Strukturni in funkcijski vidiki bioloških interakcij
|
|
|
- Εὔα Μαλαξός
- 5 χρόνια πριν
- Προβολές:
Transcript
1 Strukturni in funkcijski vidiki bioloških interakcij Urnik 30 ur predavanj + 15 ur seminarjev predavanja in seminarji: petek od do predavanj 5 seminarjev 30 ur vaj (6 x 5 ur) Obveznosti pisni izpit 60 % končne ocene seminarska naloga (pisni izdelek + predstavitev) 20 % končne ocene kolokvij iz vaj 20 % končne ocene



2 Interakcije med molekulami v bioloških sistemih Proteinske interakcije so širok pojem interakcij, ki jih proteini tvorijo z drugimi proteini, nukleinskimi kislinami in ostalimi ligandi. Študij interakcij proteinov omogoča razumevanje celičnih procesov, ki jih te interakcije posredujejo in načrtovanje novih učinkovin v medicinske namene, ki delujejo na ravni proteinov. protein-dna protein-majhna molekula (protein-ligand) protein-protein

 stični površini med obema partnerjema.")
3 Tvorba kompleksa protein-protein Pri tvorbi kompleksa pride do spremembe topilu dostopne površine obeh proteinov. Razliki med vsoto površin posameznih interakcijskih partnerjev in površine kompleksa rečemo zakopana površina in je sorazmerna (ni enaka!) stični površini med obema partnerjema. Računalniško določanje topilu dostopne površine:
4 Tvorba kompleksa protein-protein Pri tvorbi kompleksa pride do spremembe topilu dostopne površine obeh proteinov. Razliki med vsoto površin posameznih interakcijskih partnerjev in površine kompleksa rečemo zakopana površina in je sorazmerna (ni enaka!) stični površini med obema partnerjema. Pri tvorbi stične površine je pomembna komplementarnost: - geometrije - donorjev in akceptorjev vodikovih vezi - nabojev Kemijska sestava stičnih površin v proteinih: delež skupin (v %) nepolarne polarne (nenabite) polarne (nabite) površina proteinov stična površina notranjost proteinov
5 Tvorba kompleksa protein-protein Lastnosti stičnih površin: Stična površina velika ponavadi > 1100 Å 2, oz. nad > 550 Å 2 na vsakega partnerja (določitev odvisna od algoritma oz pogojev max. razdalja med interagirajočimi atomi). Zakopana površina velika je okoli 20 Å 2 na ostanek. V povprečju obsega 12% celotne površine v dimerih 17% celotne površine v trimerih 21% celotne površine v tetramerih Energija interakcije je J na Å 2 Ukrivljenost 83% do 84% interakcijskih površin je ploskih Delež sekundarne strukture 50% a-vijačnice, 20% b-ploskev
6 Tvorba kompleksa protein-protein Primerjava števila posameznih interakcij v odvisnosti od oddaljenosti med interagirajočimi skupinami, določena na podlagi vzorca 195 kompleksov protein-protein: hidrofobne H-vezi ionske interakcije vdw aromatske kation-p Kastritis & Bonvin J. R. Soc. Interface. 10
7 sive puščice.. neobstoječe poti Mehanizmi prepoznavanja med proteini lock and key induced fit dynamic fit (konformacijska selekcija) Kastritis & Bonvin J. R. Soc. Interface. 10

8 Mehanizmi prepoznavanja med proteini Tvorbo kompleksa lahko spremljajo konformacijske spremembe enega ali obeh proteinov. tioredoksin reduktaza / tioredoksin IL-1 receptor / IL-1 R antagonist Kastritis & Bonvin J. R. Soc. Interface. 10
9 Termodinamika proteinskih interakcij medmolekulske interakcije ΔG 0 = ΔH 0 TΔS 0 sprememba entropije Kastritis & Bonvin J. R. Soc. Interface. 10
10 Termodinamika proteinskih interakcij P + L k 1 k -1 PL P... akceptor L... ligand ΔG = ΔG 0 + RTlnK a v as = k 1 P L v dis = k 1 PL hitrost asociacije hitrost disociacije v ravnotežju ΔG 0 = RTlnK a v ravnotežju: v as = v dis ΔG 0 = RTlnK d k 1 P L = k 1 PL k 1 k 1 = k 1 k 1 = PL P L = K a P L PL = K d ravnotežna konstanta asociacije (enota M -1 )* ravnotežna konstanta disociacije (enota M)* K a = 1 K d t 1 = ln2 2 k 1 *v resnici so termodinamske konstante brez enot.
11 Interakcije med proteini vrste interakcij Vrste interakcij homooligomeri heterooligomeri obvezne neobvezne trajne prehodne tesne interakcije močne (sprožene) srednje močne šibke visoka afiniteta <nm nm mm mm nizka afiniteta


12 Interakcije med proteini vrste interakcij homooligomeri heterooligomeri



13 Interakcije med proteini vrste interakcij homooligomeri heterooligomeri PDB 1HHO hemoglobin heterotetramer a 2 b 2 zgradba a 2 bb ws heterotrimeren G protein (G abg ) PDB 3AH8
 same zase niso stabilne, zato je")

 srednje močne šibke PDB 1ARQ Arc represor bakteriofaga")
14 Interakcije med proteini Vrste interakcij obvezne neobvezne trajne prehodne Posamezne enote (protomeri) same zase niso stabilne, zato je tvorba kompleksa/oligomera obvezna za obstojnost proteina v celici. S tem je pogojena tudi obligatna tvorba kompleksa za opravljanje biološke vloge proteina. močne (sprožene) srednje močne šibke PDB 1ARQ Arc represor bakteriofaga P22 PDB 1HUL Interlevkin-5 (S-S vez) G ab heterodimer PDB 3AH8
 srednje močne šibke rodniin sestavljen iz dveh tandemnih ponovitev Kazalove domene prva se veže v aktivno mesto, druga pa")
15 Interakcije med proteini Vrste interakcij Ko kompleks nastane, praviloma ostane povezan dlje časa. obvezne neobvezne Interakcije protitelo-antigen, proteazeinhibitorji,.. trajne prehodne Primer: kompleks trombin-rodniin močne (sprožene) srednje močne šibke rodniin sestavljen iz dveh tandemnih ponovitev Kazalove domene prva se veže v aktivno mesto, druga pa interagira s proteinom izven njega K d = 0.2 pm k -1 = s -1 t 1/2 = 1611 s ( 27 min) PDB 1TBQ

 srednje")
16 Interakcije med proteini obvezne Vrste interakcij neobvezne Visokoafinitetne interakcije, katerih nastanek in razpad je reguliran preko različnih signalov (fosforilacija, zamenjava GTP za GDP, vezava dodatnega liganda). trajne prehodne Primer: heterotrimerni G proteini močne (sprožene) srednje močne šibke


17 Interakcije med proteini obvezne Vrste interakcij neobvezne Dinamično ravnotežje med kompleksom in prostimi proteini. Afinitete segajo od 10-8 M do 10-4 M. trajne prehodne močne (sprožene) srednje močne šibke PDB 1AVZ PDB 1HCS SH2 domene vežejo proteine, ki vsebujejo p-tyr z afiniteto okoli 10-6 M. SH3 domene vežejo motive, ki tvorijo poliprolinske vijačnice tipa 2.
18 Interakcije med proteini Nooren & Thornton EMBO J. 22(14)
19 Interakcije med proteini Nooren & Thornton EMBO J. 22(14)
20 Interakcije protein-ligand Pod pojmom ligand opisujemo manjšo (organsko) molekulo, ki se veže na vezavno mesto na proteinu. Poseben tip ligandov so substrati encimov. Dve skupini študij: - vezava naravnih ligandov študij biološkega pomena interakcije - vezava sintetičnih ligandov načrtovanje ligandov v medicinske ali raziskovalne namene, ponavadi izkoriščamo naravna (znana) vezavna mesta na proteinu. Diverziteta vezavnih mest za ligande je velika. Za načrtovanje sintetičnih ligandov je bilo razvitih veliko število eksperimentalnih in računalniških metod. Za vezavo majhnih ligandov na proteine veljajo podobne zakonitosti kot pri interakcijah protein-protein, t.j. komplementarnost: - geometrije - donorjev in akceptorjev vodikovih vezi - nabojev

21 Interakcije protein-ligand Pravilo petic Lipinskega je empirična zbirka pravil o fizikalno-kemijskih lastnostih, ki jih morajo imeti učinkovita peroralna zdravila za dobro biološko razpoložljivost: Pravila so: molska masa manj kot 500 g/mol največ 5 donorjev H-vezi največ 10 akceptorjev H-vezi log porazdelitvenega koeficienta oktanol/voda največ 5 acetilsalicilna kislina Mw = g/mol log(p) = 0.7 H-Bond Donor: 1 H-Bond Acceptor: 10 Mw = [g/mol] log(p) = 1.2 H-Bond Donor: 1 H-Bond Acceptor: 4




22 Interakcije protein-ligand Vezavna mesta za isti ligand in načini vezave liganda se med proteini lahko bistveno razlikujejo. Primer: vezava FMN na štiri različne encime. glikolat oksidaza iz špinače riboflavin kinaza iz Methanococcusa izomeraza iz Sulfolobusa fotoliaza iz Thermusa


23 Identifikacija vezavnih mest na proteinih Na proteinih je več kategorij potencialnih vezavnih žepov, vendar je težko ločiti med pravimi in nepravimi. Hajduk et al DDT. 10
). Osredotočili so se na na prispevke posameznih ostankov in njihovih atomov k interakcijam z ligandi.")
24 Interakcije protein-ligand Pred kratkim je bil objavljen članek, kjer so statistično obdelali vzorec 3295 proteinov z 9114 vezavnimi mesti (Khazanov & Carlson PLoS Comput Biol. 9(11)). Osredotočili so se na na prispevke posameznih ostankov in njihovih atomov k interakcijam z ligandi. Bolj rdeča kot je barva, večkrat atom prispeva k interakcijam.
).")
25 Interakcije protein-ligand Pred kratkim je bil objavljen članek, kjer so statistično obdelali vzorec 3295 proteinov z 9114 vezavnimi mesti (Khazanov & Carlson PLoS Comput Biol. 9(11)). Osredotočili so se na na prispevke posameznih ostankov in njihovih atomov k interakcijam z ligandi.
26 Interakcije protein-dna Temeljne lastnosti: - Specifične interakcije (branje baz) + nespecifične interakcije (ogrodje) - Konformacijske spremembe proteina in/ali DNA - Pogosto se pojavljajo nekateri DNA-vezavni motivi - Odvisne od lokalne strukture DNA - Simetrične interakcije

27 Struktura DNA



28 Struktura DNA kvadrupleksi

29 Interakcije protein-dna Rohs et al Annu Rev Biochem. 79

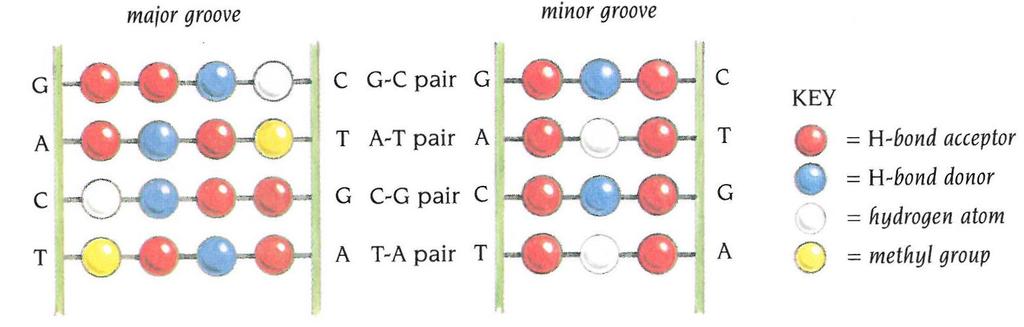
30 Struktura DNA


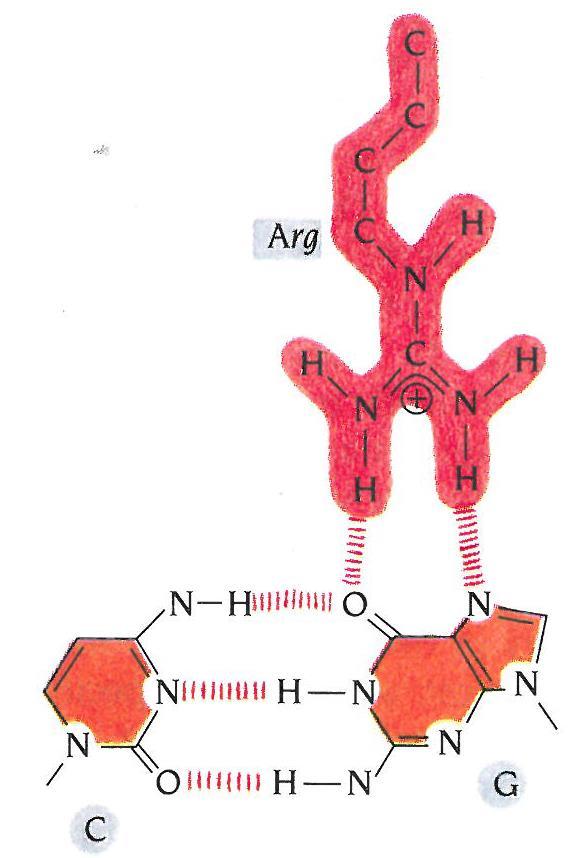
31 Interakcije protein-dna Specifične interakcije: Nespecifične interakcije:

32 Interakcije protein-dna


33 Inducirane deformacije DNA Upogib vijačnice os vijačnice je upognjena kontinuirno preko daljšega segmenta DNA Prelom vijačnice smer osi vijačnice se nenadome spremeni Rohs et al Annu Rev Biochem. 79
34 Specifično prepoznavanje DNA Kako proteini najdejo tarčno zaporedje na DNA? Drsenje difuzija v eni dimenziji (EcoRV) Skakanje vezava samo na specifično zaporedje (EcoRI) prenos med segmenti Halford & Marko Nucleic Acids Research, 32(10)

35 Specifično prepoznavanje DNA Pomen dimerizacije vezava vnaprej sestavljenih dimerov K dimer = K2 monomer geometrija dimera tandemno/ palindromno zaporedje efekt razdalje med zaporedji Georges et al. FASEB J. 24, (2010).



36 Specifično prepoznavanje DNA Pomen dimerizacije sestavljanje homo/heterodimerov na DNA kooperativnost v homooligomerih interakcije v izključujočih kombinacijah interakcije istega proteina z različnimi partnerji Georges et al. FASEB J. 24, (2010).

 v specifičnih kombinacijah.")
37 Specifično prepoznavanje DNA Pomen dimerizacije vezani transkripcijski faktorji rekrutirajo mediatorje (npr. RNApol) v specifičnih kombinacijah. Georges et al. FASEB J. 24, (2010).
38 Klasifikacija DNA-vezavnih proteinov glede na DNA-vezavne motive Pretežno a Pretežno b Mešano a/b Encimi vijačnica-zavoj-vijačnica (HTH) vijačnica-zanka-vijačnica (HLH) levcinska zadrga TATA-vezavni protein Imunoglobulinski b sendvič b-detelja motiv b-b-b cinkov prst b-pentlja-vijačnica-vijačnica ostali Kompleksne interakcije preko več DNA-vezavnih motivov/domen Proteini, ki vežejo enoverižno DNA
.")
, ki tvori stabilne dimere preko")

39 Motiv HTH Motiv HTH je najpogostejši DNA vezavni motiv pri bakterijah (in bakteriofagih). Represor Cro protein bakteriofaga l je majhen enodomenski protein (66 ostankov), ki tvori stabilne dimere preko intermolekulske b-ploskve. DNA-vezavni motiv predstavljata vijačnici 2 in 3 ter zavoj med njima. Vijačnica 3 se veže v veliki žleb DNA in ji rečemo prepoznavna vijačnica.


40 Levcinska zadrga Proteini z DNA vezavnim motivom levcinske zadrge tvorijo homo- ali heterodimere, s čimer dosežejo visoko specifičnost vezave na DNA. Primer: kvasni transkripcijski faktor GCN4

.")
41 Motiv HLH Motiv vijačnica-zanka-vijačnica je sestavljena iz bazične DNA-vezavne vijačnice, zanke in krajše vijačnice. Motiv se pogosto kombinira z levcinsko zadrgo. Člani družine so si med seboj zelo podobni (nad 2/3 identičnih ak). Ponavadi vežejo DNA zaporedja 5 -CANNTG-3. Primer: transkripcijski faktor Max



42 TATA-vezavni protein TBP je zgrajen iz dveh tandemnih enot a/b sendviča, ki tvorita kontinuirno antiparalelno beta ploskev v obliki sedla. Veže se v mali žleb DNA in povzroči veliko konformacijsko spremembo. Interakcije so pretežno hidrofobne.
43 Cinkovi prsti Cinkov prst je najbolj znan primer mešanega a/b DNA-vezavnega motiva. Vsebuje ga preko tisoč različnih transkripcijskih faktorjev. Zgradba klasičnega cinkovega prsta: Poleg Cys 2 His 2 motivov poznamo še cinkove prste s Cys 4, Cys 6,... motivi Specifične interakcije z DNA tvori prepoznavna vijačnica, ki prepozna največ 2 do 4 bazne pare. Vezava posameznega motiva cinkovega prsta na DNA je relativno šibka.


44 Cinkovi prsti Cinkov prst je najbolj znan primer mešanega a/b DNA-vezavnega motiva. Vsebuje ga preko tisoč različnih transkripcijskih faktorjev. Cinkovi prsti se vežejo na DNA v več tandemnih ponovitvah: 6 zaporednih motivov TFIIIA vezanih na DNA.


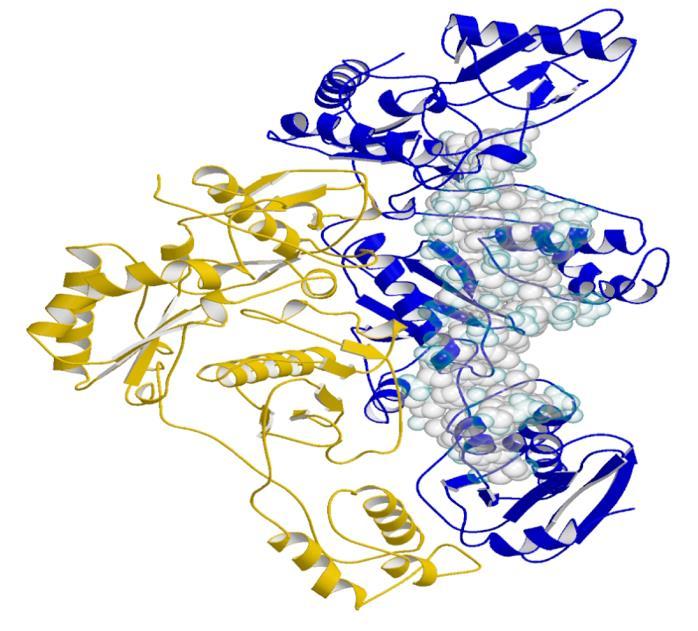
45 Encimi EcoRV DNaza I DNA topoizomeraza I EcoRI DNApol I (Bacillus) HIV RT


46 Proteini, ki vežejo enoverižno DNA Večina proteinov, ki veže enoverižno DNA, spada v družino proteinov z OB domeno (oligonukleotid/oligosaharid-vezavna domena). Primer: telomerni protein POT1 z DNA-vezavno regijo iz dveh zaporednih OB domen v kompleksu z dvema ponovitvama GGTTAG. PDB 3KJP

47 DNA kot ligand ali receptor Obravnjavanje DNA kot ligand: Pri kvantitativni karakterizaciji interakcij in vitro lahko DNA tretiramo kot ligand, ki se veže na protein. V ta namen je najlažje uporabiti sintetične oligonukleotide različnih dolžin in sestave. Obravnavanje DNA kot receptor: DNA je linearen polimer model receptorja z n (veliko) vezavnimi mesti za majhne ligande. A + nl AL n
 ali pa vežejo v")
48 DNA kot receptor Majhni ligandi se lahko interkalirajo med bazne pare (A) ali pa vežejo v mali žleb (B).


49 Glikozaminoglikani Linearni negativno nabiti polimeri iz ponavljajočih se disaharidnih enot. Tako kot DNA jih lahko tretiramo kot ligande proteinov ali pa receptorje z n vezavnimi mesti za ligande (npr. katione).


.")
50 Glikozaminoglikani Linearni negativno nabiti polimeri iz ponavljajočih se disaharidnih enot. Tako kot DNA jih lahko tretiramo kot ligande proteinov ali pa receptorje z n vezavnimi mesti za ligande (npr. katione). hondroitin sulfat dermatan sulfat heparin
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Funkcije proteinov (pogojene s strukturo)
") Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
I. OSNOVNI STRUKTURNI PRINCIPI
 I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Najpomembnejši človeški Hb
 Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks'
 Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
Najpogostejše hemoglobinopatije
 Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina
Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
Encimska kinetika govori o hitrosti encimske reakcije
 Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo
 Osnovni princip kromatografije 3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo 4. vaja Ionskoizmenjevalna ter afinitetna kromatografija Miha Pavšič komponente zmesi različne interakcije
Osnovni princip kromatografije 3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo 4. vaja Ionskoizmenjevalna ter afinitetna kromatografija Miha Pavšič komponente zmesi različne interakcije
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV. 2. predavanje: Od 1D do 3D strukture proteinov 1. del.
 IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ
 ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ α-heliks β-nagubana RAVNINA β-obrat STRUKTURNI MOTIVI STABILNOST PROTEINOV KVARTARNA ZGRADBA PROTEINOV ENA ALI VEČ KONFORMACIJ
ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ α-heliks β-nagubana RAVNINA β-obrat STRUKTURNI MOTIVI STABILNOST PROTEINOV KVARTARNA ZGRADBA PROTEINOV ENA ALI VEČ KONFORMACIJ
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
Biokemija I, 25. predavanje 1. del, , A. Videtič Paska. Proteini - splošno
 Biokemija I, 25. predavanje 1. del, 16. 4. 2012, A. Videtič Paska Proteini - splošno Razdelitev po strukturi in funkciji. Ravni proteinske strukture: - primarna in - sekundarna struktura Sinteza proteinov
Biokemija I, 25. predavanje 1. del, 16. 4. 2012, A. Videtič Paska Proteini - splošno Razdelitev po strukturi in funkciji. Ravni proteinske strukture: - primarna in - sekundarna struktura Sinteza proteinov
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 15. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
 FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
Splošno o interpolaciji
 Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Tabele termodinamskih lastnosti vode in vodne pare
 Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Jure Stojan in Marko Goličnik Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Rang & Dale's Pharmacology (7th) IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod)
 IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod)") Rang & Dale's Pharmacology (7th) IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod) Domen Sever, študent 3. letnika medicine Kazalo 2. POGLAVJE: MEHANIZMI DELOVANJA ZDRAVIL... 3 3. POGLAVJE: MEHANIZMI
Rang & Dale's Pharmacology (7th) IZBRANA POGLAVJA IZ SPLOŠNE FARMAKOLOGIJE (prevod) Domen Sever, študent 3. letnika medicine Kazalo 2. POGLAVJE: MEHANIZMI DELOVANJA ZDRAVIL... 3 3. POGLAVJE: MEHANIZMI
Neavtorizirani povzetki izbranih predavanj iz biokemije. UN študij Okolje Politehnika Nova Gorica. doc. dr. Marko Dolinar
 Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
p 1 ENTROPIJSKI ZAKON
 ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
Poglavje 10. Molekule Kovalentna vez
 Poglavje 10 Molekule Atomi se vežejo v molekule. Vezavo med atomi v molkuli posredujejo zunanji - valenčni elektroni. Pri vseh molekularnih vezeh negativni naboj elektronov posreduje med pozitinvimi ioni
Poglavje 10 Molekule Atomi se vežejo v molekule. Vezavo med atomi v molkuli posredujejo zunanji - valenčni elektroni. Pri vseh molekularnih vezeh negativni naboj elektronov posreduje med pozitinvimi ioni
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
FUNKCIJA PROTEINOV, PRIMERI FIBRILARNI PROTEINI. α KERATIN
 FUNKCIJA PROTEINOV, PRIMERI FIBRILARNI PROTEINI keratin, kolagen, fibroin, spidroin GLOBULARNI PROTEINI domenska zgradba proteinov MIOGLOBIN, HEMOGLOBIN vezava kisika zgradba konformacijske spremembe ob
FUNKCIJA PROTEINOV, PRIMERI FIBRILARNI PROTEINI keratin, kolagen, fibroin, spidroin GLOBULARNI PROTEINI domenska zgradba proteinov MIOGLOBIN, HEMOGLOBIN vezava kisika zgradba konformacijske spremembe ob
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
1. TVORBA ŠIBKEGA (SIGMATNEGA) AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ
 AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ") TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
TOPNOST, HITROST RAZTAPLJANJA
 OPNOS, HIOS AZAPLJANJA Denja: onos (oz. nasčena razona) redsavlja sanje, ko je oljene (rdn, ekoč, lnas) v ravnoežju z razono (oljenem, razoljenm v olu). - kvanavn zraz - r določen - homogena molekularna
OPNOS, HIOS AZAPLJANJA Denja: onos (oz. nasčena razona) redsavlja sanje, ko je oljene (rdn, ekoč, lnas) v ravnoežju z razono (oljenem, razoljenm v olu). - kvanavn zraz - r določen - homogena molekularna
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
ARHITEKTURA DETAJL 1, 1:10
 0.15 0.25 3.56 0.02 0.10 0.12 0.10 SESTV S2 polimer-bitumenska,dvoslojna(po),... 1.0 cm po zahtevah SIST DIN 52133 in nadstandardno, (glej opis v tehn.poročilu), npr.: PHOENIX STR/Super 5 M * GEMINI P
0.15 0.25 3.56 0.02 0.10 0.12 0.10 SESTV S2 polimer-bitumenska,dvoslojna(po),... 1.0 cm po zahtevah SIST DIN 52133 in nadstandardno, (glej opis v tehn.poročilu), npr.: PHOENIX STR/Super 5 M * GEMINI P
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
SEMINAR IZ MOLEKULSKEGA MODELIRANJA PRI FARMACEVTSKI KEMIJI III
 SEMINAR IZ MOLEKULSKEGA MODELIRANJA PRI FARMACEVTSKI KEMIJI III 1. DEL TEORETIČNE OSNOVE MOLEKULSKEGA MODELIRANJA doc. Andrej Perdih e-mail: andrej.perdih@ki.si 1. DEL: TEORETIČNE OSNOVE MOLEKULSKEGA MODELIRANJA
SEMINAR IZ MOLEKULSKEGA MODELIRANJA PRI FARMACEVTSKI KEMIJI III 1. DEL TEORETIČNE OSNOVE MOLEKULSKEGA MODELIRANJA doc. Andrej Perdih e-mail: andrej.perdih@ki.si 1. DEL: TEORETIČNE OSNOVE MOLEKULSKEGA MODELIRANJA
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * FIZIKA NAVODILA ZA OCENJEVANJE. Petek, 10. junij 2016 SPLOŠNA MATURA
 Državni izpitni center *M16141113* SPOMLADANSKI IZPITNI ROK FIZIKA NAVODILA ZA OCENJEVANJE Petek, 1. junij 16 SPLOŠNA MATURA RIC 16 M161-411-3 M161-411-3 3 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor
Državni izpitni center *M16141113* SPOMLADANSKI IZPITNI ROK FIZIKA NAVODILA ZA OCENJEVANJE Petek, 1. junij 16 SPLOŠNA MATURA RIC 16 M161-411-3 M161-411-3 3 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου...
 ΑΠΟΖΗΜΙΩΣΗ ΘΥΜΑΤΩΝ ΕΓΚΛΗΜΑΤΙΚΩΝ ΠΡΑΞΕΩΝ ΣΛΟΒΕΝΙΑ 1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου... 3 1 1. Έντυπα αιτήσεων
ΑΠΟΖΗΜΙΩΣΗ ΘΥΜΑΤΩΝ ΕΓΚΛΗΜΑΤΙΚΩΝ ΠΡΑΞΕΩΝ ΣΛΟΒΕΝΙΑ 1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου... 3 1 1. Έντυπα αιτήσεων
Iterativno reševanje sistemov linearnih enačb. Numerične metode, sistemi linearnih enačb. Numerične metode FE, 2. december 2013
 Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Funkcije več spremenljivk
 DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
Primeri: naftalen kinolin spojeni kinolin
 Primeri: naftalen kinolin spojeni kinolin 3 skupne strani 7 skupnih strani 5 skupnih strani 6 skupnih atomov 8 skupnih atomov 6 skupnih atomov orto spojen sistem orto in peri spojena sistema mostni kinolin
Primeri: naftalen kinolin spojeni kinolin 3 skupne strani 7 skupnih strani 5 skupnih strani 6 skupnih atomov 8 skupnih atomov 6 skupnih atomov orto spojen sistem orto in peri spojena sistema mostni kinolin
The Thermal Comfort Properties of Reusable and Disposable Surgical Gown Fabrics Original Scientific Paper
 24 The Thermal Comfort Properties of Surgical Gown Fabrics 1 1 2 1 2 Termofiziološke lastnosti udobnosti kirurških oblačil za enkratno in večkratno uporabo december 2008 marec 2009 Izvleček Kirurška oblačila
24 The Thermal Comfort Properties of Surgical Gown Fabrics 1 1 2 1 2 Termofiziološke lastnosti udobnosti kirurških oblačil za enkratno in večkratno uporabo december 2008 marec 2009 Izvleček Kirurška oblačila
13. Jacobijeva metoda za računanje singularnega razcepa
 13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
Frekvenčna analiza neperiodičnih signalov. Analiza signalov prof. France Mihelič
 Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Nukleinske kisline. Nukleotidi. DNA je nosilka dednih genetskih informacij.
 Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ
 GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
DISKRIMINANTNA ANALIZA
 DISKRIMINANTNA ANALIZA Z diskriminantno analizo poiščemo tako linearno kombinacijo merjenih spremenljivk, da bo maksimalno ločila vnaprej določene skupine in da bo napaka pri uvrščanju enot v skupine najmanjša.
DISKRIMINANTNA ANALIZA Z diskriminantno analizo poiščemo tako linearno kombinacijo merjenih spremenljivk, da bo maksimalno ločila vnaprej določene skupine in da bo napaka pri uvrščanju enot v skupine najmanjša.
SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV
 SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV Literatura za temo Signalne poti, ki regulirajo izražanje genov: Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH
SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV Literatura za temo Signalne poti, ki regulirajo izražanje genov: Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH
+105 C (plošče in trakovi +85 C) -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70
 -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70") KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
SPLOŠNO O BELJAKOVINAH STRUKTURA BELJAKOVIN
 SPLOŠNO O BELJAKOVINAH Beljakovine so najvažnejša sestavina vsake celice, kajti vsi življenski procesi so odvisni od njih. So makromolekularnespojine, sestavljeneiz ogljika, vodika, kisika in dušika ter
SPLOŠNO O BELJAKOVINAH Beljakovine so najvažnejša sestavina vsake celice, kajti vsi življenski procesi so odvisni od njih. So makromolekularnespojine, sestavljeneiz ogljika, vodika, kisika in dušika ter
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Prenos toplote prenos energije katerega pogojuje razlika temperatur temperatura je krajevno od točke do točke različna
 PRENOS OPOE Def. Prenos toplote prenos energije katerega pogojuje razlika temperatur temperatura je krajevno od točke do točke različna Načini prenosa toplote: PREVAJANJE (kondukcija, PRESOP (konvekcija
PRENOS OPOE Def. Prenos toplote prenos energije katerega pogojuje razlika temperatur temperatura je krajevno od točke do točke različna Načini prenosa toplote: PREVAJANJE (kondukcija, PRESOP (konvekcija
Gimnazija Krˇsko. vektorji - naloge
 Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
UPORABA POLYHIPE-BIOKOMPATIBILNIH
 UPORABA POLYHIPE-BIOKOMPATIBILNIH AKRILNIH POLIMEROV V TKIVNEM INŽENIRSTVU KOSTNIH NADOMESTKOV PoliMaT Academy TalentCamp 2012 Naloga za samostojno delo; področje C: Polimeri v medicini ddr. Matjaž DEŽELAK,
UPORABA POLYHIPE-BIOKOMPATIBILNIH AKRILNIH POLIMEROV V TKIVNEM INŽENIRSTVU KOSTNIH NADOMESTKOV PoliMaT Academy TalentCamp 2012 Naloga za samostojno delo; področje C: Polimeri v medicini ddr. Matjaž DEŽELAK,
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
IZZIVI DRUŽINSKE MEDICINE. U no gradivo zbornik seminarjev
 IZZIVI DRUŽINSKE MEDICINE Uno gradivo zbornik seminarjev študentov Medicinske fakultete Univerze v Mariboru 4. letnik 2008/2009 Uredniki: Alenka Bizjak, Viktorija Janar, Maša Krajnc, Jasmina Rehar, Mateja
IZZIVI DRUŽINSKE MEDICINE Uno gradivo zbornik seminarjev študentov Medicinske fakultete Univerze v Mariboru 4. letnik 2008/2009 Uredniki: Alenka Bizjak, Viktorija Janar, Maša Krajnc, Jasmina Rehar, Mateja
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f
 D f") IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
Kotni funkciji sinus in kosinus
 Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
Funkcije proteinov, pogojene s strukturo
 Funkcije proteinov, pogojene s strukturo Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni proteini) Oporna funkcija (strukturni proteini: keratini, kolagen)
Funkcije proteinov, pogojene s strukturo Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni proteini) Oporna funkcija (strukturni proteini: keratini, kolagen)
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Analiza'3D'struktur'makromolekul'in' modeliranje'
 Univerza'v'Ljubljani,'Fakulteta'za'kemijo'in'kemijsko'tehnologijo' Univerzitetni%študijski%program%Biokemija,%2.%letnik,%študijsko%leto%2013/2014% Analiza'3D'struktur'makromolekul'in' modeliranje' Miha%Pavšič%
Univerza'v'Ljubljani,'Fakulteta'za'kemijo'in'kemijsko'tehnologijo' Univerzitetni%študijski%program%Biokemija,%2.%letnik,%študijsko%leto%2013/2014% Analiza'3D'struktur'makromolekul'in' modeliranje' Miha%Pavšič%
APROKSIMACIJA FUNKCIJA
 APROKSIMACIJA FUNKCIJA Osnovni koncepti Gradimir V. Milovanović MF, Beograd, 14. mart 2011. APROKSIMACIJA FUNKCIJA p.1/46 Osnovni problem u TA Kako za datu funkciju f iz velikog prostora X naći jednostavnu
APROKSIMACIJA FUNKCIJA Osnovni koncepti Gradimir V. Milovanović MF, Beograd, 14. mart 2011. APROKSIMACIJA FUNKCIJA p.1/46 Osnovni problem u TA Kako za datu funkciju f iz velikog prostora X naći jednostavnu
Statistična analiza. doc. dr. Mitja Kos, mag. farm. Katedra za socialno farmacijo Univerza v Ljubljani- Fakulteta za farmacijo
 Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Vaje: Električni tokovi
 Barbara Rovšek, Bojan Golli, Ana Gostinčar Blagotinšek Vaje: Električni tokovi 1 Merjenje toka in napetosti Naloga: Izmerite tok, ki teče skozi žarnico, ter napetost na žarnici Za izvedbo vaje potrebujete
Barbara Rovšek, Bojan Golli, Ana Gostinčar Blagotinšek Vaje: Električni tokovi 1 Merjenje toka in napetosti Naloga: Izmerite tok, ki teče skozi žarnico, ter napetost na žarnici Za izvedbo vaje potrebujete
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL
 PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
Transformator. Delovanje transformatorja I. Delovanje transformatorja II
 Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
Termodinamika vlažnega zraka. stanja in spremembe
 Termodinamika vlažnega zraka stanja in spremembe Termodinamika vlažnega zraka Najpogostejši medij v sušilnih procesih konvektivnega sušenja je VLAŽEN ZRAK Obravnavamo ga kot dvokomponentno zmes Suhi zrak
Termodinamika vlažnega zraka stanja in spremembe Termodinamika vlažnega zraka Najpogostejši medij v sušilnih procesih konvektivnega sušenja je VLAŽEN ZRAK Obravnavamo ga kot dvokomponentno zmes Suhi zrak
DISKRETNA FOURIERJEVA TRANSFORMACIJA
 29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
Multivariatna analiza variance
 (MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti med več odvisnimi (številskimi) in več neodvisnimi (opisnimi) spremenljivkami. (MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti
(MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti med več odvisnimi (številskimi) in več neodvisnimi (opisnimi) spremenljivkami. (MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti
PARCIJALNI IZVODI I DIFERENCIJALI. Sama definicija parcijalnog izvoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je,
 PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda