Disanje (Respiracija)
|
|
|
- Λυδία Στεφανόπουλος
- 7 χρόνια πριν
- Προβολές:
Transcript
1 Disanje (Respiracija)
2
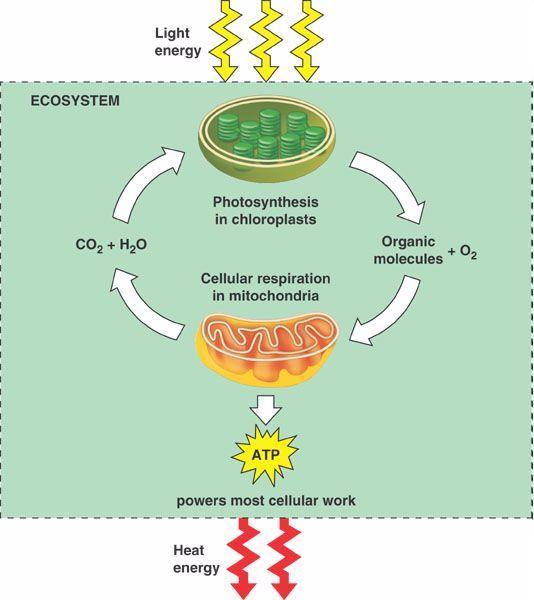
3 Osim fotosinteze, deo primarnog metabolizma biljaka je i proces ćelijskog disanja (respiracija). Dok se u fotosintezi procesima redukcije iz CO2 i vode sintetišu organska jedinjenja, u procesu disanja se energija akumulirana u hemijskim vezama tih organskih jedinjenja oslobađa njihovom oksidacijom do CO2 i vode, pri čemu je O2 krajnji akceptor elektrona i redukuje se u vodu. Oslobođena energija se skladišti privremeno u obliku ATP, koji može da je stavi na raspolaganje biljci u procesima rastenja i razvića, koji zahtevaju utrošak energije. Aerobno disanje je biološki proces u kojem se redukovana organska jedinjenja mobilišu (iz rezervnih organskih materija) i zatim kontrolisano oksiduju. Ovaj proces je sličan kod svih aerobnih eukariotskih organizama, međutim, kod biljaka postoje njegovi specifilni aspekti koji su različiti u odnosu na taj proces kod životinja.
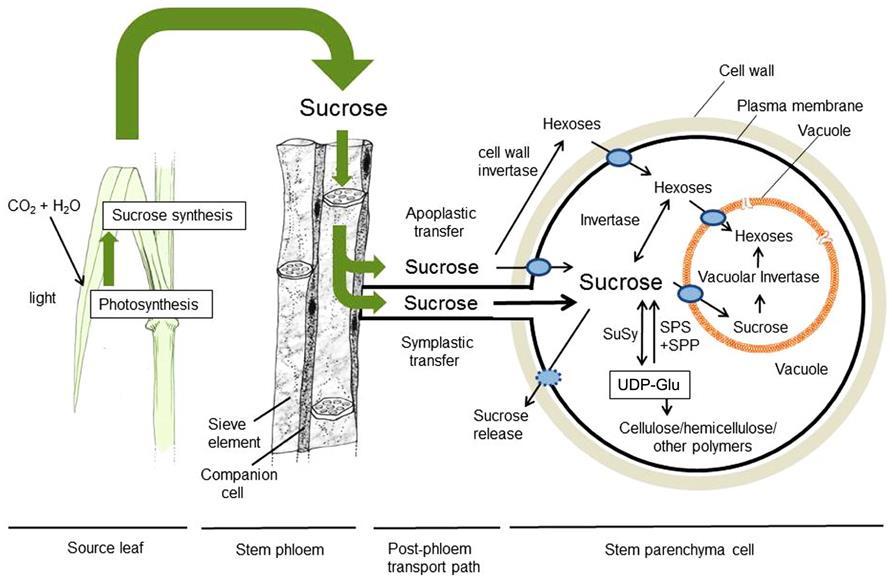
 i: saharoza (disaharid koji je sekundarni proizvod fotosinteze) heksoza-fosfati i triozafosfati (nastaju degradacijom skroba u hloroplastima ili u Kalvinovom ciklusu)")
4 Iako se glukoza najčešće navodi kao supstrat za disanje, u biljnim ćelijama redukovani ugljenik za proces disanja mogu da obezbede (tj. supstrati su za disanje) i: saharoza (disaharid koji je sekundarni proizvod fotosinteze) heksoza-fosfati i triozafosfati (nastaju degradacijom skroba u hloroplastima ili u Kalvinovom ciklusu) polimeri fruktoze (fruktani) i drugi šećeri lipidi (prvenstveno triacilgliceroli) organske kiseline i ponekad proteini.
5 Sa hemijske tačke gledišta, disanje kod biljaka može da se predstavi kao oksidacija jednog molekula saharoze (12 C- atoma) i redukcija 12 molekula O2: Potpunom oksidacijom jednog mola saharoze do CO2 i vode oslobađa se 5760 kj. Primarna uloga respiratornog metabolizma (ali ne i jedina) je kontrolisano oslobađanje slobodne energije koja se dobija oksidacijom supstrata za disanje i njegovo povezivanje sa sintezom ATP, univerzalnog prenosioca hemijske energije u živim sistemima.
6 Velika količina energije koja se oslobađa oksidacijom supstrata za disanje otpušta se postepeno, korak po korak, u većem broju uzastopnih hemijskih reakcija, kako bi se izbeglo oštećenje ćelijskih struktura. Te hemijske reakcije u respiratornom metabolizmu grupisane su u 4 procesa: Glikoliza (u citoplazmi i plastidima) Oksidativni pentozni ciklus (u citoplazmi i plastidima) Krebsov ciklus (u mitohondrijama) Oksidativna fosforilacija (u mitohondrijama) Supstrati za disanje ulaze u respiratorni proces u različitim tačkama ova 4 procesa, a pre toga moraju biti mobilisani iz rezervnih oblika organskih materija (skrob, ulja, rezervni proteini...) koje biljne ćelije deponuju. Skrob se, na primer, razlaže u semenima koja ga deponuju u amiloplastima rezervnih tkiva (endosperm) kao rezervnu materiju (semena žita), pod uticajem α-amilaze (do maltoze) i maltaze (do glukoze).
7 Energija koja se oslobodi u pojedinim koracima oksidacije tokom ova 4 procesa ne deponuje se samo u fosfatnim vezama ATP-a, već i u redukovanim piridin-nukleotidima (NAD i NADP), koji su kofaktori mnogih dehidrogenaza, i u flavin-nukleotidima (FMN i FAD). Oni su podložni reverzibilnim redukcijama i oksidacijama. U redukovanom obliku piridin-nukleotidi nose elektrone i energiju redukcije, a prilikom svoje oksidacije otpuštaju elektrone i energiju prenose na jedinjenje koje su redukovali (prenos redukujućeg ekvivalenta). Većina membrana koje okružuju organele nisu propustljive za piridin-nukleotide, tj. u svakom kompartmentu ćelije se nalazi neka količina ovih nukleotida i oni moraju biti uravnoteženi u pogledu odnosa oksidovanih i redukovanih formi da bi mogli da učestvuju u odgovarajućim metaboličkim procesima. NAD + se redukuje u glikolizi i Krebsovom ciklusu, a NADP + u fotohemijskoj fazi fotosinteze i oksidativnom pentoznom ciklusu. Najviše redukovanih kofaktora, kao i najviše ATP, nastaje u Krebsovom ciklusu. U poslednjem procesu respiratornog metabolizma, oksidativnoj fosforilaciji, redukovani kofaktori se opet oksiduju, a oslobođena energija se koristi za sintezu ATP-a.
8
9 U disanju se sintetiše 60 ATP po svakom molu oksidovane saharoze: Međutim, ne završe svi C-atomi koji uđu u respiraciju u CO2, odnosno ne iskoriste se svi za sintezu ATP. Mnogi intermedijeri respiratornog puta su početne tačke različitih biosintetskih puteva asimilacija azota, sinteza nukleotida i lipida itd.

10 Glikoliza * * Konvertuju se ugljeni hidrati u piruvat i proizvodi se manja količina ATP i NADH. Odigrava se u citoplazmi i plastidima. U početnoj fazi glikolize ugljeni hidrati se konvertuju u heksozafosfate, a ovi u trioza-fosfate. Slede reakcije u kojima se triozafosfati oksiduju i rearanžiraju dajući dva piruvata, pri čemu se konzervira energija. Glikoliza kod biljaka se odlikuje posebnom regulacijom, paralelnim glikolitičkim putem u plastidima, alternativnim enzimskim reakcijama u citoplazmi i drugačijim supstratom (saharoza umesto glukoze) i krajnjim proizvodom (osim piruvata i malat).
11
12 Alternativne glikolitičke reakcije kod biljaka PPi-zavisna fosfofrukto kinaza katalizuje istu reakciju kao i ATP-zavisna fosfofruktokinaza (ključni enzim glikolize i kod biljaka i kod životinja), ali radi i reverzibilno (za razliku od ovog drugog enzima). Nalazi se u citoplazmi svih biljnih ćelija i to u većim količinama od ATP-zavisne fosfofruktokinaze. Regulacija ovog enzima je preko fluktuacija ćelijskog metabolizma. Kao produkti glikolize kod biljaka nastaju i piruvat i malat, jer biljke imaju enzimski sistem (PEP karboksilaza i malat dehidrogenaza) koji prevodi PEP u jabučnu kiselinu. Na taj načun kod biljaka dve organske kiseline, produkti glikolize, mogu da uđu u Krebsov ciklus. Iako proces glukoneogeneze (sinteza šećera iz lipida) nije karakteristična za biljke, niti prisutan u svim biljnim ćelijama, on se odigrava u semenima onih biljnih vrsta kod kojih su rezervne materije lipidi (ulja u oleazomima). U glukoneogenezi se triacilgliceroli prevode u šećere (saharozu) koristeći neke reakcije glikolitičkog puta, ali u suprotnom smeru.
.")


