METABOLIZAM UGLJENIH HIDRATA
|
|
|
- ÍΑἰνείας Θεράπων Κουρμούλης
- 7 χρόνια πριν
- Προβολές:
Transcript
1 METABOLIZAM UGLJENIH HIDRATA
2

3

4
5
6
7

8

9
10 Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O 2. U uslovima prisustva O 2, piruvat dobijen u glikolizi se oksiduje do CO 2 u ciklusu TCA (pri čemu ATP nastaje prenosom redukcionih ekvivalenata u lancu prenosilaca elektrona i oksidativnoj fosforilaciji). U anaerobnim uslovima (ćelije bez mitohondrija, naglo smanjenje dotoka O 2 ćelijama) piruvat se prevodi u laktat, uz reoksidaciju NADH, a ATP nastaje u procesu oksidacije na nivou supstrata.
11
12 Transport glukoze u ćelije
13 GLIKOLIZA

14 Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O 2. U uslovima prisustva O 2, piruvat dobijen u glikolizi se oksiduje do CO 2 u ciklusu TCA (pri čemu ATP nastaje prenosom redukcionih ekvivalenata u lancu prenosilaca elektrona i oksidativnoj fosforilaciji). U anaerobnim uslovima (ćelije bez mitohondrija, naglo smanjenje dotoka O 2 ćelijama) piruvat se prevodi u laktat, uz reoksidaciju NADH, a ATP nastaje u procesu oksidacije na nivou supstrata.
15 Glikolizu sačinjavaju 2 faze: pripremna i faza u kojoj se dobija energija (SVAKA FAZA TRAJE PO 5 CIKLUSA). Prvih pet reakcija čine pripremnu fazu, u kojoj nastaju dva mola gliceraldehid- 3-fosfata iz jednog mola glukoze, uz utrošak 2 mola ATP-a.
16 Pripremna faza
17 (1) Fosforilacija D-glukoze: heksokinaza H OH Glukoza Heksokinaza Mg 2+ H OH H CH 2 OH H OH H O H OH Glukoza Utrošak prvog mola ATP 2- CH Ireverzibilna 2 OPO 3 reakcija CH 2 OPO 3 O H H Enzimi: heksokinaza, glukokinaza OH H OH Heksokinaza Mg 2+ O H H + ATP + ADP + H + HK Mg 2+ H OH OH H H 2- OH H OH Glukoza-6-fosfat (G6P) + ADP + H +
18 Sudbine glukozo-6-fosfata Fosforilacijom se glukoza zarobljava u ćeliji i reakcija je pod unutarćelijskim uslovima bespovratna. I druge heksoze (D-galaktoza, D-fruktoza i D- manoza), nakon fosforilacije, mogu da uđu u pripremnu fazu glikolize. Od glukozo-6-p se granaju metabolički putevi, jer je ovaj molekul prekursor praktično svakog puta glukoze : glikoliza, pentozni put, sinteza glikogena, sinteza glukuronske kiseline i aminošećera. Takođe ovaj molekuli i nastaje u nekim metaboličkim putevima (razgradnja glikogena, glukoneogeneza). Ćelijska membrana nepropusna za fosforne estre šećera, samo jetra i bubreg imaju glc-6-fosfatazu koja otpušta slobodnu glc u krv.
19 Postoji nekoliko izoenzima heksokinaze, koji katališu fosforilaciju glukoze. Većina ih ima visok afinitet za glukozu (Km<0,1 mmol/l), i inhibisani su proizvodom reakcije, glukozo-6-fosfatom Heksokinaza D (ili tip IV, glukokinaza), najzastupljeniji izoenzim u hepatocitima, ima znatno manji afinitet za glukozu od drugih heksokinaza i katališe fosforilaciju heksoza u heksozo- 6-fosfat. Ovakve karakteristike različitih izoenzima heksokinaze imaju za posledicu da: (i) porast glikemije dovodi do povećanja brzine fosforilacije glukoze u jetri (ii) u tkivima u kojima su prisutni izoenzimi heksokinaze sa velikim afinitetom za supstrat, fosforilacija glukoze postoji čak i u uslovima relativno niske koncentracije glukoze u krvi, što je od velikog značaja za ćelije koje u potpunosti zavise od glukoze kao izvora energije (mozak).
20 OH H Fruktoza-6-fosfat (F6P) (3) Fosforilacija F6P: fosfofruktokinaza Fosfofruktokinaza (PFK) Mg 2+ CH 2 OPO 3 O H H OH 2- OH OH H CH 2 OH Fruktoza-6-fosfat (F6P) + ATP PFK1 Mg 2+ CH 2 OPO 3 O H H OH 2- OH OH H CH 2 OPO 3 Fruktoza-1,6-difosfat (FDP) 2- + ADP + H + Fosfofruktokinaza (PFK) Mg 2+ Fosfofruktokinaza glavni regulatorni enzim glikolize 2- CH 2 OPO 3 O CH 2 OPO 3 2- ADP H +
21 (4) Cepanje FDP: aldolaza Dihidroksiaceton-fosfat (DHAP) HO CH 2 OPO 3 C O C H 2- Aldolaza HO CH 2 OPO 3 C O CH 2 2- H H C C OH OH CH 2 OPO 3 2- Fruktoza-1,6-difosfat (FDP) H H C C O OH CH 2 OPO 3 2- Gliceraldehid-3-fosfat (GAP)
22 (6) Oksidacija gliceraldehid-3-fosfata: gliceraldehid-fosfat dehidrogenaza ENERGETSKO JEDINJENJE O H C C H OH CH 2 OPO NAD + Pi GAPDH O H C C OPO 3 2- OH + NADH + H + CH 2 OPO 3 2- Gliceraldehid- 3-fosfat (GAP) 1,3-Difosfoglicerat (1,3-DPG) Stvaranje prvog visokoenergetskog jedinjenja, 1,3- bisfosfoglicerat, (mešoviti anhidrid karboksilne i fosforne kiseline) oksidacijom gliceraldehid-3-fosfata.
23 Faza dobijanja energije - oksidacija i fosforilacija na nivou supstrata Regeneracija NAD + : - u laktat dehidrogenaznoj reakciji, - prenošenjem redukcionih ekvivalenata u mitohondrije mehanizmima povratnog transporta.
24
25
26 Sudbine piruvata Piruvat se, u aerobnim uslovima, oksidativnom dekarboksilacijom prevodi u acetil-coa, koji ulazi u ciklus trikarboksilnih kiselina (ciklus TCA). Potpuna oksidacija glukoze do CO2 i H2O daje 29,5-31 molekul ATP-a po molekulu glukoze. U anaerobnom uslovima, piruvat se redukuje u laktat, čime se obezbeđuje oksidacija NADH+H + i dalje odvijanje glikolize

27

28 Sudbina laktata Laktat oslobođen u glikolizi prihvataju ćelije drugih tkiva (jetra, srčani i skeletni mišić) gde se oksiduje ponovo u piruvat. U jetri, piruvat je prekursor u glukoneogenezi. U ostalim tkivima, laktat se oksiduje u piruvat, koji se dalje oksiduje u ciklusu TCA.

