Τίτλος Διπλωματικής Εργασίας Ο μεταγραφικός παράγοντας Coup-TF και η διαφοροποίηση του νευρικού συστήματος στο έμβρυο του αχινού
|
|
|
- Ματταθίας Ρόκας
- 8 χρόνια πριν
- Προβολές:
Transcript
1 Τίτλος Διπλωματικής Εργασίας Ο μεταγραφικός παράγοντας Coup-TF και η διαφοροποίηση του νευρικού συστήματος στο έμβρυο του αχινού Μεταπτυχιακή Φοιτήτρια: Μπάρου Βασιλική Α.Μ: 523 Υπεύθυνος καθηγητής: Φλυτζάνης Κωνσταντίνος 1
2 Περιεχόμενα Ευχαριστήρια Παράγραφος O αχινός ως πειραματικό μοντέλο H εμβρυογένεση στον αχινό O μεταγραφικός παράγοντας COUP-TF και ο ρόλος του στην κυτταρική διαφοροποίηση και στην ανάπτυξη Tο γονίδιο COUP-TF H δομή της πρωτείνης COUP-TF Mηχανισμοί δράσης του COUP-TF H ρύθμιση του γονιδίου COUP-TF Tο γονίδιο SpCOUP-TF Η νευρογένεση στον αχινό Τα υπό μελέτη Γονίδια FoxQ Nkx Six Ο Six3 εκφράζεται νωρίς στο APD Η Λειτουργία του Six3 απαιτείται για το σχηματισμό του APD και τη διαφοροποίηση των νευρώνων Υλικά και Μέθοδοι Συλλογή γαμετών και τεχνητή γονιμοποίηση Καλλιέργεια εμβρύων του αχινού Paracentrotus lividus Απομόνωση RNA από διαφορετικά αναπτυξιακά στάδια RT-PCR Καθαρισμός των προιόντων της RT-PCR Αντίδραση λιγάσης Μετασχηματισμός Βακτηρίων Κατασκευή τρυβλίων θρεπτικού υλικού για βακτηριακές καλλιέργειες Κλωνοποίηση Mini-preps: Πέψη με ένζυμα περιορισμού PCR Καθαρισμός προιόντων PCR
3 4.14 Σήμανση των αντινοηματικών μεταγράφων Γονιδιακή καταστολή σε γονιμοποιημένα αυγά με ενέσεις με ΜΑSO Μονιμοποίηση εμβρύων για χρωμογόνο και διπλή φθορίζουσα in situ υβριδοποίηση Χρωμογόνος in situ υβριδοποίηση Διπλή φθορίζουσα in situ υβριδοποίηση (double FISH) Απομόνωση RNA (περίπου 1,5 μgr) από μικρό αριθμό control και ενεμένων εμβρύων για Q-PCR Q-PCR από RNA control και ενεμένων εμβρύων Αποτελέσματα Αποτελέσματα RT-PCR Κλωνοποίηση Πέψη με ένζυμα περιορισμού In vitro μεταγραφή Αναπτυξιακό πρότυπο έκφρασης για το γονιδιο Six Έκφραση του γονιδίου Six3 σε Coup-TF morphants Q-PCR Αναπτυξιακό πρότυπο έκφρασης για το γονιδιο FoxQ Έκφραση του γονιδίου FoxQ2 σε Coup-TF morphants Q-PCR Αναπτυξιακό πρότυπο έκφρασης για το γονιδιο Nkx Έκφραση του γονιδίου Nkx2.1 σε Coup-TF morphants Q-PCR Συζήτηση Βιβλιογραφία Παράρτημα
4 Ευχαριστήρια Παράγραφος Θέλω να ευχαριστήσω θερμά και να εκφράσω την βαθιά μου ευγνωμοσύνη στον καθηγητή μου και επιβλέποντα της διπλωματικής μου εργασίας κ. Φλυτζάνη Κωνσταντίνο για την εμπιστοσύνη που έδειξε προς το πρόσωπο μου αναθέτοντας μου τη συγκεκριμένη εργασία. Παράλληλα τον ευχαριστώ για την δυνατότητα που μου έδωσα να επεκτείνω τις γνώσεις μου πάνω σε ένα διαφορετικό πεδίο και να αγαπήσω τον «πάγκο» από την πρώτη μέρα. Ευχαριστώ ιδιαιτέρως τον κύριο Τσιρώνη Ιωάννη, ο οποίος υπήρξε πολύτιμος σύμβουλός μου και βρισκόταν πάντα εκεί όταν τον χρειαζόμουν για να με καθοδηγήσει και να με συμβουλεύσει. Καθώς και τα υπόλοιπα παιδιά του εργαστηριού για την υποστήριξη τους και την κατανόηση τους σε δύσκολες στιγμές. Μέσα από τα βάθη της καρδιάς μου ευχαριστώ την οικογένεια μου. Σε όλη την διάρκεια του μεταπτυχιακού μου αλλά και το διάστημα της υλοποίησης της παρούσας διπλωματικής εργασίας ήταν πάντα δίπλα μου και με υπομονή με ενθάρρυναν να συνεχίσω. Κλείνοντας ένα πολύ μεγάλο ευχαριστώ επίσης στον αγαπημένο μου Γκίουλη Αθανάσιο ο οποίος ήταν κάθε μέρα δίπλα μου και μου έδινε κουράγιο. Όποτε χρειαζόμουν κάποιον να μοιραστώ τα προβλήματά μου και τις δυσκολίες που μπορεί να συναντούσα ήταν πάντα εκεί και με τις πολύτιμές συμβουλές του. 4

. Εικ.1 Ο αχινός P.lividus Εικ.2 Γονιμοποιημένο αυγό αχινού υπό μικροσκοπία Τα αυγά και τα έμβρυα του αχινού αποτελούν ένα από τα πιο κοινά πειραματόζωα.")
5 1.O αχινός ως πειραματικό μοντέλο Το πειραματικό υλικό που χρησιμοποιήθηκε είναι ο Paracentrotus lividus, ο κοινός πετραχινός (εικ.1). Η συστηματική του κατάταξη φαίνεται στον πίνακα 1. Το είδος αυτό απαντάται ευρύτατα στη Μεσόγειο σε ρηχά νερά και τρέφεται με φωτόφιλα φύκη. Όπως όλα τα εχινόδερμα έτσι και ο P.lividus φέρει το πιο ξεχωριστό χαρακτηριστικό, την πεντακτινωτή συμμετρία, η οποία προκύπτει δευτερογενώς μετά από μια περίοδο αμφίπλευρης συμμετρίας (Barnes R. D. Zoology, Barnes R. S. K. The invertebrates a new synthesis). Εικ.1 Ο αχινός P.lividus Εικ.2 Γονιμοποιημένο αυγό αχινού υπό μικροσκοπία Τα αυγά και τα έμβρυα του αχινού αποτελούν ένα από τα πιο κοινά πειραματόζωα. Αυτό οφείλεται στην πληθώρα των ατόμων που είναι διαθέσιμα, στην απλή διαδικασία της εμβρυογένεσης και στο γεγονός ότι τα έμβρυα είναι διαφανή (εικ.2) και άρα δίνεται η δυνατότητα για παρατήρηση των κυτταρικών διαιρέσεων και κινήσεων και των αποτελεσμάτων των πειραματικών διαδικασιών. Σημαντική είναι επίσης η ευκολία της γονιδιακής μεταφοράς στο έμβρυο, καθώς και των βιοχημικών διαδικασιών προκειμένου να γίνει καθαρισμός μεταγραφικών παραγόντων. Ειδικά ο P.lividus χρησιμοποιήθηκε γιατί αφθονεί στις Ελληνικές θάλασσες και η συλλογή του είναι εύκολη. 5
6 Συνομοταξία Υποσυνομοταξία Ομοταξία: Υφομοταξία: Υπέρταξη: Τάξη Οικογένεια Γένος Είδος : Εχινόδερμα : Ελευθερόζωα : Εχινοειδή : Ενδοκυκλικά : Εχινόμορφα : Καμερόδοντα : Echinidae (Εχινίδες) : Paracentrotus (Παρακεντρωτός) : Paracentrotus lividus (Παρακεντρωτός ο μολυβδόχρωμος) Πίνακας H εμβρυογένεση στον αχινό H διαδικασία της εμβρυογένεσης είναι πολύ καλά μελετημένη στον αχινό. H γονιμοποίηση είναι εξωτερική και τα γονιμοποιημένα αυγά στον P.lividus περνούν από το στάδιο του βλαστιδίου, του γαστριδίου και του πλουτέα σε διάστημα δύο ημερών σε θερμοκρασία 18 0 C. Kατόπιν, τα έμβρυα μετατρέπονται σε νύμφες και μεταμορφώνονται σε νεαρούς αχινούς σε διάστημα τεσσάρων περίπου εβδομάδων από τη στιγμή της γονιμοποίησης (Davidson et al. 1982; Davidson 1986; Gilbert 1985). Άλλα είδη αχινού όπως ο Strongylocentrotus Purpuratus ολοκληρώνουν την ανάπτυξή τους με αργότερο ρυθμό, ενώ ο Lytechinus variegatus με γρηγορότερο ρυθμό μιας και ζεί σε πιο ζεστά ύδατα. Xαρακτηριστικό στοιχείο κατά την ανάπτυξη του αχινού είναι ότι μέχρι και το στάδιο του βλαστιδίου δεν πραγματοποιούνται μεταναστεύσεις κυττάρων και έτσι μπορούν να χαρτογραφηθούν και να προβλεφθούν οι αναπτυξιακές τύχες των διαφόρων εμβρυικών περιοχών (Davidson et al. 1998). Oι δύο πρώτες αυλακώσεις πραγματοποιούνται κάθετα μεταξύ τους κατά μήκος του άξονα ζ/φ, που ενώνει το ζωικό με το φυτικό πόλο και ο οποίος έχει καθοριστεί πριν τη γονιμοποίηση (Schroeder 1980a, Schroeder 1980b). Έτσι, το έμβρυο χωρίζεται σε τέσσερα ίσα τέταρτα. H τρίτη αυλάκωση πραγματοποιείται κατά μήκος του ισημερινού και διαχωρίζει ισομερώς το ζωικό από το φυτικό πόλο. H τέταρτη αυλάκωση είναι ισομερής στο ζωικό πόλο αλλά άνιση στο φυτικό. 6

. Αναλυτικά περιγράφονται στο κείμενο.")
7 F Εικ.3 Χάρτης αναπτυξιακής τύχης των κυττάρων στο έμβρυο του αχινού. Αναγνωρίζονται οι κυτταρικές σειρές στο στάδιο των 60 κυττάρων (Α), του γαστριδίου (Β, C) και και στο στάδιο του πλουτέα (D, E). Με διαφορετικά χρώματα έχουν σχεδιαστεί τα μεσομερίδια, τα μακρομερίδια και τα μικρομερίδια και με αντίστοιχα χρώματα οι περιοχές που προκύπτουν από τις τρεις αυτές κατηγορίες κυττάρων (Davidson et al. 1998). F) Στάδιο 16 κυττάρων όπου σημειώνονται τα μεσομερίδια, μακρομερίδια και μικρομερίδια. Στάδιο 60 κυττάρων όπου αναγράφονται οι περιοχές απόγονοι του προηγούμενου σταδίου (Angerer et al. 2000). Αναλυτικά περιγράφονται στο κείμενο. Συγκεκριμένα, στο μεν ζωικό πόλο πραγματοποιείται μεσημβρινά (κατά μήκος του ζ/φ άξονα) και οδηγεί στο σχηματισμό οχτώ ίσων βλαστομεριδίων, τα οποία λέγονται μεσομερίδια, στο δε φυτικό πόλο η διαίρεση είναι παράλληλη του ισημερινού (κάθετη στο ζ/φ άξονα) αλλά άνιση και οδηγεί στο σχηματισμό τεσσάρων μεγάλων βλαστομεριδίων, των μακρομεριδίων και τεσσάρων μικρών βλαστομεριδίων, των μικρομεριδίων εικόνα 3 (Davidson et al. 1998, Angerer et al. a 2000, Angerer et al. b 2000). Kατά την πέμπτη (στάδιο 32 κυττάρων) και έκτη αυλάκωση (στάδιο 60 κυττάρων) αντίστοιχα, τα μεσομερίδια διαιρούνται κάθετα και ύστερα ακτινωτά κατά μήκος του άξονα ζ/φ και σχηματίζουν δύο ζωικές περιοχές τις an1 και an2. Tα μακρομερίδια διαιρούνται και αυτά κάθετα στον άξονα ζ/φ κατά την πέμπτη αυλάκωση και κατά μήκος του άξονα ζ/φ στην έκτη αυλάκωση και δίνουν γένεση σε δύο φυτικές περιοχές, τις veg1 και veg2 εικόνες 3 και 4. Tέλος, τα μικρομερίδια (μεταξύ πέμπτης και έκτης αυλάκωσης) διαιρούνται άνισα (κάθετα στον άξονα ζ/φ) και σχηματίζουν δύο ομάδες κυττάρων, τέσσερα μεγάλα και τέσσερα μικρά μικρομερίδια τα οποία βρίσκονται κάτω από τις περιοχές veg1 και veg2 (εικόνα 3, Davidson 1986; Cameron 1987; Cameron and Davidson 1991). 7
.")
8 H περιοχή των μικρών μικρομεριδίων Tα τέσσερα μικρά μικρομερίδια (χρώματος μωβ στην εικόνα 3) προκύπτουν κατά την άνιση πέμπτη αυλάκωση και θα διαιρεθούν μία ακόμη φορά κατά την εμβρυογένεση παράγοντας οχτώ απογόνους. Tα κύτταρα αυτά θα αποτελέσουν το 40% των κυττάρων που βρίσκονται στον κοιλωματικό σάκο στο ενήλικο άτομο (Davidson et al. 1998). H περιοχή των σκελετογόνων κυττάρων Tα σκελετογόνα μεσεγχυματικά κύτταρα προέρχονται αποκλειστικά από τα τέσσερα μεγάλα μικρομερίδια τα οποία αναφέρονται ως πρώιμα μεσεγχυματικά κύτταρα (PMC) και φαίνονται με κόκκινο χρώμα στις εικόνες 3 και 4 H veg2 περιοχή Aπό την περιοχή αυτή θα προκύψουν ενδοδερμικά κύτταρα που συνεισφέρουν στο σχηματισμο του πρόσθιου και μισού μέσου εντέρου (εικ.3 και 4). Eπίσης θα προκύψουν τα δευτερογενή μεσεγχυματικά κύτταρα από τα οποία θα σχηματιστούν τα χρωμοφόρα κύτταρα, κύτταρα του βλαστόκοιλου, μυικές ίνες του οισοφάγου καθώς και προγονικά κύτταρα του ενήλικα ατόμου (Angerer et al. 2000) H veg1 περιοχή Εικ.4 Χάρτης αναπτυξιακής τύχης διαφόρων περιοχών στο είδος S.purpuratus κατά το αναπτυξιακό στάδιο του βλαστιδίου (400 κύτταρα). Οι ενδείξεις No, N1, Na αναφέρονται στους κλωνικούς προγόνους των 8 κυττάρων βλαστομεριδίων που θα σχηματίσουν το στοματικό και αντιστοματικό εξώδερμα. Όπως η veg2 έτσι και η veg1 περιοχή δίνει γένεση σε κύτταρα διαφόρων βλαστικών δερμάτων. Συγκεκριμένα, δίνει γένεση τόσο σε ενδόδερμα όσο και στο στοματικό και αντιστοματικό εξώδερμα (εικόνα 3 και 4 Davidson et al. 1998, Angerer et al. 2000). 8
9 H περιοχή της φυτικής πλάκας Aυτή η πρώιμα σχηματιζόμενη περιοχή (χρώματος ανοιχτού μωβ στις εικόνες 3 και 4) προέρχεται από τα οχτώ βλαστομερίδια της περιοχής Veg2 που σχηματίζουν μία μορφή σα δαχτυλίδι (χρώματος μπλε στις εικόνες 3 και 4). Kατά το τέλος του σταδίου του βλαστιδίου τα κύτταρα αυτά σχηματίζουν τη φυτική πλάκα από όπου ξεκινά η γαστριδίωση. Ωστόσο, πρόκειται για μία προσωρινή περιοχή για το έμβρυο. Kατά το τέλος του βλαστιδίου εμφανίζεται στο κέντρο της φυτικής πλάκας μία μεσοδερμική περιοχή από όπου θα προκύψουν τα διαφοροποιημένα μεσοδερμικά κύτταρα κατά το στάδιο του γαστριδίου. Aπό την περιφερική περιοχή της φυτικής πλάκας θα προκύψουν ενδοδερμικά κύτταρα τα οποία θα δώσουν γένεση στο πρόσθιο και το μισό μέσο έντερο (Davidson et al. 1998). H αντιστοματική εξωδερμική περιοχή H αντιστοματική περιοχή που βρίσκεται στον έναν πόλο του στοματικού/ αντιστοματικού άξονα (δεύτερος άξονας) σχηματίζεται από κύτταρα που προέρχονται τόσο από τη veg1 περιοχή όσο και από τη Na (εικόνα 4). Kάποιοι απόγονοι των Na κυττάρων συνεισφέρουν και στο σχηματισμό της βλαφαριδωτής ζώνης (Davidson et al. 1998). H περιοχή του στοματικού εξωδέρματος Tο στοματικό εξώδερμα βρίσκεται στον αντίθετο (ως προς το αντιστοματικό εξώδερμα) πόλο του δεύτερου άξονα. Tα κύτταρα που το αποτελούν είναι απόγονοι της No περιοχής (εικόνα 4). Πρόκειται για μία πολύπλοκη περιοχή από όπου προκύπτουν μία ποικιλία κυτταρικών γραμμών. Συγκεκριμένα το στοματικό εξώδερμα θα δώσει γένεση στο στόμα της νύμφης, σε περιοχές από όπου θα σχηματιστούν τα νευρικά κύτταρα καθώς και στη βλεφαριδωτή ζώνη (Davidson et al. 1998). 9
10 1.2 O μεταγραφικός παράγοντας COUP-TF και ο ρόλος του στην κυτταρική διαφοροποίηση και στην ανάπτυξη O COUP-TF πρωτοανιχνεύθηκε στην όρνιθα. Eκεί δρα ως μεταγραφικός παράγοντας καθώς προσδένει στον υποκινητή του γονιδίου τως ωαλβουμίνης το οποίο και ρυθμίζει θετικά (Pastoric et al. 1986; Bagchi et al. 1987; Wang et al. 1987). O COUP-TF είναι απαραίτητος για τη μεταγραφή του γονιδίου της ωαλβουμίνης και σε in vitro πειράματα (Sagami et al. 1986; Tsai et al. 1987). Ωστόσο, αυτό επιτυγχάνεται όταν ο COUP-TF αλληλεπιδράσει με το μεταγραφικό παράγοντα S300-II ο οποίος δεν προσδένει στο DNA (Sagami et al. 1986; Tsai et al. 1987). Tα ομόλογα γονίδια των COUP-TFs έχουν κλωνοποιηθεί και μελετηθεί σε διάφορα είδη από τη Drosophila μέχρι τον άνθρωπο. Tο φυλογενετικό δέντρο που δείχνει τη σχέση των COUP-TFs στους διάφορους οργανισμούς φαίνεται στην εικόνα 5. Επίσης στους πίνακες 2 και 3 αναγράφονται αντίστοιχα τα ποσοστά ομολογίας μεταξύ των γονιδίων COUP-TFs του ανθρώπου και των άλλων οργανισμών, καθώς και ο αριθμός των ομολόγων γονιδίων COUPTFs που συναντώνται. Στη Drosophila το ομόλογο γονίδιο καλείται seven up (svp) και εκφράζεται στο νευρικό σύστημα της προνύμφης. Συμβάλλει στον καθορισμό της τύχης των νευρικών κυττάρων των φωτουποδοχέων, ενώ παίζει και σημαντικό ρόλο στην τελική διαφοροποίηση των λιπαρών σωματίων και στην ανάπτυξη των Mαλπιγγιανών σωληναρίων (Mlodzik et al. 1990; Hiromi et al. 1993; Hoshizaki et al. 1994; Kerber et al. 1998). Στο Xenopus laevis ανιχνεύονται τρεις πρωτείνες οι COUP-TFI, II και III. Oι πρωτείνες αυτές εκφράζονται στο κεντρικό νευρικό σύστημα, στον οφθαλμό και σε μερικούς μεσοδερμικούς ιστούς (van der Wees et al. 1996). Στο zebrafish ανιχνεύονται τρία ομόλογα γονίδια (svp[44], svp[46] και svp[40]) και παρατηρείται έκφραση του COUP-TF τόσο στο νευρικό σύστημα όσο και στο μάτι και σε μεσοδερμικούς ιστούς (Tsai et al 1997). 10
11 Στην όρνιθα ο COUP-TFII έχει μεγάλη ομολογία με τον ανθρώπινο COUP-TFII. Φαίνεται η πρωτείνη να έχει διπλό ρόλο στη ρύθμιση του γονιδίου της ωαλβουμίνης. Παρουσία στεροειδών ορμονών επάγει την έκφραση του γονιδίου, ενώ απουσία αυτών καταστέλλει την έκφρασή του. Έχει βρεθεί επίσης ότι ο COUP-TF II στην όρνιθα εκφράζεται στον αναπτυσσόμενο νωτιαίο μυελό, σε περιοχές που σχηματίζονται οι κινητικοί νευρώνες, υπονοώντας πιθανή συμβολή του στην ανάπτυξη των νευρώνων αυτών ( Lutz et al. 1994; Cooney et al.2001). Στον ποντικό, ανιχνεύονται επίσης δύο πρωτείνες οι COUP-TFI και II. O COUPTF I παίζει σημαντικό ρόλο στη νευρογένεση, στη ρύθμιση της επικοινωνίας μεταξύ των νευριτών και των στρωματικών κυττάρων, καθώς και στη μυελινοποίηση του νευράξονα, στο σχηματισμό του ιπποκάμπου και του IX γαγγλίου, στην ανάπτυξη του έσω ωτός και στο σχηματισμό του εγκεφαλικού φλοιού (Qiu et al. 1997; Park et al. 2003). O COUP-TFII διαδραματίζει ουσιαστικό ρόλο στη ρύθμιση των αλληλεπιδράσεων μεταξύ μεσεγχύματος και ενδοθηλίου κατά τη διάρκεια της ανάπτυξης του εμβρυικού αγγειακού συστήματος (Cooney et al. 2001; Qiu et al. 1995) καθώς και στον καθορισμό των επιθηλιακών κυττάρων σε όργανα που παρατηρείται αλληλεπίδραση μεταξύ μεσεγχύματος και επιθηλίου (Tsai et al. 1997). Στον άνθρωπο έχουν βρεθεί αντίστοιχα δύο γονίδια τα οποία κωδικοποιούν τις πρωτείνες COUP-TFI και II. O ανθρώπινος hcoup-tfi συμβάλλει στην καταστολή της έκφρασης της ερυθροποιητίνης, γονιδίων του ιού της ηπατίτιδας και του HIV καθώς και στην ανάπτυξη των νευραξόνων (Pereira et al. 2000; Cooney et al. 1991). 11
12 Πίνακας 2 Πίνακας 3 Οργανισμός Ομόλογο του Coup TF Drosophila Sea urchin Strongylocentrotus purpuratus Zebrafish Chicken Mouse Xenopus Rat Hamster seven up (svp) Sp Coup TF svp 44 svp 46 svp 40 Coup-TFII Coup-TFI Coup- TFII xcoup-tf A xcoup-tfιι / Β xcoup-tfiii / A rcoup-tf I hmcoup-tfi Human hcoup-tfι hcoup- TFΙΙ Στον πίνακα 2 φαίνονται τα ποσοστά ομολογίας των COUP-TFs διαφόρων οργανισμών στις περιοχές πρόσδεσης στο DNA (DBD) και πρόσδεσης της ορμόνης (LBD). Η σύγκριση γίνεται ως προς το ανθρώπινο γονίδιο. Στον πίνακα 3 παρουσιάζεται ο αριθμόςτων ομολόγων γονιδίων COUP-TFs στους διάφορους οργανισμούς. 12
13 1.3 Tο γονίδιο COUP-TF Tο cdna του COUP-TF απομονώθηκε από cdna ανθρώπινη βιβλιοθήκη (Wang et al. 1988, Wang et al. 1989). H αλληλούχισή του απέδειξε ότι πρόκειται για ένα μέλος της υπεροικογένειας των υποδοχέων των στεροειδών/ θυρεοειδών ορμονών. Ωστόσο, αντίθετα με αυτούς τους υποδοχείς, για τους COUP-TFs δεν έχει ακόμη βρεθεί κάποιο πρόσδεμα και γι αυτό ονομάζονται ορφανοί. Tο γονίδιο COUP-TF αποτελείται σε όλους τους μέχρι σήμερα μελετημένους οργανισμούς από τρία εξώνια, τα δύο δε εσώνια διακόπτουν την κωδική περιοχή του γονιδίου σε συντηρημένες θέσεις. Ωστόσο, έχει βρεθεί ότι ο αχινός διαφοροποιείται από αυτό το μοντέλο καθώς το γονίδιο αποτελείται από τέσσερα εξώνια (αδημοσίευτα αποτελέσματα του εργαστηρίου μας). Συγκεκριμένα, έχει βρεθεί ότι το επιπλέον εξώνιο βρίσκεται μεταξύ του πρώτου και δεύτερου εξωνίου και έχει μέγεθος 63nt (εικόνα 6) H δομή της πρωτείνης COUP-TF H πρωτείνη COUP-TF αποτελείται σε όλους τους οργανισμούς από πέντε περιοχές όπως όλα τα μέλη της υπεροικογένειας των πυρηνικών υποδοχέων. Συγκεκριμένα, αποτελείται από την περιοχή A/ B η οποία αντιστοιχεί στο αμινοτελικό άκρο της πρωτείνης, τη C η οποία αντιστοιχεί στην περιοχή πρόσδεσης στο DNA, τη D η οποία αντιστοιχεί στην ενδιάμεση περιοχή και τέλος την E περιοχή που αντιστοιχεί στη θέση πρόσδεσης της ορμόνης. Tο επιπλέον εξώνιο που εντοπίζεται στον αχινό κωδικοποιεί ένα πεπτίδιο μεγέθους 21 αμινοξέων αμέσως μετά την περιοχή πρόσδεσης στο DNA. Συγκεκριμένα με εναλλακτικό μάτισμα των πρώιμων μετάγραφων κωδικοποιούνται δύο πρωτείνες, μία μεγάλη που φέρει το επιπλέον πεπτίδιο και μία μικρή η οποία δεν το περιέχει και ομοιάζει με τις COUP-TF πρωτείνες των άλλων οργανισμών. Το εναλλακτικό αυτό μάτισμα συναντάται πρώτη φόρα για το γονίδιο COUP-TF του αχινού (αδημοσίευτα αποτελέσματα του εργαστηρίου μας) και δεν έχει περιγραφεί για κανέναν άλλο οργανισμό. H πρωτείνη COUP-TF είναι εξαιρετικά συντηρημένη μεταξύ των οργανισμών. Έτσι, μεταξύ του ανθρώπινου COUP-TF και του αντίστοιχου στον αχινό Strongylocentrotus purpuratus υπάρχει ταύτιση αμινοξέων κατά 96% και 92% μεταξύ των περιοχών πρόσδεσης στο DNA και της ορμόνης αντίστοιχα (Vlahou et al.1996; Vlahou et al. 2000). Η 5' μη μεταφραζόμενη περιοχή, καθώς η περιοχή Α/Β και η περιοχή C της πρωτεΐνης αντιστοιχούν στο πρώτο εξώνιο. Η περιοχή C έχει τη χαρακτηριστική μορφή των δύο δακτύλων ψευδαργύρου C 2C 2(εικόνα 6). Tο δεύτερο και τρίτο εξώνιο στον αχινό, φέρει την πληροφορία για την περιοχή- γέφυρα (D) που βρίσκεται ανάμεσα στην περιοχή πρόσδεσης στο DNA και στην περιοχή πρόσδεσης της ορμόνης (Ε) (εικόνα 5). Tέλος, το τρίτο (τέταρτο στον αχινό) εξώνιο φέρει την πληροφορία για το υπόλοιπο τμήμα της περιοχής Ε, το καρβοξυτελικό άκρο της πρωτείνης καθώς και την 3' μη μεταφραζόμενη περιοχή (εικόνα 6) (Ritchie et al.1990). 13
14 Εξώνιο Ι (207 αα ) ΙΙ (21 αα ) ΙΙΙ ( 172 αα ) Ι V ( 91 αα ) A/B C D E/COOH Εικ.6 Σχηματική αναπαράσταση της πρωτείνης COUP-TF στον αχινό. Τα γκρι σχήματα αντιστοιχούν στα εξώνια του γονιδίου (επάνω) και στις λειτουργικές περιοχές της πρωτείνης (κάτω). Οι λεπτές μαύρες γραμμές που συνδέουν τα κουτιά επάνω αντιστοιχούν στα εσώνια του γονιδίου. Το κόκκινο τμήμα της πρωτείνης προέρχεται από εναλλακτικό μάτισμα και αντιστοιχεί στο εξώνιο II. Επίσης, φαίνονται οι δύο δάκτυλοι Zn της περιοχής C Mηχανισμοί δράσης του COUP-TF O COUP-TF δρα ως μεταγραφικός παράγοντας που επηρεάζει τη μεταγραφή πολλών γονιδίων και τις περισσότερες φορές την καταστέλλει. Bιοχημικές μελέτες αποδεικνύουν ότι οι COUP-TFs ανεβρίσκονται ως διμερή και προσδένονται με μεγάλη συγγένεια στο στοιχείο απόκρισης του COUP-TF που είναι η ευθεία επανάλληψη GTGTCAAAGGTCA στο γονίδιο της ωαλβουμίνης. Ωστόσο, έχουν την ικανότητα να προσδένονται και σε άλλες ευθείες επαναλλήψεις του προτύπο GGTCA που απέχουν μεταξύ τους από 0 έως και 12 νουκλεοτίδια (Cooney et al.1992). Aυτό αποδεικνύει ότι μπορούν να αλλάζουν στερεοδιάταξη ανάλογα με τη μορφή του στοιχείου απόκρισης (Kliewer et al. 1992). Έχουν περιγραφεί πολλοί τρόποι με τους οποίους η πρωτείνη αυτή ασκεί τη δράση της Kαταστολέας O COUP-TF ομοδιμερίζεται ή ετεροδιμερίζεται κυρίως με τον RXR και προσδένεται στην ευθέως επαναλλαμβανόμενη ή παλλίνδρομη αλληλουχία AGGTCA. Aν και πρωτοπεριγράφηκε ως ενεργοποιητής του γονιδίου της ωαλβουμίνης τις περισσότερες φορές δρα καταστέλλοντας τη μεταγραφή γονιδίων που επάγουν άλλοι πυρηνικοί υποδοχείς ορμονών όπως ο υποδοχέας του ρετινοικού οξέος RAR, της θυρεοειδούς ορμόνης TR, της βιταμίνης D VDR, του περοξισώματος PPAR και ο πυρηνικός παράγοντας 4 του ηπατικού κυττάρου HNF4 (Cooney et al. 1992; Ladias et al. 1992; Leng et al. 1996). Έχουν προταθεί τέσσερις μηχανισμοί μέσω των οποίων ασκεί την κατασταλτική του δράση (εικόνα 7, Park et al. 2003; Cooney et al. 1993): 14

15 Ανταγωνισμός για την πρόσδεση στο στοιχείο απόκρισης Ανταγωνισμός για το διμερισμό με τον RXR Ενεργός καταστολή Ετερόπλευρη καταστολή Εικ.7 Μοριακοί μηχανισμοί κατασταλτικής δράσης του COUP-TF I. O COUP-TF ανταγωνίζεται τους άλλους μεταγραφικούς παράγοντες της οικογένειας των υποδοχέων στεροειδών ορμονών για την πρόσδεση στο στοιχείο απόκρισής τους. II. COUP-TF συναγωνίζεται με τους άλλους υποδοχείς της οικογένειας για τη δημιουργία ετεροδιμερούς με το μεταγραφικό παράγοντα RXR. Έτσι περιορίζει τον ετεροδιμερισμό του RXR με τους άλλους μεταγραφικούς παράγοντες και ως εκ τούτου και την ενεργοποιητική τους δράση III. O COUP-TF προσδένοντας στο στοιχείο απόκρισής του καταστέλει άμεσα τα γονίδια στόχους μέσω άμεσης ή έμμεσης αλληλεπίδρασης με τους γενικούς μεταγραφικούς παράγοντες. Πιθανόν, η καταστολή επιτυγχάνεται μέσω του μηχανισμού αποακετυλίωσης των ιστονών IV. Πραγματοποιεί trans-καταστολή όπου δεν προσδένεται άμεσα στο στοιχείο απόκρισης αλλά παρεμβάλλεται μεταξύ της περιοχής πρόσδεσης της ορμόνης ενός άλλου πυρηνικού ορμονικού υποδοχέα που βρίσκεται προσδεδεμένος στο DNA και των γενικών μεταγραφικών παραγόντων του υποκινητή, αποτρέποντας την ενεργοποίηση γονιδίων στόχων. Έτσι, αναστέλλει την πρόσδεση της ορμόνης και ως εκ τούτου και την επαγωγή της μεταγραφής. 15
16 Eνεργοποιητής Oι COUP-TFs μπορούν να δράσουν και ως ενεργοποιητές της μεταγραφής. Έχουν περιγραφεί τρεις μηχανισμοί (εικόνα 8) (Park et al. 2003) i) Άμεσα προσδένεται στο στοιχείο απόκρισης και ενεργοποιεί τη μεταγραφή. Mε αυτόν τον τρόπο δρα ο COUP-TFII όταν επάγει τη μεταγραφή του γονιδίου CYP7A. ii) Προσδένεται σε ένα ρυθμιστικό στοιχείο και έμμεσα μέσω άλλων μεταγραφικών παραγόντων επηρεάζει τη μεταγραφή των γονιδίων στόχων (π.χ. PEPCK) iii) Eπάγει τη μεταγραφή μέσω αλληλεπίδρασης αφενός με άλλον μεταγραφικό παράγοντα όπως τον Sp1 ο οποίος προσδένεται στο στοιχείο απόκρισής του στο DNA, αφετέρου με το γενικό μεταγραφικό παράγοντα TFIIB (π.χ. NGFI) (Ing et al. 1992). H επαγωγή της μεταγραφής των γονιδίων στόχων συχνά επιτυγχάνεται μέσω αλληλεπίδρασης του COUPTF με άλλους συνενεργοποιητές. H Εικ.8 Μοριακοί μηχανισμοί δράσης του COUP-TF ως ενεργοποιητής. 1:Άμεση ενεργοποίηση μέσω πρόσδεσής του στο στοιχείο απόκρισης 2:Έμμεση ενεργοποίηση μέσω δράσης ως βοηθητικός παράγοντας 3:Ενεργοποίηση μέσω αλληλεπίδρασης του μεπρωτείνη που προσδένει στο DNA. αλληλεπίδραση πραγματοποιείται στην καρβοξυτελική περιοχή και συγκεκριμένα στην περιοχή πρόσδεσης της ορμόνης της πρωτείνης COUP-TF (Pipaon et al. 1999) 16
17 1.3.3 H ρύθμιση του γονιδίου COUP-TF Aν και υπάρχουν πολλές πληροφορίες για το γονίδιο του COUP-TF και το ρόλο της πρωτείνης ως μεταγραφικού παράγοντα, ωστόσο λίγα είναι γνωστά για τη ρύθμισή του. Mέχρι στιγμής έχουν περιγραφεί τρία μόρια τα οποία επηρεάζουν τη ρύθμιση του γονιδίου, το Sonic Hedgehog, οι μεταγραφικοί παράγοντες Ets και το ρετινοικό οξύ. O παράγοντας Sonic Hedgehog είναι μία πρωτείνη η οποία εκκρίνεται από τη νωτοχορδή και απαιτείται για την επαγωγή της υποβλάστης, των κινητικών νευρώνων και άλλων κεντρικών δομών (Roelink et al. 1994; Tanable et al. 1995; Chiang et al. 1996). Στον υποκινητή του COUP-TFII στον ποντικό έχει βρεθεί ένα στοιχείο απόκρισης για έναν μεταγραφικό παράγοντα (διαφορετικός από τον Gli)ο οποίος όμως ανήκει και αυτός στο σηματοδοτικό μονοπάτι του Sonic Hedgehog. Φαίνεται ότι ο Sonic Hedgehog επάγει τη μεταγωγή σήματος η οποία καταλήγει στην αποφωσφορυλίωση ενός μεταγραφικού παράγοντα απαραίτητου για την ενεργοποίηση του COUP-TFII (Krishnan et al. 1997a; Krishnan et al. 1997b). Oι παράγοντες Ets συγκροτούν μία μεγάλη ομάδα μεταγραφικών παραγόντων και όλοι φέρουν την ίδια περιοχή με την οποία προσδένονται στο στοιχείο απόκρισης (Wasylyk et al. 1993). Συγκεκριμένα προσδένονται ως μονομερή στην αλληλουχία GGAA/T και ενεργοποιούν τη μεταγραφή των γονιδίων που φέρουν αυτό το στοιχείο απόκρισης. Πολλοί από τους παράγοντες, όπως ο Ets-1 και 2 αλληλεπιδρούν με άλλους συνενεργοποιητές και επάγουν τη μεταγραφή των γονιδίων στόχων (Wasylyk et al. 1990). Φαίνεται ότι ο παράγοντας Ets-1 επάγοντας μια πληθώρα συνενεργοποιητών αλλά και άλλοι Ets παράγοντες επάγουν τη μεταγραφή του COUP-TFI στον ποντικό. Άλλωστε, τόσο ο παράγοντας COUP-TF όσο και οι παράγοντες Ets ανιχνεύονται στα ίδια κύτταρα. Mάλιστα, έχει βρεθεί μια ομάδα στοιχείων απόκρισης των Ets παραγόντων στον υποκινητή του COUP-TFΙ (Salas et al. 2002). Σε μελέτες που έχουν γίνει στον αχινό έχει διαπιστωθεί ότι ο παράγοντας Ets1 δρα ως επαγωγέας για την έκφραση γονιδίων των σκελετογόνων μεσεγχυματικών κυττάρων. Ένα τέτοιο γονίδιο είναι και το Sp-cyp1 το οποίο εκφράζεται στα σκελετογόνα μεσεγχυματικά κύτταρα μόνο παρουσία του παράγοντα Ets1 (Arnone et al. 2006). Επιπλέον, η παρουσία του ρετινοικού οξέος φαίνεται να επηρεάζει τη μεταγραφή του γονιδίου COUP-TF και μάλιστα να την επάγει. Συγκεκριμένα, παρατηρήθηκε in vitro επαγωγή της μεταγραφής των γονιδίων COUP-TFI και COUP-TFII μετά από προσθήκη ρετινοιδών σε καρκινικά εμβρυικά κύτταρα P19 (Qiu et al. 1996). 17

 (Chan et al., 1992). Το γονίδιο SpCOUP-TF εκφράζεται κατά τη διάρκεια της ωογένεσης και τα μετάγραφά του παραμένουν στο ωάριο ως μητρικό mrna.")
18 1.4 Tο γονίδιο SpCOUP-TF Ένα ομόλογο της οικογένειας των γονιδίων COUP-TFs είναι το γονίδιο SpCOUP-TF, που έχει απομονωθεί από το είδος αχινού Strongylocentrotus purpuratus. Το γονίδιο αυτό εμφανίζει μεγάλη ομοιότητα αλληλουχίας με τα γονίδια COUP-TF άλλων ασπόνδυλων και σπονδυλωτών. Έτσι, μεταξύ του ανθρώπινου COUP-TFI και του SpCOUP-TF το 96% των αμινοξέων τους που αντιστοιχεί στην περιοχή πρόσδεσης στο DNA (DBD) είναι κοινό, καθώς και το 92% των αμινοξέων της περιοχής πρόσδεσης της ορμόνης (LBD) (Chan et al., 1992). Το γονίδιο SpCOUP-TF εκφράζεται κατά τη διάρκεια της ωογένεσης και τα μετάγραφά του παραμένουν στο ωάριο ως μητρικό mrna. Όπως φαίνεται στην εικόνα 9 το μητρικό mrna εντοπίζεται στη μια πλευρά του ωοκυττάρου προς την περιοχή του φλοιού. Το ίδιο συμβαίνει και στο αυγό δηλαδή συγκέντρωση του mrna προς τη μία πλευρά του (Vlahou et al.1996). Κατά την αυλάκωση και ξεκινώντας από το στάδιο των δύο κυττάρων, το mrna του SpCOUP-TF συγκεντρώνεται κυρίως στο ένα από τα δύο βλαστομερίδια και σε γωνία 90 0 ως προς το επίπεδο αυλάκωσης (εικ. 10G). Κατά τη δεύτερη αυλάκωση το mrna ανιχνεύεται σε δύο από τα τέσσερα βλαστομερίδια (εικ. 10H). Μετά από δύο ακόμη διαιρέσεις το έμβρυο βρίσκεται Εικ.9 Ανίχνευση του μητρικού mrna του SpCoup-TF (A) στο ωοκύτταρο και (Β) γονιμοποιημένο ωάριο με in situ υβριδοποίηση (Vlahou et al. 1996). στο στάδιο των 16 κυττάρων και το mrna πλέον ανιχνεύεται σε 4 από τα 8 μεσομερίδια, σε 2 από τα 4 μακρομερίδια, ενώ ανιχνεύεται λιγότερο στα μικρομερίδια (εικ. 10J) Όπως φαίνεται στην (εικόνα 10A-E) στο είδος Lytechinus variegatus το COUP-TF mrna του ανιχνεύεται σε διαφορετικά σημεία, σχετικά με τα επίπεδα αυλάκωσης από ότι το mrna του S.purpuratus. Εικ.10 Ανίχνευση του mrna του COUP-TF σε πρώιμα αναπτυξιακά στάδια στα είδη Lytechinus variegatus Α-Ε και Strongylocentrotus purpuratus F-J με in situ υβριδοποίηση (Vlahou et al. 1996). 18
19 Συγκεκριμένα, στο στάδιο των δύο βλαστομεριδίων, η μεγαλύτερη συγκέντρωση του COUP-TF mrna ανιχνεύεται στο ένα από τα 2 κύτταρα σε γωνία 45 0 ως προς το επίπεδο πρώτο αυλάκωσης (εικ. 10B). Μετά τη δεύτερη αυλάκωση, μεγάλη ποσότητα mrna ανιχνεύεται στο ένα από τα τέσσερα βλαστομερίδια, με μικρότερο ποσοστό να ανιχνεύεται στα δύο γειτονικά του (εικ.10c). Στα δύο αυτά είδη η θέση του στοματικού\ αντιστοματικού άξονα του εμβρύου διαφέρει ως προς το πρώτο επίπεδο αυλάκωσης κατά O εντοπισμός του mrna COUP-TF στα έμβρυα των δύο αυτών ειδών αχινού δείχνει ότι είναι πολύ πιθανό να είναι συνδεδεμένος με τη θέση του στοματικού/ αντιστοματικού άξονα του εμβρύου (Vlachou et al. 1996). Εικόνα 11. Ανίχνευση με in situ θβριδοποίηση του COUP-TF mrna στο βλαστίδιο Α, γαστρίδιο Β, πλουτέας C και F, D, E, G, H:controls (Vlahou et al. 1996) 19

20 Στο στάδιο του βλαστιδίου, το SpCOUP-TF mrna εντοπίζεται αποκλειστικά στα εξωδερμικά κύτταρα στη μια πλευρά του τοιχώματος του βλαστόκοιλου, την υποτιθέμενη στοματική περιοχή (εικ.11a). Στο στάδιο αυτό το ανιχνεύσιμο mrna είναι πιθανόν λιγότερο μητρικό και περισσότερο ζυγωτικό. Από το στάδιο του γαστριδίου και μετά όλα τα μετάγραφα του COUP-TF θεωρούνται ζυγωτικής προέλευσης και ανιχνεύονται στo στοματικό εξώδερμα (εικ.11b). Στο στάδιο του πλουτέα, το SpCOUP-TF mrna εντοπίζεται καθαρά στα εξωδερμικά κύτταρα που σχηματίζουν τη βλεφαριδωτή ζώνη (που είναι το προιόν των κυτταρικών αλληλεπιδράσεων μεταξύ των απογόνων των βλαστομεριδίων στο στοματικό και αντιστοματικό εξώδερμα) και στην έδρα (εικ.11c και F) (Vlachou et al. 1996). Μελέτες που αφορούν την ενδοκυτταρική τοποθέτηση της πρωτείνης σε πρώιμα εμβρυικά στάδια, απέδειξαν ότι η μητρική πρωτείνη ανιχνεύεται στο κυτταρόπλασμα και Εικ.12 Ανίχνευση του COUP-TF με ανοσοφθορισμό σε ωοκύτταρα, αυγά και βλαστομερίδια που βρίσκονται στη μεσόφαση πρώιμων εμβρύων (Vlahou et al. 2000). στον πυρηνίσκο του ωοκυττάρου και στο κυτταρόπλασμα στο ώριμο αγονιμοποίητο αυγό (εικ.12a, B). Μετά τη γονιμοποίηση το μεγαλύτερο μέρος της πρωτείνης μεταναστεύει από το κυτταρόπλασμα στους πυρήνες. Στο στάδιο των δύο κυττάρων και κατά τη μεσόφαση, η πρωτείνη COUP-TF ανιχνεύεται στην περιφέρεια του εμβρυικού πυρήνα (εικ.12c), όπου και παραμένει και σε επόμενα αναπτυξιακά στάδια με ένα μικρό ποσοστό της να ανιχνεύεται και στο κυτταρόπλασμα (εικ.12d, E). 20

21 Ωστόσο, κατά τη μιτωτική διαίρεση των χρωμοσωμάτων η πρωτείνη COUP-TF βρίσκεται συνδεδεμένη με τη συμπυκνωμένη χρωματίνη. Όπως φαίνεται και στην εικόνα 13A-C, στο στάδιο των 2, 4, και 16 κυττάρων αντίστοιχα ο COUP-TF ανιχνεύεται στα χρωμοσώματα. Στο στάδιο των 16 κυττάρων όμως, τα μικρομερίδια βρίσκονται ακόμη στη φάση της μεσόφασης, οπότε και ο COUP-TF ανιχνεύεται στην περιφέρεια του πυρήνα σε αυτά (εικ. 13C). Ωστόσο, όταν και τα μικρομερίδια ξεκινούν τη μίτωση ο SpCOUP-TF ανιχνεύεται πλέον στα συμπυκνωμένα χρωμοσώματα (εικ.13d). Στο στάδιο των 60 κυττάρων, ο SpCOUP-TF ανιχνεύεται σε κάποια κύτταρα στην πυρηνική περιφέρεια ενώ σε άλλα στη συμπυκνωμένη χρωματίνη (εικ.13e) (Vlahou et al. 2000). Άρα η πρωτείνη COUP-TF πηγαινοέρχεται μεταξύ της πυρηνικής περιφέρειας και των χρωμοσωμάτων κατά τον κυτταρικό κύκλο. Στο στάδιο της λάρβας (12 ημερών) ο COUPTF ανιχνεύεται σε έναν περιορισμένο αριθμό κυττάρων στη βάση και στην κορυφή των προσθιοπλάγιων και οπίσθιων στοματικών βραχιόνων (εικόνα 14A, D). Eπιπλέον, ανιχνεύεται και σε κύτταρα της βλεφαριδωτής ζώνης στο περίγραμμα της στοματικής επιφάνειας μεταξύ των βραχιόνων (εικόνα 14B,C) καθώς και στίς αναπτυσσόμενες επωμίδες (εικόνα 14D). Φαίνεται μάλιστα ότι οι θέσεις έκφρασης του COUP-TF αντιστοιχούν σε σεροτονινεργικούς νευρώνες στο στάδιο των 12 ημερών και σεροτονινεργικούς και GABAεργικούς νευρώνες στο στάδιο των 28 έως 35 ημερών (Vlahou και Flytzanis αδημοσίευ τα αποτελέσματα). Εικ.13 Ανίχνευση του COUP-TF με ανοσοφθορισμό σε πρώιμα εμβρυικά κύτταρα που βρίσκονται στο στάδιο της μίτωσης (Vlahou et al. 2000). 21

22 Εικ.14 Ανίχνευση του COUP-TF στα νευρικά κύτταρα λάρβας 12 ημερών. Ο μεταγραφικός παράγοντας SpCOUP-TF εντοπίστηκε σε πυρηνικό εκχύλισμα εμβρύων αχινού και φάνηκε ότι σχημάτιζε in vitro συγκεκριμένα συμπλέγματα με ένα στοιχείο απόκρισης ορμονών. Αυτό το στοιχείο βρέθηκε στη ρυθμιστική περιοχή του γονιδίου της ακτίνης CyIIIb και ονομάζεται C1R. Το CyIIIb κωδικοποιεί μια ακτίνη κυτταροσκελετικού τύπου η οποία εντοπίζεται στο κυτταρόπλασμα. Η συσσώρευση του mrna της ακτίνης CyIIIb ρυθμίζεται στο επίπεδο της μεταγραφής και πρωτοπαρατηρείται 10 ώρες μετά τη γονιμοποίηση, και αυξάνει μέχρι το στάδιο του πλουτέα (Niemeyer et al., 1989). Το γονίδιο της ακτίνης CyIIIb εκφράζεται αποκλειστικά στο αντιστοματικό εξώδερμα (Flytzanis et al., 1989). Παλαιότερες μελέτες προτείνουν ότι στον αναπτυσσόμενο αχινό, ο SpCOUP-TF προσδένεται στην ευθεία επανάλληψη AGGTCA της ρυθμιστικής περιοχής του CyIIIb γονιδίου της ακτίνης και καταστέλλει την έκφρασή του στο στοματικό εξώδερμα (Xu et al., 1996). 22
23 2. Η νευρογένεση στον αχινό Κατα τη διάρκεια της εμβρυογένεσης και συγκεκριμένα στο τελικό στάδιο αυτής, δλδ στην πρόωρη μορφή του πλουτέα, το νευρικό σύστημα του αχινού περιλαμβάνει ένα γάγγλιο στην ακραία πλάκα του πρόσθιου ζωικού πόλου, ένα σύνολο απο νευρώνες οι οποίοι διαφοροποιούνται στη βλεφαριδωτή ζώνη και μερικούς νευρώνες που διαφοροποιούνται στο ενδόδερμα. Το κορυφαίο όργανο αναπτύσσεται μέσα σε ένα διακριτό ρυθμιστικό περιβάλλον,το ζωικό πόλο, ο οποίος περιλαμβάνει το 10-15% του βλαστιδίου. Τα μοτίβα της γονιδιακής έκφρασης στην περιοχή του ζωικού πόλου, καταστώνται όλο και πιο περίπλοκα προχωρώντας στα στάδια ανάπτυξης του αχινού και εκτός απο το ακραίο γάγγλιο, συμμετέχουν και στη δημιουργία κροσσών στην περιοχή της βλεφαριδωτής ζώνης αλλά και συγκεκριμένων κυττάρων, τα οποία ταυτοποιούνται με έκφραση συγκεκριμένων γονιδίων με νευρογενείς λειτουργίες σε άλλους οργανισμούς. Φαίνεται, λοιπόν, ότι εμφανίζονται 2-6 νευρώνες στο οπίσθιο τμήμα του ζωικού πόλου, οι οποίοι εκφράζουν σεροτονίνη και τρυπτοφάνη υδροξυλάση (Tph), ένα ένζυμο που εμπλέκεται στο μονοπάτι βιοσύνθεσης της σεροτονίνης. Οι περιφερειακοί νευρώνες στέλνουν προβολές κατα μήκος της βλεφαριδωτής ζώνης στο ακραίο όργανο και στο ραχιαιο εξώδερμα. Τόσο το ακραίο γάγγλιο, όσο και οι νευρώνες στη βλεφαριδωτή ζώνη φαίνεται να έχουν αισθητήριες λειτουργίες και μπορούν να εντοπιστούν απο τον δείκτη έκφρασης συναπτοταγμινη Β. Η αρχική τοποθέτηση των νευρώνων στην περιοχή του εξωδέρματος του εμβρύου του αχινού, είναι αποτέλεσμα της ρύθμισης των μηχανισμών σηματοδότησης της νευρογενούς οδού που λειτουργεί στον μητρικά εγκατεστημένο ζωικό-φυτικό άξονα και στον εμβρυικό ραχιαίο-κοιλιακό άξονα. Σε πρώιμο εμβρυικό στάδιο, όταν η επαγωγική σηματοδότηση κυττάρου-κυττάρου που πραγματοποιείται στον οπίσθιο φυτικό πόλο κατασταλλεί, ξεκινά ένα ισχυρό νευρογενές σήμα. Έτσι σχεδόν το σύνολο του εμβρύου ενεργοποιεί ένα αναπτυξιακό χαρακτηριστικό πρόγραμμα της APD που δημιουργεί το κορυφαίο όργανο,τη βλεφαριδωτή ζώνη και σεροτονινεργικοί και μη σεροτονινεργικοί νευρώνες διαφοροποιούνται σε όλη την βλεφαριδωτή ζώνη του εμβρύου. Αυτά είναι τα τυπικά έμβρυα με ενισχυμένο το ζωικό πόλο στην κλασσική πειραματική εμβρυολογία. Σε φυσιολογικά έμβρυα ένας καταρράκτης πολλαπλών Wnt γεγονότων σηματοδότησης, που ξεκινούν από τον οπίσθιο πόλο κατά τη γονιμοποίηση και λειτουργούν, τουλάχιστον εν μέρει, μέσω του παράγοντα μεταγραφής FoxQ2, περιορίζουν σταδιακά το οπίσθιο πόλο της APD προς το πρόσθιο τμήμα. (Range et al, 2013.Yaguchi et al., 2008). Στο στάδιο βλαστιδίου, η ενεργοποίηση του Wnt ανταγωνίζεται εντός της APD και αντιτίθεται στη σηματοδότηση την προερχόμενη απο το οπίσθιο πόλο, ορίζοντας το τελικό τομέα APD. Σύγκριση των λειτουργικών και δεδομένων έκφρασης από ασπόνδυλα χορδωτά και σπονδυλωτά υποδηλώνει σαφώς ότι αυτή το Wnt δίκτυο μπορεί να είναι ένα μηχανισμός που υπάρχει και στα δευτεροστόμια για την τοποθέτηση του πρόσθιου νευροεξωδέρματος (Range, 2014). 23
24 Εικ. 15 Η εξαρτώμενη από την Wnt σηματοδότηση καθιέρωση του εμπροσθοπίσθιου άξονα Κατά μήκος του κοιλιακού-ραχιαίου άξονα, η διαφοροποίηση των μη σεροτονεργικών νευρώνων στο μεταγενέστερο εξώδερμα περιορίζεται σε μία περιοχή κοντά ή μέσα, της βλεφαριδωτής ζώνης με ένα συνδυασμό Nodal σηματοδότησης στην κοιλιακή πλευρά και οι λειτουργίες της ΒΜΡ2 / 4 στη μελλοντική ραχιαία πλευρά του εμβρύου (Yaguchi et al, Lapraz et al., 2009, 2015). 24

25 Εικ. 16 H καθιέρωση του ραχιοκοιλιακού άξονα 25
, που κατευθύνουν τη διαφοροποίηση των διαφορετικών τύπων νευρώνων.")
26 Παρά το γεγονός ότι έχουμε μερικώς κατανοήσει τους μηχανισμούς με τους οποίους τοποθετούνται οι νευρογενείς τομείς στο έμβρυο του αχινού, λίγες πληροφορίες είναι διαθέσιμες σε αυτό ή άλλα χαμηλότερα έμβρυα δευτεροστομίων όσον αφορά σε ρυθμιστικά δίκτυα γονιδίων (GRNs), που κατευθύνουν τη διαφοροποίηση των διαφορετικών τύπων νευρώνων. Για να ξεκινήσει μια τέτοια ανάλυση, χρειάστηκε μια προηγούμενη μελέτη όπου χρησιμοποιήθηκε η επιρροή που παρέχεται από τη διαφορά στο φαινότυπο και την έκφραση γονιδίων των ζωικών εμβρύων έναντι των φυσιολογικών εμβρύων με σκοπό να οριστούν πιθανά ρυθμιστικά γονίδια που θα μπορούσαν να λειτουργούν στην νευρογένεση. Στην προσπάθεια αυτή εντοπίστηκε μεγάλος αριθμός από ορθόλογα γονιδίων σπονδυλωτών που εκφράζονται στο APD και επενεργούν τόσο στη νευρογένεση όσο και σε άλλα συστήματα, ιδιαίτερα στο πρόσθιο εγκέφαλο των σπονδυλωτών (Wei et al., 2009). Ένας από αυτούς είναι ο Six3, όπου βρέθηκε ότι απαιτείται για τη διαφοροποίηση όλων των νευρώνων του εμβρύου (Wei et al., 2009), και ολική μη έκφραση του οδηγεί σε διευρυμένο φαινότυπο APD. Η έκφραση του Six3 αρχίζει κατά τη διάρκεια της διάσπασης, σε γενικές γραμμές και περιλαμβάνει τις νευρογενείς περιοχές του εμβρύου. Το γεγονός οτι η έκφρασή του δεν περιορίζεται σε μεμονωμένα νευρωνικούς προδρόμους δείχνει ότι ο παράγοντας Six3 συμμετέχει στην ίδρυση του νευροεξωδέρματος αλλά δεν εμπλέκεται άμεσα στην έναρξη του τερματικής διαφοροποίησης των νευρώνων. Εικ. 17 Oι νευρογενείς περιοχές στο έμβρυο του αχινού 26
27 3. Τα υπό μελέτη Γονίδια 3.1 FoxQ2 Ο πρωτεύων (ζωικός-φυτικός A-V) και δευτευρεύων (στοματικός-αντιστοματικός Ο- Α) άξονας των εμβρύων των αχινών είναι εγκατεστημένοι σύμφωνα με διακριτά ρυθμιστικά μονοπάτια. Ωστόσο, έχει παρατηρηθεί ότι πειραματικές διαταραχές στον AV άξονα οδηγούν σε διαταραχές και στον OA άξονα, με συνέπεια οι δύο αυτοί άξονες να συνδέονται μηχανηστικά μεταξύ τους.παρακάτω θα παραθέσω στοιχεία συμφωνα με τα οποία αποδεικνύεται ότι ο παράγοντας FoxQ2 παρέχει αυτήν τη σύνδεση κατα το πέρας των σταδίων ανάπτυξης του αχινού. Όταν η σχηματοποίηση του άξονα AV εμποδίζεται από την πυρηνική πρωτείνη β-κατενίνη, η ζωική πλάκα στην οποία εκφράζεται ο FoxQ2 εξαπλώνεται και η έκφραση του παράγοντα nodal, ο οποίος πυροδοτεί την έναρξη του άξονα O-A, καταστέλλεται. Παραδόξως η μεταγραφή του Nodal και η διαφοροποίηση του ΟΑ διασώζονται απλά αναστέλλοντας τη μετάφραση του FoxQ2. Ως εκ τούτου ο περιορισμός του FoxQ2 στη ζωική πλάκα είναι ένα κρίσιμο στοιχείο για τη κανονική σηματοδότηση του Wnt μονοπατιού που συντονίζει και τη διαφοροποίηση των αξόνων AV και OA. Στα έμβρυα αχινού, οι πληροφορίες θέσης οι οποίες καθορίζουν το εξώδερμαμεσόδερμα-ενδόδερμα υπάρχουν κατα μήκος του πρωτεύοντος άξονα (AV) στο στάδιο του αυγού. Ο δευτερεύων (ΟΑ) είναι περισσότερο ασταθής. Αυτοί οι δύο άξονες συνδέονται μηχανικά καθώς έχει παρατηρηθεί ότι θεραπείες που διαταράσσουν τη σχηματοποίηση κατά μήκος του άξονα AV έχουν ως αποτέλεσμα και τον επηρεασμό της σχηματοποίησης του άξονα ΟΑ.Το φαινόμενο αυτό αντανακλά το γεγονός ότι η μοίρα της κυτταρικής διαφοροποίησης κατα μήκος των αξόνων απαιτεί τη είσοδο της β-κατενίνης στον πυρήνα των φυτικών βλαστομεριδίων. Ο κεντρικός ρόλος της β-κατενίνης στο ρυθμιστικό δίκτυο του ενδομεσοδέρματος (GRN) και η οποία, όπως αναφέραμε, λειτουργεί κατά μήκος του άξονα AV είναι καλά καθορισμένος. Η β-κατενίνη αρχικά εισάγεται στο πυρήνα των φυτικών βλαστομεριδίων και ενεργοποιεί το σκελετογόνο μεσέγχυμα κατά το τέλος της 4 κυτταρικής διαίρεσης. Κατά τη διάρκεια των επόμενων κυτταρικών διαίρεσεων, η β- κατενίνη εντοπίζεται διαδοχικά σε περισσότερα βλαστομερίδια του ζωικού πόλου, όπου είναι απαραίτητη και επαρκής για να ενεργοποίησει το κύμα σηματοδότησης απο το φυτικό στο ζωικό πόλο, βάσει του οποίου γίνεται και ο καθορισμός του ενδοδέρματος και μεσοδέρματος επιτυχώς. Ωστόσο, η απαραίτητη για τη β-κατενίνη πηγή που την ωθεί στη σχηματοποίηση του ΟΑ άξονα παραμένει ελειπής αν και πιστεύεται ότι συνδέεται με τη ενεργοποίηση του μεταγραφικού παράγοντα nodal. Ο μεταγραφικός παράγοντας nodal είναι επαρκής και αναγκαίος για το καθορισμό του στοματικού εξωδέρματος και στη δημιουργία ενός καταράκτη σηματοδότησης μορίων και μεταγραφικών παραγόντων, συμπεριλαμβανομένου των μεταγραφικών παραγόντων goosecoid και BMP2/4, τα οποία καθορίζουν το στοματικό και αντιστοματικό εξώδερμα αντίστοιχα. Η ενεργοποίηση του nodal ρυθμίζεται, μέσω μια διαδικασίας οξειδοαναγωγής στην οποία συμμετέχει το μητρικό μιτοχονδρικό στέλεχος. Πρόσφατη μελέτη έχει δείξει ότι cis-ρυθμιστικά στοιχεία μεσολαβούν στην 27
28 ενεργοποίηση του nodal και συνδέουν bzip μεταγραφικούς παράγοντες, οι οποίοι είναι γνωστοί από άλλα συστήματα ως οξειδοαναγωγικά ευαίσθητοι. Η μεταγραφή του παράγοντα nodal απαιτεί και την p38 MAP κινάση και υπερεκφράζεται έντονα και ραγδαία από ένα μηχανισμό θετικής ανάδρασης όπως έχει αποδείχθει και σε άλλα έμβρυα. Αν και η έκφραση του nodal στο στάδιο του μεσεγχυματικού βλαστιδίου εξαρτάται ισχυρά απο την κανονική σηματοδότηση του Wnt, η πυρηνική β-κατενίνη δε μπορεί να εντοπιστεί σε κύτταρα που μεταγράφουν nodal, οδηγώντας στην προσδοκία ότι ένα σήμα το οποίο είναι εξαρτώμενο απο τη β-κατενίνη αποστέλνεται απο τα φυτικά βλαστομερίδια προκειμένου να γίνει θετική ρύθμιση και έκφραση του nodal στα ζωικά στοματικά βλαστομερίδια. Ωστόσο, η ταυτότητα αυτού του σήματος το οποίο οδηγεί στην έκφραση του μεταγραφικού παράγοντα nodal δεν έχει προσδιοριστεί. Μολονότι, λοιπόν, οι διαταραχές στον άξονα AV επηρεάζουν τη σχηματοποίηση του ΟΑ, η αμοιβαία επίδραση δεν αποτελεί πιθανή περίπτωση. Καταστολή ή λανθασμένη έκφραση του nodal ή στόχων του σηματοδότησης όπως φαίνεται και στα σημασμένα με ραδιενέργεια έμβρυα αφήνουν άθικτο τον άξονα AV αλλάκαι τη ζωική περιοχή του εμβρύου απο την οποία προέρχεται το νευρογενές εξώδερμα. Αυτή η περιοχή είναι μοναδική διότι δεν επηρεάζεται από πειραματικές προκλήσεις οι οποίες μεταβάλλουν τη σχηματοποίηση των αξόνων AV ή ΟΑ. Επιπλέον οι περισσότεροι παράγοντες λαμβάνουν χαρακτήρα ζωικής πλάκα στην απουσία σηματοδότησης β-κατενίνης. Αυτό το εξώδερμα το οποίο είναι σημασμένο, δεν εκφράζει mrna nodal, ούτε ανταποκρίνεται σε λανθασμένη έκφραση του nodal. Αυτές οι παρατηρήσεις προτείνουν ότι η ζωική πλάκα δεν επηρεάζεται από την έκφραση ή τη λειτoυργία του nodal και ότι πρώιμοι ρυθμιστικοί παράγοντες της ζωικής πλάκας μπορούν εξίσου να επηρεάσουν τη σχηματοποίηση του άξονα ΟΑ. Το γονίδιο το οποίο κωδικοποιεί την πιο πρώιμο μεταγραφικό παράγοντα στη συγκεκριμένη περιοχή είναι το foxq2. Όπως φαίνεται και στις εικόνες, μετάγραφα του foxq2 συσσωρεύονται στο ζωικό ημισφαίριο κατα τη διάρκεια της 5 κυτταρικής διαίρεσης και εν συνεχεία εξαφανίζονται από όλο το εξώδερμα εκτός από την περιοχή η οποία διαμορφώνεται σε ζωική πλάκα κατά τη διάρκεια των επόμενων κυτταρικών διαίρεσεων, σε μια διαδικασία που απαιτεί τη λειτουργία της β-κατενίνης. Εαν αυτός ο περιορισμός αποτραπεί, τότε ο παράγοντας κυριαρχεί στο ζωικό ημισφαίριο και καταστέλλει τη μεταγραφή του nodal στο πρώιμο στοματικό εξώδερμα. Και είναι σημαντικό, καθώς η κάθαρση του Foxq2 είναι το μοναδικό β-κατενινό-εξαρτώμενο γεγονός το οποίο είναι απαραίτητο για την έκφραση του Nodal στο στοματικό εξώδερμα. Έτσι, αντί να παρέχουν μια θετική επίδραση για τη μεταγραφή του nodal, η κανονική σηματοδότηση του Wnt ρυθμίζει την έκφραση του nodal και την εδραίωση της πολικότητας του άηονα ΟΑ μέσω ενός μηχανισμού διπλού μηχανισμού καταστολής-εξάλειψη του καταστολέα του παράγοντα nodal. Προτείνεται ότι ο παράγοντας Foxq2 χρησιμεύει ως σημείο ελέγχου που συντονίζει το σωστό συγχρονισμό της ενεργοποίησης του GRN κατα μήκος του πρωτογενούς και δευτερογενούς άξονα. 28
 foxq2 and delta (vegetal) mrnas, 7th cleavage. (D) foxq2 mrna, hatching blastula (HB) stage.(e) foxq2 and z13 (formerly krl; vegetal) mrnas, hatching blastula.")
 Schematic of foxq2 expression patterns during sea urchin embryogenesis (green); magenta dot, serotonergic neurons.")
 foxq2 mrna in mesenchyme blastulae, with lateral surface view in (L).(N and O) FoxQ2 protein in mesenchyme blastulae.(p) Schematic of animal cap preparation.")
29 Εικ. 18 Macromere b- Catenin-Dependent Signals Restrict foxq2 Gene Expression to the Animal Plate (A and B) foxq2 mrna, 5th and 6th cleavage. Bar = 20 mm.(c) foxq2 and delta (vegetal) mrnas, 7th cleavage. (D) foxq2 mrna, hatching blastula (HB) stage.(e) foxq2 and z13 (formerly krl; vegetal) mrnas, hatching blastula. (F) foxq2 and nodal (oral ectoderm) mrnas, hatching blastula.(g and H) foxq2 mrna, midgastrula (MG) and prism (Pr). (I) Schematic of foxq2 expression patterns during sea urchin embryogenesis (green); magenta dot, serotonergic neurons. (J O) foxq2 expression in D-cadherin-injected (J, L, and N) and normal (K, M, and O) embryos. (J and K) DIC images of 3- day embryos.(l and M) foxq2 mrna in mesenchyme blastulae, with lateral surface view in (L).(N and O) FoxQ2 protein in mesenchyme blastulae.(p) Schematic of animal cap preparation. (Q) Schematic of micromereless embryo preparation.(r and S) FoxQ2 in animal-half embryoids. (T and U) FoxQ2 protein in micromereless embryos. Embryos are oriented with the animal pole at top and vegetal at bottom. 29
30 3.2 Nkx2.1 Ο μεταγραφικός παράγοντας Nkx2.1 ανήκει στην οικογένεια ΝΚ2, η οποία έχει προσδιορίστει στον αχινό Strongylocentrotus purpuratus, τα μετάγραφα του οποίου εντοπίζονται στην κορυφαία πλάκα του εξωδέρματος(apical plate) στο στάδιο του ώριμου βλαστιδίου και εντοπίζονται στη συγκεκριμένη περιοχή κατά τη διάρκεια τουλάχιστον 2 εβδομάδων ανάπτυξης. Μελέτη εντοπισμού πρωτεινών αποδεικνύει ότι ο παράγοντας Nkx2.1 είναι περιορισμένος στο επιθήλιο του apical plate (κορυφαία περιοχή εξωδέρματος), αλλά αποκλείεται απο τον πυρήνα των σεροτονεργικών νευρώνων. Το προφίλ έκφρασης του SpNK2.1 υπαγορεύεται μέσω δύο ξεχωριστών ρυθμιστικών συστημάτων. Αρχικά, ο SpNK2.1 περιορίζεται στην κορυφαία περιοχή (apical domain) από την επίδραση της β-κατενίνης στο ζωικο-φυτικό άξονα όπως αποδεικνύεται από μια επέκταση της έκφρασης SpNK2.1 κατά υπερέκφραση της β- κατενίνης. Ξεκινώντας από γαστριδίωση, η έκφραση στην κορυφαία πλάκα διατηρείται από τον παράγοντα SpDri. Κατάργηση του SpDri οδηγεί στην κατιούσα ρύθμιση του SpNK2.1 μετά τη γαστριδίωση, αλλά ο SpDri δεν είναι αναγκαίος για την αρχική ενεργοποίηση του SpNK2.1. Απώλεια της λειτουργίας σε πειράματα χρησιμοποιώντας SpNK2.1 ειδικά αντιπληροφοριακό μορφολινο ολιγονουκλεοτίδια και SpNK2.1 πειράματα υπερέκφραση δεν διαταράσσουν την εμβρυϊκή ανάπτυξη και δεν έχουν καμία επίδραση επί της ανάπτυξης νευρωνικών συστατικών του κορυφαίου οργάνου. Παρ 'όλα αυτά, SpNK2.1 ορίζει μια νέα πρόωρη έδαφος του έμβρυο του αχινού. Η παρουσία ενός κορυφαίου οργάνου (apical organ)είναι ένα κοινό χαρακτηριστικό ανάμεσα σε μια μεγάλη ποικιλία από θαλάσσια προνύμφες ασπόνδυλων. Αυτή η δομή αναπτύσσεται μέσα σε ένα παχυμένο επιθήλιο στο ζωικό πόλο του προνυμφικού σώματος, και ονομάζεται apical plate, και συνδέεται συχνά με μια δέσμη επιμήκων βλεφαρίδων, την apical tuft. Τοapical plate του αχινού αποτελείται από δύο συστάδες των νευρικών κυττάρων, των οποίων τα νευράξονες εξαπλωνονται πλαγίως και κατά μήκος του οπισθιο-άκρου του προφορική κουκούλα, που νευρώνει τη βλεφαριδωτή ζώνη. Τα κορυφαία όργανα πολλών ασπονδύλων προνύμφων φαίνεται να παίζουν ρόλο στην αισθητηριακή ολοκλήρωση, και έχει προταθεί ότι στη θάλασσα στον αχινό το κορυφαίο όργανο συντονίζει τη σίτιση των προνυμφών,τη μετακίνηση, και τη μεταμόρφωση. Όπως τα περισσότερα θαλάσσια ασπόνδυλα, συμπεριλαμβανομένων μαλάκια, το κορυφαίο όργανο κατά μήκος με το μεγαλύτερο μέρος της προνύμφης όπως και νευρικό σύστημα χάνεται στη μεταμόρφωση. Οι διαδικασίες καθορισμού του apical organ παραμένουν αδιευκρίνιστες. Ωστόσο μερικά ρυθμιστικά γονίδια έχουν ταυτοποιηθεί ότι εκφράζονται στο apical organ στο χορδωτό Ptychodera flava. Τα γονίδια της οικογένειας Sox, Pf-SoxB1 και Pf-SoxB2 εκφράζονται και τα δύο στο apical organ την 4 η ημέρα της προνύμφης, τόσο στην περιοχή της βλεφαριδωτής ζώνης όσο και στην περιοχή του στομαδαίου. Το ορθόλογο γονίδιο των χορδωτών στα θυλαστικά είναι ο παράγοντας Τbrain-1, ο οποίος εκφράζεται σε διαφορετικά κύτταρα στην περιοχή του εξωδέρματος στο στάδιο του γαστριδίου και τελικά περιορίζεται στο apical organ. Τελικά τα αντίγραφα του PfNK2.1, τα οποία ανήκουν στην οικογένεια ρυθμιστικών παραγόντων ΝΚ2 ανιχνεύονται στην περιοχή του εξωδέρματος στο στάδιο του γαστριδίου και τελικά εντοπίζονται σε κύτταρα του apical organ αλλά και σε κύτταρα της βλεφαριδωτής ζώνης. Βάσει αυτού 30

