Zvijanje proteinov. Zvijanje proteinov je proces, pri katerem iz razvite (denaturirane) oblike nastane zvita, funkcionalna oblika.
|
|
|
- Ζηναις Μανωλάς
- 8 χρόνια πριν
- Προβολές:
Transcript
1 Zvijanje proteinov Zvijanje proteinov je proces, pri katerem iz razvite (denaturirane) oblike nastane zvita, funkcionalna oblika. najenostavnejši model zvijanja k f denaturiran(d) nativen(n) k u K = N D = k f k u Zvijanje proteina je spontan proces, če je DG < 0. Pri 25 C ΔG 0 = ΔH 0 TΔS 0 tvorba intramolekulskih kontaktov/vezi izguba entropije zaradi urejenosti
2 Zvijanje proteinov Proste energije zvijanja (DG 0 ) nekaterih pogostejših proteinov: protein DG 0 (kj/mol) DH 0 (kj/mol) TDS 0 (kj/mol) inhibitor kimotripsina 2-27, ,3 eglin C -36, ,1 RNaza T1-37, ,6 citokrom C -37, ,9 barnaza -48, ,2 lizocim -57, ,3 kimotripsin -45, ,4 tendamistat -37, ,5
3 Zvijanje proteinov Za številne proteine velja, da se zvijajo/razvijajo v enem koraku, brez vmesnih stabilnih intermediatov. Eksperimentalno ponavadi študiramo denaturacijo proteina s spreminjanjem koncentracije denaturanta (GdmCl, urea), ph ali temperature. Signal, ki ga spremljamo, je lahko CD, intrinzična fluorescenca, FRET,...
4 Zvijanje proteinov Nekateri proteini se razvijejo v več korakih dobimo večfazni diagram. Primer: razvijanje virulenčnega faktorja pertaktina iz Bordetelle pertusis.
5 Faze zvijanja proteinov Zvijanje proteinov poteka po vnaprej določenih poteh, ki so se razvile tekom evolucije, ne pa z naključnim iskanjem po konformacijskem prostoru. Levinthalov paradoks: če bi vsak ostanek imel na voljo le 2 konformaciji, bi peptid s 100 ostanki zavzel različnih konformacij če bi naključno iskal nativno konformacijo, bi potreboval 10 miljard let. Zvijanje proteinov iz denaturirane v nativno obliko poteče v treh fazah: 1. Hitri kolaps strukture in nastanek sekundarne strukture (< ms) Gonilna sila naj bi bile hidrofobne interakcije. Nastane razrahljani klobčič (angl. molten globule). 2. Nastanek terciarne strukture (ms do s) 3. Pakiranje terciarne strukture ( kristalizacija ) v nativno obliko ( s)
6 Zgodnji dogodki zvijanja Kompaktnost strukture proteina lahko merimo z radijem giracije, tj. koren povprečnih kvadratov oddaljenosti posameznih centrov mas od središča molekule: R G = i m i x i 2 + y i 2 +z i 2 i m i protein R G (nativen) R G (denat.) razmerje SH3 domena kinaze PI3 18,6 27,5 0,7 citokrom C 17,8 32,6 0,5 lizocim 20,5 34,6 0,6 TIM 29,7 49,7 0,6 Pri hitrem kolapsu molekule nastane struktura, ki ima R G okoli 10% višji kot nativna oblika proteina razrahljani klobčič.
7 Razrahljani klobčič Razrahljani klobčič je delno strukturirano stanje med nativno in denaturirano obliko. Lahko je relativno stabilen intermediat zvijanja ali pa zgolj prehodno stanje. Ima znaten delež sekundarne strukture nativnega stanja.
.")
8 rel. fluorescenca Razrahljani klobčič Glede na nativno obliko je struktura razrahljanega klobčiča bolj ohlapna. Na tej osnovi lahko stabilne razrahljane klobčiče eksperimentalno detektiramo z uporabo fluorescenčnega barvila 8-anilino-1- naftalensulfonata (ANS). Le-ta se veže v hidrofobno notranjost klobčiča, ob čemer se intenziteta njegove fluorescence močno poveča Primer: vezava ANS na divji tip in mutant F35A stefina B. ANS 100 stfb wt stfb F35A ph
9 Primeri prehodnih stanj (razrahljanih klobčičev) inhibitor kimotripsina 2 homeodomena transkripcijskega faktorja engrailed WW domena Schaeffer in Daggett PEDS. 24

10 Energetski diagram zvijanja Energetski diagram celotnega procesa zvijanja proteina ponavadi ponazorimo v obliki lijaka.

11 Energetski diagram zvijanja Energetski diagram celotnega procesa zvijanja proteina ponavadi ponazorimo v obliki lijaka.

12 Hierarhičen model zvijanja Zvijanje se začne z interakcijami med deli, ki so si blizu v primarni strukturi in marginalno stabilni. Te lokalne strukture dalje interagirajo z ostalimi segmenti, tako da kompleksnost interakcij narašča. Hierarhičen model zvijanja l represorja: Črni krožci predstavljajo formirano vijačnico, beli pa neformirano. Linije prikazujejo interakcije med vijačnicami.


 čas")

13 Hitrost zvijanja proteinov Hitrost zvijanja je navzgor omejena s hitrostjo nastanka posameznih elementov: a-vijačnice nastanejo v času 0,1 1 ms b-lasnice nastanejo v času 1 10 ms zanke nastanejo hitreje ne omejujejo hitrosti zvijanja V splošnem je zgornja meja hitrosti N/50 ms, kjer je N št. ostankov v proteinu. PDB 1PRB PDB 2A3D albumin-vezavna domena (bakterijska) čas zvijanja 1 ms dizajniran snop treh vijačnic čas zvijanja 3 ms triptofanska kletka čas zvijanja 4,1 ms poddomena glave vilina čas zvijanja 4,3 ms
14 Hitrost zvijanja proteinov Primeri hitrosti zvijanja nekaterih proteinov: Bowman et al Curr Opin Struct Biol. 21(1)

15 Alternativne poti zvijanja Proteini se lahko iz denaturirane oblike v nativno zvijejo po eni ali več poteh. Barnaza se zvije po eni glavni poti preko enega intermediata.

16 Alternativne poti zvijanja Proteini se lahko iz denaturirane oblike v nativno zvijejo po eni ali več poteh. Lizocim se zvije po vsaj dveh različnih poteh.

17 Alternativne poti zvijanja Proteini se lahko iz denaturirane oblike v nativno zvijejo po eni ali več poteh. Na mikroskopskem nivoju zasedejo proteini med zvijanjem mnogo različnih metastabilnih konformacij. Primer: 14 najbolj stabilnih intermediatov pri ab initio zvijanju proteina NTL9 Voelz et al JACS. 132(5)

18 Tvorba disulfidov Tvorba nekaterih struktur omejuje hitrost zvijanja proteinov. Mednje spada tvorba disulfidov. Primer: zvijanje BPTI vključuje zaporedno formacijo treh disulfidnih vezi.


19 Tvorba disulfidov Tvorba nekaterih struktur omejuje hitrost zvijanja proteinov. Mednje spada tvorba disulfidov. In vivo preurejanje disulfidov katalizirajo protein disulfid izomeraze. PDB 1DSB PDB 4EKZ DsbA, protein disulfid izomeraza iz E. coli ena tioredoksinska domena (zelena) človeška protein disulfid izomeraza dve tioredoksinski domeni (modra in oranžna)
, a jih včasih najdemo v ostrih zavojih verige.")
20 Izomerizacija prolinov Ena izmed počasnih stopenj zvijanja je tudi izomerizacija prolinov. Cis-prolini so redkejši (4x manj stabilni od trans oblike), a jih včasih najdemo v ostrih zavojih verige. Cis-trans izomerizacija je počasen proces, in vivo pa ga pospešijo peptidil prolil izomeraze. človeška peptidil prolil izomeraza ciklofilin

21 Zvijanje v celici V nasprotju z in vitro poskusi je koncentracija molekul v celicah bistveno večja.

22 Šaperoni Šaperoni so molekule, ki katalizirajo zvijanje proteinov in preprečujejo agregacijo nepravilno zvitih intermediatov. Mnoge so odkrili, ker se njihovo izražanje poviša pri toplotnem stresu, zato jih pogosto imenujemo tudi proteini toplotnega šoka. Šaperonini so vrsta šaperonov v obliki dvojnega obroča, ki vežejo nepravilno zvite proteine v svoji notranjosti in katalizirajo njihovo zvijanje v nativno obliko. Vabulas et al. Cold Spring Harb Perspect Biol 2010;2:a004390



23 Šaperoni Šaperoni so molekule, ki katalizirajo zvijanje proteinov in preprečujejo agregacijo nepravilno zvitih intermediatov. Mnoge so odkrili, ker se njihovo izražanje poviša pri toplotnem stresu, zato jih pogosto imenujemo tudi proteini toplotnega šoka. Šaperonini so vrsta šaperonov v obliki dvojnega obroča, ki vežejo nepravilno zvite proteine v svoji notranjosti in katalizirajo njihovo zvijanje v nativno obliko. V E. coli poznamo šaperoninski sistem, ki ga kodirata produkta GroE operona: GroEL in GroES. Mirujoči šaperonin je sestavljen iz 14 kopij GroEL organiziranih v dva obroča okoli osrednje votline. difuzija ATP in ADP


24 Šaperoni V E. coli poznamo šaperonski sistem, ki ga kodirata produkta GroE operona: GroEL in GroES. Vsak monomer GroEL je zgrajen iz treh domen (M R 58 kda), obroča pa sta orientirane zrcalno eden na drugega.