29
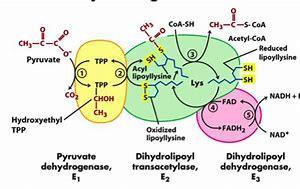

30 PIRUVAT DEHIDROGENAZNI KOMPLEKS
31 Šećer sa 5C pentoze 2,3-bisfosfoglicerat serin alanin glukoza Glukozo-6-P 1,3-bisfosfoglicerat 3-fosfoglicerat piruvat Glicerol-P MK TAG OSTALE ULOGE GLIKOLIZE Glikoliza obezbeđuje i prekursore za neke biosintetske puteve (najviše u jetri): Sinteza nukleotida Ostali šećeri (npr. UDPglukoza, manoza, sijalinska kiselina itd. Acetil-CoA Aminokiseline Triacilgliceroli TCA ciklus 2,3 bisfosfoglicerat Glutamat i druge AK
32 Regulacija enzima glikolize Enzimi heksokinaza, fosfofruktokinaza 1 i piruvat kinaza kontrolišu brzinu metaboličkog protoka u glikolizi, jer katališu reakcije koje su pod fiziološkim uslovima nepovratne (DG<0), te se smatraju regulatornim enzimima glikolize. Najvažniji regulatorni korak u glikolizi je reakcija pretvaranja fruktozo-6- fosfata u fruktozo-1,6-bisfosfat, koju katališe fosfofruktokinaza 1 (PFK 1). Aktivnost ovog enzima uglavnom je kontrolisana alosteričkim efektorima. Uopšteno uzevši, može se reći da brzina glikolize u ćeliji zavisi od: energetskog statusa ćelije (ATP, AMP, Pi) kada je nivo ATP-a visok, glikoliza je inhibirana i obrnuto; unutrašnje sredine ćelije (ph); dostupnosti alternativnih izvora energije u vidu masnih kiselina ili ketonskih tela (čijom razgradnjom se dobija acetil-coa, koji u reakciji sa oksalacetatom daje citrat) dovoljna količina supstrata ulazi u ciklus TCA i glikoliza je inhibirana; odnosa insulin/glukagon u krvi (od koga zavisi koncentracija fruktozo-2,6-bisfosfata, kao i aktivnost piruvat kinaze);
33 KREBSOV CIKLUS

34

35
36 Krebsov ciklus je amfibolički metabolički put u celini smešten u mitohondrijama Supstrat za ovaj ciklus je acetil CoA; najveći deo nastaje oksidativnom dekarboksilacijom piruvata nastalog u glikolizi pod dejstvom PDH kompleksa Nastali redukcioni ekvivalenti najvažniji izvor elektrona za lanac prenosilaca elektrona Potpunom oksidacijom 1 molekula glukoze u glikolizi, Krebsovom ciklusu i lancu prenosalaca elektrona dobija se 29,5-31 ATP, zavisno od toga kojim sistemom povratnog transporta se redukcioni ekvivalenti prebacuju iz citosola u mitohondrije.
37 Odvijanje Krebsovog ciklusa zavisi od: Odnosa NAD + /NADH unutar mitohondrija Parcijalnog pritiska O 2 Energetskog statusa ćelije (alosterička aktivacija izocitrat dehidrogenaze ADP-om) Dostupnosti jedinjenja bogatih energijom (acetil-coa, sukcinil-coa)
38 Niz međuproizvoda ovog ciklusa su početna jedinjenja u sintezi aminokiselina, hema ili steroida. Anaplerotski putevi obezbeđuju da, uprkos uključivanju u druge puteve, koncentracija međuproizvoda Krebsovog ciklusa unutar mitohondrija bude uglavnim konstantna.

39

40

41

42

43

44

45

46

47

48

49

50

51

52

53
54 OKSIDATIVNA FOSFORILACIJA
55 Oksidativna fosforilacija U eukariota u mitohondrijama. Odigrava se redukcija O2 do H2O. Donori elektrona su NADH i FADH2.
56 Najveći deo energije za svoj rast, održavanje i rad ćelija dobija oksidacijom ugljenih hidrata i lipida; ova jedinjenja su u značajnoj meri redukovana te mogu da daju veliku količinu redukcionih ekvivalenata koji se prenose do kiseonika; krajnji prozvodi oksidacije su voda i ugljen dioksid. Pri oksidaciji se oslobađa energija koja se zahvata i pretvara u hemijsku energiju koja se čuva u obliku visokoenergetskih veza.
57 Oksidacija je u ćeliji postepen proces. U prvoj fazi se redukcioni ekvivalenti odvajaju od supstrata delovanjem dehidrogenaza. U sledećoj fazi elektroni i protoni se predaju lancu prenosilica elektrona respiratornom lancu koji je smešten na unutrašnjoj membrani mitohondrije. U trećoj fazi elektroni se prenose od jednog do drugog člana respiratornog lanca do kiseonika, krajnjeg primaoca elektrona. Zahvatanje energije je ostvareno sprezanjem oslobađanja energije, koje se dešava pri prenosu elektrona sa višeg na niži potencijal, sa reakcijama u kojima se formira visokoenergetska veza u ATP-u. Celokupan proces je nazvan oksidativna fosforilacija.
58 Oksidativna fosforilacija Peter Mitchell (1961) - hemiosmotska teorija Transmembranska razlika u koncentraciji protona rezervoar za dobijanje energije iz bioloških oksidacija.
59 Dobijanje ATP-a u procesu oksidativne fosforilacije podrazumjeva učešće: davaoca elektrona (NADH ili FADH2), primaoca elektrona (O2), kao i unutrašnju membranu mitohondrija koja je nepropusna za protone, sve komponente lanca prenosilaca elektrona i ATP sintazu. Oksidativna fosforilacija je regulisana stepenom iskorišćavanja ATP-a.
60 OKSIDATIVNA FOSFORILACIJA Reakcije transfera elektrona u mitohondrijama Proteini Ugljeni hidrati Lipidi NADH+H Katabolički putevi FADH RESPIRATORNI LANAC NADPH+ H Dehidrogenaze sakupljaju elektrone iz kataboličkih puteva i prenose do univerzalnih akceptora elektrona NAD+, NADP+, FMN ili FAD.


