Ο ΡΟΛΟΣ ΤΩΝ ΑΝΟΣΟΡΡΥΘΜΙΣΤΙΚΩΝ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΣΤΗΝ ΑΝΟΣΟΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΤΗΝ ΕΞΕΛΙΞΗ ΤΗΣ HCV ΛΟΙΜΩΞΗΣ
|
|
|
- Ἀπολλωνία Μητσοτάκης
- 8 χρόνια πριν
- Προβολές:
Transcript
1 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ- ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ Β ΠΑΘΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ Γ.Π.Ν.Ν. ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΠΠΟΚΡΑΤΕΙΟ ΔΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ Ι. ΚΛΩΝΙΖΑΚΗΣ ΠΑΝΕΠΙΣΤΗΜΙΑΚΟ ΕΤΟΣ Αρ.Διατριβής 2990 Ο ΡΟΛΟΣ ΤΩΝ ΑΝΟΣΟΡΡΥΘΜΙΣΤΙΚΩΝ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΣΤΗΝ ΑΝΟΣΟΠΑΘΟΓΕΝΕΙΑ ΚΑΙ ΤΗΝ ΕΞΕΛΙΞΗ ΤΗΣ HCV ΛΟΙΜΩΞΗΣ ΜΠΕΛΛΟΥ ΑΡΙΣΤΕΑ ΛΙΑ ΙΑΤΡΟΣ ΕΙΔΙΚΗ ΠΑΘΟΛΟΓΟΣ ΔΙΔΑΚΤΟΡΙΚΗ ΔΙΑΤΡΙΒΗ ΘΕΣΣΑΛΟΝΙΚΗ 2011
2
3 Η ΤΡΙΜΕΛΗΣ ΣΥΜΒΟΥΛΕΥΤΙΚΗ ΕΠΙΤΡΟΠΗ ΜΑΡΙΑ ΡΑΠΤΟΠΟΥΛΟΥ- ΓΙΓΗ, ΟΜΟΤΙΜΗ ΚΑΘΗΓΗΤΡΙΑ ΠΑΝΑΓΙΩΤΑ ΜΠΟΥΡΑ, ΚΑΘΗΓΗΤΡΙΑ ΜΙΧΑΗΛ ΔΑΝΙΗΛΙΔΗΣ, ΚΑΘΗΓΗΤΗΣ Η ΕΠΤΑΜΕΛΗΣ ΕΞΕΤΑΣΤΙΚΗ ΕΠΙΤΡΟΠΗ ΜΑΡΙΑ ΡΑΠΤΟΠΟΥΛΟΥ- ΓΙΓΗ (ΕΠΙΒΛΕΠΟΥΣΑ) ΟΜΟΤΙΜΗ ΚΑΘΗΓΗΤΡΙΑ, ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ, ΙΑΤΡΙΚΗ ΣΧΟΛΗ, ΑΠΘ ΠΑΝΑΓΙΩΤΑ ΜΠΟΥΡΑ ΚΑΘΗΓΗΤΡΙΑ, ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ, ΙΑΤΡΙΚΗ ΣΧΟΛΗ, ΑΠΘ ΜΙΧΑΗΛ ΔΑΝΙΗΛΙΔΗΣ ΚΑΘΗΓΗΤΗΣ, ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ, ΙΑΤΡΙΚΗ ΣΧΟΛΗ, ΑΠΘ ΚΛΩΝΙΖΑΚΗΣ ΙΩΑΝΝΗΣ ΚΑΘΗΓΗΤΗΣ, ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ, ΙΑΤΡΙΚΗ ΣΧΟΛΗ, ΑΠΘ ΗΛΟΝΙΔΗΣ ΓΕΩΡΓΙΟΣ ΚΑΘΗΓΗΤΗΣ, ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ, ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΑΠΘ ΒΡΕΤΤΟΥ ΕΛΕΝΗ ΚΑΘΗΓΗΤΡΙΑ, ΤΟΜΕΑΣ ΑΝΑΤΟΜΙΑΣ ΚΑΙ ΠΑΘΟΛΟΓΙΚΗΣ ΑΝΑΤΟΜΙΚΗΣ, ΙΑΤΡΙΚΗ ΣΧΟΛΗ, ΑΠΘ ΓΟΥΛΗΣ ΙΩΑΝΝΗΣ ΕΠΙΚΟΥΡΟΣ ΚΑΘΗΓΗΤΗΣ, ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ, ΙΑΤΡΙΚΗ ΣΧΟΛΗ, ΑΠΘ «Η έγκρισης της Διδακτορικής Διατριβής υπό της Ιατρικής Σχολής του Αριστοτελείου Πανεπιστημίου Θεσσαλονίκης, δεν υποδηλοί αποδοχήν των γνωμών του συγγραφέως» (Νόμος 5343/32, αρθρ και ν. 1268/82, αρθρ. 50 8) 3
4
5 στην οικογένεια μου στα μωρά μου 5
6 6
7 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΠΡΟΕΔΡΟΣ ΤΗΣ ΣΧΟΛΗΣ ΝΙΚΟΛΑΟΣ Β. ΝΤΟΜΠΡΟΣ 7
8
9 ΠΕΡΙΕΧΟΜΕΝΑ Κυριότερες συντμήσεις 13 Πρόλογος 15 ΓΕΝΙΚΟ ΜΕΡΟΣ 19 ΜΕΡΟΣ Ι. ΗΠΑΤΙΤΙΔΑ C 21 Α. Ο ιός της ηπατίτιδας C (HCV) 21 Β. Πολλαπλασιασμός του ιού της ηπατίτιδας C. Ο κύκλος του 22 HCV Γ. Γονότυποι του HCV 23 Δ. Τρόπος μετάδοσης 23 Ε. Πρόληψη 24 ΣΤ. Διάγνωση- εργαστηριακός έλεγχος 24 Ζ. Βιοψία ήπατος 24 Η. Μη επεμβατική αξιολόγηση ηπατικής ίνωσης 25 Θ. Κλινική εικόνα και φυσική εξέλιξη της HCV λοίμωξης 25 Ι. Θεραπεία 26 ΜΕΡΟΣ ΙΙ. Η ΚΥΤΤΑΡΙΚΗ ΑΝΟΣΙΑ ΣΤΗ ΛΟΙΜΩΞΗ 33 ΜΕΡΟΣ ΙΙΙ. ΑΝΟΣΙΑΚΗ ΑΠΟΚΡΙΣΗ ΕΝΑΝΤΙ ΤΟΥ ΙΟΥ ΤΗΣ 39 ΗΠΑΤΙΤΙΔΑΣ C Α. Ανοσιακή απόκριση στην οξεία HCV λοίμωξη 41 Β. Ανοσιακή απόκριση στη χρόνια HCV λοίμωξη 42 ΜΕΡΟΣ IV. Τ ΡΥΘΜΙΣΤΙΚΑ ΛΕΜΦΟΚΥΤΤΑΡΑ 46 A. Ιστορική αναδρομή 46 B. Γονίδιο FOXP3 47 Γ. «Φυσικά αναδυόμενα» Τ ρυθμιστικά κύτταρα (ntregs, natural 49 Tregs) Δ. Επαγόμενα Tregs μέσω κλωνικής επέκτασης (itregs, 51 induced Tregs) 1.Tr1 κύτταρα 53 2.Th3 κύτταρα 54 3.CD4+CD25+ Tregs που δημιουργούνται από μετατροπή των CD4+CD25- T ενεργοποιημένων κυττάρων 54 4.CD103+ Tregs 55 5.CD8+ T ρυθμιστικά κύτταρα 55 6.Διπλά αρνητικά Tregs (CD4-CD8-, Double Negative, DN Tregs) 56 7.Επαγόμενα Tregs για θεραπευτικούς σκοπούς 57 8.Μη-Τ ρυθμιστικά κύτταρα (non-t regulatory cells) 57 Ε. Μηχανισμοί με τους οποίους τα Tregs ασκούν την 58 κατασταλτική τους δράση 9
10 1.IL Άμεση κυτταροκυτταρική επαφή 60 3.Παραγωγή ανοσοκατασταλτικών κυτταροκινών 60 4.Μεταβολική «διάσπαση» (metabolic disruption) 61 5.Έμμεσοι μηχανισμοί 62 ΣΤ. Ομοιόσταση των T ρυθμιστικών λεμφοκυττάρων 63 1.Δημιουργία ntregs στο θύμο αδένα 63 2.Μεταγραφικοί παράγοντες που είναι απαραίτητοι για τη διαφοροποίηση των Tregs Διαφοροποίηση των Τ βοηθητικών κυττάρων σε Tregs στην περιφέρεια 64 3.Κλωνική επέκταση των T ρυθμιστικών κυττάρων 64 Ζ. Ο ρόλος του TCR και η ειδικότητα των Tregs έναντι αντιγόνων 65 Η. Πώς καταφέρνουν τα κύτταρα- στόχος να ξεφεύγουν από την κατασταλτική δράση των Tregs; Θ. Στρατολόγηση των T ρυθμιστικών λεμφοκυττάρων στο σημείο της φλεγμονής 67 ΜΕΡΟΣ V. Ο ΡΟΛΟΣ ΤΩΝ T ΡΥΘΜΙΣΤΙΚΩΝ ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΣΤΗΝ HCV ΛΟΙΜΩΞΗ 69 Α. Ο ρόλος των υποπληθυσμών των T ρυθμιστικών λεμφοκυττάρων στην HCV λοίμωξη 71 1.Foxp3+ ntregs 71 2.Tr1 κύτταρα 73 3.Th3 κύτταρα 73 4.CD4+CD25+Tregs από μετατροπή των CD4+CD25-T κυττάρων 75 5.CD8+Treg κύτταρα που παράγουν IL Β. Στρατολόγηση των T ρυθμιστικών λεμφοκυττάρων στο 78 ήπαρ 65 Γ. Μηχανισμοί με τους οποίους τα T ρυθμιστικά λεμφοκύτταρα ασκούν την κατασταλτική τους δράση στην HCV λοίμωξη Δ. Ειδικότητα των Tregs έναντι αντιγόνων του ιού της ηπατίτιδας C Ε. Συνδιεγερτικά μόρια και T ρυθμιστικά λεμφοκύτταρα στην HCV λοίμωξη ΣΤ. Toll Like Receptors (TLRs) και T ρυθμιστικά λεμφοκύτταρα στην HCV λοίμωξη Ζ. T ρυθμιστικά λεμφοκύτταρα και καρκινογένεση 79 ΕΙΔΙΚΟ ΜΕΡΟΣ 85 Ι. ΥΛΙΚΟ 87 ΙΙ. ΜΕΘΟΔΟΙ 95 10
11 1.Μοριακή διάγνωση ηπατίτιδας C 95 2.Μέτρηση ALT ορού 95 3.Κυτταρομετρία ροής 95 -Προετοιμασία περιφερικού αίματος 98 -Απομόνωση λεμφοκυττάρων από το περιφερικό αίμα 100 -Μιτογόνο και in vitro καλλιέργειες περιφερικών 101 λεμφοκυττάρων -Προετοιμασία δειγμάτων λεμφοκυττάρων in vitro καλλιέργειας 101 -Ανάλυση με κυτταρομετρία ροής 102 ΙΙΙ. ΣΤΑΤΙΣΤΙΚΗ ΜΕΘΟΔΟΛΟΓΙΑ 113 IV. ΑΠΟΤΕΛΕΣΜΑΤΑ 115 Συνολικά αποτελέσματα στα τέσσερα στιγμιότυπα της μελέτης 115 Αποτελέσματα στις δύο ομάδες ασθενών (Ομάδα 1: γονότυπος 1, Ομάδα 2: γονότυπος 2 και 3) 124 V. ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ 141 Σύγκριση των αποτελεσμάτων των παραμέτρων που μελετήθηκαν σε όλους τους ασθενείς και μεταξύ των δύο 141 ομάδων ασθενών: ασθενείς με γονότυπο 1 και ασθενείς με γονότυπο 2, 3 Σύγκριση αποτελεσμάτων μεταξύ των ασθενών με γονότυπο 1 οι οποίοι απήντησαν στη θεραπεία (12/15, 80%) και των 143 ασθενών με γονότυπο 1 οι οποίοι δεν πέτυχαν (3/15, 20%) μακρά ιολογική απάντηση (SVR) Συσχετίσεις μεταβλητών (Spearman test) 152 Θηκογράμματα (Boxplots), με βάση το φύλο και το γονότυπο 163 VΙ. ΣΥΖΗΤΗΣΗ 183 VIΙ. ΣΥΜΠΕΡΑΣΜΑΤΑ 201 VIIΙ. ΠΕΡΙΛΗΨΗ 203 IΧ. SUMMARY 211 Χ. ΒΙΒΛΙΟΓΡΑΦΙΑ 219 ΠΑΡΑΡΤΗΜΑ
12
13 ΚΥΡΙΟΤΕΡΕΣ ΣΥΝΤΜΗΣΕΙΣ ανεργία: μια κατάσταση υπο-απαντητικότητας των B ή T κυττάρων σε αντιγόνα. Μετά τη διέγερση, τα ανεργικά Τ κύτταρα δεν μπορούν να παράγουν IL-2 ή να πολλαπλασιαστούν, ακόμη και παρουσία συνδιεγερτικών σημάτων ALT: αλανινική αμινοτρανσφεράση AP-1: Activator Protein-1 AST: ασπαρτική αμινοτρανσφεράση CD: Cluster Designation cdcs: συμβατικά δενδριτικά κύτταρα CHC: chronic hepatitis C (χρόνια ηπατίτιδα C) CTLA-4: Cytotoxic T-Lymphocyte-Associated Antigen 4 exhaustion: εξάντληση (κατάσταση στην οποία τα ειδικά έναντι του ιού CD8+ T κύτταρα δεν διεγείρονται από τα αντιγόνα του ιού) IFN: interferon IL: interleukin itregs: induced («επαγόμενα») T ρυθμιστικά κύτταρα kda: kilodalton LFA-3: Leucocyte Function-Associated Antigen-3 LPS: lipopolysaccharide MHC: Major Histocompatibility Complex ntregs: naive («φυσικά») T ρυθμιστικά κύτταρα NFAT: Nuclear Factor of Activated T cell NPV: negative predictive value (αρνητική προγνωστική αξία) PBMC: μονοκύτταρα του περιφερικού αίματος PD-1: Programmed Death-1 pdcs: πλασμακυτταροειδή δενδριτικά κύτταρα 13
14 PPV: positive predictive value (θετική προγνωστική αξία) συνδιέγερση ή δεύτερο σήμα: (απαιτείται προκειμένου να πολλαπλασιαστούν και να διαφοροποιηθούν τα παρθένα Τ λεμφοκύτταρα) = σύνδεση συγκεκριμένων μορίων που εκφράζονται στην επιφάνεια των ενεργοποιημένων αντιγονοπαρουσιαστικών κυττάρων με αντίστοιχους υποδοχείς στην επιφάνεια του Τ- λεμφοκυττάρου. Τα μόρια αυτά είναι γνωστά ως συνδιεγερτικά μόρια (πρώτο σήμα ή διέγερση= σύνδεση του συμπλέγματος πεπτιδίου/mhc με τον TCR) Runx πρωτεΐνες: συντομογραφία του όρου runt related proteins, πρόκειται για μια οικογένεια μεταγραφικών παραγόντων που εμπλέκονται στην ανάπτυξη του εμβρύου και την αιμοποίηση. Συνδέονται με το DNA μέσω της διατηρημένης περιοχής runt (conserved runt domain) RVR: rapid virologic response (ταχεία ιολογική απάντηση) STAT: Signal Transducer and Activator of Transcription SVR: sustained virologic response (μακροχρόνια ιολογική απάντηση) TCR: T Cell Receptor TGF-β: Transforming Growth Factor β Th: T helper cells TLR: Toll Like Receptor TNF: Tumor Necrosis Factor TRECs: η διαδικασία αναδιάταξης του TCR δημιουργεί εξωχρωμοσωματικά DNA επισώματα ή TCR excision circles, τα οποία ανιχνεύονται στα νέα Τ κύτταρα. Η παρουσία TRECs σε Τ κύτταρα τα οποία κυκλοφορούν στο περιφερικό αίμα, είναι μια ένδειξη ότι η αναδιάταξη των γονιδίων του TCR τους συνέβη πρόσφατα στο θύμο Tregs: Τ ρυθμιστικά κύτταρα 14
15 ΠΡΟΛΟΓΟΣ Η χρόνια ηπατίτιδα C είναι μια από τις πιο κοινές ιογενείς λοιμώξεις στον άνθρωπο. Στην Ευρώπη και τις Ηνωμένες Πολιτείες Αμερικής αποτελεί μια από τις πιο συχνές αιτίες κίρρωσης και ηπατοκυτταρικού καρκινώματος και μάλιστα είναι μέσα στις τρεις πρώτες αιτίες μεταμόσχευσης ήπατος, ενώ η λοίμωξη συχνά υποτροπιάζει και στο μόσχευμα. Με την ανακάλυψη των μοριακών τεχνικών έχει γίνει εφικτός ο έλεγχος όλων των παραγώγων αίματος, αλλά εξακολουθούν να υπάρχουν νέα περιστατικά με χρόνια ηπατίτιδα C, κυρίως χρήστες ενδοφλεβίων ουσιών, καθώς περίπου το 80% των ατόμων που μολύνονται εγκαθιστά χρόνια λοίμωξη και η παρασκευή εμβολίου εξακολουθεί να μην έχει καταστεί εφικτή. Επίσης, αν και η θεραπεία για τη χρόνια ηπατίτιδα C έχει εξελιχθεί την τελευταία δεκαετία, το συνολικό ποσοστό απάντησης στην θεραπεία δεν ξεπερνά το 50% των ασθενών που επιτυγχάνουν να ολοκληρώσουν την ενδεδειγμένη διάρκεια αγωγής με το συνδυασμό πεγκυλιωμένης ιντερφερόνης και ριμπαβιρίνης. Έτσι, με βάση την επιδημιολογία αλλά και τη φυσική πορεία της χρόνιας ηπατίτιδας C εκτιμάται ότι η θνητότητα και το κόστος της ιατρικής φροντίδας θα αυξηθεί τις επόμενες δεκαετίες. Είναι καλά γνωστό ότι η ισχυρή και επαρκής ανοσιακή απόκριση των CD8+ T λεμφοκυττάρων είναι απαραίτητη για την κάθαρση του ιού, ενώ στη χρόνια ηπατίτιδα C τα CD8+ T λεμφοκύτταρα έχουν «εξαντλημένο» φαινότυπο με ανεπαρκή κυτταροτοξική δράση. Όμως δεν είναι ακόμη σαφές ποιο είναι εκείνο το κομβικό σημείο της ανοσιακής απόκρισης που επιτρέπει στα CD8+ T κύτταρα να είναι αποτελεσματικά έτσι ώστε να αυτοπεριορίζουν τη λοίμωξη μόνο σε λίγους ασθενείς, ενώ στους περισσότερους οδηγεί στην μετάπτωση της λοίμωξης στη χρονιότητα. Μεταξύ των παραγόντων που έχουν προταθεί ως αίτια της λειτουργικής ανεπάρκειας των κυττάρων αυτών είναι και η δράση ενός καινούριου υποπληθυσμού των CD4+ T λεμφοκυττάρων: των T ρυθμιστικών κυττάρων. Σκοπός της παρούσας μελέτης ήταν να διερευνηθεί ο ρόλος των CD4+CD25+Foxp3+ T ρυθμιστικών κυττάρων στην ανοσοπαθογένεση της χρόνιας ηπατίτιδας C, η συμμετοχή τους στην ιστολογική βλάβη στο ήπαρ αλλά και o πιθανός τους ρόλος στην απάντηση στη θεραπεία σε ασθενείς που δεν είχαν υποβληθεί προηγουμένως σε θεραπεία. Η πρωτοτυπία της μελέτης έγκειται στο γεγονός ότι δεν έχει μελετηθεί ως σήμερα σε ασθενείς με χρόνια ηπατίτιδα C, η συσχέτιση του αριθμού των CD4+CD25+Foxp3+ T ρυθμιστικών κυττάρων στο περιφερικό αίμα 15
16 με την ιστική βλάβη και τnν in vitro καλλιέργεια των κυττάρων αυτών, σε τέσσερα διακριτά στιγμιότυπα κατά τη διάρκεια της θεραπείας, καθώς και η συσχέτισή τους με την απάντηση στη θεραπεία και με το είδος του γονότυπου του ιού. Η παρούσα μελέτη περιλαμβάνει δυο μέρη, το γενικό και το ειδικό. Στο ΓΕΝΙΚΟ ΜΕΡΟΣ, (ΜΕΡΟΣ Ι) γίνεται αρχικά ιστορική αναδρομή στην ιστορία του ιού της ηπατίτιδας C, τους τρόπους μετάδοσης και ακολούθως περιγραφή των επιδημιολογικών, ιολογικών και κλινικών χαρακτηριστικών της οξείας και χρόνιας λοίμωξης από τον ιό, ενώ περιγράφεται και η παθολογοανατομική βλάβη στο ήπαρ και το σύστημα σταδιοποίησης της, η εργαστηριακή διάγνωση, η θεραπεία και η πρόληψη. Κατόπιν (ΜΕΡΟΣ ΙΙ) αναλύονται οι μηχανισμοί κυτταρικής ανοσίας οι οποίοι εμπλέκονται στον περιορισμό της λοίμωξης και ακολούθως (ΜΕΡΟΣ ΙΙΙ) περιγράφεται η ανοσιακή απόκριση και η επιμέρους συμβολή των κυτταρικών υποπληθυσμών του ανοσιακού συστήματος στην οξεία και χρόνια λοίμωξη από τον ιό της ηπατίτιδας C, από τη στιγμή της αρχικής εισόδου του ιού έως και τον αυτοπεριορισμό της ή την εγκατάσταση της χρόνιας λοίμωξης και οι πιθανοί λόγοι μετάπτωσης της λοίμωξης σε χρονιότητα. Επίσης, (ΜΕΡΟΣ IV) γίνεται εκτενής αναφορά στον υποπληθυσμό των CD4+CD25+Foxp3+ T ρυθμιστικών κυττάρων, όσον αφορά στην ιστορία και την αναβίωση της έρευνας για αυτά, την προέλευση και την πλαστικότητα τους, τον φαινοτυπικό και λειτουργικό προσδιορισμό τους, την αλληλεπίδραση τους με άλλα κύτταρα του ανοσιακού συστήματος, τους μηχανισμούς δράσης τους και τους μηχανισμούς που ελέγχουν τη δράση τους. Τέλος, (ΜΕΡΟΣ V) γίνεται ιδιαίτερη αναφορά στις μελέτες που αφορούν στη συμμετοχή των Tregs στη χρόνια λοίμωξη από τον ιό της ηπατίτιδας C. Στο ΕΙΔΙΚΟ ΜΕΡΟΣ, αρχικά περιγράφονται το υλικό και η μεθοδολογία της μελέτης, ενώ ακολουθεί η ανάλυση, η σύγκριση των αποτελεσμάτων και η συζήτηση και η συσχέτιση αυτών με τα γνωστά έως και σήμερα δεδομένα της βιβλιογραφίας. Τέλος, αναφέρονται τα κύρια συμπεράσματα της παρούσας μελέτης, η περίληψη στην ελληνική και αγγλική γλώσσα και η βιβλιογραφία που χρησιμοποιήθηκε. Οι ασθενείς που εντάχθηκαν στη μελέτη προέρχονταν από τo Ηπατολογικό Τμήμα της Β Παθολογικής Κλινικής του Αριστοτελείου Πανεπιστημίου της Θεσσαλονίκης. Το ερευνητικό μέρος της μελέτης εκπονήθηκε στο Εργαστήριο Ανοσολογίας και Ηπατολογίας της Β Παθολογικής Κλινικής, το Εργαστήριο του Τμήματος Κλινικής 16
17 Ανοσολογίας της Β Παθολογικής Κλινικής του ΑΠΘ, καθώς και το Εργαστήριο Παθολογικής Ανατομικής του ΑΠΘ. Είναι ιδιαίτερα σημαντικό για μένα να εκφράσω τις ευχαριστίες μου στην Καθηγήτριά μου κα Μαρία Ραπτοπούλου-Γιγή, επιβλέπουσα της παρούσας διατριβής και υπεύθυνη του Ηπατολογικού Τμήματος της Β Παθολογικής Κλινικής του ΑΠΘ για την ευκαιρία που μου έδωσε να δουλέψω μαζί της, να εκπονήσω την παρούσα μελέτη αλλά και να ακολουθήσω το συναρπαστικό κόσμο της Ηπατολογίας από τα πρώτα βήματα της ειδικότητας μου στην Β Παθολογική Κλινική στην οποία διατελούσε και χρέη Διευθύντριας από το Η συμβολή της ήταν καθοριστική όχι μόνο στην επιλογή του θέματος της διατριβής αλλά και την καθοδήγηση του πρωτοκόλλου, την ώθηση για την ολοκλήρωση της και την επίβλεψη της τελικής συγγραφής. Ιδιαίτερες ευχαριστίες πρέπει να αποδώσω και στην κα Παναγιώτα Μπούρα, Καθηγήτρια και υπεύθυνη του Τμήματος Κλινικής Ανοσολογίας της Β Παθολογικής Κλινικής, μέλος της συμβουλευτικής επιτροπής, για την εξαιρετικά σημαντική της συμβολή και βοήθεια στην παρούσα μελέτη, από την εκπόνηση του πρωτοκόλλου έως και την συγγραφή της. Της χρωστάω το ότι αγάπησα την Ειδική Παθολογία από τα φοιτητικά μου χρόνια. Θερμές ευχαριστίες στον κο Μιχαήλ Δανιηλίδη, Καθηγητή, μέλος της συμβουλευτικής επιτροπής, για τις καίριες επισημάνσεις και διορθώσεις του. Θα ήταν σημαντική παράλειψη να μην αναφέρω την ουσιαστική βοήθεια, υποστήριξη και εμψύχωση του Καθηγητή κου Ιωάννη Κλωνιζάκη, Διευθυντή της Β Παθολογικής Κλινικής, καθόλη τη διάρκεια της διδακτορικής μου διατριβής. Ευχαριστώ θερμά τον συνάδελφο κο Αλέξανδρο Σαραντόπουλο για τη καίρια, ουσιαστική, αναντικατάστατη και αγόγγυστη συμβολή του σε όλη τη διάρκεια του πρωτοκόλλου της μελέτης όσον αφορά στην εκπαίδευση μου αλλά και την πραγματοποίηση του μεγαλύτερου τμήματος του πειραματικού μέρους της παρούσας μελέτης, κυρίως όσον αφορά στην κυτταρομετρία ροής και την in vitro καλλιέργεια. Ευχαριστώ πολύ την Καθηγήτρια κα Ελένη Ορφανού-Κουμερκερίδου για την ουσιαστική της συμβολή στη διενέργεια των μοριακών τεχνικών προσδιορισμού του ιικού φορτίου και του γονότυπου, αλλά και την φιλική της συμπαράσταση. Ευχαριστώ ιδιαιτέρως την Καθηγήτρια κα 17
18 Ελένη Βρεττού για την ενδελεχή μελέτη των ιστολογικών δειγμάτων των ασθενών της μελέτης και την διακριτική και συνεχή της βοήθεια στην αξιολόγηση των ιστολογικών δεδομένων των ασθενών. Ευχαριστώ ειλικρινά τους ιατρούς συνεργάτες στο Ηπατολογικό Τμήμα της Β Παθολογικής Κλινικής κα Βάνα Σούκια και κο Εμμανουήλ Σινάκο για την αναντικατάστατη βοήθεια τους στην παρακολούθηση των ασθενών της μελέτης. Χωρίς αυτούς θα ήταν αδύνατη η ολοκλήρωση της συλλογής των δειγμάτων. Σημαντική ήταν και η βοήθεια του συναδέλφου Λέκτορα Παθολογίας της Ιατρικής Σχολής Αλεξανδρούπολης, κου Παναγιώτη Σκένδρου στη διαμόρφωση των σχημάτων της παρούσας έκδοσης. Τέλος, πρέπει να ευχαριστήσω την οικογένεια μου, τον πατέρα μου κο Αντώνιο Μπέλλο, τη μητέρα μου κα Δήμητρα Μπέλλου και την αδερφή μου κα Βικτώρια Μαρία Μπέλλου για την ηθική και συναισθηματική τους στήριξη σε όλο το δύσκολο και επίπονο δρόμο της ολοκλήρωσης της παρούσας μελέτης. Στην τελευταία χρωστάω και την άρτια ολοκλήρωση της στατιστικής μελέτης. 18
19 ΓΕΝΙΚΟ ΜΕΡΟΣ 19
20
21 ΜΕΡΟΣ Ι. ΗΠΑΤΙΤΙΔΑ C Οι ιογενείς ηπατίτιδες αναγνωρίζονται σήμερα ότι αποτελούν ένα σοβαρό πρόβλημα δημόσιας υγείας και μία από τις βασικές αιτίες θνησιμότητας από λοιμώδη νοσήματα παγκόσμια. Όσον αφορά την ηπατίτιδα C, ο αριθμός των ασθενών με λοίμωξη με τον ιό της ηπατίτιδας C (HCV) ανέρχεται στα εκατομμύρια (περίπου 3% του πληθυσμού της γης), υπολογίζεται ότι 3-4 εκατομμύρια άνθρωποι μολύνονται ανά έτος, η νόσος ευθύνεται για περίπου θανάτους /έτος και ο κίνδυνος ανάπτυξης ηπατοκυτταρικού καρκινώματος αυξάνει στο 1-4% ανά έτος μετά την εγκατάσταση της κίρρωσης του ήπατος. Άλλωστε σήμερα, η κίρρωση από χρόνια HCV λοίμωξη αποτελεί την συχνότερη αιτία μεταμόσχευσης του ήπατος 1. Ο επιπολασμός της λοίμωξης ποικίλει από χώρα σε χώρα με τη μικρότερη συχνότητα να αφορά σε χώρες όπως το Ηνωμένο Βασίλειο και τη Σκανδιναβία (0,01-0,1%) και τη μεγαλύτερη την Αίγυπτο (15-20%) 2. Στην Ελλάδα, ο ακριβής επιπολασμός της HCV λοίμωξης δεν είναι γνωστός, υπολογίζεται ότι 2% του γενικού πληθυσμού πάσχει από χρόνια ηπατίτιδα C, εντούτοις φαίνεται ότι υπάρχει μεγάλη γεωγραφική διακύμανση της νόσου ( %) 3, 4, 5. Επιπλέον οι παράγοντες κινδύνου της λοίμωξης, η φυσική ιστορία και η μακρόχρονη έκβαση της, όπως και οι μεταβολές της εξέλιξής της μέσα στο χρόνο, παραμένουν άγνωστα. Ο ιός της ηπατίτιδας C είναι ένας από τους πρώτους παθογόνους για τον άνθρωπο ιούς και αναγνωρίσθηκε το 1989 χάρη στην εξέλιξη της μοριακής βιολογίας. Μεταδίδεται αιματογενώς και ευθύνεται για τη συντριπτική πλειοψηφία των περιπτώσεων ηπατίτιδας που παλαιότερα ονομάζονταν μη-α μη-β 6. Ο HCV είναι το αίτιο της οξείας και χρόνιας ηπατίτιδας C. Α. O ιός της ηπατίτιδας C (HCV) Ο HCV είναι ένας θετικός μονής έλικας RNA ιός και αποτελεί το μοναδικό μέλος του Hepacivirus της οικογένειας Flaviviridae. Το γονιδίωμα του HCV αποτελείται από μονόκλωνο RNA θετικής κατεύθυνσης, έχει μήκος 9500 νουκλεοτιδικών βάσεων με ένα και μοναδικό πλαίσιο ανάγνωσης. Του πλαισίου αυτού προηγούνται στο 5 άκρο και έπονται στο 3 άκρο αριθμός νουκλεοτιδίων που συνιστούν τις μη κωδικογραφούσες περιοχές (UTR-nontranslated regions) (Σχήμα 1). 21
22 Το πλαίσιο ανάγνωσης κωδικογράφει μια πρωτεΐνη μήκους 3010 αμινοξέων, η οποία μετά τη μετάφρασή της διαιρείται στις δομικές και μη πρωτεΐνες του ιού. Η δομική περιοχή κωδικογράφει την πρωτεΐνη του πυρηνοκαψιδίου (p22) και τις 2 γλυκοπρωτεΐνες του περιβλήματος Ε1 (gp35) και Ε2/ΝS1 (gp72). Οι μη δομικές πρωτεΐνες κωδικογράφουν τις ΝS2, NS3, NS4 και NS5. Η NS3 έχει ιδιότητες πρωτεΐνάσης με ρόλο την επεξεργασία της πολυπρωτεΐνης καθώς και ελικάσης σχετιζόμενης με τον πολλαπλασιασμό του ιού. Η ΝS5 λειτουργεί ως RNA πολυμεράση. Χαρακτηριστικό όλων των ιών οι οποίοι όπως ο HCV, πολλαπλασιάζονται μέσω μιας RNA-εξαρτώμενης RNA πολυμεράσης, είναι το σχετικό υψηλό ποσοστό εμφάνισης αυτόματων μεταλλάξεων με αποτέλεσμα να επικρατεί μια γενετική ετερογένεια. Δηλαδή σε κάθε στιγμή, ο ιός που υπάρχει σε έναν ασθενή αποτελείται από έναν πληθυσμό σχεδόν ιών (quasispecies). Η ικανότητα του ιού να μεταλλάσσεται σε ταχύ ρυθμό έχει σαν αποτέλεσμα ο ιός να επιβιώνει σε κάθε πίεση προερχόμενη είτε από την ανοσιακή απάντηση του ξενιστή είτε από τη θεραπευτική παρέμβαση. Η προκύπτουσα ετερογένεια αφορά ιδιαίτερα ορισμένες περιοχές του γονιδιώματος όπως η υπερμεταβλητή περιοχή (HVR) της E2 πρωτεΐνης. Β. Πολλαπλασιασμός του ιού της ηπατίτιδας C. Ο κύκλος ζωής του HCV Η είσοδος του ιού στο κύτταρο επιτυγχάνεται μετά τη σύνδεση του σε διάφορα μόρια που σχηματίζουν τους υποδοχείς στην επιφάνεια του κυττάρου για τον ιό : το CD81 μόριο, τον υποδοχέα Β τύπου 1, το διακυττάριο μόριο προσκόλλησης 3 μη ιντεγκρίνη (DC-SIGN) των δενδριτικών κυττάρων, το διακυττάριο μόριο προσκόλλησης 3 (ΙCAM- 3) του ήπατος/λεμφαδένων, τις πρωτεΐνες κλοντίνη (claudin) και απόφραξης (occluding) 7, 8, 9, 10 τον υποδοχέα της λιποπρωτεΐνης (LDR- R) ή τον υποδοχέα της ασιαλογλυκοπρωτεΐνης (ASGR-R) (Σχήμα 2). Η γλυκοπρωτεΐνη Ε2 του ιού θεωρείται ότι είναι υπεύθυνη για την αρχική σύνδεση του ιού στο κύτταρο ξενιστή. Μετά την ενδοκύττωση, ο HCV απελευθερώνεται από το νουκλεοκαψίδιο μέσα στο κυτταρόπλασμα και το γονιδιακό RNA μεταγράφεται άμεσα και μαζί με το RNA που έχει συνδεθεί, συνιστούν το αγγελιοφόρο RNA για τη σύνθεση της πολυπρωτεΐνης. Κατά την ΗCV λοίμωξη σχηματίζεται ένα σύμπλεγμα πολλαπλασιασμού του ιού στο οποίο συμμετέχουν ιικές πρωτεΐνες, κυτταρικά στοιχεία και νεοδημιουργηθέντα RNA γονιδιώματα. 22
23 Τελικά, ο κύκλος πολλαπλασιασμού του HCV θα μπορούσε να συνοψισθεί ως εξής (Σχήμα 3): α) διείσδυση του ιού στο κύτταρο ξενιστή και απελευθέρωση γονιδιακού RNA θετικής κατεύθυνσης (+RNA) από το ιικό σωματίδιο στο κυτταρόπλασμα. β) μεταγραφή του RNA, επεξεργασία της πολυπρωτεΐνης και σχηματισμός συμπλέγματος ρεπλικάσης που σχετίζεται με τις κυτταρικές μεμβράνες γ) χρησιμοποίηση του +RNA για τη σύνθεση του γονιδιακού RNA αρνητικής κατεύθυνσης (-) δ) παραγωγή νέων +RNA μορίων τα οποία μπορούν να χρησιμοποιηθούν για την σύνθεση RNA τα οποία χρησιμεύουν για την έκφραση πολυπρωτεΐνης ή το πακετάρισμα των προγονιδιακών ιοσωματίων ε) απελευθέρωση του ιού από το μολυσμένο κύτταρο Γ. Γονότυποι του HCV Με βάση την ποικιλομορφία της νουκλεοτιδικής ακολουθίας του HCV, διακρίνονται 6 γονότυποι του ιού (1-6), >100 διαφορετικοί υπότυποι (οι πιο συχνοί είναι οι 1 α, 1β, 2 α, 2β) και πολλά quasispecies (σχεδόν είδη) του ιού στον ίδιο ασθενή, οι οποίοι προέρχονται από διαφορετικά στελέχη, με 95% κοινή αλληλουχία νουκλεοτιδίων και προκύπτουν από αλλαγές στις υπερμεταβλητές περιοχές. Οι γονότυποι, εμφανίζουν μεταξύ τους 70-80% ομοιότητα. Η ετερογένεια των γονοτύπων προέκυψε πιθανόν από συνδυασμό της ικανότητας πολλαπλασιασμού του ιού, της «κακής» RNA πολυμεράσης, της ανοσιακής επιλογής και του τρόπου μετάδοσης και μετακίνησης πληθυσμών. Στην Ευρώπη πιο συχνός είναι ο γονότυπος 1 (60-70%) 11. Ο γονότυπος δεν φαίνεται να σχετίζεται με την φυσική εξέλιξη και της επιπλοκές της χρόνιας λοίμωξης, όμως έχει αποδειχθεί ότι καθορίζει την διάρκεια της θεραπείας καθώς είναι ο σημαντικότερος προγνωστικός παράγοντας της απάντησης. Δ. Τρόπος μετάδοσης Η ηπατίτιδα C μεταδίδεται μετά από επαφή με μολυσμένο αίμα ή προϊόντα αίματος. Σήμερα ο κυριότερος τρόπος μετάδοσης είναι η ενδοφλέβια χρήση ναρκωτικών ουσιών. Άλλοι δυνητικοί τρόποι μετάδοσης είναι: η επαγγελματική έκθεση, η σεξουαλική μετάδοση, η 23
24 ενδορινική χρήση κοκαΐνης, το τατουάζ, το τρύπημα του σώματος και η μετάδοση από τη μητέρα στο παιδί. Ο κίνδυνος της μετάδοσης μέσω των προϊόντων αίματος ή την μεταμόσχευση οργάνων, ουσιαστικά ελαχιστοποιήθηκε μετά τη δυνατότητα ανίχνευσης των antihcv αντισωμάτων στα μέσα του Σήμερα τα υψηλότερα ποσοστά HCV λοίμωξης (70-90%) αναφέρονται σε ΕΦ χρήστες ναρκωτικών ουσιών και σε ασθενείς με αιμορροφιλία οι οποίοι ελάμβαναν παράγοντες πήξεως πριν από το , 13. Ε. Πρόληψη Οι βασικές αρχές πρόληψης αφορούν αφενός στην ενημέρωση του γενικού πληθυσμού και την οργάνωση προγραμμάτων εκπαίδευσης σε ομάδες με συμπεριφορά υψηλού κινδύνου και αφετέρου στον έλεγχο του αίματος και των παραγώγων του στα Τμήματα Αιμοδοσίας με ευαίσθητες και ειδικές μεθόδους, καθώς και στον έλεγχο των υποψήφιων δοτών οργάνων σώματος και τις εγκύους προγεννητικά. Αναλυτικότερα, θα πρέπει να χρησιμοποιούνται αποστειρωμένες βελόνες και σύριγγες μιας χρήσης, να γίνεται χρήση γαντιών από το ιατρικό και νοσηλευτικό προσωπικό και να τηρούνται αυστηρά άσηπτες τεχνικές σε οδοντιάτρους, χειρουργικές αίθουσες και μονάδες τεχνητού νεφρού. ΣΤ. Διάγνωση- εργαστηριακός έλεγχος Οι διαγνωστικές εξετάσεις για τη λοίμωξη από τον ιό της ηπατίτιδας C μπορούν να διακριθούν σε δυο κατηγορίες: α) ορολογικές μέθοδοι που ανιχνεύουν αντισώματα του ιού με ανοσοενζυμικές μεθόδους (ΕΙΑ, enzyme-linked immunosorbent assay ή ELISA) και με την τεχνική RIBA δεύτερης και τρίτης γενιάς (recombinant immunoblot assay). Ο μέσος χρόνος ορομετατροπής και εμφάνισης ανιχνεύσιμου αντισώματος στον ορό μετά την μόλυνση από τον ιό είναι περίπου 10 εβδομάδες 14. β) μέθοδοι ανίχνευσης νουκλεϊκών οξέων (NAT, nucleic acid tests), πρόκειται για μοριακές μεθόδους που ανιχνεύουν ή ποσοτικοποιούν το ιικό φορτίο, δηλαδή την παρουσία ή την απουσία ιικού πολλαπλασιασμού. Ζ. Βιοψία ήπατος Η βιοψία ήπατος δεν είναι απαραίτητη για τη διάγνωση της ηπατίτιδας C, είναι όμως το gold standard για την αξιολόγηση της ιστικής βλάβης. 24
25 Προσφέρει πληροφορίες όσον αφορά στον προσδιορισμό της νεκροφλεγμονής και της ίνωσης κι έτσι την σκοπιμότητα έναρξης αγωγής αλλά και τον αποκλεισμό άλλου συνυπάρχοντα υπεύθυνου αιτίου για την αύξηση των τρανσαμινασών (π.χ. αλκοολική νόσος ήπατος, λιπώδες ήπαρ, αιμοσιδήρωση, αυτοάνοση ηπατίτιδα). Πρόκειται για αιματηρή μέθοδο με επιπλοκές σε ποσοστό 2-3% και θνητότητα 1/ Η. Μη επεμβατική αξιολόγηση της ηπατικής ίνωσης Έχουν αναπτυχθεί διάφορες μέθοδοι για την μη επεμβατική αξιολόγηση της ίνωσης στην χρόνια ηπατίτιδα C. Ένας εξ αυτών είναι οι βιολογικοί δείκτες (biomarkers) οι οποίοι διακρίνονται i) στους άμεσους, που αξιολογούν την κινητικήτης εξωκυττάριας θεμέλιας ουσίας στο ήπαρ (turnover) και ii) τους έμμεσους, οι οποίοι αντικατοπτρίζουν την ηπατική λειτουργία και τη φλεγμονή 16, 17, 18, 19, ενώ χρήσιμες έχουν αποδειχθεί και αρκετές εφαρμογές της ελαστογραφίας, δηλαδή της διαφορετικής αντίδρασης των ιστών σε μηχανικό, παλμικό ή ακουστικό ερέθισμα σε συνδυασμό με υπερήχους (Transient Elastography, Fibroscan, Echosens, Paris) αλλά και τη μαγνητική τομογραφία (MR elastography) 20, 21, 22, 23, 24. Θ. Κλινική εικόνα και φυσική εξέλιξη της HCV λοίμωξης Η φυσική ιστορία της HCV λοίμωξης είναι δύσκολο να μελετηθεί κυρίως επειδή πρόκειται για μια ασυμπτωματική λοίμωξη τόσο στην έναρξη όσο και την εξέλιξη της με αποτέλεσμα, τις περισσότερες φορές να μην είναι εφικτό να προσδιορισθεί ο ακριβής χρόνος μόλυνσης του ασθενή (Σχήμα 4). Τα δεδομένα που υπάρχουν σήμερα προέρχονται κυρίως από περιπτώσεις μετάδοσης του ιού μετά από μετάγγιση αίματος ή παραγώγων αίματος πριν το 1989, ή από ενδοφλέβια χρήση ναρκωτικών ουσιών κατά την οποία μπορεί να προσδιορισθεί ο χρόνος λοίμωξης. Ο χρόνος επώασης της HCV λοίμωξης είναι μέρες 25. Ένα ποσοστό 20-25% των ασθενών εμφανίζει κλινικά συμπτώματα όπως κόπωση, αδυναμία, ανορεξία και ίκτερο, ενώ σπάνια είναι η εμφάνιση λοίμωξης με κεραυνοβόλο ηπατική ανεπάρκεια 26. Έχει βρεθεί ότι οι ασθενείς που εμφανίζουν συμπτωματική οξεία HCV λοίμωξη και κυρίως εκείνοι με ίκτερο έχουν μικρότερο ποσοστό μετάπτωσης σε χρονιότητα 27, 28. Η οξεία ηπατίτιδα C αυτοπεριορίζεται σε ποσοστό 15-40%, ενώ σε ένα ποσοστό 60-85% δεν επιτυγχάνεται κάθαρση του ιού, το ιικό φορτίο 25
26 (HCV RNA) παραμένει θετικό για περισσότερους από έξι μήνες, οπότε η λοίμωξη μεταπίπτει σε χρονιότητα 25, 29. Το ποσοστό αυτόματης κάθαρσης του ιού και αυτοπεριορισμού της χρόνιας ηπατίτιδας C είναι εξαιρετικά μικρό (0,5%-0,74% έτη νόσου/ ασθενή) 30, 31. Η ταχύτητα της εξέλιξης της ίνωσης στη χρόνια HCV λοίμωξη και άρα η ταχύτητα εγκατάστασης της κίρρωσης φαίνεται πως δεν είναι γραμμική. Εξαρτάται από πολλούς παράγοντες όπως το φύλο (άνδρας), τη φυλή (Αφρο-αμερικανοί), την ηλικία της μόλυνσης από τον ιό (>25 ετών), την κατανάλωση αλκοόλ (>30gr/ημέρα για τους άνδρες, >20gr/ημέρα για τις γυναίκες), την επάρκεια του ανοσιακού συστήματος, την προϋπάρχουσα ιστολογική βλάβη στο ήπαρ, τις συνυπάρχουσες παθήσεις (όπως αντίσταση στην ινσουλίνη, παχυσαρκία, σακχαρώδης διαβήτης, μη αλκοολική στεατοηπατίτιδα ή μεταβολικό σύνδρομο), αλλά και την παρουσία συλλοιμώξεων από άλλους ιούς (π.χ, HBV, HDV, HIV) 32, 33, 34, 35, 36. Επιπλέον διαπιστώθηκε ότι η εμφάνιση του ηπατοκυτταρικού καρκινώματος (1-4% κατ έτος) έπεται της εγκατάσταση κίρρωσης του ήπατος 37. Όσον αφορά στα ποσοστά 3ετούς, 5ετούς και 10ετούς επιβίωσης σε αντιρροπούμενη κίρρωση έχει βρεθεί ότι είναι 96%, 91% και 79% αντίστοιχα, με πιθανότητα 5% να εμφανιστεί άρση της αντιρρόπησης τον πρώτο χρόνο. Η πιθανότητα αυτή αυξάνει σε 30% μετά από 10ετή εγκατάσταση της κίρρωσης του ήπατος. Το 5ετές ποσοστό επιβίωσης σε μη αντιρροπούμενη κίρρωση ανέρχεται στο 50% 38, 39, 40, ενώ τα ποσοστά επιβίωσης σε ηπατοκυτταρικό καρκίνο εξαρτώνται από το μέγεθος της κακοήθειας κατά τη διάγνωση. Συνολικά, φαίνεται πως η χρόνια HCV λοίμωξη έχει μικρή νοσηρότητα και θνητότητα τις πρώτες δυο δεκαετίες, αλλά κατά τη διάρκεια της τρίτης και τέταρτης δεκαετίας εμφανίζονται οι σοβαρές επιπλοκές της νόσου. Ι. Θεραπεία Όλοι οι ασθενείς με χρόνια ηπατίτιδα C και θετικό HCV RNA είναι υποψήφιοι για θεραπεία. Αντενδείξεις θεραπείας αποτελούν η μη αντιρροπούμενη κίρρωση, η προηγηθείσα μεταμόσχευση νεφρών, καρδιάς, πνευμόνων, η αυτοάνοση ηπατίτιδα ή άλλη αυτοάνοση πάθηση που επιδεινώνεται από την χορήγηση κυρίως της ιντερφερόνηςα, η θυρεοειδοπάθεια που δεν έχει ρυθμιστεί με αγωγή, έγκυες ή άτομα που δεν συμφωνούν σε επαρκή αντισυλληπτικά μέτρα, η σοβαρή συνυπάρχουσα νόσος όπως, η σοβαρή καρδιαγγειακή νόσος, η καρδιακή ανεπάρκεια, η στεφανιαία νόσος, ο αρρύθμιστος 26
27 σακχαρώδης διαβήτης, η χρόνια αποφρακτική πνευμονοπάθεια και η γνωστή υπερευαισθησία στην αγωγή. Θεραπεία της οξείας ηπατίτιδας C Η θεραπεία της οξείας ηπατίτιδας C είναι σημαντική όσον αφορά την εξέλιξη της νόσου σε χρόνια ηπατίτιδα. Ο βέλτιστος χρόνος έναρξης αγωγής με ιντερφερόνη-α είναι 8-12 εβδομάδες μετά τη μόλυνση, η ενδεδειγμένη διάρκεια αγωγής είναι οι 24 εβδομάδες και η συγχορήγηση ριμπαβιρίνης δεν είναι απαραίτητη 41, 42. Θεραπεία χρόνιας ηπατίτιδας C σε πρωτοθεραπευόμενους ασθενείς Σε όλους τους ασθενείς χορηγείται σήμερα, συνδυασμός πεγκυλιωμένης ιντερφερόνης-a και ριμπαβιρίνης (peg-ifn-a+ RIBA), εφόσον δεν υπάρχουν απόλυτες αντενδείξεις 43. Οι ασθενείς με γονότυπο 1 και 4 λαμβάνουν για 48 εβδομάδες θεραπεία συνδυασμού με peg-ifn-a + RIBA ανάλογα με το σωματικό βάρος (1000mg/ημέρα για ΣΒ<75kg και 1200mg/ημέρα για ΣΒ>75 kg), ενώ οι ασθενείς με γονότυπο 2 και 3 λαμβάνουν βραχύτερο θεραπευτικό σχήμα peg-ifn-a + RIBA διάρκειας 24 εβδομάδων, αλλά η ριμπαβιρίνη χορηγείται σε χαμηλότερη δόση (800mg/24h) 44, 45. Οι ασθενείς με γονότυπο 1 και 4 έχουν μειωμένη πιθανότητα ανταπόκρισης στη θεραπεία [μακρόχρονη ιολογική ανταπόκριση (SVR) παρατηρείται στο 30%-40% των ασθενών], σε αντίθεση με το γονότυπο 2 και 3 που τα ποσοστά ανταπόκρισης αγγίζουν το 85%. Κομβικά σημεία της αγωγής αποτελούν η τέταρτη και η δωδέκατη εβδομάδα θεραπείας. Έχει βρεθεί ότι οι ασθενείς με γονότυπο 1 που επιτυγχάνουν πλήρη πρώιμη ιολογική ανταπόκριση (EVR) έχουν πιθανότητα 60% να πετύχουν SVR, ενώ οι ασθενείς με γονότυπο 2, 3 που επιτυγχάνουν ταχεία ιολογική ανταπόκριση (RVR), έχουν 95% πιθανότητα εμφάνισης SVR 46. Τα στιγμιότυπα RVR και EVR χρησιμοποιούνται επίσης για λήψη θεραπευτικών αποφάσεων όσον αφορά τη βράχυνση ή την επιμήκυνση της διάρκειας της αγωγής 47, 48, 49 (Σχήμα 5). Εντούτοις, υπάρχουν ειδικές ομάδες ασθενών που απαιτούν εξατομίκευση και περαιτέρω μελέτη της θεραπευτικής αντιμετώπισης και αφορούν παιδιά, αιμοκαθαιρόμενους, άτομα με αιμοσφαιρινοπάθειες ή αιμορροφιλία, ενεργούς χρήστες ναρκωτικών ουσιών, ασθενείς με HIV λοίμωξη, ασθενείς με νευροψυχιατρικά προβλήματα και ασθενείς που έχουν υποβληθεί σε μεταμόσχευση ήπατος. Επιπλέον, ένα ποσοστό 10-14% των ασθενών διακόπτει την θεραπεία λόγω εμφάνισης σημαντικών παρενεργειών. Οι πιο συχνές παρενέργειες της θεραπείας είναι: το σύνδρομο που προσομοιάζει με γρίπη (flu-like 27
28 syndrome), η κόπωση, οι αιματολογικές διαταραχές (λευκοπενία, αναιμία, θρομβοπενία), τα νευροψυχιατρικά συμπτώματα (κατάθλιψη, επιθετικότητα) και η τριχόπτωση. Θεραπεία σε μη-ανταποκριθέντες σε προηγούμενα σχήματα Οι ασθενείς οι οποίοι δεν απήντησαν στην αρχική θεραπεία, είτε με μονοθεραπεία με INF-α, είτε με συνδυασμό IFN-α (πεγκυλιωμένης ή μη) και ριμπαβιρίνης, υποβάλλονται σε επαναθεραπεία με το σκεπτικό ότι δυνητικά μπορούν να ωφεληθούν από ένα νέο κύκλο θεραπείας με συνδυασμό peg-inf-α + RIBA. Το συνολικό ποσοστό επίτευξης SVR σε αυτούς τους ασθενείς είναι περίπου 20%, εάν όμως επιτευχθεί πλήρης EVR το ποσοστό SVR αγγίζει το 50% 50, 51. Σχήμα 1: Ο ιός της ηπατίτιδας C γλυκοπρωτεΐνη φακέλου Ε1 πρωτεΐνη καψιδίου C νουκλεϊκό οξύ λιπίδιο φακέλου πυρήνας RNA φάκελος εξωτερική όψη δομικά γονίδια διατομή ιικού σωματιδίου γλυκοπρωτεΐνη φακέλου Ε2 μη δομικά γονίδια 28

29 Σχήμα 2: Η σύνδεση του ιού της ηπατίτιδας C στα ηπατοκύτταρα προσαρμοσμένο από Debuisson et al, Cell Microbiol 2008;10: OCCLN σύντηξη εγκοπή επικαλυμμένη με κλαθρίνη πρώιμο ενδόσωμα απελευθέρωση γενώματος LDL-R, GAG CD81, SRB1 CLAUDIN 1, 6 ή 9 OCCLUDIN 29

30 Σχήμα 3: O κύκλος ζωής του HCV προσαρμοσμένο από Pawlotsky, Hepatology 2006;43:S σύνδεση 2. είσοδος του ιού 3. αφαίρεση του πυρήνα 4. αφαίρεση της επικάλυψης 5. μεταγραφή 6. πολλαπλασιασμός του RNA 7. συγκέντρωση 8. μεταφορά 9. απελευθέρωση του ιού από το κύτταρο 30
31 Σχήμα 4: Η φυσική εξέλιξη της HCV λοίμωξης Μόλυνση από τον ιό της ηπατίτιδας C οξεία HCV λοίμωξη (20-25% συμπτωματική, συχνότερα υποκλινική) αυτοπεριορισθείσα HCV λοίμωξη 15-40% (μη ανιχνεύσιμο HCV RNA 6 μήνες μετά) χρόνια HCV λοίμωξη 60-85% (ανιχνεύσιμο HCV RNA 6 μήνες μετά) κεραυνοβόλος οξεία ηπατική ανεπάρκεια, σπάνια κίρρωση (10-20% μετά από 20 έτη) μη αντιρροπούμενη κίρρωση (5ετής επιβίωση 50%) ΗΚΚ (1-4% ανά έτος) μεταμόσχευση ήπατος θάνατος 31
32 Σχήμα 5: Ιολογική απόκριση κατά τη διάρκεια της θεραπείας- ορισμοί πεγκυλιωμένη ιντερφερόνη και ριμπαβιρίνη υποτροπή μη ανιχνεύσιμο Rapid virologic response (RVR) (ταχεία ιολογική απάντηση) Early virologic response (EVR) (πρώιμη ιολογική απάντηση) End of treatment response (ETR) (απάντηση στο τέλος της αγωγής) Sustained virologic response (SVR) (μακροχρόνια ιολογική απάντηση) Breakthrough (διαφυγή) Relapse (υποτροπή) Null responder (ασθενής με μηδενική απάντηση στη θεραπεία, «ανθεκτικός» ) Non responder (ασθενής με μη απάντηση) Partial responder (ασθενής με μερική απάντηση) εβδομάδες μετά την έναρξη της θεραπείας Μη ανιχνεύσιμο ιικό φορτίο με ευαίσθητη ποσοτική μέθοδο την 4 η εβδομάδα αγωγής Μείωση του αρχικού ιικού φορτίου 2 δεκαδικούς λογάριθμους (partial EVR) ή μη ανιχνεύσιμο ιικό φορτίο (complete EVR) την 12 η εβδομάδα αγωγής Μη ανιχνεύσιμο ιικό φορτίο στο τέλος της αγωγής (μετά από 24 ή 48 εβδομάδες, αναλόγως του γονότυπου) Αρνητικό ιικό φορτίο 24 εβδομάδες μετά το τέλος της αγωγής Επανεμφάνιση του ιικού φορτίου κατά τη διάρκεια της αγωγής ενώ πριν είχε μειωθεί σημαντικά ή είχε γίνει μη ανιχνεύσιμο Θετικοποίηση του ιικού φορτίου μετά το τέλος της αγωγής, ενώ στο τέλος της αγωγής ήταν μη ανιχνεύσιμο Μείωση του ιικού φορτίου < 2 δεκαδικούς λογάριθμους μετά από θεραπεία 24 εβδομάδων Ανιχνεύσιμο ιικό φορτίο μετά από 24 εβδομάδες θεραπείας Μείωση ιικού φορτίου 2 δεκαδικούς λογάριθμους αλλά ανιχνεύσιμο την 24 η εβδομάδα αγωγής προσαρμοσμένο από Ghany kai Strader, Hepatology
33 ΜΕΡΟΣ ΙΙ. Η ΚΥΤΤΑΡΙΚΗ ΑΝΟΣΙΑ ΣΤΗ ΛΟΙΜΩΞΗ Οι διάφοροι παθογόνοι μικροοργανισμοί οι οποίοι δυνητικά μπορούν να προκαλέσουν λοίμωξη, θα πρέπει αρχικά να διαπεράσουν τους φυσικούς φραγμούς του δέρματος και των βλεννογόνων. Μετά την είσοδό τους, έρχονται αντιμέτωποι με την πρώτης γραμμής άμυνα του ξενιστή και ειδικότερα α) με τα φαγοκύτταρα και τα πολυμορφοπύρηνα, τα οποία φαγοκυτταρώνουν και τελικά καταστρέφουν τους μικροοργανισμούς και β) ενεργοποιούν το συμπλήρωμα, ορισμένες πρωτεΐνες του οποίου συνδέονται στην επιφάνεια των μικροοργανισμών και δρουν ως οψωνίνες, προάγοντας με τον τρόπο αυτό την φαγοκυττάρωσή τους. Η ενεργοποίηση του συμπληρώματος προάγει επίσης την τοπική φλεγμονή μέσω της απελευθέρωσης χημειοτακτικών και αγγειοδραστικών ουσιών. Στις λοιμώξεις από ιούς, ενεργοποιούνται δύο μηχανισμοί άμυνας: a)τα μολυσμένα από τον ιό κύτταρα προάγουν τη σύνθεση των IFNs και β) οι ιοί αναγνωρίζονται και καταστρέφονται. Οι μη-ειδικοί ανοσιακοί μηχανισμοί (φυσική ανοσία) παίζουν κριτικό ρόλο στα πρώιμα στάδια της λοίμωξης, καθώς η ειδική έναντι του συγκεκριμένου αντιγόνου ανοσιακή απάντηση καθυστερεί να εγκατασταθεί. Επιπλέον, οι μη-ειδικοί μηχανισμοί συνεχίζουν να δρουν καθ όλη τη διάρκεια της λοίμωξης μέχρι της αποδρομής της. Η φυσική, μη-ειδική και η ειδική (επίκτητη) ανοσιακή απάντηση αποτελούν ένα ενιαίο σύστημα άμυνας του ξενιστή στο οποίο συμμετέχουν διάφορα κύτταρα και μόρια τα οποία δρουν συνεργικά 54, 55. Οι δύο βασικοί οδοί συνεργασίας μεταξύ φυσικής και επίκτητης ανοσίας είναι: a) η φυσική ανοσιακή απάντηση σε παθογόνους μικροοργανισμούς ενεργοποιεί την επίκτητη ανοσιακή απάντηση και επηρεάζει το είδος της απάντησης και β) η ειδική ανοσιακή απάντηση χρησιμοποιεί πολλούς από τους μηχανισμούς της φυσικής ανοσίας για να περιορίσει τη λοίμωξη και επιπλέον προάγει τους μηχανισμούς άμυνας της φυσικής ανοσίας. Η επίκτητη ανοσιακή απάντηση έναντι των παθογόνων μικροοργανισμών αποτελείται από α) τη χυμική ανοσία, η οποία χρησιμοποιεί τις πρωτεΐνες που παράγονται από τα Β λεμφοκύτταρα και ονομάζονται αντισώματα προκειμένου να αναγνωρίσει τα μικροβιακά αντιγόνα, να εξουδετερώσει την μολυσματικότητα των μικροβίων χρησιμοποιώντας διάφορους μηχανισμούς. Η χυμική ανοσία αποτελεί την κύρια οδό άμυνας έναντι εξωκυττάριων μικροβίων και τοξινών μέσω της σύνδεσης των αντισωμάτων σ αυτά. Επίσης τα αντισώματα προάγουν την φαγοκυττάρωση και ενεργοποιούν την απελευθέρωση 33
34 φλεγμονωδών μεσολαβητών και β) την κυτταρική ανοσία η οποία χρησιμοποιεί τα Τ λεμφοκύτταρα για να σταματήσει ενδοκυττάριους παθογόνους μικροοργανισμούς, όπως είναι οι ιοί και ορισμένα βακτηρίδια. Περιλαμβάνει τη δράση δυο υποπληθυσμών λεμφοκυττάρων, των CD4+ T λεμφοκυττάρων (βοηθητικά, T helper cells) και των CD8+ T κυτταροτοξικών λεμφοκυττάρων (CTLs). Διαταραχή στην ανοσιακή απάντηση των Τ κυττάρων οδηγεί σε διαταραχή στην εξουδετέρωση των παθογόνων μικροοργανισμών. Τα ενδοκυττάρια παθογόνα μολύνουν πολύ γρήγορα τα κύτταρα-στόχο τους. Η ανοσιακή απάντηση έναντι αυτών από τα Τ κύτταρα εξαρτάται από την αναγνώριση των πεπτιδικών αντιγόνων τα οποία είναι συνδεδεμένα σε πρωτεΐνες του ξενιστή οι οποίες κωδικοποιούνται από γονίδια του μείζονος (MHC) συστήματος ιστοσυμβατότητας και παρουσιάζονται στην επιφάνεια των APC. Το σύστημα MHC υπάρχει σε όλα τα θηλαστικά. Στον άνθρωπο, εδράζεται στο βραχύ σκέλος του χρωμοσώματος 6 και κωδικογράφει μεμβρανικές γλυκοπρωτεΐνες στα λευκοκύτταρα (Human Leukocyte Antigens, HLA). Τα HLA αντιγόνα διακρίνονται σε τρεις τάξεις (Ι, ΙΙ, ΙΙ). Τα μόρια HLA τάξης Ι παρουσιάζουν κυρίως ενδογενή πεπτίδια στα CD8+ T λεμφοκύτταρα, τα HLA μόρια τάξης ΙΙ παρουσιάζουν κυρίως εξωγενή πεπτίδια στα CD4+ T λεμφοκύτταρα ενώ τα μόρια HLA τάξης ΙΙΙ αφορούν κυρίως σε γονίδια που έχουν σχέση με το συμπλήρωμα και τον TNF. Ως APC λειτουργούν α) τα δενδριτικά κύτταρα, τα οποία είναι υπεύθυνα για την αρχική παρουσίαση ενός αντιγόνου και την ενεργοποίηση των Τ λεμφοκυττάρων στους λεμφαδένες και β) τα μακροφάγα, τα οποία παρουσιάζουν αντιγόνα σε ενεργοποιημένα δραστικά Τ λεμφοκύτταρα. Ειδικότερα, τα αντιγόνα των παθογόνων υφίστανται ενζυματική αποδόμηση προς πεπτίδια από τα πρωτεολυτικά ένζυμα που υπάρχουν στα ενδοσωμάτια και λυσσοσωμάτια των αντιγονοπαρουσιατικών κυττάρων (APC). Συγχρόνως τάξης Ι και ΙΙ μόρια του MHC παράγονται στο ενδοπλασματικό δίκτυο, μεταφέρονται συνδεδεμένα με μία πρωτεΐνη και συνδέονται με το αντιγονικό πεπτίδιο στην σχισμή (cleft). Το όλο σύμπλεγμα μεταφέρεται στην επιφάνεια του APC όπου θα αναγνωρισθεί από τον υποδοχέα (TCR) του Τ λεμφοκυττάρου (το αντιγονικό πεπτίδιο με MHC τάξης ΙΙ από τα CD4+ κύτταρα και το συνδεδεμένο με τάξης Ι, με τα CD8+ κύτταρα. H τελική όμως ενεργοποίηση, ο πολλαπλασιασμός και η διαφοροποίηση του ήρεμου (naïve) T λεμφοκυττάρου απαιτεί 2 διαφορετικά σήματα: το ένα σήμα είναι η σύνδεση του τριμοριακού συμπλέγματος αντιγονικού πεπτιδίου, MHC και TCR (διέγερση). Το άλλο (2 ο σήμα ή συνδιέγερση) αφορά στην 34
35 σύνδεση συγκεκριμένων μορίων (συνδιεγερτικά μόρια) που εκφράζονται στην επιφάνεια του αντιγονοπαρουσιαστικού κυττάρου και στην επιφάνεια του Τ λεμφοκυττάρου. Τα συνδιεγερτικά μόρια ρυθμίζουν την έναρξη, τη διατήρηση και τον τερματισμό των ανοσιακών αποκρίσεων των Τ λεμφοκυττάρων. Η καλύτερα προσδιορισμένη οδός συνδιέγερσης είναι εκείνη που περιλαμβάνει τη σύνδεση των συνδιεγερτικών μορίων Β7-1 (CD80) και Β7-2 (CD86) των αντιγονοπαρουσιαστικών κυττάρων με τα μόρια CD28 και CTLA-4 (cytotoxic T-lymphocyte-associated antigen 4, CD152) στην επιφάνεια των Τ λεμφοκυττάρων. Άλλοι υποδοχείς που συνδέονται με συνδιεγερτικά μόρια είναι ο ICOS (Inducible Costimulator), ο PD-1 (Programmed Death-1), το OX-40, το 4-1BB αλλά και μέλη της οικογένειας του TNF. Μετά την αλληλουχία του πρώτου και του δεύτερου σήματος τα ενεργοποιημένα Τ κύτταρα εκκρίνουν κυτταροκίνες για να διευκολύνουν τον πολλαπλασιασμό τους και τις δραστικές τους λειτουργίες. Η τελική ενεργοποίηση των Τ κυττάρων επιτελείται επιπλέον με την συμμετοχή των συν-διεγερτικών μορίων (Β7-1, Β7-2) και βοηθητικών μορίων (LFA-3, ICAM-1) της επιφάνειας του APC, τα οποία συνδέονται με αντίστοιχα μόρια στην επιφάνεια του Τ λεμφοκυττάρου. Τα Τ λεμφοκύτταρα φέρουν στην επιφάνεια τους ένα μεγάλο αριθμό υποδοχέων (π.χ. TCRs, T cell receptors) με τους οποίους συνδέουν τα αντιγόνα. Ο υποδοχέας των Τ λεμφοκυττάρων (TCR) καθορίζει την αντιγονική ειδικότητα του κάθε Τ λεμφοκυττάρου και χαρακτηρίζει κάθε κλώνο Τ λεμφοκυττάρων. Αποτελείται από δυο διαμεμβρανικές πολυπεπτιδικές αλύσους με δομή ανοσοσφαιρίνης, συνήθως τις α και β, κάθε μια από τις οποίες διαθέτει μια μεταβλητή περιοχή (V) και μια σταθερή περιοχή (C). Ο TCR αναγνωρίζει τα ξένα αντιγόνα μόνο όταν το πεπτίδιο που παρουσιάζεται είναι συνδεδεμένο με ένα «ίδιο» μόριο MHC τάξεως Ι ή ΙΙ (MHC restriction, MHC περιορισμός). Όταν το αντιγόνο αναγνωρισθεί από τον TCR, πυροδοτείται το σήμα ενεργοποίησης στο Τ λεμφοκύτταρο μέσω πρωτεϊνικών τυροσινικών κινασών όπως η ZAP-70 και καταλήγει στην ενεργοποίηση συγκεκριμένων μεταγραφικών παραγόντων οι οποίοι επάγουν την έκφραση συγκεκριμένων γονιδίων. Οι κύριοι μεταγραφικοί παράγοντες που εμπλέκονται στην ενεργοποίηση των Τ λεμφοκυττάρων είναι ο NFκΒ, ο NFAT και ο AP-1 και όλοι κατά κύριο λόγο οδηγούν στην παραγωγή IL- 2, μια κυτταροκίνη που αποτελεί τον κυριότερο αυξητικό παράγοντα τόσο για τα Τ κύτταρα που την παράγουν όσο και για τα διπλανά κύτταρα, καθώς επάγει την διαφοροποίηση και τον πολλαπλασιασμό τους. 35
36 Όπως προαναφέρθηκε η κυτταρική ανοσιακή απάντηση επιτελείται από τη δράση των υποπληθυσμών των Τ λεμφοκυττάρων 1. τα κυτταροτοξικά CD8+ T λεμφοκύτταρα Τα κύτταρα αυτά είναι τα κύρια δραστικά κύτταρα έναντι ιών και άλλων ενδοκυττάριων μικροοργανισμών, έναντι των οποίων δρουν με κυτταρολυτικούς μηχανισμούς, μέσω της οδού Fas/Fas ligand, περφορίνης, granzyme κ.ά. ή με ενεργοποίηση των μακροφάγων, με κασπάσες μέσω IFN-γ, TNF-α ή λεμφοτοξίνης. Έτσι τα κύτταρα-στόχος πεθαίνουν λόγω πυροδότησης του καταρράκτη της απόπτωσης ή λύονται λόγω ώσμωσης. Η καταστροφή των μολυσμένων κυττάρων σταματάει τον κύκλο πολλαπλασιασμού του παθογόνου και το αφήνει εκτεθειμένο στα φαγοκύτταρα. 2. τα CD4+ T βοηθητικά λεμφοκύτταρα Τα CD4 T κύτταρα παίζουν κεντρικό ρόλο στο ανοσοποιητικό σύστημα και έχουν πολλαπλές δράσεις 56. Παράγουν μεγάλο εύρος κυτταροκινών και χημειοκινών μέσω των οποίων ενορχηστρώνουν τις ανοσιακές αποκρίσεις, βοηθούν τα Β κύτταρα να παράγουν αντισώματα, συμμετέχουν στην ενεργοποίηση των μακροφάγων ώστε τα τελευταία να είναι δραστικά έναντι μικροβίων και στρατολογούν ουδετερόφιλα, ηωσινόφιλα και βασεόφιλα σε θέσεις φλεγμονής. Ήδη από το 1986 είχε διατυπωθεί η άποψη ότι τα CD4+ T κύτταρα διαφέρουν ως προς τη δράση τους, ανάλογα με το προφίλ των κυτταροκινών που παράγουν 57. Σήμερα είναι γνωστό ότι κάποιοι CD4+ Τ κυτταρικοί πληθυσμοί έχουν διακριτή καταγωγή από το θύμο όπως τα naive CD4+ T κύτταρα, τα «φυσικά» ρυθμιστικά κύτταρα (naïve T regulatory cells, ntregs)όπως και τα κύτταρα φυσικοί φονείς (natural killer cells, NKT cells). Άλλοι CD4+ T κυτταρικοί πληθυσμοί αντιπροσωπεύουν εναλλακτικά μοτίβα διαφοροποίησης των naive T κυττάρων που εξέρχονται από το θύμο. Η διαφοροποίηση τους εξαρτάται από το είδος του APC κυττάρου που θα παρουσιάσει το αντιγόνο, τη φύση και τη συγκέντρωση του αντιγόνου και ιδίως τη συγγένεια του αντιγόνου με τον TCR του λεμφοκυττάρου, το είδος των συνδιεγερτικών μορίων που εμπλέκονται στη μετάδοση του δεύτερου σήματος και τις κυτταροκίνες του μικροπεριβάλλοντος στο οποίο επιτελείται η διαφοροποίηση. Τα CD4+ T βοηθητικά κύτταρα εκτός από την ετερογένεια, καταδεικνύουν και πλαστικότητα, δηλαδή κάθε υποπληθυσμός έχει την ικανότητα να αλλάζει φαινότυπο και να διαφοροποιείται σε έναν άλλο ή να υιοθετεί μικτό φαινότυπο. Η πλαστικότητα αυτή φαίνεται πως είναι 36
37 λιγότερο έντονη για τα Th1 και Th2 κύτταρα, περισσότερο για τα Tregs και ακόμη πιο έντονη για τα Th17 κύτταρα, καθώς φαίνεται ότι τα περισσότερα κύτταρα με φαινότυπο Th17 έχουν την ικανότητα να διαφοροποιούνται και προς άλλες σειρές CD4+ T κυττάρων. Ίσως αυτό σημαίνει ότι όσο πιο διαφοροποιημένος είναι ένας υποπληθυσμός, τόσο μικρότερη πλαστικότητα έχει. Ειδικότερα όσον αφορά στα Th17 τα οποία είναι και τα πιο «πλαστικά» κύτταρα, έχει βρεθεί πως κάτω από την επίδραση της IL-12 ή της IL-4 έχουν την ικανότητα να διαφοροποιούνται προς Th1 και Th2 κύτταρα αντίστοιχα. Τέλος, όσον αφορά στον υποπληθυσμό των Tregs έχει αναφερθεί πως α) κάτω από την επίδραση της IL-6 διαφοροποιούνται προς κύτταρα που παράγουν IL-17, β) κάτω από την επίδραση Th1 συνθηκών παράγουν IFN-γ, ενώ γ) η μείωση της έκφρασης του Foxp3 οδηγεί στην υιοθέτηση Th2 φαινότυπου. Βέβαια επειδή υπάρχει δυσκολία στη διάκριση των ntregs από τα itregs, δεν είναι σαφές εάν η πλαστικότητα αφορά στο σύνολό τους ή σε κάποιο υποπληθυσμό τους 58, 59. Ανάλογα με τη διαφοροποίηση, τα CD4+ T κύτταρα διακρίνονται στα: Th1 κύτταρα Τα Th1 κύτταρα αρχικά θεωρούνταν ως ένας πληθυσμός πολύ σημαντικός για την ανοσία έναντι ενδοκυττάριων μικροοργανισμών. Έχει όμως διαπιστωθεί ότι είναι υπεύθυνα και για την επαγωγή κάποιων οργανοειδικών αυτοάνοσων νοσημάτων κυρίως σε περιπτώσεις διαταραχής της ενεργοποίησης τους. Παράγουν κυρίως Ιντερφερόνη γ, η οποία έχει πολύ σημαντικό ρόλο στην ενεργοποίηση των μακροφάγων για την αύξηση της δραστικότητας τους έναντι μικροβίων, λεμφοτοξίνη α (LTa), η οποία ίσως μπορεί να χρησιμοποιηθεί ως δείκτης για την εξέλιξη της πολλαπλής σκλήρυνσης και IL-2, η οποία είναι πολύ σημαντική για την εμφάνιση μνημονικών CD4 και CD8 T κυττάρων. Th2 κύτταρα Τα Th2 κύτταρα αρχικά θεωρούνταν σημαντικά για την ανοσία έναντι πολλών εξωκυττάριων παθογόνων και την εμφάνιση αλλεργίας. Τα κύτταρα αυτά παράγουν: IL-4, η οποία αποτελεί το θετικό παλίνδρομο ερέθισμα για την Th2 διαφοροποίηση, IL-5, η οποία παίζει πολύ σημαντικό ρόλο στην στρατολόγηση των ηωσινοφίλων, IL-9, η οποία δρα στα μαστοκύτταρα και λεμφοκύτταρα, IL-10, η οποία καταστέλλει την Th1 διαφοροποίηση και τη λειτουργία των δενδριτικών κυττάρων, IL- 13, IL-25 η οποία μάλλον λειτουργεί ως εναρκτήριο ερέθισμα για τις Th2 αποκρίσεις, αυξάνοντας την παραγωγή IL-4, IL-5 και IL-13 και αμφιρεγκουλίνη (amphiregulin), η οποία εμπλέκεται στην επαγωγή του 37
38 πολλαπλασιασμού των επιθηλιακών κυττάρων και την υπεραντιδραστικότητα των αεραγωγών Th17 κύτταρα Τα Th17 αναγνωρίσθηκαν πρόσφατα και φαίνεται πως διαδραματίζουν σπουδαίο ρόλο στην προστασία έναντι μικροβίων, ιδίως εξωκυττάριων, βακτηριδίων και μυκήτων. Τα κύτταρα αυτά φαίνεται πως μεσολαβούν εν μέρει σε νοσήματα που οφείλονται σε αυτοάνοσες καταστάσεις και οι οποίες ως τώρα θεωρούνταν ότι οφείλονταν σε Th1 απαντήσεις. Τα Th17 κύτταρα παράγουν: IL-17a και IL-17f, οι οποίες είναι δυνατόν να συνεκφράζονται από το ίδιο κύτταρο, και ακριβώς επειδή χρησιμοποιούν τον ίδιο υποδοχέα (αν και με διαφορετική συγγένεια) θεωρείται πως έχουν παρόμοιες λειτουργίες, δηλαδή επάγουν φλεγμονώδεις αποκρίσεις μέσω παραγωγής προφλεγμονωδών κυτταροκινών όπως η IL-6 και στρατολογούν τα ουδετερόφιλα, η IL-21, η οποία λειτουργεί ως διεγερτικό ερέθισμα για την Th17 διαφοροποίηση (ως θετική παλίνδρομη ρύθμιση), αλλά ταυτόχρονα φαίνεται πως δρα και σε μια μεγάλη ποικιλία κυττάρων όπως τα CD8 T κύτταρα, τα B κύτταρα, τα NK κύτταρα και τα δενδριτικά κύτταρα και η IL-22, η οποία προστατεύει τα ηπατοκύτταρα σε οξεία φλεγμονή του οργάνου, αλλά η δράση της εξαρτάται σε μεγάλο βαθμό από την διέγερση των Th17 κυττάρων μέσω της IL-13. Επαγόμενα Τ ρυθμιστικά κύτταρα (adaptive, induced Tregs, itregs) Τα κύτταρα αυτά αποτελούν έναν επαγόμενο πληθυσμό κυττάρων ο οποίος ομοιάζει φαινοτυπικά και λειτουργικά με τα ntregs (naive). Τα itregs αντιπροσωπεύουν μάλλον μια κατάσταση διαφοροποίησης και υιοθέτησης ενός λειτουργικού και φαινοτυπικού προφίλ και όχι ενός ξεχωριστού κλώνου Τ κυττάρων και συμβάλλουν στην κλωνική επέκταση των ntregs. 38
39 ΜΕΡΟΣ ΙΙΙ. ΑΝΟΣΙΑΚΗ ΑΠΟΚΡΙΣΗ ΕΝΑΝΤΙ ΤΟΥ ΙΟΥ ΤΗΣ ΗΠΑΤΙΤΙΔΑΣ C Επειδή η έναρξη της HCV λοίμωξης είναι συνήθως ασυμπτωματική, οι περισσότερες πληροφορίες σχετικά με την πρώιμη κινητική του HCV προέρχονται από μελέτες σε χιμπατζήδες στις οποίες διαπιστώθηκε ότι τις πρώτες μέρες μετά την αρχική λοίμωξη, περίπου κάθε 12 ώρες το ιικό φορτίο διπλασιάζεται. Κατόπιν, ο τίτλος του ιικού φορτίου ακολουθεί μια διφασική μορφή: παρουσιάζει παροδική μείωση τις επόμενες 1-2 εβδομάδες με σύγχρονη αύξηση της ALT λόγω κάθαρσης του ιού από τα μολυσμένα ηπατοκύτταρα (με κυτταρολυτικούς και μη κυτταρολυτικούς μηχανισμούς) και στην συνέχεια ακολουθεί νέα αύξηση, μέχρι να φτάσει το ιικό φορτίο ένα πλατώ όταν πια κινητοποιηθούν ανοσιακοί μηχανισμοί ομοιόστασης. Ο μέσος χρόνος ζωής των σωματιδίων του ιού είναι 7,5 μέρες 60. Η ενδογενής παραγωγή ιντερφερονών τύπου Ι (IFNs τύπου Ι) αποτελεί την πρώτη αντιική ανοσιακή απάντηση έναντι του HCV, η οποία φαίνεται ότι ελέγχει μερικά στην αρχική φάση τον ιικό πολλαπλασιασμό. Στη συνέχεια όμως, ο ΗCV χρησιμοποιώντας διάφορους μηχανισμούς, επιτυγχάνει να περιορίσει τη δράση των IFNs. Η αντιική δράση των ιντερφερονών επιτυγχάνεται μετά από σύνδεση τους σε ειδικούς υποδοχείς στην επιφάνεια των κυττάρων. Η κύρια οδός της μεταφοράς σήματος (signaling) την οποία χρησιμοποιούν οι ιντερφερόνες και οι υποδοχείς τους επιτελείται μέσω της κινάσης της τυροσίνης, του Jak 1 και της Tyk-2, με αποτέλεσμα να ενεργοποιούνται οι μεταβιβαστές σήματος και οι ενεργοποιητές μεταγραφής (STAT1 και STAT2) προς σχηματισμό του ετεροδιμερούς STAT1/STAT2. Το σύμπλεγμα αυτό μαζί με μια πρωτεΐνη, την p-48, η οποία αποτελεί τον παράγοντα απάντησης της IFN, IRF9 (interferon response factor), σχηματίζουν τον ενεργοποιημένο από την IFN γονιδιακό παράγοντα 3 (ISGF-3) o οποίος και προάγει την μεταγραφή της IFN 61. Ο μηχανισμός της αντιικής δράσης τους δεν είναι τελείως τεκμηριωμένος, αλλά φαίνεται ότι αναστέλλουν τη μεταγραφή των ιικών mrna ή και επεμβαίνουν στη διείσδυση των ιών στα κύτταρα, στην καταστροφή του καψιδίου τους και στην εκβλάστηση και στην πρόκληση ενός αντι-ιїκού status μέσα στο κύτταρο που συνίσταται στην ενεργοποίηση ενζύμων τα οποία αποδομούν το νουκλεїνικό οξύ και αναστέλλουν την σύνθεση των ιικών πρωτεїνών, με αποτέλεσμα την αναστολή του ιικού πολλαπλασιασμού. Το τελευταίο φαίνεται ότι 39
40 επιτυγχάνεται μετά από σύνδεση της IFN σε υποδοχείς των γειτονικών κυττάρων προκαλώντας σύνθεση ορισμένων αντιικών πρωτεϊνών από αυτά, όπως είναι η πρωτεϊνική κινάση και η 2-5 ολιγοαδενυλική συνθετάση. Η πρώτη αδρανοποιεί ένα ένζυμο που είναι απαραίτητο για τη συσσώρευση των ριβοσωματίων και η δεύτερη καταλύει μία σειρά ενζύμων τα οποία διασπούν το mrna. Αποτέλεσμα είναι η εκλεκτική αναστολή της σύνθεσης των ιικών πρωτεϊνών. Κατά την λοίμωξη με τον HCV, διαπιστώθηκε ότι παράγεται το ενεργοποιημένο από την ιντερφερόνη γονίδιο (ISG)56 το οποίο αναστέλλει τη μεταγραφή του HCV. Εκτός από το ISG56, παράγονται και άλλα παρόμοια γονίδια τα οποία αναστέλλουν τον πολλαπλασιασμό του HCV παρουσία της ιντερφερόνης. Ειδικότερα στην HCV λοίμωξη, το ιικό RNA του HCV που βρίσκεται μέσα στα ηπατοκύτταρα προάγει την ενεργοποίηση δύο μορίων : α) του τύπου Toll υποδοχέα (TLR)-3 και του γονιδίου 1 του ρετινοϊκού οξέος (RIG- 1). Η ενεργοποίηση αυτών των μορίων οδηγεί σε φωσφορυλίωση του ρυθμιστικού παράγοντα της ιντερφερόνης (IRF)-3 ο οποίος μετακινείται προς τον πυρήνα του ηπατοκυττάρου και ενεργοποιεί την παραγωγή της ιντερφερόνης και με τον τρόπο αυτό δημιουργείται ένα γενικευμένο αντι-ιικό status τόσο στα μολυσμένα από τον HCV ηπατοκύτταρα όσο και στα γειτονικά ηπατοκύτταρα που δεν έχουν μολυνθεί από τον ιό. Αποτέλεσμα είναι η εκλεκτική αναστολή της σύνθεσης των ιικών πρωτεϊνών. Εντούτοις, μελέτες έδειξαν ότι ο ΗCV χρησιμοποιεί διάφορους μηχανισμούς διαφυγής στη δράση των IFN, όπως: 1) η HCVNS34A πρωτεάση της σερίνης μπλοκάρει τον μεταγραφικό παράγοντα IRF-3 (interferon response factor), ο οποίος προάγει την παραγωγή των IFNs 62 2) Η Ε2 και η NS5A του HCV αναστέλλουν τη λειτουργία της PKR (πρωτεινική κινάση) 63, 64. Η δράση της Ε2 πρωτεΐνης του HCV συνίσταται στην καταστροφή της ΡΚR επειδή η Ε2 παρουσιάζει ομολογία με τα τμήματα φωσφορυλίωσης του ενζύμου και του υποστρώματος του. Η NSSA δημιουργεί ετεροδιμερή με την PKR και με τον τρόπο αυτό αναστέλλει τη λειτουργία της. Όπως είναι γνωστό, η PKR δρα προάγοντας την απόπτωση μέσω της ενεργοποίησης της οδού της κασπάσης. Εφόσον η απόπτωση θεωρείται μία συχνή κυτταρική απάντηση στη λοίμωξη από ιούς, θα μπορούσε να θεωρηθεί ότι οι αντιαποπτωτικές ιδιότητες της NS5A, λόγω αναστολής της PHR, παίζουν κάποιο ρόλο στην παραμονή του ιού και στην εμμένουσα λοίμωξη 64 3) 40
41 Επιπλέον, διαπιστώθηκε ότι η πρωτεΐνη ΝS5Α προάγει την έκφραση της IL-8 η οποία σχετίζεται με αναστολή της δράσης της IFN. Ιδιαίτερο ρόλο στην αρχική διαφυγή του ιού από την ανοσιακή απόκριση φαίνεται πως παίζουν και τα δενδριτικά κύτταρα, τόσο τα πλασματοκυτταροειδή (plasmacytoid dendritic cells, pdcs) όσο και τα συμβατικά δενδριτικά κύτταρα (conventional dendritic cells, cdcs). Οι IFNs τύπου Ι παράγονται επίσης από τα pdcs τα οποία βρίσκονται σε ιστούς με φλεγμονή και τους επιχώριους λεμφαδένες τους. Στην HCV λοίμωξη τα κύτταρα αυτά είναι λιγότερα σε αριθμό και έχουν μικρότερη ικανότητα να παράγουν in vitro IFN-a 65, 66. Όσον αφορά τα cdcs, μελέτες έδειξαν ότι στην HCV λοίμωξη υπάρχει διαταραχή στην ωρίμανση και τη λειτουργική διαφοροποίηση των cdcs (μειωμένη παραγωγή IL-12 και αυξημένη παραγωγή IL-10 με αποτέλεσμα την κακή αντιγονοπαρουσίαση 67, 68. Α. Ανοσιακή απόκριση στην οξεία HCV λοίμωξη Ένα από τα χαρακτηριστικά της HCV λοίμωξης είναι ότι η ανοσιακή απάντηση έναντι του ιού καθυστερεί πολύ, παρά το γεγονός ότι το ο ιός πολλαπλασιάζεται πολύ γρήγορα και το ιικό φορτίο υπερδιπλασιάζεται τάχιστα. Αρχικά, ενεργοποιούνται οι μηχανισμοί φυσικής ανοσίας και επάγεται η δράση των ΙFNs. Τα ειδικά όμως έναντι του HCV T κύτταρα ανιχνεύονται καθυστερημένα, περίπου 5-9 εβδομάδες μετά τη μόλυνση, και τα ειδικά έναντι του HCV αντισώματα ανιχνεύονται μετά από 8-20 εβδομάδες 69, 70. Η καθυστερημένη αυτή ενεργοποίηση των Τ κυττάρων, πιθανολογείται ότι οφείλεται σε ελαττωματική ενεργοποίηση (defective T and B cell priming) από τον ιό, των Β και Τ λεμφοκυττάρων. Όσον αφορά την χυμική ανοσιακή απάντηση έναντι του HCV, διαπιστώνεται ότι υπολείπεται με αποτέλεσμα την καθυστερημένη εμφάνιση των αντισωμάτων. Επιπλέον φαίνεται ότι η μεγάλη μεταβλητότητα του ιού συμβάλλει στο γεγονός ότι τα αντισώματα αυτά δεν είναι εξουδετερωτικά, αν και αναφέρεται από άλλους ερευνητές ότι διαπιστώθηκε παρουσία εξουδετερωτικών αντισωμάτων τα οποία στρέφονται έναντι της υπερμεταβλητής περιοχής της Ε2 πρωτεΐνης, τα οποία επάγονται πρώιμα στην οξεία λοίμωξη και μειώνονται ή εξαφανίζονται μετά την αποδρομή της λοίμωξης 71. Όσον αφορά την Τ κυτταρική απάντηση στην οξεία HCV λοίμωξη, είναι σαφές ότι η κάθαρση του ιού επιτυγχάνεται από ειδικά έναντι του HCV 41
42 CD8+ Τ κύτταρα, καθώς η μείωση του ιικού φορτίου συμπίπτει χρονικά με την εμφάνιση τους και με την έκφραση IFN-γ στο ήπαρ 28, 69. Τα κυτταροτοξικά κύτταρα χρησιμοποιούν δύο μηχανισμούς για την κάθαρση του HCV από τα μολυσμένα ηπατοκύτταρα, α) ένα μη κυτταρολυτικό μέσω έκκρισης IFN-γ προκειμένου να κατασταλεί ο πολλαπλασιασμός του ιού και β) ένα κυτταρολυτικό μέσω της οδού Fas, της δράσης της περφορίνης και του granzyme, αλλά και μέσω της επαγόμενης απόπτωσης από συνδέτες του TNF-α (TNF-related apoptosis inducing ligand, TRAIL). Φαίνεται μάλιστα ότι η επιτυχής ανοσιακή απάντηση των CD8+ T κυττάρων προϋποθέτει μια ισχυρή και ειδική έναντι του HCV CD4+ T κυτταρική απάντηση. Τα CD8+ T κύτταρα ανιχνεύονται στο αίμα των ασθενών με οξεία HCV λοίμωξη ανεξαρτήτως της έκβασης τη λοίμωξης 72. Στην οξεία HCV λοίμωξη, τα CD8+ T κύτταρα είναι μη λειτουργικά (stunned) και χαρακτηρίζονται από μειωμένη ικανότητα πολλαπλασιασμού, μειωμένη παραγωγή IFN-γ, μειωμένη κυτταροτοξική ικανότητα και αυξημένη έκφραση PD-1. Επιπλέον τα κύτταρα αυτά είναι βραχύβια και έχουν χαρακτήρες δραστικών κυττάρων που εξαρτώνται από αντιγόνα και δεν μοιάζουν με μνημονικά Τ κύτταρα. Ο ανωτέρω φαινότυπος δεν είναι σαφές εάν επάγεται από τον ιό ή από κάποιο προϊόν του ή εάν αντιπροσωπεύει ένα προ-στάδιο της ωρίμανσης και της μετανάστευσής των CD8+ T κυττάρων στο ήπαρ. Έχει όμως διαπιστωθεί ότι όταν αναπτυχθούν ειδικές έναντι του HCV αποκρίσεις των CD4+ T κυττάρων και μειωθεί το ιικό φορτίο, τότε η δυσλειτουργία των ειδικών έναντι του HCV CD8+ T κυττάρων υποστρέφεται και αναπτύσσονται CD8+ T μνημονικά κύτταρα 69, 73. Τα ειδικά έναντι του HCV CD4+ T κύτταρα, στους ασθενείς με αυτοπεριορισθείσα HCV λοίμωξη, πολλαπλασιάζονται έντονα και παρουσιάζουν έντονη και ειδική έναντι πολλαπλών επιτόπων του HCV απόκριση με ταυτόχρονη αύξηση της IL-2 και της IFN-γ στο περιφερικό αίμα και μάλιστα ο πολλαπλασιασμός αυτός συμβαίνει κατά την αύξηση της ALT. Η απόκριση αυτή διατηρείται επί μακρόν 69, 73, 74. Αντίθετα, στους ασθενείς στους οποίους εγκαθίσταται χρόνια λοίμωξη οι ειδικές έναντι του HCV CD4+ T κυτταρικές αποκρίσεις είναι ασθενείς ή και απούσες. Β. Ανοσιακή απόκριση στη χρόνια HCV λοίμωξη 75, 76 Στη χρόνια HCV λοίμωξη, ο τίτλος των αντισωμάτων και το εύρος των αντιγονικών επιτόπων του HCV έναντι των οποίων αυτά κατευθύνονται, αυξάνει και, αν και αποτυγχάνουν να καθάρουν τον ιό, εξακολουθούν 42
43 να ασκούν πίεση στα στελέχη του ιού με αποτέλεσμα ο ιός να μεταλλάσσεται καθώς τροποποιείται η αλληλουχία των νουκλεοτιδίων του φακέλου του 77. Σε μια πρόσφατη μελέτη μάλιστα πιθανολογείται ότι ίσως σε ορισμένα άτομα, τα antihcv αντισώματα καθορίζουν και την έκβαση της λοίμωξης 78. Όσον αφορά την κυτταρική ανοσιακή απάντηση, τα CD8+ T κύτταρα αναγνωρίζουν αντιγόνα του HCV που παρουσιάζονται από μόρια HLA τάξης Ι πάνω στα αντιγονοπαρουσιαστικά κύτταρα και στα μολυσμένα ηπατοκύτταρα και η αντιιική τους δράση έναντι του HCV εκφράζεται με κυτταροτοξικότητα, πολλαπλασιασμό και έκκριση αντιιικών κυτταροκινών όπως η IFN-γ. Στη χρόνια HCV λοίμωξη οι CD8+ T κυτταρικές αποκρίσεις είναι μη λειτουργικές(stunned), ενώ και ο αριθμός αυτών των κυττάρων είναι μικρότερος με αποτέλεσμα η λειτουργία αυτών των κυττάρων και το εύρος της ανοσιακής τους απάντησης να είναι περιορισμένα 28, 79, 80. Τελικά φαίνεται να υπάρχει μια ισορροπία μεταξύ της επίθεσης του ιού και της απάντησης του ξενιστή χωρίς να επιτυγχάνεται η κάθαρση του ιού. Σε αυτό το αποτέλεσμα πιθανολογείται ότι συντείνουν ποικίλοι λόγοι όπως: α) Η πρωτοπαθής ανεπάρκεια και/ ή εξάντληση των CD8+ T κυττάρων Η προβληματική απάντηση των CD8+ T κυττάρων πιθανόν να οφείλεται σε πρωτοπαθή ανεπάρκεια της ενεργοποίησής τους (priming) ή σε πολύ γρήγορη μείωση της έντασης της απάντησής τους μετά από ενεργοποίηση λόγω εξάντλησης. Τα αίτια αυτής της κατάστασης μπορεί να είναι: το υψηλό ιικό φορτίο, η έκφραση λιγότερων TCR υποδοχέων στην επιφάνεια τους, η έκφραση ανασταλτικών υποδοχέων όπως ο PD- 1 ή και η αλληλεπίδραση με ειδικούς υποδοχείς της μεμβράνης τους (όπως ο υποδοχέας gc1qr) 81, 82, 83. β) η έλλειψη βοήθειας των CD8+ T κυττάρων από τα CD4+ T κύτταρα Στην χρόνια HCV λοίμωξη οι CD4+ T κυτταρικές αποκρίσεις είναι ασθενείς και δυσλειτουργικές (για παράδειγμα εκκρίνουν μικρότερη ποσότητα IL-2), έχουν μικρότερο εύρος και διαρκούν λίγο σε αντίθεση με την οξεία HCV λοίμωξη που αυτοπεριορίζεται όπου και παρατηρούνται έντονες και ειδικές έναντι πολλαπλών επιτόπων του HCV απαντήσεις που διατηρούνται για καιρό 84, 85. γ) η συμβολή ανασταλτικών υποδοχέων όπως ο PD-1 Ο PD-1 είναι ένα μόριο αρνητικής σηματοδότησης (negative signaling molecule) που παρεμποδίζει τις Τ κυτταρικές αποκρίσεις. Οι συνδέτες 43
44 του είναι: α) το μόριο Β7-Η1 (PD-L1), το οποίο εκφράζεται στα ενδοθηλιακά κύτταρα των κολποειδών στο ήπαρ και τα κύτταρα του Kupffer και αναστέλλει τις Τ κυτταρικές απαντήσεις 86 και β) το μόριο B7- DC (PD-L2), η έκφραση του οποίου περιορίζεται σε δενδριτικά κύτταρα και μακροφάγα, και μάλλον λειτουργεί διεγερτικά. Έτσι, φαίνεται πως το μονοπάτι PD-1/B7-H1 εμπλέκεται στη ρύθμιση των Τ κυτταρικών απαντήσεων ενδοηπατικά 87. Τα Τ κύτταρα που απομονώθηκαν από το αίμα και το ήπαρ ανθρώπων και χιμπαντζήδων με HCV λοίμωξη εκφράζουν υψηλά επίπεδα PD-1 88, 89, 90. Είναι μάλιστα πιθανόν, η πρώιμη αύξηση της έκφρασης του PD-1 να μεσολαβείται από την αλληλεπίδραση της πρωτεΐνης του πυρήνα του HCV με τον υποδοχέα gc1qr η οποία σε συνδυασμό με την συνεχή διέγερση του TCR, συμβάλει στην πρώιμη επαγωγή ανεργίας στα ειδικά έναντι του HCV κυτταροτοξικά κύτταρα (efficient escape mechanism) 90. Όμως, σε άλλες έρευνες στις οποίες διαπιστώθηκε επίσης ότι η έκφραση του PD-1 και του PDL-1 (Β7-Η1) είναι αυξημένη σε ασθενείς με χρόνια ηπατίτιδα C, αλλά και σε υγιή Τ κύτταρα τα οποία εκτέθηκαν στην πρωτεΐνη του πυρήνα του ιού, διατυπώθηκε το σκεπτικό ότι ίσως αυτή η ρύθμιση αποτελεί ένα μηχανισμό του ίδιου του ιού προκειμένου να προκαλέσει δυσλειτουργία στην ενεργοποίηση των Τ κυττάρων 87, 91. δ) Ο χρόνιος αντιγονικός ερεθισμός Είναι γνωστό ότι τα σταθερά υψηλά επίπεδα ιικών αντιγόνων οδηγούν σε χρόνια ενεργοποίηση των Τ κυττάρων που σταδιακά οδηγεί σε απώλεια της δραστικής τους λειτουργίας με πρώτη εκδήλωση τη μείωση της παραγωγής IL-2 και ακολούθως της κυτταροτοξικότητας, της παραγωγής TNF-a και IFN-γ 92. Στη χρόνια HCV λοίμωξη έχει διαπιστωθεί συσχέτιση μεταξύ της μειωμένης παραγωγής IL-2 από τα CD4+ T κύτταρα και της δυσλειτουργίας των CD8+ T κυττάρων. Τα ειδικά έναντι του HCV CD8+ T κύτταρα τα οποία παρουσιάζουν δυσλειτουργία, ως αποτέλεσμα της χρόνιας αντιγονικής διέγερσης από τον ιό, εκφράζουν σε αφθονία τον ανασταλτικό υποδοχέα PD-1 με αποτέλεσμα την αναστολή της λειτουργίας τους και την επαγωγή της απόπτωσης των κυττάρων αυτών 93, 94. ε) Αναστολή από ανασταλτικές κυτταροκίνες όπως η IL-10 Τα επίπεδα της IL-10 είναι αυξημένα στην χρόνια HCV λοίμωξη 95. Έχει διαπιστωθεί ότι η IL-10 παράγεται από μονοκύτταρα και ΝΚ κύτταρα μετά από διέγερση από πρωτεΐνες του ιού, ενώ υπάρχουν και CD8+ T κύτταρα με ρυθμιστικό φαινότυπο που παράγουν IL-10 68, 96, 97, 98, 99. Η IL-10 44
45 φαίνεται πως παρεμποδίζει την παραγωγή IFN-α, προάγει την απόπτωση των pdcs και μειώνει την ένταση των δραστικών λειτουργιών των Τ κυττάρων 66, 100. στ) οι μεταλλάξεις διαφυγής του ιού Οι μεταλλάξεις συνήθως συμβαίνουν μέσα στους πρώτους 3-4 μήνες από τη μόλυνση (πιθανώς επειδή καθυστερεί η ειδική έναντι του ιού T κυτταρική απόκριση και αυτές τείνουν να μένουν ίδιες στα κυκλοφορούντα σχεδόν είδη (quasispecies) (remain fixed within the circulating quasispecies of an individual thereafter) 101, 102. Υπάρχουν κάποια δεδομένα σύμφωνα με τα οποία η πίεση επιλογής και η εμφάνιση μεταλλάξεων επηρεάζεται από τα CD8+ T κύτταρα που αναγνωρίζουν συγκεκριμένα HLA μόρια 103, 104, 105. Επίσης, έχει διαπιστωθεί ότι η πίεση επιλογής επηρεάζεται από την παρουσία εξουδετερωτικών αντισωμάτων αλλά και από την επίδραση του ίδιου του HCV στα Τ κύτταρα καθώς φαίνεται πως επηρεάζει την σύνδεση των αντιγόνων, την επεξεργασία τους και την παρουσίαση τους, άρα και την αναγνώριση τους από τα CD8+ και CD4+ T κύτταρα 70, 101, 103. ζ) η καταστολή από πρωτεΐνες του ιού Υπάρχουν διάφορες μελέτες στις οποίες διαπιστώθηκε ότι ορισμένες πρωτεΐνες του ιού (όπως η πυρηνική πρωτεΐνη, η Ε2 κ.ά.) προκαλούν μείωση του πολλαπλασιασμού των δραστικών κυττάρων, μείωση της παραγωγής IL-2, παρεμπόδιση της ενεργοποίησης των ΝΚ κυττάρων και ελαττωματική αντιγονοπαρουσίαση 7, 68, 96. η) η διαταραχή στην ωρίμανση των CD8+ T κυττάρων Όπως διαπιστώθηκε σε αρκετές μελέτες τα CD8+ T κύτταρα εμφανίζουν ανώριμο φαινότυπο 106 με αποτέλεσμα την ελαττωμένη κυτταροτοξική δράση θ) το ηπατικό μικροπεριβάλλον το οποίο επάγει ανοχή (tolerogenic) Πιθανώς το ανοσιακό μικροπεριβάλλον στο ήπαρ νa επάγει ανοχή στα Τ κύτταρα που το διηθούν μέσω των ήδη υπαρχόντων επιθηλιακών κυττάρων στα ηπατικά κολποειδή (liver sinusoidal endothelial cells, LSEC), των κυττάρων του Kuppfer, των αστεροειδών κυττάρων και των δενδριτικών που είναι εγκατεστημένα εκεί 107, 108. ι) η δράση των Τ ρυθμιστικών κυττάρων (Tregs) 109, 110, 111,
46 ΜΕΡΟΣ ΙV. T ΡΥΘΜΙΣΤΙΚΑ ΛΕΜΦΟΚΥΤΤΑΡΑ A.Ιστορική αναδρομή Τα T ρυθμιστικά κύτταρα (T regulatory cells, Tregs) αποτελούν ένα πληθυσμό κυττάρων που πρωτο- αναφέρθηκε στη βιβλιογραφία τη δεκαετία του και η δράση τους φαινόταν ότι είναι η καταστολή της αυτοανοσίας σε φυσιολογικά πειραματόζωα 114, οπότε και χαρακτηρίστηκαν ως Τ κατασταλτικά κύτταρα. Εκείνο το χρονικό διάστημα οι ίδιοι ερευνητές εισήγαν τον όρο λοιμώδης ανοχή (infectious tolerance) για να περιγράψουν την μεταφορά της καταστολής (transfer of suppression) 115, 116. Η ύπαρξη αυτού του πληθυσμού, αργότερα, αμφισβητήθηκε επειδή τα κύτταρα αυτά δεν έφεραν ειδικούς δείκτες ώστε να μπορούν να απομονωθούν και να μελετηθούν 117, 118. Τα κύτταρα όμως αυτά με τις κατασταλτικές «ιδιότητες» προσέλκυσαν εκ νέου το ενδιαφέρον των ερευνητών και λίγο πριν το 2000 ονομάστηκαν Τ ρυθμιστικά λεμφοκύτταρα 119, 120, 121, 122. Τα πρώτα δεδομένα για τον υποπληθυσμό των CD4+ Τ ρυθμιστικών κυττάρων σε πειραματόζωα αλλά και στον άνθρωπο αφορούσαν α) την υψηλή έκφραση της a αλυσίδας του υποδοχέα της IL-2 (CD25+) 123 και β) ένα μοναδικό λειτουργικό φαινότυπο που χαρακτηρίζονταν από μειωμένη απόκριση σε in vitro αλλογενή και πολυκλωνική διέγερση του TCR υποδοχέα τους και άσκηση μη αντιγονο-ειδικής κατασταλτικής δράσης έναντι των δραστικών Τ κυττάρων μετά από επανειλημμένη διέγερση του TCR 124, 125, 126, 127. Η διαπίστωση συνεχούς και υψηλής επιφανειακής έκφρασης του CD25 στα κύτταρα αυτά, συνετέλεσε στη διάκριση και το διαχωρισμό των Tregs από τα άλλα Τ κύτταρα 126, 128, 129, στην προέλευση και ανάπτυξής τους στο θύμο και στην ανίχνευση της κατασταλτικής τους δραστηριότητας in vitro και in vivo τόσο σε πειραματόζωα όσο και στον άνθρωπο130, 131, 132, 133. Παρόλα αυτά, το επιφανειακό μόριο CD25 όχι μόνο δεν συναντάται αποκλειστικά στα Tregs αλλά και δεν εκφράζεται στην επιφάνεια όλων των Tregs 123. Επιπλέον, διαπιστώθηκε ότι όλα τα ενεργοποιημένα T κύτταρα εκφράζουν το CD25, όμως, τα Treg κύτταρα εκφράζουν τα υψηλότερα επίπεδα CD25 (CD25 high ), σε αντίθεση με τα συμβατικά CD4 T κύτταρα στα οποία η έκφραση του CD25 είναι παροδική και ασθενέστερης έντασης. Η καινοτομία στην προσέγγιση της λειτουργίας και στην απομόνωση του υποπληθυσμού των Treg κυττάρων έγινε το 2001 μετά την ανακάλυψη του γονιδίου Foxp3 στα ποντίκια, όπου διαπιστώθηκε ότι μια 46
 το IPEX, το οποίο είναι ένα Χ υπολειπόμενο φυλοσύνδετο σύνδρομο που χαρακτηρίζεται από διαταραχή της ανοσιακής ρύθμισης,")
47 σημειακή μετάλλαξη του γονιδίου Foxp3 στο Χ χρωμόσωμα οδηγεί αυτόματα σε εμφάνιση ενός σοβαρού αυτοάνοσου συνδρόμου στα άρρενα ποντίκια 134 και πως οι μεταλλάξεις στο ανθρώπειο γονίδιο FOXP3, προκαλούν δυο σοβαρά πολυοργανικά αυτοάνοσα σύνδρομα στους ανθρώπους: α) το IPEX, το οποίο είναι ένα Χ υπολειπόμενο φυλοσύνδετο σύνδρομο που χαρακτηρίζεται από διαταραχή της ανοσιακής ρύθμισης, πολυενδοκρινοπάθεια (ΣΔ τύπου 1, θυρεοειδίτιδα) και εντεροπάθεια και β) το XLAAD, το οποίο είναι ένα Χ φυλοσύνδετο σύνδρομο που χαρακτηρίζεται από αυξημένη εμφάνιση αντιδράσεων αλλεργικού τύπου (έκζεμα, ηωσινοφιλία, τροφική αλλεργία, αυξημένη IgE) και αυξημένη ευαισθησία σε λοιμώξεις και διαταραχές πήξης 119, 135, 136, 137, 138. Και τα δυο σύνδρομα ομοιάζουν με εκείνα που παρατηρήθηκαν στα ποντίκια 123, 139. B. Γονίδιο FOXP3 Το γονίδιο FOXP3 βρίσκεται στο Χ χρωμόσωμα (Xq11.23-Xq13.3), αποτελείται από 11 κωδικογράφοντα εξόνια και κωδικοποιεί μια πρωτεΐνη με μέγεθος 48kD (431 αμινοξέα) που ανήκει στην οικογένεια των μεταγραφικών παραγόντων 137, 138. Το γονίδιο FOXP3 διαθέτει πολλές περιοχές που παίζουν σπουδαίο ρόλο στη λειτουργία του και οι οποίες σχηματικά αποδίδονται στο σχήμα που ακολουθεί (Σχήμα 6) 140, 141. Σχήμα 6: Σχήμα γονιδίου F0XP3 47
48 Το γονίδιο FOXP3 εκφράζεται κυρίως στο λεμφικό ιστό, περιορίζεται στα αβ Τ κύτταρα, ενώ είναι πρακτικά μη ανιχνεύσιμο σε Β κύτταρα, γδ Τ κύτταρα, ΝΚ, μακροφάγα και δενδριτικά κύτταρα. Ο μεταγραφικός παράγοντας Foxp3 έχει υψηλή έκφραση στην ομάδα των CD4+CD25+ T κυττάρων, ενώ η έκφρασή του σε ήρεμα T κύτταρα (naive) έχει ως αποτέλεσμα να αποκτούν αυτά τα κύτταρα ρυθμιστική δραστηριότητα τόσο in vitro, όσο και in vivo. Το FOXP3 είναι ένα γονίδιο με ρόλο κλειδί για τη δημιουργία του υποπληθυσμού των Treg κυττάρων και τη διατήρηση των λειτουργικών τους χαρακτηριστικών 142, 143. Θεωρείται ότι αντιπροσωπεύει τον «κυρίαρχο ρυθμιστή» («master regulator») ή τον παράγοντα που είναι υπεύθυνος για την επιλογή του κλώνου («lineage-specification factor») των «φυσικών» Τ ρυθμιστικών κυττάρων (naive T regulatory cells, ntregs) στο θύμο. Επιπλέον, έχει πολύ μεγάλη σημασία για την ανάπτυξη των Tregs, την κλωνική τους επέκτασή και τη λειτουργικότητα τους. Είναι ενδιαφέρον ότι σε μελέτες στην οποία προκλήθηκε in vitro φλεγμονή, διαπιστώθηκε ότι η φλεγμονή οδηγεί σε αυξημένη έκφραση του Foxp3 στα naive CD4+CD25- T κύτταρα, ότι δηλαδή προκαλεί την μετατροπή αυτών των κυττάρων σε κύτταρα που έχουν, λειτουργικά και φαινοτυπικά, ρυθμιστικούς χαρακτήρες. Έτσι διαπιστώθηκε ότι τα CD4+CD25- κύτταρα σε συνθήκες in vitro πρόκλησης φλεγμονής, εμφανίζουν αυξημένη έκφραση του Foxp3, γεγονός το οποίο συμβάλλει στην έκφραση και άλλων μορίων στην επιφάνεια τους τα οποία αποτελούν δείκτες των ρυθμιστικών κυττάρων (π.χ. CD25, CTLA-4, GITR), εμφανίζουν κατασταλτική δραστηριότητα in vitro και in vivo, μειωμένη in vitro παραγωγή IL-2 και πολλαπλασιάζονται με αργούς ρυθμούς 142, 144, 145, 146,147. Επίσης, έχει βρεθεί ότι το FOXP3 ρυθμίζει ενεργά την μεταγραφή στα Treg κύτταρα μέσω στρατολόγησης ενζυματικών μορίων που λειτουργούν ως συν-διεγέρτες και/ ή ως συν-κατασταλτές 148. Η έναρξη της έκφρασης του γονιδίου ρυθμίζεται από τις Runx πρωτεΐνες 149. Οι Runx πρωτεΐνες αποτελούν μεταγραφικούς παράγοντες οι οποίοι επιπλέον παίζουν σημαντικό ρόλο στην μεταγραφική ρύθμιση του Foxp3 κατά τη διάρκεια της επαγωγής της έκφρασης αυτού στα συμβατικά CD4+ T κύτταρα, και άρα στη διαφοροποίηση τους προς Tregs, αλλά και στη διατήρηση της έκφρασής του Foxp3 σε εκείνα τα κύτταρα που είναι ήδη διαφοροποιημένα T κύτταρα με ρυθμιστικό φαινότυπο (Tregs). Η απόκτηση ρυθμιστικού φαινότυπου σημαίνει ότι τα κύτταρα είναι 48
49 ανεργικά και έχουν κατασταλτική δράση. Το γονίδιο FOXP3, φαίνεται ότι όχι μόνον είναι απολύτως απαραίτητο για την λειτουργία των Τ ρυθμιστικών κυττάρων, αλλά ότι και η ελαττωματική του λειτουργία οδηγεί σε σοβαρή διαταραχή της ανοσιακής ρύθμισης. Παλαιότερα η αναγνώριση του υποπληθυσμού των Treg ήταν εφικτή μόνο μέσω της κατάδειξης της κατασταλτικής τους δραστηριότητας. Μετά το 2003, η αναγνώριση της σχέσης του Foxp3 με τα Treg κύτταρα ήταν πρωταρχικής σημασίας για όλες τις επόμενες μελέτες καθώς επέτρεψαν την αναγνώριση αυτών των κυττάρων με μεθόδους κυτταρομετρίας ροής και ανοσοϊστοχημείας 150, 151. Σήμερα, ο μεταγραφικός παράγοντας Foxp3 θεωρείται το gold standard και αποτελεί ένα μοναδικό δείκτη για την αναγνώριση των Tregs που δημιουργούνται στο θύμο αλλά και των Tregs που δημιουργούνται στην περιφέρεια, λόγω της πλούσιας έκφρασής του σε αυτά 150. Η ανάπτυξη του μονοκλωνικού αντισώματος 151 έναντι της πρωτεΐνης του FOXP3 επέτρεψε την ενδοκυττάρια σήμανση (staining) των Tregs, οπότε και επιβεβαιώθηκε η πλούσια έκφρασή του στα ntregs 150. Σχήμα 7: Μονοκλωνικό αντίσωμα Foxp3 Γ. «Φυσικά αναδυόμενα» Τ ρυθμιστικά κύτταρα (ntregs, natural Tregs) Τα Τ ρυθμιστικά κύτταρα που δημιουργούνται στο θύμο (ntregs, naturally arising thymic-derived forkhead box P3) είναι δηλαδή FoxP3+ CD4+ CD25+ Tregs αποτελούν το 5-10% των περιφερικών CD4+ T κυττάρων 123, 128, 152, 153. Τα ntregs εξ ορισμού, έχουν δύο βασικά χαρακτηριστικά τα οποία ουσιαστικά αποτελούν και τον ορισμό του «ρυθμιστικού φαινότυπου» ή 49
50 της «ρυθμιστικής τους λειτουργίας»: α) εμφανίζουν ανεργία, δηλαδή in vitro βρίσκονται σε μια κατάσταση μειωμένης απαντητικότητας σε αντιγόνα και πολλαπλασιάζονται δύσκολα μετά από διέγερση με αυτά και β) εμφανίζουν κατασταλτική λειτουργία έναντι άλλων κυττάρων. Εντούτοις όμως άλλες μελέτες έδειξαν ότι τα ntregs όχι μόνον δεν εμφανίζουν ανεργία, αλλά έχουν μεγάλη ικανότητα να πολλαπλασιάζονται τόσο in vitro, όσο και in vivo όταν διεγερθούν in vitro με ανοσογόνα και όταν βρεθούν σε περιβάλλον με δενδριτικά κύτταρα που έχουν προηγουμένως διεγερθεί από αντιγόνο 154. Επιπλέον διαπιστώθηκε ότι τα κύτταρα αυτά in vivo, δυνητικά υφίστανται δυναμική κλωνική επέκταση ως απόκριση σε αντιγόνο και σε ομοιοστατικά σήματα 155. Τα ntregs εκφράζουν τους ίδιους αβ TCR υποδοχείς, όπως και τα συμβατικά Τ κύτταρα, αλλά αποτελούν ένα ξεχωριστό και διακριτό κλώνο κυττάρων. Προέρχονται από τα αρχέγονα κύτταρα του μυελού των οστών και αναπτύσσονται στο φλοιό του θύμου αδένα μέσα από μια διαδικασία θετικής επιλογής μετά από σύνδεση του υποδοχέα τους με ίδια αντιγόνα για τα οποία εμφανίζουν υψηλή συγγένεια 156, 157. Είναι κύτταρα τα οποία έχουν ενδογενή μακροβιότητα. Είναι πιθανόν η ισχυρή συγγένεια σύνδεσης του TCR των κυττάρων αυτών με ίδια αντιγόνα 158 να ευθύνεται για το γεγονός ότι αρχικά είναι ανεργικά και έχουν την ικανότητα να παράγουν αντι-αποπτωτικά μόρια που τα προστατεύουν από την αρνητική επιλογή στο θύμο. Ο ρόλος τους είναι να ρυθμίζουν, να ελέγχουν και να καταστέλλουν τις ανοσιακές αποκρίσεις. Τα ntregs που μεταναστεύουν από το θύμο στην περιφέρεια διατηρούν αυτό το φαινότυπο, και μάλιστα διατηρούν ακέραιη και τη λειτουργική τους ικανότητα, η οποία ασκείται με κυτταροκυτταρική επαφή μέσω μορίων που εκφράζουν στην επιφάνεια τους και είναι ανεξάρτητη από τις κυτταροκίνες. Η διατήρηση τους στην περιφέρεια υποβοηθείται από το CD28 και τους συνδέτες του (CD80 και CD86), τον TGF-β και την IL-2 η οποία λειτουργεί ως αυξητικός παράγοντας για τα ntregs και είναι πολύ σημαντική και για την κλωνική τους επέκταση 159. Τα ntregs είναι κύτταρα τα οποία χαρακτηρίζονται από την έκφραση συγκεκριμένων μορίων στην επιφάνεια τους, τα οποία συμβάλλουν και στο διαχωρισμό τους από άλλα κύτταρα. Με βάση την έκφραση αυτών των μορίων τα ntregs είναι κύτταρα τα οποία είναι CD45RO, CD45RB low CD4+, CD25 high, CD127 low είναι δηλαδή ενεργοποιημένα μνημονικά κύτταρα
51 Τα κύτταρα αυτά χαρακτηρίζονται επίσης από την σταθερά υψηλή έκφραση του μεταγραφικού παράγοντα Foxp3 ενδοκυττάρια, ο οποίος όπως έχει αναφερθεί αποτελεί δείκτη της κλωνικής τους προέλευσης και είναι ουσιαστικός για την κατασταλτική τους λειτουργία. Επιπλέον, στην επιφάνεια τους τα ntregs εκφράζουν και διάφορους άλλους δείκτες ενεργοποίησης 161, όπως: CD28, CTLA-4 (cytotoxic T lymphocyte-associated antigen 4, CD152), μέλη της υπεροικογένειας TNFR (tumor necrosis factor receptor family) όπως η GITR (glucocorticoid-induced TNF Receptor-a family-related protein 162, 163, υποδοχείς χημειοκινών 164, 165, 166, 167 οι οποίοι παίζουν ρόλο στη μετανάστευση των Tregs, Toll-like receptors (TLRs), TGF-β 168 και neuropilin-1 (Nrp-1) η οποία εκφράζεται σταθερά στα ntregs ανεξάρτητα από το αν είναι ενεργοποιημένα ή όχι, και φαίνεται ότι υπόκειται σε συνρύθμιση με το Foxp3 169, granzyme/ perforin, L-selectin (CD62L hi ) 170, lymphocyte activation gene-3 (LAG-3)(CD223) το οποίο είναι ένα μόριο MHC τάξης ΙΙ, η έκφρασή του οποίου επάγεται μετά την ενεργοποίηση των ntregs 171, 172 και τον υποδοχέα του φολικού οξέος (Folate receptor, FR4) 173 Τα ntreg κύτταρα εκφράζουν στην επιφάνεια τους και μια ποικιλία μορίων προσκόλλησης όπως το CD11a, το CD44, το CD54, και το CD103. Όλοι οι δείκτες (markers) των Tregs, συμπεριλαμβανομένων των CD25, CD127 και FoxP3, είναι μη ειδικοί ακριβώς επειδή ανιχνεύονται και στα ενεργοποιημένα λεμφοκύτταρα (CD4+ και CD8+), αλλά και πολλά άλλα ενεργοποιημένα κύτταρα του ανοσιακού συστήματος, όπως και στα αντιγονοπαρουσιαστικά κύτταρα 174, 175, 176. Το χαρακτηριστικό γνώρισμα των «φυσικά αναδυόμενων» Τreg κυττάρων (naturally arising T cells) είναι η έκφραση των προαναφερθέντων μορίων χωρίς να έχει προηγηθεί ευαισθητοποίηση από κάποιο αντιγόνο. Δ. Επαγόμενα Tregs μέσω κλωνικής επέκτασης (itregs, induced Tregs) Ορισμένοι ερευνητές υποστηρίζουν ότι όλα τα κύτταρα με Τ ρυθμιστική λειτουργία (itregs) προέρχονται από ένα και μόνο υποπληθυσμό Τ κυττάρων: τα CD4+CD25 hi Foxp3+ T κύτταρα που προέρχονται από το θύμο (thymic lineage). Είναι όμως γνωστό πως στον άνθρωπο ο θύμος υποστρέφεται στην εφηβική ηλικία. Το δεδομένο αυτό συνεπάγεται πως όλα τα Treg κύτταρα ουσιαστικά αποτελούν μια 51
52 δεξαμενή κυττάρων που έχουν μεταναστεύσει από το θύμο στην περιφέρεια και έχουν καταφέρει να αυτο-ανανεώνονται συνέχεια. Ακριβώς επειδή κάτι τέτοιο, ιδίως στον άνθρωπο, φαίνεται να είναι μάλλον απίθανο, μελέτες έδειξαν ότι σε ποντίκια, τα περιφερικά CD25- T κύτταρα στα οποία ο TCR έχει διεγερθεί παρουσία TGF-β, αποκτούν ρυθμιστικές ιδιότητες που επιβεβαιώνονται και in vitro και in vivo. Επίσης, μια πρόσφατη μελέτη έδειξε ότι στον άνθρωπο, ακόμη και σε προχωρημένη ηλικία (>70ετών), η περιφερική δεξαμενή των μνημονικών Τ κυττάρων (τα οποία είναι CD4+CD25-Foxp3-CD45RO+) περιλαμβάνει κύτταρα που είναι CD4+CD25-, τα οποία έχουν ενεργοποιηθεί από αντιγόνα, και τα οποία μετατρέπονται σε Τ ρυθμιστικά κύτταρα 177. Μάλιστα, πιθανόν το ποσοστό των Treg κυττάρων να είναι μεγαλύτερο στην μεγάλη ηλικία παρά σε άτομα νεαρής ηλικίας. Στην ίδια μελέτη διαπιστώθηκε ότι τα Treg στον άνθρωπο έχουν διπλάσιο χρόνο ζωής σε σχέση με άλλους υποπληθυσμούς κυττάρων (για παράδειγμα τα περιφερικά κλασικά μνημονικά και naive T κύτταρα), και είναι πολύ ευαίσθητα στην απόπτωση. Αυτά τα ευρήματα έθεσαν εκ νέου την υπόθεση ότι η κλωνική επέκταση του πληθυσμού των Tregs, και άρα η διαφοροποίηση και η επιβίωση των επαγόμενων Tregs, στον άνθρωπο εξαρτάται από τη συνεχή αντιγονική διέγερση και /ή την παρουσία αυξητικών παραγόντων στο μικροπεριβάλλον τους. Και ίσως τελικά αυτές οι προϋποθέσεις να είναι απαραίτητες για την ομοιόσταση του πληθυσμού των Tregs 178. Σήμερα, είναι γνωστό ότι υπάρχουν αρκετές διαφορετικές ομάδες επαγόμενων Tregs τα οποία στη βιβλιογραφία αναφέρονται ως επαγόμενα Τ ρυθμιστικά κύτταρα (adaptive ή induced Tregs, itregs) 179, 180, 181 (σχήμα 8). Τα itregs έχουν επίσης κατασταλτική λειτουργία και ασκούν αποτελεσματικό έλεγχο στις ανοσιακές αποκρίσεις έναντι ίδιων και μη αντιγόνων και εμπλέκονται σε λοιμώξεις, νεοπλασίες, αλλά και στην ανοσοκαταστολή μετά από την μεταμόσχευση. Διαφέρουν από τα ntregs επειδή η δράση τους συνήθως εξαρτάται από τις κυτταροκίνες. Ο χαρακτηρισμός των επαγόμενων πληθυσμών Τ ρυθμιστικών κυττάρων είναι ταυτόχρονα πρόβλημα και πρόκληση, καθώς εξαρτάται από: τους επιφανειακούς δείκτες που εκφράζουν, (μεταξύ των οποίων είναι το CD25, το CD127, το CTLA-4, το GITR, η neuropilin, κ.ά.) την ενδοκυττάρια έκφραση του Foxp3, τόσο όσον αφορά την έκφραση ή την μη έκφραση, αλλά και την ένταση αυτής, και τη διαφορά στο πλήρες σύμπλεγμα του Foxp3 ανάμεσα στα ntregs και τα επαγόμενα itregs, όσον αφορά στην παρουσία μεθυλιωμένων ή απομεθυλιωμένων νουκλεοτιδίων αντίστοιχα
53 την ένταση και τη σταθερότητα της κατασταλτικής τους δράσης το είδος στο οποίο μελετώνται (πειραματόζωα ή άνθρωπος) την παρουσία νόσου ή όχι Αναλυτικότερα, τα itregs διακρίνονται στους ακόλουθους υποπληθυσμούς: τα Tr1 κύτταρα, τα Th3 κύτταρα, τα CD4+CD25+ Tregs που δημιουργούνται μετά από μετατροπή των CD4+CD25- T ενεργοποιημένων κυττάρων, τα CD103+ Tregs, τα CD8+ Tregs, τα διπλά αρνητικά Tregs, τα επαγόμενα για θεραπευτικούς σκοπούς Tregs και τα μη-τ ρυθμιστικά κύτταρα. 1.Tr1 κύτταρα Τα Tr1 κύτταρα είναι CD4+ T ρυθμιστικά κύτταρα τύπου Ι, τα οποία είναι εξ ορισμού αντιγονο-ειδικά και επάγονται παρουσία της IL Τα Tr1 κύτταρα μοιάζουν λειτουργικά με τα ntregs αλλά διαφοροποιούνται από τα CD4+ επειδή χαρακτηριστικά παράγουν IL-10. Έχουν την ικανότητα να εκφράζουν CD25 μετά την ενεργοποίηση τους, όμως η έκφραση του δεν είναι υψηλή, ούτε συνεχής. Επιπλέον αν και δεν εκφράζουν Foxp3 (Foxp3-), η κατασταλτική τους δράση είναι συχνά τόσο ισχυρή όσο εκείνη των ntregs. Διακρίνονται από τα Th2 κύτταρα επειδή δεν παράγουν καθόλου IL-2 και IL-4. Επίσης, έχει διαπιστωθεί ότι αν και παράγουν IFN γ, φαίνεται ότι παράγουν αυτή την κυτταροκίνη σε μικρότερη ποσότητα από εκείνη στην οποία παράγεται από τα Th1 κύτταρα (τουλάχιστον 1 log 10 λιγότερο). Όσον αφορά στην κατασταλτική δράση τους, φαίνεται ότι οφείλεται κυρίως στην IL-10 και δευτερευόντως στον TGF-β, δηλαδή ασκείται μέσω έμμεσων μηχανισμών, αν και άλλες μελέτες υποστηρίζουν ότι η καταστολή ασκείται με κυτταροκυτταρική επαφή που είναι ανεξάρτητη από την IL Τα Tr1 κύτταρα φαίνεται πως διαδραματίζουν εξαιρετικά σημαντικό ρόλο στην ρύθμιση των ανοσιακών αποκρίσεων έναντι λοιμωδών παραγόντων, πιθανόν μέσω της άσκησης επανατροφοδοτούμενης καταστολής προκειμένου να περιοριστεί η ένταση της ανοσιακής απόκρισης και να περιοριστεί η φλεγμονώδης βλάβη 184. Ειδικότερα, καταστέλλουν τις Th1 και Th2 μεσολαβούμενες ανοσιακές αποκρίσεις έναντι παθογόνων μικροοργανισμών αλλά και έναντι καρκινικών κυττάρων και αλλοαντιγόνων. Επίσης, εμποδίζουν τον πολλαπλασιασμό των Th1 κυττάρων και την παραγωγή κυτταροκινών από αυτά, ενώ δρουν και στα δενδριτικά και μακροφάγα κύτταρα 53
54 εμποδίζοντας την παραγωγή IL-12 από αυτά, κι έτσι αναστέλλουν την παραγωγή κυτταροκινών από τα Th1 κύτταρα. Τα κύτταρα αυτά προέρχονται απόcd4+cd25- naive προγονικά Τ κύτταρα της περιφέρειας μετά από έκθεση σε αντιγόνο υπό συνθήκες συγκεκριμένης συνδιέγερσης 183, 185, 186, Th3 Τα Th3 κύτταρα είναι CD4+ κύτταρα τα οποία ονομάστηκαν T βοηθητικά (Τ helper 3) ή Th3 κύτταρα, αν και η λειτουργία τους είναι κατά κύριο λόγο κατασταλτική και όχι βοηθητική 188. Το χαρακτηριστικό τους γνώρισμα είναι η παραγωγή TGF-β 189. Είναι Τ κύτταρα που εμφανίζουν τάξης ΙΙ περιορισμό (class II-restricted) και έχουν πανομοιότυπο αβ TCR υποδοχέα με τα Th1 και Th2 κύτταρα. Η δημιουργία Th3 κυττάρων in vivo ήταν μια από τις πρώτες αναφορές κλωνικής επέκτασης των Tregs μετά από χορήγηση αντιγόνου από το στόμα, αρχικά σε μελέτες με ποντίκια. Τα Th3 κύτταρα δημιουργούνται και διεγείρονται με τρόπο που είναι μεν αντιγονοειδικός, αλλά στη συνέχεια η κατασταλτική τους δράση, μέσω έκκρισης TGF-β, δεν είναι ειδική έναντι ενός συγκεκριμένου αντιγόνου. Τα κύτταρα αυτά έχουν την ικανότητα να εκφράζουν CD25 και Foxp3 μετά την ενεργοποίηση τους. Παράγουν μεγάλη ποσότητα TGF-β, μικρή ποσότητα IL-4 και IL-10, αλλά δεν παράγουν καθόλου IFN-γ και IL-2. Έχει διαπιστωθεί ότι σε μικροπεριβάλλον με απουσία φλεγμονής, η έκκριση του TGF-β αυξάνει την έκφραση του Foxp3 στα ενεργοποιημένα Τ κύτταρα, οπότε συμβάλλει στη λειτουργική τους διαφοροποίηση (itregs) και έτσι συμβάλλει στην κλωνική επέκταση των ntregs στη περιφέρεια 190. Όσον αφορά στη δράση τους, τα κύτταρα αυτά συμβάλλουν στην παραγωγή IgA και έχουν την ικανότητα να καταστέλλουν τις Th1 και Th2 ανοσιακές αποκρίσεις. 3.CD4+CD25+ Tregs που δημιουργούνται από μετατροπή των CD4+CD25- T ενεργοποιημένων κυττάρων Αρχικά σε μελέτες που έγιναν σε ποντίκια βρέθηκε ότι κάποια από τα CD4+CD25+FoxP3+ κύτταρα προέρχονται από την περιφερική δεξαμενή των CD4+CD45RO+CD25-FoxP3- μνημονικών T κυττάρων 177, 191,
55 Ορισμένοι μάλιστα ερευνητές πιστεύουν ότι το ίδιο συμβαίνει και στον άνθρωπο 193. Είναι πιθανόν, αυτός ο υποπληθυσμός να ευθύνεται για τις διαφορές που φαίνεται πως έχουν τα ntregs που προέρχονται από το θύμο και είναι CD4+CD25+Foxp3+ σε σχέση με τα επαγόμενα T ρυθμιστικά κύτταρα (itregs) που προέρχονται από την κλωνική επέκταση των μνημονικών κυττάρων τα οποία είναι CD4+CD Άλλοι ερευνητές έχουν διαπιστώσει ότι τα αντιγονοειδικά CD4+CD25+ Tregs μπορούν να δημιουργηθούν de novo από κλωνική επέκταση των naive αλλά και των μνημονικών CD4-CD25- T προγονικών κυττάρων, μετά από διέγερση με συναφές (cognate) αντιγόνο και ασκούν την κατασταλτική τους δράση με άμεση κυτταροκυταρική επαφή CD103+ Tregs Επιπλέον, διαπιστώθηκε ένας ενδιαφέρον υποπληθυσμός των Treg κυττάρων, ο οποίος εκφράζει CD103, ένα μόριο που είναι γνωστό ως ιντεγκρίνη αεβ7 (integrin alpha Ebeta 7). Η ιντεγκρίνη αεβ7 πιθανόν να παίζει κάποιο ρόλο στην μετανάστευση και παραμονή των Tregs σε σημεία επιθηλιακής βλάβης, στην τοπική τους δράση στο επιθήλιο, αλλά ίσως δρα και ως συνδιεγερτικό μόριο. Αυτός ο υποπληθυσμός βρίσκεται κυρίως στο έντερο ή σε περιοχές φλεγμονής και ασκεί την κατασταλτική του δράση με κυτταροκυτταρική επαφή 195, 196. Συμπερασματικά φαίνεται ότι είναι ασαφές ποιο ποσοστό του συνολικού αριθμού των CD4+CD25+Foxp3+ T κυττάρων που ανιχνεύονται στο περιφερικό αίμα ενός ενήλικα ανθρώπου προέρχεται από το θύμο και ποιο ποσοστό προέρχεται από κλωνική επέκταση στην περιφέρεια. 5.CD8+ T ρυθμιστικά κύτταρα Τα CD8+ Tregs δημιουργούνται μετά την ενεργοποίηση των CD8+ T κυττάρων παρουσία συνδυασμού κυτταροκινών, δηλαδή παρουσία IL- 10 και TGF-β, και στη συνέχεια αναπτύσσουν ισχυρή κατασταλτική δράση και παράγουν με τη σειρά τους IL-10 και TGF-β, αλλά δεν έχουν κυτταροτοξική δράση 197, 198, 199. Μετά την ενεργοποίησή τους, έχουν την ικανότητα να εκφράζουν CD25. Χαρακτηρίζονται από το ότι δεν εκφράζουν CD28 (CD8+CD28-), εκφράζουν Foxp3 ενδοκυττάρια και μια πληθώρα επιφανειακών μορίων 55
56 όπως τα CTLA-4, GITR, CD62L, OX-40 και μάλιστα με την ίδια πυκνότητα όπως τα CD4+CD25+ Tregs. Όσον αφορά τη δράση των CD8+ Tregs έχει ήδη αναφερθεί ότι αναστέλλουν τον πολλαπλασιασμό των Τ κυττάρων. Έχει διαπιστωθεί ότι στοχεύουν εναντίον των αυτοαντιδρώντων CD4+ Τ κυττάρων. Επιπλέον, ορισμένοι ερευνητές υποστηρίζουν ότι τα CD8+ Tregs δύνανται να διακριθούν σε διαφορετικές «τάξεις» 181 : α) CD8+ Tregs τα οποία αναγνωρίζουν πεπτίδια που προέρχονται από αντιγόνα της κυτταρικής επιφάνειας τα οποία συσχετίζονται με κλασικά ή μη αντιγόνα MHC τάξης Ι 200 β) CD8+ Tregs τα οποία αναγνωρίζουν καθοριστές στα μόρια των ανοσοσφαιρινών και παράγουν TGF-β 201 γ) CD8+ Tregs τα οποία ομοιάζουν με τα CD4+ Tr1 κύτταρα επειδή ασκούν την κατασταλτική τους δράση κυρίως μέσω της IL-10, και δημιουργούνται από διέγερση CD8+ T κυττάρων από πλασμακυτταροειδή δενδριτικά κύτταρα παρουσία IL δ) CD8+ Tregs τα οποία χαρακτηρίζονται από υψηλή έκφραση CD122 (τη β άλυσο του υποδοχέα της IL-2) 203 ε) CD8+ Tregs τα οποία δημιουργούνται μετά από επαναλαμβανόμενη in vitro διέγερση των T κυττάρων παρουσία αλλογενών αντιγονοπαρουσιαστικών κυττάρων ή κυττάρων διεγερμένων με αντιγόνο 204 στ) CD8+ Tregs τα οποία είναι CD8+CD25+Foxp3+ και ίσως αντιπροσωπεύουν ένα πληθυσμό αντίστοιχο των ntregs, καθόσον είτε λόγω της μειωμένης έκφρασης περφορίνης και granzyme δεν έχουν κυτταρολυτική δράση ή ασκούν κατασταλτική δράση με άμεση κυτταροκυτταρική επαφή Διπλά αρνητικά Tregs (CD4-CD8-, Double Negative, DN Tregs) Τα κύτταρα αυτά αποτελούν το 1-3% των περιφερικών λεμφοκυττάρων στα τρωκτικά 206. Στον άνθρωπο, αποτελούν το 0,8-1,0% των συνολικών CD3+ T κυττάρων του περιφερικού αίματος και το 2,5% των T κυττάρων στους λεμφαδένες 207. Έχουν δράση κατασταλτική η οποία είναι δοσοεξαρτώμενη. 56
57 7.Επαγόμενα Tregs για θεραπευτικούς σκοπούς 7. Επαγόμενα Tregs για θεραπευτικούς σκοπούς Μεγάλο κομμάτι της έρευνας για τα Τ ρυθμιστικά κύτταρα έχει κινητοποιηθεί από το κίνητρο να χρησιμοποιηθούν θεραπευτικά στα πλαίσια ανοσοθεραπείας. Σε πειραματικά μοντέλα τουλάχιστον έχουν χρησιμοποιηθεί τα ntregs, μετά από αντιγονο-ειδική κλωνική επέκταση in vivo ή in vitro, για να επιτευχθεί ανοσολογική ανοχή σε μη ίδια αντιγόνα, για παράδειγμα μετά από μεταμόσχευση 208. Όμοια πρακτική έχει εφαρμοστεί σε αυτοάνοσα νοσήματα και αλλεργίες 152. Επιπλέον, όπως αναφέρεται σε ορισμένες μελέτες, έχει γίνει εφικτή η κλωνική επέκταση των Tregs σε ικανούς αριθμούς ώστε να μπορούν να χρησιμοποιηθούν θεραπευτικά 209, 210, 211, αλλά αυτό ακόμη δεν αποτελεί καθημερινή πρακτική. 8.Μη- Τ ρυθμιστικά κύτταρα (non-t regulatory cells) Τα μη-τ ρυθμιστικά κύτταρα με ρυθμιστική λειτουργία, διακρίνονται ως εξής 212, 213 : Ανοσογόνα δενδριτικά κύτταρα (tolerogenic DCs) ή μυελοειδή δενδριτικά κύτταρα (DCs) ή ρυθμιστικά δενδριτικά κύτταρα (regulatory DCs) Πλασμακυτταροειδή δενδριτικά κύτταρα (plasmacytoid DCs) Μεσεγχυματικά αρχέγονα κύτταρα (MSC) Διαφορετικά είδη αρχέγονων κυττάρων (embryonic stem cells, neural stem cells) Διάφοροι τύποι εναλλακτικά ενεργοποιημένων μακροφάγων (π.χ. α. alternatively activated macrophages, AAMΦ ή Μ2 και β. IFNγ-stimulated monocyte, IFN-γ-MdC) Κατασταλτικά κύτταρα που προέρχονται από το μυελό των οστών (Myeloid derived suppressor cells, MDSC) Τα ρυθμιστικά «μη-τ» κύτταρα έχουν μη ειδικές έναντι των αντιγόνων ανοσοκατασταλτικές ιδιότητες και συνήθως δεν επάγουν κυρίαρχη ανοχή. Οι μελέτες όμως αναφέρουν ότι συνεργάζονται με τα Τ ρυθμιστικά λεμφοκύτταρα και ενισχύουν την δράση των τελευταίων. Φαίνεται πως δρουν επάγοντας άμεσα το θάνατο του Τ κυττάρου που 57

58 αποτελεί το στόχο τους ή μέσω της αναστολής της ανάπτυξής του, και πως οι δράσεις αυτές μεσολαβούνται από ανοσοκατασταλτικά ένζυμα, κυτταροκίνες ή και κυτταροτοξικούς μηχανισμούς. Επίσης έχει βρεθεί μια διπλή αλληλεπίδραση μεταξύ των «μη-τ» ρυθμιστικών κυττάρων και των Τreg κυττάρων. Πιο συγκεκριμένα, τα πρώτα μπορούν να βοηθήσουν την κλωνική επέκταση των δεύτερων, αλλά και αντίστροφα τα Treg μπορούν να πυροδοτήσουν την ρύθμιση που ασκούν τα «μη-τ» ρυθμιστικά κύτταρα (κυρίως τα ΑΑΜΦ, Μ2). Σχήμα 8: Ενδεικτικοί υποπληθυσμοί επαγόμενων Τ ρυθμιστικών κυττάρων Ε. Μηχανισμοί με τους οποίους τα Tregs ασκούν την κατασταλτική τους δράση Οι μελέτες σχετικά με το θέμα των μηχανισμών που χρησιμοποιούν τα Tregs είναι συχνά αντικρουόμενες, εντούτοις όμως είναι πιθανόν, όχι μόνο να υπάρχουν πολλοί μηχανισμοί, αλλά και να ισχύει ένα μοντέλο στο οποίο τα Tregs ασκούν τη δράση τους με διάφορους τρόπους, οι οποίοι, σε κάποιο βαθμό, επικαλύπτονται 214, 215, 216. Τελικά, ο μηχανισμός που εμπλέκεται κάθε φορά φαίνεται ότι μάλλον εξαρτάται από τη φύση του ερεθίσματος που προκαλεί την φλεγμονή, την ποσότητα και το είδος του αντιγόνου, την κατάσταση ενεργοποίησης και τον αριθμό των κυττάρων-στόχων και βέβαια τον ιστό που εμπλέκεται στη φλεγμονώδη διαδικασία, δηλαδή από το «είδος» του «φλεγμονώδους» 58
59 μικροπεριβάλλοντος 217, 218. Ειδικότερα, έχει αναφερθεί ότι τα Treg κύτταρα μετά την αρχική τους διέγερση μέσω του TCR ασκούν τις κατασταλτικές τους λειτουργίες μέσω διάφορων μηχανισμών, για παράδειγμα μέσω: 1. IL-2 2. άμεσης κυτταροκυτταρικής επαφής 3. παραγωγής ανοσοκατασταλτικών κυτταροκινών 4. μεταβολικής διάσπασης 5. έμμεσων οδών 1.IL-2 Ιδιαίτερη σημασία για την κατασταλτική δράση των Tregs φαίνεται πως έχει η IL-2 και o υποδοχέας της. Αν και η παρουσία της IL-2 δεν είναι απαραίτητη για την δημιουργία των Tregs στο θύμο, η παρουσία της στην περιφέρεια λειτουργεί ως αυξητικός παράγοντας καθώς είναι απαραίτητη για την επιβίωση των Tregs, την διατήρηση της δραστικότητάς τους, όπως επίσης και για τη διατήρηση της μεταγραφικής τους ακεραιότητας. Μάλιστα τα Tregs ως πληθυσμός έχουν μεγαλύτερες ανάγκες σε IL-2 σε σχέση με τα Τ δραστικά κύτταρα. Η IL-2 αυξάνει την έκφραση της IL-10, του CTLA-4 και του TGF-β 219. Επίσης, η IL-2 αυξάνει την έκφραση του Foxp3 των Tregs. Ειδικότερα, όσον αφορά το Foxp3, έχει βρεθεί ότι ο συγκεκριμένος μεταγραφικός παράγοντας συνδέεται με την περιοχή του προαγωγέα της IL-2 και του CD25, και καταστέλλει την πρώτη και ενεργοποιεί τη δεύτερη περιοχή αντίστοιχα, γεγονός το οποίο σημαίνει ότι μειώνει την παραγωγή της IL-2 αλλά αυξάνει την τάση σύνδεσης των Tregs με αυτή την ιντερλευκίνη, καθώς ευοδώνει τη δημιουργία των αλύσεων του IL2R. Φαίνεται πως η μείωση της IL-2 είναι όντως, ένας από τους τρόπους δράσης των Tregs εφόσον και σε καλλιέργειες Tregs και άλλων δραστικών T κυττάρων στις οποίες προστέθηκε IL-2, διαπιστώθηκε άρση της in vitro καταστολής και ακολούθως, πολλαπλασιασμός των Τ δραστικών κυττάρων. Επιπλέον, έχει διαπιστωθεί ότι η συστηματική ανεπάρκεια της IL-2 ή του υποδοχέα της προκαλεί θανατηφόρο αυτοάνοσο ή φλεγμονώδες νόσημα 133, 220, 221. Έχει διατυπωθεί η υπόθεση ότι τα Tregs ίσως «ανταγωνίζονται» τη δράση επαγωγικών ή και κυτταροτοξικών Τ λεμφοκυττάρων δρώντας στο επίπεδο κυτταροκινών που μεταδίδουν σήματα μέσω υποδοχέων οι 59
60 οποίοι περιέχουν την κοινή γ-άλυσο (π.χ. IL-2, IL-4, IL-7), δηλαδή στερούν τις κυτταροκίνες από τα Τ δραστικά κύτταρα. Έτσι, είναι πιθανόν τα Tregs έχοντας την ικανότητα να στερούν από τα δραστικά Τ κύτταρα τον αυξητικό τους παράγοντα, τα οδηγούν πιο γρήγορα στην απόπτωση 222, 223. Το παραπάνω, δεν είναι μοναδικό προσόν των Tregs, καθώς έχουν περιγραφεί διάφορα μοντέλα T-κυτταρικού ανταγωνισμού. Επιπλέον, η κατά προτεραιότητα δέσμευση της IL-2 πιθανόν να τροποποιεί και τις δράσεις των ίδιων των Tregs, καθώς έτσι τα τελευταία παράγουν περισσότερη IL Αντίλογος στην παραπάνω υπόθεση, είναι τα ευρήματα πρόσφατης μελέτης που διαπίστωσε ότι δεν επαρκεί η μεμονωμένη εξάλειψη της IL-2 για να ασκηθεί καταστολή από τα Tregs στα δραστικά Τ κύτταρα Άμεση κυτταροκυτταρική επαφή Τα ntregs προκειμένου να ασκήσουν την κατασταλτική τους δράση απαιτούν την άμεση κυτταροκυτταρική επαφή, η οποία δεν εξαρτάται από την παρουσία ανοσοκατασταλτικών κυτταροκινών 156, 170, 178, 191. Οι κυτταρικές πρωτεΐνες που είναι υπεύθυνες για την άμεση κυτταροκυτταρική επαφή φαίνεται πως είναι ποικίλες 162, 189, 226, 227. Ειδικότερα, τα Tregs χρησιμοποιούν τα εξής μόρια: α)ctla-4 190, 228, 229, 230, 231, 232 β) TGF-β ο οποίος είναι συνδεδεμένος με τη μεμβράνη (TGF-β bound to membrane) 168, 170, 233, 234, 235 γ) LAG-3 (lymphocyte activation gene-3, CD223) 171, 236 δ) GITR (glucocorticoid-induced TNFR-a familyrelated protein) 227, 237, 238, 239 ε) PD-1(programmed death- 1, ή CD279) 228 και στ) άμεση κυτταρόλυσης για παράδειγμα μέσω granzyme και περφορίνης 240, 241, 242, Παραγωγή ανοσοκατασταλτικών κυτταροκινών Οι Vignali και συνεργάτες, το 2008 διατύπωσαν το σκεπτικό ότι τα πειραματικά μοντέλα που χρησιμοποιούνται σήμερα για τη μελέτη της άμεσης κυτταροκυτταρικής επαφής των Tregs με τα κύτταρα-στόχο δηλαδή τα Τ δραστικά κύτταρα, μάλλον εκφράζουν ένα μέτρο εγγύτητας και δεν θα πρέπει να θεωρείται δεδομένο ότι επιβεβαιώνουν την άσκηση κατασταλτικής δράσης μέσω άμεσης κυτταροκυτταρικής επαφής. Το σκεπτικό τους φαίνεται λογικό, καθώς η δράση των διαλυτών μεσολαβητών είναι πιο ισχυρή κοντά στην πηγή της παραγωγής τους. Οπότε, η εγγύτητα εξασφαλίζει και μεγάλες συγκεντρώσεις τοπικά (όπως έχει αποδειχθεί για την IL-2). Θα πρέπει επίσης να ληφθεί υπόψη και η ύπαρξη ασταθών μεσολαβητών, όπως είναι η αδενοσίνη (χρόνος 60
61 ημίσειας ζωής <10 sec), των οποίων ο εργαστηριακός προσδιορισμός είναι τεχνικά εξαιρετικά δύσκολος, αν όχι και αδύνατος με τη σημερινή τεχνολογία 244. Έτσι, ένας άλλος μηχανισμός που έχει αναφερθεί για την άσκηση της κατασταλτικής δράσης των Tregs είναι η παραγωγή κυτταροκινών με ανασταλτική δράση, όπως TGF-β 160, 223, 245, 246, 247, 248, 238, η IL , 160, 250, 251, 252, 253 και η IL Αυτός ο μηχανισμός φαίνεται πως αφορά κυρίως τα επαγόμενα itregs. 4.Μεταβολική «διάσπαση» (metabolic disruption) Πρόσφατα έχουν περιγραφεί μηχανισμοί οι οποίοι φαίνεται πως τροποποιούν τη μεταβολική κατάσταση των δραστικών Τ κυττάρωνστόχων. Έχει βρεθεί ότι τα ntregs διαθέτουν μεγάλες ποσότητες camp (cyclic adenosine monophosphatase), το οποίο είναι γνωστό ότι όχι μόνο έχει ισχυρή ανασταλτική δράση έναντι του πολλαπλασιασμού και της διαφοροποίησης των κυττάρων, αλλά και ότι μπορεί να παρεμποδίζει εκλεκτικά την έκφραση των γονιδίων που αφορούν στη σύνθεση IL-2 και IFN-γ από τα Τ κύτταρα. Η έκφραση των γονιδίων εμποδίζεται ή με μπλοκάρισμα της πρωτεϊνικής κινάσης Α του NF-κΒ ή με την ενεργοποίηση του μεταγραφικού κατασταλτή ICER (transcriptional repressor, inducible camp early repressor) 255. Ως εκ τούτου, τα Tregs είναι δυνατόν να μεταδίδουν αρνητικά σήματα αυξάνοντας τα επίπεδα του camp στα κύτταρα-στόχο με την διακυτταρική μεταφορά camp από τα ίδια σε άλλα Τ κύτταρα μέσω κενών διασύνδεσης (gap junctions) που αποτελούνται από κοννεξίνες (connexins = μεμβρανικές πρωτεΐνες). Το σκεπτικό αυτό επιβεβαιώθηκε από μελέτες στις οποίες η παρεμπόδιση του camp ή των gap junctions οδήγησε σε εξασθένηση της κατασταλτικής δράσης των Tregs 256, 257. Επίσης, η συντονισμένη και ταυτόχρονη έκφραση των εκτοενζύμων CD39 και CD73 στην επιφάνεια των Tregs δημιουργεί αδενοσίνη περικυτταρικά, η οποία ενεργοποιώντας τον υποδοχέα Α2Α της αδενοσίνης στα ενεργοποιημένα δραστικά Τ κύτταρα, τελικά τα καταστέλλει 258. Επιπλέον, η προαναφερθείσα σύνδεση ευοδώνει τη δημιουργία νέων itregs, προάγει την έκκριση TGF-β και αναστέλλει την έκφραση της IL Η αδενοσίνη έχει και επιπρόσθετες δράσεις, καθώς 61
62 ρυθμίζει την ωρίμανση των δενδριτικών κυττάρων και μάλιστα ευνοεί τη δημιουργία του ανοσογόνου φαινότυπου (tolerogenic phenotype) Έμμεσοι μηχανισμοί Τα πειραματικά μοντέλα δεν έχουν μπορέσει να αποσαφηνίσουν εάν η κατασταλτική δράση των Tregs in vivo στοχεύει έναντι των δραστικών Τ κυττάρων ή έναντι των δενδριτικών/ αντιγονοπαρουσιαστικών κυττάρων. Έτσι έχει δημιουργηθεί το ερώτημα: ποιος είναι τελικά ο πρωτεύων στόχος των κατασταλτικών μηχανισμών των Tregs; Το σκεπτικό είναι ότι θα μπορούσαν να είναι και οι δυο ομάδες κυττάρων 244. Όντως, υπάρχουν μελέτες στις οποίες διαπιστώθηκε ότι τα Tregs κατά κύριο λόγο έρχονται σε επαφή με τα δενδριτικά κύτταρα 261. Φαίνεται λοιπόν πως τα Tregs επιτυγχάνουν τις κατασταλτικές τους δράσεις και μέσω δράσης σε άλλα κύτταρα, πέραν των δραστικών Τ κυττάρων. Αυτό σημαίνει ότι έχουν την ικανότητα να αλληλεπιδρούν και με άλλα κύτταρα του ανοσιακού συστήματος. Πιο συγκεκριμένα, αλληλεπιδρούν με τα δενδριτικά κύτταρα, με τα ΝΚ κύτταρα, καθώς και με μακροφάγα και μονοπύρηνα επηρεάζοντας την ενεργοποίηση και τη λειτουργία τους 262, 263. Ειδικότερα, έχει διαπιστωθεί ότι επάγουν την παραγωγή ανοσοκατασταλτικών μορίων (όπως IDO, IL-10, TGF-β) από τα δενδριτικά κύτταρα μέσω αλληλεπίδρασης CTLA-4 και CD80 και/ ή CD86 156, 191, 264, 265, 266, 267. Επίσης σε άλλες μελέτες έχει διαπιστωθεί ότι είναι δυνατόν τα Tregs να μειώνουν την έκφραση των συνδιεγερτικών μορίων CD80, CD86 στα δενδριτικά κύτταρα, έτσι ώστε να μειώνεται η ικανότητα τους να ενεργοποιούν τα δραστικά Τ κύτταρα 268. Επιπρόσθετα, τα Tregs επιδρούν στην ωρίμανση των δενδριτικών κυττάρων 269, 270. Τέλος, η νευροπιλίνη-1 φαίνεται πως επειδή προάγει την παρατεταμένη αλληλεπίδραση των Tregs και των ανώριμων δενδριτικών κυττάρων, ίσως τελικά είναι εκείνη που τους δίνει το χρονικό πλεονέκτημα να τα τροποποιούν με τρόπο που να ευνοεί τη ρυθμιστική τους λειτουργία 271,
63 ΣΤ. Ομοιόσταση των Τ ρυθμιστικών λεμφοκυττάρων 1. Δημιουργία ntregs στο θύμο αδένα Τα ntreg κύτταρα παράγονται ως ένας ξεχωριστός κλώνος κυττάρων με ιδιαίτερα λειτουργικά χαρακτηριστικά στο θύμο, και αυτό έχει επιβεβαιωθεί σε πειραματικά μοντέλα ποντικών 149, 182, 273. Η επιλογή των CD25+ κυττάρων που έχουν ρυθμιστική λειτουργία στο θύμο, γίνεται απαραίτητα μέσω μορίων MHC τάξης ΙΙ. Πιθανόν η επιλογή των CD4+CD25+ κυττάρων μέσα στο θύμο να γίνεται με βάση μια προκαθορισμένη αναλογία αυτών σε σχέση με τα CD4+CD25- κύτταρα, καθώς τα τελευταία παράγονται λίγο νωρίτερα (χρονικά) στο θύμο. Πάντως, η διαφοροποίηση των Treg μέσα στο θύμο επηρεάζεται από πολλούς παράγοντες, όπως το είδος του αντιγόνου που παρουσιάζεται, ο χρόνος που επιλέγεται για την παρουσίαση του αντιγόνου αλλά και η συνάφεια (avidity) του αντιγόνου. Φαίνεται όμως, πως και μόνο η παρουσίαση του αντιγόνου από το επιθήλιο των κυττάρων του φλοιού του θύμου αποτελεί ένα επαρκές ερέθισμα 162. Σημαντικό ρόλο επίσης παίζει η συγγένεια (affinity) των αντιγονικών πεπτιδίων με τον TCR υποδοχέα των CD4+CD25- κυττάρων στο θύμο. Έτσι, φαίνεται πως η μικρή συγγένεια TCR και αντιγονικού πεπτιδίου οδηγεί στην θετική επιλογή των συμβατικών CD4+CD25- T κυττάρων. Αντίθετα, η υψηλή συγγένεια των αντιγονικών πεπτιδίων με τον TCR μπορεί να οδηγήσει σε δυο ενδεχόμενα: α) κατά κύριο λόγο, στην αρνητική επιλογή και την εξάλειψη των λεμφοκυττάρων τα οποία αντιδρούν με ίδια αντιγόνα (depletion), και β) υπό συνθήκες συνδιέγερσης στην δημιουργία των ntregs, που σημαίνει ότι συνδέονται συγκεκριμένα μόρια στην επιφάνεια των αντιγονοπαρουσιαστικών κυττάρων με αντίστοιχους υποδοχείς στην επιφάνεια των T λεμφοκυττάρων, έτσι ώστε να δοθεί το απαραίτητο «δεύτερο σήμα» για την δημιουργία του διακριτού κλώνου των T ρυθμιστικών κυττάρων. Ειδικότερα, θεωρείται ότι τα πρόδρομα κύτταρα των ntregs στο θύμο, εκφράζουν ένα TCR με μέση προς υψηλή συγγένεια για τα μόρια MCH τάξης ΙΙ, αλλά η συγγένεια τους είναι λίγο μικρότερη από εκείνη που θα προκαλούσε εξάλειψη του κλώνου τους. Επίσης, φαίνεται πως η σύνδεση του TCR των ntregs με το αντιγόνο υψηλής συγγένειας ταυτόχρονα αυξάνει και την έκφραση του FoxP3 γονιδίου, οπότε αποκτούν και το λειτουργικό τους φαινότυπο, δηλαδή ανεργία και κατασταλτική δράση 186. Πιθανόν στη συνδιέγερση να παίζει σημαντικό ρόλο το CD28 είτε μέσω διέγερσης της έκφρασης του Foxp3, είτε μέσω 63
64 επαγωγής άλλων γονιδίων που ευνοούν την έκφραση Foxp3, πιθανώς μάλιστα χωρίς την απαραίτητη μεσολάβηση της IL-2 274, 275. Επίσης, έχει διαπιστωθεί ότι στο θύμο, είναι απαραίτητο να είναι ακέραια τα λιπίδια της μεμβράνης των προγονικών κυττάρων των Fopx3+ Tregs αλλά και να είναι απόλυτα λειτουργικός ο υποδοχέας p56 lck του CD28 της μεμβράνης τους, έτσι ώστε να είναι εφικτό να αυξηθεί η έκφραση του Foxp3 και να διατηρηθεί επί μακρόν 276. Είναι ενδιαφέρον ότι α)τα CD4+CD25+ κύτταρα που παράγονται στο θύμο καταδεικνύουν εξαιρετικά υψηλότερο επίπεδο TREC (T cell Receptor Excision Circle), δηλαδή μεγαλύτερο αριθμό εξωχρωμοσωματικών DNA επισωμάτων που σημαίνει ότι η αναδιάταξη του TCR στο θύμο αδένα πιθανώς τους δίνει την δυνατότητα να αντισταθμίζουν την μειωμένη τους παραγωγή στην ενήλικη ζωή 277 και β)ότι η διέγερση ενός αντιγονο-ειδικού Τ κυτταρικού πληθυσμού στο θύμο από ένα ενδογενές αντιγόνο, παρουσία IL-2, οδηγεί παράλληλα ή διαδοχικά στην κλωνική επέκταση και των δραστικών κυττάρων και των Τ ρυθμιστικών κυττάρων Μεταγραφικοί παράγοντες που είναι απαραίτητοι για τη διαφοροποίηση των Τregs Διαφοροποίηση των Τ βοηθητικών σε κυττάρων Tregs στην περιφέρεια Είναι γνωστό ότι ένα Τ βοηθητικό λεμφοκύτταρο μπορεί να διαφοροποιηθεί προς διάφορους διακριτούς υποπληθυσμούς, μεταξύ των οποίων και τα Tregs. Στη διαδικασία αυτή εμπλέκονται διάφοροι μεταγραφικοί παράγοντες, όπως: α) μέλη της οικογένειας του πυρηνικού παράγοντα των ενεργοποιημένων Τ κυττάρων (Nuclear Factor of Activated T cell, NFAT), του NF-κΒ, και του ενεργοποιητή της πρωτεΐνης -1 (Activator Protein-1, AP-1) β) ο μεταγραφικός παράγοντας Foxp3 γ) το STAT5, το οποίο αποτελεί ένα μεταγραφικό παράγοντα που είναι απαραίτητος για την ανάπτυξη των Tregs, όπως επιβεβαιώθηκε από μελέτες οι οποίες έδειξαν ότι η ειδική εξάλειψη του παρεμποδίζει την ανάπτυξη τους Κλωνική επέκταση των Τ ρυθμιστικών κυττάρων Οι μελέτες έχουν δείξει ότι το ποσοστό των περιφερικών CD4 κυττάρων που είναι CD25+ είναι σταθερό σε ποντίκια που μελετήθηκαν από την ηλικία των 2 εβδομάδων έως και την ηλικία του ενός έτους. Μάλιστα, ακόμη και σε ποντίκια με θυμεκτομή, μετά την μεταφορά Tregs, 64
65 διαπιστώθηκε κλωνική επέκταση του πληθυσμού έτσι ώστε να φτάσουν το ίδιο ποσοστό που υπήρχε σε αντίστοιχης ηλικίας πειραματόζωα 280. Παρόμοια ευρήματα έθεσαν το βασικό ερώτημα κατά πόσο, ο πληθυσμός των Treg κυττάρων είναι ο ίδιος για όλη τη διάρκεια της ζωής, εάν ανανεώνεται ή εάν εμπλουτίζεται η αρχική δεξαμενή των ntregs, εάν δηλαδή υφίσταται κλωνική επέκταση από κύτταρα της περιφέρειας, έτσι ώστε ο αρχικός τους αριθμός να διατηρείται ακέραιος και πανομοιότυπος, ακόμη και σε απουσία φλεγμονής. Σήμερα, έχει αποδειχθεί ότι τα ntregs μετά την έξοδό τους από το θύμο παρουσία ίδιων αντιγόνων των ιστών πολλαπλασιάζονται μετά από διέγερση με κυτταροκίνες όπως ο TGF-β, η IL-2 και η IL-10 με σκοπό τη διατήρηση της ομοιόστασης 142, 183, 281, 282. Ζ. Ο ρόλος του TCR και η ειδικότητα των Tregs έναντι αντιγόνων Τα Tregs, όπως άλλωστε και όλα τα CD4+ T κύτταρα, διαθέτουν ένα αβ TCR με τον οποίο αναγνωρίζουν ειδικά αντιγονικά πεπτίδια με τη βοήθεια μορίων MHC τάξης II. Εντούτοις, φαίνεται ότι η λειτουργία του TCR στα Tregs διαφέρει από τα συμβατικά κύτταρα καθόσον δεν εξαρτάται από συνδιεγερτικά μόρια. Υπέρ της άποψης αυτής είναι το γεγονός ότι μεμονωμένα αντιγόνα μπορούν να πυροδοτήσουν α) τον πολλαπλασιασμό των Tregs στην περιφέρεια και β) την κατασταλτική τους δραστηριότητα έναντι των δραστικών κυττάρων, χωρίς ενδιάμεση βοήθεια από αντιγονοπαρουσιαστικά κύτταρα 156, 283. Η ενεργοποίηση των Treg είναι αντιγονο-ειδική και μάλιστα η κατασταλτική τους δραστηριότητα πυροδοτείται μετά από αντιγονοειδική διέγερση, αλλά είναι δυνατόν να επεκτείνεται και έναντι δραστικών κυττάρων με άλλη αντιγονοειδικότητα 126, 128, 273, 284, 285. Η. Πώς καταφέρνουν τα κύτταρα- στόχος να ξεφεύγουν από την κατασταλτική δράση των Tregs; Ακριβώς επειδή το φάσμα των δράσεων των Tregs είναι πολυποίκιλο, το ανοσιακό σύστημα έχει φροντίσει ώστε να υπάρχουν κάποιες δικλείδες ασφαλείας, ώστε όταν οι δράσεις τους είναι ανεπιθύμητες, να υπάρχει η δυνατότητα να περιοριστούν σε εύρος και ένταση 218, 286, 288. Αυτές οι δικλείδες αφορούν είτε στην άσκηση αρνητικής ρύθμισης στα ίδια τα Tregs είτε στην «θωράκιση» των κυττάρων-στόχων ώστε να μπορούν να αντιμετωπίσουν τα Tregs. 65
66 Η αρνητική ρύθμιση γίνεται με μια ποικιλία ερεθισμάτων, μεταξύ των οποίων περιλαμβάνεται η διέγερση των TLR υποδοχέων στα δενδριτικά κύτταρα 288, αλλά και η άμεση ρύθμιση μέσω κυτταροκινών (π.χ. IL-2, IL- 21, IL-7, IL-15, TNF-α και IL-6). Αναλυτικότερα, έχει ήδη αναφερθεί ότι η εξωγενής χορήγηση IL-2 σε in vitro μελέτες αναιρεί τον ανεργικό φαινότυπο των Tregs και αντιστρέφει την κατασταλτική τους δράση 126. Η IL-21 είναι άλλο ένα μέλος της οικογένειας της IL-2, και έχει διαπιστωθεί ότι σε πειραματικά μοντέλα σακχαρώδη διαβήτη σε ποντίκια, η υπερέκφρασή της συμβάλλει σε μικρότερη ευαλωτότητα των κυττάρωνστόχων έναντι της κατασταλτικής δράσης των Tregs κι έτσι συμβάλλει στην αποφυγή της αυτοανοσίας 289. Αντίστοιχα ευρήματα υπάρχουν και για τις IL-7 και IL-15, αλλά και για τον TNF-α 290. Όσον αφορά στην IL-6, έχει διαπιστωθεί ότι εκκρίνεται από τα ώριμα δενδριτικά κύτταρα που έχουν προηγουμένως διεγερθεί μέσω TLR, και είναι αναγκαία για τα CD4+ δραστικά κύτταρα, ώστε να καταφέρουν να υπερνικήσουν την κατασταλτική ικανότητα των CD4+CD25+Tregs 291. Συνολικά, τα σημερινά in vitro δεδομένα καταλήγουν ότι ο TNF-α και η IL-6 αποτελούν τους κυριότερους αρνητικούς ρυθμιστές της λειτουργίας των Tregs. Όσον αφορά στη «θωράκιση» των κυττάρων στόχων (δηλαδή Τ δραστικών κυττάρων και δενδριτικών), αυτή ενισχύεται με την ενίσχυση των διεγερτικών σημάτων στα Τ κύτταρα-στόχο, η οποία τα βοηθάει να ανθίστανται στη δράση των Τ ρυθμιστικών κυττάρων ή και να αποφεύγουν τελικά την καταστολή. Ειδικότερα η «θωράκιση» επιτυγχάνεται με μια πληθώρα μηχανισμών, όπως: α)με την αύξηση της έκφρασης των Β7 μορίων στην επιφάνεια τους (CD80 και CD86) 126, 128 β) με την σηματοδότηση μέσω CD28, και θετικά σήματα από μόρια της οικογένειας TNFR 163, 226, 227, 292 γ) με την απώλεια μορίων που συμμετέχουν στην ενδοκυττάρια σηματοδότηση, η οποία τελικά προσδίδει στα CD4+ T δραστικά κύτταρα, ανθεκτικότητα έναντι της κατασταλτικής δράσης των Tregs (π.χ. Cbl-b, NFATc1,c2, c3, TRAF6) 253, 293, 294,
67 Θ. Στρατολόγηση των T ρυθμιστικών λεμφοκυττάρων στο σημείο της φλεγμονής Τα ntregs δημιουργούνται στο θύμο και κατόπιν μεταναστεύουν στην περιφέρεια. Στην περιφέρεια ο πληθυσμός των Tregs επεκτείνεται κλωνικά (itregs), αλλά ακόμη δεν είναι γνωστό εάν η κλωνική επέκταση συμβαίνει μέσα στα περιφερικά όργανα του λεμφικού ιστού ή στη θέση της ιστικής φλεγμονής και αυτό ισχύει είτε η φλεγμονή αυτή προκύπτει λόγω αυτοάνοσου ή λοιμώδους νοσήματος, είτε ακόμη από νεοπλασία. Έτσι, έχουν βρεθεί Τ κύτταρα με ρυθμιστική λειτουργία και φαινότυπο στο θύμο, στο μυελό, σε περιφερικά όργανα του λεμφικού ιστού, στο περιφερικό αίμα αλλά και σε θέσεις φλεγμονής (π.χ. έντερο, ήπαρ, ενδαρθρικό υγρό, δέρμα) 120, 196, 296, 297, 298, 299, 300. Όσον αφορά στο μυελό των οστών, έχει διαπιστωθεί ότι πάνω από το ¼ των συνολικών CD4+ T κυττάρων του έχει λειτουργικούς και φαινοτυπικούς χαρακτήρες Τ ρυθμιστικών κυττάρων 296. Είναι πιθανόν ο μυελός να λειτουργεί ως ένα ενεργό όργανο του ανοσιακού συστήματος, που ρυθμίζει την κατακράτηση των Treg κυττάρων μέσα στο παρέγχυμά του, αλλά ρυθμίζει και την στοχευμένη μετακίνηση τους σε σημεία φλεγμονής. Το παραπάνω σκεπτικό υποστηρίζουν μελέτες, στις οποίες διαπιστώθηκε ότι τα Treg κύτταρα του μυελού εκφράζουν τον χημειοτακτικό υποδοχέα CXCR4 για την χημειοκίνη CXCL12. Τα Tregs όταν πρόκειται να εγκαταλείψουν το μυελό, πρώτα μειώνουν την έκφραση της CXCL12 μέσω G-CSF, κι έτσι μειώνουν την έκφραση του υποδοχέα που τα κρατά δέσμια εκεί. Αντίστροφα, διαπιστώθηκε ότι όταν τα Treg στην περιφέρεια ενεργοποιηθούν, αυξάνουν την έκφραση του CXCR4, οπότε διευκολύνουν και τη μετανάστευσή τους στο μυελό και τη σύνδεση με τον υποδοχέα CXCL Έτσι η έκφραση συγκεκριμένων υποδοχέων των χημειοκινών και η ρύθμιση της έντασης με την οποία εκφράζονται, αποτελεί το πρώτο σήμα που καθορίζει την κατεύθυνση της μετακίνησης του πληθυσμού των ntregs. Όσον αφορά στη μετανάστευση και τη συγκέντρωση των Tregs σε συγκεκριμένα ιστικά διαμερίσματα, έχει διαπιστωθεί ότι ρυθμίζεται από ποικίλα μόρια 301, όπως: α) ιντεγκρίνες Οι ιντεγκρίνες είναι σημαντικά μόρια προσκόλλησης για την in vivo μετανάστευση και στρατολόγηση των Tregs σε σημεία φλεγμονής. Ειδικότερα, έχει βρεθεί ότι τα Treg κύτταρα που δεν εκφράζουν ιντεγκρίνη και δεν εκφράζουν στην επιφάνεια τους το μόριο αε(cd103), 67
68 παρουσιάζουν υψηλή έκφραση CD62L και CCR7, και έχουν την ικανότητα να μεταναστεύουν στους λεμφαδένες όπου και καταστέλλουν κυρίως την ενεργοποίηση των naive T κυττάρων. Αντίθετα, τα Treg που εκφράζουν στην επιφάνεια τους ιντεγκρίνη [αε(cd103)(+)], εκφράζουν ασθενώς το CD62L, και έχουν την ικανότητα να μεταναστεύουν σε φλεγμαίνουσες περιοχές, όπου και καταστέλλουν την περιφερική φλεγμονή 195, 302, 303. Επίσης, έχει αναφερθεί ότι τα CD4+CD25+CD62L+ Treg κύτταρα έχουν in vivo ισχυρότερη κατασταλτική δράση, εύρημα που πιθανώς να εξηγείται επειδή μπορούν να μεταναστεύουν στους λεμφαδένες ευκολότερα σε σχέση με τα CD4+CD25+CD62L- Treg κύτταρα. Φαίνεται λοιπόν ότι η ιντεγκρίνη CD62L είναι ένα μόριο που κατευθύνει τα κύτταρα του ανοσιακού συστήματος στο λεμφικό ιστό (homing molecule) 304, 305. Επιπλέον, φαίνεται πως υπάρχει κάποια συσχέτιση μεταξύ των ιντεγκρινών και του υποπληθυσμού των itregs. Πιο συγκεκριμένα, έχει βρεθεί ότι η έκφραση της α4β7 ιντεγκρίνης στον ιστό προσελκύει τα Tr1 κύτταρα τα οποία παράγουν IL-10, ενώ η έκφραση της α4β1 ιντεγκρίνης προσελκύει τα Th3 κυττάρων τα οποία παράγουν TGF-β. Όμως, η ιντεγκρίνη α4β1 επάγεται στο ενδοθήλιο των ιστών που έχουν φλεγμονή, ενώ η α4β7 στα αγγεία των βλεννογόνων. Μπορεί λοιπόν να υποτεθεί ότι οι πληθυσμοί των Tregs μεταναστεύουν επιλεκτικά σε εκείνους τους ιστούς που όχι μόνο θα αλληλεπιδράσουν με τα αυτοαντιδρώντα T κύτταρα, αλλά και εκεί όπου θα ευνοηθεί η κλωνική επέκταση τους σε Tr1 ή Th3 κύτταρα. β) υποδοχείς χημειοκινών -ο υποδοχέας της χημειοκίνης CCR7 είναι άλλο ένα σημαντικό μόριο για την δρομολόγηση των ανοσιακών κυττάρων προς το λεμφικό ιστό. Έχει βρεθεί ότι όταν τα Treg κύτταρα κατευθύνονται στην Τ ζώνη των λεμφικών οργάνων χρησιμοποιούν τη σύνδεση CCL19/CCR7 για να διευκολυνθούν, ενώ όταν κατευθύνονται προς την Β ζώνη ο υποδοχέας CCR7 μετατρέπεται σε CXCR9, ώστε να μεταναστεύσουν προς την Β ζώνη και να συνδεθούν με τη χημειοκίνη CXCL13 που εκφράζεται εκεί 306. Τα ευρήματα αυτά αποδεικνύουν ότι τα Tregs δρουν κατασταλτικά και στις αποκρίσεις των Β κυττάρων. 68
69 ΜΕΡΟΣ V. O ΡΟΛΟΣ ΤΩΝ T ΡΥΘΜΙΣΤΙΚΩΝ ΛΕΜΦΟΚΥΤΤΤΑΡΩΝ ΣΤΗΝ HCV ΛΟΙΜΩΞΗ Όπως έχει ήδη προαναφερθεί στη βιβλιογραφία, έχουν περιγραφεί διάφοροι μηχανισμοί οι οποίοι θα μπορούσαν να ευθύνονται για την αποτυχία των CD8+ T κυττάρων να ελέγξουν την HCV λοίμωξη και ως εκ τούτου ευθύνονται για την μετάπτωση της σε χρονιότητα 75, 76, 307. Οι μηχανισμοί αυτοί είναι: γενετικοί παράγοντες του ξενιστή (HLA μόρια) οι μεταλλάξεις διαφυγής του ιού η καταστολή από παράγοντες του ιού ανασταλτικές κυτταροκίνες ανασταλτικοί συνδέτες η ανεπαρκής βοήθεια από τα CD4+ κύτταρα η ανεπαρκής διαφοροποίηση των CD8+ T κυττάρων η πρωτοπαθής ανεπάρκεια και/ ή η εξάντληση τους, και η δράση των Τ ρυθμιστικών κυττάρων (Tregs) Όσον αφορά την δράση των Τ ρυθμιστικών κυττάρων στην HCV λοίμωξη διαπιστώνονται τα εξής: Στην οξεία HCV λοίμωξη έχει διαπιστωθεί κλωνική επέκταση των Tregs και αύξηση του πληθυσμού τους 308. Τα ευρήματα αυτά επιβεβαιώθηκαν σε μια άλλη μελέτη στην οποία μελετήθηκαν 16 ασθενείς με οξεία HCV λοίμωξη από τους οποίους οι 12 είχαν γονότυπο 1. Διαπιστώθηκε αύξηση του αριθμού των Tregs αλλά και της κατασταλτικής τους δράσης στους 12 αυτούς ασθενείς οι οποίοι και εξακολούθησαν να έχουν ανιχνεύσιμο HCV RNA και 6 μήνες μετά την οξεία λοίμωξη, γεγονός το οποίο δείχνει ότι οι ασθενείς μετέπεσαν σε χρόνια νόσο. Κατά συνέπεια, φαίνεται ότι η αύξηση των Τ ρυθμιστικών κυττάρων στην έναρξη της HCV λοίμωξης με γονότυπο 1, ήταν προγνωστικός δείκτης της εγκατάστασης χρονιότητας της λοίμωξης και πιθανολογούν ότι υπάρχει κάποια επίδραση του συγκεκριμένου γονότυπου όσον αφορά την εγκατάσταση χρονιότητας μέσω της πυροδότησης των Tregs 309. Αντίθετα, άλλοι ερευνητές ανέφεραν ότι δεν διαπίστωσαν διαφορά στην έκφραση του Foxp3 και στην κατασταλτική δράση των Tregs στην αρχική φάση της οξείας λοίμωξης ανάμεσα στους ασθενείς στους οποίους η λοίμωξη μετέπεσε σε χρονιότητα και σε εκείνους στους οποίους υπήρξε αυτόματη κάθαρση του ιού. Επιπλέον διαπιστώθηκε ότι η ένταση της κατασταλτικής δράσης των Tregs 6 μήνες μετά την αποδρομή της HCV λοίμωξης ήταν σχετικά ελαττωμένη
70 Όσον αφορά στη χρόνια ηπατίτιδα C, υπάρχουν αρκετές μελέτες στις οποίες διερευνήθηκε ο παθογενετικός ρόλος των Tregs στη χρόνια νόσο και οι λειτουργικές τους μεταβολές 311. Στις μελέτες αυτές διαπιστώθηκε ότι in vitro, τα Tregs παρεμποδίζουν τον πολλαπλασιασμό, την κλωνική επέκταση των ειδικών έναντι του HCV CD8+ T κυττάρων, και την παραγωγή IFN-γ και μάλιστα με τρόπο δοσοεξαρτώμενο, γεγονός το οποίο δείχνει ότι όσο μεγαλύτερος είναι ο αριθμός των Tregs τόσο μικρότερος ο πολλαπλασιασμός των CD8+ T κυττάρων 312. Επιπλέον, ενδιαφέρον παρουσιάζουν τα ευρήματα μελετών στις οποίες διαπιστώθηκε ότι η κατασταλτική δράση των Tregs είναι συγκρίσιμου μεγέθους σε άτομα με αποδραμούσα HCV λοίμωξη και σε υγιείς αιμοδότες, αλλά είναι πιο ισχυρή σε άτομα με χρόνια HCV λοίμωξη. Το παραπάνω δεδομένο πιθανολογείται ότι οφείλεται στο γεγονός ότι ο αριθμός των CD4+CD25+ Τ κυττάρων είναι μεγαλύτερος σε ασθενείς με χρόνια ηπατίτιδα C σε σχέση με τις άλλες δυο ομάδες της μελέτης, άρα η κλωνική τους επέκταση ήταν εντονότερη 312. Τα παραπάνω ευρήματα επιβεβαιώθηκαν και σε άλλες μελέτες στις οποίες έγινε σύγκριση των ασθενών με χρόνια λοίμωξη με άτομα με αποδραμούσα λοίμωξη και με υγιή άτομα. Διαπιστώθηκε ότι υπήρχε αυξημένος αριθμός CD4+CD25+ κυττάρων στην περιφέρεια και περισσότερο στο ήπαρ, σε ασθενείς με χρόνια HCV λοίμωξη (μετά από θεραπεία ή αυτόματα) 312, 313, 314, 315, 316, 317, 318. Τα ευρήματα αυτά πιθανολογούν ότι ο μεγαλύτερος αριθμός των Tregs πιθανόν να συμβάλει στην χρόνια παραμονή του ιού 313, ενώ συγχρόνως, η κλωνική τους επέκταση προκαλεί μείωση της ιστικής βλάβης. Υπέρ του σκεπτικού αυτού είναι το γεγονός ότι υπάρχει θετική συσχέτιση του αριθμού των CD4+CD25+ Tregs και του τίτλου του ιικού φορτίου, ενώ διαπιστώθηκε αντίστροφη συσχέτιση με τη φλεγμονώδη δραστηριότητα στο ήπαρ 314. Αντίθετα, άλλοι ερευνητές διαπίστωσαν ότι δεν υπήρχε αύξηση του αριθμού των CD4+CD25+ high κυττάρων και μάλιστα ότι δεν υπήρχαν διαφορές στον αριθμό των CD4+CD25+ κυττάρων ανάμεσα σε υγιή άτομα της ομάδας ελέγχου και σε άτομα με χρόνια HCV λοίμωξη. Επιπλέον βρέθηκε ότι στα πειραματικά μοντέλα (χιμπαντζήδες) δεν υπήρχε καμία διαφορά στον αριθμό των Foxp3+CD4+CD25+ Tregs και στην ένταση της κατασταλτικής τους δράσης μεταξύ αποδραμούσας HCV λοίμωξης και χρόνιας HCV λοίμωξης 319, 320,
71 Όσον αφορά την συσχέτιση των ntregs (δηλαδή τα φυσικά αναδυόμενα Tregs τα οποία είναι CD4+CD25 high+ Foxp3+) με την δραστηριότητα της λοίμωξης, αναφέρεται αυξημένος αριθμός ntregs σε ασθενείς με χρόνια HCV λοίμωξη αλλά συγκεκριμένα στους ασθενείς που είχαν συνεχώς φυσιολογική αλανινική αμινοτρανσφεράση (ALT). Η κατασταλτική ικανότητα των ntregs ήταν πιο ισχυρή και τα κύτταρα αυτά εμφάνιζαν υψηλότερη έκφραση Foxp3 και CTLA-4, σε σχέση με τους ασθενείς που είχαν αυξημένη ALT. Επιπλέον, ο αριθμός των CD4+CD25-CD127-Foxp3+ κυττάρων ήταν μεγαλύτερος στους ασθενείς με αυξημένη ALT 322. Τα παραπάνω ευρήματα δείχνουν ότι η παρουσία των Tregs συσχετίζεται με την δραστηριότητα της χρόνιας HCV λοίμωξης και την βελτίωση της ιστικής βλάβης. Επιπλέον φαίνεται ότι υπάρχει κάποια συσχέτιση των Tregs και των CD8+ T κυττάρων. Έχει μάλιστα προταθεί ένα μοντέλο σύμφωνα με το οποίο στην οξεία HCV λοίμωξη, τα CD4+ T βοηθητικά κύτταρα συμβάλλουν στην επαγωγή και διατήρηση επαρκούς CD8+ T κυτταρικής απόκρισης έναντι του ιού. Αντίθετα στην χρόνια HCV λοίμωξη, τα Tregs καταστέλλουν τις ειδικές έναντι του ιού CD8+ T κυτταρικές αποκρίσεις, κι έτσι συμβάλλουν στην χρόνια παραμονή του ιού. Δεν έχει όμως διευκρινισθεί σε ποιο χρονικό σημείο μετά τη μόλυνση από τον ιό εμπλέκονται τα Tregs 69, 323. Α. Ο ρόλος των υποπληθυσμών των Τ ρυθμιστικών λεμφοκυττάρων στην HCV λοίμωξη 1. FoxP3+ ntregs Τα Foxp3+ ntregs έχουν συνδεθεί με τη διατήρηση της ομοιόστασης και την αυτοανοσία. Όμως, έχει αποδειχθεί ότι, με τον ανεργικό in vitro φαινότυπο τους, πολλαπλασιάζονται έντονα μετά την in vivo επαφή με συγγενή (cognate) μικροβιακά αντιγόνα. Μάλιστα, αυτά τα ntregs «διαμερισματοποιούνται» και περιορίζονται στη θέση που προκαλείται φλεγμονή από το παθογόνο και η επιβίωση τους εξαρτάται από την παρουσία αντιγόνων του παθογόνου στον ιστό 324. Όσον αφορά την χρόνια HCV λοίμωξη, η πρώτη μελέτη στην οποία απομονώθηκαν CD4+CD25+Tregs που εκφράζουν ενδοκυττάρια Foxp3 έγινε από τους Manigold και συνεργάτες και αφορούσε μελέτη σε χιμπαντζήδες. Τα κύτταρα αυτά είχαν ανεργικό φαινότυπο και κατέστειλαν τον πολλαπλασιασμό των CD4+CD25- T κυττάρων μετά 71
72 από διέγερση με μονοκλωνικό αντίσωμα έναντι του CD3. Οι ερευνητές ανέφεραν ότι τα Foxp3+CD4+CD25+ Τreg κύτταρα των πειραματόζωων με αποδραμούσα και χρόνια HCV λοίμωξη, διέφεραν από εκείνα των υγιών επειδή χαρακτηρίζονταν από αυξημένη ευαισθησία στην IL-2 και μικρότερο TREC, δεδομένα που υπονοούν ότι η HCV λοίμωξη τροποποιεί τον πληθυσμό των Tregs 319. Αργότερα διαπιστώθηκε ότι σε ασθενείς με χρόνια HCV λοίμωξη, τα Foxp3+ Tregs κύτταρα που απομονώθηκαν ήταν πανομοιότυπα με ntregs καθώς φαινοτυπικά ήταν CD45RO+ CD62L+ HLA-DR+CTLA- 4+CD127- (ήταν δηλαδή μνημονικά κύτταρα με ικανότητα να μεταναστεύουν στην περιφέρεια που εκφράζουν επιφανειακά μόρια χαρακτηριστικά του ρυθμιστικού φαινότυπου) και λειτουργικά ήταν ανεργικά χωρίς να εκφράζουν IL-10 ή TGF-β. Τα κύτταρα αυτά δεν διέφεραν από τα Foxp3+ Treg κύτταρα ατόμων χωρίς HCV λοίμωξη και ακόμη, οι κατασταλτικές τους ικανότητες ήταν πανομοιότυπες 325. Επιπλέον, διαπιστώθηκε και στο ηπατικό παρέγχυμα ασθενών με χρόνια HCV λοίμωξη η παρουσία CD4+CD25+Foxp3+ Tregs που μάλλον αντιπροσωπεύουν ntregs 317. Μετά τη διαπίστωση της παρουσίας των Foxp3+CD25+ T κυττάρων στη χρόνια HCV λοίμωξη, η επιγενετική ανάλυση επιβεβαίωσε ότι τα ειδικά έναντι του HCV Foxp3+CD25+ T κύτταρα είναι πραγματικά Tregs και όχι απλώς ενεργοποιημένα συμβατικά Τ κύτταρα που εκφράζουν Foxp3 λόγω της ενεργοποίησης τους από τον ιό καθώς τα κύτταρα αυτά εκφράζουν σταθερά φαινότυπο με Foxp3 και έχουν κατασταλτική λειτουργία. Μάλιστα, το προφίλ των γονιδίων που είναι ενεργοποιημένα και ως εκ τούτου τα μόρια που εκφράζουν τα ειδικά έναντι του HCV Foxp3+CD25+ T κύτταρα (όσον αφορά στον κυτταρικό θάνατο, την ενεργοποίηση και τον πολλαπλασιασμό, αλλά και τη ρύθμιση της μεταγραφής), τους δίνει σαφώς ένα πλεονέκτημα επιβίωσης σε σχέση με τα ειδικά έναντι του HCV δραστικά Τ κύτταρα 326. Τα ευρήματα αυτά, συνηγορούν υπέρ της άποψης ότι τα κύτταρα αυτά επηρεάζουν με κάποιο τρόπο την έκβαση της χρόνιας HCV λοίμωξης. Όπως προαναφέρθηκε, ο αριθμός των Tregs είναι αυξημένος στη χρόνια HCV λοίμωξη. Δεν είναι όμως εύκολο να διακριθεί κατά πόσο πρόκειται για ntregs ή για επαγόμενα itregs τα οποία δημιουργούνται στην περιφέρεια από τα συμβατικά CD4+ T κύτταρα μετά από έκθεση σε αντιγόνα του ιού. 72
73 2. Tr1 κύτταρα Η απομόνωση CD4+CD25+ T κυττάρων που παράγουν αυξημένη ποσότητα IL-10 από το αίμα ασθενών με χρόνια HCV λοίμωξη, θεωρήθηκε ενδεικτική ότι τα κύτταρα αυτά ήταν Tr1 κύτταρα 319. Τα Tr1 κύτταρα όμως βρίσκονταν συγκεντρωμένα κυρίως στο ήπαρ 308, 314, 327. Σημαντικό ήταν το γεγονός ότι τα Tr1 κύτταρα αναγνώριζαν τους ίδιους επίτοπους στην πρωτεΐνη του πυρήνα του ιού όπως και τα Th1 κύτταρα που παράγουν IFN-γ. Επιπλέον, σε μια άλλη μελέτη στην οποία διερευνήθηκε η κατασταλτική δράση των CD4+CD25+ Tregs ασθενών με χρόνια HCV λοίμωξη διαπιστώθηκε ότι όταν αδρανοποιήθηκε η IL-10, τα δραστικά Τ κύτταρα παρέμειναν κατεσταλμένα. Έτσι φαίνεται ότι η αναστολή της δράσης της IL-10 δεν εξουδετερώνει τα ειδικά έναντι του ιού CD4+CD25+ T κύτταρα καθώς αυτά συνεχίζουν να δρουν κατασταλτικά 312. Τα δεδομένα που υπάρχουν μέχρι σήμερα για τα Tr1 κύτταρα στην HCV λοίμωξη είναι ελλιπή και απαιτούνται περαιτέρω έρευνες για την αποσαφήνιση της δράσης αυτού του υποπληθυσμού. 3. Th3 κύτταρα Τα Τh3 κύτταρα φαίνεται ότι αποτελούν έναν υποπληθυσμό Tregs ο οποίος επάγεται κατά την HCV λοίμωξη και συμμετέχει στην εξέλιξη της χρόνιας νόσου 319. Μελέτες έδειξαν ότι οι Καυκάσιοι ασθενείς με χρόνια HCV λοίμωξη που υποβλήθηκαν σε θεραπεία με IFN-α, παρήγαγαν IL-10 και TGF-β σε ποσότητες κατώτερες του φυσιολογικού. Στην υπο-ανάλυση όμως, φάνηκε πως εκείνοι που απάντησαν στην αγωγή παρήγαγαν περισσότερο TGF-β, και λιγότερη IL-10 σε σχέση με τους μη απαντητές, γεγονός που συνηγορεί υπέρ της άποψης ότι η κλωνική επέκταση των Tregs αφορούσε κυρίως τον Th3 πληθυσμό. Τα ευρήματα αυτά δεν επιβεβαιώθηκαν στους ασθενείς με Αφρο-αμερικανική καταγωγή οι οποίοι είχαν και μεγαλύτερα ποσοστά μη απάντησης. Πάντως, είναι πιθανόν η επίδραση του ιού στην κλωνική επέκταση των Tregs και στην απάντηση στην αγωγή να επηρεάζεται και από ενδογενή χαρακτηριστικά της εθνολογικής καταγωγής των ασθενών, καθώς είναι γνωστό ότι οι Αφρο-αμερικανικής καταγωγής ασθενείς έχουν μικρότερα ποσοστά απάντησης στην θεραπεία με IFN-α
74 Στην ίδια μελέτη, διαπιστώθηκε ότι το mrna του TGF-β εκφραζόταν μόνο σε ένα μικρό ποσοστό των λευκών αιμοσφαιρίων του περιφερικού αίματος των ασθενών με χρόνια ηπατίτιδα C που δεν είχαν λάβει αγωγή και δεν διέφερε από εκείνο της ομάδας ελέγχου 329. Στο δεύτερο σκέλος αυτής της μελέτης, μετά τη θεραπεία με IFN α, διαπιστώθηκε αύξηση του αριθμού των Th3 κυττάρων που ήταν TGF-β +, εύρημα που φάνηκε να συσχετίζεται με τη μείωση των ηπατικών ενζύμων, αλλά όχι πλήρως με το ιικό φορτίο το οποίο μειώθηκε μόνο σε ένα ποσοστό 30% των ασθενών 329. Ίσως λοιπόν η θεραπεία με IFN-α να συμβάλλει στην κλωνική επέκταση του πληθυσμού των Th3 κυττάρων. Άλλοι ερευνητές ανέφεραν ότι στο 30% των ασθενών με χρόνια HCV λοίμωξη ανιχνεύονται κύτταρα CD4+ TGF-β +, ποσοστό που δεν επιβεβαιώνεται σε ασθενείς με οξεία HCV λοίμωξη 308. Οι Cabrera και συνεργάτες, ανέφεραν ότι διαπίστωσαν την παρουσία του Th3 υποπληθυσμού, καθώς τα CD4+CD25+ κύτταρα που απομόνωσαν και τα οποία είχαν φαινότυπο CD45RO high CD45RA low CD28 high CD62L high CD95 high μπορούσαν να εκκρίνουν TGF-β. Αυτά τα κύτταρα με χαρακτηριστικά μνημονικών ενεργοποιημένων κυττάρων με υψηλή ικανότητα μετανάστευσης, είχαν την ικανότητα να καταστέλλουν in vitro τον πολλαπλασιασμό των ειδικών έναντι του HCV CD4+ και CD8+ T δραστικών κυττάρων με δοσοεξαρτώμενο τρόπο και άμεση κυτταροκυτταρική επαφή 314. Επιπλέον, όταν χρησιμοποιήθηκαν εξουδετερωτικά anti-tgf-β αντισώματα διαπιστώθηκε ότι η απουσία του TGF-β είχε ανασταλτικό ρόλο στην κατασταλτική λειτουργία των CD4+CD25+ κυττάρων 314. Το εύρημα αυτό δεν επιβεβαιώθηκε από άλλες μελέτες 312. Επίσης, άλλοι ερευνητές ανέφεραν ότι τα ειδικά έναντι του HCV CD4+CD25+ high T κύτταρα παράγουν σταθερά TGF-β αλλά μόνο μικρές ποσότητες IL-10, και καθόλου IL-2 και IFN γ. Η έκκριση του TGF-β από τα ειδικά έναντι του HCV CD4+CD25+ high T κύτταρα (Th3) ήταν σημαντικά μεγαλύτερη στους ασθενείς με χρόνια ηπατίτιδα C και φυσιολογική ALT σε σχέση με τους ασθενείς με αυξημένη ALT. Επιπλέον, στην ίδια μελέτη, διαπιστώθηκε μια σημαντική αντίστροφη συσχέτιση μεταξύ της ειδικής έναντι του HCV, TGF-β απόκρισης των CD4+CD25+ high T κυττάρων και της φλεγμονής στο ήπαρ, δηλαδή όσο μεγαλύτερη ήταν η έκκριση του TGF-β τόσο μικρότερη ήταν η ηπατική φλεγμονή
75 4.CD4+CD25+Tregs από μετατροπή των CD4+CD25-T κυττάρων Όσον αφορά τα CD4+CD25+Tregs, in vitro μελέτες έδειξαν ότι ο πληθυσμός των CD4+CD25+Treg κυττάρων μπορεί να δημιουργηθεί μετά από την in vitro κλωνική επέκταση των CD4+CD25- T κυττάρων παρουσία πεπτιδίων του HCV 319. Τα επαγόμενα Tregs ήταν λειτουργικά καθώς καταδείκνυαν καταστολή της παραγωγής της IFN-γ, της επέκτασης των ειδικών έναντι του HCV Τ κυττάρων και κατέστειλαν και την απόπτωσή τους. Στην προαναφερθείσα μελέτη, τα Tregs προέρχονταν είτε από πειραματόζωα με χρόνια HCV λοίμωξη, είτε από πειραματόζωα που είχαν επιτύχει κάθαρση του ιού (αυτοπεριορισθείσα λοίμωξη). Ίσως λοιπόν τα Tregs να έχουν την δυνατότητα να διατηρούν την ικανότητα να ρυθμίζουν τις αποκρίσεις των μνημονικών Τ κυττάρων ακόμη και μετά την κάθαρση του ιού και την αποδρομή της HCV λοίμωξης. Επιπλέον, ο κλωνικά επαγόμενος πληθυσμός ήταν πιο ευαίσθητος στη δράση της IL-2 και είχε χαμηλότερο TREC, που σημαίνει ότι η HCV λοίμωξη επάγει σημαντικές αλλαγές στον πληθυσμό των Tregs. Τα χαρακτηριστικά αυτά αφορούσαν και τα Tregs σε αυτοπεριορισθείσα λοίμωξη, άρα φαίνεται ότι οι αλλαγές αυτές διατηρούνται επί μακρόν, ακόμη και μετά την κάθαρση του ιού CD8+ Treg κύτταρα που παράγουν IL-10 Η πιο κλασική μελέτη στην οποία διαπιστώθηκαν CD8+ T κυτταρικές σειρές που παράγουν IL-10 ως απόκριση σε διέγερση με αντιγόνα του HCV είναι εκείνη των Koziel και συνεργατών 331. Σε αυτή τη μελέτη, μετά τη διέγερση, διαπιστώθηκε κλωνική επέκταση των ειδικών έναντι του ιού CCR7-CD8+ Τ κύτταρα με ρυθμιστική δράση που εκκρίνουν IL-10 στο ήπαρ. Τα επαγόμενα κύτταρα είχαν την ικανότητα να καταστέλλουν in vitro τον πολλαπλασιασμό των ειδικών έναντι του HCV CD8+ T λεμφοκυττάρων που βρίσκονταν στον ηπατικό ιστό, και μάλιστα αντιγονο-ειδικά και με μηχανισμό που εξαρτάται από την IL Άλλοι ερευνητές απομόνωσαν από το περιφερικό αίμα ασθενών με HCV λοίμωξη, ειδικά έναντι του HCV CD3+CD8+CD25- κύτταρα με ρυθμιστική ικανότητα, τα οποία παράγουν IL-10 και TGF-β, αλλά η δράση τους αναστέλλεται κυρίως από το μπλοκάρισμα του TGF-β
76 Β. Στρατολόγηση των T ρυθμιστικών λεμφοκυττάρων στο ήπαρ Η στρατολόγηση των Tregs στο σημείο της βλάβης όπου υπάρχουν και τα δραστικά κύτταρα του ανοσιακού συστήματος, πιθανόν να επιτρέπει την εξέλιξη των ανοσιακών αποκρίσεων μέχρι εκείνο το σημείο στο οποίο οι αποκρίσεις τους να είναι μεν αρκετά ισχυρές για να υποστηρίξουν την άμυνα του ξενιστή, να εξασθενήσουν δηλαδή τις αποκρίσεις των Τ δραστικών κυττάρων και να βελτιώσουν την ηπατική φλεγμονή, αλλά χωρίς ταυτόχρονα να μπορούν να εξαλείψουν τον παθογόνο μικροοργανισμό, οπότε και συμβάλλουν τελικά στη μακροχρόνια εξέλιξη της νόσου και την εγκατάσταση της ίνωσης στο ήπαρ. Όπως διαπιστώθηκε, τα CD4+ και CD8+ Tregs που παράγουν IL-10 συγκεντρώνονται κυρίως στο ήπαρ 98, 317. Βέβαια, ήδη από το 2005 είχε πιθανολογηθεί ότι in vivo, τα ενεργοποιημένα Tregs συγκεντρώνονται και ασκούν την κατασταλτική τους δράση σε θέσεις στις οποίες υπάρχει και μεγάλο ιικό φορτίο, δηλαδή στο ήπαρ. Η στρατολόγηση αυτή δείχνει ότι η κατασταλτική δραστηριότητα επηρεάζει μόνο το διαμέρισμα εκείνο του οργανισμού το οποίο πρέπει να επηρεάσει, στην προκειμένη περίπτωση το ήπαρ, και επιτρέπει στις υπόλοιπες αποκρίσεις να εξελίσσονται ανεπηρέαστες. Επιπλέον, ερευνητές έδειξαν ότι σε βιοψίες ήπατος ασθενών με χρόνια ηπατίτιδα C διαπιστώθηκε ότι στις περιοχές στις οποίες είναι συγκεντρωμένα ειδικά έναντι του HCV CD8 + T κύτταρα που εκφράζουν IL-10 αλλά όχι IFN-γ, η νεκροφλεγμονώδης δραστηριότητα είναι μικρότερη ανεξάρτητα από την ηλικία, το φύλο, το ιικό φορτίο και το γονότυπο του ιού. Παρόμοια ήταν τα ευρήματα και όσον αφορά στην ίνωση, και στην απόπτωση. Αντίθετα, σε βιοψίες στις οποίες υπήρχε έντονη νεκροφλεγμονώδης δραστηριότητα, ίνωση και απόπτωση, τα CD8 T κύτταρα εξέφραζαν σε μεγαλύτερο ποσοστό IFN-γ και όχι IL-10, γεγονός το οποίο συνηγορεί υπέρ της άποψης ότι παρά το ότι τα CD8+ T κύτταρα που εκκρίνουν IL-10, δεν φαίνεται να είναι κυτταροτοξικά - αφού δεν επιτυγχάνουν τον έλεγχο του ιικού πολλαπλασιασμού-, εντούτοις επιτυγχάνουν μείωση της νεκροφλεγμονώδους δραστηριότητας και της ίνωσης και πιθανόν να είναι CD8+ Tregs 99. Εντούτοις, διαφορετικά ευρήματα διαπιστώνονται σε μια άλλη μελέτη σε βιοψίες ασθενών με χρόνια ηπατίτιδα C. Βρέθηκε ότι στα συναθροίσματα λεμφοκυττάρων στα πυλαία διαστήματα στο ήπαρ υπήρχε ιδιαίτερα μεγάλος αριθμός CD4+CD25+Tregs που εκφράζουν Foxp3, και μάλιστα με μια αναλογία 2:1 των συνολικών CD4+/FoxP3+ 76
77 κυττάρων που βρίσκονται μέσα στο ήπαρ. Τα Foxp3+ κύτταρα ήταν διεσπαρμένα και στα ηπατικά λόβια, εκεί όμως δεν υπήρχαν σε αθροίσματα (clusters) με άλλα λεμφοκύτταρα. Ο μέσος αριθμός των Foxp3+ κυττάρων και των CD4+ κυττάρων ανά διήθημα συσχετιζόταν ισχυρά με το βαθμό της πυλαίας φλεγμονής (P=0.0007), και σε μικρότερο βαθμό με το βαθμό της διαβρωτικής και της ενδολόβιας νέκρωσης (P=0.0208). Αντίθετα, δεν βρέθηκε σημαντική συσχέτιση του αριθμού των Foxp3+ κυττάρων με το στάδιο την ίνωσης. Επίσης, δεν συσχετίσθηκε το στάδιο της ίνωσης με το βαθμό της πυλαίας φλεγμονής. Ο αριθμός των CD8+ κυττάρων είχε ασθενή συσχέτιση με τον βαθμό της διαβρωτικής ή ενδολόβιας νέκρωσης (P=0.0507), αλλά είχε σημαντική συσχέτιση με τα επίπεδα της ALT και της AST στον ορό (P= και P= αντίστοιχα), δηλαδή η κυτταροτοξική δραστηριότητα αυτών των κυττάρων παίζει ρόλο στην βλάβη των ηπατικών κυττάρων. Επιπλέον η διαπίστωση της παρουσίας Foxp3+ κυττάρων με ταυτόχρονη απουσία αυξημένης έκφρασης IFN γ, επιβεβαίωσε ότι αυτά τα Foxp3+ κύτταρα ήταν στην πραγματικότητα ρυθμιστικά κύτταρα, και μάλιστα ίσως είναι ntregs. Όπως κατέληξαν οι συγγραφείς, η στρατηγική τοποθέτηση των Foxp3+ κυττάρων στο πυλαίο διάστημα τους δίνει το πλεονέκτημα της άμεσης πρόσβασης που απαιτείται προκειμένου να εμποδίσουν με κυτταροκυτταρική επαφή τον πολλαπλασιασμό των Τ δραστικών κυττάρων. Επίσης, το πυλαίο διάστημα είναι μια καίρια θέση, ώστε στα πρώιμα στάδια της μετανάστευσης και της εισόδου στο ήπαρ των ειδικών έναντι του HCV αλλά και των μη ειδικών ενεργοποιημένων λεμφοκυττάρων, να αυξάνει η πιθανότητα να συναντήσουν ένα Foxp3+ κύτταρο και να «υποκύψουν» στην καταστολή του 317. Επιπλέον διαπιστώθηκε ότι τα Tregs ναι μεν συγκεντρώνονται στο ήπαρ, αλλά η κλωνική τους επέκταση στον ιστό είναι σχετικά μικρότερη από εκείνη που παρατηρείται στον πληθυσμό των CD4+CD25+Foxp3- δραστικών T κυττάρων και όσο υψηλότερη ήταν η έκφραση του PD-1 στα Tregs τόσο μικρότερος ήταν ο πολλαπλασιασμός τους. Έτσι πιθανολογήθηκε ότι η έκφραση του PD-1 πιθανόν να έχει μεγαλύτερη αρνητική ρύθμιση στην κλωνική επέκταση των Tregs στο ήπαρ, παρά σε εκείνη των δραστικών κυττάρων. Το εύρημα αυτό επιβεβαιώθηκε όταν στα Tregs που απομονώθηκαν από το ήπαρ ασθενών με χρόνια HCV λοίμωξη διαπιστώθηκε in vitro αυξημένη κλωνική επέκταση που εξαρτώνταν από την παρουσία IL-2 και εντονότερη κατασταλτική δράση μετά από παρεμπόδιση της σύνδεσης του PD-1 και του PD-L1. Είναι ενδιαφέρον ότι η ενίσχυση της κατασταλτικής δράσης των Tregs 77
78 συσχετίσθηκε με αυξημένη έκφραση του Foxp3, αλλά και με κλινικές παραμέτρους όπως η μείωση του ιικού φορτιού και η μείωση του συνολικού βαθμού φλεγμονής στη βιοψία (HAI, High Activity Index). Συμπερασματικά φαίνεται ότι τα Tregs είναι πιο ευάλωτα στο ήπαρ επειδή το PD-1 μάλλον ελέγχει τον πολλαπλασιασμό τους, μέσω μείωσης της φωσφορυλίωσης του STAT-5 και ότι τα ενδοηπατικά Τ δραστικά κύτταρα είναι λιγότερο εκτεθειμένα στη στέρηση της IL-2 και άρα στην απόπτωση 333. Συμπερασματικά φαίνεται ότι ο πληθυσμός των Tregs είναι ένας ετερογενής πληθυσμός. Στην HCV λοίμωξη, φαίνεται ότι συνυπάρχουν ntregs και itregs, και δεν είναι εύκολο να διακριθεί το κατά πόσο ένα κύτταρο CD4+CD25+Foxp3+ προέρχεται από τα ntregs ή από τα ενεργοποιημένα έναντι του ιού CD4+CD25+ T κύτταρα. Γ. Μηχανισμοί με τους οποίους τα T ρυθμιστικά λεμφοκύτταρα ασκούν την κατασταλτική τους δράση στην HCV λοίμωξη Όσον αφορά τους μηχανισμούς δράσης των Tregs στη χρόνια HCV λοίμωξη, υπάρχουν περιορισμένα ερευνητικά δεδομένα είναι όμως σαφές ότι τα CD4+CD25+ Tregs στην χρόνια HCV λοίμωξη καταστέλλουν με ποικίλους μηχανισμούς τα ειδικά έναντι του ιού CD8+ και CD4+ κύτταρα δηλαδή καταστέλλουν τον πολλαπλασιασμό τους και την παραγωγή της IFN-γ. Όπως διαπιστώθηκε σε πολλές μελέτες, η κατασταλτική δράση των Tregs ασκούνταν με άμεση κυτταροκυτταρική επαφή και ήταν ανεξάρτητη από την παρουσία IL-10 και TGF-β. Όμως, η ένταση της καταστολής εξαρτώνταν από το μέγεθος του πληθυσμού των Tregs: όσο μεγαλύτερος ο πληθυσμός, τόσο εντονότερη η καταστολή (δοσοεξαρτώμενη) 312, 313, 314, 315, 316, 334. Παρόλα αυτά, άλλοι ερευνητές υποστηρίζουν ότι η κατασταλτική δράση των Tregs στην HCV λοίμωξη (μετρούμενη με την καταστολή της ειδικής έναντι του ιού έκκρισης IFN-γ) φαίνεται ότι ασκείται και έμμεσα μέσω της έκκρισης ανοσοκατασταλτικών κυτταροκινών και συγκεκριμένα κυρίως μέσω του TGF-β ή/ και της IL-10 και ότι στο περιφερικό αίμα ασθενών με χρόνια ηπατίτιδα C υπάρχουν αυξημένα επίπεδα της IL-10. Τα CD8+ Tregs που εκκρίνουν IL-10 συγκεντρώνονται στο ήπαρ και προφυλάσσουν από την ιστική βλάβη 99, 100, 330, 332. Με βάση τα ευρήματα των διαφόρων μελετών, φαίνεται ότι η IL-10, ανεξάρτητα από 78
79 τον είδος των κυττάρων που στοχεύει, συμβάλλει στην μείωση της ιστικής βλάβης, αλλά και στην παρεμπόδιση και στη μείωση της αποτελεσματικότητας των ανοσιακών αποκρίσεων έναντι του ιού. Συμπερασματικά, φαίνεται ότι στην χρόνια HCV λοίμωξη τα Tregs διαθέτουν πολλούς μηχανισμούς, άμεσους και έμμεσους, για να ασκήσουν την κατασταλτική τους δράση. Το αν υπερισχύει κάποιος από αυτούς ή εάν συνεργάζονται όλοι μαζί σε άλλοτε άλλο βαθμό μένει να διευκρινισθεί. Δ. Ειδικότητα των Tregs έναντι αντιγόνων του ιού της ηπατίτιδας C Έχει ήδη αναφερθεί ότι εξ ορισμού τα itregs παρουσιάζουν αντιγονοειδικότητα, ενώ τα ntregs θεωρείται ότι αναγνωρίζουν ίδια αντιγόνα καθώς αναπτύσσονται και «επιλέγονται» στο θύμο για τη διατήρηση της περιφερικής ανοχής. Όμως, έχει διαπιστωθεί ότι τα ntregs αν και στην οξεία φάση των λοιμώξεων αναγνωρίζουν ίδια αντιγόνα που απελευθερώνονται λόγω της ιστικής βλάβης, στις χρόνιες λοιμώξεις έχουν την ικανότητα να αναγνωρίζουν μικροβιακά αντιγόνα. Η πρώτη πληροφορία για Τ ρυθμιστικά κύτταρα ειδικά έναντι αντιγόνων του ιού της ηπατίτιδας C MHC τάξης Ι που είχαν την ικανότητα να ελαττώνουν τις αποκρίσεις έναντι του ιού έγινε το , και αφορούσε CD8+ T κύτταρα που παρήγαγαν IL-10, δηλαδή Tr1 κύτταρα. Η περαιτέρω επιβεβαίωση της παρατήρησης αυτής τεκμηριώθηκε μετά την απομόνωση αντιγονο-ειδικών έναντι του HCV CD8+ Tregs 98 στο ήπαρ. Επιπλέον, τα CD8+ Foxp3- Tregs από το ήπαρ και το περιφερικό αίμα ασθενών με HCV λοίμωξη που παρήγαν TGF-β ήταν επίσης ειδικά έναντι του HCV 106, 315, 332, 335. Ήδη από το 2000 είχε αναφερθεί ότι η αντιγονο-ειδικότητα διαδραματίζει κάποιο ρόλο στην κάθαρση του ιού, καθώς διαπιστώθηκε πως η κυτταροτοξική απάντηση έναντι του HCV σε ασθενείς που παρουσίασαν αυτοπεριοριζόμενη C λοίμωξη, κατευθυνόταν έναντι μεγαλύτερου αριθμού αντιγονικών επιτόπων του ιού, σε σχέση με ασθενείς στους οποίους η λοίμωξη μετέπεσε σε χρονιότητα 28. Σε μια από τις κλασικές in vitro μελέτες σε ασθενείς με αποδραμούσα C λοίμωξη αξιολογήθηκε η αντιγονο-ειδικότητα της κατασταλτικής δράσης των Tregs παρουσία διαφόρων πεπτιδίων του HCV. Διαπιστώθηκε ότι υπήρχαν έντονα ανοσογόνες περιοχές (immunogenic) στην πρωτεΐνη του πυρήνα και την NS3 πρωτεΐνη και μάλιστα ότι η ειδική έναντι του HCV 79
80 T κυτταρική απόκριση στόχευε ενάντια σε 28 από τις 36 αντιγονικές περιοχές του ιού που χρησιμοποιήθηκαν (78%). Στους ασθενείς με χρόνια HCV λοίμωξη βρέθηκε, ότι η T κυτταρική απόκριση έναντι του ιού ήταν ασθενέστερη και το αντιγονικό της φάσμα περίπου 4 φορές μικρότερο, αλλά εξακολουθούσε να αφορά τις ίδιες 8 περιοχές του γονιδιώματος του ιού που ήταν οι πιο ισχυρά ανοσογενείς. Στη μελέτη αυτή διαπιστώθηκε αυξημένος αριθμός λειτουργικών CD4+CD25+ Tregs: 7,3% στη χρόνια HCV λοίμωξη και 2,5% στην αποδραμούσα, γεγονός το οποίο ενισχύει την άποψη του μηχανισμού καταστολής των δραστικών κυττάρων. Η καταστολή των ειδικών έναντι του ιού αποκρίσεων αφορούσε στον αριθμό και τη λειτουργικότητα των δραστικών κυττάρων, αλλά όχι την ειδικότητα τους έναντι αντιγόνων του ιού 313. Η μη αντιγονο-ειδική κλωνική επέκταση των Tregs επιβεβαιώθηκε και από άλλες in vitro μελέτες, στις οποίες διαπιστώθηκε καταστολή των αποκρίσεις και έναντι άλλων ιών όπως του ιού της γρίπης και του HSV 312, 335. Όμως, υπάρχουν κάποια νεότερα δεδομένα όσον αφορά το αντιγονικό ερέθισμα το οποίο διεγείρει τα Tregs και συμβάλλει στην κλωνική τους επέκταση από μια μελέτη στην οποία χρησιμοποιήθηκαν πεπτίδια που προέρχονται από πρωτεΐνες του HCV σε καλλιέργεια PBMC HCV θετικών ασθενών. Σε αυτή τη μελέτη βρέθηκε ότι τα CD4+CD25 +high FoxP3 high Tregs επάγονται ταχέως (εύρημα που δεν αναπαραγόταν στα υγιή άτομα) και μάλιστα διευκρινίσθηκε ότι το πεπτίδιο στόχος του HCV που μπορούσε να διεγείρει τα Tregs ήταν διαφορετικό μεταξύ των ασθενών. Όμως, σε κάθε άτομο της μελέτης, μόνο ένας μικρός αριθμός πεπτιδίων ήταν ικανός να διεγείρει τα Tregs της καλλιέργειας, γεγονός που υποδηλώνει ότι ίσως υπάρχουν κυρίαρχοι επίτοποι των πεπτιδίων που αναγνωρίζουν τα Tregs προκειμένου να επεκταθούν κλωνικά 336. Στη μελέτη αυτή δεν διερευνήθηκε εάν η κατασταλτική δράση επεκτείνονταν και έναντι αντιγόνων άλλων ιών. Ε. Συνδιεγερτικά μόρια και T ρυθμιστικά λεμφοκύτταρα στην HCV λοίμωξη Η σκιαγράφηση του ρόλου των συνδιεγερτικών μορίων στη λειτουργία των Treg κυττάρων κατά τη διάρκεια της HCV λοίμωξης παραμένει αντικείμενο έρευνας. Έχει ήδη αναφερθεί ότι υπάρχουν κάποιες ενδείξεις ότι το σύστημα PD-1 είναι δυνατόν, τουλάχιστον εν μέρει, να είναι υπεύθυνο τη συμπεριφορά 80
81 των Tregs στη χρόνια HCV λοίμωξη, αν και δεν έχει ξεκαθαρίσει με ποιο τρόπο επηρεάζει την ενεργοποίηση της STAT-5 οδού 333. Επίσης, τα συνδιεγερτικά μόρια αποτελούν ένα από τους τρόπους που διαθέτουν τα Tregs για να «ελέγχουν» και άλλα κύτταρα και ειδικότερα τα δενδριτικά κύτταρα προς όφελός τους. Έχει βρεθεί ότι τα δενδριτικά κύτταρα στους ασθενείς με χρόνια ηπατίτιδα C δεν διαφέρουν από εκείνα των υγιών ατόμων ούτε σε φαινότυπο ούτε σε λειτουργία, όμως δεν μπορούν να ολοκληρώσουν τη διαδικασία διέγερσης και ωρίμανσης. Υπέρ αυτού του ευρήματος είναι το γεγονός ότι τα δενδριτικά κύτταρα μετά την απάντηση στην αντιιική θεραπεία, και την κάθαρση του HCV, έχουν φυσιολογική διέγερση και ωρίμανση 67, 68. ΣΤ. TLRs και T ρυθμιστικά λεμφοκύτταρα στην HCV λοίμωξη Οι TLRs (Toll Like Receptors) είναι οι κυτταρικοί ενεργοποιητές της φυσικής ανοσίας. Οι TLR υποδοχείς, εκτός από μικρόβια, αναγνωρίζουν και ιούς και έχει βρεθεί ότι τα ntregs επηρεάζονται άμεσα ή έμμεσα από αυτούς με αποτέλεσμα να αυξάνεται η έκφραση του Foxp3 και να ενισχύεται η κατασταλτική τους δράση 288, 337. Όσον αφορά τον ρόλο τους στην HCV λοίμωξη, μελέτες έδειξαν ότι η in vitro επανειλημμένη παρουσίαση ενός λιποπεπτιδίου του HCV από δενδριτικά κύτταρα στα CD4+ T κύτταρα, μέσω TLR2, ευοδώνει τη δράση των τελευταίων, καθώς διευκολύνει τον πολλαπλασιασμό τους και την έκκριση IFN-γ, αλλά δεν ισχύει το ίδιο και για τα CD8+ Τ κύτταρα 338. Εντούτοις δεν υπάρχουν πολλά δεδομένα για την χρόνια HCV λοίμωξη στον άνθρωπο σχετικά με τον ρόλο των TLRs στην επιβίωση και την κλωνική επέκταση των Tregs. Ζ. T ρυθμιστικά λεμφοκύτταρα και καρκινογένεση Ο ηπατοκυτταρικός καρκίνος θεωρείται ο πέμπτος συχνότερος καρκίνος παγκόσμια, με την ηπατίτιδα C να είναι μια από τις πιο συχνές αιτίες του. Καθώς έχει αποδειχθεί σε πειραματικά μοντέλα ότι τα Tregs μπορούν να καταστείλουν τις ειδικές αποκρίσεις έναντι καρκινικών όγκων, ήταν αναμενόμενο ότι ένα κομμάτι της έρευνας για τα Tregs θα στρεφόταν και προς αυτή την κατεύθυνση 339. Σε μια μελέτη στην οποία έγινε σύγκριση των αριθμών των Tregs σε άτομα με ΗΚΚ, ιογενή ηπατίτιδα και υγιείς διαπιστώθηκε ότι στο περιφερικό αίμα ασθενών με ΗΚΚ υπήρχε ο μεγαλύτερος αριθμός 81
82 Foxp3+CD4+CD25+ Tregs σε σχέση με τις τρεις ομάδες της μελέτης, και μάλιστα με φαινότυπο HLA-DR high GITR high CD45RO low. Επιπλέον, επιβεβαιώθηκε ότι τα κύτταρα αυτά καταδείκνυαν λειτουργικό ρυθμιστικό φαινότυπο, καθώς παρήγαν IL-10, ήταν ανεργικά σε διέγερση του TCR, και είχαν την ικανότητα να καταστείλουν τον πολλαπλασιασμό και την παραγωγή κυτταροκινών από κύτταρα CD4+CD25-, πιθανόν με άμεση κυτταροκυτταρική επαφή αφού η αναστολή της IL-10 δεν ανέστειλε την κατασταλτική τους δράση. Αυξημένος βρέθηκε και ο αριθμός των Tregs και στη νεοπλασματική μάζα αλλά και στο νεοπλασματικό ασκίτη, αλλά δεν φάνηκε να συσχετίζεται με το στάδιο της νεοπλασίας 321. Βέβαια η προαναφερθείσα μελέτη δεν διερεύνησε τη δράση του TGF-β ο οποίος όπως είναι γνωστό εμπλέκεται ενεργά στις αποκρίσεις έναντι καρκινικών όγκων καθώς αναστέλλει τις δράσεις των Τ δραστικών κυττάρων (έκφραση IFN-γ, περφορίνης, εξωκύττωση κοκκίων και πρόκληση κυτταρόλυσης) και επιπλέον, χρησιμοποιείται από τα Tregs στους έμμεσους μηχανισμούς άσκησης καταστολής, αλλά και για την κλωνική τους επέκταση και την επιβίωση τους 340, 341. Ενδιαφέρον προκαλεί το εύρημα μελέτης στην οποία διαπιστώθηκε ότι παρουσία ενεργοποιημένων Tregs, τα κυτταροτοξικά δραστικά κύτταρα σκοτώνουν 6,6 φορές πιο αργά τα κύτταρα-στόχο 342. Πρόσφατα, οι Kobayashi και συνεργάτες 343 ανέφεραν ότι ο αριθμός των Foxp3+ κυττάρων είναι μεγαλύτερος σε όζους ηπατοκυτταρικού καρκίνου σε σχέση με τους αναγεννητικούς όζους του κιρρωτικού ήπατος. Στην ίδια μελέτη διαπιστώθηκε ότι όσο πιο προχωρημένη ήταν η νεοπλασία, τόσο μεγαλύτερος ήταν ο αριθμός των Foxp3+ Tregs και τόσο μικρότερος ο αριθμός των CD8+ T κυττάρων. Μάλιστα ο αριθμός των FoxP3+ Tregs στο ΗΚΚ ήταν ανεξάρτητος προγνωστικός παράγοντας επιβίωσης, γεγονός το οποίο δείχνει ότι όσο μεγαλύτερος ήταν τόσο μικρότερο ήταν το ποσοστό επιβίωσης των ασθενών 344. Βέβαια, δεν είναι εύκολο να διευκρινισθεί εάν ο εμπλουτισμός με Treg είναι αποτέλεσμα του HCV ή της ανάπτυξης της νεοπλασίας, είναι όμως καλά τεκμηριωμένο ότι ηπατίτιδα C και καρκίνος συνδέονται παθογενετικά. Ο μηχανισμός με τον οποίο τα Tregs επιτυγχάνουν την καταστολή των ειδικών έναντι των καρκινικών κυττάρων αποκρίσεων, όπως προαναφέρθηκε πιθανόν να είναι η άμεση κυτταροκυτταρική επαφή ή έμμεσοι μηχανισμοί όπως η έκκριση κατασταλτικών κυτταροκινών. Υπάρχουν επιπλέον κάποια πειραματικά δεδομένα τα οποία περιγράφουν υψηλή έκφραση granzyme B στα CD4+Foxp3+ Tregs στο μικροπεριβάλλον του όγκου, και μάλιστα σε ποσοστό 5-30% 82
83 αυτών. Αυτά τα Tregs είχαν την ικανότητα να θανατώσουν NK κύτταρα και CD8+ T κύτταρα μέσω granzyme B και περφορίνης 340. Συμπερασματικά, φαίνεται πως τα T ρυθμιστικά λεμφοκύτταρα διαδραματίζουν σημαντικό ρόλο στον έλεγχο της χρόνιας λοίμωξης από τον ιό της ηπατίτιδας C και της εξέλιξής της (νεκροφλεγμονή και ίνωση στο ηπατικό παρέγχυμα, ηπατοκυτταρικό καρκίνωμα) αλλά υπάρχουν περιορισμένα ερευνητικά δεδομένα στη σημερινή βιβλιογραφία όσον αφορά στη συμμετοχή τους στην απάντηση στη θεραπεία και τη συμπεριφορά του υποπληθυσμού τους στους διαφορετικούς γονότυπους του ιού. Ως εκ τούτου, το παρόν ερευνητικό πρωτόκολλο σχεδιάστηκε με σκοπό να διερευνηθεί ο ρόλος του υποπληθυσμού των CD4+CD25+Foxp3+ T ρυθμιστικών κυττάρων σε ασθενείς με χρόνια ηπατίτιδα C και γονότυπο 1-4 που δεν είχαν προηγουμένως υποβληθεί σε θεραπεία με πεγκυλιωμένη ιντερφερόνη και ριμπαβιρίνη. Η μελέτη αφορούσε τέσσερις χρόνους, με τον πρώτο πριν την έναρξη της θεραπείας και τους υπόλοιπους κατά τη διάρκεια της, το τέλος της και τον αναμενόμενο χρόνο επίτευξης μακροχρόνιας ιολογικής απάντησης προκειμένου να διερευνηθεί ο ρόλος τους στην ανοσοπαθογένεια και την εξέλιξη της HCV λοίμωξης. 83
84
85 ΕΙΔΙΚΟ MΕΡΟΣ 85
86
87 Ι. ΥΛΙΚΟ Συνολικά μελετήθηκαν 61 άτομα με χρόνια ηπατίτιδα C, 59% των ασθενών ήταν άνδρες και 41% ήταν γυναίκες. Ο μέσος όρος ηλικίας των ασθενών ήταν τα 34 έτη. Από τους άνδρες, 56% περίπου ήταν ηλικίας 21-30, 33% μεταξύ 31 και 40 ετών και 11% είχε ηλικία μεγαλύτερη των 41 ετών. Ο μέσος όρος της ηλικίας των ανδρών που συμμετείχαν ήταν τα 31,5 έτη (γράφημα 1). Από τις γυναίκες, 40% ήταν ηλικίας 21-30, 20% μεταξύ 31 και 40 ετών και 40% είχε ηλικία μεγαλύτερη των 41 ετών. Ο μέσος όρος της ηλικίας των γυναικών που συμμετείχαν στην μελέτη ήταν τα 37,5 έτη (γράφημα 2). Γράφημα 1: Ηλικιακή κατανομή των ανδρών ασθενών της μελέτης Γράφημα 2: Ηλικιακή κατανομή των γυναικών ασθενών της μελέτης Όσον αφορά τον γονότυπο του ιού της ηπατίτιδας C, 39.3% των ασθενών είχε γονότυπο 1 (24/61), 4.9% γονότυπο 2 (3/61), 47.6% γονότυπο 3 (29/61) και 8.2% γονότυπο 4 (5/61) (γράφημα 3). 87
88 Γράφημα 3: Κατανομή των γονοτύπων στους ασθενείς της μελέτης Όλοι οι ασθενείς, ανεξάρτητα από τον τρόπο μόλυνσης από τον ιό (πίνακας 1), προέρχονταν από το Ηπατολογικό Τμήμα της Β Παθολογικής Κλινικής του ΑΠΘ. Οι ασθενείς ενημερώθηκαν για το ερευνητικό πρωτόκολλο και έδωσαν τη συγκατάθεση για τη συμμετοχή τους στη μελέτη. Ενδοφλέβια χρήση ναρκωτικών ουσιών 46/61 Μετάγγιση αίματος ή παραγώγων 5/61 Γυάλινες σύριγγες 1/61 Πιθανή ενδοοικογενειακή μετάδοση 1/61 Άγνωστος 8/61 Πίνακας 1: Τρόπος μόλυνσης των ασθενών της μελέτης από τον ιό της ηπατίτιδας C Κανένας από τους ασθενείς που συμμετείχε στη μελέτη δεν είχε λάβει στο παρελθόν θεραπεία για τη χρόνια ηπατίτιδα C. Όλοι οι ασθενείς υποβλήθηκαν σε θεραπεία με το καθιερωμένο σχήμα συνδυασμένης αγωγής με πεγκυλιωμένη ιντερφερόνη και ριμπαβιρίνη 43, 345,
89 Γονότυπος Διάρκεια Πεγκυλιωμένη αγωγής ιντερφερόνη 1, 4 48 εβδ α2α 180 μg/ εβδ Ή α2β 1.5 μg/kg/ εβδ Ριμπαβιρίνη 1000 mg/ ημέρα για ΣΒ<75 kg 1200 mg/ ημέρα για ΣΒ>75 kg 800 mg/ ημέρα για ΣΒ<65 kg 1000 mg/ ημέρα για ΣΒ kg 1200 mg/ ημέρα για ΣΒ>85 kg 2, 3 24 εβδ α2α 180 μg/ εβδ 800 mg/ ημέρα Ή α2β 1.5 μg/kg/ εβδ Πίνακας 2: Δόσεις πεγκυλιωμένης ιντερφερόνης και ριμπαβιρίνης ανάλογα με το γονότυπο του HCV Στο τέλος της θεραπείας, 59% (36/61) των ασθενών ανταποκρίθηκε στην αγωγή και πέτυχε SVR, 6.6% (4/61) δεν ανταποκρίθηκε. 34.4% των ασθενών διέκοψε την αγωγή (πίνακας 3) και ειδικότερα 8.2% διέκοψε λόγω παρενεργειών (5/61), ενώ 26.2% (16/61) δεν ολοκλήρωσε τη θεραπεία συνήθως λόγω υποτροπής στη χρήση ενδοφλεβίων ουσιών. Οι άντρες εμφάνισαν σε μεγαλύτερο ποσοστό SVR και διέκοψαν την θεραπεία λόγω παρενεργειών σε μικρότερο ποσοστό (γράφημα 3, πίνακας 4). Ωστόσο, μεγαλύτερος αριθμός ανδρών δεν ολοκλήρωσε την ενδεδειγμένη με βάση το γονότυπο διάρκεια της θεραπείας. Υποτροπή στη χρήση ενδοφλέβιων ναρκωτικών ουσιών 16/61 Ψωρίαση 1/61 Σοβαρού βαθμού αναιμία παρά τη χορήγηση ερυθροποιητίνης 1/61 Αυτοάνοση αιμολυτική αναιμία 1/61 Ιολογική διαφυγή κατά τη διάρκεια της θεραπείας 1/61 Αλλεργικό εξάνθημα 1/61 Πίνακας 3: Αιτίες διακοπής της θεραπείας στους ασθενείς της μελέτης 89
90 Γράφημα 3: Απεικόνιση της ανταπόκρισης των ασθενών της μελέτης με βάση το φύλο Τελική ανταπόκριση μη διακοπή λόγω ανταπόκριση ανταπόκριση παρενεργειών drop out Σύνολο Φύλο Άνδρας Μέτρηση % μέσα στο Φύλο 2,8% 61,1% 5,6% 30,6% 100,0% Γυναίκα Μέτρηση % μέσα στο Φύλο 12,0% 56,0% 12,0% 20,0% 100,0% Σύνολο Μέτρηση % μέσα στο Φύλο 6,6% 59,0% 8,2% 26,2% 100,0% Πίνακας 4: Ανταπόκριση των ασθενών της μελέτης με βάση το φύλο 90
91 Τα κριτήρια ένταξης στη μελέτη ήταν: 1. Θετικό αντι-hcv για >6 μήνες 2. HCV RNA (PCR) ποιοτικό θετικό με ευαίσθητη ποιοτική μέθοδο (Combas Amplicor, Roche Diagnostics, κατώτερο όριο ανίχνευσης 50 IU/ml) 3. Φυσιολογική ή αυξημένη ALT 4. Απουσία άλλης χρόνιας λοίμωξης 5. Απουσία ατομικού ή οικογενειακού ιστορικού αυτοάνοσης νόσου ή αλλεργίας 6. Απουσία συλλοίμωξης με άλλους ηπατοτρόπους ιούς (HBV, HDV) 7. Απουσία θεραπείας για τη χρόνια HCV λοίμωξη στο παρελθόν Κριτήρια αποκλεισμού 1. Άτομα <18 ετών 2. Συνυπάρχουσα σοβαρή καρδιαγγειακή πάθηση ή αιμοσφαιρινοπάθεια ή αυτοάνοσης πάθησης 3. Κύηση 4. Νέοι ενήλικες απρόθυμοι να λάβουν επαρκή αντισυλληπτικά μέτρα Το ερευνητικό πρωτόκολλο διενεργήθηκε στο Ηπατολογικό Τμήμα και στο εργαστήριο Ανοσολογίας και Ηπατολογίας του Ηπατολογικού Τμήματος, στο εργαστήριο του Τμήματος Κλινικής Ανοσολογίας της Β Παθολογικής Κλινικής και στο Εργαστήριο Παθολογικής Ανατομικής του Αριστοτελείου Πανεπιστημίου Θεσσαλονίκης. Σε βιοψία ήπατος υποβλήθηκαν 59 από τους 61ασθενείς της μελέτης με σκοπό να εκτιμηθούν α) η νεκροφλεγμονώδης δραστηριότητα της νόσου (HAI, Histological Activity Index), η οποία αξιολογήθηκε με το τροποποιημένο σύστημα Knodell 15 και β) η ίνωση, η οποία αξιολογήθηκε με τα συστήματα Metavir και Scheuer. (Πίνακας 5) Τροποποιημένο σύστημα Knodell HAI (Histological Activity Index) Ίνωση Διαβρωτική νέκρωση: , χωρίς Πυλαία φλεγμονή: 0-4 1, ελάχιστη Ενδολόβιες νεκρώσεις: 0-4 2, μέτρια ΤΟ ΑΘΡΟΙΣΜΑ ΤΩΝ ΑΝΩΤΕΡΩ ΣΤΟΙΧΕΙΩΝ 3, σημαντική ΑΞΙΟΛΟΓΕΙ ΤΗ ΦΛΕΓΜΟΝΗ, ΕΤΣΙ ΩΣΤΕ: 4, κίρρωση 0: χωρίς φλεγμονή 1-4: ελάχιστη 5-8: ήπια 9-12: μέτρια 13-18: σημαντική Πίνακας 5: Αξιολόγηση βιοψίας ήπατος με βάση το τροποποιημένο σύστημα Knodell 91
 στους ασθενείς της μελέτης.")
92 Τα χαρακτηριστικά της ιστολογικής βλάβης των ασθενών της μελέτης φαίνονται στα γραφήματα 4, 5, 6, 7, Γράφημα 4: Κατανομή της βαρύτητας του δείκτη νεκροφλεγμονώδους δραστηριότητας (ΗΑΙ) στους ασθενείς της μελέτης. Γράφημα 5: Κατανομή της βαρύτητας της διαβρωτικής νέκρωσης στους ασθενείς της μελέτης 1 Οι πλήρεις πίνακες με τα δεδομένα των μεταβλητών της μελέτης στα τέσσερα χρονικά στιγμιότυπα παρατίθενται στο Παράρτημα της διατριβής. 92

93 Γράφημα 6: Κατανομή της βαρύτητας της πυλαίας φλεγμονής στους ασθενείς της μελέτης Γράφημα 7: Κατανομή της βαρύτητας των ενδολόβιων νεκρώσεων στους ασθενείς της μελέτης 93
 αρχικά με ποιοτική μέθοδο -επειδή έχει χαμηλότερο όριο ανίχνευσης του ιικού φορτίου- και ακολούθως εάν αυτό")
94 Γράφημα 8: Κατανομή της βαρύτητας της ίνωσης στους ασθενείς της μελέτης Οι ασθενείς μελετήθηκαν σε τέσσερα στιγμιότυπα κατά τη διάρκεια της μελέτης. Σε κάθε στιγμιότυπο, οι ασθενείς υποβάλλονταν σε νέα μέτρηση της ALT και του HCV RNA (PCR) αρχικά με ποιοτική μέθοδο -επειδή έχει χαμηλότερο όριο ανίχνευσης του ιικού φορτίου- και ακολούθως εάν αυτό δεν ήταν μη ανιχνεύσιμο, με ποσοτική μέθοδο. Συγκεκριμένα: Α. Τ 0 : Πριν τη θεραπεία Β. Τ 1 : σε επιλεγμένο χρονικό στιγμιότυπο κατά τη διάρκεια της θεραπείας αναλόγως του γονότυπου, δηλαδή, -Για το γονότυπο 1 και 4: 12 η εβδομάδα, καθώς εάν ο ασθενής υπό θεραπεία επιτυγχάνει EVR (πρώιμη ιολογική απάντηση), δηλαδή έχει αρνητικό HCV RNA τη 12 η εβδομάδα της αγωγής, τότε το PPV (positive predictive value, θετική προγνωστική αξία) για την επίτευξη SVR (μακροχρόνιας ιολογικής απάντησης) είναι 60% -Για το γονότυπο 2 και 3: 4 η εβδομάδα, καθώς εάν ο ασθενής υπό θεραπεία επιτυγχάνει RVR (ταχεία ιολογική απάντηση), δηλαδή έχει HCV RNA αρνητικό την 4 η εβδομάδα αγωγής, τότε το PPV για επίτευξη SVR είναι 95% Γ. Τ 2 : στο τέλος της θεραπείας (μετά από 48 εβδομάδες για τους ασθενείς με γονότυπο 1,4 και μετά από 24 εβδομάδες για τους ασθενείς με γονότυπο 2,3) Δ. Τ 3 : 6 μήνες μετά το τέλος της θεραπείας 94
95 ΙΙ. ΜΕΘΟΔΟΙ 1. Μοριακή διάγνωση της χρόνιας ηπατίτιδας C Προσδιορισμός του γονότυπου του ιού (INNOLIPA test, BAYER Diagnostics) HCV RNA (PCR) ποιοτικό με ευαίσθητη ποιοτική μέθοδο (Combas Amplicor, Roche Diagnostics, κατώτερο όριο ανίχνευσης 50 IU/ml) HCV RNA (PCR) με ευαίσθητη ποσοτική μέτρηση (Amplicor Monitor, Roche Diagnostics, όριο ανίχνευσης <35 IU/ml) 2. Μέτρηση της ALT του ορού (αλανινική αμινοτρανσφεράση) 3. Κυτταρομετρία ροής Σε όλα τα άτομα της μελέτης, στα στιγμιότυπα της μελέτης, πραγματοποιήθηκε μέτρηση: της εκατοστιαίας αναλογίας των CD3+, CD4+ και CD8+ T λεμφοκυτταρικών υποπληθυσμών της εκατοστιαίας αναλογίας του υποδοχέα της IL-2 (IL-R2a, CD25) στον πληθυσμό των λεμφοκυττάρων, καθώς και της εκατοστιαίας συνέκφρασης CD4 και CD25 στα Τ λεμφοκύτταρα (CD4+/CD25+ διπλά θετικά, double positive) της εκατοστιαίας έκφρασης του συνδιεγερτικού μορίου CD28 στα T λεμφοκύτταρα, καθώς και της εκατοστιαίας συνέκφρασης CD4 και CD28 στα T λεμφοκύτταρα (CD4+/CD28+ διπλά θετικά, double positive) της εκατοστιαίας έκφρασης του ενδοκυττάριου μεταγραφικού παράγοντα Foxp3 στα διεγερμένα CD4+CD25+ high T λεμφοκύτταρα που αντιστοιχεί στον πληθυσμό των T ρυθμιστικών λεμφοκυττάρων (CD4+/ CD25+ high /FOXP3+) Όλες οι μετρήσεις πραγματοποιήθηκαν σε δύο διαφορετικές πειραματικές συνθήκες: α) ολικό περιφερικό αίμα (ex vivo) και β) μετά από καλλιέργεια των περιφερικών μονοπύρηνων κυττάρων (λεμφοκύτταρα, μακροφάγα) με το μιτογόνο φυτοαιμοσυγκολλητίνη (PHA). Για τις μετρήσεις των υποπληθυσμών και την έκφρασης μορίων (επιφανειακών/ μεμβρανικών και ενδοκυττάριων) χρησιμοποιήθηκε η τεχνολογία της κυτταρομετρίας ροής (Fluorescence Activated Cell Sorter, FACS). Η κυτταρομετρία ροής είναι μια μέθοδος η οποία επιτρέπει την ανίχνευση ενδοκυττάριων και εξωκυττάριων υποδοχέων ενός κυττάρου αφού προηγουμένως το κύτταρο έχει σημανθεί ενδο- ή εξω- 95
96 κυττάρια με μονοκλωνικά αντισώματα τα οποία είναι συζευγμένα με μια φθορίζουσα χρωστική. Τα κύτταρα ρέοντας σε μονήρη στήλη ισότονου διαλύματος, περνούν μέσα από σύστημα ακτίνων laser όπου, λόγω της σύνδεσής τους με φθορίζουσες χρωστικές, εκπέμπουν φθορίζουσα ακτινοβολία η οποία αναλύεται από το λογισμικό του ηλεκτρονικού υπολογιστή του κυτταρομετρητή ροής 347, 348, 349. Η κυτταρομετρία ροής επιτυγχάνει να διακρίνει αρχικά τα κύτταρα σε υποπληθυσμούς, με βάση φυσικά χαρακτηριστικά, όπως είναι το μέγεθος και η κοκκίωση τους (π.χ. πολυμορφοπύρηνα, λεμφοκύτταρα ή μονοπύρηνα). Ο διαχωρισμός των υποπληθυσμών των λευκοκυττάρων γίνεται με την ανάλυση της έντασης της αντανακλώσας ακτινοβολίας σε οριζόντιο (forward scatter, fs) και πλάγιο άξονα (90 o, side scatter, ss). Οι παράμετροι FS Και SS, δίνουν αντίστοιχα πληροφορίες για το μέγεθος και την εσωτερική δομή των κυττάρων (κοκκίωση), επιτυγχάνοντας τη διάκριση σε υποπληθυσμούς (λεμφοκύτταρα, μονοπύρηνα μακροφάγα και πολυμορφοπύρηνα). Η ανάλυση των παραμέτρων FS και SS αποδίδεται οπτικά με τα σημειακά γραφήματα κυτταροδιαγράμματος (dot plots). Η χρήση μονοκλωνικών αντισωμάτων επιτρέπει σύμφωνα με τη φθορίζουσα ακτινοβολία την αναγνώριση συγκεκριμένων αντιγόνων (CD ή άλλων μορίων) στην επιφάνεια/ μεμβράνη και το εσωτερικό των κυτταρικών υποπληθυσμών. Η φθορίζουσα ακτινοβολία των μονοκλωνικών αντισωμάτων που χρησιμοποιήθηκαν διακρίνεται σε: α) πράσινο φθορισμό με κορυφή εκπομπής τα 525 nm, ο οποίος δημιουργείται λόγω της σύνδεσης των μονοκλωνικών αντισωμάτων που έχουν επισημανθεί με ισοθειοκυανικό οξύ φλουορεσκεΐνης (Fluorescein isothiocyanate, FITC) με τα κύτταρα του δείγματος που εξετάζεται, β) πορτοκαλί φθορισμό με κορυφή εκπομπής τα 575 nm, ο οποίος δημιουργείται από τη σύνδεση των μονοκλωνικών αντισωμάτων που έχουν επισημανθεί με φυκοερυθρίνη (Phycoerythrin, PE) με τα κύτταρα του δείγματος που εξετάζεται, γ) κόκκινο φθορισμό με κορυφή εκπομπής τα 613 nm, ο οποίος εκπέμπεται από τη σύνδεση των μονοκλωνικών αντισωμάτων που έχουν επισημανθεί με φυκοερυθρίνη-texas-red -Χ (ECD) με τα κύτταρα του δείγματος που εξετάζεται. Η φθορίζουσα ακτινοβολία που ανιχνεύεται από τον κυτταρομετρητή ροής εξαρτάται από την πυκνότητα της έκφρασης του επιφανειακού αντιγόνου ή του ενδοκυττάριου μορίου. Η επεξεργασία της φθορίζουσας ακτινοβολίας παράγει τρία σήματα, το FL1 (πράσινος φθορισμός ή Forward Light 1, FITC), το FL2 (πορτοκαλί φθορισμός, 96

97 Forward Light 2, PE) και το FL3 (κόκκινος φθορισμός, Forward Light 3, ECD) τα οποία αποδίδονται οπτικά ως ιστογράμματα φθορισμού ή σημειακά γραφήματα. Η κυτταρομετρία ροής έχει διάφορες κλινικές εφαρμογές μια από τις οποίες είναι ο προσδιορισμός του ανοσοφαινότυπου, δηλαδή ο προσδιορισμός του είδους και του σταδίου διαφοροποίησης των κυττάρων με βάση τον προσδιορισμό συνδυασμού αντιγόνων στην επιφάνεια και το εσωτερικό τους. Η κυτταρομετρία ροής επιλέχθηκε για τις μετρήσεις της παρούσας μελέτης επειδή επιτυγχάνει να αναλύσει μεγάλο αριθμό κυττάρων και ταυτόχρονα να διακρίνει πολλούς υποπληθυσμούς κυττάρων. Επιπλέον, χαρακτηρίζεται από μεγάλη ταχύτητα μέτρησης (έως κύτταρα/ sec) και εξαιρετική ευαισθησία ανάλυσης (έως και 600 μόρια/ κύτταρο) σε πολύ μικρό μέγεθος δείγματος (100 μl). Για τις μετρήσεις της μελέτης χρησιμοποιήθηκε ο κυτταρομετρητής ροής EPICS XL-MCL (Beckman Coulter) του εργαστηρίου του Τμήματος Κλινικής Ανοσολογίας της Β Παθολογικής Κλινικής ο οποίος διαθέτει τη δυνατότητα τριπλού φθορισμού (Σχήμα 9). Σχήμα 9: Σχηματική απεικόνιση του τρόπου λειτουργίας του κυτταρομετρητή ροής 97
98 Σε κάθε στιγμιότυπο της μελέτης, γινόταν λήψη τριών δειγμάτων φλεβικού αίματος ανά ασθενή: α) 2 ml σε φιαλίδιο με αντιπηκτικό EDTA (ethylene-diamine-tetraacetic acid) για την ανάλυση του αιμοδιαγράμματος του ασθενή, β) 2 ml σε φιαλίδιο με αντιπηκτικό EDTA για τον άμεσο προσδιορισμό της εκατοστιαίας έκφρασης των μορίων CD3, CD4, CD8, CD25, Foxp3 ex vivo και γ) 20 ml σε αποστειρωμένο κωνικό φιαλίδιο με άμεση προσθήκη ηπαρίνης (15 IU/ ml φλεβικού αίματος) για την in vitro καλλιέργεια. Σκοπός της μέτρησης των Tregs σε καλλιέργειες λεμφοκυττάρων ήταν η έμμεση καταγραφή της λειτουργικότητας τους καθώς μετρούνταν ποσοτικά (με νέα κυτταρομετρία ροής) η δυνατότητα έκπτυξης του συγκεκριμένου πληθυσμού μετά από μη ειδική διέγερση (PHA). Κατόπιν αναζητήθηκε πιθανός συσχετισμός των ποσοτικών μετρήσεων των «επαγώμενων» Tregs με τις υπόλοιπες παραμέτρους του πρωτοκόλλου. Προετοιμασία περιφερικού αίματος Πραγματοποιούνταν λήψη 2ml φλεβικού αίματος σε αποστειρωμένο φιαλίδιο που περιείχε 2mg/ml αίματος του αντιπηκτικού EDTA. Η ιδανική σήμανση με τα μονοκλωνικά αντισώματα, απαιτεί /μl λευκοκύτταρα στο περιφερικό αίμα. Δείγματα με αριθμό λευκοκυττάρων >10000/μl αραιώνονταν με PBS (Phosphate Buffered Saline, Gibco), ενώ τα δείγματα με αριθμό λευκοκυττάρων <3000/ μl φυγοκεντρούνταν και επαναιωρούνταν, ώστε τελικά να επιτευχθούν τιμές λευκοκυττάρων μεταξύ / μl. Κατόπιν γινόταν επεξεργασία του δείγματος με 2 διαφορετικά πειραματικά πρωτόκολλα. Το πρώτο πειραματικό πρωτόκολλο αφορούσε την επεξεργασία για τη σήμανση με μονοκλωνικά αντισώματα των επιφανειακών/ μεμβρανικών υποδοχέων των λεμφοκυττάρων, ενώ το δεύτερο για τη σήμανση συνδυασμού επιφανειακών υποδοχέων και του ενδοκυττάριου FoxP3 παράγοντα στα Τ λεμφοκύτταρα. Σύμφωνα με το πρώτο πρωτόκολλο σήμανσης των επιφανειακών/ μεμβρανικών υποδοχέων, αναμιγνύονταν 100 μl αίματος, με 20 μl σεσημασμένου με FITC και 20 μl σεσημασμένου με PE μονοκλωνικού αντισώματος, σε πλαστικό σωληνάριο Falcon των 5 ml. Ακολουθούσε επώαση του δείγματος σε θερμοκρασία δωματίου (20-25 ο C), σε σκοτεινό περιβάλλον, για 15 λεπτά. Ακολούθως, γινόταν επεξεργασία του δείγματος με το λυτικό σύστημα αντιδραστηρίων Immunoprep (Beckman Coulter). To Immunoprep αποτελείται από τρία ξεχωριστά διαλύματα: Α, Β και C (πίνακας 6). 98
99 Διάλυμα Α (600 μ l) Διάλυμα Β (265 μl) Διάλυμα C (100 μl) Μυρμηκικό οξύ 1,2 Ανθρακικό οξύ Παραφορμαλδεΰδη 10 μl/ml 6 mg/ml mg/ml Χλωριούχο νάτριο 14,5 mg/ml Θειϊκό νάτριο 31,3 mg/ml Πίνακας 6: Τα διαλύματα που αποτελούν το λυτικό σύστημα αντιδραστηρίων Immunoprep Το διάλυμα Α προκαλεί λύση των ερυθρών αιμοσφαιρίων του δείγματος, το Β ρυθμίζει το ph του δείγματος (buffer) και τερματίζει τη λυτική διαδικασία, ενώ το C μονιμοποιεί τα μονοκλωνικά αντισώματα στην κυτταρική μεμβράνη του λευκοκυττάρου (fixation). Η προσθήκη των διαλυμάτων του Immunoprep, σε κάθε σωληνάριο δείγματος, γινόταν με αυτοματοποιημένη συσκευή (Q-PREP, Beckman Coulter). Συγκεκριμένα, σταθερή ποσότητα από κάθε διάλυμα προστίθετο κατά διαδοχικό τρόπο στο δείγμα (600 μl A 265 μl B 100 μl C), με ενδιάμεσες αναδεύσεις, σε κύκλο συνολικής διάρκειας 35 δευτερολέπτων. Με αυτό τον τρόπο εξασφαλίζονταν, τόσο η μικρότερη δυνατή καταστροφή, απώλεια και επιμόλυνση των κυττάρων του δείγματος, όσο και η επαναληψιμότητα στην προετοιμασία, αφού παρακάμπτονταν οι επαναλαμβανόμενες φυγοκεντρήσεις και ανασυστάσεις που απαιτούν οι κλασσικές μέθοδοι επεξεργασίας με τα λυτικά και μονιμοποιητικά διαλύματα (όπως το χλωριούχο αμμώνιο και η παραφορμαλδεϋδη 1%). Μετά την επεξεργασία με Immunoprep, το δείγμα αναλυόταν στον κυτταρομετρητή ροής είτε άμεσα, είτε εντός 24 ωρών, αφού είχε αποθηκευτεί σε σκοτεινό περιβάλλον στους 6-8 ο C. Για την προετοιμασία του δείγματος όπου θα μετρούνταν τα Tregs, εφαρμόζονταν το δεύτερο πρωτόκολλο για τη σήμανση των επιφανειακών/ μεμβρανικών υποδοχέων και του ενδοκυττάριου παράγοντα Foxp3. Αναμιγνύονταν 50 μl αίματος, με 20 μl σεσημασμένου με FITC και 20 μl σεσημασμένου με ECD μονοκλωνικού αντισώματος, σε πλαστικό σωληνάριο Falcon των 5 ml. Ακολουθούσε επώαση του δείγματος σε θερμοκρασία δωματίου (20-25 ο C), σε σκοτεινό περιβάλλον, για 15 λεπτά. Κατόπιν το δείγμα αναμιγνύονταν με 100 μl φορμαλδεΰδης (formaldehyde) που περιέχονταν στο αντιδραστήριο 1 του συστήματος αντιδραστηρίων επαγωγής διαπερατότητας λευκοκυττάρων Intraprep TM της εταιρίας Beckman coulter. Ακολουθούσε ισχυρή ανάδευση (vortex) και επώαση του δείγματος σε θερμοκρασία δωματίου (20-25 ο C), σε σκοτεινό περιβάλλον, για 15 λεπτά. Στη συνέχεια το δείγμα αραιώνονταν με 5 ml διαλύματος PBS, φυγοκεντρούνταν στις 2000 rpm για 5 λεπτά και αφαιρούνταν το υπερκείμενο, έτσι ώστε στο φιαλίδιο τελικά διατηρούνταν κύτταρα με ακέραιες και σταθεροποιημένες από τη δράση της φορμαλδεΰδης κυτταρικές μεμβράνες. Στη συνέχεια το δείγμα αναμιγνύονταν με 100 μl του αντιδραστηρίου 2 του συστήματος 99
100 Intraprep TM το οποίο περιείχε σαπονίνη (saponine) και επωάζονταν σε θερμοκρασία δωματίου (20-25 ο C), σε σκοτεινό περιβάλλον, για 5 λεπτά. Το αντιδραστήριο 2 προκαλεί αύξηση της διαπερατότητας της κυτταρικής μεμβράνης των κυττάρων του διαλύματος και λύση των ερυθρών αιμοσφαιρίων. Μετά την επώαση, το δείγμα αναμιγνύονταν με 5 μl του μονοκλωνικού αντισώματος σεσημασμένου με PE έναντι του ενδοκυττάριου μεταγραφικού παράγοντα Foxp3. Ακολουθούσε επώαση σε θερμοκρασία δωματίου (20-25 ο C), σε σκοτεινό περιβάλλον, για 15 λεπτά. Έπειτα το δείγμα επαναιωρούνταν σε 5 ml διαλύματος PBS και φυγοκεντρούνταν στις 2000 rpm για 5 λεπτά ώστε να απομακρυνθεί η περίσσεια των λυμένων ερυθρών αιμοσφαιρίων και των διαλυμάτων του συστήματος Intraprep TM. Αφαιρούνταν το υπερκείμενο και το δείγμα κυττάρων επαναιωρούνταν σε 1 ml διαλύματος PBS, προκειμένου να αναλυθεί στον κυτταρομετρητή ροής. Τα μονοκλωνικά αντισώματα που χρησιμοποιήθηκαν στη μελέτη φαίνονται στον πίνακα 7 που ακολουθεί: CD Κλώνος Φθοριόχρωμα Οίκος CD3 UCHT1 FITC Immunotech CD4 13B8.2 FITC Immunotech CD8 B9.11 PE Immunotech CD25 B PE Immunotech CD28 CD28.2 PE Immunotech CD25 B ECD Immunotech Foxp3 PCH101 PE e-bioscience Πίνακας 7: Μονοκλωνικά αντισώματα που χρησιμοποιήθηκαν στη μελέτη Απομόνωση λεμφοκυττάρων από το περιφερικό αίμα Πραγματοποιούταν λήψη 20 ml φλεβικού αίματος σε αποστειρωμένο φιαλίδιο που περιείχε ηπαρίνη (15 IU/ ml αίματος). H απομόνωση των λεμφοκυττάρων από το περιφερικό αίμα γινόταν μετά από διαχωρισμό με φυγοκέντρηση, σε βαθμίδωση ανύσματος ειδικού βάρους φικόλλης (Ficoll-Hypaque). Χρησιμοποιήθηκε η φικόλλη Histopaque (gradient d=1,077 g/ml, Sigma). Η διαδικασία πραγματοποιούνταν σε θάλαμο βιολογικής ασφάλειας (Laminar air flow hood) και είχε ως εξής: 1. Επιστίβαση 5 ml ολικού αίματος σε 5 ml φικόλλης σε κωνικό σωληνάριο Falcon των 15 ml 2. Φυγοκέντρηση σε 1500 rpm επί 30 λεπτά, σε θερμοκρασία δωματίου. 3. Συλλογή της ενδιάμεσης στιβάδας των λευκοκυττάρων και μεταφορά σε νέο σωληνάριο. 4. Προσθήκη 8 ml διαλύματος RPMI 1640 (Gibco). 5. Φυγοκέντρηση του δείγματος μετά από ήπια ανακίνηση στα 1500 rpm επί 5 λεπτά. 6. Απόρριψη του υπερκείμενου και επανάληψη της φυγοκέντρησης. 7. Επαναιώρηση του ιζήματος των μονοπυρήνων σε 1 ml διαλύματος RPMI
101 8. Μέτρηση των κυττάρων σε μικροσκόπιο με τη χρήση πλάκας Neubauer και δοκιμασία βιωσιμότητας των κυττάρων με τη μέθοδο αποκλεισμού με τη χρωστική Trypan blue (Sigma) (McAteer και Davis 1996). Αποδεκτές θεωρήθηκαν τιμές βιωσιμότητας > 85%. 9. Επαναιώρηση του ιζήματος των λευκοκυττάρων με καλλιεργητικό μέσο, ανάλογα με τον αριθμό των κυττάρων, σε τελική αραίωση 1 x 106 κύτταρα/ ml. Το καλλιεργητικό μέσο αποτελούνταν από διάλυμα RPMI 1640, εμπλουτισμένο με 10% ορό εμβρύου βοός (Foetal Calf Serum, Gibco), 2 mmol/l L-γλουταμίνη (Sigma), 100 IU/ ml πενικιλλίνη (Sigma) και 100 μg/ ml στρεπτομυκίνη (Sigma). Μιτογόνο και in vitro καλλιέργειες περιφερικών λεμφοκυττάρων Για τη μη ειδική διέγερση των T λεμφοκυττάρων επιλέχθηκε ένα κλασσικό ισχυρό μιτογόνο, η λεκτίνη φυτοαιμοσυγκολλητίνη (PHA, Seromed) 350 καθώς έχει βρεθεί ότι μετά τη διέγερση των T λεμφοκυττάρων με μιτογόνα όπως η PHA, η IL-2 η οποία είναι αυξητικός παράγοντας για τα κύτταρα αυτά, συντίθεται αρκετά γρήγορα (μέσα σε 4-12 ώρες) και μάλιστα συντίθεται κυρίως από τα CD4+ και λιγότερο από τα CD8+ Τ λεμφοκύτταρα Επίσης, η έκφραση του υποδοχέα της IL-2 (IL-2R) στα ανθρώπινα Τ-λεμφοκύτταρα που διεγείρονται με PHA, ακολουθεί άμεσα την παραγωγή της IL-2, συνεχίζεται τις πρώτες 2-3 ημέρες μετά τη διέγερση του κυττάρου, ενώ ύστερα από 6-10 ημέρες δεν ανιχνεύεται πλέον. Η βραχεία διάρκεια δράσης της IL-2 και η παροδική φύση της έκφρασης του IL-2R βοηθά τον αυτοπεριορισμό του πολλαπλασιασμού των Τ-λεμφοκυττάρων 352, 355. Στο τελικό λοιπόν εναιώρημα απομόνωσης περιφερικών λεμφοκυττάρων που περιγράφηκε παραπάνω (1 x 106 κύτταρα/ ml), προστίθετο PHA σε αναλογία 5 μg/ ml και οι καλλιέργειες με PHA επωάζονταν για 48 ώρες. Προετοιμασία δειγμάτων λεμφοκυττάρων in vitro καλλιέργειας Μετά από 48 ώρες επώασης στον ειδικό κλίβανο, η καλλιεργητική πλάκα μεταφερόταν στο θάλαμο βιολογικής ασφάλειας. Το υπερκείμενο των βοθρίων μεταφέρονταν σε σωληνάριο Falcon το οποίο φυγοκεντρούνταν και το υπερκείμενο φυλάσσονταν σε βαθειά κατάψυξη. Τα κύτταρα από τον πυθμένα των βοθρίων αποκολλούνταν προσεκτικά, με κρύο RPMI 1640 και μεταφέρονταν σε σωληνάριο Falcon. Στη συνέχεια, προστίθετο 5 ml RPMI 1640 και το δείγμα φυγοκεντρούνταν στις 1500 rpm για 5 λεπτά. Ακολουθούσε απόρριψη του υπερκείμενου και επαναιώρηση του ιζήματος των κυττάρων σε 1 ml διαλύματος RPMI Στη συνέχεια, γινόταν μέτρηση των κυττάρων σε μικροσκόπιο με τη χρήση πλάκας Neubauer και δοκιμασία βιωσιμότητας με τη μέθοδο αποκλεισμού με τη χρωστική Trypan blue. Αποδεκτές για τη μελέτη ήταν τιμές βιωσιμότητας > 85% και ιδανική συγκέντρωση θεωρούνταν τα 3-5 x 10 6 λευκοκύτταρα/ ml. Κατόπιν, σε 100 μl από το εναιώρημα γινόταν 101
 έγινε χρησιμοποιώντας τη συσχέτιση FS (μέγεθος κυττάρου) και SS (κυτταροπλασματική κοκκίωση) (εικόνα 9α).")
102 επεξεργασία σύμφωνα με το πρωτόκολλο σήμανσης των επιφανειακών και ενδοκυττάριων υποδοχέων, όπως περιγράφηκε παραπάνω και ακολουθούσε άμεσα η ανάλυση στον κυτταρομετρητή ροής. Ανάλυση με κυτταρομετρία ροής Η περίφραξη των λεμφοκυττάρων (gating) έγινε χρησιμοποιώντας τη συσχέτιση FS (μέγεθος κυττάρου) και SS (κυτταροπλασματική κοκκίωση) (εικόνα 9α). Στη συνέχεια, για τον έλεγχο της καθαρότητας της περίφραξης των λεμφοκυττάρων, χρησιμοποιήθηκε συνδυασμός μονοκλωνικών αντισωμάτων έναντι των λεμφοκυττάρων (CD45-FITC). Τα λεμφοκύτταρα ήταν CD45 εντόνως θετικά ενώ παρουσίαζαν χαμηλές τιμές FS και SS (εικόνα 9β). Η καθαρότητα του λεμφοκυτταρικού παραθύρου ήταν >97% και ο αριθμός των κυττάρων τουλάχιστον Η ανάλυση των δειγμάτων του ολικού αίματος και των διεγερμένων μετά από καλλιέργεια λεμφοκυττάρων πραγματοποιήθηκε με πρωτόκολλο κυτταρομετρίας ροής τριπλού φθορισμού. Για τη βαθμονόμηση (calibration) του κυτταρομετρητή χρησιμοποιούνταν πρότυπο (standard) σφαιριδίων φθορισμού (Flow-Check Fluorospheres, Beckman Coulter), ενώ για την εξάλειψη της αλληλοεπικάλυψης των φθοριοχρωμάτων γινόταν αντιστάθμιση FITC με PE (FL2-%FL1) και PE με FITC (FL1-%FL2) με ειδικά σφαιρίδια φθορισμού (Cyto-Comp 1, Beckman Coulter). Στις μετρήσεις που έγιναν τόσο στο περιφερικό αίμα όσο και στο εναιώρημα των καλλιεργειών, χρησιμοποιούνταν ισοτυπικό δείγμα ελέγχου (control), προκειμένου να εκτιμηθεί η μη ειδική σύνδεση των μονοκλωνικών αντισωμάτων και να τεθεί το «κατώφλι» (cut-off) μεταξύ θετικά και αρνητικά σημασμένων πληθυσμών. To ισοτυπικό δείγμα ελέγχου ορίσθηκε στο 1,5-2% (θετικός ουδός, cut-off). Εικόνα 1: Σημειακά γραφήματα (dot plots) περιφερικού αίματος. (α) περίφραξη του λεμφοκυτταρικού πληθυσμού με FS/SS συσχέτιση (β) λεμφοκυτταρική περίφραξη με συσχέτιση SS/CD45-FITC 102
, τα ιστογράμματα φθορισμού")
103 Τα ζεύγη των μονοκλωνικών αντισωμάτων που χρησιμοποιήθηκαν για τις μετρήσεις στον πληθυσμό των λεμφοκυττάρων ήταν τα εξής: Σωληνάριο 1: IgG1-FITC/IgG1-PE (ισοτυπικό δείγμα ελέγχου, control) Σωληνάριο 2: CD3-FITC/CD8-PE Σωληνάριο 3: CD4-FITC/CD25-PE Σωληνάριο 4: CD4-FITC/CD28-PE Σωληνάριο 5: CD4-FITC/CD25-ECD/FoxP3-PE Τα σημειακά γραφήματα κυτταροδιαγράμματος (dot plots), τα ιστογράμματα φθορισμού καθώς και τα περιγράμματα διάκρισης των θετικών και αρνητικών υποπληθυσμών (two colour contour plots) επεξεργάσθηκαν με το λογισμικό System II (Beckman Coulter) (εικόνες 2, α-ε και 3, α-δ) Εικόνα 2α: Ισοτυπικό δείγμα ελέγχου (IgG1-FITC/IgG1-PE) λεμφοκυττάρων περιφερικού αίματος, σε ασθενή με HCV λοίμωξη 103

104 CD3+ CD8+ CD3+/CD8+ Εικόνα 2β: Μέτρηση των CD3+ και CD8+ υποπληθυσμών του περιφερικού αίματος, στον ίδιο ασθενή 104

105 CD4+ CD25+ CD4+/CD25+ Εικόνα 2γ: Μέτρηση των CD4+ κυττάρων και της CD25 έκφρασης, στα λεμφοκύτταρα του περιφερικού αίματος, στον παραπάνω ασθενή 105
106 CD4+ CD28+ CD4+/CD28+ Εικόνα 2δ: Μέτρηση των CD4+ κυττάρων και της CD28 έκφρασης, στα λεμφοκύτταρα του περιφερικού αίματος, στον ίδιο ασθενή 106
, στα CD4+ λεμφοκύτταρα του περιφερικού αίματος, στον παραπάνω ασθενή")
107 CD4+ CD4+/CD25+ high FoxP3+ FoxP3+ FoxP3+ Εικόνα 2ε: Μέτρηση των Τ ρυθμιστικών λεμφοκυττάρων (CD4+/CD25+ high /FoxP3+), στα CD4+ λεμφοκύτταρα του περιφερικού αίματος, στον παραπάνω ασθενή 107
108 CD3+ CD8+ CD3+/CD8+ Εικόνα 3α: Μέτρηση των CD3+ και CD8+ υποπληθυσμών, μετά από καλλιέργεια με PHA, σε ασθενή με HCV λοίμωξη 108
109 CD4+ CD25+ CD4+/CD25+ Εικόνα 3β: Μέτρηση των CD4+ κυττάρων και της CD25 έκφρασης, στα λεμφοκύτταρα μετά από καλλιέργεια με PHA, στον ίδιο ασθενή 109
110 CD4+ CD28+ CD4+/CD28+ Εικόνα 3γ: Μέτρηση των CD4+ κυττάρων και της CD28 έκφρασης, στα λεμφοκύτταρα μετά από καλλιέργεια με PHA, στον παραπάνω ασθενή 110
111 CD4+/CD25+ high CD4+ FoxP3+ FoxP3+ FoxP3+ Εικόνα 3δ: Μέτρηση των Τ ρυθμιστικών λεμφοκυττάρων (CD4+/CD25+ high /FoxP3+), στα CD4+ λεμφοκύτταρα περιφερικού αίματος μετά από καλλιέργεια με PHA, στον ίδιο ασθενή 111
112
113 ΙΙΙ. ΣΤΑΤΙΣΤΙΚΗ ΜΕΘΟΔΟΛΟΓΙΑ Τα δεδομένα των πειραμάτων καταχωρήθηκαν αρχικά σε Η/Υ με τη χρήση του πακέτου επεξεργασίας φύλλων εργασίας Excel. Στη συνέχεια, εισήχθησαν στο στατιστικό πακέτο SPSS (έκδοση 18.0). Αρχικά πραγματοποιήθηκε ο έλεγχος κανονικότητας, προκειμένου να διαπιστωθεί η αναγκαιότητα της χρήσης παραμετρικών ή μη παραμετρικών στατιστικών αναλύσεων. Μετά τον έλεγχο της κανονικότητας, στην παρούσα μελέτη η πλειονότητα των μεταβλητών πλησιάζει την κανονική κατανομή σε μεγαλύτερο ή μικρότερο βαθμό. Η χρήση μη παραμετρικών στατιστικών μεθόδων προτιμήθηκε εφόσον είναι δυνατόν να χρησιμοποιηθούν τόσο σε κανονικούς όσο και σε μη κανονικούς πληθυσμούς. Συγκεκριμένα, για την περιγραφική ανάλυση του δείγματος χρησιμοποιήθηκαν οι μέσοι όροι, οι τυπικές αποκλίσεις, οι συχνότητες και τα ποσοστά εμφάνισης, αλλά και οι Πινακοειδείς Διασταυρώσεις (Cross Tabs). Ο έλεγχος των συσχετίσεων μεταξύ των μεταβλητών έγινε με το συντελεστή συσχέτισης του Spearman (Spearman Correlation Coefficient), ενώ ο έλεγχος για τις στατιστικά σημαντικές διαφορές μεταξύ δύο ανεξάρτητων δειγμάτων έγινε με τη χρήση του Mann Whitney U test (ή U test), το οποίο ελέγχει για τυχαίες μείξεις ταξινομήσεων μεταξύ των δύο δειγμάτων. Εξαίρεση στην υιοθέτηση της μη παραμετρικής στατιστικής αποτέλεσαν τα student s t για ζευγαρωτά δείγματα (paired samples student s t-test), τα οποία χρησιμοποιήθηκαν για τη σύγκριση των εξεταζόμενων μεταβλητών στα 4 χρονικά στιγμιότυπα της μελέτης. Ανεξαρτήτως στατιστικής μεθόδου, το όριο που θεωρήθηκε ως αποδεκτό επίπεδο στατιστικής σημαντικότητας ήταν το 0,05 (p<0,05). Οι γραφικές μέθοδοι που χρησιμοποιήθηκαν για την περιγραφή των μετρήσεων ήταν διάφορες: α) γραφήματα τύπου πίτας (pie charts). Σε αυτά, κάθε κομμάτι αντιπροσωπεύει και μία κατηγορία/απάντηση. Το μέγεθος του κάθε κομματιού είναι ανάλογο της συχνότητας εμφάνισης της κατηγορίας/απάντησης. β) γραφήματα ομαδοποιημένων στηλών (clustered bar charts). Τα συγκεκριμένα γραφήματα χρησιμεύουν για τη σύγκριση της συχνότητας εμφάνισης μίας κατηγορίας/απάντησης σε επιμέρους ομάδες του δείγματος. γ) γραφήματα απλών ή πολλαπλών γραμμών (simple και multiple line charts). Τα γραφήματα των απλών γραμμών χρησιμοποιήθηκαν για την καταγραφή της διαχρονικής εξέλιξης των εξεταζόμενων μεταβλητών για το σύνολο των συμμετεχόντων. Τα γραφήματα των πολλαπλών γραμμών χρησιμοποιήθηκαν για την αναπαράσταση της διαχρονικής εξέλιξης των εξεταζόμενων μεταβλητών για το σύνολο των 113
114 συμμετεχόντων, αλλά και ξεχωριστά για εκείνους με γονότυπο 1, με βάση το αν ανταποκρίθηκαν ή όχι στην αγωγή. δ) θηκογράμματα (box plots). Στα γραφήματα αυτά η έντονη μαύρη γραμμή, μέσα στα ορθογώνια πλαίσια εκφράζει τη διάμεσο τιμή (median) του δείγματος, ενώ τα άκρα των γραμμών που εκτείνονται από αυτά είναι οι μέγιστες και οι ελάχιστες τιμές. Οι μικροί κύκλοι, που παρουσιάζονται ορισμένες φορές εκτός των ορθογώνιων σχεδίων, αντιπροσωπεύουν υπερβολικά μεγάλες ή υπερβολικά μικρές μετρήσεις (outliers). Στην παρούσα μελέτη χρησιμοποιήθηκαν για την αναπαράσταση διαφορών μεταξύ των ανδρών και των γυναικών αλλά και μεταξύ των ασθενών με γονότυπο 1 και των ασθενών με γονότυπο 2 και
115 ΙV. ΑΠΟΤΕΛΕΣΜΑΤΑ Συνολικά αποτλέσματα στα τέσσερα στιγμιότυπα της μελέτης Τα συνολικά αποτελέσματα στα τέσσερα στιγμιότυπα της μελέτης (στο σύνολο των ασθενών, ανεξάρτητα από τον γονότυπο), όπως αυτά αναλύθηκαν με το student s t για ζευγαρωτά δείγματα (paired samples student s t-test), έδειξαν ότι: 1.HCV RNA Διαπιστώθηκε ότι η ποσοτική μέτρηση του αρχικού ιικού φορτίου των ασθενών, πριν από την έναρξη της θεραπείας (Τ 0 ), παρουσίασε στατιστικά σημαντικές διαφορές σε σύγκριση με όλα τα επόμενα στιγμιότυπα της μελέτης. Δηλαδή, διαπιστώθηκε μείωση του HCV RNA σε όλα τα στιγμιότυπα σε σχέση με το αρχικό. Η μεγαλύτερη μείωση στο μέσο όρο του ιικού φορτίου διαπιστώθηκε μεταξύ της τιμής του πριν την έναρξη της θεραπείας και στο τέλος της (Τ 0 - Τ 2 ), δηλαδή ήταν σημαντικά χαμηλότερο στο τέλος της θεραπείας (t=3,33, p<0,01). Η μικρότερη μείωση διαπιστώθηκε μεταξύ της τιμής του πριν την έναρξη της θεραπείας (Τ 0 ) και του πρώτου στιγμιότυπου κατά τη θεραπεία (Τ 1 w4/ w12) (t=3,85, p<0,001) (πίνακας 8, γράφημα 9). 2.ALT Οι τιμές της ALT έδειξαν στατιστικά σημαντικές διαφορές μεταξύ της αρχικής μέτρησης προ της έναρξης της θεραπείας (Τ 0 ) και όλων των επόμενων στιγμιότυπων (Τ 1, Τ 2, Τ 3 ). H αρχική τιμή της ALT ήταν σημαντικά μεγαλύτερη σε σχέση με τις τιμές κατά τη διάρκεια της μελέτης και η απόκλιση των τιμών της ALT εμφάνισε στατιστικά σημαντική συνεχή μείωση (Τ 0 - Τ 1 : t=5, 36, p<0,001, Τ 0 - Τ 3 : t= 6,24, p<0,001 και Τ 0 - Τ 2 : t=4,91,p<0,001) (πίνακας 9, γράφημα 10). 3.Αριθμός των Tregs (CD4+CD25+ high Foxp3+) στο περιφερικό αίμα Ο αρχικός αριθμός των Tregs στο περιφερικό αίμα πριν τη θεραπεία (Τ 0 ) δεν παρουσίασε στατιστικά σημαντικές διαφορές στο χρόνο Τ 1 και στο τέλος της θεραπείας (Τ 2 ), παρουσίασε όμως στατιστικά σημαντική αύξηση 6 μήνες μετά το τέλος της θεραπείας (Τ 0 -Τ 3 ) (t=-2,65, p<0,05). Επιπλέον, παρατηρήθηκε στατιστικά σημαντική διαφορά μεταξύ του χρόνου Τ 1 - Τ 3, δηλαδή αυξήθηκε μεταξύ του χρόνου Τ 1 (w4/ w12) και 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (t=3,19, p<0,01) (πίνακας 10, γράφημα 11). Δεν διαπιστώθηκαν άλλες στατιστικά σημαντικές διαφορές στον αριθμό των Tregs στα στιγμιότυπα Τ 2 - Τ
116 4.Αριθμός των Tregs στις in vitro καλλιέργειες Ο αριθμός των Tregs στις in vitro καλλιέργειες με PHA μετά από 48 ώρες δεν κατέδειξε στατιστικά σημαντικές διαφορές σε κανένα στιγμιότυπο της μελέτης (πίνακας 11, γράφημα 12). 5.Αριθμός των CD3+CD8+ T λεμφοκυττάρων στο περιφερικό αίμα Ο αριθμός των κυτταροτοξικών CD3+CD8+ Τ λεμφοκυττάρων παρουσίασε στατιστικά σημαντική διαφορά στο περιφερικό αίμα μεταξύ των χρονικών στιγμών Τ 3 και Τ 2, δηλαδή τα κύτταρα αυτά ήταν περισσότερα 6 μήνες μετά το τέλος της θεραπείας σε σχέση με το τέλος αυτής (όπου t=-2,06 με p<0,05). Δεν παρατηρήθηκαν άλλες στατιστικά σημαντικές διαφορές στον αριθμό των CD3+CD8+ T κυττάρων σε άλλα στιγμιότυπα της μελέτης (πίνακας 12, γράφημα 13). 6.Αριθμός των CD4+ T λεμφοκυττάρων στο περιφερικό αίμα Ο πληθυσμός των CD4+ Τ λεμφοκυττάρων μειώθηκε στατιστικά σημαντικά 6 μήνες μετά το τέλος της θεραπείας σε σχέση με τον αριθμό τους στο περιφερικό αίμα τη χρονική στιγμή Τ 1 (w4/ w12) κατά τη διάρκεια της θεραπείας (Τ 1 - Τ 3 : t=3,28 με p<0,01). Δεν διαπιστώθηκαν άλλες στατιστικά σημαντικές διαφορές στον αριθμό των CD4+ T κυττάρων σε άλλα στιγμιότυπα της μελέτης (πίνακας 13, γράφημα 14). 7.Αριθμός των CD4+CD25+ T λεμφοκυττάρων στο περιφερικό αίμα Ο αριθμός των CD4+CD25+ Τ λεμφοκυττάρων στο περιφερικό αίμα παρουσίασε στατιστικά σημαντική αύξηση μεταξύ των χρονικών στιγμών Τ 0 και Τ 1, δηλαδή ο αρχικός τους αριθμός (Τ 0 ) αυξήθηκε κατά τη διάρκεια της θεραπείας (Τ 1, w4/ w12) (Τ 1 - Τ 0 : t=-2,09 με p<0.05), ενώ διαπιστώθηκε πως ο αριθμός τους στο τέλος της θεραπείας (Τ 2 ) καθώς και 6 μήνες μετά (Τ 3 ) ήταν μικρότερος σε σχέση με τον αριθμό τους κατά τη διάρκεια της αγωγής την 4 η και τη 12 η εβδομάδα (Τ 1 - Τ 2 : t=2,48 με p<0,05 και Τ 1 - Τ 3 : t=2,05 με p<0,05). Δηλαδή διαπιστώθηκαν σημαντικές διακυμάνσεις του αριθμού των CD4+CD25+ T κυττάρων κατά τη διάρκεια της θεραπείας, αλλά στο τέλος της θεραπείας δεν διαπιστώθηκαν σημαντικές διαφορές σε σχέση με τον αρχικό τους αριθμό (πίνακας 14, γράφημα 15). 116
117 8.Αριθμός των CD4+CD28+ T λεμφοκυττάρων στο περιφερικό αίμα Ο αρχικός αριθμός των CD4+CD28+ T λεμφοκυττάρων στο περιφερικό αίμα πριν τη θεραπεία (Τ 0 ) ήταν στατιστικά σημαντικά μικρότερος σε σχέση με τον πληθυσμό τους κατά τη χρονική στιγμή Τ 1,δηλαδή την 4 η και τη 12 η εβδομάδα θεραπείας, αλλά και 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (Τ 0 - Τ 3 : t=2,35 με p< 0,05 και Τ 1 - Τ 3 : t=2,57 με p<0,01). Δεν διαπιστώθηκαν άλλες στατιστικά σημαντικές διαφορές σε άλλα στιγμιότυπα της μελέτης (πίνακας 15, γράφημα 16). 117
118 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο t df Σημ. HCV RNA Τ0 - HCV RNA Τ , , , , ,11 3,85 39,000 HCV RNA Τ0 - HCV RNA Τ , , , , ,86 3,33 27,003 HCV RNA Τ0 - HCV RNA Τ , , , , ,92 3,73 30,001 Πίνακας 8: Στατιστικά σημαντικές διαφορές του ιικού φορτίου (HCV RNA) των ασθενών στα χρονικά στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο t df Σημ. ALT Τ0 - ALT Τ1 55,82 65,76 10,39 34,79 76,85 5,36 39,000 ALT Τ0 - ALT Τ2 50,07 42,46 8,02 33,60 66,53 6,24 27,000 ALT Τ0 - ALT Τ3 42,54 48,21 8,65 24,86 60,23 4,91 30,000 Πίνακας 9: Στατιστικά σημαντικές διαφορές της τιμής της ALT των ασθενών στα χρονικά στιγμιότυπα της μελέτης 118
119 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο T Df Σημ. Tregs Τ0 - Tregs Τ3 -,325,648,122 -,576 -,074-2,65 27,013 Tregs Τ1 - Tregs Τ3 -,437,697,137 -,719 -,155-3,19 25,004 Πίνακας 10: Στατιστικά σημαντικές διαφορές του αριθμού των Tregs (CD4+CD25+ high Foxp3+ Τ κυττάρων) στο περιφερικό αίμα των ασθενών στα χρονικά στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο T df Σημ. Καλλιέργεια Τ0 - Καλλιέργεια Τ1 -,57 2,98,77-2,22 1,08 -,73 14,472 Καλλιέργεια Τ0 - Καλλιέργεια Τ2-2,18 3,87 1,58-6,24 1,88-1,38 5,226 Καλλιέργεια Τ1 - Καλλιέργεια Τ2-1,56 4,39 1,79-6,18 3,04 -,87 5,423 Πίνακας 11: Στατιστικά σημαντικές διαφορές του αριθμού των Tregs που αναπτύχθηκε στις in vitro καλλιέργειες μετά από 48 ώρες στα χρονικά στιγμιότυπα της μελέτης 119
120 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο T Df Σημ. CD3+CD8+ Τ2 - CD3+CD8+ Τ3-2,49 5,38 1,20-5,01,03-2,06 19,050 Πίνακας 12: Στατιστικά σημαντικές διαφορές του αριθμού των CD3+CD8+ T κυττάρων στο περιφερικό αίμα των ασθενών στα χρονικά στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο T Df Σημ. CD4+ Τ1 CD4+ Τ3 6,50 9,91 1,98 2,41 10,60 3,28 24,003 Πίνακας 13: Στατιστικά σημαντικές διαφορές του αριθμού των CD4+ T κυττάρων στο περιφερικό αίμα των ασθενών στα χρονικά στιγμιότυπα της μελέτης 120
121 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο T Df Σημ. CD4+CD25+ Τ0 - CD4+CD25+ Τ1-2,02 5,95,96-3,97 -,06-2,09 37,043 CD4+CD25+ Τ1 - CD4+CD25+ Τ2 1,09 2,24,44,18 1,99 2,48 25,020 CD4+CD25+ Τ1 - CD4+CD25+ Τ3 2,91 7,43 1,48 -,15 5,98 2,05 24,047 Πίνακας 14: Στατιστικά σημαντικές διαφορές του αριθμού των CD4+CD25+ T κυττάρων στο περιφερικό αίμα των ασθενών Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς Κατώτερο Ανώτερο T Df Σημ. CD4+CD28+ Τ0 - CD4+CD28+ Τ3 4,08 8,89 1,71,56 7,60 2,38 26,025 CD4+CD28+ Τ1 - CD4+CD28+ Τ3 5,85 10,19 2,03 1,64 10,06 2,87 24,008 Πίνακας 15: Αριθμός CD4+CD28+ T κυττάρων στο περιφερικό αίμα των ασθενών στα χρονικά στιγμιότυπα της μελέτης 121

122 Διαχρονική εξέλιξη HCV RNA , , , , , , , ,00,00 t0 T 0 t1 T 1 t2 T 2 t3 T 3 Γράφημα 9: Απεικόνιση των μετρήσεων του ιικού φορτίου των ασθενών στα χρονικά στιγμιότυπα της μελέτης 90,00 80,00 70,00 60,00 50,00 40,00 30,00 20,00 10,00 Διαχρονική εξέλιξη ALT,00 T 0 T 1 T 2 T 3 t0 t1 t2 t3 Γράφημα 10: Απεικόνιση της εξέλιξης της τιμής της ALT στους ασθενείς της μελέτης στα 4 χρονικά στιγμιότυπα 122
123 Διαχρονική εξέλιξη Tregs 3,00 2,50 2,00 1,50 1,00,50,00 t0 T 0 t1 T 1 t2t 2 Tt3 3 Γράφημα 11: Απεικόνιση της διακύμανσης του αριθμού των Tregs (CD4+CD25+ high Foxp3+ Τ κυττάρων) στους ασθενείς στα 4 στιγμιότυπα της μελέτης Διαχρονική εξέλιξη CD+ 50,00 45,00 40,00 35,00 30,00 25,00 20,00 15,00 10,00 5,00,00 Tt0 t1 t2 t3 0 T 1 T 2 T 3 CD3+CD8+ CD4+ CD4+CD25+ CD4+CD28+ Γράφημα 12: Απεικόνιση της διακύμανσης του αριθμού των CD3+CD8+, CD4+, CD4+CD25+ και CD4+CD28+ T κυτταρικών υποπληθυσμών στους ασθενείς στα 4 χρονικά στιγμιότυπα της μελέτης 123
124 Αποτελέσματα στις δύο ομάδες ασθενών (ΟΜΑΔΑ 1: γονότυπος 1- ΟΜΑΔΑ 2: γονότυπος 2 και 3) Από την τελική στατιστική ανάλυση, όσον αφορά στα αποτελέσματα των μετρήσεων των διαφόρων παραμέτρων μεταξύ των δύο ομάδων ασθενών α) με γονότυπο 1 και 4 και β) με γονότυπο 2 και 3, εξαιρέθηκαν οι ασθενείς με χρόνια ηπατίτιδα C και γονότυπο 4 λόγω της «άτυπης» συμπεριφοράς του συγκεκριμένου γονότυπου. Συγκεκριμένα, δύο ασθενείς πέτυχαν SVR, δυο διέκοψαν τη θεραπεία πριν τον πρώτο μήνα, ενώ ένας εμφάνισε ιολογική διαφυγή κατά τη διάρκεια της θεραπείας. Μετά την εξαίρεση των ασθενών με γονότυπο 4, τα νέα ποσοστά των ασθενών της μελέτης ήταν: 42,9% με γονότυπο 1 (24/56) και 57,1% με γονότυπο 2 και 3 (32/56). Οι γονότυποι 2 και 3 αποτέλεσαν μια ομάδα, επειδή είναι βιβλιογραφικά τεκμηριωμένο ότι απαιτούν την ίδια διάρκεια θεραπείας, ενώ τα ποσοστά ανταπόκρισης είναι σχεδόν παρόμοια (Γράφημα 17). Όσον αφορά την ανταπόκριση του πληθυσμού της μελέτης με βάση το γονότυπο, διαπιστώθηκε ότι οι ασθενείς με γονότυπο 2 και 3 (ΟΜΑΔΑ 2), όπως άλλωστε αναμενόταν, είχαν καλύτερη απάντηση στη θεραπεία. Για την ακρίβεια, όλοι οι ασθενείς με γονότυπο 2 και 3 που ολοκλήρωσαν τη θεραπεία (22/32) είχαν SVR, ενώ η πρώιμη διακοπή της θεραπείας λόγω παρενεργειών ήταν σημαντικά χαμηλότερη σε σχέση με τους ασθενείς με γονότυπο
125 Γράφημα 17: Απεικόνιση της ανταπόκρισης των ασθενών της μελέτης στη θεραπεία με βάση το γονότυπο Τελική ανταπόκριση μη διακοπή λόγω ανταπόκριση ανταπόκριση παρενεργειών drop out Σύνολο Γονότυποι 1 Μέτρηση % μέσα στο Γονότυποι 12,5% 50,0% 16,7% 20,8% 100,0% 2, 3 Μέτρηση % μέσα στο Γονότυποι,0% 68,8% 3,1% 28,1% 100,0% Σύνολο Μέτρηση % μέσα στο Γονότυποι 5,4% 60,7% 8,9% 25,0% 100,0% Πίνακας 16: Ανταπόκριση των ασθενών της μελέτης στη θεραπεία με βάση το γονότυπο 125
126 Τα αποτελέσματα αναλύθηκαν με βάση το γονότυπο των ασθενών στα στιγμιότυπα της μελέτης με το student s t για ζευγαρωτά δείγματα (paired samples student s t-test). ΟΜΑΔΑ 1: Ασθενείς με γονότυπο 1 (15/ 37) 1.HCV RNA Το αρχικό ιικό φορτίο πριν την έναρξη της θεραπείας (Τ 0 ) παρουσίασε στατιστικά σημαντική μείωση τη 12 η εβδομάδα αγωγής (Τ 1 ) και 6 μήνες μετά την λήξη αυτής (Τ 3 ) (Τ 0 - Τ 1 : t=2,32 με p<0,05 και Τ 0 - Τ 3 : t=2,18 με p<0,05) (πίνακας 17, γράφημα 18). 2.ALT Η αρχική τιμή της ALT πριν την έναρξη της θεραπείας (Τ 0 ) μειώθηκε στατιστικά σημαντικά την 12 η εβδομάδα αγωγής (Τ 1 ) και στο τέλος της μετά από 48 εβδομάδες (Τ 2 ) (Τ 0 - Τ 1 : t=3,68 με p<0,01 και Τ 0 - Τ 2 : t=2,60 με p<0,05) (πίνακας 18, γράφημα 19). 3.Αριθμός Tregs (CD4+CD25+ high Foxp3+ T κύτταρα) στο περιφερικό αίμα Ο αρχικός αριθμός των Tregs δεν παρουσίασε στατιστικά σημαντικές διαφορές με κανένα από τα επόμενα στιγμιότυπα. Όμως, ο αριθμός τους στο περιφερικό αίμα την 12 η εβδομάδα αγωγής (Τ 1 ) ήταν μικρότερος σε σχέση με τον πληθυσμό τους 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ), δηλαδή ο πληθυσμός τους κατά τη διάρκεια της θεραπείας (Τ 1 ) μειώθηκε σημαντικά (t=-2,26 με p<0,05), αλλά στη συνέχεια παρουσίασε αύξηση έτσι ώστε στο τέλος της θεραπείας (Τ 2 ) και 6 μήνες μετά (Τ 3 ) να μην εμφανίζει διαφορές σε σχέση με τον αρχικό τους αριθμό (πίνακας 19, γράφημα 20). 4.Αριθμός Tregs στις in vitro καλλιέργειες Ο αριθμός των Tregs δεν παρουσίασε στατιστικά σημαντικές διαφορές στις in vitro καλλιέργειες που έγιναν στα στιγμιότυπα της μελέτης. 5.Αριθμός των CD3+CD8+ T κυττάρων στο περιφερικό αίμα Ο αριθμός των κυτταροτοξικών CD3+CD8+ Τ κυττάρων δεν παρουσίασε στατιστικά σημαντικές διαφορές στα στιγμιότυπα της μελέτης (γράφημα 21). 6.Αριθμός των CD4+ T κυττάρων στο περιφερικό αίμα Ο αριθμός των CD4+ Τ λεμφοκυττάρων μειώθηκε στατιστικά σημαντικά στο τέλος της θεραπείας (μετά από 48 εβδομάδες, Τ 2 ) σε σχέση με τη 12 η εβδομάδα αγωγής (Τ 1 ), εντούτοις όμως δεν παρουσίασε σημαντικές διαφορές σε σχέση με τον αρχικό αριθμό τους (t=3,58 με p<0,01) (πίνακας 20, γράφημα 22). 126
127 7.Αριθμός των CD4+CD25+ T κυττάρων στο περιφερικό αίμα Ο αριθμός των CD4+CD25+ Τ κυττάρων δεν παρουσίασε στατιστικά σημαντικές διαφορές στα στιγμιότυπα της μελέτης (γράφημα 23). 8.Αριθμός των CD4+CD28+ T κυττάρων στο περιφερικό αίμα Ο αριθμός των CD4+CD28+ Τ λεμφοκυττάρων παρουσίασε στατιστικά σημαντικές διαφορές μόνο μεταξύ των χρονικών στιγμών Τ 1 και Τ 2, δηλαδή οι ασθενείς με γονότυπο 1 ή 4 είχαν μικρότερο αριθμό των CD4+CD28+ T λεμφοκυττάρων στο τέλος της θεραπείας, μετά από 48 εβδομάδες (Τ 2 ) σε σχέση με τη 12 η εβδομάδα αγωγής (Τ 1 ), εντούτοις δεν παρουσίασε σημαντικές διαφορές με τον αρχικό αριθμό τους (t=6,02 με p<0,001) (πίνακας 21, γράφημα 24). 127
128 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T Df Σημ. Κατώτερο Ανώτερο HCV RNA Τ0 - HCV RNA Τ , , , , ,91 2,32 17,033 HCV RNA Τ0 - HCV RNA Τ , , , , ,60 2,18 13,048 Πίνακας 17: Στατιστικά σημαντικές διαφορές του ιικού φορτίου (HCV RNA) στην ΟΜΑΔΑ 1 στα στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T Df Σημ. Κατώτερο Ανώτερο ALT Τ0 - ALT Τ1 42,55 48,99 11,54 18,19 66,92 3,68 17,002 ALT Τ0 - ALT Τ2 43,54 55,49 16,73 6,26 80,83 2,60 10,026 Πίνακας 18: Στατιστικά σημαντικές διαφορές της τιμής της ALT στους ασθενείς της ΟΜΑΔΑΣ 1 στα χρονικά στιγμιότυπα της μελέτης 128
129 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. σφάλμα Μ.Ο. Κατώτερο Ανώτερο Tregs Τ1 - Tregs Τ3 -,51,78,22-1,01 -,01-2,26 11,044 Πίνακας 19: Στατιστικά σημαντικές διαφορές του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) στο περιφερικό αίμα των ασθενών της ΟΜΑΔΑΣ 1 στα στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. 95% Διάστημα εμπιστοσύνης της διαφοράς t df Σημ. σφάλμα Μ.Ο. Κατώτερο Ανώτερο CD4+ Τ1 CD4+ Τ2 4,89 4,31 1,36 1,80 7,97 3,58 9,006 Πίνακας 20: Στατιστικά σημαντικές διαφορές του αριθμού των CD4+ Τ κυττάρων στο περιφερικό αίμα των ασθενών της ΟΜΑΔΑΣ 1 στα στιγμιότυπα της μελέτης 129
130 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. σφάλμα Μ.Ο. Κατώτερο Ανώτερο CD4+CD28+ Τ1 - CD4+CD28+ Τ2 6,09 3,19 1,01 3,80 8,37 6,02 9,000 Πίνακας 21: Στατιστικά σημαντικές διαφορές του αριθμού των CD4+CD28+ T κυττάρων στο περιφερικό αίμα των ασθενών της ΟΜΑΔΑΣ 1 στα στιγμιότυπα της μελέτης 130
131 ΟΜΑΔΑ 2: Ασθενείς με γονότυπο 2 και 3 (22/32) 1.HCV RNA Το αρχικό ιικό φορτίο πριν την έναρξη της θεραπείας (Τ 0 ) εμφάνισε στατιστικά σημαντική μείωση και στις τρεις επόμενες χρονικές στιγμές της μελέτης (Τ 1, Τ 2 και Τ 3 ) (Τ 0 - Τ 1 : t=3,37 με p<0,01, Τ 0 - Τ 2 : t=3,26 με p<0,01 και Τ 0 - Τ 3 : t=3,32 με p<0,01) (πίνακας 22, γράφημα 18). 2.ALT Η αρχική τιμή της ALT πριν την έναρξη της θεραπείας (Τ 0 ) μειώθηκε στατιστικά σημαντικά σε όλα τα επόμενα στιγμιότυπα της μελέτης (Τ 1, Τ 2 και Τ 3 ) (Τ 0 - Τ 1 : t=4,27 με p<0,001, Τ 0 - Τ 2 : t=6,81 με p<0,001 και Τ 0 - Τ 3 : t=8,29 με p<0,001) (πίνακας 23, γράφημα 19). Επιπλέον, στατιστικά σημαντική μείωση στην τιμή της ALT διαπιστώθηκε και μεταξύ της 4 ης εβδομάδας θεραπείας (Τ 1 ) και 6 μήνες μετά το τέλος της (Τ 3 ) (Τ 1 - Τ 3 : t=2,80 με p<0,05). 3.Αριθμός των Tregs (CD4+CD25+ high Foxp3+ T κύτταρα) στο περιφερικό αίμα Ο αρχικός αριθμός των Tregs πριν την έναρξη της θεραπείας (Τ 0 ) στους ασθενείς με γονότυπο 2 και 3 παρουσίασε στατιστικά σημαντική μείωση στη λήξη της θεραπείας (24 εβδομάδες μετά, Τ 2 ) (t=2,25 με p<0.05), στη συνέχεια ο αριθμός τους αυξήθηκε μεταξύ του τέλους θεραπείας (Τ 2 ) και 6 μήνες μετά το τέλος της (Τ 3 ) (Τ 2 - Τ 3 : t=-2,62 με p<0,05), χωρίς να διαφοροποιηθεί ποσοτικά από τον αρχικό και επανήλθε στα προ της θεραπείας επίπεδα (πίνακας 24, γράφημα 20). 4.Αριθμός Tregs στις in vitro καλλιέργειες Ο αριθμός των Tregs στις in vitro καλλιέργειες δεν παρουσίασε στατιστικά σημαντικές διαφορές στα στιγμιότυπα της μελέτης στους ασθενείς με γονότυπο 2 και 3. 5.Αριθμός CD3+CD8+ T κυττάρων στο περιφερικό αίμα Ο αρχικός αριθμός των κυτταροτοξικών CD3+CD8+ Τ λεμφοκυττάρων στο περιφερικό αίμα δεν παρουσίασε σημαντικές διαφορές με κανένα από τα στιγμιότυπα της μελέτης. Παρατηρήθηκε όμως ότι στο τέλος της θεραπείας (24 εβδομάδες μετά, Τ 2 ) ήταν μικρότερος σε σχέση με τον αριθμό τους 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ), δηλαδή αυξήθηκε στατιστικά σημαντικά (Τ 2 - Τ 3 : t=- 2,28, p<0,05), χωρίς όμως να μεταβληθεί από την αρχική τους τιμή (πίνακας 25, γράφημα 21). 131
132 6.Αριθμός των CD4+ T κυττάρων στο περιφερικό αίμα Ο αρχικός αριθμός των CD4+ Τ λεμφοκυττάρων στο περιφερικό αίμα δεν παρουσίασε στατιστικά σημαντικές διαφορές κατά τη διάρκεια της μελέτης, παρά το γεγονός ότι την 4 η εβδομάδα αγωγής (Τ 1 ) ήταν στατιστικά σημαντικά μεγαλύτερος σε σχέση με τον αριθμό τους 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (t=3,74 με p<0,01), τελικά όμως δεν μεταβλήθηκε η αρχική τους τιμή (πίνακας 26, γράφημα 22). 7.Αριθμός των CD4+CD25+ T κυττάρων στο περιφερικό αίμα Ο αρχικός αριθμός των CD4+CD25+ Τ κυττάρων πριν την έναρξη της θεραπείας (Τ 0 ) αυξήθηκε στατιστικά σημαντικά την 4 η εβδομάδα αγωγής (Τ 1 ) (t=-3,80 με p<0,001), αλλά στη συνέχεια μειώθηκε στατιστικά σημαντικά σε όλους τους επόμενους χρόνους της μελέτης (Τ 1 - Τ 3 : t=2,82 με p<0,05 και Τ 2 - Τ 3 : t=2,38 με p<0,05), χωρίς όμως τελικά να μεταβληθεί σε σχέση με τον αρχικό τους αριθμό (πίνακας 27, γράφημα 23). 8.Αριθμός CD4+CD28+ T κυττάρων στο περιφερικό αίμα Ο αριθμός των CD4+CD28+ T λεμφοκυττάρων παρουσίασε στατιστικά σημαντικές διαφορές σε αρκετά στιγμιότυπα. Ειδικότερα, ο αριθμός τους αυξήθηκε α) την 4 η εβδομάδα αγωγής (Τ 1 ) σε σχέση με τον αρχικό αριθμό τους (Τ 0 ) (t=-2,86 με p<0.05), β) 6 μήνες μετά τη θεραπεία (Τ 3 ) σε σχέση με τον αριθμό τους την 4 η εβδομάδα θεραπείας (Τ 1 ) και κατά τo τέλος της, μετά από 24 εβδομάδες (Τ 2 ) (Τ 1 - Τ 3 : t=3,27 με p<0,01 και Τ 2 - Τ 3 : t=2,19 με p<0,05) (πίνακας 28, γράφημα 24). 132
133 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. Κατώτερο Ανώτερο HCV RNA Τ0 - HCV RNA Τ , , , , ,754 3,37 19,003 HCV RNA Τ0 - HCV RNA Τ , , , , ,95 3,26 14,006 HCV RNA Τ0 - HCV RNA Τ , , , , ,44 3,32 14,005 Πίνακας 22: Στατιστικά σημαντικές διαφορές του ιικού φορτίου (HCV RNA) των ασθενών της ΟΜΑΔΑΣ 2 στα στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. Κατώτερο Ανώτερο ALT Τ0 - ALT Τ1 58,35 61,10 13,66 29,75 86,94 4,27 19,000 ALT Τ0 - ALT Τ2 57,93 32,93 8,50 39,69 76,16 6,81 14,000 ALT Τ0 - ALT Τ3 56,00 26,14 6,75 41,52 70,47 8,29 14,000 ALT Τ1 - ALT Τ3 10,00 12,87 3,57 2,22 17,77 2,80 12,016 Πίνακας 23: Στατιστικά σημαντικές διαφορές της τιμής της ALT των ασθενών της ΟΜΑΔΑΣ 2 στα στιγμιότυπα της μελέτης 133
134 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς t df Σημ. Κατώτερο Ανώτερο Tregs Τ0 - Tregs Τ2,23,39,10,01,46 2,25 13,042 Tregs Τ2 - Tregs Τ3 -,37,46,14 -,68 -,05-2,62 10,025 Πίνακας 24: Στατιστικά σημαντικές διαφορές του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) των ασθενών της ΟΜΑΔΑΣ 2 στα στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς t df Σημ. Κατώτερο Ανώτερο CD3+CD8+ Τ2 - CD3+CD8+ Τ3-3,01 4,38 1,32-5,96 -,07-2,28 10,046 Πίνακας 25: Αριθμός των CD3+CD8+ T κυττάρων των ασθενών της ΟΜΑΔΑΣ 2 στα στιγμιότυπα της μελέτης 134
135 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς t df Σημ. Κατώτερο Ανώτερο CD4+ Τ1 CD4+ Τ3 9,31 12,06 3,34 2,02 16,60 2,78 12,017 Πίνακας 26: Αριθμός των CD4+ T κυττάρων των ασθενών της ΟΜΑΔΑΣ 2 στα στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς t df Σημ. Κατώτερο Ανώτερο CD4+CD25+ Τ0 - CD4+CD25+ Τ1-1,30 1,53,34-2,02 -,58-3,80 19,001 CD4+CD25+ Τ1 - CD4+CD25+ Τ3 1,48 2,20,61,15 2,82 2,43 12,032 CD4+CD25+ Τ2 - CD4+CD25+ Τ3 1,08 1,50,45,07 2,08 2,38 10,
136 Πίνακας 27: Αριθμός των CD4+CD25+ T κυττάρων των ασθενών της ΟΜΑΔΑΣ 2 στα στιγμιότυπα της μελέτης Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς t df Σημ. Κατώτερο Ανώτερο CD4+CD28+ Τ0 - CD4+CD28+ Τ1-6,76 10,54 2,35-11,70-1,82-2,86 19,010 CD4+CD28+ Τ1 - CD4+CD28+ Τ3 9,31 10,26 2,84 3,11 15,51 3,27 12,007 CD4+CD28+ Τ2 - CD4+CD28+ Τ3 5,80 8,77 2,64 -,08 11,70 2,27 10,049 Πίνακας 28: Αριθμός των CD4+CD28+ T κυττάρων των ασθενών της ΟΜΑΔΑΣ 2 στα στιγμιότυπα της μελέτης 136

137 Διαχρονική εξέλιξη HCV RNA γονότυπος 1 γονότυπος 2, T 0 T 1 T 2 T 3 t0 t1 t2 t3 Γράφημα 18: Συγκριτική απεικόνιση της εξέλιξης του ιικού φορτίου στις ομάδες των ασθενών της μελέτης με βάση το γονότυπο Διαχρονική εξέλιξη ALT 90,00 80,00 70,00 60,00 50,00 40,00 γονότυπος 1 γονότυπος 2,3 30,00 20,00 10,00,00 T 0 T 1 T 2 T 3 t0 t1 t2 t3 Γράφημα 19: Συγκριτική απεικόνιση της εξέλιξης των τιμών της ALT στις ομάδες των ασθενών της μελέτης με βάση το γονότυπο 137
138 Διαχρονική εξέλιξη Tregs 5 4,5 4 3,5 3 2,5 2 1,5 1 0,5 0 t0 T 0 t1t 1 t2t 2 t3t 3 γονότυπος 1 γονότυπος 2,3 Γράφημα 20: Συγκριτική απεικόνιση της διακύμανσης του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) στις ομάδες της μελέτης με βάση το γονότυπο Διαχρονική εξέλιξη CD3+CD γονότυπος 1 γονότυπος 2, T 0 T 1 T 2 T 3 t0 t1 t2 t3 Γράφημα 21: Συγκριτική απεικόνιση της διακύμανσης του αριθμού των CD3+CD8+ T κυττάρων στις ομάδες της μελέτης με βάση το γονότυπο 138
139 Διαχρονική εξέλιξη CD γονότυπος 1 γονότυπος 2, T 0 T 1 T 2 T 3 t0 t1 t2 t3 Γράφημα 22: Συγκριτική απεικόνιση της διακύμανσης του αριθμού των CD4+ T κυττάρων στις ομάδες της μελέτης με βάση το γονότυπο Διαχρονική εξέλιξη CD4+CD T 0 T 1 T 2 T 3 t0 t1 t2 t3 γονότυπος 1 γονότυπος 2,3 Γράφημα 23: Συγκριτική απεικόνιση της διακύμανσης του αριθμού των CD4+CD25+ T κυττάρων στις ομάδες της μελέτης με βάση το γονότυπο 139
140 Διαχρονική εξέλιξη CD4+CD Tt0 0 t1 T 1 Tt2 2 t3 T 3 γονότυπος 1 γονότυπος 2,3 Γράφημα 24: Συγκριτική απεικόνιση της διακύμανσης του αριθμού των CD4+CD28+ T κυττάρων στις ομάδες της μελέτης με βάση το γονότυπο 140
141 V. ΣΤΑΤΙΣΤΙΚΗ ΑΝΑΛΥΣΗ Σύγκριση των αποτελεσμάτων των παραμέτρων που μελετήθηκαν σε όλους τους ασθενείς και μεταξύ των δύο ομάδων ασθενών: ασθενείς με γονότυπο 1 και ασθενείς με γονότυπο 2, 3. Για την αναγνώριση στατιστικά σημαντικών διαφορών στις εξεταζόμενες μεταβλητές με βάση το φύλο και το γονότυπο των ασθενών που συμμετείχαν στη μελέτη, χρησιμοποιήθηκε τo Mann-Whitney U-test. Τα αποτελέσματα της σύγκρισης έδειξαν ότι οι στατιστικά σημαντικές διαφορές που διαπιστώθηκαν με βάση το φύλο αφορούν στον αριθμό των CD4+ και των CD4+CD28+ Τ λεμφοκυττάρων στο περιφερικό αίμα στο τέλος της θεραπείας (Τ 2 ), αλλά και τον αριθμό των Tregs που αναπτύχθηκαν στην in vitro καλλιέργεια και τον αριθμό των CD3+CD8+ Τ κυττάρων την ίδια χρονική στιγμή (Τ 2 ). Πιο συγκεκριμένα, διαπιστώθηκε ότι ο αριθμός των CD4+ και CD4+CD28+ Τ λεμφοκυττάρων στο περιφερικό αίμα πριν την έναρξη της αγωγής (Τ 0 ) διέφερε σημαντικά μεταξύ ανδρών και γυναικών (επίπεδο στατιστικής σημαντικότητας α=0,01), δηλαδή οι άνδρες είχαν μικρότερους αριθμούς CD4+ και CD4+CD28+ T κυτταρικών υποπληθυσμών σε σχέση με τις γυναίκες (μέσος όρος CD4+ Τ κυττάρων: 23,90 έναντι 37,11 και μέσος όρος CD4+CD28+ Τ κυττάρων: 23,16 έναντι 35,83). Στο τέλος όμως της θεραπείας (Τ 2 ), οι άνδρες εμφάνιζαν υψηλότερο μέσο όρο στον αριθμό των CD3+CD28+ Τ κυττάρων από τις γυναίκες (15,58 έναντι 8,81 (επίπεδο στατιστικής σημαντικότητας α=0,05) (πίνακας 29). CD4+ Τ0 CD4+CD28+ Τ0 CD3+CD8+ Τ2 Mann-Whitney U 206, ,500 34,500 Z -2,926-2,850-2,084 Asymp. Sig. (2-tailed),003,004,037 Exact Sig. [2*(1-tailed Sig.)],035 Πίνακας 29: Στατιστικά σημαντικές διαφορές των μεταβλητών της μελέτης βάση του φύλου 141
142 Όσον αφορά την σύγκριση των αποτελεσμάτων των ασθενών με βάση το γονότυπο, οι μόνες σημαντικές διαφορές που διαπιστώθηκαν ήταν οι εξής: οι ασθενείς με γονότυπο 1 α) εμφάνιζαν μικρότερη τιμή ALT και μικρότερο αριθμό Tregs σε σχέση με τους ασθενείς με γονότυπο 3 στο επιλεγμένο στιγμιότυπο κατά τη διάρκεια της θεραπείας (Τ 1, 12 η εβδομάδα για τον γονότυπο 1 και 4 η εβδομάδα για τον γονότυπο 2, 3) (μέσος όρος ALT: 15,47 έναντι 23,13 και αριθμός Tregs: 14,97 έναντι 22,43 αντίστοιχα) και β) υψηλότερο ιικό φορτίο 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (17,14 έναντι 13,00) (πίνακας 30). ALT_Τ1 Tregs_Τ1 HCV_RNA_Τ3 Mann-Whitney U 107, ,500 75,000 Z -2,122-2,088-2,183 Asymp. Sig. (2- tailed) Exact Sig. [2*(1- tailed Sig.)],034,037,029,033,036,201 Πίνακας 30: Στατιστικά σημαντικές διαφορές των μεταβλητών της μελέτης βάση του γονοτύπου 142
143 Σύγκριση αποτελεσμάτων μεταξύ των ασθενών με γονότυπο 1 οι οποίοι απήντησαν στη θεραπεία (12/15, 80%) και των ασθενών με γονότυπο 1 οι οποίοι δεν πέτυχαν (3/15, 20%) μακρά ιολογική απάντηση (SVR). Η επιμέρους ανάλυση των ασθενών με γονότυπο 1 με βάση την απάντηση έγινε με το student s t για ζευγαρωτά δείγματα (paired samples student s t-test). 1.HCV RNA Το ιικό φορτίο μειώθηκε στατιστικά σημαντικά 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) σε σχέση με την αρχική του τιμή (Τ 0 ) μόνο στους ασθενείς που ανταποκρίθηκαν και πέτυχαν SVR (t=2,40 με p<0,05) (πίνακας 31, γράφημα 25). 2.ALT Η ALT μειώθηκε στατιστικά σημαντικά μόνο στους ασθενείς που ανταποκρίθηκαν στην θεραπεία, δηλαδή η ALT ήταν χαμηλότερη μετά από 12 εβδομάδες θεραπείας (Τ 1 ) και στο τέλος της (Τ 2 : μετά από 48 εβδομάδες) σε σχέση με την τιμή της πριν την έναρξη αυτής (Τ 0 ) (Τ 0 - Τ 1 : t=3,08 με p<0,05 και Τ 0 - Τ 2 : t=2,36 με p<0,05) (πίνακας 32, γράφημα 26). 3.Αριθμός Tregs (CD4+CD25+ high Foxp3+ T κύτταρα) στο περιφερικό αίμα Οι μόνες στατιστικές διαφορές των Tregs που διαπιστώθηκαν ήταν ότι οι ασθενείς με γονότυπο 1 που δεν ανταποκρίθηκαν στην αγωγή είχαν στατιστικά σημαντική αύξηση του αριθμού των Tregs στο περιφερικό αίμα 6 μήνες μετά τη λήξη της θεραπείας σε σχέση με τον αριθμό τους πριν την έναρξή της (Τ 3 - Τ 0 : t=101 με p<0,01) (πίνακας 33, γράφημα 27). 4.Αριθμός Tregs στις in vitro καλλιέργειες Δεν διαπιστώθηκαν στατιστικά σημαντικές διαφορές στον αριθμό των Tregs που αναπτύχθηκε στις in vitro καλλιέργειες ανάμεσα στους ασθενείς με γονότυπο 1 που ανταποκρίθηκαν στην θεραπεία και εκείνους που δεν πέτυχαν SVR. 5.Αριθμός CD3+CD8+ T κυττάρων στο περιφερικό αίμα Δεν διαπιστώθηκαν στατιστικά σημαντικές διαφορές στον αριθμό των CD3+CD8+ Tκυττάρων ανάμεσα στους ασθενείς με γονότυπο 1 που ανταποκρίθηκαν στην θεραπεία και εκείνους που δεν πέτυχαν SVR (γράφημα 28). 143
144 6.Αριθμός CD4+ T κυττάρων στο περιφερικό αίμα Στους ασθενείς που ανταποκρίθηκαν στη θεραπεία και πέτυχαν SVR διαπιστώθηκε ότι ο αριθμός των CD4+ Τ κυττάρων μειώθηκε στατιστικά σημαντικά στο τέλος της θεραπείας (Τ 2 ) και 6 μήνες μετά (Τ 3 ) σε σχέση με τον πληθυσμό τους την 12 η εβδομάδα της θεραπείας (Τ 1 : στο επιλεγμένο στιγμιότυπο κατά τη διάρκεια της αγωγής) (Τ 1 - Τ 2 : t=2,78 με p<0,05 και Τ 1 - Τ 3 : t=6,36 με p<0,001) (πίνακας 34, γράφημα 29). 7.Αριθμός CD4+CD25+ T κυττάρων στο περιφερικό αίμα Στους ασθενείς με γονότυπο 1 που πέτυχαν SVR, ο αριθμός των CD4+CD25+ Τ λεμφοκυττάρων αρχικά, τη 12 η εβδομάδα, είχε αυξητική τάση σε σχέση με τον πληθυσμό τους πριν την έναρξη της θεραπείας (Τ 0 ), αλλά στη συνέχεια διαπιστώθηκε ότι υπήρχε μία τάση μείωσή τους 6 μήνες μετά το τέλος της σε σχέση με τoν αριθμό τους την 12 η εβδομάδα θεραπείας (Τ 3 - Τ 1 ) όπως φαίνεται και στο γράφημα 30, χωρίς όμως να υπάρχουν στατιστικά σημαντικές διαφορές. 8.Αριθμός CD4+CD28+ T κυττάρων στο περιφερικό αίμα Ο αριθμός των CD4+CD28+ T κυττάρων μειώθηκε σε όλους τους ασθενείς με γονότυπο 1, ανεξάρτητα από την έκβαση της θεραπείας. Όμως σε εκείνους που δεν ανταποκρίθηκαν στην θεραπεία παρατηρήθηκε μείωση του αριθμού τους στο τέλος της θεραπείας (Τ 2 ) σε σχέση με τον πληθυσμό τους την 12 η εβδομάδα αγωγής (Τ 1 ) (Τ 1 - Τ 2 : t= 18, p<0.05). Στους ασθενείς που ανταποκρίθηκαν στη θεραπεία εκτός από την ίδια μείωση, διαπιστώθηκε και περαιτέρω μείωση του πληθυσμού των CD4+CD28+ T κυττάρων, 6 μήνες μετά το τέλος της θεραπείας (Τ 2 - Τ 1 : t= 4,62, p<0.001 και Τ 3 - Τ 1 : t= 3,48, p< 0.05) (πίνακας 35, γράφημα 31). 144
145 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. Κατώτερο Ανώτερο Ανταπόκριση Ανταπόκριση HCV RNA Τ , , , , ,61 2,40 10,037 HCV RNA Τ3 Πίνακας 31: Στατιστικά σημαντικές διαφορές του ιικού φορτίου των ασθενών της μελέτης με γονότυπο 1 που ανταποκρίθηκαν στη θεραπεία Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. Κατώτερο Ανώτερο ALT Τ0 ALT Τ1 51,54 55,35 16,69 14,35 88,73 3,08 10,011 ALT Τ0 ALT Τ2 48,00 60,93 20,31 1,16 94,84 2,36 8,046 Πίνακας 32: Στατιστικά σημαντικές διαφορές της τιμής της ALT στους ασθενείς με γονότυπο 1 που ανταποκρίθηκαν στη θεραπεία 145
146 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. Κατώτερο Ανώτερο Πίνακας 33: Στατιστικά σημαντικές διαφορές στον αριθμό των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) στο περιφερικό αίμα των ασθενών με γονότυπο 1 που δεν ανταποκρίθηκαν στη θεραπεία Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. Κατώτερο Ανώτερο Μη Tregs Τ0 Tregs Τ3 -,50,01,01 -,56 -,44-101,00 1,006 ανταπόκριση Ανταπόκριση CD4+ Τ1- CD4+ Τ3 4,91 2,18,77 3,08 6,73 6,36 7,000 CD4+ Τ1- CD4+ Τ2 4,80 4,87 1,72,72 8,87 2,78 7,027 Πίνακας 34: Στατιστικά σημαντικές διαφορές στον αριθμό των CD4+ T κυττάρων στο περιφερικό αίμα των ασθενών με γονότυπο 1 που ανταποκρίθηκαν στη θεραπεία 146
147 Διαφορές ανά ζεύγος Τυπική Μ.Ο. απόκλιση Τυπ. σφάλμα Μ.Ο. 95% Διάστημα εμπιστοσύνης της διαφοράς T df Σημ. Κατώτερο Ανώτερο CD4+CD28+ Τ1-7,20,56,40 2,11 12,28 18,00 1,035 CD4+CD28+ Τ2 Μη ανταπόκριση Ανταπόκριση CD4+CD28+ Τ1 - CD4+CD28+ Τ2 5,81 3,55 1,25 2,84 8,78 4,62 7,002 CD4+CD28+ Τ1 - CD4+CD28+ Τ3 4,33 3,52 1,24 1,39 7,28 3,48 7,010 Πίνακας 35: Στατιστικά σημαντικές διαφορές στον αριθμό των CD4+CD28+ T κυττάρων στο περιφερικό αίμα μεταξύ των δυο ομάδων ασθενών με γονότυπο 1 βάση της ανταπόκρισης στη θεραπεία 147
 80,00 70,00 60,00 50,00 40,00")
148 Συγκριτική εξέλιξη HCV RNA (γονότυπος 1) , , , , , ,00 HCV RNA - μη αν ταπόκριση HCV RNA_αν ταπό κριση ,00,00 T 0 t0 t1 T 1 t2 T 2 t3 T 3 Γράφημα 25: Απεικόνιση της διακύμανσης του ιικού φορτίου στις ομάδες των ασθενών με γονότυπο 1 βάση της ανταπόκρισης στη θεραπεία Συγκριτική εξέλιξη ALT (γονότυπος 1) 80,00 70,00 60,00 50,00 40,00 30,00 20,00 10,00,00 Tt0 0 t1 T 1 t2 T 2 Tt3 3 ALT αν ταπόκριση ALT μη αν ταπόκριση Γράφημα 26: Απεικόνιση της διακύμανσης της τιμής της ALT στις ομάδες των ασθενών με γονότυπο 1 βάση της ανταπόκρισης στη θεραπεία 148
 στο περιφερικό αίμα των ασθενών με γονότυπο 1 βάση της ανταπόκρισης στη θεραπεία Συγκριτική εξέλιξη CD3+CD8+ (γονότυπος 1) 35,00 30,00 25,00 20,00")
149 Συγκριτική εξέλιξη Tregs (γονότυπος 1) 2,00 1,80 1,60 1,40 1,20 1,00,80,60,40,20,00 t0 T 0 t1t 1 t2t 2 t3 T 3 tregs_αν ταπό κριση tregs_μη αν ταπόκριση Γράφημα 27: Απεικόνιση της διακύμανσης του αριθμού των Tregs (CD4+CD25+ high Foxp3 T κυττάρων) στο περιφερικό αίμα των ασθενών με γονότυπο 1 βάση της ανταπόκρισης στη θεραπεία Συγκριτική εξέλιξη CD3+CD8+ (γονότυπος 1) 35,00 30,00 25,00 20,00 15,00 CD3+CD8+_αν ταπόκριση 10,00 5,00 CD3+CD8+_μη αν ταπόκριση,00 T 0 T 1 T 2 T 3 t0 t1 t2 t3 Γράφημα 28: Απεικόνιση της διακύμανσης του αριθμού των CD3+CD8+ T κυττάρων στο περιφερικό αίμα των ασθενών με γονότυπο 1, βάση της ανταπόκρισης στη θεραπεία 149


150 Συγκριτική εξέλιξη CD4+ (γονότυπος 1) 52,00 50,00 48,00 46,00 44,00 CD4+_αν ταπ όκριση CD4+_μη αν ταπόκριση 42,00 40,00 t0 T 0 t1t 1 t2t 2 t3t 3 Γράφημα 29: Απεικόνιση της διακύμανσης του αριθμού των CD4+ T κυττάρων στο περιφερικό αίμα των ασθενών με γονότυπο 1, βάση της ανταπόκρισης στη θεραπεία Συγκριτική εξέλιξη CD4+CD25+ (γονότυπος 1) 6,00 5,00 4,00 3,00 2,00 1,00 CD4+CD25+_ αν ταπόκριση CD4+CD25+_ μη αν ταπόκριση,00 Tt0 0 t1t 1 t2 T 2 t3 T 3 Γράφημα 30: Απεικόνιση της διακύμανσης του αριθμού των CD4+CD25+ T κυττάρων στο περιφερικό αίμα των ασθενών με γονότυπο 1, βάση της ανταπόκρισης στη θεραπεία 150
151 Συγκριτική εξέλιξη CD4+CD28+ (γονότυπος 1) 49,00 48,00 47,00 46,00 45,00 44,00 43,00 42,00 41,00 40,00 39,00 38,00 T t0 0 t1 T 1 t2 T 2 t3 T 3 CD4+CD28+ _αν ταπόκριση CD4+CD28+ _μη αν ταπόκριση Γράφημα 31: Απεικόνιση της διακύμανσης του αριθμού των CD4+CD28+ T κυττάρων στο περιφερικό αίμα των ασθενών με γονότυπο 1, βάση της ανταπόκρισης στη θεραπεία 151
152 Συσχετίσεις μεταβλητών Οι συσχετίσεις μεταξύ του συνόλου των μεταβλητών που μετρήθηκαν στη μελέτη έγινε με το Spearman test, το οποίο ουσιαστικά αντικατοπτρίζει την ένταση και την κατεύθυνση (θετική ή αρνητική) της σχέσης μεταξύ δύο μεταβλητών υπό εξέταση. 1.Ηλικία Στα άτομα της μελέτης, διαπιστώθηκε ότι όσο μεγαλύτερη ήταν η ηλικία, τόσο μεγαλύτερος ήταν ο δείκτης νεκροφλεγμονώδους δραστηριότητας στον ηπατικό ιστό (ΗΑΙ) (r=0.48, p<0.01), δηλαδή η διαβρωτική νέκρωση (r=0.56, p<0.01), η πυλαία φλεγμονή (r=0.42, p<0.01), οι ενδολόβιες νεκρώσεις (r=0.26, p<0.05). Το ίδιο ισχύει και για την ίνωση (r=0.30, p<0.05). Επίσης, διαπιστώθηκε ότι η ηλικία και το ιικό φορτίο (HCV RNA) είχαν θετική συσχέτιση στο επιλεγμένο στιγμιότυπο κατά τη διάρκεια της θεραπείας (Τ 1 ) (r=0.37, p<0.05), δηλαδή όσο μεγαλύτερη ήταν η ηλικία του ασθενή τόσο μεγαλύτερο ήταν και το ιικό του φορτίο. Το εύρημα αυτό δεν υπήρξε στα άλλα στιγμιότυπα της μελέτης. 2.ΗΑΙ (δείκτης νεκροφλεγμονώδους δραστηριότητας, Histological Activity Index, ΗΑΙ) Ο δείκτης νεκροφλεγμονώδους δραστηριότητας διαπιστώθηκε ότι συσχετίζεται θετικά με την διαβρωτική νέκρωση, την πυλαία φλεγμονή και τις ενδολόβιες νεκρώσεις (r= 0.93, p<0.01, r= 0.84, p< 0.01 και r= 0.78, p< 0.01 αντίστοιχα), αλλά είχε σημαντικά εντονότερη θετική συσχέτιση με τη διαβρωτική νέκρωση. Επίσης, ο υψηλότερος ΗΑΙ διαπιστώθηκε ότι συσχετίζεται με μεγαλύτερη ίνωση (r=0.69, p<0.01). Παράλληλα, βρέθηκε ότι ο υψηλότερος HAI συσχετίζεται με υψηλότερες τιμές ALT πριν τη θεραπεία (T 0 ) (ΗΑΙ: r=0.47, p<0.01, διαβρωτική νέκρωση: r= 0.49, p<0.01, πυλαία φλεγμονή: r= 0.39, p<0.01, ενδολόβιες νεκρώσεις: r= 0.30, p<0.05). Η ίδια θετική συσχέτιση παρέμεινε μεταξύ της τιμής της ALT και του ΗΑΙ και 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (HAI: r=0.42, p<0.05, διαβρωτική νέκρωση: r=0.46, p<0.05, πυλαία φλεγμονή: r=0.42, p<0.05) αλλά δεν εξακολούθησε να υπάρχει συσχέτιση μεταξύ της τιμής της ALT (Τ 3 ) και των αρχικών ενδολόβιων νεκρώσεων της βιοψίας. 3.Ιικό φορτίο (HCV RNA) Το HCV RNA πριν την έναρξη της θεραπείας (Τ 0 ) βρέθηκε ότι συσχετίζεται με την τιμή της ALT (Τ 0 ) (r= 0.35, p<0.05). Επίσης, θετική ήταν η συσχέτιση 152
153 μεταξύ του ιικού φορτίου στους χρόνους Τ 1 και Τ 3 και 6 μήνες μετά το τέλος της θεραπείας (r= 0.67, p<0.01), αλλά και μεταξύ του HCV RNA στους χρόνους Τ 2 και Τ 3 (r= 0.50, p<0.05). Επίσης, το ιικό φορτίο 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) είχε θετική συσχέτιση με την τιμή της ALT στον ίδιο χρόνο (Τ 3 ) (r=0.52, p<0.01). 4.Αριθμός των Tregs (CD4+CD25+ high Foxp3+ T κύτταρα) στο περιφερικό αίμα Ο αριθμός των Τ ρυθμιστικών κυττάρων στο περιφερικό αίμα πριν την έναρξη της θεραπείας (Τ 0 ) δεν συσχετίστηκε με καμία από τις εξεταζόμενες παραμέτρους της μελέτης. Ωστόσο, στα επόμενα στιγμιότυπα διαπιστώθηκαν σημαντικές συσχετίσεις. Ειδικότερα, a) ο αριθμός των Tregs στο περιφερικό αίμα στο τέλος της θεραπείας (Τ 2 ): εμφάνισε θετική συσχέτιση με το τίτλο του HCV RNA στο στιγμιότυπο Τ 1 (εβδομάδα 4 ή 12 ανάλογα με τον γονότυπο, r= 0.43, p<0.05), δηλαδή οι ασθενείς οι οποίοι στον χρόνο Τ 1 είχαν υψηλότερο ιικό φορτίο, είχαν και μεγαλύτερο αριθμό Tregs στο περιφερικό αίμα στο τέλος της αγωγής (Τ 2 ). συσχετίστηκε αρνητικά με τον αριθμό των CD4+ και των CD4+CD28+ T κυττάρων στο ίδιο χρόνο (τέλος θεραπείας, Τ 2 : r=- 0.44, p<0.05 και r= -0.45, p< 0.05 αντίστοιχα), αλλά και 6 μήνες μετά το τέλος της αγωγής (Τ 3 : r= -0.49, p<0.05 και r= -0.50, p< 0.05 αντίστοιχα) είχε θετική συσχέτιση με τον αριθμό τους 6 μήνες μετά το τέλος της (Τ 3 ) (r=0,59, p<0.01) β) o αριθμός των Tregs στο περιφερικό αίμα 6 μήνες μετά το τέλος της αγωγής (Τ 3 ) είχε: αρνητική συσχέτιση με τον αριθμό των CD4+ T λεμφοκυττάρων πριν την έναρξη της θεραπείας (Τ 0 ) και κυρίως με τον αριθμό των CD4+ κυττάρων στον χρόνο Τ 1 κατά τη διάρκεια της θεραπείας (Τ 0 : r=-0.40, p<0.05 και Τ 1 : r=-0,55, p<0.01) αρνητική συσχέτιση με τον αριθμό των CD4+CD28+ T λεμφοκυττάρων στον χρόνο Τ 1 (r=-0,50, p<0.01) 153
154 θετική συσχέτιση με το ιικό φορτίο (HCV RNA) στον ίδιο χρόνο (Τ 3 ) (r= 0.47, p<0.05) θετική συσχέτιση με τον αριθμό των CD3+CD8+ Τ κυττάρων στον ίδιο χρόνο (Τ 3 ) (r= 0.43, p<0.05) αρνητική συσχέτιση με τον αριθμό των CD4+ Τ (Τ 3 ) και των CD4+CD28+ Τ κυττάρων (Τ 3 ) (CD4+: r=-0.56, p<0.01 και CD4+CD28+: r= -0.55, p< 0.01) γ) ο αριθμός των Tregs στο περιφερικό αίμα ήταν σημαντικά μεγαλύτερος στους ασθενείς με γονότυπο 2 και 3 στην 4 η εβδομάδα κατά τη διάρκεια της θεραπείας (Τ 1 ) (r=0,35, p<0,05). 5.Αριθμός των Tregs που αναπτύχθηκε στην in vitro καλλιέργεια του περιφερικού αίματος Ο αριθμός των Tregs που αναπτύχθηκε στην καλλιέργεια του περιφερικού αίματος πριν την έναρξη της θεραπείας (Τ 0 ) δεν εμφάνισε κάποια στατιστικά σημαντική συσχέτιση με κάποια από τις παραμέτρους που αξιολογήθηκαν στη μελέτη στο ίδιο χρόνο. Όμως, διαπιστώθηκε ότι ο αριθμός των Tregs που αναπτύχθηκε στην in vitro καλλιέργεια στον χρόνο Τ 1, ήταν μικρότερος στους ασθενείς που είχαν μεγαλύτερη ίνωση στη βιοψία ήπατος και μεγαλύτερο αριθμό Tregs στο περιφερικό αίμα στο τέλος της θεραπείας (Τ 2 ) (r=-0,55, p<0.05 και r=-0.81, p<0.01 αντίστοιχα). Επίσης, διαπιστώθηκε ότι στους άνδρες, στο τέλος της θεραπείας (Τ 2 ), υπήρχε η τάση ο αριθμός των Tregs να είναι μικρότερος στην in vitro καλλιέργεια (r=-0.83, p<0.05). 6.Αριθμός των κυτταροτοξικών CD3+CD8+ T λεμφοκυττάρων στο περιφερικό αίμα α) Ο αριθμός των CD3+CD8+ Τ λεμφοκυττάρων στο περιφερικό αίμα πριν την έναρξη της θεραπείας (Τ 0 ): είχε αρνητική συσχέτιση με τον πληθυσμό των CD4+, CD4+CD25+ και CD4+CD28+ Τ λεμφοκυττάρων (CD4+: r=-0.66, p<0.01, CD4+CD25+: r=-0.32, p<0.05 και CD4+CD28+: r=-0.64, p<0.01 αντίστοιχα). Όμως, στη συνέχεια στους χρόνους Τ 1, Τ 2 και Τ 3 η συσχέτιση παρέμεινε αρνητική μόνο μεταξύ των κυτταροτοξικών αρχικών, πριν από την έναρξη της θεραπείας, CD3+CD8+ T κυττάρων (Τ 0 ) και των CD4+ και CD4+CD28+ Τ υποπληθυσμών (Τ 1 : r=-0.53, p<0.01 και r=-0.59, p<0.01 αντίστοιχα, Τ 2 : r=-0.46, p<
155 και r=-0.51, p<0.01 αντίστοιχα, Τ 3 : r=-0.58, p<0.01 και r=-0.63, p<0.01 αντίστοιχα) είχε θετική συσχέτιση με τον αριθμό τους στους επόμενους χρόνους, δηλαδή όσο μεγαλύτερος ήταν ο αριθμός των κυτταροτοξικών CD3+CD8+ Τ λεμφοκυττάρων πριν την έναρξη της θεραπείας (Τ 0 ), τόσο μεγαλύτερος ήταν ο αριθμός αυτού του υποπληθυσμού και στους τρεις επόμενους χρόνους (Τ 1 : r=0.90, p<0.01, Τ 2 : r=0.76, p<0.01 και Τ 3 : r=0.87, p<0.01 αντίστοιχα). β) ο αριθμός των κυτταροτοξικών CD3+CD8+ T λεμφοκυττάρων στον χρόνο Τ 1 : είχε αρνητική συσχέτιση με τη διαβρωτική νέκρωση και την ίνωση στη βιοψία ήπατος, δηλαδή όσο μικρότερη ήταν η διαβρωτική νέκρωση και η ίνωση στη βιοψία ήπατος πριν την έναρξη της αγωγής, τόσο μεγαλύτερος ήταν ο αριθμός τους στον χρόνο Τ 1 (r=-0.40, p<0.05 και r=-0.43, p<0.01 αντίστοιχα). είχε αρνητική συσχέτιση με τον αριθμό των CD4+CD28+ Τ κυττάρων στο τέλος της θεραπείας (Τ 2 ) και 6 μήνες μετά (Τ 3 ) (Τ 2 : r=- 0.52, p<0.01, Τ 3 : r=-0.66, p<0.01). γ) Ο αριθμός των CD3+CD8+ T κυττάρων στο τέλος της θεραπείας (Τ 2 ): είχε θετική συσχέτιση με την τιμή της ALT στον ίδιο χρόνο (Τ 2 ) (r= 0.40, p< 0.05) δ) Ο πληθυσμός των CD3+CD8+ Τ κυττάρων 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ): είχε αρνητική συσχέτιση με τον αριθμό των CD4+ και των CD4+CD25+ Τ κυττάρων στον ίδιο χρόνο (Τ 3 ) (r= -0.45, p< 0.05 και r=-0.49, p< 0.01 αντίστοιχα) είχε θετική συσχέτιση με το ιικό φορτίο (HCV RNA) στον ίδιο χρόνο (Τ 3 ) (r=0.40, p<0.05) διαπιστώθηκε ότι α) οι άνδρες, στο τέλος της θεραπείας (Τ 2 ), είχαν την τάση να έχουν μικρότερο αριθμό κυτταροτοξικών CD3+CD8+ Τ λεμφοκυττάρων (r=-0.42, p<0.05) και β) οι ασθενείς με μικρότερη ηλικία είχαν την τάση να έχουν μεγαλύτερο αριθμό κυτταροτοξικών CD3+CD8+ Τ λεμφοκυττάρων κατά τη διάρκεια της θεραπείας και στο τέλος της (Τ 1 : r=-0.42, p<0.01, Τ 2 : r=-0.46, p<0.05). 155
156 7.Αριθμός των CD4+ T λεμφοκυττάρων στο περιφερικό αίμα α) Ο αριθμός των CD4+ Τ λεμφοκυττάρων στο περιφερικό αίμα, πριν την έναρξη της θεραπείας (Τ 0 ), εκτός από την προαναφερθείσα συσχέτιση με τα Tregs: βρέθηκε ότι σχετίζεται με μεγαλύτερους αριθμούς CD4+CD25+ και CD4+CD28+ Τ λεμφοκυττάρων πριν την έναρξη της θεραπείας (Τ0) (r=0.30, p<0.05 και r=0.95, p<0.01 αντίστοιχα). Στους επόμενους χρόνους της μελέτης όμως, ο αριθμός των CD4+ T λεμφοκυττάρων (Τ 0 ) βρέθηκε να εμφανίζει θετική σχέση μόνο με τον CD4+CD28+ Τ λεμφοκυτταρικό υποπληθυσμό (Τ 1 - Τ 3 ) (Τ 1 : r=0.57, p<0.01, Τ 2 : r=0.51, p<0.01, Τ 3 : r=0.72, p<0.01). είχε θετική συσχέτιση με τον αριθμό τους στους επόμενους χρόνους της μελέτης, δηλαδή όσο μεγαλύτερος ήταν ο αριθμός των CD4+ Τ κυττάρων πριν την έναρξη της θεραπείας (Τ 0 ), τόσο μεγαλύτερος ήταν ο αριθμός τους και στους υπόλοιπους χρόνους της μελέτης (Τ 1 : r=0.55, p<0.01, Τ 2 : r=0.44, p<0.05 και Τ 3 : r=0.67, p<0.01 αντίστοιχα) σχετίζεται αρνητικά με τον αριθμό των CD3+CD8+ T κυτταροτοξικών κυττάρων στον χρόνο Τ 1, στο τέλος της θεραπείας (Τ 2 ) και 6 μήνες μετά το τέλος της (Τ 3 ) (Τ 1 : r=-0.66 p<0.01, Τ 2 : r=-0.68, p<0.05, Τ 3 : r=-0.68, p<0.01) β) Ο αριθμός των CD4+ T κυττάρων του περιφερικού αίματος στον χρόνο Τ 1 είχε: αρνητική συσχέτιση με τον αριθμό των CD3+CD8+ T κυττάρων στον χρόνο Τ 1 (r=-0.39, p<0.05) αρνητική συσχέτιση με τον αριθμό των CD3+CD8+ T κυττάρων στο τέλος της θεραπείας και 6 μήνες μετά (Τ 2, Τ 3 ) (Τ2: r=-0.55, p<0.01, Τ 3 : r=-0.51, p<0.01) θετική συσχέτιση με τον αριθμό των CD4+ T κυττάρων στο τέλος της θεραπείας και 6 μήνες μετά (Τ 2, Τ 3 ) (Τ 2 : r=0.50, p<0.01, Τ 3 : r=0.68, p<0.01) θετική συσχέτιση με τον αριθμό των CD4+CD28+ Τ κυττάρων στα ίδια στιγμιότυπα (Τ 2, Τ 3 ) (Τ 2 : r=0.52, p<0.01, Τ 3 : r=0.72, p<0.01) 156
157 γ) Ο αριθμός των CD4+ T κυττάρων του περιφερικού αίματος στο τέλος της θεραπείας (Τ 2 ) είχε: αρνητική συσχέτιση με τον αριθμό των CD3+CD8+ T κυττάρων στον χρόνο Τ 1 (r= -0.47, p<0.05) και στο τέλος της θεραπείας (Τ 2 ) (r=-0.39, p<0.05) θετική συσχέτιση με τον αριθμό των CD4+CD28+ T κυττάρων στο τέλος της θεραπείας και 6 μήνες μετά (Τ 2 και Τ 3 ) (Τ 2 : r=0.97, p<0.01, Τ 3 : r=0.68, p<0.01) θετική συσχέτιση με τον αριθμό των CD4+ T κυττάρων 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (r=0.65, p<0.01) δ) Ο αριθμός των CD4+ T κυττάρων του περιφερικού αίματος 6 μήνες μετά το τέλος της αγωγής (Τ 3 ) είχε: αρνητική συσχέτιση με τον αριθμό των CD3+CD8+ T κυττάρων στον χρόνο Τ 1 (r=-0.61, p<0.01) θετική συσχέτιση με τον αριθμό των CD4+CD28+ T κυττάρων στο τέλος της θεραπείας (Τ 2 ) (r=0.70, p<0.01) Επιπρόσθετα, βρέθηκε ότι υπάρχει θετική συσχέτιση μεταξύ του HCV RNA πριν την έναρξη της θεραπείας (Τ 0 ) και του αριθμού των CD4+ Τ κυττάρων στο περιφερικό αίμα (Τ 0 ) (r=0.26, p<0.05). Τέλος, οι άνδρες ήταν πιο πιθανό να έχουν υψηλό αριθμό CD4+ T λεμφοκυττάρων πριν την έναρξη της θεραπείας (Τ 0 ) σε σχέση με τις γυναίκες (r=0.39, p<0.05). 8.Αριθμός των CD4+CD25+ T λεμφοκυττάρων στο περιφερικό αίμα α) Ο αριθμός των CD4+CD25+ Τ λεμφοκυττάρων του περιφερικού αίματος πριν την έναρξη της θεραπείας (Τ 0 ), εκτός από τις συσχετίσεις που ήδη αναφέρθηκαν, εμφανίζει: θετική συσχέτιση με τον αριθμό των CD4+CD28+ Τ λεμφοκυττάρων (r=0.36, p<0.01) την ίδια χρονική στιγμή (Τ 0 ) αρνητική συσχέτιση με τον αριθμό του CD4+CD25+ Τ υποπληθυσμού (r=-0.49, p<0.01) στον χρόνο Τ 1 αρνητική συσχέτιση με τον αριθμό των CD3+CD8+ Τ κυττάρων στο τέλος της θεραπείας (Τ 2 ) (r=-0.42, p<0.05) και 6 μήνες μετά (Τ 3 ) (r=-0.40, p<0.05) 157
158 αρνητική συσχέτιση με την τιμή της ALT στον χρόνο Τ 1 (r=-0.51, p<0.01) β) Ο αριθμός των CD4+CD25+ T κυττάρων στο περιφερικό αίμα στο τέλος της θεραπείας (Τ 2 ) είχε: θετική συσχέτιση με τον αριθμό των CD4+CD28 T κυττάρων στο τέλος της θεραπείας(τ 2 ) (r=0.56, p<0.01) θετική συσχέτιση με την τιμή της ALT στον χρόνο (Τ 1 ) (r=0.56, p<0.05) θετική συσχέτιση με τον αριθμό των CD4+ T λεμφοκυττάρων στο τέλος της θεραπείας (Τ 2 ) (r=0.51, p<0.01) γ) Ο αριθμός των CD4+CD25+ Τ κυττάρων στο περιφερικό αίμα 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) είχε: αρνητική συσχέτιση με την τιμή της ALT στον χρόνο (Τ 1 ) (r=-0.44, p<0.05) 9.Αριθμός των CD4+CD28+ T κυττάρων στο περιφερικό αίμα α) Ο αριθμός των CD4+CD28+ Τ κυττάρων του περιφερικού αίματος πριν την έναρξη της αγωγής (Τ 0 ) έχει: θετική συσχέτιση με το μέγεθος του πληθυσμού τους σε όλα τα στιγμιότυπα της μελέτης (Τ 1 : r=0.52, p<0.01, Τ 2 : r=0.51, p<0.01, Τ 3 : r=0.69, p<0.01), δηλαδή όσο περισσότερα CD4+CD28+ T κύτταρα υπήρχαν πριν τη θεραπεία, τόσο περισσότερα διαπιστώθηκαν και στα επόμενα στιγμιότυπα θετική συσχέτιση με τον αριθμό των CD4+ Τ κυττάρων επίσης σε όλα τα στιγμιότυπα της μελέτης (Τ 1 : r= 0.51, p<0.01, Τ 2 : r=0.43, p<0.05, Τ 3 : r= 0.64, p<0.01) θετική συσχέτιση με τo HCV RNA πριν την έναρξη της θεραπείας (Τ 0 ), δηλαδή όσο μεγαλύτερο ήταν το ιικό φορτίο, τόσο μεγαλύτερος ήταν και ο πληθυσμός τους (r=0.28, p<0.05) β) Ο αριθμός των CD4+CD28+ T κυττάρων του περιφερικού αίματος στον χρόνο Τ 1 διαπιστώθηκε ότι είχε: αρνητική συσχέτιση με το αριθμό των κυτταροτοξικών CD3+CD8+ T κυττάρων σε όλα τα επόμενα στιγμιότυπα της μελέτης (Τ 1, Τ 2, Τ 3 ) (Τ1: r=-0.44, p<0.01, Τ 2 : r= -0.53, p<0.01, Τ 3 : r= -0.50, p<0.05) 158
159 θετική συσχέτιση με τον αριθμό των CD4+ T κυττάρων επίσης σε όλα τα ακόλουθα στιγμιότυπα της μελέτης (Τ 1, Τ 2, Τ 3 ) (Τ 1 : r=0.88, p<0.01, Τ 2 : r= 0.88, p<0.05, Τ 3 : r= 0.67, p<0.01) θετική συσχέτιση με τον αριθμό των CD4+CD28+ T κυττάρων στο τέλος της θεραπείας και 6 μήνες μετά (Τ 2, Τ 3 ) (Τ 2 : r= 0.53, p<0.01, Τ 3 : r=0.69, p<0.01) γ) Ο πληθυσμός των CD4+CD28+ Τ κυττάρων του περιφερικού αίματος στο τέλος της θεραπείας (Τ 2 ) είχε: αρνητική συσχέτιση με τον αριθμό των CD3+CD8+ Τ κυττάρων στο τέλος της θεραπείας (Τ 2 ) (r=-0.46, p<0.05) θετική συσχέτιση με τον αριθμό των CD3+CD8+ Τ κυττάρων 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (r=0.69, p<0.01) δ) Ο αριθμός των CD4+CD28+ Τ κυττάρων 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) είχε: θετική συσχέτιση με τον αριθμό των CD4+ Τ κυττάρων 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (r= 0.94, p< 0.01) Επίσης, διαπιστώθηκε ότι οι άνδρες είναι πιο πιθανό να έχουν μεγαλύτερο αριθμό CD4+CD28+ T λεμφοκυττάρων πριν την έναρξη της θεραπείας (Τ 0 ) σε σχέση με τις γυναίκες (r=0.39, p<0.01). 10.Ασθενείς με γονότυπο 2 και 3 Οι ασθενείς με γονότυπο 2 και 3 διαπιστώθηκε ότι τείνουν να εμφανίζουν: υψηλότερη τιμή ALT και μεγαλύτερο αριθμό Tregs στο περιφερικό αίμα στον χρόνο Τ 1 (r=0.35, p<0.05, r=0.35, p<0.05) χαμηλότερο ιικό φορτίο 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) (r=-0.41, p<0.05) 159
160
161
162
163 Θηκογράμματα (Boxplots)- με βάση το φύλο Δεν υπάρχει σημαντική διαφορά ως προς τη μέση τιμή του ΗΑΙ μεταξύ ανδρών και γυναικών, αλλά στις γυναίκες το εύρος των τιμών του ΗΑΙ ήταν ελαφρώς μεγαλύτερο (γράφημα 32). Γράφημα 32: Σύγκριση της μέσης τιμής του HAI μεταξύ των ασθενών της μελέτης με βάση το φύλο Παρόμοια είναι η εικόνα και για την διαβρωτική νέκρωση, καθώς δεν φαίνεται να υπάρχει σημαντική διαφορά ως προς τη μέση τιμή της μεταξύ ανδρών και γυναικών, αλλά αυτή τη φορά οι άνδρες εμφανίζουν σημαντικά μεγαλύτερο εύρος της βαρύτητας της. Γράφημα 33: Σύγκριση της μέσης τιμής της διαβρωτικής νέκρωσης 163
164 Όσον αφορά στην πυλαία φλεγμονή και τις ενδολόβιες νεκρώσεις, δεν διαπιστώθηκαν σημαντικές διαφοροποιήσεις μεταξύ ανδρών και γυναικών, ως προς τους μέσους όρους ή το εύρος των τιμών τους. Γράφημα 34: Σύγκριση της μέσης τιμής της πυλαίας φλεγμονής Γράφημα 35: Σύγκριση της μέσης τιμής των ενδολόβιων νεκρώσεων 164
165 Εξετάζοντας τη μέση τιμή και το εύρος τιμών για τον αριθμό των Tregs στις τέσσερις μετρήσεις της μελέτης, προέκυψαν ενδιαφέρουσες αποκλίσεις μεταξύ ανδρών και γυναικών. Συγκεκριμένα, πριν την έναρξη της αγωγής, οι γυναίκες είχαν μικρότερο μέσο όρο αλλά και μικρότερο εύρος τιμών για τον πληθυσμό τους σε σχέση με τους άνδρες (Γράφημα 36). Η εικόνα αυτή, όμως, αντιστράφηκε, τουλάχιστον ως προς μέσο όρο, αμέσως μετά την έναρξη της θεραπείας (Τ 1 ), στο επιλεγμένο στιγμιότυπο 4 ή 12 εβδομάδες μετά (Γράφημα 37). Πάντως, διαπιστώθηκε αύξηση του εύρους του αριθμού των Tregs στο περιφερικό αίμα και στα δύο φύλα στη χρονική στιγμή Τ 1 σε σχέση με τη χρονική στιγμή Τ 0. T0 Γράφημα 36: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) στο περιφερικό αίμα μεταξύ ανδρών και γυναικών πριν την έναρξη της θεραπείας 165
166 T1 Γράφημα 37: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) στο περιφερικό αίμα μεταξύ ανδρών και γυναικών στο πρώτο χρονικό στιγμιότυπο της μελέτης μετά την έναρξη της θεραπείας Στο τέλος της θεραπείας (Τ 2 ), το εύρος των τιμών του μετρούμενου αριθμού των Tregs στο περιφερικό αίμα ελαχιστοποιήθηκε και στα δύο φύλα, παρότι οι γυναίκες εξακολουθούν να έχουν οριακά υψηλότερο μέσο όρο και οριακά μεγαλύτερο εύρος τιμών Tregs (Γράφημα 38). 166
167 Γράφημα 38: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) στο περιφερικό αίμα μεταξύ ανδρών και γυναικών στο τέλος της θεραπείας T3 T2 Γράφημα 38: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων) στο περιφερικό αίμα μεταξύ ανδρών και γυναικών 6 μήνες μετά το τέλος της θεραπείας 167
168 Η εικόνα μεταβλήθηκε εκ νέου σημαντικά 6 μήνες μετά τη λήξη της θεραπείας, καθώς οι γυναίκες εμφάνισαν σημαντικά χαμηλότερο μέσο όρο αλλά σημαντικά μεγαλύτερο εύρος για τον αριθμό των Tregs στο περιφερικό αίμα σε σχέση με τους άνδρες. Θηκογράμματα (Boxplots) με βάση το γονότυπο Η σύγκριση των ασθενών της μελέτης όσον αφορά το γονότυπο και την απάντηση, δεδομένου ότι όλοι οι ασθενείς με γονότυπο 2 και 3 πέτυχαν SVR ήταν εφικτή μόνο με την υποομάδα των ασθενών με γονότυπο 1 που ανταποκρίθηκαν στην αγωγή. Έτσι, όταν ομαδοποιήθηκαν οι ασθενείς της μελέτης με βάση το γονότυπο, προέκυψε ότι εκείνοι με γονότυπο 2 και 3 είχαν χαμηλότερο μέσο όρο ΗΑΙ αλλά μεγαλύτερο εύρος της βαρύτητας του ΗΑΙ σε σχέση με τους ασθενείς με γονότυπο 1. Γράφημα 39: Σύγκριση της μέσης τιμής του δείκτη νεκροφλεγμονώδους δραστηριότητας (ΗΑΙ) στη βιοψία ήπατος μεταξύ των ασθενών της μελέτης αναλόγως του γονότυπου του ιού 168
169 Η διαβρωτική νέκρωση και η πυλαία φλεγμονή, διαφέρουν μόνο όσον αφορά στο εύρος της βαρύτητας τους, δηλαδή οι ασθενείς με γονότυπο 1 έχουν μικρότερο εύρος σε σχέση με εκείνους με γονότυπο 2 και 3. Αντίθετα, η εικόνα μεταξύ των δυο ομάδων γονοτύπων είναι αρκετά ομοιογενής ως προς τις ενδολόβιες νεκρώσεις. Γράφημα 40: Σύγκριση της μέσης τιμής της νεκροφλεγμονώδους δραστηριότητας (ΗΑΙ) της βιοψίας ήπατος στους ασθενείς της μελέτης αναλόγως του γονοτύπου Γράφημα 41: Σύγκριση της μέσης τιμής της πυλαίας φλεγμονής της βιοψίας ήπατος στους ασθενείς της μελέτης αναλόγως του γονοτύπου του ιού 169
170 Γράφημα 42: Σύγκριση της μέσης τιμής των ενδολόβιων νεκρώσεων της βιοψίας ήπατος στους ασθενείς της μελέτης αναλόγως του γονοτύπου του ιού Η εξέταση του αριθμού των Tregs στο περιφερικό αίμα σε συνδυασμό με το γονότυπο αποκάλυψε ενδιαφέρουσες διαφοροποιήσεις. Συγκεκριμένα, όπως φαίνεται και στο Γράφημα 43, οι ασθενείς με γονότυπο 1 είχαν όχι μόνο μεγαλύτερο μέσο όρο Tregs πριν την έναρξη της θεραπείας αλλά και μεγαλύτερο εύρος στον αριθμό τους σε σχέση με τους ασθενείς με γονότυπο 2 και
171 T0 Γράφημα 43: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων στο περιφερικό αίμα των ασθενών της μελέτης πριν την έναρξη της θεραπείας αναλόγως του γονότυπου Στην πρώτη μέτρηση μετά την έναρξη της αγωγής, στο επιλεγμένο στιγμιότυπο την 12 η εβδομάδα, οι ασθενείς με γονότυπο 1 εξακολούθησαν να έχουν μεγαλύτερο μέσο όρο κι εύρος όσον αφορά στον αριθμό των Tregs στο περιφερικό αίμα, αλλά οι μετρήσεις ήταν κάτω από το μέσο όρο. 171
172 T1 Γράφημα 43: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων στο περιφερικό αίμα των ασθενών της μελέτης στο πρώτο χρονικό στιγμιότυπο μετά την έναρξη της θεραπείας αναλόγως του γονότυπου (w12/ w4) Στο τέλος της θεραπείας (Τ 2 ), οι ασθενείς με γονότυπο 2 και 3 εξακολούθησαν να έχουν μικρότερο μέσο όρο όσον αφορά στον αριθμό των Tregs στο περιφερικό αίμα, αλλά διαπιστώθηκε μεγαλύτερο εύρος στις μετρήσεις. 172
173 Γράφημα 44: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων στο περιφερικό αίμα των ασθενών της μελέτης στο τέλος της θεραπείας αναλόγως του γονότυπου Έξι μήνες μετά το τέλος της θεραπείας, οι ασθενείς με γονότυπο 1 εξακολούθησαν να έχουν μεγαλύτερο μέσο όρο όσον αφορά στον αριθμό των Tregs, αλλά διαπιστώθηκε εκ νέου μεγαλύτερο εύρος των μετρήσεων σε σχέση με τους ασθενείς με γονότυπο 3. T3 T2 173
174 Γράφημα 45: Σύγκριση της μέσης τιμής του αριθμού των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων στο περιφερικό αίμα των ασθενών της μελέτης 6 μήνες μετά το τέλος της θεραπείας αναλόγως του γονότυπου Στη συνέχεια, παρατίθενται τα boxplots που εξετάζουν το συνδυασμό των γονοτύπων και όλων των υπολοίπων υποπληθυσμών που μελετήθηκαν, πλην του υποπληθυσμού των CD4+CD28+ Τ κυττάρων επειδή η περαιτέρω ερμηνεία των ευρημάτων σε σχέση με αυτό τον υποπληθυσμό χωρίς προσδιορισμό του συνδέτη του θα ήταν εξαιρετικά δυσχερής. Οι συγκρίσεις που ακολουθούν αφορούν τους ασθενείς που ανταποκρίθηκαν στη θεραπεία. Όσον αφορά στον αριθμό των CD3+CD8+ Τ λεμφοκυττάρων, πριν την έναρξη της αγωγής (Τ 0 ), οι ασθενείς με γονότυπο 1 είχαν οριακά μεγαλύτερο μέσο όρο και μεγαλύτερο εύρος μετρήσεων σε σχέση με εκείνους με γονότυπο 2 και 3. T0 Γράφημα 46: Σύγκριση της μέσης τιμής του αριθμού των CD3+CD8+ T κυτταροτοξικών λεμφοκυττάρων πριν την έναρξη της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν στην θεραπεία αναλόγως του γονοτύπου του ιού Στην πρώτη μέτρηση μετά την έναρξη της αγωγής, όλοι οι ασθενείς είχαν μικρότερο μέσο όρο όσον αφορά στον αριθμό των CD3+CD8+ Τ 174
175 λεμφοκυττάρων σε σχέση με τη χρονική στιγμή Τ 0, αντίστοιχο μεταξύ τους, ενώ εκείνοι με γονότυπο 2 και 3 έχουν ελαφρά μεγαλύτερο εύρος παρατηρήσεων. T1 Γράφημα 47: Σύγκριση της μέσης τιμής του αριθμού των CD3+CD8+ T κυτταροτοξικών λεμφοκυττάρων πριν την έναρξη της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Στο τέλος της θεραπείας (Τ 2 ) οι ασθενείς με γονότυπο 2 και 3 είχαν οριακά μικρότερο αριθμό CD3+CD8+ Τ κυττάρων σε σχέση με τις προηγούμενες μετρήσεις αλλά και με τους ασθενείς με γονότυπο
176 T2 Γράφημα 48: Σύγκριση της μέσης τιμής του αριθμού των CD3+CD8+ T κυτταροτοξικών λεμφοκυττάρων στο τέλος της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Στους ασθενείς με γονότυπο 2 και 3, 6 μήνες μετά τη διακοπή της θεραπείας, ο μέσος όρος του αριθμού των CD3+CD8+ Τ κυττάρων επανήλθε στα αρχικά σχεδόν επίπεδα. Στους ασθενείς με γονότυπο 1, παρά το ότι διαπιστώθηκε αύξηση του πληθυσμού τους 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) σε σχέση με τον αριθμό τους στο τέλος της (Τ 2 ), η μέση τιμή του πληθυσμού τους ήταν μικρότερη σε σχέση με εκείνη πριν την έναρξη της θεραπείας (Τ 0 ). 176
177 Γράφημα 49: Σύγκριση της μέσης τιμής του αριθμού των CD3+CD8+ T κυτταροτοξικών λεμφοκυττάρων 6 μήνες μετά το τέλος της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν στην θεραπεία αναλόγως του γονοτύπου του ιού Ο πληθυσμός των CD4+ Τ κυττάρων στο περιφερικό αίμα πριν την έναρξη της θεραπείας (Τ 0 ) ήταν ελαφρώς μεγαλύτερος στους ασθενείς με γονότυπο 1 και παρουσίαζε μεγαλύτερο εύρος σε σχέση με τους ασθενείς με γονότυπο 2 και 3. T0 T3 177
178 Γράφημα 50: Σύγκριση της μέσης τιμής του αριθμού των CD4+ T λεμφοκυττάρων πριν την έναρξη της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Στο επιλεγμένο στιγμιότυπο κατά τη διάρκεια της θεραπείας (Τ 1 ), παρότι ο μέσος όρος του αριθμού των CD4+ Τ κυττάρων είναι σχεδόν ίδιος (αν και μεγαλύτερος από τη χρονική στιγμή Τ 0 ), ανεξαρτήτως γονότυπου, οι περισσότεροι ασθενείς με γονότυπο 2 και 3 είχαν μικρότερο πληθυσμό CD4+ T κυττάρων. CD4+ T1 Γράφημα 51: Σύγκριση της μέσης τιμής του αριθμού των CD4+ T λεμφοκυττάρων στο πρώτο χρονικό στιγμιότυπο μετά την έναρξη της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Στο τέλος της θεραπείας (Τ 2 ) ο μέσος όρος του αριθμού των CD4+ Τ κυττάρων διαπιστώθηκε ότι, ανεξαρτήτως γονότυπου, μειώνεται. Η ελάττωση του αριθμού ήταν σημαντικά μεγαλύτερη στους ασθενείς με γονότυπο
179 Γράφημα 52: Σύγκριση της μέσης τιμής του αριθμού των CD4+ T λεμφοκυττάρων στο τέλος της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Οι ασθενείς με γονότυπο 2 και 3, 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ), είχαν μικρότερο μέσο όρο κι εύρος παρατηρήσεων στον αριθμό των CD4+ Τ κυττάρων, όχι μόνο σε σχέση με τους ασθενείς με γονότυπο 1 αλλά και σε σχέση με τον αριθμό των CD4+ T κυττάρων των ίδιων πριν την έναρξη της θεραπείας (Τ 0 ). CD4+ T3 CD4+ T2 179
180 Γράφημα 53: Σύγκριση της μέσης τιμής του αριθμού των CD4+ T λεμφοκυττάρων 6 μήνες μετά το τέλος της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Οι ασθενείς με γονότυπο 1, πριν την έναρξη της θεραπείας (Τ 0 ), είχαν μεγαλύτερο μέσο όρο και μεγαλύτερο εύρος παρατηρήσεων για τον αριθμό των CD4+CD25+ Τ κυττάρων σε σχέση με τους ασθενείς με γονότυπο 2 και 3. T0 Γράφημα 54: Σύγκριση της μέσης τιμής του αριθμού των CD4+CD25+ T λεμφοκυττάρων πριν την έναρξη της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Στο επιλεγμένο στιγμιότυπο κατά τη διάρκεια της θεραπείας (Τ 1 ), η μέση τιμή του αριθμού των CD4+CD25+ Τ κυττάρων αυξήθηκε σε όλους τους ασθενείς, αλλά η μεγαλύτερη αύξηση αφορούσε στους ασθενείς με γονότυπο
181 T2 T1 Γράφημα 55: Σύγκριση της μέσης τιμής του αριθμού των CD4+CD25+ T λεμφοκυττάρων στο πρώτο στιγμιότυπο της μελέτης μετά την έναρξη της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Στους ασθενείς με γονότυπο 1 διαπιστώθηκε σημαντικότατη μείωση του μέσου όρου του αριθμού των CD4+CD25+ T κυττάρων στο τέλος της θεραπείας (Τ 2 ), σε σχέση με τον πληθυσμό τους στο επιλεγμένο στιγμιότυπο κατά τη διάρκεια της θεραπείας (Τ 1 ) και σε σχέση με τους ασθενείς με γονότυπο 2 και
182 Γράφημα 56: Σύγκριση της μέσης τιμής του αριθμού των CD4+CD25+ T λεμφοκυττάρων στο τέλος της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού Οι ασθενείς με γονότυπο 2 και 3, 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ), είχαν μικρότερο μέσο όρο στον αριθμό των CD4+CD25+ Τ κυττάρων σε σχέση με τους ασθενείς με γονότυπο 1. Ωστόσο, ο μέσος όρος του αριθμού των CD4+CD25+ Τ κυττάρων ήταν μικρότερος σε αυτό το στιγμιότυπο σε σχέση με τον πληθυσμό τους πριν από την έναρξη της αγωγής (Τ 0 ), ανεξαρτήτως γονότυπου. T3 Γράφημα 57: Σύγκριση της μέσης τιμής του αριθμού των CD4+CD25+ T λεμφοκυττάρων 6 μήνες μετά το τέλος της θεραπείας στους ασθενείς της μελέτης που ανταποκρίθηκαν αναλόγως του γονοτύπου του ιού 182
183 VI. ΣΥΖΗΤΗΣΗ Ο ιός της ηπατίτιδας C (HCV), είναι ένας μικρός RNA ιός που ανακαλύφθηκε το 1989 και σήμερα αποτελεί μια από τις πιο κοινές αιτίες χρόνιας ηπατικής νόσου παγκόσμια. Ο HCV χαρακτηρίζεται από μεγάλη ποικιλομορφία στη νουκλεοτιδική του αλληλουχία με αποτέλεσμα να διακρίνονται 6 διαφορετικοί γονότυποι αυτού (1-6), με τον πιο συχνό να είναι ο γονότυπος Επιπλέον χαρακτηρίζεται από το υψηλό ποσοστό εμφάνισης αυτόματων μεταλλάξεων με αποτέλεσμα να επικρατεί γενετική ετερογένεια. Στον ιό της ηπατίτιδας C αποδόθηκε το μεγαλύτερο ποσοστό της ηπατίτιδας που παλιότερα ονομάζονταν μη-α μη-β 6. Παρά το γεγονός ότι οι ασθενείς με χρόνια ηπατίτιδα C παραμένουν ασυμπτωματικοί για πολλά ίσως χρόνια, η λοίμωξη με τον HCV ευθύνεται για τη χρόνια ηπατική νόσο, η οποία κυμαίνεται από την χρόνια ηπατίτιδα και την κίρρωση του ήπατος έως και τον ηπατοκυτταρικό καρκίνο και αποτελεί την συχνότερη αιτία μεταμόσχευσης ήπατος στον κόσμο 1. Ο ιός μεταδίδεται κατά κύριο λόγο αιματογενώς, δηλαδή με μετάγγιση αίματος και παραγώγων αίματος, χρήση μολυσμένων ενδοφλέβιων συρίγγων, συνεδρίες αιμοκάθαρσης ή μεταμόσχευση συμπαγών οργάνων και σπανιότερα μέσω έκθεσης των βλεννογόνων σε μολυσμένο αίμα (π.χ. κατά τον τοκετό ή τη σεξουαλική πράξη). Η μετάδοση του ιού έχει περιοριστεί σε σημαντικό βαθμό μετά την ανακάλυψη του ιού το 1989 οπότε έγινε εφικτός ο ορολογικός και μοριακός έλεγχος του αίματος και των παραγώγων του, ενώ σε αυτό συνέβαλε και η βελτίωση των συνθηκών νοσηλείας και αποστείρωσης και ο υποχρεωτικός έλεγχος των ατόμων που ανήκουν σε ομάδες υψηλού κινδύνου για την λοίμωξη. Σήμερα ο υψηλότερος επιπολασμός της χρόνιας HCV λοίμωξης αφορά σε άτομα ηλικίας άνω των 40 ετών που έχουν εκτεθεί στον ιό από μολυσμένα παράγωγα αίματος και δυστυχώς αρκετοί από αυτούς τους ασθενείς εξακολουθούν να είναι αδιάγνωστοι. Όμως, τα υψηλότερα ποσοστά της HCV λοίμωξης σήμερα καταγράφονται μεταξύ των ατόμων που μολύνονται λόγω ενδοφλέβιας χρήσης ουσιών, τα οποία συνήθως είναι νεαρότερης ηλικίας 12, 13. Ο επιπολασμός της χρόνιας HCV λοίμωξης ποικίλει από χώρα σε χώρα με την παγκόσμια συχνότητα να είναι περίπου 3.0% 1, και στην Ελλάδα, παρά το γεγονός ότι ο ακριβής επιπολασμός της HCV λοίμωξης δεν είναι γνωστός, υπολογίζεται ότι κυμαίνεται περίπου στο 2.0% και εμφανίζει μεγάλη γεωγραφική διακύμανση 3, 4,
184 Έτσι αν και η συνολική μετάδοση του ιού έχει μειωθεί από το 1990 και μετά, ουσιαστικά, ο αριθμός των νέων διαγνώσεων αναμένεται να αυξηθεί τα επόμενα χρόνια λόγω της μεγαλύτερης διάρκειας των παλιών λοιμώξεων και κατά συνέπεια λόγω εμφάνισης επιπλοκών από τη χρόνια ηπατοπάθεια και της αύξησης της ενδοφλέβιας χρήσης ουσιών. Επιπλέον υπολογίζεται ότι διαχρονικά, η χρόνια ηπατίτιδα C θα επιφέρει >50% αύξηση της συχνότητας κίρρωσης, >100% αύξηση της συχνότητας της μη αντιρροπούμενης κίρρωσης, >50% αύξηση της εμφάνισης του ηπατοκυτταρικού καρκινώματος και >500% αύξηση των αναγκών για μεταμόσχευση ήπατος. Η οξεία και η χρόνια λοίμωξη από τον HCV, στη συντριπτική της πλειοψηφία, είναι ασυμπτωματική και διαδράμει υποκλινικά με συνέπεια να μην γίνεται αντιληπτή, γεγονός που δυσχεραίνει και τη μελέτη της φυσικής ιστορίας της νόσου και της εξέλιξης της. Δυστυχώς οι περισσότεροι ασθενείς εγκαθιστούν χρόνια λοίμωξη (60-85%) κυρίως λόγω της απουσίας ισχυρής ανοσολογικής απάντησης και των ιδιαίτερων χαρακτηριστικών του ιού 25, 29. Η ταχύτητα εξέλιξης της ίνωσης στο ήπαρ και η εγκατάσταση κίρρωσης είναι βραδεία αλλά δεν είναι γραμμική και εξαρτάται από πολλούς παράγοντες 32, 33, 34, 35, 36 Επιπλέον η κίρρωση του ήπατος φαίνεται ότι προάγει την εγκατάσταση ηπατοκυτταρικού καρκινώματος (1-4%) 37. Συνεπώς, συνολικά, η χρόνια HCV λοίμωξη δημιουργεί μια πολυποίκιλη και σημαντική επιβάρυνση της δημόσιας υγείας και των δαπανών της γι αυτό και όλοι οι ασθενείς με χρόνια ηπατίτιδα C είναι υποψήφιοι για θεραπεία, εκτός τους ασθενείς με απόλυτες αντενδείξεις θεραπείας 43. Η θεραπεία σήμερα συνίσταται στην χορήγηση πεγκυλιωμένης ιντερφερόνης-α και ριμπαβιρίνης 44, 45. Οι ασθενείς με γονότυπο 1 και 4 υποβάλλονται σε θεραπεία 48 εβδομάδων, ενώ οι ασθενείς με γονότυπο 2 και 3 σε θεραπεία 24 εβδομάδων και το ποσοστό απάντησης, δηλαδή επίτευξης μακράς ιολογικής απάντησης (SVR), είναι αντίστοιχα 30-40% και 85%. Εντούτοις, το συνολικό ποσοστό απάντησης στην θεραπεία δεν ξεπερνά το 50% του συνόλου των ασθενών. Στο σχεδιασμό της θεραπευτικής στρατηγικής έχει αποδειχθεί ότι είναι κρίσιμη η αρνητικοποίηση του ιικού φορτίου την τέταρτη και τη δωδέκατη εβδομάδα της θεραπείας (RVR, EVR αντίστοιχα) 46, 47, 48, 49.. Για τους παραπάνω λόγους μεγάλο κομμάτι της έρευνας εμβαθύνει στη μελέτη της ανοσοπαθογένειας της ΗCV λοίμωξης και των μηχανισμών που ευθύνονται για το μεγάλο ποσοστό εγκατάστασης χρονιότητας ή και μη απάντησης στη θεραπεία. 184
185 Όπως είναι γνωστό, οι ιοί, αφού διαπεράσουν τους φυσικούς φραγμούς έρχονται αντιμέτωποι με μηχανισμούς της φυσικής ανοσίας η οποία αποτελεί την πρώτη γραμμή άμυνας του ξενιστή. Στους μηχανισμούς αυτούς συμμετέχουν τα φαγοκύτταρα και τα πολυμορφοπύρηνα κύτταρα, το συμπλήρωμα και οι χημειοτακτικοί παράγοντες. Επιπλέον τα μολυσμένα από τον ιό κύτταρα προάγουν την παραγωγή των ιντερφερονών τύπου Ι και την επαγωγή ρυθμιστικών μορίων που ενεργοποιούν γονίδια παραγωγής ιντερφερόνης έως ότου ενεργοποιηθούν και αναλάβουν οι μηχανισμοί της επίκτητης ανοσίας την κάθαρση του παθογόνου. Η επίκτητη ανοσιακή απάντηση έναντι των ενδοκυττάριων παθογόνων, που δρουν κυτταροτοξικά ή μη, αποτελείται από τη χυμική ανοσία, η οποία χρησιμοποιεί τα Β λεμφοκύτταρα για την παραγωγή αντισωμάτων έναντι αντιγόνων και τοξινών των παθογόνων και από την κυτταρική ανοσία η οποία χρησιμοποιεί κατά κύριο λόγο τα Τ λεμφοκύτταρα. Τα τελευταία διακρίνονται σε δύο κύριους υποπληθυσμούς: τα CD8+ T κυτταροτοξικά λεμφοκύτταρα και τα CD4+ T βοηθητικά λεμφοκύτταρα. Τα βοηθητικά λεμφοκύτταρα έχουν κεντρικό ρόλο στο ανοσιακό σύστημα και είναι γνωστό ότι διακρίνονται σε πολλούς επιμέρους υποπληθυσμούς με πολλαπλές δράσεις (ncd4+, ntregs, NKT, Th1, Th2, Th17, itregs) και όπως διαφαίνεται χαρακτηρίζονται από πλαστικότητα στη λειτουργική τους διαφοροποίηση 56, 57, 58, 59. Η συνολική ανοσιακή απόκριση της φυσικής και την επίκτητης ανοσίας, σε συνδυασμό με τα ιδιαίτερα χαρακτηριστικά του κάθε ενδοκυττάριου παθογόνου και τους μηχανισμούς διαφυγής που αυτό διαθέτει, τελικά καθορίζει την έκβαση της λοίμωξης (εξουδετέρωση και κάθαρση του ιού, παραμονή ως λανθάνουσα λοίμωξη με συνοδό ύπαρξη επαρκούς αντισωματικής απάντησης, ή χρόνια λοίμωξη) 54, 55. Όσον αφορά στην HCV λοίμωξη, παρά το γεγονός ότι ενεργοποιούνται οι μηχανισμοί της φυσικής ανοσίας, ο ιός χρησιμοποιεί διάφορους μηχανισμούς διαφυγής από τη δράση των ιντερφερονών α) μπλοκάροντας τους μεταγραφικούς τους παράγοντες, β) προάγοντας την έκκριση κυτταροκινών που αναστέλλουν τη δράση τους, γ) αναστέλλοντας τις ενδοκυττάριες οδούς σηματοδότησης που οδηγούν στην παρεμπόδιση της απόπτωσης των μολυσμένων ηπατοκυττάρων και δ) μειώνοντας τον αριθμό των πλασματοκυτταροειδών δενδριτικών κυττάρων (pdc) και την ικανότητα τους να παράγουν ιντερφερόνη-α και διαταράσσοντας την ωρίμανση και την ικανότητα αντιγονοπαρουσίασης των συμβατικών δενδριτικών κυττάρων (cdc). 185
186 Το αποτέλεσμα της διαταραχής της φυσικής ανοσίας έναντι του HCV μετά τη έκθεση στον ιό, οδηγεί στην καθυστερημένη ενεργοποίηση των κυττάρων της επίκτητης ανοσίας, με αποτέλεσμα τα ειδικά έναντι του ιού T κύτταρα να ανιχνεύονται περίπου 5-9 εβδομάδες μετά, ενώ τα ειδικά έναντι του HCV αντισώματα μετά από 8-20 εβδομάδες 69, 70. Επιπλέον, λόγω της μεγάλης μεταβλητότητας του ιού τα αντισώματα -στη μεγάλη τους πλειοψηφία- δεν είναι εξουδετερωτικά 71. Στην οξεία HCV λοίμωξη, η κάθαρση του ιού επιτυγχάνεται από ειδικά έναντι του ιού CD8+ T κύτταρα 28, 69 τα οποία είτε εκκρίνουν IFN-γ για να καταστρέψουν τα μολυσμένα κύτταρα χωρίς κυτταρόλυση, είτε χρησιμοποιούν την οδό Fas, τη δράση περφορίνης και granzyme και την απόπτωση μέσω συνδετών του TNF-α για να προκαλέσουν την κυτταρόλυση των μολυσμένων κυττάρων. Επίσης έχει διαπιστωθεί ότι προκειμένου να είναι αποτελεσματικά τα CD8+ T κύτταρα, πρέπει να συνυπάρχει και μια ισχυρή και ειδική έναντι πολλαπλών επιτόπων του HCV κυτταρική απάντηση. Στην χρόνια HCV λοίμωξη, τα CD8+ T κύτταρα είναι λιγότερα σε αριθμό, δεν είναι πλήρως λειτουργικά και η ειδικότητα τους έναντι επιτόπων του ιού έχει περιορισμένο εύρος 28, 79, 80. Ο ιός φαίνεται πως επιτυγχάνει μια ισορροπία μεταξύ της ανοσιακής απόκρισης του ξενιστή, και συγκεκριμένα των ειδικών έναντι αυτού CD8+ T κυττάρων, και των μηχανισμών που χρησιμοποιεί για να διαφύγει προκειμένου να διατηρήσει τη χρονιότητα. Σε αυτό το αποτέλεσμα πιθανολογείται ότι συντείνουν πολλοί λόγοι όπως: α) Η πρωτοπαθής ανεπάρκεια και/ ή εξάντληση των CD8+ T κυττάρων β) η έλλειψη βοήθειας προς τα CD8+ T κύτταρα από τα CD4+ T κύτταρα, καθώς έχει βρεθεί ότι στη χρόνια HCV λοίμωξη οι CD4+ T κυτταρικές αποκρίσεις είναι ασθενείς και δυσλειτουργικές, έχουν μικρότερο εύρος και διαρκούν λίγο σε αντίθεση με την οξεία HCV λοίμωξη η οποία αυτοπεριορίζεται, όπου και παρατηρούνται έντονες και ειδικές έναντι πολλαπλών επιτόπων του HCV απαντήσεις που διατηρούνται για καιρό 84, 85 γ) η συμβολή ανασταλτικών υποδοχέων όπως ο PD-1, το οποίο ως μόριο αρνητικής σηματοδότησης παρεμποδίζει τις Τ κυτταρικές αποκρίσεις
187 δ) ο χρόνιος αντιγονικός ερεθισμός από τα σταθερά υψηλά επίπεδα ιικών αντιγόνων που οδηγεί σε χρόνια ενεργοποίηση των Τ κυττάρων τα οποία σταδιακά μειώνουν τη δραστική τους λειτουργία με πρώτη εκδήλωση τη μείωση της παραγωγής IL-2 και ακολούθως της κυτταροτοξικότητας, της παραγωγής TNF-a και IFN-γ ε) η αναστολή από ανασταλτικές κυτταροκίνες όπως η IL-10, τα επίπεδα της οποίας είναι αυξημένα στην χρόνια HCV λοίμωξη 66, 68,95-99, 100 στ) οι μεταλλάξεις διαφυγής του ιού, οι οποίες συμβαίνουν πιθανώς επειδή καθυστερεί η ειδική έναντι του ιού T κυτταρική απόκριση 101, 102 η οποία ευοδώνει τη μεταβλητότητα του, καθώς η πίεση επιλογής στον ιό επηρεάζεται από τα CD8+ T κύτταρα που αναγνωρίζουν συγκεκριμένα HLA μόρια 103, 104, 105, από την παρουσία εξουδετερωτικών αντισωμάτων αλλά και από την επίδραση του ίδιου του HCV στα Τ κύτταρα καθώς φαίνεται πως επηρεάζει την σύνδεση των αντιγόνων, την επεξεργασία τους και την παρουσίαση τους από τα αντιγονοπαρουσιαστικά κύτταρα, άρα και την αναγνώριση τους από τα CD8+ και CD4+ T κύτταρα 70, 101, 103. ζ) η καταστολή από πρωτεΐνες του ιού, η οποία οδηγεί σε μείωση του πολλαπλασιασμού των δραστικών κυττάρων, μείωση της παραγωγής IL-2, παρεμπόδιση της ενεργοποίησης των ΝΚ κυττάρων και ελαττωματική αντιγονοπαρουσίαση 7, 68, 96. η) η διαταραχή στην ωρίμανση των CD8+ T κυττάρων, τα οποία έχουν ανώριμο φαινότυπο 106 με αποτέλεσμα να έχουν και ελαττωμένη κυτταροτοξική δράση θ) το ηπατικό μικροπεριβάλλον το οποίο επάγει ανοχή (tolerogenic), πιθανώς μέσω των Τ κυττάρων των ηπατικών κολποειδών (liver sinusoidal endothelial cells, LSEC), των κυττάρων του Kuppfer, των αστεροειδών κυττάρων και των δενδριτικών που είναι εγκατεστημένα εκεί 107, 108. ι) η δράση των T ρυθμιστικών κυττάρων (Tregs) Τα T ρυθμιστικά κύτταρα (T regulatory cells, Tregs) είναι ένας υποπληθυσμός κυττάρων που πρωτοαναφέρθηκε στη βιβλιογραφία τη δεκαετία του 1970 από τους Gershon και Kondo 113 ως κατασταλτικά κύτταρα, αλλά μελετήθηκαν ενδελεχώς μετά το Επρόκειτο για υποπληθυσμό των CD4+ T κυττάρων που χαρακτηρίστηκε από την υψηλή έκφραση της α αλυσίδας του υποδοχέα της IL-2 (CD25+) 126, 128, 129 και ένα λειτουργικό φαινότυπο με μειωμένη απόκριση σε in vitro αλλογενή και πολυκλωνική διέγερση του TCR υποδοχέα. Τα κύτταρα 187
188 αυτά ασκούν μη αντιγονο-ειδική κατασταλτική δράση έναντι των δραστικών Τ κυττάρων μετά από επανειλημμένη διέγερση του TCR Η καινοτομία στην προσέγγιση της λειτουργίας και την απομόνωση των Tregs έγινε το 2001 μετά την ανακάλυψη του γονιδίου FOXP3 στα ποντίκια, όπου διαπιστώθηκε ότι μια σημειακή μετάλλαξη αυτού στο Χ χρωμόσωμα οδηγεί αυτόματα σε εμφάνιση ενός σοβαρού αυτοάνοσου συνδρόμου στα άρρενα ποντίκια 134 και παρομοίως, μετά από μεταλλάξεις στο ανθρώπειο γονίδιο FOXP3, προκαλούνται δυο σοβαρά πολυοργανικά αυτοάνοσα σύνδρομα στους ανθρώπους (IPEX, XLAAD) 119, Το FOXP3 είναι ένα γονίδιο με ρόλο κλειδί για τη δημιουργία του υποπληθυσμού των Treg κυττάρων και τη διατήρηση των λειτουργικών τους χαρακτηριστικών 142, 143. Θεωρείται ότι αντιπροσωπεύει τον «κυρίαρχο ρυθμιστή» («master regulator») ή τον παράγοντα που είναι υπεύθυνος για την επιλογή του κλώνου («lineage-specification factor») των «φυσικών» Τ ρυθμιστικών κυττάρων (naive T regulatory cells, ntregs) στο θύμο. Επιπλέον, έχει πολύ μεγάλη σημασία για την ανάπτυξη των Tregs, την κλωνική τους επέκταση και τη λειτουργικότητα τους. Η ανάπτυξη του μονοκλωνικού αντισώματος 151 έναντι της πρωτεΐνης που κωδικοποιεί το γονίδιο FOXP3 επέτρεψε την ενδοκυττάρια σήμανση (staining) των Tregs 150. Σήμερα είναι γνωστό πως τα Tregs αποτελούν μια ετερογενή ομάδα κυττάρων και περιλαμβάνουν α) τα «φυσικά αναδυόμενα» ή ntregs (natural Tregs) τα οποία είναι CD4+CD25+Foxp3+ Tregs και παράγονται στο θύμο 123, 128, 152, 153 και β) τα επαγόμενα Tregs (induced Tregs, itregs) 177, , 183, 188, 195, 206 τα οποία δημιουργούνται από κλωνική επέκταση στην περιφέρεια και διακρίνονται σε πολλές υπο-ομάδες κυττάρων, τα οποία είναι είτε CD4+ είτε CD8+ T κύτταρα. Το κύριο χαρακτηριστικό τους είναι η ανεργία και η κατασταλτική δράση και φαίνεται πως ασκούν αποτελεσματικό έλεγχο στις ανοσιακές αποκρίσεις έναντι ίδιων και μη αντιγόνων και εμπλέκονται σε λοιμώξεις, νεοπλασίες, αλλά και στην ανοσοκαταστολή μετά από την μεταμόσχευση. Η κατασταλτική δράση έναντι των κυττάρων- στόχων ασκείται μέσω α) της IL-2 219, , β) άμεσης κυτταρο-κυτταρικής επαφής με τα κύτταρα- στόχος μέσω μορίων όπως CTLA-4 190, 228, TGF-β συνδεδεμένο με τη μεμβράνη τους 168, 234, 235, LAG-3 171, 236, GITR 227, , PD και άμεσης κυτταρόλυσης , γ) παραγωγής ανοσοκατασταλτικών κυτταροκινών, όπως ο διαλυτός TGF-β , η IL και η IL , δ) μεταβολικής διάσπασης και ε) μέσω έμμεσων μηχανισμών
189 Τελικά, ο μηχανισμός που εμπλέκεται κάθε φορά φαίνεται ότι μάλλον εξαρτάται από τη φύση του ερεθίσματος που προκαλεί την φλεγμονή, την ποσότητα και το είδος του αντιγόνου, την κατάσταση ενεργοποίησης και τον αριθμό των κυττάρων-στόχων και βέβαια τον ιστό που εμπλέκεται στη φλεγμονώδη διαδικασία, δηλαδή από το «είδος» του «φλεγμονώδους» μικροπεριβάλλοντος 217, 218 Τα Tregs όπως έχει προαναφερθεί είναι ένα από τα πιθανολογούμενα αίτια για την αποτυχία των ειδικών έναντι του ιού CD8+ T κυττάρων να ελέγξουν αποτελεσματικά την HCV λοίμωξη 75, 76, 307. Έχει διαπιστωθεί ότι στην οξεία HCV λοίμωξη τα Tregs επεκτείνονται κλωνικά και αυξάνουν τον αριθμό και την ένταση της κατασταλτικής τους δράση, κυρίως στους ασθενείς με γονότυπο 1, όταν η λοίμωξη εξακολουθεί να είναι ενεργός 6 μήνες μετά τη μόλυνση (θετικό HCV RNA) 308 κατά συνέπεια πιθανολογείται ότι ίσως υπάρχει κάποια επίδραση του συγκεκριμένου γονότυπου στην εγκατάσταση της χρονιότητας μέσω της πυροδότησης των Tregs 309. Επίσης, έχει διαπιστωθεί ότι η ένταση της κατασταλτικής τους δράσης 6 μήνες μετά την αποδρομή της οξείας HCV λοίμωξης είναι ελαττωμένη 310. Στη χρόνια HCV λοίμωξη έχει βρεθεί ότι όσο μεγαλύτερος είναι ο αριθμός των Tregs, τόσο μικρότερος είναι ο αριθμός των CD8+ T κυττάρων και η παραγωγή IFN-γ από αυτά 312. Επιπλέον, έχει βρεθεί ότι αν και ο μεγαλύτερος αριθμός των Tregs πιθανόν να συμβάλλει στη χρόνια παραμονή του ιού 313, η κλωνική τους επέκταση προκαλεί μείωση της ιστικής βλάβης στο ήπαρ, καθώς υπάρχει θετική συσχέτιση μεταξύ του αριθμού των Tregs και του τίτλου το ιικού φορτίου αλλά αρνητική με τη φλεγμονώδη δραστηριότητα στο ήπαρ 314. Υπέρ της άποψης αυτής είναι το γεγονός ότι τα CD4+CD25 high+ Foxp3+ Tregs (ntregs) είναι περισσότερα σε ασθενείς με χρόνια HCV λοίμωξη και συνεχώς φυσιολογική ALΤ. Επίσης, υπάρχουν μελέτες που αφορούν επιμέρους υποπληθυσμούς των Tregs στη HCV λοίμωξη. Για παράδειγμα τα ntregs στη χρόνια HCV λοίμωξη α) έχουν μεγαλύτερη ευαισθησία στην IL β) έχουν βρεθεί στο ηπατικό παρέγχυμα 317 και γ) έχουν πλεονέκτημα επιβίωσης σε σχέση με τα ειδικά έναντι του HCV δραστικά Τ κύτταρα 326. Τα Tr1 κύτταρα είναι κυρίως συγκεντρωμένα στο ήπαρ 308, 314, 327 και αναγνωρίζουν τους ίδιους επιτόπους του ιού όπως και τα Th1 κύτταρα που παράγουν IFN-γ. Ο αυξημένος αριθμός Th3 κυττάρων σε ασθενείς με χρόνια HCV λοίμωξη έχει συσχετιστεί α) με την απάντηση στη θεραπεία καθώς έχει βρεθεί ότι είναι περισσότερα στους ασθενείς που απάντησαν στη 189
190 θεραπεία με IFN-a για την XHC 328 και β) με την διατήρηση ήπιας ιστολογικής φλεγμονής και με φυσιολογική ALT 330. Τα CD4+CD25+Tregs έχουν παραχθεί και in vitro παρουσία πεπτιδίων του HCV και μάλιστα με τα ίδια χαρακτηριστικά όπως τα ntregs 319, ενώ έχουν απομονωθεί και σε αναλογία 2:1 σε σχέση με τα δραστικά κύτταρα στα πυλαία διαστήματα στο ήπαρ 317. Επιπλέον τα CD4+CD25+Tregs που απομονώθηκαν από το ήπαρ ασθενών με χρόνια HCV λοίμωξη in vitro παρουσίασαν αυξημένη κλωνική επέκταση που εξαρτώνταν από την παρουσία IL-2 και εντονότερη κατασταλτική δράση μετά από παρεμπόδιση της σύνδεσης του PD-1 και του PD-L1, ενώ συσχετίστηκαν και με κλινικές παραμέτρους (HCV RNA, HAI) 333. Επιπλέον, ερευνητές υποστηρίζουν ότι η δράση των Tregs στη χρόνια HCV λοίμωξη ασκείται με άμεση κυτταροκυτταρική επαφή και μάλιστα με δοσοεξαρτώμενο τρόπο , 334, ενώ ορισμένοι ερευνητές έχουν διαπιστώσει ότι ασκείται και έμμεσα μέσω της έκκρισης ανοσοκατασταλτικών κυτταροκινών και συγκεκριμένα κυρίως μέσω του TGF-β ή/ και της IL-10 99, 100, 330, 332. Με βάση όλα τα ανωτέρω δεδομένα, σκοπός της παρούσας μελέτης ήταν να διερευνηθεί ο ρόλος των CD4+CD25+Foxp3+ T ρυθμιστικών κυττάρων στην ανοσοπαθογένεση της χρόνιας ηπατίτιδας C σε ασθενείς με χρόνια νόσο, οι οποίοι δεν είχαν προηγουμένως υποβληθεί σε θεραπεία, για όλη τη διάρκεια αυτής και έως την επίτευξη ή μη μακράς ιολογικής απάντησης. Επίσης, σκοπός ήταν να διερευνηθεί η συμμετοχή τους στην ιστολογική βλάβη στο ήπαρ. Η πρωτοτυπία της μελέτης έγκειται στο γεγονός ότι α) δεν έχει μελετηθεί ως σήμερα σε ασθενείς με χρόνια ηπατίτιδα C, η συσχέτιση του αριθμού των CD4+CD25+ high Foxp3+ T ρυθμιστικών κυττάρων στο περιφερικό αίμα με την ιστική βλάβη όπως αυτή εκφράζεται με την ίνωση και με το βαθμό της νεκροφλεγμονής (ΗΑΙ) στο ήπαρ, β) δεν έχει μελετηθεί η μεταβολή του αριθμού τους κατά τη διάρκεια της θεραπείας αλλά και κατά τον αναμενόμενο χρόνο επίτευξης μακράς ιολογικής ανταπόκρισης, γ) δεν έχει μελετηθεί η συσχέτιση των Tregs με την απάντηση στη θεραπεία ανάλογα με το είδος του γονότυπου του ιού δ) δεν έχει μελετηθεί η in vitro δυνατότητα αυτών των Tregs να εκπτύσσονται μετά από καλλιέργεια με μη ειδική διέγερση, τεχνική που προσεγγίζει έμμεσα τη λειτουργικότητα αυτών των κυττάρων και ε) δεν έχουν διερευνηθεί ταυτόχρονα οι μεταβολές των υποπληθυσμών των CD4+ βοηθητικών, CD8+ κυτταροτοξικών, CD4+CD25+ ενεργοποιημένων T βοηθητικών κυττάρων. 190
191 Για τον σκοπό αυτό μελετήθηκε η εκατοστιαία έκφραση του ενδοκυττάριου μεταγραφικού παράγοντα Foxp3 στα διεγερμένα CD4+CD25+ high T λεμφοκύτταρα που αντιστοιχεί στον πληθυσμό των T ρυθμιστικών λεμφοκυττάρων (CD4+/ CD25+ high /FOXP3+). Επιπλέον μελετήθηκαν α) η εκατοστιαία αναλογία των CD3+, CD4+ και CD8+ T λεμφοκυτταρικών υποπληθυσμών β) η εκατοστιαία αναλογία του υποδοχέα της IL-2 (IL-R2a, CD25) στον πληθυσμό των λεμφοκυττάρων καθώς και η εκατοστιαία συνέκφραση CD4 και CD25 στα Τ λεμφοκύτταρα (CD4+/CD25+ διπλά θετικά) και γ) η εκατοστιαία έκφραση του συνδιεγερτικού μορίου CD28 στα T λεμφοκύτταρα, καθώς και η εκατοστιαία συνέκφραση CD4 και CD28 στα T λεμφοκύτταρα (CD4+/CD28+ διπλά θετικά). Μετά τη λήψη φλεβικού αίματος και την απομόνωση των λεμφοκυττάρων από το περιφερικό αίμα, πραγματοποιήθηκαν μετρήσεις σε δύο διαφορετικές πειραματικές συνθήκες: α) ολικό περιφερικό αίμα (ex vivo) και β) μετά από in vitro καλλιέργεια των περιφερικών μονοπύρηνων κυττάρων (λεμφοκύτταρα, μακροφάγα) με το μιτογόνο φυτοαιμοσυγκολλητίνη (PHA). Για τις μετρήσεις των υποπληθυσμών και την έκφραση μορίων (επιφανειακών και ενδοκυττάριων) χρησιμοποιήθηκε η τεχνολογία της κυτταρομετρίας ροής, τριπλού φθορισμού. Η μέθοδος της κυτταρομετρίας ροής επιλέχθηκε επειδή επιτυγχάνει να αναλύσει μεγάλο αριθμό κυττάρων και ταυτόχρονα να διακρίνει πολλούς διαφορετικούς υποπληθυσμούς. Επιπλέον, επειδή προσφέρει τη δυνατότητα μέτρησης με μεγάλη ταχύτητα και εξαιρετική ευαισθησία ανάλυσης σε πολύ μικρό μέγεθος δείγματος. Το ενδοκυττάριο μονοκλωνικό αντίσωμα Foxp3 επιλέχθηκε επειδή έχει βρεθεί ότι η σταθερή και υψηλή έκφρασή του χαρακτηρίζει τον υποπληθυσμό των CD4+CD25+ T ρυθμιστικών κυττάρων ανεξάρτητα από το αν αυτά είναι «φυσικά αναδυόμενα» ή κλωνικά επαγόμενα στην περιφέρεια καθώς είναι ο κυρίαρχος και ουσιαστικότερος ρυθμιστής του λειτουργικού και φαινοτυπικού τους ρόλου. Συνεπώς μετά τη λήψη του περιφερικού αίματος έγινε με ειδική τεχνική η μεμβρανική (επιφανειακή) και ενδοκυττάρια χρώση των λεμφοκυττάρων των ασθενών και τελικά η μέτρηση με κυτταρομετρία ροής της εκατοστιαίας αναλογίας των CD3+, CD4+, CD8+ υποπληθυσμών, της εκατοστιαίας αναλογίας του υποδοχέα της IL-2 (IL- R2a, CD25), της εκατοστιαίας συνέκφρασης των μορίων CD4+ και CD25+, της εκατοστιαίας έκφρασης του συνδιεγερτικού μορίου CD28 και της συνέκφρασης του με το CD4+ μόριο και της εκατοστιαίας έκφρασης του ενδοκυττάριου μεταγραφικού παράγοντα Foxp3 στα διεγερμένα 191
192 CD4+CD25+ high T λεμφοκύτταρα (Tregs). Επίσης, έγινε απομόνωση και in vitro καλλιέργεια των περιφερικών μονοπύρηνων κυττάρων (λεμφοκυττάρων, μακροφάγων) με το μιτογόνο φυτοαιμοσυγκολλητίνη για την έμμεση καταγραφή της έκπτυξής τους (δηλαδή της κλωνικής τους επέκτασης) μετά από μη ειδική διέγερση για 48 ώρες. Στη μελέτη συμμετείχαν 61 ασθενείς, 59% άνδρες (μέσος όρος ηλικίας 31.5 έτη) και 41% γυναίκες (μέσος όρος ηλικίας 37.5 έτη). 39,3% των ασθενών είχε γονότυπο 1 (24/61), 4.9% γονότυπο 2 (3/61), 47.6% γονότυπο 3 (29/61) και 8.2% γονότυπο 4 (5/61). Κανένας από τους ασθενείς που συμμετείχε στη μελέτη δεν είχε λάβει στο παρελθόν θεραπεία για τη χρόνια ηπατίτιδα C. Σε βιοψία ήπατος υποβλήθηκαν οι 59/61 ασθενείς. Όλοι υποβλήθηκαν σε θεραπεία με το καθιερωμένο σχήμα συνδυασμένης αγωγής με πεγκυλιωμένη ιντερφερόνη και ριμπαβιρίνη. Οι ασθενείς με γονότυπο 1 και 4 έλαβαν θεραπεία για 48 εβδομάδες και οι ασθενείς με γονότυπο 2 και 3 έλαβαν θεραπεία για 24 εβδομάδες. Στο τέλος της θεραπείας, 59% (36/61) των ασθενών ανταποκρίθηκε στην αγωγή και πέτυχε SVR, 6.6% (4/61) δεν ανταποκρίθηκε, 8.2% διέκοψε λόγω παρενεργειών (5/61), ενώ 26.2% (16/61) δεν ολοκλήρωσε τη θεραπεία συνήθως λόγω υποτροπής στη χρήση ενδοφλεβίων ουσιών. Οι ασθενείς με γονότυπο 4 εξαιρέθηκαν λόγω της άτυπης συμπεριφοράς. Τελικά οι ασθενείς που ολοκλήρωσαν την αγωγή (37/61) διαχωρίστηκαν σε δύο ομάδες: α) ασθενείς με γονότυπο 1 (15/37) και β) ασθενείς με γονότυπο 2 και 3 (22/37). Όλες οι παράμετροι της μελέτης (HCVRNA, ALT, Tregs και οι υπόλοιποι υποπληθυσμοί των λεμφοκυττάρων) προσδιορίστηκαν σε 4 διαφορετικούς χρόνους (πριν, κατά τη διάρκεια της θεραπείας, τέλος θεραπείας και 6 μήνες μετά το τέλος της θεραπείας). Συγκεκριμένα : Α. (T 0 ): πριν τη θεραπεία Β. (T 1 ): σε επιλεγμένο στιγμιότυπο κατά τη διάρκεια της θεραπείας ανάλογα με τον γονότυπο, δηλαδή, -Για το γονότυπο 1: τη 12 η εβδομάδα, καθώς εάν ο ασθενής υπό θεραπεία επιτυγχάνει EVR (πρώιμη ιολογική απάντηση), δηλαδή αρνητικό HCV RNA τη 12 η εβδομάδα της αγωγής, τότε το PPV (positive predictive value, θετική προγνωστική αξία) για την επίτευξη SVR (μακροχρόνιας ιολογικής απάντησης) είναι 60% -Για το γονότυπο 2 και 3: την 4 η εβδομάδα, καθώς εάν ο ασθενής υπό θεραπεία επιτυγχάνει RVR (ταχεία ιολογική απάντηση), δηλαδή HCV RNA αρνητικό την 4 η εβδομάδα αγωγής, τότε το PPV για επίτευξη SVR είναι 95% Γ. (T 2 ): στο τέλος της θεραπείας (μετά από 48 εβδομάδες για τους ασθενείς με γονότυπο 1 και μετά από 24 εβδομάδες για τους ασθενείς με γονότυπο 2, 3) Δ. (T 3 ): 6 μήνες μετά το τέλος της θεραπείας 192
193 Ta αποτελέσματα της μελέτης, όπως αυτά αξιολογήθηκαν στο σύνολο των ασθενών έδειξαν ότι: Το αρχικό ιικό φορτίο (HCV RNA) παρουσίασε σημαντική ελάττωση σε όλους τους χρόνους που μελετήθηκε κατά τη διάρκεια της θεραπείας και 6 μήνες μετά το τέλος της, όπως άλλωστε αναμένονταν, με την μεγαλύτερη σημαντική ελάττωση μεταξύ της αρχικής του τιμής πριν τη θεραπεία και στο τέλος αυτής, ανεξάρτητα της επίτευξης SVR. Παράλληλα, διαπιστώθηκε σημαντική ελάττωση της ALT στους ασθενείς που είχαν αρχικά αυξημένες τιμές και επιπλέον διαπιστώθηκε ότι η ελάττωση αυτή ήταν προοδευτικά συνεχής κατά τη διάρκεια της θεραπείας αλλά και 6 μήνες μετά το τέλος της. Όσον αφορά τον αριθμό των Tregs (CD4+CD25+ high Foxp3+ T κυττάρων), δεν διαπιστώθηκαν σημαντικές διαφορές μεταξύ του αριθμού τους πριν την έναρξη της θεραπείας και στο χρόνο T 1, ούτε στο τέλος της θεραπείας (T 2 ), αλλά διαπιστώθηκε σημαντική αύξηση 6 μήνες μετά το τέλος της θεραπείας (T 0 -T 3 ). Επιπλέον, παρατηρήθηκε σημαντική αύξηση στον αριθμού τους μεταξύ του χρόνου Τ 1 και Τ 3, η οποία όμως φαίνεται ότι δεν επηρέασε το τελικό αποτέλεσμα, δηλαδή την έκβαση της θεραπείας στο σύνολο των ασθενών. Επιπλέον, ο αριθμός τους στις in vitro καλλιέργειες δεν παρουσίασε σημαντικές διακυμάνσεις. Τα ευρήματα αυτά είναι δύσκολο να εξηγηθούν ως προς τον παθογενετικό ρόλο των Tregs σε επίπεδο διακυμάνσεων του αριθμού τους ή και να συγκριθούν με τα ευρήματα άλλων μελετών καθόσον δεν υπάρχουν μελέτες παρακολούθησης της εξέλιξης των Tregs κατά τη διάρκεια της θεραπείας των ασθενών με χρόνια ηπατίτιδα C και την πιθανή συμβολή τους στην έκβαση της θεραπείας. Βέβαια όπως αναφέρεται από διάφορους ερευνητές , φαίνεται ότι τα Tregs χρησιμοποιούν διάφορους μηχανισμούς δράσης και τελικά ο μηχανισμός που εμπλέκεται κάθε φορά φαίνεται ότι μάλλον εξαρτάται από τη φύση του ερεθίσματος που προκαλεί την φλεγμονή, την ποσότητα και το είδος του αντιγόνου, την κατάσταση ενεργοποίησης και τον αριθμό των κυττάρων-στόχων και βέβαια τον ιστό που εμπλέκεται στη φλεγμονώδη διαδικασία 217, 218. Υπέρ της άποψης αυτής είναι τα ευρήματα μελετών στις οποίες έγινε σύγκριση των ασθενών με χρόνια λοίμωξη με άτομα με αποδραμούσα λοίμωξη και με υγιή άτομα και διαπιστώθηκε ότι σε εκείνους με χρόνια HCV λοίμωξη υπήρχε αυξημένος αριθμός CD4+CD25+ κυττάρων στην περιφέρεια και περισσότερο στο ήπαρ 312, 313, 314, 315, 316, 317, 318. Τα ευρήματα αυτά πιθανολογούν ότι ο μεγαλύτερος αριθμός των Tregs πιθανόν να συμβάλει στην χρόνια παραμονή του ιού 313, ενώ συγχρόνως, η κλωνική τους επέκταση προκαλεί μείωση της ιστικής βλάβης. Υπέρ του σκεπτικού αυτού είναι το γεγονός ότι υπάρχει θετική συσχέτιση του αριθμού των CD4+CD25+ Tregs και του τίτλου του ιικού φορτίου, ενώ διαπιστώθηκε αντίστροφη 193
194 συσχέτιση με τη φλεγμονώδη δραστηριότητα στο ήπαρ 314. Αντίθετα σε άλλες μελέτες διαπιστώθηκε ότι τόσο σε πειραματικά μοντέλα (χιμπαντζήδες), δεν υπήρχε καμία διαφορά στον αριθμό των Foxp3+CD4+CD25+ Tregs και στην ένταση της κατασταλτικής τους δράσης μεταξύ αποδραμούσας HCV λοίμωξης και χρόνιας HCV λοίμωξης, όσο και ανάμεσα σε υγιή άτομα της ομάδας ελέγχου και σε ασθενείς με χρόνια HCV λοίμωξη 319, 320, 321. Όσον αφορά τους υπόλοιπους υποπληθυσμούς των Τ λεμφοκυττάρων διαπιστώθηκαν τα εξής: α)ta CD3 + CD8 + Τ λεμφοκύτταρα παρουσίασαν σημαντική αύξηση του αριθμού τους μόνον 6 μήνες μετά το τέλος της θεραπείας (T 3 ). Όπως είναι γνωστό τα κύτταρα αυτά παίζουν σημαντικό ρόλο στην ανοσιακή απάντηση στην HCV λοίμωξη. Τα CD8 + T κύτταρα αναγνωρίζουν αντιγόνα του HCV που παρουσιάζονται από μόρια HLA τάξης Ι πάνω στα αντιγονοπαρουσιαστικά κύτταρα και στα μολυσμένα ηπατοκύτταρα και η αντιιική τους δράση έναντι του HCV εκφράζεται με κυτταροτοξικότητα, πολλαπλασιασμό και έκκριση αντιιικών κυτταροκινών όπως η IFN-γ. Στη χρόνια HCV λοίμωξη οι CD8 + T κυτταρικές αποκρίσεις είναι μη λειτουργικές (stunned), ενώ και ο αριθμός αυτών των κυττάρων είναι μικρότερος με αποτέλεσμα η λειτουργία τους και το εύρος της ανοσιακής τους απάντησης να είναι περιορισμένα 28, 79, 80. Τα ευρήματα της παρούσας μελέτης, δηλαδή η σημαντική αύξηση των CD8 + T κυττάρων 6 μήνες μετά το τέλος της θεραπείας, πιθανόν να οφείλεται στην επίδραση της θεραπείας και ειδικότερα στην χορήγηση της ιντερφερόνης-α, ανεξάρτητα από την απάντηση στη θεραπεία. Σύμφωνα με τα παραπάνω, είναι πιθανόν ο αριθμός τους να ελαττώνεται εκ νέου στους ασθενείς που δεν απαντούν στη θεραπεία μετά από πάροδο κάποιου χρόνου, γεγονός που θα ήταν πολύ ενδιαφέρον να μελετηθεί στα πλαίσια μιας νέας μελέτης. β) Τα CD4 + κύτταρα δεν παρουσίασαν σημαντικές διαφορές κατά τη σύγκρισή τους με τον προ της θεραπείας αριθμό, αν και ελαττώθηκαν σημαντικά 6 μήνες μετά το τέλος της θεραπείας (T 3 ) σε σχέση με τον αριθμό τους στον χρόνο T 1 (w4/ w12). Όπως αναφέρεται, στην χρόνια HCV λοίμωξη οι CD4+ T κυτταρικές αποκρίσεις είναι ασθενείς και δυσλειτουργικές, έχουν μικρότερο εύρος και διαρκούν λίγο σε αντίθεση με την οξεία HCV λοίμωξη που αυτοπεριορίζεται όπου και παρατηρούνται έντονες και ειδικές έναντι πολλαπλών επιτόπων του HCV απαντήσεις που διατηρούνται για καιρό 84, 85. Τα ευρήματα της παρούσας μελέτης, όσον αφορά τον πληθυσμό των CD4 + κυττάρων, 194
195 είναι δύσκολο να εξηγηθούν αν και φαίνεται ότι θα πρέπει να συμμετέχουν κατά τη διάρκεια της θεραπείας και η δράση τους να ελαττώνεται μετά το πέρας αυτής γ) Ο αριθμός των CD4 + CD25 + Τ λεμφοκυττάρων στο περιφερικό αίμα παρουσίασε σημαντική αύξηση μεταξύ των χρονικών στιγμών T 0 και T 1, δηλαδή ο αρχικός τους αριθμός αυξήθηκε μόνον κατά τη διάρκεια της θεραπείας, στην εβδομάδα 4 ή 12 ανάλογα με τον γονότυπο. Στο τέλος όμως της θεραπείας (T 2 ) καθώς και 6 μήνες μετά (T 3 ) δεν διαπιστώθηκαν σημαντικές διαφορές σε σχέση με τον αρχικό τους αριθμό. Τα ευρήματα αυτά δείχνουν ότι τα κύτταρα αυτά παρουσιάζουν κάποια ενεργό απάντηση δηλαδή ενεργοποιούνται αμεσότερα και πρωιμότερα σε σχέση με τους άλλους Τ κυτταρικούς υποπληθυσμούς της μελέτηςέναντι του HCV, πιθανόν μετά από ενεργοποίησή τους από την ιντερφερόνη-α για μικρό χρονικό διάστημα μετά την έναρξη της θεραπείας και επανέρχονται στον προ της θεραπείας αριθμό τους στο τέλος της θεραπείας. Ο ρόλος όμως των κυττάρων αυτών στην εξέλιξη της λοίμωξης υπό θεραπεία, δεν είναι δυνατόν να διευκρινισθεί στην παρούσα μελέτη. δ) Ο αριθμός των CD4+CD28+ T λεμφοκυττάρων στο περιφερικό αίμα παρουσίασε διακυμάνσεις, δηλαδή αυξήθηκε σημαντικά στον χρόνο T 1 (την 4 η και τη 12 η εβδομάδα θεραπείας ανάλογα με τον γονότυπο) σε σχέση με τον αρχικό αριθμό τους, ελαττώθηκε στο τέλος της θεραπείας έτσι ώστε να μην διαφέρει σημαντικά και παρουσίασε σημαντική αύξηση 6 μήνες μετά το τέλος της θεραπείας, συνηγορώντας υπέρ κάποιας δραστηριότητας των κυττάρων αυτών υπό την επίδραση της θεραπείας, χωρίς όμως η τελική συμβολή τους στην απάντηση ή μη των ασθενών στη θεραπεία να μπορεί να διευκρινισθεί στην παρούσα μελέτη και χωρίς την δυνατότητα σύγκρισης των ευρημάτων με άλλες μελέτες. Στη συνέχεια, εφόσον η απάντηση στη θεραπεία των ασθενών με χρόνια ηπατίτιδα C διαφέρει ανάλογα με τον γονότυπο του ιού, όπως άλλωστε διαπιστώθηκε και στην παρούσα μελέτη, κατά την οποία όλοι οι ασθενείς με γονότυπο 2 και 3 απήντησαν στο θεραπευτικό σχήμα επιτυγχάνοντας SVR σε αντίθεση με τους ασθενείς με γονότυπο 1, αναλύθηκαν όλοι οι παράμετροι της παρούσας μελέτης ανάλογα με τον γονότυπο, έτσι ώστε να διαπιστωθεί η πιθανή διαφορετική συμπεριφορά της ανοσιακής απάντησης και η συμβολή των Tregs και των 195
196 υποπληθυσμών των Τ λεμφοκυττάρων στην επίτευξη της απάντησης στη θεραπεία. Κατά την ανάλυση αυτή εξαιρέθηκαν οι ασθενείς με χρόνια ηπατίτιδα C και γονότυπο 4 λόγω της «άτυπης» συμπεριφοράς του συγκεκριμένου γονότυπου (5/61). Συγκεκριμένα, δύο ασθενείς πέτυχαν SVR, δυο διέκοψαν τη θεραπεία πριν τον πρώτο μήνα, ενώ ένας εμφάνισε ιολογική διαφυγή κατά τη διάρκεια της θεραπείας. Στους ασθενείς με γονότυπο 1, διαπιστώθηκε ότι ο αριθμός των Tregs ελαττώθηκε σημαντικά σε σχέση με την αρχική τους τιμή πριν από τη θεραπεία την 12 η εβδομάδα, αλλά στη συνέχεια παρουσίασε αύξηση έτσι ώστε να μην διαφέρει σημαντικά με τα προ της θεραπείας επίπεδα στο τέλος της θεραπείας και 6 μήνες μετά. Όταν όμως ο αριθμός των Tregs αναλύθηκε ανάλογα με την απάντηση στη θεραπεία, διαπιστώθηκε μία διαφοροποίηση της συμπεριφοράς των Tregs στην ομάδα των ασθενών αυτών η οποία τελικά είναι πιθανόν να επηρέασε την απάντηση στη θεραπεία. Υπέρ της άποψης αυτής είναι το γεγονός ότι τα Tregs δεν ελαττώθηκαν κατά τη διάρκεια της θεραπείας στους ασθενείς οι οποίοι δεν απήντησαν και επιπλέον παρουσίασαν σημαντική αύξηση του αριθμού τους 6 μήνες μετά το τέλος της θεραπείας (Τ 3 ) σε σχέση με την αρχικό τους αριθμό (Τ 0 ). Υπέρ του σκεπτικού αυτού συνηγορούν τα ευρήματα μελετών στις οποίες διαπιστώθηκε ότι υπάρχει θετική συσχέτιση του αριθμού των CD4+CD25+ Tregs και του τίτλου του ιικού φορτίου με αποτέλεσμα ο μεγαλύτερος αριθμός των Tregs πιθανόν να συμβάλει στην χρόνια παραμονή του ιού και την εγκατάσταση της χρόνιας λοίμωξης 312, 313. Βέβαια δεν έχει διευκρινισθεί μέχρι σήμερα εάν η αύξηση του πληθυσμού των Tregs είναι προγνωστικός δείκτης εγκατάστασης χρόνιας HCV λοίμωξης στους ασθενείς με γονότυπο 1 ή εάν αποτελεί ένα μηχανισμό που προάγει τη χρονιότητα 309. Εντούτοις όμως, τα ευρήματα αυτά δεν μπορούν να διασαφηνίσουν τους μηχανισμούς που εμπλέκονται στην διαφορετική απάντηση των ασθενών στη θεραπεία μετά την εγκατάσταση της χρόνιας HCV λοίμωξης. Όσον αφορά τα ευρήματα των in vitro καλλιεργειών των Tregs, δεν διαπιστώθηκαν σημαντικές μεταβολές στους ασθενείς με γονότυπο 1 και δεν επηρέασαν την απάντηση στη θεραπεία. Όσον αφορά την ανάλυση των υπολοίπων υποπληθυσμών των Τ λεμφοκυττάρων στους ασθενείς με γονότυπο 1 διαπιστώθηκαν τα εξής: 196
197 a) Ta CD3 + CD8 + T κύτταρα δεν παρουσίασαν σημαντικές διαφορές και ο αριθμός τους δεν διέφερε μεταξύ των ασθενών οι οποίοι απήντησαν στη θεραπεία και αυτών που δεν απήντησαν. β) Τα CD4 + T κύτταρα ελαττώθηκαν σημαντικά στο τέλος της θεραπείας σε σχέση με την τιμή τους στην 12 η εβδομάδα χωρίς όμως η ελάττωση αυτή να διαφέρει σημαντικά με τον αρχικό τους αριθμό. Εντούτοις όμως όταν ο αριθμός τους αναλύθηκε σε σχέση με την απάντηση στη θεραπεία, διαπιστώθηκε ότι οι ασθενείς οι οποίοι παρουσίασαν SVR, είχαν σημαντικά μικρότερο αριθμό CD4 + T κυττάρων στο τέλος της θεραπείας αλλά και 6 μήνες μετά, σε σχέση με την εβδομάδα 12, συνηγορώντας υπέρ μιας συμμετοχής των κυττάρων αυτών στο τελικό αποτέλεσμα της θεραπείας. γ) Τα CD4 + CD25 + T κύτταρα δεν παρουσίασαν σημαντικές διακυμάνσεις στους ασθενείς με γονότυπο 1. Εντούτοις όμως, στους ασθενείς που απήντησαν στη θεραπεία διαπιστώθηκε μία αυξητική τάση στην αρχική τους τιμή και μία τάση μείωσής τους 6 μήνες μετά το τέλος της σε σχέση με τον αριθμό τους τη 12 η εβδομάδα θεραπείας σε σύγκριση με τους ασθενείς οι οποίοι δεν είχαν τελικά SVR. δ)τα CD4 + CD28 + Τ κύτταρα παρουσίασαν σημαντικές διαφορές μεταξύ των διάφορων χρόνων μέτρησης κατά τη διάρκεια της θεραπείας, δηλαδή ελαττώθηκαν μεταξύ των χρονικών στιγμών Τ 1 και Τ 2 12 η εβδομάδα και τέλος θεραπείας- χωρίς όμως να διαφέρουν σημαντικά σε σχέση με τον αρχικό τους αριθμό. Επιπλέον όμως, στους ασθενείς οι οποίοι ανταποκρίθηκαν στη θεραπεία ο αριθμός τους ελαττώθηκε σημαντικά και μεταξύ της εβδομάδας 12 και 6 μήνες μετά το τέλος της θεραπείας. Τα ευρήματα αυτά δείχνουν ότι τα κύτταρα αυτά θα πρέπει τελικά να εμπλέκονται στην απάντηση ή μη των ασθενών στη θεραπεία ή να επηρεάζονται από τις μεταβολές των υπολοίπων Τ λεμφοκυττάρων. Στους ασθενείς με γονότυπο 2 και 3, ο αριθμός των Tregs παρουσίασε σημαντική ελάττωση στο τέλος της θεραπείας σε σχέση με τον αρχικόπριν από τη θεραπεία- αριθμό τους, στη συνέχεια όμως αυξήθηκε και 6 μήνες μετά το τέλος της θεραπείας δεν διέφερε από τον αρχικό τους αριθμό. Τα ευρήματα αυτά συνηγορούν υπέρ μιας ενεργούς συμμετοχής των Tregs κατά τη διάρκεια της θεραπείας, με αποτέλεσμα την συμβολή τους στην ελάττωση του ιικού φορτίου και την τελική επίτευξη SVR. Δεν διαπιστώθηκαν σημαντικές μεταβολές του αριθμού των Tregs στις in vitro καλλιέργειες των ασθενών με γονότυπο 2 και
198 Όσον αφορά τους υπόλοιπους υποπληθυσμούς των Τ λεμφοκυττάρων και ιδιαίτερα των CD3 + CD8 +, CD4 + και CD4 + CD25 + T λεμφοκυττάρων στους ασθενείς με γονότυπο 2 και 3, διαπιστώθηκαν ορισμένες σημαντικές μεταβολές κατά τη διάρκεια της θεραπείας χωρίς όμως τελικά οι μεταβολές αυτές να είναι σημαντικές σε σχέση με τον προ της θεραπείας αριθμό τους και χωρίς να επηρεάσουν την έκβαση της θεραπείας. Ο αριθμός όμως των CD4 + CD28 + T λεμφοκυττάρων παρουσίασε στατιστικά σημαντικές διαφορές σε αρκετά στιγμιότυπα. Ειδικότερα, ο αριθμός τους αυξήθηκε α) την 4 η εβδομάδα αγωγής σε σχέση με τον αρχικό αριθμό τους β) 6 μήνες μετά τη θεραπεία σε σχέση με τον αριθμό τους την 4 η εβδομάδα θεραπείας και του αριθμού τους στo τέλος της θεραπείας. Τα ευρήματα αυτά δείχνουν ότι τα CD4 + CD28 + T λεμφοκύτταρα συμμετέχουν ενεργά κατά τη διάρκεια της θεραπείας και πιθανόν παίζουν κριτικό ρόλο στην επίτευξη SVR. Μετά την ανάλυση των ευρημάτων και μεταβολών όλων των παραμέτρων με βάση τον γονότυπο του ιού, με σκοπό να εντοπισθούν οι παράμετροι της ανοσιακής απάντησης οι οποίοι συμβάλλουν στην διαφορετική συμπεριφορά των ασθενών στη θεραπεία, έγινε σύγκριση μεταξύ των 2 ομάδων ασθενών, δηλαδή των ασθενών με γονότυπο 1 και των ασθενών με γονότυπο 2 και 3. Τελικά, οι μόνες σημαντικές διαφορές που διαπιστώθηκαν ήταν οι εξής: οι ασθενείς με γονότυπο 1 α) εμφάνιζαν μικρότερη τιμή ALT και μικρότερο αριθμό Tregs σε σχέση με τους ασθενείς με γονότυπο 3 κατά τη διάρκεια της θεραπείας στον χρόνο Τ 1 (δηλαδή, την 12 η εβδομάδα για τον γονότυπο 1 και την 4 η εβδομάδα για τον γονότυπο 2 και 3) και β) υψηλότερο ιικό φορτίο 6 μήνες μετά το τέλος της θεραπείας, εφόσον σε όλους τους ασθενείς με γονότυπο 2 και 3 επιτεύχθηκε SVR. Τα ευρήματα αυτά είναι δύσκολο να ερμηνευθούν. Βέβαια δεν υπάρχουν άλλες μελέτες σύγκρισης των αποτελεσμάτων αυτών, αν και θα αναμένονταν ο αριθμός των Tregs να ήταν σημαντικά μικρότερος στους ασθενείς με γονότυπο 2 και 3, συμβάλλοντας έτσι στην τελική έκβαση της θεραπείας και στην επίτευξη SVR. Σε ορισμένες μελέτες μάλιστα που αφορούν τον αριθμό των Tregs και την εγκατάσταση χρόνιας HCV λοίμωξης, τα ευρήματα συνηγορούν ότι ο αυξημένος αριθμός των Tregs και κατά συνέπεια η δράση τους, οδηγούν στην εγκατάσταση χρόνιας νόσου μετά από λοίμωξη με τον HCV σε αντίθεση με τον αυτοπεριορισμό της λοίμωξης παρουσία μικρότερου αριθμού Tregs. Σε άλλες μελέτες όμως, άλλοι ερευνητές διαπίστωσαν ότι δεν υπήρχε 198
199 αύξηση του αριθμού των CD4+CD25+ high κυττάρων και μάλιστα ότι δεν υπήρχαν διαφορές στον αριθμό των CD4+CD25+ κυττάρων ανάμεσα σε υγιή άτομα της ομάδας ελέγχου και σε άτομα με χρόνια HCV λοίμωξη. Εντούτοις όλες αυτές οι μελέτες αναφέρονται στον μηχανισμό εξέλιξης της λοίμωξης με τον HCV και δεν μπορούν να συγκριθούν με την παρούσα μελέτη στην οποία οι σημαντικές μεταβολές διαπιστώθηκαν κατά τη διάρκεια της θεραπείας στην εγκαταστημένη πλέον χρόνια ηπατίτιδα C. Στη συνέχεια, έγιναν συσχετίσεις μεταξύ των διαφόρων μεταβλητών που μετρήθηκαν στην παρούσα μελέτη. Διαπιστώθηκαν οι εξής σημαντικές συσχετίσεις: α) Διαπιστώθηκε ότι όσο μεγαλύτερη ήταν η ηλικία, τόσο σημαντικά μεγαλύτερος ήταν ο δείκτης νεκροφλεγμονώδους δραστηριότητας (ΗΑΙ) και η ίνωση στον ηπατικό ιστό. β) Ο δείκτης νεκροφλεγμονώδους δραστηριότητας διαπιστώθηκε ότι συσχετίζεται θετικά με την διαβρωτική νέκρωση, την πυλαία φλεγμονή και τις ενδολόβιες νεκρώσεις, αλλά είχε σημαντικά εντονότερη θετική συσχέτιση με τη διαβρωτική νέκρωση. Επίσης, ο υψηλότερος ΗΑΙ διαπιστώθηκε ότι συσχετίζεται με μεγαλύτερη ίνωση. Παράλληλα, βρέθηκε ότι ο υψηλότερος HAI συσχετίζεται με υψηλότερες τιμές ALT πριν τη θεραπεία και η ίδια θετική συσχέτιση παρέμεινε μεταξύ της τιμής της ALT και του ΗΑΙ και 6 μήνες μετά το τέλος της θεραπείας. γ ) Ο αριθμός των Τ ρυθμιστικών κυττάρων στο περιφερικό αίμα πριν την έναρξη της θεραπείας, στο σύνολο των ασθενών και ανεξάρτητα γονότυπου, δεν συσχετίστηκε με καμία από τις εξεταζόμενες παραμέτρους της μελέτης. Ωστόσο, στα επόμενα στιγμιότυπα διαπιστώθηκαν σημαντικές συσχετίσεις. Ειδικότερα, στο τέλος της θεραπείας (Τ 2 ), οι ασθενείς οι οποίοι στον χρόνο Τ 1 (4 η εβδομάδα/ 12 η εβδομάδα) είχαν υψηλότερο ιικό φορτίο, είχαν και μεγαλύτερο αριθμό Tregs στο περιφερικό αίμα, γεγονός το οποίο συνηγορεί υπέρ της αρνητικής επίδρασης των Tregs στην καταστολή του ιού κατά τη θεραπεία. Επιπλέον, o αριθμός των Tregs στον ίδιο χρόνο (T 2 ), συσχετίστηκε αρνητικά με τον αριθμό των CD4+ και των CD4+CD28+ T κυττάρων στο τέλος της θεραπείας και 6 μήνες μετά, πιθανολογώντας ότι τα Tregs προκαλούσαν μεγαλύτερη καταστολή σε αυτούς τους υποπληθυσμούς λεμφοκυττάρων και θετικά με τον αριθμό τους 6 μήνες μετά το τέλος της θεραπείας. 199
200 Ο αριθμός των Tregs στο περιφερικό αίμα 6 μήνες μετά το τέλος της θεραπείας είχε αρνητική συσχέτιση με τον αριθμό των CD4+ T λεμφοκυττάρων πριν την έναρξη της θεραπείας και κυρίως με τον αριθμό των CD4+ κυττάρων κατά τη διάρκεια της θεραπείας (χρόνος Τ 1 ) και 6 μήνες μετά το τέλος της, αρνητική συσχέτιση με τον αριθμό των CD4+CD28+ T λεμφοκυττάρων στον χρόνο Τ 1 και 6 μήνες μετά τη θεραπεία, θετική συσχέτιση με τον αριθμό των CD3+CD8+ Τ κυττάρων 6 μήνες μετά τη θεραπεία και θετική συσχέτιση με το ιικό φορτίο στον ίδιο χρόνο. Ο αριθμός των Tregs στο περιφερικό αίμα ήταν σημαντικά μεγαλύτερος στους ασθενείς με γονότυπο 2 και 3 στην 4 η εβδομάδα κατά τη διάρκεια της θεραπείας. δ ) Ο αριθμός των Tregs που αναπτύχθηκε στην καλλιέργεια του περιφερικού αίματος στον χρόνο Τ 1 (4 η εβδομάδα/ 12 η εβδομάδα), ήταν μικρότερος στους ασθενείς που είχαν μεγαλύτερη ίνωση στη βιοψία ήπατος και μεγαλύτερο αριθμό Tregs στο περιφερικό αίμα στο τέλος της θεραπείας. Επίσης, διαπιστώθηκε ότι στους άνδρες, στο τέλος της θεραπείας, υπήρχε η τάση ο αριθμός των Tregs να είναι μικρότερος. ε ) Όσον αφορά τις συσχετίσεις των υπολοίπων Τ λεμφοκυτταρικών υποπληθυσμών μεταξύ τους, διαπιστώθηκαν διάφορες συσχετίσεις στους διαφορετικούς χρόνους της μελέτης. Οι συσχετίσεις αυτές είναι δύσκολο να ερμηνευθούν ως προς την τελική έκβαση της θεραπείας στους ασθενείς της μελέτης και την συμβολή τους ή μη στην επίτευξη μακρόχρονης ιολογικής ανταπόκρισης. 200
201 VII. ΣΥΜΠΕΡΑΣΜΑΤΑ Από την παρούσα μελέτη προέκυψαν τελικά τα εξής συμπεράσματα: 1)Το γεγονός ότι ο αριθμός των Tregs δεν παρουσίασε σημαντικές διαφορές στο τέλος της θεραπείας και παρά την αύξησή του μεταξύ του χρόνου Τ 1 και του Τ 3 (6 μήνες μετά το τέλος της θεραπείας), στο σύνολο των ασθενών, δεν επηρέασε την έκβαση της θεραπείας, δεν επιτρέπει την διεξαγωγή συμπερασμάτων ως προς τον παθογενετικό τους ρόλο κατά τη διάρκεια της θεραπείας. 2)Η σημαντική αύξηση των CD3+CD8+ Τ κυττάρων 6 μήνες μετά το τέλος της θεραπείας πιθανόν να οφείλεται στην επίδραση της θεραπείας και ειδικότερα στην χορήγηση της ιντερφερόνης-α 3)Τα CD4+ T κύτταρα ελαττώθηκαν σημαντικά 6 μήνες μετά το τέλος της θεραπείας σε σχέση με τον χρόνο Τ 1 συνηγορώντας υπέρ της συμμετοχής τους κατά τη διάρκεια της θεραπείας 4)Τα CD4+CD25+ T κύτταρα φαίνεται ότι παρουσιάζουν κάποια ενεργό δραστηριότητα έναντι του HCV αρχικά μετά την έναρξη της θεραπείας, ο ρόλος τους όμως στην εξέλιξη της θεραπείας δεν είναι σαφής 5)Τα CD4+CD28+ T κύτταρα παρουσίασαν διακυμάνσεις κατά τη θεραπεία συνηγορώντας υπέρ κάποιας δραστηριότητας των κυττάρων αυτών πιθανόν ως αποτέλεσμα της θεραπείας 6) Στους ασθενείς με γονότυπο 1, ο αριθμός των Tregs παρουσίασε μία διαφοροποίηση, δηλαδή ο αριθμός τους δεν ελαττώθηκε κατά τη διάρκεια της θεραπείας στους ασθενείς οι οποίοι δεν απήντησαν στο θεραπευτικό σχήμα και δεν πέτυχαν SVR 7) Στους ασθενείς με γονότυπο 1 οι οποίοι απήντησαν στη θεραπεία διαπιστώθηκε ότι ο αριθμός των CD4+CD28+ Τ κυττάρων ελαττώθηκε σημαντικά μεταξύ της εβδομάδας 12 και 6 μήνες μετά το τέλος της θεραπείας, συνηγορώντας ότι τα κύτταρα αυτά θα πρέπει να εμπλέκονται στην απάντηση ή μη της θεραπείας 8)Ο αριθμός των Tregs στους ασθενείς με γονότυπο 2 και 3 παρουσίασε σημαντική ελάττωση στο τέλος της θεραπείας και στη συνέχεια αυξήθηκε 6 μήνες μετά το τέλος της, συνηγορώντας υπέρ μιας ενεργού συμμετοχής των κυττάρων αυτών στην επίτευξη SVR 9)Κατά τη σύγκριση μεταξύ των δύο ομάδων ασθενών διαπιστώθηκε ότι οι ασθενείς με γονότυπο 1 εμφάνιζαν σημαντικά μικρότερο αριθμό Tregs 201
202 σε σύγκριση με τους ασθενείς με γονότυπο 3 κατά τη διάρκεια της θεραπείας στον χρόνο Τ 1 (εβδομάδα 4/ εβδομάδα 12 αντίστοιχα). Tα ευρήματα αυτά είναι δύσκολο να εκτιμηθούν αν και θα αναμένονταν ο αριθμός των Tregs να ήταν σημαντικά μικρότερος στους ασθενείς με γονότυπο 2 και 3. Το τελικό συμπέρασμα της μελέτης, όπως προκύπτει από την ανάλυση όλων των ευρημάτων, είναι ότι ο παθογενετικός ρόλος των Tregs φαίνεται να διαφέρει μεταξύ των ασθενών με γονότυπο 1 και αυτών με γονότυπο 2 και 3. Βέβαια όπως είναι γνωστό, η απάντηση στη λοίμωξη από κάποιον ιό είναι πολυπαραγοντική και η εξέλιξή της έγκειται στη συμμετοχή διαφόρων παραμέτρων της ανοσιακής απάντησης, στην παρουσία άλλων νοσημάτων, και στην γενετική διαφοροποίηση των ασθενών. Επιπλέον θα πρέπει να τονισθεί ότι η συμπεριφορά των διαφόρων υποπληθυσμών των Τ λεμφοκυττάρων στο περιφερικό αίμα δεν προδικάζει τη δράση τους στον ηπατικό ιστό όπου και εγκαθίσταται ο ιός και προκαλεί την ιστική βλάβη. Κατά συνέπεια προτείνεται η εκτίμηση της συμπεριφοράς των ανοσιακών αυτών παραμέτρων και ιδιαίτερα των Tregs, να μελετηθεί σε επίπεδο ηπατικού ιστού και να αποτελέσει το ερευνητικό πεδίο μιας άλλης μελέτης. 202
203 VIII. ΠΕΡΙΛΗΨΗ Ο ιός της ηπατίτιδας C είναι ένας μικρός RNA ιός ο οποίος αποτελεί το αίτιο της πιο συχνά αιματογενώς μεταδιδόμενης λοίμωξης και το πιο κοινό αίτιο χρόνιας ηπατικής νόσου παγκόσμιa. Είναι γνωστό ότι η παθογένεση και η έκβαση των ιογενών λοιμώξεων επηρεάζεται σημαντικά από την ανοσιακή απόκριση του ξενιστή καθώς η εκρίζωσή τους απαιτεί μια πολύπλοκη και συντονισμένη συνεργασία των μηχανισμών της φυσικής και της επίκτητης ανοσίας. Η χρόνια παραμονή του ιού της ηπατίτιδας C επιτυγχάνεται μέσω ποικίλων μηχανισμών οι οποίοι δεν είναι επαρκώς κατανοητοί. Στους μηχανισμούς αυτούς συγκαταλέγονται, η παράκαμψη των μηχανισμών της φυσικής ανοσίας από τον ίδιο τον ιό, η ανάδυση μεταλλάξεων και η δυσλειτουργία των ειδικών έναντι του ιού CD4+ και CD8+ T δραστικών κυττάρων. Σήμερα, υπάρχουν δεδομένα που υποδηλώνουν ότι τα T ρυθμιστικά κύτταρα συμβάλλουν επίσης στην καταστολή των ειδικών έναντι του ιού Τ κυτταρικών αποκρίσεων με αποτέλεσμα τη χρόνια παραμονή του, αλλά ταυτόχρονα ασκούν ευεργετική δράση περιορίζοντας την ιστική βλάβη στο ήπαρ. Τα T ρυθμιστικά κύτταρα είναι ένας ετερογενής πληθυσμός κυττάρων με ιδιαίτερα φαινοτυπικά και λειτουργικά χαρακτηριστικά ο οποίος έχει την ιδιότητα να ασκεί κατασταλτική δράση και περιλαμβάνει τα «φυσικά» αναδυόμενα Τ ρυθμιστικά κύτταρα (natural Tregs, ntregs, CD4+CD25+ high Tregs) που παράγονται στο θύμο και τα επαγόμενα Tregs (induced Tregs, itregs)που παράγονται μετά από κλωνική επέκταση στην περιφέρεια. Ο σκοπός της παρούσας μελέτης ήταν να διερευνηθεί ο ρόλος των CD4+CD25+ high Foxp3+ T ρυθμιστικών κυττάρων (Tregs) στην ανοσοπαθογένεση της χρόνιας ηπατίτιδας C σε ασθενείς που δεν είχαν προηγουμένως υποβληθεί σε θεραπεία με πεγκυλιωμένη ιντερφερόνη και ριμπαβιρίνη και στην πιθανή επίδρασή τους στο θεραπευτικό αποτέλεσμα. Στη μελέτη συμμετείχαν 61 ασθενείς, 39.3% είχε γονότυπο 1 (24/61), 4.9% γονότυπο 2 (3/61), 47.6% γονότυπο 3 (29/61) και 8.2% γονότυπο 4 (5/61). Σε βιοψία ήπατος η οποία αξιολογήθηκε με το τροποποιημένο σύστημα Knodell, υποβλήθηκαν οι 59/61 ασθενείς. Η μέτρηση του ιικού φορτίου έγινε με ευαίσθητη ποιοτική και ποσοτική μέθοδο (Combas Amplicor και Amplicor Monitor Roche Diagnostics), ενώ ο προσδιορισμός του γονότυπου έγινε με την μέθοδο INNOLIPA (Bayer Diagnostics). Όλοι οι ασθενείς πληρούσαν τα κριτήρια ένταξης στη 203
204 μελέτη, έδωσαν τη συγκατάθεση τους για να συμμετέχουν σε αυτή και υποβλήθηκαν σε θεραπεία με το καθιερωμένο σχήμα συνδυασμένης αγωγής με πεγκυλιωμένη ιντερφερόνη και ριμπαβιρίνη. Οι ασθενείς με γονότυπο 1 και 4 έλαβαν θεραπεία για 48 εβδομάδες και οι ασθενείς με γονότυπο 2 και 3 έλαβαν θεραπεία για 24 εβδομάδες. Στο τέλος της θεραπείας, 59% (36/61) των ασθενών ανταποκρίθηκε στην αγωγή και πέτυχε SVR, 6.6% (4/61) δεν ανταποκρίθηκε, 8.2% διέκοψε λόγω παρενεργειών (5/61), ενώ 26.2% (16/61) δεν ολοκλήρωσε τη θεραπεία συνήθως λόγω υποτροπής στη χρήση ενδοφλεβίων ουσιών. Οι ασθενείς με γονότυπο 4 εξαιρέθηκαν λόγω της άτυπης συμπεριφοράς του συγκεκριμένου γονότυπου. Τελικά οι ασθενείς που ολοκλήρωσαν την αγωγή (37/61) διαχωρίστηκαν σε δύο ομάδες: α) ομάδα 1: ασθενείς με γονότυπο 1 (15/37) και β) ομάδα 2: ασθενείς με γονότυπο 2 και 3 (22/37). Όλες οι παράμετροι της μελέτης (HCVRNA, ALT, Tregs και οι υπόλοιποι υποπληθυσμοί των λεμφοκυττάρων) προσδιορίστηκαν σε 4 διαφορετικούς χρόνους. Συγκεκριμένα : Α. (T 0 ): πριν τη θεραπεία Β. (T 1 ): σε επιλεγμένο στιγμιότυπο κατά τη διάρκεια της θεραπείας ανάλογα με τον γονότυπο, δηλαδή, -Για το γονότυπο 1: τη 12 η εβδομάδα, καθώς εάν ο ασθενής υπό θεραπεία επιτυγχάνει EVR (πρώιμη ιολογική απάντηση), δηλαδή αρνητικό HCV RNA τη 12 η εβδομάδα της αγωγής, τότε το PPV (positive predictive value, θετική προγνωστική αξία) για την επίτευξη SVR (μακροχρόνιας ιολογικής απάντησης) είναι 60% -Για το γονότυπο 2 και 3: την 4 η εβδομάδα, καθώς εάν ο ασθενής υπό θεραπεία επιτυγχάνει RVR (ταχεία ιολογική απάντηση), δηλαδή HCV RNA αρνητικό την 4 η εβδομάδα αγωγής, τότε το PPV για επίτευξη SVR είναι 95% Γ. (T 2 ): στο τέλος της θεραπείας (μετά από 48 εβδομάδες για τους ασθενείς με γονότυπο 1 και μετά από 24 εβδομάδες για τους ασθενείς με γονότυπο 2, 3) Δ. (T 3 ): 6 μήνες μετά το τέλος της θεραπείας. Για τη διερεύνηση του ρόλου των Tregs στη χρόνια ηπατίτιδα C μελετήθηκε η εκατοστιαία έκφραση του ενδοκυττάριου μεταγραφικού παράγοντα Foxp3 στα διεγερμένα CD4+CD25+ high T λεμφοκύτταρα ο οποίος αντιστοιχεί στον πληθυσμό των T ρυθμιστικών λεμφοκυττάρων (CD4+CD25+ high Foxp3+ Τ κυττάρων). Το ενδοκυττάριο μονοκλωνικό αντίσωμα Foxp3 επιλέχθηκε επειδή έχει βρεθεί ότι η σταθερή και υψηλή έκφρασή του χαρακτηρίζει τον υποπληθυσμό των CD4+CD25+ T ρυθμιστικών κυττάρων ανεξάρτητα από το αν αυτά είναι «φυσικά αναδυόμενα» ή κλωνικά επαγόμενα στην περιφέρεια. 204
205 Επιπλέον μελετήθηκαν α) η εκατοστιαία αναλογία των CD3+, CD4+ και CD8+ T λεμφοκυτταρικών υποπληθυσμών β) η εκατοστιαία αναλογία του υποδοχέα της IL-2 (IL-R2a, CD25) στον πληθυσμό των λεμφοκυττάρων καθώς και η εκατοστιαία συνέκφραση CD4 και CD25 στα Τ λεμφοκύτταρα (CD4+/CD25+ διπλά θετικά) και γ) η εκατοστιαία έκφραση του συνδιεγερτικού μορίου CD28 στα T λεμφοκύτταρα, καθώς και η εκατοστιαία συνέκφραση CD4 και CD28 στα T λεμφοκύτταρα (CD4+/CD28+ διπλά θετικά). Μετά τη λήψη φλεβικού αίματος και την απομόνωση των λεμφοκυττάρων από το περιφερικό αίμα, στους χρόνους της μελέτης, πραγματοποιήθηκαν μετρήσεις σε δύο διαφορετικές πειραματικές συνθήκες: α) ολικό περιφερικό αίμα (ex vivo), για τον προσδιορισμό των λεμφοκυτταρικών υποπληθυσμών στην περιφέρεια και β) μετά από in vitro καλλιέργεια των περιφερικών μονοπύρηνων κυττάρων (λεμφοκύτταρα, μακροφάγα) με το μιτογόνο φυτοαιμοσυγκολλητίνη (PHA), για τον προσδιορισμό της κλωνικής τους επέκτασης μετά από μη ειδική διέγερση. Για τις μετρήσεις των υποπληθυσμών και την έκφραση μορίων (επιφανειακών και ενδοκυττάριων) χρησιμοποιήθηκε η τεχνολογία της κυτταρομετρίας ροής, τριπλού φθορισμού (FACS, EPICS XL-MCL, Beckman Coulter) με τα μονοκλωνικά αντισώματα CD3, CD4 (FITC, Immunotech), CD4, CD8, CD25, CD28 (PE, Immunotech), CD25 (ECD, Immunotech) και Foxp3 (PE, Ebioscience). Η στατιστική ανάλυση έγινε με παραμετρικές δοκιμασίες (paired samples student s t-test) όσον αφορά τη σύγκριση των εξεταζομένων μεταβλητών στους τέσσερις χρόνους της μελέτης, και με μη παραμετρικές για την περιγραφική ανάλυση του δείγματος και τις στατιστικά σημαντικές διαφορές μεταξύ των δύο ομάδων ασθενών (Cross Tabs, Spearman Correlation Coefficient, Mann-Whitney U test). Τα αποτελέσματα στο σύνολο των ασθενών έδειξαν ότι το αρχικό ιικό φορτίο ελαττώθηκε στατιστικά σημαντικά σε όλους τους χρόνους της μελέτης, ανεξάρτητα από την επίτευξη SVR. Παράλληλα διαπιστώθηκε σημαντική, συνεχής και προοδευτική ελάττωση της ALT, επίσης ανεξάρτητα της επίτευξης SVR. Όσον αφορά στον αριθμό των Tregs δεν διαπιστώθηκαν σημαντικές διαφορές μεταξύ του αριθμού τους στους χρόνους Τ 0, Τ 1 και Τ 2 αλλά υπήρξε σημαντική αύξηση στο χρόνο Τ 3. Επίσης, διαπιστώθηκε σημαντική αύξηση μεταξύ των χρόνων Τ 1 και Τ 3. Ο αριθμός τους στις in vitro καλλιέργειες δεν παρουσίασε σημαντικές διακυμάνσεις. Τα CD3+CD8+ κυτταροτοξικά T λεμφοκύτταρα παρουσίασαν σημαντική αύξηση μόνο στο χρόνο Τ 3, ενώ τα CD4+ T βοηθητικά κύτταρα αν και παρουσίασαν αύξηση στο χρόνο Τ 1 τελικά 205
206 στο χρόνο Τ 3 μειώθηκαν ώστε ο αρχικός τους αριθμός δεν διαφοροποιήθηκε. Επίσης, ο αριθμός των CD4+CD25+ και των CD4+CD28+ T λεμφοκυττάρων αν και αυξήθηκε στο χρόνο Τ 1, στους υπόλοιπους χρόνους της μελέτης δεν διέφερε από τον αρχικό. Καθώς οι ασθενείς με χρόνια ηπατίτιδα C, ανάλογα με τον γονότυπο του ιού, απαιτούν διαφορετική χρονική διάρκεια θεραπείας και επιτυγχάνουν διαφορετικά ποσοστά SVR, τα αποτελέσματα των ασθενών που ολοκλήρωσαν την προκαθορισμένη διάρκεια θεραπείας και παρακολούθησης στη μελέτη, διαχωρίστηκαν σε δυο ομάδες: a) Oι ασθενείς της ομάδας 1 (γονότυπος 1), όταν αναλύθηκαν στο σύνολό τους, παρουσίασαν σημαντική ελάττωση του αριθμού των Tregs τη 12 η εβδομάδα (Τ 1 ), αλλά στη συνέχεια ο αριθμός τους αυξήθηκε έτσι ώστε να μη διαφέρει τελικά από τον αρχικό αριθμό τους. Όμως, οι ασθενείς με γονότυπο 1 που δεν πέτυχαν SVR, δεν παρουσίασαν μείωση του αριθμού των Tregs στο χρόνο Τ 1 και επιπλέον παρουσίασαν σημαντική αύξηση 6 μήνες μετά σε σχέση με τον αρχικό τους αριθμό. Ο αριθμός των in vitro καλλιεργειών επίσης δεν παρουσίασε σημαντικές μεταβολές, όπως και ο αριθμός των CD3+CD8+ T κυττάρων. Τα CD4+ T κύτταρα στην ομάδα 1 μειώθηκαν σημαντικά στο χρόνο Τ 2 σε σχέση με το χρόνο Τ 1, αλλά στο χρόνο Τ 3 δεν διέφεραν από τον αρχικό τους αριθμό. Όμως, στους ασθενείς με γονότυπο 1 που πέτυχαν SVR διαπιστώθηκε σημαντικά μικρότερος αριθμός CD4+ T κυττάρων στο τέλος της θεραπείας και 6 μήνες μετά σε σχέση με το χρόνο Τ 1. Τα CD4+CD25+ T κύτταρα στην ομάδα 1 συνολικά δεν παρουσίασαν σημαντικές διακυμάνσεις, όμως σε εκείνους τους ασθενείς που πέτυχαν SVR διαπιστώθηκε αρχικά μια αυξητική τάση την 12 η εβδομάδα και τελικά μια τάση μείωσης στο χρόνο Τ 3 σε σχέση με τη 12 η εβδομάδα και τον αρχικό τους αριθμό. Τα CD4+CD28+ T κύτταρα στην ομάδα 1 ελαττώθηκαν στο τέλος της θεραπείας σε σχέση με τη 12 η εβδομάδα, αλλά τελικά δεν διέφεραν από τον αρχικό τους αριθμό. Όμως, στους ασθενείς με γονότυπο 1 που πέτυχαν SVR διαπιστώθηκε μείωση του αριθμού τους στο χρόνο Τ 3 σε σχέση με τη 12 η εβδομάδα. β) Οι ασθενείς της ομάδας 2 (γονότυπος 2 και 3) που ολοκλήρωσαν τη θεραπεία πέτυχαν στο σύνολό τους SVR. Σε αυτούς ο αριθμός των Tregs παρουσίασε σημαντική ελάττωση στο τέλος της θεραπείας σε σχέση με το χρόνο Τ 0, αλλά στο χρόνο Τ 3 αυξήθηκε ώστε τελικά δεν διέφερε από τον αρχικό. Ο αριθμός των Tregs στις in vitro καλλιέργειες δεν παρουσίασε σημαντικές μεταβολές. 206
207 Οι υποπληθυσμοί των CD3+CD8+, CD4+ και CD4+CD25+ T λεμφοκυττάρων αν και παρουσίασαν ορισμένες σημαντικές μεταβολές στη διάρκεια της θεραπείας, δεν ήταν σημαντικές σε σχέση με τον προ της θεραπείας αριθμό τους. Όμως, ο αριθμός των CD4+CD28+ T λεμφοκυττάρων παρουσίασε σημαντική αύξηση την 4 η εβδομάδα σε σχέση με το χρόνο Τ 0, αλλά και στο χρόνο Τ 3 σε σχέση με την 4 η εβδομάδα. Επιπλέον, κατά την σύγκριση των δύο ομάδων ασθενών μεταξύ τους, διαπιστώθηκε ότι στην ομάδα 1 οι ασθενείς εμφάνιζαν μικρότερη τιμή ALT και μικρότερο αριθμό Tregs στο χρόνο Τ 1 και υψηλότερο ιικό φορτίο στο χρόνο Τ 3 σε σχέση με τους ασθενείς της ομάδας 2. Οι συσχετίσεις μεταξύ των διαφόρων μεταβλητών που μετρήθηκαν στην παρούσα μελέτη έδειξαν: θετική συσχέτιση ηλικίας και νεκροφλεγμονώδους δραστηριότητας (ΗΑΙ) θετική συσχέτιση ΗΑΙ και διαβρωτικής νέκρωσης, πυλαίας φλεγμονής και ενδολόβιων νεκρώσεων με την πρώτη να είναι ισχυρότερη θετική συσχέτιση ΗΑΙ και ίνωσης θετική συσχέτιση ΗΑΙ και ALT στο χρόνο Τ 0 και Τ 3 μη σημαντική συσχέτιση του αριθμού των Tregs προ θεραπείας με όλες τις λοιπές μεταβλητές θετική συσχέτιση του ιικού φορτίου στο χρόνο Τ 1 με τον αριθμό των Tregs στο χρόνο Τ 2 αρνητική συσχέτιση του αριθμού των Tregs στο τέλος της θεραπείας και του αριθμού των CD4+ και των CD4+CD28+ T κυττάρων στους χρόνους Τ 2 και Τ 3 θετική συσχέτιση του αριθμού των Tregs στο τέλος της θεραπείας με τον αριθμό τους στο χρόνο Τ 3 αρνητική συσχέτιση του αριθμού των Tregs στο χρόνο Τ 3 με τον αριθμό των CD4+ T κυττάρων στο χρόνο Τ 0, και κυρίως με τον αριθμό τους στο χρόνο Τ 1 και Τ 3 αρνητική συσχέτιση του αριθμού των Tregs στο χρόνο Τ 3 με τον αριθμό των CD4+CD28+ T λεμφοκυττάρων στο χρόνο Τ 1 και Τ 3 θετική συσχέτιση του αριθμού των Tregs στο χρόνο Τ 3 και του αριθμού των CD3+CD8+ T κυττάρων στον ίδιο χρόνο θετική συσχέτιση του αριθμού των Tregs και του ιικού φορτίου στο χρόνο Τ 3 207
208 ο αριθμός των Tregs ήταν σημαντικά μεγαλύτερος στους ασθενείς με γονότυπο 2 και 3 κατά τη διάρκεια της θεραπείας (Τ 1 ) αρνητική συσχέτιση του αριθμού των Tregs που αναπτύχθηκε στην in vitro καλλιέργεια στο χρόνο Τ 1 με την ίνωση της βιοψίας ήπατος και τον αριθμό των Tregs στο χρόνο Τ 3 οι άνδρες στο τέλος της θεραπείας είχαν την τάση να έχουν μικρότερο αριθμό Tregs Από την παρούσα μελέτη προέκυψαν τελικά τα εξής συμπεράσματα: 1)Το γεγονός ότι ο αριθμός των Tregs στο σύνολο των ασθενών, δεν παρουσίασε σημαντικές διαφορές στο τέλος της θεραπείας και παρά την αύξησή του μεταξύ του χρόνου Τ 1 και του Τ 3 (6 μήνες μετά το τέλος της θεραπείας),δεν επηρέασε την έκβαση της θεραπείας, δεν επιτρέπει την διεξαγωγή συμπερασμάτων ως προς τον παθογενετικό τους ρόλο κατά τη διάρκεια της θεραπείας. 2)Η σημαντική αύξηση των CD3 + CD8 + Τ κυττάρων 6 μήνες μετά το τέλος της θεραπείας πιθανόν να οφείλεται στην επίδραση της θεραπείας και ειδικότερα στην χορήγηση της ιντερφερόνης-α 3)Τα CD4 + T κύτταρα ελαττώθηκαν σημαντικά 6 μήνες μετά το τέλος της θεραπείας σε σχέση με τον χρόνο Τ 1 συνηγορώντας υπέρ της ενεργού συμμετοχής τους κατά τη διάρκεια της θεραπείας 4)Τα CD4 + CD25 + T κύτταρα φαίνεται ότι παρουσιάζουν κάποια ενεργό δραστηριότητα έναντι του HCV αρχικά μετά την έναρξη της θεραπείας, ο ρόλος τους όμως στην εξέλιξη της θεραπείας δεν είναι σαφής 5)Τα CD4 + CD28 + T κύτταρα παρουσίασαν διακυμάνσεις κατά τη θεραπεία συνηγορώντας υπέρ κάποιας δραστηριότητας των κυττάρων αυτών πιθανόν ως αποτέλεσμα της θεραπείας 6) Στους ασθενείς με γονότυπο 1, ο αριθμός των Tregs παρουσίασε μία διαφοροποίηση, δηλαδή ο αριθμός τους δεν ελαττώθηκε κατά τη διάρκεια της θεραπείας στους ασθενείς οι οποίοι δεν απήντησαν στο θεραπευτικό σχήμα και δεν πέτυχαν SVR 7) Στους ασθενείς με γονότυπο 1 οι οποίοι απήντησαν στη θεραπεία διαπιστώθηκε ότι ο αριθμός των CD4+CD28+ Τ κυττάρων ελαττώθηκε σημαντικά μεταξύ της εβδομάδας 12 και 6 μήνες μετά το τέλος της θεραπείας, συνηγορώντας ότι τα κύτταρα αυτά θα πρέπει να εμπλέκονται στην απάντηση ή μη της θεραπείας 208
209 8)Ο αριθμός των Tregs στους ασθενείς με γονότυπο 2 και 3 παρουσίασε σημαντική ελάττωση στο τέλος της θεραπείας και στη συνέχεια αυξήθηκε 6 μήνες μετά το τέλος της, συνηγορώντας υπέρ μιας ενεργού συμμετοχής των κυττάρων αυτών στην επίτευξη SVR 9)Κατά τη σύγκριση μεταξύ των δύο ομάδων ασθενών διαπιστώθηκε ότι οι ασθενείς με γονότυπο 1 εμφάνιζαν σημαντικά μικρότερο αριθμό Tregs σε σύγκριση με τους ασθενείς με γονότυπο 3 κατά τη διάρκεια της θεραπείας στον χρόνο Τ 1 (εβδομάδα 4/ εβδομάδα 12 αντίστοιχα). Tα ευρήματα αυτά είναι δύσκολο να εκτιμηθούν αν και θα αναμένονταν ο αριθμός των Tregs να ήταν σημαντικά μικρότερος στους ασθενείς με γονότυπο 2 και 3 οι οποίοι και όλοι επιτυγχάνουν SVR 10) Οι υπόλοιποι Τ λεμφοκυτταρικοί υποπληθυσμοί της μελέτης παρουσίασαν διάφορες συσχετίσεις στους χρόνους της μελέτης, οι οποίες είναι δύσκολο να ερμηνευθούν ως προς την τελική έκβαση της θεραπείας και τη συμβολή τους ή μη στην επίτευξη SVR Το τελικό συμπέρασμα της μελέτης, όπως προκύπτει από την ανάλυση όλων των ευρημάτων, είναι ότι ο παθογενετικός ρόλος των Tregs φαίνεται να διαφέρει μεταξύ των ασθενών με γονότυπο 1 και αυτών με γονότυπο 2 και 3. Εντούτοις όμως, δεν είναι σαφές εάν οι διακυμάνσεις του αριθμού των CD4+CD25+ high Foxp3+ Tregs στο περιφερικό αίμα μπορούν να χρησιμοποιηθούν ως προγνωστικός δείκτης της επίτευξης SVR ή όχι. Βέβαια όπως είναι γνωστό, η απάντηση στη λοίμωξη από κάποιον ιό είναι πολυπαραγοντική και η εξέλιξή της έγκειται στη συμμετοχή διαφόρων παραμέτρων της ανοσιακής απάντησης, στην παρουσία άλλων νοσημάτων, και στην γενετική διαφοροποίηση των ασθενών. Επιπλέον θα πρέπει να τονισθεί ότι η παρουσία διαφόρων υποπληθυσμών των Τ ρυθμιστικών κυττάρων δυσχεραίνει την εξαγωγή συμπερασμάτων για ένα μεμονωμένο υποπληθυσμό τους και ότι η συμπεριφορά αυτών των διαφόρων υποπληθυσμών στο περιφερικό αίμα δεν προδικάζει τη δράση τους στον ηπατικό ιστό όπου και εγκαθίσταται ο ιός και προκαλεί την ιστική βλάβη. 209
210
211 ΙΧ. SUMMARY Hepatitis C virus is a small positive sense single-stranded RNA virus and is the most important cause of chronic liver disease worldwide. A distinct and major characteristic of chronic hepatitis C, is its tendency to cause chronic liver disease, cirrhosis, liver failure and liver cancer. The immune response to viral infections comprises innate and adaptive defenses and the ability to shape the response in a virus-specific manner is depending upon communication between innate and adaptive systems. This interaction is so crucial that the adaptive response cannot occur without an innate immune system. The immunopathogenesis of chronic hepatitis is a matter of great controversy and has been suggested to involve a complex balance between several arms of the immune system. Thus, inadequate HCV specific immune reactions such as inadequate innate immune responses, insufficient or inability to maintain adaptive immune responses, viral evasion mechanisms and viral sequence variations are proposed as potential mechanisms of viral persistence. Recently, it has become evident that T regulatory cells play a role in suppressing the HCV specific antiviral cell responses favoring persistence, but at the same time preventing the host from massive liver tissue damage. T regulatory cells are a heterogeneous population with unique phenotypic and functional characteristics that have a suppressive capacity and are divided in two major subsets: naturally arising T regulatory cells (ntregs, CD4+CD25+ high Tregs) that are produced in the thymus and diverse populations of induced Tregs (itregs) that develop in the periphery after expansion. The purpose of the present study was to investigate the role of CD4+CD25+ high Foxp3+ T regulatory cells (Tregs) in the pathogenesis of chronic HCV infection and the response to treatment in naive patients. 61 patients were included in the study, 39.3% genotype 1 (24/61), 4.9% genotype 2 (3/61), 47.6% genotype 3 (29/61) and 8.2% genotype 4 (5/61). Liver biopsy was performed in 59/61 patients and was validated using the modified Knodell system. Viral load was measured with a sensitive qualitative and quantitative assay (Combas Amplicor and Amplicor Monitor, Roche Diagnostics) and genotype was assessed using INNOLIPA test (Bayer Diagnostics). The patients fulfilled the inclusion criteria, consented to participate in the study and were treated with pegylated interferon -a and ribavirin. Patients with 211
212 genotype 1 or 4 were treated for 48 weeks and patients with genotype 2 or 3 for 24 weeks. Thirty six/61 (59%) patients achieved SVR, 4/61(6.6%) did not respond to treatment, 5/61 ( 8.2%) stopped treatment due to side effects and 16/61 (26.2%) were lost to follow-up, mostly due to relapse in iv drug abuse. Patients with CHC genotype 4, were excluded from the final analysis of the results due to an atypical response to treatment. The patients that completed therapy (37/61) were divided in two groups: a) group 1, patients with CHC genotype 1 (15/37) and b) group 2, patients with genotype 2 or 3 (22/37). All study parameters (HCV RNA, ALT, Tregs, T cell subsets) were determined in 4 time-points: a. (T 0 ), prior to treatment b. (T 1 ), at a selected time-point during therapy according to CHC genotype, that is, -genotype 1: week 12, because the PPV for achieving SVR is 60% in patients with negative HCV RNA at w12 -genotype 2 and 3: week 4, because the PPV for achieving SVR is 95% in patients with negative HCV RNA at w4 c. (T 2 ): end of treatment (EOT) (48 weeks for patients with genotype 1 and 24 weeks for patients with genotype 2 or 3) d. (T 3 ): 6 months after completion of therapy (SVR) In order to investigate the role of Tregs in CHC we determined the expression of the intracellular transcription factor Foxp3 in the activated CD4+CD25+ high T lymphocytes which defines the subset of T regulatory cells (CD4+CD25+ high Foxp3+ Τ cells). The intracellular monoclonal antibody Foxp3 was selected because its constant and abundant expression characterizes the subset of CD4+CD25+ T regulatory cells, both natural and induced. In addition, we determined a) the expression of CD3+, CD4+ and CD8+ T subsets b) the a chain of the IL-2 receptor (IL-R2a, CD25) in the subset of T lymphocytes, as well as the coexpression of CD4 and CD25 on Τ lymphocytes (CD4+CD25+ double positive) and c) the expression of the costimulatory molecule CD28 on T lymphocytes as well as the coexpression of CD4 and CD28 on T lymphocytes (CD4+CD28+ double positive). Measurements at the selected time-points were performed in a) peripheral whole blood (ex vivo) and b) in vitro cultures of the 212
213 peripheral monocytes (lymphocytes, macrophages) after stimulation with phytohaemagglutinin (PHA). All the above parameters were determined using the triple colour flow cytometry (FACS, EPICS XL-MCL, Beckman Coulter) and the monoclonal antibodies CD3, CD4 (FITC, Immunotech), CD4, CD8, CD25, CD28 (PE, Immunotech), CD25 (ECD, Immunotech) and Foxp3 (PE, Ebioscience). Statistical analysis of our results was performed using a) a parametrical method (paired samples t-test) concerning the parameters in the 4- time points of the study and b) the non-parametrical methods (Cross Tabs, Spearman Correlation Coefficient and Mann-Whitney U test) for the analysis of the parameters between the 2 groups of patients. Our results showed that the initial viral load and ALT values were significantly decreased in all time-points, in all the patients, regardless of genotype or SVR achievement. The numbers of Tregs did not change significantly when Τ 1 and Τ 2, were compared to T 0, but they were significantly increased in Τ 3. A significant increase in their number was also noted from time-point Τ 1 to Τ 3. The Treg numbers in the in vitro cultures did not change significantly. The numbers of CD3+CD8+ cytotoxic T cell subset increased significantly only at Τ 3. The numbers of CD4+ T helper cells were initially significantly increased at Τ 1 but later, at time-point Τ 3, they were decreased so that they did not differ from their initial number before treatment. Similarly, although the numbers of CD4+CD25+ and CD4+CD28+ T cell subsets were both increased in T 1, they showed no significant differences at T 2 and T 3 compared to their initial number in T o. Furthermore, since the duration of treatment and SVR rates differ in the patients with CHC according to their HCV genotype, results were analyzed per patient group. In group 1, (genotype 1 patients), a significant decrease in the Tregs numbers at 12 th week of therapy was noted, but their numbers increased thereafter, so that at the end of the study, they did not differ from their initial numbers. However, when results were analyzed according to SVR achievement, the patients that did not achieve SVR did not show a decrease in their Treg numbers in Τ 1 and had a significant increase in their numbers 6 months after the end of therapy. The Treg numbers in the in vitro cultures did not change significantly. CD3+CD8+ T cells in the peripheral blood also remained unchanged. CD4+ T cells were significantly decreased at EOT in 213
214 comparison with their numbers in w12, but at Τ 3 they did not differ from their initial numbers. However, a different pattern of response was found in this patient s group in the sense that patients with SVR had a significantly smaller number of CD4+ T cells at EOT and 6 months later, in comparison with their numbers at w12. CD4+CD25+ T cells did not change significantly. However, the patients that achieved SVR showed initially a tendency to increase their CD4+CD25+ T cell numbers at w12 and a tendency to decrease them, 6 months after therapy when compared to their numbers before treatment and at week 12 th. CD4+CD28+ T cell numbers were decreased at EOT in comparison with their numbers at w12, but at the end of the study they did not differ from their initial numbers. The patients that achieved SVR had a significant decrease in CD4+CD28+ T cell numbers 6 months after therapy compared to their numbers at w12. All the patients of group 2 (genotype 2 or 3 patients) that completed therapy, achieved SVR. In group 2, Treg numbers were significantly lower at EOT in comparison with their initial numbers, but were gradually increased thereafter, so that at 6 months after the end of treatment, their numbers did not differ from those found before the initiation of treatment. Treg numbers in the in vitro cultures did not change significantly. Although the numbers of CD3+CD8+ cytotoxic, CD4+ helper and CD4+CD25+ T cells showed some statistically significant changes during therapy, at the end of the study they were not significantly different from their numbers prior to therapy. However, CD4+CD28+ T cells increased significantly at w4 compared to their initial numbers, as well as 6 months after the end of treatment in comparison with their numbers in w4. Furthermore, when patients of group 1 were compared to patients of group 2, group 1 patients had significantly lower ALT values and a smaller number of Tregs at time-point Τ 1, but higher viral load 6 months after the end of treatment. When the results were correlated in between the different parameters studied, it was shown that: age correlated positively with necroinflammatory activity in the liver biopsy (ΗΑΙ) HAI correlated positively with periportal and/or bridging necrosis, portal inflammation and intralobular degeneration, but had a stronger correlation with periportal and/or bridging necrosis ΗΑΙ correlated positively with fibrosis 214
215 ΗΑΙ correlated positively with ALT values at Τ 0 and Τ 3 Treg numbers before treatment did not correlate with any parameter Viral load at Τ 1 correlated positively with Treg numbers at EOΤ Treg numbers at EOT correlated negatively with CD4+ and CD4+CD28+ T cell numbers at EOT and Τ 3 Treg numbers at EOT correlated positively with their numbers at T 3 Treg numbers at Τ 3 correlated negatively with CD4+ T cell numbers prior to therapy, and mostly with their numbers at Τ 1 and Τ 3 Treg numbers at T 3 correlated negatively with the number of CD4+CD28+ T cells at Τ 1 and Τ 3 Tregs 6 months after the end of treatment correlated positively with CD3+CD8+ T cell numbers at the same time-point (Τ 3 ) Treg numbers at T 3 correlated positively with viral load at Τ 3 Treg numbers were significantly higher in patients with genotype 2 and 3 at Τ 1 Treg numbers at the in vitro culture at Τ 1 correlated positively with fibrosis score and with Treg numbers at Τ 3 Men at EOT had a tendency to have smaller Treg numbers than women In conclusion, the findings of the present study have shown that: 1) The fact that Treg numbers did not change significantly at EOT, although they increased between Τ 1 and Τ 3 in all patients, shows that Tregs do not probably influence the response to treatment. Thus, no conclusions as to their pathogenetic role in respect to treatment response and SVR achievement could be drawn. 2) The significant increase of CD3+CD8+ Τ cells 6 months after treatment could be possibly attributed to the influence of therapy, and mainly of pegylated interferon-a 3) The numbers of CD4 + T helper cells decreased significantly 6 months after the end of treatment in comparison with their numbers at Τ 1, a finding that could be interpreted as of their active involvement during therapy 4) CD4 + CD25 + T cells seem to have an active role against HCV in the early stages after treatment is initiated, but their role in the response to treatment is unclear 215
216 5) CD4 + CD28 + T cells showed several fluctuations in their numbers, a finding that points to the fact that they have some activity during treatment possibly initiated by pegylated interferon-a 6) In patients with genotype 1, Treg numbers showed some differentiation in between subgroups, i.e their numbers were not decreased during therapy in the group of patients that did not respond to treatment and failed to achieve SVR 7) In patients with genotype 1 that achieved SVR, CD4 + CD28 + Τ cell numbers decreased significantly 6 months after the end of treatment compared to their numbers at w12, possibly indicating that this T cell subset is somehow affecting the response to treatment. 8) Treg numbers in patients with genotype 2 or 3 (group 2) were significantly decreased at EOT and then increased 6 months later, indicating that they were actively involved in the response to treatment and the SVR achievement. 9) When the two groups of patients were compared, it was shown that group 1 had a significantly smaller number of Tregs at w12 than group 2 at w4. This finding is difficult to be interpreted in the sense that rather the opposite finding was expected since all patients with genotype 2 or 3 achieved SVR. 10) All of the T cell subsets that were determined in the four time-points of the study showed several correlations with one another, but their role concerning the response to treatment and the SVR achievement, is difficult to be clearly explained. The final conclusion of the present study, taking into consideration all the findings and correlations, is that the pathogenetic role of CD4+CD25+ high Foxp3+ T regulatory cells in the peripheral blood of naive patients with CHC seems to be different between genotype 1 and genotypes 2 or 3. However, it is not clear whether the fluctuations of Tregs in the peripheral blood during treatment could be used as a predictive marker of the response to treatment and the achievement of SVR. Furthermore, it is well known that the outcome of a viral infection is multi-factorial and that it depends on the cooperation of several mechanisms of the immune response, the co-morbidities and the genetic background of the patient. In addition, the heterogeneity of the T regulatory cell subsets makes it even more difficult to draw conclusions in the case of one subpopulation of Tregs studied and as 216
217 well as the fact that their numbers and function in the peripheral blood does not necessarily reflect their pathogenetic role and action in the liver where HCV persists and induces the tissue damage. 217
218
219 Χ. ΒΙΒΛΙΟΓΡΑΦΙΑ 1.Lavanchy D, The global burden of hepatitis C, Liver Int 2009 Jan;29 Suppl 1:74-81 ( ) 2.Fabiani FL, Epidemiology of hepatitis C virus infection, World J Gastroenterol 2007 May 7; 13(17): Gogos CA, Fouka KP, Nikiforidis G, Avgeridis K et al, Prevalence of hepatitis B and C virus infection in the general population and selected groups in the South-Western Greece, Eur J Epidemiol 2003;18(6):551-7 ( ) 4.Goritsas C, Plerou I, Agaliotis S, Spinthaki R, Mimidis K, et al, HCV infection in the general population of a Greek island: prevalence and risk factors, Hepatogastroenterology 2000 May-Jun;47(33):782-5 ( ) 5.Koulentaki M, Spanoudakis S, Kantidaki E, Drandakis P, et al, Prevalence of hepatitis B and C markers in volunteer blood donors in Crete. A 5-year study, J Viral Hepat 1999 May;6(3):243-8 ( ) 6.Choo QL, Kuo G, Weiner AJ, et al, Isolation of a cdna clone derived from a blood-borne non-a, non-b viral hepatitis genome, Science 1989 Apr 21; 244(4902): ( ) 7.Zhang J, Randall G, Higginbottom A, Monk P et al, CD81 is required for hepatitis C virus glycoprotein-mediated viral infection, J Virol 2004 Feb; 78(3): ( ) 8.Scarselli E, Ansuini H, Cerino R, Roccasecca RM et al, The human scavenger receptor class B type I is a novel candidate receptor for the hepatitis C virus, EMBO J 2002 Oct 1; 21(19): ( ) 9.Evans MJ, von Hahn T, Tscherne DM, Syder AJ et al, Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry, Nature 2007 Apr 12; 44(7137): ( ) 10.Ploss A, Evans MJ, Gaysinskaya VA, Panis M, et al, Human occluding is a hepatitis C virus entry factor required for infection of mouse cells, Nature 2009 Feb 12; 457(7231): ( ) 11.Simmonds P, Alberti A, Alter HJ, Bonino F et al, A proposed system for the nomenclature of hepatitis C viral genotypes, Hepatology, 1994;19(5): ( ) 12.Alter HJ, Seeff LB, Recovery, persistence and sequelae in hepatitis C virus infection: a perspective on long term outcome, Semin Liver Dis 2000;20(1):17-35 ( ) 219
220 13.Lauer GM, Walker BD, Hepatitis C virus infection, N Engl J Med 2001 Jul 5;345(1):41-52 ( ) 14.Gretch DR, Diagnostic tests for hepatitis C, Hepatology 1997;26(34 Suppl 1): ( ) 15.Franciscus A, HCV Diagnostic Tools: grading and staging a liver biopsy, HCSP (Hepatitis C Support Project), Version 2.2, October Castera L, Hartmann DJ, Chapel F, Guettier C et al, Serum laminin and type IV collagen are accurate markers of histologically severe alcoholic hepatitis in parients with cirrhosis, J Hepatol 2000 Mar;32(3):412-8 ( ) 17.Ichino N, Osakabe K, Nishikawa T, Sugiyama H et al, A new index for non-invasive assessment of liver fibrosis, World J Gastroenterol 2010 Oct 14;16(38): Friedrich-Rust M, Rosenberg W, Parkes J et al, Comparison of ELF, Fibrotest and Fibroscan for the non-invasine assessment of liver fibrosis, BMC Gastroenterology 2010, 10: Lee MH, Cheong JY, Um SH, Seo YS et al, Comparison of surrogate serum markers and transient elastogrphy (Fibroscan) for assessing cirrhosis in patients with chronic viral hepatitis, Dig Dis Sci 2010;55: Castera L, Foucher J, Bernard PH, Carvalho F et al, Pitfalls of liver stiffness measurement: a 5-year prospective study of examinations, Hepatology 2010, vol 51, no3, Yin M, Talwalkar JA, Glaser KJ, Manduca A et al, Assessment of hepatic fibrosis with magnetic resonance elastography, Clin Gastroenterol Hepatol 2007 Oct;5(10): ( ) 22.Huwart L, van Beers BE, MR elastography, Gastroenterol Clin Biol 2008 Sep; 32(6 Suppl 1);68-72 ( ) 23.Castera L, Verginiol J, Foucher J, Le Bail et al, Prospective comparison of transient elastography, Fibrotest, APRI, and liver biopsy for the assessment of fibrosis in chronic hepatitis C, Gastroenterology 2005 Feb;128(2): ( ) 24.Ziol M, Handra-Luca A, Kettaneh A, Christidis C et al, Noninvasive assessment of liver fibrosis by measurement of stiffness in patients with chronic hepatitis C, Hepatology 2005 Jan;41(1):48-54 ( ) 25.Alter MJ, Margolis HS, Krawczynski K, Judson FN, et al, The natural history of community-aquired hepatitis C in the United States. The Sentinel Counties Chronic non-a, non-b Hepatitis Study Team, NEJM 1992 Dec 31; 327(27): ( ) 220
221 26.Farci P, Alter HJ, Shimoda A, Govindarajan S et al, Hepatitis C virusassociated fulminant hepatic failure, NEJM 1996 Aug 29;335(9):631-4 ( ) 27.Villano SA, Vlahov D, Nelson KE, Cohn S, Thomas DL, Persistence of viremia and the importance of long-term follow-up after acute hepatitis C infection, Hepatology 1999 Mar;29(3): ( ) 28.Lechner F, Wong DK, Dunbar PR, Chapman R, Chung RT, Dohrenwend P, Robbins G, et al, Analysis of successful immune responses in persons infected with hepatitis C virus, J Exp Med 2000 May 1; 191(9): ( ) 29.Jauncey M, Micallef JM, Gilmour S et al, Clearance of hepatitis C virus after newly acquired infection in injection drug users, J Infect Dis 2004;190: Watanabe H, Saito T, Shinzawa H et al, Spontaneous elimination of serum hepatitis C virus (HCV) RNA in chronic HCV carriers: a population-based cohort study. J Med Virol 2003; 71:56-61 ( ) 31.Scott JD, McMahon BJ, Bruden D et al, High rate of spontaneous negativity for hepatitis C virus RNA after establishment of chronic infection in Alaska Natives, Clin Infect Dis 2006;42: ( ) 32.Poynard T, Bedossa P, Opolon P, Natural history of liver fibrosis progression in patients with chronic hepatitis C. The OBSVIRC, METAVIR, CLINIVIR and DOSVIRC groups, Lancet 1997 Mar 22; 349(9055): ( ) 33.Bjoro K, Froland SS, Yun Z, Samdal HH, Haaland T, Hepatitis C infection in patients with primary hypogammaglobulinemia after treatment with contaminated immune globulin, NEJM 1994 Dec 15;331(24) ( ) 34.Berenguer M, Ferrell L, Wtson J, Prieto M, Kim M et al, HCV-related fibrosis progression following liver transplantation: increase in recent years, J Hepatol 2000 Apr; 32(4): ( ) 35.Muzzi A, Leandro G, Rubbia-Brandt L, James R, Keiser O et al, Insulin resistance is associated with liver fibrosis in non-diabetic chronic hepatitis C patients, J Hepatol 2005 Jan;42(1):41-46 ( ) 36.Benhamou Y, Bochet M, Di Martino V, Charlotte F, Azria F et al, Liver fibrosis progression in human immunodeficiency virus and hepatitis C virus coinfected patients. The Multivirc Group, Hepatology 1999 Oct; 30(4): ( ) 37.Chen SL, Morgan TR, The natural History of hepatitis C virus (HCV) infection, Int J Med Sci 2006;3(2):47-52 ( ) 221
222 38.Fattovich G, Giustina G, Degos F, Tremolada F, Diodati G et al, Morbidity and mortality in compensated cirrhosis type C: a retrospective follow-up study of 384 patients, Gastroenterology, 1997 Feb; 112(2): ( ) 39.Serfaty L, Aumaitre H, Chazouilleres O, Bonnand AM et al, Determinants of outcome of compensated hepatitis C virus-related cirrhosis, Hepatology, 1998 May; 27(5): ( ) 40.Hu KQ, Tong MJ, The long-term outcomes of patients with compensated hepatitis C virus-related cirrhosis and history of parenteral exposure in the United States, Hepatology 1999 Apr; 29 (4): ( ) 41.Kamal SM, Moustafa KN, Chen J et al, Duration of peginterferon therapy in acute hepatitis C: a randomized trial, Hepatology, 2006;39: ( ) 42.Kamal SM, Fouly AE, Kamel RR et al, Peginterferon alfa-2b therapy in acute hepatitis C: impact of onset of therapy on sustained virologic response, Gastroenterology 2006;130; ( ) 43.Ghany MG, Strader DB, Thomas DL, Seeff LB, Diagnosis, management and treatment of hepatitis C: An update, Hepatology 2009 Apr;49(4): Strader DB, Wright T, Thomas DL et al, Diagnosis, management and treatment of hepatitis C, Hepatology 2004;39: ( ) 45.National Institutes of Health Consensus Development Conference Statement: Management of hepatitis C 2002 (June 10-12, 2002, Gastroenterology, 2002, 123(6): ( ) 46.Ferenci P, Fried MW, Shiffman ML, Smith CI, Marinos G et al, Predicting sustained virological responses in chronic hepatitis C patients treated with peginterferon alfa-2a (40 KD)/ribavirin, J Hepatol 2005 Sep;43(3): ( ) 47.Jensen DM, Morgan TR, Marcellin et al, Early identification of HCV genotype 1 patients responding to 24 weeks peginterferon alpha-2a (40kd)/ribavirin therapy, Hepatology 2006;43: ( ) 48.vonWagner M, Huber M, Berg et al, Peginterferon-alpha-2a (40KD) and ribavirin for 16 or 24 weeks in patients with genotype 2 or 3 chronic hepatitis C, Gastroenterology 2005;129: ( ) 49.Berg T, vonwagner M, Nasser S et al, Extended treatment duration for hepatitis C virus type 1: comparing 48 versus 72 weeks of peginterferon-alpha-2a plus riibavirin, Gastroenterology 2006;130: ( ) 222
223 50.Shiffman ML, Di Bisceglie AM, Lindsay KL, Morishima C et al, Peginterferon alfa-2a and ribavirin in patients with chronic hepatitis C who have failed prior treatment, Gastroenterology 2004 Apr;126(4): ( ) 51.Jacobson IM, Gonzalez SA, Ahmed F, Lebovics E, Min AD, et al, A randomized trial of pegylated interferon alpha-2b plus ribavirin in the retreatment of chronic hepatitis C, Am J Gastroenterol 2005 Nov;100(11): ( ) 52.Pawlotsky JM, Therapy of hepatitis C: From empiricism to eradication, Hepatology, 2006 Feb;43(2 Suppl 1):S ( ) 53.Dubuisson J, Helle F, Cocquerel L, Early steps of the hepatitis C virus life cycle, Cell Microbiol 2008 Apr;10(4):821-7 ( ) 54.Κλινική Ανοσολογία, Ραπτοπούλου- Γιγή Μ, University Studio Press, Θεσσαλονίκη Abbas ΑΚ, Lichtman ΑΗ, Pillai S, Cellular and Molecular Immunology, 6th edition, Saunders, Zhu J, Paul EW. CD4 T cells: fates, functions and faults, ASH 50 th Anniversary Review, The American Society of Hematology, Blood ; 112 (5): Mossmann TR, Cherwinski H, Bond MW, Gieldin MA, Coffman RL, Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins, J Immunol 1986 Apr 1; 136(7): ( ) 58.Zhu J, Paul WE, Heterogeneity and plasticity of T helper cells, Cell Research 2010, 20: Locksley RM, Nine lives: plasticity among T helper cell subsets, The Journal of Experimental Medicine 2009, Vol 206, no 8, (ήταν 331) 60.Dahari H, Major M, Zhang X, Mihalik K et al, Mathematical modeling of primary hepatitis C infection: noncytolytic clearance and early blockage of virion production, Gastroenterology 2005 Apr;128(4): ( ) 61.Biron CA, Interferons alpha and beta as immune regulators--a new look, Immunity 2001 Jun;14(6):661-4 ( ) 62.Foy E, Li K, Wang C, Sumpter R Jr, Ikeda M, et al, Regulation of interferon regulatory factor-3 by the hepatitis C virus serine protease, Science 2003 May 16;300(5622): ( ) 223
224 63.Gale MJ Jr, Korth MJ, Tang NM, Tan SL, Hopkins DA et al, Evidence that hepatitis C virus resistance to interferon is mediated through repression of the PKR protein kinase by the nonstructural 5A protein, Virology 1997 Apr 14;230(2): ( ) 64.Tan SL, Katze MG, How hepatitis C virus counteracts the interferon response: the jury is still out on NS5A, Virology 2001 May 25;284(1):1-12 ( ) 65.Decalf J, Fernandes S, Longman R, Ahloulay M, Audat F et al, Plasmacytoid dendritic cells initiate a complex chemokine and cytokine network and are a viable drug target in chronic HCV patients, J Exp Med 2007 Oct 1;204(10): ( ) 66.Dolganiuc A, Chang S, Kodys K, Mandrekar P, Bakis G, Cormier M, Szabo G. Hepatitis C virus (HCV) core protein-induced, monocytemediated mechanisms of reduced IFN-α and plasmacytoid dendritic cell loss in chronic HCV infection. J Immunol, 2006; 177: Auffermann-Gretzinger S, Keeffe E B, Levy S. Impaired dendritic cell maturation in patients with chronic, but not resolved, hepatitis C virus infection, Blood, 2001; 97: Dolganiuc A, Kodys K, Kopasz A, Marshall C, Do T, Romics L, Jr, Mandrekar P, Zapp M, Szabo G. Hepatitis C virus core and nonstructural protein 3 proteins induce pro- and anti-inflammatory cytokines and inhibit dendritic cell differentiation, J Immunol. 2003; 170: Thimme R, Oldach D, Chang KM, Steiger C, Ray SC, Chisari FV, Determinants of viral clearance and persistence during acute hepatitis C virus infection, J Exp Med 2001; 194(10): ( ) 70.Logvinoff C, Major ME, Oldach D, Heyward S, Talal A et al, Neutralizing antibody response during acute and chronic hepatitis C virus infection, Proc Natl Acad Sci USA 2004 Jul 6;101(27): ( ) 71.Pestka JM, Zeisel MB et al, Rapid induction of virus-neutralizing antibodies and viral clearance in a single-source outbreak of hepatitis C, Proc Natl Acad Sci USA 2007 Apr 3;104(14): ( ) 72.Kaplan DE, Sugimoto K, Newton K, Valiga ME, Ikeda F et al, Discordant role of CD4 T-cell response relative to neutralizing antibody and CD8 T-cell responses in acute hepatitis C, Gastroenterology 2007 Feb;132(2): ( ) 224
225 73.Day CL, Lauer GM, Robbins GK, McGovern B et al, Broad specificity of virus-specific CD4+ T-helper-cell responses in a resolved hepatitis C virus infection, J Virol 2002 Dec;76(24): ( ) 74.Urbani S, Amadei B, Fisicaro P, Tola D et al, Outcome of acute hepatitis C is related to virus-specific CD4 function and maturation of antiviral memory CD8 responses, Hepatology 2006 Jul;44(1): ( ) 75.Neumann-Haefelin C, Spangenberg HC, Blum HE, Thimme R, Host and viral factors contributing to CD8+ T cell failure in hepatitis C virus infection, World J Gastroenterol 2007 Sep 28; 13(36): ( ) 76.Kanto T, Hayashi N. Immunopathogenesis of Hepatitis C Virus Infection: Multifaceted Strategies Subverting Innate and Adaptive Immunity. 2006, The Japanese Society of Internal Medicine: von Hahn T, Yoon JC, Alter H, Rice CM et al, Hepatitis C virus continuously escapes from neutralizing antibody and T-cell responses during chronic infection in vivo, Gastroenterology 2007 Feb;132(2): ( ) 78.Dowd KA, Netski DM, Wang XH, Cox AL, Ray SC, Selection pressure from neutralizing antibodies drives sequence evolution during acute infection with hepatitis C virus, Gastroenterology 2009 Jun;136(7) ( ) 79.Lauer GM, Barnes E, Lucas M, Timm J, Ouchi K et al, High resolution analysis of cellular immune responses in resolved and persistent hepatitis C virus infection, Gastroenterology 2004 Sep;127(3): ( ) 80.Gruener N H, Lechner F, Jung M C, Diepolder H, Gerlach T, Lauer G, Walker B, Sullivan J, Phillips R, Pape G R, Klenerman P. Sustained dysfunction of antiviral CD8+ T lymphocytes after infection with hepatitis C virus. J Virol, 2001; 75: Moskophidis D, Lechner F, Pircher H, Zinkernagel RM, Virus persistence in acutely infected immunocompetent mice by exhaustion of antiviral cytotoxic effector T cells, Nature 1993 Apr 22;362(6422): ( ) 82.Zajac AJ, Blattman JN, Murali-Krishna K, Sourdive DJ et al, Viral immune evasion due to persistence of activated T cells without effector function, J Exp Med 1998 Dec 21;188(12): ( ) 83.Yao ZQ, Shata MT, Tricoche N, Shan MM, Brotman B et al, gc1qr expression in chimpanzees with resolved and chronic infection: potential role of HCV core/gc1qr-mediated T cell suppression in the 225
226 outcome of HCV infection, Virology 2006 Mar 15;346(2): ( ) 84.Day CL, Seth NP, Lucas M, Appel H, Gauthier L, Lauer GM et al, Ex vivo analysis of human memory CD4+ T cells specific for hepatitis C virus using MHC class II tetramers, J Clin Invest 2003; 112: ( ) 85.Semmo N, Day CL, Ward SM, Lucas M, Harcourt G, Loughry A, Klenerman P, Preferential loss of IL-2 secreting CD4+ T helper cells in chronic HCV infection, Hepatology 2005 May; 41 (5): ( ) 86.Goldberg MV, Maris CH, Hipkiss EL, Flies AS, Zhen L, Tuder RM, Grosso JF, Harris TJ, Role of PD-1 and its ligand, B7-H1, in early fate decisions of CD8 T cells, Blood 2007 July 1; 110(1): Maier H, Isogawa M, Freeman GJ, Chisari FV, PD-1: PD-L1 interactions contribute to the functional suppression of virus-specific CD8+ T lymphocytes in the liver, J Immunol 2007; 178; Urbani S, Amadei B, Tola D, Massari M, Schivazappa S, Missale G, Ferrari C. PD-1 expression in acute hepatitis C virus (HCV) infection is associated with HCV-specific CD8 exhaustion, J Virol. 2006; 80: Bowen D G, Shoukry N H, Grakoui A, Fuller M J, Cawthon A G, Dong C, Hasselschwert D L, Brasky K M, Freeman G J, Seth N P, Wucherpfennig K W, Houghton M, Walker C M. Variable patterns of programmed death-1 (PD-1) expression on fully functional memory T cells following spontaneous resolution of hepatitis C virus infection. J Virol. 2008; 82: ( ) 90.Yao Z Q, King E, Prayther D, Yin D, Moorman J. T cell dysfunction by hepatitis C virus core protein involves PD-1/PDL-1 signaling. Viral Immunol, 2007; 20: ( ) 91.Lukens JR, Cruise MW, Lassen MG, Hahn YS, Blockade of PD-1/B7-H1 interaction restores effector CD8+ T cell responses in a hepatitis C virus core murine model, J Immunol 2008; 180; Wherry EJ, Blattman JN, Murali-Krishna K, van der Most R, Ahmed R, Viral persistence alters CD8 T-cell immunodominance and tissue distribution and results in distinct stages of functional impairment, J Virol 2003 Apr;77(8): ( ) 93.Golden-Mason L, Palmer B, Klarquist J, Mengshol JA, Castelblanco N, Rosen HR, Upregulation of PD-1 expression on circulating and intrahepatic hepatitis C virus-specific CD8+ T cells associated with 226
227 reversible immune dysfunction, J Virol 2007 Sep; 81(17): ( ) 94.Radziewicz H, Ibegbu C C, Fernandez M L, Workowski K A, Obideen K, Wehbi M, Hanson H L, Steinberg J P, Masopust D, Wherry E J, Altman J D, Rouse B T, Freeman G J, Ahmed R, Grakoui A. Liver-infiltrating lymphocytes in chronic human hepatitis C virus infection display an exhausted phenotype with high levels of PD-1 and low levels of CD127 expression. J Virol, 2007; 81: Duramad O, Fearon KL, Chan JH, Kanzler H, Marshall JD, Coffman RL, Barrat FJ, IL-10 regulates plasmacytoid dendritic cell response to CpG-containing immunostimulatory sequences, Blood 2003 Dec 15; 102(13): ( ) 96.Brady MT, MacDonald AJ, Rowan AG, Milla KH, Hepatitis C virus nonstructural protein 4 suppresses TH1 responses by stimulating IL-10 production from monocytes, Eur J Immunol 2003 Dec;33(12): ( ) 97.De Maria A, Fogli M, Mazza S, Basso M, Picciotto A et al, Increased natural cytotoxicity receptor expression and relevant IL-10 production in NK cells from chronically infected viremic HCV patients, Eur J Immunol 2007 Feb;37(2): ( ) 98.Accapezzato D, Francavilla V, Paroli M, Casciaro M, Chircu L V, Cividini A, Abrignani S, Mondelli M U, Barnaba V. Hepatic expansion of a virus-specific regulatory CD8(+) T cell population in chronic hepatitis C virus infection. J Clin Invest, 2004; 113: Abel M, Sene D, Pol S, Bourliere M, Poynard T, Charlotte F, Cacoub P, Caillat-Zucman S, Intrahepatic virus-specific IL-10-producing CD8 T cells prevent liver damage during chronic hepatitis C virus infection, Hepatology 2006 Dec; 44(6): ( ) 100.Piazzolla G, Tortorella C, Schiraldi O, Antonaci S, Relationship between interferon-gamma, interleukin-10 and interleukin-12 production in chronic hepatitis C and in vitro effects of interferonalpha, J Clin Immunol 2000 Jan; 20(1): ( ) 101.Chang KM, Rehermann B, McHutchinson JG, Pasquinelli C et al, Immunological significance of cytotoxic T lymphocyte epitope variants in patients chronically infected by the hepatitis C virus, J Clin Invest 1997 Nov;100(9): ( ) 102.Cox AL, Mosbruger T, Lauer GM, Pardoll D et al, Comprehensive analyses of CD8+ T cell responses during longitudinal study of acute human hepatitis C, Hepatology 2005 Jul;42(1): ( ) 227
228 103.Timm J, Lauer GM, Kavanagh DG, Sheridan I, Kim AY et al, CD8 epitope escape and reversion in acute HCV infection, J Exp Med 2004 Dec 20;200(12): ( ) 104.Ray SC, Fanning L, Wang XH, Netski DM, Kenny-Walsh E et al, Divergent and convergent evolution after a common-source outbreak of hepatitis C virus, J Exp Med 2005 Jun 6;201(11): ( ) 105.Neumann-Haefelin C, Frick DN, Wang JJ, Pybus OG et al, Analysis of the evolutionary forces in an immunodominant CD8 epitope in hepatitis C virus at a population level, J Virol 2008 Apr;82(7): ( ) 106.Wedemeyer H, He X S, Nascimbeni M, Davis A R, Greenberg H B, Hoofnagle J H, Liang T J, Alter H, Rehermann B. Impaired effector function of hepatitis C virus-specific CD8+ T cells in chronic hepatitis C virus infection. J Immunol, 2002; 169: Crispe IN, The liver as a lymphoid organ, Annu Rev Immunol 2009;27: ( ) 108.Lau AH, Thomson AW, Dendritic cells and immune regulation in the liver, Gut 2003 Feb;52(2): ( ) 109.Rouse B T, Sarangi P P, Suvas S. Regulatory T cells in virus infections. Immunol Rev. 2006; 212: Belkaid Y. Regulatory T cells and infection: a dangerous necessity. Nat Rev Immunol, 2007; 7: Dolganiuc A, Szabo G. T cells with regulatory activity in hepatitis C virus infection: what we know and what we don t, J Leukoc Biol Sep; 84(3): Li S, Gowans EJ, Chougnet C, Plebanski M, Dittmer U, Natural regulatory T cells and persistent viral infection, J Virol Jan 2008, vol 82, no 1, pp ( ) 113.Gershon R.K., A disquisition on suppressor T cells, Transplant. Rev : 170 ( ) 114.Sakaguchi S, Fukuma K, Kuribayashi K et al. Organ-specific autoimmune diseases induced in mice by elimination of a T cell subset. I. Evidence for the active participation of T cells in natural selftolerance; deficit of a T cell subset as a possible cause of autoimmune disease. J Exp Med 1985; 161:72 87 ( ) 228
229 115.Gershon RK, Kondo K, Cell interactions in the induction of tolerance: the role of thymic lymphocytes, Immunology 1970 May; 18(5): ( ) 116.Gershon RK, Kondo K, Infectious immunological tolerance, Immunology 1971 Dec; 21(6): ( ) 117.Kapp J A, Special regulatory T-cell review: suppressors regulated but unsuppressed, Immunology 2008, 123, Germain R N, Special regulatory T-cell review: A rose by any other name: from suppressor T cells to Tregs, approbation to unbridled enthusiasm, Immunology 2008, 123, Sakaguchi S, Ono M, Setoguchi R, Yagi H, Fehrevari Z, Shimizu J, Takahashi T, Nomura T, FoxP3+CD25+CD4+ natural regulatory T cells in dominant self-tolerance and autoimmune disease. Immunol Rev 2006; 212:8-27 ( ) 120.Sakaguchi S. Naturally arising CD4+ regulatory Τ cells for immunologic self-tolerance and negative control of immune responses. Annu Rev Immunol 2004, 22: ( ) 121.Sakaguchi S, Wing K, Miyara M, Regulatory T cells- a brief history and perspective, Eur. J. Immunol : S Shevach EM, Special regulatory T cell review: How I became a T suppressor/ regulatory cell maven, Immunology 2008, 123, Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M, Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of selftolerance causes various autoimmune diseases. J Immunol 1995; 155: ( ) 124.Shevach EM, Certified Professionals: CD4+CD25+ Supressor T Cells, The Journal of Experimental Medicine, Vol 193, Number 11, Jun 4, 2001, F41-F Malek T, Robb R, Shevach EM, Identification and initial characterization of a rat monoclonal antibody reactive with the murine interleukin 2 receptor-ligand complex, Immunology, 1983, Vol 80, pp Thornton A, Shevach EM, CD4+CD25+ Immunoregulatory T cells Suppress Polyclonal T Cell Activation In Vitro by Inhibiting Interleukin 2 Production, J Exp Med 1998 July 20, Vol 188, Number 2, Jonuleit H, Schmitt E, Stassen M, et al, Identification and functional characterization of human CD4+CD25+ T cells with regulatory 229
230 properties isolated from peripheral blood, J Exp Med 2001 June 4, vol 193, no 11, Takahashi T, Kuniyasu Y, Toda M et al, Immunologic self-tolerance maintained by CD25+CD4+ naturally anergic and suppressive T cells: induction of autoimmune disease by breaking their anergic/ suppressive state, Int Immunol 1998; 10: Suri-Prayer E, Amar A, Thornton A, Shevach E, CD4+CD25+ T cells inhibit both the induction and effector function of autoreactive T cells and represent a unique lineage of immunoregulatory cells, J Immunol 1998; 160; Setoguchi R, Hori S, Takahashi T, Sakaguchi S, Homeostatic maintenance of natural Foxp3+ CD25+CD4+ regulatory T cells by interleukin (IL)-2 and induction of autoimmune disease by IL-2 neutralization, JEM 2005 March 7, 201(5): Schorle H, Holtschke T, Hunig T et al, Development and function of T cells in mice rendered interleukin-2 deficient by gene targeting, Nature : Almeida A, Legrand N, Papiernik M, et al, Homeostasis of peripheral CD4+ T cells: Il-2R alpha and Il-2 shape a population of regulatory cells that controls CD4+ T cell numbers, J Immunol : Suzuki H, Zhou Y, Kato M, et al, Normal regulatory alpha/beta T cells effectively eliminate abnormally activated T cells lacking the interleukin 2 receptor beta in vivo, J Exp Med. 1999, 190: Lahl K, Loddenkemper C, Drouin C, Freyer J, Arnason J, et al, Selective depletion of Foxp3+ regulatory T cells induces a scurfy-like disease, JEM 2007 January 22, vol 204, no 1, pp Kim JM, Rudensky A. The role of the transcription factor FoxP3 in the development of regulatory T cells. Immunol Rev 2006; 212:86-98 ( ) 136.Brunkow ME, Jeffrey EW, Hjerrild KA et al. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat Genet 2001; 27: ( ) 137.Wildin RS, Ramsdell F, Peake J et al. X-linked neonatal diabetes mellitus, enteropathy and endocrinopathy syndrome is the human equivalent of mouse scurfy. Nat Genet 2001; 27: ( ) 138.Bennett CL, Christie J, Ramsdell F et al. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused mutations of Foxp3. Nat Genet 2001; 27: 20-1 ( ) 230
231 139.Chatila TA, Blaeser F, Ho N, Lederman HM, Voulgaropoulos C, Helms C, Bowcock AM, JM2, encoding a fork head-related protein, is mutated in X-linked autoimmunity-allergic disregulation syndrome, J. Clin. Invest. 106: R75-R81 (2000) 140.van der Vliet, Nieuwenhuis EE, IPEX as a result of mutations in FOXP3, Clin Dev Immunol 2007; 2007:89017 ( ) 141.Lopes J E, Torgerson T R, Schubert L A, et al, Analysis of FOXP3 reveals multiple domains required for its function as a transcriptional repressor, The Journal of Immunology 2006, vol 177, no 5, pp ( ) 142.Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003; 299: ( ) 143.Zheng Y, Rudensky AY, Foxp3 in control of the regulatory T cell lineage. Nat Immunol 2007; 8: ( ) 144.Gavin MA, Rasmussen JP, Fontenot JD, et al. Foxp3-dependent programme of regulatory T-cell differentiation, Nature. 2007; 445: ( ) 145.Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programmes the development and function of CD4+CD25+ regulatory T cells. Nat Immunol. 2003; 4: ( ) 146.Marson A, Kretschmer K, Frampton G. M. et al, Foxp3 occupancy and regulation of key target genes during T-cell stimulation, Nature 2007, 445: Zheng Y, Josefowicz S. Z., Kas A, Chu T. T., Gavin M. A., Rudensky A. Y., Genome-wide analysis of Foxp3 target genes in developing and mature regulatory T cells, Nature 2007, 445: 936 ( ) 148.Li B, Greene M, Special regulatory T-cell review: FOXP3 biochemistry in regulatory T cells- how diverse signals regulate suppression, Immunology 2008, 123, Bruno L, Mazzarella L, Hoogenkamp M, Hertweck A, et al, Runx proteins regulate Foxp3 expression, J Exp Med 2009, vol 206, no 11, Roncador G, Brown PJ, Maestre L, et al, Analysis of FOXP3 protein expression in human CD4+CD25+ regulatory T cells at the single cell level, Eur J Immunol 2005 Jun; 35 (6): Schubert L A, Jeffery E, Zhang Y, Ramsdell F, Ziegler S F, Scurfin (FOXP3) acts as a repressor of transcription and regulates T cell 231
232 activation, J Biol Chem 2001 October 5, vol 276, no 40, pp ( ) 152.Nicolson KS, Wraith DC, Natural and induced regulatory T cells: Targets for immunotherapy of autoimmune disease and allergy, Inflamm Allergy Drug Targets September; 5(3): Shevach EH, DiPaolo Ra, Andersson J, Zhao DM, Stephens GL, Thornton AM, The lifestyle of naturally occurring CD4+CD25+FoxP3+ regulatory T cells, Immunol Rev 2006; 212:60 73 ( ) 154.Walker LSK, CD4+CD25+ Treg: divide and rule? Immunology 2004, 111, Shen S, Ding Y, Tadokoro CE, et al, Control of homeostatic proliferation by regulatory T cells, J Clin Invest 2005; 115: Raimondi G, Turner M S, Thomson A W, Morel P A. Naturally occurring regulatory T cells: recent insights in health and disease, Crit Rev Immunol. 2007; 27: Gavin M A, Torgerson T R, Houston E, DeRoos P, Ho W Y, Stray- Pedersen A, Ocheltree E L, Greenberg P D, Ochs H D, Rudensky A Y. Single-cell analysis of normal and FOXP3-mutant human T cells: FOXP3 expression without regulatory T cell development. Proc Natl Acad Sci USA. 2006; 103: Maggi E, Cosmi L, Liotta F, Romagnani P, Annunziato F, Thymic regulatory T cells, Autoimmun Rev 2005 Nov; 4(8): Liston A, Rudensky AY, Thymic development and peripheral homeostasis of regulatory T cells, Curr Opin Immunol 2007 Apr; 19(2): ( ) 160.Asseman C, Mauze S, Leach MW, Coffman RL, Powrie F, An essential role for IL-10 in the function of regulatory T cells that inhibit intestinal inflammation. J Exp Med. 1999; 190: ( ) 161.Yi H, Zhen Y, Jiang L, Zhao Y, The phenotypic characterization of naturally occurring regulatory CD4+CD25+ T cells, Cell Mol Immunol 2006; 3: ( ) 162.Valzasina B, Guiducci C, Dislich H, Killeen N, Weinberg A D, Colombo M P. Triggering of OX40 (CD134) on CD4 (+) CD25+ T cells blocks their inhibitory activity: a novel regulatory role for OX40 and its comparison with GITR. Blood, 2005; 105:
233 163.Takeda I, Ine S, Killeen N, et al, Distinct roles for the OX40-OX40 ligand interaction in regulatory and nonregulatory T cells. J Immunol 2004; 172: Kleinewietfeld M, Puentes F, Borsellino G, Battistini L, et al, CCR6 expression defines regulatory effector/ memory-like cells within the CD25+CD4+ T-cell subset, Blood 2005; 105: ( ) 165.Baecher-Allan C, Wolf E, Hafler DA, MCH class II expression identifies functionally distinct human regulatory T cells, J Immunol 2006; 176: ( ) 166.Hoffmann P, Eder R, Kunz-Schughart LA, Andreesen R, Edinger M, Large-scale in vitro expansion of polyclonal human CD4+CD25high regulatory T cells, Blood 2004; 104: ( ) 167.Wysocki CA, Jiang Q, Panoskaltsis-Mortari A, Taylor PA, McKinnon KP, Su L, et al, Critical role for CCR5 in the function of donor CD4+CD25+ regulatory T cells during acute graft-versus-host disease, Blood 2005 Nov 1; 106(9): ( ) 168.Nakamura K, Kitami A, Strober W, Cell contact-dependent immunosupression by CD4+CD25+ regulatory T cells is mediated by cell surface-bound transforming growth factor β, Journal of Experimental Medicine 2001, 194(5), ( ) 169.Bruder D, Probst-Kepper M, Westendorf AM, Geffers R, et al, Neuropilin-1: a surface marker of regulatory T cells, Eur J Immunol 2004 Mar; 34(3): ( ) 170.Shevach EM, CD4+CD25+ suppressor T cells: more questions than answers, Nat Rev Immunol, 2002, Jun; 2(6): ( ) 171.Huang CT, Workman CJ, Flies D, et al, Role of Lag-3 in regulatory T cells, Immunity Oct; 21(4): Workman CJ, Vignali DA, Negative regulation of T cell homeostasis by lymphocyte activation gene-3 (CD223), J Immunol 2005; 174: ( ) 173.Yamaguchi T, Hirota K, Nagahama K, Ohkawa K, Takahashi T, Nomura T, Sakaguchi S, Control of immune responses by antigenspecific regulatory T cells expressing the folate receptor, Immunity 2007 Jul; 27(1): ( ) 174.Fontenot JD, Rasmussen JP, Williams LM, et al, Regulatory T cell lineage specification by the forkhead transcription factor foxp3, Immunity, 2005; 22: ( ) 175.Liu W, Putnam AL, Xu-Yu Z, Szot GL, Lee MR, Zhu S, Gottlieb PA, Kapranov P, Gingeras TR, et al. CD127 expression inversely correlates 233
234 with Foxp3 and suppressive function of human CD4+ Treg cells. J Exp Med. 2006; 203 (7): ( ) 176.Seddiki N, Santner-Naman B, Martinson J, et al, Expression of interleukin (IL)-2 and Il-7 receptors discriminates between human regulatory and activated T cells, JEM 2006 July 10, vol 203, no 7, Vukmanovic-Stejic M, Zhang Y, Cook J E, Fletcher J M, McQuaid A, Masters J E, Rustin M H, Taams L S, Beverley P C, Macallan D C, Akbar A N. Human CD4+ CD25hi Foxp3+ regulatory T cells are derived by rapid turnover of memory populations in vivo. J Clin Invest. 2006; 116: Chatenoud L, Bach J F. Adaptive human regulatory T cells: myth or reality? J Clin Invest. 2006; 116: Cools N, Ponsaerts P, Van Tendeloo VFI, Berneman ZN, Regulatory T cells and human disease, Clin Dev Immunol. 2007; 2007: Jonuleit H, Schmitt E, The regulatory T cell family: distinct subsets and their interrelations, J Immunol, 2003; 171: Shevach EM, From Vanilla to 28 flavours: Multiple Varieties of T regulatory cells, Immunity 2006 Aug, 25, Lal G, Bromberg JS, Epigenetic mechanisms of regulation of Foxp3 expression, Blood 2009 Oct 29; 114(18): Barrat F J, Cua D J, Boonstra A, Richards D F, Crain C, Savelkoul H F, de Waal-Malefyt R, Coffman R L, Hawrylowicz C M, O'Garra A. In vitro generation of interleukin 10-producing regulatory CD4 (+) T cells is induced by immunosuppressive drugs and inhibited by T helper type 1 (Th1)- and Th2-inducing cytokines. J Exp Med. 2002; 195: Vieira P L, Christensen J R, Minaee S, O Neil E J, et al, IL-10-Secreting regulatory T cells do not express Foxp3 but have comparable regulatory function to naturally occurring CD4+CD25+ regulatory T cells, J Immunol 2004; 172; Kemper C, Chan AC, Green JM, Brett KA, Murphy KM, Atkinson JP, Activation of human CD4+ cells with CD3 and CD46 induces a T- regulatory cell 1 phenotype, Nature 2003l 421, Shevach EM, Regulatory/ Suppressor T cells in health and disease, Arthritis & Rheumatism, 50(9), September 2004, pp ( ) 234
235 187.Levings M, Gregori S, Tresoldi E, et al, Differentiation of Tr1 cells by immature dendritic cells requires IL-10 but no CD25+CD4+ Tr cells, Blood 2004 February 1, vol 105, no 3, pp Cerwenka A, Swain S L. TGF-β1: immunosuppressant and viability factor for T lymphocytes. Microbes Infect. 1999; 1: Tang Q, Boden E K, Henriksen K J, Bour-Jordan H, Bi M, Bluestone J A. Distinct roles of CTLA-4 and TGF-β in CD4+CD25+ regulatory T cell function. Eur J Immunol. 2004; 34: Carrier Y, Yuan J, Kuchroo VK, Weiner HL, Th3 cells in peripheral tolerance. I. Induction of Foxp3-positive regulatory T cells by Th3 cells derived from TGF-β T cell-transgenic mice, J immunol 2007; 178; Taams L S, Akbar A N. Peripheral generation and function of CD4+CD25+ regulatory T cells, Curr Top Microbiol Immunol. 2005; 293: Wang J, Ioan-Facsinay A, van der Voort E I, Huizinga T W, Toes R E. Transient expression of FOXP3 in human activated nonregulatory CD4+ T cells. Eur J Immunol. 2007; 37: Walker MR, Kasprowicz DJ, Gersuk VH, Benard A, Van Landeghen M, Buckner JH, Ziegler SF, Induction of Foxp3 and acquisition of T regulatory activity by stimulated human CD4+CD25- T cells, J Clin Invest 2003 Nov; 112(9): ( ) 194.Walker MR, Carson BD, Nepom GT, Ziegler SF, Buckner JH, De novo generation of antigen-specific CD4+CD25+ regulatory T cells from human CD4+CD25- cells, PNAS 2005 March 12; vol 102, no 11, pp Lehmann J, Huehn J, de la Rosa M, Maszyna F, Kretschmer U, Krenn V, Brunner M, Scheffold A, Hamann A. Expression of the integrin αeβ7 identifies unique subsets of CD25+ as well as CD25 regulatory T cells. Proc Natl Acad Sci USA. 2002; 99: Siegmund K, Feuerer M, Siewert C, et al, Migration matters: regulatory T cell compartmentalization determines activity in vivo, Blood 2005 November 1; 106(9): Zheng SG, Wang JH, Koss MN, Quismorio F, Gray D, Horowitz D, CD4+ and CD8+ regulatory T cells generated ex vivo with IL-2 and TGF- {beta} suppress a stimulatory graft-versus-host disease with a lupus-like syndrome, J Immunol 2004; 172;
236 198.Gray D, Hirokawa M, Horwitz D, The role of transforming growth factor β in the generation of suppression: An interaction between CD8+ T and NK cells, J Exp Med, Vol 180, Nov 1994, Gray JD, Hirokawa M, Ohtsuka K, Horowitz D, Generation of an inhibitory circuit involving CD8+ T cells, IL-2, and NK cell-derived TGF-β: Contradting effects of anti-cd2 and anti-cd3, J Immunol 1998; 160; Jiang H, Chess L, The specific regulation of immune responses by CD8+ T cells restricted by the MHC class Ib molecule, Qa-1, Annu Rev Immunol 2000; 18: ( ) 201.Hahn BH, Singh RP, La Cava A, Ebling FM, Tolerogenic treatment of lupus mice with consensus peptide induces Foxp3-expressing, apoptosis-resistance, TGFbeta-secreting CD8+ T cell suppressors, J Immunol 2005 Dec 1; 175(11): ( ) 202.Gilliet M, Liu YJ, Generation of human CD8 T regulatory cells by CD40 ligand-activated plasmacytoid dendritic cells, J Exp Med 2002 Mar 18; 195(6): ( ) 203.Rifa I M, Kawamoto Y, Nakashima I, Suzuki H, Essential roles of CD8+CD122+ regulatory T cells in the maintenance of T cell homeostasis, J Exp Med 2004 Nov; 200(9): ( ) 204.Vlad G, Cortesini R, Suciu-Foca N, Licence to heal: bidirectional interaction of antigen-specific regulatory T cells and tolerogenic APC, J Immunol 2005 May 15; 174(10): ( ) 205.Cosmi L, Liotta F, Lazzeri E, Francalanci M, Angeli B, Mazzinghi B, Santarlasci V, et al, Human CD8+CD25+ thymocytes share phenotypic and functional features with CD4+CD25+ regulatory thymocytes, Blood 2003 Dec 1;102(12): ( ) 206.Chen W, Zhou D, Torrealba JR, Waddell TK, Grant D, Zhang L, Donor lymphocyte infusion induces long-term donor-specific cardiac xenograft survival through activation of recipient double-negative regulatory T cells, J Immunol 2005 Sep 1; 175(5): ( ) 207.Fischer K, Voelkl S, Heymann J, Przybylski GK, Mondal K, Laumer M, Kunz-Schughart L, et al, Isolation and characterization of human antigen-specific TCR alpha beta+ CD4(-)CD8- double-negative regulatory T cells, Blood 2005 Apr 1; 105(7): ( ) 208.Nishimura E, Sakihama T, Setoguchi R, Tanaka K, Sakaguchi S, Induction of antigen-specific immunological tolerance by in vivo and in vitro antigen-specific expansion of naturally arising Foxp3+CD25+Cd4+ regulatory T cells, Int Immunol 2004 Aug; 16(8): ( ) 236
237 209.Tarbell KV, Yamazaki S, Olson K, Toy P, Steinman RM, CD25+CD4+ T cells, expanded with dendritic cells presenting a single autoantigenic peptide, suppress autoimmune diabetes, J Exp Med 2004; 199: ( ) 210.Tang Q, Henriksen K J, Bi M, Finger E B, et al, In vitro-expanded antigen-specific regulatory T cells suppress autoimmune diabetes, J Exp Med 2004; 199: ( ) 211.Sundstedt A, O Neil E, Nicolson KS, Wraith D, Role for IL-10 in suppression mediated by peptide-induced regulatory T cells in vivo, J. Immunol. 2003; 170; Dugast AS, Vanhove B, Immune regulation by non-lymphoid cells in transplantation, Clinical and Experimental Immunology 2009, 156: Mauri C, Carter N, Is there a feudal hierarchy amongst regulatory immune cells? More than just Tregs, Arthritis Res Ther 2009; 11(4): 237 ( ) 214.Corthay A, How do regulatory T cells work? Scandinavian Journal of Immunology 70, , 2009 Blackwell Publishing Ltd 215.Sakaguchi S, Wing K, Onishi Y, Prieto-Martin, Yamaguchi T, Regulatory T cells: how do they suppress immune responses? Int Immunology 2009 Oct; 21(10): ( ) 216.Tang Q, Bluestone JA, The Foxp3+ regulatory T cell: a jack of all trades, master of regulation, Nat Immunol 2008 Mar; 9(3): ( ) 217.Izsue A, Powrie F. Special regulatory T-cell review: regulatory T cells and the intestinal tract- patrolling the frontier, Immunology. 2008; 123: Sojka DK, Huang YH, Fowell DJ, Mechanisms of regulatory T-cell suppression- a diverse arsenal for a moving target, Immunollogy 2008, 124, Furtado GC, Curotto de Lafaille MA, et al. Interleukin 2 signaling is required for CD4 (+) regulatory T cell function. J Exp Med 2002; 196: ( ) 220.Willerford DM, Chen J, Ferry JA, Davidson L, Ma A, Alt FW, Interleukin-2 receptor alpha chain regulates the size and content of the peripheral lymphoid compartment, Immunity, 1995 Oct; 3(4): ( ) 221.Antony P.A., Paulos C.M., Ahmadzedeh, et al, Interleukin-2 dependent mechanisms of tolerance and immunity, J Immunol 2006, 176:
238 222.Barthlott T, Moncrieffe H, Veldhoen M, Atkins CJ, et al, CD25+CD4+ T cells compete with naïve CD4+ T cells for IL-2 and exploit it for the induction of IL-10, Int Immunol 2005; 17: Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ, CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivationmediated apoptosis of effector CD4+ T cells, Nat Immunol 2007; 8: ( ) 224.Ge Q, Bai A, Jones B, Eisen HN, Chen J, Competition for selfpeptide-mhc complexes and cytokines between naïve and memory CD8+ T cells expressing the same or different T cell receptors, Proc Natl Acad Sci USA 2004; 101: Oberle N, Eberhardt N, Falk CS, Krammer PH, Suri-Prayer E, Rapid suppression of cytokine transcription in human CD4+CD25+ T cells by CD4+Foxp3+ regulatory T cells: independence of IL-2 consumption, TGF-beta, and various inhibitors of TCR signaling, J Immunol 2007; 179: ( ) 226.Shimizu J, Yamazaki S, Takahashi T, Ishida Y, Sakaguchi S. Stimulation of CD25 (+) CD4 (+) regulatory T cells through GITR breaks immunological self-tolerance. Nat Immunol. 2002; 3: McHugh R S, Whitters M J, Piccirillo C A, Young D A, Shevach E M, Collins M, Byrne M C. CD4 (+) CD25 (+) immunoregulatory T cells: gene expression analysis reveals a functional role for the glucocorticoidinduced TNF receptor. Immunity, 2002; 16: ( ) 228.Fife BT, Pauken KE, Eagar TN, et al. Interactions between programmed death ligand-1 promote tolerance by blocking the T cell receptor-induced stop signal. Nat Immunol Nov; 10(11): ( ) 229.Takahashi T, Tagami T, Yamazaki S, et al, Immunologic selftolerance maintained by CD4+CD25+ regulatory T cells constitutively expressing cytotoxic T lymphocyte-associated antigen 4, Journal of Experimental Medicine 2000, vol 192, no 2, pp Read S, Malmstrom V (me umlaut, δεν ξέρω πως μπαίνουν), Powrie F, Cytotoxic T lymphocyte-associated antigen 4 plays an essential role in the function of CD25+CD4+ regulatory cells that control intestinal inflammation, J Exp Med 2000 July 17, vol 192, no 2, Sakaguchi S, Regulatory T cells: Meden Agan, Immunological Reviews 2006 Vol 212: Paust S, Lu L, McCarthy N, Cantor H, Engagement of B7 on effector T cells by regulatory T cells prevents autoimmune disease, Proceedings 238
239 of the National Academy of Sciences of the United States of America 2004, 101(28): Marie JC, Letterio JJ, Gavin M, Rudensky AY, TGF-β1 maintains suppressor function and Foxp3 expression in CD4+CD25+ regulatory T cells, JEM 2005 April 4, 201(7): Li MO, Sanjabi S, Flavell RA. Transforming growth factor-beta controls development, homeostasis, and tolerance of T cells by regulatory T cell-dependent and independent mechanisms, Immunity. 2006; 25; ( ) 235.Wan YY, Flavell RA, Regulatory T cells, transforming growth factor-β, and immune suppression, Proc Am Thorac Soc 2007 Jul; 4(3): ( ) 236.Liang B, Workman C, Lee J, Chew C, Dale BM, Colonna L, Flores M, Li N, et al, Regulatory T cells inhibit dendritic cells by LAG-3 engagement of MHC class II, J Immunol 2008; 180: ( ) 237.Shevach EM, Piccirillo CA, Thornton AM, McHugh RS, Control of T cell activation by CD4+CD25+ suppressor T cells, Immunological reviews 2001, 182: Kanamaru F, Youngnak P, Hashiguchi M, Nishioka T, Takahashi T, Sakaguchi S, Ishikawa I, Azuma M, Costimulation via glucocorticoidinduced TNF receptor in both conventional and CD15+ regulatory CD4+ T cells, J. Immunol 2004;172: Shevach EM, Stephens GL, The GITR-GITRL interaction: costimulation or contrasuppression of regulatory activity? Nat Rev Immunol 2006 Aug; 6(8): ( ) 240.Grossman WJ, Verbsky W, Tollefsen BL, Kemper C, Atkinson JP, Ley TJ, Differential expression of granzymes A and B in human cytotoxic lymphocyte subsets and T regulatory cells, Blood 2004, 104(9): Zhao DM, Thornton AM, DiPaolo RJ, Shevach EM, Activated CD4+CD25+ T cells selectively kill B lymphocytes, Blood 2006; 107: Gondek DC, Lu LF, Quezada SA, Sakaguchi S, Noelle R, Cutting edge: contact mediated suppression by CD4+CD25+ regulatory cells involves a granzyme B-dependent, perforin-independent mechanism, J Immunol 2005; 174:
240 243.Janssens W, Carlier V, Wu B et al, CD4+CD25+ T cells lyse antigenpresenting B cells by Fas- Fas igand interaction in an epitope-specific manner, J Immunol 2003; 171: Vignali DA, Collison LW, Workman CJ, How regulatory T cells work, Nat Rev Immunol 2008 Jul; 8(7): ( ) 245.Geissman F, Revy P, Regnault A, et al, TGF-beta 1 prevents the noncognate maturation of human dendritic Langerhans cells, Journal of Immunology 1999, 162(8): Chen W, Wenwen J, Hardegen N, Lei K, Li L, Marinos N, McGrady G, Wahl SM, Conversion of peripheral CD4+CD25- naïve T cells to CD4+CD25+ regulatory T cells by TGF-β induction of transcription factor Foxp3, The Journal of Experimental Medicine 2003, 198: Davidson TS, DiPaolo RJ, Andersson J, Shevach EM. Cutting edge: IL-2 is essential for TGF-beta-mediated induction of Foxp3 T regulatory cells, J Immunol. 2007; 178: Larrea E, Riezu-Boj J I, Gil-Guerrero L, Casares N, Aldabe R, Sarobe P, Civeira M P, Heeney J L, Rollier C, Verstrepen B, Wakita T, Borrás- Cuesta F, Lasarte J J, Prieto J. Upregulation of indoleamine 2,3- dioxygenase in hepatitis C virus infection. J Virol. 2007; 81: ( ) 249.Wan YY, Flavell RA. Regulatory T-cell functions are subverted and converted owing to attenuated Foxp3 expression, Nature. 2007; 445: Groux H, O Garra A, Bigler, et al. A CD4+ T cell subset inhibits antigen specific T-cell responses and prevents colitis, Nature. 1997; 389: ( ) 251.Jancovic D, Kullberg MC, Feng CG, et al. Conventional T- bet(+)foxp3(-) Th1 cells are the major source of host-protective regulatory IL-10 during intracellular protozoan infection. J Exp Med. 2007; 204: ( ) 252.Rubtsov YP, Rasmussen JP, Chi EY, et al, Regulatory T cell-derived interleukin-10 limits inflammation at environmental interfaces, Immunity. 2008; 28: ( ) 253.Moore KW, de Waal Malefyt R, Coffman RL, O Garra A. Interleukin- 10 and the interleukin-10 receptor, Annu Rev Immunol. 2001; 19: Collison LW, Workman CJ, et al. The inhibitory cytokine IL-35 contributes to regulatory T-cell function, Nature. 2007; 450: ( ) 240
241 255.Bopp T, Becker C, Klein M, Klein-Hessling S, Palmetshofer A, Serfling E, Heib V, Becker M, Kubach J, Schmitt S, Stoll S, Schild H, Staege M S, Stassen M, Jonuleit H, Schmitt E. Cyclic adenosine monophosphate is a key component of regulatory T cell-mediated suppression. J Exp Med. 2007; 204: ( ) 256.Minguet S, Huber M, Rosenkranz I, et al, Adenosine and camp are potent inhibitors of the NF-kappa B pathway downstream of immunoreceptors, Eur J Immunol 2005; 35: Bodor J, Bodorova J, Gress RE, Suppression of T cell function: a potential role for transcriptional repressor ICER, J Leukoc Biol 2000; 67: Deaglio S, Dwyer KM, Gao W, Friedman D, Usheva A, et al, Adenosine generation catalysed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression, JEM 2007 June 11, 204(6): Zarek PE, Huang CT, Lutz ER, Kowalski J, Horton MR, Linden J, et al, A2A receptor signaling promotes peripheral tolerance by inducing T- cell anergy and the generation of adaptive regulatory T cells, Blood 2008 Jan 1; 111(1): ( ) 260.Wilson JM, Ross WG, Agbai ON, Frazier R, Figler RA, Rieger J, et al, The A2B adenosine receptor impairs the maturation and immunogenicity of dendritic cells, J Immunol 2009 Apr 15; 182(8): ( ) 261.Tadokoro C E, Shakhar G, Shen S, Ding Y, Lino A C, Maraver A, Lafaille J J, Dustin M L. Regulatory T cells inhibit stable contacts between CD4+ T cells and dendritic cells in vivo. J Exp Med. 2006; 203: Taams L S, van Amelsfort J M, Tiemessen M M, Jacobs K M, de Jong E C, Akbar A N, Bijlsma J W, Lafeber F P. Modulation of monocyte/macrophage function by human CD4+CD25+ regulatory T cells. Hum Immunol. 2005; 66: Tiemessen M M, Jagger A L, Evans H G, van Herwijnen M J, John S, Taams L S. CD4+CD25+Foxp3+ regulatory T cells induce alternative activation of human monocytes/macrophages. Proc Natl Acad Sci USA. 2007; 104: Fallarino F, Grohman U, Hwang W, et al, Modulation of tryptophan catabolism by regulatory T cells, Nature Immunology 2003, vol 4, no 12, pp
242 265.Boasso A, Vaccari M. Regulatory T-cell markers, indoleamine 2, 3- dioxygenase, and virus levels in spleen and gut during progressive simian immunodeficiency virus infection, J Virol. 2007; 81(21): Munn DH, Sharma D, Lee JR, et al, Potential regulatory function of human dendritic cells expressing indoleamine 2,3-dioxygenase, Science 2002, 297(5588), pp Oderup C, Cederbom, Makowska A, Cillio C. M., Ivars F, Cytotoxic T lymphocyte antigen-4-dependent down-modulation of costimulatory molecules on dendritic cells in CD4+CD25+ regulatory T-cell-mediated suppression, Immunology 2006; 118, Cederbom L, Hall H, Ivars F, CD4+CD25+ regulatory T cells downregulate co-stimulatory molecules on antigen-presenting cells, Eur J Immunol 2000 Jun; 30(6): ( ) 269.Kryczek I, Wei S, Zou L, Zhu G, Mottram P, Xu H, Chen L, Zou W, Cutting edge: induction of B7-H4 on APCs through Il-10: novel suppressive mode for regulatory T cells, J Immunol 2006; 177: ( ) 270.Houot R, Perrot I, Garcia E, Durand I, Lebecque S. Human CD4+CD25high regulatory T cells modulate myeloid but not plasmacytoid dendritic cells activation, J Immunol. 2006; 176: Sarris M, Andersen KG, Randow F, Mayr L, Betz AG, Neuropilin-1 expression on regulatory T cells enhances their interactions with dendritic cells during antigen recognition, Immunity 2008; 28: ( ) 272.Andre S, Tough DF, Lacroix-Desmazes S, Kaveri SV, Bayry J, Surveillance of antigen-presenting cells by CD4+CD25+ regulatory T cells in autoimmunity: immunopathogenesis and therapeutic implications, Am J Pathol 2009 May; 174(5): ( ) 273.Coutinho A, Caramalho I, Seixas E, Demengeot J, Thymic commitment of regulatory T cells is a pathway of TCR-dependent selection that isolates repertoires undergoing positive or negative selection, Curr Top Microbiol Immunol 2005; 293: ( ) 274.Bour-Jordan Helene, Bluestone J A, Regulating the regulators: costimulatory signals control the homeostasis and function of regulatory T cells, Immunol Rev 2009 May; 229 (1): Tai X, Cowan M, Feigenbaum L, Singer A, CD28 costimulation of developing thymocytes induces Foxp3 expression and regulatory T cell differentiation independently of interleukin 2, Nat Immunol 2005 Feb; 6(2): ( ) 242
243 276.Nazarov-Stoica C, Surls J, Bona C, Casares S, Brumeau TD, CD28 signalling in T regulatory precursors requires p56lck and rafts integrity to stabilize the Foxp3 message, J Immunol 2009; 182; ( ) 277.Kasow KA, Chen X, Knowles J, Wichlan D, Handgretinger R, Riberdy JM, Human CD4+CD25+ regulatory T cells share equally complex and comparable repertoires with CD4+CD25- counterparts, J Immunol 2004 May 15; 172(10): ( ) 278.Knoechel B, Lohr J, Kahn E, Bluestone JA, Abbas AK, Sequential development of interleukin 2-effector and regulatory T cells in response to endogenous systemic antigen, JEM 2005 Nov 21; 202(10): Burchill MA, Yang J, et al. IL-2 receptor beta-dependent STAT5 activation is required for the development of Foxp3+ regulatory T cells, J Immunol. 2007; 178: Suri-Prayer E, Amar AZ, McHugh R, Natarajan K, Margulies DH, Shevach EM, Post-thymectomy autoimmune gastritis, fine specificity and pathogenicity of anti-h/k APTase-reactive T cells. Eur J Immunol 1999; 29: ( ) 281.Horwitz D A, Zheng S G, Gray J D, The role of the combination of IL- 2 and TGF-β or IL-10 in the generation and function of CD4+ CD25+ and CD8+ regulatory T cell subsets. J Leukoc Biol. 2003; 74: Fisson S, Darrasse- Jese G, Litvinova E, Septier F, Klatzman D, Liblau R, Salomon BL, Continuous activation of autoreactive CD4+CD25+ regulatory T cells in the steady state, J Exp Med 2003 Sep 1; 198(5): ( ) 283.Picca C C, Larkin J, III, Boesteanu A, Lerman M A, Rankin A L, Caton A J. Role of TCR specificity in CD4+ CD25+ regulatory T-cell selection. Immunol Rev. 2006; 212: Longhi MS, Hussain MJ, Kwok WW, Mieli-Vergani G et al, Autoantigen-specific regulatory T cells, a potential tool for immunetolerance reconstitution in type-2 autoimmune hepatitis, Hepatology, 2010 Oct 18 [Epub ahead of print] ( ) 285.Hu G, Liu Z, Zheng C, Zheng SG, Antigen-non-specific regulation centered on CD4+CD25+Foxp3+ Treg cells, Cell Mol Immunol 2010 Nov; 7(6): ( ) 286.Kubo T, Hatton R D, Oliver J, Liu X, Elson C O, Weaver C T. Regulatory T cell suppression and anergy are differentially regulated by proinflammatory cytokines produced by TLR-activated dendritic cells. J Immunol. 2004; 173:
244 287.Walker L S, Regulatory T cells overtuned: the effectors fight back, Immunology 2009, 126, LiuG, Zhao Y, Toll-like receptors and immune regulation: their direct and indirect modulation on regulatory CD4+CD25+ T cells, Immunology 2007; 122: King C, Olic A, Koelsch K, Sarvetnick N, Homeostatic expansion of T cells during immune insufficiency generates autoimmunity, Cell 2004; 117: ( ) 290.Valensia X, Stephens G, et al, TNF downmodulates the function of human CD4+CD25hi T-regulatory cells, Blood 2006; 108: Pasare C, Medzhitov R. Toll pathway-dependent blockade of CD4+CD25+ T cell-mediated suppression by dendritic cells. Science. 2003; 299: Stephens GL, McHugh RS, Whitters MJ, et al, Engagement of glucocorticoid-induced TNFR family-related receptor on effector T cells by its ligand mediates to suppression by CD4+CD25+ T cells. J Immunol 2004; 173: ( ) 293.Wohlfert EA, Callahan MK, Clark RB, Resistance to CD4+CD25+ regulatory T cells and TGF-beta in Cbl-b-/- mice, J Immunol 2004; 173: Bopp T, Palmetshofer A, Serfling E, et al, NFATc2 and NFATc3 transcription factors play a crucial role in suppression of CD4+ T lymphocytes by CD4+CD25+ regulatory T cells, J Exp Med 2005; 201: ( ) 295.King CG, Kobayashi T, Cejas PJ, et al, TRAF6 is a T cell-intrinsic negative regulator required for the maintenance of immune homeostasis, Nat Med 2006; 12: ( ) 296.Zou L, Barnett B, Safah H, LaRussa V F, et al, Bone marrow is a reservoir for CD4+CD25+ regulatory T cells that traffic through CXCL12/CXCR4 signals, Canser Res 2004; 64; Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, Belkaid Y, Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 Treg cells via retinoic acid, J Exp Med 2007 Aug 6; 204(8): ( ) 298.Van Amelsfort JM, Jacobs KM, Bijlsma JW, Lafeber FP et al, CD4+CD25+ regulatory T cells in rheumatoid arthritis: differences in the presence, phenotype and function between peripheral blood and synovial fluid, Arthritis Rheum 2004;50:
245 299.Mackay IR. Hepatoimmunology: A perspective. Immunol Cell Biol 2002; 80: Belkaid Y, Piccirillo C A, Mendez S, Shevach E M, Sacks D L. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature. 2002; 420: Wei S, Kryczek I, Zou W, Regulatory T-cell compartmentalization and trafficking, Blood 2006 July 15; 108(2): Huehn J, Siegmund K, Lehmann JC, et al, Developmental stage, phenotype, and migration distinguish naïve- and effector/ memory-like CD4+ regulatory T cells, J Exp Med 2004; 199; Sugiyama H, Gyulai R, Toichi E, et al, Dysfunctional blood and target tissue CD4+CD25high regulatory T cells in psoriasis: mechanism underlying unrestrained pathogenic effector T cell proliferation, J Immunol 2005; 174: Szanya V, Ermann J, Taylor C, et al, The subpopulation of CD4+CD25+ splenocytes that delays adoptive transfer of diabetes expresses L-selectin and high levels of CCR7, J Immunol 2002; 169; Ermann J, Hoffmann P, Edinger M, Dutt S, et al, Only the CD62L+ subpopulation of CD4+CD25+ regulatory T cells protects from lethal acute GVHD, Blood 2005 March 1, 105(5): Lim HW, Hillsamer P, Kim CH, Regulatory T cells can migrate to follicles upon T cell activation and suppress GC-Th cells and GC-Th celldriven B responses, J Clin Invest 2004 Dec; 114(11): Rehermann B, Hepatitis C virus versus innate and adaptive immune responses: a tale of coevolution and coexistence, J Clin Invest 2009 Jul; 119(7): ( ) 308.Ulsenheimer A, Gerlach J T, Gruener N H, Jung M C, Schirren C A, Schraut W, Zachoval R, Pape G R, Diepolder H M. Detection of functionally altered hepatitis C virus-specific CD4 T cells in acute and chronic hepatitis C. Hepatology, 2003; 37: Perrella A, Vitiello L, Atripaldi L, Conti P, Sbreglia C, Altamura S, Patarino T, Vela R, Morelli G, Bellopede P, Alone C, Racioppi L, Perrella O. Elevated CD4+/CD25+ T cell frequency and function during acute hepatitis C presage chronic evolution. Gut. 2006; 5: Smyk-Pearson S, Golden-Mason L, Klarquist J, Burton JR Jr, Tester IA, Wang CC, Culbertson N, Vandenbark AA, Rosen HR, Functional suppression by Foxp3+CD4+CD25(high) regulatory T cells during acute 245
246 hepatitis C virus infection, J Infect Dis 2008 Jan 1; 197(1):46-57 ( ) 311.Billerbeck E, Bottler T, Thimme R, Regulatory T cells in viral hepatitis, World J Gastroenterol 2007 Sep 28; 13(36): ( ) 312.Boettler T, Spangenberg H C, Neumann-Haefelin C, Panther E, Urbani S, Ferrari C, Blum H E, von Weizsäcker F, Thimme R. T cells with a CD4+CD25+ regulatory phenotype suppress in vitro proliferation of virus-specific CD8+ T cells during chronic hepatitis C virus infection. J Virol. 2005; 79: Sugimoto K, Ikeda F, Stadanlick J, Nunes F A, Alter H J, Chang K M. Suppression of HCV-specific T cells without differential hierarchy demonstrated ex vivo in persistent HCV infection. Hepatology. 2003; 38: Cabrera R, Tu Z, Xu Y, Firpi R J, Rosen H R, Liu C, Nelson D R. An immunomodulatory role for CD4 (+) CD25 (+) regulatory T lymphocytes in hepatitis C virus infection. Hepatology. 2004; 40: Rushbrook S M, Ward S M, Unitt E, Vowler S L, Lucas M, Klenerman P, Alexander G J. Regulatory T cells suppress in vitro proliferation of virusspecific CD8+ T cells during persistent hepatitis C virus infection. J Virol. 2005; 79: Yang J H, Zhang Y X, Yu R B, Su C, Sun N X. CD4+ CD25+ regulatory T cells suppress CD4+ T cell responses in patients with persistent hepatitis C virus infection. Zhonghua Nei Ke Za Zhi. 2006; 45:29 33 ( ) 317.Ward S M, Fox B C, Brown P J, Worthington J, Fox S B, Chapman R W, Fleming K A, Banham A H, Klenerman P. Quantification and localization of FOXP3+ T lymphocytes and relation to hepatic inflammation during chronic HCV infection. J Hepatol. 2007; 47: Dolganiuc A, Paek E, Kodys K, Thomas J, Szabo G, Myeloid dendritic cells of patients with chronic HCV infection induce proliferation of regulatory T lymphocytes, Gastroenterology 2008 Dec; 13(6): ( ) 319.Manigold T, Shin E C, Mizukoshi E, Mihalik K, Murthy K K, Rice C M, Piccirillo C A, Rehermann B. Foxp3+CD4+CD25+ T cells control virusspecific memory T cells in chimpanzees that recovered from hepatitis C. Blood. 2006; 107: Boyer O, Saadoun D, Abriol J, Dodille M, Piette J C, Cacoub P, Klatzmann D. CD4+CD25+ regulatory T-cell deficiency in patients with 246
247 hepatitis C-mixed cryoglobulinemia vasculitis. Blood. 2004; 103: Ormandy LA, Hillemann T, Wedemeyer H, Manns MP, Greten TF, Korangy F, Increased populations of regulatory T cells in peripheral blood of patients with hepatocellular carcinoma, Canser Res 2005 March 15; 65: Itose I, Kanto T, Kakita N, Takebe S, Inoue M, Higashitani K, Miyatake H, Sakakibara M, et al, Enhanced ability of regulatory T cells in chronic hepatitis C patients with persistently normal alanine aminotransferase levels than those with active hepatitis, J Viral Hepat 2009 Dec; 16(12): ( ) 323.Gerlach JT, Diepolder HM, Jung MC, Gruener NH, Schraut WW, Zachoval R, et al, Recurrence of hepatitis C virus after loss of virusspecific CD4(+) T-cell response in acute hepatitis C, Gastroenterology 1999 Oct; 117(4): ( ) 324.Suffia IJ, Reckling SK, Piccirillo CA, Goldzmid RS, Belkaid Y, Infected site-restricted Foxp3+ natural regulatory T cells are specific for microbial antigens, J Exp Med 2006 Mar 20; 203(3): ( ) 325.Ebinuma H, Nakamoto N, Li Y, Price D A, Gostick E, Levine B L, Tobias J, Kwok W W, Chang K M. Identification and in-vitro expansion of functional antigen-specific CD25+FoxP3+ regulatory T-cells in hepatitis C virus infection. J Virol. 2008; 82: Li S, Floess S, Hamann A, Gaudieri S, Lucas A, Hellard M, Roberts S, Paukovic M, et al, Analysis of Foxp3+ regulatory T cells that display apparent viral antigen specificity during chronic hepatitis C virus infection, PLoS Pathog 2009 Dec;5(12):e ( ) 327.MacDonald A J, Duffy M, Brady M T, McKiernan S, Hall W, Hegarty J, Curry M, Mills K H. CD4 T helper type 1 and regulatory T cells induced against the same epitopes on the core protein in hepatitis C virusinfected persons. J Infect Dis. 2002; 185: Kimball P, Elswick R K, Shiffman M. Ethnicity and cytokine production gauge response of patients with hepatitis C to interferon-α therapy. J Med Virol. 2001; 65: ( ) 329.Shapiro S, Gershtein V, Elias N, Zuckerman E, Salman N, Lahat N. mrna cytokine profile in peripheral blood cells from chronic hepatitis C virus (HCV)-infected patients: effects of interferon-α (IFN-α) treatment. Clin Exp Immunol. 1998; 114: Bolacchi F, Sinistro A, Ciaprini C, Demin F, Capozzi M, Carducci F C, Drapeau C M, Rocchi G, Bergamini A. Increased hepatitis C virus 247
248 (HCV)-specific CD4+CD25+ regulatory T lymphocytes and reduced HCV-specific CD4+ T cell response in HCV-infected patients with normal versus abnormal alanine aminotransferase levels. Clin Exp Immunol, 2006; 144: Koziel MJ, Dudley D, Afdhal N, Grakoui A, Rice CM, Choo QL, Houghton M, Walker BD, HLA class I-restricted cytotoxic T lymphocytes specific for hepatitis C virus. Identification of multiple epitopes and characterization of patterns of cytokine release, J Clin Invest 1995; 96, Alatrakchi N, Graham C S, van der Vliet H J, Sherman K E, Exley M A, Koziel M J. Hepatitis C virus (HCV)-specific CD8+ cells produce transforming growth factor β that can suppress HCV-specific T-cell responses. J Virol. 2007; 81: Franceschini D, Paroli M, Francavilla V, Videtta M, Morrone S, Labbadia G, Cerino A, et al, PD-L1 negatively regulates CD4+CD25+Foxp3+ Tregs by limiting STAT-5 phosphorylation in patients chronically infected with HCV, J Clin Invest 2009 Mar; 119(3): ( ) 334.Billerbeck E, Blum H E, Thimme R. Parallel expansion of human virusspecific FoxP3 effector memory and de novo-generated FoxP3+ regulatory CD8+ T cells upon antigen recognition in vitro. J Immunol. 2007; 179: ( ) 335.Li S, Jones K L, Woollard D J, Dromey J, Paukovics G, Plebanski M, Gowans E J. Defining target antigens for CD25+ FOXP3 + IFN-γregulatory T cells in chronic hepatitis C virus infection. Immunol Cell Biol. 2007; 85: ( ) 336.Suvas S, Kumaraguru U, Pack CD, Lee S, Rouse BT, CD4+CD25+ T cells regulate virus-specific primary and memory CD8+ T cell responses, J Exp Med 2003 Sep 15; 198(6): ( ) 337.Sutmuller R P, den Brok M H, Kramer M, Bennink E J, Toonen L W, Kullberg B J, Joosten L A, Akira S, Netea M G, Adema G J. Toll-like receptor 2 controls expansion and function of regulatory T cells. J Clin Invest. 2006; 116: Langhans B, Braunschweiger I, Nischalke H D, Nattermann J, Sauerbruch T, Spengler U. Presentation of the HCV-derived lipopeptide LP20 44 by dendritic cells enhances function of in vitro-generated CD4+ T cells via up-regulation of TLR2. Vaccine. 2006; 24: Unitt E, Rushbrook SM, Marshall A, Davies S, Gibbs P, Morris LS, Coleman N, Alexander GJ, Compromised lymphocytes infiltrate 248
249 hepatocellular carcinoma: the role of T-regulatory cells, Hepatology 2005 Apr; 41(4): ( ) 340.Cao X, Cai SF, Fehniger TA, Song J, Collins LI, Piwnica-Worms DR, Ley TJ, Granzyme B and perforin are important for regulatory T cellmediated suppression of tumor clearance, Immunity 2007 Oct; 27(4): ( ) 341.Wrzesinski SH, Wan YY, Flavell RA, Transforming growth factor-β and the immune response: Implications for anticancer therapy, Clin Cancer Res 2007 Sep 15; 13(18), pp Mempel TR, Pittet MJ, Khazaie K, Weninger W, Weissleder R, von Boehmer H, von Andrian UH, Regulatory T cells reversibly suppress cytotoxic T cell function independent of effector differentiation, Immunity 2006 Jul; 25(1) ( ) 343.Kobayashi N, Hiraoka N, Yamagami W, Ojima H, Kanai Y, Kosuge T, Nakajima A, Hirohashi S. FOXP3+ regulatory T cells affect the development and progression of hepatocarcinogenesis. Clin Cancer Res. 2007; 13: ( ) 344.Spengler U, Nattermann J. Immunopathogenesis in hepatitis C virus cirrhosis. Clin Sci (Lond). 2007; 112: NIH Consensus Statement on Management of Hepatitis C: NIH Consens State Sci Statements. 2002; 19: EASL International Consensus Conference on Hepatitis C, J Hepatol 1999; 30: Carter NP, Meyer EW, Introduction to the principles of flow cytometry. In: Ormerod MG (ed). Flow cytometry: a practical approach. 2 nd edition. New York: Oxford University Press, 1996: Μαλισιόβας Σ. Εισαγωγή στις αρχές της κυτταρομετρίας ροής. Στο: Βιβλίο Εισηγήσεων 1 ου Πανελλήνιου Συμποσίου Κυτταρομετρίας. Αθήνα: Ελληνική Εταιρεία Κυτταρομετρίας, 1999: Nunez R, Flow Cytometry: principles and instrumentation, Curr Issues Mol Biol 2001;3(2): Abbas AK, Lichtman AH, Cellular and Molecular Immunology, 5 th edition, Philadelphia: WB Saunders, Leung JC, Lai CK, Chui YL, Ho RT et al, Characterization of cytokine gene expression in CD4+ and CD8+ T cells after activation with phorbol myristate acetate and phytohaemafflutinin, Clin Exp, Immunol, 1992: 90: ( ) 249
250 352.Fan J, Nishanian P, Breen EC, McOnald M, Fahey JL, Cytokine gene expression in normal human lymphocytes in response to stimulation, Clin Diagn Lab Immunol, 1998:5: ( ) 353.Gaffen SL, Liu KD. Overview of interleukin-2 function, production and clinical applications. Cytokine. 2004; 28: ( ) 354.Acuto O, Michel F. CD28-mediated co-stimulation: a quantitative support for TCR signalling. Nat Rev Immunol. 2003; 3: ( ) 355.Hodge S, Hodge G, Flower R, Han P. Surface and intracellular interleukin-2 receptor expression on various resting and activated populations involved in cell-mediated immunity in human peripheral blood. Scand J Immunol. 2000;51:67-72 ( ) 356.Loken MR, Wells DA, Immunofluorescence of surface markers. In: Ormerod MG (ed). Flow cytometry: a practical approach. 2nd edition. New York: Oxford University Press, 1996:
251 ΠΑΡΑΡΤΗΜΑ 251
252
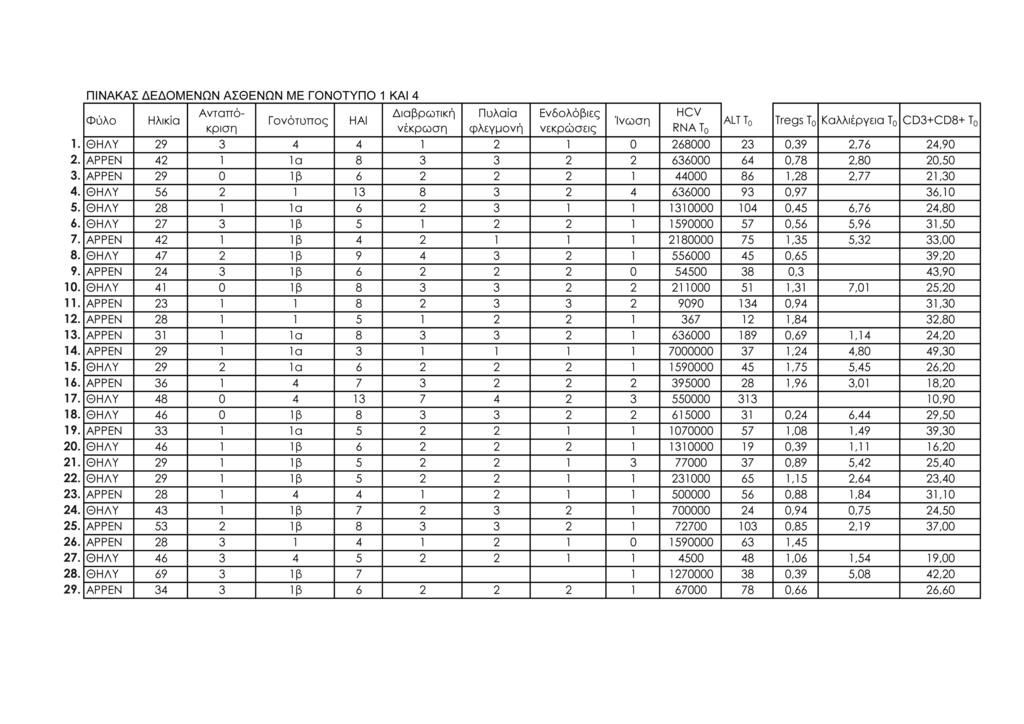
253
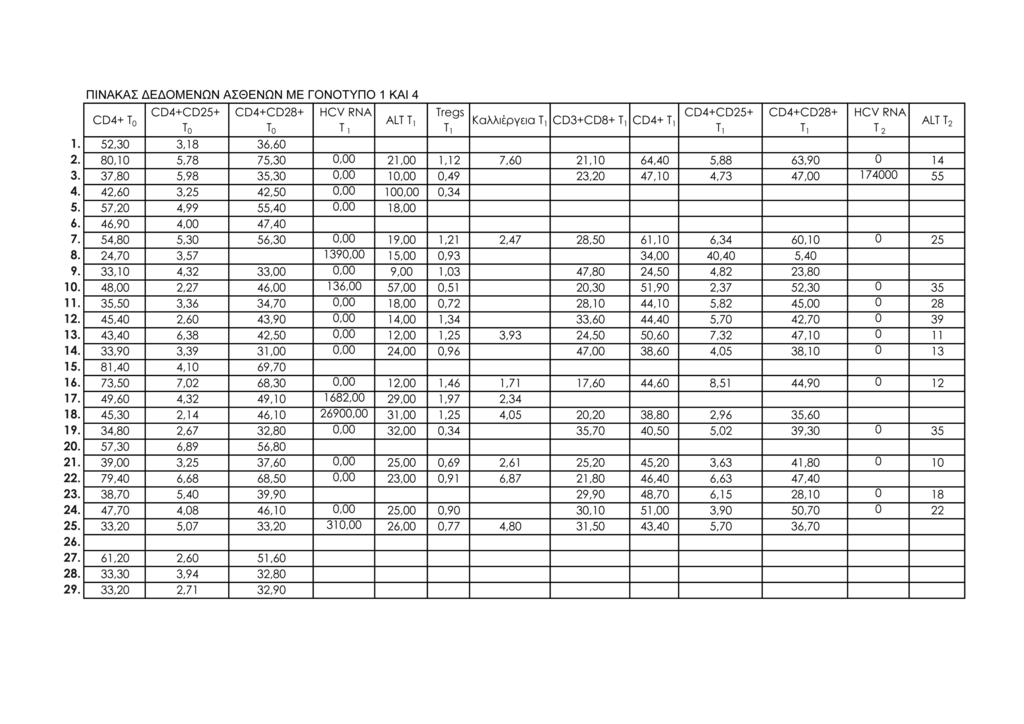
254
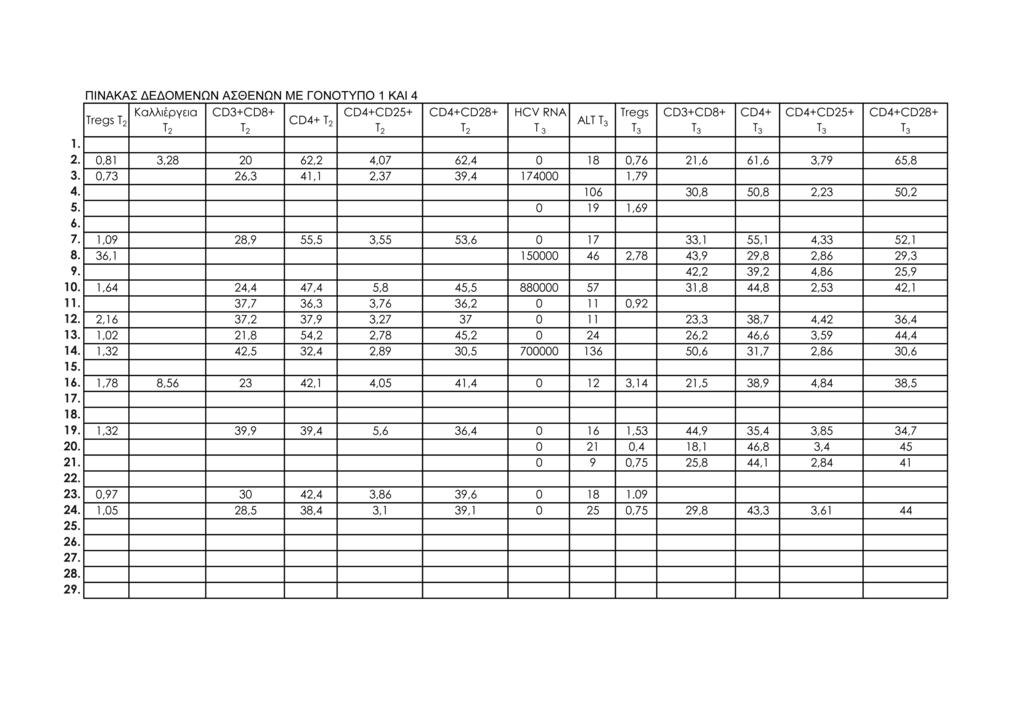
255
256
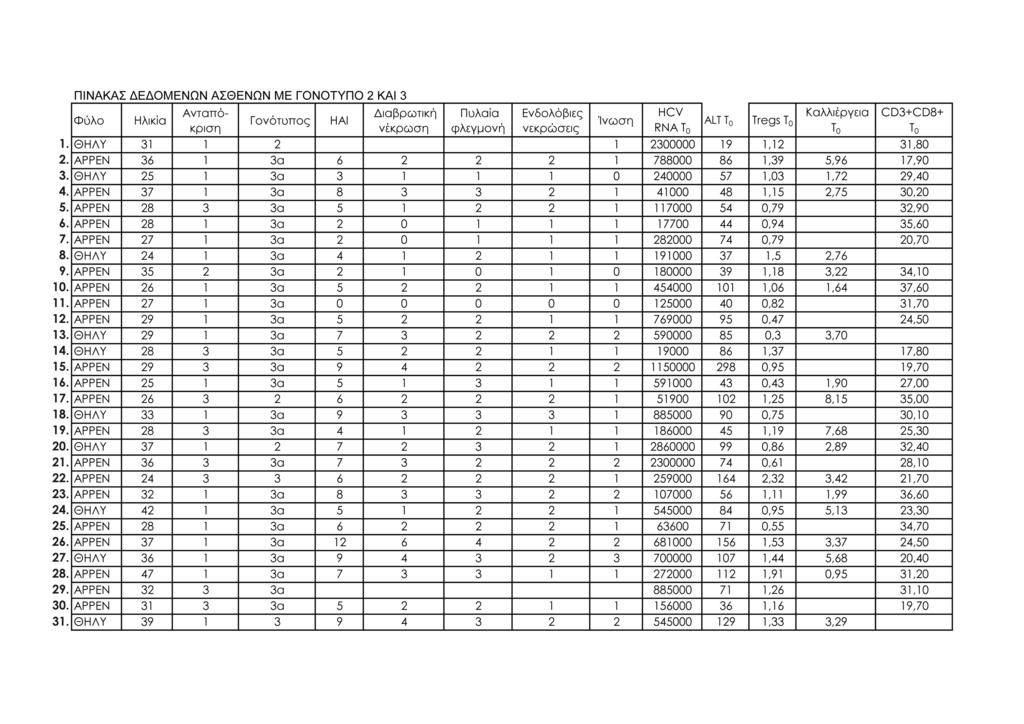
257

258
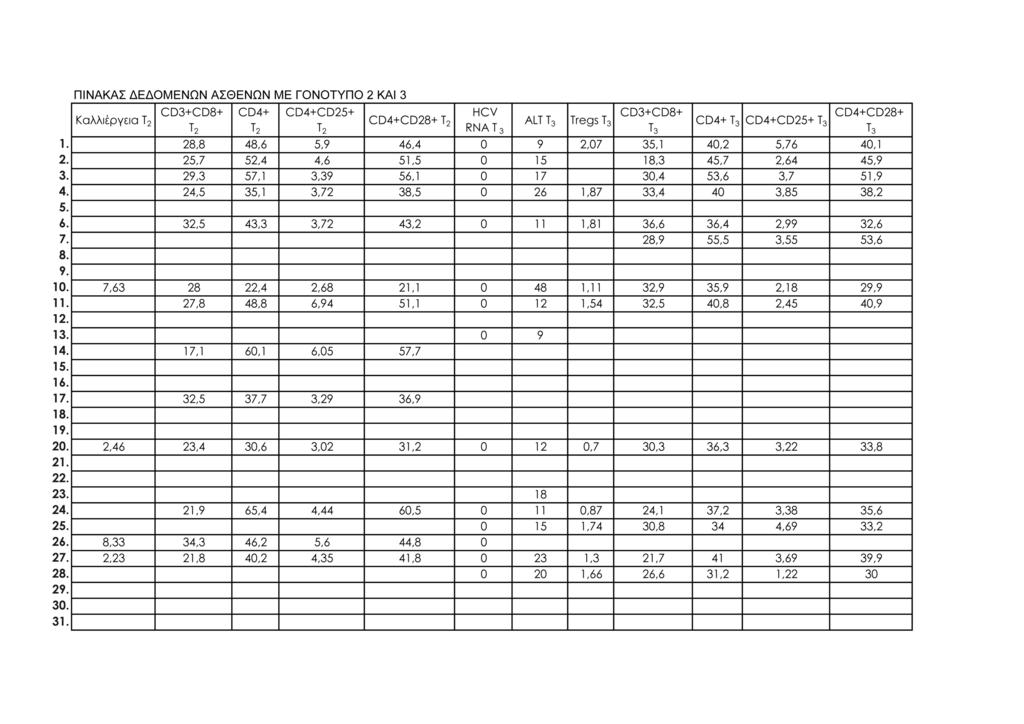
259
ΗΠΑΤΙΤΙ Α C. Ερωτήσεις-Απαντήσεις (μπορεί να υπάρχουν περισσότερες από μια σωστές απαντήσεις, οι σωστές απαντήσεις είναι με bold)
") ΗΠΑΤΙΤΙ Α C Ερωτήσεις-Απαντήσεις (μπορεί να υπάρχουν περισσότερες από μια σωστές απαντήσεις, οι σωστές απαντήσεις είναι με bold) 1) Τι είναι λάθος σχετικά με την επιδημιολογία της HCV λοίμωξης: Α. Η μέση
ΗΠΑΤΙΤΙ Α C Ερωτήσεις-Απαντήσεις (μπορεί να υπάρχουν περισσότερες από μια σωστές απαντήσεις, οι σωστές απαντήσεις είναι με bold) 1) Τι είναι λάθος σχετικά με την επιδημιολογία της HCV λοίμωξης: Α. Η μέση
ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ ΜΟΝΟΚΛΩΝΙΚΑ ΑΝΤΙΣΩΜΑΤΑ ΕΜΒΟΛΙΑ. Εργαστήριο Γενετικής, ΓΠΑ
 ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ ΜΟΝΟΚΛΩΝΙΚΑ ΑΝΤΙΣΩΜΑΤΑ ΕΜΒΟΛΙΑ Στάδια μικροβιακής λοίμωξης δημιουργία αποικίας σε εξωτερική επιφάνεια διείσδυση στον οργανισμό τοπική μόλυνση συστηματική (γενικευμένη) μόλυνση H σημασία
ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ ΜΟΝΟΚΛΩΝΙΚΑ ΑΝΤΙΣΩΜΑΤΑ ΕΜΒΟΛΙΑ Στάδια μικροβιακής λοίμωξης δημιουργία αποικίας σε εξωτερική επιφάνεια διείσδυση στον οργανισμό τοπική μόλυνση συστηματική (γενικευμένη) μόλυνση H σημασία
Μοριακή κυτταρική βιοχημεία Ανοσοποιητικό σύστημα
 Μοριακή κυτταρική βιοχημεία Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY-335 Χειμερινό εξάμηνο 2014 / 2015 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα
Μοριακή κυτταρική βιοχημεία Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY-335 Χειμερινό εξάμηνο 2014 / 2015 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα
ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ
 ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ Η πρώτη γραπτή αναφορά στο φαινόμενο της ανοσίας μπορεί να αναζητηθεί στον Θουκυδίδη, τον μεγάλο ιστορικό του Πελοποννησιακού Πολέμου Ανάπτυξη και επιβίωση o
ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ Η πρώτη γραπτή αναφορά στο φαινόμενο της ανοσίας μπορεί να αναζητηθεί στον Θουκυδίδη, τον μεγάλο ιστορικό του Πελοποννησιακού Πολέμου Ανάπτυξη και επιβίωση o
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς. Οι φάσεις των Τ κυτταρικών απαντήσεων
 ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
ΚΥΤΤΑΡΙΚΕΣ ΑΝΟΣΟΑΠΑΝΤΗΣΕΙΣ: Ενεργοποίηση των Τ κυττάρων από τους µικροοργανισµούς Οι φάσεις των Τ κυτταρικών απαντήσεων Αναγνώριση του αντιγόνου και συνδιέγερση Αναγνώριση πεπτιδίων συνδεδεµένων µε το
6. ΡΑΣΤΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΑΝΟΣΙΑΣ: Εξάλειψη των ενδοκυττάριων µικροοργανισµών
 6. ΡΑΣΤΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΑΝΟΣΙΑΣ: Εξάλειψη των ενδοκυττάριων µικροοργανισµών Οι δύο τύποι της κυτταρικής ανοσίας Η µετανάστευση των δραστικών Τ κυττάρων στις εστίες της λοίµωξης (εντόπιση
6. ΡΑΣΤΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΤΗΣ ΚΥΤΤΑΡΙΚΗΣ ΑΝΟΣΙΑΣ: Εξάλειψη των ενδοκυττάριων µικροοργανισµών Οι δύο τύποι της κυτταρικής ανοσίας Η µετανάστευση των δραστικών Τ κυττάρων στις εστίες της λοίµωξης (εντόπιση
Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I. Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ.
 Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 14/10/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Γενικά στοιχεία
Εισαγωγή στην Ανοσολογία Επίκτητη Ανοσία I Σωτήρης Ζαρογιάννης Επίκ. Καθηγητής Φυσιολογίας Εργαστήριο Φυσιολογίας Τμήμα Ιατρικής Π.Θ. 14/10/2016 Φυσιολογία Συστημάτων Ακαδημαϊκό Ετος 2016-2017 Γενικά στοιχεία
Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα
 Β λεμφοκύτταρα") Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα φυσική ή μη ειδική ανοσία δεν απαιτεί προηγούμενη έκθεση στο παθογόνο και δεν διαθέτει μνήμη. σε επίκτητη ή ειδική ανοσία χυμική ανοσία με παραγωγή
Επίκτητη Ανοσιακή Απάντηση (χυμικό σκέλος) Β λεμφοκύτταρα φυσική ή μη ειδική ανοσία δεν απαιτεί προηγούμενη έκθεση στο παθογόνο και δεν διαθέτει μνήμη. σε επίκτητη ή ειδική ανοσία χυμική ανοσία με παραγωγή
ΦΡΟΝΤΙΣΤΗΡΙΑ ΠΡΟΟΠΤΙΚΗ
 Απαντήσεις του κριτηρίου αξιολόγησης στη βιολογία γενικής παιδείας 1 ο ΚΕΦΑΛΑΙΟ ΘΕΜΑ 1 ο Να γράψετε τον αριθμό καθεμίας από τις ημιτελείς προτάσεις 1 έως και 5, και δίπλα σε αυτόν το γράμμα που αντιστοιχεί
Απαντήσεις του κριτηρίου αξιολόγησης στη βιολογία γενικής παιδείας 1 ο ΚΕΦΑΛΑΙΟ ΘΕΜΑ 1 ο Να γράψετε τον αριθμό καθεμίας από τις ημιτελείς προτάσεις 1 έως και 5, και δίπλα σε αυτόν το γράμμα που αντιστοιχεί
ΑΝΟΣΟΛΟΓΙΑ. 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ
 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ") ΑΝΟΣΟΛΟΓΙΑ 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ ΑΝΤΙΓΟΝΟ 3. Η πρόσληψη του αντιγόνου και η παρουσίασή του στα λεμφοκύτταρα 4. Η αναγνώριση του αντιγόνου. Αντιγονικοί
ΑΝΟΣΟΛΟΓΙΑ 1. Εισαγωγή (κυρίως στην επίκτητη ανοσία) 2. Φυσική ανοσία ΕΠΙΚΤΗΤΗ ΑΝΟΣΙΑ ΑΝΤΙΓΟΝΟ 3. Η πρόσληψη του αντιγόνου και η παρουσίασή του στα λεμφοκύτταρα 4. Η αναγνώριση του αντιγόνου. Αντιγονικοί
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ
 ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΕΦΑΡΜΟΓΕΣ ΤΗΣ ΒΙΟΤΕΧΝΟΛΟΓΙΑΣ ΣΤΗΝ ΙΑΤΡΙΚΗ Καθώς η επιστημονική γνώση και κατανόηση αναπτύσσονται, ο μελλοντικός σχεδιασμός βιοτεχνολογικών προϊόντων περιορίζεται μόνο από τη φαντασία μας Βιοτεχνολογία
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ / Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015
 ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015") ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ / Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015 ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Η φαγοκυττάρωση
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ / Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015 ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Η φαγοκυττάρωση
ΣΤΗΛΗ Α Αντιβιοτικό Αντισώματα ιντερφερόνες Τ- Τ- (αντιγόνα) κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera
 κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera") Α1. 1. β Βιολογία ΘΕΜΑ Α γενιικής παιιδείίας 2. γ 3. γ 4. γ 5. δ Α2. ΣΤΗΛΗ Α Αντιβιοτικό Αντισώματα ιντερφερόνες Τ- Τ- (αντιγόνα) κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera Ηπατίτιδα C + Candida albicans
Α1. 1. β Βιολογία ΘΕΜΑ Α γενιικής παιιδείίας 2. γ 3. γ 4. γ 5. δ Α2. ΣΤΗΛΗ Α Αντιβιοτικό Αντισώματα ιντερφερόνες Τ- Τ- (αντιγόνα) κυτταροτοξικά βοηθητικά Τοξίνες Vibrio cholera Ηπατίτιδα C + Candida albicans
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ. ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ
 ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
ΘΕΩΡΙΑ 3 η ΠΑΡΟΥΣΙΑΣΗ ΚΥΤΤΑΡΟΚΙΝΕΣ ή ΚΥΤΤΟΚΙΝΕΣ Dr ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ Είδαμε ότι οι ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ είναι 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις
4. Η κίρρωση του ήπατος προκαλείται εξαιτίας της αποθήκευσης στα ηπατικά κύτταρα: Πρωτεϊνών Υδατανθράκων Λιπών Αλκοόλ
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΠ Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΗΜΕΡΟΜΗΝΙΑ: 22/10/2017 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΛΑΖΑΡΑΚΗ ΝΟΤΑ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Το
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΠ Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΗΜΕΡΟΜΗΝΙΑ: 22/10/2017 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ ΛΑΖΑΡΑΚΗ ΝΟΤΑ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Το
ΠΑΡΑΓΩΓΗ ΑΝΟΣΟΡΥΘΜΙΣΤΙΚΩΝ ΚΥΤΤΑΡΟΚΙΝΩΝ ΣΤΗ ΧΡΟΝΙΑ ΗΠΑΤΙΤΙ Α C
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ ΠΑΘΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ ΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ Γ. ΗΛΟΝΙ ΗΣ ΠΑΝΕΠ. ΕΤΟΣ 2005-2006 ΑΡΙΘΜ. 2060 ΠΑΡΑΓΩΓΗ ΑΝΟΣΟΡΥΘΜΙΣΤΙΚΩΝ ΚΥΤΤΑΡΟΚΙΝΩΝ ΣΤΗ ΧΡΟΝΙΑ
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΙΑΤΡΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΠΑΘΟΛΟΓΙΑΣ ΠΑΘΟΛΟΓΙΚΗ ΚΛΙΝΙΚΗ ΙΕΥΘΥΝΤΗΣ: ΚΑΘΗΓΗΤΗΣ Γ. ΗΛΟΝΙ ΗΣ ΠΑΝΕΠ. ΕΤΟΣ 2005-2006 ΑΡΙΘΜ. 2060 ΠΑΡΑΓΩΓΗ ΑΝΟΣΟΡΥΘΜΙΣΤΙΚΩΝ ΚΥΤΤΑΡΟΚΙΝΩΝ ΣΤΗ ΧΡΟΝΙΑ
ΣΥΝΔΡΟΜΟ ΕΠΙΚΤΗΤΗΣ ΑΝΟΣΟΒΙΟΛΟΓΙΚΗΣ ΑΝΕΠΑΡΚΕΙΑΣ (AIDS)
") ΣΥΝΔΡΟΜΟ ΕΠΙΚΤΗΤΗΣ ΑΝΟΣΟΒΙΟΛΟΓΙΚΗΣ ΑΝΕΠΑΡΚΕΙΑΣ (AIDS) Στα τέλη της δεκαετίας του 1970 εμφανίστηκε μία από τις σοβαρότερες ασθένειες ανεπάρκειας του ανοσοβιολογικού συστήματος Ανοσοβιολογική ανεπάρκεια
ΣΥΝΔΡΟΜΟ ΕΠΙΚΤΗΤΗΣ ΑΝΟΣΟΒΙΟΛΟΓΙΚΗΣ ΑΝΕΠΑΡΚΕΙΑΣ (AIDS) Στα τέλη της δεκαετίας του 1970 εμφανίστηκε μία από τις σοβαρότερες ασθένειες ανεπάρκειας του ανοσοβιολογικού συστήματος Ανοσοβιολογική ανεπάρκεια
IOΓΕΝΗΣ ΗΠΑΤΙΤΙΣ Βαλεντίνη Τζιούφα-Ασημακοπούλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής ΑΠΘ
 IOΓΕΝΗΣ ΗΠΑΤΙΤΙΣ Βαλεντίνη Τζιούφα-Ασημακοπούλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής ΑΠΘ Μάρτιος 2015 Οξεία ιογενής ηπατίτιδα Αίτια Ιός της ηπατίτιδας Α (Hepatitis A Virus-HAV) Iός
IOΓΕΝΗΣ ΗΠΑΤΙΤΙΣ Βαλεντίνη Τζιούφα-Ασημακοπούλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής ΑΠΘ Μάρτιος 2015 Οξεία ιογενής ηπατίτιδα Αίτια Ιός της ηπατίτιδας Α (Hepatitis A Virus-HAV) Iός
ΘΕΜΑ 1 Ο ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 10/11/2013
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 10/11/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Οι αποικοδομητές είναι:
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΘΕΡΙΝΑ ΗΜΕΡΟΜΗΝΙΑ: 10/11/2013 ΘΕΜΑ 1 Ο Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Οι αποικοδομητές είναι:
να ταράξουν την λειτουργία των ιστών και των οργάνων του; α. τη θέση τους στο ανθρώπινο σώμα β. την γενικευμένη ή εξειδικευμένη δράση
 Ερωτήσεις κατανόησης της θεωρίας του 1 ο κεφαλαίου (συνέχεια) 1. Από τι εξαρτάται η επιβίωση του ανθρώπου και ποιοι εξωτερικοί παράγοντες θα μπορούσαν να ταράξουν την λειτουργία των ιστών και των οργάνων
Ερωτήσεις κατανόησης της θεωρίας του 1 ο κεφαλαίου (συνέχεια) 1. Από τι εξαρτάται η επιβίωση του ανθρώπου και ποιοι εξωτερικοί παράγοντες θα μπορούσαν να ταράξουν την λειτουργία των ιστών και των οργάνων
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ / Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015
 ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015") ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ / Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015 ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Η φαγοκυττάρωση
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ / Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΣΕΙΡΑ: ΗΜΕΡΟΜΗΝΙΑ: 15/11/2015 ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: Α1. Η φαγοκυττάρωση
2 Ο ΜΑΘΗΜΑ ΣΥΣΤΗΜΑ ΣΥΜΠΛΗΡΩΜΑΤΟΣ ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ
 2 Ο ΜΑΘΗΜΑ ΣΥΣΤΗΜΑ ΣΥΜΠΛΗΡΩΜΑΤΟΣ ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις μεμβράνες 2. Φυσιολογικοί φραγμοί
2 Ο ΜΑΘΗΜΑ ΣΥΣΤΗΜΑ ΣΥΜΠΛΗΡΩΜΑΤΟΣ ΠΑΠΑΓΙΑΝΝΗΣ ΔΗΜΗΤΡΗΣ ΤΜΗΜΑ ΙΑΤΡΙΚΩΝ ΕΡΓΑΣΤΗΡΙΩΝ ΤΕΙ ΘΕΣΣΑΛΙΑΣ ΜΗΧΑΝΙΣΜΟΙ ΜΗ ΕΙΔΙΚΗΣ ΑΝΟΣΙΑΣ 1. Ανατομικοί φραγμοί - Δέρμα - Βλεννώδεις μεμβράνες 2. Φυσιολογικοί φραγμοί
Βιολογία γενικής παιδείας τάξη Γ
 Βιολογία γενικής παιδείας τάξη Γ Παραδόσεις του μαθήματος Επιμέλεια: Γιάννης Αργύρης Βιολόγος M.Sc. Καθηγητής 3 ου Γεν. Λυκ. Ηλιούπολης Κεφάλαιο 1ο Άνθρωπος και υγεία 2. Μηχανισμοί Άμυνας του Ανθρώπινου
Βιολογία γενικής παιδείας τάξη Γ Παραδόσεις του μαθήματος Επιμέλεια: Γιάννης Αργύρης Βιολόγος M.Sc. Καθηγητής 3 ου Γεν. Λυκ. Ηλιούπολης Κεφάλαιο 1ο Άνθρωπος και υγεία 2. Μηχανισμοί Άμυνας του Ανθρώπινου
ΥΠΟΠΛΗΘΥΣΜΟΙ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ. Αλεξάνδρα Φλέβα Ph.D Βιολόγος Τμήμα Ανοσολογίας-Ιστοσυμβατότητας Γ.Ν. «Παπαγεωργίου»
 ΥΠΟΠΛΗΘΥΣΜΟΙ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ Αλεξάνδρα Φλέβα Ph.D Βιολόγος Τμήμα Ανοσολογίας-Ιστοσυμβατότητας Γ.Ν. «Παπαγεωργίου» Η αξιοποίηση του εύρους των δυνατοτήτων της κυτταρομετρίας ροής στην Ανοσολογία προσφέρει
ΥΠΟΠΛΗΘΥΣΜΟΙ Τ ΛΕΜΦΟΚΥΤΤΑΡΩΝ Αλεξάνδρα Φλέβα Ph.D Βιολόγος Τμήμα Ανοσολογίας-Ιστοσυμβατότητας Γ.Ν. «Παπαγεωργίου» Η αξιοποίηση του εύρους των δυνατοτήτων της κυτταρομετρίας ροής στην Ανοσολογία προσφέρει
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ
 ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ 1 ο Α. Ερωτήσεις πολλαπλής επιλογής 1. γ 2. α 3. β 4. δ 5. δ Β. Ερωτήσεις σωστού - λάθους 1. Σωστό 2. Λάθος 3. Λάθος 4. Λάθος 5. Σωστό ΘΕΜΑ
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ 1 ο Α. Ερωτήσεις πολλαπλής επιλογής 1. γ 2. α 3. β 4. δ 5. δ Β. Ερωτήσεις σωστού - λάθους 1. Σωστό 2. Λάθος 3. Λάθος 4. Λάθος 5. Σωστό ΘΕΜΑ
Ηπατίτιδα C και Σακχαρώδης διαβήτης. Όλγα Ι. Γιουλεμέ Γαστρεντερολόγος, Λέκτορας Α.Π.Θ. Α Προπ. Παθολογική Κλινική ΑΧΕΠΑ
 Ηπατίτιδα C και Σακχαρώδης διαβήτης Όλγα Ι. Γιουλεμέ Γαστρεντερολόγος, Λέκτορας Α.Π.Θ. Α Προπ. Παθολογική Κλινική ΑΧΕΠΑ HCV σοβαρό πρόβλημα δημόσιας υγείας σε παγκόσμιο επίπεδο 170 εκατομμύρια (2-3% παγκ.
Ηπατίτιδα C και Σακχαρώδης διαβήτης Όλγα Ι. Γιουλεμέ Γαστρεντερολόγος, Λέκτορας Α.Π.Θ. Α Προπ. Παθολογική Κλινική ΑΧΕΠΑ HCV σοβαρό πρόβλημα δημόσιας υγείας σε παγκόσμιο επίπεδο 170 εκατομμύρια (2-3% παγκ.
Ανοσιακή απάντηση Αικατερίνη Ταράση
 Ανοσιακή απάντηση Αικατερίνη Ταράση ιευθύντρια Τµ. Ανοσολογίας- Ιστοσυµβατότητας Γ.Ν.Α.. " Ο Ευαγγελισµός" ανοσιακή απάντηση είναι το σύνολο των πολύπλοκων διεργασιών µε τις οποίες ο οργανισµός (ξενιστής)
Ανοσιακή απάντηση Αικατερίνη Ταράση ιευθύντρια Τµ. Ανοσολογίας- Ιστοσυµβατότητας Γ.Ν.Α.. " Ο Ευαγγελισµός" ανοσιακή απάντηση είναι το σύνολο των πολύπλοκων διεργασιών µε τις οποίες ο οργανισµός (ξενιστής)
Ανοσιακή απάντηση (immune response)
") Ανοσιακή απάντηση (immune response) Το σύνολο των μηχανισμών που καθιστούν τον οργανισμό ικανό να αναγνωρίζει,να εξουδετερώνει και να απομακρύνει κάθε ξένη ουσία προς αυτόν χαρακτηρίζεται ως ανοσία. Αποτέλεσμα
Ανοσιακή απάντηση (immune response) Το σύνολο των μηχανισμών που καθιστούν τον οργανισμό ικανό να αναγνωρίζει,να εξουδετερώνει και να απομακρύνει κάθε ξένη ουσία προς αυτόν χαρακτηρίζεται ως ανοσία. Αποτέλεσμα
όλοι αναπνευστική οδός στομάχι στόμα
 κεράτινη στιβάδα περιέχει σμήγμα λιπαρά οξέα Μηχανισμοί που παρεμποδίζουν την είσοδο Δέρμα περιέχει ιδρώτας φυσιολογική μικροχλωρίδα λυσοζύμη γαλακτικό οξύ μικροοργανισμών Βλεννογόνοι όλοι αναπνευστική
κεράτινη στιβάδα περιέχει σμήγμα λιπαρά οξέα Μηχανισμοί που παρεμποδίζουν την είσοδο Δέρμα περιέχει ιδρώτας φυσιολογική μικροχλωρίδα λυσοζύμη γαλακτικό οξύ μικροοργανισμών Βλεννογόνοι όλοι αναπνευστική
Ιογενείς Ηπατίτιδες Ηπατίτιδα C. Συχνές Ερωτήσεις
 Ιογενείς Ηπατίτιδες Ηπατίτιδα C Συχνές Ερωτήσεις Tι είναι ηπατίτιδα; Το ήπαρ (συκώτι) είναι ένα ζωτικό όργανο που βρίσκεται στο δεξιό άνω τμήμα της κοιλιακής χώρας. Έχει πολλές λειτουργίες και διαδραματίζει
Ιογενείς Ηπατίτιδες Ηπατίτιδα C Συχνές Ερωτήσεις Tι είναι ηπατίτιδα; Το ήπαρ (συκώτι) είναι ένα ζωτικό όργανο που βρίσκεται στο δεξιό άνω τμήμα της κοιλιακής χώρας. Έχει πολλές λειτουργίες και διαδραματίζει
Χρόνια φλεγμονή. Βαλεντίνη Τζιούφα-Ασημακοπούλου. Νοέμβριος 2018
 Χρόνια φλεγμονή Βαλεντίνη Τζιούφα-Ασημακοπούλου Νοέμβριος 2018 Οξεία φλεγμονή Ταχεία εισβολή και λύση Εξιδρωματικά στοιχεία Πολυμορφοπύρηνα Χρόνια φλεγμονή Ύπουλη εισβολήπαρατεταμένη πορείαβραδεία λύση
Χρόνια φλεγμονή Βαλεντίνη Τζιούφα-Ασημακοπούλου Νοέμβριος 2018 Οξεία φλεγμονή Ταχεία εισβολή και λύση Εξιδρωματικά στοιχεία Πολυμορφοπύρηνα Χρόνια φλεγμονή Ύπουλη εισβολήπαρατεταμένη πορείαβραδεία λύση
Αυτοφαγία & Ανοσολογικό Σύστημα. Χαράλαμπος Μ. Μουτσόπουλος
 Αυτοφαγία & Ανοσολογικό Σύστημα Χαράλαμπος Μ. Μουτσόπουλος Απρίλιος 2018 Αυτοφαγία & Ανοσολογικό Σύστημα Ευκαρυωτικά κύτταρα: Δυο μηχανισμούς αποδόμησης και ανακύκλωσης κυτταροπλασματικών αποβλήτων Πρωτεάσες
Αυτοφαγία & Ανοσολογικό Σύστημα Χαράλαμπος Μ. Μουτσόπουλος Απρίλιος 2018 Αυτοφαγία & Ανοσολογικό Σύστημα Ευκαρυωτικά κύτταρα: Δυο μηχανισμούς αποδόμησης και ανακύκλωσης κυτταροπλασματικών αποβλήτων Πρωτεάσες
Ιοί & HPV. Ευστάθιος Α. Ράλλης. Επικ. Καθηγητής Δερματολογίας - Αφροδισιολογίας
 Ιοί & HPV Ευστάθιος Α. Ράλλης Επικ. Καθηγητής Δερματολογίας - Αφροδισιολογίας Περί ΙΩΝ Ορισμός Οι ιοί είναι ατελείς, λοιμώδεις οργανισμοί μικρότεροι σε μέγεθος από τα μικρόβια και ορατοί μόνο με το ηλεκτρονικό
Ιοί & HPV Ευστάθιος Α. Ράλλης Επικ. Καθηγητής Δερματολογίας - Αφροδισιολογίας Περί ΙΩΝ Ορισμός Οι ιοί είναι ατελείς, λοιμώδεις οργανισμοί μικρότεροι σε μέγεθος από τα μικρόβια και ορατοί μόνο με το ηλεκτρονικό
ΦΡΟΝΤΙΣΤΗΡΙΑ ΜΗΤΑΛΑΣ ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ
 ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ ΚΕΦΑΛΑΙΟ 1.3 : ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΑΝΟΣΙΑΣ ΘΕΜΑ 1 Χαρακτηρίστε τις προτάσεις ως σωστές ή λανθασµένες. 1. Οι βλεννογόνοι
ΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Γ ΛΥΚΕΙΟΥ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ ΚΕΦΑΛΑΙΟ 1.3 : ΜΗΧΑΝΙΣΜΟΙ ΑΜΥΝΑΣ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΟΡΓΑΝΙΣΜΟΥ ΒΑΣΙΚΕΣ ΑΡΧΕΣ ΑΝΟΣΙΑΣ ΘΕΜΑ 1 Χαρακτηρίστε τις προτάσεις ως σωστές ή λανθασµένες. 1. Οι βλεννογόνοι
ΔΙΑΓΩΝΙΣΜΑ ΣΤΗ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ
 23-10-11 ΘΕΡΙΝΑ ΔΙΑΓΩΝΙΣΜΑ ΣΤΗ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Όλα τα βακτήρια: Α. διαθέτουν κυτταρικό
23-10-11 ΘΕΡΙΝΑ ΔΙΑΓΩΝΙΣΜΑ ΣΤΗ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Όλα τα βακτήρια: Α. διαθέτουν κυτταρικό
Μεταβολικές ανάγκες ανοσοκυττάρων
 Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
Μεταβολικές ανάγκες ανοσοκυττάρων Στέργιος Κατσιουγιάννης PhD Μεταδιδακτορικός συνεργάτης Χαροκόπειο Πανεπιστήµιο Τµήµα Επιστήµης ιαιτολογίας και ιατροφής Μεταβολισµός και Ανοσολογία Ιστορικά το καλύτερο
ΑΝΟΣΟΒΙΟΛΟΓΙΑ. Εξεταστική Ιανουαρίου 2010
 Εξεταστική Ιανουαρίου 2010 Ποιες είναι οι διαφορές μιας πρωτογενούς από μια δευτερογενή χυμική ανοσολογική απόκριση; Περιγράψετε τους μηχανισμούς ενεργοποίησης στις δυο περιπτώσεις. ΘΕΜΑ 2 (1 μονάδα) Περιγράψετε
Εξεταστική Ιανουαρίου 2010 Ποιες είναι οι διαφορές μιας πρωτογενούς από μια δευτερογενή χυμική ανοσολογική απόκριση; Περιγράψετε τους μηχανισμούς ενεργοποίησης στις δυο περιπτώσεις. ΘΕΜΑ 2 (1 μονάδα) Περιγράψετε
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ
 23 10 2011 ΘΕΡΙΝΑ ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Όλα τα βακτήρια: Α. διαθέτουν
23 10 2011 ΘΕΡΙΝΑ ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΘΕΜΑ 1 Ο Α. Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Όλα τα βακτήρια: Α. διαθέτουν
04/11/2018 ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΝΟΤΑ ΛΑΖΑΡΑΚΗ ΘΕΜΑ Α
 ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ 04/11/2018 ΝΟΤΑ ΛΑΖΑΡΑΚΗ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Πυρηνική περιοχή διαθέτει: Α. Ο ιός της
ΑΠΑΝΤΗΣΕΙΣ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ 04/11/2018 ΝΟΤΑ ΛΑΖΑΡΑΚΗ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Πυρηνική περιοχή διαθέτει: Α. Ο ιός της
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΦΑΛΑΙΟ 1(ΥΓΕΙΑ-ΑΝΘΡΩΠΟΣ)
") ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΦΑΛΑΙΟ 1(ΥΓΕΙΑ-ΑΝΘΡΩΠΟΣ) Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη σωστή απάντηση ή στη φράση που συμπληρώνει σωστά την πρόταση: 1. Οι ιοί αποτελούνται
ΟΝΟΜΑΤΕΠΩΝΥΜΟ: ΤΜΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΦΑΛΑΙΟ 1(ΥΓΕΙΑ-ΑΝΘΡΩΠΟΣ) Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη σωστή απάντηση ή στη φράση που συμπληρώνει σωστά την πρόταση: 1. Οι ιοί αποτελούνται
ΑΡΧΕΣ ΑΝΟΣΟΛΟΓΙΑΣ ΕΙΣΑΓΩΓΗ ΣΤΟ ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ
 ΑΡΧΕΣ ΑΝΟΣΟΛΟΓΙΑΣ ΕΙΣΑΓΩΓΗ ΣΤΟ ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ Χρειάζεται η µελέτη της ανοσολογία; Εµβόλια Άµυνα κατά µικροοργανισµών Ανοσολογικές ασθένειες Αλλεργίες - υπερευαισθησίες, αυτοανοσίες, ανοσοανεπάρκειες,
ΑΡΧΕΣ ΑΝΟΣΟΛΟΓΙΑΣ ΕΙΣΑΓΩΓΗ ΣΤΟ ΑΝΟΣΟΠΟΙΗΤΙΚΟ ΣΥΣΤΗΜΑ Χρειάζεται η µελέτη της ανοσολογία; Εµβόλια Άµυνα κατά µικροοργανισµών Ανοσολογικές ασθένειες Αλλεργίες - υπερευαισθησίες, αυτοανοσίες, ανοσοανεπάρκειες,
ΕΡΩΤΗΣΕΙΣ 1 ου ΚΕΦΑΛΑΙΟΥ
 ΕΡΩΤΗΣΕΙΣ 1 ου ΚΕΦΑΛΑΙΟΥ Επιλέξτε τη σωστή απάντηση: Το τρυπανόσωμα προκαλεί α. δυσεντερία β. ελονοσία γ. ασθένεια του ύπνου δ. χολέρα Τα ενδοσπόρια σχηματίζονται από β. φυτά γ. ιούς δ. πρωτόζωα Η σύφιλη
ΕΡΩΤΗΣΕΙΣ 1 ου ΚΕΦΑΛΑΙΟΥ Επιλέξτε τη σωστή απάντηση: Το τρυπανόσωμα προκαλεί α. δυσεντερία β. ελονοσία γ. ασθένεια του ύπνου δ. χολέρα Τα ενδοσπόρια σχηματίζονται από β. φυτά γ. ιούς δ. πρωτόζωα Η σύφιλη
ΔΙΑΒΑΖΟΝΤΑΣ ΤΗ ΒΙΟΨΙΑ ΗΠΑΤΟΣ
 ΠΡΑΚΤΙΚΕΣ ΟΔΗΓΙΕΣ ΣΤΟΥΣ ΗΠΑΤΟΛΟΓΟΥΣ, ΔΙΑΒΑΖΟΝΤΑΣ ΤΗ ΒΙΟΨΙΑ ΗΠΑΤΟΣ Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Τμήματος Ιατρικής Α.Π.Θ. ΕΝΔΕΙΞΕΙΣ ΒΙΟΨΙΑΣ ΗΠΑΤΟΣ Χρόνια
ΠΡΑΚΤΙΚΕΣ ΟΔΗΓΙΕΣ ΣΤΟΥΣ ΗΠΑΤΟΛΟΓΟΥΣ, ΔΙΑΒΑΖΟΝΤΑΣ ΤΗ ΒΙΟΨΙΑ ΗΠΑΤΟΣ Πρόδρομος Χυτίρογλου Εργαστήριο Γενικής Παθολογίας και Παθολογικής Ανατομικής Τμήματος Ιατρικής Α.Π.Θ. ΕΝΔΕΙΞΕΙΣ ΒΙΟΨΙΑΣ ΗΠΑΤΟΣ Χρόνια
Μικροοργανισμοί. Οι μικροοργανισμοί διακρίνονται σε: Μύκητες Πρωτόζωα Βακτήρια Ιούς
 Μικροοργανισμοί Οι μικροοργανισμοί διακρίνονται σε: Μύκητες Πρωτόζωα Βακτήρια Ιούς Παθογόνοι μικροοργανισμοί Παθογόνοι μικροοργανισμοί ονομάζονται οι μικροοργανισμοί που χρησιμοποιούν τον άνθρωπο ως ξενιστή
Μικροοργανισμοί Οι μικροοργανισμοί διακρίνονται σε: Μύκητες Πρωτόζωα Βακτήρια Ιούς Παθογόνοι μικροοργανισμοί Παθογόνοι μικροοργανισμοί ονομάζονται οι μικροοργανισμοί που χρησιμοποιούν τον άνθρωπο ως ξενιστή
Εισαγωγή στην Ανοσολογία
 Εισαγωγή στην Ανοσολογία ρ. Γιώργος Κρασιάς Ινστιτούτο Νευρολογίας και Γενετικής Κύπρου Τµήµα Μοριακής Ιολογίας Τι είναι το Ανοσοποιητικό Σύστηµα (ΑΣ)? Το ΑΣ (Immune System) είναι ένα σύστηµα άµυνας του
Εισαγωγή στην Ανοσολογία ρ. Γιώργος Κρασιάς Ινστιτούτο Νευρολογίας και Γενετικής Κύπρου Τµήµα Μοριακής Ιολογίας Τι είναι το Ανοσοποιητικό Σύστηµα (ΑΣ)? Το ΑΣ (Immune System) είναι ένα σύστηµα άµυνας του
Η πρωτογενής ανοσοβιολογική απόκριση ενεργοποιείται κατά την πρώτη επαφή του οργανισμού με ένα αντιγόνο. Περιλαμβάνει τα εξής στάδια:
 Εικόνα 1.24: α) Σύνδεση αντισώματος - αντιγόνου, β) συμπληρωματικότητα αντισώματος - αντιγόνου, γ) ένα αντίσωμα συνδέεται με περισσότερα από ένα αντιγόνα. Στάδια ανοσοβιολογικής απόκρισης Η αντίδραση του
Εικόνα 1.24: α) Σύνδεση αντισώματος - αντιγόνου, β) συμπληρωματικότητα αντισώματος - αντιγόνου, γ) ένα αντίσωμα συνδέεται με περισσότερα από ένα αντιγόνα. Στάδια ανοσοβιολογικής απόκρισης Η αντίδραση του
ΕΦΑΡΜΟΓΕΣ ΑΣΚΗΣΕΙΣ ΣΤΟΥΣ ΜΗΧΑΝΙΣΜΟΥΣ ΑΜΥΝΑΣ
 ΕΦΑΡΜΟΓΕΣ ΑΣΚΗΣΕΙΣ ΣΤΟΥΣ ΜΗΧΑΝΙΣΜΟΥΣ ΑΜΥΝΑΣ Ερωτήσεις πολλαπλής επιλογής: επιλέξτε το γράμμα που αντιστοιχεί στη σωστή συνέχεια της πρότασης. 1. Δυσμενές χημικό περιβάλλον στην επιδερμίδα προκαλείται από:
ΕΦΑΡΜΟΓΕΣ ΑΣΚΗΣΕΙΣ ΣΤΟΥΣ ΜΗΧΑΝΙΣΜΟΥΣ ΑΜΥΝΑΣ Ερωτήσεις πολλαπλής επιλογής: επιλέξτε το γράμμα που αντιστοιχεί στη σωστή συνέχεια της πρότασης. 1. Δυσμενές χημικό περιβάλλον στην επιδερμίδα προκαλείται από:
ΕΡΩΤΗΣΕΙΣ ΣΩΣΤΟΥ - ΛΑΘΟΥΣ. ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ
 ΕΡΩΤΗΣΕΙΣ ΣΩΣΤΟΥ - ΛΑΘΟΥΣ. ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ 1. Η πήξη του αίματος συμβάλλει στην άμυνα του οργανισμού 2. Η φαγοκυττάρωση είναι αποτελεσματική μόνο έναντι των βακτηρίων 3. Οι ιντερφερόνες δρουν άμεσα
ΕΡΩΤΗΣΕΙΣ ΣΩΣΤΟΥ - ΛΑΘΟΥΣ. ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ 1. Η πήξη του αίματος συμβάλλει στην άμυνα του οργανισμού 2. Η φαγοκυττάρωση είναι αποτελεσματική μόνο έναντι των βακτηρίων 3. Οι ιντερφερόνες δρουν άμεσα
Πρόταση: καινούργιος ευρωπαϊκός ορισμός κρούσματος για ηπατίτιδα Β
 ΚΕΝΤΡΟ ΕΛΕΓΧΟΥ & ΠΡΟΛΗΨΗΣ ΝΟΣΗΜΑΤΩΝ (ΚΕ.ΕΛ.Π.ΝΟ.) ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ & ΚΟΙΝΩΝΙΚΗΣ ΑΛΛΗΛΕΓΓΥΗΣ Αλλαγή του ορισμού κρούσματος της Ηπατίτιδας Β και C Δήλωση των περιστατικών χρόνιας ηπατίτιδας Το ECDC προτείνει
ΚΕΝΤΡΟ ΕΛΕΓΧΟΥ & ΠΡΟΛΗΨΗΣ ΝΟΣΗΜΑΤΩΝ (ΚΕ.ΕΛ.Π.ΝΟ.) ΥΠΟΥΡΓΕΙΟ ΥΓΕΙΑΣ & ΚΟΙΝΩΝΙΚΗΣ ΑΛΛΗΛΕΓΓΥΗΣ Αλλαγή του ορισμού κρούσματος της Ηπατίτιδας Β και C Δήλωση των περιστατικών χρόνιας ηπατίτιδας Το ECDC προτείνει
ITP Ιδιοπαθής/ αυτοάνοση θρομβοπενική πορφύρα
 ITP Ιδιοπαθής/ αυτοάνοση θρομβοπενική πορφύρα Αθανασία Μουζάκη, Καθηγήτρια Εργαστηριακής Αιματολογίας-Αιμοδοσίας, Αιματολογικό Τμήμα, Παθολογική Κλινική, Τμήμα Ιατρικής, Παν/ο Πατρών Τα είδη της ITP είναι:
ITP Ιδιοπαθής/ αυτοάνοση θρομβοπενική πορφύρα Αθανασία Μουζάκη, Καθηγήτρια Εργαστηριακής Αιματολογίας-Αιμοδοσίας, Αιματολογικό Τμήμα, Παθολογική Κλινική, Τμήμα Ιατρικής, Παν/ο Πατρών Τα είδη της ITP είναι:
ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις:
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΑΠΟΦΟΙΤΟΙ ΗΜΕΡΟΜΗΝΙΑ: 24/01/2016 ΕΠΙΜΕΛΕΙΑ ΘΕΜΑΤΩΝ: ΝΟΤΑ ΛΑΖΑΡΑΚΗ ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΣΕΙΡΑ: ΑΠΟΦΟΙΤΟΙ ΗΜΕΡΟΜΗΝΙΑ: 24/01/2016 ΕΠΙΜΕΛΕΙΑ ΘΕΜΑΤΩΝ: ΝΟΤΑ ΛΑΖΑΡΑΚΗ ΘΕΜΑ Α Να επιλέξετε την φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες
ΠΡΟΔΡΟΜΑ ΑΠΟΤΕΛΕΣΜΑΤΑ ΠΡΟΟΠΤΙΚΗΣ ΜΕΛΕΤΗΣ
 ΔΙΑΤΑΡΑΧΕΣ ΥΠΟΠΛΗΘΥΣΜΩΝ Τ-ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΣΕ ΑΣΘΕΝΕΙΣ ΜΕ ΧΡΟΝΙΑ ΝΕΦΡΙΚΗ ΝΟΣΟ ΤΕΛΙΚΟΥ ΣΤΑΔΙΟΥ ΚΑΙ ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΩΝΕΦΡΙΚΗΣ ΚΑΘΑΡΣΗΣ ΠΡΟΔΡΟΜΑ ΑΠΟΤΕΛΕΣΜΑΤΑ ΠΡΟΟΠΤΙΚΗΣ ΜΕΛΕΤΗΣ Ε. Σαμπάνη 1, Μ. Στάγκου 1, Δ-Β.
ΔΙΑΤΑΡΑΧΕΣ ΥΠΟΠΛΗΘΥΣΜΩΝ Τ-ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΣΕ ΑΣΘΕΝΕΙΣ ΜΕ ΧΡΟΝΙΑ ΝΕΦΡΙΚΗ ΝΟΣΟ ΤΕΛΙΚΟΥ ΣΤΑΔΙΟΥ ΚΑΙ ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΩΝΕΦΡΙΚΗΣ ΚΑΘΑΡΣΗΣ ΠΡΟΔΡΟΜΑ ΑΠΟΤΕΛΕΣΜΑΤΑ ΠΡΟΟΠΤΙΚΗΣ ΜΕΛΕΤΗΣ Ε. Σαμπάνη 1, Μ. Στάγκου 1, Δ-Β.
ΚΕΦΑΛΑΙΟ 8 ο...2 I. Εφαρµογές της βιοτεχνολογίας στην ιατρική...2 ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ...7 ΝΑ ΣΥΜΠΛΗΡΩΣΕΤΕ ΤΑ ΚΕΝΑ ΜΕ ΤΗΝ ΚΑΤΑΛΛΗΛΗ ΛΕΞΗ...
 ΚΕΦΑΛΑΙΟ 8 ο ΚΕΦΑΛΑΙΟ 8 ο...2 I. Εφαρµογές της βιοτεχνολογίας στην ιατρική...2 ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ...7 ΝΑ ΣΥΜΠΛΗΡΩΣΕΤΕ ΤΑ ΚΕΝΑ ΜΕ ΤΗΝ ΚΑΤΑΛΛΗΛΗ ΛΕΞΗ...10 1 ΚΕΦΑΛΑΙΟ 8 ο I. Εφαρµογές της βιοτεχνολογίας
ΚΕΦΑΛΑΙΟ 8 ο ΚΕΦΑΛΑΙΟ 8 ο...2 I. Εφαρµογές της βιοτεχνολογίας στην ιατρική...2 ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ...7 ΝΑ ΣΥΜΠΛΗΡΩΣΕΤΕ ΤΑ ΚΕΝΑ ΜΕ ΤΗΝ ΚΑΤΑΛΛΗΛΗ ΛΕΞΗ...10 1 ΚΕΦΑΛΑΙΟ 8 ο I. Εφαρµογές της βιοτεχνολογίας
ANOΣΟΓΗΡΑΝΣΗ. Ιωάννα Οικονοµίδου. Αναπληρώτρια Καθηγήτρια Ιατρικής Πανεπιστηµίου Αθηνών
 ANOΣΟΓΗΡΑΝΣΗ Ιωάννα Οικονοµίδου Αναπληρώτρια Καθηγήτρια Ιατρικής Πανεπιστηµίου Αθηνών Το ανοσιακό σύστηµα θεωρείται αποφασιστικός παράγοντας για την διατήρηση της υγείας και την επιβίωση στους ηλικιωµένους
ANOΣΟΓΗΡΑΝΣΗ Ιωάννα Οικονοµίδου Αναπληρώτρια Καθηγήτρια Ιατρικής Πανεπιστηµίου Αθηνών Το ανοσιακό σύστηµα θεωρείται αποφασιστικός παράγοντας για την διατήρηση της υγείας και την επιβίωση στους ηλικιωµένους
ΑΝΟΣΟΛΟΓΙΑ ΘΕΩΡΙΑ 1 Ο ΜΑΘΗΜΑ
 ΑΝΟΣΟΛΟΓΙΑ ΘΕΩΡΙΑ 1 Ο ΜΑΘΗΜΑ ΑΝΟΣΙΑ Η ανοσία (α- στερητικό + νόσος) είναι η ικανότητα ενός οργανισμού να αμύνεται ενάντια σε κάποιον εξωτερικό βλαπτικό παράγοντα και να μην υφίσταται τις συνέπειές του.
ΑΝΟΣΟΛΟΓΙΑ ΘΕΩΡΙΑ 1 Ο ΜΑΘΗΜΑ ΑΝΟΣΙΑ Η ανοσία (α- στερητικό + νόσος) είναι η ικανότητα ενός οργανισμού να αμύνεται ενάντια σε κάποιον εξωτερικό βλαπτικό παράγοντα και να μην υφίσταται τις συνέπειές του.
Ανοσοποιητικό σύστημα
 Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY- 335 Χειμερινό εξάμηνο 2016 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα Η ανοσία είναι η κατάσταση προστασίας
Ανοσοποιητικό σύστημα κωδικός μαθήματος: ETY- 335 Χειμερινό εξάμηνο 2016 Μαρία Χατζηνικολαΐδου mchatzin@materials.uoc.gr Έμφυτο και προσαρμοστικό ανοσοποιητικό σύστημα Η ανοσία είναι η κατάσταση προστασίας
Ενημερώσου... γιατί. η ΗΠΑΤIΤΙΔΑ μπορεί να μη σου δώσει ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΜΕΛΕΤΗΣ ΗΠΑΤΟΣ. Αθήνα, Μάϊος 2008
 Ενημερώσου... γιατί η ΗΠΑΤIΤΙΔΑ μπορεί να μη σου δώσει μιαbευκαιρiα Αθήνα, Μάϊος 2008 ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΜΕΛΕΤΗΣ ΗΠΑΤΟΣ Ηπατίτιδα Β & Επιδημιολογία ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΜΕΛΕΤΗΣ ΗΠΑΤΟΣ ΔΙΟΙΚΗΤΙΚΟ ΣΥΜΒΟΥΛΙΟ
Ενημερώσου... γιατί η ΗΠΑΤIΤΙΔΑ μπορεί να μη σου δώσει μιαbευκαιρiα Αθήνα, Μάϊος 2008 ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΜΕΛΕΤΗΣ ΗΠΑΤΟΣ Ηπατίτιδα Β & Επιδημιολογία ΕΛΛΗΝΙΚΗ ΕΤΑΙΡΕΙΑ ΜΕΛΕΤΗΣ ΗΠΑΤΟΣ ΔΙΟΙΚΗΤΙΚΟ ΣΥΜΒΟΥΛΙΟ
4. Η κίρρωση του ήπατος προκαλείται εξαιτίας της αποθήκευσης στα ηπατικά κύτταρα: Πρωτεϊνών Υδατανθράκων Λιπών Αλκοόλ
 ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΠ Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΗΜΕΡΟΜΗΝΙΑ: 22/10/2017 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ: ΛΑΖΑΡΑΚΗ ΝΟΤΑ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Το
ΜΑΘΗΜΑ / ΤΑΞΗ : ΒΙΟΛΟΓΙΑ ΓΠ Γ ΛΥΚΕΙΟΥ (ΘΕΡΙΝΑ) ΗΜΕΡΟΜΗΝΙΑ: 22/10/2017 ΕΠΙΜΕΛΕΙΑ ΔΙΑΓΩΝΙΣΜΑΤΟΣ: ΛΑΖΑΡΑΚΗ ΝΟΤΑ ΘΕΜΑ Α Να επιλέξετε τη φράση που συμπληρώνει ορθά κάθε μία από τις ακόλουθες προτάσεις: 1. Το
ΘΕΡΑΠΕΙΑ ΤΗΣ ΗΠΑΤΙΤΙΔΑΣ C ΣΤΟΝ ΟΚΑΝΑ ΕΛΕΥΘΕΡΙΑ ΠΕΤΡΟΥΛΑΚΗ ΠΑΘΟΛΟΓΟΣ ΟΚΑΝΑ
 ΘΕΡΑΠΕΙΑ ΤΗΣ ΗΠΑΤΙΤΙΔΑΣ C ΣΤΟΝ ΟΚΑΝΑ ΕΛΕΥΘΕΡΙΑ ΠΕΤΡΟΥΛΑΚΗ ΠΑΘΟΛΟΓΟΣ ΟΚΑΝΑ Η Ηπατίτιδα C αποτελεί μία ιογενή νόσο με υψηλό φορτίο νοσηρότητας και θνητότητας Μέχρι πρόσφατα, η πλήρης ίαση των ασθενών αποτελούσε
ΘΕΡΑΠΕΙΑ ΤΗΣ ΗΠΑΤΙΤΙΔΑΣ C ΣΤΟΝ ΟΚΑΝΑ ΕΛΕΥΘΕΡΙΑ ΠΕΤΡΟΥΛΑΚΗ ΠΑΘΟΛΟΓΟΣ ΟΚΑΝΑ Η Ηπατίτιδα C αποτελεί μία ιογενή νόσο με υψηλό φορτίο νοσηρότητας και θνητότητας Μέχρι πρόσφατα, η πλήρης ίαση των ασθενών αποτελούσε
2. Τα πρωτόζωα α. δεν έχουν πυρήνα. β. είναι μονοκύτταροι ευκαρυωτικοί οργανισμοί. γ. είναι πολυκύτταρα παράσιτα. δ. είναι αυτότροφοι οργανισμοί.
 1 ΘΕΜΑΤΑ κεφ 1. Α. Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή στη φράση η οποία συμπληρώνει σωστά την ημιτελή πρόταση.
1 ΘΕΜΑΤΑ κεφ 1. Α. Να γράψετε στο τετράδιό σας τον αριθμό καθεμιάς από τις παρακάτω ημιτελείς προτάσεις και δίπλα το γράμμα που αντιστοιχεί στη λέξη ή στη φράση η οποία συμπληρώνει σωστά την ημιτελή πρόταση.
Ιογενείς Ηπατίτιδες Ηπατίτιδα Β. Συχνές Ερωτήσεις
 Ιογενείς Ηπατίτιδες Ηπατίτιδα Β Συχνές Ερωτήσεις Tι είναι ηπατίτιδα; Το ήπαρ (συκώτι) είναι ένα ζωτικό όργανο που βρίσκεται στο δεξιό άνω τμήμα της κοιλιακής χώρας. Έχει πολλές λειτουργίες και διαδραματίζει
Ιογενείς Ηπατίτιδες Ηπατίτιδα Β Συχνές Ερωτήσεις Tι είναι ηπατίτιδα; Το ήπαρ (συκώτι) είναι ένα ζωτικό όργανο που βρίσκεται στο δεξιό άνω τμήμα της κοιλιακής χώρας. Έχει πολλές λειτουργίες και διαδραματίζει
- Θεωρία- Δρ. ΠέτρουΚαρκαλούσου
 - Θεωρία- Έκδοση2008 Πρόλογος Αγαπητοίσπουδαστέςοισημειώσειςπουκρατάτεσταχέριασαςέχουνσκοπόνασας εισαγάγουνστιςβασικέςγνώσειςμιαςαπότιςσημαντικότερεςβιοιατρικέςεπιστήμες, της ανοσολογίας. Η ανοσολογίαείναιμιασχετικάνέαεπιστήμηηοποίαεμφανίστηκεστοτέλοςτου
- Θεωρία- Έκδοση2008 Πρόλογος Αγαπητοίσπουδαστέςοισημειώσειςπουκρατάτεσταχέριασαςέχουνσκοπόνασας εισαγάγουνστιςβασικέςγνώσειςμιαςαπότιςσημαντικότερεςβιοιατρικέςεπιστήμες, της ανοσολογίας. Η ανοσολογίαείναιμιασχετικάνέαεπιστήμηηοποίαεμφανίστηκεστοτέλοςτου
ΚΕΦΑΛΑΙΟ 7 Χυμικές ανοσοαπαντήσεις Ενεργοποίηση των Β λεμφοκυττάρων και παραγωγή των αντισωμάτων
 ΚΕΦΑΛΑΙΟ 7 Χυμικές ανοσοαπαντήσεις Ενεργοποίηση των Β λεμφοκυττάρων και παραγωγή των αντισωμάτων Φάσεις και τύποι των χυμικών ανοσοαπαντήσεων Διέγερση των Β λεμφοκυττάρων από το αντιγόνο Μεταβίβαση σήματος
ΚΕΦΑΛΑΙΟ 7 Χυμικές ανοσοαπαντήσεις Ενεργοποίηση των Β λεμφοκυττάρων και παραγωγή των αντισωμάτων Φάσεις και τύποι των χυμικών ανοσοαπαντήσεων Διέγερση των Β λεμφοκυττάρων από το αντιγόνο Μεταβίβαση σήματος
ΠΕΙΡΑΜΑΤΙΚΟ ΣΧΟΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΜΑΚΕΔΟΝΙΑΣ ZAΡΦΤΖΙΑΝ ΜΑΡΙΛΕΝΑ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ
 ΠΕΙΡΑΜΑΤΙΚΟ ΣΧΟΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΜΑΚΕΔΟΝΙΑΣ ZAΡΦΤΖΙΑΝ ΜΑΡΙΛΕΝΑ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΠΡΟΒΛΗΜΑΤΑ ΑΣΚΗΣΕΙΣ KEΦΑΛΑΙΟ 1ο 1. Οι παρακάτω καμπύλες αναφέρονται σε συγκεντρώσεις αντισωμάτων σε τρια
ΠΕΙΡΑΜΑΤΙΚΟ ΣΧΟΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟΥ ΜΑΚΕΔΟΝΙΑΣ ZAΡΦΤΖΙΑΝ ΜΑΡΙΛΕΝΑ ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ Γ ΛΥΚΕΙΟΥ ΠΡΟΒΛΗΜΑΤΑ ΑΣΚΗΣΕΙΣ KEΦΑΛΑΙΟ 1ο 1. Οι παρακάτω καμπύλες αναφέρονται σε συγκεντρώσεις αντισωμάτων σε τρια
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY
 ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Γενικά χαρακτηριστικά Απεικόνιση των κυτταρικών πληθυσμών με βάση: το μέγεθος την κοκκίωση το φθορισμό ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Υδροδυναμική ροή Πρόσθιος
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Γενικά χαρακτηριστικά Απεικόνιση των κυτταρικών πληθυσμών με βάση: το μέγεθος την κοκκίωση το φθορισμό ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ Υδροδυναμική ροή Πρόσθιος
Επιλεγόµενο µάθηµα:λευχαιµίες. Γεωργόπουλος Χρήστος Καρκατσούλης Μάριος Μπρίκος Νικήτας
 Επιλεγόµενο µάθηµα:λευχαιµίες Γεωργόπουλος Χρήστος Καρκατσούλης Μάριος Μπρίκος Νικήτας Ανοσολογία και καρκίνος Γενετικές και κυτταρικές αλλαγές που συµβαίνουν στον καρκίνο προσφέρουν στο ανοσιακό σύστηµα
Επιλεγόµενο µάθηµα:λευχαιµίες Γεωργόπουλος Χρήστος Καρκατσούλης Μάριος Μπρίκος Νικήτας Ανοσολογία και καρκίνος Γενετικές και κυτταρικές αλλαγές που συµβαίνουν στον καρκίνο προσφέρουν στο ανοσιακό σύστηµα
Αρχές μοριακής παθολογίας. Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ
 Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
ΝΟΣΗΛΕΙΑ ΑΣΘΕΝΟΥΣ ΜΕ HIV ΛΟΙΜΩΞΗ: ΓΝΩΣΗ, ΠΡΟΣΤΑΣΙΑ ΚΑΙ ΕΝΗΜΕΡΩΣΗ ΕΝΑΝΤΙΑ ΣΤΟΝ ΤΡΟΜΟ ΚΑΙ ΤΟΝ ΚΟΙΝΩΝΙΚΟ ΑΠΟΚΛΕΙΣΜΟ
 ΝΟΣΗΛΕΙΑ ΑΣΘΕΝΟΥΣ ΜΕ HIV ΛΟΙΜΩΞΗ: ΓΝΩΣΗ, ΠΡΟΣΤΑΣΙΑ ΚΑΙ ΕΝΗΜΕΡΩΣΗ ΕΝΑΝΤΙΑ ΣΤΟΝ ΤΡΟΜΟ ΚΑΙ ΤΟΝ ΚΟΙΝΩΝΙΚΟ ΑΠΟΚΛΕΙΣΜΟ ΤΥΜΠΑ ΕΥΔΟΚΙΑ ΜΑΝΔΕΛΗ ΕΛΕΝΗ ΕΙΣΗΓΗΤΡΙΑ: Α. ΠΑΠΟΥΤΣΗ ΟΡΙΣΜΟΙ Ορισμός AIDS: Acquired Immune
ΝΟΣΗΛΕΙΑ ΑΣΘΕΝΟΥΣ ΜΕ HIV ΛΟΙΜΩΞΗ: ΓΝΩΣΗ, ΠΡΟΣΤΑΣΙΑ ΚΑΙ ΕΝΗΜΕΡΩΣΗ ΕΝΑΝΤΙΑ ΣΤΟΝ ΤΡΟΜΟ ΚΑΙ ΤΟΝ ΚΟΙΝΩΝΙΚΟ ΑΠΟΚΛΕΙΣΜΟ ΤΥΜΠΑ ΕΥΔΟΚΙΑ ΜΑΝΔΕΛΗ ΕΛΕΝΗ ΕΙΣΗΓΗΤΡΙΑ: Α. ΠΑΠΟΥΤΣΗ ΟΡΙΣΜΟΙ Ορισμός AIDS: Acquired Immune
1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ
 1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ 1 ο Α. Στις ερωτήσεις 1-5 να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Οι ομοιοστατικοί
1 ο ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ 1 ο Α. Στις ερωτήσεις 1-5 να γράψετε στο τετράδιό σας τον αριθμό της ερώτησης και δίπλα του το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Οι ομοιοστατικοί
ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙAΣ Γ ΛΥΚΕΙΟΥ (Ιανουάριος 2014)
") ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙAΣ Γ ΛΥΚΕΙΟΥ (Ιανουάριος 2014) ΘΕΜΑ 1 ο Στις ερωτήσεις 1-5, να επιλέξετε το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Το συμπλήρωμα και η προπερδίνη είναι πρωτεΐνες
ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙAΣ Γ ΛΥΚΕΙΟΥ (Ιανουάριος 2014) ΘΕΜΑ 1 ο Στις ερωτήσεις 1-5, να επιλέξετε το γράμμα που αντιστοιχεί στη σωστή απάντηση. 1. Το συμπλήρωμα και η προπερδίνη είναι πρωτεΐνες
Νικόλαος Σιαφάκας Λέκτορας Διαγνωστικής Ιολογίας Εργαστήριο Κλινικής Μικροβιολογίας ΠΓΝ «ΑΤΤΙΚΟΝ»
 Νικόλαος Σιαφάκας Λέκτορας Διαγνωστικής Ιολογίας Εργαστήριο Κλινικής Μικροβιολογίας ΠΓΝ «ΑΤΤΙΚΟΝ» DNA RNA: ΑΝΤΙΓΡΑΦΗ, ΜΕΤΑΓΡΑΦΗ, ΜΕΤΑΦΡΑΣΗ DNA RNA: Βασικά Χαρακτηριστικά Ρόλος Κεντικό Δόγμα της Βιολογίας:
Νικόλαος Σιαφάκας Λέκτορας Διαγνωστικής Ιολογίας Εργαστήριο Κλινικής Μικροβιολογίας ΠΓΝ «ΑΤΤΙΚΟΝ» DNA RNA: ΑΝΤΙΓΡΑΦΗ, ΜΕΤΑΓΡΑΦΗ, ΜΕΤΑΦΡΑΣΗ DNA RNA: Βασικά Χαρακτηριστικά Ρόλος Κεντικό Δόγμα της Βιολογίας:
Ποιες είναι οι προϋποθέσεις που πρέπει να τηρούνται για την αποφυγή µετάδοσης ασθενειών που οφείλονται σε παθογόνους µικροοργανισµούς;
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Ποιες είναι οι προϋποθέσεις που πρέπει να τηρούνται για την αποφυγή µετάδοσης ασθενειών που οφείλονται σε παθογόνους µικροοργανισµούς; ΘΕΜΑ Β ίνεται το παρακάτω
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙ ΕΙΑΣ ΕΚΦΩΝΗΣΕΙΣ ΘΕΜΑ Α Ποιες είναι οι προϋποθέσεις που πρέπει να τηρούνται για την αποφυγή µετάδοσης ασθενειών που οφείλονται σε παθογόνους µικροοργανισµούς; ΘΕΜΑ Β ίνεται το παρακάτω
ΘΕΜΑ Α 1 δ 2 β 3 γ 4 β 5 α ΘΕΜΑ Β
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 30 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α 1 δ 2 β 3 γ
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗΣ ΗΜΕΡΗΣΙΟΥ ΚΑΙ Δ ΤΑΞΗΣ ΕΣΠΕΡΙΝΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΚΑΙ ΕΠΑΛ (ΟΜΑΔΑ Β ) ΠΑΡΑΣΚΕΥΗ 30 ΜΑΪΟΥ 2014 - ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΑΠΑΝΤΗΣΕΙΣ ΘΕΜΑ Α 1 δ 2 β 3 γ
οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά
 οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά Ανοσολογικό σύστηµα Βασικό σύστηµα του οργανισµού Λειτουργικές µονάδες του ανοσολογικού συστήµατος Οργανωµένος λεµφικός
οµή Ανοσιακού Συστήµατος Ελένη Φωτιάδου-Παππά Τµήµα Ανοσολογίας Γ.Ν. Νίκαιας-Πειραιά Ανοσολογικό σύστηµα Βασικό σύστηµα του οργανισµού Λειτουργικές µονάδες του ανοσολογικού συστήµατος Οργανωµένος λεµφικός
Γιάννης Δρακόπουλος 1. ΚΕΦ.1.3 ΘΕΜΑΤΑ (ομάδα Δ)
") Γιάννης Δρακόπουλος 1 1. ΚΕΦ.1.3 ΘΕΜΑΤΑ (ομάδα Δ) 2. Το διάγραµµα α δείχνει ότι τα αντιγόνα είναι αρχικά πολλά και εν συνεχεία ελαττώνονται διαρκώς. Δεν µεσολαβεί στάδιο αύξησης τους, επομένως δεν παρατηρείται
Γιάννης Δρακόπουλος 1 1. ΚΕΦ.1.3 ΘΕΜΑΤΑ (ομάδα Δ) 2. Το διάγραµµα α δείχνει ότι τα αντιγόνα είναι αρχικά πολλά και εν συνεχεία ελαττώνονται διαρκώς. Δεν µεσολαβεί στάδιο αύξησης τους, επομένως δεν παρατηρείται
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ 1 ου ΚΕΦΑΛΑΙΟΥ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ
 ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ 1 ου ΚΕΦΑΛΑΙΟΥ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ 1 ο A. Ερωτήσεις πολλαπλής επιλογής 1. β 2. γ 3. β 4. γ 5. δ B. Ερωτήσεις σωστού - λάθους 1. Σωστό 2. Σωστό 3. Λάθος 4. Λάθος 5.
ΑΠΑΝΤΗΣΕΙΣ ΣΤΟ ΔΙΑΓΩΝΙΣΜΑ 1 ου ΚΕΦΑΛΑΙΟΥ ΒΙΟΛΟΓΙΑΣ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΘΕΜΑ 1 ο A. Ερωτήσεις πολλαπλής επιλογής 1. β 2. γ 3. β 4. γ 5. δ B. Ερωτήσεις σωστού - λάθους 1. Σωστό 2. Σωστό 3. Λάθος 4. Λάθος 5.
ΣΥΣΧΕΤΙΣΗ ΓΕΝΕΤΙΚΩΝ ΠΟΛΥΜΟΡΦΙΣΜΩΝ ΣΤΟ ΓΟΝΙ Ι0 MYD88 ΜΕ ΤΗΝ ΑΝΑΠΤΥΞΗ ΦΥΜΑΤΙΩΣΗΣ
 ΣΥΣΧΕΤΙΣΗ ΓΕΝΕΤΙΚΩΝ ΠΟΛΥΜΟΡΦΙΣΜΩΝ ΣΤΟ ΓΟΝΙ Ι MYD88 ΜΕ ΤΗΝ ΑΝΑΠΤΥΞΗ ΦΥΜΑΤΙΩΣΗΣ Φωτεινή Μάλλη 1, Ειρήνη Γερογιάννη 1 Καλλιόπη Αγγέλου 1, Βίκυ Μολλάκη 2, Φωτεινή Μπαρδάκα 1, Κωνσταντίνος Ι. Γουργουλιάνης
ΣΥΣΧΕΤΙΣΗ ΓΕΝΕΤΙΚΩΝ ΠΟΛΥΜΟΡΦΙΣΜΩΝ ΣΤΟ ΓΟΝΙ Ι MYD88 ΜΕ ΤΗΝ ΑΝΑΠΤΥΞΗ ΦΥΜΑΤΙΩΣΗΣ Φωτεινή Μάλλη 1, Ειρήνη Γερογιάννη 1 Καλλιόπη Αγγέλου 1, Βίκυ Μολλάκη 2, Φωτεινή Μπαρδάκα 1, Κωνσταντίνος Ι. Γουργουλιάνης
ΙΪΚΟΙ ΦΟΡΕΙΣ & Γενετικά Τροποποιημένα Κύτταρα
 ΙΪΚΟΙ ΦΟΡΕΙΣ & Γενετικά Τροποποιημένα Κύτταρα ΓΕΝΙΚΑ ΠΕΡΙ ΙΩΝ 1. Οι ιοί είναι μικροοργανισμοί που προκαλούν ασθένειες σε ένα οργανισμό, μετά από μόλυνση των κυττάρων του 2. Είναι παρασιτικοί οργανισμοί
ΙΪΚΟΙ ΦΟΡΕΙΣ & Γενετικά Τροποποιημένα Κύτταρα ΓΕΝΙΚΑ ΠΕΡΙ ΙΩΝ 1. Οι ιοί είναι μικροοργανισμοί που προκαλούν ασθένειες σε ένα οργανισμό, μετά από μόλυνση των κυττάρων του 2. Είναι παρασιτικοί οργανισμοί
KΕΦΑΛΑΙΟ 1 ο : Άνθρωπος και Υγεία
 KΕΦΑΛΑΟ 1 ο : Άνθρωπος και Υγεία Α. ΕΡΩΤΗΣΕΣ ΚΛΕΣΤΟΥ ΤΥΠΟΥ Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη σωστή απάντηση ή στη φράση που συμπληρώνει σωστά την πρόταση: 1. Οι ιοί αποτελούνται από: α.
KΕΦΑΛΑΟ 1 ο : Άνθρωπος και Υγεία Α. ΕΡΩΤΗΣΕΣ ΚΛΕΣΤΟΥ ΤΥΠΟΥ Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη σωστή απάντηση ή στη φράση που συμπληρώνει σωστά την πρόταση: 1. Οι ιοί αποτελούνται από: α.
ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ. Εξειδίκευση: προϊόντα (κύτταρα ή αντισώματα) ειδικά για το αντιγόνο. Μνήμη: κύτταρα
 ειδικά για το αντιγόνο. Μνήμη: κύτταρα") ΕΙΔΙΚΗ ΑΜΥΝΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ Εξειδίκευση: προϊόντα (κύτταρα ή αντισώματα) ειδικά για το αντιγόνο Μνήμη: κύτταρα ΑΝΤΙΓΟΝΟ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΣ ΤΜΗΜΑ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΤΟΞΙΚΕΣ ΟΥΣΙΕΣ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΓΥΡΗ, ΦΑΡΜΑΚΕΥΤΙΚΕΣ
ΕΙΔΙΚΗ ΑΜΥΝΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ Εξειδίκευση: προϊόντα (κύτταρα ή αντισώματα) ειδικά για το αντιγόνο Μνήμη: κύτταρα ΑΝΤΙΓΟΝΟ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΣ ΤΜΗΜΑ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΤΟΞΙΚΕΣ ΟΥΣΙΕΣ ΜΙΚΡΟΟΡΓΑΝΙΣΜΟΥ ΓΥΡΗ, ΦΑΡΜΑΚΕΥΤΙΚΕΣ
1 Νεφρολογική κλινική Α.Π.Θ., Ιπποκράτειο Νοσοκομείο Θεσσαλονίκης. 2 Τμήμα Ανοσολογίας και Ιστοσυμβατότητας, Ιπποκράτειο Νοσοκομείο Θεσσαλονίκης
 ΔΙΑΤΑΡΑΧΕΣ ΥΠΟΠΛΗΘΥΣΜΩΝ Τ-ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΣΕ ΑΣΘΕΝΕΙΣ ΜΕ ΧΡΟΝΙΑ ΝΕΦΡΙΚΗ ΝΟΣΟ ΤΕΛΙΚΟΥ ΣΤΑΔΙΟΥ ΚΑΙ ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΩΝΕΦΡΙΚΗΣ ΚΑΘΑΡΣΗΣ ΠΡΟΔΡΟΜΑ ΑΠΟΤΕΛΕΣΜΑΤΑ ΠΡΟΟΠΤΙΚΗΣ ΜΕΛΕΤΗΣ Ε. Σαμπάνη 1, Μ. Στάγκου 1, Δ-Β.
ΔΙΑΤΑΡΑΧΕΣ ΥΠΟΠΛΗΘΥΣΜΩΝ Τ-ΛΕΜΦΟΚΥΤΤΑΡΩΝ ΣΕ ΑΣΘΕΝΕΙΣ ΜΕ ΧΡΟΝΙΑ ΝΕΦΡΙΚΗ ΝΟΣΟ ΤΕΛΙΚΟΥ ΣΤΑΔΙΟΥ ΚΑΙ ΕΠΙΔΡΑΣΗ ΤΗΣ ΕΞΩΝΕΦΡΙΚΗΣ ΚΑΘΑΡΣΗΣ ΠΡΟΔΡΟΜΑ ΑΠΟΤΕΛΕΣΜΑΤΑ ΠΡΟΟΠΤΙΚΗΣ ΜΕΛΕΤΗΣ Ε. Σαμπάνη 1, Μ. Στάγκου 1, Δ-Β.
Αρχές μοριακής παθολογίας. Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ
 Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
Αρχές μοριακής παθολογίας Α. Αρμακόλας Αν. Καθηγητής Ιατρική Σχολή ΕΚΠΑ Μοριακή Παθολογία Ανερχόμενος κλάδος της Παθολογίας Επικεντρώνεται στην μελέτη και τη διάγνωση νοσημάτων Στον καθορισμό και την πιστοποίηση
ΑΝΟΣΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΡΑΣΗΣ ΕΜΒΟΛΙΩΝ. Μαρία Χατζηστυλιανού Σιδηροπούλου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Τμήματος Ιατρικής, ΣΕΥ, ΑΠΘ
 ΑΝΟΣΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΡΑΣΗΣ ΕΜΒΟΛΙΩΝ. Μαρία Χατζηστυλιανού Σιδηροπούλου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Τμήματος Ιατρικής, ΣΕΥ, ΑΠΘ ΑΝΟΣΟΠΟΙΗΣΗ ΕΝΕΡΓΗΤΙΚΗ ΠΑΘΗΤΙΚΗ ΦΥΣΙΚΗ (ΝΟΣΗΣΗ) ΤΕΧΝΗΤΗ
ΑΝΟΣΟΛΟΓΙΚΟΙ ΜΗΧΑΝΙΣΜΟΙ ΔΡΑΣΗΣ ΕΜΒΟΛΙΩΝ. Μαρία Χατζηστυλιανού Σιδηροπούλου Ομότιμη Καθηγήτρια Παιδιατρικής Ανοσολογίας Τμήματος Ιατρικής, ΣΕΥ, ΑΠΘ ΑΝΟΣΟΠΟΙΗΣΗ ΕΝΕΡΓΗΤΙΚΗ ΠΑΘΗΤΙΚΗ ΦΥΣΙΚΗ (ΝΟΣΗΣΗ) ΤΕΧΝΗΤΗ
ΠΑΡΑΡΤΗΜΑ ΙV ΕΠΙΣΤΗΜΟΝΙΚΑ ΠΟΡΙΣΜΑΤΑ
 ΠΑΡΑΡΤΗΜΑ ΙV ΕΠΙΣΤΗΜΟΝΙΚΑ ΠΟΡΙΣΜΑΤΑ Επιστημονικά πορίσματα Λόγω των αλληλεπικαλυπτόμενων τρόπων μετάδοσης, η συλλοίμωξη με τον ιό της ηπατίτιδας В (HBV) και τον ιό της ηπατίτιδας C (HCV) δεν είναι ασυνήθιστη.
ΠΑΡΑΡΤΗΜΑ ΙV ΕΠΙΣΤΗΜΟΝΙΚΑ ΠΟΡΙΣΜΑΤΑ Επιστημονικά πορίσματα Λόγω των αλληλεπικαλυπτόμενων τρόπων μετάδοσης, η συλλοίμωξη με τον ιό της ηπατίτιδας В (HBV) και τον ιό της ηπατίτιδας C (HCV) δεν είναι ασυνήθιστη.
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY
 ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY Κυτταρομετρία Ροής Η Κυτταρομετρία Ροής είναι μια τεχνική που εκμεταλλεύεται την μέτρηση ορισμένων βιοχημικών και βιοφυσικών παραμέτρων των κυττάρων μεμονωμένα, όταν αυτά
ΚΥΤΤΑΡΟΜΕΤΡΙΑ ΡΟΗΣ FLOW CYTOMETRY Κυτταρομετρία Ροής Η Κυτταρομετρία Ροής είναι μια τεχνική που εκμεταλλεύεται την μέτρηση ορισμένων βιοχημικών και βιοφυσικών παραμέτρων των κυττάρων μεμονωμένα, όταν αυτά
Ι. Βλαχογιαννάκος, Γ. Β. Παπαθεοδωρίδης, Γ.Ν. Νταλέκος, Α. Αλεξοπούλου, Χ. Τριάντος, Ε. Χολόγκιτας, Ι. Κοσκίνας
 ΚΑΤΕΥΘΥΝΤΗΡΙΕΣ ΟΔΗΓΙΕΣ ΤΗΣ ΕΛΛΗΝΙΚΗΣ ΕΤΑΙΡΕΙΑΣ ΜΕΛΕΤΗΣ ΗΠΑΤΟΣ (ΕΕΜΗ) ΓΙΑ ΤΗΝ ΠΡΟΛΗΨΗ ΚΑΙ ΘΕΡΑΠΕΙΑ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΣΗΣ ΤΟΥ ΙΟΥ ΤΗΣ ΗΠΑΤΙΤΙΔΑΣ Β ΚΑΤΑ ΤΗ ΧΟΡΗΓΗΣΗ ΑΝΟΣΟΚΑΤΑΣΤΑΛΤΙΚΗΣ ΑΓΩΓΗΣ Ι. Βλαχογιαννάκος,
ΚΑΤΕΥΘΥΝΤΗΡΙΕΣ ΟΔΗΓΙΕΣ ΤΗΣ ΕΛΛΗΝΙΚΗΣ ΕΤΑΙΡΕΙΑΣ ΜΕΛΕΤΗΣ ΗΠΑΤΟΣ (ΕΕΜΗ) ΓΙΑ ΤΗΝ ΠΡΟΛΗΨΗ ΚΑΙ ΘΕΡΑΠΕΙΑ ΤΗΣ ΕΝΕΡΓΟΠΟΙΗΣΗΣ ΤΟΥ ΙΟΥ ΤΗΣ ΗΠΑΤΙΤΙΔΑΣ Β ΚΑΤΑ ΤΗ ΧΟΡΗΓΗΣΗ ΑΝΟΣΟΚΑΤΑΣΤΑΛΤΙΚΗΣ ΑΓΩΓΗΣ Ι. Βλαχογιαννάκος,
ΚΕΦΑΛΑΙΟ 5 ΑΝΟΣΟΑΙΜΑΤΟΛΟΓΙΑ ΑΜΕΣΗ COOMBS
 ΚΕΦΑΛΑΙΟ 5 ΑΝΟΣΟΑΙΜΑΤΟΛΟΓΙΑ ΑΜΕΣΗ COOMBS ΑΜΕΣΗ COOMBS Θεμελιώδες γνώρισμα του κάθε οργανισμού είναι ότι αναγνωρίζει τα κύτταρα των άλλων οργανισμών ως ξένα Αντιδρά με σκοπό την καταστροφή ΑΝΟΣΟΑΙΜΑΤΟΛΟΓΙΑ
ΚΕΦΑΛΑΙΟ 5 ΑΝΟΣΟΑΙΜΑΤΟΛΟΓΙΑ ΑΜΕΣΗ COOMBS ΑΜΕΣΗ COOMBS Θεμελιώδες γνώρισμα του κάθε οργανισμού είναι ότι αναγνωρίζει τα κύτταρα των άλλων οργανισμών ως ξένα Αντιδρά με σκοπό την καταστροφή ΑΝΟΣΟΑΙΜΑΤΟΛΟΓΙΑ
ΑΝΟΣΟΚΑΤΑΣΤΑΛΤΙΚΑ ΣΧΗΜΑΤΑ ΣΤΗ ΜΕΤΑΜΟΣΧΕΥΣΗ ΝΕΦΡΟΥ ΔΗΜΗΤΡΙΟΣ Σ. ΓΟΥΜΕΝΟΣ
 ΑΝΟΣΟΚΑΤΑΣΤΑΛΤΙΚΑ ΣΧΗΜΑΤΑ ΣΤΗ ΜΕΤΑΜΟΣΧΕΥΣΗ ΝΕΦΡΟΥ ΔΗΜΗΤΡΙΟΣ Σ. ΓΟΥΜΕΝΟΣ Γενικά Η πρόληψη των επεισοδίων οξείας απόρριψης και η μακροχρόνια διατήρηση του νεφρικού μοσχεύματος αποτελούν τους στόχους της
ΑΝΟΣΟΚΑΤΑΣΤΑΛΤΙΚΑ ΣΧΗΜΑΤΑ ΣΤΗ ΜΕΤΑΜΟΣΧΕΥΣΗ ΝΕΦΡΟΥ ΔΗΜΗΤΡΙΟΣ Σ. ΓΟΥΜΕΝΟΣ Γενικά Η πρόληψη των επεισοδίων οξείας απόρριψης και η μακροχρόνια διατήρηση του νεφρικού μοσχεύματος αποτελούν τους στόχους της
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΦΑΛΑΙΟ 1: ΑΝΘΡΩΠΟΣ ΚΑΙ ΥΓΕΙΑ
 ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΦΑΛΑΙΟ 1: ΑΝΘΡΩΠΟΣ ΚΑΙ ΥΓΕΙΑ 1.3, 1.4 Μηχανισμοί άμυνας του ανθρώπινου οργανισμού, AIDS ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη σωστή απάντηση:
ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ ΚΕΦΑΛΑΙΟ 1: ΑΝΘΡΩΠΟΣ ΚΑΙ ΥΓΕΙΑ 1.3, 1.4 Μηχανισμοί άμυνας του ανθρώπινου οργανισμού, AIDS ΕΡΩΤΗΣΕΙΣ ΠΟΛΛΑΠΛΗΣ ΕΠΙΛΟΓΗΣ Να βάλετε σε κύκλο το γράμμα που αντιστοιχεί στη σωστή απάντηση:
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 19 ΙΟΥΝΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ
 ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 19 ΙΟΥΝΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ (ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ) ΘΕΜΑ Α Α1. α Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1. α) Λάθος
ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ Γ ΤΑΞΗ ΗΜΕΡΗΣΙΟΥ ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΤΡΙΤΗ 19 ΙΟΥΝΙΟΥ 2018 ΕΞΕΤΑΖΟΜΕΝΟ ΜΑΘΗΜΑ: ΒΙΟΛΟΓΙΑ ΓΕΝΙΚΗΣ ΠΑΙΔΕΙΑΣ (ΕΝΔΕΙΚΤΙΚΕΣ ΑΠΑΝΤΗΣΕΙΣ) ΘΕΜΑ Α Α1. α Α2. γ Α3. α Α4. γ Α5. δ ΘΕΜΑ Β Β1. α) Λάθος
¹Γαστρεντερολογική κλινική, Πανεπιστήμιο Ιωαννίνων, Σχολή Επιστημών Υγείας, Τμήμα Ιατρικής, Παθολογικός Τομέας
 Μητσέλος, Μπαλταγιάννης, Πολίτης, Ιωάννης Γεράσιμος Δημήτριος Αθανάσιος Καββαδίας, Νικόλαος Τζαμπούρας, Δημήτριος Μπαλωμένος, Αλέξανδρος Σκαμνέλος, Κωνσταντίνος Κατσάνος, Δημήτριος Χριστοδούλου ¹Γαστρεντερολογική
Μητσέλος, Μπαλταγιάννης, Πολίτης, Ιωάννης Γεράσιμος Δημήτριος Αθανάσιος Καββαδίας, Νικόλαος Τζαμπούρας, Δημήτριος Μπαλωμένος, Αλέξανδρος Σκαμνέλος, Κωνσταντίνος Κατσάνος, Δημήτριος Χριστοδούλου ¹Γαστρεντερολογική
προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι.
 σύγχρονο Φάσµα & Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ 210 50 51 557 210 50 56 296 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 210 50 20 990 210 50 27 990 25ης Μαρτίου 74 ΠΕΤΡΟΥΠΟΛΗ
σύγχρονο Φάσµα & Group προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο Γραβιάς 85 ΚΗΠΟΥΠΟΛΗ 210 50 51 557 210 50 56 296 25ης Μαρτίου 111 ΠΕΤΡΟΥΠΟΛΗ 210 50 20 990 210 50 27 990 25ης Μαρτίου 74 ΠΕΤΡΟΥΠΟΛΗ
ΘΕΜΑ Αντιμετώπιση παθογόνων μικροοργανισμών με εμβόλια και ορούς
 ΘΕΜΑ Αντιμετώπιση παθογόνων μικροοργανισμών με εμβόλια και ορούς Ονόματα: Γωγώ Μουρίζι Έρικα Μπαλίου Τμήμα: Γ 3 Τί ονομάζουμε εμβόλια; ΕΜΒΟΛΙΑ Εμβόλια ονομάζονται τα σκευάσματα εκείνα, που χορηγούμενα
ΘΕΜΑ Αντιμετώπιση παθογόνων μικροοργανισμών με εμβόλια και ορούς Ονόματα: Γωγώ Μουρίζι Έρικα Μπαλίου Τμήμα: Γ 3 Τί ονομάζουμε εμβόλια; ΕΜΒΟΛΙΑ Εμβόλια ονομάζονται τα σκευάσματα εκείνα, που χορηγούμενα
Ερωτήµατα προς απάντηση
 Κεφ. 9. Ανοσιακή ανοχή και αυτοανοσία: Η διάκριση εαυτού-ξένου και οι διαταραχές της Ερωτήµατα προς απάντηση Πως διατηρεί το ανοσοποιητικό σύστηµα τηµη απαντητικότητα σε εαυτά αντιγόνα (=ανοχή); Ποιοι
Κεφ. 9. Ανοσιακή ανοχή και αυτοανοσία: Η διάκριση εαυτού-ξένου και οι διαταραχές της Ερωτήµατα προς απάντηση Πως διατηρεί το ανοσοποιητικό σύστηµα τηµη απαντητικότητα σε εαυτά αντιγόνα (=ανοχή); Ποιοι
Φάσμα. προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ ΓΕΝ.ΠΑΙΔΕΙΑΣ
 σύγχρονο Φάσμα προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο 25ης Μαρτίου 111 - ΠΕΤΡΟΥΠΟΛΗ - 210 50 20 990-210 50 27 990 25ης Μαρτίου 74 - ΠΕΤΡΟΥΠΟΛΗ - 210 50 50 658-210 50 60 845 Γραβιάς 85 -
σύγχρονο Φάσμα προπαρασκευή για Α.Ε.Ι. & Τ.Ε.Ι. μαθητικό φροντιστήριο 25ης Μαρτίου 111 - ΠΕΤΡΟΥΠΟΛΗ - 210 50 20 990-210 50 27 990 25ης Μαρτίου 74 - ΠΕΤΡΟΥΠΟΛΗ - 210 50 50 658-210 50 60 845 Γραβιάς 85 -
ΒΙΟΤΕΧΝΟΛΟΓΙΑ ΣΤ ΕΞΑΜΗΝΟΥ Τμήμα Ιατρικών Εργαστηρίων Διδάσκων - Δρ. Ιωάννης Δρίκος
 ΒΙΟΤΕΧΝΟΛΟΓΙΑ ΣΤ ΕΞΑΜΗΝΟΥ Τμήμα Ιατρικών Εργαστηρίων Διδάσκων - Δρ. Ιωάννης Δρίκος Επιλέξετε τη σωστή απάντηση, θέτοντας σε κύκλο το κατάλληλο γράµµα. 1. Για την παραγωγή ανθρώπινης ινσουλίνης από βακτήρια
ΒΙΟΤΕΧΝΟΛΟΓΙΑ ΣΤ ΕΞΑΜΗΝΟΥ Τμήμα Ιατρικών Εργαστηρίων Διδάσκων - Δρ. Ιωάννης Δρίκος Επιλέξετε τη σωστή απάντηση, θέτοντας σε κύκλο το κατάλληλο γράµµα. 1. Για την παραγωγή ανθρώπινης ινσουλίνης από βακτήρια
3. Η ΠΡΟΣΛΗΨΗΤΟΥ ΑΝΤΙΓΟΝΟΥ ΚΑΙ Η ΠΑΡΟΥΣΙΑΣΗ ΤΟΥ ΣΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ
 3. Η ΠΡΟΣΛΗΨΗΤΟΥ ΑΝΤΙΓΟΝΟΥ ΚΑΙ Η ΠΑΡΟΥΣΙΑΣΗ ΤΟΥ ΣΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ ΤΙ ΒΛΕΠΟΥΝ ΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ Τα αντιγόνα που αναγνωρίζονται από τα Τ λεµφοκύτταρα Πρόσληψη των πρωτεϊνικών αντιγόνων από τα αντιγονοπαρουσιαστικά
3. Η ΠΡΟΣΛΗΨΗΤΟΥ ΑΝΤΙΓΟΝΟΥ ΚΑΙ Η ΠΑΡΟΥΣΙΑΣΗ ΤΟΥ ΣΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ ΤΙ ΒΛΕΠΟΥΝ ΤΑ ΛΕΜΦΟΚΥΤΤΑΡΑ Τα αντιγόνα που αναγνωρίζονται από τα Τ λεµφοκύτταρα Πρόσληψη των πρωτεϊνικών αντιγόνων από τα αντιγονοπαρουσιαστικά
ΠΑΡΟΥΣΙΑΣΗ ΠΤΥΧΙΑΚΗΣ ΕΡΓΑΣΙΑΣ
 ΠΑΡΟΥΣΙΑΣΗ ΠΤΥΧΙΑΚΗΣ ΕΡΓΑΣΙΑΣ ΘΕΜΑ: ΙΟΓΕΝΕΙΣ ΗΠΑΤΙΤΙ ΕΣ ΚΑΙ ΝΟΣΗΛΕΥΤΙΚΕΣ ΠΑΡΕΜΒΑΣΕΙΣ ΣΠΟΥ ΑΣΤΕΣ: ΠΑΝΟΥ ΧΡΗΣΤΟΣ (Α.Μ.: 7321) ΤΟΥΜΠΟΥΛΙ ΗΣ ΚΩΝΣΤΑΝΤΙΝΟΣ (Α.Μ.:7503) ΤΙ ΕΙΝΑΙ Η ΗΠΑΤΙΤΙ Α Ηπατίτιδα σηµαίνει
ΠΑΡΟΥΣΙΑΣΗ ΠΤΥΧΙΑΚΗΣ ΕΡΓΑΣΙΑΣ ΘΕΜΑ: ΙΟΓΕΝΕΙΣ ΗΠΑΤΙΤΙ ΕΣ ΚΑΙ ΝΟΣΗΛΕΥΤΙΚΕΣ ΠΑΡΕΜΒΑΣΕΙΣ ΣΠΟΥ ΑΣΤΕΣ: ΠΑΝΟΥ ΧΡΗΣΤΟΣ (Α.Μ.: 7321) ΤΟΥΜΠΟΥΛΙ ΗΣ ΚΩΝΣΤΑΝΤΙΝΟΣ (Α.Μ.:7503) ΤΙ ΕΙΝΑΙ Η ΗΠΑΤΙΤΙ Α Ηπατίτιδα σηµαίνει
ΣΥΓΧΡΟΝΕΣ ΑΠΟΨΕΙΣ ΣΤΗΝ ΑΛΚΟΟΛΙΚΗ ΗΠΑΤΙΚΗ ΝΟΣΟ
 ΣΥΓΧΡΟΝΕΣ ΑΠΟΨΕΙΣ ΣΤΗΝ ΑΛΚΟΟΛΙΚΗ ΗΠΑΤΙΚΗ ΝΟΣΟ Ερωτήσεις (μπορεί να υπάρχουν περισσότερες από μια σωστές απαντήσεις, οι σωστές απαντήσεις είναι με bold) 1) Από τη εξαρτάται ανάπτυξη της αλκοολικής νόσου
ΣΥΓΧΡΟΝΕΣ ΑΠΟΨΕΙΣ ΣΤΗΝ ΑΛΚΟΟΛΙΚΗ ΗΠΑΤΙΚΗ ΝΟΣΟ Ερωτήσεις (μπορεί να υπάρχουν περισσότερες από μια σωστές απαντήσεις, οι σωστές απαντήσεις είναι με bold) 1) Από τη εξαρτάται ανάπτυξη της αλκοολικής νόσου
ΒΙΟΛΟΓΙΑ ΤΗΣ IDO ΚΕΡΕΝΙΔΗ ΝΟΡΑ
 ΒΙΟΛΟΓΙΑ ΤΗΣ IDO ΚΕΡΕΝΙΔΗ ΝΟΡΑ Η ύπαρξη του όγκου κινητοποιεί ανοσιακή απάντηση που περιλαμβάνει όλους τους ανοσοδραστικούς μηχανισμούς. Aυτόματη υποστροφή του καρκίνου είναι εξαιρετικά σπάνια Oι ανοσιακές
ΒΙΟΛΟΓΙΑ ΤΗΣ IDO ΚΕΡΕΝΙΔΗ ΝΟΡΑ Η ύπαρξη του όγκου κινητοποιεί ανοσιακή απάντηση που περιλαμβάνει όλους τους ανοσοδραστικούς μηχανισμούς. Aυτόματη υποστροφή του καρκίνου είναι εξαιρετικά σπάνια Oι ανοσιακές