Encimi.
|
|
|
- Ιόλη Καλάρης
- 7 χρόνια πριν
- Προβολές:
Transcript
 v produkte.")
1 Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v produkte. Aktivacijska energija je energija, ki jo morajo imeti molekule reaktantov, da se lahko pretvorijo v produkte.
 v produkte.")
2 Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v produkte. Aktivacijska energija je energija, ki jo morajo imeti molekule reaktantov, da se lahko pretvorijo v produkte.
 brez dodatkov b.")
3 Encimi - katalaza Katalaza je encim, ki ga najdemo v skoraj vseh organizmih, ki živijo v prisotnosti kisika. Vsebuje hem kot prostetično skupino. Katalizira reakcijo: 2 H 2 O 2 2 H 2 O + O 2 3 % raztopina H 2 O 2. Mehurčki, ki jih vidimo v raztopini so mehurčki nastajajočega O 2. a.) brez dodatkov b.) + Fe 3+ sol c.) + katalaza
 brez dodatkov b.")
4 Encimi - katalaza Katalaza je encim, ki ga najdemo v skoraj vseh organizmih, ki živijo v prisotnosti kisika. Vsebuje hem kot prostetično skupino. Katalizira reakcijo: 2 H 2 O 2 2 H 2 O + O 2 Prehodno stanje je kratko živeča visoko energijska, nestabilna oblika, ki se lahko pretvori v produkte ali pa nazaj v reaktante. a.) brez dodatkov b.) + Fe 3+ sol c.) + katalaza
, ki predstavljajo lokalne minimume v energetskem diagramu poteka")
5 Encimi Reakcije lahko potečejo tudi preko večih vmesnih intermediatov prehodnih metastabilnih stanj (stabilnih dlje kot s), ki predstavljajo lokalne minimume v energetskem diagramu poteka reakcije.

6 Encimi Večina encimov katalizira reakcije v obe smeri. Smer reakcije je v vsakem trenutku odvisna od razmerja koncentracij substratov in produktov. S P ΔG = ΔG 0 + RTlnK a = ΔG 0 + RTln P S
7 Encimi - lastnosti Lastnosti encima kot katalizatorja: 1. Poveča hitrost reakcije tako, da zniža aktivacijsko energijo 2. Med reakcijo se ne porabi in se trajno ne spremeni. 3. Ne vpliva na ravnotežje reakcije, ampak le na hitrost, s katero dosežemo ravnotežje. 4. Z reaktanti običajno tvori začasni kompleks in tako stabilizira prehodno stanje.
8 Encimi - lastnosti Nekateri encimi za svoje delovanje potrebujejo še neproteinsko komponento kofaktor. Kofaktorji so lahko: - kovinski ioni - koencimi šibko oz. prehodno vezane organske molekule - prostetične skupine tesno oz. stalno vezane organske molekule (npr. hem) V primerih encimov, ki za svoje delovanje potrebujejo kofaktorje, encim z vezanim kofaktorjem imenujemo holoencim, proteinsko komponento brez kofaktorja pa imenujemo apoencim.
9 Encimi poimenovanje Encime običajno imenujemo na osnovi substrata (reaktanta) reakcije, kateremu pripnemo končnico aza. tirozinaza oksidacija tirozina celulaza hidroliza celuloze nitrogenaza fiksacija dušika (pretvorbo N 2 v NH 3 ) Komisija za encime (EC) pri IUBMB teži k sistematičnem poimenovanju encimov, tako da bi vsak encim poimenovali glede na reakcijo, ki jo katalizira. Kljub temu so v uporabi še mnoga stara trivialna imena, zlasti v primerih ko je težko definirati substrate in/ali produkte reakcije ali pa so sistematična poimenovanja nerodna. Nekaj pogostih trivialnih imen: katalaza razgradnja peroksida tripsin hidroliza proteinov (peptidaza oz. proteaza) pepsin peptidaza iz želodca katepsin peptidaza iz lizosoma
, od katerih vsak naslednji bolj natančno opiše katalizirano reakcijo.")
10 Encimi - klasifikacija EC komisija vsak encim klasificira na osnovi kemijske reakcije, ki jo katalizira. Klasifikacija poteka na štirih nivojih (oblika zapisa ), od katerih vsak naslednji bolj natančno opiše katalizirano reakcijo. Primer z wikipedije:

11 Encimi razredi encimov Poznamo šest razredov encimskih reakcij:

12 Encimi oksidoreduktaze laktat dehidrogenaza

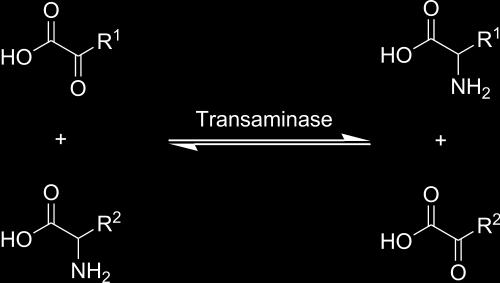
13 Encimi transferaze DNA polimeraza transaminaza
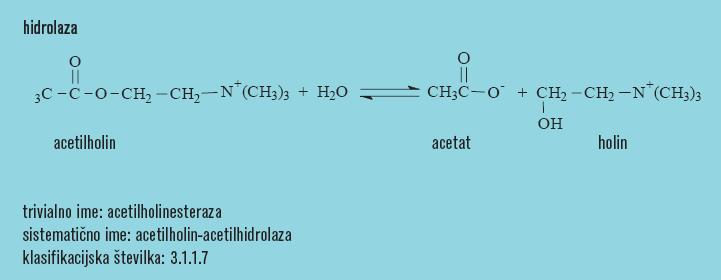
14 Encimi hidrolaze acetilholinesteraza
15 Encimi hidrolaze peptidaze (proteaze)
16 Encimi hidrolaze nukleaze (DNaze, Rnaze)

17 Encimi liaze karbonska anhidraza C enolaza +H 2 O web.campbell.edu
18 Encimi izomeraze triozafosfat izomeraza

19 Encimi ligaze piruvat karboksilaza

20 Kinetične lastnosti encimov Encimi imajo v primerjavi z nebiološkimi katalizatorji nekatere posebne kinetične lastnosti. Najenostavneje njihove kinetične lastnosti zasledujemo v sistemu, kjer uporabimo koncentracije encimov, ki so bistveno nižje od koncentracij substratov. Pod temi pogoji izmerimo začetno hitrost reakcije v 0, ki je enaka hitrosti razgradnje substratov oz. nastajanja produktov. Hitrost reakcije se z naraščanjem koncentracije substrata približuje asimptoti. Pride do nasičenja encima s substratom.


")
21 Kinetične lastnosti encimov Najenostavnejši sistem encimsko kataliziranih reakcij lahko opišemo z Michaelis-Mentenovo enačbo. Leonor Michaelis ( ) Maud Menten ( )
22 Michaelis-Mentenova enačba Najenostavnejši sistem encimsko kataliziranih reakcij lahko opišemo z Michaelis-Mentenovo enačbo. Reakcijsko shemo za ta sistem zapišemo v obliki: ES... kompleks encim-substrat Začetno hitrost reakcije opišemo z enačbo: v 0 = V max S K M + S Kjer je V max mejna hitrost, K M pa Michaelisova konstanta. K M = k 2 + k 3 k 1 K M nam pove afiniteto encima za substrat.
 po formuli: V max = k 3 E T kjer je [E] T celokupna koncentracija encima v reakcijski")
23 Michaelis-Mentenova enačba v 0 = V max S K M + S S K M v 0 = V max K M S S = K M v 0 = V max S 2 S S K M v 0 = V max S S = V max 2 = V max Iz vrednosti V max lahko določimo še vrednost pretvorbenega števila k 3 (k cat ) po formuli: V max = k 3 E T kjer je [E] T celokupna koncentracija encima v reakcijski zmesi.
. Te metode so bile statistično nezanesljive in so danes zastarele.")
24 Michaelis-Mentenova enačba v 0 = V max S K M + S 1 v 0 = K M V max 1 S + 1 V max Včasih so se za analizo encimske kinetike uporabljale metode za linearizacijo krivulj. Ena najpopularnejših je bil Lineweaver-Burkov diagram (1/v 0 v odvisnosti od 1/[S]). Te metode so bile statistično nezanesljive in so danes zastarele.


25 Primeri vrednosti K M za različne encime in njihove substrate

26 Primeri vrednosti k 3 za različne encime in njihove substrate

27 Konstanta specifičnosti Če poznamo vrednosti K M in k 3 za par encim-substrat lahko izračunamo t.i. konstanto specifičnosti, ki velja kot merilo za celokupno učinkovitost encimske katalize. konstanta specifičnosti = k 3 K M M 1 s 1 Če bi vsak trk encima in substrata privedel do pretvorbe substrata v produkt, bi bila hitrost reakcije omejena le s hitrostjo difuzije molekul v raztopini, ki je za take molekule v območju med 10 8 in 10 9 M -1 s -1. Vrednosti k 3 /K M za nekatere encime se približajo tem vrednostim. Imenujemo jih popolni encimi.

28 Značilnosti encimskih reakcij Odvisnost hitrosti reakcije od koncentracije encima

29 Značilnosti encimskih reakcij Odvisnost hitrosti reakcije od ph Tripsin v dvanajstniku Pepsin v želodcu

30 Značilnosti encimskih reakcij Odvisnost hitrosti reakcije od temperature
 2.")
31 Delovanje encimov aktivno mesto Kataliza kemijske reakcije poteka v mestu encima, ki mu rečemo aktivno mesto in ima naslednje lastnosti: 1. Je specifično prepozna le nekatere molekule substrate. Specifičnost je lahko absolutna prepozna le točno določen substrat, ali skupinska prepozna le substrate določenega tipa (npr. peptide, alkohole, itd.) 2. Aktivno mesto predstavlja malo področje v okviru celotne strukture proteina. 3. Substratne molekule se v aktivno mesto vežejo s šibkimi nekovalentnimi reverzibilnimi interakcijami. Shematski primer vezave dipeptida v aktivno mesto encima preko šibkih interakcij (prikazanih s prekinjenimi črtami). Molekula vode deluje kot nukleofil, ki hidrolozira peptidno vez.