61 Oksidativna fosforilacija se odvija u mitohondrijama Respiratorni lanac mitohondrija se sastoji od serije nosača elektrona koji djeluju sekvencijalno. Najveći broj ovih nosača su integralni proteini membrane čije prostetične grupe mogu primati ili davati jedan ili dva elektrona.
62 Respiratorni lanac čine prenosioci elektrona čiji je redosled određen rastućim elektropozitivnim potencijalom Respiratorni lanac je skup katalitičkih proteina lokalizovanih u unutrašnoj membrani mitohondrija, a čije funkcionalne grupe mogu da primaju i predaju elektrone. Ti proteini su NADH dehidrogenaza, citohrom b, citohrom c1, citohrom c, citohrom a i citohrom a3. Uz ove, respiratorni lanac čine i proteini koji sprežu oslobađanje energije koje se dešava pri promeni elektrohemijskog potencijala sa sintezom visokoenergetske veze u ATP-u. Koenzim Q, koji prenosi elektrone od flavoproteina do citohroma b takođe pripada ovom sistemu. U samom lancu elektronski nosači su poredani po rastućem elektropozitivnom potencijalu; ukupna potencijalna razlika između prvog i poslednjeg člana lanca je 1,12 V. Ova razlika u elektropozitivnosti omogućava protok elektrona uz oslobađanje energije.
63 Respiratorni lanac je organizovan u komplekse Lanac prenosilaca elektrona je organizovan sistem, i prostorno i funkcionalno. Članovi lanca prenosilaca elektrona su poredani u definisanom redosledu, prevashodno određenim njihovim redoks potencijalom od najnegativnijeg, NADHdehidrogenaze, donajpozitivnijeg, citohroma aa3. Prostorno, članovi respiratornog lanca su organizovani u funkcionalne koplekse, smeštene u unutrašnjoj membrani mitohondrija: Kompleks I NADH dehidrogenaza i Fe-S centri enzima Kompleks II sukcinat dehidrogenaza sa svojim Fe-S centrima i citohrom b Kompleks III citohromi b i c1 i specifični Fe-S centar Kompleks IV citohromi a i a3 (citohrom oksidaza) Kompleks V F0F1 ATP sintaza Ubihinon spaja kompleksa I, II i III, dok je citohrom c veza između kompleksa III i kompleksa IV.
64 Uloge pojedinih kompleksa respiratornog lanca Kompleks Sastavni delovi kompleksa Uloga u odnosu na elektrone I NADH dehodrogenaza Prima sa NADH; prenosi na CoQ II Sukcinat dehidrogenaza Ubihinon (CoQ) Prima sa sukcinata; prenosi na CoQ Prima sa I i II, prenosi na III III Citohrom bc1 kompleks Prima sa CoQ i preda je na citohrom c Uloga u odnosu na protone Ispumpava u međumembranski prostor praveći ph gradijent Ispumpava u međumembranski prostor praveći ph gradijent Citohrom c Prima sa III, prenosi na IV IV Citohrom oksidaza Prima sa citohroma c, prenosi na O2 Ispumpava u međumembranski prostor praveći ph gradijent V F0F1 ATP sintaza Omogućava ulazak u matriks mitohondrije i koristi ulazak kao pokretačku snagu za fosforilaciju
65 OKSIDATIVNA FOSFORILACIJA Respiratorni lanac Prenosioci elektrona funkcionišu u okviru multienzimskih kompleksa Kompleksi I i II katalizuju prenos elektrona do koenzima Q Kompleks III sa redukovanog koenzima Q do citohroma c Kompleks IV sa citohroma c do O2
66 Reakcije transfera elektrona u mitohondrijama Kompleks I: NADH do ubihinona NADH dehidrogenza NADH:ubihinon oksidoreduktaza 42 polipeptidna lanca FMN flavoprotein najmanje 6 FeS centara Elektroni se sa NADH+H+ prenose na koenzim Q


67 Reakcije transfera elektrona u mitohondrijama Gvožđe-sumpor proteini Gvožđe je u asocijaciji sa neorganskim sumporom ili sa atomima sumpora cisteinskih rezidua. Svi gvožđe sumpor proteini učestvuju u jedno-elektronskom transferu u kome se jedan atom gvožđa oksiduje ili redukuje. Najmanje osam FeS proteina funkcioniše u mitohondrijalnom transferu. FeS 2Fe-2S 4Fe-4S
 sa NADH i protona iz matriksa. Endergoni prenos 4 protona iz matriksa u međumembnski prostor.")
68 Reakcije transfera elektrona u mitohondrijama Kompleks I Katalizuje dva simultana i povezana procesa. Egzergoni prenos hibridnog jona (:H-) sa NADH i protona iz matriksa. Endergoni prenos 4 protona iz matriksa u međumembnski prostor. To je dakle protonska pumpa koja energiju dobija od prenosa elektrona.