31 του μοτίβου έκφρασης ο παράγοντας Nk2.1 φαίνεται να παίζει σημαντικό ρόλο ως νευρογενής παράγοντας στο στάδιο της λάρβας. Τα μετάγραφα του SpNK2.1 αρχικά εντοπίζονται στα κύτταρα του εξωδέρματος στο στάδιο του ώριμου βλαστιδίου. Αυτή η έκφραση διέπεται από αρχές σχηματοποίησης ζωικού-φυτικού πόλου, οι οποίες εξαρτώνται από τη λειτουργία της β-κατενίνης. Αργότερα ξεκινώντας η διαδικασία της γαστριδίωσης η παραμονή του SpNK2.1 στην apical plate διέπεται από ένα διαφορετικό σύνολο ρυθμιστικών παρεμβάσεων συμπεριλαμβανομένου του παράγοντα SpDri, ορθόλογο του dead ringer στον αχινό. Αν και κατάργηση του SpNK2.1 δεν οδηγεί σε ανιχνεύσιμες αναπτυξιακές ανωμαλίες, το προφίλ έκφρασης του SpNK2.1 είναι μοναδικό και προσδιορίζει μία νέα μεταγραφική περιοχή η οποία ονομάζεται apical domain, και εντοπίζεται στα πρωίμα εμβρυικά στάδια του αχινού. Εικ 19 SpNK2.1 mrna expression during embryogenesis as revealed by WMISH. (A) Lateral view of a hatched blastula (20 h pf). Transcripts are restricted to the apical plate. (B) Lateral view of mid-gastrula stage (28 h pf) embryo. (C) Lateral view of late gastrula stage (44 h pf) embryo. Note that the top of the foregut (arrow) lies beneath the ectodermal zone of expression. (D E) Oral view (D) and anal view (E) of 44-h embryo. The deceptive endodermal signal is due to overlying positively stained oral ectoderm as no signal is observed when viewed from the anal side. (F) Oral view of an early pluteus larva (72 h pf). (G)Oral view at 96 h pf.band 31
32 3.3 Six3 Δύο μεγάλα κέντρα σηματοδότησης έχει αποδειχθεί ότι ελέγχουν τη σχηματοποίηση των εμβρύων αχινού. Η κανονική σηματοδότηση του Wnt στα φυτικά βλαστομερίδια και η σηματοδότηση του Nodal στο πρώιμο στοματικό εξώδερμα είναι απαραίτητες και επαρκής προκειμένου να προκαλέσουν τη σχηματοποίηση κατα μήκος του πρωτεύοντος και δευτερεύοντος άξονα αντίστοιχα. Εδώ προσδιορίζουμε και χαρακτηρίζουμε ένα τρίτο σηματοδοτικό μονοπάτι, της περιοχής του ζωικού πόλου (Animal Pole Domain/APD) στην οποία εμπεριέχεται το νευρογενές εξώδερμα και μπορεί να αντιταχθεί στη σηματοδότηση Wnt και Nodal. Η ρυθμιστική επίδραση της APD περιορίζεται κυρίως στην περιοχή του ζωικού πόλου, αλλά φαίνεται να επηρεάζει και αρκετά άλλα κύτταρα του εμβρύου, καθώς σε περίπτωση απουσίας των Wnt και Nodal το APD επεκτείνεται σε όλο το έμβρυο.έχουν προσδιοριστεί αρκετά ρυθμιστικά γονίδια τα οποία εκφράζονται στο APD στο στάδιο του πρώιμου βλαστιδίου και φαίνεται ότι η έκφραση των περισσοτέρων απαιτεί τη λειτουργία του Six3. Επιπλέον, ο Six3 είναι απαραίτητος για την διαφοροποίηση των ποικίλων κυτταρικών τύπων του APD, συμπεριλαμβανομένου της νευρογενούς πλάκας των ζώων και του αμέσως πλευρίζοντος εξώδερματος, υποδεικνύοντας ότι λειτουργεί σε ή κοντά την κορυφή πολλών ρυθμιστικών δικτύων του APD. Η σχηματοποίηση, επομένως, του εξωδέματος στα διάφορα αναπτυξιακά στάδια του αχινού ρυθμίζεται από μια σειρά σηματοδοτικών επεισοδίων, ξεκινώντας με ένα κύμα κανονικής σηματοδότησης το οποίο προέρχεται απο τα φυρικά βλαστομερίδια του εμβρύου στο στάδιο κυττάρων και προσδιορίζει τους ιστούς του ενδομεσοδέρματος. Η Canonical σηματοδότηση Wnt είναι επίσης απαραίτητη για τη Nodal εξαρτώμενη σχηματοποίηση κατά μήκος του δευτερεύοντος άξονα αφαιρώντας ένα καταστολέα έκφρασης του Nodal, τον FoxQ2, από το πλευρικό εξώδερμα. Ο παράγοντας Nodal είναι απαραίτητος για την παραγωγή του ΒΜΡ2 / 4 το οποίο ενεργοποιεί το δίκτυο ρύθμισης για το καθορισμό του αντι-στοματικού εξωδέρματος. Ο παράγοντας είναι από τα πρώτα γονίδια το οποίο ενεργοποιείται μετά τη γονιμοποίηση και εκφράζεται στο APD στο στάδιο του βλαστιδίου. Αποδεικνύεται ότι αυτός ο παράγοντας είναι απαραίτητος για την ανάπτυξη όλων των νευρώνων, για τον ανταγωνισμό όλων των σημάτων που αναστέλλουν τη νευρική διαφοροποίηση και για την έκφραση μεγάλης πλειοψηφίας ρυθμιστικών γονιδίων που εκφράζονται πρώιμα στο APD. Επιπλέον, μη ορθή έκφραση του Six3 μπορεί να μετατρέψει σχεδόν όλα τα κύτταρα του εμβρύου στο να σχηματίσουν ένα καλά διαμορφωμένο APD. Έτσι οι Six3 εξαστώμενες ιδιότητες του APD δείχνουν ότι ο τομέας αυτός λειτουργεί ως κέντρο σηματοδότησης. 32

33 3.3.1 Ο Six3 εκφράζεται νωρίς στο APD Ο προσδιορισμός του Six3 στον αχινό είναι σαφής, επειδή η αλληλουχία του είναι πολύ υψηλά συντηρημένη στην ομοιοπεριοχή (98% ταυτόσημη με HsSix3), Επιβεβαιώθηκε σε προηγούμενες μελέτες που ότι ο Six3 εκφράζεται σε μια δυναμική πορεία στη διάρκεια της ανάπτυξης του Paracentrotus lividus ένα είδος συγγενές προς Strongylocentrotus purpuratus. Το πιο σημαντικό χαρακτηριστικό είναι ότι τα μετάγραφα του Six3 εκφράζονται στo ζωικό ημισφαίριο κατά τα τέλη της κυτταρικής διαίρεσης (Εικ. 20Α), στο APD στο στάδιο του μεταγενέστερου βλαστιδίου (Εικ. 20Β) και στη συνέχεια, σε δύο δακτυλίους (Σχ.20G,Η),έναν κοντά στην περιφέρεια του APD (Εικ. 20C-F, Η) και το άλλο στο ενδομεσόδερμα(εικ. 20C-G). Κατά τη διάρκεια της γαστριδίωσης, ο Six3 εκφράζεται σε κάποια δευτερεύοντα μεσεγχυματικά κύτταρα διάσπαρτα σε όλη το βλαστόκοιλο και στην άκρη του αρχεντέρου (Εικ. 20I, βέλος), σε κύτταρα στο ζωικό πόλο (Σχ. 20I, J) και του στοματικού εξωδέρματος (Σχ. 20J, K). Στο στάδιο πλουτέα, το Six3 RNA ανιχνεύεται σε δύο ομάδες των κυττάρων, πλευρικά του στόματος (Σχ. 20Κ, βέλη) και στη βλεφαριδωτή ζώνη (Εικ.20K). Έξι συνολικά επίπεδα mrna στο στάδιο του πρώιμου μεσεγχυματικού βλαστιδίου δεν αλλάζουν σημαντικά μετά την ένεση με Δcadherin mrna(πίνακας1).in situ υβριδοποιήσεις είναι συνεπείς με αυτό το αποτέλεσμα και δείχνουν ότι η κατανομή διαφέρει σε Δcadherin mrna-ενεμένα έμβρυα, απουσιάζει από τα φυτικά κύτταρα και διατηρώντας την ευρεία έκφραση στο ζωικό ημισφαίριο που έχει καθιερωθεί πριν από τον περιορισμό των διαδικασιών εξαρτώνται από την κανονική σηματοδότηση Wnt. Εικ 20 Whole-mount in situ hybridization for six3 mrna during development. Times as hours postfertilization are shown in the upper right corner of each image. (A) Very early blastula. (B) Hatching blastula. (C-H) Mesenchyme blastula. (I,J) Late gastrula. (K) Pluteus. All embryos are shown in lateral view, except in G and H, which illustrate vegetal pole (vv), and animal pole view (apv), respectively. Arrows in I and K mark positions of secondary mesenchyme cells and cells flanking the mouth, respectively. Scale bar:20 μm. 33

34 Για τον εντοπισμό των APD ρυθμιστικών γονιδίων, συγκρίθηκε η σχετική συγκέντρωση των μεμονωμένων mrnas σε Δcadherin mrna ενεμένα έμβρυαέναντι εμβρύων τα οποία ειχαν ενεθεί με γλεκερόλη στο στάδιο βλαστιδίου χρησιμοποιώντας μικροσυστοιχίες που αντιπροσωπεύουν όλες τις προβλέψεις γονιδίων οι οποίες βρέθηκαν στην αλληλουχία γονιδιώματος του αχινού. Τα γονίδια αυτά ταξινομούνται σε τρεις ομάδες: αυτά που εκπροσωπούνται στο μέγιστο στο μητρικής προέλευσης RNA (Εικ. 22Α), εκείνα που υπερεκφράζονται έντονα κατά τη διάρκεια των κυτταρικών διαίρεσεων (σχ. 22Β, C) και εκείνων που ρυθμίζεται αυξητικά κατά τα πρώτα στάδια γαστριδίου (Σχ. 22D). Εικ 21 Δcadherin-misexpressing embryos consist of animal pole domain (APD) tissues defined by expression of foxq2 and hbn. Whole-mount in situ hybridizations to embryos at mesenchyme blastula (A-D) and gastrulae (E-H) stages. (E,F) Glycerol-injected control. (G,H) Δcadherin-mRNA injected. (A,E,G) DIC; (B-D,F,H) Two-color fluorescence, hbn (green) and foxq2 (magenta) RNAs. Scale bar: 20 μm. 34
, 15-hour early blastula (EB), 30-hour late")
35 Εικ 22 Temporal expression profiles of genes in the provisional early APD set. Profiling methods were as described by Wei et al. (Wei et al., 2006). Values at different hours post-fertilization from 2 to 72 are shown, as a percentage of the maximum signal intensity, for each gene at 2-cell (maternal RNA), 15-hour early blastula (EB), 30-hour late mesenchyme blastula (LMB), 48-hour late gastrula (LG) and 72-hour pluteus larva (PL). Profiles are grouped according to time of earliest detectable expression: (A) maternal; (B,C) early blastula; (D) early blastula to mesenchyme blastula. The position of the dashed line represents the time of assay in Six3 morphants. Data for the six3 gene are highlighted with a red arrow. 35

36 3.3.2 Η Λειτουργία του Six3 απαιτείται για το σχηματισμό του APD και τη διαφοροποίηση των νευρώνων Για να δοκιμαστεί η λειτουργία του Six3, ενέθηκαν γονιμοποιημένα αυγά με δύο διαφορετικά Six3- μορφολινο μετάφραση αποκλεισμού, καθένα από τα οποία προκάλεσε τις ίδιες αναπτυξιακές ανωμαλίες. Έμβρυα στις 3 ημέρες παρουσίασαν στρογγυλεμένη μορφολογία (σχ. 23Α, Β) Το εξώδερμα του ζωικού πόλου δεν είχε το τυπική επιθηλιακή μορφολογία χαρακτηριστική της ζωικής πλάκας (Εικ. 23, σύγκρινε Α,Βμε C) Σε έμβρυα στα οποία απουσιάζει ο Six3, η νευρική διαφοροποίηση ανεστάλη ισχυρά, όπως προσδιορίστηκε με ανοσοχρώση για δείκτες νευρικών κανονικά εκφράζονται κατά την όψιμη gastrula και πλουτέα στάδια (εικ. 23, συγκρίνετε Α,Β με C). Η πλειοψηφία (2/3) των εμβρύων δεν περιείχε σεροτονεργικούς νευρώνες και το υπόλοιπο είχε μειωμένο αριθμό (Σχ.23D) σε σύγκριση με τον κανονικό αριθμό σε αυτό το στάδιο (3-5). Το ίδιο ήταν αληθές για όλους τους άλλους νευρώνες, και προσδιορίστηκε με τον δείκτη παν-νευρικών συναπτοταγμίνης (1e11) (Σχ. 23Α, Β), τα οποία βρίσκονται εντός του APD και της βλεφαριδωτής ζώνης των φυσιολογικών εμβρύων των 3-ημερών (Εικ. 23C). Εικ 23 Loss of Six3 results in loss of neurons and the thickened epithelium characteristic of the APD. (A,B) Three-day embryos injected with Six3-MO2 at the one-cell stage, which are typical of stronger and weaker phenotypes, respectively. (C) Normal 3-day embryo. APD in A-C indicated by brackets; 1e11 (pan-neural,magenta), serotonin (green), DAPI (nuclei, blue). (D) Numbers of embryos containing either 0, 1, 2 or 3 serotonergic neuron per embryo in Six3 morphants. At this stage, normal embryos have from 3 to 5 serotonergic neurons. (E,F) hbn mrna in normal mesenchyme blastulae (E) or in Six3 morphants (F). Scale bar: 20 μm. 36
37 4.Υλικά και Μέθοδοι 4.1 Συλλογή γαμετών και τεχνητή γονιμοποίηση 1. Ενέσεις 1ml 0,5Μ ΚCl στην περιστοματική περιοχή των ώριμων αχινών Paracentrotus lividus (περιοχή Ρίου Αχαΐας). 2. Συλλογή γαμετών (ωάρια: στο ml!, dry sperm: 1 σταγόνα αρκεί για την δημιουργία 1l καλλιέργειας!). 3. Προσθήκη γαμετών σε κατάλληλες συγκεντρώσεις σε PABA (1/4 του όγκου της καλλιέργειας, ρύθμιση ph γονιμοποίησης και μη σκλήρυνση μεμβράνης γονιμοποίησης) και ανάδευση για 10 λεπτά σε θερμοκρασία δωματίου. 4. Πέρασμα από Νitex (50 μ) για απομάκρυνση της μεμβράνης γονιμοποίησης. 5. Προσθήκη θαλασσινού νερού (3/4 του όγκου της καλλιέργειας) και αντιβιοτικού (1ml/l καλλιέργειας) στους 18ο C με ανάδευση. 4.2 Καλλιέργεια εμβρύων του αχινού Paracentrotus lividus Αναπτυξιακά στάδια : 1. Στάδιο αυγού (0 ώρες). 2. Στάδιο 16 κυττάρων (3-4 ώρες). 3. Στάδιο πρώιμου βλαστιδίου (10 ώρες). 4. Στάδιο πρώιμου γαστριδίου (20 ώρες.) 5. Στάδιο πρίσματος (25 ώρες). 6. Στάδιο πρώιμου πλουτέα (30-32 ώρες). 4.3 Απομόνωση RNA από διαφορετικά αναπτυξιακά στάδια 1. Ψυχρή φυγοκέντρηση για 5 λεπτά (full spead). 2. Aπομάκρυνση του υπερκείμενου νερού. 3. Προσθήκη 1ml φαινόλης (tripure) και ομογενοποίηση. 4. Στον πάγο για πέντε λεπτά. 5. Προσθήκη 200μl χλωροφορμίου. Vortex για 15 δευτερόλεπτα. 6. Στον πάγο για πέντε λεπτά. 7. Ψυχρή φυγοκέντρηση (12000g) για 15 λεπτά. Κρατάμε την υδατική, ανώτερη φάση. 8. Προσθήκη 500μl ισοπροπανόλης. 9. Στον πάγο για δέκα λεπτά. 10.Ψυχρή φυγοκέντρηση (12000g) για 10 λεπτά. 11.Αφαίρεση υπερκειμένου (ισοπροπανόλης). 12.Προσθήκη 75% 1ml αιθανόλης. Vortex για δευτερόλεπτα. 13.Ψυχρή φυγοκέντρηση (10000g) για 5 λεπτά. 14.Αφαιρώ υπερκειμένου (αιθανόλης). 15.Ξήρανση ιζήματος. 16.Προσθήκη 200μl RNAase free νερού και αποθήκευση στους -80ο C. 37
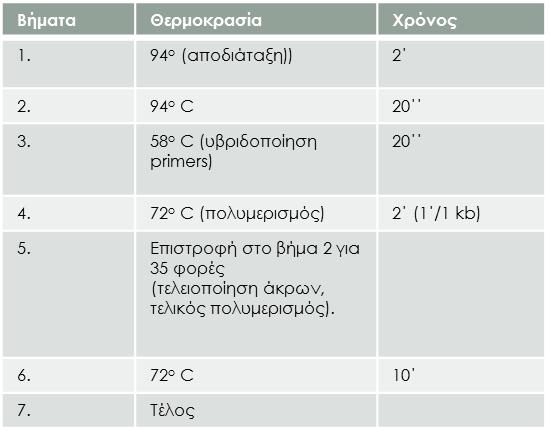
 30 5. 72 ο C (πολυμερισμός) 2 (1 /1 kb) 6. Eπιστροφή στο βήμα 3 για 39 φορές (τελειοποίηση άκρων, τελικός πολυμερισμός). 7. 72 ο C 10 8. Τέλος 38")
38 4.4 RT-PCR Η τεχνική της RT-PCR πραγματοποιήθηκε με το Kit Takara PrimeScript One Step RT- PCR kit Ver. 2. Πρόγραμμα που ακολουθούμε: Bήματα Θερμοκρασία Χρόνος ο C ο (aποδιάταξη)) ο C , 56,6, 60 ο C (υβριδοποίηση primers) ο C (πολυμερισμός) 2 (1 /1 kb) 6. Eπιστροφή στο βήμα 3 για 39 φορές (τελειοποίηση άκρων, τελικός πολυμερισμός) ο C Τέλος 38
. Φυγοκέντρηση στα 1000 g για 1 λεπτό. 3.")
.")
39 4.5 Καθαρισμός των προιόντων της RT-PCR 1. Προϊόν και RNAase free νερό: ->100μl. 2. Προσθήκη 200μl ΝΤΙ (chaotropic salt for binding of RNA in colunes). Φυγοκέντρηση στα 1000 g για 1 λεπτό. 3. Προσθήκη 700μl ΝΤ3 (ethanolic, washinh buffer). Φυγοκέντρηση στα 1000 g για 1 λεπτό. 4. Ξηρή φυγοκέντρηση στα 1000 g για 2 λεπτά. 5. Προσθήκη 50μl elution buffer (with low ionic strength conditions). Σε θερμοκρασία δωματίου για 2 λεπτά και μετά φυγοκέντρηση στα 1000 g για 1 λεπτό. 6. Αποθήκευση στους -20ο C. 4.6 Αντίδραση λιγάσης Αντιδραστήριο Overnight σε θερμοκρασία δωματίου. 39
. 6. Στον πάγο για 2 λεπτά. 7.")
40 4.7 Μετασχηματισμός Βακτηρίων 1. Eξάπλωση 80μl x-gal (ανάλογο της λακτόζης) στις plates. 2. Ξεπάγωμα στον πάγο 40μl βακτηρίων Ε. coli (Ν.Ε. Biolabs) από τους -80ο C. 3. Προσθήκη 2μl από την αντίδραση της λιγάσης. 4. Στον πάγο για 40 λεπτά. 5. Στους 42ο C για 1,5 λεπτά (heat shock). 6. Στον πάγο για 2 λεπτά. 7. Προσθήκη των βακτηρίων σε προθερμασμένο soc medium (500 μl). 8. Επώαση στους 37ο C για μία ώρα. 9. Στρώνουμε τα μετασχηματισμένα βακτήρια στις plates (25 ή 100 μl). 10.Eπώαση στους 37ο C overnight. 40
41 4.8 Κατασκευή τρυβλίων θρεπτικού υλικού για βακτηριακές καλλιέργειες Πριν τον μετασχηματισμό των δεκτικών βακτηρίων με τα προϊόντα της αντίδρασης λιγάσης, δηλαδή με τους ανασυνδυασμένους φορείς, είναι απαραίτητο να φτιαχτούν τρυβλία με κατάλληλο θρεπτικό υλικό για την ανάπτυξη των βακτηριακών καλλιεργειών. Για περίπου είκοσι plate προστίθενται σε μία κωνική φιάλητα παρακάτω: 500ml dd νερό 6,5gr nutrient E 7,5gr agar Στην συνέχεια καλύπτεται το στόμιο με αλουμινόχαρτο και αποστειρώνεται. Όταν η θερμοκρασία του θρεπτικού LB πέσει αρκετά (για να μην καταστραφεί η amp) προσθέτουμε 500μΙ αμπικίλλινη αρχικής συγκέντρωσης 100mg/ml, ώστε να υπάρχει σε τελική συγκέντρωση 100μg/μΙ. Τέλος χύνεται το θρεπτικό υλικό όσο είναι ακόμα υγρό σε τρυβλία δίπλα σε φλόγα ώστε να έχουμε στείρες συνθήκες. Αφού το θρεπτικό υλικό στερεοποιηθεί πλήρως, απλώνεται ομοιόμορφα σε κάθε τρυβλίο 80μΙ X-gal με την βοήθεια μίας πιπέτας Pasteur που έχει σχήμα Γ με την βοήθεια της φλόγας. Αυτά τα τρυβλία είναι πλέον έτοιμα για ανάπτυξη και επιλογή των μετασχηματισμένων αποικιών. 4.9 Κλωνοποίηση 1. Συλλογή των άσπρων αποικιών (blue/white screening). 2. Άγγιγμα σε μια νέα plate για την δημιουργία master plate. Επώαση overnight στους 37ο C. Διατήρηση στους -80ο C. 3. Ξέπλυμα του loop μέσα σε falcon με 5 ml LB και 5μl αμπικιλίνης για την δημιουργία υγρής καλλιέργειας. Επώαση overnight στους 37ο C. 41
. 9. Φυγοκέντρηση για 1 λεπτό στα 11000g. 10.Απόρριψη του εκλούσματος.")
42 4.10 Mini-preps: 1. Φυγοκέντρηση για 10 λεπτά στα 2600g. 2. Aφαίρεση υπερκείμενου. 3. Προσθήκη 250μl Α1 διαλύματος επαναδιάλυσης. 4. Μεταφορά σε eppendorfs. 5. Προσθήκη 250μl Α2 διαλύματος λύσης. Άμεση ανακίνηση. 6. Προσθήκη 300μl Α3 διαλύματος εξουδετέρωσης. 7. Φυγοκέντρηση για 10 λεπτά στα 11000g. 8. Μεταφορά σε κολώνα (χωρίς ίζημα). 9. Φυγοκέντρηση για 1 λεπτό στα 11000g. 10.Απόρριψη του εκλούσματος. 11.Προσθήκη 600μl Α4 διαλύματος για πλύση. 12.Ξηρή φυγοκέντρηση για 2 λεπτά στα 11000g. 13.Απόρριψη του εκλούσματος. 14.Προσθήκη 100μl mq νερού. 15.Για ένα λεπτό σε θερμοκρασία δωματίου. 16.Φυγοκέντρηση για 1 λεπτό στα 11000g. 17.Αποθήκευση στους -20ο C. Το kit είναι της MN (Nucleospin Plasmid) Πέψη με ένζυμα περιορισμού Επώαση στους 37ο C για 2 ώρες. 42

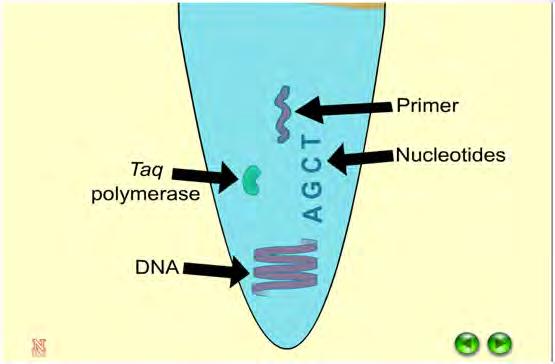
43 4.12 PCR Πρόγραμμα που ακολουθούμε: 43
44 4.13 Καθαρισμός προιόντων PCR 1. Προϊόν και RNAase free νερό 100μl. 2. Προσθήκη 200μl ΝΤΙ (chaotropic salt for binding of RNA in colunes). Φυγοκέντρηση στα 1000 g για 1 λεπτό. 3. Προσθήκη 700μl ΝΤ3 (ethanolic, washing buffer). Φυγοκέντρηση στα 1000 g για 1 λεπτό. 4. Ξηρή φυγοκέντρηση στα 1000 g για 2 λεπτά. 5. Προσθήκη 50μl elution buffer (with low ionic strength conditions). Σε θερμοκρασία δωματίου για 2 λεπτά και μετά φυγοκέντρηση στα 1000 g για 1 λεπτό. 6. Αποθήκευση στους -20ο C Σήμανση των αντινοηματικών μεταγράφων Για να εντοπισθούν οι RNA ανιχνευτές όταν θα υβριδοποιηθούν εντός των εμβρύων πρέπει να σημανθούν. Η σήμανση στην προκειμένη περίπτωση γίνεται με χημική αντίδραση και ένωση μορίων DIG και DNP απευθείας πάνω στο RNA. Στην συνέχεια τα αντιγόνα μπορεί να εντοπισθούν με αντισώματα. Όλοι οι RNA ανιχνευτές σημάνθηκαν σε αντιδράσεις που έγιναν σε PCR tubes. Τα kits που χρησιμοποιήθηκαν για την σήμανση ήταν το Label IT Digoxin Labeling Kit και το Label IT DNP Labelling Kit της εταιρείας Mirus.To πρωτόκολλο που ακολουθείται είναι το παρακάτω: 1. Σε PCR tube προστίθεται 1μg RNA υπολογίζοντας τον όγκο που απαιτείται από την μέτρηση του φωτομέτρου (~1-2μΙ). 2. Συμπληρώνεται με DEP νερό έως τελικό όγκο 8μΙ και γίνεται αποδιάταξη του RNA στους 70οC για 5 λεπτά. 3. Τοποθετούνται τα tubes στον πάγο για 2 λεπτά. 4. Προστίθενται 1μΙ 10xLabeling Buffer και 1μΙ DIG ή DNP reagent. 5. Επωάζονται τα tubes στους 37οC στον επωαστικό θάλαμο για 2 ώρες. 6. Προστίθενται 20μΙ DEP νερό. 7. Περνά το δείγμα δύο φορές από κολώνα sephadex για καθαρισμό και γίνεται συλλογή του ιχνηθετημένου ανιχνευτή. Αποδιάταξη RNA: σε 70C, για 5min -> ύστερα σε πάγο Προσθέτω: Καθαρίζω τον ιχνηθέτη με κολώνα G-50 2 φορές. Επώαση στους 37ºC, >2ώρες 44
45 4.15 Γονιδιακή καταστολή σε γονιμοποιημένα αυγά με ενέσεις με ΜΑSO Για να αποκτηθούν μονιμοποιημένα έμβρυα αχινού στο επιθυμητό στάδιο (για χρήση σε FISH σε διαφορετικά αναπτυξιακά στάδια) πρέπει πρώτα να γίνει συλλογή των γαμετών, τεχνητή γονιμοποίηση και καλλιέργεια των εμβρύων όπως αναφέρθηκε πριν. Όταν όμως πρόκειται να γίνει καταστολή του PlCoup-Tf με morpholinos και έπειτα χρωμογόνος in situ υβριδοποίηση, το πρωτόκολλο τροποποιείται σε ένα σημείο. Συγκεκριμένα η μόνη διαφορά είναι ότι μετά την συλλογή των γαμετών, στα γονιμοποιημένα ωάρια (σε 75 mi θαλασσινό νερό) με πιπέτα Pasteur προστίθενται 4 σταγόνες 0,5Μ citric acid για χημική απομάκρυνση του jelly coαt, γίνεται ανάδευση για 1 λεπτό, προστίθενται 18 σταγόνες Tris-HCl για αποκατάσταση του ph στο 8,3 για την επίτευξη γονιμοποίησης και γίνεται μηχανική αφαίρεση του jelly coαt που περιβάλλει τα ωάρια με πέρασμα από γάζα. Στην συνέχεια τα ωάρια αυτά μεταφέρονται σε μικρά πλαστικά τρυβλία για να γίνει η γονιμοποίηση αλλά και να ενεθούν με MASO (Morpholino anti-sense oligonucleotide). (Τα καπάκια έχουν επεξεργαστεί χημικά με την προσθήκη για 2 λεπτά θειικής πρωταμίνης και έχουν αποκτήσει αντίθετο φορτίο από αυτό της μεμβράνης των αυγών ώστε να συγκρατούν κολλημένα στον πάτο τα έμβρυα). Τα MASOs είναι ανάλογα νουκλεϊκών οξέων που αντί για τον δακτύλιο της πεντόζης έχουν ένα δακτύλιο morpholine. Τα MASOs διατηρούν συμπληρωματικότητα με τα νουκλεϊκά οξέα και μπορούν να προσδένονται στις συμπληρωματικές τουςαλληλουχίες χωρίς να εκκινούν μηχανισμούς καταστροφής των δίκλωνων υβριδίων. Ταυτόχρονα όμως τα MASOs είναι ανθεκτικά στους ενδογενείς μηχανισμούς καταστροφής των κυττάρων, οπότε έχουν πολύ μεγάλο χρόνο ημιζωής. Στις περισσότερες περιπτώσεις, όπως και στον Coup-TF τα MASOs είναι συμπληρωματικά με την αλληλουχία στις θέσεις έναρξης της μετάφρασης των μεταγράφων που στοχεύουν και δεν της επιτρέπουν να συμβεί. Κατ αυτόν τον τρόπο προκαλείται καταστολή (knockdown) της έκφρασης των αντίστοιχων πρωτεϊνών και των φαινοτύπων που οφείλονται σε αυτές. Τα ωάρια χωρίζονται σε 2 όμαδες, τα control και τα morphant έμβρυα: Για να αποκτηθούν control έμβρυα, τα ωάρια τοποθετούνται κάτω από το μικροσκόποιο και προστίθενται τρεις σταγόνες αραιωμένου σπέρματος και παρακολουθείται η γονιμοποίηση. Όταν εμφανιστεί η μεμβράνη γονιμοποίησης στα περισσότερα από αυτά, τότε μπαίνουν σε θάλαμο στους 18οC για να αναπτυχθούν. Για να αποκτηθούν τα morphant έμβρυα, γονιμοποιούνται με τον ίδιο τρόπο αλλά στην συνέχεια δέχονται microinjection ένα-ένα με ΜΑSO έναντι του PlCoup-TF. Στην συνέχεια και αυτά τοποθετούνται σε θάλαμο στους 18οC για να αναπτυχθούν. 45
46 Για 100μΙ injection solution: 1. 20μΙ γλυκερόλη 2. 20μΙ 1Μ PlCoup-Tf ΜΑSO (τελική συγκέντρωση 0,2mM) με αλληλουχία 5'ACTGCGATGGCGTTGAGGGAATCGAT3' 3. 60μΙ mq νερό Έπειτα ανάδευση, φιλτράρισμα και spin για 10 δευτερόλεπτα. Μετά από είκοσι με εικοσι τέσσερις ώρες που τα control έμβρυα βρίσκονται στο στάδιο του γαστριδίου και τα morphants στο στάδιο του βλαστιδίου λόγω καθυστερημένης ανάπτυξης, συλλέγονται σε πηγαδάκια σε microtiter plates. Σε κάθε ζεύγος πηγαδιών, με control και morphant έμβρυα, θα μελετηθεί η έκφραση ενός εκ των γονιδίων για τα οποία κατασκευάστηκαν ανιχνευτές Μονιμοποίηση εμβρύων για χρωμογόνο και διπλή φθορίζουσα in situ υβριδοποίηση Πρίν ξεκινήσει η χρωμογόνος in situ υβριδοποίηση στην οποία εξετάζεται η επίδραση της καταστολής του PlCoup-Tf με morpholinos στα υπό εξέταση γονίδια, πρέπει να μονιμοποιηθούν τα έμβρυα (τόσο τα control όσο και τα morphants) με τα οποία θα γίνει το πείραμα. Για να επιτευχθεί αυτό ακολουθείται η παρακάτω διαδικασία: ΠΡΟΣΟΧΗ!: Όλη η διαδικασία γίνεται κάτω από το στερεοσκόπιο για να μην ροφώνται τα έμβρυα. 1. Αφού συλλεχθούν τα έμβρυα στην microtiter plate, γεμίζεται όλο το πηγαδάκι με ASW (Artificial Sea Water). 2. Προστίθεται μία σταγόνα PFA 8% in ASW για να νεκρωθούν τα έμβρυα, να σταματήσουν να κολυμπούν και να κατακαθίσουν στον πάτο του πηγαδιού. 3. Αφαιρείται το μισό του όγκου του πηγαδιού (περίπου 150μΙ) και προστίθενται PFA 8% μέχρι να γεμίσει το πηγαδάκι. 4. Αφήνονται τα έμβρυα σε θερμοκρασία για 1 ώρα ενώ συχνά αναταράσσονται. 5. Μετά το πέρας 1 ώρας και αφού τα έμβρυα καθιζάνουν, αφαιρείται όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζεται το πηγαδάκι ξανά με ASW. 6. Αφαιρείται και πάλι όσο περισσότερο γίνεται από το υπερκείμενο και προστίθενται 100% ΜeOH έως ότου γεμίσει το πηγαδάκι. 7. Επαναλαμβάνεται 3 φορες το βήμα 6 προσέχοντας στην αναρρόφηση γιατί ο σχηματισμός αλάτων πολλές φορές εγκλωβίζει τα έμβρυα. 8. Κολλάται προστατευτικό αυτοκόλλητο πάνω στην microtiter plate και περιβάλλεται με parafilm για την αποφυγή επαφής με τον αέρα. 9. Αποθηκεύονται τα έμβρυα στους -20οC για μελλοντική χρήση. 46
47 Υλικά για την μονιμοποίηση: Aντιδραστήριο KCl Ποσότητα 0,19gr NaCl MgCl2.6H2O 7,54gr 1,73gr CaCl2.2H2O (Προστίθεται τελευταίο) 0,4gr MgSO4.7H2O 1,36gr EPPS 0,63gr Στην συνέχεια γίνεται ph μέτρηση και προσαρμογή του ph στα~8,2 με χρήση ΝaΟΗ 1Μ. PFA 8% in ASW I. Προστίθενται 4gr PFA σε falcon με 50ml νερό. II. Θερμαίνεται στους 65οC στο υδατόλουτρο για αρκετή ώρα και γίνεται συχνά Vortex. III. Όταν διαλυθεί πλήρως, αφήνεται ψυχθεί σε θερμοκρασία δωματίου. IV. Προστίθενται 1ml 1Μ ΕPPS ph 8. V. Αποθηκεύεται στους -20οC. Η παραπάνω διαδικασία ακολουθείται και για FISH για διαφορετικά αναπτυξιακά στάδια, μόνο που επειδή απαιτείται μεγαλύτερος όγκος μονιμοποιημένων εμβρύων η πορεία γίνεται σε σωληνάκια των 5ml. 47
48 4.17 Χρωμογόνος in situ υβριδοποίηση Ημέρα 1η Σημαντικό είναι να τηρούνται RNAase-free συνθήκες, να αφήνονται τα έμβρυα να καθιζάνουν πριν το επόμενο βήμα και ότι όλοι οι χειρισμοί να γίνονται σε θερμοκρασία δωματίου. 1. Αφαιρείται όσο περισσότερη μεθανόλη γίνεται (στην μεθανόλη ήταν αποθηκευμένα τα έμβρυα) και γεμίζεται το πηγαδάκι με ΜΟPS Βuffer για να ξεπλυθούν τα έμβρυα από το μονιμοποιητικό διάλυμα. H παραμονή στο συγκεκριμένο διάλυμα διαρκεί περίπου 15 λεπτά. 2. Πραγματοποιούνται άλλα δύο πλυσίματα με MOPS Βuffer αφού έχουν καθιζάνει τα έμβρυα. 3. Αφαιρείται όσο περισσότερο από το MOPS Βuffer γίνεται και προστίθεται Ηybe Βuffer με DEP νερό μέχρι να γεμίσει το πηγαδάκι για να ξεπλυθεί από το MOPS Βuffer. Τα έμβρυα στο Ηybe Βuffer είναι πολύ δυσδιάκριτα. 4. Αφαιρείται και πάλι όσο περισσότερο από το υπερκείμενο γίνεται και προστίθενται ξανά Ηybe Βuffer με DEP νερό. Σημείωση: Γίνονται πλύσεις με δύο Ηybe Βuffer με DEP νερό γιατί το δίαλυμα αυτό έχει φορμαμίδιο που είναι αποδιατακτικός παράγοντας και πρέπει να είναι σε ποσοστό~70% κατά την προυβριδοποίηση και την υβριδοποίηση. 5. Καλύπτεται η plate με αυτοκόλλητο και μπαίνει σε φούρνο στους 50οC για προυβριδοποίηση για 5 με 6 ώρες. 6. Σε eppendorf προστίθενται σε αναλογία 1:1 το προθερμασμένο (για 5 λεπτά) 2xΗybe Βuffer (δηλ. με κατάλληλη ποσότητα RNA ανιχνευτή ώστε σε τελικό όγκο 100μΙ να έχουμε συγκέντρωση~0,3-0,5ng/μι). Eπωάζονται στους 50οC για 3 μέρες. Ημέρα 2η Δεν απαιτούνται πλέον RNAase-free συνθήκες. 1. Μετά το πέρας των 3 ημερών, αφαιρούνται από το πηγαδάκι πολύ προσεκτικά 50μΙ του Ηybe Βuffer με ανιχνευτή και προστίθενται 200μΙ Ηybe Βuffer με mq νερό χωρίς t-rna. Στην συνέχεια ξανακλείνεται με αυτοκόλλητο η plate και μπαίνει στον φούρνο στους 50οC για 1 ώρα. 2. Επαναλαμβάνεται 2 φορές το βήμα 1 μόνο που αφαιρείται όσο το δυνατό περισσότερο υπερκείμενο. 3. Αφαιρείται σχεδόν όλο το υπερκείμενο και γεμίζεται το πηγαδάκι με MOPS Βuffer. Παραμένουν τα έμβρυα σε αυτό το διάλυμα για περίπου 15 λεπτά. 4. Επαναλαμβάνεται το βήμα 3 αφαιρώντας όσο μεγαλύτερη ποσότητα υπερκείμενου γίνεται. 5. Αφαιρείται όσο περισσότερο από το ΜΟPS Βuffer γίνεται, γεμίζεται το πηγαδάκι με Blocking Βuffer και επωάζονται τα έμβρυα σε αυτό για ½ με 1h. 6. Ταυτόχρονα γίνεται blocking του Αnti-Dig αντισώματος το οποίο είναι συνδεδεμένο με αλκαλική φωσφατάση (Roche) σε eppendorf με Blocking Buffer σε αναλογία 1:1500 αντίσωμα/βuffer. 48

49 7. Τελικά αφαιρείται όσο περισσότερο γίνεται από το Blocking Buffer και προστίθενται 200μΙ από το διάλυμα Blocking Buffer-αντισώματος. 8. Κλείνεται η plate με αυτοκόλλητο και παραμένει στους 4οC overnight. Hμέρα 3η 1. Ξεπλένεται το διάλυμα του αντισώματος αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας το πηγαδάκι με MOPS Βuffer. 2. Επαναλαμβάνεται το βήμα 1 4 φορές. 3. Αφαιρείται όσο περισσότερο από το MOPS Βuffer γίνεται και γεμίζεται το πηγαδάκι με AP Βuffer που προσφέρει το απαραίτητο μαγνήσιο για την δράση της φωσφατάσης. 4. Επαναλαβάνεται το βήμα 3 1 φορά. 5. Γίνονται 2 πλύσεις όπως παραπάνω με το διάλυμα AP/levamisol/Tween. Το levamisol απενεργοποιεί τις ενδογενείς φωσφατάσες των εμβρύων. 6. Αφαιρείται όσο περισσότερο γίνεται από το υπερκείμενο και προστίθενται μΙ Dye. 7. Περιβάλλεται η plate με αλουμινόχαρτο γιατί η χρωστική είναι φωτοευαίσθητη. Αφήνεται σε θερμοκρασία δωματίου. 8. Ανα διαστήματα γίνεται παρατήρηση αν τα έμβρυα έχουν βάψει. Όταν και μόνο όταν βάψουν τα control έμβρυα, σταματάται την αντίδραση. 9. Αν περάσει μεγάλο χρονικό διάστημα και δεν έχει γίνει αυτό, αλλάζεται η χρωστική (αφαιρούνται 200μΙ παλιάς και προστίθενται 200μΙ νέας χρωστικής). 10.Όταν τα έμβρυα βάψουν, αφαιρείται όση περισσότερη από την χρωστική γίνεται και γεμίζεται το πηγαδάκι με TBST/EDTA, για να σταματήσει η αντίδραση. Το EDTA ως χηλικό αντιδραστήριο δεσμεύει τα ίοντα μαγνησίου του ΑΡ που είναι απαραίτητα για την οξειδοαναγωγική αντίδραση μετατροπής της άχρωμης ουσίας σε μπλε χρωστική. 11.Επαναλαμβάνεται το βήμα 10 άλλες 2 φορές. 12.Τέλος γίνονται 2 ξεπλύματα με TBST σε όγκο 200μΙ Υλικά για την χρωμογόνο in situ υβριδοποίηση: MOPS Βuffer (για τελικό όγκο 1ml) 49


50 Hybridization (Hybe) Βuffer (για τελικό όγκο 1ml) Blocking Βuffer: Διαλύεται σε MOPS Βuffer, Ρerkin Εlmer Βlocking Reagent ώστε να δημιουργηθεί τελική συγκέντρωση του 0,5%w/v. AP buffer (για τελικό όγκο 50ml): 50

: Dye")
51 Διάλυμα AP/levamisole/Tween (για τελικό όγκο περίπου 5,2 ml): Dye (για τελικό όγκο 1ml): 51

