Jure Stojan in Marko Goličnik Medicinska fakulteta
|
|
|
- Ἀναίτις Βλαβιανός
- 7 χρόνια πριν
- Προβολές:
Transcript
1 Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do generacije 2013/14
2 Struktura predmeta: predavanja vaje seminarji Urnik Program Vaje Seminarji Kolokviji Izpit
3 MOLEKULARNA ENCIMOLOGIJA PROGRAM PREDAVANJ 2015/16 DATUM OBLIKA POUKA (P = predavanje, V = vaje) PROSTOR / četrtek P1 (MG): proteinska narava encimov, struktura, stabilnost in fleksibilnost, konformacijske spremembe, koncept aktivnega mesta, klasifikacija MF:LP / četrtek P2 (JS): encimska kataliza (kovalentna, acido-bazna, s približanjem in orientiranjem), termodinamične osnove encimske katalize, časovni potek encimske reakcije, začetna hitrost MF:LP VAJE / torek V1 (JS+MG): specifična aktivnost alkalne fosfataze razdelitev seminarjev MF:IBK vajalnica / četrtek P3 (MG): vplivi na hitrost encimske reakcije, vpliv substrata na hitrost encimske reakcije (Michaelisova kinetika), ravnotežno in stacionarno stanje MF:LP / četrtek P4 (JS): hitra kinetika in nastajanje ravnotežnih in stacionanih stanj, vrste inhibicije (reverzibilna, ireverzibilna), matematično modeliranje encimskih reakcij (holinesteraza in tubokurarin oz. eserin) MF:LP VAJE / torek V2 (JS+MG): začetna hitrost, K m, V max, k kat razdelitev seminarjev MF:IBK vajalnica K / četrtek / četrtek Integracija 1 + kolokvij 1 (MG) P5 (JS): molekulski mehanizem encimske reakcije (kimotripsin, acetilholinesteraza), alosterični pojavi, alosterija, kooperativnost, kinetika, matematične osnove, molekulski modeli, pseudokooperativnost MF:LP MF:LP VAJE / torek V3 (JS+MG): hitra kinetika in modeliranje - demonstracija MF:IBK vajalnica / četrtek P6 (JS): primeri alosteričnih encimov (PFK, CAT, ATC) MF:LP / četrtek P7 (MG): klasifikacija encimov in primeri delovanja značilnih predstavnikov posameznih encimskih razredov MF:LP K IZPIT S / četrtek / četrtek / torek / četrtek P8 (MG): uporaba encimologije v kliniki (diagnostika, terapija, encimi kot tarče zdravil) in biotehnologiji (mikroorganizmi, imobilizirani encimi, kat. protitelesa) Integracija 2 + kolokvij 2 (JS) S1 (JS+MG) S2 (JS+MG) MF:LP MF:LP MF:LP MF:LP januar 2015 izpit (1. rok) po razporedu MF:IBK februar 2015 izpit (2. rok) po razporedu MF:IBK junij 2015 izpit (3. rok) po razporedu MF:IBK september 2015 izpit (4. rok) po razporedu MF:IBK


4 KJE JE KAJ: VAJE PREDAVANJA, SEMINARJI VAJE
5 Predavanje 1: STRUKTURA PROTEINOV IN NJEN POMEN ZA DELOVANJE ENCIMOV Marko Goličnik Medicinska fakulteta GLEJ:
6 MOLEKULARNA ENCIMOLOGIJA Encimi so biološki katalizatorji: Skrbijo za dovolj veliko hitrost kemičnih reakcij. Pomembno sodelujejo pri regulaciji bioloških procesov. Omogočajo črpanje energije in njeno porabo v organizmih. V klinični biokemiji so pomembni markerji in reagenti. Po kemijski strukturi: PROTEINI in RNA (klasični encimi in ribocimi). Ribocimi so izredno pomembni pri zorenju mrna in pri sintezi proteinov Vse druge procese (reakcije) katalizirajo klasični encimi več različnih PROTEINOV (~ v človeku). ZATO MORAMO POZNATI STRUKTURO PROTEINOV!
7 PROTEINE SESTAVLJAJO AMINOKISLINE (AK) Glavne lastnosti (20 standardnih) aminokislin - klasifikacija AK - velikost - naboj - polarnost - hidrofobnost - aromatske AK - 3D-konfiguracija AK in optična aktivnost a C-atom Ta del je enak! Le po R se razlikujejo! L-AMINOKISLINA PAZI: več 100 nestandardnih AK (post-transl. modif. & D-AK)!
8 Hidrofilne (polarne) AK: - nenabit radikal nabit radikal
9 Hidrofobne (nepolarne) AK: aromatske posebne
10 PROSTA AMINOKISLINA R AMINOKISLINA (AK) R = AK radikal (stranska veriga) R 1 -COOH + H 2 N-R 2 R 1 -CONH-R 2 + H 2 O AMINOKISLINA V PROTEINU N R 3 C AK ostanek (residue) = AK H 2 O
11 Velikost AK-ostanka: Da (110 Da), a nekaj izjem! AK ostanek (residue) AK radikal (stranska veriga) dodaj 18 (H 2 O) za maso AK! odštej 56 za maso R (stranske verige)!
12 Velikost je pomembna pri zamenjavah aminokislin!
13 N R 3 C NABOJ: Odvisen od pk a vrednosti (25 C) in ph Samo R, N- and C- konci so pomembni pri proteinih Aminokislina a-cooh a-nh 3 + R Alanin Glicin Fenilalanin Serin Valin Asparaginska k Glutaminska k Histidin ! Cistein Tirosin ! Lizin Arginin
 absorbirajo UV")
14 Aromatske AK (Phe, Tyr, Trp) velike resonanca: -OH v Tyr bolj kisla polarnost: posebne interakcije (kation p) absorbirajo UV svetlobo
15 3D STRUKTURA AK DOLOČA 3D STRUKTURO PROTEINOV POMEMBNO ZA PREPOZNAVANJE: E-S, E-I, R-H, Ag-Ab rotacija linearno polarizirane svetlobe (a = [a].l.c) D,L-system Posebne funkcije! STANDARD!
16 Bliskovit in strahovito poenostavljen pregled proteinske sinteze DNA primarni transkript (pre-mrna) pre-mrna se procesira v mrna mrna je zapis za I. str. proteinov v katero se povežejo AK s peptidno vezjo (I. struktura = zaporedje AK) nekatere AK se modificirajo 1D 3D strukturo (II., III. in IV. (?) struktura) kontrola kvalitete delovanja
 Nastanek peptidne vezi: R 1 -COOH + H 2 N-R 2 R 1 -CONH-R 2")
17 Shema sinteze proteinov Transkripcija (prepis DNA mrna) nukleotidi v nukleotide pre-mrna Zorenje mrna Translacija (prevod mrna protein) Nukleotidi v aminokisline mrna Postranslacijske modifikacije AK (samo evkarionti) Nastanek peptidne vezi: R 1 -COOH + H 2 N-R 2 R 1 -CONH-R 2 + H 2 O
18 Post-translacijske modifikacije spremenijo sekvenco, ali odstranijo nezaželene dele proteina, vpeljejo nove funkcionalne skupine in imajo tudi regulatorno vlogo. Pomembne modifikacije: proteolitično procesiranje = izrezovanje delov proteinske verige spremembe N- in C-koncev glikozilacija (pripenjanje sladkorjev) pripenjanje lipidov sulfatiranje g-karboksi-glutaminska kislina hidroksilacija fosforilacija ADP-ribozilacija disulfidni mostički pripenjanje funkcionalnih skupin
19 Proteolitično procesiranje: aktivacija encimov Namen: encim postane aktiven tam kjer deluje in ne na mestu sinteze! Značilno predvsem za prebavne encime.
20 Pripenjanje lipidov: interakcija z membrano, 3D struktura
21 Pripenjanje malih funkcionalnih skupin vodi v spremembo 3D strukture in aktivnosti fosforilacija g-karboksi-glutamat metilacija sulfatiranje hidroksilacija
22 Nastanek disulfidne vezi: stabilizacija 3D-strukture
23 Primarna struktura = sekvenca aminokislin PO 4 3- peptidne vezi Ser Gly Tyr Ala Leu S G Y A L Oštevilčenje od N-konca do C-konca! Sekvenco lahko določimo neposredno iz proteina ali posredno iz mrna ali DNA.
24 S poravnavo proteinskih sekvenc določamo sorodnost med vrstami, pa tudi funkcijo neznanih proteinov.
")
25 Samosestavljanje polipeptidne verige: od primarne do terciarne (kvartarnarne) strukture


26 ŠIBKE INTERAKCIJE odboj pri dotiku van der Waalsove interakcije elektrostatske interakcije R 1 -COO - H 3 N + -R 2 vodikova vez hidrofobne interakcije ion - p interakcije (aromatske AK) CH 3 -CH 2 -CH 3 CH 3 -CH 2 -CH 3 Vse te šibke interakcije so med AK radikali, H-vezi pa tudi med CO in HN- v peptidni vezi.


27 Peptidna vez R 1 -COOH + H 2 N-R 2 R 1 -CONH-R 2 + H 2 O Delna dvojna vez, zato 6 atomov v isti ravnini! Velika večina peptidnih vezi je v trans konfiguraciji. IZJEMA: v zavojih ob Pro je približno 6% peptidnih vezi cis

28 Polipeptidna veriga v iztegnjeni konformaciji POZOR: OMEJENA GIBLJIVOST ZARADI OMEJITEV V VRTLJIVOSTI OKROG PEPTIDNE VEZI.
 Začetni položaj: Φ = Ψ")

29 Dihedralna kota Φ (fi) in Ψ (psi) Začetni položaj: Φ = Ψ = 0 stopinj Ta položaj je le teoretičen in v resnici ni mogoč!

30 C=O in N-H, ki sta povezana z vodikovo vezjo, sta udeležena v dveh različnih peptidnih vezeh 5 AK narazen. desnosučna a-vijačnica N vodikova vez med AK i in AK i+4 O H C RADIKALI (niso prikazani) MOLIJO VEN!

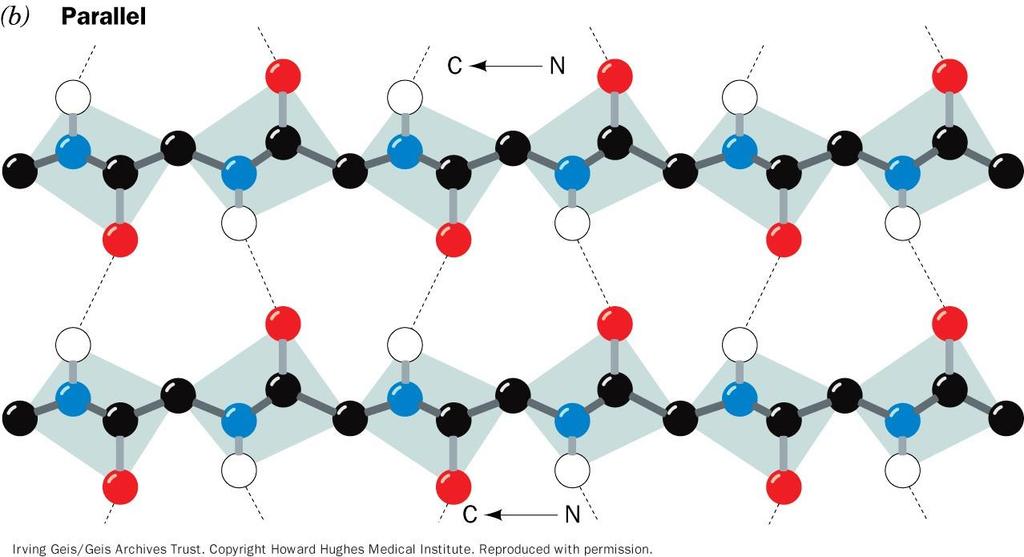
31 b struktura (antiparalelna) b struktura (paralelna)


32 Nagubani list je pogosto zvit in podprt z vijačnicami. N-t. C-t. C-t. N-t.

33 Povezave verig v nagubanem listu ANTIPARALELNA PARALELNA zavoj

34 β-zavoji RAZLIKA!

35 Zanke: Ω zanka je navadno iz 40 do 54 AK (citokrom c) Ω Ni nujno, da so vsi deli proteina v eni od teh II. struktur NEUREJENA II. STRUKTURA




36 TERCIARNA STRUKTURA GLOBULARNIH PROTEINOV Vsi encimi so globularni proteini. Pet različnih prikazov terciarne strukture mioglobina. III. struktura: zvitje v nativno konformacijo vse vrste II. strukture so običajno vključene (tudi neurejena struktura) III. strukturo vzdržujejo številne šibke interakcije med AK ostanki III. struktura je stabilna a fleksibilna poznati III. strukturo pomeni poznati položaj vseh atomov v prostoru (3D)

37 Fotografija lis na filmu po uklonu rentgenskih žarkov skozi kristal Mb. Protein moramo najprej očistiti, nato kristalizirati in skozi kristal spustiti rentgenske žarke, ki se na atomih zaradi elektronov uklonijo. Analiza uklonskih slik nam da elektronsko gostoto na določeni globini kristala. Iz tega določijo položaj vsakega atoma v proteinu.

38 Zemljevid elektronske gostote proteina z AK ostanki.

39 Struktura proteina (Src protein SH3 domena) dobljena z 2D protonsko NMR; vidijo se področja večje fleksibilnosti. Trp Phe Tyr
, ki ju lahko ločimo s prekinitvijo ene vezi v verigi, ki povezuje domeni (glej")
40 Ena podenota encima gliceraldehid-3-fosfat dehidrogenaze iz Bacillusa stearothermophilusa. POZOR: Kombinacija različnih II. struktur. Jasno se vidita dve domeni (ena rdeče in druga zelene barve), ki ju lahko ločimo s prekinitvijo ene vezi v verigi, ki povezuje domeni (glej puščico).

41 Kvartarna struktura hemoglobina.

42 Glutamin-sintetaza (Salmonella typhimurium).
")
 - POVEČANA &")
 - MULTIFUNKCIONALNI")

43 človek ZAKAJ KVARTARNA STRUKTURA? - VARNOST (Hb: 2a+2b, 2 različna gena) - ALOSTERIČNI EFEKTI (regulacija Hb, alosterični encimi) - POVEČANA & LOKALIZIRANA KONCENTRACIJA AKTIVNIH MEST (AChE) - MULTIFUNKCIONALNI PROTEINI: maščobnokislinska sintetaza: 7 aktivnih mest z različnimi funkcijami: bakterija gliva

44 Voda je integralni del proteinske strukture in pomembno prispeva k njeni stabilnosti.

45 Zvijanje proteinov preučujemo tako, da jih razvijemo (denaturiramo), nato pa opazujemo ponovno zvijanje. Denaturanti, ph, T in visok P porušijo strukturo. Proces je lahko reverzibilen.

46 Zakaj denaturacija: toplota (visoka temperatura): neposreden vnos energije ekstremni ph: ionizacija le v razvitem stanju (premik ravnotežja); elektrostatski odboji na površini proteina; porušenje ionskih interakcij. denaturanti (urea, gvanidinijev klorid): preferenčna vezava deli proteina se raje vežejo z denaturantom kot med seboj (vpliv na H-vezi in hidrofobne interakcije!)
, povsem razvit protein pa ima največjo energijo (G) in tudi največjo entropijo (S), ker je ta struktura najbolj")
47 Prerez skozi 3D energijski profil zvijanja proteina. Nativna struktura ima najmanjšo energijo (prosto entalpijo G), povsem razvit protein pa ima največjo energijo (G) in tudi največjo entropijo (S), ker je ta struktura najbolj neurejena. A POZOR, TOPILO!!! G G maksimalna, S maksimalna prispevek vezi G=H-T.S Prispevek (ne)urejenosti G minimalna, S minimalna

48 Šaperoni pomagajo proteinom pri zvijanju in skušajo ponovno zviti razvite proteine.

49 Premikanje domen v plašču šaperona pomaga pri zvijanju proteinske verige, ki je v notranjosti. Konformacijske spremembe omogoča hidroliza ATP.

 transkripcijski faktor (P.")
50 Nekateri proteini so namerno nestabilni (vsaj lokalno); to omogoča: konformacijske spremembe prilagoditve pri vezavi na druge molekule (interakcije protein - protein in protein -nukleinska kislina) transkripcijski faktor (P. Wright, Scripps)
51 Metabolično obračanje proteinov: Proteini živijo v celici različno dolgo. Npr.: razpolovni čas encimov v jetrih je od 0.2 do 150 ur. Pravilo N-konca: Razpolovni čas proteinov je povezan s strukturo N-terminalne AK: Za proteine z N-terminalnimi Met, Ser, Ala, Thr, Val ali Gly je razpolovni čas večji od 20 ur. Za proteine z N-terminalnimi Phe, Leu, Asp, Lys ali Arg je razpolovni čas 3 min ali manj. Pravilo PEST: proteini, ki imajo veliko Pro (P), Glu (E), Ser (S) ali Thr (T) se hitreje razgradijo kot drugi proteini.

