθέντα κύτταρα υφίστανται 2-3 επιπλέον διαιρέσεις καθώς αρχίζουν την μετανάστευση τους προς την αυλική επιφάνεια από την οποία αποπίπτουν 5 ημέρες αργότερα. Τα πολυδύναμα αρχέγονα κύτταρα ( stem cells ) είναι αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να πολλαπλασιάζονται, να αναπαράγουν τον εαυτό τους και μέσω του μηχανισμού διαφοροποίησης μεταναστεύουν δίνοντας όλους τους εξειδικευμένους τύπους κυττάρων του επιθηλίου του παχέος εντέρου. Είναι υπεύθυνα για την συνεχή αναγέννηση του εντερικού επιθηλίου την ομοιόσταση και την αποκατάσταση βλαβών του επιθηλίου. Οι χρόνοι ανανέωσης των επιθηλιακών κυττάρων του παχέος εντέρου είναι παρόμοιοι με εκείνους του λεπτού εντέρου δηλαδή περίπου 6 ημέρες για τα απορροφητικά και τα καλυκοειδή και έως 4 εβδομάδες για τα εντεροενδοκρινή κύτταρα. 9

10 Από τον ίδιο πληθυσμό κυττάρων (αρχέγονα επιθηλιακά κύτταρα ) προέρχονται τα εντεροενδοκρινή κύτταρα τα οποία εντοπίζονται διάσπαρτα ανάμεσα σε άλλους τύπους κυττάρων, κυρίως στο κατώτερο ήμισυ των κρυπτών. Τα νευροενδοκρινή τους κοκκία περιέχουν διάφορες ουσίες όπως, χρωμογραφίνη, σωματοστατίνη, γλυκόζη, ουσία Ρ και εντοπίζονται στο κατώτερο ήμισυ των κρυπτών. Ένας ακόμα πληθυσμός κυττάρων προέρχεται από τα αρχέγονα επιθηλιακά κύτταρα των κρυπτών, τα κύτταρα Μ, τα οποία εντοπίζονται σε περιοχές όπου υπάρχουν λεμφοζίδια. ΧΟΡΙΟ Το χόριο του παχέος εντέρου εμφανίζει κάποια επιπρόσθετα δομικά χαρακτηριστικά σε σχέση με τον υπόλοιπο πεπτικό σωλήνα. Στιβάδα κολλαγόνου πάχους έως 5μm στο φυσιολογικό ανθρώπινο κόλον και μπορεί να έχει τριπλάσιο πάχος στους υπερπλαστικούς πολύποδες του παχέος εντέρου. Συμμετέχει στην ρύθμιση της μεταφοράς ύδατος και ηλεκτρολυτών από το μεσοκυττάριο διαμέρισμα του επιθηλίου στο αγγειακό διαμέρισμα. Εκτεταμένη ανάπτυξη του ανοσοποιητικού στο κόλον, GALT, μεγάλα λεμφοζοίδια που εκτείνονται στον υποβλενογόνιο. Η ανάπτυξη του ανοσοποιητικού στο κόλον, αντανακλά το μεγάλο αριθμό και την ποικιλία μικροοργανισμών και βλαπτικών τελικών προϊόντων του μεταβολισμού. Περικρυπτικό έλυτρο ινοβλαστών το οποίο αποτελείται από ένα πληθυσμό ινοβλαστών που πολλαπλασιάζονται, διαφοροποιούνται και μεταναστεύουν προς τα πάνω παράλληλα και σύγχρονα με τα επιθηλιακά κύτταρα. Απουσία λεμφαγγείων στο χόριο μεταξύ των εντερικών αδένων, σχηματίζουν ένα δίκτυο γύρω από την βλεννογόνια μυϊκή στιβάδα όπως και στο λεπτό έντερο. Η απουσία λεμφαγγείων στο χόριο είναι σημαντική στην κατανόηση του αργού ρυθμού μετάστασης συγκεκριμένων καρκίνων του παχέος εντέρου. ΜΥΙΚΟΣ ΧΙΤΩΝΑΣ Αποτελείται από μια έσω κυκλοτερή και μία έξω επιμήκη στιβάδα λείων μυϊκών ινών. Σε αντίθεση με το λεπτό έντερο, η έξω επιμήκης στιβάδα του παχέος εντέρου είναι ασυνεχής σχηματίζοντας τρείς ταινίες, τις κολικές ταινίες. Διαπερνούν την έσω κυκλοτερή μυϊκή στιβάδα σε ακανόνιστα διαστήματα κατά μήκος και κατά την περίμετρο του παχέος εντέρου (εικόνα 1). Αυτή η ασυνέχεια του μυϊκού χιτώνα επιτρέπει στα τμήματα του παχέος εντέρου να συστέλλονται ανεξάρτητα, οδηγώντας στο σχηματισμό των κολικών κυψελών, στο τοίχωμα του παχέος εντέρου. Μεταξύ έσω και έξω στιβάδας εντοπίζεται το μυεντερικό πλέγμα του Auerbach. 10

11 Εικόνα 1.Το σχήμα απεικονίζει ορισμένα από τα κυριότερα χαρακτηριστικά των δύο μεγάλων υποδιαιρέσεων του εντέρου, και τονίζει τις ομοιότητες και τις διαφορές τους. (Dr. Thomas Caceci. ) ΟΡΟΓΟΝΟΣ Είναι το σπλαγχνικό περιτόναιο και καλύπτει πλήρως την σκωληκοειδή απόφυση, το τυφλό το εγκάρσιο και το σιγμοειδές. Αφήνει ακάλυπτο το οπίσθιο τμήμα του ανιόντος και το κατιόν. Οι επιπλοϊκές αποφύσεις είναι αναδιπλώσεις του ορογόνου χιτώνα και περιέχουν λιπώδες ιστό. 1.2 ΕΜΒΡΥΙΚΗ ΑΝΑΠΤΥΞΗ Το έντερο είναι ένα εξαιρετικά πολύπλοκο όργανο που εξυπηρετεί σημαντικές λειτουργίες όπως η πέψη, η απορρόφηση θρεπτικών συστατικών, ενδοκρινικές λειτουργίες και η ανοσία. Το παχύ έντερο προέρχεται από το μέσο και οπίσθιο αρχέγονο έντερο. Το μέσο έντερο σχηματίζει την πρωτογενή εντερική αγκύλη. Το κεφαλικό σκέλος εξελίσσεται στο περιφερικό τμήμα του δωδεκαδακτύλου μετά την εκβολή του χοληδόχου πόρου, τη νήστιδα και μέρος του ειλεού. Το ουραίο σκέλος της αγκύλης εξελίσσεται στο κατώτερο τμήμα του ειλεού, το τυφλό και την σκωληκοειδή απόφυση, το ανιόν κόλον και στα δυο πρώτα τριτημόρια του εγκάρσιου κόλου. Στην κορυφή της η πρωτογενή εντερική έλικα παραμένει προσωρινά σε ανοικτή επικοινωνία με το λεκιθικό ασκό διαμέσου του λεκιθικού πόρου χαρακτηρίζεται από ταχεία επιμήκυνση κυρίως του κεφαλικού σκέλους. Κατά συνέπεια την 6 η εβδομάδα ανάπτυξης οι εντερικές έλικες εισέρχονται στο εμβρυικό κοίλωμα του ομφάλιου λώρου (φυσιολογική ομφαλική κήλη). 11

12 Ταυτόχρονα με την ταχεία επιμήκυνση η πρωτογενείς έλικα περιστρέφεται γύρω από άξονα ο οποίος σχηματίζεται από την άνω μεσεντέρια αρτηρία. Η περιστροφή είναι αντιωρολογιακή 270 Ο (εικόνα 2) Stomach Mesenterium Parietal peritoneum 4 6 Intestinal loop Cecum Εικόνα 2 Εντερική περιστροφή κατά την εμβρυική ανάπτυξη (developed by the universities of Fribourg, Lausanne and Bern (Switzerland) with the support of the Swiss Virtual Campus ) Η καταβολή του τυφλού κάνει την εμφάνιση της όταν το έμβρυο έχει μήκος 12mm, με την μορφή μικρής κωνικής διεύρυνσης του ουραίου σκέλους της πρωτογενούς εντερικής αγκύλης και αποτελεί το τελευταίο τμήμα του εντέρου που επιστρέφει στην κοιλιακή κοιλότητα ( Langman s Medical Embryology.T Sadler ). Η σκωληκοειδής απόφυση αποτελεί το τελικό τμήμα του τυφλού. Γίνεται ορατή κατά την 8 η εβδομάδα. Στην αρχή παρουσιάζεται ως μια απλή προεξοχή του τυφλού, αλλά καθώς το τυφλό αναπτύσσεται περιστρέφεται προς την ειλεοτυφλική βαλβίδα. (Skandalakis LJet al,2004). Από το οπίσθιο έντερο προέρχεται το τελικό τμήμα του εγκάρσιου κόλου, το κατιόν κόλον, το σιγμοειδές, το ορθό και το ανώτερο τμήμα του πρωκτικού σωλήνα. Το τελικό τμήμα του οπισθίου εντέρου εκβάλει στην αμάρα, κοιλότητα η οποία επενδύεται από ενδόδερμα και επικοινωνεί απευθείας με το επιπολής εξώδερμα. Εκεί όπου έρχονται σε επαφή το ενδόδερμα με το εξώδερμα σχηματίζεται ο αμαρικός υμένας. Την 7 η εβδομάδα ο αμαρικός υμένας διαιρείται στον πρωκτικό και τον ουρογεννητικό υμένα. Ο πρωκτικός υμένας περιβάλλεται από μεσεγχυματικά ογκώματα με αποτέλεσμα την 8 η εβδομάδα να βρίσκεται στον πυθμένα ενός εξωδερμικού εντυπώματος γνωστού ως πρωκτικού βοθρίου. Την 9 η εβδομάδα ο πρωκτικός υμένας ρήγνυται. Το κατώτερο τμήμα του πρωκτικού σωλήνα έχει ενδοδερμική προέλευση και αιματώνεται από την 12
.")
13 κάτω μεσεντέρια αρτηρία ενώ το κατώτερο τριτημόριο έχει εξωδερμική προέλευση και αιματώνεται από τις αιμορροϊδικές αρτηρίες (T Sadler ). Το ώριμο εντερικό επιθήλιο αποτελείται από τις κρύπτες λάχνες και περιέχει απορροφητικά κύτταρα (εντεροκύτταρα), εκκριτικά κύτταρα (goblet, εντεροενδοκρινή, paneth, tuft) καθώς και ταχέως διαιρούμενα αρχέγονα κύτταρα (stem cells). Σε επαφή με το εντερικό επιθήλιο υπάρχουν μη ενδοδερμικής προέλευσης κύτταρα όπου σχηματίζεται το χόριο, ο υποβλεννογόνιος και ο μυϊκός χιτώνας. Η διαδικασία της γαστριδίωσης οδηγεί σε τρία κύρια βλαστικά δέρματα που συμπεριλαμβάνουν το εξώδερμα, το μεσόδερμα και το ενδόδερμα (Lawson and Pedersen et al, 1987;Lawson et al,1991; Rodaway et al,1999; Kimelman and Griffin et al,2000; Rodaway and Patient et al, 2001;Spence and Wellset al,2007; Zorn and Wells, et al,2007; Zorn and Wells, et al,2009). Με τον σχηματισμό των βλαστικών στοιβάδων το ενδόδερμα υποβάλλεται σε μια σειρά αλλαγών για να δώσει γένεση στο γαστρικό σωλήνα. Κατά την πρώιμη εμβρυϊκή ανάπτυξη, το σπλαχνικό ενδόδερμα φαίνεται ομοιογενές, είναι πολύστιβο και σηματοδοτεί τον σχηματισμό λαχνών. Στον άνθρωπο, η τελική αρχιτεκτονική του λεπτού εντέρου επιτυγχάνεται πριν από τη γέννηση και χαρακτηρίζεται από κρύπτεςλάχνες. Η μορφολογία του ώριμου παχέoς εντέρου δεν παρουσιάζει λάχνες κατά τη στιγμή της γέννησης αλλά σωληνοειδείς αδένες (κρύπτες) (εικόνα 3). Εικόνα 3 Σχηματισμός του γαστρικού σωλήνα κατά την εμβρυογένεση (T Sadler). Βασικά μοριακά μονοπάτια με καθοριστικό ρόλο στην ανάπτυξη του γαστρεντερικού αποτελούν το μονοπάτι hedgehog (Hh) στη Drosophila και στα σπονδυλωτά,(bitgood MJet al,1995) η οστική μορφογενετική πρωτεΐνη (BMP) η οποία φαίνεται να έχει σημαντικό ρόλο στην διαμόρφωση του εντέρου, το μονοπάτι Notch είναι θεμελιώδης οδός που χρησιμοποιείται από πολλά συστήματα για να κατευθύνουν την κυτταρική διαφοροποίηση, οι μεταγραφικοί παράγοντες HOΧ γονίδια που εμπλέκονται στο σχηματισμό- ανάπτυξη του συνολικού σχεδίου του σώματος, των άκρων, του ΚΝΣ και των σπλάχνων, SOX γονίδια που έχουν αναγνωριστεί ως βασικοί παράγοντες σε πολλές αναπτυξιακές διαδικασίες: καθορισμού του φύλου, νευρογένεση, διαφοροποίηση των μυών και χονδρογένεση. Τα SOX γονίδια συμμετέχουν επίσης στην παθολογική ανάπτυξη, καθώς φαίνεται να εμπλέκονται σε μονοπάτια ελέγχου του κυτταρικού πολλαπλασιασμού και στην ογκογένεση. Το σηματοδοτικό σύστημα Eph 13

14 receptors/ephrin ligands (Eph-ephrin) και το σηματοδοτικό μονοπάτι Wnt/β-catenin θεωρείται κρίσιμης σημασίας για τη διατήρηση των πολλαπλασιαζόμενων αρχέγονων κυττάρων στις κρύπτες του εντέρου (Pascal De Santa Barbara et al,2008). Μέλος της υπεροικογένειας TGF-β, ο παράγοντας Nodal απαιτείται για τον προσδιορισμό του μεσοδέρματος και του ενδοδέρματος στα σπονδυλωτά (Green and Smithet al,1990;clements et al,1999 ;Aoki et al,2002; Ben-Haim et al,2006; Hagos and Dougan et al,2007). Η έκθεση των προγονικών κυττάρων στον παράγoντα Nodal καθορίζει εάν ένα προγονικό κύτταρο θα διαφοροποιηθεί σε κύτταρο ενδοδέρματος ή μεσοδέρματος (Tremblay et al.,2000; Lowe et al., 2001; Vincent et al,2003; Weng and Stemple et al,2003;tada et al,2005 Shen et al,2007). Οδοί που εμπλέκονται στην κυτταρική διαφοροποίηση, ομοιόσταση και την απόπτωση στο επιθήλιο ενήλικων εντερικών κυττάρων είναι: 1) το μονοπάτι β-κατενίνης και TCF4 που εκφράζεται στα διαμερίσματα όπου εντοπίζονται ταχέως πολλαπλασιαζόμενα κύτταρα, τα βλαστικά κύτταρα του εντερικού επιθηλίου. Το σύμπλοκο β-catenin/tcf4 διαμεσολαβεί το WNT μονοπάτι σηματοδότησης με μεταγραφική ενεργοποίηση των γονιδίων στόχων c-myc, ΒΜΡ4, EphB2, EphB3 κα) (εικόνα 4) Εικόνα 4 Μοριακά μονοπάτια κατά την ανάπτυξη και διαφοροποίηση του εντερικού σωλήνα (Pascal De Santa Barbara et al,2008) O Shh εκφράζεται στη βάση της κρύπτης του λεπτού εντέρου και πιθανώς ρυθμίζει θετικά τον πολλαπλασιασμό των πρόδρομων κυττάρων. Ο Ihh εκφράζεται στα διαφοροποιημένα κύτταρα του κόλου και πιθανώς ρυθμίζει την ωρίμανση των εντεροκυττάρων. (Pascal De Santa Barbara et al, 2008) 14
.")
15 ΚΑΡΚΙΝΟΣ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ 1.3 ΕΠΙΔΗΜΙΟΛΟΓΙΑ Ο καρκίνος του παχέος εντέρου αποτελεί σημαντική αιτία νοσηρότητας και θνητότητας παγκοσμίως. Ευθύνεται για πάνω από 9% όλων των κρουσμάτων καρκίνου (Haggar et al, 2009). Πρόκειται για τον 3 ο συχνότερο καρκίνο στους άνδρες και 2 ο στις γυναίκες παγκοσμίως. Τα υψηλότερα ποσοστά εκτιμώνται στην Αυστραλία / Νέα Ζηλανδία και την Δυτική Ευρώπη, ενώ τα χαμηλότερα ποσοστά εκτιμώνται στην Αφρική με εξαίρεση την Ν. Αφρική και την Νότια Κεντρική Ασία (πίνακας 1).Τα ποσοστά εμφάνισης είναι σημαντικά υψηλότερα στους άνδρες σε σχέση με τις γυναίκες (1,4:1). Σημειώνονται θάνατοι από καρκίνο του παχέος εντέρου παγκοσμίως, ποσοστό 8 % του συνόλου των θανάτων από καρκίνο, καθιστώντας την τέταρτη συχνότερη αιτία θανάτου από καρκίνο (GloBocan 2008). Πίνακας 1 Περιστατικά παχέος εντέρου, συχνότητα σε σχέση με το φύλο και την γεωγραφική κατανομή Source: GLOBOCAN Η επιβίωση των ασθενών με καρκίνο παχέος εντέρου εξαρτάται σημαντικά από το στάδιο της νόσου κατά την διάγνωση και τυπικά κυμαίνεται από 90% 5 ετή επιβίωση για καρκίνο με τοπική εντόπιση, 70% για καρκίνο με τοπική επέκταση και 10% για ασθενείς με απομακρυσμένες μεταστάσεις (πίνακας 2). 15
. Σημειώνονται 608.")
16 Πίνακας 2. Πενταετή επιβίωση σε σχέση με το στάδιο της νόσου ανεξαρτήτως φύλου και φυλής. (SEER , All Races, Both Sexes by SEER Summary Stage (2000) Διάφοροι παράγοντες κινδύνου σχετίζονται με την συχνότητα εμφάνισης του καρκίνου του παχέος εντέρου όπως η ηλικία, κληρονομικοί, περιβαλλοντικοί και διατροφικοί παράγοντες. Παρατηρείται σημαντική αύξηση του ρυθμού εμφάνισης καρκίνου του παχέος εντέρου σε περιοχές που παραδοσιακά εμφάνιζαν χαμηλά ποσοστά όπως η Ισπανία και χώρες της Ανατολικής Ασίας και Ευρώπης. Τέτοιες δυσμενείς τάσεις πιστεύεται ότι αντικατοπτρίζουν έναν συνδυασμό παραγόντων, συμπεριλαμβανομένων και των αλλαγών στις διατροφικές συνήθειες, την παχυσαρκία και τον αυξημένο επιπολασμό του καπνίσματος. (de Kok IM et al,2002) Η πιθανότητα διάγνωσης αυξάνεται μετά την ηλικία των 40 ετών, ενώ παρατηρείται δραματική αύξηση μετά την ηλικία των 50. Ποσοστό 90% των περιπτώσεων ορθοκολικού καρκίνου εμφανίζονται σε άτομα ηλικίας άνω των 50, ενώ τα ποσοστά είναι κατά 50 φορές υψηλότερα σε άτομα ηλικίας 60 και 79 ετών. Στις ΗΠΑ παρατηρείται σημαντική αύξηση της εμφάνισης του καρκίνου του παχέος εντέρου σε άτομα ηλικίας ετών. Τροποποιήσιμοι παράγοντες κινδύνου για καρκίνο του παχέος εντέρου περιλαμβάνουν το κάπνισμα, η έλλειψη σωματικής άσκησης, η παχυσαρκία, η κατανάλωση κόκκινου και επεξεργασμένου κρέατος, και η υπερβολική κατανάλωση αλκοόλ. Πίνακας 3. Νέα περιστατικά, θάνατοι και 5-έτη επιβίωση.seer 9 Incidence & U.S. Mortality , All Races, Both Sexes. Rates are Age-Adjusted. 16

17 1.4 ΚΛΙΝΙΚΗ ΕΙΚΟΝΑ Ο συχνότερος ιστολογικός τύπος καρκίνου του παχέος εντέρου είναι το αδενοκαρκίνωμα. Αναφορικά με την πρωτοπαθή εντόπιση, τα δυο τρίτα των καρκίνων εντοπίζονται στο ορθό, ορθοσιγμοειδές, σιγμοειδές και το υπόλοιπο ένα τρίτο εντοπίζεται στο υπόλοιπο τμήμα του παχέος εντέρου. Στις κυριότερες κλινικές εκδηλώσεις της νόσου περιλαμβάνονται οι αλλαγές στις συνήθειες του εντέρου (44%), κοιλιακό άλγος(43%), αιμοχεσία(40%), αδυναμία(20%), αναιμία(11%) απώλεια βάρους(6%). Όλα αυτά τα συμπτώματα βρέθηκε ότι έχουν μικρή ευαισθησία (έως και κάτω του 10%) αλλά υψηλή ειδικότητα. Τα σημεία και συμπτώματα στον καρκίνο του παχέος εντέρου είναι ποικίλα και μη ειδικά. Στην πλειονότητα των περιπτώσεων η κλινική παρουσίαση της νόσου είναι υποξεία και ύπουλη. Οι ασθενείς με όγκους που εντοπίζονται στο τυφλό και δεξιό κόλον εμφανίζουν συνήθως κόπωση, αδυναμία και σιδηροπενική αναιμία λόγω της χρόνιας υποκλινικής απώλειας αίματος από το ορθό. Αντιθέτως, οι αριστεράεντοπιζόμενοι όγκοι παρουσιάζονται συχνότερα με έκδηλη αιμορραγία από το ορθό, διαταραχές κενώσεων και κοιλιακό άλγος. Σε ένα σημαντικό ποσοστό των ασθενών τα πρώτα συμπτώματα της νόσου είναι οξέα και οφείλονται σε οξεία απόφραξη ή διάτρηση του εντέρου. Το ενδεχόμενο του καρκίνου του παχέος εντέρου πρέπει να αποκλείεται σε όλους τους ασθενείς με αιμορραγία από το ορθό και σε όλους τους ασθενείς >40 ετών με σιδηροπενική αναιμία. Σε μεταστατική νόσο περί το 20% των ασθενών αναγνωρίζονται σε μη ιάσιμο στάδιο όταν υπάρχουν ήδη μεταστάσεις. Ο καρκίνος του παχέος μεθίσταται συνήθως αιματογενώς ή λεμφογενώς αν και σπάνια μπορεί να υπάρχουν περιτοναϊκές μεταστάσεις ή κατά συνέχεια ιστού οι κύριες εστίες μετάστασης είναι οι περιοχικοί λεμφαδένες, το ήπαρ, οι πνεύμονες και το περιτόναιο. Με εξαίρεση το όγκο του ορθού πρωκτού (λόγω αιμάτωσης) είναι σπανιότατη η πρώτη μετάσταση να είναι άλλη από το ήπαρ. Τα συμπτώματα είναι ανάλογα με το προσβαλλόμενο όργανο. Σπανιότερες πρώτες εκδηλώσεις είναι η τοπική διήθηση ή διάτρηση, η δημιουργία συριγγίου με κάποιο όργανο, πυρετός αγνώστου αιτιολογίας ή βακτηριαιμία από Streptococcus bovis ή Clostridium septicum (έως 25%). Ποικιλία μεθόδων χρησιμοποιούνται για την ανίχνευση και διάγνωση της νόσου, ξεκινώντας από την δακτυλική εξέταση και την εξέταση των κοπράνων για μικροσκοπική αιμορραγία. Η οριστική όμως διάγνωση τίθεται με βαριούχο υποκλυσμό και ενδοσκόπηση (ορθοσιγμοειδοσκόπηση, κολονοσκόπηση) συνοδευόμενη από βιοψία. Η εκτίμηση ολόκληρου του παχέος εντέρου στους ασθενείς με καρκίνο είναι απαραίτητη για τον αποκλεισμό της παρουσίας και άλλων όγκων. Αξονική τομογραφία και άλλες ακτινολογικές εξετάσεις πραγματοποιούνται για την εκτίμηση της έκτασης της νόσου και την παρουσία ή όχι απομακρυσμένων μεταστάσεων. Τα επίπεδα του καρκινοεμβρυϊκού αντιγόνου (CEA) στον ορό έχουν μικρή διαγνωστική σημασία λόγω χαμηλής ευαισθησίας και ειδικότητας. Ωστόσο, συμβάλλουν στην ανίχνευση μεταστατικής νόσου (υψηλά επίπεδα CEA υποδηλώνουν την παρουσία μεταστάσεων) και στην μετεγχειρητική παρακολούθηση των ασθενών για την ανάδειξη πιθανής υποτροπής της νόσου. 17
 αλλά υψηλή ειδικότητα. Τα σημεία και συμπτώματα στον καρκίνο του παχέος εντέρου είναι ποικίλα και μη ειδικά.")
18 1.5 ΑΙΤΙΟΛΟΓΙΑ-ΠΑΘΟΓΕΝΕΙΑ Η αιτιολογία του καρκίνου του παχέος εντέρου είναι πολυπαραγοντική: Περιβαλλοντικοί παράγοντες Σύμφωνα με επιδημιολογικά δεδομένα η δίαιτα σχετίζεται αιτιολογικά με την ανάπτυξη καρκινώματος του παχέος εντέρου. Η επίπτωση της νόσου είναι μεγαλύτερη σε χώρες που έχουν υιοθετήσει δυτικού τύπου δίαιτα, υψηλής δηλαδή περιεκτικότητας σε ζωικό λίπος. Από επιδημιολογικά επίσης στοιχεία προκύπτει ότι η καθιστική ζωή, το κάπνισμα και η κατανάλωση αλκοόλ αυξάνουν τον κίνδυνο της νόσου ενώ αντιθέτως προστατευτικό ρόλο φαίνεται να έχουν η φυσική δραστηριότητα, η θεραπεία υποκατάστασης με οιστρογόνα και η παρατεταμένη χρήση μη-στεροειδών αντιφλεγμονωδών φαρμάκων (ΜΣΑΦ) (Hermiston ML et al,1995). Πλήθος πράγματι επιδημιολογικών μελετών και τυχαιοποιημένων κλινικών δοκιμών δείχνουν ότι η μακρόχρονη χρήση ΜΣΑΦ συνοδεύεται από χαμηλότερη επίπτωση εμφάνισης και υποστροφή των αδενωμάτων καθώς επίσης και από χαμηλότερη επίπτωση καρκινώματος του παχέος εντέρου. Οι μηχανισμοί με τους οποίους τα ΜΣΑΦ πιθανώς αναστέλλουν την νεοπλασματική εξεργασία στο παχύ έντερο, σύμφωνα με τις μέχρι τώρα ενδείξεις, είναι η επαγωγή της απόπτωσης και η αναστολή την αγγειογένεσης. Αυξανόμενα επίσης πειραματικά δεδομένα δείχνουν ότι αυτές οι δράσεις των ΜΣΑΦ δεν οφείλονται αποκλειστικά στην αναστολή της κυκλοξυγενάσης- 2, που αναμφισβήτητα εμπλέκεται στην καρκινογένεση στο κόλον, αλλά και στην παρέμβαση σε άλλα μονοπάτια ανεξάρτητα από την οδό σύνθεσης των προσταγλανδινών (c-myc, p38, NF-kappa β, ILK) (Van de Wetering M et al,2002) Γενετική προδιάθεση Οι γενετικοί παράγοντες παίζουν πολύ σημαντικό ρόλο στην ανάπτυξη αδενοκαρκινώματος του παχέος εντέρου. Το φάσμα της γενετικής προδιάθεσης είναι ευρύ, περιλαμβάνοντας τόσο καλά καθορισμένα κληρονομικά σύνδρομα, όπως η αδενωματώδης πολυποδίαση (FAP) που οφείλεται συνήθως σε μεταλλάξεις του ογκοκατασταλτικού γονιδίου APC, το οποίο βρίσκεται στο χρωμόσωμα 5 και αποτελείται από 15 εξώνια και o κληρονομικός χωρίς πολυποδίαση καρκίνος του παχέος εντέρου (HNPCC) (non polyposis colorectal cancer), που προκαλείται κατά κύριο λόγο από μεταλλάξεις στα γονίδια MLH1, MSH2, MSH6 υπεύθυνα για την επιδιόρθωση στο DNA (MisMatch Repair genes - MMR). Μεταλλάξεις σε αυτά τα γονίδια αναλογούν στο 90% περίπου των οικογενειών (60%, 30%, 7%-10% αντιστοίχως) που έχουν κλινικά διαγνωστεί με σύνδρομο Lynch. ( Ballinger AB, et al, 2007) (εικόνα 5). Σύνδρομα με μετάλλαξη στο APC γονίδιο περιλαμβάνουν την FAP, το σύνδρομο Gardner, το σύνδρομο Turcot, και την εξασθενημένη αδενωματώδη πολυποδίαση του παχέος εντέρου (attenuated adenomatous polyposis coli AAPC). Το σύνδρομο Gardner χαρακτηρίζεται από πολυποδίαση του παχέος εντέρου (τυπική εικόνα της FAP), και συχνή παρουσία οστεωμάτων (συνηθέστερα στο κρανίο και την κάτω γνάθο), οδοντιατρικών ανωμαλιών και όγκων των μαλακών ιστών. Το σύνδρομο Turcot χαρακτηρίζεται από πολυποδίαση τυπική της FAP, μαζί με όγκους του 18
 (Hermiston ML et al,1995).")
19 κεντρικού νευρικού συστήματος (μυελοβλάστωμα). Το σύνδρομο AΑPC χαρακτηρίζεται από μικρότερο αριθμό πολυπόδων (30-35) σε σύγκριση με την κλασική FAP. Οι πολύποδες τείνουν επίσης να αναπτυχθούν σε μεγαλύτερη ηλικία (μέση ηλικία, 36 ο έτος), και τείνουν να προσβάλουν το εγγύς τμήμα του παχέος εντέρου. Το APC γονίδιο είναι ένα ογκοκατασταλτικό γονίδιο το προϊόν του οποίου ρυθμίζει έμμεσα την μεταγραφή ενός αριθμού γονιδίων κρίσιμων για τον πολλαπλασιασμό των κυττάρων, μέσω της αλληλεπίδρασης με τον παράγοντα μεταγραφής β-κατενίνης. Απώλεια του APC εμποδίζει την αποικοδόμηση της β-κατενίνης και επιτρέπει στην β- κατενίνη να συσσωρεύεται και να επάγει τον πολλαπλασιασμό των κυττάρων με επακόλουθο την ανάπτυξη των αδενωμάτων. (Nieuwenhuis MH et al,2008;schulmann Ket al,2008) O κληρονομικός χωρίς πολυποδίαση καρκίνος του παχέος εντέρου (HNPCC), είναι μια κληρονομική διαταραχή που αυξάνει τον κίνδυνο για πολλούς τύπους καρκίνου, ιδιαίτερα τον καρκίνο του παχέος εντέρου και του ορθού. Μεταλλάξεις στα γονίδια MLH1, MSH2, MSH6, PMS2 ή EpCAM αυξάνουν τον κίνδυνο ανάπτυξης του συνδρόμου Lynch. Τα γονίδια MLH1, MSH2, MSH6 και PMS2 εμπλέκονται στην επιδιόρθωση λαθών κατά την αντιγραφή του DNA. Μεταλλάξεις σε οποιοδήποτε από αυτά τα γονίδια εμποδίζουν την ορθή επιδιόρθωση λαθών αντιγραφής του DNA. Μεταλλάξεις στο γονίδιο EpCAM οδηγεί επίσης σε μειωμένη επιδιόρθωση του DNA. Το γονίδιο EpCAM βρίσκεται δίπλα στο γονίδιο MSH2 στο χρωμόσωμα 2 και ορισμένες γονιδιακές μεταλλάξεις στο EpCAM απενεργοποιούν το γονίδιο MSH2, διακόπτοντας την επιδιόρθωση του DNA και οδηγώντας σε συσσώρευση λαθών. Εικόνα 5. Απεικόνιση της εξέλιξης του καρκίνου του παχέος εντέρου σε σποραδικά και υψηλού κινδύνου γενετικά σύνδρομα. Ο όγκος ξεκινά από ένα φυσιολογικό βλαστικό κύτταρο που έχει υποστεί γενετική βλάβη με την πάροδο του χρόνου λόγω του τοπικού περιβάλλοντος και οποιαδήποτε γενετικής μετάλλαξης που έχει κληρονομήσει: FAP, η καρκινογένεση επιταχύνεται με την APC μετάλλαξη στην βλαστική σειρά, στο σύνδρομο Lynch, η εξέλιξη του όγκου επιταχύνεται λόγω της απώλειας του συστήματος DNA MMR.(Grady WM et al,2002) Στα γενετικά σύνδρομα οικογενούς πολυποδίασης περιλαμβάνεται και μια μορφή οικογενούς αδενωματώδους πολυποδίασης, η MUTYH σχετιζόμενη πολυποδίαση. Το γονίδιο MUTYH, κωδικοποιεί μια γλυκοζυλάση του DNA. Η MUTYH γλυκοζυλάση εμπλέκεται στην οξειδωτική επιδιόρθωση βλαβών του DNA. Το σύνδρομο Peutz- Jeghers, επίσης γνωστό ως κληρονομικό σύνδρομο εντερικής πολυποδίασης, είναι μία κληρονομική διαταραχή που χαρακτηρίζεται από την παρουσία αμαρτωματωδών 19

20 πολυπόδων. Η αιτία του συνδρόμου Peutz-Jeghers (PJS) στις περισσότερες περιπτώσεις (66-94%) φαίνεται να είναι μια μετάλλαξη του STK11 / LKB1 (σερίνη / θρεονίνη κινάση 11) ογκοκατασταλτικού γονιδίου, που βρίσκεται στη θέση 19p13.3. Τέλος το σύνδρομο Υπερπλαστικής πολυποδίασης είναι μια σπάνια πάθηση που έχει οριστεί από τον Παγκόσμιο Οργανισμό Υγείας ως εξής: 1. Πέντε ή περισσότεροί υπερπλαστικοί πολύποδες εγγύς στο σιγμοειδές με δύο πολύποδες μεγαλύτερους από 10 mm διάμετρο ή 2. Οποιοσδήποτε αριθμός υπερπλαστικών πολυπόδων εγγύς στο σιγμοειδές σε ένα άτομο με συγγενής πρώτου βαθμού ο οποίος έχει υπερπλαστικό σύνδρομο πολυποδίασης ή 3. Περισσότεροι από 30 υπερπλαστικοί πολύποδες με οποιοδήποτε μέγεθος σε όλη την επιφάνεια του παχέος εντέρου και του ορθού. (Stoler,M et al,2009). Αν και πιστεύεται ότι δεν εμφανίζουν κακόηθες δυναμικό έχει αποδειχθεί ότι πολύποδες του παχέος εντέρου εμφανίζουν κακοήθη εξαλλαγή. Αυτό συμβαίνει μέσω πολλαπλών μεταλλάξεων οι οποίες επηρεάζουν τον μηχανισμό επιδιόρθωσης των αταίριαστων βάσεων του DNA (DNA mismatch repair, MMR). Αυτό οδηγεί σε μικροδορυφορική αστάθεια η οποία μπορεί τελικά να οδηγήσει σε κακοήθη εξαλλαγή. Παρόλο που πιστεύεται ότι δεν εμφανίζει κακόηθες δυναμικό έχει δειχθεί ότι στη δεξιά πλευρά του παχέος εντέρου παρουσιάζεται ένα κακόηθες δυναμικό, μέσω του μηχανισμού της μικροδορυφορυκής αστάθειας Προδιαθεσικοί παράγοντες-προκαρκινικές αλλοιώσεις Οι προκαρκινικές αλλοιώσεις του καρκίνου του παχέος εντέρου είναι τα αδενώματα, σποραδικά ή στα πλαίσια οικογενούς πολυποδίασης και η Επιθηλιακή Δυσπλασία (ΕΔ) στις Ιδιοπαθείς Φλεγμονώδεις Νόσους (ΙΦΝ) (Williams G et al,2007). Η φλεγμονώδης νόσος του εντέρου αποτελεί πολύ σημαντικό αιτιολογικό παράγοντα στην ανάπτυξη καρκινώματος του παχέος εντέρου. Ο κίνδυνος αυξάνει μετά από 8-10 χρόνια νόσου και είναι υψηλότερος στους ασθενείς με πρώιμη έναρξη και εκτεταμένη νόσο (πανκολίτιδα). Τα Ιδιοπαθή Φλεγμονώδη Νοσήματα του Εντέρου (ΙΦΝΕ), περιλαμβάνουν τη νόσο Crohn και την ελκώδη κολίτιδα, οι οποίες χαρακτηρίζονται από φλεγμονή (βλάβες, εξοίδηση και αιμορραγία) του πεπτικού συστήματος. Η ελκώδης κολίτιδα προσβάλλει το βλεννογόνο του παχέος εντέρου (κόλον και ορθό) και η νόσος Crohn είναι δυνατό να προσβάλλει οποιοδήποτε τμήμα του πεπτικού σωλήνα από το στόμα μέχρι τον πρωκτό και η φλεγμονή είναι διατοιχωματική. Η επίπτωση της ΙΦΝΕ ποικίλλει στις διάφορες γεωγραφικές περιοχές. Η ελκώδης κολίτιδα και η νόσος Crohn συναντώνται και οι δύο μαζί με την μέγιστη επίπτωση στην Ευρώπη, στο Ηνωμένο Βασίλειο και την Βόρεια Αμερική. Η ηλικιακή κορύφωση της επίπτωσης της νόσου Crohn και της ελκώδους κολίτιδας εντοπίζεται μεταξύ των 15 και 30 ετών. Μία δεύτερη κορύφωση συμβαίνει μεταξύ των 60 και 80 ετών. Η αναλογία ανδρών και γυναικών για την ελκώδη κολίτιδα είναι 1:1 και για την νόσο Crohn είναι 1,1-1,8:1. 20

21 Η ακριβής αιτιολογία των δύο αυτών νόσων παραμένει άγνωστη. Σε γενετικά προδιατεθειμένα άτομα τόσο εξωγενείς παράγοντες (λοιμώδεις παράγοντες, φυσιολογική χλωρίδα του αυλού) όσο και παράγοντες του ξενιστή (λειτουργία του εντερικού επιθηλιακού φραγμού) προκαλούν ένα χρόνιο στάδιο δυσλειτουργίας της ανοσολογικής λειτουργίας του βλεννογόνου, η οποία υφίσταται περαιτέρω τροποποίηση από περιβαλλοντικούς παράγοντες όπως το κάπνισμα, η προσθήκη συντηρητικών στις τροφές, η αλλαγή στις διαιτητικές συνήθειες, ενδεχομένως το stress, καθώς και η μειονεκτική ή και λανθασμένη, για άγνωστους λόγους, αντίδραση του αμυντικού (ανοσολογικού) συστήματος του οργανισμού. Πρόσφατα, βρέθηκαν αρκετές θέσεις στο ανθρώπινο γονιδίωμα (χρωμοσώματα) που προδιαθέτουν στην εμφάνιση των νόσων αυτών, αλλά που από μόνα τους δεν είναι δυνατόν να τις προκαλέσουν. Γι αυτό χρειάζεται και η επίδραση διαφόρων περιβαλλοντικών παραγόντων. Ο κίνδυνος ανάπτυξης καρκίνου στο παχύ έντερο, περισσότερο στην ελκώδη κολίτιδα απ ότι στη νόσο Crohn, φαίνεται ότι εξαρτάται τόσο από την έκταση προσβολής του παχέος εντέρου (ολική ή εκτεταμένη κολίτιδα) όσο και από τη διάρκεια της νόσου (μεγαλύτερη από δεκαετία).της εμφάνισης του καρκίνου προηγείται, σε αρκετές περιπτώσεις, η εμφάνιση επιθηλιακής δυσπλασίας του βλεννογόνου του παχέος εντέρου (προκαρκινική κατάσταση) Αδενώματα Τα αδενώματα είναι καλοήθης επιθηλιακοί όγκοι. Η επίπτωση τους στο παχύ έντερο είναι 20-30% πριν την ηλικία των 40 ετών αυξανόμενη σε 40-50% μετά την ηλικία των 60. Γυναίκες και άνδρες προσβάλλονται εξίσου. Υπάρχει σαφής οικογενής προδιάθεση στην ανάπτυξη τους, ο κίνδυνος δε εμφάνισης καρκινώματος του παχέος εντέρου σε ασθενείς με αδένωμα είναι τετραπλάσιος σε σύγκριση με το γενικό πληθυσμό. Τα αδενώματα χαρακτηρίζονται από την παρουσία ενδοεπιθηλιακής νεοπλασίας, η οποία ιστολογικά συνίσταται σε κυτταροβρίθεια, μεγάλους υπερχρωματικούς πυρήνες, ποικίλου βαθμού στιβαδοποίηση των πυρήνων και απώλεια της πολικότητας του επιθηλίου. Αναπτύσσονται ως αποτέλεσμα γενετικών αλλαγών στα επιθηλιακά κύτταρα, με εναρκτήρια και κυριότερη την ενεργοποίηση του μονοπατιού β- catenin/tcf από τις μεταλλάξεις του γονιδίου APC, που οδηγούν σε αύξηση του επιθηλιακού πολλαπλασιασμού, μετακίνηση των πολλαπλασιαζόμενων κυττάρων από το βάθος της κρύπτης προς τον αυλό, αυξημένη διαίρεση των κρυπτών και επιθηλιακή δυσπλασία. Η επιθηλιακή δυσπλασία μπορεί να κυμαίνεται από ήπια έως τόσο σοβαρή ώστε να συνιστά καρκίνωμα in situ.(εικόνα 6) 21
22 Εικόνα 6. Γενετικές αλλαγές επιθηλιακών κύτταρων κατά την καρκινογένεση στο παχύ έντερο.reviewers: Shilpa Jain, M.D, New York University, 2 May 2012, last major update August 2010Copyright: (c) , PathologyOutlines.com, Inc. Βάσει των μακροσκοπικών τους χαρακτηριστικών τα αδενώματα διακρίνονται σε επίπεδα (flat), εμβυθιζόμενα (depressed) και υπερυψωμένα (elevated). Τα elevated αδενώματα μπορεί να φέρουν ή όχι μίσχο. Τα flat και τα depressed αναγνωρίζονται μακροσκοπικά από τις αλλαγές στην υφή και την ερυθρότητα του βλεννογόνου. Είναι προφανές ότι ο όρος αδένωμα δε αφορά μόνο σε πολυποδοειδή μορφώματα καθώς το καθοριστικό χαρακτηριστικό των αδενωμάτων είναι η ενδοεπιθηλιακή νεοπλασία. Βάσει της επιθηλιακής αρχιτεκτονικής τους τα αδενώματα διακρίνονται επιπλέον σε σωληνώδη (tubular)(εικόνα 7), θηλώδη (villous)(εικόνα 8) και σωληνοθηλώδη (tubulovillous). Τα σωληνώδη αδενώματα είναι συνήθως σφαιρικά και έμισχα ή επίπεδα και μικροσκοπικά χαρακτηρίζονται από δυσπλαστικές σωληνώδεις αδενικές δομές που καταλαμβάνουν τουλάχιστον το 80% της αυλικής επιφάνειας. Τα θηλώδη είναι συνήθως μεγαλύτερα άμισχα και μικροσκοπικά χαρακτηρίζονται από την παρουσία φυλλοειδών προεκβολών του βλεννογόνου που καλύπτονται από δυσπλαστικό αδενικό επιθήλιο και καταλαμβάνουν περισσότερο από το 80% της αυλικής επιφάνειας. Τα σωληνοθηλώδη εμφανίζουν και σωληνώδη και θηλώδη αρχιτεκτονική. Ο κίνδυνος ανάπτυξης καρκινώματος σε αδένωμα εξαρτάται από τρία ανεξάρτητα χαρακτηριστικά: το μέγεθος του αδενώματος, την ιστολογική αρχιτεκτονική και τον βαθμό της δυσπλασίας, με την μέγιστη διάμετρο του αδενώματος να αποτελεί τον κύριο παράγοντα κινδύνου. Ωστόσο, τα depressed αδενώματα παρά το μικρό τους μέγεθος (μέση διάμετρος 11χιλ) εμφανίζουν τάση εξέλιξης σε καρκινώματα. (Troussard AA, et al, 1999) 22
23 Εικόνα 7. Σωληνώδες αδένωμα αποτελούμενο από αδένια ποικίλου σχήματος με αυλό που περιβάλλονται από στρώμα. (H&E, 100; coutesy of Dr. Arnold H. Szporn, Mount Sinai School of Medicine). Εικόνα 8. Θηλώδες αδένωμα αποτελούμενο από πτυχές αδενικού επιθηλίου κάθε μία με κεντρικό ινοαγγειακό πυρήνα που περιβάλλονται από κενό χώρο (αυλός εντέρου) (H&E, 100; courtesy of Dr. Arnold H. Szporn) 23
24 ΚΕΦΑΛΑΙΟ 2 2.0ΚΑΡΚΙΝΩΜΑ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ-ΜΟΡΙΑΚΗ ΠΑΘΟΓΕΝΕΙΑ Γενετικές τροποποιήσεις που συμβάλλουν στην ανάπτυξη και εξέλιξη του καρκίνου οδηγούν σε έξι βασικά χαρακτηριστικά των καρκινικών κυττάρων (εικόνα 9) ( Hanahan D et al, 2000) Πρώτον, τα καρκινικά κύτταρα μέσω μεταλλάξεων ενεργοποιούν πρωτοογκογονίδια που επιτρέπουν στα καρκινικά κύτταρα να παράγουν τα δικά τους σήματα αύξησης-πολλαπλασιασμού, όπως ο TGF-a και ο PDGF. Δεύτερον, τα καρκινικά κύτταρα χαρακτηρίζονται από έλλειψη ευαισθησίας σε σήματα που αναστέλλουν την αύξηση, εν μέρει μέσω της διακοπής της οδού Rb. Τρίτον, τα καρκινικά κύτταρα έχουν αποκτήσει την ικανότητα να ξεφύγουν από την απόπτωση μέσω μεταλλάξεων που απενεργοποιούν ογκοκατασταλτικά γονίδια, όπως p53 ή PTEN, ή μέσω της ενεργοποίησης των οδών επιβίωσης που περιλαμβάνουν αυξητικός παράγοντας ινσουλίνης 1/2 (IGF 1/2) ή ιντερλευκίνη-3 (IL-3). Τέταρτον, τα καρκινικά κύτταρα έχουν χάσει τους μηχανισμούς που περιορίζουν τον πολλαπλασιασμό τους. Αυτό είναι αποτέλεσμα, κυρίως της ενεργοποίησης της τελομεράσης, ενός ενζύμου που διατηρεί τα τελομερή στα άκρα των χρωμοσωμάτων. Πέμπτον, τα νεοπλάσματα επάγουν την αγγειογένεση. Η διαδικασία νεοαγγείωσης επάγεται από την έκφραση προαγγειογενετικών παραγόντων όπως του αγγειακού ενδοθηλιακού αυξητικού παράγοντα (VEGF) που παράγεται από τα καρκινικά κύτταρα ή από την καταστολή μηχανισμών που αναστέλλουν την αγγειογένεση όπως η πτώση τον επιπέδων θρομβοσπονδίνης. Έκτον, κύτταρα από το πρωτοπαθές νεόπλασμα εισβάλουν σε παρακείμενους ιστούς ή σε απομακρυσμένες θέσεις και σχηματίζουν μεταστάσεις. Εικόνα 9.Σχηματική απεικόνιση των έξι βασικών χαρακτηριστικών των καρκινικών κυττάρων (Hanahan και Weinberg et al, 2000) Οι όγκοι του παχέος εντέρου εξελίσσονται μέσα από μια σειρά κλινικών και ιστοπαθολογικών σταδίων που περιλαμβάνουν την πρόοδο από δυσπλαστικές κρύπτες, σε μικρούς καλοήθεις όγκους (αδενώματα) και εν συνέχεια σε κακοήθεις όγκους (καρκίνο)( Fearon ER, et al, 1990). 24
25 Πιστεύεται ότι τα αδενώματα αναπτύσσονται από αρχέγονα κύτταρα που βρίσκονται στις βάσεις των φυσιολογικών εντερικών κρυπτών ( Bach SP et al, 2000). Οι απόγονοι των βλαστικών κυττάρων μεταναστεύουν από την κρύπτη προς τον αυλό και συνεχίζουν να διαιρούνται μέχρι να φτάσουν στο μεσαίο τμήμα της. Στη συνέχεια, τα μεταναστευτικά επιθηλιακά κύτταρα σταματούν να διαιρούνται και διαφοροποιούνται σε ώριμα κύτταρα εντερικού επιθηλίου. Όταν τα διαφοροποιημένα κύτταρα φθάσουν στην κορυφή της κρύπτης, υφίστανται απόπτωση. Αυτό το ταξίδι από τη βάση της κρύπτης στην κορυφή διαρκεί 3-6 ημέρες (Lipkin M,et al, 1963; Shorter RG et al,1964). Εάν ο λόγος γέννησης / απώλειας αυξηθεί τότε προκύπτει ένα νεόπλασμα. Όπως και με άλλους τύπους καρκίνου, η καρκινογένεση στο παχύ έντερο αποτελεί μια πολυσταδιακή διαδικασία (εικόνα10) που χαρακτηρίζεται από την αδρανοποίηση μιας ποικιλίας γονιδίων, που καταστέλλουν την ανάπτυξη των όγκων-ογκοκατασταλτικών γονιδίων(π.χ. APC, DCC, p53 ), γονιδίων που επιδιορθώνουν το DNA (π.χ.: hmsh2, hmslh1 ) και την ταυτόχρονη ενεργοποίηση πρώτο-ογκογονιδίων (π.χ. K-ras, N-Ras και c-myc ). Εικόνα10.Μοντέλο με τις κλινικοπαθολογοανατομικές αλλαγές και τις γενετικές ανωμαλίες που επάγουν την ανάπτυξη και εξέλιξη του καρκίνου του παχέος εντέρου (CRC). Το αρχικό βήμα στην ογκογένεση είναι ο σχηματισμός του αδενώματος, που συνδέεται με την απώλεια του APC. Μεγαλύτερα αδενώματα και καρκινώματα στα αρχικά στάδια αποκτούν μεταλλάξεις στο K-RAS, που ακολουθείται από την απώλεια του χρωμοσώματος 18q και Smad και μεταλλάξεις στο ΤΡ53 στο καρκίνωμα. Στον καρκίνο του παχέος εντέρου εκτός απο την Μικροδορυφορική αστάθεια (MSI +), που χαρακτηρίζεται από ανεπάρκεια του συστήματος επιδιόρθωσης, απαιτούνται και άλλες γενετικές αλλαγές όπως αυτές που περιγράφονται στην χρωμοσωμική αστάθεια (CIN) στον καρκίνο του παχέος εντέρου.(axeiw et al, 2009) Φύση της γονιδιωματικής αστάθειας Παράδειγμα γονιδιακής μετάλλαξης Σύνδρομο που σχετίζεται με μετάλλαξη βλαστικής σειράς Σχόλια Χρωμοσωμική αστάθεια Η απώλεια της λειτουργίας της APC γονιδίου Η οικογενής πολυποδίαση Σωματική APC μετάλλαξη βρέθηκε σε 85% των σποραδικών CRC. Μικροδορυφορική αστάθεια Αδρανοποίηση τους μέσω μεταλλάξεις μειώνει την δυνατότητα επιδιόρθωσης MLH1,MSH2, MSH6 και PMS2 Το σύνδρομο Lynch Σωματικές μεταλλάξεις των γονιδίων επιδιόρθωσης των αταίριαστων βάσεων βρέθηκαν στο 15% των σποραδικών CRC Ελάττωμα επιδιόρθωσης αφαίρεσης βάσεων του DNA MYH γονίδιο MYH σχετίζεται με πολυποδίαση Δεν αναγνωρίζονται σωματικές μεταλλάξεις. Πίνακας 4. Παραδείγματα της γονιδιωματικής αστάθειας στον CRC (Frontline Gastroenterol 2014.) 25
26 Σε κυτταρικό επίπεδο ο καρκίνος του παχέος εντέρου είναι αποτέλεσμα συσσώρευσης γενετικών και επιγενετικών αλλοιώσεων, που οδηγούν στην μετατροπή των φυσιολογικών επιθηλιακών κυττάρων σε κύτταρα αδενοκαρκινώματος παχέος εντέρου (πίνακας 4). Μετάλλαξη του γονιδίου APC αποτελεί από τις πρώτες χρονικά γενετικές αλλαγές και εκτιμάται ότι > 85% των όγκων παχέος εντέρου έχουν σωματικές μεταλλάξεις του APC γονιδίου ( Kinzler KW et al, 1998). Μεταλλάξεις του γονιδίου APC παρατηρούνται στο 85% των σποραδικών καρκινωμάτων καθώς και σε όλες σχεδόν τις περιπτώσεις FAP, ανευρίσκονται δε ήδη στο στάδιο των ανώμαλων εστιών στις κρύπτες (Aberrant Crypt Foci-ACF), γεγονότα ενδεικτικά ότι αποτελoύν κρίσιμο εναρκτήριο βήμα στην καρκινογένεση του παχέος εντέρου. To γονίδιο K-ras βρίσκεται στο βραχύ σκέλος του χρωμοσώματος 12 και κωδικοποιεί μια πρωτεΐνη 21Kd (p21 ras ) που συμμετέχει στην μεταγωγή σήματος μέσω G πρωτεΐνης. Το K-ras υφίσταται μετάλλαξη στα κωδικόνια 12, 13,61. Η μετάλλαξη στο K-ras οδηγεί σε ανεξέλεγκτο κυτταρικό πολλαπλασιασμό και κακοήθη εξαλλαγή (Bos J L. et al,1989). Μεταλλάξεις στο p53 παρατηρούνται καθυστερημένα στην ανάπτυξη των όγκων του παχέος εντέρου. Η απώλεια της διαμεσολαβούμενης από την πρωτεΐνη p53 απόπτωσης θεωρείται σημαντικό γεγονός για την εξαλλαγή ενός αδενώματος σε κακοήθη όγκο. Το γονίδιο για την πρωτεΐνη p53 εντοπίζεται στο βραχύ σκέλος του χρωμοσώματος 17 και είναι μεταλλαγμένο σε ποσοστό 70% των σποραδικών καρκινωμάτων του παχέος εντέρου (Baker S et al,1990). Η πρωτεΐνη p53 λειτουργεί ως μεταγραφικός παράγοντας και ελέγχει την πρόοδο του κυτταρικού κύκλου συνδεόμενος με συγκεκριμένες αλληλουχίες γονιδίων όπως p21, Bax, and Bcl-2 ως απάντηση σε κυτταρικό stess ή βλάβη του DNA( Levine A J. et al,1997). Τα επίπεδα p53 ρυθμίζονται από την λιγάση E3 ubiquitin ligase MDM2, η οποία επάγεται από το p53 και το οδηγεί σε αποικοδόμηση (May P et al,1999).(εικόνα 11) Εικόνα 11 α) Ειδικές γενετικές αλλοιώσεις προκύπτουν σε καθορισμένα στάδια κατά τη διάρκεια εξέλιξης αδενώματος-καρκινώματος το πρότυπο των αλλαγών και τα γονίδια που επηρεάζονται είναι συχνά μοναδικά σε όγκους που προκύπτουν μέσω χρωμοσωμικής αστάθειας (CIN) ή μικροδορυφορικής αστάθειας (MSI). β)αυξημένη συχνότητα των γενετικών αλλοιώσεων σε σύγκριση με το φυσιολογικό βλεννογόνο του παχέος εντέρου, η συχνότητα των οποίων αυξάνει σε προχωρημένο στάδιο καρκίνου του παχέος εντέρου. 26
27 Η απώλεια της γονιδιωματικής σταθερότητας (εικόνα 11)(πίνακας 4) φαίνεται να είναι ένα βασικό μοριακό βήμα που συμβαίνει νωρίς κατά την διαδικασία της ογκογένεσης. Τουλάχιστον δύο τύποι γονιδιωματικής αστάθειας εμπλέκονται στον καρκίνο του παχέος εντέρου: (1) μικροδορυφορική αστάθεια (MSI), (2) χρωμοσωμική αστάθεια (CIN). 2.1 Μικροδορυφορική αστάθεια (MSI) Η μικροδορυφορική αστάθεια εμφανίζεται σε ποσοστό ~ 15% των καρκίνων του παχέος εντέρου ως αποτέλεσμα αδρανοποίησης του μηχανισμού επιδιόρθωσης των αταίριαστων βάσεων (mutation mismatch repair MMR), είτε λόγω γονιδιακών μεταλλάξεων του συστήματος MMR, είτε λόγω υπερμεθυλίωσης του υποκινητή του γονιδίου MLH1 (Grand Wet al,1998). H MSI ως αποτέλεσμα της απώλειας του μηχανισμού MMR επηρεάζει κυρίως μόνο-, δι- και τρι- νουκλεοτίδια ( Eshleman Jet al,1995 ) οδηγώντας σε μεταλλάξεις γονιδίων στόχων, όπως το ογκοκατασταλτικό γονίδιο TGFBR2. Λιγότερο συχνός στόχος της μικροδορυφορικής αστάθειας είναι γονίδια όπως activin type II receptor (AVCR2 ) και ΒΑΧ. (Yamamoto H et al,1997; Yamamoto H et al,1998) Ένας όγκος χαρακτηρίζεται ως MSI-HIGH ( MSH-H ) όταν δύο ή περισσότερες από τις πέντε μικροδορυφορικές αλληλουχίες, που ορίζονται από το National Cancer Institute έχουν μεταλλαχθεί. Εάν μια εκ των μικροδορυφορικών αλληλουχιών έχει μεταλλαχθεί τότε οι όγκοι ταξινομούνται ως MSI-LOW (MSI-L). Όγκοι χωρίς μικροδορυφορικές μεταλλάξεις χαρακτηρίζονται ως σταθεροί (MSI STABLE)(Kurzawski, J. et al, 2004)(πίνακας 5) Πίνακας 5. Συστάσεις για την αξιολόγηση της MSI-H και MSI-L. Το πρότυπο του National Cancer Institute (NCI) για τον μικροδορυφορικό πίνακα περιλαμβάνει BAT25, BAT26, D2S123, D5S346 και D17S250; Ωστόσο, μπορούν να εφαρμοστούν οι ακόλουθοι περιορισμοί : Εάν μόνο επαναλήψεις δινουκλεοτιδίων είναι μεταλλαγμένες, εξετάστε ένα δεύτερο πάνελ με δείκτες μικροδορυφορικής αστάθειας με επαναλήψεις μονονουκλεοτιδίων(π.χ BAT40 και/ή MYCL) προς αποκλεισμό MSI- L. Οι επαναλήψεις δινουκλεοτιδίων είναι λιγότερο ευαίσθητες από τις επαναλήψεις σε μονονουκλεοτίδια για MSI- H, ωστόσο, παρέχουν έναν εσωτερικό έλεγχο για την πρόληψη λάθους δείγματος. Ένα πάνελ (Pentaplex quasimonomorphic) από μονομορφικές επαναλήψεις μονονουκλεοτιδίων μπορεί να είναι πιο ευαίσθητο για όγκους MSI-H σε σχέση με άλλους μικροδορυφορικούς δείκτες και μπορεί να αποτρέψει την ανάγκη για σύγκριση με φυσιολογικό ιστό. Αυτή η προσέγγιση απαιτεί τρία ή περισσότερα μεταλλαγμένα αλληλόμορφα για να δείξει MSI-H MSI-H αναφέρεται σε όγκους με αλλαγές σε δύο ή περισσότερα από τα πέντε NCI-προτεινόμενα πάνελ των μικροδορυφορικών δεικτών σε όγκους. MSI-L αναφέρεται σε όγκους με αλλαγές σε μόνο ένα από τα πέντε NCI-προτεινόμενα πάνελ των μικροδορυφορικών δεικτών σε όγκους. U Asad Uma CI J Natl Cancer Inst (2004) 96(4):
28 Το σύστημα DNA MMR θεωρείται ένα εξελικτικά συντηρημένο σύστημα το οποίο περιλαμβάνει μία σειρά πρωτεϊνών οι οποίες επιδιορθώνουν τυχόν νουκλεοτιδικές αναντιστοιχίες που έχουν διαφύγει από την DNA πολυμεράση οδηγώντας σε μετάλλαξη γονιδίων που ελέγχουν την κυτταρική ανάπτυξη επάγοντας την καρκινογένεση (πίνακας 6). Στους προκαρυωτικούς οργανισμούς, το σύστημα MMR (εικόνα 12) αποτελείται από μια οικογένεια ενζύμων που ανιχνεύουν λάθη αντιγραφής του DNA στην φάση S του κυτταρικού κύκλου, με αποτέλεσμα στην νέα συντιθέμενη έλικα να έχουμε εισαγωγή λάθος νουκλεοτιδίου. Λάθη τα οποία δεν επιδιορθώνονται από την DNA πολυμεράση κατά την διάρκεια αντιγραφής δημιουργούν προσωρινά μία αγκύλη (βρόγχο) εισαγωγής-εξάλειψης (insertion deletion loop, IDL) η οποία αναγνωρίζεται και επιδιορθώνεται από το σύστημα MMR. Αν δεν επιδιορθωθεί η βλάβη κατά την 2 η αντιγραφή η μετάλλαξη περνά στο DNA. Αλλαγή σε μία βάση προκαλεί σημειακή μετάλλαξη, ενώ η παρουσία βρόγχου εισαγωγής-εξάλειψης προκαλεί μετατόπιση του πλαισίου μεταγραφής. Το αποτέλεσμα είναι η παραγωγή μιας μη λειτουργικής πρωτεΐνης (C. Richard Boland and Ajay Goel et al,2010). Τουλάχιστον έξι γονίδια ( MSH2,MSH3, MSH6, MLH1,PMS1 και PMS2)(πίνακας 6) τα οποία κωδικοποιούν πρωτεΐνες που εμπλέκονται στο σύστημα MMR έχουν ταυτοποιηθεί τόσο στον σποραδικό καρκίνο του παχέος εντέρου όσο και στο κληρονομικό καρκίνο του παχέος εντέρου χωρίς πολυποδίαση-hnpcc (hereditary nonpolyposis colon family syndrome ). Εικόνα 12.Σχηματική παρουσίαση του συστήματος DNA MMR. Το ετεροδιμερές hmsh2-hmsh6 (hmuts) αναγνωρίζει αταίριαστες βάσεις DNA. Το ετεροδιμερές hmlh1-hpms2 (hmutl), δεσμεύεται με το hmuts και καθοδηγεί μια εξωνουκλεάση προς την απομάκρυνση των βάσεων από το πρόσφατα σχηματισμένο κλώνο DNA με την σωστή αντιστοίχιση βάσεων. Το ετεροδιμερές hmutsβ αναγνωρίζει βλάβη ενός νουκλεοτιδίου, αλλά μεγαλύτεροι βρόγχοι εισαγωγής-εξάλειψης αναγνωρίζονται από το ετεροδιμερές hmsh2-hmsh3 (hmutsβ), που ακολουθείται με δέσμευση από το hmutl. Οι ρόλοι των MLH1-PMS1 και MLH1-MLH3 στην λειτουργία του συστήματος MMR στον άνθρωπο δεν είναι απολύτως σαφείς. Όταν τα σύμπλοκα MutS δεσμεύουν το DNA, ανταλλάσσουν ADP για ATP. Για την απελευθέρωση των MMR πρωτεϊνών από το DNA, ΑΤΡ υδρολύεται προς ADP.(John P. Lynch et al,2009) 28
29 Οι πρωτεΐνες MSH2 και MSH6 ή MSH2 και MSH3 αναγνωρίζουν αταίριαστες βάσεις DNA(base base mismatches) και βρόγχοι εισαγωγής-εξάλειψης (insertion / delition loops). Τα MSH2-MSH3 ετεροδιμερή αναγνωρίζουν κυρίως insertion / delition loops. Απώλεια του MSH 6 οδηγεί σε καρκινογένεση. MMR Γονίδια Χρωμοσωμική Εντόπιση MMR partners MMR πιστότητα Σύνδρομο Lynch βλαστικές μεταλλάξεις (%) hmsh2 2p15 hmsh6, hmsh3 Πλήρης απώλεια hmsh6 2p15 hmsh2 Απλά ζεύγη βάσεων, ενιαία εισαγωγή / διαγραφή βρόχου hmsh3 5q11-q13 hmsh2 Μεγαλύτερη εισαγωγή / διαγραφή βρόχους hmlh1 3p21 hpms2 (principally), hmlh3, hpms1 Πλήρης απώλεια hpms1 2p32 hmlh1 Ασαφές 0 hpms2 7p22 hmlh1 Πλήρης απώλεια? 0 2 hmlh3 7p22 hmlh1 Ασαφές 0 Πίνακας 6. Γονίδια που εμπλέκονται στο σύστημα DNA MMR. (Lengauer Cet al, 1998). Το προφίλ MSI στο καρκίνο του παχέος εντέρου (CRC) παρέχει χρήσιμες προγνωστικές πληροφορίες για την συνολική επιβίωση των ασθενών και την ανταπόκριση στην χημειοθεραπεία που θα μπορούσαν να τροποποιήσουν τα χημειοθεραπευτικά πρωτόκολλα( Elsaleh H, et al,2000;barratt PL et al,1998 ;Barratt PL et al,2002; Ribic CMet al,2000; Elsaleh H et al,2001;chen Xet al,2002). Τα τελευταία χρόνια έχουν αναπτυχθεί μοριακοί δείκτες με προγνωστική σημασία. Μελέτες σε σποραδικά καρκινώματα παχέος εντέρου έδειξαν ότι η διατήρηση της ετεροζυγωτίας σε ένα ή περισσότερα 17p ή 18q αλληλόμορφα σε MSI- S καρκινώματα παχέος εντέρου και μεταλλάξεις στο γονίδιο type II TGF-β1 receptor σε καρκινώματα παχέος εντέρου με υψηλά επίπεδα μικροδορυφορικής αστάθειας, σχετίζονται με καλή ανταπόκριση στην χημειοθεραπεία με φθοριοουρακίλη, κυρίως για νόσο σταδίου ΙΙΙ. Πρόσφατες μελέτες έχουν δείξει επίσης ότι από την επικουρική χημειοθεραπεία με φθοριοουρακίλη επωφελήθηκαν ασθενείς με σταδίου ΙΙ ή σταδίου III, με όγκους μικροδορυφορικά σταθερούς(mss) ή όγκους που παρουσιάζουν χαμηλή συχνότητα μικροδορυφορικής αστάθειας (MSI- Low), αλλά όχι εκείνοι με καρκίνο παχέος εντέρου που παρουσιάζουν υψηλή συχνότητα μικροδορυφορικής αστάθειας (MSI-High) (Ribic CM,et al, 2003). 29
30 2.2 Χρωμοσωμική Αστάθεια (CIN, chromosomal instability) Η CIN χαρακτηρίζεται από ανευπλοειδία και αναδιατάξεις των χρωμοσωμάτων και παρατηρείται στον καρκίνο του παχέος εντέρου σε ποσοστό 80%-85% των περιπτώσεων. Περισσότερα από 100 γονίδια μπορούν να προκαλέσουν χρωμοσωμική αστάθεια στο ζυμομύκητα Saccharomyces cerevisiae. Παρά το γεγονός ότι πολλά από αυτά τα γονίδια έχουν ομόλογα και στον άνθρωπο, μόνο ένας περιορισμένος αριθμός έχει ενοχοποιηθεί σε όγκους στον άνθρωπο (πίνακας 7). Περιλαμβάνονται γονίδια που εμπλέκονται στην δομή και λειτουργία του κινητοχώρου, το σχηματισμό και την λειτουργία των μικροσωληνίσκων, την συμπύκνωση των χρωμοσωμάτων, την συνοχή των αδελφών χρωματίδων καθώς και γονίδια που ελέγχουν τον κυτταρικό κύκλο (Lengauer Cet al, 1998). Μονοπάτι Γονίδια Αλλοιώσεις των γονιδίων Διαχωρισμός των χρωμοσωμάτων BUB1? BUBR1 Σωματική μετάλλαξη hzw10? hzwilch? hrod AURKA PLK1 PTTG CENP-A? CENP-H? HEC1? INCENP APC Σωματική μετάλλαξη Γονιδιακή ενίσχυση και υπερέκφραση Υπερέκφραση του γονιδίου Υπερέκφραση του γονιδίου Υπερέκφραση του γονιδίου Σπερματικής γραμμής και σωματικές μεταλλάξεις Ρύθμιση των Τελομερών TERC Υπερέκφραση Απόκριση σε βλάβη του DNA ATM? ATR Μετάλλαξη BRCA 1/2 TP53 Mre11 Μετάλλαξη Σπερματικής γραμμής και σωματικές μεταλλάξεις Μετάλλαξη Πίνακας 7. Γονίδια που προκαλούν χρωμοσωμική αστάθεια (Lengauer Cet al, 1998). Χρωμοσωμική αστάθεια μπορεί να προκύψει από διαταραχές σε οδούς που εξασφαλίζουν τον ακριβή διαχωρισμό των χρωμοσωμάτων(πίνακας 7). Το σημείο ελέγχου της δημιουργίας της μιτωτικής ατράκτου (spindle assembly checkpoint)(εικόνα 13) αποτελεί σημαντικό μηχανισμό ελέγχου του κυτταρικού κύκλου, το οποίο εξασφαλίζει υψηλή πιστότητα στον διαχωρισμό των χρωμοσωμάτων καθυστερώντας την εκκίνηση της ανάφασης. Βλάβες σε αυτό το σημείο ελέγχου οδηγούν σε ανευπλοειδία και ανώμαλο αριθμό χρωμοσωμάτων στα θυγατρικά κύτταρα. Προϊόντα των γονιδίων MAD (mitosis arrest deficient) και BUB (budding uninhibited by benzimidazoles) λειτουργούν ως σημεία ελέγχου και μεταγωγής σήματος που ελέγχουν των διαχωρισμό των αδελφών χρωματίδων. Η ενεργοποίηση τους αναστέλλει 30
31 το σύμπλοκο APC/C(anaphase-promoting complex/c-ουβικιτίνη-πρωτεΐνη λιγάση ) και διακόπτει τον κυτταρικό κύκλο. Οι παράγοντες MAD3/BUBR1, MAD2, και BUB3 συνδέονται με το σύμπλοκο APC/C ενεργοποιούν το CDC20 δημιουργώντας ένα σύμπλοκο ελέγχου της μίτωσης και επάγουν δομικές αλλαγές στο APC/C, εμποδίζοντας τη σύνδεση και ουβικιτινίωση (Herzog F et al,2009). Η ενεργοποίηση του APC/C οδηγεί σε αποικοδόμηση την Securin και ενεργοποιεί την separases, μια κατηγορία πρωτεασών. Η Separase με τη σειρά της ρυθμίζει ένα σύμπλοκο πρωτεϊνών γνωστό ως cohesin, το οποίο δημιουργεί φυσικές συνδέσεις μεταξύ αδελφών χρωματίδων που διατηρούνται μέχρι αργά κατά την μίτωση. Λάθη σε αυτό το πολύπλοκο μηχανισμό ελέγχου προκαλεί άνισο διαχωρισμό των χρωμοσωμάτων. Εικόνα 13. Σημείο ελέγχου της μίτωσης (spindle assembly checkpoint ) Gastroenterology (2010) 138, Ένας άλλος μηχανισμός πρόκλησης χρωμοσωμικής αστάθειας (CIN), στον καρκίνο του παχέος εντέρου είναι οι διαταραχές των κεντροσωμάτων. Τα κεντροσωμάτια συντονίζουν την μίτωση αναδιοργανώνοντας τους μικροσωληνίσκους της μιτωτικής ατράκτου. Επιπλέον αριθμός κεντροσωμάτων οδηγεί στο σχηματισμό πολλαπλών ατράκτων κατά την διάρκεια της μίτωσης, με αποτέλεσμα την άνιση κατανομή των χρωμοσωμάτων. Ένα μέλος της οικογένειας AURKA (κινάση σερίνης / θρεονίνης γνωστή και ως Aurora, STK 15 / ΒΤΑΚ) ελέγχει τον κυτταρικό κύκλο ρυθμίζοντας την λειτουργία των κεντροσωματίων και τον σχηματισμό της μιτωτικής ατράκτου (Yoshifumi B et al,2009). Έρευνα σε 501 περιστατικά καρκίνου του παχέος εντέρου υποστηρίζει πιθανή συσχέτιση της κνάσεις AURKA στην εξέλιξη του καρκίνου του παχέος εντέρου μέσω μηχανισμών χρωμοσωμικής αστάθειας. Χρωμοσωμική αστάθεια μπορεί να προκύψει επίσης από βλάβη στα τελομερή που βρίσκονται σε καθένα από τα δύο άκρα ενός χρωμοσώματος. Τα τελομερή είναι DNAπρωτεϊνικά σύμπλοκα που αποτελούνται από τμήματα επαναλήψεων (TTAGGG) που προστατεύουν τα άκρα των ευκαρυωτικών χρωμοσωμάτων από την ρήξη και το σπάσιμο κατά την διάρκεια του διαχωρισμού. Σε σωματικά κύτταρα, ένα τμήμα DNA των τελομερών χάνεται μετά από κάθε κύκλο της κυτταρικής διαίρεσης, λόγω της αδυναμίας της DNA πολυμεράσης να συνθέσει πλήρως το 3'άκρο των γραμμικών χρωμοσωμάτων. Όταν ένα κρίσιμο μήκος των τελομερών χαθεί, μηχανισμοί όμοιοι με αυτούς που ενεργοποιούνται σε βλάβη του DNA ενεργοποιούν οδούς γήρανσης και κυτταρικής απόπτωσης (Maser RS et al,2002). Παρόλα αυτά εάν το p53 και Rb είναι απενεργοποιημένα τότε τα κύτταρα συνεχίζουν να διαιρούνται και εισέρχονται σε μια 31
32 φάση μαζικού κυτταρικού θανάτου (cellular crisis). Κύτταρα τα οποία καταφέρνουν να επιβιώσουν ενεργοποιούν μηχανισμούς ώστε να διατηρήσουν τα τελομερή, αυξάνοντας την έκφραση της τελομεράσης. Μελέτη σε ποντίκια με ανεπάρκεια της RNA τελομεράσης ( TERC - / - ) οδήγησε σε βράχυνση των τελομερών και αύξηση του ποσοστού δημιουργίας μικροαδενωμάτων και όγκων στο γαστρεντερικό σωλήνα. Έδειξαν επίσης ότι τα τελομερή μικραίνουν κατά το στάδιο της υψηλόβαθμης δυσπλασίας.(plentz, R et al,2003). Ομάδες ερευνητών αναφέρουν την παρουσία μικρότερων τελομερών σε ποσοστό 77% έως και 90% σε δείγματα καρκίνου του παχέος εντέρου, σε σύγκριση με τον παρακείμενο φυσιολογικό ιστό (Gertler et al,2004), καθώς και αυξημένη έκφραση της RNA πολυμεράσεις σε όγκους του παχέος εντέρου. Στα πρώτα στάδια της καρκινογένεσης η απώλεια των τελομερών οδηγεί σε CIN ενώ στα προχωρημένα στάδια της νόσου η υπερέκφραση της τελομεράσης επάγει την αθανασία των καρκινικών κυττάρων. Αυξημένο μήκος τελομερών και υπερέκφραση της τελομεράσης έχει συσχετιστεί με αυξημένο βάθος διήθησης και μετάσταση (Dukes 'C και D). Πρωτεΐνες επιδιόρθωσης του DNA επίσης εμπλέκονται στον καρκίνο στον άνθρωπο όπως ATM (ataxia telangiectasia mutated) ATR πρωτεΐνη κινάση (ataxia telangiectasia) και Rad3 που σχετίζεται με ( ATR ) πρωτεϊνική κινάση. Μεταλλάξεις σε αυτά τα γονίδια προδιαθέτουν σε συγκεκριμένα σύνδρομα, τα οποία χαρακτηρίζονται από αυξημένη προδιάθεση εμφάνισης καρκίνου, όπως η αταξία-τελαγγειεκτασία ( ATM μεταλλάξεις), το σύνδρομο Seckel ( ATR μεταλλάξεις), το σύνδρομο Li- Fraumeni ( TP53 μεταλλάξεις), και ο κληρονομικός καρκίνος μαστού-ωοθηκών ( BRCA1 και BRCA2 μεταλλάξεις). Το γονίδιο ΤΡ53 εμπλέκεται άμεσα στον καρκίνο του παχέος εντέρου στον άνθρωπο, καθώς φαίνεται να εμπλέκεται στην χρωμοσωμική αστάθεια CIN. Η απώλεια της ετεροζυγωτίας LOH (loss of heterozygosity) είναι χαρακτηριστική σε όγκους με χρωμοσωμική αστάθεια. Ένας μέσος όρος 25%-30% των αλληλίων χάνονται σε όγκους χωρίς να έχει πλήρως διευκρινιστεί ο μηχανισμός. Σε μια μελέτη απώλεια της ετεροζυγωτίας( LOH) στον καρκίνο του παχέος εντέρου, (Thiagalingam et al,2001) πραγματοποιήθηκε λεπτομερής μοριακή και κυτταρογενετική μελέτη σε 5 χρωμοσώματα (1,5, 8, 17, και 18) που χάνονται συχνότερα στον καρκίνο του παχέος εντέρου στον άνθρωπο Μοριακοί μηχανισμοί που ενεργοποιούνται από την χρωμοσωμική αστάθεια στον καρκίνο του παχέος εντέρου Αρχικό γεγονός στην καρκινογένεση του παχέος εντέρου, είναι η ενεργοποίηση του σηματοδοτικού μονοπατιού Wnt, μέσω της γενετικής διαταραχής του APC στο 5q21, η λειτουργία του οποίου είναι ογκοκατασταλτική και έχει ενοχοποιηθεί στην ανάπτυξη CIN. Σωματικές APC μεταλλάξεις παρατηρούνται σε ποσοστό 5% στις δυσπλαστικές κρύπτες, 30% -70% στα σποραδικά αδενωμάτα, και 72% στα σποραδικά καρκινώματα.(πίνακας 8) 32
33 Η φυσιολογική πρωτεΐνη APC συμμετέχει σε διάφορες κυτταρικές λειτουργίες, όπως στη ρύθμιση του κυτταρικού κύκλου, στην απόπτωση, στη κυτταρική μετανάστευση, στη κυτταρική προσκόλληση, στην οργάνωση των μικροσωληνίσκων και στην συμπύκνωση των χρωμοσωμάτων. Ωστόσο παρά το γεγονός ότι καθεμία από τις παραπάνω λειτουργίες δυνητικά σχετίζεται με την καρκινογένεση, φαίνεται ότι η ογκοκατασταλτική δράση της πρωτεΐνης APC αποδίδεται κυρίως στην ικανότητα της να ρυθμίζει τα ενδοκυττάρια επίπεδα της β-κατενίνης. Υπό φυσιολογικές συνθήκες η πρωτεΐνη APC με την βοήθεια της αξίνης και της Glycogen Synthase Kinase-3β (GSK- 3β) οδηγεί την β-κατενίνη σε αποδόμηση μέσω της οδού ουβικουιτίνηςπρωτεασώματος. Πίνακας 8: :Επικρατέστερες γενετικές μεταλλάξεις σε χρωμοσωμική αστάθεια στον καρκίνο του παχέος εντέρου Γονίδια Γενετική θέση Επίπτωση μεταλλάξεων (%) Λειτουργία Ογκογονίδια KRAS 12p Πολλαπλασιασμός, επιβίωση και μετασχηματισμός των κυττάρων. CTNNB1 3p ( 50 a ) Ρύθμιση Wnt γονιδίων που επάγουν την ανάπτυξη και εισβολή του όγκου. PIK3CA 3q26 20 Κυτταρικός πολλαπλασιασμός και επιβίωση. Ογκοκατασταλτικά γονίδια APC 5q Αναστολή Wingless/Wnt ; Ρύθμιση του κυτταροσκελετού. TP53 17p Παύση του κυτταρικού κύκλου, επαγωγή της απόπτωσης. SMAD4, SMAD2 18q Ενδοκυττάριοι μεσολαβητές TGF-β DCC 18q21 6 Κυτταρικός υποδοχέας netrin-1 Προσκόλληση κυττάρων, απόπτωση. TGF-β, transforming growth factor β.aidentified in 50% of tumors without APC mutations. (Lengauer Cet al,1998). Η πλειονότητα των μεταλλάξεων του γονιδίου APC οδηγούν σε μικρότερου μήκους πρωτεΐνη με έλλειψη των απαραίτητων περιοχών για την αποδόμηση της β-κατενίνης, με συνέπεια την αναστολή της πρωτεόλυσης και την αύξηση των ενδοκυττάριων επιπέδων της β-κατενίνης. H β-κατενίνη μετατοπίζεται στη συνέχεια στον πυρήνα όπου δεσμεύεται με τον μεταγραφικό παράγοντα TCF οδηγώντας στην ενεργοποίηση του και τη συνεχή έκφραση γονιδίων που εμπλέκονται στην ογκογένεση, όπως γονιδίων που οδηγούν σε ανεξέλεγκτο πολλαπλασιασμό (cyclin D1, c-myc), γονιδίων που εμπλέκονται στην απόπτωση (survivin, Cox-2, PPARδ) και γονιδίων που συμμετέχουν σε διαδικασίες κυτταρικής προσκόλλησης και μετανάστευσης (CD44, matrix metalloproteinase -7) (εικόνα 14). 33
34 Näthke Nature Reviews Cancer advance online publication;published 09 November 2006 Εικόνα 14.αΟι μεταλλάξεις του APC, όπως και η ενεργοποίηση του μονοπατιού Wnt αναστέλλουν την αποδόμηση της β-κατενίνης. Η β-κατενίνη στην συνέχεια εισέρχεται στον πυρήνα όπου ενεργοποιεί τον μεταγραφικό παράγοντα TCF, επάγοντας την μεταγραφή διαφόρων γονιδίων στόχων. Η β-κατενίνη αλληλεπιδρά επίσης με την Ε-κατχερίνη στις ζώνες πρόσφυσης (AJ). Το APC ρυθμίζοντας τα ελεύθερα επίπεδα της β-κατενίνης τροποποιεί την κυτταρική προσκόλληση Η πρωτεΐνη APC δεν εμπλέκεται μόνο στην ρύθμιση των ενδοκυττάριων επιπέδων της β-κατενίνης, αλλά παίζει ρόλο και σε άλλες λειτουργίες όπως η κυτταρική μετανάστευση και προσκόλληση(εικόνα 14). Συγκεκριμένα, αλληλεπιδρά με τους μικροσωληνίσκους και το σύστημα των Rac και Rho GTP-δεσμευτικών πρωτεϊνών τα οποία ως γνωστόν συμμετέχουν στα κυτταρικά γεγονότα της μετανάστευσης. Μεταλλάξεις δε του γονιδίου APC έχουν συσχετιστεί με διαταραχή της μετανάστευσης των επιθηλιακών κυττάρων κατά μήκους του άξονα των κρυπτών. Καθώς επίσης η β- κατενίνη συνδέεται με την Ε-κατχερίνη και αποτελεί σημαντικό συστατικό των ζωνών πρόσφυσης, έχει διατυπωθεί η άποψη ότι το APC ρυθμίζοντας τα ελεύθερα επίπεδα της β-κατενίνης τροποποιεί την κυτταρική προσκόλληση. Εικόνα 15.Σχηματική αναπαράσταση του πώς η απώλεια του APC και η μετέπειτα τροποποιήσεις του κυτταροσκελετού επηρεάζουν, σημαντικές διαδικασίες που ελέγχουν την ομοιόσταση-αναγέννηση του επιθηλίου του εντέρου. Näthke Nature Reviews Cancer advance online publication; published online 09 November
35 Το μονοπάτι αυτό γενικά παίζει σημαντικό ρόλο στην ανάπτυξη των σπονδυλωτών ενώ ειδικότερα εμπλέκεται και στην εμβρυογένεση και αναγέννηση του παχέος εντέρου. Το γεγονός ότι στην παθογένεια του καρκίνου του παχέος εντέρου εμπλέκονται μεταλλάξεις γονιδίων ρυθμιστών μίας οδού που παίζει ρόλο στην εμβρυογένεση και την ρύθμιση του πολλαπλασιασμού των «ενήλικων» (adult) αρχέγονων κυττάρων παρέχει ενδείξεις υπέρ των υποθέσεων ότι 1) μηχανισμοί που λειτουργούν κατά την εμβρυογένεση ενεργοποιούνται στην καρκινογένεση και 2) ο καρκίνος προέρχεται από τα αρχέγονα κύτταρα. Στην «κλασσική» αλληλουχία αδενώματος-καρκινώματος η απώλεια του APC(εικόνα 15), του καθοριστικού δηλαδή εναρκτήριου γεγονότος, ακολουθείται από μεταλλάξεις σε άλλα πρωτοογκογονίδια, όπως του K-ras και ογκοκατασταλτικά γονίδια, όπως του p53 και του DCC (Deleted in Colon Cancer), καθώς και από απώλεια διαφόρων άλλων χρωμοσωμικών τόπων τα γονίδια στόχοι των οποίων δεν είναι ακόμα γνωστά. Οι όγκοι που αναπτύσσονται μέσω αυτής της γενετικής οδού εμφανίζουν ανευπλοειδία, απώλεια ετεροζυγωτίας (LOH) και γενικά μεγάλες χρωμοσωμικές ανωμαλίες, χαρακτηρίζονται δηλαδή από χρωμοσωμική αστάθεια. Το K-ras ανήκει στην οικογένεια των RAS (K-ras, H-ras, N-ras) πρωτο-ογκογονιδίων, και δρα ως μοριακός διακόπτης on / off και κωδικοποιεί μικρές GTP-δεσμευτικές πρωτεΐνες, 21kDa οι οποίες δρουν ως μοριακοί διακόπτες που συνδέουν εξωκυττάρια σήματα με πυρηνικούς παράγοντες μεταγραφής. Το K-ras είναι μεταλλαγμένο σε ποσοστό 30% -50% στο καρκίνο του παχέος εντέρου στον άνθρωπο. Σημειακές μεταλλάξεις σε νουκλεοτίδια στα κωδικόνια 12 και 13 του εξονίου 2, και σε μικρότερο βαθμό στο κωδικόνιο 61 στο εξονίου 3, κλειδώνουν το Ras σε μια λειτουργική κατάσταση συνεχούς ενεργοποίησης. KRAS μεταλλάξεις παρατηρήθηκαν στο 60%-95% σε μη- δυσπλαστικές ή υπερπλαστικές κρύπτες, υποδεικνύοντας ότι πιθανών να αποτελούν πρόδρομες αλλοιώσεις των αδενωμάτων και του καρκίνου( Takayama, Tet al,2001;pretlow et al,2005). Το ενεργοποιημένο Ras ρυθμίζει πολλαπλές κυτταρικές λειτουργίες, όπως ο κυτταρικός κύκλος, η απόπτωση, η ανοσολογική και φλεγμονώδη απάντηση των κυττάρων. Μεταλλάξεις σε σηματοδοτικά μονοπάτια που ελέγχονται απο το Ras(εικόνα 16) έχουν συσχετιστεί με τον καρκίνο του παχέος εντέρου στον άνθρωπο πχ (Raf mitogenactivated protein kinase kinase (MEK) extracellular signal-regulated kinase (ERK, PIK3CA μεταλλάσσεται σε ποσοστό 20% στον CRCs, (Maria S. Pino et al,2010). 35
36 Εικόνα 16.Σηματοδοτικά μονοπάτια Ras.Gastroenterology , DOI: ( /j.gastro Το γονίδιο ΤΡ53 βρίσκεται στο βραχύ σκέλος του χρωμοσώματος 17 κωδικοποιεί έναν ογκοκατασταλτικό μεταγραφικό παράγοντα, 393 αμινοξέων, ο οποίος συντονίζει την απόκριση των κυττάρων στο στρες, συμπεριλαμβανομένης της βλάβης του DNA, το οξειδωτικό στρες και τα πολλαπλασιαστικά σήματα. Το ΤΡ53 επάγεται από διάφορες ογκογόνες πρωτεΐνες, όπως το c-myc, RAS, και τους αδενοϊούς Ε1Α και είναι κύριος ρυθμιστής της μεταγραφή εκατοντάδων γονιδίων που εμπλέκονται στο μεταβολισμό του DNA, την απόπτωση, τη ρύθμιση του κυτταρικού κύκλου, το γήρας, την αγγειογένεση, την ανοσοαπόκριση, την κυτταρική διαφοροποίηση, κινητικότητα και μετανάστευση. Δυσλειτουργία P53 είναι καθοριστική στην ανάπτυξη όγκων στον άνθρωπο, έχει αναφερθεί σε ποσοστό 4% -26% στα αδενώματα, και 50% -75% στον καρκίνο του παχέος εντέρου στον άνθρωπο. LOH (loss of heterozygosity) Στο 75% των CRCs έχουμε απώλεια ενός τμήματος του χρωμοσώματος 17ρ ή 18q, ή και των δύο. Το τμήμα 17ρ περιλαμβάνει το ρ53, το οποίο θεωρείται ως ένα από τα πιο σημαντικά ογκοκατασταλτικά γονίδια με μια σειρά βασικών λειτουργιών όπως η επιδιόρθωση του DNA, ο έλεγχος του κυτταρικού κύκλου και η έναρξη της απόπτωσης. Μεταλλάξεις στο p53 έχουν παρατηρηθεί σε ποσοστό 40-60% περιπτώσεων με καρκίνο παχέος εντέρου. Αλληλική απώλεια στο χρωμόσωμα 18q βρίσκεται περίπου στο 70% των CRCs και έχει συσχετιστεί με χειρότερη πρόγνωση σε στάδιο ΙΙ και ΙΙΙ. Ασθενείς σταδίου ΙΙ με 18qLOH έχουν παρόμοια κλινική πορεία με ασθενείς σταδίου ΙΙΙ, ενώ όσοι δεν παρουσιάζουν 18qLOH συμπεριφέρονται κλινικά όπως οι ασθενείς σταδίου Ι. Το τμήμα 18q περιέχει τρία ογκοκατασταλτικά γονίδια: DCC, Smad 2 και Smad 4. Το προϊόν του γονιδίου DCC είναι ένα μόριο προσκόλλησης των κυττάρων, μειωμένη έκφραση DCC μπορεί να συμβάλει στην ανάπτυξη του όγκου και την μεταστατική εξάπλωση του καρκίνου του παχέος εντέρου. Οι πρωτεΐνες Smad είναι παράγοντες μεταγραφής που εμπλέκονται στο μονοπάτι TGFβ. Αυτές οι πρωτεΐνες εμπλέκονται στην σηματοδότηση μέσω TGFβ υποδοχέα και ρυθμίζουν τη μεταγραφή γονιδίων στόχων, c-myc, CBFA1, FLRF και furin. Η μη έκφραση του γονιδίου DCC και η 18qLOH έχουν συσχετιστεί, επίσης, με την ανάπτυξη ηπατικών μεταστάσεων(kato M,et al,1993;d B Longley et al,2002) 36
37 2.3 Ο ρόλος του μικροπεριβάλοντος Πέρα από γενετικές αλλοιώσεις που παρουσιάζουν τα καρκινικά κύτταρα του παχέος εντέρου, μια δυναμική αλληλεπίδραση λαμβάνει χώρα μεταξύ των κυττάρων του καρκίνου και του μικροπεριβάλοντος επάγοντας την ανάπτυξη και εξέλιξη του καρκίνου.(tse JC, et al, 2007) Το στρώμα αποτελείται κυρίως απο ινοβλάστες και εξωκυττάρια θεμέλια ουσία (ECM). Η σημασία του στρώματος στον καρκίνο τονίζεται από ιστολογικές παρατηρήσεις που δείχνουν ότι ορισμένοι τύποι νεοπλασμάτων, όπως ο καρκίνος του παγκρέατος αποτελείται κυρίως από στρωματικά κύτταρα. Η συνομιλία μεταξύ καρκινικών κυττάρων και μικροπεριβάλλοντος (εικόνα 17) έχει δειχθεί ότι παίζει σημαντικό ρόλο στην ανάπτυξη και εξέλιξη του καρκίνου. (Mareel M et al,2003) Εικόνα 17.Ο ρόλος του στρώματος στην επαγωγή της καρκινογένεσης και μεταστάσεις.( Lance A.et al,2001) Το στρώμα του όγκου παίζει σημαντικό ρόλο στην ανάπτυξη του καρκίνου του παχέος εντέρου και αναφέρεται ως reactive stroma (Kalluri R, Zeisberg et al,2006). Παρατηρείται αύξηση του αριθμού των ινοβλαστών, αύξηση της πυκνότητας των τριχοειδών και εναπόθεση κολλαγόνου τύπου-1 και ινώδους. Τα μακροφάγα επίσης μεταναστεύουν στο αντιδραστικό στρώμα σε απόκριση της διαδικασίας επούλωσης που επάγονται από τον όγκο. Επιπλέον, αρκετές μελέτες δείχνουν ότι το αντιδραστικό στρώμα επηρεάζει την έναρξη και επαγωγή του καρκίνου μέσο της έκκρισης κυτοκινών, αυξητικών παραγόντων ή με την έκφραση των υποδοχέων τους. Για παράδειγμα, στρωματικά κύτταρα εντός του καρκίνου του παχέος εντέρου εκφράζουν υψηλά επίπεδά του υποδοχέα PDGF (PDGFR) επιδρώντας στην εξέλιξη του καρκίνου του παχέος εντέρου καθώς η αναστολή PDGFR αναστέλλει την εξέλιξη του καρκίνου. ( Kitadai Y,et al,2006). 37
38 Ινοβλάστες εντός του όγκου φέρουν μεταλλάξεις οι οποίες τους μετατρέπουν σε μυοινοβλάστες ή σε CAFs (cancer associated fibroblast). Αυτά τα κύτταρα έχουν την δυνατότητα να επάγουν τον σχηματισμό όγκου παράγοντας κυτταροκίνες όπως ο TGF-β ο οποίος ενεργοποιεί τα καρκινικά κύτταρα και προκαλεί την αποκόλληση τους από το πρωτογενές νεόπλασμα (Olumi AFet al,1999). Τα μακροφάγα είναι ένα άλλο σημαντικό είδος κυττάρων στα πρωτογενή νεοπλάσματα. Αυτά τα κύτταρα που ονομάζονται ΤΑΜs (tumor-associated macrophages) μπορεί να αποτελούν ένα μεγάλο ποσοστό της μάζας του όγκου (Pollard JW et al,2004). Η παρουσία των ΤΑΜs στους όγκους σχετίζεται με την φλεγμονώδη απάντηση στο καρκίνο και εξηγεί γιατί μερικοί καρκίνοι ανταποκρίνονται σε αντιφλεγμονώδεις παράγοντες (BalkwillFet al,2005;bingle Let al,2002). Η παρουσία των μακροφάγων ΤΑΜs εντός του όγκου συσχετίζεται επίσης με κακή πρόγνωση. Έχει προταθεί ένα μοντέλο σύμφωνα με το οποίο τα μακροφάγα επάγουν την καρκινογένεση και την μετάσταση. Διεργασίες που επάγονται από τα ΤΑΜs είναι η χρόνια φλεγμονή, η αναδιαμόρφωση της θεμέλιας ουσίας, η διήθηση, η αγγειογένεση και η μετάσταση σε απομακρυσμένες περιοχές (Condeelis J et al, 2006). Στην περίπτωση του καρκίνου του παχέος εντέρου, φαίνεται ότι τα κύτταρα ΤΑΜ επάγουν την μετάσταση μέσω της έκκρισης VEGF, προάγοντας την αγγειογένεση στην πρωτοπαθή θέση ( Barbera-Guillem Eet al, 2002). Κατά τη διάρκεια της διαδικασίας, ο φαινότυπος αυτών των κυττάρων (ΤΑΜ) είναι τροποποιημένος, καθώς συνδέεται με αυξημένη έκφραση του TGF-β1 και την έκκριση της IL-6. Αυτή η φαινοτυπική τροποποίηση μπορεί υποθετικά να συμβάλει στην περαιτέρω ενεργοποίηση της αγγειογενετικής διαδικασίας ( Popovic et al,2007) Οι όγκοι αναπτύσσουν επίσης μηχανισμούς που επιτρέπουν την διαφυγή από την άνοσο επιτήρηση. Στον καρκίνο του παχέος εντέρου έχει παρατηρηθεί αυξημένη παραγωγή του Fas lingand η σύνδεση του οποίου με κύτταρα του ανοσοποιητικού επάγει την απόπτωση και την διαφυγή από την ανοσοεπιτήρηση. Ένας άλλος μηχανισμός που αναπτύσσεται από τα καρκινικά κύτταρα είναι η αύξηση της έκφρασης υδατανθράκων στην επιφάνεια τους, εμποδίζοντας έτσι την αναγνώριση τους από κύτταρα του ανοσοποιητικού (Lu B, et al,2008). 38
39 Εικόνα 18. Ο διαχωρισμός των κυττάρων απο την πρωτοπαθή εστία και η μετάσταση αιματογενώς ή λεμφογενώς είναι μια πολυσταδιακή διαδικασία που περιλαμβάνει αλλαγές στην έκφραση πολλαπλών γονιδίων, τα οποία εμπλέκονται στην κυτταρική προσκόλληση, την επιβίωση, την χημειοταξία, αυξητικοί παράγοντες, mirnas. Στην εικόνα μπορείτε να δείτε μερικά από τα γονίδια που εμπλέκονται σε όλη αυτή τη διαδικασία(.m.e. Hern ndez- Caballe et al,2013 ) ΚΕΦΑΛΑΙΟ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ 3.1Μακροσκοπικά Χαρακτηριστικά Τα μακροσκοπικά χαρακτηριστικά των καρκινωμάτων του παχέος εντέρου ποικίλουν σημαντικά. Ο όγκος μπορεί να είναι: εξωφυτικός-πολυποδοειδής με ενδοαυλική κυρίως αύξηση, ενδοφυτικός-ελκωτικός με κυρίαρχη την ενδοτοιχωματική αύξηση, διάχυτα διηθητικός ή δακτυλιοειδής με κυκλοτερή αύξηση και απόφραξη του αυλού. Η αλληλοεπικάλυψη μεταξύ αυτών των τύπων είναι αρκετά συχνή. Τα καρκινώματα που αναπτύσσονται στο εγγύς τμήμα του παχέος εντέρου είναι συνήθως εξωφυτικά, ενώ εκείνα που αναπτύσσονται στο εγκάρσιο ή το κατιόν κόλον είναι συνήθως δακτυλιοειδή. Τα βλεννώδη αδενοκαρκινώματα εμφανίζουν συχνά περιοχές με ορατή βλέννα και τα καρκινώματα με φαινότυπο MSI-H είναι καλά περιγεγραμμένα και στο 20% των περιπτώσεων βλεννώδη (Troussard AAet al, 1999). 3.2 Ιστοπαθολογικά Χαρακτηριστικά Καθοριστικό χαρακτηριστικό του αδενοκαρκινώματος του παχέος εντέρου είναι ότι εισβάλει μέσω της υποβλεννογόνιας μυϊκής στιβάδας. Βλάβες με μορφολογικά χαρακτηριστικά αδενοκαρκινώματος που διηθούν το επιθήλιο ή διηθούν το χόριο αλλά δεν διασπούν την υποβλεννογόνιο μυϊκή στιβάδα δεν συνοδεύονται από αυξημένο κίνδυνο μεταστάσεων. Τα περισσότερα αδενοκαρκινώματα σχηματίζουν αδενικούς σχηματισμούς, οι οποίοι εμφανίζουν ποικιλότητα ως προς το μέγεθος και την διαμόρφωση των αδενικών μορφών. Σε καλά και μέτρια διαφοροποιημένο αδενοκαρκινώματα τα επιθηλιακά 39
40 κύτταρα είναι συνήθως μεγάλα και ψηλά και ο αδενικός αυλός συχνά περιέχει κυτταρικά υπολείμματα. Βλεννώδες αδενοκαρκίνωμα. Βλεννώδες αδενοκαρκίνωμα χαρακτηρίζεται ένας όγκος όταν σε ποσοστό > 50% της βλάβης αποτελείται από βλέννη. Πρόκειται για αλλοιώσεις που περιέχουν εξωκυττάρια βλέννη και κακοήθη επιθηλιακά κύτταρα, τα οποία είναι μονήρη ή σχηματίζουν σειρές ή κυψελοειδείς δομές. Οι περισσότεροι MSI- H όγκοι είναι αυτού του τύπου. Καρκίνωμα με κύτταρα Signet-ring. Χαρακτηρίζονται από την παρουσία σε ποσοστό >50% καρκινικών κυττάρων με άφθονη κυτταροπλασματική βλέννη. Τα τυπικά signet-ring κύτταρα έχουν ένα μεγάλο κυστίδιο βλέννης που γεμίζει το κυτταρόπλασμα και παρεκτοπίζει τον πυρήνα. Αυτά τα κύτταρα μπορεί να εντοπίζονται στις περιοχές της βλέννης ενός βλεννώδους αδενοκαρκινώματος ή να σχηματίζουν ομάδες που διάχυτα διηθούν το τοίχωμα με την παρουσία ελάχιστης ή καθόλου εξωκυττάριας βλέννης. Adenosquamous carcinoma (αδενοπλακώδες) Παρουσιάζουν χαρακτηριστικά τόσο πλακώδους καρκινώματος όσο και αδενοκαρκινώματος. Για να χαρακτηριστεί ένα καρκίνωμα αδενοπλακώδες θα πρέπει να εμφανίζει αρκετές εστίες πλακώδους διαφοροποίησης. Αδιαφοροποίητο καρκίνωμα. Δεν εμφανίζει καμία μορφολογική ένδειξη διαφοροποίησης πέραν αυτής ενός επιθηλιακού όγκου και έχει διάφορα ιστολογικά χαρακτηριστικά. Παρά την έλλειψη διαφοροποίησης, αυτοί οι όγκοι είναι γενετικά διακριτοί και τυπικά σχετίζονται από MSI-H. Υπάρχουν και άλλοι σπανιότεροι ιστολογικοί τύποι (μικροκυτταρικό, καρκινοσάρκωμα, γιγαντοκυτταρικό, sprindle cell, clear cell, Paneth cell-rich, stem cell, pigmented). Μedullary carcinoma ( μυελώδες καρκίνωμα) είναι ένα σπάνιος ιστολογικός τύπος που χαρακτηρίζεται από την παρουσία κακοήθων κυττάρων με κενοτοπιώδη πυρήνα, έντονο πυρηνίσκο και άφθονο ηωσινόφιλο κυτταρόπλασμα καθώς και από την παρουσία έντονης ενδοεπιθηλιακής διήθησης από λεμφοκύτταρα. Σχετίζεται με φαινότυπο MSI-H και έχει καλύτερη πρόγνωση συγκριτικά με τα κακώς διαφοροποιημένα ή αδιαφοροποίητα αδενοκαρκινώματα. 3.3Διαφοροποίηση (Grading) Η διαβάθμιση των καρκινωμάτων βασίζεται στον βαθμό σχηματισμού αδενικών δομών. Διακρίνονται σε καλώς, μετρίως και κακώς διαφοροποιημένα ή σε χαμηλού βαθμού κακοήθειας (καλώς και μετρίως διαφοροποιημένα) και υψηλού βαθμού κακοήθειας (κακώς διαφοροποιημένα και αδιαφοροποίητα) καρκινώματα. Όταν ένα καρκίνωμα εμφανίζει ετερογένεια ως προς την διαφοροποίηση του η διαβάθμιση του πρέπει να βασίζεται στο λιγότερο καλά διαφοροποιημένο στοιχείο του, πλην της περιοχής του «μετώπου» διήθησης. Το «μέτωπο» διήθησης του όγκου συχνά εμφανίζει μικρές εστίες κακής διαφοροποίησης χωρίς αυτό το στοιχείο να δικαιολογεί την ταξινόμηση του όγκου ως κακώς διαφοροποιημένου. 40
41 Για τον καθορισμό του βαθμού κακοήθειας μπορεί να χρησιμοποιηθεί το ποσοστό του όγκου που εμφανίζει αδενικές δομές. Καλώς διαφοροποιημένα (grade I) καρκινώματα εμφανίζουν αδένια σε >95% του όγκου, μετρίως διαφοροποιημένα (grade II) σε 50-95% του όγκου, κακώς διαφοροποιημένα (grade III) σε 5-50% και τα αδιαφοροποίητα καρκινώματα (grade IV) σε <5% του όγκου. Τα βλεννώδη και τα signet-ring cell καρκινώματα θεωρούνται κακώς διαφοροποιημένα ενώ τα μυελώδη με MSI-H φαίνονται αδιαφοροποίητα (εικόνα 19) (Hermiston ML et al,1995;preston SLet al,2003;powel DW,et al,1999; Van de Wetering M,et al,2002;bach SP et al, 2000;Hamilton SR et al,2002;kumar Vet al,1997; Schwartz SIet al,1999;thun MJet al,2002;alexander Net al,2002 Potter JDet al,1999; Bright-Thomas RMet al,2003;oving IMet,al,2002; Goel A, et al,2001) Α Β C D Εικόνα 19. A Καλά διαφοροποιημένο αδενοκαρκίνωμα. B Μέσης διαφοροποίησης αδενοκαρκίνωμα. C Φτωχής διαφοροποίησης αδενοκαρκίνωμα. Αυτή η αλλοίωση είναι MSI-H και δείχνει ένα αριθμό από ενδοεπιθηλιακά λεμφοκύτταρα. 3.4Σταδιοποίηση Το στάδιο περιγράφει την έκταση του καρκίνου στο σώμα. Βασίζεται στον βαθμό διήθησης του τοιχώματος του εντέρου, στην παρουσία ή όχι τοπικής επέκτασης σε γειτονικά όργανα και επιχώριους λεμφαδένες και στην παρουσία ή όχι απομακρυσμένων μεταστάσεων. Το στάδιο είναι ένας από τους σημαντικότερους παράγοντες για τον καθορισμό της πρόγνωσης των ασθενών και των θεραπευτικών επιλογών. Η σταδιοποίηση βασίζεται στα αποτελέσματα της φυσικής εξέτασης, στις βιοψίες, και στις απεικονιστικές εξετάσεις (αξονική ή μαγνητική τομογραφία, ακτινογραφίες, ΡΕΤ scan, κ.λπ.). Το συνηθέστερα χρησιμοποιούμενο σύστημα σταδιοποίησης για τον καρκίνο του παχέος εντέρου είναι η σταδιοποίηση κατά AJCC/TNM. Τα παλαιότερα συστήματα σταδιοποίησης του καρκίνου του παχέος εντέρου είναι η σταδιοποίηση κατά Dukes και Astler - Coller. Το σύστημα TNM περιγράφει 3 βασικές πληροφορίες (εικόνα 20) (Cancer Staging Manual, Seventh Edition AJCC 2010).(πίνακας 10) 41
42 Τ περιγράφει την επέκταση του όγκου στο τοίχωμα του εντέρου ή/και σε γειτονικές δομές. Tx. Ο πρωτοπαθής όγκος δεν μπορεί να εκτιμηθεί. T0. Δεν υπάρχει ένδειξη ύπαρξης πρωτοπαθούς εστίας όγκου. Tis. Καρκίνωμα in situ (ενδοεπιθηλιακό ή ενδοβλεννογόνιο καρκίνωμα). Τ1 Ο όγκος διηθεί τον υποβλεννογόνιο χιτώνα. Τ2 Ο όγκος διηθεί αλλά δεν διαπερνά τον μυϊκό χιτώνα. Τ3 Ο όγκος διηθεί τον υπορογόνο ή περικολικούς ή περιορθρικούς ιστούς. Τ4a.Ο Όγκος διεισδύει στην επιφάνεια του σπλαχνικού περιτοναίου. T4b. Ο όγκος διηθεί ή συμφύεται στερεά με γύρω ιστούς και όργανα. Περιοχικοί λεμφαδένες (Ν) N περιγράφει την έκταση της νόσου σε επιχώριους λεμφαδένες. Νx. Δεν μπορούν να αξιολογηθούν Νο:Δεν υπάρχει μετάσταση στους επιχώριους λεμφαδένες N1 Μετάσταση σε 1-3 τοπικούς λεμφαδένες. N1a Μετάσταση σε ένα περιφερικό λεμφαδένα. N1b Μετάσταση σε 2-3 περιφερικούς λεμφαδένες. N1c Εστίες καρκινικών κυττάρων στον υπορογόνιο, μεσόκολο ή στο περικολικό λίπος των εξωπεριτοναϊκών τμημάτων του παχέος εντέρου και του ορθού. N2 Μετάσταση σε 4 ή περισσότερες περιφερειακούς λεμφαδένες. Ν2a Μετάσταση σε 4-6 τοπικούς λεμφαδένες. N2b Μετάσταση σε 7 ή περισσότερους περιφερειακούς λεμφαδένες. Μ υποδεικνύει εάν ο καρκίνος έχει εξαπλωθεί (μετάσταση) σε άλλα όργανα του σώματος. Μ0 Απουσία απομακρυσμένων μεταστάσεων. Μ1 Απομακρυσμένες μεταστάσεις. Μ1a Μεταστατική νόσος σε ένα όργανο ή μία περιοχή (ήπαρ, πνεύμονες, ωοθήκες, διήθηση απομακρυσμένων λεμφαδένων). 42
43 M1b Μεταστατική νόσος σε περισσότερα από ένα όργανα ή μία περιοχή, ή περιτοναϊκές εμφυτεύσεις. Ο καρκίνος του παχέος εντέρου μπορεί να εξαπλωθεί σχεδόν οπουδήποτε στο σώμα, αλλά τα πιο κοινά σημεία μεταστάσεις είναι το ήπαρ και οι πνεύμονες. Εικόνα 20.Σταδιοποίηση και πρόγνωση για τους ασθενείς με καρκίνο του παχέος εντέρου. Πίνακας 9. Σχέση TNM σταδίου και επιβίωσης στον καρκίνο του παχέος εντέρου Στάδιο TNM 5ετής επιβίωση (%) Stage O (T1s,T1; No; Mo) >90 Stage I (T2; No; Mo) Stage II (T3,T4; No; Mo) Stage III (T2; N1,N2; Mo) Stage III (T3; N1,N2; Mo) Stage III (T4; N1,N2; Mo) Stage IV (M1) <3 43
44 Πίνακας 10. Ανατομική σταδιοποίηση κατά ΤΝΜ Σταδιοποίηση ΤΝΜ Στάδιο Τ Ν M 0 Tis N0 M0 Ι T1 N0 M0 T2 N0 M0 ΙΙΑ T3 N0 M0 ΙΙΒ T4a N0 M0 ΙΙC T4b N0 M0 IIIA T1 T2 N1/N1c M0 T1 N2a M0 IIIB T3 T4a N1/N1c M0 T2 T3 N2a M0 T1 T2 N2b M0 IIIC T4a N2a M0 T3 T4a N2b M0 T4b N1 N2 M0 IVA Κάθε Τ Κάθε Ν Μ1a IVB Κάθε Τ Κάθε Ν Μ1b 44
45 3.5 ΠΡΟΓΝΩΣΤΙΚΟΙ ΠΑΡΑΓΟΝΤΕΣ Η πρόγνωση των ασθενών με καρκίνο του παχέος εντέρου σχετίζεται με : Το βάθος της διείσδυσης του όγκου μέσω του τοιχώματος του εντέρου ( Τ ). Την παρουσία ή απουσία της συμμετοχής επιχώριων λεμφαδένων( Ν). Την παρουσία ή απουσία απομακρυσμένων μεταστάσεων ( Μ). Αγγειακή διασπορά ( V) Αυτά τα τρία χαρακτηριστικά αποτελούν τη βάση για όλα τα συστήματα σταδιοποίησης, καθορίζοντας την πρόγνωση και θεραπεία της νόσου. Προγνωστικοί δείκτες με μικρότερη ισχύ είναι η διαφοροποίηση του όγκου, η απόσταση του από το περιφερικό εγχειρητικό όριο, η επέκταση του υπολειπόμενου καρκινώματος μετά από θεραπεία (yptnm), ο ιστολογικός τύπος, ο τύπος της διηθητικής παρυφής του νεοπλάσματος και οι ιστολογικές αλλοιώσεις που υποδηλώνουν υψηλή μικροδορυφορική αστάθεια (MSΙ-Η). Η τελευταία συσχετίζεται με βελτίωση της επιβίωσης ανά συγκεκριμένο στάδιο της νόσου (Hamilton SR et al, 2010). Εντερική απόφραξη και διάτρηση του εντέρου είναι δείκτες κακής πρόγνωσης. Αυξημένα επίπεδα στον ορό του καρκινοεμβρυϊκού αντιγόνου (CEA) έχουν αρνητική προγνωστική σημασία. (Hamilton SR et al,2010) Νέοι μοριακοί δείκτες όπως η παρουσία ή όχι μικροδορυφορικής αστάθειας και η παρουσία ή όχι μεταλλάξεων στο γονίδιο Κ-RAS (Πίνακας 11) έχουν όχι μόνο προγνωστική σημασία αλλά στοχεύουν και στην πρόβλεψη με μεγαλύτερη ακρίβεια της ανταπόκριση της νόσου στην θεραπεία (Hamilton SR et al,2010). Mετάλλαξη για παράδειγμα στο KRAS αποτελεί δείκτη κακής ανταπόκρισης στην θεραπεία με αντισώματα έναντι του EGFR. 45
46 Πίνακας 11. Προγνωστικοί μοριακοί δείκτες καρκίνου του παχέος εντέρου. D B Longley, U McDermott and P G Johnston The Pharmacogenomics Journal (2002) 3.6 ΘΕΡΑΠΕΙΑ Η θεραπεία του καρκίνου του παχέος εντέρου εξαρτάται από το στάδιο της νόσου. Για τον πρώιμο καρκίνο του παχέος εντέρου η θεραπεία εκλογής είναι η χειρουργική επέμβαση. Περίπου το 95% του σταδίου Ι και 65-80% των καρκίνων του παχέος εντέρου σταδίου II είναι ιάσιμοι με χειρουργική εξαίρεση. Στο στάδιο ΙΙ μπορεί να χρειαστεί συμπληρωματική χημειοθεραπεία βάση μικροσκοπικών και μακροσκοπικών χαρακτηριστικών του όγκου κατά το χειρουργείο. Καρκίνος του ορθού, ωστόσο, μπορεί να απαιτήσει επιπλέον θεραπεία με ακτινοβολία για να ελαχιστοποιηθεί ο κίνδυνος υποτροπής. Σε προχωρημένο στάδιο (III και IV Stage) απαιτείται συνήθως ένας συνδυασμός θεραπειών, όπως: χειρουργική επέμβαση, χημειοθεραπεία και ακτινοβολία. Στο στάδιο ΙΙΙ (θετικοί επιχώριοι λεμφαδένες) απαιτείται χειρουργική εξαίρεση και συμπληρωματική χημειοθεραπεία FOLFOX (5- FU, leucovorin, και oxaliplatin) ή CapeOx (capecitabine and oxaliplatin), ενώ μπορεί να γίνει και συμπληρωματική ακτινοθεραπεία. Στο στάδιο IV ο καρκίνος έχει επεκταθεί σε γειτονικά και απομακρυσμένα όργανα όπως το ήπαρ, αλλά μπορεί επίσης να εξαπλωθεί και σε άλλα μέρη όπως οι πνεύμονες, το περιτόναιο, ή απομακρυσμένοι λεμφαδένες. Οι περισσότεροι ασθενείς με καρκίνο σταδίου IV θα αντιμετωπιστούν με χημειοθεραπεία ή / και στοχευμένες θεραπείες(εικόνα 21). Σε προχωρημένη νόσο ακτινοθεραπεία μπορεί επίσης να χρησιμοποιηθεί για την πρόληψη ή ανακούφιση από τα συμπτώματα όπως ο πόνος. 46
47 Εικόνα 21. Μοριακοί στόχοι χημειοθεραπευτικών παραγόντων, δυνητικά προγνωστικοί δείκτες. TS, thymidylate synthase; TP, thymidine phosphorylase; DPD, dihydropyrimidine dehydrogenase; 5FU, 5-fluorouracil; FdUMP, fluorodeoxyuridine monophosphate; FUMP, fluorouridine monophosphate; dtmp, deoxythymidine monophosphate; LV, leucovorin; CH2THF, 5,10-methylene tetrahydrofolate; RTX, raltitrexed; MTA, multitargeted antifolate; DHFR, dihydrofolate reductase; GARFT, glycinamide ribonucleotide formyl transferase; ERCC1, excision repair cross complementing 1 gene product; EGFR, epidermal growth factor receptor; Topo I, DNA topoisomerase I; and FTI, farnesyl transferase inhibitor. D B Longley, U McDermott and P G Johnston The Pharmacogenomics Journal (2002) Η στοχευμένη θεραπεία η οποία στοχεύει ειδικά γονίδια, πρωτεΐνες, ή το περιβάλλον του ιστού που συμβάλλει στην ανάπτυξη του καρκίνου (εικόνα 21), χρησιμοποιείται μαζί με τη χημειοθεραπεία και άλλες θεραπείες για να εμποδίσει την ανάπτυξη και την εξάπλωση των καρκινικών κυττάρων. Μια σειρά από στοχευμένες θεραπείες έχουν εγκριθεί από τον Αμερικανικό Οργανισμό Τροφίμων και Φαρμάκων (FDA) για τη θεραπεία πολλών τύπων καρκίνου. Στον καρκίνο του παχέος εντέρου έχουν αναπτυχθεί δυο τύποι στοχευμένης θεραπείας: 1) Μονοκλωνικά αντισώματα όπως cetuximab (Erbitux), panitumumab (Vectibix) που αναστέλλουν τον EGFR. Ωστόσο, περίπου 4 στους 10 καρκίνους του παχέος εντέρου έχουν μεταλλάξεις στο γονίδιο K-RAS, που καθιστούν αυτές της θεραπείες αναποτελεσματικές και 2) αντι-αγγειογενετικοί παράγοντες όπως το αντίσωμα Bevacizumab που δεσμεύεται στον VEGF και αναστέλλουν την αγγειογέννεση. 47
48 3.7 ΠΑΡΑΚΟΛΟΥΘΗΣΗ Οι οδηγίες παρακολούθησης των ασθενών με καρκίνο του παχέος εντέρου αναφέρονται στον Πίνακας 12. Parameter Ιστορικό και φυσική εξέταση. CEA CT θώρακος ESMO [JSMO](2013) Κάθε 3-6 μήνες για 3χρόνια,και μετά 6-12 μήνες στα 4 και 5 έτη. Κάθε 3-6 μήνες για 3χρόνια,και μετά 6-12 μήνες στα 4 και 5 έτη. Κάθε 6-12 μήνες για τα πρώτα 3 έτη. ASCO (2013) NCCN (2013) Κάθε 3-6μήνες για 3 έτη, μετά κάθε 6 μήνες με 5 έτη Κάθε 3 μήνες με 3 έτη Κάθε1 έτος για 3 έτη. Κάθε 3-6 μήνες για 2 έτη, μετά κάθε 6 μήνες με 5 έτη. Κάθε 3-6 μήνες για 2 έτη, μετά κάθε 6 μήνες με 5 έτη. Κάθε 1 έτος για 5 έτη. Κολονοσκόπηση CT κοιλιάς Το πρώτο 1έτος μετά το χειρ, και κάθε 3-5 έτη μετά. Το 1 έτος, μετά κάθε 5έτη, καθορίζεται απο τα ευρήματα της προηγούμενης κολονοσκόπησης. Κάθε 1 έτος για 3 έτη. Στο1 έτος, 3 έτος, και 5 έτος εάν είναι αρνητικά τα αποτελέσματα. Κάθε 6-12 μήνες για Κάθε1 έτος για 5 έτη θα πρέπει να το πρώτο 3 έτη. περιλαμβάνει την απεικόνιση της πυέλου. Πίνακας 12 Παρακολούθηση ασθενών με καρκίνο του παχέος εντέρου. ESMO = European Society of Medical Oncology; JSMO = Japanese Society of Medical Oncology; ASCO = American Society of Clinical Oncology; NCCN = National Comprehensive Cancer Network; CEA = carcinoembryonic antigen; Για ασθενείς που διατρέχουν υψηλό κίνδυνο υποτροπής (π.χ., λεμφική μετάσταση, ή χαμηλής διαφοροποίησης όγκοι)( Colon Cancer Version ; Meyerhardt JA etal, 2013) 48
49 ΚΕΦΑΛΑΙΟ 4 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ ΑΚΤΙΝΗΣ Ο κυτταροσκελετός των ευκαρυωτικών κυττάρων είναι κατασκευασμένος από νηματοειδείς πρωτεΐνες, που παρέχουν μηχανική υποστήριξη στο κύτταρο και τα κυτταροπλασματικά συστατικά του και επιτελούν σημαντικές λειτουργίες. Ο κυτταροσκελετός αποτελείται από τρεις κύριες κατηγορίες στοιχείων που διαφέρουν ως προς το μέγεθος και τη σύνθεση πρωτεϊνών. Οι μικροσωληνίσκοι είναι το μεγαλύτερο είδος νηματίων διαμέτρου περίπου 25nm και αποτελούνται από την πρωτεΐνη τουμπουλίνη. Οι μικροσωληνίσκοι είναι συνεχώς μεταβαλλόμενες δομές καθώς προστίθενται και αφαιρούνται διμερή τουμπουλίνης και στα δύο άκρα του νηματίου. Ο ρυθμός μεταβολής σε κάθε άκρο δεν είναι ίδιος, το ένα άκρο μεγαλώνει πιο γρήγορα και καλείται (+) άκρο, ενώ το άλλο άκρο είναι γνωστό ως (-) άκρο. Στα κύτταρα, το (-) άκρο των μικροσωληνίσκων είναι αγκυροβολημένο σε δομές που ονομάζονται κέντρα οργάνωσης των μικροσωληνίσκων (microtubule organizing centers -MTOCs). Οι μικροσωληνίσκοι σχηματίζουν την μιτωτική άτρακτο. Σε μη διαιρούμενα κύτταρα, τα δίκτυα των μικροσωληνίσκων παρέχουν τη βασική οργάνωση του κυτταροπλάσματος, συμπεριλαμβανομένης της θέσης των οργανιδίων. Ενδιάμεσα ινίδια, όπως υποδηλώνει το όνομά τους, είναι μεσαίου μεγέθους, με διάμετρο περίπου 10 nm. Σε αντίθεση με τα νημάτια ακτίνης και τους μικροσωληνίσκους, τα ενδιάμεσα νημάτια κατασκευάζονται από έναν αριθμό διαφορετικών πρωτεϊνικών υποομάδων. Οι λειτουργίες τους είναι κυρίως μηχανική υποστήριξη σε συνδυασμό με τους μικροσωληνίσκους. Τα νημάτια ακτίνης έχουν διάμετρο περίπου 6nm και είναι κατασκευασμένα από μια πρωτεΐνη που ονομάζεται ακτίνη. Επιτρέπουν στα κύτταρα να διατηρούν εξειδικευμένες δομές όπως οι μικρολάχνες αλλά και να κινούνται. Κάθε νημάτιο αποτελείται από μόρια σφαιρικής ακτίνης (G-ακτίνη ) που συγκροτούνται για να σχηματίζουν νηματοειδή ακτίνη (F-ακτίνη). Τα νημάτια ακτίνης παρουσιάζουν δομική πολικότητα με ένα συν και ένα πλην άκρο, αυξάνουν με την προσθήκη μονομερών ακτίνης και στα δυο άκρα, με τον ρυθμό πολυμερισμού να είναι μεγαλύτερος στο συν άκρο από το πλην. (Pollard, T et al, 2003) Τα νημάτια ακτίνης επιτελούν σημαντικές λειτουργίες όπως η ενδοκυττάρωση-φαγοκυττάρωση, η δημιουργία δομών όπως οι μικρολάχνες, η διατήρηση της κυτταρικής πολικότητας, η κυτταρική διαίρεση (κυτταροκίνησηδημιουργία συσταλτού δακτυλίου) η μεταγωγή σήματος και κυρίως η κυτταρική μετανάστευση. Η κυτταρική μετανάστευση είναι μια πολυσταδιακή διαδικασία και παίζει σημαντικό ρόλο σε πολλές φυσιολογικές και παθολογικές διαδικασίες. Κατά τη διάρκεια της εμβρυϊκής ανάπτυξης, η κυτταρική μετανάστευση απαιτείται για μορφογενετικές διεργασίες όπως η γαστριδίωση αλλά και για την μετανάστευση των κυττάρων της νευρικής ακρολοφίας. Φυσιολογικές διεργασίες όπως η ανοσολογική απάντηση και η επούλωση των ιστών συνοδεύεται από την κίνηση διαφόρων τύπου κυττάρων. Τέλος, στον καρκίνο, η ανώμαλη κυτταρική μετανάστευση είναι ένα χαρακτηριστικό γνώρισμα των μεταστατικών κυττάρων.(yarar D, et al,2001) Η κυτταρική μετανάστευση μπορεί να διαιρεθεί σε στάδια σύμφωνα με τις αλλαγές που παρατηρούνται στη μορφολογία του κυττάρου: πόλωση, δημιουργία προεξοχών στο προπορευόμενο άκρο, η προσκόλληση στο υπόστρωμα, η μετατόπιση του σώματος 49
50 του κυττάρου, και η οπισθοχώρηση του οπίσθιου τμήματος. Η φυσιολογία της κίνησης των κυττάρων είναι διαφορετική, ανάλογα με τον τύπο των κυττάρων που εμπλέκονται (Yoshida Κ et al,2006). Για παράδειγμα, ινοβλάστες και μελανοκύτταρα γενικά μεταναστεύουν σαν μεσεγχυματικά κύτταρα, μεμονωμένα κύτταρα τα οποία εμφανίζουν έντονη προσκολητική ικανότητα και πρωτεολυτική αναδιαμόρφωση της εξωκυττάριας ουσίας. Αυτά τα κύτταρα σχηματίζουν εξειδικευμένες προεξοχές της μεμβράνης, μεταξύ των οποίων είναι τα ελασματοπόδια, προεξοχές που έχουν ως βασική δομή τον κυτταροσκελετό της ακτίνης και σχηματίζονται στο οδηγό εμπρόσθιο μέρος του κινητού κυττάρου, καθώς και τα πόδια διήθησης (invadopodia), προσεκβολές της πλασματικής μεμβράνης με πρωτεολυτική δραστηριότητα που είναι υπεύθυνη για την εστιακή αποικοδόμηση των συστατικών της εξωκυττάριας θεμέλιας ουσίας (ECM) με την βοήθεια των μεταλλοπρωτεϊνασών (MMPs) ( Nabeshima K et al,2002). Επίσης, μεταξύ εξειδικευμένων προεξοχών της μεμβράνης των μεταστατικών ινοβλαστών και των μελανοκυττάρων είναι τα νηματοπόδια και τα ποδοσώματα. Κυτταρικές προσεκβολές που περιέχουν λεπτά νημάτια ακτίνης έχουν επίσης δράση αισθητήρων που ανιχνεύουν σήματα στο μικροπεριβάλλον (Wolf K et al,2003). Η ικανότητα των κακοήθων κυττάρων να μεταναστεύουν οφείλεται στην παρουσία των υποδοχέων στην επιφάνειά τους που τους επιτρέπει να ακολουθήσουν χημειοτακτικούς παράγοντες.(müller A et al,2001)(cxcr4 και CCR) Η αναδιοργάνωση του κυτταροσκελετού της ακτίνης αποτελεί βασική διεργασία κατά την κυτταρική μετανάστευση, την διήθηση και την μετάσταση και αποτελεί κύριο χαρακτηριστικό της επιθήλιο-μεσεγχυματική μετάβασης (ΕΜΤ). Η ΕΜΤ είναι μία διεργασία κατά την οποία το επιθηλιακό κύτταρο αποκτά φαινότυπο μεσεγχυματικόυ κυττάρου και χαρακτηρίζεται από την απώλεια των κυτταρικών συνδέσεων και την απόκτηση αυξημένης ικανότητας κίνησης-μετανάστευσης, η οποία σχετίζεται με την αναδιαμόρφωση του κυτταροσκελετού ακτίνες. H ΕΜΤ είναι σημαντική διεργασία που παρατηρείται κατά την εμβρυική ανάπτυξη, την επούλωση τραύματος και συμβάλλει σημαντικά και σε παθολογίες του ανθρώπου όπως η ίνωση και η εξέλιξη του καρκίνου (tumor progression).(εικόνα 22) 50
51 Εικόνα 22. Το διάγραμμα δείχνει τον κύκλο των γεγονότων κατά τη διάρκεια του οποίου τα επιθηλιακά κύτταρα μετασχηματίζονται σε μεσεγχυματικά κύτταρα και αντιστρόφως. (Jean Paul Thiery & Jonathan P. Sleeman Nature Reviews Molecular Cell Biology 7, (2006). Πιστεύεται ότι υπάρχουν τρεις υπότυποι ΕΜΤ: ΕΜΤ τύπου 1 που σχετίζεται με την εμβρυϊκή ανάπτυξη, η ΕΜΤ τύπου 2 που παρατηρείται κατά την επούλωση των πληγών, και ΕΜΤ τύπου 3 που σχετίζεται με τα καρκινικά κύτταρα (Kalluri R et al,2009). Υπάρχουν κοινές οδοί μεταξύ (εικόνα 23) ΕΜΤ τύπου 1 και της ΕΜΤ τύπου 3, συμπεριλαμβανομένης της οδού TGF-β που επάγει την φωσφορυλίωση της β- κατενίνης, η οποία με τη σειρά της ενεργοποιεί παράγοντες μεταγραφής όπως Snail (Batlle E et al,2000), αυξητικούς παράγοντες και πρωτεΐνες. Η ενεργοποίηση αυτόν των οδών οδηγεί στην απώλεια επιθηλιακών πρωτεϊνών όπως Ε-κατχερίνη, α και β- κατενίνη, Claudin, ocludin και κυτταροοκερατινών και συγχρόνως στην υπερέκφραση μεσεγχυματικών πρωτεϊνών όπως η βιμεντίνη, η ινωδονεκτίνη, μεταλλοπρωτεϊνάσες, ακτίνη και ιντεγκρίνες (Yang J et al,2008;kang Y et al, 2004). Βασικό μοριακό γεγονός κατά την ΕΜΤ είναι η απώλεια της Ε-κατχερίνης που είναι κύριο συστατικό των κυτταρικών συνδέσεων μεταξύ των επιθηλιακών κυττάρων ( Adams CL et al,1998). Εκτός από τον αυξητικό παράγοντα TGF-β, άλλoι αυξητικοί παράγοντες όπως EGF, HGF, PDGF, FGF2, TNFa και IGF, αλλά και σηματοδότηση από την εξωκυττάρια ουσία μέσω των ιντεγκρινών μπορούν να επάγουν ΕΜΤ μέσω της ενεργοποίησης καταστολέων κύριοι οι παράγοντες της οικογένειας SNAIL-της μεταγραφής του γονιδίου της Ε-καδερίνης. Στην μεταγωγή σημάτων που επάγουν ΕΜΤ εμπλέκονται διάφορες σηματοδοτικές οδοί όπως ΜΑΡΚ, ΡΙ3Κ, Wnt/ β- κατενίνης, ΝF-κΒ και Notch (N Tiwari et al,2012).(εικόνα 23) 51
52 Εικόνα 23 Μια επιλογή από τα μονοπάτια σηματοδότησης που ενεργοποιούνται κατά την EMT (επιθήλιο μεσεγχυματική μετάβαση). Jean Paul Thiery & Jonathan P. Sleeman Nature Reviews Molecular Cell Biology (2006) Ο πολυμερισμός-αποπολυμερισμός των νηματίων ακτίνης είναι μία δυναμική αυστηρά ρυθμιζόμενη διεργασία που ελέγχεται από μία ομάδα πρωτεϊνών που δεσμεύονται στην ακτίνη (ABPs, actin-binding proteins). Μονομερή σφαιρικής ακτίνης προστίθενται στα (+) άκρα ή barbed ends και αφαιρούνται από τα (-) άκρα. Έχουν περιγραφθεί τρείς μηχανισμοί δημιουργίας ελεύθερων άκρων (barbed end): (1) de novo δημιουργία σημείων έναρξης πολυμερισμού από το σύμπλοκο Arp2/3 και τις φορμίνες (formins), (2) κοπή προϋπαρχόντων νηματίων ακτίνης από τις πρωτεΐνες της οικογένειας ADF/κοφιλίνη, και (3) αφαίρεση, από το (+) άκρο προϋπαρχόντων μορίων ακτίνης (J. Condeelis et al, 2003; pollard, T. D. & Borisy et al,2003). Κατά την διάρκεια του πολυμερισμού, ΑΤΡ δεσμεύεται σε G ακτίνη και υδρολύεται προς ADP Pi σταθεροποιώντας τα νημάτια ακτίνης. Απελευθέρωση του φωσφόρου αποσταθεροποιεί τη δομή, οδηγώντας στον αποπολυμερισμό. Σηματοδότηση μέσω διαφόρων οδών μπορεί να οδηγήσει στο σχηματισμό διαφορετικών δομών της ακτίνης : (εικόνα 24) 1)Νηματοπόδια (Filopodia): πρόκειται για μακριές, λεπτές προεξοχές της κυτταρικής μεμβράνης που λειτουργούν ως αισθητήρια όργανα και επιτρέπουν στο κύτταρο να εξετάσει το μικροπεριβάλλον. Ρυθμίζονται από την Rho GTPase Cdc42 (Cramer, L et al,1997;svitkina,t et al,1999) Eπίσης παρατηρoύνται στο μεταναστευτικό μέτωπο των κυττάρων. 2)Eλασματοπόδα (Lamellipodia): είναι ευρείες προεξοχές που περιέχουν ένα διακλαδισμένο δίκτυο λεπτών νηματίων ακτίνης. Δημιουργούνται από την Rac GTPase και το WAVE / Scar σύμπλοκο και το N -WASP, οι οποίες ελέγχουν το σύμπλοκο Arp2/3, το οποίο αποτελεί τον κύριο ενεργοποιητή της έναρξης του πολυμερισμού της ακτίνης (Ponti, A et al,2004). Eχουν ένα σημαντικό ρόλο στην κυτταρική μετανάστευση επιτρέποντας στα κύτταρα να συνδέονται με το υπόστρωμα και να δημιουργούν μια δύναμη που «τραβά» το κυτταρικό σώμα προς τα εμπρός. Με τον ίδιο μηχανισμό, καρκινικά κύτταρα έρπονται στην εξωκυττάρια θεμέλια ουσία (ECM) προς τα αιμοφόρα αγγεία σε πρωτογενείς όγκους και εκτείνουν ψευδοπόδια (λειτουργικά ισοδύναμο με lamellipodia) στο μεταναστευτικό μέτωπο του όγκου (J. Condeelis et al,2003). 3)Ίνες στρες: είναι παχιές δέσμες νηματίων ακτίνης που περιέχουν πολλά αντί παράλληλα νημάτια ακτίνης. Περιέχουν μυρσίνη II, που τους προσδίδει συσταλτές ιδιότητες. 4)Τόξα (Arches): είναι μεγάλες, παχιές δέσμες νηματίων ακτίνης που παρέχουν εγκάρσια δομική υποστήριξη στο κύτταρο. Είναι προσδεδεμένα στα ραχιαία και πλευρικά τμήματα του κυττάρου και μερικές φορές καταλήγουν σε μόρια προσκόλλησης.(hotulainen P et al,2006) 52
53 Εικόνα 24. Εντός του κυττάρου, νημάτια ακτίνης μπορούν να οργανωθούν για να σχηματίσουν πολλαπλές δομές. Ίνες στρες είναι μεγάλα συγκροτήματα των νηματίων ακτίνης που μπορεί να καλύπτουν το μήκος του κυττάρου. Η παρουσία της μυοσίνης σε ίνες στρες επιτρέπει την συσταλτικότητα. Κάτω από τη μεμβράνη είναι το χαλαρά οργανωμένο δίκτυο νηματίων ακτίνης που έχει ονομαστεί φλοιώδης ακτίνη. Νημάτια ακτίνης, επίσης, μπορεί να οργανωθούν δημιουργώντας διάφορες προσεκβολές, συμπεριλαμβανομένων των ποδοσωμάτων, των ελασματοποδιων των νηματοποδίων και των μικρολάχνων. Η ακτίνη και η μυοσίνη είναι τα κύρια συστατικά του κυτταροσκελετού των κυττάρων. Παρόλο που παραδοσιακά πιστεύεται ότι λειτουργούν μόνο στο κυτταρόπλασμα, είναι ευρέως αποδεκτό ότι η ακτίνη και πολλαπλές μυοσίνες βρίσκονται και στον πυρήνα. Η ακτίνη έχει σημαντικό ρόλο στην μεταγραφή, την κίνηση προς την πυρηνική περιφέρεια και την εξαγωγή του mrna.(blaine Tet al,2004) Η πυρηνική ακτίνη φαίνεται να συμβάλει στην αναδιαμόρφωση του DNA (εικόνα 25), στην επεξεργασία του RNA, στην ρύθμιση της λειτουργίας της DNAάσης Ι. Επιπρόσθετα η ακτίνη φαίνεται να παίζει σημαντικό ρόλο στην μεταγραφή από την RNA πολυμεράση Ι, ΙΙ και ΙΙΙ. (Ricardo Uribe et al,2009; Koestler, et al,2008) 53
54 Εικόνα 25.Πυρηνική εντόπιση της ακτίνης. Ο πολυμερισμός-αποπολυμερισμός της ακτίνης είναι μια δυναμική αυστηρά ρυθμιζόμενη διεργασία που περιλαμβάνει: απομάκρυνση των πρωτεϊνών κάλυψης από το (+) άκρο, κοπή των υφιστάμενων νηματίων και δημιουργία νέων ελεύθερων (+) άκρων, de nονο σχηματισμού σημείων έναρξης του πολυμερισμού (nucleation) ακτίνης, τη φάση επιμήκυνσης και μια σταθερή κατάσταση. Κατά την φάση επιμήκυνσης προστίθεται μονομερή ακτίνης στο (+) άκρο και αυτό διευκολύνεται από πρόσθετους παράγοντες επιμήκυνσης όπως η formin. Στη σταθερή φάση η δυναμική του νήματος βρίσκεται σε κατάσταση ισορροπίας όπου μονομερή αποσυναρμολογούνται από το (-) άκρο και πολυμερίζονται στο (+) άκρο σε μια κατάσταση ισορροπίας που συντηρείται από μια κρίσιμη συγκέντρωση των μονομερών στο κυτταρόπλασμα. Αυτή η σταθερή κατάσταση συναρμολόγησης και αποσυναρμολόγησης είναι γνωστή ως «treadmilling». Ο ρυθμός αύξησης σε σταθερή κατάσταση είναι ~ 0.04 μm / min, υψηλότερο ποσοστό σε treadmilling in vivo οφείλεται σε πρωτεΐνες που αλληλεπιδρούν με νημάτια ακτίνης ή μονομερή γνωστές ως ΑΒΡs (actin binding proteins) (Pollard and Borisy et al,2003; Disanza et al,2005). Επιμήκυνση των νηματίων επιτυγχάνεται όταν ο ρυθμός προσθήκης μονομερών στο (+) άκρο είναι μεγαλύτερος από την απώλεια της ADP-ακτίνης από το (-) άκρο. (Εικόνα 26) 54
55 Εικόνα 26. Ρύθμιση treadmilling ακτίνης. 1 ) ADF συνδέεται προς την πλευρά της ADP-νημάτια ακτίνης και επάγει τον αποπολυμερισμό του μυτερού άκρου ώστε να αυξηθεί η συγκέντρωση των μονομερών ακτίνης σε σταθερή κατάσταση. 2 ) Profilin ενισχύει την ανταλλαγή ADP για την ΑΤΡ για την ανακύκλωση μονομερών ακτίνης. Σύμπλοκα προφιλίνη-ακτίνης συγκεντρώνονται αποκλειστικά στο (+) άκρο. 3) Μπλοκάροντας τα νημάτια ακτίνης στα (+)άκρα, capping πρωτεΐνες αυξάνουν τη συγκέντρωση των μονομερών ακτίνης σε σταθερή κατάσταση και επάγουν την ροή των μονομερών ακτίνης στα noncapped νημάτια τα οποία μεμονωμένα μεγαλώνουν γρηγορότερα. Η δυναμική του κυτταροσκελετού της ακτίνης ρυθμίζεται από μια μεγάλη ομάδα πρωτεϊνών ΑΒΡs (actin binding proteins)(εικόνα 27) η οποία περιλαμβάνει τις εξής κατηγορίες: 1. Πρωτεΐνες που δεσμεύονται με G ακτίνη: Ορισμένες πρωτεΐνες επάγουν την συναρμολόγηση-αποσυναρμολόγηση άμεσα, ενώ άλλες επηρεάζουν την δέσμευση ΑΤΡ, το ποσοστό της συναρμολόγησης G-ακτίνης και την κρίσιμη συγκέντρωση (Cc) στο άκρο του νήματος. Αναφέρονται πάνω από 60 οικογένειες πρωτεϊνών πρόσδεσης ακτίνης με κύριες πρωτεΐνες στα σπονδυλωτά την θυμοσίνη-β4 και την προφιλίνη. Η θυμοσίνη-β4 συνδέεται ισχυρά με ΑΤΡ-ακτίνη και αποτρέπει τη συναρμολόγηση της ακτίνης σε ίνες (Carlier et al,1998 ). Επειδή η προφιλίνη και η θυμοσίνη-β4 έχουν επικαλυπτόμενες θέσεις πρόσδεσης στην ακτίνη ( Safer D et al, 1997; Schutt CE.et al,1993 ), η προφιλίνη πρέπει να ανταγωνιστεί με την θυμοσίνη β4-κατά τη συναρμολόγηση του νήματος (Kang F et al,1999) 2. Πρωτεΐνες που διατηρούν την δεξαμενή G-ακτίνης: Οι πρωτεΐνες της κατηγορίες αυτής αποπολυμερίζουν τα νημάτια στο (-) άκρο διατηρώντας μια δεξαμενή επάγοντας την ανακύκλωση μονομερών ακτίνης που θα χρησιμοποιηθούν σε ένα καινούριο κύκλο πολυμερισμού. Σε αυτές ανήκει κυρίως η κοφιλίνη. Η κοφιλίνη δημιουργεί νέα ελεύθερα άκρα από τα οποία μπορεί να ξαναρχίσει ο πολυμερισμός, ενώ παράλληλα επάγει την ανακύκλωση μονομερών ακτίνης επάγοντας τον αποπολυμερισμό παλαιών δικτύων(condeelis J et al,2001;bamburg JR et al,2008). Η κοφιλίνη ρυθμίζεται με φωσφορυλίωση από τις LIM κινάσες, οι οποίες με τη σειρά τους ρυθμίζονται με φωσφορυλίωση από τις κινάσες ΡΑΚ και ROCK. 3. Πρωτεΐνες που διαμεσολαβούν τον πολυμερισμό της ακτίνης στο (+) άκρο: διακρίνονται σε δύο διαφορετικές ομάδες: α) επαγωγής πολυμερισμού στο (+) και β) πρωτεΐνες που καλύπτουν το (+) άκρο (capping proteins)(campellone KG, et al,2010). Στην πρώτη κατηγορία ανήκουν πρωτεΐνες που επάγουν την ταχεία αύξηση-επιμήκυνση των νηματίων π.χ οι πρωτεΐνες formins (mdia1, mdia2 και mdia3). Άλλες πρωτεΐνες εμποδίζουν την σύνδεση πρωτεϊνών κάλυψης όπως οι VASP, 55
56 Mena και Ena/VASP-like (EVL) (Bear JE et al,2009). Οι πρωτεΐνες κάλυψης αναστέλλουν την επιμήκυνση των νηματίων ακτίνης και περιλαμβάνουν τις πρωτεΐνες CapZ, adducin και γελσολίνη (Zigmond SH.et al,2004). 4. Πρωτεΐνες που επάγουν την δημιουργία σημείων έναρξης του πολυμερισμού(nucleation factors). Σε αυτήν την κατηγορία ανήκουν το σύμπλοκο Arp2/3 και πρωτεΐνες της οικογένειας WASP/WAVE. Οι παράγοντες Nucleation promoting factors (NPFs) π.χ WASP και Scar/WAVE ρυθμίζουν την διαδικασία δημιουργίας σημείων έναρξης του πολυμερισμού συνδεόμενοι ταυτόχρονα με μονομερή ακτίνης και προϋπάρχοντα νημάτια ακτίνης. Οι NPFs ανταγωνίζονται την προφιλίνη για την θέση πρόσδεσης με ελεύθερη ακτίνη, προωθώντας την ανάπτυξη του νηματίου στο ( +) άκρο.(carlier MF et al,1993;suetsugu S.et al,1998;higgs HN et al,1999; Egile C.et al,1999) 5. Πρωτεΐνες που αποκόπτουν νημάτια ακτίνης (severing proteins). Οι πρωτεΐνες αυτές με κύριο εκπρόσωπο την gelsolin και την κοφιλίνη αποκόπτουν προϋπάρχοντα νημάτια ακτίνης και δημιουργούν ελεύθερα (+) άκρα στα οποία μπορεί να γίνει επιμήκυνση. Εικόνα 27. Ομάδα πρωτεϊνών που ρυθμίζουν την δυναμική του κυτταροσκελετού της ακτίνης. Dan N. Simon November 2011 Ενώ οι περισσότερες από αυτές τις πρωτεΐνες είναι κυρίως κυτταροπλασματικές, είναι σε θέση να εισέλθoυν στον πυρήνα υπό ορισμένες συνθήκες, δηλαδή κατά τη διάρκεια του στρες, την διαφοροποίηση ή μετά από ορμονικό ερεθισμό των κυττάρων.(πίνακας 13). Επίσης κάποιες δεν περιορίζονται σε μία συγκεκριμένη λειτουργία αλλά εμφανίζουν πολλαπλές λειτουργίες, για παράδειγμα, η γελσολίνη η οποία κόβει και ταυτόχρονα καλύπτει το αγκαθωτό άκρο των νηματίων ακτίνης. 56
57 Actin-binding protein Targeting sequence Role in the nucleus Profilin NLS? pre-mrna splicing Πυρηνική εξαγωγή της ακτίνης αναστολέας myb Thymosin b4 NLS? NR effect NR CapG NLS? Η πυρηνική εντόπιση Συσχετίζεται με Gelsolin NLS? κολλαγόνο εισβολή Primary coactivator AR Ενισχύει τη PPARg, GR δράση του υποδοχέα Flightless I NLS? /NES? Secondary coactivator TR, ER a-actinins NLS Συσχέτιση με τον καρκίνο του μαστού Συσχέτιση με ιική μόλυνση Primairy coactivator Secondary coactivator Plastin NES DNA επιδιόρθωση AR, TR, ER AR, TR, ER Filamin A NLS? Απάντηση στη βλάβη του DNA καταστολή καταστολή Cofilin NLS Πυρηνική εισαγωγή της ακτίνης Intranuclear actin cofilin rods Myopodin weak NES Συσχέτιση με την εισβολή του όγκου Intranuclear actinmyopodin rods Supervillin NLS Primairy coactivator Ενίσχυση της δραστηριότητας του υποδοχέα AR GR AR PPARg, GR, ER Πίνακας 13. Εντόπιση και ρόλος ΑΒΡs στον πυρήνα Targeting and role of nuclear actin-binding proteins AR, androgen receptor; ER, estrogen receptor; GR, glucocorticoid receptor; NES, nuclear export sequence; NLS, nuclear localization sequence; NR, nuclear receptor; PPARg, peroxisome proliferator-activated receptor-g; TR, thyroid receptor. 57
58 Εικόνα 28. Οι ΑΒΡS ρυθμίζουν την δυναμική του κυτταροσκελετού ακτίνης. (Α) Treadmilling μπορεί να μεταβληθεί από profilin και ADF όπου αυξάνουν και μειώνουν το μέγεθος των νηματίων ακτίνης.(β) Νέα νημάτια ακτίνης από το ARP 2/3 σύμπλοκο, το οποίο δεσμεύει δύο μονομερή G- ακτίνης (nucleation). H Formin επάγει (nucleate) νέα νημάτια μέσω σύνδεσης G-ακτίνης και μέσω της συνεργασίας με profilin. (Γ)Οι πρωτεΐνες που διαμεσολαβούν εγκάρσιες συνδέσεις επηρεάζουν την οργάνωση των νηματίων ακτίνης σε δευτεροταγείς δομές. (Δ) Πρωτεΐνες που καλύπτουν ή αποκόπτουν τα νημάτια ακτίνης προάγουν τον πολυμερισμό/αποπολυμερισμό των νηματίων της ακτίνης. (Ε) Ο πολυμερισμός της ακτίνης μπορεί να τροποποιηθεί από γεγονότα όπως η ελεγχόμενη υδρόλυση νουκλεοτιδίων (π.χ. ΑΤΡ για ακτίνη) και αναστρέψιμες τροποποιήσεις (π.χ. φωσφορυλίωση) Mechanobiology Institute, National University of Singapore, Republic of Singapore Η απάντηση του κυττάρου σε εξωτερικά ερεθίσματα λαμβάνει χώρα μέσω ενός καταρράκτη σηματοδοτικών μορίων, που περιλαμβάνουν υποδοχείς κινάσης τυροσίνης- RTKs (receptor tyrosine kinases), υποδοχείς GPCR (G-protein coupled receptor), ιντεγκρίνες, φωσφοινοσιτίδια, οι GTPases της οικογένειας Rho- RhoΑ, CDC42 και Rac. Μεταξύ αυτών, οι GTPases RhoA, Rac και Cdc42 παίζουν κρίσιμο ρόλο στο σχηματισμό και την οργάνωση δικτύων ακτίνης (Hall et al1998; Bishop and Hall et al,2000). Η RhoA ελέγχει την συναρμολόγηση των ινών στρες. Η Rac ρυθμίζει το σχηματισμό ελασματοποδίων και η Cdc42 ελέγχει κυρίως την δημιουργία νηματοποδίων και microspikes. Κατά τη διάρκεια της κίνησης των κυττάρων, οι Rac 58
59 και Cdc42 ενισχύουν τον σχηματισμό των προεξοχών στα προπορευόμενα άκρα των κυττάρων και η RhoA επάγει την συστολή στην ουράς των κυττάρων. Αυτή η συντονισμένη αναδιοργάνωση κυτταροσκελετού επιτρέπει στα κύτταρα να κινούνται προς ένα στόχο. Οι οδοί αυτές σηματοδότησης συγκλίνουν σε πρωτεΐνες που ρυθμίζουν άμεσα την συμπεριφορά και την οργάνωση του κυτταροσκελετού της ακτίνης, συμπεριλαμβανομένης της κοφιλίνης, Arp2/3, Ena /WASP, forminins, προφιλίνη και γελσολίνη.(εικόνα 29) Εικόνα 29. Μονοπάτια ελέγχου του κυτταροσκελετού της ακτίνης (Hall,et al, 1998; Bishop and Hall, et al,2000). Μέχρι σήμερα πολλές πρωτεΐνες που ρυθμίζουν τον κυτταροσκελετό της ακτίνης έχουν ταυτοποιηθεί και έχει δειχθεί ότι υπερεκφράζονται σε διάφορους τύπους καρκίνου(e. Sahai et al,2005). Μεταξύ αυτών, είναι η πρωτεΐνη WASP (Wiskott Aldrich syndrom protein), η οικογένεια πρωτεϊνών Arp2/3, οι πρωτεΐνες της οδού LIM- κινάση/κοφιλίνη και τα μονοπάτια που ελέγχονται από την cortactin έχουν μελετηθεί εκτενώς εξαιτίας της σημασίας τους στην κυτταρική μετανάστευση και την διήθηση. 59
60 4.2WASP ( W iskot- A ldrich S yndrome) Η οικογένεια WASP αποτελείται από δύο κύριες κατηγορίες των πρωτεϊνώn, της WASP και SCAR/WAVEs. Το 1994, ταυτοποιήθηκε η πρωτεΐνη WASP ως το αίτιο του συνδρόμου W iskot- A ldrich S yndrome. Το σύνδρομο Wiskott-Aldrich (WAS) είναι μια κληρονομική ανοσολογική ανεπάρκεια, χαρακτηρίζεται από έκζεμα, αιμορραγία, και υποτροπιάζουσες λοιμώξεις. Χαρακτηρίζεται από ανεπάρκεια της κυτταρικής και χυμικής ανοσίας. Λεμφοκύτταρα και αιμοπετάλια ασθενών με WAS εμφανίζουν κυτταροσκελετικές ανωμαλίες και τα Τ λεμφοκύτταρα εμφανίζουν μειωμένη απόκριση σε διέγερση μέσω του συμπλόκου TCR-CD3. Οι πρωτεΐνες WASP ανήκουν στους παράγοντες που επάγουν την έναρξη του πολυμερισμού της ακτίνης-npfs τύπου Ι ενεργοποιώντας το σύμπλοκο Arp2/3. Οι NPFs τύπου Ι (δεσμεύουν το σύμπλοκο Arp2/3 και μονομερή ακτίνης) αποτελούνται από πέντε ομάδες πρωτεϊνών : 1. Η οικογένεια των πρωτεϊνών WASP ( W iskot- A ldrich S yndrome) και N-WASP. Οικογένεια WASP Verprolin-ομόλογες πρωτεΐνες ( WAVE ), γνωστή και ως αναστολέας του υποδοχέα camp (Scar). 2. WASP και Scar ομόλογες πρωτεΐνες (WASH). 3. WASP ομόλογη πρωτεΐνη που συνδέεται με την ακτίνη, μεμβράνες και μικροσωληνίσκους (WHAMM). 4. JMY πρωτεΐνη (junction-mediating regulatory protein) (Rohatgi R et al, 1999; Miki H et al, 2000). Το ανθρώπινο γονίδιο WASP φέρεται επί του Χ χρωμοσώματος. Οι πρωτεΐνες WASP και WAVE στον άνθρωπο αποτελούνται από 498 και 559 αμινοξέα αντίστοιχα και κωδικοποιούνται από 9 έως 12 εξώνια. Το μήκος των γονιδίων είναι σχετικά παρόμοιο, και κυμαίνεται από 67.1 kb- Ν-WASP έως 13.,2 kb - WAVE3, με εξαίρεση το WASP, το οποίο έχει μέγεθος 7,6 kb. Η περιορισμένη έκφραση των WASP σε αιμοποιητικά κύτταρα εξαρτάται από μια περιοχή 137-bp στη θέση έναρξης της μεταγραφής (Petrella A et al, 1998). Οι WASP και WAVE ομόλογες πρωτεΐνες έχουν ανακαλυφθεί σε μια ευρεία ποικιλία ευκαρυωτικών. WASP και WAVE ομόλογες πρωτεΐνες (ένα από το καθένα) έχουν ταυτοποιηθεί στο Dictyostelium discoideum (WASP και SCAR) (Bear Je et al, 1998; Myers SA et al,2005), στο Caenorhabditis elegans (WSP-1 και WVE-1) (Sawa Metal,2003 ;PatelFbetal,2008) και στη Drosophila melanogaster (WASP και SCAR) (Ben-Yaacov Set al,2001;zallen JA et al, 2002). Ο ζυμομύκητας έχει μόνο ένα WASP ομόλογο, Las17/Bee1 (Li R.et al,1997;naqvi Sn et al,1998) και δεν φαίνεται να διαθέτει WAVE. Σε αντίθεση, το φυτό Arabidopsis thaliana φαίνεται να έχει τέσσερα γονίδια WAVE, SCAR1-4 (Deeks Mjet al,2004), αλλά όχι WASP.(εικόνα 30) Δεδομένου ότι ακόμα και τα φυτά έχουν WAVE ομόλογα, η εξελικτική ιστορία των WASP και WAVE είναι πιθανό ότι ξεκινά πριν τους ευκαρυωτικούς οργανισμούς. Μαζί με την εξέλιξη του κυτταροσκελετού της ακτίνης, τα ευκαρυωτικά κύτταρα πρέπει να διαθέτουν μηχανισμό έλεγχου του πολυμερισμού της ακτίνης και της αναδιοργάνωση του κυτταροσκελετού, το οποίο πιθανώς οδήγησε στην ανάπτυξη του άξονα WASP/WAVE-Arp2/3. Οι φυλογενετικές αναλύσεις που περιλαμβάνουν τις αλληλουχίες αμινοξέων στο zebrafish έχουν δείξει ενδιαφέροντα δεδομένα για την εξέλιξη αυτών των 60
61 πρωτεϊνών στα σπονδυλωτά. Πρώτον, τόσο το WASP όσο και το Ν-WASP φαίνεται να είναι παρόν σε ένα κοινό πρόγονο ψαριών και θηλαστικών. Αυτό σημαίνει ότι το WASP πιθανόν επιτελεί εξειδικευμένη λειτουργία κατά την ανάπτυξη του ανοσοποιητικού συστήματος των σπονδυλωτών νωρίς στην εξέλιξη, καθώς είναι το πρώτο που ανευρίσκεται στα ψάρια. Δεύτερον, το WAVE είναι χωρισμένο σε τρεις διακριτές υποδιαιρέσεις, WAVE 1-3, ήδη από την εμφάνιση των σπονδυλωτών. Θεωρώντας ότι το WAVE 1 και πιθανώς το WAVE 3 εμπλέκονται στην ανάπτυξη του εγκεφάλου στα θηλαστικά (Soderling SH et al, 2007;Soderling Shet al,2003), η εμφάνιση τους θα μπορούσε να θεωρηθεί σημαντική για την ανάπτυξη του κεντρικού νευρικού συστήματος (ΚΝΣ). Εικόνα 30.Εξελικτικές σχέσεις μεταξύ της οικογένειας των πρωτεϊνών WASP και WAVE. Shusaku Kurisu -Tadaomi Takenawa et al,2009 Οι πρωτεΐνες της οικογένειας WASP και WAVE εμφανίζουν κοινή αρχιτεκτονική και αποτελούνται από μια περιοχή πλούσια σε προλίνη η οποία ακολουθείται από την περιοχή VCA που βρίσκεται στο καρβοξυτελικό άκρο και ενεργοποιεί το Arp2/3 σύμπλοκο. Η περιοχή VCA αποτελείται από τρεις επιμέρους περιοχές: (1) μία περιοχή πρόσδεσης ακτίνης (V-verprolin), γνωστή και ως WASPhomology 2 (WH2) domain), (2) μια διατηρημένη περιοχή «σύνδεσης» (Cconnecting), η οποία αποτελείται από περίπου 20 αμινοξέα και αλληλεπιδρά τόσο με το Arp2 / 3 σύμπλοκο όσο και με μονομερή ακτίνης (Marchand et al,2001) και (3) μια περιοχή (Α-acidic) που δεσμεύει το Arp2/3 σύμπλοκο.(εικόνα 31) Η περιοχή VCA είναι αναγκαία για την αποτελεσματική ενεργοποίηση του συμπλόκου Arp2/3 και λειτουργεί ως μια πλατιφόρμα για την ταυτόχρονη δέσμευση μονομερών ακτίνης και του συμπλόκου Arp2/3, φέρνοντάς τα σε άμεση γειτνίαση (Pollard TD et al,2002). Αυτό ενισχύεται περαιτέρω από την PIP2 (διφωσφορικής-4,5- φωσφατιδυλινοσιτόλη)(yarar D et al, 2002). 61
62 Εικόνα 31. Δομή των πρωτεϊνών της οικογένειας WASP και WAVE AD: autoinhibitory domain, B: basic domain, CM: Cdc42/rac interactive binding motif; PRP:proline-rich region VCA: verprolin-homology, cofilin-homology/central, and acidic domain, WH1: WASP homology 1domain.WHD/SHD: WAVE/Scar homology domain. (Marchand et al,2001;miki et al,1996; Imai et al,1999) Η αλληλουχία στο αμινοτελικό άκρο της οικογένειας WASP διαφέρει από αυτό της οικογένειας πρωτεϊνών WAVE. To αμινο τελικό άκρο της οικογένειας WASP έχει την περιοχή WH1/EVH που δεσμεύει 4,5-διφωσφορικά φωσφατιδυλινοσιτίδια (PtdIns (4,5) P2) (Miki et al,1996; Imai et al,1999). Ακολουθεί μια βασική περιοχή και μία περιοχή δεσμεύσεως GΤΡάσης (GBD/CRIB)(GTPasebinding domain/cdc42και Rac interactive binding). Η περιοχή WH1/EVH1 συνδέεται με πρωτεΐνες WIP (WASP-interacting protein) (Ramesh et al,1997) στις οποίες συμπεριλαμβάνονται οι πρωτεΐνες WIP, CR16 (κορτικοστεροειδή ) και WICH / WIRE (WIP- and CR16-homologous protein/wip-related) στα θηλαστικά (Anton IM,et al,2007; Kato M et al,2002). Στα κύτταρα, οι πρωτεΐνες WASP και Ν-WASP φαίνεται να σχηματίζουν ένα σταθερό ένα-προς-ένα σύμπλοκο με την οικογένεια πρωτεϊνών WIP (Ho Ηυ et al, 2001) οι οποίες προστατεύουν τις πρωτεΐνες WASP και Ν-WASP από την αποικοδόμηση μέσω πρωτεασών (de la Fuente MA et al, 2007;Suetsugu S et al,2007). H περιοχή GBD / CRIB είναι κρίσιμης σημασίας για τον έλεγχο των WASP και Ν- WASP. Ενεργοποιημένες μορφές του Cdc42 που εντοπίζονται στη μεμβράνη προσδένονται στην περιοχή GBD/CRIB. Η ΡΙ (4,5) Ρ 2 είναι άφθονη στην πλασματική μεμβράνη και συνδέεται προς την βασική περιοχή. Η οικογένεια τυροσινική κινάσης Src φωσφορυλιώνει υπολείμματα τυροσίνης κοντά στην περιοχή GBD/CRIB. Όλα αυτά τα γεγονότα φαίνεται να χαλαρώνουν τις ενδομοριακές αλληλεπιδράσεις μεταξύ των GBD και VCA με αποτέλεσμα την ενεργοποίηση του WASP (Takenawa et al,2007). Η πρωτεΐνη WASP αυτό-αναστέλλεται σε κατάσταση ηρεμίας εμποδίζοντας τον ανεξέλεγκτο πολυμερισμό της ακτίνης. Ενδομοριακές αλληλεπιδράσεις μεταξύ CRIB/ GBD και της περιοχής VCA διαμεσολαβούν αυτή την αυτο-αναστολή. Η περιοχή VCA απελευθερώνεται με την βοήθεια διαφόρων μορίων δέσμευσης που ανταγωνίζονται για την ίδια περιοχή δέσμευσης CRIB / GBD ή στην περιοχή γύρω από αυτή, επιτρέποντας στην περιοχή VCA να αλληλεπιδράσει και να ενεργοποιήσει το Arp2 / 3 σύμπλοκο.(εικόνα 32) 62
63 Εικόνα 32.Ν-WASP αυτο αναστέλλεται σε μια βασική κατάσταση μέσα από την αλληλεπίδραση της GBD / CRIB και της περιοχής VCA. Kurisu και Takenawa Genome Biology 2009, 10 : 226 Σε αντίθεση με το WASP, οι πρωτεΐνες SCAR/WAVE δεν αυτοαναστέλονται και αποτελούν μέρος ενός ευρύτερου μηχανισμού τεσσάρων πρωτεϊνών - PIR121, Nap1, Abi and HSPC300 (Eden et al, 2002). Οι πρωτεΐνες WASP ανήκουν στους NPFs τύπου Ι και η κύρια λειτουργία τους είναι να επάγουν τον πολυμερισμό της ακτίνης ενεργοποιώντας το σύμπλοκο Arp2/3.(εικόνα 34) Το σύμπλοκο Arp2/3 αποτελείται από εφτά υπομονάδες και έχει κρίσιμο ρόλο στην ρύθμιση της δυναμικής του κυτταροσκελετού της ακτίνης. Η ενεργοποίηση του συμπλόκου Arp2/3 αποτελεί ένα από τα σημαντικότερα βήματα για την κυτταρική κινητικότητα και τον σχηματισμό μεμβρανικών προεξοχών γνωστά ως ελασματοπόδια (Machesky Lm et al,2003). Το Arp2/3 σύμπλοκο πρέπει να ενεργοποιηθεί από πρωτεΐνες της οικογένειας Wiskott-Aldrich (WASP) παρουσία ενός νηματίου ακτίνης έτσι ώστε να δημιουργηθεί νέο θυγατρικό νημάτιο ακτίνης (Machesky et al,1999 ;Blanchoin et al,2001;zalevsky et al,2001) Οι Arp2 και Arp3 υπομονάδες συνδέονται με ΑΤΡ (Dayel et al,2001). Υδρόλυση του ΑΤΡ προωθεί την κατασκευή νηματίων ακτίνης ενώ η απελευθέρωση Pi δεν προκαλεί άμεση αποσυναρμολόγηση του νηματίου, αλλά επιτρέπει την αλληλεπίδραση με παράγοντες αποπολυμερισμού όπως η κοφιλίνη (Blanchoin και Pollard et al,1999). Η δραστηριότητα ΑΤΡάσης απαιτεί το συνδυασμό ενός προσχηματισμένου νηματίου ακτίνης, της περιοχής VCA και ενός μονομερούς ακτίνης αλλά δεν απαιτεί τον πολυμερισμό της ακτίνης. Αυτό δείχνει ότι η υδρόλυση ενεργοποιείται σχετικά νωρίς στην πυρήνωση πριν από την αντίδραση ολοκλήρωσης ενός σταθερού θυγατρικού νήματος. Κατά τον πολυμερισμό της ακτίνης υδρολύεται ΑΤΡ δημιουργώντας έτσι μια διαφορά μεταξύ της κρίσιμης συγκέντρωσης (C γ ) του ΑΤΡ δεσμευμένο στο (+) άκρο (C c = 0,06 μμ) και του ADP συνδεδεμένου στο (-) άκρο (C c = 0,6 μμ ). Σε σταθερή κατάσταση η αξία της συγκέντρωσης των μονομερών ακτίνης (C ss = 0,1 μμ) συνεπάγεται ότι το (+) άκρο επιμηκύνεται σε ισορροπία με το (-) άκρο το οποίο αποπολυμερίζεται έτσι ώστε κατά μέσο όρο το νημάτιο κινείται προς τα εμπρός και διατηρεί το ίδιο μήκος. Αυτή η διαδικασία ονομάζεται "treadmilling. Η διαδικασία αυτή ωθεί την μεμβράνη προς τα εμπρός 63
64 σχηματίζοντας τα ελασματοπόδια. (Mullins et al,1998;machesky et al,1999; Blanchoin et al,2000 ). Η ταχεία υδρόλυση του ATP στην Arp2 υπομονάδα φαίνεται να πυροδοτείται από διάφορα γεγονότα όπως : 1) δέσμευση της VCA στο Arp2/3 σύμπλοκο 2) πρόσδεση του VCA - Arp2 / 3 προς την πλευρά ενός προσχηματισμένου νηματίου 3) σύνδεση VCA - μονομερή ακτίνης στο Arp2 / 3 σύμπλοκο 4)πρόσδεση ενός δεύτερου ή τρίτου μονομερούς ακτίνης για το σχηματισμό θυγατρικού νηματίου. Το σύμπλοκο Arp2/3 επάγει in vitro την έναρξη πολυμερισμού (πυρήνωσηnucleation) της ακτίνης μετά τη δέσμευση σε προϋπάρχοντα μητρικά νημάτια υπό την παρουσία NPFs (nucleation-promoting factors) δημιουργώντας διακλαδώσεις με χαρακτηριστική γωνία 70 μεταξύ των κλάδων. Όταν στο σύμπλοκο Arp2/3 προσδένεται G -ακτίνη το αποτέλεσμα είναι να δημιουργηθεί ένα σταθερό τριμερές επάγοντας τον πολυμερισμό και την ανάπτυξη ενός νηματίου. To Arp2/3 στη συνέχεια δρα στον (-) άκρο σαν πρωτεΐνη κάλυψης επάγοντας την ταχεία ανάπτυξη του νηματίου στο (+) άκρο ( Paavilainen et al,2004). Η δομή και ο προσανατολισμός των υπομονάδων Arp2 και Arp3 εντός της κρυσταλλικής δομής του συμπλόκου υποδηλώνουν ότι αυτές οι υπομονάδες μπορούν επάγουν την πυρήνωση ενός νέου νηματίου ακτίνης, σχηματίζοντας ένα ετεροδιμερές ακτίνης που μιμείται το (+) άκρο (Robinson et al 2001). Στην κρυσταλλική δομή του μη ενεργοποιημένου συμπλόκου οι δύο υπομονάδες Arp2 και Arp3 χωρίζονται από 40 Α, έτσι ώστε ο σχηματισμός ενός διμερούς ακτίνης απαιτεί διαμορφωτική αλλαγή ( Robinson et al, 2001). Πρόσδεση του Arp2 / 3 συμπλόκου σε ένα νημάτιο και σε πρωτεΐνες της οικογένειας WASP οδηγούν τουλάχιστον εν μέρη σε αυτή την διαμορφωτική αλλαγή (Blanchoin et al, 2001; Marchand et al, 2001; Panchal et al, 2003).(εικόνα 33) Η ενεργοποίησή των πρωτεϊνών WASP επάγει μέσω της ενεργοποίησης του πολυμερισμού των νηματίων ακτίνης το σχηματισμό δομών, όπως ελασματοπόδια, νηματόδεμα και ποδοσώματα. Οι πρωτεΐνες WASP έχουν κεντρικό ρόλο στη διακίνηση μέσω της μεμβράνης, την κυτταροκίνηση καθώς και την πρόσφυση των κυττάρων στο υπόστρωμα και την κυτταρική μετανάστευση(ibarra et al, 2006). Οι πρωτεΐνες αυτές καθώς ρυθμίζουν την κυτταρική κίνηση-μετανάστευση παίζουν σημαντικό ρόλο στην διήθηση και μετάσταση των καρκινικών κυττάρων( Kurisu S et al,2005). Διαταραχή στην ρύθμιση της έκφρασης των πρωτεϊνών αυτών έχει δειχθεί σε διάφορους τύπους καρκίνου. Ωστόσο λίγα είναι γνωστά για τον ρόλο τους στον καρκίνο του παχέος εντέρου. 64