25 Šaperoni V E. coli poznamo šaperonski sistem, ki ga kodirata produkta GroE operona: GroEL in GroES. Molekule GroES (10 kda) sestavljajo heptamer. mobilna zanka
, pride do konformacijske spremembe v cis obroču, ki onemogoči vezavo GroES na drug konec GroEL.")
26 Šaperoni V E. coli poznamo šaperonski sistem, ki ga kodirata produkta GroE operona: GroEL in GroES. Ko se na GroEL veže GroES (prav tako iz sedmih podenot), pride do konformacijske spremembe v cis obroču, ki onemogoči vezavo GroES na drug konec GroEL. Vezava GroES poteče ob hidrolizi ATP v ADP. Podobno velja tudi za sproščanje GroES z GroEL. cis pozicija trans pozicija


27 Šaperoni V E. coli poznamo šaperonski sistem, ki ga kodirata produkta GroE operona: GroEL in GroES. Konformacijsko sprememba v podenotah cis obroča of vezavi GroES: PDB 1DER PDB 1AON

28 Šaperoni V E. coli poznamo šaperonski sistem, ki ga kodirata produkta GroE operona: GroEL in GroES. Konformacijsko sprememba v podenotah cis obroča of vezavi GroES: Konformacijska sprememba vsebuje premik apikalne domene za 90 glede na ekvatorialno domeno. Sprememba ima funkcijske posledice notranjost cis obroča se spremeni iz hidrofobne v hidrofilno pomembno za zvijanje proteina, ki poteka le v obroču, kjer je vezan GroES. modra brez GroES oranžna z GroES
29 Šaperoni Mehanizem delovanja šaperona: hidrofobne interakcije hidrofilne interakcije pravilno zvit protein

30 Patološka agregacija proteinov Agregacija proteinov je vzrok številnih bolezni.

31 Patološka agregacija proteinov
 je tvorba fibril sestavljenih iz b strukture t.i. cross b oz. amiloidna struktura.")
32 Patološka agregacija proteinov Skupna lastnost številnih nepovezanih patoloških stanj, povezanih z agregacijo proteinov (a zdaleč ne vseh!) je tvorba fibril sestavljenih iz b strukture t.i. cross b oz. amiloidna struktura. Strukturo fibril se lahko preučuje z rentgensko difrakcijo (določi se strukturo urejenih delov strukture b ploskev).

33 Patološka agregacija proteinov Za različne proteine so bile predpostavljene različne oblike b struktur. Amiloidom podobno zvitje lahko imajo tudi nepatološki proteini.
34 Patološka agregacija proteinov Agregacija proteinov je vzrok številnih bolezni. bolezen povezan protein anemija srpastih celic mutiran hemoglobin klasične amiloidoze lahke verige Ig, transtiretin,... pljučni emfizem mutiran a1-antitripsin Huntingtonova bolezen patološka varianta huntingtina Parkinsonova bolezen a-sinuklein Alzheimerjeva bolezen amiloid b, protein t spongiformne encefalopatije prionski protein

35 Klasične amiloidoze Zunajcelična akumulacija netopnih fibril premera Å. Izraz amiloid izvira iz dejstva, da se fibrile barvajo z jodom (kot škrob). Fibrile bogate z b strukturami, pravokotnimi na os fibrile. Primer: serumski protein transtiretin (homotetramer z maso 55 kda) PDB 3W3B PDB 3W3B dimer transtiretina tetramer transtiretina

36 Klasične amiloidoze Zunajcelična akumulacija netopnih fibril premera Å. Izraz amiloid izvira iz dejstva, da se fibrile barvajo z jodom (kot škrob). Fibrile bogate z b strukturami, pravokotnimi na os fibrile. Primer: serumski protein transtiretin (homotetramer z maso 55 kda) PDB 2M5N PDB 3W3B fibrila transtiretina tetramer transtiretina
 Fitzpatrick A W P et al.")
37 Klasične amiloidoze Primer: serumski protein transtiretin (homotetramer z maso 55 kda) Fitzpatrick A W P et al. PNAS 2013;110:

38 Huntingtonova bolezen Huntingtonova bolezen je nevrodegenerativna bolezen, ki je posledica mutacij v proteinu huntingtin 350 kda citoplazemski protein. Bolezen je avtosomna dominantna je primer poliq bolezni. PoliQ regija je podvržena polimorfizmu, zaradi katerega se št. Q ponovitev med ljudmi močno razlikuje. Če jih je manj kot 27, protein ni podvržen agregaciji. Z večanjem števila ostankov tveganje za bolezen narašča.

