REKOMBINACIJA MOLEKULA DNK
|
|
|
- Ζώνα Ελευθερίου
- 6 χρόνια πριν
- Προβολές:
Transcript

1 REKOMBINACIJA MOLEKULA DNK

2 Fenomeni odgovorni za održavanje i ekspresiju genoma
 I kod prokatiota i kod eukariota Tehnologija")
3 Svaki molekul DNK je rekombinovani molekul DNK Pojam rekombinacije Tridesete godine prošlog veka Mejoza (poslednja istraživanja i u mitozi) I kod prokatiota i kod eukariota Tehnologija rekombinovane DNK
")
4 Vrste rekombinacije DNK Homologna rekombinacija molekula DNK Rekombinacija DNK specifična za mesto Rekombinacija DNK transpozicijom Ilegitimna (nepravilna) rekombinacija


5 Rekombinacija DNK kod prokariota Horizontalni transfer Bakteriofagna infekcija Reparacija molekula DNK

6 Rekombinacije DNK kod eukariota Održanje integriteta genoma (tokom mejoze, npr.) Razmena genetičkih informacija genetički diverzitet Zaštita gen. informacija omogućava reparaciju DSB u DNK Učestvuje i u procesima regulacije ekspresije nekih gena Obezbeđuje nesmetano odvijanje replikacije pomaže da zarobljena replikaciona viljuška ponovo krene

7 HOMOLOGNA REKOMBINACIJA Fizička razmena istih ili homolognih sekvenci DNK između dva različita molekula DNK Generalna, opšta Homologna RecA zavisna (kod prokariota)
 Holidejeve strukture de novo sinteza molekula")
8 Homologna rekombinacija Fizička bliskost dva molekula DNK Homologija najmanje u 100 bp dugom regionu DNK Prekidi u jednom ili oba lanca molekula DNK Formiranje i kretanje Holidejeve strukture Razrešenje (rezolucija) Holidejeve strukture de novo sinteza molekula DNK

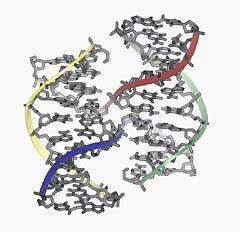
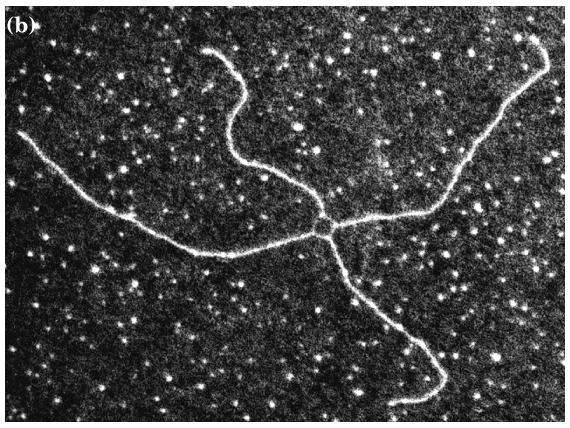

9 Pola veka istraživanja homologne rekombinacije godine otkriva je Robin Holliday ( )

10 Holidejev model homologne rekombinacije DNK Pojava jednolančanih prekida u lancima iste polarnosti u dva blisko postavljena molekula DNK predstavlja početak homologne rekombinacije

11 Holidejev model homologne rekombinacije DNK Slobodni krajevi mogu razmeniti svoje homologne partnere procesom koji se definiše kao INVAZIJA LANACA Invazija lanaca za posledicu ima formiranje mosta ili sinapse Ova strukuta se stabilizuje zatvaranjem jednolančanih prekida delovanjem enzima DNK ligaze
12 Holidejev model homologne rekombinacije DNK Dobijena hibridna struktura naziva se Holidejev rekombinovani intermedijer (Holidejeva struktura) Hibridna struktura je stabilna ali ne i statična MIGRACIJA GRANA Holidejeva veza
13 Holidejev model homologne rekombinacije DNK Migracija grana stvara kraće ili duže regione koji su u hemijskom smislu heterodupleksi a u genetičkom smislu heterozigoti


14 Holidejev model homologne rekombinacije DNK Razrešenje Holidejeve strukture Uvođenjem dva jednolančana prekida
15 Produkti homologne rekombinacije molekula DNK Dva različita Startna genotipa Patch Produkti sa umetkom Splice Splajsovani produkti
16 Delovanje endonukleaze

17 MODEL REPARACIJE DVOLANČANIH PREKIDA HOMOLOGNOM REKOMBINACIJOM DNK

18 Dvolančani prekidi DNK i njihova reparacija homolognom rekombinacijom Dvolančani prekidu u DNK molekulu česti događaji I jedan dvolančani prekid može biti letalan za E.coli Dejstvom jonizujućeg zračenja i citotoksičnih agenasa

19 Na mestu dvolančanog prekida dolazi do enzimske degradacije molekula DNK i formiranja regiona jednolančanih repova DNK ili invazivni lanci koji se završavaju slobodnom 3 OH grupom.

20 Jedan jednolančani DNK rep svojim 3 krajem napada homologni molekul dvolančane DNK Izmeštanje jednog lanca homolognog molekula DNK Kontakt oba jednolančana repa prekinutog molekula DNK sa homolognim delovima neprekinutog molekula DNK. Formiranje karakteristične strukture neprekinutog molekula DNK koja se naziva D petlja
21 Reparaciona sinteza DNK Elongacija 3 -krajeva jednočanačnih DNK repova. Matrica za DNK sintezu su homologni segmenti lanaca neprekinutog molekula DNK. DNK ligaze
22 Rekombinovani DNK intermedijer sa dve Holidejeve strukture Migracija grana je posledica raskidanja vodoničnih veza u originalnim molekulima DNK i istovremeno formiranje novih vodoničnih veza u rekombinovanom intermedijeru između lanaca dva homologna molekula DNK. Migracija grana za posledicu ima povećanje dužine segmenata DNK koji bivaju razmenjeni između homologih molekula DNK

23 Razrešenje rekombinovanog intermedijera sa dve Holidejeve strukture Odvija se uvođenjem dva jednolančana prekida Za posledicu ima stvaranje dva rekombinovana molekula DNK (dve veze sa dva mesta)
24 Razrešenje rekombinovanog intermedijera sa dve Holidejeve strukture Ako se na isti način seku obe Holidejeve veze rezultat homologne rekombinacije su produkti sa umetkom
25 Razrešenje rekombinovanog intermedijera sa dve Holidejeve strukture Ako se korišćenjem različitih mesta seku dve Holidejeve veze rezultat homologne rekombinacije su produkti sa krosingoverom splajsovani produkti

26 Kombinacija dve pojedinačne Holidejeve strukture Patch + patch = patch Splice + splice = patch Splice + patch = splice
27 Proteinska mašinerija reparacije dvolančanih prekida molekula DNK kod E.coli Specijalizovani proteini: Proteinski kompleks RecBCD promoter rekombinacije (obrada dvolančanog prekida i formiranje invazivnog lanca) Protein RecA centralni protein homologne rekombinacije (proces invazije i razmene lanaca) Proteinski kompleks RuvAB odgovoran za prepoznavanje Holidejeve veze i za proces migracije grana Protein RuvC učestvuje u razrešenju Holidejevih veza
28 Proteinska mašinerija reparacije dvolančanih prekida molekula DNK kod E.coli Pored specijalizovanih proteina u procesu homologne rekombinacije učestvuju i: DNK polimeraza DNK ligaza Topoizomeraza Proteini koji se vezuju za jednolančane molekule DNK (eng. single-strand binding protein, SSB) SSB proteini

29 Proteinski kompleks RecBCD promoter homologne rekombinacije Tri subjedinice proteinski produkti tri gena Prepoznaje dvolančani prekid u molekulu DNK odnosno njegove ravne krajeve Poseduje helikazno/nuleaznu aktivnost zavisnu od ATP Rezultat ove aktivnosti je formiranje invazivnog lanca

30 Protein RecB Dva funkcionalna domena Domen na N-kraju ima helikaznu aktivnost u smeru 3-5 Multifunkcionalan domen na C-kraju poseduje nukleaznu aktivnosti Domen na C-kraju uspostavlja interakcije sa proteinom RecA

31 Protein RecD Poseduje helikaznu aktivnost u smeru 5-3


32 Protein Rec C U svojoj strukturi poseduje izraženu petlju u vidu čiode Širi dva lanca molekula DNK i usmerava lance ka helikaznim motorima druge dve subjedinice kompleksa Prepoznaje chi mesta


33 Proteinski kompleks RecBCD promoter homologne rekombinacije Lanci DNK različitim kanalima stižu do nukleaznog centra kompleksa. Kanal kojim u nukleazni centar dospeva lanac sa 3-5 polarnošću je tako pozicioniran da je nukleazna aktivnost efikasnija na ovom lancu.
34 Proteinski kompleks RecBCD promoter homologne rekombinacije Aktivnost kompleksa RecBCD regulisanaje specifičnim sekvencama u molekulu DNK ('5-GCTGGTGG-3') koje se nazivaju chi (eng. crossover hotspot instigator, chi) ili χ mesta.
35 Prelaskom preko chi mesta dolazi do konformacione promene subjedinice RecC. Fleksibilna petlja RecC proteina zatvara kanal kojim se kreće lanac sa polarnošću 3-5. Usled toga, nukleazni centar počinje efikasnije da degradira lanac sa polarnošću 5-3. Na taj način dolazi do formiranja jednolančanog DNK repa invazivnog lanca.

36 c mesto Genoma E. coli poseduje 1009 c mesta Zaštita bakterijske od dejstva fagne DNK ili segmenata bakterijskih DNK koje ne poseduju chi mesta a koje dospevaju u bakterijsku ćeliju raznim vidovima horizontalnog transfera gena. 10x povećava frekvencu rekombinacije.

37 Proteinski kompleks RecBCD promoter homologne rekombinacije Zahvaljujući subjedinici RecB, kompleks RecBCD pomaže vezivanje proteina RecA za invazivni lanac.
38 Protein RecA Protein RecA je centralni protein homologne rekombinacije kod E. coli. Pripada proteinskoj familiji proteina za razmenu lanaca (eng. strandexchange proteins). Mutacija u proteinu RecA smanjuje učestalost rekombinacije za puta. Poseduje ATP aznu i proteaznu aktivnost. Proteni RecA vezuju se za invazivni lanac (jednolančani DNK rep) i omogućava proces invazije.

39 Filament RecA Za invazivni lanac, proteini RecA vezuju se kooperativno i na taj način formiraju filament RecA. Filament RecA sastoji se od oko 100 molekula proteina RecA koji se vezuju za 300 nukleotida dugu sekvencu DNK invazivnog lanca.

40 Filament RecA Štiti jednolančanu DNK od dejstva nukleaza i omogućava proces invazije lanaca. Vrši napad na dvolančani homologni molekul DNK. Struktura koja se sastoji od filamenta RecA i segmenta dvolančanog molekula DNK naziva se spojeni (eng. joint) molekul.

41 Filament RecA Filament RecA poseduje dva vezujuća mesta za molekule DNK. U okviru jednog nalazi se invazivni lanac i ono se naziva primarno mesto. Drugo mesto za vezivanje molekula DNK naziva se sekundarno mesto i za njega se vezuje homologni dvolančani molekul DNK.


42 Filament RecA Zahvaljujući postojanju ovih mesta u okviru filamenta RecA, tokom invazije lanaca jednolančani DNK repovi (invazivni lanci) pretražuju i pronalaze homologne regione u dvolančanom molekulu DNK.
43 Filament RecA Vezivanje dvolančanog molekula DNK za sekundarno mesto filamenta RecA je brzo, slabo, prolazno i nezavisno od sekvence. Mehanizam kojim filament RecA pretražuje dvolančani molekul DNK kako bi pronašao homologne sekvence sa sekvencom invazivnog lanca nije sasvim poznat.

44 Pretpostavka je da se baze invazivnog lanca i baze dvolančane DNK spajaju vodoničnim vezama. Energiju potrebnu za raskidanje vodoničnih veza u dvolančanom molekulu DNK obezbeđuju molekuli RecA svojom ATP-aznom aktivnošću. Raskidanjem vodoničnih veza segment dvolančanog molekula DNK privremeno se otvara i omogućava svojim bazama da formiraju vodonične veze sa bazama u jednolančanom molekulu DNK (invazivnom lancu).


45 Ispravno sparivanje 15 baza signal da je pronađena homologna sekvenca

46 Reparaciona sinteza DNK Nakon invazije lanaca i pronalaženja homologije između molekula DNK, proteini RecA disosuju, a spojeni molekul postaje mesto aktivnosti DNK polimeraza i DNK ligaza. Aktivnošću DNK polimeraze i DNK ligaze formiraju se Holidejeve veze.

47 Migracija Holidejevih veza omogućena je proteinskim kompleksom RuvAB Protein RuvA vezuje se za Holidejevu strukturu u formi tetramera i regrutuje dva molekula RuvB heksamerni protein sa ATP-aznom aktivnošću koji omogućava migraciju grana

48 U razrešenju Holidejeve strukture učestvuje protein RuvC Holidejevu strukturu prepoznaje zahvaljujući protein-protein interakciji sa kompleksomruvab. Dva molekula proteina RuvC vezuju se za kompleks RuvAB.

49 Protein RuvC Protein RuvC je endonukleaza koja uvodi jednolančane prekide u lancima DNK iste polarnosti. Jednolančane prekide uvodi posle drugog timina u sekvenci 5'-A/T-T-T-C/G -3'. Ova sekvenca se u molekulu DNK E. coli nalazi na svakih 64 nukleotida.
50 DNK ligaza još jednom DNK krajevi nastali dejstvom proteina RuvC imaju slobodnu 5 fosfatnu grupu i 3 OH grupu koje se direktno spajaju dejstvom DNK ligaze. Rekombinovani molekuli DNK međusobno se razdvajaju i time je homologna rekombinacija završena.
 - putem krosingovera predstavlja izvor genetičke varijabilnosti")
51 Homologna rekombinacija i mejoza Mejoza (dve sukcesivne deobe, redukciona i ekvaciona deoba). Homologna rekombinacija odvija se u pahitenu profaze I. Mejotička rekombinacija: - omogućava pravilno razdvajanje hormozoma (održavanje genoma) - putem krosingovera predstavlja izvor genetičke varijabilnosti (ekspresija genoma).


52 Nondisjunction Odsustvo mejotičke rekombinacije za posledicu ima nepravilno razdvajanje hromozoma. Rezultat: Povećan broj gameta bez kompletnog broja hromozma što je uzrok smanjenog fertiliteta.

53 Protein Spo11 Protein Spo11 programirano uvodi dvolančani prekid u molekul DNK tokom inicijacije mejotičke rekombinacije. Topoizomerazama sličan protein koga poseduju svi eukarioti od kvasca do čoveka. Značaj proteina Spo11 za odvijanje homologne rekombinacije i mejoze pokazan je u eksperimentima na C.elegans.
54 Protein Spo11 U formi dimera, protein Spo11 napada molekul DNK (svaki monomer napada po jedan lanac). OH grupa tirozina raskida fosfodiestarsku vezu. Generiše se kovalentna veza fosfatne grupe molekula DNK i proteina Spo11. Energija kovalentne veze može se osloboditi raskidanjem iste i iskoristiti za repariranje dvolančanog prekida u uslovima kada ćelija dobije signal za zaustavljanje procesa mejoze.

55 Protein Spo11 Seče molekul DNK na mnogim mestima sa malom selektivnošću za sekvencu. Mesta delovanja proteina SPO11 nisu nasumično raspoređena po molekulu DNK češća u regionima koji nisu čvrsto pakovani sa histonima vruća mesta (eng. hot spots) za rekombinaciju. Vremenski strogo kontrolisana aktivnost.

56 Proteinski kompleks MRX Funkcija slična funkciji kompleksa RecBCD kod E.coli. Sastoji se od tri subjedinice: Mre11, Rad 50 i Nbs1/Xrs 2. Kompleks ispoljava nukleaznu aktivnost Rezultat aktivnosti ovog kompleksa je stvaranje invazivnog lanca od najmanje 1kb.


57 Dmc1 i Rad51 Eukariotski analozi proteina RecA Dmc1 se u ćeliji eksprimira isključivo tokom mejoze Rad51 i tokom mitoze i tokom mejoze

58 Protein Dcm1 Mejotička rekombinacija dešava se između 4 kompletna dvolančana lanca DNK (svaki u po jednoj hromatidi homologih hromozoma) Dcm1 zavisna rekombinacija češće se odigrava između nesestrinskih hromatida homolognih hromozoma Mehanizam kojom protein Dcm1 promoviše rekombinaciju između nesestrinskih hromatida je nepoznat. JASNA BIOLOŠKA ULOGA

59 Brojni drugi proteini uključeni su u proces mejotičke rekombinacije. Rad52 stimuliše vezivanja jednolančane DNK repove. proteina Rad51 za Rad54 eukariotski analog RuvB (?) Veliki protein-dnk kompleksi rekombinacione fabrike. nazivaju se
 u razrešenju Holidejevih")
60 Rekombinacione fabrike Protein Mus81 je visoko konzerviran protein eukariota. Kompleks Mus81-Eme1 ima ulogu endonukleaze (resolvaze) u razrešenju Holidejevih struktura.


61 Rekombinacione fabrike Protein BRCA2 interaguje sa proteinom Rad51 i reguliše proces mejotičke rekombinacije. Poslednja istraživanja pokazuju da BRCA2 protein interaguje i sa proteinom Dmc1. Ovo otkriće, BRCA2 protein promoviše u univerzalnog regulatora rekombinacionog događaja.
62 Genska konverzija Genska konverzija je vrsta homologne rekombinacije bez krosingovera i formiranja Holidejeve veze. Zbog odsustva krosingovera naziva se i nerecipročna rekombinacija. Mehanizam kojim se odvija nerecipročna rekombinacija naziva se i sparivanje lanaca zavisno od sinteze (eng. synthesis-dependent strand annealing, SDSA).

63 Pivski kvasac
64 Lokus MAT otključana kaseta (mating type locus) Mating type geni kodiraju regulatorne proteine koji regulišu ekspresiju gena čiji proteinski produkti definišu tip ćelije. Kopije ovih gena prisutne su u svim tipovima ćelija ali se sve ne eksprimiraju. Nalaze se u okviru lokusa koji se naziva lokus MAT ili otključana kaseta.
65 Lokus MAT otključana kaseta (mating type locus) U a ćelijama u lokusu MAT prisutan gen a1. U a ćelijama u lokusu MAT prisutni geni a 1 i a 2. U diploidnim ćelijama sva tri mating-type control gena prisutna su u dva lokusa MAT.
66 Lokusi HMR i HML Utišane kasete skladište genetičkih informacija Svaka ćelija sadrži dodatne kopije kako gena a1, tako i gena a1 i a2. Ove kopije se ne eksprimiraju i nalaze se u lokusima označenim kao HML i HMR ( zaključane kasete ). Lokusi HMR i HML čuvaju genetičku informaciju neophodnu da ćelija promeni svoj tip.
67 Nerecipročna rekombinacija Lokus HML kodira utišane kompije gena a Lokus HMR kodira utišanu kopiju gena a Rekombinacija između lokusa MAT i lokusa HML, a ćelija prelazi u a ćeliju. Rekombinacija između lokusa MAT i lokusa HMR, a ćelija prelazi u a ćeliju.


68 Sparivanje lanaca zavisno od sinteze - SDSA Nerecipročna rekombinacija otpočinje uvođenjem dvolančanog prekida u okviru lokusa MAT. HO sekvencno-specifična endonukleaza. HO endonukleaza jedino prepoznaje i seče lokus MAT (nikada lokus HMR ili HML jer hromatinska struktura to ne dozvoljava).
69 Sparivanje lanaca zavisno od sinteze - SDSA Razgradnja molekula DNK u smeru 5 3 na mestu delovanja HO endonukleaze odvija se istim mehanizmom kao kod mejotičke rekombinacije dejstvom proteinskog kompleksa MRX.
70 Sparivanje lanaca zavisno od sinteze - SDSA Formirani jednolančani DNK rep iz lokusa MAT povezuje se sa proteinima Rad51 i Rad52 i napada zaključanu kasetu (dvolančanu DNK).
71 Sparivanje lanaca zavisno od sinteze - SDSA Ako je u lokusu MAT sekvenca gena a1 invazijom se napada lokus HML koji kodira utišane kopije a gena Ako je u lokusu MAT sekvenca a1 i a2 gena invazijom se napada lokus HMR koji kodira utišanu kopiju a gena
72 Sparivanje lanaca zavisno od sinteze - SDSA Nakon invazije, slobodni 3 -kraj jednolančanog DNK repa služi kao prajmer za iniciranje sinteze DNK. Matrica za ovu sintezu DNK je sekvenca zaključane kasete.
73 Sparivanje lanaca zavisno od sinteze - SDSA Suprotno mehanizmu replikacije molekula DNK, novosintetisani lanci komplementarni zaključanoj kaseti napuštaju matricu i povezuju se sa sekvenacma koje okružuju lokus MAT. Pre ove ligacije, preostali lanac DNK iz lokusa MAT biva isečen.
74 Sparivanje lanaca zavisno od sinteze - SDSA Nakon rekombinacije genetička informacija koja je u lokusu HMR nalazi se i u lokusu MAT. Informacija u lokusu MAT je zamenjena, dok je informacija u lokusu HMR ostala ista.
75 Sparivanje lanaca zavisno od sinteze - SDSA Mating-type swiching je unidirekcioni - informacija iz lokusa HML ili HMR može biti premeštena u lokus MAT, ali ne i obruto Lokus MAT je uvek recipijentni partner rekombinacije.

76 Sparivanje lanaca zavisno od sinteze SDSA

77 Sledi... Rekombinacija DNK specifična za mesto
Rekombinacija DNK TRANSPOZICIJOM
 Rekombinacija DNK TRANSPOZICIJOM Rekombinacija DNK transpozicijom Transpozicija je proces premeštanja specifičnih sekvenci molekula DNK (pokretnih genetičkih elemenata ili transpozona) sa jednog na drugo
Rekombinacija DNK TRANSPOZICIJOM Rekombinacija DNK transpozicijom Transpozicija je proces premeštanja specifičnih sekvenci molekula DNK (pokretnih genetičkih elemenata ili transpozona) sa jednog na drugo
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
Doc. dr Snežana Marković
 REPARACIJA DNK. REKOMBINACIJA DNK. Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPARACIJA OŠTEĆENIH MOLEKULA DNK Mutacija svaka promena
REPARACIJA DNK. REKOMBINACIJA DNK. Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPARACIJA OŠTEĆENIH MOLEKULA DNK Mutacija svaka promena
Regulacija ekspresije gena kod prokariota
 Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović
 DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
3.1 Granična vrednost funkcije u tački
 3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA
 NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA ANIMACIJE!!! REPLIKACIJA https://www.youtube.com/watch?v=tnkwgcfphqw TRANSKRIPCIJA https://www.youtube.com/watch?v=jqiwwjqf5d0 TRANSKRIPCIJA I TRANSLACIJA https://www.youtube.com/watch?v=-k8y0atkkai
NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA ANIMACIJE!!! REPLIKACIJA https://www.youtube.com/watch?v=tnkwgcfphqw TRANSKRIPCIJA https://www.youtube.com/watch?v=jqiwwjqf5d0 TRANSKRIPCIJA I TRANSLACIJA https://www.youtube.com/watch?v=-k8y0atkkai
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
TRANSLACIJA. Doc. dr Snežana Marković
 TRANSLACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu BIOSINTEZA PROTEINA - TRANSLACIJA U toku translacije dolazi do specifičnog
TRANSLACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu BIOSINTEZA PROTEINA - TRANSLACIJA U toku translacije dolazi do specifičnog
XI dvoqas veжbi dr Vladimir Balti. 4. Stabla
 XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
Doc. dr Snežana Marković
 REPLIKACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPLIKACIJA DNK Semikonzervativan proces Eksperimenti Mezelsona i Stala 1957.g.
REPLIKACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPLIKACIJA DNK Semikonzervativan proces Eksperimenti Mezelsona i Stala 1957.g.
PRAVA. Prava je u prostoru određena jednom svojom tačkom i vektorom paralelnim sa tom pravom ( vektor paralelnosti).
.") PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
Regulacija ekspresije gena kod eukariota
 Regulacija ekspresije gena kod eukariota Regulacija ekspresije gena Višećelijski organizmi Čovek - 250 tipova ćelija Svaka ćelija gotovo istu DNK Regulacija ekspresije gena Hijerarhija regulacije ekspresije
Regulacija ekspresije gena kod eukariota Regulacija ekspresije gena Višećelijski organizmi Čovek - 250 tipova ćelija Svaka ćelija gotovo istu DNK Regulacija ekspresije gena Hijerarhija regulacije ekspresije
18. listopada listopada / 13
 18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama.
 Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
5. Karakteristične funkcije
 5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
RIJEŠENI ZADACI I TEORIJA IZ
 RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
Zavrxni ispit iz Matematiqke analize 1
 Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Elementi spektralne teorije matrica
 Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE
 **** MLADEN SRAGA **** 011. UNIVERZALNA ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE SKUP REALNIH BROJEVA α Autor: MLADEN SRAGA Grafički urednik: BESPLATNA - WEB-VARIJANTA Tisak: M.I.M.-SRAGA
**** MLADEN SRAGA **** 011. UNIVERZALNA ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE SKUP REALNIH BROJEVA α Autor: MLADEN SRAGA Grafički urednik: BESPLATNA - WEB-VARIJANTA Tisak: M.I.M.-SRAGA
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A
, grupa A") Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE GODINE 8. RAZRED TOČNI ODGOVORI
 21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
Teorijske osnove informatike 1
 Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
Obrada signala
 Obrada signala 1 18.1.17. Greška kvantizacije Pretpostavka je da greška kvantizacije ima uniformnu raspodelu 7 6 5 4 -X m p x 1,, za x druge vrednosti x 3 x X m 1 X m = 3 x Greška kvantizacije x x x p
Obrada signala 1 18.1.17. Greška kvantizacije Pretpostavka je da greška kvantizacije ima uniformnu raspodelu 7 6 5 4 -X m p x 1,, za x druge vrednosti x 3 x X m 1 X m = 3 x Greška kvantizacije x x x p
3. razred gimnazije- opšti i prirodno-matematički smer ALKENI. Aciklični nezasićeni ugljovodonici koji imaju jednu dvostruku vezu.
 ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018.
 Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Test iz Biologije za 4. razred srednje škole R J E Š E NJ A Podgorica, 05. 05. 2018. 1 1. Spojite navedene
Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Test iz Biologije za 4. razred srednje škole R J E Š E NJ A Podgorica, 05. 05. 2018. 1 1. Spojite navedene
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
41. Jednačine koje se svode na kvadratne
 . Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
. Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
ELEKTROTEHNIČKI ODJEL
 MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
SADRŽAJ TRANSLACIJA... TRANSPORTNE RNK
 Translacija SADRŽAJ TRASLACIJA... 1 TRASPORTE RK... 2 Primarna struktura trk... 2 Sekundarna struktura trk... 3 Tercijarna struktura trk... 5 Aktivacija aminokiselina... 5 Interakcija kodon antikodon...
Translacija SADRŽAJ TRASLACIJA... 1 TRASPORTE RK... 2 Primarna struktura trk... 2 Sekundarna struktura trk... 3 Tercijarna struktura trk... 5 Aktivacija aminokiselina... 5 Interakcija kodon antikodon...
MEĐUMOLEKULSKE SILE JON-DIPOL DIPOL VODONIČNE NE VEZE DIPOL DIPOL-DIPOL DIPOL-INDUKOVANI INDUKOVANI JON-INDUKOVANI DISPERZNE SILE
 MEĐUMLEKULSKE SILE JN-DIPL VDNIČNE NE VEZE DIPL-DIPL JN-INDUKVANI DIPL DIPL-INDUKVANI INDUKVANI DIPL DISPERZNE SILE MEĐUMLEKULSKE SILE jake JNSKA VEZA (metal-nemetal) KVALENTNA VEZA (nemetal-nemetal) METALNA
MEĐUMLEKULSKE SILE JN-DIPL VDNIČNE NE VEZE DIPL-DIPL JN-INDUKVANI DIPL DIPL-INDUKVANI INDUKVANI DIPL DISPERZNE SILE MEĐUMLEKULSKE SILE jake JNSKA VEZA (metal-nemetal) KVALENTNA VEZA (nemetal-nemetal) METALNA
Ispitivanje toka i skiciranje grafika funkcija
 Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Operacije s matricama
 Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Biohemija nukleinskih kiselina. Genetska informacija
 Biohemija nukleinskih kiselina Genetska informacija deoksiribonukleinske kiseline (DNK) ribonukleinske kiseline (RNK) DNK je nosilac naslednih informacija u ćeliji, dok RNK učestvuju u prenošenju tih informacija
Biohemija nukleinskih kiselina Genetska informacija deoksiribonukleinske kiseline (DNK) ribonukleinske kiseline (RNK) DNK je nosilac naslednih informacija u ćeliji, dok RNK učestvuju u prenošenju tih informacija
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f
 D f") IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI. Doc. dr Snežana Marković
 REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REGULACIJA EKSPRESIJE
REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REGULACIJA EKSPRESIJE
NOMENKLATURA ORGANSKIH SPOJEVA. Imenovanje aromatskih ugljikovodika
 NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
numeričkih deskriptivnih mera.
 DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
Iskazna logika 3. Matematička logika u računarstvu. novembar 2012
 Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
transkripcija Matrica i enzimi Transkripcija Sličnosti između replikacije i transkripcije Razlike između replikacije i transkripcije
 Transkripcija Sinteza RNK molekula. DNK lanac služi kao matrica za sintezu RNK. Transkripcija Tip RNK molekula Ribozomalna RNK (rrnk) Male RNK (trnk; snrnk; 5S rrnk; scrnk; mirnk ) Informaciona RNK (irnk
Transkripcija Sinteza RNK molekula. DNK lanac služi kao matrica za sintezu RNK. Transkripcija Tip RNK molekula Ribozomalna RNK (rrnk) Male RNK (trnk; snrnk; 5S rrnk; scrnk; mirnk ) Informaciona RNK (irnk
Novi Sad god Broj 1 / 06 Veljko Milković Bulevar cara Lazara 56 Novi Sad. Izveštaj o merenju
 Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Reverzibilni procesi
 Reverzbln proces Reverzbln proces: proces pr koja sste nkada nje vše od beskonačno ale vrednost udaljen od ravnoteže, beskonačno ala proena spoljašnjh uslova ože vratt sste u blo koju tačku, proena ože
Reverzbln proces Reverzbln proces: proces pr koja sste nkada nje vše od beskonačno ale vrednost udaljen od ravnoteže, beskonačno ala proena spoljašnjh uslova ože vratt sste u blo koju tačku, proena ože
M086 LA 1 M106 GRP. Tema: Baza vektorskog prostora. Koordinatni sustav. Norma. CSB nejednakost
 M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
HEMIJSKA VEZA TEORIJA VALENTNE VEZE
 TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT
 METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT BEZ POLI-A REPA" Histoni su primarne proteinske komponente hromatina. Na početku se smatralo da su uglavnom uključeni u pakovanje DNK, odnosno da su važni
METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT BEZ POLI-A REPA" Histoni su primarne proteinske komponente hromatina. Na početku se smatralo da su uglavnom uključeni u pakovanje DNK, odnosno da su važni
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
LANCI & ELEMENTI ZA KAČENJE
 LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
REGULATORNI MOLEKULI RNK "SKRIVENI JEZIK RNK"
 REGULATORNI MOLEKULI RNK "SKRIVENI JEZIK RNK" Kod prokariota geni posredstvom molekula irnk kodiraju proteine koji obavljaju katalitičke, strukturne i regulatone funkcije, tako da su proteini jedini izlazni
REGULATORNI MOLEKULI RNK "SKRIVENI JEZIK RNK" Kod prokariota geni posredstvom molekula irnk kodiraju proteine koji obavljaju katalitičke, strukturne i regulatone funkcije, tako da su proteini jedini izlazni
Molekularna biologija prokariota
 Molekularna biologija prokariota I Molekularne osnove procesa: Čuvanja i prenošenja genetičke informacije u ćeliji Regulacije metabolizma - ekonomičnost ćelije i brzo prilagođavanje na uslove sredine -
Molekularna biologija prokariota I Molekularne osnove procesa: Čuvanja i prenošenja genetičke informacije u ćeliji Regulacije metabolizma - ekonomičnost ćelije i brzo prilagođavanje na uslove sredine -
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Sistemi za kontrolu kvaliteta proteina molekularni šaperoni i proteazom
 Sistemi za kontrolu kvaliteta proteina molekularni šaperoni i proteazom Kako protein nakon sinteze postaje funkcionalan? Proces ekspresije gena nije završen prevođenjem informacije sadržane u irnk u redosled
Sistemi za kontrolu kvaliteta proteina molekularni šaperoni i proteazom Kako protein nakon sinteze postaje funkcionalan? Proces ekspresije gena nije završen prevođenjem informacije sadržane u irnk u redosled
CILJNA MESTA DEJSTVA LEKOVA
 FARMACEUTSKA HEMIJA 1 CILJNA MESTA DEJSTVA LEKVA Predavač: Prof. dr Slavica Erić Ciljna mesta dejstva leka CILJNA MESTA NA MLEKULARNM NIVU: lipidi (lipidi ćelijske membrane) ugljeni hidrati (obeleživači
FARMACEUTSKA HEMIJA 1 CILJNA MESTA DEJSTVA LEKVA Predavač: Prof. dr Slavica Erić Ciljna mesta dejstva leka CILJNA MESTA NA MLEKULARNM NIVU: lipidi (lipidi ćelijske membrane) ugljeni hidrati (obeleživači
ZBIRKA POTPUNO RIJEŠENIH ZADATAKA
 **** IVANA SRAGA **** 1992.-2011. ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE POTPUNO RIJEŠENI ZADACI PO ŽUTOJ ZBIRCI INTERNA SKRIPTA CENTRA ZA PODUKU α M.I.M.-Sraga - 1992.-2011.
**** IVANA SRAGA **** 1992.-2011. ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE POTPUNO RIJEŠENI ZADACI PO ŽUTOJ ZBIRCI INTERNA SKRIPTA CENTRA ZA PODUKU α M.I.M.-Sraga - 1992.-2011.
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
Matematika 1 - vježbe. 11. prosinca 2015.
 Matematika - vježbe. prosinca 5. Stupnjevi i radijani Ako je kut φ jednak i rad, tada je veza između i 6 = Zadatak.. Izrazite u stupnjevima: a) 5 b) 7 9 c). d) 7. a) 5 9 b) 7 6 6 = = 5 c). 6 8.5 d) 7.
Matematika - vježbe. prosinca 5. Stupnjevi i radijani Ako je kut φ jednak i rad, tada je veza između i 6 = Zadatak.. Izrazite u stupnjevima: a) 5 b) 7 9 c). d) 7. a) 5 9 b) 7 6 6 = = 5 c). 6 8.5 d) 7.
KVADRATNA FUNKCIJA. Kvadratna funkcija je oblika: Kriva u ravni koja predstavlja grafik funkcije y = ax + bx + c. je parabola.
 KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI. NEUTRALNI ELEMENT GRUPOIDA.
 KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
Riješeni zadaci: Nizovi realnih brojeva
 Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
PARCIJALNI IZVODI I DIFERENCIJALI. Sama definicija parcijalnog izvoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je,
 PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
( ) ( ) 2 UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET. Zadaci za pripremu polaganja kvalifikacionog ispita iz Matematike. 1. Riješiti jednačine: 4
 ( ) 2 UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET. Zadaci za pripremu polaganja kvalifikacionog ispita iz Matematike. 1. Riješiti jednačine: 4") UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET Riješiti jednačine: a) 5 = b) ( ) 3 = c) + 3+ = 7 log3 č) = 8 + 5 ć) sin cos = d) 5cos 6cos + 3 = dž) = đ) + = 3 e) 6 log + log + log = 7 f) ( ) ( ) g) ( ) log
UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET Riješiti jednačine: a) 5 = b) ( ) 3 = c) + 3+ = 7 log3 č) = 8 + 5 ć) sin cos = d) 5cos 6cos + 3 = dž) = đ) + = 3 e) 6 log + log + log = 7 f) ( ) ( ) g) ( ) log
Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri
 Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri 1 1 Zadatak 1b Čisto savijanje - vezano dimenzionisanje Odrediti potrebnu površinu armature za presek poznatih dimenzija, pravougaonog
Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri 1 1 Zadatak 1b Čisto savijanje - vezano dimenzionisanje Odrediti potrebnu površinu armature za presek poznatih dimenzija, pravougaonog
Strukture podataka i algoritmi 1. kolokvij 16. studenog Zadatak 1
 Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Pismeni ispit iz matematike Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: ( ) + 1.
 + 1.") Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Grafičko prikazivanje atributivnih i geografskih nizova
 Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu
 Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
APROKSIMACIJA FUNKCIJA
 APROKSIMACIJA FUNKCIJA Osnovni koncepti Gradimir V. Milovanović MF, Beograd, 14. mart 2011. APROKSIMACIJA FUNKCIJA p.1/46 Osnovni problem u TA Kako za datu funkciju f iz velikog prostora X naći jednostavnu
APROKSIMACIJA FUNKCIJA Osnovni koncepti Gradimir V. Milovanović MF, Beograd, 14. mart 2011. APROKSIMACIJA FUNKCIJA p.1/46 Osnovni problem u TA Kako za datu funkciju f iz velikog prostora X naći jednostavnu
Dvanaesti praktikum iz Analize 1
 Dvaaesti praktikum iz Aalize Zlatko Lazovi 20. decembar 206.. Dokazati da fukcija f = 5 l tg + 5 ima bar jedu realu ulu. Ree e. Oblast defiisaosti fukcije je D f = k Z da postoji ula fukcije a 0, π 2.
Dvaaesti praktikum iz Aalize Zlatko Lazovi 20. decembar 206.. Dokazati da fukcija f = 5 l tg + 5 ima bar jedu realu ulu. Ree e. Oblast defiisaosti fukcije je D f = k Z da postoji ula fukcije a 0, π 2.
7 Algebarske jednadžbe
 7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
Drugi zakon termodinamike
 Drugi zakon termodinamike Uvod Drugi zakon termodinamike nije univerzalni prirodni zakon, ne važi za sve sisteme, naročito ne za neobične sisteme (mikrouslovi, svemirski uslovi). Zasnovan je na zajedničkom
Drugi zakon termodinamike Uvod Drugi zakon termodinamike nije univerzalni prirodni zakon, ne važi za sve sisteme, naročito ne za neobične sisteme (mikrouslovi, svemirski uslovi). Zasnovan je na zajedničkom
Sekundarne struktura proteina Fibrilni proteini
 Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).
![PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).](/thumbs/74/70316766.jpg "PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).") 0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
a M a A. Može se pokazati da je supremum (ako postoji) jedinstven pa uvodimo oznaku sup A.
 jedinstven pa uvodimo oznaku sup A.") 3 Infimum i supremum Definicija. Neka je A R. Kažemo da je M R supremum skupa A ako je (i) M gornja meda skupa A, tj. a M a A. (ii) M najmanja gornja meda skupa A, tj. ( ε > 0)( a A) takav da je a > M
3 Infimum i supremum Definicija. Neka je A R. Kažemo da je M R supremum skupa A ako je (i) M gornja meda skupa A, tj. a M a A. (ii) M najmanja gornja meda skupa A, tj. ( ε > 0)( a A) takav da je a > M
OSNOVI ELEKTRONIKE. Vežbe (2 časa nedeljno): mr Goran Savić
: mr Goran Savić") OSNOVI ELEKTRONIKE Vežbe (2 časa nedeljno): mr Goran Savić savic@el.etf.rs http://tnt.etf.rs/~si1oe Termin za konsultacije: četvrtak u 12h, kabinet 102 Referentni smerovi i polariteti 1. Odrediti vrednosti
OSNOVI ELEKTRONIKE Vežbe (2 časa nedeljno): mr Goran Savić savic@el.etf.rs http://tnt.etf.rs/~si1oe Termin za konsultacije: četvrtak u 12h, kabinet 102 Referentni smerovi i polariteti 1. Odrediti vrednosti
Moguća i virtuelna pomjeranja
 Dnamka sstema sa vezama Moguća vrtuelna pomjeranja f k ( r 1,..., r N, t) = 0 (k = 1, 2,..., K ) df k dt = r + t = 0 d r = r dt moguća pomjeranja zadovoljavaju uvjet: df k = d r + dt = 0. t δ r = δx +
Dnamka sstema sa vezama Moguća vrtuelna pomjeranja f k ( r 1,..., r N, t) = 0 (k = 1, 2,..., K ) df k dt = r + t = 0 d r = r dt moguća pomjeranja zadovoljavaju uvjet: df k = d r + dt = 0. t δ r = δx +
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika. Monotonost i ekstremi. Katica Jurasić. Rijeka, 2011.
 Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
1 Promjena baze vektora
 Promjena baze vektora Neka su dane dvije različite uredene baze u R n, označimo ih s A = (a, a,, a n i B = (b, b,, b n Svaki vektor v R n ima medusobno različite koordinatne zapise u bazama A i B Zapis
Promjena baze vektora Neka su dane dvije različite uredene baze u R n, označimo ih s A = (a, a,, a n i B = (b, b,, b n Svaki vektor v R n ima medusobno različite koordinatne zapise u bazama A i B Zapis
Dijagonalizacija operatora
 Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
III VEŽBA: FURIJEOVI REDOVI
 III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
POVRŠINA TANGENCIJALNO-TETIVNOG ČETVEROKUTA
 POVRŠIN TNGENIJLNO-TETIVNOG ČETVEROKUT MLEN HLP, JELOVR U mnoštvu mnogokuta zanimljiva je formula za površinu četverokuta kojemu se istoobno može upisati i opisati kružnica: gje su a, b, c, uljine stranica
POVRŠIN TNGENIJLNO-TETIVNOG ČETVEROKUT MLEN HLP, JELOVR U mnoštvu mnogokuta zanimljiva je formula za površinu četverokuta kojemu se istoobno može upisati i opisati kružnica: gje su a, b, c, uljine stranica
4. Trigonometrija pravokutnog trokuta
 4. Trigonometrij prvokutnog trokut po školskoj ziri od Dkić-Elezović 4. Trigonometrij prvokutnog trokut Formule koje koristimo u rješvnju zdtk: sin os tg tg ktet nsuprot kut hipotenuz ktet uz kut hipotenuz
4. Trigonometrij prvokutnog trokut po školskoj ziri od Dkić-Elezović 4. Trigonometrij prvokutnog trokut Formule koje koristimo u rješvnju zdtk: sin os tg tg ktet nsuprot kut hipotenuz ktet uz kut hipotenuz
BANKA PITANJA IZ BIOLOGIJE. proteini. 3. Koji se deo složenog enzima hemijski menja u toku reakcije: 4. Apoenzim i koenzim zajedno čine:
 BANKA PITANJA IZ BIOLOGIJE citologija proteini 1. α-heliks je pojam koje se odnosi na strukturu proteina. 2. Broj polipeptidnih lanaca kod složenih proteina definiše: 3. Koji se deo složenog enzima hemijski
BANKA PITANJA IZ BIOLOGIJE citologija proteini 1. α-heliks je pojam koje se odnosi na strukturu proteina. 2. Broj polipeptidnih lanaca kod složenih proteina definiše: 3. Koji se deo složenog enzima hemijski
radni nerecenzirani materijal za predavanja
 Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Kažemo da je funkcija f : a, b R u točki x 0 a, b postiže lokalni minimum ako postoji okolina O(x 0 ) broja x 0 takva da je
Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Kažemo da je funkcija f : a, b R u točki x 0 a, b postiže lokalni minimum ako postoji okolina O(x 0 ) broja x 0 takva da je
Mehanizmidejstvaenzima. Himotripsin
 Mehanizmidejstvaenzima Himotripsin Principi katalize Specifična kiselo-bazna kataliza Elektrostatska kataliza Elektrofilna kataliza Nukleofilna kataliza (kovalentna kataliza) Nukleofilna kataliza Opšta
Mehanizmidejstvaenzima Himotripsin Principi katalize Specifična kiselo-bazna kataliza Elektrostatska kataliza Elektrofilna kataliza Nukleofilna kataliza (kovalentna kataliza) Nukleofilna kataliza Opšta
KVADRATNA FUNKCIJA. Kvadratna funkcija je oblika: Kriva u ravni koja predstavlja grafik funkcije y = ax + bx + c. je parabola.
 KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije a + b + c je parabola. Najpre ćemo naučiti kako izgleda
KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije a + b + c je parabola. Najpre ćemo naučiti kako izgleda
(P.I.) PRETPOSTAVKA INDUKCIJE - pretpostavimo da tvrdnja vrijedi za n = k.
 PRETPOSTAVKA INDUKCIJE - pretpostavimo da tvrdnja vrijedi za n = k.") 1 3 Skupovi brojeva 3.1 Skup prirodnih brojeva - N N = {1, 2, 3,...} Aksiom matematičke indukcije Neka je N skup prirodnih brojeva i M podskup od N. Ako za M vrijede svojstva: 1) 1 M 2) n M (n + 1) M,
1 3 Skupovi brojeva 3.1 Skup prirodnih brojeva - N N = {1, 2, 3,...} Aksiom matematičke indukcije Neka je N skup prirodnih brojeva i M podskup od N. Ako za M vrijede svojstva: 1) 1 M 2) n M (n + 1) M,
radni nerecenzirani materijal za predavanja R(f) = {f(x) x D}
 = {f(x) x D}") Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
ТЕМПЕРАТУРА СВЕЖЕГ БЕТОНА
 ТЕМПЕРАТУРА СВЕЖЕГ БЕТОНА empertur sežeg beton menj se tokom remen i zisi od ećeg broj utijnih prmetr: Početne temperture mešine (n izsku iz mešie), emperture sredine, opote hidrtije ement, Rzmene topote
ТЕМПЕРАТУРА СВЕЖЕГ БЕТОНА empertur sežeg beton menj se tokom remen i zisi od ećeg broj utijnih prmetr: Početne temperture mešine (n izsku iz mešie), emperture sredine, opote hidrtije ement, Rzmene topote
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =
/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =") 100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
Pravilo 1. Svaki tip entiteta ER modela postaje relaciona šema sa istim imenom.
 1 Pravilo 1. Svaki tip entiteta ER modela postaje relaciona šema sa istim imenom. Pravilo 2. Svaki atribut entiteta postaje atribut relacione šeme pod istim imenom. Pravilo 3. Primarni ključ entiteta postaje
1 Pravilo 1. Svaki tip entiteta ER modela postaje relaciona šema sa istim imenom. Pravilo 2. Svaki atribut entiteta postaje atribut relacione šeme pod istim imenom. Pravilo 3. Primarni ključ entiteta postaje