Geni in regulacija njihovega prepisa
|
|
|
- Ἀλκαῖος Βασιλόπουλος
- 7 χρόνια πριν
- Προβολές:
Transcript
1 Geni in regulacija njihovega prepisa regulacija encimske aktivnosti indukcija in represija pozitivna in negativna kontrola, atenuacija globalna kontrola dvokomponentni sistemi pogovor bakterij
2 Regulacija sinteze in aktivnosti proteinov V celici ločimo dve vrsti regulacije: - regulacijo aktivnosti že sintetiziranih proteinov - regulacijo sinteze proteinov konstitutivno izraženi geni inducibilni geni
3 Regulacija encimov na različnih nivojih produkt regulacija encimske aktivnosti protein C protein D translacija transkripcija gen A gen B gen C gen D
4 Regulacija encimske aktivnosti Obstaja več mehanizmov regulacije encimske aktivnosti: aktivacija s posttranslacijsko cepitvijo proteina kovalentna modifikacija proteina (npr. dodajanje acilne skupine, AMP, ADP, fosfata, metilne skupine) deaktivacija z razgradnjo proteina mehanizem povratne zanke (pozitivna in negativna)
5 Mehanizem povratne zanke substrat intermediat 1 intermediat 2 intermediat 3 Končni produkt metabolne poti inhibitorno deluje na prvi encim v metabolni poti. Na ta način je zelo učinkovito regulirana koncentracija metabolitov v celici (npr. aminokislin). intermediat n produkt

6 Povratna zanka - alosteričen učinek


7 Regulacija transkrpicije: represija in indukcija represija encima indukcija encima


8 Genska represija (negativna kontrola) Neaktiviran represor omogoča sintezo proteina. Prisotnost korepresorja prepreči, kar negativno vpliva, na sintezo proteina.
9 Genska represija (negativna kontrola) Prisotnost represorja prepreči, negativno vpliva, na sintezo proteina. Induktor deaktivira represor, sprosti negativno kontrolo in tako omogoči sintezo proteina.


10 Genska indukcija (pozitivna kontrola) Brez induktorja ni možna vezava aktivatorskega proteina na RNA polimerazo, kar slednji onemogoča vezavo na DNA. Aktivatorski protein za razliko od represorja pozitivno vpliva na sintezo proteina.

11 Genska indukcija (pozitivna kontrola) Mesto za vezavo aktivatorja je lahko v neposredni bližini mesta za vezavo RNA polimeraze. Lahko pa je oddaljeno nekaj 100 bp od mesta za vezavo RNA polimeraze.


12 Regulacija translacije - atenuacija Višek triptofana, transkripcija zaustavljena. Regija 2 in 3 se ne moreta pariti. Pomankanje triptofana, transkripcija poteka.

13 DNA regulatorni proteini Regulacijo prepisa DNA vršijo regulatorni proteini, ki imajo specifične interakcije z regulatorno regijo tarčnega gena. Običajno so regulatorni proteini dimeri, ki se vežejo na invertirane sekvence DNA.

14 DNA regulatorni proteini cinkovi-prsti levcinska zadrga Cink interagira z dvema cisteinoma in dvema histidinoma vsaka 7. amino kislina v delu proteina, ki skrbi za interakcije med regulatornimi proteini, je levcin
15 Regulacija več genov - katabolna represija Če sta v gojišču na voljo dva vira koncentracija laktoza glukoza rast na glukozi rast na laktozi ogljika, potem običajno zasledimo preferenčno rast na enem viru ogljika, kar imenujemo diauksična rast. Represijo genov, ki so odgovorni za izrabo drugega vira ogljika imenujemo čas katabolna represija.
16 Lac operon, prototip genske regulacije RNA polimeraza cap laci P i P O lacz lacy laca mrna mrna inhibitor Z Y A O P LacI Pi LacZ LacY LacA Cap mesto vezave represorja promotor RNA polimeraze gen za represor promotor za LacI gen za beta-galaktozidazo gen za laktozno permeazo gen za tioglikozid transacetilazo gen za sintezo CAP proteina (aktivator)
17 Lac operon, ko v mediju ni prisotne laktoze RNA polimeraza inhibitor laci P i P O lacz lacy laca mrna inhibitor
18 Lac operon, ko je v mediju prisotna laktoza RNA polimeraza laci P i P O lacz lacy laca mrna mrna inhibitor Z Y A laktoza
19 Lac operator v primeru, ko sta v mediju prisotna glukoza in laktoza cap camp Cap RNA polimeraza počasen prepis brez camp-cap laci P i P O lacz lacy laca mrna mrna Cap inhibitor Z Y A glukoza camp camp Ko je v mediju prisotna glukoza je vedno tudi izrabljena. Ostali sistemi za izrabo sladkorjev morajo biti inducirani. Ko je v mediju veliko glukoze, je konc. camp nizka. Če se konc. glukoze zmanjša se poveča konc. camp, kar omgoča vezavo camp na CAP protein, le ta pa stimulira vezavo RNA polimeraze na DNA in bistveno večji prepis genov za metabolizem laktoze.
20 Dvokomponentni regulacijski sistemi ligand senzor vezava liganda P His avtofosforilacija senzorja RNA polimeraza Asp P promotor operator gen Asp P odzivni regulator Asp P His prenos fosfata iz histidina senzorja na aspartat odzivnega regulatorja
21 Nekateri dvokomponentni regulacijski sistemi v mikrobnih celicah sistem okoljski signal senzorska kinaza odzivni regulator Arc O 2 ArcB ArcA Nar NO 3-, NO - 2 NarX, NarQ NarL Ntr NH + 4 NrII NrI Pho regulon P i PhoR PhoB porinski reg. ozmotski pritisk EnvZ OmpR Evg nizka temp. EvgS EvgA Tox ph, temp., ozmoza ToxS ToxR Bvg temp., nikotinskak. BvgS BvgA
22 Quorum zaznava gostote bakterij Quorum zaznava je vrsta globalne kontrole mikroorganizmov, kjer je okoljski signal gostota bakterijske kulture. Bakterija ima encim za sintezo feromona (npr. aciliranega homoserin laktona). Feromon difundira iz celice. Njegova koncentracija postane znatna šele pri zelo gosti kulturi. celica proizvaja feromon celice začnejo agregirati celični agregat
23 Kako se bakterije pogovarjajo? Veliko signala pomeni veliko celic v okolici, čas za spremembo obnašanja! produkcija ekstracelularnih signalnih molekul - feromonov detekcija povišane celične koncentracije preko membranskih in citoplazmatskih receptorjev sprememba transkripcijskega vzorca in sprememba obnašanja
24 Kemijska narava signalnih molekul - feromonov G - bakterije G + bakterije homoserin laktoni peptidi O H N H O O 3-oxo-C6-HSL iz Vibrio fischeri H-Tyr-Ile-Asn- NH O S O O NH Asp Phe Leu Me Me Ciklični peptid iz S. aureus Druge signalne molekule: γ-butirolaktoni, quinoloni, furanoni





25 biofilm bioluminescenca sporulacija ekstracelularni encimi Kateri procesi so regulirani s quorum zaznavo? plodno telesce virulenca antibiotiki bakteriocini transfer genetskega materiala
26 Globalna kontrola sinteze proteinov Organizirani sočasni kontroli več različnih genov v organizmu pravimo globalna ali integralna kontrola. sistem signal število reguliranih genov katabolna represija ciklični AMP >300 aerobna respiracija prisotnost O 2 >50 anaerobna respiracija odsotnost O 2 70 toplotni šok temperatura 36 asimilacija dušika pomankanje NH + 4 >12 oksidativni stres oksidanti >30 SOS odgovor poškodovana DNA >20
27 Globalana kontrola signal citoplazmatska membrana signalni intermediat primarni nivo regulacija genov ekspresija induciranih genov sekundarni nivo regulacije genov
28 Mikrobna genetika mutacije homologna genska rekombinacija: transformacija transdukcija konjugacija nehomologna genska rekombinacija primerjalana genomika
29 Mutacije in mutanti Mutacija je dedna sprememba, ki se prenaša iz generacije v generacijo. Mutant je organizem, ki nosi dedno spremembo. Gene pišemo z malo črko, npr. hisc, reca. Proteine, ki nastanejo iz gena pišemo z veliko črko, npr. HisC, RecA Fenotip mutante označujemo z oznako + ali - npr. His + (organizem lahko naredi histidin) ali pa His - (organizem ne more narediti histidina).
30 Vrste mutacij Tihe mutacije: spremenjena je tretja baza kodona (ohrani se isto aminokislina) Nesmiselne mutacije: formiranje stop kodona, nekompleten protein Zamenjava aminokisline: spremenjena prva baza kodona Pogojno letalne mutante: npr. rast pri 30 o C, odsotnost rasti pri 40 o C Sprememba bralnega okvirja: popolnoma spremenjene aminokisline Revartante: povratna mutacija, ki povzroči nastanek divjega seva
31 Vrste mutacij Supresorske mutacije: vrne fenotip divjega seva (sprememba je lahko v istem genu, vendar ne na istem mestu kot revartanta, lahko je v drugem genu, ki komplementira mutiran gen, kar vrne fenotip) Delecije: izguba večjega števila nukleotidov Insercije: vgradnja večjega števila nukleotidov Translokacije: premikanje večjega dela DNA med različnimi lokacijami na kromosomu Inverzije: zamenjana orientacija DNA segmenta
32 Spontana frekvenca mutacij DNA DNA RNA protein ~ /bp ~10-5 /bp ~ 10-4 /bp Biološka ura: 10-9 mutacij/bp/leto spontana frekvenca mutacij v populaciji celic je ~ 10-6 na generacijo spontana frekvenca transpozicij je ~ 10-4 na generacijo spontana frekvenca mutacij RNA virusov je ~ 10-3 na generacijo mutatorski geni povečajo stopnjo mutacij za ~ x
33 Inducirana naključna mutageneza sredstvo učinkovanje sprememba analogi baz spremenjeno parjenje A : T T analog : G G : C HNO 2 deaminacija A in C A : T G : C NH 2 OH reakcija s C G : C A : T etilmetansulfonat dodajanje CH 3 na G G : C A : T etidijev bromid interkalacija mikroinsercije, mikrodelecije mitomicin povezava DNA vijačnic delecije UV pirimidinski dimeri delecije X-žarki prosti radikali delecije

34 Točkasta mutageneza Omogočajo zamenjavo ene same aminokisline v proteinu tako, da na mutiranem mestu v genu dodamo oligonukleotidni začetnik z nepopolnim parjenjenjem nukleotidov. Uporabljamo v biotehnologiji za pridobivanje željenih mutiranih proteinov.

35 Kasetna mutageneza Željeni del DNA lahko nadomestimo z mutiranim. Z vgradnjo genske kasete v izbrani gen na plazmidu po homologni rekombinaciji s kromosomalnim genom onsposobimo gen, obenem pa lahko vnesemo željene gene in molekularne markerje.
36 Horizontalni prenos genov Za razliko od prenosa genov iz generacije v generacijo (vertikalni prenos genov) pri horizontalnem prenosu prehaja do izmenjave (rekombinacije) genov znotraj iste generacije. Ločimo: homologno gensko rekombinacijo nehomologno gensko rekombinacijo
37 Homologna genska rekombinacija prokariontov Pri prokariontih so homologni fragmenti DNA lahko transportirani med celicami in s homologno rekombinacijo integrirani v gostiteljevo DNA. Trije najpomembnejši procesi homologne rekombinacije so: transformacija (prenos proste DNA) transdukcija (prenos DNA z virusi) konjugacija (prenos plazmidne DNA med celicami)

38 Homologna genska rekombinacija

39 Homologna genska rekombinacija

40 Rec A in SOS odgovor na DNA poškodbo
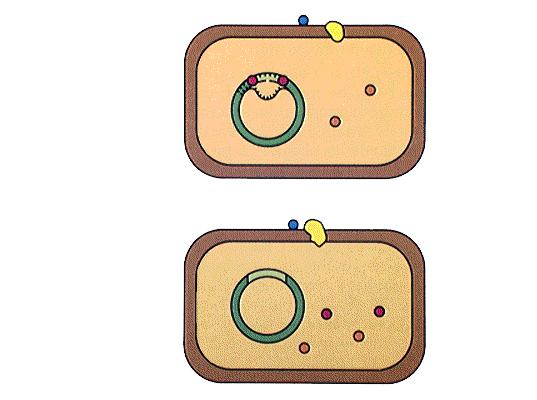
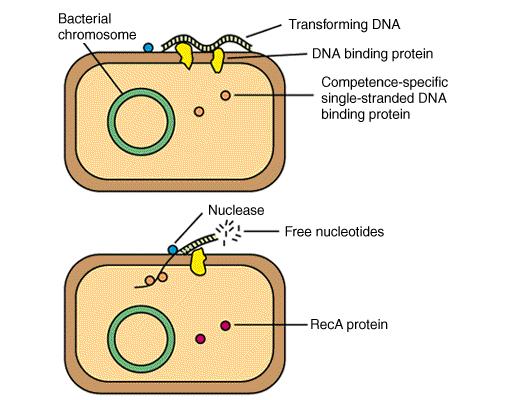
41 Genska transformacija
42 Transformacija - vnos tuje DNA NucA Nuclease? EA EC FA ATP ADP

43 Načini vnosa tuje DNA Frekvenco vnosa povečamo s Ca 2+ ioni in nizko temperaturo. DNA lahko vnesemo z elektroporacijo. Pri evkariontih DNA transformiramo z virusi, endocitozo, elektroporacijo in balističnimi izstrelki obloženimi z DNA.

44 Litični in lizogeni cikel bakteriofagov

45 Indukcija temperiranega faga
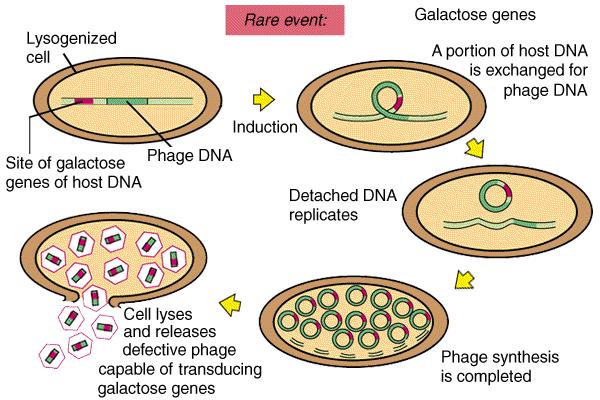
46 Specializirana transdukcija
47 Fagna konverzija bakterija fag protein ali fenotip Escherichia coli φfc3208, λ enterohemolizin, rezistenca na serum Shigella flexneri sf6, sfii O-antigen acetilaza, glukozil-transferaza Salmonella enterica Gifsy-2, superoksid dismutaza, neuraminadaza Vibrio cholerae CTXφ, VPIφ kolera toksin, TCP pilini Pseudomonas aeruginosa φctx citotoksin Clostridium botulinum C1 neurotoksin Staphylococcus aureus NA, TSST-1 enterotoksin, šok sindrom C. diphtheriae β-fag diftereja toksin
48 Plazmidi plazmid je izven kromosomska DNA podvojuje se neodvisno od kromosomske DNA. nima proteinskega plašča plazmidi imajo dvojnovijačno DNA, večinoma krožno vsebujejo od 1 do 100 kb, so supernaviti encime za replikacijo plazmid večinsko dobi od gostitelja. Število kopij varira od 1 do 100 na celico. plazmidi, ki so transportirani v celico morajo biti kompatibilni z že obstoječimi plazmidi v celici, sicer so izločeni. plazmide, ki se lahko vgradijo v kromosom, imenujemo episome
49 Vrste plazmidov rezistenčni (R) plazmidi: kodirajo zapis za odpornost na antibiotike, težke kovine, bakteriocine plazmidi s fiziološkimi funkcijami: npr. razgradnja oktanov, herbicidov, nastanek acetona, butanola, nodulacija, izraba laktoze in saharoze, produkcija pigmentov virulenčni plazmidi: imajo zapis za invazivnost: npr. koagulaze, hemolizin, eneterotoksin, K-antigen, tvorba tumorjev
 - tetracikline (tet) -streptomicin(str) - kloramfenikol")
50 R 100 (rezistenčni plazmid) Kodira zapis za rezistenco na: - živo srebro (mer) - sulfonamide (sul) - tetracikline (tet) -streptomicin(str) - kloramfenikol (clm)
51 F-plazmid gen tra orit oris inc rep funkcija transfer začetek za transfer začetek replikacije inkompatibilnost replikacija IS3, IS2, Tn/1000 (transpozoni) potencialna mesta vgradnje v gostiteljski kromosom

52 Konjugacija Informacija za konjugacijo je kodirana na plazmidu. Za konjugacijo so potrebni pili, ki skrbijo za vezavo donorske in recipientske celice, in jih imajo samo donorski sevi. Ko sta celici stabilizirani pride do fuzije obeh membran in do prehoda DNA.

53 Prenos DNA pri konjugaciji Za prenos DNA je potrebna sinteza DNA tako v donorju kot v akceptorju. Po končanem procesu ima vsaka celica eno kopijo plazmida. Akceptorska celica po ekspresiji plazmidnih genov lahko postane donorska.
54 Mobilizacija kromosoma Po integraciji F plazmida v kromosom postane tak sev Hfr ali sev z visoko sposobnostjo rekombinacije. Pri konjugaciji Hfr seva, poleg plazmida, potuje v recipientsko celico tudi del kromosoma, v katerega je vgrajen F plazmid.
55 Plazmidi in virulenčni faktorji Veliko virulenčnih faktorjev (npr. toksinov, bakteriocinov), ki omogočajo bakteriji vezavo in kolonizacijo gostitelja je kodiranih na plazmidih. Virulenčne faktorje poleg plazmidov najdemo tudi na mobilnih genskih elementih (npr. transpozoni in bakteriofagi) ter na kromosomih.
56 Transpozicija - nehomologna rekombinacija Transpozicija omogoča premikanje genov po kromosomu. Pojavlja se s frekvenco 10-5 do 10-7 na generacijo. Po kromosomu se lahko premikajo le tisti geni, ki imajo transpozabilne elemente. Ločimo: konzervativno transpozicijo (transpozon se izreže in vstavi na novo mesto) replikativno transpozicijo (transpozon se podvoji, parentalni ostane na istem mestu, hčerinski se vgradi na novo lokacijo na kromosomu).

57 Konzervativna transpozicija
58 Replikativna transpozicija
59 Mutageneza s transpozoni Če se transpozon naključno vgradi v gen ga deaktivira. Za izolacijo mutant uporabljamo transpozonske gene, ki kodirajo zapis za rezistenco na antibiotike (vgradnja pomeni rezistenco na antibiotik). Najbolj običajno uporabljamo Tn10, ki ima marker za tetraciklinsko rezistenco ali Tn5 z rezistenco na kanamicin in neomicin. gen A gen B gen C gen D IS IS transpozon
60 Integroni Nekateri transpozoni vsebujejo poleg genov za rezistenco na antibiotike še druge gene (genske kasete). Na genskih kasetah je lahko veliko genov za rezistenco na antibiotike. Najbolj poznan je Tn7 integron. inti1 P atti aadb sull inti1 P atti sull aadb integraza promotor mesto vgradnje rezist. na sulfonamide genska kaseta/rezistenca na aminoglikozilirane antibiotike
61 Kaj vemo o genomu E.coli kromosom ima bp kromosom ima bralnih okvirjev (88 % genoma), za okrog 35 % genskih produktov še vedno ne poznamo fiziološke funkcije 1 % genoma predstavljajo geni za trna 0.5 % genoma so visoko ponavljajoče se sekvence, ki ne kodirajo proteinov 10 % genoma predstavljajo regulatorne sekvence (promotorji, operatorji, začetek podvojevanja, konci podvojevanja DNA) geni so bodisi organizirani v klastre-operone (npr. hisg, hisd, hisc, hisb, hish, hisa, hisf, hisi, hise), ali pa razmetani po kromosomu (npr. arg geni) okrog 70 % genov je monocistronskih, 6 % je policistronskih
62 Kaj vemo o genomu E.coli na obeh vijačnicah je približno enako število operonov na kromosomu je 10 insercijskih elementov IS na kromosomu je do 30 % fagnih sekvenc na genomu obstaja več patogenih otokov približno 20 % celotnega genoma je E.coli pridobila s horizontalnim genskim prenosom

63 Primerjava genov pri različnih mikroorganizmih
64 Transportni sistemi Mycoplasma pneumoniae

65 Transportni sistemi Haemophilus influenzae
66 Genomika Genomika je disciplina, ki se ukvarja z mapiranjem, sekvenciranjem in analizo genoma. S primerjavo znanih sekvenc lahko določimo potrebne in zadostne pogoje za opravljanje določenega procesa. Določimo oziroma sklepamo lahko na: konzervativne in variabilne regije v genomu urejenost in grupiranje genov metabolne poti regulatorne mehanizme ortologne proteinskih družin (obstja okrog 100 ortolognih proteinskih družin, med katerimi je možen horizontalen genski prenos)
67
68 Postopki pri delu z DNA čipom populacijo celic običajno podvržemo okoljskemu faktorju in počakamo da sistem pride v stacionarno stanje ekstrahiramo mrna in jo reverzno prepišemo (mrna je hitro razgrajena, zato je potreben reverzen prepis, žal ta ni kvantitativen) fluorescentno označimo cdna (ker je vezava fluorofora odvisna od dolžine in sestave DNA ni možno kvantitativno določanje koncentracije) hibridizacija cdna na DNA čip (DNA čip je lahko narejen poljubno in vsebuje ORF ali poznane gene) posnamemo vzorec (vzbujamo z laserjem, detektiramo s CCD kamero) interpretacija dobljenega vzorca (težavna, ker hibridizacija ni kvantitativna)

69 Uporaba DNA čipov proučevanje fiziologije organizmov proučevanje ekologije organizmov odkrivanje genov diagnoza bolezni odkrivanje novih zdravil toksikološke raziskave
70 Molekularno kloniranje molekularna orodja vektorji izolacija ustreznega klona uporaba kloniranja
71 Genski inženiring Genski inženiring so postopki izolacije, manipulacije in ekspresije genetskega materiala s katerim proučujemo mehanizme genske replikacije, ekspresije in jih uporabljamo za produkcijo različnih koristnih produktov. Aplikativni uporabi genskega inženiringa pravimo tudi biotehnologija.
72 Molekularno kloniranje 1. Izbira ustreznega DNA fragmenta. DNA je lahko genomska, fragmentirana z restrikcijskimi encimi, DNA sintetizirana iz RNA, sintetiziranan s PCR ali sintetična DNA. 2. spajanje izbranega DNA fragmentov v vektor za kloniranje 3. prenos vektorja za kloniranje v izbrane gostitelje, pomnoževanje vektorja za kloniranje, ustvarjanje genske knjižnice 4. izolacija in čiščenje ustreznega klona 5. pomnoževanje izbranega klona
73 Iskanje ustreznega gena za kloniranje Gen, ki ga želimo klonirati je potrebno dobiti v relativno visokem številu (> 10 6 ), saj je vgradnja gena v vektor stohastični proces. Željeni gen za kloniranje lahko v ustrezni koncentraciji dobimo s: PCR pomnoževanjem izbranega gena iz poznane mrna sekvence z reverzno transkripcijo in PCR pomnoževanjem iz poznane proteinske sekvence s kemijsko sintezo DNA





74 Molekularna orodja - molekularne škarje Restrikcijski encimi so visokospecifične endonukleaze, ki režejo na izbranih mestih v DNA sekvenci. Za rezanje z restrikcijskimi encimi potrebujemo os simetrije okrog katere režejo restrikcijski encimi. Odrezani konci so lahko lepljivi ali pa topi. EcoRI PstI SmaI
75 Molekularna orodja - molekularno DNA lepilo DNA konce, ki jih dobimo z restrikcijskimi encimi je potrebno med seboj zlepiti ali ligirati. Zato uporabljamo encim DNA ligazo. DNA ligaza zlepi 5 fosfatni konec z 3 hidroksilnim koncem sosednjih nukleotidov.
76 Ustvarjanje rekombinantnih DNA molekul restrikcijsko mesto G A A T T C C T T A A G restrikcijsko mesto G A A T T C C T T A A G G A A T T C C T T A A lepljivi konec G G C T T A A A A T T C G A A T T C G donorska DNA G C T T A A G A A T T C C T T A A gostiteljeva DNA G donorska DNA G A A T T C C T T A A G gostiteljeva DNA
77 Ostala molekularna orodja fosfataze kinaze ssdna nukleaze DNA polimeraze RNA nukleaze RNA ligaze reverzne transkriptaze
78 Vektorji za molekularno kloniranje plazmidi bakteriofagi (M13, lambda) kozmidi (plazmidni vektorji s kohezivnimi konci iz bakteriofaga lambda) evkariontski virusi (adenovirusi, SV40, vakcinia, retrovirusi, bakulovirusi) sintetični kromosomi (HAC, YAC, BAC)
79 Specifični vektorji za molekularno kloniranje fuzijski vektorji (fuzija želejenega gena z drugim genom) ekspresijski vektorji (velika ekspresija kloniranega gena) sekrecijski vektorji (izboljšano izločanje proteinov) taksi ( shuttle ) vektorji (prenos DNA med nesorodnimi organizmi)
80 Plazmidi kot vektorji so majhni enostavna manipulacija DNA je krožna in zelo stabilna imajo neodvisno replikacijo od gostiteljevega krmosoma v celici so lahko v več kopijah prisotnost selekcijskih markerjev olajša detekcijo in selekcijo
81 Prototip plazmida za kloniranje - pbr322 je majhen, 4361 bp običajno kopij v gostitelju lahko dosežemo kopij enostavna izolacija zaradi supernavitja DNA inserti so lahko veliki do 10kb znana je njegova DNA sekvenca ima več mest za enkratno cepljenje s PstI, SalI, EcoRI, HindII, BamHI ima gen za rezistenco na ampicilin in tetraciklin, s transformacijo gre enostavno v gostitelja
82 Kloniranje s pbr322 Amp R Tc R BamHI restrikcija tuja DNA BamHI restrikcija DNA ligaze transformanta rezistentna na tetraciklin in ampicilin transformanta občutljiva na tetraciklin, rezistentna na ampicilin
83 Bakteriofag lambda kot vektor za kloniranje neesencialni del Za kloniranje ima divji sev lambde preveč restrikcijskih mest. V rekombinantnem fagu (npr. Charon 4A) donorska DNA pakiranje DNA je število restrikcijskih mest manjše. Lahko kloniramo večje fragmente kot pri plazmidih. Uspešnost transdukcije je večja kot uspešnost transformacije. infektivni fag
84 Bakteriofag M13 kot vektor za kloniranje EcoRI KpnI Smai BamHI Xbal SalI PstI...G A A T T C G A G C T C G G T A C C C G G G G A T C C T C T A G A G T C G A C C T G C A G... polilinkerska sekvenca lacz Bakteriofag M13 ima polilinkersko sekvenco (večje število enkratnih restrikcijskih mest. Polilinkerska sekvenca ne spremeni bralnega okvirja za prepis lacz gena. Če pride do vgradnje DNA segmenta se bralni okvir poruši in s tem sinteza LacZ.
85 Fuzijski vektor Spojimo, fuziramo dva gena, zaradi lažje izolacija ali detekcije izbranega genskega produkta. kontrolira Ptac promotor, ob dodatku glukoze pride do indukcije male uporaben za izolacijo fuzija inaktivira lacz mesto za začetek podvojevanja vektorja amp rezistenca na antibiotik
86 Sintetični kvasni kromosom (YAC) kot vektor za kloniranje TEL ARS CEN Not I Not I URA3 klonirana DNA TEL Kvasni kromosom, ki ga uporabljamo za kloniranje mora vsebovati: -mesto za začetek replikacije (ARS) - telomerazo (TEL) - centromero (CEN) - mesto za kloniranje (Not I) - selekcijski marker (npr. gen za uracil URA3) V YAC kromosom lahko kloniramo relativno velike segmente DNA od 250 do 1000 kb.
87 Velikost klonirane DNA pri različnih vektorjih vektor velikost (kb) plazmid < 15 bakteriofag < 90 sintetični bakterijski kromosom sintetični kvasni kromosom
88 Selekcija rekombiniranih celic Pri rekombinaciji tuje DNA dobimo tri vrste celic: celice, ki nimajo vgrajenega vektorja (npr. celice so občutljive za antibiotik) celice ki imajo vgrajen vektor, vendar nimajo želejene DNA (npr. celice rezistentne na antibiotik, brez nove fenotipske lastnosti) celice, ki imajo vgrajen vektor in željeno DNA (npr. celice rezistentne na antibiotik, imajo novo fenotipsko lastnost)
89 Selekcija ustreznega klona Željeno transformanto dobimo bodisi preko spremenjenega fenotipa ali spremenjenega genotipa: iskanje, ko je gen ekspresiran: uporabimo gostitelja, ki je mutiran za klonirani gen, kar omogoča rast samo transformantam. Alternativno uporabljamo protitelo za izbrani protein. iskanje, ko gen ni ekspresiran: uporabimo označene DNA ali RNA probe za željeni gen in naredimo hibridizacijo s klonirano DNA
90 Praktična uporaba genskega inženirstva - biotehnologija metabolni inženiring rekombinantne vakcine produkcija proteinov sesalcev transgene živali in rastline okoljska biotehnologija genska regulacija in genska terapija
91 Metabolni inženiring inkorporacija genov za večji razpon substratov na katerih organizem raste genetska sprememba metabolnih poti več produktov manj biomase
92 Proizvodnja rekombinantnih humanih proteinov protein inzulin α-1-antitripsin epidermijski rastni faktor humani rastni hormon eritropoetin faktor IX interlevkini a, b, g interlevkini urogastron funkcija uravnavanje krvnega sladkorja proteazni inhibitor rast epitelijskih celic rast celic stimulira rast krvnih celic strjevanje krvi protivirusna terapija stimulacija imunskega sistema kontrola gastrointestinalne sekrecije
93 Rekombinantne vakcine produkcija antigenov (npr. hepatitis B) uporaba spremenjenih virusov (npr. vakcinija virus za črne koze, steklino) uporaba gensko spremenjenih bakterij (npr. Salmonella typhi) produkcija antigenskih epitopov (npr. M13)
 kontrola patogenih gliv (Pseudomonas fluorescens) rezistenca na viruse utišanje genov gostitelja")
94 Uporaba rekombinantne tehnologije v kmetijstvu toleranca na herbicide rezistenca na škodljivce (Baciullus thurigiensis) kontrola patogenih gliv (Pseudomonas fluorescens) rezistenca na viruse utišanje genov gostitelja fiksacija dušika

95 Gensko spremenjena hrana
96 Nekatere od potencialnih koristi GMO povečana hranilna vrednost in obstojnost hrane povečana odpornost na škodljivce eliminacija alergenov produkcija farmacevtskih učinkovin bioremediacija toksinov in eksploziv produkcija biorazgradljive plastike zmanjševanje produkcije polutantov
97 Nekatere od potencialnih nevarnosti GMO neželjen prenos toksičnih genov med vrstami nepričakovani rezultati rekombinacije zmanjševanje biološke pestrosti navzkrižna kontaminacija med GMO in ne-gmo nezmožnost dolgoročne napovedi vpliva GMO na okolje
98 Sproščanje GMO v okolje Predno spustimo GMO v okolje moramo odgovoriti na naslednja vprašanja: ali lahko GMO preživi izven laboratorija ali se lahko razmnožuje z nemodificiranimi predstavniki iste vrste ali ima GMO selekcijsko prednost pred ostalimi organizmi v okolju ali lahko prihaja do horizontalnega prenosa genov ali se GMO lahko premika v okolju
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Kaj je DNA footprinting? Omogoča lociranje vezave proteina na DNA molekulo z ugotavljanjem katere fosfodiestrske vezi so zaščitene pred rezom Dnaze
 Ugotavljanje števila bakterij, ki tvorijo kolonije (cfu) Št kolonij zraslih na plošči faktor redčenja na plošči 10= x cfu/ml DNA fingerprinting: omogoča opazovanje genskih razlik med ljudmi na podlagi
Ugotavljanje števila bakterij, ki tvorijo kolonije (cfu) Št kolonij zraslih na plošči faktor redčenja na plošči 10= x cfu/ml DNA fingerprinting: omogoča opazovanje genskih razlik med ljudmi na podlagi
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
ODPORNOST BAKTERIJ PROTI ANTIBIOTIKOM V SLOVENIJI IN PO SVETU
 ODPORNOST BAKTERIJ PROTI ANTIBIOTIKOM V SLOVENIJI IN PO SVETU Manica Müller Premru, dr. med., spec. klin. mikrobiol. Inštitut za mikrobiologijo in imunologijo Medicinska fakulteta Univerza v Ljubljani
ODPORNOST BAKTERIJ PROTI ANTIBIOTIKOM V SLOVENIJI IN PO SVETU Manica Müller Premru, dr. med., spec. klin. mikrobiol. Inštitut za mikrobiologijo in imunologijo Medicinska fakulteta Univerza v Ljubljani
Sintezna genomika. Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi
 Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi") Sintezna genomika Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi Koncept sinteznega življenja Celic, kot so se razvile v evoluciji,
Sintezna genomika Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi Koncept sinteznega življenja Celic, kot so se razvile v evoluciji,
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
MATEMATIČNI IZRAZI V MAFIRA WIKIJU
 I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
MOLEKULARNA GENETIKA
 MOLEKULARNA GENETIKA 1 STRUKTURA DNA DNA ima več struktur: enojni, dvojni ali trojni heliks je levo- ali desnosučen, linearna ali krožna. Najpogostejša oblika je B desnosučni heliks, ki je iz dveh komplementarnih
MOLEKULARNA GENETIKA 1 STRUKTURA DNA DNA ima več struktur: enojni, dvojni ali trojni heliks je levo- ali desnosučen, linearna ali krožna. Najpogostejša oblika je B desnosučni heliks, ki je iz dveh komplementarnih
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Sinteza RNA - transkripcija
 Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
Sinteza RNA - transkripcija RNA polimeraza je encimski kompleks, ki katalizira sintezo RNA na osnovi DNA matrice sinteza RNA, ki jo usmerja DNA. Pomnoževanje poteka v 5 proti 3 smeri. Matrična veriga DNA
Genetska kontrola sinteze bjelančevina. Povijesni PaJaMo pokus Jacob-Monod model sinteze bjelančevina u prokariota
 Genetska kontrola sinteze bjelančevina Još prije otkrića genetskog koda bilo je jasno da moraju postojati načini kontroliranja broja i vrste bjelančevina koje se proizvode u stanici. Kod šumskog drveća
Genetska kontrola sinteze bjelančevina Još prije otkrića genetskog koda bilo je jasno da moraju postojati načini kontroliranja broja i vrste bjelančevina koje se proizvode u stanici. Kod šumskog drveća
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Transformator. Delovanje transformatorja I. Delovanje transformatorja II
 Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
TRANSFORMACIJA sprejem in vključitev proste DNA v genom recipienta, pri čemer celice niso v medsebojnem kontaktu Pogoji za uspešno transformacijo
 Genetika izpit: nekaj pomembnejših tem. TRANSFORMACIJA Transformacija je sprejem in vključitev proste DNA v genom recipienta, pri čemer celice niso v medsebojnem kontaktu. Prenaša se le gola DNA. Celice
Genetika izpit: nekaj pomembnejših tem. TRANSFORMACIJA Transformacija je sprejem in vključitev proste DNA v genom recipienta, pri čemer celice niso v medsebojnem kontaktu. Prenaša se le gola DNA. Celice
[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda
![[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda](/thumbs/89/99439684.jpg "[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda") kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
Molekularna diagnostika
 Molekularna diagnostika Predpogoj za uspešno zdravljenje je pravočasno detektiranje povzročiteljev - velja za ljudi, živali, rastline in okolje. Klasični postopki detekcije so vključevali odvzem okuženega
Molekularna diagnostika Predpogoj za uspešno zdravljenje je pravočasno detektiranje povzročiteljev - velja za ljudi, živali, rastline in okolje. Klasični postopki detekcije so vključevali odvzem okuženega
p 1 ENTROPIJSKI ZAKON
 ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
+105 C (plošče in trakovi +85 C) -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70
 -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70") KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
Gimnazija Krˇsko. vektorji - naloge
 Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
Neavtorizirani povzetki izbranih predavanj iz biokemije. UN študij Okolje Politehnika Nova Gorica. doc. dr. Marko Dolinar
 Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Komunikacije med mikroorganizmi v vodnih okoljih
 Letna konferenca Katedre za Biotehnologijo POMEN BIOTEHNOLOGIJE IN MIKROBIOLOGIJE ZA PRIHODNOST: VODA 18-19.1.2007 Komunikacije med mikroorganizmi v vodnih okoljih Število objav na tem področju eksponentno
Letna konferenca Katedre za Biotehnologijo POMEN BIOTEHNOLOGIJE IN MIKROBIOLOGIJE ZA PRIHODNOST: VODA 18-19.1.2007 Komunikacije med mikroorganizmi v vodnih okoljih Število objav na tem področju eksponentno
DISKRETNA FOURIERJEVA TRANSFORMACIJA
 29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
29.03.2004 Definicija DFT Outline DFT je linearna transformacija nekega vektorskega prostora dimenzije n nad obsegom K, ki ga označujemo z V K, pri čemer ima slednji lastnost, da vsebuje nek poseben element,
Regulacija ekspresije gena kod prokariota
 Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
Energetski metabolizem
 Energetski metabolizem Metabolne povezave med jetri in mišicami Corijev ciklus Glukoza-alaninski ciklus Možgani potrebe po glukozi: 120 g/dan (t.j. ca 60% ob počitku) za vzdrževanje membranskih potencialov
Energetski metabolizem Metabolne povezave med jetri in mišicami Corijev ciklus Glukoza-alaninski ciklus Možgani potrebe po glukozi: 120 g/dan (t.j. ca 60% ob počitku) za vzdrževanje membranskih potencialov
Frekvenčna analiza neperiodičnih signalov. Analiza signalov prof. France Mihelič
 Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
FARMAKOTERAPIJA BIOTEHNOLOŠKIH ZDRAVIL. Borut Štrukelj
 FARMAKOTERAPIJA BIOTEHNOLOŠKIH ZDRAVIL Borut Štrukelj Smeri razvoja biotehnologije v A) Stanje v razvitem svetu B) Perspektive OSNOVNE SMERI farmaciji 1) produkcija rekombinantnih zdravilnih učinkovin
FARMAKOTERAPIJA BIOTEHNOLOŠKIH ZDRAVIL Borut Štrukelj Smeri razvoja biotehnologije v A) Stanje v razvitem svetu B) Perspektive OSNOVNE SMERI farmaciji 1) produkcija rekombinantnih zdravilnih učinkovin
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
MOLEKULSKA GENETIKA. Osnovna zgradba polinukleotidne verige (ali kateregakoli lineranega polimera) 1
 1") nukleotidi... POLINUKLEOTIDNA VERIGA fosfodiestrske vezi Osnovna zgradba polinukleotidne verige (ali kateregakoli lineranega polimera) 1 RNK (ribonukleinska kislina) ENOJNA VIJAČNIA del molekule, ki jo
nukleotidi... POLINUKLEOTIDNA VERIGA fosfodiestrske vezi Osnovna zgradba polinukleotidne verige (ali kateregakoli lineranega polimera) 1 RNK (ribonukleinska kislina) ENOJNA VIJAČNIA del molekule, ki jo
Uravnavanje encimske aktivnosti
 Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Razmena genetičkog materijala kod prokariota
 Razmena genetičkog materijala kod prokariota Mehanizmi razmene gena kod prokariota Prenošenje dela genetičkog materijala iz ćelije donora u ćeliju recipijenta konjugacijom transformacijom transdukcijom
Razmena genetičkog materijala kod prokariota Mehanizmi razmene gena kod prokariota Prenošenje dela genetičkog materijala iz ćelije donora u ćeliju recipijenta konjugacijom transformacijom transdukcijom
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 15. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV
 SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV Literatura za temo Signalne poti, ki regulirajo izražanje genov: Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH
SIGNALNE POTI, KI REGULIRAJO IZRAŽANJE GENOV Literatura za temo Signalne poti, ki regulirajo izražanje genov: Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH
GENETIKA od dvojne vijaënice do kloniranja
 RADOVAN KOMEL GENETIKA od dvojne vijaënice do kloniranja PriroËnik za uëitelje GENETIKA od dvojne vijačnice do kloniranja Priročnik za učitelje Avtor: dr. Radovan Komel Recenzentka: Marjeta Dobravc, prof.biol.
RADOVAN KOMEL GENETIKA od dvojne vijaënice do kloniranja PriroËnik za uëitelje GENETIKA od dvojne vijačnice do kloniranja Priročnik za učitelje Avtor: dr. Radovan Komel Recenzentka: Marjeta Dobravc, prof.biol.
Do sedaj ste spoznali: Posledice mutacij. Mutacije, mutageneza, vpliv polimorfizmov/mutacij na izražanje genov
 Mutacije, mutageneza, vpliv polimorfizmov/mutacij na izražanje genov Celična biologija z genetiko LBM kozmetologija šol. leto 2012/13 Prof. dr. Irena Mlinarič-Raščan Do sedaj ste spoznali: Struktura DNA/RNA,
Mutacije, mutageneza, vpliv polimorfizmov/mutacij na izražanje genov Celična biologija z genetiko LBM kozmetologija šol. leto 2012/13 Prof. dr. Irena Mlinarič-Raščan Do sedaj ste spoznali: Struktura DNA/RNA,
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Podobnost matrik. Matematika II (FKKT Kemijsko inženirstvo) Diagonalizacija matrik
 Diagonalizacija matrik") Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Podobnost matrik Matematika II (FKKT Kemijsko inženirstvo) Matjaž Željko FKKT Kemijsko inženirstvo 14 teden (Zadnja sprememba: 23 maj 213) Matrika A R n n je podobna matriki B R n n, če obstaja obrnljiva
Funkcije več spremenljivk
 DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
DODATEK C Funkcije več spremenljivk C.1. Osnovni pojmi Funkcija n spremenljivk je predpis: f : D f R, (x 1, x 2,..., x n ) u = f (x 1, x 2,..., x n ) kjer D f R n imenujemo definicijsko območje funkcije
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
VEKTORJI. Operacije z vektorji
 VEKTORJI Vektorji so matematični objekti, s katerimi opisujemo določene fizikalne količine. V tisku jih označujemo s krepko natisnjenimi črkami (npr. a), pri pisanju pa s puščico ( a). Fizikalne količine,
VEKTORJI Vektorji so matematični objekti, s katerimi opisujemo določene fizikalne količine. V tisku jih označujemo s krepko natisnjenimi črkami (npr. a), pri pisanju pa s puščico ( a). Fizikalne količine,
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
Lastnosti molekule DNA
 Metode za odkrivanje/analizo mutacij Celična biologija z genetiko 1. letnik UŠ LBM, št. leto 2012/13 Detekcija polimorfizmov/mutacij Sprva so razlike v genomu med posamezniki ugotavljali le z opazovanjem
Metode za odkrivanje/analizo mutacij Celična biologija z genetiko 1. letnik UŠ LBM, št. leto 2012/13 Detekcija polimorfizmov/mutacij Sprva so razlike v genomu med posamezniki ugotavljali le z opazovanjem
PROTEINI POVEZANI S PRENOSOM SIGNALOV. Funkcije proteinov
 PROTEINI POVEZANI S PRENOSOM SIGNALOV Funkcije proteinov Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni proteini) Oporna funkcija (strukturni proteini,
PROTEINI POVEZANI S PRENOSOM SIGNALOV Funkcije proteinov Transport/skladiščenje določenih molekul (ligandov, npr. Hb, Mb) Uravnavanje procesov (DNA-vezavni proteini) Oporna funkcija (strukturni proteini,
Najpomembnejši človeški Hb
 Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
Najpomembnejši človeški Hb Vrsta Struktura podenot Razširjenost HbA (odrasli) α 2 β 2 97% Hb odraslih HbA 2 (odrasli) α 2 δ 2 2-3% Hb odraslih HbF (fetalni) *α 2 γ 2 Najpomembnejši Hb v 3. trimestru nosečnosti
KLONIRANJE. Lucija Smolko, Leja Antolin, Maja Jaklin
 KLONIRANJE Lucija Smolko, Leja Antolin, Maja Jaklin Kloniranje je postopek nespolnega razmnoževanja, pri katerem je novo nastala celica oz. osebek (klon) genetsko identičen izvorni celici oz. osebku. Poznamo
KLONIRANJE Lucija Smolko, Leja Antolin, Maja Jaklin Kloniranje je postopek nespolnega razmnoževanja, pri katerem je novo nastala celica oz. osebek (klon) genetsko identičen izvorni celici oz. osebku. Poznamo
Statistična analiza. doc. dr. Mitja Kos, mag. farm. Katedra za socialno farmacijo Univerza v Ljubljani- Fakulteta za farmacijo
 Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
Υγιεινή Τροφίμων. Παθογόνοι μικροοργανισμοί που σχετίζονται με τα τρόφιμα. Τροφοτοξινώσεις & Τροφολοιμώξεις
 Υγιεινή Τροφίμων Παθογόνοι μικροοργανισμοί που σχετίζονται με τα τρόφιμα Τροφοτοξινώσεις & Τροφολοιμώξεις Οι μικροοργανισμοί διακρίνονται σε παθογόνους για τον άνθρωπο και αλλοιογόνους για τα τρόφιμα Οι
Υγιεινή Τροφίμων Παθογόνοι μικροοργανισμοί που σχετίζονται με τα τρόφιμα Τροφοτοξινώσεις & Τροφολοιμώξεις Οι μικροοργανισμοί διακρίνονται σε παθογόνους για τον άνθρωπο και αλλοιογόνους για τα τρόφιμα Οι
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
Nukleinske kisline. Nukleotidi. DNA je nosilka dednih genetskih informacij.
 Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Nukleinske kisline. ribosomska informacijska prenašalna
 Nukleinske kisline Nukleinske kisline vloga pri shranjevanju, prenašanju in izražanju genetske informacije: DNA RNA proteini zgradba in delovanje celice 2 osnovni vrsti nukleinskih kislin: deoksiribonukleinska
Nukleinske kisline Nukleinske kisline vloga pri shranjevanju, prenašanju in izražanju genetske informacije: DNA RNA proteini zgradba in delovanje celice 2 osnovni vrsti nukleinskih kislin: deoksiribonukleinska
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
METABIBAΣH ΓONI IΩN ΣE BAKTHPIA BAKTHPIAKH ΣYZEYΞH
 Προκαρυωτικοί οργανισµοί = βακτήρια, κυανοβακτήρια. - µονοκύτταροι - κυκλικό χρωµόσωµα, που δεν περικλείεται µέσα σε πυρηνική µεµβράνη. - απλοειδείς - δεν σχηµατίζουν ζυγώτες - δεν εκτελούν µείωση. - προσβάλλονται
Προκαρυωτικοί οργανισµοί = βακτήρια, κυανοβακτήρια. - µονοκύτταροι - κυκλικό χρωµόσωµα, που δεν περικλείεται µέσα σε πυρηνική µεµβράνη. - απλοειδείς - δεν σχηµατίζουν ζυγώτες - δεν εκτελούν µείωση. - προσβάλλονται
Proteini. Struktura proteinov. Analiza proteinov. Proteini (proizvodnja, analiza, struktura, funkcija)
") Proteini (proizvodnja, analiza, struktura, funkcija) Celična biologija z genetiko 1. letnik UŠ LBM, Kozmetologija šol. leto 2012/13 Proteios (Gr.) = prvega reda Makromolekule polimeri aminokislin, ki so
Proteini (proizvodnja, analiza, struktura, funkcija) Celična biologija z genetiko 1. letnik UŠ LBM, Kozmetologija šol. leto 2012/13 Proteios (Gr.) = prvega reda Makromolekule polimeri aminokislin, ki so
2.1. MOLEKULARNA ABSORPCIJSKA SPEKTROMETRIJA
 2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
JEDRO (NUCLEUS PROKARYOTA(
 JEDRO (NUCLEUS) PROKARYOTA( procita) EUKARYOTA ( evcita) Nastanek: iz jedra; indirektna delitev: mitoza, mejoza-kariokineza Število: 1/ celico; 2/celico= dikariontsko stanje (Asco, - Basidiomycotina);
JEDRO (NUCLEUS) PROKARYOTA( procita) EUKARYOTA ( evcita) Nastanek: iz jedra; indirektna delitev: mitoza, mejoza-kariokineza Število: 1/ celico; 2/celico= dikariontsko stanje (Asco, - Basidiomycotina);
JEDRO (NUCLEUS PROKARYOTA(
 JEDRO (NUCLEUS) PROKARYOTA( procita) EUKARYOTA ( evcita) Nastanek: iz jedra; indirektna delitev: mitoza, mejoza-kariokineza Število: 1/ celico; 2/celico= dikariontsko stanje (Asco, - Basidiomycotina);
JEDRO (NUCLEUS) PROKARYOTA( procita) EUKARYOTA ( evcita) Nastanek: iz jedra; indirektna delitev: mitoza, mejoza-kariokineza Število: 1/ celico; 2/celico= dikariontsko stanje (Asco, - Basidiomycotina);
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks'
 Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Celični'stiki' Vrsta&povezave:'' celica.celica' celica.matriks' Povezava&in&adhezija&omogoča:'' zvezo'med'celico'in'skupnostjo'' kompartmentizacijo' prepoznavanje'in'signalizacijo' proliferacija,'diferenciacija,'migracija'
Šolsko leto 2007/08. Gregor Lakner
 Šolsko leto 2007/08 Avtorja: Veronika Vodeb Gregor Lakner Mentorica: Bojana Pevec Šmarje pri Jelšah, maj 2008 Kazalo 1. HIPOTEZE... 4 2. Mejniki v moderni biotehnologiji... 5 3. UKRIVLJENOST PROSTORA...
Šolsko leto 2007/08 Avtorja: Veronika Vodeb Gregor Lakner Mentorica: Bojana Pevec Šmarje pri Jelšah, maj 2008 Kazalo 1. HIPOTEZE... 4 2. Mejniki v moderni biotehnologiji... 5 3. UKRIVLJENOST PROSTORA...
CM707. GR Οδηγός χρήσης... 2-7. SLO Uporabniški priročnik... 8-13. CR Korisnički priručnik... 14-19. TR Kullanım Kılavuzu... 20-25
 1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
 FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
JEDRO (NUCLEUS PROKARYOTA(
 JEDRO (NUCLEUS) PROKARYOTA( procita) EUKARYOTA ( evcita) Nastanek: iz jedra; indirektna delitev: mitoza, mejoza-kariokineza Število: 1/ celico; 2/celico= dikariontsko stanje (Asco, - Basidiomycotina);
JEDRO (NUCLEUS) PROKARYOTA( procita) EUKARYOTA ( evcita) Nastanek: iz jedra; indirektna delitev: mitoza, mejoza-kariokineza Število: 1/ celico; 2/celico= dikariontsko stanje (Asco, - Basidiomycotina);
Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές:
 ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A Να επιλέξετε τη σωστή απάντηση: Α1. Το DNA απαντάται συνδεδεμένο με το RNA με ομοιοπολικό δεσμό:
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΩΝ ΣΠΟΥΔΩΝ Γ ' ΛΥΚΕΙΟΥ Τάξη: Γ Λυκείου Τμήμα: Βαθμός: Ονοματεπώνυμο: Καθηγητές: Θ Ε Μ Α A Να επιλέξετε τη σωστή απάντηση: Α1. Το DNA απαντάται συνδεδεμένο με το RNA με ομοιοπολικό δεσμό:
13. Jacobijeva metoda za računanje singularnega razcepa
 13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
Stohastični model bistabilnega stikala
 Univerza v Ljubljani Fakulteta za računalništvo in informatiko Tržaška 25 Ljubljana Seminarska naloga Stohastični model bistabilnega stikala Avtorji: Pece Adzievski, Borče Paspalovski, Aleš Uršič, Andrej
Univerza v Ljubljani Fakulteta za računalništvo in informatiko Tržaška 25 Ljubljana Seminarska naloga Stohastični model bistabilnega stikala Avtorji: Pece Adzievski, Borče Paspalovski, Aleš Uršič, Andrej
Τεχνολογία του ανασυνδυασμένου DNA
 Τεχνολογία του ανασυνδυασμένου DNA Θέση περιορισμού συμμετρική ως προς τον άξονα που περνά από το μέσο της. Στην εικόνα φαίνεται η θέση αναγνώρισης της EcoRI RI. Η αλληλουχία είναι παλίνδρο- μη: είναι
Τεχνολογία του ανασυνδυασμένου DNA Θέση περιορισμού συμμετρική ως προς τον άξονα που περνά από το μέσο της. Στην εικόνα φαίνεται η θέση αναγνώρισης της EcoRI RI. Η αλληλουχία είναι παλίνδρο- μη: είναι
Gradniki TK sistemov
 Gradniki TK sistemov renos signalov v višji rekvenčni legi Vsebina Modulacija in demodulacija Vrste analognih modulacij AM M FM rimerjava spektrov analognih moduliranih signalov Mešalniki Kdaj uporabimo
Gradniki TK sistemov renos signalov v višji rekvenčni legi Vsebina Modulacija in demodulacija Vrste analognih modulacij AM M FM rimerjava spektrov analognih moduliranih signalov Mešalniki Kdaj uporabimo
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
Viri. Nukleinske kisline. Program predavanj maj-junij DNA, RNA struktura, podvajanje, transkripcija, translacija
 Program predavanj maj-junij 2013 Datum izvajanja Vsebina predavanja / vaj 8. 5. 2013 P2 11-15 DNA, replikacija, RNA, transkripcija, translacija 15. 5. 2013 P2 11-15 Mutacije, variabilnost genoma PBL- naloge
Program predavanj maj-junij 2013 Datum izvajanja Vsebina predavanja / vaj 8. 5. 2013 P2 11-15 DNA, replikacija, RNA, transkripcija, translacija 15. 5. 2013 P2 11-15 Mutacije, variabilnost genoma PBL- naloge
A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N
 I N F O T E K N I K V o l u m e 1 5 N o. 1 J u l i 2 0 1 4 ( 61-70) A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N N o v i
I N F O T E K N I K V o l u m e 1 5 N o. 1 J u l i 2 0 1 4 ( 61-70) A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N N o v i
Rast mikrobne populacije
 Rast mikrobne populacije rast posamezne celice : rast populacije hranila fizikalno-kemijski faktorji rasti zaprt in odprt način namnoževanja merjenje rasti mikrobne populacije matematični opis rasti Rast
Rast mikrobne populacije rast posamezne celice : rast populacije hranila fizikalno-kemijski faktorji rasti zaprt in odprt način namnoževanja merjenje rasti mikrobne populacije matematični opis rasti Rast
CENTRALNI LABORATORIJ
 CENTRALNI LABORATORIJ I.ODVZEM IN POŠILJANJE VZORCEV 1 KAPILARNI ODVZEM KRVI DA DA 30min/15min 2 ODVZEM FECESA DA NE 30min/15min 3 ODVZEM URINA DA DA 30min/15min 4 POŠILJANJE BIOLOŠKIH VZORCEV ( EKSPEDIT)
CENTRALNI LABORATORIJ I.ODVZEM IN POŠILJANJE VZORCEV 1 KAPILARNI ODVZEM KRVI DA DA 30min/15min 2 ODVZEM FECESA DA NE 30min/15min 3 ODVZEM URINA DA DA 30min/15min 4 POŠILJANJE BIOLOŠKIH VZORCEV ( EKSPEDIT)
Doc. dr Snežana Marković
 REPARACIJA DNK. REKOMBINACIJA DNK. Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPARACIJA OŠTEĆENIH MOLEKULA DNK Mutacija svaka promena
REPARACIJA DNK. REKOMBINACIJA DNK. Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPARACIJA OŠTEĆENIH MOLEKULA DNK Mutacija svaka promena
OKOLJSKO NARAVOSLOVJE 2. Predavanja v študijskem letu 2. del 2012/2013
 OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju