Έκφραση και ρόλος της πρωτεογλυκάνης agrin κατά την ανάπτυξη του πρώιμου εμβρύου
|
|
|
- Θυώνη Ανδρέου
- 7 χρόνια πριν
- Προβολές:
Transcript
1 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ, ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΠΤΥΞΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ Έκφραση και ρόλος της πρωτεογλυκάνης agrin κατά την ανάπτυξη του πρώιμου εμβρύου Καπόλου Ελένη Βιολόγος Διατριβή για το Μεταπτυχιακό Δίπλωμα Ειδίκευσης στη Βιολογική Τεχνολογία Πάτρα 2008
2 Στο εξώφυλλο απεικονίζονται οι αλληλεπιδράσεις της agrin με άλλα μόρια της κυτταρικής επιφάνειας και η επαγόμενη έναρξη ποικίλων σηματοδοτικών μονοπατιών στο εσωτερικό του κυττάρου (Bezakova and Ruegg, 2003). Καθώς και πρώιμα στάδια ανάπτυξης εμβρύου όρνιθας (ΗΗ4-ΗΗ21) (Hamburger and Hamilton, 1951). 2
3 3
4 Εξεταστική Επιτροπή Νικόλας Ζάγκρης, Καθηγητής Τμήμα Βιολογίας, Πανεπιστήμιο Πατρών (επιβλέπων) Γεωργία Στεφάνου, Καθηγήτρια Τμήμα Βιολογίας, Πανεπιστήμιο Πατρών Γεώργιος Κίλιας, Αναπληρωτής Καθηγητής Τμήμα Βιολογίας, Πανεπιστήμιο Πατρών 4
5 5
6 Ευχαριστίες Θα ήθελα να ευχαριστήσω θερμά τον επιβλέποντα καθηγητή μου, καθηγητή κύριο Νικόλα Ζάγκρη για την ευκαιρία που μου έδωσε να διεκπεραιώσω την παρούσα διατριβή, καθώς και για την συνεχή καθοδήγησή του, τις γνώσεις που μου μετέδωσε και την επιστημονικότητα της κριτικής του. Θα ήθελα να εκφράσω τις ευχαριστίες μου στα μέλη της τριμελούς επιτροπής, την καθηγήτρια κυρία Γεωργία Στεφάνου και τον αναπληρωτή καθηγητή κύριο Γεώργιο Κίλια. Επίσης, ένα μεγάλο ευχαριστώ οφείλω σε όλα τα μέλη του εργαστηρίου για την βοήθεια που μου προσέφεραν κατά τη διεξαγωγή των πειραμάτων μου και ιδιαίτερα στην διδακτορική φοιτήτρια Κατερίνα Γεωργαδάκη για την πολύτιμη βοήθειά της. 6
7 7
8 Αφιερώνω αυτή τη διατριβή στην οικογένειά μου και στους φίλους μου, για την αμέριστη βοήθεια και ηθική συμπαράσταση που μου προσέφεραν κατά την πραγματοποίηση και εκπόνηση της παρούσας εργασίας. 8
9 9
10 Περιεχόμενα Εισαγωγή Η εξωκυττάρια ύλη και ο ρόλος της στην ανάπτυξη του εμβρύου Βασική μεμβράνη Πρωτεογλυκάνες Ο ρόλος των πρωτεογλυκανών Κατηγορίες πρωτεογλυκανών Πρωτεογλυκάνες θειικής ηπαράνης Σύνθεση των πρωτεογλυκανών θειικής ηπαράνης Ρόλος των πρωτεογλυκανών θειικής ηπαράνης Πρωτεογλυκάνη Agrin Δομή της agrin και μόρια που αλληλεπιδρούν με αυτήν Ισομορφές της agrin Λειτουργίες της agrin Ανάπτυξη του εμβρύου της όρνιθας Πρώιμα στάδια ανάπτυξης του εμβρύου Σχηματισμός του ενδοδέρματος και του μεσοδέρματος Νευριδίωση Προσδιορισμός των σταδίων εμβρυικής ανάπτυξης στην όρνιθα..52 Σκοπός Διατριβής
11 Υλικά και Μέθοδοι Απομόνωση/ καλλιέργεια εμβρύων Υλικά/Διαλύματα Συσκευές Παρασκευή διαλυμάτων Μεθοδολογία Μονιμοποίηση Εμβρύων Υλικά/Διαλύματα Παρασκευή διαλυμάτων Μεθοδολογία Μικροτόμηση εμβρύων Υλικα/Διαλύματα Συσκευές Μεθοδολογία.66 4.Χρώση με Alcian blue Υλικά Διαλύματα Συσκευές Μεθοδολογία Ανίχνευση πρωτεϊνών με αντισώματα-ανοσοφθορισμός Υλικά/Διαλύματα Συσκευές Παρασκευή διαλυμάτων Μεθοδολογία Έκθεση εμβρύων σε αντισώματα έναντι της agrin και καλλιέργεια 77 11
12 7 Φωτογράφηση Ανίχνευση mrna με τη μέθοδο της RT-PCR Απομόνωση εμβρύων Απομόνωση RNA Υλικά/Διαλύματα Μεθοδολογία Μέτρηση οπτικής απορρόφησης απομονωμένου RNA Υλικά/Διαλύματα Συσκευές Μεθοδολογία Σύνθεση cdna με RT-PCR Υλικά/Διαλύματα Μεθοδολογία Ηλεκτροφόρηση προϊόντων RT-PCR Υλικά/Διαλύματα Συσκευές Παρασκευή διαλυμάτων Μεθοδολογία Αποστειρώσεις. 85 Αποτελέσματα Έκφραση της πρωτεογλυκάνης agrin Έκφραση του mrna agrin Χωρο-χρονική κατανομή της πρωτεΐνης agrin Στάδιο ΗΗ3 (μέσα σταδίου γαστριδίου) Στάδιο ΗΗ4 (προχωρημένο γαστρίδιο) Στάδιο ΗΗ8 (4 σωμίτες) Στάδιο ΗΗ10 (10 σωμίτες) Στάδιο ΗΗ16 + (28-29 σωμίτες)
13 2. Αναστολή λειτουργίας της agrin Συζήτηση.108 Περιλήψεις..116 Βιβλιογραφία
14
15 ΕΙΣΑΓΩΓΗ 1. Η εξωκυττάρια ύλη και ο ρόλος της στην ανάπτυξη του εμβρύου Στους πολυκύτταρους οργανισμούς τα κύτταρα βρίσκονται σε επαφή μ ένα πολύπλοκο δίκτυο, το οποίο αποτελείται από μόρια όπως οι πρωτεογλυκάνες, οι γλυκοζαμινογλυκάνες, οι γλυκοπρωτεΐνες, το κολλαγόνο, καθώς επίσης και από αυξητικούς παράγοντες και κυτοκίνες. Όλα αυτά τα μόρια αποτελούν την εξωκυττάρια ύλη και εκκρίνονται από κύτταρα επηρεάζοντας τη συμπεριφορά τόσο των ιδίων όσο και των γειτονικών τους κυττάρων (Aumailley and Gayraud, 1998 Streuli, 1999 Kleinman et al., 2003). Η αλληλεπίδραση των μορίων αυτών, οδηγεί στο σχηματισμό δικτύων τα οποία εκτός από το δομικό ρόλο τους είναι σημαντικά στη ρύθμιση και στον έλεγχο της διαφοροποίησης των κυττάρων (Zagris, 2001). Οι διακυτταρικές αλληλεπιδράσεις καθώς και οι αλληλεπιδράσεις μεταξύ κυττάρου και εξωκυττάριας ύλης ενεργοποιούν ποικίλα μονοπάτια μεταγωγής σήματος, τροποποιούν τη ρύθμιση συγκεκριμένων γονιδίων και έχουν ως αποτέλεσμα τις διαφορετικές αποκρίσεις των κυττάρων, οι οποίες επηρεάζουν τη προσκόλληση, τη μετανάστευση, τον πολλαπλασιασμό, τη διαφοροποίηση, και την επιβίωσή τους, καθώς και τη μορφογένεση ιστών και οργάνων (Zagris, 2001). Η κυτταρική προσκόλληση προϋποθέτει εξειδίκευση και δυνατότητα ρύθμισης, ειδικότερα κατά τη μορφογένεση ιστών και οργάνων στο πρώιμο έμβρυο καθώς και σε διεργασίες αναγέννησης / ανάπλασης. Είναι σημαντική η κατάλληλη χώρο-χρονική (spatio-temporal) έκφραση πρωτεϊνών που μπορούν να επιδεικνύουν είτε προσκολλητική (adhesive) είτε απωθητική (antiadhesive) δράση. Η εξωκυττάρια ύλη αρχίζει να εμφανίζεται ήδη από το στάδιο του μοριδίου. Η δημιουργία του πλέγματος της εξωκυττάριας ύλης συμπίπτει με την έναρξη των μορφογενετικών κινήσεων της γαστριδίωσης (Zagris, 2001). Μόρια της εξωκυττάριας ουσίας αρχίζουν να ανιχνεύονται στο χώρο ανάμεσα στην επιβλάστη και την υποβλάστη (ομόλογη με τον βλαστόκοιλο των αμφιβίων) σε έμβρυα στο στάδιο του βλαστιδίου, με πιθανή προσέλευση και από τις δυο αυτές στοιβάδες κυττάρων (Zagris, 2001). 16
16 ΕΙΣΑΓΩΓΗ Συμπερασματικά, η εξωκυττάρια ύλη είναι μια δυναμική δομή που υπόκειται σε σχετικά γρήγορες ποσοτικές και ποιοτικές αλλαγές των μορίων που την δομούν. Η σύνθεση και διαφοροποίηση των εξωκυττάριων ουσιών αντανακλά τις ιδιότητες των κυττάρων που τις παράγουν και ίσως επηρεάζουν την φαινοτυπική έκφραση του προκειμένου σταδίου διαφοροποίησης (Zagris, 2001). 1.1 Βασική μεμβράνη Η βασική μεμβράνη (Εικ. 1) είναι μία εξειδικευμένη μορφή της εξωκυττάριας ύλης, αποτελούμενη από ένα πολυμερές γλυκοπρωτεϊνών, όπως η λαμινίνη, που σταθεροποιούνται μέσω της εντακτίνης και από δίκτυα κολλαγόνου ΙV εντός των οποίων παγιδεύονται πρωτεογλυκάνες με σημαντικότερη την perlecan (μαργαριτίνη). Η σύνθεση της διαφέρει από ιστό σε ιστό, ακόμα και στον ίδιο ιστό ποικίλει ανάλογα με το στάδιο ανάπτυξης. Η λειτουργική ποικιλότητα των βασικών μεμβρανών πηγάζει από την μοριακή ποικιλότητα των συστατικών της, και ιδιαίτερα από την παρουσία διαφορετικών ισομορφών κολλαγόνου ΙV και λαμινίνης (Paulsson, 1992 Timpl and Brown, 1996 Aumailley and Gayraud, 1998 Erickson and Couchman, 2000). Η βασική μεμβράνη εντοπίζεται πάντα στο κάτω μέρος του επιθηλιακού ιστού, ενώ παράλληλα απαντάται στον ενδοθηλιακό ιστό, και περιβάλλει μυϊκά, νευρικά κύτταρα (Schwann cells) και λιποκύτταρα. Συνεπώς, μπορεί να χαρακτηριστεί ως στοιβάδα επιθηλιοποίησης ιστών και επίσης διαχωρισμού του επιθηλιακού και συνδετικού ιστού (Merker, 1994). Οι σχέσεις των κυττάρων με την βασική μεμβράνη είναι ζωτικής σημασίας. Συγκεκριμένα, προσφέρει μηχανική υποστήριξη και οριοθέτηση σε κύτταρα και αναπτυσσόμενους ιστούς, ενώ παράλληλα, επηρεάζει τον κυτταρικό πολλαπλασιασμό, την πολικότητα, την προσκόλληση, τη μετανάστευση και τη διαφοροποίηση των κυττάρων. Οι ιδιότητες αυτές της βασικής μεμβράνης την κάνουν σημαντική για μία πληθώρα ζωτικών βιολογικών διεργασιών, όπως είναι η ανάπτυξη, η σταθερότητα, η αναγέννηση και η επιδιόρθωση, μετά από τραυματισμό, των ιστών (Erickson and Couchman, 2000). 17
.")
17 ΕΙΣΑΓΩΓΗ Εικόνα 1: Σχηματική αναπαράσταση της βασικής μεμβράνης και των αλληλεπιδράσεων μεταξύ μορίων που την δομούν και μορίων της επιφάνειας του κυττάρου (διαδικτυακός τόπος 1). 1.2 Πρωτεογλυκάνες Οι πρωτεογλυκάνες εκφράζονται σε όλα τα σπονδυλωτά καθώς επίσης και σε ασπόνδυλα, όπως το έντομο Drosophila melanogaster, και το νηματοειδές C. elegans (Prydz and Dalen, 2000). Εντοπίζονται σε μεγάλα ποσοστά στην εξωκυττάρια ύλη και στην κυτταρική επιφάνεια, ενώ μικρότερα ποσοστά αυτών εντοπίζονται στις ενδοκυττάριες δομές, γεγονός που αποδεικνύει την σημασία τους τόσο στην διακυτταρική επικοινωνία όσο και στις αλληλεπιδράσεις της εξωκυττάριας ύλης με το κύτταρο. Οι πρωτεογλυκάνες είναι μία οικογένεια μορίων τα οποία αποτελούνται από έναν πρωτεϊνικό κορμό και μία ή περισσότερες ομοιοπολικά συνδεδεμένες πλευρικές αλυσίδες γλυκοζαμινογλυκανών (Εικ. 2). Η σύνδεση του πρωτεϊνικού κορμού με τις γλυκοζαμινογλυκάνες γίνεται μέσω ενός γλυκοσιδικού δεσμού που ενώνει ένα κατάλοιπο σερίνης του πρωτεϊνικού κορμού της εκάστοτε πρωτεογλυκάνης, μέσω μίας ξυλόζης και δύο σακχάρων 18
18 ΕΙΣΑΓΩΓΗ γαλακτόζης, με την υπόλοιπη αλυσίδα της γλυκοζαμινογλυκάνης (Goetinck, 1991). Οι γλυκοζαμινογλυκάνες είναι γραμμικά πολυμερή από επαναλαμβανόμενους δισακχαρίτες, οι οποίοι αποτελούνται από ένα ουρικό οξύ, το D-γλυκουρονικό ή το L-ιδουρονικό οξύ και από ένα ακετυλιωμένο αμινοσάκχαρο, που μπορεί να είναι είτε Ν-ακετυλογλυκοζαμίνη είτε Ν- ακετυλογαλακτοζαμίνη. Εξαίρεση στο προηγούμενο μοτίβο του δισακχαρίτη εντοπίζεται στην η θειική κερατάνη της οποίας ο δισακχαρίτης αποτελείται από D-γαλακτόζη και Ν-ακετυλογλυκοζαμίνη (Prydz and Dalen, 2000 Frevert and Wight, 2006) (Εικ. 2). Οι επαναλαμβανόμενες αυτές μονάδες δισακχαριτών συνδέονται με γλυκοζιτικούς δεσμόυς και σχηματίζουν μεγάλα γραμμικά πολυμερή μόρια που φέρουν αρνητικό φορτίο λόγω της παρουσίας θειικών ή και καρβοξυλικών ομάδων στη δομή τους, γεγονός που συμβαίνει κατά την ενζυμική βιοσύνθεση των γλυκοζαμινογλυκανών (Goetinck, 1991). Οι κυριότερες γλυκοζαμινογλυκάνες είναι το υαλουρονικό οξύ (hyaluronan), η θειική χονδροϊτίνη (chondroitin sulphate, CS), η θειική δερματάνη (dermatan sulphate-ds), η θειική ηπαράνη/ ηπαρίνη (heparan sulphate, HS/heparin) και η θειική κερατάνη (keratan sulphate, KS). Το υαλουρονικό οξύ είναι η μόνη γλυκοζαμινογλυκάνη που δεν είναι θειικωμένη και που προσδένεται στον πρωτεϊνικό κορμό πρωτεογλυκανών μόνο μέσω συνδετικών γλυκοπρωτεϊνών (link proteins). Η πολυπλοκότητα και η ετερογένεια των πρωτεογλυκανών αποδίδεται τόσο στους περίπου 30 διαφορετικούς πρωτεϊνικούς κορμούς, όσον αφορά το μέγεθος ( kda) και τη δομή τους, όσο και στην αξιοσημείωτη ποικιλότητα που παρατηρείται στον αριθμό (1 έως >100) και στους τύπους των πλευρικά συνδεδεμένων αλυσίδων γλυκοζαμινογλυκανών (Frevert and Wight, 2006). Ωστόσο, οι πρωτεογλυκάνες έχουν τη δυνατότητα να σχηματίζουν γιγάντια σύμπλοκα με το υαλουρονικό οξύ (Εικ. 2). Σε αυτά τα γιγάντια σύμπλοκα το υαλουρονικό οξύ αποτελεί τον κορμό στον οποίο συνδέονται πλήθος πρωτεογλυκανών, μέσω της αλληλεπίδρασής τους με ειδικές συνδετικές πρωτεΐνες (Ζάγκρης, 1996). 19

19 ΕΙΣΑΓΩΓΗ Εικόνα 2: Απεικόνιση γιγαντιαίων συμπλόκων πρωτεογλυκάνης-συνδετικών πρωτεϊνών-υαλουρονικού. Από αριστερά προς τα δεξιά απεικονίζεται η συσσώρευση γιγαντιαίων μορίων πρωτεογλυκανών, όπως φαίνεται από ηλεκτρονική μικροσκοπία, η διαγραμματική απεικόνιση τμήματος των γιγαντιαίων αυτών μορίων και οι χημικοί τύποι γνωστών γλυκοζαμινογλυκανών (GAGs) (Alberts et al., 2002) Ο ρόλος των πρωτεογλυκανών Οι πρωτεογλυκάνες, λόγω της ποικιλότητας και της κατανομής τους τόσο στην εξωκυττάρια ύλη όσο και στην επιφάνεια και το εσωτερικό του κυττάρου, φαίνεται να είναι σημαντικές στις βιολογικές διαδικασίες που σχετίζονται με αλληλεπιδράσεις και επικοινωνία κυττάρου-κυττάρου και κυττάρου-εξωκυττάριας ύλης (Frevert and Sannes, 2005). Οι πρωτεογλυκάνες, φαίνεται να προσελκύουν ή να εμποδίζουν την προσκόλληση μεταξύ των κυττάρων που μεταναστεύουν. Επίσης, μπορούν να λειτουργούν ως ενδιάμεσα μόρια στις συνδέσεις μεταξύ των ιστών που απαρτίζουν ένα όργανο. Σε μερικές περιπτώσεις, οι πρωτεογλυκάνες που εκκρίνονται από έναν κυτταρικό πληθυσμό μπορεί να είναι σημαντικές για τα ίδια τα κύτταρα που τις εκκρίνουν ή για γειτονικά κύτταρα. Για παράδειγμα, στην κυτταρική μεμβράνη των αξόνων γαγγλίων ραχιαίας ρίζας, προσδένονται πρωτεογλυκάνες θειικής ηπαράνης. Απομάκρυνση των πρωτεογλυκανών 20
20 ΕΙΣΑΓΩΓΗ αυτών εμποδίζει τον πολλαπλασιασμό των προστατευτικών κυττάρων Schwann γύρω από τους άξονες (Ζάγκρης, 1996). Μία ακόμη, σημαντική λειτουργία των πρωτεογλυκανών είναι η δέσμευση αυξητικών παραγόντων, κυτοκινών και χημοκινών, μέσω των πλευρικών αλυσίδων γλυκοζαμινογλυκανών που φέρουν, αλλά και μέσω του πρωτεϊνικού κορμού τους. Η δέσμευση αυτή είναι πλέον γνωστό ότι αποτελεί μία εξειδικευμένη αλληλεπίδραση των πρωτεϊνών που προαναφέραμε με μοναδικές θέσεις δέσμευσης των γλυκοζαμινογλυκανών. Ο βαθμός εξειδίκευσης μεταξύ των αυξητικών παραγόντων και των γλυκοζαμινογλυκανών δεν είναι ακόμα γνωστός, αλλά πιστεύεται ότι υπάρχει συγκεκριμένο μοντέλο θειίκωσης για κάθε πρωτεΐνη που δεσμεύεται στην εκάστοτε αλυσίδα γλυκοσαμινογλυκανών (Frevert and Wight, 2006). Αποτέλεσμα των εξειδικευμένων αυτών αλληλεπιδράσεων είναι ο καθοδηγητικός ρόλος των πρωτεογλυκανών απέναντι στους αυξητικούς παράγοντες που μεταφέρουν, αφού τους κατευθύνουν σε ξεχωριστές θέσεις σε σχέση με το κύτταρο ανάλογα με το μοντέλο θειίκωσης της πρωτεογλυκάνης που έχουν επιλέξει να συνδεθούν. Χαρακτηριστικό παράδειγμα αποτελούν οι ινοβλαστικοί αυξητικοί παράγοντες (fibroblast growth factors) FGF-2 και -4 οι οποίοι δεσμεύονται σε αλυσίδες θειικής ηπαράνης και εντοπίζονται σε διαφορετικές ανατομικά περιοχές των πνευμόνων. Ενώ, ο FGF-2 εντοπίζεται στον ιστό των πνευμόνων και στα ενδοθηλιακά κύτταρα των αιμοφόρων αγγείων, ο FGF-4 εντοπίζεται στους πνεύμονες αλλά όχι στα ενδοθηλιακά κύτταρα. Το γεγονός αυτό σχετίζεται με το μοντέλο θειίκωσης των αλυσίδων θειικής ηπαράνης στις οποίες δεσμεύονται οι συγκεκριμένοι αυξητικοί παράγοντες, με αποτέλεσμα την διαφορετική διαμερισματοποίησή τους (Frevert and Sannes, 2005). Μία άλλη σημαντική λειτουργία των πρωτεογλυκανών που στηρίζεται και αυτή στην εξειδικευμένη αλληλεπίδραση των γλυκοζαμινογλυκανών με πρωτεϊνικά μόρια, είναι η συμβολή τους στην μεταγωγή σημάτων στο εσωτερικό του κυττάρου (Frevert and Sannes, 2005 Frevert and Wight, 2006). Το γεγονός αυτό επιτυγχάνεται με μία ποικιλία μηχανισμών ανάλογα με το είδος του σηματοδοτικού σήματος και την κυτταρική διαδικασία την οποία ρυθμίζει. Παραδείγματος χάριν, σημαντικοί αυξητικοί παράγοντες, όπως ο FGF, συνδέονται με γλυκοζαμινογλυκάνες πρωτεογλυκανών θειικής ηπαράνης διευκολύνοντας τον διμερισμό των υποδοχέων τους και κατά 21
21 ΕΙΣΑΓΩΓΗ συνέπεια την έναρξη του σηματοδοτικού μονοπατιού στο εσωτερικό του κυττάρου (Allen et al., 2001 Lin, 2004). Μία ακόμα πολύ σημαντική λειτουργία των πρωτεογλυκανών είναι η ρύθμιση της διαθεσιμότητας των αυξητικών παραγόντων αλλά και η προστασία τους από την πρωτεόλυση (Frevert and Sannes, 2005 Frevert and Wight, 2006). Οι πρωτεογλυκάνες ενεργοποιούν ποικίλα μονοπάτια μεταγωγής σήματος και επάγουν πλειοτροπικές αποκρίσεις από τα κύτταρα, συμμετέχοντας στον έλεγχο της κυτταρικής προσκόλλησης, μετανάστευσης, πολλαπλασιασμού, διαφοροποίησης και επιβίωσης των κυττάρων και στη μορφογένεση ιστών και οργάνων Κατηγορίες πρωτεογλυκανών Οι πρωτεογλυκάνες λόγω της μεγάλης ετερογένειας που παρουσιάζουν μπορούν να κατηγοριοποιηθούν είτε βάση των γλυκοζαμινογλυκανών που φέρουν στον πρωτεϊνικό τους κορμό, είτε βάση της δομής των πρωτεϊνικών τους κορμών. Στην παρούσα εργασία θα χρησιμοποιήσουμε την δεύτερη κατηγοριοποίηση, ενώ παράλληλα θα τις χωρίσουμε και με βάση την θέση (διαμερισματοποίηση) τους σε σχέση με το κύτταρο (Goetinck, 1991) (Πίνακας 1). Υπάρχουν και πρωτεογλυκάνες όπως η phosphacan και η neuroglycan που δεν ομαδοποιούνται σε κατηγορίες. Α. Εξωκυττάριες Πρωτεογλυκάνες 1. Πρωτεογλυκάνες της βασικής μεμβράνης Στην κατηγορία αυτή ανήκουν κυρίως πρωτεογλυκάνες θειικής ηπαράνης με σημαντικότερες την perlecan και την agrin. Ο πρωτεϊνικός τους κορμός φέρει τρεις θέσεις δέσμευσης γλυκοζαμινογλυκανών θειικής ηπαράνης, οι οποίες στην perlecan εντοπίζονται στο αμινοτελικό άκρο, ενώ στην agrin στην κεντρική περιοχή της. Επιπρόσθετα, τα δύο αυτά μόρια περιέχουν περιοχές πλούσιες σε κυστεϊνες, EGF-like περιοχές, καθώς και αρκετές θέσεις Ο-γλυκοζυλίωσης και Ν-γλυκοζυλίωσης. Σημαντικό χαρακτηριστικό αυτών των πρωτεογλυκανών είναι η δυνατότητα δέσμευσης 22
22 ΕΙΣΑΓΩΓΗ FGF και άλλων αυξητικών παραγόντων μέσω του πρωτεϊνικού κορμού τους αλλά και των αλυσίδων θειικής ηπαράνης. Έτσι πιστεύεται ότι οι πρωτεογλυκάνες θειικής ηπαράνης της βασικής μεμβράνης μπορούν να λειτουργούν και ως δεξαμενές αυξητικών παραγόντων (Goetinck, 1991). Στην ίδια κατηγορία ανήκει και η bamacan η οποία φέρει τρεις αλυσίδες θειικής χονδροϊτίνης και ο πρωτεϊνικός της κορμός δεν παρουσιάζει σημαντικές ομοιότητες με τις άλλες πρωτεογλυκάνες της συγκεκριμένης κατηγορίας. Χαρακτηρίζεται ωστόσο από τρείς δομές ελικοειδούς έλικας (coiled-coil), ενώ φέρει και θέσεις Ν-γλυκοζυλίωσης (Wu and Couchman, 1997). 2. Λεκτικάνες ή Υαλεκτάνες (Lecticans/Hyalectins) Η αμινοτελική περιοχή των λεκτικανών χαρακτηρίζεται από μία περιοχή δέσμευσης υαλουρονικού οξέος, η οποία περιέχει πολλαπλές περιοχές που ομοιάζουν με πλήθος πρωτεϊνών, όπως των CD44, N-CAΜ, ανοσοσφαιρινών και άλλων. Η κεντρική περιοχή, η οποία διαφέρει στο μέγεθος ανάμεσα στις λεκτικάνες, φέρει ποικίλες θέσεις δέσμευσης γλυκοζαμινογλυκανών θειικής χονδροϊτίνης και σε μικρότερο αριθμό θειικής κερατάνης. Το καρβοξυτελικό άκρο του πρωτεϊνικού κορμού των λεκτικανών αποτελείται από μία ή δύο EGR-like περιοχές, μία C-τύπου lectin-like περιοχή, μέσω της οποίας γίνεται η δέσμευση ολιγοσακχαριτών και μία συμπληρωματική ρυθμιστική περιοχή (complement control domain-ccp) η οποία εντοπίζεται σε πολλές πρωτεΐνες, όπως στην β 2 -γλυκοπρωτεΐνη, και λειτουργεί ως περιοχή αλληλεπίδρασης μεταξύ πρωτεϊνών. Κύριοι εκπρόσωποι της οικογένειας των λεκτικάνων είναι οι aggrecan, versican, neurocan και brevican. Η aggrecan είναι η πρώτη λεκτικάνη που μελετήθηκε εκτενώς και εντοπίζεται κύρια στον χόνδρο. Η neurocan, η brevican και η versican έχουν ανιχνευτεί στο κεντρικό νευρικό σύστημα (Goetinck, 1991 Bandtlow and Zimmermann, 2000 Kresse and Schönherr, 2001), αλλά οι πρωτεογλυκάνες αυτές και επίσης η aggrecan παρουσιάζουν ευρεία κατανομή (πρόσφατα αποτελέσματα του εργαστηρίου μας). 23
