ANTIGÉNY Epitopes (antigenic determinants). epitopes lymphocytes antibodies
|
|
|
- Ἡσίοδος Καραμανλής
- 6 χρόνια πριν
- Προβολές:
Transcript

1 ANTIGÉNY Epitopes (antigenic determinants). Small, specific regions on antigens, called epitopes, are bound by the antigen receptors on lymphocytes and by secreted antibodies.

2 bielkovinový nosič rôzne povrchové determinanty = organické molekuly s nízkou Mr bielkovinový nosič

3
4 ANTIGÉNY Po vniknutí cudzorodej látky imunogénu do organizmu vzniká imunitná odpoveď (IO) ako reakcia IS. Podstatou tejto reakcie je rozpoznanie vlastného a nevlastného. Imunogény sú makromolekulové látky (proteíny, polysacharidy) schopné navodiť imunitnú odpoveď voči štruktúram na svojom povrchu. Antigénový determinant (epitop) je konkrétna sekvencia aminokyselín (6-9), monosacharidov, alebo z 5-6 nukleotidov, alebo haptén. Svojou veľkosťou zodpovedá približne väzbovému miestu na protilátke. Antigény sú látky zahŕňajúce imunogény, ale aj také látky, ktoré samy o sebe nie sú schopné imunitnú odpoveď vyvolať. Tieto látky s nízkou molekulovou hmotnosťou haptény sa stávajú imunogénnymi až po spojení s makromolekulou (proteínom, polysacharidom). Všetky imunogény sú Ag, ale nie všetky Ag sú imunogénne. Odpoveď IS nevzniká na celú makromolekulu, ale iba na určitý jej úsek epitop. Na jednej imunogénnej molekule môže byť rôzny počet epitopov, pričom ich schopnosť navodiť odpoveď IS býva rôzna. Nie všetky epitopy sú rozpoznávané B- aj T-lymfocytmi, preto sa rozlišujú B-epitopy a T- epitopy.
5 ANTIGÉNY Interakciu Ag determinantu a Ab zabezpečujú hlavne nekovalentné, reverzibilné väzby (elektrostatické, vodíkové, hydrofóbne a Van der Walsove), teda sú relatívne slabé a všetky si vyžadujú tesnú blízkosť interagujúcich miest a spojenia sa prirovnávajú ku kľúču azámke. Pri reakcii Ag s Ab v živom organizme (in vivo) dochádza ku inhibícii biologickej účinnosti patogénov. Antigény sa rozdeľujú aj podľa aktivity T a B buniek v organizme a to na Ag nezávislé od týmusu (týmus independentné, Ag TI) a na antigény závislé of týmusu (týmus dependentné, Ag TD). Ag TI stimulujú B lymfocyty k tvorbe protilátok bez účasti T lymfocytov. Jedná sa väčšinou o polyméry s opakujúcimi sa antigénnymi determinantami, ktoré sú ťažko degradovateľné (LPS). Väčšina Ag patrí medzi Ag TD, kde dôležitú úlohu zohrávajú aj T lymfocyty.
6 ROZDELENIE ANTIGÉNOV Podľa funkcie: Kompletný funkčný = imunogén Nekompletný = haptén (haptein pripevniť) Podľa pôvodu: - prirodzené - syntetické - arteficiálne fyzikálnym alebo chemickým spôsobom pozmenené prirodzené Ag Podľa vzťahu k určitému organizmu: - exogénne (cudzie bb., bb. štruktúry a molekuly; mikroorganizmy, vírusy, peľ, liečivá, Ag transplantovaných tkanív) - endogénne - sú súčasťou vlastných bb. a tkanív = autoantigény autológne Ag
7 ROZDELENIE ANTIGÉNOV Vzhľadom na vzájomný vzťah 2 organizmov, medzi ktorými sa uskutočňuje prenos Ag (transplantácie tkanív): Xenogénne heterológne pochádzajú z jedinca iného druhu (prasa človek) Alogénne homológne (allos = iný) - pochádzajú z geneticky iného jedinca toho istého druhu (človek človek) Izogénne (isos = rovnaký) - pochádzajú z geneticky identického jedinca toho istého druhu (jednovajcové dvojča) Syngénne (syn = spolu) - pochádzajú z geneticky identického jedinca v rámci inbredného kmeňa (inbredný kmeň vzniká opakovaným krížením súrodencov po mnoho generácií) Autoantigénne autológne Ag vlastného organizmu organizmus ich toleruje, iba za určitých podmienok sa stávajú imnogénnymi autoimunitné ochorenia)
8 HETEROFILNÉ ANTIGÉNY - Zistené vo fylogeneticky nepríbuzných druhoch, napr. v bb. živočíchov a mikroorganizmov: Heterofilná príbuznosť medzi Ag A beta hemolytických streptokokov a Ag ľudského myokardu. Po streptokokovej infekcii sa v organizme nachádzajú anti streptokokové Ab, ktoré in vivo môžu reagovať krížovou reakciou s Ag svalových vlákien myokardu poškodenie tkaniva srdca beta hemolytické streptokoky - mikróby lyzujúce erytrocyty Krížová reakcia: Protilátka, ktorej tvorbu indukoval určitý Ag, reaguje s iným Ag, ktorý má rovnaké alebo podobné Ag determinanty.
9 Imunogén 2 základné vlastnosti: 1. Imunogénnosť schopnosť navodiť imunitnú odpoveď (vznik protilátok a senzibilizovaných Ly) 2. Špecifickosť schopnosť reagovať len s tými Ab, ktorých tvorbu daný Ag vyvolal. S ostatnými Ly a Ab zvyčajne nereaguje. Haptén ak je injektovaný sám o sebe nie je schopný vyvolať imunitnú odpoveď nie je imunogénny, môže len špecificky reagovať s už vytvorenými Ab, ale nedokáže vyvolať ich novú tvorbu. = organické molekuly s nízkou Mr Haptény a determinanty zodpovedajú za špecifickosť Ag - Ab reakcie bielkovinový nosič Imunitnú odpoveď môže vyvolať len celý komplex = imunogén
10 VLASTNOSTI IMUNOGÉNOV Fyzikálne - molekulová hmotnosť: Mr > 10 kd 10 kd slabo imunogénne, > 100 kd najsilnejšie - rozpustnosť: korpuskulárne Ag, nerozpustné koloidné Ag, rozpustné čím < rozpustnosť koloidných Ag, tým > imunogénnosť Chemické (štruktúra a degradovateľnosť) Biopolyméry bielkoviny, polysacharidy, nukleoproteíny (NK sú imunogénne len vo forme nukleoproteínov) Čisté lipidy sú len hapténmi, imunogénne sú len v komplexoch s proteínmi alebo polysacharidmi. Biologické (druhová vzdialenosť)
11 Chemické zloženie imunogénov Typ Ag polypeptidy proteíny polysacharidy glykoproteíny nukleoproteíny Príklad proteohormóny plazmatické, mikrobiálne, enzýmy kapsuly baktérií Ig, Ag krvných skupín, MHC antigény chromatín, ribozómy lipoproteíny plazmatické, bb. membrány lipopolysacharidy bb. steny G- baktérií = endotoxíny
12 VLASTNOSTI IMUNOGÉNOV Schopnosť navodiť IO nie je pri všetkých Ag rovnako silná. Sila Ag závisí najmäodmr,aleajodpočtu povrchových determinantov. Najsilnejšie Ag proteíny a polysacharidy Výnimka: želatína má napriek svojej Mr veľmi malý počet epitopov, preto je slabým imunogénom. Keď sa na želatínu naviažu haptény s nízkou Mr, preberú funkciu determinantov. Takto modifikovaná želatína môže byť imunogénna. Stupeň imunogénnosti ovplyvňuje aj degradovateľnosť molekuly imunogénu. Pri jej rozklade môže dôjsť k deštrukcii epitopov a tým k znižovaniu imunogénnosti ako aj k zmene špecifickosti. Čiastočná degradácia Ag v makrofágoch však môže mať za následok objavenie sa nových epitopov. Ak tieto nové epitopy sú v natívnej molekule skryté a teda pre receptory imunokompetentnej bunky neprístupné, môže sa po opracovaní Ag zvýšiť jeho imunogénnosť.
13 VLASTNOSTI IMUNOGÉNOV Biologické - cudzorodosť - druhová vzdialenosť - čím > fylogenetický rozdiel medzi organizmom, z ktorého Ag pochádza, a organizmom, do ktorého Ag vniká, tým > je jeho imunogénnosť (vírus človek) Aby cudzorodá látka mohla byť Ag, nestačí jej len jedna z vlastností, nato je potrebná vhodná kombinácia uvedených vlastností. Imunogény sa v prírode vyskytujú zvyčajne vo veľmi komplexnej forme, sú to najmä vírusy, mikróby, živočíšne bb. a ich súčasti) Schopnosť reagovať IO na určitý Ag sa dedí ako autozomálny dominantný znak. Kvalitu a smer IO určujú popri genóme imunizovaného jedinca a vlastnostiach Ag aj iné faktory, a to najmä miesto, kde vzniká kontakt Ag s organizmom (spôsob imunizácie) a množstvo Ag.
14 BAKTERIÁLNE Ag - sú endotoxíny G- baktérií - endotoxín = lipopolysacharidový komplex z b. steny G- baktérií (nejde o toxín pochádzajúci z vnútra baktérie, ale o látku, ktorá sa nachádza v ich povrchovej vrstve) - endotoxíny najrozšírenejšie bakteriálne Ag, s ktorými sa organizmus teplokrvných živočíchov dostáva do styku už ihneď po narodení (kolonizácia črevného traktu baktériami). Podmienkou prežitia je vytvorenie účinných Ab. - endotoxínový komplex sa uvoľňuje do okolitého prostredia vtedy, keď sa baktérie rozrušia. Endotoxín spúšťa veľké množstvo biologických reakcií: - menšie dávky spôsobujú zvýšenie telesnej T, zmeny v množstve cirkulujúcich Leu, zvýšenie glukózy v krvi a iné - ovplyvňuje cievny systém, uvoľnenie katecholamínov a glukokortikoidov, metabolizmus bb. a tkanív - menia FA a znižujú prirodzenú rezistenciu proti infekcii - väčšia dávka endotoxínov smrteľný šok
15 BAKTERIÁLNE Ag - Streptokoky A vylučujú exotoxín streptolyzín O. Protilátky proti streptolyzínu O sa volajú antistreptolyzín O (ASLO). Vysoký titer týchto protilátok (vysoké ASLO) svedčí o čerstvej streptokokovej infekcii. Najimunogénnejšia časť streptokokov A je ich povrchová zložka M- proteín. Táto bielkovina brzdí fagocytózu. Ak sa utvoria protilátky proti M- proteínu, fagocytóza sa zvýši.

16



17
18 SUPERANTIGÉNY Nie všetky antigény, ktoré sa viažu na molekuly MHC II.tr., sú viazané v peptidovom žliabku. Tým sa odlišujú od iných antigénov a patria do skupiny známej pod názvom superantigény (SAgs). SAgs sa odlišujú spôsobom väzby, čo im umožňuje stimulovať veľmi veľké množstvo T buniek, často s katastrofálnymi následkami. SAgs sú produkované mnohými odlišnými patogénmi, včítane baktérií, mykoplaziem a vírusov. SAgs sú prezentované antigén prezentujúcimi bunkami za pomoci molekúl MHC II.tr., ale nie sú viazané v peptidovom žliabku (kde sa nachádza väzbové miesto pre antigén), teda neboli spracované antigén prezentujúcimi bunkami. Sú to proteínové molekuly (hlavne bakteriálneho pôvodu, ale aj vírusov napr. besnoty), ktoré stimulujú určitý subtyp T buniek a to priamym a tesným naviazaním sa na MHC molekuly II.tr. na antigén prezentujúcich bunkách a zároveň na V β domény T-bunkových receptorov, čím sa stimuluje aktivácia T buniek. Stafylokokové enterotoxíny po požití infikovanej potravy (sú zodpovedné za otravy jedlom), s následným vznikom toxického šoku, tiež stimulujú mnoho T buniek práve týmto spôsobom.

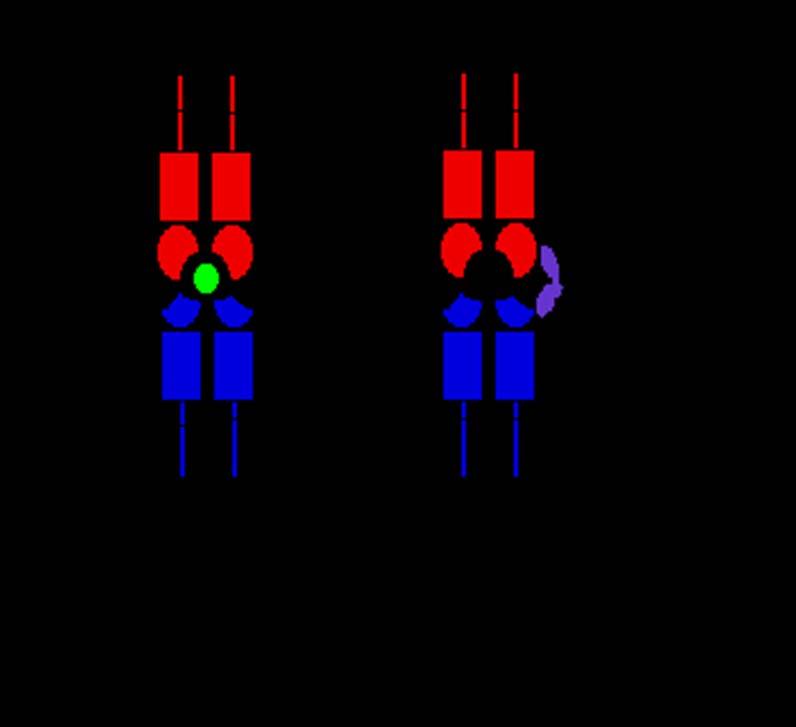

19 Cα Cβ Vα Vβ α1 β1 normal Ag Superantigen α2 β2
20 SUPERANTIGÉNY Bakteriálne toxíny, napr. Staphylococcus aureus pôsobia ako SAgs, aktivujú súčasne veľké percento T buniek s následnou sekréciou veľkého množstva cytokínov, ktoré môžu spôsobiť šokové stavy u hostiteľa. SAgs sú takým typom antigénu, ktorý aktivuje tie T bunky, ktoré vykazujú špecifický V β segment (V β 3 alebo V β 11) ako súčasť ich TCR. V β segmenty sa nachádzajú na bunkovom povrchu takmer 20% z celkovej populácie T lymfocytov, teda veľké % T buniek s rôznou antigénnou špecificitou môže byť naraz aktivovaná superantigénom. SAgs sa viažu takmer výlučne na V β oblasť TCR a nie na Vα.
21 Tumoršpecifické antigény (neoantigény, onkofetálne antigény) ich štúdium je súčasťou imunológie nádorov. Sú spravidla bežne prítomné v určitých tkanivách a telových tekutinách ľudského embrya, ale po narodení obyčajne v krátkom čase buď z organizmu úplne vymiznú, alebo sa tvoria len v extrémne nízkych koncentráciách. Pri rakovinovej premene tkaniva sa však začínajú tieto antigény vo zvýšenej miere syntetizovať. Pre túto vlastnosť sa tumoršpecifické antigény nazývajú aj onkofetálnymi. Ich biologický význam je nejasný. Predpokladá sa, že nejakým spôsobom súvisia s rýchlym rastom buniek a tkanív. V pokusoch in vitro môžu byť bunky malígnych nádorov usmrtené cytotoxickými protilátkami, alebo senzibilizovanými lymfocytmi, ktorých tvorba je indukovaná neoantigénmi, prípadne môžu byť likvidované aktivovanými makrofágmi. Predpokladá sa, že in vivo sú nádory deštruované najmä mechanizmami celulárnej imunity, protilátky majú len malú úlohu. Je otázne, prečo nádory rastú, keď máme protinádorové imunitné mechanizmy? Odpoveď nie je jednoduchá, lebo doteraz nie je presne známy mechanizmus protinádorovej imunity.
22 Tumoršpecifické antigény (neoantigény, onkofetálne antigény) Predpokladá sa, že rast nádorov in vivo umožňuje okrem špecifických vlastností nádorovej bunky aj špecifická imunologická tolerancia (lymfatický systém nerozpoznáva neoantigény ako cudzie) ako aj imunologická blokáda protilátkami. Doposiaľ je charakterizovaných niekoľko antigénov asociovaných s nádormi. Z nich sú najlepšie preskúmané karcinoembryonálny antigén a α 1 fetoproteín.
23 Karcinoembryonálny antigén Karcinoembryonálny antigén (CEA carcinoembryonic antigen) tento špecifický antigén bol objavený v r (Gold a Freedman). Výskyt: - adenokarcinóm (zhubná rakovina žliaz) v ľudskom tráviacom systéme - v orgánoch tráviaceho traktu ľudských 2 až 6 mesačných fétov. CEA molekuly sa nachádzajú na povrchu nádorových buniek, odkiaľ sa dostávajú do krvného obehu. CEA sa nachádza vo zvýšených koncentráciách v sére pacientov s karcinómom hrubého čreva a pankreasu. CEA je glykoproteín, Mr 200 kd, obsah sacharidov je viac než 50%, jeho hlavnou zložkou je N-acetylglukozamín. Stanovenie hladiny CEA v sére má účel diagnostický, ale aj prognostický a to pred chirurgickým zákrokom a po ňom. Ak zmizne CEA z cirkulácie, to znamená, že nádor bol z tela odstránený. Pretrvávanie zvýšenej hladiny CEA poukazuje na prítomnosť metastáz v organizme.
24 α 1 fetoproteín (AFP) α 1 fetoproteín (AFP) je hlavný plazmatický proteín počas vývoja ľudského fétu. Bol však dokázaný aj v sére pacientov s primárnou rakovinou pečene a aj u pacientov s teratokarcinómami (teratóm je nádor obsahujúci vlasy, zuby, kosti a iné materiály, ktoré v danej časti organizmu normálne nerastú). AFP sa syntetizuje v pečeni. Počas embryonálneho vývoja dosahuje najvyššiu koncentráciu (550 ng/ml) vo fetálnom sére v týždni gestácie. Jeho hodnota sa potom znižuje viac než stonásobne na hodnotu 2-10 ng/ml, čo je normálna hodnota u ľudí. AFP je glykoproteín s Mr 70 kd, obsahuje asi 4% sacharidov. Neboli zistené rozdiely medzi AFP z izolovaných fétov a z ľudských hepatálnych nádorov.
25 Transplantácia Už v r.1932 E.Padgett použil kožné transplantáty medzi členmi rodiny u popálených jedincov. Hoci tieto transplantáty neprežívaly dlho, dočasným prežitím umožnili reepitelizáciu, chránili pred infekciou a pred stratou telových tekutín. Už vtedy sa ukázalo, že kožné štepy blízkych členov rodiny prežívajú dlhšie než štepy náhodných darcov. V r.1937 J.B.Brown dosiahol permanentné prežívanie kože po transplantácii medzi monozygotnými dvojčatmi. Podstatu tejto transplantácie objasnil už v r Lillie, ktorý zistil, že u monozygotných dvojčiat dochádza k miešaniu krvi v placentárnej cirkulácii, čo neskôr bolo dokázané, a je to podstatou vzniku tolerancie Základným pravidlom v biológii je, že organizmus v záujme zachovania svojej jedinečnosti sa bráni pred cudzorodými bunkami a tkanivami a preto ich odvrhuje. Transplantácia je prenos živých buniek, tkanív a orgánov z jedného miesta na iné u toho istého jedinca (autotransplantácia), alebo medzi dvoma rôznymi jedincami, ktorí môžu byť geneticky totožní (jednovaječné dvojčatá), alebo medzi príslušníkmi inbredného kmeňa, resp. medzi jedincami, ktorí sa od seba antigénne líšia.

26

27
28 Transplantácia Aj napriek obrovskému technickému pokroku v medicíne a snahe lekárov pomôcť ťažko chorým ľuďom, úspešnosť transplantácií orgánov je nízka. Príčinou nie je nešikovnosť chirurgov, ale práve imunita, pretože hlavným mechanizmom neznášanlivosti tkanív darcu a príjemcu s následným odvrhnutím tkaniva je imunitná reakcia príjemcu. Naše vedomosti z oblasti regulácie imunity sú ešte nedostatočné a preto sa zatiaľ ani nedarí prekonať biologickú nezlúčivosť tkanív. Ak patria antigénne odlišní donor a recipient k rovnakému biologickému druhu, vtedy hovoríme o alogénnej transplantácii. Ak patria k inému druhu, jedná sa o xenogénnu transplantáciu. O osude transplantátu, štepu (angl. graft) rozhoduje genetický vzťah medzi darcom (donor) a príjemcom (recipient). Transplantáty prenesené medzi geneticky identickými jedincami sa prihoja, pretože je medzi nimi antigénna identita. Štepy prenesené medzi geneticky odlišnými jedincami (pri alogénnom prenose) sa neprihoja, sú odvrhnuté, vypudené. Rejekcia vypudenie je dôsledkom špecifickej imunitnej odpovede príjemcu na histokompatibilné antigény darcu. Za určitých podmienok môže byť u príjemcu navodená neschopnosť reagovať na transplantát, vzniká tzv. imunologická tolerancia.
29 Transplantácia Ak je reakcie schopný iba príjemca, dochádza k reakcii hostiteľa proti štepu, ktorá sa v angl. terminológii označuje ako host versus graft reaction (HvGR). Ak je však štep tvorený imunokompetentnými bunkami (alebo ak obsahuje určitý počet týchto buniek), reaguje štep proti hostiteľovi, reakcia sa nazýva graft versus host reaction (GvHR). Štruktúry zodpovedné za transplantačnú reakciu sú regulované histokompatibilnými génmi (gény H). Sem patria gény, ktoré určujú: 1. antigény erytrocytov 2. antigény lymfocytov 3. transplantačné antigény Uvedené delenie H génov sa opiera o metódu ich identifikácie. Antigény erytrocytov sa dokazujú predovšetkým aglutinačným testom, lymfocytové antigény hlavne lymfocytotoxickým testom a transplantačné antigény metódou tkanivových štepov (najčastejšie kožných alebo nádorových).
30 Transplantácia Podľa toho, akú intenzívnu imunitnú odpoveď vyvolávajú, rozdeľujú sa H gény na dve skupiny: 1. tie, ktoré in vivo a in vitro vyvolávajú akútnu reakciu (to sú gény hlavného histokompatibilného komplexu, MHC = major histocompatibility complex) 2. tie, ktoré spôsobujú protrahovanú chronickú reakciu (to sú gény vedľajšieho histokompatibilného komplexu, non MHC). MHC je extrémne polymorfný systém. U myší, ale aj u človeka obsahuje viac ako 50 alel, avšak pre jednotlivé non-mhc sú známe najviac 3 alely. Gény MHC sú tesne viazané s génmi, ktoré regulujú intenzitu imunitnej odpovede (Ir gény). Všeobecne platí, ža antigény MHC spôsobujú akútne vypudenie transplantovaných orgánov alebo tkanív, napr. kože.
31 Transplantačné reakcie Reakcia hostiteľa proti štepu (HvGR): Pr.: transplantovaná koža je v prvých dňoch po prenose bledá. Za 2-4 dni sa štep vaskularizuje a farba sa mení na ružovú. Rozdiel medzi aloštepom a autoštepom sa zreteľne objaví po 4-7 dňoch. Ak bol prenesený autogénny štep a ružové sfarbenie pretrváva, svedčí to o krvnom zásobení s pokračovaním epitelizácie transplantovaného miesta. Štep postupne splynie s kožou príjemcu. Iný je sled reakcií, ak sa jedná o aloštep. Ten sa stáva cyanotickým, postupne nekrotizuje a po dňoch je vypudený, odpadne. Doba, za ktorú dôjde k vypudeniu štepu je určená štyrmi základnými faktormi: 1. množstvom transplantovaného tkaniva 2. imunogénnosťou darcovho transplantačného antigénu 3. stupňom genetickej rozdielnosti medzi darcom a príjemcom 4. imunitným stavom hostiteľa U príjemcu, ktorý dostane druhý štep od toho istého darcu, doba za ktorú je štep vypudený sa skracuje na 5-7 dní. U hyperimúnnych zvierat, ktoré majú v krvi protilátky proti transplantačným antigénom darcu, prebieha odvrhnutie štepu ešte skôr, než dôjde k vaskularizácii.
32 Transplantačné reakcie Reakcia štepu proti hostiteľovi (GvHR): Táto reakcia je vyvolaná imunokompetentnými bunkami, ktoré sú implantované príjemcovi, ktorý sám imunologicky nereaguje. Areaktivita môže vzniknúť: 1. ak je príjemcom novorodený organizmus 2. ak sa dospelý príjemca stane neschopným imunologicky reagovať v dôsledku imunosupresie, alebo v dôsledku jeho patologickej imunitnej nedostatočnosti. Mechanizmus vypudenia štepu je proces, ktorý sa dá rozdeliť do troch fáz: 1. rozpoznanie 2. proliferácia spojená s diferenciáciou 3. deštrukcia Fáza rozpoznania a deštrukcie prebiehajú priamo v štepe. Proliferačná a diferenciačná fáza sa rozvíjajú v lymfatických uzlinách, ktoré drenujú lôžko štepu.
33 Transplantačné reakcie Rozpoznávacia fáza Bezprostredne po transplantácii sa začínajú antigény z darcovho tkaniva vyplavovať do krvného obehu príjemcu, alebo sa prichytia na bunkách cievneho endotelu štepu. Antigén sa tak dostáva do priameho kontaktu s bunkami imunitného systému v krvnom obehu hostiteľa. Efektorová zložka imunitnej odpovede sa skladá z lymfocytov, makrofágov a polymorfonukleárov. Každý jednotlivý typ buniek zohráva špecifickú úlohu v rozvoji imunitnej odpovede. Rozpoznanie aloantigénov je funkciou Th lymfocytov. Antigény sú rozpoznávané Th bunkami buď pri prietoku krvi štepom, alebo s prispením makrofágov, ktoré vychytávajú antigény zo štepu alebo z tkanivovej tekutiny (lymfy) a potom ich prezentujú T bunkám v lymfatickej uzline (kooperácia makrofágov s T bunkami).
34 Proliferácia a diferenciácia Ak sa lymfocyty stretli s antigénmi pri prechode štepom a rozpoznali ich ako cudzie, vracajú sa do príslušných lymfatických uzlín, tam proliferujú a stimulujú ďalšie efektorové prekurzorové bunky (T a B lymfocyty). Väčšina novo senzibilizovaných buniek potom opúšťa uzlinu a krvným obehom sa dostáva do štepu, kde buď priamo alebo s pomocou ďalších systémov (PMN, špecifické protilátky, komplement) štep zničí. Určité množstvo lymfocytov pretrváva v obehu ako tzv. pamäťové bunky. Deštrukčná fáza Existuje niekoľko možných ciest, včítane ich rôznych kombinácií: 1. aktivované zabíjačské T bunky sa viažu prostredníctvom špecifických bunkových receptorov na bunky štepu a rozrušia ich. T bunky tiež uvoľňujú faktory, napr. faktor aktivujúci migráciu makrofágov (MAF), ktoré urýchľujú infiltráciu štepu mononukleárnymi bunkami 2. aktivované B lymfocyty sa diferencujú na plazmatické bunky a produkujú špecifické protilátky proti antigénom štepu. Protilátky sa môžu priamo viazať s komplementom a aktivovať ho. Mediátory zápalu potom vo zvýšenej miere lákajú PMN na miesto štepu. Rozpadom PMN sa uvoľňujú lyzozómy a proteolytické enzýmy, ktoré spôsobia deštrukciu tkaniva štepu.
35 Spôsoby imunosupresie ožarovaním X-lúčmi injektovaním kortikosteroidov podávaním protilátok proti leukocytom recipienta
36 Nádorové bunky sa líšia od normálnych buniek z ktorých vznikli a preto by mali byť rozpoznané imunitnými mechanizmami a eliminované. Odlišnosti od normálnych buniek sú však buď príliš malé, takže sú imunitným systémom ignorované, alebo nádorové bunky majú mechanizmy, ktorými paralyzujú niektoré zbrane IS. PROTINÁDOROVÁ IMUNITA Malígna transformácia buniek dôsledok zlyhania rôznych mechanizmov regulácie delenia buniek (mutáciami v tzv. onkogénoch alebo antionkogénoch). Tieto gény v svojej nemutovanej podobe kódujú rôzne normálne signalizačné molekuly (napr. proteín-kinázy), transkripčné faktory, proteíny regulujúce adhezivitu buniek, alebo apoptózu. Mutantné formy génov môžu mať konštitutívne abnormálne zvýšenú aktivitu (produkty onkogénov), alebo zníženú (produkty anti-onkogénov). To môže mať za následok: - nekontrolovatelné delenie buniek - ich únik z normálnej lokalizácie v tkanive - disemináciu do iných tkanív a - agresívny rast v nich.
37 The presentation of the peptide fragments of the tumor cell antigens in complex with HLA class I molecules
38 Nádorové antigény Prítomnosť špecifických povrchových antigénov na nádorových bunkách je nevyhnutným predpokladom pre rozpoznanie nádorových buniek imunitným systémom. Doposiaľ identifikované dve kategórie nádorových Ag: - TSA (tumor specific antigens antigény špecifické pre nádory) -TAA (tumor associated antigens antigény asociované s nádormi) TSA sem patria proteíny, ktoré sa na normálnych bunkách nevyskytujú, napr. - komplexy MHC I.tr. s naviazanými abnormálnymi fragmentami bunkových proteínov (produkty mutovaných génov). TSA sú typické pre chemicky indukované nádory a pre niektoré leukémie. - komplexy MHC I.tr. s fragmentami proteínov onkogénnych vírusov (EBV) - abnormálne formy glykoproteínov: glykozylácia, sialylácia povrchových molekúl nádorových buniek TAA sem patria už spomínané AFP a CEA
39 Membrána normálnej bunky Membrána nádorovej bunky
40 Protinádorové mechanizmy imunity Vrodené - NK, PMN, aktivované makrofágy, komplement Adaptívne - CTL, ADCC reakcie Príčiny odolnosti nádorov voči IS - variabilita nádorových buniek (vznik mutantných foriem) - malá hustota expresie nádorových Ags ( IS ich potom ignoruje) - sialylácia povrchu nádorových buniek vedie k zamaskovaniu niektorých epitopov nádorových Ags - niektoré nádory produkujú faktory inaktivujúce T bunky - niektoré nádory exprimujú na svojom povrchu FasL - indukcia apoptózy T bb. - faktory produkované niektorými nádormi inhibujú funkcie alebo životnosť DC (napr. oxid dusnatý indukuje apoptózu DC) Terapia nádorov chirurgické odstránenie, chemoterapia, rádioterapia, imunoterapia (MoAbs, a/alebo bunkami sprostredkovanými mechanizmami).
41 Nádorové bunky sa líšia od normálnych buniek z ktorých vznikli a preto by mali byť rozpoznané imunitnými mechanizmami a eliminované. Odlišnosti od normálnych buniek sú však buď príliš malé, takže sú imunitným systémom ignorované, alebo nádorové bunky majú mechanizmy, ktorými paralyzujú niektoré zbrane IS. PROTINÁDOROVÁ IMUNITA Malígna transformácia buniek dôsledok zlyhania rôznych mechanizmov regulácie delenia buniek (mutáciami v tzv. onkogénoch alebo antionkogénoch). Tieto gény v svojej nemutovanej podobe kódujú rôzne normálne signalizačné molekuly (napr. proteín-kinázy), transkripčné faktory, proteíny regulujúce adhezivitu buniek, alebo apoptózu. Mutantné formy génov môžu mať konštitutívne abnormálne zvýšenú aktivitu (produkty onkogénov), alebo zníženú (produkty anti-onkogénov). To môže mať za následok: - nekontrolovatelné delenie buniek - ich únik z normálnej lokalizácie v tkanive - disemináciu do iných tkanív a - agresívny rast v nich.
42 Význam sacharidov pre vznik nádorov V štruktúre sacharidov rovnako ako v nukleových kyselinách a proteínoch sú uložené dôležité biologické informácie. V skutočnosti sacharidy ako nositelia informácií majú najväčšiu kapacitu, pretože majú najväčší potenciál vytvárať variabilné štruktúry. Napr. z dvoch rovnakých monosacharidov môže vzniknúť 11 rôznych disacharidov, kým dve aminokyseliny môžu vytvoriť len 1 dipeptid. Nomenklatúra rôznych sacharidov je založená na variácii cukrových podjednotiek, rozdielov väzieb medzi nimi a na prítomnosti alebo absencii rozvetvených reťazcov. Sada sacharidov nádorových buniek sa výrazne odlišuje od normálnych buniek a preto môžu byť použité ako marker. Sacharidy na povrchu nádorových buniek pomáhajú vytvárať metastázy. Nádorová bunka po penetrácii do krvného obehu cirkuluje v krvi a môže opustiť krvné cievy len cez stenu žilových kapilár.
43 V kapilárach krvných ciev nádorové bunky prichádzajú do kontaktu s E - selektínmi endotelových buniek a opúšťajú krvný obeh cez priestory medzi endotelovými bunkami. (Pozn.: E-selektíny sú špeciálne povrchové adhezívne molekuly, ktoré interagujú so sacharidmi na povrchu iných buniek.) Týmto spôsobom sa nádorová bunka dostáva do iných tkanív a orgánov, kde vytvára metastázy. Nádorová bunka potrebuje na opustenie krvného obehu 8-24 h. Len jedna z buniek, ktoré sa uvoľnili z primárneho nádoru prežije a vytvorí novú kolóniu. Nádorové bunky majú na svojom povrchu tzv. adhezívne molekuly CAM (cell adhesion molecules). Tieto molekuly sa počas tvorby metastáz viažu s CAM na bunkách orgánov a tkanív, ktoré sú im najviac podobné.
44 Čím bohatšia je sieť kapilár v orgáne, tým častejšie sa v ňom vytvárajú metastázy. To je aj vysvetlením, prečo sú najčastejšie metastázy určitých nádorov v konrétnych orgánoch a tkanivách. CAM a imunoglubulíny (Igs) sú štrukturálne homológne. Lektíny (typ proteínov schopný vytvárať rýchlo, selektívne a reverzibilne väzby so sacharidmi) zohrávajú dôležitú úlohu v raste nádoru. Sú fixované povrchovými sacharidmi na nádorových bunkách, čím ochraňujú (maskujú) nádor pred imunitným systémom. Napr. ak sú bunky melanómu opracované chemickými látkami, ktoré obsahujú laktózu, ich schopnosť vytvárať metastázy je o polovicu nižšia. Nádory sú schopné indukovať tvorbu krvných ciev (angiogenéza). Krvné cievy v sekundárnych nádoroch sú viac permeabilné, preto sekundárne nádory ľahšie metastázujú.

45 The presentation of the peptide fragments of the tumor cell antigens in complex with HLA class I molecules
46 Nádorové antigény Prítomnosť špecifických povrchových antigénov na nádorových bunkách je nevyhnutným predpokladom pre rozpoznanie nádorových buniek imunitným systémom. Doposiaľ identifikované dve kategórie nádorových Ag: - TSA (tumor specific antigens antigény špecifické pre nádory) -TAA (tumor associated antigens antigény asociované s nádormi) TSA sem patria proteíny, ktoré sa na normálnych bunkách nevyskytujú, napr. - komplexy MHC I.tr. s naviazanými abnormálnymi fragmentami bunkových proteínov (produkty mutovaných génov). TSA sú typické pre chemicky indukované nádory a pre niektoré leukémie. - komplexy MHC I.tr. s fragmentami proteínov onkogénnych vírusov (EBV) - abnormálne formy glykoproteínov: glykozylácia, sialylácia povrchových molekúl nádorových buniek TAA sem patria už spomínané AFP a CEA

47 Membrána normálnej bunky Membrána nádorovej bunky
48 Protinádorové mechanizmy imunity Vrodené - NK, PMN, aktivované makrofágy, komplement Adaptívne - CTL, ADCC reakcie Príčiny odolnosti nádorov voči IS - variabilita nádorových buniek (vznik mutantných foriem) - malá hustota expresie nádorových Ags ( IS ich potom ignoruje) - sialylácia povrchu nádorových buniek vedie k zamaskovaniu niektorých epitopov nádorových Ags - niektoré nádory produkujú faktory inaktivujúce T bunky - niektoré nádory exprimujú na svojom povrchu FasL - indukcia apoptózy T bb. - faktory produkované niektorými nádormi inhibujú funkcie alebo životnosť DC (napr. oxid dusnatý indukuje apoptózu DC) Terapia nádorov chirurgické odstránenie, chemoterapia, rádioterapia, imunoterapia (MoAbs, a/alebo bunkami sprostredkovanými mechanizmami).

49
50 Biológia rakoviny, Vanda Repiská BIOLÓGIA RAKOVINY Doc. RNDr. Vanda Repiská, PhD. Ústav lekárskej biológie a genetiky, Lekárska fakulta, Univerzita Komenského v Bratislave, Sasinkova ulica 4, I. poschodie, Bratislava, vanda.repiska@fmed.uniba.sk Rakovina vzniká nekontrolovaným množením nádorových buniek. Nádory prsníka tvarom pripomínajú kraba (nie raka = cancer), tak nepresným pomenovaním vznikol pojem rakovina. Je príčinou smrti každého piateho človeka na Zemi, pričom v tzv. rozvinutých krajinách sa, najmä u starších ľudí, postupne stáva hlavnou príčinou úmrtia. Najčastejším typom rakoviny u žien je rakovina prsníka, u mužov rakovina pľúc a hrubého čreva u oboch pohlaví. Nádorové bujnenie sa vyskytuje u všetkých mnohobunkových organizmov, zvierat aj u ľudí. Delenie buniek, rast a obnova tkanív sú veľmi presne regulovanými procesmi. Ak sa bunka, v dôsledku poruchy riadenia (deregulácie) bunkového cyklu, vymkne spod tejto prísnej kontroly môže vzniknúť nezávislá bunka, z ktorej vznikne nádor. Nádory delíme na benígne (nezhubné) a malígne (zhubné). Vznik nádoru je zvyčajne viac stupňový proces, spôsobený postupným zlyhávaním genetickej kontroly delenia buniek a nárastom ich malignity. Niektoré malígne bunky sa dokážu odpojiť a preniknúť do krvného alebo lymfatického riečiška, dostať sa do vzdialenejšieho orgánu, kde môžu vytvoriť metastázu. Tvorba metastáz je aktívny viacstupňový proces. Existujú experimentálne dôkazy, že primárny nádor už od začiatku obsahuje bunky, ktoré majú schopnosť metastázovať. Telové bunky majú svoje vlastné vnútorné biologické hodiny. Normálna bunka má určenú životnosť a stanovený počet delení. Potom zanikne prirodzenou (programovanou) smrťou (apoptóza). Pri každej replikácii DNA sa stratí krátky úsek DNA z koncov chromozómov (teloméra). Ide o sekvencie šiestich báz TTAGGG, ktoré sa v telomére opakujú priemerne asi 20 krát a pri každej replikácii sa jedno z nich nereplikuje, čím sa teloméra skracuje. Ak sa stratí aj posledné opakovanie už sa nemôže začať replikácia, bunka sa nadokáže deliť a zaniká. Zmyslom tohto mechanizmu je likvidácia starých buniek, u ktorých je zvýšená pravdepodobnosť hromadenia mutácií a môžu sa premeniť na malígne. V embryonálnych a v niektorých nádorových bunkách je aktívny enzým telomeráza, ktorý dokáže obnovovať telomerické sekvencie a tým robí bunku prakticky nesmrteľnou. Medzi najdôležitejšie faktory podporujúce vznik rakoviny patria dedičná predipozícia, vek, fajčenie, výživa, účinok onkogénnych vírusov a tzv. karcinogénne látky. Samostatné postavenie má imunitný systém, ktorý v organizme likviduje vznikajúce nádorové bunky. Zlyhanie špecifickej imunity, zvyšuje riziko vzniku nádorov, najmä zhubných, tak ako je to známe napr. o Kaposiho sarkóme u pacientov s chorobou AIDS. 1
51 Biológia rakoviny, Vanda Repiská Regulácia delenia eukaryotických buniek Delenie buniek prebieha pod dôkladnou genetickou kontrolou, je presne načasované a má definované kontrolné uzly. Ak nepokračuje správne, cyklus sa zastaví, aby sa mohli uskutočniť opravy alebo sa navodí apoptóza (programovaná smrť bunky). Význam celého procesu je v tom, aby vznikali dcérske bunky bez mutácií a aby boli likvidované rizikové bunky, ktoré by mohli zahubiť organizmus. Pre riadenie bunkového cyklu platia tri základné pravidlá: 1) normálna telová bunka sa bude deliť len po tom, ako dostane príslušný podnet z okolia, a to buď cestou endokrinnou (napr. rastovým hormónom) alebo parakrinnou (rastovými faktormi, produkovanými bunkami v okolí). Nezávisle sa delia len nádorové bunky. 2) Príkaz pre pokračovanie bunkového cyklu sa realizuje cestou tzv. signálnych dráh v bunke. Výsledkom je zvyčajne cielená expresia (transkripcia a translácia) niektorého z génov. 3) Na riadení priebehu bunkového cyklu sa podieľajú dva typy génov, tumorsupresorové gény (TSG) a protoonkogény, ktoré navzájom pôsobia. Ich vzťah sa zdá byť antagonistický, ale v skutočnosti pri každom z krokov bunkového cyklu sa nimi kódované bielkoviny (produkty) navzájom ovplyvňujú (obr. 1). Okrem týchto dvoch typov génov sa na vzniku rakoviny podieľajú aj gény opravy DNA (mutátory). bielkovina kódovaná TSG Protoonkogén proliferácia (delenie a diferenciácia buniek) Obr. 1 Interakcia tumorsupresorového génu a protoonkogénu Tumorsupresorové gény - ich normálnou funkciou je brzdiť alebo úplne zastaviť delenie buniek. Kódujú bielkoviny, ktoré buď kontrolujú niektorú etapu bunkového cyklu (napr. proteín p53), alebo riadia časovo správnu expresiu niektorých génov (napr. proteín rb1). Ich produkty teda predstavujú prirodzenú prevenciu vzniku nádorových buniek. Pri ich poruche sa zvyšuje pravdepodobnosť premeny normálnej bunky na zhubnú, pretože dôjde k vyradeniu jedného alebo viacerých kontrolných bodov v bunkovom cykle. Mutácia jednej alely zvyčajne nestačí na vznik nádorovej bunky. V ľudských nádoroch sa často vyskytuje mutovaná forma génu TP53. Tento gén kóduje bielkovinu p53. Jeho úlohou je pozastaviť začiatok replikácie DNA, čím opravné enzýmy získajú čas na opravu poškodenia DNA. Ak je poškodení príliš veľa, práve p53, navodí procesy vedúce k apoptóze. Preto sa p53 nazýva aj strážca genómu. Proteíny niektorých regulačných génov sú potrebné na replikáciu DNA. Na tieto regulačné gény sa napojí proteín rb1 (kódovaný génom RB1) a blokuje začatie replikácie. Po kontrole a oprave mutácií v DNA je rb1 uvoľnený a môže začať replikácia DNA. Ak v bunke dôjde k strate účinnosti bielkoviny, kódovanej TSG (mutáciou TSG alebo jej zablokovaním), proteíny kódované protoonkogénmi pôsobia bez kontroly a výsledkom je nebezpečenstvo deregulácie bunkového cyklu (obr. 2 ) 2

52 Biológia rakoviny, Vanda Repiská bielkovina kódovaná TSG Protoonkogén deregulácia bunkového cyklu (malignita) Obr. 2 Schéma straty účinnosti TSG a následnej deregulácie bunkového cyklu Príkladom zlyhania tumorsupresorového génu môže byť mutácia jednej alely génu TP53 (za vzniku tzv. syndrómu Li-Fraumeni), alebo mutovanie oboch alel génu RB1, ktorý vyvolá vznik retinoblastómu (nádor sietnice oka). Pri syndróme Li-Fraumeni vznikol jedinec, ktorý má v každej telovej bunke mutovanú jednu alelu TP53. Dôsledkom je, že proteín p53, ktorý produkujú jeho bunky, nie je schopný navodiť apoptózu. Následne sa v bunkách hromadia mutácie a je len otázkou času, kedy sa niektorá z nich stane malígnou. U pacientov sa vyskytujú najprv benígne a potom malígne nádory. Pri nádore sietnice v oku (retinoblastóm) je situácia trochu odlišná. Mutácia jednej alely RB1 v bunke totiž neovplyvní funkčnosť bielkoviny rb1. Sietnica je však v našom tele jednou zo štruktúr, ktoré sú najviac vystavené ultrafialovému svetlu a teda aj pravdepodobnosti, že bude gén RB1 celkom zmutovaný. Existuje dedičná a nededičná forma retinoblastómu. Nededičná forma: u zdravého človeka sa táto malignita vyskytne len ak sú v jednej bunke, postupne mutované obidve alely génu RB1. Ide zriedkavú nededičnú formu, ktorá sa prejaví zvyčajne jedným nádorom v jednom oku. Dedičná forma: naopak pri dedičnej forme, má jedinec v každej telovej bunke gén RB1 v heterozygotnom stave (dôsledok gametickej mutácie), je teda relatívne vysoká pravdepodobnosť, že v niektorej z buniek sietnice dôjde k mutácii aj druhej alely a k vzniku nádoru. Preto sa tento dedičný retinoblastóm objavuje nezriedka u detí a na oboch očiach. Príkladom straty pôsobenia TSG je zabránenie účinku bielkovín kódovaných tumorsupresorovými génmi, ako dôsledok pôsobenia onkoproteínov, po infekcii bunky niektorým DNA vírusom, napr. ľudských bradavíc (human papilloma virus - HPV). Niektoré z nich (najmä HPV 16 a 18) majú onkogénny potenciál a dávajú sa do priameho súvisu so vznikom nádorových ochorení, napr. karcinómu krčka maternice(obr. 3). Kódujú proteíny E6 a E7. Onkoproteín E6 sa viaže na p53 a E7 na Rb1 a znemožňuje ich funkciu. Obr. 3 Zhubný nádor krčka maternice (žlté šípky). Prerastanie nádorových hmôt do kanála krčku maternice (modrá šípka). Nezmenený vaječník (čierna šípka). (foto : MUDr. Michal Palkovič, PhD. Ústav patologickej anatómie LF UK v Bratislave) Protoonkogény (proliferačné gény) kódujú bielkoviny, ktoré riadia pokračovanie v bunkovom cykle. Nazývame ich onkogény, ak spôsobia dereguláciu bunkového cyklu. 3
53 Biológia rakoviny, Vanda Repiská Bielkoviny kódované protoonkogénmi sú zodpovedné za prenos signálov aktivujúcich pokračovanie v bunkovom cykle (mitogénne aktivátory) medzi bunkami, do bunky až po riadenú transkripciu určených génov. Vyskytujú sa na všetkých úrovniach riadenia bunkového cyklu (obr. 4) a tvoria signálové cesty (dráhy, kaskády) Obr. 4 Lokalizácia bielkovín kódovaných protoonkogénmi. 1. mitogénne aktivátory (endokrinná a parakrinná regulácia), 2. membránové proteíny (receptory a G proteíny), 3. plazmatické transmitery (prenášajú signál do jadra), 4. jadrové protoonkogény (transkripčné faktory, cyklíny a pod.). Pri regulácii jednotlivých procesov prebiehajúcich počas každej z etáp bunkového cyklu majú zásadný význam cyklíny. Sú to regulačné molekuly, ktoré po väzbe na príslušný enzým nazývaný cyklín-dependentná kináza (cdk) túto aktivujú. Cyklínov je 8 rodín (A až H), cyklín-dependentných kináz je tiež viacero druhov. V každej etape bunkového cyklu je prítomný iný cyklín a použije sa k nemu príslušná kináza. V adekvátnom čase je cyklín rozložený. Naopak, kinázy majú dlhšiu životnosť, ale použitý je vždy len ten typ, pre ktorý sú v jadre prítomné príslušné cyklíny. Prehľad o najdôležitejších cyklínoch a cdk pri riadení bunkového cyklu uvádza tab. 1. Tab. 1 Najdôležitejšie komplexy cyklíny, cdk a ich funkcia Cyklín cdk D 1-3 riadia 4,6 pokračovanie G 1 fázy E 2 prechod z G 1 do S fázy A 2 pokračovanie S fázy A 1 prechod z S fázy do G2 B 1 M-fáza Cyklín dependentné kinázy, po vytvorení komplexu s príslušným cyklínom, fosforylujú určitú regulačnú bielkovinu (obr. 5), čím táto sa buď aktivuje alebo inaktivuje. Preto sa tieto enzýmy nazývajú aj proteínkinázy. aktivácia cdk cyklín 4
54 Biológia rakoviny, Vanda Repiská Obr. 5 Aktívny komplex cyklín/cdk. Okrem toho existujú aj inhibítory všetkých uvedených molekúl. Súhrou všetkých zúčastnených molekúl sa zabezpečuje správne načasovanie a špecifickosť riadenia bunkového cyklu. K deregulácii bunkového cyklu a následnej malígnej transformácii bunky môže viesť aj premena protoonkogénu na onkogén (obr. 6). Stane sa to vtedy, keď ním kódovaná bielkovina zmení svoje vlastnosti (dôsledok mutácie) alebo sa jej vytvára príliš veľa (overexpresia). Obr. 6 mutácia Schéma protoonkogén onkogén deregulácia bunkového cyklu overexpresia premeny (malignita) protoonkog énu na onkogén Zmena vlastností bielkoviny kódovanej protoonkogénom je zvyčajne dôsledkom jeho mutácie. Tento proteín je potom stále aktívny a vysiela falošné podnety na pokračovanie v cykle, bez toho, aby sa uskutočnili predchádzajúce kroky v signálovej dráhe. Príkladmi sú K-ras a bcr-abl. Mutácia K-ras protoonkogénu sa vyskytuje pomerne často, napríklad u 90 % malígnych nádorov podžalúdkovej žľazy (pankreasu). V prípade bcr-abl ide o dôsledok chromozómovej prestavby (vznikne tzv. fúzny gén), ale výsledok je rovnaký proteín abl je stále aktívny a do jadra prináša príkaz na delenie (u chronickej myeloidnej leukémie). Nadmerná syntéza bielkoviny kódovanej protoonkogénom (overexpresia) vedie k deregulácii bunkového cyklu. Rýchlosť malígnej transformácie bunky je priamo úmerná nadbytku produkovaných bielkovín. Overexpresia môže byť vyvolaná dvomi spôsobmi: 1.) nadpočetnosťou (amplifikáciou) počtu alel protoonkogénu v bunke alebo 2.) poruchou riadenia transkripcie (regulačná overexpresia). 1.) Amplifikácia počtu alel môže v bunke nastať dvomi spôsobmi. a) Prvým je nerovnomerný crossing-over v profáze prvého meiotického delenia, kedy jeden z nesesterských chromatídov získa obidve alely. Ide o zriedkavý jav, ale s výnimkou protoonkogénov, sa považuje za veľmi významný pre vývoj genómu. b) Druhý spôsob amplifikácie protoonkogénov súvisí s retrovírusmi. Ide o zvláštnu skupinu RNA vírusov, ktoré sú v bunke enzýmom reverzná transkriptáza prepísané do formy 5
55 Biológia rakoviny, Vanda Repiská dvojvláknovej DNA a táto sa, ako tzv. provírus vloží (inzertuje) náhodne do niektorého z chromozómov. Prvý problém je v tom, že sa genóm retrovírusu podobá na molekulu mrna a že si môže s bunkovou mrna vymeniť časť svojej genetickej informácie (tzv. rekombinácia). Ak táto mrna vznikla transkripciou bunkového protoonkogénu (c-onc), retrovírus potom získa vírusovú kópiu tohto génu (tzv. v-onc). Druhý problém spočíva v tom, že začiatočné a koncové úseky provírusu (tzv. LTR sekvencie) veľmi silne podporujú transkripciu. Ak je v bunke potrebné transkribovať príslušný protoonkogén, sú použité všetky v jadre prítomné alely. Kým z pôvodných alel (c-onc) sa vytvorí normálny počet bielkovín, z v-onc sa (vzhľadom na účinok LTR sekvencií) vytvorí veľké množstvo danej bielkoviny. Výsledkom je náhle zlyhanie regulácie bunkového cyklu a rýchla premena normálnej bunky na malígnu, ako je to už dlhšie známe u tzv. Rousovho sarkómu alebo u niektorých akútnych leukémií. Preto označujeme retrovírusy obsahujúce v-onc ako akútne transformujúce. 2.) Regulačná overexpresia môže mať tiež dva druhy príčin. a) Prvý je dôsledkom chromozómovej mutácie, kedy došlo ku premiestneniu (translokácii) protoonkogénu na iný chromozóm, do oblasti, kde sa nachádza permanentne používaný gén. Silná transkripcia tohto génu sa potom prenesie aj na vložený protoonkogén a výsledkom je, ako napr. u Burkittovho lymfómu. b) Druhou príčinou je to, že sa pred protoonkogén inzertuje provírus z retrovírusu. Jeho LTR sekvencie zvyšujú transkripciu génu nasledujúceho za ním. Situácia síce nie je taká dramatická, ako v už uvedenom prípade, keď provírus obsahuje v-onc, ale zvýšená tvorba protoonkogénu môže spôsobiť dereguláciu bunkového cyklu. Gény opravy DNA Mutácie v DNA nastávajú pod vplyvom chemických, biologických a fyzikálnych faktorov, ale napr. aj poruchami pri jej replikácii. Aby sa tieto chyby v genetickej informácii neodovzdávali do dcérskych buniek, vyvinuli sa u všetkých organizmov, vrátane človeka, komplexné systémy zisťujúce a opravujúce mutácie v DNA. Hromadenie mutácií je vysoko rizikové, pretože môže viesť k malígnej transformácii bunky. Gény, ktoré kódujú bielkovinové systémy na zisťovanie mutácií a opravu DNA sa nazývajú mutátory. Existuje niekoľko systémov opravy DNA. Účasť génov opravy DNA na vzniku nádorov je dokázaná existenciou niektorých dedičných nádorových syndrómov. Títo pacienti majú mutované gény opravy DNA, tým nedochádza k oprave poškodenia a môže vzniknúť rakovina. Napríklad u pacientov so syndrómom Xeroderma pigmentosum je porucha génov zodpovedných za vyštiepenie poškodeného úseku tzv. excíznu reparáciu DNA, čo sa prejavuje až násobným zvýšením výskytu nádorov kože. Mutácie v génoch zodpovedajúcich za opravu chybne spárovaných báz (tzv. mismatch opravu DNA) sú príčinou dedičnej predispozície na vznik ľudského nepolypózneho kolorektálneho karcinómu (nádor hrubého čreva a konečníka, HNPCC). Spôsobuje ho mutácia v ktoromkoľvek zo šiestich známych génov. Najčastejšie sa mutácie vyskytujú v génoch MSH2 a MLH1. Ataxia teleangiektázia Typickými príznakmi sú poruchy chôdze, poruchy tvorby protilátok a s tým súvisiace respiračné infekcie a predispozícia na vznik nádorov (kože, žalúdka, maternice, prsníka...). Toto recesívne dedičné ochorenie je vyvolané mutáciami v géne ATM. Gén ATM je tumorsupresorový gén a nachádza sa na chromozóme 11. Doposiaľ sa v ňom našlo 130 rôznych mutácií. Mutácia tohto génu vedie k zvýšenej citlivosti na ionizujúce žiarenie. Nedochádza k tvorbe proteínu p53, oprava DNA je nedostatočná, čo prispieva 6
 sa radia k najčastejším nádorovým ochoreniam u žien v Slovenskej republike.")
56 Biológia rakoviny, Vanda Repiská k nahromadeniu chýb v genetickom materiáli. Mutácia v géne ATM prispieva aj k urýchleniu rakoviny prsníka. Rakovina prsníka a vaječníkov Karcinómy prsnej žľazy a vaječníkov (obr. 7 a 8) sa radia k najčastejším nádorovým ochoreniam u žien v Slovenskej republike. Výskyt oboch týchto foriem rakoviny je často geneticky podmienený, asi 5 až 10 % týchto nádorov vzniká v dôsledku dedičnej predispozície. Ochorenie je indukované predovšetkým mutáciami v dvoch génoch BRCA1 a BRCA2. Teda u nositeľa mutácie v jednom či druhom géne je zvýšené riziko vzniku rakoviny prsníka a vaječníkov. Gén BRCA2 je zodpovedný aj za vznik rakoviny prsníka u mužov. Celoživotné riziko vzniku rakoviny prsníka pre ženy v bežnej populácii je 10%, kým u žien s mutáciou v týchto génoch až 85% a rakovina sa prejaví o 10 rokov skôr. Gén BRCA1 sa nachádza na chromozóme 17 a gén BRCA2 na chromozóme 13. Gény BRCA zaraďujeme k TSG sú veľké a ich produkty sa zúčastňujú na oprave zlomov v oboch reťazcoch DNA, ktoré vznikli žiarením. Mutácia v génoch BRCA1 a BRCA2 spôsobuje stratu ich funkcie, čo vedie k zvýšenému počtu chýb pri replikácii DNA a v konečnom dôsledku k nádorovému rastu. Novo objaveným génom, ktorý súvisí s rakovinou prsníka, ako aj hrubého čreva a konečníka je CHEK2. Nachádza sa na chromozóme 22. Jeho produkt je potrebný v G2 fáze bunkového cyklu. Pri poškodení nepovoľuje vstup bunky do mitózy. Obr. 7 Karcinóm prsníka (žltá šípka) obraz získaný pri vyšetrení magnetickou rezonanciou (foto: MUDr. Katarína Rauová, Onkologický ústav Svätej Alžbety Bratislava) 7
 Rakovina prostaty Rakovina prostaty (obr.")

57 Biológia rakoviny, Vanda Repiská Obr. 8 Zhubný nádor vaječníka (žlté šípky) pozostávajúci z viacerých dutín vyplnených čírou tekutinou.. Zvyšky vaječníka (čierna šípka). (foto : MUDr. Michal Palkovič, PhD. Ústav patologickej anatómie LF UK v Bratislave) Rakovina prostaty Rakovina prostaty (obr. 9 a 10) je najčastejšie diagnostikovaná malignita u mužov po päťdesiatom roku života a druhá najčastejšia príčina smrti u mužov západnej populácie. Ročne je celosvetovo diagnostikovaných nových prípadov rakoviny prostaty. Napriek tomu sa vie len málo o genetickej predispozícii. Pozornosť sa zameriava najmä na prostatický špecifický antigén (PSA) a prostatický špecifický membránový antigén (PSM), ktoré sú potenciálnymi markermi pre odhalenie progresie a tvorby metastáz. Gén zodpovedný produkciu PSA sa nachádza na chromozóme 19, PSM na chromozóme 11. Nedávno boli objavené ďalšie dva lokusy HPC1 na chromozóme 1 a HPCX na chromozóme X ( muži ho dedia od matky) Obr. 9 Zhubný nádor prostaty (žlté šípky). Rozšírený močový mechúr (čierna šípka). (foto : MUDr. Michal Palkovič, PhD. Ústav patologickej anatómie LF UK) 8
 Apoptóza Zánik buniek je udalosť, ktorá sa v makroorganizme uskutočňuje priebežne.")
58 Biológia rakoviny, Vanda Repiská Obr. 10 Rez zhubným nádorom prostaty (žlté šípky). Močové kamene (čierna šípka). vznikajúce zabránením odtoku moču z močového mechúra. (foto : MUDr. Michal Palkovič, PhD. Ústav patologickej anatómie LF UK v Bratislave) Apoptóza Zánik buniek je udalosť, ktorá sa v makroorganizme uskutočňuje priebežne. Súvisí s neustálou prestavbou a regeneráciou tkanív, orgánov a organizmu. Má aj základný význam pri remodelačných procesoch počas embryogenézy (napr. pri oddelení sa prstov alebo spriechodnení čreva). Zároveň je spôsobom, ako sa telo chráni pred rizikom pochádzajúcim z buniek, ktoré už prekročili svoju životnosť alebo z tých, ktoré sú potenciálne nebezpečné, napríklad pre riziko ich malígnej transformácie. Najstaršie známym mechanizmom zániku buniek je patologický proces - nekróza, pri ktorej je rozpad bunky (buniek) sprevádzaný zápalovou reakciou okolia. Jedným z hlavných procesov, ktoré nekrózu vyvolávajú, je praskanie lyzozómov, uvoľnenie ich enzýmov a natrávenie bunky (tzv. autolýza)a jej okolia. Apoptóza je naopak tzv. tichá bunková smrť, ktorá nevyvoláva reakciu v okolí. Aj to je dôvod, prečo bola jej existencia zistená až pred dvomi desaťročiami. Je vyvolaná sériou procesov riadených bunkou, ktoré vedú k jej zániku (obr. 11). Preto sa nazýva aj programovaná alebo fyziologická smrť bunky. Pri apoptóze sa v bunke zastaví syntéza bielkovín, netvoria sa lyzozómy. Následne dochádza k štiepeniu jadrovej DNA na krátke úseky. Rozložený chromatín sa pripája na vnútornú stranu jadrovej membrány. Súčasne sú rozložené aj bielkoviny v cytoplazme a v organelách. Bunka sa začína scvrkávať. Apoptotický proces pokračuje vytváraním mechúrikov z jadrového obalu, ktoré obsahujú zvyšky chromatínu. Zároveň sa cytoplazmatická membrána vydúva a postupne sa odškrcujú mechúriky, ktoré obsahujú zvyšky cytoplazmy, jej obsahu a jadra. Takto vznikajú tzv. apoptotické telieska, ktoré sú v tkanive pohlcované bunkami schopnými fagocytózy, napr. makrofágmi. Materiál uložený v mechúrikoch je v tkanive recyklovaný. 9
59 Biológia rakoviny, Vanda Repiská Apoptózové telieska sú pohlcované fagocytmi a rýchlo degradované. Chromatín sa oddeľuje a tvoria sa výbežky na povrchu bunky. Vznikajú apoptózové telieska. Agregácia chromatínu na jadrovú membánu a zahustenie cytoplazmy. Bunka sa scvrkáva. V bunke sa zastavujú syntetické procesy Obr. 11 Schéma priebehu apoptózy Aktivácia (vyvolanie) apoptózy je proces, ktorým sa v bunke spúšťa séria mechanizmov a reakcií vedúcich k apoptóze. Delíme ju na dva hlavné typy, exogénnu a endogénnu. Pri exogénnej aktivácii dochádza k naviazaniu signálnej molekuly z mimobunkového priestoru na špecifický receptor na povrchu bunky. Tento spôsob je bežný napríklad pri vývoji a reakcii buniek špecifickej imunity (napr. T lymfocytov). Pri endogénnej aktivácii apoptózy vzniká podnet v bunke samotnej. Najdôležitejšia časť endogénnej aktivácie apoptózy súvisí s replikáciou DNA, teda hlavne s nahromadením neopraviteľného množstva mutácií v nej a s telomérami. Súvisia s proteínom p53 a boli opísané v časti o biológii rakoviny. Proteín p53, cez sústavu (kaskádu) pomocných proteínov, uvoľní proteín bcl2 z väzby na bielkovinový kanál bax vo vonkajšej membráne mitochondrií. Tým sa umožní, aby z priestoru medzi ich vonkajšou a vnútornou membránou mitochondrie unikol enzým cytochróm c. Tento sa viaže na bielkovinový komplex Apaf1 (apoptózu aktivujúci faktor). Spolu vytvoria apoptozóm, ktorý aktivuje proteolytický enzým kaspáza 9. Tento enzým aktivuje podriadené kaspázy (3, 6, 7), ktoré rozštiepia bunkové bielkoviny. Kaspáza 9 aktivuje aj enzýmy štiepiace DNA. tzv. DNázy, Mechanizmov spustenia a priebehu apoptózy je viac, ale ich opis je nad rámec tohto textu. 10
Matematika Funkcia viac premenných, Parciálne derivácie
 Matematika 2-01 Funkcia viac premenných, Parciálne derivácie Euklidovská metrika na množine R n všetkých usporiadaných n-íc reálnych čísel je reálna funkcia ρ: R n R n R definovaná nasledovne: Ak X = x
Matematika 2-01 Funkcia viac premenných, Parciálne derivácie Euklidovská metrika na množine R n všetkých usporiadaných n-íc reálnych čísel je reálna funkcia ρ: R n R n R definovaná nasledovne: Ak X = x
Trendy súčasnej liečby onkologických ohorení. Jozef Mardiak
 Trendy súčasnej liečby onkologických ohorení Jozef Mardiak Protinádorová liečba Poškodenie nádorovej bunky apoptózu Cytostatická liečba: nešpecifická toxicita rýchlosti proliferácie tkanív proliferácia
Trendy súčasnej liečby onkologických ohorení Jozef Mardiak Protinádorová liečba Poškodenie nádorovej bunky apoptózu Cytostatická liečba: nešpecifická toxicita rýchlosti proliferácie tkanív proliferácia
Ekvačná a kvantifikačná logika
 a kvantifikačná 3. prednáška (6. 10. 004) Prehľad 1 1 (dokončenie) ekvačných tabliel Formula A je ekvačne dokázateľná z množiny axióm T (T i A) práve vtedy, keď existuje uzavreté tablo pre cieľ A ekvačných
a kvantifikačná 3. prednáška (6. 10. 004) Prehľad 1 1 (dokončenie) ekvačných tabliel Formula A je ekvačne dokázateľná z množiny axióm T (T i A) práve vtedy, keď existuje uzavreté tablo pre cieľ A ekvačných
Obvod a obsah štvoruholníka
 Obvod a štvoruholníka D. Štyri body roviny z ktorých žiadne tri nie sú kolineárne (neležia na jednej priamke) tvoria jeden štvoruholník. Tie body (A, B, C, D) sú vrcholy štvoruholníka. strany štvoruholníka
Obvod a štvoruholníka D. Štyri body roviny z ktorých žiadne tri nie sú kolineárne (neležia na jednej priamke) tvoria jeden štvoruholník. Tie body (A, B, C, D) sú vrcholy štvoruholníka. strany štvoruholníka
(MHC = major histocompatibility complex)
") HLAVNÝ HISTOKOMPATIBILNÝ SYSTÉM = HHS (MHC = major histocompatibility complex) - HHS = MHC (major histocompatibility complex) je skupina génov, ktorých produktami sú membránové glykoproteíny (MHC molekuly
HLAVNÝ HISTOKOMPATIBILNÝ SYSTÉM = HHS (MHC = major histocompatibility complex) - HHS = MHC (major histocompatibility complex) je skupina génov, ktorých produktami sú membránové glykoproteíny (MHC molekuly
Start. Vstup r. O = 2*π*r S = π*r*r. Vystup O, S. Stop. Start. Vstup P, C V = P*C*1,19. Vystup V. Stop
 1) Vytvorte algoritmus (vývojový diagram) na výpočet obvodu kruhu. O=2xπxr ; S=πxrxr Vstup r O = 2*π*r S = π*r*r Vystup O, S 2) Vytvorte algoritmus (vývojový diagram) na výpočet celkovej ceny výrobku s
1) Vytvorte algoritmus (vývojový diagram) na výpočet obvodu kruhu. O=2xπxr ; S=πxrxr Vstup r O = 2*π*r S = π*r*r Vystup O, S 2) Vytvorte algoritmus (vývojový diagram) na výpočet celkovej ceny výrobku s
Matematika 2. časť: Analytická geometria
 Matematika 2 časť: Analytická geometria RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk Súradnicové
Matematika 2 časť: Analytická geometria RNDr. Jana Pócsová, PhD. Ústav riadenia a informatizácie výrobných procesov Fakulta BERG Technická univerzita v Košiciach e-mail: jana.pocsova@tuke.sk Súradnicové
Motivácia Denícia determinantu Výpo et determinantov Determinant sú inu matíc Vyuºitie determinantov. Determinanty. 14. decembra 2010.
 14. decembra 2010 Rie²enie sústav Plocha rovnobeºníka Objem rovnobeºnostena Rie²enie sústav Príklad a 11 x 1 + a 12 x 2 = c 1 a 21 x 1 + a 22 x 2 = c 2 Dostaneme: x 1 = c 1a 22 c 2 a 12 a 11 a 22 a 12
14. decembra 2010 Rie²enie sústav Plocha rovnobeºníka Objem rovnobeºnostena Rie²enie sústav Príklad a 11 x 1 + a 12 x 2 = c 1 a 21 x 1 + a 22 x 2 = c 2 Dostaneme: x 1 = c 1a 22 c 2 a 12 a 11 a 22 a 12
3. Striedavé prúdy. Sínusoida
 . Striedavé prúdy VZNIK: Striedavý elektrický prúd prechádza obvodom, ktorý je pripojený na zdroj striedavého napätia. Striedavé napätie vyrába synchrónny generátor, kde na koncoch rotorového vinutia sa
. Striedavé prúdy VZNIK: Striedavý elektrický prúd prechádza obvodom, ktorý je pripojený na zdroj striedavého napätia. Striedavé napätie vyrába synchrónny generátor, kde na koncoch rotorového vinutia sa
Prechod z 2D do 3D. Martin Florek 3. marca 2009
 Počítačová grafika 2 Prechod z 2D do 3D Martin Florek florek@sccg.sk FMFI UK 3. marca 2009 Prechod z 2D do 3D Čo to znamená? Ako zobraziť? Súradnicové systémy Čo to znamená? Ako zobraziť? tretia súradnica
Počítačová grafika 2 Prechod z 2D do 3D Martin Florek florek@sccg.sk FMFI UK 3. marca 2009 Prechod z 2D do 3D Čo to znamená? Ako zobraziť? Súradnicové systémy Čo to znamená? Ako zobraziť? tretia súradnica
Motivácia pojmu derivácia
 Derivácia funkcie Motivácia pojmu derivácia Zaujíma nás priemerná intenzita zmeny nejakej veličiny (dráhy, rastu populácie, veľkosti elektrického náboja, hmotnosti), vzhľadom na inú veličinu (čas, dĺžka)
Derivácia funkcie Motivácia pojmu derivácia Zaujíma nás priemerná intenzita zmeny nejakej veličiny (dráhy, rastu populácie, veľkosti elektrického náboja, hmotnosti), vzhľadom na inú veličinu (čas, dĺžka)
Cvičenie č. 4,5 Limita funkcie
 Cvičenie č. 4,5 Limita funkcie Definícia ity Limita funkcie (vlastná vo vlastnom bode) Nech funkcia f je definovaná na nejakom okolí U( ) bodu. Hovoríme, že funkcia f má v bode itu rovnú A, ak ( ε > )(
Cvičenie č. 4,5 Limita funkcie Definícia ity Limita funkcie (vlastná vo vlastnom bode) Nech funkcia f je definovaná na nejakom okolí U( ) bodu. Hovoríme, že funkcia f má v bode itu rovnú A, ak ( ε > )(
7. FUNKCIE POJEM FUNKCIE
 7. FUNKCIE POJEM FUNKCIE Funkcia f reálnej premennej je : - každé zobrazenie f v množine všetkých reálnych čísel; - množina f všetkých usporiadaných dvojíc[,y] R R pre ktorú platí: ku každému R eistuje
7. FUNKCIE POJEM FUNKCIE Funkcia f reálnej premennej je : - každé zobrazenie f v množine všetkých reálnych čísel; - množina f všetkých usporiadaných dvojíc[,y] R R pre ktorú platí: ku každému R eistuje
Matematika prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad
 Matematika 3-13. prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad Erika Škrabul áková F BERG, TU Košice 15. 12. 2015 Erika Škrabul áková (TUKE) Taylorov
Matematika 3-13. prednáška 4 Postupnosti a rady 4.5 Funkcionálne rady - mocninové rady - Taylorov rad, MacLaurinov rad Erika Škrabul áková F BERG, TU Košice 15. 12. 2015 Erika Škrabul áková (TUKE) Taylorov
IMUNITNÝ SYSTÉM výber z prednášky
 výber z prednášky Predmet: Imunológia Študijný odbor: Všeobecné lekárstvo akademický rok: 2014/2015 RNDr. Mira Horváthová, PhD. ÚKIA, LF SZU v Bratislave Imunitný systém zložitá sieť špecializovaných buniek,
výber z prednášky Predmet: Imunológia Študijný odbor: Všeobecné lekárstvo akademický rok: 2014/2015 RNDr. Mira Horváthová, PhD. ÚKIA, LF SZU v Bratislave Imunitný systém zložitá sieť špecializovaných buniek,
1. Limita, spojitost a diferenciálny počet funkcie jednej premennej
 . Limita, spojitost a diferenciálny počet funkcie jednej premennej Definícia.: Hromadný bod a R množiny A R: v každom jeho okolí leží aspoň jeden bod z množiny A, ktorý je rôzny od bodu a Zadanie množiny
. Limita, spojitost a diferenciálny počet funkcie jednej premennej Definícia.: Hromadný bod a R množiny A R: v každom jeho okolí leží aspoň jeden bod z množiny A, ktorý je rôzny od bodu a Zadanie množiny
Goniometrické rovnice a nerovnice. Základné goniometrické rovnice
 Goniometrické rovnice a nerovnice Definícia: Rovnice (nerovnice) obsahujúce neznámu x alebo výrazy s neznámou x ako argumenty jednej alebo niekoľkých goniometrických funkcií nazývame goniometrickými rovnicami
Goniometrické rovnice a nerovnice Definícia: Rovnice (nerovnice) obsahujúce neznámu x alebo výrazy s neznámou x ako argumenty jednej alebo niekoľkých goniometrických funkcií nazývame goniometrickými rovnicami
KATEDRA DOPRAVNEJ A MANIPULAČNEJ TECHNIKY Strojnícka fakulta, Žilinská Univerzita
 132 1 Absolútna chyba: ) = - skut absolútna ochýlka: ) ' = - spr. relatívna chyba: alebo Chyby (ochýlky): M systematické, M náhoné, M hrubé. Korekcia: k = spr - = - Î' pomerná korekcia: Správna honota:
132 1 Absolútna chyba: ) = - skut absolútna ochýlka: ) ' = - spr. relatívna chyba: alebo Chyby (ochýlky): M systematické, M náhoné, M hrubé. Korekcia: k = spr - = - Î' pomerná korekcia: Správna honota:
AerobTec Altis Micro
 AerobTec Altis Micro Záznamový / súťažný výškomer s telemetriou Výrobca: AerobTec, s.r.o. Pionierska 15 831 02 Bratislava www.aerobtec.com info@aerobtec.com Obsah 1.Vlastnosti... 3 2.Úvod... 3 3.Princíp
AerobTec Altis Micro Záznamový / súťažný výškomer s telemetriou Výrobca: AerobTec, s.r.o. Pionierska 15 831 02 Bratislava www.aerobtec.com info@aerobtec.com Obsah 1.Vlastnosti... 3 2.Úvod... 3 3.Princíp
Oxidačný stres v nádorových ochoreniach
 UNIVERZITA KOMENSKÉHO V BRATISLAVE LEKÁRSKA FAKULTA Ústav lekárskej chémie, biochémie a klinickej biochémie Oxidačný stres v nádorových ochoreniach Oxidačný stres Antioxidanty Prooxidanty OS chronický
UNIVERZITA KOMENSKÉHO V BRATISLAVE LEKÁRSKA FAKULTA Ústav lekárskej chémie, biochémie a klinickej biochémie Oxidačný stres v nádorových ochoreniach Oxidačný stres Antioxidanty Prooxidanty OS chronický
Mikroprostredie ako multicelulárny systém
 Mikroprostredie ako multicelulárny systém 10.1 Nádor a vnútronádorové prostredie Pri expanzii nádorov sa zvykne hovoriť o nádorovom tkanive, hoci to nie je tkanivo v zmysle histologickej klasifikácie.
Mikroprostredie ako multicelulárny systém 10.1 Nádor a vnútronádorové prostredie Pri expanzii nádorov sa zvykne hovoriť o nádorovom tkanive, hoci to nie je tkanivo v zmysle histologickej klasifikácie.
Priamkové plochy. Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava
 na nej leží potom plocha Φ je priamková. Santiago Calatrava") Priamkové plochy Priamkové plochy Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava Priamkové plochy rozdeľujeme na: Rozvinuteľné
Priamkové plochy Priamkové plochy Ak každým bodom plochy Φ prechádza aspoň jedna priamka, ktorá (celá) na nej leží potom plocha Φ je priamková. Santiago Calatrava Priamkové plochy rozdeľujeme na: Rozvinuteľné
Základné poznatky molekulovej fyziky a termodynamiky
 Základné poznatky molekulovej fyziky a termodynamiky Opakovanie učiva II. ročníka, Téma 1. A. Príprava na maturity z fyziky, 2008 Outline Molekulová fyzika 1 Molekulová fyzika Predmet Molekulovej fyziky
Základné poznatky molekulovej fyziky a termodynamiky Opakovanie učiva II. ročníka, Téma 1. A. Príprava na maturity z fyziky, 2008 Outline Molekulová fyzika 1 Molekulová fyzika Predmet Molekulovej fyziky
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy
, diferencovanie časového radu, unit root testy") Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2012/2013 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/18
Jednotkový koreň (unit root), diferencovanie časového radu, unit root testy Beáta Stehlíková Časové rady, FMFI UK, 2012/2013 Jednotkový koreň(unit root),diferencovanie časového radu, unit root testy p.1/18
ARMA modely čast 2: moving average modely (MA)
") ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2014/2015 ARMA modely časť 2: moving average modely(ma) p.1/24 V. Moving average proces prvého rádu - MA(1) ARMA modely
ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2014/2015 ARMA modely časť 2: moving average modely(ma) p.1/24 V. Moving average proces prvého rádu - MA(1) ARMA modely
IMUNITA a CHRONICKÉ OCHORENIA
 IMUNITA a CHRONICKÉ OCHORENIA I. Pre študentov študijného odboru Verejné zdravotníctvo denné štúdium II. Pre študentov študijného odboru Verejné zdravotníctvo externé štúdium RNDr. Mira Horváthová, PhD.
IMUNITA a CHRONICKÉ OCHORENIA I. Pre študentov študijného odboru Verejné zdravotníctvo denné štúdium II. Pre študentov študijného odboru Verejné zdravotníctvo externé štúdium RNDr. Mira Horváthová, PhD.
1. písomná práca z matematiky Skupina A
 1. písomná práca z matematiky Skupina A 1. Vypočítajte : a) 84º 56 + 32º 38 = b) 140º 53º 24 = c) 55º 12 : 2 = 2. Vypočítajte zvyšné uhly na obrázku : β γ α = 35 12 δ a b 3. Znázornite na číselnej osi
1. písomná práca z matematiky Skupina A 1. Vypočítajte : a) 84º 56 + 32º 38 = b) 140º 53º 24 = c) 55º 12 : 2 = 2. Vypočítajte zvyšné uhly na obrázku : β γ α = 35 12 δ a b 3. Znázornite na číselnej osi
Moderné vzdelávanie pre vedomostnú spoločnosť Projekt je spolufinancovaný zo zdrojov EÚ M A T E M A T I K A
 M A T E M A T I K A PRACOVNÝ ZOŠIT II. ROČNÍK Mgr. Agnesa Balážová Obchodná akadémia, Akademika Hronca 8, Rožňava PRACOVNÝ LIST 1 Urč typ kvadratickej rovnice : 1. x 2 3x = 0... 2. 3x 2 = - 2... 3. -4x
M A T E M A T I K A PRACOVNÝ ZOŠIT II. ROČNÍK Mgr. Agnesa Balážová Obchodná akadémia, Akademika Hronca 8, Rožňava PRACOVNÝ LIST 1 Urč typ kvadratickej rovnice : 1. x 2 3x = 0... 2. 3x 2 = - 2... 3. -4x
ARMA modely čast 2: moving average modely (MA)
") ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2011/2012 ARMA modely časť 2: moving average modely(ma) p.1/25 V. Moving average proces prvého rádu - MA(1) ARMA modely
ARMA modely čast 2: moving average modely (MA) Beáta Stehlíková Časové rady, FMFI UK, 2011/2012 ARMA modely časť 2: moving average modely(ma) p.1/25 V. Moving average proces prvého rádu - MA(1) ARMA modely
Účinky sympatikoadrenálneho systému
 Nádorové choroby Aj keď o vplyve psychosociálnych faktorov na nádorový rast uvažoval už Galén, ktorýpozoroval,že melancholické ženyvykazujúvyššiunáchylnosťnavzniknádorovprsníkaakoženyso sangvinickými povahovýmičrtami(thakerasood,2008),
Nádorové choroby Aj keď o vplyve psychosociálnych faktorov na nádorový rast uvažoval už Galén, ktorýpozoroval,že melancholické ženyvykazujúvyššiunáchylnosťnavzniknádorovprsníkaakoženyso sangvinickými povahovýmičrtami(thakerasood,2008),
13. ZÁKLADNÉ VZŤAHY MEDZI IMUNITNÝM, NERVOVÝM A ENDOKRINNÝM SYSTÉMOM
 13. ZÁKLADNÉ VZŤAHY MEDZI IMUNITNÝM, NERVOVÝM A ENDOKRINNÝM SYSTÉMOM Vzťah medzi imunitným, nervovým a endokrinným systémom bol dlho nejasný. Už dávnejšie sa dokázala tesná spolupráca medzi nervovým a
13. ZÁKLADNÉ VZŤAHY MEDZI IMUNITNÝM, NERVOVÝM A ENDOKRINNÝM SYSTÉMOM Vzťah medzi imunitným, nervovým a endokrinným systémom bol dlho nejasný. Už dávnejšie sa dokázala tesná spolupráca medzi nervovým a
HASLIM112V, HASLIM123V, HASLIM136V HASLIM112Z, HASLIM123Z, HASLIM136Z HASLIM112S, HASLIM123S, HASLIM136S
 PROUKTOVÝ LIST HKL SLIM č. sklad. karty / obj. číslo: HSLIM112V, HSLIM123V, HSLIM136V HSLIM112Z, HSLIM123Z, HSLIM136Z HSLIM112S, HSLIM123S, HSLIM136S fakturačný názov výrobku: HKL SLIMv 1,2kW HKL SLIMv
PROUKTOVÝ LIST HKL SLIM č. sklad. karty / obj. číslo: HSLIM112V, HSLIM123V, HSLIM136V HSLIM112Z, HSLIM123Z, HSLIM136Z HSLIM112S, HSLIM123S, HSLIM136S fakturačný názov výrobku: HKL SLIMv 1,2kW HKL SLIMv
REZISTORY. Rezistory (súčiastky) sú pasívne prvky. Používajú sa vo všetkých elektrických
 sú pasívne prvky. Používajú sa vo všetkých elektrických") REZISTORY Rezistory (súčiastky) sú pasívne prvky. Používajú sa vo všetkých elektrických obvodoch. Základnou vlastnosťou rezistora je jeho odpor. Odpor je fyzikálna vlastnosť, ktorá je daná štruktúrou materiálu
REZISTORY Rezistory (súčiastky) sú pasívne prvky. Používajú sa vo všetkých elektrických obvodoch. Základnou vlastnosťou rezistora je jeho odpor. Odpor je fyzikálna vlastnosť, ktorá je daná štruktúrou materiálu
Prevencia v onkológii. Úvod do problematiky
 Prevencia v onkológii Úvod do problematiky Onkológia Je oblasť medicíny zaoberajúca sa - prevenciou - diagnostikou nádorových ochorení. - liečbou Čo je rakovina? neobmedzené a nekontrolovateľné množenie
Prevencia v onkológii Úvod do problematiky Onkológia Je oblasť medicíny zaoberajúca sa - prevenciou - diagnostikou nádorových ochorení. - liečbou Čo je rakovina? neobmedzené a nekontrolovateľné množenie
Tomáš Madaras Prvočísla
 Prvočísla Tomáš Madaras 2011 Definícia Nech a Z. Čísla 1, 1, a, a sa nazývajú triviálne delitele čísla a. Cele číslo a / {0, 1, 1} sa nazýva prvočíslo, ak má iba triviálne delitele; ak má aj iné delitele,
Prvočísla Tomáš Madaras 2011 Definícia Nech a Z. Čísla 1, 1, a, a sa nazývajú triviálne delitele čísla a. Cele číslo a / {0, 1, 1} sa nazýva prvočíslo, ak má iba triviálne delitele; ak má aj iné delitele,
PATOFYZIOLÓGIA AKI. T. Baltesová. Transplantačné centrum, I.chirurgická klinika UNLP. Nefrologické sympózium, Martin
 PATOFYZIOLÓGIA AKI T. Baltesová Transplantačné centrum, I.chirurgická klinika UNLP Nefrologické sympózium, Martin 24. 26. október 2012 GFR prietok krvi obličkami (renal blood flow, RBF) efektívny cirkulujúci
PATOFYZIOLÓGIA AKI T. Baltesová Transplantačné centrum, I.chirurgická klinika UNLP Nefrologické sympózium, Martin 24. 26. október 2012 GFR prietok krvi obličkami (renal blood flow, RBF) efektívny cirkulujúci
ELEKTRICKÉ POLE. Elektrický náboj je základná vlastnosť častíc, je viazaný na častice látky a vyjadruje stav elektricky nabitých telies.
 ELEKTRICKÉ POLE 1. ELEKTRICKÝ NÁBOJ, COULOMBOV ZÁKON Skúmajme napr. trenie celuloidového pravítka látkou, hrebeň suché vlasy, mikrotén slabý prúd vody... Príčinou spomenutých javov je elektrický náboj,
ELEKTRICKÉ POLE 1. ELEKTRICKÝ NÁBOJ, COULOMBOV ZÁKON Skúmajme napr. trenie celuloidového pravítka látkou, hrebeň suché vlasy, mikrotén slabý prúd vody... Príčinou spomenutých javov je elektrický náboj,
,Zohrievanie vody indukčným varičom bez pokrievky,
 Farba skupiny: zelená Označenie úlohy:,zohrievanie vody indukčným varičom bez pokrievky, Úloha: Zistiť, ako závisí účinnosť zohrievania vody na indukčnom variči od priemeru použitého hrnca. Hypotéza: Účinnosť
Farba skupiny: zelená Označenie úlohy:,zohrievanie vody indukčným varičom bez pokrievky, Úloha: Zistiť, ako závisí účinnosť zohrievania vody na indukčnom variči od priemeru použitého hrnca. Hypotéza: Účinnosť
M6: Model Hydraulický systém dvoch zásobníkov kvapaliny s interakciou
 M6: Model Hydraulický ytém dvoch záobníkov kvapaliny interakciou Úlohy:. Zotavte matematický popi modelu Hydraulický ytém. Vytvorte imulačný model v jazyku: a. Matlab b. imulink 3. Linearizujte nelineárny
M6: Model Hydraulický ytém dvoch záobníkov kvapaliny interakciou Úlohy:. Zotavte matematický popi modelu Hydraulický ytém. Vytvorte imulačný model v jazyku: a. Matlab b. imulink 3. Linearizujte nelineárny
Nukleové kyseliny a proteosyntéza
 Nukleové kyseliny (NK) sú biomakromolekulové látky, ktoré sa spolu s bielkovinami považujú za najvýznamnejšie zložky živých sústav. V ich molekulách sa uchováva dedičná ( genetická ) informácia bunky a
Nukleové kyseliny (NK) sú biomakromolekulové látky, ktoré sa spolu s bielkovinami považujú za najvýznamnejšie zložky živých sústav. V ich molekulách sa uchováva dedičná ( genetická ) informácia bunky a
6 APLIKÁCIE FUNKCIE DVOCH PREMENNÝCH
 6 APLIKÁCIE FUNKCIE DVOCH PREMENNÝCH 6. Otázky Definujte pojem produkčná funkcia. Definujte pojem marginálny produkt. 6. Produkčná funkcia a marginálny produkt Definícia 6. Ak v ekonomickom procese počet
6 APLIKÁCIE FUNKCIE DVOCH PREMENNÝCH 6. Otázky Definujte pojem produkčná funkcia. Definujte pojem marginálny produkt. 6. Produkčná funkcia a marginálny produkt Definícia 6. Ak v ekonomickom procese počet
Podnikateľ 90 Mobilný telefón Cena 95 % 50 % 25 %
 Podnikateľ 90 Samsung S5230 Samsung C3530 Nokia C5 Samsung Shark Slider S3550 Samsung Xcover 271 T-Mobile Pulse Mini Sony Ericsson ZYLO Sony Ericsson Cedar LG GM360 Viewty Snap Nokia C3 Sony Ericsson ZYLO
Podnikateľ 90 Samsung S5230 Samsung C3530 Nokia C5 Samsung Shark Slider S3550 Samsung Xcover 271 T-Mobile Pulse Mini Sony Ericsson ZYLO Sony Ericsson Cedar LG GM360 Viewty Snap Nokia C3 Sony Ericsson ZYLO
APOPTÓZA PROGRAMOVANÁ BUNKOVÁ SMRŤ A RASTLINNÉ METABOLITY
 APOPTÓZA PROGRAMOVANÁ BUNKOVÁ SMRŤ A RASTLINNÉ METABOLITY MÁRIA FICKOVÁ a a MILAN NAGY b a Ústav experimentálnej endokrinológie SAV, Vlárska 3, 833 06 Bratislava, b Katedra farmakognózie a botaniky, Farmaceutická
APOPTÓZA PROGRAMOVANÁ BUNKOVÁ SMRŤ A RASTLINNÉ METABOLITY MÁRIA FICKOVÁ a a MILAN NAGY b a Ústav experimentálnej endokrinológie SAV, Vlárska 3, 833 06 Bratislava, b Katedra farmakognózie a botaniky, Farmaceutická
SLOVENSKO maloobchodný cenník (bez DPH)
") Hofatex UD strecha / stena - exteriér Podkrytinová izolácia vhodná aj na zaklopenie drevených rámových konštrukcií; pero a drážka EN 13171, EN 622 22 580 2500 1,45 5,7 100 145,00 3,19 829 hustota cca.
Hofatex UD strecha / stena - exteriér Podkrytinová izolácia vhodná aj na zaklopenie drevených rámových konštrukcií; pero a drážka EN 13171, EN 622 22 580 2500 1,45 5,7 100 145,00 3,19 829 hustota cca.
Forever IMMUBLEND TM
 doc. MVDr. Vojtech Lisák, CSc., MPH., člen PZ lekárov FLP SR a CR Odpovedať stručne na otázku Ako vlastne funguje naša imunita - je nesmierne ťažko. Donedávna boli vedci presvedčení, že imunitný systém
doc. MVDr. Vojtech Lisák, CSc., MPH., člen PZ lekárov FLP SR a CR Odpovedať stručne na otázku Ako vlastne funguje naša imunita - je nesmierne ťažko. Donedávna boli vedci presvedčení, že imunitný systém
Biochemické zmeny v organizme človeka v procese starnutia
 UNIVERZITA KOMENSKÉHO V BRATISLAVE JESSENIOVA LEKÁRSKA FAKULTA V MARTINE Ústav lekárskej biochémie Biochemické zmeny v organizme človeka v procese starnutia doc. Mgr. Eva Babušíková, PhD. Starnutie progresívny,
UNIVERZITA KOMENSKÉHO V BRATISLAVE JESSENIOVA LEKÁRSKA FAKULTA V MARTINE Ústav lekárskej biochémie Biochemické zmeny v organizme človeka v procese starnutia doc. Mgr. Eva Babušíková, PhD. Starnutie progresívny,
Nukleové kyseliny. Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy.
 Nukleové kyseliny Nukleové kyseliny Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy. DNA - deoxyribonukleová kyselina RNA - ribonukleová kyselina Funkcie
Nukleové kyseliny Nukleové kyseliny Nukleové kyseliny sú polymérne reťazce pozostávajúce z monomérov, ktoré sa nazývajú nukleotidy. DNA - deoxyribonukleová kyselina RNA - ribonukleová kyselina Funkcie
Gymnázium Jána Adama Raymana. Penicilín. Ročníková práca z chémie. 2005/2006 Jozef Komár 3.C
 Gymnázium Jána Adama Raymana Penicilín Ročníková práca z chémie 2005/2006 Jozef Komár 3.C Obsah 1. Úvod... 3 2. Antibiotiká... 4 3. Penicilín... 5 3.1 História... 5 3.2 Chemické zloženie... 5 3.3 Pôsobenie
Gymnázium Jána Adama Raymana Penicilín Ročníková práca z chémie 2005/2006 Jozef Komár 3.C Obsah 1. Úvod... 3 2. Antibiotiká... 4 3. Penicilín... 5 3.1 História... 5 3.2 Chemické zloženie... 5 3.3 Pôsobenie
Deliteľnosť a znaky deliteľnosti
 Deliteľnosť a znaky deliteľnosti Medzi základné pojmy v aritmetike celých čísel patrí aj pojem deliteľnosť. Najprv si povieme, čo znamená, že celé číslo a delí celé číslo b a ako to zapisujeme. Nech a
Deliteľnosť a znaky deliteľnosti Medzi základné pojmy v aritmetike celých čísel patrí aj pojem deliteľnosť. Najprv si povieme, čo znamená, že celé číslo a delí celé číslo b a ako to zapisujeme. Nech a
Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín. Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť.
 Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť. Ktoré fyzikálne jednotky zodpovedajú sústave SI: a) Dĺžka, čas,
Kontrolné otázky na kvíz z jednotiek fyzikálnych veličín Upozornenie: Umiestnenie správnej a nesprávnych odpovedí sa môže v teste meniť. Ktoré fyzikálne jednotky zodpovedajú sústave SI: a) Dĺžka, čas,
SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV KATEDRA GENETIKY A ŠĽACHTENIA RASTLÍN
 SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV KATEDRA GENETIKY A ŠĽACHTENIA RASTLÍN Prednáška z Genetiky mikroorganizmov Informačné molekuly v mikroorganizmoch
SLOVENSKÁ POĽNOHOSPODÁRSKA UNIVERZITA V NITRE FAKULTA AGROBIOLÓGIE A POTRAVINOVÝCH ZDROJOV KATEDRA GENETIKY A ŠĽACHTENIA RASTLÍN Prednáška z Genetiky mikroorganizmov Informačné molekuly v mikroorganizmoch
Vybrané kapitoly genetiky pre lesníkov
 Vybrané kapitoly genetiky pre lesníkov 1 NÁPLŇ A KLASIFIKÁCIA GENETIKY Genetika je veda o dedičnosti a premenlivosti živých organizmov. Pojem premenlivosti bol zadefinovaný v predchádzajúcej kapitole premenlivosť
Vybrané kapitoly genetiky pre lesníkov 1 NÁPLŇ A KLASIFIKÁCIA GENETIKY Genetika je veda o dedičnosti a premenlivosti živých organizmov. Pojem premenlivosti bol zadefinovaný v predchádzajúcej kapitole premenlivosť
6 Limita funkcie. 6.1 Myšlienka limity, interval bez bodu
 6 Limita funkcie 6 Myšlienka ity, interval bez bodu Intuitívna myšlienka ity je prirodzená, ale definovať presne pojem ity je značne obtiažne Nech f je funkcia a nech a je reálne číslo Čo znamená zápis
6 Limita funkcie 6 Myšlienka ity, interval bez bodu Intuitívna myšlienka ity je prirodzená, ale definovať presne pojem ity je značne obtiažne Nech f je funkcia a nech a je reálne číslo Čo znamená zápis
Chí kvadrát test dobrej zhody. Metódy riešenia úloh z pravdepodobnosti a štatistiky
 Chí kvadrát test dobrej zhody Metódy riešenia úloh z pravdepodobnosti a štatistiky www.iam.fmph.uniba.sk/institute/stehlikova Test dobrej zhody I. Chceme overiť, či naše dáta pochádzajú z konkrétneho pravdep.
Chí kvadrát test dobrej zhody Metódy riešenia úloh z pravdepodobnosti a štatistiky www.iam.fmph.uniba.sk/institute/stehlikova Test dobrej zhody I. Chceme overiť, či naše dáta pochádzajú z konkrétneho pravdep.
Klasifikácia látok LÁTKY. Zmesi. Chemické látky. rovnorodé (homogénne) rôznorodé (heterogénne)
 rôznorodé (heterogénne)") Zopakujme si : Klasifikácia látok LÁTKY Chemické látky Zmesi chemické prvky chemické zlúčeniny rovnorodé (homogénne) rôznorodé (heterogénne) Chemicky čistá látka prvok Chemická látka, zložená z atómov,
Zopakujme si : Klasifikácia látok LÁTKY Chemické látky Zmesi chemické prvky chemické zlúčeniny rovnorodé (homogénne) rôznorodé (heterogénne) Chemicky čistá látka prvok Chemická látka, zložená z atómov,
Kompilátory. Cvičenie 6: LLVM. Peter Kostolányi. 21. novembra 2017
 Kompilátory Cvičenie 6: LLVM Peter Kostolányi 21. novembra 2017 LLVM V podstate sada nástrojov pre tvorbu kompilátorov LLVM V podstate sada nástrojov pre tvorbu kompilátorov Pôvodne Low Level Virtual Machine
Kompilátory Cvičenie 6: LLVM Peter Kostolányi 21. novembra 2017 LLVM V podstate sada nástrojov pre tvorbu kompilátorov LLVM V podstate sada nástrojov pre tvorbu kompilátorov Pôvodne Low Level Virtual Machine
Metódy vol nej optimalizácie
 Metódy vol nej optimalizácie Metódy vol nej optimalizácie p. 1/28 Motivácia k metódam vol nej optimalizácie APLIKÁCIE p. 2/28 II 1. PRÍKLAD: Lineárna regresia - metóda najmenších štvorcov Na základe dostupných
Metódy vol nej optimalizácie Metódy vol nej optimalizácie p. 1/28 Motivácia k metódam vol nej optimalizácie APLIKÁCIE p. 2/28 II 1. PRÍKLAD: Lineárna regresia - metóda najmenších štvorcov Na základe dostupných
C. Kontaktný fasádny zatepľovací systém
 C. Kontaktný fasádny zatepľovací systém C.1. Tepelná izolácia penový polystyrén C.2. Tepelná izolácia minerálne dosky alebo lamely C.3. Tepelná izolácia extrudovaný polystyrén C.4. Tepelná izolácia penový
C. Kontaktný fasádny zatepľovací systém C.1. Tepelná izolácia penový polystyrén C.2. Tepelná izolácia minerálne dosky alebo lamely C.3. Tepelná izolácia extrudovaný polystyrén C.4. Tepelná izolácia penový
Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R
 Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R Ako nadprirodzené stretnutie s murárikom červenokrídlym naformátovalo môj profesijný i súkromný život... Osudové stretnutie s murárikom
Život vedca krajší od vysnívaného... s prírodou na hladine α R-P-R Ako nadprirodzené stretnutie s murárikom červenokrídlym naformátovalo môj profesijný i súkromný život... Osudové stretnutie s murárikom
PRIEMER DROTU d = 0,4-6,3 mm
 PRUŽINY PRUŽINY SKRUTNÉ PRUŽINY VIAC AKO 200 RUHOV SKRUTNÝCH PRUŽÍN PRIEMER ROTU d = 0,4-6,3 mm èíslo 3.0 22.8.2008 8:28:57 22.8.2008 8:28:58 PRUŽINY SKRUTNÉ PRUŽINY TECHNICKÉ PARAMETRE h d L S Legenda
PRUŽINY PRUŽINY SKRUTNÉ PRUŽINY VIAC AKO 200 RUHOV SKRUTNÝCH PRUŽÍN PRIEMER ROTU d = 0,4-6,3 mm èíslo 3.0 22.8.2008 8:28:57 22.8.2008 8:28:58 PRUŽINY SKRUTNÉ PRUŽINY TECHNICKÉ PARAMETRE h d L S Legenda
Zrýchľovanie vesmíru. Zrýchľovanie vesmíru. o výprave na kraj vesmíru a čo tam astronómovia objavili
 Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru
Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru a čo tam astronómovia objavili Zrýchľovanie vesmíru o výprave na kraj vesmíru
METABOLIZMUS FRUKTÓZY A GALAKTÓZY REGULÁCIA METABOLIZMU SACHARIDOV
 Katedra chémie, biochémie a biofyziky Ústav biochémie METABLIZMUS FRUKTÓZY A GALAKTÓZY METABLIZMUS GLYKGÉNU Glykogenéza Glykogenolýza REGULÁCIA METABLIZMU SACARIDV Metabolizmus fruktózy Metabolizmus fruktózy
Katedra chémie, biochémie a biofyziky Ústav biochémie METABLIZMUS FRUKTÓZY A GALAKTÓZY METABLIZMUS GLYKGÉNU Glykogenéza Glykogenolýza REGULÁCIA METABLIZMU SACARIDV Metabolizmus fruktózy Metabolizmus fruktózy
24. Základné spôsoby zobrazovania priestoru do roviny
 24. Základné spôsoby zobrazovania priestoru do roviny Voľné rovnobežné premietanie Presné metódy zobrazenia trojrozmerného priestoru do dvojrozmernej roviny skúma samostatná matematická disciplína, ktorá
24. Základné spôsoby zobrazovania priestoru do roviny Voľné rovnobežné premietanie Presné metódy zobrazenia trojrozmerného priestoru do dvojrozmernej roviny skúma samostatná matematická disciplína, ktorá
Výročná spojená konferencia Parent Project Muscular Dystrophy, Filadelfia, 12. až 14. júl 2007
 Výročná spojená konferencia Parent Project Muscular Dystrophy, Filadelfia, 12. až 14. júl 2007 Spoločná práca s cieľom skoncovať s Duchennovou svalovou dystrofiou. Viac ako 400 ľudí prišlo do Filadelfie,
Výročná spojená konferencia Parent Project Muscular Dystrophy, Filadelfia, 12. až 14. júl 2007 Spoločná práca s cieľom skoncovať s Duchennovou svalovou dystrofiou. Viac ako 400 ľudí prišlo do Filadelfie,
Príčiny SIRS Za najčastejšie uvádzané príčiny SIRS v spojení s infekciou sa považujú nasledovné patologické procesy Tab. 2.
 07. Systémová zápalová odpoveď SIRS J. Plevková V roku 1992 ACCP (American College of Chest Physicians a spoločnosť pre urgentnú medicínu definovala SIRS systémovú zápalovú odpoveď organizmu ako systémovú
07. Systémová zápalová odpoveď SIRS J. Plevková V roku 1992 ACCP (American College of Chest Physicians a spoločnosť pre urgentnú medicínu definovala SIRS systémovú zápalovú odpoveď organizmu ako systémovú
TECHNICKÁ UNIVERZITA VO ZVOLENE. Prof. Ing. Dušan Gömöry, DrSc. GENETIKA
 TECHNICKÁ UNIVERZITA VO ZVOLENE Lesnícka fakulta Katedra fytológie Prof. Ing. Dušan Gömöry, DrSc. GENETIKA 2014 prof. Ing. Dušan Gömöry, DrSc. Recenzenti: Schválené: Rektorom Technickej univerzity vo Zvolene
TECHNICKÁ UNIVERZITA VO ZVOLENE Lesnícka fakulta Katedra fytológie Prof. Ing. Dušan Gömöry, DrSc. GENETIKA 2014 prof. Ing. Dušan Gömöry, DrSc. Recenzenti: Schválené: Rektorom Technickej univerzity vo Zvolene
Odporníky. 1. Príklad1. TESLA TR
 Odporníky Úloha cvičenia: 1.Zistite technické údaje odporníkov pomocou katalógov 2.Zistite menovitú hodnotu odporníkov označených farebným kódom Schématická značka: 1. Príklad1. TESLA TR 163 200 ±1% L
Odporníky Úloha cvičenia: 1.Zistite technické údaje odporníkov pomocou katalógov 2.Zistite menovitú hodnotu odporníkov označených farebným kódom Schématická značka: 1. Príklad1. TESLA TR 163 200 ±1% L
pre I. ročník obchodných akadémií Obchodná akadémia Radlinského 1725/ Dolný Kubín samosprávny kraj
 BB iioo l ó g i a lógia pre I. ročník obchodných akadémií učebný materiál Obchodná akadémia Radlinského 1725/55 026 01 Dolný Kubín ŽILINSKÝ samosprávny kraj Z R I A Ď O VA T E Ľ Táto publikácia bola vytvorená
BB iioo l ó g i a lógia pre I. ročník obchodných akadémií učebný materiál Obchodná akadémia Radlinského 1725/55 026 01 Dolný Kubín ŽILINSKÝ samosprávny kraj Z R I A Ď O VA T E Ľ Táto publikácia bola vytvorená
MIDTERM (A) riešenia a bodovanie
 riešenia a bodovanie") MIDTERM (A) riešenia a bodovanie 1. (7b) Nech vzhl adom na štandardnú karteziánsku sústavu súradníc S 1 := O, e 1, e 2 majú bod P a vektory u, v súradnice P = [0, 1], u = e 1, v = 2 e 2. Aký predpis bude
MIDTERM (A) riešenia a bodovanie 1. (7b) Nech vzhl adom na štandardnú karteziánsku sústavu súradníc S 1 := O, e 1, e 2 majú bod P a vektory u, v súradnice P = [0, 1], u = e 1, v = 2 e 2. Aký predpis bude
Gramatická indukcia a jej využitie
 a jej využitie KAI FMFI UK 29. Marec 2010 a jej využitie Prehľad Teória formálnych jazykov 1 Teória formálnych jazykov 2 3 a jej využitie Na počiatku bolo slovo. A slovo... a jej využitie Definícia (Slovo)
a jej využitie KAI FMFI UK 29. Marec 2010 a jej využitie Prehľad Teória formálnych jazykov 1 Teória formálnych jazykov 2 3 a jej využitie Na počiatku bolo slovo. A slovo... a jej využitie Definícia (Slovo)
KATABOLIZMUS LIPIDOV BIOCHÉMIA II TÉMA 05 DOC. RNDR. MAREK SKORŠEPA, PHD.
 BIOCHÉMIA II KATEDRA CHÉMIE, FAKULTA PRÍRODNÝCH VIED, UNIVERZITA MATEJA BELA BANSKÁ BYSTRICA KATABOLIZMUS LIPIDOV TÉMA 05 DOC. RNDR. MAREK SKORŠEPA, PHD. LIPIDY AKO ZDROJ ENERGIE lipidy = tretia úrveň
BIOCHÉMIA II KATEDRA CHÉMIE, FAKULTA PRÍRODNÝCH VIED, UNIVERZITA MATEJA BELA BANSKÁ BYSTRICA KATABOLIZMUS LIPIDOV TÉMA 05 DOC. RNDR. MAREK SKORŠEPA, PHD. LIPIDY AKO ZDROJ ENERGIE lipidy = tretia úrveň
Ništiar, F., Rácz, O., Beňačka, R.: POSTAVENIE PATOFYZIOLÓGIE V ZUBNOM LEKÁRSTVE
 ÚVOD Ništiar, F., Rácz, O., Beňačka, R.: Orálna patofyziológia umožňuje lepšie pochopenie vzniku a patogenézy ochorení ústnej dutiny a tým aj kauzálnu terapiu týchto chorôb. Ich diagnostika obyčajne nie
ÚVOD Ništiar, F., Rácz, O., Beňačka, R.: Orálna patofyziológia umožňuje lepšie pochopenie vzniku a patogenézy ochorení ústnej dutiny a tým aj kauzálnu terapiu týchto chorôb. Ich diagnostika obyčajne nie
Biologická olympiáda Ročník : 47 Školský rok : 2012/2013 Kolo : Celoštátne Kategória : A Teoreticko-praktická časť.
 Biologická olympiáda Ročník : 47 Školský rok : 2012/2013 Kolo : Celoštátne Kategória : A Teoreticko-praktická časť Praktická úloha č. 1. Biochémia (49 bodov) 1. úloha: Autorské riešenia a) So zvyšujúcou
Biologická olympiáda Ročník : 47 Školský rok : 2012/2013 Kolo : Celoštátne Kategória : A Teoreticko-praktická časť Praktická úloha č. 1. Biochémia (49 bodov) 1. úloha: Autorské riešenia a) So zvyšujúcou
Hlavnou úlohou erytrocytov je prenos kyslíka. Vykonávanie tejto funkcie umožňuje erytrocytom červené krvné farbivo hemoglobín, ktorý má vo svojej mole
 LEUKÉMIA Úvod Leukémia je medzi zhubnými nádorovými ochoreniami zastúpená relatívne nízkym percentuálnym výskytom. Na Slovensku podľa najnovších štatistických údajov Národného onkologického registra SR
LEUKÉMIA Úvod Leukémia je medzi zhubnými nádorovými ochoreniami zastúpená relatívne nízkym percentuálnym výskytom. Na Slovensku podľa najnovších štatistických údajov Národného onkologického registra SR
replikačná vidlica (ori), jednosmerná-dvojsmerná replikácia
, jednosmerná-dvojsmerná replikácia") P4/1 Replikácia bakteriálneho (prokaryotického) genómu replikačná vidlica (ori), jednosmerná-dvojsmerná replikácia Dvojsmerná replikácia kruhovej chromozómovej DNA P4/2 1. Iniciácia replikácie: rozoznanie
P4/1 Replikácia bakteriálneho (prokaryotického) genómu replikačná vidlica (ori), jednosmerná-dvojsmerná replikácia Dvojsmerná replikácia kruhovej chromozómovej DNA P4/2 1. Iniciácia replikácie: rozoznanie
120 MOLEKULOVÉ MECHANIZMY PATOGENÉZY NÁDOROV
 Použité skratky ACTH adrenokortikotropný hormón ADCC antibody dependent cellular cytotoxicity, od protilátok závislá bunkami sprostredkovaná cytotoxicita AEG ALL astrocyte elevated gene akútna lymfoblastická
Použité skratky ACTH adrenokortikotropný hormón ADCC antibody dependent cellular cytotoxicity, od protilátok závislá bunkami sprostredkovaná cytotoxicita AEG ALL astrocyte elevated gene akútna lymfoblastická
Ochorenia prostaty a inkontinencia moču
 Ochorenia prostaty a inkontinencia moču Ján Breza Inkofórum 2015 Benígna prostatická hyperplázia je nezhubné zväčšenie prostaty vznikajúce v procese starnutia Histologicky ide o proliferáciu hladkého svalstva
Ochorenia prostaty a inkontinencia moču Ján Breza Inkofórum 2015 Benígna prostatická hyperplázia je nezhubné zväčšenie prostaty vznikajúce v procese starnutia Histologicky ide o proliferáciu hladkého svalstva
ŠNEKÁČI mýty o přidávání CO2 založenie akvária Poecilia reticulata REPORTÁŽE
 bulletin občianskeho združenia 2 /6.11.2006/ ŠNEKÁČI mýty o přidávání CO2 založenie akvária Poecilia reticulata REPORTÁŽE akvá ri um pr pree kre vet y, raky a krab y akva foto gr afi e Ji Jiřříí Plí š
bulletin občianskeho združenia 2 /6.11.2006/ ŠNEKÁČI mýty o přidávání CO2 založenie akvária Poecilia reticulata REPORTÁŽE akvá ri um pr pree kre vet y, raky a krab y akva foto gr afi e Ji Jiřříí Plí š
Vyhlásenie o parametroch stavebného výrobku StoPox GH 205 S
 1 / 5 Vyhlásenie o parametroch stavebného výrobku StoPox GH 205 S Identifikačný kód typu výrobku PROD2141 StoPox GH 205 S Účel použitia EN 1504-2: Výrobok slúžiaci na ochranu povrchov povrchová úprava
1 / 5 Vyhlásenie o parametroch stavebného výrobku StoPox GH 205 S Identifikačný kód typu výrobku PROD2141 StoPox GH 205 S Účel použitia EN 1504-2: Výrobok slúžiaci na ochranu povrchov povrchová úprava
ZADANIE 1_ ÚLOHA 3_Všeobecná rovinná silová sústava ZADANIE 1 _ ÚLOHA 3
 ZDNIE _ ÚLOH 3_Všeobecná rovinná silová sústv ZDNIE _ ÚLOH 3 ÚLOH 3.: Vypočítjte veľkosti rekcií vo väzbách nosník zťženého podľ obrázku 3.. Veľkosti známych síl, momentov dĺžkové rozmery sú uvedené v
ZDNIE _ ÚLOH 3_Všeobecná rovinná silová sústv ZDNIE _ ÚLOH 3 ÚLOH 3.: Vypočítjte veľkosti rekcií vo väzbách nosník zťženého podľ obrázku 3.. Veľkosti známych síl, momentov dĺžkové rozmery sú uvedené v
ROZSAH ANALÝZ A POČETNOSŤ ODBEROV VZORIEK PITNEJ VODY
 ROZSAH ANALÝZ A POČETNOSŤ ODBEROV VZORIEK PITNEJ VODY 2.1. Rozsah analýz 2.1.1. Minimálna analýza Minimálna analýza je určená na kontrolu a získavanie pravidelných informácií o stabilite zdroja pitnej
ROZSAH ANALÝZ A POČETNOSŤ ODBEROV VZORIEK PITNEJ VODY 2.1. Rozsah analýz 2.1.1. Minimálna analýza Minimálna analýza je určená na kontrolu a získavanie pravidelných informácií o stabilite zdroja pitnej
Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti rozvodu tepla
 Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti príloha č. 7 k vyhláške č. 428/2010 Názov prevádzkovateľa verejného : Spravbytkomfort a.s. Prešov Adresa: IČO: Volgogradská 88, 080 01 Prešov 31718523
Rozsah hodnotenia a spôsob výpočtu energetickej účinnosti príloha č. 7 k vyhláške č. 428/2010 Názov prevádzkovateľa verejného : Spravbytkomfort a.s. Prešov Adresa: IČO: Volgogradská 88, 080 01 Prešov 31718523
Modul pružnosti betónu
 f cm tan α = E cm 0,4f cm ε cl E = σ ε ε cul Modul pružnosti betónu α Autori: Stanislav Unčík Patrik Ševčík Modul pružnosti betónu Autori: Stanislav Unčík Patrik Ševčík Trnava 2008 Obsah 1 Úvod...7 2 Deformácie
f cm tan α = E cm 0,4f cm ε cl E = σ ε ε cul Modul pružnosti betónu α Autori: Stanislav Unčík Patrik Ševčík Modul pružnosti betónu Autori: Stanislav Unčík Patrik Ševčík Trnava 2008 Obsah 1 Úvod...7 2 Deformácie
17. ZÁPAL, HORÚČKA, STRES
 17. ZÁPAL, HORÚČKA, STRES Ništiar František a Dombrovský Peter 17.1 ZÁPAL Základné pojmy a klasifikácia Zápal (lat. inflammatio) je stereotypná obranná odpoveď organizmu, ktorú vyvoláva poškodenie alebo
17. ZÁPAL, HORÚČKA, STRES Ništiar František a Dombrovský Peter 17.1 ZÁPAL Základné pojmy a klasifikácia Zápal (lat. inflammatio) je stereotypná obranná odpoveď organizmu, ktorú vyvoláva poškodenie alebo
Neurobiológia nádorových chorôb: vplyv nervového systému na nádorový rast a tvorbu metastáz
 Neurobiológia nádorových chorôb: vplyv nervového systému na nádorový rast a tvorbu metastáz Interakcie medzi nádorovým tkanivom a nervovým systémom sú podkladom neurobiologického pohľadu na etiopatogenézu
Neurobiológia nádorových chorôb: vplyv nervového systému na nádorový rast a tvorbu metastáz Interakcie medzi nádorovým tkanivom a nervovým systémom sú podkladom neurobiologického pohľadu na etiopatogenézu
Modelovanie dynamickej podmienenej korelácie kurzov V4
 Modelovanie dynamickej podmienenej korelácie menových kurzov V4 Podnikovohospodárska fakulta so sídlom v Košiciach Ekonomická univerzita v Bratislave Cieľ a motivácia Východiská Cieľ a motivácia Cieľ Kvantifikovať
Modelovanie dynamickej podmienenej korelácie menových kurzov V4 Podnikovohospodárska fakulta so sídlom v Košiciach Ekonomická univerzita v Bratislave Cieľ a motivácia Východiská Cieľ a motivácia Cieľ Kvantifikovať
Pevné ložiská. Voľné ložiská
 SUPPORTS D EXTREMITES DE PRECISION - SUPPORT UNIT FOR BALLSCREWS LOŽISKA PRE GULIČKOVÉ SKRUTKY A TRAPÉZOVÉ SKRUTKY Výber správnej podpory konca uličkovej skrutky či trapézovej skrutky je dôležité pre správnu
SUPPORTS D EXTREMITES DE PRECISION - SUPPORT UNIT FOR BALLSCREWS LOŽISKA PRE GULIČKOVÉ SKRUTKY A TRAPÉZOVÉ SKRUTKY Výber správnej podpory konca uličkovej skrutky či trapézovej skrutky je dôležité pre správnu
Účinky glukokortikoidov
 Účinky glukokortikoidov Glukokortikoidy vykazujú mnoho priamych a nepriamych fyziologických účinkov, ktoré sprostredkúvajú stresovú reakciu, zvyšujú aktivitu iných hormónov, alebo tlmia iné zložky systémov
Účinky glukokortikoidov Glukokortikoidy vykazujú mnoho priamych a nepriamych fyziologických účinkov, ktoré sprostredkúvajú stresovú reakciu, zvyšujú aktivitu iných hormónov, alebo tlmia iné zložky systémov
OSTEOARTRITÍDA. - vek - pohlavie. Chronická degeneratívna artropatia Najčastejší typ artritídy Hlavné rizikové faktory. - obezita Prevalencia %
 OSTEOARTRITÍDA Chronická degeneratívna artropatia Najčastejší typ artritídy Hlavné rizikové faktory - vek - pohlavie - obezita Prevalencia 10 20 % Prevalencia OA Lokalizácia Rádiografická OA Symptomatická
OSTEOARTRITÍDA Chronická degeneratívna artropatia Najčastejší typ artritídy Hlavné rizikové faktory - vek - pohlavie - obezita Prevalencia 10 20 % Prevalencia OA Lokalizácia Rádiografická OA Symptomatická
Harmonizované technické špecifikácie Trieda GP - CS lv EN Pevnosť v tlaku 6 N/mm² EN Prídržnosť
 Baumit Prednástrek / Vorspritzer Vyhlásenie o parametroch č.: 01-BSK- Prednástrek / Vorspritzer 1. Jedinečný identifikačný kód typu a výrobku: Baumit Prednástrek / Vorspritzer 2. Typ, číslo výrobnej dávky
Baumit Prednástrek / Vorspritzer Vyhlásenie o parametroch č.: 01-BSK- Prednástrek / Vorspritzer 1. Jedinečný identifikačný kód typu a výrobku: Baumit Prednástrek / Vorspritzer 2. Typ, číslo výrobnej dávky
Význam železa pre ľudský organizmus. Jana Strigáčová
 Význam železa pre ľudský organizmus Jana Strigáčová MAKROMINERÁLY (7 prvkov): Ca, P, K, Mg, Na, S, Cl - sú to veľkoobjemové minerály. Podieľajú sa na stavbe organizmu, udržujú rovnováhu v tele, sú dôležité
Význam železa pre ľudský organizmus Jana Strigáčová MAKROMINERÁLY (7 prvkov): Ca, P, K, Mg, Na, S, Cl - sú to veľkoobjemové minerály. Podieľajú sa na stavbe organizmu, udržujú rovnováhu v tele, sú dôležité
Planárne a rovinné grafy
 Planárne a rovinné grafy Definícia Graf G sa nazýva planárny, ak existuje jeho nakreslenie D, v ktorom sa žiadne dve hrany nepretínajú. D sa potom nazýva rovinný graf. Planárne a rovinné grafy Definícia
Planárne a rovinné grafy Definícia Graf G sa nazýva planárny, ak existuje jeho nakreslenie D, v ktorom sa žiadne dve hrany nepretínajú. D sa potom nazýva rovinný graf. Planárne a rovinné grafy Definícia
(Úradný vestník Európskej únie L 136 z 25. mája 2012)
") Zoznam zdravotných tvrdení, ktoré možno uvádzať na potravinách podľa článku 13 ods. 3 nariadenia (ES) č. 1924/2006 v zmysle aktuálneho korigenda Korigendum k nariadeniu Komisie (EÚ) č. 432/2012 zo 16.
Zoznam zdravotných tvrdení, ktoré možno uvádzať na potravinách podľa článku 13 ods. 3 nariadenia (ES) č. 1924/2006 v zmysle aktuálneho korigenda Korigendum k nariadeniu Komisie (EÚ) č. 432/2012 zo 16.
2 Chyby a neistoty merania, zápis výsledku merania
 2 Chyby a neistoty merania, zápis výsledku merania Akej chyby sa môžeme dopustiť pri meraní na stopkách? Ako určíme ich presnosť? Základné pojmy: chyba merania, hrubé chyby, systematické chyby, náhodné
2 Chyby a neistoty merania, zápis výsledku merania Akej chyby sa môžeme dopustiť pri meraní na stopkách? Ako určíme ich presnosť? Základné pojmy: chyba merania, hrubé chyby, systematické chyby, náhodné
Kinetika fyzikálno-chemických procesov
 Kinetika fyzikálno-chemických procesov Chemická a biochemická kinetika Reálne biologické a fyzikálno-chemické procesy sú závislé na čase. Termodynamika poskytuje informácie len o možnostiach priebehu procesov,
Kinetika fyzikálno-chemických procesov Chemická a biochemická kinetika Reálne biologické a fyzikálno-chemické procesy sú závislé na čase. Termodynamika poskytuje informácie len o možnostiach priebehu procesov,
Živá hmota chemické zloženie. Biológia živočíšnej produkcie Katedra fyziológie živočíchov
 Živá hmota chemické zloženie Biológia živočíšnej produkcie Katedra fyziológie živočíchov Chemické zloženie živej hmoty Živá hmota bioplazma chemicky rôznorodá zmes látok zložitý koloidný systém Prvky v
Živá hmota chemické zloženie Biológia živočíšnej produkcie Katedra fyziológie živočíchov Chemické zloženie živej hmoty Živá hmota bioplazma chemicky rôznorodá zmes látok zložitý koloidný systém Prvky v
Vektorový priestor V : Množina prvkov (vektory), na ktorej je definované ich sčítanie a ich
, na ktorej je definované ich sčítanie a ich") Tuesday 15 th January, 2013, 19:53 Základy tenzorového počtu M.Gintner Vektorový priestor V : Množina prvkov (vektory), na ktorej je definované ich sčítanie a ich násobenie reálnym číslom tak, že platí:
Tuesday 15 th January, 2013, 19:53 Základy tenzorového počtu M.Gintner Vektorový priestor V : Množina prvkov (vektory), na ktorej je definované ich sčítanie a ich násobenie reálnym číslom tak, že platí:
LEKÁRSKA BIOLÓGIA A GENETIKA 1
 Daniel BÖHMER, Ľuboš DANIŠOVIČ, a Vanda REPISKÁ LEKÁRSKA BIOLÓGIA A GENETIKA 1 VYSOKOŠKOLSKÁ UČEBNICA PRE PREGRADUÁLNE ŠTÚDIUM NA LEKÁRSKYCH FAKULTÁCH LEKÁRSKA BIOLÓGIA A GENETIKA 1 Vysokoškolská učebnica
Daniel BÖHMER, Ľuboš DANIŠOVIČ, a Vanda REPISKÁ LEKÁRSKA BIOLÓGIA A GENETIKA 1 VYSOKOŠKOLSKÁ UČEBNICA PRE PREGRADUÁLNE ŠTÚDIUM NA LEKÁRSKYCH FAKULTÁCH LEKÁRSKA BIOLÓGIA A GENETIKA 1 Vysokoškolská učebnica
30. kapitola PATOFYZIOLÓGIA KRVI. Ján Hanáček
 30. kapitola PATOFYZIOLÓGIA KRVI Ján Hanáček Hematologický systém je najdiverzifikovanejší, najadaptabilnejší, najkoordinovanejší a najefektívnejší systém, ktorý príroda vytvorila. Je to systém vybavený
30. kapitola PATOFYZIOLÓGIA KRVI Ján Hanáček Hematologický systém je najdiverzifikovanejší, najadaptabilnejší, najkoordinovanejší a najefektívnejší systém, ktorý príroda vytvorila. Je to systém vybavený
u R Pasívne prvky R, L, C v obvode striedavého prúdu Činný odpor R Napätie zdroja sa rovná úbytku napätia na činnom odpore.
 Pasívne prvky, L, C v obvode stredavého prúdu Čnný odpor u u prebeh prúdu a napäta fázorový dagram prúdu a napäta u u /2 /2 t Napäte zdroja sa rovná úbytku napäta na čnnom odpore. Prúd je vo fáze s napätím.
Pasívne prvky, L, C v obvode stredavého prúdu Čnný odpor u u prebeh prúdu a napäta fázorový dagram prúdu a napäta u u /2 /2 t Napäte zdroja sa rovná úbytku napäta na čnnom odpore. Prúd je vo fáze s napätím.