 in nevrofibrilarnih pentelj vlaken v nevronih, zgrajenih iz proteina t")
39 Alzheimerjeva bolezen Nevrodegenerativna bolezen, pogosta pri starejših ljudeh. Povezana je s tvorbo dveh struktur senilnih plakov zunajcelični depoziti amiloida b N-končnega fragmenta amiloidnega prekurzorskega proteina (ponavadi ostanki 1-40 ali 1-42) (receptor na površini nevronov) in nevrofibrilarnih pentelj vlaken v nevronih, zgrajenih iz proteina t (stabilizira mikrotubule).
 (receptor na površini nevronov)")
40 Alzheimerjeva bolezen Nevrodegenerativna bolezen, pogosta pri starejših ljudeh. Povezana je s tvorbo dveh struktur senilnih plakov zunajcelični depoziti amiloida b N-končnega fragmenta amiloidnega prekurzorskega proteina (dolg 40 ali 42 ostankov) (receptor na površini nevronov) in nevrofibrilarnih pentelj vlaken v nevronih, zgrajenih iz proteina t (stabilizira mikrotubule). Amiloid b peptid nastane po cepitvi APP z b in g-sekretazama. a-sekretaza cepi na poziciji med obema in ščiti pred Alzheimerjevo boleznijo.
 (receptor na površini")
41 Alzheimerjeva bolezen Nevrodegenerativna bolezen, pogosta pri starejših ljudeh. Povezana je s tvorbo dveh struktur senilnih plakov zunajcelični depoziti amiloida b N-končnega fragmenta amiloidnega prekurzorskega proteina (ponavadi ostanki 1-40 ali 1-42) (receptor na površini nevronov) in nevrofibrilarnih pentelj vlaken v nevronih, zgrajenih iz proteina t (stabilizira mikrotubule). delček amiloidnega plaka Na molekulskem nivoju


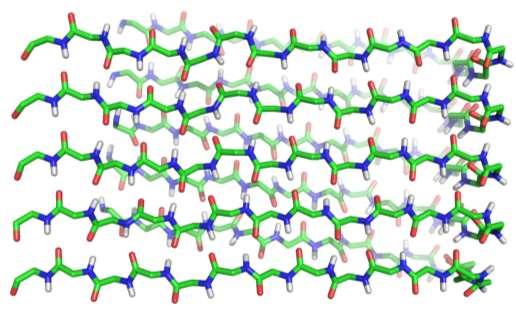
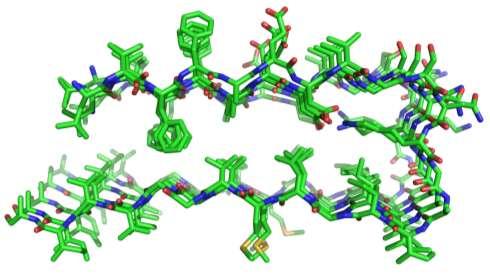
42 Alzheimerjeva bolezen Struktura cross-b ploskve amiloida b.


43 Alzheimerjeva bolezen Pri Ab je možna tvorba več morfološko različnih oblik fibril, odvisno od pogojev okolja. Tycko Annu Rev Phys Chem. 62
44 Alzheimerjeva bolezen Nevrotoksičnost amiloidnih fibiril primarno nevrotoksični naj bi bili manjši fragmenti. Cold Spring Harb Perspect Med 2012;2:a006338

45 Alzheimerjeva bolezen Regulacija in disregulacija proteina t: agregacija s parjenjem vijačnih fragmentov
 in ne vsebujejo nukleinskih kislin. Prenašajo se lahko med različimi vrstami.")
46 Prioni Prion = Protein Infection (1982, Stanley B. Prusiner) Prioni so edini kužni delci, ki so sestavljeni pretežno oz. izključno iz proteinov (patogene oblike prionskega proteina PrP) in ne vsebujejo nukleinskih kislin. Prenašajo se lahko med različimi vrstami. Prioni povzročajo bolezni, ki jih imenujemo transmisivne spongiformne encefalopatije. Pri človeku je najpogostejša Creutzfeldt-Jakobova bolezen (CJB) začne se z depresijo in težavami s spominom, nadaljuje z demenco, norostjo, nekontroliranim krčenjem mišic in konča s smrtjo znotraj 1 leta. Prisotni so tudi dedni faktorji.


47 Prioni Spongiformno možgansko tkivo
.")
48 Prionski protein Prionski protein Prp je okoli 200 ak ostankov velik protein, ki je pripet na zunanjo membrano možganskih celic. Normalna celična oblika PrP C ni nevarna, se pa iz neznanega razloga lahko pretvori v patološko obliko PrP Sc (vloga proteolize?). N-končna domena z neznano strukturo (24-105), vsebuje oktaponovitve C-končna domena pretežno iz a-vijačnic (ostanki ) Aguzzi and Falsig (2012) Nat Neurosci. 15(7)

49 Transformacija PrP Modeli transformacije in fibrilacije PrP:

50 Replikacijski cikel priona
51 Potreba po kofaktorjih Dlje časa se že preučuje možnost, ali so za toksičnost in/ali propagacijo prionov potrebni še določeni neznani kofaktorji. Definitivnih dokazov ni. Diaz-Espinoza and Soto (2012) Nat Struct Mol Biol. 19(4)
 porušenje integritete")
52 Toksičnost priona Mehanizem toksičnosti ni poznan. Ve se le, da mora za toksičnost biti PrP vezan na membrano preko GPI sidra. tvorba por interakcije s citosolnimi proteini ekscitotoksičnost (odmrtje zaradi močne vzburjenosti) porušenje integritete membrane
ZVIJANJE PROTEINOV, RAZGRADNJA IN USMERJANJE
 ZVIJANJE PROTEINOV, RAZGRADNJA IN USMERJANJE ZVIJANJE PROTEINOV Spontano Stanje odtajane kroglice Molekularni šaperoni POST-TRANSLACIJSKE MODIFIKACIJE PROTEINOV RAZGRADNJA Ubikvitin Proteasom USMERJANJE
ZVIJANJE PROTEINOV, RAZGRADNJA IN USMERJANJE ZVIJANJE PROTEINOV Spontano Stanje odtajane kroglice Molekularni šaperoni POST-TRANSLACIJSKE MODIFIKACIJE PROTEINOV RAZGRADNJA Ubikvitin Proteasom USMERJANJE
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks'
 Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 15. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
p 1 ENTROPIJSKI ZAKON
 ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
I. OSNOVNI STRUKTURNI PRINCIPI
 I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
I. OSNOVNI STRUKTURNI PRINCIPI 1. Gradnja Vsi proteini so polimeri 20 različnih amino, kislin, povezanih s peptidnimi vezmi Funkcija proteina je odvisna od 3D strukture, ki je povezana z AK sekvenco ta
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
Transformator. Delovanje transformatorja I. Delovanje transformatorja II
 Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ
 ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ α-heliks β-nagubana RAVNINA β-obrat STRUKTURNI MOTIVI STABILNOST PROTEINOV KVARTARNA ZGRADBA PROTEINOV ENA ALI VEČ KONFORMACIJ
ZGRADBA PROTEINOV SILE, KI STABILIZIRAJO 3D ZGRADBO PROTEINOV PEPTIDNA VEZ α-heliks β-nagubana RAVNINA β-obrat STRUKTURNI MOTIVI STABILNOST PROTEINOV KVARTARNA ZGRADBA PROTEINOV ENA ALI VEČ KONFORMACIJ
Biokemija I, 25. predavanje 1. del, , A. Videtič Paska. Proteini - splošno
 Biokemija I, 25. predavanje 1. del, 16. 4. 2012, A. Videtič Paska Proteini - splošno Razdelitev po strukturi in funkciji. Ravni proteinske strukture: - primarna in - sekundarna struktura Sinteza proteinov
Biokemija I, 25. predavanje 1. del, 16. 4. 2012, A. Videtič Paska Proteini - splošno Razdelitev po strukturi in funkciji. Ravni proteinske strukture: - primarna in - sekundarna struktura Sinteza proteinov
13. Jacobijeva metoda za računanje singularnega razcepa
 13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
Najpomembnejši človeški Hb
 Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
Estimation of grain boundary segregation enthalpy and its role in stable nanocrystalline alloy design
 Supplemental Material for Estimation of grain boundary segregation enthalpy and its role in stable nanocrystalline alloy design By H. A. Murdoch and C.A. Schuh Miedema model RKM model ΔH mix ΔH seg ΔH
Supplemental Material for Estimation of grain boundary segregation enthalpy and its role in stable nanocrystalline alloy design By H. A. Murdoch and C.A. Schuh Miedema model RKM model ΔH mix ΔH seg ΔH
Nevrodegeneracija. Boris Rogelj
 Nevrodegeneracija Boris Rogelj boris.rogelj@ijs.si Pregled predavanja Osnove in skupne lastnosti Alzheimerjava bolezen (AB) Parkinsonova bolezen (PB) Huntingtonova bolezen (HB) Frontotemporalna demenca
Nevrodegeneracija Boris Rogelj boris.rogelj@ijs.si Pregled predavanja Osnove in skupne lastnosti Alzheimerjava bolezen (AB) Parkinsonova bolezen (PB) Huntingtonova bolezen (HB) Frontotemporalna demenca
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
Gimnazija Krˇsko. vektorji - naloge
 Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK)
") IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
IZPIT IZ BIOKEMIJE ZA BIOLOGE (1. ROK) 25. 01. 2006. Vpisna številka: Št. točk: Procenti: Ocena: Na vprašanja 1-19 odgovoriš tako, da obkrožiš črko pred odgovorom, za katerega meniš, da je pravilen. Možnih
CM707. GR Οδηγός χρήσης... 2-7. SLO Uporabniški priročnik... 8-13. CR Korisnički priručnik... 14-19. TR Kullanım Kılavuzu... 20-25
 1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N
 I N F O T E K N I K V o l u m e 1 5 N o. 1 J u l i 2 0 1 4 ( 61-70) A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N N o v i
I N F O T E K N I K V o l u m e 1 5 N o. 1 J u l i 2 0 1 4 ( 61-70) A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N N o v i
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Funkcije proteinov (pogojene s strukturo)
") Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Funkcije proteinov (pogojene s strukturo) Oporna funkcija (strukturni proteini, npr keratini, kolagen...) Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Na pregledni skici napišite/označite ustrezne točke in paraboli. A) 12 B) 8 C) 4 D) 4 E) 8 F) 12
 12 B) 8 C) 4 D) 4 E) 8 F) 12") Predizpit, Proseminar A, 15.10.2015 1. Točki A(1, 2) in B(2, b) ležita na paraboli y = ax 2. Točka H leži na y osi in BH je pravokotna na y os. Točka C H leži na nosilki BH tako, da je HB = BC. Parabola
Predizpit, Proseminar A, 15.10.2015 1. Točki A(1, 2) in B(2, b) ležita na paraboli y = ax 2. Točka H leži na y osi in BH je pravokotna na y os. Točka C H leži na nosilki BH tako, da je HB = BC. Parabola
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
ΓΗ ΚΑΙ ΣΥΜΠΑΝ. Εικόνα 1. Φωτογραφία του γαλαξία μας (από αρχείο της NASA)
") ΓΗ ΚΑΙ ΣΥΜΠΑΝ Φύση του σύμπαντος Η γη είναι μία μονάδα μέσα στο ηλιακό μας σύστημα, το οποίο αποτελείται από τον ήλιο, τους πλανήτες μαζί με τους δορυφόρους τους, τους κομήτες, τα αστεροειδή και τους μετεωρίτες.
ΓΗ ΚΑΙ ΣΥΜΠΑΝ Φύση του σύμπαντος Η γη είναι μία μονάδα μέσα στο ηλιακό μας σύστημα, το οποίο αποτελείται από τον ήλιο, τους πλανήτες μαζί με τους δορυφόρους τους, τους κομήτες, τα αστεροειδή και τους μετεωρίτες.
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV. 2. predavanje: Od 1D do 3D strukture proteinov 1. del.
 IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
IZBIRNI PREDMET, APRIL/MAJ 2013 STRUKTURA IN FUNKCIJA PROTEINOV 2. predavanje: Od 1D do 3D strukture proteinov 1. del Matjaž Zorko E-mail: zorko@mf.uni-lj.si DOSEGLJIVO: http://ibk.mf.uni-lj.si/teaching/objave/izbir1mz.htm
Družina globinov pri človeku in bolezni.
 Družina globinov pri človeku in bolezni www.muhlenberg.edu/ Mioglobin in hemoglobin spadata v družino globinov Globinsko zvitje Mb β podenota Hb Podobnost aminokislinskega zaporedja Podobnost 3D strukture
Družina globinov pri človeku in bolezni www.muhlenberg.edu/ Mioglobin in hemoglobin spadata v družino globinov Globinsko zvitje Mb β podenota Hb Podobnost aminokislinskega zaporedja Podobnost 3D strukture
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
MATEMATIČNI IZRAZI V MAFIRA WIKIJU
 I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
DISKRETNA FOURIERJEVA TRANSFORMACIJA
 29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
1. TVORBA ŠIBKEGA (SIGMATNEGA) AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ
 AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ") TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
PROCESIRANJE SIGNALOV
 Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
Strukturni in funkcijski vidiki bioloških interakcij
 Strukturni in funkcijski vidiki bioloških interakcij Urnik 30 ur predavanj + 15 ur seminarjev predavanja in seminarji: petek od 12.00 do 15.00 10 predavanj 5 seminarjev 30 ur vaj (6 x 5 ur) Obveznosti
Strukturni in funkcijski vidiki bioloških interakcij Urnik 30 ur predavanj + 15 ur seminarjev predavanja in seminarji: petek od 12.00 do 15.00 10 predavanj 5 seminarjev 30 ur vaj (6 x 5 ur) Obveznosti
Osnove matematične analize 2016/17
 Osnove matematične analize 216/17 Neža Mramor Kosta Fakulteta za računalništvo in informatiko Univerza v Ljubljani Kaj je funkcija? Funkcija je predpis, ki vsakemu elementu x iz definicijskega območja
Osnove matematične analize 216/17 Neža Mramor Kosta Fakulteta za računalništvo in informatiko Univerza v Ljubljani Kaj je funkcija? Funkcija je predpis, ki vsakemu elementu x iz definicijskega območja
Sarò signor io sol. α α. œ œ. œ œ œ œ µ œ œ. > Bass 2. Domenico Micheli. Canzon, ottava stanza. Soprano 1. Soprano 2. Alto 1
 Sarò signor io sol Canzon, ottava stanza Domenico Micheli Soprano Soprano 2 Alto Alto 2 Α Α Sa rò si gnor io sol del mio pen sie io sol Sa rò si gnor io sol del mio pen sie io µ Tenor Α Tenor 2 Α Sa rò
Sarò signor io sol Canzon, ottava stanza Domenico Micheli Soprano Soprano 2 Alto Alto 2 Α Α Sa rò si gnor io sol del mio pen sie io sol Sa rò si gnor io sol del mio pen sie io µ Tenor Α Tenor 2 Α Sa rò
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Najpogostejše hemoglobinopatije
 Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina
Najpogostejše hemoglobinopatije Anemija srpastih celic Malarija Talasemije so napake v sestavi hemoglobina. α talasemija pomanjkanje α verige hemoglobina β-talasemija - pomanjkanje β verige hemoglobina
ΜΕΛΕΤΗ ΤΗΣ ΥΝΑΤΟΤΗΤΑΣ ΑΞΙΟΠΟΙΗΣΗΣ ΤΟΥ ΓΕΩΘΕΡΜΙΚΟΥ ΠΕ ΙΟΥ ΘΕΡΜΩΝ ΝΙΓΡΙΤΑΣ (Ν. ΣΕΡΡΩΝ)
") ελτίο της Ελληνικής Γεωλογικής Εταιρίας τοµ. XXXVI, 2004 Πρακτικά 10 ου ιεθνούς Συνεδρίου, Θεσ/νίκη Απρίλιος 2004 Bulletin of the Geological Society of Greece vol. XXXVI, 2004 Proceedings of the 10 th
ελτίο της Ελληνικής Γεωλογικής Εταιρίας τοµ. XXXVI, 2004 Πρακτικά 10 ου ιεθνούς Συνεδρίου, Θεσ/νίκη Απρίλιος 2004 Bulletin of the Geological Society of Greece vol. XXXVI, 2004 Proceedings of the 10 th
Razgradnja maščobnih kislin. Ketonska telesa H + NAD+ NADH. Pregled metabolizma MK. lipoprotein-lipaza. maščobne kisline.
 Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
- Geodetske točke in geodetske mreže
 - Geodetske točke in geodetske mreže 15 Geodetske točke in geodetske mreže Materializacija koordinatnih sistemov 2 Geodetske točke Geodetska točka je točka, označena na fizični površini Zemlje z izbrano
- Geodetske točke in geodetske mreže 15 Geodetske točke in geodetske mreže Materializacija koordinatnih sistemov 2 Geodetske točke Geodetska točka je točka, označena na fizični površini Zemlje z izbrano
Jure Stojan in Marko Goličnik Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Tabele termodinamskih lastnosti vode in vodne pare
 Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ
 GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
Podobnost matrik. Matematika II (FKKT Kemijsko inženirstvo) Diagonalizacija matrik
 Diagonalizacija matrik") Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV
 SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV Literatura za temo Signalne poti, ki regulirajo izražanje genov: Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH
SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV Literatura za temo Signalne poti, ki regulirajo izražanje genov: Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH
FUNKCIJA PROTEINOV, PRIMERI FIBRILARNI PROTEINI. α KERATIN
 FUNKCIJA PROTEINOV, PRIMERI FIBRILARNI PROTEINI keratin, kolagen, fibroin, spidroin GLOBULARNI PROTEINI domenska zgradba proteinov MIOGLOBIN, HEMOGLOBIN vezava kisika zgradba konformacijske spremembe ob
FUNKCIJA PROTEINOV, PRIMERI FIBRILARNI PROTEINI keratin, kolagen, fibroin, spidroin GLOBULARNI PROTEINI domenska zgradba proteinov MIOGLOBIN, HEMOGLOBIN vezava kisika zgradba konformacijske spremembe ob
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
Το άτομο του Υδρογόνου
 Το άτομο του Υδρογόνου Δυναμικό Coulomb Εξίσωση Schrödinger h e (, r, ) (, r, ) E (, r, ) m ψ θφ r ψ θφ = ψ θφ Συνθήκες ψ(, r θφ, ) = πεπερασμένη ψ( r ) = 0 ψ(, r θφ, ) =ψ(, r θφ+, ) π Επιτρεπτές ενέργειες
Το άτομο του Υδρογόνου Δυναμικό Coulomb Εξίσωση Schrödinger h e (, r, ) (, r, ) E (, r, ) m ψ θφ r ψ θφ = ψ θφ Συνθήκες ψ(, r θφ, ) = πεπερασμένη ψ( r ) = 0 ψ(, r θφ, ) =ψ(, r θφ+, ) π Επιτρεπτές ενέργειες
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
Logatherm WPL 14 AR T A ++ A + A B C D E F G A B C D E F G. kw kw /2013
 WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
Iterativno reševanje sistemov linearnih enačb. Numerične metode, sistemi linearnih enačb. Numerične metode FE, 2. december 2013
 Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
Nukleinske kisline. Nukleotidi. DNA je nosilka dednih genetskih informacij.
 Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL
 PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
Neavtorizirani povzetki izbranih predavanj iz biokemije. UN študij Okolje Politehnika Nova Gorica. doc. dr. Marko Dolinar
 Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba.
 1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
Νόµοςπεριοδικότητας του Moseley:Η χηµική συµπεριφορά (οι ιδιότητες) των στοιχείων είναι περιοδική συνάρτηση του ατοµικού τους αριθµού.
 των στοιχείων είναι περιοδική συνάρτηση του ατοµικού τους αριθµού.") Νόµοςπεριοδικότητας του Moseley:Η χηµική συµπεριφορά (οι ιδιότητες) των στοιχείων είναι περιοδική συνάρτηση του ατοµικού τους αριθµού. Περιοδικός πίνακας: α. Είναι µια ταξινόµηση των στοιχείων κατά αύξοντα
Νόµοςπεριοδικότητας του Moseley:Η χηµική συµπεριφορά (οι ιδιότητες) των στοιχείων είναι περιοδική συνάρτηση του ατοµικού τους αριθµού. Περιοδικός πίνακας: α. Είναι µια ταξινόµηση των στοιχείων κατά αύξοντα
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI. NEUTRALNI ELEMENT GRUPOIDA.
 KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
Kvantni delec na potencialnem skoku
 Kvantni delec na potencialnem skoku Delec, ki se giblje premo enakomerno, pride na mejo, kjer potencial naraste s potenciala 0 na potencial. Takšno potencialno funkcijo zapišemo kot 0, 0 0,0. Slika 1:
Kvantni delec na potencialnem skoku Delec, ki se giblje premo enakomerno, pride na mejo, kjer potencial naraste s potenciala 0 na potencial. Takšno potencialno funkcijo zapišemo kot 0, 0 0,0. Slika 1:
Frekvenčna analiza neperiodičnih signalov. Analiza signalov prof. France Mihelič
 Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama.
 Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Analiza'3D'struktur'makromolekul'in' modeliranje'
 Univerza'v'Ljubljani,'Fakulteta'za'kemijo'in'kemijsko'tehnologijo' Univerzitetni%študijski%program%Biokemija,%2.%letnik,%študijsko%leto%2013/2014% Analiza'3D'struktur'makromolekul'in' modeliranje' Miha%Pavšič%
Univerza'v'Ljubljani,'Fakulteta'za'kemijo'in'kemijsko'tehnologijo' Univerzitetni%študijski%program%Biokemija,%2.%letnik,%študijsko%leto%2013/2014% Analiza'3D'struktur'makromolekul'in' modeliranje' Miha%Pavšič%
Splošno o interpolaciji
 Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
SATCITANANDA. F = e E sila na naboj. = ΔW e. Rudolf Kladnik: Fizika za srednješolce 3. Svet elektronov in atomov
 Ruolf Klnik: Fizik z srenješolce Set elektrono in too Električno olje (11), gibnje elce električne olju Strn 55, nlog 1 Kolikšno netost or releteti elektron, se njego kinetičn energij oeč z 1 kev? Δ W
Ruolf Klnik: Fizik z srenješolce Set elektrono in too Električno olje (11), gibnje elce električne olju Strn 55, nlog 1 Kolikšno netost or releteti elektron, se njego kinetičn energij oeč z 1 kev? Δ W
Izpeljava Jensenove in Hölderjeve neenakosti ter neenakosti Minkowskega
 Izeljava Jensenove in Hölderjeve neenakosti ter neenakosti Minkowskega 1. Najosnovnejše o konveksnih funkcijah Definicija. Naj bo X vektorski rostor in D X konveksna množica. Funkcija ϕ: D R je konveksna,
Izeljava Jensenove in Hölderjeve neenakosti ter neenakosti Minkowskega 1. Najosnovnejše o konveksnih funkcijah Definicija. Naj bo X vektorski rostor in D X konveksna množica. Funkcija ϕ: D R je konveksna,