52 4.18 Διπλή φθορίζουσα in situ υβριδοποίηση (double FISH) Με την τεχνική αυτή επιτυγχάνεται εντοπισμός DNA ή RNA. Συγκεκριμένα, γίνεται επιλογή συμπληρωματικής RNA αλληλουχίας για ανίχνευση του mrna στον ιστό, μια μέθοδος που χρησιμοποιείται όταν η γονιδιακή ενεργότητα χρειάζεται να ελεγχθεί. Το πιο σημαντικό όμως είναι μπορεί να επιτευχθεί ταυτόχρονη ανάλυση δύο ή και περισσότερων γονιδίων στο ίδιο δείγμα, χρησιμοποιώντας τις διαφορετικές ιδιότητες διέγερσης (excitation) και εκπομπής (emission) των φθoροχρωμάτων. Αυτή η τεχνική είναι γρήγορη, ακριβής, με υψηλή αvάλυση εικόνας ακόμα και για γονίδια με χαμηλά επίδεδα έκφρασης και με μεγάλη ευελιξία στην χρήση των ψευδοχρωμάτων. Τα βελτιστοποιημένα πρωτόκολλα έχουν κάποια κοινά χαρακτηριστικά: 1. Διατήρηση της μορφολογίας των ιστών, 2. Διαπερατότητα των ιστών για τους ανιχνευτές, 3. Αποτελεσματική σύνδεση των ανιχνευτών, 4. Μείωση του μη ειδικού σήματος. Για την επίτευξη των παραπάνω, καθοριστικοί είναι οι εξής παράμετροι: η μονιμοποίηση, η διαπερατότητα, οι συνθήκες υβριδοποίησης και ο μετά την υβριδοποίηση χειρισμός. Περιληπτικά, ακολούνται τα εξής βήματα: 1. Σύνδεση του αντισώματος με το αντιγόνο (DIG ή DNP) που βρίσκεται πάνω στον αντινοηματικό ανιχνευτή. 2. Το αντίσωμα φέρει υπεροξειδάση. Το συγκεκριμένο ένζυμο όταν επωασθεί με Tyramide (TSA reagent), καταλύει την δημιουργία ελευθέρων ριζών TSA. 3. Oι ελεύθερες ρίζες TSA δημιουργούν ομοπολικούς δεσμούς με τα κατάλοιπα τυροσίνης των ενδογενών πρωτεϊνών κοντά στην υπεροξιδάση. Οι μη συνδεδεμένες ρίζες TSA δημιουργούν διμερή που απομακρύνονται με πλύσεις. Έτσι, όπου είναι εντοπισμένο το mrna, εκεί και μόνο εντοπίζεται το ένζυμο και το σήμα χωρίς να διαχεέται καθόλου. 52
53 Η διαδικασία είναι η ακόλουθη: 1η και 2η μέρα Είναι ίδια με την πορεία που ακολουθείται στην χρωμογόνο in situ υβριδοποίηση με την μόνη διαφορά ότι όλες οι διαδικασίες πραγματοποιούνται σε eppendorf και όχι σε plate και σε όγκο 200μΙ περίπου. Επίσης το Anti-Dig (Roche) είναι συνδεδεμένο με υπεροξειδάση (και όχι με αλκαλική φωσφατάση) και αραιώνεται στο Blocking Reagent της Perkin Elmer. 3η μέρα 1. Ξεπλένεται το διάλυμα του αντισώματος αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και προσθέτοντας 200μΙ MOPS Βuffer. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 2. Επαναλαμβάνεται το βήμα 1 4 φορές. 3. Ξεπλένεται το MOPS Βuffer αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και προσθέτοντας 200μΙ Αmplification Diluent (Perkin Elmer). Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 4. Ξεπλένεται το Αmplification Diluent αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας με 200μΙ Cy3 (Perkin Elmer)-Αmplification Diluent σε αναλογία 1:500i. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. Από εδώ και στο εξής διατηρούνται τα έμβρυα στο σκοτάδι. 5. Ξεπλένεται το Cy3-Αmplification Diluent αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας με 200μΙ MOPS Βuffer. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 6. Επαναλαμβάνεται το βήμα 5 4 φορές. 7. Ξεπλένεται το MOPS Βuffer αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας με 200μΙ 1% H2O2 για την απενεργοποίηση του ενζύμου. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 30 λεπτά. 8. Ξεπλένεται το διάλυμα του H2O2 αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας με 200μΙ MOPS Βuffer. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 9. Επαναλαμβάνεται το βήμα 8 4 φορές. 10.Αφαιρείται όσο περισσότερο από το ΜΟPS Βuffer γίνεται, και προστίθενται 200 μι Blocking Reagent της Perkin Elmer και επωάζονται τα έμβρυα σε αυτό για ½ με 1 ώρα. 11.Ταυτόχρονα γίνεται blocking του Αnti-DΝΡ αντισώματος το οποίο είναι συνδεδεμένο με αλκαλική φωσφατάση (Perkin Elmer) σε eppendorf με Blocking Reagent της Perkin Elmer σε αναλογία 1:1500 αντίσωμα/βuffer. 12.Τελικά αφαιρείται όσο περισσότερο γίνεται από το Blocking Buffer και προστίθενται 200μΙ από το διάλυμα Blocking Buffer-αντισώματος. 13.Παραμένουν τα έμβρυα στους 4οC overnight. 53
54 4η μέρα 1. Ξεπλένεται το διάλυμα του αντισώματος αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας με 200μΙ MOPS Βuffer. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 2. Επαναλαμβάνεται το βήμα 1 4 φορές. 3. Ξεπλένεται το MOPS Βuffer αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και προσθέτοντας 200μΙ Αmplification Diluent (Perkin Elmer). Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 4. Ξεπλένεται το Αmplification Diluent αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας με 200μΙ Cy5 (Perkin Elmer)-Αmplification Diluent σε αναλογία 1:500ii. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 5. Ξεπλένεται το Cy5-Αmplification Diluent αφαιρώντας όσο περισσότερο από το υπερκείμενο γίνεται και γεμίζοντας με 200μΙ MOPS Βuffer. Σε αυτό το διάλυμα παραμένουν τα έμβρυα για 15 λεπτά. 6. Επαναλαμβάνεται το βήμα 5 4 φορές. Σε κάθε περίπτωση μετά τις υβριδοποίσεις, τα έμβρυα διατηρούνται στο ψυγείο (εάν έχουν φθορόχρωμα, προστατευμένα από το φως) σε διάλυμα 1:1 γλυκερόλη/ MOPS Βuffer. Όταν είναι να παρατηρηθούν σε φωτονικό (μετα από χρωμογόνο υβριδοποίηση) ή σε συνεστιακό μικροσκόπιο (μετά από φθορίζουσα), σε μια καλυπτρίδα προστίθενται περίπου 15μΙ αραιωμένων εμβρύων. Στην περίπτωση της φθορίζουσας υβριδοποίσης, προστίθεται και 1μΙ DAPI (working solution) για να διακριθούν οι πυρήνες. Έπειτα καλύπτονται με καλυπτρίδα προσέχωντας να διασφαλίζεται η μορφολογία και η τρισδιάστατη δομή των εμβρύων Απομόνωση RNA (περίπου 1,5 μgr) από μικρό αριθμό control και ενεμένων εμβρύων για Q-PCR Για την ποσοτική εκτίμηση της επίδρασης της καταστολής του PlCoup-TF στα γονίδια της νευρογένεσης ήταν απαραίτητη η απομόνωση RNA από control και ενεμένα έμβρυα (20 ώρες περίπου μετά την γονιμοποίηση) και η χρήση αυτού ως υπόστρωμα για Q- PCR. Η διαδικασία που ακολουθήθηκε ήταν η παρακάτω: 1. Συλλογή των εμβρύων (από τα πλαστικά καπάκια που αναπτύσσονταν) σε eppendorf. 2. Φυγοκέντρηση για 5 λεπτά (300g) και απομάκρυνση του υπερκείμενου θαλασσινού νερού. 3. Ελαφρύ χτύπημα του eppendorf. Προσθήκη 350μΙ RLT (διαλύματος λύσης). Vortex ή πιπετάρισμα για ομογενοποίηση. 4. Προσθήκη (στο διάλυμα των λυμένων κυττάρων) ενός όγκου 70% αιθανόλης, δηλ. 350μΙ.Πιπετάρισμα και ελαφρύ χτύπημα του eppendorf. 5. Μεταφορά σε κολώνα του kit που από κάτω φέρει 2mI tube. Φυγοκέντρηση για 15 δευτερόλεπτα (8000g). Απόρριψη εκλούσματος. 54
 στην κολώνα. Φυγοκέντρηση για 15 δευτερόλεπτα (8000g). Απόρριψη εκλούσματος. 9.")
55 6. Προσθήκη 350μΙ RW1 (διαλύματος πλύσης) στην κολώνα. Φυγοκέντρηση για 15 δευτερόλεπτα (8000g). Απόρριψη εκλούσματος. 7. Προσθήκη 10μΙ DNAase I σε 70μΙ RDD (για πέψη DNA). Απαλή ανακίνηση. Προσθήκη αυτού στην κολώνα. Αναμονή σε θερμοκρασία δωματίου για 15 λεπτά. 8. Προσθήκη 350μΙ RW1 (διαλύματος πλύσης) στην κολώνα. Φυγοκέντρηση για 15 δευτερόλεπτα (8000g). Απόρριψη εκλούσματος. 9. Προσθήκη 500μΙ RPE (διαλύματος πλύσης) στην κολώνα. Φυγοκέντρηση για 15 δευτερόλεπτα (8000g). Απόρριψη εκλούσματος. 10.Προσθήκη 500μΙ 80% αιθανόλης (διαλύματος πλύσης) στην κολώνα. Φυγοκέντρηση για 2 λεπτά (8000g). Απόρριψη εκλούσματος και tube. 11.Ξηρή φυγοκέντρηση για 5 λεπτά (full spead). Απόρριψη εκλούσματος και tube. 12.Προσθήκη 25 μι DEP νερό (για έκλουση) στην κολώνα. Φυγοκέντρηση για 1 λεπτό (full spead). 13.Φύλαξη στους -80οC. H παραπάνω διαδικασία πραγματοποιήθηκε με το Qiagen RNAase Micro Kit και χρησιμοποίηθηκαν περίπου 500 έμβρυα σε κάθε ομάδα control ή ενεμένων εμβρύων για κάθε μελετηθέν γονίδιο, με 3ngr RNA/έμβρυο (ή αλλιώς 1,5μgr συνολικά) και περίπου 250 κύτταρα/βλαστίδιο (ή αλλιώς κύτταρα συνολικά) και 400 κύτταρα/γαστρίδιο (ή αλλιώς κύτταρα συνολικά) Q-PCR από RNA control και ενεμένων εμβρύων Χρησιμοποιώντας ως υπόστρωμα το απομονωμένο RNA control και ενεμένων εμβρύων, φτιάχτηκαν cdnas για μελλοντική χρήση σε QPCR πειράματα. Συγκεκριμένα ακολουθήκε η παρακάτω πορεία: 1. Φυγοκέντρηση υπό κενό των δειγμάτων έως ότου επέλθει ξήρανση. Επαναδιάλυση σε 13 μι DEP νερό με συνεχές ελαφρύ χτύπημα του eppendorf σε θερμοκρασία δωματίου για 30 λεπτά. 2. Για την αποδιάταξη των δευτερογενών δομών, απαιτείται παραμονή των δειγμάτων στους 65οC για 3 λεπτά και έπειτα στον πάγο. Αφού προστεθούν τα παρακάτω αντιδραστήρια, ακολουθείται το πρόγραμμα που περιγράφεται. 55



56 To c-dna μπορεί να χρησιμοποιηθεί ως υπόστρωμα για την Q-PCR που περιλαμβάνει τα ακόλουθα αντιδραστήρια και ακολουθεί το παρακάτω πρόγραμμα. Τα πειράματα QPCR πραγματοποίηθηκαν και σε άλλες θερμοκρασίες, όμως αυτές που παρουσιάζονται είναι οι πιο αποδοτικές. Επίσης χρησιμοιήθηκε με λιγότερη επιτυχία και ένα δεύτερο ζευγάρι primer για κάθε γονίδιο. Παρακάτων παρουσιάζονται οι αλληλουχίες των primer που χρησιμοποήθηκαν τελικά. Η SyBr Green περιέχει και τα υπόλοιπα απαραίτητα αντιδραστήρια για την αντίδραση, όπως την πολυμεράση και τα νουκλεοτίδια, και προστίθενται τελευταία για την αποφυγή του φωτός και της επίδρασης της θερμοκρασίας δωματίου στο ένζυμο. Επίσης η προστιθέμενη ποσότητα υποστρώματος εξαρτάται από την αραίωσή του και το mq Η2Ο προστίθενται αντί για το υπόστρωμα για να ελεγχεί ο πιθανός σχηματισμός παραπροϊόντών. Tα προϊόντα των αντιδράσεων στάλθηκαν για sequencing στην εταιρεία VBC μετά από καθαρισμό για απομάκρυνση της SyBr. 56

57 5 Αποτελέσματα 5.1 Αποτελέσματα RT-PCR Στόχος, λοιπόν της εργασία είναι να οδηγηθούμε σε επιτυχή παρατήρηση της έκφρασης των γονιδίων Six3, FoxQ2 και Nkx2.1 αλλά και του πως η καταστολή του ορφανού υποδοχέα Coup-TF επιδρά ή όχι στο φαινότυπο τους. Έχοντας, λοιπόν, πραγματοποιήσει απομόνωση RNA απο διαφορετικά αναπτυξιακά στάδια προχωρούμε σε αντίστροφη μεταγραφή RT PCR με κατάλληλους primers για κάθε γονίδιο σε 3 διαφορετικές θερμοκρασίες 54, 56.6 και 60 βαθμούς Κελσίου αντίστοιχα. Στη εικόνα βλέπουμε το μέγεθος των κλώνων που ενθέσαμε στο φορέα Pgem T-easy και τους κλώνους τους οποίους επιλέξαμε και στείλαμε για αλληλούχιση. Τα απότελεσματα της αλληλούχισης είναι συγκεντρωμένα στο παράρτημα και μας επαλήθευσαν την ορθότητα του πειράματος. Εικόνα 24: Αποτελέσματα RT-PCR: Οι κλώνοι ηλεκτροφορήθηκαν σε πήκτωμα αγαρόζης. Χρησιμοποίησαμε 1kb Ladder. Όπου F6: FoxQ 2 (6), N3: Nkx2.1 (3), S6: Six3 (6). 57


58 5.2 Κλωνοποίηση Εν συνεχεία αφού ενθέσαμε τους κλώνους στο φορέα Pgem T-easy πραγματοποιήθηκε μετασχηματισμός δεκτικών βακτηρίων και επιλογή των αποικιών με screening σε τρυβλία όπου υπήρχε επίστρωση X-gal. Στη συνέχεια πραγματοποίησαμε απομόνωση των πλασμιδίων ( Mini Preps ) και σε πήκτωμα αγαρόζης επιβεβαίωσαμε την ύπαρξη ενθέματος. Στις εικόνες φαίνεται η επιλογή των κλώνων οι οποιοί εστάλησαν για αλληλούχιση και επαλήθευσαν την ορθότητα του πειράματος. Εικόνα 25 και 26: Απομόνωση πλασμιδίων (mini preps) και ηλεκτροφόρηση Σε πήκτωμα αγαρόζης. Όπου F10: FoxQ 2 (10), N6: Nkx2.1 (6), S3: Six3 (3). 58
 υβριδοποιήση.")
59 5.3 Πέψη με ένζυμα περιορισμού Προχωρήσαμε σε πέψη των κλώνων με ένζυμο περιορισμού την EcoRI. και εν συνεχεία σε PCR με σκοπό τη δημιουργία ιχνηθετών για χρωμογόνο in situ υβριδοποιήση καθως και φθορίζουσα In situ (fish) υβριδοποιήση. Στην εικόνα βλέπουμε τα αποτελέσματα της PCR σε πήκτωμα αγαρόζης. Εικόνα 27: Οι επιλεγμένοι κλώνοι πολλαπλασιάστηκαν με PCR και ηλεκτροφορήθηκα σε πήκτωμα Αγαρόζης. Χρησιμοποιήσαμε 1kb Ladder. Όπου F10: FoxQ 2 (10), N6: Nkx2.1 (6), S3: Six3 (3). 59

60 5.4 In vitro μεταγραφή Με in vitro μεταγραφη δημιουργήσαμε ιχνηθετές RNA κατάλληλους με σκοπό τον εντοπισμό τους στα διάφορετικά αναπτυξιακά στάδια εμβρύων μέσω της τεχνικής In situ υβριδοποίησης και Fish υβριδοποιησης. Εικόνα 28: RNA ιχνηθέτες για τα γονίδια Nkx2.1, Six3 και FoxQ2 60
61 5.5 Αναπτυξιακό πρότυπο έκφρασης για το γονιδιο Six3 Πραγματοποιήσαμε διπλή φθορίζουσα fish υβριδοποιηση, νέα τεχνική για το εργαστήριο μας, με σκοπό να αποτυπώσουμε το πρότυπο έκφρασης των γονιδίων με φθορισμο σε 3 διαφορετικά αναπτυξιακά στάδια. Οι ιχνηθέτες που σχεδιάσαμε μας έδωσαν τη δυνατότητα καθώς δεσμεύτηκαν στις περιοχές έκφρασης των γονιδίων, δίνοντας σήμα μέσω φθορισμού. Όσον αφορά το μεταγραφικό παράγοντα Six3 παρατηρουμε ότι στο σταδιο του βλαστιδίου υπάρχει μερική και διάσπαρτη έκφραση στην περιοχή του στοματικού εξωδέρματος. Στο στάδιο του γαστριδίου η έκφραση γινεται πιο εντονη στην περιοχή ενω αρχίζει να εντοπιζεται έκφραση και στο έντερο στην περιοχή των σφικτήρων. Στο στάδιο του πλουτέα παρατηρειται έντονη έκφραση στην περιοχή του εντέρου και συγκεκριμένα στους σφικτήρες και ευρύτερα στην βλεφαριδωτή ζώνη. Ωστόσο αξίζει να σημειωθεί ότι ενώ η έκφραση του Coup TF εντοπίζεται σε όλη την περιοχή του στοματικού εξωδέρματος, η έκφραση του Six3 εντοπίζεται στην ευρύτερη περιοχή του εξωδέρματος αλλά όχι στο σημείο όπου θα δημιουργηθεί το apical organ. Παρατηρείται λοιπόν κενό στην περιοχή του ANE. 61



62 Εικόνα 29a,b,c: 29a)Συνέκφραση των Six3 και Coup-TF στο στάδιο του βλαστιδίου 29b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του Six3 στο στάδιο του βλαστιδίου 29c) Mε πράσινο φθορισμό η έκφραση του Coup-TF 62

 Με κόκκινο χρώμα φθορισμού")
 Mε")
63 Εικόνα 30a,b,c: 30a)Συνέκφραση των Six3 και Coup-TF στο στάδιο του γαστριδίου. 30b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του Six3 στο στάδιο του γαστριδίου 30c) Mε πράσινο φθορισμό η έκφραση του Coup-TF 63
 Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του Six3 στο στάδιο του")


64 Εικόνα 31a,b,c: 31a)Συνέκφραση των Six3 και Coup-TF στο στάδιο του πλουτέα. 31b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του Six3 στο στάδιο του Πλουτέα 31c) Mε πράσινο φθορισμό η έκφραση του Coup-TF. Ο λόγος που επιλέχθει να πραγματοποιηθουν πειράματα συνεντοπίσμου ήταν γιατί θέλαμε να δείξουμε και τις ακριβείς περιοχές συνέκφρασης των γονιδίων έτσι ώστε σε knock down εμβρυα Coup TF να παρατηρήσουμε τις πιθανές ή μη αλλαγές έκφραση των γονιδίων. 64


65 5.5.1 Έκφραση του γονιδίου Six3 σε Coup-TF morphants Σε knock down Coup-TF ενεμένα έμβρυα λοιπόν παρατηρήθηκε ότι ο Six3 δεν εκφράζεται. Εικόνα 32: Ενεμένα έμβρυα με αντινοηματικό CoupTF RNA (Knocked down regulation ) Ώστοσο παρότι είχαμε μια πρώτη εικόνα με φθορισμό όσον αφορά το πρότυπο έκφρασης για το γονίδιο Six3 πραγματοποίησαμε πειράματα και in situ υβριδοποίησης με AP αντισωμα ώστε να έχουμε ολοκληρωμένη εικόνα. 65


66 Γι αυτό το σκοπό πραγματοποιήσαμε ενέσεις με αντινοηματικό RNA Coup-TF σε έμβρυα και παράλληλα είχαμε control έμβρυα όπου σταματήσαμε την ανάπτυξη τους στο στάδιο του γαστριδίου. Στη συνέχεια με αντίσωμα AP καταφέραμε να παρατηρήσουμε το πρότυπο έκφρασης του Six3 τόσο στα control έμβρυα όσο και στα ενεμένα όπου ο coup ήταν σε καταστολή. Εικόνα 33: Control gastrula Six3 Στο control έμβρυο η έκφραση παρατηρείται στο στοματικό εξώδερμα και στο έντερο γύρω από την περιοχή των σφικτήρων. Εικόνα 34: Knocked down COUP-TF morphants Παρατηρούμε λοιπόν ότι η έκφραση του Six3 καταστέλλεται σε έμβρυα όπου δεν εκφράζεται και ο Coup. 66
67 5.5.2 Q-PCR Το ότι ο παράγοντας Six3 δεν εκφράζεται σε Coup-TF morphants μας επιβεβαίωσε και η ποσοτική PCR (Q-PCR). Όπως βλέπουμε στο διάγραμμα η έκφραση του γονιδίου είναι 30 φορές μικρότερη. 5.6 Αναπτυξιακό πρότυπο έκφρασης για το γονιδιο FoxQ2 Όσον αφορά το πρότυπο έκφρασης του γονιδίου FoxQ2 εντοπίζεται καθαρα στην περιοχή ΑΝΕ όπως θα δούμε σε όλα τα στάδια έκφρασης. Παρατηρουμε λοιπόν ότι ο Coup-TF(με πράσινο φθoρισμό) εκφράζεται σε όλη την περιοχή του στοματικού εξωδέρματος, ενώ ο FoxQ2 (με κοκκινο φθορισμό) μόνο στη περιοχή του ΑΝΕ και στα τρία αναπτυξιακά στάδια. Ο συνεντοπισμός στην περιοχή του ΑΝΕ με κίτρινο χρώμα επιβεβαιωνει την κοινή περιοχή έκφρασης. 67

 Με κόκκινο χρώμα φθορισμού")
Mε")
68 Εικόνα 35a,b,c: 35a)Συνέκφραση των FoxQ2 και Coup-TF στο στάδιο του βλστιδίου. 35b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του FoxQ2 στο στάδιο του βλαστιδίου 35c)Mε πράσινο φθορισμό η έκφραση του Coup-TF 68

 Με κόκκινο χρώμα φθορισμού")
 Mε")
69 Εικόνα 36a,b,c: 36a)Συνέκφραση των FoxQ2 και Coup-TF στο στάδιο του γαστριδίου. 36b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του FoxQ2 στο στάδιο του γαστριδίου 36c) Mε πράσινο φθορισμό η έκφραση του Coup- TF 69

Με κόκκινο χρώμα φθορισμού")
 Mε πράσινο")
70 Εικόνα 37a,b,c: 37a)Συνέκφραση των FoxQ2 και Coup-TF στο στάδιο του Πλουτέα. 37b)Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του FoxQ2 στο στάδιο του Πλουτέα 37c) Mε πράσινο φθορισμό η έκφραση του Coup-TF 70

71 5.6.1 Έκφραση του γονιδίου FoxQ2 σε Coup-TF morphants Σε knocked down Coup-TF morphants παρουσιάζεται ενας μερικώς φθορισμός, ωστόσο δεν είναι αρκετός για να βγάλουμε ασφαλή συμπεράσματα για το τι γίνεται με την έκφραση του FoxQ2. Εικόνα 38: Ενεμένα έμβρυα CoupTF (Knocked down regulation ) Γι αυτό το σκοπό πραγματοποιήσαμε ενέσεις με αντινοηματικό RNA Coup-TF σε έμβρυα και παράλληλα είχαμε control έμβρυα όπου σταματήσαμε την ανάπτυξη τους στο στάδιο του γαστριδίου. Στη συνέχεια με αντίσωμα AP καταφέραμε να παρατηρήσουμε το πρότυπο έκφρασης του FoxQ2 τόσο στα control έμβρυα όσο και στα ενεμένα όπου ο coup ήταν σε καταστολή. 71


72 Εικόνα 39: Control gastrula FoxQ2 Η έκφραση του FoxQ2 εντοπίζεται στην περιοχή του ΑΝΕ. Εικόνα 40: Knocked down COUP-TF morphants Παρατηρούμε λοιπόν ότι η ειδική έκφραση του FoxQ2 διατηρείται και σε ενεμένα έμβρυα. 72
73 5.6.2 Q-PCR Το ότι ο παράγοντας FoxQ2 εκφράζεται σε Coup-TF morphants μας επιβεβαίωσε και η ποσοτική PCR (Q-PCR). Όπως βλέπουμε στο διάγραμμα η έκφραση του γονιδίου είναι σχεδόν ισάξια με τα control έμβρυα. 73


Συνέκφραση των Νkx2.1 και Coup-TF στο στάδιο του βλαστιδίου.")
74 5.7 Αναπτυξιακό πρότυπο έκφρασης για το γονιδιο Nkx2.1 Η έκφραση του Nkx2.1 παρουσίαζεται διάσπαρτα στην περιοχή του ΑΝΕ στο στάδιο του βλαστιδίου. Στο στάδιο του γαστριδίου παρουσίαζεται τόσο στο ΑΝΕ όσο και στην ευρύτερη περιοχή του στοματικού εξωδέρματος, γι αυτο ο συνεντοπισμός είναι μεγαλύτερος με τον παραγοντα Coup. Στο στάδιο του πλουτέα η έκφραση φαίνεται παλι να περιορίζεται στην περιοχή του ΑΝΕ. Εικόνα 41a,b,c: 41a)Συνέκφραση των Νkx2.1 και Coup-TF στο στάδιο του βλαστιδίου. 41b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του Nkx2.1 στο στάδιο του βλαστιδίου 41c) Mε πράσινο φθορισμό η έκφραση του Coup- TF 74

 Με κόκκινο χρώμα φθορισμού")
 Mε")
75 Εικόνα 42a,b,c: 42a) Συνέκφραση των Nkx2.1 και Coup-TF στο στάδιο του γαστριδίου. 42b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του Nkx2.1 στο στάδιο του γαστριδίου 42c) Mε πράσινο φθορισμό η έκφραση του Coup- TF 75
 Mε πράσινο φθορισμό η έκφραση του Coup-TF Σε δευτέρο στάδιο κι αφού επιβεβαιώσαμε το προφιλ έκφρασης του Nkx2.")
76 Εικόνα 43a,b,c: 43a)Συνέκφραση των Nkx2.1 και Coup-TF στο στάδιο του Πλουτέα. 43b) Με κόκκινο χρώμα φθορισμού δίνεται η έκφραση του Nkx2.1 στο στάδιο του Πλουτέα 43c) Mε πράσινο φθορισμό η έκφραση του Coup-TF Σε δευτέρο στάδιο κι αφού επιβεβαιώσαμε το προφιλ έκφρασης του Nkx2.1 πραγματοποιήσαμε πειραματα με AP αντίσωμα ώστε να δούμε σε ενεμένα έμβρυα ποιο το πρότυπο έκφρασης του. 76


77 5.7.1 Έκφραση του γονιδίου Nkx2.1 σε Coup-TF morphants Σε knocked down Coup-TF ενεμένα έμβρυα παρατηρηθηκε ότι διατηρείται η έκφραση του Nkx2.1 Εικόνα 44: Control gastrula Nkx2.1 Εικόνα 45: Knocked down Coup-TF morphants 77
Λαμπρινή Καλαμπόκη Α.Μ. 302
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΕΡΕΥΝΗΤΙΚΗ ΕΡΓΑΣΙΑ ΛΕΙΤΟΥΡΓΙΚΗ ΑΝΑΛΥΣΗ ΤΗΣ ΡΥΘΜΙΣΤΙΚΗΣ ΠΕΡΙΟΧΗΣ ΤΟΥ ΓΟΝΙΔΙΟΥ COUP-TF ΣΤΟ ΕΜΒΡΥΟ ΤΟΥ ΑΧΙΝΟΥ Λαμπρινή
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΕΡΕΥΝΗΤΙΚΗ ΕΡΓΑΣΙΑ ΛΕΙΤΟΥΡΓΙΚΗ ΑΝΑΛΥΣΗ ΤΗΣ ΡΥΘΜΙΣΤΙΚΗΣ ΠΕΡΙΟΧΗΣ ΤΟΥ ΓΟΝΙΔΙΟΥ COUP-TF ΣΤΟ ΕΜΒΡΥΟ ΤΟΥ ΑΧΙΝΟΥ Λαμπρινή
Έναρξη της μεταγραφής = μείωση του ρυθμού διαίρεσης
 Αυλάκωση vs Γαστριδίωση Αυλάκωση: απανωτές μιτωτικές διαιρέσεις που οδηγούν σε λογαριθμική αύξηση του αριθμού των κυττάρων αλλά χωρίς αύξηση του συνολικού όγκου του αναπτυσσόμενου εμβρύου Έναρξη της μεταγραφής
Αυλάκωση vs Γαστριδίωση Αυλάκωση: απανωτές μιτωτικές διαιρέσεις που οδηγούν σε λογαριθμική αύξηση του αριθμού των κυττάρων αλλά χωρίς αύξηση του συνολικού όγκου του αναπτυσσόμενου εμβρύου Έναρξη της μεταγραφής
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ Ο μεταγραφικός παράγοντας Coup-TF και το γονιδιακό ρυθμιστικό δίκτυο στο νευρικό σύστημα του αχινού Paracentrotus lividus
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΓΕΝΕΤΙΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ ΚΑΙ ΑΝΑΠΤΥΞΗΣ Ο μεταγραφικός παράγοντας Coup-TF και το γονιδιακό ρυθμιστικό δίκτυο στο νευρικό σύστημα του αχινού Paracentrotus lividus
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής
 Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; ΘΩΜΑΣ ΑΠΑΝΤΗΣΗ. 2. Ποιες είναι οι κατηγορίες γονιδίων με κριτήριο το προϊόν της μεταγραφής τους;
 Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
Ο ΡΟΛΟΣ ΤΗΣ ΚΑΡΒΟΞΥ-ΤΕΛΙΚΗΣ ΕΠΕΚΤΑΣΗΣ ΤΗΣ ΕΠΙΚΡΑΤΕΙΑΣ ΠΡΟΣΔΕΣΗΣ ΣΤΟ DNA ΤΟΥ ΜΕΤΑΓΡΑΦΙΚΟΥ ΠΑΡΑΓΟΝΤΑ COUP-TF
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ Ο ΡΟΛΟΣ ΤΗΣ ΚΑΡΒΟΞΥ-ΤΕΛΙΚΗΣ ΕΠΕΚΤΑΣΗΣ ΤΗΣ ΕΠΙΚΡΑΤΕΙΑΣ ΠΡΟΣΔΕΣΗΣ ΣΤΟ DNA ΤΟΥ ΜΕΤΑΓΡΑΦΙΚΟΥ ΠΑΡΑΓΟΝΤΑ COUP-TF ΜΕΤΑΠΤΥΧΙΑΚΟ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ Ο ΡΟΛΟΣ ΤΗΣ ΚΑΡΒΟΞΥ-ΤΕΛΙΚΗΣ ΕΠΕΚΤΑΣΗΣ ΤΗΣ ΕΠΙΚΡΑΤΕΙΑΣ ΠΡΟΣΔΕΣΗΣ ΣΤΟ DNA ΤΟΥ ΜΕΤΑΓΡΑΦΙΚΟΥ ΠΑΡΑΓΟΝΤΑ COUP-TF ΜΕΤΑΠΤΥΧΙΑΚΟ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
Τμήμα Βιολογίας Πανεπιστημίου Πατρών Τομέας Γενετικής, Βιολογίας Κυττάρου και Ανάπτυξης Μεταπτυχιακό Πρόγραμμα Σπουδών
 Τμήμα Βιολογίας Πανεπιστημίου Πατρών Τομέας Γενετικής, Βιολογίας Κυττάρου και Ανάπτυξης Μεταπτυχιακό Πρόγραμμα Σπουδών Ο Coup-TF και τα προγονικά νευρικά κύτταρα στο έμβρυο του αχινού Paracentrotus lividus
Τμήμα Βιολογίας Πανεπιστημίου Πατρών Τομέας Γενετικής, Βιολογίας Κυττάρου και Ανάπτυξης Μεταπτυχιακό Πρόγραμμα Σπουδών Ο Coup-TF και τα προγονικά νευρικά κύτταρα στο έμβρυο του αχινού Paracentrotus lividus
BIOΛ 102 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ
 BIOΛ 102 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ Γονιμοποίηση Αυλάκωση Γαστριδίωση Νευριδίωση Οργανογένεση ΕΜΒΡΥΟΛΟΓΙΑ Δ. Δοκιανάκη & Μ. Παυλίδης Ηράκλειο, Νοέμβριος 2015 1 ΓΟΝΙΜΟΠΟΙΗΣΗ* 1. Επαφή, αναγνώριση & πρόσδεση
BIOΛ 102 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ Γονιμοποίηση Αυλάκωση Γαστριδίωση Νευριδίωση Οργανογένεση ΕΜΒΡΥΟΛΟΓΙΑ Δ. Δοκιανάκη & Μ. Παυλίδης Ηράκλειο, Νοέμβριος 2015 1 ΓΟΝΙΜΟΠΟΙΗΣΗ* 1. Επαφή, αναγνώριση & πρόσδεση
Αφρικανικός βάτραχος. Ψάρι ζέβρα
 Εθνικόν και Καποδιστριακόν Πανεπιστήµιον Αθηνών ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ & ΒΙΟΦΥΣΙΚΗΣ ΑΝΑΠΤΥΞΙΑΚΗ ΒΙΟΛΟΓΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΑ Επικ. Καθ. ΒασιλικήΟικονοµίδου εκέµβριος 2014 Οι 6 δηµοφιλέστεροι
Εθνικόν και Καποδιστριακόν Πανεπιστήµιον Αθηνών ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΛΟΓΙΑΣ ΚΥΤΤΑΡΟΥ & ΒΙΟΦΥΣΙΚΗΣ ΑΝΑΠΤΥΞΙΑΚΗ ΒΙΟΛΟΓΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΑ Επικ. Καθ. ΒασιλικήΟικονοµίδου εκέµβριος 2014 Οι 6 δηµοφιλέστεροι
Ζωολογία Ι. Εργαστηριακή Άσκηση : ΑΝΑΠΑΡΑΓΩΓΗ. Ενότητα 4η: Η Διαδικασία της Αναπαραγωγής
 Ζωολογία Ι Εργαστηριακή Άσκηση : ΑΝΑΠΑΡΑΓΩΓΗ Ενότητα 4η: Η Διαδικασία της Αναπαραγωγής Συγγραφείς: Α. Νικολαίδου, Μ. Βεινή Χαρίτου Διδάσκων: Σκ. Ντέντος Τμήμα ΒΙΟΛΟΓΙΑΣ, Τομέας Ζωολογίας Θαλάσσιας Βιολογίας
Ζωολογία Ι Εργαστηριακή Άσκηση : ΑΝΑΠΑΡΑΓΩΓΗ Ενότητα 4η: Η Διαδικασία της Αναπαραγωγής Συγγραφείς: Α. Νικολαίδου, Μ. Βεινή Χαρίτου Διδάσκων: Σκ. Ντέντος Τμήμα ΒΙΟΛΟΓΙΑΣ, Τομέας Ζωολογίας Θαλάσσιας Βιολογίας
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β
 ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς
 Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
ΓΟΝΙΜΟΠΟΙΗΣΗ* BIO 101 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ. Επαφή, Αναγνώριση & Πρόσδεσητου σπερματοζωαρίου στο ωάριο ΓΟΝΙΜΟΠΟΙΗΣΗ ΑΥΛΑΚΩΣΗ
 ΔΙΑΛΕΞΗ 2. BIO 101 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ ΓΟΝΙΜΟΠΟΙΗΣΗ ΑΥΛΑΚΩΣΗ ΔΙΔΑΣΚΩΝ Μιχάλης Παυλίδης (pavlidis@biology.uoc.gr) Hράκλειο, Νοέμβριος 2013 ΓΟΝΙΜΟΠΟΙΗΣΗ* 1. Επαφή, αναγνώριση & πρόσδεση σπερματοζωαρίου
ΔΙΑΛΕΞΗ 2. BIO 101 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ ΓΟΝΙΜΟΠΟΙΗΣΗ ΑΥΛΑΚΩΣΗ ΔΙΔΑΣΚΩΝ Μιχάλης Παυλίδης (pavlidis@biology.uoc.gr) Hράκλειο, Νοέμβριος 2013 ΓΟΝΙΜΟΠΟΙΗΣΗ* 1. Επαφή, αναγνώριση & πρόσδεση σπερματοζωαρίου
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
BIO 101 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ
 BIO 101 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ ΙΑΛΕΞΗ 2. ΓΟΝΙΜΟΠΟΙΗΣΗ - ΑΥΛΑΚΩΣΗ Ι ΑΣΚΩΝ Μιχάλης Παυλίδης (pavlidis@biology.uoc.gr) Hράκλειο, Νοέμβριος 2012 1 ΓΟΝΙΜΟΠΟΙΗΣΗ* 1. 2. 3. 4. Επαφή, αναγνώριση & πρόσδεση σπερματοζωαρίου
BIO 101 ΕΙΣΑΓΩΓΗ ΣΤΗ ΖΩΟΛΟΓΙΑ ΙΑΛΕΞΗ 2. ΓΟΝΙΜΟΠΟΙΗΣΗ - ΑΥΛΑΚΩΣΗ Ι ΑΣΚΩΝ Μιχάλης Παυλίδης (pavlidis@biology.uoc.gr) Hράκλειο, Νοέμβριος 2012 1 ΓΟΝΙΜΟΠΟΙΗΣΗ* 1. 2. 3. 4. Επαφή, αναγνώριση & πρόσδεση σπερματοζωαρίου
Γονιμοποίηση αναγνώριση και συνένωση ωαρίου-σπερματοζωαρίου φραγμός στην πολυσπερμία μετα μετ βολική ενεργο ενεργο ο π ίηση
 Γονιμοποίηση αναγνώριση και συνένωση ωαρίου-σπερματοζωαρίου φραγμός στην πολυσπερμία μεταβολική ενεργοποίηση του αυγού ανακατατάξεις στα συστατικά του αυγού σχηματισμός του διπλοειδή πυρήνα του ζυγωτού
Γονιμοποίηση αναγνώριση και συνένωση ωαρίου-σπερματοζωαρίου φραγμός στην πολυσπερμία μεταβολική ενεργοποίηση του αυγού ανακατατάξεις στα συστατικά του αυγού σχηματισμός του διπλοειδή πυρήνα του ζυγωτού
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΘΕΜΑ Α Α1. γ Α2. γ Α3. α Α4. β Α5. β ΘΕΜΑ B B1. B2.
 ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
Προτεινόμενες λύσεις ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16/6/17
 Πανελλήνιες 2017 Προτεινόμενες λύσεις ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16/6/17 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ ΘΕΜΑ Β Β1. Ι-Α ΙΙ-Ε ΙΙΙ-ΣΤ ΙV-Β V-Ζ VI-Γ VII-Δ Β2. Η εικόνα 1 αντιστοιχεί σε Προκαρυωτικό κύτταρο.
Πανελλήνιες 2017 Προτεινόμενες λύσεις ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16/6/17 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ ΘΕΜΑ Β Β1. Ι-Α ΙΙ-Ε ΙΙΙ-ΣΤ ΙV-Β V-Ζ VI-Γ VII-Δ Β2. Η εικόνα 1 αντιστοιχεί σε Προκαρυωτικό κύτταρο.
igenetics Mια Μεντελική προσέγγιση
 igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 19 (+ κεφάλαιο 15 Hartwell) Ρύθμιση της γονιδιακής έκφρασης σε βακτήρια και βακτηριοφάγους Ο καταστολέας του οπερονίου lac προσδεδεμένος στο DNA. igenetics 2
igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 19 (+ κεφάλαιο 15 Hartwell) Ρύθμιση της γονιδιακής έκφρασης σε βακτήρια και βακτηριοφάγους Ο καταστολέας του οπερονίου lac προσδεδεμένος στο DNA. igenetics 2
ΤΑΞΙΝΟΜΗΣΗ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ. Το πρώτο σύστημα κατάταξης των ζώων κατά τον Αριστοτέλη
 ΤΑΞΙΝΟΜΗΣΗ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ Το πρώτο σύστημα κατάταξης των ζώων κατά τον Αριστοτέλη Επόμενα συστήματα κατάταξης John Ray (1627-1705): Νέο σύστημα κατάταξης και εισαγωγή έννοιας του είδους Charles Linnaeus
ΤΑΞΙΝΟΜΗΣΗ ΖΩΙΚΩΝ ΟΡΓΑΝΙΣΜΩΝ Το πρώτο σύστημα κατάταξης των ζώων κατά τον Αριστοτέλη Επόμενα συστήματα κατάταξης John Ray (1627-1705): Νέο σύστημα κατάταξης και εισαγωγή έννοιας του είδους Charles Linnaeus
ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ. Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της...
 ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της... Γενετική Μηχανική o Περιλαμβάνει όλες τις τεχνικές με τις οποίες μπορούμε να επεμβαίνουμε στο γενετικό υλικό των οργανισμών.
ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της... Γενετική Μηχανική o Περιλαμβάνει όλες τις τεχνικές με τις οποίες μπορούμε να επεμβαίνουμε στο γενετικό υλικό των οργανισμών.
Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της
 ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΚΕΦΑΛΑΙΑ 1 ΚΑΙ 2
 ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΚΕΦΑΛΑΙΑ 1 ΚΑΙ 2 ΘΕΜΑ 1 ο Α. Στις ερωτήσεις 1-5 να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Το
ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΚΕΦΑΛΑΙΑ 1 ΚΑΙ 2 ΘΕΜΑ 1 ο Α. Στις ερωτήσεις 1-5 να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Το
ΚΕΦΑΛΑΙΟ 16. Ο κυτταρικός κύκλος. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1
 ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
ΚΕΦΑΛΑΙΟ 16 Ο κυτταρικός κύκλος Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-μια Μοριακή Προσέγγιση 1 ΕΙΚΟΝΑ 16.1 Οι φάσεις του κυτταρικού κύκλου. Ο κύκλος διαίρεσης των περισσότερων ευκαρυωτικών κυττάρων χωρίζεται
Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ ΤΩΝ ΖΩΩΝ. Αρχιτομία. Αγενής αναπαραγωγή. Παρατομία. Εκβλάστηση. Εγγενής αναπαραγωγή Διπλοφασικός κύκλος.
 Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ ΤΩΝ ΖΩΩΝ Αρχιτομία Αγενής αναπαραγωγή Παρατομία Εκβλάστηση Εγγενής αναπαραγωγή Απλοφασικός κύκλος Διπλοφασικός κύκλος Ισογαμία Ανισογαμία Ωογαμία Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ
Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ ΤΩΝ ΖΩΩΝ Αρχιτομία Αγενής αναπαραγωγή Παρατομία Εκβλάστηση Εγγενής αναπαραγωγή Απλοφασικός κύκλος Διπλοφασικός κύκλος Ισογαμία Ανισογαμία Ωογαμία Η ΑΝΑΠΑΡΑΓΩΓΗ ΚΑΙ Η ΑΝΑΠΤΥΞΗ
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ )
") Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A 1. Να επιλέξετε τη σωστή απάντηση: Α1. Το γονίδιο
γ ρ α π τ ή ε ξ έ τ α σ η σ τ ο μ ά θ η μ α ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A 1. Να επιλέξετε τη σωστή απάντηση: Α1. Το γονίδιο
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο. 3. Το DNA των μιτοχονδρίων έχει μεγαλύτερο μήκος από αυτό των χλωροπλαστών.
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
3. Σε ένα σωματικό κύτταρο ανθρώπου που βρίσκεται στη μεσόφαση πριν την αντιγραφή υπάρχουν:
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Στο οπερόνιο της λακτόζης: Α. Η πρωτεΐνη καταστολέας
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Στο οπερόνιο της λακτόζης: Α. Η πρωτεΐνη καταστολέας
Βιολογία Προσανατολισμού Γ Λυκείου Κεφάλαιο: Κεφάλαια 1,2,4 Ονοματεπώνυμο Μαθητή: Ημερομηνία: 08/12/2018 Επιδιωκόμενος Στόχος: 75/100
 Μάθημα/Τάξη: Βιολογία Προσανατολισμού Γ Λυκείου Κεφάλαιο: Κεφάλαια 1,2,4 Ονοματεπώνυμο Μαθητή: Ημερομηνία: 08/12/2018 Επιδιωκόμενος Στόχος: 75/100 ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς
Μάθημα/Τάξη: Βιολογία Προσανατολισμού Γ Λυκείου Κεφάλαιο: Κεφάλαια 1,2,4 Ονοματεπώνυμο Μαθητή: Ημερομηνία: 08/12/2018 Επιδιωκόμενος Στόχος: 75/100 ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς
Η μεταμόρφωση στη Drosophila. Δίσκοι ενηλίκου
 Η μεταμόρφωση στη Drosophila Δίσκοι ενηλίκου Ο κύκλος ζωής της Drosophila Νευροεκκριτικά κύτταρα Προθωρακικοτρόπος ορμόνη Προθωρακικός αδένας Εκδυσόνη Μεταμόρφωση Έκδυση Μεταμόρφωση Έκδυση Νεανική ορμόνη
Η μεταμόρφωση στη Drosophila Δίσκοι ενηλίκου Ο κύκλος ζωής της Drosophila Νευροεκκριτικά κύτταρα Προθωρακικοτρόπος ορμόνη Προθωρακικός αδένας Εκδυσόνη Μεταμόρφωση Έκδυση Μεταμόρφωση Έκδυση Νεανική ορμόνη
Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό
 Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
Φυσιολογία Φυτών Διαχείριση ορμονικών μορίων Τα ορμονικά μόρια και η διαχείριση τους μέσα στο φυτό Φυσιολογία Φυτών 3 ου Εξαμήνου Δ. Μπουράνης, Σ. Χωριανοπούλου 1 Φυσιολογία Φυτών Διαχείριση ορμονικών
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: 16 / 06 / 2017 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1: δ Α2:
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: 16 / 06 / 2017 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1: δ Α2:
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ
 27 Μαΐου 2016 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Ημερησίων Γενικών Λυκείων (Νέο & Παλιό Σύστημα) ΘΕΜΑ Γ Γ.1 Ο χαρακτήρας της ομάδας αίματος στον άνθρωπο
27 Μαΐου 2016 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Ημερησίων Γενικών Λυκείων (Νέο & Παλιό Σύστημα) ΘΕΜΑ Γ Γ.1 Ο χαρακτήρας της ομάδας αίματος στον άνθρωπο
ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ
 ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ 16/06/2017 ΕΠΙΜΕΛΕΙΑ: ΑΝΔΡΙΑΝΗ ΠΑΡΑΧΗ ΘΕΜΑ Α A1. Δ Α2. Δ Α3. Β Α4. Γ Α5.Α ΘΕΜΑ Β Β1) I A II E III Σ IV Β V Ζ VI Γ VII Δ Β2) ε προκαρυωτικό γιατί
ΠΑΝΕΛΛΗΝΙΕ 2017 ΑΠΑΝΣΗΕΙ ΣΟ ΜΑΘΗΜΑ ΣΗ ΒΙΟΛΟΓΙΑ ΠΡΟΑΝΑΣΟΛΙΜΟΤ 16/06/2017 ΕΠΙΜΕΛΕΙΑ: ΑΝΔΡΙΑΝΗ ΠΑΡΑΧΗ ΘΕΜΑ Α A1. Δ Α2. Δ Α3. Β Α4. Γ Α5.Α ΘΕΜΑ Β Β1) I A II E III Σ IV Β V Ζ VI Γ VII Δ Β2) ε προκαρυωτικό γιατί
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ
 ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΒΙΟΛΟΓΙΚΗ ΤΕΧΝΟΛΟΓΙΑ»
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΒΙΟΛΟΓΙΚΗ ΤΕΧΝΟΛΟΓΙΑ» ΕΡΓΑΣΤΗΡΙΟ ΜΟΡΙΑΚΗΣ ΚΑΙ ΑΝΑΠΤΥΞΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Ο PlCoup-TF και το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΒΙΟΛΟΓΙΚΗ ΤΕΧΝΟΛΟΓΙΑ» ΕΡΓΑΣΤΗΡΙΟ ΜΟΡΙΑΚΗΣ ΚΑΙ ΑΝΑΠΤΥΞΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «Ο PlCoup-TF και το
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών
 Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
ΘΕΜΑ 1 Ο ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Τα
ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Τα
ΘΕΜΑ Α ΘΕΜΑ Β ΑΠΑΝΤΗΣΕΙΣ. Α1. β. Α2. γ. Α3. δ. Α4. γ. Α5. β Β1. 5, 4, 2, 1, 3. Β2. Τα δομικά μέρη του οπερονίου της λακτόζης είναι κατά σειρά τα εξής:
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β ΘΕΜΑ
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β ΘΕΜΑ
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΟΜΑΔΑΣ ΥΓΕΙΑΣ & ΖΩΗΣ
 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΟΜΑΔΑΣ ΥΓΕΙΑΣ & ΖΩΗΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. I, Β. IV, Γ. VI, Δ. VII, Ε. ΙΙ, ΣΤ. III, Ζ. V Β2. Αντιστοιχεί σε προκαρυωτικό οργανισμό. Στους προκαρυωτικούς
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΟΜΑΔΑΣ ΥΓΕΙΑΣ & ΖΩΗΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. I, Β. IV, Γ. VI, Δ. VII, Ε. ΙΙ, ΣΤ. III, Ζ. V Β2. Αντιστοιχεί σε προκαρυωτικό οργανισμό. Στους προκαρυωτικούς
Βιολογία προσανατολισμού
 Βιολογία προσανατολισμού ΘΕΜΑ Α Στις προτάσεις από Α1-Α5 να βρείτε την σωστή απάντηση. Α1. Ένας ερευνητής απομόνωσε ένα ασυνεχές γονίδιο από το γονιδίωμα ανθρώπινων κυττάρων. Το γονίδιο συνδέθηκε με βακτηριακό
Βιολογία προσανατολισμού ΘΕΜΑ Α Στις προτάσεις από Α1-Α5 να βρείτε την σωστή απάντηση. Α1. Ένας ερευνητής απομόνωσε ένα ασυνεχές γονίδιο από το γονιδίωμα ανθρώπινων κυττάρων. Το γονίδιο συνδέθηκε με βακτηριακό
Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους.
 Τι είναι τα βλαστικά κύτταρα? Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους. Είναι σε θέση να δρουν επισκευαστικά, αναδημιουργώντας
Τι είναι τα βλαστικά κύτταρα? Αρχικά αδιαφοροποίητα κύτταρα που έχουν την ικανότητα να διαφοροποιούνται σε ιστικά εξειδικευμένους κυτταρικούς τύπους. Είναι σε θέση να δρουν επισκευαστικά, αναδημιουργώντας
3. Σχ. Βιβλίο σελ «το βακτήριο Αgrobacterium.ξένο γονίδιο» Και σελ 133 «το βακτήριο Bacillus.Βt».
 2 ο Διαγώνισμα Βιολογίας Γ Λυκείου Θέμα Α 1. Α 2. Β 3. Β 4. Α 5. C Θέμα Β 1. Σελ 40 «τα ριβοσώματα μπορούν..πρωτεινών» Και σελ 39 «ο γενετικός κώδικας είναι σχεδόν καθολικός πρωτείνη». 2. Σελ 98 «η φαινυλκετονουρία.φαινυλαλανίνης»
2 ο Διαγώνισμα Βιολογίας Γ Λυκείου Θέμα Α 1. Α 2. Β 3. Β 4. Α 5. C Θέμα Β 1. Σελ 40 «τα ριβοσώματα μπορούν..πρωτεινών» Και σελ 39 «ο γενετικός κώδικας είναι σχεδόν καθολικός πρωτείνη». 2. Σελ 98 «η φαινυλκετονουρία.φαινυλαλανίνης»
ΟΜΟΣΠΟΝΔΙΑ ΕΚΠΑΙΔΕΥΤΙΚΩΝ ΦΡΟΝΤΙΣΤΩΝ ΕΛΛΑΔΟΣ (Ο.Ε.Φ.Ε.) ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2018 A ΦΑΣΗ ΒΙΟΛΟΓΙΑ ΑΛΓΟΡΙΘΜΟΣ
 ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2018 A ΦΑΣΗ ΒΙΟΛΟΓΙΑ ΑΛΓΟΡΙΘΜΟΣ") ΤΑΞΗ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΣ: ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ Ημερομηνία: Σάββατο 13 Ιανουαρίου 2018 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς
ΤΑΞΗ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΣ: ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ Ημερομηνία: Σάββατο 13 Ιανουαρίου 2018 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. δ Α2. α Α3. α Α4. γ Α5. β. ΘΕΜΑ Β Β1. Στήλη Ι Στήλη ΙΙ 1. Γ 2. Β 3. Ε 4. Α 5. Δ
 ΘΕΜΑ Α Α1. δ Α2. α Α3. α Α4. γ Α5. β ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΡΙΤΗ 7 ΣΕΠΤΕΜΒΡΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
ΘΕΜΑ Α Α1. δ Α2. α Α3. α Α4. γ Α5. β ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΡΙΤΗ 7 ΣΕΠΤΕΜΒΡΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA
 Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA Εικόνα 24.1 Το hnrna απαντάται ως ριβονουκλεοπρωτεϊνικό σύµπλοκο µε µορφή µιας σειράς από χάντρες. Εικόνα 24.2 Το RNA τροποποιείται στον πυρήνα
Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA Εικόνα 24.1 Το hnrna απαντάται ως ριβονουκλεοπρωτεϊνικό σύµπλοκο µε µορφή µιας σειράς από χάντρες. Εικόνα 24.2 Το RNA τροποποιείται στον πυρήνα
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ 1ο 1. β 2. β 3. α 4. α 5. β
 ΘΕΜΑ 1ο 1. β 2. β 3. α 4. α 5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΡΙΤΗ 21 ΣΕΠΤΕΜΒΡΙΟΥ 2004 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ)
ΘΕΜΑ 1ο 1. β 2. β 3. α 4. α 5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΡΙΤΗ 21 ΣΕΠΤΕΜΒΡΙΟΥ 2004 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ)
3. Η μέθοδος αλυσιδωτής αντίδρασης πολυμεράσης (PCR) επιτρέπει την επιλεκτική αντιγραφή μορίων DNA, χωρίς τη μεσολάβηση ζωικών κυττάρων.
 επιτρέπει την επιλεκτική αντιγραφή μορίων DNA, χωρίς τη μεσολάβηση ζωικών κυττάρων.") ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ ΘΕΜΑ 1ο ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Σ ΗΜΕΡΗΣΙΟΥ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 3 ΙΟΥΝΙΟΥ 2003 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΣΥΝΟΛΟ ΣΕΛΙΔΩΝ: ΤΕΣΣΕΡΙΣ (4) Α. Να γράψετε τον αριθμό της
ΑΡΧΗ 1ΗΣ ΣΕΛΙΔΑΣ ΘΕΜΑ 1ο ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Σ ΗΜΕΡΗΣΙΟΥ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 3 ΙΟΥΝΙΟΥ 2003 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΣΥΝΟΛΟ ΣΕΛΙΔΩΝ: ΤΕΣΣΕΡΙΣ (4) Α. Να γράψετε τον αριθμό της
Η µελέτη της ρύθµισης της πρωτεινοσύνθεσης στο επίπεδο του Ριβοσώµατος εντοπίζεται σε τρία επίπεδα
 Η µελέτη της ρύθµισης της πρωτεινοσύνθεσης στο επίπεδο του Ριβοσώµατος εντοπίζεται σε τρία επίπεδα ΣτονΣτον ρόλο των διαφόρων οµάδων των ριβοσωµικών πρωτεινών. Κατά πόσο δηλαδή υπάρχει ετερογένεια στις
Η µελέτη της ρύθµισης της πρωτεινοσύνθεσης στο επίπεδο του Ριβοσώµατος εντοπίζεται σε τρία επίπεδα ΣτονΣτον ρόλο των διαφόρων οµάδων των ριβοσωµικών πρωτεινών. Κατά πόσο δηλαδή υπάρχει ετερογένεια στις
ΘΕΜΑ Α. 1. δ 2. δ 3. β 4. γ 5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI Δ VII Ε II ΣΤ III Ζ V Η -
 ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ 2017 ΛΥΣΕΙΣ ΘΕΜΑΤΩΝ ΕΠΙΜΕΛΕΙΑ: Π. ΚΡΥΣΤΑΛΛΙΔΗΣ 1. δ 2. δ 3. β 4. γ 5. α ΘΕΜΑ Α Β1. Α I Β IV Γ VI Δ VII Ε II ΣΤ III Ζ V Η - ΘΕΜΑ Β Β2. Η εικόνα αντιστοιχεί
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ 2017 ΛΥΣΕΙΣ ΘΕΜΑΤΩΝ ΕΠΙΜΕΛΕΙΑ: Π. ΚΡΥΣΤΑΛΛΙΔΗΣ 1. δ 2. δ 3. β 4. γ 5. α ΘΕΜΑ Α Β1. Α I Β IV Γ VI Δ VII Ε II ΣΤ III Ζ V Η - ΘΕΜΑ Β Β2. Η εικόνα αντιστοιχεί
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος. 09/04/ Μοριακή Βιολογία Κεφ. 1 Καθηγητής Δρ. Κ. Ε. Βοργιάς
 Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
ΑΠΑΝΤΗΣΕΙΣ. Β2. Η εικόνα αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς οργανισμούς το mrna αρχίζει να μεταφράζεται σε πρωτεΐνη πριν ακόμη
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 6 ΙΟΥΝΙΟΥ 207 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. δ Α2. δ Α3. β Α4. γ Α5.
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 6 ΙΟΥΝΙΟΥ 207 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. δ Α2. δ Α3. β Α4. γ Α5.
ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1.
1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1.
ΦΡΟΝΤΙΣΤΗΡΙΟ ΜΕΣΗΣ ΕΚΠΑΙΔΕΥΣΗΣ ΗΡΑΚΛΕΙΤΟΣ ΚΩΛΕΤΤΗ
 ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 ΙΟΥΝΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ε Ν Δ Ε Ι Κ Τ Ι Κ Ε Σ Α Π Α Ν Τ Η Σ Ε Ι Σ Θ Ε Μ Α Τ Ω Ν ΘΕΜΑ Α Α1-δ Α2-δ Α3-β
ΠΑΝΕΛΛΗΝΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 ΙΟΥΝΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ε Ν Δ Ε Ι Κ Τ Ι Κ Ε Σ Α Π Α Ν Τ Η Σ Ε Ι Σ Θ Ε Μ Α Τ Ω Ν ΘΕΜΑ Α Α1-δ Α2-δ Α3-β
Θετικών Σπουδών. Ενδεικτικές απαντήσεις θεμάτων
 Πανελλαδικές Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο μάθημα: Βιολογία Προσανατολισμού Θετικών Σπουδών Παρασκευή, 16 Ιουνίου 2017 Ενδεικτικές απαντήσεις θεμάτων Θέμα Α Α1. α) 3 CAT 5 β) 3 TAC 5
Πανελλαδικές Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο μάθημα: Βιολογία Προσανατολισμού Θετικών Σπουδών Παρασκευή, 16 Ιουνίου 2017 Ενδεικτικές απαντήσεις θεμάτων Θέμα Α Α1. α) 3 CAT 5 β) 3 TAC 5
ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ)
") ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ) ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό κάθε μιας από τις παρακάτω ημιτελείς προτάσεις 1-5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ) ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό κάθε μιας από τις παρακάτω ημιτελείς προτάσεις 1-5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014
 Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014 ΘΕΜΑ Α Α1. β Α2. β Α3. β Α4. β Α5. β ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ B B1. Ο όρος γονιδιακή έκφραση αναφέρεται συνήθως σε όλη τη διαδικασία με την οποία
Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014 ΘΕΜΑ Α Α1. β Α2. β Α3. β Α4. β Α5. β ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ B B1. Ο όρος γονιδιακή έκφραση αναφέρεται συνήθως σε όλη τη διαδικασία με την οποία
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
Β. ΚΑΜΙΝΕΛΛΗΣ ΒΙΟΛΟΓΙΑ. Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα).
.") ΒΙΟΛΟΓΙΑ Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα). Είδη οργανισμών Υπάρχουν δύο είδη οργανισμών: 1. Οι μονοκύτταροι, που ονομάζονται μικροοργανισμοί
ΒΙΟΛΟΓΙΑ Είναι η επιστήμη που μελετά τους ζωντανούς οργανισμούς. (Αποτελούνται από ένα ή περισσότερα κύτταρα). Είδη οργανισμών Υπάρχουν δύο είδη οργανισμών: 1. Οι μονοκύτταροι, που ονομάζονται μικροοργανισμοί
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 ΙΟΥΝΙΟΥ 2017
 Σ ε λ ί δ α 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 ΙΟΥΝΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΕΠΙΜΕΛΕΙΑ: ΛΑΜΠΡΙΝΗ ΓΑΒΡΙΗΛ ΘΕΜΑ Α Α1.
Σ ε λ ί δ α 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 ΙΟΥΝΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΕΠΙΜΕΛΕΙΑ: ΛΑΜΠΡΙΝΗ ΓΑΒΡΙΗΛ ΘΕΜΑ Α Α1.
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ
 ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ. ΘΕΜΑ Β Β1. Στήλη Ι Στήλη ΙΙ 1 Α 2 Γ 3 Α 4 Β 5 Α 6 Α 7 Γ
 ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
 ΜΑΘΗΜΑ / ΤΑΞΗ: ΒΙΟΛΟΓΙΑ ΟΠ Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΗΜΕΡΟΜΗΝΙΑ: 22/01/2017 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ: ΝΟΤΑ ΛΑΖΑΡΑΚΗ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Από
ΜΑΘΗΜΑ / ΤΑΞΗ: ΒΙΟΛΟΓΙΑ ΟΠ Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΗΜΕΡΟΜΗΝΙΑ: 22/01/2017 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ: ΝΟΤΑ ΛΑΖΑΡΑΚΗ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Από
Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού
 Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού Θέμα Α Α1) γ Α2) γ Α3) δ Α4) β Α5) β Θέμα Β Β1. Α = υδροξύλιο, Β = πρωταρχικό τμήμα, Γ = θέση έναρξης αντιγραφής, Δ = φωσφορική ομάδα, Ε = τμήμα
Ενδεικτικές απαντήσεις στα Θέματα Βιολογίας Προσανατολισμού Θέμα Α Α1) γ Α2) γ Α3) δ Α4) β Α5) β Θέμα Β Β1. Α = υδροξύλιο, Β = πρωταρχικό τμήμα, Γ = θέση έναρξης αντιγραφής, Δ = φωσφορική ομάδα, Ε = τμήμα
ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2015 Β ΦΑΣΗ ΑΠΑΝΤΗΣΕΙΣ ÏÅÖÅ
 ΤΑΞΗ: ΜΑΘΗΜΑ: 3 η ΤΑΞΗ ΕΠΑ.Λ. (Β ΟΜΑ Α) ΒΙΟΛΟΓΙΑ ΙΙ Ηµεροµηνία: Παρασκευή 19 Απριλίου 2015 ιάρκεια Εξέτασης: 3 ώρες ΘΕΜΑ Α Α1-β, Α2-γ, Α3-γ, Α4-α, Α5-α ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Β Β1. α. Με τη µέθοδο της επιλογής
ΤΑΞΗ: ΜΑΘΗΜΑ: 3 η ΤΑΞΗ ΕΠΑ.Λ. (Β ΟΜΑ Α) ΒΙΟΛΟΓΙΑ ΙΙ Ηµεροµηνία: Παρασκευή 19 Απριλίου 2015 ιάρκεια Εξέτασης: 3 ώρες ΘΕΜΑ Α Α1-β, Α2-γ, Α3-γ, Α4-α, Α5-α ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Β Β1. α. Με τη µέθοδο της επιλογής
Ημερομηνία: Πέμπτη 5 Ιανουαρίου 2016 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ
 ΑΠΟ 18/12/2016 ΕΩΣ 05/01/2017 2η ΕΞΕΤΑΣΤΙΚΗ ΠΕΡΙΟΔΟΣ ΤΑΞΗ: ΜΑΘΗΜΑ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ημερομηνία: Πέμπτη 5 Ιανουαρίου 2016 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ Στις ημιτελείς προτάσεις
ΑΠΟ 18/12/2016 ΕΩΣ 05/01/2017 2η ΕΞΕΤΑΣΤΙΚΗ ΠΕΡΙΟΔΟΣ ΤΑΞΗ: ΜΑΘΗΜΑ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ημερομηνία: Πέμπτη 5 Ιανουαρίου 2016 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ Στις ημιτελείς προτάσεις
Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Σε ένα
ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Σε ένα
Ενδεικτικές απαντήσεις
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 11 Ιουνίου 2015 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Απαντήσεις Θεμάτων Επαναληπτικών Πανελληνίων Εξετάσεων Ημερησίων & Εσπερινών Γενικών Λυκείων ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ B Β1. 1. Β 2. Γ 3. Α
11 Ιουνίου 2015 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Απαντήσεις Θεμάτων Επαναληπτικών Πανελληνίων Εξετάσεων Ημερησίων & Εσπερινών Γενικών Λυκείων ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ B Β1. 1. Β 2. Γ 3. Α
γ. δύο φορές δ. τέσσερεις φορές
 1 ο Διαγώνισμα Βιολογίας Γ Λυκείου Θέμα Α Να επιλέξετε τη σωστή απάντηση Α1. Σε ένα ανασυνδυασμένο πλασμίδιο που σχηματίστηκε με την επίδραση της EcoRI, η αλληλουχία που αναγνωρίζει η συγκεκριμένη περιοριστική
1 ο Διαγώνισμα Βιολογίας Γ Λυκείου Θέμα Α Να επιλέξετε τη σωστή απάντηση Α1. Σε ένα ανασυνδυασμένο πλασμίδιο που σχηματίστηκε με την επίδραση της EcoRI, η αλληλουχία που αναγνωρίζει η συγκεκριμένη περιοριστική
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1 ο Α. Ερωτήσεις πολλαπλής επιλογής 1. δ 2. β 3. γ 4. γ 5. β Β. Ερωτήσεις σωστού λάθους 1. Λάθος 2. Σωστό 3. Λάθος 4. Λάθος 5. Σωστό ΘΕΜΑ
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΘΕΜΑ 1 ο Α. Ερωτήσεις πολλαπλής επιλογής 1. δ 2. β 3. γ 4. γ 5. β Β. Ερωτήσεις σωστού λάθους 1. Λάθος 2. Σωστό 3. Λάθος 4. Λάθος 5. Σωστό ΘΕΜΑ
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥ ΩΝ 2017 ΑΠΑΝΤΗΣΕΙΣ ËÁÌÉÁ
 ΘΕΜΑ Α Α1: δ Α2: δ Α3: β Α4: γ Α5: α ΘΕΜΑ Β ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥ ΩΝ 2017 ΑΠΑΝΤΗΣΕΙΣ Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII - Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Αυτό
ΘΕΜΑ Α Α1: δ Α2: δ Α3: β Α4: γ Α5: α ΘΕΜΑ Β ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥ ΩΝ 2017 ΑΠΑΝΤΗΣΕΙΣ Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII - Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Αυτό
Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ÏÅÖÅ
 1 Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 ο Α. 1 - Γ 2 - Β 3-4 - Γ 5 - Β. 1 - Σ 2 - Λ 3 - Λ 4 - Λ 5 - Σ ΘΕΜΑ 2 ο ΑΠΑΝΤΗΣΕΙΣ 1. Κάθε είδος αντισώµατος που αναγνωρίζει έναν αντιγονικό καθοριστή παράγεται
1 Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 ο Α. 1 - Γ 2 - Β 3-4 - Γ 5 - Β. 1 - Σ 2 - Λ 3 - Λ 4 - Λ 5 - Σ ΘΕΜΑ 2 ο ΑΠΑΝΤΗΣΕΙΣ 1. Κάθε είδος αντισώµατος που αναγνωρίζει έναν αντιγονικό καθοριστή παράγεται
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης
 Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη
 Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
ιαγονιδιακή τεχνολογία G. Patrinos
 ιαγονιδιακή τεχνολογία Αντίστροφη γενετική Οργανισμός Γονιδίωμα ιαγονίδιο Γονίδιο Forward genetics Επαγόμενη Οργανισμός μεταλλαξογένεση Μεταλλαγμένος οργανισμός Εύρεση και μελέτη του υπεύθυνου γονιδίου
ιαγονιδιακή τεχνολογία Αντίστροφη γενετική Οργανισμός Γονιδίωμα ιαγονίδιο Γονίδιο Forward genetics Επαγόμενη Οργανισμός μεταλλαξογένεση Μεταλλαγμένος οργανισμός Εύρεση και μελέτη του υπεύθυνου γονιδίου
α) φυτοαιματογλουτίνη, κολχικίνη
 φυτοαιματογλουτίνη, κολχικίνη") ΔΙΑΓΩΝΙΣΜΑ ΚΕΦΑΛΑΙΟ 1ο 2ο και 4ο ΘΕΜΑ Α ΜΟΝΑΔΕΣ 25 Να απαντήσετε στις παρακάτω ερωτήσεις επιλέγοντας την λανθασμένη πρόταση ανάμεσα στις σωστές ή την σωστή ανάμεσα στις λανθασμένες. Α.Το καλαμπόκι (zeamays)
ΔΙΑΓΩΝΙΣΜΑ ΚΕΦΑΛΑΙΟ 1ο 2ο και 4ο ΘΕΜΑ Α ΜΟΝΑΔΕΣ 25 Να απαντήσετε στις παρακάτω ερωτήσεις επιλέγοντας την λανθασμένη πρόταση ανάμεσα στις σωστές ή την σωστή ανάμεσα στις λανθασμένες. Α.Το καλαμπόκι (zeamays)
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Θετικής Προσανατολισμού Θετικών Σπουδών, Ημ/νία: 16 Ιουνίου 2017
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Προσανατολισμού Θετικών Σπουδών, Ημ/νία: 16 Ιουνίου 2017 Απαντήσεις Θεμάτων ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Προσανατολισμού Θετικών Σπουδών, Ημ/νία: 16 Ιουνίου 2017 Απαντήσεις Θεμάτων ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ
Φάσμα& Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι.
 σύγχρονο Φάσμα& Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο 1. 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 50.27.990 50.20.990 2. 25ης Μαρτίου 74 Π. ΠΕΤΡΟΥΠΟΛΗΣ 50.50.658 50.60.845 3. Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ
σύγχρονο Φάσμα& Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο 1. 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 50.27.990 50.20.990 2. 25ης Μαρτίου 74 Π. ΠΕΤΡΟΥΠΟΛΗΣ 50.50.658 50.60.845 3. Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ
Κύτταρα πολυκύτταρων οργανισμών
 Μίτωση - Μείωση Τα ευκαρυωτικά κύτταρα διαιρούνται με δύο τρόπους: τη μίτωση και τη μείωση. Η Μίτωση είναι ο τύπος της κυτταρικής διαίρεσης που από ένα πατρικό κύτταρο καταλήγει σε δύο γενετικά πανομοιότυπα
Μίτωση - Μείωση Τα ευκαρυωτικά κύτταρα διαιρούνται με δύο τρόπους: τη μίτωση και τη μείωση. Η Μίτωση είναι ο τύπος της κυτταρικής διαίρεσης που από ένα πατρικό κύτταρο καταλήγει σε δύο γενετικά πανομοιότυπα
ΤΕΛΟΣ 1ΗΣ ΑΠΟ 5 ΣΕΛΙ ΕΣ
 ΑΡΧΗ 1ΗΣ ΣΕΛΙ ΑΣ ΝΕΟ & ΠΑΛΑΙΟ ΣΥΣΤΗΜΑ ΕΣΠΕΡΙΝΩΝ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
ΑΡΧΗ 1ΗΣ ΣΕΛΙ ΑΣ ΝΕΟ & ΠΑΛΑΙΟ ΣΥΣΤΗΜΑ ΕΣΠΕΡΙΝΩΝ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
Βιολογία προσανατολισμού
 Βιολογία προσανατολισμού Α. 1. β. 2. γ. 3. γ. 4. α. 5. δ. ΘΕΜΑ Α ΘΕΜΑ Β Β1. Σχολικό βιβλίο σελ. 131 «Το βακτήριο... στο σώμα των φυτών.» Β2.1 Ε, 2 Δ, 3 Α, 4 Β Β3. Σχολικό βιβλίο σελ. 108 «Η θερμοκρασία..
Βιολογία προσανατολισμού Α. 1. β. 2. γ. 3. γ. 4. α. 5. δ. ΘΕΜΑ Α ΘΕΜΑ Β Β1. Σχολικό βιβλίο σελ. 131 «Το βακτήριο... στο σώμα των φυτών.» Β2.1 Ε, 2 Δ, 3 Α, 4 Β Β3. Σχολικό βιβλίο σελ. 108 «Η θερμοκρασία..
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014. Απαντήσεις Θεμάτων
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
Tρίτη, 3 Ιουνίου 2003 ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ Γ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ
 Tρίτη, 3 Ιουνίου 2003 ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ Γ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 1Α. Να γράψετε τον αριθµό της καθεµιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν
Tρίτη, 3 Ιουνίου 2003 ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ Γ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 1Α. Να γράψετε τον αριθµό της καθεµιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν
ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ Ο.Ε.Φ.Ε. 2004 ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ Ο.Ε.Φ.Ε. 2004 ΘΕΜΑ 1 Ο Α. Να επιλέξετε την ορθή πρόταση: ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1. Το κωδικόνιο του mrna που κωδικοποιεί το αµινοξύ µεθειονίνη είναι α. 5 GUA
ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ Ο.Ε.Φ.Ε. 2004 ΘΕΜΑ 1 Ο Α. Να επιλέξετε την ορθή πρόταση: ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1. Το κωδικόνιο του mrna που κωδικοποιεί το αµινοξύ µεθειονίνη είναι α. 5 GUA
Malamidou A., Pantazaki A.A. *, Koliakos G., Tsolaki M.
 10th Panhellenic Conference of Alzheimer's Disease and Related Disorders (PICAD) and 2nd Mediterranean Conference of Neurodegenerative Diseases (MeCOND) Malamidou A., Pantazaki A.A. *, Koliakos G., Tsolaki
10th Panhellenic Conference of Alzheimer's Disease and Related Disorders (PICAD) and 2nd Mediterranean Conference of Neurodegenerative Diseases (MeCOND) Malamidou A., Pantazaki A.A. *, Koliakos G., Tsolaki
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη φράση που συμπληρώνει
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιο σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη φράση που συμπληρώνει
Κ Ε Φ Α Λ Α Ι Ο 25 : Το καταλυτικό RNA
 Κ Ε Φ Α Λ Α Ι Ο 25 : Το καταλυτικό RNA Εικόνα 25.1 Το αυτο-µάτισµα του πρώιµου rrna 35S της Tetrahymena thermophila µπορεί να µελετηθεί µε ηλεκτροφόρηση σε πήκτωµα. Το αποδεσµευµένο ιντρόνιο σχηµατίζει
Κ Ε Φ Α Λ Α Ι Ο 25 : Το καταλυτικό RNA Εικόνα 25.1 Το αυτο-µάτισµα του πρώιµου rrna 35S της Tetrahymena thermophila µπορεί να µελετηθεί µε ηλεκτροφόρηση σε πήκτωµα. Το αποδεσµευµένο ιντρόνιο σχηµατίζει
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ") 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ
1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ
Βιολογία Γ Γενικού Λυκείου Θετικής κατεύθυνσης. Κεφάλαιο 1α Το Γενετικό Υλικό
 Βιολογία Γ Γενικού Λυκείου Θετικής κατεύθυνσης Κεφάλαιο 1α Το Γενετικό Υλικό Το DNA είναι το γενετικό υλικό Αρχικά οι επιστήμονες θεωρούσαν ότι οι πρωτεΐνες αποτελούσαν το γενετικό υλικό των οργανισμών.
Βιολογία Γ Γενικού Λυκείου Θετικής κατεύθυνσης Κεφάλαιο 1α Το Γενετικό Υλικό Το DNA είναι το γενετικό υλικό Αρχικά οι επιστήμονες θεωρούσαν ότι οι πρωτεΐνες αποτελούσαν το γενετικό υλικό των οργανισμών.
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
σύγχρονο προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. & Group µαθητικό φροντιστήριο Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ
 σύγχρονο Φάσµα & Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. µαθητικό φροντιστήριο Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ 210 50 51 557 210 50 56 296 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 210 50 20 990 210 50 27 990 25ης Μαρτίου 74 ΠΕΤΡΟΥΠΟΛΗ
σύγχρονο Φάσµα & Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. µαθητικό φροντιστήριο Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ 210 50 51 557 210 50 56 296 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 210 50 20 990 210 50 27 990 25ης Μαρτίου 74 ΠΕΤΡΟΥΠΟΛΗ