MOLEKULARNE OSNOVE ĆELIJE. Milena Ćurčić
|
|
|
- Ξένα Καραμήτσος
- 5 χρόνια πριν
- Προβολές:
Transcript
1 MOLEKULARNE OSNOVE ĆELIJE Milena Ćurčić
2

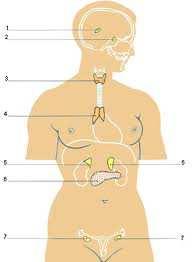
3 - RAZVIĆE EMBRIONA - OPLOĐENJE - DIFERENCIJACIJA






4 OBLIK ĆELIJA


5 - Membranski sistem ćelije Struktura ćelijske membrane
 - proteini")
 -Glikokaliks -Transport")
6 Ćelijska membrana Struktura ćelijske membrane: - lipidi (fosfolipidi, holesterol) - proteini (integralni, periferni) (transportni, receptori, enzimi, strukturni proteini) -Glikokaliks -Transport kroz ćelijsku membranu
")
7 - Bazofilna (ribozomi, RNK) - Acidofilna (mitohondrije)
8 CITOSKELET - Mikrofilamenti - intermedijalni filamenti - mikrotubuli

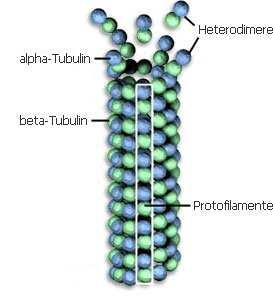

9 - Ćelijski korteks, ektoplazma
10 ĆELIJSKE ORGANELE - Nemembranske - Membranske organele




11 - Centriole - Ribozomi - diplozom - pericentriolski materijal - centrozom


12 -Endoplazmatični retikulum - Granulisani ER - sinteza proteina za sekreciju ili za potrebe same ćelije - Glatki ER - biosinteza lipida - unutarćelijski transport - sinteza holesterola, fosfolipida lipida membrane, učestvuje u metabolizmu glikogena - Sarkoplazmatični retikulum

13 - Goldžijev aparat - sinteza ugljenih hidrata - pakovanje, usmeravanje, - modifikacija proteina i lipida sintetisanih u ER - Primarni - Sekundarni - Tercijarni lizozom -Formirajuća ili cis strana i maturirajuća ili trans strana -Sadrži hidrolazne enzime za intracelularnu digestiju - autoliza - Različiti enzimi: peroksidaze, oksidaze, katalaze -U metabolizmu vodonik peroksida, detoksikaciji toksičnih supstanci

14 - Spoljašnja mitohondrijska membrana - porini - Unutrašnja mitohondrjska membrana - Kardiolipini - kontaktna mesta -Lanac za transport elektrona i kompleksi za sintezu ATP
15 - oblik, veličina, položaj jedra - Struktura jedra: jedrov omotač, nukleoplazma, nukleoskelet
16 - Hromatin - Različiti nivoi kondenzovanja lanca DNK




17 STRUKTURA NUKLEINSKIH KISELINA Nukeotid azotna baza, šećer, fosforna kiselina Nukleozid - azotna baza i šećer vezani N-glikozidnom vezom - Denaturacija DNK
18 OSNOVNA DOGMA MOLEKULARNE BIOLOGIJE - kod, kodon, antikodon
19 MEHANIZAM REPLIKACIJE KOD EUKARIOTA Replikacija je semikozervativna i semidiskontinuirana, bidirekciona Replikacija počinje istovremeno na mnogo mesta u hromozomu i završava se za nekoliko sati ARS (autonomously replicating sequences) elementi - mesta početka replikacije, oko 300 bp sa evolutivno očuvanim nizovima nukleotida; u kvascu oko 400 ARS; Replikativni počeci se nalaze u intervalu kb - genom čoveka ima rep. poč. Replikon region ARS gde započinje replikacija; replikator i inicijator Sukcesivno aktiviranje ARS Regulatorni proteini Inicijator - kompleks od šest proteina koji prepoznaje oridžin (ORC - origin recognation complex). ORC prepoznaje konzervativnu sekvencu u replikatoru kvasca (A elemenat), i manje konzervisan B region; ORC se vezuje i hidrolizuje ATP; ORC regrutuje replikacione proteine do replikatora.
 Reparacija oštećene DNK (β) Replikacija hromozomske DNK (α, δ i ε) DNK pol α od više")
 klema, egzonukleaza DNK pol δ i ε glavne replikaze DNK pol δ iseca prajmer; egzonukleazna aktivnost Replikacioni")
20 5 vrsta DNK polimeraza: α, β, γ, δ i ε (poznato još 10tak polimeraza). Replikacija mitohondrijske DNK (γ) Reparacija oštećene DNK (β) Replikacija hromozomske DNK (α, δ i ε) DNK pol α od više subjedinica, primaza, polimeraza; sinteza lanca u 5-3 smeru; nema egzonukleaznu aktivnost; + PCNA (proliferating cell nuclear antigen) klema, egzonukleaza DNK pol δ i ε glavne replikaze DNK pol δ iseca prajmer; egzonukleazna aktivnost Replikacioni faktori RFA (uloga SSB proteina) i RFC (formiranje aktivnih repl. kompleksa)
21 D N K p o l i m e r a z e b a k t e r i j e E. C o l i D N K p o l i m e r a z e e u k a r i o t a P o l I U k l a n j a n j e p oč e t n i c u, r e p e r D N K p o l α s i n t e z a p r a j m e r a z a v r e m e D N K r e p l i k a c i j e P o l I I D N K r e p e r P o l I I I R e p l i k a c i j a P o l I V D N K r e p e r, T L S ( T r a n s L e s i o n S y n t h e s i s ) p o l β o s n o v n i r e p e r e k s c i z i o n i p o l γ m i t o h o n d r ij a l n a D N K r e p l i k a c i j a i r e p e r p o l δ D N K r e p l i k a c i j a, o s n o v n i e k c i z i o n i r e p e r P o l V p o l ε T L S D N K r e p l i k a c i j a, o s n o v n i e k c i z i o n i r e p e r p o l θ D N K k r o s l i n k i n g a p o l ζ T L S r e p e r p o l λ r e p e r v e z a n z a m e j o z u p o l µ s o m a t s k e h i p e r m u t a c i j e p o l κ T L S p o l η T L S
22 Eukariotski hromozomi se replikuju samo jednom u ćelijskom ciklusu Hromozomalna DNK replikacija se dešava za vreme S faze ćelijskog ciklusa; svaki bazni par svakog hromozoma replikuje se samo jednom u toku eukariotske ćelijske deobe. Potrebni uslovi dovoljan broj replikatora mora biti aktiviran na svakom hromozomu replikatori nakon replikacije, bilo da su učestvovali u inicijaciji replikacije ili da su pasivno replicirani, moraju da ostanu inaktivni do sledeće ćelijske deobe. Replikatorska selekcija - proces identifikacije sekvence koja će inicirati replikaciju (u G1 fazi); spajanje multiproteinskog kompleksa na svakom replikatoru u genomu i formiranje pre-replikativnog kompleksa; aktivacija pojedinih ORI u S fazi (DNK rasplitanje i regrutacija DNK polimeraza). Pre-replikativni kompleks (prerc) upravlja inicijacijom replikacije kod eukariota. prepoznavanje replikatora od strane inicijatora (ORC); ORC od 6 proteina, vezuje ATP, prepoznaje replikator i regrutuje druge proteine za replikaciju. ORC regrutuje dva proteina Cdc6 i Cdt1 (punjači helikaze). ORC, Cdc6 i Cdt1 regrutuju eukariotske helikaze u viljušci (Mcm2-7 kompleks). Formiranje prerc ne mora da dovede obavezno do otvaranja originalne DNK. PreRC inicira replikaciju preko dve kinaze Cdk i Ddk; inaktivne u G1, aktivne u S fazi; aktivirane kinaze se vezuju za prerc i replikatorne proteine i fosforilišu ih; povezivanje dodatih replikacionih proteina oridžina i iniciranje replikacije. Novi proteini: tri eukariotske DNK polimeraze i brojni proteini potrebni za njihovu regrutaciju; DNK pol ε i δ se vezuju, a zatim se vezuje DNK pol α primaza; sinteza prvog RNK prajmera; punjača klizeće kleme RF-C koji vezuje klizeću klemu PCNA oko kompleksa prajmer matrica.