23 ΕΙΣΑΓΩΓΗ 3. Πρωτεογλυκάνες πλούσιες σε λευκίνη Η οικογένεια αυτή περιλαμβάνει πρωτεογλυκάνες των οποίων ο πρωτεϊνικός κορμός αποτελείται από περιοχές αμινοξέων, πλούσιες σε λευκίνη (LRPs-Leucine-rich repeats), οι οποίες επαναλαμβάνονται 7-12 φορές ανάλογα με το μόριο της πρωτεογλυκάνης. Έχει αποδειχθεί ότι υπάρχουν δύο τύποι LRPs, ο S, που αποτελείται από 21 αμινοξέα και ο Τ, που αποτελείται από 26 αμινοξέα. Οι δύο αυτοί τύποι LRPs παρουσιάζουν διαφορετική αμινοξική αλληλουχία, με συνέπεια τα κατάλοιπα λευκίνης να βρίσκονται σε διαφορετικές θέσεις (Matsushima et al., 2000). Ανάλογα λοιπόν με τον τύπο και την σειρά που εμφανίζονται τα LRPs, καθώς επίσης και με βάση το είδος και τον αριθμό των γλυκοζαμινογλυκανών που περιέχουν μπορούν να χωριστούν σε τρεις επιμέρους ομάδες. Στην ομάδα Ι, που περιέχει τις biglycan/pg-i και decorin/pg-ii, στην ομάδα ΙΙ, που περιλαμβάνει τις lumican, fibromodulin, PRELP, keratocan, και osteoadherin, και στην ομάδα ΙΙΙ, που αποτελείται από τις epiphycan/pg-lb και osteoglycin. Οι δύο πρώτες ομάδες ομοιάζουν πολύ μεταξύ τους και όλα τα μέλη τους φέρουν 12 LRPs, ενώ η τρίτη ομάδα φέρει 7 RLPs (Hocking et al., 1998 Matsushima et al., 2000). Όσον αφορά τις αλυσίδες γλυκοζαμινογλυκανών ποικίλουν ακόμα και για το ίδιο μόριο, ανάλογα με τον ιστό στον οποίο εκφράζεται αυτό. Για παράδειγμα, η decorin που εντοπίζεται στο δέρμα, τον χόνδρο και στον σκληρό χιτώνα του οφθαλμού περιέχει αλυσίδες θειικής δερματάνης, ενώ αυτή που εντοπίζεται στα οστά φέρει αλυσίδες θειικής χονδροίτίνης (Neame et al., 1989). Β. Πρωτεογλυκάνες της κυτταρικής επιφάνειας Μία άλλη κατηγορία πρωτεογλυκανών είναι αυτές που σχετίζονται με την κυτταρική επιφάνεια, είτε διαπερνώντας την κυτταρική μεμβράνη, μέσω μίας υδροφοβικής περιοχής του πρωτεϊνικού τους κορμού, είτε αγκυροβολώντας σε αυτήν μέσω της αλληλεπίδρασης του πρωτεϊνικού τους κορμού με το γλυκολιπίδιο γλυκόζυλο-φωσφατυδιλοϊνοσιτόλη (GPI) (Goetinck, 1991 Yanagishita and Hascall, 1992). Το μεγαλύτερο ποσοστό των πρωτεογλυκανών που ανήκουν σε αυτήν την κατηγορία είναι πρωτεογλυκάνες θειικής ηπαράνης. Ωστόσο, μπορεί να φέρουν και κάποιον 24
24 ΕΙΣΑΓΩΓΗ αριθμό αλυσίδων θειικής χονδροϊτίνης. Χαρακτηριστικοί εκπρώσωποι είναι οι syndecans, glypicans, τύπου ΙΙΙ υποδοχέας του TGF-β (betaglycan) και CD44 οικογένεια (Wegrowski and Maquart, 2004). Όλες, εκτός από τις glypicans, οι οποίες δεσμέυονται με το GPI, είναι διαμεμβρανικές. Οι πρωτεογλυκάνες της συγκεκριμένης κατηγορίας συναντώνται ευρέως σε πλήθος ιστών, όπως είναι τα ηπατοκύτταρα, τα επιθηλιακά κύτταρα, οι ινοβλάστες, τα στρωματικά κύτταρα του μυελού των οστών, τα ωοθηκικά κοκκιώδη κύτταρα, τα προγονικά γλοία κύτταρα και τα κύτταρα Schwann (Goetinck, 1991). Παράλληλα, οι πρωτεογλυκάνες αυτές, της κυτταρικής επιφάνειας, αλληλεπιδρούν τόσο με εξωκυττάρια όσο και με ενδοκυττάρια μόρια και εμπλέκονται λειτουργικά στις αλληλεπιδράσεις μεταξύ των κυττάρων καθώς και σε αυτές κυττάρου και εξωκυττάριας ύλης. Γ. Ενδοκυττάριες Πρωτεογλυκάνες Τα ιστιοκύτταρα (mast cells) περιέχουν ένα σχετικά υψηλό ποσοστό μικρών πρωτεογλυκανών στα εκκριτικά τους κοκκία (Stevens and Austen, 1989). Αυτές οι πρωτεογλυκάνες έχουν έναν πολύ μικρό πρωτεϊνικό κορμό που χαρακτηρίζεται από ποικίλες επαναλήψεις αλληλουχιών με κατάλοιπα σερίνης και γλυκίνης. Με βάση αυτά τα δομικά χαρακτηριστικά οι πρωτεογλυκάνες έχουν ονομαστεί serglycins. Στο πρωτεϊνικό κορμό των συγκεκριμένων πρωτεογλυκανών το μέγεθος των περιοχών που φέρουν επαναλήψεις σερίνης-γλυκίνης (Ser-Gly) ποικίλει ανάμεσα στα είδη. Οι γλυκοζαμινογλυκάνες που προσδένονται στον πρωτεϊνικό κορμό μπορεί να είναι είτε ηπαρίνη, όπως παρατηρείται στα ιστιοκύτταρα του συνδετικού ιστού, είτε υψηλά θειικωμένη μορφή θειικής χονδροϊτίνης, όπως εντοπίζονται στα ιστιοκύτταρα που προέρχονται από το μυελό των οστών. Ο τύπος της γλυκοζαμινογλυκάνης που θα συνδεθεί στον πρωτεϊνικό κορμό φαίνεται ότι ρυθμίζεται από το μικροπεριβάλλον και το στάδιο διαφοροποίησης των ιστιοκυττάρων (Steven and Austen, 1989 Goetinck, 1991). 25
25 ΕΙΣΑΓΩΓΗ Δ. Part-time Πρωτεογλυκάνες Παρόλο που η betaglycan αποτελεί, όπως αναφέραμε, μία διαμεμβρανική πρωτεογλυκάνη δεν χρειάζεται τις αλυσίδες γλυκοζαμινογλυκανών για να δράσει ως υποδοχέας. Ωστόσο, δεν είναι ξεκαθαρισμένο αν το μόριο αυτό εμφανίζεται στα κύτταρα και με τις δύο μορφές του, ταυτόχρονα. Υπάρχουν ενδείξεις ότι σε συγκεκριμένα κύτταρα εντοπίζεται και ως πρωτεογλυκάνη και ως υποδοχέας. Τέτοιου είδους πρωτεογλυκάνες αποκαλούνται part-time πρωτεογλυκάνες (Goetinck, 1991). Σε αυτήν την κατηγορία πρωτεογλυκανών ανήκουν επίσης η thrombomodulin, ο CD44, η προγονική αμυλοειδής πρωτεΐνη και τα κολλαγόνα τύπου IX, XII, XIV και XVIII (Kresse and Schönherr, 2001). ΚΑΤΗΓΟΡΙΕΣ ΠΡΩΤΕΟΓΛΥΚΑΝΩΝ Λεκτικάνες: aggrecan, versican, neurocan, brevican Εξωκυττάριες Πρωτεογλυκάνες Πρωτεογλυκάνες βασικής μεμβράνης: perlecan, agrin, bamacan Πρωτεογλυκάνες πλούσιες σε λευκίνη: biglycan, decorin, lumican, fibromodulin, PRELP, keratocan, osteoadherin, epiphycan, osteoglycin Πρωτεογλυκάνες Κυτταρικής Επιφανείας Ενδοκυττάριες Πρωτεογλυκάνες Part-time Πρωτεογλυκάνες Άλλες syndecans, glypicans, CD44, τύπου ΙΙΙ υποδοχέας του TGF-β (betaglycan) serglycins betaglycan, thrombomodulin, CD44, προγονική αμυλοειδής πρωτεΐνη, κολλαγόνα τύπου IX, XII, XIV και XVIII phosphacan, neuroglycan Πίνακας 1: Βασικές κατηγορίες των πρωτεογλυκανών σύμφωνα με την διαμερισματοποίησή τους σε σχέση με το κύτταρο και τη δομη του πρωτεϊνικού τους κορμού. 26
26 ΕΙΣΑΓΩΓΗ 2. Πρωτεογλυκάνες θειικής ηπαράνης (HSPGs) Η θειική ηπαράνη ανακαλύφθηκε πριν από περίπου 60 χρόνια σε εμπορικά παρασκευάσματα της ηπαρίνης ως λιγότερο θειικωμένη ηπαρίνη, από τους Jorpes και Gardell (Jorpes and Gardell, 1948). Είναι πλέον γνωστό ότι η αλυσίδα θειικής ηπαράνης έχει διαφορετική δομή από την ηπαρίνη και ευρύτερο φάσμα βιολογικών δραστηριοτήτων. Πρωτεογλυκάνες θειικής ηπαράνης εντοπίζονται στην κυτταρική επιφάνεια, με χαρακτηριστικούς εκπροσώπους τις syndecans, glypicans, betaglycan και CD44, ενώ επίσης ανιχνεύονται στην εξωκυττάρια ύλη, με κυριότερες τις perlecan, agrin και κολαγόνο XVIII (Εικ. 3). Μελέτες σε πειραματικά μοντέλα-οργανισμούς έχουν δείξει την σπουδαιότητα αυτών των πρωτεϊνών στην ανάπτυξη και στην ομαλή φυσιολογία ποικίλων συστημάτων του εκάστοτε οργανισμού, όπως είναι το ενδοκρινικό, το μυϊκό, το νευρικό, το αναπαραγωγικό και άλλα. Η λειτουργική ποικιλομορφία των πρωτεογλυκανών θειικής ηπαράνης οφείλεται στην ποικιλία των δεσμευτικών δυνατοτήτων των γλυκοζαμινογλυκανών θειικής ηπαράνης με άλλα μόρια, όπως για παράδειγμα με αυξητικούς παράγοντες, καθώς και στις μεταβολές της δομής των πρωτεϊνικών τους κορμών (Bishop et al., 2007). 27
27 ΕΙΣΑΓΩΓΗ Εικόνα 3: Διαγραμματική απεικόνιση πέντε αντιπροσωπευτικών πρωτεογλυκανών θειικής ηπαράνης (HSPGs). Περίπου τριάντα γονίδια κωδικοποιούν το πλήθος της συγκεκριμένης κατηγορίας πρωτεογλυκανών, η οποία περιλαμβάνει τέσσερις syndecans, έξι glypicans, μία perlecan, μία agrin και το υβρίδιο HSPG/κολλαγόνο τύπου XVIII. Επιπρόσθετα, ποικίλες ισομορφές έχουν αναφερθεί για τις perlecan, agrin και κολλαγόνο τύπου XVIII. Οι ράβδοι αντιπροσωπεύουν τον πρωτεϊνικό κορμό των πρωτεογλυκανών, με τις χαρακτηριστικές περιοχές τους, όπως περιγράφονται στο πλαίσιο και οι κυματιστές γραμμές συμβολίζουν τις αλυσίδες γλυκοζαμινογλυκανών (Iozzo, 2001). 28
28 ΕΙΣΑΓΩΓΗ 2.1 Σύνθεση των πρωτεογλυκανών θειικής ηπαράνης Τα πρόδρομα μόρια του πρωτεϊνικού κορμού κάθε πρωτεογλυκάνης θειικής ηπαράνης συντίθονται στο αδρό ενδοπλασματικό δίκτυο και στη συνέχεια μετατοπίζονται στο σύμπλεγμα Golgi, όπου συμβαίνει η βιοσύνθεση των αλυσίδων θειικής ηπαράνης. Στη διαδικασία αυτή εμπλέκεται μία ποικιλία ενζύμων, όπως, μία επιμεράση και διαφορετικές θειοτρενσφεράσες. Πολλά από τα ένζυμα αυτά υπάρχουν σε πολλαπλές ισομορφές (Perrimon and Bernfield, 2000). Όλες, εκτός από την ισομορφή 3-Ο-θειοτρανσφεράση, είναι διαμεμβρανικές πρωτεΐνες τύπου ΙΙ, δηλαδή διαπερνούν μία φορά την πλασματική μεμβράνη και το αμινοτελικό τους τμήμα βρίσκεται ενδοκυτταρικά, όπως συμβαίνει και με τις άλλες πρωτεΐνες του συμπλέγματος Golgi. Η διαδικασία της βιοσύνθεσης (Εικ. 4) των αλυσίδων θειικής ηπαράνης ξεκινά σε συγκεκριμένες θέσεις καταλοίπων σερίνης-γλυκίνης (Ser-Gly) εντός καθορισμένης αμινοξικής αλληλουχίας του πρωτεϊνικού κορμού της εκάστοτε πρωτεογλυκάνης. Το πρώτο βήμα στην διαδικασία αυτή είναι η νουκλεοτιδική (UDP) μεταφορά της ξυλόζης, στην υδροφοβική ομάδα των καταλοίπων σερίνης του πρωτεϊνικού κορμού και η δημιουργία Ο-γλυκοζυτικού δεσμού με αυτήν. Ακολουθεί η UDP μεταφορά και των άλλων σακχάρων που απαρτίζουν την τετρασακχαριτική συνδετική περιοχή, -ξυλόζη-γαλακτόζη-γαλακτόζηγλυκουρονικό οξύ- (-Xyl-Gal-Gal-GlcA-), η οποία αποτελεί εναρκτήριο σημείο για τη βιοσύνθεση τόσο της θειικής ηπαράνης όσο και της θειικής χονδροϊτίνης. Το καθοριστικό βήμα για την βιοσύνθεση αποκλειστικά αλυσίδας θειικής ηπαράνης είναι η προσθήκη ενός καταλοίπου Ν- ακετυλογλυκοζαμίνης (GlcNAc) επί του συνδετικού τετρασακχαρίτη μέσω της δράσης μίας γλυκοσυλοτρανσφεράσης. Έτσι ξεκινά η διαδοχική προσθήκη μονοσακχαριδικών καταλοίπων GlcA και GlcNAc εναλλάξ, στο άκρο της σχηματιζόμενης αλυσίδας ηπαράνης, με τη βοήθεια μίας πολυμεράσης (Stringer and Gallagher, 1997 Lindahl et al., 1998 Perrimon and Bernfield, 2000). Κατά τη διάρκεια της διαδικασίας του πολυμερισμού η αλυσίδα της θειικής ηπαράνης υφίσταται ένα πλήθος τροποποιήσεων (Lidholt and Lindahl, 1992) από μία ποικιλία ενζύμων του συμπλέγματος Golgi. Το πρώτο βήμα 29
29 ΕΙΣΑΓΩΓΗ είναι η Ν-απακετυλίωση και η Ν-θειίκωση μονάδων Ν-ακετυλογλυκοσαμίνης (GlcNAc) γεγονότα που επιτυγχάνονται μέσω της διπλής λειτουργίας του ενζύμου NDST (glucosaminyl N-deacetylase/N-sulfotransfarase). Όλες οι υπόλοιπες τροποποιήσεις λαμβάνουν χώρα εντός ή κοντά στις Ν-θειικωμένες περιοχές, καθώς τα ένζυμα φαίνεται να εξαρτώνται από την παρουσία καταλοίπων Ν-θειικής γλυκοζαμίνης (GlcNSO 3 ), την οποία αναγνωρίζουν ως υπόστρωμα (Salmivitra et al., 1996). Πρόσφατες έρευνες έχουν δείξει ότι η Ν- θειίκωση δεν αποτελεί αναγκαία προϋπόθεση για τις διαδοχικές τροποποιήσεις που συμβαίνουν (Holmborn et al., 2004). Το επόμενο ρυθμιστικό ένζυμο, η GlcA C-5 επιμεράση (C5epi) μετατρέπει το γλυκουρονικό οξύ (GlcA) σε ιδουρονικό οξύ (IdoA). Στη συνέχεια τρεις διαφορετικές αντιδράσεις Ο-θειίκωσης μπορεί να συμβούν. Αρχικά με τη δράση της 2-Ο-θειοτρανσφεράσης (2-ΟST) μεταφέρεται μία θειική ομάδα στην 2-ΟΗ θέση των καταλοίπων ιδουρονικού οξέος καθώς και σε ορισμένα κατάλοιπα γλυκουρονικού οξέος. Ακολούθως, με τον ίδιο τρόπο μέσω δράσης της 6-Ο-θειοτρανσφεράσης (6-ΟST) πραγματοποιείται θειίκωση των καταλοίπων Ν-θειική γλυκοσαμίνης και της Ν-ακετυλογλυκοσαμίνης όταν αυτή βρίσκεται κοντά με ένα κατάλοιπο Ν-θειικής γλυκοζαμινης (Lidholt and Lindahl, 1992 Stringer and Gallagher, 1997 Perrimon and Bernfield, 2000). Πρόσφατες μελέτες έχουν δείξει την ύπαρξη 6-Ο-θειικωμένων κατάλοιπα Ν- ακετυλογλυκοσαμίνης σε μη θειικωμένη θειική ηπαράνη (Holmborn et al., 2004). Σε όλα τα στάδια που αναφέραμε παραπάνω, η δράση των θειοτρανσφερασών διευκολύνεται με την συμμετοχή του συνενζύμου φωσφαδενοσίνη (PAPS), το οποίο δρα ως δότης θειικής ομάδας. Έτσι η θειοτρανσφεράση μεταφέρει την θειική αυτή ομάδα από το συνένζυμο PAPS στο αντίστοιχο σάκχαρο. 30
30 ΕΙΣΑΓΩΓΗ UDP-γλυκόζη αφυδρογονάση GlcNAc/GlcA πολυμεράση (tout velu/ext) Ν-απακετυλάση/ Ν- θειοτρανσφεράση (sulphateless) GlcA C5 επιμεράση Θειική ομάδα Συνδετική 2-, 6- και 3-Ο-θειοτρανσφεράσες περιοχή πρωτεϊνικός κορμός Εικόνα 4: Σχηματική αναπαράσταση της βιοσύνθεσης των γλυκοζαμινογλυκανών θειικής ηπαράνης. Gal, γαλακτόζη; GlcA, γλυκουρονικό οξύ; GlcNAc, Ν- ακετυλογλυκοζαμίνης; PAPS, φωσφαδενοσίνη, δότης θειικής ομάδας; UDP, νουκλεοτιδικός μεταφορέας σακχαρων; Xyl, ξυλόζη. Στις παρενθέσεις με πλάγια γράμματα αναφέρονται ορισμένα γονίδια που κωδικοποιούν τα αντίστοιχα ένζυμα (Perrimon and Bernfield, 2000). Μέσα από αυτή την πολύπλοκή βιοχημική διαδικασία προκύπτει μία πολυσακχαριδική αλυσίδα που αποτελείται ουσιαστικά από επαναλήψεις δισακχαριτών αποτελούμενων από ένα ουρονικό οξύ (D-γλυκουρονικό οξύ ή L-ιδουρονικό οξύ) και ένα κατάλοιπο D-γλυκοζαμίνης, τα οποία μπορεί να είναι Ν-θειικωμένα ή Ν-ακετυλιωμένα, και υπόκεινται σε διαφορετικές Ο- θειικώσεις σε συγκεκριμένες περιοχές. Ο μηχανισμός που καθορίζει την επέκταση της τροποποίησης της πολυσακχαριδικής αυτής αλυσίδας δεν έχει πλήρως διευκρινιστεί, αλλά βασίζεται στο επίπεδο ενεργότητας του ενζύμου Ν-απακετυλάση-Ν-θειοτρανσφεράση. Η δημιουργία συγκεκριμένων, λειτουργικά σημαντικών, αλληλουχιών θειικής ηπαράνης μπορεί να εξαρτάται 31
31 ΕΙΣΑΓΩΓΗ από την εξειδίκευση χαρακτηριστικών ενζύμων (Stringer and Gallagher, 1997 Prydz and Dalen, 2000). Η σύνθεση των αλυσίδων θειικής ηπαράνης ολοκληρώνεται σε ένα χρονικό διάστημα λεπτών, και οι πρωτεογλυκάνες που προκύπτουν οδηγούνται μέσω κυτταροπλασματικών οχημάτων στις θέσεις δράσης τους είτε στην επιφάνεια του κυττάρου (syndecans, glypicans), είτε στην βασική μεμβράνη (agrin, perlecan). Χαρακτηριστικά, οι πρωτεογλυκάνες της κυτταρικής επιφάνειας έχουν χρόνο ημιζωής 3-8 ώρες πριν αποβληθούν, είτε μέσω της δράσης πρωτεασών είτε μέσω ειδικών φωσφολιπασών για τις GPIσυνδεόμενων πρωτεογλυκανών θειικής ηπαράνης, ή μπορεί να ενδοκυτταρώνονται. Μετά την ενδοκύττωση οι πρωτεογλυκάνες θειικής ηπαράνης μπορούν να περάσουν πολλά μεταβολικά μονοπάτια, σε συνάρτηση πάντα με τον τύπο του κυττάρου στα οποία βρίσκονται (Yanagishita and Hascall, 1992). Στα κοκκιώδη κύτταρα ωοθηκών ποντικών, ενδοκυτταρωμένες GPI-συνδεόμενες πρωτεογλυκάνες θειικής ηπαράνης διαμερισματοποιούνται στα λυσοσώματα και γρήγορα αποικοδομούνται σε μονοσακχαρίτες και ανόργανο θείο. Είναι γνωστό ότι, η πλειοψηφία των πρωτεογλυκανών θειικής ηπαράνης που εγκολπόνωνται στα κοκκιώδη κύτταρα αποικοδομούνται με αργά διαδοχικά στάδια αποδίδοντας ενδιάμεσα σάκχαρα 10 και 5 kda τα οποία τελικά αποικοδομούνται στα λυσοσώματα. Τα διαφορετικά μονοπάτια αποσύνθεσης των μεμβρανικών πρωτεογλυκανών θειικής ηπαράνης δεν έχουν πλήρως αποσαφηνισθεί (Yanagishita and Hascall, 1992 Stringer and Gallagher, 1997). 2.2 Ρόλος των πρωτεογλυκανών θειικής ηπαράνης Οι πρωτεογλυκάνες θειικής ηπαράνης φαίνεται να εμπλέκονται σε πολλαπλές φυσιολογικές λειτουργίες του κυττάρου. Μελέτες έχουν δείξει την συμμετοχή των πρωτεογλυκανών θειικής ηπαράνης της εξωκυττάριας ύλης, όπως οι agrin και perlecan, στην οργάνωση και το ρόλο βασικών μεμβρανών οργάνων όπως οι πνεύμονες και το νεφρό (Bishop et al., 2007), καθώς και στην οργάνωση του αιματοεγκεφαλικού φραγμού (Kröger and Schröder, 2002). Μία όμως από τις σημαντικότερες λειτουργίες των πρωτεογλυκανών 32
32 ΕΙΣΑΓΩΓΗ θειικής ηπαράνης είναι η συμμετοχή τους στα ποικίλα αναπτυξιακά σηματοδοτικά μονοπάτια, είτε διαμέσω των πλευρικών αλυσίδων τους είτε μέσω του πρωτεϊνικού τους κορμού (Lin, 2004). Μεταλλάξεις γονιδίων που κωδικοποιούν ένζυμα σημαντικά για την βιοσύνθεση της αλυσίδας θειικής ηπαράνης οδηγούν σε μείωση ή πλήρη αναστολή της ενεργότητας σημαντικών σηματοδοτικών μονοπατιών. Για παράδειγμα, πειράματα με μεταλλάξεις στο tou velu (ttv) γονίδιο του εντόμου Drosophilla, το οποίο είναι υπεύθυνο για την κωδικοποίηση ενζύμου που μετέχει στον πολυμερισμό της αλυσίδας της θειικής ηπαράνης (Εικ. 4), έδειξαν αδυναμία των Hedgehog μορίων να μετακινηθούν από κύτταρο σε κύτταρο, με αποτέλεσμα την μειωμένη ενεργοποίηση του σηματοδοτικού μονοπατιού που επάγεται από αυτό (Perrimon and Bernfield, 2000 Lin, 2004). Παράλληλα, μεταλλάξεις και σε γονίδια που κωδικοποιούν τον πρωτεϊνικό κορμό των πρωτεογλυκανών θειικής ηπαράνης, επηρεάζουν πολλά σηματοδοτικά μονοπάτια. Για παράδειγμα, μετάλλαξη στο γονίδιο dally του εντόμου Drosophila, το οποίο κωδικοποιεί ένα από τα μέλη της οικογένειας των glypicans, οδηγεί σε φαινότυπο παρόμοιο με αυτόν που παρατηρείται απουσία της ενεργότητας του Wingless μορίου. Αυτό δείχνει ότι ο πρωτεϊνικός κορμός της glypican συμμετέχει στη μεταγωγή σήματος μέσω του Wnt μονοπατιού και τώρα είναι γνωστό ότι η πρωτεογλυκάνη glypican είναι συνυποδοχέας του wingless (Perrimon and Bernfield, 2000). Όπως γνωρίζουμε, η σηματοδότηση κατά τη διαδικασία της ανάπτυξης πρέπει να είναι αυστηρά ρυθμιζόμενη για κάθε κύτταρο ξεχωριστά έτσι ώστε να λαμβάνει τη σωστή ποσότητα σημάτων την κατάλληλη χρονική στιγμή. Η μεταγωγή σημάτων είναι μία απαραίτητη προϋπόθεση για τον καθορισμό και την διαφοροποίηση των κυττάρων, καθώς και τη μορφογένεση ιστών και οργάνων. Οι μηχανισμοί μέσω των οποίων οι πρωτεογλυκάνες θειικής ηπαράνης επηρεάζουν την ρύθμιση της μεταγωγής σήματος κατά τη διάρκεια της ανάπτυξης δεν έχουν πλήρως αποσαφηνιστεί και ίσως λειτουργούν με διαφορετικό τρόπο σε ποικίλα σηματοδοτικά συστήματα. Ωστόσο, μέχρι σημερα έχουν αναφερθεί τουλάχιστον τέσσερις μηχανισμοί με τους οποίους οι πρωτεογλυκάνες θειικής ηπαράνης μπορούν να μετέχουν στην διαδικασία της ενεργοποίησης ενός σηματοδοτικού μονοπατιού από το αντίστοιχο μόριοενεργοποιητή (Εικ. 5). Ο πρώτος μηχανισμός αφορά την διακυτταρική 33