69 Postoje tri tipa citohroma b, zatim citohromi c i c1 i citohromi a i a3. Svaki hem je udružen sa jonom Cu koji je smešten blizu Fe. Citohromi, gvožđe-sumpor proteini i koenzim Q imaju funkcionalne grupe koje mogu da primaju i otpuštaju elektrone Dva su osnovna tipa proteina koji se nalaze lancu za prenos elektrona: citohromi i proteini koji sadrže gvožđe i sumpor (Fe-S proteini). Citohromi su integralni membranski proteini, sadrže gvožđe i na osnovu apsorpcionih spektara se dele u tri grupe: a, b i c. (Izuzetak je citohrom c, koji je, uz CoQ, pokretan član lanca).
70 Citohromi su jednoelektronski prenosioci gvožđe prelazi iz +3 u +2, a bakar iz +2 u +1 stanje. Uobičajeno je svih 6 koordinativnih veza Fe popunjeno, te se O2 ne može direktno vezati za njih. Izuzetak je citohrom aa3, citohrom oksidaza, jedini član respiratornog lanca koji može da reaguje sa kiseonikom, za koji ima veoma veliki afinitet; reakcija prenosa redukcionih ekvivalenata na kiseonik je jedina nepovratna reakcija u lancu. Ove dve osobine omogućavaju neprekidni jednosmjerni prenos elektrona, čak i kada je parcijalni pritisak kiseonika mali.
71 FeS proteini sadrže gvožđe koje nije vezano za hem. U ovim molekulima, atomi gvožđa su udruženi sa atomima sumpora i u molekulima čine grupe, gvožđe-sumpor centre. Atomi gvožđa u ovim grupama prolaze kroz Fe(II) Fe(III), cikluse prenoseći redukcione ekvivalente. Niz flavin-zavisnih enzima, uključujući NADH dehidrogenazu i sukcinat dehidrogenazu, sadrže FeS centre. FeS protein
72 Koenzim Q (ubihinon; CoQ) Hinon rastvorljiv u mastima sa veoma dugim izoprenoidnim bočnim lancem (10 izoprenoidnih jedinica - CoQ10). Količinski, postoji znatan stehiometrijski višak ubihinona u odnosu na ostale članove respiratornog lanca CoQ je pokretan i sakuplja redukcione ekvivalente sa manje pokretnih flavoproteinskih kompleksa. Uloga ubihinona je da prikuplja redukcione ekvivalente ne samo sa NADH dehidrogenaze već i sa drugih flavinzavisnih dehidrogenaza (npr. Sukcinat dehidrogenaze i acil CoA dehidrogenaze masnih kiselina) i da ih predaje citohromima. Kako ubihinon prima i predaje po jedan elektron u jednom trenutku, zahvaljujući postojanju semihinonskog oblika, CoQ je tačka gdje dvoelektronski prenos postaje jednoelektronski. Pri određenim uslovima u ćeliji moguće je nagomilavanje semihinonskog oblika CoQ sa posledičnom pojavom slobodnih radikala i mogućim oštećenjima koja iz ovoga proizilaze.
73 Ubihinon preuzima i redukcione ekvivalente sa flavin zavisnih dehidrogenaza, kao što je npr. sukcinat dehidrogenaza, enzim ciklusa trikarboksilnih kiselina (kompleks II). Direktan prenos redukcionih ekvivalenata na koenzim Q vrši se i sa a- glicerofosfat dehidrogenaze (enzim koji vrši povratni transport redukcionih ekvivalenata iz citosola u mitohondrije) i acil CoA dehidrogenaze iz β-oksidacije masnih kiselina. Ubihinon predaje elektrone citohromu b kompleksa III, a citohrom c preuzima elektrone sa ovog kompleksa i prenosi ih do kompleksa IV, citohrom oksidaze, koja katališe prenos elektrona sa redukovane forme citohroma c na kiseonik, pri čemu nastaje voda: 4 cit c (2+) + 4 H + + O 2 4 cit c (3+) + 2 H 2 O Potpunom redukcijom kiseonika nastaje bezopasno jedinjenje, voda. Međutim, ukoliko je kiseonik samo delimično redukovan nastaju jedinjenja koja su veoma reaktivna i opasna za ćeliju.
74 Reakcije transfera elektrona u mitohondrijama Kompleks II Sukcinat do ubihinona Sukcinat dehidrogenaza enzim ciklusa trikarbonskih kiselina (CTK) jedini enzim CTK vezan za membranu Prenosi elektrone sa sukcinata do koenzima Q
75 Reakcije transfera elektrona u mitohondrijama Kompleks III Ubihinon do citohroma c Citohrom bc1 kompleks Ubihinon: citohrom c oksidoreduktaza Povezuje prenos elektrona sa ubihinola (QH2) na citohrom c uz vektorijalni transfer protona iz matriksa u međumembranski prostor.
 sa svoja dva hema (bh i b1). Rieske gvožđe-sumpor protein (ljubičast) sa 2Fe-2S centrom (žuto).")
76 Reakcije transfera elektrona u mitohondrijama Kompleks III Dimer sastavljen od dva identična monomera. Svaki monomer sadrži 11 različitih subjedinica. Struktura monomera Funkcionalno jezgro čine tri subjedinice. Citohrom b (zeleno) sa svoja dva hema (bh i b1). Rieske gvožđe-sumpor protein (ljubičast) sa 2Fe-2S centrom (žuto). Citohrom c1 (plavo) sa svojim hemom (c1, crveno).

77 Kompleks IV Citohrom c do O2 Citohrom oksidaza Prenosi elektrone sa citohroma c do molekularnog kiseonika dajući vodu.
.")
78 Kompleks IV Redukcija kiseonika sa 4 elektrona uključuje redoks centre koji nose po jedan elektron, i mora se odvijati bez oslobađanja nekompletno redukovanih intermedijera (slobodni radikali (vodonik peroksid i hidroksil radikal). Oni ostaju tesno vezani za kompleks do kompletne redukcije do vode.
79 Oksidativna fosforilacija Sinteza ATP-a Translokaciju protona vrše kompleksi respiratornog lanca, a sintezu ATP-a ATP sintaza. FoF1 ATP sintaza je enzim koji sintetiše ATP. FoF1 kompleks, ATP sintaza, sačinjena je od pet vrsta proteina (a,b,g, d i e), koji čine čvor i peteljku; čvor i peteljka zajedno čine F1, čiji je sastav a3b3gde. Uz to se nalazi osnova, Fo, ugrađena u unutrašnju membranu mitohondrija. U osnovi ATP sintaze nalazi se kanal kroz koji prolaze protoni. Prolazak protona daje energiju za obrtanje osnove koje se prenosi na peteljku i menja konformaciju proteina u čvoru.

80 ATP sintaza

81 Translokacija protona se dešava na tri mesta u respiratornom lancu, u kompleksima I, III i IV Smatra se da prolazak jednog para elektrona kroz cjelokupni respiratorni lanac ispumpava ukupno 10 protona. To je dovoljno za sintezu 2,5 ATP-a (deo energije se troši za izmjenu ADP:ATP između citosola i mitohondrije).

82 Kako se energija protonskog gradijenta pretvara u hemijsku vezu u ATP-u? Katalitičko mesto ATP sintaze se nalazi na spoju a i b subjedinica. b subjedinica može da bude u tri konformaciona oblika, otvorenom, opuštenom i stegnutom, što je određeno asimetričnim interakcijama sa centralno postavljenom g subjedinicom. ADP i Pi su vezani za opušteni oblik, dok se ATP nalazi vezan za stegnuti oblik b subjedinice. Pri prolasku protona kroz kanal FO komponente, ceo a3b3 skup se okrene, te stegnuto mesto postane otvoreno i ATP se otpušta; za mesto koje postaje opušteno vezuju se ADP i Pi; na mestu koje prelazi u stegnuto, ADP i Pi daju ATP. Energija je potrebna za otpuštanje ATP-a, za konformacionu promenu koja pretvara stegnutu u otvorenu formu; naime, stvaranje ATP-a od ADP-a i Pi koji su već vezani za opuštenu formu zahteva malu promenu slobodne energije, sve dok je ATP vezan za katalitičko mesto otpuštanje ATP-a zahteva utrošak energije. Makroskopski - energija je potrebna za pretvaranje ADP-a i Pi iz rastvora u ATP u rastvoru.

83 Oksidativna fosforilacija Sinteza ATP-a Pri prolasku para elektrona kroz respiratorni lanac dolazi do prebacivanja 10 H+ iz matriksa u međumembranski prostor. Za sintezu jednog molekula ATP-a potrebno je da kroz protonski kanal prođe 4 H+. NADH 10 protona 2.5 ATP FADH2 6 protona 1.5 ATP
84 Energetski bilans lanca prenosilaca elektrona Od svakog NADH+H+ u lanac prenosilaca elektrona uđu 2 elektrona, što je ekvivalent redukciji ½O2. Ukoliko se 4 protona ispumpaju na nivou kompleksa I, 4 na nivou kompleksa III i 2 protona na nivou kompleksa IV, a procenjuje se da je za sintezu ATP-a potrebno da se 4 protona vrate kroz molekul ATP sintaze, oksidacijom svakog NADH+H+ se dobiju 3 ATP-a, a od FADH2-2 ATP. To znači da se samo 30% energije dostupne oksidacijom redukovanih koenzima iskoristi za sintezu ATP-a. Deo energije se iskoristi za transport jona, a ostatak se oslobađa u vidu toplote.
85 Uloga transporta elektrona u proizvodnji toplote Ćelije mrkog masnog tkiva koriste transport elektrona za proizvodnju toplote; životinje koje hiberniraju kao i mladunčad (uključujući i čoveka) imaju relativno veću proporciju ovog specijalizovanog oblika masnog tkiva. Veća proizvodnja toplote je važna u održanju telesne temperature u novorođenčadi jer je njihova telesna površina neproporcionalno velika u odnosu na telesnu masu; u odraslih je količina mrkog masnog tkiva veoma mala. Mrko masno tkivo (boja potiče od velikog broja mitohondrija i posledično velike količine citohroma) je efikasan proizvođač toplote 400 W kg-1 u odnosu na 1 W kg-1 u tipičnom tkivu.

