Encimi v tleh. k 1 E + S ES E + P. k 3. k 2
|
|
|
- Κύρα Αθανασίου
- 5 χρόνια πριν
- Προβολές:
Transcript

1 Encimi v tleh k 1 E + S ES E + P k 2 k 3 1
 je spojina, na katero deluje encim kompleks (ES) je intermediarna struktura encima in substrata produkt (P) je spojina, ki zapusti")
2 Encimi in definicije encim (E) je protein, ki ga producira celica in deluje kot katalizator znižuje aktivacijsko energijo in na ta način pospeši hitrost reakcije substrat (S) je spojina, na katero deluje encim kompleks (ES) je intermediarna struktura encima in substrata produkt (P) je spojina, ki zapusti encim 2
3 Kinetika encimskih reakcij v i v max ½ v max K m [S] v i = začetna hitrost (mol/čas) [S] = koncentracija substrata (mol) v max = maksimalna hitrost K m = koncentracija substrata, ko je v1 = ½ v max (Michaelis Menten konstanta) 3
4 Recipročni diagram kinetike encimskih reakcij naklon = K m /v max 1/v i 1/v max -1/K m 1/S 4
5 Katalitični mehanizmi 1. Kislinsko-bazna kataliza 2. nukleofilan kataliza 3. Kataliza s kovinskimi ioni 5
6 Mehanizem delovanja encimov intramolekularne interakcije (orientacija substrata v optimalno lego) orientacija katalizatorja v optimalno lego regulacija lokalnega okolja (npr. hidrofilnost, hidrofobnost) kombinacija večih mehanizmov organske katalize (nukleofilna, kislinska bazna, kovinska kataliza) 6
7 Mehanizem delovanja encimov zniževanje aktivacijske energije pri obstoječih reakcijskih koordinatah zniževanje reakcijske energije s spremembo reakcijskih koordinat ravnotežne konstante vezava substrata na encim so v rangu od 10-2 do 10-8 M, kar je enako prosti energiji vezave v rangu od 12 do 50 kj/mol (to energijo lahko encim uporabi za reakcijo) 7
8 Mehanizem delovanja encimov K m predstavlja moč ES kompleksa in je enak (k 2 /k 1 ), večji K m pomeni šibko vezavo, majhen K m pomeni močno vezavo število kataliziranih molekul substrata (turnover number) je enako reakcijski konstanti za nastanek produkta k 3, predstavlja število molekul substrata, ki jih encim katalizira v eni sekundi, ko je encim popolnoma nasičen (v max /k 3 ), čas cikla je 1/(v max /k 3 ) kinetična učinkovitost encima (k 3 /K m ) kaže kakšno je razmerje med razpadom ES kompleksa med produkte in reaktante 8
9 Mehanizem delovanja encimov kinetična učinkovitost encima je odvisna od kemijskih reakcij na encimu in od hitrosti vezave substrata na encim vrednost k 1 je difuzijsko omejena na 10 8 do 10 9 M -1 s -1, veliko encimov ima učinkovitost, ki se približa difuzijskim omejitvam difuzijska omejitev je zmanjšana pri domenski organiziranosti encimskih kompleksov (npr. dihalna veriga) do 1000 X hitrejše reakcije s -1 encimi z velikim K m imajo nizek trurnover, vendar so izjemno učinkoviti pri nizkih koncentracijah substrata 9
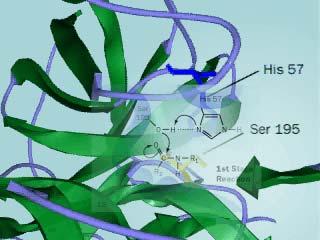

10 Primer katalitičnega mesta encima 10
11 Primer katalitičnega mehanizma encima Ser napade C v karbonilni sk.substrata tvori se tetrahedralen intermediat vezava acilne sk.. na Ser; nastane1.produkt H 2 O napade acil-encim 11

12 Primer katalitičnega mehanizma encima tvori se tetrahedralen intermediat nastane produkt Končni efekt je regeneracija encim, ki je katalizator in v reakciji niti ne nastaja niti se ne porablja. 12
13 Inhibicija encimske aktivnosti Poleg difuzije lahko na k 1 delujejo tudi inhibitorji, ki blokirajo: aktivno mesto regulatorno mesto dostop substrata do aktivnega mesta ali izstop produkta inhibitorji lahko delujejo reverzibilno ali ireverzibilno reverzibilni encimski inhibitorji vstopajo v nekovalentne interakcije, ireverzibilni tvorijo kovalentne povezave s tarčnimi skupinami na encimu 13
14 Reverzibilni inhibitorji inhibicija je večja s povečanjem koncentracije inhibitorja hitra začetna stopnja inhibicije hitrost encimske aktivnosti je znižana, vendar konstantna nekovalentne interakcije z encimom delujejo kot analog substrata - tekmovanje za specifično vezavno mesto Tipi inhibicije: - kompetitivna (konkurenčna) inhibicija - nekompetitivna inhibicija - antikompetitivna inhibicija - inhibicija mešanega tipa 14
15 Tipi reverzibilne inhibicije KOMPETITIVNI INHIBITOR NEKOMPETITIVNI INHIBITOR ANTIKOMPETITIVNI INHIBITOR INHIBITOR MEŠANEGA TIPA 15
16 Ireverzibilni inhibitorji stopnja inhibicije narašča s časom enakomerno zniževanje hitrosti encimske aktivnosti kovalentne povezave s tarčnimi skupinami na encimu - vezava z aminokislinskimi radikali v aktivnem centru inaktivacija zmanjša koncentracijo aktivnega encima dajo informacijo o funkcionalnih skupinah ali kofaktorjih encima praktično so uporabni kadar je namen uničiti encimsko aktivnost 16
17 Vezava encimov na površino Za opis adsorpcije proteinov na površine je potrebno poznati fizikalno-kemijske lastnosti adsorbenta adsorpcijske izoterme adsorpcijsko kinetiko število adsorbiranih molekul na enoto dostopne površine adsorbenta konformacijo adsorbiranega proteina 17
18 Vezava encimov na površino Sile, ki vodijo v adsorpcijo encimov na površine (predvsem neg. nabite) g ads = H ads -T. S ads Entalpijski prispevek elektrostatske interakcije van der Waalsove interakcije Entropijski prispevek hidrofobne interakcije spremembe v konformaciji proteina ionska izmenjava povečanje entropije raztopine 18
19 Sprememba katalitske aktivnosti encima zaradi adsorpcije predpostavljeno je drugačno obnašanje kot prosto v raztopini odvisno je od lege proteina na površini; lateralno premikanje odvisno od tega ali je adsorbent inertna površina (podlaga) ali aktivno sodeluje v katalizi pogosto se optimalno ph-območje pomakne višje lahko se spremeni reakcijski mehanizem 19
20 Inaktivacija encimske aktivnosti 1. Avtoklaviranje sprostijo se: org. snovi(ak, OH), Mn, N, P, S, NH 4 -N ter NO 3 -N znižajo se koncentracije topnih elementov v sledovih večina talnih encimov se inaktivira(denaturira) pri T = o C avtoklaviranje inhibira hidrolizo 20
21 Inaktivacija encimske aktivnosti 2. Sterilizacija s suhim zrakom padec koncentracije organske snovi ter porast v Mn in NH 4 -N več encimske aktivnosti v tleh se ohrani pri daljšem času sterilizacije pri nizkih T, kot pri krajši izpostavi visokim T 21

22 Inaktivacija encimske aktivnosti 3. Sterilizacija z γ-sevanjem radioaktivnih elementov- 60Co/ 137Cs povečajo se količine Mn, org. N, NH 4 -N, P celice se ne morejo več deliti, vendar so še vedno opazili biokemično aktivnost ohranise večja aktivnost encimov kot pri sterilizaciji s suhim zrakom 22
 voda v tleh in organizmih absorbira infrardeče sevanje, zato se močno poviša temperatura, to ubije")
23 Inaktivacija encimske aktivnosti 4. Mikrovalovno sevanje(2450 MHz) voda v tleh in organizmih absorbira infrardeče sevanje, zato se močno poviša temperatura, to ubije organizme postopek bolj učinkovit pri vlažnih kot pri suhih tleh močno se poveča vsebnostmn 23