33 ΕΙΣΑΓΩΓΗ μεταφορά, ή ολίσθηση του μηνυματοφόρου μορίου, μέσω των αλυσίδων θειικής ηπαράνης (Εικ. 5Α). Έτσι, οι πρωτεογλυκάνες θειικής ηπαράνης της κυτταρικής επιφάνειας μπορούν να μεσολαβούν στην κίνηση μηνυματοφόρων μορίων μεταξύ των κυττάρων, μέσω μίας ενεργητικής είτε παθητικής, εξαρτώμενη της διάχυσης, μεταφοράς. Δηλαδή, τα μόρια μπορούν να περάσουν ενεργά στα γειτονικά τους κύτταρα με ανταλλαγή προσδεμάτων μέσω των πλευρικών αλυσίδων θειικής ηπαράνης, είτε να ολισθήσουν μέσω των αλυσίδων θειικής ηπαράνης, στις οποίες δεν δεσμεύεται ισχυρά απλά τις χρησιμοποιεί ως επιταχυντή της διάχυσής τους στο κύτταρο-στόχο (Nybakken and Perrimon, 2002). Το συγκεκριμένο μοντέλο μεταφοράς έχει αποδειχθεί ότι μεσολαβεί στην μετακίνηση των μηνυματοφόρων μορίων Hedghog και Decapentaplegic, το οποίο είναι μέλος της TFG-β οικογένειας αυξητικών παραγόντων, κατά τον σχηματισμό των πτερυγίων του εντόμου Drosophila. Επίσης, είναι πιθανό οι πρωτεογλυκάνες θειικής ηπαράνης να μεσολαβούν με τον ίδιο μηχανισμό και στην μετακίνηση του Wingless σήματος, παρόλο που δεν υπάρχουν επαρκείς αποδείξεις (Lin, 2004). Σύμφωνα με τον δεύτερο μηχανισμό, οι πρωτεογλυκάνες μπορεί να συμμετέχουν στην πρόσδεση του μηνυματοφόρου μορίου στον υποδοχέα του, δηλαδή να λειτουργούν ως συνυποδοχείς (Εικ. 5Β). Ο τρίτος μηχανισμός με τον οποίο οι πρωτεογλυκάνες θειικής ηπαράνης μπορεί να μεσολαβούν στην κυτταρική σηματοδότηση είναι η προστασία των μηνυματοφόρων μορίων από πρωτεόλυση, η σταθεροποίηση σηματοδοτικών μορίων στις αλυσίδες θειικής ηπαρίνης, η αύξηση του χρόνου παραμονής τους στην κυτταρική επιφάνεια και η ρύθμιση της διαθεσιμότητας των μηνυματοφόρων μορίων στον υποδοχέα τους (Εικ. 5C) (Hacker et al., 1997 Pfeiffer et al., 2002 Lin and Perrimon, 2003 Lin, 2004). 34
34 ΕΙΣΑΓΩΓΗ Εικόνα 5: Διαγραμματική απεικόνιση των τριών πιο σημαντικών μηχανισμών με τους οποίους οι πρωτεογλυκάνες θειικής ηπαράνης (HSPGs) επηρεάζουν τη ρύθμιση της μεταγωγής μηνυματοφόρων μορίων κατά τη διάρκεια της ανάπτυξης. (Α) Μηχανισμός διακυτταρικής μεταφοράς, ή ολίσθησης μηνυματοφόρου μορίου, (Β) Μηχανισμός διμερισμού ή ολιγοδιμερισμού προσδεμάτων με τους υποδοχείς τους (C) Μηχανισμός σταθεροποίησης μυνηματοφόρου μορίου στις HS GAG (αλυσίδες θειικής ηπαράνης) και επέκταση του χρόνου δράσης του στον υποδοχέα. EGF, επιδερμικός αυξητικός παράγοντας; EFGR, υποδοχέα του EGF (Lin, 2004). 35
35 ΕΙΣΑΓΩΓΗ Σε όλους αυτούς τους μηχανισμούς το εξειδικευμένο μοντέλο θειίκωσης για κάθε θειική ηπαράνη αποτελεί σημαντικό παράγοντα. Ένα κύτταρο μπορεί, μέσω παραγωγής αλληλουχιών θειικής ηπαράνης με χαμηλή ή υψηλή συγγένεια για κάποιο μόριο, είτε να προσελκύσει είτε να απωθήσει την συγκέντρωση διαφορετικών μορίων και ως εκ τούτου να διευκολύνει τις τοπικές διαφορετικές συγκεντρώσεις. Έτσι, διαφορετικά πρότυπα θειίκωσης της θειικής ηπαράνης αποτελούν μία φυσιολογική διαδικασία κατά την ανάπτυξη μέχρι τη γήρανση. Για παράδειγμα, εξαρτώμενη της ηλικίας αύξηση στην 6-Ο-θειίκωση παρατηρείται στην θειική ηπαράνη πρωτεογλυκάνης που ανιχνεύεται στην ανθρώπινη αορτή (Feyzi et al., 1998). Διαφορετικά πρότυπα θειίκωσης της θειικής ηπαράνης παρατηρούνται σε πολλές παθολογικές καταστάσεις, όπως ο καρκίνος, η ασθένεια Alzheimer και ο διαβήτης, σε σύγκριση με τα φυσιολογικά πρότυπα. Μελέτες έχουν δείξει ότι η σύνθεση των γλυκοζαμινογλυκανών αλλάζει σε ποικίλους ιστούς στους διαβητικούς ασθενείς και στα πειραματικά διαβητικά ποντίκια. Η θειική ηπαράνη από το συκώτι των διαβητικών αρουραίων έχει λιγότερες θειικές ομάδες σε σύγκριση με αυτή από συκώτι άγριου τύπου αρουραίων (Kjellen et al., 1983) και αυτό είναι αντανάκλαση της μειωμένης ενεργότητας του NDST1 (ενζύμου που συμμετέχει στην θειίκωση της Ν-ακετυλογλυκοζαμίνης) στα ηπατοκύτταρα διαβητικών αρουραίων (Unger et al., 1991). Σε μεταλλαγμένα έμβρυα Drosophila, στα οποία ένζυμα (π.χ. NDST) που μετέχουν στην απακετυλίωση και θειίκωση της Ν-ακετυλογλυκοζαμίνης, και η αφυδρογονάση που καταλύει την UDP-γλυκόζη σε UDP-γλυκουρονικό οξύ, έχουν χάσει τη δράση τους, παρατηρήθηκε απώλεια της ενεργότητας μονοπατιών μεταγωγής σήματος που επάγονται από τον FGF (Perrimon and Bernifield, 2000 Lin, 2004). Στο αναπτυσσόμενο μυϊκό νευροεπιθήλιο, έχει παρατηρηθεί μία αλλαγή στην παραγωγή της θειικής ηπαράνης από μία δομή που προτιμά FGF2 δέσμευση και σηματοδότηση σε μία άλλη που προωθεί το FGF1 μονοπάτι την χρονική στιγμή που ξεκινά η έκφραση του FGF1 και η νευρωνική διαφοροποίηση (Nurcombe et al., 1993). Βιοχημική εξέταση της θειικής ηπαράνης, σε διαφορετικές χρονικές στιγμές ανάπτυξης του μυϊκού επιθηλίου, παρουσιάζουν διαφορετικά μοντέλα θειίκωσης, μέγεθος και αριθμός αλυσίδων θειικωμένων περιοχών. Παράλληλα, μιτογόνες αναλύσεις καλλιέργειας κυττάρων έχουν δείξει ότι ειδικές αλληλουχίες θειικής ηπαράνης 36
36 ΕΙΣΑΓΩΓΗ ευνοούν την σηματοδότηση μέσω είτε του FGF1 είτε του FGF2 αυξητικού παράγοντα (Guimond et al., 1993 Pye et al., 2000). Έχει γίνει φανερό στην βιβλιογραφία ότι διαφορετικά FGF-FGFR σύμπλοκα σε ποικίλα αναπτυξιακά στάδια απαιτούν αλληλουχίες θειικής ηπαράνης διαφορετικά θειικωμένες για να συγκροτηθούν (Allen and Rapraeger, 2003). 37
37 ΕΙΣΑΓΩΓΗ 3. Πρωτεογλυκάνη Agrin 3.1 Δομή της agrin και μόρια που αλληλεπιδρούν με αυτήν Η Agrin είναι μία πρωτεογλυκάνη θειικής ηπαράνης που εντοπίζεται κυρίως στις βασικές μεμβράνες πολλών ειδών συμπεριλαμβανομένων της όρνιθας, του ποντικού, του αρουραίου, της αγελάδας και του ανθρώπου. Ο πρωτεϊνικός της κορμός περιέχει πολυπεπτιδικές περιοχές ομόλογες με μοτίβα που εντοπίζονται και σε άλλες πρωτεΐνες της εξωκυττάριας ύλης (Rupp et al., 1991 Smith et al., 1992 Tsim et al., 1992), καθώς και περιοχές παρόμοιες με αυτές του επιδερμικού αυξητικού παράγοντα (EGF), της follistatin πρωτεΐνης και των σφαιρικών περιοχών της α-αλυσίδας της λαμινίνης. Με τη χρήση ηλεκτρονικής μικροσκοπίας αποκαλύφθηκαν οι δομικές περιοχές του πρωτεϊνικού κορμού της agrin, ο οποίος παρουσιάζεται ως ένα τμήμα μήκους περίπου 95nm και μοριακού βάρους γύρω στα 220 kda (Denzer et al., 1998). Ο πρωτεϊνικός αυτός κορμός αποτελείται από τη σφαιρική αμινοτελική περιοχή (NtA, Animo (N)-terminal agrin domain) στην αρχή της οποίας υπάρχει πεπτιδική αλληλουχία που σηματοδοτεί την έκκριση της πρωτεΐνης, την κεντρική περιοχή η οποία κυριαρχείται από περιοχές όμοιες της follistatin (follistatin-like), και από την καρβοξυτελική περιοχή αποτελούμενη από τρεις σφαιρικές περιοχές όμοιες της λαμινίνης (LG-laminin globular domain) (Bezakova and Ruegg, 2003). Στον πρωτεϊνικό κορμό της agrin φαίνεται να περιλαμβάνονται πέντε δυνητικές θέσεις Ν-συνδεδεμένων ολιγοσακχαριτών και τρεις θέσεις σύνδεσης γλυκοζαμινογλυκανών (Winzer et al., 2003). Στην Εικόνα 6 παρουσιάζεται διαγραμματική απεικόνιση της δομής της agrin και μορίων με τα οποία είναι γνωστό ότι αλληλεπιδρά. 38

38 ΕΙΣΑΓΩΓΗ Εικόνα 6: Διαγραμματική απεικόνιση της δομής της agrin και περιοχές μέσω των οποίων αλληλεπιδρά με μόρια όπως, η α-dystroglycan, οι ιντεγκρίνες, ο υποδοχέας της agrin, οι ισομορφές της λαμινίνης, τα μόρια κυτταρικής προσκόλλησης NCAM και πλήθος αυξητικών παραγόντων. EG, EGF-όμοιες περιοχές; FS, follistatin-όμοιες περιοχές; LE, laminin EGF-όμοιες περιοχές; LG, όμοιες με σφαιρικές περιοχές λαμινίνης; NtA, αμινοτελική περιοχή; SS, αρχική σηματοδοτική αλληλουχία; S/T, περιοχές πλούσιες σε σερίνη/θρεονίνη; ΤΜ, διαμεμβρανική μορφή της agrin. Α/y και B/z σημεία εναλλακτικού ματίσματος (Bezakova and Ruegg, 2003). Η αμινοτελική περιοχή, NtA περιοχή (135 αμινοξέα), της agrin φαίνεται να είναι υπεύθυνη για την δέσμευσή της στις βασικές μεμβράνες, γεγονός που επιτυγχάνεται με την αλληλεπίδραση της agrin με την γ-1 αλυσίδα του μεγάλου βραχίονα των ισομορφών της λαμινίνης (Stetefeld et al., 2001). Όμως, έχουν ανιχνευτεί και ισομορφές της agrin που δεν φέρουν την χαρακτηριστική σφαιρική NtA περιοχή, αλλά μία μικρότερη αμινοξική αλληλουχία 50 αμινοξικών καταλοίπων, στα ποντίκια (Burgess et al., 2000). Η διαφορά προκύπτει λόγω διαφορετικού σημείου έναρξης της μεταγραφής του γονιδίου της agrin, με αποτέλεσμα να προκύπτουν δύο ισομορφές όσο αφορά το αμινοτελικό άκρο της agrin. Η μία ισομορφή είναι αυτή περιγράψαμε προηγουμένως, καλείται LN (longer N-terminal) και φέρει μεγαλύτερη αμινοτελική αλληλουχία αποτελούμενη από 150 αμινοξέα τα οποία συγκροτούν την χαρακτηριστική σφαιρική NtA περιοχή των 135 αμινοξέών και την αρχική σηματοδοτική αλληλουχία (SS-signal sequence), ενώ παράλληλα φέρει και μία θέση εναλλακτικού ματίσματος αμέσως μετά την αμινοξική αλληλουχία της NtA περιοχής. Αντίθετα, η άλλη ισομορφή αποτελείται από μία μικρότερη αμινοτελική αλληλουχία, η οποία ονομάζεται SN (shorter N- terminal), και συγκροτείται από αλληλουχία 50 αμινοξέων. Η SN ισομορφή της 39
39 ΕΙΣΑΓΩΓΗ agrin, η οποία εκφράζεται αποκλειστικά στους νευρώνες, μετατρέπει την agrin από μία εκκρινόμενη, συνδεόμενη με την εξωκυττάρια ύλη πρωτεογλυκάνη, σε μία διαμεμβρανική πρωτεογλυκάνη τύπου ΙΙ, που διαπερνά την κυτταρική μεμβράνη μία μόνο φορά και έχει το αμινοτελικό άκρο κυτταροπλασματικά ενώ το καρβοξυτελικό εξωκυτταρικά (Bezakova and Ruegg, 2003 διαδικτυακός τόπος 3). Η έκφραση αυτής της διαμεμβρανικής μορφής της agrin ρυθμίζεται από ξεχωριστό υποκινητή και τα μετάγραφα που θα κωδικοποιήσουν την εν λόγω διαμεμβρανική ισομορφή είναι άφθονα στο εμβρυϊκό κεντρικό νευρικό σύστημα, και ιδιαίτερα στους αναπτυσσόμενους νευρώνες (Burgess et al., 2000 Bezakova and Ruegg, 2003). Από την παρατήρηση ότι ποντίκια, στα οποία η εκκρινόμενη ισομορφή της agrin έχει επιλεκτικά απενεργοποιηθεί, δείχνουν ακριβώς τον ίδιο φαινότυπο στην νευρομυϊκή σύναψη με τα ποντίκια που δεν εκφράζουν καθόλου την agrin, καταλήγουμε στο συμπέρασμα ότι η διαμεμβρανική ισομορφή δεν εμπλέκεται άμεσα στην δημιουργία της εν λόγω σύναψης (Burgess et al., 2000). Αντίθετα, η SN-agrin φαίνεται να διαδραματίζει συγκεκριμένους ρόλους στις αλληλεπιδράσεις μεταξύ των νευρώνων. Συμπερασματικά, και οι δύο ισομορφές της agrin είναι εκκρινόμενες αλλά η LN-agrin συγκεντρώνεται στις βασικές μεμβράνες νευρικών και μη, ιστών και δεσμεύεται σε αυτές μέσω της λαμινίνης, ενώ η SN-agrin παραμένει συνδεδεμένη αποκλειστικά με την κυτταρική επιφάνεια των νευρώνων του κεντρικού νευρικού συστήματος (Burgess et al., 2000 Bezakova and Ruegg, 2003). Ωστόσο, και στις δύο ισομορφές η αμινοξική αλληλουχία που ακολουθεί είναι ίδια, αποτελούμενη από την κεντρική και καρβοξυτελική περιοχή. Η καρβοξυτελική περιοχή του πρωτεϊνικού κορμού της agrin περιέχει όλες τις περιοχές που είναι απαραίτητες για την αλληλεπίδρασή της με υποδοχείς της κυτταρικής επιφάνειας. Για παράδειγμα, η agrin μέσω της LG3 περιοχής της επάγει τη συγκέντρωση των υποδοχέων ακετυλοχολίνης στην επιφάνεια των μυϊκών κυττάρων (Bezakova and Ruegg, 2003 Stetefeld et al., 2004). Επίσης, είναι γνωστό ότι και οι τρείς LG περιοχές συμβάλλουν στην αλληλεπίδραση της agrin με ορισμένα μόρια ιντεγκρινών, αν και συχνά η αλληλεπίδραση αυτή επιτυγχάνεται με την παρεμβολή της λαμινίνης, η οποία αλληλεπιδρά με ιντεγκρίνες καθώς και με την agrin (Εικ. 6). Η G2 περιοχή της agrin έχει τη δυνατότητα δέσμευσης ηπαρίνης που αποτελεί έναν δραστικό 40
40 ΕΙΣΑΓΩΓΗ αναστολέα της συνάθροισης των υποδοχέων ακετυλοχολίνης και ως εκ τούτου μπορεί να αναστείλει μία από τις βασικές διαδικασίες που λαμβάνουν χώρα στη νευρομυϊκή σύναψη. Η LG1 περιοχή του πρωτεϊνικού κορμού της agrin αλληλεπιδρά με την a-dystroglycan, η οποία εμφανίζεται ως ο πιο άφθονος υποδοχέας της επιφάνειας των μυϊκών κυττάρων (Hoch, 1999 Bezakova and Ruegg, 2003 Stetefeld et al., 2004). Το καρβοξυτελικό άκρο της agrin περιέχει και θέσεις εναλλακτικού ματίσματος, με αποτέλεσμα να προκύπτουν πολλαπλές ισομορφές της πρωτεογλυκάνης που ποικίλουν στα διάφορα είδη ζωικών οργανισμών. Η πλειοψηφία των ισομορφών της πρωτεογλυκάνης agrin αποτελούνται μόνο από αλυσίδες θειικής ηπαράνης, ενώ υπάρχουν και ορισμένες ισομορφές που είναι χιμαιρικές αφού φέρουν και αλυσίδες θειικής ηπαράνης αλλά και θειικής χονδροϊτίνης (Groffen et al., 1998 Winzen et al., 2003). Η σύνδεση γλυκοσαμινογλυκανών επί του πρωτεϊνικού κορμού, έχει ως αποτέλεσμα την αύξηση του συνολικού μοριακού βάρους της agrin μέχρι και 600kDa (Hoch, 1999). Οι πλευρικές αυτές αλυσίδες γλυκοσαμινογλυκανών συμβάλλουν στην αλληλεπίδραση της agrin με αυξητικούς παράγοντες, όπως οι FGF και διαμεμβρανικά μόρια γειτονικών κυττάρων όπως είναι τα NCAM (neural-cell adhesion molecule) (Bezakova and Ruegg, 2003). 3.2 Ισομορφές της agrin. Το γονίδιο της agrin έχει χαρτογραφηθεί σχετικά κοντά στο γονίδιο που κωδικοποιεί την πρωτεογλυκάνη perlecan, στην απομακρυσμένη περιοχή του μικρού βραχίονα του πρώτου χρωμοσώματος, στον άνθρωπο, ενώ στο ποντικό έχει χαρτογραφηθεί στο τέταρτο χρωμόσωμα (Iozzo, 1998). Έχουν βρεθεί τουλάχιστον τέσσερις θέσεις εναλλακτικού ματίσματος στο πρώιμο mrna (pre-mrna) με αποτέλεσμα να προκύπτουν ποικίλες ισομορφές της agrin. Η πρώτη θέση εναλλακτικού ματίσματος εντοπίζεται στην περιοχή του pre-mrna που θα κωδικοποιήσει την αμινοτελική περιοχή της agrin, με αποτέλεσμα να προκύπτουν δύο κύριες ισομορφές αυτής με διαφορετική αμινοτελική περιοχή. Όπως αναφέραμε προηγουμένως, οι ισομορφές αυτές είναι η εκκρινόμενη (LN-agrin) και η διαμεμβρανική (SN-agrin ή TM-agrin) 41
41 ΕΙΣΑΓΩΓΗ ισομορφή της agrin. Η κατανομή των mrnas που κωδικοποιούν αυτές τις ισομορφές της agrin στους διάφορους ιστούς ποικίλει ενώ, δεν έχει αποσαφηνιστεί πλήρως η διαμερισματοποίησή τους σε ορισμένους ιστούς. Υπάρχουν ιστοί όπου χαρακτηρίζονται από την παρουσία μεταγράφων που κωδικοποιούν και τις δύο ισομορφές της agrin, και άλλοι στους οποίους εντοπίζονται μετάγραφα είτε της μίας μορφής είτε της άλλης. Χαρακτηριστικά, το mrna της LN-agrin έχει ανιχνευτεί κυρίως στους πνεύμονες, τους νεφρούς, τους σκελετικούς μυς και τους κινητικούς νευρώνες. Το mrna της SN-agrin φαίνεται να κυριαρχεί στο κεντρικό νευρικό σύστημα, κυρίως στους νευρώνες και τα γαγγλιακά κύτταρα του οφθαλμού (Bezakova and Ruegg, 2003). Στην όρνιθα η LN-agrin αποτελεί την βασική ισομορφή της agrin η οποία εντοπίζεται στην βασική μεμβράνη μίας ποικιλίας ιστών (Burgess et al., 2000) Οι άλλες τρεις θέσεις εναλλακτικού ματίσματος εντοπίζονται και στις δύο ισομορφές (LN- και SN-agrin) με αποτέλεσμα να προκύπτουν διαμεμβρανικές και εκκρινόμενες ισομορφές της agrin με ένθεση ή όχι αμινοξικών αλληλουχιών. Από αυτές η πρώτη θέση καλείται x και εντοπίζεται στο pre-mrna των θηλαστικών, αλλά όχι της όρνιθας και φαίνεται να ευθύνεται για την ένθεση 3 αμινοξέων στην κεντρική περιοχή της agrin δίπλα στο SEA τμήμα αυτής. Οι άλλες δύο θέσεις εναλλακτικού ματίσματος εντοπίζονται στα θηλαστικά, με την ονομασία y, z, και στην όρνιθα, με την ονομασία Α, Β (Εικ. 6). Οι δύο αυτές θέσεις φαίνεται να είναι σημαντικές γιατί τροποποιούν το καρβοξυτελικό άκρο της agrin το οποίο είναι υπεύθυνο για πολλαπλές αλληλεπιδράσεις της με σημαντικά μόρια και υποδοχείς. Στην A/y θέση μπορεί να γίνει ένθεση 4 αμινοξέων ενώ στην θέση B/z μπορεί να γίνει ένθεση 8, 11, ή 19 αμινοξέων (Ferns et al., 1992 Rupp et al., 1992 Burgess et al., 2000). Συνεπώς, προκύπτουν πλήθος διαφορετικών ισομορφών της agrin οι οποίες διαφέρουν μόνο στην απουσία ή παρουσία μικρών παρεμβαλλόμενων αμινοξικών αλληλουχιών με λιγότερα από 20 αμινοξέα. Τα τμήματα αυτά, παρά το μικρό τους μέγεθος, επηρεάζουν ισχυρά την βιολογική ενεργότητα της πρωτεογλυκάνης. Για παράδειγμα, η LN- agrin-b/z + που φέρει οκτώ αμινοξέα στην Β/z θέση αποτελεί την ισομορφή της agrin με την ισχυρότερη δραστικότητα στην διαδικασία της νευρομυϊκής σύναψης (Hoch, 1999). Η ένθεση οκτώ αμινοξέων στην συγκεκριμένη θέση αυξάνει την 42
42 ΕΙΣΑΓΩΓΗ ενεργότητα της agrin φορές (Ferns et al., 1993), ενώ η παρουσία μίας ένθεσης έντεκα αμινοξέων στην ίδια θέση αυξάνει την ενεργότητα μόλις 50 φορές (Hoch et al., 1994a). Οι ενθέσεις και των δύο αμινοξικών αλληλουχιών, που αναφέραμε προηγουμένως, μπορούν να αλλάξουν τη στερεοδιαμόρφωση της agrin στον χώρο. Ενώ, οι ισομορφές που δεν φέρουν ένθεση στη B/z θέση (agrin-b/z - ) εντοπίζονται κυρίως σε μη νευρικούς ιστούς συμπεριλαμβανομένων του μυϊκού ιστού, του νεφρού, των αιμοφόρων αγγείων και του ανοσοποιητικού συστήματος (Bezakova and Ruegg, 2003). 3.3 Λειτουργίες της agrin Η agrin είναι μία πρωτεογλυκάνη θειικής ηπαράνης η οποία εντοπίζεται τόσο στη συναπτική βασική μεμβράνη των νευρομυϊκών συνάψεων, όπου και πρωτοανιχνεύτηκε, όσο και στις βασικές μεμβράνες του πνεύμονα, του εγκεφάλου, του νεφρού, της καρδιάς και των μυών συμβάλλοντας στην μορφολογία και στις λειτουργίες αυτών. Για παράδειγμα, στους νεφρούς η agrin μαζί με την perlecan και άλλες πρωτεΐνες, ελέγχει την επιλεκτική διαπερατότητα ουσιών, δρώντας ως ηθμός (Halfter et al., 1997 Groffen et al., 1999 Yard et al., 2001). Ενώ, είναι σημαντική για τη συναπτική διαφοροποίηση στη νευρομυϊκή σύναψη, όπου έχει μελετηθεί πολύ καλά (Gautam et al., 1995 DeChiara et al., 1996 Denzer et al., 1996 Gautam et al., 1996 Ruegg and Bixby, 1998 Hoch, 1999 Bezakova and Ruegg, 2003). Η agrin συντίθεται και απελευθερώνεται από τα μυϊκά κύτταρα και από τους κινητικούς νευρώνες και εμπεριέχεται στη βασική μεμβράνη μεταξύ των μυϊκών κυττάρων και των κινητικών νευρώνων στο σημείο της νευρομυϊκής σύναψης (Gautam et al., 1996 Denzer et al., 1997). Η agrin που αλληλεπιδρά με την dystroglycan συνεργεί στη συνάθροιση των υποδοχέων της ακετυλοχολίνης και της σχετιζόμενης με τον υποδοχέα πρωτεΐνης rapsyn στην τελική περιοχή (τελική ζώνη/τελική πλάκα) και αυτή η συνάθροιση των μορίων είναι σημαντική στη νευρομυϊκή διαβίβαση (Glass et al., 1996 Meier and Wallace, 1998 Bezakova and Markus, 2003) (Εικ. 7). 43
 στην τελική περιοχή της νευρομυϊκής σύναψης.")
