Biológia pre informatikov. Broňa Brejová Walther Flemming, 1881
|
|
|
- Φιλομήνα Δουρέντης
- 7 χρόνια πριν
- Προβολές:
Transcript

1 Biológia pre informatikov Broňa Brejová Walther Flemming,
2 Ďalšie informácie Tutoriál NCBI books Vysokoškolské učebnice molekulárnej biológie Anglická wikipédia,... Zvelebil, Baum: Understanding Bioinformatics, kap. 1 2
3 Hlavné postavy Deoxyribonukleová kyselina (DNA) Obsahuje genetickú informáciu prenášanú z generácie na generáciu. Dlhý reťazec nukleotidov z množiny {A,C,G,T} (adenín, cytozín, guanín, tymín). Informácia uložená v symbolickej, digitálnej forme. Ribonukleová kyselina (RNA) Blízka príbuzná DNA, tymín T nahradený uracylom U Proteíny (bielkoviny) Katalyzujú biochemické reakcie v bunke (enzýmy), prenášajú signály v rámci bunky/medzi bunkami, sú dôležité pre stavbu bunky a pohyb. Reťazec aminokyselín (20 rôznych aminokyselín). 3
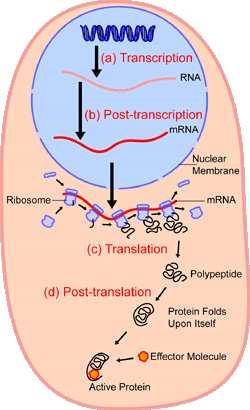
4 Aká informácia je uložená v DNA? Gény: Predpisy na tvorbu proteínov a funkčných RNA molekúl. Riadenie ich expresie: kedy a koľko sa má tvoriť. DNA: RNA: gén transkripcia do RNA replikácia RNA: RNA: spracovanie RNA proteín: translácia do proteínu regulácia 4
5 Centrálna dogma (Francis Crick 1958,1970) DNA: RNA: gén transkripcia do RNA replikácia RNA: RNA: spracovanie RNA proteín: translácia do proteínu regulácia The central dogma of molecular biology deals with the detailed residue-by-residue transfer of sequential information. It states that such information cannot be transferred back from protein to either protein or nucleic acid. 5

6 DNA, chromozómy DNA: dve komplementárne vlákna, strands (páry A-T, C-G), v opačnej orientácii (konce sa nazývajú 5 a 3 ). Napr. ACCATG je komplementárny s CATGGT. Tvar dvojitej špirály: Dvojvláknová štruktúra poskytuje redundanciu, možnosť opravy pri poškodení jedného vlákna. Pri delení bunky sa dvojvláknová DNA rozdelí a ku každému vláknu sa doplní komplement (DNA replikácia). Chromozóm: Súvislý úsek dvojvláknovej DNA a podporných proteínov. Ľudský genóm má 22 párov chromozómov plus dva pohlavné, spolu 3GB. 6

7 Technológia: sekvenovanie DNA Postup na zisťovanie poradia báz v chromozómoch genómu. Zložitý proces: chromozómy sa nasekajú na krátke kúsky, z každého sa vyrobí veľa kópií, každý sa nasekvenuje zvlášť napr. Sangerovým sekvenovaním. využíva prírodné enzýmy, napr. DNA polymerázu Bioinformatický problém: skladanie celej sekvencie z kúskov. Dostupnosť genómov umožňuje katalogizovať gény a iné funkčné úseky, hľadať podobnosti a rozdiely medzi druhmi a jedincami. 7
8 Sangerovo sekvenovanie (Sanger sequencing) Sekvenujeme AGCTAGGACT (zobrazená sprava doľava) Primer AGT + enzýmy + nukleotidy + modifikované ofarbené nukleotidy Výsledky sekvenovacej reakcie: TCAGGATCGA AGTCCTAGC TCAGGATCGA AGTCC TCAGGATCGA AGTCCTA TCAGGATCGA AGTCCTAGCT TCAGGATCGA AGTC TCAGGATCGA AGTCCT TCAGGATCGA AGTCCTAG Na géli zoradíme podľa dĺžky: AGTCCTAGCT AGTCCTAGC AGTCCTAG AGTCCTA AGTCCT AGTCC AGTC Odčítaním farieb dostaneme komplementárne vlákno: AGTCCTAGCT 8
9 9
10 PCR (polymerase chain reaction) Zvolíme si dva krátke úseky DNA (primers) PCR testuje či sú v DNA blízko seba (stovky, tisíce báz) Ak áno, namnoží úsek medzi nimi DNA primers 8 kopii 2 kopie 4 kopie 10
11 DNA: RNA: gén transkripcia do RNA replikácia RNA: RNA: spracovanie RNA proteín: translácia do proteínu regulácia 11


12 RNA Ako sa líši od DNA? obsahuje ribózu namiesto deoxyribózy obsahuje uracil namiesto tymínu (bázy A,C,G,U) jednovláknové reťazce, zvyčajne kratšie zložitá sekundárna štruktúra: spárované komplementárne úseky transferová RNA (trna) 12
13 Translácia Kodón (trojica nukleotidov) určuje 1 aminokyselinu mrna: proteín: U G G U U U G G C U C A }{{}}{{}}{{}}{{} W F G S 13
14 Genetický kód Alanine (A) GC* Cysteine (C) TGC TGT Aspartic acid (D) GAC GAT Glutamic acid (E) GAA GAG Phenylalanine (F) TTC TTT Glycine (G) GG* Histidine (H) CAC CAT Isoleucine (I) ATA ATC ATT Lysine (K) AAA AAG Leucine (L) CT* TTA TTG Methionine (M) ATG Asparagine (N) AAC AAT Proline (P) CC* Glutamine (Q) CAA CAG Arginine (R) CG* AGA AGG Serine (S) TC* AGT AGC Threonine (T) AC* Valine (V) GT* Tryptophan (W) TGG Tyrosine (Y) TAC TAT Stop codon (*) TAA TAG TGA 14
15 Proteíny Reťazce 20 rôznych aminokyselín s rôznymi chemickými vlastnosťami: Amino Acid Side chain Hydrophobic Polar Charged Alanine (A) -CH3 X Arginine (R) -(CH2)3NH-C(NH)NH2 - X basic - Asparagine (N) -CH2CONH2 - X - - Aspartic acid (D) -CH2COOH - X acidic - Cysteine (C) -CH2SH X - acidic - Glutamic acid (E) -CH2CH2COOH - X acidic - Glutamine (Q) -CH2CH2CONH2 - X - - Glycine (G) -H Histidine (H) -CH2-C3H3N2 - X weak basic Aromatic Isoleucine (I) -CH(CH3)CH2CH3 X - - Aliphatic Leucine (L) -CH2CH(CH3)2 X - - Aliphatic Lysine (K) -(CH2)4NH2 - X basic - Methionine (M) -CH2CH2SCH3 X Phenylalanine (F) -CH2C6H5 X - - Aromatic Proline (P) -CH2CH2CH2- X Serine (S) -CH2OH - X - - Threonine (T) -CH(OH)CH3 - X weak acidic - Tryptophan (W) -CH2C8H6N X - - Aromatic Tyrosine (Y) -CH2-C6H4OH X X - Aromatic Valine (V) -CH(CH3)2 X - - Aliphatic 15
16 Štruktúra proteínov Myoglobín, prvý proteín so známou štruktúrou. Proteíny sa vyskytujú poskladané v určitej stabilnej štruktúre, prípadne prechádzajú medzi niekoľkými stavmi. Hydrofóbne aminokyseliny neinteragujú s vodou, zväčša sa vyskytujú vo vnútri štrukúry. Štruktúra proteínu určuje jeho funkciu. 16
17 17
18 Regulácia expresie Bunky v rôznych tkanivách toho istého organizmu zdieľajú ten istý genóm, vyzerajú a fungujú však veľmi rôzne. Niektoré proteíny sa tvoria len za určitých okolností, alebo v premenlivom množstve. Regulácia začatia transkripcie pomocou transkripčných faktorov: DNA Transkripcne faktory RNA polymeraza Transkripcia Gen Bioinformatický problém: zisti, ktoré faktory ovplyvňujú ktorý gén, kde presne sa viažu. 18
.")
19 Technológia: microarray zname DNA vzorka 1 vzorka 2 + gen 1 gen 2 gen 3 komplementarna DNA hybridizuje oscanuj intenzitu Meranie množstva mrna prítomnej v bunke pre veľa génov naraz. Zopakujeme za rôznych podmienok, študujeme korelácie medzi génmi. Môžu byť dôsledkom spoločného regulátora (transkripčného faktoru). Bioinformatický problém: niekoľko koregulovaných génov, nájdi motív, ku ktorému sa môže viazať spoločný transkripčný faktor (motif finding) 19

20 Príklad microarray dát 20
21 Mutácie DNA V DNA občas dochádza k zmenám, mutáciám (napr. pod vplyvom prostredia, či chybou pri replikácii). Typy mutácií: substitúcia, substitution (jedna báza sa zmení na inú), inzercia, insertion (vloží sa niekoľko nových báz), delécia, deletion (vynechá sa niekoľko báz), zmeny väčšieho rozsahu (napr. translokácie). Bioinformatické problémy: Ktoré sekvencie vznikli z spoločného predka mutovaním? (hľadanie homológov, homology search) Ktoré bázy v dvoch príbuzných sekvenciách si navzájom zodpovedajú? (sequence alignment, zarovnávanie sekvencií) 21
22 Populačná genetika Mutácie sa šíria v populácii z rodičov na potomkov. Nebezpečné mutácie rýchlejšie vymiznú, prospešné sa rýchlejšie ujmú (prírodný výber, natural selection). Polymorfizmus: genetický rozdiel medzi organizmami v rámci druhu. Vedie k rozdielnosti vo fenotype, napr. výzor, dedičné choroby. Sekvenovaním viacerých jedincov toho istého druhu získame prehľad o polymorfizme. Bioinformatický problém: Izoluj polymorfizmus zodpovedný za určitý znak (napr. chorobu). 22

23 Evolúcia Vznik nových druhov (speciation): Po rozdelení populácie na viacero oddelených častí nedochádza k výmene genetického materiálu. Hromadia sa zmeny až kým nie je možné párenie: vznik nových druhov. Bioinformatický problém: Na základe dnešných sekvencií urči strom reprezentujúci vývoj druhov (fylogenetický strom, phylogenetic tree) 23
24 Prokaryotické vs. eukaryotické organizmy Prokaryoty: baktérie, jednoduché jednobunkové organizmy. Nemajú jadro (DNA priamo v cytoplazme), majú kruhový chromozóm (a prípadné kratšie plasmidy), jednoduchšia štruktúra génu atď. Eukaryoty: živočíchy, rastliny, huby, niektoré jednobunkové organizmy. Bunka obsahuje jadro s DNA, viacero organel. Mitochondrie a chloroplasty sú pohltené prokaryoty, ktoré sa stali časťou eukaryotickej bunky. Dlhší genóm v niekoľkých lineárnych chromozómoch. 24


25 Modelové organizmy Dôležité pre biologický výskum, vieme o nich viac než o príbuzných druhoch. Poznatky širšie aplikovateľné. Escherichia coli: baktéria žijúca v črevách. Jednoduchá manipulácia, delenie každých 20 min. Štúdium základných životných procesov: DNA replikácia, expresia génov, atď. Genóm s 4000 génmi, 4.6MB. Saccharomyces cerevisiae: pekárske droždie. Jednoduchý eukaryotický organizmus. Genóm s 6000 génmi, 13MB. Delenie každé 2 hodiny. Štúdium špecificky eukaryotických javov. 25
, diferenciácie buniek. Drosophila melanogaster: vínna muška. Štúdium genetiky, gény riadiace vývin jedinca.")
26 Modelové organizmy Arabidopsis thaliana: malá kvitnúca rastlina, 6-týždňový životný cyklus. Skúmanie javov špecifických pre rastliny. Caenorhabditis elegans: malý červ, nematód, žijúci v pôde. Štúdium vývinu (ontogenéza, development), diferenciácie buniek. Drosophila melanogaster: vínna muška. Štúdium genetiky, gény riadiace vývin jedinca. Stavovce: žaba Xenopus laevis (veľké, ľahko manipuľovateľné vajíčka), akvarijná ryba Danio rerio (priehľadné embryá), myš Mus musculus (existuje veľa plemien so špeciálnymi vlastnosťami). 26
27 Dostupné dáta DNA sekvencie: celé genómy, ich časti Ich anotácia: súradnice génov a iných funkčných častí Sekvencie RNA, ich štruktúra Sekvencie proteínov, ich funkcia a štruktúra Merania množstva RNA/proteínu v bunke... Dáta založené na experimetoch alebo výsledky výpočtových metód Veľa chýb (v oboch prípadoch) 27
28 Úvod do dynamického programovania, proteomika Tomáš Vinař
29 Proteomika Proteín: sekvencia pozostáva z 20 rôznych aminokyselín MGLSDGEWQLVLNVWGKVEADIPGHGQEVLIRLFKGHPETLEKFDKFKHLKSEDEMKASE DLKKHGATVLTALGGILKKKGHHEAEIKPLAQSHATKHKIPVKYLEFISECIIQVLQSKH PGDFGADAQGAMNKALELFRKDMASNYKELGFQG Z bunky sme izolovali určitý proteín, chceme zistiť jeho sekvenciu. 2

30 Proteomika: Gélová elektroforéza (gel electrophoresis) Alternatíva: kvapalinová chromatografia (liquid chromatography) 3
31 Hmotnostná spektrometria (mass spectometry) Meria pomer hmostnosť/náboj molekúl vo vzorke Používa sa na identifikáciu proteínov, napr. z 2D gélu. Proteín nasekáme enzýmom trypsín (seká na [KR]{P}) na peptidy Meriame hmostnosť kúskov, porovnáme s databázou proteínov. Tandemová hmotnostná spektrometria (MS/MS) ďalej fragmentuje každý kúsok a dosiahne podrobnejšie spektrum, ktoré obsahuje viac informácie V niektorých prípadoch tak vieme sekvenciu proteínu určiť priamo z MS/MS, bez databázy proteínov 4
32 Tandemová hmotnostná spektrometria MS/MS Štiepenie peptidu na prefixy a sufixy zdroj: Bafna and Reinert b-ióny: prefixy y-ióny: sufixy 5
33 Tandemová hmotnostná spektrometria MS/MS zdroj: Bafna and Reinert 6
34 Sekvenovanie peptidov pomocou MS/MS Pre začiatok berieme do úvahy iba b-ióny (prefixy) Všetky hmostnosti celé čísla Vstup: celková hmotnosť peptidu M, hmotnosti aminokyselín a[1],...,a[20], spektrum ako tabuľka f[0],..., f[m], ktorá hmotnosti b-iónu určí skóre podľa signálu v okolí príslušného bodu grafu Pre postupnosť aminokyselín p 1...p k nech m(p 1...p k ) je jej hmotnosť, t.j. m(p 1...p k ) = k j=1 a[p j] Výstup: postupnosť aminokyselín p 1...p k, taká, že m(p 1...p k ) = M a k i=1 f[m(p 1...p i )] je maximálna možná 7
35 Sekvenovanie peptidov pomocou MS/MS Pre peptid p = p 1,...,p k uvažujme množinu hmotností M(p), ktorá obsahuje hmostnosti všetkých jeho prefixov a sufixov M(p) = {m(p 1...p i ) i = 1...k} {m(p i...p k ) i = 1...k} Cieľ: maximalizujeme m M(p) f[m] alebo maximalizujeme m M(p),m M/2 f [m] kde f [m] = f[m]+f[m m] (pre m = M/2 vezmeme f [m] = f[m]) 8
36 Úvod do pravdepodobnosti, sekvenovanie genómov (cvičenie) Tomáš Vinař
37 G = dĺžka genómu, napr N = počet segmentov (readov), napr L = dĺžka segmentu, napr T = potrebná dĺžka prekryvu, napr. 50 2
38 Náhodne generované segmenty Usek genomu 3
39 Pokrytie jednotlivých báz
40 Počet báz s určitým pokrytím Binomicke rozdelenie Generovane data Pokrytie 5
41 Predpokladaný počet kontigov od počtu segmentov Pocet segmentov 6
42 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 0 koncov: nepokr: 274 koncov: 2 nepokr: 282 koncov: 1 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 8 koncov: 1 nepokr: 0 koncov: 0 nepokr: 12 koncov: 1 nepokr: 0 koncov: 0 nepokr: 122 koncov: 1 nepokr: 135 koncov: 1 nepokr: 111 koncov: 1 nepokr: 13 koncov: 1 nepokr: 1 koncov: 1 nepokr: 56 koncov: 1 nepokr: 265 koncov: 1 nepokr: 0 koncov: 0 nepokr: 10 koncov: 1 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 130 koncov: 1 nepokr: 217 koncov: 1 nepokr: 3 koncov: 1 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 86 koncov: 1 nepokr: 139 koncov: 2 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 76 koncov: 1 nepokr: 221 koncov: 1 nepokr: 26 koncov: 1 nepokr: 0 koncov: 0 nepokr: 1 koncov: 1 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 0 koncov: 0 nepokr: 12 koncov: 1 nepokr: 103 koncov: 2 nepokr: 0 koncov: 0 nepokr: 71 koncov: 1 nepokr: 69 koncov: 1 nepokr: 0 koncov: 0 7
43 Jadrá zarovnaní Tomáš Vinař
44 Opakovanie: Heuristické lokálne zarovnávanie, BLAST Príklad: w = 2 (začíname z jadier dĺžky 2). (V praxi sa používa w = 10 a viac.) C A G T C C T A G A C A T G T C A T A nájdi zhodné úseky 2. rozšír bez medzier 3. spoj medzerami 2
45 Senzitivita heuristického algoritmu Odhad senzitivity: Predpokladáme zarovnanie bez medzier, dĺžky L Každá pozícia je zhoda s pravdepodobnosťou p Senzitivita: f(l, p) = Pr(zarovnanie obsahuje w zhôd za sebou) 1.0 senzitivita pre w= p=0.6 p=0.7 p=0.8 p= dlzka zarovnania L 3
46 Jadrá z medzerami, spaced seeds PatternHunter [Ma, Tromp, Li 2002] Jadro s medzerami: vyžadovaná konfigurácia zhôd Príklad: match match don t care match značíme ako 1101 GTGGTGCTCTCTGACAAAGCC ATTGTTCTTAATGAGAAAGAA BLASTN jadro (11 za sebou idúcich zhôd) ekvivalentné jadru
47 Nie všetky jadrá sú rovnaké Váha jadra: počet vyžadovaných zhôd Každý krížik: senzitivita vs. čas pre jedno jadro v pravdepodobnostnom modeli Predicted false positive rate How many spurious hits? 4E-06 2E-06 0 Weight 11 Weight 9 Weight Predicted sensitivity What portion of alignments found? 5
48 Prečo sú jadrá s medzerami senzitívnejšie? Príklad: dĺžka zarovnania L = 64, pravdepodobnosť zhody p = 0.7 na každej pozícii 40 náhodných zarovnaní, výskyty jadra Sn.: 14/40, hits: Sn.: 18/40, hits: 35 6
49 Prečo sú jadrá s medzerami senzitívnejšie? Príklad: dĺžka zarovnania L = 64, pravdepodobnosť zhody p = 0.7 na každej pozícii Bez medzier S medzerami Stredná hodnota počtu výskytov v zarovnaní: = = 0.9 Pravdepodobnosť výskytu na poz. i+1 ak výskyt na i: = Výskyty často vedľa seba Výskyty nezávislejšie Senzitivita (pravdepodobnosť aspoň jedného výskytu):
50 Ďalšie hašovacie stratégie Nukleotidový BLAST: 10 zhôd za sebou Jadro s medzerami: povoľuje nezhody na 8 z 18 pozícií BLAT [Kent 2002]: povoľuje 1 nezhodu na ľub. z 11 pozícií BLASTP: 3 amino kyseliny so skóre aspoň 13 v matici BLOSUM62 Výskyt: N I R N L R Nie výskyt: A I L A I L 6+2+5= =12 Vektorové jadrá: kombinácia jadier s medzerami a BLAT/BLASTP Viaceré výskyty: začni rozširovať iba ak viac výskytov blízko seba na tej istej uhlopriečke Viaceré jadrá: zober zjednotenie výskytov 8
51 Záleží na modeli zarovnania Pravdepodobnosť zhody kolíše v rámci kodónu: Poloha v kodóne: prvá druhá tretia Pravdepodobnosť zhody: Senzitivita na testovacej vzorke exónov kódujúcich proteíny: Človek vs. Jadro Drosophila myš Optimálne pre dáta % 92% Optimalne pre kodónový model % 91% WABA [Kent, Zahler 2000] % 90% Optimálne pre i.i.d. model % 86% BLAST % 81% Najhoršie % 79% 9
52 A čo globálne zarovnania? Ukotvené zarovnanie (Anchored alignment) Nájdime lokálne zarovnania (alebo výskyty nejakého jadra) možné ukotvenia Zvoľ konzistentnú množinu ukotvení (monotónna postupnosť) Zarovnaj časti sekvencií medzi ukotveniami (pomocou dyn. prog. alebo rekurzívne ďalšim kotvením) MUMMER [Delcher 1999] GLASS [Batzoglou et al 2000] AVID [Bray et al 2003] 10
53 Ukotvené zarovnanie Modré: nezvolené ukotvenia Červené: zvolené ukotvenia Sivé: riešime dyn. prog. Čierne: globálne zarovnanie Znova protichodné vplyvy: málo spoľahlivých ukotvení dobrá kvalita, pomalé veľa slabších ukotvení rýchle (malá sivá plocha), viac chýb v ukotvení 11
54 Algoritmy pre HMM a phylohmm Tomáš Vinař
55 Opakovanie: HMM (skrytý Markovov model) model (HMM) náhodná DNA sekvencia S, náhodná anotácia A (podobné na ozajstnú DNA) Pr(S,A) pravdepodobnosť, že model vygeneruje pár (S,A) a:0.27 c:0.23 g:0.23 t: a:0.24 c:0.27 g:0.28 t:0.21 a:0.26 c:0.22 g:0.22 t:0.30 Predpokladajme, že model vždy začína v modrom stave. Pr(aca) = = Pr(aca) = =
56 Parametre HMM (označenie) Sekvencia S 1,...,S n Anotácia A 1,...,A n Parametre modelu: Prechodová pravdepodobnosť a(u,v) = Pr(A i+1 = v A i = u), Emisná pravdepodobnosť e(u,x) = Pr(S i = x A i = u), Počiatočná pravdepodobnosť π(u) = Pr(A 1 = u). a e a c g t Výsledná pravdepodobnosť: Pr(A 1,...,A n,s 1,...,S n ) = π(a 1 )e(a 1,S 1 ) n i=2 a(a i 1,A i )e(a i,s i ) 3
57 Viterbiho algoritmus Pre danú sekvenciu S nájde najpravdepodobnejšiu anotáciu A = argmax A Pr(A S) Dynamické programovanie v čase O(nm 2 ) Podproblém V[i,u]: pravdepodobnosť najpravdepodobnejšej cesty končiacej po i krokoch v stave u, pričom vygeneruje S 1 S 2...S i Rekurencia: V[1,u] = π u e u,s1 V[i,u] = max w V[i 1,w] a w,u e u,si Algoritmus: Inicializuj V[1, ] for i = 2...n (n=dĺžka reťazca) for u = 1...m (m =počet stavov) vypočítaj V[i, u] Maximálne V[n,j] je pravdepodobnosť najpravdepodobnejšej cesty 4

58 PhyloHMM: kombinácia HMM a fylogenetického stromu PhastCons: detekcia dobre zachovaných sekvencií Dva stavy: zachovaná sekv., neutrálna sekv. V každom stave generujeme celý stĺpec zarovnania Zachovaná sekvencia má kratšie hrany stromu 5
59 Dopredný algoritmus Počíta celkovú pravdepodobnosť, že vygenerujeme sekvenciu S Pr(S) = A Pr(A,S) Podproblém F[i,u]: pravdepodobnosť, že po i krokoch vygenerujeme S 1,S 2,...S i a dostaneme sa do stavu u. F[i,u] = Pr(A i = u S 1,S 2,...,S i ) = A 1,A 2,...,A i =u Pr(A 1,A 2,...,A i S 1,S 2,...,S i ) Rekurencia: F[1,u] = π u e u,s1 F[i,u] = v F[i 1,v] a v,u e u,si Celková pravdepodobnosť Pr(S) = u F[n,u] 6
60 Spätný algoritmus Obdoba dopredného algoritmu Dopredný algoritmus: F[i,u] = Pr(A i = u S 1,...,S i ) F[1,u] = π u e u,s1 F[i,u] = v F[i 1,v] a v,u e u,si Pr(S) = u F[n,u] Spätný algoritmus: B[i,u] = Pr(S i+1...,s n A i = u) 7
61 Hľadanie génov s HMM model náhodná DNA sekvencia S, náhodná anotácia A Určenie stavov a prechodov v modeli: ručne, na základe poznatkov o štruktúre génu. Trénovanie parametrov: pravdepodobnosti určíme na základe sekvencií so známymi génmi (trénovacia množina). Model zostavíme tak, aby páry (S,A) s vlastnosťami podobnými skutočným génom mali veľkú pravdepodobnosť Pr(S,A) Použitie: pre novú sekvenciu S nájdi najpravdepodobnejšiu anotáciu A = argmax A Pr(A S) Viterbiho algoritmom 8
62 Trénovanie HMM Stavový priestor + povolené prechody väčšinou ručne Parametre (pravdepodobnosti prechodu, emisie a počiatočné) automaticky z trénovacích sekvencií Čím zložitejší model a viac parametrov máme, tým potrebujeme viac trénovacích dát, aby nedošlo k preučeniu, t.j. k situácii, keď model dobre zodpovedá nejakým zvláštnostiam trénovacích dát, nie však ďalším dátam. Presnosť modelu testujeme na zvláštnych testovacích dátach, ktoré sme nepoužili na trénovanie. 9
63 Trénovanie HMM z anotovaných sekvencií Vstup: topológia modelu a niekoľko trénovacích párov S (i),a (i) Cieľ: nastaviť π u, e u,x, a u,v tak, aby i Pr(S(i),A (i) ) bola čo najväčšia Dosiahneme jednoduchým počítaním frekvencií Napr. a u,v : nájdeme všetky výskyty stavu u a zistíme, ako často za nimi ide stav v 10
64 Trénovanie HMM z neanotovaných sekvencií Vstup: topológia modelu a niekoľko trénovacích sekvencií S (i) anotácie A (i) nepoznáme Cieľ: nastaviť π u, e u,x, a u,v tak, aby i Pr(S(i) ) bola čo najväčšia Používajú sa heuristické iteratívne algoritmy, napr. Baum-Welchov, ktorý je verziou všeobecnejšieho algoritmu EM (expectation maximization). 11
65 donor0 intron0 acceptor0 donor4 intron4 acceptor4 donor5 intron5 acceptor5 donor3 intron3 acceptor3 Tvorba stavového priestoru modelu Príklad HMM na hľadanie génov translation start translation start exon intergenic exon translation stop translation stop Reverse strand Forward strand 12 donor2 intron2 acceptor2 donor1 intron1 acceptor1
66 Cvičenia pre informatikov, Zhrnutie semestra 1
67 Pravdepodobnostné modely Skryté Markovove modely (hľadanie génov, konzervovaných oblastí, fylogenetické HMM, profilové HMM) Fylogenetické stromy a substitučné modely Stochastické bezkontextové gramatiky Metóda maximálnej vierohodnosti Expectation maximization (EM) Štatistické metódy Pojem štatistickej významnosti E-value a P-value Test na pozitívny výber Linkage disequilibrium, mapovanie asociácií 2
68 Precvičenie dynamického programovania Iné Zarovnávanie sekvencií (globálne, lokálne, afínne medzery) Skryté Markovove modely (Viterbiho algoritmus) Výpočty na stromoch (úspornosť, vierohodnosť - Felsensteinov algoritmus) Hmotnostná spektrometria (MS/MS) Sekundárna štruktúra RNA Integer linear programming debruijnove grafy Zhlukovanie a klasifikácia 3
69 Ako modelovať problémy reálneho sveta Rozmyslieť si, aké máme dáta, čo by sme chceli ako výsledok Sformulovať ako informatický problém (napr. optimalizácia nejakého skóre) Pravdepodobnostné modely nám často dovolia zvoliť skórovaciu schému systematickým spôsobom Výsledný problém často NP ťažký Heuristiky, aproximačne algoritmy ILP a iné techniky na presné riešenie Nedá sa problém trochu preformulovať? Testovanie: sú výpočtové výsledky relevantné v danej doméne? (bola formulácia dostatočne realistická?) 4
70 Ďalšie predmety Strojové učenie 2-INF-150, Vinař/Petrovič (ZS, 4 hodiny, 6 kreditov) Grafové modely v strojovom učení 2-INF-238, Vinař (LS, 4 hodiny, 6 kreditov) Vybrané partie z dátových štruktúr 2-INF-237, Brejová (LS, 4 hodiny, 6 kreditov) predtým Vyhľadávanie v texte Biológia N-bCXX-055, Tomáška (ZS, 2 hodiny, 2 kredity) Všeobecná biológia N-bCXX-085, Tomáška (LS, 2 hodiny, 2 kredity) Genomika N-mCBI-303, Nosek (LS, 2 hodiny, 3 kredity) 5
Η νέα προσέγγιση στην ταχεία προγεννητική διάγνωση των χρωµοσωµατικών ανωµαλιών του εµβρύου
 Αµνιο-PCR Η νέα προσέγγιση στην ταχεία προγεννητική διάγνωση των χρωµοσωµατικών ανωµαλιών του εµβρύου Αγγελική Χατζάκη, PhD Γεωργία Χριστοπούλου, MSc Τµήµα Γενετικής και Μοριακής Βιολογίας Μαιευτήριο «ΜΗΤΕΡΑ»
Αµνιο-PCR Η νέα προσέγγιση στην ταχεία προγεννητική διάγνωση των χρωµοσωµατικών ανωµαλιών του εµβρύου Αγγελική Χατζάκη, PhD Γεωργία Χριστοπούλου, MSc Τµήµα Γενετικής και Μοριακής Βιολογίας Μαιευτήριο «ΜΗΤΕΡΑ»
Supplemental file 3. All 306 mapped IDs collected by IPA program. Supplemental file 6. The functions and main focused genes in each network.
 LIST OF SUPPLEMENTAL FILES Supplemental file 1. Primer sets used for qrt-pcr. Supplemental file 2. All 1305 differentially expressed genes. Supplemental file 3. All 306 mapped IDs collected by IPA program.
LIST OF SUPPLEMENTAL FILES Supplemental file 1. Primer sets used for qrt-pcr. Supplemental file 2. All 1305 differentially expressed genes. Supplemental file 3. All 306 mapped IDs collected by IPA program.
Supplementary Table 1. Primers used for RT-qPCR analysis of striatal and nigral tissue.
 Supplementary Table 1. Primers used for RT-qPCR analysis of striatal and nigral tissue. Gene Forward Primer (5-3 ) Reverse Primer (5-3 ) Dopaminergic Markers TH CTG GCC ATT GAT GTA CTG GA ACA CAC ATG GGA
Supplementary Table 1. Primers used for RT-qPCR analysis of striatal and nigral tissue. Gene Forward Primer (5-3 ) Reverse Primer (5-3 ) Dopaminergic Markers TH CTG GCC ATT GAT GTA CTG GA ACA CAC ATG GGA
J. Dairy Sci. 93: doi: /jds American Dairy Science Association, 2010.
 Supplementary Table 1. Primers and PCR conditions used for the amplification of the goat SCD1 cdna (PCR1 to PCR6) and three SCD1 polymorphic regions (PCR7 to PCR9) PCR Primers Sequence Position 1 Thermal
Supplementary Table 1. Primers and PCR conditions used for the amplification of the goat SCD1 cdna (PCR1 to PCR6) and three SCD1 polymorphic regions (PCR7 to PCR9) PCR Primers Sequence Position 1 Thermal
Απόφαση. Ο κ. I. Κούσκος, Οικονομικός Υπεύθυνος του Ε.Ι.Π., έχοντας υπ όψιν : Αποφασίζει. Ο Οικονομικός Υπεύθυνος του Ε.Ι.Π.
 ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΠΑΙΔΕΙΑΣ ΚΑΙ ΘΡΗΣΚΕΥΜΑΤΩΝ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΕΡΕΥΝΑΣ & ΤΕΧΝΟΛΟΓΙΑΣ Αθήνα, 27 Οκτωβρίου 2014 Αρ. Πρωτ. Ε.Ι.Π.: 3127 Αρ. Πρωτ. Διαύγειας: 1403 Θέμα: Έγκριση πρόσκληση εκδήλωσης
ΕΛΛΗΝΙΚΗ ΔΗΜΟΚΡΑΤΙΑ ΥΠΟΥΡΓΕΙΟ ΠΑΙΔΕΙΑΣ ΚΑΙ ΘΡΗΣΚΕΥΜΑΤΩΝ ΓΕΝΙΚΗ ΓΡΑΜΜΑΤΕΙΑ ΕΡΕΥΝΑΣ & ΤΕΧΝΟΛΟΓΙΑΣ Αθήνα, 27 Οκτωβρίου 2014 Αρ. Πρωτ. Ε.Ι.Π.: 3127 Αρ. Πρωτ. Διαύγειας: 1403 Θέμα: Έγκριση πρόσκληση εκδήλωσης
Matematika Funkcia viac premenných, Parciálne derivácie
 Matematika 2-01 Funkcia viac premenných, Parciálne derivácie Euklidovská metrika na množine R n všetkých usporiadaných n-íc reálnych čísel je reálna funkcia ρ: R n R n R definovaná nasledovne: Ak X = x
Matematika 2-01 Funkcia viac premenných, Parciálne derivácie Euklidovská metrika na množine R n všetkých usporiadaných n-íc reálnych čísel je reálna funkcia ρ: R n R n R definovaná nasledovne: Ak X = x
ΗΛΙΑΣΚΟΣ ΦΡΟΝΤΙΣΤΗΡΙΑ. Θετικής Κατεύθυνσης Βιολογία Γ Λυκείου ΥΠΗΡΕΣΙΕΣ ΠΑΙΔΕΙΑΣ ΥΨΗΛΟΥ ΕΠΙΠΕΔΟΥ. Επιμέλεια: ΚΩΣΤΑΣ ΓΚΑΤΖΕΛΑΚΗΣ
 ΗΛΙΑΣΚΟΣ ΦΡΟΝΤΙΣΤΗΡΙΑ ΥΠΗΡΕΣΙΕΣ ΠΑΙΔΕΙΑΣ ΥΨΗΛΟΥ ΕΠΙΠΕΔΟΥ Θετικής Κατεύθυνσης Βιολογία Γ Λυκείου Επιμέλεια: ΚΩΣΤΑΣ ΓΚΑΤΖΕΛΑΚΗΣ e-mail: info@iliaskos.gr www.iliaskos.gr 1 TO 1. µ, : i µ µ DNA ii µ DNA iii
ΗΛΙΑΣΚΟΣ ΦΡΟΝΤΙΣΤΗΡΙΑ ΥΠΗΡΕΣΙΕΣ ΠΑΙΔΕΙΑΣ ΥΨΗΛΟΥ ΕΠΙΠΕΔΟΥ Θετικής Κατεύθυνσης Βιολογία Γ Λυκείου Επιμέλεια: ΚΩΣΤΑΣ ΓΚΑΤΖΕΛΑΚΗΣ e-mail: info@iliaskos.gr www.iliaskos.gr 1 TO 1. µ, : i µ µ DNA ii µ DNA iii
4 ο ΚΕΦΑΛΑΙΟ. Γ ε ν ε τ ι κ ή
 4 ο ΚΕΦΑΛΑΙΟ Γ ε ν ε τ ι κ ή 1. Κύκλος της ζωής του κυττάρου 3ο Γελ. Ηλιούπολης επιμέλεια: Αργύρης Γιάννης 2 2. Μοριακή Γενετική i). Ροή της γενετικής πληροφορίας DNA RNA πρωτεΐνες νουκλεΐκά οξέα ή πρωτεΐνες
4 ο ΚΕΦΑΛΑΙΟ Γ ε ν ε τ ι κ ή 1. Κύκλος της ζωής του κυττάρου 3ο Γελ. Ηλιούπολης επιμέλεια: Αργύρης Γιάννης 2 2. Μοριακή Γενετική i). Ροή της γενετικής πληροφορίας DNA RNA πρωτεΐνες νουκλεΐκά οξέα ή πρωτεΐνες
Ekvačná a kvantifikačná logika
 a kvantifikačná 3. prednáška (6. 10. 004) Prehľad 1 1 (dokončenie) ekvačných tabliel Formula A je ekvačne dokázateľná z množiny axióm T (T i A) práve vtedy, keď existuje uzavreté tablo pre cieľ A ekvačných
a kvantifikačná 3. prednáška (6. 10. 004) Prehľad 1 1 (dokončenie) ekvačných tabliel Formula A je ekvačne dokázateľná z množiny axióm T (T i A) práve vtedy, keď existuje uzavreté tablo pre cieľ A ekvačných
Obvod a obsah štvoruholníka
 Obvod a štvoruholníka D. Štyri body roviny z ktorých žiadne tri nie sú kolineárne (neležia na jednej priamke) tvoria jeden štvoruholník. Tie body (A, B, C, D) sú vrcholy štvoruholníka. strany štvoruholníka
Obvod a štvoruholníka D. Štyri body roviny z ktorých žiadne tri nie sú kolineárne (neležia na jednej priamke) tvoria jeden štvoruholník. Tie body (A, B, C, D) sú vrcholy štvoruholníka. strany štvoruholníka
ΗΛΙΑΣΚΟΣ ΦΡΟΝΤΙΣΤΗΡΙΑ. Θετικής Κατεύθυνσης Βιολογία Γ Λυκείου ΥΠΗΡΕΣΙΕΣ ΠΑΙΔΕΙΑΣ ΥΨΗΛΟΥ ΕΠΙΠΕΔΟΥ. Επιμέλεια: ΘΕΟΔΟΛΙΝΤΑ ΤΕΣΤΑ
 ΗΛΙΑΣΚΟΣ ΦΡΟΝΤΙΣΤΗΡΙΑ ΥΠΗΡΕΣΙΕΣ ΠΑΙΔΕΙΑΣ ΥΨΗΛΟΥ ΕΠΙΠΕΔΟΥ Θετικής Κατεύθυνσης Βιολογία Γ Λυκείου Επιμέλεια: ΘΕΟΔΟΛΙΝΤΑ ΤΕΣΤΑ e-mail: info@iliaskos.gr www.iliaskos.gr 1 DNA Griffith (1928) : DNA 2 (Diplococcus
ΗΛΙΑΣΚΟΣ ΦΡΟΝΤΙΣΤΗΡΙΑ ΥΠΗΡΕΣΙΕΣ ΠΑΙΔΕΙΑΣ ΥΨΗΛΟΥ ΕΠΙΠΕΔΟΥ Θετικής Κατεύθυνσης Βιολογία Γ Λυκείου Επιμέλεια: ΘΕΟΔΟΛΙΝΤΑ ΤΕΣΤΑ e-mail: info@iliaskos.gr www.iliaskos.gr 1 DNA Griffith (1928) : DNA 2 (Diplococcus
Start. Vstup r. O = 2*π*r S = π*r*r. Vystup O, S. Stop. Start. Vstup P, C V = P*C*1,19. Vystup V. Stop
 1) Vytvorte algoritmus (vývojový diagram) na výpočet obvodu kruhu. O=2xπxr ; S=πxrxr Vstup r O = 2*π*r S = π*r*r Vystup O, S 2) Vytvorte algoritmus (vývojový diagram) na výpočet celkovej ceny výrobku s
1) Vytvorte algoritmus (vývojový diagram) na výpočet obvodu kruhu. O=2xπxr ; S=πxrxr Vstup r O = 2*π*r S = π*r*r Vystup O, S 2) Vytvorte algoritmus (vývojový diagram) na výpočet celkovej ceny výrobku s
Informatika pre biológov. Broňa Brejová
 Informatika pre biológov Broňa Brejová 25.9.2014 1 Formulácia problému, algoritmus Formulácia problému: jasne zadefinujeme vstupné a výstupné dáta a aký výstup očakávame pre každý vstup. Vo formulácii
Informatika pre biológov Broňa Brejová 25.9.2014 1 Formulácia problému, algoritmus Formulácia problému: jasne zadefinujeme vstupné a výstupné dáta a aký výstup očakávame pre každý vstup. Vo formulácii
DNA RNA ΝΟΥΚΛΕΙΝΙΚΑ ΟΞΕΑ. Όσα αφορούν τη δομή του DNA δόθηκαν στο κεφάλαιο οργανικές ουσίες
 ΝΟΥΚΛΕΙΝΙΚΑ ΟΞΕΑ DNA RNA Ο φορέας της γενετικής πληροφορίας (DNA), σελ. 293 320 μέχρι και την πρώτη παράγραφο. (εκτός ύλης: Ο μηχανισμός της ημισυντηρητικής αντιγραφής του DNA, σελ. 297-301, Από το 14.4
ΝΟΥΚΛΕΙΝΙΚΑ ΟΞΕΑ DNA RNA Ο φορέας της γενετικής πληροφορίας (DNA), σελ. 293 320 μέχρι και την πρώτη παράγραφο. (εκτός ύλης: Ο μηχανισμός της ημισυντηρητικής αντιγραφής του DNA, σελ. 297-301, Από το 14.4
7. FUNKCIE POJEM FUNKCIE
 7. FUNKCIE POJEM FUNKCIE Funkcia f reálnej premennej je : - každé zobrazenie f v množine všetkých reálnych čísel; - množina f všetkých usporiadaných dvojíc[,y] R R pre ktorú platí: ku každému R eistuje
7. FUNKCIE POJEM FUNKCIE Funkcia f reálnej premennej je : - každé zobrazenie f v množine všetkých reálnych čísel; - množina f všetkých usporiadaných dvojíc[,y] R R pre ktorú platí: ku každému R eistuje
DNA RNA ΝΟΥΚΛΕΙΝΙΚΑ ΟΞΕΑ. Όσα αφορούν τη δομή του DNA δόθηκαν στο κεφάλαιο οργανικές ουσίες
 Ο φορέας της γενετικής πληροφορίας (DNA), σελ. 293 320 μέχρι και την πρώτη παράγραφο. (εκτός ύλης: Ο μηχανισμός της ημισυντηρητικής αντιγραφής του DNA, σελ. 297-301, Από το 14.4 ΜΕΤΑΛΛΑΞΕΙΣ, Σύνδρομο Down
Ο φορέας της γενετικής πληροφορίας (DNA), σελ. 293 320 μέχρι και την πρώτη παράγραφο. (εκτός ύλης: Ο μηχανισμός της ημισυντηρητικής αντιγραφής του DNA, σελ. 297-301, Από το 14.4 ΜΕΤΑΛΛΑΞΕΙΣ, Σύνδρομο Down
Prechod z 2D do 3D. Martin Florek 3. marca 2009
 Počítačová grafika 2 Prechod z 2D do 3D Martin Florek florek@sccg.sk FMFI UK 3. marca 2009 Prechod z 2D do 3D Čo to znamená? Ako zobraziť? Súradnicové systémy Čo to znamená? Ako zobraziť? tretia súradnica
Počítačová grafika 2 Prechod z 2D do 3D Martin Florek florek@sccg.sk FMFI UK 3. marca 2009 Prechod z 2D do 3D Čo to znamená? Ako zobraziť? Súradnicové systémy Čo to znamená? Ako zobraziť? tretia súradnica
Nukleové kyseliny. Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy.
 Nukleové kyseliny Nukleové kyseliny Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy. DNA - deoxyribonukleová kyselina RNA - ribonukleová kyselina Funkcie
Nukleové kyseliny Nukleové kyseliny Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy. DNA - deoxyribonukleová kyselina RNA - ribonukleová kyselina Funkcie
Κεφάλαιο 2ο. Αντιγραφή, έκφραση και ρύθμιση της γενετικής πληροφορίας
 Κεφάλαιο 2ο Αντιγραφή, έκφραση και ρύθμιση της γενετικής πληροφορίας 1. Το DNA αυτοδιπλασιάζεται 3ο ε.λ. Ηλιούπολης επιμέλεια: Αργύρης Γιάννης 3 Ο μηχανισμός της αντιγραφής του DNA Ο μηχανισμός αυτοδιπλασιασμού
Κεφάλαιο 2ο Αντιγραφή, έκφραση και ρύθμιση της γενετικής πληροφορίας 1. Το DNA αυτοδιπλασιάζεται 3ο ε.λ. Ηλιούπολης επιμέλεια: Αργύρης Γιάννης 3 Ο μηχανισμός της αντιγραφής του DNA Ο μηχανισμός αυτοδιπλασιασμού
Βιομόρια, δομή κυττάρου, διαμερισματοποίηση. Ν. Κ. Μοσχονάς n_moschonas@med.upatras.gr Εργ. Γεν. Βιολογίας, Τμ. Ιατρικής Παν/μιο Πάτρας
 Βιομόρια, δομή κυττάρου, διαμερισματοποίηση Ν. Κ. Μοσχονάς n_moschonas@med.upatras.gr Εργ. Γεν. Βιολογίας, Τμ. Ιατρικής Παν/μιο Πάτρας Ποια είναι τα χαρακτηριστικά της ζωής; Θρέψη Αναπαραγωγή Εξέλιξη Ερεθιστικότητα
Βιομόρια, δομή κυττάρου, διαμερισματοποίηση Ν. Κ. Μοσχονάς n_moschonas@med.upatras.gr Εργ. Γεν. Βιολογίας, Τμ. Ιατρικής Παν/μιο Πάτρας Ποια είναι τα χαρακτηριστικά της ζωής; Θρέψη Αναπαραγωγή Εξέλιξη Ερεθιστικότητα
Chí kvadrát test dobrej zhody. Metódy riešenia úloh z pravdepodobnosti a štatistiky
 Chí kvadrát test dobrej zhody Metódy riešenia úloh z pravdepodobnosti a štatistiky www.iam.fmph.uniba.sk/institute/stehlikova Test dobrej zhody I. Chceme overiť, či naše dáta pochádzajú z konkrétneho pravdep.
Chí kvadrát test dobrej zhody Metódy riešenia úloh z pravdepodobnosti a štatistiky www.iam.fmph.uniba.sk/institute/stehlikova Test dobrej zhody I. Chceme overiť, či naše dáta pochádzajú z konkrétneho pravdep.
1. Limita, spojitost a diferenciálny počet funkcie jednej premennej
 . Limita, spojitost a diferenciálny počet funkcie jednej premennej Definícia.: Hromadný bod a R množiny A R: v každom jeho okolí leží aspoň jeden bod z množiny A, ktorý je rôzny od bodu a Zadanie množiny
. Limita, spojitost a diferenciálny počet funkcie jednej premennej Definícia.: Hromadný bod a R množiny A R: v každom jeho okolí leží aspoň jeden bod z množiny A, ktorý je rôzny od bodu a Zadanie množiny
Goniometrické rovnice a nerovnice. Základné goniometrické rovnice
 Goniometrické rovnice a nerovnice Definícia: Rovnice (nerovnice) obsahujúce neznámu x alebo výrazy s neznámou x ako argumenty jednej alebo niekoľkých goniometrických funkcií nazývame goniometrickými rovnicami
Goniometrické rovnice a nerovnice Definícia: Rovnice (nerovnice) obsahujúce neznámu x alebo výrazy s neznámou x ako argumenty jednej alebo niekoľkých goniometrických funkcií nazývame goniometrickými rovnicami
Room Temperature Ionic Liquids from 20 Natural Amino Acids. Kenta Fukumoto, Masahiro Yoshizawa, and Hiroyuki Ohno
 S1 Room Temperature Ionic Liquids from 20 Natural Amino Acids Kenta Fukumoto, Masahiro Yoshizawa, and Hiroyuki Ohno 1-Ethyl-3-methylimidazolium L-α-aminopropionic acid salt ([emim][ala]). From 1.0g (5.2mmol)
S1 Room Temperature Ionic Liquids from 20 Natural Amino Acids Kenta Fukumoto, Masahiro Yoshizawa, and Hiroyuki Ohno 1-Ethyl-3-methylimidazolium L-α-aminopropionic acid salt ([emim][ala]). From 1.0g (5.2mmol)
ΠΕΡΙΛΗΨΗ ΧΑΡΑΚΤΗΡΙΣΤΙΚΩΝ ΤΟΥ ΠΡΟΙΟΝΤΟΣ (SPC) (AMINOVEN 3.5% Glucose / Electrolytes)
 (AMINOVEN 3.5% Glucose / Electrolytes)") ΠΕΡΙΛΗΨΗ ΧΑΡΑΚΤΗΡΙΣΤΙΚΩΝ ΤΟΥ ΠΡΟΙΟΝΤΟΣ (SPC) (AMINOVEN 3.5% Glucose / Electrolytes) 1. ΕΜΠΟΡΙΚΗ ΟΝΟΜΑΣΙΑ ΤΟΥ ΦΑΡΜΑΚΕΥΤΙΚΟΥ ΠΡΟΙΟΝΤΟΣ: AMINOVEN 3.5% Glucose / Electrolytes 2. ΠΟΙΟΤΙΚΗ ΚΑΙ ΠΟΣΟΤΙΚΗ ΣΥΝΘΕΣΗ
ΠΕΡΙΛΗΨΗ ΧΑΡΑΚΤΗΡΙΣΤΙΚΩΝ ΤΟΥ ΠΡΟΙΟΝΤΟΣ (SPC) (AMINOVEN 3.5% Glucose / Electrolytes) 1. ΕΜΠΟΡΙΚΗ ΟΝΟΜΑΣΙΑ ΤΟΥ ΦΑΡΜΑΚΕΥΤΙΚΟΥ ΠΡΟΙΟΝΤΟΣ: AMINOVEN 3.5% Glucose / Electrolytes 2. ΠΟΙΟΤΙΚΗ ΚΑΙ ΠΟΣΟΤΙΚΗ ΣΥΝΘΕΣΗ
Motivácia Denícia determinantu Výpo et determinantov Determinant sú inu matíc Vyuºitie determinantov. Determinanty. 14. decembra 2010.
 14. decembra 2010 Rie²enie sústav Plocha rovnobeºníka Objem rovnobeºnostena Rie²enie sústav Príklad a 11 x 1 + a 12 x 2 = c 1 a 21 x 1 + a 22 x 2 = c 2 Dostaneme: x 1 = c 1a 22 c 2 a 12 a 11 a 22 a 12
14. decembra 2010 Rie²enie sústav Plocha rovnobeºníka Objem rovnobeºnostena Rie²enie sústav Príklad a 11 x 1 + a 12 x 2 = c 1 a 21 x 1 + a 22 x 2 = c 2 Dostaneme: x 1 = c 1a 22 c 2 a 12 a 11 a 22 a 12
Βιοπληροφορική. Πίνακες Αντικατάστασης & Οπτική Σύγκριση Αλληλουχιών. Αλέξανδρος Τζάλλας
 Βιοπληροφορική Πίνακες Αντικατάστασης & Οπτική Σύγκριση Αλληλουχιών Αλέξανδρος Τζάλλας e-mail: tzallas@teiep.gr ΤΕΙ Ηπείρου Σχολή Τεχνολογικών Εφαρμογών Τμήμα Μηχανικών Πληροφορικής ΤΕ Copyright Πανεπιστήμιο
Βιοπληροφορική Πίνακες Αντικατάστασης & Οπτική Σύγκριση Αλληλουχιών Αλέξανδρος Τζάλλας e-mail: tzallas@teiep.gr ΤΕΙ Ηπείρου Σχολή Τεχνολογικών Εφαρμογών Τμήμα Μηχανικών Πληροφορικής ΤΕ Copyright Πανεπιστήμιο
Cvičenie č. 4,5 Limita funkcie
 Cvičenie č. 4,5 Limita funkcie Definícia ity Limita funkcie (vlastná vo vlastnom bode) Nech funkcia f je definovaná na nejakom okolí U( ) bodu. Hovoríme, že funkcia f má v bode itu rovnú A, ak ( ε > )(
Cvičenie č. 4,5 Limita funkcie Definícia ity Limita funkcie (vlastná vo vlastnom bode) Nech funkcia f je definovaná na nejakom okolí U( ) bodu. Hovoríme, že funkcia f má v bode itu rovnú A, ak ( ε > )(
Βιοπληροφορική. Ενότητα 19: Υπολογιστικός Προσδιορισμός Δομής (1/3), 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου
, 2 ΔΩ. Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου") Βιοπληροφορική Ενότητα 19: Υπολογιστικός Προσδιορισμός Δομής (1/3), 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι κατανόηση της αναγκαιότητας και των εφαρμογών της υπολογιστικής
Βιοπληροφορική Ενότητα 19: Υπολογιστικός Προσδιορισμός Δομής (1/3), 2 ΔΩ Τμήμα: Βιοτεχνολογίας Όνομα καθηγητή: Τ. Θηραίου Μαθησιακοί Στόχοι κατανόηση της αναγκαιότητας και των εφαρμογών της υπολογιστικής
1. písomná práca z matematiky Skupina A
 1. písomná práca z matematiky Skupina A 1. Vypočítajte : a) 84º 56 + 32º 38 = b) 140º 53º 24 = c) 55º 12 : 2 = 2. Vypočítajte zvyšné uhly na obrázku : β γ α = 35 12 δ a b 3. Znázornite na číselnej osi
1. písomná práca z matematiky Skupina A 1. Vypočítajte : a) 84º 56 + 32º 38 = b) 140º 53º 24 = c) 55º 12 : 2 = 2. Vypočítajte zvyšné uhly na obrázku : β γ α = 35 12 δ a b 3. Znázornite na číselnej osi
PRIEMER DROTU d = 0,4-6,3 mm
 PRUŽINY PRUŽINY SKRUTNÉ PRUŽINY VIAC AKO 200 RUHOV SKRUTNÝCH PRUŽÍN PRIEMER ROTU d = 0,4-6,3 mm èíslo 3.0 22.8.2008 8:28:57 22.8.2008 8:28:58 PRUŽINY SKRUTNÉ PRUŽINY TECHNICKÉ PARAMETRE h d L S Legenda
PRUŽINY PRUŽINY SKRUTNÉ PRUŽINY VIAC AKO 200 RUHOV SKRUTNÝCH PRUŽÍN PRIEMER ROTU d = 0,4-6,3 mm èíslo 3.0 22.8.2008 8:28:57 22.8.2008 8:28:58 PRUŽINY SKRUTNÉ PRUŽINY TECHNICKÉ PARAMETRE h d L S Legenda
Nukleové kyseliny a proteosyntéza
 Nukleové kyseliny (NK) sú biomakromolekulové látky, ktoré sa spolu s bielkovinami považujú za najvýznamnejšie zložky živých sústav. V ich molekulách sa uchováva dedičná ( genetická ) informácia bunky a
Nukleové kyseliny (NK) sú biomakromolekulové látky, ktoré sa spolu s bielkovinami považujú za najvýznamnejšie zložky živých sústav. V ich molekulách sa uchováva dedičná ( genetická ) informácia bunky a
AerobTec Altis Micro
 AerobTec Altis Micro Záznamový / súťažný výškomer s telemetriou Výrobca: AerobTec, s.r.o. Pionierska 15 831 02 Bratislava www.aerobtec.com info@aerobtec.com Obsah 1.Vlastnosti... 3 2.Úvod... 3 3.Princíp
AerobTec Altis Micro Záznamový / súťažný výškomer s telemetriou Výrobca: AerobTec, s.r.o. Pionierska 15 831 02 Bratislava www.aerobtec.com info@aerobtec.com Obsah 1.Vlastnosti... 3 2.Úvod... 3 3.Princíp
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΓΕΩΠΟΝΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΤΡΟΦΙΜΩΝ ΜΑΡΙΑΣ ΦΩΤΙΟΥ ΠΤΥΧΙΟΥΧΟΥ ΓΕΩΠΟΝΟΥ
 ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΓΕΩΠΟΝΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΤΡΟΦΙΜΩΝ ΜΑΡΙΑΣ ΦΩΤΙΟΥ ΠΤΥΧΙΟΥΧΟΥ ΓΕΩΠΟΝΟΥ Συγκέντρωση των ελεύθερων αµινοξέων στο αµνιακό υγρό σε σχέση µε την εβδοµάδα
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΓΕΩΠΟΝΙΚΗ ΣΧΟΛΗ ΤΟΜΕΑΣ ΕΠΙΣΤΗΜΗΣ ΚΑΙ ΤΕΧΝΟΛΟΓΙΑΣ ΤΡΟΦΙΜΩΝ ΜΑΡΙΑΣ ΦΩΤΙΟΥ ΠΤΥΧΙΟΥΧΟΥ ΓΕΩΠΟΝΟΥ Συγκέντρωση των ελεύθερων αµινοξέων στο αµνιακό υγρό σε σχέση µε την εβδοµάδα
KATEDRA DOPRAVNEJ A MANIPULAČNEJ TECHNIKY Strojnícka fakulta, Žilinská Univerzita
 132 1 Absolútna chyba: ) = - skut absolútna ochýlka: ) ' = - spr. relatívna chyba: alebo Chyby (ochýlky): M systematické, M náhoné, M hrubé. Korekcia: k = spr - = - Î' pomerná korekcia: Správna honota:
132 1 Absolútna chyba: ) = - skut absolútna ochýlka: ) ' = - spr. relatívna chyba: alebo Chyby (ochýlky): M systematické, M náhoné, M hrubé. Korekcia: k = spr - = - Î' pomerná korekcia: Správna honota:
,Zohrievanie vody indukčným varičom bez pokrievky,
 Farba skupiny: zelená Označenie úlohy:,zohrievanie vody indukčným varičom bez pokrievky, Úloha: Zistiť, ako závisí účinnosť zohrievania vody na indukčnom variči od priemeru použitého hrnca. Hypotéza: Účinnosť
Farba skupiny: zelená Označenie úlohy:,zohrievanie vody indukčným varičom bez pokrievky, Úloha: Zistiť, ako závisí účinnosť zohrievania vody na indukčnom variči od priemeru použitého hrnca. Hypotéza: Účinnosť
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy
, diferencovanie časového radu, unit root testy") Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2012/2013 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/18
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2012/2013 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/18
ΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΓΟΝΙ ΙΑΚΗ ΕΚΦΡΑΣΗ ΚΑΙ ΣΗΜΑΤΟ ΟΤΗΣΗ
 ΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΓΟΝΙ ΙΑΚΗ ΕΚΦΡΑΣΗ ΚΑΙ ΣΗΜΑΤΟ ΟΤΗΣΗ ρ. Α. ΓΑΛΑΝΗΣ agalanis@mbg.duth.gr Figure 6.1 The Biology of Cancer ( Garland Science 2007) Figure
ΗΜΟΚΡΙΤΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΡΑΚΗΣ ΤΜΗΜΑ ΜΟΡΙΑΚΗΣ ΒΙΟΛΟΓΙΑΣ ΚΑΙ ΓΕΝΕΤΙΚΗΣ ΓΟΝΙ ΙΑΚΗ ΕΚΦΡΑΣΗ ΚΑΙ ΣΗΜΑΤΟ ΟΤΗΣΗ ρ. Α. ΓΑΛΑΝΗΣ agalanis@mbg.duth.gr Figure 6.1 The Biology of Cancer ( Garland Science 2007) Figure
Human angiogenin is a potent cytotoxin in the absence of ribonuclease inhibitor
 Human angiogenin is a potent cytotoxin in the absence of ribonuclease inhibitor SYDNEY P. THOMAS, 1 TRISH T. HOANG, 2 VALERIE T. RESSLER, 4 and RONALD T. RAINES 2,3,4 1 Graduate Program in Cell & Molecular
Human angiogenin is a potent cytotoxin in the absence of ribonuclease inhibitor SYDNEY P. THOMAS, 1 TRISH T. HOANG, 2 VALERIE T. RESSLER, 4 and RONALD T. RAINES 2,3,4 1 Graduate Program in Cell & Molecular
Kompilátory. Cvičenie 6: LLVM. Peter Kostolányi. 21. novembra 2017
 Kompilátory Cvičenie 6: LLVM Peter Kostolányi 21. novembra 2017 LLVM V podstate sada nástrojov pre tvorbu kompilátorov LLVM V podstate sada nástrojov pre tvorbu kompilátorov Pôvodne Low Level Virtual Machine
Kompilátory Cvičenie 6: LLVM Peter Kostolányi 21. novembra 2017 LLVM V podstate sada nástrojov pre tvorbu kompilátorov LLVM V podstate sada nástrojov pre tvorbu kompilátorov Pôvodne Low Level Virtual Machine
Priamkové plochy. Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava
 na nej leží potom plocha Φ je priamková. Santiago Calatrava") Priamkové plochy Priamkové plochy Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava Priamkové plochy rozdeľujeme na: Rozvinuteľné
Priamkové plochy Priamkové plochy Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava Priamkové plochy rozdeľujeme na: Rozvinuteľné
Analysis on the Ratio of Flesh Content and Nutritional. Quality of Esox lucius
 Chinese Journal of Zoology 2009,44 (3) :7075 3 ( 830000) : 6 ( Esox lucius), :6418 %,1911 %114 %,17 17138 %,7 8114 %,6136 % 45173 %,,WHOΠFAO 76135,,,,, : ; ; ; ; :Q955 :A :025023263 (2009) 03270206 Analysis
Chinese Journal of Zoology 2009,44 (3) :7075 3 ( 830000) : 6 ( Esox lucius), :6418 %,1911 %114 %,17 17138 %,7 8114 %,6136 % 45173 %,,WHOΠFAO 76135,,,,, : ; ; ; ; :Q955 :A :025023263 (2009) 03270206 Analysis
Matematika prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad
 Matematika 3-13. prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad Erika Škrabul áková F BERG, TU Košice 15. 12. 2015 Erika Škrabul áková (TUKE) Taylorov
Matematika 3-13. prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad Erika Škrabul áková F BERG, TU Košice 15. 12. 2015 Erika Škrabul áková (TUKE) Taylorov
Matematika 2. časť: Analytická geometria
 Matematika 2 časť: Analytická geometria RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk Súradnicové
Matematika 2 časť: Analytická geometria RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk Súradnicové
Απολυτήριες εξετάσεις Γ Τάξης Ημερήσιου Γενικού Λυκείου ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 18 5 2011
 Απολυτήριες εξετάσεις Γ Τάξης Ημερήσιου Γενικού Λυκείου ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 18 5 2011 ΘΕΜΑ Α 1. α 2. δ 3. γ 4. β 5. β ΘΕΜΑ Β Β1. σελ. 13: «Το 1928 ο Griffith...για το πώς γίνεται αυτό» Β2. σελ.
Απολυτήριες εξετάσεις Γ Τάξης Ημερήσιου Γενικού Λυκείου ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ 18 5 2011 ΘΕΜΑ Α 1. α 2. δ 3. γ 4. β 5. β ΘΕΜΑ Β Β1. σελ. 13: «Το 1928 ο Griffith...για το πώς γίνεται αυτό» Β2. σελ.
Gramatická indukcia a jej využitie
 a jej využitie KAI FMFI UK 29. Marec 2010 a jej využitie Prehľad Teória formálnych jazykov 1 Teória formálnych jazykov 2 3 a jej využitie Na počiatku bolo slovo. A slovo... a jej využitie Definícia (Slovo)
a jej využitie KAI FMFI UK 29. Marec 2010 a jej využitie Prehľad Teória formálnych jazykov 1 Teória formálnych jazykov 2 3 a jej využitie Na počiatku bolo slovo. A slovo... a jej využitie Definícia (Slovo)
M6: Model Hydraulický systém dvoch zásobníkov kvapaliny s interakciou
 M6: Model Hydraulický ytém dvoch záobníkov kvapaliny interakciou Úlohy:. Zotavte matematický popi modelu Hydraulický ytém. Vytvorte imulačný model v jazyku: a. Matlab b. imulink 3. Linearizujte nelineárny
M6: Model Hydraulický ytém dvoch záobníkov kvapaliny interakciou Úlohy:. Zotavte matematický popi modelu Hydraulický ytém. Vytvorte imulačný model v jazyku: a. Matlab b. imulink 3. Linearizujte nelineárny
Živá hmota chemické zloženie. Biológia živočíšnej produkcie Katedra fyziológie živočíchov
 Živá hmota chemické zloženie Biológia živočíšnej produkcie Katedra fyziológie živočíchov Chemické zloženie živej hmoty Živá hmota bioplazma chemicky rôznorodá zmes látok zložitý koloidný systém Prvky v
Živá hmota chemické zloženie Biológia živočíšnej produkcie Katedra fyziológie živočíchov Chemické zloženie živej hmoty Živá hmota bioplazma chemicky rôznorodá zmes látok zložitý koloidný systém Prvky v
MIDTERM (A) riešenia a bodovanie
 riešenia a bodovanie") MIDTERM (A) riešenia a bodovanie 1. (7b) Nech vzhl adom na štandardnú karteziánsku sústavu súradníc S 1 := O, e 1, e 2 majú bod P a vektory u, v súradnice P = [0, 1], u = e 1, v = 2 e 2. Aký predpis bude
MIDTERM (A) riešenia a bodovanie 1. (7b) Nech vzhl adom na štandardnú karteziánsku sústavu súradníc S 1 := O, e 1, e 2 majú bod P a vektory u, v súradnice P = [0, 1], u = e 1, v = 2 e 2. Aký predpis bude
Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti rozvodu tepla
 Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti príloha č. 7 k vyhláške č. 428/2010 Názov prevádzkovateľa verejného : Spravbytkomfort a.s. Prešov Adresa: IČO: Volgogradská 88, 080 01 Prešov 31718523
Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti príloha č. 7 k vyhláške č. 428/2010 Názov prevádzkovateľa verejného : Spravbytkomfort a.s. Prešov Adresa: IČO: Volgogradská 88, 080 01 Prešov 31718523
SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV KATEDRA GENETIKY A ŠĽACHTENIA RASTLÍN
 SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV KATEDRA GENETIKY A ŠĽACHTENIA RASTLÍN Prednáška z Genetiky mikroorganizmov Informačné molekuly v mikroorganizmoch
SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV KATEDRA GENETIKY A ŠĽACHTENIA RASTLÍN Prednáška z Genetiky mikroorganizmov Informačné molekuly v mikroorganizmoch
Tomáš Madaras Prvočísla
 Prvočísla Tomáš Madaras 2011 Definícia Nech a Z. Čísla 1, 1, a, a sa nazývajú triviálne delitele čísla a. Cele číslo a / {0, 1, 1} sa nazýva prvočíslo, ak má iba triviálne delitele; ak má aj iné delitele,
Prvočísla Tomáš Madaras 2011 Definícia Nech a Z. Čísla 1, 1, a, a sa nazývajú triviálne delitele čísla a. Cele číslo a / {0, 1, 1} sa nazýva prvočíslo, ak má iba triviálne delitele; ak má aj iné delitele,
Modelovanie dynamickej podmienenej korelácie kurzov V4
 Modelovanie dynamickej podmienenej korelácie menových kurzov V4 Podnikovohospodárska fakulta so sídlom v Košiciach Ekonomická univerzita v Bratislave Cieľ a motivácia Východiská Cieľ a motivácia Cieľ Kvantifikovať
Modelovanie dynamickej podmienenej korelácie menových kurzov V4 Podnikovohospodárska fakulta so sídlom v Košiciach Ekonomická univerzita v Bratislave Cieľ a motivácia Východiská Cieľ a motivácia Cieľ Kvantifikovať
ΥΠΟΔΕΙΓΜΑΤΙΚΑ ΛΥΜΕΝΕΣ ΑΣΚΗΣΕΙΣ ΚΕΦ. 2ο
 ΥΠΟΔΕΙΓΜΑΤΙΚΑ ΛΥΜΕΝΕΣ ΑΣΚΗΣΕΙΣ ΚΕΦ. 2ο 1. Δύο μόρια DΝΑ αποτελούνται το καθένα από 10.000 ζεύγη αζωτούχων βάσεων με 14 Ν. Τα μόρια μεταφέρονται σε περιβάλλον με ραδιενεργά νουκλεοτίδια που περιέχουν 15
ΥΠΟΔΕΙΓΜΑΤΙΚΑ ΛΥΜΕΝΕΣ ΑΣΚΗΣΕΙΣ ΚΕΦ. 2ο 1. Δύο μόρια DΝΑ αποτελούνται το καθένα από 10.000 ζεύγη αζωτούχων βάσεων με 14 Ν. Τα μόρια μεταφέρονται σε περιβάλλον με ραδιενεργά νουκλεοτίδια που περιέχουν 15
Planárne a rovinné grafy
 Planárne a rovinné grafy Definícia Graf G sa nazýva planárny, ak existuje jeho nakreslenie D, v ktorom sa žiadne dve hrany nepretínajú. D sa potom nazýva rovinný graf. Planárne a rovinné grafy Definícia
Planárne a rovinné grafy Definícia Graf G sa nazýva planárny, ak existuje jeho nakreslenie D, v ktorom sa žiadne dve hrany nepretínajú. D sa potom nazýva rovinný graf. Planárne a rovinné grafy Definícia
ΔΟΜΗ ΠΡΩΤΕΪΝΩΝ. Σελίδα 1 ΒΙΟΠΛΗΡΟΦΟΡΙΚΗ. Τ. Θηραίου
 ΔΟΜΗ ΠΡΩΤΕΪΝΩΝ Σελίδα 1 κύρια αλυσίδα main chain πεπτιδικός δεσμός πλευρική αλυσίδα side chain αμινομάδα amino group α άνθρακας Σελίδα 2 καρβοξυλομάδα carboxyl group Σελίδα 3 Ramachandran Plot Σελίδα 4
ΔΟΜΗ ΠΡΩΤΕΪΝΩΝ Σελίδα 1 κύρια αλυσίδα main chain πεπτιδικός δεσμός πλευρική αλυσίδα side chain αμινομάδα amino group α άνθρακας Σελίδα 2 καρβοξυλομάδα carboxyl group Σελίδα 3 Ramachandran Plot Σελίδα 4
TECHNICKÁ UNIVERZITA VO ZVOLENE. Prof. Ing. Dušan Gömöry, DrSc. GENETIKA
 TECHNICKÁ UNIVERZITA VO ZVOLENE Lesnícka fakulta Katedra fytológie Prof. Ing. Dušan Gömöry, DrSc. GENETIKA 2014 prof. Ing. Dušan Gömöry, DrSc. Recenzenti: Schválené: Rektorom Technickej univerzity vo Zvolene
TECHNICKÁ UNIVERZITA VO ZVOLENE Lesnícka fakulta Katedra fytológie Prof. Ing. Dušan Gömöry, DrSc. GENETIKA 2014 prof. Ing. Dušan Gömöry, DrSc. Recenzenti: Schválené: Rektorom Technickej univerzity vo Zvolene
Προ- & μετα-αγωνιςτικό γεφμα Για ακλθτι βάρουσ 75kg. Μαρία Χολζβα, τελειόφοιτθ τμιματοσ Επιςτιμθσ Διαιτολογίασ & Διατροφισ Χαροκοπείου Πανεπιςτθμίου
 Προ- & μετα-αγωνιςτικό γεφμα Για ακλθτι βάρουσ 75kg Μαρία Χολζβα, τελειόφοιτθ τμιματοσ Επιςτιμθσ Διαιτολογίασ & Διατροφισ Χαροκοπείου Πανεπιςτθμίου Προ-αγωνιςτικό γεφμα: ςυςτάςεισ 3-4 ϊρεσ πριν Υδατάνκρακεσ
Προ- & μετα-αγωνιςτικό γεφμα Για ακλθτι βάρουσ 75kg Μαρία Χολζβα, τελειόφοιτθ τμιματοσ Επιςτιμθσ Διαιτολογίασ & Διατροφισ Χαροκοπείου Πανεπιςτθμίου Προ-αγωνιςτικό γεφμα: ςυςτάςεισ 3-4 ϊρεσ πριν Υδατάνκρακεσ
Metódy vol nej optimalizácie
 Metódy vol nej optimalizácie Metódy vol nej optimalizácie p. 1/28 Motivácia k metódam vol nej optimalizácie APLIKÁCIE p. 2/28 II 1. PRÍKLAD: Lineárna regresia - metóda najmenších štvorcov Na základe dostupných
Metódy vol nej optimalizácie Metódy vol nej optimalizácie p. 1/28 Motivácia k metódam vol nej optimalizácie APLIKÁCIE p. 2/28 II 1. PRÍKLAD: Lineárna regresia - metóda najmenších štvorcov Na základe dostupných
Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R
 Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R Ako nadprirodzené stretnutie s murárikom červenokrídlym naformátovalo môj profesijný i súkromný život... Osudové stretnutie s murárikom
Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R Ako nadprirodzené stretnutie s murárikom červenokrídlym naformátovalo môj profesijný i súkromný život... Osudové stretnutie s murárikom
DNA. 2 η Βάση 3 η U C A G
 DNA Ο Γενετικός κώδικας θα σας είναι χρήσιμος για να απαντήσετε ορισμένες από τις ερωτήσεις που ακολουθούν 1 η Βάση U C A G 2 η Βάση 3 η U C A G Βάση UUU φαινυλανανίνη UCU σερίνη UAU τυροσίνη UGU κυστεΐνη
DNA Ο Γενετικός κώδικας θα σας είναι χρήσιμος για να απαντήσετε ορισμένες από τις ερωτήσεις που ακολουθούν 1 η Βάση U C A G 2 η Βάση 3 η U C A G Βάση UUU φαινυλανανίνη UCU σερίνη UAU τυροσίνη UGU κυστεΐνη
Εφαρμοσμένη Βιοτεχνολογία Εργαστηριακή Άσκηση Εισαγωγή στην Βιοπληροφορική
 Εφαρμοσμένη Βιοτεχνολογία Εργαστηριακή Άσκηση Εισαγωγή στην Βιοπληροφορική Δραστηριότητες 1. Εύρεση γονιδίων/πρωτεϊνών από βάσεις δεδομένων 2. Ευθυγράμμιση και σύγκριση γονιδίων/πρωτεϊνών 3. Δημιουργία
Εφαρμοσμένη Βιοτεχνολογία Εργαστηριακή Άσκηση Εισαγωγή στην Βιοπληροφορική Δραστηριότητες 1. Εύρεση γονιδίων/πρωτεϊνών από βάσεις δεδομένων 2. Ευθυγράμμιση και σύγκριση γονιδίων/πρωτεϊνών 3. Δημιουργία
KATALÓG KRUHOVÉ POTRUBIE
 H KATALÓG KRUHOVÉ POTRUBIE 0 Základné požiadavky zadávania VZT potrubia pre výrobu 1. Zadávanie do výroby v spoločnosti APIAGRA s.r.o. V digitálnej forme na tlačive F05-8.0_Rozpis_potrubia, zaslané mailom
H KATALÓG KRUHOVÉ POTRUBIE 0 Základné požiadavky zadávania VZT potrubia pre výrobu 1. Zadávanie do výroby v spoločnosti APIAGRA s.r.o. V digitálnej forme na tlačive F05-8.0_Rozpis_potrubia, zaslané mailom
Πανεπιστήμιο Δυτικής Μακεδονίας. Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών. Βιοπληροφορική
 Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 4: Πίνακες αντικατάστασης & οπτική σύγκριση αλληλουχιών Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία
Τμήμα Μηχανικών Πληροφορικής & Τηλεπικοινωνιών Βιοπληροφορική Ενότητα 4: Πίνακες αντικατάστασης & οπτική σύγκριση αλληλουχιών Αν. καθηγητής Αγγελίδης Παντελής e-mail: paggelidis@uowm.gr ΕΕΔΙΠ Μπέλλου Σοφία
Návrh vzduchotesnosti pre detaily napojení
 Výpočet lineárneho stratového súčiniteľa tepelného mosta vzťahujúceho sa k vonkajším rozmerom: Ψ e podľa STN EN ISO 10211 Návrh vzduchotesnosti pre detaily napojení Objednávateľ: Ing. Natália Voltmannová
Výpočet lineárneho stratového súčiniteľa tepelného mosta vzťahujúceho sa k vonkajším rozmerom: Ψ e podľa STN EN ISO 10211 Návrh vzduchotesnosti pre detaily napojení Objednávateľ: Ing. Natália Voltmannová
Vybrané kapitoly genetiky pre lesníkov
 Vybrané kapitoly genetiky pre lesníkov 1 NÁPLŇ A KLASIFIKÁCIA GENETIKY Genetika je veda o dedičnosti a premenlivosti živých organizmov. Pojem premenlivosti bol zadefinovaný v predchádzajúcej kapitole premenlivosť
Vybrané kapitoly genetiky pre lesníkov 1 NÁPLŇ A KLASIFIKÁCIA GENETIKY Genetika je veda o dedičnosti a premenlivosti živých organizmov. Pojem premenlivosti bol zadefinovaný v predchádzajúcej kapitole premenlivosť
x x x2 n
 Reálne symetrické matice Skalárny súčin v R n. Pripomeniem, že pre vektory u = u, u, u, v = v, v, v R platí. dĺžka vektora u je u = u + u + u,. ak sú oba vektory nenulové a zvierajú neorientovaný uhol
Reálne symetrické matice Skalárny súčin v R n. Pripomeniem, že pre vektory u = u, u, u, v = v, v, v R platí. dĺžka vektora u je u = u + u + u,. ak sú oba vektory nenulové a zvierajú neorientovaný uhol
5 -TACATGTCGCGATGCAAGTTCTAATCTCAATA CTT-3 3 -ATGTACAGCGCTACGTTCAAGATTAGAGTTAT GAA-5
 1 ( ) 18 2011 : : (5) 1 5,. 1......... 2.. DNA.. mrna.. mrna.. DNA. 3. Ti........ 4......... 1 5 2 5. T....... DNA. : 1. Griffith. 8 2.. 3. : ). ) cdna. 7 6 4. DNA : ( ) 28% (G) 28%.. 4 1.,. 2 5 3., (
1 ( ) 18 2011 : : (5) 1 5,. 1......... 2.. DNA.. mrna.. mrna.. DNA. 3. Ti........ 4......... 1 5 2 5. T....... DNA. : 1. Griffith. 8 2.. 3. : ). ) cdna. 7 6 4. DNA : ( ) 28% (G) 28%.. 4 1.,. 2 5 3., (
3. Striedavé prúdy. Sínusoida
 . Striedavé prúdy VZNIK: Striedavý elektrický prúd prechádza obvodom, ktorý je pripojený na zdroj striedavého napätia. Striedavé napätie vyrába synchrónny generátor, kde na koncoch rotorového vinutia sa
. Striedavé prúdy VZNIK: Striedavý elektrický prúd prechádza obvodom, ktorý je pripojený na zdroj striedavého napätia. Striedavé napätie vyrába synchrónny generátor, kde na koncoch rotorového vinutia sa
ARMA modely čast 2: moving average modely (MA)
") ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2014/2015 ARMA modely časť 2: moving average modely(ma) p.1/24 V. Moving average proces prvého rádu - MA(1) ARMA modely
ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2014/2015 ARMA modely časť 2: moving average modely(ma) p.1/24 V. Moving average proces prvého rádu - MA(1) ARMA modely
Πιθανοθεωρητικά µοντέλα αναπαράστασης ακολουθιών
 Πιθανοθεωρητικά µοντέλα αναπαράστασης ακολουθιών Vasilis Promponas Bioinformatics Research Laboratory Department of Biological Sciences University of Cyprus ΣΥΝΟΨΗ Εισαγωγή Αλυσίδες Markov και αλληλουχίες
Πιθανοθεωρητικά µοντέλα αναπαράστασης ακολουθιών Vasilis Promponas Bioinformatics Research Laboratory Department of Biological Sciences University of Cyprus ΣΥΝΟΨΗ Εισαγωγή Αλυσίδες Markov και αλληλουχίες
Analýza údajov. W bozóny.
 Analýza údajov W bozóny http://www.physicsmasterclasses.org/index.php 1 Identifikácia častíc https://kjende.web.cern.ch/kjende/sl/wpath_teilchenid1.htm 2 Identifikácia častíc Cvičenie 1 Na web stránke
Analýza údajov W bozóny http://www.physicsmasterclasses.org/index.php 1 Identifikácia častíc https://kjende.web.cern.ch/kjende/sl/wpath_teilchenid1.htm 2 Identifikácia častíc Cvičenie 1 Na web stránke
24. Základné spôsoby zobrazovania priestoru do roviny
 24. Základné spôsoby zobrazovania priestoru do roviny Voľné rovnobežné premietanie Presné metódy zobrazenia trojrozmerného priestoru do dvojrozmernej roviny skúma samostatná matematická disciplína, ktorá
24. Základné spôsoby zobrazovania priestoru do roviny Voľné rovnobežné premietanie Presné metódy zobrazenia trojrozmerného priestoru do dvojrozmernej roviny skúma samostatná matematická disciplína, ktorá
HASLIM112V, HASLIM123V, HASLIM136V HASLIM112Z, HASLIM123Z, HASLIM136Z HASLIM112S, HASLIM123S, HASLIM136S
 PROUKTOVÝ LIST HKL SLIM č. sklad. karty / obj. číslo: HSLIM112V, HSLIM123V, HSLIM136V HSLIM112Z, HSLIM123Z, HSLIM136Z HSLIM112S, HSLIM123S, HSLIM136S fakturačný názov výrobku: HKL SLIMv 1,2kW HKL SLIMv
PROUKTOVÝ LIST HKL SLIM č. sklad. karty / obj. číslo: HSLIM112V, HSLIM123V, HSLIM136V HSLIM112Z, HSLIM123Z, HSLIM136Z HSLIM112S, HSLIM123S, HSLIM136S fakturačný názov výrobku: HKL SLIMv 1,2kW HKL SLIMv
Improved Peptide and Protein Torsional Energetics with the OPLS-AA Force Field
 Improved Peptide and Protein Torsional Energetics with the OPLS-AA Force Field Supplementary Information Michael J. Robertson, Julian Tirado-Rives, and William L. Jorgensen* Department of Chemistry, Yale
Improved Peptide and Protein Torsional Energetics with the OPLS-AA Force Field Supplementary Information Michael J. Robertson, Julian Tirado-Rives, and William L. Jorgensen* Department of Chemistry, Yale
5 -TACATGTCGCGATGCAAGTTCTAATCTCAATA CTT-3 3 -ATGTACAGCGCTACGTTCAAGATTAGAGTTAT GAA-5
 1 ( ) 18 2011 : : (5) 1 5,. 1......... 2.. DNA.. mrna.. mrna.. DNA. 3. Ti........ 4......... 1 5 2 5. T....... DNA. : 1. Griffith. 8 2.. 3. : ). ) cdna. 7 6 4. DNA : ( ) 28% (G) 28%.. 4 1.,. 2 5 3., (
1 ( ) 18 2011 : : (5) 1 5,. 1......... 2.. DNA.. mrna.. mrna.. DNA. 3. Ti........ 4......... 1 5 2 5. T....... DNA. : 1. Griffith. 8 2.. 3. : ). ) cdna. 7 6 4. DNA : ( ) 28% (G) 28%.. 4 1.,. 2 5 3., (
Εισαγωγή στις πρωτεΐνες Δομή πρωτεϊνών Ταξινόμηση βάσει δομής Βάσεις με δομές πρωτεϊνών Ευθυγράμμιση δομών Πρόβλεψη 2D δομής Πρόβλεψη 3D δομής
 Εισαγωγή στις πρωτεΐνες Δομή πρωτεϊνών Ταξινόμηση βάσει δομής Βάσεις με δομές πρωτεϊνών Ευθυγράμμιση δομών Πρόβλεψη 2D δομής Πρόβλεψη 3D δομής Τι είναι η πρωτεΐνη Τι εννοούμε με δομή πρωτεϊνών Οικογένειες
Εισαγωγή στις πρωτεΐνες Δομή πρωτεϊνών Ταξινόμηση βάσει δομής Βάσεις με δομές πρωτεϊνών Ευθυγράμμιση δομών Πρόβλεψη 2D δομής Πρόβλεψη 3D δομής Τι είναι η πρωτεΐνη Τι εννοούμε με δομή πρωτεϊνών Οικογένειες
Vektorový priestor V : Množina prvkov (vektory), na ktorej je definované ich sčítanie a ich
, na ktorej je definované ich sčítanie a ich") Tuesday 15 th January, 2013, 19:53 Základy tenzorového počtu M.Gintner Vektorový priestor V : Množina prvkov (vektory), na ktorej je definované ich sčítanie a ich násobenie reálnym číslom tak, že platí:
Tuesday 15 th January, 2013, 19:53 Základy tenzorového počtu M.Gintner Vektorový priestor V : Množina prvkov (vektory), na ktorej je definované ich sčítanie a ich násobenie reálnym číslom tak, že platí:
Základné poznatky molekulovej fyziky a termodynamiky
 Základné poznatky molekulovej fyziky a termodynamiky Opakovanie učiva II. ročníka, Téma 1. A. Príprava na maturity z fyziky, 2008 Outline Molekulová fyzika 1 Molekulová fyzika Predmet Molekulovej fyziky
Základné poznatky molekulovej fyziky a termodynamiky Opakovanie učiva II. ročníka, Téma 1. A. Príprava na maturity z fyziky, 2008 Outline Molekulová fyzika 1 Molekulová fyzika Predmet Molekulovej fyziky
Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín. Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť.
 Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť. Ktoré fyzikálne jednotky zodpovedajú sústave SI: a) Dĺžka, čas,
Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť. Ktoré fyzikálne jednotky zodpovedajú sústave SI: a) Dĺžka, čas,
ΑΣΚΗΣΕΙΣ ΠΡΟΣ ΛΥΣΗ ΚΕΦ. 2ο
 ΑΣΚΗΣΕΙΣ ΠΡΟΣ ΛΥΣΗ ΚΕΦ. 2ο ΟΜΑΔΑ Α 1. Μόριο DΝΑ αποτελείται από 8.000 αζωτούχες βάσεις με 14 Ν. Το μόριο μεταφέρεται σε περιβάλλον με ραδιενεργά νουκλεοτίδια που περιέχουν 15 Ν και αντιγράφεται δύο φορές.
ΑΣΚΗΣΕΙΣ ΠΡΟΣ ΛΥΣΗ ΚΕΦ. 2ο ΟΜΑΔΑ Α 1. Μόριο DΝΑ αποτελείται από 8.000 αζωτούχες βάσεις με 14 Ν. Το μόριο μεταφέρεται σε περιβάλλον με ραδιενεργά νουκλεοτίδια που περιέχουν 15 Ν και αντιγράφεται δύο φορές.
Definícia parciálna derivácia funkcie podľa premennej x. Definícia parciálna derivácia funkcie podľa premennej y. Ak existuje limita.
 Teória prednáška č. 9 Deinícia parciálna deriácia nkcie podľa premennej Nech nkcia Ak eistje limita je deinoaná okolí bod [ ] lim. tak túto limit nazýame parciálno deriácio nkcie podľa premennej bode [
Teória prednáška č. 9 Deinícia parciálna deriácia nkcie podľa premennej Nech nkcia Ak eistje limita je deinoaná okolí bod [ ] lim. tak túto limit nazýame parciálno deriácio nkcie podľa premennej bode [
Genetický algoritmus
 Genetický algoritmus Vladimír Kvasnička Katedra matematiky Chemickotechnologická fakulta STU 812 35 Bratislava email: kvasnic@cvt.stuba.sk Stretnutie so študentmi Gymnázia z Groesslingovej ul., ktoré sa
Genetický algoritmus Vladimír Kvasnička Katedra matematiky Chemickotechnologická fakulta STU 812 35 Bratislava email: kvasnic@cvt.stuba.sk Stretnutie so študentmi Gymnázia z Groesslingovej ul., ktoré sa
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ
 BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΜΟΡΙΑΚΑ ΜΟΝΤΕΛΑ 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING) 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING)
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΜΟΡΙΑΚΑ ΜΟΝΤΕΛΑ 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING) 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING)
Metodicko pedagogické centrum. Národný projekt VZDELÁVANÍM PEDAGOGICKÝCH ZAMESTNANCOV K INKLÚZII MARGINALIZOVANÝCH RÓMSKYCH KOMUNÍT
 Moderné vzdelávanie pre vedomostnú spoločnosť / Projekt je spolufinancovaný zo zdrojov EÚ Kód ITMS: 26130130051 číslo zmluvy: OPV/24/2011 Metodicko pedagogické centrum Národný projekt VZDELÁVANÍM PEDAGOGICKÝCH
Moderné vzdelávanie pre vedomostnú spoločnosť / Projekt je spolufinancovaný zo zdrojov EÚ Kód ITMS: 26130130051 číslo zmluvy: OPV/24/2011 Metodicko pedagogické centrum Národný projekt VZDELÁVANÍM PEDAGOGICKÝCH
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy
, diferencovanie časového radu, unit root testy") Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2013/2014 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/27
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2013/2014 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/27
Anima-Strath. Απόλυτα φυσικό & φυτικό διατροφικό συμπλήρωμα
 Anima-Strath Απόλυτα φυσικό & φυτικό διατροφικό συμπλήρωμα Πληροφορίες Bio-Strath AG, Herrliberg - Factory Πληροφορίες Bio-Strath AG Έτος ιδρύσεως το 1961 από τον Fred Pestalozzi. Επεξεργασία στο Herrliberg
Anima-Strath Απόλυτα φυσικό & φυτικό διατροφικό συμπλήρωμα Πληροφορίες Bio-Strath AG, Herrliberg - Factory Πληροφορίες Bio-Strath AG Έτος ιδρύσεως το 1961 από τον Fred Pestalozzi. Επεξεργασία στο Herrliberg
LR(0) syntaktické analyzátory. doc. RNDr. Ľubomír Dedera
 syntaktické analyzátory. doc. RNDr. Ľubomír Dedera") LR0) syntaktické analyzátory doc. RNDr. Ľubomír Dedera Učebné otázky LR0) automat a jeho konštrukcia Konštrukcia tabuliek ACION a GOO LR0) syntaktického analyzátora LR0) syntaktický analyzátor Sám osebe
LR0) syntaktické analyzátory doc. RNDr. Ľubomír Dedera Učebné otázky LR0) automat a jeho konštrukcia Konštrukcia tabuliek ACION a GOO LR0) syntaktického analyzátora LR0) syntaktický analyzátor Sám osebe
Pevné ložiská. Voľné ložiská
 SUPPORTS D EXTREMITES DE PRECISION - SUPPORT UNIT FOR BALLSCREWS LOŽISKA PRE GULIČKOVÉ SKRUTKY A TRAPÉZOVÉ SKRUTKY Výber správnej podpory konca uličkovej skrutky či trapézovej skrutky je dôležité pre správnu
SUPPORTS D EXTREMITES DE PRECISION - SUPPORT UNIT FOR BALLSCREWS LOŽISKA PRE GULIČKOVÉ SKRUTKY A TRAPÉZOVÉ SKRUTKY Výber správnej podpory konca uličkovej skrutky či trapézovej skrutky je dôležité pre správnu
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ
 BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΜΟΡΙΑΚΑ ΜΟΝΤΕΛΑ 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING) 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING)
BIOXHMEIA, TOMOΣ I ΠANEΠIΣTHMIAKEΣ EKΔOΣEIΣ KPHTHΣ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΑΠΕΙΚΟΝΙΣΗ ΜΟΡΙΑΚΩΝ ΔΟΜΩΝ ΜΟΡΙΑΚΑ ΜΟΝΤΕΛΑ 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING) 1: ΧΩΡΟΠΛΗΡΩΤΙΚΟ ΜΟΝΤΕΛΟ (SPACE-FILLING)
Goniometrické substitúcie
 Goniometrické substitúcie Marta Kossaczká S goniometrickými funkciami ste sa už určite stretli, pravdepodobne predovšetkým v geometrii. Ich použitie tam ale zďaleka nekončí. Nazačiatoksizhrňme,čoonichvieme.Funkciesínusakosínussadajúdefinovať
Goniometrické substitúcie Marta Kossaczká S goniometrickými funkciami ste sa už určite stretli, pravdepodobne predovšetkým v geometrii. Ich použitie tam ale zďaleka nekončí. Nazačiatoksizhrňme,čoonichvieme.Funkciesínusakosínussadajúdefinovať
16. Základne rovinné útvary kružnica a kruh
 16. Základne rovinné útvary kružnica a kruh Kružnica k so stredom S a polomerom r nazývame množinou všetkých bodov X v rovine, ktoré majú od pevného bodu S konštantnú vzdialenosť /SX/ = r, kde r (patri)
16. Základne rovinné útvary kružnica a kruh Kružnica k so stredom S a polomerom r nazývame množinou všetkých bodov X v rovine, ktoré majú od pevného bodu S konštantnú vzdialenosť /SX/ = r, kde r (patri)
ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ 30-05-2012 ΑΠΑΝΤΗΣΕΙΣ
 ΘΕΜΑ Α. Α1 α Α2 γ Α3 δ Α4 β Α5 γ ΘΕΜΑ Β. ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ 30-05-2012 ΑΠΑΝΤΗΣΕΙΣ Β1. Σχ. Βιβλίο σελ. 120: «Για την επιλογή οργάνων συμβατών για μεταμόσχευση.» μέχρι «και
ΘΕΜΑ Α. Α1 α Α2 γ Α3 δ Α4 β Α5 γ ΘΕΜΑ Β. ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΚΑΤΕΥΘΥΝΣΗΣ ΠΑΝΕΛΛΑΔΙΚΕΣ ΕΞΕΤΑΣΕΙΣ 30-05-2012 ΑΠΑΝΤΗΣΕΙΣ Β1. Σχ. Βιβλίο σελ. 120: «Για την επιλογή οργάνων συμβατών για μεταμόσχευση.» μέχρι «και
Komplexné čísla, Diskrétna Fourierova transformácia 1
 Komplexné čísla, Diskrétna Fourierova transformácia Komplexné čísla C - množina všetkých komplexných čísel komplexné číslo: z = a + bi, kde a, b R, i - imaginárna jednotka i =, t.j. i =. komplexne združené
Komplexné čísla, Diskrétna Fourierova transformácia Komplexné čísla C - množina všetkých komplexných čísel komplexné číslo: z = a + bi, kde a, b R, i - imaginárna jednotka i =, t.j. i =. komplexne združené
Úvod do lineárnej algebry. Monika Molnárová Prednášky
 Úvod do lineárnej algebry Monika Molnárová Prednášky 2006 Prednášky: 3 17 marca 2006 4 24 marca 2006 c RNDr Monika Molnárová, PhD Obsah 2 Sústavy lineárnych rovníc 25 21 Riešenie sústavy lineárnych rovníc
Úvod do lineárnej algebry Monika Molnárová Prednášky 2006 Prednášky: 3 17 marca 2006 4 24 marca 2006 c RNDr Monika Molnárová, PhD Obsah 2 Sústavy lineárnych rovníc 25 21 Riešenie sústavy lineárnych rovníc
množiny F G = {t1, t2,, tn} T a pre ľubovoľný valec C so základňou B1, B2,, Bn v bodoch t1, t2,, tn, takou, že pre t G - F je Bt = E, platí PF(C) = PG
 = PG") STOCHASTICKÝ PROCES Definícia stochastického procesu Definícia 1 Nech (Ω, F, P) je pravdepodobnostný priestor a nech T je podmnožina R. Pre každé t T nech X(t, ω) je náhodná premenná definovaná na pravdepodobnostnom
STOCHASTICKÝ PROCES Definícia stochastického procesu Definícia 1 Nech (Ω, F, P) je pravdepodobnostný priestor a nech T je podmnožina R. Pre každé t T nech X(t, ω) je náhodná premenná definovaná na pravdepodobnostnom
Odporníky. 1. Príklad1. TESLA TR
 Odporníky Úloha cvičenia: 1.Zistite technické údaje odporníkov pomocou katalógov 2.Zistite menovitú hodnotu odporníkov označených farebným kódom Schématická značka: 1. Príklad1. TESLA TR 163 200 ±1% L
Odporníky Úloha cvičenia: 1.Zistite technické údaje odporníkov pomocou katalógov 2.Zistite menovitú hodnotu odporníkov označených farebným kódom Schématická značka: 1. Príklad1. TESLA TR 163 200 ±1% L
Motivácia pojmu derivácia
 Derivácia funkcie Motivácia pojmu derivácia Zaujíma nás priemerná intenzita zmeny nejakej veličiny (dráhy, rastu populácie, veľkosti elektrického náboja, hmotnosti), vzhľadom na inú veličinu (čas, dĺžka)
Derivácia funkcie Motivácia pojmu derivácia Zaujíma nás priemerná intenzita zmeny nejakej veličiny (dráhy, rastu populácie, veľkosti elektrického náboja, hmotnosti), vzhľadom na inú veličinu (čas, dĺžka)
Matematika 2. časť: Funkcia viac premenných Letný semester 2013/2014
 Matematika 2 časť: Funkcia viac premenných Letný semester 2013/2014 RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk
Matematika 2 časť: Funkcia viac premenných Letný semester 2013/2014 RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk
ΜΟΡΙΑΚΕΣ ΜΕΘΟΔΟΙ ΚΡΙΤΗΡΙΑ ΕΠΙΛΟΓΗΣ ΑΞΙΟΛΟΓΗΣΗ
 - - ό ό : Ω ί ό ί ώ.. ά ή ί ός ή ς ύ ί ί ή έ ώ ό. ή ς ά ς ής ό ός ά ς ή έ ός ή ς ί ής ί ς. ό έ ώ - ώ ή ής ή ς- ί ά ά ς ί ς. ά ί ί έ έ ά ύ ή, ό ί ά, ό ό ά έ ά ής ί ύ George Wald ή ί έ ς ί ύ ό ς ί ς ά έ
- - ό ό : Ω ί ό ί ώ.. ά ή ί ός ή ς ύ ί ί ή έ ώ ό. ή ς ά ς ής ό ός ά ς ή έ ός ή ς ί ής ί ς. ό έ ώ - ώ ή ής ή ς- ί ά ά ς ί ς. ά ί ί έ έ ά ύ ή, ό ί ά, ό ό ά έ ά ής ί ύ George Wald ή ί έ ς ί ύ ό ς ί ς ά έ