86 Ćelije mrkog masnog tkiva dobijenih iz mišićnih ćelija
87 U unutrašnjoj membrani mitohondrija u mrkom masnom tkivu se nalazi protein termogenin koji ima funkciju kanala za anjone: omogućava OH- ili Cl- jonima da brzo prolaze kroz membranu. Prenos OH- jona iz citosola u mitohondrijalni matriks ima isti efekat kao prenos H+ u suprotnom pravcu poništavanje protonskog gradijenta što znači da termogenin deluje kao supstanca koja razdvaja prenos elektrona od sinteze ATP-a, sa posledičnim oslobađanjem velike količine toplote.
88 Brus Spigelman, ćelijski biolog sa Univerziteta Harvard, načinio jako važan korak ka razumevanju razvića masnog tkiva godine Spigelman i saradnici su pokazali da se partner protein nazvan C/EBP-beta vezuje za PRDM16 protein, i da se upravo povezivanjem ova dva faktora mišićne ćelije i ćelije vezivnog tkiva pretvaraju u mrko masno tkivo.
89 Energija dobijena oksidacijom hranljivih materija se prevodi u visoko-energetske veze adenozin trifosfata (ATP) u procesu oksidativne fosforilacije. Najveći deo energije dobijen u procesu oksidacije hranljivih materija u ciklusu trikarboksilnih kiselina i drugim kataboličkim putevima se čuva u obliku redukovanih koenzima, NADH i FAD(2H). U lancu prenosilaca elektrona NADH i FAD(2H) se oksiduju, pri čemu predaju elektrone O2, koji se redukuje u H2O. Energija dobijena u redukciji O2 se koristi za fosforilaciju adenozin difosfata (ADP) u ATP dejstvom ATP sintaze (Fo F1ATPaza). Neto prinos oksidativne fosforilacije je približno 2.5 mola ATP-a po molu oksidaovanog NADH, ili 1.5 mol ATP-a po molu oksidaovanog FAD(2H).
90
91
92
93