24 Inaktivacija encimske aktivnosti 5. Sterilizacija z etilenoksidom, propilen oksidom, metilbromidom Adsorbcijaoksidanaorg.in anorg.koloidepovzroči povečano vsebnost organske snovi propilen oksid: Mn in Fe; inhibira hidrolizo etilen oksid napada proteine, NK in druge org. snovi, še posebno škodljiv je za sulfhidrilne skupine, ki jih alkilira 24
, 24 ur poveča količino organske snovi v raztopini in amonijaka poveča aktivnost")
25 Inaktivacija encimske aktivnosti 7. Preplinjevanje s kloroformom(chcl 3 ), 24 ur poveča količino organske snovi v raztopini in amonijaka poveča aktivnost arilsulfataze zmanša aktivnost: β-glukozidaze, kislih in bazičnih fosfomonoesteraz, proteaz in ureaz 25
26 Inaktivacija encimske aktivnosti 8. Inhibicija mikrobne aktivnosti z azidom(kn 3, NaN 3 ), cianidom, toluenon ali živosrebrovim kloridom azid in cianid: 1-10 %, toksična toluen: 5-10% HgCl 2 : odvisno od vzorca-500mg/kg delujejo na sulfhidrilne skupine(katalitična mesta encimov) HgCl 2 povzroči najmanj sprememb v prsti v primerjavi z drugimi postopki, deluje na celične proteine, ki vsebujejo sulfidrilne skupine azid povzroči Mn ter porast v ph, zmanjšano aktivnosti invertaze in amilaze, na fosfataze nima večjega vpliva 26
27 Inaktivacija encimske aktivnosti 9. Težke kovine: Zn in Cu sulfat inhibirajo encimske reakcije : - s tvorbo kompleksov s substratom - z vezavo na aktivno mest proteina ali - z vezavo na kompleks encim substrat inhibirajo ureaze, fosfomonoesteraze, nitratne reduktaze, amidaze 27
28 Vpliv težkih kovin na encimsko aktivnostv tleh Primer: onesnaženost z bakrom 400 mg Cu/kg zemlje: - zmanjšanje dehidrogenazne aktivnosti za 82% - zmanjšanje ureazne aktivnosti za 47% - zmanjšanje kislo fosfatazne aktivnosti za 34% - zmanjšanje alkalno fosfatazne aktivnosti za 40% 1200 mg Cu/kg zemlje: - zmanjšanje dehidrogenazne aktivnosti za 98% - zmanjšanje ureazne aktivnosti za 62% - zmanjšanje kislo fosfatazne aktivnosti za 62% - zmanjšanje alkalno fosfatazne aktivnosti za 67% 28

29 Inaktivacija encimske aktivnosti Toluen se najpogosteje uporablja pri proučevanju ekstracelularnih encimov je plazmolitičen in antiseptik učinek je odvisen od koncentracije, časa inkubacije, vsebnosti vlage v tleh in predhodnega tretiranja v krajših inkubacijskih časih inhibira sintezo encimov in prepreči asimilacijo reakcijskih produktov za zaustavitev mikrobnega podvajanja je potrebna vsaj 20% konc. inhibira aktivnost katalaz in dehidrogenaz, arilamidaz njegova plazmolitična aktivnost prispeva k aktivnosti intracelularnih encimov v tleh 29
30 Merjenje aktivnosti encimov E + S + I ES + I E + P merimo nastanek produkta (P), po dodatku substrata (S), pri kontroliranih pogojih (temperatura, EI + S ESI ph in ionska moč) če je prisoten inhibitor (I), E = encim S = substrat I = inhibitor P= produkt nastajajo poleg ES tudi ES in ESI kompleksi 30
31 Metode za kvantitativno določanje encimske reakcij SPEKTROFOTOMETRIČNE FLUORIMETRIČNE RADIOAKTIVNE TITRIMETRIČNE MANOMETRIČNE (detekcija plinskih substratov ali produktov) ELEKTRODNE (detekcija ionskih substratov ali produktov) KROMATOGRAFSKE KAPILARNA ELEKTROFOREZA 31
32 Spektrofotometrične metode kolorimetrično določanje spremembe koncentracije substrata ali produkta (neposredno ali posredno, po pretvorbi v obarvano spojino) po ekstrakciji iz vzorca tal, inkubiranega določen čas s substratom pri specifičnih pogojih (npr. T, ph ) najpogostejših načinov merjenja aktivnosti različnih hidrolaz (npr. arilsulfataz, amidaz, fosfataz, celulaz, saharaz) arilsulfataza 0SO 3 K OH + KHSO 4 NO 2 p-nitrofenil sulfat NO 2 p-nitrofenol (rumen v alkalnem) 32
33 Uporaba fluorescence uporaba različnih fluorogenih substratov (substratov s konjugiranimi spojinami, ki ob encimatski cepitvi oddajajo fluorescenco) za ugotavljanje aktivnosti različnih hidrolitičnih encimov v tleh (npr. β- glukozidaz, arilsulfataz, β-celobiaz, β-galaktozaminidaz, kislih in alkalnih fosfataz, lipaz) lipaza ROC-O- O O OH O O H 2 O + + RCOOH CH 3 CH 3 7-Hydroxy-4-methylcoumarin 7-hidroxy-4-methylumbelliferone (nefluorescenten) (močno fluorescenten) 33
34 Radioizotopske metode temeljijo na sledenju radioizotopsko označenih substratov in produktov uporaba 14 C uree za merjenje ureazne aktivnosti preko sledenja sproščenega 14 CO 2. težavo lahko predstavlja ''izotopski efekt'' (spremenjena aktivnost encima pri hidrolizi substrata s težjim izotopom, glede na običajnega) 34
35 Kaj vpliva na izmerjeno encimsko aktivnost v talnih vzorcih in pomen kontrolnih meritev 1. VPLIV TAL NA ZADRŽEVANJE MERJENIH SPOJIN: različni tipi tal imajo različno kapaciteto za zadrževanje substratov oz. produktov encimskih reakcij potrebno narediti tudi test kapacitete tal za zadrževanje različnih koncentracij merjene spojine 2. VPLIV TAL NA INTENZITETO FLUORESCENTNEGA SIGNALA talne komponente lahko vplivajo na intenziteto fluorescentnega signala, uporaba tal z znanimi koncentracijami flourescentnih produktov (npr. quench standards) 3. VPLIV ABIOTSKIH DEJAVNIKOV na NASTAJANJE MERJENIH SPOJIN: v vsakem naravnem vzorcu kemijske reakcije ne potekajo le encimsko, temveč lahko tudi spontano kemijsko, uporablja abiotske kontrole (npr. avtoklavirana zemlja, pufer, substrat) 35
36 Interpretacija izmerjene encimske aktivnosti večina testov aktivnosti talnih encimov se izvaja pri stabilnih pogojih, ki zagotavljajo optimalne hitrosti katalize (prebitek substrata, optimalna T, ph in razredčitev, ki omogoča prosto difuzijo substrata ). pri optimalnih pogojih merimo POTENCIALNO in ne DEJANSKE encimske aktivnosti v tleh koristno je narediti primerjave izmerjenih potencialnih encimskih aktivnosti z dejanskimi (npr. hidrolizo naravnih substratov, dodanih vzorcem zemlje in inkubiranih pri realističnih in situ pogojih) z zdajšnjimi metodami ne moremo ločiti med encimi, ki se nahajajo na znotraj živih ali mrtvih celic ali izven njih-prosti ali adsorbirani na anorganske koloide oz. v kompleksih s huminskimi snovmi 36
37 Primerljivost rezultatov Ključna težava, ki ovira razvoj talne encimatike, je neprimerljivost rezultatov, oziroma pomanjkanje standardiziranih metod merjenja encimske aktivnosti. 37
38 Viri encimov v tleh rastline, talne živali, mikroorganizmi encimi, ki se akumulirajo v tleh so prisotni kot: - prosti encimi (eksoencimi) - endoencimi (citoplazma, periplazma) konstitutivni encimi (pogosto v uporabi, stalno prisotni) inducibilni encimi (producirani le tedaj, ko je to nujno potrebno) 38
39 Lokacija encimov v tleh intracelualrni encimi periplazmatski encimi encimi na zunanji strani membrane encimi sproščeni med rastjo encimi v sporah, cistah, semenih, endosporah encimi na celičnem debriju in mrtvih celicah encimi sproščeni iz intaktnih ali mrtvih celic encimi v ES kompleksih adsorbirani na glino adsorbirani na humus neaktivna celica (V) Resting structure Living cells živa celica Release Release Growth/Devision Growth/Devision (iv) (iv) živa celica Living cell (ii) Secretion (i) (vii) Extracellular enzymes ekstracelularni encimi Humic acids huminske kisline (I) Death Death (iii) Lysis Lysis Humus-enzyme Humus-encim complex kompleks mrtva intaktna celica (vii) Dead intact cell (viii) Entraped enzymes (x) (vi) Lysis Dead disintegrated cell (vii) (vi) Enzyme-substrate ES-kompleks complex Clay minerali minerals glin Adsorbed enzymes (ix) Clay-enzyme Glina-encim complex kompleks mrtva desintegrirana celica 39


40 Vpliv rastlinskega pokrova na aktivnost encimov v tleh na vzorcu tal so gojili češmina, grmičasto travo in borovnice po treh mesecih so se mikrobne združbe začele močno razlikovati npr. tla poraščena z grmičasto travo so imela višje aktivnosti endocelulaze, aminopeptidaze, alkalne fosfataze in fenolne oksidaze,, ter najnižje koncentracije nitrata v tleh (rastlina( rastlina je imela večji privzem nitrata in fosfatov) 40
41 Vpliv kmetijstva na aktivnost talnih encimov encimske aktivnosti so v splošnem višje v neobdelanih tleh (naravna vegetacija, trave ali pašniki) odsotnost oranja daljša prisotnost vegetacije in obsežen koreninski sistem trav večji rizosferni efekt in večja encimska aktivnost povišano aktivnost arilsulfataze ob dodajanju organskih gnojil, povišana aktivnost fosfataz ob dodajanju fosfatnih gnojil pri stročnicah in žitih povišana aktivnosti glukozidaz in galaktozidaz intenzivne obdelave zemlje, gojenja monokultur in obsežne uporabe anorganskih gnojil povzroči znižanje aktivnosti talnih encimov 41
42 Delitev encimov glede na tip reakcije, ki jo katalizirajo klasifikacija encima oksidoreduktaze transferaze hidrolaze liaze izomeraze ligaze tip katalizirane reakcije redoks reakcije prenos funkcionalne skupine rreakcija hidrolize eliminacija funkcionalne skupine izomerizacija formiranje vezi 42
43 Glikozidaze oziroma glikozid hidrolaze So skupina encimov, ki katalizirajo hidrolizo različnih glikozidov. GLIKOZID + H 2 O sladkor + aglikan Glikozidi pa so etrom podobne spojine sladkorje. Glikozidaze običajno poimenujemo po tipu vezi, ki jo hidrolizirajo. 43
44 Glikozidaze oziroma glikozid hidrolaze α GLUKOZIDAZA (maltaza), katalizira hidrolizo α-d-glukopiranozida β- GLUKOZIDAZA (gentobioza ali celobioza), katalizira hidrolizo β-dglukopiranozida α - GALAKTOZIDAZA (melibiaza) β- GALAKTOZIDAZA (laktaza( laktaza), katalizirata hidrolizo melibioze in laktoze 44
45 β-glukozidaza hidroliza terminalnih, ne-reducirajočih β-d-glukoznih ostankov, pri katerih se sprosti β-d-glukoza celobioza + H 2 O 2 beta-d-glukoza reakcija je enosmerna encim je široko specifičen za beta-d-glukozide, inaktivira se pri 70 C 45
46 Pomembni substrati β-glukozidaze Aryl glucosides p-nitrophenyl b-d-glucoside p-nitrophenyl b-d-glucopyranoside p-nitrophenyl b-d-xyloside o-nitrophenyl b-d-glucoside Disaccharides Cellobiose (b-1,4) Gentiobiose (b-1,6) Laminaribiose (b-1,3) Sophorose (b-1,2) Salicin Phenyl-b-glucoside Alkyl b-glucosides Methyl-glucoside Hexyl-glucoside Octyl-glucoside Dodecyl-glucoside 46
47 Glukozidaze in razgradnja celuloze
48 Glukozidaze in razgradnja celuloze mineralizacija ogljika β-glukozidaza katalizira končni omejujoč korak pri razgradnji celluloze HO HOH 2 C HO O OH O-R + H 2 O β-glukozidaza HO HOH 2 C HO O OH OH + OH NO 2 p-nitrofenil-β-d-glukozid β-glukozid + p-nitrofenol 48
49 β-galaktozidaza (laktaza) Je encim, ki hidrolizira disaharid laktozo v D-glukozo in D-galaktozo. β- GALAKTOZIDAZA LAKTOZA β-d-galactopiranozil-(1,4) 4)- β-d-glukopiranoza GALAKTOZA GLUKOZA 49
50 β-fruktofuranozidaza Druga imena: ivertaza saharaza β-fruktozidaza sukraza β-h-fruktozidaza invertin maxinvert L 1000 fruktozil invertaza alkalna invertaza kisla invertaza sistematično ime: β-d-fruktofuranozid fruktohidrolaza 50
51 Saharaze maltoza, laktoza in saharoza so najbolj pogosti vodotopni disaharidi sestavljeni iz dveh monosaharidov povezanih z glikozidno vezjo eden od najbolj pogostih sladkorjev v rastlinah, mikroorganizmih in živalih je saharoza 51

52 β-fruktofuranozidaza hidroliza terminalnih nereducirajočih β-d-fruktofuranozidnih ostankov v β-d-fruktofuranozidih katalizira tudi fruktotransferazne reakcije to reakcijo lahko katalizira tudi -D-glukozidaza, ki odceplja stran terminalno glukozno enoto 52

53 Ksilanaze Za hidrolizo glavne verige sta odgovorna 2 encima: endo-β-1,4-ksilanaza cepi hrbtenico do ksilooligosaharidov β-ksilozidaza hidrolizira ksilooligosaharide do ksiloze 53
54 Aktivnost ksilanaz v odvisnosti od substrata AKTIVNOST ena enota (U) encimske aktivnosti ustreza 1 mol ksiloznega ekvivalenta sproščenega v 1 min pri pogojih: 50 mm natrijev citratni pufer, ph 5.0, 50 C vir ksilana ksilanazna aktivnost (Uml -1 ) macesen 70 oves 36 pšenični otrobi 28 breza 10 koruzni storži 9 ksiloza 3 celuloza 2 Aktivnost ekstracelularne endoksilanaze v supernatantu kulture Streptomyces sp. AMT-3 po 10 dnevih rasti pri 30 C v medijih z različnimi substrati. 54
55 Hitinaze hitin je nerazvejan polisaharid iz N-acetil-D-glukozaminskih (GlcNAc) enot povezanih z β-1,4-vezjo hitinolitsko aktivnost ima več encimov: -endohitinaza-hidroliza hitinske verige v oligomere - eksohitinaza oz. Hitobiozidaza - hidrolizira na terminalnem koncu hitinske verige ali oligomerov, nastane dimer hitobioza -1,4-β- N-acetilglukozaminidaza hidroliza kratkih oligomerov in hitobioze, nastane N-acetil-glukozamin 55
56 Encimske reakcije pri mineralizaciji dušika aminizacija amonifikacija nitrifikacija peptid NH + 4 NO - 2 NO - amino kislina 3 56
57 Mineralizacija amino kislin v tleh amino kislina amonifikacija nitrifikacija arilamidaza amino kislina amidohidrolaze npr. L-glutaminaza L-asparaginaza L-aspartaza NH 4 + NO 2 - NO 3 - Sproščanje amino kislin iz organskega materiala 57
58 Arilamidazna aktivnost NH 2 O NHCCHCH 2 CH(CH 3 ) 2 NH 2 L-leucin b-naftilamid (testni substrat) + H 2 O arilamidaza β-naftilamin + HOOC CHCH 2 CH(CH 3 ) 2 NH 2 leucin 58
59 Amidazna aktivnost RCONH 2 + H 2 O amidaza NH 3 + RCOOH katalizira hidrolizo amidov C-N vezi pri čemer nastaja amonij in ustrezna karboksilna kislina je specifična za alifatske amide prisotna v mikroorganizmih (bakterije,kvasovke in glive), živalih in rastlinah specifičnost in kinetične lastnosti amidaze v tleh so pomembne, ker so substrati tega encima (amidi) potencialna dušikova gnojila 59
60 L-asparaginazna aktivnost COOH HC NH 2 CH 2 CO COOH + H 2 O L-asparaginaza HC NH 2 + NH 3. CH 2 COOH NH 2 velik del sproščenega NH 4+ pri mineralizaciji se sprosti s hidrolizo asparagina in glutamina v naravi široko zastopan encim (mikroorganizmi, rastline) aktivnost pada z naraščajočo globino tal in z nižanjem vsebnosti org. ogljika 60
61 L-aspartazna aktivnost COOH HC NH 2 + H 2 O L-aspartaza COOH C H + NH 3 CH 2 CH COOH COOH 61
62 L-glutaminazna aktivnost COOH COOH H C NH 2 CH 2 + H 2 O L-glutaminaza H C NH 2 CH 2 + NH 3. CH 2 CH 2 CO COOH NH 2 široko zastopan v naravi, zaznali so ga pri številnih živalih, rastlinah in mikroorganizmih L-glutaminaza najbolj občutljiva amidohidrolaza na spremembe ph v tleh aktivnost encima se znižuje z globino vzorca tal, aktivnost je močno povezana s količino organskega ogljika in dušika. Močno je tudi povezana s aktivnostjo amidaze in ureaze in L-asparaginaze. Inhibitorji L-glutaminaze Ag(I), Hg(II), Sn(II), Cr(III), Ti(IV) in W(VI) 62
63 Ureazna aktivnost O NH 2 C NH 2 ureaza H 2 O CO 2 + 2NH 3 v naravi široko zastopan encim: mikrobne, rastlinske in živalske celice katalizira razgradnjo uree, dihidroksiuree in semikarbazida optimalni ph :alkalen med ph ali pa nevtralen med ph 6-7 temperaturni optimum: 60ºC, vendar vzorce tal navadno inkubiramo na 30ºC 63
64 Encimska aktivnost in mineralizacija fosforja fosfomonesteraze fosfodiesteraze trifosfomonoesteraze hidrolaze fosfatnih anhidridov hidrolazep-n vezi Izvor: večinoma mikroorganizmi in rastline, lahko ostanki živali Lokacija: znotraj živih celic, zunaj celic- prosto v tleh(nestabilne), adsorbirane na anorganske komponente tal ali v kompleksih z organskimi koloidi 64
65 Fosfatazna aktivnost O HO P O O kisle ali alkalne O monofosfataze + H 2 O HO P O + ROH O OH O P OR 1 + H 2 O fosfodiesteraze OH O P OH + R 1 OH OR 2 OR 2 65
66 Fosfomonoesteraza (FME) R-O O P O- + H 2 O FME R-OH + OH O P O- O- O- najbolje raziskana od fosfataz nizka substratna specifičnost oz. spososobnost hidrolize mnogih spojin, med katerimi najpogostejše substrate predstavljajo: inozitol- P, polifosfati, mononukleotidi in fosforilirani sladkorji glede na ph optimum delovanja, jih delimo na: - kisle: : 4-6,54 - nevtralne: : 6,5-8 - alkalne: : razmerje med kislimi in alkalnimi fosfatazami je potencialni indikator ph statusa tal 66
67 Sulfatazna aktivnost ROSO 3 _ p-nitrofenil-sulfat (testni substrat) + H 2 O arilsulfataza ROH + H + + SO p-nitrofenol (barvni produkt) 67
68 Katalaza H 2 O 2 H 2 O + ½ O 2 katalazna aktivnost v zemlji je izredno stabilna kaže se povezava katalazne aktivnosti z vsebnostjo organskega ogljika aktivnost pada z globino 68
69 Peroksidaze donor elektronov + H 2 O 2 oksidiran donor + 2H 2 O najdemo ga v rastlinah, živalih aerobnih mikroorganizmih kot donor elektronov uporablja fenole, aminofenole, organske kisline kot kofaktor se pojavlja hem ali flavin, vanadij, selen, cistein ktivnost vezana predvsem na zgornjih 5 cm huminske kisline povečajo aktivnost peroksidaz, omogočajo nastanek bolj stabilnih organskih spojin 69
70 Monooksigenaze encimi, ki inkorporirajo 1 atom O 2 v substrat, drugega pa reducirajo do H 2 O ker katalizirajo nastanek hidroksilne skupine, jih pogosto imenujemo hidroksilaze elektronski donor pri večini predstavlja NADH ali NADPH, prenos elektrona na kisik pa poteka preko flavinskega kofaktorja omogočajo izrabo ogljikovodikov nekaterim mikroorganizmom (npr. Nocardia, Pseudomonas, Mycobacterium) 70
71 Metan monooksigenaza CH 4 + O 2 CH 3 OH + H 2 O metan monooksidaza lahko deluje tudi kot oksidaza amonijaka, pri tem oba substrata tekmujeta za razpoložljiv encim amonij je za metanotrofe strupen oksidacija metana(metanotrofne bakterije) poteka na meji med oksičnim in anoksičnim delom metanotrofne bakterije za svojo rast potrebujejo metan in kisik metanotrofi vsebujejo precej sterolov, ki se nahajajo na zapletenih membranskih sistemih, ki sodelujejo pri oksidaciji metana 71
72 Amonij monooksigenaza NH 3 + O 2 + 2H+ + 2e - NH 2 OH + H 2 O encim amonij monooksidaza katalizira proces dodajanja enega kisikovega atoma na molekulo amonija (nitritacijske bakterije) pri nizkih koncentracijah amonija v zemlji, pride do inhibicije metan monooksigenaze, saj se namesto metana porablja amonij vzrok temu so soli v zemlji: ki spremenijo osmotski potencial zemlje (kar lahko vpliva na oksigenacijo metana), vplivajo na izmenjavo ionov v zemlji (kar lahko poveča dostopnost amonija) direktno vplivajo na združbo oksidatorjev metana. 72
73 Esteraze R 1 C O OR 2 R 1 C O OH + HO R 2 esteraze se od lipaz razlikujejo, ker je njihovo delovanje omejeno na kratkoverižne maščobne kisline karboksilna esteraza katalizira hidrolizo mnogih alifatskih in aromatskih estrov esteraze in druge hidrolitične encime so izolirali iz mnogih mikroorganizmov v tleh 73
74 Lipaze lipaze hidrolizirajo estre glicerola z dolgimi maščobnimi kislinami delujejo kot vmesnik med hidrofobnim substratom in hidrofilnim topilom aktivnost je visoka v emulzijah zaradi tega reakcija odstopa od Michaelis-Mentenovea modela 74
75 Dehidrogenaze XH 2 + A X + AH 2 XH 2 je pri tem organska molekula( donor vodika), A pa je pri tem akceptor vodika pomemben encim, ki ga merimo v tleh, kaže na celokupno oksidativno aktivnost celic uporabljamo tetrazolijeve soli (INT, CTC, TTC, XTT), ki so reducirane ob prisotnosti dehidrogenaz, pri tem pride do spremembe barve 75
Mehanizmi encimske katalize
 Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Mehanizmi encimske katalize Kompleks ES stabilizirajo šibke interakcije Sledi razcep ES in nastanek vezi po različnih mehanizmih, ki vključujejo prehodni nastanek kovalentnih vezi - približanje in orientacija
Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
 FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
AMILAZE. Encimi, ki hidrolizirajo ogljikove hidrate. struktura škroba
 Encimi, ki hidrolizirajo ogljikove hidrate substrati: ogljikovi hidrati (škrob, celuloza, poli in oligosaharidi) encimi: glikozidaze glikozidna vez encimska specifičnost konfiguracija glikozidne vezi kemijska
Encimi, ki hidrolizirajo ogljikove hidrate substrati: ogljikovi hidrati (škrob, celuloza, poli in oligosaharidi) encimi: glikozidaze glikozidna vez encimska specifičnost konfiguracija glikozidne vezi kemijska
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Encimi. Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije
 Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Encimi Splošne lastnosti - osnove delovanja, specifičnost, energijski vidik nekatalizirane in encimsko katalizirane reakcije Kofaktorji, koencimi in prostetične skupine Mehanizmi encimske katalize Klasifikacija
Matjaž Zorko Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Univerza v Ljubljani, Fakulteta za farmacijo, 2009 MOLEKULARNA ENCIMOLOGIJA KLASIFIKACIJA IN NOMENKLATURA ENCIMOV (s primeri) Matjaž Zorko Medicinska fakulteta GLEJ: http://ibk.mf.uni-lj.si/teaching/lab_medicina/default.html
Encimi.
 Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
Encimska kinetika govori o hitrosti encimske reakcije
 Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
Encimska kinetika govori o hitrosti encimske reakcije uvod encimska kinetika po Michaelisu in Mentenovi kinetika večsubstratnih encimskih reakcij kinetika alosteričnih encimov encimska inhibicija Kako
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Uravnavanje encimske aktivnosti
 Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Uravnavanje encimske aktivnosti Uvod Mehanizmi encimske katalize Inhibicija encimov Modulacija aktivnosti alosteričnih encimov Uravnavanje encimske aktivnosti: - uravnavanje koncentracije encimov - spreminjanje
Proizvodnja in uporaba encimov
 Kmetijska šola Grm Sevno 13 8000 Novo mesto Proizvodnja in uporaba encimov ( predmet: Biotehnologija ) Gorenja vas; 7.3.2007 Avtor: Lidija Gorenc Jožica Koračin, 3.c Mentor: Jana Goršin Fabjan Kazalo:
Kmetijska šola Grm Sevno 13 8000 Novo mesto Proizvodnja in uporaba encimov ( predmet: Biotehnologija ) Gorenja vas; 7.3.2007 Avtor: Lidija Gorenc Jožica Koračin, 3.c Mentor: Jana Goršin Fabjan Kazalo:
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
1 Uvod v biokemijo. Slika. Nekakj spoznanj s področja biokemije.
 Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
Univerza na Primorskem, Fakulteta za vede o zdravju Prehransko svetovanje - dietetika, 1. stopenjski študij Predmet: Biokemija, 1. letnik Avtorica: Doc. dr. Zala Jenko Pražnikar 1 Uvod v biokemijo Biokemijo
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
2.1. MOLEKULARNA ABSORPCIJSKA SPEKTROMETRIJA
 2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
2.1. MOLEKULARNA ABSORPCJSKA SPEKTROMETRJA Molekularna absorpcijska spektrometrija (kolorimetrija, fotometrija, spektrofotometrija) temelji na merjenju absorpcije svetlobe, ki prehaja skozi preiskovano
Talna kemija. Kaj je potrebno poznati:
 Talna kemija Kaj je potrebno poznati: splošno kemijo mol, molaren, normalnost, ekvivalent ionska jakost, aktivnost ravnotežne konstante funkcionalne skupine hidratacija, hidroliza redoks reakcije Redoks
Talna kemija Kaj je potrebno poznati: splošno kemijo mol, molaren, normalnost, ekvivalent ionska jakost, aktivnost ravnotežne konstante funkcionalne skupine hidratacija, hidroliza redoks reakcije Redoks
Teze predavanj iz mikrobne biokemije*
 Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
Tom Turk Teze predavanj iz mikrobne biokemije* Ljubljana, november 1996, oktober 2003 *slike so na CD v ppt formatu, besedilo v drobnem tisku je priporočljivo ni pa obvezno, Študentom mikrobiologije 1.
[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda
![[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda](/thumbs/89/99439684.jpg "[ E] [ ] kinetika encimske pretvorbe. razpolovni čas. ln pretvorba encimov sledi kinetiki prvega reda") kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
kinetika encimske pretvorbe pretvorba encimov sledi kinetiki prvega reda sinteza encimov sledi kinetiki nultega reda v ravnotežju de = k E s kd dt [ ] ' ' [ Et ] k s ks ' ' [ E ] k [ E ] k [ E ] = 1 e
FOTOSINTEZA Wan Hill primerjal rastlinsko fotosintezo s fotosintezo BAKTERIJ
 FOTOSINTEZA FOTOSINTEZA je proces, pri katerem s pomočjo svetlobne energijje nastajajo v živih celicah organske spojine. 1772 Priestley RASTLINA slab zrak dober zrak Rastlina s pomočjo svetlobe spreminja
FOTOSINTEZA FOTOSINTEZA je proces, pri katerem s pomočjo svetlobne energijje nastajajo v živih celicah organske spojine. 1772 Priestley RASTLINA slab zrak dober zrak Rastlina s pomočjo svetlobe spreminja
Energije in okolje 1. vaja. Entalpija pri kemijskih reakcijah
 Entalpija pri kemijskih reakcijah Pri obravnavi energijskih pretvorb pri kemijskih reakcijah uvedemo pojem entalpije, ki popisuje spreminjanje energije sistema pri konstantnem tlaku. Sistemu lahko povečamo
Entalpija pri kemijskih reakcijah Pri obravnavi energijskih pretvorb pri kemijskih reakcijah uvedemo pojem entalpije, ki popisuje spreminjanje energije sistema pri konstantnem tlaku. Sistemu lahko povečamo
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Osnovni ekološki principi
 Osnovni ekološki principi SOODVISNOST RECIKLIRANJE KOMPLEKSNOST kroženje snovi avtokatalitičnost feedback mehanizmi kroženje elementov rast in biorazgradnja PERZISTENCA Dušik 7 N dušik 14.00677 Atomsko
Osnovni ekološki principi SOODVISNOST RECIKLIRANJE KOMPLEKSNOST kroženje snovi avtokatalitičnost feedback mehanizmi kroženje elementov rast in biorazgradnja PERZISTENCA Dušik 7 N dušik 14.00677 Atomsko
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Prehrana in metabolizem
 Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
Prehrana in metabolizem Hranila Energija Kataliza in encimi Oksidacije-redukcije Prenašalci elektronov Visoko energetske spojine Fermentacija Respiracija in transport elektronov Metabolizem vsi kemični
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
ENCIMI ZGRADBA ENCIMOV NEKATERI ENCIMI IN NJIHOVI KOFAKTORJI
 Visoko specializirani proteini in mala skupina katalitičnih molekul RNA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10 16 x Zmanjšajo E a, ne vplivajo
Visoko specializirani proteini in mala skupina katalitičnih molekul RNA biološki katalizatorji Katalizirajo kemijske reakcije v živih organizmih - pospeševanje do 10 6-10 16 x Zmanjšajo E a, ne vplivajo
Ogljikovi hidrati - monosaharidi
 Ogljikovi hidrati - monosaharidi V tem poglavju se boste naučili (ponovili) - kaj so osnovne lastnosti ogljikovih hidratov, - kaj so monosaharidi, - katere so dokazne reakcije za monosaharide. Enostavni
Ogljikovi hidrati - monosaharidi V tem poglavju se boste naučili (ponovili) - kaj so osnovne lastnosti ogljikovih hidratov, - kaj so monosaharidi, - katere so dokazne reakcije za monosaharide. Enostavni
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Jure Stojan in Marko Goličnik Medicinska fakulteta
 Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
Univerza v Ljubljani, Fakulteta za farmacijo, 2015/16 Program: Laboratorijska biomedicina Predmet: MOLEKULARNA ENCIMOLOGIJA Jure Stojan in Marko Goličnik Medicinska fakulteta Prof. dr. Matjaž Zorko do
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Metabolizem oz. presnova
 Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
OKOLJSKO NARAVOSLOVJE 2. Predavanja v študijskem letu 2. del 2012/2013
 OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
OKOLJSKO NARAVOSLOVJE 2 Predavanja v študijskem letu 2. del 2012/2013 MORFOLOGIJA IN ZGRADBA BAKTERIJ Oblike bakterij Poznamo razne oblike bakterij in sicer: koki, bacili, vibrioni, spirile, aktinomicete
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
pretvarja v nestrupeno obliko, ki lahko vstopa v biosintezo nukleotidov *i) NH 4
 NH 4") 1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
1. Piruvat karboksilaza a) je aktivirana z acetil koencimom A b) je regulatorni encim glukoneogeneze c) se nahaja v citosolu d) vsebuje prostetično skupino biotin e) potrebuje za svojo aktivnost NADH *f)
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI. Razbitje celic za izolacijo intracelularnih produktov
 ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI Razbitje celic za izolacijo intracelularnih produktov Izolacija in čiščenje intracelularnega encima za uporabo v živilih Intracelularni produkti proteini (inkluzijska
ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI Razbitje celic za izolacijo intracelularnih produktov Izolacija in čiščenje intracelularnega encima za uporabo v živilih Intracelularni produkti proteini (inkluzijska
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
VODE, ODPLAKE, ODPADKI ŠTUDIJ ŽIVILSKE TEHNOLOGIJE DEZINFEKCIJA
 Slide 1 Slide 2 DEFINICIJA JE UNIČENJE MIKROORGANIZMOV, KI SO SPOSOBNI POVZROČITI BOLEZEN ESENCIALNA IN ZADNJA OVIRA PRED IZPOSTAVITVIJO LJUDI PATOGENIM MIKROORGANIZMOM, VKLJUČNO Z VIRUSI, BAKTERIJAMI,
Slide 1 Slide 2 DEFINICIJA JE UNIČENJE MIKROORGANIZMOV, KI SO SPOSOBNI POVZROČITI BOLEZEN ESENCIALNA IN ZADNJA OVIRA PRED IZPOSTAVITVIJO LJUDI PATOGENIM MIKROORGANIZMOM, VKLJUČNO Z VIRUSI, BAKTERIJAMI,
CM707. GR Οδηγός χρήσης... 2-7. SLO Uporabniški priročnik... 8-13. CR Korisnički priručnik... 14-19. TR Kullanım Kılavuzu... 20-25
 1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Ravnotežja v raztopini
 Ravnotežja v raztopini TOPILO: komponenta, ki jo je več v raztopini.v analizni kemiji uporabljamo organska in anorganska topila. Topila z veliko dielektrično konstanto (ε > 10) so polarna in ionizirajo
Ravnotežja v raztopini TOPILO: komponenta, ki jo je več v raztopini.v analizni kemiji uporabljamo organska in anorganska topila. Topila z veliko dielektrično konstanto (ε > 10) so polarna in ionizirajo
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
VPLIV REAKCIJSKIH SPREMENLJIVK NA POTEK IN HITROST MODELNE REAKCIJE NATRIJEVEGA TIOSULFATA S KLOROVODIKOVO KISLINO
 OSNOVNA ŠOLA PRIMOŽA TRUBARJA LAŠKO VPLIV REAKCIJSKIH SPREMENLJIVK NA POTEK IN HITROST MODELNE REAKCIJE NATRIJEVEGA TIOSULFATA S KLOROVODIKOVO KISLINO (RAZISKOVALNO DELO) Avtorici: Lea Lešek Povšič in
OSNOVNA ŠOLA PRIMOŽA TRUBARJA LAŠKO VPLIV REAKCIJSKIH SPREMENLJIVK NA POTEK IN HITROST MODELNE REAKCIJE NATRIJEVEGA TIOSULFATA S KLOROVODIKOVO KISLINO (RAZISKOVALNO DELO) Avtorici: Lea Lešek Povšič in
Tabele termodinamskih lastnosti vode in vodne pare
 Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
- Geodetske točke in geodetske mreže
 - Geodetske točke in geodetske mreže 15 Geodetske točke in geodetske mreže Materializacija koordinatnih sistemov 2 Geodetske točke Geodetska točka je točka, označena na fizični površini Zemlje z izbrano
- Geodetske točke in geodetske mreže 15 Geodetske točke in geodetske mreže Materializacija koordinatnih sistemov 2 Geodetske točke Geodetska točka je točka, označena na fizični površini Zemlje z izbrano
Tla in ioni. naboj mineralov. električni dvojni sloj. adsorpcija. adsorpcijske izoterme. izmenjava ionov. ravnotežje trdno - raztopina
 Tla in ioni naboj mineralov električni dvojni sloj adsorpcija adsorpcijske izoterme izmenjava ionov ravnotežje trdno raztopina Ravnotežje trdno raztopina asimilacija rastlina, mikrobi mineralizacija imobilizacija
Tla in ioni naboj mineralov električni dvojni sloj adsorpcija adsorpcijske izoterme izmenjava ionov ravnotežje trdno raztopina Ravnotežje trdno raztopina asimilacija rastlina, mikrobi mineralizacija imobilizacija
ΟΜΟΣΠΟΝ ΙΑ ΕΚΠΑΙ ΕΥΤΙΚΩΝ ΦΡΟΝΤΙΣΤΩΝ ΕΛΛΑ ΟΣ (Ο.Ε.Φ.Ε.) ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2012. Ηµεροµηνία: Τετάρτη 18 Απριλίου 2012 ΕΚΦΩΝΗΣΕΙΣ
 ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 2012. Ηµεροµηνία: Τετάρτη 18 Απριλίου 2012 ΕΚΦΩΝΗΣΕΙΣ") ΟΜΟΣΠΟΝ ΙΑ ΕΚΠΑΙ ΕΥΤΙΚΩΝ ΦΡΟΝΤΙΣΤΩΝ ΕΛΛΑ ΟΣ (Ο.Ε.Φ.Ε.) ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 0 Ε_.ΧλΘ(ε) ΤΑΞΗ: ΚΑΤΕΥΘΥΝΣΗ: ΜΑΘΗΜΑ: ΘΕΜΑ Α Β ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΧΗΜΕΙΑ Ηµεροµηνία: Τετάρτη 8 Απριλίου
ΟΜΟΣΠΟΝ ΙΑ ΕΚΠΑΙ ΕΥΤΙΚΩΝ ΦΡΟΝΤΙΣΤΩΝ ΕΛΛΑ ΟΣ (Ο.Ε.Φ.Ε.) ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ ΕΠΑΝΑΛΗΠΤΙΚΑ ΘΕΜΑΤΑ 0 Ε_.ΧλΘ(ε) ΤΑΞΗ: ΚΑΤΕΥΘΥΝΣΗ: ΜΑΘΗΜΑ: ΘΕΜΑ Α Β ΓΕΝΙΚΟΥ ΛΥΚΕΙΟΥ ΘΕΤΙΚΗ ΧΗΜΕΙΑ Ηµεροµηνία: Τετάρτη 8 Απριλίου
Izločanje zdravilnih učinkovin iz telesa:
 Izločanje zdravilnih učinkovin iz telesa: kinetični vidiki Biofarmacija s farmakokinetiko Aleš Mrhar Izločanje učinkovin Izraženo s hitrostjo in maso, dx/dt = k e U očistkom in volumnom, Cl = k e V Hitrost
Izločanje zdravilnih učinkovin iz telesa: kinetični vidiki Biofarmacija s farmakokinetiko Aleš Mrhar Izločanje učinkovin Izraženo s hitrostjo in maso, dx/dt = k e U očistkom in volumnom, Cl = k e V Hitrost
Logatherm WPL 14 AR T A ++ A + A B C D E F G A B C D E F G. kw kw /2013
 WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
WP 14 R T d 9 10 11 53 d 2015 811/2013 WP 14 R T 2015 811/2013 WP 14 R T Naslednji podatki o izdelku izpolnjujejo zahteve uredb U 811/2013, 812/2013, 813/2013 in 814/2013 o dopolnitvi smernice 2010/30/U.
Rast mikrobne populacije
 Rast mikrobne populacije rast posamezne celice : rast populacije hranila fizikalno-kemijski faktorji rasti zaprt in odprt način namnoževanja merjenje rasti mikrobne populacije matematični opis rasti Rast
Rast mikrobne populacije rast posamezne celice : rast populacije hranila fizikalno-kemijski faktorji rasti zaprt in odprt način namnoževanja merjenje rasti mikrobne populacije matematični opis rasti Rast
13. Vaja: Reakcije oksidacije in redukcije
 1. Vaja: Reakcije oksidacije in redukcije a) Osnove: Oksidacija je reakcija pri kateri posamezen element (reducent) oddaja elektrone in se pri tem oksidira (oksidacijsko število se zviša). Redukcija pa
1. Vaja: Reakcije oksidacije in redukcije a) Osnove: Oksidacija je reakcija pri kateri posamezen element (reducent) oddaja elektrone in se pri tem oksidira (oksidacijsko število se zviša). Redukcija pa
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
UVOD CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN
 = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN") CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
Vaja: Odbojnostni senzor z optičnimi vlakni. Namen vaje
 Namen vaje Spoznavanje osnovnih fiber-optičnih in optomehanskih komponent Spoznavanje načela delovanja in praktične uporabe odbojnostnega senzorja z optičnimi vlakni, Delo z merilnimi instrumenti (signal-generator,
Namen vaje Spoznavanje osnovnih fiber-optičnih in optomehanskih komponent Spoznavanje načela delovanja in praktične uporabe odbojnostnega senzorja z optičnimi vlakni, Delo z merilnimi instrumenti (signal-generator,
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - VSŠ 2005/06
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
Cefalosporini ostali β-laktami
 Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
Transformator. Delovanje transformatorja I. Delovanje transformatorja II
 Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
Transformator Transformator je naprava, ki v osnovi pretvarja napetost iz enega nivoja v drugega. Poznamo vrsto različnih izvedb transformatorjev, glede na njihovo specifičnost uporabe:. Energetski transformator.
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
Rizosfera procesi, ki vplivajo na razopložljivost mineralnih hranil. Ekofiziologija in mineralna prehrana rastlin 2005/06
 Rizosfera procesi, ki vplivajo na razopložljivost mineralnih hranil Ekofiziologija in mineralna prehrana rastlin 2005/06 Literatura Marschner H. (1995) Mineral nutrition of higher plants. Poglavje 15:
Rizosfera procesi, ki vplivajo na razopložljivost mineralnih hranil Ekofiziologija in mineralna prehrana rastlin 2005/06 Literatura Marschner H. (1995) Mineral nutrition of higher plants. Poglavje 15:
6. ΤΕΛΙΚΗ ΙΑΘΕΣΗ ΤΑΦΗ. 6.1. Γενικά
 6. ΤΕΛΙΚΗ ΙΑΘΕΣΗ ΤΑΦΗ 6.1. Γενικά Είναι γεγονός ότι ανέκαθεν ο τελικός αποδέκτης των υπολειµµάτων της κατανάλωσης και των καταλοίπων της παραγωγικής διαδικασίας υπήρξε το περιβάλλον. Στις παλιότερες κοινωνίες
6. ΤΕΛΙΚΗ ΙΑΘΕΣΗ ΤΑΦΗ 6.1. Γενικά Είναι γεγονός ότι ανέκαθεν ο τελικός αποδέκτης των υπολειµµάτων της κατανάλωσης και των καταλοίπων της παραγωγικής διαδικασίας υπήρξε το περιβάλλον. Στις παλιότερες κοινωνίες
Zakaj proučevati tla?
 Zakaj proučevati tla? medij za rast rastlin in pridelkov produkcija in absorbcija plinov medij za rast mikroorganizmov habitat za živali veliki integrator vseh delov terestričnega ekosistema vir za proučevanje
Zakaj proučevati tla? medij za rast rastlin in pridelkov produkcija in absorbcija plinov medij za rast mikroorganizmov habitat za živali veliki integrator vseh delov terestričnega ekosistema vir za proučevanje
Osnovne stehiometrijske veličine
 Osnovne stehiometrijske veličine Stehiometrija (grško: stoiheion snov, metron merilo) obravnava količinske odnose pri kemijskih reakcijah. Fizikalne veličine, s katerimi kemik najpogosteje izraža količino
Osnovne stehiometrijske veličine Stehiometrija (grško: stoiheion snov, metron merilo) obravnava količinske odnose pri kemijskih reakcijah. Fizikalne veličine, s katerimi kemik najpogosteje izraža količino
KEMIJA. Iztok Prislan Biotehniška fakulteta Oddelek za živilstvo
 KEMIJA Iztok Prislan Biotehniška fakulteta Oddelek za živilstvo Estri Najpogostejši derivati karboksilnih kislin so estri: Estri običajno nastanejo pri reakciji med kislino in alkoholom oz. fenolom (esterifikacija):
KEMIJA Iztok Prislan Biotehniška fakulteta Oddelek za živilstvo Estri Najpogostejši derivati karboksilnih kislin so estri: Estri običajno nastanejo pri reakciji med kislino in alkoholom oz. fenolom (esterifikacija):
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
K E M IČ N A V L A K N A IZ N A R AV N IH PO L IM E R O V 1. RAZDELITEV NARAVNIH POLIMEROV [1] N A R AV N A V L A K N A
![K E M IČ N A V L A K N A IZ N A R AV N IH PO L IM E R O V 1. RAZDELITEV NARAVNIH POLIMEROV [1] N A R AV N A V L A K N A](/thumbs/77/76219690.jpg "K E M IČ N A V L A K N A IZ N A R AV N IH PO L IM E R O V 1. RAZDELITEV NARAVNIH POLIMEROV [1] N A R AV N A V L A K N A") 1. RAZDELITEV NARAVNI PLIMERV [1] Na sliki 1.1 je prikazana razdelitev naravnih vlaken. N A R AV N A V L A K N A R G A N SK A A N R G A N SK A azb est SEM EN SK A STEBELN A bo m b až, k ap o k, s v i l
1. RAZDELITEV NARAVNI PLIMERV [1] Na sliki 1.1 je prikazana razdelitev naravnih vlaken. N A R AV N A V L A K N A R G A N SK A A N R G A N SK A azb est SEM EN SK A STEBELN A bo m b až, k ap o k, s v i l
Rast mikrobne populacije
 Rast mikrobne populacije rast posamezne celice : rast populacije hranila fizikalno-kemijski faktorji rasti zaprt in odprt način namnoževanja merjenje rasti mikrobne populacije matematični opis rasti Rast
Rast mikrobne populacije rast posamezne celice : rast populacije hranila fizikalno-kemijski faktorji rasti zaprt in odprt način namnoževanja merjenje rasti mikrobne populacije matematični opis rasti Rast
Homogena snov je snov, ki ima vsepovsod enake lastnosti in sestavo Heterogena snov je snov, katere sestava in lastnosti so na različnih mestih
 Homogena snov je snov, ki ima vsepovsod enake lastnosti in sestavo Heterogena snov je snov, katere sestava in lastnosti so na različnih mestih različne Postopki ločevanja zmesi:iz zmesi je mogoče ločiti
Homogena snov je snov, ki ima vsepovsod enake lastnosti in sestavo Heterogena snov je snov, katere sestava in lastnosti so na različnih mestih različne Postopki ločevanja zmesi:iz zmesi je mogoče ločiti
Če je aw manjši od 0,3 so lipidi polj izpostavljeni in zmanjšana je hidratacija. Če je aw večji od 0,5 je povečana difuzija in nabrekanje.
 ŢIVILSKA KEMIJA 1. Kaj je aktivnost vode? Definicija; katero lastnost vode opišemo s tem parametrom. Aw je parameter, s katerim kvantitativno ovrednotimo dostopnost vode v ţivilu (je merilo za hlapljivost
ŢIVILSKA KEMIJA 1. Kaj je aktivnost vode? Definicija; katero lastnost vode opišemo s tem parametrom. Aw je parameter, s katerim kvantitativno ovrednotimo dostopnost vode v ţivilu (je merilo za hlapljivost
3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo
 Osnovni princip kromatografije 3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo 4. vaja Ionskoizmenjevalna ter afinitetna kromatografija Miha Pavšič komponente zmesi različne interakcije
Osnovni princip kromatografije 3. vaja Razsoljevanje proteinov z gelsko izključitveno kromatografijo 4. vaja Ionskoizmenjevalna ter afinitetna kromatografija Miha Pavšič komponente zmesi različne interakcije
Metabolizem in energija
 Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Fazni diagram binarne tekočine
 Fazni diagram binarne tekočine Žiga Kos 5. junij 203 Binarno tekočino predstavljajo delci A in B. Ti se med seboj lahko mešajo v različnih razmerjih. V nalogi želimo izračunati fazni diagram take tekočine,
Fazni diagram binarne tekočine Žiga Kos 5. junij 203 Binarno tekočino predstavljajo delci A in B. Ti se med seboj lahko mešajo v različnih razmerjih. V nalogi želimo izračunati fazni diagram take tekočine,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Aleš Mrhar. kinetični ni vidiki. Izraženo s hitrostjo in maso, dx/dt očistkom
 Izločanje zdravilnih učinkovin u iz telesa: kinetični ni vidiki Biofarmacija s farmakokinetiko Univerzitetni program Farmacija Aleš Mrhar Izločanje učinkovinu Izraženo s hitrostjo in maso, dx/ k e U očistkom
Izločanje zdravilnih učinkovin u iz telesa: kinetični ni vidiki Biofarmacija s farmakokinetiko Univerzitetni program Farmacija Aleš Mrhar Izločanje učinkovinu Izraženo s hitrostjo in maso, dx/ k e U očistkom
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Farmacevtska kemija SOD mimetiki. doc.dr. Janez Mravljak, mag.farm.
 SD mimetiki doc.dr. Janez Mravljak, mag.farm. Priporočena literatura Pregledni članek: A. Perdih, S. Pečar: Katalitični antioksidanti kot nove zdravilne učinkovine; farm vestn 2006; 57, 24-29. doc. dr.
SD mimetiki doc.dr. Janez Mravljak, mag.farm. Priporočena literatura Pregledni članek: A. Perdih, S. Pečar: Katalitični antioksidanti kot nove zdravilne učinkovine; farm vestn 2006; 57, 24-29. doc. dr.
Nekatere interakcije v lipidnem metabolizmu
 Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
Biosinteza maščobnih kislin fosfolipidi glicerol triacilglicerol fosfatidat MK glicerol-p esterifikacija acil-oa A S L i p o g e n e z a steroidi steroidogeneza holesterol holesterogeneza glukoza piruvat
ZGRADBA ATOMA IN PERIODNI SISTEM
 ZGRADBA ATOMA IN PERIODNI SISTEM Kemijske lastnosti elementov se periodično spreminjajo z naraščajočo relativno atomsko maso oziroma kot vemo danes z naraščajočim vrstnim številom. Dmitrij I. Mendeljejev,
ZGRADBA ATOMA IN PERIODNI SISTEM Kemijske lastnosti elementov se periodično spreminjajo z naraščajočo relativno atomsko maso oziroma kot vemo danes z naraščajočim vrstnim številom. Dmitrij I. Mendeljejev,
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
VAJA 1 ALBUMINU NEZNANE KONCENTRACIJE DOLOČITE KONCENTRACIJO: Izmerite A 280 albumina v vzorcu. Vrednost absorbance vzorca naj ne preseže 1!
 VAJA 1 ALBUMINU NEZNANE KONCENTRACIJE DOLOČITE KONCENTRACIJO: Izmerite A 280 albumina v vzorcu. Vrednost absorbance vzorca naj ne preseže 1! S pomočjo standardne raztopine (raztopina albumina koncentracije
VAJA 1 ALBUMINU NEZNANE KONCENTRACIJE DOLOČITE KONCENTRACIJO: Izmerite A 280 albumina v vzorcu. Vrednost absorbance vzorca naj ne preseže 1! S pomočjo standardne raztopine (raztopina albumina koncentracije
FARMAKOKINETIKA. Hitrosti procesov Farmakokinetični ni parametri Aplikacija. Tatjana Irman Florjanc
 FARMAKOKINETIKA Hitrosti procesov Farmakokinetični ni parametri Aplikacija Tatjana Irman Florjanc Inštitut za farmakologijo in eksperimentalno toksikologijo, MF, Univerza v Ljubljani V praksi - kontrola
FARMAKOKINETIKA Hitrosti procesov Farmakokinetični ni parametri Aplikacija Tatjana Irman Florjanc Inštitut za farmakologijo in eksperimentalno toksikologijo, MF, Univerza v Ljubljani V praksi - kontrola
Razgradnja maščobnih kislin. Ketonska telesa H + NAD+ NADH. Pregled metabolizma MK. lipoprotein-lipaza. maščobne kisline.
 Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
Razgradnja maščobnih kislin Ketonska telesa Pregled metabolizma MK stradanje computing hranjeno stanje triacilgliceroli v hilomikronih in VLDL kladiščenje maščob maščobno tkivo An overview of fatty acidhormonsko
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Mehanizmi djelovanja enzima i regulacija aktivnosti enzima
 Mehanizmi djelovanja enzima i regulacija aktivnosti enzima Boris Mildner Aktivnost enzima mijenja se promjenama temperature, ph, inhibitorima te vezanjem malih molekula ili iona Temperatura povećava brzinu
Mehanizmi djelovanja enzima i regulacija aktivnosti enzima Boris Mildner Aktivnost enzima mijenja se promjenama temperature, ph, inhibitorima te vezanjem malih molekula ili iona Temperatura povećava brzinu
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - UNI
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
Heterogene ravnoteže taloženje i otapanje. u vodi u prisustvu zajedničkog iona u prisustvu kompleksirajućegreagensa pri različitim ph vrijednostima
 Heterogene ravnoteže taloženje i otapanje u vodi u prisustvu zajedničkog iona u prisustvu kompleksirajućegreagensa pri različitim ph vrijednostima Ako je BA teško topljiva sol (npr. AgCl) dodatkom
Heterogene ravnoteže taloženje i otapanje u vodi u prisustvu zajedničkog iona u prisustvu kompleksirajućegreagensa pri različitim ph vrijednostima Ako je BA teško topljiva sol (npr. AgCl) dodatkom
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
KISLINE IN BAZE ARRHENIUSOVA DEFINICIJA KISLIN IN BAZ
 6. KISLINE IN BAZE KISLINE IN BAZE ARRHENIUSOVA DEFINICIJA KISLIN IN BAZ kisline so snovi, ki v vodni raztopini disocirajo vodikove ione (H + ), baze pa snovi, ki v vodni raztopini disocirajo hidroksidne
6. KISLINE IN BAZE KISLINE IN BAZE ARRHENIUSOVA DEFINICIJA KISLIN IN BAZ kisline so snovi, ki v vodni raztopini disocirajo vodikove ione (H + ), baze pa snovi, ki v vodni raztopini disocirajo hidroksidne
LIPIDI IN PREHRANA ŠPORTNIKA. Žiga Drobnič, Filip Zupančič, 1.b
 LIPIDI IN PREHRANA ŠPORTNIKA Žiga Drobnič, Filip Zupančič, 1.b Lipide delimo na: maščobe (masti,olja) - kombinacija molekule glicerola s tremi dolgoverižnimi organskimi kislinami - maščobnimi kislinami
LIPIDI IN PREHRANA ŠPORTNIKA Žiga Drobnič, Filip Zupančič, 1.b Lipide delimo na: maščobe (masti,olja) - kombinacija molekule glicerola s tremi dolgoverižnimi organskimi kislinami - maščobnimi kislinami
POROČILO 3.VAJA DOLOČANJE REZULTANTE SIL
 POROČILO 3.VAJA DOLOČANJE REZULTANTE SIL Izdba aje: Ljubjana, 11. 1. 007, 10.00 Jan OMAHNE, 1.M Namen: 1.Preeri paraeogramsko praio za doočanje rezutante nezporedni si s skupnim prijemaiščem (grafično)..dooči
POROČILO 3.VAJA DOLOČANJE REZULTANTE SIL Izdba aje: Ljubjana, 11. 1. 007, 10.00 Jan OMAHNE, 1.M Namen: 1.Preeri paraeogramsko praio za doočanje rezutante nezporedni si s skupnim prijemaiščem (grafično)..dooči