Feb Mai Cursul 1
|
|
|
- Κάλλιστος Βαμβακάς
- 7 χρόνια πριν
- Προβολές:
Transcript
1 Feb Mai 2014 Cursul 1
2 Fluxul informaţiei genetice genă (ADN) transcriere genetică transcript primar (ARN) procesarea transcriptului ARN mesager traducerea informatiei genetice peptid ARN ribozomal ARN de transfer ARN antisens Structura Materialului Genetic ADN = acid deoxiribonucleic ARN = acid ribonucleic ADN şi ARN = un fel de polimeri Unităţile de bază (monomerii) = nucleotide, legate între ele prin legături covalente - fosfodiesterice Deci, ADN şi ARN = structuri polinucleotidice
 un rest de radical fosforic Adenina şi guanina")

3 Nucleotid = datp deoxyadenosine triphosphate bază azotată pentoză (un zahar format din 5 atomi de carbon) un rest de radical fosforic Adenina şi guanina = purine dgtp deoxyguanosine triphosphate dctp deoxycytidine triphosphate Timina şi citozina = pirimidine dttp deoxythymidine triphosphate Thymine O CH 3
 Lanţuri polinucleotidice Catene")
4 În structura ADN nucleotidele sunt legate între ele prin legături fosfodiesterice (de tip covalent) Lanţuri polinucleotidice Catene polinucleotidice
5 Majoritatea moleculelor de ADN = 2 catene polinucleotidice = ADN dublu-catenar = ADN d.c. Întotdeauna : o A de pe o catenă se împerechează cu o T de pe cealaltă catenă şi invers (o T, cu o A) o G se împerechează cu o C, şi invers Împerecherea = formarea de legături de hidrogen 2 legături de hidrogen între A şi T A T 3 legături de hidrogen între G şi C G C Cele două catene sunt legate între ele prin legături de hidrogen Cele două catene sunt complementare una cu cealaltă. Într-o moleculă de ADN d.c. numărul de A = numărul de T numărul de G = numărul de C
6 Legile lui Chargaff 1. cele 2 catene polinucleotidice sunt complementare una faţă de cealaltă Aceasta înseamnă că unei adenine de pe una din catene îi corespunde o timină pe cealaltă catenă, de care se leagă prin 2 legături de hidrogen (A = T); se spune, deci, că adenina este complementară cu timina. În mod similar, guanina este complementară cu citozina, de care se leagă prin 3 legături de hidrogen (G C). Se deduce că legăturile de hidrogen permise sunt: A = T şi G C În moleculele de acizi nucleici dublucatenare dar formate dintr-o catenă ADN şi o catenă ARN (asemenea molecule hibrid se formează de obicei în procesul de transcriere genetică), adeninelor din catena ADN le corespund în catena ARN molecule de uracil. Şi în acest caz se formează tot două legături de hidrogen: A = U. 2. într-o moleculă de acid nucleic d.c. purinele sunt în raport echimolar cu pirimidinele Această regulă este, de fapt, o consecinţă a primei legi. Astfel, dacă oricărei A de pe una din catene îi corespunde o T pe cealaltă catenă şi oricărei G îi corespunde o C, atunci numărul moleculelor de adenină este egal cu numărul celor de timină, adică A = T şi, respectiv, G = C. Prin adunarea celor două ecuaţii, se deduce: A + G = T + C. Deci, numărul purinelor dintr-o moleculă de acid nucleic d.c. este egal cu numărul pirimidinelor. Altfel spus, cele două tipuri de molecule se află în raport echimolar. 3. cele două catene polinucleotidice dintr-o moleculă de acid nucleic d.c. sunt antiparalele O catenă polinucleotidică are două capete: la un capăt se află carbonul din poziţia 5 (C5 ) al unei pentoze, iar la celălalt capăt se află carbonul din poziţia 3 (C3 ) al unei alte pentoze. În interiorul unei celule o asemenea catenă polinucleotidică este sintetizată chiar în această direcţie: 5 3. Cele 2 catene polinucleotidice ale unei molecule d.c. sunt în orientări inverse una faţă de cealaltă: capul 5 al fiecăreia corespunde cu capul 3 al celeilalte. Cele 2 catene sunt antiparalele.


7 P - fosfat Albastru - zahar Baze: ATGC
 se află la exteriorul helixului.")
 Structura schematizată a unui ADN dublu-helix.")
8 Două reprezentări ale dublului helix ADN. (a) Dublu-helix ADN de formă B. Scheletul glucido-fosforic (în gri şi marcat cu linii roşii ii) se află la exteriorul helixului. Bazele azotate se află la interior. Sunt marcate curbura majoră şi cea minoră a helixului. (b) Structura schematizată a unui ADN dublu-helix. Cele două schelete glucido-fosforice (în verde închis şi deschis) sunt în orientare inversă: 5 3, faţă de 3-5. Bazele azotate de pe cele două catene sunt prezentate în albastru şi roşu.

9 Dublul helix
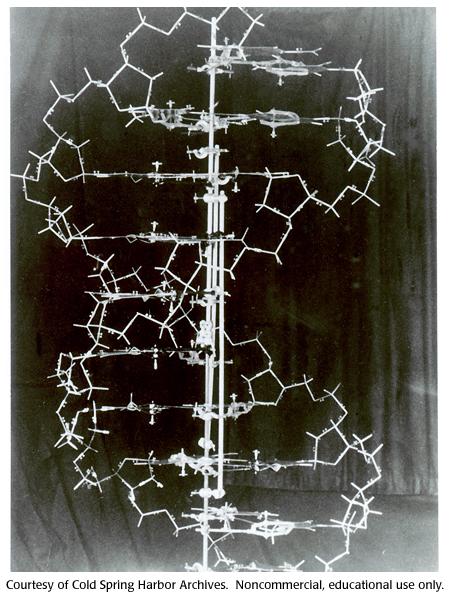
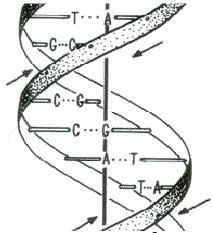
10
11 Parametri helicali n = numărul de nucleotide per tur de spiră (de elice) h = distanţa dintre perechile de nucleotide adiacente P = pasul elicei şi este distanţa traversată de-a lungul axei helixului de un tur complet de spiră (adică de o rotaţie de 360 o ). P poate fi exprimat în raport cu numărul de nucleotide dintr-un tur (n) şi cu distanţa dintre 2 nucleotide în procesul de rotaţie (h) t = unghiul de răsucire sau de rotaţie a perechile de baze azotate; acestea nu sunt perfect perpendiculare pe axa helixului (sau, altfel spus, două perechi de baze adiacente nu sunt coplanare), ci sunt înclinate cu un unghi de răsucire t care, de obicei, este 34,6 o. Aceaste este de fapt cauza pentru care cele două catene se răsucesc una în jurul celeilalte, formând o structură de dublu-helix. Valorile acestori parametri pot varia între anumite limite şi determină mai multe forme topologice ale moleculei de ADN d.c. Forma B reprezintă conformaţia cel mai des întâlnită în celule. Are un diametru de aproximativ 20 Angstrom (Å), iar distanţele dintre perechile de baze este de aproximativ 3,4 Å. În această conformaţie există o medie de 10,4 baze azotate per tur de spiră, iar unghiul de rotaţie dintre 2 baze adiacente este de +34,6º. Forma A se întâlneşte în celule în regiunile dublucatenare ale moleculelor de ARN şi în dublu-helixurile hibride ADN ARN ce se formează în procesele de transcriere genetică. Are un diametru de aproximativ 23 A, 11 baze per tur de spiră şi un unghi de +34,7º. Forma Z este mai subţire (un diametru de 18A), 12 baze per tur şi un unghi de -30º între ele. Datorită acestui lucru, formele A şi B sunt denumite de dreapta, iar forma Z este de stânga.
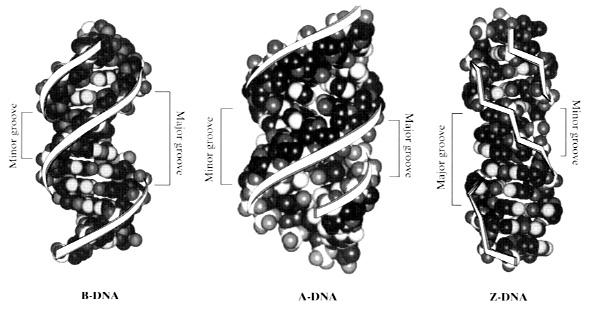
12


13 Denaturarea şi renaturarea moleculelor de ADN dublu-catenare

14 Tipuri de molecule ADN
 unele virusuri, de ex.")




15 Tipuri de molecule ADN ADN monocatenar (mc) unele virusuri, de ex. parvovirusuri ADN dublucatenar (dc) circular ADN ul unor virusuri Cromozom bacterian Plasmide bacteriene ADN mitocondrial ADN cloroplastic ADN dc circular de la virusul SV40 ADN dc circular din mitocondrii umane linear Unele virusuri, de ex. Adenovirusuri, virusuri herpes, varicella etc Cromozomi de la eucariote ADN dc linear uman in replicare
16 Replicarea ADN Toate moleculele de ADN, lineare sau circulare, se replică Dpdv chimic, replicarea = sinteza unei catene noi de ADN, folosind drept matriță o altă catenă ADN, pe bază de complementaritate (A T, G C) O moleculă ADN d.c. se desface (se desfac legăturile de hidrogen şi se separă cele 2 catene una de cealaltă) Fiecare din aceste 2 catene este folosită drept matriță: pe fiecare matriță se sintetizează o catenă nouă 1 moleculă ADN dc replicare 2 molecule ADN dc 1 moleculă m ADN dc Desfacerea legăturilor de H 2 molecule ADN mc Ataşarea area primerilor Prelungirea primerilor şi formarea catenelor noi catene parentale catene fiice
17 1 moleculă m ADN dc Desfacerea legăturilor de H 2 molecule ADN mc Ataşarea area primerilor Prelungirea primerilor şi formarea catenelor noi catene parentale catene fiice T G T T A A C T G G C A T A C A A T T G A C C G T A T G T T A A C T G G C A T A C A A T T G A C C G T A T G A C A T T A A C T G G C A T C A T T G T T A A C T G G C A T A C A A T T G A C C G T A T A A C T G G C A T C A T A C A A T T G A C C G T A A C A A T T G A C C G T A
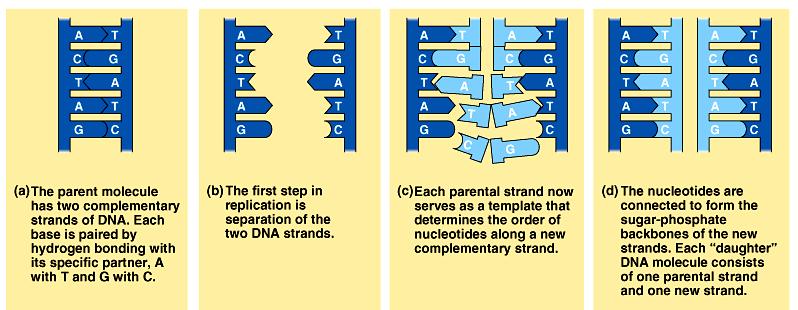
18
19
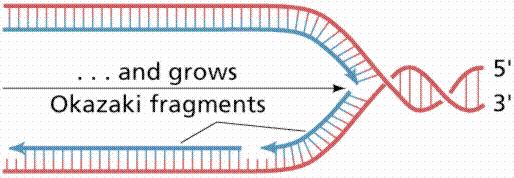
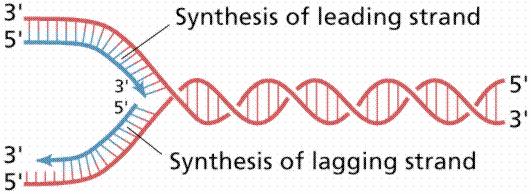
20 Replicarea ADN se desfăşoară la bifurcarea de replicare zonă unde desface dublul helix Ptr fiecare din cele 2 catene iniţiale (parentale), se sintetizează o catenă nouă, pe bază de complementaritate : - acolo unde pe catena parentală este o A, pe catena nouă vine o T, şi invers - acolo unde pe catena parentală este o G, pe catena nouă vine o C, şi invers Fiecare din catenele parenatale este folosită ca matriţă ptr sinteza unei catene noi Sinteza catenei noi se face in direcţie ie Ptr că în ADN d.c. cele 2 catene sunt în orientare inversă (una 5 3, iar cealaltă 3 5 ), sinteza celor 2 catene noi se face în direcţie opusă, dar întotdeauna în sens 5 3 La o bifurcaţie de replicare - una din catene este sintetizată continuu, de la un singur primer = catena conducătoare, leading - cealaltă catenă este sintetizată din bucăţi = fragmente Okazaki, fiecare pornind de la un primer catenă întârziată, lagging
21 1 moleculă m ADN dc P P bifurcatie de replicare Dublul helix se desface mai departe Bifurcatia de replicare se deplaseaza (spre stanga) P Atasarea unui nou primer Catena intarziata (lagging) Sinteza discontinua, de la mai multi primeri Catena conducatoare (leading) Sinteza continua, de la un singur primer Fragmente Okazaki
22 1 moleculă m ADN dc P P bifurcatie de replicare Dublul helix se desface mai departe Bifurcatia de replicare se deplaseaza (spre stanga) P Atasarea unui nou primer Catena intarziata (lagging) Sinteza discontinua, de la mai multi primeri Catena conducatoare (leading) Sinteza continua, de la un singur primer Fragmente Okazaki
23

24 Trei moduri posibile de replicare
25
26 De reținut 1 moleculă ADN dc replicare 2moleculeADNdc Fiecare catenă a moleculei parentale = matriță ptr sinteza unei catene noi Sinteza catenelor noi se face prin complementaritate cu catenele vechi: A T, G C Sinteza unei noi catene ADN începe intotdeauna de la un primer ADN polimeraza poate adăuga nucleotide pornind de la un cap 3 OH liber, dar nu poate iniția sinteza unui lanț polinucleotidic Primer = un fragment oligonucleotidic scurt, de obicei ARN O catenă nouă se sintetizează întotdeauna în direcție 5 3 Una din catenele noi este sintetizată continuu, pornind de la un singur primer Cealaltă catenă este sintetizată discontinuu, pornind de la mai mulți primeri, este formată din fragmente = fragmente Okazaki
27 Feb Mai 2014 Cursul 2
28 Enzimologia replicării ADN Pre inițiere Etape Regiunea unde începe replicarea moleculei de ADN = origine a replicării O moleculă ADN poate avea: 1 regiune ori Cromozom bacterian Plasmide bacteriene ori C ori V Pre inițiere Inițiere Elongare Terminare Cromozomul bacterian este un replicon unic
29
30 mai multe regiuni ori Cromozomi de la eucariote Cromozomii de tip eucariot sunt structuri multirepliconice

31 (după Watson, 2013, Molecular Biology of the Gene, 7th Edition)
 degete (fingers) palma / podul palmei (palm) conține situl activ, catalitic leagă 2 ioni metalici bivalenți 2 Mg 2+ sau 2 Zn")
32 Sinteza ADN este catalizată de o clasă de enzime = ADN polimeraze ADN polimerazele o mână care prinde joncțiunea/regiunea matriță primer Prin analogie, cele 3 domenii peptidice deget mare (thumb) degete (fingers) palma / podul palmei (palm) conține situl activ, catalitic leagă 2 ioni metalici bivalenți 2 Mg 2+ sau 2 Zn 2+

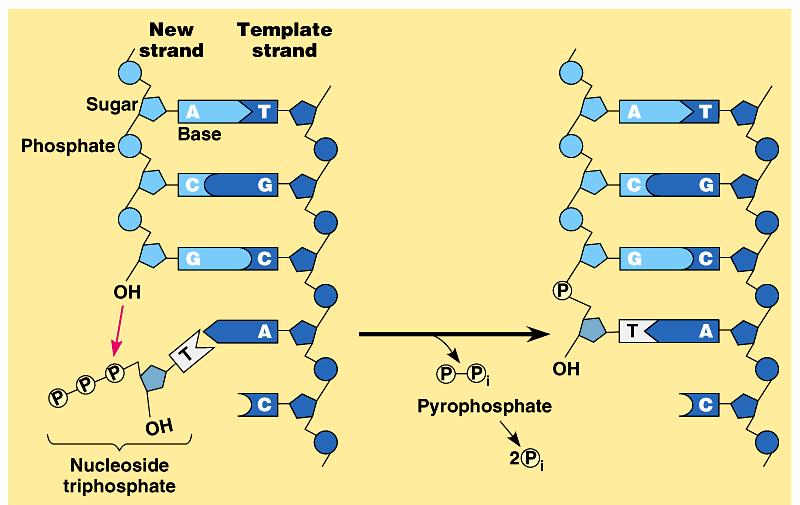
33 Ordinea evenimentelor în n sinteza ADN: Nucleotidul nou se împerechează cu următorul nucleotid liber de pe matriță Numai un nucleotid nou complementar cu cel liber de pe matriță ă se poate împerechea Domeniul palm asigură spațial complementaritatea nucleotidului nou şi nucleotidul liber de pe matriță Domeniul degete se strânge în n jurul joncțiunii catenă veche matriță catenă nouă Acum conformația enzimei poziționează ionează corect cei 2 ioni metalici Are loc reacția de formare a legăturii fosfodiesterice şi i ataşarea area nucleotidului nou la primer 1 Mg P primer primer 3 OH 5 P primer 3 O 5 P primer 3 O + dntp α P β P γ P atac nucleofilic al grupării α fosfat al dntp asupra grupării 3 O 5 P primer dntp 3 OH + β P γ P 1 Mg legătură fosfodiesterică Se redeschide domeniul degete Regiunea / joncțiunea matriță primer(+1 dntp) se deplasează cu 1 pereche de baze ADN polimeraza este gata ptr următorul ciclu,, ptr p adăugarea următorului nucleotid
34 1 Mg P primer primer 3 OH 5 P primer 3 O 5 P primer 3 O + dntp α P β P γ P atac nucleofilic al grupării α fosfat al dntp asupra grupării 3 O 5 P primer dntp 3 OH + β P γ P 1 Mg legătură fosfodiesterică
35 Ordinea evenimentelor în n sinteza ADN: Nucleotidul nou se împerechează cu următorul nucleotid liber de pe matriță Numai un nucleotid nou complementar cu cel liber de pe matriță ă se poate împerechea Domeniul degete se strânge în n jurul joncțiunii catenă veche matriță catenă nouă Acum conformația enzimei poziționează ionează corect cei 2 ioni metalici Are loc reacția de formare a legăturii fosfodiesterice şi i ataşarea area nucleotidului nou la primer Se redeschide domeniul degete Regiunea / joncțiunea matriță primer(+1 dntp) se deplasează cu 1 pereche de baze ADN polimeraza este gata ptr următorul ciclu,, ptr p adăugarea următorului nucleotid

36 La o bifurcație de replicare, ambele catene ADN sunt sintetizate împreună în n acelaşi i timp

37 Inițierea unei noi catene ADN necesită un primer Toate ADN polimerazele au nevoie de un cap 3 OH 3 liber, al unui nucleotid ADN polimerazele nu pot iniția ia sinteza de ADN nu pot ataşa a primul nucleotid Pot doar prelungi, prin p formarea unei prime legături fosfodiesterice pornind de la un cap 3 OH 3 liber Primer = fragment oligonucleotidic m.c. m ce oferă ADN polimerazei un cap 3 OH liber În n celule (in( vivo) ) primerii = fragmente ARN În anumite tehnici ( n anumite tehnici (in vitro) ) se pot utiliza şi i primeri ADN În n celule, enzima ce sintetizează primeri = PRIMAZĂ = o ARN polimerază specială Primerii = fragmente ARN m.c., de nucleotide Primaza acționează atât pe catena conducătoare,, cât şi i pe cea întârziată,, dar : Catena conducătoare necesită un singur primer,, de la el sinteza ADN mergând continuu Catena întârziată necesită mai mulți i primeri, de la fiecare sintetizându se se câte un fragment Okazaki

38 Primerii trebuie apoi îndepărtați Pentru terminarea replicării unei catene întârziate, primerii trebuie îndepărtați i din structura fragmentelor Okazaki, şi înlocuiți i cu ADN Îndepărtarea primerilor RNaza H Taie legăturile fosfodiesterice dintr o o catenă ARN, în n cadrul unui dublu helix hibrid ADN : ARN Umplerea golurilor ADN polimerază Formarea ultimei legături fosfodiesterice ADN ligază
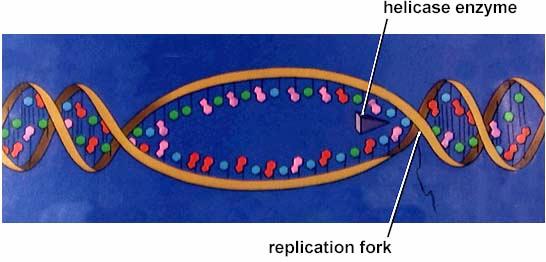
39 Separarea celor 2 catene parentale ADN helicaze La bifurcația ia de replicare, separarea celor 2 catene parentale prin ruperea legăturilor de H, este e realizată de ste realizată de ADN helicaze Proteine hexamerice cu formă de inel inelul intră pe una din cele două catene parentale şi i avansează pe ea, rupând legăturile de H dintre cele 2 catene Monocatenele ADN sunt stabilizate proteine Ssb Catenenele parentale desfăcute (denumite şi i monocatene) sunt stabilizate prin ataşare are de proteine Ssb Single stranded binding Monocatenele parentale sunt astfel disponibile ptr a fi utilizate e ca matriță în n sinteza de catene ADN noi
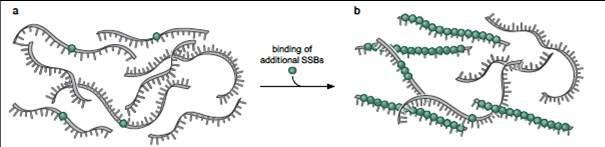
40
41 Suprarăsucirile sunt îndepărtate Topoizomeraze Desfacerea dublului helix parental şi i avansarea bifurcației iei de replicare produce suprarăsuciri în n fața a bifurcației. iei. Aceste suprarăsuciri sunt eliminate prin tăieri ale dublului helix, urmate u de derăsucire şi i re ligare Topoizomeraze Acționează pe ADN dc nereplicat, în n fața a bifurcației iei de replicare Taie una sau ambele catene ADN topoizomeraze de clasă I topoizomeraze de clasă II Rămân ataşate ate la capul tăiat şi îl l derăsucesc față ă de catenă netăiată Rămân ataşate ate la capetele tăiate şi i trec zona netăiată prin tăietură În n ambele cazuri, topoizomerazele refac legăturile pe care le au tăiat Scade gradul de răsucire a moleculei ADN parentale şi, astfel, helicaza poate desface mai departe legăturile de hidrogen dintre catenele parentale
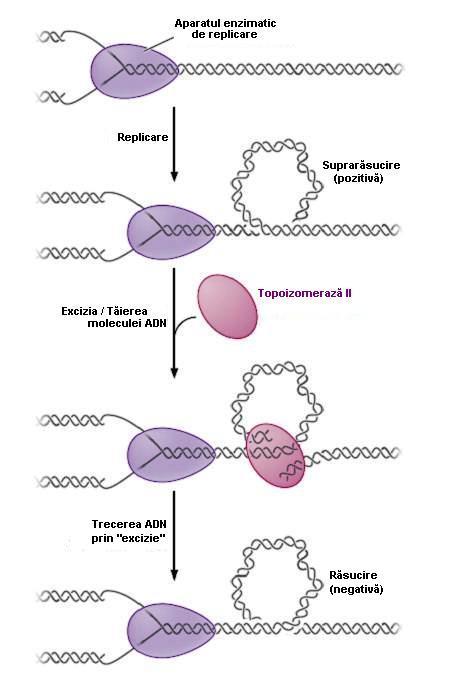
42

43 Specializarea ADN polimerazelor În n fiecare celulă există mai multe tipuri (specii moleculare) de ADN polimeraze, fiecare având un rol distinct inct Rolul central al ADN polimerazelor Replicarea corectă şi i eficientă a genomului Alte roluri Replicarea golurilor lăsate de excizarea primerilor ARN Replicarea golurilor formate în n procesele de reparare ADN Replicarea unor molecule ADN extracromozomale plasmide, ADN mitocondrial, ADN platidial Deşi i numele proteinelor este diferit de la o specie la alta, activitățile ile enzimatice desfăşurate pentru replicarea ADN cromozomal sunt practic aceleaşi i la organisme extrem de îndepărtate filogenetic bacterii, drojdii, om
44


45 Viteza ADN polimerazei este crescută de SLIDING CLAMPS Procesivitatea mare a ADN polimerazei la bifurcația ia de replicare ADN pol adaugă milioane de nucleotide fără să se desprindă de pe ADN Replicarea rapidă a cromozomului Viteza foarte mare a ADN polimerazei O proteină denumită sliding sliding clamp

46 Sliding clamps sunt poziționate de CLAMP LOADERS Proteinele sliding clamps sunt deschise şi i poziționate ionate pe molecula de ADN de către alte proteine clamp loaders Au 5 subunități şi i folosesc ATP La E.coli,, clamp loader = complexul g (γ)( În n celulele EK, clamp loader = factorul de replicare C (RF C)

47 Sinteza ADN la bifurcația de replicare La bifurcația ia de replicare, catenele conducătoare şi întârziată (leading şi lagging) ) sunt sintetizate simultan La o aceeaşi i bifurfacție ie funcționează mai multe ADN pol ie de replicare se formează un complex proteic = ADN polimeraza III holoenzimă La E.coli,, la o singură bifurcație de replicare se formează un complex proteic ADN pol III holoenzima 3 copii ADN pol III miez 1 sliding clamp loader 1 sliing clamp
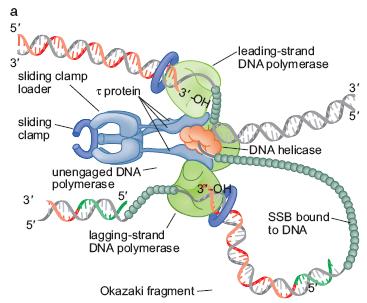
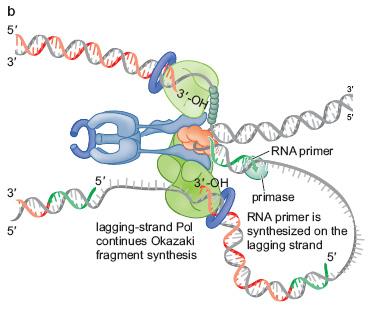
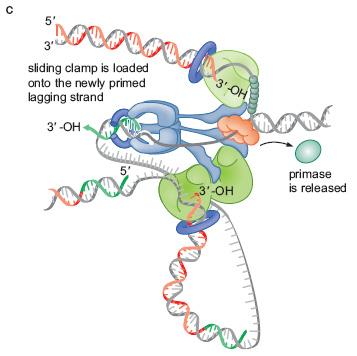
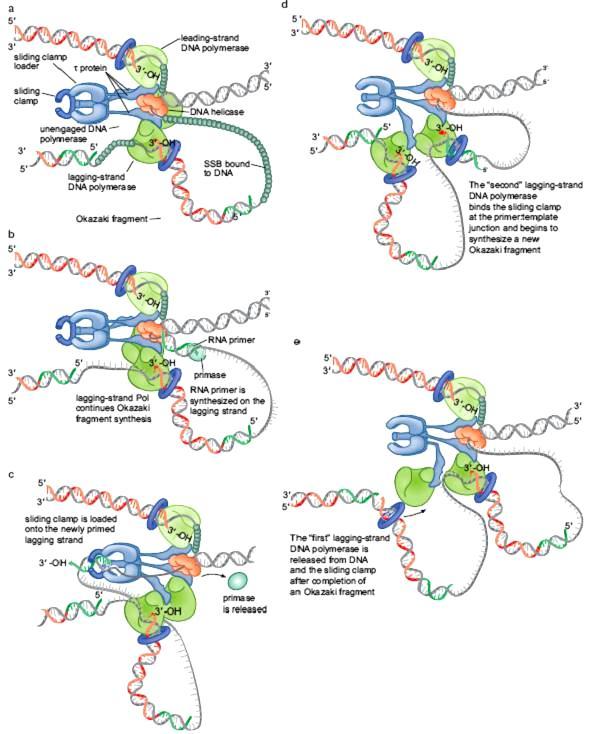
48
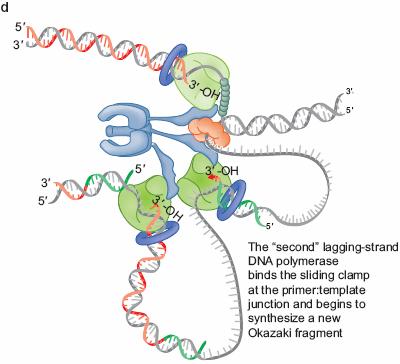
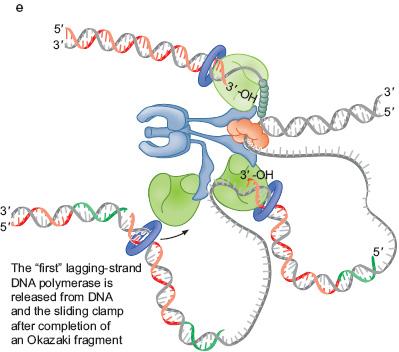
49

50 F IGUR E 9 24 Binding of the DNA helicase to DNA Pol III holoenzyme stimulates the rate of DNA strand separation. The t subunit of the sliding clamp loader interacts with both the DNA helicase and the DNA polymerase at the replication fork. (a) When this interaction occurs, the DNA helicase unwinds the DNA at approximately the same rate as the DNA polymerases replicate the DNA. (b) If the DNA helicase is not associated with DNA Pol III holoenzyme, DNA unwinding slows by 10 fold. Under these conditions, the DNA polymerases can replicate faster than the DNA helicase can separate the strands of unreplicated DNA. This allows the DNA Pol III holoenzyme to catch up to the DNA helicase and re form the replisome.
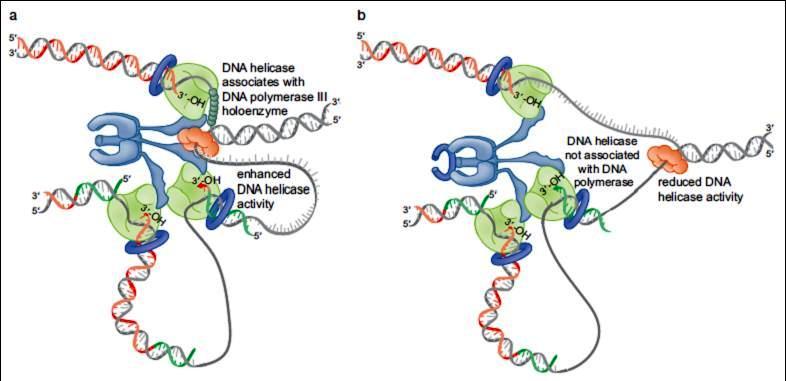
51

52 Arthur Kornberg
53 Aparat enzimatic care realizează replicarea ADN : ADN polimerază (III) holoenzima Primază = enzimă e complexă,, formată f din mai multe subunităţi i cu diverse funcţii; cea mai importantă funcţie : formarea legăturilor fosfodiesterice dintre deoxiribonucleotide Primază = o ARN polimerază,, sintetizează s scurte catene de ARN numite primeri Ligaza formează legături fosfodiesterice între nucleotide Helicază desface legăturile de hidrogen dintre cele 2 catene parentale Proteine Ssb (Single-Stranded Stranded Binding) se leagă de monocatene ADN şi i le stabilizează Topoizomeraze relaxează dublul helix,, scade s numărul de spire / kpb Topoizomeraze Nu poate porni sinteza catenei noi de novo,, ci o poate doar prelungi Are nevoie de o catenă scurtă pe care sa o prelungească - PRIMER 3 ADN pol III miez (palm, fingers, thumb) 1 sliding clamp 1 clamp loader RNază H scoate ribonucleotidele ce formează primerii ADN pol I umple golurile lăsate de RNaza H
54 Feb Mai 2014 Cursul 3
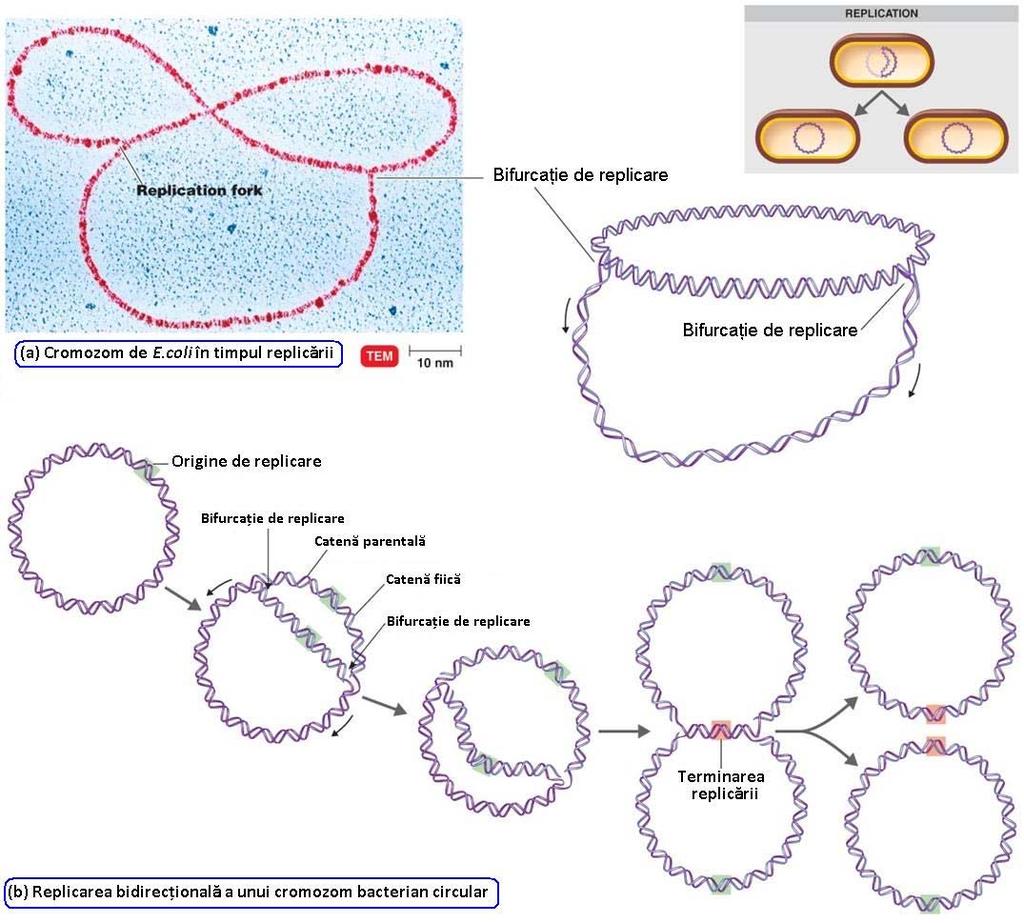
55 INIȚIEREA REPLICĂRII ADN Pre inițiere Etape Regiunea unde începe replicarea moleculei de ADN = origine a replicării O moleculă ADN poate avea: 1 regiune ori Cromozom bacterian Plasmide bacteriene ori C ori V Pre inițiere Inițiere Elongare Terminare Cromozomul bacterian este un replicon unic
56
57 mai multe regiuni ori Cromozomi de la eucariote Cromozomii de tip eucariot sunt structuri multirepliconice
58 INIȚIEREA REPLICĂRII ADN Replicarea unei molecule de ADN NU începe în n orice punct al moleculei, ci în n anumite situsuri = origini de replicare (ori) Formarea unei bifurcaţii ii de replicare necesită separarea celor 2 catene parentale Replicon = fragment de ADN replicat de la 1 origine de replicare Cromozomul de E.coli are 1 ori cromozom unirepliconic (replicon unic) Majoritatea bacteriilor au cromozomi unirepliconici Fiecare cromozom de la eucariote are mai multe regiuni ori cromozomi multirepliconici Sistemul de iniţiere iere a replicării ADN este format din 2 componente: Componenta ADN = regiunea ori, denumită şi replicator are o anumită secvenţă ă de nucleotide Componenta proteică = proteine de iniţiere, iere, denumite şi iniţiator iator, PRIMOSOM se ataşează la ori (replicator)
59 Majoritatea secvenţelor elor replicator de la diverse organisme conţin: - o regiune de legare a proteinelor de iniţiere iere - o regiune bogată în n A/T uşor de desfăcut legăturile de H de către proteinele iniţiator iator Situsul oric este în n afara acestei regiuni E.coli 5 cutii dna A (9) = regiuni 9-merice9 aici se ataşează proteina Dna A 3 regiuni 13-merice (13) bogate în n A/T aici se desface dublul helix SV40 4 situsuri P = regiuni 5-merice5 aici se ataşează proteina iniţiator iator antigenul T 2 regiuni EP (Early Palindrome) = 20 pb aici se desface dublul helix E.coli SV40 S.cerevisiae situsuri de ataşare are 5 dna A, 9-mers 4 P, 5-mers 2 A, B1 proteine iniţiator iator Dna A antigenul T ORC situsuri de desfacere 3 13-mers 2 EP (20pb) 1 B2 S.cerevisiae 2 regiuni A, B1 aici se ataşează proteina ORC 1 regiune B2 aici se ataşează helicaza aici se desface dublul helix

60 Proteinele iniţiator: iator: - se ataşează la replicator,, situs-specific: specific: E.coli 5 cutii dnaa - interacţionează ionează cu alte proteine pe care le aduc la replicator - unele desfac dublul helix în n reginile adiacente situsului de legare Principalele etape ale iniţierii ierii replicării cromozomului E.coli (a) Mai multe exemplare de proteină DnaA-ATP ATP situsurile dnaa (b) Desfacerea dublului helix în n zona situsurilor 9-mers 9 (c) Ataşarea area helicazei bacteriene Dna B, adusă a de către Dna C (complexul Dna B Dna C) (d) Deplasarea helicazei îndepărtează proteinele Dna A hill.com/sites/dl/free/ /126997/animation17.html
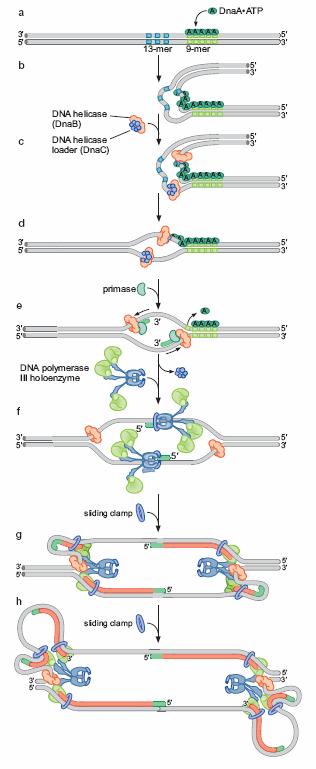
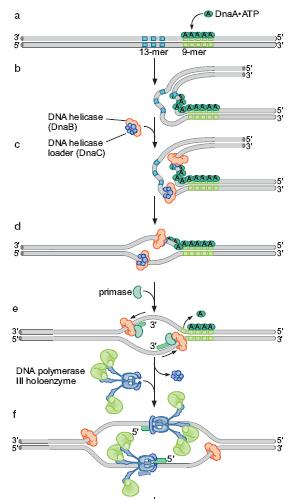
61

62 Cromozomii de la EK se replică o singură dată per ciclu celular Replicarea ADN cromozomal numai în n faza S a ciclului celular În n faza S este replicată o dată şi i doar o singură dată toată cantitatea de ADN cromozomal Originile de replicare de pe cromozomii EK sunt separate de ~ 30 kpb : crz EK mici au > 10 ori crz umani mari au mii de ori Ptr ca fiecare crt EK să fie replicat integral în n timpul fiecărei faze S, trebuie t activat un număr suficient de mare de ori (nu neapărat toate ori,, dar d dacă sunt prea puţine, atunci anumite regiuni din genom vor rămâne ne-replicate) În n mod normal, nici un ori nu poate iniţia ia 2 runde de replicare în n aceeaşi i fază S; S deci, este e inactivat până la următoarea S Un crz EK cu 5 secvenţe e replicator - mai întâi sunt activate secvenţele ele 3 şi i 5 - se formează 2 bucle de replicare 4 bifurcaţii ii de replicare bidirecţională ională - secvenţele ele ori de pe moleculele fiice sunt blocate până la următoarea rundă (X) - elongarea bifurcaţiilor iilor de replicare depăşeşte replicatorii 2 şi i 4 înainte ca aceştia să se activeze - ca urmare, 2 şi i 4 nu se mai activează,, iar refginile respective sunt replicate în n mod pasiv - în n contrast, replicatorul 1 se iniţiază iază înainte ca replicarea să ajungă în n regiunea lui Prezenţa a pe un crz a mai multor replicatori decât ar fi necesar este o redundanţă,, dar d asigură replicarea completă a fiecărui crz.
63 Încărcarea helicazei Prima etapă în n iniţierea ierea replicării la EK La EK, etapele iniţierii ierii replicării ADN se desfăşoară în n faze distincte ale ciclului celular şi în n ordine diferită faţă ă de PK Încărcarea helicazei la toţi i replicatorii în G1 (înainte de S) Activarea replicatorilor (inclusiv activarea helicazei) Asamblarea replisomului în S Încărcarea helicazei pe crz EK - proteina ORC~ATP se ataşează la replicator - se ataşează proteina Cdc6 - helicaza (Mcm2-7) este adusă de Cdt1 - helicaza este încărcată pe ADN (ca dimer) - sunt eliberate Cdc6 şi i Cdt1 Cdc6, Cdt1 sunt similare cu DnaC de la PK
64 Helicazele încărcate sunt activate de 2 protein-kinaze: CDK (cyclin-dependent dependent-kinase) ce sunt activate la intrarea în n faza S [ Protein-kinaze = proteine care ataşează ează,, covalent, c grupări fosfat la alte proteine, ţintă ] DDK (Dbf4-dependent dependent-kinase), Aceste 2 protein-kinaze sunt activate la inrarea in faza S si, fie direct, fie prin intermediul altor proteine: - desfac helicaza in monomer - ataseaza cate un monomer de helicaza pe cate o monocatena ADN - In final are loc asamblarea replisomului = complex proteic Primaza (ADN pol ) ADN pol ADN pol Clamp,, clamp loader Helicaza
65 G1 + ORC Incarcarea helicazei pe ADN S Activarea helicazei Deschiderea dublului helix Formarea replisomului


 Separarea celor 2 cromozomi = segregare Ptr că cei 2 crz sunt")
 - trage molecula")
66 TERMINAREA REPLICĂRII MOLECULELOR ADN Necesită un set de evenimente specifice Diferă de la molecule ADN CIRCULARE la molecule ADN LINEARE Terminarea replicării ADN circular cromozomul E.coli Replicarea avansează pe cele 2 bifurcaţii (replicare bidirecţională) Molecula ia forma literei greceşti THETA replicare pe model THETA La terminarea replicării, cele 2 molecule fiice ramân legate una de alta într-o structură de tip catenan Catenan = structură formată din 2 cercuri interconectate (2 zale dintr-un lanţ) Separarea celor 2 cromozomi = segregare Ptr că cei 2 crz sunt circulari: segregarea = decatenare Topoizomerază II Topoisomerase 1 and 2.flv Topoizomeraze de clasă II - taie una din cele 2 molecule (ambele catene) - trage molecula întreagă (netăiată) prin tăietură - reface molecula tăiată

 - ultimul")

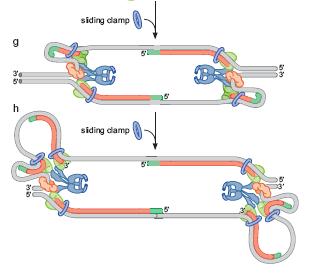
67 Terminarea replicării ADN linear - cromozomii de la eucariote 1. Deşi cromozomii d ela EK sunt molecule ADN lineare, datorită dimensiunii mari, la terminarea replicării cele 2 molecule fiice sunt înfăşurate una în jurul celeilalte Topoizomeraze de clasă II 2. Replicarea catenei întârziate ridică probleme în regiunile terminale ale cromozomilor (telomere) - ultimul primer se găseşte exact la capătul 3 al catenei parentale - după îndepărtarea acestui primer de către RNaza H, molecula rămâne cu o catenă mai lungă capul 3 -dacă procesul s-ar opri aici, atunci la fiecare rundă de replicare, cromozomii s-ar scurta semnificativ, în final cu pierdere de gene Telomeraza
68 TELOMERAZA = o ADN pol l ce nu necesită primer separat Capetele cromozomilor eucariotici = TELOMERE Repetiţii ii cap-coadă ale unei secvenţe e bogate în n TG de e ex., telomerele cromozomilor umani = repetiţii ii ale secvenţei ei 5 TTAGGG 3 Majoritatea acestor repetiţii ii sunt d.c., dar capul 3 3 al fiecărui cromozom se prelungeşte monocatenar Această structură acţionează ca o nouă origine de replicare pentru o ADN polimerază specială s - TELOMERAZA TELOMERAZA = ribonucleoproteină r = Complex de proteine, una din ele = reverstranscriptază telomerazică TERT O moleculă de ARN = ARN telomerazic TER - include o scurtă regiune complementară cu repetiţia ia telomerică La om: 5 AAUCCCAAUC 3 - ca şi i alte ADN pol, extinde capul 3 3 al unui primer - pe post de primer, foloseşte capul 3 -OH 3 al catenei parentale - pe post de catenă matriţă ă foloseşte ARN-ul propriu - astfel, extinde capul 3 3 al catenei ADN parentale - telomeraza se desprinde de pe catena ADN, se deplasează spre capul 3 şi i se reaşează ează Procesul se reia de multe ori Capul 3 -OH 3 este prelungit cu multe repetiţii ii TTAGGG Apoi telomeraza se desprinde complet ADN pol obişnuită prelungeşte capul 5 5 În n final, la EK cromozomii au capul 3 3 extins, prelungit monocatenar

