CIKLUS AZOTA I SUMPORA U PRIRODI
|
|
|
- Τηθύς Έλλη Γιάνναρης
- 5 χρόνια πριν
- Προβολές:
Transcript
1 CIKLUS AZOTA I SUMPORA U PRIRODI Asimilacija azota i sumpora zahtevaju niz biohemijskih reakcija koje spadaju u reakcije sa velikim energetskim zahtevima. Za asimilaciju azota iz nitrata (NO 3- ) potrebna je energija ekvivalentna onoj koju sadrži 12 molekula ATP po jednom atomu azota koji se asimiluje. Za proces biološke fiksacije azota (konverzija N 2 u NH 3 ), zajedno sa potonjom asimilacijom amonijaka u aminokiseline potreban je utrošak energije ekvivalentne onoj koju sadrži 16 molekula ATP, po svakom atomu azota koji se asimiluje. Za asimilaciju sulfata (SO 4 2- ) u aminokiselinu cistein troši se ekvivalent energije koji sadrži 14 molekula ATP.
2 Azot se u prirodi nalazi u više različitih oblika: kao gas N 2 (oko 78% atmosferskog vazduha) u obliku neorganskih jona NO 3- i NH 4+ ulazi u sastav zemljišta kao sastojak mnogih organskih jedinjenja rasprostranjen je u celom živom svetu.

3 Svi oblici azota se u prirodi neprekidno transformišu jedan u drugi i te promene su povezane u ciklus azota.. Ciklus započinje konverzijom atmosferskog N 2 u NH 4+ u procesu biološke fiksacije azota, pod dejstvom azotofiksatora. NH 4+ joni se se mogu usvojiti od strane korenova ili u procesu biološke oksidacije (nitrifikacije) oksidovati u NO 3-. U procesu denitrifikacije N 2 se vraća u atmosferu. U procesu amonifikacije ostaci biljaka i životinja se razlažu i oslobađa se NH 4+.
 joni.")
4 Snabdevanje azotom iz atmosfere podrazumeva raskidanje izuzetno stabilne trostruke kovalentne veze između dva atoma azota u molekulu gasovitog azota, kako bi nastali amonijum (NH 4+ ) i nitratni (NO 3- ) joni. Te hemijske reakcije su poznate pod nazivom fiksacija azota i mogu da se odvijaju i industrijskim (Haber-Bosch-ov proces proizvodnje amonijaka) i prirodnim (električna pražnjenja u atmosferi, fotohemijske reakcije i biološka fiksacija) putem.
 različite bakterije i gljivice razlažu, pri čemu se obrazuje amonijak.")
5 Više biljke dobijaju azot isključivo iz zemljišta na kome žive. Blizu 90% azota u zemljištu se nalazi u obliku organskih azotnih jedinjenja, koja u procesu mineralizacije (amonifikacije) različite bakterije i gljivice razlažu, pri čemu se obrazuje amonijak. Amonijumovi joni su vezani za anjone na površini zemljišnjih koloida. Amonijumovi joni su podložni hemosintetičkoj oksidaciji ili nitrifikaciji koju obavljaju bakterije : Nitrosomonas oksiduje NH 4+ u NO 2 - Nitrobacter oksiduje NO 2- u NO 3 -
6 Nitrat koji na taj način nastaje, sa nitratom koji potiče iz drugih izvora (veštačka đubriva i dr.), pretstavlja najvažniji oblik azota koji biljke primaju. Količina nitrata (NO 3- ) nije stabilna. Nitratni joni nisu vezani za koloide zemljišta, pa ih voda lako odnosi u dublje slojeve. Nitrat je podložan i denitrifikaciji (Micrococcus denitrificans - N 2 ) pri čemu se oslobađa u atmosferu dinitrogen (N 2 ).
7 Biološka redukcija nitrata i nitrita Azot u obliku gasa je najobilnija vrsta azota u prirodi, ali više biljke ne mogu da ga koriste. Molekul azota je veoma inertan (tri veze među atomoma) i ne stupa lako u reakciju sa drugim elementima. Ako biljka primi azot u obliku NH 4+ jona odmah ga ugrađuje u organska jedinjenja. NO 3- mora da se redukuje, a redukcija se u dva stupnja: 1. nitrat se najpre redukuje u nitrit, 2. nitrit redukuje u amonijumov jon Ove procese, koji pretstavljaju prvi korak u asimilaciji azota, obavljaju dva enzima, nitratna i nitritna reduktaza.


8 GDE SE VRŠI REDUKCIJA?

9 Redukcija nitrata može da se obavi u korenu, gde se izgrađuju aminokiseline i amidi, koji se u stablo transportuju putem ksilema. Nitratni jon može i da bude transportovan u listove, u kojima se onda obavlja redukcija i sinteza organskih jedinjenja. Pri niskoj koncentraciji nitrata (oko 1 mol m -3 ), asimilacija u korenu je znatna u odnosu na stablo. Pri većoj koncentraciji nitrata (oko 20 mol m -3 ), koja se postiže primenom đubriva, asimilacija u listovima se značajno povećava i može da prevaziđe onu u korenu.


10 Lupinus albus više od 90% apsorbovanog nitrata redukuje se u korenu. Xanthium strumarium - nitrat se transportuje u listove.
11 Redukcija nitrata u nitrit U kom ćelijskom kompartmentu se to obavlja? Koji enzim vrši redukciju?
.")

12 Redukcija nitrata se obavlja uvek u citosolu ćelija korena i lista. Enzim nitratna reduktaza (NR). Prvi stupanj u redukciji je prenos dva elektrona na nitrat, pri čemu se dobija nitrit. Donor elektrona za redukciju nitrata može biti redukovani NADPH ili NADH.

13 Pošto je nitratna reduktaza prvi enzim u asimilaciji nitrata, ona svojom aktivnošću određuje brzinu celog procesa. Izoforme NR se razlikuju po tome da li elektrone primaju samo od NADH ili i od NADH i od NADPH (ove druge su zastupljene u tkivima koja nisu zelena, tkivima korena napr.) Sekundarni aktivni transport (kotransporteri obavljaju simport protona i nitrata iz apoplasta u citoplazmu)

, citohrom b557 (hemsko gvožđe) i molibdenski kofaktor (MoCo).")
14 To je dimer, sastavljen od dva identična polipeptida, od kojih svaki ima masu od kd. Obe subjedinice nose po 3 različite prostetične grupe: flavin-adenindinukleotid (FAD), citohrom b557 (hemsko gvožđe) i molibdenski kofaktor (MoCo). Molibden vezan za pterin, jedno heterociklično jedinjenje, slično vitaminu folnoj kiselini.
15 Nitratna reduktaza se može pretstaviti kao kratak, rastvorljiv polipeptid, koji funkcioniše kao elektrontransportni lanac: NAD(P)H 2 elektrona FAD-citohrom b557 molibdopterin Redukciju nitrata inhibiraju cijanid i azid. nitrat

16 Redukcija nitrita u amonijum Redukciju nitrita obavlja enzim nitritna reduktaza (NiR). Ovaj enzim se nalazi isključivo u plastidima, i to u hloroplastima ćelija lista i u proplastidima ćelija korena. Nitrit uvek nastaje redukcijom nitrata u citoplazmi. Donor elektrona za ovu reakciju u hloroplastima je redukovani feredoksin, koji nastaje u fotosintezi svetloj fazi. U korenu elektroni potiču iz procesa disanja, pa su njihovi nosioci NADH ili NADPH.

17 Nitritna reduktaza obavlja prenos 6 elektrona na nitrit, pri čemu se ne pojavljuju međuprodukti, nego se odmah dobija amonijumov jon. NO e - + 8H + NH H 2 O Nitritna reduktaza je monomerni polipeptid od kd, čije su prostetične grupe 4Fe-4S centar i sirohem, jedno porfirinsko jedinjenje koje u centru sadrži atom gvožđa.
18 Regulacija nitratne i nitritne reduktaze Nitratna reduktaza se brzo degraduje - labilan enzim. Nitratna reduktaza je inducibilan enzim, čija aktivnost se javlja tek u prisustvu nitrata. Nitrat indukuje transkripciju gena koji kodiraju NR. Istovremeno se aktivira i sinteza drugih enzima kao NiR. Nitritna reduktaza, osim nitratom indukuje se i nitritom, koji nastaje u prvom stupnju procesa. Osim nitrata, koji je primarni signal za transkripciju gena ima i drugih faktora koji deluju na sam proces: svetlost, hormoni iz grupe citokinina, neki metaboliti.

19 Asimilacija amonijumovih jona Azot se ugrađuje u organska jedinjenja samo u obliku amonijumovih jona. Biljka može da primi amonijumov jon iz spoljašnje sredine. On takođe nastaje redukcijom nitrata u korenu ili listu. Fotorespiracija u listu C-3 biljaka je takđe izvor ovih jona. Amonijumov jona se u organska jedinjenja uvek ugrađuje preko aminokiselina. Za sintezu aminokiselina potrebno je: ketokiseline koje primaju NH 4 +, ATP i reduktanti.
 može da da dve glutaminske kiseline.")
20 U aminokseline NH 4+ ulazi preko cikličnog procesa u kome učestvuju glutaminska kiselina (glutamat) i glutamin. Amonijum grupa vezuje za glutaminsku kiselinu pri čemu nastaje glutamin, koji transaminacijom (uz prisustvo α keto glutarne kiseline) može da da dve glutaminske kiseline. Za proces su potrebni ATP i dva enzima glutamin sintetaza (GS) i glutamat sintaza (GOGAT). glutamin sintetaza (GS) i glutamat sintaza (GOGAT) čine sistem GS/GOGAT
 kao donor i jedna ketokiselina kao akceptor NH 2 grupe.")
21 Glutaminska kiselina (primarna aminakiselina) i glutamin su glavni donori NH 2 grupe za sva druga azotna jedinjenja. Većina aminokselina nastaje u procesu transaminacije u kome učestvuje jedna amnokiselina (ili amid) kao donor i jedna ketokiselina kao akceptor NH 2 grupe. Transaminaciju obavlja enzimi aminotransferaze. Od oksalacetata i glutamata nastaje aspartat. U reakciji glutaminom od aspartata se izgrađuje asparagin. Glutamat, aspartat, glutamin i asparagin su donori NH 2 grupe za sintezu drugih aminokiselina npr. alanina.
22 ASIMILACIJA SUMPORA
23 Sumpor se u zemljištu nalazi u obliku različitih mineralnih soli i organskih jedinjenja. Sumporni gasovi se takođe nalaze u atmosferi. Svi ovi oblici sumpora su dostupni biljkama i one ih mogu koristiti. Ipak je sulfatni jon najčešći oblik sumpora koji biljka apsorbuje.

24 Biološka redukcija sulfata i sulfita Sulfat se redukuje u hloroplastima ćelija lista, kao i u proplastidima ćelija korena. Kod C-4 biljaka glavno mesto redukcije su hloroplasti sare provodnog snopića. Sulfatni jon je dosta inertan i njegovoj redukciji prethodi aktivacija. Ona se sastoji u formiranju aktivnog sulfata, koji ima dva oblika: adenozin-5'-fosfosulfat (APS) i 3'-fosfoadenozin-5'- fosfosulfat (PAPS).
25 adenozin-5'-fosfosulfat (APS) 3'-fosfoadenozin-5'-fosfosulfat (PAPS) Pirofosfat (PPi) se razlaže pod dejstvom pirofosforilaze, čime se konzervira jedan deo utrošene energije. Drugu reakciju obavlja APS-kinaza: APS + ATP PAPS + ADP U redukciji je supstrat verovatno APS, a PAPS je oblik akumulacije sulfata. Redukcija APS je proces koji se uglavnom obavlja u plastidima. APS prima dva elektrona od glutationa (GSH) i dobija se sulfit. APS + 2GSH SO H + + GSSG + AMP Sulfitna reduktaza prenosi šest elektrona sa feredoksina na sulfit, dobija se sulfid (S 2- ): SO Fd red S 2- + Fd ox

26 Redukcija se sastoji od najmanje dva odvojena stupnja, za koje je potreban reduktant iz drugih metaboličkih procesa. U listu je redukcija sulfata zavisna od fotosinteze, pošto ATP i redukcioni ekvivalent potiču direktno iz svetle faze. Asimilacija sulfida podrazumeva vezivanje za serin (zamenu acetil SH grupom), pri čemu nastaje aminokiselina cistein. Cistein se konvertuje u metionin. Cistein je toksičan u većim količinama i odmah se ugrađuje u organska jedinjenja. Fotorespiracija obezbeđuje znatne količine serina (iz kojeg nastaje cistein). Asimilovani sumpor se iz listova transportuje floemom do mesta u biljci gde je intenzivna sinteza proteina (vrhovi stabla i korena, plodovi itd), uglavnom u obliku glutationa.

27 BIOLOŠKA FIKSACIJA AZOTA
28 Šta su diazotrofi?

29 Sposobnost za fiksaciju azota pripada prokariotskim mikroorganizmima iz sasvim različitih taksonomskih grupa, koje osim te sposobnosti nemaju mnogo drugih zajedničkih osobina. S obzirom na način života, oni se dele na dve osnovne grupe: 1. simbiotičke fiksatore 2. slobodne fiksatore azota


30 Simbiotički fiksatori Ovoj grupi pripadaju bakterije, aktinomicete i izvesne modrozelene alge, koje žive u simbiozi sa višim biljkama. Bakterije iz familije Rhizobiaceae (Rhizobium) prouzrokuju pojavu krtola, ili nodula, na korenima biljaka iz reda Fabales. Krtole ili nodule su organi u kojima se obavlja fiksacija azota.


, mahovina")
31 Druga grupa simbionata koji fiksiraju azot su aktinomicete, rod Frankia, koje žive u krtolama na korenu različitih dikotiledonih biljaka. Među njima je u našoj flori rasprostranjen samo Alnus (Betulaceae). Među simbiotske diazotrofe spadaju alge Nostoc i Anabaena, koje su simbionti vrsta Cycas i Ceratozamia. Simbioza sa ovim algama nalazi se i kod nekih paprati (Azolla), mahovina (Anthoceros) i kod mnogih lišajeva. Nostoc Anabaena
.")
. Obligatno anaerobne obuhvataju rod Clostridium, čija je vrsta C.")
32 Slobodne bakterije koje fiksiraju azot: Obligatno aerobne bakterije (Azotobacter), koje su dobro prilagodjene za obavljanje fiksacije N 2 i svih drugih funkcija u prisustvu normalne količine O 2 u vazduhu. Aerobne, mikroaerofilne bakterije, koje žive na vazduhu, ali fiksiraju N 2 samo pri niskoj koncentraciji O 2 (Azospirillum). Tu spada i Thiobacillus ferro-oxidans, koji je i hemosintetička bakterija, jer dobija energiju iz oksidacije fero jona u feri jon. Azotobacter Fakultativno anaerobne, koje žive sa ili bez O 2 ali N 2 fiksiraju samo u anaerobnim uslovima (neke vrste Bacillus). Obligatno anaerobne obuhvataju rod Clostridium, čija je vrsta C. pasteurianum prva uopšte pronadjena diazotrofna vrsta; ovaj rod inače pripada bakterijama koje proizvode buternu kiselinu. Bacillus
33 Nitrogenaza i redukcija N 2 N 2 + 8e - + 8H ATP 2NH 3 + H ADP + 16 Pi Za sve organizme koji fiksiraju azot, karakteristično je da enzim nitrogenaza vezuje N 2 i redukuje ga u NH 3.
34 Nitrogenaza je kompleks dva enzima. Protein I je molibdo-fero-protein (MoFe-protein), koji ima 4 subjedinice sa ukupnom masom kd. On sadrži dva atoma Mo i različit broj (do 24) ferosumpornih kompleksa. Protein II je feroprotein (Fe-protein), sastoji se od dve identične subjedinice sa masom od kd, a sadrži 4Fe-4S ferosumporni kompleks. Oba enzima su veoma osetljiva prema kiseoniku. U ćelijama u kojima se obavlja fiksacija azota moraju postojati anaerobni uslovi ili koncentacija kiseonika ne sme da pređe neke vrlo niske vrednosti.

35 Elektroni potrebni za redukciju N 2 dolaze iz različitih procesa. Prvo redukuju Fe-protein, koji ih zatim prenosi na MoFe. Za redukciju Fe-proteina potrebna je energija, koja se oslobađa hidrolizom ATP. Za jedan mol N 2 potrebno je 16 mola ATP, koji se vezuje za Fe-protein. Za korišćenje ATP-a neophodno je prisustvo jona Mg 2+.
36 Redukcija N 2 je neizbežno vezana za redukciju H +. Za jedan mol N 2 obrazuje se jedan mol H 2.. N 2 + 6e ATP + 6H + 2NH ADP + 12P i 2H + + 2e - + 4ATP H 2 + 4ADP + 4P i Redukcija azota je skup proces. Za svaki preneti elektron potrebna su 2 ATP.
37 Potrebna su tri uslova da bi došlo do redukcije: adekvatni donori elektrona ATP anaerobna sredina Kako su fiksatori azota u velikoj meri među sobom različiti, ova tri uslova ne ostvaruju se na isti način.
.")
38 Leguminoze, inficirane rizobijumom razvijaju na korenu krtole, kvržice ili nodule. Proces formiranja simbioze počinje hemotaksijom rizobija ka korenovima biljke domaćina. Ovaj lokomotorni pokret bakterija izazvan je hemijskim supstancama koje korenovi u određenoj fazi razvića biljke oslobađaju u zemljište (fenolna jedinjenja, flavonoidi i betaini). Dolazi do aktivacije nod gena koji učestvuju u biosintezi Nod faktora. Nod faktori deluju kao povratni signali, aktiviraju lektine (strukturne proteine ćelijskog zida biljaka koji imaju funkciju u prepoznavanju ćelija) u ćelijskim zidovima korenskih dlačica leguminoza, a ovi prepoznaju i olakšavaju vezivanje specifičnih rizobija koje mogu da uđu u simbiozu. Prvi vidljiv znak uvijanje vrha korenske dlačice. Vrh korena obuhvata bakterije i na tom mestu dolazi do hidrolize zida dlačice. Bakterije ulaze u ćeliju korena između ćelijskog zida i membrane.

39 Biljne ćelije luče supstace slične ćelijskom zidu, koje oko bakterija obrazuju tubularni omotač tako počinje formiranje INFEKCIONE niti. Bakterije se dele, nit se produžava u unutrašnjost korena. Infekciona nit je ispunjena matriksom od polisaharida koji verovatno luče oba simbionta. Infekcione niti i nodule se paralelno formiraju.
.")
40 Nakon 24 h dolazi i do morfoloških promena u ćelijama korena. Izvesne ćelije u primarnoj kori počinju da se dele i formira se primordija nodule. Infekciona nit se produžava do primordija i bakterije ulaze u ćelije u centru. U njima se oslobađaju infekcione niti i izlaze u citoplazmu ( na način sličan endocitozi). Po izlazku iz niti bakterije se morfološki menjaju: povećavaju se i do 40 puta, deformišu, obavijene su PERIBAKTERIODNOM membranom BAKTEROIDI. Bakteroidi su stekli status ćelijskih organela i nazivaju se SIMBIOZOMI. LEGHEMOGLOBIN pigment koji se sintetiše u ćelijama nodula i daje crvenkasu boju.

41 Razmnožavanje i diferenciranje ćelija u primordijama dovodi do formiranje krtola. Zavisno od toga da li postoji aktivan meristemski centar postoje dve vrste krtola: indeterminantne i determinantne. Indeterminantne nodule, nastaju naspram ksilemske ploče, od ćelija kore blizu endoderma. Stalno rastu, duguljastog su oblika, zadržavaju meristemsko tkivo na vrhu. Nalaze se kod leguminoza umerenog pojasa: graška, lucerke, deteline...

42 Meristem krtole Zona penetracije bakterija u ćelije Zona ekspanzije inficiranih ćelija Tkivo u kome se nalaze bakterioidi Senescentno tkivo Spoljašnja kora krtole Provodni snopić krtole Unutrašnja kora Provodni snopić korena

43 Determinantne nodule gube meristemsko tkivo nakon infekcije. Postaju deobom subepidermalnih ćelija, kojima se kasnije pridruže i ćelije pericikla.kada se pojave na površini imaju loptast oblik. Javljaju se kod biljaka tropskog porekla: soja, pasulj.. Tkivo u kome se nalaze bakterioidi (4) Senescentno tkivo (4a) Spoljašnja kora krtole (6) Provodni snopić krtole (7) Provodni snopić korena (9)

44 Redukcija N 2 u bakterioidima U TCA ciklusu redukuju se koenzimi NAD, a u respiratornom lancu bakteroida se sintetiše ATP. Energija potrebna za disanje bakterija i fiksaciju N 2, potiče iz metabolički procesa domaćina. Neposredni izvor energije su organske kiseline. Saharoza se razlaže u ćelijama nodula. Pošto su uslovi u ćelijama skoro anaerobni, monosaharidi se oksiduju putem glikolize (PEP karboksilaza vezivanjem CO 2 prizvodi oksalacetat, koji se redukuje u malat). Malat se transportuje u mitohondrije ćelija domaćina gde se u TCA ciklusu sintetišu organske kiseline (primaju NH 4+ ). Malat ili sukcinat se transportuju u bakteroid.
45 Mada je nitrogenaza osetljiva prema kiseoniku, izvesna količina kiseonika je neophodna za oksidativnu fosforilaciju i proizvodnju ATP. U isto vreme u bakteroidima koncentracija kiseonika mora biti niska. Nodule troše četiri puta više kiseonika u disanju nego korenovi. U nodulama postoji barijera za difuziju kiseonika. Unutrašnje ćelije korteksa male, zbijene, bez intercelulara sa posebnim sastavom ćelijskog zida. U ćelijskom zidu se nalazi vodeni sloj koji jako usporava difuziju kiseonika. Kada prođe ovu barijeru kiseonik difunduje u intercelularne prostore koji okružuju ćelije sa bakteroidima. U intercelularima transport kiseonika preuzima leghemoglobin (Lb). Lb ima visok afinitet prema kiseoniku. Polovina leghemoglobina je u obliku LbO 2, pa difunduje do peribakteroidne membrane i otpušta kiseonik koji se troši u disanju. Azot se u bakteroidima redukuje u obliku NH 4 + jona izlazi u citoplazmu nodule, gde se obavlja njegova asimilacija.
 leguminoze koje u")
46 Amonijak koji nastaje u nodulama je toksičan i odmah se asimiluje u organska jedinjenja koje se zatim transportuju ksilemom u listove. Na osnovu toga koja azotna jedinjenja sintetišu u nodulama postoje dva tipa leguminoza: leguminoze koje u kvržicama asimiluju amonijak u amide (uglavnom glutamin i asparagin) leguminoze koje u kvržicama asimiluju amonijak u ureide (alontoinska kiselina, alantoin i citrulin) ova jedinjenja se u listovima katabolizuju u amonijak, a zatim opet asimiluju u glutamin.
47 DISANJE Pod disanjem biljaka podrazumeva se ćelijsko disanje.

48 Disanje je proces u kome se organska materija razlaže do konačnih produkata CO 2 i H 2 O, uz oslobađanje energije. C 6 H 12 O 6 + 6O 2 6CO 2 +6H 2 O Do svih ćelija u biljci O 2 dolazi difuzijom, a CO 2 se na isti način izdvaja.
 najčešće su ugljeni hidrati (skrob, saharoza i glukoza).")
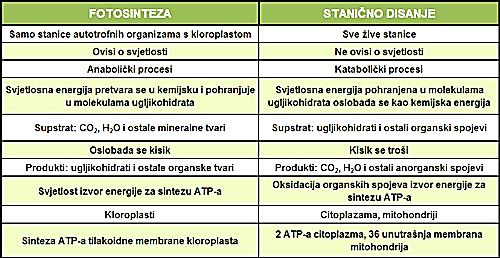
49 U procesu fotosinteze sintetišu se ugljenihidrati, dok u procesu ćelijskog disanja se oslobađa energija pohranjena u njima. Light energy ECOSYSTEM Kod biljaka kao supstrat za disanje (odnosno izvor energije) najčešće su ugljeni hidrati (skrob, saharoza i glukoza). Mogu se koristiti i lipidi (glioksalatni ciklus), organske kiseline i u manjoj meri proteini. CO 2 + H 2 O Photosynthesis in chloroplasts Cellular respiration in mitochondria Organic molecules + O 2 Prvobitno se izvrši njihovo enzimatsko razlaganje do prostijih jedinjenja koja dalje mogu biti razgrađena u procesu disanja na različitim etapama procesa. ATP powers most cellular work Figure 9.2 Heat energy
50
 Anaerobno disanje (odsustvo")
51 Dva osnovna tipa disanja: Aerobno disanje (prisustvo kiseonika) Anaerobno disanje (odsustvo kiseonika)
 koji se sastoji od amiloze i amilopektina. Sinteza primarnog skroba odvija se u hloroplastima.")
52 Da bi mogle da se oksidišu skrob i saharoza se prvo razlože do monosaharida, a zatim fosforilišu. Najrasprostranjeniji oblik rezervnih ugljenih hidrata je skrob (polimer ά-d-glukopiranoze) koji se sastoji od amiloze i amilopektina. Sinteza primarnog skroba odvija se u hloroplastima. U mnogim drugim organima postaje od saharoze koja se do njih transportuje floemom, a zatim ulazi u sve ćelije tkiva za magacioniranje (u ovim ćelijama proplastidi se razvijaju u amiloplaste, u kojima se obavlja sinteza skroba). Skrob se razgrađuje u: - amiloplastima do glukozidnih ostataka uz učešće nekoliko enzima (ά-amilaza, β- amilaza i R-enzim) - hloroplastima uz učešće skrobne fosfataze do glukozo-1-fosfata Saharoza se razlaže u citoplazmi i to: - preko invertaze do glukoze i fruktoze koje se zatim fosforilišu (da bi ušle u amiloplast) - preko saharozne sintaze i uridin difosfata (UDP) do uridin difosfoglukoze i fruktoze. Uridin difosfoglukoza prelazi u glukozo-1- fosfat. Kad se iscrpe rezerve ugljenih hidrata za disanje se kao supstrat koriste i lipidi. Lipidi se kao trigliceridi razgrađuju do masnih kiselina i glicerina (hidroliza se dešava u sferozomima). Glicerin se u citoplazmi fosforiliše i kao gliceraldehid- 3-fosfat (GAP) ulazi u glikolizu gde se oksiduje. Masne kiseline se u mitohondrijama razlažu u procesu β oksidacije i acetil-coa ulazi u Krebsov ciklus.
53 Mobilizacija ugljenih hidrata za disanje U pripremnoj fazi disanja polisaharidi se najprije razlažu na monoze, koje se zatim fosforilišu da bi mogle biti oksidovane. Krajnji produkt pripreme za disanje je fruktoza 1,6-bisfosfat (monosaharidi koji se dobiju razlaganjem drugih ugljenih hidrata se u pripremnoj fazi prvo transformišu u glukozu ili fruktozu, koje se zatim fosforilišu). Postoje dva alternativna puta u oksidativnoj degradaciji šećera glikoliza oksiduje se dva puta fosforilovana fruktoza oksidativni pentozni ciklus tzv. direktna oksidacija oksiduje se jedanput fosforilovana glukoza Korišćenje PPi za fosforilaciju čini prvu značajnu razliku između biljnih i animalnih ćelija. PPi je uzgredni produkt anabolizma i dalje se on samo razlaže na dva neorganska fosfata. Biljke mogu da koriste i ovu visokoenergetsku vezu i prištede ATP. G-1-P ulazi pomoću posebnog translokatora u amiloplast i pomoću ADP-glukozne pirofosforilaze prelazi u ADP-glukozu koja služi za sintezu skroba.

54 U aerobnom disanju ćelija koristi energiju vezanu u pruvatu, koji nastaje razgradnjom glukoze, pri čemu nastaju NADH i ATP. Disanje obuhvata tri faze u kojima glukoza oksiduje u nizu reakcija: Glikoliza Krebsov ciklus Transportni lanac elektrona Pre početka glikolize razlaže se skrob do glukoze. Zatim se glukoza aktivira (Fru-1,6-BP) i tek tada počinje glikoliza.
55 Procesi oksidacije glukoze su lokalizovani u citoplazmi, mitohondrijama i delom u plastidima (masti sferozomi i glioksizomi). U I fazi - glikolizi dolazi do razgradnje glukoze do piruvata. Glikoliza se odvija u citoplazmi. U II fazi Krebsov ciklus odvija se u matriksu mitohondrija. Tokom II faze dolazi do oksidacije acetil koenzima A do CO 2 uz redukciju NAD + u NADH. U III fazi sinteza ATP na membranama mitohondrija uz utrošak NADH. Na membranama mitohondrija generiše se gradijent protona na osnovu koga se sintetiše ATP.

56 Glikoliza Razgradnja glukoze u anaerobnom procesu, deo energije se skladišti u ATP Kod svih organizama Kod aerobnih organizama: priprema za Krebsov ciklus i oksidativnu fosforilaciju Kod anaerobnih: priprema za alkoholno i mlečnokiselinsko vrenje
57 Glikoliza Fruktoza 1,6-bifosfat se razlaže na dve trioze: gliceraldehid-3-fosfat i dihidroksiaceton aldolaza
58 Glikoliza Gliceraldehid-3-fosfat se oksiduje u 1,3-fosfoglicerinsku kiselinu Redukcija NAD u NADH+H* Gliceraldehid-3- fosfat-dehidrogenaza
59 Defosforilacija 1,3-difosfoglicerinske kiseline u fosfoenolpirogrožđanu kiselinu: Fosfatnu grupu sa 1,3-difosfoglicerinske kiseline se prenosi na ADP i nastaje 3-fosfoglicerat i ATP. SUPSTRATNA FOSFORILACIJA fosfatna grupa se direktno prenosi sa supstrata na ADP.

60 Defosforilacija fosfoenolpirogrožđane kiseline (PEP) u pirogrožđanu kiselinu; odvajanje još jedne fosfatne grupe i prenošenje na drugi ADP i sinteza drugog molekula ATP-a (supstratna fosforilacija). Piruvat kinaza I druga trioza dobijena od fruktoze takođe podleže ovim reakcijama, tako da se kao krajnji produkt glikolize dobijaju dva molekula pirogrožđane kiseline. PGK u aerobnom metabolizmu prelazi u mitohondrije i biva dalje iskorišćena za dalje dobijanje energije.

61 Energetski bilans glikolize Dva molekula ATP-a se troše u pripremi 4 molekula ATP-a se dobijaju u oksidaciji 2 molekula NAD-a se redukuju za obe trioze 2 molekula ATP-a su krajnji energetski rezultata 2 molekula NADPH+H + su produkt glikolize
62 Energetski bilans glikolize C 6 H 12 O 6 +2ATP+2NAD 2C 3 H 4 O 3 +2ATP+2NADH+2H + Od jednog molekula glukoze nastaju dva molekula pirogrožđane kiseline (PGK), dva molekula ATP-a i 2NADH Značaj glikolize: - sinteza ATP - sinteza NADH koji je bogat energijom i koristi se za: oksidaciju u mitohondrijama (sinteza ATP) kao izvor elektrona Pirogrožđana kiselina može da se koristi u procesima: fermentacije-vrenja (u anaerobnim uslovima) ciklusu trikarbonskih kiselina (u prisustvu kiseonika)
63 Na dva mesta se izbegava korišćenje adenilnih nukleotida (ADP/ATP): Enzim dehidrogenaza gliceraldehid -3-fosfata oksiduje 3-GAP pomoću NAD koenzima, bez fosforilacije ADP. Fosfoenolpiruvatna fosfataza defosforiliše PEP, odvaja neorganski fosfat bez korišćenja ADP.
64 OKSIDATIVNI PENTOZNI CIKLUS Direktna oksidacija glukoze je alternativan put za oksidaciju ugljenih hidrata. Sastoji se iz dvije faze: - oksidativne (koja je ireverzibilna) i - reverzibilne faze koja obuhvata metabolizam pentoza Ovaj proces se obavlja u citosolu i u hloroplastima u mraku (na svetlu reduktivni pentozni ciklus, a u mraku niz reverzibilnih reakcija koje predstavljaju oksidativno pentozni ciklus). U ovom procesu stvaraju se redukcioni ekvivalenti (NADPH) i pentozo fosfati koji su prekursori za ribonukleinske kiseline, a mogu se uključiti i u Kalvinov ciklus.
65 Anaerobni put pirogrožđane kiseline Dobijaju se svega dva molekula ATPa za svaki molekul glukoze. Anaerobni organizmi, bakterije, kvasci
66 Razgradnja PGK kod aerobnih organizama Odvija se u tri etape: 1. U matriksu mitohondrija obrazuje se acetil-koenzim A dekarboksilacijom piruvata uz učešće piruvat dehidrogenaze. 2. Oksidacija organskih kiselina započinje sintezom limunske kiseline (citrat) citrat sintaza. Acetil-CoA (C-2) se vezuje sa oksalsirćetnom kiselinom (C-4) i daje limunsku kiselinu (C-6). Dalje reakcije su reakcije transformacije limunske kiselina (sa tri karboksilne grupe). Odvija se oksidativna dekarboksilacija trikarbonskih kiselina 3. i oksidativna fosforilacija. 4. Transport elektrona i protona PGK 3C PGK CO2 oksalsirćetna kiselina 3NADH FADH2 + acetil - koenzim A 2C Krebsov ciklus 2CO2 koenzim A (CoA) NAD + ATP NADH limunska kiselina MITOHONDRIJA CITOPLAZMA Prvi intermedijer KC-a je limunska kiselina (ciklus limunske kisline, ciklus trikarboksilne kiseline)
67
68 Produkti Krebsovog ciklusa Dva molekula CO 2 u dva stupnja (koraci 3 i 4). Energija oslobođena u oksidacijama u 4 stupnja u obliku redukovanih koenzima NADH+H+ i FADH+H (koraci 3,4,6,8). Regenerišu se oksalsirćetna kiselina i acetil-coa koji ponovo ulaze u ciklus.
,")

69 Tri ugljenika ulaze u ciklus (kao pirogrožđana kiselina), a pojavljuje se 3 CO 2. 2 piruvata +8NAD+2FAD+2ADP+2Pi 6CO2+8NADH+2FADH+2ATP Fosforilacijom na supstratu nastaje 2 mola ATP i jedan mol glukoze, kao i redukcioni koenzimi (8NADH, 2FADH), koji ponovnm oksidacijom u završnom stupnju disanja daju ATP. Svi ugljovodonici su oksidovani u CO 2. Kod biljaka od sukcinil-coa obrazuje sukcinat uz sukcinil-coa sintetazu i sintetiše 1ATP, a ne GTP kao životinja. Kod biljaka postoji i alternativni put oksidacije C4 kiselina (malata, citrata,άketoglutarat). NAD-malat enzim učestvuje u alternativnom putu. Malat iz citoplazme ulazi u matriks mitohondrija.

70 Prenos elektrona sa redukovanih koenzima na kiseonik TERMINALNA OKSIDACIJA; a komponente koje učestvuju u prenosu elektrona čine elektron transportni lanac.
 se premeštaju u međumembranski prostor Razlika u koncentraciji protona pokreće ATP-sintetazu da sintetiše ATP u procesu oksidativne fosforilacije Usled prenošenja")
71 NADH i FADH 2 predaju elektrone elektron-transportnom lancu smeštenom u unutrašnjoj membrani mitrohondrija Krajnji primalac elektrona je O 2 : kiseonik; protoni i elektroni daju H 2 O Prilikom transporta elektrona, protoni (H + ) se premeštaju u međumembranski prostor Razlika u koncentraciji protona pokreće ATP-sintetazu da sintetiše ATP u procesu oksidativne fosforilacije Usled prenošenja elektrona kroz membranu dolazi do transporta protona kroz membranu i uspostavljanja elektrohemijskog gradijenta ET lanac biljnih mitohondrija je neosetljiv prema cijanidu i drugim inhibitorima citohrom oksidaze (IV kompleks). Elektroni ne dolaze do kompleksa III i IV već ih ubikinol oksidaza prenosi na alternativnu oksidazu koja redukuje O 2 i proizvodi vodu. Kod biljaka iz fam. Araceae cijanid čak stimuliše potrošnju kiseonika.
 se pumpaju u međumembranski prostor 4. Razlika u koncentraciji protona pokreće ATP-sintetazu da stvara ATP matriks elektron-transportni lanac 1.")
72 međumembranski prostor unutrašnja membrana Elektron-transportni lanac Oksidacijom jednog molekula NADH+H + nastaju TRI molekula ATP-a Oksidacijom jednog molekula FADH+H + nastaju DVA molekula ATP-a. 2. Protoni (H + ) se pumpaju u međumembranski prostor 4. Razlika u koncentraciji protona pokreće ATP-sintetazu da stvara ATP matriks elektron-transportni lanac 1. NADH i FADH 2 predaju elektrone elektrontransportnom lancu 3. Krajnji primalac elektrona je O 2, nastaje H 2 O ATP-sintetaza U membrani se nalazi protonska pumpa koja ima i funkciju sinteze ATP-a Ona koristi gradijent protona za sintezu ATP-a.
73 Prije svega TCA je povezan sa glikolizom i oba čine ne samo deo glavnog oksidativnog procesa u ćeliji, nego služe za proizvodnju intermedijernih jedinjenja koja su početni supstrat u drugim biosintetičkim putevima.
74 Energetski bilans glikolize, Krebsovog ciklusa i oksidativne fosforilacije Ulazni molekuli: 1 molekul glukoze ili 2 molekula trioze (PGK) 36 molekula ATP-a: 6 ATP iz glikolize 6 ATP-a iz sinteze acetil CoA (iz 2NADH+H + ) 24 ATP-a iz oksidativne fosforilacije iz NADH+H + u elektron transportnom lancu
75 Glioksilatni ciklus Predstavlja modifikaciju TCA ciklusa, obavlja se u glioksizomima i peroksizomima. I u ovom ciklusu acetil-coa se kondezuje sa oksaloacetatom u limunsku kiselinu, a zatim nastaje izolimunska kiselina. Ne dolazi do oksidativne dekarboksilacije nego se izolimunska kiselina razlaže na glioksilat-jedinjenje sa 2 C atoma, i na sukcinat. Sukcinat prelazi iz glioksizoma u mitohondriju, gdje se oksiduje u TCA ciklusu do malata. Glioksilat se kondezuje sa još jednim molekulom acetil-coa i daje jabučnu kiselinu malat (enzim malatna sintaza). Glioksalatni ciklus prelazak lipida u ugljenehidrate. Enzim malatna dehidrogenaza oksiduje malat u oksaloacetat i tako se regeneriše akceptor acetil-coa za novi ciklus. Ni jedan C se ne oksiduje do CO 2 nego se štede i koriste za dalje procese.
76 Glukoneogeneza Proces stvaranja šećera od masti zove se glukoneogeneza (biljkama su šećeri pogodniji izvor energije za rastenje i razviće). - Malat koji je nastao u mitohondriji ili glioksizomu izlazi u citoplazmu i oksiduje se u oksaloacetat. Oksaloacetat u prisustvu ATP i pod dejstvom PEP-karboksikinaze se dekarboksiliše i fosforiliše i dobija se PEP. PEP ulazi u glikolitički proces, koji protiče u obrnutom smjeru i naziva se glukoneogeneza (sve reakcije u glikolizi su reverzibilne izuzev one u kojoj PEP prelazi u piruvat-ulaskom direktno u reverznu glikolizu ovaj ireverzibilan korak je izbjegnut). Na kraju se izgrađuje se saharoza - to je način na koji se u semenima koja klijaju rezervna ulja pretvaraju u saharozu i koja se transportuje u mladu biljku i služi tek tada za sintezu gradivnog materijala i kao izvor energije. Oksidacija šećera u mladoj biljci preko TCA ciklusa ili oksidativnog pentoznog ciklusa povezuje metabolizam rezervnih lipida sa sintezom mnogih drugih jedinjenja neophodnih za rastenje. Rezervne masti se razlažu u oleazomu u ugljene hidrate, masne kiseline u glioksizomu prolaze kroz β-oksidaciju i glioksilatni ciklus; sukcinat u mitohondriji prolazi kroz TCA ciklus iz koga malat izlazi u citosol. U reverznoj glikolizi izgrađuje se prvo fruktoza bisfosfat, a zatim saharoza.
77 . Uticaj spoljašnjih činilaca na disanje Spoljašnji činioci utiču na intenzitet disanja tako što usporavaju ili ubrzavaju proces. Temperatura je najznačajniji spoljašnji faktor koji utiče na disanje jer su sve enzimske reakcije zavisne od temperature. U granicama variranja temperature od 0 do 40 o C, pri porastu temperature za 10 o C disanje ubrzava 2-3 puta. Pri visokim temperaturama iznad 45 o C pri dugotrajnom delovanju dolazi do promene na enzimima, što negativno utiče na disanje. Disanje kao i svi drugi životni procesi zavisi od prisustva vode u biljnoj ćeliji. Intenzitet disanja zavisi od hidratacije koloida protoplazme. Pri klijanju semena naglo se pojačava intenzitet disanja. U mladim listovima i organima koji sadrže dosta vode i rastu disanje je intenzivno. Disanje korena zavisi od koncentracije rastvorenog kiseonika u vodi koja se nalazi u zemljištu. Kada je zemljište prezasićeno vodom razmena gasova je mala i rastvorljivost kiseonika je mala. Koren u ovim uslovima može doći u stanje anaerobije, čime je onemogućeno disanje. Pri povećanoj koncentraciji CO 2 intenzitet disanja se smanjuje. Disanje se odvija na svetlosti i u mraku. Svetlost posredno utiče na disanje. Od svetlosti zavisi proces fotosinteze u kome se oslobađa kiseonik koji se koristi u disanju.
Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama. Hloroplast
 Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama Hloroplast Procesom ćelijskog disanja deponovana energija u šećerima erima prevodi se u ATP i druge energetske metabolite. Istovremeno se
Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama Hloroplast Procesom ćelijskog disanja deponovana energija u šećerima erima prevodi se u ATP i druge energetske metabolite. Istovremeno se
ASIMILACIJA AZOTA, SUMPORA I FOSFORA
 ASIMILACIJA AZOTA, SUMPORA I FOSFORA Više biljke, kao autotrofni organizmi, mogu da sintetišu organske supstance iz neorganskih, koje usvajaju uglavnom iz zemljišta. Inkorporacija mineralnih elemenata
ASIMILACIJA AZOTA, SUMPORA I FOSFORA Više biljke, kao autotrofni organizmi, mogu da sintetišu organske supstance iz neorganskih, koje usvajaju uglavnom iz zemljišta. Inkorporacija mineralnih elemenata
CIKLUS LIMUNSKE KISELINE (CLK)
") SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
Aminokiseline. Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina 22.12.2014
 Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Disanje (Respiracija)
") Disanje (Respiracija) Osim fotosinteze, deo primarnog metabolizma biljaka je i proces ćelijskog disanja (respiracija). Dok se u fotosintezi procesima redukcije iz CO2 i vode sintetišu organska jedinjenja,
Disanje (Respiracija) Osim fotosinteze, deo primarnog metabolizma biljaka je i proces ćelijskog disanja (respiracija). Dok se u fotosintezi procesima redukcije iz CO2 i vode sintetišu organska jedinjenja,
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
3.1 Granična vrednost funkcije u tački
 3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
METABOLIZAM UGLJENIH HIDRATA
 METABOLIZAM UGLJENIH HIDRATA 14.02.2018. Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O 2. U uslovima prisustva O 2,
METABOLIZAM UGLJENIH HIDRATA 14.02.2018. Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O 2. U uslovima prisustva O 2,
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija
 Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
Glukoneogeneza i regulacija glukoneogeneze
 Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
VODA ELEKTROLITI I ACIDO-BAZNA RAVNOTEŽA...
 SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
Ciklus limunske kiseline-2
 Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
Put pentoza fosfata. B. Mildner. Put pentoza fosfata
 Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
Kloroplasti. Fotosinteza Mitohondriji Stanično disanje
 Kloroplasti Fotosinteza Mitohondriji Stanično disanje http://en.wikipedia.org/wiki/plas tid PLASTIDI Organeli biljnih stanica i stanica algi Proizvodnja i pohranjivanje šećera i drugih molekula Pigmenti
Kloroplasti Fotosinteza Mitohondriji Stanično disanje http://en.wikipedia.org/wiki/plas tid PLASTIDI Organeli biljnih stanica i stanica algi Proizvodnja i pohranjivanje šećera i drugih molekula Pigmenti
MINERALNA ISHRANA. - Hemijski elementi se prema ulozi u biljnom organizmu mogu (uslovno) podeliti na neophodne i korisne:
 podeliti na neophodne i korisne:") MINERALNA ISHRANA - Biljke usvajaju biogene elemente iz spoljašne sredine u vidu gasova (CO 2, O 2 ), organskih molekula (malo) i mineralnih materija. - Mineralne materije učestvuju u izgradnji organskih
MINERALNA ISHRANA - Biljke usvajaju biogene elemente iz spoljašne sredine u vidu gasova (CO 2, O 2 ), organskih molekula (malo) i mineralnih materija. - Mineralne materije učestvuju u izgradnji organskih
XI dvoqas veжbi dr Vladimir Balti. 4. Stabla
 XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
PRAVA. Prava je u prostoru određena jednom svojom tačkom i vektorom paralelnim sa tom pravom ( vektor paralelnosti).
.") PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
numeričkih deskriptivnih mera.
 DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
3. razred gimnazije- opšti i prirodno-matematički smer ALKENI. Aciklični nezasićeni ugljovodonici koji imaju jednu dvostruku vezu.
 ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ISPITNA PITANJA OSNOVI BIOHEMIJE
 UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
OSNOVE METABOLIZMA MIKROORGANIZAMA. Metabolički diverzitet
 OSNOVE METABOLIZMA MIKROORGANIZAMA Metabolički diverzitet Metabolizam - obuhvata sve hemijske procese koji se dešavaju u ćeliji - anabolički (zahtevaju energiju) - katabolički (otpuštaju energiju) Energija
OSNOVE METABOLIZMA MIKROORGANIZAMA Metabolički diverzitet Metabolizam - obuhvata sve hemijske procese koji se dešavaju u ćeliji - anabolički (zahtevaju energiju) - katabolički (otpuštaju energiju) Energija
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović
 DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Iskazna logika 3. Matematička logika u računarstvu. novembar 2012
 Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Elementi spektralne teorije matrica
 Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Ispitivanje toka i skiciranje grafika funkcija
 Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Zavrxni ispit iz Matematiqke analize 1
 Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici
 Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE
 Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
Kiselo bazni indikatori
 Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
ELEKTROTEHNIČKI ODJEL
 MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
ENZIMI KOJI UČESTVUJU U "PLANT GROWTH PROMOTING" POTENCIJALU BAKTERIJA
 ENZIMI KOJI UČESTVUJU U "PLANT GROWTH PROMOTING" POTENCIJALU BAKTERIJA Klimatske promene Porast broja ljudi na planeti ZAŠTO JE OVO VAŽNO Veći zahtevi za poizvodnjom hrane, hrane za životinje, goriva posebno
ENZIMI KOJI UČESTVUJU U "PLANT GROWTH PROMOTING" POTENCIJALU BAKTERIJA Klimatske promene Porast broja ljudi na planeti ZAŠTO JE OVO VAŽNO Veći zahtevi za poizvodnjom hrane, hrane za životinje, goriva posebno
41. Jednačine koje se svode na kvadratne
 . Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
. Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
Organele života i smrti
 MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama.
 Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
M086 LA 1 M106 GRP. Tema: Baza vektorskog prostora. Koordinatni sustav. Norma. CSB nejednakost
 M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
HEMIJSKA VEZA TEORIJA VALENTNE VEZE
 TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
OKSIDACIJSKA FOSFORILACIJA
 OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
UVOD CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN
 = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN") CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
Oksidativna fosforilacija
 Oksidativna fosforilacija Oksidativna fosforilacija U eukariota u mitohondrijama. Odigrava se redukcija O2 do H2O. Donori elektrona su NADH i FADH2. Najvedi deo energije za svoj rast, održavanje i rad
Oksidativna fosforilacija Oksidativna fosforilacija U eukariota u mitohondrijama. Odigrava se redukcija O2 do H2O. Donori elektrona su NADH i FADH2. Najvedi deo energije za svoj rast, održavanje i rad
TEST PITANJA BIOHEMIJA UGLJENI HIDRATI
 TEST PITANJA BIOHEMIJA UGLJENI HIDRATI 1. Kakve osobine pokazuju monosaharidi: a) osidacione b) redukcione c) metilirajuće 2. Skrob, kao dominantan šećer u ishrani čoveka, se razlaže do disaharida u: a)
TEST PITANJA BIOHEMIJA UGLJENI HIDRATI 1. Kakve osobine pokazuju monosaharidi: a) osidacione b) redukcione c) metilirajuće 2. Skrob, kao dominantan šećer u ishrani čoveka, se razlaže do disaharida u: a)
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - UNI
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A
, grupa A") Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
III VEŽBA: FURIJEOVI REDOVI
 III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
18. listopada listopada / 13
 18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE GODINE 8. RAZRED TOČNI ODGOVORI
 21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABLIZEM GLJIKVIH HIDRATV KAK CELICA DBI GLUKZ IN STALE MNSAHARIDE? HRANA ZNTRAJCELIČNI GLIKGEN ali ŠKRB razgradnja s prebavnimi encimi GLUKZA in ostali monosaharidi fosforilitična cepitev prenos do
METABLIZEM GLJIKVIH HIDRATV KAK CELICA DBI GLUKZ IN STALE MNSAHARIDE? HRANA ZNTRAJCELIČNI GLIKGEN ali ŠKRB razgradnja s prebavnimi encimi GLUKZA in ostali monosaharidi fosforilitična cepitev prenos do
Pismeni ispit iz matematike Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: ( ) + 1.
 + 1.") Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
PARCIJALNI IZVODI I DIFERENCIJALI. Sama definicija parcijalnog izvoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je,
 PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
MEĐUMOLEKULSKE SILE JON-DIPOL DIPOL VODONIČNE NE VEZE DIPOL DIPOL-DIPOL DIPOL-INDUKOVANI INDUKOVANI JON-INDUKOVANI DISPERZNE SILE
 MEĐUMLEKULSKE SILE JN-DIPL VDNIČNE NE VEZE DIPL-DIPL JN-INDUKVANI DIPL DIPL-INDUKVANI INDUKVANI DIPL DISPERZNE SILE MEĐUMLEKULSKE SILE jake JNSKA VEZA (metal-nemetal) KVALENTNA VEZA (nemetal-nemetal) METALNA
MEĐUMLEKULSKE SILE JN-DIPL VDNIČNE NE VEZE DIPL-DIPL JN-INDUKVANI DIPL DIPL-INDUKVANI INDUKVANI DIPL DISPERZNE SILE MEĐUMLEKULSKE SILE jake JNSKA VEZA (metal-nemetal) KVALENTNA VEZA (nemetal-nemetal) METALNA
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu
 Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - VSŠ 2005/06
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
Trigonometrija 2. Adicijske formule. Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto
 Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
2 tg x ctg x 1 = =, cos 2x Zbog četvrtog kvadranta rješenje je: 2 ctg x
 Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Linearna algebra 2 prvi kolokvij,
 1 2 3 4 5 Σ jmbag smjer studija Linearna algebra 2 prvi kolokvij, 7. 11. 2012. 1. (10 bodova) Neka je dano preslikavanje s : R 2 R 2 R, s (x, y) = (Ax y), pri čemu je A: R 2 R 2 linearan operator oblika
1 2 3 4 5 Σ jmbag smjer studija Linearna algebra 2 prvi kolokvij, 7. 11. 2012. 1. (10 bodova) Neka je dano preslikavanje s : R 2 R 2 R, s (x, y) = (Ax y), pri čemu je A: R 2 R 2 linearan operator oblika
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f
 D f") IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
Konstruisanje. Dobro došli na... SREDNJA MAŠINSKA ŠKOLA NOVI SAD DEPARTMAN ZA PROJEKTOVANJE I KONSTRUISANJE
 Dobro došli na... Konstruisanje GRANIČNI I KRITIČNI NAPON slajd 2 Kritični naponi Izazivaju kritične promene oblika Delovi ne mogu ispravno da vrše funkciju Izazivaju plastične deformacije Može doći i
Dobro došli na... Konstruisanje GRANIČNI I KRITIČNI NAPON slajd 2 Kritični naponi Izazivaju kritične promene oblika Delovi ne mogu ispravno da vrše funkciju Izazivaju plastične deformacije Može doći i
Teorijske osnove informatike 1
 Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
( ) ( ) 2 UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET. Zadaci za pripremu polaganja kvalifikacionog ispita iz Matematike. 1. Riješiti jednačine: 4
 ( ) 2 UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET. Zadaci za pripremu polaganja kvalifikacionog ispita iz Matematike. 1. Riješiti jednačine: 4") UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET Riješiti jednačine: a) 5 = b) ( ) 3 = c) + 3+ = 7 log3 č) = 8 + 5 ć) sin cos = d) 5cos 6cos + 3 = dž) = đ) + = 3 e) 6 log + log + log = 7 f) ( ) ( ) g) ( ) log
UNIVERZITET U ZENICI POLITEHNIČKI FAKULTET Riješiti jednačine: a) 5 = b) ( ) 3 = c) + 3+ = 7 log3 č) = 8 + 5 ć) sin cos = d) 5cos 6cos + 3 = dž) = đ) + = 3 e) 6 log + log + log = 7 f) ( ) ( ) g) ( ) log
Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. 4/17/2013
 Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
MATEMATIKA 2. Grupa 1 Rexea zadataka. Prvi pismeni kolokvijum, Dragan ori
 MATEMATIKA 2 Prvi pismeni kolokvijum, 14.4.2016 Grupa 1 Rexea zadataka Dragan ori Zadaci i rexea 1. unkcija f : R 2 R definisana je sa xy 2 f(x, y) = x2 + y sin 3 2 x 2, (x, y) (0, 0) + y2 0, (x, y) =
MATEMATIKA 2 Prvi pismeni kolokvijum, 14.4.2016 Grupa 1 Rexea zadataka Dragan ori Zadaci i rexea 1. unkcija f : R 2 R definisana je sa xy 2 f(x, y) = x2 + y sin 3 2 x 2, (x, y) (0, 0) + y2 0, (x, y) =
Osnovni fiziološki procesi u biljkama
 Modul: Osnove agroekologije Tematska cjelina: Ekofiziologija bilja prof. dr. sc. Irena Jug Osnovni fiziološki procesi u biljkama FOTOSINTEZA Fotosinteza je u živom svijetu jedinstveni fizikalno kemijski
Modul: Osnove agroekologije Tematska cjelina: Ekofiziologija bilja prof. dr. sc. Irena Jug Osnovni fiziološki procesi u biljkama FOTOSINTEZA Fotosinteza je u živom svijetu jedinstveni fizikalno kemijski
NOMENKLATURA ORGANSKIH SPOJEVA. Imenovanje aromatskih ugljikovodika
 NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
Reverzibilni procesi
 Reverzbln proces Reverzbln proces: proces pr koja sste nkada nje vše od beskonačno ale vrednost udaljen od ravnoteže, beskonačno ala proena spoljašnjh uslova ože vratt sste u blo koju tačku, proena ože
Reverzbln proces Reverzbln proces: proces pr koja sste nkada nje vše od beskonačno ale vrednost udaljen od ravnoteže, beskonačno ala proena spoljašnjh uslova ože vratt sste u blo koju tačku, proena ože
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
Strukture podataka i algoritmi 1. kolokvij 16. studenog Zadatak 1
 Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
STRUKTURA BIOLOŠKIH MAKROMOLEKULA HEMIJSKE I BIOHEMIJSKE REAKCIJE TRANSPORT RAST I DAJE ČVRSTINU (TURGOR) VODA
 VODA") Snabdevenost biljke vodom je jedan od najvažnijih faktora koji utiče na rastenje i razviće biljaka. Od svih resursa potrebnih biljci voda je najrasprostranjenija, najobilnija i ujedno najveći ograničavajući
Snabdevenost biljke vodom je jedan od najvažnijih faktora koji utiče na rastenje i razviće biljaka. Od svih resursa potrebnih biljci voda je najrasprostranjenija, najobilnija i ujedno najveći ograničavajući
LANCI & ELEMENTI ZA KAČENJE
 LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
Fiksacija dušika i biosinteza aminokiselina
 Fiksacija dušika i biosinteza aminokiselina Boris Mildner Dušik u aminokiselinama, purinima, pirimidinima i drugim molekulama potječe od atmosferskog dušika, N 2. Biosintetski proces započinje redukcijom
Fiksacija dušika i biosinteza aminokiselina Boris Mildner Dušik u aminokiselinama, purinima, pirimidinima i drugim molekulama potječe od atmosferskog dušika, N 2. Biosintetski proces započinje redukcijom
INTEGRALNI RAČUN. Teorije, metodike i povijest infinitezimalnih računa. Lucija Mijić 17. veljače 2011.
 INTEGRALNI RAČUN Teorije, metodike i povijest infinitezimalnih računa Lucija Mijić lucija@ktf-split.hr 17. veljače 2011. Pogledajmo Predstavimo gornju sumu sa Dodamo još jedan Dobivamo pravokutnik sa Odnosno
INTEGRALNI RAČUN Teorije, metodike i povijest infinitezimalnih računa Lucija Mijić lucija@ktf-split.hr 17. veljače 2011. Pogledajmo Predstavimo gornju sumu sa Dodamo još jedan Dobivamo pravokutnik sa Odnosno
Operacije s matricama
 Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
VISOKO GOSPODARSKO UČILIŠTE U KRIŽEVCIMA
 VISOKO GOSPODARSKO UČILIŠTE U KRIŽEVCIMA Dr.sc. Marija Vukobratović Križevci, 2009. 1 UVOD U METABOLIZAM Svi živi organizmi za svoje aktivnosti (održavanje života, rast i razvoj) zahtijevaju energiju,
VISOKO GOSPODARSKO UČILIŠTE U KRIŽEVCIMA Dr.sc. Marija Vukobratović Križevci, 2009. 1 UVOD U METABOLIZAM Svi živi organizmi za svoje aktivnosti (održavanje života, rast i razvoj) zahtijevaju energiju,
7 Algebarske jednadžbe
 7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE)
") (Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
(Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
TRANSPORT JONA I ORGANSKIH JEDINJENJA KROZ MEMBRANE
 TRANSPORT JONA I ORGANSKIH JEDINJENJA KROZ MEMBRANE Transportni mehanizmi na membranama Prema potrebi za energijom i specifičnim učesnicima u transportu postoji 5 načina transporta kroz biološke membrane:
TRANSPORT JONA I ORGANSKIH JEDINJENJA KROZ MEMBRANE Transportni mehanizmi na membranama Prema potrebi za energijom i specifičnim učesnicima u transportu postoji 5 načina transporta kroz biološke membrane:
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =
/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =") 100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
5. Karakteristične funkcije
 5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
KVADRATNA FUNKCIJA. Kvadratna funkcija je oblika: Kriva u ravni koja predstavlja grafik funkcije y = ax + bx + c. je parabola.
 KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE
 **** MLADEN SRAGA **** 011. UNIVERZALNA ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE SKUP REALNIH BROJEVA α Autor: MLADEN SRAGA Grafički urednik: BESPLATNA - WEB-VARIJANTA Tisak: M.I.M.-SRAGA
**** MLADEN SRAGA **** 011. UNIVERZALNA ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE SKUP REALNIH BROJEVA α Autor: MLADEN SRAGA Grafički urednik: BESPLATNA - WEB-VARIJANTA Tisak: M.I.M.-SRAGA
METABOLIZAM PROTEINA
 METABOLIZAM PROTEINA PREGLED METABOLIZMA AMINO KISELINA Hranom unijeti proteini se razgrađuju do amino kiselina, koje se apsorbuju, prenose cirkulacijom i preuzimaju u ćelije različitih tkiva. Amino kiseline
METABOLIZAM PROTEINA PREGLED METABOLIZMA AMINO KISELINA Hranom unijeti proteini se razgrađuju do amino kiselina, koje se apsorbuju, prenose cirkulacijom i preuzimaju u ćelije različitih tkiva. Amino kiseline
REAKCIJE ELIMINACIJE
 REAKIJE ELIMINAIJE 1 . DEIDROALOGENAIJA (-X) i DEIDRATAIJA (- 2 O) su najčešći tipovi eliminacionih reakcija X Y + X Y 2 Dehidrohalogenacija (-X) X strong base + " X " X = l, Br, I 3 E 2 Mehanizam Ova
REAKIJE ELIMINAIJE 1 . DEIDROALOGENAIJA (-X) i DEIDRATAIJA (- 2 O) su najčešći tipovi eliminacionih reakcija X Y + X Y 2 Dehidrohalogenacija (-X) X strong base + " X " X = l, Br, I 3 E 2 Mehanizam Ova
Riješeni zadaci: Nizovi realnih brojeva
 Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
U unutrašnja energija H entalpija S entropija G 298. G Gibsova energija TERMOHEMIJA I TERMODINAMIKA HEMIJSKA TERMODINAMIKA
 HEMIJSKA TERMODINAMIKA Bavi se energetskim promenama pri odigravanju hemijskih reakcija. TERMODINAMIČKE FUNKCIJE STANJA U unutrašnja energija H entalpija S entropija Ako su određene na standardnom pritisku
HEMIJSKA TERMODINAMIKA Bavi se energetskim promenama pri odigravanju hemijskih reakcija. TERMODINAMIČKE FUNKCIJE STANJA U unutrašnja energija H entalpija S entropija Ako su određene na standardnom pritisku
Dijagonalizacija operatora
 Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
Pismeni ispit iz matematike GRUPA A 1. Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj, zatim naći 4 z.
 Pismeni ispit iz matematike 06 007 Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj z = + i, zatim naći z Ispitati funkciju i nacrtati grafik : = ( ) y e + 6 Izračunati integral:
Pismeni ispit iz matematike 06 007 Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj z = + i, zatim naći z Ispitati funkciju i nacrtati grafik : = ( ) y e + 6 Izračunati integral:
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem