ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΔΙΕΥΘΥΝΤΗΣ: ΓΕΩΡΓΙΟΣ ΚΩΣΤΟΠΟΥΛΟΣ
|
|
|
- Σιληνός Σαμαράς
- 6 χρόνια πριν
- Προβολές:
Transcript
1 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΔΙΕΥΘΥΝΤΗΣ: ΓΕΩΡΓΙΟΣ ΚΩΣΤΟΠΟΥΛΟΣ IN VITRO ΣΥΓΚΡΙΤΙΚΕΣ ΜΕΛΕΤΕΣ ΤΗΣ ΠΛΑΣΤΙΚΟΤΗΤΑΣ ΤΗΣ ΣΥΝΑΠΤΙΚΗΣ ΔΙΑΒΙΒΑΣΗΣ ΚΑΙ ΤΗΣ ΔΙΚΤΥΑΚΗΣ ΝΕΥΡΩΝΙΚΗΣ ΔΡΑΣΤΗΡΙΟΤΗΤΑΣ ΜΕΤΑΞΥ ΔΙΑΦΡΑΓΜΑΤΙΚΟΥ ΚΑΙ ΚΡΟΤΑΦΙΚΟΥ ΙΠΠΟΚΑΜΠΟΥ ΣΤΥΛΙΑΝΟΣ ΚΟΥΒΑΡΟΣ ΒΙΟΛΟΓΟΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΠΑΤΡΑ 2016 i
2 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΔΙΕΥΘΥΝΤΗΣ: ΓΕΩΡΓΙΟΣ ΚΩΣΤΟΠΟΥΛΟΣ IN VITRO ΣΥΓΚΡΙΤΙΚΕΣ ΜΕΛΕΤΕΣ ΤΗΣ ΠΛΑΣΤΙΚΟΤΗΤΑΣ ΤΗΣ ΣΥΝΑΠΤΙΚΗΣ ΔΙΑΒΙΒΑΣΗΣ ΚΑΙ ΤΗΣ ΔΙΚΤΥΑΚΗΣ ΝΕΥΡΩΝΙΚΗΣ ΔΡΑΣΤΗΡΙΟΤΗΤΑΣ ΜΕΤΑΞΥ ΔΙΑΦΡΑΓΜΑΤΙΚΟΥ ΚΑΙ ΚΡΟΤΑΦΙΚΟΥ ΙΠΠΟΚΑΜΠΟΥ ΣΤΥΛΙΑΝΟΣ ΚΟΥΒΑΡΟΣ ΒΙΟΛΟΓΟΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΠΑΤΡΑ 2016 ii
3 ΤΡΙΜΕΛΗΣ ΣΥΜΒΟΥΛΕΥΤΙΚΗ ΕΠΙΤΡΟΠΗ ΠΑΠΑΘΕΟΔΩΡΟΠΟΥΛΟΣ ΚΩΝΣΤΑΝΤΙΝΟΣ (ΕΠΙΒΛΕΠΩΝ)-Αναπληρωτής Καθηγητής, Επιβλέπων-Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΚΩΣΤΟΠΟΥΛΟΣ ΓΕΩΡΓΙΟΣ-Καθηγητής, Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΑΓΓΕΛΑΤΟΥ ΦΕΒΡΩΝΙΑ- Καθηγήτρια, Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΕΠΤΑΜΕΛΗΣ ΕΞΕΤΑΣΤΙΚΗ ΕΠΙΤΡΟΠΗ ΑΓΓΕΛΑΤΟΥ ΦΕΒΡΩΝΙΑ- Καθηγήτρια, Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΚΩΣΤΟΠΟΥΛΟΣ ΓΕΩΡΓΙΟΣ-Καθηγητής, Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΠΑΠΑΘΕΟΔΩΡΟΠΟΥΛΟΣ ΚΩΝΣΤΑΝΤΙΝΟΣ- Αναπληρωτής Καθηγητής, Επιβλέπων- Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΒΟΥΚΕΛΑΤΟΥ ΓΕΩΡΓΙΑ- Επίκουρη Καθηγήτρια, Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΜΗΤΣΑΚΟΥ ΑΔΑΜΑΝΤΙΑ- Καθηγήτρια, Εργαστήριο Φυσιολογίας, Τμήμα Ιατρικής, Πανεπιστήμιο Πατρών ΨΑΡΡΟΠΟΥΛΟΥ ΑΙΚΑΤΕΡΙΝΗ- Καθηγήτρια, Τμήμα Βιολογικών Εφαρμογών & Τεχνολογίας, Πανεπιστήμιο Ιωαννίνων. ΚΟΣΜΙΔΗΣ ΕΥΣΤΡΑΤΙΟΣ- Επίκουρος καθηγητής, Τμήμα Ιατρικής, Αριστοτέλειο Πανεπιστήμιο Θεσσαλονίκης. i
4 ΠΙΝΑΚΑΣ ΣΥΝΤΟΜΕΥΣΕΩΝ-ΑΚΡΩΝΥΜΙΑ A1 receptor AMPA CB1 receptor CS cell HFS IEI ICI ISI fepsp FV GABA LTP mglu receptor MUA NMDA PS PB SK channels TBS SWRs VDCC Adenosinergic 1 Receptor α-amino-3-hydroxy-5-methyl- 4-isoxazolepropionic acid Cannabinoid receptor Complex spike cell High Frequency Stimulation Inter event interval Intra cluster interval Inter spike interval Field Excitatory postsynaptic potential Fiber volley γ-aminobutyric acid Long-term potentiation Metabotropic glutamate receptor Multiunit activity N-methyl-D-aspartic acid Population Spike Primed Burst Small conductance Κ + channels Theta Burst Stimulation Sharp wave-ripples Voltage-dependent calcium channel ii
5 ΠΡΟΛΟΓΟΣ Η εργασία αυτή συνίσταται από τρεις επιμέρους μελέτες με τις οποίες προσπαθήσαμε να διερευνήσουμε: α) τους μηχανισμούς που συμμετέχουν στη ρύθμιση της διεγερτικής συναπτικής διαβίβασης (Μελέτη 1) και της μακρόχρονης πλαστικότητας (Μελέτη 2) μεταξύ ραχιαίου και κοιλιακού ιπποκάμπου και β) τις πιθανές μεταβολές που συμβαίνουν κατά το γήρας στην ιπποκάμπεια δικτυακή δραστηριότητα των οξύαιχμων κυμάτων-ριπιδισμών που παίζει κυρίαρχο ρόλο στην διεργασία της μνημονικής παγίωσης (Μελέτη 3). Το πειραματικό υλικό της παρούσας εργασίας αποτέλεσε ο ιππόκαμπος του εγκεφάλου, μία δομή που διαδραματίζει κυρίαρχο ρόλο στις ανώτερες λειτουργίες του εγκεφάλου όπως η μάθηση και η μνήμη. Η γνώση πολλών ανατομικών και φυσιολογικών στοιχείων αυτής της δομής την καθιστά ένα κατάλληλο πειραματικό μοντέλο για την διερεύνηση της συναπτικής και νευρωνικής λειτουργίας και πλαστικότητας. Η πειραματική προσέγγιση των ερωτημάτων έγινε με τη χρήση της μεθόδου in vitro διατήρησης λεπτών τομών εγκεφάλου και ηλεκτροφυσιολογικών τεχνικών σε τομές ιπποκάμπου, στις οποίες καταγράφονταν η ηλεκτρική δραστηριότητα πληθυσμού νευρώνων που είτε προκαλούνταν με ηλεκτρικό ερεθισμό είτε οργανώνονταν αυθόρμητα από τον ίδιο τον ιστό. Η εργασία αυτή συνίσταται: από το γενικό μέρος (Α) στο οποίο παρουσιάζονται τα εισαγωγικά θεωρητικά στοιχεία που συγκροτούν τη μελέτη αυτή (Α.Ι.), ο σκοπός της εργασίας (Α.ΙΙ.) και οι μεθοδολογικές αρχές της in vitro ηλεκτροφυσιολογικής τεχνικής των εξωκυττάριων καταγραφών, η πειραματική διαδικασία και το είδος της στατιστικής ανάλυσης (Α.ΙΙΙ.). από το ειδικό μέρος (Β) στο οποίο περιγράφονται τα ερωτήματα, τα αποτελέσματα, η συζήτηση και τα συμπεράσματα των επιμέρους πειραματικών μελετών (Β.Ι.1,2,3), η σύνοψη της εργασίας με τα γενικά συμπεράσματα (Β.ΙΙ), η περίληψη στην αγγλική γλώσσα (Β.ΙΙΙ) και η συνολική βιβλιογραφία (Β.ΙV). Η παρούσα διδακτορική διατριβή συν-χρηματοδοτήθηκε από την Ευρωπαϊκή Ένωση (European Social Fund ESF) και από Ελληνικό εθνικό ταμείο μέσω του επιχειρησιακού προγράμματος «Εκπαίδευση και Δια-βίου Μάθηση του Εθνικού Στρατηγικού Σχεδιασμού» (Strategic Reference Framework-NSRF) - Πρόγραμμα Χρηματοδότησης Έρευνας: Θαλής (Thales). Επενδύοντας στην κοινωνία της γνώσης μέσω του Ευρωπαϊκού Κοινωνικού Ταμείου; Αριθμός επιχορήγησης: MIS: iii
6 ΕΥΧΑΡΙΣΤΙΕΣ Σε αυτό το σημείο και με την ολοκλήρωση της διδακτορικής μου διατριβής θα ήθελα να ευχαριστήσω όλους εκείνους που συνέβαλαν στην ολοκλήρωσή της. Αρχικά, θα ήθελα να ευχαριστήσω από καρδιάς τον επιβλέποντα καθηγητή μου, τον κ. Κωνσταντίνο Παπαθεοδωρόπουλο γιατί χωρίς την πολύτιμη συμβολή του όλα αυτά τα χρόνια δεν θα ήταν δυνατή η περάτωση αυτού του έργου. Τον ευχαριστώ θερμά για τις πολύτιμες συμβουλές τόσο σε ερευνητικό όσο και σε φιλικό επίπεδο, αλλά και για την αμέριστη στήριξη του επιστημονική και οικονομική σε όλη τη διάρκεια αυτού του όμορφου ταξιδιού. Επιπλέον, θα ήθελα να ευχαριστήσω τα μέλη της τριμελούς συμβουλευτικής επιτροπής μου: τον καθηγητή κ. Γεώργιο Κωστόπουλο που με τις πολύτιμες συμβουλές του υπήρξε καθοδηγητής σε κάθε μου βήμα και ιδιαιτέρως την καθηγήτρια κ. Φεβρωνία Αγγελάτου, που είχα την τύχη να συνεργαστούμε για την ολοκλήρωση μέρους της διατριβής μου, στηρίζοντάς με σε επιστημονικό, οικονομικό αλλά και σε φιλικό επίπεδο. Ευχαριστώ επίσης την καθηγήτρια κ. Αδαμαντία Μητσάκου, την επίκουρη καθηγήτρια κ. Γεωργία Βουκελάτου, την καθηγήτρια κ. Αικατερίνη Ψαρροπούλου και τον επίκουρο καθηγητή κ. Ευστράτιο Κοσμίδη για την μεγάλη τους προθυμία να συμμετάσχουν στην εξεταστική επιτροπή. Επιπλέον, θα ήθελα να ευχαριστήσω όλη την ερευνητική ομάδα της κ. Φεβρωνίας Αγγελάτου και ιδιαιτέρως τον κ. Κωνσταντίνο Σαράντη για την ευχάριστη και αποδοτική συνεργασία μας. Επίσης ευχαριστώ όλα τα μέλη του εργαστηρίου φυσιολογίας για το φιλικό κλίμα εντός και εκτός εργαστηρίου. Τέλος, θα ήθελα να ευχαριστήσω ιδιαίτερα την οικογένεια μου, την σύντροφο μου και τους φίλους μου γιατί η αγάπη και συμπαράστασή τους σε κάθε μου βήμα, επαγγελματικό ή μη, μου δίνουν δύναμη να συνεχίσω να προσπαθώ. iv
7 Στην Οικογένεια μου Στην Κατερίνα v
8 ΠΕΡΙΕΧΟΜΕΝΑ Α. ΓΕΝΙΚΟ ΜΕΡΟΣ Ι.ΕΙΣΑΓΩΓΗ. 1 1.Γενικά περί ιπποκάμπου 1 2. Μακροσκοπική και Μικροσκοπική Οργάνωση του Ιπποκάμπου Συνδέσεις του Ιπποκάμπου Αισθητική είσοδος στον Ενδορρινικό φλοιό Διατιτραίνουσα οδός Μονοπάτι Βρυωδών Ινών Παράπλευρο συνδετικό/συνδεσμικό μονοπάτι Schaffer CA1-Υπόθεμα-Ενδορινικός φλοιός 8 3. Συναπτική Διαβίβαση Διεγερτική διαβίβαση Ανασταλτική διαβίβαση Νευρωνική Διεγερσιμότητα Συναπτική Πλαστικότητα Μακρόχρονη ενδυνάμωση Πρωτόκολλα επαγωγής της μακρόχρονης ενδυνάμωσης Ενδογενείς Ρυθμοί Ιπποκάμπου Οξύαιχμα κύματα-ριπιδισμοί Ρόλος των οξύαιχμων κυμάτων-ριπιδισμών Ιππόκαμπος και Μνήμη Διαφραγματο-κροταφική διαφοροποίηση Νευρωνική συνδεσιμότητα των δύο πόλων Γονιδιακή έκφραση και λειτουργικές διαφορές κατά μήκος του επιμήκη 30 ιπποκάμπειου άξονα Ιππόκαμπος και Γήρανση Μειωμένη είσοδος της πληροφορίας από διατιτραίνουσα οδό στην ΟΕ και της 32 CA3 περιοχή 9.2. Ισορροπία Διέγερσης/Αναστολής στον εγκέφαλο γερασμένων 33 πειραματόζωων 9.3. Μειωμένη τροποποιητική είσοδος στον γέρικο ιππόκαμπο 33 vi
9 ΙΙ. ΣΚΟΠΟΣ ΜΕΛΕΤΗΣ. 36 ΙΙΙ. ΓΕΝΙΚΕΣ ΜΕΘΟΔΟΙ Πειραματόζωα Γενικά στοιχεία των in vitro τομών ιπποκάμπου Παρασκευή Τομών Ιπποκάμπου Διαλύματα και Φαρμακολογικοί παράγοντες Εξωκυττάριες Καταγραφές Δυναμικών Πεδίου στην CA1 περιοχή του Ιπποκάμπου Δημιουργία δυναμικού πεδίου Πρόκληση Βιολογικού σήματος Ποσοτικοποίηση Δυναμικών Πεδίου Πληθυσμιακό Δυναμικό Ενέργειας Προσυναπτικών Αξόνων Διεγερτικό Μετασυναπτικό Δυναμικό Πεδίου Εμβαδόν Κυματομορφής Ορθόδρομο Πληθυσμιακό Δυναμικό Ενέργειας Μεθοδολογία Ερεθισμού Συναρτήσεις Εισόδου/εξόδου Συνάρτηση συναπτικής αποτελεσματικότητας Συνάρτηση διεγερσιμότητας Συνάρτηση μετασυναπτικής διεγερσιμότητας Σύζευξη Παλμών Παλίνδρομη αναστολή Διευκόλυνση του fepsp με σύζευξη παλμών Στατιστική Ανάλυση. 48 Β. ΕΙΔΙΚΟ ΜΕΡΟΣ I. ΑΠΟΤΕΛΕΣΜΑΤΑ ΠΕΙΡΑΜΑΤΙΚΩΝ ΜΕΛΕΤΩΝ 1. Κύριες διαφορές μεταξύ διαφραγματικού και κροταφικού πόλου στην τροποποίηση της συναπτικής διαβίβασης και νευρωνικής διεγερσιμότητας από τους υποδοχείς NMDA, mglu5 και A 2A Αδενοσίνης 1.1.Εισαγωγή Πειραματικές διαδικασίες 50 vii
10 Ζώα και προετοιμασία τομών Καταγραφές, Επεξεργασία και Ανάλυση Δεδομένων Φαρμακολογικοί παράγοντες Στατιστική Αποτελέσματα Βασικές Μετρήσεις μεταξύ DH και VH Αποκλίνουσες επιδράσεις της ενεργοποίησης των NMDARs μεταξύ DH και VH Η δράση των NMDARs δεν απαιτούν την ενεργότητα των Α1 αδενοσινεργικών υποδοχέων Συνενεργοποίηση των mglur5 και NMDARs ενισχύει τις NMDAμεσολαβούμενες δράσεις στον DH αλλά όχι στον VH Οι συνεργιστικές δράσεις των NMDARs και mglur5 εμφανίζονται και στα απομονωμένα CA1 πεδία Οι A2ARs ελέγχουν τα συνεργιστικές δράσεις των mglur5 και NMDARs στον DH αλλά όχι στον VH Τα συνεργιστικά αποτελέσματα των mglur5 και NMDARs στον DH απαιτούν τους CB1 υποδοχείς ενδοκαναβινοειδών Το μέσο ελεύθερου Μg ++ δεν επηρεάζει την συνέργεια μεταξύ mglur5 και NMDARs Οι συνεργιστικές δράσεις της mglur5-nmdar συνενεργοποίησης στην μετασυναπτική διεγερσιμότητα συνέπιπτε χρονικά με την φωσφορυλίωση των NR2B υπομονάδων (Tyr1472) στον ραχιαίο ιππόκαμπο Οι συνεργιστικές δράσεις της συνενεργοποίησης των mglur5/nmdars στην μετασυναπτική διεγερσιμότητα, στην φωσφορυλίωση του NR2B (Tyr1472) και στη συναπτική διαβίβαση, εξαρτάται από την Src οικογένεια κινασών και αδειοδοτείται από τους A2ARs Συζήτηση Συνέπειες για ραχιαίο-κοιλιακή διαφοροποίηση Ερμηνεία των δράσεων της mglu5r/nmdar συνενεργοποίησης στον DH και VH Ερμηνεία των δράσεων του NMDA στον DH και VH Συμπεράσματα 83 viii
11 2. Επαγόμενο LTP με ερεθισμό θ συχνότητας: διαφορές και ομοιότητες ανάμεσα σε συνάψεις από τον ραχιαίο και κοιλιακό ιππόκαμπο 2.1. Εισαγωγή Σκοπός Υλικά και Μέθοδοι Ζώα και προετοιμασία τομών Καταγραφές, Επεξεργασία δεδομένων και Ανάλυση Φαρμακολογικοί παράγοντες Στατιστική Αποτελέσματα Βασικές μετρήσεις σε DH και VH Το TBS επάγει μικρής διάρκειας (S-LTP) και μακράς διάρκειας LTP (L-LTP) παρόμοιου πλάτους στον DH και VH, αλλά ο DH έχει χαμηλότερα κατώφλια επαγωγής για τα δύο LTPs Οι GABABRs ελέγχουν τα διαφορετικά κατώφλια επαγωγής για το S-LTP και L-LTP μεταξύ των 2 πόλων Ο DH vs VH έχει χαμηλότερο κατώφλιο επαγωγή για NMDAR εξαρτώμενο LTP Τα L-VDCCs και mglur5 συνεισφέρουν στο 3-TBS επαγόμενο LTP εξίσου στον DH και VH Η ενίσχυση των μετασυναπτικών αποκρίσεων και η ενεργότητα των GABA B Rs κατά τη διάρκεια του TBS μπορεί να εξηγήσει τις διαφορές στο LTP μεταξύ DH και VH Συζήτηση Πιθανοί μηχανισμοί των ραχιαίο-κοιλιακών διαφορών Λειτουργικές εφαρμογές Συμπεράσματα 114 ix
12 3. Οξύαιχμα κύματα και ριπιδισμοί: επιδράσεις της γήρανσης και τροποποίηση τους μέσω NMDA υποδοχέων και L-τύπου διαύλων Ca Εισαγωγή Σκοπός Πειραματική διαδικασία Προετοιμασία τομών Ηλεκτροφυσιολογία, τύποι δραστηριότητας και ανάλυση δεδομένων Αυθόρμητη δραστηριότητα Προκλητή Δραστηριότητα Συμπεριφορική (ανοικτού πεδίου) δοκιμασία Φαρμακολογικοί παράγοντες Στατιστική Ανάλυση Αποτελέσματα Ταλάντωση Ριπιδισμού Πυροδοτήσεις συμπλόκων δυναμικών Δραστηριότητα MUA εντος των οξύαιχμων κυμάτων Γεγονότα των SWRs Επιδράσεις των φαρμάκων στα οξύαιχμα κύματα Επιδράσεις των φαρμάκων στην προκλητή δραστηριότητα Δοκιμασία ανοικτού πεδίου Συζήτηση Ερμηνεία των εξαρτώμενων από την ηλικία αλλαγών στην αυθόρμητη δραστηριότητα Ερμηνεία των φάρμακο-επαγόμενων αλλαγών στα SWRs Επιπτώσεις των αλλαγών στα SWRs στην λειτουργία της μνήμης κατά τη γήρανση Συμπεράσματα ΙΙ. ΠΕΡΙΛΗΨΗ & ΣΥΜΠΕΡΑΣΜΑΤΑ. 144 III. SUMMARY. 146 ΙV. ΒΙΒΛΙΟΓΡΑΦΙΑ 149 x







 αποτελεί μια ελκυστική")








. Εικόνα Α.Ι.")
13 Ι.ΕΙΣΑΓΩΓΗ 1. Γενικά περί ιπποκάμπου Ο ιππόκαμπος αποτελεί μία από τις καλύτερα μελετημένες περιοχές π του εγκεφαλικού φλοιού, διότι η σχετικά απλή οργάνωση των στοιβάδων των κυρίαρχων (principal) κυττάρων σε συνάφεια με την υψηλά οργανωμένη στρωματώδη κατανομή πολλών από τις εισόδους του, ενθάρρυνε τους ερευνητές στη χρήση του ως σύστημα σ μοντέλο για την μοντέρνα νευροβιολογία. Ο ιππόκαμπος, μετά την απόδειξη εμπλοκής του σε μνημονικές διαδικασίες στον άνθρωπο (Penfield et al., 1974) αποτελεί μια ελκυστική εγκεφαλική δομή για την έρευνα των διαδικασιών μνήμης και μάθησης, για την διερεύνηση της συναπτικής και νευρωνικής πλαστικότηταςς και λειτουργίας, καθώς επίσηςς και για την κατασκευή μοντέλωνν νευρωνικών δικτύων. 2. Μακροσκοπική και Μικροσκοπική Οργάνωση του ιπποκάμπου Ο Ιππόκαμπος τοποθετείται αρκετά διαφορετικά στους εγκεφάλους των τρωκτικών καιι των πρωτευόντων. Αυτό οφείλεται στον καλύτερα ανεπτυγμένο εγκεφαλικό φλοιό των πρωτευόντων, που τείνει να εξαναγκάζε ει την οδοντωτή έλικα (OE) και τον ιππόκαμπο μέσα στον κροταφικό λοβό (Εικόνα Α.Ι.1). Εικόνα Α.Ι.1: MRI εγκεφάλου αρουραίου που δείχνει την θέση του ιπποκάμπειου σχηματισμού. Α:πλάγια όψη, Β:ραχιαίαα όψη, C:πρόσθια όψη και D:πλευρική όψη. (Πηγή: Courtesy of Dr. G. Allan Johnson, Center for In Vivo Microscopy, Duke University) 1



 πάνω και")



, τις περιοχές")
, το")
,")
 και τονν")
 που")





 ευθυγραμμισμένη")
 )")





14 Ο Ιππόκαμπος είναι μια επιμηκυμένη κυλινδρική δομή με τον τ μακρύ του άξονα να επεκτείνεται σε σχήμα C από τη μέση γραμμή του εγκεφάλου ε κοντά στους διαφραγματικούς πυρήνες (πρόσθιο-ραχιαία) πάνω και πίσω από α τον θάλαμο στο έσω τμήμα του κροταφικού λοβού (οπίσθιο-κοιλιακά). Συγκροτείται από έξι διακριτές περιοχές που περιλαμβάνουν την οδοντωτή έλικα-οε (Dentatee Gyrus-DG), τις περιοχές Αμμώνιου κέρατος (Cornus ammonis, CA1, CA2, CA3), το υπόθεμα (Subiculum), το προϋπόθεμα (Presubiculum), το παραϋπόθεμα (Parasubiculum) και τονν ενδορρινικό φλοιό-εφ (Entorhinal Cortex-EC) που έχει δύο ή περισσότερες υποδιαιρέσεις (Εικόνα A.I.2). Το υπόθεμα, το προϋπόθεμα καιι το παραϋπόθεμα αναφέροντα αι συνήθωςς μαζί καιι ως υποθεματικό σύμπλοκο (Amaral et al., 1989). Εικόνα A.I.2: Απεικόνιση των τμημάτων και συνδέσεων Ιπποκάμπειου σχηματισμού: Α) σε κανονική μορφή, Β) ευθυγραμμισμένη μορφή (Πηγή: The Hippocampus Book page 38) ) Ο ιππόκαμπος είναι διαιρεμένος μέσα σε ε οριοθετημένη περιοχή: Η κυρίαρχη κυτταρική στοιβάδα ονομάζεται πυραμιδική στοιβάδα (stratum pyramidale-pcl) ), στην οποία εδράζονται τα κυτταρικά σώματα των πυραμιδικών νευρώνων. Κάτω Κ από την πυραμιδική στοιβάδα ακολουθεί μία στενή, σχετικά ακυτταρική στοιβάδα, η στοιβάδα πολυμορφικών κυττάρων (stratum oriens-so),που περιέχει τους βασικούς δενδρίτες των πυραμιδικών 2
15 κυττάρων και πολλούς τύπους ενδονευρώνων. Η ακτινωτή στοιβάδα (stratum radiatum-sr) τοποθετείται αμέσως πάνω από το πυραμιδικό στρώμα στις περιοχές CA1 και CA2. Η ακτινωτή στοιβάδα καθορίζεται ως η υπερπυραμιδική περιοχή στην οποία εδράζονται οι CA3-CA3 αντίπλευρες συνειρμικές (associational) συνδέσεις και οι CA3-CA1 παράπλευρές ίνες Schaffer. Η εξώτατη στοιβάδα του ιππόκαμπου καλείται μοριώδηςβοθριώδης στοιβάδα (stratum lacunosum-moleculare-slm) και εκεί καταλήγουν οι ίνες από τον ενδορρινικό φλοιό και από άλλες περιοχές του εγκεφάλου. Και στην ακτινωτή και στη μοριώδη-βοθριώδη στοιβάδα εδράζεται μία μεγάλη πληθώρα ενδονευρώνων (Εικόνα Α.Ι.3). Υπάρχουν αρκετοί τύποι ενδονευρώνων με τα πιο χαρακτηριστικά να είναι τα πυραμιδικά καλαθοειδή κύτταρα (Basket cells) που εδράζονται μέσα ή πλησίον της πυραμιδικής στοιβάδας και νευρώνουν το σώμα και τους εγγυς δενδριτες των πυραμιδικών κυττάρων. Αυτού του τύπου οι ενδονευρώνες ενεργοποιούνται από παλίνδρομους κλάδους των πυραμιδικών νευρώνων αλλά και από προσαγωγές ίνες, διερχόμενες από την περιοχή της στιβάδας των πολυμορφικών κυττάρων και την ακτινωτή στιβάδα, και είναι υπεύθυνοι για την ισχυρή αναστολή των πυραμιδικών κυττάρων. Ένα καλαθοειδές κύτταρο δέχεται συνάψεις περίπου από 2-15 πυραμιδικά κύτταρα και νευρώνει μέχρι 1000 πυραμιδικά κύτταρα, ασκώντας την ανασταλτική του δράση. Ένας δεύτερος ενδονευρώνας είναι το κύτταρο πολυέλαιο (Chandelier) ή αξονοαξονικό κύτταρο, που εντοπίζεται στην πυραμιδική στιβάδα και πραγματοποιεί συμμετρικές συνάψεις αποκλειστικά στην αρχική περιοχή του άξονα του πυραμιδικού κυττάρου, παρέχοντας ανασταλτικό έλεγχο στην εξερχόμενη πληροφορία από τα κύτταρα αυτά. Ένας άλλος τύπος είναι το διστοιβαδωτό κύτταρο (Bistratified cell). Αυτά τα κύτταρα έχουν ίνες που καταλήγουν και στις δενδριτικές άκανθες και κορμούς των πυραμιδικών κυττάρων. Παρόλο που οι είσοδοι αυτών των κυττάρων δεν έχουν μελετηθεί, οι δενδρίτες τους διανέμονται μέσα στη ζώνη των αντίπλευρων συνδέσεων στην CA3 περιοχή και στις παράπλευρές ίνες του Schaffer στην CA1. Συνεπώς, πιστεύεται ότι προωθούν μία πρόσσω (feed-forward) και ανάδρομη (feedback) δράση. Τέλος, υπάρχει μία ποικιλία ενδονευρώνων που εντοπίζονται στην ακτινωτή, και την μοριώδη-βοθριώδη στοιβάδα ή στο όριο αυτών. Αυτοί οι νευρώνες έχουν δενδρίτες που προσανατολίζονται οριζόντια αλλά περιστασιακά σχηματίζουν κλάδους που εκτείνονται στην πυραμιδική στοιβάδα (Lacaille et al., 1988). 3





: οι πληροφορίες που καταλήγουν")











16 Εικόνα Α.Ι.3: Μορφολογία Πυραμιδικών κυττάρων του ιππόκαμπου. Πηγή: (Ishizuka et al., 1995) 2.1. Συνδέσεις ιπποκάμπου Αισθητική είσοδος στον Ενδορρινικό φλοιό Η εισαγωγή πληροφορίας από τις νεοφλοιικές περιοχές στον ενδορρινικό φλοιό χωρίζεται σε δύο ομάδες (Εικόνες Α.Ι.4&5): οι πληροφορίες που καταλήγουν στις επιφανειακές στοιβάδες (Ι-ΙΙΙ) και αυτές που καταλήγουν ν σε βαθύτερες στοιβάδες (IV-VI). Η πρώτη κατηγορία μεταφέρει πληροφορίες στους επιφανειακά τοποθετημένους ενδορρινικούς νευρώνες, που προβάλλουν ν με την σειρά τους στην οδοντωτή έλικα, στον ιππόκαμπο και το υπόθεμα. Στα ανώτερα στρώματα καταλήγουν ν σημαντικές πληροφορίες από οσφρητικές δομές όπως, ο οσφρητικός βολβός και ο πρόσθιος οσφρητικός πυρήνας. Άλλες κρίσιμες πληροφορίες που καταλήγουν στα επιφανειακά στρώματα σ προέρχονται από τις όμορες περιρρινικές και μεταρρινικές περιοχές, που αποτελούν πολύαισθητικά κέντρα σύγκλησης που λαμβάνουν πληροφορίες από μία μεγάλη ποικιλία φλοιικώνν δομών. Ο ΕΦ προβάλλει ανάδρομαα στις οσφρητικές δομές μέσω ινών που προέρχονται κυρίως από τις στοιβάδες ΙΙ, ΙΙΙ και Va Η δεύτερη κατηγορία επιδρά κυρίως ς στους βαθύτερα τοποθετημένουςς ενδορρινικούς νευρώνες, που λαμβάνουν επεξεργασμένεςς πληροφορίες από άλλα ιπποκάμπεια πεδία και 4
17 προβάλλουν με την σειρά τους σε συγκεκριμένες φλοιικές περιοχές. Φλοιικές προβολές σε αυτά τα στρώματα προέρχονται από πολλές δομές, όπως τη νήσο του Reil και τη μέση προμετωπιαία περιοχή (μεταιχμειακό σύστημα) (Insausti et al., 1997). Πρόσθετα με τις φλοιικές πληροφορίες, ο ΕΦ δέχεται και μηνύματα από υποφλοιικές δομές. Ενώ κάποιες είσοδοι όπως οι μονοαμινεργικές και οι χολινεργικές φαίνονται ως τροποποιητικές (όπως οι ίνες από το σύμπλοκο της αμυγδαλής), ίσως να αποτελούν και επιπρόσθετες πηγές πληροφοριών. Οι σημαντικές είσοδοι από το σύμπλοκο της αμυγδαλής, που προέρχονται κυρίως από τους πλάγιους και βασικούς πυρήνες της, αποτελούν κατεξοχήν πληροφορίες για την συναισθηματική κατάσταση του οργανισμού (Pikkarainen et al., 1999). Ίνες από τον πλάγιο πυρήνα της αμυγδαλής κατανέμονται κυρίως στο βαθύ τμήμα της στοιβάδας ΙΙΙ αλλά και ανάμεσα στις νησίδες κυττάρων της στοιβάδας ΙΙ και Ι. Ίνες από τον βασικό πυρήνα καταλήγουν διαχεόμενες στις στοιβάδες ΙΙΙ-V, ενώ αυτές από τους φλοιικούς πυρήνες και τον περιαμυγδαλιδικό φλοιό στις στοιβάδες Ι και ΙΙ. Ο ΕΦ στέλνει προς τα πίσω ίνες (προερχόμενες από την στοιβάδα V) κυρίως στον βασικό πυρήνα της αμυγδαλής. Ο ΕΦ προβάλλει αμφίπλευρα στο ραβδωτό (μέσω ινών από την στοιβάδα V), κυρίως στον επικλινή πυρήνα και στα παρακείμενα τμήματα του οσφρητικού φύματος, ενώ δέχεται χολινεργική εννεύρωση κυρίως από το διάφραγμα και προβάλλει με τη σειρά του (μέσω ινών από την στοιβάδα Va και ΙΙ) πίσω σε αυτό. Επίσης δέχεται διάχυτες πληροφορίες από ποικίλες δομές του υποθαλάμου, όπως τον υπερχιασματικό πυρήνα που καταλήγει στις στοιβάδες ΙΙΙ-VI, και από την πλάγια υποθαλαμική περιοχή. Οι σημαντικότερες θαλαμικές είσοδοι προέρχονται από τον ενωτικό (reuniens) και τον κεντρικό διάμεσο πυρήνα (centralis medialis nucleus) (Van der Werf et al., 2002) ενώ δεν υπάρχουν ενδείξεις για δράση αναδραστικού συστήματος. Επιπλέον, χολινεργικές ίνες από κύτταρα της κοιλιακής καλυπτρικής περιοχής καταλήγουν κατά κύριο λόγο στην πλάγια ενδορρινική περιοχή. Οι σεροτονινεργικές ίνες προέρχονται από τους πυρήνες ραφής και καταλήγουν διάχυτα σε όλες τις στοιβάδες αλλά με μία προτίμηση στις επιφανειακές. Οι νοραδρενεργικές ίνες από τον υπομέλαινα τόπο (locus coeruleus) καταλήγουν στην στοιβάδα Ι του ΕΦ Διατιτραίνουσα οδός Η οδοντωτή έλικα λαμβάνει τις κύριες εισόδους της από τον ΕΦ μέσω της διατιτραίνουσας οδού. Στην μοριώδη στοιβάδα της ΟΕ, οι ίνες του ΕΦ σχηματίζουν συνάψεις κυρίως στις δενδριτικές άκανθες των κοκκιωδών κυττάρων, παρόλα αυτά ένας μικρός αριθμός ινών σχηματίζει συνάψεις με τους δενδριτικούς κορμούς GABAεργικών ενδονευρώνων. Η 5
18 διατιτραίνουσα οδός μπορεί να διακριθεί σε δύο μέρη βασιζόμενη στην περιοχή προέλευσής και του προτύπου κατάληξης των ινών καθώς και της εμφάνισης σε ιστοχημικές και ανοσοιστοχημικές μεθόδους. Στον αρουραίο, οι δύο υποδιαιρέσεις έχουν ονομαστεί ως πλάγια και διάμεση διατιτραίνουσα οδός προερχόμενες από πλάγιες (LEC) και διάμεσες (MEC) ενδορρινικές περιοχές, αντίστοιχα. Ίνες της πλάγιας διατιτραίνουσας οδού καταλήγουν κυρίως στο επιφανειακό 1/3 τμήμα της μοριώδους στοιβάδας της ΟΕ, ενώ ίνες της διάμεσης οδού καταλήγουν στο βαθύτερο 1/3 τμήμα της μοριώδους στοιβάδας (Εικόνες Α.Ι.4&5). Μία μεγάλη ποικιλία καλαθοειδών κυττάρων εντοπίζονται στην κοκκιώδη στοιβάδα της ΟΕ και φαίνεται να συμμετέχουν στο υπερβολικά πυκνό τελικό πλέγμα της κοκκιώδους στοιβάδας (Struble et al., 1978). Τα τελικά σημεία (συνάψεις) αυτού του πλέγματος είναι GABAεργικά και σχηματίζουν συμμετρικές ανασταλτικές συνάψεις με τα κυτταρικά σώματα και εγγύς δενδριτικούς κορμούς των κορυφαίων δενδριτών των κοκκιωδών κυττάρων. Αυτός ο πολυσυναπτικός καταρράκτης των ανασταλτικών ενδοσυνδέσεων καταδεικνύει ότι το κύκλωμα του ιππόκαμπου παρέχει έναν πολύπλοκο ανασταλτικό και από-ανασταλτικό έλεγχο της διεγερσιμότητας Μονοπάτι Βρυωδών Ινών Η ΟΕ δεν προβάλλει σε καμία άλλη περιοχή εκτός από το CA3 πεδίο του ιππόκαμπου. Οι άξονες που προβάλλουν στην CA3, οι βρυώδεις ίνες, προέρχονται αποκλειστικά από τα κοκκιώδη κύτταρα και καταλήγουν σε μία σχετικά στενή ζώνη τοποθετημένη κυρίως μόλις πάνω από την CA3 πυραμιδική στοιβάδα (Blackstad et al., 1970; Claiborne et al., 1986) που καλείται φωτεινή ή διάφανη (lucidum) στοιβάδα εξαιτίας της έλλειψης μυελίνης των βρυωδών ινών που δίνει στην στοιβάδα μία σχετικά καθαρή εμφάνιση σε φρέσκο ιστό. Μία μοναδιαία βρυώδης ίνα μπορεί να πραγματοποιήσει το πολύ 37 συνάψεις με ένα και μόνο δενδρίτη πυραμιδικού κυττάρου. Επίσης εντοπίζονται σπάνιες λεπτές ίνες που πηγάζουν από τις κύριες βρυώδεις ίνες που φαίνεται να καταλήγουν κατά προτίμηση σε GABAεργικά καλαθοειδή κύτταρα της CA3 περιοχής. Τέλος πρέπει να αναφερθεί ότι τα κοκκιώδη κύτταρα είναι διεγερτικά καθώς χρησιμοποιούν γλουταμινικό οξύ σαν νευροδιαβιβαστή. 6














 συνδεδεμένων CA3 Οι CA3")



19 Εικόνα Α.Ι.4: Βασική ανατομία του Ιπποκάμπου Η ενδογενής λειτουργική οργάνωση του ιπποκάμπου βασίζεται σε ένα τρισυναπτικό κύκλωμα με κύριο χαρακτηριστικό τις μονόδρομες προβολές μεταξύύ των περιοχών του. Η διατιτραίνουσαα πορεία, η οποία μεταφέρει πληροφορίες από τον ενδορρινικό και ι τον περιρρινικό φλοιό θεωρείται γενικά η κύρια πηγή εισαγωγής πληροφορίας στον ιππόκαμπο. Οι ίνες της διατιτραίνουσας οδού συνάπτονται με τους δενδριτες των κοκκωδών κυττάρων της οδοντωτής έλικας. Στη συνέχεια, τα κοκκώδη κύτταρα κ μέσωω των νευραξόνων τους, τις βρυώδεις ίνες, συνάπτονται με τους δενδρίτες των πυραμιδικών νευρώνων της CA3 περιοχής. Οι άξονες των κυττάρων αυτών, οι ίνες του Schaffer, άγονται προς την CA1 περιοχή όπουυ και δημιουργούν συνάψεις με τους δενδρίτες των CA1 πυραμιδικών κυττάρων. Το παραπάνω απλουστευμένο κύκλωμα αποτελείται από τρείς μονόδρομες διεγερτικές συνάψεις. Επί προσθέτως, σε αυτό το τρισυναπτικό κύκλωμα υπάρχει ένα πυκνό συνειρμικό δίκτυο (associative,, commissural network) συνδεδεμένων CA3 κυττάρων. Οι CA3 νευρώνες νευρώνονται επίσης από άμεση είσοδο από τη στοιβάδα ΙΙ του ΕΦ, ενώ οι απώτεροι δενδρίτες των CA1 κυττάρων λαμβάνουν άμεση είσοδο από την στοιβάδα ΙΙΙ του ΕΦ. Επίσηςς στον ιππόκαμπο λαμβάνουν χώρα πληθώρα αλληλεπιδράσεων μεταξύ ενδονευρώνων, οι κυριότεροι των οποίων είναι ανασταλτικοί. Για την αναλυτική περιγραφή της ανατομικής οργάνωσης του ιπποκάμπου βλέπε (Bayer, 1985; Burwell et al., 1995). Πηγή: (Neves et al., 2008) 7
20 Παράπλευρο συνδετικό/συνδεσμικό μονοπάτι Schaffer CA3 προς CA1 συνδέσεις: παράπλευρες ίνες του Schaffer Όλα τα τμήματα της CA3 και CA2 περιοχής προβάλλουν στην CA1. Στην πραγματικότητα, κάθε CA3 πυραμιδικό κύτταρο δίνει γένεση σε άξονες που διασχίζουν και το εγκάρσιο και το οβελιαίο επίπεδο στη CA1 περιοχή. Παρόλο που οι παράπλευρες ίνες του Schaffer φαίνεται να επεκτείνονται μόνο διαμέσου της ακτινωτής στοιβάδας, θα πρέπει να δοθεί έμφαση ότι και η ακτινωτή και η ακυτταρική στοιβάδα της CA1 περιοχής δέχονται πυκνή εννεύρωση από CA3 άξονες. Συνεπώς, οι ίνες Schaffer είναι υψηλά συνδεδεμένες με τους κορυφαίους δενδρίτες των CA1 κυττάρων στην ακτινωτή στοιβάδα καθώς και με τους βασικούς δενδρίτες της ακυτταρικής στοιβάδας (Εικόνες Α.Ι.3&4). Οι παχιές ίνες του Schaffer προέρχονται μόνο από τα εγγύς CA3 κύτταρα. Αυτά τα κύτταρα δίνουν γένεση σε έναν παχύ άξονα που ανέρχεται από την ακυτταρική στοιβάδα μέσα στο πιο επιφανειακό τμήμα της ακτινωτής στοιβάδας και ταξιδεύει προς το μακρινό τμήμα της CA1, όπου και συνεισφέρει πολλές ίνες. Οι άξονες από τα μακρινά CA3 κύτταρα τείνουν να είναι πιο λεπτοί και να προβάλλουν άμεσα στην εγγύς CA1, είτε μέσα στην ακυτταρική στοιβάδα είτε διαμέσου του βαθύ τμήματος της ακτινωτής στοιβάδας. Ένα πλέγμα από ένα και μόνο CA3 νευρώνα έχει 150 μέχρι 300 mm συνολικό αξονικό μήκος, στο οποίο σχηματίζονται από μέχρι συνάψεις (Li et al., 1994). CA3 προς CA3 ομόπλευρες συνδέσεις Οι συνδέσεις ενός CA3 με ένα άλλο CA3 στην ίδια πλευρά, οργανώνονται με ένα υψηλά συστηματικό τρόπο. Τα κύτταρα που εντοπίζονται στην εγγύς περιοχή της CA3 επικοινωνούν μόνο με τα κοντινά σε αυτά CA3 κύτταρα. Αμφίπλευρες συνδέσεις του Ιππόκαμπου Στον αρουραίο, τα CA3 πυραμιδικά κύτταρα δίνουν γένεση σε προβολές προς την CA3, CA2 και CA1 πεδία του αμφίπλευρου ιπποκάμπειου σχηματισμού. Όπως με τις αμφίπλευρες προβολές της ΟΕ, οι CA3 ίνες στον αμφίπλευρο ιππόκαμπο σχηματίζουν ασύμμετρες συνάψεις στις άκανθες των πυραμιδικών κυττάρων της CA3 και CA1 περιοχής αλλά επίσης καταλήγουν πάνω στους λείους δενδρίτες των ενδονευρώνων CA1-Υπόθεμα-Ενδορινικός φλοιός Η CA1 περιοχή δίνει γένεση σε δύο ενδοιπποκάμπειες συνδέσεις (προβολές). Η 1 η είναι μία τοπογραφικά οργανωμένη προβολή στο παρακείμενο υπόθεμα, ενώ η 2 η είναι 8
.")

















21 προς τα βαθειά στρώματα του ΕΦ (Εικόνεςς Α.Ι.5&6). Αυτό που είναι ε ξεκάθαρο, είναι ότι οι προβολές από την CA1 περιοχή και το υπόθεμα προς τον ΕΦ είναι και αυτές τοπογραφικά οργανωμένες. Τα κοιλιακά τμήματα της CA1 και του υποθέματος προβάλλουν κυρίωςς στα πλευρικά τμήματα του ΕΦ, ενώ τα πιο κροταφικά τμήματα προβάλλουν σε πιο διάμεσα μέρη του ΕΦ. Επιπλέον, οι προβολές από το εγγύς τμήμα της CA1 C και το μακρινό τμήμα του υποθέματος διανέμονται αποκλειστικάά στον διάμεσο ΕΦ, ενώ κύτταραα του μακρινού τμήματος της CA1 και του εγγύς υποθέματος προβάλλον κυρίως στον πλάγιο ΕΦ. Το σημαντικό σημείο για αυτές τις προς τα πίσω προβολές είναι ότι βρίσκονταιι σε ρύθμιση με τις ενδορρινικές εισόδους σε αυτές τις περιοχές. Έτσι, σε σφαιρικό σ επίπεδο, όλο το σύστημα-βρόχοτελειώνουν στον ΕΦ. Αυτή η αξιοθαύμαστη τοπογραφία επιβεβαιώνει τονν κρίσιμο ρόλο του ΕΦ όσον αφορά την είσοδο και έξοδο της πληροφορίας από τον ιπποκάμπειο σχηματισμό. είναι διαθέσιμο για την εγκατάσταση κυκλωμάτων, τα οποία αρχίζουν και Εικόνα Α.Ι.5: Σύνοψη της εγκάρσιας οργάνωσης των συνδέσεων μέσα στον σ ιπποκάμπειο σχηματισμό. Αυτή η εικόνα τονίζει την δυνατότηταα της πληροφορίας να διαχωρίζεται μέσαα στον ιπποκάμπειο σχηματισμό και τελικά να φτάνει σε διαφορετικούς παραλήπτες της ιπποκάμπειας εξόδου. (Πηγή: Hippocampus Book σελίδα 108) ). 9


.. 3.")















,")




22 Εικόνα Α.Ι.6: Σύνοψη των παράλληλων και σειριακών μονοπατιών στον ιπποκάμπειο σχηματισμό (Πηγή: Hippocampus Book page 109).. 3. Συναπτική διαβίβαση Η συναπτική διαβίβαση (synaptic transmission) μπορεί να αναλυθεί σε απελευθέρωση νευροδιαβιβαστή από το προσυναπτικο στοιχείο, διάχυση του νευροδιαβιβαστή κατά μήκος της συναπτικης σχισμής και ενεργοποίηση των μετασυναπτικών υποδοχέων. υ 3.1. Διεγερτική διαβίβαση Ο κύριος διεγερτικός νευροδιαβιβαστής στον ιππόκαμπο, καθώς και σε ολόκληρο το ΚΝΣ, είναι το γλουταμινικό οξύ, που προκαλεί την ταχεία διεγερτική διαβίβαση στις γλουταμινεργικές συνάψεις. Υπάρχουνν τρεις βασικοί τύποι ιοντοτροπικών υποδοχέων γλουταμινικού που έχουν πάρει το όνομαα τους από τους εξωγενείς αγωνιστές που τους ενεργοποιούν (Watkins et al., 1981; Nakanishi et al., 1998; Ozawa ett al., 1998): ο υποδοχέας κισκαλικού οξέος (ή AMPA), καινικού οξέος και Ν-μέθυλ-D-ασπαρτικού οξέος (NMDA). Οι υποδοχείς AMPAA και καινικού οξέος θεωρούνται υπεύθυνοι για την ταχεία συναπτική διαβίβαση, δηλαδή την ταχείαα ανάπτυξη ενός σύντομου (<1 ms) ρεύματος εισροής ιόντων νατρίου στο μετασυναπτικο στοιχείο. Αντίθεταα το μετασυναπτικο ρεύμα που προκαλείται από την ενεργοποίηση του NMDA υποδοχέα εμφανίζει ε μια πιο βραδεία ανάπτυξη και διάρκεια (20-30 ms). Οι υποδοχείς NMDA διαφέρουν από τους AMPAA σε 10
23 αρκετές σημαντικές ιδιότητες, όπως: στο εξαρτώμενο από την τάση μπλοκάρισμα του ιοντικού πόρου από το Mg ++, την υψηλή διαπερατότητα σε ιόντα ασβεστίου, την βραδεία αλλά και μεγαλύτερης διάρκειας ενεργοποίηση του καθώς και στην μεγαλύτερη χημική συγγένεια ως προς το γλουταμινικό (Dingledine et al., 1999). Με μεμβρανικό δυναμικό κοντά στο δυναμικό ηρεμίας, ο NMDA υποδοχέας παραμένει αποκλεισμένος από τα ιόντα Mg ++ (που βρίσκονται στο έξω- και ενδοκυττάριο υγρό), ακόμα και αν ο υποδοχέας έχει ενεργοποιηθεί με την πρόσδεση του γλουταμινικού. Σε καταστάσεις μετασυναπτικής εκπόλωσης, όπως συμβαίνει με την είσοδο του νατρίου από τους μετάσυναπτικούς υποδοχείς των AMPA και του καινικού οξέος, προκαλείται η απομάκρυνση του Mg ++ επιτρέποντας την διέλευση ρεύματος από τους NMDA υποδοχείς. Το ρεύμα διάμεσου των υποδοχέων αυτών αποτελείται από ιόντα νατρίου καθώς και ασβεστίου με τελικό αποτέλεσμα την εντονότερη εκπόλωση της μετασυναπτικής μεμβράνης. Και οι AMPA και οι NMDA υποδοχείς συνεντοπίζονται με πυκνότητα 100 φορές μεγαλύτερη στη σύναψη απ ότι στις εξωσυναπτικές περιοχές. Το γλουταμινικό δρα επίσης στους πιο βραδέως ενεργοποιούμενους υποδοχείς G- πρωτεΐνης, οι οποίοι δρουν μέσω 2ου αγγελιοφόρου. Αυτοί οι γλουταμινεργικοί υποδοχείς τροποποιούν την νευρωνική διεγερσιμότητα και την συναπτική διαβίβαση (τροποποιούν την απελευθέρωση του γλουταμινικού και του GABA) και αναφέρονται ως μεταβοτροπικοί υποδοχείς γλουταμινικού (mglu) (De Blasi et al., 2001). Παρουσιάζουν ευρεία κατανομή στο ΚΝΣ και παίζουν βασικούς ρόλους σε ποικίλες νευρωνικές διαδικασίες όπως τη συναπτική πλαστικότητα και τον σχηματισμό της μνήμης. Ένας mglu υποδοχέας, αποτελείται από μία μεγάλη εξωκυττάρια Ν-τελική περιοχή, μία 7-διαμεμβράνική περιοχή και ένα μεγάλο C-τελικό άκρο. Κυρίως, βασιζόμενοι στις όμοιες αλληλουχίες, αλλά και στους μηχανισμούς μεταγωγής και τις φαρμακολογικές ιδιότητες, οι mglurs έχουν κατηγοριοποιηθεί σε τρεις μεγάλες ομάδες. Η ομάδα Ι περιέχει τους mglu1 και mglu5, που ενεργοποιούν Gq πρωτεΐνες, ενώ οι υποδοχείς των ομάδων II (mglu2,3) και ΙΙΙ (mglu4,6,7,8) συζευγνύονται με άλλες G-πρωτεΐνες (Shigemoto et al., 1997). Οι mglurs εντοπίζονται κυρίως έξω από τις προ- και μετασυναπτικές μεμβράνες, καταδεικνύοντας ότι ενεργοποιούνται επιλεκτικά κατά τη διάρκεια έντονης συναπτικής ενεργοποίησης, όπως οι επαναλαμβανόμενες υψηλής συχνότητας πυροδοτήσεις, οδηγώντας σε διάχυση του νευροδιαβιβαστή στον περισυναπτικό χώρο (Baude et al., 1993; Lujan et al., 1996; Shigemoto et al., 1997). Οι υποδοχείς της ομάδας Ι ενεργοποιούνται επιλεκτικά με 3,5-διυδροφαινύλγλυκίνη (3,5-dihydroxyphenylglycine-DHPG) ή (RS)-2-χλώρο-5-υδροξυφαινυλγλυκίνη ((RS)-2-11
24 Chloro-5-hydroxyphenylglycine-CHPG) και συζευγνύονται θετικά μέσω Gq πρωτεΐνη με την φωσφολιπάση C, διεγείροντας έτσι την ενεργοποίηση της πρωτεϊνικής κινάσης C και την παραγωγή τριφωσφορικής ινοσιτόλης (IP3). Πιο συγκεκριμένα οι mglur5 εντοπίζονται σωματοδενδριτικά και η ενεργοποίησή τους οδηγεί σε ενίσχυση της νευρωνικής διεγερσιμότητας μέσω τροποποίησης του ασβεστίου (αύξηση του Ca ++ μέσω της IP3 μεσολαβούμενης απελευθέρωσης του από τις ενδοκυττάριες αποθήκες) και του καλίου. Οι υποδοχείς της ομάδας ΙΙ συνδέονται αρνητικά μέσω Gi πρωτεΐνης με την αδενυλική κυκλάση ανατέλλοντας την παραγωγή camp. Οι mglur 2,3,4,7,8 εντοπίζονται στους άξονες κοντά και μέσα στις προσυναπτικές απολήξεις και μειώνουν την απελευθέρωση του νευροδιαβιβαστή αναστέλλοντας τα τασεοελεγχόμενα κανάλια ασβεστίου και/ή επεμβαίνοντας άμεσα στην μηχανή απελευθέρωσης. Στον ιππόκαμπο, οι υποδοχείς της ομάδας ΙΙΙ προκαλούν αναστολή της διεγερτικής διαβίβασης στις ίνες του Schaffer. Ρόλος της Αδενοσίνης στην διεγερτική διαβίβαση Η αδενοσίνη είναι ένα νουκλεοτίδιο πουρίνης, το οποίο δρα ως νευροτροποποιητής σε πολλές εγκεφαλικές περιοχές, παίζοντας σημαντικό τονικό ρόλο στην δράση άλλων νευροδιαβιβαστών (Sebastiao et al., 2009). Έχει μελετηθεί ο ρόλος και η συμμετοχή της σε πολλές διαδικασίες και ασθένειες στο ΚΝΣ όπως στον ύπνο (Retey et al., 2006; Elmenhorst et al., 2007), στην επιληψία (Boison, 2010; Van Dycke et al., 2010), διαταραχή πανικού (Lam et al., 2005), άγχος (Alsene et al., 2003), στην νόσο AD (Albasanz et al., 2008), στη νόσο του Parkinson (Hurley et al., 2000), και στη σχιζοφρένεια (Uriguen et al., 2009). Η ATP που παράγεται από τον μεταβολισμό των κυττάρων καταβολίζεται ενζυμικά και οδηγεί στον σχηματισμό της αδενοσίνης. Η αδενοσίνη πιθανώς απελευθερώνεται από τους νευρώνες άμεσα, και εξωκυτταρια συσσώρευση της πραγματοποιείται σε καταστάσεις μεταβολικού στρες. Ο ρόλος της είναι διττός (Cunha, 2001) καθώς ενεργεί τόσο ως ομοιοστατική διακυτταρική αγγελιοφόρος, όσο και ως νευροτροποποιητής που ελέγχει την απελευθέρωση νευροδιαβιβαστών και τη νευρωνική διεγερσιμότητα. Η αδενοσίνη μπορεί να επηρεάσει πολλούς κυτταρικούς τύπους μέσω της ενεργοποίησης τεσσάρων υποδοχέων συζευγμένων με G πρωτεΐνες: A1, A2 A, Α2 Β και Α3. Προσυναπτικοί Α1 υποδοχείς εντοπίζονται στις διεγερτικές απολήξεις, όπου και καταστέλλουν την απελευθέρωση του γλουταμινικού μέσω G-πρωτεΐνης εξαρτώμενων 12
25 μηχανισμών (Wu et al., 1994). Η απελευθέρωση του GABA δεν επηρεάζεται σε μεγάλο βαθμό (Yoon et al., 1991; Lambert et al., 1993). Οι Α2 Α υποδοχείς βρίσκονται προσυναπτικά, σχηματίζοντας ετερομερή με τους Α1 υποδοχείς αδενοσίνης, όπου τροποποιούν την απελευθέρωση του γλουταμινικού οξέος και μετασυναπτικά, όπου σχηματίζουν ετεροδιμερή σύμπλοκα με τους mglur5 (ραβδωτό σώμα) και επομένως εμφανίζουν λειτουργική αλληλεπίδραση (Ciruela et al., 2006). Η προσυναπτική ύπαρξη των υποδοχέων σε κάποια περιοχή συνεπάγεται αναστολή της απελευθέρωσης νευροδιαβιβαστών, ενώ η μετασυναπτική εντόπιση τους συνεπάγεται υπερπόλωση του μετασυναπτικού νευρώνα μέσω της διάνοιξης ιοντικών διαύλων διαπερατών από τα ιόντα καλίου και χλωρίου. Σε περιοχές όπου οι υποδοχείς αδενοσίνης εντοπίζονται και προκαι μέτα-συναπτικά, όπως τον ιππόκαμπο, υποδεικνύεται πιθανότατα μεγαλύτερος ανασταλτικός έλεγχος της αδενοσίνης. Η αφθονία των A1 υποδοχέων αδενοσίνης στον εγκέφαλο σε συνδυασμό με την ενεργοποίηση τους από μικρές (νανομοριακές) ποσότητες αδενοσίνης δηλώνει τη σημαντικότητά τους σε κάθε κατάσταση που χρειάζεται η ανασταλτική τους δράση για να αντισταθμιστεί η υπέρμετρη λειτουργία των νευρώνων. Πολλαπλοί ρόλοι των Ιόντων Ασβεστίου Η είσοδος Ca ++ εντός του κυτταροπλάσματος των δενδριτών και των δενδριτικών ακάνθων ρυθμίζει ένα ευρύ φάσμα νευρωνικών λειτουργιών, όπως η συναπτική σηματοδότηση, η επαγωγή LTD και LTP και η ρύθμιση της γονιδιακής έκφρασης (Sabatini et al., 2001). Η συσσώρευση του Ca ++ σε αυτές τις δομές προκαλείται μέσω δυναμικών ενέργειας και συναπτικών ερεθισμάτων τα οποία προκαλούν είσοδο του Ca ++ διαμέσου πολλών τύπων ιοντικών διαύλων καθώς και απελευθέρωση από τις ενδοκυττάριες αποθήκες (Higley et al., 2008). Η συναπτικά προκαλούμενη αύξηση του κυτταροπλασματικού Ca ++ στα πυραμιδικά κύτταρα της CA1 και CA3 περιοχής του ιπποκάμπου προκύπτει από τις εξής πηγές: Από τους υποδοχείς γλουταμινικού (NMDARs και non-nmdars) που εντοπίζονται στις δενδριτικούς άξονες και άκανθες (Reid et al., 2001; Bloodgood et al., 2007), με αποτέλεσμα την εκπόλωση της μετασυναπτικής μεμβράνης. Από τους τασεοελεγχόμενους διαύλους ασβεστίου (voltage-dependent Calcium channels-vdccs), οι οποίοι εντός των δενδριτικών ακάνθων και αξόνων ανοίγουν μετά από ισχυρή συναπτικά προκαλούμενη εκπόλωση (Miyakawa et al., 1992; Regehr et al., 1992; Markram et al., 1994; Denk et al., 1995; Eilers et al., 1995; Yuste et al., 1995; Finch et al., 1998; Reid et al., 2001). Επαρκής εκπόλωση για την ενεργοποίηση των vdccs μπορεί να 13
26 προέλθει από τα αντίδρομα σωματικά παραγόμενα δυναμικά ενέργειας (back-propagated action potentials-baps) (Callaway et al., 1995; Schiller et al., 1995; Yuste et al., 1995; Svoboda et al., 1997; Helmchen et al., 1999; Koester et al., 2000; Carter et al., 2004; Nevian et al., 2004; Bloodgood et al., 2007). Πολλές μελέτες έχουν προτείνει ότι η ύπαρξη των δενδριτικών δυναμικών ενεργείας μπορεί να συνεισφέρει στην παραγωγή ειδικής ως προς τη θέση πλαστικότητας των ιπποκάμπειων συναπτικών εισόδων. Για παράδειγμα τρένα θ ρυθμού ή υψηλής συχνότητας δραστηριότητα, είναι ικανά να παράξουν δενδριτικά Ca ++ δυναμικά στους κορυφαίους δενδρίτες των CA1 πυραμιδικών νευρώνων. Το μπλοκάρισμα των NMDARs και των L-τύπου vdccs μειώνει σημαντικά και το πλάτος των Ca ++ δενδριτικών δυναμικών και την παραγωγή μακροχρόνιας ενδυνάμωσης, προτείνοντας ότι τα τοπικά Ca ++ δυναμικά παρέχουν έναν μηχανισμό επαγωγής συναπτικής πλαστικότητας απουσία παραγωγής μετασυναπτικών δυναμικών μέσω διαύλων Na + (Remy et al., 2007). Στα CA1 πυραμιδικά κύτταρα, η εισροή Ca ++ στις δενδριτικές άκανθες συμβαίνει μέσω των L-τύπου, R-τύπου και των χαμηλής ουδού Τ-τύπου vdccs (Christie et al., 1995; Magee et al., 1995; Sabatini et al., 2000), ενώ στις δενδριτικές κεφαλές φαίνεται να συμμετέχουν κυρίως οι R-τύπου με τη μικρή συνεισφορά των L-τύπου vdccs (Sabatini et al., 2000; Yasuda et al., 2003; Hoogland et al., 2004; Bloodgood et al., 2007). Τα L-τύπου vdccs έχουν δειχθεί ότι εμπλέκονται στην επαγωγή της γονιδιακής έκφρασης. Αυτοί οι υποδοχείς έχουν πολλές ιδιότητες που τους κάνουν ιδιαίτερα ταιριαστούς για τη μεταφορά του σήματος-ca ++ από το σημείο εισόδου του στη πλασματική μεμβράνη μέχρι το εσωτερικό του πυρήνα για γονιδιακή επαγωγή. Για παράδειγμα, οι L-τύπου vdccs έχουν σχετικά αργό ρυθμό απενεργοποίησης και υψηλή αγωγιμότητα για το Ca ++, επιτρέποντας την είσοδο μεγάλων ποσοτήτων ασβεστίου στους νευρώνες (Gallin et al., 1995). Πρόσθετα, αυτά τα κανάλια είναι τοποθετημένα σωματοδενδριτικά, οπότε είναι ικανά να αυξήσουν τα επίπεδα Ca ++ μέσα στο κυτταρικό σώμα σε εγγύτητα με τον πυρήνα, επιτρέποντας έτσι την αποδοτική διάδοση του σήματος εντός του πυρήνα (Westenbroek et al., 1990; Catterall, 2000). Επίσης, ένας μεγάλος αριθμός σηματοδοτικών πρωτεϊνών συσχετίζονται με τους L- τύπου vdccs, όπως η Src κιναση τυροσίνης, η φωσφατάση της καλσινευρίνης και άλλες (Greer et al., 2008). Η συνεισφορά του Ca ++ που απελευθερώνεται από τις ενδοκυττάριες αποθήκες στους δενδριτες μετά από συναπτική ενεργοποίηση είναι αμφιλεγόμενη. Σε τομές ιπποκάμπου, ισχυρή ενεργοποίηση των ινών Schaffer οδηγεί σε ενεργοποίηση των mglurs οι οποίοι στη συνέχεια μέσω της φωσφολιπασης C και της τριφωσφορικής ινοσιτόλης (IP3) επάγουν την απελευθέρωση του Ca ++ από τις ενδοκυττάριες αποθήκες και συνεισφέρουν στην 14
27 μακροπρόθεσμη ετεροσυναπτική πλαστικότητα (Dudman et al., 2007; Hong et al., 2007). Αντιθέτως, άλλες μελέτες απέτυχαν να αποδείξουν την απελευθέρωση του Ca ++ από τις ενδοκυττάριες αποθήκες ακολουθώντας μία πιο ήπια συναπτική ενεργοποίηση των ινών (Mainen et al., 1999; Yuste et al., 1999; Kovalchuk et al., 2000) Ανασταλτική διαβίβαση Ο κυριότερος ανασταλτικός νευροδιαβιβαστής στο ΚΝΣ, το GABA, συντίθεται από την αποκαρβοξυλίωση του γλουταμινικού. Η ροή της διέγερσης στον ιππόκαμπο ρυθμίζεται από τους GABAεργικούς ενδονευρώνες οι οποίοι ασκούν τις εξής δράσεις: Η απελευθέρωση GABA από αυτά τα κύτταρα προκαλεί μια ταχεία αναστολή μέσω των ιοντοτροπικών GABA A υποδοχέων που εισάγουν ιόντα Cl - (ταχύ ανασταλτικό μετασυναπτικό δυναμικό, IPSP) (Kaila, 1994). Αυτή η ταχεία GABA A ενεργοποίηση ελέγχει την συχνότητα των δυναμικών ενεργείας και αναστέλλει τα δενδριτικά Ca ++ προκαλούμενα δυναμικά. Βραδύτερες αποκρίσεις (βραδέα ανασταλτικά μετασυναπτικά δυναμικά, IPSPs, που προκαλούνται με το άνοιγμα των καναλιών καλίου) που διαρκούν από 500 έως 2000 ms, προκαλούνται από τους μεταβοτρόπους GABA B υποδοχείς. Οι προσυναπτικοί GABA B Rs είναι παρόντες στις ανασταλτικές και διεγερτικές απολήξεις, όπου και λειτουργούν ως αυτουποδοχείς και ετερουποδοχείς, αντίστοιχα. Το απελευθερωμένο GABA μπορεί να κινηθεί ανάδρομα προς τους προσυναπτικούς GABA B αυτουποδοχείς και να αναστείλει την περαιτέρω απελευθέρωση του GABA. Το GABA επίσης μπορεί να διαχυθεί στις γειτονικές διεγερτικές απολήξεις και να ενεργοποιήσει τους ετερουποδοχείς GABA B οι οποίοι αναστέλλουν την απελευθέρωση του γλουταμινικού. Η ενεργοποίηση των μετασυναπτικών GABA B Rs οδηγεί στο άνοιγμα των καναλιών GIRKs (G protein-activated inwardly rectifying potassium channels), οι οποίοι αναστέλλουν την νευρωνική δραστηριότητα μέσω συνεγγούς εκτροπής (local shunting) και παράγουν βραδέα IPSPs που υπερπολώνουν την μεμβράνη (Andrade et al., 1986; Misgeld et al., 1995; Kullmann et al., 2005; Gassmann et al., 2012). Σε γενικές γραμμές, η ηλεκτροφυσιολογική ενεργοποίηση των GABA B Rs αυτουποδοχέων απαιτεί έντονης έντασης ερεθίσματα και απελευθερωμένο GABA από πολλούς ενδονευρώνες (Scanziani, 2000; Kohl et al., 2010), που συμφωνεί με την απομακρυσμένη θέση τους σε σχέση με τα σημεία απελευθέρωσης του GABA (Kulik et al., 2006). Σε φυσιολογικές καταστάσεις, τέτοια παρατεταμένη συναπτική ενεργοποίηση συναντάται κατά τη διάρκεια πυροδοτήσεων είτε ρυθμικής φλοιικής είτε ιπποκάμπειας 15
28 δικτυακής δραστηριότητας, όταν ομάδες GABAεργικών ενδονευρώνων πυροδοτούν συγχρονισμένα (Scanziani, 2000; Kohl et al., 2010). Τα νευρογλοιακά κύτταρα αποτελούν εξαίρεση καθώς μπορούν να αποκριθούν σε μοναδιαία δυναμικά ενέργειας και να απελευθερώσουν GABA προκαλώντας IPSPs στους γειτονικούς νευρώνες (Price et al., 2008; Olah et al., 2009). Αυτή η μαζική διαβίβαση (volume transmission) είναι πολύ αποδοτική για την ενεργοποίηση των GABA B Rs με τρόπο παρακρινή και έτσι τα νευρογλοιακά κύτταρα δεν απαιτούν απαραιτήτως συναπτικές συνδέσεις με τα γειτονικά τους κύτταρα για να προκαλέσουν βραδέα IPSPs. Οι GABA B Rs εκφράζονται σχεδόν από όλους τους νευρώνες και τα γλοία κύτταρα στο ΚΝΣ και η ενεργοποίηση τους επηρεάζει πολλά νευρωνικά συστήματα και συμπεριφορικές καταστάσεις. Συνεπώς δεν μας προκαλεί έκπληξη το γεγονός ότι οι GABA B Rs έχουν εμπλακεί σε ποικιλία νευρολογικών και ψυχιατρικών ασθενειών όπως, την επιληψία, το άγχος, την κατάθλιψη, τη σχιζοφρένεια, την ιδεοψυχαναγκαστική διαταραχή, τον εθισμό και τον πόνο. Αναλυτική περιγραφή των δράσεων του GABA B R στην ανασκόπηση του Gassmann (Gassmann et al., 2012). 4. Νευρωνική Διεγερσιμότητα Νευρωνική διεγερσιμότητα καλείται η ικανότητα του νευρώνα να παράγει δυναμικά ενέργειας και εξαρτάται από την πυκνότητα και την ετοιμότητα των τασεοελεγχόμενων διαύλων Να +. Για αυτό το λόγο ο εκφυτικός κώνος έχει τη μεγαλύτερη διεγερσιμότητα αφού η πυκνότητα των διαύλων αυτών σε εκείνο το σημείο είναι αρκετά μεγάλη. Η διεγερσιμότητα επηρεάζεται και από άλλους παράγοντες όπως οι ηλεκτροτονικές ιδιότητες της μεμβράνης, οι ενδο- και εξωκυττάριες συγκεντρώσεις των ιόντων (Na +, Κ +, Cl -, Ca ++ ), καθώς και διαφορά ενδογενή και συναπτικά (ταχεία και βραδεία αναστολή) ιοντικά ρεύματα τα οποία καθορίζουν κάθε στιγμή το μεμβρανικό δυναμικό ηρεμίας και την απόσταση από το κατώφλιο (ουδό). Με τους παραπάνω τρόπους ρύθμισης, οι νευρώνες μπορούν να απαντούν ή όχι στο ίδιο ερέθισμα αλλά και να κωδικοποιούν την απάντησή τους στο ίδιο ερέθισμα με ποικίλα πρότυπα διαδοχής δυναμικών ενέργειας στον χρόνο (απλή εκφόρτιση ή υψίσυχνες ριπές δυναμικών ενέργειας). 5. Συναπτική Πλαστικότητα Η πιο συναρπαστική και σημαντική ιδιότητα του εγκεφάλου των θηλαστικών είναι η πλαστικότητα, δηλαδή η ικανότητα της εμπειρίας να τροποποιεί το νευρωνικό κύκλωμα και συνεπώς να μεταβάλλει την μελλοντική σκέψη, συμπεριφορά και συναίσθημα. Συναπτική 16
29 πλαστικότητα (synaptic plasticity) είναι η βιολογική διαδικασία κατά την οποία συγκεκριμένες μορφές συναπτικής δραστηριότητας προκαλούν αλλαγές στην συναπτική αποτελεσματικότητα (synaptic strength). Και προ- και μετασυναπτικοί μηχανισμοί συμμετέχουν στην έκφραση της συναπτικής πλαστικότητας και οι τροποποιήσεις της κατηγοριοποιούνται συνήθως βάση της χρονικής διάρκειας τους σε βραχύχρονες (παίζουν σημαντικό ρόλο στις ταχείες διαδικασίες μεταφοράς της πληροφορίας στον εγκέφαλο) και μακρόχρονες αλλαγές (συνεισφέρουν στις διαδικασίες της μάθησης και μνήμης). Oι βραχύχρονες είναι: α) η ενίσχυση (facilitation) και αποτελεί μια σύντομη (1sec) αύξηση του μετασυναπτικού δυναμικού ως συνέπεια της αυξημένης απελευθέρωσης του νευροδιαβιβαστή β) η επαύξηση (augmentation) παρόμοιο φαινόμενο με την ενίσχυση αλλά διαρκεί περισσότερα δευτερόλεπτα, γ) μετάτετανική ενδυνάμωση (postetanic potentiation), δηλαδή αύξηση της συναπτικής αποτελεσματικότητας μετά από επίδοση υψίσυχνων ηλεκτρικών ερεθισμών και διαρκεί μερικά λεπτά και δ) μετατετανική καταστολή (postetanic depression), δηλαδή μείωση της συναπτικής αποτελεσματικότητας μετά από επίδοση υψίσυχνων ηλεκτρικών ερεθισμών. Όλες αυτές οι αλλαγές της συναπτικής αποτελεσματικότητας οφείλονται κυρίως σε αλλαγές της συγκέντρωσης/διαθεσιμότητας των ιόντων ασβεστίου στην προσυναπτική απόληξη, με συνέπεια την αυξομείωση της απελευθέρωσης του νευροδιαβιβαστή. Οι μακρόχρονες αλλαγές είναι: α) η μακρόχρονη ενδυνάμωση (long term potentiation, LTP) και β) η μακρόχρονη καταστολή (long term depression, LTD) της συναπτικής αποτελεσματικότητας Μακρόχρονη ενδυνάμωση Το LTP έχει μελετηθεί ευρέως στον ιππόκαμπο (όπως στην οδοντωτή έλικα, και στις περιοχές CA1 και CA3) απ ότι κάποια άλλη περιοχή του εγκεφάλου. Ο ενθουσιασμός που περιβάλλει αυτό το φαινόμενο προέρχεται κυρίως από το γεγονός ότι: Α) ο ιππόκαμπος αποτελεί σημαντικό στοιχείο του νευρωνικού συστήματος που συμμετέχει σε ποικίλους τύπους μακρόχρονης μνήμης (Zola-Morgan et al., 1993). Β) πολλές ιδιότητες του LTP το κάνουν έναν ελκυστικό κυτταρικό μηχανισμό για αποθήκευση πληροφορίας (Kauer et al., 1988; Bliss et al., 1993). Όπως και η μνήμη, το LTP μπορεί να παραχθεί γρήγορα και να επιμηκυνθεί/ενδυναμωθεί με την επανάληψη. Επίσης, παρουσιάζει εξειδίκευση εισόδου με τρόπο ώστε να προκαλείται στις συνάψεις εκείνες που ενεργοποιούνται από προσαγωγό δραστηριότητα και όχι σε διπλανές συνάψεις στο ίδιο μετασυναπτικό κύτταρο. Αυτό το χαρακτηριστικό αυξάνει δραματικά την ικανότητα αποθήκευσης μεμονωμένων νευρώνων οι οποίοι, λόγω του ότι οι συνάψεις τους 17
30 μπορούν να τροποποιούνται ανεξάρτητα μεταξύ τους, μπορούν να συμμετέχουν στην κωδικοποίηση πολλών διαφορετικών πληροφοριών. Γ) το LTP παράγεται πολύ εύκολα σε in vitro παρασκευάσματα του ιπποκάμπου, κάνοντας το προσιτό στην πειραματική ανάλυση και Δ) το LTP έχει παρατηρηθεί σε σχεδόν όλες τις διεγερτικές συνάψεις του εγκεφάλου. Στην περιοχή CA1 in vitro, το LTP ξεκινάει μετά από 2-3 δευτερόλεπτα από τον τέτανο, κορυφώνεται στα 30 δευτερόλεπτα, και μετέπειτα μειώνεται για λεπτά όπου και καταλήγει σε μία σταθερή κατάσταση. Το πόσο διαρκεί δεν μπορεί να απαντηθεί εύκολα γιατί συνήθως παρουσιάζει μια ποικιλότητα στην αντοχή. Η διάρκεια εξαρτάται από τις παραμέτρους του τετάνου, τον τύπο του παρασκευάσματος και την περιοχή του ιπποκάμπου. Και σε τομές ιπποκάμπου και σε αναισθητοποιημένα ζώα, η μέγιστη διάρκεια περιορίζεται κυρίως από την αντοχή του παρασκευάσματος. Σε άθικτα ζώα διαρκεί από λίγες μέρες (Racine et al., 1983; Doyere et al., 1992) έως περισσότερο και από ένα χρόνο (Abraham, 2003) στην οδοντωτή έλικα. Ελάχιστες μελέτες έχουν γίνει in vivo στην CA1 περιοχή του ιπποκάμπου, αλλά έχει ειπωθεί ότι το LTP σε αυτήν την περιοχή δεν εξασθενεί (Staubli et al., 1987). Πρέπει να σημειωθεί ότι δεν μπορούν όλες οι συνάψεις να προκαλέσουν LTP με τη χρήση των διαφόρων πρωτοκόλλων, είτε εξαιτίας μη υγειών ή κατεστραμμένων κυττάρων είτε λόγω χρήσης ακατάλληλου πρωτοκόλλου για τις συγκεκριμένες συνάψεις. Για την επαγωγή του LTP απαιτούνται και οι προ- και οι μετασυναπτικοί νευρώνες την ιδία στιγμή επειδή ο μετασυναπτικός νευρώνας πρέπει να εκπολωθεί όταν το γλουταμινικό απελευθερώνεται στη συναπτική σχισμή για να αποδεσμευτεί το Mg ++ που μπλοκάρει τους NMDARs. Σαν συνέπεια της εκπόλωσης και της δέσμευσης του γλουταμινικού, θα έχουμε μέγιστη εισροή Ca ++ μέσω των NMDARs, τα οποία ενεργοποιούν ενδοκυττάρια σηματοδοτικά μονοπάτια που είναι υπεύθυνα για την αλλαγή της συναπτικής αποτελεσματικότητας Πρωτόκολλα επαγωγής της μακρόχρονης ενδυνάμωσης Το LTP μπορεί να προκληθεί από ένα ευρύ φάσμα πρωτοκόλλων ερεθισμού. Τo πιο κοινά χρησιμοποιούμενο είναι ένα τρένο παλμών (ποικίλου αριθμού) στα 100 Hz για 1 δευτερόλεπτο (High frequency stimulation-hfs). Δύο άλλα ευρέως χρησιμοποιημένα πρωτόκολλα είναι o ερεθισμός με ριπή προπαίδευσης (primed burst stimulation-pbs) και ο ερεθισμός υπό μορφή ριπής θήτα ρυθμού (Theta Burst stimulation-tbs), στα οποία το κοινό χαρακτηριστικό είναι το διάστημα των 200 ms είτε ανάμεσα στο ερέθισμα 18
31 προπαίδευσης (priming stimulus) και της σύντομης ριπής ερεθισμών όπως στο PBS (Larson et al., 1986), είτε ανάμεσα σε διαδοχικές σύντομες ριπές (συνήθως των τεσσάρων παλμών) σε ένα TBS (Rose et al., 1986). Το διάστημα των 200 ms είναι κοντά στην περιοδικότητα του θήτα ρυθμού, μίας ενδογενούς ιπποκάμπειας δραστηριότητας που παράγεται κατά την εξερευνητική κίνηση. O ερεθισμός προπαίδευσης ή η ριπή προπαίδευσης ενεργοποιεί πρόσω τους GABA ενδονευρώνες, οδηγώντας σε υπερπόλωση του πυραμιδικού κυττάρου μέσω των μετασυναπτικών GABA A και GABA B υποδοχέων (έχει αναλυθεί παραπάνω) αλλά κυρίως οδηγεί στην ενεργοποίηση των προσυναπτικών GABA B αυτό-υποδοχέων, οι οποίοι οδηγούν με τη σειρά τους στην μείωση της απελευθέρωσης του GABA που είναι βέλτιστη στα ms. Με αυτόν τον τρόπο η επόμενη σύντομη ριπή ερεθισμών προκαλεί μικρότερη υπερπόλωση της μετασυναπτικής μεμβράνης με τελικό αποτέλεσμα την ενίσχυση του ρεύματος διαμέσου των τασεοελεγχόμενων NMDA υποδοχέων. Μικρότερα πρότυπα διέγερσης αυτού του είδους είναι πιθανότερο να συμβαίνουν φυσιολογικά απ ότι τα HFS πρωτόκολλα που χρησιμοποιούνται κυρίως για λόγους συνήθειας και ευκολίας. Οι ιπποκάμπιοι πυραμιδικοί νευρώνες μπορούν και πυροδοτούν σε υψηλής συχνότητας σύντομες ριπές των 4-5 ερεθισμών, παρόλα αυτά παραμένει να αποδειχθεί το εάν το LTP επάγεται μέσω φυσιολογικών προτύπων δραστηριότητας σε ένα ελεύθερο κινούμενο πειραματόζωο (Buzsaki et al., 1987). Ο λόγος της αποτελεσματικότητας των PBS και TBS πρωτοκόλλων έναντι των HFS αναλύεται εκτενώς από (Larson & Munka, 2009). 6. Ενδογενείς Ρυθμοί του Ιπποκάμπου Στον ιππόκαμπο ενός ελεύθερα κινούμενου αρουραίου, έχουν αναγνωριστεί έξι χαρακτηριστικά EEG πρότυπα: 4 ρυθμικά και 2 μη ρυθμικά. Τα ρυθμικά πρότυπα συμπεριλαμβάνουν τον θ ρυθμό (6-12 Hz), β ρυθμό (12-30 Hz), γ ρυθμό ( Hz) και τους ριπιδισμούς ( Hz). Τα μη ρυθμικά πρότυπα συμπεριλαμβάνουν την μεγάλου μη φυσιολογικού πλάτους δραστηριότητα (Large Irregular amplitude activity, LIA) στην οποία συμπεριλαμβάνονται τα οξύαιχμα κύματα και την μικρού μη φυσιολογικού πλάτους δραστηριότητα (Small irregular amplitude activity, SIA). Κάποιες δραστηριότητες μπορεί να συνυπάρχουν (π.χ. LIA με ριπιδισμούς, θ με γ ρυθμό), ενώ άλλες φαίνεται να είναι πιο αποκλειστικές. 19
32 6.1. Οξύαιχμα κύματα-ριπιδισμοί Στην CA1 περιοχή, τα οξύαιχμα κύματα (Sharp waves-spws) συμβαίνουν πιο συχνά κατά τη διάρκεια του ύπνου βραδέων κυμάτων και κατά την ήσυχη ανάπαυση, λιγότερο συχνά κατά την πόση και κατάποση, και ελάχιστα κατά την προετοιμασία κίνησης. Φαίνεται να εμφανίζονται κατά την χαμηλή αφύπνιση και συχνά αλλά όχι πάντα αναστέλλονται από ερεθίσματα αφύπνισης. Ίσως αναπαριστούν μία κατάσταση ανάπαυσης του ιπποκάμπου σαν σύνολο ή την απουσία κάποιων όψεων της ιπποκάμπειας λειτουργίας όπως την θήτα κατάσταση. Λειτουργικά, έχει προταθεί ότι αναπαριστούν μία νευρωνική συσχέτιση με την παγίωση της μνήμης. Κάθε οξύαιχμο κύμα διαρκεί από ms με συχνότητα 0,2-5 Hz και έχει μέγιστο αρνητικό πλάτος στην ακτινωτή στοιβάδα της τάξης του 1-3,5 mv και αντιστρέφουν την πολικότητα τους στην πυραμιδική στοιβάδα. Συμβαίνουν περισσότερο ή λιγότερο συγχρονισμένα σε μεγάλες περιοχές του CA1 πεδίου του κροταφικού (κοιλιακού) ιπποκάμπου. Καταγραφές σε διαφορετικά σημεία κατά μήκος του ραχιαίο-κοιλιακού άξονα του ιππόκαμπου έχουν δείξει ότι βρίσκονται σε φάση σε όλη την έκταση (Buzsaki et al., 1992; Chrobak et al., 1996). Οι μέγιστες αρνητικές τάσεις αυτών των κυμάτων συναντώνται στην ακτινωτή στιβάδα της CA1, στο διαυγές στρώμα της CA3, στην μοριώδη στιβάδα της οδοντωτής έλικας και στη μοριώδη στιβάδα του υποθέματος. Σε κάποια ζώα θετικά οξύαιχμα κύματα έχουν καταγραφεί και στην πύλη της οδοντωτής έλικας. Το πεδίο των οξύαιχμων κυμάτων είναι σταθερά συνδεδεμένο με σύγχρονες εκφορτίσεις ενός μεγάλου νευρωνικού πληθυσμού σε όλο τον ιππόκαμπο. Η απομόνωση ενός μόνο νευρώνα στην πυραμιδική στιβάδα είναι συνήθως δύσκολη επειδή πολλοί γειτονικοί νευρώνες εκφορτίζουν κατά τη διάρκεια των οξύαιχμων κυμάτων. Όταν απομονωθούν, οι πυραμιδικοί νευρώνες δεν πυροδοτούν απαραίτητα μαζί με κάθε μοναδικό SPW, όμως η συχνότητα της εκφόρτισής τους είναι μεγαλύτερη κατά τη διάρκεια των οξύαιχμων κυμάτων παρά άλλες φορές. Ενδονευρώνες και κοκκιώδη κύτταρα αυξάνουν επίσης το βαθμό εκφόρτισής τους σε συνδυασμό με τα οξύαιχμα κύματα. Σαν κανόνας ισχύει ότι συχνά όλη η κυτταρική δραστηριότητα τερματίζεται ακολουθώντας την πληθυσμιακή έκρηξη για ms. Το μέγεθος της καταστολής είναι ανάλογο του πλάτους των πληθυσμιακών εκρήξεων. Τα οξύαιχμα κύματα δεν περιορίζονται αποκλειστικά στον ιππόκαμπο, αλλά διαδίδονται και σε άλλες περιοχές. Καταγραφές στο υπόθεμα και σε βαθειά στρώματα του ΕΦ δείχνουν ότι τα οξύαιχμα κύματα εμφανίζονται σε αυτές τις δομές περίπου την ίδια στιγμή με αυτά στον ιππόκαμπο, ενώ καταγραφές στην ΟΕ δείχνουν ότι τα κοκκιώδη 20
33 κύτταρα επηρεάζονται επίσης από την δραστηριότητα οξύαιχμων κυμάτων της CA3 περιοχής. Τα οξύαιχμα κύματα καταγράφονται ταυτοχρόνως από διαφορετικές περιοχές του ίδιου ιπποκάμπου ή και από τους δύο ιππόκαμπους (αριστερός και δεξιός) και υποδεικνύουν έναν υψηλού βαθμού συγχρονισμό κατά μήκος του επιμήκους και του εγκάρσιου άξονα του ιπποκάμπου. Μετρήσεις πλάτους από ταυτόχρονες καταγραφές ζευγών των χαρακτηριστικών των οξύαιχμων κυμάτων από CA1-CA3, και CA1-οδοντωτής έλικας δείχνουν αξιοσημείωτες συσχετίσεις (Buzsaki, 1986). Τα οξύαιχμα κύματα της οδοντωτής έλικας προτείνεται ότι αντανακλούν συγχρονισμένες διεγέρσεις από τους εγγύς δενδρίτες των κοκκιωδών κυττάρων από συνδετικά και συνδεσμικά μονοπάτια. Τα μεγάλα αρνητικά οξύαιχμα κύματα στην CA3 μπορεί να προκύπτουν από κοινές διεγέρσεις των πυραμιδικών νευρώνων διαμέσου των επαναλαμβανόμενων παράπλευρων νευραξόνων. Ο Buzsaki και οι συνεργάτες του έχουν προτείνει ότι τα οξύαιχμα κύματα παράγονται στην CA3 περιοχή και ότι αυτά που καταγράφονται στην CA1 περιοχή αποτελούν αθροισμένα διεγερτικά μετασυναπτικά δυναμικά των παράπλευρων ινών του Schaffer που προκύπτουν από την συγχρονισμένη πυροδότηση των CA3 πυραμιδικών κυττάρων (Csicsvari et al., 2000). Τελευταία αναπτύσσεται μία πιο σύνθετη θεωρία που υποστηρίζει ότι τα οξύαιχμα κύματα στην CA1 απεικονίζουν συγχρονισμένα διεγερτικά μετασυναπτικά δυναμικά των πυραμιδικών κυττάρων των παράπλευρων ινών Schaffer και της συνδετικής εισόδου, μαζί με ανασταλτικά μετασυναπτικά δυναμικά παραγόμενα από ενδονευρώνες. Οι ενδονευρώνες πιθανότατα διεγείρονται από τις παράπλευρες ίνες του Schaffer και από τους νευράξονες των πυραμιδικών νευρώνων της CA1. Άμεση διέγερση αυτών των ινών έχει ως αποτέλεσμα την δημιουργία προκλητών δυναμικών ίδιου σχήματος και έντασης. Οι ριπιδισμοί αποτελούν μια αυθόρμητη δραστηριότητα που μπορεί να εκλύεται από τα ίδια τα οξύαιχμα κύματα (Εικόνα Α.Ι.7). Για τη μεταξύ τους σχέση έχουν γίνει εκτενείς μελέτες όμως το ερώτημα της αλληλεξάρτησης οξύαιχμων κυμάτων-ριπιδισμών (Sharp waves-ripples, SWRs) παραμένει αναπάντητο. Γύρω από την αρνητική κορυφή ενός οξύαιχμου κύματος, υπάρχει μία υψηλής συχνότητας ταλάντωση μεταξύ Hz που φτάνει σε μέγιστο πλάτος περίπου μv στην CA1 πυραμιδική στοιβάδα (Buzsaki et al., 1992). Οι ριπιδισμοί έχουν ένα ιδιαίτερο σχέδιο αυθόρμητης δραστηριότητας στην ιπποκάμπεια CA1 περιοχή με συναπτικές εκρήξεις 5-15 ημιτονοειδών κυμάτων στην πυραμιδική στιβάδα σε κύκλους χρονικής διάρκειας περίπου 5 ms ( Hz). Οι ριπιδισμοί απαντώνται κατά τη διάρκεια του ύπνου βραδέων κυμάτων και πιστεύεται ότι αντιπροσωπεύουν τις αποθηκευμένες πληροφορίες που μεταφέρονται στο νεοφλοιό κατά τη 21
.")


 του ενδορρινικού φλοιού όπου")
.")

.")

34 διάρκεια της παγίωσης της μνήμης. Το κύκλωμα με το οποίο δρουν είναι ένα κλειστό κύκλωμα (Buzsaki et al., 1983; Suzuki et al., 1987). Γεννώνται στην CA3 πυραμιδική στιβάδα και μέσω του θετικού διεγερτικού κυκλώματος τωνν νευρώνωνν της περιοχής αναπτύσσουν αυθόρμητες σύγχρονες εκρήξεις δυναμικών ενέργειας και εν συνεχεία διαδίδονται στη CA1 (Draguhn et al., 2000). Οι ριπιδισμοί δεν περιορίζονται στη CA1 περιοχή. Όπως αναφέραμε παραπάνω, προκύπτουν από ταα οξύαιχμαα κύματα που δημιουργούνται στη CA3. Προς το ιπποκάμπειο μονοπάτι εξόδου, τα οξύαιχμα κύματα συνδεόμενα με τους ριπιδισμούς ταξιδεύουν μέσου της CA1 και του υποθέματος μέσα στα βαθιά στρώματα (ΙΙ, ΙV) του ενδορρινικού φλοιού όπου αποσπούνν ένα οξύαιχμο κύμα με το συνακόλουθο ριπιδισμό που καθυστερεί 5-30 ms σε σχέση μεε τη CA1 (Chrobak ett al., 1994; Chrobak et al., 1996). Παράλληλα α, τα οξύαιχμα κύματαα διεγείρουν τα κοκκιώδη κύτταρα της οδοντωτής έλικας,, πιθανώς μέσω των διεγερτικών ενδονευρώνων στην πύλη των βρυωδών κυττάρων. Έτσι, οι ριπιδισμοί εμφανίζονται και στην οδοντωτήτή έλικα (Ylinen et al., 1995; Chrobak et al., 1996). Τα οξύαιχμα κύματα με τουςς τοποθετημένους «επάνω» τους, ριπιδισμούς εμφανίζονται επίσης συχνά αμφοτερόπλευρα, όμως δεν έχει βρεθεί ακριβής συνοχή κύκλου με κύκλο μεταξύ των δύο ημισφαιρίων (Buzsaki et al., 1992; Chrobak et al., 1996). Έτσι, αρχίζουν από τη CA3 και έπειτα ταξιδεύουν σε διπλή κατεύθυνση μέσα στην οδοντωτή έλικα, καθώς επίσης και μέσω του μονοπατιού εξόδου του ιπποκάμπου CA1-υπόθεμα-ενδορρινικός φλοιός. Εικόνα Α.Ι.7: Αναπαράσταση Οξύαιχμου Κύματος-Ριπιδισμού 22
35 Το πεδίο των ριπιδισμών εμπλέκει τη δράση ενδονευρώνων και πυραμιδικών κυττάρων, που δείχνουν μια πιο σταθερή σύζευξη φάσης και υψηλότερη πιθανότητα εκφόρτισης κατά τη διάρκειά τους. Ο Buszaki πρότεινε ότι in vivo οι ριπιδισμοί γεννιούνται από μια γρήγορη συναπτική αλληλεπίδραση μεταξύ ενδονευρώνων και πυραμιδικών κυττάρων. Έχει βρεθεί ότι οι ταλαντώσεις 200 Hz δεν θα μπορούσαν να εμποδιστούν από τους ανταγωνιστές των γλουταμινεργικών ή των GABAεργικών συνάψεων. Ακόμα, διαπιστώνεται ότι η δραστηριότητα της ταλάντωσης Hz παρέμεινε ανεπηρέαστη απουσία εξωκυττάριων ιόντων ασβεστίου και έτσι δεν μπορεί να εξαρτάται από ένα κύκλωμα μόνο με χημικές συνάψεις. Προτείνεται (Draguhn et al., 1998; Traub et al., 1999) ότι οι ταλαντώσεις των 200 Hz ριπιδισμών, μπορεί να δημιουργούνται από ηλεκτρικά συνδεδεμένα δίκτυα των πυραμιδικών κυττάρων του ιπποκάμπου Ρόλος των οξύαιχμων κυμάτων-ριπιδισμών Οι δύο πόλοι του ιπποκάμπου έχει δειχθεί ότι εκτός από τις συνδέσεις διαφέρουν και σε ανώτερες γνωστικές λειτουργίες όπως στη μάθηση και στη μνήμη. Στους επίμυες έχει δειχθεί ότι ο διαφραγματικός (ραχιαίος) ιππόκαμπος παίζει σημαντικό ρόλο στις διαδικασίες της χωρικής μάθησης (Moser et al., 1998), ενώ ο κροταφικός (κοιλιακός) στην αντίληψη της εσωτερικής κατάστασης του ζώου και τις συναισθηματικές διεργασίες όπως ο φόβος και το άγχος (Hock et al., 1998). Μελέτες σε τρωκτικά έχουν αποκαλύψει ότι τα οξύαιχμα κύματα του ιππόκαμπου, συντονίζονται προσωρινά με τις νεοφλοιικές βραδείες ταλαντώσεις-άτρακτοι (Wolansky et al., 2006). Αυτή η συντονισμένη βραδέος κύματος ταλάντωση ίσως παρέχει ένα υπόστρωμα που ευνοεί τον διάλογο μεταξύ του ιππόκαμπου και του νεοφλοιού για κατοπινή σταθεροποίηση της μνήμης (memory consolidation). Εντός του ιππόκαμπου, οι υψηλής συχνότητας ταλαντώσεις γνωστές ως σύμπλοκα οξύαιχμων κυμάτων/ριπιδισμών συνδέονται με συγχρονισμένη εκφόρτιση μεγάλου αριθμού νευρώνων με πολλαπλά τμήματα του ιππόκαμπου. Το 1989, ο Buzsaki (Buzsaki, 1989) πρότεινε ότι η SPW-R δραστηριότητα ξεκινά από νευρώνες των οποίων η περιοδική συνδεσιμότητα ενδυναμώθηκε προσωρινά κατά τη διάρκεια προηγούμενης εμπειρίας όταν το πειραματόζωο ήταν ξύπνιο (awake experience). Πολλές μελέτες από τότε έχουν αποκαλύψει ότι πράγματι, ομάδες νευρώνων (π.χ. κύτταρα θέσης), που πυροδοτούν ταυτόχρονα κατά τη διάρκεια μιας συγκεκριμένης συμπεριφοράς, τείνουν να επαναλαμβάνονται κατά τη διάρκεια ενός επεισοδίου οξύαιχμου κύματος. Αυτή η δραστηριότητα που καταγράφηκε κατά τον ύπνο ή κατά τη διάρκεια ακίνητης 23
36 αγρυπνίας (awake immobility), είναι πιθανότερο να συμβαίνει κατά τη διάρκεια SPW-R επεισοδίων (Wilson et al., 1994; Kudrimoti et al., 1999). Αυτά τα κύτταρα πυροδοτούν μαζί σε υψηλές συχνότητες και θα μπορούσαν να προωθήσουν την Hebbian πλαστικότητα ( LTP, που σύμφωνα με τους περισσότερους ερευνητές αποτελεί υπόστρωμα για μακροπρόθεσμη μνήμη). Ο ύπνος μετά τη μάθηση προάγει την λειτουργική αναδιοργάνωση των μονοπατιών μνήμης με μία αυξανομένη συμμετοχή νεοφλοιικών περιοχών στην μακροπρόθεσμη αποθήκευση και ανάκληση. Τέτοια δυναμική όψη της παγίωσης της μνήμης σε συστημικό επίπεδο έχει προταθεί από πολλούς συγγραφείς και έχει επιβεβαιωθεί από μελέτες σε ζώα (Bontempi et al., 1999). Παράλληλα, αυξήσεις των γεγονότων οξύαιχμων κυμάτων/ριπιδισμών (SPW-Rs) έχουν περιγραφεί μετά από μάθηση τόσο σε ανθρώπους όσο και σε αρουραίους (Axmacher et al., 2008; Eschenko et al., 2008). Αυτά τα αποτελέσματα ενδυναμώνουν την υπόθεση ότι το σημαντικό γνώρισμα του ύπνου βραδέων κυμάτων για την παγίωση της μνήμης είναι η παρουσία των SPWRs. Επιπλέον, έχει προταθεί ότι τα SPWRs που εμφανίζονται σε συμπεριφορές αφύπνισης θα μπορούσαν να έχουν κάποια λειτουργία στην απόκτηση γνώσης (acquisition), την επεξεργασία (processing) ή στην μακροπρόθεσμη μνήμη, μαζί με τα γεγονότα που συμβαίνουν κατά τη διάρκεια του ύπνου βραδέων κυμάτων. Επιπλέον, έχει δειχθεί ότι ενώ πολλά γεγονότα παράγονται στον ιππόκαμπο, μόνο αυτά που είναι σχετικά με την παγίωση συμπεριφορικά σημαντικών πληροφοριών θα μεταφερθούν στον ΕΦ και σε περαιτέρω φλοιικές περιοχές (Axmacher et al., 2008), όπου και θα προκαλέσουν συναπτική πλαστικότητα. Τέλος, πρέπει να τονιστεί ότι υπάρχουν πολλά που πρέπει να κατανοήσουμε για τη SPW-R δραστηριότητα. Ένας πιθανός ρόλος στον σχηματισμό της μνήμης δεν πρέπει να περιοριστεί μέσα στα όρια της σημερινής επικρατούσας θεωρίας της ενδοιπποκάμπειας σταθεροποίησης ή της ιππόκαμπο-νεοφλοιικής μεταφοράς κατά την διάρκεια του ύπνου. Για παράδειγμα, τα SPWRs γεγονότα ίσως έχουν μία καθαρά ενδοιπποκάμπεια house keeping (νοικοκύρεμα) λειτουργία ή αποτελούν προϋπάρχουσα δραστηριότητα (OSAoff-line sequential activity), δηλαδή μία δραστηριότητα που ανιχνεύεται πριν το πειραματόζωο λάβει οποιαδήποτε αισθητική πληροφορία, που στρατολογείται για την κωδικοποίηση νέων μνημών (Dragoi et al., 2011). Αυτή η θεωρεία υποστηρίζεται από μία πρόσφατη ανακάλυψη ότι οι κυτταρικές ιδιότητες των πυραμιδικών κυττάρων προβλέπουν ποια κύτταρα θα ενεργοποιηθούν σε ένα καινούριο περιβάλλον (Epsztein et al., 2011). 24
37 7. Ιππόκαμπος και Μνήμη Πολλές μελέτες αποδεικνύουν τον σημαντικό ρόλο του ιπποκάμπου στην επεισοδιακή μνήμη στα ζώα και τους ανθρώπους. Ο πιο αξιοσημείωτος ασθενής ήταν ο H.M, ένας νεαρός άντρας που έπασχε από φαρμακοανθεκτική (intractable) επιληψία και υπέστη χειρουργική επέμβαση με αμφίπλευρη αφαίρεση του μέσου κροταφικού λοβού συμπεριλαμβανομένων τμημάτων των δυο ιππόκαμπων. Η επέμβαση, προκάλεσε στον Η.Μ ανικανότητα να σχηματίσει καινούριες μνήμες (εμπροσθόδρομη αμνησία, anterograde amnesia), μαζί με μία ουσιώδη αλλά όχι ολική απώλεια παλιών μνημών (οπισθόδρομη αμνησία, retrograde amnesia) (Scoville et al., 1957). Άλλες περιπτώσεις ασθενών έχουν επιβεβαιώσει ότι ο ιππόκαμπος είναι αναγκαίος για τον σχηματισμό νέων επεισοδιακών μνημών και ίσως έχει ρόλο στην μακροπρόθεσμη αποθήκευση τους. Μελέτες σε ζώα αποκάλυψαν ότι ελεγχόμενες βλάβες, φαρμακολογική απενεργοποίηση ή μοριακά Knockouts περιορισμένα αποκλειστικά και μόνο στον ιππόκαμπο είχαν σαν αποτέλεσμα την αποτυχία εκμάθησης ή την απώλεια χωρικής μνήμης (Morris et al., 1986; Tsien et al., 1996; Martin et al., 2005; Pastalkova et al., 2006). Ηλεκτροφυσιολογικές καταγραφές (Berger et al., 1983), μοριακές απεικονίσεις σε ζώα (Guzowski et al., 2001; Vazdarjanova et al., 2004) καθώς επίσης και MRI μελέτες σε ανθρώπους (Gabrieli et al., 1997; Henke et al., 1997; Maguire, 2001) παρέχουν συσχετικές αποδείξεις ότι η επεισοδιακή μνήμη και μάθηση περιλαμβάνει την ιπποκάμπεια δραστηριότητα. Μία άλλη αισθητικό/συμπεριφορική συσχέτιση της ιπποκάμπειας νευρωνικής δραστηριότητας είναι το «πεδίο θέσης». Τα πυραμιδικά κύτταρα πυροδοτούν με ρυθμούς που ποικίλουν ανάλογα με τη συγκεκριμένη τοποθεσία του αρουραίου καθώς αυτός κινείται σε ένα δεδομένο περιβάλλον. Περιοχές του περιβάλλοντος όπου ένα κύτταρο πυροδοτεί με υψηλή συχνότητα καθορίζει το πεδίο θέσης αυτού του κύτταρου και οι νευρώνες που επιδεικνύουν αυτό το πρότυπο δραστηριότητας ονομάζονται κύτταρα θέσης ή κύτταρα περιοχής (Place cells) (O'Keefe et al., 1971; O'Keefe, 1979). Η παρατήρηση των κυττάρων θέσης και η εξασθένιση μάθησης και απόδοσης στην χωρική διάκριση μετά από βλάβες του ιπποκάμπου οδήγησε στην θεωρία του χωρικού χάρτη. Αυτή η θεωρία υποστηρίζει ότι ο ιππόκαμπος συνεισφέρει στην μνήμη κωδικοποιώντας τοποθεσίες μέσα σε ένα χωροταξικό πλαίσιο. Ενώ πολλά συμπεριφορικά και ηλεκτροφυσιολογικά δεδομένα είναι σύμφωνα με την θεωρία του χωρικού χάρτη, άλλα πειραματικά αποτελέσματα καταδεικνύουν ότι τα ιπποκάμπεια κύτταρα κωδικοποιούν πληροφορίες ανεξάρτητες της τοποθεσίας και είναι αναγκαία για την μάθηση και ανάκληση άλλων μη χωρικών τύπων πληροφορίας (Winocur, 1990; Bunsey et al., 1995; Bunsey et al., 1996). 25
38 8. Διάφραγματο-κροταφική διαφοροποίηση Ο διαχωρισμός των λειτουργιών κατά μήκος του επιμήκους άξονα του ιπποκάμπου είναι μία γενικά αποδεκτή ιδέα (Small, 2002; Fanselow et al., 2010; Bannerman et al., 2014). Επί προσθέτως, πρόσφατες παρατηρούμενες διαφορές στην λειτουργικότητα του τοπικού νευρωνικού κυκλώματος ανάμεσα στους δύο πόλους του ιπποκάμπου έχουν προσελκύσει την προσοχή πολλών ερευνητών οδηγώντας στην βαθμιαία συσσώρευση αποδείξεων και στην αναδυόμενη έννοια της διαφοροποίησης του ενδογενούς νευρωνικού δικτύου ανάμεσα στον ραχιαίο (Ραχιαίος ιππόκαμπος-ρι, Dorsal hippocampus-dh) και κοιλιακό (κοιλιακός ιππόκαμπος-κι, Ventral hippocampus-vh) ιππόκαμπο. Αυτές οι παρατηρήσεις έχουν γίνει σε πολλά επίπεδα οργάνωσης όπως, στις κυτταρικές ιδιότητες (Liagkouras et al., 2008; Dougherty et al., 2012; Honigsperger et al., 2015), στους νευροχημικούς δείκτες (Gage et al., 1980; Verney et al., 1985; Sotiriou et al., 2005; Pandis et al., 2006), στη συναπτική διαβίβαση (Papatheodoropoulos et al., 2002; Petrides et al., 2007; Georgopoulos et al., 2008; Maggio et al., 2009), στη συναπτική πλαστικότητα (Papatheodoropoulos et al., 2000b; Maruki et al., 2001; Colgin et al., 2004; Maggio et al., 2007; Grigoryan et al., 2012; Kenney et al., 2013; Keralapurath et al., 2014; Pofantis et al., 2014), στα προφίλ γονιδιακής έκφρασης (Thompson et al., 2008; Dong et al., 2009), και στην δικτυακή ηλεκτρογραφική δραστηριότητα (Gilbert et al., 1985; Bragdon et al., 1986; Papatheodoropoulos et al., 2005; Sabolek et al., 2009; Mikroulis et al., 2012; Patel et al., 2012; Papatheodoropoulos, 2015). Αυτή η διαφοροποίηση αναμένεται να έχει σημαντικές επιπτώσεις για την επεξεργασία της πληροφορίας και τους λειτουργικούς ρόλους που εκτελούνται από τα δύο ιπποκάμπεια τμήματα. Η αποκάλυψη λειτουργικών διαφορών στο συναπτικό επίπεδο, συμπεριλαμβανομένων των τροποποιητικών δράσεων της πληθώρας των υποδοχέων και των αλληλεπιδράσεων τους, θα μας βοηθήσει να κατανοήσουμε το πώς τα κυκλώματα των δύο πόλων επεξεργάζονται τις πληροφορίες. Επομένως, θα είμαστε σε θέση να κάνουμε συνδέσεις μεταξύ των λειτουργιών ενός μικρής κλίμακας κυκλώματος και του μεγάλης κλίμακας λειτουργικού διαχωρισμού κατά μήκος του επιμήκους άξονα του ιπποκάμπου Νευρωνική συνδεσιμότητα των δύο πόλων Η ραχιαία CA1 περιοχή, η οποία περιέχει την μεγαλύτερη πυκνότητα των κυττάρων περιοχής, που κωδικεύουν την χωρική τοποθέτηση, στέλνει πολλές διαδοχικές, πολύσυναπτικές και πιθανόν ανάδρομες διεγερτικές προβολές στα ραχιαία τμήματα του υποθεματικού συμπλόκου (Εικόνα Α.Ι.8) (Swanson et al., 1977; van Groen et al., 1990; Witter et al., 1990; Amaral et al., 1991). Τα ραχιαία τμήματα του υποθεματικού συμπλόκου 26
39 περιέχουν τα περισσότερα κύτταρα κατεύθυνσης κεφαλής (Head cells or compass cells) που κωδικεύουν τη θέση του κεφαλιού στον χώρο (Taube et al., 1990; Taube, 2007). Οι περισσότερες φλοιικές προβολές από την ραχιαία CA1 και το ραχιαίο υπόθεμα (όχι από τα αντίστοιχα κοιλιακά) είναι προς τον οπισθιοσπλήνιο φλοιό (RSP, retrosplenial Cortex) και την πρόσθια περιοχή της έλικας του προσαγωγίου (ACA-anterior cirgulated area) στους αρουραίους (Risold et al., 1997b; Van Groen et al., 2003; Cenquizca et al., 2007), δύο περιοχές που συμμετέχουν κυρίως στην γνωστική επεξεργασία της οπτικοχωρικής πληροφορίας, στην επεξεργασία της μνήμης (Jones et al., 2005; Lavenex et al., 2006) και στην εξερεύνηση του περιβάλλοντος (χωρική πλοήγηση) στους αρουραίους (Harker et al., 2004). Είναι προφανές ότι αυτό το νευρωνικό δίκτυο που σχηματίζει ο ραχιαίος ιππόκαμπος παρέχει έναν γνωστικό χάρτη για το σύστημα πλοήγησης/κατεύθυνσης, δίνοντας την δυνατότητα στα ζώα να προσανατολιστούν και να εκτελέσουν συμπεριφορές σε ένα περιβάλλον (Taube et al., 1990; Jeffery, 2007; Taube, 2007). Επί πρόσθετα, οι ραχιαίες CA1 και CA3 περιοχές προβάλουν στον πλάγιο διαφραγματικό πυρήνα, που με τη σειρά του προβάλει στο διάμεσο διαφραγματικό σύμπλοκο (MSE) και υπερμαστικό πυρήνα (supramammillary nucleus-sum) (Risold et al., 1997a), δύο δομές που παράγουν και ελέγχουν τον ιπποκάμπειο θ ρυθμό που ενεργοποιείται κατά τη διάρκεια της εξερευνητικής κίνησης (Stewart et al., 1990; Kocsis et al., 1997). Επίσης, το ραχιαίο υπόθεμα και ο ραχιαίος πλάγιος και διάμεσος ενδορρινικός φλοιός στέλνουν μαζικές προβολές στον επικλινή πυρήνα (ACB) και στο ουραίο ή οπίσθιο κέλυφος (caudo putamen-cp) που με τη σειρά τους προβάλλουν είτε άμεσα είτε έμμεσα στην κοιλιακή καλυπτρική περιοχή (ventral tegmental area-vta) και στην μέλαινα ουσία (SNr) (Mogenson et al., 1983; Groenewegen et al., 1984; Groenewegen et al., 1996), όπου η κοιλιακή καλυπτρική περιοχή παίζει σημαντικό ρόλο στην κίνηση, ενώ οι μέλαινα ουσία στις κινήσεις προσανατολισμού των οφθαλμών, του κεφαλιού, του λαιμού και ακόμη των άνω άκρων μέσω των μαζικών προβολών τους στα βαθύτερα στρώματα του άνω διδύμιου (Hikosaka et al., 1983). Εν συντομία, το ραχιαίο σύμπλοκο ιπποκάμπου-υποθέματος σχηματίζει ένα σημαντικό φλοιικό κύκλωμα με τις οπισθιοσπλήνιες και τις πρόσθιες περιοχές της έλικας του προσαγωγίου που ολοκληρώνει γνωστικές διαδικασίες όπως η μάθηση, μνήμη, εξερεύνηση και πλοήγηση. 27








 Η κοιλιακή CA1 περιοχή διαφέρει από")

.")

 ).")






. Η εικόνα Α.Ι.")

40 Εικόνα Α.Ι.8: Σχηματική απεικόνιση που δείχνει την οργάνωση του ραχιαίου ιπποκάμπειου δικτύου ACA, πρόσθια περιοχή προσαγωγίου; ACB, επικλινής πυρήνας; ATN, πρόσθιο θαλαμικό σύμπλοκο; CP, Οπίσθιο κέλυφος; DGd, ραχιαία περιοχή της οδοντωτής έλικας; ENTl, the t πλάγιο-οπίσθια δέσμη του ενδορρινικού φλοιού; GP, ωχρά σφαίρα; LM, πλάγιος πυρήνας μαστίου; LSc, κοιλιακό τμήμα του πλάγιου septal πυρήνα; MM, ενδιάμεσος πυρήνας μαστίου; MSC, μέσο διαφραγματικό ό σύμπλοκο; PRE, προυπόθεμα; POST, μεταυπόθεμα; RSP, Οπίσθιο-σπλήνιος φλοιός; SNr, δικτυωτό τμήμα μέλαινας μ ουσίας; SUBd, ραχιαίο υπόθεμα; SUM, Υπερμαστικός πυρήνας; VTA, κοιλιακή καλυπτρική περιοχή. Πηγή: (Fanselow et al., 2010) Η κοιλιακή CA1 περιοχή διαφέρει από τηνν ραχιαία στο γεγονόςς ότι προβάλει άμεσα στον οσφρητικό βολβό και σε πολλές άλλες οσφρητικές φλοιικές περιοχές στους αρουραίους (Cenquizca et al., 2007). Τέτοιες προβολές ίσως παίζουν σημαντικό ρόλο στην εμφάνιση συμπτωμάτων που ομοιάζουν με την κατάθλιψη μετά από απώλεια της όσφρησης (Song et al., 2005) ). Επίσης, η κοιλιακή CA1 περιοχή και το κοιλιακό υπόθεμα μοιράζονται πολλές διπλής κατευθύνσεως προβολές με τους πυρήνες της αμυγδαλής που λαμβάνουν κύριεςς και συμπληρωματικές οσφρητικές αισθητικές εισόδους (Saunders et al., 1988; Petrovich et al., 2001; Cenquizca et al., 2007). Επί προσθέτως, το κοιλιακό σύμπλοκο CA1/ /υπόθεμα και οι πυρήνες της αμυγδαλής μοιράζονται διπλής κατεύθυνσης συνδέσεις με τους υποστεφανιαίους, προστεφανιαίους και ακκοκώδης νησιαίους φλοιούς (Thierry et al., 2000; Jones et al., 2005; Hoover et al., 2007; Roberts et al., 2007). Η εικόνα Α.Ι.9 δείχνει ότι αυτές οι «κοιλιακός ιππόκαμπος-υπόθεμα-αμυγδαλή-μέσος προμετωπιαίοςς φλοιός» δομές σχηματίζουν μία σειρά παράλληλων, διαχωρισμένων κατερχόμενων προβολών, είτε άμεσα είτε έμμεσα με τελικό σκοπό την νεύρωσηη του υποθαλάμου, της πρωταρχικής δομής που συμμετέχει στον έλεγχο των νευροκρινών, αυτόνομων και σωματικών κινητικών 28


















41 δραστηριοτήτων που σχετίζονται με τιςς τρεις κύριες κατηγορίες των συμπεριφορών κινήτρου που περιέχουν ισχυρά συναισθηματικά στοιχεία: κατάποση, αναπαραγωγή, φόβος και άμυνα (Kishi et al., 2000; Dong et al., 2001; Petrovich et al., 2001; 2 Dongg et al., 2006). Εικόνα Α.Ι.9: Σχηματική απεικόνιση με τις κύριες νευρωνικές συνδέσεις του τ κοιλιακούύ ιπποκάμπου ACB, επικλινής πυρήνας; AMY, περιοχές (πυρήνες) αμυγδαλής; BST, βασικός πυρήνας της τελικής ταινίας; CEA, κεντρικός πυρήνας αμυγδαλής; LSr, v, πρόσθια κοιλιακά τμήματαα του πλάγιουυ διαφραγματικού πυρήνα; MEA, μέσος πυρήνας αμυγδαλής; MPF, μέσος προμετωπιαίος φλοιός; SUBv, κοιλιακό υπόθεμα. Πηγή: (Fanselow et al., 2010) 8.2. Γονιδιακή έκφραση και λειτουργικές διαφορές κατά μήκος του τ επιμήκηη ιπποκάμπειου άξονα Έχουν αποκαλυφθεί κατά μήκος τουυ επιμήκους άξονα του ιπποκάμπου πολλαπλές διαχωρισμένες μοριακές υποπεριοχές, όπου καθεμία περιέχει ένα μοναδικό συμπλήρωμα εκφρασμένων γονιδίων (π.χ. 9 υποπεριοχές ς εντός της CA3, τρειςς κύριες υποπεριοχές εντος της οδοντωτής έλικας και της CA1) (Thompson et al.,, 2008; Dong et al., 2009; Fanselow et al., 2010). Αυτή η μοριακή διαφοροποίηση δεν είναι απλά ραχιαία-κοιλιακή. Αν συνδυάσουμε τις παραπάνω μελέτες που πραγματοποιήθηκαν σε τρειςς διαφορετικές περιοχές του ιπποκάμπου τότε καταλήγουμ με σε ένα τριμερές μοντέλο μοριακής έκφρασης. Ένα τέτοιο μοντέλοο επιβεβαιώνεται από μία αναπτυξιακή μελέτη γονιδιακής έκφρασης (O'Reilly et al., 2015). Παρόλαα αυτά, ο ακριβής αριθμός των περιοχών έκφρασης κατά 29
42 μήκος του μακρύ άξονα του ιπποκάμπου και το εάν αυτές είναι ιεραρχικά οργανωμένες παραμένουν άγνωστα. Το ενδιαφέρον είναι ότι αυτά τα πρότυπα της μοριακής έκφρασης οδηγούν σε ειδικές λειτουργικές ιδιότητες. Τα πρότυπα έκφρασης των γονιδίων που κωδικοποιούν μόρια προσκόλλησης και ιοντικά κανάλια ίσως καθορίζουν τις εγγενείς ηλεκτροφυσιολογικές ιδιότητες των διακριτών ιπποκάμπειων νευρωνικών πληθυσμών, όπως διαφορές στην νευρωνική διεγερσιμότητα (Dougherty et al., 2012) και την συναπτική πλαστικότητα (Papatheodoropoulos et al., 2000b; Maggio et al., 2007) που έχουν ανακαλυφθεί μεταξύ του ραχιαίου και κοιλιακού πόλου. Για παράδειγμα, το κανάλι HCN1 (hyperpolarizationactivated cyclic nucleotide-gated channel 1) και HCN2 που διαμεσολαβούν στα ενεργοποιούμενα μέσω μεθυπερπόλωσης ρεύματα (Ih), εκφράζονται διαφορετικά μεταξύ των δύο πόλων και είναι σημαντικά για την χωρική λειτουργία που είναι ραχιαίο-κοιλιακά διαβαθμισμένη (Garden et al., 2008; Giocomo et al., 2011; Hussaini et al., 2011). Γενικά, η έκφραση των υποδοχέων νευροδιαβιβαστών ποικίλει κατά μήκος του άξονα για την πλειοψηφία των συστημάτων νευροδιαβίβασης (Strange et al., 2014). 9. Ιππόκαμπος και γήρανση Η γήρανση του εγκεφάλου είναι ένα περίπλοκο, ετερογενές και ελάχιστα κατανοητό φαινόμενο (Kirkwood et al, 2003). Οι εγκεφαλικές αλλαγές κατά τη διάρκεια της γήρανσης φαίνεται να είναι επιλεκτικές και ειδικές ανά περιοχή (Burke and Barnes, 2006; Kelly et al, 2006; Kumar et al 2009; Burger, 2010) και κάποιες από αυτές ίσως αντιπροσωπεύουν το αποτέλεσμα αντισταθμιστικών μηχανισμών (Boric et al 2008; Kumar et al 2009; Burke and Barnes 2010). Μία συμπεριφορική ιδιότητα της γήρανσης τοy εγκεφάλου είναι η εξασθένιση του σχηματισμού νέων εμπειριών (Crook et al 1986; Burke and Mackay 1997; Balota et al 2000; Beason-Held and Horwitz, 2002), ειδικά αυτές που εξαρτώνται από τον ιππόκαμπο (Rosenzweig and Barnes, 2003). Συνεπώς, οι ηλικιωμένοι αρουραίοι ίσως εμφανίζουν εξασθενημένη ιππόκαμπο-εξαρτώμενη μνήμη (Markowska et al 1989; Gallagher and Rapp, 1997) και πιο συγκεκριμένα την βιωματική-χωρική μνήμη (Monacelli et al, 2003). Επί προσθέτως, ο ιππόκαμπος φαίνεται να υφίσταται δομικές και λειτουργικές αλλαγές κατά τη γήρανση (Rosenzweig and Barnes, 2003; Wilson et al, 2006; Oh et al, 2010). Παρόλα αυτά, δεν εμφανίζουν όλα τα ηλικιωμένα άτομα γνωστικά ελλείμματα και υπάρχει ευρεία ενδο-ατομική διαφορά στην συσχετιζόμενη με την ηλικία εξασθένιση της μνήμης τόσο στους ανθρώπους όσο και στους αρουραίους (Crook et al, 1986; Markowska et al, 1989). 30







")

 στην οδοντωτή έλικα")
 και οι ίνεςς του")





 και την διαγώνια ζώνη ζ του")
")
 προβολές")


43 Νευροβιολογικά αποτελέσματα προτείνουν ότι οι αλλαγές στον σ μέσο κροταφικό λοβό εμπλέκονται σε αυτά τα γνωστικά ελλείμματα και ότι αυτή η δομή παίζει καθοριστικό ρόλο στην επεξεργασία της μνήμης. Στην Εικόνα Α.Ι. 10 καταδεικνύονται οι βασικότερες μεταβολές στον μέσο κροταφικό λοβό. Πρόσφατες ανακαλύψεις έχουν ενισχύσει την κατανόησή των μηχανισμών που συμμετέχουν στην εξαρτώμενη με την ηλικία μείωση της μνήμης. Εικόνα Α..Ι.10: Απλοποιημένο σχεδιάγραμμα του Ιπποκάμπειου κυκλώματος και σύνοψη των νευροβιολογικών μεταβολών που συμβαίνουν στην ηλικιακό-εξαρτώμενη εξασθένιση ε της μνήμης. Το πιο τρωτό σημείο αυτού του κυκλώματος πιστεύεται ότι είναι το επίπεδο του περιρινικού φλοιού (PRC) καθώς επίσης και ο πλάγιος ενδορινικός φλοιός (LEC) και πιο συγκεκριμένα τα επιφανειακά στρώματα. Καθοδικώς,, η είσοδος της διατιτραίνουσας οδού (PP) από τον ενδορρινικό φλοιό (EF) στην οδοντωτή έλικα (DG) και την CA3 περιοχή αμβλύνεται με την ηλικία, ενώ οι βρυώδεις ίνες (MF) και οι ίνεςς του Schafferr (SC) εμφανίζονται άθικτες. Η οδοντωτή έλικα παρουσιάζει απώλεια νευρογένεσης καθώς επίσης και παρεκκλίνοντα δυναμικά, ενώ η CA3 φαίνεται ναα είναι υπερδιεγερσιμη, ενισχύοντας ε το αυτοσυνειρμικό δίκτυο παλίνδρομων παράπλευρων ινών των CA33 πυραμιδικών κυττάρων της τ περιοχής. Οι προβολέςς των ανασταλτικών ενδονευρώνων (Int) σε όλες τις ιπποκάμπειες υποπεριοχές εκφυλίζονται με τη γήρανση, συνεισφέροντας πιθανόν στην αλλαγμένη ισορροπία διέγερσης/αναστολής στον ιππόκαμπο. Η τροποποιητική χολινεργική (Ach) είσοδος από το μέσο διάφραγμαα (MS) και την διαγώνια ζώνη ζ του Broca (δεν δείχνεται) μειώνεται. Παρόμοια μειώνονται, οι ντοπαμινεργικν κές (DA) προβολές από τηνν κοιλιακή καλυπτρική περιοχή (VTA) καθώς επίσης και οι νοραδρενεργικές (ΝΕ) προβολές από την υπομέλαινα περιοχή (LC). Αλλαγές στην συναπτική πλαστικότητα και το τ ρυθμό πυροδότησης των κυττάρων θέσης συμβαίνουν επίσης. Σε γενικές γραμμές, το δίκτυο παρουσιάζει έντονες νευροβιολογικές μεταβολές που π αλλάζουνν την υπολογιστική λειτουργία του ιπποκάμπου. Οι πράσινες γραμμές και τα μαύρα έντονα κουτιά δηλώνουν μηη επαρκή δεδομένα για την εξαγωγή ολοκληρωμένων συμπερασμάτων. Πηγή: (Leal et al., 2015). 31
44 9.1. Μειωμένη είσοδος της πληροφορίας από διατιτραίνουσα οδό στην ΟΕ και της CA3 περιοχή Στους γερασμένους αρουραίους, η ΟΕ λαμβάνει περίπου κατά 1/3 λιγότερες συναπτικές συνδέσεις από τον ΕΦ (στοιβάδα ΙΙ) απ ότι στους νεαρούς. Το ποσοστό της συναπτικής μείωσης της διατιτραίνουσας οδού συσχετίζεται με τον βαθμό εξασθένισης της χωρικής μνήμης στους γέρικους αρουραίους (Geinisman et al., 1992; Smith et al., 2000). Ηλεκτροφυσιολογικές μελέτες έχουν δείξει ότι ερεθίζοντας τη διατιτραίνουσα οδό παράγεται λιγότερη διέγερση στην ΟΕ των γέρικων σε σχέση με τους νέους αρουραίους (Burke et al., 2006). Σε γηράσκοντες ανθρώπους, μετά από μετρήσεις μικροδομικών χαρακτηριστικών της λευκής ουσίας, παρατηρήθηκε μείωση της ακεραιότητας της διατιτραίνουσας οδού, η οποία συσχετίστηκε με μνημονικά ελλείμματα (Yassa et al., 2010; Yassa et al., 2011) Ισορροπία Διέγερσης/Αναστολής στον εγκέφαλο γηράσκοντων πειραματόζωων Ο Wilson και οι συνεργάτες του (Wilson et al., 2005), κατέδειξαν ότι τα κύτταρα θέσης ή περιοχής (Place cells) στην CA3 περιοχή γέρικων τρωκτικών παρουσιάζουν ανώμαλα αυξημένους ρυθμούς πυροδότησης. Πρόσφατα, ενδοκρανιακές καταγραφές σε ελευθέρα κινούμενα ζώα και εξωκυττάριες καταγραφές σε τομές ιπποκάμπου αποκάλυψαν συχνές πυροδοτήσεις δυναμικών (spikes) στην CA3 περιοχή των γέρικων ποντικιών, ενώ αυτές οι πυροδοτήσεις συνέβαιναν μόνο περιστασιακά στα νεαρά ποντίκια. Αυθόρμητα δυναμικά πεδίου με μεγάλα πλάτη που προέρχονταν από την CA3 περιοχή παρατηρηθήκαν πολύ συχνά σε τομές ιπποκάμπου γέρικων ποντικών, αλλά σπανίως σε τομές από ενήλικα (El-Hayek et al., 2013). Μελέτες fmri σε ανθρώπους έδειξαν παρόμοια αποτελέσματα υπερενεργοποίησης του OE/CA3 δικτύου (Yassa et al., 2010), η έκταση της οποίας προέβλεψε μνημονικά ελλείμματα. Ενώ είναι πιθανόν ότι πολλοί παράγοντες συμμετέχουν στις μεταβολές που προκαλούνται με την αύξηση της ηλικίας στην ισορροπία διέγερσης/αναστολής στον ιππόκαμπο, είναι ξεκάθαρο ότι η απώλεια του ανασταλτικού τόνου παίζει πολύ σημαντικό ρόλο. Πολυάριθμοι τύποι ενδονευρώνων μειώνονται σε πολλές περιοχές του ιπποκάμπου με την γήρανση (Spiegel et al., 2013) Μειωμένη τροποποιητική είσοδος στον γέρικο ιππόκαμπο Οι τροποποιητικές είσοδοι στον ιππόκαμπο μειώνονται με την γήρανση. Αυτά τα γεγονότα μπορούν να συνεισφέρουν στην ανισορροπία διέγερσης/αναστολής οδηγώντας σε απορρύθμιση του ιπποκάμπου. Παρόλο που η παραδοσιακή άποψη της τροποποιητικής 32
45 απώλειας έχει εμπλέξει την κυτταρική νευρωνική απώλεια, πιο πρόσφατα δεδομένα υποστηρίζουν ότι αυτές οι αλλαγές πιθανόν να είναι αποτέλεσμα της δενδριτικής και συναπτικής απώλειας καθώς επίσης και της νευραξονικής εκφύλισης και μείωσης της τροφικής υποστήριξης (Schliebs et al., 2011). Παρακάτω θα αναφερθούμε σε μερικά τροποιητικά συστήματα που επηρεάζονται από τη γήρανση. Το χολινεργικό σύστημα του πρόσθιου εγκεφάλου παρέχει προβολές προς τον ιππόκαμπο που πιστεύεται ότι μειώνεται με την γήρανση (Schliebs et al., 2011) και ότι παίζει σημαντικό ρόλο στην υπολογιστική ισορροπία του ιππόκαμπου. Η απουσία της φυσιολογικής τροποποίησης της ακετυλοχολίνης του ιπποκάμπου οδηγεί σε δυσλειτουργία της διακριτικής του ικανότητας μεταξύ του διαχωρισμού των μοτίβων και της επεξεργαστικής/υπολογιστικής ολοκλήρωσης της πληροφορίας (Wilson et al., 2006; Sava et al., 2008; Jacobson et al., 2013). Πολλές μελέτες έχουν αναφέρει ηλικιακό-εξαρτώμενη εκφύλιση των ντοπαμινεργικών νευρώνων της μέλαινας ουσίας/κοιλιακής καλυπτρικής περιοχής (Fearnley (Fearnley et al., 1991; Backman et al., 2006), που ίσως επηρεάζουν την ικανότητα του ιπποκάμπου να παγιώνει τις μνήμες. Πολύ συχνά αναφέρονται μειώσεις των επίπεδων ντοπαμίνης στους γέρικους αρουραίους και ανθρώπους και μελέτες που διεξήχθησαν σε υγιείς γέρικους ενήλικες προτείνουν ότι φαρμακολογική ενίσχυση της ντοπαμινεργικής ενεργότητας ενισχύει την ανάμνηση γεγονότων (Chowdhury et al., 2012), την ειδικότητα της μνήμης και την μάθηση εντοπισμού αντικειμένων (object-place) (Trossbach et al., 2014). Μείωση της εισόδου της νορεπινεφρίνης (ΝΕ) στον ιππόκαμπο (Kubanis et al., 1981) καθώς επίσης και μεταβολές των επιπέδων της περιφερικής επινεφρίνης (Sternberg et al., 1985) έχουν αναφερθεί σε γέρικα ζωικά μοντέλα. Η έκταση της εξάντλησης της ΝΕ στον ραχιαίο ιππόκαμπο σχετίζεται με εκτεταμένα μνημονικά ελλείμματα στα γέρικα ζώα (Stemmelin et al., 2000). Η εξάντληση της ΝΕ έχει συνδεθεί επίσης με εξασθενημένη μακροχρόνια ενδυνάμωση (LTP) στην CA1 περιοχή των γέρικων τρωκτικών κατά τη διάρκεια της εξαρτημένης φοβικής αντίδρασης (fear conditioning), ένα έλλειμμα που υποχωρεί μετά από χορήγηση ΝΕ στα ζώα (Luo et al., 2015). Συνεπώς, η εξάλειψη της ΝΕ φαίνεται να συνεισφέρει στην ηλικιακό-εξαρτώμενη απώλεια μνήμης και προσεγγίσεις αναπλήρωσης της ίσως είναι ευεργετικές στην αντιστροφή των ελλειμμάτων. Ενδιαφέρον παρουσιάζει το γεγονός ότι η ΟΕ είναι ο στόχος της τροποποιητικής δράσης της ΝΕ άμεσα και έμμεσα μέσω της βασεοπλευρικής αμυγδαλής (Young et al., 1979). Η ΟΕ λαμβάνει επίσης ισχυρή τροποποίηση μέσω αμέσων νοραδρενεργικών προβολών από την 33
46 υπομέλαινα περιοχή (locus coeruleus-lc) και μέσω γλουταμινεργικών προβολών από την βασεοπλευρική αμυγδαλή (BLA), η οποία νευρώνεται από νοραδρενεργικές προβολές από την LC (McGaugh, 2002). Παρόλο που η νευρογένεση συνεχίζει καθόλη τη διάρκεια της ζωής, ο ρυθμός της πιστεύεται ότι μειώνεται με την αύξηση της ηλικίας (Kuhn et al., 1996; Gould, 1999; Kempermann, 2002). Έχει προταθεί ότι η μείωση της ίσως προκαλεί τα μνημονικά ελλείμματα και ότι μπορεί να συνεισφέρει σε παθολογικές καταστάσεις όπως στη νόσο Alzheimer (Alzheimer's disease AD) (Haughey et al., 2002; Donovan et al., 2006) αλλά ο ακριβής ρόλος της νευρογένεσης στην τροποποίηση των μηχανισμών μνήμης/μάθησης δεν είναι εξακριβωμένος. Η γήρανση έχει συσχετιστεί με την μειωμένη ικανότητα να επάγει και να αποκρίνεται στους αυξητικούς παράγοντες (Smith, 1996) που είναι απαραίτητοι για την ανάπτυξη, την επιβίωση και την πλαστικότητα των νευρώνων. Ο BDNF (Brain derived growth factor) είναι ένας από τους κύριους τροφικούς παράγοντες που αλλάζει με τη γήρανση (Budni et al., 2015). Μειώσεις του παράγοντα αυτού έχει προταθεί ότι οδηγούν σε εξασθένιση της μνήμης (Tapia-Arancibia et al., 2008), μέσω μείωσης του LTP, ένα έλλειμμα που φαίνεται να είναι αντιστρεπτό αν χορηγηθεί εξωγενώς BDNF (Rex et al., 2006). Επίσης, εξάλειψη του BDNF φαίνεται να συνδέεται με συναπτική απώλεια (Schaaf et al., 2001) και τα επίπεδα του BDNF στο εγκεφαλονωτιαίο υγρό γερασμένων ανθρώπων είναι μειωμένα και σχετίζονται με χαμηλή μνημονική απόδοση (Li et al., 2009). Η εξασθένιση της μνήμης στα γέρικα ποντίκια σχετίζεται με μειωμένη ρύθμιση της ακετυλίωσης των ιστονών στον ιππόκαμπο. Μία αύξηση στην ενεργότητα αποακετυλίωσης των ιστονών λόγω της γήρανσης ίσως αποσιωπά την φυσιολογική γονιδιακή έκφραση, οδηγώντας σε μειωμένη ικανότητα σχηματισμού μνημών. Η χορήγηση αναστολέων των αποακετυλασών, που είναι γνωστό ότι ενισχύουν την ακετυλίωση των ιστονών και διευκολύνουν στην μάθηση στα τρωκτικά όταν χορηγούνται στον ιππόκαμπο (Peleg et al., 2010; Haettig et al., 2011) σχετίζεται με την ανάκαμψη της γνωστικής λειτουργίας σε ένα ζωικό μοντέλο ποντικού με AD παρόμοια νευροεκφύλιση (Fischer et al., 2007). Ηλικιακό-εξαρτώμενα ελλείμματα στην πλαστικότητα σχετίζονται με την εξασθένιση της μνήμης στα γέρικα ζώα (Barnes, 1979). Στους γέρικους αρουραίους έχει παρατηρηθεί μείωση της ικανότητας επαγωγής και διατήρησης του LTP καθώς και αυξημένη πιθανότητα μακροχρόνιας καταστολής (LTD) και αντιστροφής του LTP (Burke et al., 2006; Burke et al., 2010). Αναλύσεις DNA μικροσυστοιχιών που σύγκριναν νεαρούς, ενήλικες και γέρικους αρουραίους κατέδειξαν ότι η γονιδιακή έκφραση που είναι ειδική για την επαγωγή 34
47 του LTP έχει αλλάξει στα γέρικα ζώα και προτείνει ότι η απορύθμιση της συναπτικής πρωτεϊνικής σύνθεσης συνεισφέρει στην μείωση της διατήρησης του LTP (Ryan et al., 2015). Οι γηράσκοντες αρουραίοι φαίνεται να είναι πιο επιδεκτικοί στο LTD και την αποδυνάμωση απ ότι οι ενήλικοι (Norris et al., 1996) και το κατώφλιο επαγωγής του LTP είναι αυξημένο κάνοντας δύσκολη για τα γέρικα ζώα την κωδικοποίηση της μνήμης, ενώ από την άλλη το κατώφλιο του LTD είναι μειωμένο κάνοντας εύκολη την διαγραφή της μνήμης (Foster, 1999; Burke et al., 2010). Επίσης το NMDA-LTP και το mglur-ltp είναι σημαντικά μειωμένα στα γέρικα τρωκτικά με μνημονικά ελλείμματα. Σε γενικές γραμμές, αυτές οι αλλαγές προτείνουν ότι η πλαστικότητα είναι αλλαγμένη στον γέρικο ιππόκαμπο με τέτοιον τρόπο που εμποδίζει την μάθηση νέας πληροφορίας. ΙΙ. ΣΚΟΠΟΣ ΜΕΛΕΤΗΣ Η παρούσα διατριβή συνίσταται από τρείς επί μέρους μελέτες με τις οποίες επιχειρήθηκε α) η διερεύνηση της ρύθμισης της διεγερτικής συναπτικής διαβίβασης και της μακρόχρονης πλαστικότητάς της στην CA1 περιοχή του ιπποκάμπου συγκριτικά μεταξύ του ραχιαίου και κοιλιακού του τμήματος, και β) η διερεύνηση πιθανών μεταβολών που συμβαίνουν κατά το γήρας στην ιπποκάμπεια δικτυακή δραστηριότητα των οξύαιχμων κυμάτων-ριπιδισμών η οποία έχει κεντρικό ρόλο στην διεργασία της μνημονικής παγίωσης. Έτσι, με τις μελέτες αυτές επιδιώχθηκε α) να διασαφηνιστούν περαιτέρω οι μηχανισμοί που υπεισέρχονται στην διαφοροποίηση της λειτουργικής οργάνωσης των τοπικών νευρωνικών δικτύων μεταξύ ραχιαίου και κοιλιακού ιπποκάμπου, και β) να εντοπιστούν πιθανοί μηχανισμοί που σχετίζονται με την λειτουργία του ιπποκάμπου και μπορεί να εμπλέκονται στην μεταβολή της μνημονικής ικανότητας που συνοδεύει το εγκεφαλικό γήρας. 35
48 ΙΙΙ. ΓΕΝΙΚΕΣ ΜΕΘΟΔΟΙ 1. Πειραματόζωα Στις ακόλουθες τρεις μελέτες χρησιμοποιήθηκαν συνολικά 215 αλφικοί Wistar επίμυες ηλικίας από 2 έως 34 μηνών. Τα ζώα διατηρούνταν στο ζωοτροφείο του τμήματος Ιατρικής, σε περιβάλλον σταθερών συνθηκών θερμοκρασίας ο C, 24ωρου κύκλου φωτός (12 ώρες φως/12 ώρες σκοτάδι) και είχαν ελεύθερη πρόσβαση σε νερό και τροφή. Η μεταχείριση και οι πειραματικές διαδικασίες για όλα τα ζώα διεξήχθησαν σύμφωνα με τις Κατευθυντήριες οδηγίες για την φροντίδα και τη χρήση των πειραματόζωων εργαστηρίου του Ευρωπαϊκού Συμβουλίου Κοινοτήτων (European Communities Council Directive Guidelines (86/609/EEC, JL 358, 1, December, 12, 1987)) για τη φροντίδα και τη χρήση των πειραματόζωων και εγκρίθηκαν από την Νομαρχιακή Επιτροπή Φροντίδας και Χρήσης των πειραματόζωων ((Prefectural Animal Care and Use Committee (No: EL 13BIO04)). Αναλόγως, πάρθηκαν όλα τα μέτρα για την ελαχιστοποίηση της ταλαιπωρίας των ζώων και την μείωση του αριθμού τους στην μελέτη αυτή. 2. Γενικά στοιχεία των in vitro τομών ιπποκάμπου Η ανάπτυξη της μεθόδου in vitro διατήρησης τομών εγκεφάλου αποτέλεσε μία μεγάλη τεχνολογική πρόοδο για την έρευνα του ιπποκάμπου. Ο πρωτοπόρος αυτής της ανακάλυψης ήταν ο νευροχημικός Henry Mcllwain (Li and Mcllwain, 1957) που πραγματοποιούσε βιοχημικές μελέτες σε εγκεφαλικές τομές. Αργότερα ακολουθήσανε και οι ηλεκτροφυσιολογικές καταγραφές με την επίτευξη καταγραφών συναπτικών απαντήσεων μετά από ερεθισμό (Yamamoto et al., 1966; Richards et al., 1967; McIlwain et al., 1970) όπου τελικά εφαρμόστηκαν στις τομές ιπποκάμπου από τον Tim Bliss και τον Chris Richards (Bliss and Richards, 1971) οι οποίοι κατέδειξαν ότι τα δυναμικά πεδίου που βρέθηκαν σε άθικτα ζώα μπορούσαν να καταγραφούν και σε τομές κατά μήκος του διάφραγματο-κροταφικού άξονα του ιπποκάμπου. Οι in vitro τομές ιπποκάμπου παρουσιάζουν τα εξής πλεονεκτήματα σε σχέση με τις in vivo καταγραφές: διατήρηση των συναπτικών συνδέσεων ενός πλήρους νευρωνικού κυκλώματος μηχανική σταθερότητα που επιτρέπει τις πολύωρες καταγραφές δυνατότητα ακριβούς τοποθέτησης των ηλεκτροδίων δυνατότητα ελέγχου της χημικής σύστασης του εξωκυτταριου περιβάλλοντος όπως και άλλων συνθηκών (θερμοκρασία, ph, οξυγόνο) ελαχιστοποίηση ταλαιπωρίας των ζώων 36
49 ευκολία προσέγγισης και ταυτοποίησης των νευρώνων ευκολία παρασκευής των τομών και μεγάλου αριθμού από τον ίδιο ιππόκαμπο που ευνοεί την επαναληψιμότητα των πειραμάτων έλλειψη αναισθητικών που επηρεάζουν τις μετρήσεις Από την άλλη μεριά οι in vitro τομές παρουσιάζουν τα εξής βασικά μειονεκτήματα: απώλεια συνδέσεων με άλλες περιοχές έλλειψη ρυθμιστικών μορίων (π.χ. ορμόνες) Λαμβάνοντας υπόψη όλα τα παραπάνω αντιλαμβανόμαστε το γιατί η ανακάλυψη των in vitro τομών ιπποκάμπου αποτέλεσε την κατεξοχήν μέθοδο επιλογής των περισσοτέρων εργαστηρίων που μελετάνε ερωτήματα που αφορούν το επίπεδο οργάνωσης που εμπεριέχεται μέσα σε μία τομή Παρασκευή Τομών Ιπποκάμπου Οι τομές ιπποκάμπου από τον ραχιαίο και κοιλιακό ιππόκαμπο προετοιμάστηκαν όπως αναφέρεται προηγούμενα (Papatheodoropoulos and Kostopoulos, 200a). (Papatheodoropoulos et al., 2000a). Ειδικότερα, τα ζώα αποκεφαλίζονταν σε λαιμητόμο μετά από βαθιά αναισθησία με διαιθυλαιθέρα. Το μεγαλύτερο μέρος του κρανίου αφαιρούνταν με 3 τομές με ψαλίδι: μία κατά μήκος της μέσης οβελιαίας γραμμής και 2 κατά μήκος των δύο έσω καρωτίδων. Ο εγκέφαλος αφαιρούνταν και τοποθετούνταν σε τρυβλίο με παγωμένο (2-4 o C) τεχνητό εγκεφαλονωτιαίο υγρό (ΤΕΝΥ, ACSF). Αποχωρίζονταν η παρεγκεφαλίδα με μετωπιαία τομή στα διδύμια και διαχωρίζονταν μεταξύ τους τα δύο ημισφαίρια με νυστέρι. Στη συνέχεια μετά την προσεκτική απομάκρυνση των τμημάτων του διεγκεφάλου και κοπή της ψαλίδας αφαιρούνταν οι δύο ιππόκαμποι. Εγκάρσιες τομές μm πάχους προετοιμάστηκαν από περιοχές που εκτείνονται περισσότερο από 1 και λιγότερο από 4 mm από τον ραχιαίο (διαφραγματικό) και κοιλιακό (κροταφικό) άκρο του ιπποκάμπου με τη χρήση μικροτόμου Mcllwain. Για να διατηρήσουμε την ορθογώνιο επίπεδο κοπής κατά τη διάρκεια του κοψίματος των δύο πόλων, απαιτούταν περιστροφή του πιάτου το οποίο υποστήριζε την δομή. Από κάθε ζώο, 1-4 τομές από κάθε πόλο επιλέγονταν για πειραματισμό. Αμέσως μετά το κόψιμο, οι τομές μεταφέρονταν και διατηρούνταν σε ένα υδατόλουτρο καταγραφής τύπου διεπαφής (υγρήςαέριας φάσης) όπου διαποτίζονταν συνεχώς με ACSF (ίδιας σύστασης με το περιγραφέν 37
50 παραπάνω) και υγραίνονταν με μείγμα αερίου που περιείχε 95% Ο 2 και 5% CO 2 σε συνεχή θερμοκρασία ο C. Οι τομές αφήνονταν να ηρεμήσουν για τουλάχιστον μιάμιση ώρα μετά την προετοιμασία τους, πριν την έναρξη των καταγραφών. 3. Διαλύματα και Φαρμακολογικοί παράγοντες Σε όλες τις φάσεις του χειρισμού, ο ιστός διαβρέχονταν από τεχνητό εγκεφαλονωτιαίο υγρό (ΤΕΝΥ) συστάσεως (σε mm):124mm NaCl, 4mM KCl, 2mM MgSO4, 2mM CaCl2, 1.25mM NaH2PO4, 26mM NaHCO3 και 10mM γλυκόζη, εξισορροπημένο με 95% Ο 2 και 5% CO 2 αέριο μείγμα με ph=7,4. Χρησιμοποιήθηκαν τα ακόλουθα φάρμακα: α) ο αγωνιστής των NMDAR, το N-Methyl-D-aspartic acid (NMDA), 25μΜ, 50μΜ, β) ο ανταγωνιστής των NMDAR, το 3-((R)-2-Carboxypiperazin-4-yl)-propyl-1-phosphonic acid (CPP, 10μΜ), γ) ο επιλεκτικός αγωνιστής των mglur5 το (RS)-2-Chloro-5-hydroxyphenylglycine sodium salt (CHPG, 50μM), δ) ο επιλεκτικός ανταγωνιστής των mglur5 το 3-((2-Methyl-1,3-thiazol-4- yl)ethynyl)pyridine hydrochloride (MTEP, 200μM) ε) ο επιλεκτικός ανταγωνιστής των αδενοσινεργικών Α1 υποδοχέων το 8-Cyclopentyl-1,3- dipropylxanthine (DPCPX, 30nM), στ) ο αγωνιστής των αδενοσινεργικών Α2Α υποδοχέων το 4-[2-[[6-Amino-9-(N-ethyl-β-Dribofuranuronamidosyl)-9H-purin-2-yl]amino]ethyl]benzenepropanoic acid hydrochloride (CGS 21680, 10 nm), ζ) ο επιλεκτικός ανταγωνιστής των Α2ΑR το 4-(2-[7-Amino-2-(2-furyl)[1,2,4]triazolo[2,3- a][1,3,5]triazin-5-ylamino]ethyl)phenol (ZM , 100 nμ) η) ο επιλεκτικός ανταγωνιστής των CB1 υποδοχέων των ενδοκαναβινοειδών το 1-(2,4- Dichlorophenyl)-5-(4-iodophenyl)-4-methyl-N-4-morpholinyl-1H-pyrazole-3-carboxamide (AM 281, 5 μm). θ) ο ανταγωνιστής των GABA B R 3-[[(3,4-Dichlorophenyl)methyl]amino]propyl] diethoxymethyl) phosphinic acid (CGP 52432, 10 μm); ι) ο αποκλειστής των L-τύπου τασεο-ελεγχόμενων καναλιών, η Νιμοδιπίνη (20μΜ). κ) ο αποκλειστής των L-τύπου τασεο-ελεγχόμενων καναλιών, η Νιφεδιπίνη (20μΜ). 38
51 Τα φάρμακα αγοράστηκαν από την Tocris Cookson Ltd, UK (NMDA, CPP, CHPG, MTEP, DPCPX, CGS 21680, ZM , AM 281, CGP 52432, Νιμοδιπίνη ) καθώς επίσης και από την Sigma-Aldrich, Germany (NMDA και CHPG). Τα φάρμακα προετοιμάστηκαν αρχικά σε stock διαλύματα και έπειτα διαλύθηκαν σε ACSF και εφαρμόστηκαν στις τομές. Τα stock διαλύματα των NMDA, CPP, CHPG, MPEP και CGP πραγματοποιήθηκαν σε αποσταγμένο νερό, ενώ τα stock διαλύματα των DPCPX, ZM , AM 281, Νιφεδιπίνη και Νιμοδιπίνη σε Dimethyl-sulfoxide (DMSO) σε συγκέντρωση στην οποία όταν τα φάρμακα διαλύονταν στο ACSF εφαρμογής η τελική συγκέντρωση του DMSO ήταν μικρότερη από 0.05%. Τα stock διαλύματα στο νερό διατηρούνταν στους 4 ο C ενώ τα διαλύματα με το DMSO διαχωρίζονταν σε aliquots και διατηρούνταν στους -20 o C. Τα stock διαλύματα διαλύονταν σε ACSF στις επιθυμητές συγκεντρώσεις την ημέρα του πειράματος. 4. Εξωκυττάριες Καταγραφές Δυναμικών Πεδίου στην CA1 περιοχή του Ιπποκάμπου 4.1. Δημιουργία δυναμικού πεδίου Όταν ένας νευρώνας ενεργοποιείται συναπτικά στην περιοχή των απώτερων δενδριτών του, ο νευροδιαβιβαστής που απελευθερώνεται από τις προσυναπτικές απολήξεις προκαλεί άνοιγμα ιοντικών καναλιών της μετασυναπτικής μεμβράνης των δενδριτών και την είσοδο θετικού ιοντικού ρεύματος στον ενδοκυττάριο χώρο, προκαλώντας εκπόλωση της μεμβράνης (το σημείο αυτό καθίσταται πιο θετικό απ ότι άλλα σημεία της μεμβράνης). Σαν αποτέλεσμα το ρεύμα διατρέχει τον ενδοκυττάριο χώρο από το σημείο αυτό του υψηλού δυναμικού προς σημεία απομακρυσμένα χαμηλότερου δυναμικού (προς το σώμα) και καθώς πρέπει να κλείσει το κύκλωμα μεταξύ ενδοκυττάριου και εξωκυτταριου χώρου, εξέρχεται του νευρώνα από τα πιο απομακρυσμένα αυτά σημεία του σώματος, για να ακολουθήσει αντίθετη πορεία στο εξωκυττάριο μέσο. Το σημείο εισόδου του ρεύματος στον νευρώνα (απώτεροι δενδριτες) αποκαλείται καταβόθρα ρεύματος (current sink) ενώ αυτό της εξόδου στον εξωκυττάριο χώρο (σώμα) πηγή ρεύματος (current source) (Εικόνα Α.ΙΙΙ.1). Τα ιοντικά ρεύματα δια μέσου του εξωκυτταριου χώρου ρέουν με κατεύθυνση από τις πηγές στις καταβόθρες δημιουργώντας στιγμιαίες μεταβολές δυναμικού, τα δυναμικά πεδίου. Στη περιοχή της καταβόθρας καταγράφεται ένα αρνητικό δυναμικό, ενώ στην περιοχή της πηγής ένα θετικό, σχηματίζοντας έτσι ένα ηλεκτροστατικό δίπολο. Όταν αυτά τα δίπολα είναι παράλληλα διευθετημένα μεταξύ τους (όπως συμβαίνει με τα πυραμιδικά κύτταρα του ιπποκάμπου), τότε η άθροιση των εξωκυττάριων δυναμικών από ένα μεγάλο πληθυσμό νευρώνων δημιουργεί σχετικά μεγάλα δυναμικά πεδίου που μπορούν να καταγραφούν εξωκυτταρια. 39
 Στην παρούσα μελέτη")
 σε τομές ιπποκάμπου.")




 προκαλεί την διέγερση των νευρώνων με")
 που επικάθεται (στην")
52 Εικόνα Α.ΙΙΙ.1: Σχηματική απεικόνιση της δημιουργίας δυναμικού πεδίου (θεωρία δίπολου) Στην παρούσα μελέτη πραγματοποιήθηκαν εξωκυττάριες καταγραφές προκλητών συναπτικών δυναμικών και δυναμικών ενεργείας προερχόμενων από έναν πληθυσμό νευρώνωνν (δυναμικά πεδίου ή πληθυσμιακά δυναμικά) σε τομές ιπποκάμπου. Πιο συγκεκριμένα, ο ηλεκτρικός ερεθισμός των παράπλευρων ινών του Schaffer, προκαλεί συναπτική εκπόλωση των δενδριτών τηςς ακτινωτής στοιβάδας. Στο σημείο αυτό που αποτελεί την καταβόθρα ρεύματος, η εκπόλωση θα καταγραφεί εξωκυτταρια ως μία αρνητική απόκλιση του δυναμικού, που αποτελεί το άθροισμα των δενδριτικών εκπολώσεων ενός πληθυσμού νευρώνων και καλείται διεγερτικόό μετασυναπτικό δυναμικό πεδίου (fepsp-field excitatory postsynaptic c potential). Προκειμένου να κλείσει το κύκλωμα του δημιουργούμενου ρεύματος, αυτό θα πρέπει να διαφύγει από άλλο σημείο του νευρώνα (πηγή ρεύματος). Το σημείο αυτό είναι η περιοχή του σώματος του νευρώνα όπου καταγράφεται ως μία θετική απόκλιση του δυναμικού,, που αποτελεί κατοπτρικό είδωλο του δυναμικού που καταγράφεται στους δενδρίτες. Όταν η συναπτική εκπόλωση υπερβείί ένα συγκεκριμένο όριο (τον ουδό) προκαλεί την διέγερση των νευρώνων με την δημιουργία δυναμικών ενέργειας στην περιοχή του σώματος (εκφυτικός κώνος), που η άθροιση τους από πληθυσμό νευρώνων καταγράφεται ωςς μία αρνητική απόκλιση (καταβόθρα ρεύματος) που επικάθεται (στην κυματομορφή) του διεγερτικού μετασυναπτικού δυναμικού και καλείται πληθυσμιακό δυναμικό ενέργειας (PS-population spike) (Εικόναα Α.ΙΙΙ.2). Στην δενδριτική περιοχή το πληθυσμιακό δυναμικό ενέργειας καταγράφεται ως μία θετική απόκλιση του δυναμικού (μικρότερου μεγέθους συγκριτικά με αυτό που καταγράφεταιι στο σώμα). 40






 στην")


















53 Εικόνα Α.ΙΙΙ.2: Διάταξη των ηλεκτροδίων ερεθισμού και καταγραφής σε μία τομήή ιπποκάμπου. Η εικόνα δείχνει μία ιπποκάμπεια τομή και την τοποθέτηση των ηλεκτροδίων ερεθισμούύ και καταγραφής. Επίσης δείχνονται αντιπροσωπευτικέςς καταγραφέςς δυναμικών στο σώμα (PS) και στους δενδρίτες (fepsp) στην CA1 περιοχή μετά από ερεθισμό των ινών Schaffer. Πηγή: (Gruol et al, 2008) Πρόκληση Βιολογικού σήματος Τα ηλεκτρόδια ερεθισμού αποτελούνται από δίπολα, μονωμένα σύρματα πλατίνας/ιριδίου διαμέτρου 25μm και μεταξύ τους απόσταση 100μm σχηματίζοντας ένα ζεύγος που χρησιμοποιείται για διπολικό ερεθισμό. Τα άκρα τους που προορίζονταπ αι για ερεθισμό παραμένουν ελεύθερα μόνωσης και παράλληλα μεταξύ τους. Το σημείο ερεθισμού για όλες τις μελέτες είναι το μέσον της ακτινωτής στοιβάδας των απώτερων α δενδριτών των πυραμιδικών κυττάρων της CA1 περιοχής του ιπποκάμπου, για τον ερεθισμό των παράπλευρων ινών του Schafferr σε απόσταση μm από την στοιβάδαα των σωμάτων των πυραμιδικών νευρώνων. Το ζεύγος των ηλεκτροδίων βυθίζεται εντός της τομής κατά το ήμισυ του πάχους της. Ηλεκτρόδια καταγραφής από ίνα άνθρακα διαμέτρου 7μm (Kation Scientific, Minneapolis, USA) χρησιμοποιούνταν για την καταγραφή του βιολογικού σήματος. Σημειωτέον ότι η 41
54 απόσταση μεταξύ ηλεκτροδίων ερεθισμού και καταγραφής παραμένει σταθερή περίπου στα 500μm μεταξύ των τομών. Για την πρόκληση των βιολογικών απαντήσεων χρησιμοποιούνται ηλεκτρικοί παλμοί διάρκειας 100μs και ρυθμιζόμενης έντασης από 0.2 έως 300 μα για την εκτέλεση των συναρτήσεων εισόδου/εξόδου. Το βιολογικό σήμα που προκύπτει μετά τον ερεθισμό καταγράφεται από τα ηλεκτρόδια καταγραφής, κατευθύνεται σε προενισχυτή (ενίσχυση 100Χ), ακολούθως σε ενισχυτή (ενίσχυση 5Χ) και τέλος στο φίλτρο απομόνωσης συχνοτήτων για την απομάκρυνση συχνοτήτων και τη βελτίωση του λόγου σήμα/θόρυβος. Μετά το φίλτρο το σήμα άγεται προς: α) παλμογράφο για οπτικό έλεγχο, β) ηχεία για ακουστικό έλεγχο και γ) σε ειδικό μετατροπέα του αναλογικού σήματος σε ψηφιακό συνδεδεμένο με υπολογιστή, το CED 1401-plus interface. Τα σήματα αποκτήθηκαν με τον Neurolog ενισχυτή (Digitimer Limited, UK), band-pass φιλτραρισμένο στα 0.5Hz 2kHz, ψηφιοποιημένο στα 10kHz και αποθηκεύτηκαν στον σκληρό δίσκο του υπολογιστή με τη χρήση CED 1401-plus interface and the Signal6 λογισμικού (Cambridge Electronic Design, Cambridge, UK) για κατοπινή ανάλυση. Για την πρόσληψη, αποθήκευση και κατοπινή ανάλυση του βιολογικού σήματος σε ψηφιακή μορφή χρησιμοποιούνται τα προγράμματα Spike 2 και Signal5&6. 5. Ποσοτικοποίηση Δυναμικών Πεδίου Τα είδη των δυναμικών πεδίου που καταγράφονται στις τομές του ιπποκάμπου είναι τα εξής: 5.1. Πληθυσμιακό Δυναμικό Ενέργειας Προσυναπτικών Αξόνων- Fiber Volley, Fv. Καταγράφεται στην περιοχή των δενδριτών ως μία αρνητική απόκλιση του δυναμικού και οφείλεται στα αθροισμένα δυναμικά ενέργειας των διεγερμένων προσυναπτικών ινών Schaffer, αποτελώντας μέτρο προσυναπτικής διέγερσης. Αυξάνεται ανάλογα με την ένταση του ερεθισμού, αφού εντονότερος ερεθισμός προκαλεί τη διέγερση περισσότερων ινών. Υπολογίζεται ως η απόσταση της αρνητικής κορυφής από την οριζόντια ευθεία που ορίζει το επίπεδο του δυναμικού πριν τον ερεθισμό σε mv (Εικόνα Α.ΙΙΙ.3) Διεγερτικό Μετασυναπτικό Δυναμικό Πεδίου-Field Excitatory Postsynaptic Potential, fepsp. Το fepsp καταγράφεται τόσο στην ακτινωτή στοιβάδα των δενδριτών (όπου και δημιουργείται) όσο και στην πυραμιδική στοιβάδα (ως ανεστραμμένο είδωλο). Το fepsp αποτελεί ένα βραδύ κύμα διάρκειας ms (Εικόνα Α.ΙΙΙ.3), αποτελεί ένδειξη της έντασης της συναπτικής ενεργοποίησης και υπολογίζεται ως η μέγιστη κλίση της 42



55 ανερχόμενης φάσης του (θετικήή στην πυραμιδική στοιβάδα και αρνητική στην ακτινωτή), κλίση του EPSP (mv/ms). Στα αποτελέσματα η κλίση του fepsp, υπολογισμένη είτε στην πυραμιδική είτε στην ακτινωτή στοιβάδα, θα αναφέρεται συμβατικά με θετικό πρόσημο Εμβαδόν Κυματομορφής Το εμβαδόν της κυματομορφής (mv x ms) ορίζεται ως το εμβαδόν που περικλείεται από τη συνολική κυματομορφή του fepsp, αποτελεί μέτρο της συναπτικής ενεργοποίησης των NMDARs (Collingridge et al., 1992) και μετριέται ως η περιοχήή που οριοθετείται από την κυματομορφή του δυναμικού,, τη βασική γραμμή και την ορθογώνιαα γραμμή που διασταυρώνει την κυματομορφή στο τέλος του FV (Εικόνα Α.ΙΙΙ. 3) Ορθόδρομο Πληθυσμιακό Δυναμικό Ενέργειας-Population Spike, PS. Το ορθόδρομο PS αποτελεί μία καθυστερημένη σε σχέση με το fepsp σύντομη αλλαγή του δυναμικού, διάρκειας 2-3 ms και καταγράφεται στην πυραμιδική στοιβάδα (Εικόνα Α.ΙΙΙ.3). Αποτελεί μία μέτρηση του αριθμού των συναπτικά διεγειρόμενων νευρώνων αλλά και του συγχρονισμού διέγερσης των νευρώνων αυτών. Το PS υπολογίζεται ως η απόσταση μεταξύ της αρνητικής κορυφής και του μεσοδιαστήματος των δύο θετικών κορυφών, ύψος του PS (mv). Εικόνα Α..ΙΙΙ.3: Παραδείγματα εξωκυττάριων καταγραφών προκλητής δραστηριότητας από πυραμιδικό νευρώνα σε ακτινωτή και πυραμιδική στοιβάδα. έναν 43
56 6. Μεθοδολογία Ερεθισμού Στην παρούσα μελέτη χρησιμοποιήθηκαν δύο βασικές μέθοδοι ερεθισμού κατά τις οποίες άλλαζε είτε η ένταση του ερεθισμού ή/και το διάστημα μεταξύ των δύο διαδοχικών ερεθισμών Συναρτήσεις Εισόδου/εξόδου Οι συναρτήσεις εισόδου/εξόδου βασίζονται στη σχέση μεταξύ προσυναπτικής ενεργοποίησης και μετασυναπτικής απόκρισης. Για την κατασκευή αυτών των συναρτήσεων ερεθίζονταν κάθε 30sec οι ίνες του Schaffer με παλμούς αυξανόμενης έντασης και καταγραφόταν στην ακτινωτή (EPSP, FV) και την πυραμιδική στοιβάδα (PS) τα δυναμικά πεδίου. Όλες οι προκύπτουσες σχέσεις μπορούν να περιγραφούν διαγραμματικά και είναι οι εξής (Εικόνα Α.ΙΙΙ.4): Συνάρτηση συναπτικής αποτελεσματικότητας: fepsp/i ή fepsp/fv Αποτελεί τη συνάρτηση μεταξύ μεταβαλλόμενου ρεύματος ερεθισμού (Ι) και συναπτικής εκπόλωσης (fepsp). Η γραμμικότητα της συνάρτησης αυτής υποδηλώνει την αναλογία μεταξύ της προσυναπτικής διέγερσης και της δενδριτικής εκπόλωσης. Οι εξής δείκτες περιγράφουν την παραπάνω συνάρτηση: α) Ημιμέγιστο ρεύμα ερεθισμού (I-50 EPSP ), οριζόμενο ως το ρεύμα που αντιστοιχεί στο Ημιμέγιστο fepsp και β) Μέγιστο fepsp. Μετατόπιση της συνάρτησης προς τα αριστερά χαρακτηριζόμενη από μικρότερο I50 EPSP σημαίνει ότι απαιτείται μικρότερη ένταση ερεθισμού για την πρόκληση δεδομένης συναπτικής εκπόλωσης και συνεπώς απόδειξη ισχυρότερης συναπτικής ενεργοποίησης, δηλαδή αύξηση της συναπτικής αποτελεσματικότητας. Επίσης η προσυναπτική διέγερση εκφράζεται με τα καταγραφόμενο στην ακτινωτή στοιβάδα Fv του οποίου το ύψος είναι ανάλογο της έντασης του ρεύματος ερεθισμού. Συνεπώς η συνάρτηση fepsp/i μπορεί να αποδοθεί και ως fepsp/fv που μας παρέχει μια πιο ξεκάθαρη σχέση μεταξύ προσυναπτικής ενεργοποίησης (Fv) και μετασυναπτικής εκπόλωσης (fepsp) Συνάρτηση διεγερσιμότητας: PS/I Συνιστά σιγμοειδή συνάρτηση, που υποδηλώνει την κανονική κατανομή των νευρώνων ως προς την διεγερσιμότητα τους. Έτσι σε μικρές εντάσεις ερεθισμών, μόνο οι πιο διεγέρσιμοι νευρώνες θα φτάσουν τον ουδό διέγερσης για τη δημιουργία δυναμικών ενεργείας. Οι περισσότεροι νευρώνες πυροδοτούν εντός ενός μεγάλου εύρους ενδιάμεσων εντάσεων ερεθισμών και συνεπώς η συνάρτηση στην περιοχή αυτή θα έχει υψηλή κλίση. Οι 44
 Ημιμέγιστο ρεύμα ερεθισμού (I50 PS ), οριζόμενο ως το ρεύμα που προκαλείί το Ημιμέγιστο PS και β) ) μέγιστο PS (PS Max ), που")

 Κλίση του EPSP στο ημιμέγιστο PS (EPSPP 50 ), οριζόμενο ως η συναπτική εκπόλωση που προκαλεί το ημιμέγιστο PS και β) Μέγιστο EPSP")
57 ασυμπτωτική πορεία στις μεγάλες εντάσεις ερεθισμών οφείλεται στον μικρό αριθμό των λιγότερων διεγέρσιμων νευρώνων. Οι εξής δείκτες περιγράφουν ν την συνάρτηση αυτή: α) Ημιμέγιστο ρεύμα ερεθισμού (I50 PS ), οριζόμενο ως το ρεύμα που προκαλείί το Ημιμέγιστο PS και β) ) μέγιστο PS (PS Max ), που αποτελεί μέτρο του μέγιστου αριθμού των διεγέρσιμων νευρώνων Συνάρτηση μετασυναπτικής διεγερσιμότητας PS/fEPSP Η συνάρτηση αυτή είναι επίσης σιγμοειδής και αντιπροσωπεύει τη λειτουργική σχέση μεταξύ δενδριτικής εκπόλωσης και εκφυτικού κώνου των πυραμιδικών κυττάρων. Δηλαδή εκφράζει την ικανότητα του νευρώνα ναα πυροδοτεί δυναμικά ενέργειας με δεδομένη εκπόλωση. Η συνάρτηση αυτή περιγράφεται από τους δείκτες: : α) Κλίση του EPSP στο ημιμέγιστο PS (EPSPP 50 ), οριζόμενο ως η συναπτική εκπόλωση που προκαλεί το ημιμέγιστο PS και β) Μέγιστο EPSP (EPSPP Max ). Οι μέγιστες απαντήσεις προσδιορίζοντανν με το γεγονός ότι αύξηση της έντασης του ρεύματος ερεθισμού δεν προκαλούσε περαιτέρω αύξηση της απάντησης, ή προκαλούμε μια μικρή μείωση της απάντησης. Εικόνα Α.ΙΙΙ.4: Παραδείγματα συναρτήσεων εισόδου/εξόδου 45
58 6.2. Σύζευξη Παλμών (Ορθόδρομος Ερεθισμός) Σε αυτή τη μέθοδο ερεθισμού, δύο παλμοί επιδίδονται μέσω ενός ηλεκτροδίου με μεταβαλλόμενη χρονική απόσταση μεταξύ τους (μεσοπαλμικό διάστημα) και καταγράφεται η απάντηση από δεδομένο σημείο (στην πυραμιδική ή την ακτινωτή στοιβάδα). Διερευνάται έτσι το αποτέλεσμα (ενισχυτικό ή κατασταλτικό) που ο πρώτος ερεθισμός (conditioning stimulus inducing an unconditioned response) επιφέρει στην απάντηση που εκλύεται με τον δεύτερο ερεθισμό (conditioned or δοκιμασία response), σαν συνάρτηση της μεταξύ τους χρονικής απόστασης. Το ενισχυτικό αποτέλεσμα καλείται διευκόλυνση με σύζευξη παλμών, ενώ το κατασταλτικό αποτέλεσμα καλείται ως καταστολή με σύζευξη παλμών ή απλώς καταστολή. Το είδος και το μέγεθος της επίδρασης του πρώτου ερεθισμού στη δεύτερη απάντηση εξαρτάται: α) από τις θέσεις που επιδίδονται οι ερεθισμοί στην τομή, δηλαδή τι είδους στοιχεία διεγείρονται που σημαίνει ότι απαιτείται ο ίδιος τρόπος τοποθέτησης των ηλεκτροδίων σε κάθε τομή και β) την σχετική ένταση των δύο ερεθισμών και γ) το χρονικό διάστημα (καθυστέρηση) που μεσολαβεί των δύο ερεθισμών. Το πρωτόκολλο αυτό έχει ευρέως χρησιμοποιηθεί για τον έλεγχο τόσο της παλίνδρομης αναστολής (PS) όσο και της ενίσχυσης συζευγμένων παλμών του fepsp Παλίνδρομη αναστολή (ΠΑ) (Paired pulse Inhibition-PPI) Οι πυραμιδικοί νευρώνες του ιπποκάμπου δέχονται στην περιοχή του σώματος μια ισχυρή ανασταλτική δράση τοπικών ενδονευρώνων, οι οποίοι διεγείρονται από παλίνδρομους κλάδους των ίδιων των πυραμιδικών κυττάρων (Freund et al., 1996). Ο έλεγχος της ΠΑ υπολογίζεται μέσω ενός ορθόδρομου ερεθισμού (τον πρώτο του ζευγαριού, που προκαλεί εξάρτηση) και την παρατήρηση του ανασταλτικού αποτελέσματος στην καταστολή της απάντησης που προκαλείται από το ακόλουθο δεύτερο ερέθισμα (τεστ ερεθισμός). Το διάστημα μεταξύ των δύο παλμών ήταν 8ms στη βάση προηγούμενων παρατηρήσεων που δείχνουν μέγιστη καταστολή του PS στα διαστήματα μεταξύ 4 και 10ms (Papatheodoropoulos et al., 2002) και του γεγονότος ότι στο μικρό διάστημα υπάρχει μικρή πιθανότητα μόλυνσης της αναστολής από συναπτικά φαινόμενα διευκόλυνσης. Η δύναμη της ΠΑ ποσοτικοποιήθηκε με τον λόγο μεταξύ του PS που προκαλούνταν με τον δεύτερο ερεθισμό και αυτού με τον πρώτο ερεθισμό (PS2/PS1) με ένταση ερεθισμού ορισμένη έτσι ώστε να παράγει το 50% ή/και του 90% του PS1. Σημειωτέον ότι η απόσταση των ηλεκτροδίων ερεθισμού και καταγραφής πρέπει να είναι σταθερή στα 350μm περίπου διότι θέλουμε να αποφύγουμε την άμεση διέγερση των 46
59 ενδονευρώνων που ασκούν ανασταλτική δράση στην περιοχή του σώματος των πυραμιδικών νευρώνων από το ισχυρό ηλεκτρικό πεδίο του κοντινού ερεθισμού (Dingledine et al., 1987; Papatheodoropoulos et al., 2002) Διευκόλυνση του fepsp με σύζευξη παλμών Το πρωτόκολλο ερεθισμού συζευγμένων παλμών ερευνά την διευκόλυνση του εξωκυτταρια καταγραφόμενου διεγερτικού μετασυναπτικού δυναμικού (fepsp). Συγκεκριμένα, επιδίδονται δύο παλμοί ίδιας έντασης και υπολογίζεται ο λόγος μεταξύ της δεύτερης και πρώτης απάντησης. Κατασκευάστηκαν καμπύλες εισόδου/εξόδου αμφότερων των EPSP1 και EPSP2, μεταβάλλοντας το ρεύμα του ερεθισμού για μεσοπαλμικό διάστημα των 50 ms, στο οποίο έχει παρατηρηθεί το μεγαλύτερο ποσοστό διευκόλυνσης του fepsp (Papatheodoropoulos et al., 2002). Η διευκόλυνση με σύζευξη παλμών για τη σύγκριση μεταξύ των τομών πραγματοποιήθηκε στο 50% του μέγιστου EPSP1. 7. Στατιστική Ανάλυση Το paired t-δοκιμασία και το independent t-δοκιμασία καθώς επίσης και το μη παραμετρικό Wilcoxon δοκιμασία και Mann Whitney U-δοκιμασία χρησιμοποιήθηκαν για τις συγκρίσεις ανάμεσα στις δύο εξαρτημένες και ανεξάρτητες ομάδες τιμών, αντίστοιχα. Χρησιμοποιήθηκε επίσης η διμεταβλητή, δίπλευρη δοκιμασία συσχέτισης, με Pearson s συντελεστή r για να υπολογιστούν οι σημαντικές συσχετίσεις μεταξύ των παραμέτρων. Οι τιμές των διαφόρων παραμέτρων εκφράστηκαν Μέσος όρος + μέσο τυπικό σφάλμα (Mean+SEM) ή Μέσος όρος+μέση τυπική απόκλιση (Mean + SD) και το n σε όλο το κείμενο καταδεικνύει τον αριθμό των τομών και ζώων (τομές / ζώα) που χρησιμοποιήθηκαν στην ανάλυση. 47
60 Β.ΕΙΔΙΚΟ ΜΕΡΟΣ I. ΑΠΟΤΕΛΕΣΜΑΤΑ ΠΕΙΡΑΜΑΤΙΚΩΝ ΜΕΛΕΤΩΝ 1. Κύριες διαφορές μεταξύ κροταφικού και διαφραγματικού ιπποκάμπου στην τροποποίηση του τοπικού CA1 δικτύου από τους υποδοχείς NMDA, mglur5, αδενοσινεργικούς Α2Α και CB1 ενδοκαναβινοειδών ΕΙΣΑΓΩΓΗ Οι γλουταμινεργικοί NMDARs και οι μεταβοτροπικοί mglur5 καθώς επίσης και οι αδενοσινεργικοί υποδοχείς αποτελούν σημαντικούς τροποποιητές της νευρωνικής δραστηριότητας (Anwyl, 1999, Mukherjee and Manahan-Vaughan, 2013, Sebastiao and Ribeiro, 2014). Η οικογένεια των mglur περιλαμβάνει τρεις ομάδες υποδοχέων (Sheffler et al., 2011), με εκείνους που ανήκουν στην ομάδα Ι να έχουν ιδιαίτερα σημαντικό ρόλο στην τροποποίηση της δραστηριότητας των πυραμιδικών κυττάρων (Charpak et al., 1990, Desai and Conn, 1991, Pedarzani and Storm, 1993, Gereau and Conn, 1995b, Mannaioni et al., 1999, Mannaioni et al., 2001). Οι NMDARs (Monaghan and Cotman, 1985) και οι mglur5 είναι ιδιαίτερα άφθονοι στο CA1 πεδίο του ιπποκάμπου (Shigemoto et al., 1992, Romano et al., 1995). Σημειωτέον ότι αυτοί οι δύο υποδοχείς αλληλεπιδρούν συνεργιστικά για την ενίσχυση των NMDAR-μεσολαβούμενων αποκρίσεων (Doherty et al., 1997, Anwyl, 1999, Mannaioni et al., 2001, Tebano et al., 2005). Επίσης, πρόσφατες παρατηρήσεις έχουν καταδείξει ότι αυτή η συνεργιστική δράση των NMDARs και mglur5 βρίσκεται κάτω από τον έλεγχο των αδενοσινεργικών A2A υποδοχέων (Tebano et al., 2005). Οι Α2ΑRs έχουν ένα προεξέχοντα ρόλο στον εγκέφαλο (Cunha et al., 2008, Sebastiao and Ribeiro, 2009) και συμμετέχουν στις εξαρτώμενες από τον ιππόκαμπο διαδικασίες (Costenla et al., 2010). Σε αυτή τη μελέτη, χρησιμοποιώντας καταγραφές των τοπικών δυναμικών πεδίου από την CA1 περιοχή ιπποκάμπειων τομών ενήλικου αρουραίου δείχνουμε ότι η φαρμακολογική τροποποίηση αυτών των τριών υποδοχέων π.χ. ΝMDAR, mglur5 και A2AR, έχουν αξιοσημείωτα διαφορετικές δράσεις στην συναπτική διαβίβαση, μετασυναπτική διεγερσιμότητα και στην αναστολή με σύζευξη παλμών ανάμεσα στον DH και VH πόλο του ιπποκάμπου. Επί πρόσθετα δείξαμε ότι ένα σημαντικό μέρος αυτών των δράσεων απαιτεί την συμμετοχή των CB1 υποδοχέων των ενδοκαναβινοειδών. 48
61 1.2. ΠΕΙΡΑΜΑΤΙΚΗ ΔΙΑΔΙΚΑΣΙΑ Ζώα και προετοιμασία τομών Οι τομές ιπποκάμπου προετοιμάστηκαν από 48 ενήλικους, 2-4 μηνών αρσενικούς Wistar αρουραίους. Όλη η πειραματική μεταχείριση και διαδικασίες περιγράφονται παραπάνω στις γενικές μεθόδους (Υποκεφάλαιο Α.ΙΙΙ.2). Σημειωτέον ότι σε ένα υποσύνολο πειραμάτων, οι τομές επωάζονταν σε ένα μέσο που δεν περιείχε ιόντα μαγνησίου (ελεύθερο μαγνησίου μέσο ή Mg 2+ =0 mm) για 30 λεπτά. Επίσης, σε κάποια πειράματα, χρησιμοποιήθηκαν τομές που περιείχαν μόνο την CA1 περιοχή και όχι την CA3 μετά από αποκοπή (με νυστέρι) των δύο πεδίων με τομή των ινών του Schaffer (αναλύονται στα αποτελέσματα) Καταγραφές, Επεξεργασία και Ανάλυση Δεδομένων Στη παρούσα μελέτη καταγράφονται τα δυναμικά πεδίου που αποτελούνται από το προσυναπτικό fiber volley (Fv), το διεγερτικό μετασυναπτικό δυναμικό (fepsp) και το πληθυσμιακό δυναμικό (PS) που προκλήθηκαν με την επίδοση ηλεκτρικών παλμών όπως περιγράφεται στις γενικές μεθόδους (υποκεφάλαιο Α.ΙΙΙ.4&5&6). Οι μονοί ηλεκτρικοί παλμοί επιδίδονταν σε συχνότητα 0.033Hz. Όμως, η συχνότητα ερεθισμού αυξήθηκε στα 0.01Hz κατά τη διάρκεια των σύντομων περιόδων των φάρμακο-επαγόμενων ταχέων αλλαγών στην προκλητή απάντηση (σημειώνεται στα αποτελέσματα). Για πειραματισμό επιλέχθησαν μόνο οι τομές που παρουσίαζαν σταθερό EPSP και PS για τουλάχιστον 10 λεπτά. Καμπύλες εισόδου/εξόδου ανάμεσα στην ένταση ερεθισμού και της προκλητής απάντησης πραγματοποιήθηκαν σε συνθήκες αναφοράς (control) και κατά τη διάρκεια της εφαρμογής του φαρμάκου. Σε συνθήκες ισχυρής φάρμακο-επαγόμενης καταστολής της συναπτικής απάντησης δεν μπορούσαν να πραγματοποιηθούν καμπύλες εισόδου/εξόδου. Δεδομένου ότι το μέγεθος του Fv καταδεικνύει τον αριθμό των ενεργοποιημένων προσαγωγών ινών (Andersen et al., 1978), χρησιμοποιήσαμε την αναλογία EPSP/Fv για να ποσοτικοποιήσουμε την συναπτική αποτελεσματικότητα. Ο λόγος PS/EPSP χρησιμοποιούνταν πάντα σαν δείκτης της μετασυναπτικής διεγερσιμότητας. Η αναλογία μετρήθηκε στο 40-60% του μέγιστου EPSP και του αντίστοιχου PS. Η διακύμανση στην τιμή του EPSP οφειλόταν στο γεγονός ότι κατά τη διάρκεια των σύντομων περιόδων των ταχέων φάρμακο-επαγόμενων αλλαγών στην συναπτική απάντηση δεν ήταν εφικτή η προσαρμογή της έντασης ερεθισμού έτσι ώστε να αποκτήσουμε το 50% της απόκρισης. Παρόλα αυτά, οι συγκρίσεις της αναλογίας μεταξύ των συνθηκών control και φαρμάκου πραγματοποιούνταν πάντα στα ίδια χρονικά σημεία και χρησιμοποιώντας control EPSPs παρόμοιου πλάτους (προερχόμενα από τις καμπύλες εισόδου/εξόδου) παρά το γεγονός ότι 49
62 το EPSP μπορούσε να μειωθεί εξαιτίας της δράσης του φαρμάκου. Μετρήσαμε επίσης την διευκόλυνση του EPSP και την δύναμη της αναστολής μέσω της καταστολής του PS εφαρμόζοντας το πειραματικό πρωτόκολλο του ερεθισμού με σύζευξη παλμών όπως περιγράφεται στις γενικές μεθόδους (υποκεφάλαιο 1.6.2). Λαμβάνοντας υπόψη ότι το μέγεθος της συναπτικής διευκόλυνσης είναι αντιστρόφως σχετιζόμενο με την πιθανότητα απελευθέρωσης νευροδιαβιβαστή (Dobrunz and Stevens, 1997), χρησιμοποιήσαμε το φαινόμενο της διευκόλυνσης με σύζευξη παλμών για να αποδείξουμε το εάν συμμετέχουν προσυναπτικοί μηχανισμοί στις επιδράσεις του φαρμάκου που παρατηρήσαμε κάτω από ποικίλες φαρμακολογικές καταστάσεις. Η ισχυρή παλίνδρομη αναστολή που ελέγχει την διέγερση των ιπποκάμπειων πυραμιδικών νευρώνων (Freund and Buzsaki, 1996) μπορεί να εξεταστεί και να ποσοτικοποιηθεί με ενεργοποίηση των τοπικών δικτύων των ανασταλτικών ενδονευρώνων. Για να κάνουμε εφικτές τις συγκρίσεις ανάμεσα στις συνθήκες control και φαρμάκου, οι αναλογίες PS/EPSP και PS2/PS1 έγιναν κάτω από παρόμοιες συνθήκες μετασυναπτικής ενεργοποίησης (π.χ. EPSP ή PS1). Όταν αυτό δεν ήταν εφικτό, οι υπολογισμοί του PS/EPSP και PS2/PS1 δεν γινόντουσαν. Συνοψίζοντας, οι παράμετροι EPSP/Fv, EPSP2/EPSP1, PS/EPSP και PS2/PS1 ποσοτικοποίησαν την συναπτική αποτελεσματικότητα, την συναπτική ενίσχυση, την μετασυναπτική διεγερσιμότητα και αναστολή με σύζευξη παλμών, αντίστοιχα Φαρμακολογικοί παράγοντες Χρησιμοποιήθηκαν τα ακόλουθα φάρμακα: α) NMDA, 25μΜ (30 λεπτά), 50μΜ (20 λεπτά) β) CPP, 10μΜ (20 λεπτά) γ) CHPG, 50μM (30 λεπτά) δ) MTEP, 200μM (20 λεπτά) ε) DPCPX, 30nM (60 λεπτά) στ) CGS nm (10 λεπτά), ζ) ZM , 100 nμ (40 λεπτά) η) AM 281, 5 μm (40 λεπτά). Αναλυτική περιγραφή και τρόπος διάλυσης των φαρμάκων αναλύονται στις γενικές μεθόδους, υποκεφάλαιο Α.ΙΙΙ Στατιστική Η ανίχνευση των στατιστικά σημαντικών διαφορών έγινε με την χρήση των τομών όπως περιγράφεται στο στις γενικές μεθόδους (υποκεφάλαιο Α.ΙΙΙ.7). 50
63 1.3. ΑΠΟΤΕΛΕΣΜΑΤΑ Βασικές μετρήσεις σε DH και VH Τα δυναμικά πεδίου (Fv, EPSP και PS) προκλήθηκαν και μετρήθηκαν με ποικίλη ένταση ερεθισμού και στις DH (n=15/15) και στις VH (n=15/15) τομές (Εικόνα Β. Ι.1.1.Α- D). Αυτές οι DH και VH τομές αποκτήθηκαν από τα ίδια ζώα και συνιστούσαν ένα δείγμα του συνολικού πληθυσμού των τομών που χρησιμοποιήθηκαν σε αυτή τη μελέτη. Σε συμφωνία με προηγούμενες μελέτες (Papatheodoropoulos, 2015b) βρήκαμε ότι το Fv ήταν σημαντικά υψηλότερο στον DH σε σχέση με τον VH στις μέτριες μέχρι τις υψηλές εντάσεις ερεθισμού (Εικόνα Β.Ι.1.1.Α). Όμως, η σχέση μεταξύ προσυναπτικής ενεργοποίησης (Fv) και μετασυναπτικής εκπόλωσης (EPSP) ήταν αρκετά παρόμοια μεταξύ των δύο πόλων (Εικόνα 1Β). Επίσης, ούτε το EPSP ούτε το PS δεν διέφεραν σημαντικά ανάμεσα στον DH και VH σε καμία ένταση ερεθισμού που χρησιμοποιήθηκε (Εικόνα Β.Ι.1.1.C και ένθετο στο Β.Ι.1.1.D) επιβεβαιώνοντας προηγούμενες μελέτες (Maggio and Segal, 2007, Pofantis and Papatheodoropoulos, 2014). Κατασκευάζοντας διάγραμμα του PS έναντι του EPSP βρήκαμε ότι η μετασυναπτική διεγερσιμότητα ήταν συγκρίσιμη μεταξύ των δύο πόλων (Εικόνα Β.Ι.1.1.D). Όμως, η διευκόλυνση με σύζευξη παλμών (EPSP2/EPSP1) και η αναστολή με σύζευξη παλμών (PS2/PS1) ήταν σημαντικά ισχυρότερες στον DH (Εικόνα Β. Ι.1.1.Ε&F), επιβεβαιώνοντας προηγούμενες μελέτες (Papatheodoropoulos and Kostopoulos, 2000b, Papatheodoropoulos et al., 2002, Petrides et al., 2007). 51





.")













64 Εικόνα Β.Ι.1.1:Συγκρίσεις βασικών μετρήσεων μεταξύ DH και VH Συγκεντρωτικές καμπύλες εισόδου/εξόδου που αποτυπώνουν τις σχέσεις μεταξύ των διαφόρων παραμέτρων δείχνονται στα διαγράμματα A-D και για τον DHH (κύκλος) και για τον VH (τετράγωνο). Το ένθετο στο διάγραμμα D αποτυπώνει την σχέση μεταξύ έντασης ερεθισμού και PS. Τα γραφήματα γ E και F δείχνουν τις τιμές της διευκόλυνσης και της αναστολής με σύζευξη παλμών που ποσοτικοποιήθηκανν μέσω των λόγων EPSP2/EPSP1 και PS2/PS1, αντίστοιχα. Η διευκόλυνση με σύζευξη παλμώνν στο 50% του μέγιστου EPSP μετρήθηκε στο διάστημα 50ms μεταξύ των παλμών. Η αναστολή με σύζευξη παλμώνν μετρήθηκε σε 2 εντάσεις ερεθισμού που παρήγαγαν το 50% και το 90% του μεγίστου PS αντίστοιχα. Τα δεδομένα συλλέχτηκαν από 15 ραχιαίες και 15 κοιλιακές τομές από τα ίδια ζώα. Οι αστερίσκοι δηλώνουν στατιστικά σημαντικές διαφορές μεταξύ DH και VH στα *p<0. 05, **p<0.005 or #p< (Mann-Whitney U δοκιμασία). 52
65 Αποκλίνουσες επιδράσεις της ενεργοποίησης των NMDARs μεταξύ DH και VH Οι δράσεις του NMDA στην συναπτική διαβίβαση (EPSP/Fv), στην μετασυναπτική διεγερσιμότητα (PS/EPSP) και στην αναστολή με σύζευξη παλμών (PS2/PS1) εξετάστηκαν στις συγκεντρώσεις των 25μΜ και 50μΜ (Εικόνα Β.Ι.1.2). Αυτές οι συγκεντρώσεις ήταν παρόμοιες με εκείνες που χρησιμοποιήθηκαν από άλλους ερευνητές σε τομές ιπποκάμπου (Sah et al., 1989, Papatheodoropoulos et al., 2005, Xue et al., 2011). Σε γενικές γραμμές, οι επιπτώσεις του NMDA ήταν υψηλότερες στα 50μΜ απ ότι στα 25μΜ και στους δύο πόλους. Στα 25μΜ το φάρμακο αύξησε σημαντικά την EPSP/Fv και την EPSP2/EPSP1 αναλογία στον DH (κατά 7.37±1.9% και 10.2±2.5% αντίστοιχα, n=7/5, Wilcoxon δοκιμασία, p<0.05) αλλά όχι στον VH (n=7/4). Επίσης, στον DH η αναλογία PS2/PS1 αλλά όχι η PS/EPSP ήταν σημαντικά αυξημένη κάτω από 25μΜ NMDA (κατά 27.1±11.6%, n=7/5, Wilcoxon δοκιμασία, p<0.05). Στα 50μΜ το φάρμακο μείωσε σημαντικά την συναπτική διαβίβαση (EPSP/Fv) στον DH (κατά 18.7±2.1%, n=12/9, Wilcoxon δοκιμασία, p<0.005) και παρήγαγε μία ακόμη μεγαλύτερη μείωση στον VH (by 28.9±4.8%, n=13/7, Wilcoxon δοκιμασία, p<0.005; σύγκριση μεταξύ DH και VH με Mann-Whitney U δοκιμασία, p<0.05). Aκόμη, τα 50μΜ NMDA αύξησαν την συναπτική διευκόλυνση μόνο στον DH (κατά 7.0±2.0%, n=7/5, Wilcoxon δοκιμασία, p<0.05). Εκπληκτικό ήταν το γεγονός ότι τα 50μΜ NMDA παρήγαγαν μία έντονη αύξηση στο PS/EPSP (κατά 33.6±11.9%, n=12/9, Wilcoxon δοκιμασία, p<0.01) και PS2/PS1 (κατά 220±53%, n= 7/5, Wilcoxon δοκιμασία, p<0.05) στον DH, αλλά δεν επηρέασε αυτές τις παραμέτρους στον VH (PS/EPSP, n=13/7 και PS2/PS1, n=5/4). Επίσης, το EPSP2/EPSP1 δεν άλλαξε στον VH (n=8/5). Αυτά τα πειράματα έδειξαν ότι η ενεργοποίηση των NMDARs μείωσε τη συναπτική διαβίβαση στον VH περισσότερο από τον DH αλλά ενίσχυσε την μετασυναπτική διεγερσιμότητα και μείωσε την αναστολή μόνο στον DH. 53











 και VH (n=7/4).")

 και ζεύγος παλμών")





 Χρονοεξέλιξη της ς")

66 Εικόνα Β.Ι.1.2: Οι επιδράσεις της ενεργοποίησης των NMDARs στις ραχιαίες καιι κοιλιακές τομές τ ιπποκάμπου Α) Παραδείγματα καταγραφών πεδίου από την CA1 ακτινωτή και πυραμιδική στοιβάδα (ίχνη στο αριστερό πάνελ) που προέρχονται από μία ραχιαία και μία κοιλιακή τομή ιπποκάμπουυ πριν και κατά τη διάρκεια της εφαρμογής 25μΜ NMDA. Οι δράσειςς του ερεθισμού με σύζευξηη παλμών στοο PS δείχνονται σε καταγραφές της πυραμιδικής στοιβάδας (κατώτερα ίχνη). Το γράφημα στοο δεξιό πάνελλ αποτυπώνει την χρονοεξέλιξη της μεταβολής του EPSP/Fv κάτω από 25μΜ NMDA για τον DH D (n=7/5) και VH (n=7/4). Β) Παραδείγματα δυναμικών πεδίου από μονό παλμό (ίχνη στην πρώτη και δεύτερη σειρά) και ζεύγος παλμών (τρίτη σειρά) από μία ραχιαία και μία κοιλιακή τομή που αποτυπώνουν τηνν αντιστρεπτή ή δράση του 50μΜ NMDA. Σημειωτέον ότι το NMDA μείωσε το EPSP/Fv στον VH περισσότερο απ ότι τον DH και αύξησε το PS στον DH αλλά όχι στον VH. Ακόμη, το NMDAA αντίστρεψε την αναστολήή του PS σε έντονη διευκόλυνση στον DH αλλά όχι στον VH. C) Χρονοεξέλιξη της ς μεταβολής του EPSP/Fv που π αποτυπώνει την υψηλότερη NMDA επαγόμενη καταστολή στην συναπτική διαβίβαση στον VH (n=6/4) σε σ σύγκριση με τον DH (n= =6/4). 54
67 D) Συγκεντρωτικά αποτελέσματα των δράσεων του NMDA στην συναπτική διαβίβαση (EPSP/Fv), στην ενίσχυση με σύζευξη παλμών (EPSP2/EPSP1), στην μετασυναπτική διεγερσιμότητα (PS/EPSP) και στην αναστολή με σύζευξη παλμών (PS2/PS1) στον DH και VH (μπλε με μοτίβο και σκέτο κόκκινο, αντίστοιχα). Οι αστερίσκοι δηλώνουν τις στατιστικώς σημαντικές δράσεις του φαρμάκου στα *p<0.05 or **p<0.005 (Wilcoxon δοκιμασία). Οι διαφορές των δράσεων του φαρμάκου μεταξύ DH και VH καταδεικνύονται επίσης στα #p<0.05 (Mann-Whitney δοκιμασία). Τα artifacts σε αυτό και τα ακόλουθα σχήματα έχουν κουτσουρευτεί. Για τον DH, τα δεδομένα στα 25 μμ συλλέχτηκαν από 7 τομές/5 ζώα για όλες τις παραμέτρους, ενώ στα 50μΜ αποκτήθηκαν από 12 τομές/9 ζώα για EPSP/Fv και PS/EPSP και από 7 τομές/5 ζώα για PS2/PS1 και EPSP2/EPSP1. Για τον VH, τα δεδομένα στα 25μΜ συλλέχτηκαν από 7 τομές/4 ζώα για όλες τις παραμέτρους, ενώ στα 50μΜ αποκτήθηκαν από 13 τομές/7 ζώα για EPSP/Fv και PS/EPSP και από 5 τομές/4 ζώα για PS2/PS1 και 8 τομές/5 ζώα για το EPSP2/EPSP Η δράση των NMDARs δεν απαιτούν την ενεργότητα των Α1 αδενοσινεργικών υποδοχέων Ένας από τους προτεινόμενους μηχανισμούς που συμμετέχει στην μείωση της συναπτικής διαβίβασης που επάγεται από την ενεργοποίηση των NMDARs είναι μέσω των A1 αδενοσινεργικών υποδοχέων (A1Rs). Έτσι, η έντονη ενεργοποίηση των NMDARs προκαλεί την απελευθέρωση αδενοσίνης (Hoehn and White, 1990) η οποία μπορεί να ενεργοποιήσει τους προσυναπτικούς Α1Rs και να μειωθεί η απελευθέρωση του γλουταμινικού (Yoon and Rothman, 1991, Manzoni et al., 1994). Συνεπώς, εμείς εξετάσαμε το εάν μπλοκάρισμα των A1Rs από τον ανταγωνιστή τους το DPCPX (30nM), (Rombo et al., 2015), θα απέκλειε τις δράσεις του NMDA. Όπως φαίνεται στο Εικόνα Β.Ι.1.3, το DPCPX δεν εμπόδισε τις δράσεις του NMDA στο EPSP/Fv ούτε στον DH (n=5/5) ούτε στον VH (n=5/5). 55

 Συγκεντρωτικά δεδομένα της απουσίας δράσης με το")

.")
. 1.3.4.")




68 Εικόνα Β.Ι.1. 3: Η καταστολή της συναπτικής διαβίβασης από το NMDA δεν δ περιλαμβάνει τους A1Rs. Α) Παραδείγματα δυναμικών πεδίου από την ακτινωτή και την πυραμιδική στοιβάδα από μία ραχιαία και μία κοιλιακή τομή, Β) Συγκεντρωτικά δεδομένα της απουσίας δράσης με το μπλοκάρισμα των A1Rs από τον ανταγωνιστή DPCPX (30 nm; DH, n=5/5 and VH, n=5/5) στο κατασταλτικόό αποτέλεσμαα του NMDA στην συναπτική διαβίβαση. Β) Τα δεδομένα που προέκυψαν μόνο υπό την παρουσία του NMDA 50μΜ στην Εικόνα Β.Ι..1.2 δείχνονται και εδώ για λόγους σύγκρισης (μπάρες στα αριστερά). Οι αστερίσκοι δηλώνουν στατιστικά σημαντικές δράσεις του φαρμάκου με *p<0.05 ή **p< <0.005 (Wilcoxon δοκιμασία) Συνενεργοποίηση των mglur5 και NMDARs ενισχύει τις NMDA- τους μεσολαβούμενες δράσεις στον DH αλλά όχι στον VH. Έχει δειχθεί παλιότερα ότι η αλληλεπίδρασηη ανάμεσα στους mglur5 και NMDARs ενισχύει τις NMDA-μεσολαβούμενες αποκρίσεις (Fitzjohn et al., 1996, Mannaioni et al., 2001, Tebano et al., 2005). Έτσι, εξετάσαμε το εάν η εξωγενής ενεργοποίηση μόνο των mglur5 μπορεί ναα ενισχύσειι την δράσηη των NMDARs και έπειτα μελετήσαμε τις επιδράσεις της συνενεργοποίησης των mglur5 m και NMDARs. Εφαρμόσαμε τον επιλεκτικό αγωνιστή τωνν mglur5, το CHPG στις συγκεντρώσεις 50μΜ και 200μΜ, που ήταν παρόμοιες με εκείνες που χρησιμοποιήθηκανν σε in vitro παρασκευάσματα ιππόκαμπου ( Kotecha et al., 2003, Sarantis et al., a 2015), στον DH (n= =6/5 και n=5/3 στα 50μΜΜ και 200μM αντίστοιχα) και VH (n=4/3 και n=5/3 στα 50μΜΜ και 200μM αντίστοιχα).. Καμία από τις συγκεντρώσεις δεν είχε κάποιο αποτέλεσμα στις 56
69 παραμέτρους που μελετήθηκαν ούτε στον DH ούτε στον VH. Πιο συγκεκριμένα, οι δράσεις (αλλαγή σε ποσοστά) του 50μΜ CHPG στην EPSP/Fv, PS/EPSP και EPSP2/EPSP1 ήταν 12.7±4.8, 2.6±3.8 και 1.4±1.7 στον DH (n=6/5), και 3.2±2.1, 3.9±2.6 και 3.7±0.6 στον VH (n=4/3). Παρομοίως, οι δράσεις των 200μΜ CHPG στα EPSP/Fv, PS/EPSP, EPSP2/EPSP1 και PS2/PS1 ήταν 0.8±3.4, 6.6±7.0, 15.0±16.9 και 5.3±5.5 στον DH (n=5/3), και 7.3±3.0, 3.4±3.0, 26.8±16.4 και 7.4±4.1 στον VH (n=5/3). Έπειτα, ερευνήσαμε τις επιδράσεις της συνενεργοποίησης των mglur5 με τους NMDARs συγκριτικά ανάμεσα στον DH και VH εφαρμόζοντας ταυτόχρονα NMDA (50μΜ) και CHPG (50μΜ) σε ραχιαίες και κοιλιακές τομές. Αυτό που παρατηρήσαμε ήταν αξιοσημείωτο. Η συνενεργοποίηση των δύο αυτών υποδοχέων ενίσχυσε σημαντικά τις δράσεις της ενεργοποίησης μόνο των NMDARs στα EPSP/Fv, PS/EPSP και PS2/PS1 στον ραχιαίο αλλά όχι στον κοιλιακό ιππόκαμπο (Εικόνα Β.Ι.1.4). Πιο συγκεκριμένα, βρήκαμε ότι η συνεφαρμογή του NMDA με το CHPG στον DH παρήγαγε με συνέπεια μία αλληλουχία χρόνο-εξαρτώμενων δράσεων που αποτελούνταν από τρεις φάσεις. Η 1 η φάση ήταν σύντομη, κατά τη διάρκεια των πρώτων 3 με 5 λεπτών από την αρχή της εφαρμογής του κοκτέιλ φαρμάκων και χαρακτηριζόταν από ισχυρή μείωση της αναστολής (αύξηση του λόγου PS2/PS1 κατά 530±88%, n=11/5, Wilcoxon δοκιμασία, p<0.005) συνοδευόμενη, κατά τη διάρκεια των επόμενων δευτερολέπτων, από μία απότομη πολύ έντονη αύξηση της μετασυναπτικής διεγερσιμότητας (αύξηση του λόγου PS/EPSP κατά 641±171%, n=15/7, Wilcoxon δοκιμασία, p<0.005). Έτσι, η συνδυαστική δράση των αγωνιστών των δύο υποδοχέων στον DH υπερκέρασε κατά πολύ τις αντίστοιχες δράσεις του 50μΜ NMDA στο PS/EPSP (Mann-Whitney U δοκιμασία, p<0.001) και PS2/PS1 (Mann-Whitney U δοκιμασία, p<0.05). Σημειώνεται ότι η μεγάλη αύξηση της μετασυναπτικής διεγερσιμότητας συνέβαινε ταυτόχρονα με την βαθμιαία μείωση της διεγερτικής συναπτικής διαβίβασης (EPSP/Fv) η οποία ήταν σημαντικά μειωμένη κατά τη διάρκεια της 1 ης φάσης και στον DH (κατά 32.2±3.6%, n=19/10, Wilcoxon δοκιμασία, p<0.001) και VH (κατά 16.5±3.7%, n=10/7, Wilcoxon δοκιμασία, p<0.05). Σημειωτέον, το μειωτικό αποτέλεσμα της mglur5-nmdar συνενεργοποίησης στο EPSP/Fv ήταν σημαντικά υψηλότερο στον DH απ ότι στον VH (Mann-Whitney U δοκιμασία, p<0.05). Η φάση της έντονης αύξησης της διεγερσιμότητας και της μείωσης της αναστολής που παρατηρήθηκε στον DH ακολουθήθηκε αμέσως από μία 2 η φάση που περιείχε μία απότομη καταστολή της διεγερσιμότητας (μείωση του PS/EPSP κατά 74.4±14.4%, n=13/7, Wilcoxon δοκιμασία p<0.05) συνοδευόμενη από ανάκαμψη του PS2/PS1 σε επίπεδα control (αλλαγή 9.6±30% κοντά στα control επίπεδα, n=11/5, Wilcoxon δοκιμασία, 57
70 p>0.05). Τα αποτελέσματα αυτής της φάσης μεγιστοποιήθηκαν στα 6 με 7 λεπτά από την έναρξη της εφαρμογής του φαρμάκου. Για να αποκτήσουμε κατάλληλες μετρήσεις του PS κατά τη διάρκεια των σύντομων φάσεων των ταχέων και παροδικών αλλαγών στην μετασυναπτική διεγερσιμότητα, η συχνότητα του ερεθισμού και της καταγραφής των δυναμικών πεδίου αυξήθηκε από 0.033Hz σε 0.01Hz. Επίσης ήταν απαραίτητο να προσαρμόσουμε παροδικά την ένταση του ερεθισμού για να αποκτήσουμε το 50% του PS και να καταστίσουμε την σύγκριση με τα control δεδομένα εφικτή. Σημειωτέον, η 2 η φάση δεν παρατηρήθηκε κάτω από την ενεργοποίηση των NMDARs μόνο. Κατά τη διάρκεια της δεύτερης φάσης η αναλογία EPSP/Fv συνέχισε να μειώνεται και στον DH (46.5±4.3%, n=9/4, Wilcoxon δοκιμασία p<0.01) και στον VH (31.1±5.7%, n=9/7, Wilcoxon δοκιμασία p<0.01), με το μεγαλύτερο αποτέλεσμα να παρατηρείτε ξανά στον DH (Mann-Whitney U δοκιμασία, p<0.05). Κατά τη διάρκεια της δεύτερης φάσης της συνεφαρμογής των NMDA και CHPG (π.χ.6-7 λεπτά), ο VH δεν έδειξε σημαντική αλλαγή στην αναστολή (n=5/4), αλλά παρουσίασε σημαντική μείωση της μετασυναπτικής διεγερσιμότητας (n=5/4). Η 3 η φάση χαρακτηρίστηκε από περαιτέρω μείωση του EPSP/Fv. Πιο συγκεκριμένα, βρήκαμε ότι η απότομη μείωση του EPSP/Fv που παράχθηκε μεγιστοποιήθηκε στα λεπτά και στον DH (κατά 84.5±5.1%, n=20/10, Wilcoxon δοκιμασία, p<0.001) και στον VH (κατά 44.7±8.2%, n=12/9, Wilcoxon δοκιμασία, p<0.005). Πιο σημαντικά, το κατασταλτικό αποτέλεσμα του κοκτέιλ φαρμάκων στο EPSP/Fv στον DH υπερέβη σημαντικά αυτό που παρατηρήθηκε κάτω από την ενεργοποίηση των NMDARs μόνο (κατά 350%, Mann- Whitney δοκιμασία, p<0.001). Αντιθέτως, η μείωση του EPSP/Fv που παρατηρήθηκε στον VH ήταν παρόμοιο με αυτό της ενεργοποίησης των NMDARs μόνο (Mann-Whitney U δοκιμασία, p>0.05). Συνεπώς, το κατασταλτικό αποτέλεσμα της συνενεργοποίησης των δύο υποδοχέων στην διεγερτική συναπτική διαβίβαση ήταν σημαντικά ισχυρότερο στον DH απ ότι στον VH (Mann-Whitney U δοκιμασία, p<0.005). Κατά τη διάρκεια αυτής της τελευταίας φάσης, η μετασυναπτική διεγερσιμότητα συνέχισε να μειώνεται στον DH χωρίς σημαντική αλλαγή στην αναστολή, μέχρι που ήταν ανέφικτο να καταγράψουμε PS. Στον VH, είτε η διεγερσιμότητα είτε η αναστολή δεν παρουσίασαν καμία περαιτέρω σημαντική αλλαγή (n=5/4). Αυτά τα αποτελέσματα κατέδειξαν ότι η ενεργοποίηση των mglur5 ενδυνάμωσε τα αποτελέσματα των NMDARs στην συναπτική διαβίβαση, την μετασυναπτική διεγερσιμότητα και την αναστολή στον DH αλλά όχι στον VH. Εφαρμογή του NMDA και CHPG παρουσία του ανταγωνιστή των mglur5, το MTEP (200μΜ), (Lea et al., 2005, Sarantis et al., 2015), εμπόδισε τις συνεργιστικές δράσεις των δύο αγωνιστών (5 ραχιαίες τομές από 2 ζώα) (Εικόνα Β.Ι.1.4.D). Με παρόμοιο τρόπο το κοκτέιλ NMDA- 58



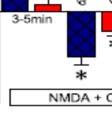





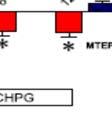























71 CHPG δεν είχε κανένα σημαντικό αποτέλεσμα σε καμία παράμετρο όταν χορηγήθηκε παρουσίαα του ανταγωνιστή των NMDARs, το CPP (5 ραχιαίες τομές από 5 ζώα), στη συγκέντρωση των 10μΜ που μπλοκάρει ταα NMDAR-μεσολαβούμενα ρεύματα στους CA1 νευρώνες (Provini et al., 1991),, (Εικόνα Β.Ι.1.4C&D) ). Εφαρμογή είτε του MTEP είτεε του CPP από μόνα τους δεν παρήγαγαν κανένα σημαντικό αποτέλεσμα σε καμία παράμετρο (n=5/2). Επίσης, το EPSP2/EPSP1 δενν άλλαξε σημαντικά κάτω απόό οποιαδήποτε φαρμακολογική κατάσταση που χρησιμοποιήθηκε σε κανένα από τους δύο πόλους (DH,( n=13/7 παρουσία NMDA-CHPG, και n=5/ /2 παρουσίαα MTEP ή CPP; C VH, n=4/4 παρουσία NMDA-CHPG). 59
72 Εικόνα Β.Ι.1.4: Συνενεργοποίηση των mglur5 και NMDARs παράγει συνεργιστικές δράσεις στην συναπτική διαβίβαση, μετασυναπτική διεγερσιμότητα και αναστολή στον DH αλλά όχι στον VH. Οι NMDARs και mglur5 ενεργοποιήθηκαν με 50μΜ NMDA και 50μΜ CHPG αντίστοιχα. Α) Σε καθένα από τα δύο πάνελ (DH και VH) δείχνονται καταγραφές δυναμικών πεδίου διευκρινίζοντας τις δράσεις του κοκτέιλ φαρμάκων στην συναπτική διαβίβαση (EPSP, πάνω σειρά), νευρωνική διεγερσιμότητα (PS, μεσαία σειρά) και αναστολή (κάτω σειρά). Δείχνονται καταγραφές από διαφορετικά χρονικά σημεία της εφαρμογής του φαρμάκου. Σημειωτέον ότι κατά τη διάρκεια των πρώτων 3-5 λεπτών της συνενεργοποίησης των δύο υποδοχέων παράχθηκε μία ισχυρή ενίσχυση στην νευρωνική διεγερσιμότητα (αποδεικνύεται με την αύξηση του PS και την εμφάνιση δευτερευόντων ) και μία μείωση της αναστολής του PS που αντιστρέφεται σε διευκόλυνση. Στα αμέσως επόμενα λεπτά (6-7) υπήρξε μία σημαντική μείωση της διεγερσιμότητας και επαναφορά της αναστολής. Πιο σημαντικά, αυτές οι διαδοχικές αλλαγές συνέβαιναν στον DH αλλά όχι στον VH. Ακόμη, τονίζεται ότι το κοκτέιλ παρήγαγε μία προοδευτική μείωση στην συναπτική διαβίβαση που ήταν μεγαλύτερη στον DH απ ότι στον VH. Β) Γράφημα της χρονοεξέλιξης του EPSP/Fv που αποτυπώνει την υψηλότερη μείωση με την εφαρμογή του κοκτέιλ στον DH απ ότι στον VH (μπλε και κόκκινο αντίστοιχα). Σημειωτέον ότι οι δράσεις της συνενεργοποίησης των 2 υποδοχέων διέφεραν τόσο εντυπωσιακά σε σχέση με τις δράσεις της ενεργοποίησης των NMDARs μόνο (Σύγκριση με το Εικόνα Β.Ι.1.2.C). C) Γράφημα χρονοεξέλιξης με την μεταβολή του EPSP/Fv που δείχνει ότι το CPP ακύρωσε το συνεργιστικό ενισχυτικό αποτέλεσμα των NMDA+CHPG στον DH. D) Γραφήματα των συγκεντρωτικών δεδομένων των δράσεων των φαρμάκων στην συναπτική διαβίβαση (EPSP/Fv), στην διευκόλυνση με σύζευξη παλμών (EPSP2/EPSP1), στην μετασυναπτική διεγερσιμότητα (PS/EPSP) και στην αναστολή (PS2/PS1) σε DH και VH. Τα αποτελέσματα πάρθηκαν από τρεις χρονικές στιγμές της εφαρμογής του κοκτέιλ έτσι ώστε να αποτυπωθούν οι παροδικές μεγάλες αλλαγές στις αναλογίες PS/EPSP και PS2/PS1. Όμως, στα γραφήματα των PS/EPSP και PS2/PS1 δεδομένα για τον DH στα λεπτά δεν παρουσιάζονται επειδή το PS δεν μπορεί να προκληθεί. Μετρήσεις κάτω από τις φαρμακολογικές καταστάσεις υπό παρουσία των ανταγωνιστών των υποδοχέων συλλέχτηκαν στα χρονικά σημεία των μεγίστων δράσεων του NMDA+CHPG κοκτέιλ (π.χ λεπτά για το EPSP/Fv και EPSP2/EPSP1 και στα 3-5 λεπτά για των PS/EPSP και PS2/PS1). Οι αστερίσκοι δηλώνουν στατιστικά σημαντικές δράσεις των φαρμάκων με *p<0.05 or **p<0.005 (Wilcoxon δοκιμασία). Οι διαφορές στις δράσεις των φαρμάκων ανάμεσα στους δύο πόλους καταδεικνύονται με #p<0.05 or ##p<0.005 (Mann- Whitney δοκιμασία). Τα δεδομένα συγκεντρώθηκαν ως εξής: i) για το EPSP/Fv και για τα τρία χρονικά σημεία, n=19/10, 9/4 και 20/10, στον DH και n=10/7, 9/7 και 12/9 στον VH. ii) για το EPSP2/EPSP1 και για τα τρία χρονικά σημεία, στον DH n=13/7, 8/4, 7/4 και στον VH n=4/4, 6/4 και 5/4 iii) για το PS/EPSP στον DH, n=15/7 (3-5 λεπτά) και n=13/7 (6-7 λεπτά); στον VH, n=10/5 (3-5 λεπτά) και n=5/4 (6-7 λεπτά και λεπτά). Για το PS2/PS1, n=11/5 (3-5 λεπτά και 6-7 λεπτά) in DH και n=5/4 (3-5 λεπτά, 6-7 λεπτά and λεπτά) στον VH. Για όλες τις παραμέτρους και για κάθε εναπομένουσα κατάσταση (MTEP, MTEP+NMDA+CHPG, CPP και CPP+NMDA+CHPG), n=5/2 (μόνο στον DH). 60


 και")





, ακολουθούμενη από μία μείωση κατά")


. Εικόνα Β.Ι.1.")


73 Οι συνεργιστικές δράσεις των NMDARs και mglur5 εμφανίζονται και στα απομονωμένα CA1 πεδία Για να εξετάσουμε το έναν η δραστηριότητα στο CA3 πεδίο ίσωςς επηρεάζει τις δράσεις των φαρμάκων στο CA1 κύκλωμα, εφαρμόσαμε CHPG (50μΜ) και NMDA (50μΜ) σε ραχιαίες τομές που περιείχανν την CA1 αλλά όχι την CA3 περιοχή. Όπως φαίνεται στην Εικόνα Β.Ι.1.5, η συνενεργοποίηση των mglur5 και των NMDARs στις CA1 μίνι τομές παρήγαγε ένα πρότυπο μεταβολών παρόμοιο μεε αυτό που παρατηρήθηκε στις άθικτες τομές του ραχιαίου ιπποκάμπου. Πιο συγκεκριμέν να, το κοκτέιλ φαρμάκων παρήγαγε μία πρώιμη, έντονη και παροδική αύξηση στο PS/EPSP κατά 932±392% (Wilcoxon δοκιμασία, p<0.05, n=6/2), ακολουθούμενη από μία μείωση κατά 80.2±19.8% (Wilcoxon δοκιμασία, p<0.01, n=6/2). Επίσης, παρήγαγε μία χρόνο-εξαρτώμενη καταστολή στην αναλογία EPSP/Fv φτάνοντας στο μέγιστο αποτέλεσμα του τ 96.5±3..5% (Wilcoxon δοκιμασία, p<0.01, n=9/2). Εικόνα Β.Ι.1.5: Δράσεις της συνενεργοποίησης των mglur5 και NMDARss στις CA1 μίνι τομές. Α) Δυναμικά πεδίου που καταγράφηκαν ταυτόχρονα από την δενδριτική και σωματική στοιβάδα (ανώτερα και χαμηλότερα ίχνη αντίστοιχα) από ραχιαία τομήή ιπποκάμπου που περιείχε το CA1 πεδίο αποσυνδεδεμένο από το CA3 με αποκοπή με νυστέρι των ινών Schaffer (CA1 μίνι-τομή),, επεξηγώντας τα αντιστρεπτές 61
74 δράσεις του κοκτέιλ στην συναπτική διαβίβαση και μετασυναπτική διεγερσιμότητα. Β) γράφημα από μια διαφορετική CA1 μίνι τομή που αποτυπώνει την αντιστρεπτή καταστολή του EPSP/Fv από το κοκτέιλ NMDA+CHPG. C) Μεταβολές που παρήχθησαν στο EPSP/Fv (n=9/2) και PS/EPSP (n=6/2) από το κοκτέιλ. Οι μετρήσεις πραγματοποιήθηκαν σε δύο διαφορετικά χρονικά σημεία. Οι αστερίσκοι καταδεικνύουν τις στατιστικώς σημαντικές διαφορές ανάμεσα στην control και φαρμακολογική συνθήκη με *p<0.05 ή **p<0.01 (Mann-Whitney U δοκιμασία) Οι A2ARs ελέγχουν τα συνεργιστικές δράσεις των mglur5 και NMDARs στον DH αλλά όχι στον VH Έχει δειχθεί ότι το ενισχυτικό αποτέλεσμα των mglur5 στην NMDA-επαγόμενη μείωση της διεγερτικής συναπτικής διαβίβασης στον ιππόκαμπο ελέγχεται από την ενδογενή επιτρεπτική δράση των A2ARs στην συνέργια των mglur5 και NMDARs (Tebano et al., 2005). Έτσι, αναρωτηθήκαμε το εάν οι ενισχυτικές δράσεις του CHPG που παρατηρήθηκαν στον DH στην διεγερτική συναπτική διαβίβαση, την μετασυναπτική διεγερσιμότητα και την αναστολή μπορούσαν να αποκλειστούν μετά το μπλοκάρισμα των A2ARs. Συνεπώς, εφαρμόσαμε 50μΜ CHPG και 50μΜ NMDA υπό την παρουσία του επιλεκτικού ανταγωνιστή των A2ARs, το ZM (100nM) (Pugliese et al., 2009). Όπως φαίνεται στην εικόνα Β.Ι.1.6, το ZM απέκλεισε τις συνεργιστικές δράσεις των mglur5 και NMDARs στην συναπτική διαβίβαση (n=12/11), την νευρωνική διεγερσιμότητα (n=7/5) και την αναστολή (n=6/4) στις ραχιαίες τομές ιπποκάμπου. Πιο συγκεκριμένα, μπλοκάρισμα των A2ARs στον DH μείωσε σημαντικά την συνεργιστική δράση του κοκτέιλ στο EPSP/Fv κατά 70% (από -84.5±5.1%, n=20/10 σε -25.4±6.0%, n=12/11, Mann- Whitney U δοκιμασία, p<0.001). Η μείωση του EPSP/Fv παρουσία του ZM έγινε παρόμοια με αυτήν που παρατηρήθηκε με την ενεργοποίηση των NMDARs μόνο. Επιπλέον, το ZM εμπόδισε τις συνεργιστικές δράσεις του κοκτέιλ στην μετασυναπτική διεγερσιμότητα και αναστολή. Πιο ειδικά, μείωσε την παροδική αύξηση του PS/EPSP κατά περισσότερο από 12 φορές (από 641±171%, n=15/7 σε 43.2±11.4%, n=7/5, Mann-Whitney U δοκιμασία, p<0.005) και περιόρισε την αύξηση του λόγου PS2/PS1 από 530±88% (n=11/5) σε 219.4±78.6% (n=6/4), (Mann-Whitney U δοκιμασία, p<0.05). Έπειτα, αναρωτηθήκαμε το εάν η απουσία της ενισχυτικής δράσης του mglur5 στις NMDA-μεσολαβούμενες δράσεις στον VH ίσως οφείλεται στην χαμηλή ενδογενή ενεργοποίηση των A2ARs σε αυτό το τμήμα του ιπποκάμπου. Εφαρμόσαμε CHPG (50μΜ) και NMDA (50μΜ) παρουσία του επιλεκτικού αγωνιστή των A2ARs το CGS (10nM) (Rodrigues et al., 2014). Όπως φαίνεται στην εικόνα 6-C&G, το CGS δεν 62
75 διευκολύνει τις συνεργιστικές δράσεις των mglur5 και NMDARs ούτε στον DH ούτε στον VH. Συνεπώς, παρουσία του CGS οι δράσεις του κοκτέιλ (CHPG+NMDA) στην PS/EPSP και PS2/PS1 ήταν παρόμοιες με αυτά που παρατηρήθηκαν χωρίς την εξωγενή ενεργοποίηση των A2ARs, π.χ. οι δράσεις του κοκτέιλ στον DH ήταν ισχυρές (αύξηση του PS/EPSP και PS2/PS1 κατά 152±38%, n=6/3 και 397±142%, n=5/3 αντίστοιχα). Στον VH οι δράσεις ήταν χαμηλότερες και μη σημαντικές (10.1±13% και 67.9±50.6% για PS/EPSP και PS2/PS1 αντίστοιχα, n=5/4). Εντούτοις, το EPSP/Fv επηρεάστηκε με παρόμοιο τρόπο από το κοκτέιλ στον DH και VH (-58.8±9.6% και -39.8±8.8% σε DH, n=6/3 και VH, n=5/4 αντίστοιχα). Το CGS από μόνο του δεν επηρέασε σημαντικά καμία από τις παραμέτρους είτε στον DΗ είτε στον VH και το EPSP2/EPSP1 δεν άλλαξε σε καμία κατάσταση είτε στον DH είτε στον VH. Αυτά τα αποτελέσματα κατέδειξαν ότι και οι A2ARs και οι mglur5 συνεισφέρουν δυναμικά στην ενίσχυση των NMDARμεσολαβούμενων δράσεων στο ραχιαίο αλλά όχι στο κοιλιακό ιπποκάμπειο κύκλωμα. 63


 Παραδείγματα δυναμικών πεδίου")
")

.")


 Χρονοεξέλιξη της μεταβολήςς του")


) εμποδίζοντας τα")





76 Εικόνα Β.Ι.1.6: Η τονική ενεργοποίηση των A2ARs στον DH αλλά όχι στονν VH ελέγχει ι τις συνεργιστικές δράσεις μεταξύ mglur5 και NMDAR. Α) Παραδείγματα δυναμικών πεδίου από την ακτινωτή (EPSP, πάνω ίχνη) και κ πυραμιδική (PS, κάτω ίχνη) στοιβάδα από μία ραχιαία και μία κοιλιακή τομή που αποτυπώνουν τις δράσεις του ανταγωνιστή των A2ARs, ZM (100nM). Σημειωτέον το μπλοκάρισμαα των A2ARs απέκλεισε τιςς δράσεις της NMDAR-mGluR5 συνέργιας (σύγκριση με τα αντίστοιχα ίχνη στην εικόνα Β.Ι.1.4.Α). Β) Χρονοεξέλιξη της μεταβολήςς του EPSP/Fv που δείχνει την ότι η συνεφαρμογή τουυ CHPG με το NMDA παρουσία ZM παρήγαγε παρόμοια μείωση στην συναπτική διαβίβαση δ στον DH (n=8/8) και στον VH (n=8/8)) εμποδίζοντας τα συνεργιστικά ενισχυτικά αποτελέσματα στον DH. C) Χρονοεξέλιξη της μεταβολής του EPSP/Fv που αποτυπώνει την απουσία αποτελέσματος του αγωνιστή των A2ARs CGS C στην δράση της συνενεργοποίησης των mglur5 με NMDARs είτεε στον DH είτε στον VH. Τα Τ δεδομένα αποκτήθηκανν από μία ραχιαίαα και μία κοιλιακή τομή. D) Δυναμικά πεδίου (ίχνη του EPSP και PS στις πάνωω και κάτω σειρές) από μία ραχιαία και μία κοιλιακή τομή που εξηγούνν τις δράσεις του κοκτέιλ παρουσία CGS (10nM). Ε- 64
77 G) Συγκεντρωτικά δεδομένα των δράσεων του ZM και CGS στην συναπτική διαβίβαση (Ε), την μετασυναπτική διεγερσιμότητα (F) και την αναστολή (G), στον DH και VH. Για λόγους σύγκρισης, τα δεδομένα που αποκτήθηκαν με τη χρήση μόνο του κοκτέιλ δείχνονται στο αριστερό μέρος των γραφημάτων. Οι αστερίσκοι δηλώνουν τις στατιστικώς σημαντικές δράσεις των φαρμάκων με *p<0.05 or **p<0.005 (Wilcoxon δοκιμασία). Οι διαφορές στις δράσεις των φαρμάκων μεταξύ DH και VH καταδεικνύονται με #p<0.05 ή ##p<0.005 (Mann-Whitney δοκιμασία). Τα δεδομένα συλλέχτηκαν ως εξής: στις «ZM» συνθήκες: n=12/11 για EPSP/Fv, n=7/5 για PS/EPSP και n=6/4 για PS2/PS1; VH, n=12/11 για EPSP/Fv και n=5/4 για PS/EPSP και PS2/PS1. Στις «CGS» συνθήκες: DH, n=6/3 για EPSP/Fv και PS/EPSP και n=5/3 για PS2/PS1; VH, n=5/4 για όλες τις παραμέτρους Τα συνεργιστικά αποτελέσματα των mglur5 και NMDARs στον DH απαιτούν τους CB1 υποδοχείς ενδοκαναβινοειδών Η ενεργοποίηση των μετασυναπτικών mglur5 ή των NMDAR ίσως οδηγεί σε απελευθέρωση των ενδοκαναβινοειδών τα οποία δρουν στους προσυναπτικούς CB1 υποδοχείς (Ohno-Shosaku et al., 2007). Ενεργοποίηση των CB1Rs μπορεί να καταστείλει την απελευθέρωση νευροδιαβιβαστή και από τις διεγερτικές και από τις ανασταλτικές απολήξεις (Chevaleyre et al., 2006, Castillo et al., 2012, Kano, 2014) και να μειώσουν την διεγερτική και ανασταλτική συναπτική διαβίβαση (Kano et al., 2009). Έτσι, οι CB1Rs θα μπορούσαν εν δυνάμει να εμπλέκονται στα συνεργιστικά αποτελέσματα των mglur5 και NMDARs. Αυτό το σκεπτικό μας ώθησε να εξετάσουμε το εάν ο αποκλεισμός των CB1Rs θα μπορούσε να επηρεάσει τις δράσεις της συνενεργοποίησης των mglur5 και NMDARs. Οπότε, εφαρμόσαμε CHPG+NMDA παρουσία του ανταγωνιστή των CB1Rs, το ΑΜ 281 (5μΜ) (Leterrier et al., 2006), σε πέντε τομές από τρία ζώα. Το Εικόνα Β.Ι.1.7 δείχνει ότι o αποκλεισμός των CB1Rs περιόρισε σημαντικά τις συνεργιστικές δράσεις του NMDA και CHPG. Πιο συγκεκριμένα, το NMDA-CHPG κοκτέιλ που εφαρμόστηκε παρουσία του ΑΜ 281 μείωσε σημαντικά το EPSP/Fv (κατά 57.8±8.1%, Wilcoxon δοκιμασία, p<0.05) και αύξησε το PS/EPSP (κατά 174.4±36.0%, Wilcoxon δοκιμασία, p<0.05) και το PS2/PS1 (κατά 169.5±89%, Wilcoxon δοκιμασία, p<0.05). Παρόλα αυτά, αυτές οι δράσεις ήταν σημαντικά χαμηλότερες από εκείνες της εφαρμογής του κοκτέιλ μόνο (σύγκριση μεταξύ των δύο καταστάσεων για όλες τις τρείς παραμέτρους Mann-Whitney U δοκιμασία, p<0.05). Επίσης, οι συνεργιστικές δράσεις των mglur5 και NMDARs παρουσία ΑΜ 281, στο EPSP/Fv και PS/EPSP, ήταν σημαντικά μεγαλύτερες σε σύγκριση με τις δράσεις του NMDA μόνου του (σύγκριση μεταξύ των δύο καταστάσεων, Mann-Whitney U δοκιμασία, p<0.005). Αντιθέτως, η δράση του κοκτέιλ στην αναστολή (PS2/PS1) παρουσία ΑΜ 281 (169.5±89%) ήταν παρόμοια με το αποτέλεσμα του NMDA μόνου του (220±53%, Βλέπε 65
78 Εικόνα Β.Ι.1.2), (Mann-Whitney U δοκιμασία, p>0.05). Σημειωτέον ότι το αποτέλεσμα του NMDA-CHPG στο EPSP/FV στον DH με το μπλοκάρισμα των CB1Rs έγινε παρόμοιο με το αποτέλεσμα των δύο αυτών φαρμάκων στον VH κάτω από κανονικές συνθήκες (χωρίς μπλοκάρισμα των CB1Rs). Θεωρητικά, οι δράσεις του μπλοκαρίσματος των CB1Rs θα μπορούσαν να ερμηνευτούν σαν να μεσολαβούν τις δράσεις των NMDARs παρά εκείνων της συνέργιας ανάμεσα στους mglur5 και NMDARs. Έτσι, για να καταλήξουμε στο εάν οι CB1Rs συμμετέχουν κατά προτίμηση σαν μεσολαβητές στις συνεργιστικές δράσεις των mglur5-nmdars ή συμμετέχουν κυρίως στις δράσεις μόνο των NMDARs, εφαρμόσαμε 50μΜ NMDA παρουσία 5μΜ ΑΜ 281. Όπως φαίνεται στην Εικόνα Β.Ι.1.7.C, κάτω από αυτές τις συνθήκες, τα αποτελέσματα της ενεργοποίησης των NMDARs ήταν παρομοια με αυτά που παράγονταν με εφαρμογή μόνο του NMDA. Πιο συγκεκριμένα, εφαρμογή του NMDA παρουσία ΑΜ 281 μείωσε σημαντικά το EPSP/Fv (κατά 26.7±6.5%, n=5/3, Wilcoxon δοκιμασία, p<0.05), και ενίσχυσε το PS/EPSP (κατά 48.8±12.9%, Wilcoxon δοκιμασία, p<0.05) και το PS2/PS1 (κατά 194.4±56.6%, Wilcoxon δοκιμασία, p<0.05). Δεδομένου ότι το κατασταλτικό αποτέλεσμα της ενεργοποίησης των NMDARs ήταν μεγαλύτερο στον VH (βλέπε Εικόνα Β.Ι.1.2), εφαρμόσαμε NMDA παρουσία ΑΜ 281 και σε κοιλιακές τομές. Παρατηρήσαμε ότι όπως στον DH, μπλοκάρισμα των CB1Rs στον VH δεν επηρέασε την δράση του NMDA στο EPSP/Fv (-36.9±6.4%, n=5/3). Σημειώνεται επίσης ότι οι CB1Rs δεν επηρέασαν την καθυστερημένη απότομη καταστολή της διεγερσιμότητας που συνοδευόταν από επαναφορά της αναστολής, στοιχεία που συνιστούσαν την 3 η φάση των NMDA-CHPG δράσεων. Έτσι, στα 6-7 λεπτά της εφαρμογής του κοκτέιλ NMDA-CHPG παρουσία ΑΜ 281, το PS/EPSP κατεστάλη κατά 74.8±5.4% (n=5/3, Wilcoxon δοκιμασία, p<0.05) και το PS2/PS1 επέστρεψε στα επίπεδα του ΑΜ 281 (0.6±35.5%, n=5/3). Αυτά τα αποτελέσματα κατέδειξαν τον σημαντικό ρόλο που παίζουν οι CB1Rs σαν μεσολαβητές των συνεργιστικών δράσεων της mglur5- NMDAR συνενεργοποίησης στον DH, χωρίς να εμπλέκονται στις δράσεις της ενεργοποίησης των NMDARs μόνο είτε στον DH είτε στον VH. Ενδιαφέρον είναι το γεγονός ότι ο αποκλεισμός των CB1Rs παρήγαγε αύξηση της βασικής διεγερτικής συναπτικής διαβίβασης στις DH αλλά όχι στις VH συνάψεις. Πιο συγκεκριμένα, εφαρμογή του ΑΜ 281 αύξησε σημαντικά το EPSP/Fv στον DH (κατά 10.9±3.8%, n=10/5, Wilcoxon δοκιμασία, p<0.05) αλλά όχι στον VH (-3.0±2.1%, n=5/3; Σύγκριση ανάμεσα στον DH και VH, Mann-Whitney U δοκιμασία, p<0.05), (Εικόνα. Β.Ι.1.7.C). Αυτός ο τονικός CB1R-εξαρτώμενος έλεγχος της συναπτικής διαβίβασης στον DH είναι συνεπής με προηγούμενα ευρήματα (Slanina and Schweitzer, 2005), ενώ η 66



 Παραδείγματα δυναμικών πεδίου από μία")

.")

.")










 και την αναστολή")
79 απουσία του στον VH είναι εμφανής όσον αφορά τον σημαντικό ρόλο παίζουν στην τροποποίηση της συναπτικής ς λειτουργίας. που οι CB1Rs Εικόνα Β.Ι.1.7: Οι συνεργιστικές δράσεις των mglur5 και NMDARs στονν DH απαιτούν τους CB1Rs. Α) Παραδείγματα δυναμικών πεδίου από μία ραχιαία τομή ιπποκάμπου που αποτυπώνει τις δράσεις της συνενεργοποίησης των mglur5 και NMDARs παρουσία του ανταγωνιστή των τ CB1Rs ΑΜ 281 (5μΜ). Β) Χρονοεξέλιξη της μεταβολής του EPSP/Fv μετά την συνεφαρμογή του CHPG-NMDA παρουσία 5μΜΜ ΑΜ 281 (n=5/3, γεμισμένα σύμβολα). Για λόγους σύγκρισης δείχνονται και οι χρονοεξελίξεις των μεταβολών του EPSP/Fv που επάγονται από την εφαρμογή μόνο του NMDA ή του NMDA+CHPG απουσία του ΑΜ 281 (ανοιχτά σύμβολα). Σημειώνεται ότι το μπλοκάρισμα των CB1Rs περιορίζει την συνεργιστική δράση δ ανάμεσα στους mglur5 και NMDARs αλλά είναι ισχυρότερη από εκείνη της δράσης της ενεργοποίησης μόνο των NMDARs. Οι τιμές σε αυτό το γράφημα υπολογίστηκαν σαν ποσοστιαία μεταβολή αυτώνν που αποκτήθηκαν κατά τη διάρκεια των τελευταίων τ 5 λεπτών της εφαρμογής του ΑΜ 281, όταν η δράση του φαρμάκου σταθεροποιήθηκε. Συνεπώς, το ενισχυτικό αποτέλεσμα του ΑΜ 2811 στην EPSP/Fv δεν φαίνεται σε αυτό το γράφημα. C) Γραφήματα των συγκεντρωτικών δεδομένων των δράσεων τωνν φαρμάκων στην συναπτική διαβίβαση (EPSP/Fv), την μετασυναπτική διεγερσιμότητα (PS/EPSP) και την αναστολή με σύζευξη παλμών (PS2/PS1). Οι αστερίσκοι δηλώνουν στατιστικά σημαντικές δράσεις με *p<0.05 (Wilcoxon δοκιμασία).. Σημαντικές διαφορές μεταξύ των φαρμακολογικών καταστάσεων τονίζονται με #p<0.05 ή 67
80 ##p<0.005 (Mann-Whitney U δοκιμασία). Τα δεδομένα συλλέχτηκαν από 5 τομές προερχόμενες από 3 ζώα. Οι συνεργιστικές δράσεις των NMDA-CHPG και του NMDA μόνου του δείχνονται επίσης για λόγους σύγκρισης. Σημειώνεται ότι η συνεργιστική δράση των mglur5-nmdar στην αναστολή (PS2/PS1) αλλά όχι της διεγερτικής διαβίβασης (EPSP/Fv) και της μετασυναπτικής διεγερσιμότητας (PS/EPSP), παρουσία ΑΜ 281, ήταν παρόμοια με αυτήν που βρέθηκε με εφαρμογή του NMDA μόνο (βλέπε επίσης στο κυρίως κείμενο) Το μέσο ελεύθερου Μg++ δεν επηρεάζει την συνέργεια μεταξύ mglur5 και NMDARs Η απουσία της συνεργιστικής δράσης των mglur5 και NMDARs στον VH μας ώθησε στο να εξετάσουμε το εάν η απαλοιφή του μπλοκαρίσματος του μαγνησίου στους NMDARs θα μπορούσε να επηρεάσει αυτή τη συνεργιστική δράση. Συνεπώς, εφαρμόσαμε NMDA και CHPG σε τομές όπου είχαν επωαστεί σε ACSF χωρίς ιόντα μαγνησίου (ελεύθερο-mg ++ μέσο, Mg 2+ =0mM). Οι τομές επωάστηκαν σε ελεύθερο Mg ++ μέσο για 30 λεπτά και τότε εφαρμόστηκε το NMDA-CHPG κοκτέιλ. Κάτω από αυτή τη κατάσταση, οι δράσεις της συνενεργοποίησης των NMDAR-mGluR5 στην διεγερτική διαβίβαση (EPSP/Fv) και στην μετασυναπτική διεγερσιμότητα (PS/EPSP) ήταν σημαντικά ενισχυμένες μόνο στον VH (Εικόνα Β.Ι.1.8). Πιο συγκεκριμένα, συνεφαρμογή του NMDA (50μΜ) και του CHPG (50μΜ) μείωσε σημαντικά την διεγερτική διαβίβαση (EPSP/Fv) και ενίσχυσε την διεγερσιμότητα (PS/EPSP) και στον DH (κατά 60.4±4.8%, και 494.0±242%, n=5/3, αντίστοιχα, Wilcoxon δοκιμασία, p<0.05) και στον VH (κατά 79.0±9.3%, και 123.8±31.0%, n=5/3, αντίστοιχα, Wilcoxon δοκιμασία, p<0.05). Στον VH αλλά όχι στον DH, αυτά τα αποτελέσματα ήταν σημαντικά μεγαλύτερα σε σύγκριση με αυτά σε κανονικό ACSF (συγκρίσεις στον VH Mann Whitney U δοκιμασία, p<0.05). Το ελεύθερο Mg ++ μέσο δεν επηρέασε σημαντικά τις δράσεις του NMDA ή του κοκτέιλ στην αναστολή (PS2/PS1) σε κανέναν πόλο. Αυτά τα αποτελέσματα του ελεύθερου Mg ++ μέσου θα μπορούσαν να ερμηνευτούν είτε ως ενίσχυση της συνέργειας μεταξύ mglur5 και NMDARs ή απλά σαν μία ενίσχυση της δραστηριότητας των NMDARs που επάγεται λόγω απουσίας των ιόντων μαγνησίου. Για να διακρίνουμε το τι πραγματικά ισχύει, εφαρμόσαμε μόνο το NMDA σε μία ομάδα τομών επωασμένων σε ελεύθερο μαγνησίου μέσον. Παρατηρήσαμε ότι οι δράσεις της ενεργοποίησης μόνο των NMDARs ήταν παρόμοιες με εκείνες της συνενεργοποίησης των NMDAR-mGluR5 (σύγκριση των 2 φαρμακολογικών καταστάσεων δείχνεται στις τελευταίες 4 μπάρες κάθε γραφήματος). Πιο συγκεκριμένα, εφαρμογή του NMDA μείωσε σημαντικά το EPSP/Fv και ενίσχυσε το PS/EPSP στον DH (κατά 53.1±4.6%, και 130.8±22.2%, n=5/3, αντίστοιχα, Wilcoxon δοκιμασία, p<0.05) και στον 68
81 VH (κατά 77.0±7.0%, και 130.0±18.3%, n=5/3, αντίστοιχα, Wilcoxon δοκιμασία, p<0.05). Αυτές οι παρατηρήσεις έδειξαν ότι αφαιρώντας τα ιόντα μαγνησίου ούτε διεγείρεται η αλληλεπίδραση μεταξύ των mglur5 και NMDAR στον VH ούτε ενισχύεται σημαντικά η συνεργιστική δράση των 2 υποδοχέων στον DH. Τέλος, εφαρμόσαμε μόνο CHPG σε 5 κοιλιακές τομές προερχόμενες από τρία ζώα, επωασμένες σε ελεύθερο Mg ++ μέσον. Καμία από τις παραμέτρους δεν παρουσίασε αξιοσημείωτη αλλαγή (EPSP/Fv, PS/EPSP και PS2/PS1 μεταβλήθηκε κατά 0.7±2.5%, 2.5±3.2% και -4.7±15.3% αντίστοιχα). Αφαιρώντας το Mg ++ από το ACSF αύξησε σημαντικά την συναπτική διαβίβαση και την μετασυναπτική διεγερσιμότητα μόνο στον DH. Πιο ειδικά, στον DH το EPSP/Fv αυξήθηκε κατά 51.7±9.3% (n=10/5, Wilcoxon δοκιμασία, p<0.005) και το PS/EPSP αυξήθηκε κατά 19.0±5.1% (n=10/5, Wilcoxon δοκιμασία, p<0.05). Αντιθέτως, στον VH το ελεύθερο Mg ++ μέσον δεν επηρέασε σημαντικά ούτε το EPSP/Fv (8.7±6.6%) ούτε το PS/EPSP (3.4±4.9%), (n=10/5). Το PS2/PS1 δεν άλλαξε σημαντικά σε κανέναν πόλο (9.9±11.9% στον DH και 22.5±10.2% στον VH). Είναι ενδιαφέρον το γεγονός ότι η αύξηση της διεγερτικής συναπτικής διαβίβασης στο ελεύθερο Mg ++ μέσον στον DH αλλά όχι στον VH είναι συνεπής με την υπόθεση ότι ο VH σε σύγκριση με τον DH παρουσιάζει υψηλότερή πιθανότητα απελευθέρωσης νευροδιαβιβαστή (Papatheodoropoulos and Kostopoulos, 2000b, Pofantis and Papatheodoropoulos, 2014). Αυτό συμβαίνει επειδή οι καταστάσεις που ενισχύουν την πιθανότητα απελευθέρωσης νευροδιαβιβαστή, όπως η αυξημένη αναλογία Ca ++ /Mg ++, θα επηρεάσει περισσότερο τις συνάψεις με σχετικά χαμηλή βασική απελευθέρωση νευροδιαβιβαστή (Wernig, 1972, Kuno and Takahashi, 1986). 69


 Συγκεντρωτικά")


")




.")

















82 Εικόνα Β.Ι.1.8: Η συνέργεια μεταξύ των mglur5 και NMDAR δεν μεταβάλλεται στοο ελεύθερο Mg++ μέσον. A-C) Συγκεντρωτικά αποτελέσματα των επιπτώσεων του ελεύθερου Mg++ μέσουυ στις δράσειςς είτε του NMDA-CHPG κοκτέιλ είτε στο NMDA D) Χρονοεξέλιξη της μεταβολής του EPSP/Fvv που επάγεται από το κοκτέιλ σε ελεύθερο Mg++ μέσον. Οι αστερίσκοι δηλώνουν τις στατιστικώς σημαντικές δράσεις με *p<0.05 or **p<0.05 (Wilcoxon δοκιμασία). Οι σημαντικές διαφορές μεταξύ των διαφορετικών φαρμακολογικών καταστάσεων ή και μεταξύ τωνν 2 πόλων καταδεικνύονται με at #p<0.05 or ##p<0.005 (Mann-Whitney U δοκιμασία). Τα δεδομένα συλλέχτηκαν από 5 ραχιαίες και 5 κοιλιακές τομές προερχόμενες από 3 ζώα. Για λόγους σύγκρισης, δείχνονται επίσης οι δράσεις των φαρμάκων σε κανονικό ACSF (μπάρες στα αριστερά), αλλά οι σημαντικές διαφορές μεταξύύ αυτών των δεδομένων δεν δείχνονται. Σημειωτέον ότι το ενισχυτικό αποτέλεσμα του ελεύθερου Mg++ μέσου στις δράσεις του κοκτέιλ στην διεγερτική διαβίβαση (EPSP/Fv) και στην μετασυναπτική διεγερσιμότητα (PS/EPSP) οφειλόταν στις ενισχυτικές δράσειςς των NMDARs. Ακόμη, προσέξτε ότι το ελεύθερο Mg++ + μέσον αύξησε τη τονική διεγερτική δ συναπτική διαβίβαση και διεγερσιμότητα στον DH αλλά όχι στον VH. 70
83 Οι συνεργιστικές δράσεις της συνενεργοποίησης μεταξύ mglur5 και NMDAR στην μετασυναπτική διεγερσιμότητα συνέπιπτε χρονικά με την φωσφορυλίωση των NR2B υπομονάδων (Tyr1472) στον ραχιαίο ιππόκαμπο Στη συνέχεια επιδιώχθηκε η διερεύνηση του μοριακού μηχανισμού μέσω του οποίου οι mglur5 αλληλεπιδρούν με τους NMDARs, ενισχύοντας με αυτό τον τρόπο τις NMDAR δράσεις στον ραχιαίο ιππόκαμπο. Αυτή η αλληλεπίδραση περιλαμβάνει πολλά στοιχεία διεργασιών ενδοκυττάριας σηματοδότησης όπως την πρωτεϊνική κινάση C (Alagarsamy et al. 1999; Mao and Wang 2002a,b) και την ERK1/2 (Yang et al. 2004). Αυτά τα Κινασόεξαρτώμενα σηματοδοτικά μονοπάτια τροποποιούν τις NMDAR-μεσολαβούμενες αποκρίσεις μέσω φωσφορυλίωσης των υπομονάδων των NMDARs (Chen and Roche 2007). Μεταξύ αυτών των σηματοδοτικών μονοπατιών, το μονοπάτι των πρωτεϊνικών κινασών της τυροσίνης, όπως των Src κινασών, και πιο συγκεκριμένα της Fyn κινάσης έχει ιδιαίτερο ενδιαφέρον, καθώς η ενεργοποίησή της οδηγεί στην ενίσχυση των NMDAR μεσολαβούμενων ρευμάτων (Wang and Salter 1994; Wang et al. 1996; Chen and Roche 2007; Rebola et al. 2008). Μία βασική περιοχή φωσφορυλίωσης της Fyn κινάσης είναι το κατάλοιπο τυροσίνης 1472 της NR2B υπομονάδας (Nakazawa et al. 2001; Takasu et al. 2002), φωσφορυλίωση της οποίας προκαλεί αναστολή της NR2B μεσολαβούμενης ενδοκύττωσης και σταθεροποίηση των NMDARs στην πλασματική μεμβράνη (Prybylowski et al. 2005; Lee 2006; Chen and Roche 2007; Lau and Zukin 2007). Αναλόγως, τα επίπεδα της φωσφορυλίωσης της Tyr1472 αυξάνονται μετά από επαγωγή μακρόχρονης ενδυνάμωσης (LTP) στον ιππόκαμπο, προτείνοντας ότι η συγκεκριμένη φωσφορυλίωση εμπλέκεται στην συναπτική πλαστικότητα (Nakazawa et al. 2001). Για αυτό το σκοπό, ερευνήσαμε τομές από το ραχιαίο τμήμα του ιπποκάμπου στις οποίες είχε απομονωθεί η CA1 περιοχή (CA1 μικροτομές) για να καθορίσουμε το εάν ενεργοποίηση των mglur5 με τον επιλεκτικό αγωνιστή τους (CHPG) θα μπορούσε να επάγει φωσφορυλίωση της NR2B υπομονάδας στο κατάλοιπο τυροσίνης 1472 (Tyr1472). Πραγματοποιήθηκαν εξωκυττάριες καταγραφές σε αυτές τις μικροτομές πριν και κατά τη διάρκεια της εφαρμογής του κοκτέιλ φαρμακολογικών παραγόντων που περιείχε τους αγωνιστές των NMDARs και mglur5, το NMDA (50μΜ) και CHPG (50μΜ), αντίστοιχα. Για να εξετάσουμε με ακρίβεια την χρονική σχέση ανάμεσα στις ηλεκτροφυσιολογικές δράσεις και της φωσφορυλίωσης του NR2B, χρησιμοποιήσαμε πέντε ομάδες με CA1 μικροτομές. Οι μικροτομές της πρώτης ομάδας διαποτίστηκαν σε μέσο με TENY (control ομάδα) ενώ οι μικροτομές των άλλων τεσσάρων ομάδων διαποτίστηκαν σε μέσο που περιείχε το κοκτέιλ των φαρμάκων NMDA-CHPG (ομάδες φαρμάκων) για προοδευτικά 71





 καιι")













84 αυξανόμενο χρόνο. Πιο συγκεκριμένα, οι CA1 μικροτομές τωνν τεσσάρωνν ομάδων με τα φάρμακα διαποτίστηκαν με κοκτέιλ φαρμάκων για 1, 2, 3-5 και 6-7 λεπτά,, αντίστοιχα. Οι μικροτομές λήφθηκαν σε συγκεκριμένη χρονική στιγμή και μεταφέρθηκαν άμεσα σε παγωμένο ισοπεντάνιο (-50 ο C) καιι αποθηκευτήκαν στους σ -80 o C έως ότου χρησιμοποιηθούν για τα πειράματα Western blotting. Τα πειράματα π μέτρησης των επιπέδων φωσφορυλίωσης πραγματοποιήθηκαν σε συνεργασία με την ερευνητική ομάδα της κ. Φεβρωνίας Αγγελάτου και πιο συγκεκριμένα από τον κ. Κωνσταντίνο Σαράντη. Οι μετρήσεις της μετασυναπτικής διεγερσιμότητας και της συναπτικής διαβίβασης αντιπροσωπεύονταν από τις αναλογίεςς PS/EPSP και EPSP/Fv, αντίστοιχα και πραγματοποιήθηκαν σε όλες τις ομάδες των μικροτομών. Όπωςς φαίνεται από την εικόνα Β.Ι.1.9, δεν ανιχνεύτηκε καμίαα σημαντική δράση κατά τη διάρκεια δ των πρώτων δύο λεπτών της εφαρμογής των φαρμάκων είτε στους δύο ηλεκτροφυσιολογικούς δείκτες (PS/EPSPP και EPSP/ /Fv) είτε στα επίπεδα φωσφορυλίωσης. 72
85 Εικόνα Β.Ι.1.9: Χρονική σύμπτωση ανάμεσα στις μοριακές και ηλεκτροφυσιολογικές δράσεις της NMDAR/mGluR5 συνέργειας. Χρονοεξέλιξη των δράσεων της συνενεργοποίησης των NMDARs και mglur5 στην: (α) μετασυναπτική διεγερσιμότητα (PS/EPSP), (b) επίπεδα φωσφορυλίωσης του NR2B (Tyr1472) και (c) συναπτική διαβίβαση (EPSP/Fv). Τα πειράματα πραγματοποιήθηκαν σε CA1 μικροτομές. Οι μετρήσεις των ηλεκτροφυσιολογικών παραμέτρων αποκτήθηκαν πριν και κατά τη διάρκεια της συνενεργοποίησης με 50μΜ NMDA και 50μΜ CHPG. Οι μετρήσεις των τριών παραμέτρων έγιναν στις ίδιες CA1 μικροτομές. Αντιπροσωπευτικά παραδείγματα των καταγραφών πεδίου από την ακτινωτή και την πυραμιδική στοιβάδα δείχνονται στην κορυφή των συγκεντρωτικών διαγραμμάτων στο PS/EPSP και EPSP/Fv, αντίστοιχα. Αντιπροσωπευτικά παραδείγματα της φωσφορυλίωσης του NR2B (Tyr1472) δείχνονται στο αντίστοιχο συγκεντρωτικό γράφημα. Η φάρμακο-επαγόμενη αύξηση στην μετασυναπτική διεγερσιμότητα εκφράστηκε με την εμφάνιση πολλαπλών PSs παρά τη μείωση της συναπτικής διαβίβασης. Αξιοσημείωτη η χρονική σύμπτωση ανάμεσα στις δράσεις της συνενεργοποίησης των NMDAR/mGluR5 στην μετασυναπτική διεγερσιμότητα και στα επίπεδα φωσφορυλίωσης των NR2B (Tyr1472). Οι αστερίσκοι δηλώνουν στατιστικά σημαντικές διαφορές σε σύγκριση με τις control τιμές στα *p < 0.05 (Wilcoxon δοκιμασία και ANOVA με Tukey post hoc δοκιμασία). Πηγή: (Sarantis et al, 2015) Με αξιοσημείωτο τρόπο, έντονες δράσεις των φαρμάκων συνέβησαν κατά τη διάρκεια των επόμενων λεπτών σε όλες τις μετρήσεις. Πιο συγκεκριμένα, παρατηρήσαμε έντονη ενίσχυση στην μετασυναπτική διεγερσιμότητα (κατά %, n=6, Wilcoxon δοκιμασία, p < 0.05) και στην ποσότητα της NR2B (Tyr1472) φωσφορυλίωσης κατά τη διάρκεια των 3-5 λεπτών της εφαρμογής των φαρμάκων (Εικόνα Β.Ι.1.9a και b) που συνέπιπταν σχεδόν χρονικά με εντυπωσιακό τρόπο. Αυτά τα φαινόμενα συνοδεύτηκαν από σημαντική μείωση της συναπτικής διαβίβασης κατά % (Wilcoxon δοκιμασία, p < 0.05, n = 6) κατά τη διάρκεια των 3-5 λεπτών της εφαρμογής των φαρμάκων (Εικόνα Β.Ι.1.9c). Η διευκόλυνση της μετασυναπτικής εκπόλωσης δεν επηρεάστηκε σημαντικά (άλλαξε κατά %, n = 5, Wilcoxon δοκιμασία, p > 0.05). Επίσης, κατά τη διάρκεια των 6-7 λεπτών της εφαρμογής των φαρμάκων η αναλογία PS/EPSP μειώθηκε εντυπωσιακά (κατά % σε σύγκριση με τις control τιμές, Wilcoxon δοκιμασία, p < 0.05, n = 6) ταυτόχρονα με την απότομη μείωση των επιπέδων της φωσφορυλίωσης (Εικόνα Β.Ι.1.9a και b). Η συναπτική διαβίβαση συνέχισε να μειώνεται προοδευτικά κατά τη διάρκεια της περιόδου των απότομων αλλαγών στην μετασυναπτική διεγερσιμότητα και φωσφορυλίωσης, φτάνοντας στη μέγιστη τιμή της στα 8-10 λεπτά της εφαρμογής των φαρμάκων. Πιο ειδικά, το EPSP/Fv μειώθηκε κατά % σε σύγκριση με τις control τιμές (Wilcoxon δοκιμασία, p < 0.05, n = 6) (Εικόνα Β.Ι.1.9c). Η διευκόλυνση του EPSP δεν άλλαξε σημαντικά (7.6 _ 13.9%, n = 4, Wilcoxon δοκιμασία, p > 0.05). Πρέπει να σημειωθεί ότι οι μετρήσεις της διεγερσιμότητας κατά τη διάρκεια της περιόδου της έντονης 73
86 καταστολής στην συναπτική διαβίβαση (8-10 λεπτά) δεν μπορούσαν να πραγματοποιηθούν, εξαιτίας αδυναμίας πρόκλησης PS, ούτε και με ισχυρό ερεθισμό (Sarantis et al, 2015) Οι συνεργιστικές δράσεις της συνενεργοποίησης των mglur5/nmdars στην μετασυναπτική διεγερσιμότητα, στην φωσφορυλίωση του NR2B (Tyr1472) και στη συναπτική διαβίβαση, εξαρτάται από την Src οικογένεια κινασών και αδειοδοτείται από τους A2ARs Όπως φαίνεται στην εικόνα Β.Ι.1.10a, ενεργοποίηση των NMDARs με 50μΜ NMDA παρήγαγε μία σημαντική αύξηση της μετασυναπτικής διεγερσιμότητας (κατά %) παρατηρούμενη στα 3-5 λεπτά της εφαρμογής του φαρμάκου (Wilcoxon δοκιμασία, p < 0.05, n = 5). Επί προσθέτως, το NMDA μείωσε σημαντικά στην διεγερτική συναπτική διαβίβαση (EPSP/Fv) παράγοντας μέγιστο αποτέλεσμα στα 8-10 λεπτά από την αρχή εφαρμογής του φαρμάκου κατά % (Wilcoxon δοκιμασία, p < 0.05, n = 6) (Εικόνα Β.Ι.1.10c). Αυτή η μείωση συνοδεύτηκε από μία μικρή αλλά σημαντική αύξηση της διευκόλυνσης του EPSP ( %, n = 5, Wilcoxon δοκιμασία, p < 0.05). Ενεργοποίηση των mglur5 με 50μΜ CHPG δεν επηρέασε ούτε το PS/EPSP (n=5, p>0.05) ούτε το EPSP/Fv (n=5, p>0.05). Όμως, συνενεργοποίηση των NMDARs και mglur5 με NMDA (50μΜ) και CHPG (50μΜ) παρήγαγε μία απότομη και πολύ έντονη αύξηση της μετασυναπτικής διεγερσιμότητας (PS/EPSP) κατά % (Wilcoxon δοκιμασία, p<0.05, n=5) κατά τη διάρκεια των πρώτων 3-5 λεπτών της εφαρμογής των φαρμάκων (Εικόνα Β.Ι.1.10a). Κατά εντυπωσιακό τρόπο, η δράσεις της συνενεργοποίησης των NMDARs και mglur5 στην μετασυναπτική διεγερσιμότητα εμφανίστηκαν παράλληλα με την μεγάλη αύξηση στα επίπεδα φωσφορυλίωσης της NR2B (Tyr1472) (Εικόνα Β.Ι.1.10.b). Επίσης, η συνενεργοποίηση των NMDARs/mGluR5 παρήγαγε μία έντονη μείωση στον λόγο EPSP/Fv κατά % (Wilcoxon δοκιμασία, p<0.01, n=9) μετρημένη στα 8-10 λεπτά από την αρχή της εφαρμογής του κοκτέιλ φαρμάκων (Εικόνα Β.Ι.1.10c). Έτσι, οι δράσεις της συνενεργοποίησης ξεπέρασαν κατά πολύ αυτές της ενεργοποίησης των NMDARs και στην μετασυναπτική διεγερσιμότητα (PS/EPSP, Wilcoxon δοκιμασία, p<0.01) και στη συναπτική διαβίβαση (EPSP/Fv, Wilcoxon δοκιμασία, p<0.01). Οι συνεργιστικές δράσεις της συνεφαρμογής των NMDA και CHPG στην μετασυναπτική διεγερσιμότητα και φωσφορυλίωση αποκλείστηκαν ολοκληρωτικά από τον επιλεκτικό ανταγωνιστή των mglur5, το MTEP (200μΜ) (Εικόνα Β.Ι.1.10a και b). Η μείωση στο 74

 που")




. Το MTEP δεν")










.")







87 EPSP/Fv κατά % (Wilcoxon δοκιμασία, p < 0.05, n = 5) που προκλήθηκε κατά τη διάρκεια της συνεφαρμογής του κοκτέιλλ (NMDA και CHPG) παρουσία MTEP ήταν παρόμοιαα με αυτήν της ενεργοποίησης μόνο των NMDARs (Εικόνα Β.Ι.1.10c). Το MTEP δεν προκάλεσε καμία σημαντική δράσηη ούτε στο PS/EPSP ούτε στο EPSP/Fv. Επί προσθέτως, ο ανταγωνιστής των NMDARs, το CPPP (10μΜ) απέκλεισε α τις δράσειςς της NMDAR/ /mglur5 συνενεργοποίησης και στο PS/EPSP ( %, n=4, μετρημένο στα 3-5 λεπτά από την εφαρμογή των φαρμάκων) και στο EPSP/ /Fv ( %, n = 4, μετρημένο στα λεπτά από την εφαρμογή των φαρμάκων (τα δεδομένα δεν δείχνονται). Εικόνα Β.Ι.1.10: Οι δράσεις της συνενεργοποίησης των NMDARs και mglur5 m στην μετασυναπτική διεγερσιμότητα, στην φωσφορυλίωση του NR2BB (Tyr1472) και στην διεγερτική συναπτική διαβίβαση υπερκέρασαν τις δράσεις της NMDAR ενεργοποίησης μόνο και απαιτούν την ενεργοποίηση των A2ARs και των Src κινασών. Δείχνονται ιστογράμματα των δράσεων των ποικίλων φαρμακολογικών καταστάσεων στα: (α) PS/EPSP, (b) φωσφορυλίωση της NR2B (Tyr1472) υπομονάδας και (c) EPSP/Fv. Παραδείγματα 75
88 καταγραφών δυναμικών πεδίου που αποκτήθηκαν πριν και κατά τη διάρκεια της εφαρμογής των φαρμάκων (αχνές και παχιές γραμμές, αντίστοιχα) δείχνονται στο πάνω μέρος των στηλών των PS/EPSP και EPSP/Fv ιστογραμμάτων. Μπάρες βαθμονόμησης για τις καταγραφές δείχνονται (στα a και c): 1mV, 5ms. Μετρήσεις των ηλεκτροφυσιολογικών παραμέτρων αποκτήθηκαν κατά τη διάρκεια της χρονικής περιόδου όταν το κοκτέιλ των NMDA-CHPG είχαν τις μέγιστες δράσεις. Σημειώνεται ότι η αύξηση της μετασυναπτικής διεγερσιμότητας (PS/EPSP) εκφράστηκε ως η αύξηση του πλάτους των PS και/ή της εμφάνισης επιπρόσθετων PSs. Οι αστερίσκοι δηλώνουν στατιστικά σημαντικές διαφορές στα *p<0.05 ή **p<0.01 (Wilcoxon δοκιμασία). Στατιστικώς σημαντικές διάφορες των φαρμακολογικών δράσεων καταδεικνύονται με την δίεση στα #p<0.05 ή ##p<0.01 (Mann Whitney U δοκιμασία και ANOVA με Tukey post hoc δοκιμασία). Αποκλεισμός των mglur5 με 200μΜ MTEP, αποκλεισμός των A2 A Rs με 1μΜ ZM και αναστολή των Src κινασών με 30μΜ PP1 απέκλεισαν τα ενδυναμωτικά αποτελέσματα της mglur5 ενεργοποίησης στις NMDAR επαγόμενες αποκρίσεις Πηγή: (Sarantis et al, 2015). Λαμβάνοντας υπόψη την συνεντόπιση των mglur5 και Α2ΑR στον ιππόκαμπο, καθώς επίσης και τον επιτρεπτικό ρόλο των Α2ΑRs στην mglur5/nmdar συνέργεια (Tebano et al. 2005), εξετάσαμε τις δράσεις του κοκτέιλ NMDA/CHPG κάτω από αποκλεισμό των A2ARs με ZM Όπως φαίνεται και στην εικόνα Β.Ι.1.10, το ZM (1μΜ) απέκλεισε εξ ολοκλήρου τις συνεργιστικές δράσεις της NMDAR/mGluR5 συνενεργοποίησης στο λόγο PS/EPSP, στη φωσφορυλίωση του NR2B (Tyr1472) και στο EPSP/Fv. Πιο συγκεκριμένα, συνενεργοποίηση των NMDARs και mglur5 με 50μΜ NMDA και 50μΜ CHPG παρουσία ZM μείωσε τα συνεργιστικά αποτελέσματα στη μετασυναπτική διεγερσιμότητα σχεδόν στα επίπεδα της ενεργοποίησης των NMDARs μόνο (Wilcoxon δοκιμασίας, p<0.05, n=6) και απέκλεισε την φωσφορυλίωση του NR2B (Tyr1472) (Εικόνα Β.Ι.1.10b). Επίσης, η συνενεργοποίηση των NMDARs/mGluR5 παρουσία ZM μείωσε τον λόγο EPSP/Fv κατά % (Wilcoxon δοκιμασία, p<0.05, n=5). Για να διευκρινίσουμε το εάν η φωσφορυλίωση των NR2B (στο κατάλοιπο Tyr1472) βρίσκεται πίσω από τις ηλεκτροφυσιολογικές δράσεις που επάγονται με το κοκτέιλ (NMDA+CHPG), εφαρμόσαμε τον επιλεκτικό αναστολέα των Src κινασών, το PP1. Βρήκαμε ότι το PP1 απέκλεισε ολοκληρωτικά τις συνεργιστικές δράσεις της συνενεργοποίησης στη μετασυναπτική διεγερσιμότητα, στην φωσφορυλίωση του NR2B (Tyr1472) και στην διεγερτική συναπτική διαβίβαση. Πιο αναλυτικά, συνεφαρμογή των NMDA (50μΜ) και CHPG (50μΜ) παρουσία του PP1 (30μΜ) σχεδόν απέκλεισε τις συνεργιστικές δράσεις της NMDAR/mGluR5 συνενεργοποίησης στο PS/EPSP και EPSP/Fv καθιστώντας αυτές παρόμοιες με εκείνες που παρατηρήθηκαν με την ενεργοποίηση των NMDARs μόνο (Wilcoxon δοκιμασία,p<0.05, n=6 και για τις δύο 76
89 παραμέτρους) και απέκλεισε την φωσφορυλίωση της NR2B (Tyr1472) υπομονάδας (Εικόνα Β.Ι.1.10b) (Sarantis et al, 2015) ΣΥΖΗΤΗΣΗ Αυτή η μελέτη καταδεικνύει με εντυπωσιακό τρόπο τις μεγάλες διαφορές ανάμεσα στον DH και VH στην αποτελεσματικότητα των NMDARs, mglu5rs και A2ARs στην τροποποίηση της συναπτικής διαβίβασης, της νευρωνικής διεγερσιμότητας και της αναστολής στο τοπικό ιπποκάμπειο μικροκύκλωμα. Τα κύρια ευρήματα ήταν: α) το μικρότερο κατασταλτικό αποτέλεσμα στην συναπτική διαβίβαση αλλά τις μεγαλύτερες ενισχυτικές δράσεις στην νευρωνική διεγερσιμότητα της ενεργοποίησης των NMDARs στον DH σε σύγκριση με τον VH, β) οι ισχυρές συνεργιστικές δράσεις των mglu5r και NMDARs που παρατηρήθηκαν στον DH αλλά όχι στον VH και γ) η τροποποιητική δράση των A2ARs στην NMDAR-mGlu5R συνέργεια στον DH αλλά όχι στον VH Ερμηνεία των δράσεων του NMDA στον DH και VH Βρήκαμε ότι η εξωγενής ενεργοποίηση των NMDARs μειώνει την συναπτική διαβίβαση και την αναστολή και αυξάνει την μετασυναπτική διεγερσιμότητα. Ποιοτικά αυτά τα αποτελέσματα είναι σύμφωνα με προηγούμενα που δείχνουν ότι η ενεργοποίηση των NMDARs καταστέλλει την διεγερτική συναπτική διαβίβαση (Chernevskaya et al., 1991, Manzoni et al., 1994, Tebano et al., 2005), ενισχύει την νευρωνική διεγερσιμότητα (Sah et al., 1989, Mor and Grossman, 2007) και μειώνει την GABAεργική αναστολή (Stelzer and Shi, 1994, Chisari et al., 2012) στο δίκτυο των πυραμιδικών νευρώνων του ιπποκάμπου. Επί πρόσθετα, βρήκαμε ότι η δράση των NMDARs στην μείωση της συναπτικής διαβίβασης ήταν ισχυρότερη στον VH απ ότι στον DH, ενώ οι δράσεις στην μετασυναπτική διεγερσιμότητα και αναστολή ήταν πολύ πιο υψηλές στον DH απ ότι στον VH. Αυτά τα αποτελέσματα καταδεικνύουν ότι διαφορετικοί μηχανισμοί βρίσκονται πίσω από τις δράσεις των NMDARs στην συναπτική διαβίβαση και τη μετασυναπτική διεγερσιμότητα με διαφορετική συνεισφορά κάθε μηχανισμού ανάμεσα στον DH και VH. Έχει δειχθεί ότι η καταστολή της συναπτικής διαβίβασης στην CA1 περιοχή από τους NMDARs μπορεί να επαχθεί μέσω αύξησης της ενεργοποίησης των A1 αδενοσινεργικών υποδοχέων λόγω αύξησης των εξωκυττάριων επιπέδων της αδενοσίνης (Chernevskaya et al., 1991, Manzoni et al., 1994). Επίσης, λαμβάνοντας υπόψη ότι η δράση των Α1 77
90 υποδοχέων είναι υψηλότερη στον DH από τον VH (Lee et al., 1983) θα περίμενε κάποιος ότι αυτή η δράση των NMDARs θα ήταν μεγαλύτερη στον DH σε σχέση με τον VH. Όμως, στην παρούσα μελέτη βρήκαμε ότι οι Α1 υποδοχείς δεν συμμετέχουν σε αυτήν την δράση των NMDARs. Ακόμη, ένα άλλο απροσδόκητο εύρημα ήταν το γεγονός ότι ενώ οι NMDAR-επαγόμενη μείωση της συναπτικής διαβίβασης στον DH συνοδευόταν από αύξηση της συναπτικής διευκόλυνσης, προτείνοντας της συμμετοχή προσυναπτικών μηχανισμών (Dobrunz and Stevens, 1997), η διευκόλυνση στον VH που παρουσίασε ακόμα μεγαλύτερη NMDAR-επαγόμενη μείωση της διαβίβασης, δεν άλλαξε σημαντικά, προτείνοντας ότι άλλοι εκτός των προσυναπτικών μηχανισμών συνεισφέρουν στην μείωση της διεγερτικής συναπτικής διαβίβασης σε αυτό το τμήμα του ιπποκάμπου. Ένας μηχανισμός μέσω του οποίου οι NMDARs μπορούν να μειώσουν την διεγερτική συναπτική διαβίβαση εμπλέκει τον πολύ πρόσφατο ανακαλυφθέν μετασυναπτικό μηχανισμό των Ca ++ - επαγόμενων SK καναλιών (Wang et al., 2014). Θα επισημάνουμε επίσης ότι χαμηλή συγκέντρωση του NMDA προκάλεσε ενίσχυση αντί για μείωση στην συναπτική διαβίβαση στον DH. Αυτή η δράση ίσως μεσολαβείτε από τους πρόσφατα ανακαλυφθέντες προσυναπτικούς NMDARs (Mameli et al., 2005, Suarez et al., 2005). Η ενισχυτική δράση των NMDARs στην νευρωνική διεγερσιμότητα μπορεί να ερμηνευτεί ευθέως με την άμεση συνεισφορά αυτών των υποδοχέων στην μετασυναπτική εκπόλωση (Forsythe and Westbrook, 1988, Andreasen et al., 1989). Ενεργοποίηση των NMDARs ενισχύει την διεγερσιμότητα των CA1 πυραμιδικών κυττάρων και επάγει αυθόρμητες εκφορτίσεις του δικτύου (Dingledine et al., 1986, Sah et al., 1989, Herreras and Jing, 1993, Mor&Grossman, 2007, Bali et al., 2014). Ένας πρόσθετος μηχανισμός που συνεισφέρει στην αυξημένη μετασυναπτική διεγερσιμότητα θα μπορούσε να περιλαμβάνει αλλαγές στην GABAεργική αναστολή. Πράγματι, ενεργοποίηση των NMDARs μπορεί να επάγει μείωση της GABAεργικής αναστολής στους ιπποκάμπειους πυραμιδικούς νευρώνες μέσω Ca ++ εξαρτώμενης αλληλεπίδρασης των NMDAR και GABA A υποδοχέων (Stelzer and Shi, 1994, Chisari et al., 2012). Η ισχυρή μείωση της αναστολής με σύζευξη παλμών που παρατηρήθηκε στον DH θα μπορούσε να προκαλείται από αυτή τη δισανασταλτική δράση των NMDARs οδηγώντας σε αύξηση της διεγερσιμότητας των πυραμιδικών κυττάρων. Όπως θα συζητηθεί πιο εκτενώς παρακάτω, ένας εναλλακτικός και πιο πρόσφατα συζητήσιμος μηχανισμός που μπορεί να εξηγήσει την κατασταλτική δράση των NMDARs στην ανασταλτική καθώς επίσης και στην διεγερτική συναπτική διαβίβαση εμπλέκει την παλίνδρομη σηματοδότηση μέσω των ενδοκαναβινοειδών στις ανασταλτικές και διεγερτικές συνάψεις (Kano et al., 2009). Όμως, εμείς δείξαμε ότι αποκλεισμός των 78
91 CB1Rs δεν επηρεάζει τις δράσεις της ενεργοποίησης μόνο των NMDARs. Ολοκληρώνοντας, μολονότι οι NMDAR-επαγόμενες δράσεις στην διεγερσιμότητα και την αναστολή μπορούν να γίνουν κατανοητές στη βάση προηγούμενων αναλυθέντων μηχανισμών, οι μηχανισμοί που συμμετέχουν στην καταστολή της διεγερτικής διαβίβασης μετά από ενεργοποίηση των NMDARs παραμένουν ακόμα ασαφείς Ερμηνεία των δράσεων της mglu5r-nmdar συνενεργοποίησης στον DH και VH. Οι ιπποκάμπιοι πυραμιδικοί νευρώνες εκφράζουν σε μεγάλο βαθμό mglu5rs (Shigemoto et al., 1992, Fotuhi et al., 1994) κυρίως τοποθετημένους μετασυναπτικά (Romano et al., 1995, Lujan et al., 1996, Mannaioni et al., 2001, Tebano et al., 2005) όπου αλληλεπιδρούν με τους NMDARs και ενισχύουν τις NMDAR-μεσολαβούμενες αποκρίσεις (Fitzjohn et al., 1996, Doherty et al., 1997, Mannaioni et al., 2001, Tebano et al., 2005). Επίσης, οι mglu5rs μειώνουν την αναστολή στους CA1 πυραμιδικούς νευρώνες (Desai and Conn, 1991, Gereau and Conn, 1995a, Mannaioni et al., 2001). Αναλόγως, εμείς βρήκαμε ότι η συνακόλουθη ενεργοποίηση των mglu5rs και των NMDARs ενισχύουν κατά πολύ τις δράσεις των NMDARs στην διεγερτική συναπτική διαβίβαση, στην μετασυναπτική διεγερσιμότητα και στην συναπτική αναστολή στον DH. Με αξιοσημείωτο τρόπο όμως, ο κοιλιακός ιππόκαμπος αποδείχτηκε αρκετά αναίσθητος στην τροποποίηση από τους mglu5rs. Τα αποτελέσματα των μοριακών πειραμάτων σε ραχιαίες CA1 μικροτομές αποκάλυψαν μία εντυπωσιακά ειδική χρόνο-εξαρτώμενη mglu5r-επαγόμενη φωσφορυλίωση των NR2B (Tyr1472) υπομονάδων των NMDARs που απαιτεί την ενεργοποίηση των Src Κινασών τυροσίνης και ταιριάζει απόλυτα με την χρονοεξέλιξη των αλλαγών της διεγερσιμότητας και της συναπτικής διαβίβασης κατά τη διάρκεια της συνενεργοποίησης των mglu5r και NMDAR (Sarantis et al., 2015) αποκλειστικά στον ραχιαίο ιππόκαμπο. Θα μπορούσε να είναι πιθανό το γεγονός να μην συμβαίνει αυτή η αλληλεπίδραση στον VH κάτω από εξωγενή ενεργοποίηση αυτών των υποδοχέων που χρησιμοποιήθηκαν στην παρούσα μελέτη. Ο τροποποιητικός ρόλος των A2ARs στις συνάψεις του ΚΝΣ έχει εδραιωθεί πολύ καλά (Cunha et al., 2008, Sebastiao and Ribeiro, 2009). Μολονότι οι A2ARs στον (ραχιαίο) ιππόκαμπο έχουν περιορισμένη κατανομή (Jarvis and Williams, 1989, Sebastiao and Ribeiro, 1992, Dixon et al., 1996), παρόλα αυτά είναι ενεργοί, παρουσιάζοντας διεγερτική δράση (Sebastiao and Ribeiro, 1992, Cunha et al., 1994, Rombo et al., 2015). Πρόσφατα έχει δειχθεί ότι η συνεργιστική αλληλεπίδραση μεταξύ των μετασυναπτικών mglu5r και 79
92 των NMDAR βρίσκεται κάτω από τον επιτρεπτικό έλεγχο των αδενοσινεργικών A2ARs (Tebano et al., 2005). Σε συμφωνία με αυτή τη μελέτη βρήκαμε ότι αποκλεισμός των A2ARs στον DH εμπόδισε τις ενισχυτικές δράσεις της συνενεργοποίησης των mglu5r και NMDARs στις NMDAR-εξαρτώμενες αποκρίσεις στις ηλεκτροφυσιολογικές παραμέτρους που μελετήθηκαν καθώς επίσης και στα επίπεδα φωσφορυλίωσης, προτείνοντας ταυτόχρονα ότι η ενδογενής τονική ενεργότητα των A2ARs ελέγχει αυτές τις συνεργιστικές δράσεις (Sarantis et al, 2015). Επίσης, η εξωγενής ενεργοποίηση των A2ARs με τον επιλεκτικό αγωνιστή τους δεν ενίσχυσε περαιτέρω τις δράσεις της συνενεργοποίησης των mglu5r/nmdar σε κανέναν από τους δύο πόλους. Αυτά τα αποτελέσματα προτείνουν ότι η ενδογενής ενεργοποίηση των A2ARs είναι αρκετή για την συνεργιστική δράση των mglu5r και NMDAR που εκφράζεται στον DH και ότι η συγκεκριμένη επιτρεπτική εμπλοκή των A2ARs στον VH είτε είναι απούσα ή πολύ μικρή για να ανιχνευτεί με τον πειραματικό σχεδιασμό αυτής της μελέτης. Παρά την έλλειψη νευροχημικών δεδομένων για τα επίπεδα των A2ARs στον VH, πρότερες παρατηρήσεις έχουν δείξει ότι υπάρχουν λειτουργικοί A2ARs στον VH και συμμετέχουν στην επιληπτογέννεση (Moschovos et al., 2012). Είναι πιθανό ότι η λειτουργική εμπλοκή των A2ARs να διαφέρει κάτω από διαφορετικές καταστάσεις ενεργοποίησης του κυκλώματος του ιπποκάμπου (Nikbakht and Stone, 2001, Sebastiao and Ribeiro, 2014). Ένας προσφάτως σημαντικός ανακαλυφθέν παίκτης στην τροποποίηση της ανασταλτικής και διεγερτικής συναπτικής διαβίβασης είναι ο CB1 υποδοχέας των ενδοκαναβινοειδών (Castillo et al., 2012, Kano, 2014). Οι προσυναπτικοί CB1Rs (Kawamura et al., 2006) εκφράζονται στον ιππόκαμπο (Herkenham et al., 1991), και μπορούν να καταστείλουν την απελευθέρωση του νευροδιαβιβαστή από τις γλουταμινεργικές και GABAεργικές απολήξεις (Schlicker and Kathmann, 2001, Wang, 2003) αφού πρώτα ενεργοποιηθούν από τα μετασυναπτικά Ca ++ -επαγόμενα ενδοκαναβινοειδή που απελευθερώνονται (Kano et al., 2009). Η μείωση στην GABAεργική διαβίβαση μπορεί στη συνέχεια να προκαλέσει ενίσχυση της μετασυναπτικής διεγερσιμότητας (Kano et al., 2009). Σε συνέπεια με αυτό το σχήμα των δράσεων των CB1Rs, βρήκαμε ότι αποκλεισμός αυτών περιέκοψε ένα τμήμα των ενισχυμένων δράσεων της συνενεργοποίησης των mglu5r με τους NMDARs στην διεγερτική διαβίβαση και την μετασυναπτική διεγερσιμότητα και ακύρωσε ολοκληρωτικά τις δράσεις συνενεργοποίησης στην αναστολή. Συνεπώς, οι συνεργιστικές δράσεις των mglu5r-nmdar στην αναστολή μπορούν να εξηγηθούν πλήρως από την ενεργοποίηση των CB1Rs στις GABAεργικές απολήξεις (Schlicker and Kathmann, 2001, Kano et al., 2009), ενώ στην διεγερτική 80
93 διαβίβαση και στην μετασυναπτική διεγερσιμότητα μπορούν μόνο εν μέρει να εξηγηθούν. Πρέπει να σημειωθεί ότι παρόλο που είναι γνωστό ότι η ενεργοποίηση των CB1R συμβαίνει μετά την ενεργοποίηση των NMDARs (Ohno-Shosaku et al., 2007) ή των mglu5rs (Ohno-Shosaku et al., 2002) και ότι κάποιο είδος αλληλεπίδρασης μεταξύ των mglu5r και των NMDARs έχει προταθεί ότι διέπουν την μακροχρόνια καταστολή της διεγερτικής διαβίβασης στην CΑ1 περιοχή (Izumi and Zorumski, 2012), αυτή είναι η πρώτη φορά, απ όσο ξέρουμε, που φαίνεται ότι CB1Rs-εξαρτώμενα βραχύχρονα αποτελέσματα συμβαίνουν μετά την συνενεργοποίηση των mglu5rs-nmdars. Η αύξηση στην μετασυναπτική διεγερσιμότητα μπορεί να είναι μία άμεση δράση της αλληλεπίδρασης των mglu5r-nmdar που οδηγεί σε ενισχυμένη NMDARμεσολαβούμενη εκπόλωση και αυξημένη πυροδότηση των CA1 πυραμιδικών νευρώνων (Mannaioni et al., 1999). Επίσης, έχει δειχθεί ότι η mglur5 εξαρτώμενη απελευθέρωση του γλουταμινικού από τα αστροκύτταρα μπορεί να ενισχύσει την μετασυναπτική διεγερσιμότητα αυξάνοντας τα NMDAR-μεσολαβούμενα ρεύματα στα CA1 πυραμιδικά κύτταρα (Angulo et al., 2004; Fellin et al., 2004; Δείτε ανασκόπηση Panatier and Robitaille, 2015). Αυτή η ρύθμιση της συναπτικης διαβίβασης από τα αστροκύτταρα θα μπορούσε εν δυνάμει να συνεισφέρει στην ενίσχυση της διεγερσιμότητας που επάγεται από την συνενεργοποίηση των mglu5r-nmdar και παραμένει μετά τον αποκλεισμό των CB1Rs. Η απουσία της CB1R-εξαρτώμενης σηματοδότησης είτε τονικής είτε επαγόμενης από την mglu5r-nmdar συνενεργοποίηση, στον VH αποτελεί μία εντυπωσιακή εξειδίκευση στον τρόπο που το τοπικό νευρωνικό κύκλωμα χειρίζεται την πληροφορία κατά μήκος του ιπποκάμπου. Στην εικόνα Β.Ι.1.11 παρουσιάζονται οι πιθανές δράσεις των ποικίλων υποδοχέων στην συναπτική διαβίβαση και νευρωνική διεγερσιμότητα που παρατηρήθηκε στον DH αλλά όχι στον VH (Kouvaros and Papatheodoropoulos, 2016) καθώς επίσης και ο μοριακός μηχανισμός της αλληλεπίδρασης των mglu5rs με τους NMDARs μέσω της φωσφορυλίωσης της NR2B (Tyr1472), (Sarantis et al., 2015). Μία ιδιαίτερη παρατήρηση στη παρούσα μελέτη ήταν η ταχυτάτη μετάβαση από την ισχυρή ενίσχυση στην βίαιη καταστολή της μετασυναπτικής διεγερσιμότητας που προκλήθηκε από την mglu5r- NMDAR συνενεργοποίηση. Αυτή η μετάβαση ήταν πολύ γρήγορη (λιγότερο από ένα λεπτό στα περισσότερα πειράματα) και θα μπορούσε να αποδοθεί στην επαγόμενη από τον αγωνιστή απευαισθητοποίηση των mglur5 (Catania et al., 1991, Gereau and Heinemann, 1998) με μία πιθανή συνεισφορά των NMDARs (Ascher and Nowak, 1988, Mayer et al., 1989, Chizhmakov et al., 1990). Είναι ενδιαφέρον το γεγονός ότι η χρονοεξέλιξη των 81


.")
























94 ταχέων αλλαγών της διεγερσιμότητας που παρατηρήθηκαν στην παρούσα μελέτη ήταν παρόμοιαα με την mglur απευαισθητοποίηση (Guerineau et al., 1997). Εικόνα Β.Ι.1.11: Σχέδιο που συνοψίζει του μηχανισμούς που βρίσκονταιι κάτω από τις συνεργιστικές δράσεις της συνενεργοποίησης των mglur5-nm MDARs. Αυτά τα αποτελέσματα παρατηρήθηκαν στον DH αλλά όχι στον VH. Η έντονη συνενεργοποίησηη των mglur5-nmdars αρχικά οδηγεί σε μία απότομηη και έντονη μείωση της αναστολής (1) που ακολουθείται αμέσως από μία απότομη και ισχυρή αύξηση της διεγερσιμότητας των CA1 πυραμιδικών νευρώνων (2). Μετά από καθυστέρηση μερικών λεπτών συμβαίνει μία ισχυρή μείωση της διεγερτικής συναπτικής διαβίβασης (3). Αυτές οι δράσεις επιτράπηκαν από την τονική ενεργοποίηση των A2ARs και εν μέρει από την ενεργοποίηση των προσυναπτικών CB1Rs οι οποίοι επάγουν μείωση και στην ανασταλτική και στην διεγερτική διαβίβαση. Αυτή η ενεργοποίηση των CB1Rs μπορεί να συμβαίνει από τα ενδοκαναβινοειδή (ecb) των οποίων η σύνθεση ίσως προκαλείτε από την mglur5-nmdars αλληλεπίδραση. Αυτή η αλληλεπίδραση μπορεί άμεσα να συνεισφέρει στην αύξηση της διεγερσιμότητας μέσω ενίσχυσης της τ NMDAR-μεσολαβούμενης εκπόλωσης,, ενώ η αλληλεπίδραση μεταξύ των NMDAR και GABAARss ίσως οδηγεί σε μείωση της αναστολής και ακολούθως σε αυξημένη διέγερση. Ενεργοποίησηη των mglur5 στα αστροκύτταρα μπορεί να επάγει απελευθέρωση του γλουταμινικού που δρώντας στους NMDARs μπορεί να οδηγήσει σε αυξημένη διεγερσιμότητα των CA1 πυραμιδικών κυττάρων (CA1 PC). Σημειωτέον ότι η ενεργοποίηση των mglur5 ενεργοποιεί τις Src κινάσες, οδηγώντας στην NR2B(Tyr1472) φωσφορυλίωση, η οποία σταθεροποιεί τους NMDARs που περιέχουν NR2B υπομονάδα στηη πλασματική μεμβράνη επάγοντας ε έντονη αύξησηη της 82
95 NMDAR εξαρτώμενης διεγερσιμότητας. Είναι ιδιαίτερης σημασίας ότι οι A2ARs αδειοδοτούν την mglur5 επαγόμενη NR2B (Tyr1472) φωσφορυλίωση (Sarantis et al, 2015) Συνέπειες για ραχιαίο-κοιλιακή διαφοροποίηση Οι NMDARs παίζουν θεμελιώδεις ρόλους στην λειτουργικότητα των νευρωνικών δικτύων και στην επεξεργασία της πληροφορίας (Daw et al., 1993, Grienberger et al., 2014) με βασική συμμετοχή τους στην συναπτική πλαστικότητα στον ιππόκαμπο και στις διαδικασίες μνήμης-μάθησης (Morris et al., 1990, Newcomer and Krystal, 2001, Collingridge et al., 2013). Όπως είναι αναμενόμενο, υπάρχουν πολλοί μηχανισμοί που συμμετέχουν στην τροποποίηση των NMDARs όπως οι mglur5 (Doherty et al., 1997, Mannaioni et al., 2001) και οι A2ARs (Tebano et al., 2005, Sebastiao and Ribeiro, 2009, 2014). Ο mglur5 συμμετέχει στα φαινόμενα της NMDAR-εξαρτώμενης συναπτικής πλαστικότητας (Rebola et al., 2010). Επί πρόσθετα, μπλοκάρισμα των A2ARs εμποδίζει το LTP στον ραχιαίο ιππόκαμπο (Fontinha et al., 2009). Η διαφορετική ραχαιοκοιλιακή τροποποίηση των NMDARs από τους mglur5 και A2ARs ίσως έχει σημαντικές επιπτώσεις στα φαινόμενα της συναπτικής πλαστικότητας, όπως η μακροχρόνια ενδυνάμωση. Αναλόγως, η προηγουμένως αναφερθείσα εντυπωσιακά χαμηλότερη ικανότητα του VH έναντι του DH να διατηρήσει την επαγωγή του NMDAR εξαρτώμενου LTP (Papatheodoropoulos and Kostopoulos, 2000a, Maruki et al., 2001, Colgin et al., 2004) ίσως σχετίζεται με την χαμηλότερη λειτουργική κατάσταση αυτών των τροποποιητικών υποδοχέων στον VH σε σχέση με τον DH. Ήταν γνωστό για πολύ καιρό ότι ο κοιλιακός ιππόκαμπος στους αρουραίους και ο αντίστοιχος πρόσθιος ιππόκαμπος στον άνθρωπο είναι το πιο ευπαθές τμήμα της δομής για επιληψία ή επιληπτογεννή δικτυακή δραστηριότητα (Andy, 1962, Brazier, 1970, Gilbert et al., 1985, Bragdon et al., 1986, Bell and Davies, 1998, Moschovos et al., 2012). Η δραστηριότητα όλων των τριών υποδοχέων NMDARs, mglur5 και A2ARs αυξάνουν την διεγερσιμότητα στον ιππόκαμπο και θα μπορούσαν εν δυνάμει να οδηγήσουν σε ανώμαλες κυτταρικές και δικτυακές εκφορτίσεις (Stasheff et al., 1989, Ireland and Abraham, 2002, Etherington and Frenguelli, 2004, Zeraati et al., 2006, Moschovos et al., 2012). Στην παρούσα μελέτη, εμείς ενεργοποιήσαμε έντονα τους NMDARs και τους mglur5 με χρήση εξωγενών αγωνιστών. Αυτή η συνθήκη ίσως μιμείται καταστάσεις υψηλής δικτυακής ενεργότητας. Αρχικά, η υψηλότερη δραστηριότητα των A2ARs και mglur5 στον DH φαίνεται ασυμβίβαστη με την αντίσταση του DH στην υπερενεργοποίηση του δικτύου. Όμως, ο DH εμφανίζει CB1R ενεργότητα μετά την mglur5-nmdar συνενεργοποίηση 83
96 που συνεισφέρει στην απότομη μείωση της διεγερτικής διαβίβασης που ακολουθεί την περίοδο της αυξημένης διεγερσιμότητας. Επιπλέον, οι CB1Rs ελέγχουν τονικά την διεγερτική συναπτική διαβίβαση στον DH καταδεικνύοντας ότι παίζουν σημαντικό ρόλο στην ρύθμιση της δικτυακής διεγερσιμότητας σε αυτό το τμήμα του ιπποκάμπου ακόμη και κάτω από φυσιολογικές συνθήκες. Επομένως οι CB1Rs στους γλουταμινεργικούς νευρώνες εμπλέκονται σε μεγάλο βαθμό στην προστασία του DH από επιληπτικές κρίσεις (Monory et al., 2006). Έτσι, σε καταστάσεις έντονης δικτυακής δραστηριότητας ο DH εμποδίζεται από το να εισέρθει σε καταστάσεις έντονων νευρωνικών εκφορτίσεων. Αντιθέτως, η απουσία της CB1R δραστηριότητας στον VH ίσως συνεισφέρει στην ευαισθησία του τμήματος αυτού σε επιληπτογενείς εκφορτίσεις. Πολλοί άλλοι μηχανισμοί ίσως συνεισφέρουν στην ευαισθησία του VH, συμπεριλαμβανομένων της υψηλής διεγερσιμότητας των πυραμιδικών νευρώνων (Dougherty et al., 2012), της χαμηλότερης συναπτικής GABAεργικής αναστολής (Papatheodoropoulos et al., 2002; Petrides et al., 2007) ή της διαφορετικής σύνθεσης των συναπτικών υποδοχέων (Sotiriou et al., 2005; Pandis et al., 2006). Παρόλα αυτά, όταν απουσιάζουν οι παράγοντες που θα μπορούσαν να αυξήσουν την διεγερσιμότητα του δικτύου, το κύκλωμα του κοιλιακού ιπποκάμπου φαίνεται να δουλεύει φυσιολογικά προτείνοντας ότι η τάση του VH για υπερδιεγερσιμότητα εξισορροπείται από προσαρμοστικές αλλαγές που μετριάζουν αυτήν την ιδιότητα. Πράγματι, πρόσφατες μελέτες προτείνουν ότι η a5gaba A R μεσολαβούμενη διαβίβαση είναι υψηλότερη στον VH απ ότι τον DH (Sotiriou et al., 2005; Pofantis and Papatheodoropoulos, 2014). Επίσης, η αρκετά χαμηλότερη ικανότητα των συνάψεων του κοιλιακού ιπποκάμπου να υφίστανται NMDAR-εξαρτώμενη μακρόχρονη ενδυνάμωση (Papatheodoropoulos and Kostopoulos, 2000a; Maruki et al., 2001; Colgin et al., 2004; Maggio and Segal, 2007) ίσως συντελεί στο να εμποδίζει την εμφάνιση επιληπτογενών δικτυακών δραστηριοτήτων δεδομένου ότι η συναπτική ενδυνάμωση ευνοεί την επιληπτογέννεση (Sutula and Steward, 1987). Έτσι, οι σημαντικές ραχιαιοκοιλιακές διαφορές στις δράσεις των NMDARs, mglur5, A2ARs και CB1Rs που παρατηρήθηκαν σε αυτή τη μελέτη θα μπορούσαν να θεωρηθούν σαν εξειδικεύσεις των τοπικών μικροκυκλωμάτων για να υποστηρίξουν αποτελεσματικά τις ειδικές λειτουργικές απαιτήσεις που αποδίδονται σε κάθε ιπποκάμπειο τμήμα. Η πρόσφατα ενδυναμωμένη ιδέα του λειτουργικού διαχωρισμού κατά μήκος του επιμήκους άξονα του ιπποκάμπου είναι ίσως εμφανώς και συνοπτικά εκφρασμένη από το δίπτυχο της αισθητικότητας και της γνωστικής λειτουργίας. Επομένως, ο DH εμπλέκεται κυρίως στην χωρική μνήμη ενώ ο VH έχει μεγαλύτερη εμπλοκή στο άγχος (Fanselow and Dong, 2010; Bast, 2011; Bannerman et al., 2014). Δεδομένης αυτής της ιδέας, το 84
97 μεγαλύτερο τροποποιητικό δυναμικό των NMDARs στον DH σε σύγκριση με τον VH, έρχεται σε συμφωνία με την ιδέα που εξηγεί ότι οι NMDARs στον ιππόκαμπο εμπλέκονται στην ολοκλήρωση γνωσιακής πληροφορίας πλαισίου και όχι στον συνειρμό μεταξύ πλαισίου και επώδυνου ερεθισμού που βασίζεται στην συγκίνηση (Fanselow and Dong, 2010) Συμπεράσματα Τα παρόντα δεδομένα καταδεικνύουν τις πολύ εμφανείς διαφορές στις δράσεις των αλληλεπιδρώντων υποδοχέων NMDARs, mglur5 και A2ARs, στην λειτουργικότητα του τοπικού νευρωνικού κυκλώματος ανάμεσα στον ραχιαίο και κοιλιακό τμήμα του ιπποκάμπου. Συνεπώς, ενεργοποίηση των NMDARs καταστέλλει την διεγερτική συναπτική διαβίβαση στον VH περισσότερο από τον DH ενώ ενισχύει την μετασυναπτική διεγερσιμότητα μόνο στον DH. Ακόμη πιο εντυπωσιακό ήταν το γεγονός ότι, η θετική τροποποίηση των NMDARs από τους mglur5, εν μέρει μέσω της NR2B (Tyr1472) φωσφορυλίωσης, ο επιτρεπτικός ρόλος των A2ARs και η συμμετοχή των CB1Rs σε αυτή τη συνέργεια ήταν ισχυρές στον DH αλλά απούσες στον VH. Θα μπορούσε να είναι ότι, κάτω από συνθήκες έντονης νευρωνικής δραστηριότητας, η εγγενή υψηλότερη διεγερσιμότητα του VH σε σύγκριση με τον DH έχει ομοιοστατικά αντισταθμιστεί από έναν προσαρμοστικό περιορισμό στην συνεισφορά κάποιων μηχανισμών που ενισχύουν την διεγερσιμότητα όπως οι mglur5 και A2ARs. Εμείς προτείνουμε ότι σε καταστάσεις υψηλής δραστηριότητας του δικτύου, ενώ η τοπική δικτυακή δραστηριότητα στον DH συντονίζεται δυναμικά από τις τροποποιητικές δράσεις των αλληλεπιδρώντων A2ARs και mglur5 στους NMDARs με τον παρεμβατικό ρόλο των CB1Rs, η συμμετοχή των NMDARs στην λειτουργία του VH είναι πιο αόριστη και στερεοτυπικά προσαρμοσμένη. Αναμένεται ότι αυτό θα έχει σημαντικές επιπτώσεις για την επεξεργασία της πληροφορίας που εκτελείται από τον DH και τον VH. 85
98 2. Επαγόμενο LTP με ερεθισμό θ συχνότητας: Διαφορές και ομοιότητες ανάμεσα σε συνάψεις από τον ραχιαίο και κοιλιακό ιππόκαμπο 2.1. Εισαγωγή Όπως αναφέρθηκε προηγουμένως (Κεφάλαιο 8) ένα ιδιαίτερα χαρακτηριστικό στοιχείο της διαφοροποίησης μεταξύ του επιμήκους άξονα του ιπποκάμπου είναι το εμφανώς διαφορετικό μέγεθος του LTP που επάγεται με υψηλής συχνότητας διέγερσης (HFS) (Papatheodoropoulos and Kostopoulos, 2000; Maruki et al., 2001; Colgin et al, 2004; Maggio and Segal, 2009-Differential modulation; Keney and Manahan-Vaughan, 2013; Pofantis and Papatheodoropoulos, 2014; Moschovos and Papatheodoropoulos, 2016). Πιο συγκεκριμένα, όπως αρχικά καταδείχτηκε (Papatheodoropoulos and Kostopoulos, 2000), ένα μονό τρένο 100 παλμών στα 100Hz επάγει LTP πολύ υψηλότερου μεγέθους στον DH απ ότι στον VH στις CA3-CA1 συνάψεις. Το LTP είναι ο πιο αποδεκτός γενικός μηχανισμός που συμμετέχει στην μνήμη και μάθηση (Martin and Morris, 2002) και εξειδικεύσεις στις παραμέτρους που αφορούν το φαινόμενο του LTP σε δεδομένες συνάψεις αναμένεται να χαρακτηρίζουν τον τρόπο με τον οποίο το συγκεκριμένο συναπτικό δίκτυο επεξεργάζεται και διατηρεί την πληροφορία. Μία σημαντική φυσιολογική παράμετρος που χαρακτηρίζει κάποιες δεδομένες συνάψεις κάτω από συγκεκριμένες συνθήκες σε μία καθορισμένη χρονική στιγμή είναι το κατώφλιο επαγωγής του LTP, το οποίο αναπαριστά την ελάχιστη ποσότητα συναπτικής ενεργοποίησης που απαιτείται για την πρόκληση μακράς διάρκειας αλλαγών στην συναπτική αποτελεσματικότητα (Lynch et al., 2013; McNaughton et al., 1978). Μία βασική λειτουργία που προτάθηκε για το κατώφλιο επαγωγής του LTP είναι η σταθεροποίηση της δικτυακής δραστηριότητας (Bliss et al., 2007). Είναι έτσι σημαντικό να καθορίσουμε το εάν υπάρχει διαφορά στο κατώφλιο του LTP ανάμεσα στα δύο ιπποκάμπεια τμήματα. Πειραματικά ο καθορισμός του κατώφλιου επαγωγής του LTP ίσως απαιτεί τη χρήση περισσότερο φυσιολογικών τύπων ερεθισμού, σε σύγκριση με την HFS. Ευρέως χρησιμοποιούμενοι τύποι ερεθισμού που υποτίθεται μιμούνται τα πρότυπα της ιπποκάμπειας δραστηριότητας είναι εκείνα τα πρότυπα που συνδυάζουν τις ριπές συμπλόκων δυναμικών (complex spike bursts) των πυραμιδικών κύτταρων (Rank, 1973) με τη συχνότητα του ρυθμού θ (5 Hz) που τροποποιεί την δραστηριότητα αυτών των κύτταρων τη στιγμή που ένας αρουραίος εξερευνεί και μαθαίνει καινούρια περιβάλλοντα (Otto, Eichenbaum, Wiener, & Wible, 1991). Αυτό το πρότυπο ερεθισμού ονομάζεται ερεθισμός υπό μορφή ριπής θήτα ρυθμού (theta-burst stimulation-tbs), (Larson et al., 86
99 1986; Larson, 2015). Γενικότερα, το TBS αποτελείται από τρένα πολλών σύντομων ριπών των τεσσάρων ή πέντε παλμών στα 100 Hz που επιδίδονται στην θ συχνότητα των 5 Hz (π.χ. με διάστημα μεταξύ των ριπών στα 200 ms). Πράγματι, το TBS χαμηλώνει το κατώφλιο της ενεργοποίησης του νευρωνικού κυκλώματος για επαγωγή LTP (Larson et al., 1986; Rose and Dunwiddie, 1986; Larson and Munkacsy, 2015). Πρόσφατα έχει προταθεί ότι η συναπτική ενίσχυση που παράγεται από ένα ζεύγος παλμών με διαπαλμικό διάστημα 200 ms (που αντιστοιχεί στη συχνότητα θ ) είναι πολύ μεγαλύτερη στις ραχιαίες απ ότι στις κοιλιακές CA1 συνάψεις (Papatheodoropoulos, 2015). Έτσι, φαίνεται ότι αυτό το είδος της βραχύχρονης πλαστικότητας είναι περισσότερο ευαίσθητη στην νευρωνική δραστηριότητα με θ συχνότητα στις DH σε σύγκριση με τις VH συνάψεις. Επίσης, στο επίπεδο του μεγαλύτερου ιπποκάμπειου νευρωνικού δικτύου καθώς και σε ολόκληρο το ζώο, ο DH σε σύγκριση με τον VH παρουσιάζει υψηλότερο πλάτος και συνάφεια με τον θ ρυθμό (Patel et al., 2012) Σκοπός Σε αυτή τη μελέτη, χρησιμοποιήσαμε μία μεγάλη γκάμα από πρωτόκολλα διέγερσης βασισμένων στην συχνότητα θ για να καθορίσουμε την ευαισθησία και την έκταση των DH και VH συνάψεων να υποστούν LTP. Βρήκαμε ότι ο DH σε σχέση με τον VH παρουσιάζει χαμηλότερα κατώφλια για βραχύχρονοo και μακρόχρονο NMDARεξαρτώμενο LTP καθώς επίσης NMDAR-ανεξάρτητο LTP με την σημαντική συμμετοχή των GABA B Rs. Άλλωστε, έπειτα από σχετικά έντονη ενεργοποίηση των προσυναπτικών ινών, οι συνάψεις και των δύο πόλων του ιπποκάμπου επιστρατεύουν τα L-VDCCs και τους mglur5 με παρόμοιο τρόπο. Λεπτομερής ανάλυση των μετασυναπτικών αποκρίσεων παρέχει στοιχεία για τους μηχανισμούς που συμμετέχουν στις ραχιαιοκοιλιακές διαφορές στην επαγωγή του LTP Υλικά και Μέθοδοι Ζώα και προετοιμασία τομών Στη παρούσα μελέτη χρησιμοποιήθηκαν 104 αρσενικοί Wistar αρουραίοι δύο έως τεσσάρων μηνών. Όλη η πειραματική μεταχείριση και διαδικασίες περιγράφονται παραπάνω στις γενικές μεθόδους (υποκεφάλαιο Α.ΙΙΙ.1&2). 87
100 Καταγραφές, Επεξεργασία δεδομένων και Ανάλυση Δυναμικά πεδίου που αποτελούνται από το προσυναπτικό fiber volley (Fv) και το διεγερτικό μετασυναπτικό δυναμικό (EPSP) κατεγράφησαν από το δενδριτικο στρώμα της ακτινωτής στοιβάδας του CA1 μετά τον ηλεκτρικό ερεθισμό των ινών του Schaffer όπως περιγράφεται στις γενικές μεθόδους (Υποκεφάλαιο Α.ΙΙΙ.4&5&6). Σε μερικά πειράματα καταγράφηκε επίσης το πληθυσμιακό δυναμικό (PS) από την πυραμιδική στοιβάδα ταυτόχρονα με τις δενδριτικές καταγραφές. Οι μονοί ηλεκτρικοί παλμοί επιδίδονταν σε συχνότητα 0.033Hz. Η ένταση ερεθισμού ρυθμίστηκε να προκαλεί το ημιμέγιστο EPSP. Μόνο οι τομές που παρουσίαζαν σταθερότητα στο FV και το EPSP για τουλάχιστον δέκα λεπτά επιλέχθησαν για περαιτέρω πειραματισμό. Καμπύλες εισόδου/εξόδου ανάμεσα στην ένταση ερεθισμού και των προκλητών απαντήσεων πραγματοποιήθηκαν και κάθε τομή. Ποσοτικοποιήσαμε τo FV, EPSP, το PS καθώς επίσης και τις αποκρίσεις των ριπών που προκληθήκαν με το TBS υπολογίζοντας το εμβαδόν (βλέπε γενικές μεθόδους). Δεδομένου ότι το μέγεθος του FV καταδεικνύει τον αριθμό των ενεργοποιούμενων προσαγωγών ινών (Andersen et al., 1978), χρησιμοποιήσαμε τον λόγο EPSP/Fv για να ποσοτικοποιήσουμε την συναπτική αποτελεσματικότητα. Για επαγωγή μακρόχρονης ενδυνάμωσης (LTP) χρησιμοποιήσαμε ερεθισμό υπό μορφή ριπής θήτα ρυθμού (TBS) αποτελούμενο από ποικίλο αριθμό ριπών και τρένων από ριπές παλμών. Πιο συγκεκριμένα, χρησιμοποιήσαμε τα ακόλουθα πειραματικά πρωτόκολλα: α) τον ερεθισμό με ριπή προπαίδευσης (PB) που αποτελείται από δύο παρόμοιες ριπές των τεσσάρων παλμών στα 100 Hz με διάστημα μεταξύ τους τα 200 ms, B) Σύντομο τρένο από πέντε ριπές που επιδίδονταν στα 5 Hz, το μισό-tbs (Half TBS, H-TBS), Γ) ένα τρένο από 10 ριπές στα 5 Hz, το ονομαζόμενο 1- TBS, δ) δύο, τρία και οκτώ τρένα των δέκα ριπών στα 5 Hz, με διάστημα μεταξύ των τρένων τα 30 sec. Μία ριπή από 4 παλμούς στα 100 Hz επιδόθηκε σε μία ομάδα τομών Φαρμακολογικοί παράγοντες Χρησιμοποιήθηκαν τα ακόλουθα φάρμακα: α) CPP 10μΜ, β) CGP μm, γ) MTEP, 200μM και δ) Νιμοδιπίνη 20μΜ. Τα φάρμακα χορηγήθηκαν για 20 (CPP και CGP 52432, MTEP) και 50 λεπτά (Νιμοδιπίνη) πριν την επίδοση του TBS και ξεπλύθηκαν 5 λεπτά μετά την επίδοση του TBS. Αναλυτική περιγραφή και τρόπος διάλυσης των φαρμάκων αναλύονται στις γενικές μεθόδους, υποκεφάλαιο Α.ΙΙΙ.3. 88

. 2.4. Αποτελέσματα 2.4.1.")
 προκλήθηκαν και μετρήθηκαν με ποικίλη ένταση ερεθισμού")
.")



101 Στατιστική Η ανίχνευση των στατιστικά σημαντικών διαφορών έγινε με την χρήση των τομών όπως περιγράφεται στο στις γενικές μεθόδους (Υποκεφάλαιο Α.ΙΙΙ.7) Αποτελέσματα Βασικές μετρήσεις σε DH και VH Τα δυναμικά πεδίου (Fv, EPSPP και PS) προκλήθηκαν και μετρήθηκαν με ποικίλη ένταση ερεθισμού και στις DH (n=20/20) και στις VH (n=20/20) τομές (Εικόνα Β.Ι. 2.1B-D). Αυτές οι DH και VH τομές αποκτήθηκαν από τα ίδια ζώα (n=20) και συνιστούσαν ν ένα δείγμα του συνολικού πληθυσμού των τομών που χρησιμοποιήθηκαν σε αυτή τη μελέτη. Σε συμφωνία με προηγούμενες μελέτες (Kouvaros and Papatheodoropoulos, 2016) βρήκαμε ότι οι σχέσεις μεταξύ του ρεύματος ερεθισμού και του EPSPP και μεταξύύ του ρεύματος και του Fv ήταν παρόμοιες στα δύο ιπποκάμπεια τμήματα (Εικόνα Β.Ι.2.1.Α). Επί πρόσθετα, παρομοια ήταν και η σχέση μεταξύ Fv καιι EPSP (Pofantis and Papatheodoropoulos, 2014). 89
102 Εικόνα Β.Ι.2.1: Χρησιμοποιούμενη μέθοδος τομής ιπποκάμπου και βασικές αποκρίσεις στον DH και VH. Α) Σχηματικό διάγραμμα της περιφέρειας του ιπποκάμπου που καταδεικνύει τον προσανατολισμό που χρησιμοποιήθηκε για το κόψιμο των τομών (λεπτές γραμμές) και την έκταση του DH και VH πόλου που κόπηκε (γραμμές με βέλη). Ο ίδιος προσανατολισμός, κάθετος στον μακρύ άξονα (διακεκομμένη γραμμή) χρησιμοποιήθηκε για την τομή των δύο ιπποκάμπειων πόλων σε τομές πάχους μm. Η περιστρεφόμενη βάση της McIlwain μικροτόμου, που βρισκόταν ο ιστός, προσαρμοζόταν για να κόψουμε εγκάρσιες τομές και από τους δύο πόλους. Β-D) Συγκεντρωτικές καμπύλες εισόδου/εξόδου ανάμεσα στην ένταση του ρεύματος ερεθισμού και του EPSP (B) ή του Fv (C) καθώς επίσης και μεταξύ του Fv και του EPSP (D) δείχνονται για τον DH (σκούρα σύμβολα) και VH (ανοιχτά σύμβολα). Τα δεδομένα συλλέχτηκαν από 20 ραχιαίες και 20 κοιλιακές τομές που αποκτήθηκαν από τα ίδια 20 ζώα Το TBS επάγει μικρής διάρκειας (S-LTP) και μακράς διάρκειας LTP (L-LTP) παρόμοιου πλάτους στον DH και VH, αλλά ο DH έχει χαμηλότερα κατώφλια επαγωγής για τα δύο LTPs Η αποτελεσματικότητα του ερεθισμού θ συχνότητας στην επαγωγή LTP μπορεί να δειχθεί από την εμφανή ικανότητα του ερεθισμού με ριπή προπαίδευσης (primed burst-pb), που περιέχει ένα ερέθισμα προπαίδευσης που προηγείται μίας τεσσάρων παλμών ριπής κατά ms, για να επάγει LTP στις CA1 συνάψεις (Larson and Lynch, 1986; Rose and Dunwiddie, 1986; Diamond et al., 1988; Corradetti et al., 1992-PS). Το ερέθισμα προπαίδευσης μπορεί να είναι είτε ένας μονός παλμός είτε μία ριπή τεσσάρων παλμών (Larson and Lynch, 1986). Στην πραγματικότητα, το PB είναι το ηπιότερο ερέθισμα που μπορεί να επάγει LTP στις CA1 συνάψεις στον ραχιαίο ιππόκαμπο (Larson and Munkacsy, 2015). Συνεπώς, χρησιμοποιήσαμε ένα PB για να καθορίσουμε το εάν μπορεί να επάγει μόνιμες αλλαγές στην διεγερτική συναπτική διαβίβαση και στις συνάψεις του κοιλιακού ιπποκάμπου. Παρατηρήσαμε λοιπόν ότι το PB παρήγαγε με συνέπεια LTP στον DH (Εικόνα B.I.2.2). Πιο συγκεκριμένα, το μέγεθος του LTP στα 60 λεπτά μετά το PB ήταν 19.44±3.86% (n=15/6, Wilcoxon δοκιμασία, p<0.05). Αυτή η παρατήρηση επιβεβαιώνεται και από προηγούμενες μελέτες που έχουν αναφέρει PB-επαγόμενο συναπτικό LTP παρόμοιου μεγέθους με τη χρήση μίας ριπής ως ερέθισμα προπαίδευσης (Davies et al., 1991; Grover et al 2009; Larson et al, 1986). Αντιθέτως, το PB δεν παρήγαγε αλλαγή της συναπτικής αποτελεσματικότητας στις κοιλιακές τομές (0.17±3.82, n=5/5; Σύγκριση μεταξύ DH και VH, Mann Whitney U δοκιμασία, p<0.005), (Εικόνα B.I.2.2). Επί πρόσθετα, παρατηρήσαμε ότι στον DH το LTP διατηρήθηκε για τουλάχιστον δύο ώρες μετά την επαγωγή του (21.64±4.25% στα 120 λεπτά μετά το PB, n=15/6, Wilcoxon δοκιμασία, p<0.005). Αυτή είναι η πρώτη φορά που δείχνεται ότι το PB πρωτόκολλο 90


 ) ούτε ο VH (2.21±3.")
 παρουσίασανν καμία σημαντική αλλαγή στο fepsp/fv στα 60 λεπτά μετά")


,")




 πριν και μετά την επίδοση του PB.")

103 μπορεί να επάγει συναπτική ενδυνάμωση που διαρκεί περισσότερο από μία ώρα. Η απροσδόκητη ικανότητα του DH να διατηρεί LTP τουλάχιστον δύο ωρώνν μας ώθησε να εξετάσουμε το εάν μία απλή ριπή τεσσάρων παλμών θα μπορούσε να επάγει χρόνιες αλλαγές στην συναπτική διαβίβαση. Όπως φαίνεται στο ένθετο γράφημα της εικόνας B.I.2.2 ούτε ο DH (9.8±7.6%, n=5/4, Wilcoxon δοκιμασία, p>0.05) ) ούτε ο VH (2.21±3.56%, n=5/3, Wilcoxon δοκιμασία,, p>0.05) παρουσίασανν καμία σημαντική αλλαγή στο fepsp/fv στα 60 λεπτά μετά την απλήή ριπή. Αυτό είναι σε συμφωνία σ μεε προηγούμενες αναφορές (Larson and Lynch, 1986; Rosee and Dunwiddie, 1986) και το γεγονός ότιι μία απλή ριπή ενεργοποιεί ένα πρόσω ανασταλτικό μετασυναπτικό δυναμικό δ τοο οποίο μειώνει το fepspp που προκαλείται από εκείνη τη ριπή (Davis et al., 1990; Nathann and Lambert, 1991; Larson and Munkacsy, 2015), αποκλείοντας με αυτόν τονν τρόπο τηνν επαγωγή LTP. Από αυτά τα αποτελέσματα συμπεράναμε ότι στον DH, το PB αναπαριστάά τον κατώφλιο ερεθισμό που απαιτείται για την επαγωγή LTP που διαρκείί τουλάχιστον δύο ώρες. Αντιθέτως, το PB δεν είναι επαρκές να επάγει LTP στον VH. Εικόνα B.I.2.2: Ο ερεθισμός με ριπή προπαίδευσης, που περιέχει δύο ριπές των τεσσάρων παλμώνν στα 100 Hz με διάστημα μεταξύ των ριπών στα 200 ms, είναι ο κατώφλιος ερεθισμός επαγωγής LTP στον DH αλλά όχι στον VH. Η χρονοεξέλιξη των αλλαγών στο fepsp/fv δείχνεται για τον DH (15/6) και τον VH (5/5) πριν και μετά την επίδοση του PB. Στον DH, το PB προκάλεσε σημαντική ενδυνάμωση της συναπτικής αποτελεσματικότητας που διατηρήθηκε σταθερή για τουλάχιστον δύο ώρες. Αντίθετα Α, το PB δεν προκάλεσε αλλαγή στον VH. Παραδείγματα καταγραφών που αποκτήθηκαν πριν (1), στα 60 λεπτά (2) ή τα 120 λεπτά (3) μετά το PB δείχνονται πάνω δεξιά και κάτω δεξιά του γραφήματος για τον DH και τον VH αντίστοιχα. Καταγραφές που αποκτήθηκαν πριν την τ επίδοση του PB είναι με γκρι. Γραμμές βαθμονόμησης: 1mV, 5ms. 91
104 Το ένθετο (πάνω αριστερό γράφημα) δείχνει ότι μία μόνο ριπή που αποτελείται από τέσσερις παλμούς των 100Hz στην ένταση του ερεθισμού δοκιμής δεν προκάλεσε LTP ούτε στον DH ούτε στον VH (πλήρη και ανοιχτά σύμβολα αντίστοιχα). Στη συνέχεια, ερευνήσαμε την αποτελεσματικότητα πρωτοκόλλων ερεθισμού που περιέχουν αυξανόμενο αριθμό ριπών που επιδίδονται με συχνότητα 5 Hz για την επαγωγή LTP στους δύο πόλους. Στην εικόνα Β.Ι.2.3Α & Β, φαίνεται ότι ερεθισμός που περιέχει πέντε ή δέκα διαδοχικά ριπές (half-tbs, H-TBS και 1-TBS αντίστοιχα) προκάλεσε LTP που μετρημένο μία ώρα μετά τον ερεθισμό είχε παρόμοιο μέγεθος και στους δύο πόλους. Πιο συγκεκριμένα, η ενδυνάμωση του fepsp/fv μετρημένο στα 60 λεπτά μετά την επίδοση του H-TBS ήταν 31.1±6.0% στον DH (n=9/6, Wilcoxon δοκιμασία, p<0.01) και 19.1±3.7% στον VH (9/6, Wicoxon δοκιμασία, p<0.01), ενώ μετά την επίδοση 1-TBS ήταν 30.7±2.6% στον DH (n=7/5, Wilcoxon δοκιμασία, p<0.05) και 16.0±4.2% στον VH (10/5, Wicoxon δοκιμασία, p<0.01). Επίσης, η ενδυνάμωση που προκλήθηκε και από τα δύο αυτά πρωτόκολλα διατηρήθηκε για τουλάχιστον 2 ώρες στον DH. Έτσι, η ενδυνάμωση του fepsp/fv στον DH μετρημένο στα 120 λεπτά μετά τον ερεθισμό ήταν 32.7±6.0% (Wilcoxon δοκιμασία, p<0.01) και 41.5±5.2% (Wilcoxon δοκιμασία, p<0.05) μετά από επίδοση H-TBS και 1-TBS αντίστοιχα. Αντιθέτως, η ενδυνάμωση που προκλήθηκε στον VH μειώθηκε στα επίπεδα ελέγχου μεταξύ της πρώτης και δεύτερης ώρας μετά τον ερεθισμό (9.7±6.1% και 6.7±4.4% αλλαγή μετρημένη στα 120 λεπτά μετά το H-TBS και 1- ΤΒS αντίστοιχα). Κατά συνέπεια, υπήρξε σημαντική διαφορά στην μεταβολή του λόγου fepsp/fv στα 120 λεπτά ανάμεσα στον DH και VH (Mann Whitney U δοκιμασία, p<0.005 για H-TBS και p<0.001 για 1-TBS). Στη βάση της διαφορετικής ικανότητας των δύο ιπποκάμπειων τμημάτων να διατηρήσουν το LTP για περισσότερο από μία ώρα ονομάσαμε την ενδυνάμωση που διήρκησε για μία ώρα ως σύντομης διάρκειας LTP (short-lasting LTP, S-LTP) και για ενδυνάμωση που διήρκησε τουλάχιστον δύο ώρες ως μακράς διάρκειας LTP (long lasting LTP, L-LTP). Έτσι, στον VH οι πέντε ριπές αναπαριστούν τον κατώφλιο ερεθισμό για την επαγωγή του S-LTP αλλά αυτός ο ερεθισμός ήταν υποκατώφλιος για επαγωγή L-LTP. Έπειτα, εξετάσαμε τις δράσεις των δύο και τριών τρένων των δέκα ριπών (2-TBS και 3- TBS αντίστοιχα). Όπως φαίνεται στην εικόνα Β.Ι.2.3C & D, τα 2-TBS και 3-TBS προκάλεσαν S-LTP και L-LTP με παρόμοιο τρόπο στους δύο πόλους. Και ότι τα 2-TBS ήταν ο κατώφλιος ερεθισμός για επαγωγή L-LTP στον VH. Ενδιαφέρον είναι το γεγονός ότι, το L-LTP διήρκησε για τουλάχιστον πέντε ώρες μετά τον ερεθισμό και στους δύο 92







, 21.9±3.")
 και 25.")
 στον DH")





. Η")


















105 πόλους. Πιο ειδικά, η ενδυνάμωση του fepsp/fv που προκλήθηκε από τα 2-TBS και μετρημένο στη μία, δύο και στιςς πέντε ώρες μετά τον ερεθισμό ήταν ή 23.4±2.9% (p<0.005), 21.9±3.7% (p<0.005) και 25.4± ±6.0% (p<0.01) στον DH (n=13/13, Wilcoxon δοκιμασία), και 33.6± ±4.1%, 32.5± ±4.4% και 28.0±5.9% στον VH (n=12/12, Wilcoxon δοκιμασία σε κάθε χρονικό σημείο, p<0.005). Η ενδυνάμωση που προκλήθηκε από τα τ 3-TBS στη μία, δύο και πέντε ώρες μετά τον ερεθισμό ήταν 66.2±9.5%, 60.7±15.2% και κ 55.2±16.4% στον DH (n=13/9, Wilcoxon δοκιμασία για κάθε χρονικό σημείο, p<0.005), p και 59.1±6.2%, 53.9±7.2% και 40.0± ±7.9% στον VH (n=7/7, Wilcoxon δοκιμασίαα για κάθε χρονικό σημείο, p<0.05). Εικόνα Β.Ι.2.3: Ο VH συγκρινόμενος με τον DH έχει υψηλότερο κατώφλιο επαγωγής για το L-LTP και παρουσιάζει διαφορετικά κατώφλιαα για το S-LTPP και L-LTP (Π.χ. ενδυνάμωση που διατηρήθηκε μέχρι 60 λεπτά και 120 λεπτά αντίστοιχα). Δείχνονται οι χρονοεξελίξεις των τ αλλαγώνν στις συναπτικές 93
106 αποτελεσματικότητες που προκλήθηκαν από το αυξανόμενο μήκος των TBS. Δείχνονται για τον DH και τον VH πόλο, ΤΒS αποτελούμενο από είτε 5 τρένα (Η-TBS, Α ), 10 τρένα (1-ΤΒS, B ) ή 3 τρένα των 10 ριπών (3-TBS, D ). Οι αριθμοί στα δεξιά κάθε γραφήματος καταδεικνύουν τον αριθμό των τομών/ζώων που χρησιμοποιήθηκαν. Δείχνονται επίσης αντιπροσωπευτικά παραδείγματα καταγραφών που προέρχονται από τα χρονικά σημεία που επισημαίνονται, στην κορυφή κάθε γραφήματος καθώς και ίχνη που αποκτήθηκαν πριν την επίδοση του ερεθισμού επαγωγής (σε γκρι χρώμα). Γραμμές βαθμονόμησης: 0.5mV, 5ms σε παραδείγματα για DH στο Α και στο D. Σε όλες τις άλλες περιπτώσεις 1mV, 5ms. Το H-TBS και 1- TBS προκάλεσαν S-LTP και L-LTP στον DH αλλά μόνο S-LTP στον VH. Στον VH το κατώφλιο επαγωγής για το L-LTP ήταν τα 2-TBS. Σημειωτέον ότι το L-LTP και στους δύο πόλους μπορούσε να διαρκέσει τουλάχιστον 5 ώρες μετά τους ερεθισμούς. Επίσης ο VH παρουσίασε διαφορετικά κατώφλια επαγωγής για το S-LTP και το L-LTP (H-TBS και 2-TBS αντίστοιχα). Επιπλέον, και το S-LTP και το L-LTP από τη στιγμή της επαγωγής τους είχαν παρόμοιο πλάτος στους δύο πόλους. Συνοψίζοντας, ο VH σε σύγκριση με τον DH εμφάνισε υψηλότερα κατώφλια ερεθισμών για επαγωγή και S-LTP και L-LTP και τα κατώφλια στον VH ήταν διαφορετικά για S-STP και L-LTP. Πιο σημαντικά, όμως, το μέγεθος των S-LTP και L-LTP από τη στιγμή που προκαλούταν η ενδυνάμωση ήταν παρόμοιο στους δύο πόλους, καταδεικνύοντας ότι το TBS είναι πιο αποτελεσματικό για επαγωγή LTP στον VH σε σύγκριση με το HFS πρωτόκολλο. Με τα παραπάνω αποτελέσματα, δείχνουμε ότι παρόλο που το μέγεθος του LTP που επάγεται με HFS είναι πολύ διαφορετικό μεταξύ DH και VH (Papatheodoropoulos and Kostopoulos, 2000a; Moruki et al., 2001; Colgin et al., 2004), σχετικά περισσότερο φυσιολογικός ερεθισμός μπορεί να επάγει LTP παρόμοιου πλάτους στους δύο πόλους Οι GABABRs ελέγχουν τα διαφορετικά κατώφλια επαγωγής για το S-LTP και L- LTP μεταξύ των 2 πόλων Έχει διαπιστωθεί ότι η αποτελεσματικότητα του ερεθισμού θ συχνότητας στην επαγωγή LTP προκύπτει από την καταστολή της πρόσω αναστολής (Pacelli et al., 1991; Davies et al., 1990; Nathan and Lambert, 1991). Πιο συγκεκριμένα, ενεργοποίηση των πρόσω GABAεργικών εισόδων οδηγεί στην ενεργοποίηση των προσυναπτικών GABA B Rs οι οποίοι καταστέλλουν την ακόλουθη απελευθέρωση GABA, μειώνοντας έτσι την μετασυναπτική αναστολή (Davies et al., 1991; Nathan and Lambert, 1991; Mott and Lewis, 1991, 1992). Αυτό το GABA B R-εξαρτώμενο δυσανασταλτικό αποτέλεσμα μεγιστοποιείται περίπου στα 5 Hz επιτρέποντας τη συγκέντρωση μετασυναπτικής εκπόλωσης αρκετής για να ενεργοποιηθούν οι NMDARs και να προκληθεί LTP (Larson and Lynch, 1988; Larson and Lynch, 1986; Larson et al., 1986; Pacelli et al., 1989; Davis et al., 1991). Συνεπώς, 94













.")




. Εικόνα Β.Ι.")






107 εμείς εξετάσαμε τις δράσεις του ανταγωνιστή των GABA B Rs CGP στην ικανότητα των TBS πρωτοκόλλων να επάγουν χρόνιες αλλαγές στον DH και VH. Παρόμοιαα με προηγούμενες παρατηρήσεις (Davis et al., 1991), αποκλεισμός των GABAA B Rs στον DH εμπόδισε την επαγωγή LTP με το PB (fepsp/fv άλλαξε μόνο κατά 1.6±2.2% % στα 60 λεπτά μετά-pb, n=6/3, Wilcoxon δοκιμασία, p>0.05; Εικόνα Β.Ι.2.4A). Επίσης, βρήκαμε ότι στον VH αποκλεισμός των GABAA B Rs εμπόδισε την επαγωγή του LTP χρησιμοποιώντας κατώφλιο ερεθισμό (Εικόνα Β.Ι.2.4C). Εικόνα Β.Ι.2.4: Οι GABABRs ελέγχουν τη διαφορά στο κατώφλιο επαγωγής του S-LTP μεταξύ DH και VH. Δείχνεται η δράση του ανταγωνιστή των GABA B Rs CGP (CGP, 101 μm) στις ενισχυτικές δράσεις του PB και των πέντε ριπών (H-TBS) στον DH ή τον VH. Αντιπροσωπευτικά παραδείγματαα καταγραφώνν που 95
108 αποκτήθηκαν τις στιγμές εκείνες που τονίζονται στα γραφήματα, δείχνονται στη κορυφή κάθε γραφήματος με ίχνη που αποκτήθηκαν πριν την επαγωγή (σε γκρι χρώμα). Οι αριθμοί εντός των παρενθέσεων καταδεικνύουν τους αριθμούς των τομών/ζώων που χρησιμοποιήθηκαν. Γραμμές βαθμονόμησης: 1mV, 5ms σε όλα τα γραφήματα. Στον DH (A-B) μπλοκάρισμα των GABA B Rs με CGP απέκλεισε το S-LTP που προκλήθηκε από τον κατώφλιο ερεθισμό (PB, Α ) και μείωσε σημαντικά το μέγεθος του S-LTP και L-LTP που προκλήθηκε με το υπερκατώφλιο H-TBS ( Β ). Στον VH, το CGP μείωσε σημαντικά το μέγεθος του S-LTP και εμπόδισε την επαγωγή του L-LTP με το Η-TBS ( C ). Η χρονοεξέλιξη ( D ) της μεταβολής του λόγου fepsp/fv στον DH μετά από αποκλεισμό των GABA B Rs δείχνεται ταυτόχρονα με την χρονοεξέλιξη που αποκτήθηκε από τον VH σε κανονικές συνθήκες ώστε να αποτυπώσουμε την αξιοσημείωτη ομοιότητα μεταξύ τους. Πιο συγκεκριμένα, η μεταβολή του fepsp/fv μετά την επίδοση του H-TBS στον VH παρουσία 10μΜ CGP ήταν 3.5±6.5% στα 60 λεπτά και -2.3±4.4% στα 120 λεπτά μετά τον ερεθισμό (n= 8/5, Wilcoxon δοκιμασία, p>0.05). Έπειτα αναρωτηθήκαμε το εάν οι GABA B Rs συνεχίζουν να εμπλέκονται στα υπερκατώφλια TBS πρωτόκολλα στον DH και VH. Παρατηρήσαμε ότι η εφαρμογή του H-TBS στον DH, παρουσία 10μΜ CGP 52432, προκάλεσε S-LTP (13.1±2.6% στα 60 λεπτά, n=7/3, Wilcoxon δοκιμασία, p<0.05) αλλά όχι L-LTP (9.9±3.8% στα 120 λεπτά, Wilcoxon δοκιμασία, n=7/3, p>0.05), (Εικόνα Β.Ι.2.4B). Επίσης το μέγεθος του S-LTP ήταν σημαντικά μικρότερο σε σύγκριση με αυτό που παρατηρήθηκε σε συνθήκες ελέγχου (Mann-Whitney U δοκιμασία, p<0.05). Είναι ενδιαφέρον το γεγονός ότι η χρονοεξέλιξη της μεταβολής του λόγου fepsp/fv που προκλήθηκε με H-TBS στον DH μετά από αποκλεισμό των GABA B Rs ήταν αρκετά όμοιο με την χρονοεξέλιξη που προκλήθηκε από το ίδιο πρωτόκολλο ερεθισμού στον VH σε ελεγχόμενες συνθήκες (Εικόνα Β.Ι.2.4D). Αυτά τα αποτελέσματα προτείνουν ότι οι GABA B Rs συνεισφέρουν σημαντικά στη διαφοροποίηση των κατωφλίων επαγωγής LTP ανάμεσα στον DH και VH κάτω από συνθήκες ενεργοποίησης με θ συχνότητα ερεθισμό. Επίσης, παρατηρήσαμε ότι αποκλεισμός των GABA B Rs δεν επηρέασε ούτε την επαγωγή ούτε το μέγεθος του S-LTP και L-LTP που προκλήθηκε στον DH με τα 2-TBS. Έτσι, παρουσία 10μΜ CGP το fepsp/fv ενδυναμώθηκε κατά 22.1±2.5% στα 60 λεπτά και κατά 24.4±3.6% στα 120 λεπτά μετά το 2-TBS (n=8/4, Wilcoxon δοκιμασία σε κάθε χρονικό σημείο, p<0.05). Αντιθέτως, παρατηρήσαμε ότι στον VH αποκλεισμός των GABA B Rs μείωσε σημαντικά το μέγεθος και του S-LTP και του L-LTP που προκλήθηκαν με 2-TBS και 3-TBS (Εικόνα Β.Ι.2.5Β&C). Πιο συγκεκριμένα, κάτω από αποκλεισμό των GABA B Rs το 2-TBS πρωτόκολλο προκάλεσε ενδυνάμωση του fepsp/fv στον VH κατά 17.4±4.2% στα 60 λεπτά (Wilcoxon δοκιμασία, p<0.05) και κατά 12.2±3.7% στα 120 λεπτά (Wilcoxon δοκιμασία, p<0.05), (Mann-Whitney U δοκιμασία ανάμεσα σε συνθήκες 96




















109 ελέγχου και μπλοκαρίσματος των GABA B BRs, p<0.05 στα 60 λεπτά λ και p<0.01 στα 120 λεπτά). Επίσης, η ενδυνάμωση που προκλήθηκε με το 3-TBS στις κοιλιακές τομές που επωάστηκαν με CGP ήταν 33.8±7.2% στα 60 λεπτά και 21.4±9.4% στα 120 λεπτά (n=6/3, Wilcoxon δοκιμασία για καθένα από τα δύο χρονικά σημεία, p<0..05). Και στους δύο χρόνους η ενδυνάμωση μετά από αποκλεισμό των GABA B Rs ήταν σημαντικά χαμηλότερη σε σύγκριση με τις συνθήκες ελέγχου (Mann-Whitney U δοκιμασία, p<0.05). Εικόνα Β.Ι.2.5: Οι GABABRs δεν συμμετέχουν στο S-LTP και το L-LTPP που επάγονται από το 2-TBS πρωτόκολλο ερεθισμού στον DH αλλά συνεισφέρει και στους δύο τύπους LTP που επάγονται από τα 2- TBS και 3-TBS στον VH. Αποκλεισμός των GABAA B Rs από το CGP δεν επηρεάζει την αποτελεσματικότητα του 2-TBS στην επαγωγή είτε S-LTP είτε ε L-LTP στις ραχιαίες τομές (A). Όμως, αποκλεισμός των GABAA B Rs 97
110 μειώνει σημαντικά το μέγεθος του S-LTP και L-LTP που επάγεται είτε από τα 2-TBS είτε από τα 3-TBS στον VH (B και C αντίστοιχα). Οι αριθμοί εντός των παρενθέσεων καταδεικνύουν τους αριθμούς των τομών/ζώων που χρησιμοποιήθηκαν. Παραδείγματα καταγραφών πάνω δεξιά σε κάθε γράφημα αποτυπώνουν την δράση του φαρμάκου στο LTP. Οι καταγραφές που αποκτήθηκαν πριν την επαγωγή του LTP δείχνονται σε γκρι χρώμα. Γραμμές βαθμονόμησης: 1mV, 5ms σε όλα τα γραφήματα. Παρατηρήστε την αύξηση του fepsp/fv κατά την εφαρμογή του CGP (A). Αυτά τα αποτελέσματα κατέδειξαν ότι οι GABA B Rs ελέγχουν την επαγωγή του LTP σε κατώφλια TBS πρωτόκολλα και συνεισφέρουν στο μέγεθος του LTP που επάγεται από τα υπερκατώφλια TBS και στους δύο πόλους. Επίσης, η δράση των GABA B Rs μειώνεται σημαντικά μετά από ισχυρή προσυναπτική ενεργοποίηση με συχνότητα θ. Εκτός από τις δράσεις του CGP στο LTP, βρήκαμε ότι αποκλεισμός των GABA B Rs προκάλεσε μία σημαντική αύξηση στην βασική συναπτική διαβίβαση στις ραχιαίες αλλά όχι στον κοιλιακές τομές. Πιο αναλυτικά, παρατηρήσαμε ότι το CGP (10μΜ) αύξησε το fepsp/fv κατά 7.0±1.4% στον DH (n=21/9, Wilcoxon δοκιμασία, p<0.001), αλλά δεν προκάλεσε καμία αξιοσημείωτη αλλαγή στον VH (0.7±1.6%, n=22/11), (Mann-Whitney U δοκιμασία μεταξύ DH και VH, p<0.005). Για να καθορίσουμε το εάν ένα LTP μεγαλύτερου μεγέθους μπορεί να προκληθεί με εντονότερα πρωτόκολλα, εφαρμόσαμε τα 8-ΤBS σε έναν πληθυσμό από DH και VH τομές. Όπως φαίνεται και στην εικόνα Β.Ι.2.6 (ένθετο γράφημα στο Α), τα 8-TBS προκάλεσαν LTP παρόμοιου πλάτους μετρούμενο στα 60 λεπτά, με εκείνο που προκάλεσε το 3-TBS και στον DH (55.8±9.2%, n=5/2, Wilcoxon δοκιμασία, p<0.05) και στον VH (45.9±13%, n=5/2, Wilcoxon δοκιμασία, p<0.05). Οι δράσεις όλων των πρωτοκόλλων ερεθισμού και του μπλοκαρίσματος των GABA B Rs στα S-LTP και L-LTP συνοψίζονται στο κύριο γράφημα της εικόνας Β.Ι.2.6. Σε γενικές γραμμές και σε συμφωνία με προηγούμενες παρατηρήσεις (Larson et al., 1986), το μέγεθος του LTP καθοριζόταν από το μήκος του TBS πρωτοκόλλου και μεγιστοποιούταν στα 3-TBS. 98






.")


 στον DH σε σύγκριση με τον VH. 2.4.")


.")



111 Εικόνα Β.Ι.2.6: Γραφήματα που συνοψίζουν τις ς δράσεις των ποικίλων TBS T πρωτοκόλλων στο S-LTP (A) και L-LTP (B) κάτω από φυσιολογικές συνθήκες και μετά από αποκλεισμό των GABABRs, στον DH και VH (διαγράμματα στα αριστερά και δεξιά αντίστοιχα). Οι απλές και με μοτίβο μπάρες αντιστοιχούν στην ενδυνάμωση που επάγεται κάτω από φυσιολογικές συνθήκες και παρουσία CGP αντίστοιχα. Οι διακεκομμένες γραμμές καταδεικνύουν το επίπεδοο στο οποίο η ενδυνάμωσηη γίνεται στατιστικά σημαντική. Το μέγεθος του LTP ήταν μεγαλύτερο για εντονότερα TBS, με το μέγιστο να επιτυγχάνεται με το πρωτόκολλοο των 8-TBS (η χρονοεξέλιξη των μεταβολών στον λόγο fepsp/fv που προκαλούνται από το 8- TBS δείχνεται στο ένθετο γράφημα). Το μέγεθος και του S-LTP και του L-LTP, από τη στιγμή που επάγεται, ήταν παρόμοιο ανάμεσα στους δύο πόλους (για στατιστικές συγκρίσεις βλέπεε στο κύριο κείμενο). Επίσης, οι GABABRs συμμετέχουν στον έλεγχο της επαγωγής του LTP νωρίτερα (π.χ. ασθενέστερα α πρωτόκολλα) στον DH σε σύγκριση με τον VH Ο DH vs VH έχει χαμηλότερο κατώφλιο επαγωγής για NMDAR εξαρτώμενο LTP Έχει προηγουμένως δειχθεί ότι το HFS πουυ περιέχει 100 παλμούςς στα 100 Hz διαφοροποιεί την ικανότητα των δύο ιπποκάμπειων τμημάτων στην επαγωγή και στο μέγεθος του LTP (Papatheodoropoulos and Kostopoulos, 2000a; Maruki et al., 2001; 2 Colgin et al., 2004; Maggio and Segal, 2007). Εδώ,, δείχνουμεε ότι τα σχετικά μεγαλύτερα TBSS που περιέχουν συγκρίσιμο αριθμό παλμών (π.χ. 2-TBS και 3-TBS με 80 και 120 παλμούς αντίστοιχα) προκάλεσαν παρόμοιο μέγεθος LTP στον DH και VH, προτείνονταπ ας ότι κάποιοι μηχανισμοί επιστρατεύονται κατά προτίμηση από το TBS για ναα αυξήσουνν την ικανότητα επαγωγής και το μέγεθος του LTP στον VH. Είναι δεδομένο ότι το LTP που επάγεται από το PB ή το TBS μέτριας διάρκειας εξαρτάται από τους NMDARss (Larson and Lynch, 1988; Larson and Munkacsy, 2015) και τους GABA B Rs ( Davis et al., 1991; Mott and Lewis, 1991; Staubli et al., 1999). 99
112 Βρήκαμε ότι οι GABA B Rs ελέγχουν την επαγωγή του LTP με κατώφλιο ερεθισμό στον DH και VH, αλλά η συνεισφορά των GABA B Rs μειώνεται με χρήση μεγαλύτερων TBS πρωτοκόλλων. Έχει επίσης δειχθεί ότι κατά τη διάρκεια της επαγωγής του LTP από σχετικά ισχυρά TBS πρωτόκολλα ίσως επιστρατεύονται, προσθετά με τους NMDARs, άλλοι μηχανισμοί, όπως τα L-τύπου τασεοελεγχόμενα κανάλια ασβεστίου (L-VDCC) και ο μεταβοτροπικός υποδοχέας 5 του γλουταμινικού (mglur5), οδηγώντας σε ένα σύνθετο LTP (Grover and Teyler, 1990; Morgan and Teyler, 2001; Raymond and Redman, 2006; Reymond, 2007). Επίσης, τα έντονα TBS μπορούν να επάγουν NMDAR-ανεξάρτητο LTP στην CA1 περιοχή (Morgan and Teyler, 2001). Συνεπώς αναρωτηθήκαμε το εάν θα μπορούσε να υπάρξει διαφορετική επιστράτευση αυτών των μηχανισμών στον DH και VH από τα TBS πρωτόκολλα που οδηγούν σε παρόμοιο μέγεθος LTP στους δύο ιπποκάμπειους πόλους. Επομένως, εξετάσαμε σε ποια έκταση οι NMDARs εμπλέκονται στην επαγωγή του LTP με τα TBS αυξανόμενου μήκους. Στην εικόνα Β.Ι.2.7 φαίνεται ότι επωάζοντας τις ραχιαίες τομές με 10μΜ CPP αποκλείστηκε ολοκληρωτικά η επαγωγή LTP με πρωτόκολλα από PB έως 2-TBS. Πιο συγκεκριμένα, μετρημένο στα 60 λεπτά μετά την επίδοση του ερεθισμού κάτω από αποκλεισμό των NMDARs, η μεταβολή του fepsp/fv ήταν - 3.5±1.9% με το PB (n=4/2), -1.0±1.4% με το H-TBS (n=4/2), 6.4±2.4% με το 1-TBS (n=6/3) και 6.9±3.8% με το 2-TBS (n=6/3). 100



")











 (Εικόνα Β. Ι.2.8A).")

, (8B).")
113 Εικόνα Β.Ι.2.7: Ερεθισμός θ συχνότητας που αποτελείται μέχρι 2-TBSS επάγει NMDAR εξαρτώμενο LTP. Οι χρονοεξελίξεις των μεταβολής του fepsp/fv που προκαλούνται στιςς ραχιαίες τομές από το H-TBS, 1-TBS και 2-TBS σε φυσιολογικές συνθήκες (Control) δείχνονται μαζί με εκείνες που αποκτήθηκαν κάτω από αποκλεισμό των NMDARs με CPP 10μΜ. Τα αντίστοιχα δεδομένα που αποκτήθηκαν α με το PB ερεθισμό δείχνονται στο ένθετο γράφημα πάνω δεξιά. Τα δεδομένα με το CPP σε κάθε πρωτόκολλο αποκτήθηκανν από τον ακόλουθο αριθμό τομών/ζώων: 4/2 στο PB,, 4/2 στο H-TBS, 6/3 στο 1-TBS και 6/3 στο 2-TBS. Παραδείγματα καταγραφών στην κορυφή των γραφημάτων επεξηγούν την κατασταλτική δράση του CPPP στο μακροχρόνια επίδραση του PB και τωνν TBS. Γραμμές βαθμονόμησης: 1mV και 5ms στο PBB και 0.5 mv και 5 ms σε όλα τα άλλα παραδείγματα. Αντιθέτως, το 3-TBS που επιδόθηκε σε ραχιαίες τομές παρουσία CPP προκαλούσε με συνέπεια σημαντική ενδυνάμωση του fepsp/fv κατά 15.2± ±2.5% (n= =6/3, Wilcoxon δοκιμασία, p<0.05) (Εικόνα Β. Ι.2.8A). Επίσης, το CPP που εφαρμόστηκε ε στις κοιλιακές τομές απέκλεισε ολοκληρωτικά την επαγωγή του LTP με το 3-TBS 3 (3.1±3.6%, n= 6/3), (Εικόνα Β.Ι.2.8B). Τα αποτελέσματα που αποκτήθηκαν απόό τον DH επιβεβαιώνουν προηγούμενες παρατηρήσεις (Morgan and Teyler, 1999). Λαμβάνοντας υπόψη ότι το LTP που προκαλείται από ισχυρό ερεθισμό και κάτω από αποκλεισμό των NMDARs μεσολαβείται από τους L-VDCCs (Morgann and Teyler, 2001), εμείς ε συνχορηγήσαμε CPP 101
114 μαζί με τον αποκλειστή των L-τύπου VDCCs, Νιμοδιπίνη (20μΜ) σε ραχιαίες τομές. Κάτω από αυτές τις συνθήκες, η επαγωγή του LTP από το 3-TBS αποκλείστηκε ολοκληρωτικά (- 6.5±3.4%, n=4/2) δείχνοντας ότι το παρατηρούμενο NMDAR-ανεξάρτητο LTP οφείλονταν στους L-VDCCs (Εικόνα Β.Ι.2.8Α). Αυτά τα αποτελέσματα κατέδειξαν ότι ο DH έχει χαμηλότερο κατώφλιο σε σύγκριση με τον VH για επαγωγή NMDAR-ανεξάρτητο LTP και ότι μεγαλύτερα TBS επάγουν σύνθετο LTP, τουλάχιστον στον DH, με τη συμμετοχή των NMDARs και των L-VDCCs. Ακόμη, αυτό το εύρημα είναι σε συμφωνία με το πρόσφατα παρατηρούμενο υψηλότερο L-VDCC-LTP που παρατηρήθηκε στον DH σε σύγκριση με τον VH στις CA3 συνειρμικές/συνδεσμικές συνάψεις (Moschovos and Papatheodoropoulos, 2016) Τα L-VDCCs και mglur5 συνεισφέρουν στο 3-TBS επαγόμενο LTP εξίσου στον DH και VH Επειδή η εκπόλωση που παρέχεται από τους NMDARs κατά τη διάρκεια σχετικά έντονων TBS ίσως οδηγεί στην ενεργοποίηση των L-VDCCs εμείς προσχωρήσαμε στο να καθορίσουμε τη συμμετοχή των L-VDCCs στο LTP στον DH κάτω από συνθήκες ενεργών NMDARs. Βρήκαμε ότι μετά από αποκλεισμό των L-VDCCs με 20μΜ Νιμοδιπίνη, τα 3- TBS προκάλεσαν σημαντική ενδυνάμωση κατά 39.1±2.8% στις ραχιαίες τομές (n=6/3, Wilcoxon δοκιμασία, p<0.05). Έπειτα χορηγήσαμε Νιμοδιπίνη σε κοιλιακές τομές και βρήκαμε ότι το 3-TBS προκάλεσε σημαντική ενδυνάμωση του fepsp/fv κατά 35.2±3.7% (n=7/3, Wilcoxon δοκιμασία, p<0.05) η οποία ήταν παρόμοια με αυτή στον DH. Επί προσθέτως, η ενδυνάμωση και στον DH και στον VH παρουσία Νιμοδιπίνης ήταν σημαντικά χαμηλότερη σε σύγκριση με αυτή που επάγεται σε κανονικές συνθήκες (Mann- Whitney U δοκιμασία, p<0.005, και για τους δύο πόλους). Παρόλο που το άθροισμα των ενδυναμώσεων που παρατηρήθηκαν στον DH μετά από αποκλεισμό των NMDARs (15.2±2.5%) ή L-VDCCs (39.1±2.8%) ήταν περίπου όμοιο με το μέγεθος της ενδυνάμωσης που προκλήθηκε σε κανονικές συνθήκες (66.2±9.5%), τα 3-TBS ίσως επιστρατεύουν τους mglur5 (Ayala et al., 2009; Raymond and Redman, 2006). Έτσι, χορηγήσαμε 3-TBS στον DH και VH παρουσία του ανταγωνιστή των mglur5 MTEP (200μΜ). Βρήκαμε ότι τα 3-TBS προκάλεσαν σημαντική ενδυνάμωση και στον DH (37.8±3.0, n=5/3, Wilcoxon δοκιμασία, p<0.05) και στον VH (35.6±7.0, n=6/3, Wilcoxon δοκιμασία, p<0.05). Επίσης, αυτή η ενδυνάμωση ήταν σημαντικά χαμηλότερη σε σύγκριση με εκείνη που παρατηρείται σε κανονικές συνθήκες και στον DH (Mann-Whitney U δοκιμασία, p<0.005) και στον VH (Mann-Whitney U δοκιμασία, p<0.005). Η Νιμοδιπίνη 102

 ούτε στον VH")


. Εικόνα Β.Ι.2.")








 και τονν VH")











115 δεν άλλαξε σημαντικά στην βασική συναπτική διαβίβαση ούτε στον DH (9.2±6.1%, n= =6/3) ούτε στον VH (-0.1±2.0%, n=7/4). Παρομοίως, το MTEP άλλαξε μόνο ελαφρά το fepsp/fv στον DH (5.9±1.0%, n=5/3) και VH (-6.3±2.4%, n=6/3). Εικόνα Β.Ι.2.8: Τo 3-TBS πρωτόκολλο προκάλεσε σύνθετο LTP με παρόμοια συνεισφορά των L-τύπου VDCCs και mglur5, και στον DH και στον VH. Επίσης, το 3-TBS προκάλεσε NMDARR ανεξάρτητο LTP στον DH αλλά όχι στον VH. Οι ποσοστιαίες μεταβολές που προκλήθηκαν στο fepsp/fv από το 3-TBS, σε φυσιολογικές συνθήκες και μετά από αποκλεισμό των NMDARs mglur5 και L-VDCC από τους αντίστοιχους αποκλειστές τους CPP, MTEP και Νιμοδιπίνη (Nimo) δείχνονται για τον DHH (A) και τονν VH (B). Κάτω από αποκλεισμό των NMDARs, το LTPP επαγόταν εύκολα στον DH D αλλά εμποδιζόταν στονν VH. Τα δεδομένα αποκτήθηκαν από τομές/ /ζώα για το Control, Nimodipine, MTEP και CPP απόό 15/9, 6/3, 5/3, και 6/3 αντίστοιχα στον DH και 7/7, 7/3,, 6/3 και 6/3 αντίστοιχα στον VH. Τα παραδείγματα π στη κορυφή κάθε γραφήματος επεξηγούν τις δράσεις των φαρμάκων στο LTP. Γραμμές βαθμονόμησης: 0.5 mv και 5 ms. Πρέπει να επισημανθεί ότι η επιλογήή του πρωτοκόλλου των 3-TBS για την εξέταση της συμμετοχής των L-VDCCs και mglur5 στο LTP βασίστηκε στο γεγονός ότι προκαλούσε LTP μέγιστου πλάτους και στους δύο πόλους και ότι ο συνολικός αριθμός των παλμών που περιείχε αυτό το πρωτόκολλο ερεθισμού ήταν συγκρίσιμο σ με εκείνο που 103
116 περιέχουν τα περισσότερα πρωτόκολλα υψηλής συχνότητας που έχουν χρησιμοποιηθεί, επιτρέποντας σε κάποιο βαθμό τη σύγκριση. Επίσης, το 3-TBS προκαλούσε NMDAR ανεξάρτητο LTP στον DH. Τα παρόντα αποτελέσματα κατέδειξαν ότι το 3-TBS επιστρατεύει L-VDCCs και mglur5, μαζί και τους NMDARs, σε παρόμοιο βαθμό και στους δύο πόλους, οδηγώντας σε ένα LTP παρόμοιου μεγέθους Η ενίσχυση των μετασυναπτικών αποκρίσεων και η ενεργότητα των GABA B Rs κατά τη διάρκεια του TBS μπορεί να εξηγήσει τις διαφορές στο LTP μεταξύ DH και VH Η επαγωγή του LTP διευκολύνεται κάτω από συνθήκες αυξημένης μετασυναπτικής εκπόλωσης (Wigstrom and Gustafsson, 1986). Όπως, αναφέρθηκε παραπάνω, κατά τη διάρκεια προσυναπτικου ερεθισμού θ συχνότητας, η ενεργοποίηση των GABA B Rs οδηγεί σε αυξημένη μετασυναπτική εκπόλωση διαμέσου της δυσαναστολής (Davies et al., 1990; Nathan et al., 1990). Έτσι, κατά τη διάρκεια του TBS, οι GABA B Rs βοηθούν την επαγωγή του LTP ενισχύοντας την μετασυναπτική εκπόλωση και ενίσχυση των NMDARs (Larson and Munkácsy, 2015). Πράγματι, έχει επανειλημμένως δειχθεί ότι οι NMDARs ενεργοποιούνται κατά τη διάρκεια του θ προτύπου ερεθισμού (Larson and Munkácsy, 2015; Larson et al., 1986; Papatheodoropoulos, 2015b). Πρόσθετα, έχει δειχθεί πρόσφατα ότι η διευκόλυνση του εμβαδού (Area) του EPSP που προκλήθηκε με ερεθισμό σύζευξης παλμών μεγιστοποιείται σε διαπαλμικό διάστημα θ προτύπου, περιλαμβάνει τους NMDARs, απαιτεί τους GABA B Rs και πιο σημαντικά, είναι ισχυρότερη στον DH σε σύγκριση με τον VH (Papatheodoropoulos, 2015b). Αυτή η διευκόλυνση είναι εμφανής σε έντονη προσυναπτική ενεργοποίηση προκαλώντας EPSP που μοιάζει με το μέγεθος της μετασυναπτικής απόκρισης που προκαλείται από τις ριπές κατά τη διάρκεια του TBS. Συνεπώς, εμείς προχωρήσαμε στην εξέταση των δράσεων του TBS στις ακόλουθες προκλητές μετασυναπτικές αποκρίσεις μετρώντας το εμβαδόν κάθε ριπής εντός των τρένων TBS, συγκριτικά ανάμεσα σε DH και VH. Παρατηρήσαμε ότι κατά τη διάρκεια του TBS σε ραχιαίες τομές υπήρξε έντονη διευκόλυνση του εμβαδού στις διαδοχικά προκαλούμενες ριπές που μεγιστοποιήθηκε στη τρίτη (65.4±3.8%) και τέταρτη (62.0±4.1%) ριπή, όπου μειωνόταν στη συνέχεια πλησιάζοντας τα κανονικά επίπεδα στη τελευταία (δέκατη) ριπή του 1 ου TBS τρένου (Εικόνα Β.Ι.2.9Α). Ένα παρόμοιο πρότυπο αλλαγών παρατηρήθηκε για το 2 ο και 3 ο τρένο TBS (π.χ. στο 2-TBS και 3-TBS πρωτόκολλο), όμως η διευκόλυνση των αντίστοιχων ριπών 104
117 ήταν σημαντικά υψηλότερη συγκρινόμενη με εκείνα του 1 ου TBS. Αντίθετα με τον DH, ο VH εμφάνισε μία περισσότερο από 3-φορές χαμηλότερη διευκόλυνση, η οποία εξαφανίστηκε μετά τη τρίτη ριπή του 1 ου TBS και τη πέμπτη ριπή του 2 ου TBS, όπου αντικαταστάθηκε από μία σταδιακά αυξανόμενη καταστολή που μεγιστοποιήθηκε στη τελευταία ριπή (-32.1±3.3% και -22.9±4.1% για το 1 ο και 2 ο TBS αντίστοιχα), (Εικόνα Β.Ι.2.9Α). Επίσης, ο VH δεν έδειξε αυξημένη διευκόλυνση των πρώτων ριπών στο 2 ο TBS, ενώ οι αποκρίσεις κατά τη διάρκεια του 3 ου TBS δεν εμφάνισαν καμία σημαντική αλλαγή. Περισσότερες λεπτομέρειες στη στατιστική ανάλυση δίνονται στα περιεχόμενα της εικόνας Β.Ι.2.9. Προφανώς, η διευκόλυνση όλων των ριπών ήταν σημαντικά υψηλότερη στον DH απ ότι στον VH. Η τεράστια αυτή διαφορά ανάμεσα στους δύο πόλους μας ώθησε στο να εξετάσουμε την σχέση μεταξύ του βαθμού της διευκόλυνση των ριπών και του μεγέθους του LTP στον DH και VH. Πιο συγκεκριμένα, μετρήσαμε την διευκόλυνση της τρίτης και τέταρτης ριπής σε κάθε TBS τρένο και την ενδυνάμωση στα 60 λεπτά μετά το TBS και χρησιμοποιήσαμε δισδιάστατη διπλής κατεύθυνσης συσχέτιση για να εξετάσουμε την πιθανή σημαντική σχέση μεταξύ των δύο παραμέτρων. Βρήκαμε ότι μία σημαντική γραμμική συσχέτιση εμφανίστηκε στον DH (r=0.39 και r=0.437 για τη 3 ο και 4 ο ριπή αντίστοιχα, p<0.05) αλλά όχι στον VH (r=0.079 και r=0.για τη 3 ο και 4 ο ριπή αντίστοιχα, p>0.05), (Εικόνα Β.Ι.2.9Β), προτείνοντας ότι η διευκόλυνση των ριπών παίζει σημαντικό ρόλο στην επαγωγή του LTP μόνο στον DH. Έπειτα, για να καθορίσουμε την συνεισφορά των NMDARs και GABA B Rs στην διευκόλυνση και καταστολή των ριπών, μετρήσαμε το εμβαδόν μετά από αποκλεισμό αυτών των υποδοχέων στα πρωτόκολλα ερεθισμού από το PB έως τα 3-TBS. Αποκλεισμός των NMDARs στον DH προκάλεσε σημαντική μείωση της διευκόλυνσης σε σχεδόν όλες τις διαδοχικές ριπές που προκλήθηκαν κατά τη διάρκεια των τριών TBS τρένων, εκτός από τις τελευταίες τέσσερις ριπές του 1 ου τρένου, τρεις από τις οποίες δεν παρουσίασαν διευκόλυνση κάτω από κανονικές συνθήκες. Περισσότερο έντονη μείωση παρατηρήθηκε στις ριπές του 2 ου και 3 ου TBS τρένου (Εικόνα Β.Ι.2.9C). Πιο συγκεκριμένα, το CPP μείωσε σημαντικά την διευκόλυνση των 2-6 ριπών του 1 ου τρένου προκαλώντας μέγιστη μείωση της διευκόλυνσης στη τρίτη ριπή κατά 25%. Επίσης, μεγαλύτερη μείωση της διευκόλυνσης παρατηρήθηκε στις ριπές των ακόλουθων TBS τρένων. Για παράδειγμα, η διευκόλυνση της τρίτης ριπής του 2 ου και 3 ου τρένου κατεστάλη κατά 55% και 65% αντίστοιχα (αναλυτική στατιστική δίνεται στα περιεχόμενα της εικόνας Β.Ι.2.9). 105

118 Εικόνα Β.Ι.2.9: Kατά τη διάρκεια του TBS, ο DH συγκρινόμενος με τον τ VH εμφάνισε υψηλότερη διευκόλυνση των μετασυναπτικών αποκρίσεων η οποία συσχετίστηκε μεε το μέγεθος του LTP και την υψηλότερηη ενεργοποίηση των GABABRs. Α) Διάγραμμα που δείχνει το βαθμό της διευκόλυνσης ή καταστολής του εμβαδού ε των διαδοχικών ριπών κατά τη διάρκεια των TBS. Οι τιμές της διευκόλυνσης των ριπών υπολογίστηκαν καιι εκφράστηκαν ως ποσοστιαία μεταβολή τους σε σχέση με το εμβαδόν ν της πρώτης ριπής του 1 ου TBS. T Τα δεδομένα συλλέχθηκαν από όλα τα πειράματα από PB έως 3-TBS. Οι αστερίσκοι και οι διέσεις δηλώνουν στατιστικά σημαντική ενίσχυση ή καταστολή στα p<0.005 και p<0.05 αντίστοιχα. Αντιπροσωπευτικά παραδείγματα της 1 ης και 3 ης ριπής του πρώτου τρένου δείχνονται με γκρι και μαύρη γραμμή αντίστοιχα. Έντονη ενίσχυση συνέβη σε σχεδόν όλες τις ριπές στον DH (εκτός από τις τελευταίες δύο του πρώτουυ τρένου και τη τελευταίαα του δευτέρου τρένου), ενώ μόνο οι δύο πρώτες ριπές του 1 ου τρένου και οι πρώτεςς πέντε του 2 ου τρένου στον VH ήταν στατιστικώς ενισχυμένες. Σημειωτέον ότι οι τελευταίες 3-4 ριπές στο 1 ο και κ 2 ο TBS τρένο στον VH ήταν στατιστικώς κατασταλμένες και ότι καμία του 3 ου τρένου στον VH δεν άλλαξε σημαντικά.. Η ενίσχυση όλων των ριπών ήταν σημαντικά υψηλότερηη στον DH σεε σύγκριση με τον VH (Mann-Whitney δοκιμασία για κάθε ριπή, p<0.005). Σύγκριση της ενίσχυσης ανάμεσα στις αντίστοιχες ριπές κατάά μήκος των TBS τρένων έδειξε έ 106
119 σημαντικές διαφορές μεταξύ του 1 ου και 2 ου TBS, p<0.05 για την ενίσχυση της δεύτερης ριπής και p<0.005 για όλες τις άλλες. Ανάμεσα στο 1 ο και 3 ο TBS, p<0.005 για όλες τις ριπές και p<0.05 για την πρώτη και τη τελευταία ριπή των δύο τρένων. Β) Η ενίσχυση των διαδοχικών ριπών κατά τη διάρκεια των TBS συσχετίστηκε με το μέγεθος του LTP στον DH αλλά όχι στον VH. Γραφήματα ανάμεσα στο βαθμό της ενίσχυσης της 3 ης (ανοιχτό σύμβολο) και της 4 ης (γεμάτο σύμβολο) ριπής στο πρώτο TBS και το μέγεθος του LTP μετρημένο στα 60 λεπτά μετά την επίδοση του TBS στον DH και VH. Το μέγεθος του LTP συσχετίστηκε γραμμικά με την ενίσχυση των ριπών στον DH (r=0.393, p<0.05 και r=0.437, p<0.05 για τη 3 η και 4 η ριπή αντίστοιχα) αλλά όχι στον VH (r=0.079, p>0.05 και r=0.072, p>0.05 για τη 3 η και 4 η ριπή αντίστοιχα). Οι γραμμές αντιπροσωπεύουν αυτό το γραμμικό ταίριασμα με τα δεδομένα. Τα δεδομένα αποκτήθηκαν από 31 τομές/24 ζώα στον DH και 36 τομες/28 ζώα στον VH. C-D) Η δράση του ανταγωνιστή των NMDARs, CPP και τους ανταγωνιστή των GABABRs, CGP (γραφήματα στα αριστερά και δεξιά αντίστοιχα) στην ενίσχυση/καταστολή του εμβαδού των ριπών που παρατηρήθηκε στον DH (C) και VH (D) δείχνονται για όλες τις ριπές σε όλα τα TBS πρωτόκολλα. Τα δεδομένα που αποκτήθηκαν σε φυσιολογικές συνθήκες και που παρουσιάζονται στο (Α) εμφανίζονται και εδώ για λόγους σύγκρισης. Οι αστερίσκοι και οι διέσεις δηλώνουν στατιστικώς σημαντική δράση του φαρμάκου με p<0.005 και p<0.05 αντίστοιχα. Παραδείγματα καταγραφών για τη 1 η και 23 η ριπή στο CPP και της 1 ης και 13 ης ριπής στο CGP52432 δείχνονται στην κορυφή των αντίστοιχων γραφημάτων με την 1 η απάντηση σε γκρι γραμμή. Προσέξτε την αυξημένη συμμετοχή των NMDARs στην ενίσχυση των ριπών του 2 ου και 3 ου TBS τρένου στον DH. Ακόμη, προσέξτε ότι το CGP κατέστειλε σημαντικά την ενίσχυση των ριπών στον DH αλλά όχι στον VH. Στον VH, το CPP ακύρωσε την διευκόλυνση των πρώτων 3-5 ριπών των τριών διαδοχικών τρένων χωρίς να επηρεάζει την διευκόλυνση των μετέπειτα (κατασταλμένων) ριπών. Αυτά τα αποτελέσματα έδειξαν ότι οι NMDARs ενεργοποιήθηκαν με σταδιακά αυξανόμενο τρόπο κατά τη διάρκεια της επίδοσης των TBS στον DH και κατέδειξαν ότι αυτή η ενεργοποίηση είναι κατά πολύ πιο έντονη στον DH απ ότι στον VH. Ακολούθως, εξετάσαμε τις δράσεις του ανταγωνιστή των GABA B Rs, CGP Παρατηρήσαμε ότι αποκλεισμός των GABA B Rs στον DH προκάλεσε ένα παρόμοιο πρότυπο αλλαγών με αυτό μετά τον αποκλεισμό των NMDARs. Πιο συγκεκριμένα, το CGP μείωσε την διευκόλυνση όλων σχεδόν των ριπών αποκρίσεων (Βλέπε εικόνα Β.Ι.2.9B και το αντίστοιχο περιεχόμενο για περισσότερα στατιστικά στοιχεία). Αυτά τα αποτελέσματα είναι σε συνέπεια με τον ρόλο των GABA B Rs στην ενεργοποίηση των NMDARs. Αντιθέτως, ο αποκλεισμός των GABA B Rs στον VH μείωσε την διευκόλυνση της 2 ης και 3 ης ριπής στο πρώτο τρένο TBS αλλά δεν επηρέασε τις άλλες αποκρίσεις σε κανένα τρένο TBS. Ενδιαφέρον είναι το γεγονός ότι το CGP προκάλεσε μία μείωση της καταστολής των τελευταίων ριπών σε κάθε τρένο (Εικόνα Β.Ι.2.9D). 107
 ειδικά")

120 Όπως περιγράψαμε παραπάνω, τα 3-TBS προκάλεσαν ένα σύνθετο LTP με τη συμμετοχή των NMDARs και L-VDCCs. Επί προσθέτως, ο DH αλλά όχι ο VH, εμφάνισε NMDAR-ανεξάρτητοεπικουρείται από τη πυροδότηση του μετασυναπτικού κυττάρου (Bi and Poo, 1998; Kelso and Brown, 1986; Wigstrom and Gustafsson, 1986) ειδικά μετά από ερεθισμό θ LTP. Έχει προταθεί ότι η επαγωγή τουυ LTP στηη CA1 περιοχή συχνότητας (Thomas et al., 1998). Επίσης, τα μεγάλα TBS ίσως προωθούν την επαγωγή του VDCC-LTP (Morgan and Teyler, 2001). Η έντονη ενίσχυση των συναπτικών αποκρίσεων που είδαμε στον DH θα μπορούσε να αυξήσει την νευρωνική ν διεγερσιμότητα προωθώντας την ενεργοποίησηη των L-VDCCs και βοηθώντας με αυτόν τον τρόπο την επαγωγή NMDAR-ανεξάρτητου L-VDCC-LTP με τα έντονα TBS. Επί προσθέτως, η ενεργοποίηση των προσυναπτικών GABA B Rs θα μπορούσε να καταστείλει την μετασυναπτική αναστολή και να ενισχύσει έτσι την διέγερση. Γι αυτό το λόγο, ποσοτικοποιήσαμε την πυροδότηση κατά τη διάρκεια των TBS μετρώντας το πλάτος των PSs που προκλήθηκαν κατά τη διάρκεια των ριπών.. Το άθροισμα του πλάτους των PSs υπολογίστηκε για κάθε ριπή και αντιπροσώπευε την απόκρισηη της ριπής. Εντυπωσιακό ήταν το γεγονός ότι ο DH και ο VH παρουσίασαν αντίθετα πρότυπα αλλαγών του PS. Πιο συγκεκριμένα, παρόλο που ο DH εμφάνισε έντονη διευκόλυνση του PS κατά μήκος όλων των TBS τρένων, η απόκριση του VH στα TBS ήταν κυρίως κατασταλτική (Εικόνα Β.Ι.2.10). Αυτά τα αποτελέσματα συμφωνούν με την υψηλότερη δισανασταλτική δράση στον DH σε σύγκριση με τον VH κατά τηη διάρκεια των TBS, προσφέροντας μία εξήγηση της υψηλότερης ικανότητας του DH να εμφανίζει L-VDCC-LTP. 108
121 Εικόνα Β.Ι.2.10: Το PS ενισχύθηκε κατά πολύ στον DH αλλά κατεστάλη στον VH κατά τη διάρκεια των TBS. Το άθροισμα του πλάτους των PSs που προκλήθηκε από κάθε ριπή στο 1 ο, 2 ο και 3 ο TBS υπολογίστηκε και εκφράστηκε σαν ποσοστιαία μεταβολή τους σε σχέση με το άθροισμα του πλάτους της πρώτης ριπής από το 1 ο TBS. Τα δεδομένα συλλέχτηκαν από 14 τομές από 14 αρουραίους στον DH και 14 τομές από 14 αρουραίους στον VH. Αντιπροσωπευτικά παραδείγματα των προκλητών PS για τη 1 η, 3 η και 9 η ριπή δείχνονται για τον DH (πάνω αριστερά) και τον VH (πάνω δεξιά). Προσέξτε ότι η έντονη ενίσχυση των PS στον DH σε όλες τις ριπές κάθε TBS τρένου και την αρκετά αντίθετη απόκριση του VH που εμφάνισε καταστολή αντί για ενίσχυση. Οι αστερίσκοι και οι διέσεις δηλώνουν σημαντικές αλλαγές των PSs των ριπών με p<0.01 και p<0.05 αντίστοιχα Συζήτηση Τα κύρια ευρήματα της παρούσας μελέτης ήταν τα εξής: η δικτυακή ενεργότητα θ συχνότητας επάγει αξιόπιστα S-LTP και L-LTP παρόμοιου μεγέθους στον DH και VH, τα κατώφλια επαγωγής και για τα δύο LTPs είναι χαμηλότερα στον DH σε σύγκριση με τον VH και επί προσθέτως, ο VH αλλά όχι ο DH παρουσιάζει διαφορετικά κατώφλια επαγωγής ανάμεσα στο S-LTP και L-LTP ο DH σε σύγκριση με τον VH παρουσιάζει πολύ υψηλότερο βαθμό διευκόλυνσης των μετασυναπτικών αποκρίσεων και υψηλότερη ενεργοποίηση των GABA B Rs κατά τη διάρκεια του ερεθισμού θ συχνότητας που μπορούν να εξηγήσουν τα χαμηλότερα κατώφλια για NMDAR-εξαρτώμενο και NMDAR-ανεξάρτητο LTP τα L-VDCCs και mglur5 συμμετέχουν στην επαγωγή του σύνθετου LTP και στους δύο πόλους με όμοιο τρόπο. Αυτή η μελέτη καταδεικνύει για πρώτη φορά ότι κάτω από συνθήκες προσυναπτικής ενεργοποίησης που μοιάζει με ένα σχετικά φυσιολογικό πρότυπο ενεργοποίησης, η ικανότητα του VH να υποστηρίζει επαγωγή και διατήρηση του LTP ενισχύεται και γίνεται παρόμοια με την ικανότητα του DH. Επιπλέον, κάτω από αυτές τις συνθήκες γίνεται εφικτό να αποκαλύψουμε ότι το κατώφλιο για επαγωγή LTP είναι διαφορετικό στους δύο πόλους. Σε αντίθεση με τον HFS (π.χ. 100 παλμοί στα 100 Hz) που επάγει LTP μεγαλύτερου μεγέθους στον DH απ ότι στον VH (Colgin et al., 2004; Maruki et al., 2001; Papatheodoropoulos and Kostopoulos, 2000), εμείς δείξαμε ότι ο ερεθισμός θ συχνότητας αποκαλύπτει την ικανότητα του VH να εμφανίζει S-LTP και L-LTP και την ίδια στιγμή καθιστά πιθανή την ανίχνευση σημαντικών διαφορών μεταξύ των δύο πόλων σχετικά με την τα κατωφλίων ενεργοποίησης που απαιτούνται για επαγωγή LTP. Βρήκαμε 109
122 ότι στον DH ακόμα και με PΒ ερεθισμό επάγεται LTP που διαρκεί τουλάχιστον δύο ώρες ενώ μακρύτερα TBS, π.χ. 2-TBS και 3-TBS επάγουν LTP που μπορεί να διατηρηθεί για τουλάχιστον πέντε ώρες. Αυτά τα αποτελέσματα επιβεβαιώνουν προηγούμενες in vivo παρατηρήσεις που έχουν δείξει ότι ένα τροποποιημένο 1-TBS που περιέχει τον ίδιο αριθμό παλμών επάγει LTP που μπορεί να διαρκέσει για εβδομάδες (Staubli and Lynch, 1987) Πιθανοί μηχανισμοί των ραχιαίο-κοιλιακών διαφορών Η επαγωγή του LTP στις CA1 συνάψεις απαιτεί την είσοδο ασβεστίου εντός του μετασυναπτικού κυτταροπλασματικού χώρου διαμέσου των NMDARs, των ενδοκυττάριων αποθηκών ή/και των L-VDCCs, ανάλογα με την δύναμη της συναπτικής ενεργοποίησης και της μετασυναπτικής διέγερσης (Malenka, 1991; Raymond, 2007). Έχει δειχθεί παλιότερα ότι το PB και το σύντομο TBS επάγουν NMDAR εξαρτώμενο LTP στον DH (Larson and Lynch, 1988). Με παρόμοιο τρόπο, βρήκαμε ότι το PB και το σύντομο TBS που αντιστοιχούσαν στους ερεθισμούς στο ή κοντά στο κατώφλιο επαγωγής του LTP ήταν διαφορετικά ανάμεσα στους δύο πόλους. Τα κρίσιμα γεγονότα που συμμετέχουν στην επιτυχή επαγωγή του LTP συμβαίνουν κατά τη διάρκεια της φάσης ερεθισμού που οδηγούν σε επαρκή μετασυναπτική εκπόλωση και ακολούθως σε επαρκή ενεργοποίηση των NMDARs και αυξημένη πιθανότητα για επαγωγή LTP (Bliss et al., 2007). Δεδομένου του σημαντικού ρόλου των NMDARs στην επαγωγή του LTP στην CA1 περιοχή (Davies and Watkins, 1981), είναι λογικό να υποθέσουμε ότι οι μηχανισμοί που ελέγχουν την ενεργοποίηση των NMDARs ίσως συνεισφέρουν σε αυτή τη διαφοροποίηση του κατώφλιου επαγωγής του LTP ανάμεσα στους δύο πόλους, χωρίς απαραιτήτως να επηρεάζουν την το μέγεθος της ενδυνάμωσης. Η επαρκής μετασυναπτική εκπόλωση που απαιτείται για την επιτυχή επαγωγή του LTP και που παράγεται κατά τη διάρκεια της φάσης επαγωγής μπορεί να επιτευχθεί διαμέσου διαδικασιών όπως της μείωσης της μετασυναπτικής αναστολής, της αύξησης της συναπτικής διευκόλυνσης και της αύξησης της μετασυναπτικής πυροδότησης. Έχει προηγουμένως αποδειχθεί, όπως και σε αυτή τη μελέτη, ότι στον DH ο κύριος μηχανισμός που συμμετέχει στην επιτυχή επαγωγή LTP μέσω κατώφλιου ερεθισμού (PB ή σύντομο TBS), είναι οι GABA B Rs (Davies et al., 1991; Mott and Lewis, 1991; Staubli and Scafidi, 1999). Βρήκαμε ότι οι GABA B Rs συμμετέχουν στην ρύθμιση του κατώφλιου επαγωγής του LTP και στους δύο πόλους, από τη στιγμή που αποκλείοντας αυτούς τους υποδοχείς αυξάνεται το μήκος του TBS που απαιτείται για να προκληθεί ένα LTP. Επιπλέον, το πρότυπο συναπτικής ενδυνάμωσης που επάγεται στον DH μέσω μόλις υπερκατώφλιου 110
123 ερεθισμού (π.χ.h-tbs) και μετά από αποκλεισμό των GABA B Rs ήταν αρκετά παρόμοιο με την ενδυνάμωση που προκλήθηκε στον VH από κατώφλιο ερεθισμό (π.χ.h-tb) και ενεργούς τους GABA B Rs, προτείνοντας ότι το κατώφλιο επαγωγής LTP στον VH είναι υψηλότερο εξαιτίας της χαμηλής ενεργότητας των GABA B Rs. Επί προσθέτως, εμείς καταδείξαμε ότι οι GABA B Rs συμμετέχουν στην τροποποίηση του μεγέθους του LTP που προκαλείται με υπερκατώφλιο ερεθισμό. Οι GABA B Rs διευκολύνουν την επαγωγή LTP από το TBS μέσω της δισανασταλτικής δράσης τους στο μετασυναπτικό στοιχείο. Πιο ειδικά, ενεργοποίηση των προσυναπτικών GABA B Rs μειώνει την απελευθέρωση του GABA και συνεπώς την μετασυναπτική αναστολή. Η μείωση της αναστολής ενισχύει την μετασυναπτικής εκπόλωση και διευκολύνει την ενεργοποίηση των NMDARs (Davies et al., 1991; Mott and Lewis, 1991). Αυτή η δράση εκφράζεται κατά τη διάρκεια του TBS, και συνεπώς μπορεί να μελετηθεί μετρώντας τις μετασυναπτικές αποκρίσεις που προκαλούνται από τους ερεθισμούς των ριπών. Σε συνέπεια με αυτές τις παρατηρήσεις, βρήκαμε ότι η ενεργότητα των GABA B Rs είναι πράγματι ενισχυμένη στον DH κατά τη διάρκεια του TBS και αυτή η ενεργότητα συνοδευόταν από ενεργοποίηση των NMDARs. Αντιθέτως, οι GABA B Rs ενεργοποιούνταν ελάχιστα στον VH. Έτσι η πιθανώς χαμηλότερη ενεργότητα των προσυναπτικών GABA B Rs στον VH δε θα μπορούσε να επιτρέψει την επαρκή ενεργοποίηση των NMDARs και την εύκολη επαγωγή LTP. Στη βάση αυτών των παρατηρήσεων συμπεράναμε ότι οι GABA B Rs συμμετέχουν σε κρίσιμο βαθμό στην διαφοροποίηση του κατώφλιου επαγωγής LTP ανάμεσα στους δύο πόλους του ιπποκάμπου. Η συναπτική διευκόλυνση κατά τη διάρκεια των ερεθισμών των ριπών ίσως συνεισφέρει σημαντικά στην αύξηση της μετασυναπτικής εκπόλωσης κατά τη διάρκεια του TBS και έτσι να διευκολύνει την ενεργοποίηση των NMDARs και την επαγωγή LTP (Larson and Lynch, 1988; Larson and Munkácsy, 2015). Συμφωνά με αυτές τις παρατηρήσεις, βρήκαμε ότι κατά τη διάρκεια των TBS οι μετασυναπτικές αποκρίσεις των ριπών ενισχύθηκαν κατά πολύ στον DH με την σημαντική συμμετοχή των NMDARs. Εντυπωσιακό είναι το γεγονός ότι η ενίσχυση των ριπών ήταν πολύ χαμηλότερη στον VH. Έτσι, υποθέσαμε ότι η διαφορά στην ενίσχυση των μετασυναπτικών αποκρίσεων κατά τη διάρκεια της φάσης επαγωγής του LTP ίσως συνεισφέρει σημαντικά στην διαφοροποίηση του LTP μεταξύ των δύο πόλων. Ένας από τους πιο αποτελεσματικούς τρόπους αύξησης της μετασυναπτικής εκπόλωσης που οδηγεί στην ενεργοποίηση των NMDARs και βοηθάει σημαντικά στην επαγωγή του LTP παρέχεται μέσω των ανάδρομων δυναμικών ενέργειας, την πυροδότηση του μετασυναπτικού κυττάρου που ίσως συμβαίνει αμέσως μετά την συναπτική 111
124 ενεργοποίηση (Magee and Johnston, 1995; Stuart and Sakmann, 1994). Επίσης, αυτός ο μηχανισμός μπορεί να μειώσει το κατώφλιο επαγωγής NMDAR-εξαρτώμενου LTP (Fuenzalida et al., 2010; Magee and Johnston, 1997). Επί προσθέτως, η εκπόλωση που παρέχεται από τον ενορχηστρωμένο συνδυασμό της συναπτικής ενεργοποίησης και της μετασυναπτικής πυροδότησης μπορεί να είναι αρκετή για να ενεργοποιήσει τους L-VDCCs και να επάγει LTP ανεξάρτητο της ενεργοποίησης των NMDARs (Cavus and Teyler, 1998; Magee and Johnston, 1995; Takahashi and Magee, 2009).Τα ανάδρομα δυναμικά ενέργειας ίσως παράγονται κατά τη διάρκεια της επαναλαμβανόμενης ενεργοποίησης που συμβαίνει με την επίδοση του TBS (Thomas et al., 1998). Βρήκαμε ότι η ικανότητα του DH να συντηρεί L-VDCC-LTP ανεξάρτητα των NMDARs με τα έντονα TBS (π.χ. 3-TBS) συνοδευόταν από ισχυρή ενίσχυση της μετασυναπτικής πυροδότησης κατά τη διάρκεια ολόκληρου του ερεθισμού θ συχνότητας. Το εντυπωσιακό εύρημα της καταστολής έναντι της ενίσχυσης της πυροδότησης στον VH μπορεί εν δυνάμει να εξηγήσει την αποτυχία του VH να εμφανίσει NMDAR-ανεξάρτητο LTP. Επίσης, από τη στιγμή που η ενίσχυση της μετασυναπτικής διέγερσης στον DH ήταν έντονη από τις πρώτες ριπές (π.χ.pb), η ραχιαιοκοιλιακή διαφορά στην πυροδότηση ίσως αντιπροσωπεύει την διαφοροποίηση του κατώφλιου επαγωγής LTP μεταξύ των δύο ιπποκάμπειων τμημάτων. Είναι αξιοσημείωτο το γεγονός ότι το μέγεθος του LTP, από τη στιγμή που επαγόταν ήταν παρόμοιο ανάμεσα στον DH και VH προτείνοντας ότι παρά τη μεγάλη διαφορά στην μετασυναπτική ενίσχυση και στην ενεργότητα των GABA B Rs κατά τη διάρκεια του ερεθισμού, η αύξηση του ενδοκυττάριου ασβεστίου που απαιτείται για επαγωγή LTP στον VH ήταν τόσο αποτελεσματική όσο στον DH. Η απελευθέρωση του ασβεστίου από το ενδοπλασματικό δίκτυο διαμέσου της ενεργοποίησης των υποδοχέων ριανοδίνης συνεισφέρει σημαντικά στην επαγωγή LTP κάτω από συγκεκριμένες συνθήκες συναπτικής ενεργότητας (Raymond, 2007). Αναλόγως, η υψηλότερη συγκέντρωση υποδοχέων ριανοδίνης στον VH σε σύγκριση με τον DH (Grigoryan and Segal, 2012) ίσως σχετίζεται με την ενισχυμένη ικανότητα του VH για LTP που προκαλείται από τα TBS. Τελικώς, σε συμφωνία με τα αποτελέσματα από προηγούμενες μελέτες (Morgan and Teyler, 2001; Raymond, 2007), βρήκαμε ότι τα μακρόσυρτα TBS επάγουν σύνθετο LTP με τη συμμετοχή των NMDARs, L-VDCCs και mglur5. Επίσης, παρόλο που μετά από αποκλεισμό των NMDARs, επαγόταν L-VDCC εξαρτώμενο LTP στον DH αλλά όχι στον VH, η συνεισφορά των L-VDCCs ήταν παρόμοια στους δύο πόλους που σημαίνει ότι η ενεργοποίησή τους στον VH απαιτεί την ενεργοποίηση των NMDARs και ότι είναι το ίδιο αποτελεσματικοί στην αύξηση του μεγέθους του LTP όπως στον DH. 112
125 Λειτουργικές εφαρμογές Το κατώφλιο επαγωγής του LTP, καθορίζοντας το πόσο εύκολα οι συναπτικές αλλαγές μπορούν να συμβούν σε ένα κύκλωμα, είναι μία σημαντική ενδογενής ιδιότητα των νευρωνικών δικτύων. Βρήκαμε ότι ο DH σε σύγκριση με τον VH ανταποκρίνεται πιο εύκολα σε ασθενή TBS πρωτόκολλα εμφανίζοντας παρατεταμένες αλλαγές στην συναπτική ενδυνάμωση. Υποθέτοντας ότι το LTP στον ιππόκαμπο αντιπροσωπεύει ή τουλάχιστον βοηθάει στην διαδικασία της μνήμης σε κυτταρικό επίπεδο (Martin and Morris, 2002), το κατώφλιο για επαγωγή LTP ίσως ομοιάζει περισσότερο με τη φυσιολογική κατάσταση της αυτόματης σύλληψης και διατήρησης μοναδιαίων γεγονότων, μία από τις ιδιότητες της ιπποκάμπειας επεισοδιακής μνήμης (Morris, 2003; Nadel and Moscovitch, 1997). Η υψηλή ευαισθησία του DH να υποστηρίζει μακράς διάρκειας συναπτικές αλλαγές με τον ελάχιστο ερεθισμό ίσως εκφράζει την υψηλότερη ικανότητα αυτού του τμήματος του ιπποκάμπου σε σύγκριση με το κοιλιακό στην απόκτηση και διατήρηση της πληροφορίας που περιέχεται σε μοναδιαία γεγονότα. Από την άλλη μεριά, παρόλο που ο VH είναι λιγότερο ευαίσθητος στην επαγωγή LTP, φαίνεται ότι μπορεί να διακρίνει με ακρίβεια εισόδους διαφορετικής δύναμης (strength) από τη στιγμή που παρουσιάζει διαφορετικά κατώφλια επαγωγής LTP ανάμεσα σε σύντομα και μεγάλα TBS τρένα. Λαμβάνοντας υπόψη ότι η μακρόχρονη μνήμη θα υποστηριζόταν καταλληλότερα από το μακροπρόθεσμο LTP (Martin and Morris, 2002), είναι λογικό να υποθέσουμε ότι η ικανότητα ενός δικτύου να ανιχνεύει σχετικά ασθενείς εισόδους και να τις διατηρεί διαμέσου σύντομης διάρκειας συναπτικές μεταβολές θα ήταν ένας κατάλληλος μηχανισμός για να υποστηρίξει την σύντομη βραχύχρονη μνήμη. Είναι ενδιαφέρον να επισημάνουμε ότι ο VH φαίνεται να εμπλέκεται στην ενεργό μνήμη σε σύγκριση με τον DH (Kesner et al., 2011; O'Neill et al., 2013) και συνεπώς θα μπορούσε να προταθεί ότι η ικανότητα του VH να διακρίνει καλύτερα τα ασθενή από τα ισχυρά πρότυπα ενεργοποίησης σε σύγκριση με τον DH ίσως είναι ένας από τους μηχανισμούς που βοηθάνε στην εμπλοκή του στην ενεργό μνήμη. Ένας σημαντικός λειτουργικός ρόλος που εκτελείται από το κατώφλιο επαγωγής LTP ίσως είναι η σταθεροποίηση της δικτυακής δραστηριότητας. Πιο συγκεκριμένα, μηχανισμοί που κρατάνε τα κατώφλια επαγωγής LTP χαμηλά αποφεύγουν τον κίνδυνο της ανεξέλεγκτης ενδυνάμωσης (Bliss et al., 2007). Θεωρητικά, εάν ένα νευρωνικό δίκτυο με σχετικά υψηλό βαθμό ενδογενούς διεγερσιμότητας έχει χαμηλό κατώφλιο για επαγωγή συναπτικής ενδυνάμωσης τότε η δραστηριότητά του θα μπορούσε εύκολα να εκτροχιαστεί. Φυσιολογικά, σε μία τέτοια κατάσταση και λαμβάνοντας υπόψη ότι το κατώφλιο είναι μία δυναμική παράμετρος που εξαρτάται από τη μέση κυτταρική ενεργότητα (Bienenstock et 113
126 al., 1982), ομοιοστατικές απαιτήσεις θα μπορούσαν να επιβάλλουν την ύπαρξη σχετικά υψηλών κατωφλίων επαγωγής LTP για να μπορέσει η δικτυακή δραστηριότητα να σταθεροποιηθεί σε ένα βέλτιστο επίπεδο εμποδίζοντας την ανεξέλεγκτη ενδυνάμωση. Πολλές μελέτες έχουν δείξει ότι το δίκτυο του VH παρουσιάζει υψηλότερη ενδογενή νευρωνική διεγερσιμότητα σε σύγκριση με τον DH (Andy, 1962; Becker et al., 1997; Bragdon et al., 1986; Brazier, 1970; Derchansky et al., 2004; Dougherty et al., 2012; Gilbert et al., 1985; Greco et al., 1994; Lee et al., 1990; Lerner-Natoli et al., 1984; Lothman and Collins, 1981; Mikroulis and Psarropoulou, 2012; Moschovos et al., 2012; Papatheodoropoulos, 2015a; Papatheodoropoulos et al., 2005; Racine et al., 1977). Έτσι, το σχετικά υψηλό κατώφλιο επαγωγής LTP του VH πόλου ίσως παρέχει στο τοπικό του δίκτυο την απαιτούμενη σταθερότητα, εξισορροπώντας την τάση για αύξηση της διέγερσης και συνεισφέροντας με αυτόν τον τρόπο στην σωστή δραστηριότητα του. Πράγματι, κάτω από φυσιολογικές συνθήκες ο κοιλιακός πόλος του ιπποκάμπου λειτουργεί τόσο αποδοτικά όσο ο ραχιαίος, διατηρώντας την ισορροπία ανάμεσα στην διέγερση /αναστολή στο δίκτυό του Συμπέρασμα Τα αποτελέσματα αυτής της μελέτης καταδεικνύουν ότι κάτω από συνθήκες ερεθισμού θ συχνότητας που πλησιάζει το φυσιολογικό πρότυπο δραστηριότητας του θ ρυθμού, το τοπικό CA1 συναπτικό κύκλωμα είναι ικανό να υποστεί S-LTP και L-LTP παρόμοιου μεγέθους και στους δύο πόλους που διαρκεί πολλές ώρες. Έτσι, κάτω από συγκεκριμένες συνθήκες της δικτυακής δραστηριότητας τα μικροκυκλώματα των δύο ιπποκάμπειων πόλων μπορούν να υποστηρίξουν με παρόμοιο τρόπο μόνιμες αλλαγές της συναπτικής πλαστικότητας. Παρόλο που τα κατώφλια επαγωγής και των δύο τύπων LTPs είναι χαμηλότερα στον DH απ ότι στον VH, ο VH μπορεί να διακρίνει με ακρίβεια εισόδους διαφορετικής δύναμης. Κάτω από έντονα πρότυπα ερεθισμού η συνεισφορά των L-VDCCs και mglur5 είναι παρόμοια μεταξύ των πόλων, παρόλο που ο DH εμφανίζει υψηλότερη ευαισθησία για L-VDCC-LTP όταν οι NMDARs έχουν αποκλειστεί. Οι μηχανισμοί που πιθανώς κρύβονται πίσω από τις ραχιαιοκοιλιακές διαφορές περιλαμβάνουν την ενεργοποίηση των GABA B Rs, την ενίσχυση της συναπτικής διευκόλυνσης και την ισχυρή αύξηση της μετασυναπτικής πυροδότησης που συμβαίνουν κατά τη διάρκεια των TBS. Υποθέσαμε ότι, η υψηλότερη ευαισθησία του DH να παρουσιάζει LTP με ασθενέστερο θ συχνότητας τύπο ερεθισμού και η ικανότητα του VH να αποκρίνεται σε 114
127 ασθενείς και ισχυρές εισόδους ίσως καταδεικνύουν την υψηλότερη εμπλοκή του DH σε μοναδιαία μνημονικά γεγονότα και τους VH στην ενεργό μνήμη αντίστοιχα. Συμπεραίνουμε ότι οι διαφορές στο LTP κατά μήκος του επιμήκους δικτύου του ιπποκάμπου αντιπροσωπεύουν προσαρμοστικές εξειδικεύσεις που βοηθούν το τοπικό κύκλωμα να επεξεργάζεται την πληροφορία με τρόπο που να επιτυγχάνει τους λειτουργικούς του στόχους. Υποθέτουμε ότι η συσσωρευμένη γνώση για τις ομοιότητες και τις διαφορές κατά μήκος ολόκληρου του φάσματος των φαινομένων της συναπτικής πλαστικότητας μεταξύ DH και VH θα μας βοηθήσει να κατανοήσουμε τη μεγάλη κλίμακα της λειτουργικής διαφοροποίησης που παρατηρείται κατά μήκος του ιπποκάμπου. 115
128 3. Οξύαιχμα κύματα και ριπιδισμοί: Επιδράσεις της γήρανσης και τροποποίηση τους μέσω NMDA υποδοχέων και L-τύπου διαύλων Ca Εισαγωγή Ο ιππόκαμπος παίζει ένα σημαντικό ρόλο στην μακροχρόνια εγκατάσταση των βιωματικών αναμνήσεων συμμετέχοντας στην διαδικασία της μνημονικής παγίωσης (Wang and Morris 2010), συντονισμένη από την επικοινωνία μεταξύ ιπποκάμπου και νεοφλοιικών κυκλωμάτων (Buzsaki 1996, Siapas and Wilson 1998, Sirota et al 2003, Battaglia et al 2004, Wierzynski et al 2009). Κατά τη διάρκεια αυτής της διαδικασίας η πληροφορία που είναι αποθηκευμένη στο ιπποκάμπειο κύκλωμα μεταφέρεται στο νεοφλοιικό κύκλωμα όπου και συνεισφέρει στην επαγωγή πλαστικών συναπτικών αλλαγών που προκαλούν μεταβολές στο κύκλωμα στο οποίο το μνημονικό περιεχόμενο ενσωματώνεται (Buzsaki 1989, Wang&Morris 2010, Inostroza&Born 2013). Συσσωρευμένες αποδείξεις προτείνουν ότι η συνεισφορά του ιπποκάμπου στην διαδικασία της παγίωσης της μνήμης πραγματοποιείται μέσω της πληθυσμιακής του δραστηριότητας που καλείται Οξύαιχμα κύματα-ριπιδισμοί (SWRs) (O Keefe and Nadel 1978, Buzsaki 1986). Πρόσφατα, καταδείχτηκε μία άμεση συσχέτιση μεταξύ των SWRs και της μνήμης/μάθησης (Eschenko et al 2008, Ramadan et al 2009, Singer and Frank 2009). Αναλόγως, η επιδόσεις της μνήμης μειώνονται όταν τα SWRs διαταράζονται (Girardeau et al 2009, Ego-Stengel and Wilson 2010). Η παραγωγή των SWRs περιλαμβάνει ενεργοποίηση της μεσολαβούμενης από GABA A R διαβίβασης (Papatheodoropoulos and Kostopoulos 2002, Wu et al 2002b, Maier et al 2003, Papatheodoropoulos 2010). Επί προσθέτως, συγκεκριμένοι τύποι ενδονευρώνων ενεργοποιούνται κατά τη διάρκεια των SWRs (Csicsvari et al 1999a, Klausberger and Somogyi 2008). Αποτελέσματα από προηγούμενες μελέτες στον ιππόκαμπο, έχουν προτείνει ότι μερικές όψεις της GABAεργικής διαβίβασης μειώνονται με την ηλικία (De Jong et al 2006, Moradi-Chameh et al 2014). Επίσης, μεταξύ πολλών συσχετιζόμενων με την ηλικία αλλαγών με αναμενόμενο αντίκτυπο στην ιπποκάμπεια λειτουργία συμβαίνουν στους N-Μεθυλ-D-ασπαρτικού οξέος υποδοχείς (NMDARs) (Magnusson et al 2010). Κάποιες σημαντικές πτυχές των SWRs όπως το πλάτος και η συχνότητα περιλαμβάνουν την ενεργοποίηση των NMDARs (Colgin et al 2005, Papatheodoropoulos 2010). Επί προσθέτως, η γήρανση συνοδεύεται από αλλαγές σε ένα άλλο κυτταρικό συστατικό που συμμετέχει στην μνήμη/μάθηση, τα L-τύπου τασεοελεγχόμενα κανάλια ασβεστίου (Lvdccs) (Moyer and Disterhoft 1994, Kumar et al 2009, Nunez-Santana et al 2013). 116
129 3.2. Σκοπός Στην παρούσα μελέτη στοχεύσαμε στη διερεύνηση των πιθανών διαφορών των SWRs μεταξύ τομών από ενήλικους και γερασμένους αρουραίους και να εξετάσουμε τις επιδράσεις των NMDARs και L-vdccs σε αυτή τη δραστηριότητα. Βρήκαμε ότι η ηλικία συνοδεύτηκε από μείωση στην ταλάντωση των ριπιδισμών και μία αυξημένη τάση παραγωγής SWRs σε μορφή ακολουθιών. Επίσης, οι αλλαγές στους ριπιδισμούς συνοδεύτηκαν από αντίστοιχες αλλαγές στην κυτταρική δραστηριότητα των συμπλόκων δυναμικών Πειραματική διαδικασία Προετοιμασία τομών Στη παρούσα μελέτη παρασκευάστηκαν εγκάρσιες τομές από ενήλικους (3-6 μηνών, gr) και ηλικιωμένους (24-34 μηνών, gr) Wistar αρουραίους. Όλη η πειραματική μεταχείριση και διαδικασίες περιγράφονται παραπάνω στις γενικές μεθόδους. Επισημαίνεται ότι χρησιμοποιήθηκαν τομές μόνο από τον κοιλιακό πόλο επειδή εμφανίζουν μεγαλύτερη πιθανότητα αυθόρμητης παραγωγής οξύαιχμων κυμάτωνριπιδισμών σε σύγκριση με τομές από την υπόλοιπη δομή (Papatheodoropoulos and Kostopoulos, 2002) Ηλεκτροφυσιολογία, τύποι δραστηριότητας και ανάλυση δεδομένων Καταγράφηκε η αυθόρμητη και προκλητή δραστηριότητα πεδίου από την πυραμιδική στοιβάδα του CA1b πεδίου με τη χρήση ηλεκτροδίων από ίνα άνθρακα. Σε μερικά πειράματα πραγματοποιήθηκαν καταγραφές και στο CA3 πεδίο. Το καταγεγραμμένο σήμα ενισχύθηκε και φιλτραρίστηκε όπως αναφέρεται στις γενικές μεθόδους (Υποκεφάλαιο Α.ΙΙΙ.4.2) Αυθόρμητη δραστηριότητα Η αυθόρμητη δραστηριότητα κατηγοριοποιήθηκε σε δικτυακή και δραστηριότητα μοναδιαίων κυττάρων (single cell or unit activity). Η δικτυακή δραστηριότητα αποτελούταν από συγχρονισμένα αργά δυναμικά πεδίου συνοδευόμενα από μια παροδική γρήγορη ταλάντωση με συχνότητα περίπου 160 Hz και μία σχετικά έντονη δραστηριότητα πολλαπλών δυναμικών από πληθυσμό κυττάρων (MUA) (Εικόνα Β.Ι.3.1Α-Β). Πρότερες in vitro μελέτες (Papatheodoropoulos and Kostopoulos, 2002, Kubota et al., 2003, Maier et al., 2003, Behrens et al., 2005, Wu et al., 2005, Papatheodoropoulos, 2010) έχουν δείξει ότι 117
130 αυτή η δραστηριότητα πεδίου μοιράζεται πολλά χαρακτηριστικά με τα in vivo οξύαιχμα κύματα-ριπιδισμούς που καταγράφονται από τον ιππόκαμπο σε συμπεριφορικές μελέτες με αρουραίους (O'Keefe and Nadel, 1978, Buzsaki et al., 1992). Συνεπώς, θα αναφερόμαστε σε αυτήν την συγχρονισμένη δραστηριότητα ως οξύαιχμα κύματα (sharp waves-spws), Ριπιδισμοί (ή ταλάντωση ριπιδισμού-ripples) και η δραστηριότητα δυναμικών ενέργειας πληθυσμού κυττάρων (multiunit activity-mua). Το σύμπλοκο συνύπαρξης αυτών των συστατικών θα αναφέρεται ως οξύαιχμα κύματα-ριπιδισμοί (Sharp waves-ripples, SWRs). Για να μετρήσουμε τις ποικίλες παραμέτρους αυτής της δραστηριότητας, την διαχωρίζουμε σε οξύαιχμα κύματα με low-pass φιλτράρισμα του αρχικού σήματος στα 40 Hz και σε ταλάντωση ριπιδισμού με band-pass στα Hz αντίστοιχα (Εικόνα Β.Ι.3.1Β). Η MUA που συνοδεύει τα SWRs αποκαλύπτεται με φιλτράρισμα της αρχικής καταγραφής στα 400 Hz-2 ΚHz. Τα οξύαιχμα κύματα ανιχνεύτηκαν στις φιλτραρισμένες καταγραφές μετά από οριοθέτηση ενός κατώφλιου επιπέδου όπου όλα τα υποτιθέμενα γεγονότα εξακριβώθηκαν μετά από πιστοποίηση μέσω οπτικού ελέγχου. Οι Ριπιδισμοί και η MUA ανιχνεύτηκαν στις φιλτραρισμένες καταγραφές μετά την οριοθέτηση κατώφλιου στις τέσσερις φορές της τυπικής απόκλισης του βασικού θορύβου που δεν εμπεριέχει SWR δραστηριότητα. Τα γεγονότα κατηγοριοποιήθηκαν σαν ριπιδισμοί μόνο όταν παρατηρήθηκαν επεισόδια τουλάχιστον δύο συνεχόμενων αρνητικών αποκλίσεων με καθυστέρηση μεταξύ τους τουλάχιστον 3 ms και όχι περισσότερο από 11 ms. Το κατώφλιο επιβεβαιώθηκε περαιτέρω και μέσω οπτικής παρατήρησης. Τα οξύαιχμα κύματα που εμφανίζονταν είτε ως μοναδιαία γεγονότα είτε με το σχηματισμό δύο ή περισσοτέρων συνεχόμενων γεγονότων ονομάζονται σύμπλεγμα ή αλληλουχίες. Το πρώτο SWR σε ένα σύμπλεγμα αναφέρεται ως πρωτεύον γεγονός, ενώ τα ακόλουθα καλούνται δευτερεύοντα γεγονότα. Τα συμπλέγματα ή αλληλουχίες των γεγονότων διαχωρίζονταν ξεκάθαρα από τα μοναδιαία γεγονότα εξαιτίας του χρονικού διαστήματος μεταξύ διαδοχικών γεγονότων σε μία αλληλουχία που ήταν σύντομο και αυστηρώς σταθερό (~100ms) σε μία δεδομένη τομή και ανάμεσα σε τομές όταν συγκρίνονταν με το διάστημα μεταξύ διακριτών επεισοδίων (αποτελούμενα από ένα απομονωμένο γεγονός ή ένα σύμπλεγμα). Η ύπαρξη των αλληλουχιών μπορεί να αποκαλυφθεί σε ένα ιστόγραμμα κατανομής των διαστημάτων μεταξύ των γεγονότων (Εικόνα Β.Ι.3.1C). Χαρακτηριστικά, αυτά τα ιστογράμματα έδειξαν δύο ευκρινώς διακριτές κορυφές μιας δίκορφης κατανομής. Από τα ιστογράμματα κατανομής κάθε τομής καθορίσαμε το εύρος των σύντομων και μεγάλων διαστημάτων και χρησιμοποιήσαμε αυτές τις μετρήσεις για να ταξινομήσουμε την δραστηριότητα σε αλληλουχίες και απομονωμένα γεγονότα. Ένα πρόσθετο κριτήριο στον καθορισμό των αλληλουχιών ήταν η συνήθως 118
131 βαθμιαία αλλαγή του πλάτους των γεγονότων μέσα σε μία αλληλουχία (εμφανές σε σχεδόν όλα τις εικόνες των SWRs). Τα γεγονότα της SWRs δραστηριότητας ποσοτικοποιήθηκαν για: 1) το πλάτος του οξύαιχμου κύματος καθορισμένο ως η διαφορά τάσης μεταξύ της θετικής κορυφής και της βασικής. Στις αλληλουχίες, τα πρωτεύοντα και τα δευτερεύοντα γεγονότα μετρήθηκαν ξεχωριστά, 2) το διάστημα μεταξύ των γεγονότων (inter-event interval, IEI) καθορίστηκε ως το χρονικό διάστημα μεταξύ διαδοχικών ξεχωριστών γεγονότων ανεξάρτητα με το εάν εμφανίζονταν ως απομονωμένα γεγονότα ή αλληλουχίες, 3) το διάστημα μεταξύ αλληλουχιών γεγονότων (Intra-cluster, ICI), καθορίστηκε ως ο μέσος όρος των διαστημάτων ανάμεσα στα διαδοχικά γεγονότα SWR με σα σε μία αλληλουχία, 4) τον αριθμό των γεγονότων των SWRs ανά λεπτό 5) τη πιθανότητα εμφάνισης αλληλουχιών που υπολογίστηκε με τον αριθμό εμφανίσεων των αλληλουχιών διαιρεμένο με τον συνολικό αριθμό των επεισοδίων. Οι ριπιδισμοί ποσοτικοποιήθηκαν για: 1) το πλάτος του γεγονότος ριπιδισμού, που καθορίστηκε ως η διαφορά τάσης μεταξύ της μέγιστης θετικής και αρνητικής τιμής κάθε ριπιδισμού 2) τη διάρκεια του γεγονότος ριπιδισμού 3) τον αριθμό των κύκλων ριπιδισμού σε ένα γεγονός ριπιδισμού υπολογισμένος ως ο αριθμός των αρνητικών αποκλίσεων μέσα στο γεγονός 4) τη συχνότητα ριπιδισμού που καθορίστηκε ως ο μέσος όρος των ενδο-ριπιδικών διαστημάτων. Οι μετρήσεις των αυθόρμητων δυναμικών έγιναν από καταγραφές που αποκτήθηκαν περίπου τρεις με πέντε ώρες μετά την τοποθέτηση των τομών στο υδατόλουτρο. Σε κάθε τομή ή πειραματική συνθήκη οι μετρήσεις των γεγονότων των SWRs σχετικά με το πλάτος και τα διαστήματα πραγματοποιήθηκαν από μία 2-λεπτη καταγραφή. Η πιθανότητα των αλληλουχιών υπολογίστηκε από μία μονόλεπτη καταγραφή. Μετρήσεις των ριπιδισμών έγιναν από περίπου τριάντα πρωτεύοντα γεγονότα. Η multiunit δραστηριότητα (MUA) μέσα στα γεγονότα των SWRs ποσοτικοποιήθηκαν μέσω του διαστήματος μεταξύ των δυναμικών. Η δραστηριότητα των μοναδιαίων δυναμικών ενέργειας οργανωμένη σε ριπές που συνέβαιναν ξεχωριστά από τα γεγονότα των SWRs κατηγοριοποιήθηκε σαν σύμπλοκα 119
132 δυναμικών (complex spikes-cs), που έχουν προηγουμένως παρατηρηθεί και περιγραφεί in vivo (Fox and Ranck, 1975; Fox and Ranck, 1981; Ranck, 1973) (Εικόνα Β.Ι.3.1D). Σύμφωνα με προηγουμένως περιγραφόμενα χαρακτηριστικά των CS, η ανίχνευση και ανάλυση των υποτιθέμενων CS ριπών στην παρούσα μελέτη έγιναν μέσω οπτικής παρατήρησης στις αρχικές καταγραφές υπακούοντας τα παρακάτω κριτήρια: Α) η δραστηριότητα των ριπών καταγράφηκε στην πυραμιδική στοιβάδα Β) οι ριπές αποτελούνταν από δύο μέχρι έξι δυναμικά Γ) το πλάτος των δυναμικών εντός της ριπής πολύ συχνά μειωνόταν από το πρώτο μέχρι το τελευταίο δυναμικό Δ) το διάστημα ανάμεσα στα δυναμικά (inter-spike interval, ISI) κυμαίνονταν από 2 έως 12 ms. Τα κριτήρια της διαδικασίας διάκρισης που χρησιμοποιήθηκαν για να συναθροιστούν τα επεισόδια των ριπών των CS κυττάρων σε διακριτές ομάδες (π.χ. κύτταρα) περιλάμβαναν το εικόνα του πρώτου και των ακόλουθων δυναμικών, το πλάτος των δυναμικών και η σταθερότητα του πλάτους του πρώτου δυναμικού από ριπή σε ριπή. Επίσης, λήφθηκε υπόψη και ο αριθμός των δυναμικών από τη στιγμή που αναφέρθηκε ότι η τάση των μεμονωμένων κυττάρων να πυροδοτούν συγκεκριμένο αριθμό δυναμικών ενέργειας μέσα σε μία ριπή είναι σχετικά σταθερή στο χρόνο (Suzuki and Smith, 1985). Από την άλλη πλευρά όμως, δεδομένου ότι οι ριπές που παράγονται από ένα δεδομένο πυραμιδικό κύτταρο ίσως αλλάζουν με την πάροδο του χρόνου (Ranck, 1973) χρησιμοποιήσαμε το κριτήριο του αριθμού των δυναμικών με προσοχή και μόνο όταν η δραστηριότητα ικανοποιούσε τα άλλα κριτήρια. Όποτε ήταν δύσκολος ο διαχωρισμός των ριπών σε διαφορετικά κύτταρα ακολουθώντας τα παραπάνω κριτήρια υποθέσαμε ότι οι διαφορετικές ριπές παράγονταν από ένα μοναδικό CS κύτταρο. Επί προσθέτως, μόνο οι ριπές με σχετικά μεγάλο πλάτος του πρώτου δυναμικού ( μv) επιλέγονταν για ανάλυση. Μετρήσεις των CS έγιναν από συνεχείς καταγραφές τουλάχιστον δέκα λεπτών. Η ποσοτικοποίηση των CS ριπών περιλάμβανε τον αριθμό των δυναμικών, τα διαστήματα μεταξύ διαδοχικών δυναμικών σε μία ριπή (inter-spike interval, ISI) καθώς επίσης και το μέσο ISI σε κάθε κύτταρο. 120
133 Προκλητή Δραστηριότητα Οι προκλητές συναπτικές απαντήσεις αποτελούμενες από διεγερτικά μετασυναπτικά δυναμικά πεδίου (fepsp) και πληθυσμιακά δυναμικά (PS) καταγράφηκαν μέσω της επίδοσης ηλεκτρικών παλμών όπως περιγράφεται αναλυτικά στις γενικές μεθόδους ( υποκεφάλαιο Α.ΙΙΙ.5). Το PS καταγραφόταν συνεχώς για να καθορίσουμε την σταθερότητα της απόκρισης. Μόνο οι τομές με σταθερές αποκρίσεις για τουλάχιστον δέκα λεπτά επιλέχθησαν για περαιτέρω πειραματισμό. Για να εξετάσουμε τα αποτελέσματα της γήρανσης και των φαρμάκων στην συναπτική δραστηριότητα, την νευρωνική διεγερσιμότητα και την GABA-εργική αναστολή κατασκευάσαμε καμπύλες εισόδου/εξόδου του fepsp και του PS. Η συναπτική αποτελεσματικότητα ποσοτικοποιήθηκε μετρώντας το fepsp σε κατώφλια ένταση ερεθισμού (fepsp thr ), την ένταση του ρεύματος ερεθισμού που απαιτήθηκε για να παραχθεί το 50% του fepsp (I 50- fepsp) και το μέγιστο fepsp (fepsp max ). Η νευρωνική διεγερσιμότητα ποσοτικοποιήθηκε μετρώντας την ένταση του ρεύματος ερεθισμού που απαιτήθηκε για να παραχθεί το 50% του PS (Ι 50-PS ), το μέγιστο PS (PS max ), τη μετασυναπτική διέγερση που απαιτήθηκε για να παραχθεί το 50% του PS (EPSP 50 ), και την αναλογία μεταξύ του 50% του PS και του αντίστοιχου fepsp (PS/fEPSP). Η δύναμη της αναστολής ποσοτικοποιήθηκε χρησιμοποιώντας το πρωτόκολλο ερεθισμού με σύζευξη παλμών όπως περιγράφεται στις γενικές μεθόδους (υποκεφάλαιο Α.ΙΙΙ.6). Η καταστολή εκφράζεται σαν μία προς τα δεξιά και προς τα κάτω μεταβολή της καμπύλης εισόδου/εξόδου του PS/I, η οποία συνεπώς πάρθηκε σαν μέτρο δύναμης της παλίνδρομης αναστολής του τοπικού δικτύου. Η προς τα δεξιά μεταβολή ποσοτικοποιήθηκε μέσω του ποσοστού αλλαγής στο I 50-PS (shift of I 50-PS ), η προς τα κάτω μεταβολή ποσοτικοποιήθηκε από την αναλογία PS2/PS1 στην ένταση ερεθισμού όπου προκλήθηκε το 50% του PS Συμπεριφορική (ανοικτού πεδίου) δοκιμασία Για να μπορέσουμε να μελετήσουμε τις κινητικές και εξερευνητικές δραστηριότητες των επίμυων, εκτελέσαμε την δοκιμασία ανοικτού πεδίου (open field δοκιμασία) (Walsh and Cummins, 1976) σε ένα υποσύνολο ενήλικων και ηλικιωμένων επίμυων. Η συσκευή αποτελούταν από μία φωτεινή κίτρινη τετράγωνη περίφραξη από κόντρα πλακέ, με διαστάσεις 90 x 90 cm με 39 cm τοίχο και το πάτωμα διαιρεμένο σε 18 x 18 cm τετραγωνικές μονάδες με χρήση μπλε λωρίδων. Τα πειραματόζωα αφήνονταν με προσοχή σε μία από τις γωνίες της συσκευής (ίδια για όλα τα ζώα) και τους επιτρεπόταν να εξερευνήσουν ελεύθερα στην περιοχή για δέκα λεπτά ενώ καταγράφονταν συνεχώς μέσω 121
134 κάμερας για κατοπινή ανάλυση. Η δοκιμασία για κάθε ζώο εκτελέστηκε εις διπλούν με μία μέρα κενό και υπολογιστήκαν για κάθε παράμετρο οι μέσοι όροι. Οι ακόλουθοι πέντε παράμετροι μετρήθηκαν σε αυτή τη δοκιμασία (Ijomone et al., 2014): α) χρόνος κίνησης (ποσοστό κίνησης επί του συνολικού χρόνου στο κουτί) β) συχνότητα κίνησης (αριθμός των διασταυρούμενων γραμμών από το ένα τετράγωνο στο άλλο όταν περνούσε τα 2/3 του ζώου) γ) συχνότητα ανασήκωσης (αριθμός φορών που το ζώο στεκόταν με τα πίσω του πόδια όρθιο στο χώρο ή ακουμπώντας τον τοίχο) δ) είσοδοι σε κεντρικά τετράγωνα (αριθμός φορών που ο επίμυς εισήρθε σε κεντρικά τετράγωνα) ε) περιποίηση (αριθμός φορών που ο επίμυς γλειφόταν ή ξυνόταν ενώ ήταν ακίνητος). Η συσκευή καθαριζόταν με 50% αιθανόλη πριν την δοκιμασία του καινούριου ζώου για να περιορίσουμε πιθανή προτίμηση λόγω οσμών από το προηγούμενο ζώο Φάρμακα Χρησιμοποιήθηκαν τα ακόλουθα φάρμακα: α) CPP, 10μΜ, (35-40 λεπτά) και β) η νιφεδιπίνη 20μΜ, (65-70 λεπτά). Αναλυτική περιγραφή και τρόπος διάλυσης των φαρμάκων αναλύονται στις γενικές μεθόδους, υποκεφάλαιο Α.ΙΙΙ.3. Οι μετρήσεις των ποικίλων παραμέτρων των δυναμικών πεδίου αποκτήθηκαν τα τελευταία 1-5 λεπτά ή 10 λεπτά πριν και κατά τη διάρκεια της εφαρμογής του φαρμάκου για την αυθόρμητη δραστηριότητα ή τα προκλητά δυναμικά, αντίστοιχα Στατιστική Ανάλυση Οι τιμές στο κείμενο εκφράζονται σαν Μέσος όρος + μέσο τυπικό σφάλμα (Mean+SEM) ή Μέσος όρος+μέση τυπική απόκλιση (Mean+SD). Οι μετρήσεις που έγιναν σε ατομικές τομές που αποκτήθηκαν από ένα δεδομένο ζώο συνενώθηκαν. Κατά μέσο όρο, από κάθε ενήλικο και γέρικο ζώο χρησιμοποιήθηκαν 1.7 και 2.0 τομές αντίστοιχα. Στα φαρμακολογικά πειράματα ο αριθμός των τομών εξισωνόταν με τον αριθμό των ζώων. Στατιστικές συγκρίσεις ανάμεσα στις δύο ηλικιακές ομάδες πραγματοποιήθηκαν χρησιμοποιώντας τον αριθμό των ζώων, μετά τον υπολογισμό της μέσης τιμής από όλες τις τομές που αποκτήθηκαν από κάθε ζώο. Παρόλα αυτά, οι συγκρίσεις σχετικές με την ηλικία για την πυροδότηση των συμπλόκων δυναμικών έγιναν με τον αριθμό των κυττάρων. Η μη παραμετρική Mann-Whitney U δοκιμασία και η δοκιμασία Wilcoxon καθώς επίσης και ο two-tailed συσχετισμός χρησιμοποιήθηκαν για την στατιστική αξιολόγηση των εξαρτημένων από την ηλικία διαφορών. 122






. Εικόνα Β.Ι.3.")







 στα 80-300Hz")
( ριπιδική ταλάντωση")

.")
135 3.4. Αποτελέσματα Τα SWRs αναπτύχτηκαν προοδευτικά σε τομές και απόό ενήλικους και γέρικους αρουραίους κατά τη διάρκεια της διατήρησής τους στο υδατόλουτρο καταγραφής.. Τα SWRs σταθεροποιήθηκαν σε πλάτος και στο ρυθμό εμφάνισής τους για περίπου δύο με τρεις ώρες μετά την τοποθέτησηη των τομών στο υδατόλουτρο και παρέμειναν αμετάβλητα τουλάχιστον για τις επόμενες τρις ώρες. (Εικόνα Β.Ι.3.1Ε). Εικόνα Β.Ι.3.1: Τύποι αυθόρμητης δραστηριότη ητας που κατεγράφησαν από τομές ιπποκάμπου Α) Παράδειγμα δραστηριότητας οξύαιχμου κύματος-ριπιδισμού από την πυραμιδική π στοιβάδα της CA1 περιοχής τομής του ιπποκάμπου που πάρθηκε απόό 26 μηνών γέρικο αρουραίο. Σημειωτέον ότι τα SWRs εμφανίζονταν σε επεισόδια αποτελούμενα από έναα ή και περισσότερα γεγονότα. Β) Δείχνεται ένα επεισόδιο που αποτελείται από τρία SWR σύμπλοκα. Φιλτράροντας την αρχική καταγραφή (πάνω ίχνος) στα Hz φανερώνεται η υψηλής συχνότητας (~160Hz)( ριπιδική ταλάντωση (μεσαίο ίχνος), ενώ φιλτράρονταςς στα 400Hz-2kH αποκαλύπτεται η multiunit δραστηριότητα (MUA) (κάτω ίχνος). Η καταγραφή πάρθηκε από 32 μηνών αρουραίο. C) Ιστόγραμμα κατανομής των διαστημάτων μεταξύ των γεγονότων γ (ΙΕΙ) αποτυπώνοντας τις δύο κορυφές που αντιστοιχούν στα σύντομα διαστήματα ανάμεσα στα διαδοχικά γεγονότα των 123
136 αλληλουχιών των SWRs και τα μεγάλα διαστήματα ανάμεσα στα επεισόδια της δραστηριότητας. Τα δεδομένα αποκτήθηκαν από δεκάλεπτη καταγραφή τομής από γέρικο αρουραίο. D) Καταγραφή από ενήλικο ζώο που καταδεικνύει την εμφάνιση μίας ριπής από ένα CS κύτταρο (*) απομονωμένο σε σχέση με τα γεγονότα των SWRs (πάνω ίχνος). Η ριπή παρουσιάζεται σε μεγαλύτερη ανάλυση ταχύτητας στο κάτω μέρος. Ε) Scatter διάγραμμα των διαστημάτων μεταξύ των γεγονότων που αποτυπώνει την σταθερότητα της αυθόρμητης δραστηριότητας στον χρόνο. Τα δεδομένα συγκεντρώθηκαν από μία τομή ενός 30-μηνών αρουραίο. Η καταγραφή ξεκίνησε τρεις ώρες μετά την τοποθέτηση της τομής στο υδατόλουτρο καταγραφής. F) Ταυτόχρονη καταγραφή SWRs από την CA3b και CA1b περιοχή, τομής από 32-μηνών αρουραίο, που διευκρινίζει τον ηγετικό ρόλο της CA3 περιοχής στην παραγωγή των SWRs. Η γραμμή τονίζει το χρονικό σημείο όπου η δραστηριότητα ξεκινάει στην CA3. G) Γράφημα Raster και ιστόγραμμα κατανομής των SWRs στην CA1b περιοχή ενεργοποιούμενα από γεγονότα στην CA3b περιοχή. Οι κουκίδες αναπαριστούν τις αυτόματα ανιχνεύσιμες κορυφές που αντιστοιχούν σε οξύαιχμα κύματα, στην low-pass φιλτραρισμένη καταγραφή. Τα δεδομένα αποκτήθηκαν από ενήλικο αρουραίο. Σημειωτέον ότι τα γεγονότα γύρω από τα 100ms μετά την ενεργοποίηση αντιστοιχούν σε δευτερεύοντα γεγονότα στις αλληλουχίες. Προηγούμενες in vivo (Buzsaki 1986, Csicsvari et al 2000) και in vitro μελέτες (Wu et al., 2005, Both et al., 2008) έχουν δείξει ότι τα SWRs στα ενήλικα ζώα ξεκινούν από την CA3 και τότε διαδίδονται στην CA1 περιοχή. Εξετάσαμε την σχέση μεταξύ των δύο ιπποκάμπειων πεδίων καταγράφοντας ταυτόχρονα από την CA3b και CA1b πεδία, με διάστημα μεταξύ των ηλεκτροδίων καταγραφής της τάξης των 2.17±0.08mm και 2.1±0.13 σε τομές από ενήλικα και γέρικα ζώα αντίστοιχα. Και στις δύο ηλικιακές ομάδες, τα SWRs συνέβαιναν αρχικά στην CA3 περιοχή και έπειτα εμφανίζονταν στην CA1 με καθυστέρηση της τάξης των 7.3±1.5ms (n=4) και 8.2±0.7 ms (n=3) σε ενήλικες και γέρικες τομές αντίστοιχα (Εικόνα Β.Ι.3.1F &G) Ταλάντωση Ριπιδισμού Πραγματοποιήσαμε λεπτομερείς μετρήσεις του πλάτους, της διάρκειας, του αριθμού και της συχνότητας των ριπιδισμών σε τομές από ενήλικους (n=33) και γέρικους (n=20) αρουραίους. Το Εικόνα Β.Ι.3.2 δείχνει ότι οι τιμές όλων των παραπάνω παραμέτρων ήταν χαμηλότερες σε τομές γέρικων ζώων. Πιο συγκεκριμένα, τομές από γέρικους αρουραίους έδειξαν μειωμένο αριθμό ριπιδισμών εντός των ριπιδικών γεγονότων (3.96±0.16 vs 4.64±0.17, αντίστοιχα Mann-Whitney U δοκιμασία, P<0.01) και μειωμένη ενδοριπιδική συχνότητα (166.7±2.9 vs 177.3±2.3 αντίστοιχα; Mann-Whitney U δοκιμασία, P<0.01). 124
 Δείχνονται οι αρχικές καταγραφέςς των SWRs από έναν ενήλικο και γέρικο αρουραίο και οι αντίστοιχες")
 Παραδείγματα φασμάτων ισχύος (power spectrum) που αποτυπώνουν την ριπιδική ταλάντωση που εμφανίζει")
 Συγκριτικά δεδομένα ποικίλων παραμέτρων της ριπιδικής ταλάντωσης μεταξύ ενηλίκων και γέρικων ζώων.")
.. 3.4.2.")




137 Εικόνα Β.Ι.3.2: Ριπιδική ταλάντωση σε ενήλικα και γέρικα ζώα. Α) Δείχνονται οι αρχικές καταγραφέςς των SWRs από έναν ενήλικο και γέρικο αρουραίο και οι αντίστοιχες band-pass φιλτραρισμένες σαρώσεις που αποκαλύπτουν την ριπιδική ταλάντωση (κατωτέρα ίχνη). Β) Παραδείγματα φασμάτων ισχύος (power spectrum) που αποτυπώνουν την ριπιδική ταλάντωση που εμφανίζει χαμηλότερηη συχνότητα σε τομές από γέρικους σε σύγκριση με τους ενηλίκους αρουραίους. C) Συγκριτικά δεδομένα ποικίλων παραμέτρων της ριπιδικής ταλάντωσης μεταξύ ενηλίκων και γέρικων ζώων. Οι αστερίσκοι υποδηλώνουνν στατιστικά σημαντικές διαφορές μεταξύ των δύο ηλικιακών η ομάδων με τιμές στα P<0.01 (Mann-Whitney U δοκιμασία) Πυροδοτήσεις συμπλόκων δυναμικών In vivo καταγραφές από την CA1 περιοχή έχουν δείξει ότιι τα πυραμιδικά κύτταρα ενεργοποιούνται εντόνως κατά τη διάρκεια των SWRs (Chrobak and Buzsaki, 1996, Csicsvari et al., 1999a) πυροδοτώντας σύντομες ριπές δυναμικών (Buzsaki, 1986, Kudrimoti et al., 1999). Εξαιτίας αυτού του τύπου πυροδότησης, τα CA1 πυραμιδικά κύτταρα ονομάζονται κύτταραα με σύμπλοκα δυναμικών (complex spikes cells) (Ranck, 1973). Στην παρούσαα μελέτη πράγματι παρατηρήθηκε καθαρή δραστηριότητα με σύμπλοκα δυναμικών να συμβαίνει κατά τη διάρκειαα των SWRs (Εικόνα Β.Ι.3.3Α). Β Επί προσθέτως, σαρώνοντας τις μακράς διάρκειας καταγραφές της αυθόρμητης δραστηριότητας ανιχνεύσαμε τη παρουσία δραστηριότηταςς από απομονωμένα κύτταρα ανάμεσα στα SWR γεγονότα σε τομές και από τις δύο ηλικιακές ομάδες (Εικόνα Β.Ι.3.3Β). Αυτή η πυροδότηση των συμπλόκων από δυναμικά παρουσίαζε πολλά από τα τυπικά χαρακτηριστικά της δραστηριότητας των συμπλόκων με δυναμικά που καταγράφεται in 125
138 vivo από τα CA1 πυραμιδικά κύτταρα (Ranck, 1973, Fox and Ranck, 1975, 1981, McNaughton et al., 1983, Suzuki and Smith, 1985, Harris et al., 2001) (δείτε επίσης τις μεθόδους Β.Ι.3.3.3). Αναλύσαμε αυτή την δραστηριότητα των κυττάρων με σύμπλοκα δυναμικών κατά τη διάρκεια της μη-ταλαντωτικής φάσης (ανάμεσα στα SWR γεγονότα), από τη στιγμή που η αναγνώριση των πυροδοτήσεων των συμπλόκων με δυναμικά κατά τη διάρκεια των SWRs δεν ήταν εφικτή. Αναλύσαμε συνολικά 52 υποτιθέμενα κύτταρα με σύμπλοκα δυναμικών από 22 ενήλικα και 30 γέρικα κύτταρα. Σύμφωνα με προηγούμενες αναφορές (Chrobak and Buzsaki, 1996, Csicsvari et al., 1999b), (Εικόνα Β.Ι.3.3C). Οι ριπές των CS κυττάρων αποτελούνται συνήθως από δύο έως πέντε δυναμικά (μέσος αριθμός των δυναμικών 3.0±0.11 δυναμικά, 52 κύτταρα) και παρουσίαζαν ένα διάστημα μεταξύ των δυναμικών (ISI) που κυμαινόταν από 2 έως 12 ms (μέσο διάστημα 7.42±0.11 ms). Η δραστηριότητα των συμπλόκων δυναμικών αποτελούνταν κυρίως από δύο διαδοχικά δυναμικά, ενώ τα σύμπλοκα δυναμικών με μεγαλύτερο αριθμό δυναμικών παρατηρήθηκαν λιγότερο συχνά (Εικόνα Β.Ι.3.3D). Παρόλα αυτά, τα CS στα ηλικιωμένα ζώα παρουσίαζαν παρόμοιες πιθανότητες εμφάνισης για δύο ή και περισσότερα δυναμικά. Ο μέσος όρος των δυναμικών ανάμεσα στις δύο ηλικιακές ομάδες ήταν σημαντικά υψηλότερος στους γέρικους απ ότι τα ενήλικους αρουραίους (3.19±0.14 vs 2.76±0.16 αντίστοιχα, Mann-Whitney U δοκιμασία και ANOVA, F=4.055, P<0.05). Επί προσθέτως, το μέσο ISI παρουσιαζόταν μεγαλύτερο στους γέρικους (8.1±0.17 ms) σε σύγκριση με τους ενήλικους (6.78±0.14ms) αρουραίους Mann-Whitney U δοκιμασία και ANOVA, F=29.9, P<0.001) (Εικόνα Β.Ι.3.3E). Τα CS από τα ηλικιωμένα ζώα εμφάνιζαν υψηλότερο εύρος μέσου ISI (από 5.5 μέχρι 7.8 ms) σε σύγκριση με τα ενήλικα (από 6.2 μέχρι 10.2 ms). Η μέση τιμή καθενός από τα τέσσερα ISI στις ριπές των συμπλόκων δυναμικών ήταν μεγαλύτερη στα κύτταρα των ηλικιωμένων αρουραίων σε σύγκριση με τους ενήλικους (Εικόνα Β.Ι.3.3Ε). 126












.")


 και το ιστόγραμμαα της")

.")





139 Εικόνα Β.Ι.3.3: Συγκρίσεις των ριπών με σύμπλοκα δυναμικών ανάμεσα σε ενήλικα και γέρικα ζώα. Α) Καταγραφή από την CA1 πυραμιδική στοιβάδα από τομή ενήλικου ζώου που π δείχνει έναα μία ριπή από ένα κύτταρο με σύμπλοκο δυναμικών να συμβαίνει κατά τη διάρκεια της ανοδικής φάσηςς ενός δευτερεύον γεγονότος (στο πλαίσιο). Αυτό αποτελεί ένα ιδιαίτερα σπάνιο παράδειγμα όπου μπορούμε εύκολα να ξεδιαλύνουμε το CS από την υπόλοιπη MUA δραστηριότητα που συμβαίνει κατά κ τη διάρκεια των SWRs. Β) Παραδείγματα καταγραφών που δείχνουν την εμφάνιση των CS ξεχωριστά από α τα SWRs που αποκτήθηκαν από ενήλικο και γέρικο ζώο. Η ριπή των τ συμπλόκου δυναμικώνν δείχνεται σε μεγέθυνση στα κατωτέρα ίχνη. C) Το πάνω ίχνος είναι μια καταγραφή από ένα γέρικο ζώο που δείχνει δύο SWRs και μία απομονωμένη ριπή ενός CS κυττάρου (αστερίσκος). Η CS ριπή και η αντίστοιχη ριπιδική ταλάντωση δείχνονται μεγεθυμένα στο κάτω αριστερό πάνελ. Το δεξιό πάνελ δείχνει έναν κύκλο μεσοποιημένου ριπιδισμού ρ (πάνω ίχνος) και το ιστόγραμμαα της κατανομής φάσης των τ συμπλόκων δυναμικών σε σχέση με την αρνητική κορυφή του ριπιδισμού (διάγραμμα στο κάτω μέρος). D) Συγκεντρωτικά δεδομένα του μέσου όρου του αριθμού των δυναμικών στις CS ριπές (αριστερό διάγραμμα) και την πιθανότητα εμφάνισης των ριπών με έναν δεδομένο αριθμό δυναμικών (δεξιόό διάγραμμα) σε ενηλίκουςς και γέρικους αρουραίους. Ε) Δείχνονται το ιστόγραμμα των διαστημάτων μεταξύ των δυναμικών για όλα τα CS κύτταρα που μελετήθηκαν (αριστερό γράφημα), το μέσο διάστημα μεταξύ των δυναμικών στις CS ριπές (μεσαίο γράφημα) και το 1 ο -5 ο διάστημα μεταξύ των δυναμικών (δεξιό γράφημα). Οι αστερίσκοι δηλώνουν τις στατιστικώς σημαντικές διαφορές μεταξύ τωνν δύο ηλικιών στα *P<0.05; ** P<0.005; *** P<0.001 (Mann-Whitney U δοκιμασία). 127





 και ήταν παρόμοια μεταξύ των ενηλίκων και")
 (Εικόνα Β.Ι.3.")

 Δείχνονται ένα γεγονός SWR και ο αντίστοιχος")



 C Ιστόγραμμα κατανομήςς των διαστημάτων μεταξύ των")
140 Δραστηριότητα MUA εντος των οξύαιχμων κυμάτων Η δραστηριότητα πολλαπλών δυναμικώνν από πληθυσμό κυττάρων (Multiunit activity- και MUA) κατά τη διάρκεια των οξύαιχμων κυμάτων αναλύθηκε σε δώδεκαα ενήλικουςς έντεκα γέρικους αρουραίους. Όπως στην περίπτωση του πυροδότησης ριπών με σύμπλοκα δυναμικών από απομονωμένα κύτταρα, η MUA συνέβαινε στις αρνητικές κορυφές της ριπιδικής ταλάντωσης (Εικόνα Β.Ι.3.4Β). Τα διαστήματα μεταξύ των δυναμικών στη MUA ήταν σύντομης διάρκειας, χαμηλότερα από 6-8 ms (Εικόνα Β.Ι.3.4C) και ήταν παρόμοια μεταξύ των ενηλίκων και γέρικων αρουραίων (2.9± ±0.135 vs 2.94±0.1 ms, αντίστοιχα) (Εικόνα Β.Ι.3.4D). Εικόνα Β.Ι.3.4: Multiunit δραστηριότητα κατάά τη διάρκεια των SWRs σε ενηλίκους και γέρικους αρουραίους. Α) Δείχνονται ένα γεγονός SWR και ο αντίστοιχος ριπιδισμός και η MUA που αποκαλύπτονται μέσω φιλτραρίσματος της καταγραφής τωνν SWRs. Β) Δείχνονται η μεσοποιημένη κυματομορφή της ριπιδικής ταλάντωσης και το ιστόγραμμα της κατανομής φάσης της MUA σε σχέση με τις αρνητικότητες της ριπιδικής ταλάντωσης. Τα δεδομένα συγκεντρώθηκαν από 15 λεπτά καταγραφής. C) C Ιστόγραμμα κατανομήςς των διαστημάτων μεταξύ των δυναμικών εντός ε της MUA μετρημένα από 15λεπτηη καταγραφή. Σημειωτέον ότι τα περισσότερα διαστήματαα κυμαίνονται κάτω των 10ms. D) Το μέσο διάστημα μεταξύ των δυναμικών κατά τη διάρκεια της MUA ήταν παρόμοιο μεταξύ των ενηλίκων και γέρικων ζώων. 128
141 Γεγονότα των SWRs Τα SWRs γεγονότα στις τομές τόσο από τους ενήλικους όσο και από τους γέρικους αρουραίους οργανώθηκαν είτε σαν μοναδιαία γεγονότα είτε σε μορφή αλληλουχιών από δύο ή περισσότερα γεγονότα (μέχρι και εφτά) (Εικόνα Β.Ι.3.1Α & Εικόνα Β.Ι.3.5Α). Το πρώτο γεγονός και τα ακόλουθα σε μία αλληλουχία ονομάζονται πρωτεύον και δευτερεύοντα γεγονότα αντίστοιχα. Και τα πρωτεύοντα και τα δευτερεύοντα γεγονότα έχουν παρόμοια πλάτη στις τομές από ενήλικους και γέρικους αρουραίους (73.5±6.6μV και 26.5±1.8μV vs 75.5±9.6μV και 27.7±3.96μV αντίστοιχα, Mann-Whitney δοκιμασία, P>0.05). Δεν βρέθηκε διαφορά στο ποσοστό εμφάνισης των SWRs εκφρασμένο σαν τον συνολικό αριθμό των SWR γεγονότων που παράγονται ανά λεπτό (167.0±5.8 vs 183.0±15.3 γεγονότα στους ενήλικους και γέρικους αρουραίους αντίστοιχα, Mann-Whitney δοκιμασία, P>0.05), στο διάστημα μεταξύ ξεχωριστών γεγονότων (ΙΕΙ) (379.7±10.6 ms vs 380.3±26.1 ms στα ενήλικα και γέρικα ζώα αντίστοιχα, Mann-Whitney δοκιμασία, P>0.05), ανάμεσα στους ενήλικες και ηλικιωμένους αρουραίους (Εικόνα Β.Ι.3.5Β). Το διάστημα μεταξύ διαδοχικών γεγονότων εντός των αλληλουχιών (intral-cluster interval (ICI)), δεν διέφερε μεταξύ των ενήλικων (107.0±2.9 ms, n=35) και των γέρικων αρουραίων (108.3±5.2 ms, n=25). Παρατηρήσαμε διαφορά στο πρότυπο παραγωγής των SWRs μεταξύ γηράσκοντων και ενήλικων αρουραίων. Τομές από γέρικα ζώα εμφάνιζαν μια τάση παραγωγής γεγονότων σε μορφή αλληλουχιών που ήταν στατιστικά σημαντικά υψηλή (Εικόνα Β.Ι.3.5Α & C) (46.0±3.1%, n=25) σε σύγκριση με τα ενήλικα ζώα (37.4±2.6%, n=38), (Mann Whitney U δοκιμασία, P<0.05). Έτσι, οι πιθανότητες εμφάνισης αλληλουχιών με τρία, τέσσερα ή και περισσότερα γεγονότα ήταν σημαντικά υψηλές στους γέρικους απ ότι τους ενήλικους αρουραίους (Εικόνα Β.Ι.3.5C). Η υψηλότερη τάση των γέρικων να εμφανίζουν μεγάλες αλληλουχίες SWRs ίσως υπονοούν ότι αντιπροσωπεύουν μία αλλαγή στον γέρικο ιππόκαμπο έτσι ώστε να εξισορροπηθεί η εξασθενημένη ριπιδική ταλάντωση. Για να εξετάσουμε το εάν αυτές οι δύο παράμετροι είναι αντιστρόφως συσχετιζόμενοι μεταξύ τους, συγκρίναμε την ικανότητα των τομών να παράγουν αλληλουχίες SWRs γεγονότων με τον αριθμό και την συχνότητα των ριπιδισμών σε κάθε δεδομένη τομή. Συγκρίνοντας την αθροιστική πιθανότητα των αλληλουχιών, δεν παρατηρήσαμε συσχετισμό μεταξύ των τιμών των παραμέτρων. Όμως, βρήκαμε μία σημαντική αντίστροφη συσχέτιση μεταξύ της συχνότητας της ριπιδικής ταλάντωσης και της τάσης των τομών να οργανώνουν αλληλουχίες με τρία ή και περισσότερα γεγονότα (r=0.45, P<0.05; one-tailed bivariate correlation). 129

 Συνεχείς καταγραφές των SWRs")









 Συγκεντρωτικά γραφήματα")
")





 Δείχνονται η πιθανότητα")
 και οι")








142 Εικόνα Β.Ι.3.5: Γεγονότα SWRs στα ενήλικα και γέρικα ζώα. Α) Συνεχείς καταγραφές των SWRs από ένα ενήλικο και ένα γέρικο ζώο (ίχνη στα αριστερά και δεξιά αντίστοιχα) ). Σημειωτέον ότι τα επεισόδια των οξύαιχμων κυμάτων στον γέρικο αρουραίοο οργανώνονται σε μεγάλες αλληλουχίες απ ότι στον ενήλικα. Β) Συγκεντρωτικά δεδομένα του πλάτους, του διαστήματος μεταξύ γεγονότων (ΙΕΙ), του ρυθμού εμφάνισης και του διαστήματος μεταξύ των αλληλουχιών (ICI) σε ενήλικα και γέρικα ζώα. Οι μετρήσεις ήταν παρόμοιες μεταξύ των 2 ηλικιών. Για διευκρινιστικούς λόγους, οι μπάρες του τυπικού σφάλματος στο μεσαίο γράφημα δείχνονται σε τυπική απόκλιση. C) Συγκεντρωτικά γραφήματα κατανομής του διαστήματος μεταξύ των γεγονότων ν (ΙΕΙ) που μετρήθηκαν σε ίσο συνολικό χρόνο καταγραφής στις δύο ηλικιακές ομάδες. Οι γηράσκοντες αρουραίοι εμφάνισαν μία έντοναα ξεχωριστή κορυφή στα μικρά διαστήματαα (σχηματισμένη από τα διαστήματα μεταξύ των γεγονότων των αλληλουχιών, α ICI) η οποία ήταν υψηλότερη από αυτήν των ενηλίκων. D) Δείχνονται η πιθανότητα εμφάνισηςς αλληλουχιώνν SWRs (αριστερό γράφημα) και οι ξεχωριστές πιθανότητες εμφάνισηςς αλληλουχιών με 2, 3, 4 και περισσότερων γεγονότων στα ενήλικα και γέρικα ζώα (δεξιό γράφημα). Οι αστερίσκοι δηλώνουν στατιστικά σημαντικέςς διαφορές μεταξύ των τιμών των δύο ηλικιών στα *P<0. 05, **P<0.01; (Mann-Whitney U δοκιμασία) Επιδράσεις των φαρμάκων στα οξύαιχμα κύματα Έχει δειχθεί από προηγούμενες μελέτες ότι οι NMDARs τροποποιούν το πλάτος των οξύαιχμων κυμάτων (Colgin et al., 2005) και παίζουν σημαντικό ρόλο στην ν οργάνωση των αλληλουχιών των SWRs γεγονότων (Papatheodoropoulos, 2010) ). Λαμβάνοντας υπόψη ότι η λειτουργία των NMDARs μεταβάλλεταιι στον γερασμένο ιππόκαμπο (Serra et al., 1994, 130
143 Magnusson et al., 2010) και ότι οι αλληλουχίες είναι μακρύτερες στα γέρικα ζώα, αναρωτηθήκαμε ποία είναι η συμμετοχή των NMDARs στις δύο ηλικιακές ομάδες. Επίσης, στους γερασμένους ιπποκάμπειους πυραμιδικούς νευρώνες, η έκφραση και ενεργότητα των L-τύπου τασεοελεγχόμενων διαύλων ασβεστίου (L-vdccs) είναι ενισχυμένη (Moyer and Disterhoft, 1994, Kumar et al., 2009b, Nunez-Santana et al., 2013). Δεδομένου ότι και οι NMDARs και L-VDCCs είναι στόχοι της διαδικασίας της γήρανσης εμείς θέσαμε ως στόχο να εκτιμήσουμε την συμμετοχή τους στην παραγωγή των SWRs, χρησιμοποιώντας φαρμακολογικούς αποκλειστές ανεξάρτητα για τους NMDARs και L-VDCCs καθώς και σε συνδυασμό. Το Εικόνα Β.Ι.3.6 δείχνει ότι ο ανταγωνιστής των NMDARs, το CPP, που εφαρμόστηκε σε τομές από ενήλικα ζώα ενίσχυσε σημαντικά το πλάτος των πρωτευόντων SWR γεγονότων (κατά 14.0±3.5%) και μείωσε την συχνότητα εμφάνισης τους (αύξησε το IEI κατά 16.3±2.8%). Επί προσθέτως, εφαρμογή του CPP μείωσε την εμφάνιση των αλληλουχιών των SWRs κατά 88±1.7%. Μπλοκάρισμα των NMDARs σε τομές από γερασμένους αρουραίους παρήγαγε παρόμοια αποτελέσματα στο πλάτος και το ΙΕΙ αλλά η μείωση στην πιθανότητα εμφάνισης αλληλουχιών (55.6±6.6%) ήταν σημαντικά χαμηλότερη από αυτή που παρατηρήθηκε στα ενήλικα ζώα (Mann-Whitney U δοκιμασία, P<0.001). Παρατηρήσαμε ότι το CPP απέκλεισε τις αλληλουχίες των SWRs σε τρείς τομές από ενήλικα ζώα αλλά σε καμία από τα γέρικα. Δεδομένου ότι η δραστηριότητα των οξύαιχμων κυμάτων αρχίζει από την CA3 περιοχή, είναι ενδιαφέρον να δούμε εάν το αποτέλεσμα του CPP που παρατηρήσαμε στην CA1 οφείλεται σε δράση του στο CA3 ή στο CA1 κύκλωμα. Συνεπώς, εξετάσαμε την δράση του CPP στο CA3 πεδίο σε εφτά τομές από ενήλικα ζώα. Όπως συνέβη και στην CA1 περιοχή, το CPP απέκλεισε την εμφάνιση των αλληλουχιών στην CA3 περιοχή κατά 93.2±1.4% (p<0.05). Το CPP δεν επηρέασε σημαντικά το ICI σε καμία από τις δύο ηλικιακές ομάδες. Αυτά τα δεδομένα κατέδειξαν ότι τα SWRs είναι λιγότερο ευαίσθητα στο μπλοκάρισμα των NMDARs στα γέρικα απ ότι τα ενήλικα ζώα. Εφαρμογή της Νιφεδιπίνης (20μΜ) παρουσίας του CPP παρήγαγε μία πρόσθετη σημαντική αύξηση στο πλάτος των SWR γεγονότων και στους ενήλικους και στους γέρικους αρουραίους (κατά 14±3% και 11±2% αντίστοιχα) χωρίς να επηρεάζουν καμία από τις άλλες παραμέτρους. Όταν η Νιφεδιπίνη χορηγήθηκε από μόνη της, το μόνο σημαντικό αποτέλεσμα που παρατηρήθηκε ήταν η αύξηση του πλάτους των SWRs στους ενήλικους αλλά όχι στα γέρικους αρουραίους (κατά 18.7±3.0%). Καμία από τις άλλες παραμέτρους των SWRs δεν επηρεάστηκε σημαντικά από την Νιφεδιπίνη. Εφαρμογή του CPP παρουσίας της Νιφεδιπίνης επηρέασε σημαντικά όλες τις παραμέτρους των οξύαιχμων κυμάτων και στα ενήλικα και στα γέρικα ζώα. Αυτές οι μεταβολές ήταν παρόμοιες με αυτές 131




























144 που παρατηρήθηκαν όταν το CPP χορηγήθηκε μόνο του τ (σύγκριση CPP Νιφεδιπίνη+CPP στο Εικόνα Β. Ι.3.6Β). με Εικόνα Β.Ι.3.6: Δράσεις του CPP και της Νιφεδιπίνης στα οξύαιχμα κύματα των ενήλικων και γέρικων ζώων. Α) Παραδείγματαα καταγραφώνν οξύαιχμων κυμάτων από ενηλίκους και γέρικους αρουραίους πριν και κατά τη διάρκεια επώασης με τον ανταγωνιστή των NMDARs, το CPP και του τ blocker των L-τύπου vdccs, την Νιφεδιπίνη. Β) Γραφήματα συγκεντρωτικών αποτελεσμάτων που δείχνουν το αποτέλεσμα του CPPP και της Νιφεδιπίνης στις διάφορες παραμέτρους των οξύαιχμων κυμάτων στα ενήλικα (γραφήματα στα αριστερά) και στα γέρικα (γραφήματα στα δεξιά) ζώα. Τέσσερις φαρμακολογικές καταστάσεις δείχνονται σε κάθε 132
145 γράφημα και ο αριθμός των ενήλικων και γέρικων ζώων που χρησιμοποιηθήκαν ήταν: CPP ( εφαρμογή μόνο του CPP, 16 ενήλικα και 14 γέρικα), CPP+NIF (εφαρμογή της νιφεδιπίνης παρουσία CPP, 11 ενήλικα και γέρικα ζώα), NIF (εφαρμογή μόνο της Νιφεδιπίνης, 21 ενήλικα και 11 γέρικα ζώα) και NIF+CPP (εφαρμογή του CPP παρουσία Νιφεδιπίνης, 12 ενήλικα και 7 γέρικα ζώα). Οι αστερίσκοι δηλώνουν τις στατιστικώς σημαντικές δράσεις των φαρμάκων στα *P<0.05; **P<0.01; ***P< Wilcoxon δοκιμασία και Mann- Whitney U δοκιμασία χρησιμοποιήθηκαν για τις συγκρίσεις των φαρμακολογικών δράσεων εντός της ηλικιακής ομάδας και ανάμεσα στις δύο ηλικίες, αντίστοιχα. Πρέπει να τονιστεί ότι τα αποτελέσματα της συνδυαστικής εφαρμογής των δύο φαρμάκων (π.χ.cpp+nif και NIF+CPP) ήταν στατιστικώς σημαντικές αλλά μόνο οι σημαντικές επιπλέον δράσεις του δεύτερου προστιθέμενου φαρμάκου έχουν σημειωθεί στα γραφήματα. Τα αποτελέσματα του CPP στα ιστογράμματα κατανομής του διαστήματος μεταξύ των γεγονότων (ΙΕΙ) στις δύο ηλικιακές ομάδες δείχνονται στο κάτω μέρος. Σημειωτέον ότι το CPP κατέστειλε ολοκληρωτικά την πρώτη κορυφή της κατανομής στα ενήλικα αλλά όχι στα γέρικα ζώα. Μπλοκάρισμα των NMDARs από το CPP παρήγαγε μία μέτρια αλλά στατιστικά σημαντική ενίσχυση της ριπιδικής ταλάντωσης. Πιο συγκεκριμένα, το CPP στα γέρικα ζώα αύξησε το πλάτος, τη διάρκεια, τον αριθμό και την συχνότητα των ριπιδισμών κατά 7.3±2.5%, 14.9±7.0%, 11.9±4.7% και 3.7±1.4% αντίστοιχα (n=13, P<0.05, Wilcoxon δοκιμασία). Στους ενήλικες αρουραίους το CPP αύξησε σημαντικά το πλάτος και τη συχνότητα της ριπιδικής ταλάντωσης κατά 12,2±4,1% και 4,1±0,5% αντίστοιχα (n=15, P<0.05, Wilcoxon δοκιμασία). Το CPP δεν επηρέασε σημαντικά καμία από τις άλλες παραμέτρους των ριπιδισμών στις τομές από ενήλικους αρουραίους. Η νιφεδιπίνη που χορηγήθηκε παρουσία του CPP δεν παρήγαγε κάποια περαιτέρω σημαντική μεταβολή ούτε στα ενήλικα ούτε στα γέρικα ζώα. Όταν όμως χορηγήθηκε μόνη της, ενίσχυσε σημαντικά το πλάτος των ριπιδισμών στα ενήλικα ζώα (κατά 11,5±3,7%) αλλά όχι στα γέρικα (25,3±17,6%), ενώ δεν προκάλεσε καμία αλλαγή στις άλλες παραμέτρους. Είναι ενδιαφέρον όμως το γεγονός ότι η Νιφεδιπίνη σχεδόν απέκλεισε εξ ολοκλήρου την δράση του CPP στους ριπιδισμούς Επιδράσεις των φαρμάκων στην προκλητή δραστηριότητα Για να εξετάσουμε τις επιδράσεις της ηλικίας στις διεγερτικές και ανασταλτικές συναπτικές ιδιότητες, καταγράψαμε προκλητά δυναμικά πεδίου με ερεθισμό του μονοπατιού των ινών του Schaffer σε 30 τομές από 12 ενήλικους αρουραίους και σε 26 τομές από 10 γερασμένους και κατασκευάζοντας καμπύλες εισόδου/εξόδου (Εικόνα Β.Ι.3.7Α). Οι μέσες τιμές για όλους τους δείκτες φαίνονται στον πίνακα Β.Ι.3.1. Κανένας από τους δείκτες που ποσοτικοποιούν την συναπτική αποτελεσματικότητα και την νευρωνική διεγερσιμότητα δεν διέφερε μεταξύ των δύο ηλικιακών ομάδων. Παρόλα αυτά, 133
146 τομές από γέρικους αρουραίους παρουσίασαν στατιστικά σημαντική μειωμένη αναστολή σε σύγκριση με τους ενήλικους. Πιο συγκεκριμένα, ο ερεθισμός με σύζευξη παλμών παρήγαγε μία σημαντική προς τα δεξιά και προς τα κάτω αλλαγή της καμπύλης PS/I στα ενήλικα (Εικόνα Β.Ι.3.7Β) όταν μετρήθηκε σαν θετική ποσοστιαία αύξηση στην I 50-PS και μείωση στην PS2/PS1 αναλογία. Στους γέρικους αρουραίους ο ερεθισμός με σύζευξη παλμών παρήγαγε μία αρνητική ποσοστιαία αλλαγή του I 50-PS, π.χ. παρήγαγε μία προς τα αριστερά αλλαγή στην καμπύλη PS/I και σημαντική αύξηση του PS2/PS1 λόγου. Για να μελετήσουμε το εάν τα L-VDCCs και οι NMDARs συμμετέχουν στις προκλητές απαντήσεις, επωάσαμε τις τομές με Νιφεδιπίνη (20μΜ) για 50 λεπτά και τότε προσθέσαμε και το CPP (10μΜ) για 30 λεπτά. Εκτελέσαμε αυτά τα πειράματα σε 12 τομές από 12 ενήλικα ζώα και σε 9 τομές από 9 γέρικα ζώα. Όπως φαίνεται στον πίνακα Β.Ι.3.1 και την εικόνα Β.Ι.3.7C, στις τομές από ενήλικα ζώα, η Νιφεδιπίνη παρήγαγε μία στατιστικά σημαντική μείωση του fepsp 50 και αύξηση του PS/fEPSP, οδηγώντας έτσι σε αύξηση της μετασυναπτικής διεγερσιμότητας. Η Νιφεδιπίνη δεν άλλαξε κανέναν από τους άλλους δείκτες στις τομές από τους ενήλικους αρουραίους παρόλο που μπορεί να παρατηρηθεί μία αυξητική τάση της αναστολής (σύγκριση Β με D στο Εικόνα Β.Ι.3.7). Στις τομές από τους γέρικους αρουραίους η νιφεδιπίνη δεν παρήγαγε καμία αλλαγή σε κανέναν δείκτη. Η χορήγηση του CPP παρήγαγε σημαντικές αλλαγές στους δείκτες που ποσοτικοποιούν την μετασυναπτική διεγερσιμότητα και αναστολή παρόμοια και στις δύο ηλικιακές ομάδες (Εικόνα Β.Ι.3.7D). Πιο συγκεκριμένα, το CPP ενίσχυσε σημαντικά την αναλογία PS/fEPSP και παρήγαγε επίσης μία έντονη αύξηση στην προς τα δεξιά μεταβολή του I50-PS, ενώ μείωσε την αναλογία PS2/PS1. Επίσης, βρήκαμε ότι οι φάρμακο επαγόμενες αλλαγές στην αναλογία PS2/PS1 και στην πιθανότητα εμφάνισης SWRs αλληλουχιών ήταν θετικά συσχετιζόμενες (two-tailed correlation δοκιμασία, r=0.587, p<0.05). Το CPP επίσης μείωσε σημαντικά το fepsp 50 σε τομές από γέρικα ζώα αλλά όχι από ενήλικα. 134





 και γέρικους (26")






 ζώο.")


.")