13 Fermentacija kod biljaka Odigrava se i kod biljaka i kod drugih organizama onda kada se nađu u hipoksičnim ili anoksičnim uslovima (kod biljaka je to kad zemljište na kojem rastu bude poplavljeno). U takvim uslovima se ne odvija Krebsov ciklus niti oksidativna fosforilacija, pa dolazi do zastoja i u glikolizi zato što je sav NAD u redukovanom obliku i ne može da se oksiduje, a u glikolizi se NAD + redukuje. Piruvat onda nastavlja da se metaboliše u alkoholnom i mlečno-kiselinskom vrenju, do etanola, odnosno laktata. U nedostatku O2 fermentacije regenerišu NAD + koji je potreban za glikolizu. Alkoholno vrenje je uobičajeno kod biljaka i dešava se u slabo aerisanim tkivima (koren, na primer), dok je mlečno-kiselinsko vrenje ređe kod biljaka. I etanol i laktat su toksični za ćelije, ali je etanol manje toksičan jer može da difunduje iz ćelije, a laktat se akumulira u ćelijama i može da utiče na kiselost citoplazme.
14 Regulacija glikolize kod biljaka Biljke se odlikuju metaboličkom fleksibilnošću za razliku od životinja, sposobne su da jedan korak u metaboličkom putu obave na više različitih načina. Glikoliza se kod biljaka odvija i u citoplazmi i u stromi plastida, pri čemu iste reakcije katalizuju različiti izoenzimi koje kodiraju geni nuklearnog genoma. Uloga glikolize u plastidima ćelija koje ne obavljaju fotosintezu i hloroplastima u odsustvu svetlosti je da doprinese razgradnji skroba i sintezi metabolita u procesu disanja, ATP-a i redukovanih koenzima. Glikolitički put koji se odvija u citoplazmi biljnih ćelija sadrži mnoge zaobilaznice ( bypass reactions ) na mestima gde se kao kosupstrati pojavljuju ATP i Pi. Te zaobilaznice angažuju pirofosfate (PPi). Na primer, PPi-zavisna fosfofruktokinaza. Kod biljaka, za razliku od životinja, ne postoje solubilne pirofosfataze (imaju pirofosfataze koje su u membrani tonoplasta i funkcionišu kao protonske pumpe), zbog čega se u citoplazmi održavaju stabilni nivoi pirofosfata. Nepovoljni uslovi (anoksija i deficijencija fosfata u zemljištu) veoma redukuju nivo adeninskih nukleotida i neorganskih fosfata u citoplazmi, što dovodi do ometanja metaboličkih procesa u kojima oni učestvuju. To nije slučaj sa pirofosfatima. Metabolička fleksibilnost omogućava biljkama da preferencijalno koriste pirofosfate kao donore energije onda kada su zalihe ATP-a i fosfata istrošene, ili kada štede ATP za druge procese.
15 Regulacija glikolize kod biljaka se ostvaruje na dva glavna mesta fosforilacija fruktozo-6-fosfata i metabolizam PEP, odnosno na nivou tri enzima: ATP-zavisne fosfofruktokinaze, PEP karboksilaze i piruvat kinaze. Dok je kod životinja primarna regulacija na nivou fosfofruktokinaze, kod biljaka je to sekundarni nivo kontrole, a primarna je regulacija na nivou metabolizma PEP. PEP inhibira ATP-zavisnu fosfofruktokinazu, dok neorganski fosfati smanjuju tu inhibiciju. To znači da aktivnost pomenutog enzima zavisi od odnosa koncentracija PEP i Pi u citoplazmi. Piruvat kinazu i PEP karboksilazu inhibiraju intermedijeri Krebsovog ciklusa, tj. organske kiseline. Ovo je bottom up regulacija. Prednost ovakve regulacije je što omogućava podešavanje sinteze piruvata nezavisno od drugih procesa kao što su Kalvinov ciklus i interkonverzija saharoza-trioza-fosfati-skrob. Postojanje dva enzima koji metabolišu PEP u dve različite organske kiseline, intermedijera Krebsovog ciklusa PEP karboksilaza može pod određenim uslovima da zaobiđe piruvat kinazu (da onemogući nastanak piruvata). Regulacija na nivou konverzije fruktozo-6-fosfata u fruktozo-1,6- bisfosfat je složena i u njoj figurira fruktozo-2,6-bisfosfat, čiji nivoi u citoplazmi variraju. F-2,6-BP inhibira ATP-zavisnu fosfofruktokinazu, a stimuliše aktivnost PPi-zavisne fosfofruktokinaze. Na taj način on u citoplazmi usmerava fluks F-6-fosfata ka glikolizi ili ka sintezi saharoze.
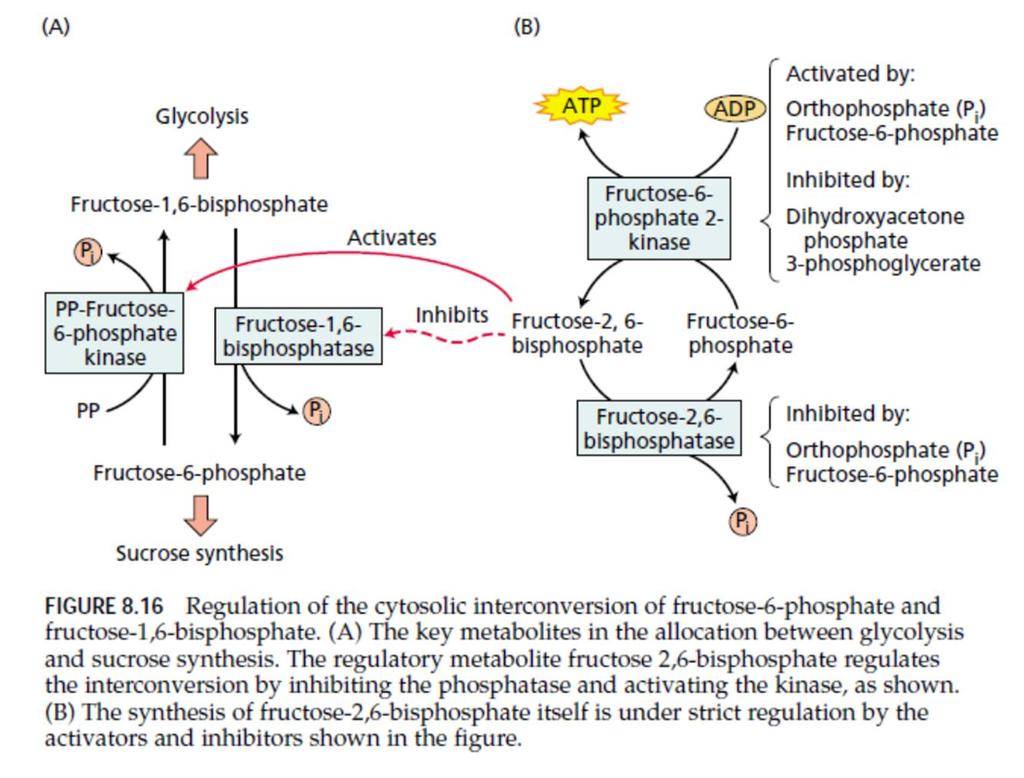
16
.")
17 Oksidativni pentozni ciklus Odigrava se u plastidima i u citoplazmi. Dominira ciklus koji se dešava u plastidima. Obradi samo 5-20% ugljenika poreklom iz heksoza (razgradnju heksoza pretežno obavlja glikoliza u većini biljnih tkiva). U početnim, oksidativnim reakcijama ovog ciklusa nastaje NADPH.
18 Uloge oksidativnog pentoznog ciklusa u metabolizmu biljaka: Produkuje redukovani NADP - NADPH učestvuje u reakcijama redukcije povezanim sa biosintezama u citoplazmi i plastidima. Doprinosi preko redukcionih ekvivalenata sintezi ATP u oksidativnoj fosforilaciji, Pošto NADPH može da oksiduje enzim NADPH dehidrogenaza (na spoljašnjoj strani unutrašnje membrane ovojnice biljnih mitohondrija). Intermedijeri (ribozo-5-fosfat i eritrozo-4-fosfat) su prekursori za sintezu riboze i dezoksiriboze (sinteza RNK i DNK), odnosno fenolnih jedinjenja (put šikimata u kojemnastaju lignin, flavonoidi i fitoaleksini). Tokom ranih faza ozelenjavanja, pre nego što listovi postanu potpuno autotrofni, oksidativni pentozni ciklus produkuje intermedijere Kalvinovog ciklusa. Regulacija oksidativnog pentoznog ciklusa obavlja se na nivou glukozo-6- fosfat dehidrogenaze, a u njoj učestvuju svetlost (samo ciklus koji se dešava u hloroplastima) i odnos NADP + /NADPH. Svetlost inhibira aktivnost pomenutog enzima (preko sistema feredoksin-tioredoksin), trioze i F-6-P se sintetišu u Kalvinovom ciklusu, a najveći deo aktivnosti oksidativnog pentoznog ciklusa odnosi se na neoksidativne reakcije (konverzije šećera) i usmeren je na produkciju pentoza.
19 U glikolizi se razgradnjom jednog molekula saharoze oslobađa manje od 25 % energije molekula saharoze, a sva ostala energija se deponuje u dva molekula piruvata. U aerobnim uslovima piruvat koji nastane u glikolizi ne metaboliše se u fermentacijama, već u Krebsovom ciklusu (ciklus limunske kiseline ili ciklus trikarboksilnih kiselina - TCA ciklus), za kojim sledi oksidativna fosforilacija (transport elektrona povezan sa sintezom ATP). U ovim procesima se dešava oksidacija ugljenikovih atoma do CO2 i redukcija O2 do vode (davaoci elektrona su redukovani koenzimi), kao i sinteza ATP-a. Oksidativni metabolizam preovladava u biljnim tkivima. Krebsov ciklus i oksidativna fosforilacija odvijaju se u matriksu mitohondrija. Mitohondrije u biljnim ćelijama imaju istu veličinu, oblik i građu kao i u animalnim ćelijama, ali je njihov broj, sa retkim izuzecima, bitno manji nego u animalnim ćelijama. Ćelije različitih biljnih tkiva sadrže različit broj mitohondrija, ali je taj broj direktno povezan sa metaboličkom aktivnošću tkiva. Na primer, ćelije zatvaračice stoma, prateće ćelije sitastih cevi i meristemske ćelije se odlikuju veoma velikim brojem mitohondrija.

 ka")

20 Mitohondrije Oblik loptast ili štapićast Veličina prečnik 0,5-1 µm, dužina do 3 µm. Okružene su ovojnicom koja se sastoji od glatke spoljašnje membrane i unutrašnje membrane koja formira veliki broj invaginacija (kriste) ka unutrašnjosti mitohondrije. Unutrašnjost mitohondrije ispunjena je matriksom. Lipidni sastav membrana uglavnom fosfolipidi (80 % fosfatidil holin i fosfatidil etanolamin). Unutrašnja membrana sadrži 50% ukupnih proteina mitohondrije.
21 Kompartmenti: Spoljašnja membrana Intermembranski prostor Unutrašnja membrana Matriks Membrane ovojnice se razlikuju po propustljivosti: Spoljašnja membrana je permeabilna za sve čestice molekulske mase manje od 10 kd propušta jone i većinu metabolita, ali NE i proteine. Unutrašnja membrana je veoma selektivna ne propušta pasivno, u matriks, jone i naelektrisane organske molekule. Mitohondrije su osmotski aktivne povećavaju volumen u hipotoničnom rastvoru. Poluautonomne su organele (kao i plastidi) sadrže ribozome (70S), RNK i DNK (cirkularna). Geni mitohondrijalnog genoma kodiraju ograničeni broj mitohondrijalnih proteina, što znači da mitohondrije ne mogu da funkcionišu bez pomoći nuklearnog genoma.
22 Mitohondrijalni genom biljaka je mnogo veći od onog kod drugih organizama. Veličina varira i između blisko srodnih vrsta. Kodira mnogo više proteina nego kod životinja, tj. oko 35 proteina, uglavnom vezanih za procese oksidativne fosforilacije (subjedinice kompleksa I, III i IV, ATP-sintaze i enzimi u biosintezi citohroma). Neki geni u mitohondrijalnoj DNK biljaka imaju introne, a važi univerzalni genetički kod bez odstupanja (kao u mitohondrijalnoj DNK životinja). Glavna kontrola ekspresije mitohondrijalnih gena kod biljaka odvija se na posttranslacionom nivou. Mitohondrije uvećavaju svoj broj deobom postojećih mitohondrija, NE biogenezom de novo.

23 Krebsov ciklus (ciklus limunske kiseline, ciklus trikarboksilnih kiselina TCA ciklus) Odvija se u matriksu mitohondrija. U Krebsov ciklus ulaze 2 piruvata, koji nastaju glikolizom iz jedne heksoze, i oni se oksiduju do CO2. Nastaju 3 CO2, 4 NADH, 1 FADH2 i 1 ATP po jednom molekulu piruvata. Piruvat dehidrogenaza je enzimski kompleks katalizuje prva 3 koraka u Krebsovom ciklusu: dekarboksilaciju, oksidaciju i konjugaciju C2 fragmenta za CoA.
.")
24 Specifičnosti Krebsovog ciklusa biljaka Reakcija koju katalizuje sukcinil-coa sintetaza kod biljaka se sintetiše ATP, a kod životinja GTP. Značaj malata i alternativni putevi oksidacije malata akumulacija malata u vakuolama mnogih biljnih ćelija (ne samo kod CAM-biljaka). Velika aktivnost enzima NAD + -malat dekarboksilaze (jabučni enzim), koja je detektovana u matriksu mitohondrija svih biljnih ćelija. Enzim katalizuje oksidativnu dekarboksilaciju malata:
25 Prisustvo ovog enzima u matriksu omogućava mitohondrijama da obavljaju različite alternativne puteve metabolizma PEP, koji nastaje u glikolizi. PEP karboksilaza u mitohondrijama prevodi PEP u malat i može da zameni nedostajuće intermedijere Krebsovog ciklusa. Na primer, α-ketoglutarat se troši za asimilaciju azota (u sintezi glutamata), što dovodi do manjka malata i usporavanja Krebsovog ciklusa, a prevazilazi se prevođenjem PEP u malat alternativnim putem, tj. ne preko Krebsovog ciklusa. U citoplazmi biljnih ćelija PEP karboksilaza i malat dehidrogenaza iz PEP sintetišu malat, koji ulazi u mitohondrije i tamo se prevodi u piruvat pod uticajem NAD + -malatne dekarboksilaze, a ovaj ulazi u Krebsov ciklus. Ovako nastali malat iz citoplazme takođe može da zameni intermedijere Krebsovog ciklusa.
26 Oksidativna fosforilacija Proces oksidativne fosforilacije odvija se u mitohondrijama i sastoji se od transporta elektrona sa kojim je povezana sinteza ATP-a. Izvor elektrona su redukovani kofaktori (NADH, NADPH i FADH2) koji su nastali u oksidacijama tokom glikolize, oksidativnog pentoznog ciklusa i Krebsovog ciklusa. Za ovaj proces potreban je O2 koji je krajnji akceptor elektrona i redukuje se do vode. Na svaki molekul saharoze, tokom glikolize u citoplazmi nastaju 4 NADH, a tokom Krebsovog ciklusa u matriksu mitohondrija 16 NADH i 4 FADH2. Svaki od ovih redukovanih koenzima može da otpusti 2 elektrona i 2 protona, tj. oksiduje se u oksidativnoj fosforilaciji. Uloga elektron-transportnog lanca je da obezbedi oksidaciju redukovanih koenzima i da energiju koja se pritom oslobodi iskoristi za generisanje elektrohemijskog gradijenta protona na unutrašnjoj membrani ovojnice mitohondrija, koji, zatim, koristi ATP sintaza za sintezu ATP-a. Elektron-transportni lanac lociran je u unutrašnjoj membrani mitohondrija. Iako je transport elektrona u disanju suštinski sličan u svim aerobnim organizmima, kod biljaka (i gljiva) lanac prenosilaca elektrona sadrži mnogobrojne NAD(P)H dehidrogenaze i alternativnu oksidazu, kojih nema u sisarskim mitohondrijama.
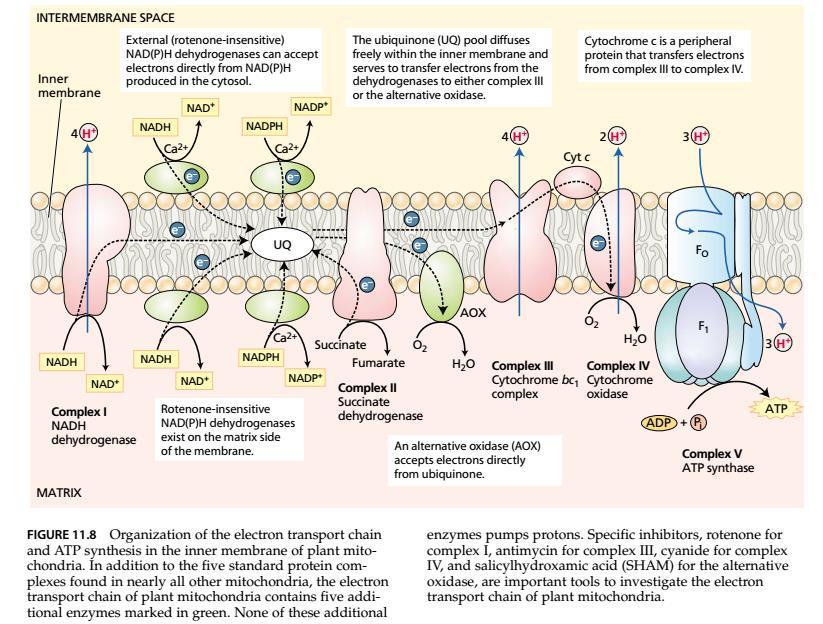
27
28 Kompleks I (NADH dehidrogenaza). Preuzima elektrone od NADH koji je nastao u Krebsovom ciklusu u matriksu, a predaje ih ubikinonu. U sklopu ovog kompleksa su i prenosioci elektrona: FMN i nekoliko fero-sumpornih centara. Za svaki elektronski par kojise prenese sa NADH na ubikinon, ovaj kompleks transportuje 4 protona iz matriksa u intermembranski prostor. Ubikinon. Mali lipofilni prenosilac elektrona i protona, koji nije čvrsto vezan ni za jedan protein i može da difunduje unutar unutrašnje membrane. Kompleks II (sukcinat dehidrogenaza). U Krebsovom ciklusu ovaj enzim koji je vezan za unutrašnju membranu oksiduje sukcinat. Prima elektrone od FADH2 i predaje ih (posredstvom nekoliko fero-sumpornih centara) ubikinonima. Ovaj kompleks ne transportuje protone. Kompleks III (kompleks citohroma bc 1 ). Preuzima elektrone od redukovanih ubikinona (ubikinola), tj. oksiduje ih, i predaje elektrone citohromu c. U sklopu kompleksa su prenosioci elektrona: fero-sumporni centri, dva citohroma b (b565 i b560) i asocirani protein citohrom c 1. Za svaki preneti elektronski par ovaj kompleks ispumpa 4 protona iz matriksa. Citohrom c. Mali protein koji je labavo vezan za spoljašnju stranu unutrašnje membrane mitohondrija. Ima funkciju mobilnog prenosioca elektrona sa kompleksa III na kompleks IV. Kompleks IV (citohrom c oksidaza, terminalna oksidaza). Sadrži 2 bakarna centra (CuA i CuB) i citohrome a i a 3. Prenosi 4 elektrona na O2, pri čemu nastaju dva molekula H2O. Ispumpava iz matriksa 2 protona po svakom prenetom elektronskom paru.
29 Strukturno i funkcinalno ubikinon je sličan plastokinonu, a citohrom bc 1 kompleksu citohroma b 6 f u fotosintetičkom elektrontransportnom lancu tilakoidnih membrana hloroplasta. Biljne mitohondrije sadrže dodatne komponente elektrontransportnog lanca, koje ne postoje u animalnim mitohondrijama. Te komponente ne transportuju protone, pa je konzervacija energije (u vidu sintetisanog ATP-a) manja kad god su one aktivne. Dodatni elektron-transportni enzimi biljnih mitohondrija su: 1. Dve NAD(P)H dehidrogenaze pričvršćene za spoljašnju stranu unutrašnje membrane mitohondrija (u kontaktu sa intermembranskim prostorom). Obe su zavisne od jona Ca 2+. One oksiduju NADH, odnosno NADPH poreklom iz citoplazme, a elektrone predaju ubikinonima. 2. NADH dehidrogenaza neosetljiva na inhibitor rotenon. Nalaze se na unutrašnjoj strani unutrašnje membrane, ka matriksu, i rade isto štoi kompleks I (oksiduju NADH poreklom iz matriksa), ali su rezistentne na rotenon. Njihova uloga je da se aktiviraju i zaobiđu ( bypass ) kompleks I kada je on preopterećen, a to je u uslovima visoke fotorespiracije. 3. NADPH dehidrogenaza na unutrašnjoh strani (ka matriksu) unutrašnje membrane. Oksiduje NADPH poreklom iz matriksa. O njoj se vrlo malo zna.
30 4. Alternativna oksidaza neosetljiva na inhibiciju cijanidom (CN - ), azidom (N3 - ) i ugljen monoksidom (CO) za razliku od citohrom c oksidaze. Redukuje kiseonik do vode, a elektrone dobija od ubikinona. Transport elektrona duž elektron-transportnog lanca odvija se niz gradijent redoks potencijala prenosilaca elektrona i povezan je sa fosforilacijom ADP i sintezom ATP koju katalizuje ATP sintaza (F0F1 ATP-sintaza, kompleks V). Broj sintetisanih molekula ATP zavisi od donora elektrona koji su raspoloživi. U elektron-transportnom lancu mitohondrija postoje tri mesta konzervacije energije (mesta gde se transportuju i protoni iz matriksa u intermembranski prostor, tj. gde je elektronski transport kuplovan sa generisanjem razlike u hemijskom potencijalu) kompleksi I, III i IV (važi i za biljke i za druge aerobne organizme). Ukoliko se kod biljaka uključi alternativna oksidaza, koja zaobilazi kompleks III i IV, konzervacija energije se smanuje, tj. broj sintetisanih ATP-a je manji. Elektroni se preusmeravaju ka alternativnoj oksidazi koja direktno redukuje O2, a pošto energija nema gde da se konzervira (nema mesta konzervacije između ubikinona i kiseonika), ona se gubi u vidu toplote. Na mestima konzervacije energije generiše se protonska motorna sila, tj. elektrohemijski gradijent protona, koji pokreće rad ATP-sintaze.
.")
31 Rezultat vektorskog transporta protona je ph matriksa oko 8 i negativan membranski potencijal unutrašnje membrane mitohondrija. Za razliku od hloroplasta, gde protonskoj motornoj sili doprinosi samo hemijska komponenta, u mitohondrijama uticaj imaju obe komponente (i hemijska i električna). Ova razlika se objašnjava time što je distribucija protona koji se vektorski transportuju iz matriksa asimetrična, a to dovodi do razlika u membranskom potencijalu.

32 Energetski bilans disanja U procesu disanja, preko glikolize i Krebsovog ciklusa, ukupno se dobija 60 ATP po jednom molu saharoze (potpuno oksidovane do CO2). To je teorijska vrednost. Veliki deo energije gubi se u obliku toplote. Međutim, čak i sa gubicima, energetski prinos disanja je mnogo veći nego u slučaju da se odvija fermentacija (u anaerobnim uslovima).
33 Kod biljaka postoji nekoliko funkcionalnih proteina koji smanjuju efikasnost konzervacije energije u oksidativnoj fosforilaciji. Smatra se da biljke to sebi mogu da priušte, jer nisu toliko limitirane izvorom energije (svetlost), koliko dostupnošću mineralnih elemenata (naročito nitrata i fosfata), pa im je važnije prilagođavanje na efikasnije korišćenje nutrijenata, nego ušteda u energiji. Koja je korist za biljke od postojanja alternativnih metaboličkih puteva u kojima se energija ne konzervira? Alternativna oksidaza cijanid-rezistentno disanje (dostiže u većini biljnih tkiva %, a u pojedinim tkivima i 100% intenziteta normalnog disanja). porast temperature (termogeneza) tokom razvića cvetova, nepostredno pred oprašivanje kod biljaka iz fam. Araceae funkcija porasta temperature cvasti je produkcija isparljivih jedinjenja koja privlače polinatore. omogućava mitohondrijama da usklade sintezu ATP i biosintezu organskih jedinjenja. Alternativni put je sigurnosni ventil u slučaju da se akumuliraju u višku supstrati koji se koriste za rastenje, kao rezerva ili za sintezu ATP-a, odnosno kada je glavni put preopterećen. Preopterećenje se dešava onda kada nivo disanja prevaziđe potrebe biljke za ATP-om (manifestuje se nedostatkom ADP u ćelijama).
, što onda izaziva oštećenja u biljnim ćelijama.")
34 funkcijau odbrani biljaka od različitih vrsta abiotičkog stresa. Abiotički stres inhibira disanje i može da dovede do nastanka ROS (zbog sporijeg protoka elektrona i nepotpune redukcije kiseonika), što onda izaziva oštećenja u biljnim ćelijama. Alternativni put preusmerava tok elektrona, alternativna oksidaza preuzima elektrone od ubikinona, sprečava da se oni nagomilaju u redukovanom obliku, što može da dovede do nastanka ROS. Na taj način ublažuje nepovoljne efekte stresa na proces disanja. Dekuplujući protein Nalazi se u unutrašnjoj membrani, povećava njenu propustljivost za protone, zbog čega se narušava elektrohemijski gradijent protona i raskida veza između transporta elektrona i fosforilacije, pa se produkuje manje ATP, a više toplote. Kod životinja ima funkciju u termogenezi. Kod biljaka ima istu funkciju kao alternativna oksidaza ublažavanje efekata stresa na disanje. NADH-dehidrogenaza neosetljiva na rotenon Služi kao zaobilaznica sa kompleks I, onda kada je on preopterećen. Kompleks I ima veći afinitet za NADH i funkcioniše dominantno pri sniženim nivoima NADH u matriksu, a to je onda kad ima dovoljno ADP. Pri sniženju količine ADP, nagomilava se NADH u matriksu i aktivira se NADH-dehidrogenaza. Pošto ona ne transportuje protone i nije mesto konzervacije energije, posledica njenog rada je manja količina sintetisanog ATP-a.


35 Regulacija respiracije Obavlja se u sve tri faze ADP i Pi su ključni regulatori brzine glikolize, Krebsovog ciklusa i oksidativne fosforilacije Najbolje okarakterisano mesto regulacije Krebsovog ciklusa je kompleks piruvat dehidrogenaze. bottom up regulacija


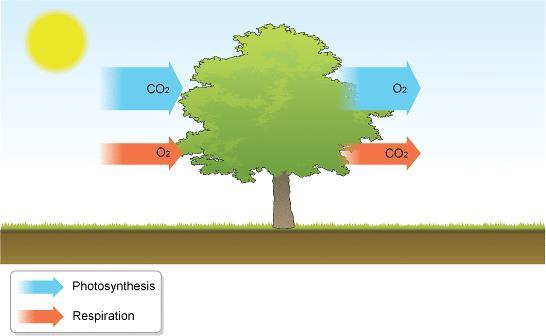
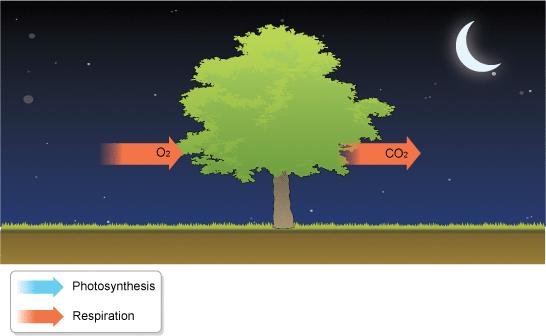
36 Respiracija in vivo Biljke istovremeno i fotosintetišu i dišu na svetlosti. U mraku samo dišu. Na svetlosti se intenzitet disanja smanjuje u listovima, ali mitohondrije i dalje najvećim delom snabdevaju citoplazmu ATP-om. U disanju se izgubi 30-60% u fotosintezi asimiliranog ugljenika. Različita biljna tkiva i organi karakterišu se različitim intenzitetom disanja. Disanje direktno zavisi od metaboličke aktivnosti ćelija. Na intenzitet disanja utiče i stadijum razvića, tj. starost. Sa starošću biljaka, opada intenzitet disanja, osim u slučaju klimakteričnih plodova kod kojih prilikom sazrevanja dolazi do porasta disanja (uglavnom na račun alternativnog puta), a pod kontrolom hormona etilena.
37 Na intenzitet disanja utiču sledeći faktori spoljašnje sredine: Kiseonik, O2 krajnji akceptor elektrona u disanju. Utiče na smanjenje intenziteta disanja ako je snabdevanje njime smanjeno (hipoksični uslovi, korenovi koji rastu na poplavljenom zemljištu). Uglavnom nije ograničavajući faktor zbog dobro razvijenih puteva (aerenhim, intercelulari, lenticele) u biljkama, kojima može u gasovitom stanju da dođe do svih tkiva. Problem može da predstavlja samo njegova mala koncentracija u vodenim rastvorima (rastvorljivost se smanjuje sa porastom temperature) i sporija difuzija u vodenim rastvorima (hidroponi). Temperatura sa porastom temperature ubrzava se disanje (u opsegu od 0 do 30 C). Na svakih 10 C u tom opsegu, intenzitet disanja se udvostruči. Intenzitet disanja dostiže plato na C, a na višim temperaturama brzo opada. Na niskim temperaturama (ispod 5 C) disanje je usporeno (koristise za dugotrajno skladištenje plodova i krtola). Koncentracija CO2 visoke koncentracije imaju ograničen direktni inhibitorni efekat na disanje usporavaju disanje na nivou Krebsovog ciklusa, ali se broj mitohondrija povećava.
38 Procesi koji sačinjavaju metabolički put respiracije ne snabdevaju biljne ćelije samo redukovanim kofaktorima i energijom (ATP), već predstavljaju i izvor prekursora za različite biosintetske procese.
Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama. Hloroplast
 Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama Hloroplast Procesom ćelijskog disanja deponovana energija u šećerima erima prevodi se u ATP i druge energetske metabolite. Istovremeno se
Svetlosna energija absorbuje se hlorofilima u biljnim ćelijama Hloroplast Procesom ćelijskog disanja deponovana energija u šećerima erima prevodi se u ATP i druge energetske metabolite. Istovremeno se
METABOLIZAM UGLJENIH HIDRATA
 METABOLIZAM UGLJENIH HIDRATA 14.02.2018. Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O 2. U uslovima prisustva O 2,
METABOLIZAM UGLJENIH HIDRATA 14.02.2018. Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O 2. U uslovima prisustva O 2,
CIKLUS LIMUNSKE KISELINE (CLK)
") SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
SVEUČILIŠTE U ZAGREBU FAKULTET KEMIJSKOG INŽENJERSTVA I TEHNOLOGIJE CIKLUS LIMUNSKE KISELINE (CLK) Doc. dr. sc. Dragana Vuk Metabolička sudbina piruvata 1. Oksidacijska dekarboksilacija piruvata 2. Ciklus
3.1 Granična vrednost funkcije u tački
 3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
3 Granična vrednost i neprekidnost funkcija 2 3 Granična vrednost i neprekidnost funkcija 3. Granična vrednost funkcije u tački Neka je funkcija f(x) definisana u tačkama x za koje je 0 < x x 0 < r, ili
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET SIGNALI I SISTEMI. Zbirka zadataka
 UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
UNIVERZITET U NIŠU ELEKTRONSKI FAKULTET Goran Stančić SIGNALI I SISTEMI Zbirka zadataka NIŠ, 014. Sadržaj 1 Konvolucija Literatura 11 Indeks pojmova 11 3 4 Sadržaj 1 Konvolucija Zadatak 1. Odrediti konvoluciju
Put pentoza fosfata. B. Mildner. Put pentoza fosfata
 Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
Put pentoza fosfata B. Mildner Put pentoza fosfata Svrha ovog puta je: A) da se stanici omogući dovoljno NADPH, koji služi kao reducens u biosintetskim reakcijama kao i u zaštiti stanica od kisikovih radikala.
Oksidativna fosforilacija
 Oksidativna fosforilacija Oksidativna fosforilacija U eukariota u mitohondrijama. Odigrava se redukcija O2 do H2O. Donori elektrona su NADH i FADH2. Najvedi deo energije za svoj rast, održavanje i rad
Oksidativna fosforilacija Oksidativna fosforilacija U eukariota u mitohondrijama. Odigrava se redukcija O2 do H2O. Donori elektrona su NADH i FADH2. Najvedi deo energije za svoj rast, održavanje i rad
Organele života i smrti
 MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
MITOHONDRIJE Organele života i smrti OTKRIĆE MITOHONDRIJA 1857. Albert Kolliker uređeni nizovi granula u mišićnim ćelijama 1893. Richard Altman bioblasti vrsta bakterija? 1. menjaju oblik 2. umnožavaju
Glukoneogeneza i regulacija glukoneogeneze
 Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
Glukoneogeneza i regulacija glukoneogeneze Boris Mildner Glukoneogeneza Sinteza ugljikohidrata iz jednostavnih preteča Put od fosfoenolpiruvata do glukoza 6-fosfata zajednički je za mnoge preteče ugljikohidrata.
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović
 DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
DISKRETNA MATEMATIKA - PREDAVANJE 7 - Jovanka Pantović Novi Sad April 17, 2018 1 / 22 Teorija grafova April 17, 2018 2 / 22 Definicija Graf je ure dena trojka G = (V, G, ψ), gde je (i) V konačan skup čvorova,
CIKLUS AZOTA I SUMPORA U PRIRODI
 CIKLUS AZOTA I SUMPORA U PRIRODI Asimilacija azota i sumpora zahtevaju niz biohemijskih reakcija koje spadaju u reakcije sa velikim energetskim zahtevima. Za asimilaciju azota iz nitrata (NO 3- ) potrebna
CIKLUS AZOTA I SUMPORA U PRIRODI Asimilacija azota i sumpora zahtevaju niz biohemijskih reakcija koje spadaju u reakcije sa velikim energetskim zahtevima. Za asimilaciju azota iz nitrata (NO 3- ) potrebna
PRAVA. Prava je u prostoru određena jednom svojom tačkom i vektorom paralelnim sa tom pravom ( vektor paralelnosti).
.") PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
PRAVA Prava je kao i ravan osnovni geometrijski ojam i ne definiše se. Prava je u rostoru određena jednom svojom tačkom i vektorom aralelnim sa tom ravom ( vektor aralelnosti). M ( x, y, z ) 3 Posmatrajmo
SEKUNDARNE VEZE međumolekulske veze
 PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
PRIMARNE VEZE hemijske veze među atomima SEKUNDARNE VEZE međumolekulske veze - Slabije od primarnih - Elektrostatičkog karaktera - Imaju veliki uticaj na svojstva supstanci: - agregatno stanje - temperatura
Eliminacijski zadatak iz Matematike 1 za kemičare
 Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Za mnoge reakcije vrijedi Arrheniusova jednadžba, koja opisuje vezu koeficijenta brzine reakcije i temperature: K = Ae Ea/(RT ). - T termodinamička temperatura (u K), - R = 8, 3145 J K 1 mol 1 opća plinska
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija
 Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
Mitohondriji i kloroplasti Stanično disanje Fotosinteza Evolucija metaboličkih reakcija MITOHONDRIJI -u svim eukariotskim stanicama -njihov broj ovisi o metaboličkoj aktivnosti stanice (nekoliko stotina
numeričkih deskriptivnih mera.
 DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
DESKRIPTIVNA STATISTIKA Numeričku seriju podataka opisujemo pomoću Numeričku seriju podataka opisujemo pomoću numeričkih deskriptivnih mera. Pokazatelji centralne tendencije Aritmetička sredina, Medijana,
Osnovni primer. (Z, +,,, 0, 1) je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju
 je komutativan prsten sa jedinicom: množenje je distributivno prema sabiranju") RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
RAČUN OSTATAKA 1 1 Prsten celih brojeva Z := N + {} N + = {, 3, 2, 1,, 1, 2, 3,...} Osnovni primer. (Z, +,,,, 1) je komutativan prsten sa jedinicom: sabiranje (S1) asocijativnost x + (y + z) = (x + y)
Ispitivanje toka i skiciranje grafika funkcija
 Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Ispitivanje toka i skiciranje grafika funkcija Za skiciranje grafika funkcije potrebno je ispitati svako od sledećih svojstava: Oblast definisanosti: D f = { R f R}. Parnost, neparnost, periodičnost. 3
Elementi spektralne teorije matrica
 Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Elementi spektralne teorije matrica Neka je X konačno dimenzionalan vektorski prostor nad poljem K i neka je A : X X linearni operator. Definicija. Skalar λ K i nenula vektor u X se nazivaju sopstvena
Ciklus limunske kiseline-2
 Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
Ciklus limunske kiseline-2 Boris Mildner Katabolizam proteina, masti i ugljikohidrata u tri faze staničnog disanja. Faza 1.: oksidacija masnih kiselina, masti i ugljikohidrata kako bi nastao acetil-coa.
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15
 1 / 15") MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
MATRICE I DETERMINANTE - formule i zadaci - (Matrice i determinante) 1 / 15 Matrice - osnovni pojmovi (Matrice i determinante) 2 / 15 (Matrice i determinante) 2 / 15 Matrice - osnovni pojmovi Matrica reda
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI)
") IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
IZRAČUNAVANJE POKAZATELJA NAČINA RADA NAČINA RADA (ISKORIŠĆENOSTI KAPACITETA, STEPENA OTVORENOSTI RADNIH MESTA I NIVOA ORGANIZOVANOSTI) Izračunavanje pokazatelja načina rada OTVORENOG RM RASPOLOŽIVO RADNO
S t r a n a 1. 1.Povezati jonsku jačinu rastvora: a) MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:
 MgCl 2 b) Al 2 (SO 4 ) 3 sa njihovim molalitetima, m. za so tipa: M p X q. pa je jonska jačina:") S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
S t r a n a 1 1.Povezati jonsku jačinu rastvora: a MgCl b Al (SO 4 3 sa njihovim molalitetima, m za so tipa: M p X q pa je jonska jačina:. Izračunati mase; akno 3 bba(no 3 koje bi trebalo dodati, 0,110
Iskazna logika 3. Matematička logika u računarstvu. novembar 2012
 Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
Iskazna logika 3 Matematička logika u računarstvu Department of Mathematics and Informatics, Faculty of Science,, Serbia novembar 2012 Deduktivni sistemi 1 Definicija Deduktivni sistem (ili formalna teorija)
SISTEMI NELINEARNIH JEDNAČINA
 SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
SISTEMI NELINEARNIH JEDNAČINA April, 2013 Razni zapisi sistema Skalarni oblik: Vektorski oblik: F = f 1 f n f 1 (x 1,, x n ) = 0 f n (x 1,, x n ) = 0, x = (1) F(x) = 0, (2) x 1 0, 0 = x n 0 Definicije
OKSIDACIJSKA FOSFORILACIJA
 OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
OKSIDACIJSKA FOSFORILACIJA Sinteza ATP B. Mildner & M. Kekez 2012. 1. Što od navedenog nije dio Mittchelove kemiosmotske hipoteze? a) Dio energije koji se dobiva transportom elektrona koristi se za dobivanje
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici
 Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
Metabolizam Uvod u metabolizam - procesi izgradnje i razgradnje u živoj stanici Izv. prof. dr. sc. Lidija Šver jelokupnost svih kemijskih pretvorbi u stanici ili organizmu Pretvorba (transformacija) tvari
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - VSŠ 2005/06
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Substrat: škrob saharoza fruktani drugi sladkorji lipidi organske kisline proteini
DIHANJE. Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo. Agronomija - UNI
 DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
DIHANJE Univerza v Ljubljani Biotehniška fakulteta Oddelek za agronomijo Agronomija - UNI Fotosinteza + Dihanje + Svetlobno dihanje Dihanje Fotosinteza 6CO 2 + 6H 2 O C 6 H 12 O 6 + 6O 2 Fotosintetski
41. Jednačine koje se svode na kvadratne
 . Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
. Jednačine koje se svode na kvadrane Simerične recipročne) jednačine Jednačine oblika a n b n c n... c b a nazivamo simerične jednačine, zbog simeričnosi koeficijenaa koeficijeni uz jednaki). k i n k
Kloroplasti. Fotosinteza Mitohondriji Stanično disanje
 Kloroplasti Fotosinteza Mitohondriji Stanično disanje http://en.wikipedia.org/wiki/plas tid PLASTIDI Organeli biljnih stanica i stanica algi Proizvodnja i pohranjivanje šećera i drugih molekula Pigmenti
Kloroplasti Fotosinteza Mitohondriji Stanično disanje http://en.wikipedia.org/wiki/plas tid PLASTIDI Organeli biljnih stanica i stanica algi Proizvodnja i pohranjivanje šećera i drugih molekula Pigmenti
OSNOVE METABOLIZMA MIKROORGANIZAMA. Metabolički diverzitet
 OSNOVE METABOLIZMA MIKROORGANIZAMA Metabolički diverzitet Metabolizam - obuhvata sve hemijske procese koji se dešavaju u ćeliji - anabolički (zahtevaju energiju) - katabolički (otpuštaju energiju) Energija
OSNOVE METABOLIZMA MIKROORGANIZAMA Metabolički diverzitet Metabolizam - obuhvata sve hemijske procese koji se dešavaju u ćeliji - anabolički (zahtevaju energiju) - katabolički (otpuštaju energiju) Energija
M086 LA 1 M106 GRP. Tema: Baza vektorskog prostora. Koordinatni sustav. Norma. CSB nejednakost
 M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
M086 LA 1 M106 GRP Tema: CSB nejednakost. 19. 10. 2017. predavač: Rudolf Scitovski, Darija Marković asistent: Darija Brajković, Katarina Vincetić P 1 www.fizika.unios.hr/grpua/ 1 Baza vektorskog prostora.
XI dvoqas veжbi dr Vladimir Balti. 4. Stabla
 XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
XI dvoqas veжbi dr Vladimir Balti 4. Stabla Teorijski uvod Teorijski uvod Definicija 5.7.1. Stablo je povezan graf bez kontura. Definicija 5.7.1. Stablo je povezan graf bez kontura. Primer 5.7.1. Sva stabla
Aminokiseline. Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina 22.12.2014
 Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
Anabolizam azotnihjedinjenja: Biosinteza aminokiselina, glutationa i biološki aktivnih amina Predavanja iz opšte biohemije Školska 2014/2015. godina Aminokiseline 1 Metabolizam aminokiselina Proteini iz
ISPITNA PITANJA OSNOVI BIOHEMIJE
 UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
UNIVERZITET PRIVREDNA AKADEMIJA, NOVI SAD STOMATOLOŠKI FAKULTET PANČEVO ISPITNA PITANJA OSNOVI BIOHEMIJE Prof. dr Esma R. Isenović 1. Biohemija kao nauka, zadaci izučavanja i discipline 1. Koja je definicija
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A
, grupa A") Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Kontrolni zadatak (Tačka, prava, ravan, diedar, poliedar, ortogonalna projekcija), grupa A Ime i prezime: 1. Prikazane su tačke A, B i C i prave a,b i c. Upiši simbole Î, Ï, Ì ili Ë tako da dobijeni iskazi
Teorijske osnove informatike 1
 Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Teorijske osnove informatike 1 9. oktobar 2014. () Teorijske osnove informatike 1 9. oktobar 2014. 1 / 17 Funkcije Veze me du skupovima uspostavljamo skupovima koje nazivamo funkcijama. Neformalno, funkcija
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama.
 Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
Apsolutno neprekidne raspodele Raspodele apsolutno neprekidnih sluqajnih promenljivih nazivaju se apsolutno neprekidnim raspodelama. a b Verovatno a da sluqajna promenljiva X uzima vrednost iz intervala
PITANJA ZA USMENI ISPIT IZ BIOHEMIJE
 PITANJA ZA USMENI ISPIT IZ BIOHEMIJE PROTEINI STRUKTURA I FUNKCIJE 1. Struktura proteina nivoi organizacije molekula 2. Proteini koji transportuju kiseonik hemoglobin i mioglobin ENZIMI 1. Opšte osobine
PITANJA ZA USMENI ISPIT IZ BIOHEMIJE PROTEINI STRUKTURA I FUNKCIJE 1. Struktura proteina nivoi organizacije molekula 2. Proteini koji transportuju kiseonik hemoglobin i mioglobin ENZIMI 1. Opšte osobine
METABOLIZEM OGLJIKOVIH HIDRATOV
 METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
METABOLIZEM OGLJIKOVIH HIDRATOV KAKO CELICA DOBI GLUKOZO IN OSTALE MONOSAHARIDE? HRANA ZNOTRAJCELIČNI GLIKOGEN ali ŠKROB razgradnja s prebavnimi encimi GLUKOZA in ostali monosaharidi fosforilitična cepitev
ELEKTROTEHNIČKI ODJEL
 MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
MATEMATIKA. Neka je S skup svih živućih državljana Republike Hrvatske..04., a f preslikavanje koje svakom elementu skupa S pridružuje njegov horoskopski znak (bez podznaka). a) Pokažite da je f funkcija,
Operacije s matricama
 Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Linearna algebra I Operacije s matricama Korolar 3.1.5. Množenje matrica u vektorskom prostoru M n (F) ima sljedeća svojstva: (1) A(B + C) = AB + AC, A, B, C M n (F); (2) (A + B)C = AC + BC, A, B, C M
Kaskadna kompenzacija SAU
 Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
Kaskadna kompenzacija SAU U inženjerskoj praksi, naročito u sistemima regulacije elektromotornih pogona i tehnoloških procesa, veoma često se primenjuje metoda kaskadne kompenzacije, u čijoj osnovi su
PARCIJALNI IZVODI I DIFERENCIJALI. Sama definicija parcijalnog izvoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je,
 PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
PARCIJALNI IZVODI I DIFERENCIJALI Sama definicija parcijalnog ivoda i diferencijala je malo teža, mi se njome ovde nećemo baviti a vi ćete je, naravno, naučiti onako kako vaš profesor ahteva. Mi ćemo probati
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE
 Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
Ciklus limunske kiseline-1 KOMPLEKS PIRUVAT DEHIDROGENAZE Boris Mildner Citratni ciklus /Krebsov ciklus Piruvat koji nastaje glikolizom, umjesto da se reducira u laktat, odnosno u etanol, dalje se oksidira
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu
 Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
Zadaci sa prethodnih prijemnih ispita iz matematike na Beogradskom univerzitetu Trigonometrijske jednačine i nejednačine. Zadaci koji se rade bez upotrebe trigonometrijskih formula. 00. FF cos x sin x
ISPITNA PITANJA ZA USMENI DIO ISPITA
 UNIVERZITET CRNE GORE MEDICINSKI FAKULTET MEDICINSKA BIOHEMIJA ISPITNA PITANJA ZA USMENI DIO ISPITA STUDIJSKI PROGRAM MEDICINA I ENZIMOLOGIJA 1. Opšte osobine enzima i struktura molekula enzima 2. Izoenzimi.
UNIVERZITET CRNE GORE MEDICINSKI FAKULTET MEDICINSKA BIOHEMIJA ISPITNA PITANJA ZA USMENI DIO ISPITA STUDIJSKI PROGRAM MEDICINA I ENZIMOLOGIJA 1. Opšte osobine enzima i struktura molekula enzima 2. Izoenzimi.
Glukoneogeneza. Glukoneogeneza. Glukoneogeneza. poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih.
 poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
poteka v jetrih in ledvični skorji, v citoplazmi in delno v mitohondrijih. Izhodne spojine:, laktat, in drugi intermediati cikla TKK glukogene aminokisline, glicerol Kaj pa maščobne kisline? Ireverzibilne
18. listopada listopada / 13
 18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
18. listopada 2016. 18. listopada 2016. 1 / 13 Neprekidne funkcije Važnu klasu funkcija tvore neprekidne funkcije. To su funkcije f kod kojih mala promjena u nezavisnoj varijabli x uzrokuje malu promjenu
FTN Novi Sad Katedra za motore i vozila. Teorija kretanja drumskih vozila Vučno-dinamičke performanse vozila: MAKSIMALNA BRZINA
 : MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
: MAKSIMALNA BRZINA Maksimalna brzina kretanja F O (N) F OI i m =i I i m =i II F Oid Princip određivanja v MAX : Drugi Njutnov zakon Dokle god je: F O > ΣF otp vozilo ubrzava Kada postane: F O = ΣF otp
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP
 Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
Nastanek NADH in NADPH Prenos elektronov in nastanek ATP Glavne metabolične poti glukoze Glikoliza (Embden Meyerhofova metabolna pot) Fosfoglukonatna (pentozafosfatna) pot: nekatere živali Katabolizem
VODA ELEKTROLITI I ACIDO-BAZNA RAVNOTEŽA...
 SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
SADRŽAJ UVOD 1 1. BIOHEMIJA ĆELIJE... 1-1 1.1 UVOD... 1-2 1.2 ĆELIJA KAO OSNOVNA ŽIVA JEDINICA TELA... 1-2 1.3 VANĆELIJSKA TEČNOST UNUTRAŠNJA OKOLINA... 1-2 1.4 BIOELEMENTI I BIOMOLEKULI... 1-3 1.5 ĆELIJA
TRANSPORT JONA I ORGANSKIH JEDINJENJA KROZ MEMBRANE
 TRANSPORT JONA I ORGANSKIH JEDINJENJA KROZ MEMBRANE Transportni mehanizmi na membranama Prema potrebi za energijom i specifičnim učesnicima u transportu postoji 5 načina transporta kroz biološke membrane:
TRANSPORT JONA I ORGANSKIH JEDINJENJA KROZ MEMBRANE Transportni mehanizmi na membranama Prema potrebi za energijom i specifičnim učesnicima u transportu postoji 5 načina transporta kroz biološke membrane:
5. Karakteristične funkcije
 5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
5. Karakteristične funkcije Profesor Milan Merkle emerkle@etf.rs milanmerkle.etf.rs Verovatnoća i Statistika-proleće 2018 Milan Merkle Karakteristične funkcije ETF Beograd 1 / 10 Definicija Karakteristična
KVADRATNA FUNKCIJA. Kvadratna funkcija je oblika: Kriva u ravni koja predstavlja grafik funkcije y = ax + bx + c. je parabola.
 KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
KVADRATNA FUNKCIJA Kvadratna funkcija je oblika: = a + b + c Gde je R, a 0 i a, b i c su realni brojevi. Kriva u ravni koja predstavlja grafik funkcije = a + b + c je parabola. Najpre ćemo naučiti kako
PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).
![PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).](/thumbs/74/70316766.jpg "PID: Domen P je glavnoidealski [PID] akko svaki ideal u P je glavni (generisan jednim elementom; oblika ap := {ab b P }, za neko a P ).") 0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
0.1 Faktorizacija: ID, ED, PID, ND, FD, UFD Definicija. Najava pojmova: [ID], [ED], [PID], [ND], [FD] i [UFD]. ID: Komutativan prsten P, sa jedinicom 1 0, je integralni domen [ID] oblast celih), ili samo
IZVODI ZADACI ( IV deo) Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo
 Rešenje: Najpre ćemo logaritmovati ovu jednakost sa ln ( to beše prirodni logaritam za osnovu e) a zatim ćemo") IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
IZVODI ZADACI ( IV deo) LOGARITAMSKI IZVOD Logariamskim izvodom funkcije f(), gde je >0 i, nazivamo izvod logarima e funkcije, o jes: (ln ) f ( ) f ( ) Primer. Nadji izvod funkcije Najpre ćemo logarimovai
Osnovne veličine, jedinice i izračunavanja u hemiji
 Osnovne veličine, jedinice i izračunavanja u hemiji Pregled pojmova veličina i njihovih jedinica koje se koriste pri osnovnim izračunavanjima u hemiji dat je u Tabeli 1. Tabela 1. Veličine i njihove jedinice
Osnovne veličine, jedinice i izračunavanja u hemiji Pregled pojmova veličina i njihovih jedinica koje se koriste pri osnovnim izračunavanjima u hemiji dat je u Tabeli 1. Tabela 1. Veličine i njihove jedinice
HEMIJSKA VEZA TEORIJA VALENTNE VEZE
 TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
TEORIJA VALENTNE VEZE Kovalentna veza nastaje preklapanjem atomskih orbitala valentnih elektrona, pri čemu je region preklapanja između dva jezgra okupiran parom elektrona. - Nastalu kovalentnu vezu opisuje
ASIMILACIJA AZOTA, SUMPORA I FOSFORA
 ASIMILACIJA AZOTA, SUMPORA I FOSFORA Više biljke, kao autotrofni organizmi, mogu da sintetišu organske supstance iz neorganskih, koje usvajaju uglavnom iz zemljišta. Inkorporacija mineralnih elemenata
ASIMILACIJA AZOTA, SUMPORA I FOSFORA Više biljke, kao autotrofni organizmi, mogu da sintetišu organske supstance iz neorganskih, koje usvajaju uglavnom iz zemljišta. Inkorporacija mineralnih elemenata
MEĐUMOLEKULSKE SILE JON-DIPOL DIPOL VODONIČNE NE VEZE DIPOL DIPOL-DIPOL DIPOL-INDUKOVANI INDUKOVANI JON-INDUKOVANI DISPERZNE SILE
 MEĐUMLEKULSKE SILE JN-DIPL VDNIČNE NE VEZE DIPL-DIPL JN-INDUKVANI DIPL DIPL-INDUKVANI INDUKVANI DIPL DISPERZNE SILE MEĐUMLEKULSKE SILE jake JNSKA VEZA (metal-nemetal) KVALENTNA VEZA (nemetal-nemetal) METALNA
MEĐUMLEKULSKE SILE JN-DIPL VDNIČNE NE VEZE DIPL-DIPL JN-INDUKVANI DIPL DIPL-INDUKVANI INDUKVANI DIPL DISPERZNE SILE MEĐUMLEKULSKE SILE jake JNSKA VEZA (metal-nemetal) KVALENTNA VEZA (nemetal-nemetal) METALNA
SEMINAR IZ KOLEGIJA ANALITIČKA KEMIJA I. Studij Primijenjena kemija
 SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
SEMINAR IZ OLEGIJA ANALITIČA EMIJA I Studij Primijenjena kemija 1. 0,1 mola NaOH je dodano 1 litri čiste vode. Izračunajte ph tako nastale otopine. NaOH 0,1 M NaOH Na OH Jak elektrolit!!! Disoira potpuno!!!
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =
/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med =") 100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
100g maslaca: 751kcal = 20g : E maslac E maslac = (751 x 20)/100 E maslac = 150,2kcal 100g med: 320kcal = 30g : E med E med = (320 x 30)/100 E med = 96kcal 100g mleko: 49kcal = 250g : E mleko E mleko =
III VEŽBA: FURIJEOVI REDOVI
 III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
III VEŽBA: URIJEOVI REDOVI 3.1. eorijska osnova Posmatrajmo neki vremenski kontinualan signal x(t) na intervalu definisati: t + t t. ada se može X [ k ] = 1 t + t x ( t ) e j 2 π kf t dt, gde je f = 1/.
STRUKTURA BIOLOŠKIH MAKROMOLEKULA HEMIJSKE I BIOHEMIJSKE REAKCIJE TRANSPORT RAST I DAJE ČVRSTINU (TURGOR) VODA
 VODA") Snabdevenost biljke vodom je jedan od najvažnijih faktora koji utiče na rastenje i razviće biljaka. Od svih resursa potrebnih biljci voda je najrasprostranjenija, najobilnija i ujedno najveći ograničavajući
Snabdevenost biljke vodom je jedan od najvažnijih faktora koji utiče na rastenje i razviće biljaka. Od svih resursa potrebnih biljci voda je najrasprostranjenija, najobilnija i ujedno najveći ograničavajući
TEST PITANJA BIOHEMIJA UGLJENI HIDRATI
 TEST PITANJA BIOHEMIJA UGLJENI HIDRATI 1. Kakve osobine pokazuju monosaharidi: a) osidacione b) redukcione c) metilirajuće 2. Skrob, kao dominantan šećer u ishrani čoveka, se razlaže do disaharida u: a)
TEST PITANJA BIOHEMIJA UGLJENI HIDRATI 1. Kakve osobine pokazuju monosaharidi: a) osidacione b) redukcione c) metilirajuće 2. Skrob, kao dominantan šećer u ishrani čoveka, se razlaže do disaharida u: a)
I.13. Koliki je napon između neke tačke A čiji je potencijal 5 V i referentne tačke u odnosu na koju se taj potencijal računa?
 TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
TET I.1. Šta je Kulonova sila? elektrostatička sila magnetna sila c) gravitaciona sila I.. Šta je elektrostatička sila? sila kojom međusobno eluju naelektrisanja u mirovanju sila kojom eluju naelektrisanja
NOMENKLATURA ORGANSKIH SPOJEVA. Imenovanje aromatskih ugljikovodika
 NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
NOMENKLATURA ORGANSKIH SPOJEVA Imenovanje aromatskih ugljikovodika benzen metilbenzen (toluen) 1,2-dimetilbenzen (o-ksilen) 1,3-dimetilbenzen (m-ksilen) 1,4-dimetilbenzen (p-ksilen) fenilna grupa 2-fenilheptan
3. razred gimnazije- opšti i prirodno-matematički smer ALKENI. Aciklični nezasićeni ugljovodonici koji imaju jednu dvostruku vezu.
 ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
ALKENI Acikliči ezasićei ugljovodoici koji imaju jedu dvostruku vezu. 2 4 2 2 2 (etile) viil grupa 3 6 2 3 2 2 prope (propile) alil grupa 4 8 2 2 3 3 3 2 3 3 1-bute 2-bute 2-metilprope 5 10 2 2 2 2 3 2
Riješeni zadaci: Nizovi realnih brojeva
 Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Riješei zadaci: Nizovi realih brojeva Nizovi, aritmetički iz, geometrijski iz Fukciju a : N R azivamo beskoači) iz realih brojeva i ozačavamo s a 1, a,..., a,... ili a ), pri čemu je a = a). Aritmetički
Verovatnoća i Statistika I deo Teorija verovatnoće (zadaci) Beleške dr Bobana Marinkovića
 Beleške dr Bobana Marinkovića") Verovatnoća i Statistika I deo Teorija verovatnoće zadaci Beleške dr Bobana Marinkovića Iz skupa, 2,, 00} bira se na slučajan način 5 brojeva Odrediti skup elementarnih dogadjaja ako se brojevi biraju
Verovatnoća i Statistika I deo Teorija verovatnoće zadaci Beleške dr Bobana Marinkovića Iz skupa, 2,, 00} bira se na slučajan način 5 brojeva Odrediti skup elementarnih dogadjaja ako se brojevi biraju
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f
 D f") IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f IspitivaƬe funkcija: 1. Oblast definisanosti funkcije (ili domen funkcije) D f 2. Nule i znak funkcije; presek sa y-osom IspitivaƬe
Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. 4/17/2013
 Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
Metabolizam Svi živi organizmi imaju potrebu za konstantnim prilivom energije kako bi održali ćelijsku strukturu i rast. Kemotrofni organizmi; dobivaju slobodnu energiju gj oksidacijom hranjivih tvari
2 tg x ctg x 1 = =, cos 2x Zbog četvrtog kvadranta rješenje je: 2 ctg x
 Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
Zadatak (Darjan, medicinska škola) Izračunaj vrijednosti trigonometrijskih funkcija broja ako je 6 sin =,,. 6 Rješenje Ponovimo trigonometrijske funkcije dvostrukog kuta! Za argument vrijede sljedeće formule:
INTELIGENTNO UPRAVLJANJE
 INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
INTELIGENTNO UPRAVLJANJE Fuzzy sistemi zaključivanja Vanr.prof. Dr. Lejla Banjanović-Mehmedović Mehmedović 1 Osnovni elementi fuzzy sistema zaključivanja Fazifikacija Baza znanja Baze podataka Baze pravila
Zavrxni ispit iz Matematiqke analize 1
 Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
Građevinski fakultet Univerziteta u Beogradu 3.2.2016. Zavrxni ispit iz Matematiqke analize 1 Prezime i ime: Broj indeksa: 1. Definisati Koxijev niz. Dati primer niza koji nije Koxijev. 2. Dat je red n=1
VISOKO GOSPODARSKO UČILIŠTE U KRIŽEVCIMA
 VISOKO GOSPODARSKO UČILIŠTE U KRIŽEVCIMA Dr.sc. Marija Vukobratović Križevci, 2009. 1 UVOD U METABOLIZAM Svi živi organizmi za svoje aktivnosti (održavanje života, rast i razvoj) zahtijevaju energiju,
VISOKO GOSPODARSKO UČILIŠTE U KRIŽEVCIMA Dr.sc. Marija Vukobratović Križevci, 2009. 1 UVOD U METABOLIZAM Svi živi organizmi za svoje aktivnosti (održavanje života, rast i razvoj) zahtijevaju energiju,
U unutrašnja energija H entalpija S entropija G 298. G Gibsova energija TERMOHEMIJA I TERMODINAMIKA HEMIJSKA TERMODINAMIKA
 HEMIJSKA TERMODINAMIKA Bavi se energetskim promenama pri odigravanju hemijskih reakcija. TERMODINAMIČKE FUNKCIJE STANJA U unutrašnja energija H entalpija S entropija Ako su određene na standardnom pritisku
HEMIJSKA TERMODINAMIKA Bavi se energetskim promenama pri odigravanju hemijskih reakcija. TERMODINAMIČKE FUNKCIJE STANJA U unutrašnja energija H entalpija S entropija Ako su određene na standardnom pritisku
UVOD CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN
 = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN") CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
CIKLUS CITRONSKE KISLINE (CCK) = KREBSOV CIKLUS = CIKLUS TRIKARBOKSILNIH KISLIN Glavne metabolične poti oglj. hidratov pri rastlinah in živalih GLIKOGEN, ŠKROB Riboza 5-fosfat + NADPH+H + katabolizem fosfoglukonatna
Dvanaesti praktikum iz Analize 1
 Dvaaesti praktikum iz Aalize Zlatko Lazovi 20. decembar 206.. Dokazati da fukcija f = 5 l tg + 5 ima bar jedu realu ulu. Ree e. Oblast defiisaosti fukcije je D f = k Z da postoji ula fukcije a 0, π 2.
Dvaaesti praktikum iz Aalize Zlatko Lazovi 20. decembar 206.. Dokazati da fukcija f = 5 l tg + 5 ima bar jedu realu ulu. Ree e. Oblast defiisaosti fukcije je D f = k Z da postoji ula fukcije a 0, π 2.
OM2 V3 Ime i prezime: Index br: I SAVIJANJE SILAMA TANKOZIDNIH ŠTAPOVA
 OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
OM V me i preime: nde br: 1.0.01. 0.0.01. SAVJANJE SLAMA TANKOZDNH ŠTAPOVA A. TANKOZDN ŠTAPOV PROZVOLJNOG OTVORENOG POPREČNOG PRESEKA Preposavka: Smičući napon je konsanan po debljini ida (duž pravca upravnog
Osnovne teoreme diferencijalnog računa
 Osnovne teoreme diferencijalnog računa Teorema Rolova) Neka je funkcija f definisana na [a, b], pri čemu važi f je neprekidna na [a, b], f je diferencijabilna na a, b) i fa) fb). Tada postoji ξ a, b) tako
Osnovne teoreme diferencijalnog računa Teorema Rolova) Neka je funkcija f definisana na [a, b], pri čemu važi f je neprekidna na [a, b], f je diferencijabilna na a, b) i fa) fb). Tada postoji ξ a, b) tako
PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE)
") (Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
(Enegane) List: PARNA POSTROJENJA ZA KOMBINIRANU PROIZVODNJU ELEKTRIČNE I TOPLINSKE ENERGIJE (ENERGANE) Na mjestima gdje se istovremeno troši električna i toplinska energija, ekonomičan način opskrbe energijom
OSNOVI ELEKTRONIKE. Vežbe (2 časa nedeljno): mr Goran Savić
: mr Goran Savić") OSNOVI ELEKTRONIKE Vežbe (2 časa nedeljno): mr Goran Savić savic@el.etf.rs http://tnt.etf.rs/~si1oe Termin za konsultacije: četvrtak u 12h, kabinet 102 Referentni smerovi i polariteti 1. Odrediti vrednosti
OSNOVI ELEKTRONIKE Vežbe (2 časa nedeljno): mr Goran Savić savic@el.etf.rs http://tnt.etf.rs/~si1oe Termin za konsultacije: četvrtak u 12h, kabinet 102 Referentni smerovi i polariteti 1. Odrediti vrednosti
Pismeni ispit iz matematike Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: ( ) + 1.
 + 1.") Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
Pismeni ispit iz matematike 0 008 GRUPA A Riješiti sistem jednačina i diskutovati rješenja sistema u zavisnosti od parametra: λ + z = Ispitati funkciju i nacrtati njen grafik: + ( λ ) + z = e Izračunati
RIJEŠENI ZADACI I TEORIJA IZ
 RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
RIJEŠENI ZADACI I TEORIJA IZ LOGARITAMSKA FUNKCIJA SVOJSTVA LOGARITAMSKE FUNKCIJE OSNOVE TRIGONOMETRIJE PRAVOKUTNOG TROKUTA - DEFINICIJA TRIGONOMETRIJSKIH FUNKCIJA - VRIJEDNOSTI TRIGONOMETRIJSKIH FUNKCIJA
1 Afina geometrija. 1.1 Afini prostor. Definicija 1.1. Pod afinim prostorom nad poljem K podrazumevamo. A - skup taqaka
 1 Afina geometrija 11 Afini prostor Definicija 11 Pod afinim prostorom nad poljem K podrazumevamo svaku uređenu trojku (A, V, +): A - skup taqaka V - vektorski prostor nad poljem K + : A V A - preslikavanje
1 Afina geometrija 11 Afini prostor Definicija 11 Pod afinim prostorom nad poljem K podrazumevamo svaku uređenu trojku (A, V, +): A - skup taqaka V - vektorski prostor nad poljem K + : A V A - preslikavanje
Dijagonalizacija operatora
 Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
Dijagonalizacija operatora Problem: Može li se odrediti baza u kojoj zadani operator ima dijagonalnu matricu? Ova problem je povezan sa sljedećim pojmovima: 1 Karakteristični polinom operatora f 2 Vlastite
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI. NEUTRALNI ELEMENT GRUPOIDA.
 KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
KOMUTATIVNI I ASOCIJATIVNI GRUPOIDI NEUTRALNI ELEMENT GRUPOIDA 1 Grupoid (G, ) je asocijativa akko važi ( x, y, z G) x (y z) = (x y) z Grupoid (G, ) je komutativa akko važi ( x, y G) x y = y x Asocijativa
Kiselo bazni indikatori
 Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
Kiselo bazni indikatori Slabe kiseline ili baze koje imaju različite boje nejonizovanog i jonizovanog oblika u rastvoru Primer: slaba kiselina HIn(aq) H + (aq) + In (aq) nejonizovani oblik jonizovani oblik
Trigonometrija 2. Adicijske formule. Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto
 Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
Trigonometrija Adicijske formule Formule dvostrukog kuta Formule polovičnog kuta Pretvaranje sume(razlike u produkt i obrnuto Razumijevanje postupka izrade složenijeg matematičkog problema iz osnova trigonometrije
Seminar 13.b. Glikogen GLIKOGEN. B. Mildner
 Seminar 13.b Glikogen B. Mildner GLIKOGEN 1 Glikogen Nereducirani kraj Glikogen je jako dostupni skladišni oblik glukoze; kao i jako velik, razgranat polimer; Glukozne jedinice su povezane α-1,4-glikozidnim
Seminar 13.b Glikogen B. Mildner GLIKOGEN 1 Glikogen Nereducirani kraj Glikogen je jako dostupni skladišni oblik glukoze; kao i jako velik, razgranat polimer; Glukozne jedinice su povezane α-1,4-glikozidnim
Grafičko prikazivanje atributivnih i geografskih nizova
 Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
Grafičko prikazivanje atributivnih i geografskih nizova Biserka Draščić Ban Pomorski fakultet u Rijeci 17. veljače 2011. Grafičko prikazivanje atributivnih nizova Atributivni nizovi prikazuju se grafički
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika. Monotonost i ekstremi. Katica Jurasić. Rijeka, 2011.
 Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
Veleučilište u Rijeci Stručni studij sigurnosti na radu Akad. god. 2011/2012. Matematika Monotonost i ekstremi Katica Jurasić Rijeka, 2011. Ishodi učenja - predavanja Na kraju ovog predavanja moći ćete:,
Računarska grafika. Rasterizacija linije
 Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
Računarska grafika Osnovni inkrementalni algoritam Drugi naziv u literaturi digitalni diferencijalni analizator (DDA) Pretpostavke (privremena ograničenja koja se mogu otkloniti jednostavnim uopštavanjem
IZVODI ZADACI (I deo)
") IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
IZVODI ZADACI (I deo) Najpre da se podsetimo tablice i osnovnih pravila:. C`=0. `=. ( )`= 4. ( n )`=n n-. (a )`=a lna 6. (e )`=e 7. (log a )`= 8. (ln)`= ` ln a (>0) 9. = ( 0) 0. `= (>0) (ovde je >0 i a
Cauchyjev teorem. Postoji više dokaza ovog teorema, a najjednostvniji je uz pomoć Greenove formule: dxdy. int C i Cauchy Riemannovih uvjeta.
 auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,
auchyjev teorem Neka je f-ja f (z) analitička u jednostruko (prosto) povezanoj oblasti G, i neka je zatvorena kontura koja čitava leži u toj oblasti. Tada je f (z)dz = 0. Postoji više dokaza ovog teorema,