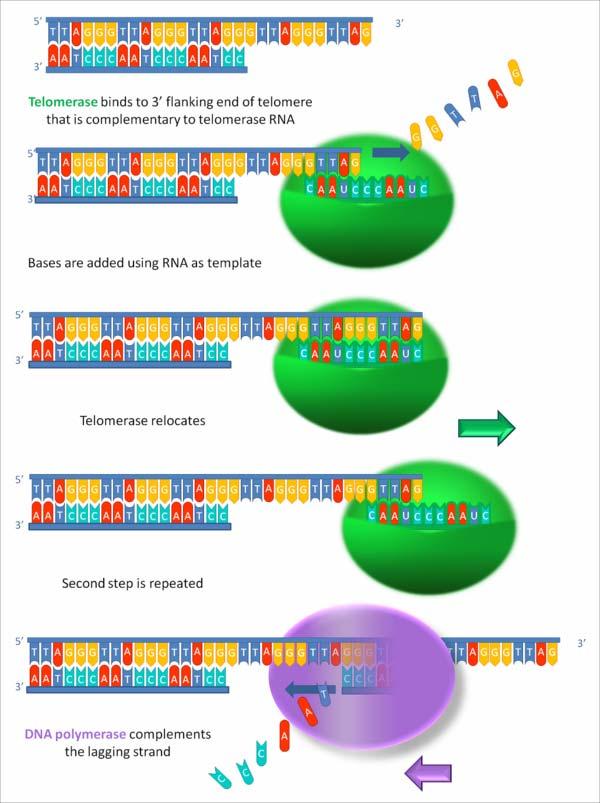
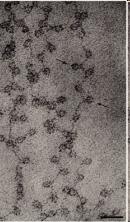
69 MBOG 8 Action of telomerase
70 Ordinea evenimentelor în n replicarea telomerelor - Telomeraza (T) se aşează a cu ARN-ul propriu (TER) în n zona de complementariatte a monocatenei ADN de la capul 3 -OH3 - extinde această monocatenă ADN,, folosind drept primer capul 3 -OH 3 al acesteia drept matriţă propriul ARN - T repetă de multe ori acest ciclu Extinde capul 3 3 cu multe repetiţii ii [ TTAGGG ] - apoi T se desprinde complet - capul 5 5 este prelungit, pe modelul unei catene întârziate, de către o ADN pol obişnuită În n final, la EK cromozomii au capul 3 3 extins, prelungit monocatenar
71 REPLICAREA ADN recapitulare I. INIŢIEREA IEREA ori 1 ori pe crz bacterian crz unirepliconic n ori pe crz EK crz multirepliconici Replicon = secvenţă ă ADN replicată de la un ori Secvenţele ele ADN din ori au funcţie de replicator situsuri de legare a proteinelor iniţiator iator + Proteine iniţiator iator = PRIMOSOM regiunile A/T, uşor de desfăcut Aici se formează bucla de replicare PK Complexul proteic primosom Proteina DnaA se ataşează la cutiile dnaa Distorsionarea ADN în n zona de legare Desfacerea dublului helix în n regiunile A/T PK Complexul proteic de iniţiere iere Proteina ORC~ATP se ataşează la replicator + proteine CDC ( - DnaC) Încărcarea helicazei (MCM)) pe ADN G1 + proteine CDK Activarea helicazei S Încărcarea helicazei (DnaB)) pe ADN de către DnaC Desfacerea dublului helix Formarea buclei de replicare cu cele 2 bifurcaţii ii Asamblarea replisomului Formarea buclei de replicare cu cele 2 bifurcaţii ii Asamblarea replisomului
72 II. ELONGAREA REPLISOM Primază Helicază ADN polimerază Proteine ajutătoare Sliding Clamp,, Clamp C loader RNază Ligază PK ADN pol III holoenzima 3 copii ADN pol III miez 1 sliding clamp 1 clamp loader Catena ADN nouă este sintetizată în n direcţie ie Catena ADN matriţă ă este citită în n direcţie ie inversă Catena ADN nouă este sintetizată prin complementaritate cu cea vechev La o bifurcaţie ie de replicare Una din catenele noi = catena c conducătoare,, in direcţia ia de deplasare a bifurcaţiei iei Cealaltă catenă nouă = catena întârziată, în n direcţie ie inversă
73 Funcţii Principala proteină de iniţiere a replicării Desfacerea legăturilor de H - Helicază Stabilizarea monocatenelor ADN DnaA DnaB Ssb Procariote ORC MCM RPA Eucariote Sinteză de primeri Primază = o ARN polimerază specială Menţine ADN pol pe ADN şi îi creşte procesivitatea Încarcă sliding clamp pe ADN ADN polimeraza replicativă Procesarea fragmentelor Okazaki (a) Eliminarea primerilor (b) Umplerea golurilor prin sinteză ADN (c) Ligarea fragmentelor Okazaki Derăsucirea ADN parental în faţa buclei de replicare Derăsucirea moleculelor d.c. fiice Terminarea replicării PK segregarea celor 2 crz bacterieni EK terminarea replicării la telomere DnaG Sliding Clamp ( -Clamp) Clamp Loader (subunităţi: ) ADN pol III miez RNaza H ADN pol I ADN ligaza H Topoizomerază de clasă I Topoizomerază de clasă II (ADN girază) Topo cls I, Topo cls II Topoizomerază cls II ADN pol PCNA RFC ADN pol pe catena întârziată ADN pol pe cat.conducătoare RNaza H Dna2, FEN1 ADN ligaza I Topoizomerază de clasă I Topoizomerază de clasă II Topo cls I, Topo cls II Telomeraza
74 III. TERMINAREA PK PK Cromozomul bacterian = ADN dc CIRCULAR Cromozomii eucariotelor = ADN dc LINEAR Terminarea replicării 2 molecule ADN dc circulare interconectate Terminarea replicării capul 3 3 al moleculei parentale este extins Desfacerea celor 2 molecule TOPO cls II TELOMERAZA (a) Prelungeşte capul 3 3 cu multe repetiţii ii TTAGGG - Primer = capul 3 3 al catenei vechi - Matriţă = ARN telomerazic (TER) Acest proces de reverstranscriere se repetă de multe ori (b) Se desprinde complet de pe ADN (c) ADN pol prelungeşte capul 5 5 (sinteză prin fragmente Okazaki) În n final, cromozomii au capul 3 3 extins, prelungit monocatenar
75 Alfabet grecesc Literă mare Literă mică Se citeşte Literă mare Literă mică Se citeşte alfa niu beta ksi gama omicron delta pi epsilon rho (ro) zeta sigma eta tau theta ipsilon iota phi (fi) kappa chi (hi) lambda psi miu omega
76 Feb Mai 2014 Cursul 4
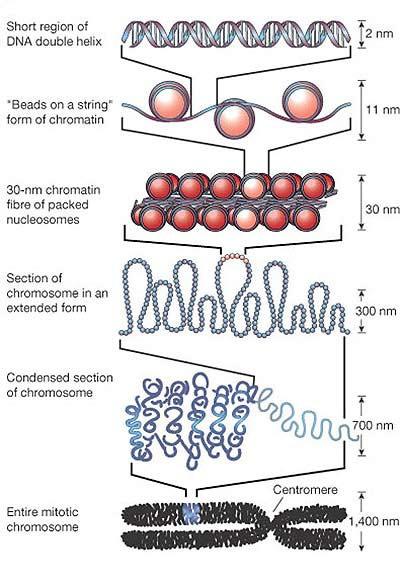
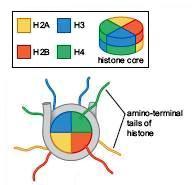
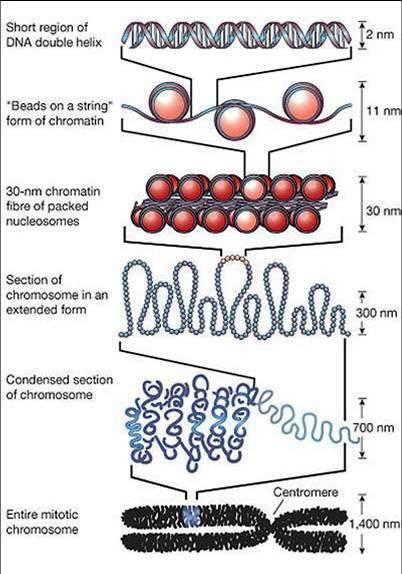
77 De la ADN la cromozomi În n celule, moleculele de ADN sunt asociate cu proteine Fiecare moleculă ADN + proteinele asociate = cromozom, şi i la PK, şi i la EK Cuvântul cromozom : chroma = culoare, soma = corp cromozom = corp colorat Împachetarea ADN ului în n cromozomi îndeplineşte mai multe funcții : ADN ul celular are dimensiune mai mare decât celula ptr a încăpea în n celulă,, ADN ul trebuie compactat Împachetarea ADN ului şi i asocierea cu proteine protejează față ă de alterări ADN ul nud este destul de instabil în n celule, spre deosebire de cromozomi, care sunt mult mai stabili Doar în n formă compactată în n cromozomi, ADN ul poate fi transmis eficient şi i egal la cele 2 celule fiice Crz conferă o organizare globală fiecărei molecule de ADN,, organizare o ce conferă şi i accesibilitate către ADN ½ din masa unui crz EK = proteine Cromatină = regiuni din ADN asociate cu proteine Majoritatea proteinelor asociate cu moleculele de ADN = proteine cu g.m. g Mică,, bazice HISTONE Alte proteine asociate cu ADN : NON HISTONE reglează replicarea,, repararea, recobinarea şi i transcrierea Primul rol improtant al proteinelor asociate cu ADN, în n mod special al histonelor, este compactarea moleculelor de ADN De ex: grosimea medie a unei pb = 3,4 Å 1 celulă c umană conține 3 x 10 9 pb / set haploid, ~ Å ~ 1 m 1 cel. are în n nucleu ( φ =10 15mm) 15mm) 2 m de ADN
78 Fiecare celulă are un anumit număr, specie-specific, de cromozomi PK Nu există membrană nucleară nu există nucleu Majoritatea PK au 1 cromozom = 1 moleculă de ADN d.c. d circular Cromozomul bacterian nucleoid ADN extracromozomal: PK pot avea şi i plasmide (molecule ADN dc, de obicei circulare) Organismele se înmulțesc prin diviziune celulară simplă fisiune binară EK Majoritatea celulelor EK sunt DIPLOIDE: : au câte 2 exemplare din fiecare crz, dar cele 2 exemplare nu sunt s identice Număr par de cromozomi în n nucleu (2n),( fiecare crz = 1 moleculă ADN d.c. d linear ADN extracromozomal: în n mitocondrii, cloroplaste câte molecule ADNdc circular ADN mt Diploidia asigură înmulțirea de tip sexuat a organismelor EK gameții au ½ nr.crz Organismele EK (majoritatea) se înmulțesc prin reproducere sexuată (câte 1 exemplar din fiecare tip de crz) Celule 2n meioză Gameți n fecundație unirea a 2 gameți Celule 2n





79 Cromozomul la procariote c r o m o so m b a cter ia n p l i er e c i r ca 5 0 d o m e n i i p e r c r o m o so m b a cter ia n REP su p r a r a su ci r e REP d o m en i i su p r a sp i r a l i z a t e n ega tiv A D N gir a za
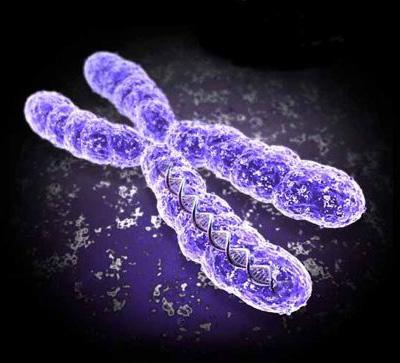
80 Cromozomul la eucariote
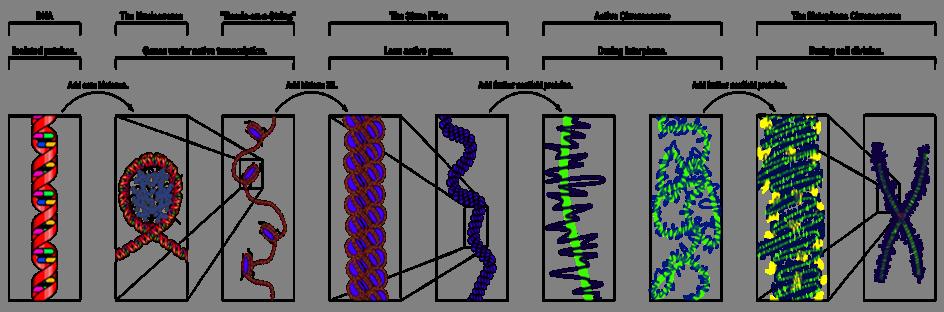
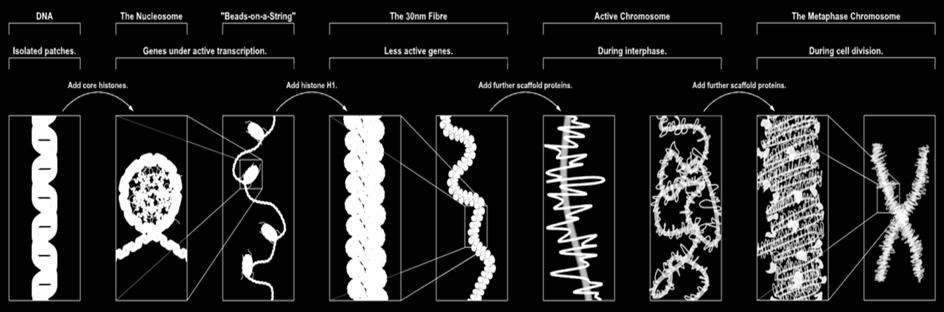
81
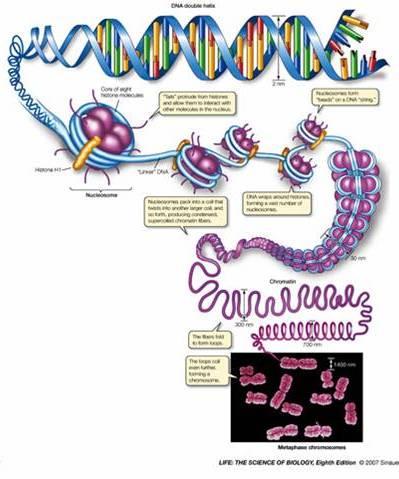
82
83 SPECIE Nr.total crz Nr.seturi haploide Nr.crz. / set haploid Forma crz Dimensiune genom [Mpb] P R O C A R I O T E Escherichia coli circular 4,6 Staphylococcus aureus circular 2,8 Vibrio cholerae circular Agrobacterium tumefaciens C, 1 L 5,7 E U C A R I O T E Saccharomyces cerevisiae drojdia de bere 32 1 sau 2 16 linear 12,1 Caenorhabditis elegans linear 97 Arabidopsis thaliana linear 125 Allium cepa linear Pisum sativum mazăre linear 4500 Drosophila melanogaster linear 180 Mus musculus linear 2600 Felis catus pisica domestică Canis familiaris câine linear linear Sus scrofa porc domestic linear 2700 Ovis aries oaie linear 2700 Bos taurus vaca domestică linear 2900 Equus caballus cal linear 2600 Gorilla gorilla linear 2900 Pan troglodytes linear 3300 Homo sapiens linear 3200

84 Organismele mai complexe au densitate genică scăzută
 Asamblarea/împachetarea mpachetarea în n nucleosomi = prima etapă şi i esențială în n compactarea ADN ului")
85 NUCLEOZOMUL Nucleosomes Are the Building Blocks of Chromosomes În n celulele EK, majoritatea cantității ii ADN este împachetat în n NUCLEOSOMI Nucleosom = miez de 8 protenie histonice + regiunea ADN înfăşurată în n jurul lor ADN ul dintre 2 nucleosomi adiacenți i = ADN linker (de legătură) Asamblarea/împachetarea mpachetarea în n nucleosomi = prima etapă şi i esențială în n compactarea ADN ului Prin asamblare/împachetare mpachetare în n nucleosomi, ADN ul se scurtează de 6 ori ADN înfăşurat în n jurul unui octamer histonic ~ 147 pb, de 1,7 ori Aspect de mărgele pe un fir H2A H2B H3 H4
 11 15 15 KDa H2 H4 H4 21 KDa H1 5 tipuri: H1, H2A, H2B, H3, H4 histonele miez 2 x [ H2A, H2B, H3, H4 ] = MIEZUL NUCLEOSOMULUI 1 x H1 1 se leagă la ADN ul linker, dintre 2")
86 Histonele = proteine mici, încărcate pozitiv Histonele = cele mai abundente proteine asociate cu ADN la EK proteine mici, încărcate pozitiv (conținut ridicat în n AA încărcat pozitiv) KDa H2 H4 H4 21 KDa H1 5 tipuri: H1, H2A, H2B, H3, H4 histonele miez 2 x [ H2A, H2B, H3, H4 ] = MIEZUL NUCLEOSOMULUI 1 x H1 1 se leagă la ADN ul linker, dintre 2 nucleosomi

87 Miezul proteic al nucleosomului are formă de disc În n soluție, cele c 4 tipuri t de histone se asamblează astfel: H2A + H2B heterodimer H2A H2B H2B H3 + H4 heterodimer H3 H4 H4 2 heterodimeri H3 H4 H4 tetramer H3 H4 H4


![H2A dimer 2[H2A] 2 x H2B dimer 2[H2B] 2.](/docs-images/69/61506101/images/88-2.jpg "Inițierea ierea nucleosomului 1 tetramer 4[H3 H4]")
![H4] se ataşează la ADN 3.](/docs-images/69/61506101/images/88-3.jpg "Terminarea formării nucleosomului dimeri 2[H2a]")
88 H2A H2B H3 H4 Asamblarea unui nucleosom este realizată într o o anumită ordine: 1. Dimerizarea histonelor H3 + H4 heterodimer [H3 H4] H4] 2 x [H3 H4] H4] tetramer 4[H3 H4] H4] 2 x H2A dimer 2[H2A] 2 x H2B dimer 2[H2B] 2. Inițierea ierea nucleosomului 1 tetramer 4[H3 H4] H4] se ataşează la ADN 3. Terminarea formării nucleosomului dimeri 2[H2a] şi i 2[H2B] se ataşează la acest complex NUCLEOSOM
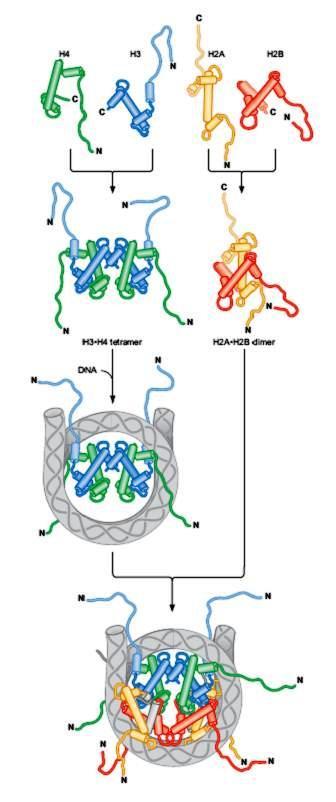
89

90 STRUCTURA CROMATINEI Eucromatina şi i Heterocromatina De foarte multe ori, cromozomii nu se colorează uniform: Eucromatina se colorează mai slab este mai activă ca rată de transcriere Heterocromatina se colorează mai intens este mai puțin activă ca rată de transcriere Histona H1 seleagă la ADN linker dintre nucleosomi Odată formați i nucleosomii, următarea etapă în împachetarea moleculelor de ADN = legarea histonei H1 H1 = o proteină mică, încărcată pozitiv,, se leagă la ADN ul dintre nucleosomi Se leagă la ADN în n 2 zone: o zonă este exact în n afara unui nucleosom o zonă este regiunea centrală de înfăşurare a ADN în n jurul histonelor miez În n acest mod, H1 1 stabilizează s asocierea ADN ului cu proteinele nucleosomale şi i deci, stabilizează structura nucleosomului



91 Adăugarea histonei H1 H un nivel superior de compactare a ADN ului

 ) ai a fibra de 30 nm cu structură ZIGZAG Într un")
92 Următoarea etapă de compactare = FIBRA DE 30 nm Eviden 2 modele m de structură a fibrei de 30 nm Modelul SOLENOID Evidențiată iată şi in vitro şi in vivo Se formează un superhelix ce conține 6 nucleosomi n per spiră ADN ul linker se găseşte spre interiorul superhelixului, dar NU trece prin axul central l al acestuia Zonele de intrare şi ieşire a ADN din nucleosomi NU sunt accesibile Lungimea ADN ului linker variază (distanța dintre 2 nucleosomi) Modelul ZIGZAG Se formează un superhelix ce conține 4 nucleosomi n per spiră ADN ul linker TRECE prin axul central al superhelixului Zonele de intrare şi ieşire a ADN din nucleosomi nu sunt mai accesibile Regiunile cu linker mai lung (deci cu densitate mai mică de nucleosomi) ) ai a fibra de 30 nm cu structură ZIGZAG Într un acelaşi i nucleu se pot întâlni în n simultan ambele forme
 interacționează ionează cu nucleosomii")
93 Regiunile amino terminale ale histonelor sunt necesare ptr formarea fibrei 30 nm Cozile amino terminale ale histonelor nucleosomale (H2A, H2B, H3, H4) interacționează ionează cu nucleosomii adiacenți
 2 clase de")
94 Următoarea etapă în n compactarea ADN = formarea de bucle mari Formarea nucleosomilor + formarea fibrei 30 nm compactarea ADN cu scurtare pînă p la de 40 de ori Este încă insuficient este necesară compactarea mai puternică a ADN Model: Fibra 30nm formează bucle mari,, de kpb, ținute de o structură proteică denumită NUCLEAR SCAFFOLD (traducere ad literam: : schelă s nucleară) 2 clase de proteine: Topo II: : la baza fiecărei bucle,, menține forma buclată; ; buclele sunt topoizomeri Proteine SMC (Structural Maintenance of Chromosome) Se formează fibra de cromatină = structură s de bază a cromozomilor

95 Chromosome Structure Changes as Eukaryotic Cells Divide Gradul de condensare a materialului genetic variază în n timpul unui ciclu celular Interfază condensare mai redusă a materialului genetic,, permite transcriere, replicare, reparare După replicare,, moleculele fiice: asamblare nucleosomi formarea fibrei 30nm coeziunea cromatidelor surori complex proteic denumit coezină În n interfază nu se văd cromozomii ca entități i distincte citologic; sunt vizibile doar blocuri cromatice cromatină, unele colorate mai slab eucromatină,, altele a colorate mai intens heterocromatină


96 Profază continuă condensarea cromatinei (deci, a materialului genetic) formarea cromozomilor ca entități vizibile la microscop (procesul se mai numeşte şi condensarea cromozomilor ) La sfârşitul diviziunii celulare reîncepe procesul de decondensare a cromozomilor, pâna la gradul de fibră de cromatină
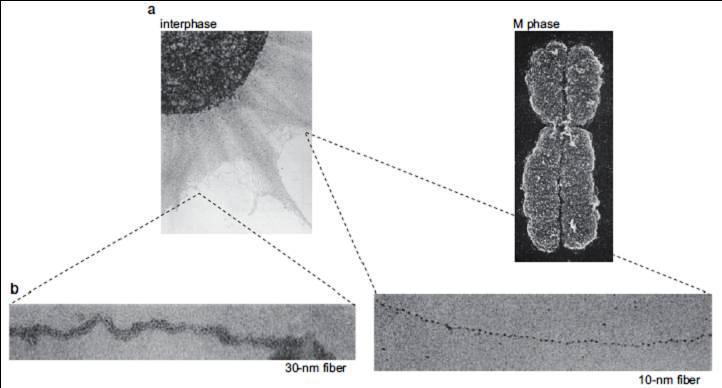
97

98 DEZASAMBLAREA / ASAMBLAREA NUCLEOSOMILOR în replicarea ADN Nucleosomii sunt dezasamblați i pe molecula parentală, în n fața a bifurcației iei de replicare asamblați i pe moleculele fiice, în n ordinea anterior prezentată Tetramerii H3 H4 H4 vechi trec ca atare, fie la una din moleculele noi, fie f la cealaltă Dimerii H2A şi i H2B vechi nu trec direct la moleculele noi, ci intâi în n soluție.
99
Planul determinat de normală şi un punct Ecuaţia generală Plane paralele Unghi diedru Planul determinat de 3 puncte necoliniare
 1 Planul în spaţiu Ecuaţia generală Plane paralele Unghi diedru 2 Ecuaţia generală Plane paralele Unghi diedru Fie reperul R(O, i, j, k ) în spaţiu. Numim normala a unui plan, un vector perpendicular pe
1 Planul în spaţiu Ecuaţia generală Plane paralele Unghi diedru 2 Ecuaţia generală Plane paralele Unghi diedru Fie reperul R(O, i, j, k ) în spaţiu. Numim normala a unui plan, un vector perpendicular pe
MARCAREA REZISTOARELOR
 1.2. MARCAREA REZISTOARELOR 1.2.1 MARCARE DIRECTĂ PRIN COD ALFANUMERIC. Acest cod este format din una sau mai multe cifre şi o literă. Litera poate fi plasată după grupul de cifre (situaţie în care valoarea
1.2. MARCAREA REZISTOARELOR 1.2.1 MARCARE DIRECTĂ PRIN COD ALFANUMERIC. Acest cod este format din una sau mai multe cifre şi o literă. Litera poate fi plasată după grupul de cifre (situaţie în care valoarea
(a) se numeşte derivata parţială a funcţiei f în raport cu variabila x i în punctul a.
 se numeşte derivata parţială a funcţiei f în raport cu variabila x i în punctul a.") Definiţie Spunem că: i) funcţia f are derivată parţială în punctul a în raport cu variabila i dacă funcţia de o variabilă ( ) are derivată în punctul a în sens obişnuit (ca funcţie reală de o variabilă
Definiţie Spunem că: i) funcţia f are derivată parţială în punctul a în raport cu variabila i dacă funcţia de o variabilă ( ) are derivată în punctul a în sens obişnuit (ca funcţie reală de o variabilă
5. FUNCŢII IMPLICITE. EXTREME CONDIŢIONATE.
 5 Eerciţii reolvate 5 UNCŢII IMPLICITE EXTREME CONDIŢIONATE Eerciţiul 5 Să se determine şi dacă () este o funcţie definită implicit de ecuaţia ( + ) ( + ) + Soluţie ie ( ) ( + ) ( + ) + ( )R Evident este
5 Eerciţii reolvate 5 UNCŢII IMPLICITE EXTREME CONDIŢIONATE Eerciţiul 5 Să se determine şi dacă () este o funcţie definită implicit de ecuaţia ( + ) ( + ) + Soluţie ie ( ) ( + ) ( + ) + ( )R Evident este
REPLICAREA ŞI REPARAREA ADN
 1 Capitolul 7 7 REPLICAREA ŞI REPARAREA ADN Replicarea ADN este procesul molecular prin care se realizează copierea exactă a moleculelor de ADN (a secvenţei nucleotidice). Datorită replicării are loc transmiterea
1 Capitolul 7 7 REPLICAREA ŞI REPARAREA ADN Replicarea ADN este procesul molecular prin care se realizează copierea exactă a moleculelor de ADN (a secvenţei nucleotidice). Datorită replicării are loc transmiterea
Metode iterative pentru probleme neliniare - contractii
 Metode iterative pentru probleme neliniare - contractii Problemele neliniare sunt in general rezolvate prin metode iterative si analiza convergentei acestor metode este o problema importanta. 1 Contractii
Metode iterative pentru probleme neliniare - contractii Problemele neliniare sunt in general rezolvate prin metode iterative si analiza convergentei acestor metode este o problema importanta. 1 Contractii
Seminariile Capitolul X. Integrale Curbilinii: Serii Laurent şi Teorema Reziduurilor
 Facultatea de Matematică Calcul Integral şi Elemente de Analiă Complexă, Semestrul I Lector dr. Lucian MATICIUC Seminariile 9 20 Capitolul X. Integrale Curbilinii: Serii Laurent şi Teorema Reiduurilor.
Facultatea de Matematică Calcul Integral şi Elemente de Analiă Complexă, Semestrul I Lector dr. Lucian MATICIUC Seminariile 9 20 Capitolul X. Integrale Curbilinii: Serii Laurent şi Teorema Reiduurilor.
Curs 10 Funcţii reale de mai multe variabile reale. Limite şi continuitate.
 Curs 10 Funcţii reale de mai multe variabile reale. Limite şi continuitate. Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie p, q N. Fie funcţia f : D R p R q. Avem următoarele
Curs 10 Funcţii reale de mai multe variabile reale. Limite şi continuitate. Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie p, q N. Fie funcţia f : D R p R q. Avem următoarele
 4. CIRCUITE LOGICE ELEMENTRE 4.. CIRCUITE LOGICE CU COMPONENTE DISCRETE 4.. PORŢI LOGICE ELEMENTRE CU COMPONENTE PSIVE Componente electronice pasive sunt componente care nu au capacitatea de a amplifica
4. CIRCUITE LOGICE ELEMENTRE 4.. CIRCUITE LOGICE CU COMPONENTE DISCRETE 4.. PORŢI LOGICE ELEMENTRE CU COMPONENTE PSIVE Componente electronice pasive sunt componente care nu au capacitatea de a amplifica
Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor X) functia f 1
 functia f 1") Functii definitie proprietati grafic functii elementare A. Definitii proprietatile functiilor. Fiind date doua multimi X si Y spunem ca am definit o functie (aplicatie) pe X cu valori in Y daca fiecarui
Functii definitie proprietati grafic functii elementare A. Definitii proprietatile functiilor. Fiind date doua multimi X si Y spunem ca am definit o functie (aplicatie) pe X cu valori in Y daca fiecarui
DISTANŢA DINTRE DOUĂ DREPTE NECOPLANARE
 DISTANŢA DINTRE DOUĂ DREPTE NECOPLANARE ABSTRACT. Materialul prezintă o modalitate de a afla distanţa dintre două drepte necoplanare folosind volumul tetraedrului. Lecţia se adresează clasei a VIII-a Data:
DISTANŢA DINTRE DOUĂ DREPTE NECOPLANARE ABSTRACT. Materialul prezintă o modalitate de a afla distanţa dintre două drepte necoplanare folosind volumul tetraedrului. Lecţia se adresează clasei a VIII-a Data:
a n (ζ z 0 ) n. n=1 se numeste partea principala iar seria a n (z z 0 ) n se numeste partea
 n. n=1 se numeste partea principala iar seria a n (z z 0 ) n se numeste partea") Serii Laurent Definitie. Se numeste serie Laurent o serie de forma Seria n= (z z 0 ) n regulata (tayloriana) = (z z n= 0 ) + n se numeste partea principala iar seria se numeste partea Sa presupunem ca,
Serii Laurent Definitie. Se numeste serie Laurent o serie de forma Seria n= (z z 0 ) n regulata (tayloriana) = (z z n= 0 ) + n se numeste partea principala iar seria se numeste partea Sa presupunem ca,
Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor
 Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor. Fiind date doua multimi si spunem ca am definit o functie (aplicatie) pe cu valori in daca fiecarui element
Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor. Fiind date doua multimi si spunem ca am definit o functie (aplicatie) pe cu valori in daca fiecarui element
Analiza în curent continuu a schemelor electronice Eugenie Posdărăscu - DCE SEM 1 electronica.geniu.ro
 Analiza în curent continuu a schemelor electronice Eugenie Posdărăscu - DCE SEM Seminar S ANALA ÎN CUENT CONTNUU A SCHEMELO ELECTONCE S. ntroducere Pentru a analiza în curent continuu o schemă electronică,
Analiza în curent continuu a schemelor electronice Eugenie Posdărăscu - DCE SEM Seminar S ANALA ÎN CUENT CONTNUU A SCHEMELO ELECTONCE S. ntroducere Pentru a analiza în curent continuu o schemă electronică,
Curs 1 Şiruri de numere reale
 Bibliografie G. Chiorescu, Analiză matematică. Teorie şi probleme. Calcul diferenţial, Editura PIM, Iaşi, 2006. R. Luca-Tudorache, Analiză matematică, Editura Tehnopress, Iaşi, 2005. M. Nicolescu, N. Roşculeţ,
Bibliografie G. Chiorescu, Analiză matematică. Teorie şi probleme. Calcul diferenţial, Editura PIM, Iaşi, 2006. R. Luca-Tudorache, Analiză matematică, Editura Tehnopress, Iaşi, 2005. M. Nicolescu, N. Roşculeţ,
Aplicaţii ale principiului I al termodinamicii la gazul ideal
 Aplicaţii ale principiului I al termodinamicii la gazul ideal Principiul I al termodinamicii exprimă legea conservării şi energiei dintr-o formă în alta şi se exprimă prin relaţia: ΔUQ-L, unde: ΔU-variaţia
Aplicaţii ale principiului I al termodinamicii la gazul ideal Principiul I al termodinamicii exprimă legea conservării şi energiei dintr-o formă în alta şi se exprimă prin relaţia: ΔUQ-L, unde: ΔU-variaţia
5.4. MULTIPLEXOARE A 0 A 1 A 2
 5.4. MULTIPLEXOARE Multiplexoarele (MUX) sunt circuite logice combinaţionale cu m intrări şi o singură ieşire, care permit transferul datelor de la una din intrări spre ieşirea unică. Selecţia intrării
5.4. MULTIPLEXOARE Multiplexoarele (MUX) sunt circuite logice combinaţionale cu m intrări şi o singură ieşire, care permit transferul datelor de la una din intrări spre ieşirea unică. Selecţia intrării
Laborator 11. Mulţimi Julia. Temă
 Laborator 11 Mulţimi Julia. Temă 1. Clasa JuliaGreen. Să considerăm clasa JuliaGreen dată de exemplu la curs pentru metoda locului final şi să schimbăm numărul de iteraţii nriter = 100 în nriter = 101.
Laborator 11 Mulţimi Julia. Temă 1. Clasa JuliaGreen. Să considerăm clasa JuliaGreen dată de exemplu la curs pentru metoda locului final şi să schimbăm numărul de iteraţii nriter = 100 în nriter = 101.
Curs 14 Funcţii implicite. Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi"
 Curs 14 Funcţii implicite Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie F : D R 2 R o funcţie de două variabile şi fie ecuaţia F (x, y) = 0. (1) Problemă În ce condiţii ecuaţia
Curs 14 Funcţii implicite Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie F : D R 2 R o funcţie de două variabile şi fie ecuaţia F (x, y) = 0. (1) Problemă În ce condiţii ecuaţia
5.5. REZOLVAREA CIRCUITELOR CU TRANZISTOARE BIPOLARE
 5.5. A CIRCUITELOR CU TRANZISTOARE BIPOLARE PROBLEMA 1. În circuitul din figura 5.54 se cunosc valorile: μa a. Valoarea intensității curentului de colector I C. b. Valoarea tensiunii bază-emitor U BE.
5.5. A CIRCUITELOR CU TRANZISTOARE BIPOLARE PROBLEMA 1. În circuitul din figura 5.54 se cunosc valorile: μa a. Valoarea intensității curentului de colector I C. b. Valoarea tensiunii bază-emitor U BE.
Integrala nedefinită (primitive)
") nedefinita nedefinită (primitive) nedefinita 2 nedefinita februarie 20 nedefinita.tabelul primitivelor Definiţia Fie f : J R, J R un interval. Funcţia F : J R se numeşte primitivă sau antiderivată a funcţiei
nedefinita nedefinită (primitive) nedefinita 2 nedefinita februarie 20 nedefinita.tabelul primitivelor Definiţia Fie f : J R, J R un interval. Funcţia F : J R se numeşte primitivă sau antiderivată a funcţiei
riptografie şi Securitate
 riptografie şi Securitate - Prelegerea 12 - Scheme de criptare CCA sigure Adela Georgescu, Ruxandra F. Olimid Facultatea de Matematică şi Informatică Universitatea din Bucureşti Cuprins 1. Schemă de criptare
riptografie şi Securitate - Prelegerea 12 - Scheme de criptare CCA sigure Adela Georgescu, Ruxandra F. Olimid Facultatea de Matematică şi Informatică Universitatea din Bucureşti Cuprins 1. Schemă de criptare
Curs 4 Nucleul. Acizi nucleici. Notiuni de genetica moleculara
 Biologie Celulara si Histologie Curs 4 Nucleul. Acizi nucleici. Notiuni de genetica moleculara Cuprins Nucleul celular structura Acizi nucleici Transcriptia, translatia informatiei genetice; sinteza de
Biologie Celulara si Histologie Curs 4 Nucleul. Acizi nucleici. Notiuni de genetica moleculara Cuprins Nucleul celular structura Acizi nucleici Transcriptia, translatia informatiei genetice; sinteza de
III. Serii absolut convergente. Serii semiconvergente. ii) semiconvergentă dacă este convergentă iar seria modulelor divergentă.
 semiconvergentă dacă este convergentă iar seria modulelor divergentă.") III. Serii absolut convergente. Serii semiconvergente. Definiţie. O serie a n se numeşte: i) absolut convergentă dacă seria modulelor a n este convergentă; ii) semiconvergentă dacă este convergentă iar
III. Serii absolut convergente. Serii semiconvergente. Definiţie. O serie a n se numeşte: i) absolut convergentă dacă seria modulelor a n este convergentă; ii) semiconvergentă dacă este convergentă iar
a. 11 % b. 12 % c. 13 % d. 14 %
 1. Un motor termic funcţionează după ciclul termodinamic reprezentat în sistemul de coordonate V-T în figura alăturată. Motorul termic utilizează ca substanţă de lucru un mol de gaz ideal având exponentul
1. Un motor termic funcţionează după ciclul termodinamic reprezentat în sistemul de coordonate V-T în figura alăturată. Motorul termic utilizează ca substanţă de lucru un mol de gaz ideal având exponentul
Activitatea A5. Introducerea unor module specifice de pregătire a studenţilor în vederea asigurării de şanse egale
 POSDRU/156/1.2/G/138821 Investeşte în oameni! FONDUL SOCIAL EUROPEAN Programul Operaţional Sectorial pentru Dezvoltarea Resurselor Umane 2007 2013 Axa prioritară nr. 1 Educaţiaşiformareaprofesionalăînsprijinulcreşteriieconomiceşidezvoltăriisocietăţiibazatepecunoaştere
POSDRU/156/1.2/G/138821 Investeşte în oameni! FONDUL SOCIAL EUROPEAN Programul Operaţional Sectorial pentru Dezvoltarea Resurselor Umane 2007 2013 Axa prioritară nr. 1 Educaţiaşiformareaprofesionalăînsprijinulcreşteriieconomiceşidezvoltăriisocietăţiibazatepecunoaştere
Curs 4 Serii de numere reale
 Curs 4 Serii de numere reale Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Criteriul rădăcinii sau Criteriul lui Cauchy Teoremă (Criteriul rădăcinii) Fie x n o serie cu termeni
Curs 4 Serii de numere reale Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Criteriul rădăcinii sau Criteriul lui Cauchy Teoremă (Criteriul rădăcinii) Fie x n o serie cu termeni
V.7. Condiţii necesare de optimalitate cazul funcţiilor diferenţiabile
 Metode de Optimizare Curs V.7. Condiţii necesare de optimalitate cazul funcţiilor diferenţiabile Propoziţie 7. (Fritz-John). Fie X o submulţime deschisă a lui R n, f:x R o funcţie de clasă C şi ϕ = (ϕ,ϕ
Metode de Optimizare Curs V.7. Condiţii necesare de optimalitate cazul funcţiilor diferenţiabile Propoziţie 7. (Fritz-John). Fie X o submulţime deschisă a lui R n, f:x R o funcţie de clasă C şi ϕ = (ϕ,ϕ
1.7. AMPLIFICATOARE DE PUTERE ÎN CLASA A ŞI AB
 1.7. AMLFCATOARE DE UTERE ÎN CLASA A Ş AB 1.7.1 Amplificatoare în clasa A La amplificatoarele din clasa A, forma de undă a tensiunii de ieşire este aceeaşi ca a tensiunii de intrare, deci întreg semnalul
1.7. AMLFCATOARE DE UTERE ÎN CLASA A Ş AB 1.7.1 Amplificatoare în clasa A La amplificatoarele din clasa A, forma de undă a tensiunii de ieşire este aceeaşi ca a tensiunii de intrare, deci întreg semnalul
Subiecte Clasa a VIII-a
 Subiecte lasa a VIII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate pe foaia de raspuns in dreptul
Subiecte lasa a VIII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate pe foaia de raspuns in dreptul
COLEGIUL NATIONAL CONSTANTIN CARABELLA TARGOVISTE. CONCURSUL JUDETEAN DE MATEMATICA CEZAR IVANESCU Editia a VI-a 26 februarie 2005.
 SUBIECTUL Editia a VI-a 6 februarie 005 CLASA a V-a Fie A = x N 005 x 007 si B = y N y 003 005 3 3 a) Specificati cel mai mic element al multimii A si cel mai mare element al multimii B. b)stabiliti care
SUBIECTUL Editia a VI-a 6 februarie 005 CLASA a V-a Fie A = x N 005 x 007 si B = y N y 003 005 3 3 a) Specificati cel mai mic element al multimii A si cel mai mare element al multimii B. b)stabiliti care
R R, f ( x) = x 7x+ 6. Determinați distanța dintre punctele de. B=, unde x și y sunt numere reale.
 = x 7x+ 6. Determinați distanța dintre punctele de. B=, unde x și y sunt numere reale.") 5p Determinați primul termen al progresiei geometrice ( b n ) n, știind că b 5 = 48 și b 8 = 84 5p Se consideră funcția f : intersecție a graficului funcției f cu aa O R R, f ( ) = 7+ 6 Determinați distanța
5p Determinați primul termen al progresiei geometrice ( b n ) n, știind că b 5 = 48 și b 8 = 84 5p Se consideră funcția f : intersecție a graficului funcției f cu aa O R R, f ( ) = 7+ 6 Determinați distanța
Esalonul Redus pe Linii (ERL). Subspatii.
. Subspatii.") Seminarul 1 Esalonul Redus pe Linii (ERL). Subspatii. 1.1 Breviar teoretic 1.1.1 Esalonul Redus pe Linii (ERL) Definitia 1. O matrice A L R mxn este in forma de Esalon Redus pe Linii (ERL), daca indeplineste
Seminarul 1 Esalonul Redus pe Linii (ERL). Subspatii. 1.1 Breviar teoretic 1.1.1 Esalonul Redus pe Linii (ERL) Definitia 1. O matrice A L R mxn este in forma de Esalon Redus pe Linii (ERL), daca indeplineste
Asupra unei inegalităţi date la barajul OBMJ 2006
 Asupra unei inegalităţi date la barajul OBMJ 006 Mircea Lascu şi Cezar Lupu La cel de-al cincilea baraj de Juniori din data de 0 mai 006 a fost dată următoarea inegalitate: Fie x, y, z trei numere reale
Asupra unei inegalităţi date la barajul OBMJ 006 Mircea Lascu şi Cezar Lupu La cel de-al cincilea baraj de Juniori din data de 0 mai 006 a fost dată următoarea inegalitate: Fie x, y, z trei numere reale
Curs 5 Structura acizilor nucleici
 BIOINFORMTICĂ PLICTĂ ÎN BIOLOI STRUCTURLĂ 3/28/2018 Curs V - Structura acizilor nucleici 1 29.03.2018 Curs 5 Structura acizilor nucleici Structura chimică a DN-ului - în 1869 Friedrich Miescher izolează
BIOINFORMTICĂ PLICTĂ ÎN BIOLOI STRUCTURLĂ 3/28/2018 Curs V - Structura acizilor nucleici 1 29.03.2018 Curs 5 Structura acizilor nucleici Structura chimică a DN-ului - în 1869 Friedrich Miescher izolează
Criptosisteme cu cheie publică III
 Criptosisteme cu cheie publică III Anul II Aprilie 2017 Problema rucsacului ( knapsack problem ) Considerăm un număr natural V > 0 şi o mulţime finită de numere naturale pozitive {v 0, v 1,..., v k 1 }.
Criptosisteme cu cheie publică III Anul II Aprilie 2017 Problema rucsacului ( knapsack problem ) Considerăm un număr natural V > 0 şi o mulţime finită de numere naturale pozitive {v 0, v 1,..., v k 1 }.
I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare.
 Capitolul 3 COMPUŞI ORGANICI MONOFUNCŢIONALI 3.2.ACIZI CARBOXILICI TEST 3.2.3. I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Reacţia dintre
Capitolul 3 COMPUŞI ORGANICI MONOFUNCŢIONALI 3.2.ACIZI CARBOXILICI TEST 3.2.3. I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Reacţia dintre
Capitolul 2 - HIDROCARBURI 2.5.ARENE
 Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.2 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Radicalul C 6 H 5 - se numeşte fenil. ( fenil/
Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.2 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Radicalul C 6 H 5 - se numeşte fenil. ( fenil/
4. Elemente de biologie celulară şi moleculară
 4. Elemente de biologie celulară şi moleculară O caracteristică esenţială a materiei vii este reprezentată de structura sa celulară, alături de încă două proprietăţi fundamentale: metabolismul şi reproducerea.
4. Elemente de biologie celulară şi moleculară O caracteristică esenţială a materiei vii este reprezentată de structura sa celulară, alături de încă două proprietăţi fundamentale: metabolismul şi reproducerea.
Ecuaţia generală Probleme de tangenţă Sfera prin 4 puncte necoplanare. Elipsoidul Hiperboloizi Paraboloizi Conul Cilindrul. 1 Sfera.
 pe ecuaţii generale 1 Sfera Ecuaţia generală Probleme de tangenţă 2 pe ecuaţii generale Sfera pe ecuaţii generale Ecuaţia generală Probleme de tangenţă Numim sferă locul geometric al punctelor din spaţiu
pe ecuaţii generale 1 Sfera Ecuaţia generală Probleme de tangenţă 2 pe ecuaţii generale Sfera pe ecuaţii generale Ecuaţia generală Probleme de tangenţă Numim sferă locul geometric al punctelor din spaţiu
Teoria mecanic-cuantică a legăturii chimice - continuare. Hibridizarea orbitalilor
 Cursul 10 Teoria mecanic-cuantică a legăturii chimice - continuare Hibridizarea orbitalilor Orbital atomic = regiunea din jurul nucleului în care poate fi localizat 1 e - izolat, aflat într-o anumită stare
Cursul 10 Teoria mecanic-cuantică a legăturii chimice - continuare Hibridizarea orbitalilor Orbital atomic = regiunea din jurul nucleului în care poate fi localizat 1 e - izolat, aflat într-o anumită stare
Activitatea A5. Introducerea unor module specifice de pregătire a studenților în vederea asigurării de șanse egale
 Investește în oameni! FONDUL SOCIAL EUROPEAN Programul Operațional Sectorial pentru Dezvoltarea Resurselor Umane 2007 2013 Axa prioritară nr. 1 Educația și formarea profesională în sprijinul creșterii
Investește în oameni! FONDUL SOCIAL EUROPEAN Programul Operațional Sectorial pentru Dezvoltarea Resurselor Umane 2007 2013 Axa prioritară nr. 1 Educația și formarea profesională în sprijinul creșterii
Sisteme diferenţiale liniare de ordinul 1
 1 Metoda eliminării 2 Cazul valorilor proprii reale Cazul valorilor proprii nereale 3 Catedra de Matematică 2011 Forma generală a unui sistem liniar Considerăm sistemul y 1 (x) = a 11y 1 (x) + a 12 y 2
1 Metoda eliminării 2 Cazul valorilor proprii reale Cazul valorilor proprii nereale 3 Catedra de Matematică 2011 Forma generală a unui sistem liniar Considerăm sistemul y 1 (x) = a 11y 1 (x) + a 12 y 2
Metode de interpolare bazate pe diferenţe divizate
 Metode de interpolare bazate pe diferenţe divizate Radu Trîmbiţaş 4 octombrie 2005 1 Forma Newton a polinomului de interpolare Lagrange Algoritmul nostru se bazează pe forma Newton a polinomului de interpolare
Metode de interpolare bazate pe diferenţe divizate Radu Trîmbiţaş 4 octombrie 2005 1 Forma Newton a polinomului de interpolare Lagrange Algoritmul nostru se bazează pe forma Newton a polinomului de interpolare
Geometrie computationala 2. Preliminarii geometrice
 Platformă de e-learning și curriculă e-content pentru învățământul superior tehnic Geometrie computationala 2. Preliminarii geometrice Preliminarii geometrice Spatiu Euclidean: E d Spatiu de d-tupluri,
Platformă de e-learning și curriculă e-content pentru învățământul superior tehnic Geometrie computationala 2. Preliminarii geometrice Preliminarii geometrice Spatiu Euclidean: E d Spatiu de d-tupluri,
V O. = v I v stabilizator
 Stabilizatoare de tensiune continuă Un stabilizator de tensiune este un circuit electronic care păstrează (aproape) constantă tensiunea de ieșire la variaţia între anumite limite a tensiunii de intrare,
Stabilizatoare de tensiune continuă Un stabilizator de tensiune este un circuit electronic care păstrează (aproape) constantă tensiunea de ieșire la variaţia între anumite limite a tensiunii de intrare,
Capitolul 2 - HIDROCARBURI 2.3.ALCHINE
 Capitolul 2 - HIDROCARBURI 2.3.ALCHINE TEST 2.3.3 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Acetilena poate participa la reacţii de
Capitolul 2 - HIDROCARBURI 2.3.ALCHINE TEST 2.3.3 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Acetilena poate participa la reacţii de
 10. STABILIZATOAE DE TENSIUNE 10.1 STABILIZATOAE DE TENSIUNE CU TANZISTOAE BIPOLAE Stabilizatorul de tensiune cu tranzistor compară în permanenţă valoare tensiunii de ieşire (stabilizate) cu tensiunea
10. STABILIZATOAE DE TENSIUNE 10.1 STABILIZATOAE DE TENSIUNE CU TANZISTOAE BIPOLAE Stabilizatorul de tensiune cu tranzistor compară în permanenţă valoare tensiunii de ieşire (stabilizate) cu tensiunea
2. Circuite logice 2.4. Decodoare. Multiplexoare. Copyright Paul GASNER
 2. Circuite logice 2.4. Decodoare. Multiplexoare Copyright Paul GASNER Definiţii Un decodor pe n bits are n intrări şi 2 n ieşiri; cele n intrări reprezintă un număr binar care determină în mod unic care
2. Circuite logice 2.4. Decodoare. Multiplexoare Copyright Paul GASNER Definiţii Un decodor pe n bits are n intrări şi 2 n ieşiri; cele n intrări reprezintă un număr binar care determină în mod unic care
RECOMBINAREA GENETICĂ
 14 RECOMBINAREA GENETICĂ Prin recombinare genetică se defineşte fenomenul producerii unor combinaţii genetice noi prin rearanjarea, reasortarea sau redistribuţia materialului genetic cuprins în două unităţi
14 RECOMBINAREA GENETICĂ Prin recombinare genetică se defineşte fenomenul producerii unor combinaţii genetice noi prin rearanjarea, reasortarea sau redistribuţia materialului genetic cuprins în două unităţi
BIOCHIMIA ACIZILOR NUCLEICI
 BIOCHIMIA ACIZILOR NUCLEICI Baze pirimidinice: pirimidina; citozina (2-oxo-4-amino-pirimidina); uracilul (2,4-dioxo-pirimidina); timina (5-metil-2,4-dioxo-pirimidina). Baze purinice: purina (imidazolpirimidină);
BIOCHIMIA ACIZILOR NUCLEICI Baze pirimidinice: pirimidina; citozina (2-oxo-4-amino-pirimidina); uracilul (2,4-dioxo-pirimidina); timina (5-metil-2,4-dioxo-pirimidina). Baze purinice: purina (imidazolpirimidină);
Profesor Blaga Mirela-Gabriela DREAPTA
 DREAPTA Fie punctele A ( xa, ya ), B ( xb, yb ), C ( xc, yc ) şi D ( xd, yd ) în planul xoy. 1)Distanţa AB = (x x ) + (y y ) Ex. Fie punctele A( 1, -3) şi B( -2, 5). Calculaţi distanţa AB. AB = ( 2 1)
DREAPTA Fie punctele A ( xa, ya ), B ( xb, yb ), C ( xc, yc ) şi D ( xd, yd ) în planul xoy. 1)Distanţa AB = (x x ) + (y y ) Ex. Fie punctele A( 1, -3) şi B( -2, 5). Calculaţi distanţa AB. AB = ( 2 1)
Fig Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36].
![Fig Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36].](/thumbs/72/67648751.jpg "Fig Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36].") Componente şi circuite pasive Fig.3.85. Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36]. Fig.3.86. Rezistenţa serie echivalentă pierderilor în funcţie
Componente şi circuite pasive Fig.3.85. Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36]. Fig.3.86. Rezistenţa serie echivalentă pierderilor în funcţie
BARAJ DE JUNIORI,,Euclid Cipru, 28 mai 2012 (barajul 3)
") BARAJ DE JUNIORI,,Euclid Cipru, 8 mi 0 (brjul ) Problem Arătţi că dcă, b, c sunt numere rele cre verifică + b + c =, tunci re loc ineglitte xy + yz + zx Problem Fie şi b numere nturle nenule Dcă numărul
BARAJ DE JUNIORI,,Euclid Cipru, 8 mi 0 (brjul ) Problem Arătţi că dcă, b, c sunt numere rele cre verifică + b + c =, tunci re loc ineglitte xy + yz + zx Problem Fie şi b numere nturle nenule Dcă numărul
CROMOZOMUL LA ORGANISME 3 PRO- SI EUCARIOTE
 CROMOZOMUL LA ORGANISME 3 PRO- SI EUCARIOTE 3.1 CROMOZOMUL BACTERIAN 3.1.1 Dimensiune Imensa majoritate a bacteriilor deţin ca material genetic esenţial o moleculă ADN dublu catenar circular covalent închis,
CROMOZOMUL LA ORGANISME 3 PRO- SI EUCARIOTE 3.1 CROMOZOMUL BACTERIAN 3.1.1 Dimensiune Imensa majoritate a bacteriilor deţin ca material genetic esenţial o moleculă ADN dublu catenar circular covalent închis,
Definiţia generală Cazul 1. Elipsa şi hiperbola Cercul Cazul 2. Parabola Reprezentari parametrice ale conicelor Tangente la conice
 1 Conice pe ecuaţii reduse 2 Conice pe ecuaţii reduse Definiţie Numim conica locul geometric al punctelor din plan pentru care raportul distantelor la un punct fix F şi la o dreaptă fixă (D) este o constantă
1 Conice pe ecuaţii reduse 2 Conice pe ecuaţii reduse Definiţie Numim conica locul geometric al punctelor din plan pentru care raportul distantelor la un punct fix F şi la o dreaptă fixă (D) este o constantă
5.1. Noţiuni introductive
 ursul 13 aitolul 5. Soluţii 5.1. oţiuni introductive Soluţiile = aestecuri oogene de două sau ai ulte substanţe / coonente, ale căror articule nu se ot seara rin filtrare sau centrifugare. oonente: - Mediul
ursul 13 aitolul 5. Soluţii 5.1. oţiuni introductive Soluţiile = aestecuri oogene de două sau ai ulte substanţe / coonente, ale căror articule nu se ot seara rin filtrare sau centrifugare. oonente: - Mediul
Capitolul ASAMBLAREA LAGĂRELOR LECŢIA 25
 Capitolul ASAMBLAREA LAGĂRELOR LECŢIA 25 LAGĂRELE CU ALUNECARE!" 25.1.Caracteristici.Părţi componente.materiale.!" 25.2.Funcţionarea lagărelor cu alunecare.! 25.1.Caracteristici.Părţi componente.materiale.
Capitolul ASAMBLAREA LAGĂRELOR LECŢIA 25 LAGĂRELE CU ALUNECARE!" 25.1.Caracteristici.Părţi componente.materiale.!" 25.2.Funcţionarea lagărelor cu alunecare.! 25.1.Caracteristici.Părţi componente.materiale.
Subiecte Clasa a VII-a
 lasa a VII Lumina Math Intrebari Subiecte lasa a VII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate
lasa a VII Lumina Math Intrebari Subiecte lasa a VII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate
Conice. Lect. dr. Constantin-Cosmin Todea. U.T. Cluj-Napoca
 Conice Lect. dr. Constantin-Cosmin Todea U.T. Cluj-Napoca Definiţie: Se numeşte curbă algebrică plană mulţimea punctelor din plan de ecuaţie implicită de forma (C) : F (x, y) = 0 în care funcţia F este
Conice Lect. dr. Constantin-Cosmin Todea U.T. Cluj-Napoca Definiţie: Se numeşte curbă algebrică plană mulţimea punctelor din plan de ecuaţie implicită de forma (C) : F (x, y) = 0 în care funcţia F este
Proiectarea filtrelor prin metoda pierderilor de inserţie
 FITRE DE MIROUNDE Proiectarea filtrelor prin metoda pierderilor de inserţie P R Puterea disponibila de la sursa Puterea livrata sarcinii P inc P Γ ( ) Γ I lo P R ( ) ( ) M ( ) ( ) M N P R M N ( ) ( ) Tipuri
FITRE DE MIROUNDE Proiectarea filtrelor prin metoda pierderilor de inserţie P R Puterea disponibila de la sursa Puterea livrata sarcinii P inc P Γ ( ) Γ I lo P R ( ) ( ) M ( ) ( ) M N P R M N ( ) ( ) Tipuri
Capitolul 2 - HIDROCARBURI 2.4.ALCADIENE
 Capitolul 2 - HIDROCARBURI 2.4.ALCADIENE TEST 2.4.1 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. Rezolvare: 1. Alcadienele sunt hidrocarburi
Capitolul 2 - HIDROCARBURI 2.4.ALCADIENE TEST 2.4.1 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. Rezolvare: 1. Alcadienele sunt hidrocarburi
Problema a II - a (10 puncte) Diferite circuite electrice
 Diferite circuite electrice") Olimpiada de Fizică - Etapa pe judeţ 15 ianuarie 211 XI Problema a II - a (1 puncte) Diferite circuite electrice A. Un elev utilizează o sursă de tensiune (1), o cutie cu rezistenţe (2), un întrerupător
Olimpiada de Fizică - Etapa pe judeţ 15 ianuarie 211 XI Problema a II - a (1 puncte) Diferite circuite electrice A. Un elev utilizează o sursă de tensiune (1), o cutie cu rezistenţe (2), un întrerupător
RĂSPUNS Modulul de rezistenţă este o caracteristică geometrică a secţiunii transversale, scrisă faţă de una dintre axele de inerţie principale:,
 REZISTENTA MATERIALELOR 1. Ce este modulul de rezistenţă? Exemplificaţi pentru o secţiune dreptunghiulară, respectiv dublu T. RĂSPUNS Modulul de rezistenţă este o caracteristică geometrică a secţiunii
REZISTENTA MATERIALELOR 1. Ce este modulul de rezistenţă? Exemplificaţi pentru o secţiune dreptunghiulară, respectiv dublu T. RĂSPUNS Modulul de rezistenţă este o caracteristică geometrică a secţiunii
Fiziologia fibrei miocardice
 Fiziologia fibrei miocardice CELULA MIOCARDICĂ = celulă excitabilă având ca şi proprietate specifică contractilitatea Fenomene electrice ale celulei miocardice Fenomene mecanice ale celulei miocardice
Fiziologia fibrei miocardice CELULA MIOCARDICĂ = celulă excitabilă având ca şi proprietate specifică contractilitatea Fenomene electrice ale celulei miocardice Fenomene mecanice ale celulei miocardice
(N) joncţiunea BC. polarizată invers I E = I C + I B. Figura 5.13 Prezentarea funcţionării tranzistorului NPN
 joncţiunea BC. polarizată invers I E = I C + I B. Figura 5.13 Prezentarea funcţionării tranzistorului NPN") 5.1.3 FUNŢONAREA TRANZSTORULU POLAR Un tranzistor bipolar funcţionează corect, dacă joncţiunea bază-emitor este polarizată direct cu o tensiune mai mare decât tensiunea de prag, iar joncţiunea bază-colector
5.1.3 FUNŢONAREA TRANZSTORULU POLAR Un tranzistor bipolar funcţionează corect, dacă joncţiunea bază-emitor este polarizată direct cu o tensiune mai mare decât tensiunea de prag, iar joncţiunea bază-colector
Ι Ο Λ Ο Γ Ι Μ Ο - Α Π Ο Λ Ο Γ Ι Μ Ο Μ Η Ν Ο Γ Δ Κ Δ Μ Β Ρ Ι Ο Υ 2 0 1 5
 Μ Ρ : 0 9 / 0 1 / 2 0 1 6 Ρ. Ρ Ω. : 7 Λ Γ Μ - Λ Γ Μ Μ Η Γ Δ Κ Δ Μ Β Ρ Υ 2 0 1 5 Δ Γ Ρ Ϋ Λ Γ Θ Δ ΚΔ Μ Β Δ Β Ω Θ Δ Δ Ρ Υ Θ Δ 0111 Χ / Γ Δ Θ Μ Θ Δ Ρ Ω Κ - - - 0112 Χ / Γ Λ Ρ Γ Κ Δ 2 3. 2 1 3. 0 0 0, 0 0-2
Μ Ρ : 0 9 / 0 1 / 2 0 1 6 Ρ. Ρ Ω. : 7 Λ Γ Μ - Λ Γ Μ Μ Η Γ Δ Κ Δ Μ Β Ρ Υ 2 0 1 5 Δ Γ Ρ Ϋ Λ Γ Θ Δ ΚΔ Μ Β Δ Β Ω Θ Δ Δ Ρ Υ Θ Δ 0111 Χ / Γ Δ Θ Μ Θ Δ Ρ Ω Κ - - - 0112 Χ / Γ Λ Ρ Γ Κ Δ 2 3. 2 1 3. 0 0 0, 0 0-2
SERII NUMERICE. Definiţia 3.1. Fie (a n ) n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0
 n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0") SERII NUMERICE Definiţia 3.1. Fie ( ) n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0 şirul definit prin: s n0 = 0, s n0 +1 = 0 + 0 +1, s n0 +2 = 0 + 0 +1 + 0 +2,.......................................
SERII NUMERICE Definiţia 3.1. Fie ( ) n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0 şirul definit prin: s n0 = 0, s n0 +1 = 0 + 0 +1, s n0 +2 = 0 + 0 +1 + 0 +2,.......................................
Seminar 5 Analiza stabilității sistemelor liniare
 Seminar 5 Analiza stabilității sistemelor liniare Noțiuni teoretice Criteriul Hurwitz de analiză a stabilității sistemelor liniare În cazul sistemelor liniare, stabilitatea este o condiție de localizare
Seminar 5 Analiza stabilității sistemelor liniare Noțiuni teoretice Criteriul Hurwitz de analiză a stabilității sistemelor liniare În cazul sistemelor liniare, stabilitatea este o condiție de localizare
TEHNOLOGIA ADN RECOMBINANT
 11 TEHNOLOGIA ADN RECOMBINANT Descifrând enigmele structurii genelor, savanţii au început să se preocupe de izolarea şi sinteza lor. În urmă cu 30 de ani cei care credeau în reuşita acestor deziderate
11 TEHNOLOGIA ADN RECOMBINANT Descifrând enigmele structurii genelor, savanţii au început să se preocupe de izolarea şi sinteza lor. În urmă cu 30 de ani cei care credeau în reuşita acestor deziderate
2. Circuite logice 2.5. Sumatoare şi multiplicatoare. Copyright Paul GASNER
 2. Circuite logice 2.5. Sumatoare şi multiplicatoare Copyright Paul GASNER Adunarea în sistemul binar Adunarea se poate efectua în mod identic ca la adunarea obişnuită cu cifre arabe în sistemul zecimal
2. Circuite logice 2.5. Sumatoare şi multiplicatoare Copyright Paul GASNER Adunarea în sistemul binar Adunarea se poate efectua în mod identic ca la adunarea obişnuită cu cifre arabe în sistemul zecimal
* K. toate K. circuitului. portile. Considerând această sumă pentru toate rezistoarele 2. = sl I K I K. toate rez. Pentru o bobină: U * toate I K K 1
 FNCȚ DE ENERGE Fie un n-port care conține numai elemente paive de circuit: rezitoare dipolare, condenatoare dipolare și bobine cuplate. Conform teoremei lui Tellegen n * = * toate toate laturile portile
FNCȚ DE ENERGE Fie un n-port care conține numai elemente paive de circuit: rezitoare dipolare, condenatoare dipolare și bobine cuplate. Conform teoremei lui Tellegen n * = * toate toate laturile portile
Valori limită privind SO2, NOx şi emisiile de praf rezultate din operarea LPC în funcţie de diferite tipuri de combustibili
 Anexa 2.6.2-1 SO2, NOx şi de praf rezultate din operarea LPC în funcţie de diferite tipuri de combustibili de bioxid de sulf combustibil solid (mg/nm 3 ), conţinut de O 2 de 6% în gazele de ardere, pentru
Anexa 2.6.2-1 SO2, NOx şi de praf rezultate din operarea LPC în funcţie de diferite tipuri de combustibili de bioxid de sulf combustibil solid (mg/nm 3 ), conţinut de O 2 de 6% în gazele de ardere, pentru
Vectori liberi Produs scalar Produs vectorial Produsul mixt. 1 Vectori liberi. 2 Produs scalar. 3 Produs vectorial. 4 Produsul mixt.
 liberi 1 liberi 2 3 4 Segment orientat liberi Fie S spaţiul geometric tridimensional cu axiomele lui Euclid. Orice pereche de puncte din S, notată (A, B) se numeşte segment orientat. Dacă A B, atunci direcţia
liberi 1 liberi 2 3 4 Segment orientat liberi Fie S spaţiul geometric tridimensional cu axiomele lui Euclid. Orice pereche de puncte din S, notată (A, B) se numeşte segment orientat. Dacă A B, atunci direcţia
Reflexia şi refracţia luminii.
 Reflexia şi refracţia luminii. 1. Cu cat se deplaseaza o raza care cade sub unghiul i =30 pe o placa plan-paralela de grosime e = 8,0 mm si indicele de refractie n = 1,50, pe care o traverseaza? Caz particular
Reflexia şi refracţia luminii. 1. Cu cat se deplaseaza o raza care cade sub unghiul i =30 pe o placa plan-paralela de grosime e = 8,0 mm si indicele de refractie n = 1,50, pe care o traverseaza? Caz particular
Capitolul 2 - HIDROCARBURI 2.5.ARENE
 Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.3 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Sulfonarea benzenului este o reacţie ireversibilă.
Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.3 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Sulfonarea benzenului este o reacţie ireversibilă.
III. Reprezentarea informaţiei în sistemele de calcul
 Metode Numerice Curs 3 III. Reprezentarea informaţiei în sistemele de calcul III.1. Reprezentarea internă a numerelor întregi III. 1.1. Reprezentarea internă a numerelor întregi fără semn (pozitive) Reprezentarea
Metode Numerice Curs 3 III. Reprezentarea informaţiei în sistemele de calcul III.1. Reprezentarea internă a numerelor întregi III. 1.1. Reprezentarea internă a numerelor întregi fără semn (pozitive) Reprezentarea
Biochimia moleculelor informaţionale 14. XII XII Metabolismul ADN-ului
 Biochimia moleculelor informaţionale 14. XII. 2017 1730-1800 21. XII. 2017 1100-1500 Metabolismul ADN-ului Experimente ce indică localizarea și natura materialului genetic 1. Experimentele lui Hammerling
Biochimia moleculelor informaţionale 14. XII. 2017 1730-1800 21. XII. 2017 1100-1500 Metabolismul ADN-ului Experimente ce indică localizarea și natura materialului genetic 1. Experimentele lui Hammerling
Tranzistoare bipolare şi cu efect de câmp
 apitolul 3 apitolul 3 26. Pentru circuitul de polarizare din fig. 26 se cunosc: = 5, = 5, = 2KΩ, = 5KΩ, iar pentru tranzistor se cunosc următorii parametrii: β = 200, 0 = 0, μa, = 0,6. a) ă se determine
apitolul 3 apitolul 3 26. Pentru circuitul de polarizare din fig. 26 se cunosc: = 5, = 5, = 2KΩ, = 5KΩ, iar pentru tranzistor se cunosc următorii parametrii: β = 200, 0 = 0, μa, = 0,6. a) ă se determine
REACŢII DE ADIŢIE NUCLEOFILĂ (AN-REACŢII) (ALDEHIDE ŞI CETONE)
 (ALDEHIDE ŞI CETONE)") EAŢII DE ADIŢIE NULEFILĂ (AN-EAŢII) (ALDEIDE ŞI ETNE) ompușii organici care conțin grupa carbonil se numesc compuși carbonilici și se clasifică în: Aldehide etone ALDEIDE: Formula generală: 3 Metanal(formaldehida
EAŢII DE ADIŢIE NULEFILĂ (AN-EAŢII) (ALDEIDE ŞI ETNE) ompușii organici care conțin grupa carbonil se numesc compuși carbonilici și se clasifică în: Aldehide etone ALDEIDE: Formula generală: 3 Metanal(formaldehida
SEMINAR 14. Funcţii de mai multe variabile (continuare) ( = 1 z(x,y) x = 0. x = f. x + f. y = f. = x. = 1 y. y = x ( y = = 0
 ( = 1 z(x,y) x = 0. x = f. x + f. y = f. = x. = 1 y. y = x ( y = = 0") Facultatea de Hidrotehnică, Geodezie şi Ingineria Mediului Matematici Superioare, Semestrul I, Lector dr. Lucian MATICIUC SEMINAR 4 Funcţii de mai multe variabile continuare). Să se arate că funcţia z,
Facultatea de Hidrotehnică, Geodezie şi Ingineria Mediului Matematici Superioare, Semestrul I, Lector dr. Lucian MATICIUC SEMINAR 4 Funcţii de mai multe variabile continuare). Să se arate că funcţia z,
z a + c 0 + c 1 (z a)
") 1 Serii Laurent (continuare) Teorema 1.1 Fie D C un domeniu, a D şi f : D \ {a} C o funcţie olomorfă. Punctul a este pol multiplu de ordin p al lui f dacă şi numai dacă dezvoltarea în serie Laurent a funcţiei
1 Serii Laurent (continuare) Teorema 1.1 Fie D C un domeniu, a D şi f : D \ {a} C o funcţie olomorfă. Punctul a este pol multiplu de ordin p al lui f dacă şi numai dacă dezvoltarea în serie Laurent a funcţiei
FLUXURI MAXIME ÎN REŢELE DE TRANSPORT. x 4
 FLUXURI MAXIME ÎN REŢELE DE TRANSPORT Se numeşte reţea de transport un graf în care fiecărui arc îi este asociat capacitatea arcului şi în care eistă un singur punct de intrare şi un singur punct de ieşire.
FLUXURI MAXIME ÎN REŢELE DE TRANSPORT Se numeşte reţea de transport un graf în care fiecărui arc îi este asociat capacitatea arcului şi în care eistă un singur punct de intrare şi un singur punct de ieşire.
2.1 Sfera. (EGS) ecuaţie care poartă denumirea de ecuaţia generală asferei. (EGS) reprezintă osferă cu centrul în punctul. 2 + p 2
 ecuaţie care poartă denumirea de ecuaţia generală asferei. (EGS) reprezintă osferă cu centrul în punctul. 2 + p 2") .1 Sfera Definitia 1.1 Se numeşte sferă mulţimea tuturor punctelor din spaţiu pentru care distanţa la u punct fi numit centrul sferei este egalăcuunnumăr numit raza sferei. Fie centrul sferei C (a, b,
.1 Sfera Definitia 1.1 Se numeşte sferă mulţimea tuturor punctelor din spaţiu pentru care distanţa la u punct fi numit centrul sferei este egalăcuunnumăr numit raza sferei. Fie centrul sferei C (a, b,
2CP Electropompe centrifugale cu turbina dubla
 2CP Electropompe centrifugale cu turbina dubla DOMENIUL DE UTILIZARE Capacitate de până la 450 l/min (27 m³/h) Inaltimea de pompare până la 112 m LIMITELE DE UTILIZARE Inaltimea de aspiratie manometrică
2CP Electropompe centrifugale cu turbina dubla DOMENIUL DE UTILIZARE Capacitate de până la 450 l/min (27 m³/h) Inaltimea de pompare până la 112 m LIMITELE DE UTILIZARE Inaltimea de aspiratie manometrică
Capitolul 14. Asamblari prin pene
 Capitolul 14 Asamblari prin pene T.14.1. Momentul de torsiune este transmis de la arbore la butuc prin intermediul unei pene paralele (figura 14.1). De care din cotele indicate depinde tensiunea superficiala
Capitolul 14 Asamblari prin pene T.14.1. Momentul de torsiune este transmis de la arbore la butuc prin intermediul unei pene paralele (figura 14.1). De care din cotele indicate depinde tensiunea superficiala
ΥΧΡΩΜΑ ΜΟΛΥΒΙΑ. «Γ λ υ κ ό κ α λ ο κ α ι ρ ά κ ι» της Γ ω γ ώ ς Α γ γ ε λ ο π ο ύ λ ο υ
 ΤΑ Π ΥΧΡΩΜΑ ΜΟΛΥΒΙΑ Εφη μ ε ρ ί δ α τ ο υ τ μ ή μ α τ ο ς Β τ ο υ 1 9 ου Δ η μ ο τ ι κ ο ύ σ χ ο λ ε ί ο υ Η ρ α κ λ ε ί ο υ Α ρ ι θ μ ό ς φ ύ λ λ ο υ 1 Ι ο ύ ν ι ο ς 2 0 1 5 «Γ λ υ κ ό κ α λ ο κ α ι ρ
ΤΑ Π ΥΧΡΩΜΑ ΜΟΛΥΒΙΑ Εφη μ ε ρ ί δ α τ ο υ τ μ ή μ α τ ο ς Β τ ο υ 1 9 ου Δ η μ ο τ ι κ ο ύ σ χ ο λ ε ί ο υ Η ρ α κ λ ε ί ο υ Α ρ ι θ μ ό ς φ ύ λ λ ο υ 1 Ι ο ύ ν ι ο ς 2 0 1 5 «Γ λ υ κ ό κ α λ ο κ α ι ρ
Transformata Laplace
 Tranformata Laplace Tranformata Laplace generalizează ideea tranformatei Fourier in tot planul complex Pt un emnal x(t) pectrul au tranformata Fourier ete t ( ω) X = xte dt Pt acelaşi emnal x(t) e poate
Tranformata Laplace Tranformata Laplace generalizează ideea tranformatei Fourier in tot planul complex Pt un emnal x(t) pectrul au tranformata Fourier ete t ( ω) X = xte dt Pt acelaşi emnal x(t) e poate
OBIECTIVE NUCLEUL. Caracteristici generale. pag. 1. Curs 3 histologie (Nucleul. Diviziunea celulară), anul I, sem. 1, 2017/2018
, anul I, sem. 1, 2017/2018") pag. 1 OBIECTIVE - familiarizarea studenților cu principalele caracteristici ale nucleului, structura și funcțiile acestuia; - definirea ciclului celular și a punctelor de restricție ca elemente importante
pag. 1 OBIECTIVE - familiarizarea studenților cu principalele caracteristici ale nucleului, structura și funcțiile acestuia; - definirea ciclului celular și a punctelor de restricție ca elemente importante
Capitolul 4 PROPRIETĂŢI TOPOLOGICE ŞI DE NUMĂRARE ALE LUI R. 4.1 Proprietăţi topologice ale lui R Puncte de acumulare
 Capitolul 4 PROPRIETĂŢI TOPOLOGICE ŞI DE NUMĂRARE ALE LUI R În cele ce urmează, vom studia unele proprietăţi ale mulţimilor din R. Astfel, vom caracteriza locul" unui punct în cadrul unei mulţimi (în limba
Capitolul 4 PROPRIETĂŢI TOPOLOGICE ŞI DE NUMĂRARE ALE LUI R În cele ce urmează, vom studia unele proprietăţi ale mulţimilor din R. Astfel, vom caracteriza locul" unui punct în cadrul unei mulţimi (în limba
MULTIMEA NUMERELOR REALE
 www.webmteinfo.com cu noi totul pre mi usor MULTIMEA NUMERELOR REALE office@ webmteinfo.com 1.1 Rdcin ptrt unui numr nturl ptrt perfect Ptrtul unui numr rtionl este totdeun pozitiv su zero (dic nenegtiv).
www.webmteinfo.com cu noi totul pre mi usor MULTIMEA NUMERELOR REALE office@ webmteinfo.com 1.1 Rdcin ptrt unui numr nturl ptrt perfect Ptrtul unui numr rtionl este totdeun pozitiv su zero (dic nenegtiv).
STRUCTURA BIOLOGICĂ. FORME DE REPREZENTARE 2.1. SECVENŢE DE AMINOACIZI: Sorana D. BOLBOACĂ
 STRUCTURA BIOLOGICĂ. 2.1. SECVENŢE DE AMINOACIZI: FORME DE REPREZENTARE Sorana D. BOLBOACĂ Despre 2 Informaţia genetică: Definiţie Forme de prezentare Genomul Codul genetic Aminoacizi: Definiţie Clasificare
STRUCTURA BIOLOGICĂ. 2.1. SECVENŢE DE AMINOACIZI: FORME DE REPREZENTARE Sorana D. BOLBOACĂ Despre 2 Informaţia genetică: Definiţie Forme de prezentare Genomul Codul genetic Aminoacizi: Definiţie Clasificare
a. 0,1; 0,1; 0,1; b. 1, ; 5, ; 8, ; c. 4,87; 6,15; 8,04; d. 7; 7; 7; e. 9,74; 12,30;1 6,08.
 1. În argentometrie, metoda Mohr: a. foloseşte ca indicator cromatul de potasiu, care formeazǎ la punctul de echivalenţă un precipitat colorat roşu-cărămiziu; b. foloseşte ca indicator fluoresceina, care
1. În argentometrie, metoda Mohr: a. foloseşte ca indicator cromatul de potasiu, care formeazǎ la punctul de echivalenţă un precipitat colorat roşu-cărămiziu; b. foloseşte ca indicator fluoresceina, care
Să se arate că n este număr par. Dan Nedeianu
 Primul test de selecție pentru juniori I. Să se determine numerele prime p, q, r cu proprietatea că 1 p + 1 q + 1 r 1. Fie ABCD un patrulater convex cu m( BCD) = 10, m( CBA) = 45, m( CBD) = 15 și m( CAB)
Primul test de selecție pentru juniori I. Să se determine numerele prime p, q, r cu proprietatea că 1 p + 1 q + 1 r 1. Fie ABCD un patrulater convex cu m( BCD) = 10, m( CBA) = 45, m( CBD) = 15 și m( CAB)
Repere pentru pregătirea către totalizarea II la Biologia Moleculară 1. Definiţi noţiunile
 Repere pentru pregătirea către totalizarea II la Biologia Moleculară 1. Definiţi noţiunile ADN-polimerază Aminoacid Anticodon Apoptoza ARN-polimeraza I ARN-polimeraza II ARN-polimeraza III CAP-are Catena
Repere pentru pregătirea către totalizarea II la Biologia Moleculară 1. Definiţi noţiunile ADN-polimerază Aminoacid Anticodon Apoptoza ARN-polimeraza I ARN-polimeraza II ARN-polimeraza III CAP-are Catena
7. Fie ABCD un patrulater inscriptibil. Un cerc care trece prin A şi B intersectează
 TEMĂ 1 1. În triunghiul ABC, fie D (BC) astfel încât AB + BD = AC + CD. Demonstraţi că dacă punctele B, C şi centrele de greutate ale triunghiurilor ABD şi ACD sunt conciclice, atunci AB = AC. India 2014
TEMĂ 1 1. În triunghiul ABC, fie D (BC) astfel încât AB + BD = AC + CD. Demonstraţi că dacă punctele B, C şi centrele de greutate ale triunghiurilor ABD şi ACD sunt conciclice, atunci AB = AC. India 2014
Algebra si Geometrie Seminar 9
 Algebra si Geometrie Seminar 9 Decembrie 017 ii Equations are just the boring part of mathematics. I attempt to see things in terms of geometry. Stephen Hawking 9 Dreapta si planul in spatiu 1 Notiuni
Algebra si Geometrie Seminar 9 Decembrie 017 ii Equations are just the boring part of mathematics. I attempt to see things in terms of geometry. Stephen Hawking 9 Dreapta si planul in spatiu 1 Notiuni
2. Sisteme de forţe concurente...1 Cuprins...1 Introducere Aspecte teoretice Aplicaţii rezolvate...3
 SEMINAR 2 SISTEME DE FRŢE CNCURENTE CUPRINS 2. Sisteme de forţe concurente...1 Cuprins...1 Introducere...1 2.1. Aspecte teoretice...2 2.2. Aplicaţii rezolvate...3 2. Sisteme de forţe concurente În acest
SEMINAR 2 SISTEME DE FRŢE CNCURENTE CUPRINS 2. Sisteme de forţe concurente...1 Cuprins...1 Introducere...1 2.1. Aspecte teoretice...2 2.2. Aplicaţii rezolvate...3 2. Sisteme de forţe concurente În acest
Examen AG. Student:... Grupa:... ianuarie 2011
 Problema 1. Pentru ce valori ale lui n,m N (n,m 1) graful K n,m este eulerian? Problema 2. Să se construiască o funcţie care să recunoască un graf P 3 -free. La intrare aceasta va primi un graf G = ({1,...,n},E)
Problema 1. Pentru ce valori ale lui n,m N (n,m 1) graful K n,m este eulerian? Problema 2. Să se construiască o funcţie care să recunoască un graf P 3 -free. La intrare aceasta va primi un graf G = ({1,...,n},E)
Biochimia moleculelor informaţionale. 11.I Metabolismul ADN-ului 3 Degradarea ADN-ului si a nucleotidelor
 Biochimia moleculelor informaţionale 11.I. 2018-1100-1500 Metabolismul ADN-ului 3 Degradarea ADN-ului si a nucleotidelor Catabolismul ADN-ului Structura şi mecanismele de reparaţie fac ca molecula de ADN
Biochimia moleculelor informaţionale 11.I. 2018-1100-1500 Metabolismul ADN-ului 3 Degradarea ADN-ului si a nucleotidelor Catabolismul ADN-ului Structura şi mecanismele de reparaţie fac ca molecula de ADN