65 Εικόνα 33. Το συγκρότημα ARP2/3 βρίσκεται αρχικά σε μια ανενεργή, ανοικτή διαμόρφωση. Δέσμευση του WCA (Wiskott Aldrich syndrome protein (WASP)-homology-2, central, acidic) προωθεί μια διαμόρφωση, η οποία λαμβάνει χώρα κατά την πρόσδεση της WCA-ακτίνη ARP2/3 μητρικό νήμα, κατά προτίμηση κοντά στο (+)(barbed end). (Βήμα 2) ΑΤΡ υδρολύεται στην ARP2 ταυτόχρονα με την πυρήνωση του θυγατρικού νηματίου. Η WCA διασπάται, αν και το έναυσμα για αυτό είναι άγνωστο. (Βήμα 3) Φωσφορικό απελευθερώνεται από ARP2. Μητρικά και θυγατρικά νημάτια επιμηκύνονται κατά την υδρόλυση του ΑΤΡ και απελευθερώνεται Ρι.(Στάδιο 4) Η απελευθέρωση Pi από την ARP2 και η γήρανση του νήματος αποδυναμώνει την αλληλεπίδραση μεταξύ ARP2/3 και θυγατρικού ή και μητρικού νηματίου (βήμα 5), που επιτρέπει την αποσυναρμολόγηση και την απελευθέρωση του ARP2/3 (Βήμα 6) νουκλεοτιδική ανταλλαγή ARP2 (και, ενδεχομένως, σε ARP3) εμφανίζεται και ο κύκλος αρχίζει ξανά. Σχήμα 34. Η ενεργοποίηση συμπλόκου Arp2/3 εξαρτάται από την άμεση αλληλεπίδραση με παράγοντες προαγωγής σχηματισμού θέσεων έναρξης πολυμερισμού (NPFs) τύπου Ι, οι οποίοι περιέχουν την περιοχή VCA. Οι NPFs ρυθμίζονται από φωσφατιδυλινοσιτίδα (PtdIns), πρωτεΐνες με δραστηριότητα GΤΡασών καθώς και από κινάσες. Πρόσφατα ανακαλύφθηκαν ARP2/3 αναστολείς, συμπεριλαμβανομένων των παραγόντων GMFs (glia maturation factors), coronins, κοφιλίνη, gadkin και την πρωτεΐνη PICK1. Οι ανασταλτικοί παράγοντες υπόκεινται επίσης σε ρύθμιση από διάφορες οδούς σηματοδότησης.( Nature Reviews Molecular Cell Biology (2013). 65
66 4.3 LIM ΚΙΝΑΣΕΣ (LIMK1 και LIMK2) Η οικογένεια LIM κινασών αποτελεί μια υποκατηγορία κινασών σερίνης / θρεονίνης που περιλαμβάνει δύο πρωτεΐνες, την LIMK1 και την LIMK2. Οι κινάσες παίζουν κεντρικό ρόλο στα ενδοκυττάρια συστήματα μεταγωγής σήματος που εμπλέκονται στον πολλαπλασιασμό των κυττάρων, τη διαφοροποίηση, τον μεταβολισμό, και άλλες αποκρίσεις σε εξωτερικά ερεθίσματα.(cohen Pet al,1992; Hunter T.et al,1995). Κύρια δράση και των δύο πρωτεϊνών της οικογένειας LIMK είναι να ρυθμίζουν το πολυμερισμό-αποπολυμερισμό του κυτταροσκελετού ακτίνης φωσφορυλιώνοντας στην Ser-3 και αδρανοποιώντας τις πρωτεΐνες της οικογένειας ADF / κοφιλίνη. Οι πρωτεΐνες της οικογένειας αυτής με κύρια την κοφιλίνη ανήκουν στις ABPs και παίζουν σημαντικό ρόλο στην ρύθμιση της δυναμικής του κυτταροσκελετού ακτίνης. H κοφιλίνη παίζει σημαντικό ρόλο στη διατήρηση και την επέκταση μεμβρανικών προεξοχών κίνησης όπως τα ελασματοπόδια που βρίσκονται στην αιχμή των μεταναστευτικών κυττάρων (Bailly Jones et al,2003; Dawe et al, 2003). Είναι ευρέως διαδεδομένη (πρωτεΐνη kda) έχοντας σημαντικό ρόλο στην κυτταρική μετανάστευση, την κυτοκίνηση, την ενδοκυττάρωση κα. (Carlier et al, 1999). Πρωτεΐνες της οικογένειας ADF/cofilin από διαφορετικούς οργανισμούς παρουσιάζουν υψηλό βαθμό ομολογίας στην αλληλουχία και ο γενικός μηχανισμός δράσης των διαφόρων ADF/cofilins είναι συντηρημένη. Η τρισδιάστατη δομή προσδιορίστηκε με κρυσταλλογραφία ακτίνων-χ και δείχνει πέντε κεντρικά βήταφύλλα πλαισιώνονται από τρις έως τέσσερις άλφα-έλικες. (Carlier, M.-F et al,1999). Η κοφιλίνη επάγει τον πολυμερισμό της ακτίνης με δύο μηχανισμούς 1) αποκόπτοντας σχηματισμένα νημάτια ακτίνης δημιουργώντας νέα ελεύθερα άκρα και 2) αυξάνει το ρυθμό αποπολυμερισμού στο (-) άκρο, διατηρώντας έτσι μια σταθερή δεξαμενή μονομερών ακτίνης (Condeelis, J et al, 2001). Η κοφιλίνη αδρανοποιείται από τις LIM κινάσες (LΙΜΚ) μέσω της φωσφορυλίωσης στη Ser-3 (Arber et al,1998;yang et al,1998;toshima et al,2001). Αυτή η φωσφορυλίωση αναστέλλει την αλληλεπίδραση της κοφιλίνης με νημάτια και μονομερή ακτίνης. H ανενεργή κοφιλίνη (P-κοφιλίνη) αποφωσφορυλιώνεται και ενεργοποιείται από μια οικογένεια φωσφατασών τις Slingshot (SSH) (Niwa et al, 2002; Ohta et al, 2003)(εικόνα 35) Εικόνα 35 Σχηματική απεικόνιση, ADF / κοφιλίνη και ρυθμιστών. 66
67 Μέχρι σήμερα ομόλογα LIM κινάσης έχουν ταυτοποιηθεί σε μία ποικιλία ειδών όπως στον άνθρωπο (Mizuno K,et al,1994; Nunoue K,et al,1995;okano Iet al,1995), τα ποντίκια (αρχικά ονομάστηκε Kiz-1) (Bernard O et al,1994), τους αρουραίους (Nunoue K,et al,1995) και τα κοτόπουλα (Koshimizu U,et al,1997). Σχετικά ομόλογα LIMK1 εκφράζονται επίσης σε είδη με πιο μακρινή συγγένεια όπως Xenopus laevis (XLIMK Takahashi Tet al,1997), Drosophila melanogaster (DLIMK, Ohashi K, et al,2000) και στα Αφρικανικά κουνούπια Anopheles gambiae (Εικόνα 36). LIMK2 BOS LIMK2 CANIS LIMK2 RATTUS LIMK2 MUS LIMK2 HOMO LIMK ANOPHELES LIMK2 GALLUS LIMK2 DANIO LIMK1 DROSOPHILA LIMK1 DANIO LIMK1 CANIS LIMK1 HOMO LIMK1 PAN LIMK1MUS LIMK1 RATTUS LIMK1 XENOPUS LIMK1 GALLUS Εικόνα36. Φυλογενετικό δενδρόγραμμα 1 LIM K 1- LIMK- 2 Υπάρχουν περίπου 40 γνωστές πρωτεΐνες LIM στα ευκαρυωτικά, που ονομάστηκαν έτσι για τις LIM περιοχές που περιέχουν. Οι LIM περιοχές είναι εξαιρετικά διατηρημένες δομές πλούσιες σε κυστεΐνη που περιέχουν 2 δακτυλίους ψευδαργύρου. Μολονότι οι δακτύλιοι ψευδαργύρου λειτουργούν με δέσμευση στο DNA ή RNA στις LIM-κινάσες πιθανώς διαμεσολαβούν πρωτεϊνικές αλληλεπιδράσεις. Οι LIM κινάση-1 και LIM κινάση-2 ανήκουν σε μια υποοικογένεια πρωτεϊνών LIM με ένα μοναδικό συνδυασμό από 2 Ν-τελικά μοτίβα LIM και μια περιοχή πρωτεΐνης κινάσης στο καρβοξυτελικό άκρο. Εμφανίζουν 50% ομολογία στην αλληλουχία αμινοξέων παρά το γεγονός ότι τα γονίδια βρίσκονται σε διαφορετικά χρωμοσώματα.(bernard et al,1996;ikebe et al,1997). Πιο συγκεκριμένα οι αλληλουχίες στις περιοχές LIM, ΡDΖ, και στην περιοχή της κινάσης εμφανίζουν ομοιότητα 50%, 45% και 70%, αντιστοίχως. Οι κινάσες που συνδέονται πιο στενά με την LIMK1 και LIMK2 είναι οι TESK1 και TESK2 (εκφράζονται στους όρχεις), στις οποίες η περιοχή της κινάσης έχει περίπου 50% ομοιότητα με την περιοχή κινάσης της LIMK1. 67

68 Το γονίδιo LIMK1 βρίσκεται στο μακρύ (q) βραχίονα του χρωμοσώματος 7 στην θέση 11,23. Συγκεκριμένα από το ζεύγος βάσεων έως ζεύγος βάσεων στο χρωμόσωμα 7.(εικόνα 37) Εικόνα 37. Σχηματική αναπαράσταση τoυ γονιδιακού τόπου της LIMK1 στο χρωμόσωμα 7 Atlas of Genetics and Cytogenetics in Oncology and Haematology Jean-Loup Huret, INSERM U 935, GeneticsUniversity Hospital of Poitiers, F Poitiers, France Το γονίδιο της LIMK1 αποτελείται από 16 εξώνια. Τα εξόνια 2-4 κωδικοποιούν δύο LIM περιοχές. Οι LIM περιοχές είναι εξαιρετικά διατηρημένες δομές πλούσιες σε κυστεΐνη που περιέχουν 2 δακτυλίους ψευδαργύρου. Τα εξώνια 4-6 κωδικοποιούν την περιοχή ΡDΖ και τα εξώνια 8-16 κωδικοποιούν την περιοχή κινάσης σερίνης /θρεονίνης. Σε μία παραλλαγή του ματίσματος το ιντρόνιο 7 επεκτείνεται σε 61 επιπλέον βάσεις στο 5 'άκρο του εξώνιου 8, προκαλώντας την μετατόπιση του πλαισίου μεταγραφής και πρόωρο τερματισμό της μεταγραφής. Το γονίδιο της LIMK2 βρίσκεται στο μακρύ (q) βραχίονα του χρωμοσώματος 22, 22q12. (εικόνα 38) Η έκφραση του mrna της LIM K2 (3,7-kb) ανιχνεύθηκε σε ιστούς, με υψηλότερα επίπεδα έκφρασης στον πλακούντα, και μέτρια επίπεδα στο ήπαρ, πνεύμονα, νεφρό και πάγκρεας. Εμφανίζει ιδιότητες κινάσης σερίνης/θρεονίνης και φωσφορυλίωνει την βασική πρωτεΐνη της μυελίνης (ΜΒΡ) in vitro. Εικόνα 38. Σχηματική παράσταση του γονιδίου της LIMK2,στο μακρύ (q)βραχίονα του χρωμοσώματος 22 68
Συνέντευξη με τον Παθολόγο - Ογκολόγο, Στυλιανό Γιασσά
 Συνέντευξη με τον Παθολόγο - Ογκολόγο, Στυλιανό Γιασσά Ο καρκίνος του παχέος εντέρου ορθού αποτελεί το δεύτερο πιο συχνό καρκίνο σε γυναίκες και άνδρες και αντιπροσωπεύει το 13% όλων των καρκίνων. Στην
Συνέντευξη με τον Παθολόγο - Ογκολόγο, Στυλιανό Γιασσά Ο καρκίνος του παχέος εντέρου ορθού αποτελεί το δεύτερο πιο συχνό καρκίνο σε γυναίκες και άνδρες και αντιπροσωπεύει το 13% όλων των καρκίνων. Στην
ΠΡΟΚΑΡΚΙΝΙΚΕΣ ΑΛΛΟΙΩΣΕΙΣ ΣΤΟΜΑΧΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ
 ΠΡΟΚΑΡΚΙΝΙΚΕΣ ΑΛΛΟΙΩΣΕΙΣ ΣΤΟΜΑΧΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΕΛΛΗΝΙΚΗ ΠΑΘΟΛΟΓΟAΝΑΤΟΜΙΚΗ ΕΤΑΙΡΕΙΑ ΟΜΑΔΑ ΠΕΠΤΙΚΟΥ ΑΘΗΝΑ 15 Δεκεμβρίου 2004 Mαρία Δαιμονάκου- Βατοπούλου Αναπληρώτρια Διευθύντρια Σισμανόγλειο ΓΠΝΑ Kαρκινογέννεση
ΠΡΟΚΑΡΚΙΝΙΚΕΣ ΑΛΛΟΙΩΣΕΙΣ ΣΤΟΜΑΧΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΕΛΛΗΝΙΚΗ ΠΑΘΟΛΟΓΟAΝΑΤΟΜΙΚΗ ΕΤΑΙΡΕΙΑ ΟΜΑΔΑ ΠΕΠΤΙΚΟΥ ΑΘΗΝΑ 15 Δεκεμβρίου 2004 Mαρία Δαιμονάκου- Βατοπούλου Αναπληρώτρια Διευθύντρια Σισμανόγλειο ΓΠΝΑ Kαρκινογέννεση
Εμβρυολογία πεπτικού συστήματος
 Εμβρυολογία πεπτικού συστήματος Υπατία Δούση-Αναγνωστοπούλου Εργαστήριο Ιστολογίας και Εμβρυολογίας AΡΧΕΓΟΝΟ ΕΝΤΕΡΟ : - πρόσθιο έντερο, - μέσο έντερο, - οπίσθιο έντερο. `σχηματίζεται την 4η εβδομάδα (από
Εμβρυολογία πεπτικού συστήματος Υπατία Δούση-Αναγνωστοπούλου Εργαστήριο Ιστολογίας και Εμβρυολογίας AΡΧΕΓΟΝΟ ΕΝΤΕΡΟ : - πρόσθιο έντερο, - μέσο έντερο, - οπίσθιο έντερο. `σχηματίζεται την 4η εβδομάδα (από
ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ
 ΕΛΛΗΝΙΚΗ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΙΚΗ ΕΤΑΙΡΕΙΑ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΝΩΣΗ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΩΝ ΕΛΛΑΔΑΣ ΕΛΛΗΝΙΚΟ ΙΔΡΥΜΑ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΙΑΣ ΚΑΙ ΔΙΑΤΡΟΦΗΣ ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΑΘΗΝΑ 2008 ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ
ΕΛΛΗΝΙΚΗ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΙΚΗ ΕΤΑΙΡΕΙΑ ΕΠΑΓΓΕΛΜΑΤΙΚΗ ΕΝΩΣΗ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΩΝ ΕΛΛΑΔΑΣ ΕΛΛΗΝΙΚΟ ΙΔΡΥΜΑ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΙΑΣ ΚΑΙ ΔΙΑΤΡΟΦΗΣ ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΑΘΗΝΑ 2008 ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ. Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη
 Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΗ ΕΚΤΙΜΗΣΗ ΑΣΘΕΝΩΝ ΜΕ Ι.Φ.Ν.Ε. Αξιολόγηση της ιστολογικής απάντησης Αντιπαράθεση Παθολογοανατόμου-Γαστρεντερολόγου
 ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΗ ΕΚΤΙΜΗΣΗ ΑΣΘΕΝΩΝ ΜΕ Ι.Φ.Ν.Ε. Αξιολόγηση της ιστολογικής απάντησης Αντιπαράθεση Παθολογοανατόμου-Γαστρεντερολόγου Χ. Σπηλιάδη Παθολογοανατόμος ΕΛΚΩΔΗΣ ΚΟΛΙΤΙΔΑ ΝΟΣΟΣ CROHN «ΑΠΡΟΣΔΙΟΡΙΣΤΗ»
ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΗ ΕΚΤΙΜΗΣΗ ΑΣΘΕΝΩΝ ΜΕ Ι.Φ.Ν.Ε. Αξιολόγηση της ιστολογικής απάντησης Αντιπαράθεση Παθολογοανατόμου-Γαστρεντερολόγου Χ. Σπηλιάδη Παθολογοανατόμος ΕΛΚΩΔΗΣ ΚΟΛΙΤΙΔΑ ΝΟΣΟΣ CROHN «ΑΠΡΟΣΔΙΟΡΙΣΤΗ»
ΕΘΝΙΚΗ ΕΠΙΤΡΟΠΗ ΠΡΟΛΗΨΗΣ ΚΑΡΚΙΝΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ
 ΕΘΝΙΚΗ ΕΠΙΤΡΟΠΗ ΠΡΟΛΗΨΗΣ ΚΑΡΚΙΝΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΑΘΗΝΑ 2015 ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ Τι είναι και πόσο συχνός είναι ο καρκίνος του παχέος εντέρου;
ΕΘΝΙΚΗ ΕΠΙΤΡΟΠΗ ΠΡΟΛΗΨΗΣ ΚΑΡΚΙΝΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΑΘΗΝΑ 2015 ΠΡΟΛΗΨΗ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ Τι είναι και πόσο συχνός είναι ο καρκίνος του παχέος εντέρου;
Η σημασία του προληπτικού ελέγχου για τον ΚΠΕ. Όλγα Ι Γιουλεμέ Επίκουρη Καθηγήτρια Γαστρεντερολογίας ΑΠΘ Β Προπ. Παθολογική Κλινική ΓΠΝΘ ΙΠΠΟΚΡΑΤΕΙΟ
 Η σημασία του προληπτικού ελέγχου για τον ΚΠΕ Όλγα Ι Γιουλεμέ Επίκουρη Καθηγήτρια Γαστρεντερολογίας ΑΠΘ Β Προπ. Παθολογική Κλινική ΓΠΝΘ ΙΠΠΟΚΡΑΤΕΙΟ καρκίνος παχέος εντέρου (ΚΠΕ) Ευρώπη: 2 η αιτία θανάτου
Η σημασία του προληπτικού ελέγχου για τον ΚΠΕ Όλγα Ι Γιουλεμέ Επίκουρη Καθηγήτρια Γαστρεντερολογίας ΑΠΘ Β Προπ. Παθολογική Κλινική ΓΠΝΘ ΙΠΠΟΚΡΑΤΕΙΟ καρκίνος παχέος εντέρου (ΚΠΕ) Ευρώπη: 2 η αιτία θανάτου
Πρόληψη κληρονομικού καρκίνου παχέος εντέρου. Μιχάλης Οικονόμου Γαστρεντερολόγος
 Πρόληψη κληρονομικού καρκίνου παχέος εντέρου Μιχάλης Οικονόμου Γαστρεντερολόγος Καρκίνος Παχέος Εντέρου (ΚΠΕ) ΚΠΕ είναι ο 3 ος συχνότερος καρκίνος ΚΠΕ είναι η 2 η αιτία θανάτου από καρκίνο Μείωση αριθμών
Πρόληψη κληρονομικού καρκίνου παχέος εντέρου Μιχάλης Οικονόμου Γαστρεντερολόγος Καρκίνος Παχέος Εντέρου (ΚΠΕ) ΚΠΕ είναι ο 3 ος συχνότερος καρκίνος ΚΠΕ είναι η 2 η αιτία θανάτου από καρκίνο Μείωση αριθμών
Παράγοντες κινδύνου για τον καρκίνο του Παγκρέατος. Στέργιος Δελακίδης Γαστρεντερολόγος
 Παράγοντες κινδύνου για τον καρκίνο του Παγκρέατος Στέργιος Δελακίδης Γαστρεντερολόγος Καρκίνος Παγκρέατος: πρόγνωση Κακή πρόγνωση Συνολική πενταετής επιβίωση 6%.
Παράγοντες κινδύνου για τον καρκίνο του Παγκρέατος Στέργιος Δελακίδης Γαστρεντερολόγος Καρκίνος Παγκρέατος: πρόγνωση Κακή πρόγνωση Συνολική πενταετής επιβίωση 6%.
Δεκαπεντάλεπτη προετοιμασία του φοιτητή ιατρικής για το μάθημα του καρκίνου του όρχη βασικές γνώσεις :
 Δεκαπεντάλεπτη προετοιμασία του φοιτητή ιατρικής για το μάθημα του καρκίνου του όρχη βασικές γνώσεις : Οι όρχεις αποτελούν κομμάτι του αναπαραγωγικού συστήματος (παραγωγή σπερματοζωάριων) του άνδρα αλλά
Δεκαπεντάλεπτη προετοιμασία του φοιτητή ιατρικής για το μάθημα του καρκίνου του όρχη βασικές γνώσεις : Οι όρχεις αποτελούν κομμάτι του αναπαραγωγικού συστήματος (παραγωγή σπερματοζωάριων) του άνδρα αλλά
Καρκίνος του παχέος εντέρου
 Καρκίνος τι Πρ έπ ει να ''ξέρω... του Π. Πετρίδη* Παχύ έντερο Το παχύ έντερο είναι το τελευταίο τμήμα του πεπτικού συστήματος και αποτελεί τη συνέχεια του λεπτού εντέρου. Αποτελείται από 6 τμήματα που
Καρκίνος τι Πρ έπ ει να ''ξέρω... του Π. Πετρίδη* Παχύ έντερο Το παχύ έντερο είναι το τελευταίο τμήμα του πεπτικού συστήματος και αποτελεί τη συνέχεια του λεπτού εντέρου. Αποτελείται από 6 τμήματα που
αμινοξύ. Η αλλαγή αυτή έχει ελάχιστη επίδραση στη στερεοδιάταξη και τη λειτουργικότητα της πρωτεϊνης. Επιβλαβής
 Κεφάλαιο 6: ΜΕΤΑΛΛΑΞΕΙΣ -ΘΕΩΡΙΑ- Μεταλλάξεις είναι οι αλλαγές που συμβαίνουν στο γενετικό υλικό ενός οργανισμού, τόσο σε γονιδιακό επίπεδο (γονιδιακές μεταλλάξεις) όσο και σε χρωμοσωμικό επίπεδο (χρωμοσωμικές
Κεφάλαιο 6: ΜΕΤΑΛΛΑΞΕΙΣ -ΘΕΩΡΙΑ- Μεταλλάξεις είναι οι αλλαγές που συμβαίνουν στο γενετικό υλικό ενός οργανισμού, τόσο σε γονιδιακό επίπεδο (γονιδιακές μεταλλάξεις) όσο και σε χρωμοσωμικό επίπεδο (χρωμοσωμικές
Μήπως έχω Σκληρόδερµα;
 Μήπως έχω Σκληρόδερµα; Για να πληροφορηθώ µýëïò ôçò Σπάνιος ναι... Μόνος όχι Η Πανελλήνια Ένωση Σπανίων Παθήσεων (Π.Ε.Σ.ΠΑ) είναι ο μόνος φορέας, μη κερδοσκοπικό σωματείο, συλλόγων ασθενών σπανίων παθήσεων
Μήπως έχω Σκληρόδερµα; Για να πληροφορηθώ µýëïò ôçò Σπάνιος ναι... Μόνος όχι Η Πανελλήνια Ένωση Σπανίων Παθήσεων (Π.Ε.Σ.ΠΑ) είναι ο μόνος φορέας, μη κερδοσκοπικό σωματείο, συλλόγων ασθενών σπανίων παθήσεων
ΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΝΕΥΜΟΝΑ
 ΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΝΕΥΜΟΝΑ Χρ.Τσοµπανίδου Παθολογοανατόµος Συντονίστρια Διευθύντρια Νοσοκοµείο Θεσ/νίκης «Άγιος Δηµήτριος» προέρχονται από το φυσιολογικό επιθήλιο µε την
ΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΝΕΥΜΟΝΑ Χρ.Τσοµπανίδου Παθολογοανατόµος Συντονίστρια Διευθύντρια Νοσοκοµείο Θεσ/νίκης «Άγιος Δηµήτριος» προέρχονται από το φυσιολογικό επιθήλιο µε την
φυσιολογικό δέρμα - 1
 φυσιολογικό δέρμα -1 Επιδερμίδα (επιθήλιο, εξωδερμική προέλευση) Α Α Α Μ θηλώδες χόριο (επιπολής) ακανθωτή στιβάδα βασική στιβάδα χόριο Μ = Μελανοκύτταρο (νευροεξωδερμική προέλευση, νευρική ακρολοφία)
φυσιολογικό δέρμα -1 Επιδερμίδα (επιθήλιο, εξωδερμική προέλευση) Α Α Α Μ θηλώδες χόριο (επιπολής) ακανθωτή στιβάδα βασική στιβάδα χόριο Μ = Μελανοκύτταρο (νευροεξωδερμική προέλευση, νευρική ακρολοφία)
ΠΡΟΚΑΡΚΙΝΩΜΑΤΩΔΕΙΣ ΑΛΛΟΙΩΣΕΙΣ ΤΟΥ ΠΑΓΚΡΕΑΤΟΣ
 ΠΡΟΚΑΡΚΙΝΩΜΑΤΩΔΕΙΣ ΑΛΛΟΙΩΣΕΙΣ ΤΟΥ ΠΑΓΚΡΕΑΤΟΣ Ελένη Σπανίδου-Καρβούνη Καρβούνη, FRCPC Παθολογοανατόμος Αρεταίειο Νοσοκομείο Προκαρκινωματώδεις αλλοιώσεις του παγκρέατος Παγκρεατική Ενδοεπιθηλιακή Νεοπλασία
ΠΡΟΚΑΡΚΙΝΩΜΑΤΩΔΕΙΣ ΑΛΛΟΙΩΣΕΙΣ ΤΟΥ ΠΑΓΚΡΕΑΤΟΣ Ελένη Σπανίδου-Καρβούνη Καρβούνη, FRCPC Παθολογοανατόμος Αρεταίειο Νοσοκομείο Προκαρκινωματώδεις αλλοιώσεις του παγκρέατος Παγκρεατική Ενδοεπιθηλιακή Νεοπλασία
Η ρομποτική χειρουργική στον καρκίνο του παχέος εντέρου - Ο Δρόμος για την Θεραπεία Τρίτη, 21 Σεπτέμβριος :49
 Συνέντευξη με τον Χειρουργό, Νικόλαο Τσαγκούλη Ο καρκίνος του παχέος εντέρου είναι ο τρίτος πιο συχνά εμφανιζόμενος καρκίνος σε άνδρες και γυναίκες. Παρόλα αυτά η θνησιμότητα από καρκίνο του παχέος εντέρου
Συνέντευξη με τον Χειρουργό, Νικόλαο Τσαγκούλη Ο καρκίνος του παχέος εντέρου είναι ο τρίτος πιο συχνά εμφανιζόμενος καρκίνος σε άνδρες και γυναίκες. Παρόλα αυτά η θνησιμότητα από καρκίνο του παχέος εντέρου
VIN (Ενδοεπιθηλιακή νεοπλασία αιδοίου)
") VIN (Ενδοεπιθηλιακή νεοπλασία αιδοίου) Διαφορές μεταξύ τραχήλου και αιδοίου Κύτταρο-στόχος: κυρίως πλακώδες δεν υπάρχει εκτεθειμένος πολυδύναμος πληθυσμός, όπως στον τράχηλο Σχετιζόμενοι HPV: κυριαρχούν
VIN (Ενδοεπιθηλιακή νεοπλασία αιδοίου) Διαφορές μεταξύ τραχήλου και αιδοίου Κύτταρο-στόχος: κυρίως πλακώδες δεν υπάρχει εκτεθειμένος πολυδύναμος πληθυσμός, όπως στον τράχηλο Σχετιζόμενοι HPV: κυριαρχούν
Section A: Είδος επέμβασης
 Section A: Είδος επέμβασης A1. Είδος Επέμβασης Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple ), μερική παγκρεατεκτομή Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple), ολική παγκρεατεκτομή PPPD, μερική παγκρεατεκτομή
Section A: Είδος επέμβασης A1. Είδος Επέμβασης Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple ), μερική παγκρεατεκτομή Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple), ολική παγκρεατεκτομή PPPD, μερική παγκρεατεκτομή
Εργασία για το μάθημα της βιολογίας Υπεύθυνος Καθηγητής : Dr Κεραμάρης Κων/νος Συντελεστές : Αϊναλάκης Πέτρος Γ 1 Κυρίκος Κυριάκος Γ 1
 Εργασία για το μάθημα της βιολογίας Υπεύθυνος Καθηγητής : Dr Κεραμάρης Κων/νος Συντελεστές : Αϊναλάκης Πέτρος Γ 1 Κυρίκος Κυριάκος Γ 1 Καρκίνος..1 Γενικά χαρακτηριστικά καρκινικών κυττάρων...2 Ανάλυση
Εργασία για το μάθημα της βιολογίας Υπεύθυνος Καθηγητής : Dr Κεραμάρης Κων/νος Συντελεστές : Αϊναλάκης Πέτρος Γ 1 Κυρίκος Κυριάκος Γ 1 Καρκίνος..1 Γενικά χαρακτηριστικά καρκινικών κυττάρων...2 Ανάλυση
Λίγα λόγια για τους καρκίνους, τη γενετική τους βάση και διερεύνηση
 Λίγα λόγια για τους καρκίνους, τη γενετική τους βάση και διερεύνηση Ο καρκίνος είναι μια ιδιαίτερα ετερογενής νόσος, που προκύπτει από συσσώρευση μεταλλάξεων του DNA. Η αιτιολογία του καρκίνου είναι πολυπαραγοντική,
Λίγα λόγια για τους καρκίνους, τη γενετική τους βάση και διερεύνηση Ο καρκίνος είναι μια ιδιαίτερα ετερογενής νόσος, που προκύπτει από συσσώρευση μεταλλάξεων του DNA. Η αιτιολογία του καρκίνου είναι πολυπαραγοντική,
ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΕΤΟΣ ΚΑΡΚΙΝΟΣ ΚΑΙ ΜΕΤΑΛΛΑΞΕΙΣ
 ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΕΤΟΣ 2012-2013 ΚΑΡΚΙΝΟΣ ΚΑΙ ΜΕΤΑΛΛΑΞΕΙΣ Τι είναι ο καρκίνος ; Ο Καρκίνος είναι ένα από τα σοβαρότερα προβλήματα υγείας που παρατηρούνται σήμερα στις αναπτυγμένες χώρες. Οι στατιστικές
ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΕΤΟΣ 2012-2013 ΚΑΡΚΙΝΟΣ ΚΑΙ ΜΕΤΑΛΛΑΞΕΙΣ Τι είναι ο καρκίνος ; Ο Καρκίνος είναι ένα από τα σοβαρότερα προβλήματα υγείας που παρατηρούνται σήμερα στις αναπτυγμένες χώρες. Οι στατιστικές
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4
 ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
ΙΦΝΕ (ΕΚ, ν.crohn, απροσδιόριστη) Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες
 Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες") ΦΛΕΓΜΟΝΩΔΗ ΝΟΣΗΜΑΤΑ Π.Ε ΙΦΝΕ (ΕΚ, ν.crohn, απροσδιόριστη) Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες (ΜΣΑΦ κ.ά) Ισχαιμική Μετακτινική
ΦΛΕΓΜΟΝΩΔΗ ΝΟΣΗΜΑΤΑ Π.Ε ΙΦΝΕ (ΕΚ, ν.crohn, απροσδιόριστη) Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες (ΜΣΑΦ κ.ά) Ισχαιμική Μετακτινική
ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ. «Ιατρική Χημεία: Σχεδιασμός και Ανάπτυξη Φαρμακευτικών Προϊόντων»
 Τμήματα Χημείας και Ιατρικής ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «Ιατρική Χημεία: Σχεδιασμός και Ανάπτυξη Φαρμακευτικών Προϊόντων» Δείκτες επιθήλιο-μεσεγχυματικής
Τμήματα Χημείας και Ιατρικής ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΔΙΑΤΜΗΜΑΤΙΚΟ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «Ιατρική Χημεία: Σχεδιασμός και Ανάπτυξη Φαρμακευτικών Προϊόντων» Δείκτες επιθήλιο-μεσεγχυματικής
igenetics Mια Μεντελική προσέγγιση
 igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 22 (+κεφ. 17 Hartwell) Γενετική του καρκίνου Η πρωτεΐνη p53 προσδένεται στο DNA. 2 ΕΙΚΟΝΑ 22.1 Μαστογραφία που απεικονίζει έναν όγκο. Όγκος 3 Κύρια σημεία: Καρκίνος
igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 22 (+κεφ. 17 Hartwell) Γενετική του καρκίνου Η πρωτεΐνη p53 προσδένεται στο DNA. 2 ΕΙΚΟΝΑ 22.1 Μαστογραφία που απεικονίζει έναν όγκο. Όγκος 3 Κύρια σημεία: Καρκίνος
Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης. Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010
 ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Ι) ΗΠΑΤΕΚΤΟΜΗ ΓΙΑ ΕΝ ΟΗΠΑΤΙΚΟ ΧΟΛΑΓΓΕΙΟΚΑΡΚΙΝΩΜΑ: ΜΑΚΡΟΣΚΟΠΙΚΗ ΕΞΕΤΑΣΗ:
 ΗΠΑΤΕΚΤΟΜΗ ΓΙΑ ΕΝ ΟΗΠΑΤΙΚΟ ΧΟΛΑΓΓΕΙΟΚΑΡΚΙΝΩΜΑ: ΜΑΚΡΟΣΚΟΠΙΚΗ ΕΞΕΤΑΣΗ:") ΧΕΙΡΟΥΡΓΙΚΑ ΠΑΡΑΣΚΕΥΑΣΜΑΤΑ ΕΚΤΟΜΗΣ ΧΟΛΑΓΓΕΙΟΚΑΡΚΙΝΩΜΑΤΟΣ Β. Τζιούφα, Αναπληρώτρια Καθηγήτρια Ιατρικής Σχολής Εργαστηρίου Παθολογικής Ανατοµικής, Αριστοτέλειο Πανεπιστήµιο Θεσσαλονίκης Ι) ΗΠΑΤΕΚΤΟΜΗ ΓΙΑ
ΧΕΙΡΟΥΡΓΙΚΑ ΠΑΡΑΣΚΕΥΑΣΜΑΤΑ ΕΚΤΟΜΗΣ ΧΟΛΑΓΓΕΙΟΚΑΡΚΙΝΩΜΑΤΟΣ Β. Τζιούφα, Αναπληρώτρια Καθηγήτρια Ιατρικής Σχολής Εργαστηρίου Παθολογικής Ανατοµικής, Αριστοτέλειο Πανεπιστήµιο Θεσσαλονίκης Ι) ΗΠΑΤΕΚΤΟΜΗ ΓΙΑ
ΓΕΝΙΚΑ ΣΤΟΙΧΕΙΑ ΓΙΑ ΤΗΝ ΚΥΚΛΟΦΟΡΙΑ. Σχηµατική απεικόνιση της µεγάλης και της µικρής κυκλοφορίας
 ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ Ι ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιµος Π. Βανδώρος ΑΙΜΟΦΟΡΑ ΑΓΓΕΙΑ ΑΡΤΗΡΙΕΣ - ΦΛΕΒΕΣ - ΤΡΙΧΟΕΙ Η 1 ΓΕΝΙΚΑ ΣΤΟΙΧΕΙΑ ΓΙΑ ΤΗΝ ΚΥΚΛΟΦΟΡΙΑ Μεγάλη και µικρή κυκλοφορία Σχηµατική
ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΑΝΑΤΟΜΙΑ Ι ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιµος Π. Βανδώρος ΑΙΜΟΦΟΡΑ ΑΓΓΕΙΑ ΑΡΤΗΡΙΕΣ - ΦΛΕΒΕΣ - ΤΡΙΧΟΕΙ Η 1 ΓΕΝΙΚΑ ΣΤΟΙΧΕΙΑ ΓΙΑ ΤΗΝ ΚΥΚΛΟΦΟΡΙΑ Μεγάλη και µικρή κυκλοφορία Σχηµατική
Σύμφωνα με τον παγκόσμιο οργανισμό υγείας, κάθε χρόνο υπάρχουν 1.38 εκατομμύρια καινούρια περιστατικά και περίπου 458 000 θάνατοι από τον καρκίνο του
 1 Σύμφωνα με τον παγκόσμιο οργανισμό υγείας, κάθε χρόνο υπάρχουν 1.38 εκατομμύρια καινούρια περιστατικά και περίπου 458 000 θάνατοι από τον καρκίνο του μαστού. Ο καρκίνος του μαστού είναι με μεγάλη διαφορά
1 Σύμφωνα με τον παγκόσμιο οργανισμό υγείας, κάθε χρόνο υπάρχουν 1.38 εκατομμύρια καινούρια περιστατικά και περίπου 458 000 θάνατοι από τον καρκίνο του μαστού. Ο καρκίνος του μαστού είναι με μεγάλη διαφορά
TI NEOTEΡΟ ΣΤΟΥΣ ΝΕΥΡΟΕΝΔΟΚΡΙΝΕΙΣ ΟΓΚΟΥΣ ΤΟ 2015. Ορισμοί-Κατάταξη-Ιστολογία. Δ Ροντογιάννη
 TI NEOTEΡΟ ΣΤΟΥΣ ΝΕΥΡΟΕΝΔΟΚΡΙΝΕΙΣ ΟΓΚΟΥΣ ΤΟ 2015 Ορισμοί-Κατάταξη-Ιστολογία Δ Ροντογιάννη Νευροενδοκρινή νεοπλάσματα Ταξινόμηση Βιολογικοί δείκτες Μοριακή Παθολογία Νευροενδοκρινή νεοπλάσματα Ταξινόμηση
TI NEOTEΡΟ ΣΤΟΥΣ ΝΕΥΡΟΕΝΔΟΚΡΙΝΕΙΣ ΟΓΚΟΥΣ ΤΟ 2015 Ορισμοί-Κατάταξη-Ιστολογία Δ Ροντογιάννη Νευροενδοκρινή νεοπλάσματα Ταξινόμηση Βιολογικοί δείκτες Μοριακή Παθολογία Νευροενδοκρινή νεοπλάσματα Ταξινόμηση
Section A: ΕΠΙΔΗΜΙΟΛΟΓΙΚΑ ΣΤΟΙΧΕΙΑ
 ΣΤΟΙΧΕΙΑ ΑΣΘΕΝΟΥΣ ΜΕ ΚΑΡΚΙΝΟ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΟΡΘΟΥ ΠΡΟΕΓΧΕΙΡΗΤΙΚΗ ΕΚΤΙΜΗΣΗ Section A: ΕΠΙΔΗΜΙΟΛΟΓΙΚΑ ΣΤΟΙΧΕΙΑ A1. Εθνικότητα Ελληνική Άλλη Εθνικότητα Άλλη Εθνικότητα A2. Τωρινό Επάγγελμα A3. Έτη στο τωρινό
ΣΤΟΙΧΕΙΑ ΑΣΘΕΝΟΥΣ ΜΕ ΚΑΡΚΙΝΟ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ ΟΡΘΟΥ ΠΡΟΕΓΧΕΙΡΗΤΙΚΗ ΕΚΤΙΜΗΣΗ Section A: ΕΠΙΔΗΜΙΟΛΟΓΙΚΑ ΣΤΟΙΧΕΙΑ A1. Εθνικότητα Ελληνική Άλλη Εθνικότητα Άλλη Εθνικότητα A2. Τωρινό Επάγγελμα A3. Έτη στο τωρινό
Ατυπία Υπερπλασία- Δυσπλασία. Κίττυ Παυλάκη
 Ατυπία Υπερπλασία- Δυσπλασία Κίττυ Παυλάκη Jeanne Calment Κάπνιζε µέχρι τα 117 Πέθανε στα 122 Η σωστή λειτουργία των οργανισµών απαιτεί τη δυνατότητα προσαρµογής των κυττάρων και κατά συνέπεια και των
Ατυπία Υπερπλασία- Δυσπλασία Κίττυ Παυλάκη Jeanne Calment Κάπνιζε µέχρι τα 117 Πέθανε στα 122 Η σωστή λειτουργία των οργανισµών απαιτεί τη δυνατότητα προσαρµογής των κυττάρων και κατά συνέπεια και των
Βιοψία εκτομής (εκπυρήνιση) Μερική παγκρεατεκτομή, σώμα-ουρά παγκρέατος. Μερική παγκρεατεκτομή, ουρά παγκρέατος. Δεν καθορίζεται.
 Μερική παγκρεατεκτομή, σώμα-ουρά παγκρέατος. Μερική παγκρεατεκτομή, ουρά παγκρέατος. Δεν καθορίζεται.") Section A: Είδος επέμβασης A1. Βιοψία εκτομής (εκπυρήνιση) Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple ), μερική παγκρεατεκτομή Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple), ολική παγκρεατεκτομή Μερική παγκρεατεκτομή,
Section A: Είδος επέμβασης A1. Βιοψία εκτομής (εκπυρήνιση) Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple ), μερική παγκρεατεκτομή Παγκρεατο12δακτυλεκτομή(επέμβαση Whipple), ολική παγκρεατεκτομή Μερική παγκρεατεκτομή,
Κεφάλαιο 6: Μεταλλάξεις
 Κεφάλαιο 6: Μεταλλάξεις ΕΛΕΓΧΟΣ ΓΝΩΣΕΩΝ 1. Τι ονομάζονται μεταλλάξεις και ποια τα κυριότερα είδη τους; 2. Ποιες οι διαφορές μεταξύ γονιδιακών και χρωμοσωμικών μεταλλάξεων; 3. Οι μεταλλάξεις στα σωματικά
Κεφάλαιο 6: Μεταλλάξεις ΕΛΕΓΧΟΣ ΓΝΩΣΕΩΝ 1. Τι ονομάζονται μεταλλάξεις και ποια τα κυριότερα είδη τους; 2. Ποιες οι διαφορές μεταξύ γονιδιακών και χρωμοσωμικών μεταλλάξεων; 3. Οι μεταλλάξεις στα σωματικά
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟ ΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ. ΝΙΚΟΛΑΟΣ ΚΑΒΑΝΤΖΑΣ Επίκουρος Καθηγητής
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟ ΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ ΝΙΚΟΛΑΟΣ ΚΑΒΑΝΤΖΑΣ Επίκουρος Καθηγητής ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ Επιφυσιοβλάστωµα Επιφυσιοκύττωµα Ογκος
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟ ΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ ΝΙΚΟΛΑΟΣ ΚΑΒΑΝΤΖΑΣ Επίκουρος Καθηγητής ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ Επιφυσιοβλάστωµα Επιφυσιοκύττωµα Ογκος
Ανι χνευση μεταλλα ξεων στον καρκι νο και εξατομικευμε νη θεραπει α
 Ανι χνευση μεταλλα ξεων στον καρκι νο και εξατομικευμε νη θεραπει α Περικλής Μακρυθανα σης, MD, PhD, FMH, PD Κλινικός γενετιστής Κύριος Ερευνητής Ίδρυμα Ιατροβιολογικών Ερευνών της Ακαδημι ας Αθηνών 4o
Ανι χνευση μεταλλα ξεων στον καρκι νο και εξατομικευμε νη θεραπει α Περικλής Μακρυθανα σης, MD, PhD, FMH, PD Κλινικός γενετιστής Κύριος Ερευνητής Ίδρυμα Ιατροβιολογικών Ερευνών της Ακαδημι ας Αθηνών 4o
Βιολογία ΘΕΜΑ Α ΘΕΜΑ B
 Βιολογία προσανατολισμού Α. 1. β 2. γ 3. δ 4. γ 5. δ ΘΕΜΑ Α B1. 4,1,2,6,8,3,5,7 ΘΕΜΑ B B2. Σχολικό βιβλίο σελ. 103 Η γενετική καθοδήγηση είναι.υγιών απογόνων. Σχολικό βιβλίο σελ. 103 Παρ ότι γενετική καθοδήγηση
Βιολογία προσανατολισμού Α. 1. β 2. γ 3. δ 4. γ 5. δ ΘΕΜΑ Α B1. 4,1,2,6,8,3,5,7 ΘΕΜΑ B B2. Σχολικό βιβλίο σελ. 103 Η γενετική καθοδήγηση είναι.υγιών απογόνων. Σχολικό βιβλίο σελ. 103 Παρ ότι γενετική καθοδήγηση
ΚΑΤΑΛΟΓΟΣ ΑΣΘΕΝΕΙΩΝ ΠΟΥ ΑΝΑΦΕΡΟΝΤΑΙ ΣΤΟ ΣΧΟΛΙΚΟ ΕΓΧΕΙΡΙΔΙΟ
 ΑΣΘΕΝΕΙΕΣ ΚΑΤΑΛΟΓΟΣ ΑΣΘΕΝΕΙΩΝ ΠΟΥ ΑΝΑΦΕΡΟΝΤΑΙ ΣΤΟ ΣΧΟΛΙΚΟ ΕΓΧΕΙΡΙΔΙΟ ΤΡΟΠΟΣ ΚΛΗΡΟΝΟΜΙΚΟΤΗΤΑΣ ΣΥΝΕΠΕΙΕΣ ΕΙΔΟΣ ΜΕΤΑΛΛΑΞΗΣ Οικογενή υπερχοληστερολαιμία Αυτοσωμική επικρατής κληρονομικότητα Σχετίζεται με αυξημένο
ΑΣΘΕΝΕΙΕΣ ΚΑΤΑΛΟΓΟΣ ΑΣΘΕΝΕΙΩΝ ΠΟΥ ΑΝΑΦΕΡΟΝΤΑΙ ΣΤΟ ΣΧΟΛΙΚΟ ΕΓΧΕΙΡΙΔΙΟ ΤΡΟΠΟΣ ΚΛΗΡΟΝΟΜΙΚΟΤΗΤΑΣ ΣΥΝΕΠΕΙΕΣ ΕΙΔΟΣ ΜΕΤΑΛΛΑΞΗΣ Οικογενή υπερχοληστερολαιμία Αυτοσωμική επικρατής κληρονομικότητα Σχετίζεται με αυξημένο
Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία
 ΣΥΓΧΡΟΝΕΣ ΜΕΘΟΔΟΛΟΓΙΕΣ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ - ΓΕΝΕΤΙΚΗΣ Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία ΠΑΠΑΝΙΚΟΛΑΟΥ ΕΛΕΝΗ, Ph.D. Λέκτορας Εργαστήριο Βιολογίας, Ιατρική Σχολή Αθηνών
ΣΥΓΧΡΟΝΕΣ ΜΕΘΟΔΟΛΟΓΙΕΣ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ - ΓΕΝΕΤΙΚΗΣ Σύγχρονες μεθοδολογίες μοριακής βιολογίας και γενετικής στη γυναικολογία ΠΑΠΑΝΙΚΟΛΑΟΥ ΕΛΕΝΗ, Ph.D. Λέκτορας Εργαστήριο Βιολογίας, Ιατρική Σχολή Αθηνών
Γαστρεντερικές ορμόνες, νεύρωση & αιμάτωση. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Γαστρεντερικές ορμόνες, νεύρωση & αιμάτωση Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 02/12/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Οικογένεια
Γαστρεντερικές ορμόνες, νεύρωση & αιμάτωση Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 02/12/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Οικογένεια
ÐÁÍÅÐÉÓÔÇÌÉÏ ÉÙÁÍÍÉÍÙÍ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ÔÏÌÅÁÓ ΜΟΡΦΟΛΟΓΙΚΟΣ - ÊËÉÍÉÊÏÅÑÃÁÓÔÇÑÉÁÊÏÓ ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΑΣ
 ÐÁÍÅÐÉÓÔÇÌÉÏ ÉÙÁÍÍÉÍÙÍ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ÔÏÌÅÁÓ ΜΟΡΦΟΛΟΓΙΚΟΣ - ÊËÉÍÉÊÏÅÑÃÁÓÔÇÑÉÁÊÏÓ ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΑΣ ΜΕΛΕΤΗ ΤΗΣ ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗΣ ΕΚΦΡΑΣΗΣ ΤΩΝ ΓΟΝΙΔΙΩΝ ΕΠΙΔΙΟΡΘΩΣΗΣ ΚΑΙ
ÐÁÍÅÐÉÓÔÇÌÉÏ ÉÙÁÍÍÉÍÙÍ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ÔÏÌÅÁÓ ΜΟΡΦΟΛΟΓΙΚΟΣ - ÊËÉÍÉÊÏÅÑÃÁÓÔÇÑÉÁÊÏÓ ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΑΣ ΜΕΛΕΤΗ ΤΗΣ ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗΣ ΕΚΦΡΑΣΗΣ ΤΩΝ ΓΟΝΙΔΙΩΝ ΕΠΙΔΙΟΡΘΩΣΗΣ ΚΑΙ
Εργασία του μαθητή Θ. Σιδηρόπουλου :καρκίνος και μεταλλάξεις. Τμήμα Γ 2
 ΚΑΡΚΙΝΟΣ: Ο Καρκίνος είναι ένα από τα σοβαρότερα προβλήματα υγείας που παρατηρούνται σήμερα στις αναπτυγμένες χώρες. Οι στατιστικές δείχνουν ότι αποτελεί τη δεύτερη πιο συχνή αιτία θανάτου μετά τις καρδιοπάθειες.
ΚΑΡΚΙΝΟΣ: Ο Καρκίνος είναι ένα από τα σοβαρότερα προβλήματα υγείας που παρατηρούνται σήμερα στις αναπτυγμένες χώρες. Οι στατιστικές δείχνουν ότι αποτελεί τη δεύτερη πιο συχνή αιτία θανάτου μετά τις καρδιοπάθειες.
Τα οφέλη της λαπαροσκοπικής χολοκυστεκτομής στην πράξη - Ο Δρόμος για την Θεραπεία Δευτέρα, 27 Δεκέμβριος :17
 Απαντά ο κ. Θεμιστοκλής Ευκαρπίδης, Γενικός Χειρουργός Ίσως λίγοι από εμάς να είμαστε ενημερωμένοι για τη σπουδαία εργασία που εκτελεί στο σώμα μας, ένα από τα όργανα του, η χοληδόχος κύστη. Εκεί μέσα
Απαντά ο κ. Θεμιστοκλής Ευκαρπίδης, Γενικός Χειρουργός Ίσως λίγοι από εμάς να είμαστε ενημερωμένοι για τη σπουδαία εργασία που εκτελεί στο σώμα μας, ένα από τα όργανα του, η χοληδόχος κύστη. Εκεί μέσα
ΗPV και ενδοεπιθηλιακές αλλοιώσεις κατωτέρου γεννητικού συστήματος. Ε.Κωστοπούλου Λέκτορας Παθολογικής Ανατομικής Πανεπιστήμιο Θεσσαλίας
 ΗPV και ενδοεπιθηλιακές αλλοιώσεις κατωτέρου γεννητικού συστήματος Ε.Κωστοπούλου Λέκτορας Παθολογικής Ανατομικής Πανεπιστήμιο Θεσσαλίας H.M. Παράγοντες κινδύνου καρκινώματος τραχήλου Ηλικία κατά την πρώτη
ΗPV και ενδοεπιθηλιακές αλλοιώσεις κατωτέρου γεννητικού συστήματος Ε.Κωστοπούλου Λέκτορας Παθολογικής Ανατομικής Πανεπιστήμιο Θεσσαλίας H.M. Παράγοντες κινδύνου καρκινώματος τραχήλου Ηλικία κατά την πρώτη
ΥΠΕΡΠΛΑΣΙΑ ΤΟΥ ΕΝΔΟΜΗΤΡΙΟΥ ΚΑΛΠΑΚΤΣΙΔΟΥ ΒΑΚΙΑΝΗ ΜΑΧΗ
 ΥΠΕΡΠΛΑΣΙΑ ΤΟΥ ΕΝΔΟΜΗΤΡΙΟΥ ΚΑΛΠΑΚΤΣΙΔΟΥ ΒΑΚΙΑΝΗ ΜΑΧΗ Ανωορρηκτικοί κύκλοι Εξωγενή οιστρογόνα Σύνδρομο πολυκυστικών ωοθηκών Παχυσαρκία Όγκοι ωοθηκών Εφηβία Παραγωγική ηλικία Κλιμακτήριο εμμηνόπαυση ΥΠΕΡΠΛΑΣΙΑ
ΥΠΕΡΠΛΑΣΙΑ ΤΟΥ ΕΝΔΟΜΗΤΡΙΟΥ ΚΑΛΠΑΚΤΣΙΔΟΥ ΒΑΚΙΑΝΗ ΜΑΧΗ Ανωορρηκτικοί κύκλοι Εξωγενή οιστρογόνα Σύνδρομο πολυκυστικών ωοθηκών Παχυσαρκία Όγκοι ωοθηκών Εφηβία Παραγωγική ηλικία Κλιμακτήριο εμμηνόπαυση ΥΠΕΡΠΛΑΣΙΑ
Το σύστημα τελομερών/τελομεράσης στις χρόνιες φλεγμονώδεις διαταραχές
 Το σύστημα τελομερών/τελομεράσης στις χρόνιες φλεγμονώδεις διαταραχές Βιβλιογραφική Ανασκόπηση Κορδίνας Βασίλειος Μοριακός Βιολόγος και Γενετιστής Ειδικευόμενος Παθολόγος Γενικό Νοσοκομείο Νίκαιας-Πειραιά
Το σύστημα τελομερών/τελομεράσης στις χρόνιες φλεγμονώδεις διαταραχές Βιβλιογραφική Ανασκόπηση Κορδίνας Βασίλειος Μοριακός Βιολόγος και Γενετιστής Ειδικευόμενος Παθολόγος Γενικό Νοσοκομείο Νίκαιας-Πειραιά
Όρχεις -Χειρισμός παρασκευάσματος -Εισαγωγή στους όγκους. Α.. Κιζιρίδου, Αναπ. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.
 Όρχεις -Χειρισμός παρασκευάσματος -Εισαγωγή στους όγκους Α.. Κιζιρίδου, Αναπ. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.Θεαγένειο Περιγραφική Ανατομική 2 αδένες με σχήμα δαμάσκηνου διαστ.5χ3χ2.5 Κρέμονται
Όρχεις -Χειρισμός παρασκευάσματος -Εισαγωγή στους όγκους Α.. Κιζιρίδου, Αναπ. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.Θεαγένειο Περιγραφική Ανατομική 2 αδένες με σχήμα δαμάσκηνου διαστ.5χ3χ2.5 Κρέμονται
 ΙΑΦΟΡΙΚΗ ΙΑΓΝΩΣΗ ΚΑΡΑΤΖΑΣ ΠΑΝΤΕΛΗΣ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΙΚΟ ΤΜΗΜΑ ΓΝΑ ΕΥΑΓΓΕΛΙΣΜΟΣ ΣΥΝΟΨΗ ΠΕΡΙΣΤΑΤΙΚΟΥ Άνδρας 59 ετών ΑΥ υσλιπιδαιµία Γνωστό λέµφωµα κυττάρων µανδύα ελεύθερος νόσου από το 2010 ιαταραχές κενώσεων
ΙΑΦΟΡΙΚΗ ΙΑΓΝΩΣΗ ΚΑΡΑΤΖΑΣ ΠΑΝΤΕΛΗΣ ΓΑΣΤΡΕΝΤΕΡΟΛΟΓΙΚΟ ΤΜΗΜΑ ΓΝΑ ΕΥΑΓΓΕΛΙΣΜΟΣ ΣΥΝΟΨΗ ΠΕΡΙΣΤΑΤΙΚΟΥ Άνδρας 59 ετών ΑΥ υσλιπιδαιµία Γνωστό λέµφωµα κυττάρων µανδύα ελεύθερος νόσου από το 2010 ιαταραχές κενώσεων
Πρώτη εμφάνιση βαριάς κολίτιδας χωρίς ιστορικό ΙΦΝΕ. Πως μπορεί να βοηθήσει ο Παθολογοανατόμος. Μαρία Σωτηροπούλου Νοσοκομείο «Αλεξάνδρα»
 Πρώτη εμφάνιση βαριάς κολίτιδας χωρίς ιστορικό ΙΦΝΕ. Πως μπορεί να βοηθήσει ο Παθολογοανατόμος Μαρία Σωτηροπούλου Νοσοκομείο «Αλεξάνδρα» Το ερώτημα: Είναι ΙΦΝΕ ή κάποιο άλλο είδος κολίτιδας που εξηγεί
Πρώτη εμφάνιση βαριάς κολίτιδας χωρίς ιστορικό ΙΦΝΕ. Πως μπορεί να βοηθήσει ο Παθολογοανατόμος Μαρία Σωτηροπούλου Νοσοκομείο «Αλεξάνδρα» Το ερώτημα: Είναι ΙΦΝΕ ή κάποιο άλλο είδος κολίτιδας που εξηγεί
Κυτταρική Βιολογία. Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
Η παγκόσμια οργάνωση υγείας συνιστά την κατάταξη σε 4 βασικούς ιστολογικούς τύπους: καρκίνωμα μεταβατικού επιθηλίου, καρκίνωμα πλακώδους
 ΚΑΡΚΙΝΟΣ ΟΥΡΟΔΟΧΟΥ ΚΥΣΤΕΩΣ ΣΥΧΝΟΤΗΤΑ ΚΑΡΚΙΝΟΥ ΟΥΡΟΔΟΧΟΥ ΚΥΣΤΕΩΣ. Είναι ο τέταρτος πιο συχνός καρκίνος της στον Δυτικό κόσμο. Η μέση ηλικία κατά την διάγνωση είναι τα 60 με 65 έτη.είναι πιο συχνός στους
ΚΑΡΚΙΝΟΣ ΟΥΡΟΔΟΧΟΥ ΚΥΣΤΕΩΣ ΣΥΧΝΟΤΗΤΑ ΚΑΡΚΙΝΟΥ ΟΥΡΟΔΟΧΟΥ ΚΥΣΤΕΩΣ. Είναι ο τέταρτος πιο συχνός καρκίνος της στον Δυτικό κόσμο. Η μέση ηλικία κατά την διάγνωση είναι τα 60 με 65 έτη.είναι πιο συχνός στους
ΜΕΤΑΜΟΣΧΕΥΣΗ ΝΕΦΡΟΥ. Λειτουργία των νεφρών. Συμπτώματα της χρόνιας νεφρικής ανεπάρκειας
 ΜΕΤΑΜΟΣΧΕΥΣΗ ΝΕΦΡΟΥ Η χρόνια νεφρική ανεπάρκεια είναι η προοδευτική, μη αναστρέψιμη μείωση της νεφρικής λειτουργίας, η οποία προκαλείται από βλάβη του νεφρού ποικίλης αιτιολογίας. Η χρόνια νεφρική ανεπάρκεια
ΜΕΤΑΜΟΣΧΕΥΣΗ ΝΕΦΡΟΥ Η χρόνια νεφρική ανεπάρκεια είναι η προοδευτική, μη αναστρέψιμη μείωση της νεφρικής λειτουργίας, η οποία προκαλείται από βλάβη του νεφρού ποικίλης αιτιολογίας. Η χρόνια νεφρική ανεπάρκεια
Σήμερα η παγκόσμια ημέρα κατά του καρκίνου: Στατιστικά-πρόληψη
 Σήμερα η παγκόσμια ημέρα κατά του καρκίνου: Στατιστικά-πρόληψη Η Παγκόσμια Ημέρα κατά του Καρκίνου καθιερώθηκε με πρωτοβουλία της Διεθνούς Ένωσης κατά του Καρκίνου και πραγματοποιείται κάθε χρόνο στις
Σήμερα η παγκόσμια ημέρα κατά του καρκίνου: Στατιστικά-πρόληψη Η Παγκόσμια Ημέρα κατά του Καρκίνου καθιερώθηκε με πρωτοβουλία της Διεθνούς Ένωσης κατά του Καρκίνου και πραγματοποιείται κάθε χρόνο στις
ΚΑΡΚΙΝΟ του ΠΡΟΣΤΑΤΗ. Πως να προλάβετε τον ΔΙΑΓΝΩΣΗ Η ΕΓΚΑΙΡΗ ΣΩΖΕΙ ΖΩΕΣ. Επιστημονική Επιμέλεια Ελληνική Ουρολογική Εταιρία
 Η ΕΓΚΑΙΡΗ ΔΙΑΓΝΩΣΗ ΣΩΖΕΙ ΖΩΕΣ Πως να προλάβετε τον ΚΑΡΚΙΝΟ του ΠΡΟΣΤΑΤΗ με ΤΗΝ ΕΥΓΕΝΙΚΗ Χορηγία Επιστημονική Επιμέλεια Ελληνική Ουρολογική Εταιρία με ΤΗ ΣΥΝΕΡΓΑΣΙΑ «Αυτές οι πληροφορίες προορίζονται για
Η ΕΓΚΑΙΡΗ ΔΙΑΓΝΩΣΗ ΣΩΖΕΙ ΖΩΕΣ Πως να προλάβετε τον ΚΑΡΚΙΝΟ του ΠΡΟΣΤΑΤΗ με ΤΗΝ ΕΥΓΕΝΙΚΗ Χορηγία Επιστημονική Επιμέλεια Ελληνική Ουρολογική Εταιρία με ΤΗ ΣΥΝΕΡΓΑΣΙΑ «Αυτές οι πληροφορίες προορίζονται για
Εικόνες ζωής. Στάση πρόληψης.
 Εικόνες ζωής. Στάση πρόληψης. Αν είστε 55-70 ετών ελάτε και εσείς για ΔΩΡΕΑΝ προληπτικές εξετάσεις για τον καρκίνο παχέος εντέρου, από 1-31 Μαρτίου σε Αθήνα-Πειραιά, Θεσσαλονίκη, Πάτρα και Ηράκλειο. Πληροφορίες
Εικόνες ζωής. Στάση πρόληψης. Αν είστε 55-70 ετών ελάτε και εσείς για ΔΩΡΕΑΝ προληπτικές εξετάσεις για τον καρκίνο παχέος εντέρου, από 1-31 Μαρτίου σε Αθήνα-Πειραιά, Θεσσαλονίκη, Πάτρα και Ηράκλειο. Πληροφορίες
ΣΑΚΧΑΡΩΔΗΣ ΔΙΑΒΗΤΗΣ και ΚΑΡΚΙΝΟΣ: συνύπαρξη ή αιτιολογική σχέση;
 ΣΑΚΧΑΡΩΔΗΣ ΔΙΑΒΗΤΗΣ και ΚΑΡΚΙΝΟΣ: συνύπαρξη ή αιτιολογική σχέση; Δ. Καραγιάννη, Β. Κουρκούμπας, Δ. Μπαλτζής, Γ. Κοτρώνης, Ε. Κιντιράκη, Χ.Τρακατέλλη, Α. Παυλίδου, Μ. Σιών Γ Παθολογική Κλινική ΑΠΘ, ΓΠΝΘ
ΣΑΚΧΑΡΩΔΗΣ ΔΙΑΒΗΤΗΣ και ΚΑΡΚΙΝΟΣ: συνύπαρξη ή αιτιολογική σχέση; Δ. Καραγιάννη, Β. Κουρκούμπας, Δ. Μπαλτζής, Γ. Κοτρώνης, Ε. Κιντιράκη, Χ.Τρακατέλλη, Α. Παυλίδου, Μ. Σιών Γ Παθολογική Κλινική ΑΠΘ, ΓΠΝΘ
Δυσκοίλιο, θεωρούμε ένα άτομο όταν εμφανίζει δύο τουλάχιστον από τα παρακάτω προβλήματα για 3 ή περισσότερους μήνες:
 Δυσκοιλιότητα. Η δυσκοιλιότητα αποτελεί ένα από τα πιο συχνά συμπτώματα του κατώτερου πεπτικού σωλήνα, επηρεάζοντας κάθε χρόνο ένα μεγάλο αριθμό ασθενών, κυρίως των χωρών του δυτικού κόσμου. Εικόνα 1.
Δυσκοιλιότητα. Η δυσκοιλιότητα αποτελεί ένα από τα πιο συχνά συμπτώματα του κατώτερου πεπτικού σωλήνα, επηρεάζοντας κάθε χρόνο ένα μεγάλο αριθμό ασθενών, κυρίως των χωρών του δυτικού κόσμου. Εικόνα 1.
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου
 Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Στα χρωμοσώματα ενός ανθρώπινου σωματικού κυττάρου στο στάδιο της μετάφασης της μίτωσης υπάρχουν: Α. 23 μόρια DNA Β. 92 μόρια DNA Γ. 46 μόρια DNA Δ.
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Στα χρωμοσώματα ενός ανθρώπινου σωματικού κυττάρου στο στάδιο της μετάφασης της μίτωσης υπάρχουν: Α. 23 μόρια DNA Β. 92 μόρια DNA Γ. 46 μόρια DNA Δ.
Αιμοσφαιρίνες. Αιμοσφαιρίνη Συμβολισμός Σύσταση A HbA α 2 β 2 F HbF α 2 γ 2 A 2 HbA 2 α 2 δ 2 s. Σύγκριση γονιδιακών και χρωμοσωμικών μεταλλάξεων
 A. ΔΙΑΓΡΑΜΜΑ Αιμοσφαιρίνες Αιμοσφαιρίνη Συμβολισμός Σύσταση A HbA α 2 β 2 F HbF α 2 γ 2 A 2 HbA 2 α 2 δ 2 S HbS s α 2 β 2 Σύγκριση γονιδιακών και χρωμοσωμικών μεταλλάξεων ΓΟΝΙΔΙΑΚΕΣ Αλλαγή σε αζωτούχες
A. ΔΙΑΓΡΑΜΜΑ Αιμοσφαιρίνες Αιμοσφαιρίνη Συμβολισμός Σύσταση A HbA α 2 β 2 F HbF α 2 γ 2 A 2 HbA 2 α 2 δ 2 S HbS s α 2 β 2 Σύγκριση γονιδιακών και χρωμοσωμικών μεταλλάξεων ΓΟΝΙΔΙΑΚΕΣ Αλλαγή σε αζωτούχες
ΗPV και Καρκίνος Δέρµατος. Ηλέκτρα Νικολαΐδου Επ. Καθηγήτρια Δερµατολογίας ΕΚΠΑ Νοσ. «Α. Συγγρός»
 ΗPV και Καρκίνος Δέρµατος Ηλέκτρα Νικολαΐδου Επ. Καθηγήτρια Δερµατολογίας ΕΚΠΑ Νοσ. «Α. Συγγρός» ΓΕΝΗ HPV Α γένος: βλεννογόνοι αιτιολογική συσχέτιση µε καρκίνο τραχήλου µήτρας, πρωκτού, αιδοίου, πέους
ΗPV και Καρκίνος Δέρµατος Ηλέκτρα Νικολαΐδου Επ. Καθηγήτρια Δερµατολογίας ΕΚΠΑ Νοσ. «Α. Συγγρός» ΓΕΝΗ HPV Α γένος: βλεννογόνοι αιτιολογική συσχέτιση µε καρκίνο τραχήλου µήτρας, πρωκτού, αιδοίου, πέους
ΓΕΝΙΚΟ ΝΟΣΟΚΟΜΕΙΟ ΡΟΔΟΥ «ΑΝΔΡΕΑΣ ΠΑΠΑΝΔΡΕΟΥ» ΚΑΡΚΙΝΟΣ ΤΗΣ ΟΥΡΟΔΟΧΟΥ ΚΥΣΤΗΣ
 ΓΕΝΙΚΟ ΝΟΣΟΚΟΜΕΙΟ ΡΟΔΟΥ «ΑΝΔΡΕΑΣ ΠΑΠΑΝΔΡΕΟΥ» ΚΑΡΚΙΝΟΣ ΤΗΣ ΟΥΡΟΔΟΧΟΥ ΚΥΣΤΗΣ ΟΥΡΟΔΟΧΟΣ ΚΥΣΤΗ Κοίλο μυώδες όργανο που υποδέχεται, αποθηκεύει και αποβάλλει τα ούρα Μέρη της κύστης Κορυφή Σώμα Πυθμένας Ανατομικά
ΓΕΝΙΚΟ ΝΟΣΟΚΟΜΕΙΟ ΡΟΔΟΥ «ΑΝΔΡΕΑΣ ΠΑΠΑΝΔΡΕΟΥ» ΚΑΡΚΙΝΟΣ ΤΗΣ ΟΥΡΟΔΟΧΟΥ ΚΥΣΤΗΣ ΟΥΡΟΔΟΧΟΣ ΚΥΣΤΗ Κοίλο μυώδες όργανο που υποδέχεται, αποθηκεύει και αποβάλλει τα ούρα Μέρη της κύστης Κορυφή Σώμα Πυθμένας Ανατομικά
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ (PATHOLOGY) Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ.
 Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ.") ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ (PATHOLOGY) Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ. Παθολογική Ανατομική εξέταση των δομικών αλλοιώσεων
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ (PATHOLOGY) Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ. Παθολογική Ανατομική εξέταση των δομικών αλλοιώσεων
Το συγκεκριμένο θέμα παρουσιάστηκε ως ανακοίνωση στο ΠΑΓΚΥΠΡΙΟ ΣΥΝΕ ΡΙΟ ΧΕΙΡΟΥΡΓΙΚΗΣ, ΗILTON CYPRUS.
 edtime - Σταδιοποίηση Καρκίνου Παχέος Εντέρου Written by Παπασταματίου Μιλτιάδης, Νικολόπουλος Δημήτριος Παρασκευή, 15 Ιανουάριος 2010 There are no translations available Παπασταματίου Μιλτιάδης, MD, PhD,
edtime - Σταδιοποίηση Καρκίνου Παχέος Εντέρου Written by Παπασταματίου Μιλτιάδης, Νικολόπουλος Δημήτριος Παρασκευή, 15 Ιανουάριος 2010 There are no translations available Παπασταματίου Μιλτιάδης, MD, PhD,
Συστηματικός ερυθηματώδης λύκος: το πρότυπο των αυτόάνοσων ρευματικών νοσημάτων
 Συστηματικός ερυθηματώδης λύκος: το πρότυπο των αυτόάνοσων ρευματικών νοσημάτων Φ.Ν. Σκοπούλη Καθηγήτρια τον Χαροκόπειου Πανεπιστημίου Αθηνών συστηματικός ερυθηματώδης λύκος θεωρείται η κορωνίδα των αυτοάνοσων
Συστηματικός ερυθηματώδης λύκος: το πρότυπο των αυτόάνοσων ρευματικών νοσημάτων Φ.Ν. Σκοπούλη Καθηγήτρια τον Χαροκόπειου Πανεπιστημίου Αθηνών συστηματικός ερυθηματώδης λύκος θεωρείται η κορωνίδα των αυτοάνοσων
ΚΑΛΟΗΘΕΙΣ ΚΥΣΤΙΚΕΣ ΠΑΘΗΣΕΙΣ ΤΟΥ ΝΕΦΡΟΥ ΑΘΑΝΑΣΙΟΣ ΚΟΡΔΕΛΑΣ, ΠΑΘΟΛΟΓΟΝΑΤΟΜΟΣ ΕΠΙΜΕΛΗΤΗΣ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΟΥ ΕΡΓΑΣΤΗΡΙΟΥ ΝΑΥΤΙΚΟΥ ΝΟΣΟΚΟΜΕΙΟΥ ΑΘΗΝΩΝ
 ΚΑΛΟΗΘΕΙΣ ΚΥΣΤΙΚΕΣ ΠΑΘΗΣΕΙΣ ΤΟΥ ΝΕΦΡΟΥ ΑΘΑΝΑΣΙΟΣ ΚΟΡΔΕΛΑΣ, ΠΑΘΟΛΟΓΟΝΑΤΟΜΟΣ ΕΠΙΜΕΛΗΤΗΣ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΟΥ ΕΡΓΑΣΤΗΡΙΟΥ ΝΑΥΤΙΚΟΥ ΝΟΣΟΚΟΜΕΙΟΥ ΑΘΗΝΩΝ Ταξινόμηση κυστικής νόσου του νεφρού 1. Απλή νεφρική κύστη
ΚΑΛΟΗΘΕΙΣ ΚΥΣΤΙΚΕΣ ΠΑΘΗΣΕΙΣ ΤΟΥ ΝΕΦΡΟΥ ΑΘΑΝΑΣΙΟΣ ΚΟΡΔΕΛΑΣ, ΠΑΘΟΛΟΓΟΝΑΤΟΜΟΣ ΕΠΙΜΕΛΗΤΗΣ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΟΥ ΕΡΓΑΣΤΗΡΙΟΥ ΝΑΥΤΙΚΟΥ ΝΟΣΟΚΟΜΕΙΟΥ ΑΘΗΝΩΝ Ταξινόμηση κυστικής νόσου του νεφρού 1. Απλή νεφρική κύστη
ΕΜΒΡΥΟΛΟΓΙΑ ΗΠΑΤΟΣ ΧΟΛΗΔΟΧΟΥ ΚΥΣΤΕΩΣ ΠΑΓΚΡΕΑΤΟΣ
 ΕΜΒΡΥΟΛΟΓΙΑ ΗΠΑΤΟΣ ΧΟΛΗΔΟΧΟΥ ΚΥΣΤΕΩΣ ΠΑΓΚΡΕΑΤΟΣ Ανάπτυξη ήπατος Ηπατικό εκκόλπωμα: προσεκβολή της ενδοδερματικής επένδυσης του προσθίου εντέρου ανάπτυξη ηπατικών δοκίδων και καταβολών του συστήματος χοληφόρων
ΕΜΒΡΥΟΛΟΓΙΑ ΗΠΑΤΟΣ ΧΟΛΗΔΟΧΟΥ ΚΥΣΤΕΩΣ ΠΑΓΚΡΕΑΤΟΣ Ανάπτυξη ήπατος Ηπατικό εκκόλπωμα: προσεκβολή της ενδοδερματικής επένδυσης του προσθίου εντέρου ανάπτυξη ηπατικών δοκίδων και καταβολών του συστήματος χοληφόρων
ΜΑΘΗΜΑΤΑ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ 6. Α.Παπαχατζοπούλου
 ΜΑΘΗΜΑΤΑ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ 6 ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΙΑΤΡΙΚΗΣ ΧΗΜΕΙΑΣ Α.Παπαχατζοπούλου 2016-2017 Η ΓΕΝΕΤΙΚΗ ΤΟΥ ΚΑΡΚΙΝΟΥ Ταξινόμηση των γενετικών νοσημάτων Πολυπαραγοντικά + environment Μονογονιδιακά Χρωμοσωμικές
ΜΑΘΗΜΑΤΑ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ 6 ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΙΑΤΡΙΚΗΣ ΧΗΜΕΙΑΣ Α.Παπαχατζοπούλου 2016-2017 Η ΓΕΝΕΤΙΚΗ ΤΟΥ ΚΑΡΚΙΝΟΥ Ταξινόμηση των γενετικών νοσημάτων Πολυπαραγοντικά + environment Μονογονιδιακά Χρωμοσωμικές
Επιδημιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία Πνεύμονα. Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής
 Επιδημιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία Πνεύμονα Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Επιδημιολογικά στοιχεία καρκίνου του πνεύμονα Ο καρκίνος
Επιδημιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία Πνεύμονα Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Επιδημιολογικά στοιχεία καρκίνου του πνεύμονα Ο καρκίνος
Ακανθοκυτταρικό καρκίνωμα κατωτέρου γεννητικού συστήματος θήλεος. Μαρία Σωτηροπούλου Νοσοκομείο Αλεξάνδρα
 Ακανθοκυτταρικό καρκίνωμα κατωτέρου γεννητικού συστήματος θήλεος Μαρία Σωτηροπούλου Νοσοκομείο Αλεξάνδρα Ακανθοκυτταρικό καρκίνωμα αιδοίου Σπάνιο, 1,5 ανά 100 000 γυναίκες στις ΗΠΑ Τρειςομάδεςγυναικών:
Ακανθοκυτταρικό καρκίνωμα κατωτέρου γεννητικού συστήματος θήλεος Μαρία Σωτηροπούλου Νοσοκομείο Αλεξάνδρα Ακανθοκυτταρικό καρκίνωμα αιδοίου Σπάνιο, 1,5 ανά 100 000 γυναίκες στις ΗΠΑ Τρειςομάδεςγυναικών:
ΠΑΘΗΣΕΙΣ ΜΑΖΙΚΟΥ ΑΔΕΝΑ. Τριανταφυλλιά Κολέτσα Λέκτορας
 ΠΑΘΗΣΕΙΣ ΜΑΖΙΚΟΥ ΑΔΕΝΑ Τριανταφυλλιά Κολέτσα Λέκτορας ΕΓΠΠΑ, Α.Π.Θ. Μαστός Λοβοί: εκβάλουν στη θηλή με γαλακτοφόρο πόρο. Διακλαδιζόμενοι πόροι κατάληξη-λοβιακές λοβιακές μονάδες. Λοβιακή μονάδα: αποτελείται
ΠΑΘΗΣΕΙΣ ΜΑΖΙΚΟΥ ΑΔΕΝΑ Τριανταφυλλιά Κολέτσα Λέκτορας ΕΓΠΠΑ, Α.Π.Θ. Μαστός Λοβοί: εκβάλουν στη θηλή με γαλακτοφόρο πόρο. Διακλαδιζόμενοι πόροι κατάληξη-λοβιακές λοβιακές μονάδες. Λοβιακή μονάδα: αποτελείται
Η ενημέρωση ζευγαριών σχετικά με τον κίνδυνο που διατρέχουν να αποκτήσουν ένα παιδί με μία συγγενή ανωμαλία ή μία γενετική διαταραχή και τις επιλογές
 Η ενημέρωση ζευγαριών σχετικά με τον κίνδυνο που διατρέχουν να αποκτήσουν ένα παιδί με μία συγγενή ανωμαλία ή μία γενετική διαταραχή και τις επιλογές που έχουν όσον αφορά στην διαχείριση του κινδύνου Προχωρημένη
Η ενημέρωση ζευγαριών σχετικά με τον κίνδυνο που διατρέχουν να αποκτήσουν ένα παιδί με μία συγγενή ανωμαλία ή μία γενετική διαταραχή και τις επιλογές που έχουν όσον αφορά στην διαχείριση του κινδύνου Προχωρημένη
ΕΓΚΟΛΕΑΣΜΟΣ ΛΕΠΤΟΥ ΕΝΤΕΡΟΥ
 Χειρουργική Κλινική Γ.Ν. Χαλκίδας Ο εγκολεασμός αποτελεί το συχνότερο αίτιο εντερικής απόφραξης και το δεύτερο αίτιο οξείας κοιλίας στα παιδιά. Αντιθέτως, στους ενήλικες εμφανίζεται σε ποσοστό μικρότερο
Χειρουργική Κλινική Γ.Ν. Χαλκίδας Ο εγκολεασμός αποτελεί το συχνότερο αίτιο εντερικής απόφραξης και το δεύτερο αίτιο οξείας κοιλίας στα παιδιά. Αντιθέτως, στους ενήλικες εμφανίζεται σε ποσοστό μικρότερο
ΜΟΡΙΑΚΗ ΒΑΣΗ ΓΕΝΕΤΙΚΩΝ ΑΣΘΕΝΕΙΩΝ ΘΕΜΑ: ΠΟΛΛΑΠΛΗ ΝΕΥΡΟΙΝΩΜΑΤΩΣΗ. Ομάδα ΑΡΕΑΛΗ ΑΝΤΩΝΙΑ ΜΠΑΗ ΙΩΑΝΝΑ ΠΑΤΕΡΑΚΗ ΕΛΕΝΗ
 ΜΟΡΙΑΚΗ ΒΑΣΗ ΓΕΝΕΤΙΚΩΝ ΑΣΘΕΝΕΙΩΝ ΘΕΜΑ: ΠΟΛΛΑΠΛΗ ΝΕΥΡΟΙΝΩΜΑΤΩΣΗ Ομάδα ΑΡΕΑΛΗ ΑΝΤΩΝΙΑ ΜΠΑΗ ΙΩΑΝΝΑ ΠΑΤΕΡΑΚΗ ΕΛΕΝΗ ΑΠΡΙΛΙΟΣ 2015 ΣΚΟΠΟΣ Σκοπός της εργασίας αυτής είναι η παρουσίαση των χαρακτηριστικών της
ΜΟΡΙΑΚΗ ΒΑΣΗ ΓΕΝΕΤΙΚΩΝ ΑΣΘΕΝΕΙΩΝ ΘΕΜΑ: ΠΟΛΛΑΠΛΗ ΝΕΥΡΟΙΝΩΜΑΤΩΣΗ Ομάδα ΑΡΕΑΛΗ ΑΝΤΩΝΙΑ ΜΠΑΗ ΙΩΑΝΝΑ ΠΑΤΕΡΑΚΗ ΕΛΕΝΗ ΑΠΡΙΛΙΟΣ 2015 ΣΚΟΠΟΣ Σκοπός της εργασίας αυτής είναι η παρουσίαση των χαρακτηριστικών της
 94 95 96 97 98 Ο ρόλος της αγγειογένεσης στη μετάσταση Η νεοαγγείωση αποτελεί ένα απαραίτητο τμήμα της ογκογόνου διεργασίας που διασφαλίζει τη γρηγορότερη και ανεμπόδιστη ανάπτυξη του όγκου. Η λειτουργική
94 95 96 97 98 Ο ρόλος της αγγειογένεσης στη μετάσταση Η νεοαγγείωση αποτελεί ένα απαραίτητο τμήμα της ογκογόνου διεργασίας που διασφαλίζει τη γρηγορότερη και ανεμπόδιστη ανάπτυξη του όγκου. Η λειτουργική
Ο ρόλος της ΕΘΟ. στην αναγέννηση. & την επανόρθωση
 Ο ρόλος της ΕΘΟ στην αναγέννηση & την επανόρθωση Νοvo E & Parola M. Fibrogenesis & Tissue Repair 2008, 1:5 Χρόνια παγκρεατίτιδα Ιστολογία παγκρεατικού καρκινώµατος Αδενοκαρκίνωµα εξ εκφορητικών
Ο ρόλος της ΕΘΟ στην αναγέννηση & την επανόρθωση Νοvo E & Parola M. Fibrogenesis & Tissue Repair 2008, 1:5 Χρόνια παγκρεατίτιδα Ιστολογία παγκρεατικού καρκινώµατος Αδενοκαρκίνωµα εξ εκφορητικών
ΠΑΡΟΥΣΙΑΣΗ ΠΕΡΙΣΤΑΤΙΚΟΥ
 ΠΑΡΟΥΣΙΑΣΗ ΠΕΡΙΣΤΑΤΙΚΟΥ Άνδρας ηλικίας 47 ετών προσέρχεται λόγω πνευματουρίας και απώλειας κοπράνων από την ουρήθρα, από δεκαημέρου. Ο ασθενής αναφέρει γριππώδη συνδρομή που προηγήθηκε της παρούσης κατάστασης.
ΠΑΡΟΥΣΙΑΣΗ ΠΕΡΙΣΤΑΤΙΚΟΥ Άνδρας ηλικίας 47 ετών προσέρχεται λόγω πνευματουρίας και απώλειας κοπράνων από την ουρήθρα, από δεκαημέρου. Ο ασθενής αναφέρει γριππώδη συνδρομή που προηγήθηκε της παρούσης κατάστασης.
Πολύποδες χοληδόχου κύστης: Τι είναι και ποιά η αντιμετώπιση τους.
 Πολύποδες χοληδόχου κύστης: Τι είναι και ποιά η αντιμετώπιση τους. Οι πολύποδες της χοληδόχου κύστης είναι ένα σύνηθες εύρημα στις διαγνωστικές εξετάσεις στην πλειονότητα των ασθενών. Οι πολύποδες στην
Πολύποδες χοληδόχου κύστης: Τι είναι και ποιά η αντιμετώπιση τους. Οι πολύποδες της χοληδόχου κύστης είναι ένα σύνηθες εύρημα στις διαγνωστικές εξετάσεις στην πλειονότητα των ασθενών. Οι πολύποδες στην
Ιδιαιτερότητες χειρουργικής αντιμετώπισης ΝΕΤ εντέρου
 Ιδιαιτερότητες χειρουργικής αντιμετώπισης ΝΕΤ εντέρου Γεώργιος Χ. Σωτηρόπουλος, MD, PhD, FACS, FEBS Καθηγητής Χειρουργικής Πανεπιστημίου Essen Αν. Καθηγητής Χειρουργικής ΕΚΠΑ Αθήνα 11.11.2017 Αντιμετώπιση
Ιδιαιτερότητες χειρουργικής αντιμετώπισης ΝΕΤ εντέρου Γεώργιος Χ. Σωτηρόπουλος, MD, PhD, FACS, FEBS Καθηγητής Χειρουργικής Πανεπιστημίου Essen Αν. Καθηγητής Χειρουργικής ΕΚΠΑ Αθήνα 11.11.2017 Αντιμετώπιση
ΓΕΝΙΚΕΣ ΑΡΧΕΣ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΑΣ ΚΥΤΤΑΡΙΚΗ ΠΡΟΣΑΡΜΟΓΗ
 ΓΕΝΙΚΕΣ ΑΡΧΕΣ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΑΣ ΚΥΤΤΑΡΙΚΗ ΠΡΟΣΑΡΜΟΓΗ ΕΥΑΓΓΕΛΙΑ ΠΙΠΕΡΗ ΕΠΙΚΟΥΡΗ ΚΑΘΗΓΗΤΡΙΑ Εργαστήριο Στοματολογίας ΕΚΠΑ ΚΥΤΤΑΡΙΚΕΣ ΑΝΤΙΔΡΑΣΕΙΣ ΣΕ ΚΑΤΑΣΤΑΣΕΙΣ STRESS ΚΑΙ ΚΥΤΤΑΡΙΚΑ ΕΡΕΘΙΣΜΑΤΑ ΟΜΟΙΟΣΤΑΣΗ:
ΓΕΝΙΚΕΣ ΑΡΧΕΣ ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΑΣ ΚΥΤΤΑΡΙΚΗ ΠΡΟΣΑΡΜΟΓΗ ΕΥΑΓΓΕΛΙΑ ΠΙΠΕΡΗ ΕΠΙΚΟΥΡΗ ΚΑΘΗΓΗΤΡΙΑ Εργαστήριο Στοματολογίας ΕΚΠΑ ΚΥΤΤΑΡΙΚΕΣ ΑΝΤΙΔΡΑΣΕΙΣ ΣΕ ΚΑΤΑΣΤΑΣΕΙΣ STRESS ΚΑΙ ΚΥΤΤΑΡΙΚΑ ΕΡΕΘΙΣΜΑΤΑ ΟΜΟΙΟΣΤΑΣΗ:
ΜΟΡΙΑΚΗ ΒΙΟΛΟΓΙΑ ΚΑΙ ΓΕΝΕΤΙΚΗ ΤΟΥ ΝΕΦΡΟΒΛΑΣΤΩΜΑΤΟΣ (ΟΓΚΟΥ ΤΟΥ WILMS) Σπυριδάκης Ιωάννης 2, Καζάκης Ι 2, Δογραματζής Κωνσταντίνος 1, Κοσμάς Νικόλαος 1,
 Σπυριδάκης Ιωάννης 2, Καζάκης Ι 2, Δογραματζής Κωνσταντίνος 1, Κοσμάς Νικόλαος 1,") 1 Ιατρική Σχολή ΑΠΘ, Στρατιωτική Σχολή Αξιωματικών Σωμάτων, 2Β Πανεπιστημιακή Κλινική Χειρουργικής Παίδων ΑΠΘ, Γ.Ν Παπαγεωργίου (Διευθυντής: Καθηγητής Αντώνιος Φιλιππόπουλος) Εισαγωγή Ο όγκος του Wilms
1 Ιατρική Σχολή ΑΠΘ, Στρατιωτική Σχολή Αξιωματικών Σωμάτων, 2Β Πανεπιστημιακή Κλινική Χειρουργικής Παίδων ΑΠΘ, Γ.Ν Παπαγεωργίου (Διευθυντής: Καθηγητής Αντώνιος Φιλιππόπουλος) Εισαγωγή Ο όγκος του Wilms
3. Το σχεδιάγραμμα παρουσιάζει τομή ανθρώπινου πεπτικού συστήματος.
 ΠΕΠΤΙΚΟ 1. Α. Να γράψετε τα είδη των δοντιών Α, Β, Γ, Δ και τα μέρη του δοντιού Ε Μ. Β. Πόσα δόντια έχει ένα παιδί 3 χρόνων; Γ. Ποιοι αδένες αφήνουν το έκκριμά τους στη στοματική κοιλότητα και ποιο το
ΠΕΠΤΙΚΟ 1. Α. Να γράψετε τα είδη των δοντιών Α, Β, Γ, Δ και τα μέρη του δοντιού Ε Μ. Β. Πόσα δόντια έχει ένα παιδί 3 χρόνων; Γ. Ποιοι αδένες αφήνουν το έκκριμά τους στη στοματική κοιλότητα και ποιο το
Εφαρμοσμένη Διατροφική Ιατρική
 Γλωσσάρι για το Μάθημα της Διατροφικής Ιατρικής Λιπαρά οξέα: περιέχουν μακριές αλυσίδες μορίων που αποτελούν σχεδόν όλο το σύμπλεγμα λιπιδίων τόσο για τα ζωικά όσο και για τα φυτικά λίπη. Αν αποκοπούν
Γλωσσάρι για το Μάθημα της Διατροφικής Ιατρικής Λιπαρά οξέα: περιέχουν μακριές αλυσίδες μορίων που αποτελούν σχεδόν όλο το σύμπλεγμα λιπιδίων τόσο για τα ζωικά όσο και για τα φυτικά λίπη. Αν αποκοπούν
ΟΓΚΟΛΟΓΙΑ - ΡΑΔΙΟΒΙΟΛΟΓΙΑ
 ΟΓΚΟΛΟΓΙΑ - ΡΑΔΙΟΒΙΟΛΟΓΙΑ Βασικές γνώσεις I SBN 960-372-069-0 ΠΕΡΙΕΧΟΜΕΝΑ ΠΑΡΙΣ Α. ΚΟΣΜΙΔΗΣ ΓΕΩΡΓΙΟΣ ΤΣΑΚΙΡΗΣ Μ Ε Ρ Ο Σ Ι ΟΓΚΟΛΟΓΙΑ Κεφάλαιο 1 ΕΙΣΑΓΩΓΗ ΣΤΟΝ ΚΑΡΚΙΝΟ... 3 Το καρκινικό κύτταρο... 3 Κυτταρικός
ΟΓΚΟΛΟΓΙΑ - ΡΑΔΙΟΒΙΟΛΟΓΙΑ Βασικές γνώσεις I SBN 960-372-069-0 ΠΕΡΙΕΧΟΜΕΝΑ ΠΑΡΙΣ Α. ΚΟΣΜΙΔΗΣ ΓΕΩΡΓΙΟΣ ΤΣΑΚΙΡΗΣ Μ Ε Ρ Ο Σ Ι ΟΓΚΟΛΟΓΙΑ Κεφάλαιο 1 ΕΙΣΑΓΩΓΗ ΣΤΟΝ ΚΑΡΚΙΝΟ... 3 Το καρκινικό κύτταρο... 3 Κυτταρικός
ΧΕΙΡΟΥΡΓΙΚΗ ΠΛΟΗΓΗΣΗΣ
 ΧΕΙΡΟΥΡΓΙΚΗ ΠΛΟΗΓΗΣΗΣ Μέχρι πρόσφατα, παρ όλες τις δυνατότητες που έχουμε στο οπλοστάσιο μας για να απεικονίζουμε όγκους στο ήπαρ και στο πάγκρεας, λόγω της έλλειψης λεπτομερούς απεικονίσεως αλλά και ουσιαστικής
ΧΕΙΡΟΥΡΓΙΚΗ ΠΛΟΗΓΗΣΗΣ Μέχρι πρόσφατα, παρ όλες τις δυνατότητες που έχουμε στο οπλοστάσιο μας για να απεικονίζουμε όγκους στο ήπαρ και στο πάγκρεας, λόγω της έλλειψης λεπτομερούς απεικονίσεως αλλά και ουσιαστικής
 Περιβαλλοντικά Καρκινογόνα Κυρίως τρεις τύποι: Χηµικά (µόλυνση, κάπνισµα, αµίαντος) Φυσικά (ιονίζουσα ακτινοβολία, µή- ιονίζουσα???) Βιολογικά (ιοί π.χ.. HPV) Καρκίνος παχέος εντέρου ΓΕΝΕΤΙΚΟΣ
Περιβαλλοντικά Καρκινογόνα Κυρίως τρεις τύποι: Χηµικά (µόλυνση, κάπνισµα, αµίαντος) Φυσικά (ιονίζουσα ακτινοβολία, µή- ιονίζουσα???) Βιολογικά (ιοί π.χ.. HPV) Καρκίνος παχέος εντέρου ΓΕΝΕΤΙΚΟΣ
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ
 ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Περιγραφή της περίπτωσης Άνδρας 43 ετών εισάγεται σε κλινική λόγω επιγαστραλγίας. Μετά από έλεγχο ετέθη η διάγνωση του πεπτικού
ΚΛΙΝΙΚΗ ΠΕΡΙΠΤΩΣΗ Αναστολή αντλίας πρωτονίων ΦΥΣΙΟΛΟΓΙΑ ΚΥΤΤΑΡΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Περιγραφή της περίπτωσης Άνδρας 43 ετών εισάγεται σε κλινική λόγω επιγαστραλγίας. Μετά από έλεγχο ετέθη η διάγνωση του πεπτικού
Μοριακή βιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία πνεύμονα. Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής
 Μοριακή βιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία πνεύμονα Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Εισαγωγή Ο καρκίνος του πνεύμονα παρουσιάζει άφθονες
Μοριακή βιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία πνεύμονα Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Εισαγωγή Ο καρκίνος του πνεύμονα παρουσιάζει άφθονες
ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ Διακρίνονται σε: - Πρωτογενή και - Δευτερογενή Πρωτογενή είναι τα όργανα στα οποία γίνεται η ωρίμανση των κυττάρων του ανοσοποιητικού: - Θύμος
ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΟΡΓΑΝΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ Διακρίνονται σε: - Πρωτογενή και - Δευτερογενή Πρωτογενή είναι τα όργανα στα οποία γίνεται η ωρίμανση των κυττάρων του ανοσοποιητικού: - Θύμος
Φλεγμονή. Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ
 Φλεγμονή Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ Μη ειδική ανοσολογική άμυνα ΑΝΑΤΟΜΙΚΟΙ ΦΡΑΓΜΟΙ Φυσικοί: δέρμα, βλεννογόνοι, βλέννα, βήχας Χημικοί: λυσοζύμη, αντιμικροβιακά
Φλεγμονή Α. Χατζηγεωργίου Επίκουρος Καθηγητής Φυσιολογίας Ιατρικής Σχολής ΕΚΠΑ Μη ειδική ανοσολογική άμυνα ΑΝΑΤΟΜΙΚΟΙ ΦΡΑΓΜΟΙ Φυσικοί: δέρμα, βλεννογόνοι, βλέννα, βήχας Χημικοί: λυσοζύμη, αντιμικροβιακά
ΣΤΟΙΧΕΙΑ ΑΝΑΤΟΜΙΑΣ ΤΗΣ ΜΗΤΡΑΣ
 ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιµος Π. Βανδώρος ΣΤΟΙΧΕΙΑ ΑΝΑΤΟΜΙΑΣ ΤΗΣ ΜΗΤΡΑΣ Η µήτρα (εικόνα 1) είναι κοίλο µυώδες όργανο µήκους περίπου 8 cm που προέρχεται από την συνένωση
ΤΕΙ ΠΑΤΡΑΣ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΥΠΕΥΘΥΝΟΣ ΚΑΘΗΓΗΤΗΣ : Γεράσιµος Π. Βανδώρος ΣΤΟΙΧΕΙΑ ΑΝΑΤΟΜΙΑΣ ΤΗΣ ΜΗΤΡΑΣ Η µήτρα (εικόνα 1) είναι κοίλο µυώδες όργανο µήκους περίπου 8 cm που προέρχεται από την συνένωση
ΝΕΟΠΛΑΣΜΑΤΑ ΚΕΝΤΡΙΚΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Εργαστήριο Γενικής Παθολογίας και Παθολογικικής Ανατομικής
 ΝΕΟΠΛΑΣΜΑΤΑ ΚΕΝΤΡΙΚΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Εργαστήριο Γενικής Παθολογίας και Παθολογικικής Ανατομικής Φυσιολογική εγκεφαλική ουσία Νεοπλάσματα ΚΝΣ ανάλογα με την προέλευσή τους σε πρωτοπαθή (2/3) μεταστατικά
ΝΕΟΠΛΑΣΜΑΤΑ ΚΕΝΤΡΙΚΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Εργαστήριο Γενικής Παθολογίας και Παθολογικικής Ανατομικής Φυσιολογική εγκεφαλική ουσία Νεοπλάσματα ΚΝΣ ανάλογα με την προέλευσή τους σε πρωτοπαθή (2/3) μεταστατικά
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου
 Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Η αναλογία Α+G/T+C στο γενετικό υλικό ενός ιού είναι ίση με 2/3. Ο ιός μπορεί να είναι: α. ο φάγος λ. β. ο ιός της πολιομυελίτιδας. γ. φορέας κλωνοποίησης
Βιολογία Ο.Π. Θετικών Σπουδών Γ' Λυκείου ΘΕΜΑ Α Α1. Η αναλογία Α+G/T+C στο γενετικό υλικό ενός ιού είναι ίση με 2/3. Ο ιός μπορεί να είναι: α. ο φάγος λ. β. ο ιός της πολιομυελίτιδας. γ. φορέας κλωνοποίησης
ΣΠΥΡΙΔΩΝ ΧΡΙΣΤΟΔΟΥΛΟΥ
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΠΑΝΕΠΙΣΤΗΜΙΑΚΟ ΓΕΝΙΚΟ ΝΟΣΟΚΟΜΕΙΟ «ΑΤΤΙΚΟΝ» Δ ΧΕΙΡΟΥΡΓΙΚΗ ΚΛΙΝΙΚΗ ΑΝΑΛΥΣΗ ΤΗΣ ΕΚΦΡΑΣΗΣ ΚΑΙ ΚΛΙΝΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΤΟΥ ΓΟΝΙΔΙΟΥ
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΠΑΝΕΠΙΣΤΗΜΙΑΚΟ ΓΕΝΙΚΟ ΝΟΣΟΚΟΜΕΙΟ «ΑΤΤΙΚΟΝ» Δ ΧΕΙΡΟΥΡΓΙΚΗ ΚΛΙΝΙΚΗ ΑΝΑΛΥΣΗ ΤΗΣ ΕΚΦΡΑΣΗΣ ΚΑΙ ΚΛΙΝΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΤΟΥ ΓΟΝΙΔΙΟΥ
Στην περισσότερο επιτυχημένη αντιμετώπιση του καρκίνου έχει συμβάλλει σημαντικά η ανακά-λυψη και εφαρμογή των καρκινι-κών δεικτών.
 Όλες μαζί οι μορφές καρκίνου αποτελούν, παγκοσμίως τη δεύτερη αιτία θανάτου μετά από τα καρδιαγγειακά νοσήματα. Τα κρούσματα συνεχώς αυξάνονται και σε πολλές αναπτυγμένες χώρες αποτελεί την πρώτη αιτία
Όλες μαζί οι μορφές καρκίνου αποτελούν, παγκοσμίως τη δεύτερη αιτία θανάτου μετά από τα καρδιαγγειακά νοσήματα. Τα κρούσματα συνεχώς αυξάνονται και σε πολλές αναπτυγμένες χώρες αποτελεί την πρώτη αιτία
ΔΙΑΒΑΖΟΝΤΑΣ ΤΗ ΒΙΟΨΙΑ ΗΠΑΤΟΣ
 ΠΡΑΚΤΙΚΕΣ ΟΔΗΓΙΕΣ ΣΤΟΥΣ ΗΠΑΤΟΛΟΓΟΥΣ, ΔΙΑΒΑΖΟΝΤΑΣ ΤΗ ΒΙΟΨΙΑ ΗΠΑΤΟΣ Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Τμήματος Ιατρικής Α.Π.Θ. ΕΝΔΕΙΞΕΙΣ ΒΙΟΨΙΑΣ ΗΠΑΤΟΣ Χρόνια
ΠΡΑΚΤΙΚΕΣ ΟΔΗΓΙΕΣ ΣΤΟΥΣ ΗΠΑΤΟΛΟΓΟΥΣ, ΔΙΑΒΑΖΟΝΤΑΣ ΤΗ ΒΙΟΨΙΑ ΗΠΑΤΟΣ Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Τμήματος Ιατρικής Α.Π.Θ. ΕΝΔΕΙΞΕΙΣ ΒΙΟΨΙΑΣ ΗΠΑΤΟΣ Χρόνια
Kυτταρική$Bιολογία$ Πολυκυτταρική+οργάνωση+και+ καρκίνος+ ΔIAΛEΞΕΙΣ*15*&*16! (18!&!21/5/2012)! Δρ.$Xρήστος$Παναγιωτίδης,$Τμήμα$Φαρμακευτικής$Α.Π.Θ.
! Δρ.$Xρήστος$Παναγιωτίδης,$Τμήμα$Φαρμακευτικής$Α.Π.Θ.") Kυτταρική$Bιολογία$ ΔIAΛEΞΕΙΣ*15*&*16! (18!&!21/5/2012)! Πολυκυτταρική+οργάνωση+και+ καρκίνος+ Ας+ξαναθυμηθούμε+για+λίγο+ τη+κυτταρική+θεωρία+ Η*κυτταρική*θεωρία*! OΛOI!OI!ZΩNTANOI!OPΓANIΣMOI! AΠOTEΛOYNTAI!AΠO!KYTTAPA!H!AΠO!
Kυτταρική$Bιολογία$ ΔIAΛEΞΕΙΣ*15*&*16! (18!&!21/5/2012)! Πολυκυτταρική+οργάνωση+και+ καρκίνος+ Ας+ξαναθυμηθούμε+για+λίγο+ τη+κυτταρική+θεωρία+ Η*κυτταρική*θεωρία*! OΛOI!OI!ZΩNTANOI!OPΓANIΣMOI! AΠOTEΛOYNTAI!AΠO!KYTTAPA!H!AΠO!