Molekularna biotehnologija: encimi, pretvorba biomase
|
|
|
- Χριστός Μαρκόπουλος
- 6 χρόνια πριν
- Προβολές:
Transcript
1 Molekularna biotehnologija: encimi, pretvorba biomase
 je povpraševanja po encimih za razgradnjo polisaharidov do mono- in disaharidov ( karbohidraza ), sledijo proteaze (27 %), lipaze, polimeraze in nukleaze.")
2 Biotehnološka proizvodnja encimov Svetovno tržišče za encime ocenjujejo na 4,4 mrd USD (2013) in naj bi se širil 8 % letno, tako da naj bi leta 2020 dosegel skoraj 7,7 mrd USD. Največ (45 %) je povpraševanja po encimih za razgradnjo polisaharidov do mono- in disaharidov ( karbohidraza ), sledijo proteaze (27 %), lipaze, polimeraze in nukleaze. Največji potrošnik encimov je živilska industrija, naraščala pa bo poraba proteaz za pripravo močnih krmil. Poraba za pripravo biogoriv in detergentov naj se ne bi bistveno spreminjala. Največji proizvajalec encimov so ZDA (37 %), najhitreje pa bo v naslednjem obdobju rasla proizvodnja v Azijsko-Pacifiški regiji. Svetovno tržišče danes obvladujejo trije proizvajalci Novozymes (47 %), Danisco (DuPont; 21 %) in DSM (6 %), ki skupaj pokrivajo >70 % potreb po encimih. Polovica proizvedenih encimov izhaja iz GSO. 2013
.")
3 Encimi v pekarski industriji: polisaharidi v kruhu Izdelki iz moke, kot je na primer kruh, imajo sestavo in izgled, ki ju v veliki meri določajo encimi. Ti izhajajo iz moke same, pa tudi iz kvasovk (pri kvašenem testu). Ker je zanašanje na naravne encime negotovo, v žitne izdelke dodajajo encime v točno določenih koncentracijah, ki omogočajo hitrejše in bolj ponovljive postopke, hkrati pa je (ob vse strožjih predpisih) zaradi encimov v izdelku lahko manj aditivov. Škrob, glavna sestavina izdelkov iz žita, je sestavljen iz nerazvejane amiloze (do 6000 glukoznih enot, povezanih z alfa-1,4 vezmi) in močno razvejanega amilopektina (10-60 glukoznih enot v verigi, razvejitve dolge enot, vezanih preko alfa-1,6 vezi skupaj do 2 M glukoznih enot). Ta je substrat za več encimov: : (a) α-amilaze, (b) amiloglukozidaze; (c) β-amilaze; (d) izoamilaze in pululanaze. Razen škroba moka vsebuje še nekatere druge polisaharide, od katerih so za lastnosti testa predvsem pomembni arabinoksilani in arabinogalaktani, ki lahko zadržujejo vodo in s tem stabilizirajo testo, povečujejo pa tudi njegovo viskoznost in povečajo prepustnost za pline. S tem povečujejo volumen kruha in podaljšujejo rok trajanja.
 in polimerne glutenine (iztegnjene verige, povezane s S-S; 80 kda nekaj M kda).")
4 Za lastnosti testa so najpomembnejši proteini iz skupine glutenov, ki testu dajejo elastičnost in omogočajo zadrževanje plinov v njem. Glede na njihovo topnost v 70-odstotnem etanolu glutene razdelimo na monomerne gliadine (globularni, kda) in polimerne glutenine (iztegnjene verige, povezane s S-S; 80 kda nekaj M kda). Gliadini zagotavljajo plastičnost, viskoznost in vlečljivost, glutenini pa elastičnost in povezovano moč. Moke se med seboj razlikujejo po razmerju med različnimi gluteni in dolžini ter strukturi polimerov. Encimi v pekarski industriji: proteini v kruhu
5
6 Priprava kruha in pomen dodanih encimov Med mešanjem, vzhajanjem in pečenjem poteče več fizikalnih, kemijskih in biokemijskih sprememb, ki jih še vedno ne razumemo v celoti: ekspanzija volumna, evaporacija vode, nastanek porozne strukture, denaturacija proteinov, želatiniziranje škroba, razvoj skroje, reakcije porjavenja. Za vzhajanje uporabljamo kvasovke S. cerevisiae, pa tudi nekatere druge vrste, za kislo testo pa laktobacile. Dodatek encimov omogoča standardizacijo postopka, vpliva pa na reološke lastnosti testa, zadrževanje plinov, mehkost drobtin in zmanjšuje nastajanje akrilamidov (asparaginaza) med peko. Encime dodajajo ločeno, ali pa kot predpripravljeno mešanico. V fazi mešanja najpogosteje dodajajo alfaamilaze, ki krajšajo polisaharidne verige do oligosaharidov in dekstrinov (razvejani). Encimi pridejo iz slada ali pa jih pridobivajo iz različnih mikroorganizmov. Produkt delovanja betaamilaze je med drugim maltoza, ki je uporabna kot vir ogljika za kvasovke, ostali produkti pa predstavljajo sestavino kruha. Odvisno od temperaturne stabilnosti encimov lahko dosežemo tudi podaljšanje življenjske dobe kruha (najpogosteje uporabljajo srednje obstojno maltogeno alfaamilazo iz bakterije Bacillus stearothermophillus). Za razgradnjo arabinoznih polimerov uporabljajo že ~40 let mikrobne ksilanaze, ki optimizirajo reološke lastnosti testa in okus kruha. Lipaze uporabljajo ~20 let in trenutno uporabljajo inženirske encime 3. generacije. Posledica je podaljšan rok uporabe, bolj mehko testo in kasneje kruh, poveča volumen štruc in naredi debelejšo skorjo. Uporabljajo tudi lipoksigenaze, predvsem iz sojine moke. Produkti teh encimov so razni radikali, ki oksidirajo pigmente in -SH skupine v (poli)peptidih. Hkrati pride do beljenja moke. Proteaze v moki so večinoma aspartatne proteaze in karboksipeptidaze, ki so aktivne v kislem. Proteaze dodajajo za zmanjševanje čvrstosti testa, skrajšanje časa mešanja, zagotavljajo enotnost mase in izboljšujejo okus. Uporabljajo glivne proteaze, predvsem pri visokoglutenskih mokah. Razen tega preprečujejo krčenje gotovih izdelkov po tem, ko so pečeni. Glukoza oksidaza odstranjuje glukozo in s tem podaljšuje rok uporabe, hkrati pa odstranjuje kisik, nastali peroksid pa deluje protimikrobno. Ugotovili so tudi vpliv na reološke lastnosti, verjetno preko vpliva na zamreženost proteinov preko S-S. Uporabljajo tudi transglutaminaze, lakazo.
