Celične strukture. mikroskopija. površina celic. celična stena. citoplazemska membrana. citoplazma. spore in sporulacija
|
|
|
- Βασίλης Στεφανόπουλος
- 6 χρόνια πριν
- Προβολές:
Transcript
1 Celične strukture mikroskopija površina celic celična stena citoplazemska membrana citoplazma spore in sporulacija
2 Mikroskopija mikrobiologija v svetlem polju mikroskopija v temnem polju stereomikroskopija fazno kontrastna mikroskopija Nomarski mikroskopija polarizacijska mikroskopija mikroskopija v bližnjem polju mikroskopija na atomsko silo fluorescenčna mikroskopija konfokalna fluorescenčna mikroskopija elektronska transmisijaksa mikroskopija vrstična elektronska mikroskopija

3 Mikroskopija v svetlem polju Za kvaliteto mikroskopa je najpomembnejša ločljivost, sposobnost ločevanja dveh točk. S povečevanjem objekta ne dosežemo veliko, če je ločljivost slaba. Ločljivost, D, je odvisna od: -aperture objektiva, α - lomnega količnika, n - valovne dolžine svetlobe, λ D = 0.61λ / (n x sin α) Zaradi tehničnih omejitev glede α, λ in n je meja ločljivosti pri svetlobni mikroskopiji okoli 0.2 µm.

4 Video svetlobna mikroskopija
5 Merjenje dimenzij s svetlobnim mikroskopom
6 Štetje mikroorganizmov s svetlobnim mikroskopom Število dobimo tako, da: preštejemo število celic upoštevamo redčitev vzorca upoštevamo volumen števne komore

7 Kontrastiranje

8 Barvanje po Gramu

9 Barvanje po Gramu G+ temno vijolično G- rožnato Barvo vedno primerjamo z ustrezno kontrolo (poznana G+ in G- bakterija, ki je bila enako pripravljena kot neznani vzorec.
10 Druga barvanja negativno barvanje (nigrozin, indijsko črno, kristal vijolično, metilensko modro) acidorezistentno barvanje (za barvanje mikobakterij) barvanje endospor barvanje cist (npr. ciste pri Azotobacter sp.) barvanje kapsule barvanje flagel barvanje citoplazmatskih vključkov (polifosfat, poli-β-hidroksibutirat, glikogen, kisli polisaharidi) barvanje nukleoida barvanje spirohet,rikecij, legionel, mikoplazm

11 Običajne oblike mikrobnih celic

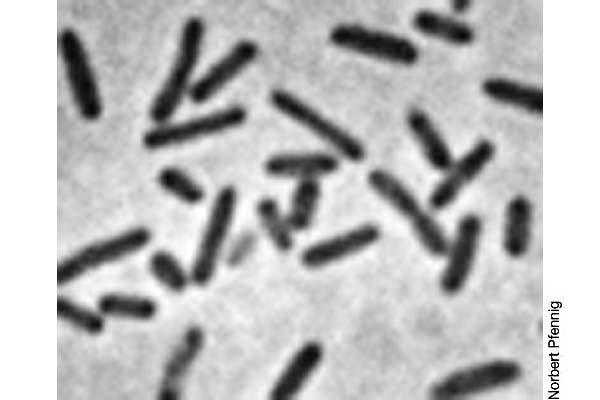



12 Oblike mikrobnih celic
13 Oblike kokov diplokoki streptokoki stafilokoki tetrade sarcine


14 Stereo mikroskopija Guignardia bidwellii, črna grozna gniloba

15 Mikroskopija v temnem polju Arachnoidiscus ehrenbergi, diatomeja



16 DIC (differential interference contrast) ali Nomarski mikroskopija Sordaria fimicola, plodno telesce z askusi
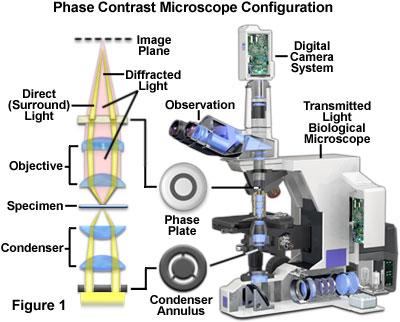

17 Fazno kontrastna mikroskopija


18 Polarizacijska mikroskopija a sferulit b polikarbonat c DNA

19 Epi-fluorescenčni mikroskop Olympus BX51

20 LSCM - laserska vrstična konfokalna mikroskopija


21 Dekonvolucijska mikroskopija


22 Fluorofori za G+ in G- bakterije Pseudomonas aeruginosa zeleno Bacillus cereus rumeno Pseudomonas aeruginosa zeleno Bacilus cereus oranžno


23 Fluorofori za ločevanje živih in mrtvih celic Micrococcus luteus žive celice zelene mrtve celice modre Micrococcus luteus in Bacilus cereus žive celice zelene mrtve celice rdeče

24 Fluorofori za določanje spor membrane rdeče DNA modro zeleno fuzijski zeleni protein
")
25 Fluorofori za določanje virusov Večje rumene pike celice, manjše zelene pike virusi (Syber Green-1)

26 Fluorofori na osnovi 16S rrna 16S rrna specifična barvila za dve vrsti bakterij na globokomorskem črvu



27 Fluorofori za določanje celičnih struktur aktin rdeče membrane zeleno jedro modro tubulin kromosomi

28 Konjugacija ciliate Spolna delitev ciliate Tetrahymena thermophila tubulin rdeče makro in mikrojedra zeleno

29 Fluorofori za vakuole in celične stene Sacharomyces cerevisae vakuola rdeča celična stena modra Sacharomyces cerevisae lumen vakuole moder

30 Fluorofori za ER in Golgijev aparat Endoplazmatski retikulum rdeč vakuole zelene Golgijev aparat rumen jedro zeleno tubulin rdeč


31 Fluorofori za mitohondrije mitohondriji rdeče aktin zeleno jedro modro lizosomi modro mitohondriji rdeče

32 Aktivnost alkalne fosfataze v celici aktivna mesta v celici, kjer deluje alkalna fosfataza so zelena jedro so modro obarvana

33 Fluorescenčna vizualizacija lipidnih domen Domenska struktura velikih lipidnih veziklov izgrajenih iz DPPC/DLPC mešanice.

34 TEM - transmisijska elektronska mikroskopija Osnovni principi so enaki kot pri svetlobni mikroskopiji. Namesto svetlobe uporabljamo snop elektronov in elektromagnetne leče. Zaradi absrpcije elektronov v zraku je potrebno delati v vakumu. Vir elektronov je vroča katoda z napetostjo kv, ki pospeši elektrone proti anodi. Elektrone detektiramo na fluorescenčnem zaslonu. ločljivost je ~ 0.1 nm potrebna posebna priprava vzorca


35 Priprava vzorca za TEM Ultramikroroton: vzorec fiksiramo v glutaraldehidu ali ozmijevem tetroksidu, ga posušimo v etanolu ali acetonu in ga nato damo v tekočo plastiko, ki jo s toploto polimeriziramo. Polimerizirano plastiko in vzorec razrežemo na ultratanke rezine ~ 0.1 µm, ki jih naložimo na mikroskopirne mrežice. Freze-fracture: vzorce hitro zamrznemo v tekočem dušiku in jih nato z ostrim nožem lomimo. Odlomljene dele naparimo s težkimi kovinami.
, z elektroni vrstično preizkujemo vzorec in detektiramo sekundarne elektrone, z obdelavo podatkov lahko dobimo navidezno 3D sliko.")
36 SEM - vrstična elektronska mikroskopija omogoča opazovanje površine vzorca vzorec je potrebno predhodno obdelati s težkimi kovinami (npr. platino), z elektroni vrstično preizkujemo vzorec in detektiramo sekundarne elektrone, z obdelavo podatkov lahko dobimo navidezno 3D sliko.
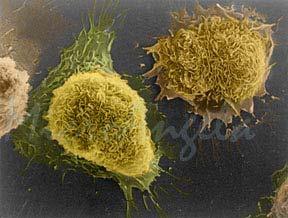


37 SEM slike


38 NSOM (Near Field Scanning Optical Microscopy)


39 AFM - mikroskopija na atomsko silo AFM bacteriorodopsina iz Holobacterium salinarium

40 Velikost mikrobnih celic
41 Zakaj so prokariontske celice manjše od evkariontskih? Z zmanjšanjem celice se povečuje razmerje med volumnom in površino. To pomeni, da imajo manjše celice večjo relativno površino. Npr.pri krogli je razmerje med površino S = 4πr 2 in volumnom V = 4/3πr 3 : pri r = 1 je S/V = 3 pri r = 2 je S/V = 1.5 pri r = 5 je S/V = 0.6 pri r = 10 je S/V = 0.3
42 Strukture, ki jih najdemo na površini bički fimbriji fibrile pili invazosomi kodri S-plast kapsule

43 Flagele - bički peritrihne flagele polarne flagele lopotrihne flagele lopotrihne flagele Flagela je izgrajena iz proteina flagelina, ki tvori dolgo votlo cev.

44 Struktura bakterijskega bička
45 Delovanje kemotaktičnega motorja CheR CH 3 CheW CheA P CheY P CH3 CheB P CheY CheY P CheZ


46 Taksa - gibanje bakterij v gradientu Poznamo več vrst takse: -kemotaksa -aerotaksa -magnetotaksa - fototaksa - ozmotaksa
 tanjši in številčnejši od flagel izgrajeni iz fimbrilinov in pilinov, ki tvorijo vijačnico pomagajo pri adsorpciji bakterij na")
47 Fimbriji strukturno podobni flagelam, vendar niso pomembne za gibanje so manjši (0.5 do 10 µm) tanjši in številčnejši od flagel izgrajeni iz fimbrilinov in pilinov, ki tvorijo vijačnico pomagajo pri adsorpciji bakterij na površino (adhezini) pomembni pri patogenezi in nastanku biofilmov
48 Fibrile krajše (do 0.5 µm), tanše in številčnejše od fimbrijev izgrajene iz večjih proteinov pomembne pri adsorpciji

49 Pili daljši in debelejši od fimbrij običajno jih je le nekaj (1 do 10) na celico specifični receptorji za viruse pomembni pri konjugaciji pomembni pri adheziji na površino
50 Curli - kodri zavite kodrasne strukture na površii celic sestavljene iz polisaharidov in proteinov najdemo samo pri E.coli in Salmonelli omogočajo vezavo na epitelijske celice gostitelja ali pa na fibronektin in plazminogen
51 Ivazosomi površinske proteinske strukture pri Salmonelli typhimurium podobni so čepkom, 3x debelejše od flagel in 10x krajše od flagel do 100 na celico po internalizaciji bakterije v celice gostitelja se struktura invazosoma razgradi

52 S-plast S-plast najdemo na površini nekaterih bakterijskih in arhejskih celic in ima heksagonalno ali tetragonalno simetrijo. S-plast je sestavljena iz ene vrste proteinov oziroma glikoproteinov in se samo-sestavi. Z ostalo celico je povezana preko nekovalentnih vezi. Integriteta S-plasti je vezana na prisotnost divalentnih kationov.
53 Kapsula Veliko mikroorganizmov na svojo površini izloča sluzast material, predvsem polisaharide in proteine. Kapsula ni trdno vezana na površino celice. Kapsula ima več funkcij: vezava na površino vezava na gostitelja zaloga za vodo in hranila zaščita pred infekcijo tvori matrkis pri nastanku biofilma močno antigena, pomembna pri virulenci
54 Centrifugiranje Diferencialno centrifugiranje: Centrifugiramo pri različnih pospeških oziroma različno dolgo npr. 10 min pri 5000 x g sedimentiramo intaktne celice 20 min prii x g fragmenti stene in večji membranski fragmenti 60 min pri x g sedimentiramo membranske vezikle in ribosome Ravnotežno gradientno centrifugiranje: Strukture s podobno velikostjo vendar različno gostoto lahko ločimo v saharoznem gradientu (lažje strukture) ali CsCl gradientu (težje strukture). v= d 2 (p p = 18η p l)g d je premer delca, p p je gostota delca, p l je gostota tekočine, g je zemeljski pospešek, η je viskoznost tekočine
55 Frakcioniranje celičnih komponent Mehansko razbijanje celic: francoska preša (1000 atm) ultrasonikacija zamrzovanje in odtajanje mikrovalovna pečica Kemijska liza celic: detergenti ozmotska liza segrevanje alkalna liza
56 Celična stena bakterij

57 Osnovna sladkorja peptidoglikana N-acetilglukozamin N-acetil muramična kislina

58 Osnovna enota peptidoglikana N-acetilglukozamin N-acetilmuramična kislina
59 Povezava glikanskih verig pri G+ in G- bakterijah M-G-M-G-M-G-M-G G- M-G-M-G-M-G-M-G G+ L-Ala L-Ala D-Glu D-Glu DAP D-Ala D-Ala DAP D-Glu D-Ala [Gly] 5 L-Lys D-Ala [Gly] 5 D-Ala L-Lys D-Glu D-Ala [Gly] 5 M-G-M-G-M-G-M-G M-G-M-G-M-G-M-G
60 Razlike med peptidoglikani preko 100 kemijsko različnih peptidoglikanov G- bakterije imajo DAP (diaminopimelično kislino), G+ bakterije imajo namesto DAP običajno lizin. različne povezave med tetrapeptidi različna sestava tetrapeptidov (npr. glicin, treonin, serin in aspartat). V celični steni ne najdemo razvejanih aminokislin, aromatskih aminokislin, žveplo vsebujočih aminokislin, histidina, arginina in prolina.
61 Sinteza peptidoglikana Dva temeljna problema: kako pridejo sestavni deli v periplazmo kako poteka sinteza peptidoglikana v periplazmi brez ATP Sinteza peptidoglikana poteka v več stopnjah: - sinteza pentapeptida in derivatizacija UDP sladkorjev - prenos pentapeptida in derivatiziranih sladkorjev preko membrane z lipidnim prenašalcem baktoprenolom - polimerizacija osnovne enote peptidoglikana v periplazmi - transpeptidacija
62 Nastanek in derivatizacija UDP sladkorjev fruktoza-6-p glukozamin-6-p N-acetilglukozamin-6-P N- acetilglukozamin-1-p UDP-N-acetilglukozamin UDP-Nacetilglukozamin-3-enolpiruvileter UDP-N-acetilmuramična kislina UDP-muramil-pentapeptid Nastanek UDP-muramil pentapeptida in UDP-N-acilgkukozamina je vezana na citoplazmo.
63 Prenos UDP-sladkorjev preko membrane citoplazma UDP-M citoplazmatska membrana C 55 -P periplazma UMP UDP-G M - PP - C 55 C 55 -PP - M -G -M -G -M -G C 55 - PP - M - G G -M -PP -C 55 UDP

64 Rast peptidoglikana Komponente celične stene so dodajajo v obstoječo celično steno v katero zareže avtolizin zareže, kar omogoča dodajanje nove osnovne enote peptidoglikana. Zaradi tega na mestu brazgotina dodajanja nastane brazgotina. V kolikor avtolizin zareže, nov celični material pa ni dodan, lahko pride do avtolize.
65 Transpeptidacija - rast peptidoglikana Reakcijo katalizirata karboksipeptidaza in transpeptidaza, ki sta pomembni tarči za delovanje antibiotikov.
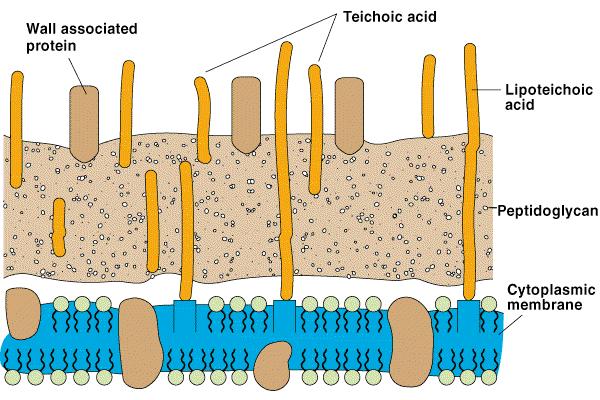
66 G+ bakterijska stena
67 Tehojske in tehuronske kisline O O -O-P-O-CH 2 -CH -CH 2 -O-P-O- O - O O - R n R=alanin glicerol fosfat tehojska kislina O O R=alanin GlcNac R -O-P-O-CH 2 -CH -CH 2 -O-P-O- O - OH O - n glicerol fosfat tehojska kislina O O R=glukoza -O -P -O -CH 2 -CH -CH -CH -CH 2 -O -P -O - O - O OH OH R O - n ribitol fosfat tehuronska kislina - ManNacUA - D-Glu - - D-GluUA - GalNac - tehuronska kislina z N-acetilmanuronsko kislino tehuronska kislina z N-acetilgalaktozaminom
68 Lipotehojske kisline polimeri glicerol fosfata, ki je substituiran z D-alaninom in sladkorji kovalentno vezane na lipide vzdržujejo strukturo celične stene vezavno mesto za površinske proteine vzdržujejo negativen naboj celične površine antigeni, povzročajo vnetni odziv
69 Proteini vezani na površino večinoma so kovaletno vezani na stranske verige aminokislin v peptidoglikanu, ostali interagirajo s tehojsko in lipotehojsko kislino pomembni pri adheziji pomembni pri razgradnji polimerov pomembni pri indukciji citokinov
")
70 M - protein streptokokov A štrli do 60 nm izven membrane. omogoča serotipizacijo streptokokov A (obstaja preko 80 različnih tipov) pomemben virulenčni dejavnik
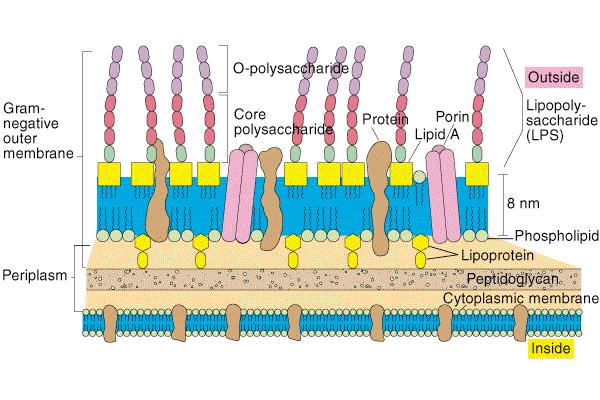
71 G- bakterijska stena
72 Periplazma Komponente periplazme: - oligosaharidi (ozmotska regulacija) - proteini, ki vežejo topljence (npr. transport sladkorjev, amino kislin, železo) - citokormi (npr. citokrom c) - hidrolitični encimi (npr. alkalna fosfataza, glikozidaze) - detoksificirajoči agensi (npr. β-laktamaza)
73 Lipoprotein Zunanja membrana gram negativnih bakterij je povezana s peptidoglikanom preko lipoproteina z nekovalentnimi vezmi, kar omogoča selektivno odstranitev zunanje membrane.
74 Nastanek protoplastov Peptidoglikan lahko razgradimo z lizocimom, ki cepi 1-4 β-glikozidno vez. Ob dodatku EDTA se zunanja membrana loči. Celice brez celične stene imenujemo protoplasti. Velikokrat pa celične stene ne moremo popolnoma odstraniti in dobimo sferoplaste. Nekatere bakterije (npr. mikoplazme) lahko preživijo brez celične stene, vendar imajo v svoji plazmatski membrani povečano koncentracijo sterolov, ki omogočajo vzdrževanje integritete celične membrane.
75 Zunanja membrana porin Notranji monosloj je sestavljen predvsem iz fosfolipidov, medtem, ko je zunanji monosloj sestavljen predvsem iz lipopolisaharidov LPS. Relativno propustna membrana.

76 Porin Porin neselektivno prepušča molekule, ki so manjše od 600 Da, najdemo ga samo v zunanji membrani gram negativnih bakterij. V celici je več različnih porinov, npr. E.coli ima: OmpC porin ima najmanjše pore, difuzijska bariera OmpF porin ima večje pore, olajšan transport PhoE porin je selektivna pora za transport fosfata
77 LPS - lipopolisaharid O - antigen sredica lipid A P KDO P Glu- Nac Gal Hep P KDO GlcN Glu Gal Glu Hep Hep KDO GlcN n P P GlcN KDO Hep Glu Gal GluNac glukozamin ketodeoksikonat heptoza glukoza galaktoza N-acetilglukozamin
78 Sinteza LPS Sinteza lipida A: fruktoza-6-p glukozamin-6-p N-acetilglukozamin-6-P N-acetilglukozamin-1-P UDP-N-acetilglukozamin UDP-3-monomiristiol- GlcNac UDP-2,3-dimiristoilglukozamin 1-6 tetramiristoil disaharid-1-p tetramiristoil-disaharid-1,4-difosfat lipid A Sinteza sredice: na lipid A se ena za drugo vezejo ketodeoksonat, heptoza, glukoza, galaktoza in N-acetilglukozamin. Ko so na lipid A vezani vsi sladkorji sredice se lipid A skupaj s sredico prenese v periplazmatski prostor, kjer se dodajo sladkorji O antigena. Sinteza O-antigena: sinteza poteka ločeno od sinteza lipida A in sredice. Sinteza poteka z baktoprenolom podobno kot pri sintezi peptidoglikana. O- antigen raste tako, da baktoprenol s sladkorjem iizpodrine baktoprenol, ki ima vezano veriogo sladkorjev O-antigena.
79 Sinteza LPS citoplazmatska membrana sredica Lipid A Lipid A sredica -O -O -O -O -O O C 55 -PP C 55 -PP -O -O -O -O -O C 55 -PP -O -O -O -O O - PP - C 55 C 55 - PP - O

80 Pseudopeptidoglikan (arheje) Bistvena razlika med bakterijskim in arhejskim peptidoglikanom je: N-acetiltalosaminuronska kislina namesto N-acetilmuramične kisline 1-3 β-glikozidna vez namesto 1-4 β-glikozidne vezi nekatere arheje sploh nimajo pseudopeptidoglikana in imajo le polisaharide ali proteine (S-plast)

81 Biološka membrana
82 Funkcije biološke membrane transport snovi v celico in iz celice energijske transformacije signaliziranje gibanje
83 Gradnja biološke membrane Lipidi: predstavljajo semipermeabilno pregrado in matriks v katerem so vgrajeni proteini. Proteini: skrbijo za encimske reakcije v membrani, omogočajo strukturne povezave s citoskeletom, mediirajo energijske transformacije, sprejemajo in prevajajo kemijske signale. Ogljikovi hidrati: v povezavi z lipidi in proteini skrbijo za sprejem signala, tvorijo zaščito pred fizikalno-kemijskim stresom in so rezervoar za hranila.
84 Običajne fosfolipidne glave NH 3 + -CH 2 -C-COO - H -CH 2 -CH 2 -NH+ 3 fosfatidil serin, PS fosfatidil etanolamin, PE -CH 2 -CH -CH 2 OH OH fosfatidil glicerol, PG O difosfatidil glicerol (kardiolipin), CL -CH 2 -CH -CH 2 -O-P-CH 2 -CH -CH 2 OH OH O- OH
85 Običajne maščobne kisline nasičene maščobne kisline C atmov: 1, 2, 3, 4, 6, 10, 16, 18 (-C -C -C -) mono nenasičene maščobne kisline C atomov: 4, 12, 14, 16, 18 (- C - C = C - C -) di nenasičene maščobne kisline C atomov: 6, 18 (- C = C - C = C -) tri nenasičene maščobne kisline C atomov: 18 (- C = C - C = C - C = C -) tetra nenasičene maščobne kisline C atomov: 20 (- C = C - C = C - C = C - C = C -) penta nenasičene maščobne kisline C atomov: 20 (- C = C - C = C - C = C - C = C - C = C -)
86 Običajne maščobne kisline Hidroksialkanojske kisline C atmov: 12, 18 (- CH 2 -CH -COOH) OH Keto in ciklo maščobne kisline C atmov: 5, 19 (-C -C -C -) (-C -C -C -C -) O CH 2 Razvejane maščobne kisline C atomov: 5, 10, 16, 19, 20 CH 3 CH 2 CH 3 CH - (CH 2 )n - COOH CH - (CH 2 )n - COOH CH 3 CH 3

87 Kemijske vezi, ki jih najdemo pri lipidih etrske vezi samo pri arhejah estrske vezi pri bakterijah in evkariontih izoprenska enota samo pri arhejah

88 Dinamika lipidnih molekul porazdelitev fosfatne skupine vzdolž lipidnega dvosloja verjetnost

89 Dinamika lipidnih molekul



90 Polimorfizem lipidov invezna bikontinuirana kubična faza Pn3m inverzna heksagonalna faza H II Multilamelarni lipidni dvosloj

91 Struktura in topologija integralnih membranskih proteinov Dve najbolj običajni strukturi membranskih proteinov α-heliks β-naguban list
92 Topologija perifernega peptida v lipidnem dvosloju
93 Intracitoplazemske membrane Veliko prokariontov ima v citoplazmi močno nagubane membrane, ki so povezane s citoplazmatsko membrano in imajo posebno fiziološko funkcijo. Takšne membrane najdemo pri: metanotrofih fiksatorjih dušika nitrifikatorjih fototrofih
94 Lipidno-proteinske interakcije Za pravilno funkcioniranje biološke membrane je pomembna: sestava lipidov in proteinov ter njihovo razmerje in selektivnost konfiguracija lipidnih acilnih verig ter njihova fluidnost ukrivljenost lipidnega dvosloja interakcija lipidnih polarnih glav s proteini struktura proteina topologija proteina agregacijski status proteinov hidrofobno neujemanje proteina in lipidnega dvosloja
95 Transport snovi skozi membrano Vsa hranila morajo skozi citoplazmatsko membrano. Za nekatere npr. O 2, CO 2, NH 3 in H 2 O velja, da hitro difundirajo skozi membrano. Nižje maščobne kisline lahko prosto prehajajo membrano (višje maščobne kisline potrebujejo transporterje). Za vse ostale spojine velja, da je njihova difuzija skozi membrano močno omejena in je potreben aktivni transport.
96 Difuzija snovi skozi membrano Membrana je različno permeabilna za snovi. Permeabilnost (P) za: vodo ~ 10-3 cm/s glicerol ~ 10-5 cm/s glukozo ~ 10-7 cm/s Cl - ~ cm/s Na + ~ cm/s Tok snovi skozi membrano je J = c P S kjer je c razlika v koncentraciji, P je permeabilnost, S je površina membrane. Za glukozo velja, da bi bil pri razliki v koncentracije glukoze znotraj in zunaj celice 10-4 mol/cm 3 in permeabilnostjo = 10-7 cm/s pretok skozi 1 cm 2 membrane ~10-11 mol/sek oziroma 6 x 10 4 molekul na sekundo skozi 1µm 2.


97 Difuzija vode skozi akvaporine

98 Kinetika transporta topljencev skozi membrano Z difuzijo poteka transport počasi. Z aktivnim transportom poteka prenos hitro vendar prihaja pri višjih koncentracijah substrata do saturiranosti transporterja. Koncentracijo substrata pri kateri je hitrost transporta enaka 1/2 max hitrosti imenujemo K m ali afinitetna konstanta za substrat.
99 Aktivni transport snovi skozi membrano Različno velike spojine potujejo skozi membrano na različne načine: majhni topljenci (npr. ioni, monomeri) s permeazami in transporterji veliki topljenci (npr. proteini) s transporterji delci s fagocitozo
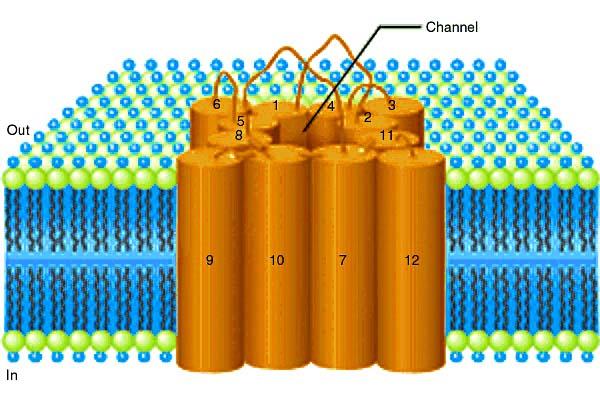
100 Struktura transporterjev
101 Nastanek gradientov preko membrane Snov vedno potuje vzdolž gradienta. Kontinuiran nastanek gradientov H +, K +, Na + ali H 2 PO 4- ionov preko membrane omogočajo metabolne reakcije s pomočjo: svetlobe redoks reakcij kotransporta hidrolize ATP Zmanjševanje gradienta ionov preko membrane spremlja sproščanje energije.
102 Energetika prenosa naboja preko membrane Sprememba proste energije zaradi transporta spojine preko membrane je G = -RT ln c in /c out Sprememba proste energije zaradi prenosa naboja preko membrane je G = nfv pri ravnotežnih pogojih velja nfv -RT ln c in /c out = 0 oziroma, električni potencial preko membrane, ko ni toka elektronov je V = RT/nF ln c in /c out V primeru, da je koncentracija K + v celici 140 mm, zunaj pa 5 mm se pri sobne temperaturi razvije 89 mv potenciala.

103 Sekundarni transportni sistemi Sekundarne transportne sisteme poganjajo sklopitve s primarnimi transportnimi sistemi, ki generirajo gradient ionov preko membrane. -simporterji - antiporterji
104 Energetika sekundarnega transporta simport elektronevtralnega topljenca s protonom [H + ]/[S] p = -60 log [S in ]/[S out ] H + S simport monovalentnega aniona s protonom H + ([H + ]/[S - ] - 1) -[H + ]/[S - ] 60 ph = -60 log [S - in ]/[S- out ] S - antiport kationa s protonom H + ([H + ]/[S + ] - 1) -[H + ]/[S + ] 60 ph = -60 log [S + in ]/[S+ out ] S -
105 Transport s simportom in antiportom simport antiport H + /glicin H + /galaktoza H + /Ca 2+ H + /histidin H + /laktoza H + /CaHPO 4 H + /lizin H + /fosfat H + /K + H + /fenilalanin Na +, H + /glutamat H + /Na + H + /laktat Na + /prolin K + /CH 3 NH 4 H + /piruvat Na + /melobioza H 2 PO 4- /organski anioni H + /sukcinat Mg 2+,H + /citrat H 2 PO 4- / heksoza-6-p H + /glukonat H + /arabinoza H 2 PO 4- / glicerol-3-p H 2 PO 4- / fosfoenolpiruvat

106 Transportni sistemi Haemophilus influenzae

107 Fosforilacija sladkorja po prehodu skozi membrano Sisetem za prenos sladkorjev je izgrajen iz 24 proteinov. Fosfatna skupina prihaja iz fosfoenolpiruvata. Glukoza-6P vstopa direktno v glikolizo.

108 ABC transportni sistemi pri G- bakterijah ABC je akronim za ATPbinding cassette in je sestavljen iz 3 delov. Pri prokariontih poznamo več kot 200 različnih ABC transporterjev. Za vse ABC transporterje je značilna visoka konstanta za vezavo substrata v periplazmi.

109 Transport železa, železo-kelirajoč agensi (hidroksamat) hidrokamat Fe 3+ Fe 3+ - hidrokamat citoplazemska membrana Fe 3+ - hidrokamat Fe 3+ -hidrokamat hidrokamat Fe 2+ redukcija porfirin hem
110 Transporterji za ostale ione ion Ca 2+ Fe 3+ Cu 2+ Ni 2+ Zn 2+ Co 2+ transporter fosfoprotein siderogfori/citrat/transferin caeruloplazmin albumin/histidin albumin vitamin B12
111 Transport proteinov skozi membrano Obstaja več sistemov za transport proteinov skozi membrano: Sec sistem tip I tip II tip III tip IV tip V šaperoni zunanja membrana notranja membrana šaperoni tip II tip IV tip V tip I Sec Tat tip III
112 Tip I sistem Tol C Hyl D Hyl B ADP + P i ATP Pul E
113 Tip III sistem TARČNA CELICA gostiteljska membrana Yop B, D zunanja membrana Lcr D / Ysc notranja membrana ADP + P i ATP
114 Sec sistem SecD, F SecY, E, G FtsY SecB SRP ribosom
115 Tat sistem Tat A, B, E citoplazemska membrana
116 Tip II sistem Pul D Pul C, F - O SecD, F SecY, E, G Pul E
117 Citoplazma inkluzijska telesca in granule ribosomi plinski vezikli nukleoid citosol znotrajcelične membrane poli hidroksi butirati polifosfati polisilfudi elementarno žveplo glikogen karboksisom, klorosom magnetosom metanotrofi nitrifikatorji fototrofi fiksatorji dušika

118 Granule elementarnega žvepla Veliko mikroorganizmov lahko oksidira H 2 S in tiosulfat. Pri tem lahko pride do akumulacije elementarnega žvepla v obliki granul. Granule se večajo dokler je na voljo reducirana oblika žvepla. Ko reducirane oblike zmanjka pride do porabe granul in oksidacije žvepla do sulfata.
119 Polifosfat Veliko mikroorganizmov akumulira rezerve anorganskega fosfata v obliki granul polifosfata. Anorganski fosfat je pomemben pri celični energetiki in signaliziranju. Polifosfat v celici lahko pobarvamo z bazičnimi barvili.
120 Poli-β-hidroksialkaonati Inkluzijska telesca iz poli-β-hidroksibutirata so izgrajena iz več podenot poli-β-hidroksibutirata povezanih z esterskimi vezmi. Dolžina monomerov je različna od C4 (butirat) do C18. Poli-β-hidroksialkaonate producirajo bakterije in arheje. Poli-β hidroksialkaonate uporabljamo za izdelavo biorazgradljive plastike. O CH 3 O CH 3 O CH 3 C O CH C CH C CH CH 2 O CH 2 O CH 2
121 Glikogen V glikogenu so glukozni ostanki povezani z α-1,4 in α-1,6 glikozidno vezjo. Granule glikogena so običajno majhne. Glikogen se podobno kot poli-β-hidroksalkanoati uporablja kot energijska rezerva in kot vir ogljika.
122 Karboksisom Avtotrofne bakterije, ki uporabljajo CO 2 kot edini ali glavni vir ogljika imajo velika (do 100 nm) poliedrska inkluzijska telesca, v katerih se nahaja ribuloza-bifosfat (glavni encim Calvinovega cikla). Take vključke najdemo pri: nitrifikatorjih žveplo oksidirajočih bakterijah cijanobakterijah

123 Klorosom Zelene žveplove in nežveplove bakterije imajo elipsoidno inkluzijsko telesce, ki ga obdaja membrana. Klorosom je pomemben pri fotosintezi.

124 Magnetotaktične bakterije in magnetosomi Magnetosomi so intracelularni kristali magnetita Fe 3 O 4. Magnetosom daje celici permanenten magnetni dipol, kar omogoča bakteriji, da se orientira v magnetnem polju. Magnetosomi so obdani s fosfolipidno membrano in glikoproteini.


125 Plinski vezikli Omogočajo celici lebdenje v vodnem stoplu, izgrajeni iz proteinov ~ 2 nm debeli, kar omogoča rigidnost. V veziklih je sestava atmosfere enaka kot v citoplazmi, napolnjeni vezikli so od GvpA 5 do 20 krat lažji od ostalega celičnega materiala. GvpC

126 Ribosomi Ribosomi so ribonukleoproteinski delci, kjer poteka sinteza proteinov. So ~ 30 nm veliki, sestavljeni iz velike in male podenote. Njihovo število se spreminja glede na metabolni status celice.

127 Nukleoid Nukleoid je mesto sinteze DNA in RNA. Nahaja se v središču bakterij. Hitro rastoče bakterije odprta struktura supernavita struktura imajo lahko več nukleoidov. Bazični proteini ovijajo DNA in skrbijo za njeno supernavitje.
128 Citosol Citosol je tekoči del citoplazme. V citosolu so raztopljeni minerali, monomeri, intermediati metabolizma, nukleinske kisline, oligosaharidi in vodotopni proteini. Med proteini v citosolu ločimo: samostojne proteine encimske komplekse: npr. piruvat dehidrogenaza (sestavljen iz 3 različnih encimov in 50 proteinov), α-ketoglutarat (3 različni encimi in 48 različnih proteinov, maščobno kislinska sintetaza (7 različnih encimov)
129 Neugodni pogoji za rast Mikrobne celice imajo izjemno sposobnost, da ostanejo žive v pogojih, ki ne dovoljujejo rasti in začno zelo hitro rasti, ko ponovno nastopijo ugodni pogoji. Mikrobne celice se na neugodne pogoje v okolju prilagodijo tako da: zmanjšajo metabolno aktivnost agregirajo s sosednjimi celicami spremenijo morfologijo sporulirajo
130 Morfološke spremembe stradajočih celic Zaradi stradanja se spremeni morfologija celic glede na rastoče celice. Celice postanejo: manjše okrogle spremenijo se razmerja med subcelularni kompartmenti (kondenzirana citoplazma, povečana periplazma) poveča se hidrofobnost površine celic (agregacija) spremeni se sestave membrane (povečanje nasičenosti lipidov) spremeni se struktura celične stene prihaja do kondenzacije kromosomov in plazmidov
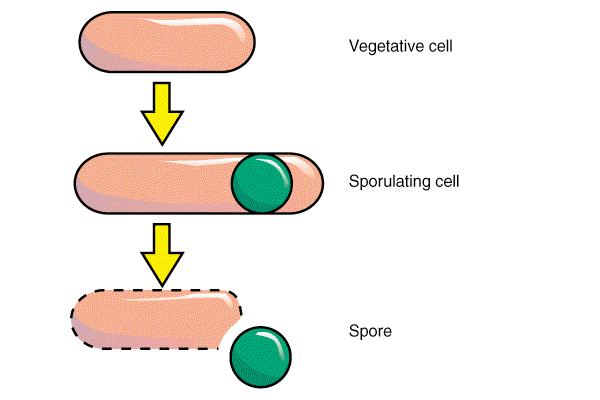
131 Spore in sporulacija

132 Nastanek spore I

133 Nastanek spore II

134 Nastanek spore III

135 Struktura evkariontske celice
136 Evkariontska celica Obnovi znanje o strukturi evkariotske celice, ki si ga pridobil pri predmetu Biologija celice: sestava evkariontske membrane organeli in njihova funkcija struktura citoskeleta struktura znotrajceličnega vezikularnega transporta struktura signalnih poti pri evkariontskih celicah struktura evkariontskega genetskega aparata
137 Primerjava prokarionti : evkarionti struktura/funkcija prokarionti evkarionti skupine bakterije, arheje alge, glive, protozoji, rastline, živali velikostni rang < 2 µm > 2 µm celična organizacija enocelični enocelični, večcelični jedrna membrana nima ima diferenciacija nediferencirani diferencirani število kromosomov 1 > 1 gibanje kromosomov citoplazmatska m. delitveno vreteno jedrce nima ima DNA nima histonov histoni
138 Primerjava prokarionti : evkarionti struktura/funkcija prokarionti evkarionti introni redki običajni plazmidi prisotni odsotni telomeraza nima ima spolni ciklus redek, nepopolen običajen, popolen celični cikel nima ima transkr./translacija v istem kompartmentu ločeno operon kontrola več encimov kontrola enega encima - regulacija transkripcije običajna neobičajna + regulacija transkripcije neobičajna običajna mrna policistrinska monocistronska
139 Primerjava prokarionti : evkarionti struktura/funkcija prokarionti evkarionti modifikacija mrna redka poli A in metilacija rrna 5S, 16S, 23S 5S, 5.8S, 18S, 28S mala p. ribosoma 21 proteinov 30 proteinov velika p. ribosoma 34 proteinov 50 proteinov poliproteini niso prisotni lahko prisotni posttranslacijske mod. neobičajne običajne glikokaliks prisoten, debel tanjši flagele rotirajoče nerotirajoče flagele-sestava flagelin mikrotubuli neflagelarno gibanje drsenje ameboidno
140 Primerjava prokarionti : evkarionti struktura/funkcija prokarionti evkarionti fimbrije pili prisotni odsotni plazmatska m. nima sterolov ima sterole plazmatska m. nima ogljikohidratov ima notranje membrane samo nekateri običajne fagocitoza odsotna prisotna mesto respiracije citoplazmatska m. mitohondriji mesto fotosinteze citoplazmatska m. kloroplasti organeli nima ima vakuole redke običajne mikrotubuli nima prisotni
141 Primerjava prokarionti : evkarionti struktura/funkcija prokarionti evkarionti zračni vezikli lahko ima nima metabolizem aeroben/anaeroben aeroben eksosopre pri nekaterih običajne pri glivah endospore lahko prisotne nima rezervne snovi žveplo, polifosfati škrob glikogen polibutirati
142 Pomembna gesla ABC transporter periplazma kristalografija kemotaksa glikokaliks fluorofor endospora peritrihen koki flagela lopotrihe bacili plinski vezikli fototaksa pili G- in G+ bakterije PHB fimbrije transport skupin protoplast LPS sferoplast magnetosom TEM nukleoid SEM organeli LSCM peptidoglikan AFM
Uvod v mikrobiologijo
 Uvod v mikrobiologijo Predmet: Mikrobiologija Predavanja: 10 ur Seminarske vaje: 10 ur Laboratorijske vaje: 10 ur Študijske obveznosti: izpit Učitelj: Prof. dr. David Stopar email: david.stopar@bf.uni-lj.si
Uvod v mikrobiologijo Predmet: Mikrobiologija Predavanja: 10 ur Seminarske vaje: 10 ur Laboratorijske vaje: 10 ur Študijske obveznosti: izpit Učitelj: Prof. dr. David Stopar email: david.stopar@bf.uni-lj.si
Prve raziskave. Diagram prvega modela membrane.
 Celična membrana imajo jo vse celice omogoča fizično in kemično osamitev od okolja ohranja stabilne razmere znotraj celice tanka dvoslojna plast, obdaja citoplazmo, celično jedro ali organele 7-10 nm debela
Celična membrana imajo jo vse celice omogoča fizično in kemično osamitev od okolja ohranja stabilne razmere znotraj celice tanka dvoslojna plast, obdaja citoplazmo, celično jedro ali organele 7-10 nm debela
Osnovna zgradba prokariontske celice
 Osnovna zgradba prokariontske celice Citoplazemska membrana Celična stena Citoplazma Ribosomi Vključki - Inkluzije Nukleotidi Citoplazma Na citoplazmo so vezane funkcije saj v njej potekajo naloge celične
Osnovna zgradba prokariontske celice Citoplazemska membrana Celična stena Citoplazma Ribosomi Vključki - Inkluzije Nukleotidi Citoplazma Na citoplazmo so vezane funkcije saj v njej potekajo naloge celične
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
TRANSPORT RAZTOPIN. Agronomija - UNI
 TRANSPORT RAZTOPIN Agronomija - UNI Transport na celičnem nivoju oz. transport preko membrane je osnova za transport na višjih nivojih (tkiva). Pomen biološki membran (plazmalema, tonoplast,...) homeostaza
TRANSPORT RAZTOPIN Agronomija - UNI Transport na celičnem nivoju oz. transport preko membrane je osnova za transport na višjih nivojih (tkiva). Pomen biološki membran (plazmalema, tonoplast,...) homeostaza
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - VSŠ 2005/06
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
ZGODOVINA MIKROBIOLOGIJE
 ZGODOVINA MIKROBIOLOGIJE Prvi mikroskopi so omogočili prva opazovanja mikroorganizmov. Robert Hook je na usnju prvi videl plesni in je začetnik celične teorije. Antonie van Leuwenhoek je sestavil prvi
ZGODOVINA MIKROBIOLOGIJE Prvi mikroskopi so omogočili prva opazovanja mikroorganizmov. Robert Hook je na usnju prvi videl plesni in je začetnik celične teorije. Antonie van Leuwenhoek je sestavil prvi
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
Svetlobni mikroskop. Princip delovanja Pomembna kakovost leč
 Mikroskopija Steklena krogla napolnjena z vodo - prva povečevalna naprava - Plinij prvo stoletje Antonij van Leeuwenhoek (1632 1723) izdelal leče v velikosti bucikine glave (eritrocite, bakterije) Zaharias
Mikroskopija Steklena krogla napolnjena z vodo - prva povečevalna naprava - Plinij prvo stoletje Antonij van Leeuwenhoek (1632 1723) izdelal leče v velikosti bucikine glave (eritrocite, bakterije) Zaharias
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
Metabolizem in energija
 Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem oz. presnova
 Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
6 ogljikovih atomov: HEKSOZE (npr. glukoza, fruktoza, galaktoza) Ločimo dve vrsti glukoze: α glukoza in. β glukoza, ki se
 Ločimo dve vrsti glukoze: α glukoza in. β glukoza, ki se") OGLJIKOVI HIDRATI Monosaharidi enostavni sladkorji Spojine C, H, O v razmerju 1:2:1 3 ogljikovi atomi: TRIOZE 5 ogljikovih atomov: PENTOZE (npr. riboza, deoksiriboza) 6 ogljikovih atomov: HEKSOZE (npr.
OGLJIKOVI HIDRATI Monosaharidi enostavni sladkorji Spojine C, H, O v razmerju 1:2:1 3 ogljikovi atomi: TRIOZE 5 ogljikovih atomov: PENTOZE (npr. riboza, deoksiriboza) 6 ogljikovih atomov: HEKSOZE (npr.
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
Razgradnja maščobnih kislin. Ketonska telesa H + NAD+ NADH. Pregled metabolizma MK. lipoprotein-lipaza. maščobne kisline.
 Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - UNI
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
TRANSPORT RAZTOPIN. Agronomija - UNI 2005/06
 TRANSPORT RAZTOPIN Transport na celičnem nivoju oz. transport preko membrane je osnova za transport na višjih nivojih (tkiva). Pomen biološki membran (plazmalema, tonoplast,...) homeostaza rastlinske celice
TRANSPORT RAZTOPIN Transport na celičnem nivoju oz. transport preko membrane je osnova za transport na višjih nivojih (tkiva). Pomen biološki membran (plazmalema, tonoplast,...) homeostaza rastlinske celice
UVOD CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN
 = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN") CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Biološke membrane in transport Model tekočega mozaika, asimetričnost, omogočeno lateralno gibanje lipidov in proteinov, oligosahardine verige lipidov
 Biološke membrane in transport Model tekočega mozaika, asimetričnost, omogočeno lateralno gibanje lipidov in proteinov, oligosahardine verige lipidov in proteinov na zunanji strani Sestava in struktura
Biološke membrane in transport Model tekočega mozaika, asimetričnost, omogočeno lateralno gibanje lipidov in proteinov, oligosahardine verige lipidov in proteinov na zunanji strani Sestava in struktura
PRIROČNIK ZA UČENJE PRI PREDMETU OSNOVE MIKROBIOLOGIJE (1. LETNIK - BOLONJSKI PROGRAM PRI PROF. NINI GUNDE CIMERMAN)
") PRIROČNIK ZA UČENJE PRI PREDMETU OSNOVE MIKROBIOLOGIJE (1. LETNIK - BOLONJSKI PROGRAM PRI PROF. NINI GUNDE CIMERMAN) Priročnik je napisan v obliki kratkih povzetkov in zajema snov 1. letnika smeri Biologija
PRIROČNIK ZA UČENJE PRI PREDMETU OSNOVE MIKROBIOLOGIJE (1. LETNIK - BOLONJSKI PROGRAM PRI PROF. NINI GUNDE CIMERMAN) Priročnik je napisan v obliki kratkih povzetkov in zajema snov 1. letnika smeri Biologija
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
BIOLOGIJA RASTLINSKE CELICE Teze predavanj. Marina DERMASTIA
 BIOLOGIJA RASTLINSKE CELICE Teze predavanj Marina DERMASTIA UVOD 5 KEMIČNA SESTAVA RASTLINSKE CELICE 6 ATOMI IN MOLEKULE 6 VELIKE MOLEKULE: POLIMERI IN NJIHOVI MONOMERI 6 OGLJIKOVI HIDRATI 7 STRUKTURNI
BIOLOGIJA RASTLINSKE CELICE Teze predavanj Marina DERMASTIA UVOD 5 KEMIČNA SESTAVA RASTLINSKE CELICE 6 ATOMI IN MOLEKULE 6 VELIKE MOLEKULE: POLIMERI IN NJIHOVI MONOMERI 6 OGLJIKOVI HIDRATI 7 STRUKTURNI
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
SINTEZA SAHAROZE IN ŠKROBA
 SINTEZA SAHAROZE IN ŠKROBA Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Stroma kloroplasta Škrob (primarni ali asimilacijski) Calvinov cikel Sladkor (trioza) Sladkor (trioza) Pi Sladkor
SINTEZA SAHAROZE IN ŠKROBA Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Stroma kloroplasta Škrob (primarni ali asimilacijski) Calvinov cikel Sladkor (trioza) Sladkor (trioza) Pi Sladkor
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko
 Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Univerzitetni študij GOZDARSTVA Predmet BOTANIKA S FIZIOLOGIJO
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Univerzitetni študij GOZDARSTVA Predmet BOTANIKA S FIZIOLOGIJO
FOTOSINTEZA Wan Hill primerjal rastlinsko fotosintezo s fotosintezo BAKTERIJ
 FOTOSINTEZA FOTOSINTEZA je proces, pri katerem s pomočjo svetlobne energijje nastajajo v živih celicah organske spojine. 1772 Priestley RASTLINA slab zrak dober zrak Rastlina s pomočjo svetlobe spreminja
FOTOSINTEZA FOTOSINTEZA je proces, pri katerem s pomočjo svetlobne energijje nastajajo v živih celicah organske spojine. 1772 Priestley RASTLINA slab zrak dober zrak Rastlina s pomočjo svetlobe spreminja
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
OKOLJSKO NARAVOSLOVJE 2. Predavanja v študijskem letu 2. del 2012/2013
 OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda
![[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda](/thumbs/89/99439684.jpg "[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda") kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
Biologija celice. ločljivost: oko 0'2 mm,svetlobni mikroskop 0'2 μm, elektronski 0'2 ηm
 Biologija celice Živa bitja: - so kemijsko kompleksna in visoko organizirana (celična zgradba) - imajo presnovo (metabolizem), uporabljajo in pretvarjajo energijo o disimilacija (katabolizem)-razgradnji
Biologija celice Živa bitja: - so kemijsko kompleksna in visoko organizirana (celična zgradba) - imajo presnovo (metabolizem), uporabljajo in pretvarjajo energijo o disimilacija (katabolizem)-razgradnji
Funkcije proteinov (pogojene s strukturo)
") Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje molekul (ligandov, npr. O2 v Hb, Mb) Uravnavanje - DNA-vezavni proteini
Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje molekul (ligandov, npr. O2 v Hb, Mb) Uravnavanje - DNA-vezavni proteini
BIOLOGIJA CELICE TEZE PREDAVANJ ZA 1. LETNIK ŠTUDENTOV BIOKEMIJE
 BIOLOGIJA 1 BIOLOGIJA CELICE TEZE PREDAVANJ ZA 1. LETNIK ŠTUDENTOV BIOKEMIJE MARINA DERMASTIA BIOLOGIJA 2 OBLIKA IN VELIKOST RASTLINSKE CELICE 3 JEDRO 3 VAKUOLA 5 PLASTIDI 5 Proplastidi 6 Amiloplasti 6
BIOLOGIJA 1 BIOLOGIJA CELICE TEZE PREDAVANJ ZA 1. LETNIK ŠTUDENTOV BIOKEMIJE MARINA DERMASTIA BIOLOGIJA 2 OBLIKA IN VELIKOST RASTLINSKE CELICE 3 JEDRO 3 VAKUOLA 5 PLASTIDI 5 Proplastidi 6 Amiloplasti 6
Nekatere interakcije v lipidnem metabolizmu
 Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
+105 C (plošče in trakovi +85 C) -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70
 -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70") KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Transformator. Delovanje transformatorja I. Delovanje transformatorja II
 Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks'
 Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Matjaž Zorko Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
Aminokiseline. Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina 22.12.2014
 Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
Statistična analiza. doc. dr. Mitja Kos, mag. farm. Katedra za socialno farmacijo Univerza v Ljubljani- Fakulteta za farmacijo
 Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV. 2. predavanje: Od 1D do 3D strukture proteinov 1. del.
 IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
Kvantni delec na potencialnem skoku
 Kvantni delec na potencialnem skoku Delec, ki se giblje premo enakomerno, pride na mejo, kjer potencial naraste s potenciala 0 na potencial. Takšno potencialno funkcijo zapišemo kot 0, 0 0,0. Slika 1:
Kvantni delec na potencialnem skoku Delec, ki se giblje premo enakomerno, pride na mejo, kjer potencial naraste s potenciala 0 na potencial. Takšno potencialno funkcijo zapišemo kot 0, 0 0,0. Slika 1:
Neavtorizirani povzetki izbranih predavanj iz biokemije. UN študij Okolje Politehnika Nova Gorica. doc. dr. Marko Dolinar
 Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI. Membranski separacijski procesi: diafiltracija, elektrodializa, reverzna osmoza, pervaporacija
 ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI Membranski separacijski procesi: diafiltracija, elektrodializa, reverzna osmoza, pervaporacija Membranski separacijski procesi v biotehnologiji proces mikrofiltracija
ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI Membranski separacijski procesi: diafiltracija, elektrodializa, reverzna osmoza, pervaporacija Membranski separacijski procesi v biotehnologiji proces mikrofiltracija
AMILAZE. Encimi, ki hidrolizirajo ogljikove hidrate. struktura škroba
 Encimi, ki hidrolizirajo ogljikove hidrate substrati: ogljikovi hidrati (škrob, celuloza, poli in oligosaharidi) encimi: glikozidaze glikozidna vez encimska specifičnost konfiguracija glikozidne vezi kemijska
Encimi, ki hidrolizirajo ogljikove hidrate substrati: ogljikovi hidrati (škrob, celuloza, poli in oligosaharidi) encimi: glikozidaze glikozidna vez encimska specifičnost konfiguracija glikozidne vezi kemijska
Biokemija I, 17. predavanje-1. del, , T. Režen
 Biokemija I, 17. predavanje-1. del, 27. 3. 2012, T. Režen Vrste polisaharidov Homopolisaharidi enake monosaharidne enote nerazvejani razvejani Heteropolisaharidi različne monosaharidne enote Dve vrs7 Več
Biokemija I, 17. predavanje-1. del, 27. 3. 2012, T. Režen Vrste polisaharidov Homopolisaharidi enake monosaharidne enote nerazvejani razvejani Heteropolisaharidi različne monosaharidne enote Dve vrs7 Več
1.MIKROORGANIZMI KOT CELICA
 1.MIKROORGANIZMI KOT CELICA -celica je samostojna enota, izolirana od ostalih s cel.membrano in mogoče cel.st.(nek.bakterije in protozoi). Cel.membrana je bariera,ki loči cel.od okolja. Jedro oz.jedrna
1.MIKROORGANIZMI KOT CELICA -celica je samostojna enota, izolirana od ostalih s cel.membrano in mogoče cel.st.(nek.bakterije in protozoi). Cel.membrana je bariera,ki loči cel.od okolja. Jedro oz.jedrna
Nukleinske kisline. Nukleotidi. DNA je nosilka dednih genetskih informacij.
 Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
