Preiskovanje knjižnic
|
|
|
- Ἡρακλῆς Χρηστόπουλος
- 7 χρόνια πριν
- Προβολές:
Transcript

1 Preiskovanje knjižnic bakterijske kolonije prepivnamo na membrano, bakterije liziramo, DNA denaturiramo in fiksiramo 0,5 M NaOH/nevtralizacija/proteinaza K, spiranje; zapekanje pri 80 C, hibridiziramo z označeno sondo [metoda po Grunsteinu in Hognessu (1975)]
 denaturirano tarčno DNA vežemo na membrano inkubiramo z označeno sondo spiramo pri pogojih, ki onemogočajo nespecifične")
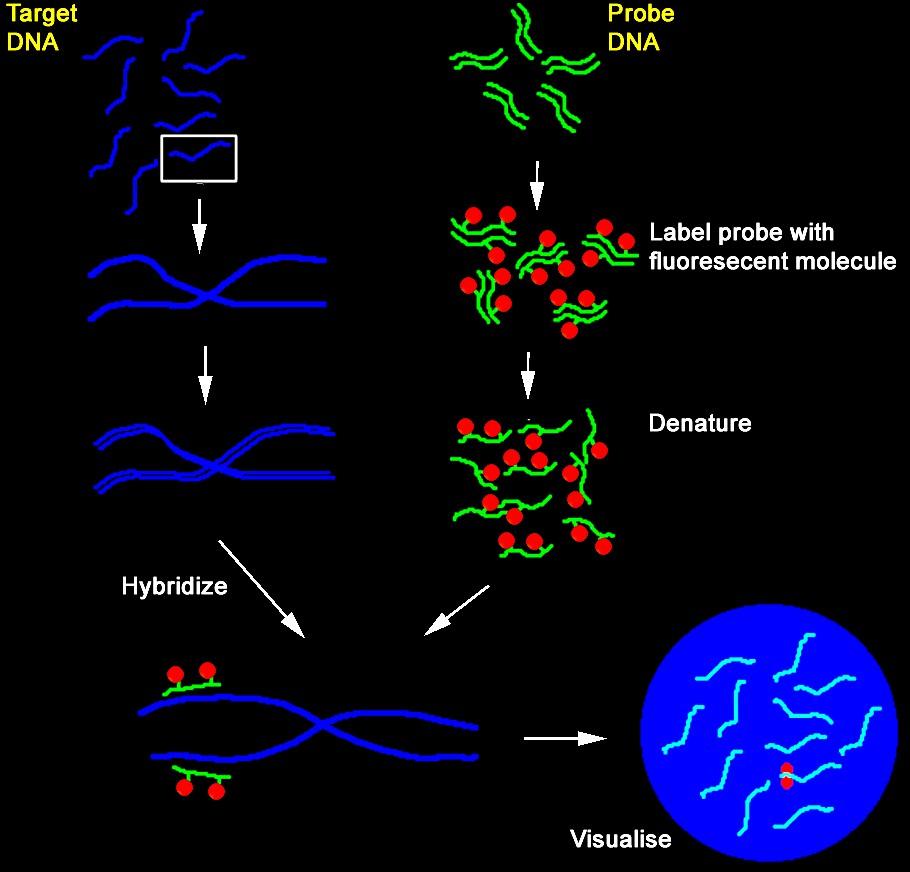
2 Hibridizacija Prileganje identičnih ali podobnih zaporedij nukleinskih kislin preko nastanka stabilnih baznih parov. [Stabilni bazni pari: >80 % identičnost na odseku ~50 bp.] sonda reagira s tarčnim zaporedjem v obliki ssdna (dsdna + NaOH -> ssdna) denaturirano tarčno DNA vežemo na membrano inkubiramo z označeno sondo spiramo pri pogojih, ki onemogočajo nespecifične interakcije Sonde so večinoma dolge b (! izjeme) Značilnosti hibridizacijske reakcije: - specifičen proces (do vezave pride samo na mestu ujemanja baz) - prisotnost nespecifičnih tarč ne vpliva na interakcijo med sondo in specifično tarčo Uporaba: detektiranje specifične DNA v knjižnicah detektiranje specifičnega fragmenta po elektroforezi določanje relativne količine specifične mrna v vzorcih hibridizacija in situ

.")
3 Načini prenašanja na membrano Nukleinske kisline običajno prenašamo na membrano z elektroforeznega gela, na katerem smo opravili ločevanje po velikosti. Ξ Hibiridizacija v raztopini je z eksperimentalnega stališča nesmiselna (izjeme: PCR, mol. svetila). Prenos lahko dosežemo na več načinov: kapilarni prenos: elektro-prenos:


 hibridiziranje v")

4 Načini prenašanja /2 suhi prenos - gel namočimo v pufru, odcedimo in nanj pritisnemo membrano kontaktni prenos - enako, brez predhodnega namakanja gela vakuumski prenos - prenos pospešimo z vakuumom (zadošča 1 h) hibridiziranje v gelu je možno, a je precej neučinkovito mokri prenos polsuhi prenos točkovni prenos vakuumski prenos
 - manjša občutljivost kot najlonske membrane, DNA na NC ne moremo pritrditi")


5 Prenašalne membrane vrste membran: - nitrocelulozne membrane (samo za kolorimetrično ali radioaktivno detekcijo) - manjša občutljivost kot najlonske membrane, DNA na NC ne moremo pritrditi z UV-svetlobo - pozitivno nabite najlonske membrane (tudi za kemiluminiscenco*) bolj obstojne membrane, večja občutljivost - nenabite najlonske membrane (manjša kapaciteta vezave kot nabite membrane) * za kemiluminiscenčno detekcijo je potrebna hidrofobna membrana, zato NC membrane niso primerne Lastnosti membran so lahko odvisne od proizvajalca in šarže (npr. gostota naboja previsoka povzroča visoka ozadja).




, tako da se")
6 Delo s prenašalnimi membranami /1 DNA in RNA imata veliko afiniteto do nitroceluloznih in nabitih najlonskih membran pred prenosom DNA z agaroze je treba DNA denaturirati (močne alkalne raztopine: npr. 0,5 M NaOH), tako da se na membrano veže kot ssdna

 nukleacija - tvorba kratkega niza sparjenih baz (vsaj 3) - počasi b) zapiranje -")
7 Delo s prenašalnimi membranami /2 nezasedena mesta na membrani je treba blokirati, da se tja ne veže sonda; blokiramo z nespecifično DNA (npr. telečja, lososova) dodamo sondo v molarnem prebitku in inkubiramo več ur, da se ustvarijo hibridne molekule proces hibridizacije je dvostopenjski: a) nukleacija - tvorba kratkega niza sparjenih baz (vsaj 3) - počasi b) zapiranje - podaljševanje sparjene regije - hitro speremo nevezano sondo, da zmanjšamo ozadje; postopek večkrat ponovimo
 tipične razmere med hibridizacijo: 6x SSC, 0,2 % SDS, 1x Denhardtova raztopina (0,02 % BSA, 0,02 % fikol, 0,02 %")
8 Delo s prenašalnimi membranami /3 s spreminjanjem pogojev spiranja lahko detektiramo bolj ali manj podobne molekule (spiranje pri višji temperaturi večja specifičnost signala) detektiranje signala: avtoradiografija ali neradioaktivni načini (kemične ali encimske kromogene reakcije) tipične razmere med hibridizacijo: 6x SSC, 0,2 % SDS, 1x Denhardtova raztopina (0,02 % BSA, 0,02 % fikol, 0,02 % polivinilpirolidon) ali 1 % mleko sonda: ng/ml 65 C, h tipična raztopina za spiranje: 3x SSC, 0,2 % SDS 1x SSC, 0,2 % SDS (SSC = 150 mm NaCl, 15 mm Na-citrat ph 7)
 Fiksiranje z UV: vezava je preko T, ki se kovalentno vežejo na membrano.")
9 Fiksiranje DNA na membrane UV-crosslinker (30-60 s, RT) zapekanje v vakuumu (80 C 2 h) - NC/nekoč inkubiranje v alkalnem: samo za poz. nabite membrane (4-6 h, RT) Fiksiranje z UV: vezava je preko T, ki se kovalentno vežejo na membrano. Najti je treba optimalne pogoje vezave (dovolj čvrsto, a ne preko vseh T, da so ti še na voljo za parjenje s sondo).
, ki jo ločimo na denaturirajočih agaroznih")
: detektiramo specifična zaporedja DNA.")

10 Tipi prenosov northern: analiziramo RNA (10-20 µg/nanos), ki jo ločimo na denaturirajočih agaroznih gelih (v prisotnosti glioksala ali formaldehida) Southern (1975): detektiramo specifična zaporedja DNA. Genomske fragmente ločujemo po velikosti na agaroznih gelih - osnova RFLP in podobnih metod. TIBS 25, (2000)
11
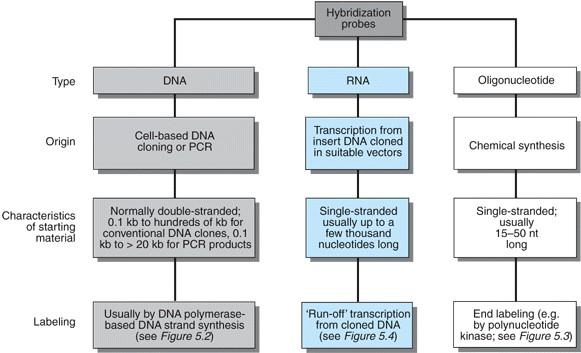
12 Sonde Sonde so označene ssdna ali RNA, ki so vsaj delno komplementarne zaporedju, ki ga iščemo. Uporabljamo jih pri tehnikah prenosa (blotting) in pri študijah celic in situ. vrste označevanja: detekcija: radioaktivno (32P, 35S, 125I, 3H) avtoradiografija, števci biotin preko avidina/steptavidina encimi (alk.fosfataza, hrenova peroksidaza) test aktivnosti kemoluminiscenca luminometer, film fluorescenca fluorimetri, mikroskopija antigeni protitelesa
13 Sonde /2 Načini pritrjevanja oznak na DNA (RNA): - pomikanje zareze (nick translation) - podaljševanje začetnega oligonukleotida (primer extension) - RNA-polimeraza - označevanje koncev - direktno označevanje - označevanje s PCR Sondo lahko pripravimo na osnovi zaporedja iz sorodnega organizma (heterologne sonde) ali pa jih sintetiziramo na osnovi znanega aminokislinskega zaporedja proteina, za katerega iščemo zapis.
14 Sonde /3 Priprava označene sonde s pomikom zareze (Rigby et al., 1977) 1.) osnova reakcije sta DNA, ki jo želiš označiti, in DNA-polimeraza 2.) v reakcijo dodaj dntp (1 označen) in vnesi zareze v DNA [DNaza I] 3.) dodaj polimerazo, ki bo zareze zapolnjevala z dntp (5 3 ) 4.) polimeraza cepi vezi pred sabo [eksonukleazna aktivnost 5 3 ] in vgrajuje nove nukleotide vključno z označenim dntp 5.) DNA denaturiramo in ločimo označeno sondo od preostanka DNA
15 Sonde /4 Priprava sonde po metodi z naključnimi začetnimi oligonukleotidi (random primer) oz. metodi s podaljševanjem oligonukleotidov (Feinberg & Vogelstein, 1983). Začetni oligonukl.: 6-10 b.

16 Sonde /5 Priprava sonde po metodi z označevanjem koncev (end labelling): Običajno označimo 5 -konec DNA/RNA tako, da dodamo označevalno skupino s pomočjo delovanja polinukleotid kinaze (PNK). Vendar tako lahko vgradimo samo 1 označevalno skupino, kar pomeni manjšo specifično aktivnost pri detekciji. Najprej s CIP odstranimo fosfatne skupine, nato s T4-PNK dodamo na 5 -konec označen fosfat 32P.
17 Sonde /6 Priprava sonde s PCR: Priprava z RNA-polimerazo: Vgradnja označevalca v repe s terminalno transferazo:
18 Sonde /6 Priprava sonde s PCR: Priprava z RNA-polimerazo: Vgradnja označevalca v repe s terminalno transferazo: Direktno označevanje: DNA pri 85 C inkubiramo 30 minut s cisplatinovo linkersko molekulo, na katero je že vezana oznaka. Vezava poteče na N7 gvanozina in citozina. DIG Chem-Link, Roche
19 Sonde: neradioaktivno označevanje
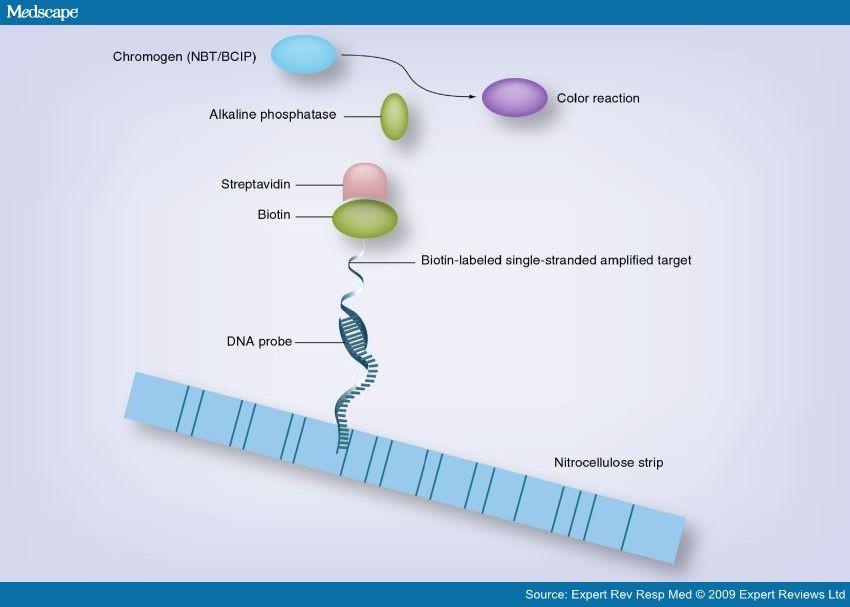
20 Označevanje in detekcija /1 kemično označena sonda: po prenosu membrano inkubiramo s protitelesi proti digoksigeninu, ta pa so konjugirana z reporterskim encimom (npr. alkalno fosfatazo) DETEKCIJA 1: dodamo substrata NTB (nitro blue tetrazolium) in BCIP (5-bromo-4-kloro-3-indolilfosfat), ki ju fosfataza razgradi in nastane netopno modro barvilo na mestu vezanih protiteles DETEKCIJA 2: dodamo kemoluminiscenčni substrat za alkalno fosfatazo (CSPD) in detektiramo sproščeno svetlobo (luminometer ali film)

21 Označevanje in detekcija /2 Iste membrane z vzorci lahko zapored uporabimo z dvema različno označenima sondama (npr. digoksigenin in fluorescein). Iste membrane lahko uporabimo večkrat tudi v primeru, da po končani detekciji sondo speremo z imobilizirane tarčne DNA. Metode spiranja so odvisne od vrste označevanja, v osnovi pa gre za inkubiranje v 0,2 M NaOH, 0,1 % SDS (2 x 15 min).

 fosforotioat reagira z 5-jodoacetamid-fluoresceinom (5-IAF), kar povzroči vezavo fluoresceina na žveplov atom tako označen")
22 Označevanje in detekcija /3 Označevanje s fluoresceinom: označimo lahko katerikoli oligonukleotid - naročimo markiranje ob sintezi ali oznako vnesemo v laboratoriju: 1.) s T4-PNK na 5 -konec DNA uvedemo fosforotioatno skupino (uporabimo reagent adenozin-5 -O-3-tiotrifosfat) 2.) fosforotioat reagira z 5-jodoacetamid-fluoresceinom (5-IAF), kar povzroči vezavo fluoresceina na žveplov atom tako označen oligonukleotid uporabimo za označevanje sond, npr. s PCR DETEKCIJA: detektiramo s posebno aparaturo (npr. FlouroImager)
23 Označevanje in detekcija /4 Označevanje preko biotina omogoča podobne načine detekcije kot so bili opisani pri digoksigeninu (kemiluminiscenca ali kolorimetrično detektiranje preko kompleksa streptavidin - alkalna fosfataza). Biotin smo vezali na sondo kemično preko psoralena ali že v postopku sinteze. Primerjava radioaktivnih in neradioaktivnih označevalcev: radioaktivnost manj reagentov, standardizirani postopki, podaljševanje ekspozicije močnejši signali alternativni postopki: trajnost reagentov (ni upada aktivnosti izvora), ni nevarnosti za okolje in delavce, kratki časi eksponiranja, možnost barvnih reakcij Na tržišču je več deset kompletov reagentov za različne načine označevanja in detekcije. Najprej se moramo odločiti, ali bomo označevali radioaktivno ali ne, nato pa po katerem postopku bomo označevali sondo. Kupiti je mogoče tudi membrane za prenos northern z že ločenimi vzorci mrna iz posameznih normalnih in patoloških tkiv.
24
25 Označevanje in detekcija: FISH
26
27 Določanje nukleotidnega zaporedja DNA Osnovni metodi: 1) Maxam & Gilbert (1977): kemična cepitev dsdna 2) Sanger (1977): zaključevanje verige pri sintezi s ssdna Pri obeh osnovnih metodah uporabljamo radioaktivno označevanje (pri novejših izvedbah Sangerjeve metode v avtomatskih postopkih so oznake fluorescenčne). Pri obeh pristopih analiziramo fragmente DNA. Prekinitve vnesemo kemično. Prekinitve vnašamo na specifičnih mestih, ki omogočajo ločevanje med 4 bazami. V začetni stopnji moramo izolirati DNA, ki jo želimo analizirati, v čisti obliki, nato pa izvedemo reakcije, ki vodijo do vnosa markerja in vnosa prekinitev. Sledi ločevanje fragmentov po velikosti (PAGE; avtomatizirani postopki: kapilarna elektroforeza - CE) Pri fluorescenčnem označevanju ločimo: označevanje preko začetnega oligonukleotida (dye primer chemistry) označevanje preko terminatorja (dye terminator chemistry)
: G z dimetilsulfatom C+T s hidrazinom A+G z mravlj.")
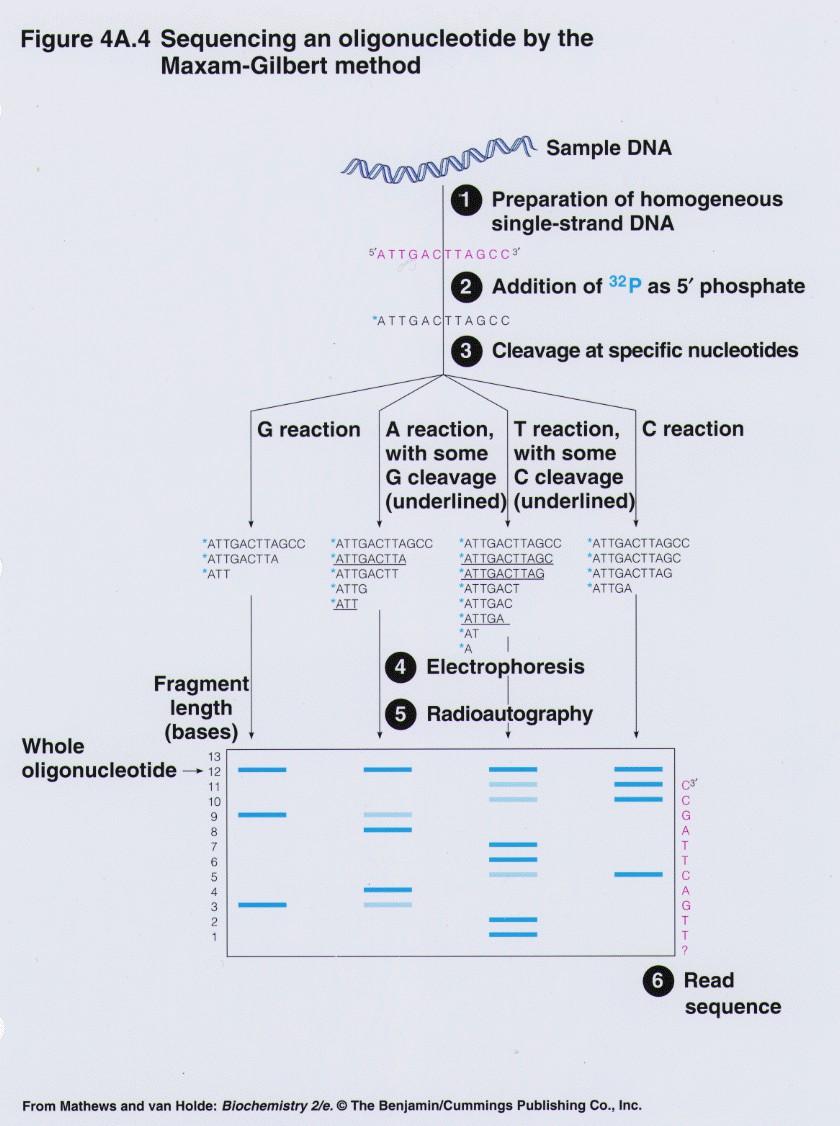
28 Metoda po Maxamu in Gilbertu Postopek: * dsdna označimo na 5 -koncu s 32P-ATP * 2 verigi ločimo pri 90 C v DMSO, nanesemo na PAGE in nato izoliramo posamezni verigi * ssdna razdelimo na 4 alikvote in obdelamo ločeno (modifikacija baz): G z dimetilsulfatom C+T s hidrazinom A+G z mravlj. kislino C s hidrazinom v alkalnem Na mestu modificiranih baz s piperidinom v kislem prekinemo verigo fragmente ločimo s PAGE. Razcep ni kvantitativen, zato dobimo veliko število radioaktivno označenih produktov.
29
 dodamo encim, dntp in označeni nukleotid (označevalna reakcija) dodamo terminatorje")
30 Določanje zaporedja po dideoksi-metodi (1) izoliramo dsdna in jo pretvorimo v ssdna obliko (denaturacija) vežemo začetne oligonukleotide (15-20 b) dodamo encim, dntp in označeni nukleotid (označevalna reakcija) dodamo terminatorje (ddntp)

31 Določanje zaporedja po dideoksi-metodi (2) zaustavimo reakcijo ločimo nastale fragmente po velikosti (elektroforeza na 7 % PAG + 7 M urea)
")

32 Določanje zaporedja po dideoksi-metodi (3) dideoksinukleotidi kot terminatorji v polimerazni reakciji:
33 Avtomatski sekvenatorji Dva različna pristopa: - vsako bazo označimo s svojim barvilom in analiziramo v 1 nanosu (4 reakcije = 1 zaporedje hkrati); detektor ločeno zazna posamezne signale in program generira izpis - uporabljamo samo 1 barvilo in vsako bazo nanesemo ločeno (4 nanosi za 1 zaporedje); detektor prebere vsakega od 4 signalov ločeno, program integrira signale in generira izpis Barvila so fluorescenčna (emisija v vidnem ali IR delu spektra) ali radioaktivna. Fragmente ločujemo s klasično PAGE ali s kapilarno elektroforezo. Aparatura ima ločevalni del, detektorski del in kontrolni/integracijski del. Problemi: prekrivajoči spektri barvil, sekundarne strukture in kompresije Postopek: priprava matrične DNA (plazmid, fag, ) sekvenčna reakcija (vgradnja barvila med polimerazno reakcijo) čiščenje označene DNA nanos na elektroforezni nosilec ločevanje na nosilcu detekcija in interpretacija signalov (surovi signal odštevanje ozadja, mapiranje signalov, oblikovanje izpisa)

 Ločevanje pri 10-30 kv.")
, zanesljivost")
34 Naprave na osnovi kapilarne elektroforeze Proizvajalec ABI (Applied Biosystems) Kapilare so običajno iz silikata, dolge cm in imajo notranji premer ~50 µm; nanos nl. Za delo z velikim številom vzorcev so na voljo večkapilarni sistemi z vzporedno detekcijo. Uporabljamo komercialna barvila (Big-Dye; ABI) Ločevanje pri kv. Prednosti sistema: ni potrebno pripravljati PA-gelov, ni gibljivih delov. Slabosti: potrošni material je sorazmerno drag (kapilare, reagenti), zanesljivost določenega zaporedja je ~98,5 %.

35 Kemizem Big-Dye Označeni so terminatorji ( vedno lahko uporabimo enake)
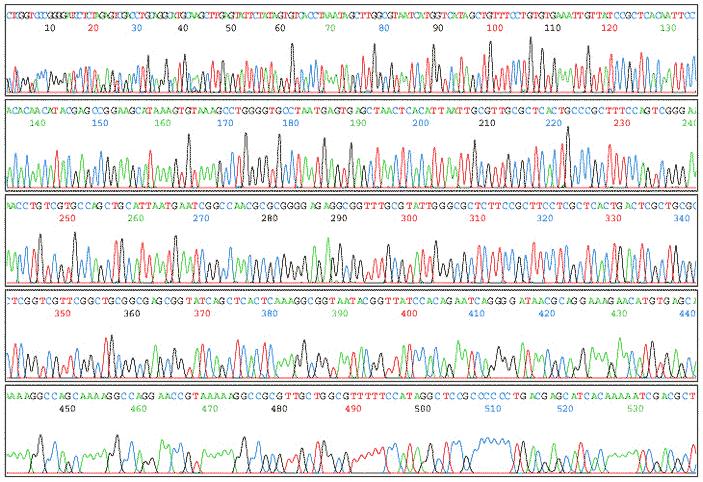
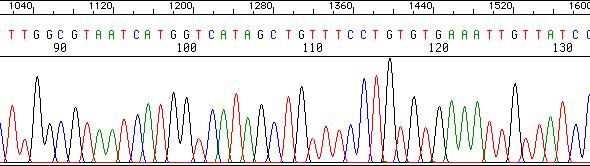
36 Sekvenatorji na osnovi CE (3) Izpis:
Diferencialna enačba, v kateri nastopata neznana funkcija in njen odvod v prvi potenci
 Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Linearna diferencialna enačba reda Diferencialna enačba v kateri nastopata neznana funkcija in njen odvod v prvi potenci d f + p= se imenuje linearna diferencialna enačba V primeru ko je f 0 se zgornja
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 22. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 22. oktober 2013 Kdaj je zaporedje {a n } konvergentno, smo definirali s pomočjo limite zaporedja. Večkrat pa je dobro vedeti,
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 5. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 5. december 2013 Primer Odvajajmo funkcijo f(x) = x x. Diferencial funkcije Spomnimo se, da je funkcija f odvedljiva v točki
Funkcijske vrste. Matematika 2. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 2. april Gregor Dolinar Matematika 2
 Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
Matematika 2 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 2. april 2014 Funkcijske vrste Spomnimo se, kaj je to številska vrsta. Dano imamo neko zaporedje realnih števil a 1, a 2, a
KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK
 1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
1 / 24 KODE ZA ODKRIVANJE IN ODPRAVLJANJE NAPAK Štefko Miklavič Univerza na Primorskem MARS, Avgust 2008 Phoenix 2 / 24 Phoenix 3 / 24 Phoenix 4 / 24 Črtna koda 5 / 24 Črtna koda - kontrolni bit 6 / 24
Tretja vaja iz matematike 1
 Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Tretja vaja iz matematike Andrej Perne Ljubljana, 00/07 kompleksna števila Polarni zapis kompleksnega števila z = x + iy): z = rcos ϕ + i sin ϕ) = re iϕ Opomba: Velja Eulerjeva formula: e iϕ = cos ϕ +
Molekularna diagnostika
 Molekularna diagnostika Predpogoj za uspešno zdravljenje je pravočasno detektiranje povzročiteljev - velja za ljudi, živali, rastline in okolje. Klasični postopki detekcije so vključevali odvzem okuženega
Molekularna diagnostika Predpogoj za uspešno zdravljenje je pravočasno detektiranje povzročiteljev - velja za ljudi, živali, rastline in okolje. Klasični postopki detekcije so vključevali odvzem okuženega
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 21. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 21. november 2013 Hiperbolične funkcije Hiperbolični sinus sinhx = ex e x 2 20 10 3 2 1 1 2 3 10 20 hiperbolični kosinus coshx
Booleova algebra. Izjave in Booleove spremenljivke
 Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
Izjave in Booleove spremenljivke vsako izjavo obravnavamo kot spremenljivko če je izjava resnična (pravilna), ima ta spremenljivka vrednost 1, če je neresnična (nepravilna), pa vrednost 0 pravimo, da gre
8. Diskretni LTI sistemi
 8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
8. Diskreti LI sistemi. Naloga Določite odziv diskretega LI sistema s podaim odzivom a eoti impulz, a podai vhodi sigal. h[] x[] - - 5 6 7 - - 5 6 7 LI sistem se a vsak eoti impulz δ[] a vhodu odzove z
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 14. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 14. november 2013 Kvadratni koren polinoma Funkcijo oblike f(x) = p(x), kjer je p polinom, imenujemo kvadratni koren polinoma
NEPARAMETRIČNI TESTI. pregledovanje tabel hi-kvadrat test. as. dr. Nino RODE
 NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
NEPARAMETRIČNI TESTI pregledovanje tabel hi-kvadrat test as. dr. Nino RODE Parametrični in neparametrični testi S pomočjo z-testa in t-testa preizkušamo domneve o parametrih na vzorcih izračunamo statistike,
Frekvenčna analiza neperiodičnih signalov. Analiza signalov prof. France Mihelič
 Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Frekvenčna analiza neperiodičnih signalov Analiza signalov prof. France Mihelič Vpliv postopka daljšanja periode na spekter periodičnega signala Opazujmo družino sodih periodičnih pravokotnih impulzov
Poglavje 7. Poglavje 7. Poglavje 7. Regulacijski sistemi. Regulacijski sistemi. Slika 7. 1: Normirana blokovna shema regulacije EM
 Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Slika 7. 1: Normirana blokovna shema regulacije EM Fakulteta za elektrotehniko 1 Slika 7. 2: Principielna shema regulacije AM v KSP Fakulteta za elektrotehniko 2 Slika 7. 3: Merjenje komponent fluksa s
Proteini. Struktura proteinov. Analiza proteinov. Proteini (proizvodnja, analiza, struktura, funkcija)
") Proteini (proizvodnja, analiza, struktura, funkcija) Celična biologija z genetiko 1. letnik UŠ LBM, Kozmetologija šol. leto 2012/13 Proteios (Gr.) = prvega reda Makromolekule polimeri aminokislin, ki so
Proteini (proizvodnja, analiza, struktura, funkcija) Celična biologija z genetiko 1. letnik UŠ LBM, Kozmetologija šol. leto 2012/13 Proteios (Gr.) = prvega reda Makromolekule polimeri aminokislin, ki so
SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK
 SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
SKUPNE PORAZDELITVE SKUPNE PORAZDELITVE VEČ SLUČAJNIH SPREMENLJIVK Kovaec vržemo trikrat. Z ozačimo število grbov ri rvem metu ( ali ), z Y a skuo število grbov (,, ali 3). Kako sta sremelivki i Y odvisi
vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov. 6. vaja Kvan*ta*vno določanje proteinov
 28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
28. 3. 11 UV- spektrofotometrija Biuretska metoda Absorbanca pri λ=28 nm (A28) UV- spektrofotometrija Biuretska metoda vstopni žarek intenziteta I Lowrijeva metoda Bradfordova metoda Bradfordova metoda
13. Jacobijeva metoda za računanje singularnega razcepa
 13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
13. Jacobijeva metoda za računanje singularnega razcepa Bor Plestenjak NLA 25. maj 2010 Bor Plestenjak (NLA) 13. Jacobijeva metoda za računanje singularnega razcepa 25. maj 2010 1 / 12 Enostranska Jacobijeva
Odvod. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 10. december Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 10. december 2013 Izrek (Rolleov izrek) Naj bo f : [a,b] R odvedljiva funkcija in naj bo f(a) = f(b). Potem obstaja vsaj ena
Lastnosti molekule DNA
 Metode za odkrivanje/analizo mutacij Celična biologija z genetiko 1. letnik UŠ LBM, št. leto 2012/13 Detekcija polimorfizmov/mutacij Sprva so razlike v genomu med posamezniki ugotavljali le z opazovanjem
Metode za odkrivanje/analizo mutacij Celična biologija z genetiko 1. letnik UŠ LBM, št. leto 2012/13 Detekcija polimorfizmov/mutacij Sprva so razlike v genomu med posamezniki ugotavljali le z opazovanjem
Gradniki TK sistemov
 Gradniki TK sistemov renos signalov v višji rekvenčni legi Vsebina Modulacija in demodulacija Vrste analognih modulacij AM M FM rimerjava spektrov analognih moduliranih signalov Mešalniki Kdaj uporabimo
Gradniki TK sistemov renos signalov v višji rekvenčni legi Vsebina Modulacija in demodulacija Vrste analognih modulacij AM M FM rimerjava spektrov analognih moduliranih signalov Mešalniki Kdaj uporabimo
Kotne in krožne funkcije
 Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kotne in krožne funkcije Kotne funkcije v pravokotnem trikotniku Avtor: Rok Kralj, 4.a Gimnazija Vič, 009/10 β a c γ b α sin = a c cos= b c tan = a b cot = b a Sinus kota je razmerje kotu nasprotne katete
Kontrolne karte uporabljamo za sprotno spremljanje kakovosti izdelka, ki ga izdelujemo v proizvodnem procesu.
 Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
Kontrolne karte KONTROLNE KARTE Kontrolne karte uporablamo za sprotno spremlane kakovosti izdelka, ki ga izdeluemo v proizvodnem procesu. Izvaamo stalno vzorčene izdelkov, npr. vsako uro, vsake 4 ure.
1. Definicijsko območje, zaloga vrednosti. 2. Naraščanje in padanje, ekstremi. 3. Ukrivljenost. 4. Trend na robu definicijskega območja
 ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
ZNAČILNOSTI FUNKCIJ ZNAČILNOSTI FUNKCIJE, KI SO RAZVIDNE IZ GRAFA. Deinicijsko območje, zaloga vrednosti. Naraščanje in padanje, ekstremi 3. Ukrivljenost 4. Trend na robu deinicijskega območja 5. Periodičnost
PONOVITEV SNOVI ZA 4. TEST
 PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
PONOVITEV SNOVI ZA 4. TEST 1. * 2. *Galvanski člen z napetostjo 1,5 V požene naboj 40 As. Koliko električnega dela opravi? 3. ** Na uporniku je padec napetosti 25 V. Upornik prejme 750 J dela v 5 minutah.
Numerično reševanje. diferencialnih enačb II
 Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Numerčno reševanje dferencaln enačb I Dferencalne enačbe al ssteme dferencaln enačb rešujemo numerčno z več razlogov:. Ne znamo j rešt analtčno.. Posamezn del dferencalne enačbe podan tabelarčno. 3. Podatke
Zaporedja. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 15. oktober Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 15. oktober 2013 Oglejmo si, kako množimo dve kompleksni števili, dani v polarni obliki. Naj bo z 1 = r 1 (cosϕ 1 +isinϕ 1 )
Funkcije. Matematika 1. Gregor Dolinar. Fakulteta za elektrotehniko Univerza v Ljubljani. 12. november Gregor Dolinar Matematika 1
 Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
Matematika 1 Gregor Dolinar Fakulteta za elektrotehniko Univerza v Ljubljani 12. november 2013 Graf funkcije f : D R, D R, je množica Γ(f) = {(x,f(x)) : x D} R R, torej podmnožica ravnine R 2. Grafi funkcij,
1. Trikotniki hitrosti
 . Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
. Trikotniki hitrosti. Z radialno črpalko želimo črpati vodo pri pogojih okolice z nazivnim pretokom 0 m 3 /h. Notranji premer rotorja je 4 cm, zunanji premer 8 cm, širina rotorja pa je,5 cm. Frekvenca
vaja Izolacija kromosomske DNA iz vranice in hiperkromni efekt. DNA RNA Protein. ime deoksirbonukleinska kislina ribonukleinska kislina
 transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
transkripcija translacija Protein 12. vaja Izolacija kromosomske iz vranice in hiperkromni efekt sladkorji deoksiriboza riboza glavna funkcija dolgoročno shranjevanje genetskih informacij prenos informacij
*M * Osnovna in višja raven MATEMATIKA NAVODILA ZA OCENJEVANJE. Sobota, 4. junij 2011 SPOMLADANSKI IZPITNI ROK. Državni izpitni center
 Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
Državni izpitni center *M40* Osnovna in višja raven MATEMATIKA SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sobota, 4. junij 0 SPLOŠNA MATURA RIC 0 M-40-- IZPITNA POLA OSNOVNA IN VIŠJA RAVEN 0. Skupaj:
GEL ELEKTROFOREZA. Seminar pri predmetu Molekularna Biofizika. Avtorica: Tjaša Parkelj
 GEL ELEKTROFOREZA Seminar pri predmetu Molekularna Biofizika Avtorica: Tjaša Parkelj Povzetek: V tem seminarju bom predstavila fizikalno ozadje elektroforeze. Začela bom z opisom gibanja nabitega delca
GEL ELEKTROFOREZA Seminar pri predmetu Molekularna Biofizika Avtorica: Tjaša Parkelj Povzetek: V tem seminarju bom predstavila fizikalno ozadje elektroforeze. Začela bom z opisom gibanja nabitega delca
Iterativno reševanje sistemov linearnih enačb. Numerične metode, sistemi linearnih enačb. Numerične metode FE, 2. december 2013
 Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
Numerične metode, sistemi linearnih enačb B. Jurčič Zlobec Numerične metode FE, 2. december 2013 1 Vsebina 1 z n neznankami. a i1 x 1 + a i2 x 2 + + a in = b i i = 1,..., n V matrični obliki zapišemo:
MATEMATIČNI IZRAZI V MAFIRA WIKIJU
 I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
I FAKULTETA ZA MATEMATIKO IN FIZIKO Jadranska cesta 19 1000 Ljubljan Ljubljana, 25. marec 2011 MATEMATIČNI IZRAZI V MAFIRA WIKIJU KOMUNICIRANJE V MATEMATIKI Darja Celcer II KAZALO: 1 VSTAVLJANJE MATEMATIČNIH
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
IZPIT IZ ANALIZE II Maribor,
 Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
Maribor, 05. 02. 200. (a) Naj bo f : [0, 2] R odvedljiva funkcija z lastnostjo f() = f(2). Dokaži, da obstaja tak c (0, ), da je f (c) = 2f (2c). (b) Naj bo f(x) = 3x 3 4x 2 + 2x +. Poišči tak c (0, ),
p 1 ENTROPIJSKI ZAKON
 ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
ENROPIJSKI ZAKON REERZIBILNA srememba: moža je obrjea srememba reko eakih vmesih staj kot rvota srememba. Po obeh sremembah e sme biti obeih trajih srememb v bližji i dalji okolici. IREERZIBILNA srememba:
Delovna točka in napajalna vezja bipolarnih tranzistorjev
 KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
KOM L: - Komnikacijska elektronika Delovna točka in napajalna vezja bipolarnih tranzistorjev. Določite izraz za kolektorski tok in napetost napajalnega vezja z enim virom in napetostnim delilnikom na vhod.
ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI. Membranski separacijski procesi: diafiltracija, elektrodializa, reverzna osmoza, pervaporacija
 ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI Membranski separacijski procesi: diafiltracija, elektrodializa, reverzna osmoza, pervaporacija Membranski separacijski procesi v biotehnologiji proces mikrofiltracija
ZAKLJUČNI PROCESI V BIOTEHNOLOGIJI Membranski separacijski procesi: diafiltracija, elektrodializa, reverzna osmoza, pervaporacija Membranski separacijski procesi v biotehnologiji proces mikrofiltracija
matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):
![matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):](/thumbs/87/96579490.jpg "matrike A = [a ij ] m,n αa 11 αa 12 αa 1n αa 21 αa 22 αa 2n αa m1 αa m2 αa mn se števanje po komponentah (matriki morata biti enakih dimenzij):") 4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
4 vaja iz Matematike 2 (VSŠ) avtorica: Melita Hajdinjak datum: Ljubljana, 2009 matrike Matrika dimenzije m n je pravokotna tabela m n števil, ki ima m vrstic in n stolpcev: a 11 a 12 a 1n a 21 a 22 a 2n
Imunofluorescencija. vizualizacija molekula protutijela obilježenih fluorokromom vezanih za antigene na stanicama ili tkivnim preparatima
 Imunofluorescencija 1944. - Robert Coons protutijela se mogu označiti molekulama koje imaju sposobnost fluorescencije fluorokromi - apsorbiraju svjetlost jedna valne duljine (ekscitacija), a emitiraju
Imunofluorescencija 1944. - Robert Coons protutijela se mogu označiti molekulama koje imaju sposobnost fluorescencije fluorokromi - apsorbiraju svjetlost jedna valne duljine (ekscitacija), a emitiraju
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * NAVODILA ZA OCENJEVANJE. Sreda, 3. junij 2015 SPLOŠNA MATURA
 Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
Državni izpitni center *M15143113* SPOMLADANSKI IZPITNI ROK NAVODILA ZA OCENJEVANJE Sreda, 3. junij 2015 SPLOŠNA MATURA RIC 2015 M151-431-1-3 2 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor Naloga Odgovor
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
1. TVORBA ŠIBKEGA (SIGMATNEGA) AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ
 AORISTA: Največ grških glagolov ima tako imenovani šibki (sigmatni) aorist. Osnova se tvori s. γραψ") TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
TVORBA AORISTA: Grški aorist (dovršnik) izraža dovršno dejanje; v indikativu izraža poleg dovršnosti tudi preteklost. Za razliko od prezenta ima aorist posebne aktivne, medialne in pasivne oblike. Pri
Integralni račun. Nedoločeni integral in integracijske metrode. 1. Izračunaj naslednje nedoločene integrale: (a) dx. (b) x 3 +3+x 2 dx, (c) (d)
 dx. (b) x 3 +3+x 2 dx, (c) (d)") Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Integralni račun Nedoločeni integral in integracijske metrode. Izračunaj naslednje nedoločene integrale: d 3 +3+ 2 d, (f) (g) (h) (i) (j) (k) (l) + 3 4d, 3 +e +3d, 2 +4+4 d, 3 2 2 + 4 d, d, 6 2 +4 d, 2
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
Nukleinske kisline. Nukleotidi. DNA je nosilka dednih genetskih informacij.
 Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Nukleinske kisline DNA je nosilka dednih genetskih informacij. RNA je posrednik, ki omogoča sintezo proteinov na osnovi zapisa na DNA. Nukleotidi Nukleinske kisline so polimeri nukleotidov. OH... RNA ribonukleinska
Katedra za farmacevtsko kemijo. Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks. 25/11/2010 Vaje iz Farmacevtske kemije 3 1
 Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Katedra za farmacevtsko kemijo Sinteza mimetika encima SOD 2. stopnja: Mn 3+ ali Cu 2+ salen kompleks 25/11/2010 Vaje iz Farmacevtske kemije 3 1 Sinteza kompleksa [Mn 3+ (salen)oac] Zakaj uporabljamo brezvodni
Analiza'3D'struktur'makromolekul'in' modeliranje'
 Univerza'v'Ljubljani,'Fakulteta'za'kemijo'in'kemijsko'tehnologijo' Univerzitetni%študijski%program%Biokemija,%2.%letnik,%študijsko%leto%2013/2014% Analiza'3D'struktur'makromolekul'in' modeliranje' Miha%Pavšič%
Univerza'v'Ljubljani,'Fakulteta'za'kemijo'in'kemijsko'tehnologijo' Univerzitetni%študijski%program%Biokemija,%2.%letnik,%študijsko%leto%2013/2014% Analiza'3D'struktur'makromolekul'in' modeliranje' Miha%Pavšič%
Osnove elektrotehnike uvod
 Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
Osnove elektrotehnike uvod Uvod V nadaljevanju navedena vprašanja so prevod testnih vprašanj, ki sem jih našel na omenjeni spletni strani. Vprašanja zajemajo temeljna znanja opredeljenega strokovnega področja.
1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου...
 ΑΠΟΖΗΜΙΩΣΗ ΘΥΜΑΤΩΝ ΕΓΚΛΗΜΑΤΙΚΩΝ ΠΡΑΞΕΩΝ ΣΛΟΒΕΝΙΑ 1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου... 3 1 1. Έντυπα αιτήσεων
ΑΠΟΖΗΜΙΩΣΗ ΘΥΜΑΤΩΝ ΕΓΚΛΗΜΑΤΙΚΩΝ ΠΡΑΞΕΩΝ ΣΛΟΒΕΝΙΑ 1. Έντυπα αιτήσεων αποζημίωσης... 2 1.1. Αξίωση αποζημίωσης... 2 1.1.1. Έντυπο... 2 1.1.2. Πίνακας μεταφράσεων των όρων του εντύπου... 3 1 1. Έντυπα αιτήσεων
13. Vaja: Reakcije oksidacije in redukcije
 1. Vaja: Reakcije oksidacije in redukcije a) Osnove: Oksidacija je reakcija pri kateri posamezen element (reducent) oddaja elektrone in se pri tem oksidira (oksidacijsko število se zviša). Redukcija pa
1. Vaja: Reakcije oksidacije in redukcije a) Osnove: Oksidacija je reakcija pri kateri posamezen element (reducent) oddaja elektrone in se pri tem oksidira (oksidacijsko število se zviša). Redukcija pa
Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba.
 1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
1. Osnovni pojmi Enačba, v kateri poleg neznane funkcije neodvisnih spremenljivk ter konstant nastopajo tudi njeni odvodi, se imenuje diferencialna enačba. Primer 1.1: Diferencialne enačbe so izrazi: y
Gimnazija Krˇsko. vektorji - naloge
 Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
Vektorji Naloge 1. V koordinatnem sistemu so podane točke A(3, 4), B(0, 2), C( 3, 2). a) Izračunaj dolžino krajevnega vektorja točke A. (2) b) Izračunaj kot med vektorjema r A in r C. (4) c) Izrazi vektor
BIOKEMIJA IN MOLEKULARNA BIOLOGIJA
 Katja Repnik, Uroš Potočnik BIOKEMIJA IN MOLEKULARNA BIOLOGIJA Navodila za laboratorijske vaje Maribor, 2013/2014 DELO IN VARNOST V LABORATORIJU Osnovna pravila varnega dela v laboratoriju - V laboratorij
Katja Repnik, Uroš Potočnik BIOKEMIJA IN MOLEKULARNA BIOLOGIJA Navodila za laboratorijske vaje Maribor, 2013/2014 DELO IN VARNOST V LABORATORIJU Osnovna pravila varnega dela v laboratoriju - V laboratorij
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
Tabele termodinamskih lastnosti vode in vodne pare
 Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
Univerza v Ljubljani Fakulteta za strojništvo Laboratorij za termoenergetiko Tabele termodinamskih lastnosti vode in vodne pare po modelu IAPWS IF-97 izračunano z XSteam Excel v2.6 Magnus Holmgren, xsteam.sourceforge.net
ARHITEKTURA DETAJL 1, 1:10
 0.15 0.25 3.56 0.02 0.10 0.12 0.10 SESTV S2 polimer-bitumenska,dvoslojna(po),... 1.0 cm po zahtevah SIST DIN 52133 in nadstandardno, (glej opis v tehn.poročilu), npr.: PHOENIX STR/Super 5 M * GEMINI P
0.15 0.25 3.56 0.02 0.10 0.12 0.10 SESTV S2 polimer-bitumenska,dvoslojna(po),... 1.0 cm po zahtevah SIST DIN 52133 in nadstandardno, (glej opis v tehn.poročilu), npr.: PHOENIX STR/Super 5 M * GEMINI P
Matematika 1. Gabrijel Tomšič Bojan Orel Neža Mramor Kosta
 Matematika Gabrijel Tomšič Bojan Orel Neža Mramor Kosta 6. november 200 Poglavje 2 Zaporedja in številske vrste 2. Zaporedja 2.. Uvod Definicija 2... Zaporedje (a n ) = a, a 2,..., a n,... je predpis,
Matematika Gabrijel Tomšič Bojan Orel Neža Mramor Kosta 6. november 200 Poglavje 2 Zaporedja in številske vrste 2. Zaporedja 2.. Uvod Definicija 2... Zaporedje (a n ) = a, a 2,..., a n,... je predpis,
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna hitrost
 FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
FFA: Laboratorijska medicina, Molekularna encimologija, 2010/2011 3.predavanje Jure Stojan 2. predavanje termodinamične osnove, encimske katalize encimska kataliza časovni potek encimske reakcije začetna
A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N
 I N F O T E K N I K V o l u m e 1 5 N o. 1 J u l i 2 0 1 4 ( 61-70) A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N N o v i
I N F O T E K N I K V o l u m e 1 5 N o. 1 J u l i 2 0 1 4 ( 61-70) A N A L I S I S K U A L I T A S A I R D I K A L I M A N T A N S E L A T A N S E B A G A I B A H A N C A M P U R A N B E T O N N o v i
DNA in RNA: zgradba in vloga. Velika predavalnica IJS,
 DNA in RNA: zgradba in vloga Velika predavalnica IJS, 10. 4. 2014 Nukleinske kisline Shranjevanje, prenašanje in izražanje genetske informacije. Dve vrsti nukleinskih kislin: deoksiribonukleinska kislina
DNA in RNA: zgradba in vloga Velika predavalnica IJS, 10. 4. 2014 Nukleinske kisline Shranjevanje, prenašanje in izražanje genetske informacije. Dve vrsti nukleinskih kislin: deoksiribonukleinska kislina
Kaj je DNA footprinting? Omogoča lociranje vezave proteina na DNA molekulo z ugotavljanjem katere fosfodiestrske vezi so zaščitene pred rezom Dnaze
 Ugotavljanje števila bakterij, ki tvorijo kolonije (cfu) Št kolonij zraslih na plošči faktor redčenja na plošči 10= x cfu/ml DNA fingerprinting: omogoča opazovanje genskih razlik med ljudmi na podlagi
Ugotavljanje števila bakterij, ki tvorijo kolonije (cfu) Št kolonij zraslih na plošči faktor redčenja na plošči 10= x cfu/ml DNA fingerprinting: omogoča opazovanje genskih razlik med ljudmi na podlagi
Sintezna genomika. Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi
 Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi") Sintezna genomika Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi Koncept sinteznega življenja Celic, kot so se razvile v evoluciji,
Sintezna genomika Ponovno zapisovanje (Rewriting) Preoblikovanje kode (Refactoring) Transplantacija genoma Sintezni genomi Sintezni organizmi Koncept sinteznega življenja Celic, kot so se razvile v evoluciji,
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Matematika 1. Gregor Dolinar. 2. januar Fakulteta za elektrotehniko Univerza v Ljubljani. Gregor Dolinar Matematika 1
 Mtemtik 1 Gregor Dolinr Fkultet z elektrotehniko Univerz v Ljubljni 2. jnur 2014 Gregor Dolinr Mtemtik 1 Izrek (Izrek o povprečni vrednosti) Nj bo m ntnčn spodnj mej in M ntnčn zgornj mej integrbilne funkcije
Mtemtik 1 Gregor Dolinr Fkultet z elektrotehniko Univerz v Ljubljni 2. jnur 2014 Gregor Dolinr Mtemtik 1 Izrek (Izrek o povprečni vrednosti) Nj bo m ntnčn spodnj mej in M ntnčn zgornj mej integrbilne funkcije
Splošno o interpolaciji
 Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Splošno o interpolaciji J.Kozak Numerične metode II (FM) 2011-2012 1 / 18 O funkciji f poznamo ali hočemo uporabiti le posamezne podatke, na primer vrednosti r i = f (x i ) v danih točkah x i Izberemo
Neavtorizirani povzetki izbranih predavanj iz biokemije. UN študij Okolje Politehnika Nova Gorica. doc. dr. Marko Dolinar
 Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
Neavtorizirani povzetki izbranih predavanj iz biokemije UN študij Okolje Politehnika Nova Gorica doc. dr. Marko Dolinar obravnavane teme v študijskem letu 2003/2004: Pretok genetskih informacij (5. poglavje)
svari Real-time RT-PCR RSV
 19 39-43 12 Real-time RS svarireal-time RSV subgroup HMPV RSV N Real-time 100 100 Real-time RSV subgroup RSV HMPV F Real-time 55.8 95.5 genotype A2 B1 TaqMan svari InfV RS RSV HMPV HRV PIV RSV HMPV RSV
19 39-43 12 Real-time RS svarireal-time RSV subgroup HMPV RSV N Real-time 100 100 Real-time RSV subgroup RSV HMPV F Real-time 55.8 95.5 genotype A2 B1 TaqMan svari InfV RS RSV HMPV HRV PIV RSV HMPV RSV
CM707. GR Οδηγός χρήσης... 2-7. SLO Uporabniški priročnik... 8-13. CR Korisnički priručnik... 14-19. TR Kullanım Kılavuzu... 20-25
 1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
1 2 3 4 5 6 7 OFFMANAUTO CM707 GR Οδηγός χρήσης... 2-7 SLO Uporabniški priročnik... 8-13 CR Korisnički priručnik... 14-19 TR Kullanım Kılavuzu... 20-25 ENG User Guide... 26-31 GR CM707 ΟΔΗΓΟΣ ΧΡΗΣΗΣ Περιγραφή
Tehnologija rekombinantne DNA
 Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Tehnologija rekombinantne DNA Pod pojmom tehnologija rekombinantne DNA razumemo nabor tehnik za manipulacijo nukleinskih kislin, ki se uporabljajo v laboratorijih v različne namene. Rekombinantna DNA je
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar
 Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Pripravili: Ana Bernard in Eva Srečnik Dopolnil: Matic Dolinar Študijsko leto 2011/2012 KAZALO: 1. UVOD V BIOKEMIJO 2. PRENOS BIOLOŠKIH INFORMACIJ: CELIČNA KOMUNIKACIJA 3. BIOLOŠKE MOLEKULE V VODI 4. AMINOKISLINE,
Spoznajmo sedaj definicijo in nekaj osnovnih primerov zaporedij števil.
 Zaporedja števil V matematiki in fiziki pogosto operiramo s približnimi vrednostmi neke količine. Pri numeričnemu računanju lahko npr. število π aproksimiramo s števili, ki imajo samo končno mnogo neničelnih
Zaporedja števil V matematiki in fiziki pogosto operiramo s približnimi vrednostmi neke količine. Pri numeričnemu računanju lahko npr. število π aproksimiramo s števili, ki imajo samo končno mnogo neničelnih
+105 C (plošče in trakovi +85 C) -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70
 -50 C ( C)* * Za temperature pod C se posvetujte z našo tehnično službo. ϑ m *20 *40 +70") KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
KAIFLEX ST Tehnični podatki Material Izjemno fleksibilna zaprtocelična izolacija, fleksibilna elastomerna pena (FEF) Opis Uporaba Temperaturno območje Toplotna prevodnost W/(m K ) pri različnih srednjih
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Državni izpitni center SPOMLADANSKI IZPITNI ROK *M * FIZIKA NAVODILA ZA OCENJEVANJE. Petek, 10. junij 2016 SPLOŠNA MATURA
 Državni izpitni center *M16141113* SPOMLADANSKI IZPITNI ROK FIZIKA NAVODILA ZA OCENJEVANJE Petek, 1. junij 16 SPLOŠNA MATURA RIC 16 M161-411-3 M161-411-3 3 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor
Državni izpitni center *M16141113* SPOMLADANSKI IZPITNI ROK FIZIKA NAVODILA ZA OCENJEVANJE Petek, 1. junij 16 SPLOŠNA MATURA RIC 16 M161-411-3 M161-411-3 3 IZPITNA POLA 1 Naloga Odgovor Naloga Odgovor
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ
 GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
GR ΠΡΙΤΣΙΝΑΔΟΡΟΣ ΛΑΔΙΟΥ ΑΕΡΟΣ ΓΙΑ ΠΡΙΤΣΙΝΙΑ M4/M12 ΟΔΗΓΙΕΣ ΧΡΗΣΗΣ - ΑΝΤΑΛΛΑΚΤΙΚΑ H OLJLAJNYOMÁSÚ SZEGECSELŐ M4/M12 SZEGECSEKHEZ HASZNÁLATI UTASÍTÁS - ALKATRÉSZEK SLO OLJNO-PNEVMATSKI KOVIČAR ZA ZAKOVICE
- Geodetske točke in geodetske mreže
 - Geodetske točke in geodetske mreže 15 Geodetske točke in geodetske mreže Materializacija koordinatnih sistemov 2 Geodetske točke Geodetska točka je točka, označena na fizični površini Zemlje z izbrano
- Geodetske točke in geodetske mreže 15 Geodetske točke in geodetske mreže Materializacija koordinatnih sistemov 2 Geodetske točke Geodetska točka je točka, označena na fizični površini Zemlje z izbrano
MOLEKULARNA GENETIKA
 MOLEKULARNA GENETIKA 1 STRUKTURA DNA DNA ima več struktur: enojni, dvojni ali trojni heliks je levo- ali desnosučen, linearna ali krožna. Najpogostejša oblika je B desnosučni heliks, ki je iz dveh komplementarnih
MOLEKULARNA GENETIKA 1 STRUKTURA DNA DNA ima več struktur: enojni, dvojni ali trojni heliks je levo- ali desnosučen, linearna ali krožna. Najpogostejša oblika je B desnosučni heliks, ki je iz dveh komplementarnih
Encimi.
 Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
Encimi Encimi so biološki katalizatorji, ki pospešijo hitrost kemijskih reakcij v bioloških sistemih. Delujejo tako, da znižajo aktivacijsko energijo za pretvorbo reaktantov (imenujemo jih substrati) v
PROCESIRANJE SIGNALOV
 Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
Rešive pisega izpia PROCESIRANJE SIGNALOV Daum: 7... aloga Kolikša je ampliuda reje harmoske kompoee arisaega periodičega sigala? f() - -3 - - 3 Rešiev: Časova fukcija a iervalu ( /,/) je lieara fukcija:
VEKTORJI. Operacije z vektorji
 VEKTORJI Vektorji so matematični objekti, s katerimi opisujemo določene fizikalne količine. V tisku jih označujemo s krepko natisnjenimi črkami (npr. a), pri pisanju pa s puščico ( a). Fizikalne količine,
VEKTORJI Vektorji so matematični objekti, s katerimi opisujemo določene fizikalne količine. V tisku jih označujemo s krepko natisnjenimi črkami (npr. a), pri pisanju pa s puščico ( a). Fizikalne količine,
2 tg x ctg x 1 = =, cos 2x Zbog četvrtog kvadranta rješenje je: 2 ctg x
 Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Kotni funkciji sinus in kosinus
 Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
Kotni funkciji sinus in kosinus Oznake: sinus kota x označujemo z oznako sin x, kosinus kota x označujemo z oznako cos x, DEFINICIJA V PRAVOKOTNEM TRIKOTNIKU: Kotna funkcija sinus je definirana kot razmerje
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL
 PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
PROTITELESA IN IMUNSKI ODGOVOR BIOKEMIJA ČUTIL Povzetek dveh predavanj pri predmetu Biokemija študijski program Kemija 2008/2009 doc. dr. Marko Dolinar Pred vami je del gradiva, ki ga pripravljam za naslednje
ENCIMI V ORGANIZIRANIH SISTEMIH
 ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
ENCIMI V ORGANIZIRANIH SISTEMIH EC klasifikacija: po reakcijah, ki jih katalizira Encimski sistemi: so poteini z več kot eno encimsko akevnostjo RazvrsEtev po organiziranose: muleencimski protein proteini
Τα αναλώσιμα μαζί με τα τεχνικά χαρακτηριστικά τους περιγράφονται αναλυτικά ανά ομάδα:
 Στα πλαίσια του έργου ΠΡΟΪΟΝΤΑ ΜΕΙΩΜΕΝΟΥ ΚΙΝΔΥΝΟΥ ΓΙΑ ΤΗ ΝΙΚΟΤΙΝΗ: ΣΥΓΚΡΙΤΙΚΕΣ ΜΕΛΕΤΕΣ ΕΠΙΔΡΑΣΗΣ ΣΤΟΝ ΑΝΑΠΝΕΥΣΤΙΚΟ & ΛΙΠΩΔΗ ΙΣΤΟ, ΟΠΣ 5006015, Φ.Κ. 80534 με επιστημονικό υπεύθυνο τον Αναπληρωτή καθηγητή
Στα πλαίσια του έργου ΠΡΟΪΟΝΤΑ ΜΕΙΩΜΕΝΟΥ ΚΙΝΔΥΝΟΥ ΓΙΑ ΤΗ ΝΙΚΟΤΙΝΗ: ΣΥΓΚΡΙΤΙΚΕΣ ΜΕΛΕΤΕΣ ΕΠΙΔΡΑΣΗΣ ΣΤΟΝ ΑΝΑΠΝΕΥΣΤΙΚΟ & ΛΙΠΩΔΗ ΙΣΤΟ, ΟΠΣ 5006015, Φ.Κ. 80534 με επιστημονικό υπεύθυνο τον Αναπληρωτή καθηγητή
8. Navadne diferencialne enačbe
 8. Navadne diferencialne enačbe 8.1. Začetni problem prvega reda Iščemo funkcijo y(x), ki zadošča diferencialni enačbi y = f(x, y) in začetnemu pogoju y(x 0 ) = y 0, kjer je f dana dovolj gladka funkcija
8. Navadne diferencialne enačbe 8.1. Začetni problem prvega reda Iščemo funkcijo y(x), ki zadošča diferencialni enačbi y = f(x, y) in začetnemu pogoju y(x 0 ) = y 0, kjer je f dana dovolj gladka funkcija
Vaja: Odbojnostni senzor z optičnimi vlakni. Namen vaje
 Namen vaje Spoznavanje osnovnih fiber-optičnih in optomehanskih komponent Spoznavanje načela delovanja in praktične uporabe odbojnostnega senzorja z optičnimi vlakni, Delo z merilnimi instrumenti (signal-generator,
Namen vaje Spoznavanje osnovnih fiber-optičnih in optomehanskih komponent Spoznavanje načela delovanja in praktične uporabe odbojnostnega senzorja z optičnimi vlakni, Delo z merilnimi instrumenti (signal-generator,
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
Določanje strukture. mikrobne združbe
 Določanje strukture mikrobne združbe 1 Določanje strukture mikrobne združbe mikroskopija gojenje (fiziološka različnost) FAME, PLFA imunološki testi tipizacije (fagotipizacija, ribotipizacija) analiza
Določanje strukture mikrobne združbe 1 Določanje strukture mikrobne združbe mikroskopija gojenje (fiziološka različnost) FAME, PLFA imunološki testi tipizacije (fagotipizacija, ribotipizacija) analiza
Statistična analiza. doc. dr. Mitja Kos, mag. farm. Katedra za socialno farmacijo Univerza v Ljubljani- Fakulteta za farmacijo
 Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Statistična analiza opisnih spremenljivk doc. dr. Mitja Kos, mag. arm. Katedra za socialno armacijo Univerza v Ljubljani- Fakulteta za armacijo Statistični znaki Proučevane spremenljivke: statistični znaki
Cefalosporini ostali β-laktami
 Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
Cefalosporini ostali β-laktami doc.dr. Marko Anderluh 12. januar 2012 Vir cefalosporinov Cephalosporium acremonium Cefalosporin C Enaka tarča kot pri penicilinih Podoben mehanizem delovanja Cefalosporini
POROČILO 3.VAJA DOLOČANJE REZULTANTE SIL
 POROČILO 3.VAJA DOLOČANJE REZULTANTE SIL Izdba aje: Ljubjana, 11. 1. 007, 10.00 Jan OMAHNE, 1.M Namen: 1.Preeri paraeogramsko praio za doočanje rezutante nezporedni si s skupnim prijemaiščem (grafično)..dooči
POROČILO 3.VAJA DOLOČANJE REZULTANTE SIL Izdba aje: Ljubjana, 11. 1. 007, 10.00 Jan OMAHNE, 1.M Namen: 1.Preeri paraeogramsko praio za doočanje rezutante nezporedni si s skupnim prijemaiščem (grafično)..dooči
Multivariatna analiza variance
 (MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti med več odvisnimi (številskimi) in več neodvisnimi (opisnimi) spremenljivkami. (MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti
(MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti med več odvisnimi (številskimi) in več neodvisnimi (opisnimi) spremenljivkami. (MANOVA) MANOVA je multivariatna metoda za proučevanje odvisnosti