7
8

9 Rekombinantna lipaza Maščobne madeže lahko odstranimo s povišano temperaturo in alkalnimi detergenti, kar pa skrajšuje življenjsko dobo tkanin in je energetsko potratno. Alternativo predstavlja encimska razgradnja, pri čemer bi lahko izhajali iz bakterijske lipaze Pseudomonas alcaligenes. Bakterija ga proizvaja zelo malo, rekombinantnega pa ga ni bilo mogoče prekomerno izraziti, čeprav so preizkusili različne bakterijske in glivne ekspresijske sisteme. Predvideli so, da je za težavo odgovorna odsotnost nekega drugega proteina, ki bi lahko pripomogel k izločanju ali k stabilizaciji lipaze. Vsekakor je bilo treba poiskati najprej zapis za lipazo in potem še za morebitne druge proteine. Pot do gena je potekala preko izoliranega encima. Na osnovi N- končnega ak zaporedja so pripravili sondo in pri presejanju genomske knjižnice P. alcaligenes našli klon, ki je vseboval celoten gen za lipazo (lipa) in del sosednjega gena (lipb). Ta klon so uporabili za novo sondo in z njo pregledali drugo gensko knjižnico. Tako so našli še preostanek gena lipb in oba dela združili. Sestavljeni produkt so preko ekspresijskega vektorja širokega spektra vnesli v P. alcaligenes. Ob uporabi vektorja z majhnim številom kopij je bila aktivnost lipaze 4-5x večja kot pri wt ne glede na vsebnost lipb. Z vektorjem z velikim številom kopij je bila aktivnost 20x večja kot pri wt v odsotnosti lipb, v prisotnosti lipb pa 35x večja. Sekrecijski vektorji omogočajo prenos rekombinatnega proteina v gojišče, od koder ga je lažje izolirati kot iz celic za praške zadošča koncentiranje gojišča brez celic. V 10m 3 reaktorju je bila proizvodnja manjša od pričakovane zaradi nastajanja povečane konc. CO 2. S spremembo pogojev obratovanja se je zmanjšala konc. CO 2, s tem pa se je povečala vsebnost lipaze.
, iz mikroorganizmov z zelo različnimi pogoji rasti za optimalno")


10 Rekombinante restrikcijske endonukleaze Komercialno dostopnih je nad 600 različnih restriktaz (236 različnih specifičnosti), iz mikroorganizmov z zelo različnimi pogoji rasti za optimalno proizvodnjo. Veliko genov za restriktaze so zato klonirali in jih izrazili v večjem obsegu v E. coli. Pri tem pa je treba preprečiti, da bi tuja restriktaza razgradila gostiteljsko DNA. Običajno to dosežejo s koekspresijo gena za metilacijo DNA. Pri tem je treba zagotoviti izražanje metiltransferaze pred izražanjem restriktaze. Microbiol. Mol. Biol. Rev. 77(1), (2013)

11 Rekombinanti PstI Primer: PstI (Providencia stuartii) v E. coli. Pripravili so genomsko knjižnico P. stuartii v E. coli. Transformante so rasle v tekočem gojišču, nato pa so jih inficirali z bakteriofagom l. Celice, ki so izražale funkcionalni zapis za restriktazo, so preživele, ostale pa je fag liziral. Odporne bakterije so namnožili, jih lizirali in preverili specifičnost restriktaze ter metilazno aktivnost. V opisanem poskusu so identificirali en sam klon z vključkom dolžine ~4 kb, ki je vseboval gen za restriktazo in metilazo na istem operonu, ki je vključeval skupni promotor. Proizvodnja v E. coli je bila ~10x večja kot v P. stuartii, restriktazno aktivnost so dokazali v periplazmi, metilazno pa v citoplazmi.
.")
12 Rekombinante restrikcijske endonukleaze /2 Alternativna pot za iskanje genov za metilazo in restriktazo: Iz seva, za katerega so vedeli, da proizvaja restriktazo, so pripravili genomsko knjižnico v E. coli, pri tem pa so uporabili vektor, ki vsebuje prepoznavno zaporedje za to restriktazo (predpostavljamo, da bo na vključku tudi gen za metilazo in da imata oba gena lastni promotor). Iz kolonij so izolirali plazmidno DNA in jo rezali s predhodno izolirano restriktazo (pbr322 vsebuje 8 prepoznavnih mest za DdeI), s tako obdelano plazmidno DNA pa so transformirali bakterijske celice. Metilaza bi morala zaščititi plazmidno DNA pred rezanjem, zato bi plazmid ostal nerazrezan, s takim pa se bakterije zlahka transformirajo. Zraslim transformantam določijo le še restriktazno aktivnost (da imajo metilazno, že vedo). Tak pristop je uporaben v vseh tistih primerih, ko sta gena za metilazo in restriktazo en ob drugem, to pa je v večini primerov.
.")
13 Pretvorba sestavin biomase: škrob Škrob je pogosta zaloga energije pri rastlinah in jo sestavlja mešanica amiloze (linearni homopolimer) in amilopektina (razvejani homopolimer D-glukoze). Dolžina / razvejanost je različna, odvisna od vira in starosti škroba. Iz škroba npr. lahko v biotehnološkem postopku proizvedemo fruktozo, lahko pa tudi alkohol (primer biogoriva). Pri pretvorbi v prvi stopnji uporabljajo a-amilazo, nato pa glukoamilazo in glukoza izomerazo. Ti trije encimi po vrednosti predstavljajo 30 % vseh industrijsko uporabljenih encimov. Pretvorba škroba vključuje encimske in neencimske reakcije. Pogosta surovina je koruzno zrnje, ki vsebuje 40 % škroba. Encim a-amilaza cepi vezi tako v amilozi kot v amilopektinu, kar privede do nastanka glukoze, maltoze, maltotrioze in dekstrinov (pri cepitvi amilopektina razvejanih dekstrinov). Vir a-amilaze je običajno Bacillus amyloliquefaciens. Včasih uporabijo dodatno še b-amilazo, ki odceplja disaharide s koncev verig. Glukoamilaza sicer cepi a-1,3, a-1,4 in a-1,6 vezi, a a-1,4 manj učinkovito kot a-amilaza. Uporabljajo jo npr. za lahka suha piva, ker hidrolizira razvejane dekstrine. Industrijski encim pridobivajo predvsem iz glive Aspergillus niger.

14 Postopek pretvorbe škroba Zrnje zmeljejo, nato ga želatinizirajo s kuhanjem pod povišanim tlakom s tem se izpostavi površina škroba za lažjo encimsko razgradnjo. Ko se želatinasta masa ohladi na C, dodajo a-amilazo, ki cepi a-1,4 vezi in s tem hidrolitično skrajša verige, to pa povzroči utekočinjenje mase. Dodajo glukoamilazo, s katero poteče popolna hidroliza polisaharida: saharificirana masa, ki jo predstavlja večinoma glukoza. Glukozo lahko encimsko pretvorijo v fruktozo (glukoza izomeraza), ali pa poteče alkoholna fermentacija, s katero v anaerobnih pogojih nastane etanol. Proizvodnja fruktoze po zgornjem postopku je cenejša od proizvodnje sladkorja (saharoza), zato saharozo kot sladilo v živilski industriji nadomeščajo s fruktozo visokofruktozni (koruzni) sirup, ki pa sicer vsebuje še okrog 50 % glukoze. Znana so kodirajoča zaporedja iz več mikroorganizmov, ki omogočajo pripravo rekombinantnih organizmov (največkrat kvasovk) z izboljšanimi lastnostmi. Z mutacijami v genu za glukozaizomerazo so ob zamenjavi 2 ak, ki sodelujeta pri vezavi substrata, uspeli dobiti seve, ki so bistveno bolj učinkoviti pri biosintezi fruktoze.

15 Možnosti za cenejšo pretvorbo škroba Pri proizvodnji fruktoze in alkohola največji strošek predstavlja cena encimov, ki jih uporabijo samo enkrat. Potrebni so novi pristopi za pripravo cenenih encimov. Možne rešitve bi bile: - prekomerno izražanje rekombinantnih encimov v mikrobih, ki hitro rastejo na cenenih substratih; - priprava izboljšane a-amilaze, ki bi bila aktivna pri visoki temperaturi (80 90 C), s čimer bi odpadli stroški hlajenja mase, postopek pretvorbe pa bi se pospešil; - priprava takih encimov, da bi imeli temperaturni in ph-optimum enak, tako da bi lahko utekočinjenje in saharifikacija potekla pri enakih pogojih; - razvoj encimov, ki bi lahko razgrajevali škrob, ne da bi bila potreba stopnja želatinizacije; - razvoj fermentacijskega mikrobnega seva, ki bi izločal glukoamilazo in je s tem ne bi bilo treba dodajati. Gen za a-amilazo so izolirali iz različnih organizmov, npr. B. amyloliquefaciens in bolj termofilne B. stearothermophylus idr. Genomsko knjižnico bacilusa so pripravili v B. subtilis, ki nima gena za a-amilazo, ter preverili, ali kateri od klonov proizvaja in izloča a-amilazo. Gojišče s škrobom, na katerem so zrasle transformante, so izpostavili hlapom joda. Ker so kolonije, ki so proizvajale encim, razgradile škrob, so na gojišču opazili neobarvan kolobar okrog kolonije. Iz Aspergillus awamori so izolirali cdna za glukoamilazo in jo vstavili v vektor za S. cerevisiae pod kontrolo kvasovkinega promotorja za enolazo (ENO1). Kvasovke so hkrati pretvarjale oligosaharide in izvajale fermentacijo, tako kot so pričakovali, a učinkovitost ni bila velika.
; - izražanje cdna je bilo nizko; -")
16 Problemi pri izražanju glukoamilazne cdna v S. cerevisiae za sočasno pretvorbo oligosaharidov in alkoholno fermentacijo: - sev mora biti odporen na visoko koncentracijo etanola (laboratorijski sevi niso); - izražanje cdna je bilo nizko; - izgubljanje plazmidov iz celic v odsotnosti selekcijskega pritiska. Rešitve: - uporaba seva iz pivovarniške industrije, ki je odporen na povišane koncentracije etanola; - delecija 175 bp iz promotorske regije ENO1 (delovalo kot negativni regulatorski element); - zamenjava ori z regijo za homologno rekombinacijo integrativni vektor. Tako so pridobili dva nova seva, ki sta bila po svojih lastnostih boljša od naravne amilolitične kvasovke S. diastaticus in boljša od izhodiščnih sevov S. cerevisiae. Poskusili so tudi pripraviti seve A. niger s povečanim številom kopij gena za glukoamilazo. Ugotovili so, da izražanje ni bilo povezano s številom vnesenih kopij, pač pa predvsem z mestom vstavitve ene ali več dodatnih kopij.

17 Kvasovke za slajše vino Sladkost vina je do neke mere odvisna od vsebnosti glicerola in znaša 4-9 g/l. Da bi dobili lažja in slajša vina, so pripravili sev S. cerevisae, ki prekomerno izraža gen GPD1, ki zapisuje glicerol-3-fosfat dehidrogenazo. Med alkoholno fermentacijo se prebiteh NADH z GPD1 pretvori v NAD+, hkrati pa isti encim pretvarja dihidroksiaceton v glicerol-3-fosfat, tega pa glicerol-3-fosfataza pretvori v glicerol. Konstrukte so vstavili v 6 različnih sevov kvasovk za vinarstvo in s tem pridobili v vinu glicerol namesto 7-9 g/l g/l, hkrati pa se je vsebnost alkohola znižala z 9,5-10 % na 8,5-9 %. Vsebnost acetata se ni spremenila.
 za glukozo kot za ksilozo, zato se ksiloza na encim veže tesneje kot glukoza in se pretvarja hitreje.")

18 Izboljšana proizvodnja fruktoze Glukoza izomeraza je pravzaprav ksiloza/glukoza izomeraza. Ob pretvorbi (5C) ksiloze v ksilulozo v stranski reakciji pretvarja (6C) glukozo v fruktozo. Encim ima za nižjo vrednost k cat in višjo K m (vezavno konstanto) za glukozo kot za ksilozo, zato se ksiloza na encim veže tesneje kot glukoza in se pretvarja hitreje. Glukoza izomeraza je citoplazemski encim, zato ga je težje izolirati v čisti obliki kot encime, ki se izločajo iz celic, hkrati pa je proizvodnja dražja. Zato bi bilo smiselno uporabiti ta encim večkrat, kar je možno, če je encim v pretvorbenem postopku imobiliziran na trden nosilec, kar tudi stabilizira encim. Izomerizacijska reakcija je reverzibilna, vsebnost frukoze pa je odvisna od temperature. V industrijskih postopkih je ta običajno 60 C. Učinkovitost bi bilo mogoče izboljšati s pripravo bolj temperaturno obstojnih encimov s višjim temperaturnim optimumom. Tak encim najdemo pri bakteriji Thermus thermophilus (T opt =95 C), a ga celice proizvedejo zelo malo, zato so gen zanj prestavili v različne vektorje in ga izrazili v E. coli oz. Bacillus brevis. V enem od primerov je bilo izražanje 1000-krat večje kot v termofilni bakteriji, iz katere izhaja gen.

19 Izboljšana glukoza izomeraza Substratno sprecifičnost so izboljšali z usmerjeno mutagenezo gena iz bakterije Clostridim thermosulfurogenes, pri katerem so najprej mutirali eno od dveh ak, ki sodelujeta pri vezavi substrata in s tem dosegli do 2,6x večjo katalitično aktivnost na glukozo in do 7x manjšo aktivnost na ksilozo. Če so zamenjali obe ak, je bila aktivnost na glukozo 5,7x večja, na ksilozo pa 4,5x manjša. Ker je v izhodišču encim 17-krat bolj aktiven na ksilozo, je zdaj 1,5-krat bolj reaktiven na glukozo.
 poteka drugače kot pri kvasovki (2 mol ATP/1 mol glukoze).")
20 Zymomonas kot alternativa za Saccharomyces Zymomonas mobilis je gramnegativna bakterija, ki lahko fermentira glukozo, fruktozo in saharozo ter proizvaja dokaj visoke koncentracije etanola. Pri tem ne raste intenzivno in torej ne porablja veliko substrata za lastno rast. Energetski metabolizem (1 mol ATP/1 mol glukoze) poteka drugače kot pri kvasovki (2 mol ATP/1 mol glukoze). Zymomonas uporabljajo ponekod v tropih za pripravo alkoholnih pijač (npr. pulque iz agave v Mehiki; 3-5 % alkohola). Hitrost sinteze etanola je večja kot pri kvasovki, zato bi bila dobra alternativa, vendar ima nekatere slabosti: - uporablja manj različnih virov ogljika; - vektorji za te bakterije niso dovolj stabilni; - bakterije so odporne proti številnim antibiotikom, ki jih uporabljamo v genski tehnologiji.
.")


21 Optimizacija sevov bakterije Zymomonas V bakterijo Zymomonas mobilis so vstavili gene, ki omogočajo razgradnjo laktoze, škroba, celuloze, ksiloze in celobioze. Ti geni so se izrazili, niso pa omogočali, da bi bakterija določen vir ogljika izrabljala kot edinega. Izjema je ksiloza (npr. odpadki pri izdelavi celuloze). Za rast na ksilozi so v bakterije vstavili gena za glukoza/ksiloza izomerazo in ksilulokinazo pod kontrolo močnega konstitutivnega promotorja gap. Nastajale so fosforilirane pentoze, njihova pretvorba pa je bila počasna. Zato so vstavili še dodaten operon, ki je omogočal porabo pentoz (gena za transketolazo in transaldolazo pod kontrolo gena za enolazo). Pri pretvorbi zelenih rastlin je najpogostejša pentoza arabinoza. Zato so v Z. mobilis vstavili gene za uporabo arabinoze: ribulokinazo, arabinoza izomerazo, ribuloza-5- fosfat-4-epimerazo iz E. coli pod kontrolo promotorja gap ter oba gena za pretvorbo pentoz. Alternativa bi bila, da pa bi iz Z. mobilis izolirali gene, ki omogočajo sintezo etanola in jih prestavili v E. coli, ki je sama sposobna pretvarjati in uporabljati tako ksilozo kot arabinozo. Ključno je, kako preusmeriti piruvat v sintezo alkohola. Uporabili so gena za piruvat dekarboksilazo in alkohol dehidrogenazo iz Z. mobilis. Enako so naredili z bakterijo Klebsiella oxytoca.
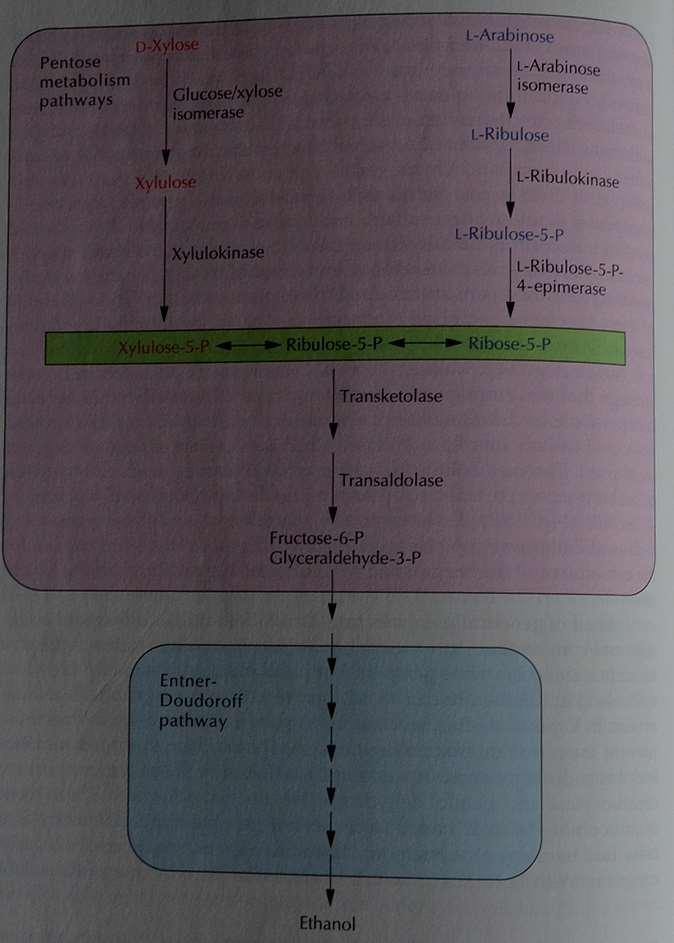
22

23 Silažna fermentacija Trave in deteljo za krmo je treba shraniti za zimo, kar je mogoče z naravnimi mlečnokislinskimi bakterijami, ki rastlinski material pretvarjajo v mlečno in ocetno kislino, zaradi znižanja ph pa ne pride do kvarjenja silaže. Ker je včasih naravnih mlečnokislinskih bakterij malo, jih je smiselno dodajati največkrat je to Lactobacillus plantarum. Naravni sevi so precej neučinkoviti v primerih, ko je vsebnost vodotopnih ogljikovih hidratov nizka. Zato so pripravili GS bakterije, ki sintetizirajo a-amilazo iz L. amylovorus (ki pa ne omogoča siliranja). Gen so s homologno rekombinacijo vstavili v genom na mesto gena za hidrolazo cbh tega bakterija ne rabi, če raste na silažnem rastlinskem materialu (rabi pa ga, če se naseli v živalskih prebavilih).
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
AMILAZE. Encimi, ki hidrolizirajo ogljikove hidrate. struktura škroba
 Encimi, ki hidrolizirajo ogljikove hidrate substrati: ogljikovi hidrati (škrob, celuloza, poli in oligosaharidi) encimi: glikozidaze glikozidna vez encimska specifičnost konfiguracija glikozidne vezi kemijska
Encimi, ki hidrolizirajo ogljikove hidrate substrati: ogljikovi hidrati (škrob, celuloza, poli in oligosaharidi) encimi: glikozidaze glikozidna vez encimska specifičnost konfiguracija glikozidne vezi kemijska
Sintezna genomika. Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi
 Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi") Sintezna genomika Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi Koncept sinteznega življenja Celic, kot so se razvile v evoluciji,
Sintezna genomika Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi Koncept sinteznega življenja Celic, kot so se razvile v evoluciji,
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
Proizvodnja in uporaba encimov
 Kmetijska šola Grm Sevno 13 8000 Novo mesto Proizvodnja in uporaba encimov ( predmet: Biotehnologija ) Gorenja vas; 7.3.2007 Avtor: Lidija Gorenc Jožica Koračin, 3.c Mentor: Jana Goršin Fabjan Kazalo:
Kmetijska šola Grm Sevno 13 8000 Novo mesto Proizvodnja in uporaba encimov ( predmet: Biotehnologija ) Gorenja vas; 7.3.2007 Avtor: Lidija Gorenc Jožica Koračin, 3.c Mentor: Jana Goršin Fabjan Kazalo:
Gimnazija Krˇsko. vektorji - naloge
 Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
ENCIMI PRI PROCESIRANJU ŠKROBA. gelatinizacija škroba nabrekanje, viskozna suspenzija. raztapljanje škroba delna hidroliza, izguba viskoznosti
 ENCIMI PRI PROCESIRANJU ŠKROBA gelatinizacija škroba nabrekanje, viskozna suspenzija raztapljanje škroba delna hidroliza, izguba viskoznosti saharifikacija škroba nastanek glukoze in maltoze škrobne granule
ENCIMI PRI PROCESIRANJU ŠKROBA gelatinizacija škroba nabrekanje, viskozna suspenzija raztapljanje škroba delna hidroliza, izguba viskoznosti saharifikacija škroba nastanek glukoze in maltoze škrobne granule
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου...
 ΑΠΟΖΗΜΙΩΣΗ ΘΥΜΑΤΩΝ ΕΓΚΛΗΜΑΤΙΚΩΝ ΠΡΑΞΕΩΝ ΣΛΟΒΕΝΙΑ 1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου... 3 1 1. Έντυπα αιτήσεων
ΑΠΟΖΗΜΙΩΣΗ ΘΥΜΑΤΩΝ ΕΓΚΛΗΜΑΤΙΚΩΝ ΠΡΑΞΕΩΝ ΣΛΟΒΕΝΙΑ 1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου... 3 1 1. Έντυπα αιτήσεων
+105 C (plošče in trakovi +85 C) -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70
 -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70") KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
 FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
FOTOSINTEZA Wan Hill primerjal rastlinsko fotosintezo s fotosintezo BAKTERIJ
 FOTOSINTEZA FOTOSINTEZA je proces, pri katerem s pomočjo svetlobne energijje nastajajo v živih celicah organske spojine. 1772 Priestley RASTLINA slab zrak dober zrak Rastlina s pomočjo svetlobe spreminja
FOTOSINTEZA FOTOSINTEZA je proces, pri katerem s pomočjo svetlobne energijje nastajajo v živih celicah organske spojine. 1772 Priestley RASTLINA slab zrak dober zrak Rastlina s pomočjo svetlobe spreminja
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
Tabele termodinamskih lastnosti vode in vodne pare
 Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Ogljikovi hidrati - monosaharidi
 Ogljikovi hidrati - monosaharidi V tem poglavju se boste naučili (ponovili) - kaj so osnovne lastnosti ogljikovih hidratov, - kaj so monosaharidi, - katere so dokazne reakcije za monosaharide. Enostavni
Ogljikovi hidrati - monosaharidi V tem poglavju se boste naučili (ponovili) - kaj so osnovne lastnosti ogljikovih hidratov, - kaj so monosaharidi, - katere so dokazne reakcije za monosaharide. Enostavni
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Molekularna biotehnologija: rekombinantna zdravila
 Molekularna biotehnologija: rekombinantna zdravila Proteinska zdravila Pred razvojem tehnologije rekombinantne DNA je bilo proteinskih zdravil zelo malo, bila so slabo okarakterizirana in draga. Danes
Molekularna biotehnologija: rekombinantna zdravila Proteinska zdravila Pred razvojem tehnologije rekombinantne DNA je bilo proteinskih zdravil zelo malo, bila so slabo okarakterizirana in draga. Danes
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABLIZEM GLJIKVIH HIDRATV KAK CELICA DBI GLUKZ IN STALE MNSAHARIDE? HRANA ZNTRAJCELIČNI GLIKGEN ali ŠKRB razgradnja s prebavnimi encimi GLUKZA in ostali monosaharidi fosforilitična cepitev prenos do
METABLIZEM GLJIKVIH HIDRATV KAK CELICA DBI GLUKZ IN STALE MNSAHARIDE? HRANA ZNTRAJCELIČNI GLIKGEN ali ŠKRB razgradnja s prebavnimi encimi GLUKZA in ostali monosaharidi fosforilitična cepitev prenos do
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 15. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matjaž Zorko Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
13. Jacobijeva metoda za računanje singularnega razcepa
 13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
p 1 ENTROPIJSKI ZAKON
 ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
Frekvenčna analiza neperiodičnih signalov. Analiza signalov prof. France Mihelič
 Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Statistična analiza. doc. dr. Mitja Kos, mag. farm. Katedra za socialno farmacijo Univerza v Ljubljani- Fakulteta za farmacijo
 Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Encimska kinetika govori o hitrosti encimske reakcije
 Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Metabolizem oz. presnova
 Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Splošno o interpolaciji
 Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
VEKTORJI. Operacije z vektorji
 VEKTORJI Vektorji so matematični objekti, s katerimi opisujemo določene fizikalne količine. V tisku jih označujemo s krepko natisnjenimi črkami (npr. a), pri pisanju pa s puščico ( a). Fizikalne količine,
VEKTORJI Vektorji so matematični objekti, s katerimi opisujemo določene fizikalne količine. V tisku jih označujemo s krepko natisnjenimi črkami (npr. a), pri pisanju pa s puščico ( a). Fizikalne količine,
Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba.
 1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
Iterativno reševanje sistemov linearnih enačb. Numerične metode, sistemi linearnih enačb. Numerične metode FE, 2. december 2013
 Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
ENCIMI PRI PRODUKCIJI ŽGANIH PIJAČ. Alkohol v medicini in industriji
 ENCIMI PRI PRODUKCIJI ŽGANIH PIJAČ Alkoholne žgane pijače maraska calvados slivovica tekila viski burbon vodka (višnje) (jabolka) (slive) (agava) (ječmen) (koruza) (rž, krompir) Alkohol v medicini in industriji
ENCIMI PRI PRODUKCIJI ŽGANIH PIJAČ Alkoholne žgane pijače maraska calvados slivovica tekila viski burbon vodka (višnje) (jabolka) (slive) (agava) (ječmen) (koruza) (rž, krompir) Alkohol v medicini in industriji
Kaj je DNA footprinting? Omogoča lociranje vezave proteina na DNA molekulo z ugotavljanjem katere fosfodiestrske vezi so zaščitene pred rezom Dnaze
 Ugotavljanje števila bakterij, ki tvorijo kolonije (cfu) Št kolonij zraslih na plošči faktor redčenja na plošči 10= x cfu/ml DNA fingerprinting: omogoča opazovanje genskih razlik med ljudmi na podlagi
Ugotavljanje števila bakterij, ki tvorijo kolonije (cfu) Št kolonij zraslih na plošči faktor redčenja na plošči 10= x cfu/ml DNA fingerprinting: omogoča opazovanje genskih razlik med ljudmi na podlagi
Mehanizmi encimske katalize
 Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Geni in regulacija njihovega prepisa
 Geni in regulacija njihovega prepisa regulacija encimske aktivnosti indukcija in represija pozitivna in negativna kontrola, atenuacija globalna kontrola dvokomponentni sistemi pogovor bakterij Regulacija
Geni in regulacija njihovega prepisa regulacija encimske aktivnosti indukcija in represija pozitivna in negativna kontrola, atenuacija globalna kontrola dvokomponentni sistemi pogovor bakterij Regulacija
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
1. TVORBA ŠIBKEGA (SIGMATNEGA) AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ
 AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ") TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
ODPORNOST BAKTERIJ PROTI ANTIBIOTIKOM V SLOVENIJI IN PO SVETU
 ODPORNOST BAKTERIJ PROTI ANTIBIOTIKOM V SLOVENIJI IN PO SVETU Manica Müller Premru, dr. med., spec. klin. mikrobiol. Inštitut za mikrobiologijo in imunologijo Medicinska fakulteta Univerza v Ljubljani
ODPORNOST BAKTERIJ PROTI ANTIBIOTIKOM V SLOVENIJI IN PO SVETU Manica Müller Premru, dr. med., spec. klin. mikrobiol. Inštitut za mikrobiologijo in imunologijo Medicinska fakulteta Univerza v Ljubljani
Logatherm WPL 14 AR T A ++ A + A B C D E F G A B C D E F G. kw kw /2013
 WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
UVOD CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN
 = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN") CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
Transformator. Delovanje transformatorja I. Delovanje transformatorja II
 Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Podobnost matrik. Matematika II (FKKT Kemijsko inženirstvo) Diagonalizacija matrik
 Diagonalizacija matrik") Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
MATEMATIČNI IZRAZI V MAFIRA WIKIJU
 I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
Če je aw manjši od 0,3 so lipidi polj izpostavljeni in zmanjšana je hidratacija. Če je aw večji od 0,5 je povečana difuzija in nabrekanje.
 ŢIVILSKA KEMIJA 1. Kaj je aktivnost vode? Definicija; katero lastnost vode opišemo s tem parametrom. Aw je parameter, s katerim kvantitativno ovrednotimo dostopnost vode v ţivilu (je merilo za hlapljivost
ŢIVILSKA KEMIJA 1. Kaj je aktivnost vode? Definicija; katero lastnost vode opišemo s tem parametrom. Aw je parameter, s katerim kvantitativno ovrednotimo dostopnost vode v ţivilu (je merilo za hlapljivost
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
ARHITEKTURA DETAJL 1, 1:10
 0.15 0.25 3.56 0.02 0.10 0.12 0.10 SESTV S2 polimer-bitumenska,dvoslojna(po),... 1.0 cm po zahtevah SIST DIN 52133 in nadstandardno, (glej opis v tehn.poročilu), npr.: PHOENIX STR/Super 5 M * GEMINI P
0.15 0.25 3.56 0.02 0.10 0.12 0.10 SESTV S2 polimer-bitumenska,dvoslojna(po),... 1.0 cm po zahtevah SIST DIN 52133 in nadstandardno, (glej opis v tehn.poročilu), npr.: PHOENIX STR/Super 5 M * GEMINI P
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Funkcije več spremenljivk
 DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
K E M IČ N A V L A K N A IZ N A R AV N IH PO L IM E R O V 1. RAZDELITEV NARAVNIH POLIMEROV [1] N A R AV N A V L A K N A
![K E M IČ N A V L A K N A IZ N A R AV N IH PO L IM E R O V 1. RAZDELITEV NARAVNIH POLIMEROV [1] N A R AV N A V L A K N A](/thumbs/77/76219690.jpg "K E M IČ N A V L A K N A IZ N A R AV N IH PO L IM E R O V 1. RAZDELITEV NARAVNIH POLIMEROV [1] N A R AV N A V L A K N A") 1. RAZDELITEV NARAVNI PLIMERV [1] Na sliki 1.1 je prikazana razdelitev naravnih vlaken. N A R AV N A V L A K N A R G A N SK A A N R G A N SK A azb est SEM EN SK A STEBELN A bo m b až, k ap o k, s v i l
1. RAZDELITEV NARAVNI PLIMERV [1] Na sliki 1.1 je prikazana razdelitev naravnih vlaken. N A R AV N A V L A K N A R G A N SK A A N R G A N SK A azb est SEM EN SK A STEBELN A bo m b až, k ap o k, s v i l
Teze predavanj iz mikrobne biokemije*
 Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
Reševanje sistema linearnih
 Poglavje III Reševanje sistema linearnih enačb V tem kratkem poglavju bomo obravnavali zelo uporabno in zato pomembno temo linearne algebre eševanje sistemov linearnih enačb. Spoznali bomo Gaussovo (natančneje
Poglavje III Reševanje sistema linearnih enačb V tem kratkem poglavju bomo obravnavali zelo uporabno in zato pomembno temo linearne algebre eševanje sistemov linearnih enačb. Spoznali bomo Gaussovo (natančneje
CM707. GR Οδηγός χρήσης... 2-7. SLO Uporabniški priročnik... 8-13. CR Korisnički priručnik... 14-19. TR Kullanım Kılavuzu... 20-25
 1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ
 GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
Matematika 1. Gregor Dolinar. 2. januar Fakulteta za elektrotehniko Univerza v Ljubljani. Gregor Dolinar Matematika 1
 Mtemtik 1 Gregor Dolinr Fkultet z elektrotehniko Univerz v Ljubljni 2. jnur 2014 Gregor Dolinr Mtemtik 1 Izrek (Izrek o povprečni vrednosti) Nj bo m ntnčn spodnj mej in M ntnčn zgornj mej integrbilne funkcije
Mtemtik 1 Gregor Dolinr Fkultet z elektrotehniko Univerz v Ljubljni 2. jnur 2014 Gregor Dolinr Mtemtik 1 Izrek (Izrek o povprečni vrednosti) Nj bo m ntnčn spodnj mej in M ntnčn zgornj mej integrbilne funkcije
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
Metabolizem in energija
 Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
PROCESIRANJE SIGNALOV
 Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
Encimi.
 Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
UPOTREBA ENZIMA U OBRADI HRANE
 UPTREBA ENZIMA U BRADI HRANE PDRUČJA PREHRAMBENE INDUSTRIJE U KJIMA SE KRISTE ENZIMSKI PRCESI BRADA PEKARSKIH PRIZVDA BRADA VĆA I VĆNIH SKVA UPTREBA ENZIMA U PIVARSTVU BRADA MLIJEČNIH PRIZVDA BRADA MESA
UPTREBA ENZIMA U BRADI HRANE PDRUČJA PREHRAMBENE INDUSTRIJE U KJIMA SE KRISTE ENZIMSKI PRCESI BRADA PEKARSKIH PRIZVDA BRADA VĆA I VĆNIH SKVA UPTREBA ENZIMA U PIVARSTVU BRADA MLIJEČNIH PRIZVDA BRADA MESA
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
Effect of Fibre Fineness on Colour and Reflectance Value of Dyed Filament Polyester Fabrics after Abrasion Process Izvirni znanstveni članek
 Učinek finosti filamentov na barvne vrednosti in odbojnost svetlobe 8 Učinek finosti filamentov na barvne vrednosti in odbojnost svetlobe barvanih poliestrskih filamentnih tkanin po drgnjenju July November
Učinek finosti filamentov na barvne vrednosti in odbojnost svetlobe 8 Učinek finosti filamentov na barvne vrednosti in odbojnost svetlobe barvanih poliestrskih filamentnih tkanin po drgnjenju July November
OKOLJSKO NARAVOSLOVJE 2. Predavanja v študijskem letu 2. del 2012/2013
 OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
Rešitve nalog. Biologija. za gimnazije
 Rešitve nalog Biologija za gimnazije 1 Značilnosti živega (str. 8) 1. Citoplazma, celična membrana, metabolizem (celici lastni presnovni mehanizmi). 2. a) velikost Virusi so zelo majhni, v splošnem 50-
Rešitve nalog Biologija za gimnazije 1 Značilnosti živega (str. 8) 1. Citoplazma, celična membrana, metabolizem (celici lastni presnovni mehanizmi). 2. a) velikost Virusi so zelo majhni, v splošnem 50-
pretvarja v nestrupeno obliko, ki lahko vstopa v biosintezo nukleotidov *i) NH 4
 NH 4") 1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
DISKRETNA FOURIERJEVA TRANSFORMACIJA
 29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Kotni funkciji sinus in kosinus
 Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
Matematika 1. Gabrijel Tomšič Bojan Orel Neža Mramor Kosta
 Matematika Gabrijel Tomšič Bojan Orel Neža Mramor Kosta 6. november 200 Poglavje 2 Zaporedja in številske vrste 2. Zaporedja 2.. Uvod Definicija 2... Zaporedje (a n ) = a, a 2,..., a n,... je predpis,
Matematika Gabrijel Tomšič Bojan Orel Neža Mramor Kosta 6. november 200 Poglavje 2 Zaporedja in številske vrste 2. Zaporedja 2.. Uvod Definicija 2... Zaporedje (a n ) = a, a 2,..., a n,... je predpis,