ΠΑΡΑΣΚΕΥΗ ΔΗΜΟΡΑΓΚΑ Πτυχιούχος Μηχανικός Πληροφορικής ΑΤΕΙΘ
|
|
|
- Γοργοφόνη Γκόφας
- 7 χρόνια πριν
- Προβολές:
Transcript


1 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ Ανάλυση Μικροσυστοιχιών mirna και mrna του Πολυσταδιακού Μοντέλου Δερματικής Καρκινογένεσης του Ποντικιού ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΠΑΡΑΣΚΕΥΗ ΔΗΜΟΡΑΓΚΑ Πτυχιούχος Μηχανικός Πληροφορικής ΑΤΕΙΘ Επιβλέπων: Κωνσταντίνος E. Βοργιάς Καθηγητής Βιοχημείας ΕΚΠΑ ΑΘΗΝΑ, Μάρτιος 2015
2 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ Ανάλυση Μικροσυστοιχιών mirna και mrna του Πολυσταδιακού Μοντέλου Δερματικής Καρκινογένεσης του Ποντικιού ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ ΠΑΡΑΣΚΕΥΗ ΔΗΜΟΡΑΓΚΑ Πτυχιούχος Μηχανικός Πληροφορικής ΑΤΕΙΘ Τριμελής Εξεταστική Επιτροπή Υπογραφή 1. Κ. E. Βοργιάς Καθηγητής Βιοχημείας ΕΚΠΑ 2. Π. Μπάγκος Αναπ. Καθ. Βιοπλ. και Βιοστ. Πανεπιστήμιο Θεσσαλίας 3. B. Α. Οικονομίδου Επικ. Καθ. Βιοφυσικής - Μοριακής Βιοφυσικής ΕΚΠΑ ΑΘΗΝΑ, Μάρτιος 2015
3 i Στο σύζυγο μου, Παναγιώτη Δ. Κουρδή, για όλα όσα μου προσέφερε...
4 Η έγκριση της μεταπτυχιακής εργασίας από το Τμήμα Βιολογίας του Εθνικού & Καποδιστριακού Πανεπιστημίου Αθηνών δεν υποδηλώνει αποδοχή των γνωμών του συγγραφέα. ii (Ν. 5343/1932, Άρθρο 202)
5 Ευχαριστίες Η παρούσα διπλωματική εργασία εκπονήθηκε στα πλαίσια του Μεταπτυχιακού Διπλώματος Ειδίκευσης, «Βιοπληροφική», του τμήματος Βιολογίας του Εθνικού και Καποδιστριακού Πανεπιστημίου Αθηνών υπό την επίβλεψη του Καθ. Κων/νου Βοργιά. Θα ήθελα να ευχαριστήσω τον Δρ. Ιωάννη Μιχαλόπουλο, Ειδικού Λειτ. Επιστήμ. Β, ΙΙΒΕΑΑ για την εμπιστοσύνη που έδειξε στο πρόσωπό μου με την ανάθεση ενός επιστημονικά επίκαιρου και απαιτητικού θέματος Βιοπληροφορικής. Η συνεχής καθοδήγηση και στήριξη του, σε συνδυασμό με την άρτια επιστημονική του κατάρτιση, αποτέλεσαν καθοριστικό παράγοντα για την επιτυχή έκβαση της παρούσας διπλωματικής εργασίας. Επίσης, ένα μεγάλο ευχαριστώ οφείλω στο Δρ. Βασίλειο Ζουμπουρλή Ερευνητή Α' EIE, ο οποίος μου εμπιστεύτηκε τα πειραματικά δεδομένα του. Τέλος θα ήθελα να ευχαριστήσω έναν προς ένα όλους τους διδάσκοντες του M.Δ.Ε. της Βιοπληροφορικής του ΕΚΠΑ, καθώς οι γνώσεις που αποκόμισα μέσω του υψηλού επιπέδου διδακτικού τους έργου είναι πολύτιμες και αποτελούν σημαντική παρακαταθήκη για την περαιτέρω επιστημονική μου εξέλιξη. Ευχαριστώ επίσης τους Αναπ. κ. Καθ. Παντελή Μπάγκο, Παν. Θεσσαλίας, και Επίκ. Καθ. κ. Βασιλική Οικονομίδου που δέχτηκαν να συμμετάσχουν στην τριμελή εξεταστική επιτροπή. Τέλος ευχαριστώ τον μέχρι πρότινος διευθυντή του Μ.Δ.Ε. και νυν ομότιμο καθηγητή Σταύρο Ι. Χαμόδρακα, για την ευκαιρία φοίτησης που μου έδωσε και τη συνολική προσφορά του στην εκπαίδευσή μου. i
6 Περίληψη Τα micrornas είναι μικρά, μη-κωδικά RNAs, τα οποία λειτουργούν ως μετα-μεταγραφικοί ρυθμιστές εκατοντάδων mrna στόχων. Δεδομένου ότι η γονιδιακή έκφραση απορυθμίζεται σε πολλούς τύπους καρκίνου, αυτά αποτελούν πιθανούς βιοδείκτες πρόβλεψης, πρόγνωσης και διάγνωσης, καθώς και θεραπευτικούς στόχους επόμενης γενιάς. Η συμμετοχή των mirnas στο NMSC (Non-Melanoma Skin Carcinogenesis) τύπο καρκίνου, ο οποίος παρουσιάζει αυξανόμενη συχνότητα δεν έχει μελετηθεί εκτενώς. Αξίζει να σημειωθεί επίσης η απουσία μιας ολοκληρωμένης μελέτης των mirnas από την αρχικοποίηση, την προαγωγή μέχρι την εξέλιξη του καρκίνου. Για το σκοπό αυτό, χρησιμοποιήθηκε το πολυσταδιακό μοντέλο δερματικής καρκινογένεσης στον ποντικό, για τον εντοπισμό mirnas τα οποία συμμετέχουν καθ όλη τη διάρκεια της δερματικής καρκινογένεσης. Οι κυτταρικές σειρές που αποτελούν αυτό το μοντέλο υπεβλήθησαν σε ανάλυση γονιδιακής έκφρασης mirna μικροσυστοιχιών, βιοπληροφορική ανάλυση, επαλήθευση με q-pcr και θεραπεία με mirna ρυθμιστές. Δείξαμε ότι μεταξύ όλων των mirnas η έκφραση των οποίων απορυθμίζεται, μόνο τα μέλη της οικογένειας του mir-200 και το mir-205 εμφανίζουν ένα συνεχές πρότυπο στο οποίο η έκφραση αυτών μειώνεται καθ όλη τη διάρκεια των σταδίων της προαγωγής και της εξέλιξης. Γονίδια η έκφραση των οποίων πιθανόν να ρυθμίζεται ταυτόχρονα από δύο μέλη της οικογένειας αυτής, είναι τα Akt3, ROCK2 και Rap1a,. Συμπεραίνεται ότι η απορρύθμιση της έκφρασης αυτής της ομάδας mirnas εμπλέκεται σταθερά σε όλα τα στάδια της δερματικής καρκινογένεσης, αυξάνοντας την έκφραση ενός συνόλου mrna στόχων οι οποίοι συμμετέχουν στο μονοπάτι της κυτταρικής πρόσφυσης και το οποίο ευνοεί την εξέλιξη του καρκίνου. Λέξεις κλειδιά: mir-200, mir-205, AKT3, ROCK2, Rap1a, δερματική καρκινογένεση στον ποντικό ii
7 Abstract MicroRNAs are small, non-coding RNAs which act as post-transcriptional regulators of hundreds of target mrnas. Given that their expression is deregulated in several cancer types, they represent potential diagnostic, prognostic and predictive biomarkers, and also next-generation therapeutic targets. The involvement of mirnas in non-melanoma skin cancer, a type of cancer with increasing prevalence, is not extensively studied. Notably, a comprehensive characterization of the mirnas that are progressively altered throughout initiation, promotion and progression stages is missing. To this end, we exploited the multistage mouse skin carcinogenesis model, to identify mirnas consistently implicated in skin carcinogenesis. The cell lines comprising this model were subjected to mirna expression profiling using microarrays, followed by bioinformatics analysis and validation with q-pcr and treatment with mirna modulators. We showed that among all deregulated mirnas in our system, only a group consisting of the mir- 200 family members and mir-205 displays a continuous pattern of downregulation throughout promotion and progression stages. Genes putatively co-regulated by at least two members of this basic group of mirnas, such as Akt3, ROCK2 and Rap1a. We conclude that deregulation of this mirna group is consistently involved in all stages of oncogenic transformation of skin cells by increasing the expression of a set of crucial, co-regulated, focal adhesion-involved targets that favor cancer progression. Key words: mir-200, mir-205, AKT3, ROCK2, Rap1a, mouse skin carcinogenesis iii
8 Εισαγωγή Η συχνότητα εμφάνισης του καρκίνου του δέρματος αυξάνει παγκοσμίως και παρά το γεγονός ότι ο NMSC τύπος είναι σπάνιος, η ελλιπής πρόγνωση του και η συμβατική θεραπεία είναι αναποτελεσματική σε μεταστατικά στάδια. Επιπλέον, o NMSC μακροπρόθεσμα προκαλεί παρενέργειες στη μεταμόσχευση των οργάνων, τονίζοντας την ανάγκη για καλύτερη πρόληψη και διαχείριση. Συνεπώς, η διαλεύκανση των μοριακών μηχανισμών που διέπουν την καρκινογένεση του δέρματος είναι ζωτικής σημασίας. Ένα καλά τεκμηριωμένο σύστημα κυτταρικών σειρών για τη διερεύνηση του δερματικού καρκίνου είναι το πολυσταδιακό μοντέλο δερματικής καρκινογένεσης στον ποντικό. Αυτό αναπτύχθηκε στο εργαστήριο του Καθ. A. Balmain με την επαγωγή χημικής καρκινογένεσης στην επιδερμίδα του ποντικού χρήση DΜΒΑ (7,12-διμεθυλοβενζο [α] ανθρακένιο). Οι κυτταρικές σειρές οι οποίες παράγονται από τους αναπτυσσόμενους όγκους στο δέρμα, ταξινομούνται σε τέσσερις βασικές ομάδες: στα αθανατοποιημένα μη καρκινογόνα κερατινοκύτταρα (C5N και H- rass null); στα καλοήθη θηλώματα (P1, P6, MSP5); στα κακοήθη πλακώδη κύτταρα (PDV, PDVC57, Β9, Ε4) και στα πιο επιθετικά ατρακτοειδή καρκινικά κύτταρα (D3, H11, A5, CarB, CarC). H κυτταρική σειρά PDVC57 αποτελεί την κλωνική επέκταση της PDV ενώ οι Α5 και D3 προέρχονται από τον ίδιο αρχικό όγκο Β9 ο οποίος αντιπροσωπεύει την εξέλιξη στα πιο επιθετικά στάδια. Συνολικά τα ζεύγη PDV:PDVC57, Β9:A5 και B9:D3 προσομοιώνουν την κλωνική επέκταση των καρκινικών κυττάρων στο δέρμα του ποντικού και επομένως χρησιμεύουν στην εξέταση των βασικών αλλοιώσεων κατά την εξέλιξη και τη EMT (Εpithelial to Mesenchymal Transition) μετάβαση. Το μοντέλο αυτό είναι ιδανικό για τη σταδιακή ανάλυση μοριακών γεγονότων καίριας σημασίας στην ογκογένεση του δέρματος. Το σημαντικότερο είναι πως τα πιο σημαντικά ευρήματα αυτού του μοντέλου δεν περιορίζονται στον καρκίνο του δέρματος, αλλά σχεδόν σε όλους τους επιθηλιακούς τύπους καρκίνου. Μέχρι στιγμής, το μοντέλο αυτό έχει χρησιμοποιηθεί επιτυχώς για την παρακολούθηση κομβικών μεταβολών στα mrnas και των πρωτεϊνών που κωδικοποιούν, αλλά ποτέ για τον έλεγχο όλων των μεταβολών μικρών μη-κωδικών RNAs, όπως αυτών των mirnas σε όλα τα στάδια του. iv
9 Tην τελευταία δεκαετία, η κατανόηση των μονοπατιών των μοριακών μηχανισμών που ακολουθεί ο καρκίνος, θεωρήθηκε επανάσταση λόγω της ανακάλυψης του ρόλου των mirnas σε αυτόν. Τα micrornas, η κατηγορία των μικρών αυτών μη κωδικών μορίων RNA, ρυθμίζουν την έκφραση πολλών κωδικών γονιδίων μέσω της πρόσδεσης τους στο 3 'UTRs των mrna στόχων τους, προκλύοντας είτε την υποβάθμιση ή την αναστολή της λειτουργίας του mrna στόχου. Τα mirnas συχνά λειτουργούν είτε ως ογκογονίδια ή καταστολείς όγκων η έκφραση των οποίων συχνά απορυθμίζεται στον καρκίνο. Ο προσδιορισμός των mirnas που συμμετέχουν στον καρκίνο του ανθρώπου καθώς και των στόχων αυτών, αποτελούν επί του παρόντος αντικείμενα εντατικής έρευνας παρά τον περιορισμένο όγκο διαθέσιμων πληροφοριών για την επίδραση σε όλα τα διαδοχικά στάδια της ογκογένεσης. Η αποκλίνουσα έκφραση των mirnas σε διαφορετικά στάδια στο NMSC έχει μελετηθεί μέχρι στιγμής με τη σύγκριση της γονιδιακής έκφρασης καρκινικών βιοψιών δέρματος έναντι φυσιολογικών. Το πολυσταδιακό μοντέλο που χρησιμοποιούμε είναι ένα κατάλληλο σύστημα για τον έλεγχο των αλλαγών στα διαφορετικά στάδια. Η αξιοποίηση του μοντέλου αυτού σε συνδυασμό με τη χρήση μικροσυστοιχιών και κατόπιν βιοπληροφορικής ανάλυσης επιχειρεί να διαλευκάνει την έκφραση των mirnas σε μια σειρά διαδοχικών σταδίων της δερματικής καρκινογένεσης δίνοντας έμφαση στα πιο επιθετικά στάδια. Η ανάλυση αυτή έδειξε πως η μεγαλύτερη μείωση της έκφρασης των mirναs στα πιο επιθετικά στάδια της δερματικής καρκινογένεσης του ποντικού βρέθηκε στα μέλη της οικογένειας mir-200 και mir-205 και πως πιθανόν να χρειάζονται τουλάχιστον δύο από αυτά για τη ρύθμιση της έκφρασης των γονιδίων Akt3, ROCK2 and Rap1a. Όσον αφορά τις υπολογιστικές μεθόδους που χρησιμοποιήθηκαν για την εξαγωγή των παραπάνω συμπερασμάτων πρέπει να ειπωθεί πώς: i) η ανάλυση μικροσυστοιχιών mirnas της Exiqon μας επιτρέπει να εντοπίσουμε mirnas που εκφράζονται διαφορικά ii) ο συνδυασμός των mirnas που εκφράζονται διαφορικά σε συνδυασμό με γονίδια από βάσεις δεδομένων μας επιτρέπει να εντοπίσουμε γονίδια στόχους των mirnas που θα έπρεπε να εκφράζονται διαφορικά βάσει της διαφορικής έκφρασης των mirnas, iii) η σύγκριση των συμπερασμάτων του (ii) με τη σύγκριση γονιδίων στόχων μικροσυστοιχιών εξονίων που όντως εκφράζονται διαφορικά μας δίνει την τομή ενός συνόλου με τα γονίδια που θα έπρεπε και όντως εκφράζονται v
10 διαφορικά. Τέλος, καθορίζονται οι βιολογικές λειτουργίες στις οποίες συμμετέχουν αυτά τα γονίδια, μέσα από τον εντοπισμό τυχόν υπερεκπροσωπήσεων βιολογικών διεργασιών με τη χρήση δεδομένων Γονιδιακής Οντολογίας. Για την ανάλυση της υπερεκπροσώπησης των βιολογικών διεργασιών χρησιμοποιείται ανάλυση υπεργεωμετρικής κατανομής. vi
11 Περιεχόμενα vii
12 Ευχαριστίες i Περίληψη... ii Abstract.... iii Εισαγωγή iv Περιεχόμενα.... vii Κεφάλαιο 1 ο mirnas Εισαγωγή Ιστορική αναδρομή Ονοματολογία Βιογένεση Kανονική Βιογένεση των micrornas Bιογένεση ανεξάρτητη του ενζύμου Drosha Mirtrons mirnas παραγόμενα από snornas Μη-κανονική βιογένεση των micrornas mirnas παραγόμενα από shrnas mirnas παραγόμενα από trnas MHV68 (murine γ-herpesvirus 68) mirnas mirnas παραγόμενα από sirnas Βιογένεση μη-εξαρτώμενη από τo ένζυμο Dicer Antisense mirna (mirna*) Πρόσδεση των mirnas στους mrna στόχους Νουκλεοτιδική αλληλουχία ενός mirna Χαρακτηριστικά των περιοχών στόχων Στερεοδιάταξη των περιοχών στόχων των mirnas Παράγοντες αποτελεσματικότητας των mirnas Επίπεδα συγκέντρωσης των mirs και των στόχων τους Πρωτεΐνες πρόσδεσης στο RNA Competing endogenous RNAs (cernas) Circular RNAs (cirnas). 18 viii
13 1.8 Μη κανονικές λειτουργίες των mirnas Ενεργοποίηση εκκινητή Ενεργοποίηση μετάφρασης μέσω πρόσδεσης στο στόχο Συνδέτες υποδοχέων Αναγνώριση mirnas και εκκινητών τους Βιβλιογραφία. 23 Κεφάλαιο 2 ο Τεχνολογία Μικροσυστοιχιών Ανάλυση γονιδιακής έκφρασης Ιστορικά στοιχεία Συσχετισμός ανάλυσης γονιδιακής έκφρασης με άλλες βιολογικές πληροφορίες Πρόβλεψη περιοχών πρόσδεσης Πρόβλεψη αλληλεπίδρασης και λειτουργίας πρωτεϊνών Τύποι μικροσυστοιχιών Επίπεδες (δισδιάστατες) μικροσυστοιχίες Μη-επίπεδες (τρισδιάστατες) μικροσυστοιχίες Πειραματική διαδικασία Επιλογή ανιχνευτών Ακινητοποίηση ανιχνευτών Απομόνωση βιολογικού υλικού Ιχνηθέτηση του στόχου Μονοχρωματικές μικροσυστοιχίες Διχρωματικές μικροσυστοιχίες Πειραματικός σχεδιασμός Δημιουργία αντιγράφων Ανιχνευτές ελέγχου Κοινό δείγμα αναφοράς Διχρωματικές μικροσυστοιχίες: Dye-swapping Υβριδοποίηση Σάρωση Πλατφόρμες μικροσυστοιχιών In situ σύνθεση ολιγονουκλεοτιδικών μικροσυστοιχιών cdna μικροσυστοιχίες εκτύπωσης. 47 ix
14 2.9.3 Ολιγονουκλεοτιδικές μικροσυστοιχίες εκτύπωσης Βιβλιογραφία. 49 Κεφάλαιο 3 ο Ανάλυση Πρωτογενών Δεδομένων Επεξεργασία πρωτογενών δεδομένων μικροσυστοιχιών Μέθοδοι για την ανάλυση χαμηλού επιπέδου Διόρθωση υποβάθρου Κανονικοποίηση Κανονικοποίηση εντός των μικροσυστοιχιών Composite loess κανονικοποίηση Κανονικοποίηση μεταξύ των μικροσυστοιχιών Αquantile κανονικοποίηση Έλεγχος Στατιστικών Υποθέσεων Ανάλυση Διακύμανσης κατά ένα Παράγοντα (One-way ANOVA) Παραδοχές στις οποίες βασίζεται η Aνάλυση Διακύμανσης Αποφάσεις ελέγχου Post-hoc ανάλυση Μέθοδοι ανάλυσης υψηλού επιπέδου Συντελεστής συσχέτισης Pearson Correlation Coefficient (PCC) False Discovery Rate (FDR) Υπεργεωμετρική κατανομή Βιβλιογραφία 71 Κεφάλαιο 4 ο Μέθοδοι Περιγραφή βιολογικών δειγμάτων Exiqon mircury LNA TM microrna array kit Περιγραφή ανιχνευτών mircury LNA TM microrna Array Exiqon s mircury LNA TM Hi-Power Labelling Kit Πειραματικός σχεδιασμός Αρχεία πρωτογενών δεδομένων Ανάλυση χαμηλού επιπέδου Εγκατάσταση R και εγκατάσταση Limma/Bioconductor Ανάγνωση πρωτογενών δεδομένων. 81 x
15 4.6.3 Διόρθωση υποβάθρου Οπτική αναπαράσταση γονιδιακής έκφρασης (ΜΑplots) Κανονικοποίηση εντός των μικροσυστοιχιών Κανονικοποίηση μεταξύ των μικροσυστοιχιών Ανάλυση υψηλού επιπέδου Ανάλυση διακύμανσης και εύρεση διαφορικά εκφρασμένων mirnas Παρουσίαση αποτελεσμάτων σε βάση δεδομένων Προσδιορισμός λειτουργικών γονιδίων στόχων mirnas Γονίδια ειδικού ενδιαφέροντος Ανάλυση του ρόλου των γονιδίων στόχων των mirs με όρους γονιδιακής οντολογίας Βιβλιογραφία. 106 Κεφάλαιο 5 ο Αποτελέσματα και Συζήτηση Αποτελέσματα H Προοδευτική μείωση της έκφρασης των μελών της οικογένειας mir-200 και του mir-205 κατά την εξέλιξη της δερματικής καρκινογένεσης στον ποντικό Λειτουργική συντήρηση των μελών της mir-200 οικογενείας και του mir Παράλληλη υπερέκφραση των στόχων της οικογενείας mir-200 και του mir-205 στη δερματική καρκινογένεση Διαφορική έκφραση mirnas κατά τη μετάβαση από πλακώδη κακοήθη σε αναπλαστικά καρκινώματα του δέρματος Συζήτηση Βιβλιογραφία. 121 Παράρτημα Ι 123 I.1 Έγρωμα ΜΑ plots μη κανονικοποιημένων δεδομένων. 124 Ι.2 Έγχρωμα ΜΑplots μετά την κανονικοποίηση εντός των μικροσυστοιχιών.131 Ι.3 Ασπρόμαυρα ΜΑplots μετά την εντός μικροσυστοιχιών κανονικοποίηση - ομαδική παρουσίαση. 138 I.4 Boxplots μετά την κανονικοποίηση εντός των μικροσυστοιχιών I.5 ΜΑ plots μετά την κανονικοποίηση μεταξύ των μικροσυστοιχιών Ι.6 Ασπρόμαυρα ΜΑplots μετά την μεταξύ μικροσυστοιχιών κανονικοποίηση ομαδική παρουσίαση 148 I.7 Boxplots μετά την κανονικοποίηση μεταξύ των μικροσυστοιχιών I.8 Eντολές δημιουργίας πινάκων SQL xi
16 Κεφάλαιο 1 ο mirnas 1
17 Κεφάλαιο 1. MicroRNAs 1.1 Εισαγωγή Τα micrornas (mirnas ή mirs), είναι υψηλά συντηρημένα μικρά μη-κωδικά μονόκλωνου RNA, μήκους ~22 νουκλεοτιδικών βάσεων τα οποία εντοπίζονται σε φυτά, ζώα και μερικούς ιούς. Εκτός της μεθυλίωσης του DNA και της τροποποίησης των ιστονών (Zheng et al., 2013), τα mirnas ρυθμίζουν την αυτο-ανανέωση, τη διαφοροποίηση και τη διαίρεση των κυττάρων, μέσω της μετα-μεταγραφικής σίγησης της γονιδιακής έκφρασης. Η σίγηση της γονιδιακής έκφρασης επιτυγχάνεται είτε μέσω της παρεμπόδισης της πρωτεϊνικής έκφρασης (μετάφρασης) είτε μέσω της αποδόμησης συγκεκριμένων μεταγράφων RNAs (mrnas). Έχει βρεθεί πως ο βαθμός της συμπληρωματικότητας των mirnas με τους mrna στόχους καθορίζει τον τρόπο με τον οποίο γίνεται η σίγηση της έκφρασης. Τα mirnas των φυτών παρουσιάζουν εκτεταμένη ή πλήρη συμπληρωματικότητα ενώ τα ζωικά mirnas παρουσιάζουν μικρή συμπληρωματικότητα με τους στόχους τους έκτασης ~7 nt (Axtell et al., 2011; Rhoades et al., 2002). Εάν το mirna με το mrna στόχο είναι πλήρως συμπληρωματικά τότε γίνεται η αποδόμηση του mrna, διαφορετικά παρεμποδίζεται η πρωτεϊνοσύνθεση (Hutvagner and Zamore, 2002). Ωστόσο οι περιοχές στόχοι με πλήρη συμπληρωματικότητα είναι πιο ευαίσθητες συγκριτικά με αυτές που παρουσιάζουν ατελή συμπληρωματικότητα (Okamura et al., 2008) πιθανόν εξαιτίας της μεγαλύτερης ισχύς που παρουσιάζει η αποκοπή από την AGO2 σε σχέση με την AGO1 (Forstemann et al., 2007). Ένα mirna μπορεί να ρυθμίζει την έκφραση ακόμη και διακοσίων mrna στόχων. Αυτοί μπορεί να είναι μεταγραφικοί και εκκριτικοί παράγοντες, υποδοχείς ή και κυτταρικοί διαβιβαστές. Δυνητικά, λοιπόν τα mirnas έχουν επίδραση σε πολλά διαφορετικά γενετικά μονοπάτια. Συνεπώς, η απορρύθμισή τους πιθανόν να οδηγεί σε πολλές ασθένειες, συμπεριλαμβανομένου και του καρκίνου. Σύμφωνα με την 20 η έκδοση της βάσης mirbase ( (Kozomara and Griffiths-Jones, 2011), έχουν σχολιασθεί περισσότερα από 1100 ποντικίσια και 1800 ανθρώπινα mirnas. Στον άνθρωπο, παρά το γεγονός ότι μόνο το 3% των γονιδίων του ανθρώπου κωδικοποιεί mirnas, τα εν λόγω mirnas ρυθμίζουν περίπου το 50% των γονιδίων 2
18 που κωδικοποιούν πρωτεΐνες, ωστόσο ένα μέρος αυτών είναι πιθανό να είναι μην έχουν προσδιοριστεί σωστά ως mirs (Chiang et al., 2010). 1.2 Ιστορική αναδρομή Το πρώτο microrna (mirna) ανακαλύφθηκε το 1993 στο νηματώδη σκώληκα Caenorhabditis elegans από τους Victor Ambros, Rosalind Lee και Rhonda Feinbaum (Lee et al., 1993) και ονομάστηκε lin-4. Βρέθηκε ότι η πρωτεΐνη LIN-14 ρυθμιζόταν από ένα μικρό RNA που κωδικοποιείτο από το γονίδιο lin-4. Το πρόδρομο lin-4 mirna αποτελείτο από 61 νουκλεοτίδια ενώ στην ώριμή του μορφή από 22 και είναι μερικώς συμπληρωματικό με το 3 UTR του lin-14 mrna. Το δεύτερο mirna let-7 το οποίο προσδιορίστηκε το 2000, καταστέλλει την έκφραση των lin-14, lin-28, lin-41, lin-42 και daf-12 γονιδίων και είναι συντηρημένο σε πολλά είδη (Pasquinelli et al., 2000; Reinhart et al., 2000). 1.3 Ονοματολογία Ένα πρόδρομο microrna (precursor mirna ή pre-mirna) συμβολίζεται με το πρόθεμα mir το οποίο ακολουθείτο από μία παύλα και έναν αριθμό, με τον τελευταίο να υποδεικνύει τη χρονική σειρά της ονομασίας του. Παραδείγματος χάριν, το mir-123 έχει ονομαστεί και είναι πιθανό να έχει ανακαλυφθεί πριν από το mir-456. Ένα ώριμο mirna συμβολίζεται με τo πρόθεμα mir. Τα mirnas με σχεδόν ταυτόσημες ακολουθίες πλην ενός ή δύο νουκλεοτιδίων συμβολίζονται με ένα επιπλέον πεζό γράμμα, π.χ. mir-133a και mir-133b. Τα πρόδρομα mirnas από τα οποία προκύπτουν ταυτόσημα mirnas αλλά εντοπίζονται σε διαφορετικές τοποθεσίες του γονιδιώματος συμβολίζονται με μία παύλα και ένα αριθμητικό επίθεμα επιπλέον. Για παράδειγμα τα mirnas hsa-mir-133a-1 και hsa-mir-133a-2 παράγουν και τα δύο το hsa-mir- 133a mirna. To είδος προέλευσης του mirna συμβολίζεται με ένα πρόθεμα τριών γραμμάτων. Π.χ. το hsa-mir-133a είναι ένα ανθρώπινο mirna. Δύο mirnas που προέρχονται από τα αντίθετα άκρα του ίδιου pre-mirna συμβολίζονται επιπροσθέτως με τα επιθέματα 3p και 5p. 3
19 1.4 Βιογένεση Τα mirnas προέρχονται τόσο από τη μεταγραφή μη-κωδικών DNA περιοχών, όσο και από την αντιγραφή των ήδη υπαρχόντων mirnas. Πιο αναλυτικά περίπου το 40% όλων των mirnas κωδικοποιούνται από εσώνια κωδικών γονιδίων, ενώ περίπου το 10% από εσώνια mrna γονιδίων που δεν κωδικοποιούν πρωτεΐνες. Επιπλέον περίπου 10% είναι τα mirnas προέρχονται από εξώνια mrna γονιδίων που δεν κωδικοποιούν πρωτεΐνες (Rodriguez et al., 2004) ή ακόμη και από το 3 UTR mrna γονιδίων Κανονική Βιογένεση των micrornas Τα ώριμα mirnas αποτελούν μετάγραφα μόρια, μεγαλύτερων μεταγράφων τμημάτων, αρχικών mirnas (primary mirnas ή pri-mirναs), με τα περισσότερα mirnas να μεταγράφονται από την RNA πολυμεράση ΙΙ και μόνο κάποια από την RNA πολυμεράση ΙΙΙ (Corcoran et al., 2009). Η πλειονότητα των mirnas μεταγράφεται από τη συρραφή των εσωνίων στο pre-mrna έχοντας όπως τα περισσότερα αντίγραφα της RNA II πολυμεράσης ένα κάλυπτρο στην 5 και μία 3 poly(α) αμετάφραστη περιοχή. H συρραφή των εσωνίων οδηγεί στο σχηματισμό ενός pri-mir το οποίο κατόπιν μετατρέπεται σε πολλά ώριμα mirs (Saini et al., 2007). Μέσα στη δομή των primirs σχηματίζονται μία ή περισσότερες δομές φουρκέτας οι οποίες χαρακτηρίζονται από ατελή συμπληρωματικότητα ~30 bp (Han et al., 2006). Στα ζώα, η ωρίμανση των mirnas υφίσταται σε δύο στάδια, με το πρώτο να λαμβάνει χώρα στον πυρήνα και το δεύτερο στο κυτταρόπλασμα (Miyoshi et al., 2010) και κυρίως με την παρεμπόδιση της μετάφρασης των mrna μεταγράφων (Lee et al., 2004b). Μέσα στον πυρήνα, οι δομές φουρκέτας του pri-mir αναγνωρίζονται και αποκόπτονται από ένα πρωτεϊνικό σύμπλοκο με κύρια συστατικά τη Drosha ριβονουκλεάση (Rnase III) και από τη DGCR8 (Pasha στα ασπόνδυλα) η οποία είναι μία πρωτεΐνη η οποία προσδένεται σε δίκλωνο RNA (double stranded RNA-binding domain (dsrbd)). Η DGCR8 αναγνωρίζει τα άκρα της φουρκέτας και καθοδηγεί τη Drosha στην αποκοπή του pri-mir ~11 bp ή κατά ~1 έλικα (Han et al., 2006) από το «κοτσάνι» της φουρκέτας, παράγοντας μία μικρότερη φουρκέτα μήκους ~60-70 nt (Han et al., 2006). Πάραυτα, έχουν παρατηρηθεί στη Drosophila και φουρκέτες διπλάσιου μεγέθους 4
20 (Ruby et al., 2007). Η νέα μικρότερη φουρκέτα που προκύπτει ονομάζεται pre-mir και διαθέτει μία προεξοχή 2 nt στο 3 άκρο. Από ένα pri-mir μπορούν να προκύψουν μέχρι και 6 pre-mirs. H 3 προεξοχή που άφησε το ένζυμο Drosha αναγνωρίζεται από την exportin-5 και το pre-mir μεταφέρεται στο κυτταρόπλασμα μέσω του RAN-GTP μηχανισμού. H υδρόλυση του προσδεδεμένου στη RAN πρωτεΐνη, GTP στο κυτταρόπλασμα, αποτελεί το έναυσμα για τη διάσπαση του συμπλόκου, επιτρέποντας στο pre-mir να προσδεθεί μέσου του 3 άκρου (Lund and Dahlberg, 2006) στη δίκλωνη ριβονουκλεάση ΙΙΙ, τη Dicer. Η Dicer η οποία είναι προσδεδεμένη με τις dsrbp, TRBP, PACT και Loquacious (στη Δροσόφιλα) πρωτεΐνες αποκόπτει το βρόχο του pre-mir κατά ~2 στροφές έλικας, απελευθερώνοντας ένα δίκλωνο mirna : mirna* ~22nt και το οποίο διαθέτει προεξοχές 2 nt και στα δύο 3 άκρα. Από τους δύο κλώνους το mirna προκύπτει από αυτόν (οδηγός κλώνος) του οποίου το 5 άκρο διαθέτει τη μικρότερη ελεύθερη ενέργεια βάσει της συμπληρωματικότητας των βάσεων (Khvorova et al., 2003). Το mirna προσδένεται με ένα μέλος της Argonaute (Ago) οικογένειας πρωτεϊνών για να σχηματίσουν τον πυρήνα του RNA συμπλόκου που επάγει τη γονιδιακή σίγηση (mirna-induced Silencing Complex RISC ή mirisc) (Miyoshi et al., 2010). Ο κλώνος (mirna*) αποδομείται, χωρίς να αποκλείεται κάποιες φορές να είναι βιώσιμος. Με ανάλυση βαθιάς αλληλούχισης έχει δειχθεί ότι ένα μεγάλο πλήθος mirnas* συνδέεται με της οικογενείας AGO. Στις μύγες, ενώ τα mirnas προσδένονται στην AGO1, παρατηρείται ορισμένες φορές τα mirnas και mirnas* να συνδέονται με σε διαφορετικές AGO πρωτεΐνες, όπως στην AGO1 και AGO2 αντίστοιχα (Seitz et al., 2008). Oι πρωτεΐνες Αργοναύτες περιέχουν δύο περιοχές πρόσδεσης, τις PAZ και PIWI οι οποίες προσδένονται στο 3 και 5 άκρο αντίστοιχα των mirnas προσανατολίζοντας τα για αλληλεπίδραση με τους mrna στόχους. Έχει βρεθεί πως αυτές οι πρωτεΐνες εμπλέκονται σε διάφορους καρκίνους. Για παράδειγμα στον άνθρωπο, τα γονίδια Αργοναύτες ago1, ago2 και ago4 τα οποία βρίσκονται ομαδοποιημένα στο χρωμόσωμα 1 παρουσιάζουν ελλείψεις σε καρκίνους του νεφρού, ενώ το γονίδιο Αργοναύτης hiw-1 που εκφράζεται στους όρχεις ίσως στην ομαλή λειτουργία του ρυθμίζει των πολλαπλασιασμό των γεννητικών κυττάρων και λειτουργεί ως ογκογονίδιο κατά την απορρύθμισή του. Το γονίδιο αυτό έχει βρεθεί μεταλλαγμένο σε 12 από 19 καρκίνους των όρχεων. 5
21 Tέλος το mirna καθοδηγεί το mirisc σύμπλοκο στις συμπληρωματικές περιοχές των mrna στόχων, συνήθως στο 3 UTR (Εικόνα 1.1) (Ameres et al., 2007). Στα φυτά ωστόσο, αυτή σε αυτή τη διαδικασία δεν παίρνουν μέρος δύο ένζυμα, ένα μέσα και ένα έξω από τον πυρήνα, αλλά μόνο το DL1 (Dicer-Like 1) ομόλογο του Dicer, το οποίο εκφράζεται στον πυρήνα των φυτικών κυττάρων. Στη συνέχεια το δίκλωνο mirna : mirna* μεταφέρεται εκτός πυρήνα από μια πρωτεΐνη ομόλογη της Exportin 5, την HST (Hasty) και από εκεί το ώριμο mirna ενσωματώνεται στο RISC (Lelandais-Briere et al., 2010). Εικόνα 1.1: Βιογένεση mirna (Doxakis, 2013). 6
22 1.4.2 Βιογένεση ανεξάρτητη του ενζύμου Drosha Mirtrons Ένα εναλλακτικό μονοπάτι βιογένεσης των mirnas στο οποίο δε λαμβάνει μέρος το Drosha ένζυμο, είναι το mirton μονοπάτι, το οποίο πιθανό να προκύπτει εξαιτίας μεταλλάξεων στη Δροσόφιλα, ή στο Drosha/DGCR8 σύμπλοκο. Τα mirtrons είναι μικρά RNAs τα οποία αναγνωρίστηκαν πρώτα στη μύγα και το σκώληκα, ενώ παρόμοιες περιοχές αναγνωρίστηκαν αργότερα στον ποντικό και στο ρύζι (Zhu et al., 2008). Στη Δροσόφιλα το μήκος των εσωνίων έχει μέγιστο μήκος ~60 nt, όσο δηλαδή περίπου το μήκος των pre-mirnas. Αναλύσεις βαθιάς αλληλούχισης σε μικρά RNAs Δροσόφιλας, έδειξαν ότι μικρά RNAs τα οποία προέρχονται από μικρά εσώνια ~60 nt όσο δηλαδή και το μέγεθος των premirnas, έχουν δυνατότητα σχηματισμού φουρκέτας (Lee et al., 1993). Το αρχικό προϊόν της συρραφής αυτών των εσωνίων δεν είναι γραμμικό αλλά δημιουργεί μια θηλειά (σαν φουρκέτα) πάνω στο 5 UTR του οποίο συνδέεται το 3 άκρο. Μέσω του Ldbr (Lariat debranching) ενζύμου επιτυγχάνεται το άνοιγμα της θηλειάς, δίπλωμα της σε μορφή pre-mirna και μετέπειτα μεταφορά στο κυτταρόπλασμα από την Exportin-5, αποκοπή από την Dicer και σύνδεση με την οικογένεια πρωτεϊνών Ago (Okamura et al., 2007). Το κοτσάνι των πρόδρομων mirtrons έχει μικρότερο μήκος από τα mirnas λόγω έλλειψης της ελικοειδούς στροφής στο κοτσάνι η οποία προσλαμβάνει το Dicer/DGCR8 σύμπλοκο και για αυτό έχει προταθεί πως η συρραφή υποκαθιστά την αποκοπή της Drosha. Για το λόγο αυτό, θεωρείται κατά κάποιο τρόπο ότι πολλά pre-mirnas μετάγραφα είναι ταυτόχρονα και pri-mirnas (Εικόνα 1.2Β). Τα mirtrons, αποτελούν ένα μικρό μέρος των mirnas τόσο στα θηλαστικά καθώς όσο και στη Δροσόφιλα, επειδή μόνο κάποια από αυτά τα μικρά εσώνια σχηματίζουν δομές φουρκέτας στο μέγεθος των mirnas (Berezikov et al., 2007). O εντοπισμός μη-δομημένων περιοχών τόσο στο 5 UTR όσο και στο 3 άκρο των φουρκετών των pre-mirnas στα mirtrons σημαίνει την παραγωγή ενός ενδιάμεσου προϊόντος. H αποκοπή αυτών των ουρών στο 3 άκρο γίνεται στον πυρήνα από το σύμπλοκο exosome. Αντιστοίχως στο 5 η αποκοπή της ουράς δεν είναι γνωστό από ποια ένζυμα γίνεται (Εικόνα 1.2C). 7
23 Εικόνα 1.2: Σχηματική επισκόπηση βιογένεσης των mirtrons mirnas παραγόμενα από snornas Πρόδρομα mirnas προκύπτουν επίσης από μικρά πυρηνικά RNAs τα οποία είναι εξελικτικά συντηρημένα με μήκος ~ nt. Από αναλύσεις βαθιάς αλληλούχισης μικρών RNAs συνδεδεμένα με τις Ago1 και Ago2 πρωτεΐνες προσδιορίστηκαν mirna-like μόρια τα οποία πρέρχονται από συγκεκριμένα snornas, η δευτεροταγής δομή των οποίων περιλαμβάνει δύο pre-mirna-like δομές φουρκέτας συνδεδεμένες μεταξύ τους (Ender et al., 2008). Συνεπώς κάποια snornas αποτελούν ταυτόχρονα τμήμα πυρηνικών ριβονουκλεοπρωτεϊνών αλλά και 8
24 πηγή σχηματισμού mirnas. Ερωτήματα προκύπτουν για το πώς τα υπό κανονικές συνθήκες snornps τα οποία βρίσκονται στον πυρήνα, μεταφέρονται στο κυτταρόπλασμα για περαιτέρω επεξεργασία από τη Dicer και πως αυτή προσδένεται πάνω σε αυτά στα οποία υπάρχουν ήδη προσδεδεμένες πολλές πρωτεΐνες. MiRNAs που προέρχονται από πυρηνικά RNAs εντοπίζονται σε θηλαστικά (π.χ. άνθρωπος) και στο πρωτόζωο Giardia lambia Μη-κανονική βιογένεση των micrornas mirnas παραγόμενα από shrnas Έχει προσδιοριστεί πως ένα από τα shrnas παράγει μετάγραφο του οποίου η δευτεροταγή δομή περιέχει δύο pre-mirna-like φουρκέτες από τις οποίες μόνο η μία παράγει το mir-1980 (Doxakis, 2013) mirnas παραγόμενα από trnas Σε αντίθεση με τα περισσότερα trnas τα οποία των οποίων τα pri-trnas μετάγραφα σχηματίζουν δευτεροταγείς δομές τριφυλλιού, μπορεί να προκύψουν pri-trnas μετάγραφα με μία εναλλακτική δευτεροταγή δομή, μιας φουρκέτας μήκους 110 nt. Συνεπώς ένα pri-trna μετάγραφο μπορεί να έχει διπλή λειτουργία, να μετατρέπεται σε πρόδρομο trna ή σε πρόδρομο mirna (Doxakis, 2013) MHV68 (murine γ-herpesvirus 68) mirnas Στον ποντικό τα mirnas που κωδικοποιούνται από το παθογόνο γ-herpesvirus 68 (MHV68), μεταγράφονται από την RNA III πολυμεράση με αποτέλεσμα το pri-mirna να αποτελείται από μία pre-mirna φουρκέτα και ένα trna. Εδώ τα pri-mirnas δεν επεξεργάζονται από τη Drosha αλλά από το ένζυμο trnase Z στον πυρήνα. To ένζυμο αυτό αποκόπτει το trna στο 3 άκρο του, απελευθερώνοντας το pre-mirna το οποίο κατόπιν υφίσταται επεξεργασία από τη Dicer προκειμένου να επέλθει η ωρίμανση του. 9
25 mirnas παραγόμενα από sirnas Τα dsrnas και τα pre-mirnas είναι μικρά RNAs με διαφορετική δομή. Ωστόσο η επεξεργασία τους σε mirnas και sirnas αντιστοίχως γίνεται από την ίδια οικογένεια πρωτεϊνών τη Dicer. Συγκεκριμένα η Dicer-1 και η AGO1 συμμετέχει στην παρεμπόδιση της μετάφρασης του στόχου του mirna ενώ η Dicer-2 και η AGO2 στην αποδόμηση του mrna αν υπάρχει πλήρης συμπληρωματικότητα. Αντιθέτων στην περίπτωση ενός sirna η συμπληρωματικότητα είναι συνήθως πλήρης και το mrna μόνο αποδομείται (Holen et al., 2002; Parrish et al., 2000). Ένα υποσύνολο των sirnas παράγεται από επιμήκη RNA μετάγραφα με αποτέλεσμα την ύπαρξη επιμηκών φουρκετών με ατελή συμπληρωματικότητα σε πολλά σημεία. Η μετατροπή σε ώριμο sirna γίνεται με τη συμμετοχή του συμπλόκου Dicer2-Loqs-PD. Το σύμπλοκο Dicer-2-Loqs-PD επεξεργάζεται το πρόδρομο sirna αλλά δεν μπορεί να επεξεργαστεί την τελική φουρκέτα που προκύπτει. Σε αυτό το σημείο η τελική φουρκέτα αναγνωρίζεται ως πρόδρομο mirna και στη συνέχεια επεξεργάζεται από το σύμπλοκο Dicer1-Loqs PA/PB. Τα mirnas που προκύπτουν με αυτή τη διαδικασία ονομάζονται αδέλφια των sirnas ή semirnas Βιογένεση μη-εξαρτώμενη από το ένζυμο Dicer To pre-mir-451 το οποίο παράγεται από τη Drosha προσδένεται στην Αgo2 χωρίς τη μεσολάβηση του ενζύμου Dicer. Αυτό έρχεται σε αντίθεση με την αποκοπή από τη Dicer και την πρόσδεση στο RISC σύμπλοκο (Chendrimada et al., 2005). Αντ αυτού το ρόλο της Dicer στο κυτταρόπλασμα αναλαμβάνει η Ago2 το pre-mir-451 ενώ μια κυτταρική νουκλεάση αναλαμβάνει την περαιτέρω ωρίμανσή του. Τα trnas διακρίνονται σε δύο τάξεις, σε αυτά που η μετατροπή τους σε μικρά RNAs μεγέθους mirnas είναι εξαρτώμενη από τη Dicer και σε αυτά που είναι ανεξάρτητα της Dicer αλλά εξαρτώνται από τη trnase Z. Στη δεύτερη περίπτωση τα πρόδρομα trnas αποκόπτονται στο 5 άκρο τους από την RNase P και στο 3 άκρο τους από τη trnase Z. Επειδή αυτό που προκύπτει δε συνάδει με τον ακριβή ορισμό του mirna ονομάζονται trfs (trna-derived RNA Fragments) (Lee et al., 2009) (Εικόνα 1.3). 10
. 1.")
26 Εικόνα 1.3: Μονοπάτια βιογένεσης των mirnas (Miyoshi et al., 2010). 1.5 Antisense mirna (mirna*) Παρά τη μεταγραφή των pre-mirnas (ή των mirtrons) από το mrna σε ίσες ποσότητες κλώνων, η συσσώρευση των mirnas και mirnas* κλώνων σε κατάσταση δυναμικής ισορροπίας (steady state) είναι ασύμμετρη (Lau et al., 2001). Παρόλο που τα mirnas* είναι σπάνια, κάποια είναι σχετικώς άφθονα καθώς επίσης έχουν παρατηρηθεί περιπτώσεις mirnas* με μεγαλύτερη αφθονία στο κύτταρο συγκριτικά με τα mirnas. Ανάμεσα σε είδη Δροσόφιλας παρατηρούνται απόλυτα συντηρημένα mirnas*. Το γεγονός αυτό καταδεικνύει ότι μεγαλύτερος βαθμός συντήρησης των νουκλεοτιδίων των mirnas* συνεπάγεται και μεγαλύτερη συγκέντρωση ενός mirna* σε κατάσταση δυναμικής ισορροπίας (Okamura et al., 2008). Αυτή η συσχέτιση ανάμεσα στην εξελικτική πίεση και το 11
27 επίπεδο έκφρασης των mirnas* καθώς και το γεγονός ότι ~40% των mirna* νουκλεοτιδικών ακολουθιών δεν αποκλίνει ανάμεσα σε ένα μεγάλο εύρος διαφορετικών ειδών, δε συμβαδίζει με την ιδέα ότι τα mirnas* είναι μόνο φορείς κλώνοι προκειμένου να διατηρήσουν το συμπληρωματικό τους κλώνο σε δομή φουρκέτας. Δύναται λοιπόν να αντικατοπτρίζει τη χρήση των mirnas ως ρυθμιστές της έκφρασης των στόχων τους (Okamura et al., 2008). H λειτουργικότητα των mirnas* εκτείνεται και στα mirtrons (Okamura et al., 2008), ενώ η παρουσία μόνο mirnas* στο συνολικό RNA δε σημαίνει απαραιτήτως ότι έχουν και κάποιο ρυθμιστικό ρόλο αλλά πιθανώς να αποδομούνται με πιο αργούς ρυθμούς από τα υπόλοιπα. Όμοια με ότι συμβαίνει στα mirnas, τα 5 και 3 άκρα των mirnas* είναι περισσότερο συντηρημένα συγκριτικά με τις περιοχές στο εσωτερικό τους και επιπλέον το 5 άκρο είναι ελαφρώς πιο συντηρημένο από το 3. Αν ο στόχος των mirnas* ήταν η συμμετοχή του μόνο στη διαδικασία της ωρίμανσης του mirna θα προσδοκούσε κανείς το 3 άκρο να είναι περισσότερο συντηρημένο συγκριτικά με το 5 καθώς ζευγαρώνει με τη φύτρα του mirna. Παρόλο που τα mirnas* είναι λιγότερο συντηρημένα έναντι των mirnas, παρατηρούνται σε αυτά μοτίβα η νουκλεοτιδική απόκλιση των οποίων είναι σύμφωνη με τη ρυθμιστική τους ενεργότητα. Tέλος το πλήθος των στόχων των mirnas* είναι μικρότερο από αυτόν των mirnas. 1.6 Πρόσδεση των mirnas στους mrna στόχους Μέχρι στιγμής, δεν είναι απολύτως κατανοητοί ποιοι είναι οι παράγοντες οι οποίοι καθορίζουν ποια mrnas αποτελούν στόχο ποιων mirnas ή ποιος είναι ο μηχανισμός ο οποίος καταστέλλει τη μετάφραση ή τη σταθερότητα των mrnas Νουκλεοτιδική αλληλουχία ενός mirna Μελέτες στο μεταγράφωμα και στο πρωτέωμα έχουν δείξει πως ο κύριος παράγοντας πρόσδεσης ενός mir στο mrna στόχο είναι η πλήρης ή σχεδόν πλήρης συμπληρωματικότητα των βάσεων κατά Watson-Crick στην περιοχή 2-7 ή 2-8 του 5 άκρου του mir (Brennecke et al., 2005), κάτι το οποίο έχει επιβεβαιωθεί πειραματικά από μελέτες κρυσταλλογραφίας στο Ago- 12
28 mir σύμπλοκο. Η 2-7 ή 2-8 περιοχή ονομάζεται φύτρα (seed) και παρουσιάζει ελικοειδή στερεοδιάταξη (Elkayam et al., 2012; Wang et al., 2008). Υπάρχουν τέσσερις τέτοιοι τύποι περιοχών στην περιοχή της φύτρας, ένα εξαμερές, δύο επταμερή και ένα οκταμερές. Το εξαμερές εμφανίζει πλήρη συμπληρωματικότητα με την περιοχή της φύτρας (mirna 2-7 nt). Το ένα επταμερές περιλαμβάνει τα νουκλεοτίδια 2-8 του mirna ενώ το άλλο τα νουκλεοτίδια 1-7 με το πρώτο να είναι μια αδενίνη. Τέλος το οκταμερές περιλαμβάνει τα νουκλεοτίδια 1-8 με το πρώτο να είναι επίσης μια αδενίνη. H ελλιπής συμπληρωματικότητα στην περιοχή της φύτρας μπορεί να αντισταθμιστεί από ένα επταμερές ή οκταμερές κατά Watson-Crick ζευγάρωμα των βάσεων στις θέσεις του 3 άκρου ενός mirna. Ειδικότερα το τετραμερές στις θέσεις έχει συσχετιστεί με υποέκφραση του mirna στόχου (Grimson et al., 2007). Η παρουσία εκτενούς συμπληρωματικότητας του 5 UTR άκρου δεν εγγυάται σε κάποιες περιπτώσεις τη σίγαση του στόχου του mir (Shin et al., 2010). Από αναλύσεις στον εγκέφαλο του ποντικού έχει διαπιστωθεί η ύπαρξη G:U wobble ζεύγη βάσεων ακόμη και στο κέντρο της πρόσδεσης του στόχου με τη φύτρα (Chi et al., 2012). Καταλήγουμε στο συμπέρασμα πως η περιοχή της φύτρας δεν είναι απαραίτητη ούτε επαρκής για τη σίγαση της έκφρασης του στόχου. Η έλλειψη συμπληρωματικότητας σε αυτή, δεν εμποδίζει την ύπαρξη λειτουργικών mirs τα οποία μπορεί να διαθέτουν συνεχή νουκλεοτιδικά ζεύγη βάσεων στην κέντρο τους χωρίς την ύπαρξη περιοχών με συμπληρωματικότητα σε κανένα από τα δύο άκρα όπως και έχει διαπιστωθεί. Σίγαση της έκφρασης μπορεί να επιτευχθεί επιπλέον όταν υπάρχει συμπληρωματικότητα mir γονιδίου στόχου σε περιοχές που δεν ανήκουν σε καμία από τις παραπάνω κατηγορίες. Τα mirnas μπορούν να ταξινομηθούν σε οικογένειες με βάση την ομολογία στο 5' άκρο τους, αν και δεν είναι ακόμη γνωστό αν τα μέλη της ίδιας οικογένειας ρυθμίζουν παρόμοιες βιολογικές διαδικασίες Χαρακτηριστικά των περιοχών στόχων Επιπλέον κριτήριο για τον προσδιορισμό των στόχων των mirs με μεγαλύτερη ακρίβεια, αποτελεί η διαπίστωση από τον Grimson της παρουσίας περιοχών πλούσιων σε AU 13
29 δινουκλεοτίδια, κοντά στην περιοχή πρόσδεσης του στόχου ή μακριά από το κέντρο μεγάλων 3 UTRs (Grimson et al., 2007). H παρουσία αυτών των AU δινουκλεοτιδίων υποδηλώνει την ύπαρξη υψηλά συντηρημένων περιοχών (Grimson et al., 2007). Βέλτιστη καταστολή της έκφρασης φαίνεται να επιτυγχάνεται όταν δύο περιοχές οι οποίες αποτελούν στόχο των mirs απέχουν nt (Hon and Zhang, 2007). Ωστόσο mrnas με περιοχές στόχων οι οποίες απέχουν περισσότερο μεταξύ τους φαίνεται να παρουσιάζουν χαμηλή πρωτεϊνική έκφραση. Η ύπαρξη περισσοτέρων του ενός περιοχών στόχων, δεν εγγυάται και τη μεγαλύτερη αποσταθεροποίηση αυτού. Παρόλα αυτά, έχουν παρατηρηθεί περιπτώσεις όπου ο εντοπισμός δύο περιοχών στόχων σε κοντινή απόσταση λειτουργεί συνεργατικά προσφέροντας μεγαλύτερη αποσταθεροποίηση απ ότι κάθε περιοχή ατομικά, ωστόσο, είναι πιο πιθανό δύο υψηλά συντηρημένες περιοχές να βρίσκονται σε μεγαλύτερη μεταξύ τους απόσταση. Από τα παραπάνω, προκύπτει πως συνεργατική λειτουργία των mirnas συνεπάγεται και ευκολότερη αποσταθεροποίηση του mrna στόχου σε μικρές αλλαγές της έκφρασης των mirnas (Grimson et al., 2007). Οι περιοχές στόχων των mirs τείνουν να είναι λιγότερο συντηρημένες στα πρώτα ~15 nt μετά το κωδικόνιο λήξης πιθανόν για να μην εκτοπίσουν το mirisc σύμπλοκο (Grimson et al., 2007). Τέλος, έχει αναφερθεί πως στην περίπτωση της καταστολής της έκφρασης και όταν οι περιοχές στόχοι εντοπίζονται σε κωδικές περιοχές αντί στο 3 UTR των mrna στόχων φαίνεται να επιτυγχάνεται γρηγορότερη καταστολή αυτής Στερεοδιάταξη των περιοχών στόχων των mirnas Πολλές περιοχές στόχων των mirs στην αρχή και το τέλος του 3 UTR είναι μη-δομημένες. Τα μισά κατά προσέγγιση ανθρώπινα γονίδια παράγουν μετάγραφα με μεταβλητό μήκος του 3 UTR. Παρατηρείται λοιπόν ότι σε γονίδια με υψηλή έκφραση το 3 UTR είναι μικρότερο σε μήκος και άρα με μικρότερο πλήθος περιοχών στόχων των mirs συγκριτικά με ισόμορφά με μεγαλύτερο μήκος 3 UTR (Ji et al., 2009). Στον πίνακα 1.1 περιλαμβάνονται πολλά από τα διαθέσιμα εργαλεία πρόβλεψης στόχων mirnas. 14
30 1.7 Παράγοντες αποτελεσματικότητας των mirnas Επίπεδα συγκέντρωσης των mirs και των στόχων τους Η αποτελεσματικότητα της καταστολής της έκφρασης εξαρτάται επιπρόσθετα από τα επίπεδα συγκέντρωσης του (α) RNA στόχου, (β) του mirna και (γ) του mirisc συμπλόκου. Χαμηλά επίπεδα συγκέντρωσης των mirnas στο κύτταρο ίσως αποτυγχάνουν να ρυθμίσουν την έκφραση των mrna στόχων τους από μόνα τους, για αυτό στην περίπτωση χαμηλής συγκέντρωσης ενός mirna αυτό μπορεί να ρυθμίσει την έκφρασης ενός στόχου σε συνεργασία με άλλα mirnas (Paschou and Doxakis, 2012) 15
31 TargetScan miranda PicTar (συμπεριλαμβανομένου του dorina) RNA-hybrid DIANA-microT 3.0 ΕΙΜΜο microcosm Targets mirecords PITA Πίνακας 1.1: Κύρια εργαλεία πρόβλεψης στόχων mirnas. Εργαλεία πρόβλεψης στόχων mirnas Βάση πρόβλεψης συντηρημένων στόχων σε 5 σπονδυλωτά. Εύρεση στόχων βάσει κανόνων θέσης και συντήρησης μεταξύ διαφορετικών ειδών. Λεπτομέρειες στοίχισης του 3 UTR. Εύρεση του βέλτιστου δυνατού στόχου βάσει της ελάχιστης ενέργειας υβριδισμού. Υπολογισμός πολλών παραμέτρων από κάθε mirna ταυτόχρονα, συνδυασμός πληροφοριών από συντηρημένα και μη mirnas. Προσδιορισμός mirna στόχων με ανάλυση γενετικών μονοπατιών και βάσει εξελικτικής συντήρησης. Πρόβλεψη mirna στόχων ανάμεσα σε πολλά είδη. Βάση δεδομένων πειραματικά προσδιορισμένων mirna στόχων σε 9 είδη ζώων. Πρόβλεψη στόχων βάσει της προσβασιμότητας στο UTR _prediction.html starbase Βάση δεδομένων, πρόβλεψη στόχων από συνδυασμό 5 αλγορίθμων mirtar Προσδιορισμός βιολογικών λειτουργιών και συσχετίσεων μεταξύ γνωστών mirnas και κωδικών γονιδίων psrnatarget Πρόβλεψη στόχων μικρών RNAs (και mirnas) φυτών. MicroInspector Πρόβλεψη mrna στόχων πολλών ειδών οργανισμών. mirwip Υπολογισμός mrna στόχων βάσει ελεύθερης ενέργειας mirna στόχου, δομική προσβασιμότητα του στόχου και τοπολογίας των βάσεων της φύτρας του mirna. 16
32 17
33 1.7.2 Πρωτεΐνες πρόσδεσης στο RNA Η πρόσδεση των mirnas σε mrnas με μοτίβα πλούσια σε U προκαλεί την αποσταθεροποίηση τους ενώ η πρόσδεση σε AU μοτίβα προκαλεί τη σταθεροποίηση τους (Jacobsen et al., 2010; Larsson et al., 2010). Ωστόσο οι αλλαγές στη γονιδιακή έκφραση δεν εξηγείται πλήρως μόνο από την πρόσδεση των mirnas στα mrnas στόχους. Οι πρωτεΐνες πρόσδεσης στο RNA (RNA binding proteins - RBPs) ρυθμίζουν καίριες λειτουργίες της γονιδιακής έκφρασης όπως η συρραφή και η μετάφραση, μέσω της πρόσδεσης τους σε περιοχές στο 5 και 3 UTR των mrnas που είναι πλούσια σε U και AU νουκλεοτίδια. Πρόσφατα βρέθηκε μία συσχέτιση μεταξύ των mirs και των RBPs. Στο παράδειγμα της HuR πρωτεΐνης όταν οι περιοχές πρόσδεσης αυτής και ενός mirna αλληλεπικαλύπτονται η έκφραση του μεταγράφου ρυθμίζεται κατά προτίμηση από την HuR ενώ όταν δεν υπάρχει αλληλοεπικάλυψη η έκφραση του μεταγράφου ρυθμίζεται από το mir (Abdelmohsen et al., 2008; Guo et al., 2009; Lebedeva et al., 2011) Competing endogenous RNAs (cernas) Πρόσφατα διαπιστώθηκε ένας νέος μετα-μεταγραφικός ρόλος των RNAs, σύμφωνα με τον οποίο ο ρόλος των RNA στόχων δεν είναι παθητικός αποτελώντας μόνο υπόστρωμα των mirs. Τα RNAs που αποτελούν στόχο των ίδιων mirs συναγωνίζονται μεταξύ τους για το ποιο θα προσλάβει ένα συγκεκριμένο microrna. Με αυτό τον έμμεσο τρόπο τα RNAs ρυθμίζουν το ένα την έκφραση του άλλου (Cesana and Daley, 2013; Salmena et al., 2011) Circular RNAs (cirnas) Επιπλέον κάποια κυκλικά RNAs (circrnas) τα οποία σχηματίζονται με ομοιοπολικό δεσμό των 3 και 5 UTRs είναι ανθεκτικά στη δράση των mirs που προσδένονται πάνω τους. Παράδειγμα αποτελεί το cirs-7 το οποίο καταστέλλει τη δράση του mir-7 και δεν αποσταθεροποιείται με την πρόσδεση του (Memczak et al., 2013) (Εικόνα 1.4). 18
34 Εικόνα 1.4: Παράγοντες που επηρεάζουν τη διαθεσιμότητα των mirnas (Doxakis, 2013). 1.8 Μη κανονικές λειτουργίες των mirnas Από ένα μικρό πλήθος μελετών συμπεραίνεται ότι τα mirnas εκτός από κατασταλτικό χαρακτήρα μπορούν να ενεργοποιήσουν την έκφραση ενός γονιδίου Ενεργοποίηση εκκινητή Παλαιότερες μελέτες έχουν δείξει πως η εφαρμογή μικρών εξωγενών δίκλωνων RNAs τα οποία είναι συμπληρωματικά στους εκκινητές των γονιδίων ενεργοποιούν τη γονιδιακή έκφραση (RNA activation - RNAa) (Janowski et al., 2007; Li et al., 2006). To ίδιο ακριβώς φαίνεται να ισχύει με ορισμένα mirnas όπως το mir-373 του οποίου στόχοι έχουν βρεθεί στην περιοχή του εκκινητή των E-cadherin CSDC2 (Cold Shock Domain-Containing protein) (Place et al., 2008). Ομοίως το mir-205 το οποίο εντοπίζεται στον εκκινητή των ογκοκατασταλτικών γονιδίων ιντερλευκίνης, IL- 24 και IL-32 ενεργοποιεί την έκφραση τους (Majid et al., 2010). 19
35 1.8.2 Ενεργοποίηση μετάφρασης μέσω πρόσδεσης στο στόχο Σε ορισμένες περιπτώσεις το mirisc σύμπλοκο ενεργοποιεί την έκφραση μέσω της πρόσδεσης ορισμένων mirnas στο 5 και 3 UTR. Στον εγκέφαλο η πρωτεΐνη RIP140 (Receptor-Interacting protein 140) αποτελεί στόχο του mir-346 (Tsai et al., 2009) Συνδέτες υποδοχέων Οι TLR7 στον ποντικό και TLR8 στον άνθρωπο διαμεμβρανικοί υποδοχείς - μέλη της οικογένειας πρωτεϊνών TLR (Toll-Like Receptor) αναγνωρίζουν φυσιολογικά και προσδένουν πάνω τους μονόκλωνα ιικά RNAs μήκους ~20 nt (Heil et al., 2004; Lund et al., 2004). Όταν τα mirnas εκκρίνονται σε κυστίδια και καθώς έχουν παρόμοιο μέγεθος έχει διαπιστωθεί ότι μπορούν να αποτελέσουν TLR7/8 συνδέτες. Πιο αναλυτικά τα mir-21 και mir29a μπορεί να αποτελέσουν συνδέτες για τους TLR7/8 και ενεργοποιώντας την προαγωγή της μετάστασης ως απόκριση σε μια φλεγμονή (Fabbri et al., 2012). 1.9 Αναγνώριση mirnas και εκκινητών τους O μηχανισμός μεταγραφής των γονιδίων από τα οποία προκύπτουν τα mirnas παραμένει άγνωστος μέχρι σήμερα. Συνεπώς είναι απαραίτητος ο ακριβής προσδιορισμός των περιοχών των εκκινητών προκειμένου να προσδιοριστούν οι περιοχές έναρξης της μεταγραφής (transcriptional start sites - TSSs) των mirna γονιδίων. Από τη στιγμή που τα περισσότερα mirnas γονίδια μεταγράφονται από την RNA πολυμεράση II τα μοντέλα πρόβλεψης των εκκινητών βασίζονται στα χαρακτηριστικά μεταγραφής της RNA πολυμεράσης ΙΙ (Lee et al., 2004a). Αν και πολλές υπολογιστικές μέθοδοι έχουν επιτύχει υψηλές επιδόσεις στην αναγνώριση εκκινητών των γονιδίων με τη χρήση μεθόδων μηχανικής μάθησης (Megraw et al., 2009; Sonnenburg et al., 2006; Wang et al., 2009) δεν ισχύει το ίδιο και στο πεδίο της πρόβλεψης των εκκινητών των mirnas. H πρόβλεψη των εκκινητών είναι δύσκολη εξαιτίας της ετερογένειας με την οποία παράγονται τα mirnas. 20
36 Τα mirnas διακρίνονται σε διαγονιδιακά (intergenic) όταν εντοπίζονται ανάμεσα σε γονίδια που έχουν σχολιασθεί και ενδογονιδιακά (intragenic), όταν εντοπίζονται στα εξώνια (εξωνικά mirnas) ή τα εσώνια (εσωνικά mirnas) των γονιδίων (Εικόνα 1.5b,c). Ωστόσο κάποια mirnas εντοπίζονται στα σύνορα εξωνίων και εσωνίων (Rodriguez et al., 2004). Τα γονίδια τα οποία περιέχουν mirnas στα εσώνια τους ονομάζονται γονίδια ξενιστές (host). Στα θηλαστικά ~50% των mirnas είναι ενδογονιδιακά (He et al., 2012). Στον άνθρωπο σχεδόν το 50% των mirnas είναι διαγονιδιακά. Σχεδόν το άλλο 50% είναι εσωνικά mirnas και πολύ λίγα είναι εξωνικά (Hinske et al., 2010). Επιπλέον το 20% των mirnas έχει ως στόχο το ίδιο το γονίδιο ξενιστή, ίσως και κάποια γειτονικά γονίδια του ξενιστή. Τα γονίδια ξενιστές των mirnas είναι τρεις φορές μεγαλύτερα συγκριτικά με τα συνήθη γονίδια, περιέχοντας περισσότερο επιμήκη στο 5 UTR και περιέχουν σε αυτό περισσότερο του 50% των εσωνίων. Το ποσοστό των εσωνίων που βρίσκονται κοντά στο 5 UTR ανέρχεται στο 60%. Επιπλέον το 3 άκρο των mirnas γονιδίων ξενιστών είναι 40% μεγαλύτερο (Hinske et al., 2010). Από τη μελέτη των περιοχών πρόσδεσης 49 εξωνικών mirnas με 51 mrnas έχει αποδειχθεί ότι παρά το μικρό πλήθος των γονιδίων στόχων τα εξωνικά mirnas προσδένονται στις κωδικές περιοχές και στο 3 UTR αλλά όχι στο 5 UTR των mrna γονιδίων στόχων τους. Ένα mirna μπορεί να έχει περισσότερους από έναν εκκινητές. Οι εκκινητές των mirnas χαρακτηρίζονται ως ενδογενείς όταν αναφερόμαστε στους εσωνικούς εκκινητές και τους εκκινητές των γονιδίων ξενιστών, ενώ ως υβριδικοί εκκινητές χαρακτηρίζονται όσοι εντοπίζονται σε διαγονιδιακές περιοχές πριν των ενδογονιδιακών mirnas και δεν είναι ξεκάθαρο αν ανήκουν στο mirna ή όχι. Tα εσωνικά mirnas δε συμμεταγράφονται απαραίτητα μαζί με το γονίδιο ξενιστή τους (Εικόνα 1.5a) και έχει βρεθεί ότι το 62% των σχολιασμένων εσωνικών mirnas έχουν το δικό τους (εσωνικό) εκκινητή (Marsico et al., 2013), ενώ παλαιότερες μελέτες υπολόγισαν αυτό το ποσοστό σε 30-35% (Monteys et al., 2010; Ozsolak et al., 2008). Πρόσφατες μελέτες έχουν δείξει ότι αυτοί οι εναλλακτικοί εσωνικοί εκκινητές μπορεί να σχετίζονται με ασθένειες όπως ο καρκίνος (Relle et al., 2014). Κάθε τύπος εκκινητών έχει και διαφορετικά χαρακτηριστικά. Για παράδειγμα οι νησίδες CpG χρησιμοποιούνται για την αναγνώριση των εκκινητών. Οι μεγάλοι εκκινητές συσχετίζονται με τις νησίδες CpG και οι εκκινητές των γονιδίων ξενιστών είναι πιο μεγάλοι συγκριτικά με τους διαγονιδιακούς και εσωνικούς εκκινητές. 21
37 Τα συντηρημένα ( παλιά ) mirnas στα σπονδυλωτά είναι πιο πιθανό να έχουν τους δικούς τους εκκινητές σε σχέση με τα λιγότερο συντηρημένα ( νεαρά ) mirnas τα οποία εξαρτώνται από τον εκκινητή του γονιδίου ξενιστή. Οι εσωνικοί εκκινητές ρυθμίζονται από διαφορετικούς μεταγραφικούς παράγοντες σε σχέση με τους διαγονιδιακούς και τους εκκινητές του γονιδίου ξενιστή υποδηλώνοντας και έναν διαφορετικό μηχανισμό εξέλιξης. Είναι συνήθως μικρότεροι των 100 bp και χρησιμοποιούνται σε μερικούς ιστούς περισσότερο από ότι σε άλλους. Επίσης οι εσωνικοί εκκινητές έχουν μεγαλύτερη συνάφεια πρόσδεσης στο πλαίσιο TATA συγκριτικά με τους διαγονιδιακούς και τους εκκινητές του ξενιστή. Εικόνα 1.5: Γονιδιακή οργάνωση των mirnas. (a) Ανεξάρτητοι εκκινητές για τη μεταγραφή των mir-1 1 και mir-133a-2, οι οποίοι ρυθμίζονται από τους μεταγραφικούς παράγοντες SRF and MyoD. (b), (c) Ενδογενείς εκκινητές. H γονιδιακή περιοχής της mirna ακολουθίας απεικονίζει με μπλε το pre-mirna και με κόκκινο το ώριμο mirna (Zhao and Srivastava, 2007). 22
38 1.10 Βιβλιογραφία Abdelmohsen, K., Srikantan, S., Kuwano, Y., and Gorospe, M. (2008). mir-519 reduces cell proliferation by lowering RNA-binding protein HuR levels. Proc Natl Acad Sci U S A 105, Ameres, S.L., Martinez, J., and Schroeder, R. (2007). Molecular basis for target RNA recognition and cleavage by human RISC. Cell 130, Axtell, M.J., Westholm, J.O., and Lai, E.C. (2011). Vive la difference: biogenesis and evolution of micrornas in plants and animals. Genome Biol 12, 221. Berezikov, E., Chung, W.J., Willis, J., Cuppen, E., and Lai, E.C. (2007). Mammalian mirtron genes. Mol Cell 28, Brennecke, J., Stark, A., Russell, R.B., and Cohen, S.M. (2005). Principles of microrna-target recognition. PLoS Biol 3, e85. Cesana, M., and Daley, G.Q. (2013). Deciphering the rules of cerna networks. Proc Natl Acad Sci U S A 110, Chendrimada, T.P., Gregory, R.I., Kumaraswamy, E., Norman, J., Cooch, N., Nishikura, K., and Shiekhattar, R. (2005). TRBP recruits the Dicer complex to Ago2 for microrna processing and gene silencing. Nature 436, Chi, S.W., Hannon, G.J., and Darnell, R.B. (2012). An alternative mode of microrna target recognition. Nat Struct Mol Biol 19, Chiang, H.R., Schoenfeld, L.W., Ruby, J.G., Auyeung, V.C., Spies, N., Baek, D., Johnston, W.K., Russ, C., Luo, S., Babiarz, J.E., et al. (2010). Mammalian micrornas: experimental evaluation of novel and previously annotated genes. Genes Dev 24, Corcoran, D.L., Pandit, K.V., Gordon, B., Bhattacharjee, A., Kaminski, N., and Benos, P.V. (2009). Features of mammalian microrna promoters emerge from polymerase II chromatin immunoprecipitation data. PLoS One 4, e5279. Doxakis, E. (2013). Principles of mirna-target regulation in metazoan models. Int J Mol Sci 14, Elkayam, E., Kuhn, C.D., Tocilj, A., Haase, A.D., Greene, E.M., Hannon, G.J., and Joshua-Tor, L. (2012). The structure of human argonaute-2 in complex with mir-20a. Cell 150, Fabbri, M., Paone, A., Calore, F., Galli, R., Gaudio, E., Santhanam, R., Lovat, F., Fadda, P., Mao, C., Nuovo, G.J., et al. (2012). MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc Natl Acad Sci U S A 109, E Forstemann, K., Horwich, M.D., Wee, L., Tomari, Y., and Zamore, P.D. (2007). Drosophila micrornas are sorted into functionally distinct argonaute complexes after production by dicer-1. Cell 130, Grimson, A., Farh, K.K., Johnston, W.K., Garrett-Engele, P., Lim, L.P., and Bartel, D.P. (2007). MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol Cell 27, Guo, X., Wu, Y., and Hartley, R.S. (2009). MicroRNA-125a represses cell growth by targeting HuR in breast cancer. RNA Biol 6, Han, J., Lee, Y., Yeom, K.H., Nam, J.W., Heo, I., Rhee, J.K., Sohn, S.Y., Cho, Y., Zhang, B.T., and Kim, V.N. (2006). Molecular basis for the recognition of primary micrornas by the Drosha-DGCR8 complex. Cell 125, He, C., Li, Z., Chen, P., Huang, H., Hurst, L.D., and Chen, J. (2012). Young intragenic mirnas are less coexpressed with host genes than old ones: implications of mirna-host gene coevolution. Nucleic Acids Res 40, Heil, F., Hemmi, H., Hochrein, H., Ampenberger, F., Kirschning, C., Akira, S., Lipford, G., Wagner, H., and Bauer, S. (2004). Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 303,
39 Hinske, L.C., Galante, P.A., Kuo, W.P., and Ohno-Machado, L. (2010). A potential role for intragenic mirnas on their hosts' interactome. BMC Genomics 11, 533. Holen, T., Amarzguioui, M., Wiiger, M.T., Babaie, E., and Prydz, H. (2002). Positional effects of short interfering RNAs targeting the human coagulation trigger Tissue Factor. Nucleic Acids Res 30, Hon, L.S., and Zhang, Z. (2007). The roles of binding site arrangement and combinatorial targeting in microrna repression of gene expression. Genome Biol 8, R166. Hutvagner, G., and Zamore, P.D. (2002). A microrna in a multiple-turnover RNAi enzyme complex. Science 297, Jacobsen, A., Wen, J., Marks, D.S., and Krogh, A. (2010). Signatures of RNA binding proteins globally coupled to effective microrna target sites. Genome Res 20, Janowski, B.A., Younger, S.T., Hardy, D.B., Ram, R., Huffman, K.E., and Corey, D.R. (2007). Activating gene expression in mammalian cells with promoter-targeted duplex RNAs. Nat Chem Biol 3, Ji, Z., Lee, J.Y., Pan, Z., Jiang, B., and Tian, B. (2009). Progressive lengthening of 3' untranslated regions of mrnas by alternative polyadenylation during mouse embryonic development. Proc Natl Acad Sci U S A 106, Khvorova, A., Reynolds, A., and Jayasena, S.D. (2003). Functional sirnas and mirnas exhibit strand bias. Cell 115, Kozomara, A., and Griffiths-Jones, S. (2011). mirbase: integrating microrna annotation and deepsequencing data. Nucleic Acids Res 39, D Larsson, E., Sander, C., and Marks, D. (2010). mrna turnover rate limits sirna and microrna efficacy. Mol Syst Biol 6, 433. Lau, N.C., Lim, L.P., Weinstein, E.G., and Bartel, D.P. (2001). An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science 294, Lebedeva, S., Jens, M., Theil, K., Schwanhausser, B., Selbach, M., Landthaler, M., and Rajewsky, N. (2011). Transcriptome-wide analysis of regulatory interactions of the RNA-binding protein HuR. Mol Cell 43, Lee, R.C., Feinbaum, R.L., and Ambros, V. (1993). The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 75, Lee, Y., Kim, M., Han, J., Yeom, K.H., Lee, S., Baek, S.H., and Kim, V.N. (2004a). MicroRNA genes are transcribed by RNA polymerase II. EMBO J 23, Lee, Y.S., Nakahara, K., Pham, J.W., Kim, K., He, Z., Sontheimer, E.J., and Carthew, R.W. (2004b). Distinct roles for Drosophila Dicer-1 and Dicer-2 in the sirna/mirna silencing pathways. Cell 117, Lee, Y.S., Shibata, Y., Malhotra, A., and Dutta, A. (2009). A novel class of small RNAs: trna-derived RNA fragments (trfs). Genes Dev 23, Lelandais-Briere, C., Sorin, C., Declerck, M., Benslimane, A., Crespi, M., and Hartmann, C. (2010). Small RNA diversity in plants and its impact in development. Curr Genomics 11, Li, L.C., Okino, S.T., Zhao, H., Pookot, D., Place, R.F., Urakami, S., Enokida, H., and Dahiya, R. (2006). Small dsrnas induce transcriptional activation in human cells. Proc Natl Acad Sci U S A 103, Lund, E., and Dahlberg, J.E. (2006). Substrate selectivity of exportin 5 and Dicer in the biogenesis of micrornas. Cold Spring Harb Symp Quant Biol 71, Lund, J.M., Alexopoulou, L., Sato, A., Karow, M., Adams, N.C., Gale, N.W., Iwasaki, A., and Flavell, R.A. (2004). Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc Natl Acad Sci U S A 101, Majid, S., Dar, A.A., Saini, S., Yamamura, S., Hirata, H., Tanaka, Y., Deng, G., and Dahiya, R. (2010). MicroRNA-205-directed transcriptional activation of tumor suppressor genes in prostate cancer. Cancer 116,
40 Marsico, A., Huska, M.R., Lasserre, J., Hu, H., Vucicevic, D., Musahl, A., Orom, U.A., and Vingron, M. (2013). PROmiRNA: a new mirna promoter recognition method uncovers the complex regulation of intronic mirnas. Genome Biol 14, R84. Megraw, M., Pereira, F., Jensen, S.T., Ohler, U., and Hatzigeorgiou, A.G. (2009). A transcription factor affinity-based code for mammalian transcription initiation. Genome Res 19, Memczak, S., Jens, M., Elefsinioti, A., Torti, F., Krueger, J., Rybak, A., Maier, L., Mackowiak, S.D., Gregersen, L.H., Munschauer, M., et al. (2013). Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 495, Miyoshi, K., Miyoshi, T., and Siomi, H. (2010). Many ways to generate microrna-like small RNAs: noncanonical pathways for microrna production. Mol Genet Genomics 284, Monteys, A.M., Spengler, R.M., Wan, J., Tecedor, L., Lennox, K.A., Xing, Y., and Davidson, B.L. (2010). Structure and activity of putative intronic mirna promoters. RNA 16, Okamura, K., Hagen, J.W., Duan, H., Tyler, D.M., and Lai, E.C. (2007). The mirtron pathway generates microrna-class regulatory RNAs in Drosophila. Cell 130, Okamura, K., Phillips, M.D., Tyler, D.M., Duan, H., Chou, Y.T., and Lai, E.C. (2008). The regulatory activity of microrna* species has substantial influence on microrna and 3' UTR evolution. Nat Struct Mol Biol 15, Ozsolak, F., Poling, L.L., Wang, Z., Liu, H., Liu, X.S., Roeder, R.G., Zhang, X., Song, J.S., and Fisher, D.E. (2008). Chromatin structure analyses identify mirna promoters. Genes Dev 22, Parrish, S., Fleenor, J., Xu, S., Mello, C., and Fire, A. (2000). Functional anatomy of a dsrna trigger: differential requirement for the two trigger strands in RNA interference. Mol Cell 6, Paschou, M., and Doxakis, E. (2012). Neurofibromin 1 is a mirna target in neurons. PLoS One 7, e Pasquinelli, A.E., Reinhart, B.J., Slack, F., Martindale, M.Q., Kuroda, M.I., Maller, B., Hayward, D.C., Ball, E.E., Degnan, B., Muller, P., et al. (2000). Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 408, Place, R.F., Li, L.C., Pookot, D., Noonan, E.J., and Dahiya, R. (2008). MicroRNA-373 induces expression of genes with complementary promoter sequences. Proc Natl Acad Sci U S A 105, Reinhart, B.J., Slack, F.J., Basson, M., Pasquinelli, A.E., Bettinger, J.C., Rougvie, A.E., Horvitz, H.R., and Ruvkun, G. (2000). The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 403, Relle, M., Becker, M., Meyer, R.G., Stassen, M., and Schwarting, A. (2014). Intronic promoters and their noncoding transcripts: a new source of cancer-associated genes. Mol Carcinog 53, Rhoades, M.W., Reinhart, B.J., Lim, L.P., Burge, C.B., Bartel, B., and Bartel, D.P. (2002). Prediction of plant microrna targets. Cell 110, Rodriguez, A., Griffiths-Jones, S., Ashurst, J.L., and Bradley, A. (2004). Identification of mammalian microrna host genes and transcription units. Genome Res 14, Ruby, J.G., Stark, A., Johnston, W.K., Kellis, M., Bartel, D.P., and Lai, E.C. (2007). Evolution, biogenesis, expression, and target predictions of a substantially expanded set of Drosophila micrornas. Genome Res 17, Saini, H.K., Griffiths-Jones, S., and Enright, A.J. (2007). Genomic analysis of human microrna transcripts. Proc Natl Acad Sci U S A 104, Salmena, L., Poliseno, L., Tay, Y., Kats, L., and Pandolfi, P.P. (2011). A cerna hypothesis: the Rosetta Stone of a hidden RNA language? Cell 146, Seitz, H., Ghildiyal, M., and Zamore, P.D. (2008). Argonaute loading improves the 5' precision of both MicroRNAs and their mirna* strands in flies. Curr Biol 18, Shin, C., Nam, J.W., Farh, K.K., Chiang, H.R., Shkumatava, A., and Bartel, D.P. (2010). Expanding the microrna targeting code: functional sites with centered pairing. Mol Cell 38,
41 Sonnenburg, S., Zien, A., and Ratsch, G. (2006). ARTS: accurate recognition of transcription starts in human. Bioinformatics 22, e Tsai, N.P., Lin, Y.L., and Wei, L.N. (2009). MicroRNA mir-346 targets the 5'-untranslated region of receptorinteracting protein 140 (RIP140) mrna and up-regulates its protein expression. Biochem J 424, Wang, X., Xuan, Z., Zhao, X., Li, Y., and Zhang, M.Q. (2009). High-resolution human core-promoter prediction with CoreBoost_HM. Genome Res 19, Wang, Y., Sheng, G., Juranek, S., Tuschl, T., and Patel, D.J. (2008). Structure of the guide-strand-containing argonaute silencing complex. Nature 456, Zhao, Y., and Srivastava, D. (2007). A developmental view of microrna function. Trends Biochem Sci 32, Zheng, H., Fu, R., Wang, J.T., Liu, Q., Chen, H., and Jiang, S.W. (2013). Advances in the Techniques for the Prediction of microrna Targets. Int J Mol Sci 14, Zhu, Q.H., Spriggs, A., Matthew, L., Fan, L., Kennedy, G., Gubler, F., and Helliwell, C. (2008). A diverse set of micrornas and microrna-like small RNAs in developing rice grains. Genome Res 18,
42 Κεφάλαιο 2 ο Τεχνολογία Μικροσυστοιχιών 27
43 2.1 Ανάλυση γονιδιακής έκφρασης Η λειτουργική γονιδιωματική, αποτελεί πεδίο της μοριακής βιολογίας η οποία εν αντιθέσει με τη γονιδιωματική που ασχολείται με την αλληλούχιση του DNA, επιχειρεί την απάντηση ερωτημάτων σχετικών με τη μεταγραφή των γονιδίων, τη μετάφραση και την αλληλεπίδραση των πρωτεϊνών. Χαρακτηριστικό της λειτουργικής γονιδιωματικής είναι ότι χρησιμοποιεί μεθόδους υψηλής απόδοσης για την ανάλυση πληροφοριών μεγάλων δεδομένων που παράγονται από την ποσοτικοποίηση βιολογικών δραστηριοτήτων πολλών πειραμάτων. Με τον τρόπο αυτό γίνονται κατανοητές οι λειτουργίες των γονιδίων και των πρωτεϊνών καθώς και η αλληλεπίδραση των τελευταίων αλλά και η σχέση του γονιδιώματος του οργανισμού και του φαινοτύπου του. Μια τέτοιου είδους μεγάλης κλίμακας ανάλυση για τον έλεγχο της διακύμανσης της έκφρασης των γονιδίων, του RNA και των πρωτεϊνών στο χρόνο μέσω της στιγμιαίας απεικόνισης της μεταγραφικής δραστηριότητας του βιολογικού δείγματος (Conlon et al., 2003) είναι η τεχνολογία των μικροσυστοιχιών. Αυτή, σε αντίθεση με τα παραδοσιακά εργαλεία μοριακής βιολογίας τα οποία επιτρέπουν τη μελέτη μεμονωμένων βιολογικών λειτουργιών λίγων συσχετιζόμενων γονιδίων, πρωτεϊνών ή κυτταρικών μονοπατιών, η ανάλυση δεδομένων μικροσυστοιχιών χρησιμοποιεί υπολογιστικά και στατιστικά εργαλεία για αναλύσεις μεγάλης κλίμακας. 2.2 Ιστορικά στοιχεία Η τεχνολογία των μικροσυστοιχιών γεννήθηκε τον Οκτώβριο του 1995 (Schena et al., 1995), ενώ προπομπός αυτής υπήρξε η τεχνολογία των μακροσυστοιχιών σε μεμβράνες. 2.3 Συσχετισμός ανάλυσης γονιδιακής έκφρασης με άλλες βιολογικές πληροφορίες Στην παράγραφο αυτή απαριθμούνται επιπλέον βιολογικά ερωτήματα στα οποία μπορεί να απαντήσει η ανάλυση της γονιδιακής έκφρασης. 28
44 2.3.1 Πρόβλεψη περιοχών πρόσδεσης Γονίδια με όμοια προφίλ γονιδιακής έκφρασης ρυθμίζονται από τους ίδιους μεταγραφικούς παράγοντες και συνεπώς έχουν ίδιες περιοχές πρόσδεσης αυτών. Η διαδικασία που ακολουθείται είναι: i. Εύρεση των γονιδίων που συν εκφράζονται. ii. iii. Εξαγωγή ακολουθιών εκκινητών των συν εκφραζόμενων γονιδίων. Προσδιορισμός pattern ακολουθιών που υπερ εκπροσωπούνται Πρόβλεψη αλληλεπίδρασης και λειτουργίας πρωτεϊνών Η ενσωμάτωση πληροφοριών εξελικτικής συντήρησης (evolutionary conservation) των πρωτεϊνών σε δεδομένα γονιδιακής έκφρασης, χρησιμοποιούνται για την πρόβλεψη της πρωτεϊνικής λειτουργίας, αλληλεπίδρασης. Πρωτεΐνες που προκύπτουν από όμοια προφίλ γονιδιακής έκφρασης είναι πιθανό να αλληλεπιδρούν (Conlon et al., 2003; Ge et al., 2001), ενώ πρωτεΐνες τα γονίδια των οποίων είναι συντηρημένα και συνεκφράζονται είναι πιθανό να αποτελούν μέρος του ίδιου μονοπατιού και συνεπώς είναι δυνατή η πρόβλεψη της λειτουργίας των γονιδίων τους. Η διαδικασία που ακολουθείται είναι: i. Προσδιορισμός γονιδίων που συν εκφράζονται σε δύο οργανισμούς. ii. iii. Προσδιορισμός συντηρημένων (ορθόλογων) πρωτεϊνών. Εύρεση στιγμιοτύπων όπου οι συντηρημένες (ορθόλογες) πρωτεΐνες συνεκφράζονται και στους δύο οργανισμούς 2.4 Τύποι μικροσυστοιχιών Επίπεδες (δισδιάστατες) μικροσυστοιχίες Η διάταξη μιας μικροσυστοιχίας DNA (slide ή gene chip ή DNA chip) είναι συνήθως επίπεδη και κατασκευασμένη από γυαλί. Κυρίαρχο συστατικό του γυαλιού είναι το διοξείδιο του πυριτίου 29
45 (SiO 2 ), ενώ συχνά σε αυτό προστίθενται οξείδια μετάλλων προκειμένου να αυξηθεί το σημείο τήξης του. Το γυαλί αποτελεί το πιο διαδεδομένο υλικό κατασκευής μικροσυστοιχιών για τους ακόλουθους λόγους: Είναι διαφανές (Cheung et al., 1999) Παρουσιάζει χαμηλό φθορισμό (Cheung et al., 1999) Είναι ανθεκτικό σε υψηλές θερμοκρασίες (Cheung et al., 1999) Έχει χαμηλό κόστος (Cheung et al., 1999) Είναι χημικά αδρανές (π.χ. δεν αλλάζει ιδιότητες κατά την επαφή με το νερό) (Dufva et al., 2004) Τροποποιείται εύκολα από σιλάνια (Dufva et al., 2004; Zammatteo et al., 2000) Δεν είναι πορώδες. Η ιδιότητα αυτή αποτρέπει τη διείσδυση του διαλύματος των προς υβριδισμό στόχων με αποτέλεσμα να υβριδίζονται απευθείας με τους ανιχνευτές χωρίς το φαινόμενο της εσωτερικής διάχυσης (Southern et al., 1999). Άλλα υλικά τα οποία χρησιμοποιούνται στην κατασκευή μικροσυστοιχιών είναι το διαφανές πλαστικό ή κάποιο πολυμερές (π.χ. συντηγμένη σίλικα) με μαύρη επίστρωση προκειμένου ο αυτοφθορισμός υποβάθρου να είναι ο ελάχιστος (Venkatasubbarao, 2004) Μη επίπεδες (τρισδιάστατες) μικροσυστοιχίες Αν και οι επίπεδες μικροσυστοιχίες είναι αυτές που χρησιμοποιούνται κατά κόρον, οι εταιρίες προσφέρουν εναλλακτικές τύπους μικροσυστοιχιών όπως αυτές που φιλοξενούν κάποιο δίκτυο μικρο καναλιών στο πορώδες υπόστρωμα τους ή αυτές των τρισδιάστατων μικροσυστοιχιών μικρο σφαιριδίων (micro beads) της Luminex. O σχεδιασμός αυτός στοχεύει στην αύξηση του λόγου της διαθέσιμης επιφάνειας προς το διαθέσιμο όγκο υβριδισμού παρέχοντας έτσι μικροσυστοιχίες ακόμη υψηλότερης πυκνότητας 1 (Venkatasubbarao, 2004). Στις μικροσυστοιχίες μικροσφαιριδίων, ένας ανιχνευτής ακινητοποιείται στην επιφάνεια ενός φθορίζοντος μικροσφαιριδίου και κατόπιν υβριδίζεται με το σημασμένο στόχο. Ο υβριδισμός λαμβάνει χώρα μέσα σε διάλυμα και είναι πιο αποτελεσματικός από τον υβριδισμό στερεάς φάσης. Η χρήση μικρο σφαιριδίων με πολλές διαφορετικές φθορίζουσες χρωστικές δημιουργεί 30

46 πολλούς διαφορετικούς μοναδικούς συνδυασμούς μετά τον υβριδισμό με το στόχο και ο οποίος προσδιορίζεται μονοσήμαντα μέσα στο διάλυμα (εικόνα 2.1). 1 Εικόνα 2.1: Υβριδισμός πολλαπλών στόχων με τη χρήση μονοσήμαντα σημασμένων μικρο σφαιριδίων 2.5 Πειραματική διαδικασία Επιλογή ανιχνευτών Κάθε chip περιέχει χιλιάδες spots και σε κάθε ένα από αυτά βρίσκονται ακινητοποιημένα πολλά πανομοιότυπα συνθετικά μονόκλωνα μόρια DNA (ssdna) τα οποία ονομάζονται ανιχνευτές (probes). To σύνολο των ανιχνευτών ενός spot ονομάζεται probe set οι οποίοι αντιστοιχούν στο ίδιο γονίδιο καλύπτοντας μια περιοχή ~600nt αυτού (εικόνα 2.2). Άλλα βιομόρια το οποία μπορούν να ακινητοποιηθούν πάνω σε ένα chip είναι ολιγονουκλεοτίδια, RNA, προϊόντα PCR, καθώς και πεπτίδια, πρωτεΐνες, λιπίδια ή υδατάνθρακες (Venkatasubbarao, 2004). 1 H πυκνότητα της μικροσυστοιχίας καθορίζεται από τη διάμετρο του spot και για τον υπολογισμό της πρέπει να είναι γνωστή η απόσταση μεταξύ των κέντρων δύο spot. Η διάμετρος ενός spot κυμαίνεται από nm έως nm. 31


47 Εικόνα 2.2: Κάθε slide μικροσυστοιχίας διαθέτει χιλιάδες spots. Κάθε spot περιέχει πολλαπλά αντίγραφα ανιχνευτών που αντιστοιχούν στο ίδιο πάντα γονίδιο Ακινητοποίηση ανιχνευτών Η αποτελεσματικότητα των μικροσυστοιχιών εξαρτάται κυρίως από την αλληλουχία των ανιχνευτών και από τον τρόπο με τον οποίοι αυτοί είναι ακινητοποιημένοι στην επιφάνεια (Zammatteo et al., 2000), οι οποίοι μπορεί να είναι: i) Ακινητοποίηση ανιχνευτών βάσει αλληλεπιδράσεων συνάφειας. Στην περίπτωση των πρωτεϊνικών μικροσυστοιχιών, αυτές σημαίνονται με βιοτίνη και εν συνεχεία επισυνάπτονται σε παραγωγοποιημένες επιφάνειες στρεπταβιδίνης ή σημαίνονται με πολυϊστιδίνη και επισυνάπτονται σε επιφάνειες με χηλικό νικέλιο (εικόνα 2.3) (Venkatasubbarao, 2004). ii) Ιοντικές αλληλεπιδράσεις. Η ακινητοποίηση του DNA ανιχνευτή γίνεται με υπεριώδη ακτινοβόληση και την αλληλεπίδραση των καταλοίπων θυμιδίνης του DNA και των θετικά φορτισμένων αμινομάδων των πλακιδίων (Joos et al., 1997). Ωστόσο με τη 32
. H επίστρωση της μικροσυστοιχίας με σιλάνια (Schena et al.")
48 μέθοδο αυτή, οι περιοχές πρόσδεσης του DNA και το πλήθος αυτών δεν προσδιορίζονται με ακρίβεια γιατί εξαρτάται κάθε φορά από την ακολουθία και το μήκος της (εικόνα 2.4). Για τους λόγους αυτούς προτιμάται η ακινητοποίηση του ανιχνευτή να γίνεται μέσω των άκρων του (εικόνα 2.5). H επίστρωση της μικροσυστοιχίας με σιλάνια (Schena et al., 1995) και οι υδρόφοβες αλληλεπιδράσεις (Allemand et al., 1997) κάτω από συνθήκες υψηλών θερμοκρασιών ή υψηλής αλατότητας. Για τους λόγους αυτούς η διαδικασία της σύζευξης θα πρέπει να είναι γρήγορη και ομοιοπολική (Zammatteo et al., 2000). iii) Ομοιοπολικοί δεσμοί. Μετά από σύγκριση πολλών τρόπων ομοιοπολικής πρόσδεσης του DNA (σύζευξη αμινομάδων, καρβοξυλομάδων, φωσφορυλιωμένου DNA πάνω σε καρβοξυλο ή αμινοτροποποιημένη μικροσυστοιχία και αμινοτροποποιημένου DNA πάνω σε αλδεϋδοτροποποιημένο γυαλί), η σύζευξη αμινοτροποποιημένου DNA ανιχνευτή σε αλδεϋδοτροποποιημένη επιφάνεια είναι προτιμότερη (εικόνα 2.6) (Zammatteo et al., 2000). Εικόνα 2.3: Ακινητοποίηση πρωτεϊνικών ανιχνευτών βάσει συνάφειας. 33



49 Εικόνα 2.4: Ακινητοποίηση DNA ανιχνευτή καθ όλο το μήκος του. Εικόνα 2.5: Ακινητοποίηση DNA ανιχνευτή μέσω των άκρων του Εικόνα 2.6: Ομοιοπολική πρόσδεση ανιχνευτών στην αλδεϋδοτροποποιημένη επιφάνεια της μικροσυστοιχίας. 34
50 To φιλμ αγαρόζης προτείνεται σαν ένας πιο απλοποιημένος τρόπος επίστρωσης απ ευθείας στη μη τροποποιημένη επιφάνεια των μικροσυστοιχιών, σε αντίθεση με την επίστρωση με σιλάνια η οποία αποδεικνύεται λιγότερο αποτελεσματική (χωρίς να αποκλείεται επίστρωση και σε τροποποιημένη επιφάνεια) (Dufva et al., 2004). Η επιφάνεια του γυαλιού, όντας επίπεδη, θέτει στερικούς περιορισμούς κατά τον υβριδισμό οι οποίοι αντιμετωπίζονται με χρήση δενδριμερών προσδετών αυξάνοντας τη χωρητικότητα πρόσδεσης των ανιχνευτών στο γυαλί (Beier and Hoheisel, 1999; Benters et al., 2002; Le Berre et al., 2003). Τα δενδριμερή ανυψώνουν επίσης τους DNA ανιχνευτές από την επιφάνεια βελτιώνοντας έτσι την αποτελεσματικότητα του υβριδισμού (Shchepinov et al., 1997). Το φιλμ αγαρόζης δρα επιπλέον σαν δενδριμερές και η χρήση του έχει σαν αποτέλεσμα υψηλότερα επίπεδα υβριδισμού και άρα υψηλότερο σήμα φθορισμού καθώς και σχετικά χαμηλό σήμα υποβάθρου σε σχέση με την αλδεϋδοτροποποιημένη επιφάνεια του γυαλιού (Dufva et al., 2004). Η ακινητοποίηση αμινοτροποποιημένων ή μη DNA ανιχνευτών σε αλδεϋδοτροποποιημένο ή μη αντιστοίχως φιλμ αγαρόζης έδειξε να είναι πιο αποτελεσματική όταν γίνεται με υπεριώδη ακτινοβόληση συγκριτικά με την ολονύκτια επώαση (Dufva et al., 2004). Επίσης ο υβριδισμός με ολονύκτια επώαση φαίνεται να εξαρτάται σε μεγάλο βαθμό από την παρουσία αλδεϋδομάδων στο φιλμ αγαρόζης (Dufva et al., 2004). Τέλος δεν παρατηρείται διαφορά στο σήμα υβριδισμού μεταξύ τροποποιημένου ή μη DNA ανιχνευτή (Dufva et al., 2004). Επιπλέον παράγοντες οι οποίοι συμβάλουν στην αποτελεσματική ακινητοποίηση των ανιχνευτών και κατά συνέπεια τη λήψη ενός ικανοποιητικού σήματος του υβριδισμού είναι: Το μήκος του ανιχνευτή. Το μεγάλο μήκος cdna ανιχνευτών επιτρέπει αποτελεσματικότερο υβριδισμό με το στόχο γι αυτό και οι μέθοδοι απευθείας εναπόθεσης των ανιχνευτών στα DNA chips πλεονεκτούν έναντι της in situ σύνθεσης (Zammatteo et al., 2000). Η πυκνότητα των DNA ανιχνευτών στην επιφάνεια (Venkatasubbarao, 2004). Μικρή πυκνότητα ανιχνευτών στο spot έχει ως αποτέλεσμα και χαμηλό σήμα υβριδισμού. Ωστόσο η σχέση πυκνότητας και σήματος υβριδισμού δεν είναι γραμμική. Υπάρχει ένα 35
51 όριο πάνω από το οποίο η αύξηση της πυκνότητας των ανιχνευτών στην επιφάνεια δε δίνει απαραίτητα και μεγαλύτερο σήμα υβριδισμού. Αυτό θα συμβεί όταν παρουσιαστούν στερικές παρεμποδίσεις μεταξύ των ακινητοποιημένων ανιχνευτών με συνέπεια τον περιορισμό της πρόσβασης στο DNA στόχο (Shchepinov et al., 1997). Συνεπώς προτείνεται το μήκος του άκρου του ανιχνευτή να είναι αρκετά μεγάλο ώστε να αποφεύγονται αυτού του είδους οι παρεμποδίσεις. Η κινητική της πρόσδεσης. Ο όγκος του διαλύματος που περιέχει τους ανιχνευτές και εκτυπώνεται από τις ακίδες στην επιφάνεια της μικροσυστοιχίας είναι πολύ μικρός, με αποτέλεσμα να εξατμίζεται γρήγορα. Συνεπώς είναι απαραίτητο η σύζευξη να γίνεται γρήγορα (Zammatteo et al., 2000). Η συγκέντρωση των μορίων στόχων στο διάλυμα. Το φθορίζον σήμα κάθε spot είναι ανάλογο με το σημασμένο DNA που εγχέεται στο συγκεκριμένο spot. Για να είναι ανιχνεύσιμο το σήμα θεωρείται ότι ο λόγος του σήματος προς το υπόβαθρο πρέπει να είναι μεγαλύτερος του 2 (Zammatteo et al., 2000). Η ελάχιστη μη ειδική πρόσδεση (Venkatasubbarao, 2004). Ο μειωμένος αυτοφθορισμός των ανιχνευτών (Venkatasubbarao, 2004) Απομόνωση βιολογικού υλικού Για τη διεξαγωγή ενός πειράματος ανάλυσης μικροσυστοιχιών, απαιτείται εξαγωγή και προετοιμασία του DNA. Οι διαθέσιμοι μέθοδοι απόκτησης DNA είναι: i) Πολλαπλασιασμός DNA που εξάγεται από τον πυρήνα ή τα μιτοχόνδρια με αλυσιδωτή αντίδραση πολυμεράσης (Polymerase Chain Reaction PCR). ii) Αντίστροφη μεταγραφή του mrna σε cdna με τη χρήση του ενζύμου της αντίστροφης μεταγραφάσης. iii) Χημική σύνθεση DNA. To βιολογικό δείγμα συλλέγονται από ιστούς οι οποίοι έχουν αναπτυχθεί σε διαφορετικές συνθήκες με αποτέλεσμα κάποιοι από αυτούς να είναι υγιείς και κάποιοι να νοσούν. Το δείγμα 36
52 το οποίοι προέρχεται από υγιή κύτταρα ή ιστούς αποτελεί το δείγμα αναφοράς (reference sample) ενώ αυτό που προέρχεται από κύτταρα ή ιστούς που νοσούν αποτελεί το δείγμα ελέγχου (control sample) Ιχνηθέτηση του στόχου Η σήμανση του στόχου μπορεί να γίνει είτε άμεσα είτε έμμεσα. Στην άμεση σήμανση φθορίζοντα μόρια προσδένονται απευθείας στο στόχο με χημικό ή ενζυμικό μέσο. Στην έμμεση απαιτείται ένα ενδιάμεσο μόριο ως γέφυρα μεταξύ του στόχου και του φθορίζοντος μορίου Μονοχρωματικές μικροσυστοιχίες Τα πειράματα μικροσυστοιχιών χαρακτηρίζονται ως μονοχρωματικά (single channel) όταν και τα δύο δείγματα σημαίνονται με την ίδια φθορίζουσα χρωστική. Για τη σήμανση των μονοχρωματικών μικροσυστοιχιών. Για τη σήμανση αυτών μπορεί να χρησιμοποιηθεί τόσο πράσινη (Cy3) όσο και κόκκινη (Cy5) χρωστική. Ωστόσο χρησιμοποιείται κυρίως η Cy3 λόγω της μεγαλύτερης σταθερότητας της. Επειδή δεν είναι δυνατή ταυτόχρονη χρήση δύο δειγμάτων σημασμένων με διαφορετικές φθορίζουσες χρωστικές, απαιτούνται δύο slides προκειμένου να συγκριθεί ένα δείγμα υπό μελέτη και ένα δείγμα αναφοράς. Χαρακτηριστικό παράδειγμα μονοχρωματικών μικροσυστοιχιών είναι οι Affymetrix GeneChip (εικόνα 2.7) Διχρωματικές μικροσυστοιχίες Τα πειράματα μικροσυστοιχιών χαρακτηρίζονται ως διχρωματικά (dual channel) όταν κάθε δείγμα σημαίνεται με φθορίζουσα χρωστική διαφορετικού χρώματος. Συνήθως το δείγμα ελέγχου σημαίνεται με Cy5 και το δείγμα αναφοράς με Cy3 χρωστική. Παράδειγμα διχρωματικών μικροσυστοιχιών είναι αυτές της Exiqon (εικόνα 2.8). 37

53 Εικόνα 2.7: Κατασκευή μονοχρωματικών μικροσυστοιχιών 38

54 Εικόνα 2.8: Κατασκευή διχρωματικών μικροσυστοιχιών 2.6 Πειραματικός σχεδιασμός Δημιουργία αντιγράφων Οι αριθμητικές τιμές που λαμβάνονται από ένα πείραμα μικροσυστοιχιών, παρουσιάζουν διακύμανση η οποία μπορεί να οφείλεται : i) Σε τεχνικούς παράγοντες. Η σήμανση των ακολουθιών που ανήκουν στο ίδιο γονίδιο ποτέ δεν είναι όμοια ακόμη και αν βρίσκονται στο ίδιο μείγμα καθώς μπορεί να επηρεαστεί από παράγοντες όπως τη μη σταθερή θερμοκρασία τη συγκέντρωση των ανιχνευτών και την υγρασία που επικρατεί κατά την εκτύπωση στη μικροσυστοιχία. Για τη μετρίαση της διακύμανσης που προκύπτει από τέτοιες ενδείκνυται η δημιουργία τεχνικών αντιγράφων. 39
55 ii) Στη διακύμανση εντός του βιολογικού δείγματος. H διακύμανση είναι αναπόφευκτη ακόμη και σε βιολογικά δείγματα γενετικά όμοιων οργανισμών, εξαιτίας των διαφορετικών περιβαλλοντικών παραγόντων, όπως η διατροφή, ο μεταβολισμός, η ηλικία, ποια ώρα της ημέρας θα ληφθεί το δείγμα, κ.λπ. Επιπροσθέτως ανεξαρτήτως εάν η λήψη του βιολογικού δείγματος γίνει από ολόκληρο τον οργανισμό, από έναν ιστό μόνο ή πρόκειται για μια καλλιέργεια κυτταρικής σειράς, διαφορετικές μέθοδοι εξαγωγής RNA και ο διαφορετικοί μέθοδοι πολλαπλασιασμού του μπορούν να εισάγουν επιπλέον διακύμανση. H ύπαρξη αντιγράφων αποτελεί θεμέλιο λίθο στη στατιστική συμπερασματολογία (Bolstad et al., 2004) με αυτά να διακρίνονται σε δύο κατηγορίες: i) Τεχνικά αντίγραφα. Η δημιουργία spot αντιγράφων ή αντιγράφων μικροσυστοιχιών από τον ίδιο βιολογικό ιστό ενός οργανισμού που λαμβάνεται σε διαφορετικές χρονικές στιγμές. ii) Βιολογικά αντίγραφα. Βιολογικά αντίγραφα θεωρείται το mrna που εξάγεται από δύο ή περισσότερους οργανισμούς (π.χ. ποντίκια). Η διαθεσιμότητα βιολογικών αντιγράφων κρίνεται πιο σημαντική καθώς τα τεχνικά αντίγραφα συνήθως δίνουν παρόμοιες τιμές με αυτές του αρχικού υβριδισμού ή προκύπτουν χρήσιμα συμπεράσματα που αφορούν το ίδιο άτομο (Bolstad et al., 2004). Στην πλειονότητα της η όποια διακύμανση που παρουσιάζεται στα πειράματα των μικροσυστοιχιών οφείλεται σε βιολογικούς και όχι τεχνικούς παράγοντες (Yauk et al., 2004) και για το λόγο αυτό είναι πιο σημαντικό ο σχεδιασμός ενός πειράματος μικροσυστοιχιών να περιλαμβάνει βιολογικά αντίγραφα έναντι των τεχνικών αντιγράφων, ωστόσο ο συνδυασμός βιολογικών και τεχνικών αντιγράφων είναι συχνά επιθυμητός. Τα βιολογικά αντίγραφα είναι κατάλληλα όταν θέλουμε να οδηγηθούμε σε συμπεράσματα για το σύνολο του πληθυσμού, ενώ τα τεχνικά αντίγραφα είναι προτιμητέο να χρησιμοποιούνται όταν θέλουμε να εξάγουμε συμπεράσματα για ένα συγκεκριμένο οργανισμό υπό τις συγκεκριμένες συνθήκες στις οποίες διεξάχθηκε το πείραμα. 40
56 Αντίγραφα κάθε probe set εντοπίζονται σε διακριτές περιοχές της μικροσυστοιχίας. To πλήθος των αντιγράφων που απαιτούνται εξαρτάται από πολλούς παράγοντες. Εάν η ανάλυση στοχεύει στην εύρεση μεγάλων διαφορών (μεγαλύτερες από 2 fold), τότε τρία αντίγραφα σε κάθε κατάσταση συνήθως επαρκούν. Για την εύρεση μικρότερων διαφορών ή σχεδόν όλων των μεγάλων τότε προτείνεται η χρήση πέντε αντιγράφων του δείγματος; χρήση τεσσάρων ή περισσότερων δίνουν λογικούς υπολογισμούς Ανιχνευτές ελέγχου Εκτός των ανιχνευτών που αντιπροσωπεύουν το υπό διερεύνηση γονίδιο, μια μικροσυστοιχία διαθέτει επιπλέον ανιχνευτές σε προκαθορισμένες θέσεις. Παράδειγμα αυτών αποτελούν οι: i) Αρνητικοί ανιχνευτές ελέγχου (negative control probes). Μετά τον υβριδισμό, δεν αναμένεται από τα αρνητικά controls κάποιο σήμα φθορισμού και επομένως η οποιαδήποτε παρουσία σήματος σε αυτό τον τύπο ανιχνευτών αποτελεί ένδειξη προσμίξεων στο σημασμένο στόχο, ατελή υβριδισμό ή ότι οι συνθήκες πλύσης της μικροσυστοιχίας δεν ήταν αρκετά αυστηρές. Οι ανιχνευτές αυτοί είναι μερικής συμπληρωματικότητας. ii) Θετικοί ανιχνευτές ελέγχου (positive control probes). Αυτοί οι ανιχνευτές λειτουργούν το αντίθετο, αφού αναμένεται να έχουν προβλέψιμο φθορίζον σήμα μετά τον υβριδισμό και χρησιμοποιούνται για την κανονικοποίηση της έντασης του σήματος υβριδισμού των μικροσυστοιχιών. Τα θετικά controls είναι ανιχνευτές πλήρους συμπληρωματικότητας οι οποίοι χαρακτηρίζονται από σταθερή έκφραση σε όλα τα δείγματα και βοηθούν στην εύρεση του προσανατολισμού της μικροσυστοιχίας ή μεταγενέστερα στην κανονικοποίηση των δεδομένων που προκύπτουν από την ανάλυση μικροσυστοιχιών. Τα θετικά controls υβριδίζονται με τα λεγόμενα γονίδια συντήρησης (housekeeping genes). Πρόκειται για ενδογενή γονίδια τα οποία εκφράζονται σε όλα τα κύτταρα ενός οργανισμού υπό κανονικές και παθολογικές συνθήκες αφού απαιτούνται για τη συντήρηση βασικών λειτουργιών του κυττάρου. Η επιλογή τέτοιων ανιχνευτών 41
57 αμφισβητείται δεδομένου ότι είναι δύσκολο να προσδιοριστούν γονίδια των οποίων η έκφραση είναι σταθερή κατά τη διάρκεια όλων των πειραματικών συνθηκών (Suzuki et al., 2000; Thellin et al., 1999; Warrington et al., 2000). Μία πιθανή μέθοδος είναι η χρήση ανιχνευτών, γνωστών για τη σταθερή τους έκφραση από προηγούμενα πειράματα μικροσυστοιχιών. Ωστόσο η σταθερή έκφραση αυτών των γονιδίων είναι υψηλή και δεν είναι αντιπροσωπευτική των γονιδίων που μας ενδιαφέρουν και τα οποία έχουν χαμηλότερη έκφραση με αποτέλεσμα να εισάγεται κάποιο σφάλμα έντασης (intensity bias). Πιο κατάλληλη λύση θεωρείται η χρήση εξωγενών spike in ανιχνευτών. Πρόκειται για ακολουθίες οι οποίες ανήκουν σε διαφορετικό από τον εξεταζόμενο οργανισμό ή για τελείως συνθετικούς ανιχνευτές. Αυτές οι ακολουθίες είναι μετάγραφα RNA σε γνωστή ποσότητα και τα οποία αναμιγνύονται σε διαφορετικές συγκεντρώσεις στο προς μελέτη δείγμα (Zhu et al., 2008) και κατόπιν υβριδίζονται με τα θετικά control. Το σημαντικό χαρακτηριστικό αυτών των ανιχνευτών ελέγχου είναι ότι δεν έχουν σημαντική ομολογία με άλλες ακολουθίες στο υπό εξέταση δείγμα αλλά έχουν ίδιες γενικές ιδιότητες υβριδισμού με τους υπόλοιπους ανιχνευτές. Δόκιμο είναι πολλαπλά αντίγραφα των ανιχνευτών ελέγχου να εξαπλώνονται σε όλο το εύρος μιας μικροσυστοιχίας Κοινό δείγμα αναφοράς Στην περίπτωση των δύο ή τριών δειγμάτων είναι εφικτό να γίνουν όλοι οι δυνατοί κατά ζεύγη συγκρίσεις απευθείας. Ωστόσο όταν το πλήθος δειγμάτων είναι μεγαλύτερο ίσες ποσότητες όλων των προς ανάλυση δειγμάτων σχηματίζουν το κοινό δείγμα αναφοράς (common reference) (εικόνα 2.9). Αυτό αναμιγνύεται κάθε φορά μαζί με ένα από τα προς εξέταση δείγματα και όλο μαζί εγχέεται στη μικροσυστοιχία. Σε αυτού του είδους τον πειραματικό σχεδιασμό κάθε δείγμα συγκρίνεται με έμμεσο τρόπο με όλα τα υπόλοιπα. 42

58 Εικόνα 2.9: Κοινό δείγμα αναφοράς τεσσάρων βιολογικών δειγμάτων Διχρωματικές μικροσυστοιχίες: Dye swapping Ένας τρόπος για να αντιμετωπιστεί η έλλειψη αντιγράφων ή η πιθανή αναποτελεσματική σήμανση των ανιχνευτών με τις χρωστικές είναι το dye swapping όπου τα ίδια δείγματα συγκρίνονται δύο φορές μετά από σήμανση και με τις δύο χρωστικές. Η μέθοδος αυτή μπορεί είναι σημαντική όταν ενδιαφέρει η σύγκριση του δείγματος ελέγχου με το δείγμα αναφοράς και μπορεί να εφαρμοστεί και με χρήση βιολογικών αντιγράφων. 2.7 Υβριδοποίηση Στη συνέχεια της πειραματικής διαδικασίας τα δύο cdna δείγματα αναμειγνύονται (αν πρόκειται για διχρωματικά) σε ίσες ποσότητες σε διάλυμα το οποίο περιέχει άλατα για τη ρύθμιση του ph και κατόπιν εγχέονται πάνω στη μικροσυστοιχία. Στο στάδιο αυτό οι μονόκλωνοι DNA ανιχνευτές μήκους βάσεων προσδένονται τη δημιουργία υδρογονικών δεσμών με τα μονόκλωνα cdna μόρια στόχους σε συγκεκριμένα spots της μικροσυστοιχίας. Η υβριδοποίηση διαρκεί ~2 18 ώρες, σε θερμοκρασία περίπου 45 ο C. Η αποτελεσματικότητα της υβριδοποίησης εξαρτάται από πολλούς παράγοντες όπως: i) Tη σύσταση της νουκλεοτιδικής αλληλουχίας ii) Tο μήκος των ανιχνευτών και των στόχων iii) Tη θερμοκρασία υβριδοποίησης. iv) Tη δευτεροταγή δομή v) Tη συγκέντρωση αλάτων vi) Tο ph και άλλους παράγοντες 43
59 Αυξημένες τιμές θερμοκρασίας αποτρέπουν τη δημιουργία δευτεροταγών δομών σε ανιχνευτές και στόχους ενώ το ph προτείνεται να κυμαίνεται μεταξύ (ουδέτερο) ώστε να σχηματίζονται ευκολότερα δεσμοί υδρογόνου. Η σύσταση της νουκλεοτιδικής αλληλουχίας είναι σημαντική διότι κάθε νουκλεοτιδικό ζεύγος G:C περιέχει τρεις δεσμούς υδρογόνου σε σχέση με τα ζεύγη A:T. Συνεπώς μεγαλύτερο ποσοστό ζευγών G:C αυξάνει τη συγγένεια της υβριδοποίησης. Σημαντικός ανασταλτικός παράγοντας για την υβριδοποίηση είναι και το μέγεθος του στόχου, καθώς όταν είναι μεγάλο συμβαίνει αναδίπλωση και εμφανίζονται επίσης δευτεροταγείς δομές παρεμποδίζοντας την. 2.8 Σάρωση Μετά τον υβριδισμό, ακτίνες laser διεγείρουν τις φθορίζουσες χρωστικές οι οποίες ανιχνεύονται σε διαφορετικό μήκος κύματος (550nm για τη Cy3 και 649nm για τη Cy5) σαρώνοντας κάθε ένα από τα pixels κάθε spot της μικροσυστοιχίας. Η τιμή της έντασης του σήματος που παράγεται κατά τη διέγερση των χρωστικών είναι ανάλογη της ποσότητας των μορίων που έχουν υβριδιστεί. Οι επίπεδες μικροσυστοιχίες υποστηρίζουν ακριβή σάρωση από την οποία προκύπτει ένα 16 bit TIFF αρχείο το οποίο απεικονίζει τη σχετική έκφραση (και των δύο δειγμάτων) κάθε γονιδίου. Το πλήθος των διαφορετικών εντάσεων που μπορεί να πάρει ένα 16 bitt αρχείο (από 0 έως αφού = 65535) διαφορετικών επιπέδων έντασης. Aν ένα γονίδιο εκφράζεται περισσότερο στο δείγμα ελέγχου από ό,τι στο δείγμα αναφοράς, τότε αντίστοιχο spot θα είναι πράσινο. Εάν το γονίδιο αυτό εκφράζεται το ίδιο και στα δύο δείγματα η απόχρωση του spot θα είναι κίτρινη ενώ στην περίπτωση που δεν εκφράζεται σε κανένα δείγμα το spot είναι μαύρο. Οι σημερινοί σαρωτές μπορούν να ανιχνεύσουν έως 11 αποχρώσεις. Δύο είναι οι μέθοδοι για τον υπολογισμό της έντασης του σήματος του spot μετά τη σάρωση. Η πρώτη περιλαμβάνει τον υπολογισμό της έντασης για ένα σταθερό εμβαδόν γύρω από το κέντρο του κάθε spot. Η μέθοδος αυτή έχει μικρότερο υπολογιστικό κόστος αλλά δυσχεραίνει τη διάκριση της έντασης των pixels, σε ένταση υποβάθρου και στην πραγματική ένταση του σήματος. Η δεύτερη μέθοδος, περιλαμβάνει τον αυστηρό καθορισμό των ορίων του spot και τον 44
60 υπολογισμό του σήματος βάσει όλων τον pixels εντός των ορίων αυτών. Ο υπολογισμός βάσει αυτής της μεθόδου, είναι ακριβέστερος αλλά υπολογιστικά πιο χρονοβόρος. 2.9 Πλατφόρμες μικροσυστοιχιών In situ σύνθεση ολιγονουκλεοτιδικών μικροσυστοιχιών Η μέθοδος αυτή αναπτύχθηκε αρχικά από την Affymetrix. Πρόκειται για την επί τόπου σύνθεση ολιγονουκλεοτιδίων μήκους έως 25 βάσεων στην επιφάνεια του πλακιδίου κάνοντας χρήση μεθόδων φωτολιθογραφίας και συνδυαστικής χημείας (Pease et al., 1994). Η φωτολιθογραφία αποτελεί μια φωτο κατευθυνόμενη μέθοδο κατά την οποία χρησιμοποιούνται ειδικές μάσκες οι οποίες επιτρέπουν την έκθεση στην υπεριώδη ακτινοβολία μόνο συγκεκριμένων περιοχών κάθε φορά. Οι ολιγονουκλεοτιδικοί ανιχνευτές συντίθενται σε βηματισμό μίας βάσης τη φορά. Εν συνεχεία με χημική σύζευξη στο 5 άκρο του ανιχνευτή, τοποθετούνται νέες βάσεις πάνω στις προηγούμενες. Μια χημική ομάδα (MeNPOC μεθυλονιτροπιπερονυλοκαρβονύλιο) στο 5 άκρο του ανιχνευτή είναι αυτή που εμποδίζει τον πολυμερισμό του DNA εκτός και αν εκτεθεί σε υπεριώδες φως (εικόνα 2.10). Η χημική αυτή ομάδα υπάρχει σε κάθε προστιθέμενη βάση. Κάθε γονίδιο αντιπροσωπεύεται συνήθως από ζεύγη ολιγονουκλεοτιδίων. Κάθε ζεύγος περιλαμβάνει μία perfect match (PM) ακολουθία με πλήρη συμπληρωματικότητα ως προς το γονίδιο και μία mismatch (MM) ακολουθία η οποία διαφέρει μόνο ως προς το κεντρικό νουκλεοτίδιο (13 η βάση). Γενικά ένας PM ανιχνευτής κουβαλά περισσότερη πληροφορία αλλά αυτό δε συμβαίνει πάντα εξαιτίας της διασταυρούμενης υβριδοποίησης. Γι αυτό προτάθηκε από την Affymetrix η χρήση του ΜΜ ανιχνευτή ως control για τον PM. Εκτός της διασταυρούμενης υβριδοποίησης η μη ειδική πρόσδεση του MM επιτρέπει στην ανίχνευση και άλλων λανθανουσών σημάτων (τα οποία κατόπιν αφαιρούνται από το PM) αλλά και στην ποσοτικοποίηση των mrnas που εκφράζονται ασθενώς στο δείγμα (εικόνα 2.11). 45


61 Εικόνα 2.10: Μέθοδος in situ σύνθεσης ολιγονουκλεοτιδίων με τη μέθοδο της φωτολιθογραφίας Εικόνα 2.11: Τεχνολογία Affymetrix GeneChip: Ολιγονουκλεοτιδικό PM:ΜΜ ζεύγος. Σήμερα είναι εφικτή η κατασκευή μικροσυστοιχιών ολιγονουκλεοτιδίων μήκους βάσεων ή και περισσότερων (Barrett and Kawasaki, 2003). Η τεχνική αυτή επιτρέπει την κατασκευή μικροσυστοιχιών υψηλής πυκνότητας ακόμη και spots. Ιδανικό μήκος για τους ολιγονουκλεοτιδικούς ανιχνευτές θεωρούνται όταν είναι 70 nt, καθώς όταν είναι μικρότερο παρουσιάζει περιορισμένη ειδικότητα υβριδοποίησης ενώ μήκη μεγαλύτερα των 70 nt αυξάνουν τον κίνδυνο της διασταυρούμενης υβριδοποίησης. 46
62 2.9.2 cdna μικροσυστοιχίες εκτύπωσης Οι cdna μικροσυστοιχίες εκτύπωσης (spotted) προκύπτουν με μηχανική μικρο εναπόθεση ίσων ποσοτήτων αλληλουχιών cdna μήκους kb. Οι cdna στόχοι αποτελούν προϊόντα πολλαπλασιασμού της αλυσιδωτής αντίδρασης πολυμεράσης (PCR) (Pease et al., 1994). Το πλεονέκτημα της τεχνολογίας αυτής είναι έγκειται στο μεγάλο μήκος των ανιχνευτών και άρα μεγαλύτερη απόδοση του υβριδισμού. Επιπλέον πλεονέκτημα θεωρείται η δυνατότητα τοποθέτησης στις μικροσυστοιχίες άγνωστων αλληλουχιών έτσι ώστε να δημιουργηθούν προφίλ γονιδιακής έκφρασης για οργανισμούς των οποίων η αλληλουχία των γονιδίων τους είναι περιορισμένη Ολιγονουκλεοτιδικές μικροσυστοιχίες εκτύπωσης Η διαφορά ολιγονουκλεοτιδικών μικροσυστοιχιών εκτύπωσης από αυτών των cdna μικροσυστοιχιών εκτύπωσης είναι ότι χρησιμοποιούνται συνθετικοί ολιγονουκλεοτιδικοί ανιχνευτές έναντι των cdna αλληλουχιών. Όταν οι επιθυμητές αλληλουχίες βάσεων είναι γνωστές, τότε υπάρχει η δυνατότητα σχεδίασης και σύνθεσης ολιγονουκλεοτιδικών ανιχνευτών με τη βοήθεια ειδικής συσκευής. Η εναπόθεση των ολιγονουκλεοτιδίων στην επιφάνεια του πλακιδίου μετά τη σύνθεση τους μπορεί να γίνει από τα ίδια ρομπότ που χρησιμοποιούνται στις τυπωμένες cdna μικροσυστοιχίες. H απευθείας εκτύπωση κατηγοριοποιείται σε: i) Εκτύπωση με επαφή. Σε αυτή την περίπτωση η επιφάνεια έρχεται σε επαφή με τις ακίδες κατά την εναπόθεση των ανιχνευτών απελευθερώνοντας σταγονίδια όγκου 50pl 100 nl σε spots με διάμετρο που κυμαίνεται από μm. Μειονέκτημα αυτής της μεθόδου θεωρείται η έλλειψη αντοχής των ακίδων εξαιτίας των κτυπημάτων πάνω στην επιφάνεια καθώς και η πιθανότητα καταστροφής της επίστρωσης της μικροσυστοιχίας. ii) Εκτύπωση χωρίς επαφή. Ακολουθείται η ίδια αρχή με τους εκτυπωτές ψεκασμού (inkjet) με χορήγηση σταγονιδίων όγκου εκατοντάδων picoliters (10 12 liter). Μειονέκτημα αυτής της μεθόδου είναι η διάχυση σταγονιδίων εκτός των ορίων του spot. 47
63 Οι ολιγονουκλεοτιδικοί ανιχνευτές παρέχουν κάποια σημαντικά πλεονεκτήματα έναντι των cdna ανιχνευτών τα οποία είναι: a) H απευθείας σύνθεση τους από βιβλιοθήκες αλληλουχιών και όχι μέσω PCR όπως τα cdnas b) Δυνατότητα ελέγχου του ακριβούς τμήματος του γονιδίου θα υβριδοποιηθεί c) Οι ολιγονουκλεοτιδικοί ανιχνευτές μπορούν να σχεδιαστούν με βάση μοναδικές περιοχές συγκεκριμένου γονιδίου ώστε να αποφευχθεί κάποια ενδεχόμενη διασταυρούμενη υβριδοποίηση (cross hybridization), όπως θα μπορούσε να συμβεί στην περίπτωση των τυπωμένων cdna μικροσυστοιχιών. Η διασταυρούμενη υβριδοποίηση συνήθως παρατηρείται όταν υπάρχει ομολογία >70%. d) Σημαντικό πλεονέκτημα των in situ μικροσυστοιχιών είναι ότι εγγυώνται υψηλή καθαρότητα εξαιτίας των καταλοίπων που πιθανόν να αφήσει το αναποτελεσματικό πλύσιμο των εκτυπωτικών συσκευών (spotter). e) Τα ολιγονουκλεοτίδια όντας μονόκλωνα και όχι δίκλωνα όπως τα cdnas δεν απαιτούν περαιτέρω επεξεργασία πριν την υβριδοποίηση, ούτε αναδιπλώνονται όπως τα cdnas. 48
64 2.10 Βιβλιογραφία Allemand, J.F., Bensimon, D., Jullien, L., Bensimon, A., and Croquette, V. (1997). ph dependent specific binding and combing of DNA. Biophys J 73, Barrett, J.C., and Kawasaki, E.S. (2003). Microarrays: the use of oligonucleotides and cdna for the analysis of gene expression. Drug Discov Today 8, Beier, M., and Hoheisel, J.D. (1999). Versatile derivatisation of solid support media for covalent bonding on DNA microchips. Nucleic Acids Res 27, Benters, R., Niemeyer, C.M., Drutschmann, D., Blohm, D., and Wohrle, D. (2002). DNA microarrays with PAMAM dendritic linker systems. Nucleic Acids Res 30, E10. Bolstad, B.M., Collin, F., Simpson, K.M., Irizarry, R.A., and Speed, T.P. (2004). Experimental design and lowlevel analysis of microarray data. Int Rev Neurobiol 60, Cheung, V.G., Morley, M., Aguilar, F., Massimi, A., Kucherlapati, R., and Childs, G. (1999). Making and reading microarrays. Nat Genet 21, Conlon, E.M., Liu, X.S., Lieb, J.D., and Liu, J.S. (2003). Integrating regulatory motif discovery and genomewide expression analysis. Proc Natl Acad Sci U S A 100, Dufva, M., Petronis, S., Jensen, L.B., Krag, C., and Christensen, C.B. (2004). Characterization of an inexpensive, nontoxic, and highly sensitive microarray substrate. Biotechniques 37, , 294, 296. Ge, H., Liu, Z., Church, G.M., and Vidal, M. (2001). Correlation between transcriptome and interactome mapping data from Saccharomyces cerevisiae. Nat Genet 29, Joos, B., Kuster, H., and Cone, R. (1997). Covalent attachment of hybridizable oligonucleotides to glass supports. Anal Biochem 247, Le Berre, V., Trevisiol, E., Dagkessamanskaia, A., Sokol, S., Caminade, A.M., Majoral, J.P., Meunier, B., and Francois, J. (2003). Dendrimeric coating of glass slides for sensitive DNA microarrays analysis. Nucleic Acids Res 31, e88. Pease, A.C., Solas, D., Sullivan, E.J., Cronin, M.T., Holmes, C.P., and Fodor, S.P. (1994). Light generated oligonucleotide arrays for rapid DNA sequence analysis. Proc Natl Acad Sci U S A 91, Schena, M., Shalon, D., Davis, R.W., and Brown, P.O. (1995). Quantitative monitoring of gene expression patterns with a complementary DNA microarray. Science 270, Shchepinov, M.S., Case Green, S.C., and Southern, E.M. (1997). Steric factors influencing hybridisation of nucleic acids to oligonucleotide arrays. Nucleic Acids Res 25, Southern, E., Mir, K., and Shchepinov, M. (1999). Molecular interactions on microarrays. Nat Genet 21, 5 9. Suzuki, T., Higgins, P.J., and Crawford, D.R. (2000). Control selection for RNA quantitation. Biotechniques 29, Thellin, O., Zorzi, W., Lakaye, B., De Borman, B., Coumans, B., Hennen, G., Grisar, T., Igout, A., and Heinen, E. (1999). Housekeeping genes as internal standards: use and limits. J Biotechnol 75, Venkatasubbarao, S. (2004). Microarrays status and prospects. Trends Biotechnol 22, Warrington, J.A., Nair, A., Mahadevappa, M., and Tsyganskaya, M. (2000). Comparison of human adult and fetal expression and identification of 535 housekeeping/maintenance genes. Physiol Genomics 2, Yauk, C.L., Berndt, M.L., Williams, A., and Douglas, G.R. (2004). Comprehensive comparison of six microarray technologies. Nucleic Acids Res 32, e124. Zammatteo, N., Jeanmart, L., Hamels, S., Courtois, S., Louette, P., Hevesi, L., and Remacle, J. (2000). Comparison between different strategies of covalent attachment of DNA to glass surfaces to build DNA microarrays. Anal Biochem 280, Zhu, J., He, F., Hu, S., and Yu, J. (2008). On the nature of human housekeeping genes. Trends Genet 24,
65 Κεφάλαιο 3 ο Επεξεργασία Πρωτογενών Δεδομένων 50
66 3.1 Επεξεργασία πρωτογενών δεδομένων μικροσυστοιχιών Σε ένα πείραμα διχρωματικών μικροσυστοιχιών οι κύριες ποσότητες που προκύπτουν μετά την κατάτμηση και τη διόρθωση υποβάθρου είναι για κάθε γονίδιο, ένα ζεύγος τιμών εντάσεων R (R=Cy5) και G (G=Cy3). Ο λόγος R/G δίνει μία εικόνα της αλλαγής της έκφρασης αλλά χειρίζεται διαφορετικά τα γονίδια που υπερεκφράζονται και αυτά που υποεκφράζονται. Παραδείγματος χάριν για τιμές έκφρασης R=4 και G=2 η έκφραση ενός γονιδίου διπλασιάζεται, ο λόγος R/G είναι 2 ενώ όταν το γονίδιο υπο εκφράζεται για R=2 και G=4 είναι 0.5. Γι αυτό ο λόγος R/G εκφράζεται με βάση το log 2. O λόγος log 2 (R/G) χειρίζεται συμμετρικά την υπερέκφραση και την υποέκφραση των γονιδίων, παράδειγμα log 2 (2)=1, log 2 (1/2)= 1 και log 2 (4)=2, log 2 (1/4)= 2 (Quackenbush, 2002)., Η οπτική απεικόνιση των (R,G) δεδομένων ονομάζεται ΜΑ plot, όπου Μ=log2(R/G) και A= log 2 (RXG). Στην περίπτωση που ο υβριδισμός είναι ιδανικός οι τιμές του M σε ένα ΜΑ plot θα έπρεπε να είναι ομοιόμορφα κατανεμημένες γύρω από το 0 κατά μήκος όλων των Α τιμών έντασης. Μιας και στην πράξη αυτό σπάνια συμβαίνει, τα MA plots είναι χρήσιμα στον προσδιορισμό των προβληματικών spot. 3.2 Μέθοδοι για την ανάλυση χαμηλού επιπέδου Το λογισμικό ανάλυσης εικόνων που προκύπτει από ένα πείραμα μικροσυστοιχιών δίνει τα πρωτογενή δεδομένα τα οποία πρέπει να μετατραπούν σε μετρήσεις γονιδιακής έκφρασης. Αυτού του είδους η ανάλυση ονομάζεται ανάλυση χαμηλού επιπέδου Διόρθωση υποβάθρου Στα πρωτογενή δεδομένα μικροσυστοιχιών, απαιτείται διόρθωση υποβάθρου προκειμένου να αφαιρεθεί η επίδραση του φθορισμού υποβάθρου. Ο φθορισμός υποβάθρου προκύπτει εντός της μικροσυστοιχίας λόγω της μη ειδικής πρόσδεσης ή της χωρικής ανομοιογένειας που προκαλείται από κατάλοιπα που έχουν μείνει στην επιφάνεια της μικροσυστοιχίας μετά το 51
67 στάδιο της πλύσης ή από το θόρυβο που εισάγει το scanner. Άλλες πηγές θορύβου, είναι ορατές μεταξύ των μικροσυστοιχιών και μπορεί να προκύψουν από γδαρσίματα ή άλλα κατάλοιπα (σκόνη, σάλιο) σε κάποιες από τις μικροσυστοιχίες. Τα περισσότερα προγράμματα ανάλυσης εικόνας υπολογίζουν ως σήμα υποβάθρου τη διάμεσο ή τη μέση τιμής της τοπικής έντασης του περιβάλλοντος χώρου κάθε spot. Συνήθως προτιμάται η διάμεσος έναντι της μέσης τιμής αυτών καθώς επηρεάζεται λιγότερο από τις ακραίες τιμές. Αφαιρώντας το σήμα υποβάθρου (R b ) από το πραγματικό σήμα (R f ) η ένταση που προκύπτει είναι αντικειμενική (unbiased) αλλά με μη επιθυμητές στατιστικές ιδιότητες λόγω της απουσίας πολλών λογαριθμικών λόγων, εξαιτίας των αρνητικών εντάσεων που προκύπτουν όταν ο φθορισμός υποβάθρου είναι μεγαλύτερος, καθώς και λόγω της μεγάλης μεταβλητότητας στους λογαριθμικούς λόγους χαμηλής έντασης (Ritchie et al., 2007). H διόρθωση υποβάθρου δε βοηθάει μόνο στην ευκολότερη ανίχνευση των διαφορικά εκφρασμένων γονιδίων αλλά και στο ότι η μη εφαρμογή της ελλοχεύει υποτίμηση του πλήθους των λογαριθμικών λόγων των spikein γονιδίων (Yang, Buckley, & Speed, 2001). Η συνήθης θεώρηση για τη διόρθωση υποβάθρου των διχρωματικών μικροσυστοιχιών είναι πως τα Rb και Gb σήματα υποβάθρου, αποτελούν πρόσθετα των πραγματικών R και G σημάτων, ισχύει δηλαδή R=R f R b και G=G f G b. H μέθοδος η οποία επιλέχθηκε στην παρούσα διπλωματική για τη διόρθωση υποβάθρου είναι η normexp+offset της συνάρτησης backgroundcorrect() του πακέτου Limma. H normexp, βασίζεται στο δημοφιλή RMA (Robust Multi Array Analysis) αλγόριθμο ο οποίος χρησιμοποιεί ένα μοντέλο συνέλιξης (convolution model) το οποίο μοντελοποιεί τις παρατηρηθείσες εντάσεις των pixel ως άθροισμα δύο ανεξάρτητων τυχαίων μεταβλητών, μίας κανονικά κατανεμημένης μεταβλητής η οποία αναπαριστά το σήμα υποβάθρου και μίας εκθετικά κατανεμημένης μεταβλητής η οποία αναπαριστά το σήμα φθορισμού. Ο λόγος για τον οποίο η ένταση υποβάθρου περιγράφεται από μία κανονική κατανομή είναι διότι η κανονική κατανομή περιγράφει ικανοποιητικά τυχαίους αριθμούς, όπως τυχαία είναι και τα σφάλματα που επηρεάζουν την τιμή της έντασης υποβάθρου. 52
68 To μοντέλο αυτό προσαρμόζει τις παρατηρηθείσες εντάσεις αφαιρώντας τις εντάσεις υποβάθρου με τη χρήση μιας μονοτονικής συνάρτησης έτσι ώστε οι διορθωμένες τιμές να είναι θετικές. Η normexp θέτει για το κόκκινο κανάλι R f = R b + B + S, όπου S η πραγματική ένταση του σήματος και B η ένταση υποβάθρου που δεν υπολογίζεται από την R b. Το σήμα S ακολουθεί την εκθετική κατανομή με μέση τιμή α, ενώ το B την κανονική κατανομή με μέση τιμή μ και διακύμανση σ 2. Από Χ = R f R b το μοντέλο γίνεται: Χ = Β + S όπου, Χ η παρατηρηθείσα ένταση και η κοινή αθροιστική συνάρτηση κατανομής f B,S (b, s; μ, σ, α) = exp( )φ(b; μ, σ2 ) Ο υπολογισμός των τριών παραμέτρων μ, σ, α γίνεται με μία από τις διαθέσιμες μεθόδους της normexp, mle, saddle, rma, rma75. Η normexp εφαρμόζεται σε κάθε ένα από τα R και G κανάλια, οι εντάσεις των οποίων μετά μετατρέπονται στους λογαριθμικούς λόγους M = log 2 (R/G) και Α = ½log 2 (RG) για κάθε spot. Ο αλγόριθμος διόρθωσης υποβάθρου normerxp βελτιώνεται περεταίρω με την εισαγωγή ενός offset k πριν τη λογαρίθμηση των εντάσεων, έτσι ώστε ο λογαριθμικός λόγος να είναι M = log 2 [(R+k)/(G+k)]. Σκοπός του offset είναι η απομάκρυνση των διορθωμένων τιμών έντασης από το 0. Η τιμή του offset επιλέγεται με βάση το MA plot και πλεονεκτεί στο ότι εφαρμόζεται σε εκ των προτέρων θετικές τιμές της έντασης, επιτρέποντας τη σταθεροποίηση της διακύμανσης των M τιμών (που είναι πιο εμφανής στις χαμηλές Α τιμές) χωρίς την ανάγκη επίτευξης θετικών τιμών ταυτόχρονα. 53
69 3.2.2 Κανονικοποίηση Η κανονικοποίηση των εντάσεων των μικροσυστοιχιών απαιτείται για την εξομάλυνση της συστηματικής διακύμανσης, δηλαδή της διακύμανσης που οφείλεται σε τεχνολογικούς παράγοντες παρά στις διαφορές των βιολογικών δειγμάτων ή στις διαφορές μεταξύ των τυπωμένων ανιχνευτών. Στόχος της κανονικοποίησης είναι η μετρίαση της συστηματικής διακύμανσης ώστε οι βιολογικές διαφορές να εντοπίζονται πιο εύκολα. Τέτοιου είδους διακυμάνσεις μπορεί να οφείλονται: i. Στη διακύμανση των πειραματικών συνθηκών οι οποίες πιθανόν να δώσουν διαφορετικές εκφράσεις μεταξύ των αντιγράφων ενός γονιδίου. ii. Στη μη αποτελεσματική σήμανση των ανιχνευτών με την πράσινη ή την κόκκινη χρωστική, εξαιτίας του ότι οι φυσικοχημικές ιδιότητες του μπορεί να επηρεάζονται από το φως, τη θερμοκρασία κ.λπ. iii. Στην ύπαρξη ατελειών στην επιφάνεια της μικροσυστοιχίας, σκόνης, κ.λπ. Στη μη κανονική εναπόθεση των πολλαπλασιασμένων με PCR, cdnas από τις ακίδες (print tips) οι οποίες μπορεί να διαφέρουν ελαφρώς στο μήκος τους ή στο άνοιγμα τους ή μετά από πολλές ώρες εκτύπωσης να μην έχουν την ίδια απόδοση. iv. Στη χρήση διαφορετικών ρυθμίσεων στο σαρωτή (π.χ. όριο ανίχνευσης). v. Στη θέση της μικροσυστοιχίας στο χώρο. Για τους παραπάνω λόγους είναι απαραίτητη η κανονικοποίηση τόσο εντός των ίδιων των μικροσυστοιχιών όσο και μεταξύ αυτών. Στην περίπτωση των διχρωματικών μικροσυστοιχιών, συνήθως οι περισσότεροι μέθοδοι κανονικοποίησης εφαρμόζονται στους λογαριθμικούς λόγους της έντασης του κόκκινου προς το πράσινο κανάλι και η κανονικοποίηση στο ένα κανάλι πρέπει να γίνεται με βάση την ένταση του άλλου καναλιού αφού πρώτα έχει προηγηθεί η διόρθωση υποβάθρου. Η ποιότητα της κανονικοποίησης ελέγχεται από το ΜΑ plot. 54
70 Κανονικοποίηση εντός των μικροσυστοιχιών Composite loess κανονικοποίηση Στην παρούσα διπλωματική και για την κανονικοποίηση εντός των μικροσυστοιχιών, επιλέχθηκε η μέθοδος τοπικής παλινδρόμησης Composite loess, προκειμένου να διορθωθούν τυχόν αποκλίσεις που προκύπτουν λόγω των διαφορετικών χρωστικών. Η κανονικοποίηση loess βασίζεται στη θεώρηση ότι η πλειονότητα των γονιδίων δεν εκφράζεται διαφορικά (Yang et al., 2002) και η composite εκδοχή αυτής, χρησιμοποιείται όταν υπάρχουν διαθέσιμα spots ανιχνευτών ελέγχου οι οποίοι όπως είναι γνωστό δεν εκφράζονται διαφορικά. Οι εντάσεις αυτών των spots ελέγχου θα πρέπει να καλύπτουν το μεγαλύτερο δυνατό εύρος εντάσεων (Smyth & Speed, 2003). Για ένα δεδομένο υπόπλεγμα, η composite loess κανονικοποίηση, είναι μια σταθμισμένη μέση τιμή της καμπύλης loess που βασίζεται σε όλα τα γονίδια του υποπλέγματος και των ανιχνευτών ελέγχου. Η προσαρμογή της καμπύλης μέσω των spots ελέγχου, εξασφαλίζει την αντικειμενικότητα της κανονικοποίησης, δηλαδή πως η καμπύλη δεν διαμορφώνεται από γονίδια που εκφράζονται διαφορικά. Πιο συγκεκριμένα, τα βάρη εξαρτώνται από το πλήθος των γονιδίων σε διαφορετικά επίπεδα έκφρασης Α. Δηλαδή, η καμπύλη κανονικοποίησης για τις χαμηλές log 2 A τιμές εντάσεων η βασίζεται σε όλα τα γονίδια που αντιστοιχούν στο αντίστοιχο εύρος έντασης κάθε, ενώ για τις υψηλότερες log 2 A τιμές εντάσεων η βασίζεται στους ανιχνευτές ελέγχου (Yang et al., 2002) Κανονικοποίηση μεταξύ των μικροσυστοιχιών Η κανονικοποίηση μεταξύ των μικροσυστοιχιών βασίζεται στην υπόθεση ότι οι διακυμάνσεις των κατανομών μεταξύ των μικροσυστοιχιών είναι αποτέλεσμα των πειραματικών συνθηκών και όχι κάποιας βιολογικής μεταβλητότητας. 55
71 Αquantile κανονικοποίηση H κανονικοποίηση ποσοστημορίου (quantile) τροποποιεί τις εντάσεις των ανιχνευτών έτσι ώστε οι κατανομές των εντάσεων των ανιχνευτών μεταξύ των διαφορετικών πλακιδίων μικροσυστοιχιών να είναι ίσες. Στο παράδειγμα που ακολουθεί φαίνεται η διαδικασία με την οποία κανονικοποιούνται τρεις ομάδες δεδομένων με τέσσερα γονίδια η κάθε μία: Σε κάθε πίνακα ανατίθεται ένας αριθμός κατάταξης της έντασης των γονιδίων από i iv: Οι τιμές κάθε πίνακα ταξινομούνται κατά αύξουσα τιμή Κατόπιν γίνεται εύρεση της μέσης τιμής κάθε γραμμής των τριών πινάκων: Στον πίνακα κατάταξης που σχεδιάστηκε νωρίτερα γίνεται ανάθεση των μέσω τιμών που υπολογίστηκαν με την ανάλογη κατάταξη: 56
72 Μετά την κανονικοποίηση με ποσοστημόρια οι μετρήσεις κάθε μικροσυστοιχίας έχουν ίσια μέση τιμή ίση με 0 και τυπική απόκλιση ίση με 1 (εικόνα 3.1). Στην παρούσα ανάλυση χρησιμοποιήθηκε η Aquantile μέθοδος κανονικοποίησης η οποία διασφαλίζει πως οι A τιμές έχουν την ίδια τιμή μεταξύ των διαφορετικών πλακιδίων των μικροσυστοιχιών ενώ διατηρώντας τις M τιμές στα ίδια επίπεδα. Εικόνα 3.1: Boxplot κανονικοποίησης ποσοστημορίου Έλεγχος Στατιστικών Υποθέσεων Σε πολλά προβλήματα δεν ενδιαφέρει η εκτίμηση της τιμής μιας παραμέτρου, αλλά η διαπίστωση αν η παράμετρος είναι μικρότερη ή μεγαλύτερη από μια δεδομένη τιμή που έχει φυσική σημασία για το πρόβλημα. 57
73 Ανάλυση Διακύμανσης κατά ένα Παράγοντα (One way ANOVA) Η ανάλυση διακύμανσης χρησιμοποιεί ένα F test για τον έλεγχο της ύπαρξης στατιστικά σημαντικής διαφοράς μεταξύ των μέσων τιμών των πειραμάτων. Το γονίδιο αποτελεί την εξαρτημένη συνεχή μεταβλητή (ή παράγοντα) και γι αυτό η ανάλυση ονομάζεται μονομεταβλητή (univariate) ενώ το πείραμα μικροσυστοιχιών την ανεξάρτητη κατηγορική μεταβλητή. Λόγω της μίας ανεξάρτητης μεταβλητής, η ανάλυση διακύμανσης χαρακτηρίζεται one way. Η ανάλυση διακύμανσης διαφέρει από το t test ως προς το ότι μπορεί να συγκρίνει την ισότητα τριών ή περισσότερων μέσων τιμών τη φορά, σε αντίθεση με το student s t test του οποίου η ανεξάρτητη μεταβλητή μπορεί να έχει δύο μόνο επίπεδα. Ο τυχαίος ορισμός επιπέδων της ανεξάρτητης μεταβλητής ανάμεσα στα άλλα επίπεδα ονομάζει το μοντέλο της ανάλυσης τυχαίο (random), ενώ σε περίπτωση επιλογής ορισμένου ενδιαφέροντος επιπέδων ή όλων, το μοντέλο της ανάλυσης ονομάζεται σταθερό (fixed) Παραδοχές στις οποίες βασίζεται η Aνάλυση Διακύμανσης H ανάλυση διακύμανσης αποτελεί παραμετρικό έλεγχο. Στους παραμετρικούς ελέγχους χρησιμοποιούνται ποσοτικά δεδομένα και εφαρμόζονται υπό τις ακόλουθες προϋποθέσεις. i. Ανεξαρτησία των παρατηρήσεων. Η τιμή μιας παρατήρησης της ανεξάρτητης μεταβλητής δε θα πρέπει να παρέχει καμία ένδειξη για την τιμή κάποιας άλλης παρατήρησης μεταξύ των διαφορετικών πειραμάτων. Ανεξαρτησία των παρατηρήσεων απαιτείται και εντός της ομάδας. Παραβίαση αυτής της παραδοχής συμβαίνει όταν παρατηρήσεις σχετίζονται χρονικά μεταξύ τους. Στην περίπτωση αυτή κατάλληλο θεωρείται το blocked one way ANOVA test. ii. Ομοιογένεια της διακύμανσης. Η ύπαρξη ή όχι ακραίων τιμών εξετάζεται με το Levene s και το Bartlett s test. H επίδραση της ανομοιογένειας της διακύμανσης μετριάζεται όταν κάθε πείραμα αποτελείται από ίσο πλήθος παρατηρήσεων (balanced design). Ως ίσοι χαρακτηρίζονται πληθυσμοί των οποίων οι παρατηρήσεις δε μπορούν να διαφέρουν 58
74 περισσότερο από 1 ½. Εάν τα δεδομένα δεν ικανοποιούν αυτή την παραδοχή της ομοιογένειας αναλύονται με Welch Anova και με το Games Howell ή το Dunnet s C posthoc test αντί του Tukey. iii. Κανονικότητα. Οι τιμές της εξαρτημένης μεταβλητής θεωρείται ότι ακολουθούν την κανονική κατανομή και ο έλεγχος της παραδοχής αυτής ελέγχεται με το Shapiro Wilk test. Καθ ότι όμως αυτό είναι πολύ συντηρητικό, η κανονικότητα είναι ελέγχεται εναλλακτικά και γραφικά με τη χρήση boxplots και Q Q plots. Ύπαρξη ακραίων τιμών στα boxplots και μια μη γραμμική σχέση στα Q Q plots εγείρει ανησυχία για την παραδοχή της κανονικότητας. Υπάρχουν πολλοί τρόποι χειρισμού δεδομένων που παραβιάζουν την παραδοχή της κανονικοποίησης όπως η κανονικοποίηση τους ή η διαγραφή κάποιων εκ των δεδομένων. Το αποτέλεσμα των παραβιάσεων των παραπάνω παραδοχών εξαρτάται από μέγεθος της παραβίασης. Η επίδραση της ανομοιογένειας της διακύμανσης και της μη κανονικότητας των δεδομένων είναι εντονότερη στους μικρότερους πληθυσμούς ή σε πληθυσμούς με άνισες παρατηρήσεις. Αν οι παραδοχές της κανονικότητας και της ομοιογένειας της διακύμανσης παραβιάζονται προτείνεται η χρήση του μη παραμετρικού Kruskal Wallis test αντί της ανάλυσης διακύμανσης (Πίνακας 3.1). Τα μη παραμετρικά tests ενδείκνυνται όταν οι παραπάνω προϋποθέσεις δεν ικανοποιούνται και επιπλέον όταν υπάρχουν περισσότερα από πέντε αντίγραφα της εξαρτημένης μεταβλητής ανά κάθε πείραμα ενώ ο έλεγχος ΑΝΟVA όπως και το t test ενδείκνυται στην περίπτωση μικρού αριθμού αντιγράφων. Στον πίνακα φαίνεται εν συντομία o κατάλληλος τύπος ελέγχου της ανάλυσης διακύμανσης σε κάθε περίπτωση. 59
75 Πίνακας 3.1: Κριτήρια επιλογής του κατάλληλου τεστ Ανάλυσης διακύμανσης. Τύπος test Ανάλυση 2 ομάδων Ανάλυση 3 ομάδων Παραμετρικό Student s t test ANOVA (ίσες διακυμάνσεις) Παραμετρικό Welch t test Welch ANOVA (ανίσες διακυμάνσεις) Μη παραμετρικό Wilcoxon Mann Whitney test Kruskal Wallis test Στο ακόλουθο παράδειγμα περιγράφεται αναλυτικά ο τρόπος ελέγχου της Ανάλυσης Διακύμανσης κατά ένα Παράγοντα. Έστω ότι έχουμε τέσσερα αντίγραφα του γονιδίου mir και τρία πειράματα μικροσυστοιχιών η έκφραση των οποίων σε κάθε πείραμα δίνεται στον πίνακα 3.2. Πίνακας 3.2: Παράδειγμα δεδομένων προς Ανάλυση διακύμανσης. Πειράματα Γονίδιο Slide1 Slide2 Slide3 mir a mir b mir c mir d Αποφάσεις ελέγχου Για να απαντήσουμε στα παραπάνω ερωτήματα αρχικά διατυπώνουμε τη μηδενική υπόθεση Η 0 η οποία ορίζει πως σε όλα τα πειράματα δεν υπάρχει στατιστικά σημαντική διαφορά της μέσης έκφρασης του γονιδίου ως H 0 : μ Slide1 = μ Slide2 = μ Slide3 και ως εναλλακτική υπόθεση H 1, ότι τουλάχιστον μία μέση τιμή της έκφρασης του γονιδίου διαφέρει μεταξύ των άλλων μέσων τιμών των πειραμάτων. 60
76 (α) μ Slide1 =μ Slide2 =μ Slide3 (β) μ Slide1 = μ Slide2 μ Slide3 ή μ Slide1 μ Slide2 μ Slide3 Εικόνα 3.2: (α) Ορισμός μηδενικής υπόθεσης: Δεν υπάρχει στατιστικά σημαντική διαφορά της μέσης τιμής έκφρασης στα πειράματα (β) Ορισμός εναλλακτικής υπόθεσης: Τουλάχιστον μία μέση τιμή έκφρασης διαφέρει σημαντικά μεταξύ των πειραμάτων. 61

77 Κατόπιν προσδιορίζεται το πλήθος των βαθμών ελευθερίας μεταξύ και εντός των ομάδων για να προσδιορίσουμε την κρίσιμη F τιμή. Οι βαθμοί ελευθερίας εκφράζουν το πλήθος των ελάχιστων ανεξάρτητων πληροφοριών προκειμένου να προσδιοριστεί μια άλλη πληροφορία. Για παράδειγμα αν γνωρίζουμε ότι το άθροισμα n μεταβλητών είναι 0 πρέπει να γνωρίζουμε τις n 1 για να προσδιοριστεί η τελευταία τιμή της μεταβλητής. df BETWEEN = k 1 = 3 1 = 2 df WITHIN = k(n 1)= N k = 12 3 = 9 Όπου k το πλήθος των πειραμάτων μικροσυστοιχιών στην περίπτωση μας, n το πλήθος των παρατηρήσεων για κάθε πείραμα και N το συνολικό πλήθος παρατηρήσεων. Οι συνολικοί βαθμοί ελευθερίας είναι: df TOTAL = df BETWEEN + df WITHIN = 2+9 = 11 O αριθμός των βαθμών ελευθερίας μεταξύ των ομάδων, είναι σύμφωνα με τον πίνακα της F κατανομής και για επίπεδο σημαντικότητας α=.05 (βλέπε πίνακα 3.3) ίσο με τον αριθμό των βαθμών ελευθερίας του αριθμητή, ενώ ο αριθμός των βαθμών ελευθερίας εντός των ομάδων είναι σύμφωνα με τον πίνακα της κατανομής ίσος με τον αριθμό των βαθμών ελευθερίας του παρονομαστή. Άρα από τη διασταύρωση της στήλης 2 με τη γραμμή 9 του πίνακα ο συνολικός αριθμός βαθμών ελευθερίας είναι F CRITICAL = Πίνακας 3.3: F κρίσιμες τιμές για επίπεδο σημαντικότητας
78 Για κάθε συνδυασμό βαθμών ελευθερίας του αριθμητή και του παρονομαστή, υπάρχει μια διαφορετική F κατανομή η οποία έχει μεγαλύτερο εύρος για μικρούς βαθμούς ελευθερίας. i) Υπολογισμός Αθροίσματος Τετραγώνων H ολική μεταβλητότητα που υπάρχει στα δεδομένα διασπάται σε επιμέρους συνιστώσες που οφείλονται σε διαφορετικές πηγές προέλευσης. Ο όρος μεταβλητότητα αναφέρεται στο άθροισμα των τετραγώνων των αποκλίσεων των παρατηρήσεων από τη μέση τιμή τους που για συντομία καλείται άθροισμα τετραγώνων. Αρχικά υπολογίζεται ο καθολικός μέσος όρος ο οποίος ισούται με: x = = 5.92 Η ανάλυση διακύμανσης διχοτομεί τη συνολική διακύμανση των δεδομένων (Sum of Squares Total SST) στην εσωτερική διακύμανση κάθε ομάδας ή πειράματος (Within groups Sum of Squares SS W ) και στη διακύμανση που υπάρχει μεταξύ των ομάδων (Between groups Sum of Squares SS B ). ii) Ολικό άθροισμα τετραγώνων Αρχικά υπολογίζεται το ολικό άθροισμα των τετραγώνων των αποκλίσεων των παρατηρήσεων από τον καθολικό μέσο. x x = (8 5.92) 2 + (7 5.92) 2 + (8 5.92) 2 + (9 5.92) 2 + (6 5.92) 2 + (6 5.92) 2 + (8 5.92) 2 + (7 5.92) 2 + (3 5.92) 2 + (4 5.92) 2 + (4 5.92) 2 + (3 5.92) 2 =
79 iii) Άθροισμα τετραγώνων εντός των ομάδων ή άθροισμα σφάλματος Εκφράζει τη μεταβλητότητα κάθε τιμής γύρω από το μέσο όρο της ομάδας που ανήκει. Υπολογίζεται αρχικά το άθροισμα των τετραγωνισμένων αποκλίσεων των παρατηρήσεων από τον δειγματικό μέσο κάθε ομάδας και κατόπιν αθροίζονται τα επιμέρους αποτελέσματα για όλες τις ομάδες. Η μέση έκφραση του γονιδίου για κάθε πείραμα είναι x 1 = 8, x 2= 6.75 και x 3 = 3. SS WITHIN = n x x = (8 8) 2 + (7 8) 2 + (8 8) 2 + (9 8) 2 + (6 6.75) 2 + (6 6.75) 2 + (8 6.75) 2 + (7 6.75) 2 + (3 3) 2 + (4 3) 2 + (4 3) 2 + (3 3) 2 = Όπου: i= μέλος ενός επιπέδου (πειράματος) j= αριθμός επιπέδου nj = πλήθος παρατηρήσεων σε ένα συγκεκριμένο επίπεδο x ij = παρατήρηση i, j x = μέση τιμή του επιπέδου j iv) Άθροισμα τετραγώνων μεταξύ των ομάδων Εκφράζει τη μεταβλητότητα εξαιτίας της ανεξάρτητης μεταβλητής. Εξετάζει τις διαφορές των μέσων όρων κάθε ομάδας υπολογίζοντας τη μεταβλητότητα (ή τετραγωνική απόκλιση) του κάθε μέσου γύρω από τον καθολικό μέσο και κατόπιν πολλαπλασιάζει το αποτέλεσμα με το πλήθος των παρατηρήσεων της ομάδας. Ορίζεται από τον τύπο: SS BETWEEN = x x Ωστόσο υπολογίζεται εύκολα αν γνωρίζουμε το άθροισμα τετραγώνων μέσα στις ομάδες και το ολικό άθροισμα τετραγώνων. SS BETWEEN = SS TOTAL SS WITHIN =

80 Η μετατροπή του αθροίσματος των τετραγώνων σε μέσα τετράγωνα (mean squares) μέσω της διαίρεσης τους με τους βαθμούς ελευθερίας επιτρέπει να καταλήξουμε σε συμπέρασμα για το αν υπάρχει σημαντικά στατιστική διαφορά μεταξύ των διαφορετικών πειραμάτων μικροσυστοιχιών. Από τον υπολογισμό της συστηματικής διαφοράς που οφείλεται στον παράγοντα: MS BETWEEN = =. = και τον υπολογισμός της τυχαίας μη συστηματική διαφοράς: MS WITHIN = =. = 1.75 Προκύπτει: F = =.. = Οι F τιμές είναι πάντα θετικές γιατί η κατανομή (η οποία είναι όμοια με την χ 2 ) είναι θετική λοξή και καθ ότι F CRITICAL < F απορρίπτεται η μηδενική υπόθεση. Γενικότερα λοιπόν αν F 1 δεν παρατηρούνται διαφορές στους μέσους όρους των ομάδων και η Η 0 απορρίπτεται (εικόνα 3.3). Περιοχή Μη Απόρριψης 90% F 0.95 = 1/F CRITICAL F 0.05 ή F CRITICAL = 4.26 Εικόνα 3.3: Περιοχή απόρριψης και αποδοχής της μηδενικής υπόθεσης. 65
81 3.2.4 Post hoc ανάλυση Σε ενδεχόμενη απόρριψη της μηδενικής υπόθεσης, γεννιέται ένα ακόμη πιο σημαντικό ερώτημα, αυτού της εύρεσης ποιοι είναι η στατιστικά σημαντικές διαφορές των μέσων τιμών μεταξύ όλων των δυνατών πειραματικών συνδυασμών (Slide1 Slide2, Slide2 Slide3, Slide1 Slide3). Η χρήση του λόγου F δεν απαντά σε αυτό το ερώτημα. Επιπλέον κατά τη σύγκριση δύο μέσων τιμών η πιθανότητα να πάρουμε ένα ψευδώς θετικό (false positive) αποτέλεσμα και να απορρίψουμε τη μηδενική υπόθεση όταν αυτή αληθεύει ονομάζεται σφάλμα Τύπου Ι (ή σφάλμα απόρριψης). Η πιθανότητα να διαπραχθεί ένα σφάλμα τύπου Ι καθορίζεται από το προκαθορισμένο επίπεδο σημαντικότητας του ελέγχου (τιμή α). Εάν πραγματοποιηθούν δηλαδή κατ επανάληψη έλεγχοι της υπόθεσης διατηρώντας το επίπεδο σημαντικότητας στο 0.05, θα απορριφθεί λανθασμένα η μηδενική υπόθεση, ενώ είναι αληθής. Για τους παραπάνω λόγους εφαρμόζονται post hoc αναλύσεις και στο συγκεκριμένο παράδειγμα το Tukey s (Honestly Significance Difference HSD) test πολλαπλών συγκρίσεων. Oυσιαστικά πρόκειται για μια σειρά επαναλαμβανόμενων t test με αυστηρότερη πιθανότητα αποδοχής ώστε να μην αυξάνεται το σφάλμα Τύπου Ι. Με τον τρόπο αυτόν επιτυγχάνεται η διεξαγωγή πολλαπλών συγκρίσεων διατηρώντας το σφάλμα Τύπου Ι σε ένα προκαθορισμένο α επίπεδο για το σύνολο των συγκρίσεων. Στη στατιστική των πολλαπλών κατά ζεύγη συγκρίσεων των μέσων τιμών συναντώνται δύο περιπτώσεις σφάλματος Τύπου Ι: i. Comparison wise Error Rate (CER) ή Family wise Error Rate σφάλμα, ορίζεται ως ο λόγος των Type I σφαλμάτων προς το συνολικό αριθμό των κατά ζεύγη συγκρίσεων. H πιθανότητα CER σφάλματος είναι μικρότερη όταν όταν το επίπεδο σημαντικότητας είναι α=0.10 αντί 0.5 και αν η εσφαλμένη απόρριψη ενός σφάλματος σε ένα πείραμα δεν επηρρεάζει τα άλλα συμπεράσματα στο πείραμα, τότε ο υπολογισμός του CER είναι πιο σημαντικός. ii. Experimentwise Error Rate (EER) σφάλμα και εκφράζει το λόγο των πειραμάτων στα οποία εμφανίζονται περισσότερα του ενός Type I σφάλματα προς το συνολικό αριθμό πειραμάτων. Συνήθως για έως 10 συγκρίσεις εφαρμόζεται το κριτήριο του Bonferroni τo οποίο ελέγχει για CER και σπανιότερα το κριτήριο του Tukey ή του Scheffe. 66
82 Το EER σφάλμα είναι δύσκολο να υπολογιστεί στην περίπτωση του τα σφάλματα δεν είναι ανεξάρτητα, αλλά είναι δυνατό να υπολογιστεί ένα ανώτατο όριο για το EER θεωρώντας ότι η πιθανότητα σφάλματος α σε μία σύγκριση είναι ανεξάρτητη από τις άλλες συγκρίσεις. Ένας από τους τρόπους με το οποίο συμβολίζεται το experimentwise σφάλμα είναι ως: α e, όπου α e = 1 (1 α) c To Τukey τεστ ελέγχει το maximum experiment wise error rate (MEER) τύπο σφάλματος. Το επίπεδο σημαντικότητας α ορίζεται για κάθε σύγκριση έτσι ώστε το ενδεχόμενο των false positives στο σύνολο των c κατά ζεύγη συγκρίσεων (c=k(k 1)/2) το α e διατηρείται στο μέγιστο επίπεδο του 0.05 και η μεταξύ τους σχέση ορίζεται ως: α 1 1 H εύρεση των ζευγών των μέσων τιμών οι τιμές των οποίων διαφέρουν σημαντικά γίνεται με τη σύγκριση της απόλυτης διαφοράς τους με την HSD (Honestly Significance Difference) κρίσιμη τιμή η οποία υπολογίζεται από τον ακόλουθο τύπο: q, όπου q η τιμή του πίνακα της Q κατανομής Από τον πίνακα αυτό για προκύπτει ότι q=3.95 (βλ. πίνακα 3.4) και άρα HSD = 1.31 Συγκρίνοντας τώρα την απόλυτη διαφορά των μέσων τιμών x 1, x 2 και x 3 των τριών πειραμάτων με τη HSD τιμή προκύπτει: x 1 x 2 = = 1.25 < HSD = 1.31 x 1 x 3 = 8 3 = 5 > HSD = 1.31 x 2 x 3 = = 3.75 > HSD = 1.31 Συμπεραίνεται ότι στατιστικά σημαντική διαφορά υπάρχει μεταξύ του 1 ου και του 3 ου πειράματος καθώς και μεταξύ 2 ου και 3 ου. 67
83 Πίνακας 3.4: Studentized Range q πίνακας 3.3 Μέθοδοι ανάλυσης υψηλού επιπέδου Η διατύπωση συμπερασμάτων βιολογικής φύσεως όπως η γονιδιακή λειτουργία, ανάλυση μονοπατιού (pathway analysis), αλλαγές στη γονιδιακή έκφραση στο χρόνο, κ.λπ., εμπίπτουν στην κατηγορία της ανάλυσης υψηλού επιπέδου. Μια σύντομη περιγραφή των μεθόδων που χρησιμοποιούνται για το σκοπό αυτό στην παρούσα διπλωματική παρατίθενται παρακάτω Συντελεστής συσχέτισης Pearson Correlation Coefficient (PCC) O συντελεστής συσχέτισης Pearson Correlation Coefficient (r value) (Pearson, 1895)αποτελεί μέτρο του βαθμού της γραμμικής συσχέτισης μεταξύ των τιμών δύο ομάδων Χ, Υ. Το πεδίο ορισμού της r είναι: 1 r 1 Θετικές r τιμές υποδηλώνουν θετική γραμμική συσχέτιση των τιμών, ενώ όσο πιο αρνητικές είναι τόσο πιο αρνητική είναι και η συσχέτισή τους. Για r=0 δεν υπάρχει καμία γραμμική συσχέτιση των τιμών. Παρατίθεται το ακόλουθο παράδειγμα υπολογισμού της r value. Για το σύνολο των τιμών των ομάδων Χ και Υ που βλέπουμε στον πίνακα 3.5 υπολογίζεται: 68
84 Πίνακας 3.5: Χ Υ ΧΥ Χ 2 Υ Σ ΣΧ ΣΥ ΣΧΥ Χ 2 Υ 2 r= False Discovery Rate (FDR) Η μέθοδος του ρυθμού ψευδών ανακαλύψεων (False Discovery Rate FDR) (Benjamini Hochberg, 1995) είναι εναλλακτική της FWER μεθόδου, για την προσαρμογή της τιμής του p value. Σε παραδοσιακές τεχνικές όπως η διόρθωση Bonferroni (FWER) όταν ο αριθμός των τεστ ανέρχεται σε χιλιάδες, αυτές γίνονται πολύ συντηρητικές, με την έννοια ότι μειώνοντας τον αριθμό των ψευδοθετικών ευρημάτων μειώνεται ταυτόχρονα και ο αριθμός των αληθών ευρημάτων. Η ΒΗ FDR, είναι λιγότερο συντηρητική, γι αυτό έχει μεγαλύτερη ικανότητα εύρεσης πραγματικά αληθών αποτελεσμάτων. Παρακάτω, παρατίθεται αναλυτικό παράδειγμα της διαδικασίας με την οποία γίνεται η διόρθωση FDR στις τιμές των p values (πίνακας 3.6): Oι p values τιμές, ταξινομούνται κατά αύξουσα σειρά. Θέτουμε k, τη σειρά στην οποία βρίσκεται η ταξινομημένη τιμή του p value και Ν=5 το πλήθος των p values. Για κάθε p value υπολογίζεται το N p value/k. Θέτουμε μια προσωρινή τιμή tmp=1 για το μεγαλύτερο p value. Ξεκινώντας από την προσωρινή τιμή 1, η διορθωμένη τιμή του, adj p value, είναι η μικρότερη τιμή μεταξύ του γινομένου N p value/k και της προσωρινής τιμής. Μετά το τέλος κάθε διόρθωσης, προσωρινή τιμή γίνεται η διορθωμένη. 69
85 Πίνακας 3.6: p value Ascending p value N p value / k tmp ΒΗ adj. p value Υπεργεωμετρική κατανομή Η διαφορική έκφραση των γονιδίων από μόνη της δε δίνει πολλές πληροφορίες, γι αυτό συνδυάζουμε όρους γονιδιακής οντολογίας (Gene Ontology GO) (Ashburner et al., 2000) και τη βάση δεδομένων KEGG (Kyoto Encyclopedia of Genes and Genomes) (Du et al., 2014), για την κατηγοριοποίηση των γονιδίων σε ομάδες που θα μας δώσουν τις πληροφορίες που χρειαζόμαστε. Οι όροι γονιδιακής οντολογίας περιγράφουν λειτουργίες γονιδίων οι οποίες μπορεί να συμβαίνουν σε διαφορετικό βαθμό εξαιτίας διαφορετικών συνθηκών. Σε κάθε γονιδιακό όρο, ανατίθεται ένα σύνολο γονιδίων. Η βάση δεδομένων KEGG ταξινομεί τα γονίδια, ανάλογα με το βιολογικό μονοπάτι στο οποίο ανήκουν. Η πιο κοινή μέθοδος για την εύρεση της υπερεκπροσώπησης ή μη ενός όρου γονιδιακής οντολογίας, είναι η υπεργεωμετρική κατανομή. Η υπεργεωμετρική κατανομή είναι μια διακριτή συνάρτηση κατανομής τυχαίας μεταβλητής. Περιγράφει ένα τυχαίο πείραμα με δυο πιθανά αποτελέσματα (επιτυχία αποτυχία) σε πεπερασμένο πληθυσμό που επαναλαμβάνεται n φορές χωρίς επανατοποθέτηση. Παρακάτω παρατίθεται ένα παράδειγμα για την κατανόηση της υπεργεωμετρικής κατανομής: Aπό ένα σύνολο 50 γονιδίων, τα 20 σχετίζονται με το ErbB μονοπάτι σήμανσης και τα 30 όχι. Αν επιλεχθούν τυχαία 10 γονίδια ποια η πιθανότητα ακριβώς 5 γονίδια να σχετίζονται με το ErbB; Όπου Ν=50, Μ=20, n=10, k=5 και άρα P(5)=
86 3.4 Βιβλιογραφία Ashburner, M., Ball, C. A., Blake, J. A., Botstein, D., Butler, H., Cherry, J. M.,... Sherlock, G. (2000). Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet, 25(1), doi: /75556 Du, J., Yuan, Z., Ma, Z., Song, J., Xie, X., & Chen, Y. (2014). KEGG PATH: Kyoto encyclopedia of genes and genomes based pathway analysis using a path analysis model. Mol Biosyst, 10(9), doi: /c4mb00287c Pearson, K. (1895). Notes on regression and inheritance in the case of two parents. Proceedings of the Royal Society of London, Quackenbush, J. (2002). Microarray data normalization and transformation. Nat Genet, 32 Suppl, doi: /ng1032 Ritchie, M. E., Silver, J., Oshlack, A., Holmes, M., Diyagama, D., Holloway, A., & Smyth, G. K. (2007). A comparison of background correction methods for two colour microarrays. Bioinformatics, 23(20), doi: /bioinformatics/btm412 Smyth, G. K., & Speed, T. (2003). Normalization of cdna microarray data. Methods, 31(4), Yang, Y. H., Buckley, M. J., & Speed, T. P. (2001). Analysis of cdna microarray images. Brief Bioinform, 2(4), Yang, Y. H., Dudoit, S., Luu, P., Lin, D. M., Peng, V., Ngai, J., & Speed, T. P. (2002). Normalization for cdna microarray data: a robust composite method addressing single and multiple slide systematic variation. Nucleic Acids Res, 30(4), e15. 71
87 Κεφάλαιο 4 ο Μέθοδοι 72
88 4.1 Περιγραφή βιολογικών δειγμάτων Το σύνολο των βιολογικών δειγμάτων που χρησιμοποιήθηκε στην παρούσα ανάλυση μικροσυστοιχιών mirnas, διακρίνεται σε δεκατέσσερις κυτταρικές σειρές σύμφωνα με το τεχνητό πολυσταδιακό μοντέλο δερματικής καρκινογένεσης που αναπτύχθηκε στο εργαστήριο του Dr. A. Balmain. Όλες οι κυτταρικές σειρές πλην της C5N προέρχονται από όγκους αρχικοποιημένους in vivo που αναπαριστούν διαφορετικά στάδια της δερματικής καρκινογένεσης, μετά από δερματική επάλειψη με τον κυκλικό αρωματικό υδρογονάνθρακα DMBA (7,12 διμεθυλοβενζο [α] ανθρακένιο). Τα στάδια αυτά κατατάσσονται στις εξής ομάδες: Aθανατοποιημένα (immortalized) κύτταρα. Καλοήθη θηλώματα (benign papillomas). Κακοήθη πλακώδη κύτταρα (malignant squamous cells). Κακοήθη ατρακτοκυτταρικά (malignant spindle cells). Στον πίνακα 4.1 απαριθμούνται οι προς σύγκριση δεκατέσσερις κυτταρικές σειρές της παρούσας ανάλυσης μικροσυστοιχιών. Αναφέρεται ότι η κυτταρική σειρά PDVC57 αναπαριστά την κλωνική επέκταση της PDV, ενώ οι Β9, A5 και D3 κυτταρικές σειρές προέρχονται από τον ίδιο αρχικό όγκο αναπαριστώντας την εξέλιξη στα πιο επιθετικά στάδια (Zoumpourlis et al., 2003). Πίνακας 4.1: Κυτταρικές σειρές και κατηγοριοποίηση τους σε ομάδες Slide Κυτταρική σειρά 1 C5N 2 H ras null 3 P1 4 P6 5 MSP5 6 PDV 7 PDVC57 8 B9 9 E4 10 A5 11 D3 12 H11 13 CarB 14 CarC Κατηγοριοποίηση κυτταρικών σειρών Αθανατοποιημένα κύτταρα Καλοήθη θηλώματα Κακοήθη πλακώδη κύτταρα Κακοήθη ατρακτοκυτταρικά 73
 η οποία είναι σχεδιασμένη τόσο για διχρωματική ανάλυση Hy3 TM /Hy5 ΤΜ όσο και για μονοχρωματική ανάλυση διαφορικής έκφρασης. Όπως απεικονίζεται στην εικόνα 4.")
 αποτελείται από 18 γραμμές και 19 στήλες και άρα 18x19 θα είναι το πλήθος των spots σε κάθε block. Εικόνα 4.1: Διαστάσεις mircury LNA TM microrna μικροσυστοιχίας Εικόνα 4.")
89 4.2 Exiqon mircury LNA TM microrna array kit Για την ανάλυση της γονιδιακής έκφρασης των mirnas χρησιμοποιήθηκε η πλατφόρμα μικροσυστοιχιών Exiqon mircury LNA TM microrna array kit (6 th generation με αριθμό προϊόντος ) η οποία είναι σχεδιασμένη τόσο για διχρωματική ανάλυση Hy3 TM /Hy5 ΤΜ όσο και για μονοχρωματική ανάλυση διαφορικής έκφρασης. Όπως απεικονίζεται στην εικόνα 4.1 οι μικροσυστοιχίες αυτού του τύπου αποτελούνται από 36 επιμέρους blocks με spots διαμέτρου 100 μm και απόσταση μεταξύ αυτών 215 μm. Κάθε ένα από αυτά τα 36 blocks (εικόνα 4.2) αποτελείται από 18 γραμμές και 19 στήλες και άρα 18x19 θα είναι το πλήθος των spots σε κάθε block. Εικόνα 4.1: Διαστάσεις mircury LNA TM microrna μικροσυστοιχίας Εικόνα 4.2: Διάταξη mircury LNA TM microrna μικροσυστοιχίας 74
90 4.2.1 Περιγραφή ανιχνευτών mircury LNA TM microrna Array H πλατφόρμα της Exiqon ξεχωρίζει εξαιτίας των βελτιωμένων με απροσπέλαστο RNA (LNA TM ) ανιχνευτών. Τα DNA ή RNA ολιγονουκλεοτίδια που είναι τροποποιημένα με LNA ΤΜ παρουσιάζουν: Αυξημένη συνάφεια και εξειδίκευση (ανίχνευση μονονουκλεοτιδικής αναντιστοιχίας). Πρωτοφανή θερμική σταθερότητα κατά τον υβριδισμό τους με DNA ή RNA. Ίδια επίπεδα θερμικής σταθερότητας όταν το μήκος τους είναι μικρότερο των παραδοσιακών DNA ή RNA ανιχνευτών. Παρόμοια εξειδίκευση με όλες τις ακολουθίες ανεξαρτήτου του GC περιεχομένου τους μεταβάλλοντας το περιεχόμενο των ανιχνευτών σε LNA και άρα να ανίχνευση όλων των mirnas. Κάθε slide περιέχει probes με 4 αντίγραφα ανιχνευτών το κάθε ένα. Οι ανιχνευτές αυτοί που είναι συμπληρωματικοί όλων των mirnas του ανθρώπου, του ποντικού και του αρουραίου, είναι διαθέσιμοι από τη βάση δεδομένων mirnas mirbase 19.0 (Kozomara and Griffiths Jones, 2011). Επιπλέον περιέχει ανιχνευτές ανθρώπινων mirnas που δεν περιέχονται στη mirbase. Στο σύνολο αυτών των ανιχνευτών περιέχονται επιπλέον θετικοί και αρνητικοί ανιχνευτές ελέγχου (πίνακας 4.2), spike in ανιχνευτές (πίνακας 4.3), ανώνυμα spots των οποίων η αλληλουχία αποτελεί πνευματική ιδιοκτησία της Exiqon καθώς και κενά spots όπου δεν υπάρχει ανιχνευτής σε αυτά (εικόνα 4.3). Κάθε ένα από τα 36 blocks διαθέτει 5 ανιχνευτές (ένα σε κάθε γωνία και δύο στην κάτω δεξιά γωνία) οι οποίοι σημαίνονται απευθείας με Hy3 TM. Ο ρόλος αυτών των 180 συνολικά ανιχνευτών περιορίζεται στη διατήρηση του σωστού προσανατολισμού της μικροσυστοιχίας (εικόνα 4.4). 75


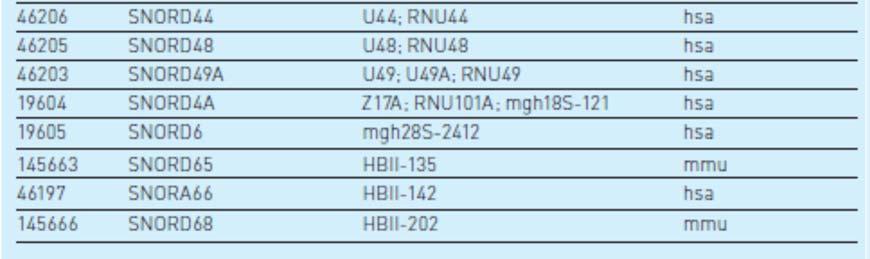
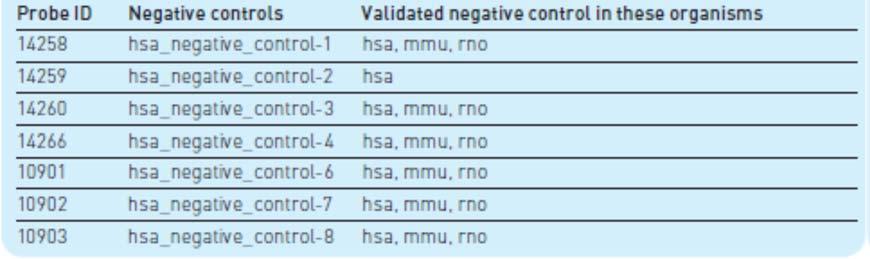
91 Πίνακας 4.2: Λίστα θετικών και αρνητικών ανιχνευτών ελέγχου 76




92 Πίνακας 4.3: Λίστα ανιχνευτών ελέγχου spike in mirna kit v2 και τα ID s αυτών Εικόνα 4.3: Κενά spots μικροσυστοιχιών 77

93 Εικόνα 4.4: Οι θέσεις των spots που καταλαμβάνουν οι spike in ανιχνευτές επί των blocks. 4.3 Exiqon s mircury LNA TM Hi Power Labelling Kit Σε κάθε ένα από τα δεκατέσσερα πλακίδια μικροσυστοιχιών, το προς εξέταση δείγμα σημαίνεται με χρωστική Hy3 TM και το δείγμα αναφοράς με Hy5 TM. Οι χρωστικές αυτές είναι φασματοσκοπικά ισοδύναμες με τις Cy3 και Cy5. H σήμανση των ανιχνευτών γίνεται με το Hi Power Labelling Kit της Exiqon, το οποίο δίνει διπλάσια ένταση σήματος σε σχέση με το standard labelling kit. Αυτό πρακτικά σημαίνει την ανίχνευση περισσότερων mirnas με την ίδια ποσότητα δείγματος RNA ή ότι μπορεί να χρησιμοποιηθεί σχεδόν η μισή ποσότητα δείγματος για την ανίχνευση mirnas χωρίς να υποβαθμίζεται η ποιότητα των αποτελεσμάτων του πειράματος. 4.4 Πειραματικός σχεδιασμός Ο πειραματικός σχεδιασμός που ακολουθείται από την παρούσα ανάλυση είναι βάσει ενός κοινού δείγματος αναφοράς (common reference) αφού τα βιολογικά προς σύγκριση δείγματα είναι πολλά. 4.5 Αρχεία πρωτογενών δεδομένων Από τη σάρωση των δεκατεσσάρων μικροσυστοιχιών προκύπτει πλήθος 16 bit TIFF αρχείων, τα σχετικά επίπεδα έκφρασης των οποίων ποσοτικοποιούνται με το λογισμικό ImaGene 9 78
94 (Biodiscovery) που προσφέρεται από την Exiqon. Το ImaGene 9 υπολογίζει τη μέση τιμή και τη διάμεσο του σήματος, και του σήματος υποβάθρου για κάθε spot κάθε καναλιού. Από αυτό παράγοντα είκοσι οκτώ αρχεία (πίνακας 4.4), δεκατέσσερα για κάθε κανάλι. Επιπλέον των σχετικών τιμών έκφρασης τα αρχεία αυτά περιέχουν και άλλες πληροφορίες, όπως το πλήθος των blocks της μικροσυστοιχίας και το πλήθος των spot κάθε block. Πίνακας 4.4: Παραγωγή αρχείων πρωτογενών δεδομένων από το ImaGene 9 (Biodiscovery) Φθορίζουσα χρωστική Hy3 Hy5 Αρχείο πρωτογενών δεδομένων 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 1_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt 0_Exiqon_ _S01.txt Κυτταρική σειρά C5N H ras null P1 P6 MSP5 PDV PDVC57 B9 E4 A5 D3 H11 CarB CarC C5N H ras null P1 P6 MSP5 PDV PDVC57 B9 E4 A5 D3 H11 CarB CarC Ομάδα A B C D A B C D 79
95 4.6 Ανάλυση χαμηλού επιπέδου Στο στάδιο αυτό επεξεργαζόμαστε τα πρωτογενή δεδομένα, ώστε να έχουν την κατάλληλη μορφή για τη στατιστική ανάλυση που πρέπει να ακολουθηθεί. Αναλυτικότερα τα βήματα που ακολουθούνται είναι: Εγκατάσταση R και εγκατάσταση Limma/Bioconductor Η ανάλυση των πρωτογενών δεδομένων έγινε με τη γλώσσα στατιστικού προγραμματισμού R η οποία είναι διαθέσιμη στη διεύθυνση: project.org/index.html. Έπειτα απαιτείται η εγκατάσταση της σουίτας λογισμικών Bioconductor (Gentleman et al., 2004), χρήσει της εντολής R: source(" Το Bioconductor επιτρέπει την εγκατάσταση επιμέρους πακέτων η υλοποίηση και διάθεση των οποίων γίνεται από διάφορες ερευνητικές ομάδες ανά τον κόσμο. Κάθε ένα από τα πακέτα αυτά προορίζεται για διαφορετικές στατιστικές αναλύσεις, ή αναλύσεις που υλοποιούνται με διαφορετική μεθοδολογία. Για την ανάλυση χαμηλού επιπέδου χρησιμοποιήθηκε το πακέτο Limma (Ritchie et al., 2015). Η εγκατάσταση αυτού γίνεται στο τερματικό της R με την εντολή: bioclite("limma") Εναλλακτικά το Limma εγκαθίσταται επιλέγοντας Packages Install package(s) από τη γραφική διεπαφή της R. Κάθε φορά πριν τη χρήση του πακέτου Limma απαιτείται η φόρτωση του, ακόμη και αν αυτό ήδη εγκατεστημένο: library(limma) 80
 Εν συνεχεία, διαβάζεται το αρχείο Targets.txt.")
96 4.6.2 Ανάγνωση πρωτογενών δεδομένων Αρχικά απαιτείται ο ορισμός του path του φακέλου εργασίας στον οποίο βρίσκονται τα πρωτογενή δεδομένα προς ανάλυση. setwd("c:/users/ /Raw data files") Εν συνεχεία, διαβάζεται το αρχείο Targets.txt. Πρόκειται για ένα tab delimited αρχείο το οποίο περιγράφει ποιο από τα 28 αρχεία των πρωτογενών δεδομένων που προκύπτουν από την ανάλυση των TIFF εικόνων με το ImaGene, συσχετίζεται με ποια φθορίζουσα χρωστική (εικόνα 4.5). Το αρχείο μπορεί να ονομαστεί με οποιοδήποτε όνομα αλλά το προκαθορισμένο όνομα που διαβάζει η συνάρτηση readtargets() είναι Targets.txt. Διαφορετικά, ορίζεται το όνομά του μέσα, ως παράμετρος της συνάρτησης: targets < readtargets() Από την ανάγνωση του Targets.txt δημιουργείται το targets data frame, η έξοδος του οποίου φαίνεται στην εικόνα 4.6. Εικόνα 4.5: Μορφή αρχείου Targets.txt 81

 με την εντολή: files < targets[,c(\"filenamecy5\",\"filenamecy3\")] ή files <")
97 Εικόνα 4.6: Targets data frame έξοδος Targets.txt στο τερματικό Από το targets data frame, κρατώντας μόνο τις δύο τελευταίες στήλες, δημιουργείται ένα character vector (εικόνα 4.7) με την εντολή: files < targets[,c("filenamecy5","filenamecy3")] ή files < targets[,c(2:3)] Εικόνα 4.7: Έξοδος data frame files στο τερματικό 82
98 Οι εντάσεις σήματος και υποβάθρου (και άλλες πληροφορίες), διαβάζονται από τα αρχεία που προκύπτουν με το λογισμικό ανάλυσης των εικόνων ImaGene και αποθηκεύονται σε αντικείμενα της RG λίστας η οποία δημιουργείται με τη μέθοδο read.maimages(), ως εξής: RG < read.maimages(files, source="imagene", path=" C:/Users/ /Raw data files ") Από τoν ιστότοπο της Exiqon, επιλέγουμε το πλέον πρόσφατο tab delimited αρχείο GAL (GenePix Array List) το οποίο περιέχει τα IDs κάθε ανιχνευτή και πληροφορίες σχετικά με τη σειρά, τη στήλη και το block στο οποίο ανήκουν. Το περιεχόμενο του αρχείου αυτού αντικαθιστά το RG$genes data frame του αντικειμένου RG που είχε οριστεί με την προηγούμενη εντολή, διότι το ID ενός γονιδίου πιθανόν να αλλάξει σε μια νεότερη έκδοση της mirbase: RG$genes< readgal() Το πιο πρόσφατο GAL αρχείο γίνεται διαθέσιμο στη διεύθυνση: downloads επιλέγοντας τα φίλτρα που ορίζονται στην εικόνα 4.8. Εικόνα 4.8: Επιλογή παραμέτρων για τη λήψη του κατάλληλου GAL αρχείου Διόρθωση υποβάθρου Η μέθοδος διόρθωσης υποβάθρου που εφαρμόζεται στην παρούσα ανάλυση είναι η normexp+offset (Ritchie et al., 2007; Shi et al., 2010), επιλέγοντας εμπειρικά offset=10. RG < backgroundcorrect(rg, method="normexp", offset=10) 83
99 Οπτική αναπαράσταση γονιδιακής έκφρασης (ΜΑplots) Η γραφική απεικόνιση των εντάσεων μετά τη διόρθωση υποβάθρου, μπορεί να γίνει με έγχρωμα MA plots, στα οποία κάθε τύπος ανιχνευτή έχει διαφορετικό χρώμα, ώστε να διακρίνονται μεταξύ τους. Αρχικά απαιτείται η δημιουργία ενός data frame στο οποίο τα probe sets κατηγοριοποιούνται ανάλογα με τον τύπο τους και τα IDs τους στις επιμέρους ομάδες: 5S_rRNA, u6b, mmu let, mmu mir, mghv mir, mcmv mir, Hy3, spike_control, blank, negative, empty, snord και RNU, μέσω pattern matching κανονικών εκφράσεων. Σε κάθε μία από αυτές, ορίζεται και ένα διαφορετικό χρώμα με το οποίο θα απεικονίζονται τα spots στα MAplots. types < data.frame(spottype=c("5s_rrna","u6b","mmu let","mmu mir","mghv mir","mcmvmir","hy3","spike_control","blank","negative","empty","snord","rnu"),id=c("46202","46207","*","*","* ","*","13138","*","*","*","0","*","*"),Name=c("5S_rRNA","U6B","mmu let *","mmu mir *","mghv mir *","mcmv mir *","Hy3","spike_control_*","","hsa_negative_control_*","Empty","SNOR*","RNU*"),col=c("orange","cya n","black","black","brown","blue","green","red","yellow","magenta","pink","purple","grey")) To data frame που δημιουργήθηκε, αποτελείται από τα vectors SpotType, ID, Name και col, όπως φαίνεται στην εντολή που προηγείται αλλά και σε αυτή που έπεται: objects(types) H έξοδος της οποίας είναι: [1] "col" "ID" "Name" "SpotType" RG$genes$Status< controlstatus(types,rg) Το data frame RG$genes που δημιουργείται με τη μέθοδο controlstatus(), μεταξύ άλλων περιέχει ένα vector (RG$genes$Status) στο οποίο ορίζεται ο τύπος κάθε ανιχνευτή (εικόνα 4.9). Επίσης, περιέχει ένα vector με το ID του κάθε ανιχνευτή. 84
 (βλ. Παράρτημα I.1), με την ακόλουθη εντολή: plotma(rg, array=i) για i=1,2,3,,14) 4.6.")
100 Εικόνα 4.9: Προσδιορισμός πληθυσμού κάθε τύπου ανιχνευτών στη μικροσυστοιχία από το RG$genes$Status chatacter vector. Τα έγχρωμα ΜΑ plots προκύπτουν για κάθε μία από τις δεκατέσσερεις μικροσυστοιχίες (array=1, array=2,, array=14) (βλ. Παράρτημα I.1), με την ακόλουθη εντολή: plotma(rg, array=i) για i=1,2,3,,14) Κανονικοποίηση εντός των μικροσυστοιχιών Μετά τη διόρθωση υποβάθρου, ακολουθεί η κανονικοποίηση των τιμών των εντάσεων εντός των δεκατεσσάρων μικροσυστοιχιών, με τη μέθοδο normalizewithinarrays. Για την κανονικοποίηση, η normalizewithinarrays απαιτεί τον ορισμό του data frame layout, τα vectors του οποίου ορίζουν τις διαστάσεις και το πλήθος των grids της μικροσυστοιχίας. layout < list(ngrid.r=rg$printer$ngrid.r, ngrid.c=rg$printer$ngrid.c, nspot.r=rg$printer$nspot.r, nspot.c=rg$printer$nspot.c) H μέθοδος κανονικοποίησης που επιλέχτηκε στην παρούσα ανάλυση, είναι η composite loess (Oshlack et al., 2007) η οποία βασίζεται στους spike controls ανιχνευτές. To σύνολο των spike controls το παίρνουμε με τη μέθοδο grep. H grep ψάχνει όλα τα στοιχεία του vector RG$genes για το pattern spike_control : 85
101 SC < grep("spike_control", RG$genes[,"Name"]) ΜA < normalizewithinarrays(rg, layout, method="composite", controlspots=sc) Ομοίως, μπορούμε να δούμε το αποτέλεσμα της κανονικοποίησης με έγχρωμα MA plots (βλ. Παράρτημα Ι.2), με την ίδια όπως προηγουμένως εντολή: plotma(rg, array=i) για i=1,2,3,,14) Εφικτή είναι η δημιουργία όλων των ΜΑ plots ταυτοχρόνως σε μικρότερη κλίμακα με μία εντολή (βλ. Παράρτημα Ι.3): plotma3by2(rg) Επίσης η ποιότητα της κανονικοποίησης αναπαρίσταται γραφικά με boxplots (βλ. Παράρτημα Ι.4) Κανονικοποίηση μεταξύ των μικροσυστοιχιών Την κανονικοποίηση εντός των μικροσυστοιχιών ακολουθεί η κανονικοποίηση μεταξύ των μικροσυστοιχιών. Η μέθοδος που εφαρμόζεται στην παρούσα ανάλυση είναι η Aquantile (Bolstad et al., 2003; Smyth and Speed, 2003), χωρίς να ληφθούν υπ όψιν τα spots των αρνητικών controls, spike in, blank, Empty και Hy3 ανιχνευτών που ορίζονται και στην εντολή που ακολουθεί: MA < MA[!( (MA$genes$Name =="Empty") (MA$genes$Name =="Hy3") (substr(ma$genes$name,1,12)=="hsa_negative") (substr(ma$genes$name,1,14)=="spike_control_") (MA$genes$Name=="") ),] MA< normalizebetweenarrays(ma, method="aquantile") plotma(rg, array=i) για i=1,2,3,,14) plotma3by2(rg) 86
102 Η μέθοδος write.csv εξάγει ένα comma separated αρχείο το οποίο περιέχει κανονικοποιημένες τις τιμές των εντάσεων για 4412 ανιχνευτές στα 14 slides. Επίσης, περιέχει πληροφορίες για τη θέση των ανιχνευτών στα πλέγματα. write.csv(ma, "MA.csv", row.names=false) H μέθοδος readtable διαβάζει ένα αρχείο σε μορφή πίνακα και δημιουργεί ένα data frame από αυτό. H μεταβλητή header παίρνει τη λογική τιμή TRUE, διαφορετικά η R προσθέτει μία επιπλέον γραμμή, δίνοντας ένα όνομα σε κάθε στήλη. MAtable < read.table("ma.csv", header=true, ",") Από το data frame ΜAtable δημιουργείται ένα νέο data frame το οποίο περιλαμβάνει μόνο τα ονόματα των probe sets και τις κανονικοποιημένες εντάσεις τους. MAnova < MAtable[5:19] Κατόπιν, ταξινομούμε το data frame αλφαβητικά ως προς τα probe sets. ordered_probes < MAnova[order(MAnova$Name),] Από την εξαγωγή ενός αρχείου μορφής csv, φαίνεται η μέχρι τώρα μορφοποίηση των κανονικοποιημένων δεδομένων. write.csv(ordered_probes, "ordered_probes.csv", row.names=false) Η κανονικοποίηση μεταξύ των μικροσυστοιχιών αναπαρίσταται γραφικά στο Παράρτημα Ι.5, Ι.6, Ι.7 με ΜΑplots και boxplots αντιστοίχως. 4.7 Ανάλυση υψηλού επιπέδου Για την εύρεση της διαφορικής έκφρασης των mirnas χρησιμοποιείται το λογισμικό SPSS: 01.ibm.com/software/analytics/spss/ το οποίο απαιτεί την εισαγωγή των κανονικοποιημένων δεδομένων σε longform format. 87
103 Με τον ίδιο τρόπο που εγκαταστάθηκε το πακέτο limma, εγκαθίσταται και φορτώνεται το πακέτο reshape2 (Wickham, 2007): source(" bioclite("reshape2") library(reshape2) Από το αρχείο ordered_probes.csv, δημιουργείται το παρακάτω data frame: data.r < read.table("c:/users/ /Raw data files/ordered_probes.csv", header=t, sep=",", na.strings="na", dec=".") Κατόπιν με τη συνάρτηση melt του πακέτου reshape τα δεδομένα διαμορφώνονται υπό την μορφή των εξής τριών στηλών: Name, η οποία αναφέρεται στο όνομα του ανιχνευτή. Variable, η οποία περιγράφει τα ονόματα των δεκατεσσάρων μικροσυστοιχιών. Value, η οποία εκφράζει την κανονικοποιημένη τιμή έντασης του ανιχνευτή. H στήλη με την ονομασία variable μπορεί να αλλάξει στην επιθυμητή ονομασία μέσω της παραμέτρου variable_name. longform < melt(data.r,id=c("name"), variable_name="array_slide", na.rm=true) Τέλος, γίνεται αλφαβητική ταξινόμηση των ανιχνευτών. longform < longform[order(longform$name),] Η εξ ορισμού τιμή της παραμέτρου na.rm είναι FALSE. Με τον ορισμό της σε TRUE αποφεύγεται πιθανή εμφάνιση επιπλέον κελιών με NA (missing values) τιμές. na.rm=true, αν παραλειφθεί εμφανίζει επιπλέον κελιά με τιμή ΝΑ (missing values) write.csv(longform, "longform.csv", row.names=false) 1. 1 Στο longform.csv αντικαθίσταται το όνομα του γονιδίου mmu mir 466a 3p/mmu mir 466b 3p/mmu mir 466c 3p/mmu mir 466e 3p/mmu mir 466p 3p με mmu mir 466p 3p, διότι στην ανάλυση που ακολουθεί το μέγιστο εύρος ενός String είναι 64 χαρακτήρες, διαφορετικά προκύπτει σφάλμα Επίσης, αλλάζουμε τη μεταβλητή Name σε mir_name. 88
. O παράγοντας αυτός είναι η ανεξάρτητη μεταβλητή array_slide.")
104 4.7.1 Ανάλυση διακύμανσης και εύρεση διαφορικά εκφρασμένων mirnas Για τη διερεύνηση της ύπαρξης στατιστικά σημαντικής διαφοράς μεταξύ των μέσων τιμών των εντάσεων κάθε γονιδίου στις δεκατέσσερις κυτταρικές σειρές, απαιτείται η εφαρμογή ανάλυσης διακύμανσης κατά ένα παράγοντα (Analysis of Variance ANOVA). O παράγοντας αυτός είναι η ανεξάρτητη μεταβλητή array_slide. H μορφή των δεδομένων στο αρχείο longform.csv ομοιάζει με το σχεδιασμό για δύο ανεξάρτητες μεταβλητές ( array_slide, mir_name ), ο οποίος απαιτεί την ανάλυση διακύμανσης κατά δύο παράγοντες. Προκειμένου να εφαρμοστεί ανάλυση διακύμανσης κατά ένα παράγοντα για κάθε σύνολο ανιχνευτών (probe set) επί των 4412 ανιχνευτών, δημιουργούμε υποσύνολα επιλέγοντας από τη διεπαφή του SPSS: Data Split File Organize output by groups Groups Based on: mir_name (εικόνα 4.10) Κατόπιν επιλέγεται: Analyze General Linear Model Dependent Variable: value & Fixed Factor(s): mir_name, array_slide (εικόνα 4.11). Tέλος από την ίδια διεπαφή, επιλέγουμε (εικόνα 4.11): Options Tukey Post Hoc Tests for: array_slide (εικόνα 4.12). Εικόνα 4.10: Εφαρμογή ANOVA σε κάθε ένα σύνολο ανιχνευτών (probe set). 89


105 Εικόνα 4.11: Ορισμός εξαρτημένων και ανεξάρτητων μεταβλητών στο SPSS. Εικόνα 4.12: Επιλογή του τεστ πολλαπλών συγκρίσεων Tukey για τη μεταβλητή array_slide. 90
106 Τα αποτελέσματα της ανάλυσης αποθηκεύονται στο SPSS_output.spv αρχείο και μετέπειτα εξάγονται σε αρχείο μορφής excel (SPSS_output.xlsx) με ταυτόχρονη μορφοποίηση των δεδομένων της πέμπτης στήλης σε scientific με δύο δεκαδικά ψηφία (δεξί κλικ format cells). Εν συνεχεία, τα δεδομένα της ανάλυσης εξάγονται εκ νέου στο SPSS_output.txt, ένα ιδιότυπο tab delimited αρχείο. Από αυτό, ενδιαφέρουν οι πληροφορίες (I) array_slide, Mean Difference (I J) και Sig. από τις οποίες φαίνεται η μέση διαφορά έκφρασης κάθε συνόλου ανιχνευτών (mir) στις 91 ανά ζεύγη συγκρίσεις των 14 κυτταρικών σειρών και το p value αυτών. Τα δεδομένα αυτά είναι διαθέσιμα στο αρχείο ExiqonmiRs.txt μετά από επεξεργασία του SPSS_output.txt με το 14_cell_lines.php script Παρουσίαση αποτελεσμάτων σε βάση δεδομένων Τα αποτελέσματα της παρούσας ανάλυσης είναι διαθέσιμα στη σχεσιακή MySQL βάση δεδομένων ενός PHP based ιστότοπου στη διεύθυνση στιγμιότυπο της αρχικής σελίδας της οποίας απεικονίζεται στην εικόνα Στο σύνδεσμο Differential Expression Reports απαριθμούνται τα 30 mirnas με τη μεγαλύτερη διαφορά μέσης έκφρασης μεταξύ όλων των δυνατών συνδυασμών ζευγών των 14 πειραμάτων. Ο χρήστης μπορεί να εφαρμόσει ή όχι διόρθωση FDR και να επιλέξει το cutoff του επιπέδου σημαντικότητας (p value = 0.05 ή 0.01 ή 0.01) (εικόνα 4.14 και πίνακας 4.5). 91


107 Εικόνα 4.13: Διεπαφή της αρχικής σελίδας της php/mysql βάσης δεδομένων. Εικόνα 4.14: Εύρεση διαφορικής έκφρασης των επιθυμητών ζευγών κυτταρικών σειρών. 92
 απεικονίζεται γραφικά το πρότυπο έκφρασής του στο σύνολο των δεκατεσσάρων κυτταρικών σειρών (εικόνα 4.15).")
108 Πίνακας 4.5: Διαφορική έκφραση των top 30 mirnas για το ζεύγος κυτταρικών σειρών PDV:PDVC57. Επιλέγοντας οποιοδήποτε mir (πίνακας 4.5) απεικονίζεται γραφικά το πρότυπο έκφρασής του στο σύνολο των δεκατεσσάρων κυτταρικών σειρών (εικόνα 4.15). Παράλληλα με διπλό κλικ στο γράφημα αυτό, προκύπτει ο πίνακας των ακριβών μέσων τιμών έκφρασης για το συγκεκριμένο mir (πίνακας 4.6). 93

109 Εικόνα 4.15: Πρότυπο έκφρασης mmu mir 205 5p στις δεκατέσσερεις κυτταρικές σειρές. Πίνακας 4.6: Μέση τιμή έκφρασης του mmu mir 205 5p στο σύνολο των 14 κυτταρικών σειρών. Στο σύνδεσμο List of mirs, επίσης εφαρμόζοντας ή όχι διόρθωση FDR και επιλέγοντας το επιθυμητό cutoff (0.05 ή 0.01 ή 0.01) απαριθμούνται όλα τα γονίδια που ικανοποιούν το 94
110 κριτήριο του cutoff, ταξινομημένα βάσει της απολύτου τιμής της μέγιστης τυπικής απόκλισης μεταξύ των δεκατεσσάρων κυτταρικών σειρών. Ένα τμήμα αυτών, με βάση τα κριτήρια της εικόνας 4.16 απεικονίζεται στον πίνακα 4.7. Για την εξαγωγή των μέχρι τώρα παραπάνω συμπερασμάτων απαιτείται η δημιουργία στη βάση δεδομένων των πινάκων ExiqonmiRs και mirexp όπως φαίνεται και από το διάγραμμα EER (Enhanced Entity Relationship) της εικόνας Εικόνα 4.16: Επιλογή φίλτρων για την εύρεση των mirnas με την ταξινόμηση των mirnas με διακύμανση 95

111 Πίνακα 4.7: mirnas με τη μέγιστη διακύμανση στο σύνολο των κατά ζεύγη συγκρίσεων των 14 κυτταρικών σειρών. 96
112 Εικόνα 4.17: EER διάγραμμα βάσης δεδομένων 97
 τους.")

113 4.7.2 Προσδιορισμός λειτουργικών γονιδίων στόχων mirnas Από την καρτέλα Download της mirbase (Kozomara and Griffiths Jones, 2011), ελήφθη το αρχείο mature.fa (εικόνα 4.17). Από αυτό το FASTA αρχείο, ενδιαφέρουν τα ονόματα των 1908 ώριμων mirna ανιχνευτών του ποντικού και τα MIMAT ids (ΜiRBase Μature mirna ids) τους. Τα δεδομένα αυτά αποθηκεύονται στο mirmimat αρχείο και μεταφορτώνονται με τη δημιουργία πίνακα στη βάση δεδομένων, όπως φαίνεται στο διάγραμμα EER. Εικόνα 4.17: mirbase: Λήψη όλων των αλληλουχιών mirnas σε FASTA μορφή. Ο ιστότοπος (Betel et al., 2008) ο οποίος συντηρείται από το κέντρο υπολογιστικής Βιολογίας του Memorial Sloan Kettering Cancer Center (MSKCC), αποτελεί πηγή στόχων micrornas για ζώα όπως ο άνθρωπος, ο ποντικός, ο αρουραίος, η μύγα, κ.λπ. Από την καρτέλα Downloads της έκδοσης Αυγούστου 2010 αυτού, ελήφθη το mouse_predictions_s_c_aug2010.txt tab delimited αρχείο με καλό mirsvr score 2 (εικόνα 4.18), 2 Κάθε προβλεφθείσα περιοχή στόχου χαρακτηρίζεται από μία πιθανότητα απορρύθμισης της έκφρασης της (mirsvr score). To score αυτό εκφράζει πόσο ισχυρή είναι η θερμοδυναμική αλληλεπίδραση της με το mirna. To mirsvr προκύπτει από ένα μοντέλο γραμμικής παλινδρόμησης το οποίο υπολογίζει το σταθμισμένο άθροισμα λαμβάνοντας υπ όψιν: i) τις συντηρημένες ακολουθίες (ένα mirna θεωρείται συντηρημένο αν είναι καταχωρημένο στον πίνακα mir_family_info_table του TargetScan γι αυτό δεν 98
 και τα ids των γονιδίων της Entrez (gene_id) (Maglott et al.")
114 το οποίο περιλαμβάνει τις περιοχές των γονιδίων, η έκφραση των οποίων ελέγχεται πιθανώς από κάποιο mirna που ανήκει στον οργανισμό του ποντικού. Από το αρχείο αυτό, ενδιαφέρουν τα MIMAT ids των mirnas (#mirbase_acc) και τα ids των γονιδίων της Entrez (gene_id) (Maglott et al., 2011) που στοχεύουν αυτά, τα οποία αποθηκεύονται στο αρχείο MIMATEntrez.txt και μεταφορτώνονται στη βάση δεδομένων με τη δημιουργία του αντίστοιχου πίνακα (βλ. ΕΕR). Εικόνα 4.18: microrna.org: Περιοχές στόχοι των mirnas στον ποντικό. Για τις δεκατέσσερεις κυτταρικές σειρές που μας ενδιαφέρουν, μας παραχωρήθηκαν από τον Καθ. κ. Balmain, δεδομένα μικροσυστοιχιών Mouse Genome Array [Mouse430_2] chip της Affymetrix των γονιδίων που κωδικοποιούν πρωτεΐνες. Τα δεδομένα αυτά είναι δευτερογενή δεδομένα έκφρασης που έχουν προέλθει από κανονικοποίηση RMA (Irizarry et al., 2003) και έχουν ενσωματωθεί στη βάση δεδομένων, έτσι ώστε εκτός από τα πρότυπα έκφρασης των mirnas, η βάση δεδομένων να παρέχει τα πρότυπα έκφρασης των πιθανών λειτουργικών στόχων αυτών. Ο πλήρης πίνακας (full table) που περιγράφει τα σύνολα ανιχνευτών (probe sets) υπάρχει κάποιο προκαθορισμένο cutoff ανά τις διάφορες εκδόσεις), ii) το μήκος του UTR, iii) τη θέση της περιοχής στόχου σε αυτό, iv) το conservation score, v) τη σύνθεση γύρω από την περιοχή πρόσδεσης του στόχου σε A/U και vi) την προσβασιμότητα εξαιτίας των δευτεροταγών δομών. Η παρούσα έκδοση περιλαμβάνει περιοχές στόχους με μήκος φύτρας μεγαλύτερο ή ίσο από αυτό του εξαμερούς και mirsvr
115 της συγκεκριμένης πλατφόρμας μικροσυστοιχιών, της Affymetrix, είναι διαθέσιμος από τo δημόσιο αποθετήριο δεδομένων μικροσυστοιχιών NCBI GEO (Gene Expression Omnibus) (Barrett et al., 2013), στη διεύθυνση: Στο αρχείο Mouse Oncogenesis Cell Lines.xlsx, περιλαμβάνονται οι γονιδιακές εκφράσεις για διάφορες κυτταρικές σειρές. Τα Affymetrix GeneChip arrays βοηθούν στη γρήγορη ανίχνευση της παρουσίας συγκεκριμένων γονιδίων στο βιολογικό δείγμα. Από το αρχείο αυτό, επιλέγονται τα ονόματα των γονιδίων (Gene Symbol) με τα ID τους (Probe ID) και οι εκφράσεις τους στις 14 κυτταρικές σειρές για τη δημιουργία του Affymetrix.txt και μετέπειτα η longform μορφή του (geneexp.txt). Τέλος μεταφορτώνεται στη βάση ο ομώνυμος πίνακας (geneexp). Για διαφορετικά είδη οργανισμών, τo Gene Symbol προκύπτει από διαφορετικές πηγές. Η Affymetrix για τον άνθρωπο, το ποντίκι και τον αρουραίο χρησιμοποιεί τις εγγραφές της UniGene (Schuler, 1997; Wheeler et al., 2005) Εν συνεχεία, από την καρτέλα Technical Documentation του ιστότοπου επιλέγοντας Mouse430_2 Annotations, CSV format, Release 34 (21 MB, 10/24/13), λαμβάνεται το αρχείο Mouse430_2.na34.annot σε csv μορφή. Από αυτό, επιλέγονται τα δεδομένα των στηλών Probe Set (στήλη Α), Gene Title (στήλη Ν), Gene Symbol (στήλη Ο) και MGI Name (στήλη AB), απορρίπτοντας τα δεδομένα των πρώτων εικοσιτριών γραμμών. Με αυτά τα δεδομένα, δημιουργείται στη βάση δεδομένων, ο πίνακας probesetmgi. To MGI Name αποτελεί προσδιοριστικό της Mouse Genome Informatics (MGI) βάσης (Eppig et al., 2012). Επιπροσθέτως, από το ίδιο αρχείο, ένας δεύτερος πίνακας δημιουργείται στη βάση δεδομένων με την ονομασία probesetentrez και από τις στήλες Probe Set και Entrez Gene (στήλη Ο). Για κάθε ζεύγος mirna στόχου, υπολογίζεται ο συντελεστής συσχέτισης Pearson (r value) (Pearson, 1895) μεταξύ του mirna και του στόχου. Με βάση το r value ταξινομούνται με αύξουσα σημαντικότητα τα πρότυπα έκφρασης των mirnas με τα πρότυπα έκφρασης των εξονικών στόχων της Affymetrix που έχουν τη μεγαλύτερη αρνητική συσχέτιση. Από το r value υπολογίζεται και το p value (Kendall et al., 1977), ενώ εξαιτίας των πολλαπλών πιθανών στόχων ενός mirna, εφαρμόστηκε διόρθωση πολλαπλών ερωτημάτων FDR (Benjamini and Hochberg, 100

116 1995) για τον υπολογισμό του adjusted p value και την εύρεση των στατιστικά σημαντικών περιπτώσεων αυτών. Από τους υπολογισμούς αυτούς δημιουργήθηκε ο πίνακας PCC ώστε να βρεθούν εν τέλει τα γονίδια η έκφραση των οποίων ρυθμίζεται αρνητικά από τα mirnas. Με τη δημιουργία των επιπλέον βοηθητικών πινάκων βάσεων δεδομένων mir, mirsplit, MIMAT, Entrez, geneexp, CLorder και mir_probeset επιτυγχάνεται, όπως είναι ορατό και από το διάγραμμα EER, η διασύνδεση όλων των παραπάνω πινάκων που έχουν περιγραφεί. Όλες οι εντολές MySQL και οι εντολές δημιουργίας πινάκων της βάσης δεδομένων βρίσκονται στο Παράρτημα Ι Γονίδια ειδικού ενδιαφέροντος Στον τρίτο σύνδεσμο της βάσης δεδομένων Specific genes of interest είναι διαθέσιμα τα πρότυπα έκφρασης των πιθανών στόχων των mirnas στα στάδια της προαγωγής και της εξέλιξης του καρκίνου (εικόνα 4.19) (Zoumpourlis et al., 2003). H λίστα αυτών εμφανίζεται επιλέγοντας ακολούθως το σύνδεσμο Target probe set list. Ομοίως με πριν, με διπλό κλικ πάνω στο γράφημα κάθε στόχου εμφανίζεται μια λίστα με την έκφραση του στόχου σε κάθε κυτταρική σειρά. Εικόνα 4.19: Συγκεκριμένες γονιδιακές μεταλλάξεις σε κάθε στάδιο της καρκινογένεσης. 101
117 4.7.4 Ανάλυση του ρόλου των γονιδίων στόχων των mirs με όρους γονιδιακής οντολογίας Οι λίστες γονιδίων στόχων των mirnas, μπορούν να αναλυθούν περαιτέρω για την εύρεση υπερεκπροσώπισης όρων γονιδιακής οντολογίας (Gene Ontology) (Ashburner et al., 2000) ή μονοπατιών KEGG (Kyoto Encyclopedia for Genes and Genomes) (Du et al., 2014), με τη χρήση του web tool Webgestalt (Wang et al., 2013), όπου η ανάλυση υπεργεωμετρικής κατανομής (Chvátal, 1979), σε συνδυασμό με FDR, προσδιορίζει τις βιολογικές διεργασίες τις οποίες επηρεάζουν τα υπό μελέτη mirnas. Για την κατανόηση των όσων προαναφέρθηκαν, θα περιγράψουμε αναλυτικά τη διαδικασία που ακολουθήθηκε, με παράδειγμα το mir 141 3p (MIMAT ): Βρίσκουμε όλα τα Entrez (User) IDs των στόχων του mir. Κατόπιν στο σύνδεσμο του Webgestalt ακολουθούμε τα εξής βήματα: 1) Eπιλέγουμε START 2) Select the organism of interest > mmusculus (από το drop down menu) 3) Select gene ID type mmusculus_entrezgene (από το drop down menu) 4) Στο Upload gene list κάνουμε επικόλληση 5181 Entrez User IDs ή φορτώνουμε του mmumir 141 3p ή φορτώνουμε κάποιο αρχείο. 5) Enter Στο σημείο αυτό προκύπτει πως από τους 5181 User IDs στόχους του mmu mir141 3p με τους οποίους τροφοδοτήσαμε το Webgestalt, 5160 από αυτούς αντιστοιχούν σε 3756 μοναδικά Entrez Gene IDs και άρα γονίδια. Τα υπόλοιπα 21 User IDs είτε αντιστοιχούν σε περισσότερα του ενός Entrez Gene IDs, είτε δεν αντιστοιχούν σε κανένα (εικόνα 4.20). 102
 Enrichment analysis KEGG Analysis 7) Select Reference Set for Enrichment Analysis mmusculus_mouse430_2 8) Statistical Method Hypergeometric 9) Multiple Test Adjustment BH (Benjamin Hochberg) 10)")
118 Εικόνα 4.20: Πληροφορίες αντιστοίχισης Entrez User IDs σε μοναδικά Entrez Gene IDs. 6) Enrichment analysis KEGG Analysis 7) Select Reference Set for Enrichment Analysis mmusculus_mouse430_2 8) Statistical Method Hypergeometric 9) Multiple Test Adjustment BH (Benjamin Hochberg) 10) Significance Level ) Minimum number of genes for a Category 2 12) Run Enrichment Analysis 13) View Results Από την έως τώρα ανάλυση προκύπτουν τα κυτταρικά μονοπάτια στα οποία συμμετέχουν κάποια από τα γονίδια τα οποία στοχεύει το mmu mir 141 3p, με πρώτα να εμφανίζονται τα πιο πιθανά μονοπάτια. Ας εξηγήσουμε πιο αναλυτικά τι σημαίνουν τα στατιστικά αριθμητικά δεδομένα για ένα από αυτά. 103
119 Πίνακας 4.8: Επεξήγηση στατιστικών ευρημάτων για το μονοπάτι σήμανσης ErbB στο οποίο συμμετέχουν γονίδια στόχοι του mmu mir 141 3p. Pathway name #Gene Entrez Gene Statistics C=85; ErbB signaling pathway O=34; E=15.78; R=2.15; rawp=3.35e 06 adjp= H μεταβλητή Category C=85 παριστάνει τον αριθμό των γονιδίων (Entrez Gene IDs) τα οποία σχετίζονται με το μονοπάτι σήμανσης ErbB, στο σύνολο αναφοράς mmusculus_affymouse_430_2 (τα γονίδια, την έκφραση των οποίων μελετά το Mouse Genome Array chip). Η Observed Ο=34 παριστάνει το πλήθος των γονιδίων που σχετίζονται με το συγκεκριμένο μονοπάτι στο σύνολο της λίστας των μοναδικών Gene IDs που εξετάσαμε (3756) και που αποτελούν στόχους του mmu mir141 3p. Ωστόσο, αν το δείγμα ήταν αντιπροσωπευτικό, αυτά θα έπρεπε να ισούνται με τη μεταβλητή Expected Ε= Η μεταβλητή Ratio of enrichment R=2.15, προκύπτει ως εξής: R=O/E=34/15.8=2.15 και εκφράζει το μέγεθος της υπερεκπροσώπησης. Με βάση τα μοναδικά Gene IDs, C, E και την απλή μέθοδο των τριών, υπολογίζουμε πως το σύνολο αναφοράς περιέχει γονίδια που περιέχει Entrez γονίδια. Θέλοντας να δούμε κατά πόσο το σύνολο των 3756 Gene IDs είναι αντιπροσωπευτικό, δηλαδή η όποια υπερεκπροσώπηση προέκυψε κατά τύχη, υπολογίζουμε την πιθανότητα της μηδενικής υπόθεσης H null : Aν στα γονίδια, τα 85 σχετίζονται με το ErbB μονοπάτι, υπολογίζουμε την πιθανότητα raw P στο υποσύνολο των 3756 γονιδίων, τουλάχιστον τα 34 να σχετίζονται με το μονοπάτι από τύχη (δηλαδή το δείγμα να είναι αντιπροσωπευτικό), με τη μέθοδο της υπεργεωμετρικής κατανομής (Chvátal, 1979). Η μεταβλητή raw P=3.35e 06, σημαίνει ότι η πιθανότητα να είναι το δείγμα αντιπροσωπευτικό, είναι Tέλος, επειδή εξετάζουμε πολλά κυτταρικά μονοπάτια, εφαρμόζεται διόρθωση πολλαπλών συγκρίσεων FDR, και υπολογίζεται η μεταβλητή της τροποποιημένης p value, adjp= Ορίζοντας adjp cut 104

120 off=0.05, σημαίνει ότι το 5% των κυτταρικών μονοπατιών που φαίνεται να ρυθμίζει η αλλαγή της έκφρασης του mmu mir141 3p είναι ψευδώς θετικά. Στον πίνακα 4.9, φαίνονται τα κυτταρικά μονοπάτια τα οποία επηρεάζουν τα mmu mir205 5p, mmu mir200b 3p, mmu mir200c 3p, mmu mir429 3p και mmu mir141 3p τα οποία είναι ιδιαίτερου ενδιαφέροντος, όπως θα αναπτυχθεί στο 5 ο κεφάλαιο, μέσω της ρύθμισης της έκφρασης των γονιδίων στόχων τους. Πίνακας 4.9: ΚΕGG μονοπάτια τα οποία επηρεάζει η αλλαγή της έκφρασης των συγκεκριμένων mirs. 105
121 4.8 Βιβλιογραφία Ashburner, M., Ball, C.A., Blake, J.A., Botstein, D., Butler, H., Cherry, J.M., Davis, A.P., Dolinski, K., Dwight, S.S., Eppig, J.T., et al. (2000). Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 25, Barrett, T., Wilhite, S.E., Ledoux, P., Evangelista, C., Kim, I.F., Tomashevsky, M., Marshall, K.A., Phillippy, K.H., Sherman, P.M., Holko, M., et al. (2013). NCBI GEO: archive for functional genomics data sets update. Nucleic Acids Res 41, D Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Statist Soc Ser B 57, Betel, D., Wilson, M., Gabow, A., Marks, D.S., and Sander, C. (2008). The microrna.org resource: targets and expression. Nucleic Acids Res 36, D Bolstad, B.M., Irizarry, R.A., Astrand, M., and Speed, T.P. (2003). A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 19, Chvátal, V. (1979). The tail of the hypergeometric distribution. Discrete Math 25, Du, J., Yuan, Z., Ma, Z., Song, J., Xie, X., and Chen, Y. (2014). KEGG PATH: Kyoto encyclopedia of genes and genomes based pathway analysis using a path analysis model. Mol Biosyst 10, Eppig, J.T., Blake, J.A., Bult, C.J., Kadin, J.A., Richardson, J.E., and Mouse Genome Database, G. (2012). The Mouse Genome Database (MGD): comprehensive resource for genetics and genomics of the laboratory mouse. Nucleic Acids Res 40, D Gentleman, R.C., Carey, V.J., Bates, D.M., Bolstad, B., Dettling, M., Dudoit, S., Ellis, B., Gautier, L., Ge, Y., Gentry, J., et al. (2004). Bioconductor: open software development for computational biology and bioinformatics. Genome Biol 5, R80. Irizarry, R.A., Hobbs, B., Collin, F., Beazer Barclay, Y.D., Antonellis, K.J., Scherf, U., and Speed, T.P. (2003). Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 4, Kendall, M., Stuart, A., and Ord, J. (1977). The Advanced Theory of Statistics., 4th edn (London: Charles Griffin). Kozomara, A., and Griffiths Jones, S. (2011). mirbase: integrating microrna annotation and deepsequencing data. Nucleic Acids Res 39, D Maglott, D., Ostell, J., Pruitt, K.D., and Tatusova, T. (2011). Entrez Gene: gene centered information at NCBI. Nucleic Acids Res 39, D Oshlack, A., Emslie, D., Corcoran, L.M., and Smyth, G.K. (2007). Normalization of boutique two color microarrays with a high proportion of differentially expressed probes. Genome Biol 8, R2. Pearson, K. (1895). Notes on regression and inheritance in the case of two parents. Proceedings of the Royal Society of London Ritchie, M.E., Phipson, B., Wu, D., Hu, Y., Law, C.W., Shi, W., and Smyth, G.K. (2015). limma powers differential expression analyses for RNA sequencing and microarray studies. Nucleic Acids Res. Ritchie, M.E., Silver, J., Oshlack, A., Holmes, M., Diyagama, D., Holloway, A., and Smyth, G.K. (2007). A comparison of background correction methods for two colour microarrays. Bioinformatics 23, Schuler, G.D. (1997). Pieces of the puzzle: expressed sequence tags and the catalog of human genes. J Mol Med (Berl) 75, Shi, W., Oshlack, A., and Smyth, G.K. (2010). Optimizing the noise versus bias trade off for Illumina whole genome expression BeadChips. Nucleic Acids Res 38, e204. Smyth, G.K., and Speed, T. (2003). Normalization of cdna microarray data. Methods 31, Wang, J., Duncan, D., Shi, Z., and Zhang, B. (2013). WEB based GEne SeT AnaLysis Toolkit (WebGestalt): update Nucleic Acids Res 41, W
122 Wheeler, D.L., Barrett, T., Benson, D.A., Bryant, S.H., Canese, K., Church, D.M., DiCuccio, M., Edgar, R., Federhen, S., Helmberg, W., et al. (2005). Database resources of the National Center for Biotechnology Information. Nucleic Acids Res 33, D Wickham, H. (2007). Reshaping Data with the {reshape} Package. Journal of Statistical Software 21, Zoumpourlis, V., Solakidi, S., Papathoma, A., and Papaevangeliou, D. (2003). Alterations in signal transduction pathways implicated in tumour progression during multistage mouse skin carcinogenesis. Carcinogenesis 24,
123 Κεφάλαιο 5 ο Αποτελέσματα και Συζήτηση 108
124 5.1 Αποτελέσματα Προοδευτική μείωση της έκφρασης των μελών της οικογένειας mir-200 και του mir-205 κατά την εξέλιξη της δερματικής καρκινογένεσης στον ποντικό Η επιλογή των mirnas με τη μεγαλύτερη διαφορική έκφραση έγινε βάσει των επιπλέον ακόλουθων κριτηρίων: i. Επιλογή από τη βάση δεδομένων των mirnas με τη μεγαλύτερη τυπική απόκλιση της σχετικής έκφρασης μεταξύ των δεκατεσσάρων κυτταρικών σειρών. ii. iii. iv. Προσδιορισμός ενός συνόλου mirnas των οποίων η έκφραση αλλάζει αντί μεμονωμένων mirnas, έχει προταθεί ότι έχει ογκογόνα ή ογκοκατασταλτική έκβαση (Iorio and Croce, 2012). Επιλογή των mirnas που συνεκφράζονται (είτε αυξάνει ή έκφραση όλων είτε μειώνεται) σε όλες κυτταρικές σειρές (εικόνα 5.1 και 5.2). Επιλογή των mirnas που παρουσιάζουν ένα μονοπάτι συνεχούς υποέκφρασης ή συνεχούς υπερέκφρασης κατά τα στάδια της καρκινογένεσης, πιστεύεται ότι συμβάλει καθοριστικά στη δερματική καρκινογένεση (εικόνα 5.2). Εικόνα 5.1: Ομαδοποίηση (clustering) των περισσότερο διαφορικά εκφρασμένων mirnas. Τα έξι mirnas που προσδιορίστηκαν με τη μεγαλύτερη διαφορική έκφραση ομαδοποιούνται στο ίδιο κλαδί ενός δέντρου. Αυτό αποτελεί ένδειξη συνέκφρασης των έξι mirnas στις δεκατέσσερεις κυτταρικές σειρές του μοντέλου της δερματικής καρκινογένεσης στον ποντικό. 109
125 Εικόνα 5.2: Σχετική έκφραση του mir-205-5p και της mir-200 οικογένειας mirnas στις δεκατέσσερεις κυτταρικές σειρές. 110
126 Τα mirnas που πληρούν τα παραπάνω κριτήρια είναι τα: mir-200a-3p mir-200b-3p mir-200c-3p mir-141-3p mir-429-3p mir-205-5p. Tα πρώτα πέντε mirnas συνιστούν την οικογένεια mir-200. Τα έξι αυτά mirnas (τα οποία στο εξής χάριν συντομίας θα αποκαλούνται MOS MiRNAs Of Study) υποεκφράζονται κατά την τεχνητή δερματική καρκινογένεση στον ποντικό και ειδικά κατά τη μετάβαση από τα πλακώδη κύτταρα στους πιο επιθετικούς ατρακτοκυτταρικούς φαινοτύπους Παράλληλη υπερέκφραση των στόχων της οικογενείας mir-200 και του mir-205 στη δερματική καρκινογένεση Όπως προαναφέρθηκε η αλλαγή της έκφρασης των mirnas κατά ομάδες, μπορεί να έχει ογκογόνα ή ογκοκατασταλτική έκβαση (Iorio and Croce, 2012; Lu et al., 2005). Επίσης, η ρύθμιση ενός γονιδιακού στόχου από ένα σύνολο mirnas φαίνεται να είναι η αφορμή ο στόχος αυτός να συμμετέχει στη ρύθμιση της λειτουργίας πολλαπλά κυτταρικά που συμμετέχουν στην ογκογένεση (Wang et al., 2014b). Με βάση τα παραπάνω, ευρέθησαν πιθανοί στόχοι οι οποίοι ρυθμίζονται τουλάχιστον από δύο MOS καθώς και τα μονοπάτια που υπερεκπροσωπούνται και στα οποία συμμετέχουν αυτοί. Οι προβλεφθέντες στόχοι απαριθμούνται στον πίνακα 5.1 ενώ τα KEGG μονοπάτια τα οποία συν-ρυθμίζονται από τα MOS απεικονίζονται στην εικόνα 5.3. Μεταξύ των προβλεθέντων στόχων, τρείς από αυτούς ο akt3 (πιθανός στόχος των mir-200b, mir-200c και mir-429),ο rock2 (πιθανός στόχος όλων των μελών της οικογενείας mir-200) και ο rap1a (πιθανός στόχος των mir-200b, mir-200c, mir-429 and mir-205) συμμετέχουν στη ρύθμιση του μονοπατιού της κυτταρικής πρόσφυσης (focal adhesion) (πίνακας 5.1), το οποίο συσχετίζεται με επιθετικούς φαινοτύπους (Hanahan and Weinberg, 2011). Σε αυτούς τους τρείς στόχους, η μεγαλύτερη αρνητική συσχέτισης της έκφρασης παρατηρείται βάσει του συντελεστή συσχέτισης Pearson, στα ζεύγη: i) mir-200c - akt3 (εικόνα 5.4a), ii) mir rock2 (εικόνα 5.4(b)) 111

127 και iii) mir rap1a (εικόνα 5.4(c)). Πίνακας 5.1: Υπερεκπροσώπηση KEGG μονοπατιών. 112

128 Εικόνα 5.3: Στατιστκά σημαντική υπερ-εκπροσώπιση των KEGG μονοπατιών. 113
129 (a) (b) (c) Εικόνα 5.4: Συσχέτιση έκφρασης των a) mir-200c-3p, b) mir-141-3p, c) mir-205-5p και των στόχων τους. Στο διάγραμμα φαίνεται η αρνητική συσχέτιση των log 2 τιμών έκφρασης αυτών 114
130 5.1.3 Διαφορική έκφραση mirnas κατά τη μετάβαση από πλακώδη κακοήθη σε αναπλαστικά καρκινώματα του δέρματος Για τον προσδιορισμό περισσότερων mirnas των οποίων η έκφραση αλλάζει σημαντικά στα πιο επιθετικά στάδια της δερματικής καρκινογένεσης στον ποντικό, εξετάζονται τα 30 mirnas με τη μεγαλύτερη διαφορική έκφραση μεταξύ των PDV και PDVC57 κυτταρικών σειρών και με cut-off για Mean Difference = 1.5 ως προς τη διαφορική τους έκφραση. Κατά τη μετάβαση στην PDVC57 κυτταρική σειρά η οποία εκφράζει τη μετάβαση στα αναπλαστικά καρκινώματα (Caulin et al., 1993) παρατηρείται σημαντική υποέκφραση mir-708-5p και σημαντική υπερέκφραση του mir-181a-5p (πίνακας 5.2). Πίνακας 5.2: Λίστα των top-30 διαφορικά εκφρασμένων mirnas για το PDV:PDVC57 ζεύγος κυτταρικών σειρών. mirna Mean Difference: PDV-PDVC57 p-value mmu-mir-205-5p mmu-mir-708-5p mmu-mir-181a-5p mmu-mir-712-5p mmu-mir mmu-mir-22-3p mmu-mir-365-3p mmu-mir-193a-3p mmu-mir p mmu-mir-665-3p mmu-mir-25-5p mmu-mir-147-3p mmu-mir-222-5p mmu-mir-146a-5p mmu-mir-350-3p mmu-mir-672-5p mmu-mir-218-5p mmu-mir mmu-mir-344e-3p mmu-mir-500-3p mmu-mir-7a-5p mmu-mir-185-3p mmu-mir p mmu-mir-196a-5p mmu-mir-362-3p
131 mmu-mir p mmu-mir-532-5p mmu-mir-10b-5p mmu-mir-125b-1-3p mmu-mir-668-3p Διαφορική έκφραση mirnas κατά τη μετάβαση από πλακώδη σε ατρακτοκυτταρικά κακοήθη καρκινώματα Ομοίως στον πίνακα 5.3 απαριθμούνται τα 30 mirnas με τη μεγαλύτερη διαφορική έκφραση στο B9:D3 ζεύγος κυτταρικών σειρών ενώ στον πίνακα 5.4 τα mirnas για το B9:A5 ζεύγος κυτταρικών σειρών. Και τα δύο αυτά ζεύγη κυτταρικών σειρών, προσομοιώνουν τη μετάβαση από τα μη-διηθητικά (non-invasive) πλακώδη κακοήθη στα διηθητικά ατρακτοκυτταρικά κακοήθη καρκινώματα (Oft et al., 2002). Παρατηρείται πως τα mirnas που εκφράζονται διαφορικά τόσο στο B9:D3 όσο και στο B9:A5 ζεύγος κυτταρικών σειρών (με τιμή cut-off Mean Difference =1.5) είναι τα: mir-143-3p, mir- 181d-5p και mir-181a-5p. Πιο συγκεκριμένα παρατηρείται υποέκφραση αυτών, κατά μετάβαση από τις μη-διηθητικές στις διηθητικές κυτταρικές σειρές. Πίνακας 5.3: Λίστα των top-30 διαφορικά εκφρασμένων mirnas για το B9:A5 ζεύγος κυτταρικών σειρών. mirna Mean Difference: B9-A5 p-value mmu-mir-199a-5p mmu-mir-205-5p mmu-mir-143-3p mmu-mir-199a-3p/mmu-mir-199b-3p mmu-mir-34c-5p mmu-mir-200c-3p mmu-mir-141-3p mmu-mir-34b-5p mmu-mir-199b-5p mmu-mir-200a-3p mmu-mir-214-3p mmu-mir-200b-3p mmu-mir-429-3p mmu-mir-350-3p mmu-mir-34a-5p mmu-mir-146a-5p
132 mmu-mir-181d-5p mmu-mir-708-5p mmu-mir-181a-5p mmu-mir-9-5p mmu-mir-195a-5p mmu-mir-214-5p mmu-mir-34c-3p mmu-mir-667-3p mmu-mir-145a-5p mmu-mir-540-3p mmu-mir-322-5p mmu-mir-582-5p mmu-let-7i-5p mmu-mir-378a-3p Πίνακας 5.4: Λίστα των top-30 διαφορικά εκφρασμένων mirnas για το B9:D3 ζεύγος κυτταρικών σειρών. mirna Mean Difference: B9-D3 p-value mmu-mir-205-5p mmu-mir-200c-3p mmu-mir-141-3p mmu-mir-200a-3p mmu-mir-200b-3p mmu-mir-429-3p mcmv-mir-m88-1-3p mmu-mir-142-5p mmu-mir-218-5p mmu-mir-210-3p mmu-mir-143-3p mmu-mir-193a-3p mmu-mir-181d-5p mmu-mir-181a-5p mmu-mir-205-3p mmu-mir-378a-3p mmu-mir-99a-5p mmu-mir-33-5p mmu-mir-350-3p mmu-mir mmu-mir-21a-3p mmu-mir-138-5p mmu-mir-155-5p mmu-mir-125b-5p mmu-mir-335-5p
133 mmu-mir-203-3p SNORD mmu-mir-322-5p mmu-mir-125b-1-3p mmu-mir-467b-5p Συζήτηση Εξαιτίας της θνησιμότητας του NMSC (Non Melanoma Skin Carcinogenesis) στην περίπτωση μη έγκαιρης διάγνωσης, θα πρέπει, το τρέχον θεραπευτικό οπλοστάσιο της ασθένειας αυτής να εμπλουτιστεί. Οι θεραπείες που βασίζονται στα mirnas έχουν σηματοδοτήσει νέους ορίζοντες στη διαχείριση του καρκίνου. Tα ευνοϊκά χαρακτηριστικά των mirnas, όπως η ικανότητα τους να λειτουργούν ως κύριοι ρυθμιστές του γονιδιώματος και η μη αναφορά δυσμενών συμβάντων, τα καθιστούν ελκυστικά ώστε να αποτελέσουν την επόμενη γενιά θεραπευτικών στόχων. Δοκιμαζόμενα φάρμακα που βασίζονται στη ρύθμιση της έκφρασης των mirnas εισχώρησαν γρήγορα σε αντικαρκινικές κλινικές δοκιμές (Li et al., 2014), ενώ οι αιτήσεις ευρεσιτεχνίας στις Η.Π.Α. που σχετίζονται με τα mirναs έχουν αυξηθεί δραματικά την τελευταία δεκαετία (van Rooij et al., 2012). Ένα mirna από μόνο του μπορεί να μην είναι ικανό για την έκβαση ενός ογκογόνου ή ογκοκατασταλτικού αποτελέσματος και μπορεί να δρα συνεργατικά με ένα επιπλέον σύνολο mirnas. Επιπλέον πρέπει να ληφθεί υπ όψη ότι υπάρχουν mirnas με μία yin-yang συμπεριφορά στον καρκίνο (So et al., 2013). Για το σκοπό αυτό, αναζητήθηκαν ομάδες mirnas οι οποίες φαίνεται να απορυθμίζουν συνεργατικά την έκφραση των NMSC έναντι των φυσιολογικών κυττάρων. Η προσέγγιση αυτή αναμένεται να διαχωρίσει καλύτερα τα mirnas τα οποία έχουν κυρίαρχο ρόλο στη NMSC από αυτά τα οποία η έκφραση τους απορυθμίζεται τυχαία και συνεπώς είναι αμφιλεγόμενα. H συντονισμένη απώλεια έκφρασης των μελών της οικογενείας mir-200 και στο mir-205 είναι κομβικής σημασίας στην κακοήθη μετατροπή πολλών τύπων καρκίνου επιθηλιακής προέλευσης όπως αυτών του πνεύμονα (Tellez et al., 2011), της ουροδόχου κύστης (Wiklund et al., 2011) και του στήθος (Gregory et al., 2008) και πιθανώς προαπαιτούμενη καθολικά. Η ταυτόχρονη υποέκφραση των mir-200s και του mir-205, στους πιο επιθετικούς φαινοτύπους και δεδομένου ότι τα γονίδια ξενιστές αυτών των mirnas δε βρίσκονται όλα στον ίδια χρωμοσωμικό τόπο ούτε 118
134 στον ποντικό ούτε στον άνθρωπο (Korpal et al., 2008) δύναται να είναι οφείλεται στη συλλογική τους δράση. Η δράση μιας ομάδας mirnas πιθανόν να έχει αντι-ογκογόνο αποτέλεσμα εν αντιθέσει με την ατομική δράση των mirnas της ομάδας αυτής η οποία μπορεί να έχει διαφορετικές εκβάσεις. Συνεπώς η επαναφορά της έκφρασης των mir-200s και mir-205 φαίνεται να είναι μια πολλά υποσχόμενη στρατηγική για την παρεμπόδιση της εξέλιξης και της μετάστασης πολλών καρκινικών τύπων συμπεριλαμβανομένου και του NMSC. Πράγματι στοιχεία που αποδεικνύουν ότι η επαναφορά της έκφρασης των mir-200s αποτρέπει τη μετάσταση στον καρκίνο των πνευμόνων και των ηπατοκυττάρων (Ding et al., 2012) ενθαρρύνουν περαιτέρω προσπάθειες προς αυτή την κατεύθυνση. Αξίζει να αναφερθεί πως ο ρόλος των mir-200s στην ΕΜΤ (Epithelial to Mesenchymal Transition) αναστολή είναι καλά τεκμηριωμένος και σταθερός (Feng et al., 2014), σε αντίθεση με το mir- 205 το οποίο πότε καταστέλλει και πότε προάγει την ογκογένεση (συμπεριφορά yin-yang) (Qin et al., 2013). Αυτό ίσως οφείλεται στο ότι οι αντίστοιχες μελέτες επικεντρώνονται μόνο στο mir- 205 και όχι στην εξέταση της έκφρασης και άλλων mirnas μαζί. Αυτή η συμπεριφορά πιθανόν να μπορεί να εξηγηθεί από mirnas η έκφραση των οποίων συν-απορυθμίζεται με το mir-205. Σε μελλοντικές μελέτες προτείνεται ένας πληρέστερος προσδιορισμός του ρόλου του mir-205 ως προς τους πιο επιθετικούς φαινοτύπους με συγκριτικές μελέτες μαζί με τα υποεκφραζόμενα mir-200s στο ζεύγος PDV:PDVC57 έναντι των Β9: D3 και Β9: Α5 ζευγών κυτταρικών σειρών. Αναφορικά με τα γονίδια η ρύθμιση της έκφρασης των οποίων απαιτεί δύο MOS, παρατηρείται μια αρνητική συσχέτιση μεταξύ αυτών των mirnas και των στόχων που σχετίζονται με το μονοπάτι κυτταρικής πρόσφυσης (Akt3, Rock2, Rap1a), με αυτά να υπερεκφράζονται όταν ή έκφραση των MOS φθίνει. H Akt3, Ser/Thr κινάση, εμπλέκεται τόσο στην προαγωγή (Cheung et al., 2008) όσο και στην εξέλιξη (Stahl et al., 2004) του μελανώματος. Η Rock2 (Rho-associated protein kinase 2) είναι ένα ακόμη παράδειγμα κινάσης Ser/Thr που απαιτείται για την ανάπτυξη και τη διήθηση σε μεγάλα καρκινικά κύτταρα των πνευμόνων. Όσο για το Rap1a (Ras-related πρωτεΐνη 1a), είναι μία μικρή Ras-like GTPάση η οποία πρόσφατα αναφέρθηκε πως προάγει την κυτταρική μετανάστευση (migration) και τη διήθηση των καρκινικών κυττάρων (Chen et al., 2013; Wang et al., 2012). 119
135 Από την υποέκφραση του mir-708-5p με την υπερέκφραση του mir-181a-5p στο ζεύγος PDV:PDVC57 και η υπερέκφραση των mir-143-3p, mir-181d-5p και mir-181a-5p κατά τη μετάβαση από τα πλακώδη κύτταρα σε πιο επιθετικούς αναπλαστικούς/ατρακτοκυτταρικούς φαινοτύπους, διαπιστώνεται πως το mir-181a-5p αποτελεί κοινό παρονομαστή σε αυτή την εξέλιξη. Πρόσφατες μελέτες αναφέρουν επίσης ότι η υπερέκφραση του mir-181a προάγει την εξέλιξη του καρκίνου των ωοθηκών, ενώ το mir-708 καταστέλλει τη μετάσταση του καρκίνου του μαστού (Ryu et al., 2013). Από την άλλη πλευρά ο ρόλος των λιγότερο μελετημένων mir- 181d και mir-143 στην εξέλιξη του καρκίνου είναι προς το παρόν αντιφατικός: το mir-181d υποεκφράζεται στα γλοιώματα (Wang et al., 2014a), αλλά υποεκφράζεται στα καρκινικά ηπατοκύτταρα (Ji et al., 2011). Ομοίως, το mir-143 είναι ογκογόνο στον καρκίνο των οστών (Zhang et al., 2010) και ενώ στον καρκίνο του παχέος εντέρου όχι (Kitade and Akao, 2010). Με βάση την τρέχουσα εμπειρία από κλινικές δοκιμές, ποσοστό μεγαλύτερο του 90% των προκλινικά επιτυχόντων mirna στόχων αποτυγχάνουν σε μεταγενέστερες δοκιμές. Δεδομένης της πληθώρας των mirnas που προτείνονται ως αντικαρκινικοί στόχοι με βάση τις προκλινικές δοκιμές, απαιτείται διάκριση των πραγματικά λειτουργικών αντικαρκινικών mirna στόχων. Εισαγωγή καινοτόμων προσεγγίσεων και επιπλέον αυστηρών κριτηρίων καταλληλότητας (π.χ. λειτουργική συντήρηση στα είδη και ιστούς, σταθερό ρόλο σε όλα τα στάδια της ογκογένεσης κ.λπ) σε προκλινικό επίπεδο, θα μπορούσε να οδηγήσει σε σοφότερη επιλογή των στόχων και ενδεχομένως στην αύξηση του ποσοστού επιτυχίας των κλινικών δοκιμών. 120
136 5.3 Βιβλιογραφία Caulin, C., Bauluz, C., Gandarillas, A., Cano, A., and Quintanilla, M. (1993). Changes in keratin expression during malignant progression of transformed mouse epidermal keratinocytes. Exp Cell Res 204, Chen, C.H., Chuang, H.C., Huang, C.C., Fang, F.M., Huang, H.Y., Tsai, H.T., Su, L.J., Shiu, L.Y., Leu, S., and Chien, C.Y. (2013). Overexpression of Rap-1A indicates a poor prognosis for oral cavity squamous cell carcinoma and promotes tumor cell invasion via Aurora-A modulation. Am J Pathol 182, Cheung, M., Sharma, A., Madhunapantula, S.V., and Robertson, G.P. (2008). Akt3 and mutant V600E B- Raf cooperate to promote early melanoma development. Cancer Res 68, Ding, W., Dang, H., You, H., Steinway, S., Takahashi, Y., Wang, H.G., Liao, J., Stiles, B., Albert, R., and Rountree, C.B. (2012). mir-200b restoration and DNA methyltransferase inhibitor block lung metastasis of mesenchymal-phenotype hepatocellular carcinoma. Oncogenesis 1, e15. Federhen, S. (2012). The NCBI Taxonomy database. Nucleic Acids Res 40, D Feng, X., Wang, Z., Fillmore, R., and Xi, Y. (2014). MiR-200, a new star mirna in human cancer. Cancer Lett 344, Gregory, P.A., Bert, A.G., Paterson, E.L., Barry, S.C., Tsykin, A., Farshid, G., Vadas, M.A., Khew-Goodall, Y., and Goodall, G.J. (2008). The mir-200 family and mir-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat Cell Biol 10, Hanahan, D., and Weinberg, R.A. (2011). Hallmarks of cancer: the next generation. Cell 144, Iorio, M.V., and Croce, C.M. (2012). microrna involvement in human cancer. Carcinogenesis 33, Ji, J., Yamashita, T., and Wang, X.W. (2011). Wnt/beta-catenin signaling activates microrna-181 expression in hepatocellular carcinoma. Cell Biosci 1, 4. Kitade, Y., and Akao, Y. (2010). MicroRNAs and their therapeutic potential for human diseases: micrornas, mir-143 and -145, function as anti-oncomirs and the application of chemically modified mir-143 as an anti-cancer drug. J Pharmacol Sci 114, Korpal, M., Lee, E.S., Hu, G., and Kang, Y. (2008). The mir-200 family inhibits epithelial-mesenchymal transition and cancer cell migration by direct targeting of E-cadherin transcriptional repressors ZEB1 and ZEB2. J Biol Chem 283, Li, X.J., Ren, Z.J., and Tang, J.H. (2014). MicroRNA-34a: a potential therapeutic target in human cancer. Cell Death Dis 5, e1327. Lu, J., Getz, G., Miska, E.A., Alvarez-Saavedra, E., Lamb, J., Peck, D., Sweet-Cordero, A., Ebert, B.L., Mak, R.H., Ferrando, A.A., et al. (2005). MicroRNA expression profiles classify human cancers. Nature 435, Oft, M., Akhurst, R.J., and Balmain, A. (2002). Metastasis is driven by sequential elevation of H-ras and Smad2 levels. Nat Cell Biol 4, Qin, A.Y., Zhang, X.W., Liu, L., Yu, J.P., Li, H., Wang, S.Z., Ren, X.B., and Cao, S. (2013). MiR-205 in cancer: an angel or a devil? Eur J Cell Biol 92, Ryu, S., McDonnell, K., Choi, H., Gao, D., Hahn, M., Joshi, N., Park, S.M., Catena, R., Do, Y., Brazin, J., et al. (2013). Suppression of mirna-708 by polycomb group promotes metastases by calcium-induced cell migration. Cancer Cell 23, So, A.Y., Zhao, J.L., and Baltimore, D. (2013). The Yin and Yang of micrornas: leukemia and immunity. Immunol Rev 253, Stahl, J.M., Sharma, A., Cheung, M., Zimmerman, M., Cheng, J.Q., Bosenberg, M.W., Kester, M., Sandirasegarane, L., and Robertson, G.P. (2004). Deregulated Akt3 activity promotes development of malignant melanoma. Cancer Res 64, Tellez, C.S., Juri, D.E., Do, K., Bernauer, A.M., Thomas, C.L., Damiani, L.A., Tessema, M., Leng, S., and Belinsky, S.A. (2011). EMT and stem cell-like properties associated with mir-205 and mir-200 epigenetic 121
137 silencing are early manifestations during carcinogen-induced transformation of human lung epithelial cells. Cancer Res 71, van Rooij, E., Purcell, A.L., and Levin, A.A. (2012). Developing microrna therapeutics. Circ Res 110, Wang, K., Li, J., Guo, H., Xu, X., Xiong, G., Guan, X., Liu, B., Li, J., Chen, X., Yang, K., et al. (2012). MiR-196a binding-site SNP regulates RAP1A expression contributing to esophageal squamous cell carcinoma risk and metastasis. Carcinogenesis 33, Wang, L., Shi, Z.M., Jiang, C.F., Liu, X., Chen, Q.D., Qian, X., Li, D.M., Ge, X., Wang, X.F., Liu, L.Z., et al. (2014a). MiR-143 acts as a tumor suppressor by targeting N-RAS and enhances temozolomide-induced apoptosis in glioma. Oncotarget 5, Wang, L.J., Chou, Y.F., Chen, P.R., Su, B., Hsu, Y.C., Chang, C.H., and Lee, J.W. (2014b). Differential mirna expression in repeated recurrence of nasopharyngeal carcinoma. Cancer Lett 344, Wiklund, E.D., Bramsen, J.B., Hulf, T., Dyrskjot, L., Ramanathan, R., Hansen, T.B., Villadsen, S.B., Gao, S., Ostenfeld, M.S., Borre, M., et al. (2011). Coordinated epigenetic repression of the mir-200 family and mir-205 in invasive bladder cancer. Int J Cancer 128, Zhang, H., Cai, X., Wang, Y., Tang, H., Tong, D., and Ji, F. (2010). microrna-143, down-regulated in osteosarcoma, promotes apoptosis and suppresses tumorigenicity by targeting Bcl-2. Oncol Rep 24,
138 Παράρτημα Ι 123
139 I.1 Έγχρωμα ΜΑ plots μη κανονικοποιημένων δεδομένων 124
140 125
141 126
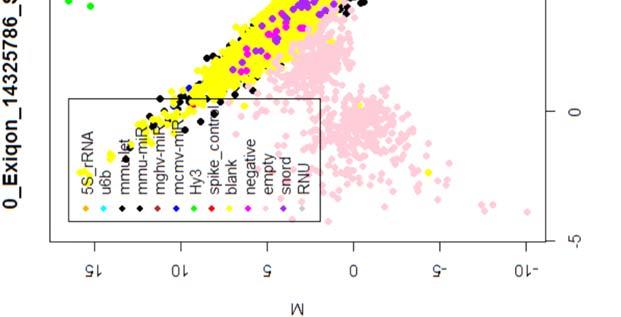
142 127
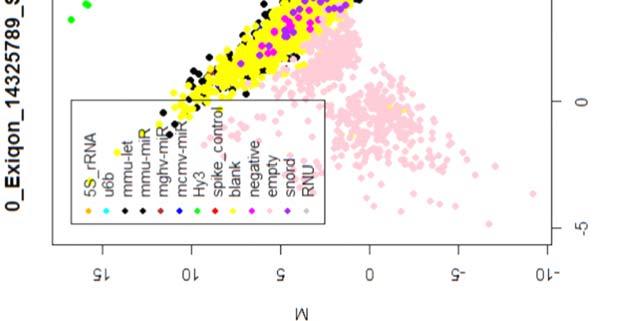
143 128
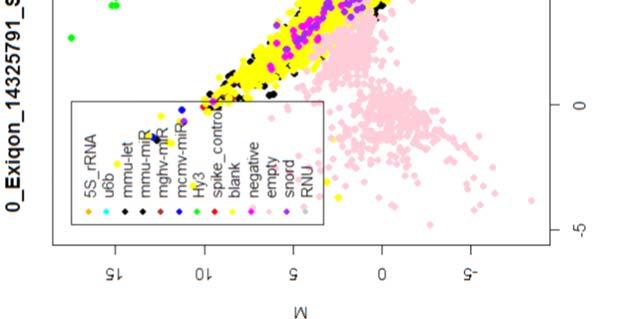
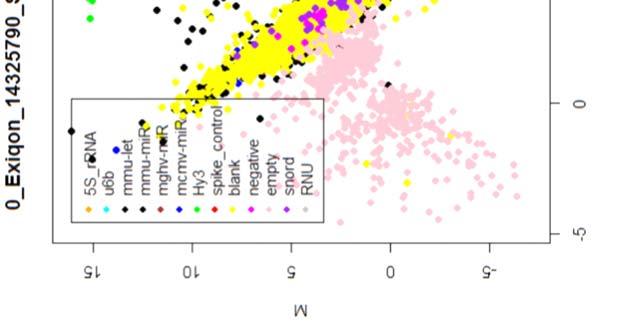
144 129
145 130


146 Ι.2 Έγχρωμα ΜΑplots μετά την κανονικοποίηση εντός των μικροσυστοιχιών 131
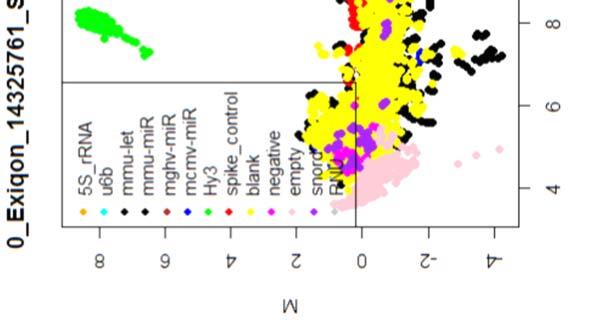
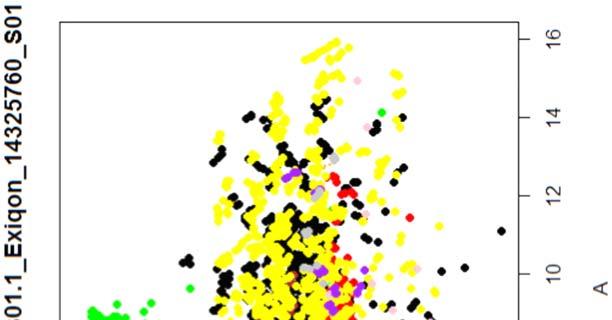
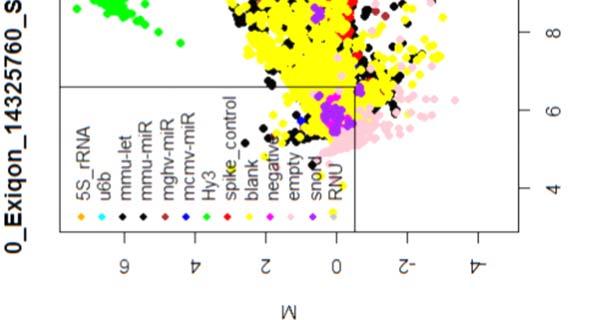
147 132
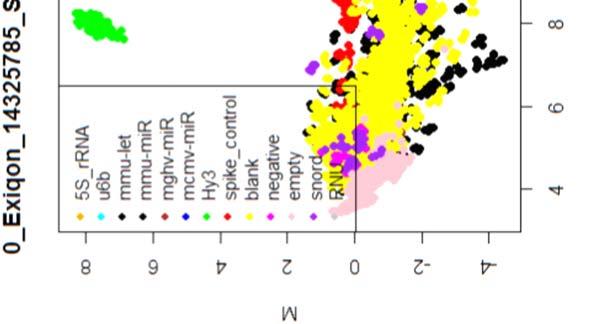
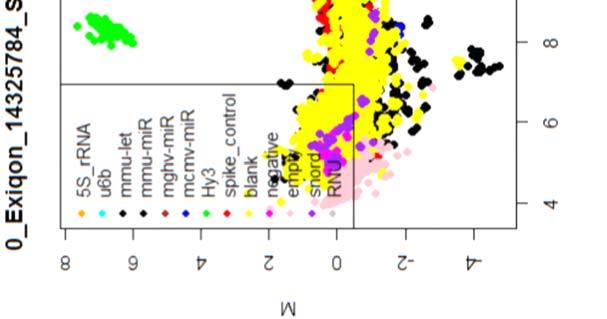
148 133
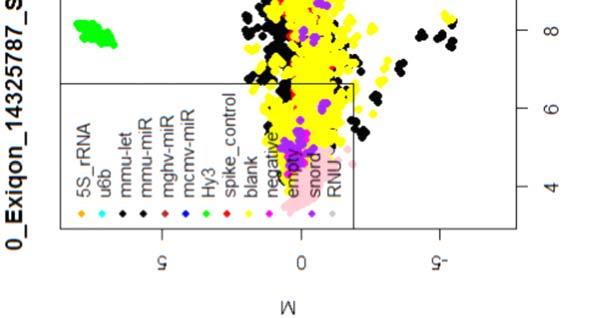
149 134
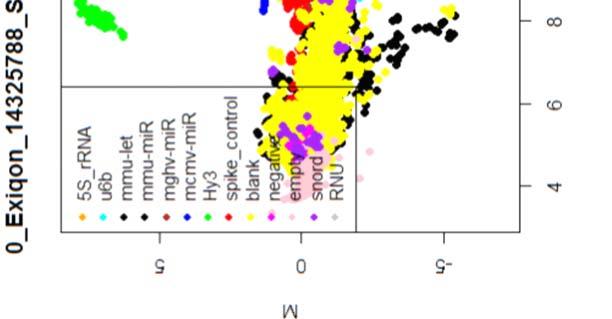
150 135
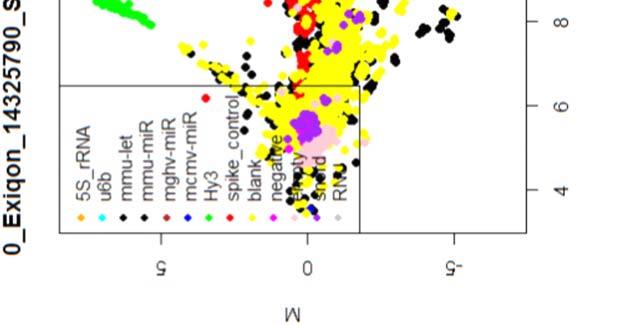
151 136
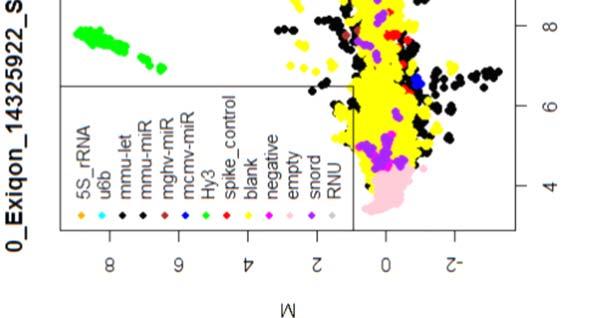
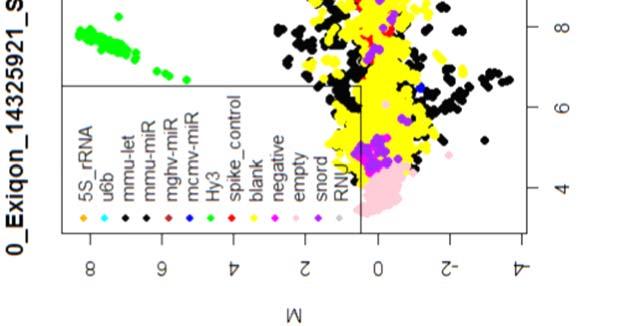
152 137
Εισαγωγή στα microrna Δ. ΣΙΔΕΡΗΣ
 Εισαγωγή στα microrna Δ. ΣΙΔΕΡΗΣ Non-Coding RNA: Παλαιότερα γνωστό ως «JUNK» NC-RNAs: Αποτελούν την πλειοψηφία των μεταγράφων σε σύνθετα γονιδιώματα Non-Coding RNA: Κλειδί στην πολυπλοκότητα των ευκαρυωτικών?
Εισαγωγή στα microrna Δ. ΣΙΔΕΡΗΣ Non-Coding RNA: Παλαιότερα γνωστό ως «JUNK» NC-RNAs: Αποτελούν την πλειοψηφία των μεταγράφων σε σύνθετα γονιδιώματα Non-Coding RNA: Κλειδί στην πολυπλοκότητα των ευκαρυωτικών?
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10
 Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ-ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής Βιολογίας Εργαστήριο Φαρμακολογίας, Τομέας Φαρμακογνωσίας/Φαρμακολογίας
Μοριακή Bιολογία ΔIAΛEΞΕΙΣ 9 & 10 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ-ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Χρήστος Παναγιωτίδης, Ph.D. Καθηγητής Κυτταρικής/Μοριακής Βιολογίας Εργαστήριο Φαρμακολογίας, Τομέας Φαρμακογνωσίας/Φαρμακολογίας
ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ- ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ
 MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞH 9 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ- ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής TO BAΣIKO ΔOΓMA THΣ MOPIAKHΣ BIOΛOΓIAΣ ΣE EYKAPYΩTIKA
MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞH 9 ΩΡΙΜΑΝΣΗ ΤΟΥ RNA, ΙΝΤΡΟΝΙΑ/ΕΞΟΝΙΑ & ΜΕΤΑ- ΜΕΤΑΓΡΑΦΙΚΗ ΡΥΘΜΙΣΗ Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής TO BAΣIKO ΔOΓMA THΣ MOPIAKHΣ BIOΛOΓIAΣ ΣE EYKAPYΩTIKA
1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; ΘΩΜΑΣ ΑΠΑΝΤΗΣΗ. 2. Ποιες είναι οι κατηγορίες γονιδίων με κριτήριο το προϊόν της μεταγραφής τους;
 Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
Βιολογία Γ Ενιαίου Λυκείου / Θετική Κατεύθυνση κεφαλαιο 2ο: αντιγραφη, εκφραση και ρυθμιση τησ ΓενετικηΣ ΠληροφοριαΣ 1. Πού πραγματοποιούνται η αντιγραφή και η μεταγραφή; Ευκαρυωτικά κύτταρα: στον πυρήνα,
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ
 ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΕΡΩΤΗΣΕΙΣ ΚΑΤΑΝΟΗΣΗΣ ΚΕΦΑΛΑΙΟ 2 ο 1. Με ποιο μηχανισμό αντιγράφεται το DNA σύμφωνα με τους Watson και Crick; 2. Ένα κύτταρο που περιέχει ένα μόνο χρωμόσωμα τοποθετείται σε θρεπτικό υλικό που περιέχει ραδιενεργό
ΘΕΜΑ Α Α1. γ Α2. γ Α3. α Α4. β Α5. β ΘΕΜΑ B B1. B2.
 ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
ΘΕΜΑ Α Α1. γ (το πριμόσωμα) Α2. γ (οι υποκινητές και οι μεταγραφικοί παράγοντες κάθε γονιδίου) Α3. α (μεταφέρει ένα συγκεκριμένο αμινοξύ στο ριβόσωμα) Α4. β (αποδιάταξη των δύο συμπληρωματικών αλυσίδων)
Μοριακή Βιολογία. Ενότητα # (5): Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής
: Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής") ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (5): Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (5): Ωρίμανση του RNA, ιντρόνια/εξώνια και μεταγραφική ρύθμιση Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
ΕΙΚΟΝΑ 2.4 ΣΤΑΔΙΑ ΜΕΤΑΦΡΑΣΗΣ σ ε λ ί δ α 1 ΕΙΚΟΝΑ 4.2β ΕΡΩΤΗΣΕΙΣ 1. Να συμπληρώσετε τα κενά πλαίσια της εικόνας με την κατάλληλη λέξη ή φράση 2. Να γράψετε τον προσανατολισμό της μετακίνησης του ριβοσώματος
Γονιδιωματική. G. Patrinos
 Γονιδιωματική Η μεταγονιδιωματική εποχή... Σημαντικότερα επιτεύγματα POST GENOME ERA Ολοκλήρωση της αποκρυπτογράφησης της αλληλουχίας των γονιδιωμάτων πολλών οργανισμών. Προτύπωση μεθοδολογιών για προσδιορισμό
Γονιδιωματική Η μεταγονιδιωματική εποχή... Σημαντικότερα επιτεύγματα POST GENOME ERA Ολοκλήρωση της αποκρυπτογράφησης της αλληλουχίας των γονιδιωμάτων πολλών οργανισμών. Προτύπωση μεθοδολογιών για προσδιορισμό
ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ
 14.1 ΕΙΣΑΓΩΓΗ") 1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
1 ΕΝΟΤΗΤΑ 14: Ο ΦΟΡΕΑΣ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ (DNA) 14.1 ΕΙΣΑΓΩΓΗ Οι δύο πολυνουκλεοτιδικές αλυσίδες του DNA αποτελούνται από νουκλεοτίδια τα οποία ενώνονται με φωσφοδιεστερικούς δεσμούς. Πιο συγκεκριμένα
Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014
 Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014 ΘΕΜΑ Α Α1. β Α2. β Α3. β Α4. β Α5. β ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ B B1. Ο όρος γονιδιακή έκφραση αναφέρεται συνήθως σε όλη τη διαδικασία με την οποία
Διαγώνισμα Βιολογίας στα Κεφάλαια 1 έως 4 ΚΥΡΙΑΚΗ 7 ΔΕΚΕΜΒΡΙΟΥ 2014 ΘΕΜΑ Α Α1. β Α2. β Α3. β Α4. β Α5. β ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ B B1. Ο όρος γονιδιακή έκφραση αναφέρεται συνήθως σε όλη τη διαδικασία με την οποία
Ανασυνδυασμένο DNA. Richard M. Myers Jan A. Witkowski. James D. Watson Amy A. Caudy. Γονίδια και Γονιδιώματα Μία Συνοπτική Παρουσίαση
 Ανασυνδυασμένο DNA Γονίδια και Γονιδιώματα Μία Συνοπτική Παρουσίαση James D. Watson Amy A. Caudy Richard M. Myers Jan A. Witkowski Κεφάλαιο 9 Η παρεμβολή RNA ρυθμίζει τη λειτουργία των γονιδίων 2 Κλασσική
Ανασυνδυασμένο DNA Γονίδια και Γονιδιώματα Μία Συνοπτική Παρουσίαση James D. Watson Amy A. Caudy Richard M. Myers Jan A. Witkowski Κεφάλαιο 9 Η παρεμβολή RNA ρυθμίζει τη λειτουργία των γονιδίων 2 Κλασσική
Κεφάλαιο 4: Ανασυνδυασμένο DNA
 Κεφάλαιο 4: Ανασυνδυασμένο DNA 1. Η ανάπτυξη της γενετικής μηχανικής επέτρεψε: α. την κατανόηση των μηχανισμών αντιγραφής του γενετικού υλικού β. την απομόνωση των πλασμιδίων από τα βακτήρια γ. την πραγματοποίηση
Κεφάλαιο 4: Ανασυνδυασμένο DNA 1. Η ανάπτυξη της γενετικής μηχανικής επέτρεψε: α. την κατανόηση των μηχανισμών αντιγραφής του γενετικού υλικού β. την απομόνωση των πλασμιδίων από τα βακτήρια γ. την πραγματοποίηση
ΒΙΟΛΟΓΙΑ Γ ΛΥΚΕΙΟΥ, ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
EIKONA 2.1 Ημισυντηρητικός μηχανισμός αντιγραφής του DNA 1. Να γράψετε τα ένζυμα που (α) προκαλούν ξετύλιγμα των αλυσίδων του αρχικού (μητρικού μορίου) DNA και (β) συνθέτουν τις νέες αλυσίδες του DNA.
ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα
 Εργασία στη Βιολογία ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Η ροή της πληροφορίας για το σχηματισμό των πρωτεϊνών, προϋποθέτει τη μεταφορά της από το DNA στο RNA (ΜΕΤΑΓΡΑΦΗ).
Εργασία στη Βιολογία ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Περετσή Χριστίνα Πιτσικάλη Παναγιώτα ΜΕΤΑΓΡΑΦΗ ΤΟΥ DNA Η ροή της πληροφορίας για το σχηματισμό των πρωτεϊνών, προϋποθέτει τη μεταφορά της από το DNA στο RNA (ΜΕΤΑΓΡΑΦΗ).
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ. ΘΕΜΑ Β Β1. Στήλη Ι Στήλη ΙΙ 1 Α 2 Γ 3 Α 4 Β 5 Α 6 Α 7 Γ
 ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΠΑΡΑΣΚΕΥΗ 27 ΜΑΪΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ ΣΥΣΤΗΜΑ) ΒΙΟΛΟΓΙΑ
ΘΕΜΑ 1 Ο ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Τα
ΜΑΘΗΜΑ / ΤΑΞΗ : ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 01/12/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Τα
Δοµή και ιδιότητες του DNA. 09/04/ Μοριακή Βιολογία Κεφ. 1 Καθηγητής Δρ. Κ. Ε. Βοργιάς
 Δοµή και ιδιότητες του DNA 09/04/2014 1 09/04/2014 2 Τόσο τα νεκρά (µε θερµική επεξεργασία) βακτήρια S όσο και τα ζωντανά βακτήρια R δεν µπορούν να θανατώσουν ποντικούς. Όµως, η ταυτόχρονη µόλυνση µε αυτά
Δοµή και ιδιότητες του DNA 09/04/2014 1 09/04/2014 2 Τόσο τα νεκρά (µε θερµική επεξεργασία) βακτήρια S όσο και τα ζωντανά βακτήρια R δεν µπορούν να θανατώσουν ποντικούς. Όµως, η ταυτόχρονη µόλυνση µε αυτά
ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ. Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της...
 ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της... Γενετική Μηχανική o Περιλαμβάνει όλες τις τεχνικές με τις οποίες μπορούμε να επεμβαίνουμε στο γενετικό υλικό των οργανισμών.
ΓΕΝΕΤΙΚΗ ΜΗΧΑΝΙΚΗ Η τεχνολογία του ανασυνδυασμένου DNA και οι εφαρμογές της... Γενετική Μηχανική o Περιλαμβάνει όλες τις τεχνικές με τις οποίες μπορούμε να επεμβαίνουμε στο γενετικό υλικό των οργανισμών.
Β. Σελ 60 σχολικού: «Η αποµόνωση του συνολικού έως και σελ 61 από µία cdna βιβλιοθήκη.». Γ. ι ι α α α ι α α ι α α α! " # $ % & ' ( ) ( ) ( * % + α ι α
 ( ) ( * % + α ι α") ! THΛ: 270727 222594 THΛ: 919113 949422 Απαντήσεις: " # $ % & ' 1=γ, 2=β, 3=γ, 4=β, 5=δ. " # $ % ( ' εδοµένα από την ανάλυση του ποσοστού των βάσεων σε µόρια DNA από διαφορετικούς οργανισµούς έδειχναν
! THΛ: 270727 222594 THΛ: 919113 949422 Απαντήσεις: " # $ % & ' 1=γ, 2=β, 3=γ, 4=β, 5=δ. " # $ % ( ' εδοµένα από την ανάλυση του ποσοστού των βάσεων σε µόρια DNA από διαφορετικούς οργανισµούς έδειχναν
ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ)
") ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ) ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό κάθε μιας από τις παρακάτω ημιτελείς προτάσεις 1-5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
ΑΠΑΝΤΗΣΕΙΣ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (Β ΛΥΚΕΙΟΥ) ΘΕΜΑ Α Να γράψετε στο τετράδιο σας τον αριθμό κάθε μιας από τις παρακάτω ημιτελείς προτάσεις 1-5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA
 Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA Εικόνα 24.1 Το hnrna απαντάται ως ριβονουκλεοπρωτεϊνικό σύµπλοκο µε µορφή µιας σειράς από χάντρες. Εικόνα 24.2 Το RNA τροποποιείται στον πυρήνα
Κ Ε Φ Α Λ Α Ι Ο 24 : Το µάτισµα και η επεξεργασία του RNA Εικόνα 24.1 Το hnrna απαντάται ως ριβονουκλεοπρωτεϊνικό σύµπλοκο µε µορφή µιας σειράς από χάντρες. Εικόνα 24.2 Το RNA τροποποιείται στον πυρήνα
Ανασυνδυασµένο DNA. Richard M. Myers Jan A. Witkowski. James D. Watson Amy A. Caudy. Γονίδια και Γονιδιώµατα Μία Συνοπτική Παρουσίαση
 Ανασυνδυασµένο DNA Γονίδια και Γονιδιώµατα Μία Συνοπτική Παρουσίαση James D. Watson Amy A. Caudy Richard M. Myers Jan A. Witkowski Κεφάλαιο 9 Η παρεµβολή RNA ρυθµίζει τη λειτουργία των γονιδίων 2 Κλασσική
Ανασυνδυασµένο DNA Γονίδια και Γονιδιώµατα Μία Συνοπτική Παρουσίαση James D. Watson Amy A. Caudy Richard M. Myers Jan A. Witkowski Κεφάλαιο 9 Η παρεµβολή RNA ρυθµίζει τη λειτουργία των γονιδίων 2 Κλασσική
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών
 Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
Κεφ. 4 DNA, RNA και η ροή των γενετικών πληροφοριών Η οικογενειακή ομοιότητα, οφείλεται στα κοινά γονίδια. Τα γονίδια πρέπει να εκφραστούν για να έχουν αποτέλεσμα, και η έκφραση αυτή ρυθμίζεται από πρωτεΐνες.
5 GTG CAC CTG ACT CCT GAG GAG 3 3 CAC GTG GAC TGA GGA CTC CTC 5
 Βιολογία Κατεύθυνσης Γ Λυκείου Απαντήσεις διαγωνίσματος στο Κεφάλαιο 4 ο ΘΕΜΑ Α Α1. β Α2. β Α3. γ Α4. β Α5. β ΘΕΜΑ B B1. Ο κλώνος είναι μια ομάδα πανομοιότυπων μορίων, κυττάρων, ή οργανισμών. B2. Η υβριδοποίηση
Βιολογία Κατεύθυνσης Γ Λυκείου Απαντήσεις διαγωνίσματος στο Κεφάλαιο 4 ο ΘΕΜΑ Α Α1. β Α2. β Α3. γ Α4. β Α5. β ΘΕΜΑ B B1. Ο κλώνος είναι μια ομάδα πανομοιότυπων μορίων, κυττάρων, ή οργανισμών. B2. Η υβριδοποίηση
ΧΡΗΣΤΟΣ ΚΑΚΑΒΑΣ 1 ΚΑΘΗΓΗΤΗΣ ΒΙΟΛΟΓΟΣ Μ.Δ.Ε
 ΚΕΦΑΛΑΙΟ 2 ον. ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ ΤΙ ΠΡΕΠΕΙ ΝΑ ΞΕΡΩ. 1. Τη δομή της δίκλωνης έλικας πάρα πολύ καλά. 2. Τους δεσμούς υδρογόνου μεταξύ των συμπληρωματικών βάσεων και την επίπτωσή
ΚΕΦΑΛΑΙΟ 2 ον. ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ ΤΙ ΠΡΕΠΕΙ ΝΑ ΞΕΡΩ. 1. Τη δομή της δίκλωνης έλικας πάρα πολύ καλά. 2. Τους δεσμούς υδρογόνου μεταξύ των συμπληρωματικών βάσεων και την επίπτωσή
NATIONAL AND KAPODISTRIAN UNIVERSITY OF ATHENS SCHOOL OF SCIENCE FACULTY OF INFORMATICS AND TELECOMMUNICATIONS
 NATIONAL AND KAPODISTRIAN UNIVERSITY OF ATHENS SCHOOL OF SCIENCE FACULTY OF INFORMATICS AND TELECOMMUNICATIONS INTERDICIPLINARY POSTGRADUATE PROGRAMME "INFORMATION TECHNOLOGIES IN MEDICINE AND BIOLOGY"
NATIONAL AND KAPODISTRIAN UNIVERSITY OF ATHENS SCHOOL OF SCIENCE FACULTY OF INFORMATICS AND TELECOMMUNICATIONS INTERDICIPLINARY POSTGRADUATE PROGRAMME "INFORMATION TECHNOLOGIES IN MEDICINE AND BIOLOGY"
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής
 Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
Κ Ε Φ Α Λ Α Ι Ο 22 : Η ενεργοποίηση της µεταγραφής Εικόνα 22.1 Η γονιδιακή έκφραση ελέγχεται κυρίως κατά την έναρξη της µεταγραφής και σπάνια στα επόµενα στάδια της γονιδιακής έκφρασης, παρόλο που ο έλεγχος
ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 2016
 ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 2016 ΘΕΜΑ Α Α1 Β Α2 Β Α3 Δ Α4 Γ Α5 Γ ΘΕΜΑ Β Β1 1. Α 2. Γ 3. Α 4. Β 5. Α 6. Α 7. Γ Β2 ΣΕΛ.24 σχολ.βιβ. «Κάθε φυσιολογικό µεταφασικό.. Η απεικόνιση αυτή αποτελεί
ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΒΙΟΛΟΓΙΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 2016 ΘΕΜΑ Α Α1 Β Α2 Β Α3 Δ Α4 Γ Α5 Γ ΘΕΜΑ Β Β1 1. Α 2. Γ 3. Α 4. Β 5. Α 6. Α 7. Γ Β2 ΣΕΛ.24 σχολ.βιβ. «Κάθε φυσιολογικό µεταφασικό.. Η απεικόνιση αυτή αποτελεί
Η παρεμβολή RNA, RNAi
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΤΜΗΜΑ ΒΙΟΧΗΜΕΙΑΣ ΚΑΙ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Η ΡΥΘΜΙΣΗ ΤΗΣ ΓΟΝΙΔΙΑΚΗΣ ΕΚΦΡΑΣΗΣ στους ευκαρυωτικούς οργανισμούς Μέρος Β Η παρεμβολή RNA, RNAi Νικόλαος Μπαλατσός RNA interference, RNAi: Specific
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΤΜΗΜΑ ΒΙΟΧΗΜΕΙΑΣ ΚΑΙ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ Η ΡΥΘΜΙΣΗ ΤΗΣ ΓΟΝΙΔΙΑΚΗΣ ΕΚΦΡΑΣΗΣ στους ευκαρυωτικούς οργανισμούς Μέρος Β Η παρεμβολή RNA, RNAi Νικόλαος Μπαλατσός RNA interference, RNAi: Specific
Κεντρικό δόγμα της βιολογίας
 Κεντρικό δόγμα της βιολογίας DNA RNA Πρωτεΐνη Μεταγραφή Σύνθεση (μονόκλωνου) RNA από ένα δίκλωνο μόριο DNA κυρίως με τη βοήθεια του ενζύμου RNA πολυμεράση Το προϊόν της μεταγραφής ονομάζεται πρωτογενές
Κεντρικό δόγμα της βιολογίας DNA RNA Πρωτεΐνη Μεταγραφή Σύνθεση (μονόκλωνου) RNA από ένα δίκλωνο μόριο DNA κυρίως με τη βοήθεια του ενζύμου RNA πολυμεράση Το προϊόν της μεταγραφής ονομάζεται πρωτογενές
Βιολογία ΘΕΜΑ Α ΘΕΜΑ B
 Βιολογία προσανατολισμού Α. 1. β 2. γ 3. δ 4. γ 5. δ ΘΕΜΑ Α B1. 4,1,2,6,8,3,5,7 ΘΕΜΑ B B2. Σχολικό βιβλίο σελ. 103 Η γενετική καθοδήγηση είναι.υγιών απογόνων. Σχολικό βιβλίο σελ. 103 Παρ ότι γενετική καθοδήγηση
Βιολογία προσανατολισμού Α. 1. β 2. γ 3. δ 4. γ 5. δ ΘΕΜΑ Α B1. 4,1,2,6,8,3,5,7 ΘΕΜΑ B B2. Σχολικό βιβλίο σελ. 103 Η γενετική καθοδήγηση είναι.υγιών απογόνων. Σχολικό βιβλίο σελ. 103 Παρ ότι γενετική καθοδήγηση
Β1. Β2. ΘΕΜΑ 2ο 1. 2.
 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 20 ΣΕΠΤΕΜΒΡΙΟΥ 2002 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ) ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο Α1.
1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 20 ΣΕΠΤΕΜΒΡΙΟΥ 2002 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: BΙΟΛΟΓΙΑ (ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ) ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο Α1.
Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ
 Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΝΤΙΓΡΑΦΗ ΜΕΤΑΓΡΑΦΗ Γίνεται σύνθεση DNA. Γίνεται σύνθεση RNA. Εξασφαλίζεται η διαιώνιση της γενετικής
Ι. ΘΕΩΡΙΑ ΠΙΝΑΚΑΣ 2.1: ΣΥΓΚΡΙΤΙΚΟΣ ΠΙΝΑΚΑΣ ΑΝΤΙΓΡΑΦΗΣ-ΜΕΤΑΓΡΑΦΗΣ ΣΤΟΝ ΠΥΡΗΝΑ ΤΩΝ ΕΥΚΑΡΥΩΤΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΝΤΙΓΡΑΦΗ ΜΕΤΑΓΡΑΦΗ Γίνεται σύνθεση DNA. Γίνεται σύνθεση RNA. Εξασφαλίζεται η διαιώνιση της γενετικής
ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ & ΕΠΑΛ (ΟΜΑΔΑ Β ) ΝΕΟ & ΠΑΛΑΙΟ ΣΥΣΤΗΜΑ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 27/05/2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΟΠ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1:
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ & ΕΠΑΛ (ΟΜΑΔΑ Β ) ΝΕΟ & ΠΑΛΑΙΟ ΣΥΣΤΗΜΑ ΗΜΕΡΟΜΗΝΙΑ ΕΞΕΤΑΣΗΣ: 27/05/2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΟΠ ΠΡΟΤΕΙΝΟΜΕΝΕΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑΤΩΝ Θέμα Α Α1:
ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 02/12/2012 ΑΠΑΝΤΗΣΕΙΣ
 ΤΣΙΜΙΣΚΗ &ΚΑΡΟΛΟΥ ΝΤΗΛ ΓΩΝΙΑ THΛ: 270727 222594 ΑΡΤΑΚΗΣ 12 - Κ. ΤΟΥΜΠΑ THΛ: 919113 949422 ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 02/12/2012 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1 ο Α. Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη
ΤΣΙΜΙΣΚΗ &ΚΑΡΟΛΟΥ ΝΤΗΛ ΓΩΝΙΑ THΛ: 270727 222594 ΑΡΤΑΚΗΣ 12 - Κ. ΤΟΥΜΠΑ THΛ: 919113 949422 ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 02/12/2012 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1 ο Α. Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ )
") Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
Κεφάλαιο 10 ΤΟ ΟΠΕΡΟΝΙΟ (σελ. 387-417) Ένα ρυθμιστικό γονίδιο κωδικοποιεί μια πρωτεΐνη που δρα σε μια θέση-στόχο πάνω στο DNA και ρυθμίζει την έκφραση ενός άλλου γονιδίου. Στον αρνητικό έλεγχο, μία trans-δραστική
Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Σε ένα
ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ 04/11/2018 Νότα Λαζαράκη Αλέξανδρος Παπαγιαννακόπουλος ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Σε ένα
ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1.
1 ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΕΜΠΤΗ 11 ΙΟΥΝΙΟΥ 2015 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1.
Εξόρυξη Γνώσης από Βιολογικά εδομένα
 Παρουσίαση Διπλωματικής Εργασίας Εξόρυξη Γνώσης από Βιολογικά εδομένα Καρυπίδης Γεώργιος (Μ27/03) Επιβλέπων Καθηγητής: Ιωάννης Βλαχάβας MIS Πανεπιστήμιο Μακεδονίας Φεβρουάριος 2005 Εξόρυξη Γνώσης από Βιολογικά
Παρουσίαση Διπλωματικής Εργασίας Εξόρυξη Γνώσης από Βιολογικά εδομένα Καρυπίδης Γεώργιος (Μ27/03) Επιβλέπων Καθηγητής: Ιωάννης Βλαχάβας MIS Πανεπιστήμιο Μακεδονίας Φεβρουάριος 2005 Εξόρυξη Γνώσης από Βιολογικά
Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά;
 ΒΙΟΛΟΓΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ 12 26/10/2016 Κεφάλαιο 3 Α μέρος Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά; Ποια είναι η δομή
ΒΙΟΛΟΓΙΚΗ ΑΝΘΡΩΠΟΛΟΓΙΑ 12 26/10/2016 Κεφάλαιο 3 Α μέρος Εισαγωγή στη Γενετική και στη Γονιδιωματική Τι είναι η κληρονομικότητα, και πώς μεταβιβάζεται η πληροφορία από γενιά σε γενιά; Ποια είναι η δομή
ΛΥΣΗ ΤΗΣ ΑΣΚΗΣΗΣ ΕΠΑΝΑΛΗΨΗΣ
 ΛΥΣΗ ΤΗΣ ΑΣΚΗΣΗΣ ΕΠΑΝΑΛΗΨΗΣ 1. Ο γενετικός κώδικας είναι ένας κώδικας αντιστοίχισης των κωδικονίων του mrna με αμινοξέα στην πολυπεπτιδική αλυσίδα. Σύμφωνα με αυτόν η 3 μετάφραση όλων των mrna αρχίζει
ΛΥΣΗ ΤΗΣ ΑΣΚΗΣΗΣ ΕΠΑΝΑΛΗΨΗΣ 1. Ο γενετικός κώδικας είναι ένας κώδικας αντιστοίχισης των κωδικονίων του mrna με αμινοξέα στην πολυπεπτιδική αλυσίδα. Σύμφωνα με αυτόν η 3 μετάφραση όλων των mrna αρχίζει
Τηλ: Ανδρέου Δημητρίου 81 & Ακριτών 26 -ΚΑΛΟΓΡΕΖΑ
 ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ- ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ (Ιανουάριος 2014) 1 ο ΘΕΜΑ Απαντήστε στις παρακάτω ερωτήσεις πολλαπλής επιλογής. Μία απάντηση είναι η σωστή. 1. Υβριδοποίηση: Α. Είναι ιδιότητα του DNA
ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ- ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ (Ιανουάριος 2014) 1 ο ΘΕΜΑ Απαντήστε στις παρακάτω ερωτήσεις πολλαπλής επιλογής. Μία απάντηση είναι η σωστή. 1. Υβριδοποίηση: Α. Είναι ιδιότητα του DNA
Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ. Ημερομηνία: Κυριακή 23 Οκτωβρίου 2016 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ
 ΤΑΞΗ: ΜΑΘΗΜΑ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Ημερομηνία: Κυριακή 23 Οκτωβρίου 2016 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Στις ημιτελείς προτάσεις Α1 Α4 να γράψετε στο
ΤΑΞΗ: ΜΑΘΗΜΑ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Ημερομηνία: Κυριακή 23 Οκτωβρίου 2016 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Στις ημιτελείς προτάσεις Α1 Α4 να γράψετε στο
ΜΕΛΕΤΗ ΑΝΘΕΚΤΙΚΟΤΗΤΑΣ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΣΤΗ ΘΕΡΑΠΕΙΑ
 ΕΡΕΥΝΗΤΙΚΗ ΜΟΝΑΔΑ: ΜΟΡΙΑΚΗ ΒΙΟΛΟΓΙΑ ΥΠΕΥΘΥΝΟΣ: ΤΣΕΖΟΥ ΑΣΠΑΣΙΑ ΜΕΛΕΤΗ ΑΝΘΕΚΤΙΚΟΤΗΤΑΣ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΣΤΗ ΘΕΡΑΠΕΙΑ ΠΑΠΑΝΙΚΟΛΑΟΥ Βασίλης, PhD Λάρισα, 2012 ΚΑΡΚΙΝΟΣ ΜΑΣΤΟΥ Αποτελεί την πιο κοινή μορφή
ΕΡΕΥΝΗΤΙΚΗ ΜΟΝΑΔΑ: ΜΟΡΙΑΚΗ ΒΙΟΛΟΓΙΑ ΥΠΕΥΘΥΝΟΣ: ΤΣΕΖΟΥ ΑΣΠΑΣΙΑ ΜΕΛΕΤΗ ΑΝΘΕΚΤΙΚΟΤΗΤΑΣ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΣΤΗ ΘΕΡΑΠΕΙΑ ΠΑΠΑΝΙΚΟΛΑΟΥ Βασίλης, PhD Λάρισα, 2012 ΚΑΡΚΙΝΟΣ ΜΑΣΤΟΥ Αποτελεί την πιο κοινή μορφή
Μόρια-κλειδιά των ζωντανών οργανισμών καθώς περιέχουν την γενετική πληροφορία Νουκλεϊκά οξέα:
 Μόρια-κλειδιά των ζωντανών οργανισμών καθώς περιέχουν την γενετική πληροφορία 1. Δεοξυριβονουκλεϊκό οξύ (DNA) Νουκλεϊκά οξέα: Φορέας της γενετικής πληροφορίας 2. Ριβονουκλεϊκό οξύ (RNA) Συμμετοχή στην
Μόρια-κλειδιά των ζωντανών οργανισμών καθώς περιέχουν την γενετική πληροφορία 1. Δεοξυριβονουκλεϊκό οξύ (DNA) Νουκλεϊκά οξέα: Φορέας της γενετικής πληροφορίας 2. Ριβονουκλεϊκό οξύ (RNA) Συμμετοχή στην
Φάσμα group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι.
 σύγχρονο Φάσμα group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. µαθητικό φροντιστήριο Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ 50.51.557 50.56.296 25ης Μαρτίου 74 ΠΛ.ΠΕΤΡΟΥΠΟΛΗΣ 50.50.658 50.60.845 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 50.27.990
σύγχρονο Φάσμα group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. µαθητικό φροντιστήριο Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ 50.51.557 50.56.296 25ης Μαρτίου 74 ΠΛ.ΠΕΤΡΟΥΠΟΛΗΣ 50.50.658 50.60.845 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 50.27.990
ΟΜΟΣΠΟΝΔΙΑ ΕΚΠΑΙΔΕΥΤΙΚΩΝ ΦΡΟΝΤΙΣΤΩΝ ΕΛΛΑΔΟΣ (Ο.Ε.Φ.Ε.) ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2018 A ΦΑΣΗ ΒΙΟΛΟΓΙΑ ΑΛΓΟΡΙΘΜΟΣ
 ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2018 A ΦΑΣΗ ΒΙΟΛΟΓΙΑ ΑΛΓΟΡΙΘΜΟΣ") ΤΑΞΗ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΣ: ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ Ημερομηνία: Σάββατο 13 Ιανουαρίου 2018 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς
ΤΑΞΗ: Γ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΣ: ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ Ημερομηνία: Σάββατο 13 Ιανουαρίου 2018 Διάρκεια Εξέτασης: 3 ώρες ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς
Δοµή και ιδιότητες του DNA. 23/02/ Μοριακή Βιολογία Κεφ. 1 Καθηγητής Δρ. Κ. Ε. Βοργιάς
 Δοµή και ιδιότητες του DNA 23/02/2014 1 23/02/2014 2 Τόσο τα νεκρά (µε θερµική επεξεργασία) βακτήρια S όσο και τα ζωντανά βακτήρια R δεν µπορούν να θανατώσουν ποντικούς. Όµως, η ταυτόχρονη µόλυνση µε αυτά
Δοµή και ιδιότητες του DNA 23/02/2014 1 23/02/2014 2 Τόσο τα νεκρά (µε θερµική επεξεργασία) βακτήρια S όσο και τα ζωντανά βακτήρια R δεν µπορούν να θανατώσουν ποντικούς. Όµως, η ταυτόχρονη µόλυνση µε αυτά
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1ο Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις 1 έως 5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή τη φράση, η οποία
Μηχανισμοί Μεταμεταγραφικού Ελέγχου
 Μηχανισμοί Μεταμεταγραφικού Ελέγχου Δημήτρης Λ. Κοντογιάννης, PhD Ερευνητής Β, Ινστιτούτο Ανοσολογίας ΕΕΠΑ Αθήνα 6-2-2010 Crowne-Plaza Μεταμεταγραφικοί Μηχανισμοί Χρήσης- Τα Βασικά... ΕΕΠΑ Αθήνα 6-2-2010
Μηχανισμοί Μεταμεταγραφικού Ελέγχου Δημήτρης Λ. Κοντογιάννης, PhD Ερευνητής Β, Ινστιτούτο Ανοσολογίας ΕΕΠΑ Αθήνα 6-2-2010 Crowne-Plaza Μεταμεταγραφικοί Μηχανισμοί Χρήσης- Τα Βασικά... ΕΕΠΑ Αθήνα 6-2-2010
Ενδεικτικές απαντήσεις βιολογίας κατεύθυνσης 2014
 Θέμα Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β Θέμα Β ΑΓ.ΚΩΝΣΤΑΝΤΙΝΟΥ 11 -- ΠΕΙΡΑΙΑΣ -- 18532 -- ΤΗΛ. 210-4224752, 4223687 Ενδεικτικές απαντήσεις βιολογίας κατεύθυνσης 2014 Β1. 4 2 1 6 3 5 Β2. α. DNA πολυμεράση
Θέμα Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β Θέμα Β ΑΓ.ΚΩΝΣΤΑΝΤΙΝΟΥ 11 -- ΠΕΙΡΑΙΑΣ -- 18532 -- ΤΗΛ. 210-4224752, 4223687 Ενδεικτικές απαντήσεις βιολογίας κατεύθυνσης 2014 Β1. 4 2 1 6 3 5 Β2. α. DNA πολυμεράση
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης
 Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Εξέλιξη και ανθρώπινος πολιτισμός: Η ρύθμιση του γονιδίου της λακτάσης Η διατήρηση του ενζύμου της λακτάσης στους ενήλικες είναι ένα παράδειγμα πρόσφατης εξέλιξης στον άνθρωπο. Μας δείχνει επίσης πώς μεταλλαγές
Το mir-29a ως βιοδείκτης στην υπερτροφική μυοκαρδιοπάθεια. Δ. Ντέλιος 1,2
 Το mir-29a ως βιοδείκτης στην υπερτροφική μυοκαρδιοπάθεια Δ. Ντέλιος 1,2 Υπερτροφική μυοκαρδιοπάθεια - 1:500 στο γενικό πληθυσμό Πρωτεΐνες του σαρκομεριδίου ΜΥΗ7, MYBPC3, TNNI3, TNNT2, TPM1, MYL3, MYL2
Το mir-29a ως βιοδείκτης στην υπερτροφική μυοκαρδιοπάθεια Δ. Ντέλιος 1,2 Υπερτροφική μυοκαρδιοπάθεια - 1:500 στο γενικό πληθυσμό Πρωτεΐνες του σαρκομεριδίου ΜΥΗ7, MYBPC3, TNNI3, TNNT2, TPM1, MYL3, MYL2
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΤΗΣ Γ' ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ. ΟΙ ΛΥΣΕΙΣ ΤΩΝ ΘΕΜΑΤΩΝ ΑΠΟ ΤΟΝ ΚΑΘΗΓΗΤΗ κύριο ΤΡΙΓΚΑ ΓΕΩΡΓΙΟ του ΦΡΟΝΤΙΣΤΗΡΙΟΥ
 ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΤΗΣ Γ' ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΙ ΛΥΣΕΙΣ ΤΩΝ ΘΕΜΑΤΩΝ ΑΠΟ ΤΟΝ ΚΑΘΗΓΗΤΗ κύριο ΤΡΙΓΚΑ ΓΕΩΡΓΙΟ του ΦΡΟΝΤΙΣΤΗΡΙΟΥ www.orionidef.gr ΘΕΜΑ A Α1. α Α2. δ Α3. γ Α4. β Α5. β ΘΕΜΑ
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ ΤΗΣ Γ' ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΙ ΛΥΣΕΙΣ ΤΩΝ ΘΕΜΑΤΩΝ ΑΠΟ ΤΟΝ ΚΑΘΗΓΗΤΗ κύριο ΤΡΙΓΚΑ ΓΕΩΡΓΙΟ του ΦΡΟΝΤΙΣΤΗΡΙΟΥ www.orionidef.gr ΘΕΜΑ A Α1. α Α2. δ Α3. γ Α4. β Α5. β ΘΕΜΑ
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη
 Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
Χρωμοσώματα και ανθρώπινο γονιδίωμα Πεφάνη Δάφνη 12.02.2019 Νουκλεoτίδια-Δομικοί λίθοι του DNA H διπλή έλικα του DNAχωροπληρωτικό μοντέλο To ευκαρυωτικό DNA οργανώνεται σε χρωμοσώματα Τα χρωμοσώματα περιέχουν
Οργάνωση των NA σε ιούς. 09/04/ Μοριακή Βιολογία Κεφ. 1 Καθηγητής Δρ. Κ. Ε. Βοργιάς
 Οργάνωση των NA σε ιούς 09/04/2014 1 Η οργάνωση των ιών γίνεται µε βάση το εύρος των ξενιστών που µπορούν να µολύνουν και συµπεριφέρονται σαν παράσιτα: Μόνο βακτήρια: βακτηριοφάγος ή φάγος (phage) Ζωϊκά
Οργάνωση των NA σε ιούς 09/04/2014 1 Η οργάνωση των ιών γίνεται µε βάση το εύρος των ξενιστών που µπορούν να µολύνουν και συµπεριφέρονται σαν παράσιτα: Μόνο βακτήρια: βακτηριοφάγος ή φάγος (phage) Ζωϊκά
ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ Α Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις Α1 έως Α5 και δίπλα το γράμμα που αντιστοιχεί στη λέξη
ΘΕΜΑ Α ΘΕΜΑ Β ΑΠΑΝΤΗΣΕΙΣ. Α1. β. Α2. γ. Α3. δ. Α4. γ. Α5. β Β1. 5, 4, 2, 1, 3. Β2. Τα δομικά μέρη του οπερονίου της λακτόζης είναι κατά σειρά τα εξής:
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β ΘΕΜΑ
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΔΕΥΤΕΡΑ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β ΘΕΜΑ
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ
 ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
ΑΠΟΛΥΤΗΡΙΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 27 ΜΑΪΟΥ 2008 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 1 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ 1ο 1. β 2. δ 3. β 4. δ 5. β ΘΕΜΑ 2ο 1. Σχολικό βιβλίο, σελ.
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος. 09/04/ Μοριακή Βιολογία Κεφ. 1 Καθηγητής Δρ. Κ. Ε. Βοργιάς
 Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
Δοµή και ιδιότητες του DNA σε επίπεδο χρωµατίνηςνουκλεοσώµατος 09/04/2014 1 09/04/2014 2 Η καθαρά δοµική εικόνα της χρωµατίνης µας παρέχει µόνο µια στατική περιγραφή της. Δυναµική εικόνα της χρωµατίνης
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2014
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2014 ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ Β Β1. Η σειρά των βημάτων που οδηγούν στην κατασκευή καρυότυπου είναι: 4, 2, 1, 6, 3, 5 Β2. α.
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΣΤΑ ΘΕΜΑΤΑ ΕΞΕΤΑΣΕΩΝ 2014 ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ Β Β1. Η σειρά των βημάτων που οδηγούν στην κατασκευή καρυότυπου είναι: 4, 2, 1, 6, 3, 5 Β2. α.
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ") 1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ
1 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β) ΤΕΤΑΡΤΗ 4 ΙΟΥΝΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. γ Α3. β Α4. γ Α5. β ΘΕΜΑ
ΒΙΟΛΟΓΙΑ Ο.Π. ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ. Να σημειώσετε το γράμμα που συμπληρώνει κατάλληλα τη φράση:
 Κανάρη 36, Δάφνη Τηλ. 210 9713934 & 210 9769376 ΔΙΑΓΩΝΙΣΜΑ ΠΡΟΣΟΜΟΙΩΣΗΣ ΒΙΟΛΟΓΙΑ Ο.Π. ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΘΕΜΑ Α Να σημειώσετε το γράμμα που συμπληρώνει κατάλληλα τη φράση: Α1. Το βακτήριο Agrobacterium tumefaciens
Κανάρη 36, Δάφνη Τηλ. 210 9713934 & 210 9769376 ΔΙΑΓΩΝΙΣΜΑ ΠΡΟΣΟΜΟΙΩΣΗΣ ΒΙΟΛΟΓΙΑ Ο.Π. ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ ΘΕΜΑ Α Να σημειώσετε το γράμμα που συμπληρώνει κατάλληλα τη φράση: Α1. Το βακτήριο Agrobacterium tumefaciens
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων. Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014. Απαντήσεις Θεμάτων
 Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
Πανελλήνιες Εξετάσεις Ημερήσιων Γενικών Λυκείων Εξεταζόμενο Μάθημα: Βιολογία Θετικής Κατεύθυνσης, Ημ/νία: 04 Ιουνίου 2014 Απαντήσεις Θεμάτων ΘΕΜΑ Α A1. Τα πλασμίδια είναι: δ. κυκλικά δίκλωνα μόρια DNA
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ
 11 Ιουνίου 2015 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Απαντήσεις Θεμάτων Επαναληπτικών Πανελληνίων Εξετάσεων Ημερησίων & Εσπερινών Γενικών Λυκείων ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ B Β1. 1. Β 2. Γ 3. Α
11 Ιουνίου 2015 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Απαντήσεις Θεμάτων Επαναληπτικών Πανελληνίων Εξετάσεων Ημερησίων & Εσπερινών Γενικών Λυκείων ΘΕΜΑ Α Α1. β Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ B Β1. 1. Β 2. Γ 3. Α
ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ
 16 Ιουνίου 2017 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Εσπερινών Λυκείων Γενικών ΘΕΜΑ Α Α.1 δ Α.2 δ Α.3 β Α.4 γ Α.5 α ΘΕΜΑ B B.1 I. A II. E III. ΣΤ IV.
16 Ιουνίου 2017 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Εσπερινών Λυκείων Γενικών ΘΕΜΑ Α Α.1 δ Α.2 δ Α.3 β Α.4 γ Α.5 α ΘΕΜΑ B B.1 I. A II. E III. ΣΤ IV.
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β
 ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
ΘΕΜΑ Α Α1. β Α2. γ Α3. δ Α4. γ Α5. β 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 10 ΣΕΠΤΕΜΒΡΙΟΥ 2014 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ:
Ογκογόνα Μικρό - RNAs (oncomirs): micrornas με Ρόλο σιον Καρκίνο
: micrornas με Ρόλο σιον Καρκίνο") Ογκογόνα Μικρό - RNAs (oncomirs): micrornas με Ρόλο σιον Καρκίνο Στέλλα Λογοθέτη, Αθηνά Γιαρίκα, Δημήτριος Μήκος, Σπύρος Βλαχόπουλος και Βασίλειος Ζουμπουρλής Μονάδα Βιοϊατρικών Εφαρμογών, Ινστιτούτο Βιολογικών
Ογκογόνα Μικρό - RNAs (oncomirs): micrornas με Ρόλο σιον Καρκίνο Στέλλα Λογοθέτη, Αθηνά Γιαρίκα, Δημήτριος Μήκος, Σπύρος Βλαχόπουλος και Βασίλειος Ζουμπουρλής Μονάδα Βιοϊατρικών Εφαρμογών, Ινστιτούτο Βιολογικών
ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΕΠΛ 450 ΥΠΟΛΟΓΙΣΤΙΚΗ ΒΙΟΛΟΓΙΑ. Παύλος Αντωνίου
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΕΠΛ 450 ΥΠΟΛΟΓΙΣΤΙΚΗ ΒΙΟΛΟΓΙΑ Παύλος Αντωνίου Με μια ματιά: Εισαγωγή στη Βιολογία Ευθυγράμμιση Ακολουθιών Αναζήτηση ομοίων ακολουθιών από βάσεις δεδομενων Φυλογενετική πρόβλεψη Πρόβλεψη
ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΕΠΛ 450 ΥΠΟΛΟΓΙΣΤΙΚΗ ΒΙΟΛΟΓΙΑ Παύλος Αντωνίου Με μια ματιά: Εισαγωγή στη Βιολογία Ευθυγράμμιση Ακολουθιών Αναζήτηση ομοίων ακολουθιών από βάσεις δεδομενων Φυλογενετική πρόβλεψη Πρόβλεψη
Αθήνα, 18/5/2011 ΠΑΝΕΛΛΗΝΙΑ ΕΝΩΣΗ ΒΙΟΕΠΙΣΤΗΜΟΝΩΝ
 Αθήνα, 18/5/2011 ΠΑΝΕΛΛΗΝΙΑ ΕΝΩΣΗ ΒΙΟΕΠΙΣΤΗΜΟΝΩΝ Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Θετικής Κατεύθυνσης των Ημερησίων Γενικών Λυκείων. Η Επιτροπή Παιδείας
Αθήνα, 18/5/2011 ΠΑΝΕΛΛΗΝΙΑ ΕΝΩΣΗ ΒΙΟΕΠΙΣΤΗΜΟΝΩΝ Σας αποστέλλουμε τις προτεινόμενες απαντήσεις που αφορούν τα θέματα της Βιολογίας Θετικής Κατεύθυνσης των Ημερησίων Γενικών Λυκείων. Η Επιτροπή Παιδείας
Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΑΠΑΝΤΗΣΕΙΣ ÏÅÖÅ
 1 Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 ο 1 γ 2 δ 3 β 4 α 5 γ ΘΕΜΑ 2 ο ΑΠΑΝΤΗΣΕΙΣ Μονάδες 25 (5Χ5) Α. ιαγονιδιακά ζώα ονοµάζονται εκείνα στα οποία το γενετικό τους υλικό έχει τροποποιηθεί µε την
1 Γ' ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΚΑΤΕΥΘΥΝΣΗ ΒΙΟΛΟΓΙΑ ΘΕΜΑ 1 ο 1 γ 2 δ 3 β 4 α 5 γ ΘΕΜΑ 2 ο ΑΠΑΝΤΗΣΕΙΣ Μονάδες 25 (5Χ5) Α. ιαγονιδιακά ζώα ονοµάζονται εκείνα στα οποία το γενετικό τους υλικό έχει τροποποιηθεί µε την
) 4 x 10 5 ) 2 x 10 5
 4 x 10 5 ) 2 x 10 5") 1 & ( ) 27 2016 - : ( ) ( ) : (5) 1 5,,,. 1.. DNA. DNA. DNA. RNA. 2.. 46... DNA 1,5 x 10 9. 3. Dolly. DNA.. 1. DNA. 4. (ADA),. AIDS.... 1 5 2 & 5. Ti. Agrobacterium tumefaciens. T 2. DNA.. 1. N,,,. 1.
1 & ( ) 27 2016 - : ( ) ( ) : (5) 1 5,,,. 1.. DNA. DNA. DNA. RNA. 2.. 46... DNA 1,5 x 10 9. 3. Dolly. DNA.. 1. DNA. 4. (ADA),. AIDS.... 1 5 2 & 5. Ti. Agrobacterium tumefaciens. T 2. DNA.. 1. N,,,. 1.
Βιολογία προσανατολισμού
 Βιολογία προσανατολισμού ΘΕΜΑ Α Στις προτάσεις από Α1-Α5 να βρείτε την σωστή απάντηση. Α1. Ένας ερευνητής απομόνωσε ένα ασυνεχές γονίδιο από το γονιδίωμα ανθρώπινων κυττάρων. Το γονίδιο συνδέθηκε με βακτηριακό
Βιολογία προσανατολισμού ΘΕΜΑ Α Στις προτάσεις από Α1-Α5 να βρείτε την σωστή απάντηση. Α1. Ένας ερευνητής απομόνωσε ένα ασυνεχές γονίδιο από το γονιδίωμα ανθρώπινων κυττάρων. Το γονίδιο συνδέθηκε με βακτηριακό
Το 1998 άρχισε να εγκαθιδρύεται η. αντεπιδρούν µε το mrna και να
 Το 1998 άρχισε να εγκαθιδρύεται η αντίληψη ότι τα δίκλωνα RNA µόρια µπορούν να αντεπιδρούν µε τα γονίδια και να τα αδρανοποιούν ή µπορούν να αντεπιδρούν µε το mrna και να προωθούν την αποικοδόµηση του
Το 1998 άρχισε να εγκαθιδρύεται η αντίληψη ότι τα δίκλωνα RNA µόρια µπορούν να αντεπιδρούν µε τα γονίδια και να τα αδρανοποιούν ή µπορούν να αντεπιδρούν µε το mrna και να προωθούν την αποικοδόµηση του
Βιοπληροψορική, συσιημική βιολογία και εξατομικευμένη θεραπεία
 Βιοπληροψορική, συσιημική βιολογία και εξατομικευμένη θεραπεία Φραγκίσκος Κολίσης Καθηγητής Βιοτεχνολογίας, Σχολή Χημικών Μηχανικών ΕΜΠ, Διευθυντής Ινστιτούτου Βιολογικών Ερευνών και Βιοτεχνολογίας, EIE
Βιοπληροψορική, συσιημική βιολογία και εξατομικευμένη θεραπεία Φραγκίσκος Κολίσης Καθηγητής Βιοτεχνολογίας, Σχολή Χημικών Μηχανικών ΕΜΠ, Διευθυντής Ινστιτούτου Βιολογικών Ερευνών και Βιοτεχνολογίας, EIE
Ενδεικτικές απαντήσεις
 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΠΟΥ ΥΠΗΡΕΤΟΥΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΠΑΡΑΣΚΕΥΗ 8 ΣΕΠΤΕΜΒΡΙΟΥ 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Ενδεικτικές απαντήσεις
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΠΑΡΑΣΚΕΥΗ 16 IOYNIOY 2017 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α I Β IV Γ VI
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ
 ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
ΠΑΝΕΛΛAΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ 16-06-2017 ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Α. φωσφορική ομάδα (Ι) E. υδροξύλιο (II) Β. mrna
ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ
 27 Μαΐου 2016 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Ημερησίων Γενικών Λυκείων (Νέο & Παλιό Σύστημα) ΘΕΜΑ Γ Γ.1 Ο χαρακτήρας της ομάδας αίματος στον άνθρωπο
27 Μαΐου 2016 ΒΙΟΛΟΓΙΑ ΟΜΑΔΑΣ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Απαντήσεις Θεμάτων Πανελλαδικών Εξετάσεων Ημερησίων Γενικών Λυκείων (Νέο & Παλιό Σύστημα) ΘΕΜΑ Γ Γ.1 Ο χαρακτήρας της ομάδας αίματος στον άνθρωπο
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
Μοριακή βάση του καρκίνου της ουροδόχου κύστεως
 Πανεπιστήμιο Αθηνών Τμήμα Βιολογίας Τομέας Βιοχημείας & Μοριακής Βιολογίας Μοριακή βάση του καρκίνου της ουροδόχου κύστεως Μαργαρίτης Αυγέρης, Ανδρέας Σκορίλας Τομέας Βιοχημείας και Μοριακής Βιολογίας,
Πανεπιστήμιο Αθηνών Τμήμα Βιολογίας Τομέας Βιοχημείας & Μοριακής Βιολογίας Μοριακή βάση του καρκίνου της ουροδόχου κύστεως Μαργαρίτης Αυγέρης, Ανδρέας Σκορίλας Τομέας Βιοχημείας και Μοριακής Βιολογίας,
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ Κυριακή 9 Μαρτίου 2014
 ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ Κυριακή 9 Μαρτίου 2014 ΘΕΜΑ Α Α1. β, Α2. δ, Α3. γ, Α4. δ, Α5. γ. ΘΕΜΑ Β Β1. Σχολικό βιβλίο σελ. 90-91: «Το παράδειγμα της δρεπανοκυτταρικής
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ Κυριακή 9 Μαρτίου 2014 ΘΕΜΑ Α Α1. β, Α2. δ, Α3. γ, Α4. δ, Α5. γ. ΘΕΜΑ Β Β1. Σχολικό βιβλίο σελ. 90-91: «Το παράδειγμα της δρεπανοκυτταρικής
Οργανοτυπικές Καλλιέργειες
 Οργανοτυπικές Καλλιέργειες Καθηγητής-Διευθυντής Βασίλης Γοργούλης Επ. Καθηγητής Ιωάννης Πατέρας Εργαστήριο Ιστολογίας και Εμβρυολογίας Ιατρική Σχολή Εθνικό και Καποδιστριακό Πανεπιστήμιο Αθηνών Αθήνα,
Οργανοτυπικές Καλλιέργειες Καθηγητής-Διευθυντής Βασίλης Γοργούλης Επ. Καθηγητής Ιωάννης Πατέρας Εργαστήριο Ιστολογίας και Εμβρυολογίας Ιατρική Σχολή Εθνικό και Καποδιστριακό Πανεπιστήμιο Αθηνών Αθήνα,
Βιολογία ομάδας προσανατολισμού θετικών σπουδών. Πανελλαδικές εξετάσεις
 Βιολογία ομάδας προσανατολισμού θετικών σπουδών Πανελλαδικές εξετάσεις 2015-2016 ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ ΘΕΜΑ Β Β1. 1 Α 2 Γ 3 Α 4 Β 5 Α 6 Α 7 Γ Β2. Καρυότυπος ονομάζεται η απεικόνιση των μεταφασικών
Βιολογία ομάδας προσανατολισμού θετικών σπουδών Πανελλαδικές εξετάσεις 2015-2016 ΘΕΜΑ Α Α1. β Α2. β Α3. δ Α4. γ Α5. γ ΘΕΜΑ Β Β1. 1 Α 2 Γ 3 Α 4 Β 5 Α 6 Α 7 Γ Β2. Καρυότυπος ονομάζεται η απεικόνιση των μεταφασικών
ΑΠΑΝΤΗΣΕΙΣ. Β2. Η εικόνα αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς οργανισμούς το mrna αρχίζει να μεταφράζεται σε πρωτεΐνη πριν ακόμη
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 6 ΙΟΥΝΙΟΥ 207 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. δ Α2. δ Α3. β Α4. γ Α5.
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 6 ΙΟΥΝΙΟΥ 207 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α Α. δ Α2. δ Α3. β Α4. γ Α5.
ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ
 ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ ΠΑΝΕΛΛΗΝΙΩΝ 2017 ΘΕΜΑ Α Α1. δ Α2. δ Α3. β Α4. γ Α5. α ΘΕΜΑ Β Β1. Ι Α, ΙΙ Ε, ΙΙΙ ΣΤ, ΙV Β, V Ζ, VII Γ, VII Δ Β2. Η εικόνα 1 αντιστοιχεί σε προκαρυωτικό κύτταρο. Στους προκαρυωτικούς
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς
 Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
Κεφάλαιο 20 Η ρύθμιση της γονιδιακής έκφρασης στους ευκαρυωτικούς οργανισμούς Πυρίνας ανθρώπινου μεσοφασικού κυττάρου στον οποίο παρατηρούμε, με ανοσοφθορισμό, τη διάστικτη κατανομή της απακετυλάσης των
Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της
 ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΚΕΦ. 2 ο ΕΡΩΤΗΣΕΙΣ ΚΡΙΣΕΩΣ Ποιες είναι οι ομοιότητες και οι διαφορές μεταξύ της αντιγραφής και της μεταγραφής; Διαφορές Αντιγραφή Μεταγραφή 1. Διατηρείται και μεταβιβάζεται η 1. Μεταβιβάζεται η γενετική
ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 16-2-2014 ΘΕΜΑ 1 ο Α. Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη σωστή απάντηση. (Μονάδες 25)
") ΤΣΙΜΙΣΚΗ &ΚΑΡΟΛΟΥ ΝΤΗΛ ΓΩΝΙΑ THΛ: 270727 222594 ΑΡΤΑΚΗΣ 12 - Κ. ΤΟΥΜΠΑ THΛ: 919113 949422 ΕΠΩΝΥΜΟ:... ΟΝΟΜΑ:... ΤΜΗΜΑ:... ΗΜΕΡΟΜΗΝΙΑ:... ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 16-2-2014 ΘΕΜΑ 1 ο Α. Να βάλετε σε
ΤΣΙΜΙΣΚΗ &ΚΑΡΟΛΟΥ ΝΤΗΛ ΓΩΝΙΑ THΛ: 270727 222594 ΑΡΤΑΚΗΣ 12 - Κ. ΤΟΥΜΠΑ THΛ: 919113 949422 ΕΠΩΝΥΜΟ:... ΟΝΟΜΑ:... ΤΜΗΜΑ:... ΗΜΕΡΟΜΗΝΙΑ:... ΒΙΟΛΟΓΙΑ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ 16-2-2014 ΘΕΜΑ 1 ο Α. Να βάλετε σε
ΒΙΟΤΕΧΝΟΛΟΓΙΑ ΣΤ ΕΞΑΜΗΝΟΥ Τμήμα Ιατρικών Εργαστηρίων Τ.Ε.Ι. Αθήνας
 ΒΙΟΤΕΧΝΟΛΟΓΙΑ ΣΤ ΕΞΑΜΗΝΟΥ Τμήμα Ιατρικών Εργαστηρίων Τ.Ε.Ι. Αθήνας Μάθημα 15 ο Νανοσωματίδια για γονιδιακή μεταφορά Διδάσκων Δρ. Ιωάννης Δρίκος Απόφοιτος Ιατρικής Σχολής Ιωαννίνων (ΠΙ) Απόφοιτος Βιολογίας,
ΒΙΟΤΕΧΝΟΛΟΓΙΑ ΣΤ ΕΞΑΜΗΝΟΥ Τμήμα Ιατρικών Εργαστηρίων Τ.Ε.Ι. Αθήνας Μάθημα 15 ο Νανοσωματίδια για γονιδιακή μεταφορά Διδάσκων Δρ. Ιωάννης Δρίκος Απόφοιτος Ιατρικής Σχολής Ιωαννίνων (ΠΙ) Απόφοιτος Βιολογίας,
ΘΕΜΑΤΑ : ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Γ ΛΥΚΕΙΟΥ ΕΞΕΤΑΖΟΜΕΝΗ ΥΛΗ: ΚΕΦ /12/2017
 ΘΕΜΑΤΑ : ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Γ ΛΥΚΕΙΟΥ ΕΞΕΤΑΖΟΜΕΝΗ ΥΛΗ: ΚΕΦ 1-2-4 03/12/2017 ΘΕΜΑ A Α. Να επιλέξετε την ορθή πρόταση στα παρακάτω: Α1. Βασική μονάδα οργάνωσης της χρωματίνης αποτελεί το α. νουκλεοτίδιο
ΘΕΜΑΤΑ : ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ Γ ΛΥΚΕΙΟΥ ΕΞΕΤΑΖΟΜΕΝΗ ΥΛΗ: ΚΕΦ 1-2-4 03/12/2017 ΘΕΜΑ A Α. Να επιλέξετε την ορθή πρόταση στα παρακάτω: Α1. Βασική μονάδα οργάνωσης της χρωματίνης αποτελεί το α. νουκλεοτίδιο
Μεταπτυχιακή Εργασία
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΜΗΧΑΝΙΚΩΝ ΥΠΟΛΟΓΙΣΤΩΝ ΕΠΙΣΚΟΠΗΣΗ - ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΥΠΟΛΟΓΙΣΤΙΚΩΝ ΠΡΟΓΡΑΜΜΑΤΩΝ ΠΟΥ ΕΝΤΟΠΙΖΟΥΝ ΣΤΟΧΟΥΣ ΤΩΝ ΜΙΚΡΩΝ RNA Μεταπτυχιακή
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΠΟΛΥΤΕΧΝΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΜΗΧΑΝΙΚΩΝ ΥΠΟΛΟΓΙΣΤΩΝ ΕΠΙΣΚΟΠΗΣΗ - ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΥΠΟΛΟΓΙΣΤΙΚΩΝ ΠΡΟΓΡΑΜΜΑΤΩΝ ΠΟΥ ΕΝΤΟΠΙΖΟΥΝ ΣΤΟΧΟΥΣ ΤΩΝ ΜΙΚΡΩΝ RNA Μεταπτυχιακή
igenetics Mια Μεντελική προσέγγιση
 igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 22 (+κεφ. 17 Hartwell) Γενετική του καρκίνου Η πρωτεΐνη p53 προσδένεται στο DNA. 2 ΕΙΚΟΝΑ 22.1 Μαστογραφία που απεικονίζει έναν όγκο. Όγκος 3 Κύρια σημεία: Καρκίνος
igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 22 (+κεφ. 17 Hartwell) Γενετική του καρκίνου Η πρωτεΐνη p53 προσδένεται στο DNA. 2 ΕΙΚΟΝΑ 22.1 Μαστογραφία που απεικονίζει έναν όγκο. Όγκος 3 Κύρια σημεία: Καρκίνος
ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ
 ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ Θέμα 1 ο : Να επιλέξετε τη σωστή απάντηση: 1. Το σύμπλοκο έναρξης της πρωτεϊνοσύνθεσης δεν περιλαμβάνει α. το mrna β. τη μεγάλη ριβοσωμική υπομονάδα γ.
ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ Θέμα 1 ο : Να επιλέξετε τη σωστή απάντηση: 1. Το σύμπλοκο έναρξης της πρωτεϊνοσύνθεσης δεν περιλαμβάνει α. το mrna β. τη μεγάλη ριβοσωμική υπομονάδα γ.
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ 1ο 1. γ 2. γ 3. β 4. α 5. δ
 ΘΕΜΑ 1ο 1. γ 2. γ 3. β 4. α 5. δ 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 9 ΣΕΠΤΕΜΒΡΙΟΥ 2009 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ: ΒΙΟΛΟΓΙΑ
ΘΕΜΑ 1ο 1. γ 2. γ 3. β 4. α 5. δ 1 ΕΙΣΑΓΩΓΙΚΕΣ ΕΞΕΤΑΣΕΙΣ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΤΟΥ ΕΞΩΤΕΡΙΚΟΥ ΚΑΙ ΤΕΚΝΩΝ ΕΛΛΗΝΩΝ ΥΠΑΛΛΗΛΩΝ ΣΤΟ ΕΞΩΤΕΡΙΚΟ ΤΕΤΑΡΤΗ 9 ΣΕΠΤΕΜΒΡΙΟΥ 2009 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ: ΒΙΟΛΟΓΙΑ
ΑΠΑΝΤΗΣΕΙΣ. ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ
 ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
ΘΕΜΑ Α A1. β Α2. γ Α3. γ Α4. α Α5. δ ΕΠΑΝΑΛΗΠΤΙΚΕΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΤΕΤΑΡΤΗ 15 ΙΟΥΝΙΟΥ 2016 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΠΡΟΣΑΝΑΤΟΛΙΣΜΟΥ (ΝΕΟ
Επιδίωρθωση Βλαβών στο DNA Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Εργαστήριο Βιολογίας
 Επιδίωρθωση Βλαβών στο DNA 18.02.2019 Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Εργαστήριο Βιολογίας Πηγές Βλαβών στο DNA 1. Ενδογενείς βλάβες: Προκαλούνται κατά τη διάρκεια κυτταρικών διαδικασιών όπως η αντιγραφή
Επιδίωρθωση Βλαβών στο DNA 18.02.2019 Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Εργαστήριο Βιολογίας Πηγές Βλαβών στο DNA 1. Ενδογενείς βλάβες: Προκαλούνται κατά τη διάρκεια κυτταρικών διαδικασιών όπως η αντιγραφή
Δοµή και ιδιότητες του DNA
 Δοµή και ιδιότητες του DNA Βακτηριακό χρωµόσωµα ευκαρυωτικό χρωµόσωµα και χρωµατίνη 28/02/2014 1 Tο βακτηριακό γονιδίωµα περιέχεται σε ένα κυκλικό DNA µήκους 1300 µm εντός του βακτηριακού κυττάρου που
Δοµή και ιδιότητες του DNA Βακτηριακό χρωµόσωµα ευκαρυωτικό χρωµόσωµα και χρωµατίνη 28/02/2014 1 Tο βακτηριακό γονιδίωµα περιέχεται σε ένα κυκλικό DNA µήκους 1300 µm εντός του βακτηριακού κυττάρου που