43 ΕΙΣΑΓΩΓΗ Εικόνα 7: Διαγραμματική απεικόνιση της αλληλεπίδρασης της agrin με την α- dystroglycan του DAPC (dystrophin-associated protein complex) συμπλόκου και η επακόλουθη συνάθροιση της rapsyn και των υποδοχέων ακετυλοχολίνης (AChRs) στην τελική περιοχή της νευρομυϊκής σύναψης. Το DAPC σύμπλοκο δρα ως υποδοχέας της λαμινίνης, της agrin και της perlacan και η βασική του λειτουργία είναι η σύνδεση της εξωκυττάριας ύλης με τον κυτταροσκελετό του μυϊκού κυττάρου και η έμμεση εμπλοκή του στη διαδικασία συνάθροισης AChRs στη μετασυναπτική μεμβράνη της νευρομυϊκής σύναψης, μέσω αλληλεπίδρασης της rapsyn με την dystroglycan. DB, dystrobrevin; DG, dystroglycans; R, rapsyn; SG, sacroglycans; SP, sarcospan; SYN, syntrophin (Chamberlain, 1999). Είναι γνωστή η ικανότητα των ιντεγκρινών να μεταβιβάζουν σινιάλα από τα μόρια της εξωκυττάριας ύλης στο εσωτερικό του κύτταρου (Hynes, 1992, 1996). Επιπλέον, η αφθονία των ιντεγκρινών τόσο στο έμβρυο όσο και στον ενήλικα οργανισμό, καθώς και η μεσολάβησή τους στις περισσότερες επιδράσεις της λαμινίνης, η οποία με τη σειρά της αλληλεπιδρά με την agrin, (Rupp et al., 1991 Ruegg et al., 1992 Mercurio, 1995 Engvall and Wewer, 1996), αποτελούν σημαντικές ενδείξεις για να ερευνηθεί η πιθανότητα των ιντεγκρινών να παίζουν κάποιο ρόλο στο σηματοδοτικό μονοπάτι της agrin (Εικ. 8). Οι τρεις LG δομές της καρβοξυτελικής περιοχής της agrin εμπλέκονται στην αλληλεπίδρασή της με μόρια ιντεγκρινών. Πράγματι, κύτταρα που φέρουν τις αvβ1 και αvβ3 ιντεγκρίνες φαίνεται να συνδέονται με την διαλυτή μορφή της agrin (LN-agrin). Για παράδειγμα, η αναστολή της 44
44 ΕΙΣΑΓΩΓΗ λειτουργίας ιντεγκρινών που περιέχουν τις αλυσίδες β1 ή αv, μέσω χρήσης αντισωμάτων έναντι αυτών, αναστέλλει την επαγόμενη από την agrin συνάθροιση υποδοχέων ακετυλοχολίνης (AChRs) (Martin and Sanes, 1997). Παράλληλα, από βιβλιογραφικά δεδομένα γνωρίζουμε ότι η agrin φαίνεται να συντονίζει χωροχρονικά ποικίλα σηματοδοτικά μονοπάτια που εκκινούνται από την ενεργοποίηση υποδοχέων όπως ο MuSK (musclespedigic kinase), τα διάφορα μέλη των ErbBs υποδοχέων, οι ιντεγκρίνες και οι SFKs κινάσες (Scr family kinases). Τα σηματοδοτικά αυτά μονοπάτια που διαμορφώνουν τον κυτταροσκελετό, επιτρέπουν την ενσωμάτωση διαφορετικών σημάτων και ενεργοποιούν τη μεταγραφή συγκεκριμένων γονιδίων (Bezakova and Ruegg, 2003). Εικόνα 8: Σχηματική αναπαράσταση της αλληλεπίδρασης της agrin με μόρια της κυτταρικής επιφάνειας και της εξωκυττάριας ύλης και τα σηματοδοτικά μονοπάτια που ενεργοποιούνται από τις αλληλεπιδράσεις αυτές (Bezakova and Ruegg, 2003). Συζητείται στη βιβλιογραφία ότι η agrin συνεργεί όχι μόνο στο σχηματισμό της νευρομυϊκής σύναψης, αλλά ότι μπορεί ο ρόλος της να είναι επίσης σημαντικός στο σχηματισμό ενδονευρικών συνάψεων. Αποτελέσματα 45
45 ΕΙΣΑΓΩΓΗ που συνηγορούν υπέρ της υπόθεσης αυτής είναι τα εξής: η agrin ανιχνεύεται σε υψηλά επίπεδα στις συναπτικές σχισμές των ενδονευρωνικών συνάψεων (Mann and Kröger, 1996 Koulen et al., 1999), ενώ έχει ανιχνευτεί υψηλή συγκεντρωση της agrin εντός των συναπτοσωμάτων νευρώνων του κεντρικού νευρικού συστήματος (Böse et al., 2000) καθώς και η έκκριση της συγκεκριμένης πρωτεογλυκάνης σε καλλιέργειες ιστών του κεντρικού νευρικού συστήματος (Mann and Kröger, 1996). Η agrin εκφράζεται σε υψηλά επίπεδα στους νευρώνες του κεντρικού νευρικού συστήματος, πριν τον σχηματισμό συνάψεων και συγκεκριμένα τη χρονική στιγμή της επιμήκυνσης αξόνων προς τα κύτταρα στόχους (Halfter et al., 1997 Koulen et al., 1999 Serpinskaya et al., 1999 Neumann et al., 2001). Το γεγονός αυτό υποδηλώνει ότι η agrin μπορεί να παίζει σημαντικό ρόλο κατά την διαδικασία της επιμήκυνσης τόσο των αξόνων όσο και των δενδριτών. Πειράματα αναστολής της έκφρασης της agrin μέσω αντινοηματικών ολιγονουκλεοτιδίων ή αντισωμάτων έναντι αυτής, έδειξαν μείωση του ρυθμού ανάπτυξης των διακλαδώσεων των δενδριτών, αλλά δεν παρατηρήθηκε αλλαγή της επιμήκυνσης του άξονα (Böse et al., 2000). Η agrin εκφράζεται τόσο στους νευρώνες όσο και στα γλοία κύτταρα στο κεντρικό νευρικό σύστημα και αυτό υποδεικνύει ότι η agrin μπορεί να έχει επιπρόσθετες λειτουργίες που δεν σχετίζονται μόνο με το σχηματισμό συνάψεων. Είναι τώρα γνωστό ότι η agrin συμμετέχει στην ενεργοποίηση και τον πολλαπλασιασμό των Τ-κυττάρων του ανοσοποιητικού συστήματος (Bezakova and Ruegg, 2003). Παράλληλα, η agrin φαίνεται να εμπλέκεται και στην αιτιολογία της νευροεκφυλιστικής νόσου Alzheimer. Μελέτες έχουν δείξει ότι η agrin μπορεί να δεσμευτεί σε β-αμυλοειδή κυρίως μέσω των πλευρικών αλυσίδων γλυκοσαμινογλυκανών που φέρει στον πρωτεϊνικό της κορμό, με αποτέλεσμα να συσσωρεύεται στις αμυλοειδείς πλάκες που παρατηρούνται στον εγκέφαλο ασθενών με Alzheimer (Cotman et al., 2000). Στο πρώιμο έμβρυο zebrafish, αναστολή της έκφρασης της agrin προκάλεσε ανωμαλίες στον οφθαλμό, στον ακουστικό πόρο, στους σωμίτες και στο όριο μεταξύ του μεσεγκεφάλου και οπίσθεγκεφάλου (Kim et al., 2007). 46
46 ΕΙΣΑΓΩΓΗ 4. Ανάπτυξη του εμβρύου της όρνιθας 4.1 Πρώιμα στάδια ανάπτυξης του εμβρύου Στο ζυγωτό της όρνιθας (τηλολέκιθα ωάρια), οι αλλεπάλληλες αυλακώσεις που είναι μεροβλαστικές και δεν επεκτείνονται στο λεκιθικό κυτταρόπλασμα σχηματίζουν ένα δίσκο στον ζωικό πόλο, τo βλαστόδισκο (blastodisk). Το πρώιμο έμβρυο στο ανεπώαστο γονιμοποιημένο αυγό όρνιθας είναι ένας δίσκος περίπου 2mm σε διάμετρο, απαρτίζεται από περίπου κύτταρα, ονομάζεται βλαστόδερμα (blastoderm) και είναι στο στάδιο του μοριδίου (Ζάγκρης, 1996). Στο στάδιο αυτό το βλαστόδερμα μπορεί να διαχωριστεί σε δύο διακριτές περιοχές, την κεντρική διάφανη περιοχή (area pellucida) η οποία περικλείεται από την περιφερειακή αδιάφανη περιοχή (area opaca). Η διάφανη περιοχή του βλαστοδέρματος υποδιαιρείται στην εσώτερη κεντρική περιοχή (central area) και στην περιθωριακή ζώνη (marginal zone), ένα δαχτυλίδι γύρω από την κεντρική περιοχή (Ζάγκρης, 1996). Η περιθωριακή ζώνη στο οπίσθιο άκρο του εμβρύου (posterior marginal zone / οπίσθια περιθωριακή ζώνη) είναι η ομόλογη περιοχή του κέντρου του Nieuwkoop στα αμφίβια. Η διάφανη περιοχή του βλαστοδέρματος θα σχηματίσει σχεδόν εξ ολοκλήρου το έμβρυο (Gilbert, 2003). Στο βλαστόδερμα, τα κύτταρα είναι μικρά και απόλυτα καθορισμένα στη διάφανη περιοχή ενώ στην περιφέρειά του, στην αδιάφανη περιοχή αρκετά κύτταρα είναι ακόμη ανοικτά προς τη λέκιθο. Στο στάδιο αυτό κύτταρα της επιβλάστης με τη διεργασία της πολυδιείσδυσης (polyingression) και επίσης κύτταρα κάτω (deep cells) από την οπίσθια περιθωριακή ζώνη μεταναστεύουν οριζόντια με κατεύθυνση προς τα εμπρός, ενώνονται και σχηματίζουν μια ομοιόμορφη στοιβάδα, την υποβλάστη (hypoblast). Το έμβρυο στο στάδιο στο οποίο έχει ολοκληρωθεί ο σχηματισμός της υποβλάστης, αποτελείται από δυο στοιβάδες, την υποβλάστη και την επιβλάστη, που συγκρατόνται μαζί στο περιθώριο της αδιάφανης περιοχής, και ο χώρος μεταξύ των δύο στοιβάδων ονομάζεται βλαστόκοιλος (Gilbert, 2003). Το έμβρυο είναι στο στάδιο του βλαστιδίου (πεπλατυσμένο βλαστίδιο) και η υποβλάστη αντιπροσωπεύει το φυτικό ενώ η επιβλάστη το ζωικό πόλο (Ζάγκρης, 1996). 47
47 ΕΙΣΑΓΩΓΗ Ο εμβρυϊκός άξονας σχηματίζεται εξ ολοκλήρου από την επιβλάστη, όπου έχουν την αρχή τους και οι τρεις βλαστικές στοιβάδες, εξώδερμα, μεσόδερμα και ενδόδερμα. Η υποβλάστη δεν λαμβάνει μέρος στο σχηματισμό των εμβρυϊκών δομών (Rosenquist, 1972), αλλά είναι γνωστό ότι συνεργεί στην επαγωγή της πρωτογενούς αύλακας στην επιβλάστη, σταθεροποιεί τον εμβρυϊκό άξονα, και συμμετέχει στο σχηματισμό έξω-εμβρυϊκών υμένων (Ζάγκρης, 1996 Gilbert, 2003). Προς το τέλος του σχηματισμού της υποβλάστης ένα συσσώρευμα κυττάρων, που ονομάζεται Koller s sickle ( δρεπάνι του Koller / ονομάζεται «δρεπάνι» λόγω της χαρακτηριστικής μορφολογίας του) και βρίσκεται στη μέση θέση του πίσω άκρου της κεντρικής περιοχής της επιβλάστης (ακριβώς πρόσθια της οπίσθιας περιθωριακής ζώνης), αρχίζει να μεταναστεύει με κατεύθυνση προς τα εμπρός έως ότου φθάσει το μέγιστο μήκος του (περίπου 1,88mm) και ονομάζεται πρωτογενής γραμμή (primitive streak). Στην μετανάστευση αυτή έχουν συμμετάσχει και κύτταρα της οπίσθιας περιθωριακής ζώνης που δομούν το πίσω μέρος της πρωτογενούς γραμμής. Στο ραχιαίο εμπρόσθιο μέρος της πρωτογενούς γραμμής αρχίζει να σχηματίζεται μια αύλακα (εσοχή primitive groove), η οποία εκτείνεται βαθμιαία προς τα πίσω σε όλο το μήκος της πρωτογενούς γραμμής που τώρα ονομάζεται πρωτογενής αύλακα. Η πρωτογενής αύλακα, ομόλογη του βλαστοπόρου στα αμφίβια, λειτουργεί ως άνοιγμα μέσω του οποίου κύτταρα εγκολπώνονται μέσα στο βλαστόκοιλο. Ενώ επιμηκύνεται η πρωτογενής αύλακα, κύτταρα από την επιβλάστη εγκολπώνονται στο εσωτερικό της σε όλη την έκτασή της και σχηματίζουν τη μεσοβλάστη (μεσόδερμα) και μια βαθύτερη στοιβάδα, την ενδοβλάστη (οριστική υποβλάστη-ενδόδερμα) (Ζάγκρης, 1996). Στο πρόσθιο άκρο της, η πρωτογενή αύλακα καταλήγει στο κόμβο του Hensen (Hensen s node), μια πύκνωση κυττάρων που έχει σχηματιστεί από τα κύτταρα που πρώτα έχουν να μεταναστεύουν από το Koller s sickle (Gilbert, 2006). Τα πρώτα κύτταρα που εγκολπώνονται μέσα στον κόμβο του Hensen καθορίζονται ως ενδόδερμα και μεσόδερμα της κεφαλής και ακολουθούνται από κύτταρα που θα σχηματίσουν τη νωτοχορδή (pre-chordal plate). Ο κόμβος του Hensen είναι ο οργανωτής του εμβρύου και είναι 48
48 ΕΙΣΑΓΩΓΗ ομόλογη δομή του ραχιαίου χείλους του βλαστοπόρου του οργανωτή του εμβρύου των αμφιβίων (Ζάγκρης, 1996). Τα κύτταρα που μεταναστεύουν από τα πλευρικά τμήματα της πρωτογενούς αύλακας θα σχηματίσουν την πλειοψηφία των ένδο- και μεσοδερμικών ιστών (Schoenwolf et al., 1992). 4.2 Σχηματισμός του ενδοδέρματος και του μεσοδέρματος Έχοντας φθάσει το μέγιστο μήκος της, η πρωτογενής αύλακα σμικρύνεται βαθμιαία με οπισθοδρόμηση του κόμβου του Hensen, ενώ στη θέση της σχηματίζονται διαδοχικά η νωτοχορδή, η κεφαλική πτυχή και ο νευρικός σωλήνας. Όταν έχει αρχίσει να σχηματίζεται η νωτοχορδή (αξονικό μεσόδερμα/ axial mesoderm), η κύρια μάζα του μεσοδέρματος συνεχίζει να αυξάνεται και να απλώνεται κάτω από το εξώδερμα στην περιφέρεια της πρωτογενούς αύλακας. Το μεσόδερμα που διατάσσεται εκατέρωθεν της νωτοχορδής ονομάζεται παρα-αξονικό (paraxial) μεσόδερμα. Διάτμηση του παρα-αξονικού μεσοδέρματος δημιουργεί πλάκες που ονομάζονται σωμίτες, οι οποίες αργότερα θα σχηματίσουν τα οστά και τους μύες της ράχης και τους μύες των άκρων καθώς και της γλώσσας. Πιο πλευρικά, το μεσόδερμα αποχωρίζεται σε δυο πέταλα, το ένα πλησιέστερα προς το εξώδερμα (σωματικό) και το άλλο προς το ενδόδερμα (σπλαχνικό). Μεταξύ τους σχηματίζεται μια κοιλότητα, η σωματική κοιλότητα. Η κοιλότητα αυτή δεν σχηματίζεται ταυτόχρονα σε όλο το μήκος του μεσοδέρματος, αλλά το πρώτο σημείο αυτής που σχηματίζεται είναι το σημείο στο οποίο θα σχηματιστεί η καρδιά (Ζάγκρης, 1996). Το σπλαχνικό μεσόδερμα, εκατέρωθεν του εμβρυϊκού άξονα στο επίπεδο της μελλοντικής καρδιάς του εμβρύου, σχηματίζει συστάδες κυττάρων (heart primordia/καταβολές της καρδιάς ή προκαρδιακά κύτταρα) που μεταναστεύουν προς τον εμβρυϊκό άξονα, συνδυάζονται και σχηματίζουν τον καρδιακό σωλήνα (heart tube). Ο καρδιακός σωλήνας αποτελείται από ένα εξωτερικό τοίχωμα, το μυοκάρδιο, και ένα εσωτερικό μονοστοίβαδο τοίχωμα, το ενδοκάρδιο και θα σχηματίσει την καρδιά. 49
49 ΕΙΣΑΓΩΓΗ Από τα παραπάνω γίνεται κατανοητό ότι, ως συνέπεια της αλληλουχείας με την οποία δίνονται οι διεργασίες για τον σχηματισμό του μεσοδέρματος και της νωτοχορδής, στα έμβρυα των πτηνών υπάρχει μία διακριτή προσθοπίσθια κλίση αναπτυξιακής ωριμότητας (Gilbert, 2003). Έτσι, ενώ στο πρόσθιο τμήμα του εμβρύου έχει ήδη ξεκινήσει η οργανογένεση, στο οπίσθιο επιτελούνται ακόμα οι μορφογενετικές κινήσεις της γαστριδίωσης (Darnell et al., 1999). 4.3 Νευριδίωση Το μεσόδερμα και ενδόδερμα της κεφαλής και η νωτοχορδή, οι κυτταρικοί πληθυσμοί που έχουν προέλθει από τον οργανωτή, επάγουν το υπερκείμενό τους εξώδερμα να καθοριστεί ως νευροεξώδερμα (νευρική πλάκα) που σχηματίζεται από το 50% των εξωδερμικών κυττάρων του εμβρύου. Μία μπαταρία από μόρια που συντίθενται και εκκρίνονται από τα κύτταρα του οργανωτή συνεργούν στον καθορισμό και τη διαμερισματοποίηση του νευροεξωδέρματος (Gilbert, 2006). Η νευρική πλάκα έχει σχήμα αχλαδιού, είναι πεπλατυσμένη εμπρόσθια, όπου θα σχηματίσει τον εγκέφαλο και στενότερη οπίσθια, όπου θα σχηματίσει το νωτιαίο μυελό. Αφού επαχθεί η νευρική πλάκα, παρατηρούνται αλλαγές στη μορφολογία των κυττάρων της. Τα κύτταρα στα άκρα της νευρικής πλάκας αρχίζουν να επιμηκύνονται, ενώ τα γειτονικά τους επιθηλιακά κύτταρα διατηρούν το πεπλατυσμένο σχήμα τους (Ζάγκρης, 1996). Καθώς συνεχίζεται η ανάπτυξη του εμβρύου, τα άκρα της νευρικής πλάκας ανασηκώνονται σχηματίζοντας τις νευρικές πτυχές (neural folds). Καθώς η νευρική πλάκα μοιάζει με αύλακα, το έμβρυο αρχίζει να επιμηκύνεται προσθοπίσθια κατά μήκος της αύλακας. Οι νευρικές πτυχές συνεχίζουν να αυξάνουν σε ύψος, συγκλίνουν η μία προς την άλλη έως ότου τα άκρα τους συναντώνται και συνεννοούνται σχηματίζοντας ένα κοίλο σωλήνα, το νευρικό σωλήνα (neural tube). Πλευρικά, το επιδερμικό εξώδερμα ενώνεται πάνω από το νευρικό σωλήνα, ο οποίος αποχωρίζεται από το εξώδερμα και καταβυθίζεται μέσα στο μεσόδερμα. Από το νευρικό σωλήνα σχηματίζεται όλο το κεντρικό νευρικό σύστημα (Ζάγκρης, 1996). 50
50 ΕΙΣΑΓΩΓΗ Οι νευρικές πτυχές δεν συνεννοούνται ταυτόχρονα, αλλά αυτό γίνεται σταδιακά αρχίζοντας από το πρόσθιο προς το οπίσθιο άκρο του νευρικού σωλήνα. Το κλείσιμο ολόκληρου του νευρικού σωλήνα έχει τελειώσει πριν αποχωρισθεί από το επιδερμικό εξώδερμα. Τα διάφορα τμήματα του εγκεφάλου (προσθεγκέφαλος, μεσεγκέφαλος, οπισθεγκέφαλος) καθώς επίσης και ο νωτιαίος μυελός πιο οπίσθια, σχηματίζονται από το νευρικό σωλήνα. Στο πλάι και κατά μήκος της σύμφυσης των νευρικών πτυχών, καθώς έχει κλείσει ο νευρικός σωλήνας, ένας κυτταρικός πληθυσμός που μοιάζει με «τσαμπιά σταφύλια», οι νευρικές κρηπίδες ή νευρική ακρολοφία (neural crest), αρχίζει να μεταναστεύει σε διάφορα σημεία του εμβρύου. Από τα κύτταρα των νευρικών κρηπίδων σχηματίζεται όλο το περιφερικό νευρικό σύστημα. Μια πολυπληθής ομάδα των κυττάρων των νευρικών κρηπίδων εξαπλώνεται κάτω από το εξώδερμα και σχηματίζει τα χρωματοφόρα κύτταρα του δέρματος και επίσης συμμετέχει στην παραγωγή τριχών και νυχιών. Μερικά γίνονται μεσένχυμα και συνεισφέρουν στο συνδετικό ιστό. Άλλα γίνονται χονδρίτης ιστός της κάτω σιαγόνας, οστά του κρανίου, γάγγλια των κρανιακών και νωτιαίων νευρώνων, και προστατευτικά κύτταρα Schwann των νευρώνων (Ζάγκρης, 1996). 51
51 ΕΙΣΑΓΩΓΗ 5. Προσδιορισμός των σταδίων εμβρυικής ανάπτυξης στην όρνιθα Τα στάδια ανάπτυξης των εμβρύων της όρνιθας συνήθως προσδιορίζονται με βάση την κλίμακα που προτάθηκε από τους ερευνητές Hamburger και Hamilton (Hamburger and Hamilton, 1951). Σύμφωνα με την κλίμακα αυτή, τα στάδια ορίζονται με τους αραβικούς αριθμούς 1-46 (π.χ. ΗΗ 4 = στάδιο 4 κατά Hamburger και Hamilton 1951) και τα οποία περιλαμβάνουν και τις 21 ημέρες ανάπτυξης του εμβρύου. Για τα πρώιμα έμβρυα, χρησιμοποιείται συχνά μια άλλη κλίμακα που προτάθηκε από τους Eyal-Giladi και Kochav και η οποία επιτρέπει τον προσδιορισμό των πρώιμων σταδίων με μεγαλύτερη ακρίβεια (Eyal-Giladi and Kochav, 1976). Στην κλίμακα αυτή, χρησιμοποιούνται οι λατινικοί αριθμοί I-XIV, οι οποίοι προσδιορίζουν τα στάδια από το πρώιμο μορίδιο (στάδιο Ι), μέχρι το προχωρημένο βλαστίδιο (στάδιο XIV). Τα στάδια αυτά προηγούνται του σταδίου ΗΗ 2, των Hamburger and Hamilton (1951). 52
52 53 ΕΙΣΑΓΩΓΗ
53
54 ΣΚΟΠΟΣ ΔΙΑΤΡΙΒΗΣ Η εξωκυττάρια ουσία αποτελεί ένα πολύπλοκο και δυναμικό δίκτυο μορίων, ιδιαίτερα σημαντικό για την επιτέλεση των αναπτυξιακών διαδικασιών στους πολυκύτταρους οργανισμούς. Η αλληλεπίδραση των μορίων αυτών με υποδοχείς της κυτταρικής επιφανείας έχει ως αποτέλεσμα τη ρύθμιση βασικών λειτουργιών των κυττάρων, όπως είναι η προσκόλληση, η κατευθυνόμενη μετανάστευση, ο πολλαπλασιασμός, η επιβίωση, η διαφοροποίηση τους και η οργάνωσή τους σε ιστούς και όργανα. Το έμβρυο της όρνιθας αποτελεί ιδανικό σύστημα μελέτης των διαδικασιών αυτών. Είναι πολύ πιο εύκολα προσιτό σε σχέση με τα έμβρυα θηλαστικών που αρχίζουν να εμφυτεύονται στη μήτρα με την έναρξη του σταδίου του βλαστιδίου, γεγονός που τα κάνει απρόσιτα για τη μελέτη του καθορισμού και διαφοροποίησης κυττάρων και τη μορφογένεση των ιστών και οργάνων. Είναι σημαντικό ότι η ανάπτυξη του πρώιμου εμβρύου της όρνιθας είναι εξαιρετικά παρόμοια με αυτή των θηλαστικών έτσι ώστε το έμβρυο όρνιθας να θεωρείται ως σύστημα πρότυπο (Εικ. 9). Είναι ενδιαφέρον ότι το έμβρυο όρνιθας στο στάδιο ΗΗ20 (Εικ. 10) παρουσιάζεται ως πρότυπο της ανάπτυξης των ανωτέρων σπονδυλωτών συμπεριλαμβανομένου και του εμβρύου του ανθρώπου- το έμβρυο όρνιθας στο στάδιο ΗΗ20 είναι ομόλογο του εμβρύου του ανθρώπου 5 εβδομάδων. Στη διαδικασία αυτή της μορφογένεσης ιστών και οργάνων στο πρώιμο έμβρυο συνεργεί η πρωτεογλυκάνη agrin ως συστατικό της εξωκυττάριας ύλης. Η agrin είναι πρωτεογλυκάνη θειικής ηπαράνης που αλληλεπιδρά με την λαμινίνη καθώς και με άλλες πρωτεΐνες που βρίσκονται στην κυτταρική επιφάνεια, όπως οι ιντεγκρίνες και η α-dystroglycan. Το γεγονός αυτό, σε συνδυασμό με την συμμετοχή της agrin σε αλληλεπιδράσεις με αυξητικούς παράγοντες, όπως για παράδειγμα με τον FGF, υποδεικνύει την σημασία της συγκεκριμένης πρωτεογλυκάνης στις διακυτταρικές και στις κυττάρουεξωκυττάριας ύλης αλληλεπιδράσεις. Ο ρόλος και η έκφραση της agrin κατά τη διάρκεια των μορφογενετικών διαδικασιών κατά την ανάπτυξη του εμβρύου, δεν έχει μελετηθεί πολύ καλά, ενώ ελάχιστα είναι γνωστά για τα πρώιμα στάδια της εμβρυογένεσης. Σκοπός της παρούσας διατριβής, είναι η μελέτη της έκφρασης και του ρόλου της agrin στο πρώιμο έμβρυο. Συγκεκριμένα, θα μελετήσουμε πότε αρχίζει να εκφράζεται το mrna της agrin, σε ποιούς κυτταρικούς πληθυσμούς 56

55 ΣΚΟΠΟΣ ΔΙΑΤΡΙΒΗΣ εκφράζεται η πρωτεογλυκάνη agrin και ποιος είναι ο ρόλος της κατά την ανάπτυξη πρώιμου εμβρύου όρνιθας. Εικόνα 9: Σχηματική αναπαράσταση των εμβρύων, ψαριού, σαλαμάνδρας, χελώνας, όρνιθας, μοσχαριού και ανθρώπου. Διακρίνεται η καταπληκτική ομοιότητα στα πρώιμα στάδια ανάπτυξης μεταξύ αυτών των οργανισμών. Διακριτικά χαρακτηριστικά εμφανίζονται αργότερα στην ανάπτυξη (διαδικτυακός τόπος 2). Εικόνα 10: Έμβρυο όρνιθας σταδίου ΗΗ20 (Hamburger and Hamilton, 1951). 57
56
57 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 1. Απομόνωση/ καλλιέργεια εμβρύων 1.1 Υλικά/Διαλύματα Διάλυμα Ringer Σταγονόμετρο Γυάλινη πιπέτα (2ml) Πιπέτες Pasteur Μικροβελόνες Χαρτί, βαμβάκι Τρυβλία petri Λαβίδα Φρέσκα Μικρά τρυβλία petri Κουτάλι γονιμοποιημένα αυγά Γυάλινα δαχτυλίδια Ψαλίδι Πλαστικό δοχείο Πουαρ Πορσελάνινη κάψα απορριμάτων 1.2 Συσκευές Θάλαμος επώασης MEMMERT (36-37 o C) Στερεοσκόπιο Stemi 2000-C-Zeiss Συσκευή πηγής φωτός (υγρών κρυστάλλων) Schott KL 1500LCD Φωτογραφικό σύστημα με ψηφιακή κάμερα για φωτογράφιση μέσω στερεοσκοπίου 1.3 Παρασκευή διαλυμάτων Διάλυμα Ringer buffer 10x (stock, πυκνό διάλυμα) NaCl 72,5 g KCl 3,7 g Διαλύονται σε 1 l dh 2 O CaCl 2 1,8 g Στην απομόνωση και την καλλιέργεια εμβρύων χρησιμοποιείται διάλυμα Ringer που προκύπτει μετά από αραίωση του πυκνού διαλύματος Ringer σε dh 2 O με αναλογία 1:9 (100ml Ringer stock : 900ml dh 2 O). 60
58 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 1.4 Μεθοδολογία Ρίχνουμε λίγο Ringer στην κάψα, σπάμε το αυγό και το ρίχνουμε μέσα στην κάψα απαλά έτσι ώστε να μην σχιστεί η λεκιθική μεμβράνη ή ακόμα και το ίδιο το έμβρυο. Το έμβρυο διακρίνεται συνήθως σαν ένας μικρός άσπρος κύκλος επί του κρόκου. Χρησιμοποιούμε το κουτάλι για να ακινητοποιήσουμε τον κρόκο και με τη βοήθεια του ψαλιδιού κόβουμε τη μεμβράνη γύρω από το έμβρυο με γρήγορες και προσεκτικές κινήσεις. Αφού το κόψουμε, με τη βοήθεια της λαβίδας και του κουταλιού απομακρύνουμε το έμβρυο από τον κρόκο και το μεταφέρουμε σε τρυβλίο petri (γεμάτο κατά ¾ περίπου με Ringer) με κοντά τοιχώματα. Επίσης, με την λαβίδα μεταφέρουμε τη λεκιθική μεμβράνη σε ένα petri με ψηλά τοιχώματα. Η λεκιθική μεμβράνη θα χρησιμοποιηθεί -στην περίπτωση της καλλιέργειας- για την κατασκευή της σχεδίας στην επιφάνεια της οποίας θα απλώσουμε το έμβρυο. Επιπλέον, κατά το σπάσιμο του αυγού απομακρύνουμε την αλβουμίνη και τη φυλάσσουμε σε ένα ξεχωριστό petri. Το petri με το έμβρυο τοποθετείται κάτω από το στερεοσκόπιο όπου το έμβρυο καθαρίζεται από τη λέκιθο (από τη πλευρά της υποβλάστης) με τη βοήθεια δύο μικροβελόνων και με λεπτούς χειρισμούς ώστε να μη σκισθεί. Η διαδικασία αυτή γίνεται χρησιμοποιώντας το στερεοσκόπιο. Είναι απαραίτητο να αφαιρεθεί όσο το δυνατόν περισσότερη λέκιθος, επειδή η λέκιθος εμποδίζει μηχανικά τις μορφογενετικές κινήσεις και τη μορφογένεση του εμβρύου. Το καθαρισμένο έμβρυο πρέπει να δείχνει σχεδόν διαφανές. Το καθαρισμένο έμβρυο μεταφέρεται με σταγονόμετρο σε καθαρό petri με Ringer*. Η λεκιθική μεμβράνη καθαρίζεται από τη λέκιθο και τη χάλαζα με τη βοήθεια της λαβίδας, με προσοχή ώστε να μην δημιουργήσουμε τρύπες. Αναδιπλώνουμε τη λεκιθική μεμβράνη γύρω από ένα γυάλινο δαχτυλίδι, ολοκληρώνοντας έτσι την κατασκευή σχεδίας από τη λεκιθική μεμβράνη. Μεταφέρουμε ένα από τα καθαρισμένα έμβρυα στη σχεδία με σταγονόμετρο. Εναποθέτουμε το έμβρυο με την επιβλάστη προς τα κάτω στη σχεδία και αφαιρώντας το Ringer προσπαθούμε να το ακινητοποιήσουμε στο κέντρο της σχεδίας. Μόλις απομακρυνθεί το Ringer, το έμβρυο προσκολλάται στην επιφάνεια της σχεδίας. Η σχεδία μεταφέρεται σε μικρό petri καλλιέργειας που περιέχει 61
59 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ περίπου 1,2 ml αλβουμίνη η οποία αποτελεί το θρεπτικό μέσο της καλλιέργειας. Στη συνέχεια το μικρό petri τοποθετείται σε μεγαλύτερο petri με βαμβάκι καλά νοτισμένο με dh 2 Ο και το έμβρυο επωάζεται σε επωαστικό θάλαμο στους 37,5 o C. * Στην περίπτωση που θέλουμε να απομονώσουμε έμβρυα, για παράδειγμα για την απομόνωση RNA ή πρωτεϊνών, τα συγκεντρώνουμε σε ένα σωληνάκι eppendorf και τα αποθηκεύουμε στον καταψύκτη (-40 o C). 62
60 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 2. Μονιμοποίηση εμβρύων 2.1 Υλικα/Διαλύματα Διάλυμα Ringer Διάλυμα Carnoy Αιθανόλη 100% (ethanol, C 2 H 5 OH) N-amyl-acetate (C 7 H 14 O 2 ) Καθαρή βενζίνη (thiophene free C 6 H 6 ) Βενζίνη-παραφίνη (1:1) Καθαρή παραφίνη Ζωτική χρωστική (άνθρακας ή καρμίνη) Χλωροφόρμιο (chloroform, CHCl 3 ) Οξικό οξύ (glacial acetic acid, CH 3 COOH) Πιπέτες Pasteur petri κλασσικού τύπου petri με χωρίσματα Μικροβελόνα Ψαλίδι. Μικρή μεταλλική σπάτουλα.μεταλλικές λαβίδες.μεταλλικά καλούπια Νυστέρι 2.2 Παρασκευή διαλυμάτων Μονιμοποιητικό διάλυμα Carnoy (Carnoy s Fixative, Formula B, Humason 1972): Αναλογία ml Αιθανόλη 100% (C 2 H 5 OH) Χλωροφόρμιο (CHCl 3 ) 3 30 Οξικό οξύ (CH 3 COOH) Mεθοδολογία Μετά την καλλιέργεια τοποθετoύμε τη σχεδία με το έμβρυο, με τη βοήθεια λαβίδας, σε ένα petri με κοντό τοίχωμα το οποίο περιέχει μικρή ποσότητα Ringer (για να μην υπάρχει κίνδυνος επαναδίπλωσης της σχεδίας με Ringer και καταστροφής του εμβρύου) Με την άκρη μιας μικροβελόνας συλλέγουμε μικρή ποσότητα ζωτικής χρωστικής (carmin) και σημαδεύουμε τον προσανατολισμό του 63
61 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ εμβρυϊκού άξονα. Με τον τρόπο αυτό καθίσταται ορατός ο προσανατολισμός του εμβρυϊκού άξονα και μετά την έγκλιση του εμβρύου σε καλούπι παραφίνης. Η σήμανση γίνεται πάνω στη λεκιθική μεμβράνη με μια ευθεία ή μικρή καμπύλη γραμμή κάθετη στον εμβρυϊκό άξονα. Τοποθετούμε τη σχεδία με το σημασμένο έμβρυο στο διάλυμα Carnoy για περίπου 20 λεπτά στους 4 ο C. Το διάλυμα Carnoy ρίχνεται προσεκτικά μέσα στη σχεδία με τη βοήθεια πιπέτας. Λαμβάνεται μνεία έτσι ώστε η μεταφορά της σχεδίας να γίνεται με κλίση προς τη μεριά της χρωστικής και όχι προς τη μεριά του εμβρύου, ώστε να αποφευχθεί διάχυση της χρωστικής μέσα στην εμβρυϊκή περιοχή και κυρίως στον εμβρυϊκό άξονα. Τα έμβρυα αφυδατώνεται περνώντας τα διαδοχικά από τα εξής διαλύματα: 100% αιθανόλη 2 φορές, 5 λεπτά κάθε φορά N-αμυλικό-οξικό άλας (N-amyl-acetate) 3 φορές, 5 λεπτά κάθε φορά Στο τρίτο διάλυμα Ν-αμυλο-οξικό άλας τα έμβρυα μπορούν να μείνουν για μήνες. Στο στάδιο αυτό το δαχτυλίδι απομακρύνεται από τη σχεδία και το εκάστοτε έμβρυο τοποθετείται σε ένα petri με κοντό τοίχωμα που περιέχει το τρίτο άμυλο-οξικό άλας. Με νυστέρι και με τη βοήθεια μικροβελόνας κόβεται συμμετρικά η λεκιθική μεμβράνη με το έμβρυο σε μικρή απόσταση από τη χρωστική και την περιοχή του εμβρύου. Στις επόμενες εμβαπτίσεις του εμβρύου στα υπόλοιπα διαλύματα, το έμβρυο μεταφέρεται με τη βοήθεια μεταλλικής σπάτουλας. Ακολουθεί η διαπότιση του εμβρύου στα παρακάτω διαλύματα για τους αντίστοιχους χρόνους, όπως περιγράφονται: καθαρή βενζίνη 3 φορές, 20 λεπτά κάθε φορά βενζίνη-παραφίνη (1:1) 1 ώρα στους 60 ο C καθαρή παραφίνη 1 ώρα στους 60 ο C καθαρή παραφίνη 1 ώρα στους 60 ο C Σε μεταλλικά καλούπια τοποθετούμε ρευστή παραφίνη (60 ο C). Τοποθετούμε το κάθε έμβρυο στη ρευστή παραφίνη με γρήγορες κινήσεις και με τη βοήθεια μικροβελόνας του δίνουμέ τον επιθυμητό 64
62 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ προσανατολισμό, λαμβάνοντας υπόψη και τη σήμανση με τη χρωστική που είχε προηγηθεί, για να καταστεί γνωστός ο προσανατολισμός του εμβρυϊκού αξονα. Στο εμπρόσθιο μέρος του κάθε καλουπιού βυθίζουμε ένα χαρτάκι με τα στοιχεία του εκάστοτε εμβρύου. Αφήνουμε την παραφίνη να στερεοποιηθεί. Αποχωρίζουμε το μπλοκ παραφίνης από το καλούπι και το λεπταίνουμε με τη βοήθεια νυστεριού, έτσι ώστε το μπλοκ της παραφίνης που εγκλείει το έμβρυο να αποκτήσει ένα λεπτό παραλληλόγραμμο σχήμα. 65
63 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 3. Μικροτόμηση εμβρύων 3.1 Υλικα/Διαλύματα Διάλυμα συγκόλλησης Ξυλόλη 70% Απιονισμένο H 2 O Μεταλλική σπάτουλα Καλυπτρίδες Μικροβελόνες Αντικειμενοφόροι Διάλυμα συγκόλλησης Αναλογία ml Αλβουμίνη αυγού όρνιθας 1 10 Γλυκερόλη (CH 2 OHCHOHCH 2 OH) 1 10 dh 2 O ½ 5 Τα υλικά ανακατεύονται καλά για λεπτά. Το διάλυμα συγκόλλησης διατηρείται στο ψυγείο. 3.2 Συσκευές Κλίβανος MEMMERT Μικροτόμος Θερμαντική πλάκα Bioblock scientific model No C (slide warmer) Θερμαινόμενο υδατόλουτρο (40 ο C) Θερμαντική λυχνία Μικροσκόπιο 3.3 Μεθοδολογία Κολλάμε το μπλοκ παραφίνης πάνω σε μεταλλική βάση. Αυτό επιτυγχάνεται με την πυράκτωση μίας σπάτουλας στη φλόγα της λυχνίας, τοποθέτησή της στη βάση του μπλοκ έτσι ώστε να λιώσει η 66
64 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ παραφίνη στο σημείο αυτό και ακολουθεί η άμεση τοποθέτηση του μπλοκ πάνω στην πυρακτωμένη θέση. Αφήνουμε την παραφίνη να στερεοποιηθεί τουλάχιστον 1 ώρα πριν αρχίσει η μικροτόμηση. Eν τω μεταξύ, οι αντικειμενοφόροι καθαρίζονται προσεκτικά με καθαρή αιθανόλη και στη συνέχεια επιστρώνονται με διάλυμα συγκόλλησης. Προσανατολίζουμε το μπλοκ παραφίνης πάνω από το μαχαίρι του μικροτόμου προσεκτικά (το μπλοκ παράλληλα στο μαχαίρι). Αρχίζουμε τη μικροτόμηση οι τομές έχουν πάχος 7μm. Η κορδέλα παραφίνης (ομάδα 8-9 τομών) όπως κόβεται μεταφέρεται προσεκτικά με σπάτουλα ή/και πινέλο σε υδατόλουτρο στους 40 ο C για να ισιώσει και κατόπιν προσλαμβάνεται σε αντικειμενοφόρο πλάκα που είναι επιστρωμένη με μια σταγόνα διαλύματος συγκόλλησης. Οι αντικειμενοφόροι τοποθετούνται σε θερμαντική πλάκα (περίπου 40 ο C) για να κολλήσουν οι τομές για τουλάχιστον μισή ώρα. Η μικροτόμηση ολοκληρώνεται αφού εξασφαλιστεί -με τη βοήθεια μικροσκοπίου- ότι δεν υπάρχει άλλος ιστός πάνω στις τομές. Τοποθετούμε τις αντικειμενοφόρους σε γυάλινα δοχεία (Coplin jars) και κατόπιν στον κλίβανο αποπαραφινοποίησης (60 ο C) για να λιώσει η περίσσεια της παραφίνης. Παραμένουν εκεί για τουλάχιστον 48 ώρες. 67
65 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 4. Χρώση με Αlcian blue 4.1 Υλικα Pyrex tank για τη χρώση Ογκομετρικοί κύλινδροι του 1l Χωνιά Διηθητικό χαρτί 4.2 Διαλύματα Διάλυμα χρωστικής Alcian Blue 8 GX, ph 2,5 Alcian Blue 8 GX (C.I ) ΑπιονισμdH 2 O Οξικό οξύ (acetic acid, CH 3 COOH) 1 g 100 ml 3 ml Διάλυμα χρωστικής Nuclear Fast Red (Kernechtrot) Nuclear Fast Red (Kernechtrot) 5% θειικό αλουμίνιο [aluminum sulfate, Al 2 (SO 4 ) 2.18H 2 O] 0,1 g 100 ml Διαλύουμε τη χρωστική ζεσταίνοντας (περίπου 45 ο C) το διάλυμα. Αφήνουμε το διάλυμα να φθάσει σε θερμοκρασία δωματίου και περνάμε το διάλυμα από φίλτρο. Προσθέτουμε 1 κρύσταλλο θυμόλη (thymol) για να αποφευχθεί η ανάπτυξη μυκήτων. Κόλλα ENTELLAN neu für mikroskopie (Merck) Αιθανόλη 70%, 95% και 100% Ξυλόλη Οξικό οξύ 3% 68
66 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 4.3 Συσκευές Κλίβανος MEMMERT 4.4 Μεθοδολογία Παίρνουμε ανά 2 αντικειμενοφόρους (1, 3, 5, 7 ) και τις τοποθετούμε στο θάλαμο στους 60 ο C για αποπαραφινοποίηση για 48 ώρες τουλάχιστον, ο χρόνος αυτός είναι αρκετός για να γίνει αποπαραφινοποίηση αλλά προληπτικά μπορούν να μείνουν για μεγαλύτερο χρονικό διάστημα. Τοποθετούμε τις αντικειμενοφόρους με τις τομές σε δοχεία Coplin και τις περνάμε από τα ακόλουθα διαλύματα: Ενυδάτωση τομών Ξυλόλη (έχουμε τοποθετήσει δοχείο Coplin με ξυλόλη στους 60 ο C να ζεσταθεί 20 λεπτά πριν τη χρήση της) Αιθανόλη 100% Αιθανόλη 95% Αιθανόλη 75% Η 2 Ο βρύσης (Το νερό πρέπει να τρέχει απαλά στα τοιχώματα του δοχείου και όχι πάνω στις τομές) 10 λεπτά 1 λεπτό 1 λεπτό 1 λεπτό 5 λεπτά Οξικό οξύ 3% (αναδεύεται πριν από τη χρήση). 3 λεπτά Διάλυμα χρωστικής Alcian Blue (χρειάζεται φιλτράρισμα πριν από κάθε χρήση) Η 2 Ο βρύσης (Το νερό πρέπει να τρέχει απαλά στα τοιχώματα του δοχείου και όχι πάνω στις τομές) 30 λεπτά 10 λεπτά Διάλυμα χρωστικής Nuclear Fast Red (χρειάζεται φιλτράρισμα πριν από κάθε χρήση) Γρήγορο ξέπλυμα των αντικειμενοφόρων με dh 2 O 5 λεπτά 2 φορές / 1λεπτό 69
67 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Αφυδάτωση τομών Αιθανόλη 70 % Αιθανόλη 95 % Αιθανόλη 100 % Ξυλόλη 1 λεπτό 1 λεπτό 1 λεπτό. 1 λεπτό Αφήνουμε τις αντικειμενοφόρους να στεγνώσουν, περίπου ένα λεπτό. Στη συνέχεια, με μια γυάλινη ράβδο τοποθετούμε 2 σταγόνες κόλλα ENTELLAN neu πάνω σε κάθε αντικειμενοφόρο. Με κλίση αφήνουμε την καλυπτρίδα να πέσει πάνω στις τομές απαλά η κόλλα απλώνεται. Προσοχή να μην δημιουργηθούν φυσαλίδες. Σημείωση 1: Η χρωστική Alcian blue ανιχνεύει τις σουλφολιωμένες και μη πρωτεογλυκάνες και η nuclear fast red τους πυρήνες και είναι ορατοί οι ιστοί και τα όργανα του εμβρύου. Με βάση αυτή την εικόνα γίνεται η επιλογή των αντικειμενοφόρων που θα χρησιμοποιηθούν για την επιτέλεση πειράματος, για παράδειγμα, αν πρόκειται για πείραμα ανοσοφθορισμού, όπου λαμβάνονται οι γειτονικές, από τις χρωσμένες, αντικειμενοφόροι στο επίπεδο του εμβρυϊκού άξονα που επιθυμείται (π.χ. οφθαλμός, καρδιά). 70
68 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 5. Ανίχνευση πρωτεϊνών με αντισώματα- Ανοσοφθορισμός 5.1 Υλικά/Διαλύματα Μονοκλωνικό αντίσωμα (40μg/ml, stock) ειδικό για την agrin (κλώνος 6D2, Hybridoma bank, Iowa, USA). Για κάθε αντικειμενοφόρο, το αντίσωμα αραιώθηκε έως την τελική συγκέντρωση 10μg/ml σε τελικό όγκο 500μl. Διαλύτης του αντισώματος : PBS * που περιέχει 0,1% Triton X-100 και 1% BSA βλέπετε διαλύματα πιο κάτω. Fluorescein Goat F Anti-Mouse (δευτερογενές αντίσωμα με πρόσδεμα FITC, Santa Cruz, USA), stock concentration 200μg/0,5ml. Για κάθε αντικειμενοφόρο, το αντίσωμα αραιώθηκε έως την τελική συγκέντρωση 5μg/ml και σε τελικό όγκο 800μl (V FITC/stock =10μl και V δ/τη PBS =790μl). PAP PEN BSA (bovine serum albumin) Διάλυμα PBS (Phosphate Buffered Saline) Διάλυμα PBS που περιέχει 0,1% Τriton X-100 Διαλύτης (PBS που περιέχει 0,1% Triton X-100 και 1% BSA) για πολυκλωνικά ή καθαρισμένα μονοκλωνικά αντισώματα Γλυκερόλη (Glycerol) σε συγκεντρώσεις 10%, 20%, 40%, 80%, 100% σε PBS Ξυλόλη (Xylene) Αιθανόλη 100%, Αιθανόλη 95%, Αιθανόλη 75% dh 2 O Πιπέτες Pasteur Ειδικά γυάλινα δοχεία (coplin jars) Μεγάλο Petri επιστρωμένο με διηθητικό χαρτί νοτισμένο με dh 2 O Διάλυμα MOWIOL 4-88 Καλυπτρίδες Ποτήρια ζέσεως * Σε όλες της διαδικασίες χρησιμοποιήσαμε PBS 1x 71
69 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 5.2 Συσκευές Κλίβανος MEMMERT Επωαστικός θάλαμος Θερμαινόμενο ανακινούμενο υδατόλουτρο Gallenkamp Μικροσκόπιο και φωτογραφικό σύστημα Sony με ψηφιακή κάμερα 5.3 Παρασκευή διαλυμάτων Διάλυμα Phosphate Buffered Saline(PBS) 10x (STOCK, πυκνό διάλυμα) Molarity M r Αναλογίες Na 2 HPO 4 16mM 141,96 22,7 g NaH 2 PO 4 H 2 O 4mM 137,99 5,5 g NaCl 140mM 58,44 81,8 g NaN 3 0,2 g Διαλύουμε τα συστατικά που αναγράφονται στον παραπάνω πίνακα σε dh 2 O ώστε να φτάσει σε τελικό όγκο 1000ml. Διάλυμα Phosphate Buffered Saline(PBS) 1x Από το πυκνό διάλυμα (stock) παίρνουμε 100ml και το φέρνουμε σε τελικό όγκο 1000ml με την προσθήκη dh 2 O. Διάλυμα PBS (1x), 0,1% Triton X-100, BSA 1% ΒSA. Σε 500ml διαλύματος PBS (1x) προσθέτω 0,5ml Triton X-100 και 5g Διάλυμα Mowiol g γλυκερόλης (glycerol, analytical grade) τοποθετούνται σε ποτήρι ζέσεως των 50 ml. Στη συνέχεια, 2,4 g Mowiol 4-88 προστίθενται και 72
70 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ αναδεύονται για 1 ώρα, ώστε να αναμειχθεί το Μowiol με την γλυκερόλη. Συμπληρώνουμε με 6 ml dh 2 O και γίνεται ανάδευση για 2 ώρες σε θερμοκρασία δωματίου. Προσθέτουμε 12 ml από 0,2Μ Tris-HCl buffer (διαλύουμε 2,42 g Tris σε 100 ml dh 2 O, ρυθμίζουμε το ph στο 8,5 με HCl). Το διάλυμα τοποθετείται σε υδατόλουτρο 50 ο C για 10 λεπτά αναδεύοντας περιστασιακά για να διαλύσουμε το Mowiol. To διάλυμα φυγοκεντρείται σε 5000 g (6300 rpm) για 15 λεπτά και εν συνεχεία το μοιράζουμε σε σωληνάκια eppendorf. Διατηρείται στους -20 ο C για τουλάχιστον 12 μήνες. Μετά την απόψυξη το διάλυμα είναι σταθερό για ένα μήνα σε θερμοκρασία δωματίου. 5.4 Μεθοδολογία Αποπαραφινοποίηση Αρχικά επιλέγονται οι αντικειμενοφόροι κάθε εμβρύου που θα χρησιμοποιηθούν. Οι αντικειμενοφόροι επιλέγονται με βάση τις χρωσμένες που παρατηρούνται μικροσκοπικά. Συνήθως χρησιμοποιούνται δύο αντικειμενοφόροι ανά πείραμα. Τοποθετούμε τις αντικειμενοφόρους με τις τομές στο θάλαμο αποπαραφινοποίησης (56-60 C) για τουλάχιστον 48 ώρες. Τοποθετούμε τις αντικειμενοφόρους σε ξυλόλη (προθερμασμένη στους 60 C, έτσι ώστε η θερμοκρασία της να εξισορροπεί τη θερμοκρασία των τομών και η αποπαραφινοποίηση να είναι πιο εύκολη). Ενυδάτωση Οι αντικειμενοφόροι τοποθετούνται στα εξής διαλύματα : 100% αιθανόλη 1 λεπτό 95% αιθανόλη 1 λεπτό 75% αιθανόλη 1 λεπτό τρεχούμενο Η 2 Ο 5 λεπτά d Η 2 Ο 2-3 φορές (απομάκρυνση αλάτων) 73
71 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Τοποθετούμε τις αντικειμενοφόρους σε PBS, 2 φορές για 5 λεπτά κάθε φορά. Στεγνώνουμε τις αντικειμενοφόρους (σκουπίζουμε απαλά πλάγια και κάτω) και τα τοποθετούμε σε μαύρο πλαίσιο, ώστε να είναι ορατές οι τομές. Χωρίζουμε τις τομές στην αντικειμενοφόρο σε πειραματικές (πιο πολλές) και σε μάρτυρα (2-3). Φτιάχνουμε ένα περίγραμμα με το ειδικό στυλό PAP PEN, έτσι ώστε να περικλείει όλες τις πειραματικές τομές. Το PAP PEN είναι μια υδρόφοβη ουσία και εμποδίζει τη διάχυση των διαλυμάτων μεταξύ των πειραματικών τομών και των τομών του μάρτυρα. Επωάζουμε σε PBS στο οποίο έχει προστεθεί 0,1% Triton X-100 και 1% BSA για 30 λεπτά στους 37 C. To Triton X-100 είναι απαλό (nonionic) απορρυπαντικό, χαλαρώνει τα κύτταρα, δεν σπάει τις μεμβράνες και επιτρέπει τη είσοδο του αντισώματος στο εσωτερικό των κυττάρων. Οι επωάσεις γίνονται σε μεγάλο τρυβλίο petri με υγρασία που τοποθετείται στο θάλαμο καλλιέργειας (37,5 ο C) Σκουπίζουμε με χαρτί τις αντικειμενοφόρους και ανάμεσα στις τομές, προσεκτικά, ώστε να αφαιρεθεί το περίσσιο του PBS για να μην αλλάξει η συγκέντρωση του αντισώματος που ακολουθεί. Προσθέτουμε στις τομές το πρωτογενές αντίσωμα (μονοκλωνικό αντίσωμα έναντι της agrin, κλώνος 6D2). Το αντίσωμα έχει διαλυθεί σε PBS που περιέχει 0,1% Triton X-100 και 1% BSA σε τελική συγκέντρωση 10μg/ml. Χρησιμοποιούμε 500 μl από το διάλυμα με το πρωτογενές αντίσωμα για κάθε αντικειμενοφόρο. Οι αντικειμενοφόροι που φέρουν τις τομές με το πρωτεογενές αντίσωμα τοποθετούνται σε μεγάλο τρυβλίο petri με νοτισμένο φίλτρο για υγρασία, που τοποθετείται στο θάλαμο καλλιέργειας στους 37,5 C. Η επώαση διαρκεί overnight, με κατάλληλη υγρασία, όπως περιγράφουμε. (Μπορούμε να το επωάσουμε και για 2h εάν η πρωτεΐνη που ανιχνεύουμε εκφράζεται σε υψηλά επίπεδα). 74
72 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Μετά την επώαση τοποθετούμε τις αντικειμενοφόρους σε δοχείο Coplin και ξεπλένουμε τις τομές με PBS και 0,1% Triton X φορές για 5 λεπτά κάθε φορά για να απομακρύνουμε το αντίσωμα που δεν έχει προσκολληθεί στο μόριο μας. Στη συνέχεια ξεπλένουμε με PBS με ανακίνηση, 1 ώρα μέσα σε υδατόλουτρο, 37 C. Σπάμε τη 1 ώρα σε 10 λεπτά, 15 λεπτά, 15 λεπτά, 20 λεπτά και κάθε φορά αλλάζουμε PBS. Σκουπίζουμε τις αντικειμενοφόρους προσεκτικά από κάτω και πάνω γύρω από τις τομές. Όλες οι διαδικασίες που ακολουθούν γίνονται στο απόλυτο σκοτάδι ή με φωτισμό μειωμένο στο ελάχιστο. Στις τομές τοποθετούμε προσεκτικά το δευτερογενές αντίσωμα Fluorescein Goat F Anti-Mouse στο απόλυτο σκοτάδι. Τοποθετούμε τις αντικειμενοφόρους σε μεγάλο τρυβλίο petri με υγρό φίλτρο για υγρασία - το τρυβλίο petri είναι καλυμμένο με αλουμινόχαρτο για απόλυτο σκοτάδι- και επωάζουμε για 1 ώρα στους 37,5 o C. Το δευτερογενές αντίσωμα του φθορισμού έχει διαλυθεί σε PBS και η τελική συγκέντρωση του είναι 5 μg/ml. Όταν το αντίσωμα είναι παλιό χρησιμοποιούμε διπλάσια (10μg/ml) ή τριπλάσια (15μg/ml) τελική συγκέντρωση. Χρησιμοποιούμε 800 μl από το διάλυμα με το δευτερογενές αντίσωμα για κάθε αντικειμενοφόρο. Ξεπλένουμε 1 ώρα με PBS μέσα σε υδατόλουτρο όπως πιο πάνω. Χωρίζουμε τα πλυσίματα σε 10 λεπτά 15 λεπτά, 15 λεπτά, 20 λεπτά και κάθε φορά αλλάζουμε PBS. Σκουπίζουμε την αντικειμενοφόρο κάτω και πάνω γύρω από τις τομές προσεκτικά δίχως να στεγνώσουν οι τομές. Προσθέτουμε στις τομές διαλύματα γλυκερόλης σε PBS στο σκοτάδι ως εξής : 10% γλυκερόλη σε PBS 15 λεπτά 20% γλυκερόλη σε PBS 15 λεπτά 40% γλυκερόλη σε PBS 15 λεπτά 80% γλυκερόλη σε PBS 15 λεπτά 100% γλυκερόλη σε PBS 15 λεπτά Αδειάζουμε και σκουπίζουμε καλά 75
73 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Προσθέτουμε Mowiol 4-88 (2-3 σταγόνες) σε όλες τις τομές για τη διατήρηση του φθορισμού και αμέσως τοποθετούμε καλυπτρίδα με προσοχή ώστε να μη δημιουργηθούν φυσαλίδες. Βάζουμε τις τομές σε ειδική θήκη, σε οριζόντια θέση στο σκοτάδι μέχρι τη φωτογράφιση. Σημείωση 1: Από τη στιγμή που θα χρησιμοποιηθεί το δευτερογενές αντίσωμα FITC όλες οι διαδικασίες γίνονται στο σκοτάδι. Όλα τα δοχεία που χρησιμοποιούνται είναι καλυμμένα ερμητικά καλά με αλουμινόχαρτο. Σημείωση 2: Μετά το τέλος του πειράματος οι αντικειμενοφόροι πρέπει να παραμένουν σε απολύτως οριζόντια θέση επειδή το Mowiol δεν στερεώνει μόνιμα την καλυπτρίδα στην αντικειμενοφόρο και τυχόν μετατόπισή του καταστρέφει τις τομές. Το Mowiol 4-88 χρησιμεύει για τη διατήρηση του φθορισμού για μεγάλο χρονικό διάστημα. Η φωτογράφιση των τομών γίνεται το συντομότερο δυνατό γιατί ο φθορισμός μειώνεται με το χρόνο. 76
74 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 6. Έκθεση εμβρύων σε αντισώματα έναντι της agrin και καλλιέργεια Στα πειράματα αυτά χρησιμοποιήσαμε μονοκλωνικά αντισώματα έναντι της agrin για να μελετήσουμε το ρόλο του μορίου αυτού κατά την ανάπτυξη του πρώιμου εμβρύου. Έμβρυα όρνιθας, στα στάδια ΗΗ3-4 (στάδιο γαστριδίου), καθαρίζονται από τη λέκιθο, όπως περιγράφεται πιο πάνω, και τοποθετούνται σε αντικειμενοφόρο με λακουβίτσα (Arthur Thomas Culture Slide) χωρητικότητας 500μl. Προστίθεται το μονοκλωνικό αντίσωμα έναντι της agrin (κλώνος 6D2) με τελική συγκέντρωση 1,2μg/ml σε 500μl τελικό όγκο διαλύματος Ringer. Συγκεκριμένα, σε 485μl Ringer προστέθηκαν 15μl του αντισώματος από το stock των 40μg/ml. Η αντικειμενοφόρος με τα έμβρυα τοποθετείται σε μεγάλο petri που είναι επιστρωμένο με νοτισμένο διηθητικό χαρτί για υγρασία και που τοποθετείται στο θάλαμο καλλιέργειας. Τα έμβρυα επωάζονται με το αντίσωμα για 2 ώρες στους 37,5 ο C. Στο τέλος της επώασης τα έμβρυα ξεπλένονται με διάλυμα Ringer προσεκτικά, τοποθετούνται σε σχεδίες λεκιθικής μεμβράνης και καλλιεργούνται όπως έχει περιγραφεί πιο πάνω. Η αναπτυξιακή πορεία των εμβρύων παρακολουθείται και καταγράφεται ανά τακτά χρονικά διαστήματα. Ακολουθεί φωτογράφιση των εμβρύων στο στερεοσκόπιο. Η μονιμοποίηση και η μικροτόμηση των εμβρύων καθώς και η χρώση των τομών πραγματοποιούνται όπως έχει περιγραφεί παραπάνω. 7. Φωτογράφιση Η φωτογράφιση των τομών έγινε με μικροσκόπιο MicroST-25 (Zeiss) και ψηφιακή camera ExwaveHAD (SONY) και το πρόγραμμα Studio DC10plus (Pinnacle Systems). 77
75 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 8. Ανίχνευση mrna με τη μέθοδο της RT-PCR 8.1 Απομόνωση εμβρύων Η διαδικασία αυτή είναι ίδια με αυτή που περιγράψαμε κατά το πρώτο στάδιο της καλλιέργειας εμβρύου. Με τη διαφορά ότι εδώ απομονώνουμε τα έμβρυα από τη λέκιθο, τα καθαρίζουμε με τη βοήθεια των μικροβελόνων, τα τοποθετούμε σε ένα σωληνάκι eppendorf και τα αποθηκεύουμε στον καταψύκτη μέχρι την ώρα της επεξεργασίας τους. 8.2 Aπομόνωση RNA Υλικά/Διαλύματα Qiagen RNeasy Mini Kit (Qiagen) Σωληνάκια eppendorf 1,5 και 2ml Σύριγγα με μικροβελόνα διαμέτρου 23 gauge Πιπέτες Pasteur Φυγόκεντρος β-μερκαπτοαιθανόλη 70% αιθανόλη RNase-free Η 2 Ο Μεθοδολογία Απομονώνουμε ολικό RNA από έμβρυα όρνιθας σε διαφορετικά στάδια ανάπτυξης με τη βοήθεια του Qiagen RNeasy Mini Kit (Qiagen). Σύμφωνα με το πρωτόκολλο της εταιρείας, ομογενοποιούμε τα έμβρυα (μόλις τα βγάλουμε από τον καταψύκτη) σε 600μl RLT buffer (προμηθεύεται από την εταιρεία και περιέχει guanidine-thiocyanate και απενεργοποιεί αμέσως τις RNases) στο οποίο προσθέτουμε 6μl β-μερκαπτοαιθανόλης. Το ομογενοποίημα το παιρνάμε από σύριγγα απαλλαγμένη από RNase, στην οποία είναι προσαρτημένη μικροβελόνα διαμέτρου 23 gauge (Sambrook et al., 1989), για καλύτερη ομογενοποίηση του υλικού. Με φυγοκέντρηση του ομογενοποίηματος για 3 min (14000rpm, maximum speed), απομακρύνονται οι πυρήνες και οι μεμβράνες που δεν έχουν ομογενοποιηθεί. Το υπερκείμενο 78
76 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ μεταφέρεται με πιπέτα Pasteur σε καινούριο σωληνάκι eppendorf και προστίθενται σ αυτό 600μl 70% αιθανόλη (η αιθανόλη παρέχει ευνοϊκές συνθήκες για εκλεκτική πρόσδεση του RNA στη RNeasy μεμβράνη) και ανακατεύεται χωρίς καθυστέρηση, γιατί η αιθανόλη προκαλεί κατακρήμνιση. Στη συνέχεια, το παραπάνω μείγμα μεταφέρεται στη RNeasy mini στήλη με πιπέτα Pasteur, φυγοκεντρείται για 15sec (10.000rpm). Απομακρύνεται το διάλυμα που έχει περάσει μέσα από τη μεμβράνη. Ακολουθούν τρία ξεπλύματα της μεμβράνης, ένα με 700μl RW1 buffer και δύο με 500μl RPE buffer. Ανάμεσα στα ξεπλύματα γίνεται φυγοκέντρηση για 15 sec (10.000rpm). Τελικό βήμα είναι ο διαχωρισμός με εκχύλιση του καθαρού RNA (40μl RNasefree Η 2 Ο, φυγοκέντρηση για 1min / rpm). Για μεγαλύτερη συγκέντρωση του τελικού προϊόντος το τελευταίο βήμα επαναλαμβάνεται για δεύτερη φορά με την ίδια ποσότητα RNase-free Η 2 Ο. 8.3 Μέτρηση οπτικής απορρόφησης απομονωμένου RNA Υλικά/Διαλύματα 100% αιθανόλη Ποτήρια ζέσεως 0,2% DEPC(Diethylpyrocarbonate) -H 2 O Πιπέτες Σωληνάκια eppendorf 1,5ml Κυβέτα 6Q Συσκευές Φωτόμετρο Μεθοδολογία Προετοιμάζεται το φωτόμετρο. Παρασκευάζουμε τα προς φωτομέτρηση δείγματα: α) 3μl RNA και 597μl DEPC-H 2 O (δείγμα) και β) 600μl DEPC-H 2 O (τυφλό). Μηδενίζεται το φωτόμετρο με το τυφλό στα 260nm και φωτομετρούμε στην συνέχεια το δείγμα (OD Δ260) ). Στη συνέχεια, φωτομετρούμε ξανά το τυφλό και το δείγμα στα 280nm (OD Τ280 και OD Δ280 ). Από τις τιμές των 79
77 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ απορροφήσεων θα εκτιμηθεί η ποσότητα του RNA που θα χρησιμοποιηθεί στην μέθοδο της RT-PCR. Υπολογίζεται η συγκέντρωση του RNA που απομονώθηκε μέσω του τύπου: C RNA = ( OD Δ260 x 40 x 200) / 1000 μg/μl, όπου OD Δ260 η απορρόφηση του δείγματος στα 260 nm. Στη συνέχεια, με την απλή μέθοδο των τριών υπολογίζονται τα μl του διαλύματος που πρέπει να χρησιμοποιήθουν ώστε σε αυτό να περιέχονται 2μg RNA. 8.4 Σύνθεση cdna με RT-PCR Υλικά/Διαλύματα Qiagen OneStep RT-PCR kit (Qiagen) Πιπέτα Σωληνάκι eppendorf 1,5ml Σωληνάκια για PCR Μεθοδολογία Συνθέσαμε cdna για την agrin όρνιθας με RT-PCR σύμφωνα με τις οδηγίες του Qiagen OneStep RT-PCR kit (Qiagen). Για κάθε PCR αντίδραση χρησιμοποιήθηκαν 2μg RNA. Oι primers της agrin όρνιθας που χρησιμοποιήθηκαν (τελική συγκέντρωση 0.6μΜ) είναι: sense που καλύπτει τα 6531 ως 6550bp 5 -GTC TTA TGC CTG TTT GCT GG-3 και antisense που καλύπτει τα 6807 ως 6824bp 5 - GGA CCC AAT CCC ATA CCC -3 δίνοντας ένα προϊόν στα 294bp (βάσεις , chick). Σχεδιάσαμε τους primers με τη χρήση του Primer Premier software (PREMIER Biosoft International) από την αλληλουχία cdna της agrin (Tsim et al., 1992). Οι συνθήκες που χρησιμοποιήθηκαν για την ανίχνευση της agrin εμφανίζονται στον πίνακα: 80
78 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ AGRIN Χρόνος Θερμοκρασία Αντίστροφη μεταγραφή (Reverse transcription) 30 min 50ºC Εναρκτήρια PCR αντίδραση (initial PCR activation) 15 min 95ºC 3 βήματα/κύκλο Αποδιάταξη (Denaturation) 30 sec 94ºC Επαναδιάταξη (Annealing) 1 min 55ºC Επιμήκυνση (Extension) 1min 72ºC Αριθμός κύκλων 35 (και 45) Τελική επιμήκυνση (Final extension) 10min 72ºC Ως μάρτυρας χρησιμοποιήθηκε η β-ακτίνη (β-actin) που εκφράζεται σε όλους τους οργανισμούς. Οι primers της β-ακτίνης είναι: sense 5 -CGG TAT TGT CAC CAA CTG G-3 και antisense 5 - TGG CTA CAT ACA TGG CTG G -3 δίνοντας ένα προϊόν στα 187 bp. Οι primers για την β-ακτίνη είναι όπως έχουν περιγραφεί στη βιβλιογραφία (Corpuz et al., 2000). Οι συνθήκες που χρησιμοποιήθηκαν για την β-actin παρουσιάζονται στον πίνακα: β-actin Χρόνος Θερμοκρασία Αντίστροφη μεταγραφή (Reverse transcription) 30 min 50ºC Εναρκτήρια PCR αντίδραση (initial PCR activation) 15 min 95ºC 3 βήματα/κύκλο Αποδιάταξη (Denaturation) 30 sec 94ºC Επαναδιάταξη (Annealing) 1 min 55ºC Επιμήκυνση (Extension) 1min 72ºC Αριθμός κύκλων 35 (και 45) Τελική επιμήκυνση (Final extension) 10min 72ºC 81
79 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Τα δείγματα που χρησιμοποιήθηκαν για PCR παρασκευάζονταν σύμφωνα με τον ακόλουθο πίνακα: Volume/reaction Volume/reaction Master Mix Agrin Master mix β-actin Agrin β-actin control control 5x buffer 10μl 10μl 5x buffer 10μl 10μl dntps 2μl 2μl dntps 2μl 2μl Sag1 3μl 3μl Q-solution 10μl 10μl ASAg1 3μl 3μl SAC 3μl 3μl Enzyme mix 2μl 2μl ASAC 3μl 3μl H 2 O? 30μl Enzyme mix 2μl 2μl RNA? _ H 2 O? 20μl RNA? _ Total Volume 50μl 50μl 50μl 50μl Σημείωση 1: Η ποσότητα του διαλύματος RNA ποικίλει ανάλογα με την συγκέντρωση που έχει προκύψει από την απομόνωση του RNA. 8.5 Ηλεκτροφόρηση προϊόντων RT-PCR Υλικά/Διαλύματα 40% Acr/bis (Acrylamide/N,N- Βρωμιούχο αιθίδιο (ethidium Methylendiacrylamide) bromide 5mg/ml) Ουρία (Urea) Ποτήρια ζέσεως 1xTBE (Tris/Borate/EDTA) Ογκομετρικός κύλινδρος 10xTBE Πιπέτες 10% APS (ammonium persulfate) Σύριγγα Αποστειρωμένο ddh 2 O Σωληνάκι eppendorf 1,5ml TEMED (Tetramethylethylenediamine) 0,2μm Nalgene φίλτρο (filter) Sample buffer 82
80 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Συσκευές Υδατόλουτρο Συσκευή ηλεκτροφόρησης UV transilluminator (UVP BioDoc It System) Παρασκευή διαλυμάτων Διάλυμα 40% Acr/bis Acrylamide 9,5g Bis 0,5g Διαλύονται σε 25ml ddh 2 O Διάλυμα 10xTBE (stock) Tris 109,02g Boric Acid 55,647g Διαλύονται σε 1l ddh 2 O EDTA 9,31g (EDTA ethylenediaminetetraacetic acid) Χρησιμοποιείται διάλυμα 1xΤΒΕ, που προκύπτει μετά από αραίωση του stock ΤΒΕ(10x) σε ddh 2 O, με αναλογία 1:9. Συνεπώς, για να φτιάξουμε 1200ml TBE(1x) που θα χρειαστούμε αραιώνουμε 120ml TBE(10x) σε 1080ml ddh 2 O. Διάλυμα 10% APS (ammonium persulfate) (1000μl) Διαλύονται 0,1g APS σε 1000μl ddh 2 O. Διαλυμα βρωμιούχο αιθίδιο (ethidium bromide) (5mg/ml) Αναμιγνύονται 500μl βρωμιούχου αιθίδιου 10mg/ml σε 500μl ddh 2 O. 83
81 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ Διάλυμα sample buffer Bromophenol blue 0,0125g Xylene cyanol FF 0,0125g Διαλύονται σε 5ml deionized formamide Glycerol 2,5ml EDTA 0,5M 10μl Μεθοδολογία Προετοιμασία 5% πηκτώματος πολυακρυλαμιδίου (0,75mm πάχος) Ανακατεύουμε 1,875ml 40% Acr/bis, 6,3g Urea, 1,5ml TBE(10x) και 5ml αποστειρωμένο ddh 2 O, μέχρι να διαλυθούν πλήρως και ογκομετρούμε. Συμπληρώνουμε αποστειρωμένο ddh 2 O ως τελικού όγκου 15ml και ανακατεύουμε ξανά το διάλυμα, το οποίο στη συνέχεια φιλτράρουμε μέσω 0,2μm φίλτρου (Nalgene) και προσθέτουμε 9μl TEMED και 90μl 10% APS. Ανακατεύουμε γρήγορα το διάλυμα και το ρίχνουμε με πιπέτα Pasteur στην σχισμή μεταξύ δύο γυάλινων πλακών. Τοποθετούμε το ειδικό πλαστικό χτενάκι για να σχηματιστούν τα πηγαδάκια φορτώματος και αφήνουμε το διάλυμα να πήξει. Αφού σταθεροποιηθεί το πήκτωμα βγάζουμε το χτενάκι και τα πηγαδάκια ξεπλένονται (από την ουρία) με ΤΒΕ(1Χ). Επεξεργασία δειγμάτων Για την β-actin χρησιμοποιούμε 3μl προϊόντος από τη PCR προσθέτοντας 0,8μl sample buffer. Για το πειραματικό δείγμα (agrin) χρησιμοποιούμε 10μl προϊόντος από την PCR και προσθέτουμε 1μl sample buffer. Θερμαίνουμε τα δείγματα για 2-3min στους 65ºC, σε υδατόλουτρο. Ακολουθεί σύντομη φυγοκέντρηση και τα δείγματα φορτώνονται στο πήκτωμα. Χρησιμοποιούμε 6μl marker (Fermentas SMO331, bp). Γεμίζουμε το δοχείο της συσκευής με TBE(1x) και ηλεκτροφορούμε στα 40Volt για περίπου 2,5 ώρες. Στη συνέχεια γίνεται χρώση με 20μl βρωμιούχο αιθίδιο σε 120ml TBE(1x) για min. Ακολουθεί φωτογράφιση του πηκτώματος ηλεκτροφόρησης σε UV transilluminator (UVP BioDoc It System). 84
82 ΥΛΙΚΑ & ΜΕΘΟΔΟΙ 9. Αποστειρώσεις Ο γυάλινος εξοπλισμός (petri, γυάλινα δαχτυλίδια, σταγονόμετρα κλπ), καθώς και τα μεταλλικά αντικείμενα (ψαλίδια, λαβίδες κ.α.) αποστειρώνονται με ξηρή αποστείρωση σε κλίβανο στους 160 ο C για 1 ½ ώρες. Τα διαλύματα, ο γυάλινος εξοπλισμός με μεταλλικά εξαρτήματα ή με screw caps και οτιδήποτε περιέχει λάστιχο, πολυπροπυλένιο ή teflon αποστειρώνεται στο αυτόκαυστο σε πίεση 25ppm (1atm) και σε θερμοκρασία περίπου 130 ο C για 30 λεπτά. 85
83
84 ΑΠΟΤΕΛΕΣΜΑΤΑ 1. Έκφραση της πρωτεογλυκάνης agrin 1.1 Έκφραση του mrna agrin Μελετήσαμε την έκφραση του mrna agrin στα στάδια XI-XII (μορίδιο), ΧΙΙΙ - (προχωρημένο βλαστίδιο), HH1-2 (έναρξη σταδίου γαστριδίου), ΗΗ3 (μέσα σταδίου γαστριδίου), HH4-5 (προχωρημένο γαστρίδιο/πρώιμο νευρίδιο), ΗΗ8-9 (4-7 σωμίτες) και ΗΗ9-10 (7-10 σωμίτες) στο πρώιμο έμβρυο της όρνιθας (Εικ. 11). Χρησιμοποιήσαμε τη μέθοδος της RT-PCR (reverse transcriptase polymerase chain reaction) για να ανιχνεύσουμε το mrna της agrin (Εικ. 11), και να προσδιορίσουμε το αναπτυξιακό στάδιο που αρχίζει να εκφράζεται το γονίδιο της agrin και εάν ρυθμίζεται αναπτυξιακά κατά την ανάπτυξη του εμβρύου της όρνιθας. Τα αποτελέσματά μας έδειξαν ότι το mrna της agrin δεν εκφράζεται στο στάδιο του μοριδίου, όταν το έμβρυο είναι ιστολογικά απλό. Το mrna agrin ανιχνεύτηκε σε πολύ χαμηλά επίπεδα στο στάδιο μετά τα μέσα του σταδίου του βλαστιδίου (XIII - ) όταν έχει σχεδόν ολοκληρωθεί ο σχηματισμός της υποβλάστης. Η έκφραση του mrna agrin παραμένει στα ίδια επίπεδα στο στάδιο ΗΗ1-2 που σηματοδοτεί την έναρξη των μορφογενετικών κινήσεων της γαστριδίωσης. Στα στάδια XIII - και ΗΗ1-2, το mrna agrin ανιχνεύτηκε όταν πραγματοποιήσαμε την αντίδραση της PCR για 45 κύκλους όχι όμως για 35 κύκλους όπως ανιχνεύτηκε στα επόμενα στάδια ανάπτυξης του εμβρύου. Παρατηρήσαμε ένταση έκφρασης του mrna agrin στα μέσα του σταδίου του γαστριδίου (ΗΗ3), ενώ στο στάδιο του προχωρημένου γαστριδίου/πρώιμου νευριδίου (ΗΗ4-5) η ένταση έκφρασης του γονιδίου της agrin έχει μειωθεί σημαντικά. Τα μέσα του σταδίου του γαστριδίου χαρακτηρίζονται από την ένταση των μορφογενετικών κινήσεων των κυττάρων μέσα και κάτω από την πρωτογενή αύλακα, η οποία στο στάδιο αυτό εκτείνεται από το οπίσθιο μέρος μέχρι πιο πάνω από το κέντρο της διάφανης περιοχής της επιβλάστης. Στο στάδιο ΗΗ4-5, η πρωτογενής αύλακα, έχοντας φτάσει στο μέγιστο μήκος της (1,88mm, Spratt, 1946), αρχίζει να υποχωρεί προς τα πίσω, ενώ στη θέση της αρχίζει να σχηματίζεται 88
85 ΑΠΟΤΕΛΕΣΜΑΤΑ η νωτοχορδή από το μεσόδερμα, ενώ έχει επαχθεί η νευρική πλάκα (neural plate). Στο στάδιο των 4-7 σωμιτών (ΗΗ8-9) παρατηρήσαμε ένταση έκφρασης του mrna της agrin και μείωση των επιπέδων του στο στάδιο των 7-10 σωμιτών (ΗΗ9-10). Στα στάδια ΗΗ8-9 και ΗΗ9-10 έχει ήδη αρχίσει η οργανογένεση καθώς ενδοδερμικά κύτταρα εκατέρωθεν του κεντρικού σημείου του εμβρυϊκού άξονα συγκλίνουν και σχηματίζουν το κάτω τοίχωμα του εντέρου, τα προκαρδιακά κύτταρα (προκαρδιακά πεδία/pre-cardiac fields) συγκλίνουν και σχηματίζουν τον καρδιακό σωλήνα και αρχίζει η διαμερισματοποίηση του εγκεφάλου και ο σχηματισμός των οπτικών κυστιδίων. Οι ζώνες που πήραμε μετά την ηλεκτροφόρηση των cdnas ήταν στο αναμενόμενο μέγεθος, στα 294bp για την agrin και στα 187bp για την β- ακτίνη. Η ζώνη μεγέθους 370bp πάνω από τη ζώνη της agrin θα μπορούσε να είναι μία από τις ισομορφές της. 89
86 ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα 11: Εκφραση του γονιδίου της agrin σε διαφορετικά στάδια στο πρώιμο έμβρυο όρνιθας, όπως ανιχνεύθηκε με τη μέθοδο της RT- PCR. Ηλεκτροφόρηση των προϊόντων της RT-PCR μεγέθους 294bp για την agrin και 187bp για την β-ακτίνη, που έχουν παραχθεί με ειδικούς primers. Χρώση πηκτώματος με βρωμιούχο αιθίδιο (ethidium bromide). Ολικό RNA απομονώθηκε από έμβρυα σε διάφορα αναπτυξιακά στάδια ανάπτυξης (μορίδιο (XI-XII), προχωρημένο βλαστίδιο (XIII-), πρώιμο γαστρίδιο (HH1-2), μέσα σταδίου γαστριδίου (HH3), πρώιμο νευρίδιο (HH4-5), 4-7 (HH8-9) και 7-10 (HH9-10) σωμίτες) και χρησιμοποιήθηκε ως εκμαγείο (template). axis, εμβρυϊκός άξονας; area vasculosa, αιμοτοποιητική περιοχή. 90
 (Εικ. 12Α), ΗΗ4 (προχωρημένο γαστρίδιο) (Εικ. 13Α), ΗΗ8 (4 σωμίτες) (Εικ. 14Α), ΗΗ10 (10 σωμίτες) (Εικ.")
87 ΑΠΟΤΕΛΕΣΜΑΤΑ 1.2 Χωρο-χρονική κατανομή της πρωτεΐνης agrin Μελετήσαμε τη χωροχρονική κατανομή της agrin στα πρώιμα στάδια ανάπτυξης του εμβρύου της όρνιθας, με τη μέθοδο του ανοσοφθορισμού. Απομονώσαμε έμβρυα από τα στάδια ΗΗ3 (μέσα σταδίου γαστριδίου) (Εικ. 12Α), ΗΗ4 (προχωρημένο γαστρίδιο) (Εικ. 13Α), ΗΗ8 (4 σωμίτες) (Εικ. 14Α), ΗΗ10 (10 σωμίτες) (Εικ. 15Α) και ΗΗ16 + (28-29 σωμίτες) (Εικ. 16Α), τα επωάσαμε με πρωτογενές μονοκλωνικό αντίσωμα έναντι της agrin (κλώνος 6D2) και ακολούθως με δευτερογενές αντίσωμα που φέρει τη φθορίζουσα ουσία φθορεσίνη (FITC) Στάδιο ΗΗ3 (μέσα σταδίου γαστριδίου) Στο στάδιο ΗΗ3 πολύ αχνός φθορισμός ανιχνεύτηκε μόνο στα μεσεγχυματικά κύτταρα μετά την εγκόλπωσή τους μέσα από την πρωτογενή αύλακα (Εικ. 12). B Α Β Εικόνα 12: (Α) Μορφολογία εμβρύου στο στάδιο ΗΗ3 (Β) Εγκάρσια τομή (7μm) εμβρύου στο στάδιο ΗΗ3 στο επίπεδο (κόκκινης γράμμης) που φαίνεται στο σχήμα (Α) και ανίχνευση της agrin με τη μέθοδο του ανοσοφθορισμού. en, ενδόδερμα; ep, επιβλάστη; m, μεσόδερμα; ps, πρωτογενής αύλακα; v, λεκιθική μεμβράνη. 91
 Στο στάδιο του προχωρημένου γαστριδίου (ΗΗ4) παρατηρήσαμε ένταση φθορισμού στην επιβλάστη όπως και στην πρωτογενή αύλακα και ιδιαίτερη ένταση φθορισμού στα")
88 ΑΠΟΤΕΛΕΣΜΑΤΑ Στάδιο ΗΗ4 (προχωρημένο γαστρίδιο) Στο στάδιο του προχωρημένου γαστριδίου (ΗΗ4) παρατηρήσαμε ένταση φθορισμού στην επιβλάστη όπως και στην πρωτογενή αύλακα και ιδιαίτερη ένταση φθορισμού στα κύτταρα που έχουν μεταναστεύσει έξω από την πρωτογενή αύλακα και αρχίζουν να σχηματίζουν το μεσόδερμα και το ενδόδερμα (Εικ. 13). Α Εικόνα 13: (Α) Μορφολογία του εμβρύου στο στάδιο ΗΗ4. (Β) Εγκάρσια τομή (7μm) εμβρύου στο στάδιο ΗΗ4, όπως φαίνεται στο (Α) και ανίχνευση της agrin με τη μέθοδο του ανοσοφθορισμού. en, ενδόδερμα; ep, επιβλάστη; m, μεσόδερμα; ps, πρωτογενής αύλακα; y, ενδοδερμικά κύτταρα φορτωμένα με λέκιθο (yolk laden cells). Β 92
 Στο στάδιο του πρώιμου νευριδίου (ΗΗ8) ανιχνεύσαμε ένταση φθορισμού στη νευρική πλάκα και ιδιαίτερα στην κεντρική κοίλη περιοχή της νευρικής πλάκας (neural groove).")
89 ΑΠΟΤΕΛΕΣΜΑΤΑ Στάδιο ΗΗ8 (4 σωμίτες) Στο στάδιο του πρώιμου νευριδίου (ΗΗ8) ανιχνεύσαμε ένταση φθορισμού στη νευρική πλάκα και ιδιαίτερα στην κεντρική κοίλη περιοχή της νευρικής πλάκας (neural groove). Στο μεσόδερμα και το ενδόδερμα παρατηρείται έκφραση της agrin αλλά με λιγότερη ένταση, σε σύγκριση με αυτήν στην νευρική πλάκα (Εικ. 14). Α Εικόνα 14: (Α) Μορφολογία του εμβρύου στο στάδιο ΗΗ8. (Β) Εγκάρσια τομή (7μm) εμβρύου στο στάδιο ΗΗ8, όπως φαίνεται στο (Α) και ανίχνευση της agrin με τη μέθοδο του ανοσοφθορισμού. e, εξώδερμα; en, ενδόδερμα; m, μεσόδερμα; nf, νευρικές πτυχές; ng, αύλακας νευροεπιθηλίου (neural groove); y, ενδοδερμικά κύτταρα φορτωμένα με λέκιθο (yolk laden cells). Β 93
90 ΑΠΟΤΕΛΕΣΜΑΤΑ Στάδιο ΗΗ10 (10 σωμίτες) Κατά την οργανογένεση στα έμβρυα στο στάδιο ΗΗ10 παρατηρήσαμε ένταση φθορισμού στο νευρικό σωλήνα καθώς και στη μεσοκυττάρια ουσία στην κοιλότητα του νευρικού σωλήνα και στα κύτταρα των νευρικών κρηπίδων που αρχίζουν να μεταναστεύουν (Εικ. 15). Ανιχνεύσαμε ένταση έκφρασης στο εσωτερικό τοίχωμα ολόκληρου του πεπτικού σωλήνα, με αξιοσημείωτη ένταση φθορισμού στην κεντρική περιοχή του πάνω τοιχώματος αυτού (Εικ. 15Β, C). Ήταν ενδιαφέρον να παρατηρήσουμε τη διαφορική ένταση φθορισμού της πρωτεΐνης agrin κατά τον σχηματισμό του εντέρου και του καρδιακού σωλήνα (Εικ. 15Β-D). Αρχικά, παρατηρήθηκε έντονη έκφραση της agrin στα προκαρδιακά κύτταρα (heart primordia) και επίσης στο ενδόδερμα που θα σχηματίσει το κάτω τοίχωμα του πεπτικού σωλήνα (Εικ. 15Β). Στη συνέχεια της οργανογένεσης, εξαιρετικά υψηλά επίπεδα φθορισμού παρατηρήθηκαν στο σημείο συρραφής του κάτω τοιχώματος του εντέρου και επίσης στο σημείο της σύνδεσης των προκαρδιακών κυττάρων (Εικ. 15C). Μετά τον σχηματισμό του εντέρου παρατηρήσαμε ένταση έκφρασης της agrin στην εσωτερική επιφάνεια του κάτω τοιχώματος και στην κεντρική περιοχή του πάνω τοιχώματος του εντέρου (Εικ. 15D). Στην καρδία ανιχνεύσαμε έκφραση της agrin σε ολόκληρη την καρδιά (Εικ. 15D), όχι όμως με την ένταση με την οποία ανιχνεύτηκε κατά τη σύνδεση των προκαρδιακών κυττάρων (heart fields). Με τομή στο επίπεδο των σωμιτών (εικόνα 15Ε), η οποία ορίζεται από την κόκκινη γραμμή στην Εικόνα 15Α (τομή b), παρατηρήσαμε φθορισμό κυρίως στον νευρικό σωλήνα και στα κύτταρα των νευρικών κρηπίδων που μεταναστεύουν από αυτόν. Ενώ, λίγο χαμηλότερη ένταση φθορισμού παρατηρήσαμε στους σωμίτες, στο μεσόδερμα και στο ενδόδερμα. Στο στάδιο αυτό παρατηρήσαμε έντονο φθορισμό στον ποικίλο αριθμό συστάδων αιμοκυττάρων (blood islands) (Εικ. 15F). 94
 Εγκάρσια τομή (7μm) στο επίπεδο της καρδιάς (κόκκινη γραμμή στην εικόνα Α, τομή a) και (Ε,F) εγκάρσια τομή στο επίπεδο των σωμιτών (κόκκινη γραμμή στην εικόνα Α, τομή b) εμβρύου στο στάδιο ΗΗ10.")
91 ΑΠΟΤΕΛΕΣΜΑΤΑ a b Α Εικόνα 15: (Α) Μορφολογία του εμβρύου στο στάδιο ΗΗ10. (Β-D) Εγκάρσια τομή (7μm) στο επίπεδο της καρδιάς (κόκκινη γραμμή στην εικόνα Α, τομή a) και (Ε,F) εγκάρσια τομή στο επίπεδο των σωμιτών (κόκκινη γραμμή στην εικόνα Α, τομή b) εμβρύου στο στάδιο ΗΗ10. Ανίχνευση της agrin με τη μέθοδο του ανοσοφθορισμού κατά την οργανογένεση. a, ραχιαία αορτή; bl, νησίδες αίματος; dm, δερμομυοτόμος; e, εξώδερμα; em, μεσοκυττάρια ουσία; en, ενδόδερμα; g, πεπτικός σωλήνας; hp, προκαρδιακά κύτταρα; in, συρραφή τοιχώματος εντέρου (fusion of margins of anterior intestinal portal); m, μεσόδερμα; mc, μυοκάρδιο; n, νωτοχορδή; nc, κύτταρα νευρικών κρηπίδων; nt, νευρικός σωλήνας; sc, σκληροτόμος; sm, σωματοπλευρά (somatopleure); sp, σπλαχνοπλευρά (splanchnopleure) 95
92 ΑΠΟΤΕΛΕΣΜΑΤΑ Β C D 96
93 ΑΠΟΤΕΛΕΣΜΑΤΑ Ε F 97
94 ΑΠΟΤΕΛΕΣΜΑΤΑ Στάδιο ΗΗ16 + (28-29 σωμίτες) Στα έμβρυα στο στάδιο των σωμιτών (Εικ. 16Α) φθορισμός της agrin ανιχνεύτηκε στα τοιχώματα του μυελεγκεφάλου, στο μυοτόμο, στα τοιχώματα των φαρυγγικών και των αορτικών τόξων, στη μεσοκυττάρια ουσία εντός της κοιλότητας του εντέρου, στα τοιχώματα του διεγκεφάλου και στην μεσοκυττάρια ουσία εντός της κοιλότητας αυτού, στον υπό ανάπτυξη οφθαλμό και στα κύτταρα των νευρικών κρηπίδων που μεταναστεύουν προς τον οφθαλμό, μετέχοντας στον σχηματισμό του κερατοειδούς χιτώνα. Στον οφθαλμό ισχυρή έκφραση της agrin ανιχνεύτηκε στο φακό, το χρωματοφόρο και τον νευρικό αμφιβληστροειδή και επίσης στο εξώδερμα που θα σχηματίσει τον κερατοειδή χιτώνα και με τη συμβολή των κυττάρων των νευρικών κρηπίδων. Επιπλέον, ισχυρή ένταση παρατηρήσαμε στην προέκταση του διεγκεφάλου (infandibulum) που αλληλεπιδρά με το στομόδαιο για να σχηματίσουν αργότερα στην ανάπτυξη τους δύο λοβούς της υπόφυσης, στην μεσοκυττάρια ουσία της σωματικής κοιλότητας, καθώς και στο εξώδερμα και το άμνιο. Μικρότερη ένταση παρατηρήσαμε επίσης στη νωτοχορδή (Eικ. 16Β). Στο ίδιο έμβρυο στο επίπεδο της καρδιάς (Εικ. 16C), έντονος φθορισμός της agrin ανιχνεύτηκε στο νευρικό σωλήνα, στα κύτταρα των νευρικών κρηπίδων πλευρικά αυτού, στο μυοτόμιο, αλλά όχι στο δερμοτόμιο και σκληροτόμιο του σωμίτη, στα τοιχώματα της ραχιαίας αορτής και στα αιματοκύτταρα εντός αυτής, στα τοιχώματα καθώς και στη μεσοκυττάρια ουσία της κοιλότητας του εντέρου. Ένταση φθορισμού παρατηρήσαμε επίσης τόσο στο πολυστίβαδο μυοκάρδιο όσο και στο μονοστίβαδο ενδοκάρδιο, καθώς και στο εξώδερμα και το άμνιο. 98
 Εγκάρσιες τομές (7μm) στο επίπεδο του εγκεφάλου και της καρδιάς, αντίστοιχα (κόκκινες γραμμές στην εικόνα Α, τομή a και b, αντίστοιχα) εμβρύου στο στάδιο ΗΗ16 +.")
95 ΑΠΟΤΕΛΕΣΜΑΤΑ Α a b Εικόνα 16: (Α) Μορφολογία του εμβρύου στο στάδιο ΗΗ16 +. (Β, C) Εγκάρσιες τομές (7μm) στο επίπεδο του εγκεφάλου και της καρδιάς, αντίστοιχα (κόκκινες γραμμές στην εικόνα Α, τομή a και b, αντίστοιχα) εμβρύου στο στάδιο ΗΗ16 +. Ανίχνευση της agrin με τη μέθοδο του ανοσοφθορισμού. a, ραχιαία αορτή; aa, αορτικά τόξα; am, άμνιο; ca, ενδοκάρδιο; d, διεγκέφαλος; dc, ραχιαίο μεσοκάρδιο; dm, δερμομυοτόμος; dt, δερμοτόμιο; e, εξώδερμα; em, μεσοκυττάρια ουσία; g, πεπτικός σωλήνας; ln, φακός; mc, μυοκάρδιο; mp, μυελεγκέφαλος; mt, μυοτόμος n, νωτοχορδή; nc, κύτταρα νευρικών κρηπίδων; nt, νευρικός σωλήνας; p, φαρυγγικά τόξα; pl, χρωματοφόρος αμφιβληστροειδής; sc: σκληροτόμος; sl: νευρικός αμφιβληστροειδής; sr, κερατοειδής (sclera). Β 99

96 ΑΠΟΤΕΛΕΣΜΑΤΑ C 100
97 ΑΠΟΤΕΛΕΣΜΑΤΑ 2. Αναστολή λειτουργίας της agrin Έμβρυα στα στάδια ΗΗ3-4 (στάδιο γαστριδίου) επωάστηκαν με μονοκλωνικά αντισώματα έναντι της agrin (κλώνος 6D2) για 2 ώρες στους 37,5º C, με σκοπό να ανασταλεί η λειτουργία της agrin. Μετά την απομάκρυνση του αντισώματος, τα έμβρυα ξεπλύθηκαν σε διάλυμα Ringer και τοποθετήθηκαν σε σχεδίες από λεκιθική μεμβράνη, με την επιβλάστη σε επαφή με τη σχεδία. Ακολούθησε καλλιέργεια σε αλβουμίνη για περίπου 44 ώρες στους 37,5º C, μονιμοποίηση των εμβρύων και μικροτόμηση σε διαδοχικές τομές των 7μm. Παράλληλα, άλλα έμβρυα, χωρίς να έχουν επωαστεί με αντίσωμα, καλλιεργήθηκαν όπως τα παραπάνω πειραματικά έμβρυα σε αλβουμίνη για περίπου 44 ώρες στους 37,5º C. Τα έμβρυα αυτά αποτελούν τους μάρτυρες, ώστε να γίνεται η σύγκριση με τα πειραματικά έμβρυα. Στην Εικόνα 17Α και Β παρουσιάζονται ένα από τα πειραματικά έμβρυα (Β) και ένα από τα έμβρυα μάρτυρες (Α). Σε εγκάρσια τομή (τομή a) του εμβρύου (Β) στο επίπεδο του εγκεφάλου (Εικ. 17D), η βασική μεμβράνη ειδικότερα στο εσωτερικό τοίχωμα του εγκεφάλου εμφανίστηκε αποδιοργανωμένη και τα κύτταρα σε ορισμένα σημεία έδειξαν μεσεγχυματικά χαρακτηριστικά. Το έντερο εμφανίστηκε τελείως αποδιοργανωμένο και το πάνω τοίχωμά του είχε χάσει τη συνοχή του. Είναι επίσης ενδιαφέρον ότι το ενδόδερμα δεν είχε επιθηλιοποιηθεί και δημιούργησε χαρακτηριστικές εκχυμώσεις. Τομή στο ίδιο επίπεδο (τομή a) εμβρύου μάρτυρα (Α) παρουσιάζεται στην Εικόνα 17C, για σύγκριση. Στην Εικόνα 17G απεικονίζεται εγκάρσια τομή του πειραματικού εμβρύου (Β) στο επίπεδο της καρδιάς (τομή b). Τα κύτταρα του νευρικού σωλήνα δεν ήταν πλήρως επιθηλιοποιημένα και η βασική μεμβράνη στην περιφέρεια του νευρικού σωλήνα ήταν αποδιοργανωμένη. Μια πολύ ενδιαφέρουσα παρατήρηση ήταν ότι ενώ τα κύτταρα των νευρικών κρηπίδων έχουν εξέλθει από τη ραχιαία επιφάνεια του νευρικού σωλήνα δεν είχαν τη δυνατότητα να μεταναστεύσουν, αλλά είναι ακόμα σε επαφή με τα τοιχώματα του νευρικού σωλήνα. Στην Εικόνα 17Ι παρουσιάζονται ο νευρικός σωλήνας και τα κύτταρα των νευρικών κρηπίδων σε μεγέθυνση. Παρατηρήσαμε επίσης την πλήρη 101
98 ΑΠΟΤΕΛΕΣΜΑΤΑ απουσία του πάνω τοιχώματος του εντέρου και το αποδιοργανωμένο κάτω τοίχωμά του που μπορεί να είναι αποτέλεσμα της αποδιοργανωμένης βασικής μεμβράνης του. Τα προ-καρδιακά κύτταρα έχουν μεταναστεύσει να σχηματίσουν τον καρδιακό σωλήνα, δεν είναι επιθηλιοποιημένα και δείχνουν μεσεγχυματικά χαρακτηριστικά, ενώ η βασική μεμβράνη είναι αποδιοργανωμένη. Τα τοιχώματα του καρδιακού σωλήνα έχουν καταρρεύσει και τα κύτταρα του ραχιαίου μεσοκάρδιου που συγκρατούν την καρδιά δεν είναι επιθηλιοποιημένα. Στην Εικόνα 17H παρουσιάζονται η καρδιά και το κάτω τοίχωμα του εντέρου σε μεγέθυνση. Το ενδόδερμα εμφάνισε χαρακτηριστικές εκχυμώσεις αδυνατώντας να επιθηλιοποιηθεί, όπως το αναφέραμε και πιο πάνω. Τομή του εμβρύου μάρτυρα (Α) στο ίδιο επίπεδο (τομή b) παρουσιάζεται στις Εικόνες 17E, F, για σύγκριση. Εγκάρσια τομή στο επίπεδο των σωμιτών (τομή c) του πειραματικού εμβρύου (Β) έδειξε ότι οι σωμίτες δεν είχαν επιθηλιοποιηθεί και τα σωμιτικά κύτταρα δεν περιορίζονταν από τη χαρακτηριστική βασική μεμβράνη, αλλά εμφάνιζαν μεσεγχυματικά χαρακτηριστικά. Το ενδόδερμα έδειξε χαρακτηριστικές εκχυμώσεις, όπως παρουσιάσαμε πιο πάνω, και αδυναμία επιθηλιοποίησης (Εικ. 17L). Στις Εικόνες 17Μ και Ν παρουσιάζονται ένας σωμίτης και εκχύμωση μη επιθηλιοποιημένου ενδοδέρματος, αντίστοιχα, σε μεγέθυνση. Τομή του εμβρύου μάρτυρα (Α) στο ίδιο επίπεδο (τομή c) παρουσιάζεται στις Εικόνες 24J, K, για σύγκριση. Στην εικόνα 17Ρ που απεικονίζεται μια εγκάρσια τομή (τομή d) στο οπίσθιο μέρος του πειραματικού εμβρύου (Β) παρατηρήσαμε ότι η εξωεμβρυϊκή αιματοποιητική περιοχή (area vasculosa) του εμβρύου ήταν αποδιοργανωμένη. Το ενδόδερμα δεν ήταν επιθηλιοποιημένο και έδειξε χαρακτηριστικές εκχυμώσεις. Τομή του εμβρύου μάρτυρα (Α) στο ίδιο επίπεδο (τομή d) παρουσιάζεται στην Εικόνα 17Ο, για σύγκριση. 102

99 ΑΠΟΤΕΛΕΣΜΑΤΑ Εικόνα 17: (A) και (B) Έμβρυα απομονώθηκαν στο στάδιο ΗΗ3-4, επωάστηκαν απουσία (Α) ή παρουσία (Β) μονοκλωνικού αντισώματος έναντι της agrin για 2h στους 37,5 ο C και καλλιεργήθηκαν σε αλβουμίνη για 44h. (C) και (D) Εγκάρσια τομή των εμβρύων (Α) και (Β), αντίστοιχα, στο επίπεδο του εγκεφάλου (τομή a) (Ε) και (G) εγκάρσια τομή των εμβρύων (Α) και (Β), αντίστοιχα, στο επίπεδο της καρδιάς (τομή b), (F) και (Η, Ι) λεπτομέρεια των εικόνων (Ε) και (G), αντίστοιχα, δείχνουν νευρικό σωλήνα, κύτταρα νευρικών κρηπίδων και καρδιά. (J) και (L) εγκάρσια τομή των εμβρύων (Α) και (Β), αντίστοιχα, στο επίπεδο των σωμιτών (τομή c) (Κ) και (Μ, Ν) λεπτομέρεια εικόνων (J) και (L), αντίστοιχα, δείχνει σωμίτες και ενδόδερμα. (Ο) και (Ρ) εγκάρσια τομή των εμβρύων (Α) και (Β), αντίστοιχα, στο οπίσθιο μέρος τους (τομή d), δείχνει την αιμοποιητική περιοχή (area vasculosa). bl: νησίδες αίματος, bm: βασική μεμβράνη (basement membrane) ca: ενδοκάρδιο, dc: ραχιαίο μεσοκάρδιο, e: εξώδερμα, en: ενδόδερμα, g: πεπτικός σωλήνας, m: μεσόδερμα, mc: μυοκαρδιο, ms: μεσεγκέφαλος, n: νωτοχορδή, nc: κύτταρα νευρικών κρηπίδων, np: νευρική πλάκα, nt: νευρικός σωλήνας, s: σωμίτης, v: λεκιθική μεμβράνη, vv: αγγείο λεκιθικού ασκού (viteline vessel) 103
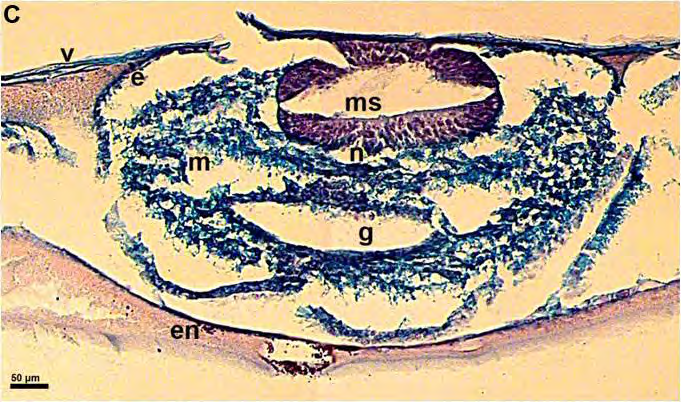
100 ΑΠΟΤΕΛΕΣΜΑΤΑ 104
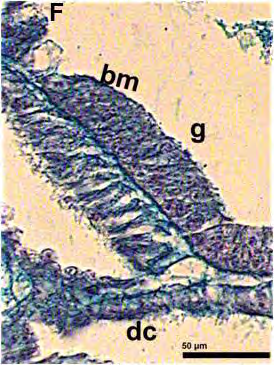
101 ΑΠΟΤΕΛΕΣΜΑΤΑ 105
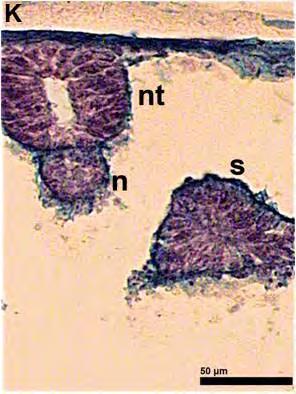
102 ΑΠΟΤΕΛΕΣΜΑΤΑ 106
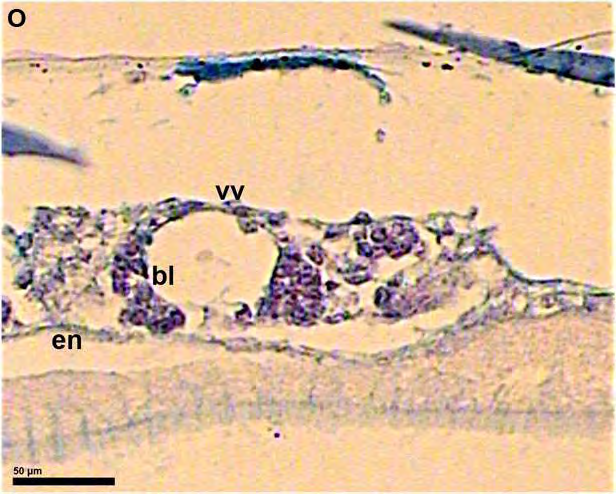
103 ΑΠΟΤΕΛΕΣΜΑΤΑ 107
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ)
") Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
Εξωκυττάριο στρώμα (ΕΣ) 1 Παραδείγματα εξωκυτταρικού στρώματος. Στιβάδες επιθηλιακών κυττάρων στηρίζονται σε μια λεπτή στιβάδα εξωκυτταρικού στρώματος που ονομάζεται βασικός υμένας. Κάτω από τον βασικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ & ΑΝΑΠΤΥΞΗΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ & ΑΝΑΠΤΥΞΗΣ Έκφραση και ρόλος των πρωτεογλυκανών CD44 και versican κατά την ανάπτυξη του πρώιμου εμβρύου ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ & ΑΝΑΠΤΥΞΗΣ Έκφραση και ρόλος των πρωτεογλυκανών CD44 και versican κατά την ανάπτυξη του πρώιμου εμβρύου ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ
ΚΕΦΑΛΑΙΟ 11 Υδατάνθρακες
 ΚΕΦΑΛΑΙΟ 11 Υδατάνθρακες Οι υδατάνθρακες είναι σημαντικά καύσιμα μόρια, αλλά παίζουν και πολλούς άλλους βιοχημικούς ρόλους, συμπεριλαμβανομένης της προστασίας ενάντια σε δυνάμεις με υψηλή κρούση. Ο χόνδρος
ΚΕΦΑΛΑΙΟ 11 Υδατάνθρακες Οι υδατάνθρακες είναι σημαντικά καύσιμα μόρια, αλλά παίζουν και πολλούς άλλους βιοχημικούς ρόλους, συμπεριλαμβανομένης της προστασίας ενάντια σε δυνάμεις με υψηλή κρούση. Ο χόνδρος
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016)
") Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (29/2 & 2/3/2016) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
Συστήματα επικοινωνίας Ανθρωπίνου σώματος. ενδοκρινολογικό νευρικό σύστημα
 Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Κύτταρο Το κύτταρο αποτελείται από μέρη τα οποία έχουν συγκεκριμένη δομή και επιτελούν μία συγκεκριμένη λειτουργία στην όλη οργάνωση του κυττάρου. Δομή κυτταροπλασματικής μεμβράνης Συστήματα επικοινωνίας
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΗ 4 (6/3/2013)
") Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
Kυτταρική Bιολογία ΔIAΛEΞΗ 4 (6/3/2013) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές Φωσφολιπιδική μεμβράνη
και χρειάζεται μέσα στο ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. ενζύμων κύτταρο τρόπους
 Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
Για να εξασφαλιστεί η σωστή και αρμονική έκφραση των ενζύμων μέσα στο κύτταρο χρειάζεται ρύθμιση εναρμόνιση των διαφόρων ενζυμικών δραστηριοτήτων. και Η εναρμόνιση αυτή επιτυγχάνεται με διάφορους τρόπους
Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών
 Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
Kυτταρική Bιολογία ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ ΔIAΛEΞΕΙΣ 4 & 5 (3/3 & 6/3/2017)
") Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (3/3 & 6/3/2017) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
Kυτταρική Bιολογία ΔIAΛEΞΕΙΣ 4 & 5 (3/3 & 6/3/2017) ΒΙΟΛΟΓΙΚΕΣ ΜΕΜΒΡΑΝΕΣ, ΜΕΜΒΡΑΝΙΚΑ ΔΙΑΜΕΡΙΣΜΑΤΑ & ΔΙΑΛΟΓΗ ΠΡΩΤΕΪΝΩΝ Οι λιπιδικές διπλοστιβάδες λειτουργούν ως φραγμοί Νερό Υδρόφιλες φωσφολιπιδικές κεφαλές
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής
 Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ
 ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
ΚΕΦΑΛΑΙΟ 9 ο ΣΤΡΑΤΗΓΙΚΕΣ ΚΑΤΑΛΥΣΗΣ Είδαμε τους μηχανισμούς με τους οποίους καταλύονται οι χημικές/βιολογικές αντιδράσεις (θα επανέλθουμε αν έχουμε χρόνο) Θα εξετάσουμε δύο παραδείγματα ενζύμων και του
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β
 ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης. Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010
 ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
Κυτταρική Βιολογία. Ενότητα 13 : Πολυκυτταρική οργάνωση και διακυτταρικές συνδέσεις. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 13 : Πολυκυτταρική οργάνωση και διακυτταρικές συνδέσεις Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 13 : Πολυκυτταρική οργάνωση και διακυτταρικές συνδέσεις Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
Η κυτταρική µετατόπιση των πρωτεϊνών
 9-1 Κεφάλαιο 9 Η κυτταρική µετατόπιση των πρωτεϊνών Εισαγωγή Στο κύτταρο η έκφραση των πρωτεϊνών γίνεται από µόνο ένα τύπο ριβοσώµατος (εκτός των µιτοχονδριακών και των χλωροπλαστικών που µοιάζουν µε αυτά
9-1 Κεφάλαιο 9 Η κυτταρική µετατόπιση των πρωτεϊνών Εισαγωγή Στο κύτταρο η έκφραση των πρωτεϊνών γίνεται από µόνο ένα τύπο ριβοσώµατος (εκτός των µιτοχονδριακών και των χλωροπλαστικών που µοιάζουν µε αυτά
ΚΕΦΑΛΑΙΟ 15. Κυτταρική ρύθμιση. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΚΕΦΑΛΑΙΟ 15 Κυτταρική ρύθμιση Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 15.1 Μηχανισμοί διακυτταρικής σηματοδότησης. Η διακυτταρική σηματοδότηση μπορεί να συμβαίνει είτε απευθείας
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: 16 / 06 / 2017 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1: δ Α2:
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: 16 / 06 / 2017 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1: δ Α2:
Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά;
 ΒΙΟΛΟΓΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ 12 26/10/2016 Κεφάλαιο 3 Α μέρος Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά; Ποια είναι η δομή
ΒΙΟΛΟΓΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ 12 26/10/2016 Κεφάλαιο 3 Α μέρος Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά; Ποια είναι η δομή
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ. Πετρολιάγκης Σταμάτης Τμήμα Γ4
 ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Πετρολιάγκης Σταμάτης Τμήμα Γ4 ΕΝΝΟΙΑ ΤΗΣ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Η κυτταρική μεμβράνη ή πλασματική μεμβράνη είναι η εξωτερική μεμβράνη που περιβάλλει το κύτταρο
ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Πετρολιάγκης Σταμάτης Τμήμα Γ4 ΕΝΝΟΙΑ ΤΗΣ ΠΛΑΣΜΑΤΙΚΗΣ ΜΕΜΒΡΑΝΗΣ Η κυτταρική μεμβράνη ή πλασματική μεμβράνη είναι η εξωτερική μεμβράνη που περιβάλλει το κύτταρο
Ενδεικτικές απαντήσεις βιολογίας κατεύθυνσης 2014
 Θέμα Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β Θέμα Β ΑΓ.ΚΩΝΣΤΑΝΤΙΝΟΥ 11 -- ΠΕΙΡΑΙΑΣ -- 18532 -- ΤΗΛ. 210-4224752, 4223687 Ενδεικτικές απαντήσεις βιολογίας κατεύθυνσης 2014 Β1. 4 2 1 6 3 5 Β2. α. DNA πολυμεράση
Θέμα Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β Θέμα Β ΑΓ.ΚΩΝΣΤΑΝΤΙΝΟΥ 11 -- ΠΕΙΡΑΙΑΣ -- 18532 -- ΤΗΛ. 210-4224752, 4223687 Ενδεικτικές απαντήσεις βιολογίας κατεύθυνσης 2014 Β1. 4 2 1 6 3 5 Β2. α. DNA πολυμεράση
Προτεινόμενες λύσεις ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16/6/17
 Πανελλήνιες 2017 Προτεινόμενες λύσεις ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16/6/17 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ ΘΕΜΑ Β Β1. Ι-Α ΙΙ-Ε ΙΙΙ-ΣΤ ΙV-Β V-Ζ VI-Γ VII-Δ Β2. Η εικόνα 1 αντιστοιχεί σε Προκαρυωτικό κύτταρο.
Πανελλήνιες 2017 Προτεινόμενες λύσεις ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16/6/17 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ ΘΕΜΑ Β Β1. Ι-Α ΙΙ-Ε ΙΙΙ-ΣΤ ΙV-Β V-Ζ VI-Γ VII-Δ Β2. Η εικόνα 1 αντιστοιχεί σε Προκαρυωτικό κύτταρο.
Θέματα πριν τις εξετάσεις. Καλό διάβασμα Καλή επιτυχία
 Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
Θέματα πριν τις εξετάσεις Καλό διάβασμα Καλή επιτυχία 2013-2014 Θέματα πολλαπλής επιλογής Μετουσίωση είναι το φαινόμενο α. κατά το οποίο συνδέονται δύο αμινοξέα για τον σχηματισμό μιας πρωτεΐνης β. κατά
Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό
 Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία. Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ
 Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014. Απαντήσεις Θεμάτων
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ
 ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ 16/06/2017 ΕΠΙΜΕΛΕΙΑ: ΑΝΔΡΙΑΝΗ ΠΑΡΑΧΗ ΘΕΜΑ Α A1. Δ Α2. Δ Α3. Β Α4. Γ Α5.Α ΘΕΜΑ Β Β1) I A II E III Σ IV Β V Ζ VI Γ VII Δ Β2) ε προκαρυωτικό γιατί
ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ 16/06/2017 ΕΠΙΜΕΛΕΙΑ: ΑΝΔΡΙΑΝΗ ΠΑΡΑΧΗ ΘΕΜΑ Α A1. Δ Α2. Δ Α3. Β Α4. Γ Α5.Α ΘΕΜΑ Β Β1) I A II E III Σ IV Β V Ζ VI Γ VII Δ Β2) ε προκαρυωτικό γιατί
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. γ Α2. α Α3. δ Α4. β Α5. α
 ΘΕΜΑ Α Α1. γ Α2. α Α3. δ Α4. β Α5. α 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 5 ΣΕΠΤΕΜΒΡΙΟΥ 2012 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
ΘΕΜΑ Α Α1. γ Α2. α Α3. δ Α4. β Α5. α 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 5 ΣΕΠΤΕΜΒΡΙΟΥ 2012 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
Kυτταρική Bιολογία. Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος ΔIAΛEΞΗ 20 (9/5/2017) Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.
 Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.") Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
ΘΕΜΑ Α ΘΕΜΑ Β ΑΠΑΝΤΗΣΕΙΣ. Α1. β. Α2. γ. Α3. δ. Α4. γ. Α5. β Β1. 5, 4, 2, 1, 3. Β2. Τα δομικά μέρη του οπερονίου της λακτόζης είναι κατά σειρά τα εξής:
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β ΘΕΜΑ
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β ΘΕΜΑ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ") 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ
1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4
 ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
ΜΕΤΑΛΛΑΞΕΙΣ ΚΑΙ ΚΑΡΚΙΝΟΣ ΕΡΓΑΣΙΑ ΣΤΟ ΜΑΘΗΜΑ ΤΗΣ ΒΙΟΛΟΓΙΑΣ ΟΝΟΜΑ:ΕΥΑΓΓΕΛΙΑ ΕΠΙΘΕΤΟ:ΠΡΙΦΤΗ ΤΑΞΗ:Γ ΤΜΗΜΑ:4 Τι εννοούμε με τον όρο μεταλλάξεις; Το γενετικό υλικό μπορεί να υποστεί αλλαγές με πολλούς διαφορετικούς
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ
 ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
Βιολογία Γενικής Παιδείας Β Λυκείου
 Απρίλιος Μάιος 12 Βιολογία Γενικής Παιδείας Β Λυκείου Βιολογία Γενικής Παιδείας Β Λυκείου (Ερωτήσεις που παρουσιάζουν ενδιαφέρον) 1. Τι είναι τα βιομόρια και ποια είναι τα βασικά χαρακτηριστικά τους; Βιομόρια
Απρίλιος Μάιος 12 Βιολογία Γενικής Παιδείας Β Λυκείου Βιολογία Γενικής Παιδείας Β Λυκείου (Ερωτήσεις που παρουσιάζουν ενδιαφέρον) 1. Τι είναι τα βιομόρια και ποια είναι τα βασικά χαρακτηριστικά τους; Βιομόρια
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία. Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ
 Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
Στηρικτικά Κύτταρα και Εξωκυττάρια Ουσία Κοτσίνας Αθανάσιος Επικ. Καθηγητής Εργ. Ιστολογίας-Εμβρυολογίας Ιατρική Σχολή - ΕΚΠΑ Συνδετικός Ιστός - Ορισμός Παρέχει το: Υποστηρικτικό και Συνδετικό πλαίσιο
ΚΕΦΑΛΑΙΟ 10. Στρατηγικές ρύθμισης
 ΚΕΦΑΛΑΙΟ 10. Στρατηγικές ρύθμισης Oι μεταβολικές πορείες, όπως και η κυκλοφοριακή κίνηση ρυθμίζονται από σήματα. Η CTP, το τελικό προϊόν μιας πορείας πολλών βημάτων, ελέγχει τη ροή των αντιδράσεων σύνθεσής
ΚΕΦΑΛΑΙΟ 10. Στρατηγικές ρύθμισης Oι μεταβολικές πορείες, όπως και η κυκλοφοριακή κίνηση ρυθμίζονται από σήματα. Η CTP, το τελικό προϊόν μιας πορείας πολλών βημάτων, ελέγχει τη ροή των αντιδράσεων σύνθεσής
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ. ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ
 ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
Κυτταρική Βιολογία. Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
BΑΣΙΚΕΣ ΑΡΧΕΣ ΒΙΟΧΗΜΕΙΑΣ ΕΝΕΡΓΕΙΑ ΚΑΙ ΜΕΤΑΒΟΛΙΣΜΟΣ ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΩΝ ΚΥΤΤΑΡΩΝ
 BΑΣΙΚΕΣ ΑΡΧΕΣ ΒΙΟΧΗΜΕΙΑΣ ΕΝΕΡΓΕΙΑ ΚΑΙ ΜΕΤΑΒΟΛΙΣΜΟΣ ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΩΝ ΚΥΤΤΑΡΩΝ 1. ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΩΝ ΚΥΤΤΑΡΩΝ 2. BΑΣΙΚΕΣ ΑΡΧΕΣ ΒΙΟΧΗΜΕΙΑΣ Ι. ΑΤΟΜΑ ΚΑΙ ΜΟΡΙΑ ΙΙ. ΧΗΜΙΚΟΙ ΔΕΣΜΟΙ ΙΙΙ. ΜΑΚΡΟΜΟΡΙΑ ΣΤΑ ΚΥΤΤΑΡΑ
BΑΣΙΚΕΣ ΑΡΧΕΣ ΒΙΟΧΗΜΕΙΑΣ ΕΝΕΡΓΕΙΑ ΚΑΙ ΜΕΤΑΒΟΛΙΣΜΟΣ ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΩΝ ΚΥΤΤΑΡΩΝ 1. ΧΗΜΙΚΗ ΣΥΣΤΑΣΗ ΤΩΝ ΚΥΤΤΑΡΩΝ 2. BΑΣΙΚΕΣ ΑΡΧΕΣ ΒΙΟΧΗΜΕΙΑΣ Ι. ΑΤΟΜΑ ΚΑΙ ΜΟΡΙΑ ΙΙ. ΧΗΜΙΚΟΙ ΔΕΣΜΟΙ ΙΙΙ. ΜΑΚΡΟΜΟΡΙΑ ΣΤΑ ΚΥΤΤΑΡΑ
IΣTOΛOΓIA. Tα δείγµατα του βιολογικού υλικού λαµβάνονται µε > βελόνες ενδοσκοπικούς σωλήνες εύκαµπτους καθετήρες
 IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
αποτελούν το 96% κ.β Ποικιλία λειτουργιών
 ΧΗΜΙΚΑ ΣΤΟΙΧΕΙΑ ΠΟΥ ΣΥΝΘΕΤΟΥΝ ΤΟΥΣ ΟΡΓΑΝΙΣΜΟΥΣ 92 στοιχεία στο φλοιό της Γης 27 απαραίτητα για τη ζωή H, Ο, Ν, C αποτελούν το 96% κ.β S, Ca, P, Cl, K, Na, Mg αποτελούν το 4% κ.β. Fe, I Ιχνοστοιχεία αποτελούν
ΧΗΜΙΚΑ ΣΤΟΙΧΕΙΑ ΠΟΥ ΣΥΝΘΕΤΟΥΝ ΤΟΥΣ ΟΡΓΑΝΙΣΜΟΥΣ 92 στοιχεία στο φλοιό της Γης 27 απαραίτητα για τη ζωή H, Ο, Ν, C αποτελούν το 96% κ.β S, Ca, P, Cl, K, Na, Mg αποτελούν το 4% κ.β. Fe, I Ιχνοστοιχεία αποτελούν
ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ. (Γενετικό γονιδιακής έκφρασης) Μαντώ Κυριακού 2015
 Μαντώ Κυριακού 2015") ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ (Γενετικό υλικό των βακτηρίων ρύθμιση της γονιδιακής έκφρασης) Μαντώ Κυριακού 2015 Γενετικό υλικό των βακτηρίων Αποτελείται από ένα μόριο DNA σε υπερελιγμένη μορφή και τα άκρα του
ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ (Γενετικό υλικό των βακτηρίων ρύθμιση της γονιδιακής έκφρασης) Μαντώ Κυριακού 2015 Γενετικό υλικό των βακτηρίων Αποτελείται από ένα μόριο DNA σε υπερελιγμένη μορφή και τα άκρα του
«ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ»
 «ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ» Τι είναι οι πρωτεΐνες; Από τι αποτελούνται; Ποιος είναι ο βιολογικός του ρόλος; Ας ρίξουμε μία ματιά σε όλα αυτά τα ερωτήματα που μας απασχολούν ΚΕΦΑΛΑΙΟ 1:
«ΠΡΩΤΕΪΝΕΣ: ΧΗΜΙΚΗ ΔΟΜΗ ΚΑΙ ΒΙΟΛΟΓΙΚΟΣ ΡΟΛΟΣ» Τι είναι οι πρωτεΐνες; Από τι αποτελούνται; Ποιος είναι ο βιολογικός του ρόλος; Ας ρίξουμε μία ματιά σε όλα αυτά τα ερωτήματα που μας απασχολούν ΚΕΦΑΛΑΙΟ 1:
Βιολογία Β Λυκείου θέματα
 Ι. Οι υδατάνθρακες διακρίνονται σε μονοσακχαρίτες, δισακχαρίτες και πολυσακχαρίτες. α) Να αναφέρετε από δύο παραδείγματα μονοσακχαριτών, δισακχαριτών και πολυσακχαριτών. (6μ) β) Σε ένα κύτταρο συναντώνται
Ι. Οι υδατάνθρακες διακρίνονται σε μονοσακχαρίτες, δισακχαρίτες και πολυσακχαρίτες. α) Να αναφέρετε από δύο παραδείγματα μονοσακχαριτών, δισακχαριτών και πολυσακχαριτών. (6μ) β) Σε ένα κύτταρο συναντώνται
τα βιβλία των επιτυχιών
 Τα βιβλία των Εκδόσεων Πουκαμισάς συμπυκνώνουν την πολύχρονη διδακτική εμπειρία των συγγραφέων μας και αποτελούν το βασικό εκπαιδευτικό υλικό που χρησιμοποιούν οι μαθητές των φροντιστηρίων μας. Μέσα από
Τα βιβλία των Εκδόσεων Πουκαμισάς συμπυκνώνουν την πολύχρονη διδακτική εμπειρία των συγγραφέων μας και αποτελούν το βασικό εκπαιδευτικό υλικό που χρησιμοποιούν οι μαθητές των φροντιστηρίων μας. Μέσα από
ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ
 ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ ΠPΩTEINEΣ Οι πρωτεΐνες παίζουν σημαντικό ρόλο σε όλες σχεδόν τις βιολογικές διεργασίες. H σημασία τους φαίνεται στις παρακάτω περιπτώσεις: 1. Κατάλυση (πχ. ένζυμα) 2. Μεταφορά
ΔΟΜΗ ΚΑΙ ΔΡΑΣΗ ΠΡΩΤΕΙΝΩΝ ΠPΩTEINEΣ Οι πρωτεΐνες παίζουν σημαντικό ρόλο σε όλες σχεδόν τις βιολογικές διεργασίες. H σημασία τους φαίνεται στις παρακάτω περιπτώσεις: 1. Κατάλυση (πχ. ένζυμα) 2. Μεταφορά
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1
 Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
Η ΔΟΜΗ ΚΑΙ Η ΛΕΙΤΟΥΡΓΙΑ ΤΟΥ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΥ ΚΥΡΙΑΚΗ ΒΑΣΙΛΙΚΟΥ Γ1 ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ Τρισδιάστατο δίκτυο που αποτελείται από μικροσωληνίσκους, μικροϊνίδια και ενδιάμεσα ινίδια. Οι νηματοειδείς πρωτεΐνες του
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
Δομικές κατηγορίες πρωτεϊνών
 3-1 Κεφάλαι ο Δομικές κατηγορίες πρωτεϊνών 3.1. α-δομές πρωτεϊνών Οι α-έλικες είναι δομικά στοιχεία που μπορούν να σχηματίσουν πολλές κατηγορίες στερεοδομών και με πολλές διαφορετικές λειτουργίες. Εκτός
3-1 Κεφάλαι ο Δομικές κατηγορίες πρωτεϊνών 3.1. α-δομές πρωτεϊνών Οι α-έλικες είναι δομικά στοιχεία που μπορούν να σχηματίσουν πολλές κατηγορίες στερεοδομών και με πολλές διαφορετικές λειτουργίες. Εκτός
ΘΕΜΑ Α. 1. δ 2. δ 3. β 4. γ 5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI Δ VII Ε II ΣΤ III Ζ V Η -
 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ 2017 ΛΥΣΕΙΣ ΘΕΜΑΤΩΝ ΕΠΙΜΕΛΕΙΑ: Π. ΚΡΥΣΤΑΛΛΙΔΗΣ 1. δ 2. δ 3. β 4. γ 5. α ΘΕΜΑ Α Β1. Α I Β IV Γ VI Δ VII Ε II ΣΤ III Ζ V Η - ΘΕΜΑ Β Β2. Η εικόνα αντιστοιχεί
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ 2017 ΛΥΣΕΙΣ ΘΕΜΑΤΩΝ ΕΠΙΜΕΛΕΙΑ: Π. ΚΡΥΣΤΑΛΛΙΔΗΣ 1. δ 2. δ 3. β 4. γ 5. α ΘΕΜΑ Α Β1. Α I Β IV Γ VI Δ VII Ε II ΣΤ III Ζ V Η - ΘΕΜΑ Β Β2. Η εικόνα αντιστοιχεί
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10
 Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ-ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής Βιολογίας Εργαστήριο Φαρμακολογίας, Τομέας Φαρμακογνωσίας/Φαρμακολογίας
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ-ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής Βιολογίας Εργαστήριο Φαρμακολογίας, Τομέας Φαρμακογνωσίας/Φαρμακολογίας
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014. Απαντήσεις Θεμάτων
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A 1. Να επιλέξετε τη σωστή απάντηση: Α1. Το γονίδιο
γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A 1. Να επιλέξετε τη σωστή απάντηση: Α1. Το γονίδιο
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Βασικοί μηχανισμοί προσαρμογής
 ΣΥΓΚΡΙΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 23-24, 18/4/2016 Π.Παπαζαφείρη Βασικοί μηχανισμοί προσαρμογής Προσαρμογή σε μοριακό και γονιδιακό επίπεδο Επίπεδα ελέγχου 1. Πρωτεïνική δράση 2. Πρωτεïνοσύνθεση 3. Ρύθμιση της
ΣΥΓΚΡΙΤΙΚΗ ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 23-24, 18/4/2016 Π.Παπαζαφείρη Βασικοί μηχανισμοί προσαρμογής Προσαρμογή σε μοριακό και γονιδιακό επίπεδο Επίπεδα ελέγχου 1. Πρωτεïνική δράση 2. Πρωτεïνοσύνθεση 3. Ρύθμιση της
ΘΕΜΑ Α Α1. γ Α2. γ Α3. α Α4. β Α5. β ΘΕΜΑ B B1. B2.
 ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ
 ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
ΕΡΑΣΜΕΙΟΣ ΕΛΛΗΝΟΓΕΡΜΑΝΙΚΗ ΣΧΟΛΗ Ιδιωτικό Γενικό Λύκειο Όνομα: Ημερομηνία:./04/2014 ΤΑΞΗ : A Λυκείου ΕΡΩΤΗΣΕΙΣ ΓΙΑ ΤΟ 1 ο ΘΕΜΑ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΕΡΩΤΗΣΕΙΣ ΚΕΦΑΛΑΙΟ 11: Ενδοκρινείς αδένες ΒΙΟΛΟΓΙΑ Α ΛΥΚΕΙΟΥ
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014. Ενδεικτικές απαντήσεις Θέµα Β
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 Θέµα Α Α1. δ Α2. γ Α3. β A4. γ A5. β Ενδεικτικές απαντήσεις Θέµα Β Β1. Η σειρά των βημάτων που οδηγούν στην κατασκευή καρυοτύπου
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 Θέµα Α Α1. δ Α2. γ Α3. β A4. γ A5. β Ενδεικτικές απαντήσεις Θέµα Β Β1. Η σειρά των βημάτων που οδηγούν στην κατασκευή καρυοτύπου
ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ
 27 Μαΐου 2016 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Ημερησίων Γενικών Λυκείων (Νέο & Παλιό Σύστημα) ΘΕΜΑ Γ Γ.1 Ο χαρακτήρας της ομάδας αίματος στον άνθρωπο
27 Μαΐου 2016 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Ημερησίων Γενικών Λυκείων (Νέο & Παλιό Σύστημα) ΘΕΜΑ Γ Γ.1 Ο χαρακτήρας της ομάδας αίματος στον άνθρωπο
ΤΟ DNA ΚΑΙ RNA. Θανος Εξαρχου Γ1
 ΤΟ DNA ΚΑΙ RNA Θανος Εξαρχου Γ1 ΤΟ DNA Το δε(σ)οξυριβο(ζο)νουκλεϊ(νι)κό οξu είναι νουκλεϊκό οξύ που περιέχει τις γενετικές πληροφορίες που καθορίζουν τη βιολογική ανάπτυξη όλων των κυτταρικών μορφών ζωής
ΤΟ DNA ΚΑΙ RNA Θανος Εξαρχου Γ1 ΤΟ DNA Το δε(σ)οξυριβο(ζο)νουκλεϊ(νι)κό οξu είναι νουκλεϊκό οξύ που περιέχει τις γενετικές πληροφορίες που καθορίζουν τη βιολογική ανάπτυξη όλων των κυτταρικών μορφών ζωής
ΦΡΟΝΤΙΣΤΗΡΙΟ ΜΕΣΗΣ ΕΚΠΑΙΔΕΥΣΗΣ ΗΡΑΚΛΕΙΤΟΣ ΚΩΛΕΤΤΗ
 ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 ΙΟΥΝΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ε Ν Δ Ε Ι Κ Τ Ι Κ Ε Σ Α Π Α Ν Τ Η Σ Ε Ι Σ Θ Ε Μ Α Τ Ω Ν ΘΕΜΑ Α Α1-δ Α2-δ Α3-β
ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 ΙΟΥΝΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ε Ν Δ Ε Ι Κ Τ Ι Κ Ε Σ Α Π Α Ν Τ Η Σ Ε Ι Σ Θ Ε Μ Α Τ Ω Ν ΘΕΜΑ Α Α1-δ Α2-δ Α3-β
Κ Ε Φ Α Λ Α Ι Ο 21 : Υποκινητές και Ενισχυτές
 Κ Ε Φ Α Λ Α Ι Ο 21 : Υποκινητές και Ενισχυτές Εικόνα 21.1 Ένα τυπικό γονίδιο που µεταγράφεται από την RNA πολυµεράση ΙΙ έχει έναν υποκινητή ο οποίος εκτείνεται ανοδικά από τη θέση έναρξης της µεταγραφής.
Κ Ε Φ Α Λ Α Ι Ο 21 : Υποκινητές και Ενισχυτές Εικόνα 21.1 Ένα τυπικό γονίδιο που µεταγράφεται από την RNA πολυµεράση ΙΙ έχει έναν υποκινητή ο οποίος εκτείνεται ανοδικά από τη θέση έναρξης της µεταγραφής.
ΜΕΣΟΘΕΡΑΠΕΙΑ ΕΝΕΣΙΜΗ ΤΟΠΙΚΗ ΘΕΡΑΠΕΙΑ
 ΜΕΣΟΘΕΡΑΠΕΙΑ ΕΝΕΣΙΜΗ ΤΟΠΙΚΗ ΘΕΡΑΠΕΙΑ ΜΕΣΟΘΕΡΑΠΕΙΑ Αυτό σημαίνει ότι χρησιμοποιούμε μόνο ενέσιμα φάρμακα και μόνο στο σημείο που πάσχει. ΜΕΣΟΘΕΡΑΠΕΙΑ Ξεκίνησε στη λογική του γιατί να μη χορηγήσω ένα αντιφλεγμονώδες
ΜΕΣΟΘΕΡΑΠΕΙΑ ΕΝΕΣΙΜΗ ΤΟΠΙΚΗ ΘΕΡΑΠΕΙΑ ΜΕΣΟΘΕΡΑΠΕΙΑ Αυτό σημαίνει ότι χρησιμοποιούμε μόνο ενέσιμα φάρμακα και μόνο στο σημείο που πάσχει. ΜΕΣΟΘΕΡΑΠΕΙΑ Ξεκίνησε στη λογική του γιατί να μη χορηγήσω ένα αντιφλεγμονώδες
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ )
") Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
Έκφραση και ρόλος της aggrecan και perlecan στο πρώιµο έµβρυο
 ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ, ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΠΤΥΞΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ Έκφραση και ρόλος της aggrecan και perlecan στο πρώιµο έµβρυο ΝΙΚΟΛΙΤΣΑ ΣΟΥΛΙΝΤΖΗ
ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ, ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ ΕΡΓΑΣΤΗΡΙΟ ΑΝΑΠΤΥΞΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ Έκφραση και ρόλος της aggrecan και perlecan στο πρώιµο έµβρυο ΝΙΚΟΛΙΤΣΑ ΣΟΥΛΙΝΤΖΗ
Malamidou A., Pantazaki A.A. *, Koliakos G., Tsolaki M.
 10th Panhellenic Conference of Alzheimer's Disease and Related Disorders (PICAD) and 2nd Mediterranean Conference of Neurodegenerative Diseases (MeCOND) Malamidou A., Pantazaki A.A. *, Koliakos G., Tsolaki
10th Panhellenic Conference of Alzheimer's Disease and Related Disorders (PICAD) and 2nd Mediterranean Conference of Neurodegenerative Diseases (MeCOND) Malamidou A., Pantazaki A.A. *, Koliakos G., Tsolaki
ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ 2014
 ΦΡΟΝΤΙΣΤΗΡΙΑ ΜΕΣΗΣ ΕΚΠΑΙΔΕΥΣΗΣ : - ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ 2014 ΘΕΜΑ A Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ B Β1. 4 2 1 6 3 5 Β2. α. DNA πολυμεράση β. Πριμόσωμα γ. DNA δεσμάση δ. DNA
ΦΡΟΝΤΙΣΤΗΡΙΑ ΜΕΣΗΣ ΕΚΠΑΙΔΕΥΣΗΣ : - ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ 2014 ΘΕΜΑ A Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ B Β1. 4 2 1 6 3 5 Β2. α. DNA πολυμεράση β. Πριμόσωμα γ. DNA δεσμάση δ. DNA
Πανελλαδικές εξετάσεις 2017
 Πανελλαδικές εξετάσεις 2017 Ενδεικτικές απαντήσεις στο μάθημα «ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ» Θέμα Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α Θέμα Β Β1 I: Α, ΙΙ: E, III: ΣΤ, ΙV: B, V:Z, VI: Γ, VII: Δ Η έννοια πυρηνική
Πανελλαδικές εξετάσεις 2017 Ενδεικτικές απαντήσεις στο μάθημα «ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ» Θέμα Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α Θέμα Β Β1 I: Α, ΙΙ: E, III: ΣΤ, ΙV: B, V:Z, VI: Γ, VII: Δ Η έννοια πυρηνική
ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ
 14.1 ΕΙΣΑΓΩΓΗ") 1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
Κυτταρική Βιολογία. Ενότητα 08 : Βιολογικές μεμβράνες, μεμβρανικά διαμερίσματα, μεταφορά πρωτεϊνών Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 08 : Βιολογικές μεμβράνες, μεμβρανικά διαμερίσματα, μεταφορά πρωτεϊνών Παναγιωτίδης Χρήστος Φαρμακευτικής ΑΠΘ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 08 : Βιολογικές μεμβράνες, μεμβρανικά διαμερίσματα, μεταφορά πρωτεϊνών Παναγιωτίδης Χρήστος Φαρμακευτικής ΑΠΘ
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς
 Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα
 Εργασία στη Βιολογία ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Η ροή της πληροφορίας για το σχηματισμό των πρωτεϊνών, προϋποθέτει τη μεταφορά της από το DNA στο RNA (ΜΕΤΑΓΡΑΦΗ).
Εργασία στη Βιολογία ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Η ροή της πληροφορίας για το σχηματισμό των πρωτεϊνών, προϋποθέτει τη μεταφορά της από το DNA στο RNA (ΜΕΤΑΓΡΑΦΗ).
ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ. Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της...
 ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της... Γενετική Μηχανική o Περιλαμβάνει όλες τις τεχνικές με τις οποίες μπορούμε να επεμβαίνουμε στο γενετικό υλικό των οργανισμών.
ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της... Γενετική Μηχανική o Περιλαμβάνει όλες τις τεχνικές με τις οποίες μπορούμε να επεμβαίνουμε στο γενετικό υλικό των οργανισμών.
Ρόλος Υδατανθρακών. Μονοσακχαρίτες Ολιγοσακχαρίτες, Πολυσακχαρίτες
 Ρόλος Υδατανθρακών Καύσιµα, αποθήκες ενέργειας, µεταβολικά ενδιάµεσα Δοµικά στοιχεία RNA, DNA, κυτταρικού τοιχώµατος φυτών & βακτηρίων Μεσολαβητές κυτταρικών αλληλεπιδράσεων Μονοσακχαρίτες Ολιγοσακχαρίτες,
Ρόλος Υδατανθρακών Καύσιµα, αποθήκες ενέργειας, µεταβολικά ενδιάµεσα Δοµικά στοιχεία RNA, DNA, κυτταρικού τοιχώµατος φυτών & βακτηρίων Μεσολαβητές κυτταρικών αλληλεπιδράσεων Μονοσακχαρίτες Ολιγοσακχαρίτες,
ΒΙΟΧΗΜΕΙΑ Ι. ΚΕΦΑΛΑΙΟ 2 ο Βιοχημική εξέλιξη
 ΒΙΟΧΗΜΕΙΑ Ι ΚΕΦΑΛΑΙΟ 2 ο Βιοχημική εξέλιξη ΣΥΝΔΕΣΗ ΜΕ ΤΑ ΠΡΟΗΓΟΥΜΕΝΑ Τι είναι ΒΙΟΧΗΜΕΙΑ DNA ΠΡΩΤΕΙΝΕΣ ΑΛΛΑ ΣΥΝΔΕΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΧΗΜΕΙΑΣ (Δεσμοί, ενέργεια, δομή) ΕΞΕΛΙΞΗ ΤΗΣ ΖΩΗΣ Υπάρχει μια συνεχή εξελικτική
ΒΙΟΧΗΜΕΙΑ Ι ΚΕΦΑΛΑΙΟ 2 ο Βιοχημική εξέλιξη ΣΥΝΔΕΣΗ ΜΕ ΤΑ ΠΡΟΗΓΟΥΜΕΝΑ Τι είναι ΒΙΟΧΗΜΕΙΑ DNA ΠΡΩΤΕΙΝΕΣ ΑΛΛΑ ΣΥΝΔΕΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΧΗΜΕΙΑΣ (Δεσμοί, ενέργεια, δομή) ΕΞΕΛΙΞΗ ΤΗΣ ΖΩΗΣ Υπάρχει μια συνεχή εξελικτική
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης
 Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Μόρια κυτταρικής πρόσφυσης
 Μόρια κυτταρικής πρόσφυσης 1 Ομαδες μοριων κυτταρικης προσφυσης Καντχερίνες -CAMs Σελεκτίνες ΙντεγκρίνεςCAMs Σελεκτίνες Ιντεγκρίνες Συνδέσεις μεταξύ πρωτεινών με ομοιοφιλικό είτε με ετεροφιλικό τρόπο Κυτταρικές
Μόρια κυτταρικής πρόσφυσης 1 Ομαδες μοριων κυτταρικης προσφυσης Καντχερίνες -CAMs Σελεκτίνες ΙντεγκρίνεςCAMs Σελεκτίνες Ιντεγκρίνες Συνδέσεις μεταξύ πρωτεινών με ομοιοφιλικό είτε με ετεροφιλικό τρόπο Κυτταρικές
ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΚΕΦΑΛΑΙΑ 1 ΚΑΙ 2
 ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΚΕΦΑΛΑΙΑ 1 ΚΑΙ 2 ΘΕΜΑ 1 ο Α. Στις ερωτήσεις 1-5 να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Το
ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΚΕΦΑΛΑΙΑ 1 ΚΑΙ 2 ΘΕΜΑ 1 ο Α. Στις ερωτήσεις 1-5 να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Το
ΑΠΑΝΤΗΣΗ ΤΗΣ ΣΥΝΔΥΑΣΤΙΚΗΣ ΑΣΚΗΣΗΣ
 ΑΠΑΝΤΗΣΗ ΤΗΣ ΣΥΝΔΥΑΣΤΙΚΗΣ ΑΣΚΗΣΗΣ 1. Το γενεαλογικό δένδρο είναι η διαγραμματική απεικόνιση των μελών μιας οικογένειας για πολλές γενιές, στην οποία αναπαριστώνται οι γάμοι, η σειρά των γεννήσεων, το φύλο
ΑΠΑΝΤΗΣΗ ΤΗΣ ΣΥΝΔΥΑΣΤΙΚΗΣ ΑΣΚΗΣΗΣ 1. Το γενεαλογικό δένδρο είναι η διαγραμματική απεικόνιση των μελών μιας οικογένειας για πολλές γενιές, στην οποία αναπαριστώνται οι γάμοι, η σειρά των γεννήσεων, το φύλο
Κεφάλαιο 4: Ανασυνδυασμένο DNA
 Κεφάλαιο 4: Ανασυνδυασμένο DNA 1. Η ανάπτυξη της γενετικής μηχανικής επέτρεψε: α. την κατανόηση των μηχανισμών αντιγραφής του γενετικού υλικού β. την απομόνωση των πλασμιδίων από τα βακτήρια γ. την πραγματοποίηση
Κεφάλαιο 4: Ανασυνδυασμένο DNA 1. Η ανάπτυξη της γενετικής μηχανικής επέτρεψε: α. την κατανόηση των μηχανισμών αντιγραφής του γενετικού υλικού β. την απομόνωση των πλασμιδίων από τα βακτήρια γ. την πραγματοποίηση
ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΔΕΥΤΕΡΑ 18 ΙΟΥΝΙΟΥ 2012 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΔΕΥΤΕΡΑ 18 ΙΟΥΝΙΟΥ 2012 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. δ Α4. β Α5. α 2 β 5 γ 6 δ 1 ε 3 ΑΠΑΝΤΗΣΕΙΣ
1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΔΕΥΤΕΡΑ 18 ΙΟΥΝΙΟΥ 2012 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. δ Α4. β Α5. α 2 β 5 γ 6 δ 1 ε 3 ΑΠΑΝΤΗΣΕΙΣ
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2014
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2014 ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ Β Β1. Η σειρά των βημάτων που οδηγούν στην κατασκευή καρυότυπου είναι: 4, 2, 1, 6, 3, 5 Β2. α.
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2014 ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ Β Β1. Η σειρά των βημάτων που οδηγούν στην κατασκευή καρυότυπου είναι: 4, 2, 1, 6, 3, 5 Β2. α.
φροντιστήρια Απαντήσεις Βιολογίας Γ λυκείου Προσανατολισμός Θετικών Σπουδών
 Απαντήσεις Βιολογίας Γ λυκείου Προσανατολισμός Θετικών Σπουδών Θέμα Α Α1. α. 2 β. 1 γ. 4 δ. 1 ε. 2 Θέμα Β Β1. α. 1-Γ, 2-Γ, 3-Β, 4-Β, 5-Α, 6-B, 7-Α β. 1. Σύμπλοκο έναρξης πρωτεϊνοσύνθεσης: Το σύμπλοκο που
Απαντήσεις Βιολογίας Γ λυκείου Προσανατολισμός Θετικών Σπουδών Θέμα Α Α1. α. 2 β. 1 γ. 4 δ. 1 ε. 2 Θέμα Β Β1. α. 1-Γ, 2-Γ, 3-Β, 4-Β, 5-Α, 6-B, 7-Α β. 1. Σύμπλοκο έναρξης πρωτεϊνοσύνθεσης: Το σύμπλοκο που
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ Έκφραση και ρόλος των πρωτεογλυκανών neurocan και phosphacan κατά την ανάπτυξη του πρώϊμου εμβρύου Neurocan Phosphacan
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ, ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ Έκφραση και ρόλος των πρωτεογλυκανών neurocan και phosphacan κατά την ανάπτυξη του πρώϊμου εμβρύου Neurocan Phosphacan
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών
 Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ
 Επαναληπτικό διαγώνισμα ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΚΕΦΑΛΑΙΑ 1-9 ΗΜΕΡΟΜΗΝΙΑ 1/3/2015 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ Σημείωση: η διπλή αρίθμηση στις σελίδες αφορά την παλιά και τη νέα έκδοση του σχολικού βιβλίου
Επαναληπτικό διαγώνισμα ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΚΕΦΑΛΑΙΑ 1-9 ΗΜΕΡΟΜΗΝΙΑ 1/3/2015 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ Σημείωση: η διπλή αρίθμηση στις σελίδες αφορά την παλιά και τη νέα έκδοση του σχολικού βιβλίου
Β. ΚΑΜΙΝΕΛΛΗΣ ΒΙΟΛΟΓΙΑ. Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα).
.") ΒΙΟΛΟΓΙΑ Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα). Είδη οργανισμών Υπάρχουν δύο είδη οργανισμών: 1. Οι μονοκύτταροι, που ονομάζονται μικροοργανισμοί
ΒΙΟΛΟΓΙΑ Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα). Είδη οργανισμών Υπάρχουν δύο είδη οργανισμών: 1. Οι μονοκύτταροι, που ονομάζονται μικροοργανισμοί
ΠΕΤΡΟΣ ΔΕΔΕΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΣΙ ΠΕΡΙΛΗΨΗ
 ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΣΙ ΠΕΡΙΛΗΨΗ Το οστό αποτελεί ένα ιδανικό περιβάλλον για μετάσταση, καθώς η συνεχής και δυναμική ανάπλασή τον παρέχει μια γόνιμη βάση για την παλιννόστηση και τον πολλαπλασιασμό των καρκινικών
ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΣΙ ΠΕΡΙΛΗΨΗ Το οστό αποτελεί ένα ιδανικό περιβάλλον για μετάσταση, καθώς η συνεχής και δυναμική ανάπλασή τον παρέχει μια γόνιμη βάση για την παλιννόστηση και τον πολλαπλασιασμό των καρκινικών
Α2. Το αντικωδικόνιο είναι τριπλέτα νουκλεοτιδίων του α. mrna β. snrna γ. trna δ. rrna Μονάδες 5
 ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και δίπλα στο γράμμα που αντιστοιχεί στη λέξη ή στη φράση, η οποία συμπληρώνει
ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και δίπλα στο γράμμα που αντιστοιχεί στη λέξη ή στη φράση, η οποία συμπληρώνει
ΘΕΜΑ 1 Ο ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Τα
ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Τα