94
95
96

97

98

99

100

101

102

103

104

105

106

107

108

109

110

111

112

113

114

115

116

117

118

119

120

121
122

123
Oksidativna fosforilacija
 Oksidativna fosforilacija Oksidativna fosforilacija U eukariota u mitohondrijama. Odigrava se redukcija O2 do H2O. Donori elektrona su NADH i FADH2. Najvedi deo energije za svoj rast, održavanje i rad
Oksidativna fosforilacija Oksidativna fosforilacija U eukariota u mitohondrijama. Odigrava se redukcija O2 do H2O. Donori elektrona su NADH i FADH2. Najvedi deo energije za svoj rast, održavanje i rad
Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama. Hloroplast
 Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama Hloroplast Procesom ćelijskog disanja deponovana energija u šećerima erima prevodi se u ATP i druge energetske metabolite. Istovremeno se
Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama Hloroplast Procesom ćelijskog disanja deponovana energija u šećerima erima prevodi se u ATP i druge energetske metabolite. Istovremeno se
CIKLUS LIMUNSKE KISELINE (CLK)
") SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
Disanje (Respiracija)
") Disanje (Respiracija) Osim fotosinteze, deo primarnog metabolizma biljaka je i proces ćelijskog disanja (respiracija). Dok se u fotosintezi procesima redukcije iz CO2 i vode sintetišu organska jedinjenja,
Disanje (Respiracija) Osim fotosinteze, deo primarnog metabolizma biljaka je i proces ćelijskog disanja (respiracija). Dok se u fotosintezi procesima redukcije iz CO2 i vode sintetišu organska jedinjenja,
Glukoneogeneza i regulacija glukoneogeneze
 Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
Aminokiseline. Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina 22.12.2014
 Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Put pentoza fosfata. B. Mildner. Put pentoza fosfata
 Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija
 Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Ciklus limunske kiseline-2
 Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
ISPITNA PITANJA OSNOVI BIOHEMIJE
 UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
VODA ELEKTROLITI I ACIDO-BAZNA RAVNOTEŽA...
 SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
OKSIDACIJSKA FOSFORILACIJA
 OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici
 Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
numeričkih deskriptivnih mera.
 DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
TEST PITANJA BIOHEMIJA UGLJENI HIDRATI
 TEST PITANJA BIOHEMIJA UGLJENI HIDRATI 1. Kakve osobine pokazuju monosaharidi: a) osidacione b) redukcione c) metilirajuće 2. Skrob, kao dominantan šećer u ishrani čoveka, se razlaže do disaharida u: a)
TEST PITANJA BIOHEMIJA UGLJENI HIDRATI 1. Kakve osobine pokazuju monosaharidi: a) osidacione b) redukcione c) metilirajuće 2. Skrob, kao dominantan šećer u ishrani čoveka, se razlaže do disaharida u: a)
HEMIJSKA VEZA TEORIJA VALENTNE VEZE
 TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
CIKLUS AZOTA I SUMPORA U PRIRODI
 CIKLUS AZOTA I SUMPORA U PRIRODI Asimilacija azota i sumpora zahtevaju niz biohemijskih reakcija koje spadaju u reakcije sa velikim energetskim zahtevima. Za asimilaciju azota iz nitrata (NO 3- ) potrebna
CIKLUS AZOTA I SUMPORA U PRIRODI Asimilacija azota i sumpora zahtevaju niz biohemijskih reakcija koje spadaju u reakcije sa velikim energetskim zahtevima. Za asimilaciju azota iz nitrata (NO 3- ) potrebna
Organele života i smrti
 MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
PITANJA ZA USMENI ISPIT IZ BIOHEMIJE
 PITANJA ZA USMENI ISPIT IZ BIOHEMIJE PROTEINI STRUKTURA I FUNKCIJE 1. Struktura proteina nivoi organizacije molekula 2. Proteini koji transportuju kiseonik hemoglobin i mioglobin ENZIMI 1. Opšte osobine
PITANJA ZA USMENI ISPIT IZ BIOHEMIJE PROTEINI STRUKTURA I FUNKCIJE 1. Struktura proteina nivoi organizacije molekula 2. Proteini koji transportuju kiseonik hemoglobin i mioglobin ENZIMI 1. Opšte osobine
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE
 Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
Kiselo bazni indikatori
 Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
ISPITNA PITANJA ZA USMENI DIO ISPITA
 UNIVERZITET CRNE GORE MEDICINSKI FAKULTET MEDICINSKA BIOHEMIJA ISPITNA PITANJA ZA USMENI DIO ISPITA STUDIJSKI PROGRAM MEDICINA I ENZIMOLOGIJA 1. Opšte osobine enzima i struktura molekula enzima 2. Izoenzimi.
UNIVERZITET CRNE GORE MEDICINSKI FAKULTET MEDICINSKA BIOHEMIJA ISPITNA PITANJA ZA USMENI DIO ISPITA STUDIJSKI PROGRAM MEDICINA I ENZIMOLOGIJA 1. Opšte osobine enzima i struktura molekula enzima 2. Izoenzimi.
[ C][ D] [ A][ B] Integracija metabolizma. Metabolički putevi koji omogućuju život izuzetno su složeni i međusobno isprepleteni
![[ C][ D] [ A][ B] Integracija metabolizma. Metabolički putevi koji omogućuju život izuzetno su složeni i međusobno isprepleteni](/thumbs/66/55930464.jpg "[ C][ D] [ A][ B] Integracija metabolizma. Metabolički putevi koji omogućuju život izuzetno su složeni i međusobno isprepleteni") Metabolički putevi koji omogućuju život izuzetno su složeni i međusobno isprepleteni Integracija metabolizma Mitohondriji u štapićima Svi metabolički putevi moraju udovoljiti dvama uvjetima 1. individualne
Metabolički putevi koji omogućuju život izuzetno su složeni i međusobno isprepleteni Integracija metabolizma Mitohondriji u štapićima Svi metabolički putevi moraju udovoljiti dvama uvjetima 1. individualne
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović
 DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
Kloroplasti. Fotosinteza Mitohondriji Stanično disanje
 Kloroplasti Fotosinteza Mitohondriji Stanično disanje http://en.wikipedia.org/wiki/plas tid PLASTIDI Organeli biljnih stanica i stanica algi Proizvodnja i pohranjivanje šećera i drugih molekula Pigmenti
Kloroplasti Fotosinteza Mitohondriji Stanično disanje http://en.wikipedia.org/wiki/plas tid PLASTIDI Organeli biljnih stanica i stanica algi Proizvodnja i pohranjivanje šećera i drugih molekula Pigmenti
3.1 Granična vrednost funkcije u tački
 3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3. razred gimnazije- opšti i prirodno-matematički smer ALKENI. Aciklični nezasićeni ugljovodonici koji imaju jednu dvostruku vezu.
 ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
PRAVA. Prava je u prostoru određena jednom svojom tačkom i vektorom paralelnim sa tom pravom ( vektor paralelnosti).
.") PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
BIOREGULATORI 13.XII.2016.
 BIREGULATRI 13.XII.2016. Bioregulatori Mnogi biohemijski procesi u organizmu zavise od strukture i funkcije specifičnih molekula koji se nalaze u ćeliji. eki od njih su nazvani bioregulatorima. To su najčešće
BIREGULATRI 13.XII.2016. Bioregulatori Mnogi biohemijski procesi u organizmu zavise od strukture i funkcije specifičnih molekula koji se nalaze u ćeliji. eki od njih su nazvani bioregulatorima. To su najčešće
Ispitivanje toka i skiciranje grafika funkcija
 Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABLIZEM GLJIKVIH HIDRATV KAK CELICA DBI GLUKZ IN STALE MNSAHARIDE? HRANA ZNTRAJCELIČNI GLIKGEN ali ŠKRB razgradnja s prebavnimi encimi GLUKZA in ostali monosaharidi fosforilitična cepitev prenos do
METABLIZEM GLJIKVIH HIDRATV KAK CELICA DBI GLUKZ IN STALE MNSAHARIDE? HRANA ZNTRAJCELIČNI GLIKGEN ali ŠKRB razgradnja s prebavnimi encimi GLUKZA in ostali monosaharidi fosforilitična cepitev prenos do
KATABOLIZAM UGLJENIH HIDRATA
 KATABOLIZAM UGLJENIH HIDRATA 20.02.2018. SINTEZA I RAZGRADNJA GLIKOGENA Glikogen je homopolimer glukoze, oblik u kojem se ugljeni hidrati čuvaju u životinja. Čuvanjem glukoze u obliku glikogena
KATABOLIZAM UGLJENIH HIDRATA 20.02.2018. SINTEZA I RAZGRADNJA GLIKOGENA Glikogen je homopolimer glukoze, oblik u kojem se ugljeni hidrati čuvaju u životinja. Čuvanjem glukoze u obliku glikogena
7 Algebarske jednadžbe
 7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
7 Algebarske jednadžbe 7.1 Nultočke polinoma Skup svih polinoma nad skupom kompleksnih brojeva označavamo sa C[x]. Definicija. Nultočka polinoma f C[x] je svaki kompleksni broj α takav da je f(α) = 0.
Seminar 13.b. Glikogen GLIKOGEN. B. Mildner
 Seminar 13.b Glikogen B. Mildner GLIKOGEN 1 Glikogen Nereducirani kraj Glikogen je jako dostupni skladišni oblik glukoze; kao i jako velik, razgranat polimer; Glukozne jedinice su povezane α-1,4-glikozidnim
Seminar 13.b Glikogen B. Mildner GLIKOGEN 1 Glikogen Nereducirani kraj Glikogen je jako dostupni skladišni oblik glukoze; kao i jako velik, razgranat polimer; Glukozne jedinice su povezane α-1,4-glikozidnim
OSNOVE METABOLIZMA MIKROORGANIZAMA. Metabolički diverzitet
 OSNOVE METABOLIZMA MIKROORGANIZAMA Metabolički diverzitet Metabolizam - obuhvata sve hemijske procese koji se dešavaju u ćeliji - anabolički (zahtevaju energiju) - katabolički (otpuštaju energiju) Energija
OSNOVE METABOLIZMA MIKROORGANIZAMA Metabolički diverzitet Metabolizam - obuhvata sve hemijske procese koji se dešavaju u ćeliji - anabolički (zahtevaju energiju) - katabolički (otpuštaju energiju) Energija
UVOD CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN
 = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN") CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Elementi spektralne teorije matrica
 Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
XI dvoqas veжbi dr Vladimir Balti. 4. Stabla
 XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
Iskazna logika 3. Matematička logika u računarstvu. novembar 2012
 Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Bioregulatori bioregulatorima. Koenzimi Vitamini
 BIREGULATRI Bioregulatori Mnogi biohemijski procesi u organizmu zavise od strukture i funkcije specifičnih molekula koji se nalaze u ćeliji. eki od njih su nazvani bioregulatorima. To su najčešće organski
BIREGULATRI Bioregulatori Mnogi biohemijski procesi u organizmu zavise od strukture i funkcije specifičnih molekula koji se nalaze u ćeliji. eki od njih su nazvani bioregulatorima. To su najčešće organski
Ugljikohidrati i glikoliza
 Ugljikohidrati i glikoliza Seminar 11b 1 1. Suspenzija stanica kvasca uzgajana je u anaerobnim uvjetima te se glukoza fermentirala u etanol i O 2. Ako se želi promatrati količina 14 O 2, na kojem mjestu
Ugljikohidrati i glikoliza Seminar 11b 1 1. Suspenzija stanica kvasca uzgajana je u anaerobnim uvjetima te se glukoza fermentirala u etanol i O 2. Ako se želi promatrati količina 14 O 2, na kojem mjestu
Operacije s matricama
 Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Elektrotehnički fakultet univerziteta u Beogradu 17.maj Odsek za Softversko inžinjerstvo
 Elektrotehnički fakultet univerziteta u Beogradu 7.maj 009. Odsek za Softversko inžinjerstvo Performanse računarskih sistema Drugi kolokvijum Predmetni nastavnik: dr Jelica Protić (35) a) (0) Posmatra
Elektrotehnički fakultet univerziteta u Beogradu 7.maj 009. Odsek za Softversko inžinjerstvo Performanse računarskih sistema Drugi kolokvijum Predmetni nastavnik: dr Jelica Protić (35) a) (0) Posmatra
Teorijske osnove informatike 1
 Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
Glikoliza. Metabolične poti. Organizacija metaboličnih poti Ciklična pot (intermediati se reciklirajo) Kako poteka oksidacija goriv v našem organizmu?
 Kako poteka oksidacija goriv v našem organizmu?") Glikoliza Sežig v bombnem kalorimetru organizacija in osnove uravnavanja metaboličnih poti aerobna glikoliza anaerobna glikoliza vstop drugih sladkorjev v glikolizo glicerolfosfatni in malat-aspartatni
Glikoliza Sežig v bombnem kalorimetru organizacija in osnove uravnavanja metaboličnih poti aerobna glikoliza anaerobna glikoliza vstop drugih sladkorjev v glikolizo glicerolfosfatni in malat-aspartatni
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
STVARANJE VEZE C-C POMO]U ORGANOBORANA
![STVARANJE VEZE C-C POMO]U ORGANOBORANA](/thumbs/55/37829094.jpg "STVARANJE VEZE C-C POMO]U ORGANOBORANA") STVAAJE VEZE C-C PM]U GAAA 2 6 rojne i raznovrsne reakcije * idroborovanje alkena i reakcije alkil-borana 3, Et 2 (ili TF ili diglim) Ar δ δ 2 2 3 * cis-adicija "suprotno" Markovnikov-ljevom pravilu *
STVAAJE VEZE C-C PM]U GAAA 2 6 rojne i raznovrsne reakcije * idroborovanje alkena i reakcije alkil-borana 3, Et 2 (ili TF ili diglim) Ar δ δ 2 2 3 * cis-adicija "suprotno" Markovnikov-ljevom pravilu *
Novi Sad god Broj 1 / 06 Veljko Milković Bulevar cara Lazara 56 Novi Sad. Izveštaj o merenju
 Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
Broj 1 / 06 Dana 2.06.2014. godine izmereno je vreme zaustavljanja elektromotora koji je radio u praznom hodu. Iz gradske mreže 230 V, 50 Hz napajan je monofazni asinhroni motor sa dva brusna kamena. Kada
Pismeni ispit iz matematike Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: ( ) + 1.
 + 1.") Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
PARCIJALNI IZVODI I DIFERENCIJALI. Sama definicija parcijalnog izvoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je,
 PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner
 Boris Mildner") Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner Čestice glikogena u hepatocitima. Glikogen, čestice za pohranjivanje glukoze, u hepatocitima se nalazi u citoplazmi blizu tubula endoplazmatske
Glikogen (razgradnja, sinteza i njihova regulacija) Boris Mildner Čestice glikogena u hepatocitima. Glikogen, čestice za pohranjivanje glukoze, u hepatocitima se nalazi u citoplazmi blizu tubula endoplazmatske
Trigonometrija 2. Adicijske formule. Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto
 Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE)
") (Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
(Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu
 Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
18. listopada listopada / 13
 18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
OSNOVI ELEKTRONIKE. Vežbe (2 časa nedeljno): mr Goran Savić
: mr Goran Savić") OSNOVI ELEKTRONIKE Vežbe (2 časa nedeljno): mr Goran Savić savic@el.etf.rs http://tnt.etf.rs/~si1oe Termin za konsultacije: četvrtak u 12h, kabinet 102 Referentni smerovi i polariteti 1. Odrediti vrednosti
OSNOVI ELEKTRONIKE Vežbe (2 časa nedeljno): mr Goran Savić savic@el.etf.rs http://tnt.etf.rs/~si1oe Termin za konsultacije: četvrtak u 12h, kabinet 102 Referentni smerovi i polariteti 1. Odrediti vrednosti
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama.
 Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =
/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =") 100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
Pošto pretvaramo iz veće u manju mjernu jedinicu broj 2.5 množimo s 1000,
 PRERAČUNAVANJE MJERNIH JEDINICA PRIMJERI, OSNOVNE PRETVORBE, POTENCIJE I ZNANSTVENI ZAPIS, PREFIKSKI, ZADACI S RJEŠENJIMA Primjeri: 1. 2.5 m = mm Pretvaramo iz veće u manju mjernu jedinicu. 1 m ima dm,
PRERAČUNAVANJE MJERNIH JEDINICA PRIMJERI, OSNOVNE PRETVORBE, POTENCIJE I ZNANSTVENI ZAPIS, PREFIKSKI, ZADACI S RJEŠENJIMA Primjeri: 1. 2.5 m = mm Pretvaramo iz veće u manju mjernu jedinicu. 1 m ima dm,
Riješeni zadaci: Nizovi realnih brojeva
 Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Verovatnoća i Statistika I deo Teorija verovatnoće (zadaci) Beleške dr Bobana Marinkovića
 Beleške dr Bobana Marinkovića") Verovatnoća i Statistika I deo Teorija verovatnoće zadaci Beleške dr Bobana Marinkovića Iz skupa, 2,, 00} bira se na slučajan način 5 brojeva Odrediti skup elementarnih dogadjaja ako se brojevi biraju
Verovatnoća i Statistika I deo Teorija verovatnoće zadaci Beleške dr Bobana Marinkovića Iz skupa, 2,, 00} bira se na slučajan način 5 brojeva Odrediti skup elementarnih dogadjaja ako se brojevi biraju
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - UNI
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
OBRTNA TELA. Vladimir Marinkov OBRTNA TELA VALJAK
 OBRTNA TELA VALJAK P = 2B + M B = r 2 π M = 2rπH V = BH 1. Zapremina pravog valjka je 240π, a njegova visina 15. Izračunati površinu valjka. Rešenje: P = 152π 2. Površina valjka je 112π, a odnos poluprečnika
OBRTNA TELA VALJAK P = 2B + M B = r 2 π M = 2rπH V = BH 1. Zapremina pravog valjka je 240π, a njegova visina 15. Izračunati površinu valjka. Rešenje: P = 152π 2. Površina valjka je 112π, a odnos poluprečnika
RAD, SNAGA I ENERGIJA
 RAD, SNAGA I ENERGIJA SADRŢAJ 1. MEHANIĈKI RAD SILE 2. SNAGA 3. MEHANIĈKA ENERGIJA a) Kinetiĉka energija b) Potencijalna energija c) Ukupna energija d) Rad kao mera za promenu energije 4. ZAKON ODRŢANJA
RAD, SNAGA I ENERGIJA SADRŢAJ 1. MEHANIĈKI RAD SILE 2. SNAGA 3. MEHANIĈKA ENERGIJA a) Kinetiĉka energija b) Potencijalna energija c) Ukupna energija d) Rad kao mera za promenu energije 4. ZAKON ODRŢANJA
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
ELEKTROTEHNIČKI ODJEL
 MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. 4/17/2013
 Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
Osnovne veličine, jedinice i izračunavanja u hemiji
 Osnovne veličine, jedinice i izračunavanja u hemiji Pregled pojmova veličina i njihovih jedinica koje se koriste pri osnovnim izračunavanjima u hemiji dat je u Tabeli 1. Tabela 1. Veličine i njihove jedinice
Osnovne veličine, jedinice i izračunavanja u hemiji Pregled pojmova veličina i njihovih jedinica koje se koriste pri osnovnim izračunavanjima u hemiji dat je u Tabeli 1. Tabela 1. Veličine i njihove jedinice
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE GODINE 8. RAZRED TOČNI ODGOVORI
 21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
21. ŠKOLSKO/OPĆINSKO/GRADSKO NATJECANJE IZ GEOGRAFIJE 2014. GODINE 8. RAZRED TOČNI ODGOVORI Bodovanje za sve zadatke: - boduju se samo točni odgovori - dodatne upute navedene su za pojedine skupine zadataka
Zavrxni ispit iz Matematiqke analize 1
 Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
M086 LA 1 M106 GRP. Tema: Baza vektorskog prostora. Koordinatni sustav. Norma. CSB nejednakost
 M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
Cauchyjev teorem. Postoji više dokaza ovog teorema, a najjednostvniji je uz pomoć Greenove formule: dxdy. int C i Cauchy Riemannovih uvjeta.
 auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,
auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,
Prirodno-matematički fakultet Društvo matematičara I fizičara Crne Gore
 Prirodno-matematički fakultet Društvo matematičara I fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Rješenja zadataka iz HEMIJE za IX razred osnovne škole 1. Koju zapreminu, pri standardnim uslovima, zauzimaju
Prirodno-matematički fakultet Društvo matematičara I fizičara Crne Gore OLIMPIJADA ZNANJA 2018. Rješenja zadataka iz HEMIJE za IX razred osnovne škole 1. Koju zapreminu, pri standardnim uslovima, zauzimaju
41. Jednačine koje se svode na kvadratne
 . Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
. Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE
 **** MLADEN SRAGA **** 011. UNIVERZALNA ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE SKUP REALNIH BROJEVA α Autor: MLADEN SRAGA Grafički urednik: BESPLATNA - WEB-VARIJANTA Tisak: M.I.M.-SRAGA
**** MLADEN SRAGA **** 011. UNIVERZALNA ZBIRKA POTPUNO RIJEŠENIH ZADATAKA PRIRUČNIK ZA SAMOSTALNO UČENJE SKUP REALNIH BROJEVA α Autor: MLADEN SRAGA Grafički urednik: BESPLATNA - WEB-VARIJANTA Tisak: M.I.M.-SRAGA
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A
, grupa A") Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - VSŠ 2005/06
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
Svakodnevni poslovi našeg tela
 Poglavlje 2 Svakodnevni poslovi našeg tela U ovom poglavlju Razmatranje svakodnevnih automatskih funkcija tela Šta se događa u svakoj ćeliji Značaj homeostaze Izgradnja i održavanje delova tela O vo poglavlje
Poglavlje 2 Svakodnevni poslovi našeg tela U ovom poglavlju Razmatranje svakodnevnih automatskih funkcija tela Šta se događa u svakoj ćeliji Značaj homeostaze Izgradnja i održavanje delova tela O vo poglavlje
OM2 V3 Ime i prezime: Index br: I SAVIJANJE SILAMA TANKOZIDNIH ŠTAPOVA
 OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
- pravac n je zadan s točkom T(2,0) i koeficijentom smjera k=2. (30 bodova)
 i koeficijentom smjera k=2. (30 bodova)") MEHANIKA 1 1. KOLOKVIJ 04/2008. grupa I 1. Zadane su dvije sile F i. Sila F = 4i + 6j [ N]. Sila je zadana s veličinom = i leži na pravcu koji s koordinatnom osi x zatvara kut od 30 (sve komponente sile
MEHANIKA 1 1. KOLOKVIJ 04/2008. grupa I 1. Zadane su dvije sile F i. Sila F = 4i + 6j [ N]. Sila je zadana s veličinom = i leži na pravcu koji s koordinatnom osi x zatvara kut od 30 (sve komponente sile
Pismeni ispit iz matematike GRUPA A 1. Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj, zatim naći 4 z.
 Pismeni ispit iz matematike 06 007 Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj z = + i, zatim naći z Ispitati funkciju i nacrtati grafik : = ( ) y e + 6 Izračunati integral:
Pismeni ispit iz matematike 06 007 Napisati u trigonometrijskom i eksponencijalnom obliku kompleksni broj z = + i, zatim naći z Ispitati funkciju i nacrtati grafik : = ( ) y e + 6 Izračunati integral:
Metabolizem oz. presnova
 Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Metabolizem oz. presnova Metabolne poti Metabolizem (presnova) Vsota vseh kemijskih reakcij v organizmu, njihova koordinacija, regulacija in energetske potrebe Substrati se pretvarjajo v produkte preko
Vodik. dr.sc. M. Cetina, doc. Tekstilno-tehnološki fakultet, Zavod za primijenjenu kemiju
 Vodik Najzastupljeniji element u svemiru (maseni udio iznosi 90 %) i sastavni dio Zvijezda. Na Zemlji je po masenom udjelu deseti element po zastupljenosti. Zemljina gravitacija premalena je da zadrži
Vodik Najzastupljeniji element u svemiru (maseni udio iznosi 90 %) i sastavni dio Zvijezda. Na Zemlji je po masenom udjelu deseti element po zastupljenosti. Zemljina gravitacija premalena je da zadrži
Metabolizem in energija
 Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,
Metabolizem in energija . Metabolizem Vsota vseh kemijskih reakcij v organizmu, njihovo uravnavanje, in vse energetske spremembe ki potekajo v organizmih. Metabolizem vključuje različne metabolične poti,