ΜΟΡΦΟΛΟΓΙΚΗ, ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΚΑΙ ΜΟΡΙΑΚΗ ΔΙΕΡΕΥΝΗΣΗ ΤΩΝ ΑΝΑΠΛΑΣΤΙΚΩΝ ΛΕΜΦΩΜΑΤΩΝ (ALCL)
|
|
|
- Ωσαννά Αλεξανδρίδης
- 7 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ANATOMIKHΣ ΚΑΙ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΕΡΓΑΣΤΗΡΙΟ ΓΕΝΙΚΗΣ ΠΑΘΟΛΟΓΙΑΣ ΚΑΙ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΔΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ ΓΕΩΡΓΙΟΣ Σ. ΚΑΡΚΑΒΕΛΑΣ ΠΑΝΕΠ. ΕΤΟΣ ΑΡΙΘΜ ΜΟΡΦΟΛΟΓΙΚΗ, ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΗ ΚΑΙ ΜΟΡΙΑΚΗ ΔΙΕΡΕΥΝΗΣΗ ΤΩΝ ΑΝΑΠΛΑΣΤΙΚΩΝ ΛΕΜΦΩΜΑΤΩΝ (ALCL) ΜΑΤΘΑΙΟΣ Θ. ΜΠΟΜΠΟΣ ΙΑΤΡΟΣ-ΠΑΘΟΛΟΓΟΑΝΑΤΟΜΟΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΥΠΟΒΛΗΘΗΚΕ ΣTΗΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΥ ΑΡΙΣΤΟΤΕΛΕΙΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΘΕΣΣΑΛΟΝΙΚΗΣ Η παρούσα ερευνητική εργασία χρηματοδοτήθηκε από το Έργο 4902 της Επιτροπής Ερευνών Α.Π.Θ.» ΘΕΣΣΑΛΟΝΙΚΗ
2 Η ΤΡΙΜΕΛΗΣ ΕΠΙΤΡΟΠΗ ΙΩΑΝΝΗΣ ΚΩΣΤΟΠΟΥΛΟΣ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΒΑΣΙΛΙΚΗ ΚΑΛΟΥΤΣΗ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΡΙΑ ΒΑΣΙΛΙΚΗ ΚΩΤΟΥΛΑ, ΕΠΙΚ. ΚΑΘΗΓΗΤΗΣ Η ΕΠΤΑΜΕΛΗΣ ΕΠΙΤΡΟΠΗ ΙΩΑΝΝΗΣ ΚΩΣΤΟΠΟΥΛΟΣ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΗΣ ΒΑΣΙΛΙΚΗ ΚΑΛΟΥΤΣΗ, ΑΝΑΠΛ. ΚΑΘΗΓΗΤΡΙΑ ΒΑΣΙΛΙΚΗ ΚΩΤΟΥΛΑ, ΕΠΙΚ. ΚΑΘΗΓΗΤΡΙΑ ΕΛΕΝΗ ΒΡΕΤΤΟΥ, ΚΑΘΗΓΗΤΡΙΑ ΓΕΩΡΓΙΟΣ ΚΑΡΚΑΒΕΛΑΣ, ΚΑΘΗΓΗΤΗΣ ΙΩΑΝΝΗΣ ΚΛΩΝΙΖΑΚΗΣ, ΚΑΘΗΓΗΤΗΣ ΓΕΩΡΓΙΟΣ ΦΟΥΝΤΖΗΛΑΣ, ΚΑΘΗΓΗΤΗΣ «Η έγκριση της διδακτορικής διατριβής από την Ιατρική Σχολή του Αριστοτελείου Πανεπιστημίου δεν υποδηλώνει την αποδοχή των γνωμών του συγγραφέα». (Νόμος 5343/32, αρθρ και ν. 1268/82, αρθρ. 50 8). 2
3 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΡΟΕΔΡΟΣ ΤΗΣ ΣΧΟΛΗΣ ΝΙΚΟΛΑΟΣ ΝΤΟΜΠΡΟΣ 3
4 4
5 Στη σύζυγό μου Άννα Στην κόρη μου Θεοδοσία Στους δασκάλους μου 5
6 6
7 ΠΕΡΙΕΧΟΜΕΝΑ ΣΥΝΤΜΗΣΕΙΣ...11 ΠΡΟΛΟΓΟΣ...13 ΓΕΝΙΚΟ ΜΕΡΟΣ ΟΡΙΣΜΟΣ...19 ΙΣΤΟΡΙΚΑ ΣΤΟΙΧΕΙΑ...19 ΕΠΙΔΗΜΙΟΛΟΓΙΑ...23 ΕΝΤΟΠΙΣΗ...23 ΑΙΤΙΟΛΟΓΙΑ...24 ΚΛΙΝΙΚΗ ΕΙΚΟΝΑ...24 ΜΟΡΦΟΛΟΓΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ...25 ΑΝΟΣΟΦΑΙΝΟΤΥΠΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΟΥ ΑΛΜΚ...30 ΜΟΡΙΑΚΗ ΠΑΘΟΛΟΓΙΑ ΤΟΥ ΑΛΜΚ...32 TNFRSF8/CD30/Ki ALK...33 Cyclin D Fascin...45 htert...47 MUC ΚΛΙΝΙΚΗ ΤΑΞΙΝΟΜΗΣΗ...51 ΕΡΓΑΣΤΗΡΙΑΚΑ ΕΥΡΗΜΑΤΑ...52 ΣΤΑΔΙΟΠΟΙΗΣΗ...52 ΠΡΟΓΝΩΣΤΙΚΟΙ ΚΑΙ ΠΡΟΒΛΕΠΤΙΚΟΙ ΔΕΙΚΤΕΣ...53 ΘΕΡΑΠΕΙΑ...53 ΕΙΔΙΚΟ ΜΕΡΟΣ ΥΛΙΚΟ ΚΑΙ ΜΕΘΟΔΟΙ...59 ΑΣΘΕΝΕΙΣ...59 ΥΛΙΚΟ...59 ΕΝΤΟΠΙΣΗ...59 ΚΛΙΝΙΚΗ ΤΑΞΙΝΟΜΗΣΗ...60 ΙΣΤΟΛΟΓΙΚΗ ΤΑΞΙΝΟΜΗΣΗ...60 ΑΝΟΣΟΦΑΙΝΟΤΥΠΟΣ
8 ΜΕΘΟΔΟΙ ΕΠΙΛΟΓΗ ΥΛΙΚΟΥ...62 ΔΗΜΙΟΥΡΓΙΑ ΙΣΤΙΚΩΝ ΜΙΚΡΟΣΥΣΤΟΙΧΙΩΝ (TMA)...62 PCR...63 TCR-γ...64 RT-PCR...65 t(2;5)(p23;q35)...65 htert...66 Bdel ΚΑΙ Bplus ΜΕΤΑΓΡΑΦΑ ΤΗΣ HTERT...66 CYCLIN D FISH...67 LSI ALK PROBE...68 CEP 2 PROBE...70 CYCLIN D1/CEP11 PROBE...70 FICTION...71 ISH...72 mrna ΤΗΣ CYCLIN D mrna των ισομορφών της htert...74 ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΕΣ ΤΕΧΝΙΚΕΣ...76 ΑΠΛΗ ΑΝΟΣΟΪΣΤΟΧΗΜΕΙΑ...76 ΔΙΠΛΗ ΑΝΟΣΟΪΣΤΟΧΗΜΕΙΑ...77 ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΑΝΟΣΟΧΡΩΣΕΩΝ...77 ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ...78 ΑΠΟΤΕΛΕΣΜΑΤΑ ΓΕΝΙΚΑ...79 TCR-γ...79 ΕΚΦΡΑΣΗ ΤΗΣ ΧΙΜΑΙΡΙΚΗΣ ALK ΠΡΩΤΕΪΝΗΣ ΚΑΙ ΑΝΑΛΥΣΗ ΤΩΝ ΓΟΝΙΔΙΑΚΩΝ ΜΕΤΑΘΕΣΕΩΝ ΜΕ ΤΙΣ ΜΟΡΙΑΚΕΣ ΤΕΧΝΙΚΕΣ...80 ALK...80 t(2;5)(p23;q35) p Συσχέτιση των μεθόδων ανίχνευσης της ALK...82 CEP Cyclin D
9 Cyclin D1 πρωτεΐνη...82 mrna της cyclin D CCND1 γονίδιο...84 FICTION...84 htert...84 ISH για τους Bplus και Bdel ανιχνευτές...85 htert πρωτεΐνη...86 RT-PCR για ανίχνευση των ισομορφών της htert...86 RT-PCR για τα Bdel και Bplus μετάγραφα...87 ΑΙΧ ΓΙΑ ΤΙΣ ΠΡΩΤΕΪΝΕΣ FASCIN ΚΑΙ MUC FASCIN...87 MUC Συσχέτιση της έκφρασης των Fascin, htert και MUC ΠΙΝΑΚΕΣ, ΓΡΑΦΗΜΑΤΑ...89 ΕΙΚΟΝΕΣ ΣΥΖΗΤΗΣΗ ΣΥΜΠΕΡΑΣΜΑΤΑ ΠΕΡΙΛΗΨΗ SUMMARY ΒΙΒΛΙΟΓΡΑΦΙΑ
10 10
11 ΣΥΝΤΜΗΣΕΙΣ ΑΙΧ: ΑΛΜΚ: ΠΔ-ΑΛΜΚ: Σ-ΑΛΜΚ: ΜΗΛ: ALK: Ανοσοϊστοχημεία Αναπλαστικό Λέμφωμα από Μεγάλα Κύτταρα Πρωτοπαθές Δερματικό ΑΛΜΚ Συστηματικό ΑΛΜΚ Μη-Hodgkin λέμφωμα Anaplastic lymphoma kinase / Κινάση του Αναπλαστικού λεμφώματος CEP PROBE: FICTION: Centromeric Probe / Κεντρομεριδιακός ανιχνευτής / ιχνηθέτης Fluorescence immunophenotyping and interphase cytogenetics as a tool for the investigation of neoplasms / Φθορίζουσα ανοφαινοτυπική και γονιδιακή ανάλυση FISH: Fluorescence In Situ Hybridization / Φθορίζων εντοπισμένος/επιτόπιος υβριδισμός ISH: htert: LSI PROBE: In Situ Hybridization / Επιτόπιος υβριδισμός human Telomerase Reverse Transcriptase locus specific probes / ειδικοί ανιχνευτές τμημάτων χρωμοσωμάτων NPM: PCR: Nucleophosmin Polymerase Chain Reaction / Αλυσιδωτή αντίδραση πολυμεράσης) RT-PCR: TCR-γ: ΤΜΑ: Reverse Transcription PCR / PCR ανάστροφης μεταγραφής Τ-cell receptor γ / Τ-κυτταρικός υποδοχέας γ Tissue Microarrays / Ιστικές μικροσυστοιχίες 11
12 12
13 ΠΡΟΛΟΓΟΣ Τα μη-hodgkin λεμφώματα (ΜΗΛ) αποτελούν σπάνια κακοήθη νεοπλάσματα του αιμολεμφικού συστήματος με περίπου, νέες παγκοσμίως περιπτώσεις [1]. Διακρίνονται, σε λιγότερο ή περισσότερο διακριτές ανοσολογικές ομάδες, με βάση τα δύο βασικά λεμφοκυτταρικά συστήματα (Β και Τ). Η διάκριση των διαφόρων υποκατηγοριών των Β- και Τ-λεμφωμάτων γίνεται με βάση τα μορφολογικά, τα ιστοχημικά, τα ανοσοϊστοχημικά, τα ανοσολογικά και πρόσφατα τα μοριακά κριτήρια. Το αναπλαστικό λέμφωμα από μεγάλα κύτταρα (ΑΛΜΚ) / anaplastic large cell lymphoma (ALCL) αποτελεί μία σχετικά νέα κατηγορία των Τ-κυτταρικής αρχής λεμφωμάτων. Είναι ένας σπάνιος τύπος λεμφώματος που προσβάλει συνήθως παιδιά και μεσήλικες. Εμφανίσθηκε για πρώτη φορά στην αναθεωρημένη κατάταξη του Κιέλου το 1988 με την ονομασία «ALCL». Στην παραπάνω ταξινόμηση, με την παραπάνω ονομασία, ανήκαν το συστηματικό αναπλαστικό λέμφωμα από μεγάλα κύτταρα (Σ-ΑΛΜΚ) και το πρωτοπαθές δερματικό αναπλαστικό λέμφωμα από μεγάλα κύτταρα (ΠΔ-ΑΛΜΚ). Το τελευταίο, αποτέλεσε ξεχωριστή κατηγορία των Τ- ΜΗΛ μόλις το 1994, στην REAL (Revised European-American Lymphoma classification) ταξινόμηση, όπου και διαχωρίσθηκε από το Σ-ΑΛΜΚ λόγω των ιδιαίτερων βιολογικών του χαρακτηριστικών. Οι ραγδαίες εξελίξεις στον χώρο της βιολογίας, της ιατρικής τεχνολογίας και της ιατρικής επιστήμης γενικότερα, τις τελευταίες δεκαετίες, συνέδραμαν σε ανακαλύψεις που σχετίζονται με την παθοφυσιολογία, την βιολογική συμπεριφορά, την αντιμετώπιση των ΑΛΜΚ και οδήγησαν στον περαιτέρω χαρακτηρισμό των ΑΛΜΚ. Η ανεύρεση της μετάθεσης t(2;5)(p23;q35) μεταξύ της κινάσης του αναπλαστικού λεμφώματος (anaplastic lymphoma kinase/alk) και της νουκλεοφωσμίνης (nucleophosmin/npm) στα μέσα της δεκαετίας του 80 και η περαιτέρω έρευνα, οδήγησε στην αξιολόγηση της, ως της βασικής γενετικής ανωμαλίας σε αυτό τον τύπο των νεοπλασμάτων. Η ανάλυση των δεδομένων κλινικών μελετών επιβίωσης καθώς και των πολυάριθμων κλινικοπαθολογοανατομικών μελετών έδειξαν ότι τα ΑΛΜΚ που εκφράζουν την κινάση του αναπλαστικού λεμφώματος (ΑΛΜΚ, ALK+) αποτελούν μία κατηγορία λεμφωμάτων με ξεχωριστή βιολογική συμπεριφορά και πρόσφατα η νεώτερη κατάταξη της Π.Ο.Υ. τα διαχώρισε από τα ΑΛΜΚ, ALK-. 13
14 Τα ΑΛΜΚ, μεταξύ των άλλων τους χαρακτηριστικών, εμφανίζουν αυξημένο ρυθμό κυτταρικής διαίρεσης, ελαττωμένη απόπτωση (ιδιαίτερα τα ALK+) και σημαντικά αυξημένο μεταστατικό δυναμικό. Στα βασικά μόρια που συμμετέχουν στον έλεγχο του κυτταρικού κύκλου, μέσω σύνθετων μηχανισμών, ανήκει και η πρωτεΐνη κυκλίνη D1 (Cyclin D1) η οποία παίζει σημαντικό ρόλο στη παθογένεση του Β-κυτταρικής αρχής λεμφώματος του μανδύα, στο πλασματοκυτταρικό μυέλωμα και σε αρκετούς τύπους επιθηλιακών και μη νεοπλασιών. Στο ΑΛΜΚ ο ρόλος της πρωτεΐνης Cyclin D1 δεν έχει διευκρινησθεί πλήρως και οι λίγες μελέτες εμφανίζουν αντικροούμενα αποτελέσματα. Σημαντικότατο ρόλο στην κυτταρική διαίρεση και ευρύτερα στην καρκινογένεση διαδραματίζει και η καταλυτική υπομονάδα της τελομεράσης (htert-human Telomerase Reverse Transcriptase), ένα ένζυμο με δράση αντίστροφης μεταγραφάσης, η οποία σε συνέργεια με άλλα ένζυμα οδηγεί σε σταθεροποίηση των τελομερών, που αποτελούν το μιτωτικό ρολόϊ του κυττάρου. Νεώτερες μελέτες αναδεικνύουν επίδραση της htert σε μηχανισμούς που σχετίζονται με την ρύθμιση της αύξησης και επιβίωσης των νεοπλασματικών κυττάρων. Ενεργότητα της htert παρατηρείται σε σπάνιους τύπους σωματικών κυττάρων και στην πλειονότητα των κακοήθων νεοπλασμάτων. Για τα ΑΛΜΚ δεν υπάρχουν βιβλιογραφικές αναφορές. Τα ΑΛΜΚ μεταξύ των πολυάριθμων μορίων που τα χαρακτηρίζουν, εκφράζουν και πρωτεΐνες που εμπλέκονται στην κυτταρική επικοινωνία, στην διήθηση και μετανάστευση. Στις παραπάνω ανήκουν και οι πρωτεΐνες Fascin και MUC-1. Ο σκοπός της παρούσας μελέτης ήταν να διερευνηθούν σε μια σειρά 45 περιπτώσεων ΑΛΜΚ, Τ-Null-κυτταρικής αρχής, από 42 ασθενείς το προφίλ των γενετικών / μοριακών αλλαγών σε σχέση με κλασσικές παραμέτρους που μελετώνται στα ΑΛΜΚ. Ειδικότερα μελετήθηκαν με μεθόδους ανοσοϊστοχημείας (ΑΙΧ) η ALK πρωτεΐνη, ανάστροφης αλυσιδωτής αντίδρασης πολυμεράσης (RT-PCR) η μετάθεση t(2;5)(p23;q35) και φθορίζοντος υβριδισμό in situ (FISH) οι μεταθέσεις και χρωμοσωμικές ανωμαλίες του ALK γονιδίου. Διερευνήθηκε επίσης, με PCR, η παρουσία ανασυνδυασμού του Τ-κυτταρικού υποδοχέα τύπου γ (TCR-γ, Τ-cell receptor γ). Επιμέρους στόχους αποτέλεσαν η διερερεύνηση της έκφρασης των ισομορφών (mrna) και της πρωτεΐνης htert, η διερεύνηση του γονιδιακού προφίλ του CCND1 γονιδίου, της έκφρασης του mrna της cyclin D1 και της πρωτεΐνης Cyclin D1 με ΑΙΧ, καθώς και η ανίχνευση της πρωτεϊνικής έκφρασης των Fascin και MUC-1 (EMA, CD227) με ΑΙΧ. Τα Bplus και Bdel εναλλακτικά ματισμένα 14
15 μετάγραφα της htert μελετήθηκαν με ενδοεργαστηριακά παρασκευασμένους ανιχνευτές χρησιμοποιώντας τις τεχνικές RT-PCR και ISH, ενώ η htert πρωτεΐνη με ΑΙΧ. Η Cyclin D1 διερευνήθηκε σε επίπεδο πρωτεΐνης με τη χρήση δύο ειδικών μονοκλωνικών αντισωμάτων με τις μεθόδους τις απλής και διπλής ΑΙΧ, σε επίπεδο mrna, χρησιμοποιώντας ενδοεργαστηριακά παρασκευασμένους ανιχνευτές για τις ISH και RT-PCR και το CCND1 γονίδιο με εμπορικά διαθέσιμο ιχνηθέτη FISH. Επιπρόσθετα, στην περίπτωση της μελέτης του CCND1, εφαρμόσθηκε η μέθοδος της ταυτόχρονης φθορίζουσας ανοφαινοτυπικής και γονιδιακής ανάλυσης (FICTION). Οι ανοσοϊστοχημικές και in situ τεχνικές πραγματοποιήθηκαν στην πλειονότητα των περιπτώσεων σε ιστικές μικροσυστοιχίες (ΤΜΑ), μία σύγχρονη τεχνική, η οποία επέτρεψε την σημαντική μείωση του κόστους, του χρόνου, και του χρησιμοποιούμενου βιοπτικού υλικού της μελέτης. Τα αποτελέσματα συσχετίσθηκαν με γνωστές κλινικοπαθολογικές παραμέτρους. Στο σημείο αυτό αισθάνομαι την ανάγκη να εκφράσω την μεγάλη μου ευγνωμοσύνη στον επιβλέποντα της παρούσας μελέτης, Αναπληρωτή Καθηγητή κ. Ιωάννη Κωστόπουλο, για την συνεχή καθοδήγηση, την ουσιαστική βοήθεια στην διάρκεια της εκπόνησης του εργαστηριακού τμήματος της μελέτης, την συμπαράσταση και την πολύτιμη βοήθεια που μου προσέφερε στην διάρκεια της συγγραφής της μελέτης. Την Αναπληρώτρια Καθηγήτρια κ. Βασιλική Καλούτση ευχαριστώ ιδιαίτερα για την καθοδήγηση, τις υποδείξεις και την συμπαράσταση της. Ευχαριστώ θερμά την Επίκουρο Καθηγήτρια κ. Βασιλική Κωτούλα-Δημητριάδου. Η έμπρακτη συμβολή της στη μοριακή μελέτη των ΑΛΜΚ με πολύπλοκες μεθόδους και τεχνικές, η πολύπλευρη και αμέριστη βοήθεια της από την αρχή έως το τέλος της μελέτης ήταν καθοριστικές στην εκπόνηση της διατριβής αυτής. Βαθύτατα ευχαριστώ τον τέως Διευθυντή του Εργαστηρίου Παθολογικής Ανατομικής Α.Π.Θ., Καθηγητή Παθολογικής Ανατομικής κ. Κωνσταντίνο Παπαδημητρίου για την ανάθεση της παρούσας διατριβής, που μου έδωσε την δυνατότητα να ασχοληθώ με ένα τόσο ενδιαφέρον θέμα. Η καθοδήγηση του, η βοήθεια και συμπαράσταση που μου προσέφερε στην διάρκεια της επίβλεψης του, ήταν ανεκτίμητες. Τον νυν Διευθυντή, Καθηγητή κ. Γεώργιο Καρκαβέλα, την Καθηγήτρια κ. Ελένη Βρεττού, τον Καθηγητή κ. Γεώργιο Φούντζηλα και τον Καθηγητή ΑΤΕΙΘ κ. Ιωάννη Χατζημπούγια για την συμπαράσταση τους και τις παροτρύνσεις τους. 15
16 Ιδιαίτερες ευχαριστίες απευθύνω στον Αναπληρωτή Καθηγητή, Ιατρικής Σχολής Α.Π.Θ. κ. Ιωάννη Κουφογιάννη και την κ. Βουρλή Γεωργία, στατιστικολόγο, για την πολύτιμη προσφορά τους στην ανάλυση και επιβεβαίωση της ορθότητας της στατιστικής ανάλυσης της μελέτης. Επίσης θέλω να ευχαριστήσω θερμά τις παρασκευάστριες και τεχνολόγους του Εργαστηρίου Παθολογικής Ανατομικής Α.Π.Θ. κκ. Αναστασία Τσατσαρόνη, Αικατερίνη Θεολόγου, Γεωργία Ιωσηφίδου, Αφροδίτη Παγώνη και Μαρία Μητσικάρη για την συνεισφορά τους στην εφαρμογή των τεχνικών της μελέτης. Θα ήταν παράλειψη να μην ευχαριστήσω τους συναδέλφους παθολογοανατόμους κ. Σωτήριο Μπαρμπάνη για την βοήθεια στην συλλογή του υλικού και κ. Στέφανο Μηλιά για την βοήθεια στη δημιουργία των ιστικών μικροστοιχιών. Την κ. Ελένη Νικολακάκη, Επίκουρο Καθηγήτρια στο Εγαστήριο Βιολογικής Χημείας του Χημικού Τμήματος Α.Π.Θ., ευχαριστώ ιδιαίτερα για την δημιουργία των κλώνων που χρησιμοποιήθηκαν για την παρασκευή των in situ ανιχνευτών. Τέλος, θέλω να ευχαριστήσω από καρδιάς την σύζυγο μου για την ηθική συμπαράσταση και για τις αυξημένες ευθύνες που επωμίσθηκε στη διάρκεια της εκπόνησης της μελέτης. 16
17 ΓΕΝΙΚΟ ΜΕΡΟΣ 17
18 18
19 ΟΡΙΣΜΟΣ Το συστηματικό αναπλαστικό λέμφωμα από μεγάλα κύτταρα (Σ-ΑΛΜΚ) είναι ένα Τ-κυτταρικής αρχής λέμφωμα που αποτελείται συνήθως από μεγάλα λεμφοειδή κύτταρα με άφθονο κυτταρόπλασμα και πλειόμορφους, συχνά δίκην σέλας ή εμβρύου πυρήνες. Τα νεοπλασματικά κύτταρα εκφράζουν στην πλειονότητα τους το αντιγόνο CD30/Ki-1 και σε πολλές περιπτώσεις κυτταροτοξικές πρωτεΐνες [2]. Μεγάλο ποσοστό των Σ-ΑΛΜΚ εκφράζουν την κινάση του αναπλαστικού λεμφώματος (ALK-anaplastic lymphoma kinase), αν και αρνητικές στην ALK περιπτώσεις, έχουν συμπεριληφθεί στην ως εν λόγω κατηγορία. Το πρωτοπαθές δερματικό αναπλαστικό λέμφωμα από μεγάλα κύτταρα (ΠΔ- ΑΛΜΚ) είναι επίσης ένα Τ-κυτταρικής αρχής λέμφωμα του δέρματος που χαρακτηρίζεται και αυτό από την παρουσία μεγάλων νεοπλασματικών λεμφοειδών κυττάρων με κυτταρολογικά χαρακτηριστικά παρόμοια με εκείνα του Σ-ΑΛΜΚ. Το αντιγόνο CD30/Ki-1 εκφράζεται επίσης στην πλειονότητα των νεοπλασματικών λεμφοειδών κυττάρων, ενώ η ALK πρωτεΐνη είναι θετική σε σπανιότατες περιπτώσεις [2, 3]. Το ΠΔ-ΑΛΜΚ μαζί με την λεμφωματοειδή βλατίδωση ανήκουν στις λεγόμενες πρωτοπαθείς δερματικές CD30+ Τ-κυτταρικής αρχής λεμφοϋπερπλασίες (T-cell lymphoproliferative disorders, T-LPDs) [2-5]. ΙΣΤΟΡΙΚΑ ΣΤΟΙΧΕΙΑ Το ΑΛΜΚ αποτελεί μία σχετικά νέα κατηγορία λεμφωμάτων (Εικόνα 1, σελ. 20). Η ανάδειξη του σε ξεχωριστή κατηγορία λεμφωμάτων βασίστηκε στην ανακάλυψη το 1982 του Ki-1 αντιγόνου [6, 7] το οποίο εκφραζόταν από τα νεοπλασματικά κύτταρα της νόσου Hodgkin και προσδιορίσθηκε στο σύστημα ταξινόμησης CD (Cluster Designation) ως CD30 to 1986 [8]. Tο 1985, μία ομάδα αιμοπαθολογοανατόμων από πέντε διαφορετικές Ευρωπαϊκές χώρες ταυτοποίησε μία νέα κατηγορία λεμφωμάτων την οποία και ονόμασε αναπλαστικό λέμφωμα από μεγάλα κύτταρα ή αναπλαστικό μεγαλοκυτταρικό λέμφωμα (anaplastic large cell lymphoma-alcl) [9]. Η ομάδα αυτή παρουσίασε ένα σύνολο 45 λεμφωμάτων με ειδικά ιστολογικά χαρακτηριστικά και θετικότητα σε συγκεκριμένο αντιγόνο. Ήταν όλα μεγαλοκυτταρικά νεοπλάσματα με κυτταρική μορφολογία που κυμαινόταν μεταξύ ανοσοβλαστικών και μεγάλων αναπλαστικών, με άφθονο κυτταρόπλασμα, 19
20 κυτταρικών μορφών. Πολλές περιπτώσεις από τις 45 είχαν διαγνωσθεί ως κακοήθεις ιστιοκυτταρικοί όγκοι, αναπλαστικά (μεταστατικά) καρκινώματα, κακοήθη μελανώματα, σεμινώματα, ή ακόμα και ως κακοήθη ινώδη ιστιοκυτώματα [9-11]. Τα παραπάνω νεοπλάσματα διηθούσαν κυρίως την παραφλοιώδη περιοχή του λεμφαδένα (συχνά με διατήρηση των βλαστικών κέντρων), τους περιφερικούς λεμφόκολπους, ενώ συχνά αναγνωρίζονταν και εστίες νέκρωσης. Σε όλες τις περιπτώσεις, το σύνολο των νεοπλασματικών κυττάρων, εμφάνιζε χαρακτηριστική θετικότητα στο αντιγόνο Ki-1/CD30 και συχνά ήταν θετικά στους Β ή Τ-λεμφοκυτταρικούς δείκτες. Τα λεμφώματα αυτά, μαζί με την προσθήκη του ΠΔ-ΑΛΜΚ (C-ALCL, cuataneous anaplastic large cell lymphoma) αποτέλεσαν κατηγορία T-μη Hodgkin λεμφωμάτων (Τ-ΜΗΛ) για πρώτη φορά στην αναθεωρημένη κατάταξη του Κιέλου το 1988 κάτω από την ίδια ονομασία: αναπλαστικό λέμφωμα από μεγάλα κύτταρα. Ιδιαίτερα το ΠΔ-ΑΛΜΚ/C-ALCL, αποτέλεσε ξεχωριστή κατηγορία των Τ-ΜΗΛ μόλις το 1994, στην REAL ταξινόμηση, όπου και πρωτοεμφανίσθηκε λόγω των ιδιαίτερων βιολογικών του χαρακτηριστικών σε σχέση με το Σ-ΑΛΜΚ. Ουσιαστικά η κατάταξη αυτή είναι περισσότερο κλινική γιατί και οι δύο τύποι πιθανολογείται ότι αποτελούν το ίδιο λέμφωμα με διαφορετική πρωτοπαθή εντόπιση, το Σ-ΑΛΜΚ μπορεί να εμφανισθεί σε διάφορα όργανα και ιστούς, ενώ το ΠΔ-ΑΛΜΚ περιορίζεται στο δέρμα, χωρίς να μπορεί να εντοπισθεί άλλη πρωτοπαθής εστία την στιγμή της διάγνωσης ή και έξι μήνες μετά την διάγνωση. Στα τέλη της δεκαετίας του 80 ανακαλύφθηκε η μετάθεση t(2;5)(p23;q35) [12-14], η οποία αργότερα και ταυτοποίηθηκε. Ειδικότερα, νεότερες μελέτες έδειξαν ότι η παραπάνω μετάθεση αφορούσε την κινάση του αναπλαστικού λεμφώματος (anaplastic lymphoma kinase/alk) και την νουκλεοφωσμίνη (nucleophosmin/npm) [15]. Η ανεύρεση της οδήγησε στην περαιτέρω αξιολόγηση της ως της βασικής γενετικής ανωμαλίας σε αυτό τον τύπο των νεοπλασμάτων και η ογκογόνος δράση της επιβεβαιώθηκε σε πειραματικά μοντέλα [16]. Πολυκλωνικά και αργότερα μονοκλωνικά αντισώματα χρησιμοποιήθηκαν για να ταυτοποιήσουν και να χαρακτηρίσουν τα ΑΛΜΚ [17, 18]. Στη συνέχεια, ακολούθησε μία άνθηση της μελέτης της ALK στα ΑΛΜΚ και σε άλλες νεοπλασίες η οποία οδήγησε στην ανακάλυψη έως σήμερα συνολικά 14 χρωμοσωμικών ανωμαλίων στα οποία εμπλέκεται το γονίδιο της ALK [19]. Το 2001, στην ταξινόμηση της WHO τα Β- κυτταρικής αρχής λεμφώματα με αναπλαστική μορφολογία και έκφραση της ALK διαχωρίζονται από τα ΑΛΜΚ Τ-null κυτταρικής αρχής και κατατάσσονται στα Β- 20
21 λεμφώματα διαχύτου τύπου από μεγάλα κύτταρα (DLBCL) [2]. Μελέτες γονιδιακού προφίλ (gene expression profile) συντελέσαν στην περαιτέρω αξιολόγηση και ταυτοποίηση των ΑΛΜΚ [20, 21], ενώ επιβεβαιώθηκε και η αρχή του εξαρτώμενου από την ALK φαινότυπου (ALK addiction) των ΑΛΜΚ, ALK+ με την χρήση small hairpin RNA (shrna) [22]. Πρόσφατα έχουν παρουσιασθεί μικρά μόρια ως αναστολείς της ογκογόνου δράσης της ALK με ελπιδοφόρα αποτελέσματα και θεραπευτικές δράσεις σε πειραματικά μοντέλα [23]. Επιπρόσθετα, έχει δοκιμασθεί και η θεραπευτική χρήση του anti-cd30 mab σε μελέτη φάσης Ι/ΙΙ (NCT ) που αφορά παιδιατρικούς ασθενείς καθώς και σε μελέτες φάσης ΙΙ με δερματικό (NCT ) και συστηματικό (NCT ) ΑΛΜΚ. Τα αποτελέσματα των παραπάνω μελετών αναμένονται να αναδείξουν μελλοντικούς δρόμους στοχεύουσας θεραπείας. Πολύ πρόσφατα, η ανάλυση των πολυάριθμων κλινικών μελετών επιβίωσης καθώς και των παθολογοανατομικών μελετών, έδειξαν ότι τα ΑΛΜΚ που εκφράζουν την κινάση του αναπλαστικού λεμφώματος (ΑΛΜΚ, ALK+) αποτελούν μία ειδική κατηγορία λεμφωμάτων και εντάχθηκαν ως αυτόνομη κατηγορία στη νεώτερη κατάταξη της Π.Ο.Υ., όπου και διαχωρίσθηκαν από τα ΑΛΜΚ, ALK- [5]. Η εισαγωγή σύγχρονων μεθόδων και τεχνικών (γονιδιωμική, πρωτεϊνωμική) για την περαιτέρω ταυτοποίηση και διερεύνηση της παθογένεσης των ΑΛΜΚ, η ανεύρεση νέων μορίων αναστολέων της ογκογόνου δράσης μορίων κλειδιών και η χορήγηση αποτελεσματικότερων και λιγότερα τοξικών θεραπευτικών σχημάτων, προμηνύουν ένα συναρπαστικό και πολλά υποσχόμενο μέλλον, για τους ασθενείς με ΑΛΜΚ. 21

22 Εικόνα 1. Το χρονοδιάγραμμα των σημαντικότερων ανακαλύψεων στον τομέα των ΑΛΜΚ [τροποποιημένο από Chiarle et al., 2008 (19)]. 22
23 ΕΠΙΔΗΜΙΟΛΟΓΙΑ Το Σ-ΑΛΜΚ αποτελεί ένα σχετικά σπάνιο νεόπλασμα. Αφορά περίπου το 2-8% των ΜΗΛ στους ενήλικες και περίπου το 4-8% των διάχυτων λεμφωμάτων από μεγάλα κύτταρα [2, 24, 25]. Στα παιδιά αποτελεί το 10-30% των ΜΗΛ [2]. Εκτιμάται ότι 1000 περίπου νέες περιπτώσεις εμφανίζονται στις Η.Π.Α. κάθε χρόνο, ενώ σε 15 χώρες της Ευρωπαϊκής Ένωσης ο αριθμός αυτών υπολογίζεται στις Η καμπύλη εμφάνισης της νόσου σε σχέση με την ηλικία παρουσιάζει δύο κορυφές προσβολής, τις 2 η και 3 η δεκαετίες και σε μικρότερο βαθμό τις 5 η και 6 η δεκαετίες [26]. Προσβάλλει συχνότερα τους άρρενες. Το ΑΛΜΚ, ALK+ αποτελεί περίπου το 3% των ΜΗΛ των ενηλίκων και το 10-20% των παιδικών λεμφωμάτων [3]. Είναι πιο συχνό σε νεαρές ηλικίες έως τα 35 έτη και εμφανίζεται συχνότερα στους άρρενες σε σχέση με τα θήλεα (1,5/1). Το ΑΛΜΚ, ALK- προσβάλλει συχνότερα άρρενες ενήλικες (40-65 έτη), αν και σπάνια έχουν αναφερθεί περιπτώσεις εμφάνισης της νόσου και σε μικρότερες ή μεγαλύτερες ηλικίες [3, 27]. Το ΠΔ-ΑΛΜΚ αφορά περίπου το 9% των δερματικών λεμφωμάτων και το 25% των πρωτοπαθών δερματικών Τ-ΜΗΛ [2, 28]. Η νόσος προσβάλλει συνήθως ενήλικες / ηλικιωμένους, με μέση ηλικία εμφάνισης τα 60 έτη, ενώ είναι σπάνιο στα παιδιά [4, 5]. Η αναλογία προσβολής ανδρών/γυναικών είναι 2-3:1. ΕΝΤΟΠΙΣΗ Το ΑΛΜΚ, ALK+ μπορεί ταυτόχρονα να προσβάλλει λεμφαδένες και εξωλεμφαδενικούς ιστούς. Οι συχνότερες εξωλεμφαδενικές εντοπίσεις αφορούν το δέρμα, τα οστά, τα μαλακά μόρια, τους πνεύμονες και το ήπαρ [5]. Η διήθηση του ΓΕΣ και του ΚΝΣ είναι σπάνια, όπως επίσης και η υπεζοκωτική εντόπιση της νόσου. Η διήθηση του μυελού των οστών είναι στις περισσότερες (>90%) περιπτώσεις εμφανής μορφολογικά στην κλασσική χρώση της Αιματοξυλίνης-Εωσίνης και κυμαίνεται έως το 17% των περιπτώσεων [5, 29, 30]. Το παραπάνω ποσοστό, φαίνεται ότι δεν αυξάνεται σημαντικά με τη χρήση ειδικών αντισωμάτων (με ΑΙΧ) για την ανίχνευση των νεοπλασματικών λεμφοειδών κυττάρων [30], ενώ με την χρήση μοριακών μεθόδων (RT-PCR και RΤ-Q-PCR για NPM και ALK) το ποσοστό διήθησης στις ελάχιστες υπάρχουσες μελέτες κυμαίνεται από 47,5-61% [31, 32]. 23
24 Το ΑΛΜΚ, ALK- μπορεί επίσης, αν και σπανιότερα σε σχέση με το ALK+, να προσβάλλει ταυτόχρονα λεμφαδένες και εξωλεμφαδενικούς ιστούς, με συχνότερες εντοπίσεις τα οστά, τα μαλακά μόρια και το δέρμα [5]. Το ΠΔ-ΑΛΜΚ εμφανίζεται συχνότερα στις περιοχές του θώρακα, του προσώπου, τα άκρα και στους γλουτούς [4, 5, 33]. ΑΙΤΙΟΛΟΓΙΑ Η ανάπτυξη του ΑΛΜΚ έως σήμερα δεν κατέστη δυνατόν να αποδοθεί σε κάποιο αίτιο ή να συσχετισθεί με περιβαλλοντικούς, γενετικούς, οικογενείς παράγοντες. Πολλές μελέτες ασχολήθηκαν με την τυχόν παρουσία σχετιζόμενης ιογενούς λοίμωξης και ειδικότερα λοίμωξης από τον ιό Epstein Barr (EBV) [34]. Φαίνεται ότι η λοίμωξη από τον EBV απουσιάζει ή είναι σπανιότατη σε ασθενείς με ΑΛΜΚ. Στο Σ-ΑΛΜΚ (ALK+ και ALK-) θεωρείται ότι τα νεοπλασματικά κύτταρα προέρχονται από ώριμα ενεργοποιημένα κυτταροτοξικά Τ-λεμφοκύτταρα, ενώ στο ΠΔ-ΑΛΜΚ ο νεοπλασματικός πληθυσμός αποτελείται από ενεργοποιημένα, συχνά με κυτταροτοξικό ανοσοφαινότυπο, Τ-λεμφοκύτταρα του δέρματος [5]. ΚΛΙΝΙΚΗ ΕΙΚΟΝΑ Η πλειονότητα των ασθενών με Σ-ΑΛΜΚ (50-70%), ALK+ ή ALKδιαγιγνώσκεται σε προχωρημένο στάδιο της νόσου (III-IV) [2, 5, 26]. Σε αυτό το στάδιο ανευρίσκονται διογκωμένοι περιφερικοί ή/και κοιλιακοί λεμφαδένες, ενώ συχνά παρατηρείται εξωλεμφαδενική επέκταση και διήθηση του μυελού των οστών [5]. Οι ασθενείς εμφανίζουν συχνά Β-συμπτώματα (75%) και ειδικότερα υψηλό πυρετό [5]. Στα ΠΔ-ΑΛΜΚ η εμφάνιση της νόσου ποικίλει. Η νόσος εμφανίζεται συχνά ως μονήρες ασυμπτωματικό οζίδιο ή ογκίδιο και σπανιότερα με τη μορφή βλατίδας, συχνά με συνοδό εξέλκωση [5, 35]. Πολλαπλή προσβολή παρατηρείται στο 20% των ασθενών. Συχνά παρατηρείται αυτόματη μερική ή πλήρης υποστροφή της νόσου, όπως και στη λεμφωματοειδή βλατίδωση. Εξωδερματική διασπορά της νόσου παρατηρείται σε περίπου 10% των ασθενών, με συχνότερη προσβολή των επιχώριων λεμφαδένων [5, 36]. 24
25 ΜΟΡΦΟΛΟΓΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ Το χαρακτηριστικό μορφολογικό γνώρισμα του ΑΛΜΚ είναι η παρουσία σε όλες τις περιπτώσεις χαρακτηριστικών μεγάλου μεγέθους κυττάρων με έκκεντρους δίκην σέλας, εμβρύου ή νεφρού πυρήνες συχνά με ηωσινόφιλη περιοχή πλησίον του πυρήνα. Τα ως άνω κύτταρα επειδή αναγνωρίζονται σε διαφόρου βαθμού ποσόστωση σε όλους τους ιστολογικούς υποτύπους του ΑΛΜΚ ονομάζονται στην αγγλοσαξονική βιβλιογραφία hallmark cells. Παρόμοιους μορφολογικούς χαρακτήρες εμφανίζουν συχνά και τα μικρότερου μεγέθους νεοπλασματικά λεμφοειδή κύτταρα και η παρουσία αυτών δύναται σε πολλές περιπτώσεις να οδηγήσει στη σωστή διάγνωση [37]. Σε αρκετές περιπτώσεις, μικρός αριθμός νεοπλασματικών κυττάρων εμφανίζει ενδοπυρηνικά έγκλειστα. Αυτά δεν είναι πραγματικά έγκλειστα αλλά αναδιπλώσεις της πυρηνικής μεμβράνης και συχνά αναφέρονται ως doughnut cells, κύτταρα δίκην «λουκουμά» [2]. To ΑΛΜΚ, ALK+ χαρακτηρίζεται από την παρουσία συνήθως μεγάλου μεγέθους hallmark κυττάρων, αν και μικρότερα κύτταρα με παρόμοιους μορφολογικούς χαρακτήρες μπορεί να παρατηρηθούν [37]. Πολυπύρηνα κύτταρα με διάταξη των πυρήνων δίκην «στεφανιού» είναι δυνατόν να παρατηρηθούν, όπως επίσης και κύτταρα με μορφολογία παρόμοια με τα κύτταρα Reed-Sternberg της νόσου Hodgkin. Οι λεμφαδενικές/εξωλεμφαδενικές διηθήσεις είναι συχνά διάχυτες και αποτελούνται από ποικίλου μεγέθους νεοπλασματικές ομάδες χωρίς σαφή οργανοειδή διάταξη. Όταν παρατηρείται μερική λεμφαδενική διήθηση, το νεόπλασμα χαρακτηριστικά διηθεί τους λεμφόκολπους, δίνοντας την εντύπωση μεταστατικού καρκινώματος. Το ΑΛΜΚ, ALK- εμφανίζει χαμηλότερου βαθμού διαφοροποίηση σε σχέση με το ΑΛΜΚ, ALK+ και παρόμοια μορφολογικά χαρακτηριστικά. Συχνά διηθεί διάχυτα λεμφαδένες ή εξωλεμφαδενικούς ιστούς με τη μορφή πολλαπλών, συχνά συρρέουσων ομάδων. Παρόμοια με το ALK+, η μερική λεμφαδενική διήθηση δίνει την εντύπωση καρκινωματωδών μεταστάσεων, ενώ όχι σπάνια παρατηρείται σκλήρυνση του στρώματος και εωσινοφιλική συνοδός διήθηση. Τα νεοπλασματικά κύτταρα του ΑΛΜΚ, ALK- τείνουν να είναι ελαφρώς μεγαλύτερα και πιο πλειόμορφα από αυτά του ALK+ και/ή εμφανίζουν αυξημένο λόγο πυρήνα/κυτταροπλάσματος [2, 5, 38-40]. Στην παραπάνω περίπτωση πρέπει να 25
26 αποκλεισθεί η πιθανότητα διήθησης από περιφερικό Τ-κυτταρικής αρχής λέμφωμα μη ειδικού τύπου (PTCL, NOS). Στο ΠΔ-ΑΛΜΚ μικροσκοπικά παρατηρείται διάχυτη, οζοειδής, από μεγάλα νεοπλασματικά κύτταρα, διήθηση του δέρματος και σπανιότερα του υποδορίου λιπώδους ιστού συνήθως χωρίς επιδερμοτροπισμό. Στην πλειονότητα των περιπτώσεων, τα λεμφοειδή κύτταρα εμφανίζουν τα μορφολογικά χαρακτηριστικά του Σ-ΑΛΜΚ. Μιτώσεις και αποπτωτικά κύτταρα ανευρίσκονται συχνά. Αγγειακή διήθηση μπορεί να παρατηρηθεί. Σπανιότερα (20-25%) των περιπτώσεων, τα νεοπλασματικά λεμφοειδή κύτταρα δεν εμφανίζουν σαφείς αναπλαστικούς μορφολογικούς χαρακτήρες, ενώ δύναται να παρατηρηθούν, συχνά στην περιφέρεια του όγκου λεμφοκυτταρικές αντιδραστικές αθροίσεις [4, 41]. Στις περιπτώσεις με εξέλκωση της επιδερμίδας η ιστολογική εικόνα προσομοιάζει με αυτή της λεμφωματοειδούς βλατίδωσης. Παρατηρείται πυκνή, μικτή κυτταρική διήθηση από αντιδραστικά Τ-λεμφοκύτταρα, ιστιοκύτταρα, πολυμοφροπύρηνα λευκοκύτταρα και σπάνια CD30+ κύτταρα [5]. Σπανιότατα έχουν αναφερθεί και περιπτώσεις που προσομοιάζουν ιστολογικά με πυογόνο κοκκίωμα, λόγω τις πυκνής ουδετεροφιλικής φλεγμονής και αποτελούν διαφοροδιαγνωστικό πρόβλημα [42]. Έχουν αναφερθεί στην διεθνή βιβλιογραφία αρκετοί ιστολογικοί τύποι του ΑΛΜΚ. Οι περισσότεροι απαντούνται και στους δύο τύπους των Σ-ΑΛΜΚ καθώς και στα ΠΔ-ΑΛΜΚ (Πίνακας 1, σελ. 29). Στο 10% των περιπτώσεων, οι ασθενείς δύναται να εμφανίσουν παραπάνω του ενός ιστολογικού τύπου [37]. Επιπροσθέτως, στις υποτροπές της νόσου, τα ιστολογικά χαρακτηριστικά του λεμφώματος μπορεί να είναι διαφορετικά από εκείνα της πρώτης προσβολής. Στα ΑΛΜΚ, ALK- δεν έχει ως τώρα περιγραφεί ο μικροκυτταρικός τύπος [5]. Ο κοινός ή κλασσικός τύπος (80%) διακρίνεται σε δύο υποτύπους: τον πλειόμορφο και τον μονόμορφο κλασσικό τύπο [43]. Ο πρώτος αποτελείται από πλειόμορφα μεγάλα λεμφοειδή κύτταρα με άφθονο κυτταρόπλασμα, πτυχωτούς οδοντωτούς πυρήνες με μερικώς διάχυτη χρωματίνη και εμφανή πυρήνια. Τα χαρακτηριστικά hallmark cells κύτταρα υπάρχουν σε μεγάλους αριθμούς, ενώ σπάνια παρατηρούνται και γιγαντοκυτταρικές μορφές. Χαρακτηριστικά, τα μεγάλου μεγέθους κύτταρα εμφανίζουν περιαγγειακή κατανομή. Ο μονόμορφος υπότυπος χαρακτηρίζεται απο σχετικά ομοιόμορφο κυτταρικό πληθυσμό που αποτελείται από μεσαίου και μεγάλου μεγέθους λεμφοειδή κύτταρα, υποστρόγγυλου συνήθως σχήματος και μικρού βαθμού πυρηνική ατυπία. Τα hallmark cells υπάρχουν σε 26
27 μικρούς αριθμούς. Τέλος υπάρχουν και περιπτώσεις που χαρακτηρίζονται ως μικτές, στις οποίες ο νεοπλασματικός πληθυσμός αποτελείται από πλειόμορφα μεγάλα λεμφοειδή κύτταρα με άφθονο κυτταρόπλασμα και μεσαίου και μεγάλου μεγέθους λεμφοειδή κύτταρα, υποστρόγγυλου συνήθως σχήματος και μικρού βαθμού πυρηνική ατυπία με περίπου ισότιμη κατανομή [5, 44]. Ο μικροκυτταρικός τύπος (6%) αποτελείται κυρίως από μικρού και μέτριου μεγέθους νεοπλασματικά λεμφοειδή κύτταρα με ανώμαλους/ακανόνιστους πυρήνες και μικρό αριθμό hallmark cells θετικών στο CD30 αντιγόνο. Τα μικρού μεγέθους νεοπλασματικά κύτταρα είναι αρνητικά ή εκφράζουν ήπια το CD30 και στην πλειονότητα των περιπτώσεων εκφράζουν την ALK πρωτεΐνη. Ο υπότυπος αυτός προσομοιάζει ιστολογικά με το PTCL (NOS) ΜΗΛ και παρατηρείται μόνο στα συστηματικά ALK+ και ΠΔ-ΑΛΜΚ. Ειδικό εύρημα, αποτελεί η παρουσία μεγάλων λεμφοειδών κυττάρων CD30+ με περιαγγειακή κατανομή. Στις περισσότερες περιπτώσεις, τα κύτταρα αυτά διηθούν και το τοίχωμα των αγγείων [2, 45]. Στο ¼ των περιπτώσεων, μετά από πάροδο μηνών, ο παραπάνω τύπος μπορεί να μεταπέσει στον μονόμορφο ή στον κοινό τύπο [44]. Αυτή η μετατροπή συνοδεύεται από ραγδαία επιδείνωση της κλινικής πορείας, με το 75% των ασθενών να καταλήγουν σε διάστημα κάτω του ενός έτους. Ο λεμφοϊστιοκυτταρικός τύπος (5%) χαρακτηρίζεται από την παρουσία μεγάλου αριθμού αντιδραστικών ιστιοκυττάρων τα οποία αναμιγνύονται με τα νεοπλασματικά κύτταρα. Κάποια από τα ιστιοκύτταρα εμφανίζουν πλασματοκυτταροειδείς χαρακτήρες και σπανιότερα στοιχεία ερυθροφαγοκυττάρωσης. Τα νεοπλασματικά λεμφοειδή κύτταρα είναι μικρού ή μέσου μεγέθους και ανευρίσκονται κυρίως ως ομάδες γύρω από τα αγγεία του λεμφαδένα [2, 43, 46]. O τύπος πλούσιος σε γιγαντοκύτταρα (5-8%) χαρακτηρίζεται από την παρουσία πολυάριθμων πλειόμορφων μεγάλου μεγέθους συχνά πολυπύρηνων νεοπλασματικών κυττάρων. Περίπου το 1/3 των περιπτώσεων εκφράζουν την ALK πρωτεΐνη [40]. Ο τύπος από κύτταρα δίκην «σφραγιστήρος δακτυλίου» (<1%) αποτελεί εξαιρετικά σπάνιο φαινότυπο των ΑΛΜΚ. Τα λεμφοειδή κύτταρα συχνά εμφανίζουν κυτταροπλασμικά κενοτόπια, αρνητικά για παρουσία βλέννης, ενώ ο πυρήνας είναι συχνά απωθημένος στην περιφέρεια και ελλειπτικού σχήματος. Μεταξύ των παραπάνω κυττάρων ανευρίσκονται και μεσαίου και μεγάλου μεγέθους νεοπλασματικά κύτταρα με υποστρόγγυλους, ωοειδείς ή ακανόνιστους πυρήνες, συχνά με εμφανή πυρήνια. Τα κενοτόπι συχνά εμφανίζουν μερική ή εκτεταμένη 27
28 θετικότητα σε ειδικούς ανοσοϊστοχημικούς δείκτες (CD30, CD15, CD25) [47], η οποία ερμηνεύεται ως αποτέλεσμα της εσωτερίκευσης επιφανειακών αντιγόνων και της παρουσίας τους στα κυστίδια της συσκευής Golgi [48]. Ο σαρκωματοειδής τύπος (<1%) είναι ιδιαίτερα σπάνιος [5, 44]. Τα νεοπλασματικά κύτταρα είναι μέσου μεγέθους, ατρακτόμορφου ή αστεροειδούς σχήματος και εκφράζουν το CD30 αντιγόνο και σπανιότερα άλλους λεμφοκυτταρικούς δείκτες. Το στρώμα είναι συχνά μυξοειδές και οιδηματώδες. Λόγω του ατρακτόμορφου σχήματος των νεοπλασματικών κυττάρων, των αλλοιώσεων του στρώματος και της ανώμαλης αντιγονικής έκφρασης (vimentin, SMA) η διάγνωση είναι ιδιαίτερα δύσκολη και συχνά απαιτείται πληθώρα ανοσοϊστοχημικών δεικτών για την ακριβή διάγνωση και τον αποκλεισμό ενός κακοήθους σαρκώματος [49]. Ο φλεγμονώδης (ηωσινοφιλικός) τύπος (πυογόνο ΑΛΜΚ) (<1%) παρατηρείται σε όλους τους τύπους του ΑΛΜΚ και χαρακτηρίζεται από την παρουσία πυκνών ηωσινοφιλικών αθροίσεων, σε εστιακή ή διάχυτη κατανομή, μεταξύ των νεοπλασματικών λεμφοειδών κυττάρων [50, 51]. Σπάνια είναι δυνατόν να παρατηρηθούν και πολυμορφοπυρηνικά αποστημάτια [42]. Στον αντιδραστικό πληθυσμό συμμετέχουν επίσης ουδετερόφιλα πολυμορφοπύρηνα, Β- και Τ- λεμφοκύτταρα και επιθηλιοειδή ιστιοκύτταρα. Ο υποκυτταρικός τύπος/hypocellular (<1%) αποτελεί σπανιότατο τύπο του Σ-ΑΛΜΚ και του ΠΔ-ΑΛΜΚ [5, 44, 52, 53]. Η υποκυτταρική εμφάνιση του νεοπλάσματος οφείλεται στην χαλαρή διάταξη των κυττάρων στο έντονα οιδηματώδες και κατά θέσεις ινομυξοειδές στρώμα, πλούσιο σε φλεβίδια [52]. Ο λεμφοειδής πληθυσμός αποτελείται από κυρίως μικρού, μέσου και σπάνια μεγάλου μεγέθους κύτταρα. Σπάνια παρατηρούνται και ατρακτόμορφα κύτταρα κυρίως σε δεσμιδωτή διάταξη. Ο Hodgkin-like τύπος παρατηρείται μόνο στο Σ-ΑΛΜΚ και εμφανίζει μορφολογικά χαρακτηριστικά παρόμοια με τον τύπο της οζώδους σκλήρυνσης της νόσου Hodgkin [5]. Στον ως άνω τύπο, παρατηρούνται νεοπλασματικοί όζοι από πλειόμορφα, αναπλαστικά κύτταρα, αρκετά απο τα οποία εμφανίζουν μορφολογία κυττάρων Reed-Sternberg. Στο υπόστρωμα ανευρίσκεται πυκνός λεμφοκοκκιωματώδης ιστός, καθώς και ινώδεις ταινίες που σχηματίζουν ατελείς ή πλήρεις όζους. Επιπρόσθετα, αναγνωρίζεται και πάχυνση της λεμφαδενικής κάψας [43]. 28
29 Πίνακας 1. Ιστολογικοί τύποι του ΑΛΜΚ [2, 5, 42, 44, 52] Ιστολογικός τύπος ALK+ ALK- ΠΔ-ΑΛΜΚ Κοινός ή κλασσικός Πλειόμορφος υπότυπος Μονόμορφος υπότυπος Μικτός υπότυπος Μικροκυτταρικός Λεμφοϊστιοκυτταρικός Γιγαντοκυτταρικός Με κύτταρα δίκην «σφραγιστήρος δακτυλίου» Σαρκωματοειδής Φλεγμονώδης (ηωσινοφιλικός) Υποκυτταρικός Hodgkin-like 29
30 ΑΝΟΣΟΦΑΙΝΟΤΥΠΙΚΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΟΥ ΑΛΜΚ Τα κύτταρα του ΑΛΜΚ και συχνότερα των ΑΛΜΚ, ALK+ είναι θετικά στις περισσότερες περιπτώσεις στο επιθηλιακό μεμβρανικό αντιγόνο (EMA-epithelial membrane antigen/muc1- mucin 1, cell surface associated, CD227) [2, 54, 55]. Το κοινό λευκοκυτταρικό αντιγόνο (LCA-leukocyte common antigen) εκφράζεται επίσης σε μεγάλο ποσοστό στα ΑΛΜΚ [2, 54]. Υψηλό ποσοστό των ΑΛΜΚ εμφανίζει θετικότητα σε έναν ή περισσότερους Τ-δείκτες. Ωστόσο έχουν αναφερθεί και περιπτώσεις ΑΛΜΚ που δεν εκφράζουν κανένα Τ-κυτταρικό δείκτη και μνημονεύονται ως ΑΛΜΚ χωρίς ειδικό ανοσοφαινότυπο ( null cell ALCL) [2]. Στα ως άνω λεμφώματα ανευρίσκονται με μοριακές τεχνικές σε μεγάλο ποσοστό αναδιατάξεις του T-κυτταρικού υποδοχέα (TCR, T-cell receptor rearrangement) [56, 57]. Επειδή όμως έως και σήμερα δεν έχει παρατηρηθεί κάποια διακριτή μορφολογική, κλινική και μοριακή διαφορά μεταξύ των δύο παραπάνω τύπων, του ΑΛΜΚ Τ-κυτταρικής αρχής και του ΑΛΜΚ χωρίς ειδικό ανοσοφαινότυπο, θεωρούνται ως μία οντότητα και κατατάσσονται σύμφωνα με την τελευταία ταξινόμηση γενικώς στα ΑΛΜΚ [2, 5, 37]. Το CD3 αντιγόνο, που αποτελεί τον πιο διαδεδομένο Τ-κυτταρικό δείκτη, είναι αρνητικό σε ποσοστό >50% των ΑΛΜΚ περιπτώσεων ενώ, στα ΑΛΜΚ, ALK- το ποσοστό είναι ακόμα μεγαλύτερο [2]. Τα αντιγόνα CD5 και CD7 είναι επίσης αρνητικά, όπως επίσης και το CD8 εκτός σπάνιων εξαιρέσεων. Σε αντιδιαστολή, τα CD2 και CD4 αντιγόνα είναι θετικά σε ένα σημαντικό ποσοστό των ΑΛΜΚ. Τα νεοπλασματικά κύτταρα του ΑΛΜΚ εκφράζουν επίσης σε υψηλά ποσοστά αντιγόνα έναντι κυτταροτοξικών πρωτεϊνών που περιέχονται στα ενδοκυττάρια κυστίδια όπως οι ΤΙΑ-1, granzyme B, perforin [57, 58], ενώ σε μικρό αριθμό περιπτώσεων εκφράζουν επίσης και το αντιγόνο CD56 [55]. Το CD15 αντιγόνο είναι θετικό στο 20% των περιπτώσεων, σε λίγα κύτταρα και κυρίως στα ΑΛΜΚ, ALK- [55]. Σε τομές ψυκτικού μικροτόμου τα νεοπλασματικά κύτταρα είναι εντόνως θετικά στο CD25 αντιγόνο. Tόσο το Τ-κυτταρικής αρχής ΑΛΜΚ όσο και το ΑΛΜΚ χωρίς ειδικό ανοσοφαινότυπο εκφράζουν σε ποσοστό >50% την πρωτεΐνη FASCIN [59, 60], όπως επίσης και τη γλυκοπρωτεΐνη MUC-1/DF3 [61]. Έχουν περιγραφεί σημαντικές διαφορές μεταξύ των ΑΛΜΚ, ALK+ και ΑΛΜΚ, ALK- ως προς την έκφραση μορίων που ελέγχουν την απόπτωση. Ειδικότερα τα ΑΛΜΚ, ALK+ δεν εκφράζουν τον αντι-αποπτωτικό δείκτη BCL2, και 30
31 είναι σπανιότατα θετικά στον BCL-X L. Σε αντίθεση, τα ΑΛΜΚ, ALK- είναι θετικά σε ποσοστό άνω του 50% στο BCL2 και άνω του 40% στο BCL-X L [62-65]. Σημαντική διαφορά επίσης παρατηρείται μεταξύ των δύο ομάδων και ως προς την έκφραση της ενεργού κασπάσης (ac-3) και της ανενεργού proc-3 κασπάσης, μορίων που ελέγχουν επίσης την απόπτωση [65]. Οι αποπτωτικοί δείκτες BAX και BCL-X S εκφράζονται συχνότερα στα ALK+ απ ότι στα ALK- [63]. Η διαφορά της έκφρασης των αποπτωτικών δεικτών επιβεβαιώνεται και από τη μέθοδο TUNNEL, η οποία δείχνει σημαντικές διαφορές (3.4% vs 1.1%, p=0.0002) μεταξύ των ALK+ και ALKλεμφωμάτων [63]. Η μιτωτική δραστηριότητα των Σ-ΑΛΜΚ, όπως αυτή μετριέται με τον δείκτη κυτταρικού πολλαπλασιασμού Ki-67/ΜΙΒ-1, είναι ιδιαίτερα αυξημένη και στους δύο τύπους των Σ-ΑΛΜΚ, χωρίς σημαντικές στατιστικά διαφορές (μέσος όρος 75%, διακύμανση από 20,5% έως 94,6%) [63]. Τα νεοπλασματικά κύτταρα του ΠΔ-ΑΛΜΚ είναι στην πλειονότητα τους θετικά στο CD30, συνήθως θετικά στο CD4 αντιγόνο, και σε ποσοστό >70% στα αντιγόνα έναντι των πρωτεϊνών των κυτταροτοξικών κοκκίων (granzyme B, TIA-1, perforin) [66, 67]. Συχνά δεν εκφράζουν άλλους Τ-δείκτες (CD2, CD5, CD3), ενώ οι περιπτώσεις ΠΔ-ΑΛΜΚ χωρίς ειδικό ανοσοφαινότυπο είναι σπάνιες [2]. Τα ΠΔ- ΑΛΜΚ, σε ελάχιστες περιπτώσεις εκφράζουν την γλυκοπρωτεΐνη EMA [35], ενώ το δερματικό λεμφοκυτταρικό αντιγόνο (CLA-cutaneous lymphocyte antigen) ή επίτοπος HECA-452, ανευρίσκεται σε >50% των περιπτώσεων [66]. 31
32 ΜΟΡΙΑΚΗ ΠΑΘΟΛΟΓΙΑ ΤΟΥ ΑΛΜΚ TNFRSF8/CD30/Ki-1 Η αναγνώριση του ΑΛΜΚ ως ειδικού τύπου των ΜΗΛ στηρίχθηκε κυρίως στην ανοσοϊστοχημική έκφραση από τα νεοπλασματικά λεμφοειδή κύτταρα του αντιγόνου Ki-1 που ανακαλύφθηκε το 1982 από τους Stein και συν. [6, 7] και προσδιορίσθηκε στο σύστημα ταξινόμησης CD (Cluster Designation) ως CD30 to 1986 [8]. Περαιτέρω βιοχημικές και μοριακές μελέτες [68, 69] έδειξαν ότι το εν λόγω μόριο κωδικοποιείται από το αντίστοιχο γονίδιο που εδράζεται στο χρωμόσωμα 1 (1p36). Το μόριο Ki-1 είναι μία διαμεμβρανική πρωτεΐνη με μοριακό βάρος 120 kd και ανήκει στην ομάδα των υποδοχέων του αυξητικού παράγοντα των νεύρων/ παράγοντα της νέκρωσης των όγκων (NGF-nerve growth factor/tnf - tumor necrosis factor) και αναφέρεται από την HUGO Gene Nomenclature Committee ως: TNFRSF8 (tumor necrosis factor receptor superfamily, member 8). Αρχικά η παρουσία του εν λόγω αντιγόνου ανιχνεύθηκε σε κυτταρικές σειρές της νόσου Hodgkin (L428) όπου και εκφραζόταν από τα Hodgkin και Reed-Sternberg κύτταρα καθώς και σε μικρό αριθμό μεγάλων κυττάρων (λεμφοβλαστών) που εδράζονταν γύρω από τα βλαστικά κέντρα σε αντιδραστικές λεμφαδενίτιδες [6, 69, 70]. Περαιτέρω μελέτες έδειξαν ότι το μονοκλωνικό αντίσωμα έναντι του αντιγόνου Ki-1 (κλώνος Ber-H2) εκφράζεται από ενεργοποιημένα Β και Τ-κύτταρα καθώς και από ενεργοποιημένα ιστιοκύτταρα [71, 72]. Επίσης εκφράζεται σε διάφορες νεοπλασίες του λεμφικού ιστού όπως σε πρωτοπαθή Β-κυτταρικής αρχής λεμφώματα των σωματικών κοιλοτήτων, σε Β-λεμφώματα με πλασματοκυτταροειδή διαφοροποίηση και σε θυμικά λεμφώματα από μεγάλα Β-κύτταρα [11, 73]. Επιπροσθέτως, η πλειονότητα των διάχυτων Β-λεμφωμάτων από μεγάλα κύτταρα με «αναπλαστική μορφολογία» εμφανίζουν εστιακή θετικότητα στο εν λόγω αντιγόνο, ενώ περιπτώσεις με μη «αναπλαστική μορφολογία» σπάνια εκφράζουν το Ki-1/CD30 [2]. Τα νεοπλασματικά λεμφοειδή κύτταρα του ΑΛΜΚ εκφράζουν στην πλειονότητα τους (>90%) το CD30 στην κυτταρική μεμβράνη καθώς και στη περιοχή που εδράζεται η συσκευή του Golgi. Εντονότερη χρωστική αντίδραση παρατηρείται στα μεγάλου μεγέθους νεοπλασματικά κύτταρα, ενώ στα μικρότερα η έκφραση του CD30 είναι ήπια ή αρνητική [2, 37]. 32
33 Η έκφραση του CD30 στα νεοπλασματικά κύτταρα του ΑΛΜΚ συνδέεται με την παθογένεση του λεμφώματος. Πρόσφατα ευρήματα, σε κυτταρικές σειρές Σ- ΑΛΜΚ, ALK+ και ALK-, δείχνουν ότι η CD30 ελεγχόμενη σηματοδότηση επηρεάζει βασικούς κυτταρικούς μηχανισμούς [74]. Μηχανισμούς που επηρεάζουν την μεταγραφή γονιδίων που σχετίζονται με την επαγωγή ενδοκυττάριων σημάτων, την έκφραση κυτοκινών, της ανοσολογικής απάντησης και της απόπτωσης (Εικόνα 2, σελ. 37). Μεγάλος αριθμός αυτών των γονιδίων ρυθμίζονται από τον πυρηνικό παράγοντα κβ (Nuclear Factor κb, NF-κB). Στα κύτταρα του ΑΛΜΚ εμπλέκονται και οι δύο κυτταρικές οδοί ενεργοποίησης του NF-κB, η κανονική και η εναλλακτική (canonical & alternative NF-κB pathway) [74, 75]. ALK Η ραγδαία πρόοδος της μοριακής βιολογίας τόσο σε θεωρητικό όσο και σε τεχνικό επίπεδο έχει οδηγήσει σε σημαντικές ανακαλύψεις σε όλους σχεδόν τους τομείς της σύγχρονης ιατρικής. Στο ΑΛΜΚ, οι εξελιγμένες μοριακές τεχνικές και οι νέες γνώσεις σε υποκυτταρικό επίπεδο, οδήγησαν σε μεγάλο βαθμό στην κατανόηση της παθογένειας της νόσου και στην μοριακή ταυτοποίηση της (Εικόνα 1, σελ. 22). Ειδικότερα, στα μέσα της δεκαετίας του 80 αναφέρθηκαν για πρώτη φορά στη βιβλιογραφία περιστατικά «κακοήθων ινωδών ιστιοκυτωμάτων», τα οποία αποδείχθηκαν στη συνέχεια ότι ήταν ΑΛΜΚ, και έφεραν μία χαρακτηριστική μετάθεση την: t(2;5)(p23;q35) [76, 77]. Περαιτέρω μελέτες έδειξαν ότι αυτή η μετάθεση ήταν χαρακτηριστική για αυτό τον τύπο των λεμφωμάτων [14, 78]. Σημείο αναφοράς στη μοριακή διερεύνηση του ΑΛΜΚ αποτέλεσε η κλωνοποίηση του γονιδίου της «κινάσης του αναπλαστικού λεμφώματος» (ALK) από την ομάδα του Morris το 1994 [79]. Οι Morris και συν. [79] περιέγραψαν την μετάθεση τμήματος του 2p23 όπου εδράζεται το γονίδιο της ALK, στο τελικό άκρο του χρωμοσώματος 5 και συγκεκριμένα στη θέση 5q35 όπου κωδικοποιείται μία πυρηνική πρωτεΐνη, η νουκλεοφωσμίνη (nucleophosmin, NPM). Η NPM (Β23) ανακαλύφθηκε στα τέλη της δεκαετίας του 70 ως μία RNA συνδεδεμένη πυρηνική φωσφοπρωτεΐνη με μοριακό βάρος D [80, 81], που λειτουργεί ως μεταφορέας νεοσυντεθημένων ριβονουκλεοπρωτεΐνων από το κυτταρόπλασμα και ειδικότερα από τα ριβοσώματα στον πυρήνα [82, 83, 84, 85]. Η ALK είναι μία γλυκοπρωτεΐνη, ένας διαμεμβρανικός υποδοχέας, του τύπου της τυροσινικής κινάσης (RTK-receptor tyrosine kinase), μέλος της ομάδας των 33
34 υποδοχέων της ινσουλίνης με μοριακό βάρος 200 kd με μήκος cdna 6226 bp και αποτελείται απο 1620 αμινοξέα [79, 86-88]. Η φυσιολογική (wild-type) ALK πρωτεΐνη είναι γενικώς ανενεργή, παίζει όμως σημαντικό ρόλο στην ενδομήτρια ανάπτυξη του νευρικού συστήματος, όπου και εκφράζεται από ολιγάριθμα νευρικά στοιχεία, όπως γαγλιακά κύτταρα, πυρήνες στην περιοχή του θάλαμου και υποθάλαμου, σπάνια ενδοθηλιακά κύτταρα και περικύτταρα [87, 88, 89]. Επίσης η ALK πρωτεΐνη έχει ανιχνευθεί και σε κυτταρικές σειρές νεοπλασμάτων νευροεκτοδερμικής προέλευσης (νευροβλάστωμα, γλοιοβλάστωμα, ES/PNET, μελάνωμα, μικροκυτταρικό καρκίνωμα, ραβδομυοσάρκωμα) [90, 91]. Έχει αναφερθεί από τoυς Cessna και συν. [92] ότι η ALK πρωτεΐνη εκφράζεται σε κακοήθεις όγκους προερχόμενους από τα περιφερικά έλυτρα των νεύρων (MPNST-malignant peripheral nerve sheath tumors), σε μικρό ποσοστό λειομυοσαρκωμάτων και κακοήθων ινωδών ιστιοκυτωμάτων, σε φλεγμονώδεις μυοϊνοβλαστικούς όγκους (inflammatory myofibroblastic tumors-imt) και σε περιπτώσεις ραβδομυοσαρκωμάτων. Ανοσοϊστοχημική έκφραση της ALK παρατηρήθηκε και σε άλλους όγκους των μαλακών μορίων όπως το λιποσάρκωμα, το συνοβιακό σάρκωμα και το εξωσκελετικό οστεοσάρκωμα [93]. Η παρουσία της NPM-ALK μετάθεσης αρχικά περιγράφηκε με τη μέθοδο Southern blot [94] και ακολούθησε η ανίχνευση της με τη μέθοδο RT-PCR [94-98], με υβριδισμό in situ [99], και με διπλού φάσματος φθορίζων in situ υβριδισμό [100]. Η ευκολία και η πληθώρα μεθόδων και τεχνικών οδήγησε σε άνθηση της μελέτης της ALK στα ΑΛΜΚ και σε άλλες νεοπλασίες με αποτέλεσμα την ανακάλυψη έως σήμερα συνολικά 14 χρωμοσωμικών ανωμαλιών στις οποίες εμπλέκεται το γονίδιο της ALK [19]. Σε όλες τις μεταθέσεις, όπως και στην κλασσική, το αποτέλεσμα είναι η ενεργοποίηση του τμήματος της ALK και η έκφραση «χιμαιρικών» πρωτεϊνών με αποτέλεσμα την κακοήθη εξαλλαγή. Το αποτέ-λεσμα της μετάθεσης t(2;5)(p23;q35) είναι η δημιουργία ενός νέου «χιμαιρικού» γονιδίου, του NPM-ALK, στο οποίο ο εκκινητής και τα 4 πρώτα εξόνια προέρχονται από το NPM γονίδιο, ενώ η υπόλοιπη αλληλουχία, από το τμήμα του γονιδίου της ALK που κωδικοποιεί το κυτταροπλασμικό τμήμα της ALK πρωτεΐνης [84]. Το προϊόν αυτού του γονιδίου είναι μια νέα πρωτεΐνη, η NPM-ALK, μοριακού βάρους 80 kd, που περιλαμβάνει στο μόριο της το πολυμερές τμήμα της NPM και την ενδοκυττάρια περιοχή της ALK (το τμήμα της τυροσινικής κινάσης). Πειραματικά, τόσο in vitro όσο και in vivo, έχει αποδειχθεί ότι η NPM-ALK δύναται να προκαλέσει ογκογένεση [101, 102, 103]. Η 34
35 δράση αυτή της NPM-ALK «χιμαιρικής πρωτεΐνης» επιβεβαιώθηκε σε πειραματικά μοντέλα in vitro όπου οδήγησε σε κακοήθη εξαλλαγή των NIH-3T3 και Fr3T3 κυτταρικών σειρών ινοβλαστικών κυττάρων τρωκτικών [101] και in vivo στην γένεση κακοήθων Β-κυτταρικής αρχής λεμφωμάτων σε πειραματικά πρότυπα ακτινοβοληθέντων BALB/cByJ ποντικών [16]. Σημαντική πρόοδος στη διάγνωση και μελέτη του ΑΛΜΚ προήλθε από την εμφάνιση μονοκλωνικών και πολυκλωνικών αντισωμάτων έναντι του επίτοπου της ALK πρωτεΐνης και της «χιμαιρικής» NPM-ALK πρωτεΐνης [17, 18]. Ακολούθησαν μεγάλες πολυκεντρικές κλινικοεργαστηριακές μελέτες οι οποίες έδειξαν ότι η έκφραση της NPM-ALK σχετιζόταν με επιμέρους μορφολογικά, κλινικά και βιολογικά χαρακτηριστικά του ΑΛΜΚ [17, 104, 105]. Λόγω της ειδικής μοριακής δομής της, η NPM-ALK πρωτεΐνη ανιχνεύεται ανοσοϊστοχημικά στο κυτταρόπλασμα ή/και στον πυρήνα/πυρήνιο. Οι άλλες μεταθέσεις που έχουν παρατηρηθεί στο ΑΛΜΚ εμφανίζουν διαφορετική κυτταρική εντόπιση (κυτταροπλασμική ή κυτταροπλασμική/κοκκώδης, μεμβρανική, περιπυρηνική). Πρόσφατα έχουν δημιουργηθεί και αντισώματα που στοχεύουν φωσφορυλιωμένες μορφές της ALK και της NPM. Εκτός από την παραπάνω μετάθεση, έχουν αναφερθεί σε ποσοστό περίπου 20% των ΣΑΛΜΚ και άλλες μεταθέσεις που εμπλέκουν το γονίδιο της ALK (Πίνακας 2) [5, 19, 79, 90, ], οι οποίες αναφέρονται στον Πίνακα 2 (επόμενη σελίδα). 35
36 Πίνακας 2. Μεταθέσεις και «χιμαιρικές» πρωτεΐνες στις οποίες εμπλέκεται η ALK και έχουν ανιχνευθεί στα ΑΛΜΚ. Χρωμοσωμική μετάθεση t(2;5)(p23;q35) Εμπλεκόμενη Πρωτεΐνη Nucleophosmin (NPM) t(1;2)(q25;p23) Tropomyosin 3 (TPM3) t(2;3)(p23;q21) TRK-fused gene (TFG) TFGXlong TFGlong TFshort % Χιμαιρική πρωτεΐνη (KDa) Εντόπιση NPM ALK (80) Π/ΠΡ/Κ Π/Κ Νεόπλασμα ALK+ ALCL & ALK+ DLBCL TPM3 ALK (104) ΔΚ ALK+ ALCL & IMT 2 TFG ALK (85-113) ΔΚ ALK+ ALCL TFG ALK (113) ΔΚ TFG ALK (97) ΔΚ TFG ALK (85) ΔΚ inv(2)(p23;q35) ATIC 2 ATIC ALK (96) ΚΚ ALK+ ALCL & IMT t(2;17)(p23;q23) Clathrin heavy 2 CLTC1 ALK (250) ΚΚ ALK+ ALCL, chainlike 1 (CLTC1) IMT & ALK+ DLBCL t(2;x)(p23;q11 12) Moesin (MSN) <1 MSN ALK (125) Μ ALK+ ALCL t(2;19)(p23;p13) Tropomyosin 4 <1 TPM4 ALK (95 ΔΚ ALK+ ALCL (TPM4) 105) & IMT t(2;17)(p23;q25) ALO17 <1 ALO17 ALK (ΑΓ) Κ ALK+ ALCL t(2;22)(p23;q11.2) Non-muscle myosin <1 MYH9 ALK (220) ΔΚ ALK+ ALCL heavy chain (MYH9) ins(3 ALK)(4q22 24) Άγνωστη <1 Άγνωστη ΚΚ ALK+ DLBCL Επεξηγήσεις: ΑΓ: άγνωστη, ΔΚ: διάχυτη κυτταροπλασμική, Κ: κυτταρόπλασμα, ΚΚ: κοκκώδης κυτταροπλασμική, Μ: μεμβρανική, Π: πυρήνας, ΠΡ: περιπυρηνική. ALCL: anaplastic large cell lymphoma, ALK: anaplastic lymphoma kinase, ALO17: ALK lymphoma oligomerization partner on chromosome 17, ATIC: 5-aminoimidazole-4-carboxamide ribonucleotide formyltransferase/imp cyclohydrolase, DLBCL: diffuse large B-cell lymphoma, IMT: inflammatory myofibroblastic tumor. Επίδραση της NPM-ALK στην ογκογένεση Θεωρείται ότι στα ΑΛΜΚ, ALK+ η έκφραση του CD30 ελέγχεται από την ALK μέσω της φωσφωρυλίωσης των μορίων μεταγωγής σήματος STAT3 και 5 (signal transducer and activator of transcription 3 και 5) και της υπερέκφρασης της JUNB (jun B proto-oncogene) πρωτεΐνης λόγω της ενεργοποίησης των εξωκυττάριων κινασών μεταγωγής σήματος ERK1 και 2 (extracellular signal-regulated kinase 1 και 2) (Εικόνα 2, σελ. 39). Η φωσφωρυλίωση των STAT3 και 5 και η ενεργοποίηση του συμπλέγματος AP1- JUNB οδηγεί σε ενεργοποίηση της μεταγραφής του CD30 γονιδίου. Η «χιμαιρική» NPM-ALK πρωτεΐνη παρεμβάλλεται επίσης στην CD30 ελεγχόμενη σηματοδότηση και ενεργοποίηση του NF-κB μέσω της αναστολής ένωσης του TRAF2 (tumour necrosis factor receptor-associated factor 2) στον CD30 εξαιτίας του διμερισμού της με την wild-type (WT) NPM. Αυτή η δέσμευση του CD30 έχει ως αποτέλεσμα την αποδόμηση του TRAF2 και την φωσφορυλίωση της 36
37 BCL3 (B-cell CLL/lymphoma 3) που δρά ως ενεργοποιητής της μεταγραφής λόγω ένωσης της με τον NF-κB (Εικόνα 2). Επιπρόσθετα, στα ΑΛΜΚ, ALK+ έχει παρατηρηθεί ενεργοποίηση του JAK3, μορίου που αποτελεί τον φυσιολογικό ενεργοποιητή του STAT3, σε κυτταρικές σειρές και πρωτοπαθείς όγκους [116]. Ο JAK3 συνδέεται επίσης απευθείας με την NPM-ALK και η αναστολή του ελλαττώνει την τυροσινική της δράση [116]. Πρόσφατα έχει αναφερθεί και αυτοκρινής παραγωγή από τα ΑΛΜΚ, ALK+ κύτταρα της IL-9, η οποία αποτελεί προς τα πάνω (upstream) διαμορφωτή της JAK3/STAT3 οδού σηματοδότησης [117]. Η κυτταρική οδός σηματοδότησης PI3K/Akt είναι επίσης ενεργοποιημένη στα ΑΛΜΚ, ALK+. Ειδικότερα, η ενεργοποίηση της Akt σε κυτταρική σειρά λεμφώματος (BaF3) οδηγεί στη φωσφωρυλίωση και έξοδο από τον πυρήνα του μεταγραφικού παράγοντα FOXO3a (FKHRL1) [118]. Η παραπάνω δράση έχει ως αποτέλεσμα την ενεργοποίηση της Cyclin D2 και την αδρανοποίηση των p27 και Bim-1 [118]. Η ενεργοποίηση της Akt δρά και αντιαποπτωτικά, απομονώντας την προαπτωτική πρωτεΐνη Bad και προάγοντας την ένωση της με τις πρωτεΐνες, με αποτέλεσμα την αύξηση του ελέυθερου Bcl-X L [119]. Σε πειραματικά μοντέλα έχει φανεί ότι η ενεργοποιημένη Akt αναστέλλει την ενεργότητα της κασπάσης 9, ενώ αντίθετα αυξάνει την ενεργότητα του NF-κB μέσω επίδρασης στην αποδόμηση του αναστολέα του NF-κB, ΙκΒ και προάγει την ενεργότητα της κινάσης τύπου σερίνης/θρεονίνης mtor [120]. Η παραπάνω κυτταρική οδός επηρεάζει και την πρόοδο του κυτταρικού κύκλου, λόγω ελλάτωσης (downregulation) του κυκλινοεξαρτόμενου αναστολέα p27 [121, 122]. Η ενεργοποίηση της φωσφολιπάσης PLC-γ λόγω της αλληλεπίδρασης της με τους υποδοχείς της τυροσινικής κινάσης (στην προκειμένη περίπτωση την NPM- ALK) οδηγεί σε υδρόλυση της phophatidylinositol-4,5-biphosphate (PIP2) και την μετατροπή της στην διαλυτή inositol-1,4,5-triphosphate (IP3) και στην μεμβρανική diacylglycerol (DAG). Τα παραπάνω μόρια λειτουργούν ως δευτερεύοντες ενδοκυττάριοι σηματοδότες, απελευθερώνοντας μόρια ασβεστίου από το ενδοπλασματικό δίκτυο (IP3) και ενεργοποιώντας την κινάση PKC. Η ογκογόνος δράση NPM-ALK έχει αποδειχθεί σε πειραματικά μοντέλα ότι επάγεται και μέσω της αλληλεπίδρασης της με το Ras [123]. Η NPM-ALK οδηγεί σε ενεργοποίηση του Ras και την φωσφορυλίωση των εξωκυττάριων μιτογόνων κινασών της ομάδας MAPK [124]. Στα παραπάνω κύτταρα, η NPM-ALK λειτουργεί ως υπόστρωμα σύνδεσης υποκείμενων πρωτεϊνών προσαρμογής (downstream adaptors), 37
38 scaffolding πρωτεϊνών με SRC homology 2 (SH2) και υπόστρωμα στο οποίο οι αυτοφωσφορυλιωμένες περιοχές λειτουργούν ως θέσεις φωσφορυλίωσης των παραπάνω μορίων που τελικά οδηγούν στην ενεργοποίηση της Ras-ERK κυτταρικής οδού σηματοδότησης [101]. Οι SH2 domain-containing transforming protein (SHC), η insulin receptor substrate 1 (IRS1) και ο growth factor receptor-bound protein 2 (GRB2) είναι οι συχνότερα εμπλεκόμενες πρωτεΐνες στην RTK σηματοδότηση που έχει ως αποτέλεσμα τον κυτταρικό πολλαπλασιασμό μέσω της επαγωγής μιτογόνων σημάτων από την NPM-ALK. Η NPM-ALK επίσης επάγει τη έκφραση του μεταγραφικού παράγοντα Myc [125]. Η NPM-ALK επίσης φωσφωρυλιώνει την JUN N-terminal kinase (JNK, MAPK8) καθώς και υποκείμενα μόρια, όπως την JUN με αποτέλεσμα την επαγωγή της ενεργότητας του AP1 μεταγραφικού παράγοντα. Αυτή η δράση έχει ως αποτέλεσμα την απώλεια ελέγχου της κυτταρικής διαίρεσης και του κυτταρικού πολλαπλασιασμού λόγω αναστολής του p21 και της επαγωγής της Cyclin D3 και Cyclin A [126]. Εικόνα 2. Μοριακές οδοί ενεργοποίησης στα ΑΛΜΚ. Στα ΑΛΜΚ και ιδιαίτερα στα ALK+ έχει αναφερθεί ένα πολύπλοκο δίκτυο επιδράσεων μεταξύ της NPM-ALK και μορίων που επιδρούν/επάγουν πολυάριθμες κυτταρικές οδούς με τελικό αποτέλεσμα την επιβίωση των νεοπλασματικών κυττάρων. Επίσης στην ίδια εικόνα παρουσιάζεται και μέρος των επιδράσεων του CD30 στην παθογένεση των ΑΛΜΚ. Επεξηγήσεις: ALK: anaplastic lymphoma kinase, AP1: activator protein 1, BAD: Bcl-2 antagonist of cell death, BCL2 : B-cell CLL/lymphoma 2, BCL3 : B-cell CLL/lymphoma 3, Bcl-X L : BCL2-like 1 protein, BIM: BCL2-like 11, DAG: diacylglycerol, ELF4EBP1: eukaryotic initiation factor 4E-binding protein 1, ERK1/2: Extracellular signal-regulated kinase 1/2, FOXO3a: Forkhead box protein O3, GRB2: growth factor receptor-bound protein 2, IκΒ: Inhibitor of nuclear factor kappa-b kinase subunit beta, IL: interleukin, IRS1: insulin receptor substrate 1, JAK: Janus kinase, JNK: c-jun N-terminal kinase, JUN: transcription factor AP-1, MCL1: Induced myeloid leukemia cell differentiation protein Mcl-1, mtor: mammalian target of rapamycin, MYC: Myc proto-oncogene protein, NF-kB: nuclear factor kappa-b, NPM: nucleophosmin, PIP2: phosphatidylinositol-4,5-bisphosphate, PIP3: phosphatidylinositol-4,5-triphosphate, PKC: protein kinase C, PLC-γ: Phospholipase C-γ, PTEN: phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase and dual-specificity protein phosphatase PTEN, p70s6k: phospho S6 kinase, RAS: rat sarcoma, SHC: SRC homology 2 domain-containing, SRC: Schmidt-Ruppin A-2, SOS: son of sevenless, SHP: tyrosine-protein phosphatase non-receptor type 11, STAT: Signal transducer and activator of transcription, S6RP: S6 riboprotein, TRAF: tumour necrosis factor receptor-associated factor 2, WT: wild-type. 38
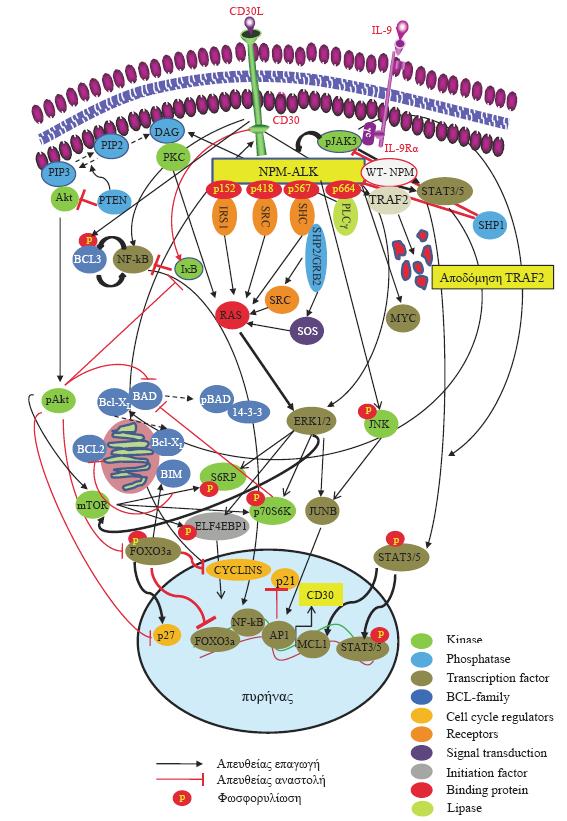
39 39
40 Η SHP2 ενεργοποιημένη από την NPM-ALK επιδρά στην ανάπτυξη των ΑΛΜΚ μέσω της επαγωγής της φωσφορυλίωσης των ERK1 (MAPK3) και ERK2 (MAPK1), πιθανότατα ως αποτελέσμα της ένωσης με το σύμπλεγμα GRB2 SOS1 [127]. Παράλληλα η σίγαση (silencing) της SHP2 αναστέλλει την φωσφορυλίωση της NPM-ALK στη θέση Tyr416 και επιδρά στον κυτταρικό πολλαπλασιασμό, όπως έχει φανεί και με την δράση αναστολέων της οικογένειας των SRC κινασών καθώς και με την χρήση ειδικών SRC sirna [128]. Σε αντίθεση με την SHP2, η απώλεια της SHP1 (PTPN6, protein tyrosine phosphatase, non-receptor type 6) έχει παρατηρηθεί στο ~50% των ΑΛΜΚ [129]. Η SHP1 λειτουργεί ως ογκοκατασταλτικό γονίδιο και η απώλεια της, συνήθως σχετίζεται με μη ελεγχόμενο πολλαπλασιασμό των κυττάρων, λόγω ενεργοποίησης της JAK3-STAT3 κυτταρικής οδού. Έχει δειχθεί ότι η αποκατάσταση της έκφρασης του SHP1 στα ΑΛΜΚ οδηγεί στην αποφωσφωρυλίωση της NPM-ALK και την μερική της αποδόμηση στο πρωτεάσωμα, με αποτέλεσμα την επίσης αποφωσφωρυλίωση των JAK3 και STAT3 και σταθεροποίηση της κυτταρικής ανάπτυξης [130]. Σημαντικό ρόλο στα ΑΛΜΚ διαδραματίζει και η κινάση mtor (mammalian target of rapamycin, FRAP1) και οι υποκείμενες αυτού πρωτεΐνες, ribosomal protein S6 kinase (p70s6k), S6 ribosomal protein (S6RP) και eukaryotic initiation factor 4Ebinding protein 1 (EIF4EBP1). Οι παραπάνω πρωτεΐνες είναι υποκείμενα δραστικά μόρια των PI3K-Akt και Ras-ERK κυτταρικών οδών σηματοδότησης [131]. Ωστόσο στα ΑΛΜΚ φαίνεται ότι η NPM-ALK-μεσολαβούμενη mtor επαγωγή γίνεται κυρίως μέσω της Ras-ERK κυτταρικής οδού [131]. Ο αναστολέας της mtor, rapamycin, επιδρά έντονα στην κυτταρική αύξηση και οδηγεί σε απόπτωση τα νεοπλασματικά κύτταρα του ΑΛΜΚ [131]. 40
41 Cyclin D1 Η ανάπτυξη και αύξηση του κυττάρου γίνεται μέσω μίας καλά ρυθμιζόμενης διαδικασίας, τον κυτταρικό κύκλο, ο οποίος διασφαλίζει τον πλήρη διπλασιασμό του DNA πριν από την κυτταρική διαίρεση. Ο κυτταρικός κύκλος ελέγχεται από ένα πολύπλοκο σύστημα πρωτεϊνών, με βασικότερους «πρωταγωνιστές» τις κυκλίνες και τις κυκλινοεξαρτώμενες κινάσες. Το κύτταρο αντιδρά σε διάφορα αυξητικά και περιβαλλοντικά ερεθίσματα κατά την διάρκεια του κύκλου. Ανασταλτικό ρόλο σε διάφορα σημεία ελέγχου του κυτταρικού κύκλου παίζουν διάφορα εξωτερικά ερεθίσματα, ογκογονίδια (p53, Rb1) και παράγοντες που χαρακτηρίζονται ως αναστολείς των κυκλινοεξαρτώμενων κινασών. Οι κυκλινοεξαρτώμενες κινάσες (cyclin-dependent kinases- CDKs) αποτελούν μία ομάδα πρωτεϊνοκινασών (CDK1 έως CDK6) η ενεργότητα των οποίων αυξομειώνεται στη διάρκεια του κύκλου και ελέγχεται κυρίως από τις πρωτεΐνες του κυτταρικού κύκλου, τις κυκλίνες [132]. Πoλυάριθμες κυκλίνες έχουν έως τώρα αναγνωρισθεί (A-M, T) στα ευκαρυωτικά κύτταρα. Οι κυκλίνες τις ομάδας D (D1, D2 και D3) κωδικοποιούνται από διαφορετικά γονίδια και εκφράζονται σε όλα τα πολλαπλασιαζόμενα κύτταρα [133]. Ειδικότερα, η κυκλίνη D1 (Cyclin D1) είναι μία πυρηνική πρωτεΐνη, μοριακού βάρους 33,7 kd και μεγέθους 295 αμινοξέων, η οποία κωδικοποιείται από το γονίδιο CCND1, το οποίο αναφέρεται και ως PRAD1, λόγω της συχνής παρουσίας μετάθεσης του εν λόγω γονιδίου με το γονίδιο της παραθυρεοειδικής ορμόνης (PTH) στα καλοήθη αδενώματα των παραθυρεοειδών σωματίων. Το CCND1 γονίδιο εδράζεται στο χρωμόσωμα 11, περιοχή 11q13, bp Το CCND1 γονίδιο περιέχει 5 εξόνια [134]. Το εν λόγω γονίδιο θεωρείται ότι λειτουργεί ως ογκογονίδιο όπως έχει αποδειχθεί πειραματικά. Εμφανίζει ενίσχυση στο 15% των πρωτοπαθών καρκινωμάτων του μαστού, ενώ υπερέκρφαση του mrna στο 30%, υπαινίσσοντας ότι συμπληρωματικοί μηχανισμοί ως προς την γονιδιακή ενίσχυση πιθανότατα οδηγούν στην ως άνω υπερέκφραση, όπως η επαγωγή άλλων ογκογόνων σημάτων, σωματικές μεταλλάξεις και μεταθέσεις του CCND1 γονιδίου, απώλεια συνδετών της κυκλίνης D1 (pocket proteins), έκφραση άλλου ογκογονιδίου ή συνδυασμός των παραπάνω [135, 136]. Ο συχνός μονονουκλεοτιδικός πολυμορφισμός τύπου A/G (A870G) στο ιντρόνιο 4 του CCND1 γονιδίου οδηγεί στην δημιουργία 2 διαφορετικών mrna 41
42 μεταγράφων (ισομορφές a και b/ cyclin D1a και cyclin D1b), που κωδικοποιούν δύο διαφορετικές δομικά και από άποψη μεγέθους πρωτεΐνες, τις cyclin D1a και cyclin D1b. Η cyclin D1b πρωτεΐνη, που κωδικοποιείται από την ισομορφή β και εμφανίζει αλλαγές στα τελευταία 55 αμινοξέα από το καρβοξυλικό άκρο (C-terminus), έχει αναφερθεί ότι συναντάται εκλεκτικά σε κάποιους τύπους καρκινωμάτων (παχέος εντέρου, πνεύμονα, οισοφάγου) και εμφανίζει αυξημένη ογκογόνο δράση λόγω της συνεχούς παραμονής της στον πυρήνα. Η παραμονή της στον πυρήνα έχει αποδειχθεί ότι οφείλεται στην μη σύνδεση της με τις πρωτεϊνες GSK-3β (Glycogen Synthase Kinase-3β) και CRM1 που λειτουργούν ως μεταφορείς (προσδέτες) της Cyclin D1 κατά την έξοδο της από τον πυρήνα στο κυτταρόπλασμα όπου και συντελείται η αποδόμηση της [137]. Η κυκλίνη D1 διαδραματίζει σημαντικότατο ρόλο στην ρύθμιση του κυτταρικού κύκλου. Η σύνδεση της κυκλίνης D1 με τα καταλυτικά ένζυμα Cdk4 και Cdk6 οδηγεί σε φωσφορυλίωση του προϊόντος του γονιδίου του ρετινοβλαστώματος (Rb) (Εικόνα 3, σελ. 44). Η φωσφορυλιωμένη Rb (prb) μορφή δεν μπορεί πλέον να συνδεθεί με τον μεταγραφικό παράγοντα E2F-DP-1, με τελικό αποτέλεσμα την μετάβαση του κυττάρου από την G1 στην S φάση του κυτταρικού κύκλου. Κύτταρα τα οποία στερούνται της λειτουργικής prb μορφής εμφανίζουν ιδιαίτερα χαμηλά επίπεδα κυκλίνης D1 και του συμπλέγματος κυκλίνης D1- Cdk4. Τα παραπάνω στοιχεία, δείχνουν, ότι η φωσφορυλιωμένη Rb συμμετέχει στη διέγερση της μεταγραφής της κυκλίνης D1. Σε πειραματικά μοντέλα ποντικών με λέμφωμα που υπερεκφράζουν τις κυκλίνες D, η prb είναι υπερφοσφωρυλιωμένη σε σχέση με την prb των κυττάρων που δεν υπερεκφράζουν τις D κυκλίνες. Η ένωση στο σύμπλεγμα κυκλίνης D1/Cdk4 μορίων της οικογένειας των CDK αναστολέων Cip/Kip, οδηγεί σε ενεργοποίηση του συμπλέγματος κυκλίνης Ε/Cdk2 με αποτελέσμα της πρόοδο του κυτταρικού κύκλου. Αξίξει να σημειωθεί εδώ, ότι η ένωση των Cip/Kip πρωτεϊνών με την κυκλίνη D1, δεν αποτελεί ανασταλτικό παράγοντα στην δράση της κυκλίνης D1. Το αντίθετο, διεκολύνει την σύνδεση της κυκλίνης D1 με την Cdk4 και διασφαλίζει την παραμονή της στον πυρήνα, αναστέλλοντας της έξοδο της από αυτόν. Αυξητικοί παράγοντες επάγουν την κυκλίνη D1 κυρίως μέσω των Ras εξαρτώμενων κυτταρικών οδών σηματοδότησης και της ενεργοποίησης μορίων της ομάδας των μιτογόνο-ενεργοποιημένων πρωτεϊνοκινασο-κινασών (mitogen-activated protein kinase-kinases, MEK1 και 2) καθώς και των εξωκυττάριων πρωτεϊνοκινασών ρυθμιστών σηματοδότησης (extracellular signal-regulated protein kinases, ERKs) 42
43 [138]. Η πρωτεόλυση της κυκλίνης D1 και η ρύθμιση της ενδοκυττάριας κυκλοφορίας της επίσης επάγονται από αυξητικούς παράγοντες. Η απανταχού κυτταρική εντόπιση της κυκλίνης D1 (polyubiquitination) και έξοδος της από τον πυρήνα εξαρτάται από την φωσφορυλίωση της πρωτεΐνης στην θέση Thr-286 από την GSK-3β [139]. Αυξητικοί παράγοντες, σήματα των οποίων μεταφέρονται μέσω των Ras, PI3K και Akt οδών σηματοδότησης ρυθμίζουν και εδώ την ενεργότητα και την κυτταρική εντόπιση της GSK-3β. Η φωσφορυλίωση της GSK-3β στη σερίνη 9 από την Akt, έχει ως αποτέλεσμα την δραστική μείωση της καλυτικής επίδρασης της GSK-3β στην κυκλίνη D1 που τελικά οδηγεί στην ενδοπυρηνική της συσσώρευση στην G1 φάση του κυτταρικού κύκλου, οδηγώντας έτσι μέσω της επίδρασης της στο Rb στην πρόοδο του κύκλου. Υπερέκφραση της «κανονικής» πρωτεΐνης κυκλίνης D1 (cyclin D1a) έχει αναφερθεί σε πληθώρα νεοπλασίων, όπως του μαστού, οισοφάγου, παγκρέατος, εντέρου, πνεύμονα, ουροδόχου κύστεως, προστάτη, ενδομητρίου, κεφαλής και τραχήλου, εξωτραχήλου μήτρας, όπως επίσης και σε αστροκυττώματα, σαρκώματα και λεμφώματα [134, 140, 141]. Περίπου το 50% των καρκινωμάτων του μαστού εμφανίζουν υπερέκφραση της κυκλίνης D1, που δεν σχετίζεται πάντα με ενίσχυση του γονιδίου ή με παρουσία στεροειδών υποδοχέων. Στα καρκινώματα κεφαλής και τραχήλου, ανιχνεύεται συχνά υπερέκφραση της κυκλίνης D1, σε πολλές περιπτώσεις με ταυτόχρονη ενίσχυση του γονιδίου. Στο καρκίνωμα του παχέος εντέρου η υπερέκφραση της κυκλίνης D1 παρατηρείται στα αρχόμενα στάδια της καρκινογένεσης. Παραδόξως, η υπερέκφραση της «κανονικής» πρωτεΐνης κυκλίνης D1 από μόνη της δεν οδηγεί σε κακοήθη εξαλλαγή [142]. Στα λεμφώματα η υπερέκφραση της πρωτεΐνης κυκλίνης D1 λόγω της μετάθεσης t(11;14)(q13;q32) έχει αναφερθεί στο λέμφωμα από κύτταρα του μανδύα, στο πολλαπλούν μυέλωμα (ΠΜ), καθώς και σε σπάνιες περιπτώσεις Β- λεμφοκυτταρικής λευχαιμίας/λεμφώματος και προλεμφοκυτταρικής λευχαμίας [140]. Στα ΑΛΜΚ ελάχιστες αναφορές υπάρχουν για ανίχνευση της πρωτεΐνης ή του mrna της κυκλίνης D1. 43
 και το απελευθερώνει από την ένωση του με το σύμπλεγμα E2F/DP-1.")
44 Εικόνα 3. Η επίδραση της κυκλίνης D1 στον κυτταρικό κύκλο. Η κυκλίνη D1 συνδεόμενη με τις CDK4&6 σχηματίζει ένα ολοένζυμο το οποίο φωσφορυλιώνει το προϊόν του γονιδίου του ρετινοβλαστώματος (Rb) και το απελευθερώνει από την ένωση του με το σύμπλεγμα E2F/DP-1. Με αυτό τον τρόπο οι E2F/DP-1 μεταγραφικοί παράγοντες προωθούν τον κυτταρικό κύκλο πέρα από το περιοριστικό σημείο (restriction point) και από την G1 στην S φάση. Επεξηγήσεις: cdk4 & cdk6: cyclin-depedent kinases 4 & 6, DP-1: Transcription factor Dp-1, Rb: retinoblastoma. 44
45 Fascin Οι πρωτεΐνες Fascins παρατηρούνται τόσο στα σπονδυλωτά όσο και τα ασπόνδυλα ζώα. Ονομάσθηκαν έτσι λόγω τις ικανότητας τους να ενώνονται στενά και σταθερά με την F-actin δημιουργώντας δεμάτια (από την λατινική λέξη fasciculus: δεμάτιο, δέσμη). Υπάρχουν 3 Fascins: η γενική Fascin ή Fascin-1, η Fascin του αμφιβληστροειδή χιτώνα ή Fascin-2 και η ορχική Fascin ή Fascin-3. [143]. Το γονίδιο FSCN1 βρίσκεται στο χρωμόσωμα 7q22 και κωδικοποιεί την αποτελούμενη από 493 αμινοξέα Fascin-1, το γονίδιο της Fascin-2 στο 17q25 και της Fascin-3 στο 7q31. Στην παρούσα μελέτη, ο όρος Fascin θα χρησιμοποιηθεί για την Fascin-1. Το μόριο της Fascin εκτός από την σύνδεση με την F-actin, η οποία γίνεται μέσω 2 περιοχών σύνδεσης, η πρώτη πλησίον του αμινοτελικού άκρου (αμινοξέα 33-47) και η δεύτερη στις θέσεις μεταξύ των αμινοξέων , αποτελεί υπόστρωμα για την πρωτεΐνη kinase C alpha (PKCα), ενώ αλληλεπιδρά και με το κυτταροπλασμικό τμήμα της p75ntr (p75 neurotrophin receptor) [144, 145]. Η φωσφορυλίωση της Fascin στην θέση Ser-39 αναστέλλει την ικανότητα σύνδεσης της με την F-actin. In vitro και in vivo πειράματα σε κυτταρικές σειρές έδειξαν ότι η Fascin σχηματίζει καλά οργανωμένα δεμάτια, σε παράλληλη διάταξη, με την F-actin. Τα παραπάνω δεμάτια είναι σημαντικά δομικά στοιχεία του κυτταροσκελετού, τα οποία στηρίζουν τον σχηματισμό των μεμβρανικών προσεκβολών, απαραίτητων στην κίνηση και μετανάστευση των κυττάρων. Ανάλογα με τον τύπο των κυττάρων οι παραπάνω προσεκβολές είναι εστιακές (filopodia), ή εκτεταμένες προβολές της κυτταρικής μεμβράνης (lamellipodia), μικροπροσεκβολές (microspikes) ή επιμήκεις προσεκβολές (δενδρίτες). Η Fascin εκφράζεται κυρίως σε στοιχεία του αναπτυσσόμενου νευρικού συστήματος, ενδοθηλιακά κύτταρα, ινοβλάστες, ώριμα δενδριτικά κύτταρα και απουσιάζει από τους περισσότερους επιθηλιακούς ιστούς [146, 147]. Ήπια έκφραση της έχει αναφερθεί στα κύτταρα της βασικής στιβάδας των πλακωδών επιθηλίων, όπως του δέρματος και του οισοφάγου. Σε αντίθεση με τους φυσιολογικούς ιστούς, υπερέκφραση της Fascin παρατηρείται σε πολυάριθμες νεοπλασίες, όπως καρκινώματα (μαστός, παχύ έντερο, πάγκρεας, οισοφάγος, πνεύμονας, οωθήκες, στόμαχος, δέρμα, μήτρα, νεφρό, λάρυγγας, ουροδόχος κύστη, προστάτης, ενδοκρινικοί όγκοι του ΓΕΣ), σαρκώματα (αγγειοσάρκωμα, λιποσάρκωμα, λειομυοσάρκωμα, κακόηθες ινώδες ιστιοκύττωμα, στρωματικά σαρκώματα του 45
46 ενδομητρίου), νεοπλασίες της γλοίας (αστροκυττώματα, γλοιοβλαστώματα) και του λεμφοαιμοποιητικού ιστού [143, 145, ]. Η Fascin απουσιάζει από τα λεμφοκύτταρα. Υπερέκφραση της Fascin έχει παρατηρηθεί στα HeLa (Henrietta Lacks) κύτταρα, EBV-μετασχηματισμένα Β- λεμφοκύτταρα, στα Reed-Sternberg (RS) κύτταρα της νόσου Hodgkin, σε περιπτώσεις λεμφωματοειδούς βλατίδωσης και σε δερματικά και συστηματικά ΑΛΜΚ [154, 155]. Αν και πιθανολογείται ότι υπάρχει ένας κοινός μηχανισμός που εξηγεί αυτή την υπερέκφραση της Fascin στις παραπάνω νεοπλασίες, θωρείται πιθανότερο ότι αυτή η υπερέκφραση είναι ειδική για κάθε ιστό (tissue specific). Η τελευταία θεωρία ενισχύεται από την παρουσίαση ερευνητικών ευρημάτων με μεγάλες αποκλίσεις ως προς την ανεύρεση της Fascin σε διάφορες νεοπλασίες. Ειδικότερα, τα παγκρεατικά διηθητικά καρκινώματα υπερεκφράζουν σε πάνω από 95% των περιπτώσεων την Fascin και τα σταδίου Ι μη-μικροκυτταρικά καρκινώματα του πνεύμονα στο 89%. Σε άλλα καρκινώματα, η υπερέκφραση της Fascin είναι λιγότερο συχνή. Στο γαστρικό καρκίνωμα η υπερέκφραση της Fascin είναι 12,5% στα ενδοβλεννογόνια καρκινώματα και αυξάνεται στο 53,3% στα στάδιου Τ4 διηθητικά καρκινώματα. Στο καρκίνωμα του μαστού η υπερέκφραση της Fascin κυμαίνεται μεταξύ 16 και 25% και αυτή η υπερέκφραση σχετίζεται με πιο επιθετική βιολογική συμπεριφορά, αρνητικούς οιστρογονικούς και προγεστερονικούς υποδοχείς και μείωση τόσο της ελεύθερης νόσου επιβίωσης, όσο της συνολικής επιβίωσης [145, 156]. Τα έως τώρα ερευνητικά δεδομένα συνηγορούν για την εμπλοκή μεταφραστικών και/ή μετα-μεταφραστικών μηχανισμών, καθώς και διαταραχών της συνοχής (σταθερότητας/ευστάθειας) του mrna που οδηγούν στην υπερέκφραση της Fascin και σπανιότερα, γονιδιακών διαταραχών του FSCN1 γονιδίου. Η σημαντική αύξηση της έκφρασης των ESTs (expressed sequence tag) δηλ. μικρών sub-sequence του cdna μετάγραφου της Fascin, έχουν παρατηρηθεί σε καρκινώματα του πνεύμονα, παγκρέατος, προστάτη και παχέος εντέρου [145]. Αύξηση των τιμών του mrna της Fascin ως προς το φυσιολογικό επιθήλιο, έχει αναφερθεί σε μελέτη με τη χρήση γονιδιακών μικροσυστοιχιών (gene microarray) στο παγκρεατικό αδενοκαρκίνωμα και με semi-quantitative RT-PCR στο οισοφαγικό καρκίνωμα [145]. 46
47 htert (human Telomerase Reverse Transcriptase) Τα άκρα των χρωμοσωμάτων του ανθρώπου αποτελούνται από επαναλαμβανόμενες εξαμερείς TTAGGG νουκλεοτιδικές αλληλουχίες, τα καλούμενα τελομερή τα οποία παρουσιάζουν προοδευτική ελάττωση του μήκους τους κατά την διάρκεια της S φάσης του κυτταρικού κύκλου. Η μείωση του μήκους των τελομερών είναι ευθέως ανάλογη με την ικανότητα για κυτταρική διαίρεση και λειτουργεί ως το «μιτωτικό ρολόϊ» του κυττάρου. Όταν το μήκος των τελομερών προσεγγίσει ένα ελάχιστο κρίσιμο όριο, πυροδοτείται μια σειρά μοριακών γεγονότων, που σηματοδοτούν τη μόνιμη λήξη του αναδιπλασιασμού του DNA στο συγκεκριμένο κύτταρο, το οποίο μπαίνει στο στάδιο της αναπαραγωγικής γήρανσης (replicative senescence). Η τελομεράση αποτελεί ένα σύνθετο ριβονουκλεοπρωτεϊνικό σύμπλοκο με ενζυμική δράση ειδικής ανάστροφης μεταγραφάσης που επιμηκύνει τα τελομερή προσθέτοντας στο 3 τους άκρο (GGGTTA)n νουκλεοτιδικές αλληλουχίες [157]. Η ικανότητα της τελομεράσης να προσθέτει GGGTTA αλληλουχίες στα άκρα των τελομερών, αποτελεί δείκτη ενεργότητας του ενζύμου και η μέτρηση της γίνεται με την στηριζόμενη στην αλυσιδωτή αντίδραση πολυμεράσης μέθοδο TRAP (Telomeric Repeat Amplification Protocol) [158]. Η ενεργότητα της τελομεράσης ανιχνεύεται σε χαμηλά επίπεδα σε σπάνιους τύπους σωματικών κυττάρων (λεμφοκύτταρα). Εξαίρεση αποτελούν οι εμβρυϊκοί ιστοί, τα γεννητικά κύτταρα και τα μητρικά κύτταρα (stem cells). Αποτελείται από δύο απαραίτητες για την λειτουργία της υπομονάδες, την RNA υπομονάδα ή htr (human Telomerase RNA) και την καταλυτική υπομονάδα ή htert. Η htert αποτελεί τον πρωτεϊνικό πυρήνα της τελομεράσης και εμφανίζει δράση ειδικής ανάστροφης μεταγραφάσης. Το γονίδιο που την κωδικοποιεί εντοπίζεται στο χρωμόσωμα 5 (5p15.33). Η πρωτεΐνη αποτελείται από 1132 αμινοξέα, με μοριακό βάρος 127kDa. Σύμφωνα με τους Moriarty και συν. [159] η htert πρωτεΐνη εμφανίζει ένα αμινο-τελικό-(n)-rna-καταλυτικό τμήμα (RID1), ακολουθούμενο από την περιοχή διασύνδεσης (linker region), δεύτερο RNAκαταλυτικό τμήμα (RID2), τμήμα με δράση ανάστροφης μεταγραφάσης και C-τελικό άκρο. Η htert απουσιάζει από τα περισσότερα φυσιολογικά κύτταρα, ενώ υπερεκφράζεται στην πλειονότητα των καρκινωμάτων [160]. 47
48 Η ρύθμιση της ενεργότητας της τελομεράσης γίνεται κυρίως μέσω μηχανισμών που στοχεύουν στην καταλυτική υπομονάδα του ενζύμου και επιδρούν σε προ-μεταγραφικό, μεταγραφικό, μετα-μεταγραφικό και μετα-μεταφραστικό επίπεδο. Α. ΠΡΟ-ΜΕΤΑΓΡΑΦΙΚΟ ΕΠΙΠΕΔΟ Η μεθυλίωση του προαγωγέα καθώς και των ιστονών θεωρούνται οι σημαντικότεροι μηχανισμοί καταστολής της γονιδιακής έκφρασης. Σε αντίθεση, η απομεθυλίωση ενεργοποιεί την γονιδιακή μεταγραφή. Η απομεθυλίωση του DNA προκαλεί την κινητοποίηση της histone acetyltransferase (HAT) με συνέπεια την μεταγραφή της htert [161]. Β. ΜΕΤΑΓΡΑΦΙΚΟ ΕΠΙΠΕΔΟ Η ρύθμιση της μεταγραφής του γονιδίου της htert αποτελεί τον κύριο μηχανισμό για την ρύθμιση της ενεργότητας της τελομεράσης στις νεοπλασίες. Πολυάριθμοι παράγοντες έχουν έως τώρα αναγνωρισθεί που επηρεάζουν θετικά ή αρνητικά την μεταγραφή του συγκεκριμένου γονιδίου όταν δεσμεύονται σε ειδικές θέσεις του προαγωγέα της htert. Οι παράγοντες αυτοί είναι πρωτεΐνες που προάγουν την μεταγραφή (c-myc, Sp1, HIF-1, AP2, ER, Ets) ή ογκοκατασταλτικές πρωτεΐνες (p53, WT1, Menin, Rb, p21 Waf-1, TGF-β) [161]. Γ. ΜΕΤΑ-ΜΕΤΑΓΡΑΦΙΚΟ ΕΠΙΠΕΔΟ Η παραπάνω ρύθμιση της htert, μέσω του εναλλακτικού τρόπoυ ματίσματος (alternative splicing) του αρχικού της mrna, οδηγεί στην δημιουργία εναλλακτικών ματισμένων μορφών (splice variants). Δεκατρείς συνολικά ματισμένες μορφές έχουν περιγραφεί [162]. Τρείς προκαλούνται από την αποκοπή εξονίων (deletion variants), ενώ οι 10 υπόλοιπες προκύπτουν από την προσθήκη ιντρονικών ακολουθιών (insertion variants) [162, 163]. Οι πιο καλά μελετημένες εναλλακτικά ματισμένες μορφές είναι οι: Adel που προκύπτει από την αποκοπή 36 bp από την αρχή του έκτου εξονίου, η Bdel που σχηματίζεται από την αποκοπή 183 bp που αντιστοιχούν στα εξόνια 7 και 8 και η Adel/Bdel, από την οποία λείπουν και οι δύο παραπάνω περιοχές. Η Adel μορφή, φαίνεται να αποτελεί αρνητικό ρυθμιστή της τελομεράσης [164], ενώ η Bdel δεν επιδρά στην ενεργότητα της τελομεράσης. Η Adel/Bdel ματισμένη μορφή έχει ανιχνευθεί σε ποσοστό εως 90% σε διάφορες κυτταρικές σειρές [165] και πιθανότατα επιδρά στην ενεργότητα της τελομεράσης [166]. Η Adel/Bplus μορφή, όταν υπερεκφράζεται, εμφανίζει ρυθμιστική δράση δρώντας ως 48
49 αναστολέας της ενεργότητας της τελομεράσης τόσο σε φυσιολογικούς ιστούς, όσο και σε νεοπλασίες [166]. Δ. ΜΕΤΑ-ΜΕΤΑΦΡΑΣΤΙΚΟ ΕΠΙΠΕΔΟ Η φωσφορυλίωση της htert από διάφορες κινάσες (Akt, κινάση C) οδηγεί στην αύξηση της δραστηριότητας της τελομεράσης με σύγχρονη μετατόπιση της htert απο το κυτταρόπλασμα στον πυρήνα. Εξαίρεση αποτελεί η φωσφορυλίωση της από την c-abl κινάση η οποία οδηγεί σε αδρανοποίηση της καταλυτικής της δράσης. Τα λεμφοκύτταρα, με εξαίρεση τα παρθένα Β-λεμφοκύτταρα των βλαστικών κέντρων, εκφράζουν συχνά την htert. Οι νεοπλασίες του λεμφοαιμοποιητικού ιστού εμφανίζουν ενεργότητα της τελομεράσης και έκφραση της htert. Υψηλές τιμές ενεργότητας της τελομεράσης έχουν παρατηρηθεί σε υποομάδες λευχαιμιών και στο πολλαπλούν μυέλωμα (ΠΜ). Η υπερέκφραση της ενεργότητας της τελομεράσης, σε συνδυασμό με τη μείωση των τελομερών, εμφανίζονται ως δείκτες πτωχής πρόγνωσης στο ΠΜ [167] και την Τ-κυτταρικής αρχής λευχαιμία των ενηλίκων [168]. MUC-1 (Mucin-1, Polymorphic epithelial mucin) Οι βλεννοπρωτεΐνες (Mucins) αποτελούν οικογένεια μεγάλου μοριακού βάρους (>200 kda) γλυκοπρωτεϊνών στις οποίες η ολιγοσακχαριτική ομάδα είναι συνδεδεμένη στο OH- της πλευρικής αλυσίδας μίας σερίνης ή θρεονίνης ή υδροξυλυσίνης με Ο-γλυκοσιδικό δεσμό [169]. Δεκαέξι mucins (MUC-1-16) έχουν έως τώρα περιγραφεί. Η σύνθεση τους γίνεται κυρίως από τα επιθηλιακά κύτταρα και κατατάσσονται σε εκκριτικού και μεμβρανικού τύπου. Η MUC-1 γλυκοπρωτεΐνη [γνωστή και ως EMA (epithelial membrane antigen/επιθηλιακό μεμβρανικό αντιγόνο), breast carcinoma-associated antigen DF3, CD227] είναι διαμεμβρανική πρωτεΐνη, μεγέθους 1255 αμινοξέων και κωδικοποιείται από το ομόνυμο γονίδιο που εδράζεται στο 1q21. Στο μόριο της παρατηρείται ένα μεγάλου μεγέθους εξωκυττάριο τμήμα, αποτελούμενο κυρίως από μία περιοχή τυχαίων επαναλήψεων (tandem repeats) των 20 αμινοξέων, διαμεμβρανικό τμήμα (28 αμινοξέα) και ενδοκυττάριο τμήμα μεγέθους 72 αμινοξέων (Εικόνα 4, σελ. 51). Μέσω του εναλλακτικού τρόπoυ ματίσματος (alternative splicing) του αρχικού της mrna, η MUC-1 εμφανίζει 12 ματισμένες μορφές. 49
50 Αποτελείται από την α και β υπομονάδες. Η α-υπομονάδα εμφανίζει συγκολλητικές (adhesive) ιδιότητες και επιδρά θετικά ή αρνητικά στην κυτταρική συνοχή. Πειραματικά δεδομένα δείχνουν ότι δημιουργεί προστατευτικό στρώμα έναντι βακτηρίων και βλαπτικών για το κύτταρο ενζύμων. Η β-υπομονάδα περιέχει C-τελικό τμήμα και εμπλέκεται στην μεταγωγή σήματος, μέσω φωσφορυλίωσης και πρωτεϊνικής αλληλεπίδρασης στις ERK, SRC και NF-kB οδούς σηματοδότησης. Στα ενεργοποιημένα Τ-λεμφοκύτταρα επηρεάζει άμεσα ή έμμεσα την Ras/MAPK οδό σηματοδότησης. Πιστεύεται ότι δρά ως λιγάση για την ICAM-1 (intercellular adhesion molecule-1) όταν εκφράζεται στα Τ-λεμφοκύτταρα και ενδοθηλιακά κύτταρα [170]. Η ένωση της με τον KLF4 στον PE21 προαγωγέα της P53 προάγει την εξέλιξη των όγκων μέσω επίδρασης στην P53-ελεγχόμενη μεταγραφή [171, 172]. Εκφράζεται στο κυτταρόπλασμα και/ή μεμβράνη των κυττάρων. Στα επιθηλιακά κύτταρα της αεροφόρου οδού, του μαστού και του ενδομητρίου εμφανίζει αυλική εντόπιση. Ανιχνεύεται στα ενεργοποιημένα και αδρανή Τ-λεμφοκύτταρα και σε επιθηλιακές και μη νεοπλασίες. Στις νεοπλασίες, η έκφραση της MUC-1 είναι υψηλή, δεν εμφανίζει όμως την χαρακτηριστική πολικότητα στις κορυφαίες επιφάνειες του κυττάρου όπως συμβαίνει στα φυσιολογικά επιθήλια. Υπάρχουν τουλάχιστον 56 διαφορετικά μονοκλωνικά αντισώματα που συνδέονται με ειδικούς επιτόπους του μορίου της MUC-1. Η MUC-1 εκφράζεται συχνά στα ΑΛΜΚ (30-60%), συχνότερα στα ALK+ [28, 61] και λιγότερο συχνά, περίπου στο 30% των ΠΔ-ΑΛΜΚ [35]. Η ανεύρεση της πρωτεΐνης, σύμφωνα με τα ευρήματα των Rassidakis και συν. αποτελεί ειδικό και ανεξάρτητο προγνωστικό δείκτη σε ασθενείς με ΑΛΜΚ και ειδικότερα της ομάδας ασθενών με ALK- νεοπλασματικό ανοσοφαινότυπο [61]. 50
 περιοχή σημειώνεται με μπλέ χρώμα και τα εναπομείναντα τμήματα του μορίου με καφέ.")
51 Εικόνα 4. Η δομή της MUC-1. Το κυτταροπλασμικό τμήμα με τα 72 αμινοξέα είναι χρωματισμένο με βυσσινί. Το διαμεμβρανικό τμήμα (28 αμινοξέα) εμφανίζεται πράσινο. Η VNTR (variable number of tandem repeats) περιοχή σημειώνεται με μπλέ χρώμα και τα εναπομείναντα τμήματα του μορίου με καφέ. Η σκιαγραφημένη περιοχή αντιπροσωπεύει τους πολυάριθμους Ο-γλυκοσιδικούς δεσμούς στην VNTR περιοχή. ΚΛΙΝΙΚΗ ΤΑΞΙΝΟΜΗΣΗ Το ΑΛΜΚ κλινικά διακρίνεται σε: α) Πρωτοπαθές (De novo): - Πρωτοπαθές του δέρματος (ΠΔ-ΑΛΜΚ, C-ALCL) - Συστηματικό ΑΛΜΚ (λεμφαδενικό με ή χωρίς εξωλεμφαδενική επέκταση) Η ανοσοϊστοχημική έκφραση της ALK από τα κύτταρα του ΑΛΜΚ διαχωρίζει πλέον δύο κλινικοπαθολογικές οντότητες τα: ΑΛΜΚ, ALK+ και ΑΛΜΚ, ALK-, οι οποίες εμφανίζουν διαφορετικά βιολογικά και κλινικά χαρακτηριστικά. - Σχετιζόμενο με HIV-λοίμωξη β) Δευτεροπαθές: Θεωρείται το ΑΛΜΚ το οποίο εμφανίζεται ως μετάπτωση προηγούμενης νόσου όπως οι παρακάτω: - Λεμφωματοειδής βλατίδωση - Νόσος Hodgkin 51
52 ΕΡΓΑΣΤΗΡΙΑΚΑ ΕΥΡΗΜΑΤΑ Η LDH ορού είναι συνήθως υψηλή όπως επίσης και η β 2 μικροσφαιρίνη [61, 104]. Αναιμία εμφανίζει περίπου το 40% των ασθενών. Στις σπάνιες αναφερόμενες περιπτώσεις πυογόνου ΑΛΜΚ έχει αναφερθεί περιφερική ηωσινοφιλία ή ουδετεροφιλία [42, 50, 51]. ΣΤΑΔΙΟΠΟΙΗΣΗ Η σταδιοποίηση των ασθενών γίνεται με το σύστημα Ann Arbor [173] ή το Διεθνή Προγνωστικό Δείκτη (International Prognostic Index-IPI) [174]. Στα παιδιά εφαρμόζεται το St Jude σύστημα σταδιοποίησης των ΜΗΛ [175]. Στο Ann Arbor σύστημα σταδιοποίησης χρησιμοποιούνται δύο ταξινομήσεις, η κλινική (Πίνακας 2, σελ. 55) και η παθολογοανατομική (Πίνακας 3, σελ. 55). Στο παραπάνω σύστημα χρησιμοποιούνται επίσης οι υποδιαιρέσεις Α, Β, Ε και Χ που υποδηλώνουν απουσία (Α) ή παρουσία (Β) γενικών συμπτωμάτων, εξωλεμφαδενική επέκταση (Ε), και ευρεία/εκτεταμένη (bulky) λεμφαδενική νόσος μεγέθους >10 εκ ή διεύρυνση >1/3 του μεσοθωρακίου (Χ). Η Β-ταξινόμηση γίνεται σε ασθενείς που: α) εμφανίζουν ανεξήγητη απώλεια βάρους πάνω από το 10% του βάρους σώματος τους τελευταίους 6 μήνες, β) ανεξήγητη πυρετική κίνηση πάνω από τους 38 ο C, γ) νυχτερινές εφιδρώσεις. Ο IPI αποτελεί το άθροισμα επιμέρους κλινικών παραμέτρων και αποτελεί ένα προβλεπτικό μοντέλο επιβίωσης των ασθενών που έχουν προσβληθεί από επιθετικό ή μη τύπο ΜΗΛ και βασίζεται σε επιμέρους κλινικά χαρακτηριστικά προ θεραπείας (Πίνακας 4, σελ. 56). Για την σταδιοποίηση της νόσου στην παιδική ηλικία έχουν προταθεί διαφορετικά συστήματα, απο τα οποία το πιο δημοφιλές είναι το μοντέλο του St. Jude Children's Research Hospital [175]. Το παραπάνω σύστημα διαχωρίζει τους ασθενείς σε τέσσερα στάδια σύμφωνα με την έκταση προσβολής (Πίνακας 5, σελ. 56). 52
53 ΠΡΟΓΝΩΣΤΙΚΟΙ ΚΑΙ ΠΡΟΒΛΕΠΤΙΚΟΙ ΔΕΙΚΤΕΣ Ο IPI στα ΑΛΜΚ, ΑΛΚ+ δεν εμφανίζει την σημαντικότητα όπως σε άλλου τύπου λεμφώματα, έχει όμως κάποια προβλεπτική αξία [5]. Η παρουσία της ALK αποτελεί σημαντικότατο προβλεπτικό δείκτη επιβίωσης. Η συνολική 5-ετής επιβίωση παιδιατρικών ασθενών με ΑΛΜΚ, ΑΛΚ+ πλησιάζει το 80%, ενώ σε ασθενείς με ΑΛΜΚ, ΑΛΚ- το 48%. Σε ενήλικες, το ποσοστό της 5-ετούς επιβίωσης ασθενών με ΑΛΜΚ, ΑΛΚ+ είναι στο 70% και των ΑΛΜΚ, ΑΛΚ- στο 49%, παραμένει όμως η διαφορά στατιστικά σημαντική [176]. Ο τύπος των μεταθέσεων δεν σχετίζεται με σημαντικές διαφορές ως προς την επιβίωση. Υποτροπές της νόσου εμφανίζονται στο 30% των περιπτώσεων, συχνά όμως οι ασθενείς παραμένουν ευαίσθητοι στην χημειοθεραπεία. Τα ΑΛΜΚ, ΑΛΚ- αν και αποτελούν την επιθετικότερη μορφή, εμφανίζουν καλύτερη πρόγνωση σε σχέση με τα PTCL (NOS) (49% έναντι 32% ως προς την συνολική 5-ετή επιβίωση) και 36% έναντι 20% ως προς την FFS-failure-free survival δηλ. την επιβίωση από την στιγμή της διάγνωσης έως την πρώτη επανεμφάνιση της νόσου, ή της υποτροπής μετά από απάντηση στη θεραπεία ή του θανάτου από οποιαδήποτε αιτία [176, 177]. Η 10-ετής επιβίωση των ασθενών με ΠΔ-ΑΛΜΚ πλησιάζει το 90% [5]. Η προσβολή επιχώριων λεμφαδένων δεν επηρεάζει την επιβίωση και είναι παρόμοια με τη πρωτοπαθή δερματική νόσο. Η αυτόματη υποχώρηση της νόσου, σε συνδυασμό με την ηλικία <60 έτη σχετίζεται με καλύτερη πρόγνωση, ενώ η εξωδερματική επέκταση και η μεγάλη ηλικία με μειωμένη επιβίωση [178]. Η μορφολογία του νεοπλάσματος δεν αποτελεί προγνωστικό δείκτη. ΘΕΡΑΠΕΙΑ Η θεραπευτική προσέγγιση του ΑΛΜΚ απαιτεί προσεκτική εκτίμηση όλων των κλινικών και εργαστηριακών ευρημάτων. Η θεραπεία εκλογής στις περισσότερες περιπτώσεις είναι η χημειοθεραπεία (ΧΜΘ). Η θεραπευτική ακτινοβόληση περιορίζεται σε ασθενείς με εντοπιζόμενη νόσο, όπως τα ΠΔ-ΑΛΜΚ χωρίς εξωδερματική επέκταση [179]. Οι θεραπευτικές στρατηγικές ποικίλουν: μόνο-χμθ, συνδυασμένη ΧΜΘ, μόνο-ακτινοθεραπεία, συνδυασμένη ΧΜΘ με ακτινοβολία, ΧΜΘ υψηλών δόσεων ακολουθούμενη από αυτόλογη μεταμόσχευση μυελού των 53
54 οστών ή βλαστικών κυττάρων, θεραπεία διάσωσης και θεραπεία συντήρησης [180]. Τα θεραπευτικά σχήματα βασίζονται συνήθως στις ανθρακυκλίνες. Σχήμα πρώτης γραμμής ΧΜΘ αποτελεί η CHOP (cyclophosphamide, hydroxy-daunorubicin, vincristine, prednisone). Επίσης χρησιμοποιούνται CHOP-like σχήματα (COP- BLAM: cyclophosphamide, vincristine, procarbazine, bleomycin, adriamycin, methylprednisolone, ή HOAP-Bleo: hydroxy-daunorubicin, vincristine, cytosinearabinoside, prednisolone, bleomycin). Μελλοντικές θεραπευτικές προσεγγίσεις. Σε πρόσφατες κλινικές μελέτες (ClinicalTrials.gov) έχει χρησιμοποιηθεί η ανοσοθεραπεία με το μονοκλωνικό anti- CD30 αντίσωμα, όπως το SGN-30 [181]. Η χρήση μικρών μορίων που δρούν ως αναστολείς της ALK έχουν επίσης δοκιμασθεί με ελπιδοφόρα αποτελέσματα [fused pyrrolocarbazole (FP)-derived kinase inhibitors και το NVP-TAE684, (5-chloro-2,4-diaminophenylpyrimidine) [19]. Ο χορηγούμενος per os αναστολέας του MET και της ALK (PF ) εμφανίζει σημαντική αντιογκογόνο δράση σε κυτταρικές σειρές ΑΛΜΚ, ALK+ (Karpas299 και SU-DHL- 1) λόγω αναστολής της κυτταρικής διαίρεσης και επαγωγής της απόπτωσης [182]. Επιπρόσθετα, λόγω της πλήρους εξάρτησης των κυττάρων του ΑΛΜΚ, ALK+ από τις τυροσινικές δράσεις της ALK, το ως άνω μόριο αποτελεί ιδανικό στόχο εμβολιασμού. 54
55 Πίνακας 2. Κλινική σταδιοποίηση (CS - clinical classification) των ασθενών με λέμφωμα σύμφωνα με το σύστημα Ann Arbor. Στάδιο Ι Προσβολή μίας λεμφαδενικής ομάδας (Ι) Ενός εξωλεμφαδενικού οργάνου ή τμήματος (ΙΕ) Προσβολή 2 ή περισσότερων λεμφαδενικών ομάδων στην ίδια πλευρά του διαφράγματος (ΙΙ) Στάδιο ΙI Τμηματική προσβολή εξωλεμφαδενικού οργάνου ή τμήματος με ταυτόχρονη προσβολή ενός ή περισσότερων λεμφαδενικών ομάδων στην ίδια πλευρά του διαφράγματος (ΙΙΕ) Προσβολή λεμφαδενικών ομάδων αμφοτερόπλευρα του διαφράγματος (ΙΙΙ) Στάδιο ΙIΙ - Η παραπάνω προσβολή μπορεί να συνοδεύεται από τμηματική διήθηση εξωλεμφαδενικού οργάνου ή τμήματος (ΙΙΙΕ), ή από προσβολή του σπλήνα (ΙΙΙS), ή και τα δύο (IIIES) Διάχυτη προσβολή ενός ή περισσοτέρων εξωλεμφαδενικών οργάνων Στάδιο ΙV και ιστών με ή χωρίς συνοδό λεμφαδενική προσβολή Υποσημείωση: Στα εξωλεμφαδενικά όργανα ανήκουν όλα, εκτός από τα παρακάτω: λεμφαδένες, σπλήνας, θύμος, σκωληκοειδής απόφυση, πλάκες Peyer, δακτύλιος του Waldeyer. Πίνακας 3. Παθολογοανατομική σταδιοποίηση (PS - pathological classification) των ασθενών με λέμφωμα σύμφωνα με το σύστημα Ann Arbor. Ν+ ή Ν- Παρουσία διήθησης λεμφαδένα (Ν+), απουσία διήθησης (Ν-) Η+ ή Η- Παρουσία διήθησης του ήπατος (Η+), απουσία διήθησης (Η-) S+ ή S- Παρουσία διήθησης του σπλήνα (S+), απουσία διήθησης (S-) μετά από σπληνεκτομή L+ ή L- Παρουσία διήθησης του πνεύμονα (L+), απουσία διήθησης (L-) M+ ή Μ- Ρ+ ή Ρ- Παρουσία διήθησης του μυελού των οστών (Μ+), απουσία διήθησης (με επίχρισμα ή βιοψία) (Μ-) Παρουσία διήθησης του υπεζοκώτα (Ρ+), απουσία διήθησης (Ρ-) (κυτταρολογικά ή ιστολογικά) Ο+ ή Ο- Παρουσία διήθησης οστού (Ο+), απουσία διήθησης (Ο-) D+ ή D- Παρουσία διήθησης του δέρματος (D+), απουσία διήθησης (D-) 55
56 Πίνακας 4. Σταδιοποίηση ασθενών σύμφωνα με το Διεθνή Προγνωστικό Δείκτη (International Prognostic Index-IPI). Κλινικά χαρακτηριστικά Βαθμός Ηλικία 60 1 LDH ορού πάνω από τις φυσιολογικές τιμές (>1) 1 Performance status 2 1 Στάδιο νόσου ΙΙΙ - ΙV 1 Πολλαπλή εξωλεμφαδενική προσβολή 1 Διεθνής Προγνωστικός Δείκτης Ομάδες επικινδυνότητας σύμφωνα με τον IPI Ομάδα χαμηλού κινδύνου 0-1 Ομάδα χαμηλού ενδιάμεσου κινδύνου 2 Ομάδα ενδιάμεσου- υψηλού κινδύνου 3 Ομάδα υψηλού κινδύνου 4-5 Πίνακας 5. Κλινική σταδιοποίηση της νόσου στην παιδική ηλικία σύμφωνα με το σύστημα St. Jude. Στάδιο Ι Στάδιο ΙI Στάδιο ΙIΙ Στάδιο ΙV Προσβολή μίας λεμφαδενικής ομάδας Μονήρης όγκος εκτός της κοιλίας και του υπεζωκότα Μονήρης όγκος με προσβολή μίας επιχώριας λεμφαδενικής ομάδας Δύο ή περισσότεροι όγκοι ή λεμφαδενικές ομάδες στην ίδια πλευρά του διαφράγματος Πρωτοπαθής όγκος του ΓΕΣ με ή χωρίς προσβολή των επιχώριων λεμφαδένων Πολλαπλοί όγκοι Προσβολή λεμφαδενικών ομάδων αμφοτερόπλευρα του διαφράγματος Πρωτοπαθής υπεζωκοτικός ή ενδοκοιλιακός όγκος Περισπονδυλικός όγκος ή όγκος που διηθεί την σκληρά μήννιγγα Προσβολή του ΚΝΣ Προσβολή του μυελού των οστών (ιστολογικά διήθηση > 5% του μυελού) 56
57 ΕΙΔΙΚΟ ΜΕΡΟΣ ΕΙΔΙΚΟ ΜΕΡΟΣ 57
58 58
59 ΥΛΙΚΟ ΚΑΙ ΜΕΘΟΔΟΙ ΑΣΘΕΝΕΙΣ Στη μελέτη συμπεριλήφθησαν 42 ασθενείς με αναπλαστικό λέμφωμα από μεγάλα κύτταρα (ΑΛΜΚ), που επιλέχθηκαν από τα αρχεία του Εργαστηρίου Γενικής Παθολογίας και Παθολογικής Ανατομικής του Α.Π.Θ. για το χρονικό διάστημα Η διάγνωση του ΑΛΜΚ στηρίχθηκε στα ιστολογικά, ανοσοϊστοχημικά και σε αρκετές περιπτώσεις στα κλινικά ευρήματα. Όλες οι περιπτώσεις επανεξετάσθηκαν και ταξινομήθηκαν σύμφωνα με τις τελευταίες οδηγίες της Π.Ο.Υ. [2]. Σε αρκετές περιπτώσεις έγινε και πρόσθετος ανοσοϊστοχημικός έλεγχος. Επιπρόσθετα σε 3 ασθενείς υπήρχε η δυνατότητα παρακολούθησης της εξέλιξης της νόσου λόγω βιοψιών από υποτροπές της νόσου, από άλλες εστίες ή μεταστάσεις. Ειδικότερα, σε 2 ασθενείς παρατηρήθηκε υποτροπή της νόσου στο ίδιο όργανο (δέρμα και λεμφαδένα) και σε 1 ασθενή υποτροπή σε άλλο όργανο. Η χρονική διάρκεια των υποτροπών της νόσου κυμαινόταν από 3 έως 84 μήνες. ΥΛΙΚΟ Το υλικό που χρησιμοποιήθηκε αφορούσε σε 36 ασθενείς, κύβους παραφίνης ιστολογικών παρασκευασμάτων που είχαν αποσταλεί στο Εργαστήριο για συμβουλευτική εκτίμηση και σε 6 ασθενείς, βιοψίες εντός διαλύματος φορμόλης, που μονιμοποιήθηκαν και εγκλείσθηκαν σε παραφίνη στο Εργαστήριο σύμφωνα με τα υπάρχοντα Πρωτόκολλα για το χειρισμό του χειρουργικού υλικού. Σε 2 ασθενείς υπήρχε παραληφθείς κύβος παραφίνης καθώς και προγενέστερο ή μεταγενέστερο μονιμοποιημένο στο Εργαστήριο χειρουργικό βιοπτικό υλικό και σε 1 ασθενή υλικό από 2 η βιοψία από άλλη ανατομική θέση. Συνολικά, 45 βιοψίες/υλικό από 42 ασθενείς συμπεριλήφθησαν στη μελέτη. ΕΝΤΟΠΙΣΗ Η συχνότερη πρωτοπαθής εστία εμφάνισης της νόσου ήταν οι λεμφαδένες σε 22/42 περιπτώσεις (52%). Από τις εξωλεμφαδενικές εντοπίσεις (20/42, 48%), το δέρμα (12/42, 28%) αποτέλεσε τη συχνότερη και ακολούθησαν τα μαλακά μόρια 59
60 (4/42, 9,5%). Άλλες εντοπίσεις ήταν τα οστά, ο πνεύμονας, ο σπλήνας και ο στόμαχος. Ταυτόχρονη λεμφαδενική και εξωλεμφαδενική εντόπιση (δέρμα) παρατηρήθηκε σε δύο μόνον περιπτώσεις. ΚΛΙΝΙΚΗ ΤΑΞΙΝΟΜΗΣΗ Με βάση τα κλινικά στοιχεία 34/42 (81%) περιπτώσεις ταξινομήθηκαν ως Σ- ΑΛΜΚ και 8/42 (19%) περιπτώσεις ως ΠΔ-ΑΛΜΚ. ΙΣΤΟΛΟΓΙΚΗ ΤΑΞΙΝΟΜΗΣΗ Η ιστολογική ταξινόμηση έγινε σύμφωνα με τα αναθεωρημένα κριτήρια της Π.Ο.Υ. [2]. Σε ολιγάριθμες περιπτώσεις παρατηρήθηκαν και γιγάντια νεοπλασματικά κύτταρα, ενώ συχνά αναγνωρίσθηκαν και κύτταρα με ηωσινόφιλη περιοχή πλησίον του πυρήνα. Σε αρκετές περιπτώσεις ανευρέθησαν και μικροί αριθμοί νεοπλασματικών κυττάρων με ενδοπυρηνικά έγκλειστα καθώς και διπύρηνες μορφές, δίκην κυττάρων Reed-Sternberg. Οι μιτώσεις ήταν συχνές σε αριθμό. ΑΝΟΣΟΦΑΙΝΟΤΥΠΟΣ Η αναγνώριση του ΑΛΜΚ ως ειδικού τύπου των ΜΗΛ στηρίζεται κυρίως στην ανοσοϊστοχημική έκφραση από τα νεοπλασματικά λεμφοειδή κύτταρα του αντιγόνου CD30/Ki-1. Στη μελέτη μας, σε όλες τις περιπτώσεις, τα νεοπλασματικά κύτταρα εμφάνιζαν έντονη χρωστική αντίδραση στην κυτταρική μεμβράνη. Σε αρκετά νεοπλασματικά κύτταρα, κυρίως μεγάλου μεγέθους, παρατηρήθηκε θετικότητα και στη περιοχή που εδράζεται η συσκευή του Golgi. Για την ανοσοϊστοχημική ταυτοποίηση των ΑΛΜΚ στα δύο βασικά λεμφοκυτταρικά συστήματα (Β-Τ) έγιναν για διαγνωστικούς σκοπούς σε όλες τις περιπτώσεις οι ανοσοχρώσεις για Β-λεμφοκύτταρα CD20 (Dako, Glostrup, Denmark) και CD45RΑ (Dako), ενώ σε 10 περιπτώσεις για τον Β-λεμφοκυτταρικό δείκτη CD79a (Dako). Για τα Τ-λεμφοκύτταρα διενεργήθηκε ανοσοϊστοχημεία για τα αντιγόνα CD45RΟ (Dako), CD3 (Dako) και CD43 (Novocastra, Newcastle, U.K.). Σε αρκετές αρχειακές περιπτώσεις οι ανοσοχρώσεις για τα αντιγόνα CD8, CD15, CD45, EMA, CD68 (Dako), CD4, CD5 (Novocastra) and Ki67 (clone MIB1, Dako) ήταν 60
61 επίσης προσβάσιμες και χρησιμοποιήθηκαν για την ανοσοφαινοτυπική ταυτοποίηση των ΑΛΜΚ. Επίσης για διαγνωστικούς και προβλεπτικούς σκοπούς σε 20/42 περιπτώσεις (από το 2000) διενεργήθηκε και ανοσοϊστοχημεία για το αντιγόνο ALK1 (Dako), και σε λίγες περιπτώσεις PCR για τον TCR-γ και RT-PCR για την μετάθεση t(2;5)(p23;q35). 61
62 ΜΕΘΟΔΟΙ ΕΠΙΛΟΓΗ ΥΛΙΚΟΥ Από όλους τους κύβους παραφίνης κάθε περιστατικού εξετάσθηκαν οι τομές Αιματοξυλίνης/Εωσίνης (Η/Ε), η ανοσοχρώση για το αντιγόνο CD30/Ki-1 καθώς και όπου ύπηρχε, η χρώση για την ALK1. Σε ολιγάριθμες περιπτώσεις λόγω σημαντικής απόκλισης του αρχικού με το εναπομείναν υλικό στους κύβους παραφίνης κόπηκαν νέες τομές οι οποίες και χρωματίσθηκαν με Η/Ε. Στις παραπάνω περιπτώσεις έγινε και νέα ανοχρώση για το αντιγόνο CD30/Ki-1. Επίσης σε 25 περιπτώσεις διενεργήθηκε η ανοσοχρώση ALK1 (CD246) (Πίνακας 6, σελ. 74). ΔΗΜΙΟΥΡΓΙΑ ΙΣΤΙΚΩΝ ΜΙΚΡΟΣΥΣΤΟΙΧΙΩΝ (ΤΜΑ) Για την εφαρμογή των ανοσοϊστοχημικών και των επιμέρους in situ τεχνικών δημιουργήθηκαν νέοι κύβοι παραφίνης με μικροσυστοιχίες ιστικών κυλίνδρων (ΤΜΑ-tissue microarrays) από συνολικά 49 κύβους παραφίνης με ΑΛΜΚ που αφορούσαν 38 ασθενείς. Για την δημιουργία των ΤΜΑ κύβων παραφίνης χρησιμοποιήθηκε το ειδικό χειροκίνητο υψηλής ακρίβειας μηχάνημα (TMA Model I, Beecher Instruments, Sun Prairie, WI, U.S.A.) που διαθέτει το Εργαστήριο σύμφωνα με τις οδηγίες του κατασκευαστή. Ελήφθησαν 5 τουλάχιστον ιστικοί κύλινδροι ανά περίπτωση διαμέτρου 0,6 χιλ. και εγκλείσθηκαν σε 2 νέους κύβους παραφίνης μεγάλου μεγέθους. Μετά την δημιουργία των ΤΜΑ, οι κύβοι, τοποθετήθηκαν σε κλίβανο στους 60 0 C για προκειμένου να ομογενοποιηθεί η παραφίνη και να αποκατασταθεί η συνοχή και οι μικρο-ρωγμές της παραφίνης. Όπως έχει αναφερθεί στην βιβλιογραφία [183, 184] με τη χρησιμοποίηση τουλάχιστον 3 ιστικών κυλίνδρων διαμέτρου 0,6 χιλ. είναι δυνατή η αντιπροσώπευση του >95% της ολόκληρης ιστικής τομής, ενώ στην περίπτωση της τοποθέτησης 5 ιστικών κυλίνδρων το ποσοστό ξεπερνά το 99% [183]. Οι τομές Η/Ε και η ανοχρώση για το αντιγόνο CD30/Ki-1 χρησιμοποιήθηκαν ως οδηγοί των εγκλεισθέντων σε παραφίνη βιοψιών. Ειδικότερα στα παραπάνω πλακίδια σημειώθηκαν με μαρκαδόρο ή/και με ειδική μικροσκοπική σφραγίδα όλες οι νεοπλασματικές θέσεις. Σε 5 περιπτώσεις το υλικό ήταν επαρκές για την τοποθέτηση 10 ιστικών κυλίνδρων, ενώ σε άλλες 5 τοποθετήθηκαν και 5 επιπλέον κύλινδροι από 62
63 μετάχρονη βιοψία ή άλλη εντόπιση. Η απόσταση μεταξύ των ιστικών κυλίνδρων ήταν 1,0 χιλ. Στους ΤΜΑ κύβους εγκλείσθηκαν επίσης και ιστικοί κύλινδροι από ιστούς ως δείκτες αξιοπιστίας (μάρτυρες) των ειδικών τεχνικών (IHC, FISH, ISH). Ειδικότερα, τοποθετήθηκαν 10 ιστικοί κύλινδροι ανά ΤΜΑ από 5 περιπτώσεις λεμφωμάτων από κύτταρα του μανδύα (Mantle cell lymphoma, MCL), 4 ιστικοί κύλινδροι ανά ΤΜΑ από 2 περιπτώσεις καρκινωμάτων παχέος εντέρου και 5 ιστικοί κύλινδροι ανά ΤΜΑ από 1 περίπτωση λοβιακού καρκινώματος του μαστού. Οι θέσεις των ιστικών κυλίνδρων στους 2 ΤΜΑ κύβους καταγράφησαν σε αρχείο Excel (2 worksheets). Το παραπάνω αρχείο χρησιμοποιήθηκε ως βάση για την χαρτογράφηση και ταξινόμηση των περιπτώσεων καθώς και για την καταγραφή των αποτελεσμάτων ανοσοχρώσεων και των in situ μεθόδων. Στις υπόλοιπες περιπτώσεις, όπου ήταν δυνατόν, η ανοσοϊστοχημεία και οι in situ μέθοδοι έγιναν σε ξεχωριστές για κάθε περίπτωση τομές παραφίνης. Αχρωμάτιστες τομές παραφίνης πάχους 5-10μm κόπηκαν από τους κύβους παραφίνης και τοποθετήθηκαν σε μη φορτισμένα πλακίδια. Στη συνέχεια και με την Η/Ε ως οδηγό επιλέχθησαν οι νεοπλασματικές περιοχές για την χειροκίνητη μικροεκτομή (microdissection) τομών και τοποθετήθηκαν σε αποστειρωμένα φιαλίδια Eppendorf. Η εκχύλιση του ολικού DNA και RNA από τα δείγματα της μελέτης έγινε σύμφωνα με γνωστά πρωτόκολλα μετά από ολονύκτια επώαση του υλικού σε διάλυμα πρωτεινάσης Κ και στη συνέχεια στο αντιδραστήριο TRIZOL-LS (Invitrogen, Paisley, UK). PCR Η Αλυσιδωτή αντίδραση πολυμεράσης, γνωστή και με το ακρωνύμιο PCR (Polymerase Chain Reaction) αποτελεί μια μη-κυτταρική μέθοδο κλωνοποίησης του DNA και η εμφάνιση της στα μέσα της δεκαετίας του 80 [185, 186] οδήγησε σε επανάσταση στους τομείς της μοριακής βιολογίας και γενετικής. Είναι σύντομη, όχι δαπανηρή, εύκολη στην εφαρμογή της και ιδιαίτερα ευαίσθητη ως τεχνική. Οι διάφορες παραλλαγές της έχουν επιτρέψει την ανάλυση ποικίλων δειγμάτων και ιστών καθώς και τη δυνατότητα ελέγχου πολυάριθμων γενετικών ανωμαλίων. Η εξαγωγή DNA θεωρείται ιδανική όταν γίνεται από φρέσκο ιστό ή κυτταρικά εναιωρήματα. Η αποθήκευση του ιστού στη βαθιά κατάψυξη (-70 0 C) αποτελεί τον πιο συχνό τρόπο αποθήκευσης δειγμάτων και ουσιαστικά δεν επηρεάζει την ποιότητα των 63
64 πυρηνικών οξέων. Η εξαγωγή DNA είναι δυνατή και από ιστούς μονιμοποιημένους σε φορμόλη και μάλιστα με ικανοποιητικά αποτελέσματα. Σημαντικό ρόλο στην ποιότητα του DNA παίζουν ο τρόπος και χρόνος μονιμοποιήσης του ιστού καθώς και το εφαρμοζόμενο πρωτόκολλο εξαγωγής του DNA [187, 188]. Πρόσφατα, έχουν δημοσιευθεί πρωτόκολλα τα οποία επιτρέπουν την εξαγωγή και ενίσχυση του DNA σχεδόν σε όλες τις περιπτώσεις μονιμοποιημένων σε φορμόλη ιστών, ακόμα και για μικρού μεγέθους αλληλουχίες νουκλεϊνικών οξέων ( bp) [189, 190]. Στην παρούσα μελέτη χρησιμοποιήθηκε η κλασσική PCR μέθοδος καθώς και παραλλαγές αυτής όπως η PCR ανάστροφης μεταγραφής (Reverse Transcription PCR/RT-PCR) και η πολλαπλή (multiplex) PCR. Στην RT-PCR, το υπό εξέταση DNA δεν είναι γενωμικό, αλλά προέρχεται απο ανάστροφη μεταγραφή του εξαχθέντος RNA (cdna), επιτρέποντας έτσι μελέτη της γονιδιακής έκφρασης, ενώ στην multiplex-pcr, δίνεται η δυνατότητα στόχευσης δύο ή περισσοτέρων αλληλουχιών με την ταυτόχρονη χρησιμοποίηση διαφορετικών ζευγών από εκκινητές. Ειδικότερα, στην μελέτη χρησιμοποιήθηκε η κλασσική PCR για την ανίχνευση της παρουσίας ανασυνδυασμού του Τ-κυτταρικού υποδοχέα τύπου γ (Τcell receptor γ, TCR-γ) ενώ η RT-PCR για α) την ανίχνευση της χρωμοσωμιακής μετάθεσης t(2;5)(p23;q35), β) την έκφραση της htert, γ) την ανίχνευση των Bplus και Bdel μεταγράφων της htert και δ) του mrna της cyclin D1. TCR-γ Εφαρμόσθηκε το τυποποιημένο πρωτόκολλο PCR του Εργαστηρίου για τον TCR-γ κατά Algara και συν. [191]. Σε όλα τα δείγματα χρησιμοποιήθηκαν θετικοί και αρνητικοί μάρτυρες. 64
65 RT-PCR t(2;5)(p23;q35). Η multiplex PCR εφαρμόσθηκε για την ταυτόχρονη ανίχνευση ολιγονουκλεοτιδικών αλληλουχιών ειδικών για το χιμαιρικό NPM/ALK μεταγραφικό προϊόν και το NPM μετάγραφο σύμφωνα με τους Wellmann και συν. [98]. Οι εκκινητές που χρησιμοποιήθηκαν για την ειδική ανάστροφη μεταγραφή της ALK ή της NPM ήταν οι παρακάτω και αποτέλεσαν τους 3 εκκινητές: 3 -ALK, 5 CGA GGT GCG GAG CTT GCT CAG C 3 και της NPM: 3 NPM, 5 GCT ACC ACC TCC AGG GGC AGA 3. Ο παρακάτω 5 NPM εκκινητής χρησιμοποιήθηκε για την ενίσχυση τόσο του NPM-ALK cdna όσο και του NPM cdna: 5 -NPM, 5 TCC CTT GGG GGC TTT GAA ATA ACA CC 3. Ως μάρτυρας της ειδικότητας του ενισχυμένου προϊόντος χρησιμοποιήθηκε ο germline 3 NPM εκκινητής: 5 AGC ACT TAG TAG CTG TGG AGG AAG 3 και ως μάρτυρας για την ποιότητα του RNA και του συμπληρωματικού DNA η ενίσχυση του τμήματος του μετάγραφου της c-erbb2. Για τη σύνθεση του μονόκλωνου cdna, το μισό απο το εξαχθέν RNA επωάσθηκε στους 70 C για 10 λεπτά με τα 3 ALK και 3 NPM ολιγονουκλεοτίδια σε diethylpyrocarbonate (DEPC). Η ανάστροφη μεταγραφή έγινε με επώαση σε Superscript Reverse Transcriptase (Life Technologies) σε θερμοκρασία δωματίου για 10 λεπτά, ακολούθησε επώαση στους 42 C για 50 λεπτά και τέλος επώαση στους 70 C για 15 λεπτά. Στη συνέχεια προστέθηκαν στο μίγμα οι εκκινητές (3' ALK ή 3' NPM και 5 NPM), η Taq πολυμεράση όπως και διάλυμα PCR (PCR buffer), ακολούθησε αποδιάταξη εκκινητών στους 94 C για 4 λεπτά και στη συνέχεια επώαση του μίγματος σε θερμικό κυκλοποιητή για 35 κύκλους ως εξής: 1 λεπτό υβριδισμός (annealing) στους 60 C, 1 λεπτό extension στους 72 C και 1 λεπτό αποδιάταξη (denaturation) στους 94 C. Το ενισχυμένο χιμαιρικό NPM/ALK μεταγραφικό προϊόν είχε μεγέθος 177 ζεύγη βάσεων (bp). Όλα τα PCR προϊόντα ανιχνεύθηκαν με ηλεκτροφόρηση σε πηκτή αγαρόζης 2% και χρώση με βρωμιούχο αζίδιο. Η ευαισθησία της μεθόδου δίνει την δυνατότητα ανίχνευσης, 1 έως 5 κυττάρων με μετάθεση, σε 10 5 κυττάρων. 65
66 htert Σχεδιάστηκε και χρησιμοποιήθηκε πρωτόκολλο duplex-pcr για την ταυτόχρονη ανίχνευση μεταγράφων της νουκλεοφωσμίνης (NPM) και της htert [192]. Το προϊόν της NPM είναι 186 ζεύγη βάσεων (νουκλεοτίδια στο M της Genbank) και το προϊόν της htert 95 ζεύγη βάσεων (μεταξύ 10 και 11 εξονίου του αντίστοιχου mrna [192]. Τα προϊόντα της αντίδρασης ανιχνεύθηκαν με ηλεκτροφόρηση σε πηκτή αγαρόζης 4%. Bdel και Bplus μετάγραφα της htert Η ανίχνευση των Bdel και Bplus μεταγράφων της htert έγινε σε μηνεοπλασματικούς λεμφαδένες. Η μέθοδος αυτή χρησιμοποιήθηκε για την επαλήθευση της έκφρασης των htert μεταγράφων σε φυσιολογικούς λεμφαδένες. Η εξαγωγή του RNA έγινε από κατεψυγμένους ιστούς μη-νεοπλασματικών λεμφαδένων επίσης με την χρήση του TRIZOL-LS. Το εξαχθέν RNA χρησιμοποιήθηκε για την ανάστροφη μεταγραφή σύμφωνα με γνωστό πρωτόκολλο [193]. Οι εκκινητές που χρησιμοποιήθηκαν για τα Bdel (203 bp) και Bplus (385 bp) προϊόντα ήταν οι παρακάτω: forward: 5 -AGCATCATCAAACCCCAGAAC-3 ; reverse: 5 - GCCGAATCCCCGCAAACAGC-3. Η Touchdown PCR του μίγματος που ακολούθησε έγινε για 30 κύκλους ως εξής: 68 C (2 κύκλοι), 66 C (2 κύκλοι) και 63 C (26 κύκλοι). Τα προϊόντα της αντίδρασης ανιχνεύθηκαν με ηλεκτροφόρηση σε πηκτή αγαρόζης 4% κάτω από υπεριώδη φωτισμό. CYCLIN D1 Εφαρμόσθηκε πρωτόκολλο duplex RT-PCR. Αναλύθηκαν 8 περιπτώσεις ΑΛΜΚ παράλληλα με 3 δείγματα περιφερικού αίματος που χρησιμοποιήθηκαν ως αρνητικοί μάρτυρες. Η εξαγωγή του RNA έγινε επίσης με την χρήση του TRIZOL-LS σύμφωνα με τα εφαρμοζόμενα πρωτόκολλα του Εργαστήριου, ενώ η RT-PCR σύμφωνα με τους Ronchetti και συν. [194]. Οι παρακάτω εκκινητές χρησιμοποιήθηκαν για την ανίχνευση του mrna της cyclin D1, forward, 5 - AACAGATCATCCGCAAACAC-3 ; reverse, 5 -TCACACTTGATCACTCTGGA-3. Οι συνθήκες επώασης ήταν οι εξής: 1 κύκλος στους 94 C για 3 λεπτά, 58 C για 1 66
67 λεπτό, 72 C για 1 λεπτό, 29 κύκλοι στους 94 C για 30 δευτερόλεπτα, 60 C για 30 δευτερόλεπτα και 72 C για 30 δευτερόλεπτα. Το ενισχυμένο προϊόν, μεγέθους 196 ζευγών βάσεων, ανιχνεύθηκε με ηλεκτροφόρηση σε πηκτή αγαρόζης 2% και χρώση με βρωμιούχο αζίδιο. Ως γονίδιο μάρτυρας για την ποιότητα του RNA και του cdna, χρησιμοποιήθηκε η ενίσχυση του τμήματος του μετάγραφου της γλυκεραλδεΰδης-3- φωσφορικής-δεϋδρογενάσης (GAPDH) και ειδικότερα οι παρακάτω εκκινητές: forward, 5 - ACTGGCGTCTTCACCACCAT -3 ; reverse, 5 - GGGATGATGTTCTGGAGAGC -3. FISH O φθορίζων εντοπισμένος/επιτόπιος υβριδισμός (Fluorescence In Situ Hybridization, FISH) είναι μία κυτταρογενετική-μοριακή τεχνική, η οποία χρησιμοποιεί DNA ή RNA ιχνηθέτες/ανιχνευτές επισημασμένους με μόρια αναφοράς, τα οποία με τη σειρά τους φθορίζουν. Τα σήματα των ιχνηθετών γίνονται ορατά με το μικροσκόπιο φθορισμού χρησιμοποιώντας ειδικά φίλτρα για τον ακριβή διαχωρισμό των φθοριοχρωμάτων. Η χρήση ειδικών μηχανών λήψης (CCD) στα μικροσκόπια φθορισμού προσδίδει στην τεχνική FISH μεγάλη ευαισθησία, αλλά και δυνατότητες ποσοτικής μέτρησης των εικόνων φθορισμού και ανίχνευσης σημάτων σε μήκη κύματος πέραν του ορατού για το ανθρώπινο μάτι φάσματος. Επιπρόσθετα με τη χρησιμοποιήση ειδικών λογισμικών προγραμμάτων είναι δυνατή η αποθήκευση των εικόνων FISH σε ηλεκτρονική μορφή και η περαιτέρω ανάλυση τους. Τα νεότερης γενιάς λογισμικά προγράμματα δίνουν την ευχέρεια στον χρήστη της βελτιστοποίησης των σημάτων φθορισμού, λόγω μείωσης της έντασης του υπόβαθρου (backround noise) και της δυνατότητας πολλαπλών επιπέδων λήψης, καθώς και της αυτόματης ανάγνωσης των σημάτων φθορισμού. Με τη χρήση της FISH είναι δυνατός ο προσδιορισμός του αριθμού και της τοποθεσίας συγκεκριμένων ακολουθιών DNA σε κύτταρα. Ειδικότερα με τη DNA- FISH είναι δυνατή η παρατήρηση μεμονωμένων γονιδίων, τμημάτων ή ολόκληρων χρωμοσωμάτων, η αποσαφήνηση ισορροπημένων ή όχι μεταθέσεων, η αναγνώριση χρωματοσωματικών διπλοποιήσεων και ανασυνδυασμών. Η ευαισθησία της κλασσικής τεχνικής FISH είναι ανώτερη της κλασσικής χρώσης των χρωμοσωμάτων με Giemsa (καρυότυπος) και της συγκριτικής υβριδοποίησης γενώματος (Comparative Genome Hybridization, CGH), ενώ οι παραλλαγές της FISH, η TSA- 67
68 FISH (Tyramide signal amplification) [195] και η νεότερη two-pass TSA-FISH [196] συγκρίσημες των μεθόδων PCR για την διερεύνηση ελάχιστου μεγέθους μοριακών διαταραχών (<1 ζεύγος βάσεων). Οι τύποι ιχνηθετών που αποτελούν και το βασικότερο στοιχείο της μεθοδολογίας FISH διακρίνονται σε: ιχνηθέτες DNA επαναλαμβανόμενων ακολουθιών (repetitive DNA probes), σύνθετους ιχνηθέτες DNA (complex DNA probes) ή εξειδικευμένους γονιδιακούς ιχνηθέτες [gene specific probes, locus specific probes (LSI)] και ιχνηθέτες ολόκληρων χρωμοσωμάτων (whole chromosome probes). Στη μελέτη μας χρησιμοποιήθηκαν ιχνηθέτες DNA επαναλαμβανόμενων ακολουθιών και εξειδικευμένοι γονιδιακοί ιχνηθέτες. Η μέθοδος FISH εφαρμόσθηκε σε τομές παραφίνης πάχους 4,5μm οι οποίες είχαν τοποθετηθεί σε θετικά φορτισμένα αντικειμενοφόρα πλακίδια. LSI ALK PROBE Το ΑΛΜΚ εμφανίζει μεταθέσεις που περιλαμβάνουν πάντα τμήμα του ALK γονιδίου (περιοχή 2p23) με άλλες χρωμοσωμικές περιοχές. Η πιο συχνή μετάθεση είναι η t(2;5)(p23;q35) μεταξύ του ALK και του NPM γονιδίου. Το σύμπλοκο ιχνηθετών LSI ALK dual color, break apart rearrangement probe (Vysis, USA) περιέχει δύο ιχνηθέτες σημασμένους με διαφορετικό φθοριόχρωμα οι οποίοι συνδέονται ειδικά σε τμήματα του ALK γονιδίου (περιοχή 2p23) που βρίσκονται αμφοτερόπλευρα του σημείου διάσπασης του γονιδίου (ALK gene breakpoint region). Ειδικότερα ο σημαινόμενος με Spectrum Orange (ερυθρό σήμα) ιχνηθέτης, μεγέθους περίπου 250 kb, συνδέεται με ομόλογες νουκλεοτιδικές αλληλουχίες στο σημείο διάσπασης του γονιδίου (ALK gene breakpoint region) και προς το τελομερικό άκρο του γονιδίου (3 άκρο). Ο άλλος ιχνηθέτης, μεγέθους περίπου 300 kb, είναι σημασμένος με Spectrum Green (πράσινο σήμα) και ενώνεται ειδικά στο άλλο σημείο διάσπασης, προς το κεντρομερικό τμήμα του γονιδίου (5 άκρο) (Εικόνα 5, σελ. 69). Με τον τρόπο αυτό είναι δυνατή η ανίχνευση της διάσπασης του γονιδίου, στοιχείο αποδεικτικό της μετάθεσης του γονιδιακού τμήματος 2p23, χωρίς όμως την ακριβή ταυτοποίηση αυτής της μετάθεσης. Σε φυσιολογικά κύτταρα παρατηρούνται στους πυρήνες 2 ζεύγη πράσινων/ερυθρών (κίτρινων) σημάτων (2RG ή 2Υ/F). Σε περίπτωση παρουσίας της μετάθεσης t(2;5) ή άλλης μετάθεσης της περιοχής 2p23 θα παρατηρηθεί συνήθως 1 ερυθρό, 1 πράσινο 68

69 και 1 κίτρινο σήμα ή ζευγος ερυθρού/πράσινου σήματος (1R1G1F ή 1R1G1GR). Στην περίπτωση αυτή, το ερυθρό μεμονωμένο σήμα, αντιστοιχεί στο τμήμα της περιοχής 2p23 που έχει μετατεθεί σε άλλο χρωμόσωμα, το πράσινο σήμα στο υπόλοιπο αμετάθετο τμήμα του ALK γονιδίου και το κίτρινο ή πράσινο/ερυθρό σήμα στο μη διεσπασμένο τμήμα του ALK γονιδίου. Παραλλαγές της παραπάνω εμφάνισης των σημάτων έχουν παρατηρηθεί, και αυτές συνήθως είναι του τύπου 2G1R1F και 2G2R1F. Η μέθοδος FISH πραγματοποιήθηκε σύμφωνα με τις οδηγίες του κατασκευαστή του ιχνηθέτη (Vysis, USA), με μικρές αλλαγές. Ειδικότερα, η αποπαραφίνωση των τομών έγινε με ολονύκτια επώαση στους 60 ο C την οποία ακολούθησε εμβάπτιση σε ξυλόλη (2x5 λεπτά) και αφυδάτωση σε 100% αλκοόλη (2x5 λεπτά). Οι τομές στη συνέχεια αφέθηκαν να στεγνώσουν σε θερμοκρασία δωματίου. Ακολούθησε προκατεργασία του ιστού με NaSCN (80 C για 30 λεπτά) και πέψη με πρωτεάση (37 C για 10 λεπτά). Η αποδιάταξη του DNA έγινε με εμβύθιση των τομών σε φορμαμίδη 70%, ph 7,5, στους 72 C για 5 λεπτά. Ακολούθησε ο υβριδισμός ο οποίος έγινε με ολονύκτια επώαση των τομών με 10μL του ιχνηθέτη στους 37 C. Την επόμενη ημέρα ακολούθησε μεθυβριδική ξέπλυση με διάλυμα SSC και 0,3% NP-40 σε θερμοκρασία δωματίου για 5 λεπτά και εμβύθιση των τομών στο ίδιο διάλυμα στους 72 C για 7 λεπτά. Επακολούθησε στέγνωμα των τομών, αντίχρωση με DAPI, στέγνωμα στον αέρα και κάλυψη. Η αξιολόγηση του υβριδισμού έγινε σε μικροσκόπιο Zeiss (Axioskop 2 plus HBO 100) το οποίο φέρει υψηλής ποιότητας αντικειμενικούς φακούς, επιλεγμένα φίλτρα (EX BP360/51 για την DAPI, EX BP485/17 για το FITC/spectrum green και EX BP560/18 για τη rhodamine/spectrum orange) καθώς και ψηφιακή κάμερα. Η ανάλυση των ψηφιακών εικόνων έγινε με τη βοήθεια ειδικού προγράμματος (FISH Imager TM METASYSTEMS, Germany). Ως φυσιολογικό σημείο αποκοπής (normal cutoff) θεωρήθηκε το 4,5% των νεοπλασματικών πυρήνων [197]. Εικόνα 5. Απεικόνιση του LSI ALK ιχνηθέτη. 69
70 CEP 2 PROBE Με τους CEP (Centromeric Probes) ιχνηθέτες οι οποίοι ανήκουν στην κατηγορία των ιχνηθετών επαναλαμβανόμενων ακολουθιών είναι δυνατή η διερεύνηση των αριθμητικών ανωμαλιών (ανευσωμίας) επιμέρους χρωμοσωμάτων. Ο ιχνηθέτης αποτελείται από επαναλαμβανόμενες νουκλεοτιδικές αλληλουχίες μεγέθους 171 bp που βρίσκονται στο τμήμα που αποτελεί το α-δορυφορικό DNA του χρωμοσώματος, στην περιοχή του κεντρομεριδίου. Ο υβριδισμός εφαρμόσθηκε σε τομές παραφίνης χρησιμοποιώντας τον CEP 2 (D2Z1, 2p11.1-q11.1) SpectrumOrange Probe (Vysis), (Εικόνα 6), σύμφωνα με τις οδηγίες του κατασκευαστή. Ο παραπάνω ιχνηθέτης εμφανίζει συχνά προβλήματα διασταυρούμενου υβριδισμού με κεντρομεριδιακές θέσεις άλλων χρωμοσωμάτων [198], όπως και άλλοι CEP ιχνηθέτες [199]. Τα φθορίζοντα (ερυθρά) υβριδικά σήματα μετρήθηκαν σε 100 μη επικαλυπτόμενους νεοπλασματικούς πυρήνες.. Ως πολυσωμία του CEP 2 θεωρήθηκε η καταγραφή σε >10% των καταμετρημένων νεοπλασματικών πυρήνωνν >2 ερυθρών σημάτων, ενώ ως μονοσωμία η παρουσία σε >40% των καταμετρημένων νεοπλασματικών πυρήνων <2 ερυθρών σημάτων. Η αξιολόγηση του υβριδισμού έγινε στο παραπάνω περιγραφέν σύστημα μικροσκοπίου με το άναλογο λογισμικό. Εικόνα 6. Απεικόνιση του CEP 2 (D2Z1) ιχνηθέτη. CYCLIN D1/CEP11 PROBE Με τον παραπάνω σύνθετο (διπλό) ιχνηθέτη είναι δυνατή η διερεύνησηη της γονιδιακής κατάστασης του CCND1 και η ταυτόχρονη παρακολούθηση των αριθμητικών ανωμαλιών του χρωμοσώματος