23
 Cdk aktivnost inhibira formiranje novih prerc.")

24 Regulacija replikacije kod Eukariota Regulacija formiranja i aktivacije prerc putem ciklin zavisne kinaze Cdks; (i) Cdks su potrebne za aktivaciju prerc da bi započela DNK replikacija; (ii) Cdk aktivnost inhibira formiranje novih prerc. Visoka Cdk aktivnost je potrebna za postojanje prerc kompleksa za otpočinjanje replikacije, ali ne dozvoljava formiranje prerc. Niska Cdk aktivnost je stimulativna za formiranje novih prerc ali je neadekvatna da otpočne DNK replikaciju inicijacijom novoformiranih prerc. Čvrsta povezanost između prerc uloge, Cdk nivoa i faze ćelijskog ciklusa - eukariotski genom se replikuje samo jednom u ćelijskom ciklusu.

25 Replikacija DNK u telomerama Uklanjanje prajmera sa 3 kraja matrice za lanac koji zaostaje ne može da se sintetiše 5 kraj 3 krajevi linearnih molekula DNK su jednolančani (12-16 bp) i oko 1000 umnožaka kratkog nukleotidnog niza sa mnogo G Telomeraza ribonukleoproteinski kompleks; RNK sadrži segment komplementaran nizu u telomeri i taj segment služi kao matrica za sintezu 3 krajeva lanaca DNK (proces sličan reversnoj transkripciji)
26 TRANSKRIPCIJA KOD EUKARIOTA Modifikacija (obrada primarnih transkripata) RNK Transkripcija i translacija vremenski i prostorno odvojeni procesi
 RNK pol I transkribuje velike rrnk (18S, 28S i 5.")

27 U jedrima eukariota: RNK pol I, RNK pol II i RNK pol III (po 2 velike i 12 malih subjedinica) RNK pol u organelama RNK pol II transkribuje strukturne gene; RNK pol kvasca (RPB1-10; RPB1 sadrži CTD rep bogat OH grupama na Ser) RNK pol I transkribuje velike rrnk (18S, 28S i 5.8S) RNK pol III transkribuje trnk, malu rrnk (5S) i snrnk Različiti promotori za RNK pol Transkripcioni faktori specifično prepoznaju nizove nukleotida u promotoru, vezuju RNK pol za promotor, tačna inicijacija transkripcije Promotori za RNK pol I i II su uzvodno od mesta početka transkripcije, a za RNK pol III unutar strukturnih gena
28 Хумане РНК полимеразе Полимеразе Локација Продукт РНК пол I Нуклеолус 18S, 28S, 5.8S ррнк РНК пол II Нуклеоплазма хнрнк/ирнк У1, У2, У4, У5 снрнк РНК пол III Нуклеоплазма трнк 5S РНК, У6 снрнк, 7SL РНК Мит РНК пол Митохондрије Све мит РНК
 CCAAT blok na -70 do -90 nukleotida Četiri elementa konsenzusne sekvence u sastavu osnovnog promotora za Pol li BRE element prepoznaje TFIIB faktor TATA element prepoznaje")
29 PROMOTORI ZA RNK POL II Konstitutivni geni sadrže GC blok Geni koji se eksprimiraju samo u nekim tkivima najčešće ne sadrže GC blok, već imaju TATA blok na -20 do 30 nukleotida (tačnost inicijacije i efikasnost transkripcije) CCAAT blok na -70 do -90 nukleotida Četiri elementa konsenzusne sekvence u sastavu osnovnog promotora za Pol li BRE element prepoznaje TFIIB faktor TATA element prepoznaje TBP Inicijator (Inr) - prepoznaju TAF proteini TFIID kompleksa. Nizvodni promotorski element DPE - prepoznaju TAF proteini TFIID kompleksa. Pojačivači, prigušivači i insulatori
30 PROMOTORI RNK POL I Strukturno specificni za vrstu organizma rrnk geni u više kopija Dva niza nukleotida +6 do -31 nukleotid -187 do -107 nukleotid

31 PROMOTORI ZA RNK POL III Unutar strukturnih gena 5S geni promotor u okviru +40 do +80 nukleotida Geni za trnk A i B blok od po 10-tak nukleotida

 omogućavajući")

32 Za inicijaciju transkripcije neophodno je otvaranje hromatina hipersenzitivna mesta DNK se u toku transkripcije namotava oko nukleozoma (kao oko kalema) omogućavajući pristup RNK pol Destabilizacija nukleozoma ispred RNK pol, a njihovo formiranje iza enzima


33 Transkripcija kod eukariota Inicijacija Osnovni transkripcioni faktori (TFIIB, D, E, F, H, A) Preinicijacioni kompleks RNK pol II čiji CTD nije fosforilisan može da inicira transkripciju. Elongacija Elongacija transkripcije nakon fosforilacije CTD Elongacioni faktori TEFb stimuliše elongaciju; poseduje kinaznu aktivnost, stimuliše elongaciju preko fosforilacije CTD repa Pol II. hspt5 regrutuje enzime za dodavnje 5 kape TAT-SF1 protein regrutuje komponente splajsozoma. Terminacija Specifični nizovi nukleotida ili specifične sekundarne strukture Signal za adenilaciju AAUAAA, transkripcija se nastavlja i do 1000 nukleotida nakon toga PoliA polimeraza dodaje polia rep Nakon terminacije transkripcije fosfataze recikliraju RNK pol II u nefosforilisani oblik.
 Komunikacija TF preko aktivacionih domena Insulatori")

34 Regulacija inicijacije transkripcije na tkivno specifičan način Geni se eksprimiraju tkivno- i vremenski-specifično Regulatorni i inducibilni TF vezuju se za pojačivače Ubrzavanje ili usporavanje transkripcije Hormoni preko svojih receptora deluju kao TF preko elemenata odgovora (RES) Komunikacija TF preko aktivacionih domena Insulatori - segmenti u DNK koji funkcionalno izdvajaju pojedine gene ili grupe gena; ograničavaju delovanje pojačivača ili utišivača i sprečavaju da se njihov uticaj proširi van željenih regiona. Membranski receptori - tirozinske kinaze EGF Dimerizacija Autofosforilacija P P P P P P Signalni put kroz ćeliju Jedro Aktivacija transkripcionih faktora
35
36 PRODUKTI TRANSKRIPCIJE RNK POLIMERAZOM II Heterogene nuklearne RNK (hnrnk) Ribonukleoproteinski kompleksi (RNP) Male jedarne RNK (snrnk ili U-RNK) Proteini hnrnk ili irnk OBRADA TRANSKRIPATA KOVALENTNIM MODIFIKACIJAMA Elongacija i terminacija transkripcije i obrada irnk su međusobno povezani i koordinisani procesi. Svi enzimi uključeni u obradu RNK reaguju sa CTD repom Pol II, zamenjuju inicijatorne transkripcione faktore sa enzimima koji dodaju 5 kapu i vrše splajsovanje irnk, faktorima za poliadenilaciju i kidanje irnk. 5 kapa, pozitivno naelektrisana, bitna za vezivanje za ribozom Na 5 kraj hnrnk dodaje se 7-metil-guanozin (5-5 trifosfatnim mostom vezan za prvi ribonukleotid) Metilacija 2 OH grupe prve (nekad i druge) riboze u nizu Metilacija N 6 atoma purinskog prstena ako je prva baza adenin Na 3 kraju 3 -poli(a) rep Na hnrnk poli(a)polimeraza dodaje A nukleotida Signal za poliadenilaciju je AAUAAA Endonukleaza hidrolizuje prim.tr. 15 nukl. nizvodno od signala za poliadenilaciju, a zatim deluje poli(a)polimeraza uz ATP

37
")
38 OBRADA TRANSKRIPATA ISECANJEM INTRONA hnrnk je nestabilna i samo mali deo izbegne degradaciju (oko 5%) Obrada hnrnk do irnk traje oko 30 min Introni i egzoni Obrada transkripata isecanjem introna (RNA splicing) u jedru
,")


39 Dužina introna od 100 do 1000 nukleotida GU na 5 kraju (donori), a AG na 3 kraju (akceptori) Komplementarni snrnk u snrnp česticama Formiranje RNK-RNK hibrida Prekidanje lanca u granicama introna i spajanje egzona
40 snrnk (U-RNK) U1-RNK su komplementarne graničnim nizovima introna U2-RNK su komplementarne nizovima unutar introna U4-RNK su komplementarne signalu za poliadenilaciju U5-RNK su komplementarne 3 krajevima introna snrnp (U1-RNP, U2-RNP, U4-RNP,..) Za obradu hnrnk Za transport RNK iz jedra u citoplazmu Pakovanje ribozoma u nukleolusu Splajsozomi (50-60S) od hnrnk, U-RNK i proteina Prva faza: presecanje veze između I i E i formiranje 2,5 -fosfodiestarske veze između G (5 kraja I) i A (u okviru I) omča Druga faza: iskrajanje I i spajanje E; energija tranesterifikacije Proteini koji imaju ključnu ulogu u regulaciji splajsovanja i ulogu aktivatora u regulaciji alterantivnog splajsovanja su familija SR proteina Proteini SR familije poseduju dva domena: Domen preko koga se vezuju za RNK sekvencu (eng. RNA recognition motif, RRM). Domen bogat argininskim i serinskim ostacima (RS motiv) preko koga interaguju sa komponentama splajsozoma, regrutujući ih na susedno splice mesto. Proteini represori alternativnog splajsovanja Transport zrelih irnk kroz jedrove pore u citoplazmu Proteini koji markiraju zrelu irnk su SR proteini (ostaju vezani za zrelu irnk) i proteini koji se specifično vezuju samo za egzon-egzon granice.
41

42 Introni genetiča fleksibilnost ćelije Alternativna obrada transkripata Intra- i intermolekulske obrade transkripata Zrela irnk može sadržati sve egzone ili može biti alterantivno obrađena Preskakanjem egzona Produžavanjem egzona Zadržavanjem sekvence introna Alterantivnim korišćenjem dva egzona - zrela irnk je smeša dve različite irnk. Editovanje RNK - proces u kome se menjaju sekvence na nivou informacione RNK (irnk). Mesto-specifičnom deaminacija C ili A Insercijom ili delecijom U uz pomoć vodič RNK molekula (grna).
43 TRANSLACIJA - AKTIVACIJA AMINOKISELINA (AK) Aminoacil-tRNK sintetaze specifične za AK

