Struktura nukleoida E. coli. a) u izolovanom nespiralizovanom stanju. b) hromozom podeljen u nekoliko petlji povezanih malim RNK molekulima.
|
|
|
- Τιτάνος Κορωναίος
- 5 χρόνια πριν
- Προβολές:
Transcript
1 PRINCIPI GENETIKE 8 Euhromatin i heterohromatin. Struktura i funkcija hromozoma. Genetičke osnove deobe ćelija. Ponašanje hromozoma u mitozi i mejozi. Vanjedarno nasleđivanje. Organizacija genoma mitohondrija
2 Struktura nukleoida E. coli. a) u izolovanom nespiralizovanom stanju. b) hromozom podeljen u nekoliko petlji povezanih malim RNK molekulima. c) svaka petlja se dodatno superspiralizuje. d) tretman spiralizovanog genoma endonukleazom dovodi do relaksacije DNK samo u tretiranim petljama e) tretman RNA-zom raspliće samo delove povezane malim RNK molekulima.
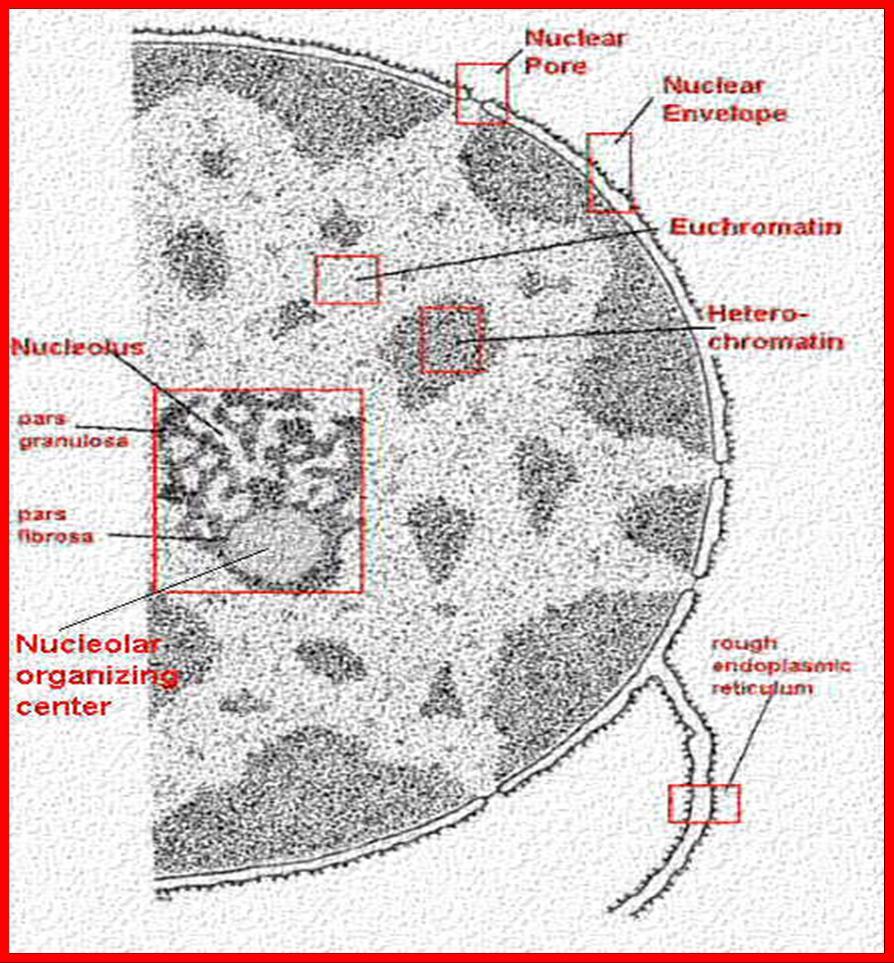
3 NUCLEUS

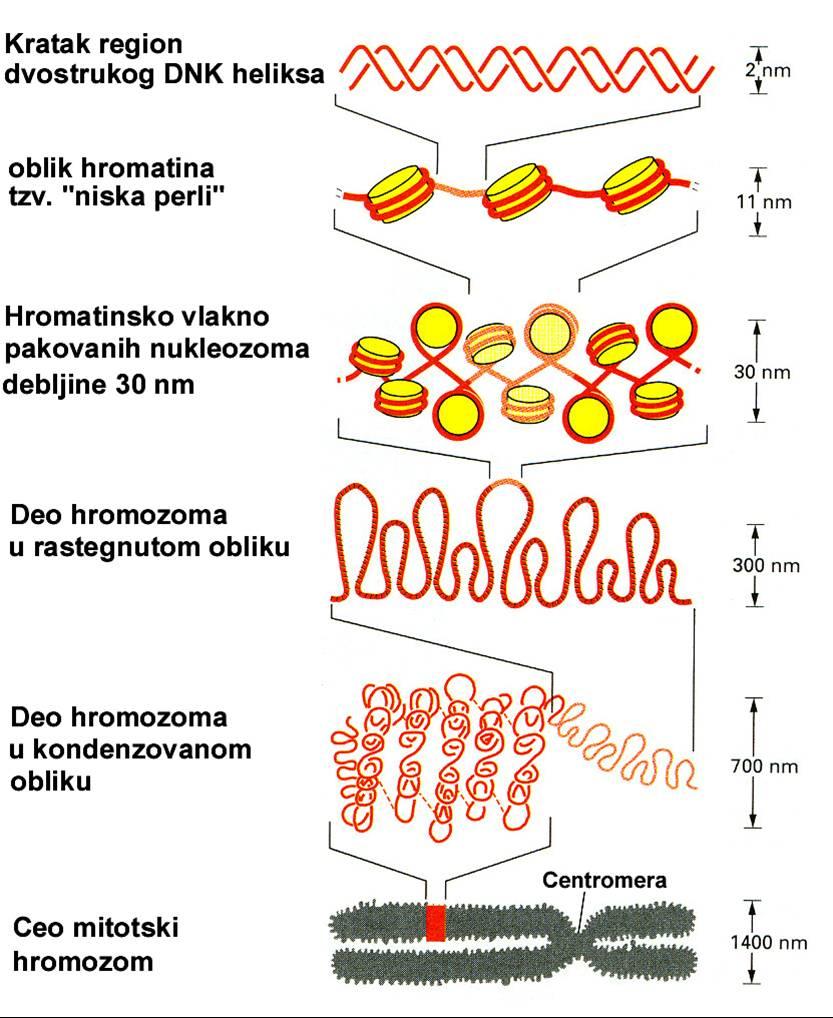
4 Nivoi pakovanja nasledne informacije u ćeliji
5 Šematska ilustracija strukture nukleozomskog čvorića Svaki nukleozomski čvorić koji se sastoji od histonskog oktamera je obmotan sa DNK koju čini 146 baznih parova. Ovaj proteinski histonski oktamer čini po dva elementa histona H2A, H2B, H3 i H4. Svaki histon se sastoji od amino kiselina. Svaki histonski oktamer vezan je za susedni slobodnim delom vezujuće DNK.
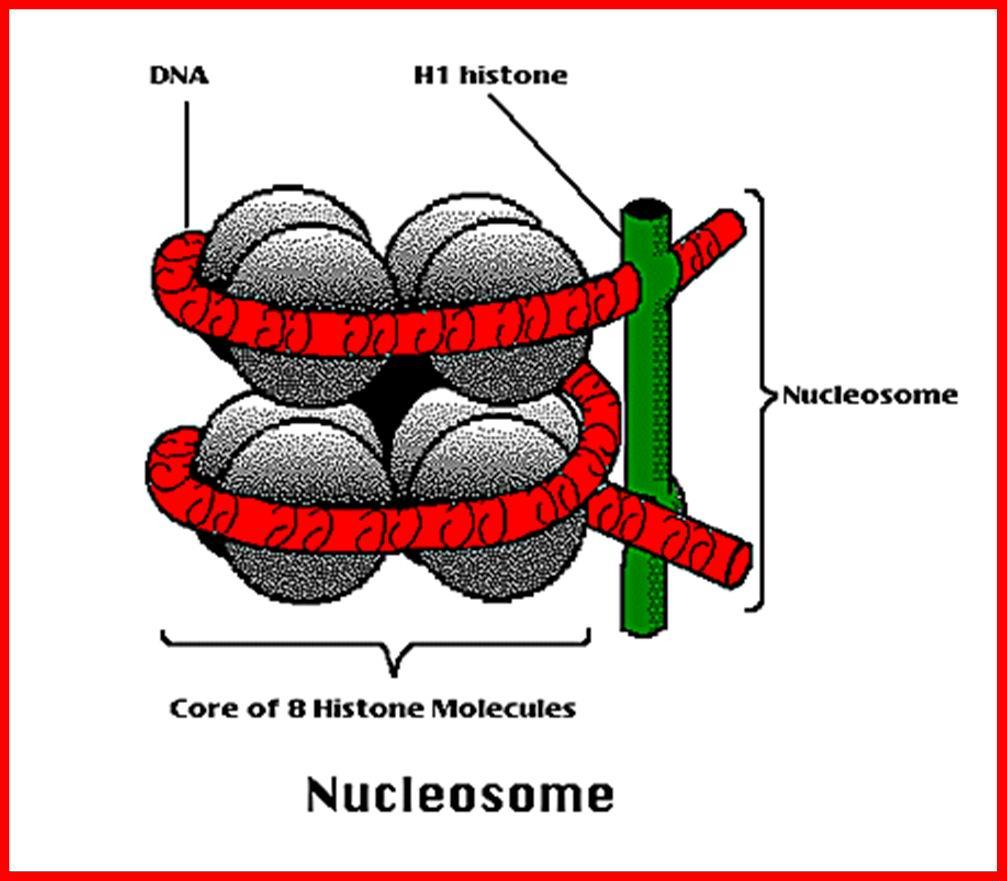
6 Građa nukleozoma
7 Hromatin je kompleks DNK, proteina i malih količina RNK. Hemijske analize izolovanog hromatina govore da se sastoji iz DNK i proteina uz male količine RNK. Proteini su grupisani u dve velike klase: histoni i nehistonski hromozomski proteini. Histoni imaju glavnu ulogu u strukturiranju hromatina. Prisutni su u hromatinu svih viših eukariota u količinama ekvivalentnim količini DNK (približno isti težinski odnos). Histoni obuhvataju 5 različitih proteina: H1, H2a, H2b, H3 i H4, prisutnih skoro u svim tipovima eukariotskih ćelija uz malobrojne izuzetke (u spermatozoidima se umesto histona nalaze mali bazni proteini tzv. Protamini). Histoni su bazni proteini, bogati aminokiselinama lizinom i argininom (sadrže 20-30% arginina i lizina). Pozitivno naelektrisani ostaci baznih aminokiselina histona mogu lako da interreaguju sa kiselim fosfatnim grupama u DNK. Histoni kompleksuju sa DNK i formiraju posebne strukture nukleozome. Svaki nukleozom sadrži po četiri molekula histona H2a, H2b, H3 i H4 (oktamer histona) oko kojih se nalazi DNK
8 Građa nukleozoma.

9 Struktura hromozoma
10 Lokalni region hromozoma se može ponašati kao posebna strukturna jedinica zahvaljujući specijalnoj ulogzi koju imaju histoni H1 u povezivanju nukleozomskih oktamera. Otpuštanjem histona H1, region se može naglo dekondenzovati po primanju informacije od strane nekog spoljašnjeg faktora. Smatra se da se dekondenzacija ovog tipa dešava pri procesu aktivacije gena.
11 Histoni pomažu pakovanju DNK u hromozome; kod čoveka se nalazi skoro 2 m DNK u diploidnom jedru. Otuda se postavlja pitanje kako je DNK upakovana u hromozome. Danas je izvesno da se u svakom hromozomu nalazi isključivo jedan linearni molekul DNK, koji se pruža od jednog kraja hromozoma preko centromere do drugog kraja hromozoma. Najveći hromozom u humanom genomu ima DNK dužine oko 85 mm (85000μm). Ovaj DNK molekul upakuje se u metafazni hromozom dijametra 0,5 μm i dužine oko 10 μm. To znači da se od golog molekula DNK do hromozoma ostvaruje kondenazcija od oko10000 puta. Nasuprot histonima, nehistonski proteini obuhvataju frakciju sastavljenu iz velikog broja jako heterogenih proteina. Sastav nehistonskih proteina jako varira ne samo između različitih vrsta, već i između različitih ćelija unutar istog organizma. Među nehistonskim proteinima treba, pored ostalog, tražiti regulatore genske aktivnosti.
12 Građa nukleozoma.
13 Pojam heterohromatina i euhromatina HETEROHROMATIN više kondenzovan, tamno obojen a) KONSTITUTIVNI DEO KONSTITUCIJE - centromerni i telomerni regioni b) FAKULTATIVNI inaktivirani X hromozom, Barovo telo EUHROMATIN svetliji, manje kondenzovan, aktivan
14 NUCLEUS
15 X hromozom konstutivni heterohromatin Centromerni heterohromatin euhromatin Humani kariotip (2n = 46, XY) obojen tehnikom G-traka
16 Genetičke analize ukazuju da je heterohromatin uglavnom genetički neaktivan. Većina do danas okarakterisanih eukariotskih gena locirana je u euhromatinskim regionima hromozoma. Štaviše, heterohromatin je, po pravilu, bogat visoko repetitivnim, tandemski raspoređenim DNK sekvencama. Molekularno biološke analize eukariotskoh hromatina pokazale su da postoje tri osnovna tipa DNK sekvenci: - visoko repetitivni nizovi (satelitska DNK) sastavljeni od višestruko ponovljenih kratkih nizova nukleotida, najčešće različitog baznog sastava od ukupne DNK. Prisutni su u svim, ili skoro svim, hromozomima, a sama funkcija ovih nizova još uvek nije dovoljno pojašnjena. - intermedijarna (srednje repetitivna) DNK obuhvata DNK sekvence sa znatno manjim brojem kopija u genomu - u poređenju sa visokorepetitivnom DNK. Pored ostalog, u srednje ponovljene nizove spadaju geni za rrnk, geni za trnk, za histone, α i β globine, tubuline, aktine itd.-
17 - unikalni nizovi ( jedinstvene sekvence) pokazuju veliku raznovrsnost i obuhvataju najveći broj strukturnih gena. Geni se mogu javiti samo u jednoj kopiji (engl. single copy ) ili u svega 2-3 kopije u genomu. Tipovi DNK sekvenci otkriveni su pre svega zahvaljujući proučavanju kinetike renaturacije DNK. Naime, rastavljanje lanaca DNK ostvareno dejstvom visokih temperatura ili hemijskih agenasa označeno je kao denaturacija DNK. Suprotan proces - ponovno uspostavljanje dvolančanog heliksa do kojeg dolazi po uklanjanju agenasa za denaturaciju, naziva se renaturacija DNK. Stepen renaturacije uveliko zavisi od koncentracije DNK u rastvoru i kompeksnosti DNK sekvence. Stoga je proces renaturacije kod visokorepetitivnih DNK najbrži i njaefikasniji, dok se renaturacija unikalnih nizova ostvaruje najsporije. Konačno, treba istaći da eukariotski hromozomi poseduju posebne strukture na svojim krajevima, tzv. telomere.
18 Već decenijama se zna da telomerni regioni imaju jedinstvene karakteristike. Barbara McClintock je pokazala da se odstranjivanjem telomera formiraju lepljivi krajevi hromozoma koji imaju tendenciju da se međusobno spajaju. Nasuprot tome, prirodni krajevi normalnih hromozoma stabilni su i nemaju tendenciju ka međusobnom spajanju. Danas se zna da postoji specifičan enzim telomeraza (transferaza telomera), sposoban da dodaje telomere na krajeve hromozoma. Funkcija samih telomera verovatno se sastoji u: - stabilizaciji hromozoma usled sprečavanja egzonukleaza da vrše degradaciju linearnih DNK s kraja hromozoma - sprečavanju fuzionisanja krajeva hromozoma sa drugim DNK molekulima - olakšavanju replikacije krajeva linearnih DNK. Naime, da nema telomera krajevi hromozoma bi se posle svakog ciklusa replikacije malo skraćivali - zato što na samom kraju lanca sa diskontinuiranom replikacijom ( lagging strand ) ne bi postojala DNK koja omogućuje sintezu RNK početnice sa slobodnim 3 -OH krajem.
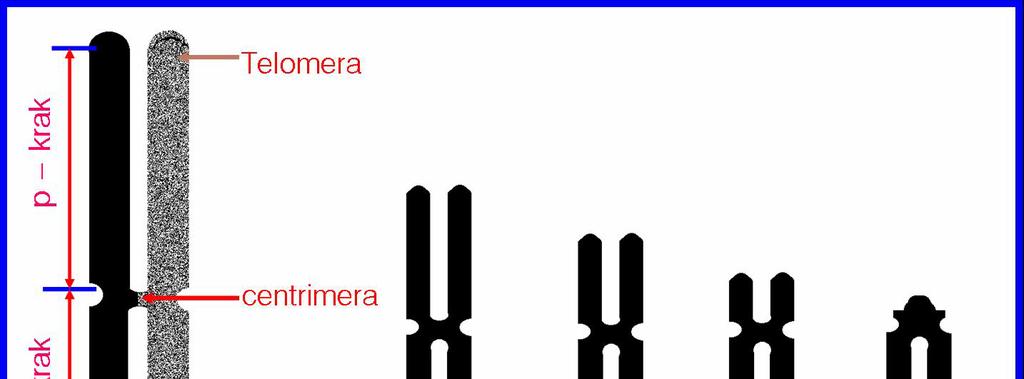
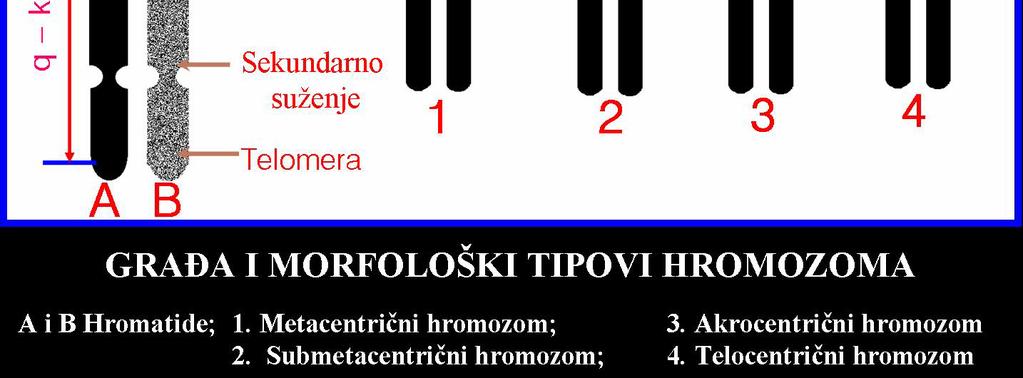
19

20 B A C A. K A R I O T I P, B. K A R I O G R A M, C. I D I O G R A M

21 Četkasti hromozomi u ovocitima vodozemaca posmatrani pod svetlosnim

22 Politeni hromozomi pljuvačnih žlezda Drosophila melanogaster posmatrani pod svetlosnim mikroskopom
23 Kariogram psa 2n=78,XY
24 Kariogram mačke 2n=38,XY
25 Kariogram svinje 2n=38,XY


26 Kariogram govečeta 2n=60,XX

27 Kariotip bika obojen tehnikom G-traka, 2n = 60, XY.

28 Kariogram jarca 2n=60,XY
29 Kariogram ovna 2n=54,XY
30 Kariogram kobile 2n=64,XX
31 Kariogram kokoši 2n=18,ZW
32 Kariogram radilice medonosne pčele 2n=32,XX


33 Normalni kariotip svinje (2n=38, XX)
34 FUNKCIJA HROMOZOMA Poređenje kariotipova dve srodne vrste jelena: A. Muntacus reevesi i B. Muntiacus muntiak. (2n=46,XX/XY ) (2n=7,XY 1 Y 2 )
35 Ne postoji direktna korelacija između stepena filogentskog razvoja i broja hromozoma. Mada se kod srodnih vrsta može da javi sličan broj hromozoma, poznati su primeri da broj hromozoma znatno razlikuje kod veoma srodnih vrsta. Poređenjem broja hromozoma dve veoma srodne vrste jelena, utvrđeno je da vrsta Muntiacus munitak iz Indije ima samo dva para autozoma i tri polna hromozoma, dok vrsta Muntiacus reevesi iz Kine ima 22 autozoma i dva polna hromozoma Pojava blokova sa identičnim rasporedom gena u kariotipovima srodnih vrsta naziva se sintenijom hromozoma. Poređenjem genoma miša i čoveka pronađeno je ne samo da ove dve vrste imaju veliki broj zajedničkih gena, već veliki blokovi u genomu miša i čoveka sadrže isti redosled gena. Sintenija hromozoma svedoči o preuređivanjima (rearanžmanima) blokova gena tokom evolucije genoma. Hromozomi svojim ponašanjem tokom ćelijskog ciklusa omogućavaju ispoljavanje genetičkog materijala potrebno za ostvarivanje fenotipa ćelije, kao i ravnomernu raspodelu genetičkog materijala u ćerke ćelije pri ćelijskoj deobi
36 U periodu između dve ćelijske deobe (interfaza) hromozomi su u relativno dekondenzovanom stanju, tako da su omogućeni procesi transkripcije i translacije. Sintezom specifičnih proteina ostvaruje se karakterističan izgled i funkcionisanje ćelije. Pored toga, u interfazi dolazi do replikacije DNK, što predstavlja jedan od osnovnih preduslova da se ćelija kasnije podeli. Nakon replikacije svaki hromozom ima dve sestrinske hromatide, a svaka od njih sadrži isti molekul DNK. Za vreme deobe ćelije, hromozomi se prevode u maksimalno kondenzovan oblik pri čemu svaki hromozom sadrži dva identična molekula DNK u svojim sestrinskim hromatidama. Visok stepen kondenzacije DNK i proteina u metafaznim hromozomima neophodan je radi ravnomerne podele naslednog materijala. Velika količina DNK u eukariotskim genomima stvorila je potrebu za evolutivnim postankom kompleksno građenih hromozoma i ćeljskog ciklusa.

37 Šematski prikaz ćelijskog ciklusa

38 Faze ćelijskog ciklusa tipične ćelije sisara

39 Šematski prikaz osnovnih faza deobe životinjske ćelije
40 Šematski prikaz deobnog vretena kod životinjskih ćelija
 ćelijski zid; b) citoplazma; c) jedro; 1")
41 b c Mitoza somatskih ćelija korenčića crnog luka a 1. INTERFAZA a) ćelijski zid; b) citoplazma; c) jedro; 1
 hromatin 2")
42 d Mitoza somatskih ćelija korenčića crnog luka 2. PROFAZA d) hromatin 2
 hromozomi u ekvatorijalnoj ravni; f) vlakna")
43 e Mitoza somatskih ćelija korenčića crnog luka 3. METAFAZA e) hromozomi u ekvatorijalnoj ravni; f) vlakna deobnog vretena; f 3
 interhromozomska vlakna deobnog vretena; h) homologi")
44 h g Mitoza somatskih ćelija korenčića crnog luka 4. ANAFAZA g) interhromozomska vlakna deobnog vretena; h) homologi hromozomi u razdvajanju; 4

45 i Mitoza somatskih ćelija korenčića crnog luka 5. TELOFAZA i) fragmoblast 5
 ćelijska membrana; b) citoplazma; c)")
46 b a c Mitoza somatskih ćelija embriona pastrmke 1. INTERFAZA a) ćelijska membrana; b) citoplazma; c) jedro; 1

47 d Mitoza somatskih ćelija embriona pastrmke 2. PROFAZA d) hromatin 2
 hromozomi u ekvatorijalnoj ravni; f)")
48 e Mitoza somatskih e ćelija embriona pastrmke 3. METAFAZA e) hromozomi u ekvatorijalnoj ravni; f) aster; f 3
 interhromozomska vlakna deobnog vretena; h) homologi")
49 g Mitoza somatskih ćelija embriona pastrmke h 4. ANAFAZA g) interhromozomska vlakna deobnog vretena; h) homologi hromozomi u razdvajanju; 4

50 i Mitoza somatskih ćelija embriona pastrmke 5. TELOFAZA i) konstrikciona brazda 5

51 Ploidnost ćelije tokom mejoze i mitoze kod diploidne vrste organizama

52 Sinaptonemalni kompleks elektromikrografija Šematski prikaz mejoze I. Šematski prikaz Sinaptonemalnog kompleksa
")
")
53 a) b) Sinaptonemalni kompleks: a) elektronska mikrografija; b) šematski prikaz Diploten Pahiten
54 Shematski prikaz spermatogeneze (levo) i ovogeneze (desno) kod sisara
55 Faze gametogeneze

56 Uporedni prikaz spermatogeneze i oogeneze

57 Uporedni prikaz mejoze i mitoze

58 Poređenje mitoze i mejoze.
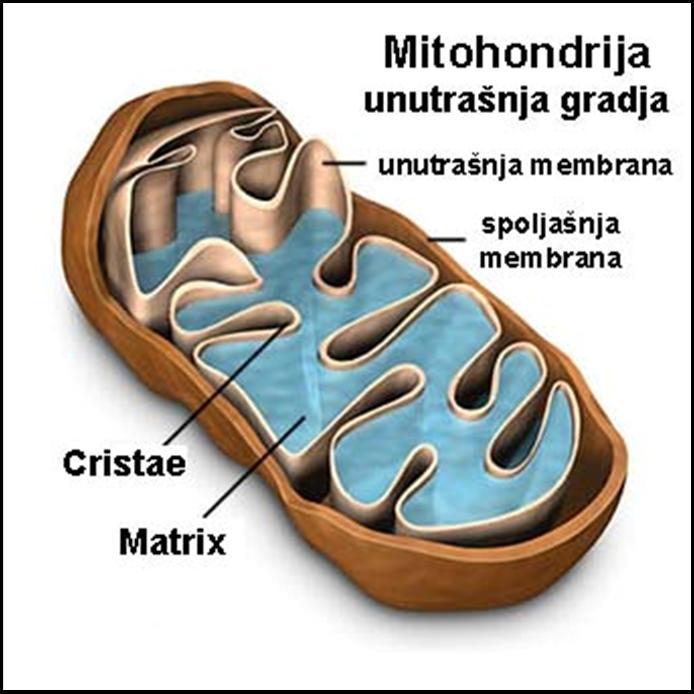

59 Vanjedarno (ekstranuklearno) nasleđivanje
60 Pod vanjedarnim (ekstranuklearnim) nasleđivanjem podrazumeva se ne- Mendelovski način distribucije fenotipskih osobina kod potomaka determinisanih genima u mitohondrijama ili plastidima. Izuzetno, neke bakterije i virusi, takođe, mogu predstavljati materijalnu osonvu vanjedarnih genetičkih elemeanta. Osnovni kriterijumi po kojima se može utvrditi da je reč o vanjedarnom nasleđivanju su: -razlike u fenotipskim odnosima pri recipročnim ukrštanjima. Pri recipročnim ukrštanjima sparuju se ženke soja A sa mužjacima soja B, a zatim mužjaci soja A sa ženkama soja B. Pod uslovom da se isključi nasleđivanje vezano za polne hromozome (odnosno u slučaju autozomalnih gena), razlike u fenotipskim odnosima pri recipročnim ukrštanjima ukazuju da jedan roditelj (najčešće ženka) ima veći uticaj na ispoljavanje osobine od drugog roditelja (mužjaka). -ženski gamet obično sadrži znatno više citoplazme od muškog gameta, te se može očekivati da utiče na odstupanje od Mendelovih pravila. Organele i simbionti u citoplazmi mogu da se izoluju i analiziraju i ukoliko se vanjedarna DNK dovede u vezu sa prenošenjem praćenih osobina, smatra se da je dokazano vanjedarno nasleđivanje.
61 - hromozomalni geni imaju svoje mesto na hromozomima u odnosu na ostale gene. Ukoliko se ne može ustanoviti vezanost sa nekim od hromozomalnih gena, osnovano se pretpostavlja da on pripada vanjedarnim genetičkim sistemima. -odsustvo Mendelovskog načina segregacije gena i nepostojanje tipičnih Mendelovih odnosa pri pojedinim tipovima ukrštanja navodi na sumnju da je reć o vanjedarnom nasleđivanju, - ukoliko se nakon eksperimenta zamene jedara, osobine prenose nezavisno od jedarnih gena, postoje jake indicije da su praćene osobine determinisane vanjedarnim genima.
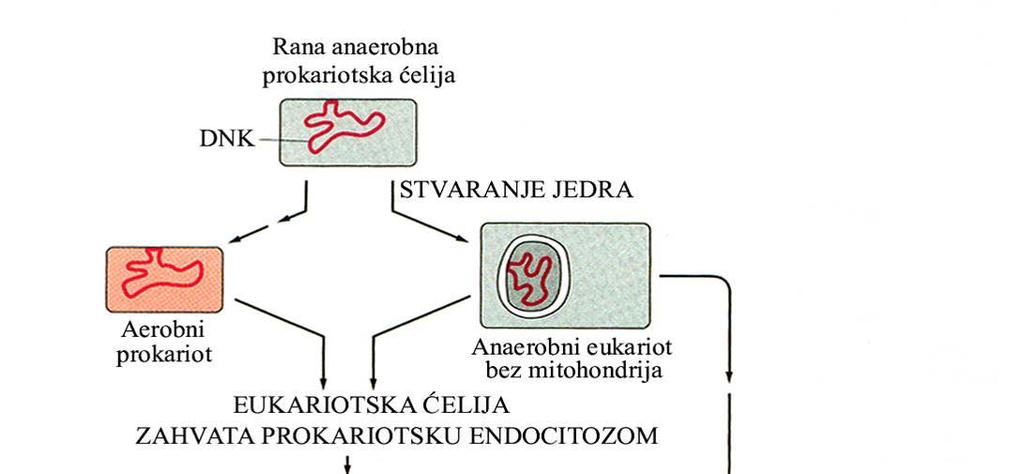
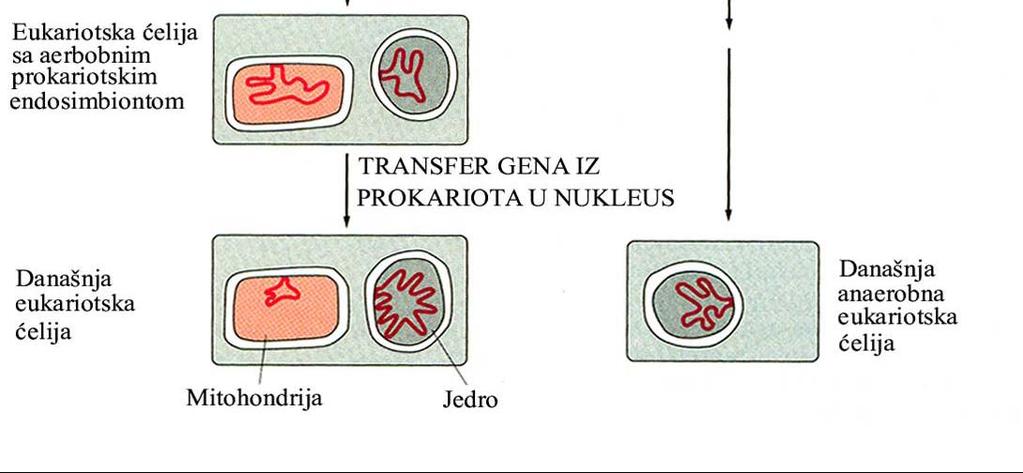
62 Evolutivni put nastanka mitohondrija
63 - Mitohondrijalna DNK (mtdnk) nalazi se u nekoliko kopija u matriksu mitohondrija gde je obično raspoređena u nekoliko grupacija (nukleoidi). Nukleoidi su pričvršćeni za unutrašnju membranu mitohondrija. Po svojoj strukturnoj organizaciji nukleotidi su znatno bliži bakterijskim DNK, a ne eukariotskom hromatinu. Kao i kod bakterija, u sastavu mitohondrijalnih nukleoida nema histona. Veličina mtdnk slična je virusnim DNK i kreće se u rasponu od oko 6000 bp (Plasmodium falciparum, izazivač malarije kod čoveka) do preko bp kod nekih kopnenih biljaka. Kao i kod bakterija, molekuli mtdnk su većinom cirkularni, mada su primećene linearne mtdnk. Kod sisara je mitohondrijalni genom predstavljen cirkularnom DNK od oko bp (manje od 0,001% od veličine jedarnog genoma). Približno ista veličina mtdnk kao kod sisara, konstatovana je kod nekih jako udaljenih vrsta kao što su Drosophila i morski ježevi.
64 Relativna količina mtdnk u nekim tipovima eukariotskih ćelija Organizam Tkivo ili tip ćelija DNK molekula po organeli Organela po ćeliji % mtdnk od ukupne ćelijske DNK Pacov Jetra Kvasac* Vegetativna ćel Žaba Jajna ćelija
65 U ćelijama sisara mtdnk zauzima manje od 1% ukupne ćelijske DNK (Tabela 1). Međutim, u jajniim ćelijama vodozemaca, gde su potrebne velike rezverve mitohondrija za ranu embirogenezu, mtdnk zauzima čak 99% ukupne ćelijske DNK. Zahvaljujući relativno maloj veličini humane mtdnk, celokupna sekvenca od nukleotidnih parova određena je još godine. Primećene su neke neobične karakteristike humane mtdnk kojih nema ne samo u jedarnom genomu, već isto tako ni kod DNK bakterija i hloroplasta. Za razliku od ostalih genoma, u humanoj mtdnk skoro svaki nukleotid je deo kodirajućih sekvenci (za protein ili za rrnk), tako da je količlina regulatornih DNK sekvenci svedena na minimum. Pored toga, postoje svega 22 trnk koje učestvuju u sintezi proteina u humanim mihohondrijama (u hloroplastima ima bar 30 različitih trnk). Stoga su normalna pravila sparivanja kodon-antikodon u mitohondrijama fleksibilna. Naime, veći broj trnk prepoznaje samo prva dva kodona, dok na trećem kodonu postoji kolebljivost. Na taj način, sinteza proteina moguća je sa manjim brojem trnk nego u drugim genomima.

66 Opšta organizacija humane mitohondrjalne DNK
67 - Konačno, možda najmarkantnija razlika, je da 4 od ukupno 64 kodona u sastavu humane mtdnk imaju drugačije značenje (odstupanje od univerzalnosti genetičkog koda) Kodon Univerzalni kod sisari Mitohondrijalni kodovi* beskičme njaci kvasci biljke UGA STOP Trp Trp Trp STOP AUA Ile Met Met Met Ile CUA Leu Leu Leu Thr Leu AGA ili AGG Arg STOP Ser Arg Arg * kodovi koji odstupaju od univerzalnosti predstavljeni belom bojom
68 Univerzalnost genetičkog koda predstavlja jedan od osnovnih dokaza da svi organizmi potiču od istih predaka (monofiletsko poreklo života na Zemlji). Smatra se da usled neobično malog broja proteina kodiranih mtdnk, mogu da se tolerišu povremene promene u značenjima kodona, dok bi takva promena u velikom genomu ugrozila funkciju mnogih proteina i doprinela uništavanju ćelije. Ovu pretpostavku podržavaju nalazi da odstupanja od univerzalnosti genetičkog koda nisu ista u različitim gupama organizama. Interesanto je da se u mtdnk zamena nukleotida tokom evolucije javlja oko 10 puta brže u poređenju sa jedarnom DNK. Uzrok ove pojave može se tražiti u smanjenoj vernosti replikacije i/ili nedovoljno efikasnoj reparaciji mtdnk. Relativno velika brzina evolucije mitohondrijalne DNK omogućava da se poređenjem nukleotidnih sekvenci mtdnk kod bliskih vrsta odrede njihovi filogenetski odnosi.
69 MATERINSKIO NASLEĐIVANJE Pri materinskom nasleđivanju fenotip potomaka određen je jedarnim genima prisutnim u jajnoj ćeliji. Prema tome, za razliku od biparentalnog nasleđivanja u kome su fenotipske osobine potomaka određene jedarnim genima oba roditelja, pri materinskom nasleđivanju dolazi do prepisivanja (transkripcija) nasledne informacije ženskog gameta i akumulacije genskog produkta (irnk ili protein) u jajnoj ćeliji. Nakon oplođenja, akumulirani genski produkti tokom ranog razvića utiču na ispoljavanje fenotipskih osobina kod potomaka. Materinsko nasleđivanje može da se identifikuje na osnovu različitih fenotipskih odnosa kod potomaka iz recipročnih ukrštanja. Opisani su primeri trajne promene fenotipa pod uticajem gena majke, kao što je oblik uvijanja ljušture kod puža Limnaea peregra.
70 Jedinke genotipa dd imaju sinistralne (uvijene na levo) ljušture, dok se kod ostalih (genotipovi DD i Dd) javljaju dekstralne (na desno uvijene) ljušture. S obzirom da su ovi puževi hermafroditi mogu se izvršiti različiti oblici ukrštanja i samooplođenja. Pri recipročnim ukrštanjima rezultat zavisi od genotipa ženskog roditelja. Ženke genotipa DD ili Dd produkuju isključivo potomke sa dekstralnom ljušturom, dok su potomci dd ženki uvek sa sinistralnom ljušturom. Interesantno je da oblik ljušture zavisi samo od genotipa ženke, nezavisno od njenog fenotipa. Istraživanja ranog razvića Limnaea pokazala su da orijentacija deobnog vretena u prvoj deobi nakon oplođenja odrećuje pravac uvrtanja ljušture. Orijentacija deobnog vretena zavisna od dejstva materinskih gena koji se ispoljavaju još tokom ovogeneze. Ustanovljeno je da DD ili Dd ženke imaju funkcionalan genski produkt D alela koji se akumulira u citoplazmi jajne ćelije. Čak i u slučaju kada ovocit kod ovih ženki nakon mejoze sadrži genski alel d i oplodi se spermatozoidom sa genskim alelom d, potomci su genotipa dd ali će ipak imati dekstralne ljušture.
71 Nasleđivanje oblika ljušture kod Limnaea peregra
3.1 Granična vrednost funkcije u tački
 3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović
 DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
numeričkih deskriptivnih mera.
 DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
41. Jednačine koje se svode na kvadratne
 . Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
. Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA
 NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA ANIMACIJE!!! REPLIKACIJA https://www.youtube.com/watch?v=tnkwgcfphqw TRANSKRIPCIJA https://www.youtube.com/watch?v=jqiwwjqf5d0 TRANSKRIPCIJA I TRANSLACIJA https://www.youtube.com/watch?v=-k8y0atkkai
NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA ANIMACIJE!!! REPLIKACIJA https://www.youtube.com/watch?v=tnkwgcfphqw TRANSKRIPCIJA https://www.youtube.com/watch?v=jqiwwjqf5d0 TRANSKRIPCIJA I TRANSLACIJA https://www.youtube.com/watch?v=-k8y0atkkai
Iskazna logika 3. Matematička logika u računarstvu. novembar 2012
 Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Zavrxni ispit iz Matematiqke analize 1
 Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A
, grupa A") Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Ispitivanje toka i skiciranje grafika funkcija
 Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT
 METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT BEZ POLI-A REPA" Histoni su primarne proteinske komponente hromatina. Na početku se smatralo da su uglavnom uključeni u pakovanje DNK, odnosno da su važni
METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT BEZ POLI-A REPA" Histoni su primarne proteinske komponente hromatina. Na početku se smatralo da su uglavnom uključeni u pakovanje DNK, odnosno da su važni
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama.
 Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Konstruisanje. Dobro došli na... SREDNJA MAŠINSKA ŠKOLA NOVI SAD DEPARTMAN ZA PROJEKTOVANJE I KONSTRUISANJE
 Dobro došli na... Konstruisanje GRANIČNI I KRITIČNI NAPON slajd 2 Kritični naponi Izazivaju kritične promene oblika Delovi ne mogu ispravno da vrše funkciju Izazivaju plastične deformacije Može doći i
Dobro došli na... Konstruisanje GRANIČNI I KRITIČNI NAPON slajd 2 Kritični naponi Izazivaju kritične promene oblika Delovi ne mogu ispravno da vrše funkciju Izazivaju plastične deformacije Može doći i
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
5. Karakteristične funkcije
 5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
18. listopada listopada / 13
 18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
APROKSIMACIJA FUNKCIJA
 APROKSIMACIJA FUNKCIJA Osnovni koncepti Gradimir V. Milovanović MF, Beograd, 14. mart 2011. APROKSIMACIJA FUNKCIJA p.1/46 Osnovni problem u TA Kako za datu funkciju f iz velikog prostora X naći jednostavnu
APROKSIMACIJA FUNKCIJA Osnovni koncepti Gradimir V. Milovanović MF, Beograd, 14. mart 2011. APROKSIMACIJA FUNKCIJA p.1/46 Osnovni problem u TA Kako za datu funkciju f iz velikog prostora X naći jednostavnu
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
Teorijske osnove informatike 1
 Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018.
 Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Test iz Biologije za 4. razred srednje škole R J E Š E NJ A Podgorica, 05. 05. 2018. 1 1. Spojite navedene
Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Test iz Biologije za 4. razred srednje škole R J E Š E NJ A Podgorica, 05. 05. 2018. 1 1. Spojite navedene
XI dvoqas veжbi dr Vladimir Balti. 4. Stabla
 XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
BANKA PITANJA IZ BIOLOGIJE. proteini. 3. Koji se deo složenog enzima hemijski menja u toku reakcije: 4. Apoenzim i koenzim zajedno čine:
 BANKA PITANJA IZ BIOLOGIJE citologija proteini 1. α-heliks je pojam koje se odnosi na strukturu proteina. 2. Broj polipeptidnih lanaca kod složenih proteina definiše: 3. Koji se deo složenog enzima hemijski
BANKA PITANJA IZ BIOLOGIJE citologija proteini 1. α-heliks je pojam koje se odnosi na strukturu proteina. 2. Broj polipeptidnih lanaca kod složenih proteina definiše: 3. Koji se deo složenog enzima hemijski
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f
 D f") IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
Operacije s matricama
 Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
M086 LA 1 M106 GRP. Tema: Baza vektorskog prostora. Koordinatni sustav. Norma. CSB nejednakost
 M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
PRAVA. Prava je u prostoru određena jednom svojom tačkom i vektorom paralelnim sa tom pravom ( vektor paralelnosti).
.") PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
ELEKTROTEHNIČKI ODJEL
 MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
Funkcije dviju varjabli (zadaci za vježbu)
") Funkcije dviju varjabli (zadaci za vježbu) Vidosava Šimić 22. prosinca 2009. Domena funkcije dvije varijable Ako je zadano pridruživanje (x, y) z = f(x, y), onda se skup D = {(x, y) ; f(x, y) R} R 2 naziva
Funkcije dviju varjabli (zadaci za vježbu) Vidosava Šimić 22. prosinca 2009. Domena funkcije dvije varijable Ako je zadano pridruživanje (x, y) z = f(x, y), onda se skup D = {(x, y) ; f(x, y) R} R 2 naziva
3. razred gimnazije- opšti i prirodno-matematički smer ALKENI. Aciklični nezasićeni ugljovodonici koji imaju jednu dvostruku vezu.
 ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
Novi Sad god Broj 1 / 06 Veljko Milković Bulevar cara Lazara 56 Novi Sad. Izveštaj o merenju
 Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
2 tg x ctg x 1 = =, cos 2x Zbog četvrtog kvadranta rješenje je: 2 ctg x
 Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI. Doc. dr Snežana Marković
 REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REGULACIJA EKSPRESIJE
REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REGULACIJA EKSPRESIJE
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
PARCIJALNI IZVODI I DIFERENCIJALI. Sama definicija parcijalnog izvoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je,
 PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu
 Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
radni nerecenzirani materijal za predavanja R(f) = {f(x) x D}
 = {f(x) x D}") Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
Grafičko prikazivanje atributivnih i geografskih nizova
 Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
Elementi spektralne teorije matrica
 Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE GODINE 8. RAZRED TOČNI ODGOVORI
 21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
7 Algebarske jednadžbe
 7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
NUKLEUS. 1) STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi)
 STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi)") NUKLEUS 1) STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi) Saznanja o ćelijama do kojih se došlo posredstvom novih
NUKLEUS 1) STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi) Saznanja o ćelijama do kojih se došlo posredstvom novih
Cauchyjev teorem. Postoji više dokaza ovog teorema, a najjednostvniji je uz pomoć Greenove formule: dxdy. int C i Cauchy Riemannovih uvjeta.
 auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,
auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,
RIJEŠENI ZADACI I TEORIJA IZ
 RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
Uvod u neparametarske testove
 Str. 148 Uvod u neparametarske testove Predavač: Dr Mirko Savić savicmirko@ef.uns.ac.rs www.ef.uns.ac.rs Hi-kvadrat testovi c Str. 149 Koristi se za upoređivanje dve serije frekvencija. Vrste c testa:
Str. 148 Uvod u neparametarske testove Predavač: Dr Mirko Savić savicmirko@ef.uns.ac.rs www.ef.uns.ac.rs Hi-kvadrat testovi c Str. 149 Koristi se za upoređivanje dve serije frekvencija. Vrste c testa:
Strukture podataka i algoritmi 1. kolokvij 16. studenog Zadatak 1
 Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Obrada signala
 Obrada signala 1 18.1.17. Greška kvantizacije Pretpostavka je da greška kvantizacije ima uniformnu raspodelu 7 6 5 4 -X m p x 1,, za x druge vrednosti x 3 x X m 1 X m = 3 x Greška kvantizacije x x x p
Obrada signala 1 18.1.17. Greška kvantizacije Pretpostavka je da greška kvantizacije ima uniformnu raspodelu 7 6 5 4 -X m p x 1,, za x druge vrednosti x 3 x X m 1 X m = 3 x Greška kvantizacije x x x p
HEMIJSKA VEZA TEORIJA VALENTNE VEZE
 TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
Elektrotehnički fakultet univerziteta u Beogradu 17.maj Odsek za Softversko inžinjerstvo
 Elektrotehnički fakultet univerziteta u Beogradu 7.maj 009. Odsek za Softversko inžinjerstvo Performanse računarskih sistema Drugi kolokvijum Predmetni nastavnik: dr Jelica Protić (35) a) (0) Posmatra
Elektrotehnički fakultet univerziteta u Beogradu 7.maj 009. Odsek za Softversko inžinjerstvo Performanse računarskih sistema Drugi kolokvijum Predmetni nastavnik: dr Jelica Protić (35) a) (0) Posmatra
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika. Monotonost i ekstremi. Katica Jurasić. Rijeka, 2011.
 Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri
 Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri 1 1 Zadatak 1b Čisto savijanje - vezano dimenzionisanje Odrediti potrebnu površinu armature za presek poznatih dimenzija, pravougaonog
Betonske konstrukcije 1 - vežbe 3 - Veliki ekscentricitet -Dodatni primeri 1 1 Zadatak 1b Čisto savijanje - vezano dimenzionisanje Odrediti potrebnu površinu armature za presek poznatih dimenzija, pravougaonog
Trigonometrija 2. Adicijske formule. Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto
 Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).
![PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).](/thumbs/74/70316766.jpg "PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).") 0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
NOMENKLATURA ORGANSKIH SPOJEVA. Imenovanje aromatskih ugljikovodika
 NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
Sume kvadrata. mn = (ax + by) 2 + (ay bx) 2.
 2 + (ay bx) 2.") Sume kvadrata Koji se prirodni brojevi mogu prikazati kao zbroj kvadrata dva cijela broja? Propozicija 1. Ako su brojevi m i n sume dva kvadrata, onda je i njihov produkt m n takoder suma dva kvadrata.
Sume kvadrata Koji se prirodni brojevi mogu prikazati kao zbroj kvadrata dva cijela broja? Propozicija 1. Ako su brojevi m i n sume dva kvadrata, onda je i njihov produkt m n takoder suma dva kvadrata.
Sekundarne struktura proteina Fibrilni proteini
 Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
Riješeni zadaci: Nizovi realnih brojeva
 Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
MATEMATIKA 2. Grupa 1 Rexea zadataka. Prvi pismeni kolokvijum, Dragan ori
 MATEMATIKA 2 Prvi pismeni kolokvijum, 14.4.2016 Grupa 1 Rexea zadataka Dragan ori Zadaci i rexea 1. unkcija f : R 2 R definisana je sa xy 2 f(x, y) = x2 + y sin 3 2 x 2, (x, y) (0, 0) + y2 0, (x, y) =
MATEMATIKA 2 Prvi pismeni kolokvijum, 14.4.2016 Grupa 1 Rexea zadataka Dragan ori Zadaci i rexea 1. unkcija f : R 2 R definisana je sa xy 2 f(x, y) = x2 + y sin 3 2 x 2, (x, y) (0, 0) + y2 0, (x, y) =
Verovatnoća i Statistika I deo Teorija verovatnoće (zadaci) Beleške dr Bobana Marinkovića
 Beleške dr Bobana Marinkovića") Verovatnoća i Statistika I deo Teorija verovatnoće zadaci Beleške dr Bobana Marinkovića Iz skupa, 2,, 00} bira se na slučajan način 5 brojeva Odrediti skup elementarnih dogadjaja ako se brojevi biraju
Verovatnoća i Statistika I deo Teorija verovatnoće zadaci Beleške dr Bobana Marinkovića Iz skupa, 2,, 00} bira se na slučajan način 5 brojeva Odrediti skup elementarnih dogadjaja ako se brojevi biraju
OM2 V3 Ime i prezime: Index br: I SAVIJANJE SILAMA TANKOZIDNIH ŠTAPOVA
 OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
KVADRATNA FUNKCIJA. Kvadratna funkcija je oblika: Kriva u ravni koja predstavlja grafik funkcije y = ax + bx + c. je parabola.
 KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
III VEŽBA: FURIJEOVI REDOVI
 III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
Biohemija nukleinskih kiselina. Genetska informacija
 Biohemija nukleinskih kiselina Genetska informacija deoksiribonukleinske kiseline (DNK) ribonukleinske kiseline (RNK) DNK je nosilac naslednih informacija u ćeliji, dok RNK učestvuju u prenošenju tih informacija
Biohemija nukleinskih kiselina Genetska informacija deoksiribonukleinske kiseline (DNK) ribonukleinske kiseline (RNK) DNK je nosilac naslednih informacija u ćeliji, dok RNK učestvuju u prenošenju tih informacija
Pismeni ispit iz matematike Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: ( ) + 1.
 + 1.") Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
10. STABILNOST KOSINA
 MEHANIKA TLA: Stabilnot koina 101 10. STABILNOST KOSINA 10.1 Metode proračuna koina Problem analize tabilnoti zemljanih maa vodi e na određivanje odnoa između rapoložive mičuće čvrtoće i proečnog mičućeg
MEHANIKA TLA: Stabilnot koina 101 10. STABILNOST KOSINA 10.1 Metode proračuna koina Problem analize tabilnoti zemljanih maa vodi e na određivanje odnoa između rapoložive mičuće čvrtoće i proečnog mičućeg
radni nerecenzirani materijal za predavanja
 Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Kažemo da je funkcija f : a, b R u točki x 0 a, b postiže lokalni minimum ako postoji okolina O(x 0 ) broja x 0 takva da je
Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Kažemo da je funkcija f : a, b R u točki x 0 a, b postiže lokalni minimum ako postoji okolina O(x 0 ) broja x 0 takva da je
Pravilo 1. Svaki tip entiteta ER modela postaje relaciona šema sa istim imenom.
 1 Pravilo 1. Svaki tip entiteta ER modela postaje relaciona šema sa istim imenom. Pravilo 2. Svaki atribut entiteta postaje atribut relacione šeme pod istim imenom. Pravilo 3. Primarni ključ entiteta postaje
1 Pravilo 1. Svaki tip entiteta ER modela postaje relaciona šema sa istim imenom. Pravilo 2. Svaki atribut entiteta postaje atribut relacione šeme pod istim imenom. Pravilo 3. Primarni ključ entiteta postaje
Reverzibilni procesi
 Reverzbln proces Reverzbln proces: proces pr koja sste nkada nje vše od beskonačno ale vrednost udaljen od ravnoteže, beskonačno ala proena spoljašnjh uslova ože vratt sste u blo koju tačku, proena ože
Reverzbln proces Reverzbln proces: proces pr koja sste nkada nje vše od beskonačno ale vrednost udaljen od ravnoteže, beskonačno ala proena spoljašnjh uslova ože vratt sste u blo koju tačku, proena ože
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =
/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =") 100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
Regulacija ekspresije gena kod eukariota
 Regulacija ekspresije gena kod eukariota Regulacija ekspresije gena Višećelijski organizmi Čovek - 250 tipova ćelija Svaka ćelija gotovo istu DNK Regulacija ekspresije gena Hijerarhija regulacije ekspresije
Regulacija ekspresije gena kod eukariota Regulacija ekspresije gena Višećelijski organizmi Čovek - 250 tipova ćelija Svaka ćelija gotovo istu DNK Regulacija ekspresije gena Hijerarhija regulacije ekspresije
4.7. Zadaci Formalizam diferenciranja (teorija na stranama ) 343. Znajući izvod funkcije x arctg x, odrediti izvod funkcije x arcctg x.
 343. Znajući izvod funkcije x arctg x, odrediti izvod funkcije x arcctg x.") 4.7. ZADACI 87 4.7. Zadaci 4.7.. Formalizam diferenciranja teorija na stranama 4-46) 340. Znajući izvod funkcije arcsin, odrediti izvod funkcije arccos. Rešenje. Polazeći od jednakosti arcsin + arccos
4.7. ZADACI 87 4.7. Zadaci 4.7.. Formalizam diferenciranja teorija na stranama 4-46) 340. Znajući izvod funkcije arcsin, odrediti izvod funkcije arccos. Rešenje. Polazeći od jednakosti arcsin + arccos
INTEGRALNI RAČUN. Teorije, metodike i povijest infinitezimalnih računa. Lucija Mijić 17. veljače 2011.
 INTEGRALNI RAČUN Teorije, metodike i povijest infinitezimalnih računa Lucija Mijić lucija@ktf-split.hr 17. veljače 2011. Pogledajmo Predstavimo gornju sumu sa Dodamo još jedan Dobivamo pravokutnik sa Odnosno
INTEGRALNI RAČUN Teorije, metodike i povijest infinitezimalnih računa Lucija Mijić lucija@ktf-split.hr 17. veljače 2011. Pogledajmo Predstavimo gornju sumu sa Dodamo još jedan Dobivamo pravokutnik sa Odnosno
Antene. Srednja snaga EM zračenja se dobija na osnovu intenziteta fluksa Pointingovog vektora kroz sferu. Gustina snage EM zračenja:
 Anene Transformacija EM alasa u elekrični signal i obrnuo Osnovne karakerisike anena su: dijagram zračenja, dobiak (Gain), radna učesanos, ulazna impedansa,, polarizacija, efikasnos, masa i veličina, opornos
Anene Transformacija EM alasa u elekrični signal i obrnuo Osnovne karakerisike anena su: dijagram zračenja, dobiak (Gain), radna učesanos, ulazna impedansa,, polarizacija, efikasnos, masa i veličina, opornos
TEST PITANJA IZ BIOLOGIJE
 TEHNOLOŠKI 33 FAKULTETZVORNIK TEST PITANJA IZ BIOLOGIJE 2015. Prof. dr Milan Kulić Mr Nikolina Elez Burnjaković BIOLOGIJA ĆELIJE 1. Biološka disciplina koja se bavi proučavanjem organizacije ćelije se
TEHNOLOŠKI 33 FAKULTETZVORNIK TEST PITANJA IZ BIOLOGIJE 2015. Prof. dr Milan Kulić Mr Nikolina Elez Burnjaković BIOLOGIJA ĆELIJE 1. Biološka disciplina koja se bavi proučavanjem organizacije ćelije se
Linearna algebra 2 prvi kolokvij,
 1 2 3 4 5 Σ jmbag smjer studija Linearna algebra 2 prvi kolokvij, 7. 11. 2012. 1. (10 bodova) Neka je dano preslikavanje s : R 2 R 2 R, s (x, y) = (Ax y), pri čemu je A: R 2 R 2 linearan operator oblika
1 2 3 4 5 Σ jmbag smjer studija Linearna algebra 2 prvi kolokvij, 7. 11. 2012. 1. (10 bodova) Neka je dano preslikavanje s : R 2 R 2 R, s (x, y) = (Ax y), pri čemu je A: R 2 R 2 linearan operator oblika
LANCI & ELEMENTI ZA KAČENJE
 LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
konst. Električni otpor
 Sveučilište J. J. Strossmayera u sijeku Elektrotehnički fakultet sijek Stručni studij Električni otpor hmov zakon Pri protjecanju struje kroz vodič pojavljuje se otpor. Georg Simon hm je ustanovio ovisnost
Sveučilište J. J. Strossmayera u sijeku Elektrotehnički fakultet sijek Stručni studij Električni otpor hmov zakon Pri protjecanju struje kroz vodič pojavljuje se otpor. Georg Simon hm je ustanovio ovisnost
Osnovne teoreme diferencijalnog računa
 Osnovne teoreme diferencijalnog računa Teorema Rolova) Neka je funkcija f definisana na [a, b], pri čemu važi f je neprekidna na [a, b], f je diferencijabilna na a, b) i fa) fb). Tada postoji ξ a, b) tako
Osnovne teoreme diferencijalnog računa Teorema Rolova) Neka je funkcija f definisana na [a, b], pri čemu važi f je neprekidna na [a, b], f je diferencijabilna na a, b) i fa) fb). Tada postoji ξ a, b) tako
OBRTNA TELA. Vladimir Marinkov OBRTNA TELA VALJAK
 OBRTNA TELA VALJAK P = 2B + M B = r 2 π M = 2rπH V = BH 1. Zapremina pravog valjka je 240π, a njegova visina 15. Izračunati površinu valjka. Rešenje: P = 152π 2. Površina valjka je 112π, a odnos poluprečnika
OBRTNA TELA VALJAK P = 2B + M B = r 2 π M = 2rπH V = BH 1. Zapremina pravog valjka je 240π, a njegova visina 15. Izračunati površinu valjka. Rešenje: P = 152π 2. Površina valjka je 112π, a odnos poluprečnika
Inženjerska grafika geometrijskih oblika (5. predavanje, tema1)
") Inženjerska grafika geometrijskih oblika (5. predavanje, tema1) Prva godina studija Mašinskog fakulteta u Nišu Predavač: Dr Predrag Rajković Mart 19, 2013 5. predavanje, tema 1 Simetrija (Symmetry) Simetrija
Inženjerska grafika geometrijskih oblika (5. predavanje, tema1) Prva godina studija Mašinskog fakulteta u Nišu Predavač: Dr Predrag Rajković Mart 19, 2013 5. predavanje, tema 1 Simetrija (Symmetry) Simetrija
Linearna algebra 2 prvi kolokvij,
 Linearna algebra 2 prvi kolokvij, 27.. 20.. Za koji cijeli broj t je funkcija f : R 4 R 4 R definirana s f(x, y) = x y (t + )x 2 y 2 + x y (t 2 + t)x 4 y 4, x = (x, x 2, x, x 4 ), y = (y, y 2, y, y 4 )
Linearna algebra 2 prvi kolokvij, 27.. 20.. Za koji cijeli broj t je funkcija f : R 4 R 4 R definirana s f(x, y) = x y (t + )x 2 y 2 + x y (t 2 + t)x 4 y 4, x = (x, x 2, x, x 4 ), y = (y, y 2, y, y 4 )
Riješeni zadaci: Limes funkcije. Neprekidnost
 Riješeni zadaci: Limes funkcije. Neprekidnost Limes funkcije Neka je 0 [a, b] i f : D R, gdje je D = [a, b] ili D = [a, b] \ { 0 }. Kažemo da je es funkcije f u točki 0 jednak L i pišemo f ) = L, ako za
Riješeni zadaci: Limes funkcije. Neprekidnost Limes funkcije Neka je 0 [a, b] i f : D R, gdje je D = [a, b] ili D = [a, b] \ { 0 }. Kažemo da je es funkcije f u točki 0 jednak L i pišemo f ) = L, ako za
Regulacija ekspresije gena kod prokariota
 Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
Rekombinacija DNK TRANSPOZICIJOM
 Rekombinacija DNK TRANSPOZICIJOM Rekombinacija DNK transpozicijom Transpozicija je proces premeštanja specifičnih sekvenci molekula DNK (pokretnih genetičkih elemenata ili transpozona) sa jednog na drugo
Rekombinacija DNK TRANSPOZICIJOM Rekombinacija DNK transpozicijom Transpozicija je proces premeštanja specifičnih sekvenci molekula DNK (pokretnih genetičkih elemenata ili transpozona) sa jednog na drugo
Osnovne karakteristike 3-D strukture molekula DNK i RNK
 Osnovne karakteristike 3-D strukture molekula DNK i RNK Rendgenska strukturna analiza (vlakana) DNK Watson-Crickov model (B) DNK Zašto dvostruki heliks? Polimorfizam DNK: kanonske (standardne/prosečne)
Osnovne karakteristike 3-D strukture molekula DNK i RNK Rendgenska strukturna analiza (vlakana) DNK Watson-Crickov model (B) DNK Zašto dvostruki heliks? Polimorfizam DNK: kanonske (standardne/prosečne)
a M a A. Može se pokazati da je supremum (ako postoji) jedinstven pa uvodimo oznaku sup A.
 jedinstven pa uvodimo oznaku sup A.") 3 Infimum i supremum Definicija. Neka je A R. Kažemo da je M R supremum skupa A ako je (i) M gornja meda skupa A, tj. a M a A. (ii) M najmanja gornja meda skupa A, tj. ( ε > 0)( a A) takav da je a > M
3 Infimum i supremum Definicija. Neka je A R. Kažemo da je M R supremum skupa A ako je (i) M gornja meda skupa A, tj. a M a A. (ii) M najmanja gornja meda skupa A, tj. ( ε > 0)( a A) takav da je a > M
Matematika 1 - vježbe. 11. prosinca 2015.
 Matematika - vježbe. prosinca 5. Stupnjevi i radijani Ako je kut φ jednak i rad, tada je veza između i 6 = Zadatak.. Izrazite u stupnjevima: a) 5 b) 7 9 c). d) 7. a) 5 9 b) 7 6 6 = = 5 c). 6 8.5 d) 7.
Matematika - vježbe. prosinca 5. Stupnjevi i radijani Ako je kut φ jednak i rad, tada je veza između i 6 = Zadatak.. Izrazite u stupnjevima: a) 5 b) 7 9 c). d) 7. a) 5 9 b) 7 6 6 = = 5 c). 6 8.5 d) 7.
Periodičke izmjenične veličine
 EHNČK FAKULE SVEUČLŠA U RJEC Zavod za elekroenergeiku Sudij: Preddiploski sručni sudij elekroehnike Kolegij: Osnove elekroehnike Nosielj kolegija: Branka Dobraš Periodičke izjenične veličine Osnove elekroehnike
EHNČK FAKULE SVEUČLŠA U RJEC Zavod za elekroenergeiku Sudij: Preddiploski sručni sudij elekroehnike Kolegij: Osnove elekroehnike Nosielj kolegija: Branka Dobraš Periodičke izjenične veličine Osnove elekroehnike