32 Delovanje encimov vezava substrata v aktivno mesto Model ključa in ključavnice Model inducirane prilagoditve Resources/cfb/images/07aa.gif Aktivno mesto je že samo po sebi komplementarno strukturi substrata. Aktivno mesto se prilagodi obliki substrata.

33 Delovanje encimov vezava substrata v aktivno mesto Model analoga prehodnega stanja Predpostavlja da substrat ob vezavi v aktivno mesto preide v strukturo prehodnega stanja, ki se stabilizira preko interakcij z encimom.
34 Delovanje encimov mehanizmi encimske katalize Kislinsko-bazna hidroliza funkcionalne skupine ostankov v aktivnem mestu delujejo kot kisline oz. baze pri prenosu protonov. kisline - donorji H + (-NH3 + in -COOH skupine) baze - akceptorji H + (-NH2 in -COO - skupine) Primer: s kislino katalizirana hidroliza amidne (peptidne) vezi
 sodeluje pri prenosu")
35 Delovanje encimov mehanizmi encimske katalize Kataliza s kovinskim ionom: a) pomaga pravilno orientirati substrat b) polarizira vez, kjer bo potekla reakcija ali stabilizira negativni naboj intermediata c) sodeluje pri prenosu elektronov
36 Delovanje encimov mehanizmi encimske katalize Kovalentna kataliza nukleofilna skupina na encimu s substratom tvori kovalentno povezan intermediat. Primer: delovanje serinskih peptidaz
37 Encimi - lastnosti Nekateri encimi za svoje delovanje potrebujejo še neproteinsko komponento kofaktor. Kofaktorji so lahko: - kovinski ioni - koencimi šibko oz. prehodno vezane organske molekule - prostetične skupine tesno oz. stalno vezane organske molekule (npr. hem) V primerih encimov, ki za svoje delovanje potrebujejo kofaktorje, encim z vezanim kofaktorjem imenujemo holoencim, proteinsko komponento brez kofaktorja pa imenujemo apoencim.

38 Kofaktorji kovinski ioni

 NADP + (nikotinamid adenine")

39 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: NAD + (nikotinamid adenine dinukleotid) NADP + (nikotinamid adenine dinukleotid fosfat) Sodelujeta v redoks reakcijah NAD + pretežno v katabolnih, NADP + pretežno v anabolnih. Sintetizirata se iz niacina (nikotinamid).

 FMN (flavin mononukleotid) Sodelujeta v reakcijah oksidacije.")
40 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: FAD (flavin adenine dinukleotid) FMN (flavin mononukleotid) Sodelujeta v reakcijah oksidacije. Sintetizirata se iz riboflavina (vitamin B 2 ).

41 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: koencim A Deluje kot prenašalec vmesnih produktov v razgradnji maščobnih kislin in glukoze. Za njegovo biosintezo je potreben pantotenat (vitamin B5). pantotenat


42 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: tetrahidrofolat Sodeluje pri reakcijah prenosa metilnih skupin pomemben zlasti pri sintezi DNA in RNA ter v mehanizmih preprečevanja poškodb DNA. Sintetizira se iz folne kisline. Zadosten vnos folne kisline je zlasti pomemben pri nosečnicah. Zaviralci biosinteze tetrahidrofolata (metotreksat) se uporabljajo za zdravljenje raka ter za sprožanje splava pri nosečnicah (npr. pri izvenmaternični nosečnosti). folna kislina

 L-askorbinska kislina je pomemben antioksidant in koencim v reakcijah")
43 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: L-askorbinska kislina (vitamin C) L-askorbinska kislina je pomemben antioksidant in koencim v reakcijah hidroksilacije prolina in lizina v kolagenu. Njeno pomanjkanje povzroča skorbut.


44 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: biotin (vitamin B 7, vitamin H, koencim R) Sodeluje pri reakcijah karboksilacije (aktivacija in prenos CO 2. piridoksal fosfat Sodeluje pri prenosu in amino skupin. Je aktivna oblika vitamina B 6 (skupno ime za piridoksal, piridoksamin, piridoksin).

. Sintetizira se iz tiamina (vitamina B 1 ).")
45 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: tiamin pirofosfat Sodeluje pri reakcijah dekarboksilacije (odstranitev CO 2 ). Sintetizira se iz tiamina (vitamina B 1 ). tiamin lipojska kislina Sodeluje pri reakcijah prenosa acilnih skupin. Prekurzor ni vitamin (sintetiziramo jo sami).


46 Kofaktorji organske molekule Večino kofaktorjev ljudje sintetiziramo iz prekurzorjev, ki jih moramo zaužiti s hrano vitaminov. Nekaj primerov pogostejših kofaktorjev ter njihovih prekurzorjev: kobalamin Sodeluje pri prenosu metilnih skupin. Ena ključnih vlog je regeneracija THF. Drugo ime za vse različne oblike je vitamin B 12. Osnovna struktura je korinski obroč. Zaužijemo ga s hrano živalskega izvora ali prehranskimi nadomestki. Pomanjkanje kobalamina se ponavadi izrazi kot pomanjkanje folata. Uporablja se za zdravljenje zastrupitev s cianidom. bakterije evkarionti polsintetičen
47 Inhibicija encimov Inhibitorji so molekule, ki se vežejo na encim in s tem zavrejo njegovo delovanje. V osnovi inhibitorje razdelimo na ireverzibilne in reverzibilne. Ireverzibilni inhibitorji (inaktivatorji) tvorijo kovalentno ali zelo močno nekovalentno vez z encimom in ga s tem trajno onesposobijo. Primer: diisopropil fluorofosfat je živčni strup, ki deluje kot ireverzibilni inhibitor encima acetilholinesteraza (desno). Aspirin (acetilsalicilna kislina, spodaj) deluje kot ireverzibilni inhibitor encima ciklooksigenaza.


48 Reverzibilni inhibitorji Reverzibilni inhibitorji se vežejo na encim, a lahko od njega tudi oddisociirajo. Prisotno je ravnotežje med neinhibirano in inhibirano obliko encima. mešani

49 Kompetitivni inhibitorji Najpogostejša vrsta inhibitorjev v naravi so kompetitivni inhibitorji, ki se vežejo v aktivno mesto encima in s tem preprečijo vezavo substrata. Reakcijska shema: K i Afiniteto inhibitorja do encima podajamo s konstanto inhibicije K i.
50 Kompetitivni inhibitorji Najpogostejša vrsta inhibitorjev v naravi so kompetitivni inhibitorji, ki se vežejo v aktivno mesto encima in s tem preprečijo vezavo substrata. Delovanje inhibitorjev preučujemo tako, da merimo in primerjamo hitrost v prisotnosti in odsotnosti inhibitorja. v = Vnav S K m nav + S K m nav = K m 1 + I K i V = V nav
51 Kompetitivni inhibitorji Najpogostejša vrsta inhibitorjev v naravi so kompetitivni inhibitorji, ki se vežejo v aktivno mesto encima in s tem preprečijo vezavo substrata. Primer: sukcinat dehidrogenazo inhibirajo analogi substrata malonat, oksalat in pirofosfat. inhibitorji reakcija sukcinat dehidrogenaze
52 Kompetitivni inhibitorji Kot kompetitivne inhibitorje pogosto srečamo analoge prehodnega stanja tj. molekule, ki imajo strukturo podobno prehodnem stanju, a jih encim ne more kemično pretvoriti.


53 Kompetitivni inhibitorji Najpogostejša vrsta inhibitorjev v naravi so kompetitivni inhibitorji, ki se vežejo v aktivno mesto encima in s tem preprečijo vezavo substrata. Terapevtski primer: ritonavir inhibitor proteaze HIV-1 struktura ritonavira kristalna struktura HIV-1 proteaze z vezanim ritonavirom



54 Kompetitivni inhibitorji Najpogostejša vrsta inhibitorjev v naravi so kompetitivni inhibitorji, ki se vežejo v aktivno mesto encima in s tem preprečijo vezavo substrata. Terapevtski primer: captopril inhibitor angiotenzin pretvarjajočega encima tarča za zdravljenje povišanega krvnega tlaka struktura captoprila kristalna struktura ACE in captoprila (levo v prikazu cartoon, desno molekulska površina)


55 Kompetitivni inhibitorji Kot kompetitivne inhibitorje lahko srečamo tudi proteine makromolekulski inhibitorji. stefin serpin katepsin tripsin inhibitor stefin A vezan v aktivno mesto lizosomske proteaze iz družine katepsinov inhibitor serpin vezan v aktivno mesto tripsina

56 Reverzibilni inhibitorji Preostali dve obliki inhibicije, akompetitivna in mešana, sta v naravi redki. K m nav = K m 1 + I K i K i V nav = V 1 + I K i

57 Reverzibilni inhibitorji Preostali dve obliki inhibicije, akompetitivna in mešana, sta v naravi redki. mešani K m nav = K m 1 + I K ic 1 + I K iu V nav = V 1 + I K iu K ic K iu Posebni primer: K ic = K iu K m nav = K m V nav = nekompetitivni inhibitor V 1 + I K iu

58 Alosterični encimi Alosterija (allos gr. drug, stereos gr. mesto) pomeni prenos informacije med dvema mestoma (ali več mesti) v proteinu. Alosterična regulacija je pogost mehanizem uravnavanja delovanja encimov. Alosterijo so v začetku opisali v oligomernih proteinih. Podobno kot pri vezavi kisika na hemoglobin se pri oligomernih encimih alosterija lahko pokaže kot kooperativnost pri vezavi substrata. Gre za komunikacijo med aktivnimi (katalitičnimi) mesti. Oligomerni encimi, ki kažejo znake kooperativnosti, se ne obnašajo skladno z Michaelis-Mentenovo kinetiko. Krivulje odvisnosti hitrosti od koncentracije substrata so sigmoidne.
 na alosterična mesta lahko poviša aktivnost encima (pozitivni regulatorji) ali jo zniža (negativni regulatorji).")
59 Alosterični encimi Podobno kot v hemoglobinu so tudi v oligomernih encimih lahko poleg aktivnih mest prisotna tudi regulatorna (alosterična) mesta. Vezava modulatorjev (oz. efektorjev) na alosterična mesta lahko poviša aktivnost encima (pozitivni regulatorji) ali jo zniža (negativni regulatorji). Vpliva lahko na afiniteto encima do substrata in/ali katalitično hitrost encima.

 in regulatorne (b) podenote")
60 Alosterični encimi Alosterični encimi niso nujno sestavljeni iz večih podenot s katalitičnimi lastnostmi. dimer katalitične (a) in regulatorne (b) podenote monomer z aktivnim in alosteričnim mestom
 obstaja v dveh konformacijskih oblikah: oblika T (tense) je manj aktivna (ali neaktivna) od oblike R (relaxed).")
61 Modeli alosteričnega uravnavanja Najenostavnejši modeli alosteričnega uravnavanja predpostavljajo, da vsaka podenota alosteričnih encimov (oz. proteinov) obstaja v dveh konformacijskih oblikah: oblika T (tense) je manj aktivna (ali neaktivna) od oblike R (relaxed). Modeli so bili v osnovi postavljeni za pojasnjevanje alosterije v oligomernih encimih, lahko pa jih analogno uporabimo tudi za monomerne sisteme. Model MWC (Monod, Wyman, Changeux; 1965)
 obstaja v dveh konformacijskih oblikah: oblika T (tense) je manj aktivna (ali neaktivna) od oblike R (relaxed).")
62 Modeli alosteričnega uravnavanja Najenostavnejši modeli alosteričnega uravnavanja predpostavljajo, da vsaka podenota alosteričnih encimov (oz. proteinov) obstaja v dveh konformacijskih oblikah: oblika T (tense) je manj aktivna (ali neaktivna) od oblike R (relaxed). Modeli so bili v osnovi postavljeni za pojasnjevanje alosterije v oligomernih encimih, lahko pa jih analogno uporabimo tudi za monomerne sisteme. Model KNF sekvenčni model (Koshland, Nemethy, Filmer; 1966)
63 Regulacija s kovalentnimi modifikacijami V naravi je delovanje encimov često uravnavano s kovalentnimi modifikacijami.
64 Regulacija s kovalentnimi modifikacijami Eden najpogostejših načinov znotrajceličnega uravnavanja je fosforilacija. Pri tem procesu sodelujejo kinaze (dodajajo fosfatno skupino) in fosfataze (odstranjujejo fosfatno skupino). Primer: regulacija encimov v sintezi glikogena.

65 Regulacija s proteolitičnim procesiranjem Nekateri encimi se sintetizirajo kot neaktivni cimogeni in se s proteolitično cepitvijo pretvorijo v aktivno obliko. Primer: aktivacija kimotripsinogena.


66 Regulacija s proteolitičnim procesiranjem Nekateri encimi se sintetizirajo kot neaktivni cimogeni in se s proteolitično cepitvijo pretvorijo v aktivno obliko. kimotripsinogen (N-končen propeptid je rdeč, izrezana ostanka 14 in 15 rumena, izrezana ostanka 147 in 148 zelena) prokatepsin K
 LDH je tetramer, sestavljen iz dveh vrst podenot, M in H, ki sta produkta različnih genov.")
67 Regulacija z izoencimi Izoencimi so različne oblike med seboj podobnih si encimov, ki katalizirajo isto reakcijo, a imajo različne kinetične lastnosti. Primer: laktat dehidrogenaza (LDH) LDH je tetramer, sestavljen iz dveh vrst podenot, M in H, ki sta produkta različnih genov. Oligomerna sestava se med organi razlikuje: V skeletnih mišicah M 4 V srčni mišici H 4 V drugih tkivih zmesi 5 oblik (M 4, M 3 H, M 2 H 2, MH 3, H 4 ), katere lahko ločimo z elektroforezo (desno).


68 Abcimi Abcimi oz. katalitična protitelesa so protitelesa s sposobnostjo kataliziranja kemijskih reakcij. antigen katalitičnega protitelesa je analog prehodnega stanja reakcije. Protitelo veže substrat in povzroči njegov prehod v prehodno stanje. V primerjavi z naravnimi encimi so bistveno slabši katalizatorji.



69 Ribocimi Ribocimi so katalitične RNA molekule.najbolj znan je ribocim hammerhead. Ribocime najdemo pri: samoizrezujočih elementih RNA procesiranju molekul RNA (ribonukleaza P) sintezi proteinov
Encimska kinetika govori o hitrosti encimske reakcije
 Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Mehanizmi encimske katalize
 Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Uravnavanje encimske aktivnosti
 Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
 FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
Matjaž Zorko Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
ENCIMI ZGRADBA ENCIMOV NEKATERI ENCIMI IN NJIHOVI KOFAKTORJI
 Visoko specializirani proteini in mala skupina katalitičnih molekul RNA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10 16 x Zmanjšajo E a, ne vplivajo
Visoko specializirani proteini in mala skupina katalitičnih molekul RNA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10 16 x Zmanjšajo E a, ne vplivajo
Prehrana in metabolizem
 Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
pretvarja v nestrupeno obliko, ki lahko vstopa v biosintezo nukleotidov *i) NH 4
 NH 4") 1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
Encimi kot tarče učinkovin
 Encimi kot tarče učinkovin doc. dr. Janez Ilaš 1. december 2011 1 Uvod Biokemija 2. letnik Predavanje št. 5: Encimi uvod 2011 encimi so KATALIZATORJI globularni proteini, ki za svoje nemoteno delovanje
Encimi kot tarče učinkovin doc. dr. Janez Ilaš 1. december 2011 1 Uvod Biokemija 2. letnik Predavanje št. 5: Encimi uvod 2011 encimi so KATALIZATORJI globularni proteini, ki za svoje nemoteno delovanje
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Nekatere interakcije v lipidnem metabolizmu
 Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
Jure Stojan in Marko Goličnik Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 15. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
ZGRADBA ENCIMOV ENCIMI ENCIMI REAKCIJE, KINETIKA IN INHIBICIJA. Aktivni samo v nativni konformaciji. M = Da. Enostavni in sestavljeni
 EIMI REAKIJE, KIETIKA I IIBIIJA Visoko specializirani proteini in mala skupina katalitičnih molekul RA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10
EIMI REAKIJE, KIETIKA I IIBIIJA Visoko specializirani proteini in mala skupina katalitičnih molekul RA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda
![[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda](/thumbs/89/99439684.jpg "[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda") kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Proizvodnja in uporaba encimov
 Kmetijska šola Grm Sevno 13 8000 Novo mesto Proizvodnja in uporaba encimov ( predmet: Biotehnologija ) Gorenja vas; 7.3.2007 Avtor: Lidija Gorenc Jožica Koračin, 3.c Mentor: Jana Goršin Fabjan Kazalo:
Kmetijska šola Grm Sevno 13 8000 Novo mesto Proizvodnja in uporaba encimov ( predmet: Biotehnologija ) Gorenja vas; 7.3.2007 Avtor: Lidija Gorenc Jožica Koračin, 3.c Mentor: Jana Goršin Fabjan Kazalo:
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Funkcije proteinov (pogojene s strukturo)
") Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
Encimi v tleh. k 1 E + S ES E + P. k 3. k 2
 Encimi v tleh k 1 E + S ES E + P k 2 k 3 1 Encimi in definicije encim (E) je protein, ki ga producira celica in deluje kot katalizator znižuje aktivacijsko energijo in na ta način pospeši hitrost reakcije
Encimi v tleh k 1 E + S ES E + P k 2 k 3 1 Encimi in definicije encim (E) je protein, ki ga producira celica in deluje kot katalizator znižuje aktivacijsko energijo in na ta način pospeši hitrost reakcije
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
Energije in okolje 1. vaja. Entalpija pri kemijskih reakcijah
 Entalpija pri kemijskih reakcijah Pri obravnavi energijskih pretvorb pri kemijskih reakcijah uvedemo pojem entalpije, ki popisuje spreminjanje energije sistema pri konstantnem tlaku. Sistemu lahko povečamo
Entalpija pri kemijskih reakcijah Pri obravnavi energijskih pretvorb pri kemijskih reakcijah uvedemo pojem entalpije, ki popisuje spreminjanje energije sistema pri konstantnem tlaku. Sistemu lahko povečamo
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
Metabolizem oz. presnova
 Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
KOENCIMI IN VITAMINI REGULACIJA ENCIMSKE AKTIVNOSTI KOENCIMI
 REGULAIJA EIMSKE AKTIVSTI Regulacija na genetskem nivoju ormoni Prisotnost koencimov in esencialnih kovinskih ionov Kovalentne modifikacije Izoencimi Proteolitska aktivacija Alosterične interakcije KEIMI
REGULAIJA EIMSKE AKTIVSTI Regulacija na genetskem nivoju ormoni Prisotnost koencimov in esencialnih kovinskih ionov Kovalentne modifikacije Izoencimi Proteolitska aktivacija Alosterične interakcije KEIMI
Razgradnja maščobnih kislin. Ketonska telesa H + NAD+ NADH. Pregled metabolizma MK. lipoprotein-lipaza. maščobne kisline.
 Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Tabele termodinamskih lastnosti vode in vodne pare
 Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Fazni diagram binarne tekočine
 Fazni diagram binarne tekočine Žiga Kos 5. junij 203 Binarno tekočino predstavljajo delci A in B. Ti se med seboj lahko mešajo v različnih razmerjih. V nalogi želimo izračunati fazni diagram take tekočine,
Fazni diagram binarne tekočine Žiga Kos 5. junij 203 Binarno tekočino predstavljajo delci A in B. Ti se med seboj lahko mešajo v različnih razmerjih. V nalogi želimo izračunati fazni diagram take tekočine,
13. Jacobijeva metoda za računanje singularnega razcepa
 13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
UVOD CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN
 = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN") CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK)
") IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Teze predavanj iz mikrobne biokemije*
 Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba.
 1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
Splošno o interpolaciji
 Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
VPLIV REAKCIJSKIH SPREMENLJIVK NA POTEK IN HITROST MODELNE REAKCIJE NATRIJEVEGA TIOSULFATA S KLOROVODIKOVO KISLINO
 OSNOVNA ŠOLA PRIMOŽA TRUBARJA LAŠKO VPLIV REAKCIJSKIH SPREMENLJIVK NA POTEK IN HITROST MODELNE REAKCIJE NATRIJEVEGA TIOSULFATA S KLOROVODIKOVO KISLINO (RAZISKOVALNO DELO) Avtorici: Lea Lešek Povšič in
OSNOVNA ŠOLA PRIMOŽA TRUBARJA LAŠKO VPLIV REAKCIJSKIH SPREMENLJIVK NA POTEK IN HITROST MODELNE REAKCIJE NATRIJEVEGA TIOSULFATA S KLOROVODIKOVO KISLINO (RAZISKOVALNO DELO) Avtorici: Lea Lešek Povšič in
Metabolizem in energija
 Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
CIKLUS LIMUNSKE KISELINE (CLK)
") SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
Ravnotežja v raztopini
 Ravnotežja v raztopini TOPILO: komponenta, ki jo je več v raztopini.v analizni kemiji uporabljamo organska in anorganska topila. Topila z veliko dielektrično konstanto (ε > 10) so polarna in ionizirajo
Ravnotežja v raztopini TOPILO: komponenta, ki jo je več v raztopini.v analizni kemiji uporabljamo organska in anorganska topila. Topila z veliko dielektrično konstanto (ε > 10) so polarna in ionizirajo
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Gimnazija Krˇsko. vektorji - naloge
 Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks'
 Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
1. TVORBA ŠIBKEGA (SIGMATNEGA) AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ
 AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ") TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
Mehanizmi djelovanja enzima i regulacija aktivnosti enzima
 Mehanizmi djelovanja enzima i regulacija aktivnosti enzima Boris Mildner Aktivnost enzima mijenja se promjenama temperature, ph, inhibitorima te vezanjem malih molekula ili iona Temperatura povećava brzinu
Mehanizmi djelovanja enzima i regulacija aktivnosti enzima Boris Mildner Aktivnost enzima mijenja se promjenama temperature, ph, inhibitorima te vezanjem malih molekula ili iona Temperatura povećava brzinu
Logatherm WPL 14 AR T A ++ A + A B C D E F G A B C D E F G. kw kw /2013
 WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
p 1 ENTROPIJSKI ZAKON
 ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
MATEMATIČNI IZRAZI V MAFIRA WIKIJU
 I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
Iterativno reševanje sistemov linearnih enačb. Numerične metode, sistemi linearnih enačb. Numerične metode FE, 2. december 2013
 Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Primeri: naftalen kinolin spojeni kinolin
 Primeri: naftalen kinolin spojeni kinolin 3 skupne strani 7 skupnih strani 5 skupnih strani 6 skupnih atomov 8 skupnih atomov 6 skupnih atomov orto spojen sistem orto in peri spojena sistema mostni kinolin
Primeri: naftalen kinolin spojeni kinolin 3 skupne strani 7 skupnih strani 5 skupnih strani 6 skupnih atomov 8 skupnih atomov 6 skupnih atomov orto spojen sistem orto in peri spojena sistema mostni kinolin
Talna kemija. Kaj je potrebno poznati:
 Talna kemija Kaj je potrebno poznati: splošno kemijo mol, molaren, normalnost, ekvivalent ionska jakost, aktivnost ravnotežne konstante funkcionalne skupine hidratacija, hidroliza redoks reakcije Redoks
Talna kemija Kaj je potrebno poznati: splošno kemijo mol, molaren, normalnost, ekvivalent ionska jakost, aktivnost ravnotežne konstante funkcionalne skupine hidratacija, hidroliza redoks reakcije Redoks
VAJA 1 ALBUMINU NEZNANE KONCENTRACIJE DOLOČITE KONCENTRACIJO: Izmerite A 280 albumina v vzorcu. Vrednost absorbance vzorca naj ne preseže 1!
 VAJA 1 ALBUMINU NEZNANE KONCENTRACIJE DOLOČITE KONCENTRACIJO: Izmerite A 280 albumina v vzorcu. Vrednost absorbance vzorca naj ne preseže 1! S pomočjo standardne raztopine (raztopina albumina koncentracije
VAJA 1 ALBUMINU NEZNANE KONCENTRACIJE DOLOČITE KONCENTRACIJO: Izmerite A 280 albumina v vzorcu. Vrednost absorbance vzorca naj ne preseže 1! S pomočjo standardne raztopine (raztopina albumina koncentracije
CM707. GR Οδηγός χρήσης... 2-7. SLO Uporabniški priročnik... 8-13. CR Korisnički priručnik... 14-19. TR Kullanım Kılavuzu... 20-25
 1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
OKOLJSKO NARAVOSLOVJE 2. Predavanja v študijskem letu 2. del 2012/2013
 OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
Funkcije več spremenljivk
 DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
VODA ELEKTROLITI I ACIDO-BAZNA RAVNOTEŽA...
 SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
Podobnost matrik. Matematika II (FKKT Kemijsko inženirstvo) Diagonalizacija matrik
 Diagonalizacija matrik") Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Frekvenčna analiza neperiodičnih signalov. Analiza signalov prof. France Mihelič
 Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Homogena snov je snov, ki ima vsepovsod enake lastnosti in sestavo Heterogena snov je snov, katere sestava in lastnosti so na različnih mestih
 Homogena snov je snov, ki ima vsepovsod enake lastnosti in sestavo Heterogena snov je snov, katere sestava in lastnosti so na različnih mestih različne Postopki ločevanja zmesi:iz zmesi je mogoče ločiti
Homogena snov je snov, ki ima vsepovsod enake lastnosti in sestavo Heterogena snov je snov, katere sestava in lastnosti so na različnih mestih različne Postopki ločevanja zmesi:iz zmesi je mogoče ločiti
Kotni funkciji sinus in kosinus
 Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
Ta uredba je v celoti zavezujoča in se neposredno uporablja v vseh državah članicah.
 L 314/36 Uradni list Evropske unije 1.12.2009 UREDBA KOMISIJE (ES) št. 1170/2009 z dne 30. novembra 2009 o spremembi Direktive 2002/46/ES Evropskega parlamenta in Sveta in Uredbe (ES) št. 1925/2006 Evropskega
L 314/36 Uradni list Evropske unije 1.12.2009 UREDBA KOMISIJE (ES) št. 1170/2009 z dne 30. novembra 2009 o spremembi Direktive 2002/46/ES Evropskega parlamenta in Sveta in Uredbe (ES) št. 1925/2006 Evropskega
TOPNOST, HITROST RAZTAPLJANJA
 OPNOS, HIOS AZAPLJANJA Denja: onos (oz. nasčena razona) redsavlja sanje, ko je oljene (rdn, ekoč, lnas) v ravnoežju z razono (oljenem, razoljenm v olu). - kvanavn zraz - r določen - homogena molekularna
OPNOS, HIOS AZAPLJANJA Denja: onos (oz. nasčena razona) redsavlja sanje, ko je oljene (rdn, ekoč, lnas) v ravnoežju z razono (oljenem, razoljenm v olu). - kvanavn zraz - r določen - homogena molekularna
Raztopine. Raztopine. Elektroliti. Elektrolit je substanca, ki pri raztapljanju (v vodi) daje ione. A a B b aa b+ + bb a-
 daje ione. A a B b aa b+ + bb a-") Raztopine Mnoge analizne metode temeljijo na opazovanju ravnotežnih sistemov, ki se vzpostavijo v raztopinah. Najpogosteje uporabljeno topilo je voda! RAZTOPINE: topljenec topilo (voda) (Enote za koncentracije!)
Raztopine Mnoge analizne metode temeljijo na opazovanju ravnotežnih sistemov, ki se vzpostavijo v raztopinah. Najpogosteje uporabljeno topilo je voda! RAZTOPINE: topljenec topilo (voda) (Enote za koncentracije!)
I. OSNOVNI STRUKTURNI PRINCIPI
 I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
DISKRETNA FOURIERJEVA TRANSFORMACIJA
 29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
PROCESIRANJE SIGNALOV
 Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
Statistična analiza. doc. dr. Mitja Kos, mag. farm. Katedra za socialno farmacijo Univerza v Ljubljani- Fakulteta za farmacijo
 Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
vezani ekstremi funkcij
 11. vaja iz Matematike 2 (UNI) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 ekstremi funkcij več spremenljivk nadaljevanje vezani ekstremi funkcij Dana je funkcija f(x, y). Zanimajo nas ekstremi nad
11. vaja iz Matematike 2 (UNI) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 ekstremi funkcij več spremenljivk nadaljevanje vezani ekstremi funkcij Dana je funkcija f(x, y). Zanimajo nas ekstremi nad
Vaje: Električni tokovi
 Barbara Rovšek, Bojan Golli, Ana Gostinčar Blagotinšek Vaje: Električni tokovi 1 Merjenje toka in napetosti Naloga: Izmerite tok, ki teče skozi žarnico, ter napetost na žarnici Za izvedbo vaje potrebujete
Barbara Rovšek, Bojan Golli, Ana Gostinčar Blagotinšek Vaje: Električni tokovi 1 Merjenje toka in napetosti Naloga: Izmerite tok, ki teče skozi žarnico, ter napetost na žarnici Za izvedbo vaje potrebujete