dogma centrala a biologiei moleculare
|
|
|
- Ῥεβέκκα Λειβαδάς
- 8 χρόνια πριν
- Προβολές:
Transcript
1 Sinteza proteinelor Este activitatea fundamentala a fiecarei celule din organism Ea reprezinta expresia informatiei genetice din ADN si este un fenomen care se desfasoara intr-un singur sens: ADN ARN proteine Datorita universalitatii sale, acest proces este cunoscut sub denumirea de dogma centrala a biologiei moleculare

2 Dogma centrala a biologiei moleculare Descrie procesul format din doua etape, transcriptia si translatia prin care se produce fluxul informatiei genetice din ADN genic pana la proteina nousintetizata.
3 Fluxul informatiei genetice In realitate, fluxul informatiei genetice nu este in totalitate unidirectional Exista secvente de ADN-retrotranspozoni, care sunt transcrise mai intai in ARN si apoi sub actiunea unei revers transcriptaze sunt copiate sub forma de ADNc care se insera aleatoriu in ADN Exemplu de virus ARN care se insera prin reverstranscriptie in genomul ADN este vs. HIV.
4 Fluxul informatiei genetice - Etape I. Transferul informatiei genetice din ADN-ul cromozomial in citoplasma celulei. Acest transfer este realizat prin sinteza moleculelor de ARN mesager care, conform principiului complementaritatii bazelor azotate, respecta riguros structura primara a ADN-ului; II. Formarea nisei polizom unde se asambleaza specific aminoacizii in lanturile polipeptidice(pe baza ARNm,ARNt si ARNr)care duc la edificarea unei proteine ale carei structura si functie sunt specifice fiecarui individ si fiecarei specii; III. Materializarea informatiei genetice sub forma de molecule proteice, enzimatice, imunologice; IV. Finalizarea caracterelor anatomo-structurale. Realizate ontogenetic, controlate genetic si influentate de factorii de mediu, caracterele morfologice sunt particulare fiecarui individ si fiecarei specii.
5 Teoria semantica a fluxului informatiei genetice Biopolimerii celulari pot fi clasificati dupa gradul semnificatiei genetice si participarea la edificarea caracterului(zuckernandl si Pauling). Semantida primara - ADN-ul cromozomial are drept caracteristica dispunerea liniara a informatiei genetice - detine informatia primara a genelor; Semantida secundara ARN-ul celular are rolul de mesager intre nucleu si citoplasma - copiaza informatia genetica din ADN, folosita apoi in citoplasma pentru ordonarea aminoacizilor din structura polipeptidului; Proteinele, formate din lanturi polipeptidice sunt considerate semantide tertiare; Molecule episemantice - Glucidele si lipidele se sintetizeaza sub controlul unor enzime specifice; ele ca produs final nu exprima in totalitate informatia obtinuta din primele trei tipuri de semantide.
6 CODUL GENETIC Intelegerea fluxului de informatie genetica cere cunoasterea modului particular in care sunt codificate mesajele in structura ADN-ului cromozomial; Informatia genetica este inregistrata in moleculele acizilor nucleici sub forma unui cod genetic. Codul genetic este determinat de ordinea bazelor azotate in structura primara a ADN-ului. Orice informatie genetica este reprezentata conventional prin semne simboluri (A, G, C, T/U); Totalitatea semnelor conventionale care servesc la obtinerea unui mesaj genetic reprezinta alfabetul genetic;
7 CODUL GENETIC GENERALITATI 4 simboluri(a, G, C, T/U) = 20 aminoacizi? Watson si Crick au introdus in 1961 notiunea de codon unitatea de functie a codului genetic, format dintr-o succesiune de baze azotate, necesare pentru codificarea unui aminoacid din structura proteinelor; Initial s-a crezut ca unitatea de functie este reprezentata de un singur nucleotid, adica codonul univoc. Daca codonul ar specifica strict un aminoacid, numarul codonilor obtinuti ar fi de 4 x 1 = 4, deci ar fi specificati numai 4 aminoacizi, iar restul de 16 ar ramane nespecificati; Acelasi rationament poate fi aplicat si in cazul codonului biunivoc 4 x 4 = 16 codoni, respectiv 16 aminoacizi, admitand faptul ca ordinea bazelor modifica tipul de aminoacid; Nierenberg si Matthaei(1961) au demonstrat ca, dimensional, codonul este triunivoc, iar fiecare aminoacid este codificat de o tripleta, succesiunea a trei nucleotide adiacente; Prin aceasta asociere numarul combinatiilor posibile este de 4 x 4 x 4 = 64, care acopera si depaseste numarul aminoacizilor din structura proteinelor.
8 Descifrarea codului genetic Codul genetic a fost descris de catre Robert Holley, Har Gobind Khorana si Marshall Nirenberg in 1965 si a insemnat inceputul erei geneticii moleculare Codul a fost descifrat experimental folosind polinucleotide sintetice cu secventa cunoscuta de ARNm si analizand secventa de aminoacizi a proteinei sintetizate artificial De exemplu, ARNm sintetic al poliuracilului a produs un polipeptid alcatuit numai din fenilalanina, deci tripleta UUU codifica fenilalanina. Premiul Nobel pentru chimie in 1968

9 CODUL GENETIC Structura codului genetic
10 CARACTERISTICILE CODULUI GENETIC Unitatea de functie a codului genetic este codonul. El este format din secventa liniara a trei nucleotide; Reprezinta un sistem de corespondenta intre informatia din structura ADN-ului si un anumit aminoacid
11 CARACTERISTICILE CODULUI GENETIC Codul genetic este degenerat. Numarul combinatiilor posibile este 4³ = 64 codoni. Fata de numarul aminoacizilor avem un surplus de 44 de codoni. Dintre aceste triplete, multe participa la codificarea aceluiasi aminoacid. De exemplu, leucina este codificata de 6 codoni diferiti: UUA, UUG, CUU, CUC, CUA si CUG, iar arginina de 6 codoni: CGU, CGC, CGA, CGG, AGA si AGG. Acestia sunt codoni sinonimi iar codul genetic este degenerat, adica un aminoacid este specificat de mai multi codoni. Acest aspect se numeste redundanta informationala; Desi este un termen peiorativ, caracterul degenerat al codului genetic este un avantaj informational deoarece unele mutatii genice ce produc codoni sinonimi nu modifica proteina codificata de gena(mutatii silentioase sau izo-semantice) Aceasta degenerare este un sistem de protectie impotriva efectului mutatiilor.
12 CARACTERISTICILE CODULUI GENETIC Codul genetic inscris in molecula ADN-ului este recunoscut in structura ARN-ului mesager prin complementaritatea bazelor: C» G, G» C, T» A, A» U. Codul genetic nu este ambiguu. Un codon dat recunoaste un singur tip de aminoacid. Codificarea este inepuizabila. De exemplu, secventa de 10 dezoxiribonucleotide realizeaza 1 milion de combinatii sau fraze diferite, iar dintr-o secventa de 15 dezoxiribonucleotide pot rezulta cateva sute de milioane de combinatii diferite.
13 CARACTERISTICILE CODULUI GENETIC Mesajele au punctuatie. Semnele de punctuatie sunt reprezentate de un codon start sau initiator AUG, si 3 codoni denumiti nonsens(uaa, UAG, UGA) care semnifica oprirea sintezei proteice. Codonul initiator AUG(cu care se incepe cadrul deschis de lectura) specifica metionina. Codonii stop semnaleaza incheierea sintezei si eliberarea polipeptidului format(prin fixarea unor factori de eliberare).
14 CARACTERISTICILE CODULUI GENETIC Codoni start alternativi pot fi GUG sau UUG; acestia in mod obisnuit reprezinta valina si leucina dar in prezenta unor factori initiatori ai translatiei sunt tradusi drept metionina si formil-metionina Ceilalti 61 de codoni sunt codoni sens care semnifica cei 20 de aminoacizi.
15 CARACTERISTICILE CODULUI GENETIC Intre codoni nu exista semne de separare sau virgule. Citirea mesajului se face in flux continuu pana la codonul terminator. Intre codoni nu raman dezoxi-ribonucleotide izolate, neintegrate in componenta unui triplet functional. Codul genetic nu accepta suprapunere. Fiecare codon poseda bazele proprii, care nu participa in acelasi timp la structurarea altui codon.
16 CARACTERISTICILE CODULUI GENETIC Codul genetic este universal. Aceeiasi codoni codifica aceiasi aminoacizi la marea majoritate a genelor la animale, plante si micoorganisme, iar semnalele START si STOP sunt universale si ele. Exceptii de la universalitate ale codului genetic Acestea se refera mai ales la desemnarea unuia sau doi codoni terminatori drept codoni codificatori la nivelul: Genelor mitocondriale Aminoacizilor nestandardizati
17 Exceptii de la universalitatea codului genetic- Mitocondrii Gene mitocondriale La animale si microorganisme codonul UGA codifica triptofan Experiment: ARNm mitocondrial impreuna cu celelalte componente ale complexului citozolic de sinteza proteica(aminoacizi, enzime, ARNt, ribozomi) acesta nu va fi translat in proteine, deoarece codonul UGA codifica triptofan si nu oprirea sintezei lantului polipeptidic. Sinteza proteica se opreste acolo unde ar fi trebuit sa fie inserat aa. trp. La majoritatea mitocondriilor animale codonul AUA codifica metionina si nu izoleucina; Toate vertebratele au mitocondrii ce folosesc codonii AGA(arginina) si AGG(arginina) drept codoni terminatori ai lantului polipeptidic; Mitocondriile de drojdie folosesc toti codonii care incep cu CU pentru desemnarea treoninei in locul leucinei(cuu, CUC, CUA, CUG).
18 Exceptii de la universalitatea codului genetic-aminoacizii nestandardizati Aminoacizi nestandardizati Marea majoritate a proteinelor este asamblata din cei 20 de aminoacizi, chiar daca unii pot fi alterati prin fosforilare uneori; Cu toate acestea, au fost identificate doua cazuri in care un aminoacid care nu face parte dintre cei douazeci este inserat printr-un ARNt in lantul polipeptidic: Selenocisteina(aa 21) Acest aminoacid este codificat prin UGA. UGA este utilizat si ca terminator al lantului, dar complexul ribozomal este capabil sa discearna cand trebuie folosit ca selenocisteina, respectiv secventa terminator. Aceasta utilizare diferita a codonului a fost semnalata la Archea, eubacteria. La om s-au evidentiat 25 de proteine diferite continand seleniu. Pirolizina(aa 22) La cateva specii de Archea si de bacterii acest aminoacid este codificat prin UAG, dar poate actiona ca un codon stop si opreste si sinteza lantului proteic.
19 CARACTERISTICILE CODULUI GENETIC Codul genetic se modifica prin mutatie. Principalele tipuri de mutatii, limitate la un singur cistron constau din: insertia repetitiva, substitutia sau deletia unei baze din structura ADN-ului Daca intr-o celula prin mutatie se suprima sau se insera 1-2 nucleotide (sau un alt numar care nu este multiplu de 3) se produce o decalare a cadrului de lectura al genei care duce la aparitia altui codon. Toti codonii vor fi diferiti si se va sintetiza o proteina in care toti aminoacizii vor fi diferiti de la locul in care s-a produs mutatia(numita frameshift)


20 Mutatiile in codul genetic
21 Mutatiile in codul genetic Repetitii trinucleotidice Boala Huntington: insertii CAG care adauga serii de glutamina Ataxia Friedreich: repetitii GAA in primul intron Atrofia dentatorubral-pallidoluysiana (DRPLA), produsa prin amplificarea unor secvente CAG ce produce poliglutamina in proteina atrofina-1 Boala Kennedy, produsa prin amplificarea trinucleotidica a secventei CAG in primul exon al genei receptorului androgenic Sindromul X Fragil: insertii CGG pe cromozomul X; practic acestea nu sunt legate de arginina deoarece mutatia este localizata in regiunea 5 netranslata; CTG in distrofia miotrofica ; Ataxia spinocerebelara, cu repetitii CAG
22 Mutatiile in codul genetic Mutatii missense Anemia Sickle-cell: GAG se transforma in GTG in gena beta a hemoglobinei; APC: varianta missense GAC la GTC Cancer prostatic: mutatie missense GCC la ACC in gena steroid 5 alfa-reductaza Mutatii nonsens: β-thalassemia (β-globin gene) Boala McArdle s glicogenoza prin mutatie nonsens in gena miofosforilazei Deletia: Fibroza chistica deletia UUU
23 CARACTERISTICILE CODULUI GENETIC Amplificarea codului genetic Din 2001 au fost creati 40 de aminoacizi de sinteza H. Murakami si M. Sisido au emis teoria codonului format din 4 si 5 baze Steven A. Benner a construit al 65-lea codon functional(in vivo).
24 SINTEZA DE PROTEINE SI POLIPEPTIDE poate fi impartita in 2 etape Transcriptia informatiei genetice de la ADN catre moleculele de ARNm; Translatia/ Traductia informatiei genetice intr-un lant polipeptidic.
25 TRANSCRIPTIA INFORMATIEI GENETICE Prin asamblarea ribonucleotidelor in structura ARN-m, respectandu-se principiul complementaritatii bazelor se reproduce fidel, in negativ, informatia genetica din segmentul cistronului de ADN. Transcriptia ARN-m are loc in 3 etape: Initierea; Elongatia; Terminarea.
26 Diferente între replicare si transcriere Transcrierea, spre deosebire de replicare,care angajeaza simultan întregul cromozom, vizeaza numai anumite portiuni de ADN; Transcrierea are loc ori de cite ori este necesara o anumita proteina, deci nu este un eveniment unic, spre deosebire de replicare, care este un eveniment unic în viata celulei.
27 Cum poate ADN-ul sa codifice informatia cu privire la sinteza proteinelor? Ideea ca ARN-ul actioneaza ca intermediar a fost sugerata de urmatoarele constatari experimentale realizate la eucariote: 1.ADN-ul este în principal asociat cu cromozomii si se gaseste în nucleu, în timp ce ribozomii au sediul sintezei proteinelor în citoplasma; 2.ARN este sintetizat în nucleu; 3.ARN migreaza în citoplasma, unde sunt sintetizate proteinele; 4.Cantitatea de ARN este în general direct proportionala cu cantitatea de proteine din celula.
28 ARN polimeraza Sinteza ARN implica existenta unei enzime Enzima capabila sa sintetizeze ARN se numeste ARNpolimeraza(ARNpol), mai exact ARNpolimeraza-ADN-dependenta; ARN-P pentru a functiona necesita: 1.ADN dublu catenar 2.Cele 4 ribonucleozide 5 -trifosforilate NTP(ATP, GTP, CTP, UTP) 3.Mg2+.
29 Comparatie ARN-pol si ADN-pol Asemanari între ARN-polimeraza si ADN-polimeraza Sensul sintezei este întotdeauna 5-3. Necesita o matrita, 4 nucleotide si Mg2+. Diferente între ADN-polimeraza si ARN-polimeraza Spre deosebire de ADN-pol, ARNpol: -nu necesita primer pentru a initia sinteza -are o fidelitate mai mica (1 eroare la baze): face mai multe erori decat ADN-pol (dar eroarea persista doar o generatie de proteine, nu toata viata celulei, ca în cazul ADN-pol)
30 Tipuri de ARN-polimeraze Procariote Toate cele 3 tipuri de ARN sunt transcrise de aceeasi ARN-polimeraza Eucariote Fiecare tip de ARN este transcris de catre un tip diferit de ARN-polimeraza ARN-pol I(localizata în nucleol): ARNr 45 S ARN-poI II(localizata în nucleoplasma): ARNm, ARNsn ARN-pol III(localizata în nucleoplasma): ARNt si ARN 5S, ARNsn.
31 FAZA DE INITIERE A TRANSCRIPTIEI Incepe cu o secventa particulara de ADN denumita situs promotor, unde se ataseaza enzima ARN-polimeraza- ADN-dependenta, care initiaza transcriptia; ARN-polimeraza sintetizeaza acid nucleic in directia 5 la 3, iar citirea matritei de ADN se face in directia 3 la 5 ; La acest nivel se gaseste si o subunitate proteica denumita factor sigma, care are rolul de a mentine conformatia de dublu helix necesara recunoasterii situsului promotor; In momentul in care complexul ARN-polimeraza/factor sigma recunoaste corect promotorul, factorul sigma se disociaza de ARN polimeraza si enzima incepe sa despiralizeze helixul de ADN, formand o regiune de dezoxiribonucleotide neimperecheate, care serveste drept matrita pentru sinteza ARN-ului.
32 Initiation
33 Faza de initiere a transcriptiei ARN polimeraza de la E. coli este formata din 6 subunitati polipeptidice: 2 subunitati alfa; 1 beta; 1 beta prim; 1 omega Subunitatea sigma-asigura legarea specifica a ARN-pol la anumite secvente de ADN numite promotor fata de care creste afinitatea ARNpol si scade afinitatea enzimei pentru catena non-matrita; Primele 5 subunitati sunt strans unite in miezul enzimei Enzima miez se poate lega nespecific la ADN; Holoenzima ARN polimeraza recunoaste secventa initiala a unei gene, regiunea de start, care este cunoscuta sub denumirea de regiunea promotor; Informatia continuta de promotor face enzima ARN polimeraza capabila sa recunoasca precis punctul initial de la care incepe transcriptia;
34 ARN polimeraza - Rol subunitati α-leaga ADN(secvente reglatorii), proteine, ARN-pol; β-catalitic(leaga NTP si formeaza legaturile fosfatdiesterice; β - leaga ADN; σ recunoaste promotorul; Ω asambleaza ARN-pol. Beta si beta prim formeaza impreuna centrul activ al enzimei.
35 Structura promotorului Este alcatuit din regiunea de recunoastere situata in pozitia - 35 pb fata de situsul initial de transcriptie(tss). Are structura TTGACA si la nivelul ei se ataseaza ARN polimeraza. Aceasta structura se refera la catena superioara care nu va fi transcrisa. Polimeraza paraseste secventa -35 si se deplaseaza in aval spre o alta regiune conservata. Aceasta este a doua secventa de consens(tataat) si este situata in pozitia -10 fata de tss. Este denumita PRIBNOW BOX(PB)
36 Faza de initiere a transcriptiei Distanta dintre PB si secventa -35 este de aproximativ 16-18b; PB este regiunea unde polimeraza se ataseaza ferm de ADN si se considera ca este pozitia in care helixul de ADN se deschide pentru a permite imperecherea de recunoastere intre matrita de ADN si ARNm; Atasarea ARN-polimerazei este conditionata de o serie de factori de transcriptie; Regiunea deschisa se intinde de la PB pana la cateva pozitii inaintea punctului +1(tss); Numai una dintre cele doua catene de ADN este folosita drept matrita, catena numita antisens(3 5 ); Se obisnuieste sa se descrie secventa genica de pe catena sens; Transcriptul de ARN(5 3 ) va fi complementar ca directie si structura( cu exceptia U care inlocuieste T) cu catena sens; Primul nucleotid al transcriptului de ARN este de obicei o purina(a sau G) situata la capatul 5 ; Aceasta inseamna ca pe catena antisens a secventei de ADN care va fi transcrisa se va gasi la capatul 3 o pirimidina(t sauc).
37 Faza de initiere a transcriptiei Initierea transcriptiei se incheie odata cu formarea primei legaturi fosfat diesterice intre NTP+1 si NTP+2
38 Complexul de elongare-elementul cheie al elongarii lantului polinucleotidic Complexul de elongare cuprinde: -ARNpol (enzima miez, fara subunitatea σ) -ADN matrita, -ARN în crestere -topoizomeraza
39 Faza de elongatie In timp ce polimeraza se deplaseaza de-a lungul catenei antisens, pe lantul de ARN sunt adaugate resturi de ribonucleozid monofosfati(amp, CMP, GMP, sau UMP) la capatul 3 ; Aceste nucleotide rezulta din clivarea precursorilor ribonucleozid trifosfati(atp, CTP, GTP si UTP) de o molecula de pirofosfat(ppi); Aceasta inseamna ca la capatul 5 al transcriptului ARN se va gasi un nucleotid initiator diferit, cu un grup trifosfat la 5 ; Produsul transcriptiei este totdeauna un ARN(ARNt, ARNm, ARNsn); Pe masura ce lantul de ARN se elongheaza, enzima despiralizeaza ADN-ul, ataseaza ribonucleozidmonofosfati si elibereaza difosfatul;

40 Faza de elongatie
41 Faza de elongatie Subunitatea sigma se detaseaza de pe restul enzimei dupa ce s-au format 6-10 legaturi fosfodiesterice de NTP si se poate atasa la un alt complex polimerazic; Regiunea deschisa a ADN-ului se extinde pe o portiune de aprox. 15pb, deoarece dublul helix de ADN se reface in spatele enzimei sub actiunea topoizomerazei; Fiecare transcript in crestere va avea un capat 3 liber la care se poate atasa capatul 5 al unui nou nucleotid
42 Faza de elongatie
43 Etapa de terminare a transcriptiei Se realizeaza prin doua mecanisme: Ro dependent Ro independent Mecanismul ro dependent Ro este o helicaza ATP-dependenta care are rolul de a desface duplexul ADN-ARN; ARN-pol intalneste o zona bogata in GC si incetineste transcrierea; Proteina ro ajunge ARN-pol si disociaza ARN de ADN.
44 Etapa de terminare a transcriptiei Mecanismul ro independent: Situsurile ter sunt bogate in secvente palindromice GC cu structura simetrica in oglinda urmate de regiuni bogate in secvente AT(aprox. 6-8 resturi A) pe catena codificatoare); Secventa inversata, prin transcriere, genereaza un ARN in ac de par(prin formarea unei regiuni dicatenare), care se termina cu numeroase secvente uridilice(complementare cozii poli-a); ARN-pol se opreste cand ajunge la un situs ter; Pauza permite ARN sa formeze un ac de par; Acul de par cu coada poli-u distruge legaturile spatiale intre ARN si ARN-pol; Complexul de elongare este astfel disociat si elongarea se termina
45 TERMINATOR SEQUENCE
46 Transcriptia la eucariote Diferita fata de procariote Eucariotele au 3 ARN-polimeraze diferite, in vreme ce procariotele au decat una Eucariotele nu sintetizeaza ARNm policistronic; ARNm eucariot este prelucrat inainte de a fi utilizat pentru sinteza proteinelor.
47 Transcriptia la eucariote ARN polimeraza I (nucleol) ARNr 20S si ARNr 50S;); ARN polimeraza II sintetizeaza ARNm codificat in toate genele structurale si ARNsn; ARN polimeraza III - ARNt, ARNsn si ARNr 5S
48 Transcriptia la eucariote Cele trei polimeraze nu sunt capabile sa recunoasca si sa se lege de secvente specifice din promotori; Ele au nevoie de factorii de transcriptie, de care depind in totalitate; FT sunt proteine care recunosc specific secventele promotorilor si initiaza transcriptia; Majoritatea promotorilor care interactioneaza cu ARN polimeraza II contin o secventa conservata numita caseta HOGNESS( TATA box).

49 Transcriptia la eucariote-structura promotorului Este localizat in pozitia -25pb in amonte fata de tss(+1) si are secventa de consens 5 -TATAAA- 3 ; Este inconjurat de secvente GC; Factorii de transcriptie care se leaga in vecinatatea casetei TATA se atasaza pe rand; Primul care se atasaza este TF IID, cunoscut sub denumirea de proteina TATA deoarece recunoaste secventa TATA; Dupa atasarea TF IID este permisa atasarea TF A, TF B, ARNpolimeraza si TF IIE
50
51 Transcriptia la eucariote Secventa TATA determina situsul exact pentru initierea transcriptiei, dar nu este singurul; Alte doua secvente sunt caseta CAAT si caseta GC. Acestea au rol in modificarea expresiei genice prin controlul transcriptiei; I.CAAT box are secventa de consens GGCCAATCT si este localizata in pozitia -80 in amonte fata de +1. Mutatiile in CAAT reduc nivelul transcriptiei de la promotor in aval; Mutatiile in TATA box reduc partial activitatea transcriptionala, dar altereaza major situsul initial de transcriptie.
52 Transcriptia la eucariote II.GC box cuprinde 2 casete, fiecare cu secventa de consens GGGCGG; Una dintre casete este situata in amonte si cealalta in aval fata de caseta CAAT; TATA box si CG box interactioneaza cu cativa factori de transcriptie bine definiti; CAAT box este recunoscuta de diferite molecule proteice, unele dintre ele facilitand transcriptia; Secventele enhancer sunt situsuri de legare pentru o mare varietate de FT. Ele au rolul de a stimula rata transcriptiei.

53 Transcriptia la eucariote
54 Maturarea ARNm precursor Transcriptele primare de ARN formate sufera un proces de maturare nucleara care fac ARN disponibil si util translatiei 2 procese: Modificarea extremitatilor ARNm precursor cresterea stabilitatii(protectie la actiunea endonucleazelor), facilitarea transportului in citoplasma, recunoasterea si fixarea la subunitatea 40S a ribozomului Procesarea ARNm precursor-matisarea
55 Prelucrarea extremitatilor transcriptului primar Precoce in transcriptie este adaugat un nucleotid neobisnuit 7-metilguanilat la capatul 5 al ARN-pm, numit cap(capac, boneta, calota); Acesta are rolul de atasare a ribozomilor pentru translatie; In momentul in care transcriptia este aproape completa, la capatul 3 este atasata o serie de nucleotide cu adenina, numita coada poli-a; La aceasta coada se adauga proteina PABP(polyA binding protein)

56 Procesarea ARNm precursor Rolul sau este de a transporta ARN-m in afara nucleului si de a-l stabiliza impotriva degradarii in citoplasma; Dupa transcriptia ARN-pm, regiunile non-codante sunt excizate iar regiunile codante sunt unite prin complexe de ribonucleoproteine numite spliceozomi pentru a forma ARN-m matur, care trece apoi in citoplasma pentru a fi translat in proteine.
57 Matisarea ARNm Termen care semnifica innadirea a doua capete Se descriu urmatoarele Secvente de semnal 5 GT(GU in ARN) situs donor 3 AG situs acceptor Situsul de conexiune sau de legatura(branch site) cu nucleotidul A
 si formarea unei structuri lasou/bucla 3.")
58 Matisarea ARNm Etape 1. Sectionarea jonctiunii exon intron 5 GU 2. Atasarea nucleotidului terminal G la nucleotidul A al situsului de bransare(transesterificare) si formarea unei structuri lasou/bucla 3. Sectionarea intronului la jonctiunea 3 AG, indepartarea lui si unirea segmentelor de ARN exonic. Reactiile sunt mediate printr-un complex ARNproteine(spliceozomi) format din 5 tipuri de ARNsn(U1,U2,U4,U5,U6) si proteine
59 Matisarea alternativa Ordinea exonilor in ADN si transcriptul primar sunt pastrate in ARNm matur Exista, cu toate acestea, o flexibilitate in utilizarea exonilor, in sensul ca in celule diferite vor fi retinuti in ARNm numai o parte din exoni, restul fiind eliminati odata cu intronii Matisare alternativa Aceasta duce la tipuri diferite de ARN din aceeasi gena, deci tipuri diferite de polipeptide Aceste proteine pot fi inrudite(izoforme) ca in cazul mielinei sau troponinei C sau complet diferite(de exemplu gena calcitoninei produce in celulele C tiroidiene precursorul calcitoninei iar in creier precursorul unui neuromediator CGRP.
60 TRANSLATIA/TRADUCTIA INFORMATIEI GENETICE Elementele implicate in realizarea translatiei sunt: ARN-r ARN-t Aminoacizi Enzime Donatorii de energie
61 Elementele implicate in realizarea translatiei: ARN-t, aminoacizi In timpul translatiei ARN-t specifici fixeaza aminoacizii specifici, ii transfera pe ribozomi si ii insera intr-o pozitie specifica, in functie de mesajul din ARN-m; Cuplarea si transportul specific se fac in prezenta ATP-ului si a aminoacil-arn-t-sintetaza specifica; Activarea aminoacidului si transportul sau specific se desfasoara in doua etape:
62 Elementele implicate in realizarea translatiei: Etapele activarii aminoacidului - prima etapa corespunde activarii aminoacidului: R-CH-COOH + Enzima + ATP = R-CH-CO-O-AMP-E + PP NH2 NH2 aminoacid aminoacid activat - in etapa a doua are loc cuplarea aminoacidului activat cu ARN-t: R-CH-CO-O-AMP-E + ARN-t = R-CH-CO-O-ARN-t + AMP + E NH2 aminoacil-arn-t
63 Aminoacid activation
64 Elementele implicate in realizarea translatiei: ARN-t Acest proces este efectuat de catre portiunea anticodon a moleculelor de ARN-t complementare cu secventa de codoni de pe ARN-m; ARN-t este o molecula tridimensionala cu forma de frunza de trifoi inversata, cu lungime de aprox. 70 nucleotide;
65 Elementele implicate in realizarea translatiei: ARN-t La capatul 3 se poate atasa un aminoacid specific; La capatul opus se gaseste bucla anticodon, care contine o serie de trei baze complementare cu codonul de pe ARN-m, numit capatul de citire; Bucla DHU dihidrouridina recunoaste si leaga aminoacil- ARNt sintetaza Bucla TΨC contine pseudouridina( Ψ )si ribotimina(t) asigura legarea de subunitatea 50S a ribozomului

66 Elementele implicate in realizarea translatiei: ARN-r, ribozomi Ribozomii, descrisi de E. Pallade in 1953, au un diametru de Å, fiind formati din doua subunitati inegale, cuplate intre ele prin legaturi stabilizate de ionii de Mg²+;
67 Elementele implicate in realizarea translatiei: ARN-r, ribozomi La bacterii sunt formati din doua subunitati, diferite prin constanta de sedimentare: marea subunitate are 50S(ARNr 23S, 5S, 34 proteine) iar mica subunitate are constanta 30S(ARNr 16S, 21 tipuri de proteine); La om, subunitatile sunt 60S(ARNr 5S, 5,8S, 28S, 46 tipuri de proteine) si 40S(ARNr 18S, 33 tipuri de proteine). Fiecare subunitate este constituita din complexe de proteine(40%) si ARN-r(60%); Ribozomii au 4 situsuri functionale: Pe subunitatea mica situsul de fixare al extremitatii 5 a ARNm Pe subunitatea mare situsul P(peptidil) si A(aminoacil) unde se fixeaza moleculele de ARNt incarcate cu aminoacizi specifici si situsul de iesire E (exit) O molecula de ARN-m poate fi explorata de mai multi ribozomi, formand poliribozomul sau polizomul; Fixarea ARN-m de ribozomi se face la la subunitatea de 40S.
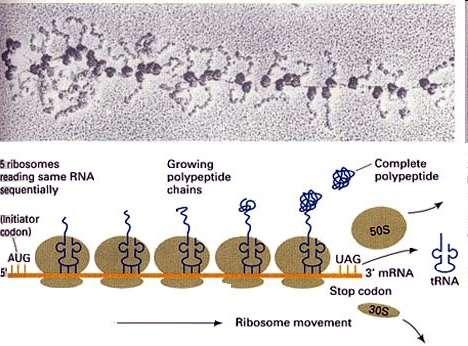
68 Polyribosome
69 TRANSLATIA/TRADUCTIA INFORMATIEI GENETICE FAZA DE INITIERE Pentru a initia translatia o unitate ribozomala de 30 S se leaga la o secventa scurta de ARNm numita situsul de legatura ribozomal; Secventa incepe intotdeauna cu semnalul AUG, denumit codon initiator; Anticodonul complementar de pe ARN-t va fi UAC si transporta specific numai metionina; Aceasta cuplare se realizeaza in prezenta a doi factori de initiere de natura proteica(fc si Fb);
70 TRANSLATIA/TRADUCTIA INFORMATIEI GENETICE FAZA DE INITIERE Subunitatea ribozomala de 50S se ataseaza apoi la complexul de initiere, iar factorii de initiere se desprind si se formeaza ribozomul de 70S; Se formeaza astfel capul de citire, favorizat de un alt factor de initiere Fa, in prezenta unei molecule de GTP; Subunitatea mare are doua situsuri alaturate: situsul aminoacil-a si situsul peptidil-p; Situsul P este primul ocupat de ARN-t adaptor.
71 TRANSLATIA/TRADUCTIA INFORMATIEI GENETICE FAZA DE ELONGATIE Soseste al doilea ARN-t adaptor, care transporta un aminoacid care care se fixeaza de ribozom in situsul A; In prezenta peptidiltransferazei din ribozom se stabileste o legatura peptidica intre cei doi aminoacizi; Dupa legarea aminoacizilor, ARN-t initiator paraseste situsul P ribozomal prin situsul de iesire E; Ribozomul avanseaza trei baze pe secventa ARN-m;

72 TRANSLATIA/TRADUCTIA INFORMATIEI GENETICE FAZA DE ELONGATIE Prin avansare, al doilea ARN-t trece pe situsul P; Situsul A este eliberat prin avansare, cuprinzand codonul urmator din secventa ARN-m; Soseste al treilea ARN-t adaptor, care ocupa situsul A; Se formeaza o noua legatura peptidica intre aminoacizii 2 si 3, dupa care ARN-t paraseste ribozomul; Lantul polipeptidic in crestere strabate un tunel in interiorul subunitatii 50S;

73 TRANSLATIA/TRADUCTIA INFORMATIEI GENETICE FAZA DE TERMINARE Acest proces continua de nenumarate ori in directia 3-5 pana cand ribozomul atinge un codon stop(uaa, UAG, UGA); Factorii eliberatori elibereaza lantul polipeptidic de pe ARN-t, iar cele doua subunitati ribozomale vor fi reciclate; In timpul fazei de elongatie proteina ia o forma functionala tridimensionala;
74 TRANSLATIA/TRADUCTIA INFORMATIEI GENETICE Acest mecanism asigura adaugarea ordonata si repetata a aminoacizilor pana la formarea lantului polipeptidic; Legarea aminoacizilor nu se face la intamplare, ci conform ordonarii codonilor din structura ARN-m, care asigura astfel specificitatea structurala si functionala a polipeptidului; Sinteza unui polipeptid in celula se desfasoara foarte rapid; de exemplu, pentru constituirea lantului polipeptidic al hemoglobinei cu 150 de aminoacizi este necesar un timp de aprox. 9 minute.
75 REGLAREA SINTEZEI PROTEICE Mecanismele de reglare au fost studiate cel mai bine la bacterii. Exemplul clasic: Modelul operonului descris de catre Francois Jacob si Jacques Monod la E.coli premiul Nobel Ei au demonstrat experimental si teoretic ca sinteza proteinelor nu este constanta in timp ci variaza in functie de necesitatile celulei
76 REGLAREA SINTEZEI PROTEICE Mecanismele de reglare au la baza actiunea unor enzime si au fost descrise pentru procariote si eucariote; Enzimele care intervin in reglarea sintezei proteice pot fi controlate prin 2 mecanisme: 1. Controlul genetic al sintezei enzimei; 2. Controlul activitatii enzimatice(inhibitie de feedback)
77 REGLAREA SINTEZEI PROTEICE A). CONTROL GENETIC Controlul genetic al sintezei enzimatice se refera la controlul transcriptiei ARN-m necesar pentru sinteza unei enzime; Acest control se realizeaza prin intermediul proteinelor reglatoare care se pot atasa la ADN si pot bloca sau amplifica functia ARN-polimerazei; Proteinele reglatoare pot functiona ca: -Represori(control genetic negativ) - Corepresori(operonul trp); - Inductori(operonul lac); -Activatori(control genetic pozitiv).
78 REGLAREA SINTEZEI PROTEICE 1). CONTROL GENETIC NEGATIV Represorii sunt proteine reglatoare care blocheaza transcriptia ARN-m; Represorii se leaga la o portiune de ADN numita operator care se gaseste in aval de promotor; Legarea proteinei reglatoare la operator blocheaza trecerea ARNpolimerazei de operator si transcrierea secventei de gene structurale ale enzimei control genetic negativ; Represorii sunt proteine alosterice care au un situs de legare pentru o molecula specifica numita: - corepresor- care modifica forma proteinei represor astfel incat ea sa se poata cupla cu operatorul si sa blocheze transcriptia; - inductor- altereaza forma proteinei reglatoare, blocheza cuplarea cu operatorul si permite transcriptia.
79 REGLAREA SINTEZEI PROTEICE PRIN COREPRESOR Corepresor - modifica forma proteinei represor astfel incat ea sa se poata cupla cu operatorul si sa blocheze transcriptia Este produsul final al sintezei proteice
80 REGLAREA SINTEZEI PROTEICE PRIN COREPRESOR Operonul represibil in absenta corepresorului (operon Trp) Pasul 1: Gena reglatoare codifica o proteina represoare inactiva; Pasul 2: Proteina represor inactivata nu se poate lega de regiunea operator a operonului;
 este capabila acum sa se lege de regiunea promotor a operonului; Pasul 4: ARN polimeraza este capabila acum sa transcrie cele")
81 REGLAREA SINTEZEI PROTEICE PRIN COREPRESOR Un operon represibil in absenta corepresorului (operon Trp) Pasul 3: deoarece proteina represor inactiva nu este capabila sa se ataseze de regiunea operator, ARN polimeraza (enzima responsabila pentru transcrierea genei) este capabila acum sa se lege de regiunea promotor a operonului; Pasul 4: ARN polimeraza este capabila acum sa transcrie cele 5 gene ce codifica enzima in ARNm; Pasul 5: prin transcrierea acestor gene sunt sintetizate cele 5 enzime necesare bacteriei pentru sinteza aminoacidului triptofan.

82 REGLAREA SINTEZEI PROTEICE PRIN COREPRESOR Un operon represibil in prezenta unui corepresor(operon Trp) Pasul 1: Gena reglatoare codifica o proteina represor inactiva; Pasul 2: Daca corepresorul triptofan este prezent, el se leaga de proteina represor inactiva; Pasul 3: Legarea corepresorului produce activarea proteinei represor inactive; Pasul 4: Proteina represor activata se leaga apoi la regiunea operator a operonului;
 nu mai este capabila sa se lege la regiunea promotoare a operonului; Pasul 6: Daca ARN polimeraza nu se mai leaga la regiunea promotoare, cele 5 gene")
83 REGLAREA SINTEZEI PROTEICE PRIN COREPRESOR Un operon represibil in prezenta unui corepresor (operonul Trp) Pasul 5: Odata cu legarea proteinei represor active la regiunea operatoare, ARN polimeraza (enzima responsabila pentru transcrierea genelor) nu mai este capabila sa se lege la regiunea promotoare a operonului; Pasul 6: Daca ARN polimeraza nu se mai leaga la regiunea promotoare, cele 5 gene enzimatice nu vor mai fi transcrise in ARNm; Pasul 7: In lipsa transcriptiei celor 5 gene, cele 5 enzime necesare pentru ca bacteria sa sintetizeze aminoacidul triptofan nu mai sunt sintetizate.
84 REGLAREA SINTEZEI PROTEICE PRIN INDUCTOR Inductor- altereaza forma proteinei reglatoare, blocheza cuplarea cu operatorul si permite transcriptia Este substanta care trebuie metabolizata
85 The lactose (lac) operon Operonul lac este necesar pentru transportul si metabolizarea lactozei la E.coli
86 REGLAREA SINTEZEI PROTEICE PRIN INDUCTOR Un operon inductibil in absenta inductorului (operonul lactoza) Pasul 1: Gena reglatoare codifica o proteina represor activa; Pasul 2: Proteina represor se leaga apoi la regiunea operator a operonului;
 nu sunt transcrise in ARNm; Pasul 5: In lipsa transcriptiei")
87 REGLAREA SINTEZEI PROTEICE PRIN INDUCTOR Un operon inductibil in absenta inductorului (operonul Lac) Pasul 3: ARN polimeraza nu se poate lega de regiunea promotor a operonului cind proteina represor activa este legata de regiunea operator; Pasul 4: Daca ARN polimeraza nu se leaga la regiunea promotor genele celor 3 enzime (Z, Y si A) nu sunt transcrise in ARNm; Pasul 5: In lipsa transcriptiei celor 3 gene enzimatice, cele 3 enzime necesare pentru utilizarea lactozei de catre bacterie nu sunt sintetizate.
88 REGLAREA SINTEZEI PROTEICE PRIN INDUCTOR Un operon inductibil in prezenta inductorului (operonul Lac) Pasul 1: Gena reglatoare codifica o proteina represor activa; Pasul 2: Lactoza, molecula inductoare se leaga de proteina represor activa; Pasul 3: Legarea inductorului inactiveaza proteina represor; Pasul 4: Proteina represor inactiva nu se mai poate lega la regiunea operatoare a operonului;
89 REGLAREA SINTEZEI PROTEICE PRIN INDUCTOR Un operon inductibil in prezenta inductorului(operonul Lac) Pasul 5: Deoarece proteina represor inactiva nu este capabila sa se lege la regiunea operatoare, ARN polimeraza va fi din nou capabila sa se lege la regiunea promotor a operonului; Pasul 6: ARN polimeraza este acum capabila sa transcrie cele 3 gene enzimatice (Z, Y si A) in ARNm; Pasul 7: Odata cu transcrierea acestor gene sunt sintetizate cele 3 enzime necesare bacteriei pentru a utiliza lactoza.
90 REGLAREA SINTEZEI PROTEICE 2). CONTROL GENETIC POZITIV Activatorii sunt proteine reglatoare care promoveaza sau initiaza transcriptia; Activatorii controleaza gene care au un promotor la care ARN-polimeraza nu se poate atasa; Activatorul este o proteina allosterica care se poate lega de situsul activator de legatura; ARN-polimeraza nu se va putea lega de promotor si nu va putea transcrie genele; Legarea unei proteine inductor la activator ii va modifica forma si aceasta se va atasa la situsul activator de legatura; ARN-polimeraza se leaga de promotor si initiaza transcriptia control genetic pozitiv.

91 REGLAREA SINTEZEI PROTEICE CONTROL GENETIC POZITIV O proteina activatoare in absenta inductorului Gena reglatoare produce o proteina activatoare inactiva; Proteina activatoare nu se poate lega de situsul de legare activator; ARN polimeraza nu se poate lega la promotor; Gena X nu se transcrie iar enzima nu se sintetizeaza.

92 REGLAREA SINTEZEI PROTEICE O proteina activatoare in prezenta inductorului pasul 1 Gena reglatoare produce o proteina activatoare inactiva; Inductorul se leaga la activator; Legarea inductorului produce modificarea formei activatorului; Activatorul se poate lega la situsul activator de reglare.

93 REGLAREA SINTEZEI PROTEICE O proteina activatoare in prezenta inductorului pas 2 Legarea activatorului evidentiaza situsul de legare al ARN polimerazei la promotor; ARN polimeraza se leaga la promotor si este capabila sa transcrie gena X in ARN-m; Sinteza enzimei.
94 REGLAREA SINTEZEI PROTEICE B). CONTROLUL ACTIVITATII ENZIMATICE (Inhibitie prin feed-back) Inhibitie non-competitiva; Inhibitie competitiva.

95 REGLAREA SINTEZEI PROTEICE Inhibitia noncompetitiva prin enzime allosterice Produsul final (inhibitor) al caii metabolice se combina cu situsul allosteric al enzimei, aceasta altereaza situsul activ al enzimei astfel incit ea nu se mai poate lega la substratul initial al caii; Aceasta blocheaza sinteza produsului final.

96 REGLAREA SINTEZEI PROTEICE Inhibitia competitiva a activitatii enzimatice Produsul final (inhibitor) al caii metabolice se leaga la situsul activ al primei enzime a caii; Ca rezultat enzima nu se mai poate lega la substratul initiator al caii metabolice.
97 Exista diferente majore intre reglarea sintezei la eucariote si procariote Eucariotele au histone si cromozomii sunt stans infasurati in cromatina, iar procariotele au proteine legate direct de ADN care se poate exprima mult mai usor; Procariotele nu au introni iar fiecare gena are propria sa regiune reglatoare; Eucariotele folosesc factori de transcriptie si regiuni promotoare pentru reglarea controlata a fiecarei gene; Eucariotele au introni; ca urmare se produce procesarea ARNm, care este folosit mai ales in reglarea expresiei genice la eucariote.
98 Reglarea expresiei genice se produce la mai multe niveluri CONTROL TRANSCRIPTIONAL CONTROL POSTTRANSCRIPTIONAL
99 CONTROL TRANSCRIPTIONAL Punctul primar de control este reglarea activitatii ARN-polimerazei II Activitatea ei este controlata de: Elemente cis-reglatoare Factori de transcriptie transreglatori Controlul la distanta al expresiei genice prin enhancers sau silencers Reglare ca raspuns la semnalele extracelulare liganzi Selectarea promotorilor pentru gene care au mai multi promotori
100 CONTROL POSTTRANSCRIPTIONAL Sunt mecanisme secundare care controleaza expresia genica dupa transcriptie. Acestea cuprind: Controlul procesarii ARN; Controlul translational; Controlul degradarii mrna; Controlul activitatii proteice;
101 I. Controlul procesarii ARN Este limitat la eucariote si determina cum si cand este splitat sau procesat transcriptul primar pentru a forma ARNm; De exemplu, unele transcripturi de ARN produc diferite tipuri de ARNm din aceeasi gena, in diferite tipuri de celule numai prin folosirea selectiva a intronilor-matisare alternativa
102 II. Controlul translational Determina care ARNm va fi translat in proteine si cand; De exemplu, translatia poate fi inhibata de ARNm cuplat cu diferite proteine specifice sau stimulata de secvente nucleotidice speciale de la capatul ARNm, care faciliteaza cuplarea ribozomilor.
103 II.Controlul translational Se realizeaza prin urmatoarele mecanisme: Adaugarea secventei cap la extremitatea 5 a ARNm cu rolul de a-l proteja de actiunea 5 a exonucleazelor si de atasare a ribozomilor. Procesul de splicing Adaugarea cozii poli-a(poliadenilare) la capatul3 actioneaza protector impotriva exonucleazelor si stimuleaza viteza translatiei; Procesul de editare al ARNm este un mecanism evolutiv prin care se modifica structura ARNm in timpul procesului de maturare(modificari nucleozidice, deaminare, editare la virusuri)
104 III. Controlul degradarii ARNm Se realizeaza prin stabilizarea sau destabilizarea selectiva unor tipuri diferite de ARNm. O proteina care este necesara in cantitati mari pe o perioada lunga de timp are un ARNm foarte stabil. De exemplu, ARNm al β globinei are un timp de injumatatire de 10 ore(timp de injumatatire timpul necesar pentru ca 50% dintr molecule sa fie degradate).alte proteine sunt necesare numai tranzitor si au un ARNm cu timp de injumatatire de cateva minute.
105 IV. Controlul activitatii proteice Presupune activarea si inactivarea selectiva, modificarea moleculelor proteice specifice dintro celula sau un anumit tip de celula, influentand cand si cum actioneaza o proteina. De exemplu - unele proteine sunt necesare pentru evenimente legate de dezvoltare si actioneaza numai in anumite celule sau intr-un moment specific al dezvoltarii(hbe, HbF). Aceste proteine produc efecte profunde asupra altor celule si trebuie sa fie inactivate imediat dupa ce au actionat, deoarece pot produce anomalii de dezvoltare.
Controlul expresiei genice
 Controlul expresiei genice REGLAREA EXPRESIEI GENICE!!! Alegerea genei pentru transcripţie!!! Activarea secvenţei de ADN!!! Controlul activităţii ARN-polimerazei II!!! Controlul cantităţii şi calităţii
Controlul expresiei genice REGLAREA EXPRESIEI GENICE!!! Alegerea genei pentru transcripţie!!! Activarea secvenţei de ADN!!! Controlul activităţii ARN-polimerazei II!!! Controlul cantităţii şi calităţii
(a) se numeşte derivata parţială a funcţiei f în raport cu variabila x i în punctul a.
 se numeşte derivata parţială a funcţiei f în raport cu variabila x i în punctul a.") Definiţie Spunem că: i) funcţia f are derivată parţială în punctul a în raport cu variabila i dacă funcţia de o variabilă ( ) are derivată în punctul a în sens obişnuit (ca funcţie reală de o variabilă
Definiţie Spunem că: i) funcţia f are derivată parţială în punctul a în raport cu variabila i dacă funcţia de o variabilă ( ) are derivată în punctul a în sens obişnuit (ca funcţie reală de o variabilă
Metode iterative pentru probleme neliniare - contractii
 Metode iterative pentru probleme neliniare - contractii Problemele neliniare sunt in general rezolvate prin metode iterative si analiza convergentei acestor metode este o problema importanta. 1 Contractii
Metode iterative pentru probleme neliniare - contractii Problemele neliniare sunt in general rezolvate prin metode iterative si analiza convergentei acestor metode este o problema importanta. 1 Contractii
Repere pentru pregătirea către totalizarea II la Biologia Moleculară 1. Definiţi noţiunile
 Repere pentru pregătirea către totalizarea II la Biologia Moleculară 1. Definiţi noţiunile ADN-polimerază Aminoacid Anticodon Apoptoza ARN-polimeraza I ARN-polimeraza II ARN-polimeraza III CAP-are Catena
Repere pentru pregătirea către totalizarea II la Biologia Moleculară 1. Definiţi noţiunile ADN-polimerază Aminoacid Anticodon Apoptoza ARN-polimeraza I ARN-polimeraza II ARN-polimeraza III CAP-are Catena
4. Elemente de biologie celulară şi moleculară
 4. Elemente de biologie celulară şi moleculară O caracteristică esenţială a materiei vii este reprezentată de structura sa celulară, alături de încă două proprietăţi fundamentale: metabolismul şi reproducerea.
4. Elemente de biologie celulară şi moleculară O caracteristică esenţială a materiei vii este reprezentată de structura sa celulară, alături de încă două proprietăţi fundamentale: metabolismul şi reproducerea.
Planul determinat de normală şi un punct Ecuaţia generală Plane paralele Unghi diedru Planul determinat de 3 puncte necoliniare
 1 Planul în spaţiu Ecuaţia generală Plane paralele Unghi diedru 2 Ecuaţia generală Plane paralele Unghi diedru Fie reperul R(O, i, j, k ) în spaţiu. Numim normala a unui plan, un vector perpendicular pe
1 Planul în spaţiu Ecuaţia generală Plane paralele Unghi diedru 2 Ecuaţia generală Plane paralele Unghi diedru Fie reperul R(O, i, j, k ) în spaţiu. Numim normala a unui plan, un vector perpendicular pe
Curs 10 Funcţii reale de mai multe variabile reale. Limite şi continuitate.
 Curs 10 Funcţii reale de mai multe variabile reale. Limite şi continuitate. Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie p, q N. Fie funcţia f : D R p R q. Avem următoarele
Curs 10 Funcţii reale de mai multe variabile reale. Limite şi continuitate. Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie p, q N. Fie funcţia f : D R p R q. Avem următoarele
Aplicaţii ale principiului I al termodinamicii la gazul ideal
 Aplicaţii ale principiului I al termodinamicii la gazul ideal Principiul I al termodinamicii exprimă legea conservării şi energiei dintr-o formă în alta şi se exprimă prin relaţia: ΔUQ-L, unde: ΔU-variaţia
Aplicaţii ale principiului I al termodinamicii la gazul ideal Principiul I al termodinamicii exprimă legea conservării şi energiei dintr-o formă în alta şi se exprimă prin relaţia: ΔUQ-L, unde: ΔU-variaţia
5.4. MULTIPLEXOARE A 0 A 1 A 2
 5.4. MULTIPLEXOARE Multiplexoarele (MUX) sunt circuite logice combinaţionale cu m intrări şi o singură ieşire, care permit transferul datelor de la una din intrări spre ieşirea unică. Selecţia intrării
5.4. MULTIPLEXOARE Multiplexoarele (MUX) sunt circuite logice combinaţionale cu m intrări şi o singură ieşire, care permit transferul datelor de la una din intrări spre ieşirea unică. Selecţia intrării
TRANSCRIPŢIA GENETICĂ
 TRANSCRIPŢIA GENETICĂ principii, etape, ARN polimeraze, promotori şi terminatori, factori de transcripţie (NOTE DE CURS BM3/BM4) - procesarea ARNm la eucariote (NOTE DE CURS BM4) Atât celulele procariote
TRANSCRIPŢIA GENETICĂ principii, etape, ARN polimeraze, promotori şi terminatori, factori de transcripţie (NOTE DE CURS BM3/BM4) - procesarea ARNm la eucariote (NOTE DE CURS BM4) Atât celulele procariote
 4. CIRCUITE LOGICE ELEMENTRE 4.. CIRCUITE LOGICE CU COMPONENTE DISCRETE 4.. PORŢI LOGICE ELEMENTRE CU COMPONENTE PSIVE Componente electronice pasive sunt componente care nu au capacitatea de a amplifica
4. CIRCUITE LOGICE ELEMENTRE 4.. CIRCUITE LOGICE CU COMPONENTE DISCRETE 4.. PORŢI LOGICE ELEMENTRE CU COMPONENTE PSIVE Componente electronice pasive sunt componente care nu au capacitatea de a amplifica
V.7. Condiţii necesare de optimalitate cazul funcţiilor diferenţiabile
 Metode de Optimizare Curs V.7. Condiţii necesare de optimalitate cazul funcţiilor diferenţiabile Propoziţie 7. (Fritz-John). Fie X o submulţime deschisă a lui R n, f:x R o funcţie de clasă C şi ϕ = (ϕ,ϕ
Metode de Optimizare Curs V.7. Condiţii necesare de optimalitate cazul funcţiilor diferenţiabile Propoziţie 7. (Fritz-John). Fie X o submulţime deschisă a lui R n, f:x R o funcţie de clasă C şi ϕ = (ϕ,ϕ
BIOCHIMIA ACIZILOR NUCLEICI
 BIOCHIMIA ACIZILOR NUCLEICI Baze pirimidinice: pirimidina; citozina (2-oxo-4-amino-pirimidina); uracilul (2,4-dioxo-pirimidina); timina (5-metil-2,4-dioxo-pirimidina). Baze purinice: purina (imidazolpirimidină);
BIOCHIMIA ACIZILOR NUCLEICI Baze pirimidinice: pirimidina; citozina (2-oxo-4-amino-pirimidina); uracilul (2,4-dioxo-pirimidina); timina (5-metil-2,4-dioxo-pirimidina). Baze purinice: purina (imidazolpirimidină);
 10. STABILIZATOAE DE TENSIUNE 10.1 STABILIZATOAE DE TENSIUNE CU TANZISTOAE BIPOLAE Stabilizatorul de tensiune cu tranzistor compară în permanenţă valoare tensiunii de ieşire (stabilizate) cu tensiunea
10. STABILIZATOAE DE TENSIUNE 10.1 STABILIZATOAE DE TENSIUNE CU TANZISTOAE BIPOLAE Stabilizatorul de tensiune cu tranzistor compară în permanenţă valoare tensiunii de ieşire (stabilizate) cu tensiunea
5. FUNCŢII IMPLICITE. EXTREME CONDIŢIONATE.
 5 Eerciţii reolvate 5 UNCŢII IMPLICITE EXTREME CONDIŢIONATE Eerciţiul 5 Să se determine şi dacă () este o funcţie definită implicit de ecuaţia ( + ) ( + ) + Soluţie ie ( ) ( + ) ( + ) + ( )R Evident este
5 Eerciţii reolvate 5 UNCŢII IMPLICITE EXTREME CONDIŢIONATE Eerciţiul 5 Să se determine şi dacă () este o funcţie definită implicit de ecuaţia ( + ) ( + ) + Soluţie ie ( ) ( + ) ( + ) + ( )R Evident este
Catedra Biologie moleculară şi Genetică umană
 Catedra Biologie moleculară şi Genetică umană biologiemoleculara.usmf.md acces: bmgu 1 Tema 1 Macromoleculele = biopolimerii Tipuri Structură Biogeneză Proprietăţi Funcţii Localizare Interacţiuni 2 Functiile
Catedra Biologie moleculară şi Genetică umană biologiemoleculara.usmf.md acces: bmgu 1 Tema 1 Macromoleculele = biopolimerii Tipuri Structură Biogeneză Proprietăţi Funcţii Localizare Interacţiuni 2 Functiile
Biochimia moleculelor informaţionale. 11.I Metabolismul ADN-ului 3 Degradarea ADN-ului si a nucleotidelor
 Biochimia moleculelor informaţionale 11.I. 2018-1100-1500 Metabolismul ADN-ului 3 Degradarea ADN-ului si a nucleotidelor Catabolismul ADN-ului Structura şi mecanismele de reparaţie fac ca molecula de ADN
Biochimia moleculelor informaţionale 11.I. 2018-1100-1500 Metabolismul ADN-ului 3 Degradarea ADN-ului si a nucleotidelor Catabolismul ADN-ului Structura şi mecanismele de reparaţie fac ca molecula de ADN
DISTANŢA DINTRE DOUĂ DREPTE NECOPLANARE
 DISTANŢA DINTRE DOUĂ DREPTE NECOPLANARE ABSTRACT. Materialul prezintă o modalitate de a afla distanţa dintre două drepte necoplanare folosind volumul tetraedrului. Lecţia se adresează clasei a VIII-a Data:
DISTANŢA DINTRE DOUĂ DREPTE NECOPLANARE ABSTRACT. Materialul prezintă o modalitate de a afla distanţa dintre două drepte necoplanare folosind volumul tetraedrului. Lecţia se adresează clasei a VIII-a Data:
Curs 4 Serii de numere reale
 Curs 4 Serii de numere reale Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Criteriul rădăcinii sau Criteriul lui Cauchy Teoremă (Criteriul rădăcinii) Fie x n o serie cu termeni
Curs 4 Serii de numere reale Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Criteriul rădăcinii sau Criteriul lui Cauchy Teoremă (Criteriul rădăcinii) Fie x n o serie cu termeni
Curs 14 Funcţii implicite. Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi"
 Curs 14 Funcţii implicite Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie F : D R 2 R o funcţie de două variabile şi fie ecuaţia F (x, y) = 0. (1) Problemă În ce condiţii ecuaţia
Curs 14 Funcţii implicite Facultatea de Hidrotehnică Universitatea Tehnică "Gh. Asachi" Iaşi 2014 Fie F : D R 2 R o funcţie de două variabile şi fie ecuaţia F (x, y) = 0. (1) Problemă În ce condiţii ecuaţia
Seminariile Capitolul X. Integrale Curbilinii: Serii Laurent şi Teorema Reziduurilor
 Facultatea de Matematică Calcul Integral şi Elemente de Analiă Complexă, Semestrul I Lector dr. Lucian MATICIUC Seminariile 9 20 Capitolul X. Integrale Curbilinii: Serii Laurent şi Teorema Reiduurilor.
Facultatea de Matematică Calcul Integral şi Elemente de Analiă Complexă, Semestrul I Lector dr. Lucian MATICIUC Seminariile 9 20 Capitolul X. Integrale Curbilinii: Serii Laurent şi Teorema Reiduurilor.
Analiza în curent continuu a schemelor electronice Eugenie Posdărăscu - DCE SEM 1 electronica.geniu.ro
 Analiza în curent continuu a schemelor electronice Eugenie Posdărăscu - DCE SEM Seminar S ANALA ÎN CUENT CONTNUU A SCHEMELO ELECTONCE S. ntroducere Pentru a analiza în curent continuu o schemă electronică,
Analiza în curent continuu a schemelor electronice Eugenie Posdărăscu - DCE SEM Seminar S ANALA ÎN CUENT CONTNUU A SCHEMELO ELECTONCE S. ntroducere Pentru a analiza în curent continuu o schemă electronică,
a n (ζ z 0 ) n. n=1 se numeste partea principala iar seria a n (z z 0 ) n se numeste partea
 n. n=1 se numeste partea principala iar seria a n (z z 0 ) n se numeste partea") Serii Laurent Definitie. Se numeste serie Laurent o serie de forma Seria n= (z z 0 ) n regulata (tayloriana) = (z z n= 0 ) + n se numeste partea principala iar seria se numeste partea Sa presupunem ca,
Serii Laurent Definitie. Se numeste serie Laurent o serie de forma Seria n= (z z 0 ) n regulata (tayloriana) = (z z n= 0 ) + n se numeste partea principala iar seria se numeste partea Sa presupunem ca,
a. 11 % b. 12 % c. 13 % d. 14 %
 1. Un motor termic funcţionează după ciclul termodinamic reprezentat în sistemul de coordonate V-T în figura alăturată. Motorul termic utilizează ca substanţă de lucru un mol de gaz ideal având exponentul
1. Un motor termic funcţionează după ciclul termodinamic reprezentat în sistemul de coordonate V-T în figura alăturată. Motorul termic utilizează ca substanţă de lucru un mol de gaz ideal având exponentul
III. Serii absolut convergente. Serii semiconvergente. ii) semiconvergentă dacă este convergentă iar seria modulelor divergentă.
 semiconvergentă dacă este convergentă iar seria modulelor divergentă.") III. Serii absolut convergente. Serii semiconvergente. Definiţie. O serie a n se numeşte: i) absolut convergentă dacă seria modulelor a n este convergentă; ii) semiconvergentă dacă este convergentă iar
III. Serii absolut convergente. Serii semiconvergente. Definiţie. O serie a n se numeşte: i) absolut convergentă dacă seria modulelor a n este convergentă; ii) semiconvergentă dacă este convergentă iar
Esalonul Redus pe Linii (ERL). Subspatii.
. Subspatii.") Seminarul 1 Esalonul Redus pe Linii (ERL). Subspatii. 1.1 Breviar teoretic 1.1.1 Esalonul Redus pe Linii (ERL) Definitia 1. O matrice A L R mxn este in forma de Esalon Redus pe Linii (ERL), daca indeplineste
Seminarul 1 Esalonul Redus pe Linii (ERL). Subspatii. 1.1 Breviar teoretic 1.1.1 Esalonul Redus pe Linii (ERL) Definitia 1. O matrice A L R mxn este in forma de Esalon Redus pe Linii (ERL), daca indeplineste
Curs 1 Şiruri de numere reale
 Bibliografie G. Chiorescu, Analiză matematică. Teorie şi probleme. Calcul diferenţial, Editura PIM, Iaşi, 2006. R. Luca-Tudorache, Analiză matematică, Editura Tehnopress, Iaşi, 2005. M. Nicolescu, N. Roşculeţ,
Bibliografie G. Chiorescu, Analiză matematică. Teorie şi probleme. Calcul diferenţial, Editura PIM, Iaşi, 2006. R. Luca-Tudorache, Analiză matematică, Editura Tehnopress, Iaşi, 2005. M. Nicolescu, N. Roşculeţ,
MARCAREA REZISTOARELOR
 1.2. MARCAREA REZISTOARELOR 1.2.1 MARCARE DIRECTĂ PRIN COD ALFANUMERIC. Acest cod este format din una sau mai multe cifre şi o literă. Litera poate fi plasată după grupul de cifre (situaţie în care valoarea
1.2. MARCAREA REZISTOARELOR 1.2.1 MARCARE DIRECTĂ PRIN COD ALFANUMERIC. Acest cod este format din una sau mai multe cifre şi o literă. Litera poate fi plasată după grupul de cifre (situaţie în care valoarea
Metode de interpolare bazate pe diferenţe divizate
 Metode de interpolare bazate pe diferenţe divizate Radu Trîmbiţaş 4 octombrie 2005 1 Forma Newton a polinomului de interpolare Lagrange Algoritmul nostru se bazează pe forma Newton a polinomului de interpolare
Metode de interpolare bazate pe diferenţe divizate Radu Trîmbiţaş 4 octombrie 2005 1 Forma Newton a polinomului de interpolare Lagrange Algoritmul nostru se bazează pe forma Newton a polinomului de interpolare
Curs 4 Nucleul. Acizi nucleici. Notiuni de genetica moleculara
 Biologie Celulara si Histologie Curs 4 Nucleul. Acizi nucleici. Notiuni de genetica moleculara Cuprins Nucleul celular structura Acizi nucleici Transcriptia, translatia informatiei genetice; sinteza de
Biologie Celulara si Histologie Curs 4 Nucleul. Acizi nucleici. Notiuni de genetica moleculara Cuprins Nucleul celular structura Acizi nucleici Transcriptia, translatia informatiei genetice; sinteza de
Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor X) functia f 1
 functia f 1") Functii definitie proprietati grafic functii elementare A. Definitii proprietatile functiilor. Fiind date doua multimi X si Y spunem ca am definit o functie (aplicatie) pe X cu valori in Y daca fiecarui
Functii definitie proprietati grafic functii elementare A. Definitii proprietatile functiilor. Fiind date doua multimi X si Y spunem ca am definit o functie (aplicatie) pe X cu valori in Y daca fiecarui
riptografie şi Securitate
 riptografie şi Securitate - Prelegerea 12 - Scheme de criptare CCA sigure Adela Georgescu, Ruxandra F. Olimid Facultatea de Matematică şi Informatică Universitatea din Bucureşti Cuprins 1. Schemă de criptare
riptografie şi Securitate - Prelegerea 12 - Scheme de criptare CCA sigure Adela Georgescu, Ruxandra F. Olimid Facultatea de Matematică şi Informatică Universitatea din Bucureşti Cuprins 1. Schemă de criptare
Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor
 Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor. Fiind date doua multimi si spunem ca am definit o functie (aplicatie) pe cu valori in daca fiecarui element
Functii definitie, proprietati, grafic, functii elementare A. Definitii, proprietatile functiilor. Fiind date doua multimi si spunem ca am definit o functie (aplicatie) pe cu valori in daca fiecarui element
Asupra unei inegalităţi date la barajul OBMJ 2006
 Asupra unei inegalităţi date la barajul OBMJ 006 Mircea Lascu şi Cezar Lupu La cel de-al cincilea baraj de Juniori din data de 0 mai 006 a fost dată următoarea inegalitate: Fie x, y, z trei numere reale
Asupra unei inegalităţi date la barajul OBMJ 006 Mircea Lascu şi Cezar Lupu La cel de-al cincilea baraj de Juniori din data de 0 mai 006 a fost dată următoarea inegalitate: Fie x, y, z trei numere reale
1.7. AMPLIFICATOARE DE PUTERE ÎN CLASA A ŞI AB
 1.7. AMLFCATOARE DE UTERE ÎN CLASA A Ş AB 1.7.1 Amplificatoare în clasa A La amplificatoarele din clasa A, forma de undă a tensiunii de ieşire este aceeaşi ca a tensiunii de intrare, deci întreg semnalul
1.7. AMLFCATOARE DE UTERE ÎN CLASA A Ş AB 1.7.1 Amplificatoare în clasa A La amplificatoarele din clasa A, forma de undă a tensiunii de ieşire este aceeaşi ca a tensiunii de intrare, deci întreg semnalul
Seminar 5 Analiza stabilității sistemelor liniare
 Seminar 5 Analiza stabilității sistemelor liniare Noțiuni teoretice Criteriul Hurwitz de analiză a stabilității sistemelor liniare În cazul sistemelor liniare, stabilitatea este o condiție de localizare
Seminar 5 Analiza stabilității sistemelor liniare Noțiuni teoretice Criteriul Hurwitz de analiză a stabilității sistemelor liniare În cazul sistemelor liniare, stabilitatea este o condiție de localizare
Sisteme diferenţiale liniare de ordinul 1
 1 Metoda eliminării 2 Cazul valorilor proprii reale Cazul valorilor proprii nereale 3 Catedra de Matematică 2011 Forma generală a unui sistem liniar Considerăm sistemul y 1 (x) = a 11y 1 (x) + a 12 y 2
1 Metoda eliminării 2 Cazul valorilor proprii reale Cazul valorilor proprii nereale 3 Catedra de Matematică 2011 Forma generală a unui sistem liniar Considerăm sistemul y 1 (x) = a 11y 1 (x) + a 12 y 2
V O. = v I v stabilizator
 Stabilizatoare de tensiune continuă Un stabilizator de tensiune este un circuit electronic care păstrează (aproape) constantă tensiunea de ieșire la variaţia între anumite limite a tensiunii de intrare,
Stabilizatoare de tensiune continuă Un stabilizator de tensiune este un circuit electronic care păstrează (aproape) constantă tensiunea de ieșire la variaţia între anumite limite a tensiunii de intrare,
5.5. REZOLVAREA CIRCUITELOR CU TRANZISTOARE BIPOLARE
 5.5. A CIRCUITELOR CU TRANZISTOARE BIPOLARE PROBLEMA 1. În circuitul din figura 5.54 se cunosc valorile: μa a. Valoarea intensității curentului de colector I C. b. Valoarea tensiunii bază-emitor U BE.
5.5. A CIRCUITELOR CU TRANZISTOARE BIPOLARE PROBLEMA 1. În circuitul din figura 5.54 se cunosc valorile: μa a. Valoarea intensității curentului de colector I C. b. Valoarea tensiunii bază-emitor U BE.
SERII NUMERICE. Definiţia 3.1. Fie (a n ) n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0
 n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0") SERII NUMERICE Definiţia 3.1. Fie ( ) n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0 şirul definit prin: s n0 = 0, s n0 +1 = 0 + 0 +1, s n0 +2 = 0 + 0 +1 + 0 +2,.......................................
SERII NUMERICE Definiţia 3.1. Fie ( ) n n0 (n 0 IN) un şir de numere reale şi (s n ) n n0 şirul definit prin: s n0 = 0, s n0 +1 = 0 + 0 +1, s n0 +2 = 0 + 0 +1 + 0 +2,.......................................
Laborator 11. Mulţimi Julia. Temă
 Laborator 11 Mulţimi Julia. Temă 1. Clasa JuliaGreen. Să considerăm clasa JuliaGreen dată de exemplu la curs pentru metoda locului final şi să schimbăm numărul de iteraţii nriter = 100 în nriter = 101.
Laborator 11 Mulţimi Julia. Temă 1. Clasa JuliaGreen. Să considerăm clasa JuliaGreen dată de exemplu la curs pentru metoda locului final şi să schimbăm numărul de iteraţii nriter = 100 în nriter = 101.
Integrala nedefinită (primitive)
") nedefinita nedefinită (primitive) nedefinita 2 nedefinita februarie 20 nedefinita.tabelul primitivelor Definiţia Fie f : J R, J R un interval. Funcţia F : J R se numeşte primitivă sau antiderivată a funcţiei
nedefinita nedefinită (primitive) nedefinita 2 nedefinita februarie 20 nedefinita.tabelul primitivelor Definiţia Fie f : J R, J R un interval. Funcţia F : J R se numeşte primitivă sau antiderivată a funcţiei
Criptosisteme cu cheie publică III
 Criptosisteme cu cheie publică III Anul II Aprilie 2017 Problema rucsacului ( knapsack problem ) Considerăm un număr natural V > 0 şi o mulţime finită de numere naturale pozitive {v 0, v 1,..., v k 1 }.
Criptosisteme cu cheie publică III Anul II Aprilie 2017 Problema rucsacului ( knapsack problem ) Considerăm un număr natural V > 0 şi o mulţime finită de numere naturale pozitive {v 0, v 1,..., v k 1 }.
R R, f ( x) = x 7x+ 6. Determinați distanța dintre punctele de. B=, unde x și y sunt numere reale.
 = x 7x+ 6. Determinați distanța dintre punctele de. B=, unde x și y sunt numere reale.") 5p Determinați primul termen al progresiei geometrice ( b n ) n, știind că b 5 = 48 și b 8 = 84 5p Se consideră funcția f : intersecție a graficului funcției f cu aa O R R, f ( ) = 7+ 6 Determinați distanța
5p Determinați primul termen al progresiei geometrice ( b n ) n, știind că b 5 = 48 și b 8 = 84 5p Se consideră funcția f : intersecție a graficului funcției f cu aa O R R, f ( ) = 7+ 6 Determinați distanța
Subiecte Clasa a VII-a
 lasa a VII Lumina Math Intrebari Subiecte lasa a VII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate
lasa a VII Lumina Math Intrebari Subiecte lasa a VII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate
Fiziologia fibrei miocardice
 Fiziologia fibrei miocardice CELULA MIOCARDICĂ = celulă excitabilă având ca şi proprietate specifică contractilitatea Fenomene electrice ale celulei miocardice Fenomene mecanice ale celulei miocardice
Fiziologia fibrei miocardice CELULA MIOCARDICĂ = celulă excitabilă având ca şi proprietate specifică contractilitatea Fenomene electrice ale celulei miocardice Fenomene mecanice ale celulei miocardice
Definiţia generală Cazul 1. Elipsa şi hiperbola Cercul Cazul 2. Parabola Reprezentari parametrice ale conicelor Tangente la conice
 1 Conice pe ecuaţii reduse 2 Conice pe ecuaţii reduse Definiţie Numim conica locul geometric al punctelor din plan pentru care raportul distantelor la un punct fix F şi la o dreaptă fixă (D) este o constantă
1 Conice pe ecuaţii reduse 2 Conice pe ecuaţii reduse Definiţie Numim conica locul geometric al punctelor din plan pentru care raportul distantelor la un punct fix F şi la o dreaptă fixă (D) este o constantă
SEMINAR 14. Funcţii de mai multe variabile (continuare) ( = 1 z(x,y) x = 0. x = f. x + f. y = f. = x. = 1 y. y = x ( y = = 0
 ( = 1 z(x,y) x = 0. x = f. x + f. y = f. = x. = 1 y. y = x ( y = = 0") Facultatea de Hidrotehnică, Geodezie şi Ingineria Mediului Matematici Superioare, Semestrul I, Lector dr. Lucian MATICIUC SEMINAR 4 Funcţii de mai multe variabile continuare). Să se arate că funcţia z,
Facultatea de Hidrotehnică, Geodezie şi Ingineria Mediului Matematici Superioare, Semestrul I, Lector dr. Lucian MATICIUC SEMINAR 4 Funcţii de mai multe variabile continuare). Să se arate că funcţia z,
Structura genei 1. Structura genei
 Structura genei 1 Structura genei Gena (din gr. genos descendentă) reprezintă unitatea elementară a eredităńii. Pentru a specifica particularităńile eredităńii G. Mendel (1865) a folosit termenul de factor
Structura genei 1 Structura genei Gena (din gr. genos descendentă) reprezintă unitatea elementară a eredităńii. Pentru a specifica particularităńile eredităńii G. Mendel (1865) a folosit termenul de factor
REACŢII DE ADIŢIE NUCLEOFILĂ (AN-REACŢII) (ALDEHIDE ŞI CETONE)
 (ALDEHIDE ŞI CETONE)") EAŢII DE ADIŢIE NULEFILĂ (AN-EAŢII) (ALDEIDE ŞI ETNE) ompușii organici care conțin grupa carbonil se numesc compuși carbonilici și se clasifică în: Aldehide etone ALDEIDE: Formula generală: 3 Metanal(formaldehida
EAŢII DE ADIŢIE NULEFILĂ (AN-EAŢII) (ALDEIDE ŞI ETNE) ompușii organici care conțin grupa carbonil se numesc compuși carbonilici și se clasifică în: Aldehide etone ALDEIDE: Formula generală: 3 Metanal(formaldehida
COLEGIUL NATIONAL CONSTANTIN CARABELLA TARGOVISTE. CONCURSUL JUDETEAN DE MATEMATICA CEZAR IVANESCU Editia a VI-a 26 februarie 2005.
 SUBIECTUL Editia a VI-a 6 februarie 005 CLASA a V-a Fie A = x N 005 x 007 si B = y N y 003 005 3 3 a) Specificati cel mai mic element al multimii A si cel mai mare element al multimii B. b)stabiliti care
SUBIECTUL Editia a VI-a 6 februarie 005 CLASA a V-a Fie A = x N 005 x 007 si B = y N y 003 005 3 3 a) Specificati cel mai mic element al multimii A si cel mai mare element al multimii B. b)stabiliti care
Lucrare. Varianta aprilie I 1 Definiţi noţiunile de număr prim şi număr ireductibil. Soluţie. Vezi Curs 6 Definiţiile 1 şi 2. sau p b.
 Lucrare Soluţii 28 aprilie 2015 Varianta 1 I 1 Definiţi noţiunile de număr prim şi număr ireductibil. Soluţie. Vezi Curs 6 Definiţiile 1 şi 2 Definiţie. Numărul întreg p se numeşte număr prim dacă p 0,
Lucrare Soluţii 28 aprilie 2015 Varianta 1 I 1 Definiţi noţiunile de număr prim şi număr ireductibil. Soluţie. Vezi Curs 6 Definiţiile 1 şi 2 Definiţie. Numărul întreg p se numeşte număr prim dacă p 0,
Capitolul 4-COMPUŞI ORGANICI CU ACŢIUNE BIOLOGICĂ-
 Capitolul 4 COMPUŞI ORGANICI CU ACŢIUNE BIOLOGICĂ 4.1.ZAHARIDE.PROTEINE. TEST 4.1.2. I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. Rezolvare
Capitolul 4 COMPUŞI ORGANICI CU ACŢIUNE BIOLOGICĂ 4.1.ZAHARIDE.PROTEINE. TEST 4.1.2. I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. Rezolvare
I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare.
 Capitolul 3 COMPUŞI ORGANICI MONOFUNCŢIONALI 3.2.ACIZI CARBOXILICI TEST 3.2.3. I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Reacţia dintre
Capitolul 3 COMPUŞI ORGANICI MONOFUNCŢIONALI 3.2.ACIZI CARBOXILICI TEST 3.2.3. I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Reacţia dintre
RĂSPUNS Modulul de rezistenţă este o caracteristică geometrică a secţiunii transversale, scrisă faţă de una dintre axele de inerţie principale:,
 REZISTENTA MATERIALELOR 1. Ce este modulul de rezistenţă? Exemplificaţi pentru o secţiune dreptunghiulară, respectiv dublu T. RĂSPUNS Modulul de rezistenţă este o caracteristică geometrică a secţiunii
REZISTENTA MATERIALELOR 1. Ce este modulul de rezistenţă? Exemplificaţi pentru o secţiune dreptunghiulară, respectiv dublu T. RĂSPUNS Modulul de rezistenţă este o caracteristică geometrică a secţiunii
STRUCTURA BIOLOGICĂ. FORME DE REPREZENTARE 2.1. SECVENŢE DE AMINOACIZI: Sorana D. BOLBOACĂ
 STRUCTURA BIOLOGICĂ. 2.1. SECVENŢE DE AMINOACIZI: FORME DE REPREZENTARE Sorana D. BOLBOACĂ Despre 2 Informaţia genetică: Definiţie Forme de prezentare Genomul Codul genetic Aminoacizi: Definiţie Clasificare
STRUCTURA BIOLOGICĂ. 2.1. SECVENŢE DE AMINOACIZI: FORME DE REPREZENTARE Sorana D. BOLBOACĂ Despre 2 Informaţia genetică: Definiţie Forme de prezentare Genomul Codul genetic Aminoacizi: Definiţie Clasificare
Fig Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36].
![Fig Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36].](/thumbs/72/67648751.jpg "Fig Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36].") Componente şi circuite pasive Fig.3.85. Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36]. Fig.3.86. Rezistenţa serie echivalentă pierderilor în funcţie
Componente şi circuite pasive Fig.3.85. Impedanţa condensatoarelor electrolitice SMD cu Al cu electrolit semiuscat în funcţie de frecvenţă [36]. Fig.3.86. Rezistenţa serie echivalentă pierderilor în funcţie
Problema a II - a (10 puncte) Diferite circuite electrice
 Diferite circuite electrice") Olimpiada de Fizică - Etapa pe judeţ 15 ianuarie 211 XI Problema a II - a (1 puncte) Diferite circuite electrice A. Un elev utilizează o sursă de tensiune (1), o cutie cu rezistenţe (2), un întrerupător
Olimpiada de Fizică - Etapa pe judeţ 15 ianuarie 211 XI Problema a II - a (1 puncte) Diferite circuite electrice A. Un elev utilizează o sursă de tensiune (1), o cutie cu rezistenţe (2), un întrerupător
a. Caracteristicile mecanice a motorului de c.c. cu excitaţie independentă (sau derivaţie)
") Caracteristica mecanică defineşte dependenţa n=f(m) în condiţiile I e =ct., U=ct. Pentru determinarea ei vom defini, mai întâi caracteristicile: 1. de sarcină, numită şi caracteristica externă a motorului
Caracteristica mecanică defineşte dependenţa n=f(m) în condiţiile I e =ct., U=ct. Pentru determinarea ei vom defini, mai întâi caracteristicile: 1. de sarcină, numită şi caracteristica externă a motorului
Să se arate că n este număr par. Dan Nedeianu
 Primul test de selecție pentru juniori I. Să se determine numerele prime p, q, r cu proprietatea că 1 p + 1 q + 1 r 1. Fie ABCD un patrulater convex cu m( BCD) = 10, m( CBA) = 45, m( CBD) = 15 și m( CAB)
Primul test de selecție pentru juniori I. Să se determine numerele prime p, q, r cu proprietatea că 1 p + 1 q + 1 r 1. Fie ABCD un patrulater convex cu m( BCD) = 10, m( CBA) = 45, m( CBD) = 15 și m( CAB)
4. Măsurarea tensiunilor şi a curenţilor electrici. Voltmetre electronice analogice
 4. Măsurarea tensiunilor şi a curenţilor electrici oltmetre electronice analogice oltmetre de curent continuu Ampl.c.c. x FTJ Protectie Atenuator calibrat Atenuatorul calibrat divizor rezistiv R in const.
4. Măsurarea tensiunilor şi a curenţilor electrici oltmetre electronice analogice oltmetre de curent continuu Ampl.c.c. x FTJ Protectie Atenuator calibrat Atenuatorul calibrat divizor rezistiv R in const.
2.1 Sfera. (EGS) ecuaţie care poartă denumirea de ecuaţia generală asferei. (EGS) reprezintă osferă cu centrul în punctul. 2 + p 2
 ecuaţie care poartă denumirea de ecuaţia generală asferei. (EGS) reprezintă osferă cu centrul în punctul. 2 + p 2") .1 Sfera Definitia 1.1 Se numeşte sferă mulţimea tuturor punctelor din spaţiu pentru care distanţa la u punct fi numit centrul sferei este egalăcuunnumăr numit raza sferei. Fie centrul sferei C (a, b,
.1 Sfera Definitia 1.1 Se numeşte sferă mulţimea tuturor punctelor din spaţiu pentru care distanţa la u punct fi numit centrul sferei este egalăcuunnumăr numit raza sferei. Fie centrul sferei C (a, b,
SEMINARUL 3. Cap. II Serii de numere reale. asociat seriei. (3n 5)(3n 2) + 1. (3n 2)(3n+1) (3n 2) (3n + 1) = a
(3n 2) + 1. (3n 2)(3n+1) (3n 2) (3n + 1) = a") Capitolul II: Serii de umere reale. Lect. dr. Lucia Maticiuc Facultatea de Hidrotehică, Geodezie şi Igieria Mediului Matematici Superioare, Semestrul I, Lector dr. Lucia MATICIUC SEMINARUL 3. Cap. II Serii
Capitolul II: Serii de umere reale. Lect. dr. Lucia Maticiuc Facultatea de Hidrotehică, Geodezie şi Igieria Mediului Matematici Superioare, Semestrul I, Lector dr. Lucia MATICIUC SEMINARUL 3. Cap. II Serii
REPLICAREA ŞI REPARAREA ADN
 1 Capitolul 7 7 REPLICAREA ŞI REPARAREA ADN Replicarea ADN este procesul molecular prin care se realizează copierea exactă a moleculelor de ADN (a secvenţei nucleotidice). Datorită replicării are loc transmiterea
1 Capitolul 7 7 REPLICAREA ŞI REPARAREA ADN Replicarea ADN este procesul molecular prin care se realizează copierea exactă a moleculelor de ADN (a secvenţei nucleotidice). Datorită replicării are loc transmiterea
TEHNOLOGIA ADN RECOMBINANT
 11 TEHNOLOGIA ADN RECOMBINANT Descifrând enigmele structurii genelor, savanţii au început să se preocupe de izolarea şi sinteza lor. În urmă cu 30 de ani cei care credeau în reuşita acestor deziderate
11 TEHNOLOGIA ADN RECOMBINANT Descifrând enigmele structurii genelor, savanţii au început să se preocupe de izolarea şi sinteza lor. În urmă cu 30 de ani cei care credeau în reuşita acestor deziderate
5.1. Noţiuni introductive
 ursul 13 aitolul 5. Soluţii 5.1. oţiuni introductive Soluţiile = aestecuri oogene de două sau ai ulte substanţe / coonente, ale căror articule nu se ot seara rin filtrare sau centrifugare. oonente: - Mediul
ursul 13 aitolul 5. Soluţii 5.1. oţiuni introductive Soluţiile = aestecuri oogene de două sau ai ulte substanţe / coonente, ale căror articule nu se ot seara rin filtrare sau centrifugare. oonente: - Mediul
2. Circuite logice 2.2. Diagrame Karnaugh. Copyright Paul GASNER 1
 2. Circuite logice 2.2. Diagrame Karnaugh Copyright Paul GASNER Diagrame Karnaugh Tehnică de simplificare a unei expresii în sumă minimă de produse (minimal sum of products MSP): Există un număr minim
2. Circuite logice 2.2. Diagrame Karnaugh Copyright Paul GASNER Diagrame Karnaugh Tehnică de simplificare a unei expresii în sumă minimă de produse (minimal sum of products MSP): Există un număr minim
Capitolul ASAMBLAREA LAGĂRELOR LECŢIA 25
 Capitolul ASAMBLAREA LAGĂRELOR LECŢIA 25 LAGĂRELE CU ALUNECARE!" 25.1.Caracteristici.Părţi componente.materiale.!" 25.2.Funcţionarea lagărelor cu alunecare.! 25.1.Caracteristici.Părţi componente.materiale.
Capitolul ASAMBLAREA LAGĂRELOR LECŢIA 25 LAGĂRELE CU ALUNECARE!" 25.1.Caracteristici.Părţi componente.materiale.!" 25.2.Funcţionarea lagărelor cu alunecare.! 25.1.Caracteristici.Părţi componente.materiale.
Catedra Biologie moleculară şi Genetică umană
 Catedra Biologie moleculară şi Genetică umană www.bimogeum.ucoz.com Cursul Biologie moleculară Semestrul I Prelegeri 34 ore Lucrări practice 51 ore două totalizări (lucrare scrisă + teste la calculator)
Catedra Biologie moleculară şi Genetică umană www.bimogeum.ucoz.com Cursul Biologie moleculară Semestrul I Prelegeri 34 ore Lucrări practice 51 ore două totalizări (lucrare scrisă + teste la calculator)
Capitolul 2 - HIDROCARBURI 2.4.ALCADIENE
 Capitolul 2 - HIDROCARBURI 2.4.ALCADIENE TEST 2.4.1 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. Rezolvare: 1. Alcadienele sunt hidrocarburi
Capitolul 2 - HIDROCARBURI 2.4.ALCADIENE TEST 2.4.1 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. Rezolvare: 1. Alcadienele sunt hidrocarburi
Curs 2 DIODE. CIRCUITE DR
 Curs 2 OE. CRCUTE R E CUPRN tructură. imbol Relația curent-tensiune Regimuri de funcționare Punct static de funcționare Parametrii diodei Modelul cu cădere de tensiune constantă Analiza circuitelor cu
Curs 2 OE. CRCUTE R E CUPRN tructură. imbol Relația curent-tensiune Regimuri de funcționare Punct static de funcționare Parametrii diodei Modelul cu cădere de tensiune constantă Analiza circuitelor cu
FLUXURI MAXIME ÎN REŢELE DE TRANSPORT. x 4
 FLUXURI MAXIME ÎN REŢELE DE TRANSPORT Se numeşte reţea de transport un graf în care fiecărui arc îi este asociat capacitatea arcului şi în care eistă un singur punct de intrare şi un singur punct de ieşire.
FLUXURI MAXIME ÎN REŢELE DE TRANSPORT Se numeşte reţea de transport un graf în care fiecărui arc îi este asociat capacitatea arcului şi în care eistă un singur punct de intrare şi un singur punct de ieşire.
Proiectarea filtrelor prin metoda pierderilor de inserţie
 FITRE DE MIROUNDE Proiectarea filtrelor prin metoda pierderilor de inserţie P R Puterea disponibila de la sursa Puterea livrata sarcinii P inc P Γ ( ) Γ I lo P R ( ) ( ) M ( ) ( ) M N P R M N ( ) ( ) Tipuri
FITRE DE MIROUNDE Proiectarea filtrelor prin metoda pierderilor de inserţie P R Puterea disponibila de la sursa Puterea livrata sarcinii P inc P Γ ( ) Γ I lo P R ( ) ( ) M ( ) ( ) M N P R M N ( ) ( ) Tipuri
Examen AG. Student:... Grupa:... ianuarie 2011
 Problema 1. Pentru ce valori ale lui n,m N (n,m 1) graful K n,m este eulerian? Problema 2. Să se construiască o funcţie care să recunoască un graf P 3 -free. La intrare aceasta va primi un graf G = ({1,...,n},E)
Problema 1. Pentru ce valori ale lui n,m N (n,m 1) graful K n,m este eulerian? Problema 2. Să se construiască o funcţie care să recunoască un graf P 3 -free. La intrare aceasta va primi un graf G = ({1,...,n},E)
z a + c 0 + c 1 (z a)
") 1 Serii Laurent (continuare) Teorema 1.1 Fie D C un domeniu, a D şi f : D \ {a} C o funcţie olomorfă. Punctul a este pol multiplu de ordin p al lui f dacă şi numai dacă dezvoltarea în serie Laurent a funcţiei
1 Serii Laurent (continuare) Teorema 1.1 Fie D C un domeniu, a D şi f : D \ {a} C o funcţie olomorfă. Punctul a este pol multiplu de ordin p al lui f dacă şi numai dacă dezvoltarea în serie Laurent a funcţiei
Capitolul 2 - HIDROCARBURI 2.5.ARENE
 Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.2 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Radicalul C 6 H 5 - se numeşte fenil. ( fenil/
Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.2 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Radicalul C 6 H 5 - se numeşte fenil. ( fenil/
7. RETELE ELECTRICE TRIFAZATE 7.1. RETELE ELECTRICE TRIFAZATE IN REGIM PERMANENT SINUSOIDAL
 7. RETEE EECTRICE TRIFAZATE 7.. RETEE EECTRICE TRIFAZATE IN REGIM PERMANENT SINSOIDA 7... Retea trifazata. Sistem trifazat de tensiuni si curenti Ansamblul format din m circuite electrice monofazate in
7. RETEE EECTRICE TRIFAZATE 7.. RETEE EECTRICE TRIFAZATE IN REGIM PERMANENT SINSOIDA 7... Retea trifazata. Sistem trifazat de tensiuni si curenti Ansamblul format din m circuite electrice monofazate in
CIRCUITE INTEGRATE MONOLITICE DE MICROUNDE. MMIC Monolithic Microwave Integrated Circuit
 CIRCUITE INTEGRATE MONOLITICE DE MICROUNDE MMIC Monolithic Microwave Integrated Circuit CUPRINS 1. Avantajele si limitarile MMIC 2. Modelarea dispozitivelor active 3. Calculul timpului de viata al MMIC
CIRCUITE INTEGRATE MONOLITICE DE MICROUNDE MMIC Monolithic Microwave Integrated Circuit CUPRINS 1. Avantajele si limitarile MMIC 2. Modelarea dispozitivelor active 3. Calculul timpului de viata al MMIC
a. 0,1; 0,1; 0,1; b. 1, ; 5, ; 8, ; c. 4,87; 6,15; 8,04; d. 7; 7; 7; e. 9,74; 12,30;1 6,08.
 1. În argentometrie, metoda Mohr: a. foloseşte ca indicator cromatul de potasiu, care formeazǎ la punctul de echivalenţă un precipitat colorat roşu-cărămiziu; b. foloseşte ca indicator fluoresceina, care
1. În argentometrie, metoda Mohr: a. foloseşte ca indicator cromatul de potasiu, care formeazǎ la punctul de echivalenţă un precipitat colorat roşu-cărămiziu; b. foloseşte ca indicator fluoresceina, care
Activitatea A5. Introducerea unor module specifice de pregătire a studenţilor în vederea asigurării de şanse egale
 POSDRU/156/1.2/G/138821 Investeşte în oameni! FONDUL SOCIAL EUROPEAN Programul Operaţional Sectorial pentru Dezvoltarea Resurselor Umane 2007 2013 Axa prioritară nr. 1 Educaţiaşiformareaprofesionalăînsprijinulcreşteriieconomiceşidezvoltăriisocietăţiibazatepecunoaştere
POSDRU/156/1.2/G/138821 Investeşte în oameni! FONDUL SOCIAL EUROPEAN Programul Operaţional Sectorial pentru Dezvoltarea Resurselor Umane 2007 2013 Axa prioritară nr. 1 Educaţiaşiformareaprofesionalăînsprijinulcreşteriieconomiceşidezvoltăriisocietăţiibazatepecunoaştere
Spatii liniare. Exemple Subspaţiu liniar Acoperire (înfăşurătoare) liniară. Mulţime infinită liniar independentă
 liniară. Mulţime infinită liniar independentă") Noţiunea de spaţiu liniar 1 Noţiunea de spaţiu liniar Exemple Subspaţiu liniar Acoperire (înfăşurătoare) liniară 2 Mulţime infinită liniar independentă 3 Schimbarea coordonatelor unui vector la o schimbare
Noţiunea de spaţiu liniar 1 Noţiunea de spaţiu liniar Exemple Subspaţiu liniar Acoperire (înfăşurătoare) liniară 2 Mulţime infinită liniar independentă 3 Schimbarea coordonatelor unui vector la o schimbare
CURS XI XII SINTEZĂ. 1 Algebra vectorială a vectorilor liberi
 Lect. dr. Facultatea de Electronică, Telecomunicaţii şi Tehnologia Informaţiei Algebră, Semestrul I, Lector dr. Lucian MATICIUC http://math.etti.tuiasi.ro/maticiuc/ CURS XI XII SINTEZĂ 1 Algebra vectorială
Lect. dr. Facultatea de Electronică, Telecomunicaţii şi Tehnologia Informaţiei Algebră, Semestrul I, Lector dr. Lucian MATICIUC http://math.etti.tuiasi.ro/maticiuc/ CURS XI XII SINTEZĂ 1 Algebra vectorială
Capitolul 2 - HIDROCARBURI 2.5.ARENE
 Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.3 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Sulfonarea benzenului este o reacţie ireversibilă.
Capitolul 2 - HIDROCARBURI 2.5.ARENE TEST 2.5.3 I. Scrie cuvântul / cuvintele dintre paranteze care completează corect fiecare dintre afirmaţiile următoare. 1. Sulfonarea benzenului este o reacţie ireversibilă.
Capitolul 4 Amplificatoare elementare
 Capitolul 4 mplificatoare elementare 4.. Etaje de amplificare cu un tranzistor 4... Etajul emitor comun V CC C B B C C L L o ( // ) V gm C i rπ // B // o L // C // L B ro i B E C E 4... Etajul colector
Capitolul 4 mplificatoare elementare 4.. Etaje de amplificare cu un tranzistor 4... Etajul emitor comun V CC C B B C C L L o ( // ) V gm C i rπ // B // o L // C // L B ro i B E C E 4... Etajul colector
CURS 11: ALGEBRĂ Spaţii liniare euclidiene. Produs scalar real. Spaţiu euclidian. Produs scalar complex. Spaţiu unitar. Noţiunea de normă.
 Sala: 2103 Decembrie 2014 Conf. univ. dr.: Dragoş-Pătru Covei CURS 11: ALGEBRĂ Specializarea: C.E., I.E., S.P.E. Nota: Acest curs nu a fost supus unui proces riguros de recenzare pentru a fi oficial publicat.
Sala: 2103 Decembrie 2014 Conf. univ. dr.: Dragoş-Pătru Covei CURS 11: ALGEBRĂ Specializarea: C.E., I.E., S.P.E. Nota: Acest curs nu a fost supus unui proces riguros de recenzare pentru a fi oficial publicat.
2. Circuite logice 2.4. Decodoare. Multiplexoare. Copyright Paul GASNER
 2. Circuite logice 2.4. Decodoare. Multiplexoare Copyright Paul GASNER Definiţii Un decodor pe n bits are n intrări şi 2 n ieşiri; cele n intrări reprezintă un număr binar care determină în mod unic care
2. Circuite logice 2.4. Decodoare. Multiplexoare Copyright Paul GASNER Definiţii Un decodor pe n bits are n intrări şi 2 n ieşiri; cele n intrări reprezintă un număr binar care determină în mod unic care
3.2. Enzime utilizate în tehnologia ADN recombinant
 Genetica microorganismelor şi inginerie genetică microbiană Note de curs şi tehnici de laborator 3.2. Enzime utilizate în tehnologia ADN recombinant Enzimele de restricţie Introducere Sistemele de restricţie
Genetica microorganismelor şi inginerie genetică microbiană Note de curs şi tehnici de laborator 3.2. Enzime utilizate în tehnologia ADN recombinant Enzimele de restricţie Introducere Sistemele de restricţie
Repere pentru pregătirea către examen la Biologia Moleculară Repere pentru pregătirea către examen la Biologia Moleculară:
 Repere pentru pregătirea către examen la Biologia Moleculară: Repetați pentru examen toate temele de la 1 la 14 (conspect prelegeri, caiet LP, manual, site); + reperele la totalizarea I, II, III; + subiectele
Repere pentru pregătirea către examen la Biologia Moleculară: Repetați pentru examen toate temele de la 1 la 14 (conspect prelegeri, caiet LP, manual, site); + reperele la totalizarea I, II, III; + subiectele
Teoria mecanic-cuantică a legăturii chimice - continuare. Hibridizarea orbitalilor
 Cursul 10 Teoria mecanic-cuantică a legăturii chimice - continuare Hibridizarea orbitalilor Orbital atomic = regiunea din jurul nucleului în care poate fi localizat 1 e - izolat, aflat într-o anumită stare
Cursul 10 Teoria mecanic-cuantică a legăturii chimice - continuare Hibridizarea orbitalilor Orbital atomic = regiunea din jurul nucleului în care poate fi localizat 1 e - izolat, aflat într-o anumită stare
2. Sisteme de forţe concurente...1 Cuprins...1 Introducere Aspecte teoretice Aplicaţii rezolvate...3
 SEMINAR 2 SISTEME DE FRŢE CNCURENTE CUPRINS 2. Sisteme de forţe concurente...1 Cuprins...1 Introducere...1 2.1. Aspecte teoretice...2 2.2. Aplicaţii rezolvate...3 2. Sisteme de forţe concurente În acest
SEMINAR 2 SISTEME DE FRŢE CNCURENTE CUPRINS 2. Sisteme de forţe concurente...1 Cuprins...1 Introducere...1 2.1. Aspecte teoretice...2 2.2. Aplicaţii rezolvate...3 2. Sisteme de forţe concurente În acest
III. Reprezentarea informaţiei în sistemele de calcul
 Metode Numerice Curs 3 III. Reprezentarea informaţiei în sistemele de calcul III.1. Reprezentarea internă a numerelor întregi III. 1.1. Reprezentarea internă a numerelor întregi fără semn (pozitive) Reprezentarea
Metode Numerice Curs 3 III. Reprezentarea informaţiei în sistemele de calcul III.1. Reprezentarea internă a numerelor întregi III. 1.1. Reprezentarea internă a numerelor întregi fără semn (pozitive) Reprezentarea
Subiecte Clasa a VIII-a
 Subiecte lasa a VIII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate pe foaia de raspuns in dreptul
Subiecte lasa a VIII-a (40 de intrebari) Puteti folosi spatiile goale ca ciorna. Nu este de ajuns sa alegeti raspunsul corect pe brosura de subiecte, ele trebuie completate pe foaia de raspuns in dreptul
Geometrie computationala 2. Preliminarii geometrice
 Platformă de e-learning și curriculă e-content pentru învățământul superior tehnic Geometrie computationala 2. Preliminarii geometrice Preliminarii geometrice Spatiu Euclidean: E d Spatiu de d-tupluri,
Platformă de e-learning și curriculă e-content pentru învățământul superior tehnic Geometrie computationala 2. Preliminarii geometrice Preliminarii geometrice Spatiu Euclidean: E d Spatiu de d-tupluri,
BARAJ DE JUNIORI,,Euclid Cipru, 28 mai 2012 (barajul 3)
") BARAJ DE JUNIORI,,Euclid Cipru, 8 mi 0 (brjul ) Problem Arătţi că dcă, b, c sunt numere rele cre verifică + b + c =, tunci re loc ineglitte xy + yz + zx Problem Fie şi b numere nturle nenule Dcă numărul
BARAJ DE JUNIORI,,Euclid Cipru, 8 mi 0 (brjul ) Problem Arătţi că dcă, b, c sunt numere rele cre verifică + b + c =, tunci re loc ineglitte xy + yz + zx Problem Fie şi b numere nturle nenule Dcă numărul
Algebra si Geometrie Seminar 9
 Algebra si Geometrie Seminar 9 Decembrie 017 ii Equations are just the boring part of mathematics. I attempt to see things in terms of geometry. Stephen Hawking 9 Dreapta si planul in spatiu 1 Notiuni
Algebra si Geometrie Seminar 9 Decembrie 017 ii Equations are just the boring part of mathematics. I attempt to see things in terms of geometry. Stephen Hawking 9 Dreapta si planul in spatiu 1 Notiuni
Câmp de probabilitate II
 1 Sistem complet de evenimente 2 Schema lui Poisson Schema lui Bernoulli (a bilei revenite) Schema hipergeometrică (a bilei neîntoarsă) 3 4 Sistem complet de evenimente Definiţia 1.1 O familie de evenimente
1 Sistem complet de evenimente 2 Schema lui Poisson Schema lui Bernoulli (a bilei revenite) Schema hipergeometrică (a bilei neîntoarsă) 3 4 Sistem complet de evenimente Definiţia 1.1 O familie de evenimente
EDITURA PARALELA 45 MATEMATICĂ DE EXCELENŢĂ. Clasa a X-a Ediţia a II-a, revizuită. pentru concursuri, olimpiade şi centre de excelenţă
 Coordonatori DANA HEUBERGER NICOLAE MUŞUROIA Nicolae Muşuroia Gheorghe Boroica Vasile Pop Dana Heuberger Florin Bojor MATEMATICĂ DE EXCELENŢĂ pentru concursuri, olimpiade şi centre de excelenţă Clasa a
Coordonatori DANA HEUBERGER NICOLAE MUŞUROIA Nicolae Muşuroia Gheorghe Boroica Vasile Pop Dana Heuberger Florin Bojor MATEMATICĂ DE EXCELENŢĂ pentru concursuri, olimpiade şi centre de excelenţă Clasa a
Aparate de măsurat. Măsurări electronice Rezumatul cursului 2. MEE - prof. dr. ing. Ioan D. Oltean 1
 Aparate de măsurat Măsurări electronice Rezumatul cursului 2 MEE - prof. dr. ing. Ioan D. Oltean 1 1. Aparate cu instrument magnetoelectric 2. Ampermetre şi voltmetre 3. Ohmetre cu instrument magnetoelectric
Aparate de măsurat Măsurări electronice Rezumatul cursului 2 MEE - prof. dr. ing. Ioan D. Oltean 1 1. Aparate cu instrument magnetoelectric 2. Ampermetre şi voltmetre 3. Ohmetre cu instrument magnetoelectric
Ακαδημαϊκός Λόγος Κύριο Μέρος
 - Επίδειξη Συμφωνίας În linii mari sunt de acord cu...deoarece... Επίδειξη γενικής συμφωνίας με άποψη άλλου Cineva este de acord cu...deoarece... Επίδειξη γενικής συμφωνίας με άποψη άλλου D'une façon générale,
- Επίδειξη Συμφωνίας În linii mari sunt de acord cu...deoarece... Επίδειξη γενικής συμφωνίας με άποψη άλλου Cineva este de acord cu...deoarece... Επίδειξη γενικής συμφωνίας με άποψη άλλου D'une façon générale,
Codificatorul SN74148 este un codificator zecimal-bcd de trei biţi (fig ). Figura Codificatorul integrat SN74148
. Figura Codificatorul integrat SN74148") 5.2. CODIFICATOAE Codificatoarele (CD) sunt circuite logice combinaţionale cu n intrări şi m ieşiri care furnizează la ieşire un cod de m biţi atunci când numai una din cele n intrări este activă. De regulă
5.2. CODIFICATOAE Codificatoarele (CD) sunt circuite logice combinaţionale cu n intrări şi m ieşiri care furnizează la ieşire un cod de m biţi atunci când numai una din cele n intrări este activă. De regulă