52 Selektivno razgradnjo proteinov uravnavajo znotrajcelični in zunajcelični signali. Eden od njih je vezava ubikvitina (majhen protein) na za razgranjo namenjen (predvsem delno razvit) protein. Razgradnja ubikvitiniranih proteinov poteka v proteasomih. signalna sekvenca Primarna struktura za razgradnjo namenjenega proteina H 2 N veriga ubikvitinov COO Veriga 4 ali več ubikvitinov usmeri protein v razgradnjo. ubikvitin PDB 1TBE


53 a b b a 20 S Proteasom (kvasovka) v zaprtem stanju dva pogleda PDB 1JD2 Proteasom je velik kompleks. Jedro ima 4 obroče (2 α in 2 β), vsak je iz 7 proteinov, znotraj pa je votlina kjer se razgrajujejo ubikvitirani proteini. Po 3 podenote v β obročih so proteolitični encimi.
54 Kontrola kvalitete proteinske sinteze Proteasomi prepoznajo in razgrajujejo poli-ubikvitinirane proteine, ki so celici lastni. Nekateri celici lastni proteini se le mono-ubikvitinirajo; ti verjetno doživijo razgradnjo v lizosomih. Lizosomi so organeli, ki vsebujejo proteolitične encime (katepsine); namenjeni so razgradnji predvsem izvenceličnih proteinov (v celico pridejo z endocitozo). Oba sistema sta potrebna za odstranjevanje izrabljenih in napačno zvitih ali razvitih proteinov. Okrog 50 % vseh sintetiziranih proteinov se takoj razgradi! VSE KAR SMO POVEDALI O ZGRADBI, ZVIJANJU IN RAZGRADNJI PROTEINOV VELJA SEVEDA TUDI ZA ENCIME IN JIM OMOGOČA NJIHOVO DELOVANJE!
55 Kako encimi delujejo? nekatalizirana reakcija S P substrat produkt encimsko katalizirana reakcija Med katalizo encim E specifično veže substrat S, ga pomaga spremeniti v produkt P, encim pa se nespremenjen odcepi. Tako se lahko ponovno in večkrat uporabi. Med tem procesom se tvori kratko živeči kompleks ES (in drugi kompleksi, npr. EP). E + S ES EP E + P E encim S substrat, P produkt ES kompleks encim-substrat EP kompleks encim-produkt POZOR, nista aktivirana kompleksa!
56 Encimi so bio-katalizatorji, ki zmanjšajo aktivacijsko energijo reakcije (S 1 + S 2 )

57 S P E + S ES EP E + P DG o ΔG o = ΔG o = - RT ln K ΔG # < ΔG # v > v

58 Substrat se veže na aktivno mesto, ki je navadno le manjši del pravilno 3D-oblikovane encimske molekule. Tu vidimo pomen III. In IV. proteinske strukture encima. Neaktivni del encima je veliko večji od aktivnega mesta, a je potreben, da se aktivno mesto pravilno oblikuje in za mnoge druge funkcije, med drugim za kompenzacijo konformacijskih sprememb med katalizo in za interakcije z drugimi molekulami, s katerimi se omogoča in regulira aktivnost (koencimi, modulatorji in drugi proteini). Encim predstavlja specifično okolje za molekulo substrata.

59 Nastanek kompleksa encim substrat omogoča geometrijska in fizikalnokemijska komplementarnost encima in substrata. Skupine na substratu in v aktivnem mestu encima si ustrezajo tako, da se lahko medsebojno povežejo. Koncept aktivnega mesta Fischerjev koncept: popolno ustrezanje substrata in encima. To ni realistično, ker bi to stabiliziralo osnovno stanje substrata in ta bi teže prešel v prehodno stanje.

60 Za razlikovanje enantiomerov mora imeti encim vsaj tri specifična mesta za vezavo substrata. R
 ENCIM VODA (S 2 ) SUBSTRAT (S 1 ) S 1 + S 2 P 1 + P 2 Koshlandov koncept: encim le delno ustreza substratu, a se oba prilagodita pri tem encim vsili substratu konformacijo prehodnega stanja v")
61 Peptidaza veže substrat (dipeptid) s tremi šibkimi vezmi, vsili substratu novo konformacijo, kar destabilizira peptidno vez (prehodno stanje!) ENCIM VODA (S 2 ) SUBSTRAT (S 1 ) S 1 + S 2 P 1 + P 2 Koshlandov koncept: encim le delno ustreza substratu, a se oba prilagodita pri tem encim vsili substratu konformacijo prehodnega stanja v kateri se ta najbolje veže na encim in se zaradi tega laže (in tudi hitreje) pretvori v produkt!

62 V aktivnem mestu pridejo zaradi III. strukture skupaj AK, ki so navadno v I.strukturi med seboj precej oddaljene. Struktura tripsina Katalitična triada Namen: aktivacija Ser (OH) N 1 7 His 57 Asp 102 Ser C
63 KOENCIMI IN PROSTETIČNE SKUPINE (nastanejo iz vitaminov), ZAGOTOVIJO DODATNE FUNKCIONALNE SKUPINE PREKURZOR (VITAMIN) SKUPINA PRENOSA KOENCIM/PROST. SK. tiamin (vitamin B1) aldehidna skupina tiamin pirofosfat riboflavin (vitamin B2) elektroni in protoni flavin mononukleotid FMN, FAD nikotinska kislina (niacin) hidridni ion (H - ) nikotinamid adenin dinukleotid NAD pantotenska kislina in druge molekule acilne skupine koencim A piridoksin (vitamin B6) amino skupine piridoksal fosfat vitamin B12 H-atomi in alkilne skupine (ciano)kobalamin biotin CO 2 (-COOH) biocitin folna kislina skupine z enim C-atomom tetrahidrofolna kislina lipojska kislina elektroni in acilne skupine lipojska kislina
64 KOENCIMI Se sprehajajo med encimi in nanje niso trajno vezani Na enem E sprejmejo neko skupino ali molekulo, na drugem jo oddajo Tako povezujejo različne encime v kompleksne večstopenjske procese in prenašajo skupine ali molekule med različnimi substrati A-x CoE KOFAKTORJI x = skupina, ki se prenaša B-x A-x PROSTETIČNE SKUPINE So trajno vezani na določen encim, ki ga nikoli ne zapustijo Na istem E sprejmejo neko skupino ali molekulo od enega S in jo oddajo drugemu S Tudi povezujejo večstopenjske procese, a običajno sodelujejo z omejenim številom substratov PSk E B-x E 1 E 2 A CoE-x B A PSk-x E B Kofaktorji so lahko tudi le ustrezni kovinski ioni (cink, magnezij, baker itd) APOENCIM predstavlja le proteinski del, HOLOENCIM pa celoten aktivni encim; tj. proteinski del + kofaktor.
65 PREKURZOR (VITAMIN) SKUPINA PRENOSA KOENCIM pantotenska kislina in druge molekule acilne skupine koencim A R-COOH + HS- R-CO-S- Vir: vsaka hrana; dnevna priporočena količina: 4-7 mg. Pomanjkanja niso nikoli opazili, s težavo se ga da umetno izzvati v strogo nadzorovanih eksperimentalnih razmerah.
66 PREKURZOR (VITAMIN) SKUPINA PRENOSA KOENCIM (PROST. SK.) riboflavin (vitamin B2) elektroni in protoni FMN, FAD! 2 e - in 2 H +! Vir: meso, jetra, jajca, mleko; dnevna priporočena količina: 1,5 mg Pomanjkanje: prizadete sluznice in koža, anemija - posebno pri alkoholikih in pri novorojenčkih s hiperbilirubinemijo, ki so pod osvetljevalno terapijo (razgradnja bilirubina in riboflavina).
67 PREKURZOR (VITAMIN) SKUPINA PRENOSA KOENCIM nikotinska kislina (niacin) hidridni ion (H - ) NAD -OH 2 e - in 1 H + = H - = hidridni ion! Vir: meso, jetra, kvas, semena; dnevna priporočena količina: mg (tudi neuč. sinteza iz Trp). Pomanjkanje (koruza!): pelagra - prizadeta je koža, v resnejši obliki pa dermatitis, diareja in demenca (lahko tudi ireverzibilna). V razvitem svetu redka.
68 PREKURZOR (VITAMIN) SKUPINA PRENOSA PROSTETIČNA SKUPINA biotin karboksilna skupina biocitin (kovalentno vezan na Lys)
69 2. predavanje termodinamske in kinetične osnove katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV. 2. predavanje: Od 1D do 3D strukture proteinov 1. del.
 IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
 FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
Encimska kinetika govori o hitrosti encimske reakcije
 Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Encimi.
 Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Mehanizmi encimske katalize
 Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Matjaž Zorko Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Biokemija I, 25. predavanje 1. del, , A. Videtič Paska. Proteini - splošno
 Biokemija I, 25. predavanje 1. del, 16. 4. 2012, A. Videtič Paska Proteini - splošno Razdelitev po strukturi in funkciji. Ravni proteinske strukture: - primarna in - sekundarna struktura Sinteza proteinov
Biokemija I, 25. predavanje 1. del, 16. 4. 2012, A. Videtič Paska Proteini - splošno Razdelitev po strukturi in funkciji. Ravni proteinske strukture: - primarna in - sekundarna struktura Sinteza proteinov
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda
![[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda](/thumbs/89/99439684.jpg "[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda") kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ
 ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ α-heliks β-nagubana RAVNINA β-obrat STRUKTURNI MOTIVI STABILNOST PROTEINOV KVARTARNA ZGRADBA PROTEINOV ENA ALI VEČ KONFORMACIJ
ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ α-heliks β-nagubana RAVNINA β-obrat STRUKTURNI MOTIVI STABILNOST PROTEINOV KVARTARNA ZGRADBA PROTEINOV ENA ALI VEČ KONFORMACIJ
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
STRUKTURNE LASTNOSTI AMINOKISLIN
 AMINOKISLINE Amino-karboksilne kisline (izjema: prolin, iminokarboksilna kislina) Vloga aminokislin: 1. Gradniki proteinov 2. Vir energije 3. Izhodišče za sintezo drugih pomembnih biomolekul (nukleinske
AMINOKISLINE Amino-karboksilne kisline (izjema: prolin, iminokarboksilna kislina) Vloga aminokislin: 1. Gradniki proteinov 2. Vir energije 3. Izhodišče za sintezo drugih pomembnih biomolekul (nukleinske
Uravnavanje encimske aktivnosti
 Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
ZVIJANJE PROTEINOV, RAZGRADNJA IN USMERJANJE
 ZVIJANJE PROTEINOV, RAZGRADNJA IN USMERJANJE ZVIJANJE PROTEINOV Spontano Stanje odtajane kroglice Molekularni šaperoni POST-TRANSLACIJSKE MODIFIKACIJE PROTEINOV RAZGRADNJA Ubikvitin Proteasom USMERJANJE
ZVIJANJE PROTEINOV, RAZGRADNJA IN USMERJANJE ZVIJANJE PROTEINOV Spontano Stanje odtajane kroglice Molekularni šaperoni POST-TRANSLACIJSKE MODIFIKACIJE PROTEINOV RAZGRADNJA Ubikvitin Proteasom USMERJANJE
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
3/25/2016. Hemijske komponente ćelije
 Hemijske komponente ćelije Molekuli u ćeliji Najbitniji molekuli u ćeliji su poznati. Putevi sinteze i razgradnje su poznati za većinu ćelijskih konstituenata. Hemijska energija pokreće biosintezu. Organizacija
Hemijske komponente ćelije Molekuli u ćeliji Najbitniji molekuli u ćeliji su poznati. Putevi sinteze i razgradnje su poznati za većinu ćelijskih konstituenata. Hemijska energija pokreće biosintezu. Organizacija
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
Aminokiseline. Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina 22.12.2014
 Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK)
") IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
pretvarja v nestrupeno obliko, ki lahko vstopa v biosintezo nukleotidov *i) NH 4
 NH 4") 1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
CILJNA MESTA DEJSTVA LEKOVA
 FARMACEUTSKA HEMIJA 1 CILJNA MESTA DEJSTVA LEKVA Predavač: Prof. dr Slavica Erić Ciljna mesta dejstva leka CILJNA MESTA NA MLEKULARNM NIVU: lipidi (lipidi ćelijske membrane) ugljeni hidrati (obeleživači
FARMACEUTSKA HEMIJA 1 CILJNA MESTA DEJSTVA LEKVA Predavač: Prof. dr Slavica Erić Ciljna mesta dejstva leka CILJNA MESTA NA MLEKULARNM NIVU: lipidi (lipidi ćelijske membrane) ugljeni hidrati (obeleživači
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
Funkcije proteinov (pogojene s strukturo)
") Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 15. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
I. OSNOVNI STRUKTURNI PRINCIPI
 I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Nekatere interakcije v lipidnem metabolizmu
 Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Prehrana in metabolizem
 Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
ÂÓÈÎ ÁÈ ÙÔ K ÙÙ ÚÔ 1 Ô KÂÊ Ï ÈÔ 1.1 E Ë Î ÙÙ ÚˆÓ 1.1.1 ÚÔÎ Ú ˆÙÈÎ Î ÙÙ Ú
 11 1 Ô KÂÊ Ï ÈÔ ÂÓÈÎ ÁÈ ÙÔ K ÙÙ ÚÔ 1.1 E Ë Î ÙÙ ÚˆÓ Στο κεφάλαιο αυτό θα αναφερθούμε σύντομα στο κύτταρο, τα είδη (ευκαρυωτικά και προκαρυωτικά) και γενικά στα διάφορα στοιχεία του, όπως πυρήνα, κυτταρόπλασμα
11 1 Ô KÂÊ Ï ÈÔ ÂÓÈÎ ÁÈ ÙÔ K ÙÙ ÚÔ 1.1 E Ë Î ÙÙ ÚˆÓ Στο κεφάλαιο αυτό θα αναφερθούμε σύντομα στο κύτταρο, τα είδη (ευκαρυωτικά και προκαρυωτικά) και γενικά στα διάφορα στοιχεία του, όπως πυρήνα, κυτταρόπλασμα
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
ENCIMI ZGRADBA ENCIMOV NEKATERI ENCIMI IN NJIHOVI KOFAKTORJI
 Visoko specializirani proteini in mala skupina katalitičnih molekul RNA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10 16 x Zmanjšajo E a, ne vplivajo
Visoko specializirani proteini in mala skupina katalitičnih molekul RNA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10 16 x Zmanjšajo E a, ne vplivajo
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
1. TVORBA ŠIBKEGA (SIGMATNEGA) AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ
 AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ") TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
Sekundarne struktura proteina Fibrilni proteini
 Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Malgorzata Korycka-Machala, Marcin Nowosielski, Aneta Kuron, Sebastian Rykowski, Agnieszka Olejniczak, Marcin Hoffmann and Jaroslaw Dziadek
 Molecules 2017, 21, 154; doi:10.3390/molecules22010154 Supplementary Materials: Naphthalimides Selectively Inhibit the Activity of Bacterial, Replicative DNA Ligases and Display Bactericidal Effect against
Molecules 2017, 21, 154; doi:10.3390/molecules22010154 Supplementary Materials: Naphthalimides Selectively Inhibit the Activity of Bacterial, Replicative DNA Ligases and Display Bactericidal Effect against
Nukleinske kisline. ribosomska informacijska prenašalna
 Nukleinske kisline Nukleinske kisline vloga pri shranjevanju, prenašanju in izražanju genetske informacije: DNA RNA proteini zgradba in delovanje celice 2 osnovni vrsti nukleinskih kislin: deoksiribonukleinska
Nukleinske kisline Nukleinske kisline vloga pri shranjevanju, prenašanju in izražanju genetske informacije: DNA RNA proteini zgradba in delovanje celice 2 osnovni vrsti nukleinskih kislin: deoksiribonukleinska
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Sinteza RNA - transkripcija
 Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks'
 Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Družina globinov pri človeku in bolezni.
 Družina globinov pri človeku in bolezni www.muhlenberg.edu/ Mioglobin in hemoglobin spadata v družino globinov Globinsko zvitje Mb β podenota Hb Podobnost aminokislinskega zaporedja Podobnost 3D strukture
Družina globinov pri človeku in bolezni www.muhlenberg.edu/ Mioglobin in hemoglobin spadata v družino globinov Globinsko zvitje Mb β podenota Hb Podobnost aminokislinskega zaporedja Podobnost 3D strukture
ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ
 ΜΙΝΟΠΕΤΡΟΣ ΚΩΝΣΤΑΝΤΙΝΟΣ ΦΥΣΙΚΟΣ - Ρ/Η ΚΑΘΗΓΗΤΗΣ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΥΠΕΥΘΥΝΟΣ ΣΕΦΕ 2 ου ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΠΕΡΑΜΑΤΟΣ ΕΡΓΑΣΤΗΡΙΑΚΗ ΑΣΚΗΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ
ΜΙΝΟΠΕΤΡΟΣ ΚΩΝΣΤΑΝΤΙΝΟΣ ΦΥΣΙΚΟΣ - Ρ/Η ΚΑΘΗΓΗΤΗΣ ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΥΠΕΥΘΥΝΟΣ ΣΕΦΕ 2 ου ΕΝΙΑΙΟΥ ΛΥΚΕΙΟΥ ΠΕΡΑΜΑΤΟΣ ΕΡΓΑΣΤΗΡΙΑΚΗ ΑΣΚΗΣΗ ΒΙΟΛΟΓΙΑΣ ΚΑΤΕΥΘΥΝΣΗΣ Γ ΛΥΚΕΙΟΥ ΑΝΤΙΓΡΑΦΗ ΚΑΙ ΕΚΦΡΑΣΗ ΤΗΣ ΓΕΝΕΤΙΚΗΣ ΠΛΗΡΟΦΟΡΙΑΣ
-NH 3. Degradación de aminoácidos. 1) Eliminación del NH 3. 2) Degradación de esqueletos carbonados. Ac. grasos c. cetónicos glucosa.
 Eliminación del NH 3. 2) Degradación de esqueletos carbonados. Ac. grasos c. cetónicos glucosa.") Degradación de aminoácidos 1) Eliminación del NH 3 interfiere polarización/despolariación nerviosa compite transportadores metales alcalinos -NH 3 + 2) Degradación de esqueletos carbonados Ac. grasos c.
Degradación de aminoácidos 1) Eliminación del NH 3 interfiere polarización/despolariación nerviosa compite transportadores metales alcalinos -NH 3 + 2) Degradación de esqueletos carbonados Ac. grasos c.
SPLOŠNO O BELJAKOVINAH STRUKTURA BELJAKOVIN
 SPLOŠNO O BELJAKOVINAH Beljakovine so najvažnejša sestavina vsake celice, kajti vsi življenski procesi so odvisni od njih. So makromolekularnespojine, sestavljeneiz ogljika, vodika, kisika in dušika ter
SPLOŠNO O BELJAKOVINAH Beljakovine so najvažnejša sestavina vsake celice, kajti vsi življenski procesi so odvisni od njih. So makromolekularnespojine, sestavljeneiz ogljika, vodika, kisika in dušika ter
Poglavje 10. Molekule Kovalentna vez
 Poglavje 10 Molekule Atomi se vežejo v molekule. Vezavo med atomi v molkuli posredujejo zunanji - valenčni elektroni. Pri vseh molekularnih vezeh negativni naboj elektronov posreduje med pozitinvimi ioni
Poglavje 10 Molekule Atomi se vežejo v molekule. Vezavo med atomi v molkuli posredujejo zunanji - valenčni elektroni. Pri vseh molekularnih vezeh negativni naboj elektronov posreduje med pozitinvimi ioni
Splošno o interpolaciji
 Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Transformator. Delovanje transformatorja I. Delovanje transformatorja II
 Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Cefalosporini ostali β-laktami
 Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
2.1. MOLEKULARNA ABSORPCIJSKA SPEKTROMETRIJA
 2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
Nukleinske kisline. Nukleotidi. DNA je nosilka dednih genetskih informacij.
 Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Tabele termodinamskih lastnosti vode in vodne pare
 Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
Iterativno reševanje sistemov linearnih enačb. Numerične metode, sistemi linearnih enačb. Numerične metode FE, 2. december 2013
 Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
4 ο ΚΕΦΑΛΑΙΟ. Γ ε ν ε τ ι κ ή
 4 ο ΚΕΦΑΛΑΙΟ Γ ε ν ε τ ι κ ή 1. Κύκλος της ζωής του κυττάρου 3ο Γελ. Ηλιούπολης επιμέλεια: Αργύρης Γιάννης 2 2. Μοριακή Γενετική i). Ροή της γενετικής πληροφορίας DNA RNA πρωτεΐνες νουκλεΐκά οξέα ή πρωτεΐνες
4 ο ΚΕΦΑΛΑΙΟ Γ ε ν ε τ ι κ ή 1. Κύκλος της ζωής του κυττάρου 3ο Γελ. Ηλιούπολης επιμέλεια: Αργύρης Γιάννης 2 2. Μοριακή Γενετική i). Ροή της γενετικής πληροφορίας DNA RNA πρωτεΐνες νουκλεΐκά οξέα ή πρωτεΐνες
Podobnost matrik. Matematika II (FKKT Kemijsko inženirstvo) Diagonalizacija matrik
 Diagonalizacija matrik") Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Neavtorizirani povzetki izbranih predavanj iz biokemije. UN študij Okolje Politehnika Nova Gorica. doc. dr. Marko Dolinar
 Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * FIZIKA NAVODILA ZA OCENJEVANJE. Petek, 10. junij 2016 SPLOŠNA MATURA
 Državni izpitni center *M16141113* SPOMLADANSKI IZPITNI ROK FIZIKA NAVODILA ZA OCENJEVANJE Petek, 1. junij 16 SPLOŠNA MATURA RIC 16 M161-411-3 M161-411-3 3 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor
Državni izpitni center *M16141113* SPOMLADANSKI IZPITNI ROK FIZIKA NAVODILA ZA OCENJEVANJE Petek, 1. junij 16 SPLOŠNA MATURA RIC 16 M161-411-3 M161-411-3 3 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
Gimnazija Krˇsko. vektorji - naloge
 Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
ZGRADBA ENCIMOV ENCIMI ENCIMI REAKCIJE, KINETIKA IN INHIBICIJA. Aktivni samo v nativni konformaciji. M = Da. Enostavni in sestavljeni
 EIMI REAKIJE, KIETIKA I IIBIIJA Visoko specializirani proteini in mala skupina katalitičnih molekul RA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10
EIMI REAKIJE, KIETIKA I IIBIIJA Visoko specializirani proteini in mala skupina katalitičnih molekul RA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10
Ζεύγη βάσεων ΓΕΝΕΤΙΚΗ. Γουανίνη Κυτοσίνη. 4α. Λειτουργία γενετικού υλικού. Φωσφοδιεστερικός δεσμός
 εύγη βάσεων Αδενίνη Θυμίνη Γουανίνη Κυτοσίνη ΓΕΝΕΤΙΚΗ Φωσφοδιεστερικός δεσμός 4α. Λειτουργία γενετικού υλικού 1 ΛΕΙΤΟΥΡΓΙΑ ΓΕΝΕΤΙΚΟΥ ΥΛΙΚΟΥ Αντιγραφή (διπλασιασμός) DNA: DNA DNA Έκφραση γενετικής πληροφορίας:
εύγη βάσεων Αδενίνη Θυμίνη Γουανίνη Κυτοσίνη ΓΕΝΕΤΙΚΗ Φωσφοδιεστερικός δεσμός 4α. Λειτουργία γενετικού υλικού 1 ΛΕΙΤΟΥΡΓΙΑ ΓΕΝΕΤΙΚΟΥ ΥΛΙΚΟΥ Αντιγραφή (διπλασιασμός) DNA: DNA DNA Έκφραση γενετικής πληροφορίας:
DNA in RNA: zgradba in vloga. Velika predavalnica IJS,
 DNA in RNA: zgradba in vloga Velika predavalnica IJS, 10. 4. 2014 Nukleinske kisline Shranjevanje, prenašanje in izražanje genetske informacije. Dve vrsti nukleinskih kislin: deoksiribonukleinska kislina
DNA in RNA: zgradba in vloga Velika predavalnica IJS, 10. 4. 2014 Nukleinske kisline Shranjevanje, prenašanje in izražanje genetske informacije. Dve vrsti nukleinskih kislin: deoksiribonukleinska kislina
Metabolizem in energija
 Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Logatherm WPL 14 AR T A ++ A + A B C D E F G A B C D E F G. kw kw /2013
 WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
Najpomembnejši človeški Hb
 Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
6 ogljikovih atomov: HEKSOZE (npr. glukoza, fruktoza, galaktoza) Ločimo dve vrsti glukoze: α glukoza in. β glukoza, ki se
 Ločimo dve vrsti glukoze: α glukoza in. β glukoza, ki se") OGLJIKOVI HIDRATI Monosaharidi enostavni sladkorji Spojine C, H, O v razmerju 1:2:1 3 ogljikovi atomi: TRIOZE 5 ogljikovih atomov: PENTOZE (npr. riboza, deoksiriboza) 6 ogljikovih atomov: HEKSOZE (npr.
OGLJIKOVI HIDRATI Monosaharidi enostavni sladkorji Spojine C, H, O v razmerju 1:2:1 3 ogljikovi atomi: TRIOZE 5 ogljikovih atomov: PENTOZE (npr. riboza, deoksiriboza) 6 ogljikovih atomov: HEKSOZE (npr.
Definiranje okolja mikroorganizmov
 Definiranje okolja mikroorganizmov David Stopar Novi sad 2007 Izr. prof. dr. David Stopar Univerza v Ljubljani, BF Oddelek za živilstvo Katedra za mikrobiologijo Večna pot 111 Email: david.stopar@bf.uni-lj.si
Definiranje okolja mikroorganizmov David Stopar Novi sad 2007 Izr. prof. dr. David Stopar Univerza v Ljubljani, BF Oddelek za živilstvo Katedra za mikrobiologijo Večna pot 111 Email: david.stopar@bf.uni-lj.si
Strukturni in funkcijski vidiki bioloških interakcij
 Strukturni in funkcijski vidiki bioloških interakcij Urnik 30 ur predavanj + 15 ur seminarjev predavanja in seminarji: petek od 12.00 do 15.00 10 predavanj 5 seminarjev 30 ur vaj (6 x 5 ur) Obveznosti
Strukturni in funkcijski vidiki bioloških interakcij Urnik 30 ur predavanj + 15 ur seminarjev predavanja in seminarji: petek od 12.00 do 15.00 10 predavanj 5 seminarjev 30 ur vaj (6 x 5 ur) Obveznosti
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo
 Osnovni princip kromatografije 3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo 4. vaja Ionskoizmenjevalna ter afinitetna kromatografija Miha Pavšič komponente zmesi različne interakcije
Osnovni princip kromatografije 3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo 4. vaja Ionskoizmenjevalna ter afinitetna kromatografija Miha Pavšič komponente zmesi različne interakcije
Najpogostejše hemoglobinopatije
 Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina
Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina