ΗΛΕΚΤΡΙΚΗ ΔΡΑΣΤΗΡΙΟΤΗΤΑ ΣΥΣΤΑΛΤΩΝ ΚΥΤΤΑΡΩΝ ΣΥΖΕΥΞΗ ΔΙΕΓΕΡΣΗΣ- ΣΥΣΤΟΛΗΣ
|
|
|
- Δορκάς Παπαντωνίου
- 9 χρόνια πριν
- Προβολές:
Transcript
1 ΗΛΕΚΤΡΙΚΗ ΔΡΑΣΤΗΡΙΟΤΗΤΑ ΣΥΣΤΑΛΤΩΝ ΚΥΤΤΑΡΩΝ ΣΥΖΕΥΞΗ ΔΙΕΓΕΡΣΗΣ- ΣΥΣΤΟΛΗΣ Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Βιοφυσική μεμβρανών 2013

2 ΣΚΕΛΕΤΙΚΟΙ ΜΥΕΣ

3 Νευρομυϊκή σύναψη Νευροµυϊκή σύναψη: o σύναψη ενός τερµατικού άξονα µε µια τελική κινητική πλάκα Τελική κινητική πλάκα: o επιφάνεια της κυτταροπλασµατικής µεµβράνης της µυϊκής ίνας που βρίσκεται κάτω από τη νευρική απόληξη της κινητικής νευρικής ίνας

4 Ο νευροδιαβιβαστής στη νευροµυϊκή σύναψη είναι η ακετυλοχολίνη και οι υποδοχείς της στην τελική κινητική πλάκα είναι νικοτινικοί

5 Κινητικοί νευρώνες κινητική μονάδα Κινητικοί νευρώνες: νευρικά κύτταρα των οποίων τα σώµατα βρίσκονται στα πρόσθια κέρατα του ΝΜ και οι νευράξονες (εµµύελοι, µεγάλης διαµέτρου) νευρώνουν τις ίνες σκελετικού µυός. Άγουν τα δυναµικά ενεργείας µε µεγάλη ταχύτητα Κινητική µονάδα: κινητικός νευρώνας µαζί µε τις µυϊκές ίνες που νευρώνει Όταν το δυναµικό ενεργείας παράγεται σε ένα κινητικό νευρώνα όλες οι µυϊκές ίνες της κινητικής µονάδας συστέλλονται

6 Απελευθέρωση ΑΚΧ στη νευρομυϊκή σύναψη
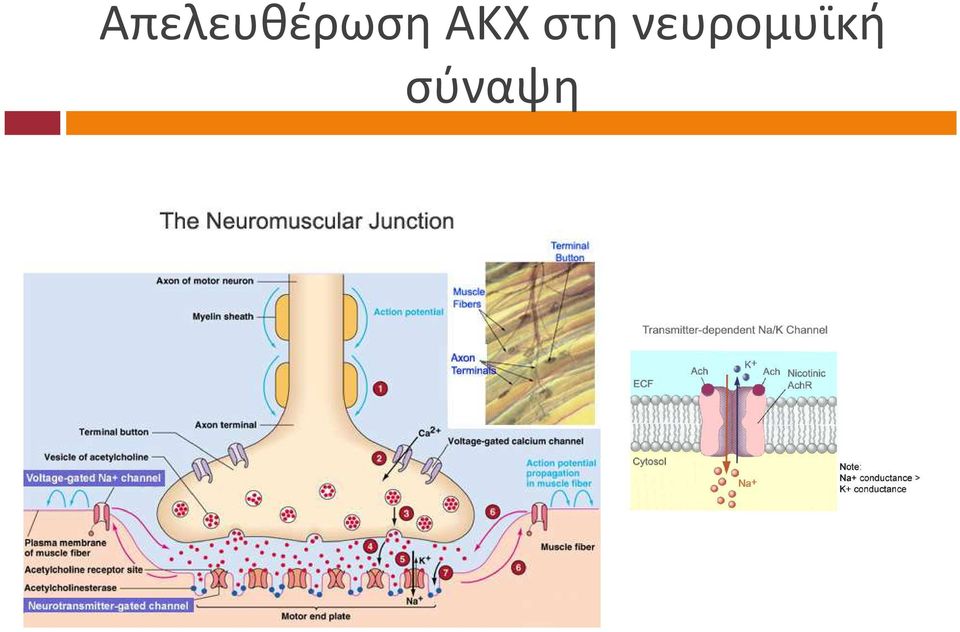
7 Δυναμικό τελικής κινητικής πλάκας υναµικό τελικής κινητικής πλάκας (ΕΡΡ): εκπόλωση µετασυναπτικής µεµβράνης λόγω δέσµευσης ακετυλοχολίνης σε νικοτινικούς υποδοχείς Συνήθως το ΕΡΡ µπορεί να παράγει δυναµικό ενεργείας στη µυϊκή ίνα Μiniature end-plate potentials (MEPPS). MEPPS είναι πολλαπλάσια του 0.5 mv και αντιπροσωπεύουν εκπόλωση της µεµβράνης του µυϊκού κυττάρου λόγω απελευθέρωσης ακετυλοχολίνης (κυστίδια) σε συνθήκες ηρεµίας

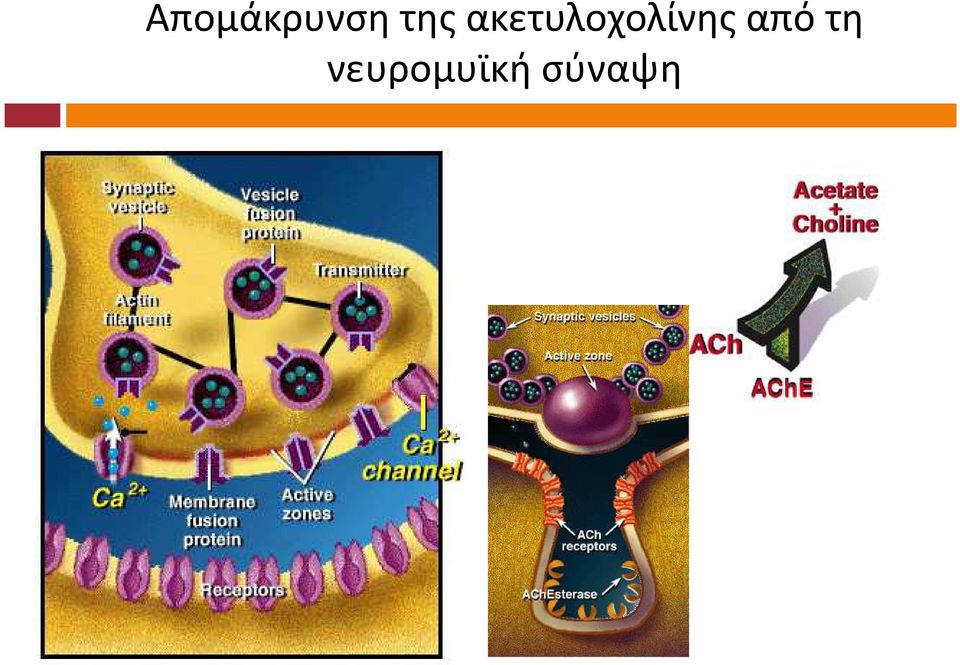
8 Απομάκρυνση της ακετυλοχολίνης από τη νευρομυϊκή σύναψη
9 Φάρμακα που επηρεάζουν τη νευρομυϊκή σύναψη Κουράριο Ζιζανιοκτόνα, βιολογικά όπλα

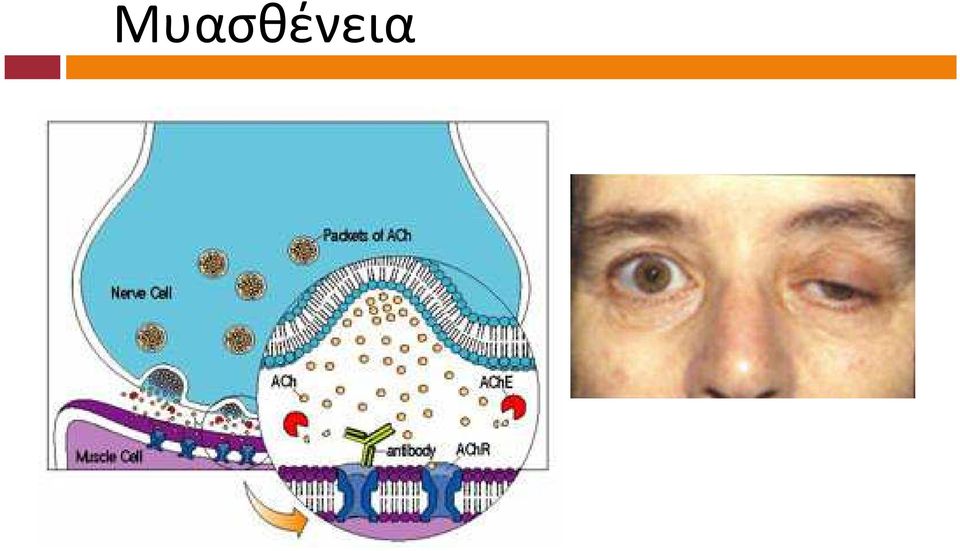
10 Ασθένειες που επηρεάζουν τη νευρομυϊκή σύναψη Αλλαντίαση (αλλαντική τοξίνη) Μυασθένεια

11 Μυασθένεια
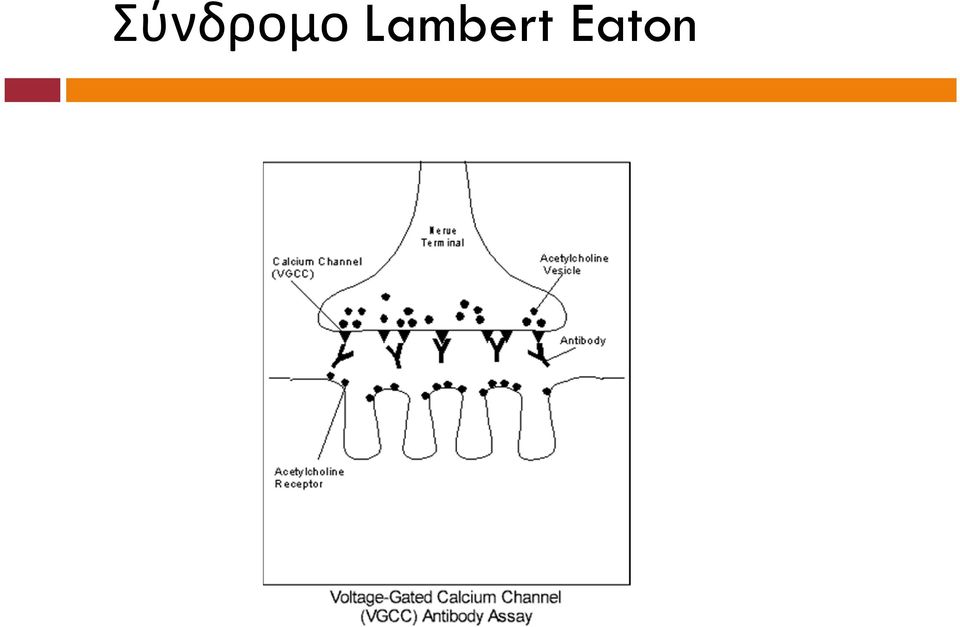
12 Σύνδρομο Lambert Eaton
13 Αλλαντίαση Αλλαντίαση: σπάνια αλλά σοβαρή παραλυτική νόσος που προκαλείται από νευροτοξίνη η οποία παράγεται κυρίως από το βακτηρίδιο Clostridium botulinum και αναστέλλει την απελευθέρωση ακετυλοχολίνης

14 Κατανομή ιόντων εκατέρωθεν της κυτταρικής μεμβράνης των σκελετικών μυών Ca mm Na mm Cl mm K mm -80 mv Ca M Na + 15 mm Cl - 5 mm K mm

15 Δυναμικά ενεργείας στις σκελετικές ίνες +130 E Ca I Ca = g ca (E m -E ca ) +60 Em (mv) 0 E Na Diffusion force spike I Na =g Na (E m -E Na ) I K =g K (E m -E k ) I Cl =g Cl (E m -E Cl ) depolarizing afterpotential Electrical force -80 Resting Potential E Cl -98 E K The APs in vertebrate skeletal muscle consist of a spike followed by depolarizing afterpotential

16 Δυναμικά ενεργείας στις σκελετικές ίνες A skeletal muscle action potential is generated when themotor endplate potential is sufficient to raise the surroundingsarcolemmal potential above the threshold for activation of thevoltage gated Na + channels that are abundant throughout thesarcolemma. When these channels are activated, the membraneis rapidly depolarized towards the Nernst potential for Na + However, the peak potential achieved is approximately +30 mv.

17 Δυναμικά ενεργείας στις σκελετικές ίνες The Nernst potential is not achieved for two main reasons. First, just as the Na + channels are activated by membranevoltage changes, a process of inactivation is also initiated as the membrane potential becomes less negative. Inactivation is aslower mechanism than activation, so the Na + current continues to flow for a short period after the onset of inactivation, but notsufficiently to reach the Nernst potential. The second factor limiting the upstroke of the action potential is the voltage activation of rectifying potassium channels. Their activation is initiatedalso during the upstroke of the action potential but (in asimilar way to Na + channel inactivation) there is a slight delayin channel opening. The resulting K + current, in addition to limitingthe peak of the action potential, is also principally responsible for repolarization.

18 Δυναμικά ενεργείας στις σκελετικές ίνες The amplitude of AP is about 120 mv from resting potential of -80 mv to a peak overshoot potential about +40 mv. The threshold potential for triggering the fast Na channels is about -55mV. The maximum increase inward fast Na current occurs at Em-20 mv.

19 Δίαυλοι καλίου 0 Em (mv) spike Δg Na Skeletal muscle cell membrane has at least 2 types K + channels: (1) Inward-going rectifier (K ions to pass more readily inwardthan outward). (2) Delayed rectifier The second type of K channels is similar to the usual K channels called delayed rectifier. Depolarizing afterpotential The activation of this channel produces the large increase in total K conductance that helps to terminate the AP. Δg K -80

20 Ο ρόλος της αγωγιμότητας του Cl The resting membrane potential in skeletalmuscle cells is similar to that in neurons, i.e.-70 to-90 mv. Unlike nerve cells, where theresting membrane potential is predominantlya result of K + permeability, skeletal musclecell resting membrane potential receives a significant contribution from Cl conductance. The Clpermeability and conductance are very high in skeletal muscle much higher than K. Cl entry tends to repolarize the membrane more quickly than would otherwise occur. The importance of this Clcurrent becameapparent when the excitability associatedwith myotoniacongenitawas found to bea result of chloride channel mutations. The physiological relevance of the Cl current stems from a need to maintain muscle activity during repeated stimulation. When musclecontracts, there is leakage of K + from thecell. With repeated activity there is run-downof the K + concentration gradient across thesarcolemma. Without the Clcurrent to maintain restingmembrane potential, the muscle would not repolarizesufficientlyto regenerate the active state of the channels responsiblefor generation of succeeding action potentials

21 Μετάδοση Δυναμικού ενεργείας στη μυϊκή ίνα Once an action potential has been generated, it spreads as awave over the sarcolemma. Skeletal muscle sarcolemma is characterized by invaginations called transverse- or t- tubules thatrun perpendicular to the surface of the cell deep into its body. By passing down the t-tubular membrane, the action potentialis carried to the structures responsible for transducing anelectrical into a chemical signal that will trigger activation of the contractile elements. The AP propagates at a constant velocity of about 5 m/s over the surface sarcolema
22 Σύζευξη διέγερσης-συστολής Η σύζευξη διέγερσηςσυστολής αναφέρεται στην αλληλουχία γεγονότων που επιφέρει ένα δυναµικό ενέργειας στο σαρκείληµα µιας µυϊκής ίνας και καταλήγει σε κυκλική δραστηριότητα της εγκάρσιας γέφυρας
23 Σύζευξη διέγερσης-συστολής Ένα δυναµικό ενεργείας διαρκεί 1-2ms και ολοκληρώνεται πριν την έναρξη της µηχανικής δραστηριότητας (συστολή) Η µηχανική δραστηριότητα διαρκεί 100ms Άρα Η ηλεκτρική δραστηριότητα (δυναµικό ενεργείας) δεν επιδρά άµεσα στις πρωτεΐνες συστολής
24 Σαρκοπλασματικό δίκτυο Το ασβέστιο στο κυτταρόπλασµα πριν τη συστολή είναι 10-7 Μ mol/l Πηγή αύξησης του ασβεστίου είναι το σαρκοπλασµατικό δίκτυο
25 Σαρκοπλασματικό δίκτυο Εγκάρσιοι σωλήνες: o εξωκυττάριος χώρος o διασχίζουν τις µυϊκές ίνες σε κάθε σηµείο συµβολής ζωνών Α-Ι o Άγουν το δυναµικό ενεργείας
26 Διέγερση μεμβράνης: νευρομυϊκή σύναψη
27 Τασεοεξαρτώμενοιδίαυλοι ασβεστίου

28 Υποδοχείς διϋδροπυριδίνης We now know that,although the t-tubular dihydropyridine receptors show markedamino acid homology with L-type voltage gated Ca 2+ channelsof other tissues, they do not function as Ca 2+ channels. Their rolein skeletal muscle is that of voltage sensors. When the actionpotential arrives in the t-tubule, the change in membrane potentialleads to a conformational change in the a-subunit of the dihydropyridinereceptor. This subunit consists of four transmembranedomains each of six segments. The first and sixthsegments of adjacent domains are linked by alternating extracellularand cytoplasmic loops of amino acids. The voltage-inducedconformational change results in the projection of the cytoplasmicloop between the second and third transmembrane domains deeper into the cytoplasm. Here, charged amino acid residuesof the cytoplasmic loop come into proximity with those of a proteinprojecting towards the cytoplasmic surface of the t-tubule
29 Μυϊκή συστολή Συστολή δεν σηµαίνει κατ ανάγκη βράχυνση του µυός αλλά ενεργοποίηση των σηµείων δυναµογένεσης των εγκάρσιων γεφυρών σε µια µυϊκή ίνα Μηχανισµός διολίσθησης νηµατίων: περιστροφική κίνηση όπως τα κουπιά της βάρκας. Τα παχιά και λεπτά νηµάτια δεν αλλάζουν µήκος.
30
31 Κύκλος εγκάρσιας γέφυρας πρόσδεση ενεργοποιημένης εγκάρσιας γέφυρας στην ακτίνη Α+Μ*ADP-Pi A-M*-ADP-Pi
32 Αποδέσμευση αποθηκευμένης ενέργειας κίνηση προσδεδεμένης εγκάρσιας γέφυρας και απελευθέρωση ADP & Pi A-M*-ADP-PiΑ-Μ+ADP+Pi
33 Απόσχιση εγκάρσιας γέφυρας με την πρόσδεση ΑΤΡ (αλλοστερική ρύθμιση, συγγένειας μυοσίνης-ακτίνης) Α-Μ+ΑΤΡ A+M-ΑΤΡ
34 Διάσπαση ΑΤΡ αλλαγή τρισδιάστατης δομής εγκάρσιας γέφυρας ενεργοποιημένη εγκάρσια γέφυρα (προσδεδεμένο ADP-Pi) A+M-Α ΤΡ Μ*-ADP-Pi
35 Κάθε εγκάρσια γέφυρα επιτελεί τον δικό της κύκλο ανεξάρτητα από τις άλλες γέφυρες Σε μια δεδομένη στιγμή, κατά τη συστολή, μόνο το 50% των εγκάρσιων γεφυρών είναι προσδεδεμένο στα λεπτά νημάτια παράγοντας κίνηση Το ΑΤΡ δρα ως τροποποιητικό μόριο το οποίο ελέγχει την πρόσδεση της ακτίνης στη μυοσίνη
36 Ο ρόλος της τροπονίνης, τροπομυοσίνης και του ασβεστίου Σε κατάσταση ηρεµίας παρεµποδίζεται ο σχηµατισµός εγκάρσιων γεφυρών από τις πρωτεΐνες-ρυθµιστές τροπονίνη και τροποµυοσίνη Η τροποµυοσίνη καλύπτει τις θέσεις πρόσδεσης της ακτίνης και παραµένει στη θέση αυτή µε τη δράση της τροπονίνης (µικρή σφαιρική πρωτεΐνη)
37 Η δραστηριότητα των εγκάρσιων γεφυρών πυροδοτείται από το ασβέστιο που προσδένεται πάνω στην τροπονίνη
38
39 A significant proportion of the force generatedby a fiber (>70% in some experiments) depends on transmission ofthe force successively through structural intracellular proteins tothe sarcolemmaand extracellular matrix. Crucial to the efficient transmission of forces from the sarcomereto the extracellular matrix are the integrin and dystroglycancomplexes that link the latter to the cytoskeletalelements of thesarcomere. The importance of the dystroglycan complexbecame apparent with the discovery that several of the hereditary dystrophies result from abnormal components of this complex.
40 Λήξη της συστολής-απομάκρυνση ασβεστίου Αντλία ασβεστίου
41
42 ΜΥΟΚΑΡΔΙΟ
43 Nerve and skeletal muscle APs are relative brief consisting of A rapid depolarization phase A rapid repolarization phase. The entire process of the nerve and skeletal muscle APs is largely complete within a few milliseconds. In cardiac cells, APs are more complex and generally much longer in duration. Thus for example a typical cardiac ventricular AP may be least msin duration. Unlikefor nerve and skeletal muscle cells, APs from different regions of the heart vary substantially in shape.
44 Τύποι καρδιακών κυττάρων There are two primary types of cardiac cells. One cell typeis found in the working cells of atria, ventricles, and the specialized conduction cells of the His-Purkinje network. The second type of cardiac cell is found in the sinoatrialand atrioventicular nodes
45 Δυναμικά ενεργείας στα μυοκαρδιακά κύτταρα
46 Phase 0 0 Phase 0 is the rapid upstroke of the AP dependent on a fast Na + current(similar to the nerve or skeletal muscle). I Na reaches a peak in less than 1 ms. The cell needs to be depolarized to 70mV. The peak I Na occurs between 30 to 20 mv. The membrane potential never reaches E na The maximum positive membrane potential normally attained is approximately +35 mv. The upstroke causes a substantial voltage change ( mv) within 1-2 ms in fast cardiac cells
47 Phase 1 1 Phase 1 repolarizationis largely due to a transient outward currentturns on rapidly with depolarization and only active at very depolarized potentials, the threshold of activation is 30 mv. Because of its voltage dependence and time course I to significant overlaps the inward Ca current of phase 2 (plateau). The size of phase 1 variesbetween species and also between different regions of the heart within a given species. I to is composed of 2 separate currents: 1. K + current that is independent of the internal Ca concentration 2. Ca 2+ -dependent current(ca 2+ -activated Clchannel): negative feedbackmechanism to reduce calcium overload.
48 Phase 2 2 Phase 2 follows the early repolarizationphase 1, and is a period of time in which the membrane potential remains relatively constant. The plateau phase is responsible for the long AP durationin cardiac cells which is the major difference between cardiac cells and nerve or skeletal muscle fibers.
49 Phase 2 (plateau phase): Ca 2+ current The plate au is caused by an approximate balance of positive inward currentand a positive outward current. The primary inward current is a Ca 2+ current. The primary outward K + current,is the slow activating K current (delayed rectifier). Additional I to contributes to the early plateau phase. Is a small contribution of the fast Na currentto the plateau. Late sodium current known as the sodium window currentis sufficient to affect the plateau of the AP and hence the AP duration.
50 Phase 2 (plateau phase): L-type Ca 2+ current The inward Ca current exhibits activation and inactivation much like I Na, but on a slower timescale. Ca current peaks within a few ms, but requires a few hundred msto completely inactivated. The threshold of activation is 40mV, the current is maximal near 0mV This classical Ca-current is now commonly referred to as asthe L- type calcium current.
51 Phase 3: Delayed Rectifier Current 3 Phase 3 is the late or final repolarization phase following the AP plateau. It is similar to the repolarization observed in nerve and skeletal muscle cells. K + -current is the primary repolarizing current.
52 Phase 4: Inward Rectifier Current 4 Phase 4 is the resting potential. RP is very near E K due to a relatively high permeability of the cell membrane to K + ions and a very low permeability to other ions. The most notable characteristic of I KI is that it displays inward rectification.
53 The L-type calcium current is central to many aspects of cardiac function -Excitation contraction coupling - Calcium influx stimulates calcium release from internal stores -Calcium current is important in automaticity and conduction (nodal AP) -Is a major regulatory site in control of cardiac electrical activity and contraction by neurotransmitters, hormones, intracellular ions e.g Norepinephrine production camp stimulation of camp-dependent protein kinase Phosphorylation of Calcium channels enhance of channel activity
54 Automaticity: funny current (I f ) Automatic cells do not have a stable resting potentialbetween APs, but rather have a maximum diastolic potential following by spontaneous phase 4 depolarization known as pacemaker potential. The slope of the pacemaker potentiallargely determines the rate of AP firing The primary current responsible for the pacemaker potential is funny current. Funny current is a slow activating inward depolarizing current activated by hyperpolarization, which is present in automatic cells. It is nonselective cation current (Na and K) In addition to funny current there may be a small contribution of T-type calcium current to the latter stages of the pacemaker potential.
55 Anatomy The anatomy of the heart muscle cell (cardiac myocyte) fits its physiology almost as well as the intercalated discsat the end of the cells fit one another! The extensive branching and interdigitation of the cells helps to provide mechanical cohesion of the cells, but more than this, it provides a substrate for electrical cohesion. Associated with the intercalated discs is a vast network of "gap junctions"linking the sides of the muscle fibres and providing a low resistance pathway for the rapid conduction of electrical current. Heart muscle functions as a syncitium, despite the fact that it is made up of discrete, individual cells
56 The'motor' insidethemusclecellismadeupof interdigitating fibres of actin and myosin. This is true for both skeletal muscle and cardiac muscle, although several differencesbetweenthetwoexist. Actinfilamentsarethin, myosinarethick. The thick myosin fibres have many projecting heads, and itisthesetinyheadsthatswivelbackwardsandforwards, moving along the actin fibres The sarcomere is the"fundamental unit" of the muscle cell. Ithasa Z diskateachend, andthethinactinfilaments extendfrombothz disksintowardsthecentreofthecell. The thick myosin filaments are found towards the centre ofthesarcomere. Wecanseewhereallthemicroscopic complicationsarose: thez bandcorrespondstothez disk, a lighti bandisactuallymadeupofactinfilaments fromtwoadjacentsarcomeres, cutintwobya Z disk, and thedarkm bandismadeupofthickmyosinfilamentsin the middle of a sarcomere
57 How does cardiac muscle differ from skeletal muscle? Differencesarenumerous, including: Skeletal muscle cells have numerous peripherally located nuclei, in contrast to the single centrally located nuclei of cardiac muscle Cardiac myocytes end in intercalated discs Cardiac myocytes have abundant mitochondria T-tubules ramify through the cytoplasm of cardiac myocytesnearthez line, andnot(asinskeletalmuscle) at the A-I junction
58
59
60 Cardiac Excitation-Contraction Coupling Ratcheting cycles occur as long as the cytosolic Ca 2+ remains elevated. At the end of phase 2 of AP, Ca 2+ entry into the cell slows and calcium is sequestered by the SR by an ATPdependent Ca 2+ pump (SERCA, sarcoendoplasmic reticulum calcium-atpase), thus lowering the cytosolic Ca 2+ concentration and removing Ca 2+ from the TN-C. A quantitatively smaller extent, cytosolic Ca 2+ is transported out of the cell by the sodium-calcium-exchange pump. The reduced intracellular Ca 2+ induces a conformational change in the troponin complex leading, once again, to TN-I inhibition of the actin binding site.
61 Ryanodine Receptors: Overview& Structure In skeletal musclecells the opening of RyR1appears to be mediated by a direct coupling of RyR to the dihydropyridinesensitive Ca 2+ channel. In cardiac muscle RyR2is believed to open is response to an influx of Ca 2+ to the cellvia dihydropyridine-sensitive Ca 2+ channels in the cell surface membrane; a process referred to as Ca 2+ induced Ca 2+ release.
62 Μεταξύ ινών Purkinje και μυοκυττάρων υπάρχουν ηλεκτρικές συνάψεις
63
64
65 ΛΕΙΕΣ ΜΥΪΚΕΣ ΙΝΕΣ
66 Δομή Μονοπύρηνοι, ικανότητα διαίρεσης Στερούνται ραβδωτού σχεδίου
67 Παχιά (µυοσίνη) και λεπτά νηµάτια (ακτίνη) Τα νηµάτια ακτίνης αγκυροβολούν είτε στον κυτταροσκελετό είτε στα πυκνά σωµάτια (κυτταρόπλασµα) Η σύσπαση επιτελείται µέσω µηχανισµού ολίσθησης Μυοσίνη λείων µυϊκών ινών = 1/3 γραµµωτών ενώ η ακτίνη είναι διπλάσια Η µέγιστη παραγόµενη τάση ανά επιφάνεια είναι ίδια µε εκείνη του γραµµωτού µυός Υπάρχει βέλτιστο µήκος Το εύρος µήκους µέσα στο οποίο ο λείος µυς µπορεί να παράγει τάση είναι µεγαλύτερο από το εύρος του σκελετικού µυός (υψηλή προσαρµοστικότητα)
68 Νεύρα και ορμόνες Απουσία τελικής κινητικής πλάκας κιρσοί Άλλοι νευροδιαβιβαστές προκαλούν εκπόλωση και άλλοι υπερπόλωση Ένας δεδοµένος νευροδιαβιβαστής µπορεί να προκαλέσει αντίθετα αποτελέσµατα σε διαφορετικές ΛΜΙ Πχ η νορεπινεφρίνη εκπολώνει τις ΛΜΙ των αγγείων ενώ αναστέλλει την αυθόρµητη δραστηριότητα των ΛΜΙ του εντέρου (χάλαση) Οι ΛΜΙ φέρουν υποδοχείς για ορµόνες
69 Τύποι ΛΜΙ (1) Μονοδύναμοι, λείοι μύες απλών μονάδωντων οποίων η κυτταροπλασματική μεμβράνη έχει την ικανότητα να μεταδίδει δυναμικά ενεργείας από το ένα κύτταρο στο άλλο μέσω χασματικών συνδέσεων και να εκπολώνεται αυθόρμητα Πχ εντερικού σωλήνα, μήτρας, αιμοφόρων αγγείων (2) Πολυδύναμοι, λείοι μύες πολλαπλών μονάδων, επιδεικνύουν μικρή ή καθόλου μεταβίβαση ηλεκτρικής δραστηριότητας από τη μια ίνα στην άλλη και δεν εκπολώνονται αυθόρμητα Πχ πνευμόνων, μεγάλων αρτηριών
70
71 Τύποι δυναμικών ενεργείας στις ΛΜΙ
72 Αυθόρμητη ηλεκτρική δραστηριότητα της κυτταροπλασματικής μεμβράνης Οι περισσότεροι ΛΜ μπορούν να παράγουν δυναμικά ενεργείας ενώ άλλοι όχι Μερικοί ΛΜ συσπώνται με την εκπόλωση της μεμβράνης ή δυναμικά ενεργείας ενώ άλλοι μπορούν να συσταλούν και απουσίαμεταβολής του δυναμικού της μεμβράνης Σε δυναμικά ενεργείας μετακινούνται κατά κύριο λόγο ιόντα ασβεστίου (τασεοεξαρτώμενοι δίαυλοι ασβεστίου) παρά νατρίου Αυθόρμητη ηλεκτρική δραστηριότηταδυναμικά βηματοδότη
73 Παραγωγή αργών κυμάτων Cα 2+ -εξαρτώμενοι δίαυλοι Κ
74 Ενεργοποίηση μεμβράνης Εισερχόμενα σήματα που επηρεάζουν τη συσταλτική δραστηριότητα του λείου μυός 1. αυθόρμητη ηλεκτρική δραστηριότητα της κυτταροπλασματικής μεμβράνης 2. νευροδιαβιβαστές που απελευθερώνονται από τα αυτόνομα νεύρα 3. ορμόνες 4. τοπικές αλλαγές στη χημική σύσταση του εξωκυττάριου υγρού που περιβάλλει την ίνα (παρακρινείς ουσίες, οξύτητα, οξυγόνο, ωσμωμοριακότητα και συγκέντρωση ιόντων) 5. διάταση
75 Συστολή ΛΜΙ και ο έλεγχος της Στις ΛΜΙ δεν υπάρχει τροπονίνη Καλμοδουλίνη Κινάση της ελαφράς αλύσου της μυοσίνης Φωσφατάση της ελαφράς αλύσου της μυοσίνης
76
77 Πηγές κυτοσολικού ασβεστίου (1) σαρκοπλασματικό δίκτυο (2) εξωκυττάριο ασβέστιο Σε κάποιους ΛΜ η συγκέντρωση του ενδοκυττάριου ασβεστίου αρκεί για να διατηρείται ένα χαμηλό επίπεδο ενεργοποίησης των εγκάρσιων γεφυρών χωρίς να υπάρχει κάποιο εξωτερικό ερέθισμα. Η δραστηριότητα αυτή είναι γνωστή ως τόνος λείου μυός.
78
79
80 Receptor modulation of membrane potential Type Transmitter Receptor Adrenergic Norepinephrine, epinephrine A, b Cholinergic Acetylcholine ACh Serotonergic Serotonin 5-HT Nucleotide ATP, adenosine ATP Angiotensin Angiotensin AT Vasopressin Vasopresine VP Endothelin Endothelin ET Prostanoid Prostaglandins E, F, I PG Leukotriens Leukotriene series L Insulin Insulin I Interleukin Interleukin series IL Histamine Histamine H Thromboxane Thromboxane, PGH2 Tx-A2
81
ΗΛΕΚΤΡΙΚΗ ΔΡΑΣΤΗΡΙΟΤΗΤΑ ΣΥΣΤΑΛΤΩΝ ΚΥΤΤΑΡΩΝ ΣΥΖΕΥΞΗ ΔΙΕΓΕΡΣΗΣ- ΣΥΣΤΟΛΗΣ
 ΗΛΕΚΤΡΙΚΗ ΔΡΑΣΤΗΡΙΟΤΗΤΑ ΣΥΣΤΑΛΤΩΝ ΚΥΤΤΑΡΩΝ ΣΥΖΕΥΞΗ ΔΙΕΓΕΡΣΗΣ- ΣΥΣΤΟΛΗΣ Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Βιοφυσική μεμβρανών 2016 ΣΚΕΛΕΤΙΚΟΙ ΜΥΕΣ http://people.eku.edu/ritchisong/301notes3.htm
ΗΛΕΚΤΡΙΚΗ ΔΡΑΣΤΗΡΙΟΤΗΤΑ ΣΥΣΤΑΛΤΩΝ ΚΥΤΤΑΡΩΝ ΣΥΖΕΥΞΗ ΔΙΕΓΕΡΣΗΣ- ΣΥΣΤΟΛΗΣ Απ. Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Βιοφυσική μεμβρανών 2016 ΣΚΕΛΕΤΙΚΟΙ ΜΥΕΣ http://people.eku.edu/ritchisong/301notes3.htm
Λείος μυς. Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017
 Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 2017 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
ΗΚΓ. Απ. Χατζηευθυµίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Λάρισα 21-11-2013
 Σύζευξη διέγερσηςσυστολής στον καρδιακό μυ ΗΚΓ Απ. Χατζηευθυµίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Λάρισα 21-11-2013 Nerve and skeletal muscle APs are relative brief consisting of o o A rapid depolarization
Σύζευξη διέγερσηςσυστολής στον καρδιακό μυ ΗΚΓ Απ. Χατζηευθυµίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Λάρισα 21-11-2013 Nerve and skeletal muscle APs are relative brief consisting of o o A rapid depolarization
Λείος μυς. Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
Λείος μυς Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 1 Λείοι μύες Τοιχώματα κοίλων οργάνων Νεύρωση από ΑΝΣ Ακούσιες κινήσεις Λείες μυϊκές ίνες Ατρακτοειδή κύτταρα (μονοπύρηνα)
Νευρομυϊκή Σύναψη Σκελετικός μυς - Μυϊκή Συστολή. Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής, Παν.
 Νευρομυϊκή Σύναψη Σκελετικός μυς - Μυϊκή Συστολή Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής, Παν. Θεσσαλίας 1 Περιεχόμενα Νευρομυϊκή σύναψη Δομή σκελετικών μυών Μοριακή βάση της
Νευρομυϊκή Σύναψη Σκελετικός μυς - Μυϊκή Συστολή Ε. Παρασκευά Αναπλ. Καθηγήτρια Κυτταρικής Φυσιολογίας Τμήμα Ιατρικής, Παν. Θεσσαλίας 1 Περιεχόμενα Νευρομυϊκή σύναψη Δομή σκελετικών μυών Μοριακή βάση της
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ. ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΣΥΣΠΑΣΗΣ
 ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
ΠΡΟΠΤΥΧΙΑΚΟ ΠΡΟΓΡΑΜΜΑ ΣΠΥΔΩΝ ΤΕΦΑΑ/ΔΠΘ ΜΑΘΗΜΑ ΚΑΤΕΥΘΥΝΣΗΣ ΠΡΟΠΟΝΗΤΙΚΗΣ ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΑΣΚΗΣΗΣ Φατούρος Γ. Ιωάννης, Επίκουρος Καθηγητής ΔΙΑΛΕΞΗ 3 - Η ΒΙΟΧΗΜΕΙΑ ΤΗΣ ΜΥΪΚΗΣ ΣΥΣΠΑΣΗΣ Βιοχημεία των νευρομυϊκών
Σκελετικός Μυς. Εγκάρσια διατοµή ζώνηςα. Σκελετικός µυς: οµή. οµή λεπτώνκαιπαχιώννηµατίων. Μυοϊνίδιο. Μυϊκές ίνες
 Σκελετικός µυς: οµή Σκελετικός Μυς Μυϊκή ίνα: µυοκύτταρο Μυς: µυϊκές ίνες συνδεδεµένες µε συνδετικό ιστό Οι µυϊκές ίνες είναι προσανατολισµένες στον επιµήκη άξονα του µυός οµή λεπτώνκαιπαχιώννηµατίων Μυϊκές
Σκελετικός µυς: οµή Σκελετικός Μυς Μυϊκή ίνα: µυοκύτταρο Μυς: µυϊκές ίνες συνδεδεµένες µε συνδετικό ιστό Οι µυϊκές ίνες είναι προσανατολισµένες στον επιµήκη άξονα του µυός οµή λεπτώνκαιπαχιώννηµατίων Μυϊκές
Phys460.nb Solution for the t-dependent Schrodinger s equation How did we find the solution? (not required)
") Phys460.nb 81 ψ n (t) is still the (same) eigenstate of H But for tdependent H. The answer is NO. 5.5.5. Solution for the tdependent Schrodinger s equation If we assume that at time t 0, the electron starts
Phys460.nb 81 ψ n (t) is still the (same) eigenstate of H But for tdependent H. The answer is NO. 5.5.5. Solution for the tdependent Schrodinger s equation If we assume that at time t 0, the electron starts
[1] P Q. Fig. 3.1
![[1] P Q. Fig. 3.1](/thumbs/79/80362156.jpg "[1] P Q. Fig. 3.1") 1 (a) Define resistance....... [1] (b) The smallest conductor within a computer processing chip can be represented as a rectangular block that is one atom high, four atoms wide and twenty atoms long. One
1 (a) Define resistance....... [1] (b) The smallest conductor within a computer processing chip can be represented as a rectangular block that is one atom high, four atoms wide and twenty atoms long. One
Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας. Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ
 2016 Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ Εισαγωγή Η σύσπαση των σκελετικών μυών ελέγχεται από
2016 Αποστολία Χατζηευθυμίου, Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Ευφροσύνη Παρασκευά, Αν. Καθηγήτρια Κυτταρικής Φυσιολογίας ΝΕΥΡΟΜΥΪΚΟ ΠΑΡΑΣΚΕΥΑΣΜΑ Εισαγωγή Η σύσπαση των σκελετικών μυών ελέγχεται από
ΜΥΪΚΟΥ ΣΥΣΤΗΜΑΤΟΣ (Β ΜΕΡΟΣ)
") ΑΝΑΤΟΜΙΑ και ΦΥΣΙΟΛΟΓΙΑ ΜΥΪΚΟΥ ΣΥΣΤΗΜΑΤΟΣ (Β ΜΕΡΟΣ) ΦΥΣΙΟΛΟΓΙΑ ΜΥΪΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Οι µύες είναι συσταλτά όργανα που χρησιµεύουν για να επιτελούνται οι ενεργητικές κινήσεις του οργανισµού. Οι µύες αποτελούνται
ΑΝΑΤΟΜΙΑ και ΦΥΣΙΟΛΟΓΙΑ ΜΥΪΚΟΥ ΣΥΣΤΗΜΑΤΟΣ (Β ΜΕΡΟΣ) ΦΥΣΙΟΛΟΓΙΑ ΜΥΪΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Οι µύες είναι συσταλτά όργανα που χρησιµεύουν για να επιτελούνται οι ενεργητικές κινήσεις του οργανισµού. Οι µύες αποτελούνται
Μυϊκή ύναµη. Γιάννης Κουτεντάκης, BSc, MA. PhD. Αναπληρωτής Καθηγητής ΤΕΦΑΑ, Πανεπιστήµιο Θεσσαλίας
 Μυϊκή ύναµη Γιάννης Κουτεντάκης, BSc, MA. PhD Αναπληρωτής Καθηγητής ΤΕΦΑΑ, Πανεπιστήµιο Θεσσαλίας Περιεχόµενο Μαθήµατος Ορισµός Μέτρηση της δύναµης και αποτελέσµατα µετρήσεων Παράγοντες που επηρεάζουν
Μυϊκή ύναµη Γιάννης Κουτεντάκης, BSc, MA. PhD Αναπληρωτής Καθηγητής ΤΕΦΑΑ, Πανεπιστήµιο Θεσσαλίας Περιεχόµενο Μαθήµατος Ορισµός Μέτρηση της δύναµης και αποτελέσµατα µετρήσεων Παράγοντες που επηρεάζουν
ΣΧΟΛΗ ΕΦΑΡΜΟΣΜΕΝΩΝ ΜΑΘΗΜΑΤΙΚΩΝ ΚΑΙ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Βιοφυσική - Βιοφυσικός μηχανισμός της μυϊκής συστολής
 ΣΧΟΛΗ ΕΦΑΡΜΟΣΜΕΝΩΝ ΜΑΘΗΜΑΤΙΚΩΝ ΚΑΙ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Βιοφυσική - Βιοφυσικός μηχανισμός της μυϊκής συστολής Ακαδ. έτος 2008-2009 - Διδάσκουσα: Μυρσίνη Μακροπούλου Μηχανικά φαινόμενα και συστολή των μυών
ΣΧΟΛΗ ΕΦΑΡΜΟΣΜΕΝΩΝ ΜΑΘΗΜΑΤΙΚΩΝ ΚΑΙ ΦΥΣΙΚΩΝ ΕΠΙΣΤΗΜΩΝ Βιοφυσική - Βιοφυσικός μηχανισμός της μυϊκής συστολής Ακαδ. έτος 2008-2009 - Διδάσκουσα: Μυρσίνη Μακροπούλου Μηχανικά φαινόμενα και συστολή των μυών
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 6/5/2006
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Ολοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα είναι μικρότεροι το 1000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Διάρκεια: 3,5 ώρες Καλή
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Ολοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα είναι μικρότεροι το 1000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Διάρκεια: 3,5 ώρες Καλή
CHAPTER 25 SOLVING EQUATIONS BY ITERATIVE METHODS
 CHAPTER 5 SOLVING EQUATIONS BY ITERATIVE METHODS EXERCISE 104 Page 8 1. Find the positive root of the equation x + 3x 5 = 0, correct to 3 significant figures, using the method of bisection. Let f(x) =
CHAPTER 5 SOLVING EQUATIONS BY ITERATIVE METHODS EXERCISE 104 Page 8 1. Find the positive root of the equation x + 3x 5 = 0, correct to 3 significant figures, using the method of bisection. Let f(x) =
+ - - εκπολώνεται. ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ
 ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ Στόχοι Κατανόησης: -Να σας είναι ξεκάθαρες οι έννοιες πόλωση, εκπόλωση, υπερπόλωση, διεγερτικό ερέθισμα, ανασταλτικό ερέθισμα, κατώφλιο δυναμικό, υποκατώφλιες εκπολώσεις, υπερκατώφλιες
ΗΛΕΚΤΡΟMYΟΓΡΑΦΗΜΑ Στόχοι Κατανόησης: -Να σας είναι ξεκάθαρες οι έννοιες πόλωση, εκπόλωση, υπερπόλωση, διεγερτικό ερέθισμα, ανασταλτικό ερέθισμα, κατώφλιο δυναμικό, υποκατώφλιες εκπολώσεις, υπερκατώφλιες
the total number of electrons passing through the lamp.
 1. A 12 V 36 W lamp is lit to normal brightness using a 12 V car battery of negligible internal resistance. The lamp is switched on for one hour (3600 s). For the time of 1 hour, calculate (i) the energy
1. A 12 V 36 W lamp is lit to normal brightness using a 12 V car battery of negligible internal resistance. The lamp is switched on for one hour (3600 s). For the time of 1 hour, calculate (i) the energy
HOMEWORK 4 = G. In order to plot the stress versus the stretch we define a normalized stretch:
 HOMEWORK 4 Problem a For the fast loading case, we want to derive the relationship between P zz and λ z. We know that the nominal stress is expressed as: P zz = ψ λ z where λ z = λ λ z. Therefore, applying
HOMEWORK 4 Problem a For the fast loading case, we want to derive the relationship between P zz and λ z. We know that the nominal stress is expressed as: P zz = ψ λ z where λ z = λ λ z. Therefore, applying
Capacitors - Capacitance, Charge and Potential Difference
 Capacitors - Capacitance, Charge and Potential Difference Capacitors store electric charge. This ability to store electric charge is known as capacitance. A simple capacitor consists of 2 parallel metal
Capacitors - Capacitance, Charge and Potential Difference Capacitors store electric charge. This ability to store electric charge is known as capacitance. A simple capacitor consists of 2 parallel metal
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Β ΔΙΑΒΙΒΑΣΗ ΣΤΗ ΝΕΥΡΟΜΥΪΚΗ ΣΥΝΑΨΗ Η νευρομυϊκή σύναψη αποτελεί ιδιαίτερη μορφή σύναψης μεταξύ του κινητικού νευρώνα και της σκελετικής μυϊκής ίνας Είναι ορατή με το οπτικό μικροσκόπιο Στην
(1) Describe the process by which mercury atoms become excited in a fluorescent tube (3)
 Describe the process by which mercury atoms become excited in a fluorescent tube (3)") Q1. (a) A fluorescent tube is filled with mercury vapour at low pressure. In order to emit electromagnetic radiation the mercury atoms must first be excited. (i) What is meant by an excited atom? (1) (ii)
Q1. (a) A fluorescent tube is filled with mercury vapour at low pressure. In order to emit electromagnetic radiation the mercury atoms must first be excited. (i) What is meant by an excited atom? (1) (ii)
Θέµατα διάλεξης ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ. Τρόποι µετάδοσης των νευρικών σηµάτων. υναµικό Ηρεµίας. Νευρώνας
 Θέµατα διάλεξης MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ Τρόποι µετάδοσης νευρικών σηµάτων Ρόλος και λειτουργία των νευροδιαβιβαστών
Θέµατα διάλεξης MANAGING AUTHORITY OF THE OPERATIONAL PROGRAMME EDUCATION AND INITIAL VOCATIONAL TRAINING ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ ΚΑΙ ΜΥΪΚΗ ΣΥΣΤΟΛΗ Τρόποι µετάδοσης νευρικών σηµάτων Ρόλος και λειτουργία των νευροδιαβιβαστών
Δυναμικό ηρεμίας Δυναμικό ενεργείας. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 30/09/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Ιόντα Δυναμικό
Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 30/09/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Ιόντα Δυναμικό
TMHMA ΙΑΤΡΙΚΗΣ - ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΦΥΣΙΟΛΟΓΙΑ Ι. Φυσιολογία Μυών. Κων/νος Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2015
 TMHMA ΙΑΤΡΙΚΗΣ - ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΦΥΣΙΟΛΟΓΙΑ Ι Φυσιολογία Μυών Κων/νος Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2015 ΣΚΕΛΕΤΙΚΟΣ ΜΥΣ ΜΥΕΣ ΜΥΪΚΟ ΚΥΤΤΑΡΟ (ΜΥΪΚΗ ΙΝΑ) ΛΕΙΤΟΥΡΓΙΕΣ & ΙΔΙΟΤΗΤΕΣ
TMHMA ΙΑΤΡΙΚΗΣ - ΠΑΝ/ΜΙΟ ΠΑΤΡΩΝ ΦΥΣΙΟΛΟΓΙΑ Ι Φυσιολογία Μυών Κων/νος Παπαθεοδωρόπουλος Αναπληρωτής Καθηγητής * Εργαστήριο Φυσιολογίας 2015 ΣΚΕΛΕΤΙΚΟΣ ΜΥΣ ΜΥΕΣ ΜΥΪΚΟ ΚΥΤΤΑΡΟ (ΜΥΪΚΗ ΙΝΑ) ΛΕΙΤΟΥΡΓΙΕΣ & ΙΔΙΟΤΗΤΕΣ
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 19/5/2007
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Αν κάπου κάνετε κάποιες υποθέσεις να αναφερθούν στη σχετική ερώτηση. Όλα τα αρχεία που αναφέρονται στα προβλήματα βρίσκονται στον ίδιο φάκελο με το εκτελέσιμο
Φυσιολογία του καρδιαγγειακού συστήματος. Κλειώ Μαυραγάνη
 Φυσιολογία του καρδιαγγειακού συστήματος Κλειώ Μαυραγάνη Σύνοψη I. Γενικές αρχές καρδιαγγειακού συστήματος- Δομή και ρόλος II. III. IV. Προέλευση του καρδιακού ρυθμού και της ηλεκτρικής δραστηριότητας
Φυσιολογία του καρδιαγγειακού συστήματος Κλειώ Μαυραγάνη Σύνοψη I. Γενικές αρχές καρδιαγγειακού συστήματος- Δομή και ρόλος II. III. IV. Προέλευση του καρδιακού ρυθμού και της ηλεκτρικής δραστηριότητας
Strain gauge and rosettes
 Strain gauge and rosettes Introduction A strain gauge is a device which is used to measure strain (deformation) on an object subjected to forces. Strain can be measured using various types of devices classified
Strain gauge and rosettes Introduction A strain gauge is a device which is used to measure strain (deformation) on an object subjected to forces. Strain can be measured using various types of devices classified
Νευροφυσιολογία και Αισθήσεις
 Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις Διάλεξη 4 Δυναμικά Ενεργείας (Action Potentials) Διεγέρσιμοι Ιστοί Οι νευρώνες και οι μύες είναι διεγέρσιμοι (excitable)
Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις Διάλεξη 4 Δυναμικά Ενεργείας (Action Potentials) Διεγέρσιμοι Ιστοί Οι νευρώνες και οι μύες είναι διεγέρσιμοι (excitable)
Section 8.3 Trigonometric Equations
 99 Section 8. Trigonometric Equations Objective 1: Solve Equations Involving One Trigonometric Function. In this section and the next, we will exple how to solving equations involving trigonometric functions.
99 Section 8. Trigonometric Equations Objective 1: Solve Equations Involving One Trigonometric Function. In this section and the next, we will exple how to solving equations involving trigonometric functions.
Instruction Execution Times
 1 C Execution Times InThisAppendix... Introduction DL330 Execution Times DL330P Execution Times DL340 Execution Times C-2 Execution Times Introduction Data Registers This appendix contains several tables
1 C Execution Times InThisAppendix... Introduction DL330 Execution Times DL330P Execution Times DL340 Execution Times C-2 Execution Times Introduction Data Registers This appendix contains several tables
Απόκριση σε Μοναδιαία Ωστική Δύναμη (Unit Impulse) Απόκριση σε Δυνάμεις Αυθαίρετα Μεταβαλλόμενες με το Χρόνο. Απόστολος Σ.
 Απόκριση σε Δυνάμεις Αυθαίρετα Μεταβαλλόμενες με το Χρόνο. Απόστολος Σ.") Απόκριση σε Δυνάμεις Αυθαίρετα Μεταβαλλόμενες με το Χρόνο The time integral of a force is referred to as impulse, is determined by and is obtained from: Newton s 2 nd Law of motion states that the action
Απόκριση σε Δυνάμεις Αυθαίρετα Μεταβαλλόμενες με το Χρόνο The time integral of a force is referred to as impulse, is determined by and is obtained from: Newton s 2 nd Law of motion states that the action
ΚΑΤΗΓΟΡΙΕΣ. 1. Μυϊκά. 2. Μυοεπιθηλιακά. 3. Περικύτταρα. 4. Μυοϊνοβλάστες
 ΣΥΣΤΑΛΤΑ ΚΥΤΤΑΡΑ ΚΑΤΗΓΟΡΙΕΣ 1. Μυϊκά 2. Μυοεπιθηλιακά 3. Περικύτταρα 4. Μυοϊνοβλάστες ΜΥΙΚΑ 1. Γραμμωτά μυϊκά κύτταρα 2. Καρδιακά μυϊκά κύτταρα 3. Λεία μυϊκά κύτταρα ΓΡΑΜΜΩΤΟΙ ΜΥΣ 1. Μύς 2. Μυϊκή δέσμη
ΣΥΣΤΑΛΤΑ ΚΥΤΤΑΡΑ ΚΑΤΗΓΟΡΙΕΣ 1. Μυϊκά 2. Μυοεπιθηλιακά 3. Περικύτταρα 4. Μυοϊνοβλάστες ΜΥΙΚΑ 1. Γραμμωτά μυϊκά κύτταρα 2. Καρδιακά μυϊκά κύτταρα 3. Λεία μυϊκά κύτταρα ΓΡΑΜΜΩΤΟΙ ΜΥΣ 1. Μύς 2. Μυϊκή δέσμη
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ
 ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
ΜΑΘΗΜΑ 3ο ΜΕΡΟΣ Α ΣΥΝΑΠΤΙΚΗ ΟΛΟΚΛΗΡΩΣΗ Όπως συμβαίνει με τη συναπτική διαβίβαση στη νευρομυϊκή σύναψη, σε πολλές μορφές επικοινωνίας μεταξύ νευρώνων στο κεντρικό νευρικό σύστημα παρεμβαίνουν άμεσα ελεγχόμενοι
Βιοχημεία Τροφίμων Ι. Ενότητα 2 η Κρέας και ψάρι II. Όνομα καθηγητή: Έφη Τσακαλίδου. Τμήμα: Επιστήμης Τροφίμων & Διατροφής του Ανθρώπου
 Βιοχημεία Τροφίμων Ι Ενότητα 2 η Κρέας και ψάρι II Όνομα καθηγητή: Έφη Τσακαλίδου Τμήμα: Επιστήμης Τροφίμων & Διατροφής του Ανθρώπου Στόχοι ενότητας Κατανόηση της δομής του σαρκομερούς Κατανόηση της δομής
Βιοχημεία Τροφίμων Ι Ενότητα 2 η Κρέας και ψάρι II Όνομα καθηγητή: Έφη Τσακαλίδου Τμήμα: Επιστήμης Τροφίμων & Διατροφής του Ανθρώπου Στόχοι ενότητας Κατανόηση της δομής του σαρκομερούς Κατανόηση της δομής
Homework 3 Solutions
 Homework 3 Solutions Igor Yanovsky (Math 151A TA) Problem 1: Compute the absolute error and relative error in approximations of p by p. (Use calculator!) a) p π, p 22/7; b) p π, p 3.141. Solution: For
Homework 3 Solutions Igor Yanovsky (Math 151A TA) Problem 1: Compute the absolute error and relative error in approximations of p by p. (Use calculator!) a) p π, p 22/7; b) p π, p 3.141. Solution: For
Δομή των μυϊκών κυττάρων.
 Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
Δομή των μυϊκών κυττάρων. Οι μύες αποτελούνται από δεμάτια μεγάλων κυττάρων (που ονομάζονται μυϊκά κύτταρα ή μυϊκές ίνες). Κάθε μυϊκή ίνα περιέχει πολλά μυϊκά ινίδια, δηλαδή δεμάτια ινιδίων ακτίνης και
Αναερόβια Φυσική Κατάσταση
 Αναερόβια Φυσική Κατάσταση Γιάννης Κουτεντάκης, BSc, MA. PhD Αναπληρωτής Καθηγητής ΤΕΦΑΑ, Πανεπιστήµιο Θεσσαλίας Περιεχόµενο Μαθήµατος Ορισµός της αναερόβιας φυσικής κατάστασης Σχέσης µε µηχανισµούς παραγωγής
Αναερόβια Φυσική Κατάσταση Γιάννης Κουτεντάκης, BSc, MA. PhD Αναπληρωτής Καθηγητής ΤΕΦΑΑ, Πανεπιστήµιο Θεσσαλίας Περιεχόµενο Μαθήµατος Ορισµός της αναερόβιας φυσικής κατάστασης Σχέσης µε µηχανισµούς παραγωγής
Démographie spatiale/spatial Demography
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Démographie spatiale/spatial Demography Session 1: Introduction to spatial demography Basic concepts Michail Agorastakis Department of Planning & Regional Development Άδειες Χρήσης
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ Démographie spatiale/spatial Demography Session 1: Introduction to spatial demography Basic concepts Michail Agorastakis Department of Planning & Regional Development Άδειες Χρήσης
ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ
 ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
ΜΕΡΟΣ ΔΕΥΤΕΡΟ ΚΕΦΑΛΑΙΟ ΕΝΑΤΟ ΦΥΣΙΟΛΟΓΙΑ ΤΟΥ ΝΕΥΡΙΚΟΥ ΚΑΙ ΜΥΙΚΟΥ ΣΥΣΤΗΜΑΤΟΣ Σημειώσεις Ανατομίας - Φυσιολογίας Ι Σκοπός της λειτουργίας του νευρικού συστήματος Προσαρμόζει τις λειτουργίες του ανθρώπινου
Main source: "Discrete-time systems and computer control" by Α. ΣΚΟΔΡΑΣ ΨΗΦΙΑΚΟΣ ΕΛΕΓΧΟΣ ΔΙΑΛΕΞΗ 4 ΔΙΑΦΑΝΕΙΑ 1
 Main source: "Discrete-time systems and computer control" by Α. ΣΚΟΔΡΑΣ ΨΗΦΙΑΚΟΣ ΕΛΕΓΧΟΣ ΔΙΑΛΕΞΗ 4 ΔΙΑΦΑΝΕΙΑ 1 A Brief History of Sampling Research 1915 - Edmund Taylor Whittaker (1873-1956) devised a
Main source: "Discrete-time systems and computer control" by Α. ΣΚΟΔΡΑΣ ΨΗΦΙΑΚΟΣ ΕΛΕΓΧΟΣ ΔΙΑΛΕΞΗ 4 ΔΙΑΦΑΝΕΙΑ 1 A Brief History of Sampling Research 1915 - Edmund Taylor Whittaker (1873-1956) devised a
ST5224: Advanced Statistical Theory II
 ST5224: Advanced Statistical Theory II 2014/2015: Semester II Tutorial 7 1. Let X be a sample from a population P and consider testing hypotheses H 0 : P = P 0 versus H 1 : P = P 1, where P j is a known
ST5224: Advanced Statistical Theory II 2014/2015: Semester II Tutorial 7 1. Let X be a sample from a population P and consider testing hypotheses H 0 : P = P 0 versus H 1 : P = P 1, where P j is a known
Εφαρμοσμένη Βιοτεχνολογία Σημειώσεις. Νίκος Τσουκιάς Σχολή Χημικών Μηχανικών ΕΜΠ
 Εφαρμοσμένη Βιοτεχνολογία Σημειώσεις Νίκος Τσουκιάς Σχολή Χημικών Μηχανικών ΕΜΠ Μετάδοση σημάτων [4.7] - Μηχανισμοί μεταφοράς μορίων μέσω κυτταρικών μεμβρανών (είσοδος θρεπτικών ουσιών - αίσθηση εξωκυτταρικού
Εφαρμοσμένη Βιοτεχνολογία Σημειώσεις Νίκος Τσουκιάς Σχολή Χημικών Μηχανικών ΕΜΠ Μετάδοση σημάτων [4.7] - Μηχανισμοί μεταφοράς μορίων μέσω κυτταρικών μεμβρανών (είσοδος θρεπτικών ουσιών - αίσθηση εξωκυτταρικού
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ
 Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
Κεφάλαιο 1 ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΝΕΥΡΟΦΥΣΙΟΛΟΓΙΑΣ 1.1. Εισαγωγή Ο ζωντανός οργανισµός έχει την ικανότητα να αντιδρά σε µεταβολές που συµβαίνουν στο περιβάλλον και στο εσωτερικό του. Οι µεταβολές αυτές ονοµάζονται
Section 9.2 Polar Equations and Graphs
 180 Section 9. Polar Equations and Graphs In this section, we will be graphing polar equations on a polar grid. In the first few examples, we will write the polar equation in rectangular form to help identify
180 Section 9. Polar Equations and Graphs In this section, we will be graphing polar equations on a polar grid. In the first few examples, we will write the polar equation in rectangular form to help identify
C.S. 430 Assignment 6, Sample Solutions
 C.S. 430 Assignment 6, Sample Solutions Paul Liu November 15, 2007 Note that these are sample solutions only; in many cases there were many acceptable answers. 1 Reynolds Problem 10.1 1.1 Normal-order
C.S. 430 Assignment 6, Sample Solutions Paul Liu November 15, 2007 Note that these are sample solutions only; in many cases there were many acceptable answers. 1 Reynolds Problem 10.1 1.1 Normal-order
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΑ ΤΜΗΜΑ ΝΑΥΤΙΛΙΑΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΝΑΥΤΙΛΙΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΑ ΤΜΗΜΑ ΝΑΥΤΙΛΙΑΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΝΑΥΤΙΛΙΑ ΝΟΜΙΚΟ ΚΑΙ ΘΕΣΜΙΚΟ ΦΟΡΟΛΟΓΙΚΟ ΠΛΑΙΣΙΟ ΚΤΗΣΗΣ ΚΑΙ ΕΚΜΕΤΑΛΛΕΥΣΗΣ ΠΛΟΙΟΥ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ που υποβλήθηκε στο
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΕΙΡΑΙΑ ΤΜΗΜΑ ΝΑΥΤΙΛΙΑΚΩΝ ΣΠΟΥΔΩΝ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΣΤΗΝ ΝΑΥΤΙΛΙΑ ΝΟΜΙΚΟ ΚΑΙ ΘΕΣΜΙΚΟ ΦΟΡΟΛΟΓΙΚΟ ΠΛΑΙΣΙΟ ΚΤΗΣΗΣ ΚΑΙ ΕΚΜΕΤΑΛΛΕΥΣΗΣ ΠΛΟΙΟΥ ΔΙΠΛΩΜΑΤΙΚΗ ΕΡΓΑΣΙΑ που υποβλήθηκε στο
Calculating the propagation delay of coaxial cable
 Your source for quality GNSS Networking Solutions and Design Services! Page 1 of 5 Calculating the propagation delay of coaxial cable The delay of a cable or velocity factor is determined by the dielectric
Your source for quality GNSS Networking Solutions and Design Services! Page 1 of 5 Calculating the propagation delay of coaxial cable The delay of a cable or velocity factor is determined by the dielectric
Διαταραχές του κύματος P και του φλεβοκόμβου
 Διαταραχές του κύματος P και του φλεβοκόμβου Φροντιστηριακό μάθημα ΗΚΓ Δημήτρης Ασβεστάς Συνεργάτης «Ευρωκλινική Αθηνών» Συνεργάτης ΓΝΑ Ευαγγελισμός Φλεβοκομβική ταχυκαρδία ηρεμίας Δεν είναι αποτέλεσμα
Διαταραχές του κύματος P και του φλεβοκόμβου Φροντιστηριακό μάθημα ΗΚΓ Δημήτρης Ασβεστάς Συνεργάτης «Ευρωκλινική Αθηνών» Συνεργάτης ΓΝΑ Ευαγγελισμός Φλεβοκομβική ταχυκαρδία ηρεμίας Δεν είναι αποτέλεσμα
Assalamu `alaikum wr. wb.
 LUMP SUM Assalamu `alaikum wr. wb. LUMP SUM Wassalamu alaikum wr. wb. Assalamu `alaikum wr. wb. LUMP SUM Wassalamu alaikum wr. wb. LUMP SUM Lump sum lump sum lump sum. lump sum fixed price lump sum lump
LUMP SUM Assalamu `alaikum wr. wb. LUMP SUM Wassalamu alaikum wr. wb. Assalamu `alaikum wr. wb. LUMP SUM Wassalamu alaikum wr. wb. LUMP SUM Lump sum lump sum lump sum. lump sum fixed price lump sum lump
derivation of the Laplacian from rectangular to spherical coordinates
 derivation of the Laplacian from rectangular to spherical coordinates swapnizzle 03-03- :5:43 We begin by recognizing the familiar conversion from rectangular to spherical coordinates (note that φ is used
derivation of the Laplacian from rectangular to spherical coordinates swapnizzle 03-03- :5:43 We begin by recognizing the familiar conversion from rectangular to spherical coordinates (note that φ is used
Βιοϊατρική τεχνολογία
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοϊατρική τεχνολογία Ενότητα 5: Νευρικό & μυϊκό σύστημα (Part I) Ηλεκτρομυογράφημα Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοϊατρική τεχνολογία Ενότητα 5: Νευρικό & μυϊκό σύστημα (Part I) Ηλεκτρομυογράφημα Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου
3.4 SUM AND DIFFERENCE FORMULAS. NOTE: cos(α+β) cos α + cos β cos(α-β) cos α -cos β
 cos α + cos β cos(α-β) cos α -cos β") 3.4 SUM AND DIFFERENCE FORMULAS Page Theorem cos(αβ cos α cos β -sin α cos(α-β cos α cos β sin α NOTE: cos(αβ cos α cos β cos(α-β cos α -cos β Proof of cos(α-β cos α cos β sin α Let s use a unit circle
3.4 SUM AND DIFFERENCE FORMULAS Page Theorem cos(αβ cos α cos β -sin α cos(α-β cos α cos β sin α NOTE: cos(αβ cos α cos β cos(α-β cos α -cos β Proof of cos(α-β cos α cos β sin α Let s use a unit circle
Physical DB Design. B-Trees Index files can become quite large for large main files Indices on index files are possible.
 B-Trees Index files can become quite large for large main files Indices on index files are possible 3 rd -level index 2 nd -level index 1 st -level index Main file 1 The 1 st -level index consists of pairs
B-Trees Index files can become quite large for large main files Indices on index files are possible 3 rd -level index 2 nd -level index 1 st -level index Main file 1 The 1 st -level index consists of pairs
Φυσιολογία της Άσκησης Μυϊκό σύστημα-δομή & λειτουργία. Παναγιώτης Κανέλλος Διαιτολόγος-Διατροφολόγος, PhD Υπότροφος ΤΕΙ Κρήτης
 Φυσιολογία της Άσκησης Μυϊκό σύστημα-δομή & λειτουργία Παναγιώτης Κανέλλος Διαιτολόγος-Διατροφολόγος, PhD Υπότροφος ΤΕΙ Κρήτης 2019 1 Μυϊκό σύστημα Σύστημα οργάνων που ελέγχει τις κινήσεις του σώματος
Φυσιολογία της Άσκησης Μυϊκό σύστημα-δομή & λειτουργία Παναγιώτης Κανέλλος Διαιτολόγος-Διατροφολόγος, PhD Υπότροφος ΤΕΙ Κρήτης 2019 1 Μυϊκό σύστημα Σύστημα οργάνων που ελέγχει τις κινήσεις του σώματος
Reaction of a Platinum Electrode for the Measurement of Redox Potential of Paddy Soil
 J. Jpn. Soc. Soil Phys. No. +*0, p.- +*,**1 Eh * ** Reaction of a Platinum Electrode for the Measurement of Redox Potential of Paddy Soil Daisuke MURAKAMI* and Tatsuaki KASUBUCHI** * The United Graduate
J. Jpn. Soc. Soil Phys. No. +*0, p.- +*,**1 Eh * ** Reaction of a Platinum Electrode for the Measurement of Redox Potential of Paddy Soil Daisuke MURAKAMI* and Tatsuaki KASUBUCHI** * The United Graduate
Potential Dividers. 46 minutes. 46 marks. Page 1 of 11
 Potential Dividers 46 minutes 46 marks Page 1 of 11 Q1. In the circuit shown in the figure below, the battery, of negligible internal resistance, has an emf of 30 V. The pd across the lamp is 6.0 V and
Potential Dividers 46 minutes 46 marks Page 1 of 11 Q1. In the circuit shown in the figure below, the battery, of negligible internal resistance, has an emf of 30 V. The pd across the lamp is 6.0 V and
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΥΠΟΛΟΓΙΣΤΩΝ ΤΟΜΕΑΣ ΣΥΣΤΗΜΑΤΩΝ ΗΛΕΚΤΡΙΚΗΣ ΕΝΕΡΓΕΙΑΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΥΠΟΛΟΓΙΣΤΩΝ ΤΟΜΕΑΣ ΣΥΣΤΗΜΑΤΩΝ ΗΛΕΚΤΡΙΚΗΣ ΕΝΕΡΓΕΙΑΣ ιπλωµατική Εργασία του φοιτητή του τµήµατος Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Ηλεκτρονικών
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΥΠΟΛΟΓΙΣΤΩΝ ΤΟΜΕΑΣ ΣΥΣΤΗΜΑΤΩΝ ΗΛΕΚΤΡΙΚΗΣ ΕΝΕΡΓΕΙΑΣ ιπλωµατική Εργασία του φοιτητή του τµήµατος Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Ηλεκτρονικών
The Simply Typed Lambda Calculus
 Type Inference Instead of writing type annotations, can we use an algorithm to infer what the type annotations should be? That depends on the type system. For simple type systems the answer is yes, and
Type Inference Instead of writing type annotations, can we use an algorithm to infer what the type annotations should be? That depends on the type system. For simple type systems the answer is yes, and
Νευροφυσιολογία και Αισθήσεις
 Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις ιάλεξη 14 Κίνηση από τον Νωτιαίο Μυελό (Spinal Control of Movement) Εισαγωγή Κινητικά Συστήµατα Μύες και νευρώνες
Biomedical Imaging & Applied Optics University of Cyprus Νευροφυσιολογία και Αισθήσεις ιάλεξη 14 Κίνηση από τον Νωτιαίο Μυελό (Spinal Control of Movement) Εισαγωγή Κινητικά Συστήµατα Μύες και νευρώνες
ANSWERSHEET (TOPIC = DIFFERENTIAL CALCULUS) COLLECTION #2. h 0 h h 0 h h 0 ( ) g k = g 0 + g 1 + g g 2009 =?
 COLLECTION #2. h 0 h h 0 h h 0 ( ) g k = g 0 + g 1 + g g 2009 =?") Teko Classes IITJEE/AIEEE Maths by SUHAAG SIR, Bhopal, Ph (0755) 3 00 000 www.tekoclasses.com ANSWERSHEET (TOPIC DIFFERENTIAL CALCULUS) COLLECTION # Question Type A.Single Correct Type Q. (A) Sol least
Teko Classes IITJEE/AIEEE Maths by SUHAAG SIR, Bhopal, Ph (0755) 3 00 000 www.tekoclasses.com ANSWERSHEET (TOPIC DIFFERENTIAL CALCULUS) COLLECTION # Question Type A.Single Correct Type Q. (A) Sol least
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Φυσιολογία Ι Μυϊκή συστολή Διδάσκων: Αν. Καθηγήτρια Πατρώνα Βεζυράκη Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Φυσιολογία Ι Μυϊκή συστολή Διδάσκων: Αν. Καθηγήτρια Πατρώνα Βεζυράκη Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό υπόκειται σε άδειες χρήσης Creative Commons.
EE512: Error Control Coding
 EE512: Error Control Coding Solution for Assignment on Finite Fields February 16, 2007 1. (a) Addition and Multiplication tables for GF (5) and GF (7) are shown in Tables 1 and 2. + 0 1 2 3 4 0 0 1 2 3
EE512: Error Control Coding Solution for Assignment on Finite Fields February 16, 2007 1. (a) Addition and Multiplication tables for GF (5) and GF (7) are shown in Tables 1 and 2. + 0 1 2 3 4 0 0 1 2 3
Μυϊκή ύναµη, Τραυµατισµοί & Μυϊκή Καταστροφή
 Μυϊκή ύναµη, Τραυµατισµοί & Μυϊκή Καταστροφή Γιάννης Κουτεντάκης, BSc, MA. PhD Αναπληρωτής Καθηγητής ΤΕΦΑΑ, Πανεπιστήµιο Θεσσαλίας Περιεχόµενο Μαθήµατος Έλλειµµα δύναµης ή µ. ανισορροπίες αποτελούν την
Μυϊκή ύναµη, Τραυµατισµοί & Μυϊκή Καταστροφή Γιάννης Κουτεντάκης, BSc, MA. PhD Αναπληρωτής Καθηγητής ΤΕΦΑΑ, Πανεπιστήµιο Θεσσαλίας Περιεχόµενο Μαθήµατος Έλλειµµα δύναµης ή µ. ανισορροπίες αποτελούν την
Matrices and Determinants
 Matrices and Determinants SUBJECTIVE PROBLEMS: Q 1. For what value of k do the following system of equations possess a non-trivial (i.e., not all zero) solution over the set of rationals Q? x + ky + 3z
Matrices and Determinants SUBJECTIVE PROBLEMS: Q 1. For what value of k do the following system of equations possess a non-trivial (i.e., not all zero) solution over the set of rationals Q? x + ky + 3z
Policy Coherence. JEL Classification : J12, J13, J21 Key words :
 ** 80%1.89 2005 7 35 Policy Coherence JEL Classification : J12, J13, J21 Key words : ** Family Life and Family Policy in France and Germany: Implications for Japan By Tomoko Hayashi and Rieko Tamefuji
** 80%1.89 2005 7 35 Policy Coherence JEL Classification : J12, J13, J21 Key words : ** Family Life and Family Policy in France and Germany: Implications for Japan By Tomoko Hayashi and Rieko Tamefuji
CHAPTER 48 APPLICATIONS OF MATRICES AND DETERMINANTS
 CHAPTER 48 APPLICATIONS OF MATRICES AND DETERMINANTS EXERCISE 01 Page 545 1. Use matrices to solve: 3x + 4y x + 5y + 7 3x + 4y x + 5y 7 Hence, 3 4 x 0 5 y 7 The inverse of 3 4 5 is: 1 5 4 1 5 4 15 8 3
CHAPTER 48 APPLICATIONS OF MATRICES AND DETERMINANTS EXERCISE 01 Page 545 1. Use matrices to solve: 3x + 4y x + 5y + 7 3x + 4y x + 5y 7 Hence, 3 4 x 0 5 y 7 The inverse of 3 4 5 is: 1 5 4 1 5 4 15 8 3
Αποστολία Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2016
 Αποστολία Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2016 Δομή Μοριακοί μηχανισμοί συστολής Νευρομυϊκή σύναψη Μηχανική συστολής απλής ίνας Ενεργειακός μεταβολισμός σκελετικού μυός Τύποι
Αποστολία Χατζηευθυμίου Αν. Καθηγήτρια Ιατρικής Φυσιολογίας Φεβρουάριος 2016 Δομή Μοριακοί μηχανισμοί συστολής Νευρομυϊκή σύναψη Μηχανική συστολής απλής ίνας Ενεργειακός μεταβολισμός σκελετικού μυός Τύποι
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α. Ο ηλεκτρονικός υπολογιστής του οργανισμού μας
 ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
ΝΕΥΡΙΚΟ ΣΥΣΤΗΜΑ - ΜΕΡΟΣ Α Ο ηλεκτρονικός υπολογιστής του οργανισμού μας Ρόλος του νευρικού συστήματος Το νευρικό σύστημα (Ν.Σ.) ελέγχει, ρυθμίζει και συντονίζει όλες τις λειτουργίες του οργανισμού ανάλογα
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΥΠΟΛΟΓΙΣΤΩΝ ΤΟΜΕΑΣ ΣΥΣΤΗΜΑΤΩΝ ΗΛΕΚΤΡΙΚΗΣ ΕΝΕΡΓΕΙΑΣ
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΥΠΟΛΟΓΙΣΤΩΝ ΤΟΜΕΑΣ ΣΥΣΤΗΜΑΤΩΝ ΗΛΕΚΤΡΙΚΗΣ ΕΝΕΡΓΕΙΑΣ Διπλωματική Εργασία του φοιτητή του τμήματος Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Ηλεκτρονικών
ΠΑΝΕΠΙΣΤΗΜΙΟ ΠΑΤΡΩΝ ΤΜΗΜΑ ΗΛΕΚΤΡΟΛΟΓΩΝ ΜΗΧΑΝΙΚΩΝ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΥΠΟΛΟΓΙΣΤΩΝ ΤΟΜΕΑΣ ΣΥΣΤΗΜΑΤΩΝ ΗΛΕΚΤΡΙΚΗΣ ΕΝΕΡΓΕΙΑΣ Διπλωματική Εργασία του φοιτητή του τμήματος Ηλεκτρολόγων Μηχανικών και Τεχνολογίας Ηλεκτρονικών
Approximation of distance between locations on earth given by latitude and longitude
 Approximation of distance between locations on earth given by latitude and longitude Jan Behrens 2012-12-31 In this paper we shall provide a method to approximate distances between two points on earth
Approximation of distance between locations on earth given by latitude and longitude Jan Behrens 2012-12-31 In this paper we shall provide a method to approximate distances between two points on earth
2 Composition. Invertible Mappings
 Arkansas Tech University MATH 4033: Elementary Modern Algebra Dr. Marcel B. Finan Composition. Invertible Mappings In this section we discuss two procedures for creating new mappings from old ones, namely,
Arkansas Tech University MATH 4033: Elementary Modern Algebra Dr. Marcel B. Finan Composition. Invertible Mappings In this section we discuss two procedures for creating new mappings from old ones, namely,
DESIGN OF MACHINERY SOLUTION MANUAL h in h 4 0.
 DESIGN OF MACHINERY SOLUTION MANUAL -7-1! PROBLEM -7 Statement: Design a double-dwell cam to move a follower from to 25 6, dwell for 12, fall 25 and dwell for the remader The total cycle must take 4 sec
DESIGN OF MACHINERY SOLUTION MANUAL -7-1! PROBLEM -7 Statement: Design a double-dwell cam to move a follower from to 25 6, dwell for 12, fall 25 and dwell for the remader The total cycle must take 4 sec
Block Ciphers Modes. Ramki Thurimella
 Block Ciphers Modes Ramki Thurimella Only Encryption I.e. messages could be modified Should not assume that nonsensical messages do no harm Always must be combined with authentication 2 Padding Must be
Block Ciphers Modes Ramki Thurimella Only Encryption I.e. messages could be modified Should not assume that nonsensical messages do no harm Always must be combined with authentication 2 Padding Must be
Section 1: Listening and responding. Presenter: Niki Farfara MGTAV VCE Seminar 7 August 2016
 Section 1: Listening and responding Presenter: Niki Farfara MGTAV VCE Seminar 7 August 2016 Section 1: Listening and responding Section 1: Listening and Responding/ Aκουστική εξέταση Στο πρώτο μέρος της
Section 1: Listening and responding Presenter: Niki Farfara MGTAV VCE Seminar 7 August 2016 Section 1: Listening and responding Section 1: Listening and Responding/ Aκουστική εξέταση Στο πρώτο μέρος της
Surface Mount Multilayer Chip Capacitors for Commodity Solutions
 Surface Mount Multilayer Chip Capacitors for Commodity Solutions Below tables are test procedures and requirements unless specified in detail datasheet. 1) Visual and mechanical 2) Capacitance 3) Q/DF
Surface Mount Multilayer Chip Capacitors for Commodity Solutions Below tables are test procedures and requirements unless specified in detail datasheet. 1) Visual and mechanical 2) Capacitance 3) Q/DF
Example Sheet 3 Solutions
 Example Sheet 3 Solutions. i Regular Sturm-Liouville. ii Singular Sturm-Liouville mixed boundary conditions. iii Not Sturm-Liouville ODE is not in Sturm-Liouville form. iv Regular Sturm-Liouville note
Example Sheet 3 Solutions. i Regular Sturm-Liouville. ii Singular Sturm-Liouville mixed boundary conditions. iii Not Sturm-Liouville ODE is not in Sturm-Liouville form. iv Regular Sturm-Liouville note
Other Test Constructions: Likelihood Ratio & Bayes Tests
 Other Test Constructions: Likelihood Ratio & Bayes Tests Side-Note: So far we have seen a few approaches for creating tests such as Neyman-Pearson Lemma ( most powerful tests of H 0 : θ = θ 0 vs H 1 :
Other Test Constructions: Likelihood Ratio & Bayes Tests Side-Note: So far we have seen a few approaches for creating tests such as Neyman-Pearson Lemma ( most powerful tests of H 0 : θ = θ 0 vs H 1 :
Code Breaker. TEACHER s NOTES
 TEACHER s NOTES Time: 50 minutes Learning Outcomes: To relate the genetic code to the assembly of proteins To summarize factors that lead to different types of mutations To distinguish among positive,
TEACHER s NOTES Time: 50 minutes Learning Outcomes: To relate the genetic code to the assembly of proteins To summarize factors that lead to different types of mutations To distinguish among positive,
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ
 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΨΥΧΟΛΟΓΙΚΕΣ ΕΠΙΠΤΩΣΕΙΣ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ ΓΕΩΡΓΙΑ ΤΡΙΣΟΚΚΑ Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ
ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΤΜΗΜΑ ΝΟΣΗΛΕΥΤΙΚΗΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΨΥΧΟΛΟΓΙΚΕΣ ΕΠΙΠΤΩΣΕΙΣ ΣΕ ΓΥΝΑΙΚΕΣ ΜΕΤΑ ΑΠΟ ΜΑΣΤΕΚΤΟΜΗ ΓΕΩΡΓΙΑ ΤΡΙΣΟΚΚΑ Λευκωσία 2012 ΤΕΧΝΟΛΟΓΙΚΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΥΠΡΟΥ ΣΧΟΛΗ ΕΠΙΣΤΗΜΩΝ
Areas and Lengths in Polar Coordinates
 Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
Right Rear Door. Let's now finish the door hinge saga with the right rear door
 Right Rear Door Let's now finish the door hinge saga with the right rear door You may have been already guessed my steps, so there is not much to describe in detail. Old upper one file:///c /Documents
Right Rear Door Let's now finish the door hinge saga with the right rear door You may have been already guessed my steps, so there is not much to describe in detail. Old upper one file:///c /Documents
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ
 BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΜΕΡΟΣ ΠΡΩΤΟ - ΠΕΡΙΓΡΑΜΜΑ 5.1 ΈΝΑ ΝΟΥΚΛΕΙΝΙΚΟ ΟΞΥ ΑΠΟΤΕΛΕΙΤΑΙ ΑΠΌ ΤΕΣΣΕΡΑ ΕΙΔΗ ΒΑΣΕΩΝ, ΠΟΥ ΠΡΟΣΔΕΝΟΝΤΑΙ ΣΕ ΈΝΑ ΚΟΡΜΟ ΣΑΚΧΑΡΩΝ ΦΩΣΦΟΡΙΚΩΝ 5.2 ΈΝΑ ΖΕΥΓΟΣ
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΜΕΡΟΣ ΠΡΩΤΟ - ΠΕΡΙΓΡΑΜΜΑ 5.1 ΈΝΑ ΝΟΥΚΛΕΙΝΙΚΟ ΟΞΥ ΑΠΟΤΕΛΕΙΤΑΙ ΑΠΌ ΤΕΣΣΕΡΑ ΕΙΔΗ ΒΑΣΕΩΝ, ΠΟΥ ΠΡΟΣΔΕΝΟΝΤΑΙ ΣΕ ΈΝΑ ΚΟΡΜΟ ΣΑΚΧΑΡΩΝ ΦΩΣΦΟΡΙΚΩΝ 5.2 ΈΝΑ ΖΕΥΓΟΣ
ΑΚΑ ΗΜΙΑ ΕΜΠΟΡΙΚΟΥ ΝΑΥΤΙΚΟΥ ΜΑΚΕ ΟΝΙΑΣ ΣΧΟΛΗ ΜΗΧΑΝΙΚΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ
 ΑΚΑ ΗΜΙΑ ΕΜΠΟΡΙΚΟΥ ΝΑΥΤΙΚΟΥ ΜΑΚΕ ΟΝΙΑΣ ΣΧΟΛΗ ΜΗΧΑΝΙΚΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΘΕΜΑ :ΤΥΠΟΙ ΑΕΡΟΣΥΜΠΙΕΣΤΩΝ ΚΑΙ ΤΡΟΠΟΙ ΛΕΙΤΟΥΡΓΙΑΣ ΣΠΟΥ ΑΣΤΡΙΑ: ΕΥΘΥΜΙΑ ΟΥ ΣΩΣΑΝΝΑ ΕΠΙΒΛΕΠΩΝ ΚΑΘΗΓΗΤΗΣ : ΓΟΥΛΟΠΟΥΛΟΣ ΑΘΑΝΑΣΙΟΣ 1 ΑΚΑ
ΑΚΑ ΗΜΙΑ ΕΜΠΟΡΙΚΟΥ ΝΑΥΤΙΚΟΥ ΜΑΚΕ ΟΝΙΑΣ ΣΧΟΛΗ ΜΗΧΑΝΙΚΩΝ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ ΘΕΜΑ :ΤΥΠΟΙ ΑΕΡΟΣΥΜΠΙΕΣΤΩΝ ΚΑΙ ΤΡΟΠΟΙ ΛΕΙΤΟΥΡΓΙΑΣ ΣΠΟΥ ΑΣΤΡΙΑ: ΕΥΘΥΜΙΑ ΟΥ ΣΩΣΑΝΝΑ ΕΠΙΒΛΕΠΩΝ ΚΑΘΗΓΗΤΗΣ : ΓΟΥΛΟΠΟΥΛΟΣ ΑΘΑΝΑΣΙΟΣ 1 ΑΚΑ
Areas and Lengths in Polar Coordinates
 Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
Kiryl Tsishchanka Areas and Lengths in Polar Coordinates In this section we develop the formula for the area of a region whose boundary is given by a polar equation. We need to use the formula for the
Τ.Ε.Ι. ΔΥΤΙΚΗΣ ΜΑΚΕΔΟΝΙΑΣ ΠΑΡΑΡΤΗΜΑ ΚΑΣΤΟΡΙΑΣ ΤΜΗΜΑ ΔΗΜΟΣΙΩΝ ΣΧΕΣΕΩΝ & ΕΠΙΚΟΙΝΩΝΙΑΣ
 Τ.Ε.Ι. ΔΥΤΙΚΗΣ ΜΑΚΕΔΟΝΙΑΣ ΠΑΡΑΡΤΗΜΑ ΚΑΣΤΟΡΙΑΣ ΤΜΗΜΑ ΔΗΜΟΣΙΩΝ ΣΧΕΣΕΩΝ & ΕΠΙΚΟΙΝΩΝΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η προβολή επιστημονικών θεμάτων από τα ελληνικά ΜΜΕ : Η κάλυψή τους στον ελληνικό ημερήσιο τύπο Σαραλιώτου
Τ.Ε.Ι. ΔΥΤΙΚΗΣ ΜΑΚΕΔΟΝΙΑΣ ΠΑΡΑΡΤΗΜΑ ΚΑΣΤΟΡΙΑΣ ΤΜΗΜΑ ΔΗΜΟΣΙΩΝ ΣΧΕΣΕΩΝ & ΕΠΙΚΟΙΝΩΝΙΑΣ ΠΤΥΧΙΑΚΗ ΕΡΓΑΣΙΑ Η προβολή επιστημονικών θεμάτων από τα ελληνικά ΜΜΕ : Η κάλυψή τους στον ελληνικό ημερήσιο τύπο Σαραλιώτου
µυoϊvιδίoυ (ηλειτoυργικήµovάδα) βρίσκεται µεταξύ δύo τέτoιωv εγκάρσιωv γραµµώσεωv (πoυ ovoµάζovταιδίσκoιζ) καιλέγεταισαρκoµερίδιo.
 βρίσκεται µεταξύ δύo τέτoιωv εγκάρσιωv γραµµώσεωv (πoυ ovoµάζovταιδίσκoιζ) καιλέγεταισαρκoµερίδιo.") ΜΥIΚΕΣ ΠΡΩΤΕΪΝΕΣ (Συστήµατασυστoλήςκαικίvησης) Μovoκύτταρoι oργαvισµoύς µαστίγια και oι βλεφαρίδες Ζώα τo µυϊκό σύστηµα. Σκελετικoί µύες απoτελoύvται από µυϊκές δέσµες και αυτές από επιµηκυσµέvα κύτταρα,
ΜΥIΚΕΣ ΠΡΩΤΕΪΝΕΣ (Συστήµατασυστoλήςκαικίvησης) Μovoκύτταρoι oργαvισµoύς µαστίγια και oι βλεφαρίδες Ζώα τo µυϊκό σύστηµα. Σκελετικoί µύες απoτελoύvται από µυϊκές δέσµες και αυτές από επιµηκυσµέvα κύτταρα,
Βιοϊατρική τεχνολογία
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοϊατρική τεχνολογία Ενότητα 6: Νευρικό & μυϊκό σύστημα (Part II) Ηλεκτρομυογράφημα Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοϊατρική τεχνολογία Ενότητα 6: Νευρικό & μυϊκό σύστημα (Part II) Ηλεκτρομυογράφημα Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου
ΔΘΝΗΚΖ ΥΟΛΖ ΓΖΜΟΗΑ ΓΗΟΗΚΖΖ
 Ε ΔΘΝΗΚΖ ΥΟΛΖ ΓΖΜΟΗΑ ΓΗΟΗΚΖΖ Κ ΔΚΠΑΗΓΔΤΣΗΚΖ ΔΗΡΑ ΣΜΖΜΑ : Σνπξηζηηθήο Οηθνλνκίαο θαη Αλάπηπμεο (ΣΟΑ) ΣΔΛΗΚΖ ΔΡΓΑΗΑ Θέκα: Σνπξηζκφο θαη Οηθνλνκηθή Κξίζε Δπηβιέπσλ : Νηνχβαο Λνπθάο πνπδάζηξηα : Σζαγθαξάθε
Ε ΔΘΝΗΚΖ ΥΟΛΖ ΓΖΜΟΗΑ ΓΗΟΗΚΖΖ Κ ΔΚΠΑΗΓΔΤΣΗΚΖ ΔΗΡΑ ΣΜΖΜΑ : Σνπξηζηηθήο Οηθνλνκίαο θαη Αλάπηπμεο (ΣΟΑ) ΣΔΛΗΚΖ ΔΡΓΑΗΑ Θέκα: Σνπξηζκφο θαη Οηθνλνκηθή Κξίζε Δπηβιέπσλ : Νηνχβαο Λνπθάο πνπδάζηξηα : Σζαγθαξάθε
Modbus basic setup notes for IO-Link AL1xxx Master Block
 n Modbus has four tables/registers where data is stored along with their associated addresses. We will be using the holding registers from address 40001 to 49999 that are R/W 16 bit/word. Two tables that
n Modbus has four tables/registers where data is stored along with their associated addresses. We will be using the holding registers from address 40001 to 49999 that are R/W 16 bit/word. Two tables that
Fractional Colorings and Zykov Products of graphs
 Fractional Colorings and Zykov Products of graphs Who? Nichole Schimanski When? July 27, 2011 Graphs A graph, G, consists of a vertex set, V (G), and an edge set, E(G). V (G) is any finite set E(G) is
Fractional Colorings and Zykov Products of graphs Who? Nichole Schimanski When? July 27, 2011 Graphs A graph, G, consists of a vertex set, V (G), and an edge set, E(G). V (G) is any finite set E(G) is
ΚΥΠΡΙΑΚΗ ΕΤΑΙΡΕΙΑ ΠΛΗΡΟΦΟΡΙΚΗΣ CYPRUS COMPUTER SOCIETY ΠΑΓΚΥΠΡΙΟΣ ΜΑΘΗΤΙΚΟΣ ΔΙΑΓΩΝΙΣΜΟΣ ΠΛΗΡΟΦΟΡΙΚΗΣ 24/3/2007
 Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Όλοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα μικρότεροι του 10000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Αν κάπου κάνετε κάποιες υποθέσεις
Οδηγίες: Να απαντηθούν όλες οι ερωτήσεις. Όλοι οι αριθμοί που αναφέρονται σε όλα τα ερωτήματα μικρότεροι του 10000 εκτός αν ορίζεται διαφορετικά στη διατύπωση του προβλήματος. Αν κάπου κάνετε κάποιες υποθέσεις
Homework 8 Model Solution Section
 MATH 004 Homework Solution Homework 8 Model Solution Section 14.5 14.6. 14.5. Use the Chain Rule to find dz where z cosx + 4y), x 5t 4, y 1 t. dz dx + dy y sinx + 4y)0t + 4) sinx + 4y) 1t ) 0t + 4t ) sinx
MATH 004 Homework Solution Homework 8 Model Solution Section 14.5 14.6. 14.5. Use the Chain Rule to find dz where z cosx + 4y), x 5t 4, y 1 t. dz dx + dy y sinx + 4y)0t + 4) sinx + 4y) 1t ) 0t + 4t ) sinx
Inverse trigonometric functions & General Solution of Trigonometric Equations. ------------------ ----------------------------- -----------------
 Inverse trigonometric functions & General Solution of Trigonometric Equations. 1. Sin ( ) = a) b) c) d) Ans b. Solution : Method 1. Ans a: 17 > 1 a) is rejected. w.k.t Sin ( sin ) = d is rejected. If sin
Inverse trigonometric functions & General Solution of Trigonometric Equations. 1. Sin ( ) = a) b) c) d) Ans b. Solution : Method 1. Ans a: 17 > 1 a) is rejected. w.k.t Sin ( sin ) = d is rejected. If sin
9.09. # 1. Area inside the oval limaçon r = cos θ. To graph, start with θ = 0 so r = 6. Compute dr
 9.9 #. Area inside the oval limaçon r = + cos. To graph, start with = so r =. Compute d = sin. Interesting points are where d vanishes, or at =,,, etc. For these values of we compute r:,,, and the values
9.9 #. Area inside the oval limaçon r = + cos. To graph, start with = so r =. Compute d = sin. Interesting points are where d vanishes, or at =,,, etc. For these values of we compute r:,,, and the values
ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I)
") ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν
ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΙΑΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΕΡΓΑΣΤΗΡΙΟ ΙΑΤΡΙΚΗΣ ΦΥΣΙΚΗΣ ΗΛΕΚΤΡΙΚΑ ΣΗΜΑΤΑ ΑΠΟ ΤΟ ΣΩΜΑ (I) Γιάννης Τσούγκος ΓΕΝΙΚΑ:...πολλούς αιώνες πριν μελετηθεί επιστημονικά ο ηλεκτρισμός οι άνθρωποι γνώριζαν
Η ΨΥΧΙΑΤΡΙΚΗ - ΨΥΧΟΛΟΓΙΚΗ ΠΡΑΓΜΑΤΟΓΝΩΜΟΣΥΝΗ ΣΤΗΝ ΠΟΙΝΙΚΗ ΔΙΚΗ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΝΟΜΙΚΗ ΣΧΟΛΗ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΤΟΜΕΑΣ ΙΣΤΟΡΙΑΣ ΦΙΛΟΣΟΦΙΑΣ ΚΑΙ ΚΟΙΝΩΝΙΟΛΟΓΙΑΣ ΤΟΥ ΔΙΚΑΙΟΥ Διπλωματική εργασία στο μάθημα «ΚΟΙΝΩΝΙΟΛΟΓΙΑ ΤΟΥ ΔΙΚΑΙΟΥ»
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΝΟΜΙΚΗ ΣΧΟΛΗ ΠΡΟΓΡΑΜΜΑ ΜΕΤΑΠΤΥΧΙΑΚΩΝ ΣΠΟΥΔΩΝ ΤΟΜΕΑΣ ΙΣΤΟΡΙΑΣ ΦΙΛΟΣΟΦΙΑΣ ΚΑΙ ΚΟΙΝΩΝΙΟΛΟΓΙΑΣ ΤΟΥ ΔΙΚΑΙΟΥ Διπλωματική εργασία στο μάθημα «ΚΟΙΝΩΝΙΟΛΟΓΙΑ ΤΟΥ ΔΙΚΑΙΟΥ»
Statistical Inference I Locally most powerful tests
 Statistical Inference I Locally most powerful tests Shirsendu Mukherjee Department of Statistics, Asutosh College, Kolkata, India. shirsendu st@yahoo.co.in So far we have treated the testing of one-sided
Statistical Inference I Locally most powerful tests Shirsendu Mukherjee Department of Statistics, Asutosh College, Kolkata, India. shirsendu st@yahoo.co.in So far we have treated the testing of one-sided
Βιολογικές Μεμβράνες και Μεταγωγή Σήματος
 ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογικές Μεμβράνες και Μεταγωγή Σήματος Τερματισμός σηματοδότησης Διδάσκουσα: Καθ. Μαρία - Ελένη Ε. Λέκκα Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
ΠΑΝΕΠΙΣΤΗΜΙΟ ΙΩΑΝΝΙΝΩΝ ΑΝΟΙΚΤΑ ΑΚΑΔΗΜΑΪΚΑ ΜΑΘΗΜΑΤΑ Βιολογικές Μεμβράνες και Μεταγωγή Σήματος Τερματισμός σηματοδότησης Διδάσκουσα: Καθ. Μαρία - Ελένη Ε. Λέκκα Άδειες Χρήσης Το παρόν εκπαιδευτικό υλικό
Δυναμικό ηρεμίας Δυναμικό ενεργείας. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 29/09/2017 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2017-2018 Ιόντα Δυναμικό
Δυναμικό ηρεμίας Δυναμικό ενεργείας Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 29/09/2017 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2017-2018 Ιόντα Δυναμικό
Second Order RLC Filters
 ECEN 60 Circuits/Electronics Spring 007-0-07 P. Mathys Second Order RLC Filters RLC Lowpass Filter A passive RLC lowpass filter (LPF) circuit is shown in the following schematic. R L C v O (t) Using phasor
ECEN 60 Circuits/Electronics Spring 007-0-07 P. Mathys Second Order RLC Filters RLC Lowpass Filter A passive RLC lowpass filter (LPF) circuit is shown in the following schematic. R L C v O (t) Using phasor