Podstata fotosyntézy
|
|
|
- Οἰνεύς Σκλαβούνος
- 7 χρόνια πριν
- Προβολές:
Transcript
1 Fotosyntéza Plastidy Asimilačné pigmenty a absorpcia svetla Komplexy tylakoidov (fotosystém II, I, komplex cyt b6-f, ATP syntáza) Elektrónový a protónový transport Fotofosforylácia Fixácia CO2 a syntéza cukrov (Calvinov cyklus, fotorespirácia, syndróm C3, Hatchov a Slackov cyklus, syndróm C4, syndróm CAM) Syntéza a odbúravanie sacharózy a translokácie asimilátov floémom, škrob. Ekofyziológia fotosyntézy. Difúzne procesy. Energetická účinnosť. Faktory prostredia (žiarenie, CO2, teplota a ostatné faktory).
2 Podstata fotosyntézy
3
4
5

6 10-12 s 10-9 s
7
8 PS 1 plastokyanín/ cytochróm c 6 na feredoxín oxidoreduktáza Cyt b 6 f plastohydrochinón na plastokyanín/ cytochróm c 6 oxidoreduktáza PS 2 voda-plastochinón oxidoreduktáza

9 Redukcia plastochinónu v PSII

10 Prenos elektrónov a protónov v tylakoidnej membráne 4hν 4 H + stróma + 2PQ + 2 H 2 O 2PQH 2 + O H + lumen
11 Fotofosforylácia chloroplastová ATP syntáza
12 Calvinov cyklus
13
14
15 Regeneračná fáza Calvinovho cyklu

16 Rubisko katalyzuje karboxyláciu alebo oxygenáciu
17 Fotorespirácia
18 Fotorespirácia
19
20 C 3 stredne produktívne, mierne pásmo C 4 vysoko produktívne, tropické a subtropické pásmo CAM málo produktívne, púšte
21 C4
22 C4
23 CAM
24 Syntéza sacharózy
25 amylóza
26 amylopektín
27
28 Ekofyziológia fotosyntézy Faktory prostredia žiarenie CO 2 teplota
29 Priečny rez rôzne osvetleným listom (Fagus silvatica)
30 Listová mozaika
31 Svetelný tok a ožiarenie
32
33
34
35
36 CO2
37
38 Teplota

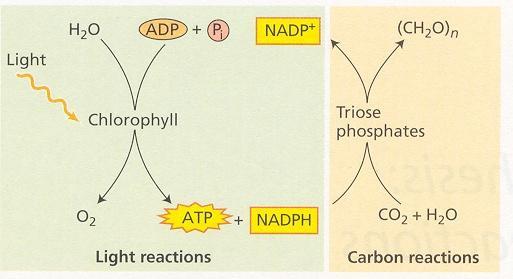
39 FOTOSYNTÉZA Život na Zemi nevyhnutne závisí na energii slnečného žiarenia. Fotosyntéza má základný význam, lebo je v podstate jediným mechanizmom vstupu energie do živých organizmov. Fotosyntetizujúce systémy premieňajú energiu svetla na energiu chemických väzieb v organických látkach. Prvé fotosyntetizujúce baktérie sa objavili pred 3000 miliónmi rokov. Sinice k ním pribudli asi pred 2000 miliónmi rokov a napokon prvé cievnaté suchozemské rastliny sa objavili pred asi 400 miliónmi rokov. Značná časť energetických zdrojov na Zemi je výsledkom fotosyntézy. Popri recentnej biomase sú to aj zásoby fosílnych palív. V súčasnosti fotosyntetizujúce organizmy produkujú okolo ton hmoty ročne. Schopnosť fotosyntetizovať majú len primárni producenti, rastlinné organizmy obsahujúce chlorofyl a a baktérie obsahujúce baktériochlorofyly. Orgánom pre fotosyntézy cievnatých rastlín je list. Najviac aktívnych fotosyntetizujúcich buniek je v mezofylovom pletive listov. Jeho bunky obsahujú chloroplasty, v ktorých prebieha fotosyntéza. Podstatou fotosyntézy sú procesy premeny energie. V tylakoidných membránach chloroplastov, kde prebiehajú svetelné reakcie, chlorofyl excitovaný svetlom vyvolá oddelenie oxidujúcej a redukujúcej jednotky. Časť energie absorbovaných fotónov sa využije na oxidáciu vody spojenú s výdajom O 2 do atmosféry. Ďalej sa pomocou tejto energie prenášajú elektróny v redoxných reakciách cez sériu látok, ktoré pôsobia ako elektrónové donory a akceptory. Konečným akceptorom elektrónov je NADP +. Časť svetelnej energie spôsobí vytvorenie protónového gradientu cez tylakoidnú membránu a využije sa na syntézu makroergických väzieb ATP. Výsledné produkty svetelných reakcií, energeticky bohaté látky, reduktant NADPH a ATP sa v uhlíkových reakciách v stróme spotrebujú na redukciu CO 2 a syntézu väzby C-C- v cukroch a ďalších organických látkach. Obr. 1 Svetelné a uhlíkové reakcie fotosyntézy 15

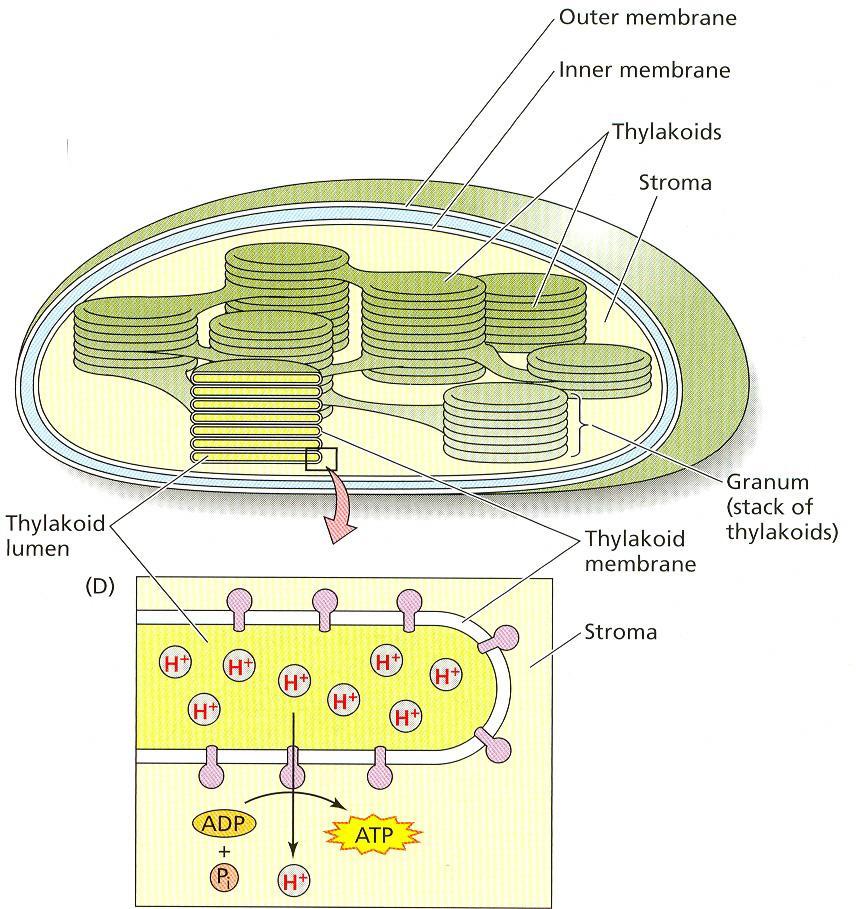
40 Chloroplasty Fotosyntetickým aparátom v prokaryotických bunkách fotosyntetizujúcich baktérií a siníc sú špecializované membrány neohraničené od základnej cytoplazmy. Eukaryotické rastlinné organizmy majú bunkové organely plastidy, ktoré od cytoplazmy oddeľuje povrchová membrána. Nové plastidy vznikajú výlučne delením už existujúcich plastidov. Pri pohlavnom rozmnožovaní sa kontinuita plastidov zabezpečuje prenosom cez vajcovú bunku. V meristematických bunkách sú bezfarebné proplastidy malých rozmerov. Z nich sa diferencujú chloroplasty. Limitujúcim faktorom vývinu chloroplastov je svetlo. Rôzne druhy rastlín majú chloroplasty rôznych tvarov a veľkostí. Semiautonómny charakter chloroplastov spôsobujú DNA, RNA a ribozómy, ktoré sú zastúpené v stróme. Plastidové gény kódujú niektoré proteíny, ale väčšina chloroplastových proteínov je kódovaná jadrovými génmi. Ultraštrukúra funkčných chloroplastov sa vyznačuje rozsiahlym membránovým systémom. Každý chloroplast je obklopený povrchovou membránou, ktorá kontroluje prenos molekúl dovnútra a von. Medzi jej dvomi vrstvami je periplastidový priestor. Povrchová membrána neobsahuje chlorofyly, ale sú v nej karotenoidy s ochrannou funkciou. Vnútro chloroplastov tvorí amorfná gélovitá stróma, ktorá je bohatá na enzýmy. Obr. 2 Štruktúra chloroplastu Membránový systém chloroplastov tvoria tylakoidy, ktoré majú tvar plochých vrecúšok. Tylakoidy uložené tesne vedľa seba tvoria gránum. Funkčne sú odlíšené časti tylakoidných membrán v gránach, ktoré sú v kontakte s inými a časti membrán ktoré sú v kontakte so strómou. Vnútorný priestor tylakoidov, ich lúmen, je vyplnený vodou a rozpustenými soľami a má úlohu pri tvorbe ATP. V tylakoidnej membráne sú uložené chlorofyly a ďalšie asimilačné pigmenty ako aj zložky reťazca prenosu elektrónov. Chlorofyly a karotenoidy sú nekovalentnými väzbami viazané na molekuly bielkovín. V tylakoidných lipidoch sú hojne zastúpené nenasýtené mastné kyseliny, čo spôsobuje, že tylakoidná membrána je neobvykle fluidná a niektoré zložky elektróntransportného reťazca sú v nej pohyblivé. 16
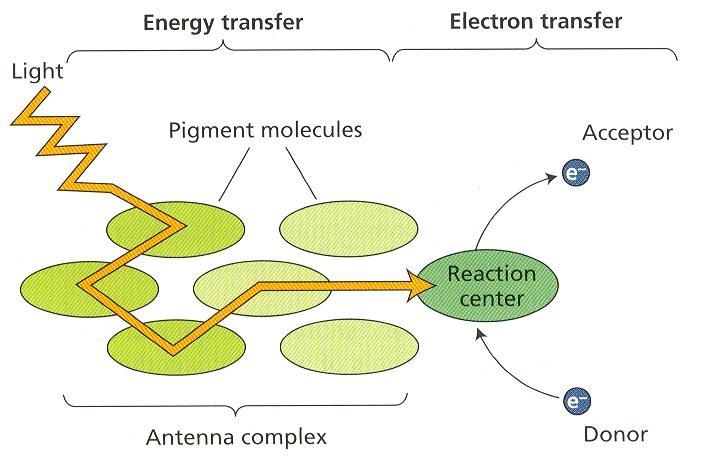
41 Asimilačné pigmenty Väčšina pigmentov absorbujúcich svetlo vo fotosyntéze sú zberače svetla. Chlorofyly a karotenoidy sú molekuly, ktoré absorbujú svetelné kvantá, prenášajú absorbovanú energiu na iné molekuly, až do reakčného centra a znova sa vrátia do základného stavu schopného ďalšej absorbcie. Vo vyšších rastlinách sú zastúpené dva chlorofyly, a a b. Ich molekula má časť cyklickú, ktorou je porfyrín pozostávajúci zo štyroch pyrolových heterocyklov a centrálne viazaného atómu horčíka. Esterovou väzbou je na porfyrín naviazaný lineárny diterpénový alkohol, fytol. Táto nepolárna časť molekuly chlorofylu sa javí ako kotva do hydrofóbnej časti membránového prostredia. Nahradením horčíka dvomi atómami vodíka v molekule chlorofylu vzniká feofytín. Pyrolové jadrá porfyrínu majú konjugovaný -elektrónový systém v dôsledku striedania jednoduchých a dvojitých väzieb. Tento systém sa môže vzbudiť relatívne nízkymi dávkami energie. Dôsledkom toho je schopnosť chlorofylu absorbovať kvantá dlhovlnného viditeľného svetla. Chlorofyl sa nám javí ako zelený, lebo absorbuje svetlo v červenej a modrej časti spektra, zelené sa odráža alebo prepúšťa. Obr. 3 Chlorofyl a a b Delokalizácia π elektrónov v porfyrínovej časti molekuly. Karotenoidy sú žltooranžové izoprenoidné pigmenty, ktoré absorbujú v modrej a fialovej časti spektra. Karotény sú uhľovodíky, v chloroplastoch sú najviac zastúpené -karoténom. Xantofyly obsahujú v molekule aj kyslík, v listoch je najhojnejší luteín. 17
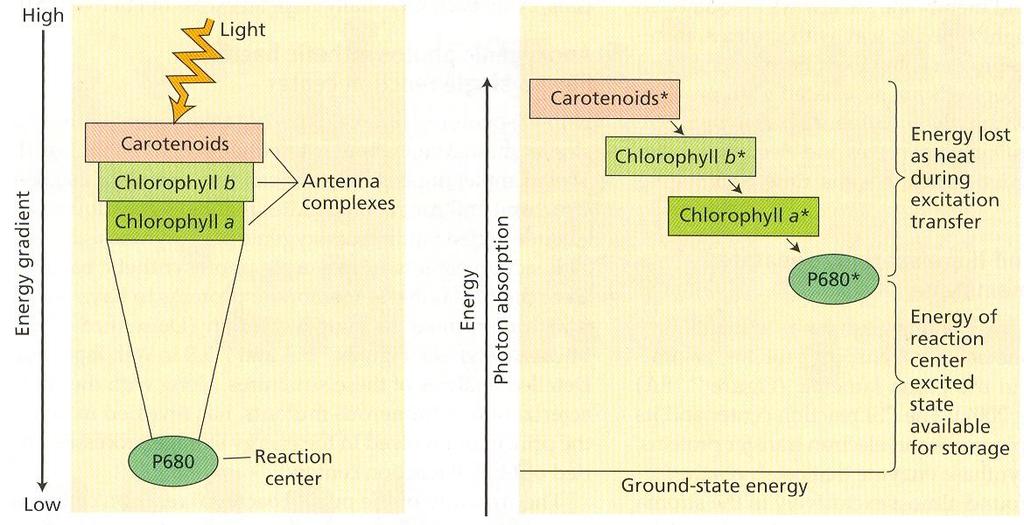
42 luteín Podobne ako chlorofyly väčšina karotenoidov účinne prenáša excitačnú energiu a prispieva k fotosyntéze. Karotenoidy majú aj ochrannú funkciu pri vysokých hodnotách ožiarenia. Mechanizmom xantofylového cyklu chránia lipoidné molekuly pred deštrukciou singletovým kyslíkom. Po osvetlení violaxantín (diepoxidový xantofyl) sa v deepoxidačnej reakcii mení cez anteroxantín (monoepoxid) na zeaxantín (bez epoxi- skupín). Za šera alebo v tme sa zeaxantín naspäť epoxiduje na violaxantín. Svetelnú deepoxidačnú reakciu katalyzuje enzým na lúmenovej strane membrány, ktorý má optimum ph pri 5,2 a je neaktívny pri ph okolo 7. Do reakcie vstupuje askorbát a redukovaný glutation. Epoxidácia prebieha na strómalnej strane tylakoidnej membrány v podmienkach osvetlenej strómy pri ph 7,5, katalyzuje ju oxygenáza so zmiešanou funkciou. Do reakcie vstupuje NADPH, vzniká voda a NADP +, ktorý môže prijímať ďalšie elektróny. Aktivita xantofylového cyklu závisí od svetlom indukovaného protónového gradientu cez tylakoidnú membránu, vzhľadom na lokalizáciu v dvoch kompartmentoch a rozdielne hodnoty optima ph. Hlavnou úlohou xantofylového cyklu je pravdepodobne regulovať pomer NADPH ku ATP, znížiť možnosť poškodenia membrány svetlom a optimalizovať podmienky pre funkciu membrány. Absorpcia svetla Hnacou silou fotosyntézy je svetlo, časť energie žiarenia, ktoré je viditeľné pre ľudské oči. Z celého elektromagnetického spektra svetlo predstavuje len malú oblasť. Svetlo má dvojaký charakter, vlnový a časticový. Fotóny majú energiu kvánt nepriamo úmernú vlnovej dĺžke. Kvantá fotónov modrých vlnových dĺžok sú bohatšie na energiu ako fotóny dlhších červených vlnových dĺžok. Základným princípom vo fotochémii je, že molekula môže absorbovať súčasne iba jeden fotón a excitovať sa môže iba jeden elektrón. Získaná energia umožňuje preskok elektrónu zo základného do excitovaného orbitu, teda na energeticky vyššiu hladinu. Väčšia hodnota energie kvanta umožňuje dosiahnutie vyššieho excitačného stavu, ale jeho životnosť je kratšia ako pri nižšom excitačnom stave. Absorbciou modrého svetla sa excituje chlorofyl do vyššieho excitačného stavu (2. singlet, životnosť s). Ak sa časť energie odovzdá ako teplo elektrón môže prejsť do nižšieho excitačného stavu (1. singlet, životnosť 10-9 s). Absorbciou červeného svetla sa dosiahne excitácia do 1. singletového stavu. Dlhšie trvanie toho stavu umožňuje niekoľko ciest pre použitie voľnej energie. 18

43 Obr. 4 Absorpcia a emisia svetla chlorofylmi. (A) diagram hladín energie; (B) spektrá absorpcie a fluorescencie Emisiou fotónu fluorescenciou sa môže elektrón vrátiť do základného stavu. Vlnová dĺžka fluorescencie bude dlhšia asi o 10 nm, lebo časť energie sa premení na teplo. Druhou možnosťou je premena excitačnej energie na teplo bez emisie fotónu. Tretí proces je fotochemický, absorbovaná energia sa prenesie fluorescenčným rezonančným prenosom na inú susediacu pigmentovú molekulu. Migráciou energie z molekuly na molekulu sa umožňuje sústredenie svetelnej energie v chlorofyle reakčného centra (P680 alebo P700). Tu dochádza k oddeleniu kladného a záporného náboja a tým ku konzervovaniu energie do chemických väzieb. Modré svetlo je vo fotosyntéze vždy menej účinné ako červené svetlo. Pri excitácii chlorofylu modrým svetlom sú nevyhnutné energetické straty výdajom tepla. Pri absorbcii fotónu červeného svetla sa excitačná energia využije na prenos bez strát. Grafické zobrazenie relatívnej absorbancie rôznych vlnových dĺžok svetla látkou sa volá absorpčné spektrum. Absorpčné spektrá chlorofylov ukazujú, že oba chlorofyly absorbujú v modrej a červenej oblasti spektra a že absorbujú veľmi málo zeleného svetla. Ak sa porovná účinok rôznych vlnových dĺžok na rýchlosť fotosyntézy tak, že sa nedosiahne úroveň nasýtenia, získa sa akčné spektrum. Porovnanie absorbčných spektier jednotlivých pigmentov a akčného spektra prekvapuje v tom, že vo fotosyntéze sa značne využíva aj zelené svetlo. Príčinou tohoto faktu je mnohonásobne opakované odrážanie zeleného svetla od chloroplastu ku chloroplastu v 19
44 liste. S každým odrazom sa malé percento týchto vlnových dĺžok absorbuje až nakoniec sa absorbuje značná časť zeleného svetla. Chlorofylové molekuly absorbujú niekoľko fotónov každú sekundu. Efektívnosť využitia enzýmov fotosyntézy sa dosahuje veľkým počtom pigmentov, ktoré dodávajú energiu do reakčného centra. Vysoko špecifické nekovalentné väzby molekúl chlorofylov s bielkovinami spôsobujú modifikáciu ich absorbčných spektier. Červené absorbčné maximum sa posúva o 10 až 30 nm k dlhším hodnotám vlnových dĺžok. In vivo existuje v tylakoidnej membráne niekoľko foriem chlorofylu a líšiacich sa absorpčnými maximami. Červené svetlo s vlnovými dĺžkami nad 690 nm sa chloroplastami výrazne absorbuje, ale vo fotosyntéze je málo účinné. Zistilo sa, že ak sa chloroplasty osvetlili súčasne svetlom kratších vlnových dĺžok a aj dlhovlnným červeným svetlom, intenzita fotosyntézy bola väčšia, ako jej suma ak sa obe osvetlenia aplikovali zvlášť. Toto zvýšenie fotosyntézy sa dá vysvetliť iba existenciou dvoch oddelených skupín pigmentov, fotosystémov. Fotosystém II (PSII) absorbuje vlnové dĺžky kratšie než 690 nm a sériovo kooperujúci fotosystém I (PSI) vlnové dĺžky do 700 nm. Hlavné komplexy tylakoidov V tylakoidnej membrány sú lokalizované štyri hlavné proteínové komplexy fotosyntézy (fotosystém II, komplex cytochrómov b 6 -f, fotosystém I a ATP syntáza). Obr. 5 Proteínové komplexy prenosu elektrónov a protónov v tylakoidnej membráne 20
45 Fotosystém II (voda-plastochinón oxidoreduktáza). Jadrový komplex fotosystému II pozostáva zo šiestich polypeptidov, ktoré sú navzájom pospájané nekovalentnými väzbami a reakčného centra P 680. Dva z týchto polypeptidov (D1 a D2) priamo viažu P680 a chinóny potrebné na oxidáciu vody. S jadrovým komplexom sú prepojené aj tri periférne polypeptidy. Pigment P680 je chlorofyl a s absorpčným maximom 680 nm. Predpokladá sa, že polypeptidy viažu Ca 2+ a Cl -, potrebné pre fotolýzu vody. Okrem polypeptidov jadrový komplex obsahuje 40 molekúl chlorofylu a, niekoľko molekúl -karoténu, membránové lipidy (väčšinou galaktolipidy), štyri mangánové ióny, jeden nekovalentne viazaný Fe, jeden alebo viac Ca 2+, niekoľko Cl -, dve molekuly plastochinónu a dve molekuly feofytínu. Svetlozberný komplex fotosystému II (LHCII) zachytáva svetelnú energiu pomocou 250 molekúl chlorofylu a a b (zastúpené sú v približne rovnakom počte) a mnohými molekulami xantofylov. Každá molekula pigmentu je viazaná s integrálnym proteínom. Na jednu molekulu proteínu sa viaže 10 chlorofylov a 2 alebo 3 xantofyly. Komplexy PSII sú prítomné najmä v častiach tylakoidov grán, ktoré nie sú v kontakte so strómou. Celková funkcia PSII je pomocou svetelnej energie redukovať plastochinón (PQ na PQH 2 ) za využitia elektrónov z vody, pričom sa potrebné protóny odoberajú zo strómy. Cytochrómový komplex b 6 -f. Tento komplex pozostáva zo štyroch odlišných integrálnych polypeptidov, tri z nich obsahujú Fe 3+, ktoré sa počas toku elektrónov redukuje na Fe 2+ a potom späť oxiduje. Prvé dva sú cytochróm b 6 a cytochróm f. Každý cytochróm obsahuje železo v hémovej prostetickej skupine. Tretí je proteín s dvomi nehémovými atómami železa, ktoré sú spojené s dvomi nebielkovinovými atómami síry a tiež s dvomi atómami síry cysteínov v proteíne (2Fe - 2S proteín). Štvrtý polypeptid neobsahuje železo a nie je známa jeho funkcia. Komplex cyt b 6 - f je zhruba rovnako zastúpený v tylakoidoch grán a strómy. Jeho hlavnou funkciou je preniesť elektróny z PSI na PSII. Uskutočňuje sa to oxidáciou PQH 2 a redukciou plastokyaninu. Zároveň sa protóny uvoľňujú do lúmenu tylakoidu. Plastokyanin je malý proteín, dobre pohyblivý v tylakoidnej membráne. Pozostáva zo štyroch polypeptidov, každý ma po atóme medi koordinovanom s dvomi atómami dusíka histidínu a dvomi atómami síry cysteínu. V oxidovanom stave plastokyanin je modrý. Fotosystém I. Jadrový komplex PSI obsahuje 11 odlišných polypeptidov. Jeden z nich je polypeptid centra P700. Pigmentom P700 je chlorofyl a. Dva najväčšie polypeptidy (označené Ia a Ib) viažu reakčné centrum s inými polypeptidmi a viažu tiež 50 až 100 molekúl chlorofylu a, niekoľko molekúl -karoténu a tri prenášače elektrónov (A 0, A i a X). A o je chlorofyl a, A 1 je naftochinón fylochinón a X je 4Fe - 4S skupina. FeS skupiny sa zistili aj na ďalších dvoch polypeptidoch. Na polypeptid svetlozberného komplexu I (LHCI) sa viaže okolo 100 anténových molekúl chlorofylu a a b (v pomere 4:1). LHCI obklopuje jadrový komplex. Fotosystém I je lokalizovaný v tylakoidoch strómy a úsekoch tylakoidov grán, ktoré sú orientované do strómy. PSI funguje ako na svetle závislý systém oxidácie redukovaného plastokyaninu, ktorý prenáša elektróny na ferredoxin. 21

46 Ferredoxin je malý periférny Fe-S proteín pripojený voľne na strómatickej strane tylakoidov. Syntetizuje sa na cytoplazmatických ribozómoch. Môže byť donórom elektrónov aj pre sulfitreduktázu a nitritreduktázu. ATP syntáza. Konečným komplexom v tylakoidoch je skupina deviatich polypeptidov, ktorá mení ADP a P i na ATP. Nachádza sa len v nestlačených úsekoch tylakoidov grán a tylakoidoch strómy, tesne vedľa PSI. Má dve hlavné časti, stopkovú (CF o ), ktorá prebieha z lúmenu cez tylakoidnú membránu do strómy a sférickú (CF 1 ), ktorá leží v stróme. ATP syntáza spája tvorbu ATP s transportom protónov z lúmenu cez tylakoidnú membránu do strómy. Mechanizmus elekrónového a protónového transportu Po absorbcii fotónov vhodných vlnových dĺžok v pigmentoch P680 a P700 v reakčných centrách fotosystémov dochádza k ich elektrónovej excitácii do singletového stavu. Každý z excitovaných pigmentov P * 680 a P * 700 prenáša v primárnej reakcii elektrón v priebehu 20 nanosekúnd na molekulu reakčného partnera, na eletrónový akceptor. Takto sa energia žiarenia využije na intermolekulový prenos elektrónu, pričom energia fotónu sa viaže do formy redoxnej energie. V ďalších sekundárnych reakciách sa oddelí náboj na protiľahlych stranách tylakoidnej membrány za asi 200 pikosekúnd. Obr. 6 Cesty toku elektrónov a protónov v tylakoidnej membráne Transport elektrónov a protónov sa uskutočňuje kaskádou oxidačno - redukčných reakcií pomocou štyroch komplexov (PSII, cyt b 6 -f, PSI a ATP syntáza). Tieto komlexy sú v tylakoidnej membráne vektorovo orientované. Voda sa oxiduje na O 2 v lúmene tylakoidu, NADP + sa redukuje na NADPH na strómalnej strane tylakoidnej membrány, ATP sa uvoľňuje do strómy v spojení s pohybom H + z lúmenu do strómy. 22
47 Fotolýza vody Elektróny z H 2 O odoberá komplex výdaja O 2 fotosystému II. Uvoľnenie jednej molekuly O 2 vyžaduje oxidáciu dvoch H 2 O a premiestnenie štyroch elektrónov. Predpokladá sa, že prebieha jednostupňová oxidácia dvoch molekúl vody, ktorej podmienkou je naakumulovanie štyroch kladných nábojov. Pre vytvorenie štvornásobného kladného náboja, sa predpokladá účasť rôznych oxidačných stupňov mangánu (Mn 2+, Mn 3+, Mn 4+ ) naviazaných na polypeptidy D 1 a D 2 na lúmenovej strane tylakoidnej membrány, kde sa O 2 uvoľňuje. Pri oxidácii H 2 O majú úlohu Cl - a Ca 2+. Prvou látkou, ktorá získa elektróny uvoľnené oxidáciou H 2 O je aminokyselina tyrozín v polypeptide D1. Elektróny prenáša po jednom na P P680 odovzdá elektrón feofytínu, ktorý ho ďalej prenesie na plastochinón Q A pevne viazaný na D2. Z neho sa odovzdá elektrón na inú molekulu plastochinónu (Q B ) voľne viazanú na polypeptid D1. Úplná redukcia plastochinónu vyžaduje dva elektróny a súčasne dva H +. Protóny pochádzajú zo strómy a keď sa neskôr PQH 2 oxiduje, uvoľňujú sa do lúmenu tylakoidu. Redukovaný PQ B je pohyblivý a prenáša elektróny na komplex cyt b 6 -f. Na D1 sa potom viaže iná molekula PQ B, ktorá prenesie dva elektróny z druhej molekuly H 2 O a zároveň odoberie protóny zo strómy. Obr. 7 Z schéma fotosyntézy 23
48 Komplex cyt b 6 -f môže prijať elektróny každého mobilného redukovaného plastochinónu. Elektróny sa prenášajú po jednom na Fe 3+, ktoré sa redukuje na Fe 2+. Fe 3+ je buď v Fe-S proteíne alebo cytochróme b 6. Zároveň z každého PQH 2 sa dva H + uložia do lúmenu tylakoidu. Každý elektrón z Fe 2+ v cyt b 6 alebo FeS proteíne sa prenesie na cyt f, v ktorom sa redukuje Fe 3+ na Fe 2+. Cyt f dodá elektrón na Cu 2+ v plastokyanine, ktorý redukuje na Cu +. Mobilný plastokyanový prenášač dodáva elektróny do komplexu fotosystému I. Prvou molekulou, ktorá prijíma elektróny z Cu + v plastokyanine je P Každý pohyb elektrónu z P700 na akceptor A o vyžaduje dodanie energie zo svetlozberného systému LHCI. Z A o sa elektrón prenesie na A 1 a potom na železo v rôznych 4Fe-4S proteínoch v jadre komplexu fotosystému I. Napokon príjme elektrón mobilný ferredoxin a prenesie ho pomocou ferredoxin - NADP + reduktázy na NADP +. Reakcie poháňané svetlom, ktorými sa lineárne prenášajú elektróny z H 2 O cez tylakoidnú membránu umožnia v stróme vznik NADPH. Na redukciu jednej molekuly NADP + sú potrebné dva elektróny z H 2 O. Súčasne sa pri oxidácii každej molekuly H 2 O uložia do lúmenu tylakoidu štyri H + : dva z oxidácie vody a dva z PQH 2. Pri výdaji jednej molekuly O 2 sa akumuluje 8 H +. Fotofosforylácia Časť energie fotónu zachytenej pigmentami fotosyntetického aparátu sa chemiosmotickým mechanizmom viaže do ATP. Asymetrická podstata tylakoidnej membrány a jednosmerný tok protónov počas transportu elektrónov vytvoria gradient H +. Rozdiely v koncentráciach protónov sú ekvivalentné gradientu ph, stróma sa stane alkalickou (ph 8) a lúmen tylakoidov kyslým (ph 5). Transport elektrónov a protónová pumpa vyvolajú tiež rozdiel v elektrických potenciáloch cez tylakoidnú membránu. Protónový chemický potenciál a transmembránový elektrický potenciál predstavujú zásobu voľnej energie, ktorá sa označuje spolu ako protónom motivovaná sila. Lipidová membrána tylakoidov je prakticky nepriepustná pre H +. Protóny sa môžu prenášať späť do strómy kontrolovaným spôsobom cez komplex ATP syntázy. Protóny sa z lúmenu tylakoidu transportujú rotujúcim c polypeptidom a vypudia do strómy. Tento enzým (CF1) využije protónom motivovanú silu pri vzniku makroergických väzieb počas syntézy ATP z ADP a P i. Na tvorbu jednej molekuly ATP je potrebné transportovať 3 protóny. Obr. 8 Chloroplastová ATP syntáza 24
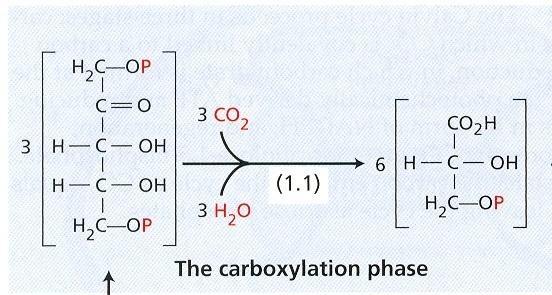
49 Cesty transportu elektrónov a fotofosforylácia Fotofosforylácia môže byť spojená s dvojakými cestami toku elektrónov. Dosiaľ popísaný lineárny transport elektrónov z vody na NADP + sa označuje ako necyklický. Pri necyklickej fotofosforylácii 8 H + umožňuje tvorbu 3 ATP. ATP sa môže tvoriť aj v spojení s cyklickým transportom elektrónov. Ak je necyklický tok elektrónov zablokovaný, napríklad nedostatkom oxidovaných NADP +, môže prebiehať cyklický transport elektrónov. Elektróny z PSI alebo veľmi pravdepodobne z ferredoxínu sa vrátia späť na plastochinón cez cytochrómy. Energia sa využije iba na prenos H + do lúmenu tylakoidu. Fotosystém II sa sa na cyklickom transporte nepodieľa. Význam necyklickej fotofosforylácie in vivo nie je jasný. Vo fyziologicky intaktných pletivách môže byť dôležitá vtedy, keď necyklický transport je pomalý, napríklad pri nedostatku CO 2 a slabom svetle. Metabolizmus uhlíka - fixácia CO 2 a syntéza sacharidov Svetelná energia uložená do makroergických väzieb NADPH a ATP predstavuje redukčnú silu, ktorá sa využíva na redukciu CO 2 po jeho naviazaní ako karboxylovej skupiny do organických látok. NADPH je reduktantom v týchto reakciách, ATP iba podporuje redukciu. Prvým stabilným produktom po naviazaní CO 2 je buď kyselina 3-fosfoglycerová (Calvinov cyklus) alebo organické kyseliny pozostávajúce zo štyroch kyslíkov (Hatchov a Slackov cyklus). Calvinov cyklus V tomto cykle je akceptorom CO 2 ribulozo-1,5-bisfosfát a prvým stabilným produktom po fixácií kyselina 3-fosfoglycerová. Reakcie Calvinovho cyklu prebiehajú v stróme chloroplastov, kde sú lokalizované enzýmy, ktoré ich katalyzujú. Cyklus má tri fázy: 1. karboxylačnú, 2. redukčnú a 3. regeneračnú. Obr. 9 Calvinov cyklus 25
. Na jedného človeka pripadá 20 kg. Rubisko tvorí 40 % z celkových rozpustných proteínov väčšiny listov.")
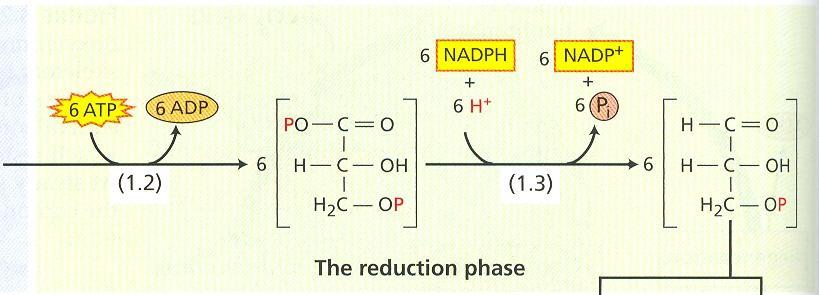
50 Prvú, karboxylačnú reakciu katalyzuje ribulóza 1,5-bisfosfát karboxyláza-oxygenáza (rubisko). Rubisko je enzým zodpovedný za fixáciu 200 biliónov ton CO 2 ročne. Je to kvantitatívne najvýznamnejší enzým na svete (10 7 ton). Na jedného človeka pripadá 20 kg. Rubisko tvorí 40 % z celkových rozpustných proteínov väčšiny listov. Aktívny enzým sa skladá 8 jednotiek. Každá jednotka sa tvorí z dvoch podjednotiek, menšia (14 kd) je kódovaná jadrovým genómom, väčšia (53 kd) chloroplastovým genómom. Expresia génov pre menšiu podjednotku je regulovaná svetlom cez fytochróm a ďalšie proteíny. Rubisko katalyzuje reakciu ribulóza-1,5-bisfosfátu a CO 2. Očakávanú šesťuhlíkovú látku sa nepodarilo zistiť ani ako medziprodukt. Vznikajú dve molekuly kyseliny 3-fosfoglycerovej. Preto rastliny, ktoré primárne viažu CO 2 Calvinovým cyklom sa označujú ako C-3 rastliny. V redukčnej fáze sa karboxylová skupina kyseliny 3-fosfoglycerovej redukuje na aldehydovú skupinu v 3-fosfoglyceraldehyde. Reakcia neprebieha priamo, najprv za spotreby ATP dochádza ku fosforylácii na kyselinu 1,3-bisfosfoglycerovú, až potom sa spotrebuje NADPH na redukciu. Celkom na redukciu 1 molekuly CO 2 sú potrebné 3 ATP a 2 NADPH. Tretia molekula ATP sa spotrebuje v regeneračnej fáze. Obr. 10 Karboxylačná a redukčná fáza fotosyntézy 1.1 ribulóza 1,5-bisfosfát karboxyláza/oxygenáza (rubisko); fosfoglycerátkináza; 1.3 NADP-glyceraldehyd-3-fosfátdehydrogenáza 1,5-bisfosfát sa regeneruje cez viaceré cukrové medziprodukty so 4, 5, 6 a 7 uhlíkmi. Tretí ATP je potrebný na fosforyláciu ribulóza-5-fosfátu. V troch opakovaniach cyklu sa fixujú tri molekuly CO 2 a vytvorí sa jeden 3- fosfoglyceraldehyd. Niektoré molekuly 3-fosfoglyceraldehydu sa využijú v chloroplastoch na tvorbu asimilačného škrobu, ktorý je bezprostredným produktom fotosyntézy. Ďalšie molekuly 3- fosfoglyceraldehydu sa transportujú von z chloroplastu antiportovým prenášačovým systémom výmenou za P i. Iné molekuly 3-PGA sa menia na dihydroxyacetón fosfát a môžu sa tým istým antiportovým prenášačovým systémom vyniesť von z chloroplastu, do cytosólu. Tento transportný systém udržiava konštantné množstvo fosfátu v chloroplastoch. Triozafosfáty v cytosóle sa využijú na tvorbu sacharózy, polysacharidov bunkovej steny a stoviek ďalších látok. 26
51 Obr. 11 Regeneračná fáza Calvinovho cyklu. Fotorespirácia V tme dýchajú listy C-3 rastlín rýchlosťou, ktorá je približne šestinou rýchlosti fotosyntézy. Na svetle je rýchlosť dýchania vyššia, často dva až trikrát ako v tme. V poľných podmienkach sa dýchaním môže uvoľniť tretina až štvrtina CO 2, ktorý sa viazal fotosyntézou. Dýchanie v osvetlených orgánoch prebieha dvomi mechanizmami: tmavým lokalizovaným v cytosóle a mitochondriách a fotorespiráciou (kooperujú chloroplasty, peroxizómy a mitochondrie). Na rozdiel od mitochondriálnej respirácie, ktorá prebieha na svetle aj v tme, fotorespirácia je svetlom stimulovaný príjem O 2 a výdaj CO 2. Obr. 12 Karboxylácia a oxygenácia ribulóza-1,5-bisfosfátu enzýmom rubisko 27

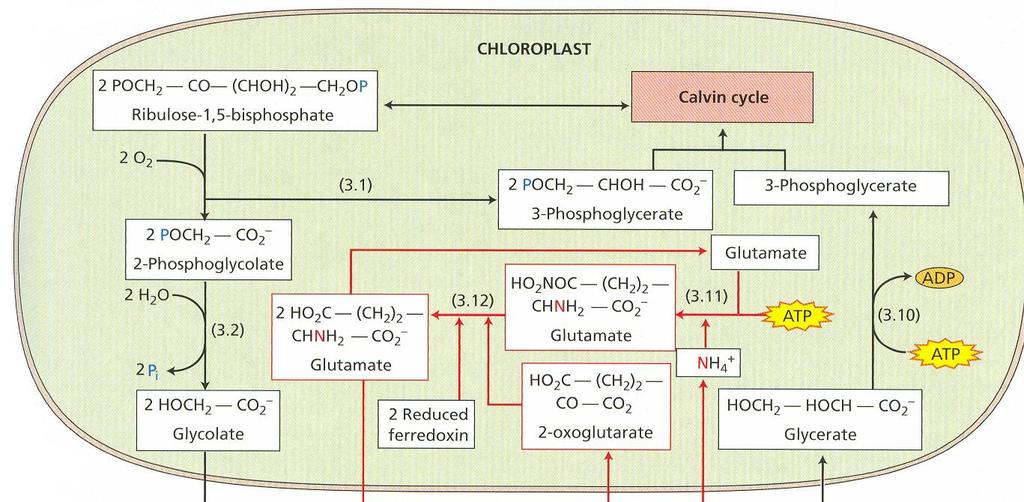
52 Obr. 13 Fotorespirácia C 2 oxidatívny cyklus fotosyntézy 28
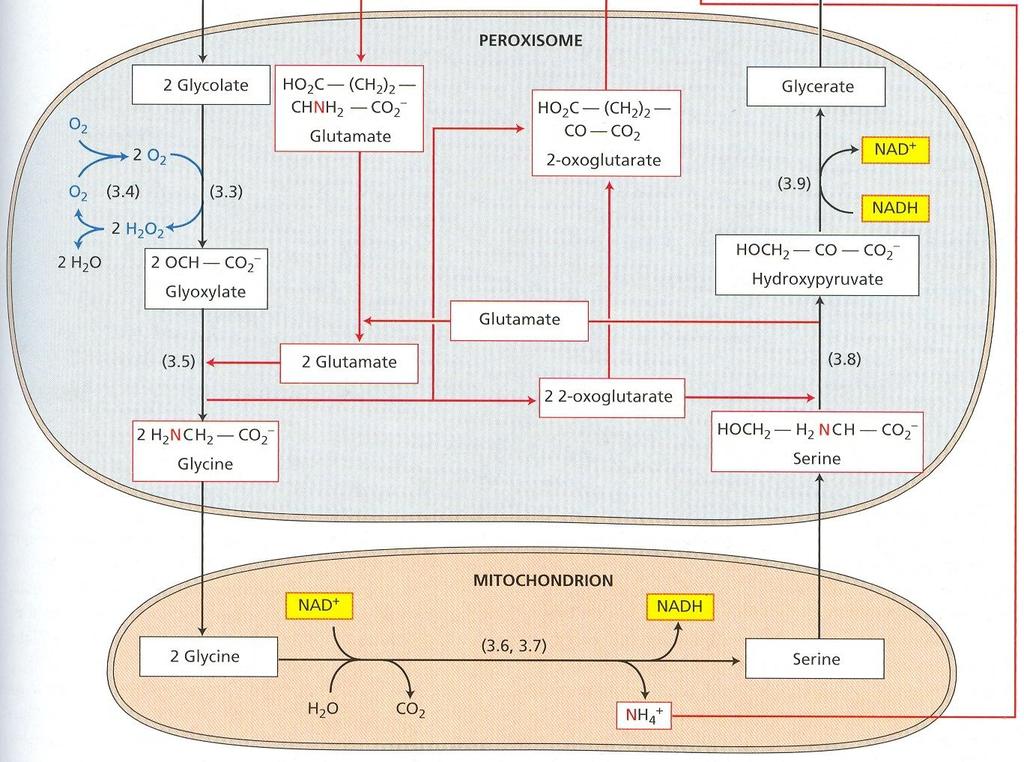
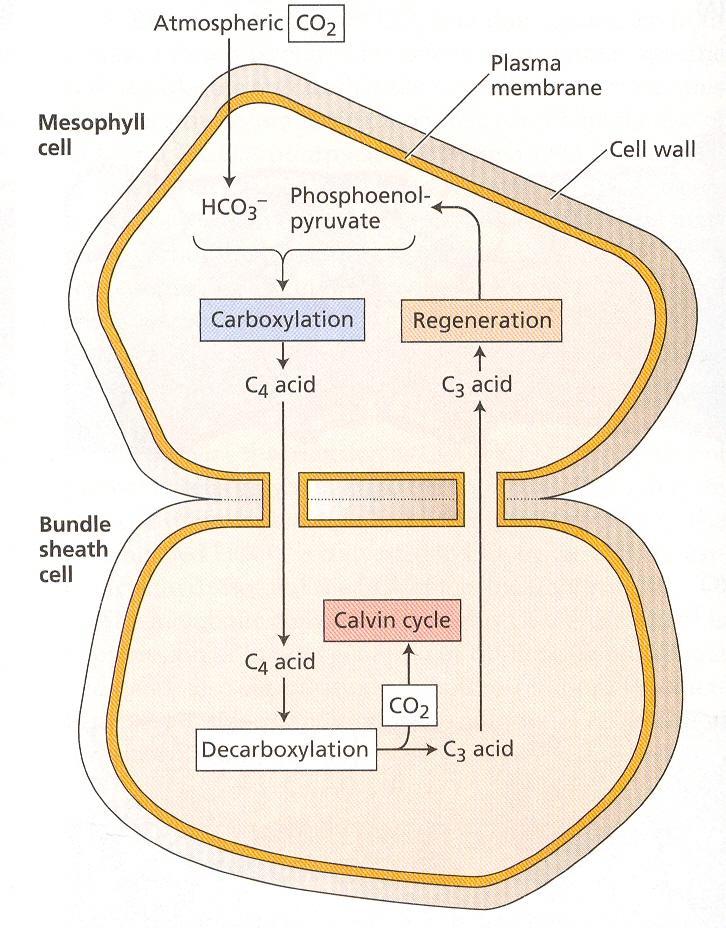
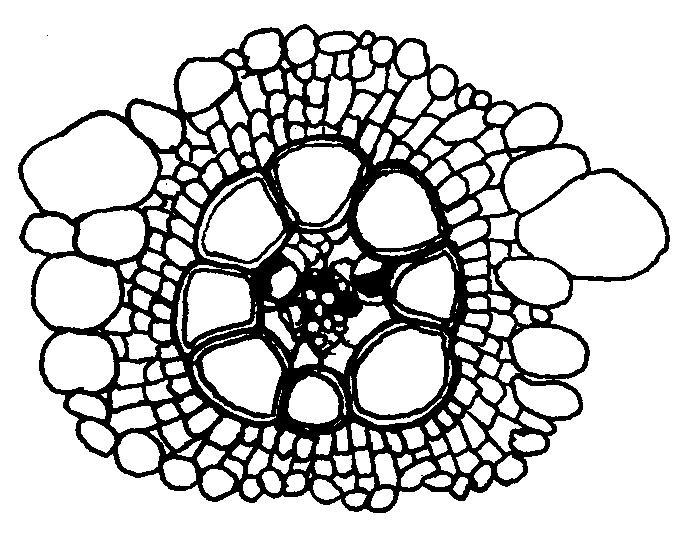
53 Fotosyntéza a fotorespirácia sú navzájom súvisiace procesy. Pre objasnenie mechanizmu fotorespirácie bolo dôležité zistenie, že rubisko má zároveň oxygenázovú aktivitu. Ribulóza-1,5- bisfosfát karboxyláza/oxygenáza má iba jedno aktívne centrum, na ktorom prebieha reakcia s ribulóza-1,5-bisfosfátom a CO 2 alebo O 2, ktoré sú v kompetitívnom vzťahu. Pri rovnakej koncentrácií CO 2 a O 2 rubisko 80 krát rýchlejšie katalyzuje fixáciu CO 2 ako oxygenáciu. V reakcii s O 2 (obr. 13) sa ribulóza-1,5-bisfosfát štiepi na kyselinu fosfoglykolovú a kyselinu 3- fosfoglycerovú. Z kyseliny fosfoglykolovej špecifická fosfatáza odštiepi P i ešte v chloroplastoch. Kyselina glykolová sa transportuje do peroxizómov. Peroxizómy sú druhom mikroteliesok, ktoré sú prítomné len vo fotosyntetických pletivách. Sú v priamom kontakte s chloroplastami. V nich sa kyselina glykolová oxiduje na kyselinu glyoxálovú, elektróny sa prenášajú na O 2 pričom vzniká H 2 O 2. Dve molekuly H 2 O 2 sa rozložia katalázou na H 2 O a O 2. Kyselina glyoxálová sa transamináciou konvertuje na aminokyselinu glycín, ktorý sa transportuje do mitochondrií. V nich dve molekuly glycínu reagujú za vzniku aminokyseliny serín, pričom sa odštiepi CO 2 a NH 4 +. Serín sa transportuje do peroxizómov, kde transamináciou vzniká hydroxypyruvát, ten sa redukuje na glycerát a transportuje do chloroplastov. Fotorespirácia znižuje čistú fotosyntézu a v konečnom dôsledku rýchlosť rastu. Jednou z možnosti vysvetlenia jej existovania je, že sa takto odstraňuje prebytok ATP a NADPH pri veľmi vysokých hodnotách ožiarenia. Cesta C-4 dikarboxylových kyselín - Hatchov a Slackov cyklus V listoch cukrovej trstiny, a neskôr aj v iných druhoch, sa zistili ako prvé produkty fixácie CO 2 organické kyseliny (oxaloacetát, malát a aspartát). Fixáciu CO 2 v tomto cykle katalyzuje fosfoenolpyruvát karboxyláza lokalizovaná v cytoplazme. CO 2 sa viaže na fosfoenolpyruvát za vzniku oxaloacetátu a P i. Oxaloacetát v ďalšej reakcii vyžadujúcej NADPH, za katalytického pôsobenia malátdehydrogenázy sa mení na malát. Malátdehydrogenáza je lokalizovaná v stróme chloroplastov, to znamená, že oxaloacetát sa musí transportovať do chloroplastu. Antiportový systém, ktorý to zabezpečuje zároveň vynáša málat von z chloroplastu. Oxaloacetát v cytozole sa môže transaminačnou reakciou meniť na aspartát. Amínoskupina z alanínu nahradí kyslík v α- oxoskupine, zároveň vznikne pyruvát. Postupne sa zistilo, že aj rastliny C-4 majú všetky enzýmy Calvinovho cyklu. Fixácia CO 2 v nich prebieha dvakrát. Z hľadiska evolúcie sa za pôvodné, vývojové staršie typy považujú C-3, z nich sa neskôr vyvinuli C-4 ako adaptácia na sucho a veľkú intenzitu žiarenia. Prítomnosť C-4 cyklu nie je taxonomicky viazaná. V rámci jedného rodu sa môžu vyskytnúť druhy C-3 aj C-4. Priestorová kompartmentácia fixácie CO 2 Pre rastliny C-4 je význačný vencovitý typ anatomickej stavby listov. Okolo pošiev cievných zväzkov je nápadná jedna alebo dve vrstvy hrubostenných buniek s veľkým obsahom chlorofylu, 29
54 ktorá oddeľuje cievne zväzky od mezofylu. V C-3 rastlinách takéto rozdiely nie sú. V bunkách pošiev cievnych zväzkov C-4 rastlín je veľa chloroplastov, ktoré často obsahujú temer všetok listový škrob. V mezofylových bunkách škrob je málo zastúpený. Malát a aspartát sa tvoria v mezofylových bunkách. 3-fosfoglyceraldehyd, sacharóza a škrob sa tvoria hlavne v bunkách pošiev cievnych zväzkov. Obr. 14 C 4 fotosyntetický cyklus v periférnych bunkách listu (mezofyl) a cievnej oblasti. Priestorová kompartmentácia je charakteristická aj pre lokalizáciu enzýmov. Rubisko a ďalšie enzýmy Calvinovho cyklu sú iba v bunkách pošiev cievnych zväzkov. Fosfoenolpyruvát karboxyláza sa vyskytuje hlavne v mezofylových bunkých. CO 2 po vstupe do listu cez prieduchy najprv preniká do mezofylových buniek, kde sa viaže Hatch - Slackovým cyklom ako karboxylová skupina malátu alebo aspartátu. Tieto C-4 organické kyseliny sa rýchle prenášajú do buniek pošiev cievnych zväzkov. V nich sa dekarboxylujú, uvoľnený CO 2 sa fixuje rubiskom do 3-fosfoglycerátu. Pyruvát a alanin ako produkty dekarboxylácie sa vrátia do mezofylových buniek. Možno konštatovať, že tvorba organických kyselín v mezofyle bez čistého zisku energie slúži na zachytávanie CO 2 a prenášanie do buniek pošiev cievnych zväzkov, kde sa tvoria asimiláty. Skupina C-4 rastlín nie je homogénna, podľa enzýmov a látok, ktoré sa transportujú medzi bunkami sa zaraďujú do troch skupín. 30
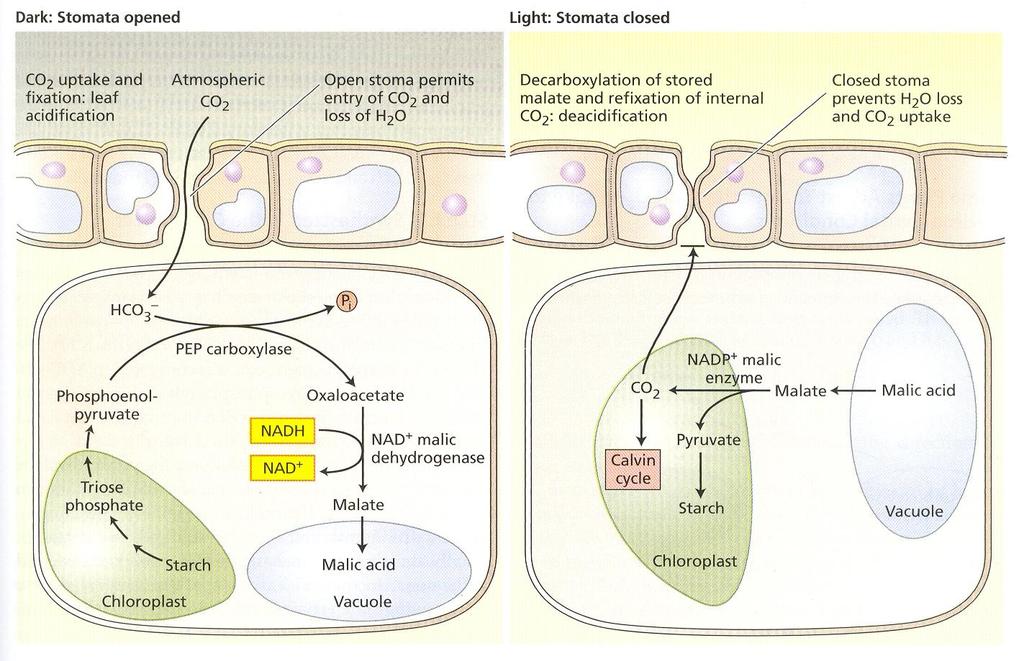
55 1. typ NADP-ME Rastliny, v ktorých sa prevažne syntetizuje malát. Enzým chloroplastov mezofylu malátdehydrogenáza využíva ako zdroj elektrónov koenzým NADPH. Ten istý koenzým vyžaduje aj dekarboxylačný jablčný enzým buniek pošiev cievnych zväzkov. Chloroplasty v bunkách pošiev cievnych zväzkov majú vretenovitý tvar, sú umiestnené pozdĺž vonkajšej steny buniek. Nemajú vyvinuté typické grána, ale vyznačujú sa veľkým počtom škrobových zŕn. Patrí tu kukurica (Zea mays), cukrová trstina (Saccharum officinarum), cirok sudánsky (Sorghum sudanense). 2. typ NAD-ME Aspartátové rastliny s veľkým množstvom NADH jablčného enzýmu v mitochondriách buniek pošiev cievnych zväzkov, kde sa dekarboxyluje a v chloroplastoch fixuje Calvinovým cyklom. Do mezofylových buniek sa transportuje alanín. V bunkách pošiev cievnych zväzkov je veľa mitochondrií. Chloroplasty sú uložené centripetálne a majú vyvinuté grána. 3. typ PCK Patria tu aspartátové rastliny s veľkým obsahom fosfoenolpyruvát karboxykinázy v cytozole buniek pošiev cievnych zväzkov. Aspartát z mezofylových buniek sa transportuje do buniek pošiev cievnych zväzkov, kde sa transamináciou mení na oxaloacetát a ten sa PCK dekarboxyluje. Do mezofylových buniek sa transportuje alanín. Chloroplasty sú rozložené pozdĺž vonkajšej steny parenchymatickej pošvy buniek a majú vyvinuté grána. Patria tu Panicum maximum, Chloris gayana, Sporobolis fimbriatus. Syndrom CAM Metabolizmus organických kyselín rastlín zistený najprv v čeľadi Crassulaceae je adaptáciou na extrémne suché stanovištia a vysoké intenzity žiarenia. Rastliny z tejto čeľade sa označujú ako sukulenty. Majú hrubé listy s pomerne malým pomerom povrchu k objemu. Hrubá kutikula výrazne znižuje rýchlosť transpirácie. Obvykle nemajú dobre vyvinutý palisádový parenchým, ale fotosyntéza prebieha v špongiovom mezofyle. Jeho bunky majú veľké vakuoly. Metabolizmus CAM sa zistil v stovkách druhov 26 čeľadí vyšších rastlín. Podstatnou črtou rastlín CAM je časová kompartmentácia fotosyntézy. V noci pri nízkych teplotách sú prieduchy otvorené, CO 2 sa asimiluje karboxyláciou fosfoenolpyruvátu, ktorý sa získava odbúraním škrobu. Fosfoenolpyruvátkarboxyláza je lokalizovaná v cytozole. Malát sa akumuluje vo vakuole. Ráno je bunková šťava rastlín CAM kyslá (ph 4). Cez deň sú prieduchy uzavreté. Malát sa transportuje do cytozolu a dekarboxyluje. Uvoľnený CO 2 sa refixuje rubiskom do triozofosfátov. 31
56 Obr. 15 Časová kompartmentácia fotosyntézy (CAM). Porovnanie rastlín typu C-3, C-4 a CAM Prevažujúce podmienky na stanovištiach C - 3 mierne klimatické podmienky C - 4 sucho, vysoká intenzita slnečného žiarenia, nízka vzdušná vlhkosť CAM arídne podmienky, sucho, vysoká intenzita slnečného žiarenia, vysoké denné a nízke nočné teploty Anatomická stavba listu C - 3 mezofyl väčšinou rozlíšený na hubový a palisádový parenchým C - 4 mezofyl s parenchymatickou pošvou okolo cievnych zväzkov CAM bunkový mezofyl s veľkými vakuolami Štruktúra chloroplastov C - 3 majú grána, škrobové zrná a plastoglobuly, pomer chlorofylu a:b - 3:1 32
57 C - 4 dimorfizmus chloroplastov v bunkách mezofylu (granárne) a pošvách cievnych zväzkov (granárne alebo agranárne), pomer chlorofylu a:b - 5:1 CAM chloroplasty s menším počtom grán, s mnohými škrobovými zrnami a plastoglobulami, pomer chlorofylu a:b - 3:1 Karboxylačné enzýmy C - 3 rubisko C - 4 fosfoenolpyruvát karboxyláza a rubisko, oddelená lokalizácia CAM fosfoenolpyruvát karboxyláza a rubisko, časové oddelenie Minimálna spotreba ATP a NADPH pre fixáciu 1 molekuly CO 2 C ATP + 2 NADPH C ATP + 2 NADPH CAM 5 ATP + 2 NADPH Strata vody pri fixácii 1g CO 2 C g C g CAM g Vplyv intenzity žiarenia C - 3 saturácia pri asi tretine z maximálneho ožiarenia C - 4 ani pri maximálnom ožiarení nedochádza k saturácii CAM protichodné údaje Fotorespirácia C - 3 do 40 % fotosyntézy C - 4 nedetekovateľná CAM nedetekovateľná Rozdeľovanie produktov asimilácie C - 3 pomalé C - 4 rýchle CAM variabilné Produkcia sušiny C - 3 stredná C - 4 veľká CAM malá Syntéza sacharózy a polysacharidov Na konci každej z troch hlavných ciest, ktorými sa fixuje CO 2, z triozofosfátov sa syntetizujú dva základné produkty disacharid sacharóza a polysacharid škrob. Glukóza a fruktóza, ani iné voľné hexózy sa vo fotosyntetických bunkách obvykle neakumulujú. V listoch niektorých tráv namiesto škrobu sa syntetizujú fruktozány. Syntéza oligosacharidov a polysacharidov sú kompetitívne procesy. 33
58 Syntéza sacharózy Sacharóza je základnou transportnou formou sacharidov v rastline. V rýchle rastúcich rastlinách je prevažujúcim konečným produktom fotosyntézy, je zdrojom energie. Ekonomicky významná je jej akumulácia v koreňoch cukrovej repy a v stonkách cukrovej trstiny. Sacharóza sa syntetizuje v cytozole. 3-fosfoglyceraldehyd a dihydroxyacetón fosfát sa v cytozole za katalytického pôsobenia fruktóza-1,6-bisfosfát aldolázy menia na fruktóza-1,6- bisfosfát. Fosfatázou sa odštiepi P i za vzniku fruktóza-6-fosfátu, ktorý sa izomerázou mení na glukóza-6-fosfát a v ďalšej reakcie katalyzovanej fosfoglukomutázou na glukóza-1-fosfát. Molekula glukóza-1-fosfátu sa pyrofosforylázou aktivuje reakciou s UTP za vzniku uridíndifosfoglukózy. Za katalytického pôsobenia enzýmu sacharóza-fosfát syntáza UDPG reaguje s fruktózo-6-fosfátom, pričom vzniká sacharóza-6-fosfát. Sacharóza-fosfát fosfatázou sa odštiepi fosfát. Na syntézu jednej molekuly sacharózy je potrebná energia vo forme 1 ATP. Sacharóza sa uskladňuje vo vakuole. Pri mobilizácii sa zásobná sacharóza hydrolyzuje kyslou invertázou vakuoly (optimum ph je 5,0) a hexózy sa uvoľňujú do cytoplazmy. Nosič je v tonoplaste ten istý pre oba smery a je energeticky nezávislý, preto sa riadi relatívnými koncentráciami. Sacharóza sa transportuje z miest syntézy (zdroja) do sinkov (napr. meristémov). Pri transporte na krátke vzdialenosti sa uplatňuje apoplastická aj symplastická cesta. Transport na dlhé vzdialenosti sa uskutočňuje sitkovicami. Pohyb je možný v oboch smeroch, poháňaný je koncentračným gradientom. Sacharóza sa potom štiepi buď alkalickou invertázou (optimum ph je 7,5) v cytoplazme alebo kyslou invertázou v bunkovej stene a vakuole. Obr. 16 Syntéza sacharózy v cytosole. 34
59 Syntéza a odbúravanie škrobu V chloroplastoch cez deň sa syntetizuje nestály asimilačný škrob. Časť viazaného uhlíka sa môže využiť pre metabolizmus fotosyntetizujúcej bunky. Odbúrava sa prednostne účinkom exoenzýmu fosforylázy, ktorý fosforolyticky odštiepuje glukóza-1-fosfát. Fosfátový translokátor je schopný transportovať maltózu a glukóza-1-fosfát von z chloroplastu. V noci sa asimilačný škrob mobilizuje a transportuje do sinkov. Transportné formy sú dočasne vo vakuole. Najvýznamnejším činiteľom regulácie distribúcie asimilátov je turgorový tlak v sitkoviciach. Synchronizáciu medzi zdrojom a sinkom riadia aj ďalšie faktory, hormóny (nepriamym vplyvom), sacharóza, draslík a fosfát. Škrob je hlavným zásobným sacharidom väčšiny rastlín. Napríklad v bunkách dozrievajúcich semien sa menia proplastidy na amyloplasty. Tie nefotosyntetizujú, ale sa špecializujú na zhromažďovanie zásobného škrobu pre vyvíjajúce sa semeno. V amyloplastoch zásobných pletív sa na dobu niekoľkých mesiacov ukladá škrob vo forme granúl nerozpustných vo vode. Tvorí sa zo sacharózy alebo iných transportných foriem. Sacharóza sa v cytoplazme štiepi alkalickou invertázou a ako glukóza-6-fosfát transportuje do plastidov. Pri syntéze škrobu do reakcie vstupuje ADP- a UDP-glukóza. Škrob má dva komponenty, amylózu - lineárne reťazce molekúl glukózy viazaných 1,4- -D-glukozidovými väzbami a amylopektín, kde rozvetvenie molekúl je 1,6- -D-glukozidovými väzbami. Tvorbu 1,6 rozvetvenia v amylopektíne katalyzuje Q-enzým. Rozštiepi sa väzba 1,4- -D a časť reťazca sa prenesie na iný glukozylový zbytok za vzniku 1,6- - D väzby. Reakcia môže prebiehať intramolekulove alebo intermolekulove. Zásobný škrob pozostáva z 20 až 25 % amylózy a 75 až 80 % amylopektínu. R-enzým hydrolyzuje 1,6 rozvetvenie, vzniká dextrán. a -amyláza štiepia 1,4- -D glukozidové väzby za vzniku maltózy. Fosforyláza škrobu štiepi maltózu a fosforyluje glukózu. amylóza 35
60 amylopektín Obr. 17 Štruktúra zložiek škrobu Translokácie floémom Transport na dlhé vzdialenosti v rastlinách prebieha v cievach xylému (vzostupný transpiračný prúd) a floému (asimilačný tok). Floém je pletivo, ktorým sa translokujú produkty fotosyntézy z dospelých listov (zo zdroja) do sinkov - miest rastu a zásob (mladé listy, plody, korene. Floémom sa však redistribuje aj voda, hormóny a ďalšie látky. Smer transportu vo floéme nie je vo vzťahu ku gravitácii. Vodivú funkciu floému majú sitkové bunky väčšiny nižších a nahosemenných rastlín a články sitkovíc krytosemenných rastlín, ktoré tvoria sitkovice spolu so sprievodnými bunkami, lykovým parenchýmom a sklerenchymatickými lykovými vláknami. Floémové bunky sú prepojené plazmodezmami prechádzajúcimi prederaveniami stien. Na priečnych stenách článkov sitkovíc sú dokonalejšie sitkové platničky, ktoré tvoria vzájomné prepojenie susediacich živých protoplastov. Dospelé články sitkovíc majú unikátnu stavbu medzi živými bunkami. Nemajú jadro, tonoplast ani ďalšie štruktúry: mikrofilamenty, mikrotubuly, Golgiho telieska a ribozómy. Steny nie sú lignifikované. Floémový cytozol je bohatý na P-proteín, ktorý tvorí slizové telieska. Plastidy obsahujú buď škrob (S-typ) buď proteín (P-typ). Medzi plazmatickou membránou a bunkovou stenou v póre sitka sa ukladá kalóza. Je to - 1,3-glukán syntetizovaný enzýmom plazmalemy. Syntáza kalózy je uložená vektorovo, substrát získava z cytozolovej strany, produkt sa ukladá na bunkovú stenu. Kalóza sa syntetizuje ako odpoveď na poranenie alebo iný stres a za normálnych podmienok ako príprava na dormanciu. Uložením kalózy sa účinne uzavrie poškodený článok sitkovice a oddelí od intaktného pletiva. Po obnovení článku sitkovice sa kalóza odstráni. 36

61 Každý článok sitkovice je plazmodezmami prepojený so sprievodnými bunkami. Tie zabezpečujú metabolické funkcie ako je proteosyntéza, dodávanie ATP a v listoch majú tiež úlohu v transporte asimilátov z produkujúcich buniek. Obyčajné sprievodné bunky majú dobre vyvinuté chloroplasty s tylakoidmi a bunkovú stenu s hladkým vnútorným povrchom. Transferové bunky majú početné výrastky bunkových stien zasahujúce do vnútra bunky, čím sa zväčšuje povrchová plocha plazmalemy. Predpokladá sa, že sú adaptované pre príjem solútov z apoplastu. Intermediárne sprievodné bunky majú početné plazmodezmy, ktorými sa spájajú s okolitými, zvlášť s bunkami pošiev cievnych zväzkov. Floémom sa transportuje voda a solúty, ktorými sú najmä cukry. Sacharóza sa zistila v koncentráciách 0,3 až 0,9 M. Ďalej sa transportujú oligosacharidy, cukrové alkoholy manitol a sorbitol. Netransportujú sa redukujúce cukry. Z dusíkatých látok sa transportujú aminokyseliny glutamát, aspartát, amidy glutamín a asparagín. Druhy s nodulmi fixujúcimi dusík transportujú ureidy (alantoin). Všetky hormóny (auxín, giberelíny, cytokiníny, kyselina abscisová a ACC) ako aj nukleotidové fosfáty a proteíny (proteínové kinázy, tioredoxín ubikvitín a i.) sa zistili vo floéme. Z anorganických solútov sa pohybujú vo floéme draslík, horčík, fosfát a chlorid. Obr. 18 Translokácie floémom 37

62 Ekofyziológia fotosyntézy Počas evolúcie sa rastliny dedične adaptovali štruktúrou a metabolizmom na rôzne podmienky pre fotosyntézu (C 3, C 4, CAM, heliofyty, sciofyty, efeméry a pod.). Dostupnosť minerálnych živín K, N a ďalších prvkov pre dobrú funkciu fotosyntetického aparátu je úzko spätá so zásobovaním vodou. P - makroergické väzby, Fe - cytochrómy, feredoxín, Cu - plastocyanín, Mn a Cl - fotolýza vody. Pre priebeh fotosyntézy je dôležitý obsah chlorofylu. Priemerný obsah chlorofylu je 0,5 g.m -2 listovej plochy. Syntéza chlorofylu prebieha len na svetle. V tme listy obsahujú nezelené etioplasty. Intenzita fotosyntézy počas ontogenézy sa mení, optimálna je v štádiu fotosynteticky dospelého, plne aktívneho listu. Fotosyntéza v gradiente listovej inzercie stúpa a potom nasleduje klesanie. Časové faktory. Cirkadiánne rytmy deň/noc; poludňajšia depresia fotosyntézy; zmeny počas vegetačnej sezóny. Fotosyntézu limitujú aj kapacity pre hromadenie asimilátov (sinky). Patologické faktory a stres tiež znižujú rýchlosť fotosyntézy. Rastliny v poraste - distribúcia žiarenia, CO 2 a pod., vnútro druhová konkurencia, medzidruhová konkurencia v zápoji. Svetlo V priebehu ontogenézy sa rastliny aklimujú na podmienky prostredia. Napríklad v anatomickej stavbe listu, mohutnosť listového mezofylu závisí od toho či sa vyvíja na svetle, alebo v tieni. Obr. 19 Anatomická stavba listov buka lesného (Fagus silvatica) rastúcich v rôznom svetelnom prostredí 38
. Ak rovnaké množstvá svetla dopadajú pod rôznym uhlom, ožiarenosť bude ovplyvnená uhlom dopadu [(B) = (A) x kosínus α]. Obr.")
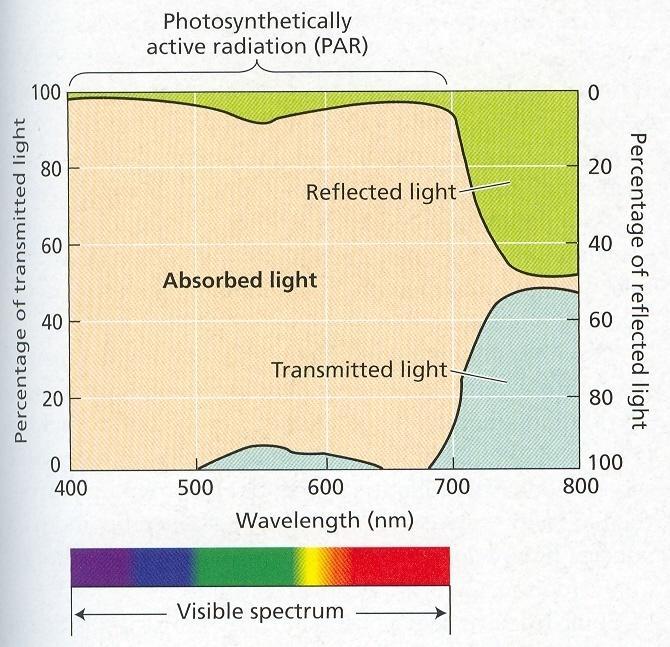
63 Listová mozaika, rozloženie listových čepelí tak aby absorbovali čo najviac z dopadajúceho žiarenia. Obr. 20 Listová mozaika Svetlo dopadajúce na povrch rastlín je tok fotónov. Ožiarenosť je množstvo energie dopadajúce na plochý sensor (W m2). Ak rovnaké množstvá svetla dopadajú pod rôznym uhlom, ožiarenosť bude ovplyvnená uhlom dopadu [(B) = (A) x kosínus α]. Obr. 21 Svetelný tok a ožiarenie Kvalita - FAR, fotosynteticky aktívna radiácia, nm. 39 až 48 % z dopadajúceho žiarenia je FAR, listy absorbujú 1 až 5 %, ostatné odrážajú. 39


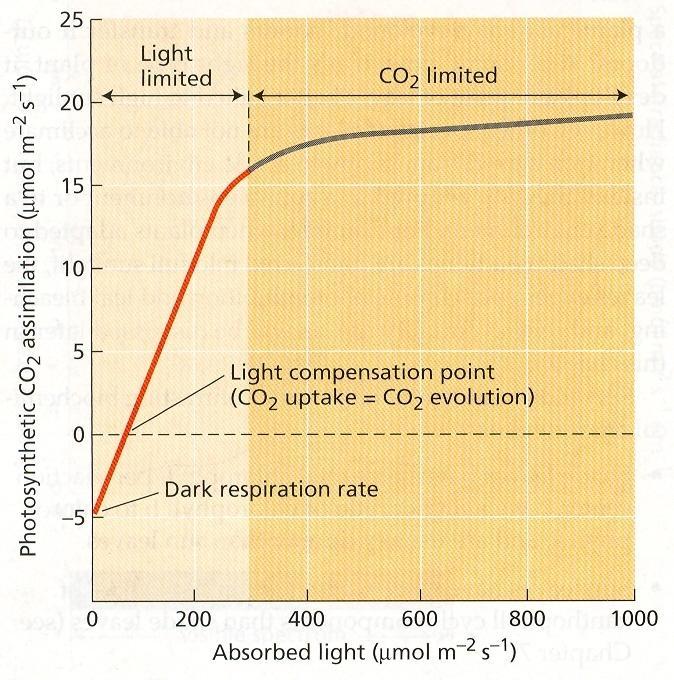
64 Obr. 22 Absorpcia, prepúšťanie a odrážanie podľa kvality svetla Kvantita - pri veľmi nízkej intenzite žiarenia prevažuje dýchanie nad fotosyntézou. Kompenzačný svetelný bod je taká intenzita svetla, pri ktorej je fotosyntéza a dýchanie, spotreba a výdaj CO 2 v rovnováhe. Ďalším zvyšovaním intenzity žiarenia sa zvyšuje výkon fotosyntézy až po bod svetelného nasýtenia. Zvyšovanie kvantity radiácie nad bod svetelného nasýtenia nemá vplyv na rýchlosť fotosyntézy. Za týchto podmienok iné faktory limitujú fotosyntézu. Veľmi vysoké hodnoty žiarenia majú inhibičný efekt. Konkrétne hodnoty sú druhovo špecifické. Heliofyty - dedične adaptované na plné osvetlenie, sciofyty - dedične adaptované na rast v tieni. Obr. 23 Odpoveď fotosyntézy na svetlo v C 3 rastlinách 40
 Obr.")
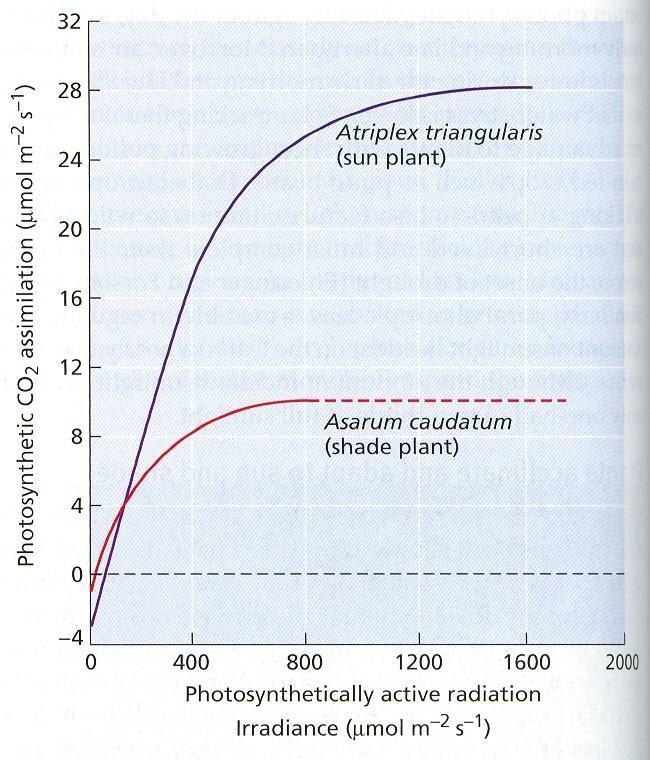
65 a sciofyty (Asarum caudatum) Obr. 24 Heliofyty (Atriplex triangularis) Obr. 25 Svetlo dopadajúce na povrch Zeme a účinnosť fotosyntézy 41
, pre submerzné rastliny je to hlavná forma príjmu anorganického uhlíka.")

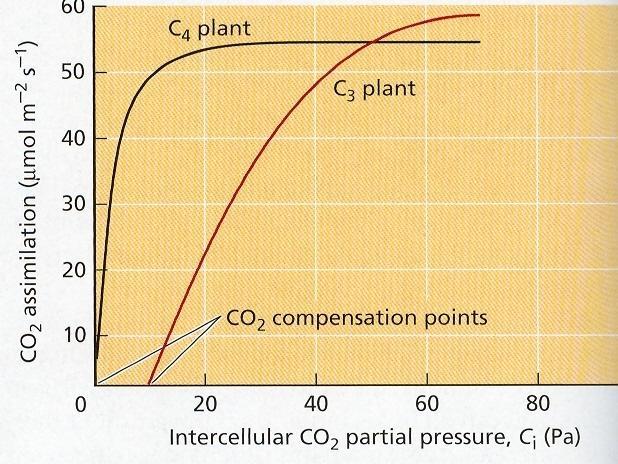
66 CO 2 Bunkový metabolizmus uhlíka spája s vonkajším prostredím výmena plynov. Oxid uhličitý je substrátom fotosyntézy. V atmosfére je 0,02 až 0,03 % CO 2. V plynnej forme vstupuje do listového mezofylu cez prieduchy. Rastliny prijímajú aj bikarbonátový anión koreňmi (HCO 3 - ), pre submerzné rastliny je to hlavná forma príjmu anorganického uhlíka. Zvyšovanie koncentrácie CO 2 do 3 až 5 % v atmosfére pozitívne ovplyvňuje fotosyntézu, ak nie sú iné faktory limitujúce. Obr. 26 Difúzne odpory pri vstupe CO 2 do listového mezofylu C 3 rastliny pri nižších koncentráciách CO 2 majú nižšiu rýchlosť fotosyntézy ako C 4, pri vysokých koncentráciách naopak. Dvojitá fixácia CO 2 C 4 rastlín je efektívnym mechanizmom, preto sa fotosyntéza začína už pri veľmi nízkych koncentráciách CO 2 v pletivách týchto rastlín aj pri uzavretých prieduchoch. Obr. 27 Kompenzácia a saturácia fotosyntézy CO 2 v C 3 a C 4 rastlinách 42

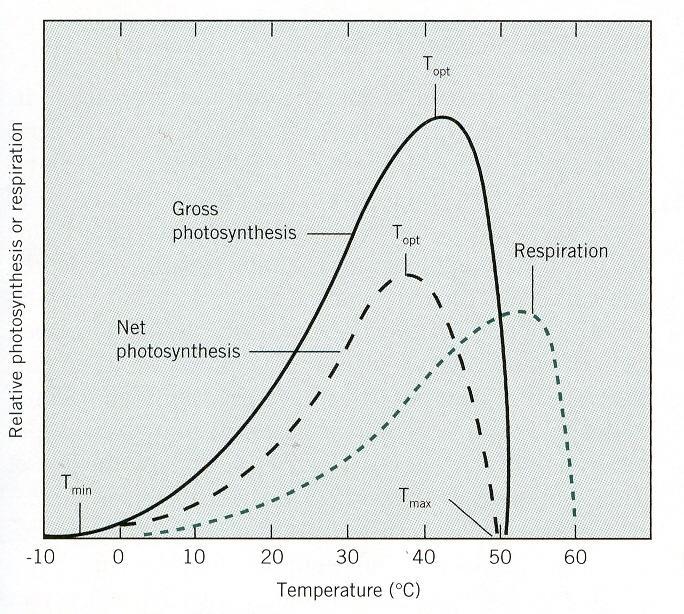
67 V C 3 rastlinách sú straty fotosyntézy fotorespiráciou. Nedostatok atmosférickej vlhkosti môže znížiť fotosyntézu najmä v spojení so suchým teplým vetrom, ktorý silne zvyšuje transpiráciu, spôsobuje zatváranie prieduchov a tým obmedzuje difúziu CO 2 do listových pletív. Zvyšovanie koncentrácie CO 2 v atmosfére sa zistilo dlhodobým meraním. Teplota Teplota neovplyvňuje fotochemické reakcie, ale má rozhodujúci vplyv na enzymatické reakcie. Zvýšenie rýchlosti spôsobené zvýšením teploty o 10 C udáva teplotný koeficient Q 10, ktorý je takmer konštantný (2 až 3). Rozsah teplôt, pri ktorých čistá fotosyntéza dosahuje viac ako 90 % z možného maxima možno považovať za optimum. Postupne narastajúca rýchlosť dýchania čistú fotosyntézu znižuje. Mnohé C 4 rastliny majú teplotné optimum medzi 30 až 40 C, niekedy až do 50 C. C 3 rastliny majú teplotné optimum v širokom rozsahu teplôt, v závislosti od ekologickej charakteristiky rastlinného druhu. Byliny výslnných stanovíšť a stromy teplých oblastí dosahujú najvyššiu fotosyntézu pri 25 až 30 C. Obr. 28 Vplyv teploty na rýchlosť fotosyntézy a dýchania 43
Antiport membránový transport, pri ktorom sa prenášajú dva rôzne ióny (malé molekuly) v navzájom opačnom smere. Apoplast neživý voľný priestor
 v navzájom opačnom smere. Apoplast neživý voľný priestor") Výkladový slovník ABA pozri kyselina abscisová Acetyl-CoA acetyl-koenzým A, je molekula dôležitá pre metabolizmus buniek. Jeho hlavnou funkciou je prenos atómov uhlíka do Krebsovho cyklu, počas ktorého
Výkladový slovník ABA pozri kyselina abscisová Acetyl-CoA acetyl-koenzým A, je molekula dôležitá pre metabolizmus buniek. Jeho hlavnou funkciou je prenos atómov uhlíka do Krebsovho cyklu, počas ktorého
Cytológia. Veda o bunke
 Cytológia Veda o bunke Bunková teória 1665 Cellulae- korok Bunková teória 1838 Zhrnutie bunkovej teórie: 1.základnou štruktúrnou a funkčnou jednotkou živého organizmu je bunka 2.nie je podstatný rozdiel
Cytológia Veda o bunke Bunková teória 1665 Cellulae- korok Bunková teória 1838 Zhrnutie bunkovej teórie: 1.základnou štruktúrnou a funkčnou jednotkou živého organizmu je bunka 2.nie je podstatný rozdiel
Obvod a obsah štvoruholníka
 Obvod a štvoruholníka D. Štyri body roviny z ktorých žiadne tri nie sú kolineárne (neležia na jednej priamke) tvoria jeden štvoruholník. Tie body (A, B, C, D) sú vrcholy štvoruholníka. strany štvoruholníka
Obvod a štvoruholníka D. Štyri body roviny z ktorých žiadne tri nie sú kolineárne (neležia na jednej priamke) tvoria jeden štvoruholník. Tie body (A, B, C, D) sú vrcholy štvoruholníka. strany štvoruholníka
ELEKTRICKÉ POLE. Elektrický náboj je základná vlastnosť častíc, je viazaný na častice látky a vyjadruje stav elektricky nabitých telies.
 ELEKTRICKÉ POLE 1. ELEKTRICKÝ NÁBOJ, COULOMBOV ZÁKON Skúmajme napr. trenie celuloidového pravítka látkou, hrebeň suché vlasy, mikrotén slabý prúd vody... Príčinou spomenutých javov je elektrický náboj,
ELEKTRICKÉ POLE 1. ELEKTRICKÝ NÁBOJ, COULOMBOV ZÁKON Skúmajme napr. trenie celuloidového pravítka látkou, hrebeň suché vlasy, mikrotén slabý prúd vody... Príčinou spomenutých javov je elektrický náboj,
Cytológia. Veda o bunke
 Cytológia Veda o bunke Bunková teória 1665 Cellulae- korok Cytológia Súvisí s vývojom a zdokonalením mikroskopu 1610- G. Galilei zostrojil 1. mikroskop 1838- Schleiden, Schwann- formovanie bunkovej teórie:
Cytológia Veda o bunke Bunková teória 1665 Cellulae- korok Cytológia Súvisí s vývojom a zdokonalením mikroskopu 1610- G. Galilei zostrojil 1. mikroskop 1838- Schleiden, Schwann- formovanie bunkovej teórie:
Matematika Funkcia viac premenných, Parciálne derivácie
 Matematika 2-01 Funkcia viac premenných, Parciálne derivácie Euklidovská metrika na množine R n všetkých usporiadaných n-íc reálnych čísel je reálna funkcia ρ: R n R n R definovaná nasledovne: Ak X = x
Matematika 2-01 Funkcia viac premenných, Parciálne derivácie Euklidovská metrika na množine R n všetkých usporiadaných n-íc reálnych čísel je reálna funkcia ρ: R n R n R definovaná nasledovne: Ak X = x
3. Striedavé prúdy. Sínusoida
 . Striedavé prúdy VZNIK: Striedavý elektrický prúd prechádza obvodom, ktorý je pripojený na zdroj striedavého napätia. Striedavé napätie vyrába synchrónny generátor, kde na koncoch rotorového vinutia sa
. Striedavé prúdy VZNIK: Striedavý elektrický prúd prechádza obvodom, ktorý je pripojený na zdroj striedavého napätia. Striedavé napätie vyrába synchrónny generátor, kde na koncoch rotorového vinutia sa
Ekvačná a kvantifikačná logika
 a kvantifikačná 3. prednáška (6. 10. 004) Prehľad 1 1 (dokončenie) ekvačných tabliel Formula A je ekvačne dokázateľná z množiny axióm T (T i A) práve vtedy, keď existuje uzavreté tablo pre cieľ A ekvačných
a kvantifikačná 3. prednáška (6. 10. 004) Prehľad 1 1 (dokončenie) ekvačných tabliel Formula A je ekvačne dokázateľná z množiny axióm T (T i A) práve vtedy, keď existuje uzavreté tablo pre cieľ A ekvačných
METABOLIZMUS FRUKTÓZY A GALAKTÓZY REGULÁCIA METABOLIZMU SACHARIDOV
 Katedra chémie, biochémie a biofyziky Ústav biochémie METABLIZMUS FRUKTÓZY A GALAKTÓZY METABLIZMUS GLYKGÉNU Glykogenéza Glykogenolýza REGULÁCIA METABLIZMU SACARIDV Metabolizmus fruktózy Metabolizmus fruktózy
Katedra chémie, biochémie a biofyziky Ústav biochémie METABLIZMUS FRUKTÓZY A GALAKTÓZY METABLIZMUS GLYKGÉNU Glykogenéza Glykogenolýza REGULÁCIA METABLIZMU SACARIDV Metabolizmus fruktózy Metabolizmus fruktózy
Klasifikácia látok LÁTKY. Zmesi. Chemické látky. rovnorodé (homogénne) rôznorodé (heterogénne)
 rôznorodé (heterogénne)") Zopakujme si : Klasifikácia látok LÁTKY Chemické látky Zmesi chemické prvky chemické zlúčeniny rovnorodé (homogénne) rôznorodé (heterogénne) Chemicky čistá látka prvok Chemická látka, zložená z atómov,
Zopakujme si : Klasifikácia látok LÁTKY Chemické látky Zmesi chemické prvky chemické zlúčeniny rovnorodé (homogénne) rôznorodé (heterogénne) Chemicky čistá látka prvok Chemická látka, zložená z atómov,
16. Reakcia glukóza + ATP glukóza-6-fosfát + ADP a) predstavuje reakciu aktivácie glukózy pred jej vstupom do glykolýzy
 predstavuje reakciu aktivácie glukózy pred jej vstupom do glykolýzy") Sacharidy 1. Hlavné látky, v podobe ktorých sa privádza glukóza do organizmu: a) sú monosacharidy napr. glukóza b) sú polysacharidy napr. celulóza prítomná hlavne v ovocí c) sú polysacharidy obsahujúce
Sacharidy 1. Hlavné látky, v podobe ktorých sa privádza glukóza do organizmu: a) sú monosacharidy napr. glukóza b) sú polysacharidy napr. celulóza prítomná hlavne v ovocí c) sú polysacharidy obsahujúce
1. Hlavné látky, v podobe ktorých sa privádza glukóza do organizmu:
 1. Hlavné látky, v podobe ktorých sa privádza glukóza do organizmu: a. sú monosacharidy napr. glukóza b. sú polysacharidy napr. celulóza prítomná hlavne v ovocí c. sú polysacharidy obsahujúce 1,4-glykozidovú
1. Hlavné látky, v podobe ktorých sa privádza glukóza do organizmu: a. sú monosacharidy napr. glukóza b. sú polysacharidy napr. celulóza prítomná hlavne v ovocí c. sú polysacharidy obsahujúce 1,4-glykozidovú
Citrátový cyklus a dýchací reťazec. Kristína Tomášiková
 Citrátový cyklus a dýchací reťazec Kristína Tomášiková 15.3.2012 2 Hans Adolf Krebs (1900 1981) študoval oxidáciu živín, medziprodukty metabolismu, vznik močoviny v pečeni u cicavcov, syntézu kyseliny
Citrátový cyklus a dýchací reťazec Kristína Tomášiková 15.3.2012 2 Hans Adolf Krebs (1900 1981) študoval oxidáciu živín, medziprodukty metabolismu, vznik močoviny v pečeni u cicavcov, syntézu kyseliny
Priamkové plochy. Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava
 na nej leží potom plocha Φ je priamková. Santiago Calatrava") Priamkové plochy Priamkové plochy Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava Priamkové plochy rozdeľujeme na: Rozvinuteľné
Priamkové plochy Priamkové plochy Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava Priamkové plochy rozdeľujeme na: Rozvinuteľné
KATABOLIZMUS LIPIDOV BIOCHÉMIA II TÉMA 05 DOC. RNDR. MAREK SKORŠEPA, PHD.
 BIOCHÉMIA II KATEDRA CHÉMIE, FAKULTA PRÍRODNÝCH VIED, UNIVERZITA MATEJA BELA BANSKÁ BYSTRICA KATABOLIZMUS LIPIDOV TÉMA 05 DOC. RNDR. MAREK SKORŠEPA, PHD. LIPIDY AKO ZDROJ ENERGIE lipidy = tretia úrveň
BIOCHÉMIA II KATEDRA CHÉMIE, FAKULTA PRÍRODNÝCH VIED, UNIVERZITA MATEJA BELA BANSKÁ BYSTRICA KATABOLIZMUS LIPIDOV TÉMA 05 DOC. RNDR. MAREK SKORŠEPA, PHD. LIPIDY AKO ZDROJ ENERGIE lipidy = tretia úrveň
AerobTec Altis Micro
 AerobTec Altis Micro Záznamový / súťažný výškomer s telemetriou Výrobca: AerobTec, s.r.o. Pionierska 15 831 02 Bratislava www.aerobtec.com info@aerobtec.com Obsah 1.Vlastnosti... 3 2.Úvod... 3 3.Princíp
AerobTec Altis Micro Záznamový / súťažný výškomer s telemetriou Výrobca: AerobTec, s.r.o. Pionierska 15 831 02 Bratislava www.aerobtec.com info@aerobtec.com Obsah 1.Vlastnosti... 3 2.Úvod... 3 3.Princíp
Start. Vstup r. O = 2*π*r S = π*r*r. Vystup O, S. Stop. Start. Vstup P, C V = P*C*1,19. Vystup V. Stop
 1) Vytvorte algoritmus (vývojový diagram) na výpočet obvodu kruhu. O=2xπxr ; S=πxrxr Vstup r O = 2*π*r S = π*r*r Vystup O, S 2) Vytvorte algoritmus (vývojový diagram) na výpočet celkovej ceny výrobku s
1) Vytvorte algoritmus (vývojový diagram) na výpočet obvodu kruhu. O=2xπxr ; S=πxrxr Vstup r O = 2*π*r S = π*r*r Vystup O, S 2) Vytvorte algoritmus (vývojový diagram) na výpočet celkovej ceny výrobku s
Zrýchľovanie vesmíru. Zrýchľovanie vesmíru. o výprave na kraj vesmíru a čo tam astronómovia objavili
 Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru
Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru
REZISTORY. Rezistory (súčiastky) sú pasívne prvky. Používajú sa vo všetkých elektrických
 sú pasívne prvky. Používajú sa vo všetkých elektrických") REZISTORY Rezistory (súčiastky) sú pasívne prvky. Používajú sa vo všetkých elektrických obvodoch. Základnou vlastnosťou rezistora je jeho odpor. Odpor je fyzikálna vlastnosť, ktorá je daná štruktúrou materiálu
REZISTORY Rezistory (súčiastky) sú pasívne prvky. Používajú sa vo všetkých elektrických obvodoch. Základnou vlastnosťou rezistora je jeho odpor. Odpor je fyzikálna vlastnosť, ktorá je daná štruktúrou materiálu
METABOLIZMUS A VÝŽIVA MIKROORGANIZMOV
 6. METABOLIZMUS A VÝŽIVA MIKROORGANIZMOV Bunka je základnou stavebnou a funkčnou jednotkou všetkých žijúcich organizmov. Organizmus môže byť tvorený len jednou bunkou ako napríklad baktérie (jednobunkové
6. METABOLIZMUS A VÝŽIVA MIKROORGANIZMOV Bunka je základnou stavebnou a funkčnou jednotkou všetkých žijúcich organizmov. Organizmus môže byť tvorený len jednou bunkou ako napríklad baktérie (jednobunkové
Motivácia Denícia determinantu Výpo et determinantov Determinant sú inu matíc Vyuºitie determinantov. Determinanty. 14. decembra 2010.
 14. decembra 2010 Rie²enie sústav Plocha rovnobeºníka Objem rovnobeºnostena Rie²enie sústav Príklad a 11 x 1 + a 12 x 2 = c 1 a 21 x 1 + a 22 x 2 = c 2 Dostaneme: x 1 = c 1a 22 c 2 a 12 a 11 a 22 a 12
14. decembra 2010 Rie²enie sústav Plocha rovnobeºníka Objem rovnobeºnostena Rie²enie sústav Príklad a 11 x 1 + a 12 x 2 = c 1 a 21 x 1 + a 22 x 2 = c 2 Dostaneme: x 1 = c 1a 22 c 2 a 12 a 11 a 22 a 12
Návrh vzduchotesnosti pre detaily napojení
 Výpočet lineárneho stratového súčiniteľa tepelného mosta vzťahujúceho sa k vonkajším rozmerom: Ψ e podľa STN EN ISO 10211 Návrh vzduchotesnosti pre detaily napojení Objednávateľ: Ing. Natália Voltmannová
Výpočet lineárneho stratového súčiniteľa tepelného mosta vzťahujúceho sa k vonkajším rozmerom: Ψ e podľa STN EN ISO 10211 Návrh vzduchotesnosti pre detaily napojení Objednávateľ: Ing. Natália Voltmannová
7. FUNKCIE POJEM FUNKCIE
 7. FUNKCIE POJEM FUNKCIE Funkcia f reálnej premennej je : - každé zobrazenie f v množine všetkých reálnych čísel; - množina f všetkých usporiadaných dvojíc[,y] R R pre ktorú platí: ku každému R eistuje
7. FUNKCIE POJEM FUNKCIE Funkcia f reálnej premennej je : - každé zobrazenie f v množine všetkých reálnych čísel; - množina f všetkých usporiadaných dvojíc[,y] R R pre ktorú platí: ku každému R eistuje
Základné poznatky molekulovej fyziky a termodynamiky
 Základné poznatky molekulovej fyziky a termodynamiky Opakovanie učiva II. ročníka, Téma 1. A. Príprava na maturity z fyziky, 2008 Outline Molekulová fyzika 1 Molekulová fyzika Predmet Molekulovej fyziky
Základné poznatky molekulovej fyziky a termodynamiky Opakovanie učiva II. ročníka, Téma 1. A. Príprava na maturity z fyziky, 2008 Outline Molekulová fyzika 1 Molekulová fyzika Predmet Molekulovej fyziky
1. písomná práca z matematiky Skupina A
 1. písomná práca z matematiky Skupina A 1. Vypočítajte : a) 84º 56 + 32º 38 = b) 140º 53º 24 = c) 55º 12 : 2 = 2. Vypočítajte zvyšné uhly na obrázku : β γ α = 35 12 δ a b 3. Znázornite na číselnej osi
1. písomná práca z matematiky Skupina A 1. Vypočítajte : a) 84º 56 + 32º 38 = b) 140º 53º 24 = c) 55º 12 : 2 = 2. Vypočítajte zvyšné uhly na obrázku : β γ α = 35 12 δ a b 3. Znázornite na číselnej osi
Prechod z 2D do 3D. Martin Florek 3. marca 2009
 Počítačová grafika 2 Prechod z 2D do 3D Martin Florek florek@sccg.sk FMFI UK 3. marca 2009 Prechod z 2D do 3D Čo to znamená? Ako zobraziť? Súradnicové systémy Čo to znamená? Ako zobraziť? tretia súradnica
Počítačová grafika 2 Prechod z 2D do 3D Martin Florek florek@sccg.sk FMFI UK 3. marca 2009 Prechod z 2D do 3D Čo to znamená? Ako zobraziť? Súradnicové systémy Čo to znamená? Ako zobraziť? tretia súradnica
Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín. Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť.
 Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť. Ktoré fyzikálne jednotky zodpovedajú sústave SI: a) Dĺžka, čas,
Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť. Ktoré fyzikálne jednotky zodpovedajú sústave SI: a) Dĺžka, čas,
M O N I T O R 2002 pilotné testovanie maturantov MONITOR Chémia. 2. časť. Realizácia projektu: EXAM, Bratislava. (2002) Štátny pedagogický ústav
 Štátny pedagogický ústav") M O N I T O R 2002 pilotné testovanie maturantov MONITOR 2002 Chémia 2. časť Odborný garant projektu: Realizácia projektu: Štátny pedagogický ústav, Bratislava EXAM, Bratislava 1 MONITOR 2002 Voda je jedna
M O N I T O R 2002 pilotné testovanie maturantov MONITOR 2002 Chémia 2. časť Odborný garant projektu: Realizácia projektu: Štátny pedagogický ústav, Bratislava EXAM, Bratislava 1 MONITOR 2002 Voda je jedna
Goniometrické rovnice a nerovnice. Základné goniometrické rovnice
 Goniometrické rovnice a nerovnice Definícia: Rovnice (nerovnice) obsahujúce neznámu x alebo výrazy s neznámou x ako argumenty jednej alebo niekoľkých goniometrických funkcií nazývame goniometrickými rovnicami
Goniometrické rovnice a nerovnice Definícia: Rovnice (nerovnice) obsahujúce neznámu x alebo výrazy s neznámou x ako argumenty jednej alebo niekoľkých goniometrických funkcií nazývame goniometrickými rovnicami
Stavba atómového jadra
 Objavy stavby jadra: 1. H. BECQUEREL (1852 1908) objavil prenikavé žiarenie vysielané zlúčeninami prvku uránu. 2. Pomocou žiarenia α objavil Rutherford so svojimi spolupracovníkmi atómové jadro. Žiarenie
Objavy stavby jadra: 1. H. BECQUEREL (1852 1908) objavil prenikavé žiarenie vysielané zlúčeninami prvku uránu. 2. Pomocou žiarenia α objavil Rutherford so svojimi spolupracovníkmi atómové jadro. Žiarenie
Motivácia pojmu derivácia
 Derivácia funkcie Motivácia pojmu derivácia Zaujíma nás priemerná intenzita zmeny nejakej veličiny (dráhy, rastu populácie, veľkosti elektrického náboja, hmotnosti), vzhľadom na inú veličinu (čas, dĺžka)
Derivácia funkcie Motivácia pojmu derivácia Zaujíma nás priemerná intenzita zmeny nejakej veličiny (dráhy, rastu populácie, veľkosti elektrického náboja, hmotnosti), vzhľadom na inú veličinu (čas, dĺžka)
1. Limita, spojitost a diferenciálny počet funkcie jednej premennej
 . Limita, spojitost a diferenciálny počet funkcie jednej premennej Definícia.: Hromadný bod a R množiny A R: v každom jeho okolí leží aspoň jeden bod z množiny A, ktorý je rôzny od bodu a Zadanie množiny
. Limita, spojitost a diferenciálny počet funkcie jednej premennej Definícia.: Hromadný bod a R množiny A R: v každom jeho okolí leží aspoň jeden bod z množiny A, ktorý je rôzny od bodu a Zadanie množiny
,Zohrievanie vody indukčným varičom bez pokrievky,
 Farba skupiny: zelená Označenie úlohy:,zohrievanie vody indukčným varičom bez pokrievky, Úloha: Zistiť, ako závisí účinnosť zohrievania vody na indukčnom variči od priemeru použitého hrnca. Hypotéza: Účinnosť
Farba skupiny: zelená Označenie úlohy:,zohrievanie vody indukčným varičom bez pokrievky, Úloha: Zistiť, ako závisí účinnosť zohrievania vody na indukčnom variči od priemeru použitého hrnca. Hypotéza: Účinnosť
Gazometrické merania a modelovanie A-Ci kriviek (manuál)
") Gazometrické merania a modelovanie A-Ci kriviek (manuál) 1 Obsah 1. Základy merania výmeny plynov... 3 1.1. Príjem a výdaj CO 2 rastlinou... 3 1.2. Princíp merania výmeny plynov gazometrickým systémom...
Gazometrické merania a modelovanie A-Ci kriviek (manuál) 1 Obsah 1. Základy merania výmeny plynov... 3 1.1. Príjem a výdaj CO 2 rastlinou... 3 1.2. Princíp merania výmeny plynov gazometrickým systémom...
Monitoring mikrobiálnych pomerov pôdy na kalamitných plochách Tatier
 Monitoring mikrobiálnych pomerov pôdy na kalamitných plochách Tatier Erika Gömöryová Technická univerzita vo Zvolene, Lesnícka fakulta T. G.Masaryka 24, SK960 53 Zvolen email: gomoryova@tuzvo.sk TANAP:
Monitoring mikrobiálnych pomerov pôdy na kalamitných plochách Tatier Erika Gömöryová Technická univerzita vo Zvolene, Lesnícka fakulta T. G.Masaryka 24, SK960 53 Zvolen email: gomoryova@tuzvo.sk TANAP:
Pevné ložiská. Voľné ložiská
 SUPPORTS D EXTREMITES DE PRECISION - SUPPORT UNIT FOR BALLSCREWS LOŽISKA PRE GULIČKOVÉ SKRUTKY A TRAPÉZOVÉ SKRUTKY Výber správnej podpory konca uličkovej skrutky či trapézovej skrutky je dôležité pre správnu
SUPPORTS D EXTREMITES DE PRECISION - SUPPORT UNIT FOR BALLSCREWS LOŽISKA PRE GULIČKOVÉ SKRUTKY A TRAPÉZOVÉ SKRUTKY Výber správnej podpory konca uličkovej skrutky či trapézovej skrutky je dôležité pre správnu
Kontrolné otázky z jednotiek fyzikálnych veličín
 Verzia zo dňa 6. 9. 008. Kontrolné otázky z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej odpovede sa môže v kontrolnom teste meniť. Takisto aj znenie nesprávnych odpovedí. Uvedomte si
Verzia zo dňa 6. 9. 008. Kontrolné otázky z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej odpovede sa môže v kontrolnom teste meniť. Takisto aj znenie nesprávnych odpovedí. Uvedomte si
KATEDRA DOPRAVNEJ A MANIPULAČNEJ TECHNIKY Strojnícka fakulta, Žilinská Univerzita
 132 1 Absolútna chyba: ) = - skut absolútna ochýlka: ) ' = - spr. relatívna chyba: alebo Chyby (ochýlky): M systematické, M náhoné, M hrubé. Korekcia: k = spr - = - Î' pomerná korekcia: Správna honota:
132 1 Absolútna chyba: ) = - skut absolútna ochýlka: ) ' = - spr. relatívna chyba: alebo Chyby (ochýlky): M systematické, M náhoné, M hrubé. Korekcia: k = spr - = - Î' pomerná korekcia: Správna honota:
2.2 Rádioaktivita izotopy stabilita ich atómových jadier rádioaktivita žiarenie jadrové
 2.2 Rádioaktivita Koniec 19. storočia bol bohatý na významné objavy vo fyzike a chémii, ktoré poskytli základy na vybudovanie moderných predstáv o zložení atómu. Medzi najvýznamnejšie objavy patrí objavenie
2.2 Rádioaktivita Koniec 19. storočia bol bohatý na významné objavy vo fyzike a chémii, ktoré poskytli základy na vybudovanie moderných predstáv o zložení atómu. Medzi najvýznamnejšie objavy patrí objavenie
100626HTS01. 8 kw. 7 kw. 8 kw
 alpha intec 100626HTS01 L 8SplitHT 8 7 44 54 8 alpha intec 100626HTS01 L 8SplitHT Souprava (tepelná čerpadla a kombivané ohřívače s tepelným čerpadlem) Sezonní energetická účinst vytápění tepelného čerpadla
alpha intec 100626HTS01 L 8SplitHT 8 7 44 54 8 alpha intec 100626HTS01 L 8SplitHT Souprava (tepelná čerpadla a kombivané ohřívače s tepelným čerpadlem) Sezonní energetická účinst vytápění tepelného čerpadla
ZADANIE 1_ ÚLOHA 3_Všeobecná rovinná silová sústava ZADANIE 1 _ ÚLOHA 3
 ZDNIE _ ÚLOH 3_Všeobecná rovinná silová sústv ZDNIE _ ÚLOH 3 ÚLOH 3.: Vypočítjte veľkosti rekcií vo väzbách nosník zťženého podľ obrázku 3.. Veľkosti známych síl, momentov dĺžkové rozmery sú uvedené v
ZDNIE _ ÚLOH 3_Všeobecná rovinná silová sústv ZDNIE _ ÚLOH 3 ÚLOH 3.: Vypočítjte veľkosti rekcií vo väzbách nosník zťženého podľ obrázku 3.. Veľkosti známych síl, momentov dĺžkové rozmery sú uvedené v
Biochémia hraničná vedná disciplína. Chemické deje. Podstata základných životných procesov Metabolizmus, rast, pohyb, dráždivosť, rozmnožovanie
 BIOCHÉMIA Biochémia hraničná vedná disciplína Chemické deje Podstata základných životných procesov Metabolizmus, rast, pohyb, dráždivosť, rozmnožovanie Biochémia vedy o športe 30. roky 20. storočia Laktát,
BIOCHÉMIA Biochémia hraničná vedná disciplína Chemické deje Podstata základných životných procesov Metabolizmus, rast, pohyb, dráždivosť, rozmnožovanie Biochémia vedy o športe 30. roky 20. storočia Laktát,
SLOVENSKO maloobchodný cenník (bez DPH)
") Hofatex UD strecha / stena - exteriér Podkrytinová izolácia vhodná aj na zaklopenie drevených rámových konštrukcií; pero a drážka EN 13171, EN 622 22 580 2500 1,45 5,7 100 145,00 3,19 829 hustota cca.
Hofatex UD strecha / stena - exteriér Podkrytinová izolácia vhodná aj na zaklopenie drevených rámových konštrukcií; pero a drážka EN 13171, EN 622 22 580 2500 1,45 5,7 100 145,00 3,19 829 hustota cca.
Zmeny potravín počas skladovania a prepravy
 Zmeny potravín počas skladovania a prepravy 541P308 Hygiena distribúcie a predaja potravín (POVINNÝ PREDMET) ZIMNÝ SEM ESTER AKADEMICKÝ ROK 2016/2017 Ing. Peter Zajác, PhD. 1 Osnova 1. Zloženie potravín
Zmeny potravín počas skladovania a prepravy 541P308 Hygiena distribúcie a predaja potravín (POVINNÝ PREDMET) ZIMNÝ SEM ESTER AKADEMICKÝ ROK 2016/2017 Ing. Peter Zajác, PhD. 1 Osnova 1. Zloženie potravín
Zateplite fasádu! Zabezpečte, aby Vám neuniklo teplo cez fasádu
 Zateplite fasádu! Zabezpečte, aby Vám neuniklo teplo cez fasádu Austrotherm GrPS 70 F Austrotherm GrPS 70 F Reflex Austrotherm Resolution Fasáda Austrotherm XPS TOP P Austrotherm XPS Premium 30 SF Austrotherm
Zateplite fasádu! Zabezpečte, aby Vám neuniklo teplo cez fasádu Austrotherm GrPS 70 F Austrotherm GrPS 70 F Reflex Austrotherm Resolution Fasáda Austrotherm XPS TOP P Austrotherm XPS Premium 30 SF Austrotherm
M6: Model Hydraulický systém dvoch zásobníkov kvapaliny s interakciou
 M6: Model Hydraulický ytém dvoch záobníkov kvapaliny interakciou Úlohy:. Zotavte matematický popi modelu Hydraulický ytém. Vytvorte imulačný model v jazyku: a. Matlab b. imulink 3. Linearizujte nelineárny
M6: Model Hydraulický ytém dvoch záobníkov kvapaliny interakciou Úlohy:. Zotavte matematický popi modelu Hydraulický ytém. Vytvorte imulačný model v jazyku: a. Matlab b. imulink 3. Linearizujte nelineárny
ÚLOHA MIKROORGANIZMOV V PREMENÁCH LÁTOK
 8. ÚLOHA MIKROORGANIZMOV V PREMENÁCH LÁTOK Kolobeh hmoty a energie je v ekosystémoch podmienkou života všetkých organizmov. Autotrofné organizmy syntetizujú z jednoduchých minerálnych látok látky organické,
8. ÚLOHA MIKROORGANIZMOV V PREMENÁCH LÁTOK Kolobeh hmoty a energie je v ekosystémoch podmienkou života všetkých organizmov. Autotrofné organizmy syntetizujú z jednoduchých minerálnych látok látky organické,
HASLIM112V, HASLIM123V, HASLIM136V HASLIM112Z, HASLIM123Z, HASLIM136Z HASLIM112S, HASLIM123S, HASLIM136S
 PROUKTOVÝ LIST HKL SLIM č. sklad. karty / obj. číslo: HSLIM112V, HSLIM123V, HSLIM136V HSLIM112Z, HSLIM123Z, HSLIM136Z HSLIM112S, HSLIM123S, HSLIM136S fakturačný názov výrobku: HKL SLIMv 1,2kW HKL SLIMv
PROUKTOVÝ LIST HKL SLIM č. sklad. karty / obj. číslo: HSLIM112V, HSLIM123V, HSLIM136V HSLIM112Z, HSLIM123Z, HSLIM136Z HSLIM112S, HSLIM123S, HSLIM136S fakturačný názov výrobku: HKL SLIMv 1,2kW HKL SLIMv
Nukleové kyseliny. Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy.
 Nukleové kyseliny Nukleové kyseliny Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy. DNA - deoxyribonukleová kyselina RNA - ribonukleová kyselina Funkcie
Nukleové kyseliny Nukleové kyseliny Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy. DNA - deoxyribonukleová kyselina RNA - ribonukleová kyselina Funkcie
Matematika 2. časť: Analytická geometria
 Matematika 2 časť: Analytická geometria RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk Súradnicové
Matematika 2 časť: Analytická geometria RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk Súradnicové
difúzne otvorené drevovláknité izolačné dosky - ochrana nie len pred chladom...
 (TYP M) izolačná doska určená na vonkajšiu fasádu (spoj P+D) ρ = 230 kg/m3 λ d = 0,046 W/kg.K 590 1300 40 56 42,95 10,09 590 1300 60 38 29,15 15,14 590 1300 80 28 21,48 20,18 590 1300 100 22 16,87 25,23
(TYP M) izolačná doska určená na vonkajšiu fasádu (spoj P+D) ρ = 230 kg/m3 λ d = 0,046 W/kg.K 590 1300 40 56 42,95 10,09 590 1300 60 38 29,15 15,14 590 1300 80 28 21,48 20,18 590 1300 100 22 16,87 25,23
Termodynamika. Doplnkové materiály k prednáškam z Fyziky I pre SjF Dušan PUDIŠ (2008)
") ermodynamika nútorná energia lynov,. veta termodynamická, Izochorický dej, Izotermický dej, Izobarický dej, diabatický dej, Práca lynu ri termodynamických rocesoch, arnotov cyklus, Entroia Dolnkové materiály
ermodynamika nútorná energia lynov,. veta termodynamická, Izochorický dej, Izotermický dej, Izobarický dej, diabatický dej, Práca lynu ri termodynamických rocesoch, arnotov cyklus, Entroia Dolnkové materiály
Moderné vzdelávanie pre vedomostnú spoločnosť Projekt je spolufinancovaný zo zdrojov EÚ M A T E M A T I K A
 M A T E M A T I K A PRACOVNÝ ZOŠIT II. ROČNÍK Mgr. Agnesa Balážová Obchodná akadémia, Akademika Hronca 8, Rožňava PRACOVNÝ LIST 1 Urč typ kvadratickej rovnice : 1. x 2 3x = 0... 2. 3x 2 = - 2... 3. -4x
M A T E M A T I K A PRACOVNÝ ZOŠIT II. ROČNÍK Mgr. Agnesa Balážová Obchodná akadémia, Akademika Hronca 8, Rožňava PRACOVNÝ LIST 1 Urč typ kvadratickej rovnice : 1. x 2 3x = 0... 2. 3x 2 = - 2... 3. -4x
KATEDRA BUNKOVEJ A MOLEKULÁRNEJ BIOLÓGIE LIEČIV
 KATEDRA BUNKOVEJ A MOLEKULÁRNEJ BIOLÓGIE LIEČIV Predmet: BIOCHÉMIA, rok 2016/2017 Forma výučby: prednášky, semináre, cvičenia Ročník: 2. Celkový počet hodín: 98, LS Rozsah hodín v týždni: prednášky 3 semináre
KATEDRA BUNKOVEJ A MOLEKULÁRNEJ BIOLÓGIE LIEČIV Predmet: BIOCHÉMIA, rok 2016/2017 Forma výučby: prednášky, semináre, cvičenia Ročník: 2. Celkový počet hodín: 98, LS Rozsah hodín v týždni: prednášky 3 semináre
Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti rozvodu tepla
 Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti príloha č. 7 k vyhláške č. 428/2010 Názov prevádzkovateľa verejného : Spravbytkomfort a.s. Prešov Adresa: IČO: Volgogradská 88, 080 01 Prešov 31718523
Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti príloha č. 7 k vyhláške č. 428/2010 Názov prevádzkovateľa verejného : Spravbytkomfort a.s. Prešov Adresa: IČO: Volgogradská 88, 080 01 Prešov 31718523
Živá hmota chemické zloženie. Biológia živočíšnej produkcie Katedra fyziológie živočíchov
 Živá hmota chemické zloženie Biológia živočíšnej produkcie Katedra fyziológie živočíchov Chemické zloženie živej hmoty Živá hmota bioplazma chemicky rôznorodá zmes látok zložitý koloidný systém Prvky v
Živá hmota chemické zloženie Biológia živočíšnej produkcie Katedra fyziológie živočíchov Chemické zloženie živej hmoty Živá hmota bioplazma chemicky rôznorodá zmes látok zložitý koloidný systém Prvky v
Odporníky. 1. Príklad1. TESLA TR
 Odporníky Úloha cvičenia: 1.Zistite technické údaje odporníkov pomocou katalógov 2.Zistite menovitú hodnotu odporníkov označených farebným kódom Schématická značka: 1. Príklad1. TESLA TR 163 200 ±1% L
Odporníky Úloha cvičenia: 1.Zistite technické údaje odporníkov pomocou katalógov 2.Zistite menovitú hodnotu odporníkov označených farebným kódom Schématická značka: 1. Príklad1. TESLA TR 163 200 ±1% L
Margita Vajsáblová. ρ priemetňa, s smer premietania. Súradnicová sústava (O, x, y, z ) (O a, x a, y a, z a )
 (O a, x a, y a, z a )") Mrgit Váblová Váblová, M: Dekriptívn geometri pre GK 101 Zákldné pom v onometrii Váblová, M: Dekriptívn geometri pre GK 102 Definíci 1: onometri e rovnobežné premietnie bodov Ε 3 polu prvouhlým úrdnicovým
Mrgit Váblová Váblová, M: Dekriptívn geometri pre GK 101 Zákldné pom v onometrii Váblová, M: Dekriptívn geometri pre GK 102 Definíci 1: onometri e rovnobežné premietnie bodov Ε 3 polu prvouhlým úrdnicovým
C. Kontaktný fasádny zatepľovací systém
 C. Kontaktný fasádny zatepľovací systém C.1. Tepelná izolácia penový polystyrén C.2. Tepelná izolácia minerálne dosky alebo lamely C.3. Tepelná izolácia extrudovaný polystyrén C.4. Tepelná izolácia penový
C. Kontaktný fasádny zatepľovací systém C.1. Tepelná izolácia penový polystyrén C.2. Tepelná izolácia minerálne dosky alebo lamely C.3. Tepelná izolácia extrudovaný polystyrén C.4. Tepelná izolácia penový
Matematika prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad
 Matematika 3-13. prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad Erika Škrabul áková F BERG, TU Košice 15. 12. 2015 Erika Škrabul áková (TUKE) Taylorov
Matematika 3-13. prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad Erika Škrabul áková F BERG, TU Košice 15. 12. 2015 Erika Škrabul áková (TUKE) Taylorov
VYŠETROVANIE VONKAJŠIEHO FOTOELEKTRICKÉHO JAVU A URČENIE PLANCKOVEJ KONŠTANTY
 45 VYŠETROVANE VONKAJŠEHO FOTOELEKTRCKÉHO JAV A RČENE PLANCKOVEJ KONŠTANTY doc. RNDr. Drahoslav Vajda, CSc. Teoretický úvod: Vonkajší fotoelektrický jav je veľmi presvedčivým dôkazom kvantovej povahy elektromagnetického
45 VYŠETROVANE VONKAJŠEHO FOTOELEKTRCKÉHO JAV A RČENE PLANCKOVEJ KONŠTANTY doc. RNDr. Drahoslav Vajda, CSc. Teoretický úvod: Vonkajší fotoelektrický jav je veľmi presvedčivým dôkazom kvantovej povahy elektromagnetického
Praktikum z biochémie 2. vydanie, Sedlák, Danko, Varhač, Paulíková, Podhradský, 2007
 Praktikum z biochémie 2. vydanie, Sedlák, Danko, arhač, Paulíková, Podhradský, 2007 5 ENZÝMY Enzýmy (z gréckeho enzymon = v droždí), katalyzátory v biologických systémoch, sú pozoruhodné molekulové prístroje,
Praktikum z biochémie 2. vydanie, Sedlák, Danko, arhač, Paulíková, Podhradský, 2007 5 ENZÝMY Enzýmy (z gréckeho enzymon = v droždí), katalyzátory v biologických systémoch, sú pozoruhodné molekulové prístroje,
CHÉMIA PRE BIOLÓGOV ŠTUDIJNÝ TEXT
 CHÉMIA PRE BIOLÓGOV ŠTUDIJNÝ TEXT Mária Linkešová, Ivona Paveleková CHÉMIA AKO PRÍRODNÁ VEDA Chémia je prírodná veda, ktorá študuje štruktúru atómov, molekúl a látok z nich utvorených, sleduje ich vlastnosti
CHÉMIA PRE BIOLÓGOV ŠTUDIJNÝ TEXT Mária Linkešová, Ivona Paveleková CHÉMIA AKO PRÍRODNÁ VEDA Chémia je prírodná veda, ktorá študuje štruktúru atómov, molekúl a látok z nich utvorených, sleduje ich vlastnosti
CHÉMIA Ing. Iveta Bruončová
 Výpočet hmotnostného zlomku, látkovej koncentrácie, výpočty zamerané na zloženie roztokov CHÉMIA Ing. Iveta Bruončová Moderné vzdelávanie pre vedomostnú spoločnosť/projekt je spolufinancovaný zo zdrojov
Výpočet hmotnostného zlomku, látkovej koncentrácie, výpočty zamerané na zloženie roztokov CHÉMIA Ing. Iveta Bruončová Moderné vzdelávanie pre vedomostnú spoločnosť/projekt je spolufinancovaný zo zdrojov
ŽIACKE AKTIVITY POZOROVANIE ABSORPČNÉHO A FLUORESCENČNÉHO SPEKTRA CHLOROFYLU RASTLÍN
 ŽIACKE AKTIVITY POZOROVANIE ABSORPČNÉHO A FLUORESCENČNÉHO SPEKTRA CHLOROFYLU RASTLÍN Jana Útla Fakulta matematiky, fyziky a informatiky Univerzity Komenského v Bratislave Abstrakt: Príspevok sa zaoberá
ŽIACKE AKTIVITY POZOROVANIE ABSORPČNÉHO A FLUORESCENČNÉHO SPEKTRA CHLOROFYLU RASTLÍN Jana Útla Fakulta matematiky, fyziky a informatiky Univerzity Komenského v Bratislave Abstrakt: Príspevok sa zaoberá
CHÉMIA A ŽIVOTNÉ PROSTREDIE
 CHÉMIA A ŽIVOTNÉ PROSTREDIE Mária Orolínová Trnavská univerzita v Trnave Pedagogická fakulta 2009 Mária Orolínová Recenzenti: Vydala: doc. Ing. Maroš Soldán, CSc. Ing. Viera Peterková, PhD. Trnavská univerzita
CHÉMIA A ŽIVOTNÉ PROSTREDIE Mária Orolínová Trnavská univerzita v Trnave Pedagogická fakulta 2009 Mária Orolínová Recenzenti: Vydala: doc. Ing. Maroš Soldán, CSc. Ing. Viera Peterková, PhD. Trnavská univerzita
alu OKNÁ, ZA KTORÝMI BÝVA POHODA DREVENÉ OKNÁ A DVERE Profil Mirador Alu 783 Drevohliníkové okno s priznaným okenným krídlom.
 DREVENÉ OKNÁ A DVERE m i r a d o r 783 OKNÁ, ZA KTORÝMI BÝVA POHODA EXTERIÉROVÁ Profil Mirador Alu 783 Drevohliníkové okno s priznaným okenným krídlom. Je najviac používané drevohliníkové okno, ktoré je
DREVENÉ OKNÁ A DVERE m i r a d o r 783 OKNÁ, ZA KTORÝMI BÝVA POHODA EXTERIÉROVÁ Profil Mirador Alu 783 Drevohliníkové okno s priznaným okenným krídlom. Je najviac používané drevohliníkové okno, ktoré je
Cvičenie č. 4,5 Limita funkcie
 Cvičenie č. 4,5 Limita funkcie Definícia ity Limita funkcie (vlastná vo vlastnom bode) Nech funkcia f je definovaná na nejakom okolí U( ) bodu. Hovoríme, že funkcia f má v bode itu rovnú A, ak ( ε > )(
Cvičenie č. 4,5 Limita funkcie Definícia ity Limita funkcie (vlastná vo vlastnom bode) Nech funkcia f je definovaná na nejakom okolí U( ) bodu. Hovoríme, že funkcia f má v bode itu rovnú A, ak ( ε > )(
Podnikateľ 90 Mobilný telefón Cena 95 % 50 % 25 %
 Podnikateľ 90 Samsung S5230 Samsung C3530 Nokia C5 Samsung Shark Slider S3550 Samsung Xcover 271 T-Mobile Pulse Mini Sony Ericsson ZYLO Sony Ericsson Cedar LG GM360 Viewty Snap Nokia C3 Sony Ericsson ZYLO
Podnikateľ 90 Samsung S5230 Samsung C3530 Nokia C5 Samsung Shark Slider S3550 Samsung Xcover 271 T-Mobile Pulse Mini Sony Ericsson ZYLO Sony Ericsson Cedar LG GM360 Viewty Snap Nokia C3 Sony Ericsson ZYLO
METABOLIZMUS prednáška z fyziológie pre študentov zubného lekárstva
 METABOLIZMUS prednáška z fyziológie pre študentov zubného lekárstva Katarína Babinská, MD, PhD, MSc., Fyziologický ústav LFUK, Bratislava 2016 palivo funkcia produkty spaľovania potrava palivo pre organizmus
METABOLIZMUS prednáška z fyziológie pre študentov zubného lekárstva Katarína Babinská, MD, PhD, MSc., Fyziologický ústav LFUK, Bratislava 2016 palivo funkcia produkty spaľovania potrava palivo pre organizmus
kovalentná väzba - Lewisov model
 Modely chemickej väzby klasické elektrostatické úvahy kovalentná väzba Lewisov model Geometria, VSEPR kvantovomechanické model hybridných orbitalov teória molekulových orbitalov teória valenčných väzieb
Modely chemickej väzby klasické elektrostatické úvahy kovalentná väzba Lewisov model Geometria, VSEPR kvantovomechanické model hybridných orbitalov teória molekulových orbitalov teória valenčných väzieb
ARMA modely čast 2: moving average modely (MA)
") ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2014/2015 ARMA modely časť 2: moving average modely(ma) p.1/24 V. Moving average proces prvého rádu - MA(1) ARMA modely
ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2014/2015 ARMA modely časť 2: moving average modely(ma) p.1/24 V. Moving average proces prvého rádu - MA(1) ARMA modely
Návod na vyplnenie hárku. Test 100
 Návod na vyplnenie hárku Súčasťou testu je priložený snímací hárok. Na ten budete zaznamenávať odpovede na otázky testu. Hárok je ohraničený veľkým čiernym rámom. Vpisujte len do vnútornej oblasti tohoto
Návod na vyplnenie hárku Súčasťou testu je priložený snímací hárok. Na ten budete zaznamenávať odpovede na otázky testu. Hárok je ohraničený veľkým čiernym rámom. Vpisujte len do vnútornej oblasti tohoto
Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R
 Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R Ako nadprirodzené stretnutie s murárikom červenokrídlym naformátovalo môj profesijný i súkromný život... Osudové stretnutie s murárikom
Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R Ako nadprirodzené stretnutie s murárikom červenokrídlym naformátovalo môj profesijný i súkromný život... Osudové stretnutie s murárikom
Chemická väzba 1. R O Č N Í K SŠ
 Chemická väzba 1. R O Č N Í K SŠ Atómy nemajú radi samotu o Iba vzácne plyny sú radi sami o Vo všetkých ostatných látkach sú atómy spájané pomocou chemických väzieb Prečo sa atómy zlučujú? Atómy sa zlučujú,
Chemická väzba 1. R O Č N Í K SŠ Atómy nemajú radi samotu o Iba vzácne plyny sú radi sami o Vo všetkých ostatných látkach sú atómy spájané pomocou chemických väzieb Prečo sa atómy zlučujú? Atómy sa zlučujú,
Vyhlásenie o parametroch stavebného výrobku StoPox GH 205 S
 1 / 5 Vyhlásenie o parametroch stavebného výrobku StoPox GH 205 S Identifikačný kód typu výrobku PROD2141 StoPox GH 205 S Účel použitia EN 1504-2: Výrobok slúžiaci na ochranu povrchov povrchová úprava
1 / 5 Vyhlásenie o parametroch stavebného výrobku StoPox GH 205 S Identifikačný kód typu výrobku PROD2141 StoPox GH 205 S Účel použitia EN 1504-2: Výrobok slúžiaci na ochranu povrchov povrchová úprava
Ebook. TESTY Biológia Príprava na maturitu a prijímacie skúšky na vysokú školu.
 Ebook Všetky práva sú vyhradené. Nijaká časť tejto publikácie sa nesmie reprodukovať mechanicky, elektronicky, fotokopírovaním alebo iným spôsobom rozširovania bez predchádzajúceho súhlasu majiteľov autorských
Ebook Všetky práva sú vyhradené. Nijaká časť tejto publikácie sa nesmie reprodukovať mechanicky, elektronicky, fotokopírovaním alebo iným spôsobom rozširovania bez predchádzajúceho súhlasu majiteľov autorských
KATALÓG KRUHOVÉ POTRUBIE
 H KATALÓG KRUHOVÉ POTRUBIE 0 Základné požiadavky zadávania VZT potrubia pre výrobu 1. Zadávanie do výroby v spoločnosti APIAGRA s.r.o. V digitálnej forme na tlačive F05-8.0_Rozpis_potrubia, zaslané mailom
H KATALÓG KRUHOVÉ POTRUBIE 0 Základné požiadavky zadávania VZT potrubia pre výrobu 1. Zadávanie do výroby v spoločnosti APIAGRA s.r.o. V digitálnej forme na tlačive F05-8.0_Rozpis_potrubia, zaslané mailom
PRIEMER DROTU d = 0,4-6,3 mm
 PRUŽINY PRUŽINY SKRUTNÉ PRUŽINY VIAC AKO 200 RUHOV SKRUTNÝCH PRUŽÍN PRIEMER ROTU d = 0,4-6,3 mm èíslo 3.0 22.8.2008 8:28:57 22.8.2008 8:28:58 PRUŽINY SKRUTNÉ PRUŽINY TECHNICKÉ PARAMETRE h d L S Legenda
PRUŽINY PRUŽINY SKRUTNÉ PRUŽINY VIAC AKO 200 RUHOV SKRUTNÝCH PRUŽÍN PRIEMER ROTU d = 0,4-6,3 mm èíslo 3.0 22.8.2008 8:28:57 22.8.2008 8:28:58 PRUŽINY SKRUTNÉ PRUŽINY TECHNICKÉ PARAMETRE h d L S Legenda
KAGEDA AUTORIZOVANÝ DISTRIBÚTOR PRE SLOVENSKÚ REPUBLIKU
 DVOJEXCENTRICKÁ KLAPKA je uzatváracia alebo regulačná armatúra pre rozvody vody, horúcej vody, plynov a pary. Všetky klapky vyhovujú smernici PED 97/ 23/EY a sú tiež vyrábané pre výbušné prostredie podľa
DVOJEXCENTRICKÁ KLAPKA je uzatváracia alebo regulačná armatúra pre rozvody vody, horúcej vody, plynov a pary. Všetky klapky vyhovujú smernici PED 97/ 23/EY a sú tiež vyrábané pre výbušné prostredie podľa
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy
, diferencovanie časového radu, unit root testy") Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2012/2013 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/18
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2012/2013 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/18
Meranie na jednofázovom transformátore
 Fakulta elektrotechniky a informatiky TU v Košiciach Katedra elektrotechniky a mechatroniky Meranie na jednofázovom transformátore Návod na cvičenia z predmetu Elektrotechnika Meno a priezvisko :..........................
Fakulta elektrotechniky a informatiky TU v Košiciach Katedra elektrotechniky a mechatroniky Meranie na jednofázovom transformátore Návod na cvičenia z predmetu Elektrotechnika Meno a priezvisko :..........................
SLOVENSKÁ KOMISIA CHEMICKEJ OLYMPIÁDY CHEMICKÁ OLYMPIÁDA. 54. ročník, školský rok 2017/2018 Kategória B. Krajské kolo RIEŠENIE A HODNOTENIE
 SLVENSKÁ KMISIA CHEMICKEJ LYMPIÁDY CHEMICKÁ LYMPIÁDA 54. ročník, školský rok 2017/2018 Kategória B Krajské kolo RIEŠENIE A HDNTENIE TERETICKÝCH A PRAKTICKÝCH ÚLH RIEŠENIE A HDNTENIE ÚLH Z VŠEBECNEJ A ANRGANICKEJ
SLVENSKÁ KMISIA CHEMICKEJ LYMPIÁDY CHEMICKÁ LYMPIÁDA 54. ročník, školský rok 2017/2018 Kategória B Krajské kolo RIEŠENIE A HDNTENIE TERETICKÝCH A PRAKTICKÝCH ÚLH RIEŠENIE A HDNTENIE ÚLH Z VŠEBECNEJ A ANRGANICKEJ
(kvalitatívna, kvantitatívna).
.") 3. FUNKČNÁ ANALÝZA (kvalitatívna, kvantitatívna). Inštrumentálne analytické metódy: Infračervená a Ramanova spektrometria. UV/VIS molekulová absorpčná spektrometria. Röntgenová spektrometria. Spektrálne
3. FUNKČNÁ ANALÝZA (kvalitatívna, kvantitatívna). Inštrumentálne analytické metódy: Infračervená a Ramanova spektrometria. UV/VIS molekulová absorpčná spektrometria. Röntgenová spektrometria. Spektrálne
u R Pasívne prvky R, L, C v obvode striedavého prúdu Činný odpor R Napätie zdroja sa rovná úbytku napätia na činnom odpore.
 Pasívne prvky, L, C v obvode stredavého prúdu Čnný odpor u u prebeh prúdu a napäta fázorový dagram prúdu a napäta u u /2 /2 t Napäte zdroja sa rovná úbytku napäta na čnnom odpore. Prúd je vo fáze s napätím.
Pasívne prvky, L, C v obvode stredavého prúdu Čnný odpor u u prebeh prúdu a napäta fázorový dagram prúdu a napäta u u /2 /2 t Napäte zdroja sa rovná úbytku napäta na čnnom odpore. Prúd je vo fáze s napätím.
Inkrementy na výpočet chemických posunov protónov >C=CH substituovaných alkénov
 Inkrementy na výpočet chemických posunov protónov >C=CH substituovaných alkénov Substituent X z gem z cis z trans H 0 0 0 Alkyl 0.45-0.22-0.28 Aryl 1.38 0.36-0.07 CH 2 -Hal 0.70 0.11-0.04 CH 2 -O 0.64-0.01-0.02
Inkrementy na výpočet chemických posunov protónov >C=CH substituovaných alkénov Substituent X z gem z cis z trans H 0 0 0 Alkyl 0.45-0.22-0.28 Aryl 1.38 0.36-0.07 CH 2 -Hal 0.70 0.11-0.04 CH 2 -O 0.64-0.01-0.02
2.2 Elektrónový obal atómu
 2.2 Elektrónový obal atómu Chemické vlastnosti prvkov závisia od usporiadania elektrónov v elektrónových obaloch ich atómov, presnejšie od počtu elektrónov vo valenčnej vrstve atómov. Poznatky o usporiadaní
2.2 Elektrónový obal atómu Chemické vlastnosti prvkov závisia od usporiadania elektrónov v elektrónových obaloch ich atómov, presnejšie od počtu elektrónov vo valenčnej vrstve atómov. Poznatky o usporiadaní
Modul pružnosti betónu
 f cm tan α = E cm 0,4f cm ε cl E = σ ε ε cul Modul pružnosti betónu α Autori: Stanislav Unčík Patrik Ševčík Modul pružnosti betónu Autori: Stanislav Unčík Patrik Ševčík Trnava 2008 Obsah 1 Úvod...7 2 Deformácie
f cm tan α = E cm 0,4f cm ε cl E = σ ε ε cul Modul pružnosti betónu α Autori: Stanislav Unčík Patrik Ševčík Modul pružnosti betónu Autori: Stanislav Unčík Patrik Ševčík Trnava 2008 Obsah 1 Úvod...7 2 Deformácie
Biochemické zmeny v organizme človeka v procese starnutia
 UNIVERZITA KOMENSKÉHO V BRATISLAVE JESSENIOVA LEKÁRSKA FAKULTA V MARTINE Ústav lekárskej biochémie Biochemické zmeny v organizme človeka v procese starnutia doc. Mgr. Eva Babušíková, PhD. Starnutie progresívny,
UNIVERZITA KOMENSKÉHO V BRATISLAVE JESSENIOVA LEKÁRSKA FAKULTA V MARTINE Ústav lekárskej biochémie Biochemické zmeny v organizme človeka v procese starnutia doc. Mgr. Eva Babušíková, PhD. Starnutie progresívny,
ARMA modely čast 2: moving average modely (MA)
") ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2011/2012 ARMA modely časť 2: moving average modely(ma) p.1/25 V. Moving average proces prvého rádu - MA(1) ARMA modely
ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2011/2012 ARMA modely časť 2: moving average modely(ma) p.1/25 V. Moving average proces prvého rádu - MA(1) ARMA modely
Příloha č. 1 etiketa. Nutrilon Nenatal 0
 Příloha č. 1 etiketa Nutrilon Nenatal 0 Čelní strana Logo Nutrilon + štít ve štítu text: Speciální výživa pro nedonošené děti a děti s nízkou porodní hmotností / Špeciálna výživa pre nedonosené deti a
Příloha č. 1 etiketa Nutrilon Nenatal 0 Čelní strana Logo Nutrilon + štít ve štítu text: Speciální výživa pro nedonošené děti a děti s nízkou porodní hmotností / Špeciálna výživa pre nedonosené deti a
ZÁKLADNÉ POJMY. Svetlo. Svetlo ako vlnenie, vlnová dĺžka
 ÚVOD Laser sa v dnešnej dobe využíva v rôznych oblastiach ľudskej činnosti, vo vede, technike, strojárenstve, biológii, geodézii, holografii, medicíne atď. Za štvrťstoročie sa stal z laboratórneho systému
ÚVOD Laser sa v dnešnej dobe využíva v rôznych oblastiach ľudskej činnosti, vo vede, technike, strojárenstve, biológii, geodézii, holografii, medicíne atď. Za štvrťstoročie sa stal z laboratórneho systému
24. Základné spôsoby zobrazovania priestoru do roviny
 24. Základné spôsoby zobrazovania priestoru do roviny Voľné rovnobežné premietanie Presné metódy zobrazenia trojrozmerného priestoru do dvojrozmernej roviny skúma samostatná matematická disciplína, ktorá
24. Základné spôsoby zobrazovania priestoru do roviny Voľné rovnobežné premietanie Presné metódy zobrazenia trojrozmerného priestoru do dvojrozmernej roviny skúma samostatná matematická disciplína, ktorá
6. ΤΕΛΙΚΗ ΙΑΘΕΣΗ ΤΑΦΗ. 6.1. Γενικά
 6. ΤΕΛΙΚΗ ΙΑΘΕΣΗ ΤΑΦΗ 6.1. Γενικά Είναι γεγονός ότι ανέκαθεν ο τελικός αποδέκτης των υπολειµµάτων της κατανάλωσης και των καταλοίπων της παραγωγικής διαδικασίας υπήρξε το περιβάλλον. Στις παλιότερες κοινωνίες
6. ΤΕΛΙΚΗ ΙΑΘΕΣΗ ΤΑΦΗ 6.1. Γενικά Είναι γεγονός ότι ανέκαθεν ο τελικός αποδέκτης των υπολειµµάτων της κατανάλωσης και των καταλοίπων της παραγωγικής διαδικασίας υπήρξε το περιβάλλον. Στις παλιότερες κοινωνίες
Elektromagnetické žiarenie a jeho spektrum
 Elektromagnetické žiarenie a jeho spektrum Elektromagnetické žiarenie je prenos energie v podobe elektromagnetického vlnenia. Elektromagnetické vlnenie alebo elektromagnetická vlna je lokálne vzniknutá
Elektromagnetické žiarenie a jeho spektrum Elektromagnetické žiarenie je prenos energie v podobe elektromagnetického vlnenia. Elektromagnetické vlnenie alebo elektromagnetická vlna je lokálne vzniknutá
BIOCHÉMIA II KATEDRA CHÉMIE, FAKULTA PRÍRODNÝCH VIED, UNIVERZITA MATEJA BELA BANSKÁ BYSTRICA CITRÁTOVÝ CYKLUS TÉMA 03 DOC. RNDR. MAREK SKORŠEPA, PHD.
 BIOCHÉMIA II KATEDRA CHÉMIE, FAKULTA PRÍRODNÝCH VIED, UNIVERZITA MATEJA BELA BANSKÁ BYSTRICA CITRÁTOVÝ CYKLUS TÉMA 03 DOC. RNDR. MAREK SKORŠEPA, PHD. POSTAVENIE CITRÁTOVÉHO CYKLU V METABOLIZME 3 centrálne
BIOCHÉMIA II KATEDRA CHÉMIE, FAKULTA PRÍRODNÝCH VIED, UNIVERZITA MATEJA BELA BANSKÁ BYSTRICA CITRÁTOVÝ CYKLUS TÉMA 03 DOC. RNDR. MAREK SKORŠEPA, PHD. POSTAVENIE CITRÁTOVÉHO CYKLU V METABOLIZME 3 centrálne
Modelovanie dynamickej podmienenej korelácie kurzov V4
 Modelovanie dynamickej podmienenej korelácie menových kurzov V4 Podnikovohospodárska fakulta so sídlom v Košiciach Ekonomická univerzita v Bratislave Cieľ a motivácia Východiská Cieľ a motivácia Cieľ Kvantifikovať
Modelovanie dynamickej podmienenej korelácie menových kurzov V4 Podnikovohospodárska fakulta so sídlom v Košiciach Ekonomická univerzita v Bratislave Cieľ a motivácia Východiská Cieľ a motivácia Cieľ Kvantifikovať
Pilota600mmrez1. N Rd = N Rd = M Rd = V Ed = N Rd = M y M Rd = M y. M Rd = N 0.
 Bc. Martin Vozár Návrh výstuže do pilót Diplomová práca 8x24.00 kr. 50.0 Pilota600mmrez1 Typ prvku: nosník Prostředí: X0 Beton:C20/25 f ck = 20.0 MPa; f ct = 2.2 MPa; E cm = 30000.0 MPa Ocelpodélná:B500
Bc. Martin Vozár Návrh výstuže do pilót Diplomová práca 8x24.00 kr. 50.0 Pilota600mmrez1 Typ prvku: nosník Prostředí: X0 Beton:C20/25 f ck = 20.0 MPa; f ct = 2.2 MPa; E cm = 30000.0 MPa Ocelpodélná:B500
Rozsah akreditácie 1/5. Príloha zo dňa k osvedčeniu o akreditácii č. K-003
 Rozsah akreditácie 1/5 Názov akreditovaného subjektu: U. S. Steel Košice, s.r.o. Oddelenie Metrológia a, Vstupný areál U. S. Steel, 044 54 Košice Rozsah akreditácie Oddelenia Metrológia a : Laboratórium
Rozsah akreditácie 1/5 Názov akreditovaného subjektu: U. S. Steel Košice, s.r.o. Oddelenie Metrológia a, Vstupný areál U. S. Steel, 044 54 Košice Rozsah akreditácie Oddelenia Metrológia a : Laboratórium
1 Prevod miestneho stredného slnečného času LMT 1 na iný miestny stredný slnečný čas LMT 2
 1 Prevod miestneho stredného slnečného času LMT 1 na iný miestny stredný slnečný čas LMT 2 Rozdiel LMT medzi dvoma miestami sa rovná rozdielu ich zemepisných dĺžok. Pre prevod miestnych časov platí, že
1 Prevod miestneho stredného slnečného času LMT 1 na iný miestny stredný slnečný čas LMT 2 Rozdiel LMT medzi dvoma miestami sa rovná rozdielu ich zemepisných dĺžok. Pre prevod miestnych časov platí, že