147 Εικόνα Β.Ι.3.7: Συγκρίσεις των προκλητών δυναμικών μεταξύ ενήλικων και κ γέρικων ζώων. Α) Συγκεντρωτικές καμπύλες εισόδου/εξόδου για τις παραμέτρους fepsp/i και PS/Ι δείχνονται στα αριστερά και μεσαία διαγράμματαα για ενήλικους (19 τομές/ /12 ζώα) και γέρικους (26 τομές/10 ζώα) αρουραίους. Το scatter διάγραμμα στα δεξιά δείχνει τη σχέση μεταξύ PS και fepsp για όλες τις τομές πουυ μελετήθηκαν. Β) Καμπύλες εισόδου/εξόδου που αποτυπώνουν το κατασταλτικό αποτέλεσμα του ερεθισμού με σύζευξη παλμών πάνω στο PS2 σε τομές από ένα ενήλικο (αριστερά) και ένα γέρικο (δεξιά) ζώο. Τα βέλη καταδεικνύουν τις τιμές του I50-PS που αντιστοιχεί στις καμπύλες για το PS1 και PS2 (βέλος με διάστικτη και συμπαγή γραμμή,, αντίστοιχα). Παραδείγματα καταγραφών δείχνονται στα ένθετα. Μπάρες βαθμονόμησης: 1mV, 5ms στα ενήλικα και 0.5mVV και 5ms στα γέρικα ζώα. Προσέξτε ότιι η καταστολή του PS2, εκφρασμένη σαν προς τα δεξιά και προς τα κάτω μεταβολή στην αντίστοιχη καμπύλη, είναι απούσα στην τομή από γέρικο ζώο. C) Παραδείγματα καμπυλών εισόδου/εξόδου από ξεχωριστές τομές που δείχνουν τα αποτελέσματα της Νιφεδιπίνης καιι του CPP στην μετασυναπτική διεγερσιμότητα στα ενήλικα και γέρικα 135
148 ζώα. Τα βέλη καταδεικνύουν τις τιμές του fepsp για τις 3 καμπύλες (η τιμή στην control καμπύλη δείχνεται με το διάστικτο βέλος). Σημειωτέον ότι η νιφεδιπίνη παρήγαγε μία προς τα αριστερά αλλαγή του EPSP50 (έτσι αυξάνεται η μετασυναπτική διεγερσιμότητα) στην τομή από το ενήλικο αλλά όχι το γέρικο ζώο. D) Παραδείγματα PS/I καμπυλών για το PS1 και PS2 που δείχνουν το ανασταλτικό αποτέλεσμα της διέγερσης με σύζευξη παλμών πριν και κατά τη διάρκεια της διαδοχικής εφαρμογής της νιφεδιπίνης και του CPP. Σημειωτέον ότι η καταστολή του PS2 (βέλη) ήταν υψηλότερη υπό παρουσία CPP παρά στις control συνθήκες (γεμισμένα και ανοιχτά σύμβολα αντίστοιχα) και στα ενήλικα και στα γέρικα ζώα. Τα παραδείγματα που δείχνονται στο B, C και D αποκτήθηκαν από διαφορετικά πειράματα. Συναπτική Αποτελεσματικότητα Νευρωνική Διεγερσιμότητα Αναστολή fepsp thr I 50-EPSP fepsp max I 50-PS PS max fepsp 50 PS/fEPSP Shift of I 50-PS PS2/PS1 Ενήλικα Control 0.34±0.04 (12) 134.8± ±0.5 (12) 102.8±11.9 (12) 3.7±0.4 (12) 0.83±0.11 (12) 2.4±0.2 (12) 11.0±4.9% (#) (12) 0.88±0.06 % (12) (12) Nife -9.7±5.4% 3.8±3.3% - 5.5±4.9% 0.36±3.6% -8.0±2.1% 9.4±4.2% * 4.1±19.3% - (12) 7.1±4.1% ** 5.8±3.2% Γέρικα + CPP 7.5±10.3% 1.3±1.8% 6.5±7.8% -4.2±3.8% 5.9±3.0% -3.5±3.1% 10.9±2.9% * 138.3±29.3%** - (12) 18.3±3.7 % ** Control 0.5± ± ± ± ± ± ±0.1 (10) 4 (10) (10) (10) (10) (10) (10) -6.6±4.0% * 1.11±0.05 (9) % * (9) Nife 4.4±13.1% 0.06± ±3.9% 11.4±7% -7.1±5.7% 18.5±11.1% 23.7±15.5% 0.8±1.5% (9) % 3.5±5.5% + CPP -0.7±8.4% 1.1±1.5% ±2.1% 1.1±2.8% -6.6±4.1% * 11.7±4.3% * 128.5±57.5%** - (9) 3.4±3.9% 13.7±2.1 % ** Πίνακας Β.Ι.3.1: Προκλητά δυναμικά Οι τιμές στις παρενθέσεις αντιπροσωπεύουν τον αριθμό των πειραματόζωων. Οι αστερίσκοι που εμφανίζονται στην σειρά Old-Control δηλώνουν τις στατιστικά σημαντικές διαφορές μεταξύ των γέρικων ομάδων με τη χρήση Mann-Whitney δοκιμασίας. Οι αστερίσκοι που εμφανίζονται στις σειρές Nifedipine και CPP δηλώνουν στατιστικά σημαντικές διαφορές μεταξύ Control vs Nifedipine και Nifedipine vs CPP με Wilcoxon δοκιμασία. Σημαντικές διαφορές παρατηρήθηκαν στα: * p<0.05 και **p<0.005, αντίστοιχα. Το # δηλώνει σημαντικό αποτέλεσμα του PPS στο I50-PS (Wilcoxon δοκιμασία, P<0.05) Δοκιμασία ανοικτού πεδίου Η κίνηση είναι ένα από τα σημαντικά συστατικά της εξερεύνησης στις δοκιμασίες της μάθησης και μνήμης (Jeltsch et al., 2001; Flores-Abreu et al., 2014). Η χωρική μνήμη και η κίνηση σε ένα ανοικτό πεδίο επηρεάζονται από την δυσλειτουργία του ιπποκάμπου (Jeltsch et al, 2001; Bardgett et al, 2006) και συν-τροποποιούνται από ένα εμπλουτισμένο περιβάλλον (Mohammed et al, 1990; Iuvone et al, 1996). Η κινητική δραστηριότητα μπορεί να χρησιμοποιηθεί σαν ένα μέτρο για να προβλέψουμε την απόδοση της μνήμης σε μία 136





149 δοκιμασίαα ανοικτού πεδίου. Η κινητική δραστηριότητα ίσως αλλάζει με τηνν ηλικία (Welzl et al, 1988) επηρεάζοντας την ικανότητα των ζώων για μάθηση και μνήμη. Για να εξετάσουμε το εάν οι μεταβολές στην αυθόρμητη δραστηριότητα που παρατηρήθηκαν in vitro ίσως συσχετίζονται με πιθανές ηλικιακό-εξαρτώμενες αλλαγές στην κινητική συμπεριφορά, εκτελέσαμε την δοκιμασία ανοικτού πεδίου σε έναα υποσύνολο ενήλικωνν και γέρικων ζώων πριν την εξέταση της νευρωνικής δραστηριότητας στις τομές τους. Μετρήσαμε 6 διαφορετικούς δείκτες ποσοτικοποίησης της συμπεριφοράς των ζώων. Όπως δείχνεται στον πίνακα Β.Ι.3.2, κανένας από αυτούς του δείκτες δεν διέφερε σημαντικά ανάμεσα στις δύο ηλικιακές ομάδες. Ενδιαφέρον βέβαια ήταν το γεγονόςς ότι ο ρυθμός εμφάνισης SWR γεγονότων ήταν αντιστρεπτά συσχετιζόμενος με μ τον χρόνο κίνησης στα ενήλικα ζώα (r=-0. 63, P<0.05) αλλά γραμμικά συσχετιζόμενος στα γηράσκοντα ζώα (r=0.83, P<0.005). Επίσης, δεν ανιχνεύσαμε διαφορές ανάμεσα στα άτομα σε καμία παράμετρο ούτε στα ενήλικα ούτε στα γέρικα ζώα. Πίνακας Β..Ι.3.2. Μετρήσεις παραμέτρων δοκιμασίας ανοικτού πεδίου 3.5. Συζήτηση Η παρούσα μελέτη δείχνει ότι το ιπποκάμπειο κύκλωμα στους γέρικους αρουραίους εμφανίζει εξασθενημένη ριπιδική ταλάντωση και αυξημένηη πιθανότητα οργάνωσης μεγάλων αλληλουχιών SWRs γεγονότων.. Η μειωμένη συχνότητα των ριπιδισμών (π.χ. αυξημένο ενδο-ριπιδικό διάστημα) σε τομές από γέρικουςς αρουραίους συνέβαινε παράλληλα με το αυξημένο διάστημα μεταξύ των δυναμικών στις ριπέςς με σύμπλοκα δυναμικών υπονοώντας ότι οι αλλαγές στην πυροδότηση των πυραμιδικών κυττάρων ίσως ί συνεισφέρουν στην αλλαγμένη ταλάντωση του δικτύου. Επίσης, η συμμετοχή των NMDARs στην παραγωγή των SWRs αλληλουχιών μειώθηκε στις τομέςς από γέρικους αρουραίους. 137
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
ΑΣΚΗΣΗ: MΕΤΡΗΣΗ ΜΕΤΑΣΥΝΑΠΤΙΚΩΝ ΔΥΝΑΜΙΚΩΝ ΣΕ ΤΟΜΕΣ ΙΠΠΟΚΑΜΠΟΥ ΑΡΟΥΡΑΙΟΥ.
 ΑΣΚΗΣΗ: MΕΤΡΗΣΗ ΜΕΤΑΣΥΝΑΠΤΙΚΩΝ ΔΥΝΑΜΙΚΩΝ ΣΕ ΤΟΜΕΣ ΙΠΠΟΚΑΜΠΟΥ ΑΡΟΥΡΑΙΟΥ. Η μετάδοση σημάτων μεταξύ των νευρώνων επιτυγχάνεται μέσω χημικών συνάψεων. Κατά τη χημική συναπτική διαβίβαση απελευθερώνεται από
ΑΣΚΗΣΗ: MΕΤΡΗΣΗ ΜΕΤΑΣΥΝΑΠΤΙΚΩΝ ΔΥΝΑΜΙΚΩΝ ΣΕ ΤΟΜΕΣ ΙΠΠΟΚΑΜΠΟΥ ΑΡΟΥΡΑΙΟΥ. Η μετάδοση σημάτων μεταξύ των νευρώνων επιτυγχάνεται μέσω χημικών συνάψεων. Κατά τη χημική συναπτική διαβίβαση απελευθερώνεται από
Νευροβιολογία των Μνημονικών Λειτουργιών
 Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 3: Ιππόκαμπος και Μνήμη Στυλιανός Κούβαρος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας Μνημονικά Συστήματα Ιππόκαμπος και Μνήμη Πειραματική Μεθοδολογία
Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 3: Ιππόκαμπος και Μνήμη Στυλιανός Κούβαρος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας Μνημονικά Συστήματα Ιππόκαμπος και Μνήμη Πειραματική Μεθοδολογία
Η βιολογία της μάθησης και της μνήμης: Μακρόχρονη ενδυνάμωση/αποδυνάμωση
 Η βιολογία της μάθησης και της μνήμης: Μακρόχρονη ενδυνάμωση/αποδυνάμωση 1 Τι λέμε μνήμη? Η ικανότητα να καταγράφουμε, να αποθηκεύουμε και να ανακαλούμε πληροφορίες Είναι ένας πολύπλοκος συνδυασμός υποσυστημάτων
Η βιολογία της μάθησης και της μνήμης: Μακρόχρονη ενδυνάμωση/αποδυνάμωση 1 Τι λέμε μνήμη? Η ικανότητα να καταγράφουμε, να αποθηκεύουμε και να ανακαλούμε πληροφορίες Είναι ένας πολύπλοκος συνδυασμός υποσυστημάτων
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΜΕ ΤΙΤΛΟ
 ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΜΕ ΤΙΤΛΟ ΠΑΡΑΓΟΝΤΕΣ ΕΞΕΙΔΙΚΕΥΣΗΣ ΤΟΥ ΕΝΔΟΓΕΝΟΥΣ ΝΕΥΡΩΝΙΚΟΥ ΔΙΚΤΥΟΥ ΙΠΠΟΚΑΜΠΟΥ ΕΠΙΜΥΟΣ ΒΑΣΙΛΕΙΟΣ ΠΑΠΑΛΕΩΝΙΔΟΠΟΥΛΟΣ ΒΙΟΛΟΓΟΣ
ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΜΕ ΤΙΤΛΟ ΠΑΡΑΓΟΝΤΕΣ ΕΞΕΙΔΙΚΕΥΣΗΣ ΤΟΥ ΕΝΔΟΓΕΝΟΥΣ ΝΕΥΡΩΝΙΚΟΥ ΔΙΚΤΥΟΥ ΙΠΠΟΚΑΜΠΟΥ ΕΠΙΜΥΟΣ ΒΑΣΙΛΕΙΟΣ ΠΑΠΑΛΕΩΝΙΔΟΠΟΥΛΟΣ ΒΙΟΛΟΓΟΣ
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ Ορίζουμε ως διαβιβαστή μια ουσία που απελευθερώνεται από έναν νευρώνα σε μια σύναψη και που επηρεάζει ένα άλλο κύτταρο, είτε έναν νευρώνα είτε ένα κύτταρο
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Γ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ Ορίζουμε ως διαβιβαστή μια ουσία που απελευθερώνεται από έναν νευρώνα σε μια σύναψη και που επηρεάζει ένα άλλο κύτταρο, είτε έναν νευρώνα είτε ένα κύτταρο
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ
 ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΑΠΕΛΕΥΘΕΡΩΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ ΥΠΟΔΟΧΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ 1 Ηλεκτρικές και Χημικές Συνάψεις 2 Ηλεκτρικές
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΕΣ ΑΠΕΛΕΥΘΕΡΩΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ ΥΠΟΔΟΧΕΙΣ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΩΝ 1 Ηλεκτρικές και Χημικές Συνάψεις 2 Ηλεκτρικές
Γνωστική-Πειραµατική Ψυχολογία
 Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Μηχανισµοί της ΣΔ à Βάση διεργασιών όπως η αντίληψη, µάθηση, εκούσια κίνηση. 10.000 à Μέσος νευρώνας à 1000 (1011 1014). 2 θεµελιώδεις µηχανισµοί ΣΔς:
Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Μηχανισµοί της ΣΔ à Βάση διεργασιών όπως η αντίληψη, µάθηση, εκούσια κίνηση. 10.000 à Μέσος νευρώνας à 1000 (1011 1014). 2 θεµελιώδεις µηχανισµοί ΣΔς:
Συστήματα αισθήσεων. Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
 Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας 2018 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας 2018 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Συστήματα αισθήσεων. Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
 Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Συστήματα αισθήσεων Αισθητικοί υποδοχείς Νευρικές αισθητικές οδοί Συνειρμικός φλοιός και διαδικασία αντίληψης Πρωτοταγής αισθητική κωδικοποίηση
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α. Ο ηλεκτρονικός υπολογιστής του οργανισμού μας
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ
 ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Νευρικό Σύστημα 2 Ανατομία ΝΣ ΝΣ ΚΝΣ (εγκέφαλος + ΝΜ) ΠΝΣ Προσαγωγό (αισθητικό) Απαγωγό Κινητικοί νευρώνες ΑΝΣ Συμπαθητικό Παρασυμπαθητικό 3 Οι διαταραχές της λειτουργίας
ΕΙΣΑΓΩΓΗ ΣΤΟ ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Νευρικό Σύστημα 2 Ανατομία ΝΣ ΝΣ ΚΝΣ (εγκέφαλος + ΝΜ) ΠΝΣ Προσαγωγό (αισθητικό) Απαγωγό Κινητικοί νευρώνες ΑΝΣ Συμπαθητικό Παρασυμπαθητικό 3 Οι διαταραχές της λειτουργίας
Απ. Χατζηευθυμίου Αν Καθηγήτρια Ιατρικής Φυσιολογίας
 Απ. Χατζηευθυμίου Αν Καθηγήτρια Ιατρικής Φυσιολογίας Το 80% περίπου της γεύσης του φαγητού παρέχεται στην πραγματικότητα από την αίσθηση της όσφρησης. Η μυρωδιά μιας ουσίας σχετίζεται άμεσα με τη χημική
Απ. Χατζηευθυμίου Αν Καθηγήτρια Ιατρικής Φυσιολογίας Το 80% περίπου της γεύσης του φαγητού παρέχεται στην πραγματικότητα από την αίσθηση της όσφρησης. Η μυρωδιά μιας ουσίας σχετίζεται άμεσα με τη χημική
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ
 Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
Βιοδυναμικά: Ασθενή ηλεκτρικά ρεύματα τα οποία παράγονται στους ιστούς των ζωντανών οργανισμών κατά τις βιολογικές λειτουργίες.
 Bιοηλεκτρισμός To νευρικό σύστημα Το νευρικό κύτταρο Ηλεκτρικά δυναμικά στον άξονα Δυναμικά δράσης Ο άξονας ως ηλεκτρικό καλώδιο Διάδοση των δυναμικών δράσης Δυναμικά δράσεις στους μύες Δυναμικά επιφανείας
Bιοηλεκτρισμός To νευρικό σύστημα Το νευρικό κύτταρο Ηλεκτρικά δυναμικά στον άξονα Δυναμικά δράσης Ο άξονας ως ηλεκτρικό καλώδιο Διάδοση των δυναμικών δράσης Δυναμικά δράσεις στους μύες Δυναμικά επιφανείας
ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 7-8/ Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ
 ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 7-8/02-03-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ Νευροδιαβιβαστές α) παραγωγή, είδη β) αναγνώριση γ) σηματοδότηση Σε πρώτη προσέγγιση, ο ορισμός του διαβιβαστή είναι: Διαβιβαστής είναι μια ουσία που
ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 7-8/02-03-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ Νευροδιαβιβαστές α) παραγωγή, είδη β) αναγνώριση γ) σηματοδότηση Σε πρώτη προσέγγιση, ο ορισμός του διαβιβαστή είναι: Διαβιβαστής είναι μια ουσία που
Δυναμικό ηρεμίας Δυναμικό ενεργείας. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 30/09/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Ιόντα Δυναμικό
Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 30/09/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Ιόντα Δυναμικό
ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I)
") ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν
Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός. Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας
 Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Θάλαμος Ο θάλαμος: «πύλη προς τον εγκέφαλο» Είναι μια μεγάλη συλλογή νευρώνων στο διεγκέφαλο Παίρνει
Θάλαμος, Φλοιός του Εγκεφάλου & Δικτυωτός Σχηματισμός Α. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Θάλαμος Ο θάλαμος: «πύλη προς τον εγκέφαλο» Είναι μια μεγάλη συλλογή νευρώνων στο διεγκέφαλο Παίρνει
Β. Να επιλέξετε την ορθή απάντηση αναγράφοντας στον πίνακα της ακόλουθης
 Ονοματεπώνυμο:.. Βαθμός: Ωριαία γραπτή εξέταση Α Τετραμήνου στη Βιολογία [Κεφ. 9 ο, σελ. 153-158] Α. Να χαρακτηρίσετε τις ακόλουθες προτάσεις με το γράμμα Ο, εφόσον είναι ορθές, ή με το γράμμα Λ, αν είναι
Ονοματεπώνυμο:.. Βαθμός: Ωριαία γραπτή εξέταση Α Τετραμήνου στη Βιολογία [Κεφ. 9 ο, σελ. 153-158] Α. Να χαρακτηρίσετε τις ακόλουθες προτάσεις με το γράμμα Ο, εφόσον είναι ορθές, ή με το γράμμα Λ, αν είναι
Μνήμη και μάθηση. Συμπεριφορικό επίπεδο Κυτταρικό επίπεδο. Παρασκευή, 5 Απριλίου 13
 Μνήμη και μάθηση Συμπεριφορικό επίπεδο Κυτταρικό επίπεδο Κατηγορίες μνήμης Βραχύχρονη/Μακρόχρονη μνήνη Έκδηλη μνήμη (ιππόκαμπος/φλοιός) Άδηλη μνήμη (αμυγδαλή, παρεγκεφαλίδα) Συνειρμική Μη συνειρμική Διαδικαστική
Μνήμη και μάθηση Συμπεριφορικό επίπεδο Κυτταρικό επίπεδο Κατηγορίες μνήμης Βραχύχρονη/Μακρόχρονη μνήνη Έκδηλη μνήμη (ιππόκαμπος/φλοιός) Άδηλη μνήμη (αμυγδαλή, παρεγκεφαλίδα) Συνειρμική Μη συνειρμική Διαδικαστική
Συνάψεις Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2018
 Συνάψεις Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2018 Σύναψη Σύναψη είναι μια ανατομικά εξειδικευμένη σύνδεση μεταξύ δύο νευρώνων. Σύναψη Στη σύναψη η ηλεκτρική δραστηριότητα
Συνάψεις Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2018 Σύναψη Σύναψη είναι μια ανατομικά εξειδικευμένη σύνδεση μεταξύ δύο νευρώνων. Σύναψη Στη σύναψη η ηλεκτρική δραστηριότητα
Σκοπός του μαθήματος είναι ο συνδυασμός των θεωρητικών και ποσοτικών τεχνικών με τις αντίστοιχες περιγραφικές. Κεφάλαιο 1: περιγράφονται οι βασικές
 Εισαγωγή Ασχολείται με τη μελέτη των ηλεκτρικών, η λ ε κ τ ρ ο μ α γ ν η τ ι κ ώ ν κ α ι μ α γ ν η τ ι κ ώ ν φαινομένων που εμφανίζονται στους βιολογικούς ιστούς. Το αντικείμενο του εμβιοηλεκτρομαγνητισμού
Εισαγωγή Ασχολείται με τη μελέτη των ηλεκτρικών, η λ ε κ τ ρ ο μ α γ ν η τ ι κ ώ ν κ α ι μ α γ ν η τ ι κ ώ ν φαινομένων που εμφανίζονται στους βιολογικούς ιστούς. Το αντικείμενο του εμβιοηλεκτρομαγνητισμού
Λείος μυς. Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017
 Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ. Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.
 2310-999185 (γραμματεία) anogian@auth.") ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Σύνοψη των όσων εξετάσαμε για τους ιοντικούς διαύλους: 1. Διαπερνούν
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Σύνοψη των όσων εξετάσαμε για τους ιοντικούς διαύλους: 1. Διαπερνούν
Εγκέφαλος και Έλεγχος της Κίνησης
 Παρεγκεφαλίδα Εγκέφαλος και Έλεγχος της Κίνησης 3 4 Γενικά Cerebellum = Little brain 10% όγκου εγκεφάλου, >50% του συνόλου των νευρώνων του εγκεφάλου Κανονικότητα στη διάταξη των νευρώνων και τις διασυνδέσεις
Παρεγκεφαλίδα Εγκέφαλος και Έλεγχος της Κίνησης 3 4 Γενικά Cerebellum = Little brain 10% όγκου εγκεφάλου, >50% του συνόλου των νευρώνων του εγκεφάλου Κανονικότητα στη διάταξη των νευρώνων και τις διασυνδέσεις
Σύναψη µεταξύ της απόληξης του νευράξονα ενός νευρώνα και του δενδρίτη ενός άλλου νευρώνα.
 ΟΙ ΝΕΥΡΩΝΕΣ ΕΠΙΚΟΙΝΩΝΟΥΝ ΜΕΣΩ ΤΗΣ ΣΥΝΑΨΗΣ Άντα Μητσάκου Αναπληρώτρια Καθηγήτρια, Ιατρική Σχολή, Πανεπιστήµιο Πατρών Γνωρίζουµε ότι είµαστε ικανοί να εκτελούµε σύνθετες νοητικές διεργασίες εξαιτίας της
ΟΙ ΝΕΥΡΩΝΕΣ ΕΠΙΚΟΙΝΩΝΟΥΝ ΜΕΣΩ ΤΗΣ ΣΥΝΑΨΗΣ Άντα Μητσάκου Αναπληρώτρια Καθηγήτρια, Ιατρική Σχολή, Πανεπιστήµιο Πατρών Γνωρίζουµε ότι είµαστε ικανοί να εκτελούµε σύνθετες νοητικές διεργασίες εξαιτίας της
Βιολογία Α Λυκείου Κεφ. 9. Νευρικό Σύστημα. Δομή και λειτουργία των νευρικών κυττάρων
 Βιολογία Α Λυκείου Κεφ. 9 Νευρικό Σύστημα Δομή και λειτουργία των νευρικών κυττάρων Νευρικό Σύστημα Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων φροντίζουν να διατηρείται σταθερό το εσωτερικό
Βιολογία Α Λυκείου Κεφ. 9 Νευρικό Σύστημα Δομή και λειτουργία των νευρικών κυττάρων Νευρικό Σύστημα Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων φροντίζουν να διατηρείται σταθερό το εσωτερικό
+ - - εκπολώνεται. ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ
 ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ Στόχοι Κατανόησης: -Να σας είναι ξεκάθαρες οι έννοιες πόλωση, εκπόλωση, υπερπόλωση, διεγερτικό ερέθισμα, ανασταλτικό ερέθισμα, κατώφλιο δυναμικό, υποκατώφλιες εκπολώσεις, υπερκατώφλιες
ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ Στόχοι Κατανόησης: -Να σας είναι ξεκάθαρες οι έννοιες πόλωση, εκπόλωση, υπερπόλωση, διεγερτικό ερέθισμα, ανασταλτικό ερέθισμα, κατώφλιο δυναμικό, υποκατώφλιες εκπολώσεις, υπερκατώφλιες
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων. Μεταιχµιακό Σύστηµα
 Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Μεταιχµιακό Σύστηµα Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά Γάγγλια µεταιχµιακό (στεφανιαίο) σύστηµα διάµεσος εγκέφαλος
Εσωτερική Κατασκευή των Εγκεφαλικών Ηµισφαιρίων Μεταιχµιακό Σύστηµα Στο εσωτερικό των ηµισφαιρίων υπάρχου πλάγιες κοιλίες λευκή ουσία Βασικά Γάγγλια µεταιχµιακό (στεφανιαίο) σύστηµα διάµεσος εγκέφαλος
ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΣΤΙΣ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ. Διπλωματική Εργασία
 ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΣΤΙΣ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ Διπλωματική Εργασία «Φαρμακολογικές επιδράσεις επί των in vitro Οξέων Κυμάτων σε λεπτές τομές ιπποκάμπου επίμυος» Γιαννόπουλος Παναγιώτης Α.Μ.
ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΣΤΙΣ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ Διπλωματική Εργασία «Φαρμακολογικές επιδράσεις επί των in vitro Οξέων Κυμάτων σε λεπτές τομές ιπποκάμπου επίμυος» Γιαννόπουλος Παναγιώτης Α.Μ.
Συναπτική ολοκλήρωση. Η διαδικασία της άθροισης όλων των εισερχόμενων διεγερτικών και ανασταλτικών σημάτων σε ένα νευρώνα. Τετάρτη, 20 Μαρτίου 13
 Συναπτική ολοκλήρωση Η διαδικασία της άθροισης όλων των εισερχόμενων διεγερτικών και ανασταλτικών σημάτων σε ένα νευρώνα http://www.mpg.de/13795/learning_memory_perception?print=yes 2 Τοποθεσία συνάψεων
Συναπτική ολοκλήρωση Η διαδικασία της άθροισης όλων των εισερχόμενων διεγερτικών και ανασταλτικών σημάτων σε ένα νευρώνα http://www.mpg.de/13795/learning_memory_perception?print=yes 2 Τοποθεσία συνάψεων
Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας. Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ
 2016 Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ Εισαγωγή Η σύσπαση των σκελετικών μυών ελέγχεται από
2016 Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ Εισαγωγή Η σύσπαση των σκελετικών μυών ελέγχεται από
ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ
 ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ Δρ. Κυριακή Σιδηροπούλου Λέκτορας Νευροφυσιολογίας Γραφείο: Γ316δ ΤΗΛ: 28103940871 (γραφείο) E- MAIL: sidirop@imbb.forth.gr Εισαγωγή Σιδηροπούλου - Νευροβιολογία 1 Δομή μαθήματος
ΒΙΟ492: ΝΕΥΡΟΒΙΟΛΟΓΙΑ Δρ. Κυριακή Σιδηροπούλου Λέκτορας Νευροφυσιολογίας Γραφείο: Γ316δ ΤΗΛ: 28103940871 (γραφείο) E- MAIL: sidirop@imbb.forth.gr Εισαγωγή Σιδηροπούλου - Νευροβιολογία 1 Δομή μαθήματος
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ Συγκριτική μελέτη των φαρμακολογικών ιδιοτήτων του συμπλόκου του υποδοχέα GABA A /Βενζοδιαζεπινών μεταξύ του διαφραγματικού
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ Συγκριτική μελέτη των φαρμακολογικών ιδιοτήτων του συμπλόκου του υποδοχέα GABA A /Βενζοδιαζεπινών μεταξύ του διαφραγματικού
Κεφάλαιο 1: Βασικές αρχές ανατοµίας και οργάνωσης του νευρικού συστήµατος
 Κεφάλαιο 1: Βασικές αρχές ανατοµίας και οργάνωσης του νευρικού συστήµατος Περίληψη Σε αυτό το πρώτο κεφάλαιο, θα παρουσιαστεί η βασική οργάνωση του νευρικού συστήµατος, δηλαδή ο διαχωρισµός σε κεντρικό
Κεφάλαιο 1: Βασικές αρχές ανατοµίας και οργάνωσης του νευρικού συστήµατος Περίληψη Σε αυτό το πρώτο κεφάλαιο, θα παρουσιαστεί η βασική οργάνωση του νευρικού συστήµατος, δηλαδή ο διαχωρισµός σε κεντρικό
Τεχνική και μεθοδολογία της ηλεκτροεγκεφαλογραφικής καταγραφής Το μηχάνημα που χρησιμοποιείται για τη λήψη του ΗΕΓ ονομάζεται
 ΗΛΕΚΤΡΟΕΓΚΕΦΑΛΟΓΡΑΦΗΜΑ (ΗΕΓ) Ιστορικά στοιχεία Οι πρώτοι ερευνητές που διαπίστωσαν με τη βοήθεια γαλβανόμετρου την ύπαρξη ηλεκτρικής δραστηριότητας στον εγκέφαλο κουνελιών ήταν ο Άγγλος βιολόγος Caton
ΗΛΕΚΤΡΟΕΓΚΕΦΑΛΟΓΡΑΦΗΜΑ (ΗΕΓ) Ιστορικά στοιχεία Οι πρώτοι ερευνητές που διαπίστωσαν με τη βοήθεια γαλβανόμετρου την ύπαρξη ηλεκτρικής δραστηριότητας στον εγκέφαλο κουνελιών ήταν ο Άγγλος βιολόγος Caton
Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών
 Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
Χηµική Μεταβίβαση Σήµατος Ηλίας Ηλιόπουλος Εργαστήριο Γενετικής, Τµήµα Γεωπονικής Βιοτεχνολογίας, Γεωπονικό Πανεπιστήµιο Αθηνών 1 Η Επικοινωνία στα Ζωϊκά Κύτταρα 1. Δίκτυα εξωκυτταρικών και ενδοκυτταρικών
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ. Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας (προσωπικό) (γραμματεία)
 (γραμματεία)") ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Αρχές της ηλεκτρικής διακυτταρικής επικοινωνίας Ή πως το νευρικό
ΓΕΝΙΚΗ ΦΥΣΙΟΛΟΓΙΑ Γιώργος Ανωγειανάκις Εργαστήριο Πειραματικής Φυσιολογίας 2310-999054 (προσωπικό) 2310-999185 (γραμματεία) anogian@auth.gr Αρχές της ηλεκτρικής διακυτταρικής επικοινωνίας Ή πως το νευρικό
ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΕΦΑΡΜΟΓΕΣ ΣΤΙΣ ΒΙΟΪΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ»
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΕΦΑΡΜΟΓΕΣ ΣΤΙΣ ΒΙΟΪΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ "Η ΣΥΝΕΡΓΙΣΤΙΚΗ ΔΡΑΣΗ ΤΩΝ mglur5
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΕΦΑΡΜΟΓΕΣ ΣΤΙΣ ΒΙΟΪΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ "Η ΣΥΝΕΡΓΙΣΤΙΚΗ ΔΡΑΣΗ ΤΩΝ mglur5
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ. ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΣΥΣΠΑΣΗΣ
 ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ. Μεμβρανική Βιοφυσική. Δίαυλοι: απο το γονίδιο στην εξέλιξη Διδάσκων: Λεκ. Χαράλαμπος Λαμπρακάκης
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μεμβρανική Βιοφυσική Δίαυλοι: απο το γονίδιο στην εξέλιξη Διδάσκων: Λεκ. Χαράλαμπος Λαμπρακάκης Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μεμβρανική Βιοφυσική Δίαυλοι: απο το γονίδιο στην εξέλιξη Διδάσκων: Λεκ. Χαράλαμπος Λαμπρακάκης Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε
«In vivo και in vitro μελέτες της φυσιολογίας και της φαρμακολογίας της GABA-εργικής συναπτικής αναστολής στον εγκέφαλο μυών και επίμυων.
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΑΝΘΡΩΠΟΥ ΚΑΙ ΖΩΩΝ «In vivo και in vitro μελέτες της φυσιολογίας και της φαρμακολογίας της GABA-εργικής συναπτικής αναστολής
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΑΝΘΡΩΠΟΥ ΚΑΙ ΖΩΩΝ «In vivo και in vitro μελέτες της φυσιολογίας και της φαρμακολογίας της GABA-εργικής συναπτικής αναστολής
Θέµατα διάλεξης ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ. Τρόποι µετάδοσης των νευρικών σηµάτων. υναµικό Ηρεµίας. Νευρώνας
 Θέµατα διάλεξης MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ Τρόποι µετάδοσης νευρικών σηµάτων Ρόλος και λειτουργία των νευροδιαβιβαστών
Θέµατα διάλεξης MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ Τρόποι µετάδοσης νευρικών σηµάτων Ρόλος και λειτουργία των νευροδιαβιβαστών
ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ
 ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 5-6/29-02-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ 1. ΕΙΣΑΓΩΓΗ 2. ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 3. ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 4. ΚΕΝΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΡΥΘΜΙΣΗ 1. ΕΙΣΑΓΩΓΗ διαβίβαση στο νευρικό
ΦΥΣΙΟΛΟΓΙΑ ΖΩΩΝ 5-6/29-02-2016 Π.Παπαζαφείρη ΣΥΝΑΨΕΙΣ 1. ΕΙΣΑΓΩΓΗ 2. ΗΛΕΚΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 3. ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ:ΔΟΜΗ ΛΕΙΤΟΥΡΓΙΑ 4. ΚΕΝΤΡΙΚΕΣ ΣΥΝΑΨΕΙΣ ΡΥΘΜΙΣΗ 1. ΕΙΣΑΓΩΓΗ διαβίβαση στο νευρικό
MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΟΡΜΟΝΕΣ ΚΑΙ ΑΣΚΗΣΗ. ΘΑΝΑΣΗΣ ΤΖΙΑΜΟΥΡΤΑΣ, Ph.D., C.S.C.
 MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΟΡΜΟΝΕΣ ΚΑΙ ΑΣΚΗΣΗ ΘΑΝΑΣΗΣ ΤΖΙΑΜΟΥΡΤΑΣ, Ph.D., C.S.C.S Αδένες Έκκρισης Ορμονών Υπόφυση Θυρεοειδής Αδένας Παραθυροειδείς
MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΟΡΜΟΝΕΣ ΚΑΙ ΑΣΚΗΣΗ ΘΑΝΑΣΗΣ ΤΖΙΑΜΟΥΡΤΑΣ, Ph.D., C.S.C.S Αδένες Έκκρισης Ορμονών Υπόφυση Θυρεοειδής Αδένας Παραθυροειδείς
Διάλεξη 5, Κ. Ψαρροπούλου
 Διάλεξη 5, 2014 Κ. Ψαρροπούλου Διεγερτικά αμινοξέα Γλουταμικό Ασπαρτικό Ρόλος που βρίσκεται Tο γλουταμικό (glutamic acid, glutamate) είναι ο κύριος διεγερτικός ΝΔ του ΚΝΣ, και απελευθερώνεται τόσο από
Διάλεξη 5, 2014 Κ. Ψαρροπούλου Διεγερτικά αμινοξέα Γλουταμικό Ασπαρτικό Ρόλος που βρίσκεται Tο γλουταμικό (glutamic acid, glutamate) είναι ο κύριος διεγερτικός ΝΔ του ΚΝΣ, και απελευθερώνεται τόσο από
Μελέτη της επίδρασης των α5gaba A υποδοχέων στη συναπτική πλαστικότητα μεταξύ ραχιαίου & κοιλιακού ιπποκάμπου
 Μελέτη της επίδρασης των α5gaba A υποδοχέων στη συναπτική πλαστικότητα μεταξύ ραχιαίου & κοιλιακού ιπποκάμπου Μεταπτυχιακό Πρόγραμμα Σπουδών: Εισαγωγή στις Βασικές Ιατρικές Επιστήμες Διπλωματική εργασία
Μελέτη της επίδρασης των α5gaba A υποδοχέων στη συναπτική πλαστικότητα μεταξύ ραχιαίου & κοιλιακού ιπποκάμπου Μεταπτυχιακό Πρόγραμμα Σπουδών: Εισαγωγή στις Βασικές Ιατρικές Επιστήμες Διπλωματική εργασία
Βασικά γάγγλια. Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας
 Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί η πρόθεση δράσης
Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί η πρόθεση δράσης
Νευροδιαβιβαστές και συμπεριφορά
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Νευροδιαβιβαστές και συμπεριφορά Νευροβιολογία της λήψης τροφής Διδάσκοντες: Καθ. Α. Ψαρροπούλου, Λεκ. Χ. Λαμπρακάκης Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Νευροδιαβιβαστές και συμπεριφορά Νευροβιολογία της λήψης τροφής Διδάσκοντες: Καθ. Α. Ψαρροπούλου, Λεκ. Χ. Λαμπρακάκης Άδειες Χρήσης Το παρόν εκπαιδευτικό
Μεθοδολογική προσέγγιση της Βιοηλεκτρικής βάσης του νευρικού ενεργού δυναμικού
 Μεθοδολογική προσέγγιση της Βιοηλεκτρικής βάσης του νευρικού ενεργού δυναμικού ΕΥΣΤΡΑΤΙΟΣ ΚΟΣΜΙΔΗΣ, Ph.D. ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ Ο εγκέφαλος και οι νευρώνες Προσυναπτικά Μετασυναπτικά Ένας
Μεθοδολογική προσέγγιση της Βιοηλεκτρικής βάσης του νευρικού ενεργού δυναμικού ΕΥΣΤΡΑΤΙΟΣ ΚΟΣΜΙΔΗΣ, Ph.D. ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ Ο εγκέφαλος και οι νευρώνες Προσυναπτικά Μετασυναπτικά Ένας
Κυτταροαρχιτεκτονική Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή
 Κυτταροαρχιτεκτονική Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή Τελικός Εγκ Εγκεφαλικά ηµισφαίρια Διάµεσος εγκ & Βασικά γάγγλια Διαίρεση του ΚΝΣ Στέλεχος του εγκέφαλου Μέσος εγκ Γέφυρα Προµήκης
Κυτταροαρχιτεκτονική Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή Τελικός Εγκ Εγκεφαλικά ηµισφαίρια Διάµεσος εγκ & Βασικά γάγγλια Διαίρεση του ΚΝΣ Στέλεχος του εγκέφαλου Μέσος εγκ Γέφυρα Προµήκης
«ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ» Λειτουργία Νευρικού Συστήματος
 ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ «ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ» Λειτουργία Νευρικού Συστήματος Κώστας Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2013 Ομοιόσταση Ορισμός: Το σύνολο των φυσιολογικών
ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ «ΝΕΥΡΟΕΠΙΣΤΗΜΕΣ» Λειτουργία Νευρικού Συστήματος Κώστας Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2013 Ομοιόσταση Ορισμός: Το σύνολο των φυσιολογικών
Δυναμικό ηρεμίας Δυναμικό ενεργείας. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 29/09/2017 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2017-2018 Ιόντα Δυναμικό
Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 29/09/2017 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2017-2018 Ιόντα Δυναμικό
ΜΑΘΗΜΑ 7ο ΜΕΡΟΣ Α Η ΔΟΜΗ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ
 ΜΑΘΗΜΑ 7ο ΜΕΡΟΣ Α Η ΔΟΜΗ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Η ΛΕΥΚΗ ΟΥΣΙΑ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ Η λευκή ουσία συντίθεται από εμύελες νευρικές ίνες διαφόρων διαμέτρων και νευρογλοία Οι νευρικές ίνες κατατάσσονται
ΜΑΘΗΜΑ 7ο ΜΕΡΟΣ Α Η ΔΟΜΗ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ Η ΛΕΥΚΗ ΟΥΣΙΑ ΤΩΝ ΗΜΙΣΦΑΙΡΙΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ Η λευκή ουσία συντίθεται από εμύελες νευρικές ίνες διαφόρων διαμέτρων και νευρογλοία Οι νευρικές ίνες κατατάσσονται
Η ΚΥΤΤΑΡΙΚΗ ΜΕΜΒΡΑΝΗ
 ΗΛΕΚΤΡΟΦΥΣΙΟΛΟΓΙΑ Η ΚΥΤΤΑΡΙΚΗ ΜΕΜΒΡΑΝΗ Ο ρόλος της κυτταρικής μεμβράνης Φαινόμενα μεταφοράς Διάχυση Φαινόμενα μικρο-διαπερατότητας της κυτταρικής μεμβράνης Ασύμμετρη κατανομή ιόντων Ενεργός μεταφορά Ενεργός
ΗΛΕΚΤΡΟΦΥΣΙΟΛΟΓΙΑ Η ΚΥΤΤΑΡΙΚΗ ΜΕΜΒΡΑΝΗ Ο ρόλος της κυτταρικής μεμβράνης Φαινόμενα μεταφοράς Διάχυση Φαινόμενα μικρο-διαπερατότητας της κυτταρικής μεμβράνης Ασύμμετρη κατανομή ιόντων Ενεργός μεταφορά Ενεργός
Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος
 MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞΕΙΣ 10-12 Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής
MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞΕΙΣ 10-12 Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής
Βασικά γάγγλια. Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017
 Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί
Βασικά γάγγλια Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Μάρτιος 2017 Ιεραρχία κινητικού ελέγχου ΠΡΟΘΕΣΗ Αναμετάδοση της πληροφορίας Εξειδίκευση της θέσης και της κίνησης για να εκτελεστεί
ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ
 ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Εγκέφαλος Μεγάλη αιµάτωση, πολύ σηµαντική για την λειτουργία του Επικοινωνία µε το περιβάλλον Χρησιµοποιεί το 20% του Ο 2 και ως πηγή ενέργειας γλυκόζη Στις χειρουργικές επεµβάσεις
ΚΕΝΤΡΙΚΟ ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Εγκέφαλος Μεγάλη αιµάτωση, πολύ σηµαντική για την λειτουργία του Επικοινωνία µε το περιβάλλον Χρησιµοποιεί το 20% του Ο 2 και ως πηγή ενέργειας γλυκόζη Στις χειρουργικές επεµβάσεις
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ. Κορνηλία Πουλοπούλου Αναπληρώτρια Καθ. Νευροφυσιολογίας Νευρολογική Κλινκή
 ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ Κορνηλία Πουλοπούλου Αναπληρώτρια Καθ. Νευροφυσιολογίας Νευρολογική Κλινκή ΣΥΝΑΨΕΙΣ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ Εξυπηρετεί την επικοινωνία ανάμεσα στα νευρικά κύτταρα
ΦΥΣΙΟΛΟΓΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ Κορνηλία Πουλοπούλου Αναπληρώτρια Καθ. Νευροφυσιολογίας Νευρολογική Κλινκή ΣΥΝΑΨΕΙΣ ΣΥΝΑΠΤΙΚΗ ΔΙΑΒΙΒΑΣΗ Εξυπηρετεί την επικοινωνία ανάμεσα στα νευρικά κύτταρα
Κεφάλαιο 8 Λειτουργίες µνήµης και µάθησης
 Κεφάλαιο 8 Λειτουργίες µνήµης και µάθησης Σύνοψη Σε αυτό το κεφάλαιο, θα παρουσιαστεί η νευροβιολογική βάση των διαδικασιών µνήµης. Στο πρώτο µέρος του κεφαλαίου, θα γίνει µια εισαγωγή στους διαφορετικούς
Κεφάλαιο 8 Λειτουργίες µνήµης και µάθησης Σύνοψη Σε αυτό το κεφάλαιο, θα παρουσιαστεί η νευροβιολογική βάση των διαδικασιών µνήµης. Στο πρώτο µέρος του κεφαλαίου, θα γίνει µια εισαγωγή στους διαφορετικούς
Νευροβιολογία των Μνημονικών Λειτουργιών
 Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 6: Μακρόχρονη Συναπτική Πλαστικότητα Κωνσταντίνος Παπαθεοδωρόπουλος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας (1/2) Γενική επισκόπηση των φαινομένων
Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 6: Μακρόχρονη Συναπτική Πλαστικότητα Κωνσταντίνος Παπαθεοδωρόπουλος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας (1/2) Γενική επισκόπηση των φαινομένων
ΦΑΡΜΑΚΟ ΥΝΑΜΙΚΗ ΙΙ. Γενικές έννοιες (Θεωρία υποδοχέων - Αγωνιστής ανταγωνιστής) Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων
 Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων") ΦΑΡΜΑΚΟ ΥΝΑΜΙΚΗ ΙΙ Γενικές έννοιες (Θεωρία υποδοχέων - Αγωνιστής ανταγωνιστής) Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων Υποδοχείς (φαρµάκων) και ενδοκυττάριες σηµατοδοτικές πορείες - Συστήµατα Μεταγωγής
ΦΑΡΜΑΚΟ ΥΝΑΜΙΚΗ ΙΙ Γενικές έννοιες (Θεωρία υποδοχέων - Αγωνιστής ανταγωνιστής) Σηµεία ράσης Μοριακοί Μηχανισµοί ράσης Φαρµάκων Υποδοχείς (φαρµάκων) και ενδοκυττάριες σηµατοδοτικές πορείες - Συστήµατα Μεταγωγής
Οι Κυριότερες Νευρικές Οδοί
 Οι Κυριότερες Νευρικές Οδοί Κατιόντα (φυγόκεντρα) δεµάτια Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή φυσιολογικά δεµάτια (κατά τον επιµήκη άξονα) έχουν κοινή έκφυση πορεία απόληξη λειτουργία
Οι Κυριότερες Νευρικές Οδοί Κατιόντα (φυγόκεντρα) δεµάτια Ελίζαµπεθ Τζόνσον Εργαστήριο Ανατοµίας Ιατρική Σχολή φυσιολογικά δεµάτια (κατά τον επιµήκη άξονα) έχουν κοινή έκφυση πορεία απόληξη λειτουργία
9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΔΟΜΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΝΕΥΡΙΚΩΝ. Νευρώνες
 9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος (ομοιόσταση), ελέγχοντας και συντονίζοντας τις λειτουργίες των
9. ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ Το νευρικό σύστημα μαζί με το σύστημα των ενδοκρινών αδένων συμβάλλουν στη διατήρηση σταθερού εσωτερικού περιβάλλοντος (ομοιόσταση), ελέγχοντας και συντονίζοντας τις λειτουργίες των
Ιππόκαμπος: Ανατομία, Φυσιολογία, Παθολογία
 Ιππόκαμπος: Ανατομία, Φυσιολογία, Παθολογία Άννα Σιατούνη Νευρολόγος Εφαρμοσμένη Νευροανατομία ΙΙΙ: Εν τω βάθει δομές του εγκεφάλου Πρόγραμμα Μεταπτυχιακών Σπουδών «ΕΦΑΡΜΟΣΜΕΝΗ ΝΕΥΡΟΑΝΑΤΟΜΙΑ» Αρχαιοφλοιός
Ιππόκαμπος: Ανατομία, Φυσιολογία, Παθολογία Άννα Σιατούνη Νευρολόγος Εφαρμοσμένη Νευροανατομία ΙΙΙ: Εν τω βάθει δομές του εγκεφάλου Πρόγραμμα Μεταπτυχιακών Σπουδών «ΕΦΑΡΜΟΣΜΕΝΗ ΝΕΥΡΟΑΝΑΤΟΜΙΑ» Αρχαιοφλοιός
«ΚΑΤΑΝΟΜΗ GABA ΝΕΥΡΩΝΩΝ ΣΤΗ ΡΑΧΙΑΙΑ ΚΑΙ ΚΟΙΛΙΑΚΗ ΜΟΙΡΑ ΤΟΥ ΙΠΠΟΚΑΜΠΟΥ»
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΚΛΙΝΙΚΕΣ ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ» ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ «ΚΑΤΑΝΟΜΗ GABA ΝΕΥΡΩΝΩΝ ΣΤΗ ΡΑΧΙΑΙΑ ΚΑΙ ΚΟΙΛΙΑΚΗ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΚΛΙΝΙΚΕΣ ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ» ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ «ΚΑΤΑΝΟΜΗ GABA ΝΕΥΡΩΝΩΝ ΣΤΗ ΡΑΧΙΑΙΑ ΚΑΙ ΚΟΙΛΙΑΚΗ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
32. Πλαστικότητα Ορισμός και Γενικές Παρατηρήσεις
 32. Πλαστικότητα Σύνοψη Η έννοια της πλαστικότητας είναι μια έννοια ευρύτατης χρήσης και εφαρμογής στο πεδίο της Νευροεπιστήμης και περιλαμβάνει φαινόμενα σε όλα τα επίπεδα οργάνωσης του νευρικού συστήματος
32. Πλαστικότητα Σύνοψη Η έννοια της πλαστικότητας είναι μια έννοια ευρύτατης χρήσης και εφαρμογής στο πεδίο της Νευροεπιστήμης και περιλαμβάνει φαινόμενα σε όλα τα επίπεδα οργάνωσης του νευρικού συστήματος
Αμυγδαλή (ΑΜΥ)* Ι. Εισαγωγή ΙΙ. Ανατομική οργάνωση
* Ι. Εισαγωγή ΙΙ. Ανατομική οργάνωση") Αμυγδαλή (ΑΜΥ)* Ι. Εισαγωγή Η αμυγδαλή είναι μια σύνθετη δομή που εμπλέκεται σε μια πληθώρα φυσιολογικών συμπεριφορικών λειτουργιών αλλά και ψυχιατρικών καταστάσεων. Για πολύ καιρό η αμυγδαλή αποτελούσε
Αμυγδαλή (ΑΜΥ)* Ι. Εισαγωγή Η αμυγδαλή είναι μια σύνθετη δομή που εμπλέκεται σε μια πληθώρα φυσιολογικών συμπεριφορικών λειτουργιών αλλά και ψυχιατρικών καταστάσεων. Για πολύ καιρό η αμυγδαλή αποτελούσε
Ηλεκτροφυσιολογία της Κυτταρικής Μεμβράνης
 ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ - ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΦΥΣΙΟΛΟΓΙΑ Ι Ηλεκτροφυσιολογία της Κυτταρικής Μεμβράνης Κων/νος Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2017 Βοηθήματα: Αρχές Φυσιολογίας: Berne
ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ - ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΦΥΣΙΟΛΟΓΙΑ Ι Ηλεκτροφυσιολογία της Κυτταρικής Μεμβράνης Κων/νος Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2017 Βοηθήματα: Αρχές Φυσιολογίας: Berne
2. Μεμβρανικά δυναμικά του νευρικού κυττάρου
 2. Μεμβρανικά δυναμικά του νευρικού κυττάρου Στόχοι κατανόησης: Διαφορά δυναμικού της κυτταρικής μεμβράνης ενός νευρικού κυττάρου: Τί είναι; Πώς δημιουργείται; Ποιά είδη διαφοράς δυναμικού της μεμβράνης
2. Μεμβρανικά δυναμικά του νευρικού κυττάρου Στόχοι κατανόησης: Διαφορά δυναμικού της κυτταρικής μεμβράνης ενός νευρικού κυττάρου: Τί είναι; Πώς δημιουργείται; Ποιά είδη διαφοράς δυναμικού της μεμβράνης
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 11-13 OΔΟΙ ΚΑΙ ΜΗΧΑΝΙΣΜΟΙ ΕΥΚΑΡΥΩΤΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής
«ΑΛΛΗΛΕΠΙΔΡΑΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΙΚΩΝ ΣΥΣΤΗΜΑΤΩΝ ΑΔΕΝΟΣΙΝΗΣ ΚΑΙ ΓΛΟΥΤΑΜΙΝΙΚΟΥ ΟΞΕΟΣ ΣΕ ΔΙΑΔΙΚΑΣΙΕΣ ΝΕΥΡΩΝΙΚΗΣ ΠΛΑΣΤΙΚΟΤΗΤΑΣ ΣΤΟΝ ΙΠΠΟΚΑΜΠΟ ΕΠΙΜΥΟΣ»
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΕΦΑΡΜΟΓΕΣ ΣΤΙΣ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «ΑΛΛΗΛΕΠΙΔΡΑΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΙΚΩΝ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΕΦΑΡΜΟΓΕΣ ΣΤΙΣ ΒΑΣΙΚΕΣ ΙΑΤΡΙΚΕΣ ΕΠΙΣΤΗΜΕΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ «ΑΛΛΗΛΕΠΙΔΡΑΣΗ ΝΕΥΡΟΔΙΑΒΙΒΑΣΤΙΚΩΝ
ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ
 K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ κατάσταση ετοιμότητος του μυός ενός βαθμού μόνιμης σύσπασης που διατηρούν οι μύες στην ηρεμία αποτελεί
K. I. Boυμβουράκης Αν. Καθηγητής Νευρολογίας Β Νευρολογική Κλινική Πανεπιστημίου Αθηνών Π.Γ.Ν. ΑΤΤΙΚΟΝ κατάσταση ετοιμότητος του μυός ενός βαθμού μόνιμης σύσπασης που διατηρούν οι μύες στην ηρεμία αποτελεί
Νευροβιολογία των Μνημονικών Λειτουργιών
 Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 2: Κατηγοριοποιήσεις Μάθησης & Μνήμης Κωνσταντίνος Παπαθεοδωρόπουλος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας Ανάλυση των γενικών αξόνων που
Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 2: Κατηγοριοποιήσεις Μάθησης & Μνήμης Κωνσταντίνος Παπαθεοδωρόπουλος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας Ανάλυση των γενικών αξόνων που
Νευροφυσιολογία και Αισθήσεις
 Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις Διάλεξη 5 Μοντέλο Hodgkin-Huxley (Hodgkin-Huxley Model) Απόκριση στην Έγχυση Ρεύματος 2 Hodgin και Huxley Οι Sir Alan
Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις Διάλεξη 5 Μοντέλο Hodgkin-Huxley (Hodgkin-Huxley Model) Απόκριση στην Έγχυση Ρεύματος 2 Hodgin και Huxley Οι Sir Alan
Τεχνητά Νευρωνικά Δίκτυα. Τσιριγώτης Γεώργιος Τμήμα Μηχανικών Πληροφορικής ΤΕΙ Ανατολικής Μακεδονίας & Θράκης
 Τεχνητά Τσιριγώτης Γεώργιος Τμήμα Μηχανικών Πληροφορικής ΤΕΙ Ανατολικής Μακεδονίας & Θράκης Ο Βιολογικός Νευρώνας Δενδρίτες Συνάψεις Πυρήνας (Σώμα) Άξονας 2 Ο Βιολογικός Νευρώνας 3 Βασικά Χαρακτηριστικά
Τεχνητά Τσιριγώτης Γεώργιος Τμήμα Μηχανικών Πληροφορικής ΤΕΙ Ανατολικής Μακεδονίας & Θράκης Ο Βιολογικός Νευρώνας Δενδρίτες Συνάψεις Πυρήνας (Σώμα) Άξονας 2 Ο Βιολογικός Νευρώνας 3 Βασικά Χαρακτηριστικά
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
Νευροβιολογία των Μνημονικών Λειτουργιών
 Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 3: Ενεργός Μνήμη - Μη Συνειρμική Μάθηση & Μνήμη - Κλασσική Εξαρτημένη Μάθηση Κωνσταντίνος Παπαθεοδωρόπουλος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί
Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 3: Ενεργός Μνήμη - Μη Συνειρμική Μάθηση & Μνήμη - Κλασσική Εξαρτημένη Μάθηση Κωνσταντίνος Παπαθεοδωρόπουλος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ. Βιολογία A λυκείου. Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος:
 ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ Βιολογία A λυκείου Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος: 2013-2014 Ένα αισθητικό σύστημα στα σπονδυλωτά αποτελείται από τρία βασικά μέρη: 1. Τους αισθητικούς υποδοχείς,
ΔΑΜΔΑΣ ΙΩΑΝΝΗΣ Βιολογία A λυκείου Υπεύθυνη καθηγήτρια: Μαριλένα Ζαρφτζιάν Σχολικό έτος: 2013-2014 Ένα αισθητικό σύστημα στα σπονδυλωτά αποτελείται από τρία βασικά μέρη: 1. Τους αισθητικούς υποδοχείς,
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΑΝΑΡΤΗΤΕΑ ΣΤΟ ΔΙΑΔΙΚΤΥΟ
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΑΣ, ΥΠΟΔΟΜΩΝ, ΝΑΥΤΙΛΙΑΣ ΚΑΙ ΤΟΥΡΙΣΜΟΥ ΕΙΔΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΙΑΧΕΙΡΙΣΗΣ ΤΟΜΕΑΚΩΝ ΕΠ ΤΟΥ ΕΚΤ ΕΙΔΙΚΗ ΥΠΗΡΕΣΙΑ ΔΙΑΧΕΙΡΙΣΗΣ Ε.Π. "ΑΝΑΠΤΥΞΗ ΑΝΘΡΩΠΙΝΟΥ ΔΥΝΑΜΙΚΟΥ, ΕΚΠΑΙΔΕΥΣΗ
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΟΙΚΟΝΟΜΙΑΣ, ΥΠΟΔΟΜΩΝ, ΝΑΥΤΙΛΙΑΣ ΚΑΙ ΤΟΥΡΙΣΜΟΥ ΕΙΔΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΔΙΑΧΕΙΡΙΣΗΣ ΤΟΜΕΑΚΩΝ ΕΠ ΤΟΥ ΕΚΤ ΕΙΔΙΚΗ ΥΠΗΡΕΣΙΑ ΔΙΑΧΕΙΡΙΣΗΣ Ε.Π. "ΑΝΑΠΤΥΞΗ ΑΝΘΡΩΠΙΝΟΥ ΔΥΝΑΜΙΚΟΥ, ΕΚΠΑΙΔΕΥΣΗ
Γνωστική-Πειραµατική Ψυχολογία
 Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Γνωστική λειτουργία & φλοιός. Γνωστική λειτουργία & φλοιός. Γνωστικές λειτουργίες à επεξεργασία πληροφοριών από διαφορετικές περιοχές (µεγαλύτερη ( αποκλειστική)
Γνωστική-Πειραµατική Ψυχολογία ΕΑΡΙΝΟ ΕΞΑΜΗΝΟ 2018 Γνωστική λειτουργία & φλοιός. Γνωστική λειτουργία & φλοιός. Γνωστικές λειτουργίες à επεξεργασία πληροφοριών από διαφορετικές περιοχές (µεγαλύτερη ( αποκλειστική)
Οπτική οδός. Έξω γονατώδες σώµα. Οπτική ακτινοβολία
 Όραση Γ Όραση Οπτική οδός Έξω γονατώδες σώµα Οπτική ακτινοβολία Οπτικό χίασµα: Οι ίνες από το ρινικό ηµιµόριο περνούν στην αντίπλευρη οπτική οδό ενώ τα κροταφικά ηµιµόρια δεν χιάζονται. Εποµένως κάθε οπτική
Όραση Γ Όραση Οπτική οδός Έξω γονατώδες σώµα Οπτική ακτινοβολία Οπτικό χίασµα: Οι ίνες από το ρινικό ηµιµόριο περνούν στην αντίπλευρη οπτική οδό ενώ τα κροταφικά ηµιµόρια δεν χιάζονται. Εποµένως κάθε οπτική
Νευροβιολογία των Μνημονικών Λειτουργιών
 Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 4: Μνήμη και Συναπτική Πλαστικότητα Στυλιανός Κούβαρος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας Ιστορικά στοιχεία περί συναπτικής Πλαστικότητας.
Νευροβιολογία των Μνημονικών Λειτουργιών Ενότητα 4: Μνήμη και Συναπτική Πλαστικότητα Στυλιανός Κούβαρος Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Σκοποί ενότητας Ιστορικά στοιχεία περί συναπτικής Πλαστικότητας.
ΣΥΓΚΡΙΣΗ ΤΗΣ ΙΕΓΕΡΣΙΜΟΤΗΤΑΣ ΝΕΥΡΏΝΩΝ ΤΟΥ ΡΑΧΙΑΙΟΥ ΚΑΙ ΚΟΙΛΙΑΚΟΥ ΙΠΠΟΚΑΜΠΟΥ ΠΟΥ ΕΧΟΥΝ ΑΝΑΓΝΩΡΙΣΘΕΙ ΜΟΡΦΟΛΟΓΙΑΚΑ ΕΥΑΓΓΕΛΟΣ ΚΟΥΒΑΡΑΣ ΒΙΟΛΟΓΟΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΦΑΡΜΑΚΟΛΟΓΙΑΣ ιευθυντής: Καθηγητής Ν. Σακελλαρίδης 1 ΣΥΓΚΡΙΣΗ ΤΗΣ ΙΕΓΕΡΣΙΜΟΤΗΤΑΣ ΝΕΥΡΏΝΩΝ ΤΟΥ ΡΑΧΙΑΙΟΥ ΚΑΙ ΚΟΙΛΙΑΚΟΥ ΙΠΠΟΚΑΜΠΟΥ ΠΟΥ ΕΧΟΥΝ ΑΝΑΓΝΩΡΙΣΘΕΙ ΜΟΡΦΟΛΟΓΙΑΚΑ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΦΑΡΜΑΚΟΛΟΓΙΑΣ ιευθυντής: Καθηγητής Ν. Σακελλαρίδης 1 ΣΥΓΚΡΙΣΗ ΤΗΣ ΙΕΓΕΡΣΙΜΟΤΗΤΑΣ ΝΕΥΡΏΝΩΝ ΤΟΥ ΡΑΧΙΑΙΟΥ ΚΑΙ ΚΟΙΛΙΑΚΟΥ ΙΠΠΟΚΑΜΠΟΥ ΠΟΥ ΕΧΟΥΝ ΑΝΑΓΝΩΡΙΣΘΕΙ ΜΟΡΦΟΛΟΓΙΑΚΑ
ΣΧΟΛΗ ΙΑΤΡΙΚΗΣ ΦΥΣΙΟΛΟΓΙΑ Ι * ΕΡΓΑΣΤΗΡΙΑΚΗ ΦΡΟΝΤΙΣΤΗΡΙΑΚΗ ΑΣΚΗΣΗ: ΔΥΝΑΜΙΚΟ ΕΝΕΡΓΕΙΑΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΦΥΣΙΟΛΟΓΙΑ Ι * ΕΡΓΑΣΤΗΡΙΑΚΗ ΦΡΟΝΤΙΣΤΗΡΙΑΚΗ ΑΣΚΗΣΗ: ΔΥΝΑΜΙΚΟ ΕΝΕΡΓΕΙΑΣ Υπεύθυνος: Κων/νος Παπαθεοδωρόπουλος, Αναπληρωτής καθηγητής ΑΣΚΗΣΗ 1. ΕΞΑΣΚΗΣΗ
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΙΑΤΡΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΦΥΣΙΟΛΟΓΙΑΣ ΦΥΣΙΟΛΟΓΙΑ Ι * ΕΡΓΑΣΤΗΡΙΑΚΗ ΦΡΟΝΤΙΣΤΗΡΙΑΚΗ ΑΣΚΗΣΗ: ΔΥΝΑΜΙΚΟ ΕΝΕΡΓΕΙΑΣ Υπεύθυνος: Κων/νος Παπαθεοδωρόπουλος, Αναπληρωτής καθηγητής ΑΣΚΗΣΗ 1. ΕΞΑΣΚΗΣΗ
Εγκέφαλος και Έλεγχος της Κίνησης. 4 διακριτά υποσυστήματα που αλληλεπιδρούν
 Τα Βασικά Γάγγλια Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης
Τα Βασικά Γάγγλια Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης 4 διακριτά υποσυστήματα που αλληλεπιδρούν Εγκέφαλος και Έλεγχος της Κίνησης
ΑΣΚΗΣΕΙΣ ΦΥΣΙΟΛΟΓΙΑΣ ΙΙΙ:
 ΑΣΚΗΣΕΙΣ ΦΥΣΙΟΛΟΓΙΑΣ ΙΙΙ: Εργαστήριο Φυσιολογίας Τµήµα Ιατρικής Πανεπιστηµίου Θεσσαλίας Λάρισα 2010 ΗΛΕΚΤΡΟΕΓΚΕΦΑΛΟΓΡΑΦΗΜΑ ΥΠΝΟΣ 2 ΗΛΕΚΤΡΟΕΓΚΕΦΑΛΟΓΡΑΦΗΜΑ Ιστορικά στοιχεία Οι πρώτοι ερευνητές που διαπίστωσαν
ΑΣΚΗΣΕΙΣ ΦΥΣΙΟΛΟΓΙΑΣ ΙΙΙ: Εργαστήριο Φυσιολογίας Τµήµα Ιατρικής Πανεπιστηµίου Θεσσαλίας Λάρισα 2010 ΗΛΕΚΤΡΟΕΓΚΕΦΑΛΟΓΡΑΦΗΜΑ ΥΠΝΟΣ 2 ΗΛΕΚΤΡΟΕΓΚΕΦΑΛΟΓΡΑΦΗΜΑ Ιστορικά στοιχεία Οι πρώτοι ερευνητές που διαπίστωσαν
ηδιεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς
 Μάθηση και Μνήµη Μάθηση Μνήµη Μάθηση: Μνήµη: ηδιεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς ηαποθήκευσητωνπληροφοριώναυτών η ικανότητα να
Μάθηση και Μνήµη Μάθηση Μνήµη Μάθηση: Μνήµη: ηδιεργασία µε την οποία αποκτούµε γνώσεις για τον κόσµο η οποία θα οδηγήσει στην τροποποίηση µιας συµπεριφοράς ηαποθήκευσητωνπληροφοριώναυτών η ικανότητα να
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ Εργαστήριο Φυσιολογίας Καθηγητής: Ηλίας Κούβελας
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ Εργαστήριο Φυσιολογίας Καθηγητής: Ηλίας Κούβελας Συγκριτική Μελέτη της Έκφρασης των Υποµονάδων του GABA A Υποδοχέα και των Πρώιµων Γονιδίων c-fos
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤΜΗΜΑ ΙΑΤΡΙΚΗΣ Εργαστήριο Φυσιολογίας Καθηγητής: Ηλίας Κούβελας Συγκριτική Μελέτη της Έκφρασης των Υποµονάδων του GABA A Υποδοχέα και των Πρώιµων Γονιδίων c-fos
Ν Ε Υ Ρ Ο Ψ Υ Χ Ο Λ Ο Γ Ι Κ Ο Ι Π Α Ρ Α Γ Ο Ν Τ Ε Σ Π Ο Υ Ε Π Η Ρ Ε Α Ζ Ο Υ Ν Τ Η Μ Α Θ Η Σ Η
 Ν Ε Υ Ρ Ο Ψ Υ Χ Ο Λ Ο Γ Ι Κ Ο Ι Π Α Ρ Α Γ Ο Ν Τ Ε Σ Π Ο Υ Ε Π Η Ρ Ε Α Ζ Ο Υ Ν Τ Η Μ Α Θ Η Σ Η 2 ΤΙ ΕΙΝΑΙ ΣΥΝΑΙΣΘΗΜΑ; Μηχανισμός επικοινωνίας που διατηρούν την κοινωνική τάξη / δομή Μαθημένη συμπεριφορά
Ν Ε Υ Ρ Ο Ψ Υ Χ Ο Λ Ο Γ Ι Κ Ο Ι Π Α Ρ Α Γ Ο Ν Τ Ε Σ Π Ο Υ Ε Π Η Ρ Ε Α Ζ Ο Υ Ν Τ Η Μ Α Θ Η Σ Η 2 ΤΙ ΕΙΝΑΙ ΣΥΝΑΙΣΘΗΜΑ; Μηχανισμός επικοινωνίας που διατηρούν την κοινωνική τάξη / δομή Μαθημένη συμπεριφορά
Αισθητικά συστήματα. Σωματοαισθητικό σύστημα. Όραση Ακοή/Αίσθηση ισορροπίας Γεύση Όσφρηση. Αφή Ιδιοδεκτικότητα Πόνος Θερμοκρασία
 Αισθητικά συστήματα Σωματοαισθητικό σύστημα Αφή Ιδιοδεκτικότητα Πόνος Θερμοκρασία Όραση Ακοή/Αίσθηση ισορροπίας Γεύση Όσφρηση Σιδηροπούλου - Νευροβιολογία!1 1 Δομή αισθητικών συστημάτων Υποδοχέας Πρωτοταγής
Αισθητικά συστήματα Σωματοαισθητικό σύστημα Αφή Ιδιοδεκτικότητα Πόνος Θερμοκρασία Όραση Ακοή/Αίσθηση ισορροπίας Γεύση Όσφρηση Σιδηροπούλου - Νευροβιολογία!1 1 Δομή αισθητικών συστημάτων Υποδοχέας Πρωτοταγής
ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΚΛΙΝΙΚΕΣ ΕΦΑΡΜΟΓΕΣ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤMHMA ΙΑΤΡΙΚΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΚΛΙΝΙΚΕΣ ΕΦΑΡΜΟΓΕΣ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Ανοσοϊστοχημική χρώση τομών ιπποκάμπου
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ ΥΓΕΙΑΣ ΤMHMA ΙΑΤΡΙΚΗΣ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ «ΚΛΙΝΙΚΕΣ ΕΦΑΡΜΟΓΕΣ ΜΟΡΙΑΚΗΣ ΙΑΤΡΙΚΗΣ» ΜΕΤΑΠΤΥΧΙΑΚΗ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ Ανοσοϊστοχημική χρώση τομών ιπποκάμπου
ΜΑΘΗΜΑ 10ο ΜΕΡΟΣ Γ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ
 ΜΑΘΗΜΑ 10ο ΜΕΡΟΣ Γ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ Πρόσφατες εργασίες έχουν αποδείξει ότι στη μνήμη παρεμβαίνουν πολλές περιοχές του εγκεφάλου Παρ όλα αυτά, υπάρχουν διαφορετικοί τύποι μνήμης και ορισμένες
ΜΑΘΗΜΑ 10ο ΜΕΡΟΣ Γ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ ΜΑΘΗΣΗ ΚΑΙ ΜΝΗΜΗ Πρόσφατες εργασίες έχουν αποδείξει ότι στη μνήμη παρεμβαίνουν πολλές περιοχές του εγκεφάλου Παρ όλα αυτά, υπάρχουν διαφορετικοί τύποι μνήμης και ορισμένες