71 Για τη μέθοδο FISH χρησιμοποιήθηκε το εμπορικά διαθέσιμο σύμπλεγμα ιχνηθετών LSI TM Cyclin D1 (11q13) Spectrum Orange/CEP11 Spectrum Green Probe (Vysis). Ο υβριδισμός εφαρμόσθηκε σε τομές παραφίνης σύμφωνα με τις οδηγίες του κατασκευαστή. Ο Cyclin D1 (11q13) Spectrum Orange (ερυθρό σήμα) ιχνηθέτης (Εικόνα 7) έχει μέγεθος περίπου 300 kb και σημαίνει ολόκληρο το γονίδιο της κυκλίνης D1 (CCND1) που βρίσκεται στο μακρύ σκέλος του χρ. 11 (τμήμα 11q13), ενώ ο CEP11 ιχνηθέτης που είναι σημασμένος με Spectrum Green (πράσινο σήμα), χρησιμοποιείται για την ανίχνευση του α-δορυφορικού DNA που αντιστοιχεί στην περιοχή του κεντρομεριδίου του χρωμοσώματος 11 (11p11.11-q11). Τα φθορίζοντα υβριδικά σήματα μετρήθηκαν σε 100 μη επικαλυπτόμενους νεοπλασματικούς πυρήνες. Υπολογίσθηκε ο μέσος όρος σημάτων για το CCND1 γονίδιο, το κεντρομερίδιο 11 (CEP11) καθώς και ο λόγος CCND1/CEP11. Περιπτώσεις με λόγο CCND1/CEP11>2 θεωρήθηκαν ότι εμφανίζουν γονιδιακή ενίσχυση [200]. Ως επαύξηση (πολυσωμία) του CCND1 γονιδίου θεωρήθηκε η καταγραφή σε >20% των καταμετρημένων νεοπλασματικών πυρήνων >2 ερυθρών και πράσινων σημάτων [201]. Εικόνα 7. Απεικόνιση του LSI Cyclin D1 ιχνηθέτη. FICTION Η φθορίζουσα ανοσοφαινοτυπική και κυτταρογενετική ταυτοποίηση των νεοπλασματικών κυττάρων (fluorescence immunophenotyping and interphase cytogenetics as a tool for the investigation of neoplasms) αποτελεί μία νέα τεχνική [202], η οποία συνδυάζει την ταυτόχρονη ανοσοφαινοτυπική ταυτοποίηση νεοπλασματικών κυττάρων με φθορίζοντα αντίσωματα και τον φθορίζοντα in situ υβριδισμό (FISH). Οι FISH ιχνηθέτες μπορεί να είναι κεντρομεριδιακοί (CEP probes), γονιδιακοί (gene specific probes) και ολοχρωμοσωματικοί (whole 71
72 chromosome probes). Παραλλαγή της μεθόδου αποτελεί η M-FICTION η οποία επιτρέπει την ανοσοφαινοτυπική ταυτοποίηση σε συνδυασμό με την χρήση πολλαπλών FISH ιχνηθετών [203, 204]. Οι δυνατότητες της μεθόδου είναι πολλές, τόσο σε ερευνητικό, όσο σε διαγνωστικό επίπεδο. Στη μελέτη μας, η μέθοδος FICTION, εφαρφόσθηκε σε περίπτωση Σ-ΑΛΜΚ, ΑLK+, που εμφάνιζε έντονη θετικότητα στο αντιγόνο CD30, ανευπλοειδία του χρωμοσώματος 11 και επαύξηση του CCND1 γονιδίου με την μέθοδο FISH. Η τεχνική εφαρμόσθηκε σε τομές πάχους 4 μm. Η επώαση, αποπαραφίνωση, αφυδάτωση, αντιγονική ανάκτηση και επώαση των ιστικών τομών για το αντιγόνο CD30 έγιναν με βάση το χρησιμοποιούμενο στο Εργαστήριο πρωτόκολλο. Την επόμενη ημέρα ακολουθήθηκε το πρωτόκολλο των Martinez-Ramirez και συν. με μικρές αλλαγές [204]. Ειδικότερα, εφαρμόσθηκε έκπλυση των τομών με διάλυμα PBS και επώαση των τομών για 20 λεπτά με το δευτερογενές αντίσωμα NCL-SAM- FITC (Novocasta, αραίωση 1:20 σε PBS). Το παραπάνω πολυκλωνικό αντίσωμα προβάτου είναι σημασμένο με φλουοροσκεΐνη και συνδέεται ειδικά με αντισώματα ποντικού κλάσης IgG (όπως το CD30). Ακολούθησε έκπλυση με διάλυμα PBS, κάλυψη των τομών με γλυκεραλδεΰδη και έλεγχος του φθορισμού στο μικροσκόπιο. Οι τομές με ικανοποιητικό ανοσοφθορισμό χρησιμοποιήθηκαν περαιτέρω για την διενέργεια της FISH με τον ιχνηθέτη LSI Cyclin D1/CEP11 όπως περιγράφηκε παραπάνω. Η αξιολόγηση και μελέτη της FICTION έγινε στο σύστημα φθορισμού του Εργαστηρίου. ISH Ο επιτόπιος υβριδισμός (ISH) αποτελεί τεχνική της μοριακής βιολογίας [ ] που επιτρέπει την ανίχνευση ακολουθιών πυρηνικών οξέων σε κύτταρα που έχουν στερεωθεί πάνω σε αντικειμενοφόρους πλάκες μικροσκοπίου. Η τεχνική αυτή επιτρέπει τον καθορισμό της θέσης διαφόρων ακολουθιών πυρηνικών οξέων πάνω σε μεσοφασικά χρωμοσώματα ή μεσοφασικούς πυρήνες καθώς και στην ανίχνευση και κατανομή του κυτταρικού RNA. Ο ISH μπορεί να χρησιμοποιηθεί ως ερευνητική και διαγνωστική μέθοδος. Οι ανιχνευτές μπορεί να είναι DNA ή RNA αλληλουχίες ολιγονουκλεοτιδίων. Συχνότερα χρησιμοποιούνται οι ριβονουκλεοτιδικοί ανιχνευτές (ριβοανιχνευτές). Για κάθε mrna που ανιχνεύεται χρησιμοποιούνται δύο ριβοανιχνευτές, σε χωριστές αντιδράσεις. Ο antisense ανιχνευτής στοχεύει το 72
73 επιλεγμένο τμήμα mrna και ο sense ανιχνευτής, που είναι συμπληρωματικός ως προς τον antisense, αποτελεί τον αρνητικό μάρτυρα της αντίδρασης. Για την οπτική απεικόνηση της αντίδρασης χρησιμοποιείται κυρίως η μη ραδιοϊσοτοπική σήμανση του συμπλέγματος με βιοτίνη, διγοξυγενίνη, φλουοροσκεΐνη, αλκαλική φωσφατάση ή βρωμοδεοξυουριδίνη και στη συνέχεια επώαση με χρωμογόνο. Στην παρούσα μελέτη χρησιμοποιήθηκαν sense και antisense ριβοανιχνευτές για την ανίχνευση α) τμήματος του mrna της cyclin D1 και β) τμημάτων του mrna της htert. Οι υβριδισμοί πραγματοποιήθηκαν σε αντικειμενοφόρα θετικά φορτισμένα πλακίδια ιστικών τομών παραφίνης πάχους 2 μm. mrna ΤΗΣ CYCLIN D1 Ριβονουκλεοτιδικοί (sense και antisense) ανιχνευτές παρασκευασμένοι στο Εργαστήριο με in vitro μεταγραφή (DIG RNA labeling kit, SP6/T7; Roche Hellas, GR) χρησιμοποιώντας ως υπόστρωμα προυπάρχοντα RT-PCR προϊόντα και σημασμένοι με διγοξυγενίνη (Digoxigenin [DIG]-labeled riboprobes) εφαρμόσθηκαν για την ανίχνευση του mrna της cyclin D1. Το προϊόν της ενίσχυσης, μεγέθους 196 ζευγών βάσεων, είχε κλωνοποιηθεί με TA-cloning σε p-gem-t πλασμίδια (p-gem- T easy kit; Promega, U.K.). Οι κλωνοποιημένες αλληλουχίες επιβεβαιώθηκαν με ανάλυση αλληλουχίας (sequencing). Οι αλληλουχίες ενισχύθηκαν με puc primers (Promega), ώστε να περιέχονται τα τμήματα SP6 και T7 και ακολούθησε in vitro μεταγραφή. Οι ιστικές τομές μετά από τα στάδια της επώασης, αποπαραφίνωσης, πρωτεόλυσης και ακετυλίωσης προϋβριδοποιήθηκαν σε διάλυμα 50% απιονισμένης φορμαμίδης στους 48 C για 2 ώρες. Ακολούθησε το στάδιο αποδιάταξης των ριβονουκλεοτιδικών ανιχνευτών με επώαση των πλακιδίων στους 70 C για 10 λεπτά και στη συνέχεια το στάδιο του υβριδισμού με επώαση στους 42 C για 24 ώρες (overnight). Το στάδιο εμφάνισης πραγματοποιήθηκε με έκπλυση σε διάλυμα 0,2 SSC στους 55 C για 30 λεπτά, επώαση με δεσμευτικό διάλυμα για 1 ώρα, επώαση με το σύμπλοκο αντί-διγοξυγενίνη-αλκαλική φωσφατάση για 2 ώρες και έκπλυση σε διάλυμα TRIS/NaCl/Tween 20. Το σύμπλοκο οπτικοποιήθηκε με την τοποθέτηση του χρωμογόνου nitroblue tetrazolium/5-bromo-4-chloro-3-indolyl-phosphate (NBT/BCIP, Roche) στους 38 C για 30 λεπτά, έκπλυση με απεσταγμένο νερό και κάλυψη με γλυκερόλη χωρίς αντίχρωση (counterstain). 73
74 Παράλληλα με τον παραπάνω υβριδισμό, διενεργήθηκε και υβριδισμός σε ιστικές τομές χωρίς την χρήση των ειδικών ριβοανιχνευτών, ο οποίος και αποτέλεσε τον εξωτερικό αρνητικό μάρτυρα της μεθόδου. Ως θετικός μάρτυρας, για την παρουσία ανιχνεύσιμου mrna, χρησιμοποιήθηκε το ολιγονουκλεοτίδιο Poly-T (BioGenex, U.S.A.) που εφαρμόσθηκε παράλληλα με τους sense και antisense ανιχνευτές στις ιστικές τομές. Επιπρόσθετα, θετικούς εσωτερικούς μάρτυρες του υβριδισμού, αποτέλεσαν στις ιστικές τομές των ΤΜΑ, οι περιπτώσεις, με γνωστή θετικότητα στην κυκλίνη D1, λεμφωμάτων από κύτταρα του μανδύα. Μόνο οι περιπτώσεις που εμφάνιζαν θετικότητα για το Poly-T αξιολογήθηκαν και συμπεριλήφθησαν στην μελέτη. Ως θετική αντίδραση υβριδισμού θεωρήθηκε η παρουσία περιπυρηνικής κυανόμαυρης κοκκώδους χρώσης στα νεοπλασματικά κύτταρα. Για την ακριβέστερη αναγνώριση των θετικών νεοπλασματικών κυττάρων έγινε σύγκριση των πλακιδίων του υβριδισμού με την αντίστοιχη Η&Ε και την ανοσοχρώση για το CD30. mrna των ισομορφών της htert Σχετικά με την ανίχνευση του mrna της htert χρησιμοποιήθηκαν για τον υβριδισμό, σε ιστικές τομές πάχους 2 μm, 4 ριβονουκλεοτιδικοί ανιχνευτές (Βplus sense [FS] και antisense [FT], Bdel sense [BS] και antisense [BT]) οι οποίοι παρασκευάσθηκαν στο Εργαστήριο με in vitro μεταγραφή και ταυτόχρονη σήμανση με διγοξυγενίνη. Ως υπόστρωμα, χρησιμοποιήθηκαν υπάρχοντα PCR προϊόντα για τις αλληλουχίες (AF ) του mrna της htert, τα οποία είχαν κλωνοποιηθεί με TA-cloning σε p-gem-t πλασμίδια (Promega) [193]. Παράλληλα, ως εξωτερικοί αρνητικοί μάρτυρες της μεθόδου, πραγματοποιήθηκαν και υβριδισμοί σε ιστικές τομές χωρίς την χρήση των ειδικών ριβοανιχνευτών, καθώς και έλεγχος του mrna με την χρήση του ολιγονουκλεοτιδίου Poly-T (θετικός μάρτυρας). Μόνο οι περιπτώσεις με θετικό mrna αξιολογήθηκαν και συμπεριλήφθησαν στην μελέτη. Για την διασφάλιση της αναπαραγωγιμότητας των αποτελεσμάτων οι υβριδισμοί πραγματοποιήθηκαν σε τρείς ξεχωριστούς κύκλους. Η παρουσία περιπυρηνικής κυανόμαυρης κοκκώδους χρώσης στα νεοπλασματικά κύτταρα θεωρήθηκε ως θετική αντίδραση υβριδισμού και η αναγνώριση των νεοπλασματικών κυττάρων έγινε με τη σύγκριση των πλακιδίων του υβριδισμού με την αντίστοιχη Η/Ε και την ανοσοχρώση 74

75 για το CD30. Θετική κυτταροπλασμική χρώση παρατηρήθηκε και σε μικρό αριθμό αντιδραστικών λεμφοκυττάρων, η οποία λόγω της μικρής ποσότητας του κυτταροπλάσματος των λεμφοκυττάρων, εμφάνιζε περιπυρηνική εντόπιση. Ο Bplus antisense ανιχνευτής χρησιμοποιήθηκε για την σήμανση του ολικού mrna της htert, το οποίο είναι υπεύθυνο για την ενεργότητα της τελομεράσης. Επίσης, ο παραπάνω ριβοανιχνευτής ανιχνεύει και κάποιες από τις εναλλακτικά ματισμένες μορφές της htert (splice variants), οι οποίες παράγονται σε ελάχιστες ποσότητες (Εικόνα 7). Ο Bdel antisense ριβοανιχνευτής ανιχνεύει την Bdel εναλλακτικά ματισμένη μορφή, η οποία προκύπτει από την αποκοπή 182 bp που αντιστοιχούν στα εξόνια 7 και 8 της htert. Η μορφή αυτή της htert δεν εμφανίζει ικανότητα ανάστροφης μεταγραφής και συνεπώς δεν εμφανίζει ενεργότητα της τελομεράσης (Εικόνα 7). Εικόνα 7. Απεικόνιση των ριβοανιχνευτών Bplus και Bdel της htert. Ο Bplus antisense ανιχνευτής ανιχνεύει το ολικό mrna της htert, ενώ Bdel antisense ριβοανιχνευτής, την αντίστοιχη εναλλακτικά ματισμένη μορφή της htert, η οποία προκύπτει από την αποκοπή αλληλουχιών που αντιστοιχούν στα εξόνια 7 και 8 [Τροποποιημένο από Kotoula V et al., 2004 (193)]. 75
76 ΑΝΟΣΟΪΣΤΟΧΗΜΙΚΕΣ ΤΕΧΝΙΚΕΣ Διενεργήθηκε: Α) Απλή Ανοσοϊστοχημεία έναντι πέντε αντιγόνων (Simple IHC ) και B) Διπλή ανοσοϊστοχημεία (Doublestain IHC) με αντισώματα έναντι 2 κλώνων της κυκλίνης D1 (Cyclin D1 κλώνοι SP4 ή DCS-6) και έναντι του CD30 αντιγόνου Η ανοσοϊστοχημική διερεύνηση και για τις δύο τεχνικές εφαρμόσθηκε σε τομές πάχους 2 μm, από τους δύο ΤΜΑ κύβους παραφίνης καθώς και από ολόκληρες τομές, όπου ήταν απαραίτητο. Οι ιστικές τομές τοποθετήθηκαν για επώαση σε κλίβανο στους 60 C και ακολούθησε αποπαραφίνωση, αφυδάτωση και εμβάπτιση των ιστικών τομών σε διάλυμα υπεροξειδίου του υδρογόνου (Η 2 Ο 2 ) σε μεθανόλη 3%. Για την διασφάλιση της αναπαραγωγιμότητας των αποτελεσμάτων οι ανοσοχρώσεις πραγματοποιήθηκαν σε διαφορετικούς κύκλους για κάθε αντίσωμα. Παράλληλα, σε κάθε κύκλο, μαζί με τις υπό μελέτη ιστικές τομές διενεργήθηκαν και ανοσοχρώσεις με θετικούς και αρνητικούς μάρτυρες. ΑΠΛΗ ΑΝΟΣΟΪΣΤΟΧΗΜΕΙΑ Μετά τα παραπάνω στάδια ακολούθησαν τα στάδια της αντιγονικής ανάκτησης, της επώασης των αντισωμάτων και της δέσμευσης των ανοσοσυμπλεγμάτων με ειδικούς αντιορούς (Πίνακας 6). Ως χρωμογόνο χρησιμοποιήθηκε η 3, 3 - διαμινοβενζιδίνη (DAB) και ως αντίχρωση η αιματοξυλίνη Mayer. Πίνακας 6. Ανάλυση των αντισωμάτων και ανοσοϊστοχημικών μεθόδων που εφαρμόσθηκαν στην παρούσα μελέτη. Αντίσωμα Κλώνος/Εταιρεία ΑΑ/Τ/ΡΔ Aραίωση Ε ΣΑ/Εταιρεία ALK1 (m, mab) ALK1 (1) KO/MW, 21 /TBS 1:30 O/Ν StrepABC (1)* Cyclin D1 (r, mab) SP4 (2) TR/ST, /TBS 1:45 30 Envision+ TM (1)** Cyclin D1 (m, mab) DCS-6 (1) ER2/HP, 20 /PBS 1: Bond Polymer (4) *** Fascin (m, mab) IM29 (3) KO/MW, 21 /TBS 1:100 O/Ν StrepABC (1)* htert (m, mab) 44F12 (3) TRª/WB, 30 /TBS 1:100 O/Ν Envision+ TM (1)** MUC-1 (m, mab) Ma695 (3) KO/MW, 21 /TBS 1: StrepABC (1)* 76
77 Επεξηγήσεις: ΑΑ: Αντιγονική ανάκτηση, Ε: επώαση, KO: Κιτρικό οξύ, ph 6.0, ΡΔ: Ρυθμιστικό διάλυμα, ΣΑ: Σύστημα ανίχνευσης, ER2: ethylenediaminetetraacetate, ph8.8, HP: Hot plate, m: mouse, mab: monoclonal antibody, MW: Microwave, O/N, overnight, PBS: phosphate buffered saline, r: rabitt, ST: Steamer, Τ: Χρόνος αντιγονικής ανάκτησης, TBS: tris buffered saline, TR: ethylenediaminetetraacetate, ph9.0, TRª: ethylenediaminetetraacetate, ph6.2, WB: Water Bath. (1): Dako, DK; (2): Spring Bioscience, U.S.A.; (3): Novocastra, U.K.; (4): Vision Biosystems, AUS. * Τα στάδια της 1ης ημέρας πραγματοποιήθηκαν manually, ενώ τα στάδια της 2ης ημέρας (μετά την επώαση του αντισώματος έως και την έκπλυση μετά την αντίχρωση, έγιναν στο αυτόματο μηχάνημα ανοσοχρώσεων i6000 Biogenex, U.S.A. ** Όλα τα στάδια της μεθόδου πραγματοποιήθηκαν manually*** Όλα τα στάδια της μεθόδου, έως και την έκπλυση μετά την αντίχρωση, έγιναν στο αυτόματο μηχάνημα ανοσοχρώσεων Bond Max autostainer (Vision Biosystems, AUS) ΔΙΠΛΗ ΑΝΟΣΟΪΣΤΟΧΗΜΕΙΑ Για την διενέργεια της διπλής ανοσοϊστοχημείας χρησιμοποιήθηκε το σύστημα ανίχνευσης Envision G/2 System/AP (Dako). Όλα τα στάδια της μεθόδου καθώς και το στάδιο της χρωμογόνου αντίδρασης με την 3, 3 - διαμινοβενζιδίνη έγιναν όπως περιγράφονται στον Πίνακα 6. Στη συνέχεια, μετά την εμφάνιση του ανοσοσυμπλέγματος (Cyclin D1 SP4 ή DCS-6) ακολούθησε η επώαση των τομών με το αντίσωμα CD30 (1:50, κλώνος Ber-H2, Dako) για 30 λεπτά. Μετά την έκπλυση των τομών με ρυθμιστικό διάλυμα TBS (3x5 λεπτά), οι τομές επωάσθηκαν από 30 λεπτά με τους αντιορούς Rabbit/Mouse (LINK) και AP Enzyme (ENHANCER) αντίστοιχα. Ακολούθησε επώαση των ιστικών τομών με το διάλυμα ερυθρού χρωμογόνου (Permanent Red Chromogen) μέχρι να γίνει εμφανής η αντίδραση (16 λεπτά). Ακολούθησε έκπλυση σε τρεχούμενο νερό, αντίχρωση με αιματοξυλίνη Mayer, έκπλυση σε τρεχούμενο νερό, στέγνωμα σε θερμοκρασία δωματίου και κάλυψη με βάλσαμο του Καναδά. ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΑΝΟΣΟΧΡΩΣΕΩΝ Η αξιολόγηση της ανοσοϊστοχημικής έκφρασης των υπό μελέτη πρωτεϊνών έγινε σύμφωνα με αντίστοιχες παλαιότερες μελέτες. Αναλυτικότερα για κάθε πρωτεΐνη εφαρμόσθηκαν τα παρακάτω πρωτόκολλα: Περιπτώσεις με θετικότητα ακόμα και ελάχιστων νεοπλασματικών κυττάρων, ανεξάρτητα της χρωστικής έντασης και κυτταρικής εντόπισης της ανοσοχρώσης ALK1 αξιολογήθηκαν ως θετικές [104]. 77
78 Οι ανοσοχρώσεις και για τους δύο κλώνους της κυκλίνης D1 αξιολογήθηκαν ως θετικές όταν παρουσίαζαν, οποιουδήποτε βαθμού χρωστικής έντασης πυρηνική ή/και κυτταροπλασμική χρώση, τουλάχιστον στο 10% των νεοπλασματικών κυττάρων [208, 209]. Ως θετικές στην πρωτεΐνη Fascin, καταμετρήθηκαν οι περιπτώσεις που εμφάνιζαν οποιουδήποτε βαθμού χρωστικής έντασης κυτταροπλασμική χρώση, σε ποσοστό >10% των νεοπλασματικών κυττάρων [154]. Η πρωτεΐνη htert αξιολογήθηκε ως θετική όταν, οποιουδήποτε βαθμού χρωστικής έντασης, κυρίως πυρηνική ή/και σπανιότερα κυτταροπλασμική χρώση, παρατηρήθηκε σε ποσοστό >10% των νεοπλασματικών κυττάρων [210]. Η έκφραση της πρωτεΐνης MUC-1 θεωρήθηκε ως θετική στις περιπτώσεις που αναγνωρίσθηκε τουλάχιστον ήπιου βαθμού κυτταροπλασμική χρώση ποσοστό >10% των νεοπλασματικών κυττάρων [61]. ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ Η στατιστική ανάλυση έγινε με τη χρήση του προγράμματος SPSS (Statistical Package for Social Science, version 11.5, SPSS Inc, Chicago, IL, U.S.A.) για το λειτουργικό σύστημα Windows XP (Microsoft, U.S.A.). Οι μέσες τιμές των διαφόρων μεταβλητών με συνεχή δεδομένα αναλύθηκαν με τη χρήση του x 2 και Fisher exact test. Το kappa test χρησιμοποιήθηκε για την μελέτη των εξαρτημένων μεταβλητών, ενώ για την σύγκριση των ποιοτικών ISH αποτελεσμάτων το likelihood ratio (LR). Τιμή του P value = <0,05 θεωρήθηκε στατιστικά σημαντική, ενώ τιμή του P value = 0,05 οριακής στατιστικής σημασίας. Η συμφωνία μεταξύ των μεταβλητών (kappa agreement) έγινε σύμφωνα με της κατευθυντήριες οδηγίες των Landis και Koch [211] και ειδικότερα: κ=0, ασυμφωνία; κ =>0 - <0,20, ήπια συμφωνία; κ => < 0.40, επαρκής συμφωνία; κ= > <0.60, μέτρια συμφωνία; κ=> <0.80, σημαντική συμφωνία και κ >0.80 έως 1.00, σχεδόν τέλεια συμφωνία. 78
79 ΑΠΟΤΕΛΕΣΜΑΤΑ ΓΕΝΙΚΑ Η μελέτη συμπεριέλαβε 42 ασθενείς, 30 άρρενες και 12 θήλεα (αναλογία Α:Θ = 2,5:1), με μέση ηλικία τα 54,3 έτη, διακύμανση από 8-76 έτη, με σταθερή απόκλιση (±SD) τα 17,65 έτη. Έξι από τους 42 ασθενείς εμφάνισαν την νόσο σε ηλικία 35 έτη [104]. Η πλειονότητα των πρωτοπαθών συστηματικών ΑΛΜΚ (30/34, 88,2%) με βάση τα μορφολογικά κριτήρια κατατάχθηκαν στον κοινό τύπο (Εικόνα 8Α). Σε τέσσερις περιπτώσεις (# 9, 24, 26, 27) ανευρέθησαν πολυάριθμα ιστιοκύτταρα μεταξύ των νεοπλασματικών κυττάρων. Οι παραπάνω περιπτώσεις χαρακτηρίστηκαν ως λεμφοϊστιοκυτταρικός τύπος του ΑΛΜΚ. Σε τέσσερις επίσης περιπτώσεις παρατηρήθηκαν νεοπλασματικές εστίες νέκρωσης. Στα πρωτοπαθή ΑΛΜΚ του δέρματος (Εικόνα 8Β) παρατηρήθηκε σε 3/8 περιπτώσεις επιδερμοτροπισμός, σε μία περίπτωση εξέλκωση της επιδερμίδας και σε μία περίπτωση διήθηση του επιθηλίου των τριχοθυλάκων. Συχνά, η εξεργασία επεκτεινόταν στον υποκείμενο υποδόριο λιπώδη ιστό. Μεταξύ των νεοπλασματικών κυττάρων αναγνωρίσθηκαν μικροί αριθμοί πλασματοκυττάρων και ηωσινόφιλων πολυμορφοπυρήνων λευκοκυττάρων. Όλα τα ΠΔ-ΑΛΜΚ ανήκαν στον κοινό τύπο. Τα νεοπλασματικά λεμφοειδή κύτταρα σε όλες τις περιπτώσεις (42/42) ήταν θετικά στο CD30/Ki1 αντιγόνο (Εικόνα 9Α, Β) και αρνητικά στις ανοσοχρώσεις για Β-δείκτες CD20 (42/42) και CD45RO (42/42) και CD79α (10/10). Σε 34/42 (81%) περιπτώσεις παρατηρήθηκε θετική ανοσοαντίδραση στα νεοπλασματικά κύτταρα για το CD45RA αντιγόνο, σε 22/42 (52%) περιπτώσεις για το αντιγόνο CD3 και σε 35/39 (90%) περιπτώσεις για το CD43. Σε 1/42 περιπτώσεις δεν παρατηρήθηκε θετικότητα για τους Τ-κυτταρικούς δείκτες (CD45RA, CD3, CD43, CD4, CD5, CD8) και αυτή θεωρήθηκε ως Σ-ΑΛΜΚ χωρίς ειδικό ανοσοφαινότυπο (null-cell ALCL). Τα κλινικά στοιχεία των ασθενών, τα ιστολογικά και τα γενικά ανοσοϊστοχημικά ευρήματα αναγράφονται στον Πίνακα 7. TCR-γ Η ποιότητα του εξαχθέντος DNA ήταν ικανοποιητική σε 37/42 περιπτώσεις. Μονοκλωνικός πληθυσμός Τ-κυτταρικής αρχής παρατηρήθηκε σε 22 από τα 37 (59%) ΑΛΜΚ, 18/29 Σ-ΑΛΜΚ και 4/8 ΠΔ-ΑΛΜΚ (Πίνακας 8), (Εικόνα 10). 79
80 ΕΚΦΡΑΣΗ ΤΗΣ ΧΙΜΑΙΡΙΚΗΣ ALK ΠΡΩΤΕΪΝΗΣ ΚΑΙ ΑΝΑΛΥΣΗ ΤΩΝ ΓΟΝΙΔΙΑΚΩΝ ΜΕΤΑΘΕΣΕΩΝ ΜΕ ΤΙΣ ΜΟΡΙΑΚΕΣ ΤΕΧΝΙΚΕΣ ALK Ο ανοσοϊστοχημικός έλεγχος με το ALK1 αντιγόνο έδειξε θετικότητα σε 13/42 (31%) περιπτώσεις οι οποίες χαρακτηρίσθηκαν ως ΑΛΜΚ, ALK+, ενώ οι υπόλοιπες ως ΑΛΜΚ, ALK- (Πίνακας 8). Ειδικότερα, σε 8/13 ΑΛΜΚ, ALK+ παρατηρήθηκε παρουσία της ανοσοχρώσης στον πυρήνα και το κυτταρόπλασμα των νεοπλασματικών κυττάρων (Εικόνα 11Α) ενώ σε 5/13 ΑΛΜΚ, ALK+ κυτταροπλασμική χρώση (Εικόνα 11Β). Τα αποτελέσματα της χρώσης ALK1 ήταν ταυτόσημα μεταξύ πρώτης βιοψίας και υποτροπής ή επανεμφάνισης της νόσου (Πίνακας 8). Η ανίχνευση της ALK1 ήταν σημαντικά συχνότερη σε μικρότερες ηλικίες ( 35 έτη) σε σύγκριση με μεγαλύτερες ηλικίες (> 35 έτη), (6/6 vs 7/21, Fisher's Exact Test p=0,001) (Γράφημα 1). Σε καμία περίπτωση ΠΔ-ΑΛΜΚ (0/8), δεν παρατηρήθηκε έκφραση της ALK1, η οποία ανευρέθη σε 13/34 Σ-ΑΛΜΚ (p=0,035) (Γράφημα 2). t(2;5)(p23;q35) Σε όλες τις υπό εξέταση περιπτώσεις (42/42, 100%), η ποιότητα του RNA, ήταν ικανοποιητική. Σε 11/42 (26%) ΑΛΜΚ ανιχνεύθηκε το χιμαιρικό μετάγραφο NPM-ALK (Εικόνα 12), (Πίνακας 8). Τα αποτελέσματα της RT-PCR ήταν ταυτόσημα (θετικά ή αρνητικά) με αυτά της ΑΙΧ για την ALK1 πρωτεΐνη σε 34/42 περιπτώσεις (concordance: 81%, kappa=0,535, Fisher's Exact Test p=0,001) (Πίνακας 9Α). Από τις υπόλοιπες 8 περιπτώσεις, 3 ήταν θετικές με την RT-PCR και αρνητικές με την ΑΙΧ και οι άλλες 5 αρνητικές με RT-PCR και θετικές με ΑΙΧ. Η συμφωνία ως προς τις θετικές περιπτώσεις ήταν 61,5% (8/13, kappa=0,535, Fisher's Exact Test p=0,001). Αξίζει να σημειωθεί εδώ ότι οι 5 αρνητικές με RT-PCR περιπτώσεις εμφάνιζαν κυτταροπλασμική έκφραση της ALK1 πρωτεΐνης η οποία παρατηρείται σε μεταθέσεις εκτός της t(2;5) και η ανίχνευση τους ήταν αδύνατη με τη χρήση των ιχνηθετών της μελέτης. Με την απάλειψη των παραπάνω περιπτώσεων από την ανάλυση συμφωνίας (kappa agreement) μεταξύ των δύο μεταβλητών η συμφωνία εκτοξεύεται στο 100% για τις θετικές περιπτώσεις (kappa=0,840, Fisher's Exact Test p<0,001) και στο 92% για το σύνολο των περιπτώσεων (34/37), (Πίνακας 80
81 9Β). Η ανίχνευση της NPM-ALK ήταν σημαντικά συχνότερη σε μικρότερες ηλικίες ( 35 έτη) σε σύγκριση με μεγαλύτερες ηλικίες (> 35 έτη), (6/6 vs 5/31, Fisher's Exact Test p<0,001) (Γράφημα 1). Ενα ΠΔ-ΑΛΜΚ, (1/8) ήταν θετικό για την παρουσία της NPM-ALK, η οποία ανευρέθη σε 10/34 Σ-ΑΛΜΚ, (p=0,32) (Γράφημα 2). 2p23(FISH) Σε 18 από τις 42 περιπτώσεις (43%) των ΑΛΜΚ παρατηρήθηκαν διαχωριζόμενα φθορίζοντα υβριδικά σήματα ενδεικτικά της t(2;5) ή άλλης μετάθεσης του 3 άκρου της περιοχής 2p23 σε ποσοστό κυττάρων πάνω από την καθορισμένη οριακή τιμή ( 4,5%) (Πίνακας 8). Στις περιπτώσεις αυτές παρατηρήθηκαν νεοπλασματικοί πυρήνες με 1 ερυθρό, 1 πράσινο και 1 κίτρινο σήμα ή 1 ζεύγος πράσινου/ερυθρού σήματος (1R1G1F ή 1R1G1RG), αποδεικτικό στοιχείο μετάθεσης του 2p23 (Εικόνα 12Α). Σε 10/42 (24%) περιπτώσεις, καταμετρήθηκαν σε >10% των κυττάρων, επιπλέον 3-5 πράσινα σήματα (5 τμήματα του 2p23) συνήθως ως ζεύγη πράσινων/ερυθρών σημάτων και σπανιότερα ως μεμονωμένα σήματα (Εικόνα 12Β). Τα επιπλέον αυτά 5 τμήματα του 2p23 καταμετρήθηκαν σε 4 FISH+ και 6 FISHπεριπτώσεις (Πίνακας 8), σε ισάριθμα λεμφαδενικά και εξωλεμφαδενικά Σ-ΑΛΜΚ. Στους πυρήνες των φυσιολογικών λεμφοκυττάρων παρατηρήθηκαν κυρίως 2 ζεύγη πράσινων/ερυθρών (κίτρινων) σημάτων (2RG ή 2F). Σε σπάνια κύτταρα (1-3%), ανευρέθησαν 1 ερυθρό, 1 πράσινο και 1 κίτρινο σήμα ή 1 ζεύγος πράσινου/ερυθρού σήματος. Τα αποτελέσματα της FISH ήταν ταυτόσημα (θετικά ή αρνητικά) με αυτά της ΑΙΧ για την ALK1 σε 37/42 (88%) περιπτώσεις (kappa=0,748, Fisher's Exact Test p<0,001), ενώ και οι 13 ALK+ περιπτώσεις ήταν FISH θετικές (concordance: 100%) (Πίνακας 9Γ). Οι υπόλοιπες 5 περιπτώσεις, αφορούσαν 4 συστηματικά ΑΛΜΚ, ALK- και 1 ΠΔ-ΑΛΜΚ, όλες θετικές με την FISH και αρνητικές με την ΑΙΧ. Το ΠΔ- ΑΛΜΚ ήταν επίσης θετικό στην NPM-ALK. Δεκαεπτά από τα 34 Σ-ΑΛΜΚ (50%) και 1/8 ΠΔ-ΑΛΜΚ εμφάνισαν την 2p23 μετάθεση (p=0,054), (Γράφημα 2). Η ανίχνευση της μετάθεσης του 2p23 ήταν σημαντικά συχνότερη στις ηλικίες 35 έτη σε σύγκριση με ηλικίες > 35 έτη, (6/6 vs 12/36, Fisher's Exact Test p=0,004) (Γράφημα 1). Τα αποτελέσματα ως προς την μετάθεση του 2p23 στις 3 συνολικά 81
82 περιπτώσεις με υποτροπή ή επανεμφάνιση της νόσου ήταν ταυτόσημα με εκείνα της πρώτης βιοψίας (Πίνακας 8). Συσχέτιση των μεθόδων ανίχνευσης της ALK Στην στατιστική ανάλυση αναδείχτηκαν σημαντικές συσχετίσεις μεταξύ των μεθόδων της RT-PCR με τη FISH (kappa=0,642, p<0,001) (Πίνακας 9Δ) και συνολικά των μοριακών μεθόδων (RT-PCR και FISH) με την έκφραση της ALK1 (kappa=0,704, p<0,001) (Πίνακας 9Ε). Η ταύτιση των αποτελεσμάτων των μοριακών τεχνικών (RT-PCR, FISH) με την ALK1 πρωτεΐνη ήταν απόλυτη (100%, 13/13 περιπτώσεις) όσον αφορούσε τις θετικές περιπτώσεις. Η παρουσία του NPM-ALK μετάγραφου και της μετάθεσης του 2p23 με τις παραπάνω μοριακές τεχνικές εμφάνισε στατιστικά σημαντική διαφορά, ως προς την ηλικία εμφάνισης της νόσου ( 35 έτη vs >35 έτη, Fisher s exact test, p=0,005), (Γράφημα 1) και την κλινική ταξινόμηση των ΑΛΜΚ (συστηματικά vs πρωτοπαθή δερματικά, p=0,039), (Γράφημα 2). CEP2 Με τον ιχνηθέτη για το κεντρομερίδιο του χρωμοσώματος 2 (CEP2), αναδείχθηκαν 12 από τα 42 (29%) ΑΛΜΚ (5 ALK+) με ανευσωμία, και ειδικότερα πολυπλοειδία (Πίνακας 8). Τα φθορίζοντα ερυθρά σήματα στις παραπάνω περιπτώσεις που ανευρέθησαν σε ποσοστό κυττάρων >10% ήταν αυξημένα, με ανώτερο αριθμό τα 5 σήματα (Εικόνα 13). Η πολυπλοειδία του χρ.2 παρατηρήθηκε σε 11 Σ-ΑΛΜΚ (6 λεμφαδενικά και 5 εξωλεμφαδικά) και 1 ΠΔ-ΑΛΜΚ. Η συμφωνία της πλοειδίας του χρ.2 με τους CEP2 και LSI ALK ιχνηθέτες ήταν υψηλή (kappa=0,877, p<0,001), (Γράφημα 3). Cyclin D1 Cyclin D1 πρωτεΐνη Τα νεοπλασματικά λεμφοειδή κύτταρα, σε όλες τις περιπτώσεις (0/42), δεν εξέφραζαν κανένα από τους κλώνους της πρωτεΐνης κυκλίνης D1, ενώ πυρηνική 82
83 θετικότητα παρατηρήθηκε στα ενδοθηλιακά κύτταρα, τους ινοβλάστες και τα βασικάπαραβασικά ακανθοκύτταρα της επιδερμίδας (Εικόνα 15Α-Δ, 16 Γ, Δ). Σε μικρό αριθμό μη-νεοπλασματικών κυττάρων παρατηρήθηκε μέτριας έντασης κυτταροπλασμική χρώση με τον κλώνο DCS-6. Σε σπάνιες ΑΛΜΚ περιπτώσεις παρατηρήθηκαν διάσπαρτα, μεταξύ των νεοπλασματικών κυττάρων, μέσου μεγέθους κύτταρα με ωοειδής πυρήνες και μέτρια ποσότητα κυτταροπλάσματος τα οποία εμφάνιζαν πυρηνική χρώση. Τα παραπάνω κύτταρα, πιθανότατα ιστιοκύτταρα, όπως αποδείχθηκε με την διπλή ΑΙΧ, είταν αρνητικά στο αντιγόνο CD30. Τα νεοπλασματικά λεμφοειδή κύτταρα του λεμφώματος από κύτταρα του μανδύα (MCL) εμφάνισαν έντονη πυρηνική θετικότητα και με τους δύο κλώνους της κυκλίνης D1 (Εικόνα 16Α, Β). Τα αποτελέσματα της ΑΙΧ και με τους δύο κλώνους της πρωτεΐνης κυκλίνης D1 παρουσιάζονται στον Πίνακα 10. mrna της cyclin D1 Σαράντα-μια από τις 42 υπό εξέταση περιπτώσεις εμφάνισαν ικανοποιητική ποιότητα του poly-t ανιχνευτή (Εικόνα 17Α) (Πίνακας 10). Η παρουσία κυανόμαυρης περιπυρηνικής χρώσης, η οποία αντιστοιχούσε στο mrna της cyclin D1, παρατηρήθηκε σε 29/41 (71%) περιπτώσεις (Πίνακας 10). Η ένταση της χρώσης αξιολογήθηκε ως ήπια (+), μέτρια (++) και έντονη (+++). Θετικότητα παρατηρήθηκε μόνο στα νεοπλασματικά λεμφοειδή κύτταρα, ενώ τα αντιδραστικά λεμφοκύτταρα ήταν αρνητικά (Εικόνα 17Β, Γ). Σε σπάνιες περιπτώσεις, μικροί αριθμοί ινοβλαστών εμφάνισαν ήπια θετικότητα. Χρωστικό σήμα δεν παρατηρήθηκε με τον sense ανιχνευτή (Εικόνα 17Δ) καθώς και στις περιπτώσεις που επωάσθηκαν απουσία ανιχνευτών. Οι εσωτερικοί μάρτυρες της μεθόδου (MCL), εμφάνισαν έντονη έκφραση του mrna της cyclin D1. Το 79% (26/33) των Σ-ΑΛΜΚ και 3/8 (37.5%) ΠΔ-ΑΛΜΚ εξέφραζαν το mrna της cyclin D1 (Fisher's Exact Test p=0,034) (Πίνακας 11). Ταυτόσημα αποτελέσματα παρατηρήθηκαν και στις περιπτώσεις ασθενών με παραπάνω από 1 βιοψίες (Πίνακας 10). Υψηλό ποσοστό ΑΛΜΚ, ALK+ περιπτώσεων με τις μοριακές μεθόδους (15/18, 83%) εξέφραζαν το mrna της cyclin D1, ενώ η ομάδα των ΑΛΜΚ, ALK- το 61% (14/23), (p=0,12) (Πίνακας 11). Η παρουσία του mrna της cyclin D1 διασταυρώθηκε με τη μέθοδο της RT- PCR σε 8 ΑΛΜΚ περιπτώσεις (Εικόνα 18). Τα αποτελέσματα του ISH και της RT- PCR ήταν ταυτόσημα σε όλες τις εξεταζόμενες περιπτώσεις (7/7 +, 1/1 -). 83
84 CCND1 γονίδιο Η μέθοδος FISH ήταν επιτυχής σε 38/42 περιπτώσεις. Σε καμία περίπτωση δεν παρατηρήθηκε ενίσχυση του γονιδίου της CCND1. Αντίθετα, καταμετρήθηκαν αυξημένοι αριθμοί αντιγράφων του CCND1 γονιδίου και του κεντρομεριδίου του χρωμοσώματος 11 (D11Z1) σε 15/38 (39%) περιπτώσεις, 14 Σ-ΑΛΜΚ (6/12 ALK+, 8/19 ALK-) και 1/7 ΠΔ-ΑΛΜΚ (Πίνακας 10, 11). Στις παραπάνω περιπτώσεις, 3 έως 5 φωτεινά ερυθρά και πράσινα σήματα καταμετρήθηκαν στους νεοπλασματικούς πυρήνες (Εικόνα 19). Οι ως άνω αριθμητικές ανωμαλίες δεν ανέδειξαν στατιστικά σημαντικές συσχετίσεις με την παρουσία των μοριακών διαταραχών του ALK γονιδίου (μεταθέσεις, χιμαιρικό μετάγραφο), την έκφραση της ALK1, την κλινική και ιστολογική ταξινόμηση των ΑΛΜΚ (Πίνακας 11). FICTION Τα νεοπλασματικά λεμφοειδή κύτταρα εμφάνισαν έντονη μεμβρανική χρώση με το αντιγόνο CD30 (Εικόνα 20Α, πράσινο φθοριόχρωμα, μονό φίλτρο FITC). Με την χρήση και των 3 φίλτρων φθορισμού, παρατηρήθηκαν στους νεοπλασματικούς πυρήνες 3 έως 5 σήματα για κάθε ιχνηθέτη, γεγονός αποδεικτικό επαύξησης των CCND1 και D11Z1 (α-δορυφορικό DNA χρ. 11) χρωμοσωμικών περιοχών (Εικόνα 20Β). htert Η αξιολόγηση της ISH και της ΑΙΧ για την htert στα ΑΛΜΚ με την μέθοδο των ιστικών μικροσυστοιχιών ήταν δυνατή εξαιτίας της προγενέστερης πιλοτικής εφαρμογής των παραπάνω μεθόδων σε ιστικές τομές μη-νεοπλασματικών λεμφαδένων (Εικόνα 21), όπου μελετήθηκε και αξιολογήθηκε η κατανομή των htert θετικών λεμφοκυττάρων. Γενικά, τα νεοπλασματικά κύτταρα ήταν εμφανή λόγω του μεγάλου μεγέθους τους και την ομοιογενή κατανομή τους στους ιστούς. Σε αντίθεση ως προς την σχεδόν ισότιμη κατανομή των θετικών κυττάρων στους Bplus και Bdel ανιχνευτές στους μη-νεοπλασματικούς λεμφαδένες, σπάνια ΑΛΜΚ εμφάνισαν αυτή την εικόνα (Εικόνες 22Α-Γ, Ζ). Με την παραπάνω μέθοδο το mrna των εναλλακτικά ματισμένων μορφών Bplus και Bdel της htert 84
85 ανιχνεύθηκε σε όλους τους μη νεοπλασματικούς λεμφαδένες (Εικόνες 21Α, Β, Δ, Ε, Η, Θ). Ειδικότερα, έντονη έκφραση τους παρατηρήθηκε στα λεμφοκύτταρα των βλαστικών κέντρων και Τ-ζωνών καθώς και στα λεμφοκύτταρα των περιφερικών λεμφόκολπων. Στις άλλες λεμφαδενικές περιοχές (λεμφοζιδιακός μανδύας, περιφέρεια των λεμφοζιδίων) σπάνια λεμφοκύτταραρα ήταν επίσης θετικά. Συνήθως, τα λεμφοκύτταρα, ήταν θετικά και για τους δύο ριβοανιχνευτές (Εικόνες 21B, E). ISH για τους Bplus και Bdel ανιχνευτές Τα αποτελέσματα της ISH παρουσιάζονται στον Πίνακα 12. Τα κυριότερα ευρήματα είναι: 1) Συνολικά, το mrna της htert παρατηρήθηκε σε 27 από τις 38 (71%) ΑΛΜΚ περιπτώσεις που εξετάσθηκαν, με μεγαλύτερη συχνότητα στην ομάδα των ΑΛΜΚ, ALK+ (12/13, 92%) σε σχέση με την ομάδα των ΑΛΜΚ, ALK- (11/17, 65%) (x 2, p=0,077, LR=0,062), (Γράφημα 4) και στην ομάδα των Σ-ΑΛΜΚ (23/30, 77%) έναντι των ΠΔ-ΑΛΜΚ (4/8, 50%), (x 2, p=0,14), (Γράφημα 4). Στις αρνητικές περιπτώσεις δεν παρατηρήθηκε εμφανής χρώση των λεμφοειδών κυττάρων και στους δύο ριβοανιχνευτές (Εικόνες 22 Ξ, Ο). Οι παραπάνω περιπτώσεις ήταν επίσης αρνητικές και στην αντίστοιχη πρωτεΐνη. Αξίζει εδώ να σημειωθεί, ότι στις ως άνω περιπτώσεις, τα μη-νεοπλασματικά λεμφοκύτταρα ήταν θετικά τόσο στους Bplus και Bdel ανιχνευτές, όσο και στην πρωτεΐνη (Εικόνες 22 Ξ-Π). 2) Η πλειονότητα των ΑΛΜΚ θετικών για την htert περιπτώσεων ήταν κυρίως θετική για τον Bplus ανιχνευτή και σπανιότερα για τον Bdel ανιχνευτή. Ειδικότερα, έκφραση του mrna των Bplus και Bdel παρατηρήθηκε σε 24/38 (63%) και 15/38 (39,5%) ΑΛΜΚ περιπτώσεις αντίστοιχα (Πίνακας 12). Τα νεοπλασματικά θετικά κύτταρα ήταν εμφανώς περισσότερα στον Bplus ανιχνευτή, εκτός από 3 περιπτώσεις, οι οποίες ήταν θετικές μόνο στον Bdel ανιχνευτή. Στον εν λόγω ανιχνευτή, τα νεοπλασματικά θετικά κύτταρα ήταν σπάνια (Εικόνα 22 Β, Λ) ή τελείως αρνητικά (Εικόνα 22 Δ, Η, Ο). 3) Παρουσία του Bplus mrna παρατηρήθηκε σε 11/13 (85%) ΑΛΜΚ, ALK+ και 9/17 ΑΛΜΚ, ALK- (52%), (x 2, p=0,048, LR=0,040) και σε 14/18 (78%) ΑΛΜΚ, ALK+ περιπτώσεις με τις μοριακές τεχνικές έναντι 10/20 (50%) ΑΛΜΚ, ALK- (p=0,076) (Γράφημα 5). 85
86 4) Έκφραση του Bdel mrna ανευρέθη σε 9/13 (69%) ΑΛΜΚ, ALK+ και 4/17 ΑΛΜΚ, ALK- (x 2, p=0,012, LR=0,025), (Γράφημα 6). 5) Με βάση την ομαδοποίηση των αποτελεσμάτων του υβριδισμού σε τέσσερις ομάδες: Bplus+/Bdel-, Bplus-/Bdel+, Bplus-/Bdel- και Bplus+/Bdel+ παρατηρήθηκε διαφορά στο προφίλ έκφρασης μεταξύ των παραπάνω ομάδων. Η ομάδα Bplus+/Bdel- εκφραζόταν ελαφρώς υψηλότερα στα ΑΛΜΚ, ALK+ σε σχέση με τα ΑΛΜΚ, ALK- (27% vs 7%), (Fisher s exact test, p=0,029, LR=0,042), (Γράφημα 7) και στην ομάδα των Σ-ΑΛΜΚ (23/30, 77%) έναντι των ΠΔ-ΑΛΜΚ (4/8, 50%), (Fisher's Exact Test, p=ns) (Γράφημα 7). htert πρωτεΐνη Τριάντα οκτώ περιπτώσεις μελετήθηκαν ανοσοϊστοχημικά για την πρωτεΐνη htert (Πίνακας 12). Έκφραση της htert παρατηρήθηκε στον πυρήνα των νεοπλασματικών κυττάρων του λεμφώματος σε 27/38 (71%) περιπτώσεις με την μορφή πολλαπλών εμφανών κουκκίδων καθώς και διάχυτα (Εικόνα 22Θ). Κυτταροπλασμική κατά θέσεις εντόπιση της htert παρατηρήθηκε σε πέντε θετικές περιπτώσεις, όλες ΑΛΜΚ, ALK+. Στα μη-νεοπλασματικά λεμφοκύτταρα παρατηρήθηκε μεμονωμένη πυρηνική εντόπιση της πρωτεΐνης (Εικόνα 21Ι). Η έκφραση της htert ήταν συχνότερη στην ομάδα των ΑΛΜΚ, ALK+ (12/13, 92%) σε σύγκριση με την ομάδα των ΑΛΜΚ, ALK- (11/17, 65%) (x 2, p=0,077) και στα Σ- ΑΛΜΚ έναντι των ΠΔ-ΑΛΜΚ (77% vs 50%, x 2, p=0,13), (Γράφημα 8). Όλες οι θετικές περιπτώσεις για το htert mrna, συμπεριλαμβανομένων και αυτών με το προφίλ Bplus-/Bdel+ ήταν θετικές για την htert πρωτεΐνη (Fisher's Exact Test, p<0,001). Εν κατακλείδι, τα θετικά νεοπλασματικά κύτταρα για το mrna των εναλλακτικά ματισμένων μορφών Bplus και Bdel της htert ήταν επίσης θετικά και για την htert πρωτεΐνη. RT-PCR για ανίχνευση των ισομορφών της htert Με την duplex RT-PCR όλα τα δείγματα των ΑΛΜΚ (42/42, 100%) ήταν θετικά για την έκφραση της htert (Εικόνα 23Α). 86
87 RT-PCR για τα Bdel και Bplus μετάγραφα Μετάγραφα των Bplus και Bdel εναλλακτικά ματισμένων μορφών της htert ανιχνεύθηκαν και στους 3 αντιδραστικούς λεμφαδένες (Εικόνα 23Β). ΑΙΧ ΓΙΑ ΤΙΣ ΠΡΩΤΕΪΝΕΣ FASCIN ΚΑΙ MUC-1 FASCIN Η πρωτεΐνη Fascin ανιχνεύθηκε σε 35/42 (83%) περιπτώσεις (Πίνακας 13). Στις περιπτώσεις αυτές τα νεοπλασματικά λεμφοειδή κύτταρα εμφάνιζαν από ήπια έως έντονη κυτταροπλασμική θετικότητα, η οποία σε 29/36 περιπτώσεις αφορούσε πάνω από το 50% των κυττάρων (Εικόνα 24). Έκφραση της πρωτεΐνης παρατηρήθηκε επίσης στα ενδοθηλιακά (Εικόνα 24) και δενδριτικά κύτταρα τα οποία και αποτελούσαν τους εσωτερικούς θετικούς μάρτυρες της ανοσοχρώσης. Η Fascin ανιχνεύθηκε στο σύνολο των ΑΛΜΚ, ALK+ (13/13, 100%) και σε 12/13 περιπτώσεις σε >50% των νεοπλασματικών κυττάρων. Παρατηρήθηκε στατιστικά σημαντική διαφορά της έκφρασης της Fascin μεταξύ των ΑΛΜΚ, ALK+ (100%) και ΑΛΜΚ, ALK- (71%), (x 2, p=0,040, Fisher's Exact Test p=0,040), (Γράφημα 9). Το σύνολο των εξωλεμφαδενικών (12/12, 100%) και το 73% (16/22) των λεμφαδενικών ΑΛΜΚ περιπτώσεων ήταν θετικά στην παραπάνω πρωτεΐνη (x 2, p=0,046), (Γράφημα 9). Στα δερματικά ΑΛΜΚ, το 57% των περιπτώσεων εξέφραζε σε >50% των νεοπλασματικών κυττάρων την παραπάνω πρωτεΐνη, ποσοστό όμως σημαντικά χαμηλότερο από το 89% των Σ-ΑΛΜΚ (x 2, p=0,044), (Γράφημα 9). MUC-1 Κυτταροπλασμική έκφραση της πρωτεΐνης MUC-1 παρατηρήθηκε σε 15/42 (36%) περιπτώσεις (Πίνακας 13), με κυρίως εστιακή κατανομή (13/15 περιπτώσεις) (Εικόνα 25). Διαπιστώθηκε σημαντικά υψηλότερη έκφραση της MUC-1 στις ΑΛΜΚ, ALK+ (8/13, 61,5%) περιπτώσεις σε σύγκριση με τις ΑΛΜΚ, ALK- (5/21, 24%) περιπτώσεις (Γράφημα 10), (x 2, p=0,028). Η απώλεια έκφρασης της MUC-1 ήταν σημαντικά συχνότερη σε μεγαλύτερες ηλικίες (> 35 έτη) σε σύγκριση με μικρότερες ηλικίες ( 35 έτη), (26/36 vs 1/6, Fisher's Exact Test p=0,016) (Γράφημα 11). 87
88 Συσχέτιση της έκφρασης των Fascin, htert και MUC-1 Οι δεκαπέντε θετικές περιπτώσεις στην MUC-1 εξέφραζαν επίσης την Fascin (x 2, p=0,031) καθώς και την htert (x 2, p=0,001), (Γράφημα 12 & 13). Ο ανοσοφαινότυπος Fascin+ / MUC-1+ / htert+ ήταν πιο συχνός στα ΑΛΜΚ, ALK+ (8/13) σε σύγκριση με τα ΑΛΜΚ, ALK- (5/17) (x 2, p=0,13) και στις ηλικίες 35 έτη σε σύγκριση με > 35 έτη (5/6 vs 10/32, x 2, p=0,17). 88
89 ΠΙΝΑΚΕΣ & ΓΡΑΦΗΜΑΤΑ 89
90 90
91 Πίνακας 7. Κλινικοπαθολογικά και ανοσοφαινοτυπικά χαρακτηριστικά των περιπτώσεων της μελέτης (n=45). Α.Π Η Φ ΕΝΤ. ΚΛ. ΤΑΞ. ΑΝΦ CD30 CD3 CD43 CD45 RO CD Θ ΛΦ Σ-ΑΛΜΚ T Α ΔΕΡΜΑ Σ-ΑΛΜΚ T Α ΜΜ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Θ ΜΜ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T * 32 Α ΛΦ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Θ ΛΦ Σ-ΑΛΜΚ T Α ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T + + ΔΕ Θ ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T Θ Λ-Δ Σ-ΑΛΜΚ T # 47 Θ Λ-Δ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΔΕΡΜΑ Σ-ΑΛΜΚ T Θ ΛΦ Σ-ΑΛΜΚ T Θ ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T Α Λ-Δ Σ-ΑΛΜΚ T * 56 Α Λ-Δ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T + + ΔΕ Α ΠΝ. Σ-ΑΛΜΚ T Θ ΛΦ Σ-ΑΛΜΚ T + - ΔΕ Α ΣΠΛ. Σ-ΑΛΜΚ NULL Α ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T Α ΣΤ. Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΟΣΤΟ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Θ ΛΦ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Θ ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T Α ΜΜ Σ-ΑΛΜΚ T Α ΛΦ Σ-ΑΛΜΚ T Α ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T Θ ΛΦ Σ-ΑΛΜΚ T Α ΜΜ Σ-ΑΛΜΚ T Α ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T Θ ΛΦ Σ-ΑΛΜΚ T Α ΔΕΡΜΑ ΠΔ-ΑΛΜΚ T CD45 RA 91
92 Επεξηγήσεις: Α: άρρεν, ΑΝΦ: Ανοσοφαινότυπος, Α.Π: αριθμός περίπτωσης, ΔΕ: δεν έγινε, ΕΝΤ: εντόπιση, Θ: θήλυ, Η: ηλικία, ΚΛ. ΤΑΞ.: κλινική ταξινόμηση, Λ-Δ: λεμφαδένας-δέρμα, ΛΦ: λεμφαδένας, ΜΜ: μαλακά μόρια, ΠΔ-ΑΛΜΚ: πρωτοπαθές δερματικό ΑΛΜΚ, ΠΝ: πνεύμονας, Σ- ΑΛΜΚ: συστηματικό ΑΛΜΚ, ΣΤ: στόμαχος, ΣΠΛ: σπλήνας, Φ: φύλο. * επανεμφάνιση, # υποτροπή. 92
93 Πίνακας 8. Ανάλυση των αποτελεσμάτων της μελέτης ως προς την ανίχνευση της χιμαιρικής ALK πρωτεΐνης, του χιμαιρικού μετάγραφου NPM-ALK [t(2;5)(p23q21)] με RT-PCR, της μετάθεσης του τμήματος 2p23 με FISH, της επαύξησης των 2p23 και CEP2 χρωμοσωμικών τμημάτων επίσης με FISH καθώς και του T-cell receptor γ (TCR-γ) με PCR, (n=45). Α.Π 2p23 CEP2 ALK1 TCR-γ t(2;5)(p23q35) 2p23 status status (ΑΙΧ) (PCR) (RT-PCR) (FISH) (FISH) (FISH) 1 Π/Κ Φ Φ 2 Π/Κ Φ Φ 3 Π/Κ E E 4 Π/Κ Φ Φ 5 Π/Κ E E 6 Π/Κ Φ Φ 6* Π/Κ - ΔΑ + Φ Φ 7 Κ E E E E 9 Κ Φ E Φ Φ Φ Φ Φ Φ Φ Φ 14 Κ Φ Φ 14# Κ + ΔΕ + Φ Φ Φ Φ 16 - ΔΑ - - Φ Φ Φ Φ 18 - ΔΑ - - Φ Φ Φ Φ Φ Φ 20* - ΔΑ ΔΕ - Φ Φ 21 Κ Φ Φ Φ E Π/Κ E Φ Φ Φ Φ Φ E E Φ E 28 Κ E Φ E E 93
94 Φ Φ 31 - ΔΑ - - Φ Φ Φ Φ E E Φ Φ 35 Π/Κ Φ Φ 36 - ΔΑ - - Φ Φ E E Φ Φ Φ Φ Φ Φ 41 - ΔΑ - - Φ E Φ Φ Επεξηγήσεις: Α.Π: αριθμός περίπτωσης, ΔΑ: δεν απέδωσε, ΔΕ: δεν έγινε, E: επαύξηση, Κ: κυτταρόπλασμα, Π/Κ: πυρήνας και κυτταρόπλασμα, Φ: φυσιολογικό. * επανεμφάνιση, # υποτροπή 94




. * αφορά 34")





.")
95 Γράφημα 1. Παρουσία της ALK χιμαιρικής πρωτεΐνης με ΑΙΧ, της NPM-ALK με RT-PCRαποτελεσμάτων των μοριακών μεθόδων ανίχνευσης της ALK, σε σχέση με την ηλικία εμφάνισης της νόσου (n= 42). * αφορά 34 περιπτώσεις Σ-ΑΛΜΚ. ** Ο διαχωρισμός των ηλικιακών ομάδων έγινε σύμφωνα με τους Falini και συν. της μετάθεσης του τμήματος 2p23 με FISH και συνολικά των [104]. Γράφημα 2. Παρουσία της ALK χιμαιρικής πρωτεΐνης με ΑΙΧ, της NPM-ALK με RT-PCRανίχνευσης της ALK, σε σχέση με την κλινική ταξινόμησηη των ΑΛΜΚ της μελέτης (n=42). της μετάθεσης του τμήματος 2p23 με FISH και των μοριακών μεθόδων 95

96 Γράφημα 3. Σ υμφωνία ως προς την ανίχνευση της πολυπλοειδίας του χρωμοσώματος 2 με τον κεντρομεριδιακό ιχνηθέτη (CEP2) και τον διπλό ιχνηθέτη αποκοπής του 2p23 με FISH (kappa=0,877) (n=42). 96
97 Πίνακας 9. Συσχέτιση συμφωνίας (kappa agreement) των μοριακών μεθόδων ανίχνευσης της t(2;5), της μετάθεσης του τμήματος 2p23 με FISH και της ALK1 πρωτεΐνης. Α. Συσχέτιση της ανοσοϊστοχημικής έκφρασης (ΑΙΧ) της ALK1 με την παρουσία του χιμαιρικού μετάγραφου NPM-ALK [t(2;5)(p23q35)] με RT-PCR (n=42). ALK1 (ΑΙΧ) ΣΥΝΟΛΟ - + ALK (RT-PCR) ΣΥΝΟΛΟ Συμφωνία (concordance) θετικών περιπτώσεων (+): 8/13 (81%) Συμφωνία όλων των περιπτώσεων: (26+8)/42= 34/42 (61,5%) Kappa for agreement= 0,535, Fisher's Exact Test p=0,001 Β. Συσχέτιση της ALK1 με την ανίχνευση της μετάθεσης του τμήματος 2p23 με τη FISH (n=42). ALK (ΑΙΧ) ΣΥΝΟΛΟ - + ALK (FISH) ΣΥΝΟΛΟ Συμφωνία θετικών περιπτώσεων (+): 13/13 (100%) Συμφωνία όλων των περιπτώσεων: (24+13)/42= 37/42 (88%) Kappa for agreement= 0,748, Fisher's Exact Test p<0,001 Γ. Συσχέτιση της πυρηνοκυτταροπλασμικής εντόπισης της ALK1 πρωτεΐνης (που υποδηλώνει την παρουσία της NPM-ALK χιμαιρικής πρωτεΐνης) με την παρουσία του χιμαιρικού μετάγραφου NPM-ALK [t(2;5)(p23q35)] με RT-PCR (n=37). ALK1 (ΑΙΧ) ΣΥΝΟΛΟ - + NPM/ALK ΣΥΝΟΛΟ Συμφωνία (concordance) θετικών περιπτώσεων (+): 8/8 (100%) Συμφωνία όλων των περιπτώσεων: (26+8)/37= 34/37 (91,8%) Kappa for agreement= 0,840, Fisher's Exact Test p<0,001 97
98 Δ. Συσχέτιση της RT-PCR με τη FISH (n=42). ALK (FISH) ΣΥΝΟΛΟ - + ALK (RT-PCR) ΣΥΝΟΛΟ Συμφωνία θετικών περιπτώσεων (+): 11/18 (61%) Συμφωνία όλων των περιπτώσεων: (24+11)/42= 35/42 (83%) Kappa for agreement= 0,642, Fisher's Exact Test p<0,001 Ε. Συσχέτιση συνολικά των ευρημάτων της RT-PCR και της FISH με την ΑΙΧ έκφραση της ALK1 (n=42). ALK1 (AIX) ΣΥΝΟΛΟ - + ALK (RT-PCR + FISH) ΣΥΝΟΛΟ Συμφωνία θετικών περιπτώσεων (+): 13/13 (100%) Συμφωνία όλων των περιπτώσεων: (23+13)/42= 36/42 (86%) Kappa for agreement= 0,704, Fisher's Exact Test p<0,001 98
99 Πίνακας 10. Ανάλυση των αποτελεσμάτων της μελέτης της Cyclin D1 σε επίπεδο πρωτεΐνης, mrna και γονιδίου σε 45 βιοψίες από 42 ασθενείς. Μέθοδος Ανίχνευσης Μέθοδος Ανίχνευσης ΑΙΧ ISH FISH ΑΙΧ ISH FISH Α.Π Cyclin Cyclin Cyclin Cyclin Cyclin 11/11q13 Cyclin 11/11q13 D1 D1 poly-t Α.Π D1 D1 poly-t D1 (CCND1) D1 (CCND1) (SP1) (DCS-6) (SP1) (DCS-6) Δ ΔΑ Δ Π Π Π Δ Δ Π Δ Δ Δ 6* Δ Δ Π Π Π Π Δ Π Π Δ Δ ΔΑ ΔΑ Δ Δ Δ Δ Δ Π 14# Δ ΔΑ Δ Π Δ Δ Π Δ Π Δ Δ ΔΑ Δ Π 20* Δ Επεξηγήσεις: ΑΙΧ: ανοσοϊστοχημεία, Α.Π: αριθμός περίπτωσης, ΔΑ: δεν απέδωσε, Δ: δισωμία, Π: πολυσωμία. * επανεμφάνιση, # υποτροπή. 99
100 Πίνακας 11. Έκφραση του mrna της Cyclin D1 και της πολυσωμίας του 11q13 (CCND1 γονίδιο) σε σχέση με την ανοσοϊστοχημική (ΑΙΧ) έκφραση της χιμαιρικής ALK, τις μοριακές διαταραχές του ALK γονιδίου, την κλινική ταξινόμηση και την πρωτοπαθή εντόπιση της νόσου στις ΑΛΜΚ περιπτώσεις της μελέτης. Cyclin D1 mrna, ISH (n=41) 11q13 status (CCND1 γονίδιο), FISH (n=38) Cyclin D1+ Cyclin D1- Πολυσωμία Δισωμία ALK (ΑΙΧ)* ALK θετικά ALK αρνητικά P 0.51 a 0.67 b ALK (FISH & PCR) Θετικές περιπτώσεις Αρνητικές περιπτώσεις P Κλινική Ταξινόμηση Συστηματικά ΑΛΜΚ Πρωτοπαθή δερματικά ΑΛΜΚ P Πρωτοπαθής εντόπιση** Λεμφαδενικά Εξωλελφαδενικά P 0.69 c 0.98 d * Αφορά μόνο τα Σ-ΑΛΜΚ: a: 33 περιπτώσεις; b: 31 περιπτώσεις. ** Αφορά μόνο τα Σ-ΑΛΜΚ: c: 33 περιπτώσεις; d: 31 περιπτώσεις. 100
101 Πίνακας 12. Ανάλυση των αποτελεσμάτων της έκφρασης της htert με τις μεθόδους της PCR, ISH και ΑΙΧ στις 42 περιπτώσεις ΑΛΜΚ της μελέτης. Α.Π duplex PCR ISH, Bplus ISH, Bdel htert (AIX) Α.Π duplex PCR ISH, Bplus ISH, Bdel htert (AIX) Κ Κ * ΔΕ ΔΕ ΔΕ Κ Κ ΔΑ ΔΑ ΔΑ ΔΑ Κ # ΔΕ ΔΕ ΔΕ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ ΔΑ * ΔΕ ΔΕ ΔΕ ΔΑ Επεξηγήσεις: Α.Π: αριθμός περίπτωσης, ΔΑ: δεν απέδωσε, ΔΕ: δεν έγινε Κ: κυτταρόπλασμα. * επανεμφάνιση, # υποτροπή. 101
102 Γράφημα 4. Εκφραση του mrna της htertt σε 38 ΑΛΜΚ (Σ-ΑΛΜΚ και ΑΛΜΚ, ΑΛΜΚ, ALK+ και ΑΛΜΚ, ALK-). ΠΔ- Γράφημα 5. Εκφραση της μορφής Bplus της htert σε 38 ΑΛΜΚ (ΑΛΜΚ, ALK+ και ΑΛΜΚ, ALK- σύμφωνα με την ΑΙΧ έκφραση της ALK1, του μεταγράφου NPM- ALK με RT-PCR και της μετάθεσης του τμήματος του 2p23 με FISH). 102


103 Γράφημα 6. Εκφραση της μορφής Bdel της htert σε 38 ΑΛΜΚ (Σ-ΑΛΜΚ ALK+ και ALK- με ΑΙΧ). Γράφημα 7. Εκφραση των μορφών Bplus και Bdel της htert σε 38 ΑΛΜΚ (Σ- ΑΛΜΚ, ΠΔ-ΑΛΜΚ και στα Σ-ΑΛΜΚ, ALK+ και Σ-ΑΛΜΚ, ALK- σύμφωνα με την έκφραση της ALK1). Επεξηγήσεις: ΘΘ: θετικές περιπτώσεις στους Bplus και Bdel mrna s ανιχνευτές, ΘΑ: θετικές περιπτώσεις στον Bplus/αρνητικές περιπτώσεις στον Bdel, ΑΘ: αρνητικές περιπτώσεις στον Bplus/θετικές περιπτώσεις στον Bdel, ΑΑ: αρνητικές περιπτώσεις και στους δύο ανιχνευτές. 103
104 Γράφημα 8. Εκφραση της htert πρωτεΐνης σε 38 ΑΛΜΚ (Σ-ΑΛΜΚ, ΠΔ-ΑΛΜΚ και στα ΑΛΜΚ, ALK+ και ΑΛΜΚ, ALK- σύμφωνα με την έκφραση της ALK1). 104
105 Πίνακας 13. Ανοσοϊστοχημική έκφραση των Fascin και MUC-1 πρωτεϊνών σε 45 βιοψίες από 42 ασθενείς με ΑΛΜΚ. Α.Π Fascin MUC-1 Α.Π Fascin MUC * # * + - Επεξηγήσεις: Α.Π: αριθμός περίπτωσης. * επανεμφάνιση, # υποτροπή. 105



, ΕΞ.")
.")

106 Γράφημα 9. Έκφραση της πρωτεΐνης Fascin στις 42 ΑΛΜΚ περιπτώσεις της μελέτης σε σχέση με την κλινική ταξινόμηση,, την έκφραση της ALK1 και την λεμφαδενική ή εξωλεμφαδενική εντόπιση. Επεξηγήσεις: ΛΦ-ΑΛΜΚ: λεμφαδενικά ΑΛΜΚ (22 περιπτώσεις), ΕΞ.ΛΦ-ΑΛΜΚ: εξωλεμφαδενικά ΑΛΜΚ (12 περιπτώσεις). * έκφραση της Fascin σε >50% των νεοπλασματικών κυττάρων. Γράφημα 10. Έκφραση της πρωτεΐνης MUC-1 στις 42 μελέτης σε σχέση με την έκφραση της ALK1. ΑΛΜΚ περιπτώσεις της 106
107 Γράφημα 11. Έκφραση της πρωτεΐνης MUC-1 στις 42 ΑΛΜΚ περιπτώσεις της μελέτης σε σχέση με την ηλικία εμφάνισης της νόσου. Ο διαχωρισμός των ηλικιακών ομάδων έγινε σύμφωνα με τους Falini και συν. [ 104]. Γράφημα 12. Συσχέτιση της συνέκφρασης των πρωτεϊνών MUC-1 και Fascin στις 42 ΑΛΜΚ περιπτώσεις της μελέτης. Επεξηγήσεις: ΘΘ: θετικές περιπτώσεις και στις δύο πρωτεΐνες, ΘΑ: θετικές περιπτώσεις στην MUC-1 /αρνητικές περιπτώσεις στην Fascin, ΑΘ: αρνητικές περιπτώσεις στην MUC-1/θετικές περιπτώσεις στην Fascin, ΑΑ: αρνητικές περιπτώσεις και στις δύο πρωτεΐνες. 107
108 Γράφημα 13. Συσχέτιση της συνέκφρασης των πρωτεϊνώνν MUC-1 και htert στις 42 ΑΛΜΚ περιπτώσεις της μελέτης. Επεξηγήσεις: ΘΘ: θετικές περιπτώσεις και στις δύο πρωτεΐνες, ΘΑ: θετικές περιπτώσεις στην MUC-1 /αρνητικές περιπτώσεις στην htert, ΑΘ: αρνητικές περιπτώσεις στην MUC-1/θετικές περιπτώσεις στην htert, ΑΑ: αρνητικές περιπτώσεις και στις δύο πρωτεΐνες. 108
109 ΕΙΚΟΝΕΣ 109
110 110



, x 100 (Β).")



. # Β.")

111 Α Α. Β. Β Εικ κόνα 8. Η&Ε Ε. Α. ΑΛΜΚ, ALK+ A (περ.. #1). Β. ΠΔ-ΑΛΜΚ Π Κ χωρίς επιιδερμοτροπιισμό, με εκτεεταμένη διή ήθηση του χορίου χ (περ.. #38). Η&Ε Ε x 200 (Α), x 100 (Β).. Α Α. Β. Β Εικ κόνα 9.Ανοσ σοϊστοχημεεία για το CD30/Ki1 C αντιγόνο. α Α. Θετική αννοσοαντίδραση των νεοπλασμα ατικών λεμ μφοειδών κκυττάρων ΑΛΜΚ, ALK+ + στο CD300/Ki1 (περ. #23). # Β. ΠΔ Δ-ΑΛΜΚ με επιδερμοττροπισμό ο οπ ποίος αναδυυκνύεται μεε την ανοσοοχρώση για το CD30/K Ki1 (περ. #113). Α, Β x
 # και πολυκλωνικόςς (περρ. #28) στονν TCR-γ πλλυθυσμός.")

. x 200. εντόπ πιση (περ. #1). Β. Εικ κόνα 12. RT-PCR.")

112 Εικ κόνα 10. PC CR για τονν TCR-γ. Μονοκλωνι Μ ικός (περ. #8) # και πολυκλωνικόςς (περρ. #28) στονν TCR-γ πλλυθυσμός. L= L κλίμακα (ladder) ανα α 100bp DN NA. Α. Β. κόνα 11. Ανοσοϊστοχημ μεία για τη ην ALK1. Εικ Α. Έντονη κυτταροπλλασμική και κ πυρηνιική κυττταροπλασμιική εντόπισ ση (περ. #7). x 200. εντόπ πιση (περ. #1). Β. Εικ κόνα 12. RT-PCR. Ανίχνευση η της NP PM/ALK, t(2;5)(p23;;q35) στηνν περίίπτωση τηςς εικόνας 11Α. Αριστεερά στην στήλη L απ πεικονίζετα αι ο θετικόςς μάρρτυρας. L= κλίμακα κ (laadder) ανα 100bp 1 DNA A. 112

 με παρουσία 1")
,")




113 Α. Β. Εικόνα 13. FISH με τη χρήση του LSI ALK (Anaplastic Lymphoma Kinase) Dual Color, Break Apart Rearrangement Probe. Α. Νεοπλασματικοί πυρήνες (βέλη) με παρουσία 1 ερυθρού, 1 πράσινου και 1 κίτρινου σήματος ή 1 ζεύγους πράσινου/ερυθρού σήματος (1R1G1F ή 1R1G1RG), αποδεικτικό στοιχείο μετάθεσης του 2p23 (περ. #1). Β. Νεοπλασματικοί ανευπλοειδικοί πυρήνες με 5 ζεύγη πράσινου/ /ερυθρού σήματος (βέλος) και πέντε 5 τμήματα του 2p23 και μετάθεση του 2p23 (κεφαλή βέλους) (περ. #3). Α x 1000, Β zoom Εικόνα 14. FISH με τη χρήση του κεντρομεριδιακού ιχνηθέτη (D2Z1, 2p11.1- q11.1). Νεοπλασματικοί πυρήνες με παρουσία 3 ερυθρών σημάτων (βέλη), τεσσάρων ερυθρών σημάτων (κεφαλή βέλους) και πέντε ερυθρών σημάτων (2 κεφαλές βέλους), (περ. #3). x

.")
 και")

114 Α. Β. Γ. Δ. Εικόνα 15. Διπλή ανοσοϊστοχημεία για τα CD30/Cyclin D1, clone SP4 αντιγόνα (A, B) και CD30/Cyclin D1 clone DCS-6 αντιγόνα (Γ, Δ). Και τα δύο αντισώματα έναντι της Cyclin D1 παρουσίασαν έντονη πυρηνική θετικότητα στα ενδοθηλιακά (βέλη) και επιδερμιδικά κύτταρα (κεφαλές βέλους). Το CD30 αντιγόνο απεικονίστηκε χρησιμοποιώντας την αλκαλική φωσφατάση ως χρωμογόνο (περ #3 και #13 αντίστοιχα). Α - Δ x 200. (Bobos M et al., 2009, in press, ref. 212) 114


, ενώ τα ενδοθηλιακά κύτταρα (βέλη) χρησιμοποιήθηκαν ως")

115 Α. Β. Γ. Δ. Εικόνα 16. Απλή ανοσοϊστοχημεία για την Cyclin D1. Οι MCL περιπτώσεις παρουσίασαν έντονη πυρηνική θετικότητα και στα δύο αντι- Cyclin D1 McAbs (Α: SP4 κλώνος Β: κλώνος DCS-6). Οι ΑΛΜΚ περιπτώσεις ήταν αρνητικές για την Cyclin D1 και με τα δύο mab (περ. #3), ενώ τα ενδοθηλιακά κύτταρα (βέλη) χρησιμοποιήθηκαν ως εσωτερικοί θετικοί μάρτυρες (Γ: SP4 κλώνος Δ: κλώνος DCS-6). Α - Δ: x 200. (Bobos M et al., 2009, in press, ref. 212) MCL: mantle cell lymphoma (λέμφωμα από κύτταρα του μανδύα) 115
 έντονα θετικό σήμα υβριδισμού για mrna (x 200). Β, Γ.")

.")
 Εικόνα 18.")
 και αρνητικών (στήλη 8) ΑΛΜΚ περιπτώσεων.")
116 Α. Β. Γ. Δ. Εικόνα 17. ISH για το mrna της cyclin D1. A. ΑΛΜΚ (περ. #3) έντονα θετικό σήμα υβριδισμού για mrna (x 200). Β, Γ. Η ίδια περίπτωση εμφανίζει έντονη κυτταροπλασματική/περιπυρηνική κυανόμαυρη χρώση, ενδεικτική της παρουσίας του mrna της cyclin D1 στα νεοπλασματικά λεμφοειδή κύτταρα (Β x 200, Γ x 400) με τον antisense ανιχνευτή Δ. Υβριδοποίηση του sense ανιχνευτή της cyclin D1(περ. #3). Κανένα σήμα δεν ανιχνεύθηκε στα κύτταρα του λεμφώματος (x 200). (Bobos M et al., 2009, in press, ref. 212) Εικόνα 18. duplex RT-PCR για την cyclin D1. Παρουσίαση των θετικών (στήλες 1-7) και αρνητικών (στήλη 8) ΑΛΜΚ περιπτώσεων. PB = περιφερικό δείγμα αίματος, αρνητικό για την cyclin D1. L= κλίμακα (ladder) ανα 100bp DNA. (Bobos M et al., 2009, in press, ref. 212) 116
 και του χρωμοσώματος 11 (πράσινα σήματα). x 1000.")

, αναγνωρίζονται 4 αντίγραφα του")
117 Εικόνα 19. FISH με τους LSI Cyclin D1 (11q13) / CEP11 ιχνηθέτες. Αντιπροσωπευτική εικόνα της περίπτωσης #3 που παρουσιάζει τρία αντίγραφα του γονιδίου CCND1 (κόκκινα σήματα) και του χρωμοσώματος 11 (πράσινα σήματα). x (Bobos M et al., 2009, in press, ref. 212) A. B. Εικόνα 20. FICTION με το CD30 αντίσωμα και τον διπλό ιχνηθέτη CCND1/CEP11. Α. Ίδια περίπτωση με αυτή της εικόνας 19. Νεοπλασματικό κύτταρο που φωτογραφίζεται χρησιμοποιώντας το φίλτρο FITC/spectrum green. Το κύτταρο εκφράζει το CD30 αντιγόνο (πράσινο χρώμα, βέλη) και εμφανίζει στον πυρήνα τέσσερα αντίγραφα του χρωμοσώματος 11 (πράσινα σήματα, κεφαλή βέλους) Β. Στον πυρήνα του ίδιου κυττάρου, που φωτογραφίζεται εδώ με τρία φίλτρα (DAPI, FITC/spectrum green, rhodamine/ spectrum orange), αναγνωρίζονται 4 αντίγραφα του χρωμοσώματος 11 και 5 αντίγραφα του CCND1 γονιδίου Η μεμβράνη του κυττάρου είναι πράσινη (βέλη) με τον έμμεσο ανοσοφθορισμό για CD30. Α x 1000, Β zoom. (Bobos M et al., 2009, in press, ref. 212) 117
118 Α Β Γ Δ Ε Ζ Α Η Θ Ι Εικ κόνα 21. ISH H για τους Bplus, Bdeel, sense κα αι antisencee ανιχνευτέςς της hter RT. Έκφ φραση τωνν εναλλακττικά ματισ σμένων mr RNA μορφών της ht TERT σε μη νεοπ πλασματικοούς λεμφα αδένες. Περιοχές Π από α το ίδιο βλασ στικό κένντρο παρρουσιάζοντα αι μετά την υβριδοποίη ηση με τουςς Bplus antisence (Α, Β Β), Bplus seense (Γ),, Bdel antisense (Δ, Ε) Ε και Bdeel sense (Ζ Ζ) ανιχνευτέές. Σημειώσ στε ότι πολλλά (βέλλη) αλλά όχχι όλα (κεφ φαλή βέλουυς) κύτταρα α εκφράζουνν ταυτόχροονα και τα δύο mrn NAs. Η περιφερειακ π κή, ενεργοπ ποιημένη Τ ζώνη Τ απ πό τον ίδιο λεμφαδδένα απεικονίζεται με μ τους Bpllus antisensse (Η) και Bdel B antisennse ανιχνευττές (Θ), καθθώς επίσ σης και με το τ αντίσωμα α NCL-hTE ERT (Ι). Β, Ε x 400, A, Γ, Δ, Ζ-Ι xx200. (Koto oula V ett al., 2006, ref r 164) 118
119 Εικ κόνα 22. ht TERT ISH (Bplus ( και Bdel ισομο ορφές) και ΑΙΧ σε ΑΛ ΛΜΚ. Ανττιπροσωπευτικά παραδείγματα τωνν htert mrnas m και της πρωτεϊννικής έκφρα ασης σε ΑΛΜΚ. Στην Σ περίπττωση #1 παρατηρούντται ισάριθμ μα θετικά νεοπλασμα ατικά λεμφοειδή κύττταρα στουςς Bplus (Α)) και Bdel ανιχνευτές α (Β), με δια αφορετική όμως ό κατανομή (πλα αίσια). Περρίπτωση #31 θετική στον σ Bplus ανιχνευτή (Γ), αρνη ητική στονν Bdel (Δ Δ). Σημειώ ώστε ότι, τα θετικά ά μη-νεοπλλασματικά λεμφοκύττταρα παρραμένουν θεετικά στον Bdel B ανιχνεευτή (Δ). Πεερίπτωση #33. Η πλειοννότητα των Ki-1 K θετιικών κυττάρων (Ε) είνναι θετικά στον Bpluss ανιχνευτή ή (Ζ), αρνητικά στον Bdel B (Η) ανιχνευτή,, ενώ τα πεερισσότερα λεμφοειδή κύτταρα είναι ε θετικά ά στην ΑΙΧ Χ για 119
120 την htert (Θ). Περίπτωση #7. Εδώ, τα θετικά στο Ki-1 κύτταρα (Ι) δεν εκφράζουν τον Bplus (Κ), ενώ είναι προφανώς λιγότερα τα θετικά κύτταρα στον Bdel (Λ). Στα πλαίσια παρουσιάζεται η αντίστοιχη περιοχή στην ΑΙΧ (Μ), όπου παρατηρείται κυτταροπλασματική χρώση σε ολιγάριθμα νεοπλασματικά κύτταρα (βέλη). Περίπτωση #21. Τα Ki-1 θετικά κύτταρα (Ν) είναι αρνητικά στους Bplus (Ξ) και Bdel (Ο) ανιχνευτές, καθώς και στην πρωτεΐνη (Π). Σε αντίθεση, τα γύρω λεμφοκύτταρα είναι θετικά για τα htert mrnas (Ξ, Ο) και στην πρωτεΐνη (Π). Μ x 400, Α-Λ, Ν-Π x200. (Kotoula V et al., 2006, ref 164) Εικόνα 23. htert έκφραση με duplex RT-PCRR σε ΑΛΜΚ και μηνεοπλασματικούς λεμφαδένες. Α. Όλες οι παρουσιαζόμενες ΑΛΜΚ περιπτώσεις είναι θετικές για το htert μετάγραφο. Β.. Σύγχρονη ενίσχυση των Bplus και Bdel μεταγράφων σε μη μηνεοπλασματικούς λεμφαδένες. NPM: nucleophosmin ως εσωτερικός μάρτυρας,, L= κλίμακα (ladder) ανα 100bp DNA. (Kotoula V et al., 2006, ref 164) 120
.")
. x 200 Εικόνα 25. Ανοσοϊστοχημεία για τη MUC-1.")
121 Εικόνα 24. Ανοσοϊστοχημεία για τη Fascin. Έντονη κυτταροπλασμική θετικότητα των κυττάρων του λεμφώματος (περ. #39). Σημειώνεται ότι, τα ενδοθηλιακά κύτταρα που αποτελούν εσωτερικούς μάρτυρες της τεχνικής, είναι επίσης θετικά (βέλος). x 200 Εικόνα 25. Ανοσοϊστοχημεία για τη MUC-1. Διάσπαρτα νεοπλασματικά κύτταρα με κυτταροπλασμική εντόπιση της πρωτεΐνης (περ. #5). x
Λεμφώματα πνεύμονα (συνέχεια) Πηνελόπη Κορκολοπούλου Επ. Καθηγήτρια Εργαστήριο Παθολογικής Ανατομικής Πανεπιστημίου Αθηνών
 Πηνελόπη Κορκολοπούλου Επ. Καθηγήτρια Εργαστήριο Παθολογικής Ανατομικής Πανεπιστημίου Αθηνών") Λεμφώματα πνεύμονα (συνέχεια) Πηνελόπη Κορκολοπούλου Επ. Καθηγήτρια Εργαστήριο Παθολογικής Ανατομικής Πανεπιστημίου Αθηνών Αγγειοκεντρική ανοσοϋπερπλαστική νόσος / λέμφωμα πνεύμονος ΕΒV B T/NΚ Λεμφωματώδης
Λεμφώματα πνεύμονα (συνέχεια) Πηνελόπη Κορκολοπούλου Επ. Καθηγήτρια Εργαστήριο Παθολογικής Ανατομικής Πανεπιστημίου Αθηνών Αγγειοκεντρική ανοσοϋπερπλαστική νόσος / λέμφωμα πνεύμονος ΕΒV B T/NΚ Λεμφωματώδης
ΠΑΡΟΥΣΙΑΣΗ ΠΕΡΙΣΤΑΤΙΚΟΥ
 ΠΑΡΟΥΣΙΑΣΗ ΠΕΡΙΣΤΑΤΙΚΟΥ ΠΟΥΛΙΑΝΙΤΗΣ ΝΙΚΟΛΑΟΣ ΕΙΔΙΚΕΥΟΜΕΝΟΣ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΓΝΑ Ο ΕΥΑΓΓΕΛΙΣΜΟΣ ΙΣΤΟΡΙΚΟ Βιοψίες κατά χρονολογική σειρά παραλαβής: 1. Βιοψία 2 ης μοίρας δωδεκαδακτύλου με αριθμό πρωτοκόλλου
ΠΑΡΟΥΣΙΑΣΗ ΠΕΡΙΣΤΑΤΙΚΟΥ ΠΟΥΛΙΑΝΙΤΗΣ ΝΙΚΟΛΑΟΣ ΕΙΔΙΚΕΥΟΜΕΝΟΣ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΓΝΑ Ο ΕΥΑΓΓΕΛΙΣΜΟΣ ΙΣΤΟΡΙΚΟ Βιοψίες κατά χρονολογική σειρά παραλαβής: 1. Βιοψία 2 ης μοίρας δωδεκαδακτύλου με αριθμό πρωτοκόλλου
ΛΕΜΦΩΜΑ HODGKIN. Τριανταφυλλιά Κολέτσα Λέκτορας Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής, ΑΠΘ
 ΛΕΜΦΩΜΑ HODGKIN Τριανταφυλλιά Κολέτσα Λέκτορας Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής, ΑΠΘ Ταξινόμηση λεμφωμάτων- Βασικές αρχές Στην ταξινόμηση WHO: Λαμβάνονται υπόψη μορφολογικά, ανοσοϊστοχημικά,
ΛΕΜΦΩΜΑ HODGKIN Τριανταφυλλιά Κολέτσα Λέκτορας Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής, ΑΠΘ Ταξινόμηση λεμφωμάτων- Βασικές αρχές Στην ταξινόμηση WHO: Λαμβάνονται υπόψη μορφολογικά, ανοσοϊστοχημικά,
ΟΓΚΟΙ ΤΗΣ ΑΣΤΡΟΚΥΤΤΑΡΙΚΗΣ ΓΛΟΙΑΣ. Θωµάς Ζαραµπούκας. Αν. Καθ. Παθολογικής Ανατοµικής. Ιατρική Σχολή Α.Π.Θ.
 ΟΓΚΟΙ ΤΗΣ ΑΣΤΡΟΚΥΤΤΑΡΙΚΗΣ ΓΛΟΙΑΣ Θωµάς Ζαραµπούκας Αν. Καθ. Παθολογικής Ανατοµικής Ιατρική Σχολή Α.Π.Θ. Όγκοι της Αστροκυτταρικής Γλοίας Πιλοκυτταρικό αστροκύτωµα Πιλοµυξοειδές αστροκύτωµα Πλειόµορφο ξανθοαστροκύτωµα
ΟΓΚΟΙ ΤΗΣ ΑΣΤΡΟΚΥΤΤΑΡΙΚΗΣ ΓΛΟΙΑΣ Θωµάς Ζαραµπούκας Αν. Καθ. Παθολογικής Ανατοµικής Ιατρική Σχολή Α.Π.Θ. Όγκοι της Αστροκυτταρικής Γλοίας Πιλοκυτταρικό αστροκύτωµα Πιλοµυξοειδές αστροκύτωµα Πλειόµορφο ξανθοαστροκύτωµα
ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ. Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ.
 ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Λεμφαδενικά Εξωλεμφαδενικά Λέμφωμα Hodgkin Μη Hodgkin λεμφώματα Β-κυτταρικής
ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Λεμφαδενικά Εξωλεμφαδενικά Λέμφωμα Hodgkin Μη Hodgkin λεμφώματα Β-κυτταρικής
ΕΙΔΙΚΗ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ
 ΕΙΔΙΚΗ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. 5o ΦΡΟΝΤΙΣΤΗΡΙΟ ΛΕΜΦΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ Διδάσκοντες: Αγγελική Χέβα Παθολογοανατόμος, ΠΓΝΘ «Γ. Παπανικολάου» Επιστημονικός
ΕΙΔΙΚΗ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. 5o ΦΡΟΝΤΙΣΤΗΡΙΟ ΛΕΜΦΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ Διδάσκοντες: Αγγελική Χέβα Παθολογοανατόμος, ΠΓΝΘ «Γ. Παπανικολάου» Επιστημονικός
TI NEOTEΡΟ ΣΤΟΥΣ ΝΕΥΡΟΕΝΔΟΚΡΙΝΕΙΣ ΟΓΚΟΥΣ ΤΟ 2015. Ορισμοί-Κατάταξη-Ιστολογία. Δ Ροντογιάννη
 TI NEOTEΡΟ ΣΤΟΥΣ ΝΕΥΡΟΕΝΔΟΚΡΙΝΕΙΣ ΟΓΚΟΥΣ ΤΟ 2015 Ορισμοί-Κατάταξη-Ιστολογία Δ Ροντογιάννη Νευροενδοκρινή νεοπλάσματα Ταξινόμηση Βιολογικοί δείκτες Μοριακή Παθολογία Νευροενδοκρινή νεοπλάσματα Ταξινόμηση
TI NEOTEΡΟ ΣΤΟΥΣ ΝΕΥΡΟΕΝΔΟΚΡΙΝΕΙΣ ΟΓΚΟΥΣ ΤΟ 2015 Ορισμοί-Κατάταξη-Ιστολογία Δ Ροντογιάννη Νευροενδοκρινή νεοπλάσματα Ταξινόμηση Βιολογικοί δείκτες Μοριακή Παθολογία Νευροενδοκρινή νεοπλάσματα Ταξινόμηση
Ιδιαίτερες νοσολογικές οντότητες στα λεμφώματα υψηλού βαθμού κακοηθείας
 Ιδιαίτερες νοσολογικές οντότητες στα λεμφώματα υψηλού βαθμού κακοηθείας Απαρτιωμένη διδασκαλία στην Αιματολογία 2015 Αργύρης Σ. Συμεωνίδης Οντότητες μη Hodgkin λεμφωμάτων που θα αναφερθούν Πρωτοπαθές Β-λέμφωμα
Ιδιαίτερες νοσολογικές οντότητες στα λεμφώματα υψηλού βαθμού κακοηθείας Απαρτιωμένη διδασκαλία στην Αιματολογία 2015 Αργύρης Σ. Συμεωνίδης Οντότητες μη Hodgkin λεμφωμάτων που θα αναφερθούν Πρωτοπαθές Β-λέμφωμα
ΛΕΜΦΑ ΕΝΙΚΟ ΛΕΜΦΩΜΑ ΟΡΙΑΚΗΣ ΖΩΝΗΣ ΙΣΤΟΛΟΓΙΚΗ ΕΙΚΟΝΑ
 ΛΕΜΦΑ ΕΝΙΚΟ ΛΕΜΦΩΜΑ ΟΡΙΑΚΗΣ ΖΩΝΗΣ ΙΣΤΟΛΟΓΙΚΗ ΕΙΚΟΝΑ Μικρά ή µεσαίου µεγέθους λεµφοειδή κύτταρα Πυρήνες στρογγυλοί ή ανωµάλου σχήµατος (µονοκυτταροειδείς ή σαν κεντροκύτταρα) ιαυγές κυτταρόπλασµα ΛΕΜΦΑ
ΛΕΜΦΑ ΕΝΙΚΟ ΛΕΜΦΩΜΑ ΟΡΙΑΚΗΣ ΖΩΝΗΣ ΙΣΤΟΛΟΓΙΚΗ ΕΙΚΟΝΑ Μικρά ή µεσαίου µεγέθους λεµφοειδή κύτταρα Πυρήνες στρογγυλοί ή ανωµάλου σχήµατος (µονοκυτταροειδείς ή σαν κεντροκύτταρα) ιαυγές κυτταρόπλασµα ΛΕΜΦΑ
ΛΕΜΦΙΚΟΣ ΙΣΤΟΣ. Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ.
 ΛΕΜΦΙΚΟΣ ΙΣΤΟΣ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΛΕΜΦΑΔΕΝΕΣ ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα CD20/L26
ΛΕΜΦΙΚΟΣ ΙΣΤΟΣ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΛΕΜΦΑΔΕΝΕΣ ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα CD20/L26
Ιστολογικά χαρακτηριστικά χαρακτηριστικά ΚΟΕΝ Ποικίλη µορφολογία Ατρακτόµορφα κύτταρα µε ποικίλο ηωσινόφιλο κυτταρόπλασµα,κυµατοειδείς πυρήνες Τύπου
 Ιστολογικά χαρακτηριστικά χαρακτηριστικά ΚΟΕΝ Ποικίλη µορφολογία Ατρακτόµορφα κύτταρα µε ποικίλο ηωσινόφιλο κυτταρόπλασµα,κυµατοειδείς πυρήνες Τύπου ινοσαρκώµατος εσµιδωτό πρότυπα ανάπτυξης Ιστολογικά
Ιστολογικά χαρακτηριστικά χαρακτηριστικά ΚΟΕΝ Ποικίλη µορφολογία Ατρακτόµορφα κύτταρα µε ποικίλο ηωσινόφιλο κυτταρόπλασµα,κυµατοειδείς πυρήνες Τύπου ινοσαρκώµατος εσµιδωτό πρότυπα ανάπτυξης Ιστολογικά
Ιδιαίτερες νοσολογικές οντότητες στα λεμφώματα υψηλού βαθμού κακοηθείας
 Ιδιαίτερες νοσολογικές οντότητες στα λεμφώματα υψηλού βαθμού κακοηθείας Απαρτιωμένη διδασκαλία στην Αιματολογία 2013 Αργύρης Σ. Συμεωνίδης Οντότητες μη Hodgkin λεμφωμάτων που θα αναφερθούν Πρωτοπαθές Β-λέμφωμα
Ιδιαίτερες νοσολογικές οντότητες στα λεμφώματα υψηλού βαθμού κακοηθείας Απαρτιωμένη διδασκαλία στην Αιματολογία 2013 Αργύρης Σ. Συμεωνίδης Οντότητες μη Hodgkin λεμφωμάτων που θα αναφερθούν Πρωτοπαθές Β-λέμφωμα
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟ ΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ. ΝΙΚΟΛΑΟΣ ΚΑΒΑΝΤΖΑΣ Επίκουρος Καθηγητής
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟ ΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ ΝΙΚΟΛΑΟΣ ΚΑΒΑΝΤΖΑΣ Επίκουρος Καθηγητής ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ Επιφυσιοβλάστωµα Επιφυσιοκύττωµα Ογκος
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟ ΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ ΝΙΚΟΛΑΟΣ ΚΑΒΑΝΤΖΑΣ Επίκουρος Καθηγητής ΟΓΚΟΙ ΕΠΙΦΥΣΗΣ Επιφυσιοβλάστωµα Επιφυσιοκύττωµα Ογκος
Μοριακή βιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία πνεύμονα. Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής
 Μοριακή βιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία πνεύμονα Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Εισαγωγή Ο καρκίνος του πνεύμονα παρουσιάζει άφθονες
Μοριακή βιολογία καρκίνου του πνεύμονα Ενότητα 1: Ογκολογία πνεύμονα Κυριάκος Καρκούλιας, Επίκουρος Καθηγητής Σχολή Επιστημών Υγείας Τμήμα Ιατρικής Εισαγωγή Ο καρκίνος του πνεύμονα παρουσιάζει άφθονες
Παθολογοανατομική προσέγγιση διάγνωση. Σχολιασμός Δημ. Σαμπαζιώτης, Ειδικευόμενος
 Παθολογοανατομική προσέγγιση διάγνωση Σχολιασμός Δημ. Σαμπαζιώτης, Ειδικευόμενος Παρασκευή Οστεομυελικής βιοψίας (ΟΜΒ) Επαρκής διαγνωστική ΟΜΒ πρέπει να είναι τουλάχιστον 1.5εκ, με απουσία crush artifact
Παθολογοανατομική προσέγγιση διάγνωση Σχολιασμός Δημ. Σαμπαζιώτης, Ειδικευόμενος Παρασκευή Οστεομυελικής βιοψίας (ΟΜΒ) Επαρκής διαγνωστική ΟΜΒ πρέπει να είναι τουλάχιστον 1.5εκ, με απουσία crush artifact
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογία ΙI Κυτταρική Επικοινωνία Διδάσκοντες: Σ. Γεωργάτος, Θ. Τζαβάρας, Π. Κούκλης, Χ. Αγγελίδης Υπεύθυνος μαθήματος: Σ. Γεωργάτος Άδειες Χρήσης Το
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΧΗΜΕΙΑΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΧΗΜΕΙΑΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΛΙΝΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΚΑΙ ΜΕΛΕΤΗ ΙΣΟΜΟΡΦΩΝ ΤΗΣ L-DOPA ΑΠΟΚΑΡΒΟΞΥΛΑΣΗΣ ΚΑΙ ΤΟΥ
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΧΗΜΕΙΑΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΛΙΝΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΚΑΙ ΜΕΛΕΤΗ ΙΣΟΜΟΡΦΩΝ ΤΗΣ L-DOPA ΑΠΟΚΑΡΒΟΞΥΛΑΣΗΣ ΚΑΙ ΤΟΥ
Iστοπαθολογία νευροενδοκρινικών όγκων πνεύμονος. Δ Ροντογιάννη
 Iστοπαθολογία νευροενδοκρινικών όγκων πνεύμονος Δ Ροντογιάννη Νευρώνες ΚΝΣ C Κύτταρα Θυρεοειδούς Κύτταρα παραθυρεοειδών αδένων Μεμονωμένα κύτταρα βρογχικού βλεννογόνου και βλεννογόνουγεσ ΔΙΑΧΥΤΟ ΝΕΥΡΟΕΝΔΟΚΡΙΝΙΚΟ
Iστοπαθολογία νευροενδοκρινικών όγκων πνεύμονος Δ Ροντογιάννη Νευρώνες ΚΝΣ C Κύτταρα Θυρεοειδούς Κύτταρα παραθυρεοειδών αδένων Μεμονωμένα κύτταρα βρογχικού βλεννογόνου και βλεννογόνουγεσ ΔΙΑΧΥΤΟ ΝΕΥΡΟΕΝΔΟΚΡΙΝΙΚΟ
Η Κυτταρογενετική στις αιματολογικές κακοήθειες
 Εργαστήριο Υγειοφυσικής & Περιβαλλοντικής Υγείας, ΙΠΤ-Α, Ε.Κ.Ε.Φ.Ε. «Δημόκριτος» Η Κυτταρογενετική στις αιματολογικές κακοήθειες Μανωλά Καλλιόπη, Ph.D Ερευνήτρια Γ Κυτταρογενετική Κλάδος της Γενετικής
Εργαστήριο Υγειοφυσικής & Περιβαλλοντικής Υγείας, ΙΠΤ-Α, Ε.Κ.Ε.Φ.Ε. «Δημόκριτος» Η Κυτταρογενετική στις αιματολογικές κακοήθειες Μανωλά Καλλιόπη, Ph.D Ερευνήτρια Γ Κυτταρογενετική Κλάδος της Γενετικής
Μελέτη έκφρασης των αντιγόνων των οικογενειών Survivin και MAGE-A στον καρκίνο του πνεύμονα
 Μονάδα Ανοσολογίας Καρκίνου Εργαστήριο Ανοσολογίας-Ιστοσυμβατότητας Πανεπιστημιακό Νοσοκομείο Λάρισας Τμήμα Ιατρικής Πανεπιστήμιο Θεσσαλίας Μελέτη έκφρασης των αντιγόνων των οικογενειών Survivin και MAGE-A
Μονάδα Ανοσολογίας Καρκίνου Εργαστήριο Ανοσολογίας-Ιστοσυμβατότητας Πανεπιστημιακό Νοσοκομείο Λάρισας Τμήμα Ιατρικής Πανεπιστήμιο Θεσσαλίας Μελέτη έκφρασης των αντιγόνων των οικογενειών Survivin και MAGE-A
Συχνότητα. Άντρες Γυναίκες 5 1. Νεαρής και μέσης ηλικίας
 Η αιτιολογία της πάθησης είναι άγνωστη, αν και έχει μεγάλη σχέση με το κάπνισμα καθώς το 90% των ασθενών είναι ενεργείς καπνιστές Συχνότητα Άντρες Γυναίκες 5 1 Νεαρής και μέσης ηλικίας Στο 60% των περιπτώσεων
Η αιτιολογία της πάθησης είναι άγνωστη, αν και έχει μεγάλη σχέση με το κάπνισμα καθώς το 90% των ασθενών είναι ενεργείς καπνιστές Συχνότητα Άντρες Γυναίκες 5 1 Νεαρής και μέσης ηλικίας Στο 60% των περιπτώσεων
ΛΕΜΦΙΚΟΣ ΙΣΤΟΣ - ΛΕΜΦΑΔΕΝΙΤΙΔΕΣ. Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ.
 ΛΕΜΦΙΚΟΣ ΙΣΤΟΣ - ΛΕΜΦΑΔΕΝΙΤΙΔΕΣ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΛΕΜΦΑΔΕΝΕΣ ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά
ΛΕΜΦΙΚΟΣ ΙΣΤΟΣ - ΛΕΜΦΑΔΕΝΙΤΙΔΕΣ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΛΕΜΦΑΔΕΝΕΣ ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά κέντρα ΛΕΜΦΑΔΕΝΕΣ Βλαστικά
Όγκοι των Όρχεων. Α. Κιζιρίδου, Αναπ.. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.Θεαγένειο
 Όγκοι των Όρχεων Α. Κιζιρίδου, Αναπ.. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.Θεαγένειο ΙΣΤΟΛΟΓΙΚΗ ΤΑΞΙΝΟΜΗΣΗ ΤΩΝ ΟΓΚΩΝ ΤΟΥ ΟΡΧΕΩΣ ΑΠΟ ΓΕΝΝΗΤΙΚΑ ΚΥΤΤΑΡΑ (WHO 2004) Όγκοι ενός ιστολογικού τύπου (αμιγείς
Όγκοι των Όρχεων Α. Κιζιρίδου, Αναπ.. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.Θεαγένειο ΙΣΤΟΛΟΓΙΚΗ ΤΑΞΙΝΟΜΗΣΗ ΤΩΝ ΟΓΚΩΝ ΤΟΥ ΟΡΧΕΩΣ ΑΠΟ ΓΕΝΝΗΤΙΚΑ ΚΥΤΤΑΡΑ (WHO 2004) Όγκοι ενός ιστολογικού τύπου (αμιγείς
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ. ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ
 ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
ΗPV και ενδοεπιθηλιακές αλλοιώσεις κατωτέρου γεννητικού συστήματος. Ε.Κωστοπούλου Λέκτορας Παθολογικής Ανατομικής Πανεπιστήμιο Θεσσαλίας
 ΗPV και ενδοεπιθηλιακές αλλοιώσεις κατωτέρου γεννητικού συστήματος Ε.Κωστοπούλου Λέκτορας Παθολογικής Ανατομικής Πανεπιστήμιο Θεσσαλίας H.M. Παράγοντες κινδύνου καρκινώματος τραχήλου Ηλικία κατά την πρώτη
ΗPV και ενδοεπιθηλιακές αλλοιώσεις κατωτέρου γεννητικού συστήματος Ε.Κωστοπούλου Λέκτορας Παθολογικής Ανατομικής Πανεπιστήμιο Θεσσαλίας H.M. Παράγοντες κινδύνου καρκινώματος τραχήλου Ηλικία κατά την πρώτη
Γονίδια και καρκινογένεση
 Γονίδια και καρκινογένεση Σύνολο 40.000 γονίδια Υπό έλεγχο Φυσιολογική ανάπτυξη Εκτός ελέγχου Καρκινογένεση Αποτελεί μια μη ισοτοπική τεχνική μοριακού υβριδισμού Επιτρέπει τη διατήρηση της ιστικής, κυτταρικής
Γονίδια και καρκινογένεση Σύνολο 40.000 γονίδια Υπό έλεγχο Φυσιολογική ανάπτυξη Εκτός ελέγχου Καρκινογένεση Αποτελεί μια μη ισοτοπική τεχνική μοριακού υβριδισμού Επιτρέπει τη διατήρηση της ιστικής, κυτταρικής
ΜΕΛΕΤΗ ΚΑΙ ΚΛΙΝΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΤΟΥ ΝΕΟΥ ΑΠΟΠΤΩΤΙΚΟΥ ΓΟΝΙΔΙΟΥ BCL2L12 ΚΑΙ ΤΗΣ L-DOPA ΑΠΟΚΑΡΒΟΞΥΛΑΣΗΣ ΣΤΟΝ ΚΑΡΚΙΝΟ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΧΗΜΕΙΑΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΜΕΛΕΤΗ ΚΑΙ ΚΛΙΝΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΤΟΥ ΝΕΟΥ ΑΠΟΠΤΩΤΙΚΟΥ ΓΟΝΙΔΙΟΥ BCL2L12 ΚΑΙ ΤΗΣ
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΣΧΟΛΗ ΘΕΤΙΚΩΝ ΕΠΙΣΤΗΜΩΝ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ ΤΟΜΕΑΣ ΒΙΟΧΗΜΕΙΑΣ ΚΑΙ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΜΕΛΕΤΗ ΚΑΙ ΚΛΙΝΙΚΗ ΑΞΙΟΛΟΓΗΣΗ ΤΟΥ ΝΕΟΥ ΑΠΟΠΤΩΤΙΚΟΥ ΓΟΝΙΔΙΟΥ BCL2L12 ΚΑΙ ΤΗΣ
Αρχές μοριακής παθολογίας. Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ
 Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
ΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΝΕΥΜΟΝΑ
 ΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΝΕΥΜΟΝΑ Χρ.Τσοµπανίδου Παθολογοανατόµος Συντονίστρια Διευθύντρια Νοσοκοµείο Θεσ/νίκης «Άγιος Δηµήτριος» προέρχονται από το φυσιολογικό επιθήλιο µε την
ΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΙΣΤΟΛΟΓΙΚΟΙ ΤΥΠΟΙ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΠΝΕΥΜΟΝΑ Χρ.Τσοµπανίδου Παθολογοανατόµος Συντονίστρια Διευθύντρια Νοσοκοµείο Θεσ/νίκης «Άγιος Δηµήτριος» προέρχονται από το φυσιολογικό επιθήλιο µε την
Εφαρμογές τεχνολογιών Μοριακής Βιολογίας στην Γενετική
 Εφαρμογές τεχνολογιών Μοριακής Βιολογίας στην Γενετική Πεφάνη Δάφνη 06.03.2019 Επίκουρη καθηγήτρια Εργαστήριο Βιολογίας Τεχνικές μοριακής βιολογίας Επιτρέπουν την μελέτη της δομής του DNA και της έκφρασης
Εφαρμογές τεχνολογιών Μοριακής Βιολογίας στην Γενετική Πεφάνη Δάφνη 06.03.2019 Επίκουρη καθηγήτρια Εργαστήριο Βιολογίας Τεχνικές μοριακής βιολογίας Επιτρέπουν την μελέτη της δομής του DNA και της έκφρασης
Συχνότητα και παράγοντες κινδύνου
 Συχνότητα και παράγοντες κινδύνου Είδος µεταµοσχευµένου οργάνου (σε συµπαγές όργανο 1,4-2 %, καρδιά 1,8-9,8 %, καρδιά µε πνεύµονα 4,6-12,5 % και πάγκρεας µε άλλο όργανο µέχρι 32% ) Ηλικία του λήπτη (συχνότερα
Συχνότητα και παράγοντες κινδύνου Είδος µεταµοσχευµένου οργάνου (σε συµπαγές όργανο 1,4-2 %, καρδιά 1,8-9,8 %, καρδιά µε πνεύµονα 4,6-12,5 % και πάγκρεας µε άλλο όργανο µέχρι 32% ) Ηλικία του λήπτη (συχνότερα
Ο ρόλος της ΕΘΟ. στην αναγέννηση. & την επανόρθωση
 Ο ρόλος της ΕΘΟ στην αναγέννηση & την επανόρθωση Νοvo E & Parola M. Fibrogenesis & Tissue Repair 2008, 1:5 Χρόνια παγκρεατίτιδα Ιστολογία παγκρεατικού καρκινώµατος Αδενοκαρκίνωµα εξ εκφορητικών
Ο ρόλος της ΕΘΟ στην αναγέννηση & την επανόρθωση Νοvo E & Parola M. Fibrogenesis & Tissue Repair 2008, 1:5 Χρόνια παγκρεατίτιδα Ιστολογία παγκρεατικού καρκινώµατος Αδενοκαρκίνωµα εξ εκφορητικών
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος
 Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
Έλεγχος κυτταρικού κύκλου-απόπτωση Πεφάνη Δάφνη Επίκουρη καθηγήτρια, Ιατρική σχολή ΕΚΠΑ Μιχαλακοπούλου 176, 1 ος όροφος Κυτταρικός κύκλος Φάσεις του κυτταρικού κύκλου G1:Αύξηση του κυττάρου και προετοιμασία
ΗPV και Καρκίνος Δέρµατος. Ηλέκτρα Νικολαΐδου Επ. Καθηγήτρια Δερµατολογίας ΕΚΠΑ Νοσ. «Α. Συγγρός»
 ΗPV και Καρκίνος Δέρµατος Ηλέκτρα Νικολαΐδου Επ. Καθηγήτρια Δερµατολογίας ΕΚΠΑ Νοσ. «Α. Συγγρός» ΓΕΝΗ HPV Α γένος: βλεννογόνοι αιτιολογική συσχέτιση µε καρκίνο τραχήλου µήτρας, πρωκτού, αιδοίου, πέους
ΗPV και Καρκίνος Δέρµατος Ηλέκτρα Νικολαΐδου Επ. Καθηγήτρια Δερµατολογίας ΕΚΠΑ Νοσ. «Α. Συγγρός» ΓΕΝΗ HPV Α γένος: βλεννογόνοι αιτιολογική συσχέτιση µε καρκίνο τραχήλου µήτρας, πρωκτού, αιδοίου, πέους
Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης. Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010
 ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
ΠΕΡΙΛΗΨΗ Υποψήφιος διδάκτορας: Καββαδάς Παναγιώτης Έτος ολοκλήρωσης διδακτορικής διατριβής: 2010 Μελέτη τοπ ρόλοπ της ιντεγκρινοσπνδεόμενης κινάσης στην πνεπμονική ίνσση, Διδακτορική Διατριβή, Πανεπιστήμιο
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ. Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη
 Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
Γυμνάσιο Κερατέας ΚΑΡΚΙΝΟΣ & ΜΕΤΑΛΛΑΞΕΙΣ Αναστασία Σουλαχάκη Κωνσταντίνα Πρίφτη 2013 ΠΕΡΙΕΧΟΜΕΝΑ : Ορολογία και λίγα λόγια για τον καρκίνο Χαρακτηριστικά του καρκίνου Μεταλλάξεις Μεταλλάξεις και καρκίνος
ΛΕΜΦΩΜΑΤΑ ΟΡΙΑΚΗΣ ΖΩΝΗΣ
 ΛΕΜΦΩΜΑΤΑ ΟΡΙΑΚΗΣ ΖΩΝΗΣ Πηνελόπη Κορκολοπούλου Αν. Καθηγήτρια Παθολογικής Ανατοµικής Παν/µίου Αθηνών 1 Β-ΛΕΜΦΩΜΑ ΑΠO ΤΟ ΚΥΤΤΑΡΟ ΤΗΣ ΟΡΙΑΚΗΣ ΖΩΝΗΣ (MZL) (WHO 2001) Tρεις τύποι λεµφωµάτων τα οποία αποτελούν
ΛΕΜΦΩΜΑΤΑ ΟΡΙΑΚΗΣ ΖΩΝΗΣ Πηνελόπη Κορκολοπούλου Αν. Καθηγήτρια Παθολογικής Ανατοµικής Παν/µίου Αθηνών 1 Β-ΛΕΜΦΩΜΑ ΑΠO ΤΟ ΚΥΤΤΑΡΟ ΤΗΣ ΟΡΙΑΚΗΣ ΖΩΝΗΣ (MZL) (WHO 2001) Tρεις τύποι λεµφωµάτων τα οποία αποτελούν
IΣTOΛOΓIA. Tα δείγµατα του βιολογικού υλικού λαµβάνονται µε > βελόνες ενδοσκοπικούς σωλήνες εύκαµπτους καθετήρες
 IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
IΣTOΛOΓIA H ιστολογία κλάδος της ιατρικής που µελετά > υφή βιολογικού υλικού και τους τρόπους που τα επιµέρους συστατικά στοιχεία σχετίζονται µεταξύ τους δοµικά & λειτουργικά Tα δείγµατα του βιολογικού
ΑΙΜΟΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΑ
 ΑΙΜΟΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΑ Η συµβολή της µοριακής ανάλυσης Eλισάβετ Οικονοµάκη Βιολόγος Αιµοπαθολογοανατοµικό Εργαστήριο ΠΓΝΑ > ΜΟΡΙΑΚΕΣ ΜΕΘΟ ΟΙ (Μη µορφολογικές) Αλυσιδωτή Αντίδραση Πολυµεράσης
ΑΙΜΟΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΑ Η συµβολή της µοριακής ανάλυσης Eλισάβετ Οικονοµάκη Βιολόγος Αιµοπαθολογοανατοµικό Εργαστήριο ΠΓΝΑ > ΜΟΡΙΑΚΕΣ ΜΕΘΟ ΟΙ (Μη µορφολογικές) Αλυσιδωτή Αντίδραση Πολυµεράσης
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΝΕΟΠΛΑΣΜΑΤΑ ΚΕΝΤΡΙΚΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Εργαστήριο Γενικής Παθολογίας και Παθολογικικής Ανατομικής
 ΝΕΟΠΛΑΣΜΑΤΑ ΚΕΝΤΡΙΚΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Εργαστήριο Γενικής Παθολογίας και Παθολογικικής Ανατομικής Φυσιολογική εγκεφαλική ουσία Νεοπλάσματα ΚΝΣ ανάλογα με την προέλευσή τους σε πρωτοπαθή (2/3) μεταστατικά
ΝΕΟΠΛΑΣΜΑΤΑ ΚΕΝΤΡΙΚΟΥ ΝΕΥΡΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Εργαστήριο Γενικής Παθολογίας και Παθολογικικής Ανατομικής Φυσιολογική εγκεφαλική ουσία Νεοπλάσματα ΚΝΣ ανάλογα με την προέλευσή τους σε πρωτοπαθή (2/3) μεταστατικά
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY
 ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Γενικά χαρακτηριστικά Απεικόνιση των κυτταρικών πληθυσμών με βάση: το μέγεθος την κοκκίωση το φθορισμό ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Υδροδυναμική ροή Πρόσθιος
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Γενικά χαρακτηριστικά Απεικόνιση των κυτταρικών πληθυσμών με βάση: το μέγεθος την κοκκίωση το φθορισμό ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Υδροδυναμική ροή Πρόσθιος
DEPARTMENT OF IMMUNOLOGY & HISTOCOMPATIBILITY UNIVERSITY OF THESSALY
 Μαμάρα Α. (1), Καλαλά Φ. (1), Λιαδάκη Κ. (1), Παπαδούλης Ν. (2), Μανδαλά Ε. (3), Παπαδάκης Ε. (4), Γιαννακούλας N. (5), Παλασοπούλου M. (5), Λαφιωνιάτης Σ. (6), Κιουμή A. (4), Κοραντζής I. (4), Καρτάσης
Μαμάρα Α. (1), Καλαλά Φ. (1), Λιαδάκη Κ. (1), Παπαδούλης Ν. (2), Μανδαλά Ε. (3), Παπαδάκης Ε. (4), Γιαννακούλας N. (5), Παλασοπούλου M. (5), Λαφιωνιάτης Σ. (6), Κιουμή A. (4), Κοραντζής I. (4), Καρτάσης
Kυτταρική Bιολογία. Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος ΔIAΛEΞΗ 20 (9/5/2017) Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.
 Δρ. Xρήστος Παναγιωτίδης, Τμήμα Φαρμακευτικής Α.Π.Θ.") Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
Kυτταρική Bιολογία ΔIAΛEΞΗ 20 (9/5/2017) Απόπτωση, ή Προγραμματισμένος Κυτταρικός Θάνατος Τι είναι απόπτωση; Απόπτωση είναι ο προγραμματισμένος κυτταρικός θάνατος Η καταστροφή του κυττάρου γίνεται «ήπια»
ΘΕΡΑΠΕΥΤΙΚΗ ΑΝΤΙΜΕΤΩΠΙΣΗ ΟΓΚΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ. Β. ΠΑΝΑΓΙΩΤΟΠΟΥΛΟΣ Λέκτορας Νευροχειρουργικής Π.Γ.Ν.Π.
 ΘΕΡΑΠΕΥΤΙΚΗ ΑΝΤΙΜΕΤΩΠΙΣΗ ΟΓΚΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ Β. ΠΑΝΑΓΙΩΤΟΠΟΥΛΟΣ Λέκτορας Νευροχειρουργικής Π.Γ.Ν.Π. ΓΛΟΙΩΜΑΤΑ ΕΓΚΕΦΑΛΟΥ ΓΛΟΙΩΜΑΤΑ ΕΓΚΕΦΑΛΟΥ ΠΙΛΟΚΥΤΤΑΡΙΚΟ ΑΣΤΡΟΚΥΤΩΜΑ (WHO I) ΑΣΤΡΟΚΥΤΩΜΑ (WHO II) ΑΝΑΠΛΑΣΤΙΚΟ
ΘΕΡΑΠΕΥΤΙΚΗ ΑΝΤΙΜΕΤΩΠΙΣΗ ΟΓΚΩΝ ΤΟΥ ΕΓΚΕΦΑΛΟΥ Β. ΠΑΝΑΓΙΩΤΟΠΟΥΛΟΣ Λέκτορας Νευροχειρουργικής Π.Γ.Ν.Π. ΓΛΟΙΩΜΑΤΑ ΕΓΚΕΦΑΛΟΥ ΓΛΟΙΩΜΑΤΑ ΕΓΚΕΦΑΛΟΥ ΠΙΛΟΚΥΤΤΑΡΙΚΟ ΑΣΤΡΟΚΥΤΩΜΑ (WHO I) ΑΣΤΡΟΚΥΤΩΜΑ (WHO II) ΑΝΑΠΛΑΣΤΙΚΟ
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY
 ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY Κυτταρομετρία Ροής Η Κυτταρομετρία Ροής είναι μια τεχνική που εκμεταλλεύεται την μέτρηση ορισμένων βιοχημικών και βιοφυσικών παραμέτρων των κυττάρων μεμονωμένα, όταν αυτά
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY Κυτταρομετρία Ροής Η Κυτταρομετρία Ροής είναι μια τεχνική που εκμεταλλεύεται την μέτρηση ορισμένων βιοχημικών και βιοφυσικών παραμέτρων των κυττάρων μεμονωμένα, όταν αυτά
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΕΠΗΡΜΕΝΟ ΔΕΡΜΑΤΟΪΝΟΣΑΡΚΩΜΑ AΓΓΕΛΙΚΗ ΜΠΕΦΟΝ ΕΠΙΜΕΛΗΤΡΙΑ Α ΔΕΡΜΑΤΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ ΕΣΥ ΝΟΣΟΚΟΜΕΙΟ ΑΝΔΡΕΑΣ ΣΥΓΓΡΟΣ
 ΕΠΗΡΜΕΝΟ ΔΕΡΜΑΤΟΪΝΟΣΑΡΚΩΜΑ AΓΓΕΛΙΚΗ ΜΠΕΦΟΝ ΕΠΙΜΕΛΗΤΡΙΑ Α ΔΕΡΜΑΤΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ ΕΣΥ ΝΟΣΟΚΟΜΕΙΟ ΑΝΔΡΕΑΣ ΣΥΓΓΡΟΣ ΕΠΗΡΜΕΝΟ ΔΕΡΜΑΤΟΪΝΟΣΑΡΚΩΜΑ DERMATOFIBROSARCOMA PROTUBERANS DFSP 1924: «προοδευτικό και υποτροπιάζον
ΕΠΗΡΜΕΝΟ ΔΕΡΜΑΤΟΪΝΟΣΑΡΚΩΜΑ AΓΓΕΛΙΚΗ ΜΠΕΦΟΝ ΕΠΙΜΕΛΗΤΡΙΑ Α ΔΕΡΜΑΤΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ ΕΣΥ ΝΟΣΟΚΟΜΕΙΟ ΑΝΔΡΕΑΣ ΣΥΓΓΡΟΣ ΕΠΗΡΜΕΝΟ ΔΕΡΜΑΤΟΪΝΟΣΑΡΚΩΜΑ DERMATOFIBROSARCOMA PROTUBERANS DFSP 1924: «προοδευτικό και υποτροπιάζον
Όρχεις -Χειρισμός παρασκευάσματος -Εισαγωγή στους όγκους. Α.. Κιζιρίδου, Αναπ. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.
 Όρχεις -Χειρισμός παρασκευάσματος -Εισαγωγή στους όγκους Α.. Κιζιρίδου, Αναπ. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.Θεαγένειο Περιγραφική Ανατομική 2 αδένες με σχήμα δαμάσκηνου διαστ.5χ3χ2.5 Κρέμονται
Όρχεις -Χειρισμός παρασκευάσματος -Εισαγωγή στους όγκους Α.. Κιζιρίδου, Αναπ. Διευθύντρια Παθολογοανατομικού Τμήματος A.Ν.Θ.Θεαγένειο Περιγραφική Ανατομική 2 αδένες με σχήμα δαμάσκηνου διαστ.5χ3χ2.5 Κρέμονται
 94 95 96 97 98 Ο ρόλος της αγγειογένεσης στη μετάσταση Η νεοαγγείωση αποτελεί ένα απαραίτητο τμήμα της ογκογόνου διεργασίας που διασφαλίζει τη γρηγορότερη και ανεμπόδιστη ανάπτυξη του όγκου. Η λειτουργική
94 95 96 97 98 Ο ρόλος της αγγειογένεσης στη μετάσταση Η νεοαγγείωση αποτελεί ένα απαραίτητο τμήμα της ογκογόνου διεργασίας που διασφαλίζει τη γρηγορότερη και ανεμπόδιστη ανάπτυξη του όγκου. Η λειτουργική
Αρχές μοριακής παθολογίας. Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ
 Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
Ενότητα 2011: Λεμφώματα
 Κλινικοεργαστηριακή Εκπαίδευση στην Αιματολογία Ενότητα 2011: Λεμφώματα 23-24 Σεπτεμβρίου 2011 Ηράκλειο, Κρήτη Διοργάνωση Ίδρυμα Ελληνικής Αιματολογικής Εταιρείας και Αιματολογική Κλινική Ιατρικής Σχολής
Κλινικοεργαστηριακή Εκπαίδευση στην Αιματολογία Ενότητα 2011: Λεμφώματα 23-24 Σεπτεμβρίου 2011 Ηράκλειο, Κρήτη Διοργάνωση Ίδρυμα Ελληνικής Αιματολογικής Εταιρείας και Αιματολογική Κλινική Ιατρικής Σχολής
igenetics Mια Μεντελική προσέγγιση
 igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 22 (+κεφ. 17 Hartwell) Γενετική του καρκίνου Η πρωτεΐνη p53 προσδένεται στο DNA. 2 ΕΙΚΟΝΑ 22.1 Μαστογραφία που απεικονίζει έναν όγκο. Όγκος 3 Κύρια σημεία: Καρκίνος
igenetics Mια Μεντελική προσέγγιση Κεφάλαιο 22 (+κεφ. 17 Hartwell) Γενετική του καρκίνου Η πρωτεΐνη p53 προσδένεται στο DNA. 2 ΕΙΚΟΝΑ 22.1 Μαστογραφία που απεικονίζει έναν όγκο. Όγκος 3 Κύρια σημεία: Καρκίνος
Παθολογοανατομικό εργαστήριο Κυριακόπουλος Γεώργιος Εδ Ειδικευόμενος ιατρός
 Παθολογοανατομικό εργαστήριο Κυριακόπουλος Γεώργιος Εδ Ειδικευόμενος ιατρός ΒΙΟΨΙΑ ΝΕΦΡΟΥ Σπειράματα λειτουργικά, με φυσιολογική αρχιτεκτονική και κυτταροβρίθεια ΒΙΟΨΙΑ ΝΕΦΡΟΥ Εικόνα Διάμεσης και Σωληναριακής
Παθολογοανατομικό εργαστήριο Κυριακόπουλος Γεώργιος Εδ Ειδικευόμενος ιατρός ΒΙΟΨΙΑ ΝΕΦΡΟΥ Σπειράματα λειτουργικά, με φυσιολογική αρχιτεκτονική και κυτταροβρίθεια ΒΙΟΨΙΑ ΝΕΦΡΟΥ Εικόνα Διάμεσης και Σωληναριακής
ΝΟΣΟΣ MAREK (MAREK s DISEASE)
") econteplusproject Organic.Edunet ΝΟΣΟΣ MAREK (MAREK s DISEASE) Δρ. Ευτυχία Ξυλούρη Φραγκιαδάκη Κτηνίατρος Υγιεινολόγος, Αναπλ. Καθηγήτρια Υγιεινής Αγρ. Ζώων, Τμήμα Επιστήμης Ζωικής Παραγωγής και Υδατοκαλλιεργειών,
econteplusproject Organic.Edunet ΝΟΣΟΣ MAREK (MAREK s DISEASE) Δρ. Ευτυχία Ξυλούρη Φραγκιαδάκη Κτηνίατρος Υγιεινολόγος, Αναπλ. Καθηγήτρια Υγιεινής Αγρ. Ζώων, Τμήμα Επιστήμης Ζωικής Παραγωγής και Υδατοκαλλιεργειών,
Λόγοι έκδοσης γνώμης για τον χαρακτηρισμό φαρμακευτικού προϊόντος ως ορφανού
 Παράρτημα 1 Λόγοι έκδοσης γνώμης για τον χαρακτηρισμό φαρμακευτικού προϊόντος ως ορφανού Η Επιτροπή Ορφανών Φαρμάκων (COMP), έχοντας εξετάσει την αίτηση, κατέληξε στα ακόλουθα: Σύμφωνα με το άρθρο 3 παράγραφος
Παράρτημα 1 Λόγοι έκδοσης γνώμης για τον χαρακτηρισμό φαρμακευτικού προϊόντος ως ορφανού Η Επιτροπή Ορφανών Φαρμάκων (COMP), έχοντας εξετάσει την αίτηση, κατέληξε στα ακόλουθα: Σύμφωνα με το άρθρο 3 παράγραφος
Πέτρος Καρακίτσος. Καθηγητής Εργαστήριο Διαγνωστικής Κυτταρολογίας Ιατρικής Σχολής Παμεπιστημίου Αθηνών Πανεπιστημιακό Γενικό Νοσοκομείο «ΑΤΤΙΚΟΝ»
 Πέτρος Καρακίτσος Καθηγητής Εργαστήριο Διαγνωστικής Κυτταρολογίας Ιατρικής Σχολής Παμεπιστημίου Αθηνών Πανεπιστημιακό Γενικό Νοσοκομείο «ΑΤΤΙΚΟΝ» Αναπαραγωγιμότητα Αντικειμενοποίηση Εφαρμογή τεχνικών:
Πέτρος Καρακίτσος Καθηγητής Εργαστήριο Διαγνωστικής Κυτταρολογίας Ιατρικής Σχολής Παμεπιστημίου Αθηνών Πανεπιστημιακό Γενικό Νοσοκομείο «ΑΤΤΙΚΟΝ» Αναπαραγωγιμότητα Αντικειμενοποίηση Εφαρμογή τεχνικών:
ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ. Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ.
 ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Λεμφαδενικά Εξωλεμφαδενικά Λέμφωμα Hodgkin Μη Hodgkin λεμφώματα Β-κυτταρικής
ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Ιωάννης Κωστόπουλος Καθηγητής Εργαστήριο Γεν. Παθολογίας και Παθολογικής Ανατομικής Α.Π.Θ. ΚΑΚΟΗΘΗ ΛΕΜΦΩΜΑΤΑ Λεμφαδενικά Εξωλεμφαδενικά Λέμφωμα Hodgkin Μη Hodgkin λεμφώματα Β-κυτταρικής
ΣΧΗΜΑΤΙΣΜΟΣ ΕΠΑΓΩΜΕΝΩΝ ΠΟΛΥΔΥΝΑΜΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΙΚΟΣ ΕΠΑΝΑΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ
 ΣΧΗΜΑΤΙΣΜΟΣ ΕΠΑΓΩΜΕΝΩΝ ΠΟΛΥΔΥΝΑΜΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΙΚΟΣ ΕΠΑΝΑΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ Υπάρχουν Βλαστικά κύτταρα με διαφορετικές ιδιότητες: Τα Πολυδύναμα - Pluripotent Εμβρυονικά Βλαστικά κύτταρα - Embryonic Stem
ΣΧΗΜΑΤΙΣΜΟΣ ΕΠΑΓΩΜΕΝΩΝ ΠΟΛΥΔΥΝΑΜΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΙΚΟΣ ΕΠΑΝΑΠΡΟΓΡΑΜΜΑΤΙΣΜΟΣ Υπάρχουν Βλαστικά κύτταρα με διαφορετικές ιδιότητες: Τα Πολυδύναμα - Pluripotent Εμβρυονικά Βλαστικά κύτταρα - Embryonic Stem
Μοριακή Βιολογία. Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής
: Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής") ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Μοριακή Βιολογία Ενότητα # (6): Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος Παναγιωτίδης Χρήστος Άδειες Χρήσης Το παρόν εκπαιδευτικό
Μεσεγχυματογενή Νεοπλάσματα (μικροσκοπική εξέταση) Γιώργος Αγρογιάννης Αναπλ. Καθηγητής
 Γιώργος Αγρογιάννης Αναπλ. Καθηγητής") 1 ο ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ Διευθυντής: Καθηγητής Α. Λάζαρης 4 η Εργαστηριακή Άσκηση Μεσεγχυματογενή Νεοπλάσματα (μικροσκοπική εξέταση) Γιώργος Αγρογιάννης Αναπλ.
1 ο ΕΡΓΑΣΤΗΡΙΟ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ Διευθυντής: Καθηγητής Α. Λάζαρης 4 η Εργαστηριακή Άσκηση Μεσεγχυματογενή Νεοπλάσματα (μικροσκοπική εξέταση) Γιώργος Αγρογιάννης Αναπλ.
ΛΕΜΦΩΜΑ ΗΟDGKIN. Αλεξάνδρα Κουράκλη-Συμεωνίδου Απαρτιωμένη διδασκαλία 2015
 ΛΕΜΦΩΜΑ ΗΟDGKIN Αλεξάνδρα Κουράκλη-Συμεωνίδου Απαρτιωμένη διδασκαλία 2015 ΛΕΜΦΩΜΑ ΗΟDGKIN Επιδημιολογία - Αιτιοπαθογένεια Δικόρυφη καμπύλη εμφάνισης κορυφές: 15-40 έτη, >60 έτη : = Μικρή υπεροχή (1.5-2.0/1)
ΛΕΜΦΩΜΑ ΗΟDGKIN Αλεξάνδρα Κουράκλη-Συμεωνίδου Απαρτιωμένη διδασκαλία 2015 ΛΕΜΦΩΜΑ ΗΟDGKIN Επιδημιολογία - Αιτιοπαθογένεια Δικόρυφη καμπύλη εμφάνισης κορυφές: 15-40 έτη, >60 έτη : = Μικρή υπεροχή (1.5-2.0/1)
ΝΕΟΠΛΑΣΜΑΤΑ ΔΕΡΜΑΤΟΣ. Ι. Γ. Παναγιωτίδης Β Εργαστήριο Παθολογικής Ανατομικής Ιατρική Σχολή Εθνικού & Καποδιστριακού Παν/μίου Αθηνών
 ΝΕΟΠΛΑΣΜΑΤΑ ΔΕΡΜΑΤΟΣ Ι. Γ. Παναγιωτίδης Β Εργαστήριο Παθολογικής Ανατομικής Ιατρική Σχολή Εθνικού & Καποδιστριακού Παν/μίου Αθηνών ΕΙΔΗ ΑΚΤΙΝΟΒΟΛΙΑΣ ΒΛΑΒΕΣ DNA ΑΠΟ ΥΠΕΡΙΩΔΗ ΑΚΤΙΝΟΒΟΛΙΑ ΥΠΕΡΙΩΔΗΣ ΑΚΤΙΝΟΒΟΛΙΑ
ΝΕΟΠΛΑΣΜΑΤΑ ΔΕΡΜΑΤΟΣ Ι. Γ. Παναγιωτίδης Β Εργαστήριο Παθολογικής Ανατομικής Ιατρική Σχολή Εθνικού & Καποδιστριακού Παν/μίου Αθηνών ΕΙΔΗ ΑΚΤΙΝΟΒΟΛΙΑΣ ΒΛΑΒΕΣ DNA ΑΠΟ ΥΠΕΡΙΩΔΗ ΑΚΤΙΝΟΒΟΛΙΑ ΥΠΕΡΙΩΔΗΣ ΑΚΤΙΝΟΒΟΛΙΑ
Είδη κυτταρικών βλαβών ή εκφυλίσεων
 Είδη κυτταρικών βλαβών ή εκφυλίσεων Αναστρέψιμες κυτταρικές βλάβες κυρίως στο κυτταρόπλασμα Α. Θολερή εξοίδηση Β. Υδρωπική εκφύλιση Γ. Λιπώδης εκφύλιση Θολερή εξοίδηση ή κυτταρική εξοίδηση Η πρώτη μορφολογική
Είδη κυτταρικών βλαβών ή εκφυλίσεων Αναστρέψιμες κυτταρικές βλάβες κυρίως στο κυτταρόπλασμα Α. Θολερή εξοίδηση Β. Υδρωπική εκφύλιση Γ. Λιπώδης εκφύλιση Θολερή εξοίδηση ή κυτταρική εξοίδηση Η πρώτη μορφολογική
ΠΝΕΥΜΟΝΙΚΗ ΑΓΓΕΙΪΤΙΣ ΚΑΙ ΚΟΚΚΙΩΜΑΤΩΣΗ
 ΠΝΕΥΜΟΝΙΚΗ ΑΓΓΕΙΪΤΙΣ ΚΑΙ ΚΟΚΚΙΩΜΑΤΩΣΗ Πνευμονική αγγειίτις και κοκκιωμάτωση είναι ένας περιγραφικός όρος που χαρακτηρίζεται από κυτταρική διήθηση του τοιχώματος των αγγείων (αγγειίτις) με καταστροφή και
ΠΝΕΥΜΟΝΙΚΗ ΑΓΓΕΙΪΤΙΣ ΚΑΙ ΚΟΚΚΙΩΜΑΤΩΣΗ Πνευμονική αγγειίτις και κοκκιωμάτωση είναι ένας περιγραφικός όρος που χαρακτηρίζεται από κυτταρική διήθηση του τοιχώματος των αγγείων (αγγειίτις) με καταστροφή και
Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος
 MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞΕΙΣ 10-12 Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής
MOPIAKH BIOΛOΓIA ΦAPMAKEYTIKHΣ ΔIAΛEΞΕΙΣ 10-12 Oδοί και μηχανισμοί ευκαρυωτικής μεταγωγής σήματος (Πως γίνονται αντιληπτά τα μηνύματα και πως δίδονται οι απαντήσεις) Δρ. Xρήστος Παναγιωτίδης, Tµήµα Φαρµακευτικής
Κυτταρική Βιολογία. Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος. Παναγιωτίδης Χρήστος Τμήμα Φαρμακευτικής ΑΠΘ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΙΚΑ ΜΑΘΗΜΑΤΑ Κυτταρική Βιολογία Ενότητα 12 : Απόπτωση ή Προγραμματισμένος κυτταρικός θάνατος Παναγιωτίδης Χρήστος ΑΠΘ Άδειες Χρήσης Το παρόν εκπαιδευτικό
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ
 ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
ΜΟΝΟΠΑΤΙΑ ΕΝΔΟΚΥΤΤΑΡΙΚΗΣ ΜΕΤΑΓΩΓΗΣ ΣΗΜΑΤΟΣ Το ένζυμο Αδενυλική κυκλάση, υπεύθυνο για τη βιοσύνθεση του camp. Το camp είναι ένα παράδειγμα μορίου «αγγελιοφόρου» καθοδικά των G πρωτεινών Αύξηση του camp
οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά
 οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά Ανοσολογικό σύστηµα Βασικό σύστηµα του οργανισµού Λειτουργικές µονάδες του ανοσολογικού συστήµατος Οργανωµένος λεµφικός
οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά Ανοσολογικό σύστηµα Βασικό σύστηµα του οργανισµού Λειτουργικές µονάδες του ανοσολογικού συστήµατος Οργανωµένος λεµφικός
Μεταβολικές ανάγκες ανοσοκυττάρων
 Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
ΙΕΡΕΥΝΗΣΗ ΤΟΥ ΑΞΟΝΑ ΤΕΛΟΜΕΡΑΣΗΣ/ΤΕΛΟΜΕΡΩΝ ΣΕ ΜΕΣΕΓΧΥΜΑΤΙΚΑ ΚΥΤΤΑΡΑ ΜΥΕΛΟΥ ΤΩΝ ΟΣΤΩΝ ΑΣΘΕΝΩΝ ΜΕ ΡΕΥΜΑΤΟΕΙ Η ΠΝΕΥΜΟΝΑ
 ΙΕΡΕΥΝΗΣΗ ΤΟΥ ΑΞΟΝΑ ΤΕΛΟΜΕΡΑΣΗΣ/ΤΕΛΟΜΕΡΩΝ ΣΕ ΜΕΣΕΓΧΥΜΑΤΙΚΑ ΚΥΤΤΑΡΑ ΜΥΕΛΟΥ ΤΩΝ ΟΣΤΩΝ ΑΣΘΕΝΩΝ ΜΕ ΡΕΥΜΑΤΟΕΙ Η ΠΝΕΥΜΟΝΑ ΓΙΩΡΓΟΣ Α. ΜΑΡΓΑΡΙΤΟΠΟΥΛΟΣ 1,2, ΑΘΑΝΑΣΙΑ ΠΡΟΚΛΟΥ 1,2, ΚΩΣΤΑΣ ΚΑΡΑΓΙΑΝΝΗΣ 2, ΙΣΜΗΝΙ ΛΑΣΗΘΙΩΤΑΚΗ
ΙΕΡΕΥΝΗΣΗ ΤΟΥ ΑΞΟΝΑ ΤΕΛΟΜΕΡΑΣΗΣ/ΤΕΛΟΜΕΡΩΝ ΣΕ ΜΕΣΕΓΧΥΜΑΤΙΚΑ ΚΥΤΤΑΡΑ ΜΥΕΛΟΥ ΤΩΝ ΟΣΤΩΝ ΑΣΘΕΝΩΝ ΜΕ ΡΕΥΜΑΤΟΕΙ Η ΠΝΕΥΜΟΝΑ ΓΙΩΡΓΟΣ Α. ΜΑΡΓΑΡΙΤΟΠΟΥΛΟΣ 1,2, ΑΘΑΝΑΣΙΑ ΠΡΟΚΛΟΥ 1,2, ΚΩΣΤΑΣ ΚΑΡΑΓΙΑΝΝΗΣ 2, ΙΣΜΗΝΙ ΛΑΣΗΘΙΩΤΑΚΗ
Καραπέτσας Θανάσης. Διπλωματική Εργασία:
 Διαπανεπιστημιακό Πρόγραμμα Μεταπτυχιακών Σπουδών «Κλινική Φαρμακολογία & Θεραπευτική» Επιβλέπων: Δρ. Αλ. Γαλάνης, Λέκτορας Μορ. Βιολογίας, «Σχεδιασμός Ειδικών Πεπτιδίων Αναστολέων της Αλληλεπίδρασης του
Διαπανεπιστημιακό Πρόγραμμα Μεταπτυχιακών Σπουδών «Κλινική Φαρμακολογία & Θεραπευτική» Επιβλέπων: Δρ. Αλ. Γαλάνης, Λέκτορας Μορ. Βιολογίας, «Σχεδιασμός Ειδικών Πεπτιδίων Αναστολέων της Αλληλεπίδρασης του
Λέµφωµα Hodgkin. Βλαχάκη Ευθυµία Επίκουρη Καθηγήτρια Αιµατολογίας-Αιµοσφαιρινοπαθειών ΑΠΘ
 Λέµφωµα Hodgkin Βλαχάκη Ευθυµία Επίκουρη Καθηγήτρια Αιµατολογίας-Αιµοσφαιρινοπαθειών ΑΠΘ Λεµφώµατα Κακοήθη νεοπλάσµατα του λεµφικού ιστού. Προσβάλλουν κυρίως τους λεµφαδένες, το σπλήνα και το µυελό των
Λέµφωµα Hodgkin Βλαχάκη Ευθυµία Επίκουρη Καθηγήτρια Αιµατολογίας-Αιµοσφαιρινοπαθειών ΑΠΘ Λεµφώµατα Κακοήθη νεοπλάσµατα του λεµφικού ιστού. Προσβάλλουν κυρίως τους λεµφαδένες, το σπλήνα και το µυελό των
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ (PATHOLOGY) Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ.
 Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ.") ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ (PATHOLOGY) Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ. Παθολογική Ανατομική εξέταση των δομικών αλλοιώσεων
ΕΙΣΑΓΩΓΗ ΣΤΗΝ ΠΑΘΟΛΟΓΙΚΗ ΑΝΑΤΟΜΙΚΗ (PATHOLOGY) Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Tμήματος Ιατρικής Α.Π.Θ. Παθολογική Ανατομική εξέταση των δομικών αλλοιώσεων
Νικόλαος Σιαφάκας Λέκτορας Διαγνωστικής Ιολογίας Εργαστήριο Κλινικής Μικροβιολογίας ΠΓΝ «ΑΤΤΙΚΟΝ»
 Νικόλαος Σιαφάκας Λέκτορας Διαγνωστικής Ιολογίας Εργαστήριο Κλινικής Μικροβιολογίας ΠΓΝ «ΑΤΤΙΚΟΝ» DNA RNA: ΑΝΤΙΓΡΑΦΗ, ΜΕΤΑΓΡΑΦΗ, ΜΕΤΑΦΡΑΣΗ DNA RNA: Βασικά Χαρακτηριστικά Ρόλος Κεντικό Δόγμα της Βιολογίας:
Νικόλαος Σιαφάκας Λέκτορας Διαγνωστικής Ιολογίας Εργαστήριο Κλινικής Μικροβιολογίας ΠΓΝ «ΑΤΤΙΚΟΝ» DNA RNA: ΑΝΤΙΓΡΑΦΗ, ΜΕΤΑΓΡΑΦΗ, ΜΕΤΑΦΡΑΣΗ DNA RNA: Βασικά Χαρακτηριστικά Ρόλος Κεντικό Δόγμα της Βιολογίας:
ΟΓΚΟΛΟΓΙΑ - ΡΑΔΙΟΒΙΟΛΟΓΙΑ
 ΟΓΚΟΛΟΓΙΑ - ΡΑΔΙΟΒΙΟΛΟΓΙΑ Βασικές γνώσεις I SBN 960-372-069-0 ΠΕΡΙΕΧΟΜΕΝΑ ΠΑΡΙΣ Α. ΚΟΣΜΙΔΗΣ ΓΕΩΡΓΙΟΣ ΤΣΑΚΙΡΗΣ Μ Ε Ρ Ο Σ Ι ΟΓΚΟΛΟΓΙΑ Κεφάλαιο 1 ΕΙΣΑΓΩΓΗ ΣΤΟΝ ΚΑΡΚΙΝΟ... 3 Το καρκινικό κύτταρο... 3 Κυτταρικός
ΟΓΚΟΛΟΓΙΑ - ΡΑΔΙΟΒΙΟΛΟΓΙΑ Βασικές γνώσεις I SBN 960-372-069-0 ΠΕΡΙΕΧΟΜΕΝΑ ΠΑΡΙΣ Α. ΚΟΣΜΙΔΗΣ ΓΕΩΡΓΙΟΣ ΤΣΑΚΙΡΗΣ Μ Ε Ρ Ο Σ Ι ΟΓΚΟΛΟΓΙΑ Κεφάλαιο 1 ΕΙΣΑΓΩΓΗ ΣΤΟΝ ΚΑΡΚΙΝΟ... 3 Το καρκινικό κύτταρο... 3 Κυτταρικός
Ανι χνευση μεταλλα ξεων στον καρκι νο και εξατομικευμε νη θεραπει α
 Ανι χνευση μεταλλα ξεων στον καρκι νο και εξατομικευμε νη θεραπει α Περικλής Μακρυθανα σης, MD, PhD, FMH, PD Κλινικός γενετιστής Κύριος Ερευνητής Ίδρυμα Ιατροβιολογικών Ερευνών της Ακαδημι ας Αθηνών 4o
Ανι χνευση μεταλλα ξεων στον καρκι νο και εξατομικευμε νη θεραπει α Περικλής Μακρυθανα σης, MD, PhD, FMH, PD Κλινικός γενετιστής Κύριος Ερευνητής Ίδρυμα Ιατροβιολογικών Ερευνών της Ακαδημι ας Αθηνών 4o
Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 14/10/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Γενικά στοιχεία
Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 14/10/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Γενικά στοιχεία
Aιμοφαγοκυτταρικό Σύνδρομο ή Αιμοφαγοκυτταρική Λεμφοϊστιοκυττάρωση HLH
 Aιμοφαγοκυτταρικό Σύνδρομο ή Αιμοφαγοκυτταρική Λεμφοϊστιοκυττάρωση HLH Aιμοφαγοκυτταρικό Σύνδρομο ή Αιμοφαγοκυτταρική Λεμφοϊστιοκυττάρωση HLH Υπερφλεγμονώδες σύνδρομο Aιμοφαγοκυτταρικό Σύνδρομο ή Αιμοφαγοκυτταρική
Aιμοφαγοκυτταρικό Σύνδρομο ή Αιμοφαγοκυτταρική Λεμφοϊστιοκυττάρωση HLH Aιμοφαγοκυτταρικό Σύνδρομο ή Αιμοφαγοκυτταρική Λεμφοϊστιοκυττάρωση HLH Υπερφλεγμονώδες σύνδρομο Aιμοφαγοκυτταρικό Σύνδρομο ή Αιμοφαγοκυτταρική
φυσιολογικό δέρμα - 1
 φυσιολογικό δέρμα -1 Επιδερμίδα (επιθήλιο, εξωδερμική προέλευση) Α Α Α Μ θηλώδες χόριο (επιπολής) ακανθωτή στιβάδα βασική στιβάδα χόριο Μ = Μελανοκύτταρο (νευροεξωδερμική προέλευση, νευρική ακρολοφία)
φυσιολογικό δέρμα -1 Επιδερμίδα (επιθήλιο, εξωδερμική προέλευση) Α Α Α Μ θηλώδες χόριο (επιπολής) ακανθωτή στιβάδα βασική στιβάδα χόριο Μ = Μελανοκύτταρο (νευροεξωδερμική προέλευση, νευρική ακρολοφία)
ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. :
 ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. : 2010336 ΕΡΓΑΣΙΑ ΓΙΑ ΤΟ Μ.Π.Σ. ΜΕΤΑΒΟΛΙΚΑ ΝΟΣΗΜΑΤΑ ΟΣΤΩΝ ΚΥΤΟΚΙΝΕΣ ΠΑΡΑΓΟΜΕΝΕΣ ΑΠΟ ΤΑ Τ-ΚΥΤΤΑΡΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΠΟΥ ΕΜΠΛΕΚΟΝΤΑΙ ΣΤΗΝ ΟΣΤΕΟΚΛΑΣΤΟΓΕΝΕΣΗ
ΚΕΧΑΓΙΑΣ ΒΑΣΙΛΕΙΟΣ ΑΘΗΝΑ, 16/11/10 A.M. : 2010336 ΕΡΓΑΣΙΑ ΓΙΑ ΤΟ Μ.Π.Σ. ΜΕΤΑΒΟΛΙΚΑ ΝΟΣΗΜΑΤΑ ΟΣΤΩΝ ΚΥΤΟΚΙΝΕΣ ΠΑΡΑΓΟΜΕΝΕΣ ΑΠΟ ΤΑ Τ-ΚΥΤΤΑΡΑ ΤΟΥ ΑΝΟΣΟΠΟΙΗΤΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ ΠΟΥ ΕΜΠΛΕΚΟΝΤΑΙ ΣΤΗΝ ΟΣΤΕΟΚΛΑΣΤΟΓΕΝΕΣΗ
Μοριακή βάση του καρκίνου της ουροδόχου κύστεως
 Πανεπιστήμιο Αθηνών Τμήμα Βιολογίας Τομέας Βιοχημείας & Μοριακής Βιολογίας Μοριακή βάση του καρκίνου της ουροδόχου κύστεως Μαργαρίτης Αυγέρης, Ανδρέας Σκορίλας Τομέας Βιοχημείας και Μοριακής Βιολογίας,
Πανεπιστήμιο Αθηνών Τμήμα Βιολογίας Τομέας Βιοχημείας & Μοριακής Βιολογίας Μοριακή βάση του καρκίνου της ουροδόχου κύστεως Μαργαρίτης Αυγέρης, Ανδρέας Σκορίλας Τομέας Βιοχημείας και Μοριακής Βιολογίας,
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ
 ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ: «ΝΕΟΠΛΑΣΜΑΤΙΚΗ ΝΟΣΟΣ ΣΤΟΝ ΑΝΘΡΩΠΟ: ΣΥΓΧΡΟΝΗ ΚΛΙΝΙΚΟΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΗ ΠΡΟΣΕΓΓΙΣΗ ΚΑΙ
ΕΘΝΙΚΟ ΚΑΙ ΚΑΠΟΔΙΣΤΡΙΑΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΑΘΗΝΩΝ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΜΗΜΑ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ ΜΕΤΑΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΟΥΔΩΝ: «ΝΕΟΠΛΑΣΜΑΤΙΚΗ ΝΟΣΟΣ ΣΤΟΝ ΑΝΘΡΩΠΟ: ΣΥΓΧΡΟΝΗ ΚΛΙΝΙΚΟΠΑΘΟΛΟΓΟΑΝΑΤΟΜΙΚΗ ΠΡΟΣΕΓΓΙΣΗ ΚΑΙ
ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΕΤΟΣ ΚΑΡΚΙΝΟΣ ΚΑΙ ΜΕΤΑΛΛΑΞΕΙΣ
 ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΕΤΟΣ 2012-2013 ΚΑΡΚΙΝΟΣ ΚΑΙ ΜΕΤΑΛΛΑΞΕΙΣ Τι είναι ο καρκίνος ; Ο Καρκίνος είναι ένα από τα σοβαρότερα προβλήματα υγείας που παρατηρούνται σήμερα στις αναπτυγμένες χώρες. Οι στατιστικές
ΓΥΜΝΑΣΙΟ ΚΕΡΑΤΕΑΣ ΕΤΟΣ 2012-2013 ΚΑΡΚΙΝΟΣ ΚΑΙ ΜΕΤΑΛΛΑΞΕΙΣ Τι είναι ο καρκίνος ; Ο Καρκίνος είναι ένα από τα σοβαρότερα προβλήματα υγείας που παρατηρούνται σήμερα στις αναπτυγμένες χώρες. Οι στατιστικές
Πανεπιστήμιο Θεσσαλίας Τμήμα Ιατρικής Εργαστήριο Ακτινολογίας Ιατρικής Απεικόνισης
 Πανεπιστήμιο Θεσσαλίας Τμήμα Ιατρικής Εργαστήριο Ακτινολογίας Ιατρικής Απεικόνισης Διδάσκοντες Ιωάννης Β. Φεζουλίδης Καθηγητής Μαριάννα Βλυχού Αναπλ. Καθηγήτρια Έφη Καψαλάκη Αναπλ. Καθηγήτρια Αικατερίνη
Πανεπιστήμιο Θεσσαλίας Τμήμα Ιατρικής Εργαστήριο Ακτινολογίας Ιατρικής Απεικόνισης Διδάσκοντες Ιωάννης Β. Φεζουλίδης Καθηγητής Μαριάννα Βλυχού Αναπλ. Καθηγήτρια Έφη Καψαλάκη Αναπλ. Καθηγήτρια Αικατερίνη
Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα
 Β λεμφοκύτταρα") Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα φυσική ή μη ειδική ανοσία δεν απαιτεί προηγούμενη έκθεση στο παθογόνο και δεν διαθέτει μνήμη. σε επίκτητη ή ειδική ανοσία χυμική ανοσία με παραγωγή
Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα φυσική ή μη ειδική ανοσία δεν απαιτεί προηγούμενη έκθεση στο παθογόνο και δεν διαθέτει μνήμη. σε επίκτητη ή ειδική ανοσία χυμική ανοσία με παραγωγή
Δεκαπεντάλεπτη προετοιμασία του φοιτητή ιατρικής για το μάθημα του καρκίνου του όρχη βασικές γνώσεις :
 Δεκαπεντάλεπτη προετοιμασία του φοιτητή ιατρικής για το μάθημα του καρκίνου του όρχη βασικές γνώσεις : Οι όρχεις αποτελούν κομμάτι του αναπαραγωγικού συστήματος (παραγωγή σπερματοζωάριων) του άνδρα αλλά
Δεκαπεντάλεπτη προετοιμασία του φοιτητή ιατρικής για το μάθημα του καρκίνου του όρχη βασικές γνώσεις : Οι όρχεις αποτελούν κομμάτι του αναπαραγωγικού συστήματος (παραγωγή σπερματοζωάριων) του άνδρα αλλά
VIN (Ενδοεπιθηλιακή νεοπλασία αιδοίου)
") VIN (Ενδοεπιθηλιακή νεοπλασία αιδοίου) Διαφορές μεταξύ τραχήλου και αιδοίου Κύτταρο-στόχος: κυρίως πλακώδες δεν υπάρχει εκτεθειμένος πολυδύναμος πληθυσμός, όπως στον τράχηλο Σχετιζόμενοι HPV: κυριαρχούν
VIN (Ενδοεπιθηλιακή νεοπλασία αιδοίου) Διαφορές μεταξύ τραχήλου και αιδοίου Κύτταρο-στόχος: κυρίως πλακώδες δεν υπάρχει εκτεθειμένος πολυδύναμος πληθυσμός, όπως στον τράχηλο Σχετιζόμενοι HPV: κυριαρχούν
Στεργίου Ιωάννης Ά ΠΡΟΠΑΙΔΕΥΤΙΚΗ ΠΑΘΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ. Ά ΕΡΓΑΣΤΗΡΙΟ ΜΙΚΡΟΒΙΟΛΟΓΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΑΠΘ Πέμπτη 12 Νοεμβρίου ο Συνέδριο ΔΕΒΕ
 ΜΕΛΕΤΗ ΕΠΙΠΕΔΩΝ stnf arii ΚΛΙΝΙΚΗ Στεργίου Ιωάννης Ά ΠΡΟΠΑΙΔΕΥΤΙΚΗ ΠΑΘΟΛΟΓΙΚΗ Ά ΕΡΓΑΣΤΗΡΙΟ ΜΙΚΡΟΒΙΟΛΟΓΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΑΠΘ Πέμπτη 12 Νοεμβρίου 2009 23 ο Συνέδριο ΔΕΒΕ Σκοπός της μελέτης Έλεγχος της πιθανής
ΜΕΛΕΤΗ ΕΠΙΠΕΔΩΝ stnf arii ΚΛΙΝΙΚΗ Στεργίου Ιωάννης Ά ΠΡΟΠΑΙΔΕΥΤΙΚΗ ΠΑΘΟΛΟΓΙΚΗ Ά ΕΡΓΑΣΤΗΡΙΟ ΜΙΚΡΟΒΙΟΛΟΓΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΑΠΘ Πέμπτη 12 Νοεμβρίου 2009 23 ο Συνέδριο ΔΕΒΕ Σκοπός της μελέτης Έλεγχος της πιθανής
Η έκφραση του μεταγραφικού παράγοντα GATA 4 σε ισχαιμικές αλλοιώσεις του μυοκαρδίου σαν ισχυρός προγνωστικός παράγοντας.
 Η έκφραση του μεταγραφικού παράγοντα GATA 4 σε ισχαιμικές αλλοιώσεις του μυοκαρδίου σαν ισχυρός προγνωστικός παράγοντας. Α. Τσίπης 1, Α.Μ. Αθανασιάδου 1, Ε. Πέτρου 2, Δ. Μηλιόπουλος 2, Α. Γκουζιούτα 2,
Η έκφραση του μεταγραφικού παράγοντα GATA 4 σε ισχαιμικές αλλοιώσεις του μυοκαρδίου σαν ισχυρός προγνωστικός παράγοντας. Α. Τσίπης 1, Α.Μ. Αθανασιάδου 1, Ε. Πέτρου 2, Δ. Μηλιόπουλος 2, Α. Γκουζιούτα 2,
ΙΦΝΕ (ΕΚ, ν.crohn, απροσδιόριστη) Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες
 Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες") ΦΛΕΓΜΟΝΩΔΗ ΝΟΣΗΜΑΤΑ Π.Ε ΙΦΝΕ (ΕΚ, ν.crohn, απροσδιόριστη) Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες (ΜΣΑΦ κ.ά) Ισχαιμική Μετακτινική
ΦΛΕΓΜΟΝΩΔΗ ΝΟΣΗΜΑΤΑ Π.Ε ΙΦΝΕ (ΕΚ, ν.crohn, απροσδιόριστη) Συνήθεις λοιμώδεις, παρατεταμένες συστηματικές, αφροδισιακές-παρασιτικές, ιογενείς λοιμώξεις Φάρμακα και τοξίνες (ΜΣΑΦ κ.ά) Ισχαιμική Μετακτινική
ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΧΗΜΕΙΟΘΕΡΑΠΕΙΑΣ ΝΕΟΠΛΑΣΜΑΤΙΚΩΝ ΝΟΣΩΝ. Αργύρης Συμεωνίδης Καθηγητής Αιματολογίας Πανεπιστημίου Πατρών
 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΧΗΜΕΙΟΘΕΡΑΠΕΙΑΣ ΝΕΟΠΛΑΣΜΑΤΙΚΩΝ ΝΟΣΩΝ Αργύρης Συμεωνίδης Καθηγητής Αιματολογίας Πανεπιστημίου Πατρών Προσέγγιση του ασθενούς με νεοπλασματική νόσο Ι. Θέματα που αφορούν το είδος της νόσου
ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΧΗΜΕΙΟΘΕΡΑΠΕΙΑΣ ΝΕΟΠΛΑΣΜΑΤΙΚΩΝ ΝΟΣΩΝ Αργύρης Συμεωνίδης Καθηγητής Αιματολογίας Πανεπιστημίου Πατρών Προσέγγιση του ασθενούς με νεοπλασματική νόσο Ι. Θέματα που αφορούν το είδος της νόσου
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς. Οι φάσεις των Τ κυτταρικών απαντήσεων
 ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
6 o ΕΚΠΑΙΔΕΥΤΙΚΟ ΣΕΜΙΝΑΡΙΟ
 6 o ΕΚΠΑΙΔΕΥΤΙΚΟ ΣΕΜΙΝΑΡΙΟ ΜΟΡΙΑΚΗΣ ΟΓΚΟΛΟΓΙΑΣ & ΣΤΟΧΕΥΟΥΣΑΣ ΘΕΡΑΠΕΙΑΣ ΓΙΑ ΤΟΝ ΚΛΙΝΙΚΟ ΟΓΚΟΛΟΓΟ ΜΕΤΣΟΒΟ 10-12 Φεβρουαρίου, 2012 Συνεδριακό Κέντρο Διάσελο Υπό την αιγίδα της ΕΟΠΕ Υπό την αιγίδα του Ελληνικού
6 o ΕΚΠΑΙΔΕΥΤΙΚΟ ΣΕΜΙΝΑΡΙΟ ΜΟΡΙΑΚΗΣ ΟΓΚΟΛΟΓΙΑΣ & ΣΤΟΧΕΥΟΥΣΑΣ ΘΕΡΑΠΕΙΑΣ ΓΙΑ ΤΟΝ ΚΛΙΝΙΚΟ ΟΓΚΟΛΟΓΟ ΜΕΤΣΟΒΟ 10-12 Φεβρουαρίου, 2012 Συνεδριακό Κέντρο Διάσελο Υπό την αιγίδα της ΕΟΠΕ Υπό την αιγίδα του Ελληνικού
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο. 3. Το DNA των μιτοχονδρίων έχει μεγαλύτερο μήκος από αυτό των χλωροπλαστών.
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΘΕΜΑ 1 Ο Α. Να γράψετε τον αριθμό της καθεμιάς από τις παρακάτω προτάσεις 1-5 και δίπλα του τη λέξη Σωστό, αν η πρόταση είναι σωστή, ή Λάθος, αν η πρόταση
Η κυτταρομετρία ροής στα Β λεμφοϋπερπλαστικά νοσήματα. Ελένη Καψάλη Επίκουρη Καθηγήτρια Αιματολογίας
 Η κυτταρομετρία ροής στα Β λεμφοϋπερπλαστικά νοσήματα Ελένη Καψάλη Επίκουρη Καθηγήτρια Αιματολογίας Κατάταξη λεμφικών νεοπλασμάτων Τα νεοπλάσματα των Β, Τ, ΝΚ λεμφοκυττάρων είναι όγκοι από ανώριμα και
Η κυτταρομετρία ροής στα Β λεμφοϋπερπλαστικά νοσήματα Ελένη Καψάλη Επίκουρη Καθηγήτρια Αιματολογίας Κατάταξη λεμφικών νεοπλασμάτων Τα νεοπλάσματα των Β, Τ, ΝΚ λεμφοκυττάρων είναι όγκοι από ανώριμα και
ΙΪΚΟΙ ΦΟΡΕΙΣ & Γενετικά Τροποποιημένα Κύτταρα
 ΙΪΚΟΙ ΦΟΡΕΙΣ & Γενετικά Τροποποιημένα Κύτταρα ΓΕΝΙΚΑ ΠΕΡΙ ΙΩΝ 1. Οι ιοί είναι μικροοργανισμοί που προκαλούν ασθένειες σε ένα οργανισμό, μετά από μόλυνση των κυττάρων του 2. Είναι παρασιτικοί οργανισμοί
ΙΪΚΟΙ ΦΟΡΕΙΣ & Γενετικά Τροποποιημένα Κύτταρα ΓΕΝΙΚΑ ΠΕΡΙ ΙΩΝ 1. Οι ιοί είναι μικροοργανισμοί που προκαλούν ασθένειες σε ένα οργανισμό, μετά από μόλυνση των κυττάρων του 2. Είναι παρασιτικοί οργανισμοί
ΚΑΡΚΙΝΟΣ ΤΟΥ ΛΑΡΥΓΓΑ
 ΚΩΝΣΤΑΝΤΙΝΟΣ ΒΛΑΧΤΣΗΣ ΙΑΤΡΟΣ ΩΤΟΡΙΝΟΛΑΡΥΓΓΟΛΟΓΟΣ Ι ΑΚΤΟΡΑΣ ΑΡΙΣΤΟΤΕΛΕΙΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΘΕΣΣΑΛΟΝΙΚΗΣ ΚΑΡΚΙΝΟΣ ΤΟΥ ΛΑΡΥΓΓΑ ΟΓΚΟΓΕΝΕΣΗ ΚΑΙ ΣΥΓΧΡΟΝΟΙ ΜΟΡΙΑΚΟΙ ΠΡΟΓΝΩΣΤΙΚΟΙ ΕΙΚΤΕΣ ΘΕΣΣΑΛΟΝΙΚΗ 2006 3 Κωνσταντίνος
ΚΩΝΣΤΑΝΤΙΝΟΣ ΒΛΑΧΤΣΗΣ ΙΑΤΡΟΣ ΩΤΟΡΙΝΟΛΑΡΥΓΓΟΛΟΓΟΣ Ι ΑΚΤΟΡΑΣ ΑΡΙΣΤΟΤΕΛΕΙΟΥ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΘΕΣΣΑΛΟΝΙΚΗΣ ΚΑΡΚΙΝΟΣ ΤΟΥ ΛΑΡΥΓΓΑ ΟΓΚΟΓΕΝΕΣΗ ΚΑΙ ΣΥΓΧΡΟΝΟΙ ΜΟΡΙΑΚΟΙ ΠΡΟΓΝΩΣΤΙΚΟΙ ΕΙΚΤΕΣ ΘΕΣΣΑΛΟΝΙΚΗ 2006 3 Κωνσταντίνος