DOLOČANJE)ENCIMSKE)AKTIVNOSTI)V)KLINIČNE)NAMENE)
ENCIMSKE)AKTIVNOSTI)V)KLINIČNE)NAMENE)") DLČANJEENCIMSKEAKTIVNSTIVKLINIČNENAMENE 20encimovseru=nskopregledujevkliniki 1954sougotovilipovezanostsrčnegainfarktainpovišanekonc. aspartataminotransferazevserumu danesnarapolagovelikoabzapreciznodoločanjekoncproteinov
DLČANJEENCIMSKEAKTIVNSTIVKLINIČNENAMENE 20encimovseru=nskopregledujevkliniki 1954sougotovilipovezanostsrčnegainfarktainpovišanekonc. aspartataminotransferazevserumu danesnarapolagovelikoabzapreciznodoločanjekoncproteinov
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
Rešitve nalog. Biologija. za gimnazije
 Rešitve nalog Biologija za gimnazije 1 Značilnosti živega (str. 8) 1. Citoplazma, celična membrana, metabolizem (celici lastni presnovni mehanizmi). 2. a) velikost Virusi so zelo majhni, v splošnem 50-
Rešitve nalog Biologija za gimnazije 1 Značilnosti živega (str. 8) 1. Citoplazma, celična membrana, metabolizem (celici lastni presnovni mehanizmi). 2. a) velikost Virusi so zelo majhni, v splošnem 50-
LIPIDI IN LIPIDNE MEMBRANE
 LIPIDI IN LIPIDNE MEMBRANE Maščobne kisline Sestavljeni lipidi (mašč.kisline + alkohol) Triacilgliceroli Glicerofosfatidi Sfingolipidi Voski Enostavni lipidi (vsi so izoprenski derivati) Terpeni Steroidi
LIPIDI IN LIPIDNE MEMBRANE Maščobne kisline Sestavljeni lipidi (mašč.kisline + alkohol) Triacilgliceroli Glicerofosfatidi Sfingolipidi Voski Enostavni lipidi (vsi so izoprenski derivati) Terpeni Steroidi
Jerneja Čučnik Mikroskopiranje in tipi celic Gimnazija Celje Center Mikroskopiranje in tipi celic
 Ime in priimek: Jerneja Čučnik Razred: 4.b Šola: Gimnazija Celje Center Mentor: Saša ogrizek, prof. Datum izvedbe vaje: 24.9.2009 1 1. UVOD Mikroskop je instrument za preučevanje predmetov, ki so premajhni,
Ime in priimek: Jerneja Čučnik Razred: 4.b Šola: Gimnazija Celje Center Mentor: Saša ogrizek, prof. Datum izvedbe vaje: 24.9.2009 1 1. UVOD Mikroskop je instrument za preučevanje predmetov, ki so premajhni,
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko
 Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Univerzitetni študij KMETIJSTVO:Agronomija Predmet BOTANIKA
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Univerzitetni študij KMETIJSTVO:Agronomija Predmet BOTANIKA
Biologija rastlinske celice
 Barbara Vilhar Biologija rastlinske celice Teorija za vaje Predmet Splošna botanika Pedagoška fakulteta, 1. letnik Interno študijsko gradivo Univerza v Ljubljani, Biotehniška fakulteta Ljubljana, 2006
Barbara Vilhar Biologija rastlinske celice Teorija za vaje Predmet Splošna botanika Pedagoška fakulteta, 1. letnik Interno študijsko gradivo Univerza v Ljubljani, Biotehniška fakulteta Ljubljana, 2006
Univerza v Ljubljani,, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko
 Univerza v Ljubljani,, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Univerzitetni študij Kmetijstvo-Zootehnika Predmet BOTANIKA
Univerza v Ljubljani,, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Univerzitetni študij Kmetijstvo-Zootehnika Predmet BOTANIKA
pretvarja v nestrupeno obliko, ki lahko vstopa v biosintezo nukleotidov *i) NH 4
 NH 4") 1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
HIGIENA IN MIKROBIOLOGIJA
 HIGIENA IN MIKROBIOLOGIJA Romina Bernard univ. dipl. biologinja in prof. biologije Romina Bernard, gradivo za HMB 1 KAZALO: UVOD VIRUSI BAKTERIJE GLIVE PARAZITSKE ŽIVALI IMUNOLOGIJA HIGIENA IN UNIČEVANJE
HIGIENA IN MIKROBIOLOGIJA Romina Bernard univ. dipl. biologinja in prof. biologije Romina Bernard, gradivo za HMB 1 KAZALO: UVOD VIRUSI BAKTERIJE GLIVE PARAZITSKE ŽIVALI IMUNOLOGIJA HIGIENA IN UNIČEVANJE
BIOLOGIJA CELICE. Uredil Primož Pirih Društvo študentov biologije
 Društvo študentov biologije Stran 1/44 http://dsb.biologija.org BIOLOGIJA CELICE Zapiski s predavanj pri predmetu Biologija celice za študente biologije. Predavatelji Jezernik, Strle, Pšeničnik. Zadnja
Društvo študentov biologije Stran 1/44 http://dsb.biologija.org BIOLOGIJA CELICE Zapiski s predavanj pri predmetu Biologija celice za študente biologije. Predavatelji Jezernik, Strle, Pšeničnik. Zadnja
Cefalosporini ostali β-laktami
 Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
CENTRALNI LABORATORIJ
 CENTRALNI LABORATORIJ I.ODVZEM IN POŠILJANJE VZORCEV 1 KAPILARNI ODVZEM KRVI DA DA 30min/15min 2 ODVZEM FECESA DA NE 30min/15min 3 ODVZEM URINA DA DA 30min/15min 4 POŠILJANJE BIOLOŠKIH VZORCEV ( EKSPEDIT)
CENTRALNI LABORATORIJ I.ODVZEM IN POŠILJANJE VZORCEV 1 KAPILARNI ODVZEM KRVI DA DA 30min/15min 2 ODVZEM FECESA DA NE 30min/15min 3 ODVZEM URINA DA DA 30min/15min 4 POŠILJANJE BIOLOŠKIH VZORCEV ( EKSPEDIT)
2.1. MOLEKULARNA ABSORPCIJSKA SPEKTROMETRIJA
 2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
Fotosinteza. 1. Sinteza NADPH+H + in ATP 2. Fiksacija CO 2
 Fotosinteza 1. Sinteza NADPH+H + in ATP 2. Fiksacija CO 2 Oris 1 Fotoreaktivnost klorofila 2 Z-shema fotosinteze 3 Svetlobno-gnana ATP-sinteza - Fotofosforilacija 4 Fiksacija ogljikovega dioksida 5 Calvin-Bensonov
Fotosinteza 1. Sinteza NADPH+H + in ATP 2. Fiksacija CO 2 Oris 1 Fotoreaktivnost klorofila 2 Z-shema fotosinteze 3 Svetlobno-gnana ATP-sinteza - Fotofosforilacija 4 Fiksacija ogljikovega dioksida 5 Calvin-Bensonov
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK)
") IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
2. Membranski proteini značilnosti zgradbe, vrste in različne naloge proteinov.
 1. Celična membrana - zgradba plazemske membrane 1. Sestava fosfolipidnega dvosloja (P in E sloj membranskih dvoslojev). Plazemska membrana je zgrajena iz lipidov in proteinov. Lipidi dajejo celici osnovno
1. Celična membrana - zgradba plazemske membrane 1. Sestava fosfolipidnega dvosloja (P in E sloj membranskih dvoslojev). Plazemska membrana je zgrajena iz lipidov in proteinov. Lipidi dajejo celici osnovno
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
Sinteza RNA - transkripcija
 Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
Definiranje okolja mikroorganizmov
 Definiranje okolja mikroorganizmov David Stopar Novi sad 2007 Izr. prof. dr. David Stopar Univerza v Ljubljani, BF Oddelek za živilstvo Katedra za mikrobiologijo Večna pot 111 Email: david.stopar@bf.uni-lj.si
Definiranje okolja mikroorganizmov David Stopar Novi sad 2007 Izr. prof. dr. David Stopar Univerza v Ljubljani, BF Oddelek za živilstvo Katedra za mikrobiologijo Večna pot 111 Email: david.stopar@bf.uni-lj.si
Ogljikovi hidrati - monosaharidi
 Ogljikovi hidrati - monosaharidi V tem poglavju se boste naučili (ponovili) - kaj so osnovne lastnosti ogljikovih hidratov, - kaj so monosaharidi, - katere so dokazne reakcije za monosaharide. Enostavni
Ogljikovi hidrati - monosaharidi V tem poglavju se boste naučili (ponovili) - kaj so osnovne lastnosti ogljikovih hidratov, - kaj so monosaharidi, - katere so dokazne reakcije za monosaharide. Enostavni
Teze predavanj iz mikrobne biokemije*
 Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko
 Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Visokošolski strokovni študij GOZDARSTVA Predmet BOTANIKA
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za agronomijo Katedra za aplikativno botaniko, ekologijo, fiziologijo rastlin in informatiko Visokošolski strokovni študij GOZDARSTVA Predmet BOTANIKA
Prehrana in metabolizem
 Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
MIKROSKOP IN MIKROSKOPIRANJE
 Gimnazija Murska Sobota POROČILO K LABORATORIJSKI VAJI MIKROSKOP IN MIKROSKOPIRANJE Sandra Gorčan, 4.c prof. Edita Vučak Murska Sobota,8.10.2003 UVOD: Mikroskop je naprava, ki služi za gledanje mikroskopsko
Gimnazija Murska Sobota POROČILO K LABORATORIJSKI VAJI MIKROSKOP IN MIKROSKOPIRANJE Sandra Gorčan, 4.c prof. Edita Vučak Murska Sobota,8.10.2003 UVOD: Mikroskop je naprava, ki služi za gledanje mikroskopsko