44
45
 Ternarni kompleks (eif-2, GTP, Met-tRNK i Met ) Mala subjedinica ribozoma (40S) sa eif-3 irnk, eif-4, uz hidrolizu")
46 INICIJACIJA TRANSLACIJE KOD EUKARIOTA Start signal je AUG Metionin u inicijatorskoj trnk (Met-tRNK i Met ) nije formilovan Inicijacioni kompleks Binarni kompleks (eif-2 i GTP) Ternarni kompleks (eif-2, GTP, Met-tRNK i Met ) Mala subjedinica ribozoma (40S) sa eif-3 irnk, eif-4, uz hidrolizu ATP
 u blizini start kodona i inicijatorska trnk se vezuje za kompletno P mesto, uz hidrolizu")
47 Mala (40S) subjedinica se vezuje pomoću eif-4 za irnk (za 5 kapu) pomera se duž čeonog niza do prvog start kodona Prepoznavanje AUG kodona omogućeno je sparivanjem sa antikodonom inicijatorske trnk Velika subjedinica se vezuje kada se oslobode eif-2 i eif-3 uz pomoć eif-5 Stavranje kompletnog ribozoma (80S) u blizini start kodona i inicijatorska trnk se vezuje za kompletno P mesto, uz hidrolizu GTP
48 INICIJACIONI FAKTORI EUKARIOTA eif-1, eif-2, eif-3, eif-4a, eif-4b učestvuju u formiranju 40S inicijacionog kompleksa eif-5 - uklanja prethodne faktore eif4c povezuje ribozomske subjedinice eif-3 stimuliše disocijaciju ribozoma eif-6 vezivanjem za 60S sprečava reasocijaciju ribozoma
49 TERMINACIJA TRANSLACIJE Stop kodon (UAA, UAG, UGA) se nađe u A mestu, za koje se vezuje terminacioni faktor Prokarioti: RF-1, RF-2, RF-3 RF-1 prepoznaje stop kodone UAA i UAG RF-2 prepoznaje stop kodone UAA i UGA RF-3 vezuje GTP i stimuliše aktivnost prva dva faktora Eukarioti: erf Pri vezivanju terminacionih faktora stimulisana je hidrolitička aktivnost peptidil transferaze (hidroliza veze između trnk i sintetisanog polipeptidnog lanca) Hidrolizom GTP dobija se energija za oslobađanje terminacionih faktora Izbacivanje trnk, disocijacija irnk sa ribozoma


50
51 PRATIOCI PROTEINA Obrada polipeptidnog lanca biološki aktivan oblik Nativna konformacija, kovalentne modifikacije, lokalizacija u ćeliji, udruživanje sa drugim polipeptidnim lancima Pratioci proteina - PROTEINI TOPLOTNOG STRESA (heat shock proteins - HSP) Sprečavaju agregaciju i pogrešno savijanje novosintetisanih proteina Omogućuju uju transport kroz biološke membrane Omogućuju udruživanje polipeptida u oligomere Omogućuju disagregaciju, odvijanje i ponovno savijanje u nativni oblik Protein u odvijenom (O), ispravno savijenom (S) i pogrešno savijenom obliku (A/D) HSP imaju ATPaznu i odvijajuću aktivnost i stimulišu prelaz polipeptidnih lanaca iz A/D u O i O u S oblik HSP70 (monomeri, vezuju se za proteine u toku sinteze ) i HSP60 (oligomeri, vezuju se za nepravilno savijene ili dentaurisane proteine)





52 Endoplazmatski retikulum granulirani i negranulirani Goldžijev kompleks Proteini kao gradivni elementi ćelije, proteini enzimi, proteini organela, proteini za export Proteinsko pakovanje u ER (glikozilacija, stvaranje S-S mostova) Signalna sekvenca polipeptida sa amino kraja, hidrofobna Proteozomi ili lizozomi razgradnja nepravilno savijenih proteina (izloženi hidrofobni segmenti; ubikvitin)



53

54
55 POSTRANSLACIONE MODIFIKACIJE PROTEINA Kovalentne modifikacije: Proteolitičko uklanjanje vodećeg metionina (formilmetionina) Ograničena proteoliza (preproteini proteini; tripsin, himotripsin) Odstranjivanje signalnih peptida na NH 2 kraju (transmembranski i sekretorni proteini) pomoću peptidaza Digestija poliproteina pomoću proteaza (hormoni) Modifikacije aminokiselinskih bočnih grupa (fosforilacije, acetilacije, glikozilacije, hidroksilacije, metilacije, ADP-ribozilacije, i sl.) regulatorni značaj Ne modifikuju se: Ala, Gly, Ile, Leu, Met, Val Vezivanje koenzima (biotin, piridoksal fosfat)
TRANSLACIJA. Doc. dr Snežana Marković
 TRANSLACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu BIOSINTEZA PROTEINA - TRANSLACIJA U toku translacije dolazi do specifičnog
TRANSLACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu BIOSINTEZA PROTEINA - TRANSLACIJA U toku translacije dolazi do specifičnog
Doc. dr Snežana Marković
 TRANSKRIPCIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu RIBONUKLEINSKE KISELINE VRSTE I FUNKCIJE RNK irnk trnk rrnk snrnk male nuklearne
TRANSKRIPCIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu RIBONUKLEINSKE KISELINE VRSTE I FUNKCIJE RNK irnk trnk rrnk snrnk male nuklearne
Doc. dr Snežana Marković
 REPLIKACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPLIKACIJA DNK Semikonzervativan proces Eksperimenti Mezelsona i Stala 1957.g.
REPLIKACIJA Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPLIKACIJA DNK Semikonzervativan proces Eksperimenti Mezelsona i Stala 1957.g.
REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI. Doc. dr Snežana Marković
 REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REGULACIJA EKSPRESIJE
REGULACIJA EKSPRESIJE GENA KOD EUKARIOTA TRANSKRIPCIONI FAKTORI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REGULACIJA EKSPRESIJE
NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA
 NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA ANIMACIJE!!! REPLIKACIJA https://www.youtube.com/watch?v=tnkwgcfphqw TRANSKRIPCIJA https://www.youtube.com/watch?v=jqiwwjqf5d0 TRANSKRIPCIJA I TRANSLACIJA https://www.youtube.com/watch?v=-k8y0atkkai
NIVOI ORGANIZACIJE I EKSPRESIJE GENOMA ANIMACIJE!!! REPLIKACIJA https://www.youtube.com/watch?v=tnkwgcfphqw TRANSKRIPCIJA https://www.youtube.com/watch?v=jqiwwjqf5d0 TRANSKRIPCIJA I TRANSLACIJA https://www.youtube.com/watch?v=-k8y0atkkai
transkripcija Matrica i enzimi Transkripcija Sličnosti između replikacije i transkripcije Razlike između replikacije i transkripcije
 Transkripcija Sinteza RNK molekula. DNK lanac služi kao matrica za sintezu RNK. Transkripcija Tip RNK molekula Ribozomalna RNK (rrnk) Male RNK (trnk; snrnk; 5S rrnk; scrnk; mirnk ) Informaciona RNK (irnk
Transkripcija Sinteza RNK molekula. DNK lanac služi kao matrica za sintezu RNK. Transkripcija Tip RNK molekula Ribozomalna RNK (rrnk) Male RNK (trnk; snrnk; 5S rrnk; scrnk; mirnk ) Informaciona RNK (irnk
Biohemija nukleinskih kiselina. Genetska informacija
 Biohemija nukleinskih kiselina Genetska informacija deoksiribonukleinske kiseline (DNK) ribonukleinske kiseline (RNK) DNK je nosilac naslednih informacija u ćeliji, dok RNK učestvuju u prenošenju tih informacija
Biohemija nukleinskih kiselina Genetska informacija deoksiribonukleinske kiseline (DNK) ribonukleinske kiseline (RNK) DNK je nosilac naslednih informacija u ćeliji, dok RNK učestvuju u prenošenju tih informacija
Molekularna biologija prokariota
 Molekularna biologija prokariota I Molekularne osnove procesa: Čuvanja i prenošenja genetičke informacije u ćeliji Regulacije metabolizma - ekonomičnost ćelije i brzo prilagođavanje na uslove sredine -
Molekularna biologija prokariota I Molekularne osnove procesa: Čuvanja i prenošenja genetičke informacije u ćeliji Regulacije metabolizma - ekonomičnost ćelije i brzo prilagođavanje na uslove sredine -
Regulacija ekspresije gena kod eukariota
 Regulacija ekspresije gena kod eukariota Regulacija ekspresije gena Višećelijski organizmi Čovek - 250 tipova ćelija Svaka ćelija gotovo istu DNK Regulacija ekspresije gena Hijerarhija regulacije ekspresije
Regulacija ekspresije gena kod eukariota Regulacija ekspresije gena Višećelijski organizmi Čovek - 250 tipova ćelija Svaka ćelija gotovo istu DNK Regulacija ekspresije gena Hijerarhija regulacije ekspresije
Regulacija ekspresije gena kod prokariota
 Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
Regulacija ekspresije gena kod prokariota Bakterije Jednoćelijski organizmi koji nemaju jedro i druge organele. Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT
 METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT BEZ POLI-A REPA" Histoni su primarne proteinske komponente hromatina. Na početku se smatralo da su uglavnom uključeni u pakovanje DNK, odnosno da su važni
METABOLIZAM I REGULACIJA HISTONSKIH IRNK "ŽIVOT BEZ POLI-A REPA" Histoni su primarne proteinske komponente hromatina. Na početku se smatralo da su uglavnom uključeni u pakovanje DNK, odnosno da su važni
Rekombinacija DNK TRANSPOZICIJOM
 Rekombinacija DNK TRANSPOZICIJOM Rekombinacija DNK transpozicijom Transpozicija je proces premeštanja specifičnih sekvenci molekula DNK (pokretnih genetičkih elemenata ili transpozona) sa jednog na drugo
Rekombinacija DNK TRANSPOZICIJOM Rekombinacija DNK transpozicijom Transpozicija je proces premeštanja specifičnih sekvenci molekula DNK (pokretnih genetičkih elemenata ili transpozona) sa jednog na drugo
PROTEINI. Doc. dr Snežana Marković
 PROTEINI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu Proteos prvi, najvažniji. 20 proteinogenih L- aminokiselina (AK). AK na istom
PROTEINI Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu Proteos prvi, najvažniji. 20 proteinogenih L- aminokiselina (AK). AK na istom
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
Transkripcija u eukariota
 Transkripcija u eukariota 1 Transkripcija u eukariota osnovni principi isti kao kod prokariota razlika: veći broj RNA-polimeraza relativno složene kontrolne sekvence Primarni transkript: - u prokariota
Transkripcija u eukariota 1 Transkripcija u eukariota osnovni principi isti kao kod prokariota razlika: veći broj RNA-polimeraza relativno složene kontrolne sekvence Primarni transkript: - u prokariota
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
3.1 Granična vrednost funkcije u tački
 3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018.
 Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Test iz Biologije za 4. razred srednje škole R J E Š E NJ A Podgorica, 05. 05. 2018. 1 1. Spojite navedene
Prirodno-matematički fakultet Društvo matematičara i fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Test iz Biologije za 4. razred srednje škole R J E Š E NJ A Podgorica, 05. 05. 2018. 1 1. Spojite navedene
SADRŽAJ TRANSLACIJA... TRANSPORTNE RNK
 Translacija SADRŽAJ TRASLACIJA... 1 TRASPORTE RK... 2 Primarna struktura trk... 2 Sekundarna struktura trk... 3 Tercijarna struktura trk... 5 Aktivacija aminokiselina... 5 Interakcija kodon antikodon...
Translacija SADRŽAJ TRASLACIJA... 1 TRASPORTE RK... 2 Primarna struktura trk... 2 Sekundarna struktura trk... 3 Tercijarna struktura trk... 5 Aktivacija aminokiselina... 5 Interakcija kodon antikodon...
3/25/2016. Hemijske komponente ćelije
 Hemijske komponente ćelije Molekuli u ćeliji Najbitniji molekuli u ćeliji su poznati. Putevi sinteze i razgradnje su poznati za većinu ćelijskih konstituenata. Hemijska energija pokreće biosintezu. Organizacija
Hemijske komponente ćelije Molekuli u ćeliji Najbitniji molekuli u ćeliji su poznati. Putevi sinteze i razgradnje su poznati za većinu ćelijskih konstituenata. Hemijska energija pokreće biosintezu. Organizacija
BANKA PITANJA IZ BIOLOGIJE. proteini. 3. Koji se deo složenog enzima hemijski menja u toku reakcije: 4. Apoenzim i koenzim zajedno čine:
 BANKA PITANJA IZ BIOLOGIJE citologija proteini 1. α-heliks je pojam koje se odnosi na strukturu proteina. 2. Broj polipeptidnih lanaca kod složenih proteina definiše: 3. Koji se deo složenog enzima hemijski
BANKA PITANJA IZ BIOLOGIJE citologija proteini 1. α-heliks je pojam koje se odnosi na strukturu proteina. 2. Broj polipeptidnih lanaca kod složenih proteina definiše: 3. Koji se deo složenog enzima hemijski
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
numeričkih deskriptivnih mera.
 DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
ISPITNA PITANJA ZA USMENI DIO ISPITA
 UNIVERZITET CRNE GORE MEDICINSKI FAKULTET MEDICINSKA BIOHEMIJA ISPITNA PITANJA ZA USMENI DIO ISPITA STUDIJSKI PROGRAM MEDICINA I ENZIMOLOGIJA 1. Opšte osobine enzima i struktura molekula enzima 2. Izoenzimi.
UNIVERZITET CRNE GORE MEDICINSKI FAKULTET MEDICINSKA BIOHEMIJA ISPITNA PITANJA ZA USMENI DIO ISPITA STUDIJSKI PROGRAM MEDICINA I ENZIMOLOGIJA 1. Opšte osobine enzima i struktura molekula enzima 2. Izoenzimi.
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović
 DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
ISPITNA PITANJA OSNOVI BIOHEMIJE
 UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
18. listopada listopada / 13
 18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
41. Jednačine koje se svode na kvadratne
 . Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
. Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
Organele života i smrti
 MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
NOMENKLATURA ORGANSKIH SPOJEVA. Imenovanje aromatskih ugljikovodika
 NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
Iskazna logika 3. Matematička logika u računarstvu. novembar 2012
 Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
REKOMBINACIJA MOLEKULA DNK
 REKOMBINACIJA MOLEKULA DNK Fenomeni odgovorni za održavanje i ekspresiju genoma Svaki molekul DNK je rekombinovani molekul DNK Pojam rekombinacije Tridesete godine prošlog veka Mejoza (poslednja istraživanja
REKOMBINACIJA MOLEKULA DNK Fenomeni odgovorni za održavanje i ekspresiju genoma Svaki molekul DNK je rekombinovani molekul DNK Pojam rekombinacije Tridesete godine prošlog veka Mejoza (poslednja istraživanja
Sistemi za kontrolu kvaliteta proteina molekularni šaperoni i proteazom
 Sistemi za kontrolu kvaliteta proteina molekularni šaperoni i proteazom Kako protein nakon sinteze postaje funkcionalan? Proces ekspresije gena nije završen prevođenjem informacije sadržane u irnk u redosled
Sistemi za kontrolu kvaliteta proteina molekularni šaperoni i proteazom Kako protein nakon sinteze postaje funkcionalan? Proces ekspresije gena nije završen prevođenjem informacije sadržane u irnk u redosled
Elementi spektralne teorije matrica
 Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Riješeni zadaci: Nizovi realnih brojeva
 Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
CILJNA MESTA DEJSTVA LEKOVA
 FARMACEUTSKA HEMIJA 1 CILJNA MESTA DEJSTVA LEKVA Predavač: Prof. dr Slavica Erić Ciljna mesta dejstva leka CILJNA MESTA NA MLEKULARNM NIVU: lipidi (lipidi ćelijske membrane) ugljeni hidrati (obeleživači
FARMACEUTSKA HEMIJA 1 CILJNA MESTA DEJSTVA LEKVA Predavač: Prof. dr Slavica Erić Ciljna mesta dejstva leka CILJNA MESTA NA MLEKULARNM NIVU: lipidi (lipidi ćelijske membrane) ugljeni hidrati (obeleživači
PITANJA ZA USMENI ISPIT IZ BIOHEMIJE
 PITANJA ZA USMENI ISPIT IZ BIOHEMIJE PROTEINI STRUKTURA I FUNKCIJE 1. Struktura proteina nivoi organizacije molekula 2. Proteini koji transportuju kiseonik hemoglobin i mioglobin ENZIMI 1. Opšte osobine
PITANJA ZA USMENI ISPIT IZ BIOHEMIJE PROTEINI STRUKTURA I FUNKCIJE 1. Struktura proteina nivoi organizacije molekula 2. Proteini koji transportuju kiseonik hemoglobin i mioglobin ENZIMI 1. Opšte osobine
Osnovne karakteristike 3-D strukture molekula DNK i RNK
 Osnovne karakteristike 3-D strukture molekula DNK i RNK Rendgenska strukturna analiza (vlakana) DNK Watson-Crickov model (B) DNK Zašto dvostruki heliks? Polimorfizam DNK: kanonske (standardne/prosečne)
Osnovne karakteristike 3-D strukture molekula DNK i RNK Rendgenska strukturna analiza (vlakana) DNK Watson-Crickov model (B) DNK Zašto dvostruki heliks? Polimorfizam DNK: kanonske (standardne/prosečne)
PRAVA. Prava je u prostoru određena jednom svojom tačkom i vektorom paralelnim sa tom pravom ( vektor paralelnosti).
.") PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
Konstruisanje. Dobro došli na... SREDNJA MAŠINSKA ŠKOLA NOVI SAD DEPARTMAN ZA PROJEKTOVANJE I KONSTRUISANJE
 Dobro došli na... Konstruisanje GRANIČNI I KRITIČNI NAPON slajd 2 Kritični naponi Izazivaju kritične promene oblika Delovi ne mogu ispravno da vrše funkciju Izazivaju plastične deformacije Može doći i
Dobro došli na... Konstruisanje GRANIČNI I KRITIČNI NAPON slajd 2 Kritični naponi Izazivaju kritične promene oblika Delovi ne mogu ispravno da vrše funkciju Izazivaju plastične deformacije Može doći i
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE GODINE 8. RAZRED TOČNI ODGOVORI
 21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
Pismeni ispit iz matematike Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: ( ) + 1.
 + 1.") Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
3. razred gimnazije- opšti i prirodno-matematički smer ALKENI. Aciklični nezasićeni ugljovodonici koji imaju jednu dvostruku vezu.
 ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
Zavrxni ispit iz Matematiqke analize 1
 Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Sekundarne struktura proteina Fibrilni proteini
 Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
Sekundarne struktura proteina Fibrilni proteini Nivoi strukture proteina (strukturna hijerarhija) proteina Nivoi strukture proteina Primarna struktura Sekundarna struktura Super-sekundarna struktura Tercijarnastruktura
XI dvoqas veжbi dr Vladimir Balti. 4. Stabla
 XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
Biosintetski procesi na ER-u i GA-u i mehanizmi vezikularnog transporta
 Biosintetski procesi na ER-u i GA-u i mehanizmi vezikularnog transporta Intracelularne membrane 1 Endoplazmatični retikulum ER je mreža cevčica i cisterni okruženih membranom ER formira membrana koja od
Biosintetski procesi na ER-u i GA-u i mehanizmi vezikularnog transporta Intracelularne membrane 1 Endoplazmatični retikulum ER je mreža cevčica i cisterni okruženih membranom ER formira membrana koja od
ELEKTROTEHNIČKI ODJEL
 MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A
, grupa A") Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
NUKLEUS. 1) STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi)
 STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi)") NUKLEUS 1) STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi) Saznanja o ćelijama do kojih se došlo posredstvom novih
NUKLEUS 1) STRUKTURA (hromozomske teritorije, nukleusne organele) 2) FUNKCIJA (formiranje nukleusnih prekursora za sintezu proteina u citoplazmi) Saznanja o ćelijama do kojih se došlo posredstvom novih
HEMIJSKA VEZA TEORIJA VALENTNE VEZE
 TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
Operacije s matricama
 Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
M086 LA 1 M106 GRP. Tema: Baza vektorskog prostora. Koordinatni sustav. Norma. CSB nejednakost
 M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
Ćelijska signalizacija
 Ćelijska signalizacija Interakcije organizma i sredine posredovane su signalima Primanje signala iz okoline, odgovor na signale i odašiljanje signala u okolinu osobinu živih organizama Posebno važno i
Ćelijska signalizacija Interakcije organizma i sredine posredovane su signalima Primanje signala iz okoline, odgovor na signale i odašiljanje signala u okolinu osobinu živih organizama Posebno važno i
Doc. dr Snežana Marković
 REPARACIJA DNK. REKOMBINACIJA DNK. Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPARACIJA OŠTEĆENIH MOLEKULA DNK Mutacija svaka promena
REPARACIJA DNK. REKOMBINACIJA DNK. Doc. dr Snežana Marković Institut za biologiju i ekologiju Prirodno-matematički fakultet Univerzitet u Kragujevcu REPARACIJA OŠTEĆENIH MOLEKULA DNK Mutacija svaka promena
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu
 Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
RECEPTORI KAO CILJNA MESTA DEJSTVA LEKOVA. Kako se prenose informacije u organizmu? Predavač: Doc. dr Slavica Erić FARMACEUTSKA HEMIJA 1
 FARMACEUTSKA HEMIJA 1 RECEPTORI KAO CILJNA MESTA DEJSTVA LEKOVA Predavač: Doc. dr Slavica Erić Kako se prenose informacije u organizmu? receptori imaju ulogu prenosioca poruka većina receptora se nalazi
FARMACEUTSKA HEMIJA 1 RECEPTORI KAO CILJNA MESTA DEJSTVA LEKOVA Predavač: Doc. dr Slavica Erić Kako se prenose informacije u organizmu? receptori imaju ulogu prenosioca poruka većina receptora se nalazi
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f
 D f") IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =
/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =") 100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
Biologija ćelije CITOSKELET
 Biologija ćelije CITOSKELET Kompleksna mreža proteinskih filamenata 3 osnovna elementa: 1. Mikrofilamenti (prečnika oko 7 nm) od proteina aktina 2. Intermedijarni filamenti (oko 8-11 nm) - od 6 glavnih
Biologija ćelije CITOSKELET Kompleksna mreža proteinskih filamenata 3 osnovna elementa: 1. Mikrofilamenti (prečnika oko 7 nm) od proteina aktina 2. Intermedijarni filamenti (oko 8-11 nm) - od 6 glavnih
Mehanizmidejstvaenzima. Himotripsin
 Mehanizmidejstvaenzima Himotripsin Principi katalize Specifična kiselo-bazna kataliza Elektrostatska kataliza Elektrofilna kataliza Nukleofilna kataliza (kovalentna kataliza) Nukleofilna kataliza Opšta
Mehanizmidejstvaenzima Himotripsin Principi katalize Specifična kiselo-bazna kataliza Elektrostatska kataliza Elektrofilna kataliza Nukleofilna kataliza (kovalentna kataliza) Nukleofilna kataliza Opšta
ENZIMI KAO CILJNA MESTA DEJSTVA LEKOVA. Enzimi kao ciljna mesta dejstva lekova
 FARMACEUTSKA HEMIJA 1 ENZIMI KAO CILJNA MESTA DEJSTVA LEKOVA Predavač: Prof. dr Slavica Erić Enzimi kao ciljna mesta dejstva lekova -enzimi učestvuju u hemijskoj reakciji ali pri tome ostaju nepromenjeni
FARMACEUTSKA HEMIJA 1 ENZIMI KAO CILJNA MESTA DEJSTVA LEKOVA Predavač: Prof. dr Slavica Erić Enzimi kao ciljna mesta dejstva lekova -enzimi učestvuju u hemijskoj reakciji ali pri tome ostaju nepromenjeni
2 tg x ctg x 1 = =, cos 2x Zbog četvrtog kvadranta rješenje je: 2 ctg x
 Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
radni nerecenzirani materijal za predavanja R(f) = {f(x) x D}
 = {f(x) x D}") Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
Matematika 1 Funkcije radni nerecenzirani materijal za predavanja Definicija 1. Neka su D i K bilo koja dva neprazna skupa. Postupak f koji svakom elementu x D pridružuje točno jedan element y K zovemo funkcija
1. Sistemi za kontrolu kvaliteta proteina, molekularni šaperoni i proteazom
 1. Sistemi za kontrolu kvaliteta proteina, molekularni šaperoni i proteazom Proces ekspresije gena koji kodiraju proteine nije završen prevođenjem informacije sadržane u irnk u redosled aminokiselina polipeptidnog
1. Sistemi za kontrolu kvaliteta proteina, molekularni šaperoni i proteazom Proces ekspresije gena koji kodiraju proteine nije završen prevođenjem informacije sadržane u irnk u redosled aminokiselina polipeptidnog
Ispitivanje toka i skiciranje grafika funkcija
 Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Dijagonalizacija operatora
 Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
Teorijske osnove informatike 1
 Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Aminokiseline. Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina 22.12.2014
 Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
VODA ELEKTROLITI I ACIDO-BAZNA RAVNOTEŽA...
 SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
LANCI & ELEMENTI ZA KAČENJE
 LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
LANCI & ELEMENTI ZA KAČENJE 0 4 0 1 Lanci za vešanje tereta prema standardu MSZ EN 818-2 Lanci su izuzetno pogodni za obavljanje zahtevnih operacija prenošenja tereta. Opseg radne temperature se kreće
REGULATORNI MOLEKULI RNK "SKRIVENI JEZIK RNK"
 REGULATORNI MOLEKULI RNK "SKRIVENI JEZIK RNK" Kod prokariota geni posredstvom molekula irnk kodiraju proteine koji obavljaju katalitičke, strukturne i regulatone funkcije, tako da su proteini jedini izlazni
REGULATORNI MOLEKULI RNK "SKRIVENI JEZIK RNK" Kod prokariota geni posredstvom molekula irnk kodiraju proteine koji obavljaju katalitičke, strukturne i regulatone funkcije, tako da su proteini jedini izlazni
Strukture podataka i algoritmi 1. kolokvij 16. studenog Zadatak 1
 Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
Strukture podataka i algoritmi 1. kolokvij Na kolokviju je dozvoljeno koristiti samo pribor za pisanje i službeni šalabahter. Predajete samo papire koje ste dobili. Rezultati i uvid u kolokvije: ponedjeljak,
III VEŽBA: FURIJEOVI REDOVI
 III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
PRILOG. Tab. 1.a. Dozvoljena trajna opterećenja bakarnih pravougaonih profila u(a) za θ at =35 C i θ=30 C, (θ tdt =65 C)
 za θ at =35 C i θ=30 C, (θ tdt =65 C)") PRILOG Tab. 1.a. Dozvoljena trajna opterećenja bakarnih pravougaonih profila u(a) za θ at =35 C i θ=30 C, (θ tdt =65 C) Tab 3. Vrednosti sačinilaca α i β za tipične konstrukcije SN-sabirnica Tab 4. Minimalni
PRILOG Tab. 1.a. Dozvoljena trajna opterećenja bakarnih pravougaonih profila u(a) za θ at =35 C i θ=30 C, (θ tdt =65 C) Tab 3. Vrednosti sačinilaca α i β za tipične konstrukcije SN-sabirnica Tab 4. Minimalni
1892. Ivanovski, filtrabilni uzročnik mozaične bolesti duvana Bejerink, potvrdio eksperimente Ivanovskog Lefler i Froš, filtrabilni
 1892. Ivanovski, filtrabilni uzročnik mozaične bolesti duvana 1898. Bejerink, potvrdio eksperimente Ivanovskog 1898. Lefler i Froš, filtrabilni uzročnik slinavke i šapa 1909. Landštajner i Poper, filtrabilni
1892. Ivanovski, filtrabilni uzročnik mozaične bolesti duvana 1898. Bejerink, potvrdio eksperimente Ivanovskog 1898. Lefler i Froš, filtrabilni uzročnik slinavke i šapa 1909. Landštajner i Poper, filtrabilni
CITO T SKE K L E ET E
 CITOSKELET ULOGE CITOSKELETNIH ELEMENATA ćelije gajene u kulturi aktinski filamenti mikrotubule intermedijarni filamenti enterocit specifičnost organizacija STRUKTURA -PRATEĆI PROTEINI FUNKCIJA debljina
CITOSKELET ULOGE CITOSKELETNIH ELEMENATA ćelije gajene u kulturi aktinski filamenti mikrotubule intermedijarni filamenti enterocit specifičnost organizacija STRUKTURA -PRATEĆI PROTEINI FUNKCIJA debljina
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI. NEUTRALNI ELEMENT GRUPOIDA.
 KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
Cauchyjev teorem. Postoji više dokaza ovog teorema, a najjednostvniji je uz pomoć Greenove formule: dxdy. int C i Cauchy Riemannovih uvjeta.
 auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,
auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,
PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).
![PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).](/thumbs/74/70316766.jpg "PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).") 0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
Metode molekularne biologije
 Metode molekularne biologije Metode za analizu hromozoma Hromozom χρώμα (hroma) - boja i σώμα (soma) telo Sposobnost bojenja hromozoma je bila osnov na kojoj su se bazirale pionirske metode za analizu
Metode molekularne biologije Metode za analizu hromozoma Hromozom χρώμα (hroma) - boja i σώμα (soma) telo Sposobnost bojenja hromozoma je bila osnov na kojoj su se bazirale pionirske metode za analizu
ZBIRKA POTPUNO RIJEŠENIH ZADATAKA
 **** IVANA SRAGA **** 1992.-2011. ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE POTPUNO RIJEŠENI ZADACI PO ŽUTOJ ZBIRCI INTERNA SKRIPTA CENTRA ZA PODUKU α M.I.M.-Sraga - 1992.-2011.
**** IVANA SRAGA **** 1992.-2011. ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE POTPUNO RIJEŠENI ZADACI PO ŽUTOJ ZBIRCI INTERNA SKRIPTA CENTRA ZA PODUKU α M.I.M.-Sraga - 1992.-2011.
Numerička matematika 2. kolokvij (1. srpnja 2009.)
") Numerička matematika 2. kolokvij (1. srpnja 29.) Zadatak 1 (1 bodova.) Teorijsko pitanje. (A) Neka je G R m n, uz m n, pravokutna matrica koja ima puni rang po stupcima, tj. rang(g) = n. (a) Napišite puni
Numerička matematika 2. kolokvij (1. srpnja 29.) Zadatak 1 (1 bodova.) Teorijsko pitanje. (A) Neka je G R m n, uz m n, pravokutna matrica koja ima puni rang po stupcima, tj. rang(g) = n. (a) Napišite puni
Genetska kontrola sinteze bjelančevina. Povijesni PaJaMo pokus Jacob-Monod model sinteze bjelančevina u prokariota
 Genetska kontrola sinteze bjelančevina Još prije otkrića genetskog koda bilo je jasno da moraju postojati načini kontroliranja broja i vrste bjelančevina koje se proizvode u stanici. Kod šumskog drveća
Genetska kontrola sinteze bjelančevina Još prije otkrića genetskog koda bilo je jasno da moraju postojati načini kontroliranja broja i vrste bjelančevina koje se proizvode u stanici. Kod šumskog drveća
Novi Sad god Broj 1 / 06 Veljko Milković Bulevar cara Lazara 56 Novi Sad. Izveštaj o merenju
 Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
OM2 V3 Ime i prezime: Index br: I SAVIJANJE SILAMA TANKOZIDNIH ŠTAPOVA
 OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
Biohemija proteina i nukleinskih kiselina
 Biohemija proteina i nukleinskih kiselina Biohemija proteina i nukleinskih kiselina Predavanja: Profesor Vesna Niketić vniketic@chem.bg.ac.rs Docent Natalija Polović polovicn@chem.bg.ac.rs Vežbe: Dr Natalija
Biohemija proteina i nukleinskih kiselina Biohemija proteina i nukleinskih kiselina Predavanja: Profesor Vesna Niketić vniketic@chem.bg.ac.rs Docent Natalija Polović polovicn@chem.bg.ac.rs Vežbe: Dr Natalija
Obrada signala
 Obrada signala 1 18.1.17. Greška kvantizacije Pretpostavka je da greška kvantizacije ima uniformnu raspodelu 7 6 5 4 -X m p x 1,, za x druge vrednosti x 3 x X m 1 X m = 3 x Greška kvantizacije x x x p
Obrada signala 1 18.1.17. Greška kvantizacije Pretpostavka je da greška kvantizacije ima uniformnu raspodelu 7 6 5 4 -X m p x 1,, za x druge vrednosti x 3 x X m 1 X m = 3 x Greška kvantizacije x x x p
RIJEŠENI ZADACI I TEORIJA IZ
 RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA